Перекрестная ссылка на родственные заявки
[1] Настоящая заявка заявляет приоритет предварительной заявки на патент США № 61/898387, поданной 31 октября 2013 г., полное содержание которой включено в данный документ посредством ссылки.
Область техники
[2] Гемопоэтические стволовые клетки/клетки-предшественники (HSPC) и/или не-Т эффекторные клетки генетически модифицируют для экспрессии (i) внеклеточного компонента, содержащего лигандсвязывающий домен, который связывает клеточный маркер, преимущественно экспрессируемый на нежелательной клетке; и (ii) внутриклеточного компонента, содержащего эффекторный домен. Помимо других применений, модифицированные клетки можно вводить пациентам для нацеливания на нежелательные раковые клетки без необходимости проверки иммунологической совместимости перед введением.
Уровень техники
[3] В области генетической инженерии Т-клеток иммунной системы был сделан существенный прогресс в отношении нацеливания и уничтожения нежелательных типов клеток, таких как раковые клетки. Например, были генетически сконструированы Т-клетки для экспрессии молекул, содержащих внеклеточные компоненты, которые связывают конкретные антигены-мишени, и внутриклеточные компоненты, которые управляют действиями Т-клеток после связывания внеклеточного компонента с антигеном-мишенью. Например, внеклеточный компонент можно сконструировать так, чтобы он связывал антигены-мишени, находящиеся на раковых клетках, а после связывания внутриклеточный компонент направлял бы Т-клетку для уничтожения связанной раковой клетки. Примеры таких молекул включают генетически сконструированные рецепторы Т-клеток (РТК) и химерные антигенные рецепторы (ХАР).
[4] Хотя генетически сконструированные Т-клетки имеют существенное преимущество в отношении способности нацеливаться и уничтожать нежелательные типы клеток, они требуют проверки иммунологической совместимости в случае каждого конкретного субъекта перед тем, как их можно будет применять в схеме лечения. После нахождения соответствующего донора (или получения Т-клеток от нуждающегося в лечении субъекта), клетки необходимо модифицировать и размножить перед тем, как их можно будет применять к субъекту. Этот длительный и дорогостоящий процесс в некоторых случаях может приводить к летальным задержкам в лечении.
Сущность изобретения
[5] В настоящем изобретении предложены генетически модифицированные стволовые клетки, которые можно вводить в качестве терапевтического средства без необходимости проверки иммунологической совместимости в случае конкретных субъектов. Таким образом, эти модифицированные стволовые клетки можно использовать как «готовые» лекарственные средства, устраняя задержки и расходы в лечении, связанные с нахождением донора и последующей модификацией и размножением клеток. Модифицированные стволовые клетки можно вводить сами по себе или в комбинации с различными другими видами лечения для достижения многочисленных лечебных целей. В конкретных вариантах реализации изобретения перед введением модифицированные стволовые клетки дифференцируют в модифицированные не-Т эффекторные клетки.
[6] Конкретнее, гемопоэтические стволовые клетки/клетки-предшественники (HSPC) генетически модифицируют для экспрессии молекул, содержащих внеклеточный компонент, который связывает конкретные клеточные маркеры, находящиеся преимущественно на нежелательных типах клеток, и внутриклеточный компонент, который управляет действиями генетически модифицированной клетки после связывания внеклеточным компонентом клеточного маркера. Например, внеклеточный компонент можно сконструировать так, чтобы он связывал клеточные маркеры, находящиеся преимущественно на раковых клетках, а после связывания внутриклеточный компонент направлял бы генетически модифицированную клетку для уничтожения связанной раковой клетки. Примеры таких молекул включают генетически сконструированные рецепторы Т-клеток (РТК), химерные антигенные рецепторы (ХАР) и другие молекулы, раскрытые в данном документе. В конкретных вариантах реализации изобретения перед введением модифицированные HSPC можно дифференцировать в не-Т эффекторные клетки.
Краткое описание графических материалов
[7] ФИГ. 1. Нуклеотидная последовательность анти-CD19 химерного рецептора с коротким спейсером GMCSFRss-CD19scFv-IgG4hinge-CD28tm-41BB-Zeta-T2A-EGFRt.
[8] ФИГ 2. Аминокислотная последовательность GMCSFRss-CD19scFv-IgG4hinge-CD28tm-41BB-Zeta-T2A-EGFRt.
[9] ФИГ. 3A и 3B. ФИГ. 3A иллюстрируют карту участков нуклеотидных и аминокислотных последовательностей ZXR-014. ФИГ. 3B иллюстрирует типовые последовательности праймеров.
[10] ФИГ. 4. Аминокислотная последовательность и карта участков Uniprot P0861 IgG4-Fc.
[11] ФИГ. 5. Аминокислотная последовательность и карта участков Uniprot P10747 CD28.
[12] ФИГ. 6. Аминокислотная последовательность и карта участков Uniprot Q07011 4-1BB.
[13] ФИГ. 7. Аминокислотная последовательность и карта участков Uniprot P20963 человека CD3ζ изоформы 3.
[14] ФИГ. 8. Типовые последовательности шарнирной области.
[15] ФИГ. 9. Последовательность ХАР R12 с длинным спейсером: PJ_R12-CH2-CH3-41BB-Z-T2A-tEGFR.
[16] ФИГ. 10. Последовательность Leader_R12-Hinge-CH2-CH3-CD28tm/41BB-Z-T2A-tEGFR.
[17] ФИГ. 11. Последовательность ХАР R12 со средним спейсером: PJ_R12-CH3-41BB-Z-T2A-tEGFR.
[18] ФИГ. 12. Последовательность Leader _R12-Hinge-CH3-CD28tm/41BB-Z-T2A-tEGFR.
[19] ФИГ. 13. Последовательность ХАР R12 с коротким спейсером: PJ_R12-Hinge-41BB-Z-T2A-tEGFR.
[20] ФИГ. 14. Последовательность Leader_R12-CD28tm/41BB-Z-T2A-tEGFR.
[21] ФИГ. 15. Последовательность ХАР R11 с длинным спейсером: PJ_R11-CH2-CH3-41BB-Z-T2A-tEGFR.
[22] ФИГ. 16. Последовательность Leader_R11-Hinge-CH2-CH3-CD28tm/41BB-Z-T2A-tEGFR.
[23] ФИГ. 17. Последовательность ХАР R11 со средним спейсером: PJ_R11-CH3-41BB-Z-T2A-tEGFR.
[24] ФИГ. 18. Последовательность Leader_R11-Hinge-CH3-CD28tm/41BB-Z-T2A-tEGFR.
[25] ФИГ. 19. Последовательность ХАР R11 с коротким спейсером: PJ_R11-41BB-Z-T2A-tEGFR.
[26] ФИГ. 20. Последовательность Leader_R11-Hinge-CD28tm/41BB-Z-T2A-tEGFR.
[27] ФИГ. 21. Типовые спейсерные последовательности.
[28] ФИГ. 22. Последовательность конструкции Her2 с коротким спейсером GMCSFss-Her2scFv-IgG4hinge-CD28tm-41BB-Zeta-T2A-EGFRt.
[29] ФИГ. 23. Последовательность конструкции Her2 со средним спейсером.
[30] ФИГ. 24. Последовательность конструкции Her2 с длинным спейсером.
[31] ФИГ. 25. Библиотека спейсерных последовательностей. Конструировали плазмидную библиотеку, которая содержит кодон-оптимизированные последовательности ДНК, которые кодируют внеклеточные компоненты, включая части шарнирной области IgG4, шарнирную область IgG4, соединенную с доменами CH2 и CH3, или шарнирную область IgG4, соединенную с доменом CH3. Любую последовательность scFV (VH и VL) можно клонировать 5' к последовательностям, кодируемым этой библиотекой вариабельных спейсерных доменов. Спейсерные домены в свою очередь связаны с трансмембранными и внутриклеточными сигнальными доменами CD28 и с CD3 ζ. Последовательность T2A в векторе отделяет химерный рецептор от селектируемого маркера, кодирующего усеченный рецептор человеческого эпидермального фактора роста (EGFR).
[32] ФИГ. 26A и 26B. Конструкция химерных рецепторов ROR1 с изменяемой длиной спейсера и полученных из 2A2 и R12 scFV с разной аффинностью. (ФИГ. 26A) Конструкция лентивирусных трансгенных вставок, кодирующих панель химерных рецепторов ROR1, содержащих 2A2 scFV, полученный из IgG4-Fc спейсер «Hinge-CH2-CH3» (длинный спейсер, 229 AA), «Hinge-CH3» (средний, 119 AA) или только «Hinge» (короткий, 12 AA), и сигнальный модуль CD3ζ и CD28. Каждая кассета химерного рецептора содержит усеченный маркер EGFR, кодируемый ниже элемента T2A. (ФИГ. 26B) Лентивирусные трансгенные вставки, кодирующие ROR1-специфические химерные рецепторы, полученные из R12 и 2A2 scFV, с коротким IgG4-Fc «Hinge» спейсером (12 AA) и сигнальным модулем, содержащим CD28 или 4-1BB и CD3ζ, соответственно (всего 4 конструкции).
[33] ФИГ. 27A и 27B. ФИГ. 27A) Изображение расположения эпитопа Fab герцептина на проксимальном эпитопе мембраны опухолевой клетки на человеческом HER2, ФИГ. 27B) Структура вариантов ХАР scFv герцептина со спейсерами разной длины в виде -T2A- связанных белков с карбоксильным маркерным трансмембранным белком EGFRt.
[34] ФИГ. 28. Векторы CD19-химерных рецепторов. Конструкция лентивирусных трансгенных вставок, кодирующих панель CD19-специфических химерных рецепторов, которые отличаются длиной внеклеточного спейсера и внутриклеточной костимуляцией. Каждый химерный рецептор кодировал CD19-специфический одноцепочечный вариабельный фрагмент, полученный из FMC63 mAb в ориентации VL-VH, полученный из IgG4 спейсерный домен Hinge-CH2-CH3 (длинный спейсер, 229 AA) или только Hinge (короткий спейсер, 12 AA), а также сигнальный модуль, содержащий CD3ζ с CD28 или 4-1BB поодиночке или в тандеме. Каждая кассета химерного рецептора содержит усеченный маркер EGFR, кодируемый ниже отщепляемого элемента 2A.
[35] ФИГ. 29A и 29B. Типовые СИН (самоинактивирующиеся) лентивирусные плазмиды. ФИГ. 29A иллюстрирует СИН CD19-специфический scFvFc-CD3ζCD28 ХАР и лентивирусную плазмиду huEGFRt. ФИГ. 29B иллюстрирует СИН CD19-специфический scFv-4-1BBCD3ζ ХАР и лентивирусную плазмиду huEGFRt.
[36] ФИГ. 30A и 30B. Экспрессия EGFR как маркер эффективности трансдукции/стабильности генной экспрессии в процентах (ФИГ. 30A) и абсолютном значении (ФИГ. 30B). HSPC культивировали на Delta, как описывалось ранее. На день +3 клетки трансдуцировали, используя ХАР scFvFc-CD3ζCD28 и вектор huEGFRt при MOI 3 в присутствии протамина сульфата и подвергали спинфекции. На протяжении курса культивирования измеряли трансгенную экспрессию проточным методом, используя эрбитукс, который связывается с меткой EGFRt. В указанные культуры на день +7 добавляли облученные ЛКЛ в соотношении 1:1.
[37] ФИГ. 31. Клетки ПК CD34+, культивируемые с лигандом Notch, подвергали трансдукции лентивирусом на день +3 при MOI 3, используя ХАР scFvFc-CD3ζCD28 и вектор huEGFRt. На 7 день в указанные культуры добавляли ЛКЛ в соотношении 1:1 (трансдуцированные (■), трансдуцированные с ЛКЛ (X), нетрансдуцированные (большей степенью не видные за линией ■), нетрансдуцированные с ЛКЛ (▲)). Кратность размножения CD34 повышалась при добавлении ЛКЛ на фоне общей кратности размножения TNC.
[38] ФИГ. 32. День 14, MOI 3, применение ХАР scFv-4-1BB/CD3ζ и вектора huEGFRt для трансдукции с и без ЛКЛ. Добавление ЛКЛ на день +7 не стимулировало пролиферацию экспрессирующих ХАР HSPC или их потомства, что видно по аналогичному популяционному распределению в культуре с и без ЛКЛ.
[39] ФИГ. 33. Культуральный фенотип на конечном этапе. HSPC культивировали на Delta, как описывалось ранее. Указанные культуры на день +3 трансдуцировали лентивирусом при MOI 3 для экспрессии ХАР scFv-4-1BB/CD3ζ и huEGFRt. Кроме того в указанные культуры на день +7 добавляли облученные ЛКЛ в соотношении 1:1. На 14 день культуры анализировали методом проточной цитометрии. Не было выявлено существенных различий между трансдуцированными и нетрансдуцированными культурами. Аналогично, не было выявлено различий между общей популяцией клеток и клетками EGFRt+, что позволяет предположить, что конструкция ХАР равномерно распределена между подгруппами.
[40] ФИГ. 34. Функциональный анализ ХАР scFvFc-CD3ζCD28 и вектора huEGFRt. Через 14 дней культивирования на Delta клетки забирали с Delta, помещали в среду RPMI, дополненную IL-2 и IL-15 на дополнительную неделю, чтобы получить популяцию NK.
[41] ФИГ. 35. Анализ с радиоактивным хромом для целевой клетки K562 (x и ● ) или ЛКЛ (▲ и ♦) с применением NK-эффекторных клеток, полученных из клеток ПК CD34+, размноженных с лигандом Notch и трансдуцированных для экспрессии CD19-специфического ХАР scFvFc-CD3ζCD28 и huEGFRt (● и ♦) или нетрансдуцированных (▲ и x). Зрелые NK-клетки были получены посредством дополнительной недели в культуре с RPMI, IL-2 и IL-15.
[42] ФИГ. 36. У мышей, получавших трансдуцированные ХАР scFv-4-1BB/CD3ζ и вектором huEGFRt клетки, наблюдали нарушенное приживление CD19, что демонстрирует анти-CD19 действие, которое зависело от экспрессии трансгена.
[43] ФИГ. 37. Мыши NOG, получавшие клетки из культур, которые были трансдуцированы лентивирусом, кодирующим ХАР scFv-4-1BB/CD3ζ и huEGFRt, и демонстрировали значительную экспрессию EGFRt и сниженное приживление CD19.
Подробное описание изобретения
[44] В области генетической инженерии Т-клеток иммунной системы был сделан существенный прогресс в отношении нацеливания и уничтожения нежелательных типов клеток, таких как раковые клетки. Например, были генетически сконструированы Т-клетки для экспрессии молекул, содержащих внеклеточный компонент, который связывает конкретные антигены-мишени, и внутриклеточный компонент, который управляет действиями Т-клеток после связывания внеклеточного компонента с антигеном-мишенью. Например, внеклеточный компонент можно сконструировать так, чтобы он связывал антигены-мишени, находящиеся преимущественно на раковых клетках, а после связывания внутриклеточный компонент направлял бы Т-клетку для уничтожения связанной раковой клетки. Примеры таких молекул включают генетически сконструированные рецепторы Т-клеток (РТК) и химерные антигенные рецепторы (ХАР).
[45] Хотя генетически сконструированные Т-клетки имеют существенное преимущество в отношении способности нацеливаться и уничтожать нежелательные типы клеток, они требуют проверки иммунологической совместимости в случае каждого конкретного субъекта перед тем, как их можно будет применять в схеме лечения. После нахождения соответствующего донора (или получения Т-клеток от нуждающегося в лечении субъекта), клетки необходимо модифицировать и размножить перед тем, как их можно будет применять к субъекту. Этот длительный и дорогостоящий процесс в некоторых случаях может приводить к летальным задержкам в лечении.
[46] В настоящем изобретении предложены генетически модифицированные стволовые клетки, которые можно вводить в качестве терапевтического средства без необходимости проверки иммунологической совместимости в случае конкретных субъектов. Таким образом, эти модифицированные стволовые клетки можно использовать как «готовые» лекарственные средства, устраняя задержки и расходы в лечении, связанные с нахождением донора и последующими модификацией и размножением клеток. Модифицированные стволовые клетки можно вводить сами по себе или в комбинации с различными другими видами лечения для достижения многочисленных лечебных целей. В конкретных вариантах реализации изобретения перед введением модифицированные стволовые клетки могут быть дифференцированы в модифицированные не-Т эффекторные клетки.
[47] Конкретнее, гемопоэтические стволовые клетки/клетки-предшественники (HSPC) генетически модифицируют для экспрессии молекул, содержащих внеклеточный компонент, который связывает конкретные клеточные маркеры, и внутриклеточный компонент, который управляет действиями генетически модифицированной клетки после связывания внеклеточным компонентом клеточного маркера. Например, внеклеточный компонент можно сконструировать так, чтобы он связывал клеточные маркеры, находящиеся преимущественно на раковых клетках, а после связывания внутриклеточный компонент направлял бы генетически модифицированную клетку для уничтожения связанной раковой клетки. Примеры таких молекул включают генетически сконструированные рецепторы Т-клеток (РТК), химерные антигенные рецепторы (ХАР) и другие молекулы, раскрытые в данном документе. Перед введением HSPC можно дифференцировать в не-Т эффекторные клетки.
[48] В качестве примера применения конкретного варианта реализации изобретения можно привести трансплантацию пуповинной крови (ТПК), которая является стандартным видом лечения рецидивной детской острой лимфобластической лейкемии (ALL), когда невозможно найти подходящего совместимого донора. Это особенно актуально для пациентов миноритарного или смешанного этнического происхождения (и 30% представителей белой европеоидной расы), для которых вероятность нахождения подходящего донора очень мала.
[49] Способность ТПК излечивать ALL и обеспечивать продолжительную ремиссию частично связана с эффектом «трансплантат против лейкемии» (ТПЛ). Однако до сих пор уровень рецидивов ALL после ТПК составляет около 40% (Smith et al., Biol Blood Marrow Transplant, 2009. 15(9): p. 1086-93; Tomblyn et al., J Clin Oncol, 2009. 27(22): p. 3634-41), а общая выживаемость ограничена смертностью как вследствие рецидива, так и вследствие лечения, включая болезнь «трансплантат против хозяина» (БТПХ). Раскрытые в данном документе композиции и препараты могут усиливать эффект ТПЛ, не повышая уровень БТПХ. Эту стратегию можно осуществить клинически, применяя ex vivo размножение HSPC пуповинной крови (ПК) путем активации эндогенного пути сигнализации Notch при помощи лиганда Notch, что приводит к более чем 100-кратному увеличению количества клеток CD34+. В клинических условиях эти размноженные HSPC можно инфузировать вместе с необработанной единицей, что приводит к временному приживлению размноженных HSPC с потомством, полученным от размноженной единицы, в то время как долгосрочное приживление в конечном итоге обеспечивает необработанная единица.
[50] Размноженные при помощи лиганда Notch HSPC ПК восприимчивы к генетическим модификациям с применением векторов, которые экспрессируют CD19-специфические ХАР. Учитывая преимущества системы размножения ПК при помощи лиганда Notch, ТПЛ можно прививать в ТПК путем генетической модификации размноженных HSPC для экспрессии CD19 ХАР, в результате чего приживленные миелоидные и лимфоидные эффекторные клетки распознают и лизируют остаточные клетки лейкемии.
[51] Далее заявляемое изобретение будет описано более широко.
[52] Гемопоэтические стволовые клетки/клетки-предшественники или HSPC относятся к гемопоэтическим стволовым клеткам и/или гемопоэтическим клеткам-предшественникам. HSPC могут самообновляться или могут дифференцироваться в (i) миелоидные клетки-предшественники, которые в конечном итоге приводят к образованию моноцитов и макрофагов, нейтрофилов, базофилов, эозинофилов, эритроцитов, мегакариоцитов/тромбоцитов или дендритных клеток; или (ii) лимфоидные клетки-предшественники, которые в конечном итоге приводят к образованию Т-клеток, В-клеток и лимфоцит-подобных клеток, называемых естественными клетками-киллерами (NK-клетками). В отношении общих сведений по гемопоэзу и дифференцировке HSPC смотрите главу 17, Differentiated Cells and the Maintenance of Tissues, Alberts et al., 1989, Molecular Biology of the Cell, 2nd Ed., Garland Publishing, New York, NY; главу 2 из Regenerative Medicine, Department of Health and Human Services, August 5, 2006, и главу 5 из Hematopoietic Stem Cells, 2009, Stem Cell Information, Department of Health and Human Services.
[53] HSPC могут быть позитивными в отношении специфического маркера, экспрессируемого в HSPC на повышенных уровнях по сравнению с другими типами гемопоэтических клеток. Например, такие маркеры включают CD34, CD43, CD45RO, CD45RA, CD59, CD90, CD109, CD117, CD133, CD166, HLA DR или их комбинацию. HSPC также могут быть негативными в отношении экспрессируемого маркера по сравнению с другими типами гемопоэтических клеток. Например, такие маркеры включают Lin, CD38 или их комбинацию. Предпочтительно HSPC представляют собой клетки CD34+.
[54] Источники HSPC включают пуповинную кровь, плацентарную кровь и периферическую кровь (смотрите патенты США № 5004681; 7399633; и 7147626; Craddock et al., 1997, Blood 90(12):4779-4788; Jin et al., 2008, Journal of Translational Medicine 6:39; Pelus, 2008, Curr. Opin. Hematol. 15(4):285-292; Papayannopoulou et al., 1998, Blood 91(7):2231-2239; Tricot et al., 2008, Haematologica 93(11):1739-1742; и Weaver et al., 2001, Bone Marrow Transplantation 27(2):S23-S29). Способы, касающиеся сбора, антикоагуляции и обработки и т.д. образцов крови, хорошо известны в данной области техники. Смотрите, например, Alsever et al., 1941, N.Y. St. J. Med. 41:126; De Gowin, et al., 1940, J. Am. Med. Ass. 114:850; Smith, et al., 1959, J. Thorac. Cardiovasc. Surg. 38:573; Rous and Turner, 1916, J. Exp. Med. 23:219; и Hum, 1968, Storage of Blood, Academic Press, New York, pp. 26-160. Источники HSPC также включают костный мозг (смотрите Kodo et al., 1984, J. Clin Invest. 73:1377-1384), эмбриональные клетки, клетки, полученные из тканей аорты-гонад-мезонефроса, лимфу, печень, вилочковую железу и селезенку от подходящих по возрасту доноров. Все собранные образцы HSPC можно исследовать в отношении нежелательных компонентов и отбраковывать, обрабатывать или применять в соответствии с общепринятыми текущими стандартами в данное время.
[55] HSPC можно собирать и выделять из образца, используя любой подходящий метод. Подходящие процедуры сбора и выделения включают разделение в магнитном поле; сортировку флуоресцентно-активированных клеток (FACS; Williams et al., 1985, J. Immunol. 135:1004; Lu et al., 1986, Blood 68(1):126-133); аффинную хроматографию; использование цитотоксических агентов, присоединенных к моноклональному антителу или применяемых в сочетании с моноклональным антителом, например, комплемента и цитотоксинов; «пэннинг» с применением антитела, иммобилизованного на твердой матрице (Broxmeyer et al., 1984, J. Clin. Invest. 73:939-953); селективную агглютинацию с применением лектина, такого как соя (Reisner et al., 1980, Proc. Natl. Acad. Sci. U.S.A. 77:1164); и т.д.
[56] В конкретных вариантах реализации изобретения образец HSPC (например, свежую дозу пуповинной крови) можно обрабатывать для селекции/обогащения в отношении клеток CD34+, используя анти-CD34 антитела, прямым или непрямым образом конъюгированные с магнитными частицами, и магнитный клеточный сепаратор, например, систему для разделения клеток CliniMACS® (Miltenyi Biotec, Bergisch Gladbach, Germany). Смотрите также раздел 5.4.1.1 патента США № 7399633, в котором описано обогащение CD34+ HSPC от 1-2% нормальной клеточной популяции костного мозга до 50-80% популяции.
[57] Аналогично, HSPC, экспрессирующие CD43, CD45RO, CD45RA, CD59, CD90, CD109, CD117, CD133, CD166, HLA DR или их комбинацию, можно обогащать, используя антитела против этих антигенов. В патенте США № 5877299 описаны дополнительные подходящие гемопоэтические антигены, которые можно применять для выделения, сбора и обогащения клеток HSPC из образцов.
[58] После выделения и/или обогащения HSPC можно размножать с целью увеличения числа HSPC. Способы выделения и/или размножения описаны, например, в патентах США № 7399633 и 5004681; публикации патента США № 2010/0183564; международных публикациях патентов № (WO) WO2006/047569; WO2007/095594; WO 2011/127470; и WO 2011/127472; Vamum-Finney et al., 1993, Blood 101:1784-1789; Delaney et al., 2005, Blood 106:2693-2699; Ohishi et al., 2002, J. Clin. Invest. 110:1165-1174; Delaney et al., 2010, Nature Med. 16(2): 232-236; и главе 2 из Regenerative Medicine, Department of Health and Human Services, August 2006, а также в приведенных в них ссылках. Каждый из упоминаемых способов сбора, выделения и размножения можно применять в конкретных вариантах реализации изобретения.
[59] Предпочтительные способы размножения HSPC включают размножение HSPC с агонистом Notch. В отношении информации, касающейся размножения HSPC с использованием агонистов Notch, смотрите разделы 5.1 и 5.3 патента США № 7399633; патенты США № 5780300; 5648464; 5849869; и 5856441; WO 1992/119734; Schlondorfiand Blobel, 1999, J. Cell Sci. 112:3603-3617; Olkkonen and Stenmark, 1997, Int. Rev. Cytol. 176:1-85; Kopan et al., 2009, Cell 137:216-233; Rebay et al., 1991, Cell 67:687-699 и Jarriault et al., 1998, Mol. Cell. Biol. 18:7423-7431. В конкретных вариантах реализации изобретения агонист Notch иммобилизуют во время размножения.
[60] Агонисты Notch включают любое соединение, которое связывается или каким-либо другим образом взаимодействует с белками Notch или другими белками пути Notch так, чтобы стимулировать активность пути Notch. Типовыми агонистами Notch являются внеклеточные связывающие лиганды Delta и Serrate (например, Jagged), RBP JϰI супрессор Hairless, Deltex, Fringe или их фрагменты, которые стимулируют активацию пути Notch. Нуклеотидные и аминокислотные последовательности представителей семейства Delta и представителей семейства Serrate были выделены из нескольких видов и описаны, например, в WO 1993/12141; WO 1996/27610; WO 1997/01571; и Gray et al., 1999, Am. J. Path. 154:785-794.
[61] В конкретных вариантах реализации изобретения агонист Notch представляет собой Delta1ext-IgG. В конкретных вариантах реализации изобретения Delta1ext-IgG иммобилизуют на твердой подложке в концентрации между 0,2 и 20 мкг/мл, между 1,25 и 10 мкг/мл или между 2 и 6 мкг/мл.
[62] В конкретных вариантах реализации изобретения во время размножения HSPC культивируют в присутствии агониста Notch и антагониста арил-гидрокарбонового рецептора. Агонист Notch может быть иммобилизован, а антагонист арил-гидрокарбонового рецептора может находиться в жидкости, контактирующей с клетками.
[63] Как понятно специалисту в данной области техники, дополнительные условия культивирования могут включать размножение в присутствии одного или более факторов роста, таких как: ангиопоэтин-подобные белки (Angptl, например, Angptl2, Angptl3, Angptl7, Angpt15 и Mfap4); эритропоэтин; фактор роста фибробластов-1 (FGF-1); лиганд Flt-3 (Flt-3L); гранулоцитарный колониестимулирующий фактор (G-CSF); гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF); инсулиновый фактор роста-2 (IFG-2); интерлейкин-3 (IL-3); интерлейкин-6 (IL-6); интерлейкин-7 (IL-7); интерлейкин-11 (IL-11); фактор стволовых клеток (SCF; также известный как лиганд c-kit или фактор роста тучных клеток); тромбопоэтин (TPO); и их аналоги (при этом аналоги включают любые структурные варианты факторов роста, имеющие биологическую активность фактора роста природного происхождения; смотрите, например, WO 2007/1145227 и публикацию патента США № 2010/0183564).
[64] В конкретных вариантах реализации изобретения количество или концентрация факторов роста, подходящие для размножения HSPC, представляют собой количество или концентрацию, достаточные для стимуляции пролиферации HSPC, но практически без дифференцировки HSPC. Также популяции клеток размножают преимущественно до достижения достаточного числа клеток, чтобы обеспечить по меньшей мере одну инфузию субъекту-человеку, как правило, составляющего от 104 клеток/кг до 109 клеток/кг.
[65] Количество или концентрация факторов роста, подходящие для размножения HSPC, зависят от активности препаратов факторов роста и видового соответствия между факторами роста и HSPC и т.д. В общем случае, когда фактор(ы) роста и HSPC получены от одного вида, общее количество фактора роста в культуральной среде находится в диапазоне от 1 нг/мл до 5 мкг/мл, от 5 нг/мл до 1 мкг/мл или от 5 нг/мл до 250 нг/мл. В дополнительных вариантах реализации изобретения количество фактора роста может находиться в диапазоне 5-1000 или 50-100 нг/мл.
[66] В конкретных вариантах реализации изобретения вышеуказанные факторы роста присутствуют в культуральных условиях для размножения HSPC в следующих концентрациях: 25-300 нг/мл SCF, 25-300 нг/мл Flt-3L, 25-100 нг/мл TPO, 25-100 нг/мл IL-6 и 10 нг/мл IL-3. В более конкретных вариантах реализации изобретения можно использовать 50, 100 или 200 нг/мл SCF; 50, 100 или 200 нг/мл Flt-3L; 50 или 100 нг/мл TPO; 50 или 100 нг/мл IL-6; и 10 нг/мл IL-3.
[67] В конкретных вариантах реализации изобретения HSPC можно размножать, подвергая HSPC воздействию иммобилизованного агониста Notch и 50 нг/мл или 100 нг/мл SCF; иммобилизованного агониста Notch и 50 нг/мл или 100 нг/мл каждого из Flt-3L, IL-6, TPO и SCF; или иммобилизованного агониста Notch и 50 нг/мл или 100 нг/мл каждого из Flt-3L, IL-6, TPO и SCF и 10 нг/мл IL-11 или IL-3.
[68] HSPC можно размножать на планшете для тканевого культивирования, с которым связан белок внеклеточного матрикса, такой как фибронектин (FN) или его фрагмент (например, CH-296 (Dao et. al., 1998, Blood 92(12):4612-21)) или ретронектин® (фрагмент рекомбинантного человеческого фибронектина; (Clontech Laboratories, Inc., Madison, WI).
[69] В конкретном варианте реализации изобретения способы размножения HSPC включают культивирование выделенных HSPC ex vivo на твердой поверхности, покрытой иммобилизованными Delta1ext-IgG и CH-296 и четырьмя или более факторами роста, выбранными из IL-6, TPO, Flt-3L, CSF и IL-3; что позволяет получить образец с размноженными HSPC.
[70] В конкретных вариантах реализации размножения HSPC клетки культивируют на пластиковом планшете для тканевого культивирования, содержащем иммобилизованные лиганд Delta и фибронектин и 25 нг/мл или 100 нг/мл (или любого значения между этими величинами) и предпочтительно 50 нг/мл каждого из SCF и TPO. В конкретных вариантах реализации размножения HSPC клетки культивируют на пластиковом планшете для тканевого культивирования, содержащем иммобилизованные лиганд Delta и фибронектин, в присутствии 25 нг/мл или 100 нг/мл (или любого значения между этими величинами) и предпочтительно 50 нг/мл каждого из SCF и Flt-3L. В конкретных вариантах реализации размножения HSPC клетки культивируют на пластиковом планшете для тканевого культивирования, содержащем иммобилизованные лиганд Delta и фибронектин и 25 нг/мл или 100 нг/мл (или любое значение между этими величинами) и предпочтительно 50 нг/мл каждого из SCF, Flt-3L и TPO. В конкретных вариантах реализации размножения HSPC клетки культивируют на пластиковом планшете для тканевого культивирования, содержащем иммобилизованные лиганд Delta и фибронектин и 25 нг/мл или 100 нг/мл (или любое значение между этими величинами) и предпочтительно 50 нг/мл каждого из SCF, Flt-3L, TPO и IL-6. В конкретных вариантах реализации изобретения HSPC дополнительно культивируют в присутствии от 5 до 15 нг/мл и предпочтительно 10 нг/мл IL-3. В конкретных вариантах реализации изобретения HSPC дополнительно культивируют в присутствии от 5 до 15 нг/мл и предпочтительно 10 нг/мл GM-CSF. В конкретных вариантах реализации изобретения один или более из применяемых факторов роста не является GM-SCF или IL-7. В конкретных альтернативных вариантах реализации изобретения фибронектин удаляют из планшетов для тканевого культивирования или замещают другим белком внеклеточного матрикса. Дополнительные способы и подробности, касающиеся размножения HSPC, можно найти в WO 2013/086436.
[71] В конкретных вариантах реализации изобретения процентное содержание клеток CD34+ в размноженном образце HSPC, полученном описанными способами, превышает процентное содержание клеток CD34+ в выделенных HSPC до размножения. В отношении дополнительной информации касательно подходящих условий культивирования смотрите патент США № 7399633; публикацию патента США № 2010/0183564; и Freshney Culture of Animal Cells, Wiley-Liss, Inc., New York, NY (1994)).
[72] Модифицированные HSPC. В конкретных вариантах реализации изобретения HSPC модифицируют для экспрессии молекул, содержащих внеклеточный компонент и внутриклеточный компонент. Внеклеточный и внутриклеточный компоненты могут быть связаны напрямую или посредством спейсерной области, трансмембранного домена, последовательности метки и/или линкерной последовательности.
[73] Внеклеточные компоненты. Внеклеточные компоненты включают по меньшей мере один лигандсвязывающий домен (далее называемый связывающим доменом). Связывающий домен сконструирован для нацеливания модифицированной клетки на конкретный нежелательный тип клеток путем связывания клеточного маркера, который находится преимущественно на нежелательном типе клеток.
[74] Клеточные маркеры. В конкретных вариантах реализации изобретения клеточные маркеры преимущественно экспрессируются нежелательными клетками, такими как нежелательные раковые клетки. «Преимущественно экспрессируемый» означает, что клеточный маркер находится на нежелательном типе клеток на более высоких уровнях по сравнению с другими, не являющимися мишенями клетками. Разница в уровне экспрессии является достаточно существенной для того, чтобы с медицинской точки зрения введение клетки, которая будет нацеливаться и уничтожать нежелательную клетку на основании наличия маркера, превышает риск параллельного уничтожения других, не являющихся мишенями клеток, которые также могут в меньшей степени экспрессировать маркер. В некоторых случаях клеточный маркер экспрессируется только нежелательным типом клеток. В других случаях клеточный маркер экспрессируется на нежелательном типе клеток по меньшей мере на 25%, 35%, 45%, 55%, 65%, 75%, 85%, 95%, 96%, 97%, 98%, 99% или 100% больше, чем на не являющихся мишенями клетках. Типовые нежелательные раковые клетки включают раковые клетки рака надпочечников, рака мочевого пузыря, рака крови, рака костей, рака головного мозга, рака молочной железы, карциномы, рака шейки матки, рака толстой кишки, колоректального рака, рака тела матки, рака уха, носа и горла (ENT), рака эндометрия, рака пищевода, рака желудочно-кишечного тракта, рака головы и шеи, болезни Ходжкина, рака кишечника, рака почек, рака гортани, лейкемии, рака печени, рака лимфатических узлов, лимфомы, рака легких, меланомы, мезотелиомы, миеломы, рака носоглотки, нейробластомы, неходжкинской лимфомы, рака полости рта, рака яичников, рака поджелудочной железы, рака полового члена, рака глотки, рака простаты, рака прямой кишки, саркомы, семиномы, рака кожи, рака желудка, тератомы, рака яичка, рака щитовидной железы, рака матки, вагинального рака, сосудистой опухоли и их метастазов.
[75] Путем включения во внеклеточный компонент связывающего домена, который связывает ассоциированный(е) клеточный(е) маркер(ы), можно нацеливаться на следующие конкретные виды рака:
[76] Не ограничивая вышеуказанное, клеточные маркеры также включают A33; BAGE; Bcl-2; β-катенин; B7H4; BTLA; CA125; CA19-9; CD5; CD19; CD20; CD21; CD22; CD33; CD37; CD44v6; CD45; CD123; CEA; CEACAM6; c-Met; CS-1; циклин B1; DAGE; EBNA; EGFR; эфрин B2; ErbB2; ErbB3; ErbB4; EphA2; рецептор эстрогена; FAP; ферритин; α-фетопротеин (AFP); FLT1; FLT4; фолатсвязывающий белок; Frizzled; GAGE; G250; GD-2; GHRHR; GHR; GM2; gp75; gp100 (Pmel 17); gp130; HLA; HER-2/neu; HPV E6; HPV E7; hTERT; HVEM; IGF1R; IL6R; KDR; Ki-67; LIFRβ; LRP; LRP5; LTβR; мезотелин; OSMRβ; p53; PD1; PD-L1; PD-L2; PRAME; рецептор прогестерона; PSA; PSMA; PTCH1; MAGE; MART; мезотелин; MUC; MUC1; MUM-1-B; myc; NYESO-1; RANK; ras; Robo1; RORl; сурвивин; TCRα; TCRβ; тенасцин; TGFBR1; TGFBR2;TLR7; TLR9; TNFR1; TNFR2; TNFRSF4; TWEAK-R; TSTA тирозиназа; VEGF; и WT1.
[77] Клеточные маркеры конкретных раковых клеток включают:
[78] Нежелательные клетки и клеточные маркеры не ограничены раковыми клетками и раковыми клеточными маркерами, но также могут включать, например, инфицированные вирусом клетки, такие как те, которые экспрессируют поверхностный антиген вируса гепатита В.
[79] Связывающие домены. Связывающие домены включают любое вещество, которое связывается с клеточным маркером с образованием комплекса. Примеры связывающих доменов включают лиганды клеточных маркеров, лиганды рецепторов, антитела, пептиды, пептидные аптамеры, рецепторы (например, рецепторы Т-клеток) и их комбинации.
[80] Антитела являются одним из примеров связывающих доменов и включают цельные антитела или связывающие фрагменты антител, например, Fv, Fab, Fab', F(ab')2, Fc, и их одноцепочечные (sc) формы или фрагменты, которые специфически связываются с клеточным маркером. Дополнительные примеры включают грабатела на основе scFv и растворимые VH-доменные антитела. Эти антитела образуют связывающие области с использованием только вариабельных областей тяжелой цепи. Смотрите, например, Jespers et al., Nat. Biotechnol. 22:1161, 2004; Cortez-Retamozo et al., Cancer Res. 64:2853, 2004; Baral et al., Nature Med. 12:580, 2006; и Barthelemy et al., J. Biol. Chem. 283:3639, 2008).
[81] Антитела или антигенсвязывающие фрагменты могут включать все или часть поликлональных антител, моноклональных антител, человеческих антител, гуманизированных антител, синтетических антител, химерных антител, биспецифических антител, минител и линейных антител.
[82] Антитела человеческого происхождения или гуманизированные антитела обладают сниженной иммуногенностью или неиммуногенны в организме человека и содержат меньшее количество неиммуногенных эпитопов по сравнению с нечеловеческими антителами. Антитела или их фрагменты в общем случае выбирают так, чтобы они имели сниженный уровень антигенности или были неантигенны в организме субъекта-человека.
[83] Антитела, которые специфически связывают конкретный клеточный маркер, можно получить, используя методы получения моноклональных антител, методы фагового дисплея, методы генерации человеческих или гуманизированных антител или методы с использованием трансгенных животных или растений, сконструированных для выработки антител, как известно специалистам в данной области техники (смотрите, например, патенты США № 6291161 и 6291158). Библиотеки фагового дисплея для частично или полностью синтетических антител являются общедоступными, и можно проводить их скрининг в отношении антитела или его фрагмента, который может связываться с клеточным маркером. Например, связывающие домены можно определить, проводя скрининг фаговой библиотеки Fab в отношении фрагментов Fab, которые специфически связываются с представляющим интерес клеточным маркером (смотрите Hoet et al., Nat. Biotechnol. 23:344, 2005). Также доступны библиотеки фагового дисплея для человеческих антител. Кроме того, для получения связывающих доменов можно применять традиционные стратегии разработки гибридом, используя представляющий интерес клеточный маркер в качестве иммуногена в удобных системах (например, мышах, мышах HuMAb ® (GenPharm Int'l. Inc., Mountain View, CA), мышах TC® (Kirin Pharma Co. Ltd., Tokyo, JP), мышах KM ® (Medarex, Inc., Princeton, NJ), ламах, курах, крысах, хомяках, кроликах и т.д.). В конкретных вариантах реализации изобретения антитела специфически связываются с клеточным маркером, экспрессируемым преимущественно конкретным нежелательным типом клеток, и не вступают в перекрестную реакцию с неспецифическими компонентами или неродственными мишенями. После выявления аминокислотную последовательность антитела и генную последовательность, кодирующую антитело, можно выделять и/или определять.
[84] Альтернативный источник связывающих доменов включает последовательности, которые кодируют случайные пептидные библиотеки, или последовательности, которые кодируют сконструированное разнообразие аминокислот в петлевых областях альтернативных не принадлежащих антителам каркасов, такие как scTCR (смотрите, например, Lake et al., Int. Immunol.11:745, 1999; Maynard et al., J. Immunol. Methods 306:51, 2005; патент США № 8361794), домены фибриногена (смотрите, например, Weisel et al., Science 230:1388, 1985), домены Кунитца (смотрите, например, патент США № 6423498), сконструированные белки с анкириновым повтором (DARPins; Binz et al., J. Mol. Biol. 332:489, 2003 и Binz et al., Nat. Biotechnol. 22:575, 2004), фибронектинсвязывающие домены (аднектины или монотела; Richards et al., J. Mol. Biol. 326:1475, 2003; Parker et al., Protein Eng. Des. Selec. 18:435, 2005 и Hackel et al. (2008) J. Mol. Biol. 381:1238-1252), минимизированные белки с цистеиновыми узлами (Vita et al., 1995, Proc. Nat′l. Acad. Sci. (USA) 92:6404-6408; Martin et al., 2002, Nat. Biotechnol. 21:71, 2002 и Huang et al. (2005) Structure 13:755, 2005), домены с тетратрикопептидным повтором (Main et al., Structure 11:497, 2003 и Cortajarena et al., ACS Chem. Biol. 3:161, 2008), домены с лейцин-богатым повтором (Stumpp et al., J. Mol. Biol. 332:471, 2003), домены липокалина (смотрите, например, WO 2006/095164, Beste et al., Proc. Nat′l. Acad. Sci. (USA) 96:1898, 1999 и Schönfeld et al., Proc. Nat′l. Acad. Sci. (USA) 106:8198, 2009), V-подобные домены (смотрите, например, публикацию заявки на патент США № 2007/0065431), домены лектина С-типа (Zelensky and Gready, FEBS J. 272:6179, 2005; Beavil et al., Proc. Nat′l. Acad. Sci. (USA) 89:753, 1992 и Sato et al., Proc. Nat′l. Acad. Sci. (USA) 100:7779, 2003), mAb2 или Fcab™ (смотрите, например, WO 2007/098934 и WO 2006/072620), белки с армадилло повтором (смотрите, например, Madhurantakam et al., Protein Sci. 21: 1015, 2012; WO 2009/040338), аффилин (Ebersbach et al., J. Mol. Biol. 372: 172, 2007), аффитело, авимеры, ноттинсы, финомеры, атримеры, ассоциированный с цитотоксическими Т-лимфоцитами белок-4 (Weidle et al., Cancer Gen. Proteo. 10:155, 2013) или подобные соединения (Nord et al., Protein Eng. 8:601, 1995; Nord et al., Nat. Biotechnol. 15:772, 1997; Nord et al., Euro. J. Biochem. 268:4269, 2001; Binz et al., Nat. Biotechnol. 23:1257, 2005; Boersma and Plückthun, Curr. Opin. Biotechnol. 22:849, 2011).
[85] В конкретных вариантах реализации изобретения связывающий домен представляет собой одноцепочечный рецептор Т-клеток (scTCR), включая цепи Vα/β и Cα/β (например, Vα Cα, Vβ Cβ, Vα Vβ), или включая пары Vα Cα, Vβ-Cβ, Vα-Vβ, специфические в отношении представляющего интерес клеточного маркера (например, комплекса пептид-ГКГС).
[86] Пептидные аптамеры содержат пептидную петлю (которая является специфической в отношении клеточного маркера), присоединенную с обоих концов к белковому каркасу. Это двойное структурное ограничение повышает аффинность связывания пептидных аптамеров до уровней, сравнимых с антителами. Длина вариабельной петли составляет, как правило, от 8 до 20 аминокислот, а каркасом может служить любой белок, который является стабильным, растворимым, небольшим и нетоксичным. Отбор пептидного аптамера можно проводить, используя разные системы, такие как дрожжевая двухгибридная система (например, дрожжевая двухгибридная система Gal4) или система ловушки для взаимодействий LexA.
[87] В конкретных вариантах реализации изобретения связывающий домен может представлять собой антитело, которое связывает клеточный маркер CD19. В конкретных вариантах реализации изобретения связывающий домен представляет собой одноцепочечный фрагмент Fv (scFv), который содержит VH- и VL-области, специфические в отношении CD19. В конкретных вариантах реализации изобретения VH- и VL-области являются человеческими. Типовые VH- и VL-области включают сегменты анти CD19 специфического моноклонального антитела FMC63. В конкретных вариантах реализации изобретения scFV является человеческим или гуманизированным и содержит вариабельную легкую цепь, содержащую последовательность CDRL1 RASQDISKYLN (SEQ ID NO. 108), последовательность CDRL2 SRLHSGV (SEQ ID NO. 111) и последовательность CDRL3 GNTLPYTFG (SEQ ID NO. 104). В других вариантах реализации изобретения scFV является человеческим или гуманизированным ScFv, содержащим вариабельную тяжелую цепь, содержащую последовательность CDRH1 DYGVS (SEQ ID NO. 103), последовательность CDRH2 VIWGSETTYYNSALKS (SEQ ID NO. 114) и последовательность CDRH3 YAMDYWG (SEQ ID NO. 115).
[88] Генная последовательность, кодирующая связывающий домен, приведена на ФИГ. 1 в виде scFv из антитела, которое специфически связывает CD19, такого как FMC63. Генная последовательность, кодирующая гибкий линкер, содержащая аминокислоты GSTSGSGKPGSGEGSTKG (SEQ ID NO.30), разделяет цепи VH и VL в scFV. Аминокислотная последовательность scFv, включая линкер, приведена на ФИГ. 2 (SEQ ID NO.34). Известны другие нацеленные на CD19 антитела, такие как SJ25C1 (Bejcek et al. Cancer Res 2005, PMID 7538901) и HD37 (Pezutto et al. JI 1987, PMID 2437199). SEQ ID NO. 10 представляет собой последовательность ДНК анти-CD19 scFv (VH-VL), а SEQ ID NO. 9 представляет собой аминокислотную последовательность анти-CD19 scFv (VH-VL).
[89] В конкретных вариантах реализации изобретения связывающий домен связывает клеточный маркер ROR1. В конкретных вариантах реализации изобретения scFV является человеческим или гуманизированным scFv, содержащим вариабельную легкую цепь, содержащую последовательность CDRL1 ASGFDFSAYYM (SEQ ID NO. 101), последовательность CDRL2 TIYPSSG (SEQ ID NO. 112) и последовательность CDRL3 ADRATYFCA (SEQ ID NO. 100). В конкретных вариантах реализации изобретения scFV является человеческим или гуманизированным scFv, содержащим вариабельную тяжелую цепь, содержащую последовательность CDRH1 DTIDWY (SEQ ID NO. 102), последовательность CDRH2 VQSDGSYTKRPGVPDR (SEQ ID NO. 113) и последовательность CDRH3 YIGGYVFG (SEQ ID NO. 117).
[90] В конкретных вариантах реализации изобретения связывающий домен связывает клеточный маркер ROR1. В конкретных вариантах реализации изобретения scFV является человеческим или гуманизированным scFv, содержащим вариабельную легкую цепь, содержащую последовательность CDRL1 SGSDINDYPIS (SEQ ID NO. 109), последовательность CDRL2 INSGGST (SEQ ID NO. 105) и последовательность CDRL3 YFCARGYS (SEQ ID NO. 116). В конкретных вариантах реализации изобретения scFV является человеческим или гуманизированным ScFv, содержащим вариабельную тяжелую цепь, содержащую последовательность CDRH1 SNLAW (SEQ ID NO. 110), последовательность CDRH2 RASNLASGVPSRFSGS (SEQ ID NO. 107) последовательность CDRH3 NVSYRTSF (SEQ ID NO. 106). Специалистам в данной области техники известно большое количество дополнительных антител, специфических в отношении ROR1.
[91] В конкретных вариантах реализации изобретения связывающий домен связывает клеточный маркер Her2. Специалистам в данной области техники известно большое количество антител, специфических в отношении Her2, последовательность, связывание эпитопов и аффинность которых легко оценить. В конкретных вариантах реализации изобретения связывающий домен содержит последовательность scFV из антитела герцептин. В конкретных вариантах реализации изобретения связывающий домен содержит человеческий или гуманизированный ScFv, содержащий вариабельную легкую цепь, содержащую последовательность CDRL1, последовательность CDRL2 и последовательность CDRL3 антитела герцептин. В конкретных вариантах реализации изобретения scFV является человеческим или гуманизированным ScFv, содержащим вариабельную тяжелую цепь, содержащую последовательность CDRH1, последовательность CDRH2 и последовательность CDRH3 антитела герцептин. Последовательности CDR можно легко определить по аминокислотной последовательности герцептина. Типовая генная последовательность, кодирующая домен, связывающий лиганд Her2, приведена в SEQ ID NO. 39 и 40.
[92] В конкретных вариантах реализации изобретения области CDR находятся в пределах областей антитела согласно нумерации Кабата следующим образом: для легкой цепи: CDRL1 представляют аминокислоты 24-34; CDRL2 представляют аминокислоты 50-56; CDRL3 представляют аминокислоты 89-97, и для тяжелой цепи: CDRH1 представляют аминокислоты 31-35; CDRH2 представляют аминокислоты 50-65; и CDRH3 представляют аминокислоты 95-102.
[93] Другие антитела хорошо известны и коммерчески доступны. Например, анти-PSMA и анти-PSCA антитела доступны от Abcam plc (ab66912 и ab15168, соответственно). Мезотелин и антитела WT1 доступны от Santa Cruz Biotechnology, Inc. Анти-CD20 антитела, такие как ритуксимаб (торговые наименования ритуксан, мабтера и зитукс), были разработаны IDEC Pharmaceuticals.
[94] Внутриклеточные компоненты. Внутриклеточные компоненты экспрессируемых молекул могут содержать эффекторные домены. Эффекторные домены способны передавать функциональные сигналы в клетку. В конкретных вариантах реализации изобретения эффекторный домен прямым или непрямым образом стимулирует клеточный ответ путем ассоциации с одним или более другими белками, которые прямо стимулируют клеточный ответ. Эффекторные домены могут обеспечивать активацию по меньшей мере одной функции модифицированной клетки после связывания с клеточным маркером, экспрессируемым на нежелательной клетке. Активация модифицированной клетки может включать одно или более из дифференцировки, пролиферации и/или активации или другие эффекторные функции.
[95] Эффекторный домен может содержать один, два, три или более рецепторных сигнальных доменов (например, цитоплазматические сигнальные последовательности), костимулирующие домены или их комбинации. Типовые эффекторные домены включают сигнальные и стимулирующие домены, выбранные из: 4-1BB, CARD11, CD3 гамма, CD3 дельта, CD3 эпсилон, CD3ζ, CD27, CD28, CD79A, CD79B, DAP10, FcRα, FcRβ, FcRγ, Fyn, HVEM, ICOS, LAG3, LAT, Lck, LRP, NKG2D, NOTCH1, pTα, PTCH2, OX40, ROR2, Ryk, SLAMF1, Slp76, TCRα, TCRβ, TRIM, Wnt, Zap70 или любую их комбинацию.
[96] Основные цитоплазматические сигнальные последовательности, которые действуют стимулирующим образом, могут содержать сигнальные мотивы, известные как мотивы активации рецепторов на основе тирозина или iTAM. Примеры iTAM, содержащих основные цитоплазматические сигнальные последовательности, включают полученные из CD3γ, CD3δ, CD3ε, CD3ζ, CD5, CD22, CD66d, CD79a, CD79b и FeR гамма. В конкретных вариантах реализации изобретения в вариантах CD3ζ сохранены по меньшей мере одна, две, три или все области ITAM, как показано на ФИГ. 7.
[97] В конкретных вариантах реализации изобретения эффекторный домен содержит цитоплазматическую часть, которая связывается с цитоплазматическим сигнальным белком, при этом цитоплазматический сигнальный белок представляет собой рецептор лимфоцитов или его сигнальный домен - белок, содержащий несколько ITAM, костимулирующий домен или любую их комбинацию.
[98] Примеры внутриклеточных сигнальных доменов включают цитоплазматические последовательности цепи CD3ζ и/или корецепторы, которые вместе инициируют передачу сигнала после взаимодействия со связывающим доменом.
[99] В конкретных вариантах реализации изобретения внутриклеточный сигнальный домен молекулы, экспрессируемой модифицированной клеткой, можно сконструировать так, чтобы он содержал внутриклеточный сигнальный домен в комбинации с любым другим необходимым цитоплазматическим доменом(ами). Например, внутриклеточный сигнальный домен может содержать внутриклеточный сигнальный домен и костимулирующий домен, такой как костимулирующая сигнальная область.
[100] Костимулирующая сигнальная область относится к части молекулы, включая внутриклеточный домен костимулирующего домена. Костимулирующий домен представляет собой молекулу клеточной поверхности, отличную от связывающего домена экспрессируемого клеточного маркера, которая может быть необходима для ответа лимфоцитов на связывание клеточного маркера. Примеры таких молекул включают CD27, CD28, 4-1BB (CD 137), OX40, CD30, CD40, лимфоцитарный функциональный антиген-1 (LFA-1), CD2, CD7, LIGHT, NKG2C, B7-H3 и лиганд, который специфически связывается с CD83.
[101] В конкретных вариантах реализации изобретения аминокислотная последовательность внутриклеточного сигнального домена, содержащая вариант CD3ζ и часть внутриклеточного сигнального домена 4-1BB, соответствует приведенной на ФИГ. 2. Типовая генная последовательность приведена на ФИГ. 1 (SEQ ID NO.16; SEQ ID NO.1).
[102] В конкретных вариантах реализации изобретения внутриклеточный сигнальный домен содержит (i) весь или часть сигнального домена CD3ζ, (ii) весь или часть сигнального домена CD28, (iii) весь или часть сигнального домена 4-1BB или (iv) весь или часть сигнального домена CD3ζ, CD28 и/или 4-1BB.
[103] Последовательности внутриклеточных сигнальных доменов экспрессируемой молекулы могут быть связаны друг с другом в произвольном или определенном порядке. Необязательно, связь может быть образована коротким олиго- или белковым линкером, предпочтительно длиной от 2 до 10 аминокислот.
[104] Спейсерные области. В конкретных вариантах реализации изобретения спейсерная область расположена между связывающим доменом и внутриклеточным компонентом экспрессируемой молекулы. В конкретных вариантах реализации изобретения спейсерная область является частью внеклеточного компонента экспрессируемой молекулы.
[105] Длину спейсерной области можно подбирать для определенных клеточных маркеров на нежелательных клетках, чтобы оптимизировать распознавание и уничтожение нежелательной клетки. В конкретных вариантах реализации изобретения длину спейсерной области можно подбирать на основании расположения эпитопа клеточного маркера, аффинности связывающего домена к этому эпитопу и/или способности модифицированных клеток, экспрессирующих молекулу, пролиферировать in vitro и/или in vivo в ответ на распознавание клеточного маркера.
[106] Как правило, спейсерная область находится между связывающим доменом и трансмембранным доменом экспрессируемой молекулы. Спейсерные области могут обеспечивать гибкость связывающих доменов и высокие уровни экспрессии в модифицированных клетках. В конкретных вариантах реализации изобретения спейсерная область содержит по меньшей мере от 10 до 250 аминокислот, по меньшей мере от 10 до 200 аминокислот, по меньшей мере от 10 до 150 аминокислот, по меньшей мере от 10 до 100 аминокислот, по меньшей мере от 10 до 50 аминокислот или по меньшей мере от 10 до 25 аминокислот. В дополнительных вариантах реализации изобретения спейсерная область содержит 250 аминокислот или менее; 200 аминокислот или менее, 150 аминокислот или менее; 100 аминокислот или менее; 50 аминокислот или менее; 40 аминокислот или менее; 30 аминокислот или менее; 20 аминокислот или менее; или 10 аминокислот или менее.
[107] В конкретных вариантах реализации изобретения спейсерные области могут быть получены из шарнирной области иммуноглобулин-подобной молекулы, например, всей или части шарнирной области человеческого lgG1, IgG2, IgG3 или IgG4. Шарнирные области можно модифицировать, чтобы избежать нежелательных структурных взаимодействий, таких как димеризация. В конкретных вариантах реализации изобретения всю или часть шарнирной области можно комбинировать с одним или более доменами константной области иммуноглобулина. Например, часть шарнирной области можно комбинировать со всем или частью домена CH2 или CH3. В конкретных вариантах реализации изобретения спейсерная область не содержит 47-48-аминокислотную последовательность шарнирной области из CD8α.
[108] В конкретных вариантах реализации изобретения спейсерная область выбрана из группы, содержащей последовательность шарнирной области из IgG1, IgG2, lgG3 и lgG4 в комбинации со всей или частью области CH2; всей или частью области CH3; или всей или частью области CH2 и всей или частью области CH3.
[109] В конкретных вариантах реализации изобретения короткая спейсерная область содержит 12 аминокислот или менее и содержит всю или часть последовательности шарнирной области IgG4 (например, белок, кодируемый SEQ ID NO.50), спейсер средней длины содержит 119 аминокислот или менее и содержит всю или часть последовательности шарнирной области IgG4 и область CH3 (например, SEQ ID NO.52), а длинный спейсер содержит 229 аминокислот или менее и содержит всю или часть последовательности шарнирной области IgG4, область CH2 и область CH3 (например, SEQ ID NO.50).
[110] В конкретных вариантах реализации изобретения, когда связывающий домен связывается с частью клеточного маркера, которая находится очень близко к мембране нежелательной клетки, выбирают длинный спейсер (например, длиной 229 аминокислот или менее, но более чем 119 аминокислот). Очень близко к мембране нежелательной клетки означает в пределах первых 100 внеклеточных аминокислот клеточного маркера.
[111] В конкретных вариантах реализации изобретения, когда связывающий домен связывается с частью клеточного маркера, которая находится далеко от мембраны нежелательной клетки, выбирают средний или короткий спейсер (например, длиной 119 аминокислот или менее или 12 аминокислот или менее).
[112] Специалисту в данной области техники понятно, что определить, находится ли связывающая часть клеточного маркера близко или далеко от мембраны, можно путем моделирования трехмерных структур или на основании анализа кристаллической структуры.
[113] В конкретном варианте реализации изобретения экспрессируемая молекула содержит связывающий домен, содержащий scFV, который связывается с эпитопом ROR1, расположенным в мембране далеко от домена Ig/Frizzled, и спейсер длиной 15 аминокислот или менее. В конкретных вариантах реализации изобретения экспрессируемая молекула содержит связывающий домен, содержащий scFV, который связывает эпитоп ROR1, расположенный в мембране близко к домену Kringle, и спейсер, длина которого превышает 15 аминокислот. В конкретных вариантах реализации изобретения экспрессируемая молекула содержит связывающий домен, содержащий scFV, который связывает CD19, и спейсер длиной 15 аминокислот или менее.
[114] В конкретных вариантах реализации изобретения, когда связывающий домен содержит (i) вариабельную легкую цепь, содержащую последовательность CDRL1 RASQDISKYLN (SEQ ID NO. 108), последовательность CDRL2 SRLHSGV (SEQ ID NO. 111) и последовательность CDRL3 GNTLPYTFG (SEQ ID NO. 104), и вариабельную тяжелую цепь, содержащую последовательность CDRH1 DYGVS (SEQ ID NO. 103), последовательность CDRH2 VIWGSETTYYNSALKS (SEQ ID NO. 114) и последовательность CDRH3 YAMDYWG (SEQ ID NO. 115), или (ii) вариабельную легкую цепь, содержащую последовательность CDRL1 ASGFDFSAYYM (SEQ ID NO. 101), последовательность CDRL2 TIYPSSG (SEQ ID NO. 112) и последовательность CDRL3 ADRATYFCA (SEQ ID NO. 100), и вариабельную тяжелую цепь, содержащую последовательность CDRH1 DTIDWY (SEQ ID NO. 102), последовательность CDRH2 VQSDGSYTKRPGVPDR (SEQ ID NO. 113) и последовательность CDRH3 YIGGYVFG (SEQ ID NO. 117), спейсер может содержать 12 аминокислот или менее и в более конкретном варианте реализации изобретения может содержать SEQ ID NO.47.
[115] В конкретных вариантах реализации изобретения, когда связывающий домен содержит (i) вариабельную легкую цепь, содержащую последовательность CDRL1 SGSDINDYPIS (SEQ ID NO. 109), последовательность CDRL2 INSGGST (SEQ ID NO. 105) и последовательность CDRL3 YFCARGYS (SEQ ID NO. 116), и вариабельную тяжелую цепь, содержащую последовательность CDRH1 SNLAW (SEQ ID NO. 110), последовательность CDRH2 RASNLASGVPSRFSGS (SEQ ID NO. 107) и последовательность CDRH3 NVSYRTSF (SEQ ID NO. 106), или (ii) вариабельную легкую цепь, содержащую последовательность CDRL1, последовательность CDRL2 и последовательность CDRL3 антитела герцептин, и вариабельную тяжелую цепь, содержащую последовательность CDRH1, последовательность CDRH2 и последовательность CDRH3 антитела герцептин спейсер может содержать 229 аминокислот или менее и в более конкретном варианте реализации изобретения может содержать SEQ ID NO.61.
[116] Трансмембранные домены. Раскрытые в данном документе экспрессируемые молекулы также могут содержать трансмембранный домен, по меньшей мере часть которого расположена между внеклеточным компонентом и внутриклеточным компонентом. Трансмембранный домен может заякоривать экспрессируемую молекулу в мембране модифицированной клетки. Трансмембранный домен может быть получен из природного и/или синтетического источника. В случае природного источника трансмембранный домен может быть получен из любого мембраносвязанного или трансмембранного белка. Трансмембранные домены могут содержать по меньшей мере трансмембранную(ые) область(и) альфа-, бета- или зета-цепи рецептора Т-клеток CD28, CD3, CD45, CD4, CD5, CD9, CD16, CD22; CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154. Трансмембранные домены могут включать приведенные на ФИГ. 2 и ФИГ. 6.
[117] В конкретных вариантах реализации изобретения трансмембранный домен содержит аминокислотную последовательность трансмембранного домена CD28, как показано на ФИГ. 2, или аминокислотную последовательность трансмембранного домена CD4. Типовая генная последовательность, кодирующая трансмембранный домен CD28, показана на ФИГ. 1 (SEQ ID NO.12). SEQ ID NO.118 является типовой генной последовательностью, кодирующей трансмембранный домен CD4.
[118] Последовательности меток. В конкретных вариантах реализации изобретения экспрессируемая молекула дополнительно содержит последовательность метки. Последовательность метки может обеспечивать выявление и/или отбор трансдуцированных клеток. Можно применять большое количество разных последовательностей меток. Позитивные последовательности селектируемых меток могут кодироваться геном, который после внесения в модифицированную клетку экспрессирует доминантный фенотип, позволяющий проводить позитивную селекцию клеток, несущих этот ген. Гены такого типа известны в данной области техники и включают ген гигромицин-B-фосфотрансферазы (hph), который придает устойчивость к гигромицину B, ген аминогликозид фосфотрансферазы (neo или aph) из Tn5, который кодирует устойчивость к антибиотику G418, ген дигидрофолатредуктазы (DHFR), ген аденозиндезаминазы (ADA) и ген мультилекарственной резистентности (MDR). В конкретных вариантах реализации изобретения последовательность метки представляет собой усеченный EGFR, как показано на ФИГ. 2. типовая генная последовательность, кодирующая усеченный EGFR, показана на ФИГ. 1. (SEQ ID NO.27).
[119] В конкретных вариантах реализации изобретения в модифицированные HSPC можно вносить функциональные гены для проведения негативной селекции in vivo. «Негативная селекция» означает, что вводимая клетка может быть уничтожена в результате изменения in vivo состояния пациента. Негативный селектируемый фенотип может быть результатом внесения гена, который придает чувствительность к водимому агенту. Негативные селектируемые гены известны в данной области техники и включают: ген тимидинкиназы вируса простого герпеса типа I (HSV-I TK), который придает чувствительность к ганцикловиру; ген клеточной гипоксантин-фосфорибозилтрансферазы (HPRT), ген клеточной аденин-фосфорибозилтрансферазы (APRT) и бактериальной цитозиндезаминазы. В отношении дополнительного вспомогательного описания негативной селекции смотрите Lupton S.D. et. al., Mol. and Cell Biol., 11:6 (1991); Riddell et al., Human Gene Therapy 3:319-338 (1992); WO 1992/008796 и WO 1994/028143 и патент США № 6040177 параграфы 14-17).
[120] Конструкцию конкретных молекул, предназначенных для экспрессии модифицированными клетками, можно подбирать в зависимости от типа целевого клеточного маркера, аффинности связывающего домена в отношении клеточного маркера, гибкости, необходимой для связывающего клеточный маркер домена, и/или внутриклеточного сигнального домена. В конкретных вариантах реализации изобретения на in vitro и in in vivo моделях тестируют большое количество конструкций, чтобы определить способность модифицированных клеток размножаться в культуре и/или уничтожать нежелательные клетки. В конкретных вариантах реализации изобретения выбирают молекулу, которая обеспечивает способность по меньшей мере 30% модифицированных эффекторов (например, дифференцированных модифицированных HSPC) пролиферировать на протяжении по меньшей мере двух поколений in vitro и/или в течение 72 часов после внесения in vivo. В конкретных вариантах реализации изобретения не выбирают молекулу, которая приводит к тому, что более 50% клеток в организме иммунодефицитных мышей подвергаются индуцированной активацией гибели клеток (AICD) в течение 72 часов in vivo и не способны снижать количество опухолевых клеток.
[121] В нижеследующем описании приведены более конкретные примеры экспрессируемых молекул и связанных с ними векторов.
[122] «Химерный антигенный рецептор» или «ХАР» относится к синтетически сконструированному рецептору, содержащему связывающий домен, который связывается с клеточным маркером, преимущественно ассоциированным с нежелательной клеткой, который соединен с эффекторным доменом. Связывающий домен и эффекторный домен могут быть соединены посредством спейсерного домена, трансмембранного домена, последовательности метки и/или линкерной последовательности.
[123] В конкретных вариантах реализации изобретения ROR1-специфические и CD19-специфические ХАР можно сконструировать, используя сегменты цепи VL и VH, принадлежащие 2A2, R12 и R11 mAh (ROR1) и FMC63 mAb (CD19). Последовательности вариабельных областей R11 и R12 приведены в Yang et al, Plos One 6(6):e21018, June 15, 2011. Каждый scFV может быть соединен при помощи белка (G4S)3 (SEQ ID NO.60) со спейсерным доменом, полученным из IgG4-Fc (База данных Uniprot: P0186l, SEQ ID NO.92), содержащим любые последовательности из «Hinge-CH2-CH3» (229 AA, SEQ ID NO.61), «Hinge-CH3» (119 AA, SEQ ID NO. 52) или только «Hinge» (12 AA, SEQ. ID NO:47) (ФИГ. 1). Все спейсеры могут содержать замену S→P внутри домена «Hinge» в позиции 108 нативного белка IgG4-Fc и могут быть соединены с трансмембранным доменом 27 AA человеческого CD28 (Uniprot: P10747, SEQ ID NO.93) и с сигнальным модулем эффекторного домена, включая (i) цитоплазматический домен 41 AA человеческого CD28 с заменой LL→GG в позициях 186-187 нативного белка CD28 (SEQ ID NO.93) или (ii) цитоплазматический домен 42 AA человеческого 4-1BB (Uniprot: Q07011, SEQ ID NO. 95), каждый из которых может быть соединен с цитоплазматическим доменом 112 AA изоформы 3 человеческого CD3ζ (Uniprot: P20963, SEQ ID NO.94). Конструкция кодирует рибосомный элемент пропуска T2A (SEQ ID NO.88)) и последовательность tEGFR (SEQ ID NO.27) ниже химерного рецептора. Можно синтезировать кодон-оптимизированные генные последовательности, кодирующие каждый трансген (Life Technologies), и клонировать их в лентивирусный вектор epHIV7, используя сайты рестрикции Nhel и Not1. Лентивирусный вектор epHIV7 можно получить из вектора pHIV7 путем замещения промотора цитомегаловируса pHIV7 промотором EF-1. ROR1-химерный рецептор, CD19-химерный рецептор или tEGFR-кодирующие лентивирусы можно получать в клетках 293T, используя пакующие векторы pCHGP-2, pCMV-Rev2 и pCMV-G и трансфекционный реагент Calphos® (Clontech).
[124] HER2-специфические химерные рецепторы можно сконструировать, используя сегменты цепей VL и VH HER2-специфического mAb, которое распознает проксимальный мембранный эпитоп на HER2 (ФИГ. 12A), а scFV могут быть соединены с внеклеточными спейсерными доменами, содержащими шарнирную область/CH2/CH3 IgG4, шарнирную область/CH3 IgG4 и только шарнирную область IgG4, и с трансмембранным доменом CD28, сигнальными доменами 4-1BB и CD3ζ (ФИГ. 12B).
[125] Как указано, каждый химерный рецептор CD19 может содержать одноцепочечный вариабельный фрагмент, соответствующий последовательности CD19-специфического mAb FMC63 (scFv: VL-VH), спейсер, полученный из IgG4-Fc, содержащий домен «Hinge-CH2-CH3» (229 AA, длинный спейсер) или только домен «Hinge» (12 AA, короткий спейсер), и сигнальный модуль CD3ζ с близкорасположенными к мембране костимулирующими доменами CD28 или 4-1BB, как поодиночке, так и в тандеме (ФИГ. 13A). Трансгенная кассета может содержать усеченный EGFR (tEGFR) ниже гена химерного рецептора и отделяться отщепляемым элементом T2A, чтобы служить в качестве последовательности метки для трансдукции, отбора и in vivo отслеживания модифицированных при помощи химерного рецептора клеток.
[126] Как понятно специалисту в данной области техники, модифицированные HSPC можно получать рекомбинантным методом путем внесения рекомбинантной генной последовательности в HSPC. Описание генетически сконструированных HSPC можно найти в разделе 5.1 патента США № 7399633. Ген, экспрессия которого необходима в модифицированной клетке, вносят в HSPC таким образом, чтобы он мог экспрессироваться клетками и/или их потомством.
[127] Необходимые гены можно вносить в HSPC любым известным в данной области техники способом, включая трансфекцию, электропорацию, микроинъекцию, липофекцию, опосредованную фосфатом кальция трансфекцию, инфицирование вирусным вектором или вектором бактериофага, содержащим генные последовательности, клеточное слияние, опосредованный хромосомами перенос генов, опосредованный микроклетками перенос генов, слияние шеропластов и т.д. В данной области техники известны многочисленные способы внесения в клетки чужеродных генов (смотрите, например, Loeffler and Behr, 1993, Meth. Enzymol. 217:599-618; Cohen et al., 1993, Meth. Enzymol. 217:618-644; Cline, 1985, Pharmac. Ther. 29:69-92), которые можно применять при условии, что не происходит нарушения необходимого развития и физиологических функций клеток-реципиентов. Способ должен обеспечивать стабильный перенос гена в клетку так, чтобы ген экспрессировался клеткой и предпочтительно наследовался и экспрессировался потомством клетки. Как указано, в конкретных вариантах реализации изобретения способ переноса включает перенос в клетки последовательности селектируемой метки. Затем клетки подвергают селекции, чтобы выделить те клетки, которые приняли и экспрессируют перенесенный ген.
[128] Термин «ген» относится к последовательности нуклеиновой кислоты (взаимозаменяемо употребляемой с полинуклеотидной или нуклеотидной последовательностью), которая кодирует молекулу, содержащую внеклеточный компонент и внутриклеточный компонент, как описано в данном документе. Это определение включает различные полиморфизмы последовательностей, мутации и/или варианты последовательностей, при этом такие изменения существенно не влияют на функцию кодируемой молекулы. Термин «ген» может включать не только кодирующие последовательности, но также регуляторные области, такие как промоторы, энхансеры и терминационные области. Этот термин может дополнительно включать все интроны и другие последовательности ДНК, сплайсированные из транскрипта мРНК, вместе с вариантами, получающимися в результате наличия альтернативных участков сплайсинга. Генные последовательности, кодирующие молекулу, может представлять ДНК или РНК, которая управляет экспрессией молекулы. Эти нуклеотидные последовательности могут представлять собой последовательность цепи ДНК, которая транскрибируется в РНК, или последовательность РНК, которая транслируется в белок. Нуклеотидные последовательности включают как полноразмерные нуклеотидные последовательности, так и не полноразмерные последовательности, полученные из полноразмерного белка. Последовательности также могут включать вырожденные кодоны нативной последовательности или последовательности, которые можно вносить, чтобы обеспечить предпочтительное использование кодонов в конкретном типе клеток. Как понятно специалисту в данной области техники, в тексте описания приведены ссылки на части полных генных последовательностей.
[129] Генную последовательность, кодирующую связывающий домен, эффекторный домен, спейсерную область, трансмембранный домен, последовательность метки, линкерную последовательность или любую другую описанную в данном документе белковую или пептидную последовательность, легко можно получить при помощи синтетических или рекомбинантных способов из соответствующей аминокислотной последовательности. В вариантах реализации изобретения генная последовательность, кодирующая любую из этих последовательностей, также может содержать один или более участков расщепления рестрикционными ферментами в 5' и/или 3' концах кодирующей последовательности, чтобы обеспечить легкое вырезание и замещение генной последовательности, кодирующей последовательность, другой генной последовательностью, кодирующей отличную последовательность. В вариантах реализации изобретения генная последовательность, кодирующая последовательности, может быть кодон-оптимизированной для экспрессии в клетках млекопитающих.
[130] «Кодирование» относится к способности конкретных последовательностей нуклеотидов в гене, таких как кДНК или мРНК, служить в качестве матриц для синтеза других макромолекул, таких как определенные последовательности аминокислот. Таким образом, ген кодирует белок, если транскрипция и трансляция мРНК, соответствующей этому гену, приводит к выработке белка в клетке или другой биологической системе. «Генная последовательность, кодирующая белок» включает все нуклеотидные последовательности, которые являются вырожденными версиями друг друга, и которые кодируют одинаковую аминокислотную последовательность или одинаковые аминокислотные последовательности, имеющие в значительной степени сходную форму и функцию.
[131] Полинуклеотидные генные последовательности, кодирующие более одной части экспрессируемой молекулы, могут быть функционально связаны друг с другом и соответствующими регуляторными последовательностями. Например, может существовать функциональная связь между регуляторной последовательностью и гетерологичной нуклеотидной последовательностью, что приводит к экспрессии последней. В другом примере первая нуклеотидная последовательность может быть функционально связана со второй нуклеотидной последовательностью, если первая нуклеотидная последовательность размещена в функциональной взаимосвязи со второй нуклеотидной последовательностью. Например, промотор функционально связан с кодирующей последовательностью, если промотор влияет на транскрипцию или экспрессию кодирующей последовательности. В общем случае функционально связанные последовательности ДНК являются смежными и, в случае необходимости или целесообразности, соединяют кодирующие области в одной рамке считывания.
[132] Можно использовать ретровирусные векторы (смотрите Miller et al., 1993, Meth. Enzymol. 217:581-599). В таких вариантах реализации изобретения предназначенный для экспрессии ген клонируют в ретровирусный вектор для доставки его в HSPC. В конкретных вариантах реализации изобретения ретровирусный вектор содержит все цис-действующие последовательности, необходимые для упаковки и интеграции вирусного генома, т.е. (a) длинный концевой повтор (ДКП) или его части с каждого конца вектора; (b) участки связывания праймера для синтеза негативной и позитивной цепи ДНК; и (c) пакующий сигнал, необходимый для внесения геномной РНК в вирионы. Больше подробностей в отношении ретровирусных векторов можно найти в Boesen et al., 1994, Biotherapy 6:291-302; Clowes et al., 1994, J. Clin. Invest. 93:644-651; Kiem et al., 1994, Blood 83:1467-1473; Salmons and Gunzberg, 1993, Human Gene Therapy 4:129-141; и Grossman and Wilson, 1993, Curr. Opin. in Genetics and Devel. 3:110-114. Также можно использовать аденовирусы, аденоассоциированные вирусы (ААВ) и альфа-вирусы. Смотрите Kozarsky and Wilson, 1993, Current Opinion in Genetics and Development 3:499-503, Rosenfeld et al., 1991, Science 252:431-434; Rosenfeld et al., 1992, Cell 68:143-155; Mastrangeli et al., 1993, J. Clin. Invest. 91:225-234; Walsh et al., 1993, Proc. Soc. Exp. Bioi. Med. 204:289-300; и Lundstrom, 1999, J. Recept. Signal Transduct. Res. 19: 673-686. Другие способы доставки генов включают применение искусственных хромосом млекопитающих (Vos, 1998, Curr. Op. Genet. Dev. 8:351-359); липосом (Tarahovsky and Ivanitsky, 1998, Biochemistry (Mosc) 63:607-618); рибозимов (Branch and Klotman, 1998, Exp. Nephrol. 6:78-83); и трехспиральной ДНК (Chan and Glazer, 1997, J. Mol. Med. 75:267-282).
[133] Дополнительные варианты реализации изобретения включают последовательности, обладающие 70% идентичности последовательностей; 80% идентичности последовательностей; 81% идентичности последовательностей; 82% идентичности последовательностей; 83% идентичности последовательностей; 84% идентичности последовательностей; 85% идентичности последовательностей; 86% идентичности последовательностей; 87% идентичности последовательностей; 88% идентичности последовательностей; 89% идентичности последовательностей; 90% идентичности последовательностей; 91% идентичности последовательностей; 92% идентичности последовательностей; 93% идентичности последовательностей; 94% идентичности последовательностей; 95% идентичности последовательностей; 96% идентичности последовательностей; 97% идентичности последовательностей; 98% идентичности последовательностей; или 99% идентичности последовательностей с любой генной, белковой или пептидной последовательностью, раскрытой в данном документе.
[134] « % идентичности последовательностей» относится к взаимосвязи между двумя или более последовательностями, определяемой путем сравнения последовательностей. В данной области техники «идентичность» также обозначает степень родства между белковыми последовательностями, определяемую по совпадению между участками таких последовательностей. «Идентичность» (часто называемую «сходством») можно легко рассчитать при помощи известных способов, включая описанные в: Computational Molecular Biology (Lesk, A. M., ed.) Oxford University Press, NY (1988); Biocomputing: Informatics and Genome Projects (Smith, D. W., ed.) Academic Press, NY (1994); Computer Analysis of Sequence Data, Part I (Griffin, A. M., and Griffin, H. G., eds.) Humana Press, NJ (1994); Sequence Analysis in Molecular Biology (Von Heijne, G., ed.) Academic Press (1987); и Sequence Analysis Primer (Gribskov, M. and Devereux, J., eds.) Oxford University Press, NY (1992). Предпочтительные способы определения идентичности последовательностей разработаны так, чтобы получать наилучшее совпадение между исследуемыми последовательностями. Способы определения идентичности и сходства последовательностей представлены в общедоступных компьютерных программах. Выравнивание последовательностей и расчет процента идентичности можно проводить, используя программу Megalign из биоинформационного вычислительного пакета LASERGENE (DNASTAR, Inc., Madison, Wisconsin). Множественное выравнивание последовательностей также можно проводить, используя метод выравнивания Clustal (Higgins and Sharp CABIOS, 5, 151-153 (1989) с установленными по умолчанию параметрами (штраф за открытие гэпа=10, штраф за длину гэпа=10). Соответствующие программы также включают пакет программ GCG (Wisconsin Package Version 9.0, Genetics Computer Group (GCG), Madison, Wisconsin); BLASTP, BLASTN, BLASTX (Altschul, et al., J. Mol. Biol. 215:403-410 (1990); DNASTAR (DNASTAR, Inc., Madison, Wisconsin); и программу FASTA, в которой используется алгоритм Смита-Уотермана (Pearson, Comput. Methods Genome Res., [Proc. Int. Symp.] (1994), Meeting Date 1992, 111-20. Editor(s): Suhai, Sandor. Publisher: Plenum, New York, N.Y. В контексте настоящего изобретения следует понимать, что в случае применения для анализа программного обеспечения для анализа последовательностей, результаты анализа основаны на «установленных по умолчанию величинах» соответствующей программы. «Установленные по умолчанию величины» означают любую группу величин или параметров, которые изначально загружаются с программным обеспечением при первом использовании.
[135] Не ограничивая вышесказанного, белки и пептиды, обладающие идентичностью последовательностей с раскрытыми в данном документе последовательностями, включают их варианты и D-замещенные аналоги.
[136] «Варианты» раскрытых в данном документе последовательностей включают последовательности, содержащие одну или более добавок, делеций, стоп-позиций или замен по сравнению с раскрытой в данном документе последовательностью.
[137] Аминокислотная замена может быть консервативной или неконсервативной заменой. Раскрытые в данном документе варианты белковых или пептидных последовательностей могут включать те, которые содержат одну или более аминокислотных замен. «Консервативная замена» включает замену, соответствующую одной из следующих групп консервативных замен: Группа 1: аланин (Ala или A), глицин (Gly или G), Ser, Thr; Группа 2: аспарагиновая кислота (Asp или D), Glu; Группа 3: аспарагин (Asn или N), глутамин (Gln или Q); Группа 4: Arg, лизин (Lys или K), гистидин (His или H); Группа 5: Ile, лейцин (Leu или L), метионин (Met или M), валин (Val или V); и Группа 6: Phe, Tyr, Trp.
[138] Кроме того, аминокислоты могут быть отнесены к группам консервативных замен на основании сходной функции, химической структуры или состава (например, кислотные, основные, алифатические, ароматические, серосодержащие). Например, алифатическая группа может включать в целях замены Gly, Ala, Val, Leu и Ile. Другие группы, содержащие аминокислоты, которые считаются консервативными заменами по отношению друг к другу, включают: серосодержащие: Met и Cys; кислотные: Asp, Glu, Asn и Gln; малые алифатические, неполярные или слабополярные остатки: Ala, Ser, Thr, Pro и Gly; полярные, отрицательно заряженные остатки и их амиды: Asp, Asn, Glu, и Gln; полярные, положительно заряженные остатки: His, Arg и Lys; крупные алифатические, неполярные остатки: Met, Leu, Ile, Val и Cys; и крупные ароматические остатки: Phe, Tyr и Trp. Дополнительную информацию можно найти в Creighton (1984) Proteins, W.H. Freeman and Company.
[139] «D-замещенные аналоги» включают раскрытые в данном документе белки или пептиды, в которых одна или более L-аминокислот замещены одной или более D-аминокислотами. D-аминокислота может принадлежать тому же самому аминокислотному типу, который обнаруживается в контрольной последовательности, или может являться отличной аминокислотой. Соответственно, D-аналоги также могут являться вариантами.
[140] Не ограничивая вышесказанного и исключительно в качестве примера:
[141] В конкретных вариантах реализации изобретения связывающий домен содержит последовательность, которая имеет по меньшей мере 80%; 81%; 82%; 83%; 84%; 85%; 86%; 87%; 88%; 89%; 90%; 91%; 92%; 93%; 94%; 95%; 96%; 97%; 98%; или 99% идентичности последовательностей с аминокислотной последовательностью раскрытой в данном документе вариабельной области легкой цепи (VL) или вариабельной области тяжелой цепи (VH) или ними двумя, при этом каждая CDR не содержит изменений или содержит не более одного, двух или трех изменений по сравнению с моноклональным антителом или его фрагментом, которое специфически связывает представляющий интерес клеточный маркер.
[142] В конкретных вариантах реализации изобретения связывающий домен содержит последовательность, которая имеет по меньшей мере 80%; 81%; 82%; 83%; 84%; 85%; 86%; 87%; 88%; 89%; 90%; 91%; 92%; 93%; 94%; 95%; 96%; 97%; 98%; или 99% идентичности последовательностей с аминокислотной последовательностью РТК Vα, Vβ, Cα или Cβ, при этом каждая CDR не содержит изменений или содержит не более одного, двух или трех изменений по сравнению с РТК или его фрагментом, который специфически связывается с представляющим интерес клеточным маркером.
[143] В конкретных вариантах реализации изобретения связывающий домен области Vα, Vβ, Cα или Cβ может быть получен из или на основе Vα, Vβ, Cα или Cβ известного РТК (например, высокоаффинного РТК) и содержать одну или более (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) вставок, одну или более (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) делеций, одну или более (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) аминокислотных замен (например, консервативных аминокислотных замен или неконсервативных аминокислотных замен) или комбинацию вышеуказанных изменений по сравнению с Vα, Vβ, Cα или Cβ известного РТК. Вставка, делеция или замена может находиться в любом месте области Vα, Vβ, Cα или Cβ, включая амино- или карбокси-конец, или оба конца этих областей, при условии, что каждая CDR не содержит изменений или содержит не более одного, двух или трех изменений, а связывающий домен, содержащий модифицированную область Vα, Vβ, Cα или Cβ, способен специфически связывать свою мишень с аффинностью, сходной с диким типом.
[144] В конкретных вариантах реализации изобретения VH- или VL-область связывающего домена могут быть получены из или на основе VH или VL известного моноклонального антитела и могут по отдельности или вместе содержать одну или более (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) вставок, одну или более (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) делеций, одну или более (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) аминокислотных замен (например, консервативных аминокислотных замен или неконсервативных аминокислотных замен) или комбинацию вышеуказанных изменений по сравнению с VH или VL известного моноклонального антитела. Вставка, делеция или замена может находиться в любом месте VH- или VL-области, включая амино- или карбокси-конец, или оба конца этих областей, при условии, что каждая CDR не содержит изменений или содержит не более одного, двух или трех изменений, а связывающий домен, содержащий модифицированную VH- или VL-область, способен специфически связывать свою мишень с аффинностью, сходной с диким типом.
[145] В конкретных вариантах реализации изобретения связывающий домен содержит последовательность, которая имеет по меньшей мере 80%; 81%; 82%; 83%; 84%; 85%; 86%; 87%; 88%; 89%; 90%; 91%; 92%; 93%; 94%; 95%; 96%; 97%; 98%; или 99% идентичности последовательностей с соответствующим доменом из (i) scFv для FMC63 (ii) scFv для R12; (iii) scFv для R11; или (iv) scFv для герцептина.
[146] В конкретных вариантах реализации изобретения внутриклеточный сигнальный домен может иметь по меньшей мере 80%; 81%; 82%; 83%; 84%; 85%; 86%; 87%; 88%; 89%; 90%; 91%; 92%; 93%; 94%; 95%; 96%; 97%; 98%; или 99% идентичности последовательностей с CD3ζ, имеющим последовательность, приведенную на ФИГ. 2.
[147] В конкретных вариантах реализации изобретения костимулирующий сигнальный домен может иметь по меньшей мере 80%; 81%; 82%; 83%; 84%; 85%; 86%; 87%; 88%; 89%; 90%; 91%; 92%; 93%; 94%; 95%; 96%; 97%; 98%; или 99% идентичности последовательностей с внутриклеточным доменом CD28, показанным на ФИГ. 5, или с 4-1BB, имеющим последовательность, приведенную на ФИГ. 2. В конкретных вариантах реализации изобретения вариант внутриклеточного домена CD28 содержит аминокислотную замену в позициях 186-187, в которой LL замещены GG.
[148] В конкретных вариантах реализации изобретения трансмембранный домен может быть выбран или модифицирован при помощи аминокислотной замены (аминокислотных замен), чтобы избежать связывания таких доменов с трансмембранными доменами таких же или отличных поверхностных мембранных белков, чтобы минимизировать взаимодействия с другими участниками рецепторного комплекса. В дополнительных конкретных вариантах реализации изобретения синтетические или вариантные трансмембранные домены содержат преимущественно гидрофобные остатки, такие как лейцин и валин. Вариантные трансмембранные домены предпочтительно имеют индекс гидрофобности, составляющий по меньшей мере 50, согласно расчетам Kyte Doolittle. В конкретных вариантах реализации изобретения трансмембранный домен может иметь по меньшей мере 80%; 81%; 82%; 83%; 84%; 85%; 86%; 87%; 88%; 89%; 90%; 91%; 92%; 93%; 94%; 95%; 96%; 97%; 98%; или 99% идентичности последовательностей с последовательностью с ФИГ. 2 или 6.
[149] Также включены белки и пептиды, обладающие такой же функциональной способностью, что и те, которые явным образом раскрыты в данном документе.
[150] В случае если они явным образом не приведены в данном документе, для выявления родственных и соответствующих белковых и пептидных последовательностей, а также генных последовательностей, кодирующих такие белки и пептиды, можно использовать информацию по последовательностям из общедоступных баз данных и знания специалистов в данной области техники.
[151] Дифференцировка. В конкретных вариантах реализации изобретения модифицированные HSPC перед введением субъекту дифференцируют в модифицированные не-Т эффекторные клетки. В случае необходимости дифференцировки модифицированных HSPC, HSPC можно подвергать воздействию одного или более факторов роста, которые стимулируют дифференцировку в не-Т эффекторные клетки. Факторы роста и условия клеточного культивирования, которые стимулируют дифференцировку, известны в данной области техники (смотрите, например, патент США № 7399633 раздел 5.2 и раздел 5.5). Например, можно использовать SCF в комбинации с GM-SCF или IL-7 для дифференцировки HSPC в миелоидные стволовые клетки/клетки-предшественники или лимфоидные стволовые клетки/клетки-предшественники, соответственно. В конкретных вариантах реализации изобретения HSPC можно дифференцировать в лимфоидную стволовую клетку/клетку-предшественника, подвергая HSPC воздействию 100 нг/мл каждого из SCF и GM-SCF или IL-7. В конкретных вариантах реализации изобретения для стимуляции дифференцировки HSPC используют антагонист рецептора ретиноевой кислоты (RAR) или предпочтительно полностью транс-ретиноевую кислоту (ATRA). Дифференцировку в естественные клетки-киллеры можно осуществить, подвергая культивируемые HSPC воздействию среды RPMI, дополненной человеческой сывороткой, IL-2 в концентрации 50 Е/мл и IL-15 в концентрации 500 нг/мл. В дополнительных вариантах реализации изобретения среда RPMI также может быть дополнена L-глутамином.
[152] В конкретных вариантах реализации изобретения модифицированные HSPC могут быть дифференцированы в не-Т эффекторные клетки, включая естественные клетки-киллеры (NK) или нейтрофилы. NK-клетки выполняют две основные функции: (i) распознавания и уничтожения опухолевых клеток и других инфицированных вирусом клеток; и (ii) регуляции врожденных и адаптивных иммунных ответов путем секреции CCL3, CCL4, CCL5 и/или хемокинов XCL1 или цитокинов, таких как гранулоцитарно-макрофагальный колониестимулирующий фактор, фактор некроза опухолей-α или IFN-γ. Нейтрофилы в общем случае циркулируют в кровотоке, пока не попадают к участками воспаления, где они нацеливаются и разрушают аберрантные типы клеток.
[153] Композиции и составы. Клетки и модифицированные клетки можно готовить в виде композиций и/или составов для введения субъекту. Композиция относится к клетке и/или модифицированной клетке, приготовленной с фармацевтически приемлемым носителем для введения субъекту. Состав относится по меньшей мере к двум типам клеток в фармацевтически приемлемом носителе (далее - носителе) для введения субъекту.
[154] На разных этапах приготовления композиции или состава может быть необходимо или целесообразно проводить криоконсервацию клетки. Термины «замороженный/заморозка» или «криоконсервированный/криоконсервация» можно употреблять взаимозаменяемо.
[155] Как известно специалисту в данной области техники, заморозка клеток может быть разрушительной (смотрите Mazur, P., 1977, Cryobiology 14:251-272), но существует большое количество процедур, способных предотвратить такое повреждение. Например, повреждение можно предотвратить путем (a) применения криопротекторного агента, (b) регуляции скорости заморозки и/или (c) хранения при достаточно низкой температуре для минимизации разрушительных реакций. Типовые криопротекторные агенты включают диметилсульфоксид (ДМСО) (Lovelock and Bishop, 1959, Nature 183:1394-1395; Ashwood-Smith, 1961, Nature 190:1204-1205), глицерол, поливинилпирролидин (Rinfret, 1960, Ann. N.Y. Acad. Sci. 85:576), полиэтиленгликоль (Sloviter and Ravdin, 1962, Nature 196:548), альбумин, декстран, сахарозу, этиленгликоль, i-эритритол, D-рибитол, D-маннитол (Rowe et al., 1962, Fed. Proc. 21:157), D-сорбитол, i-инозитол, D-лактозу, хлорид холина (Bender et al.., 1960, J. Appl. Physiol. 15:520), аминокислоты (Phan The Tran and Bender, 1960, Exp. Cell Res. 20:651), метанол, ацетамид, моноацетат глицерина (Lovelock, 1954, Biochem. J. 56:265) и неорганические соли (Phan The Tran and Bender, 1960, Proc. Soc. Exp. Biol. Med. 104:388; Phan The Tran and Bender, 1961, in Radiobiology, Proceedings of the Third Australian Conference on Radiobiology, Ilbery ed., Butterworth, London, p. 59). В конкретных вариантах реализации изобретения можно применять ДМСО. Добавление плазмы (например, до концентрации 20-25%) может усиливать защитное действие ДМСО. После добавления ДМСО клетки можно хранить при 0° C до заморозки, так как концентрация ДМСО, составляющая 1%, может быть токсичной при температуре выше 4° C.
[156] Для криоконсервации клеток может иметь критическое значение медленная регулируемая скорость охлаждения (Rapatz et al., 1968, Cryobiology 5(1): 18-25), а разные типы клеток характеризуются разными оптимальными скоростями охлаждения (смотрите, например, Rowe and Rinfret, 1962, Blood 20:636; Rowe, 1966, Cryobiology 3(1):12-18; Lewis, et al., 1967, Transfusion 7(1):17-32; и Mazur, 1970, Science 168:939- 949 в отношении влияния скорости охлаждения на выживаемость стволовых клеток и их трансплантационный потенциал). Фаза нагревания или плавления, когда вода превращается в лед, должна быть минимальной. Процедуру охлаждения можно проводить, используя, например, программируемое устройство для заморозки или процедуру с применением метаноловой бани. Программируемые устройства для заморозки позволяют определить оптимальную скорость охлаждения и облегчают стандартное охлаждение с возможностью дальнейшего восстановления.
[157] В конкретных вариантах реализации изобретения предварительно обработанные ДМСО клетки могут быть предварительно охлаждены на льду и перенесены на планшет, содержащий охлажденный метанол, который, в свою очередь, помещают в механический рефрижератор (например, Harris или Revco) при -80° C. Измерения, проводимые при помощи термопары, показывают, что предпочтительная скорость охлаждения может составлять от 1° до 3° C/минуту. По меньшей мере через два часа образцы охлаждаются до температуры -80° C, после чего их можно помещать непосредственно в жидкой азот (-196° C).
[158] После полной заморозки клетки можно быстро переносить в криогенный резервуар для длительного хранения. В предпочтительном варианте реализации изобретения образцы могут криогенно храниться в жидком азоте (-196° C) или паре (-1° C). Такое хранение облегчается доступностью высокоэффективных рефрижераторов с охлаждением жидким азотом.
[159] Дополнительные аспекты и процедуры обработки, криоконсервации и долгосрочного хранения клеток можно найти в следующих типовых ссылках патенты США № 4199022; 3753357; и 4559298; Gorin, 1986, Clinics In Haematology 15(1):19-48; Bone-Marrow Conservation, Culture and Transplantation, Proceedings of a Panel, Moscow, July 22-26, 1968, International Atomic Energy Agency, Vienna, pp. 107-186; Livesey and Linner, 1987, Nature 327:255; Linner et al., 1986, J. Histochem. Cytochem. 34(9):1123-1135; Simione, 1992, J. Parenter. Sci. Technol. 46(6):226-32).
[160] После криоконсервации замороженные клетки можно размораживать для применения в соответствии со способами, известными специалистам в данной области техники. Замороженные клетки предпочтительно быстро размораживают и охлаждают непосредственно после размораживания. В конкретных вариантах реализации изобретения сосуд, содержащий замороженные клетки, можно до горлышка погружать в теплую водяную баню; легкое вращение гарантирует смешивание клеточной суспензии во время размораживания и увеличивает перенос тепла от теплой воды к внутренней ледяной массе. Сразу после того, как лед полностью растаял, сосуд можно немедленно переносить на лед.
[161] В конкретных вариантах реализации изобретения можно применять способы для предотвращения агрегации клеток во время размораживания. Типовые способы включают: добавление до и/или после заморозки ДНКазы (Spitzer et al., 1980, Cancer 45:3075-3085), низкомолекулярного декстрана и цитрата, гидроксиэтилового крахмала (Stiff et al., 1983, Cryobiology 20:17-24) и т.д.
[162] Специалисту в данной области техники понятно, что в случае применения криопротекторного агента, который токсичен для человека, его следует удалять перед терапевтическим применением. ДМСО не обладает сильной токсичностью.
[1] Типовые носители и способы введения клеток описаны на страницах 14-15 публикации патента США № 2010/0183564. Дополнительные фармацевтические носители описаны в Remington: The Science and Practice of Pharmacy, 21st Edition, David B. Troy, ed., Lippicott Williams & Wilkins (2005).
[163] В конкретных вариантах реализации изобретения клетки можно собирать из культуральной среды и промывать и добавлять в носитель в терапевтически эффективном количестве. Типовые носители включают солевой раствор, забуференный солевой раствор, физиологический солевой раствор, воду, раствор Хенкса, раствор Рингера, Nonnosol-R (Abbott Labs), Plasma-Lyte A® (Baxter Laboratories, Inc., Morton Grove, IL), глицерин, этанол и их комбинации.
[164] В конкретных вариантах реализации изобретения носители могут быть дополнены человеческим сывороточным альбумином (ЧСА) или другими компонентами человеческой сыворотки или фетальной бычьей сывороткой. В конкретных вариантах реализации изобретения носитель для инфузии содержит забуференный солевой раствор с 5% РЧА (раствором человеческого альбумина) или декстрозой. Дополнительные изотонические агенты включают многоатомные сахарные спирты, включая трехатомные или высшие сахарные спирты, такие как глицерин, эритритол, арабитол, ксилитол, сорбитол или маннитол.
[165] Носители могут включать буферные агенты, такие как цитратные буферы, сукцинатные буферы, тартратные буферы, фумаратные буферы, глюконатные буферы, оксалатные буферы, лактатные буферы, ацетатные буферы, фосфатные буферы, гистидиновые буферы и/или соли триметиламинов.
[166] Стабилизаторы относятся к большой категории вспомогательных веществ, функция которых может варьироваться от объемообразующего агента до добавки, которая помогает препятствовать адгезии клеток к стенкам контейнера. Типичные стабилизаторы могут включать многоатомные сахарные спирты; аминокислоты, такие как аргинин, лизин, глицин, глутамин, аспарагин, гистидин, аланин, орнитин, L-лейцин, 2-фенилаланин, глутаминовая кислота и треонин; органические сахара или сахарные спирты, такие как лактоза, трегалоза, стахиоза, маннитол, сорбитол, ксилитол, рибитол, миоинозитол, галактитол, глицерин и циклитолы, такие как инозитол; ПЭГ; аминокислотные полимеры; серосодержащие восстановительные агенты, такие как мочевина, глутатион, тиоктовая кислота, тиогликолят натрия, тиоглицерин, альфа-монотиоглицерин и тиосульфат натрия; низкомолекулярные полипептиды (т.е. <10 остатков); белки, такие как ЧСА, бычий сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; моносахариды, такие как ксилоза, манноза, фруктоза и глюкоза; дисахариды, такие как лактоза, мальтоза и сахароза; трисахариды, такие как раффиноза, и полисахариды, такие как декстран.
[2] В случае необходимости или целесообразности композиции или составы могут содержать местный анестетик, такой как лидокаин, для облегчения боли в месте инъекции.
[167] Типовые консерванты включают фенол, бензиловый спирт, мета-крезол, метилпарабен, пропилпарабен, октадецилдиметилбензил хлорид аммония, бензалкония галиды, гексаметония хлорид, алкилпарабены, такие как метилпарабен или пропилпарабен, катехол, резорцин, циклогексанол и 3-пентанол.
[168] Терапевтически эффективное количество клеток в композициях или составах может быть больше чем 102 клеток, больше чем 103 клеток, больше чем 104 клеток, больше чем 105 клеток, больше чем 106 клеток, больше чем 107 клеток, больше чем 108 клеток, больше чем 109 клеток, больше чем 1010 клеток или больше чем 1011.
[169] В раскрытых в данном документе композициях и составах клетки обычно находятся в объеме, составляющем литр или меньше, 500 мл или меньше, 250 мл или меньше или 100 мл или меньше. Следовательно, плотность вводимых клеток, как правило, превышает 104 клеток/мл, 107 клеток/мл или 108 клеток/мл.
[170] Как указано, композиции содержат один тип клеток (например, модифицированные HSPC или модифицированные эффекторы). Составы могут содержать HSPC, модифицированные HSPC и/или модифицированные эффекторы (такие как модифицированные NK-клетки) в комбинации. В конкретных вариантах реализации изобретения смешивают комбинации модифицированных HSPC и модифицированных эффекторов с одним и тем же связывающим доменом. В других вариантах реализации изобретения смешивают модифицированные HSPC и модифицированные эффекторы разных связывающих доменов. Аналогично, все другие аспекты экспрессируемой молекулы (например, компоненты эффекторного домена, спейсерные области и т.д.) могут быть одинаковыми или разными в различных комбинациях модифицированных HSPC и модифицированных эффекторов в составе. Кроме того, в состав могут быть вместе включены модифицированные HSPC, экспрессирующие разные молекулы или их компоненты, и в состав могут быть вместе включены модифицированные эффекторы, экспрессирующие разные молекулы или их компоненты. В конкретных вариантах реализации изобретения состав может содержать по меньшей мере две модифицированные HSPC, экспрессирующие разные молекулы, и по меньшей мере два модифицированных эффектора, экспрессирующих разные молекулы.
[171] HSPC, модифицированные HSPC и модифицированные эффекторы можно смешивать в разных соотношениях, например, соотношении 1:1:1, соотношении 2:1:1, соотношении 1:2:1, соотношении 1:1:2, соотношении 5:1:1, соотношении 1:5:1, соотношении 1:1:5, соотношении 10:1:1, соотношении 1:10:1, соотношении 1:1:10, соотношении 2:2:1, соотношении 1:2:2, соотношении 2:1:2, соотношении 5:5:1, соотношении 1:5:5, соотношении 5:1:5, соотношении 10:10:1, соотношении 1:10:10, соотношении 10:1:10 и т.д. Эти соотношения также можно применять к количеству клеток, экспрессирующих одинаковые или разные молекулярные компоненты. Если смешивают только два типа клеток или только 2 комбинации экспрессируемых молекулярных компонентов включены в состав, соотношение может включать любую 2-числовую комбинацию, которую можно получить из 3-числовых комбинаций, приведенных выше. В вариантах реализации изобретения комбинированные популяции клеток исследуют в отношении эффективности и/или клеточной пролиферации in vitro и/или in vivo, и выбирают то соотношение клеток, которое обеспечивает необходимую эффективность и/или клеточную пролиферацию.
[172] Раскрытые в данном документе композиции и составы можно готовить для введения путем, например, инъекции, инфузии, перфузии или лаважа. Дополнительно можно получать композиции и составы для инъекции в костный мозг, внутривенной, интрадермальной, внутриартериальной, внутриузловой, внутрилимфатической, внутрибрюшинной, внутриочаговой, внутрипростатической, внутривагинальной, внутрипрямокишечной, местной, интратекальной, внутриопухолевой, внутримышечной, внутрипузырной и/или подкожной инъекции.
[173] Наборы. Наборы могут содержать один или более контейнеров, содержащих одну или более клеток, композиций или составов, описанных в данном документе. В конкретных вариантах реализации изобретения наборы могут содержать один или более контейнеров, содержащих одну или более клеток, композиций или составов и/или композиции, предназначенные для применения в комбинации с другими клетками, композициями или составами. К такому контейнеру(ам) может прилагаться вкладыш в форме, установленной государственным органом, регулирующим производство, использование или продажу фармацевтических средств или биологических продуктов, в котором отражено разрешение со стороны указанного органа на производство, использование или продажу для применения на людях. Во вкладыше может быть указано, что предлагаемые клетки, композиции или составы можно вводить субъекту без проверки иммунологической совместимости. Наборы могут дополнительно содержать инструкции по применению набора, например, инструкции по приготовлению клеток, композиций и/или составов для введения; надлежащей утилизации отходов; и тому подобное. Инструкции могут иметь форму печатных инструкций, прилагаемых к набору, или инструкции могут быть напечатаны на самом наборе. Инструкции могут иметь форму листка-вкладыша, проспекта, брошюры, CD-Rom или машиночитаемого носителя, или могут содержать ссылки на инструкции в удаленной локации, такой как вебсайт. В конкретных вариантах реализации изобретения наборы также могут содержать некоторые или все необходимые медицинские принадлежности для эффективного применения набора, такие как шприцы, ампулы, капельница, маска, безыгольное устройство для переливания жидкостей, инъекционный колпачок, губки, стерильные липкие пластыри, Chloraprep, перчатки и тому подобное. Содержание описанных в данном документе наборов может меняться.
[174] Способы применения. Раскрытые в данном документе способы включают лечение субъектов (людей, животных ветеринарного значения (собак, кошек, рептилий, птиц и т.д.), сельскохозяйственных животных (лошадей, крупный рогатый скот, коз, свиней, курей и т.д.) и подопытных животных (обезьян, крыс, мышей, рыб и т.д.) раскрытыми в данном документе клетками. Лечение субъектов включает доставку терапевтически эффективного количества. Терапевтически эффективное количество включает то, которое обеспечивает эффективное количество, профилактическое лечение и/или терапевтическое лечение.
[175] «Эффективное количество» представляет собой количество клеток, необходимое, чтобы в организме субъекта произошло желательное физиологическое изменение. Эффективное количество часто вводят в исследовательских целях. Раскрытое в данном документе эффективное количество выполняет одно или более действий из: (i) обеспечивает нормальный состав крови путем снижения иммунодефицита, панцитопении, нейтропении и/или лейкопении (например, восполняя популяцию клеток иммунной системы) и (ii) обладает противораковым действием.
[176] «Профилактическое лечение» включает лечение, применяемое к субъекту, который не обнаруживает признаков или симптомов нуждающегося в лечении патологического состояния или обнаруживает только ранние признаки или симптомы нуждающегося в лечении патологического состояния, так, что лечение применяется в целях уменьшения, предотвращения или снижения риска развития патологического состояния. Таким образом, профилактическое лечение выполняет функцию превентивного лечения патологического состояния.
[177] «Терапевтическое лечение» включает лечение, применяемое к субъекту, который обнаруживает признаки или симптомы патологического состояния, и применяется к субъекту с целью снижения тяжести или прогрессирования патологического состояния.
[178] Фактическое количество вводимой конкретному субъекту дозы может быть определено врачом, ветеринаром или исследователем на основании параметров, таких как физические и физиологические факторы, включая, например, цель; массу тела; тип патологического состояния; тяжесть патологического состояния; возможные связанные явления, если они известны; предыдущие или сопутствующие терапевтические вмешательства; идиопатию субъекта; и путь введения. Кроме того, необязательно, можно использовать вспомогательные методы in vitro и in vivo анализа для определения оптимальных диапазонов дозировок.
[179] Предназначенное для введения терапевтически эффективное количество может включать более чем 102 клеток, более чем 103 клеток, более чем 104 клеток, более чем 105 клеток, более чем 106 клеток, более чем 107 клеток, более чем 108 клеток, более чем 109 клеток, более чем 1010 клеток или более чем 1011.
[180] Как указано, раскрытые в данном документе композиции и составы можно вводить, например, путем инъекции, инфузии, перфузии или лаважа и, конкретнее, их можно вводить одним или более путями из инъекций в костный мозг, внутривенных, интрадермальных, внутриартериальных, внутриузловых, внутрилимфатических, внутрибрюшинных, внутриочаговых, внутрипростатических, внутривагинальных, внутрипрямокишечных, местных, интратекальных, внутриопухолевых, внутримышечных, внутрипузырных и/или подкожных инфузий и/или болюсных инъекций.
[181] Применение немодифицированных HSPC описано в разделе 5.6.1 патента США № 7399633 и WO 2013/086436. HSPC и модифицированные HSPC можно вводить с одной и той же целью или в разных целях. Обычные цели включают обеспечение гемопоэтической функции нуждающемуся в этом субъекту; и/или лечение одного или более явлений из иммунодефицита, панцитопении, нейтропении и/или лейкопении (включая циклическую нейтропению и идиопатическую нейтропению) (вместе называемые «целями»). HSPC и модифицированные HSPC можно вводить субъектам, которые имеют пониженный уровень кровяных телец или подвержены риску снижения уровня кровяных телец по сравнению со стандартным уровнем кровяных телец. В конкретных вариантах реализации изобретения субъект имеет анемию или подвержен риску развития анемии.
[182] Лечение в определенных целях может быть необходимым при прохождении интенсивной химиотерапии, включая применение одного или более из алкилирующих агентов, Ara-C, азатиоприна, карбоплатина, цисплатина, хлорамбуцила, клофарабина, циклофосфамида, ифосфамида, мехлоретамина, меркаптопурина, оксалиплатина, таксанов и алкалоидов барвинка (например, винкристина, винбластина, винорелбина и виндезина).
[183] Лечение в определенных целях также может быть необходимым при миелоаблативном режиме при трансплантациии гемопоэтических клеток (ТГК). В конкретных вариантах реализации изобретения HSPC и/или модифицированные HSPC вводят донору костного мозга, при риске истощения костного мозга или снижения или ограничения уровня кровяных телец. Введение можно проводить до и/или после получения костного мозга. HSPC и/или модифицированные HSPC также можно вводить реципиенту трансплантата костного мозга.
[184] Лечение в определенных целях также может быть необходимым при воздействии острого ионизирующего облучения и/или воздействии других лекарственных препаратов, которые могут приводить к супрессии костного мозга или гемопоэтическому дефициту, включая антибиотики, пенициллин, ганцикловир, дауномицин, сульфамидные лекарственные препараты, фенотиазоны, транквилизаторы, мепробамат, анальгетики, аминопирин, дипирон, противосудорожные препараты, фенитоин, карбамазепин, антитиреоиды, пропилтиоурацил, метимазол и диуретики.
[185] Лечение в определенных целях также может быть необходимым при вирусных (например, ВИЧI, ВИЧII, ВТКЛЧVI, ВТКЛЧVII, ВТКЛЧVIII), микробных или паразитарних инфекциях и/или в результате лечения болезни почек или почечной недостаточности, например, диализа. Лечение HSPC и/или модифицированными HSPC также может оказывать благоприятное влияние при различных иммунодефицитах, например, при снижении количества Т- и/или В-лимфоцитов, или иммунных нарушениях, например, ревматоидном артрите. Иммунодефицит также может быть результатом прохождения другого медицинского лечения.
[186] HSPC и/или модифицированные HSPC также можно применять для лечения апластической анемии, синдрома Чедиак-Хигаши, системной красной волчанки (СКВ), лейкемии, миелодиспластического синдрома, миелофиброза или тромбоцитопении. Тяжелая тромбоцитопения может быть результатом генетических дефектов, таких как анемия Фанкони, синдромов Вискотта-Олдрича или Мея-Хегглина. Приобретенная тромбоцитопения может быть связана с ауто- или алло-антителами, как в случае тромбоцитопенической пурпуры, системной красной волчанки, гемолитической анемии или несовместимости в системе мать-плод. Кроме того, к тромбоцитопении могут приводить спленомегалия, синдром диссеминированного внутрисосудистого свертывания, тромботическая тромбоцитопеническая пурпура, инфекция и/или искусственные клапаны сердца. Также тромбоцитопения может быть связана с поражение костного мозга карциномой, лимфомой, лейкемией или фиброзом.
[187] В конкретных вариантах реализации изобретения у субъекта наблюдается потеря крови вследствие, например, травмы, или субъект подвержен риску потери крови. В конкретных вариантах реализации изобретения у субъекта наблюдается истощение костного мозга, связанное, например, с наследственным, генетическим или приобретенным синдромом, характеризующимся потерей костного мозга или истощением костного мозга. В конкретных вариантах реализации изобретения субъект нуждается в гемопоэзе.
[188] Как было указано в отношении доноров костного мозга, введение HSPC или модифицированных HSPC субъекту может происходить в любое время на протяжении лечения, которое лечащий врач посчитает уместным. В качестве неограничивающих примеров, HSPC и/или модифицированные HSPC можно вводить субъекту, например, до, одновременно или после химиотерапии, лучевой терапии или пересадки костного мозга. HSPC и/или модифицированные HSPC могут эффективно обеспечивать приживление согласно анализу, проводимому через 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 дней (или более или менее чем 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 дней); 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 недель (или более или менее чем 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 недель); 1; 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 месяцев (или более или менее чем 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 месяцев); или 1, 2, 3, 4, 5 лет (или более или менее чем 1, 2, 3, 4, 5 лет) после введения субъекту HSPC и/или модифицированных HSPC. В конкретных вариантах реализации изобретения HSPC и/или модифицированные HSPC эффективно обеспечивают приживление согласно анализу, проводимому через 10 дней, 2 недели, 3 недели, 4 недели, 6 недель или 13 недель после введения субъекту HSPC и/или ХАР-HSPC.
[189] HSPC, модифицированные HSPC и модифицированные эффекторы. HSPC, модифицированные HSPC и модифицированные эффекторы можно вводить в разных целях в рамках схемы лечения. Применение HSPC и модифицированных HSPC, чтобы обеспечить нормальный состав крови, и модифицированных HSPC и модифицированных эффекторов, чтобы обеспечить эффект «трансплантат против лейкемии» при лечении ALL, описано выше. Подобные подходы можно применять для обеспечения нормального состава крови и нацеливания на нежелательные раковые клетки, а также в качестве вспомогательного лечения при химиотерапии или облучении.
[190] Типовые виды рака, которые можно лечить модифицированными HSPC и модифицированными эффекторами, включают рак надпочечников, рак мочевого пузыря, рак крови, рак костей, рак головного мозга, рак молочной железы, карциному, рак шейки матки, рак толстой кишки, колоректальный рак, рак тела матки, рак уха, носа и горла (ENT), рак эндометрия, рак пищевода, рак желудочно-кишечного тракта, рак головы и шеи, болезнь Ходжкина, рак кишечника, рак почек, рак гортани, лейкемию, рак печени, рак лимфатических узлов, лимфому, рак легких, меланому, мезотелиому, миелому, рак носоглотки, нейробластому, неходжкинскую лимфому, рак полости рта, рак яичников, рак поджелудочной железы, рак полового члена, рак глотки, рак простаты, рак прямой кишки, саркому, семиному, рак кожи, рак желудка, тератому, рак яичка, рак щитовидной железы, рак матки, вагинальный рак, сосудистые опухоли и их метастазы.
[191] В контексте рака терапевтически эффективное количество обладает противораковым эффектом. Противораковый эффект можно оценить, наблюдая снижение количества опухолевых клеток, снижение количества метастазов, уменьшение объема опухоли, увеличение прогнозируемой продолжительности жизни, индукцию апоптоза раковых клеток, индукцию гибели раковых клеток, подавление пролифиреции раковых клеток, подавление роста опухоли, предотвращение возникновения метастазов, увеличение продолжительности жизни субъекта и/или снижение вероятности рецидива или повторного возникновения рака после лечения.
[192] В контексте обеспечения нормального состава крови терапевтически эффективное количество лечит иммунодефицит, панцитопению, нейтропению и/или лейкопению, повышая количество необходимых клеток в системе кровообращения субъекта. Повышение количества необходимых клеток в системе кровообращения субъекта может восстанавливать популяцию иммунной системы субъекта, увеличивая количество клеток иммунной системы и/или клеток-предсшественников иммунной системы.
[193] В конкретных вариантах реализации изобретения, в которых применяют модифицированные HSPC и модифицированные эффекторы, можно проводить характеристику раковых клеток субъекта в отношении наличия клеточных маркеров. Связывающий домен, экспрессируемый модифицированными HSPC или модифицированными эффекторами, можно выбирать на основании характеристики клеточного маркера. В конкретных вариантах реализации изобретения ранее полученные модифицированные HSPC и модифицированные эффекторы выбирают для лечения субъекта на основании их способности связывать клеточный маркер, экспрессируемый преимущественно на конкретных раковых клетках субъекта.
[194] В случае лечения рака раскрытые композиции и составы также могут содержать плазмидную ДНК, несущую один или более противораковых генов, выбранных из p53, RB, BRCA1, E1A, bcl-2, MDR-1, p21, p16, bax, bcl-xs, E2F, IGF-I VEGF, ангиостатина, онкостатина, эндостатина, GM-CSF, IL-12, IL-2, IL-4, IL-7, IFN-γ, TNF-α и/или HSV-tk. Композиции и составы также могут содержать или водиться в комбинации с одним или более противоопухолевыми лекарственными препаратами, включая адриамицин, ангиостатин, азатиоприн, блеомицин, бусульфан, камптотецин, карбоплатин, кармустин, хлорамбуцил, хлорметамин, хлорхиноксалин сульфонамид, цисплатин, циклофосфамид, циклоплатам, цитарабин, дакарбазин, дактиномицин, даунорубицин, дидокс, доксорубицин, эндостатин, энлоплатин, эстрамустин, этопозид, эстрамустинфосфат, флуцитозин, фтордезоксиуридин, фторурацил, нитрат галлия, гидроксимочевину, идоксуридин, интерфероны, интерлейкины, лейпролид, лобаплатин, ломустин, манномустин, мехлоретамин, мехлорэтамин оксид, мелфалан, меркаптопурин, метотрексат, митрамицин, митобронитол, митомицин, микофеноловую кислоту, нокодазол, онкостатин, оксалиплатин, паклитаксел, пентамустин, комплекс платина-триамин, пликамицин, преднизолон, преднизон, прокарбазин, ингибиторы протеинкиназы С, пуромицин, семустин, ингибиторы сигнальной трансдукции, спироплатин, стрептозотоцин, ингибиторы стромелизина, таксол, тегафур, ингибиторы теломеразы, тенипозид, талидомид, тиамиприн, тиогуанин, тиотепу, тиамиприн, третамин, триазиквон, трифосфамид, ингибиторы тирозинкиназы, урамустин, видарабин, винбластин, алкалоиды барвинка, винкристин, виндезин, ворозол, зениплатин, цениплатин или циностатин.
[195] Модифицированные HSPC и модифицированные эффекторы. Модифицированные HSPC и/или модифицированные эффекторы можно применять без HSPC, когда лечение для обеспечения гемопоэтической функции или для лечения иммунодефицита; панцитопении; нейтропении и/или лейкопении не является нужным или необходимым.
[196] Как понятно специалисту в данной области техники, животные модели разных видов нарушений крови и рака хорошо известны и могут быть использованы для оценки эффективности конкретных подходов лечения, если это необходимо или целесообразно.
[197] Нижеприведенные Примеры и Типовые варианты реализации изобретения включены, чтобы продемонстрировать конкретные варианты реализации изобретения. Специалистам в данной области техники следует понимать, что в свете настоящего изобретения в конкретные раскрытые в данном документе варианты реализации можно вносить большое количество изменений и получать при этом сходный или аналогичный результат, не отступая от сущности и объема изобретения.
[198] Типовые варианты реализации изобретения.
1. Гемопоэтическая стволовая клетка-предшественник (HSPC) CD34+, генетически модифицированная, чтобы экспрессировать (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает CD19; (ii) внутриклеточный компонент, содержащий эффекторный домен, содержащий цитоплазматический домен CD28 или 4-1BB; (iii) спейсерную область, содержащую шарнирную область человеческого IgG4; и (iv) человеческий трансмембранный домен CD4 или CD28.
2. HSPC по варианту реализации изобретения 1, отличающаяся тем, что лигандсвязывающий домен представляет собой одноцепочечный фрагмент Fv (scFv), содержащий последовательность CDRL1 RASQDISKYLN (SEQ ID NO. 108), последовательность CDRL2 SRLHSGV (SEQ ID NO. 111), последовательность CDRL3 GNTLPYTFG (SEQ ID NO. 104), последовательность CDRH1 DYGVS (SEQ ID NO. 103), последовательность CDRH2 VIWGSETTYYNSALKS (SEQ ID NO. 114) и последовательность CDRH3 YAMDYWG (SEQ ID NO. 115).
3. HSPC по вариантам реализации изобретения 1 или 2, отличающаяся тем, что спейсерная область состоит из 12 аминокислот или менее.
4. HSPC по любому из вариантов реализации изобретения 1-3, отличающаяся тем, что спейсерная область содержит SEQ ID NO. 47.
5. Не-Т эффекторная клетка, генетически модифицированная, чтобы экспрессировать (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает CD19; (ii) внутриклеточный компонент, содержащий эффекторный домен, содержащий цитоплазматический домен CD28 или 4-1BB; (iii) спейсерную область, содержащую шарнирную область человеческого IgG4; и (iv) человеческий трансмембранный домен CD4 или CD28.
6. Не-Т эффекторная клетка по варианту реализации изобретения 5, отличающаяся тем, что лигандсвязывающий домен представляет собой одноцепочечный фрагмент Fv (scFv), содержащий последовательность CDRL1 RASQDISKYLN (SEQ ID NO. 108), последовательность CDRL2 SRLHSGV (SEQ ID NO. 111), последовательность CDRL3 GNTLPYTFG (SEQ ID NO. 104), последовательность CDRH1 DYGVS (SEQ ID NO. 103), последовательность CDRH2 VIWGSETTYYNSALKS (SEQ ID NO. 114) и последовательность CDRH3 YAMDYWG (SEQ ID NO. 115).
7. Не-Т эффекторная клетка по вариантам реализации изобретения 5 или 6, отличающаяся тем, что спейсерная область состоит из 12 аминокислот или менее.
8. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 5-7, отличающаяся тем, что спейсерная область содержит SEQ ID NO. 47.
9. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 5-8, отличающаяся тем, что не-Т эффекторная клетка является естественной клеткой-киллером.
10. Гемопоэтическая стволовая клетка-предшественник (HSPC), генетически модифицированная, чтобы экспрессировать химерный антигенный рецептор (ХАР) с SEQ ID NO. 34, 53, 54, 55, 56, 57 или 58.
11. HSPC по варианту реализации изобретения 10, отличающаяся тем, что HSPC представляет собой CD34+.
12. Не-Т эффекторная клетка, генетически модифицированная, чтобы экспрессировать ХАР с SEQ ID NO. 34, 53, 54, 55, 56, 57 или 58.
13. Не-Т эффекторная клетка по варианту реализации изобретения 12, отличающаяся тем, что не-Т эффекторная клетка является естественной клеткой-киллером.
14. HSPC, генетически модифицированная, чтобы экспрессировать (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает клеточный маркер, экспрессируемый преимущественно на нежелательной клетке; и (ii) внутриклеточный компонент, содержащий эффекторный домен.
15. HSPC по варианту реализации изобретения 14, отличающаяся тем, что лигандсвязывающий домен является фрагментом антитела.
16. HSPC по вариантам реализации изобретения 14 или 15, отличающаяся тем, что лигандсвязывающий домен представляет собой одноцепочечный вариабельный фрагмент антитела.
17. HSPC по любому из вариантов реализации изобретения 14-16, отличающаяся тем, что лигандсвязывающий домен связывает CD19.
18. HSPC по любому из вариантов реализации изобретения 14-17, отличающаяся тем, что лигандсвязывающий домен представляет собой scFv, содержащий последовательность CDRL1 RASQDISKYLN (SEQ ID NO. 108), последовательность CDRL2 SRLHSGV (SEQ ID NO. 111), последовательность CDRL3 GNTLPYTFG (SEQ ID NO. 104), последовательность CDRH1 DYGVS (SEQ ID NO. 103), последовательность CDRH2 VTWGSETTYYNSALKS (SEQ ID NO. 114) и последовательность CDRH3 YAMDYWG (SEQ ID NO. 115).
19. HSPC по варианту реализации изобретения 18, отличающаяся тем, что HSPC также генетически модифицирована, чтобы экспрессировать спейсерную область, состоящую из 12 аминокислот или менее.
20. HSPC по варианту реализации изобретения 19, отличающаяся тем, что спейсерная область содержит SEQ ID NO. 47.
21. HSPC по любому из вариантов реализации изобретения 14-16, отличающаяся тем, что лигандсвязывающий домен связывает ROR1.
22. HSPC по любому из вариантов реализации изобретения 14-16 или 21, отличающаяся тем, что лигандсвязывающий домен представляет собой scFv, содержащий последовательность CDRL1 ASGFDFSAYYM (SEQ ID NO. 101), последовательность CDRL2 TIYPSSG (SEQ ID NO. 112), последовательность CDRL3 ADRATYFCA (SEQ ID NO. 100), последовательность CDRH1 DTIDWY (SEQ ID NO. 102), последовательность CDRH2 VQSDGSYTKRPGVPDR (SEQ ID NO. 113) и последовательность CDRH3 YIGGYVFG (SEQ ID NO. 117).
23. HSPC по любому из вариантов реализации изобретения 14-16 или 21, отличающаяся тем, что лигандсвязывающий домен представляет собой scFv, содержащий последовательность CDRL1 SGSDINDYPIS (SEQ ID NO. 109), последовательность CDRL2 INSGGST (SEQ ID NO. 105), последовательность CDRL3 YFCARGYS (SEQ ID NO. 116), последовательность CDRH1 SNLAW (SEQ ID NO. 110), последовательность CDRH2 RASNLASGVPSRFSGS (SEQ ID NO. 107) и последовательность CDRH3 NVSYRTSF (SEQ ID NO. 106).
24. HSPC по варианту реализации изобретения 23, отличающаяся тем, что HSPC также генетически модифицирована, чтобы экспрессировать спейсерную область, состоящую из 229 аминокислот или менее.
25. HSPC по варианту реализации изобретения 24, отличающаяся тем, что спейсерная область содержит SEQ ID NO. 61.
26. HSPC по любому из вариантов реализации изобретения 14-16, отличающаяся тем, что лигандсвязывающий домен связывает PSMA, PSCA, мезотелин, CD20, WT1 или Her2.
27. HSPC по любому из вариантов реализации изобретения 14-26, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий один или более сигнальных и/или стимулирующих доменов, выбранных из: сигнальных и/или стимулирующих доменов 4-1BB, CARD11, CD3 γ, CD3δ, CD3ε, CD3ζ, CD27, CD28, CD79A, CD79B, DAP10, FcRα, FcRβ, FcRγ, Fyn, HVEM, ICOS, LAG3, LAT, Lck, LRP, NKG2D, NOTCH1, pTα, PTCH2, OX40, ROR2, Ryk, SLAMF1, Slp76, TCRα, TCRβ, TRIM, Wnt и Zap70.
28. HSPC по любому из вариантов реализации изобретения 14-27, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий внутриклеточный сигнальный домен CD3ζ, CD28ζ или 4-1BB.
29. HSPC по любому из вариантов реализации изобретения 14-28, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий один или более костимулирующих доменов, выбранных из: костимулирующих доменов CD27, CD28, 4-1BB, OX40, CD30, CD40, лимфоцитарного функционального антигена-1 (LFA-1), CD2, CD7, LIGHT, NKG2C или B7-H3.
30. HSPC по любому из вариантов реализации изобретения 14-29, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий внутриклеточный сигнальный домен, содержащий (i) весь или часть сигнального домена CD3ζ, (ii) весь или часть сигнального домена CD28, (iii) весь или часть сигнального домена 4-1BB или (iv) весь или часть сигнального домена CD3ζ, CD28 и/или 4-1BB.
31. HSPC по любому из вариантов реализации изобретения 14-30, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий вариант CD3ζ и/или часть внутриклеточного сигнального домена 4-1BB.
32. HSPC по любому из вариантов реализации изобретения 14-18, 21-23 или 26-31, отличающаяся тем, что HSPC также генетически модифицирована, чтобы экспрессировать спейсерную область.
33. HSPC по варианту реализации изобретения 32, отличающаяся тем, что спейсерная область содержит часть шарнирной области человеческого антитела.
34. HSPC по варианту реализации изобретения 32 или 33, отличающаяся тем, что спейсерная область содержит шарнирную область и по меньшей мере одну другую часть Fc-домена человеческого антитела, выбранного из CH1, CH2, CH3 или их комбинаций.
35. HSPC по варианту реализации изобретения 32 или 33, отличающаяся тем, что спейсерная область содержит Fc-домен и шарнирную область тяжелой цепи человеческого IgG4.
36. HSPC по варианту реализации изобретения 32, отличающаяся тем, что длина спейсерной области выбрана из 12 аминокислот или менее, 119 аминокислот или менее или 229 аминокислот или менее.
37. HSPC по варианту реализации изобретения 32, отличающаяся тем, что спейсерная область представляет собой SEQ ID NO.47, SEQ ID NO.52 или SEQ ID NO.61.
38. HSPC по любому из вариантов реализации изобретения 14-37, отличающаяся тем, что HSPC также генетически модифицирована, чтобы экспрессировать трансмембранный домен.
39. HSPC по варианту реализации изобретения 38, отличающаяся тем, что трансмембранный домен является трансмембранным доменом CD28 или трансмембранным доменом CD4.
40. HSPC по любому из вариантов реализации изобретения 14-39, отличающаяся тем, что внеклеточный компонент дополнительно содержит последовательность метки.
41. HSPC по варианту реализации изобретения 40, отличающаяся тем, что последовательность метки представляет собой EGFR, в котором отсутствует внутриклеточный сигнальный домен.
42. HSPC по любому из вариантов реализации изобретения 14-41, отличающаяся тем, что HSPC представляет собой CD34+.
43. Не-Т эффекторная клетка, генетически модифицированная, чтобы экспрессировать (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает клеточный маркер на нежелательной клетке; и (ii) внутриклеточный компонент, содержащий эффекторный домен.
44. Не-Т эффекторная клетка по варианту реализации изобретения 43, отличающаяся тем, что лигандсвязывающий домен является фрагментом антитела.
45. Не-Т эффекторная клетка по варианту реализации изобретения 43 или 44, отличающаяся тем, что лигандсвязывающий домен представляет собой одноцепочечный вариабельный фрагмент антитела.
46. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-45, отличающаяся тем, что лигандсвязывающий домен связывает CD19.
47. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-46, отличающаяся тем, что лигандсвязывающий домен представляет собой scFv, содержащий последовательность CDRL1 RASQDISKYLN (SEQ ID NO. 108), последовательность CDRL2 SRLHSGV (SEQ ID NO. 111), последовательность CDRL3 GNTLPYTFG (SEQ ID NO. 104), последовательность CDRH1 DYGVS (SEQ ID NO. 103), последовательность CDRH2 VIWGSETTYYNSALKS (SEQ ID NO. 114) и последовательность CDRH3 YAMDYWG (SEQ ID NO. 115).
48. Не-Т эффекторная клетка по варианту реализации изобретения 47, отличающаяся тем, что не-Т эффекторная клетка также генетически модифицирована, чтобы экспрессировать спейсерную область, состоящую из 12 аминокислот или менее.
49. Не-Т эффекторная клетка по варианту реализации изобретения 48, отличающаяся тем, что спейсерная область содержит SEQ ID NO. 47.
50. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-45, отличающаяся тем, что лигандсвязывающий домен связывает ROR1.
51. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-45 или 50, отличающаяся тем, что лигандсвязывающий домен представляет собой scFv, содержащий последовательность CDRL1 ASGFDFSAYYM (SEQ ID NO. 101), последовательность CDRL2 TIYPSSG (SEQ ID NO. 112), последовательность CDRL3 ADRATYFCA (SEQ ID NO. 100), последовательность CDRH1 DTIDWY (SEQ ID NO. 102), последовательность CDRH2 VQSDGSYTKRPGVPDR (SEQ ID NO. 113) и последовательность CDRH3 YIGGYVFG (SEQ ID NO. 117).
52. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-45 или 50, отличающаяся тем, что лигандсвязывающий домен представляет собой одноцепочечный фрагмент Fv (scFv), содержащий последовательность CDRL1 SGSDINDYPIS (SEQ ID NO. 109), последовательность CDRL2 INSGGST (SEQ ID NO. 105), последовательность CDRL3 YFCARGYS (SEQ ID NO. 116), последовательность CDRH1 SNLAW (SEQ ID NO. 110), последовательность CDRH2 RASNLASGVPSRFSGS (SEQ ID NO. 107) и последовательность CDRH3 NVSYRTSF (SEQ ID NO. 106).
53. Не-Т эффекторная клетка по варианту реализации изобретения 52, отличающаяся тем, что не-Т эффекторная клетка также генетически модифицирована, чтобы экспрессировать спейсерную область, состоящую из 229 аминокислот или менее.
54. Не-Т эффекторная клетка по варианту реализации изобретения 53, отличающаяся тем, что спейсерная область содержит SEQ ID NO. 61.
55. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-45, отличающаяся тем, что лигандсвязывающий домен связывает PSMA, PSCA, мезотелин, CD20, WT1 или Her2.
56. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-55, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий один или более сигнальных и/или стимулирующих доменов, выбранных из: сигнальных и/или стимулирующих доменов 4-1BB, CARD11, CD3γ, CD3δ, CD3ε, CD3ζ, CD27, CD28, CD79A, CD79B, DAP10, FcRα, FcRβ, FcRγ, Fyn, HVEM, ICOS, LAG3, LAT, Lck, LRP, NKG2D, NOTCH1, pTα, PTCH2, OX40, ROR2, Ryk, SLAMF1, Slp76, TCRα, TCRβ, TRIM, Wnt и Zap70.
57. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-56, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий внутриклеточный сигнальный домен CD3ζ, CD28ζ или 4-1BB.
58. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-57, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий один или более костимулирующих доменов, выбранных из: костимулирующих доменов CD27, CD28, 4-1BB, OX40, CD30, CD40, LFA-1, CD2, CD7, LIGHT, NKG2C или B7-H3.
59. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-58, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий внутриклеточный сигнальный домен, содержащий (i) весь или часть сигнального домена CD3ζ, (ii) весь или часть сигнального домена CD28, (iii) весь или часть сигнального домена 4-1BB или (iv) весь или часть сигнального домена CD3ζ, CD28 и/или 4-1BB.
60. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-59, отличающаяся тем, что внутриклеточный компонент содержит эффекторный домен, содержащий вариант CD3ζ и/или часть внутриклеточного сигнального домена 4-1BB.
61. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-47, 50-52 или 55-60, генетически модифицированная, чтобы экспрессировать спейсерную область.
62. Не-Т эффекторная клетка по варианту реализации изобретения 61, отличающаяся тем, что спейсерная область содержит часть шарнирной области человеческого антитела.
63. Не-Т эффекторная клетка по варианту реализации изобретения 61 или 62, отличающаяся тем, что спейсерная область содержит шарнирную область и по меньшей мере одну другую часть Fc-домена человеческого антитела, выбранного из CH1, CH2, CH3 или их комбинаций.
64. Не-Т эффекторная клетка по варианту реализации изобретения 61 или 62, отличающаяся тем, что спейсерная область содержит Fc-домен и шарнирную область тяжелой цепи человеческого IgG4.
65. Не-Т эффекторная клетка по варианту реализации изобретения 61, отличающаяся тем, что длина спейсерной области выбрана из 12 аминокислот или менее, 119 аминокислот или менее или 229 аминокислот или менее.
66. Не-Т эффекторная клетка по варианту реализации изобретения 61, отличающаяся тем, что спейсерная область представляет собой SEQ ID NO.47, SEQ ID NO.52 или SEQ ID NO.61.
67. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-66, отличающаяся тем, что не-Т эффекторная клетка также генетически модифицирована, чтобы экспрессировать трансмембранный домен.
68. Не-Т эффекторная клетка по варианту реализации изобретения 67, отличающаяся тем, что трансмембранный домен является трансмембранным доменом CD28 или трансмембранным доменом CD4.
69. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-68, отличающаяся тем, что внеклеточный компонент дополнительно содержит последовательность метки.
70. Не-Т эффекторная клетка по варианту реализации изобретения 69, отличающаяся тем, что последовательность метки представляет собой EGFR, в котором отсутствует внутриклеточный сигнальный домен.
71. Не-Т эффекторная клетка по любому из вариантов реализации изобретения 43-70, отличающаяся тем, что не-Т эффекторная клетка является естественной клеткой-киллером.
72. Композиция, содержащая генетически модифицированную HSPC по любому из вариантов реализации изобретения 1-4, 10, 11 или 14-42.
73. Композиция, содержащая не-Т эффекторную клетку по любому из вариантов реализации изобретения 5-9, 12, 13 или 43-71.
74. Композиция по варианту реализации изобретения 72 или 73, полученная для инфузии или инъекции.
75. Состав, содержащий HSPC и генетически модифицированную HSPC по любому из вариантов реализации изобретения 1-4, 10, 11 или 14-42.
76. Состав, содержащий HSPC и генетически модифицированную не-Т эффекторную клетку по любому из вариантов реализации изобретения 5-9, 12, 13 или 43-71.
77. Состав, содержащий генетически модифицированную HSPC по любому из вариантов реализации изобретения 1-4, 10, 11 или 14-42 и не-Т эффекторную клетку по любому из вариантов реализации изобретения 5-9, 12, 13 или 43-71.
78. Состав по варианту реализации изобретения 77, дополнительно содержащий HSPC.
79. Состав по любому из вариантов реализации изобретения 75-78, полученный для инфузии или инъекции.
80. Набор, содержащий композиции по любому из вариантов реализации изобретения 72-74, отличающийся тем, что набор содержит инструкции, указывающие, что композиции или составы можно вводить субъекту без проверки иммунологической совместимости.
81. Набор, содержащий составы по любому из вариантов реализации изобретения 75-79, отличающийся тем, что набор содержит инструкции, указывающие, что композиции или составы можно вводить субъекту без проверки иммунологической совместимости.
82. Набор, содержащий композиции по любому из вариантов реализации изобретения 72-74 и составы по любому из вариантов реализации изобретения 75-79, отличающийся тем, что набор содержит инструкции, указывающие, что композиции или составы можно вводить субъекту без проверки иммунологической совместимости.
83. Способ восстановления популяции иммунной системы нуждающегося в этом субъекта и нацеливания на нежелательные раковые клетки в организме субъекта, включающий введение терапевтически эффективного количества генетически модифицированных HSPC, при этом генетически модифицированные HSPC экспрессируют (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает клеточный маркер, который экспрессируется преимущественно на нежелательных раковых клетках, и (ii) внутриклеточный компонент, содержащий эффекторный домен, восстанавливая, таким образом, популяцию иммунной системы субъекта и осуществляя нацеливание на нежелательные раковые клетки.
84. Способ по варианту реализации изобретения 83, дополнительно включающий введение генетически модифицированных не-Т эффекторных клеток, при этом генетически модифицированные не-Т эффекторные клетки экспрессируют (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает клеточный маркер, который экспрессируется преимущественно на нежелательных раковых клетках, и (ii) внутриклеточный компонент, содержащий эффекторный домен.
85. Способ по варианту реализации изобретения 83 или 84, дополнительно включающий введение HSPC.
86. Способ по любому из вариантов реализации изобретения 83-85, отличающийся тем, что перед введением не требуется проверки иммунологической совместимости с субъектом.
87. Способ по любому из вариантов реализации изобретения 83-86, отличающийся тем, что клеточный маркер представляет собой CD19, ROR1, PSMA, PSCA, мезотелин, CD20, WT1 или Her2.
88. Способ по любому из вариантов реализации изобретения 83-87, отличающийся тем, что восстановление популяции необходимо на основании воздействия миелоаблативного режима при трансплантации гемопоэтических клеток (ТГК), а нежелательные раковые клетки представляют собой клетки острой лимфобластической лейкемии, экспрессирующие CD19.
89. Способ по любому из вариантов реализации изобретения 83-88, отличающийся тем, что субъект является педиатрическим пациентом с рецидивом острой лимфобластической лейкемии.
90. Способ нацеливания на нежелательные раковые клетки в организме субъекта, включающий выявление по меньшей мере одного клеточного маркера, экспрессируемого преимущественно на раковой клетке из организма субъекта; введение субъекту терапевтически эффективного количества генетически модифицированных не-Т эффекторных клеток, при этом генетически модифицированные не-Т эффекторные клетки экспрессируют (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает преимущественно экспрессируемый клеточный маркер, и (ii) внутриклеточный компонент, содержащий эффекторный домен.
91. Способ по варианту реализации изобретения 90, дополнительно включающий введение субъекту генетически модифицированных HSPC, при этом генетически модифицированные HSPC экспрессируют (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает преимущественно экспрессируемый клеточный маркер, и (ii) внутриклеточный компонент, содержащий эффекторный домен.
92. Способ нацеливания на нежелательные раковые клетки в организме субъекта, включающий выявление по меньшей мере одного клеточного маркера, экспрессируемого преимущественно на раковой клетке из организма субъекта; введение субъекту генетически модифицированных HSPC, при этом генетически модифицированные HSPC экспрессируют (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает преимущественно экспрессируемый клеточный маркер, и (ii) внутриклеточный компонент, содержащий эффекторный домен.
93. Способ по любому из вариантов реализации изобретения 90-92, дополнительно включающий лечение иммунодефицита, панцитопении, нейтропении и/или лейкопении у субъекта путем введения субъекту терапевтически эффективного количества HSPC.
94. Способ по варианту реализации изобретения 93, отличающийся тем, что иммунодефицит, панцитопения, нейтропения и/или лейкопения является результатом химиотерапии, лучевой терапии и/или миелоаблативного режима при ТГК.
95. Способ по любому из вариантов реализации изобретения 90-94, отличающийся тем, что клеточный маркер представляет собой CD19, ROR1, PSMA, PSCA, мезотелин, CD20, WT1 или Her2.
96. Способ по любому из вариантов реализации изобретения 90-95, отличающийся тем, что перед введением не требуется проверки иммунологической совместимости с субъектом.
97. Способ по любому из вариантов реализации изобретения 90-96, отличающийся тем, что нежелательные раковые клетки представляют собой клетки острой лимфобластической лейкемии, экспрессирующие CD19.
98. Способ по любому из вариантов реализации изобретения 90-97, отличающийся тем, что субъект является педиатрическим пациентом с рецидивом острой лимфобластической лейкемии.
99. Способ восстановления популяции иммунной системы нуждающегося в этом субъекта, включающий введение субъекту терапевтически эффективного количества HSPC и/или генетически модифицированных HSPC, восстанавливая, таким образом, популяцию иммунной системы субъекта.
100. Способ по варианту реализации изобретения 99, отличающийся тем, что восстановление популяции необходимо на основании наличия одного или более из иммунодефицита, панцитопении, нейтропении или лейкопении.
101. Способ по варианту реализации изобретения 99 или 100, отличающийся тем, что восстановление популяции необходимо на основании наличия одного или более из вирусной инфекции, микробной инфекции, паразитарной инфекции, болезни почек и/или почечной недостаточности.
102. Способ по любому из вариантов реализации изобретения 99-101, отличающийся тем, что восстановление популяции необходимо на основании воздействия режима химиотерапии, миелоаблативного режима при ТГК и/или острого ионизирующего излучения.
103. Способ по любому из вариантов реализации изобретения 99-102, отличающийся тем, что восстановление популяции необходимо на основании воздействия лекарственных препаратов, которые приводят к супрессии костного мозга или гемопоэтическому дефициту.
104. Способ по любому из вариантов реализации изобретения 99-103, отличающийся тем, что восстановление популяции необходимо на основании воздействия пенициллина, ганцикловира, дауномицина, мепробамата, аминопирина, дипирона, фенитоина, карбамазепина, пропилтиоурацила и/или метимазола.
105. Способ по любому из вариантов реализации изобретения 99-104, отличающийся тем, что восстановление популяции необходимо на основании воздействия диализа.
106. Способ по любому из вариантов реализации изобретения 99-105, дополнительно включающий нацеливание на нежелательные раковые клетки в организме субъекта путем введения генетически модифицированных HSPC и/или генетически модифицированных не-Т эффекторных клеток, при этом генетически модифицированные HSPC и/или генетически модифицированные не-Т эффекторные клетки экспрессируют (i) внеклеточный компонент, содержащий лигандсвязывающий домен, который связывает клеточный маркер, который, как известно, экспрессируется преимущественно на раковых клетках в организме субъекта, и (ii) внутриклеточный компонент, содержащий эффекторный домен.
107. Способ по варианту реализации изобретения 106, отличающийся тем, что раковые клетки принадлежат раку надпочечников, раку мочевого пузыря, раку крови, раку костей, раку головного мозга, раку молочной железы, карциноме, раку шейки матки, раку толстой кишки, колоректальному раку, раку тела матки, раку уха, носа и горла (ENT), раку эндометрия, раку пищевода, раку желудочно-кишечного тракта, раку головы и шеи, болезни Ходжкина, раку кишечника, раку почек, раку гортани, лейкемии, раку печени, раку лимфатических узлов, лимфоме, раку легких, меланоме, мезотелиоме, миеломе, раку носоглотки, нейробластоме, неходжкинской лимфоме, раку полости рта, раку яичников, раку поджелудочной железы, раку полового члена, раку глотки, раку простаты, раку прямой кишки, саркоме, семиноме, раку кожи, раку желудка, тератоме, раку яичка, раку щитовидной железы, раку матки, вагинальному раку, сосудистой опухоли и их метастазам.
108. Способ по варианту реализации изобретения 106 или 107, отличающийся тем, что клеточный(е) маркер(ы) выбран(ы) из A33; BAGE; Bcl-2; β-катенина; B7H4; BTLA; CA125; CA19-9; CD5; CD19; CD20; CD21; CD22; CD33; CD37; CD44v6; CD45; CD123; CEA; CEACAM6; c-Met; CS-1; циклина B1; DAGE; EBNA; EGFR; эфрина B2; ErbB2; ErbB3; ErbB4; EphA2; рецептора эстрогена; FAP; ферритина; α-фетопротеина (AFP); FLT1; FLT4; фолатсвязывающего белка; Frizzled; GAGE; G250; GD-2; GHRHR; GHR; GM2; gp75; gp100 (Pmel 17); gp130; HLA; HER-2/neu; HPV E6; HPV E7; hTERT; HVEM; IGF1R; IL6R; KDR; Ki-67; LIFRβ; LRP; LRP5; LTβR; мезотелина; OSMRβ; p53; PD1; PD-L1; PD-L2; PRAME; рецептора прогестерона; PSA; PSMA; PTCH1; MAGE; MART; мезотелина; MUC; MUC1; MUM-1-B; myc; NYESO-1; RANK; ras; Robo1; RORl; сурвивина; TCRα; TCRβ; тенасцина; TGFBR1; TGFBR2;TLR7; TLR9; TNFR1; TNFR2; TNFRSF4; TWEAK-R; TSTA тирозиназы; VEGF; и WT1.
109. Способ по любому из вариантов реализации изобретения 106-108, отличающийся тем, что рак представляет собой лейкемию/лимфому, а клеточный(е) маркер(ы) представляет(ют) собой один или более из CD19, CD20, CD22, ROR1, CD33 и WT-1; отличающийся тем, что рак представляет собой множественную миелому, а клеточный маркер представляет собой BCMA; отличающийся тем, что рак представляет собой рак простаты, а клеточный(е) маркер(ы) представляет(ют) собой один или более из PSMA, WT1, PSCA и SV40 T; отличающийся тем, что рак представляет собой рак молочной железы, а клеточный(е) маркер(ы) представляет(ют) собой один или более из HER2, ERBB2 и ROR1; что рак представляет собой рак стволовых клеток, а клеточный маркер представляет собой CD133; отличающийся тем, что рак представляет собой рак яичников, а клеточный(е) маркер(ы) представляет(ют) собой один или более из L1-CAM, MUC-CD, фолатного рецептора, Lewis Y, ROR1, мезотелина и WT-1; отличающийся тем, что рак представляет собой мезотелиому, а клеточный маркер представляет собой мезотелин; отличающийся тем, что рак представляет собой почечно-клеточную карциному, а клеточный маркер представляет собой CAIX; отличающийся тем, что рак представляет собой меланому, а клеточный маркер представляет собой GD2; отличающийся тем, что рак представляет собой рак поджелудочной железы, а клеточный(е) маркер(ы) представляет(ют) собой один или более из мезотелина, CEA, CD24 и ROR1; или отличающийся тем, что рак представляет собой рак легких, а клеточный маркер представляет собой ROR1.
110. Способ по любому из вариантов реализации изобретения 106-109, отличающийся тем, что рак представляет собой острую лимфобластическую лейкемию, а пациент является педиатрическим пациентом.
111. Способ по любому из вариантов реализации изобретения 106-110, отличающийся тем, что перед введением не требуется проверка иммунологической совместимости с субъектом.
112. Способ нацеливания на клетки, преимущественно экспрессирующие CD19, для их разрушения, включающий введение нуждающемуся в этом субъекту терапевтически эффективного количества генетически модифицированных HSPC и/или генетически модифицированных не-Т эффекторных клеток, при этом генетически модифицированные клетки экспрессируют (i) внеклеточный компонент, содержащий CD19-лигандсвязывающий домен, и (ii) внутриклеточный компонент, содержащий эффекторный домен, осуществляя, таким образом, нацеливание и разрушение клеток, преимущественно экспрессирующих CD19.
113. Способ по варианту реализации изобретения 112, дополнительно включающий лечение иммунодефицита, панцитопении, нейтропении и/или лейкопении у субъекта путем введения субъекту терапевтически эффективного количества HSPC.
114. Способ по варианту реализации изобретения 113, отличающаяся тем, что отличающийся тем, что иммунодефицит, панцитопения, нейтропения и/или лейкопения является результатом химиотерапии, лучевой терапии и/или миелоаблативного режима при ТГК.
115. Способ по любому из вариантов реализации изобретения 112-114, отличающийся тем, что перед введением не требуется проверки иммунологической совместимости с субъектом.
116. Способ по любому из вариантов реализации изобретения 112-115, отличающийся тем, что клетки, преимущественно экспрессирующие CD19, представляют собой клетки острой лимфобластической лейкемии.
117. Способ по любому из вариантов реализации изобретения 112-116, отличающийся тем, что субъект является педиатрическим пациентом с рецидивом острой лимфобластической лейкемии.
[199] Пример 1. Осуществляли конструирование и цГМФ-получение двух лентивирусных векторов третьего поколения для координированной экспрессии CD19-ХАР и селекционной/суицидной конструкции huEGFRt. В обоих случаях в условиях цГМФ разрабатывали псевдотипированный СИН (самоинактивирующийся) лентивирусный вектор на основе вируса везикулярного стоматита G (VSV-G), который кодирует специфический в отношении CD19 ХАР и huEGFRt, который представляет собой усеченный человеческий белок EGFR, который не содержит внутриклеточный сигнальный домен. Специфический в отношении CD19 ХАР scFvFc-CD3ζCD28 и вектор huEGFRt содержит гибридный 5' ДКП, в котором область U3 замещена промотором ЦМВ, и 3' ДКП, в котором цис-действующие регуляторные последовательности полностью удалены из области U3. В результате как 5', так и 3' ДКП инактивируются при выработке и интеграции провируса в хромосому. CD19-ХАР содержит человеческую лидерную последовательность GMCSFRα-цепи, VL- и VH-последовательности, полученные из CD19-специфического мышиного IgG1mAb (FMC63), Fc и шарнирные области тяжелой цепи человеческого IgG4, трансмембранную область человеческого CD28 и цитоплазматический домен CD3ζ и CD28. Эту конструкцию клонировали в модифицированный pHIV7, в котором промотор ЦМВ был заменен человеческим альфа-промотором EF-1 (Фигура 29A). Вектор позволяет получить приблизительно 1:1 экспрессию CD19-ХАР и huEGFRt посредством применения элемента T2A. Второй - CD19-специфический фрагмент ХАР scFv-4-1BB/CD3ζ - кодирует N-концевой лидерный пептид сигнальной последовательности альфа-цепи рецептора человеческого GMCSF для управления поверхностной экспрессией, CD19-специфический scFv, полученный из мышиного моноклонального антитела IgG1 (FMC63), человеческую шарнирную область IgG4 и человеческую трансмембранную область CD28 и костимулирующий элемент 4-1BB с цитоплазматическим хвостом человеческого CD3ζ (Фигура 29B). Это вектор также позволяет получить приблизительно 1:1 экспрессию CD19-ХАР и huEGFRt посредством применения элемента T2A.
[200] Экспрессия huEGFRt обеспечивает наличие второго маркера клеточной поверхности, который облегчает измерение эффективности трансдукции. Биотинилированный эрбитукс связывается с huEGFRt, экспрессируемым на клеточной поверхности, и его можно метить флуорохромом для анализа методом проточной цитометрии. Кроме того, его можно использовать в качестве суицидного гена в клинических условиях при лечении эрбитуксом. Также генерировали сходный вектор с уЗФБ вместо ХАР.
[201] Пример 2. Notch-опосредуемое ex vivo размножение HSPC ПК является клинически утвержденным продуктом клеточной терапии, который хорошо переносится, может употребляться в виде готового продукта без проверки HLA-совместимости и обеспечивает временное миелоидное приживление как при ТГК, так и при интенсивной химиотерапии. Готовые к применению размноженные единицы инфузировали >85 субъектам, при этом не наблюдали серьезных побочных явлений за исключением одной аллергической реакции, связанной с ДМСО. Кроме того, через 180 дней при ТГК и 14 дней после инфузии при химиотерапии устойчивое приживление не наблюдалось.
[202] Способы. Единицы пуповинной крови/плацентарной крови получали от людей при рождении. Собранную кровь смешивали с антикоагулянтом для предотвращения свертывания и хранили. Перед планируемой инициацией размножаемых культур емкости для тканевого культивирования сначала покрывали на протяжении ночи при 4 °C или на протяжении минимум 2 часов при 37 °C Delta1ext-lgG в концентрации 2,5 мкг/мл и ретронектином® (фрагментом рекомбинантного человеческого фибронектина) (Clontech Laboratories, Inc., Madison, Wl) в концентрации 5 мкг/мл в фосфатно-солевом буферном растворе (PBS). Затем пробирки промывали PBS и блокировали PBS-2% человеческим сывороточным альбумином (ЧСА). Для единицы свежей пуповинной крови проводили лизис эритроцитов и обрабатывали для отбора клеток CD34+ при помощи системы для разделения клеток autoMACS® (Miltenyi Biotec GmbH, Gladbach, Germany). После обогащения процентное содержание клеток CD34+ в образце возрастало по сравнению с процентным содержанием клеток CD34+ в образце до обогащения. Обогащенную фракцию клеток CD34+ пересуспендировали в конечной культуральной среде, которая состоит из бессывороточной среды для размножения STEMSPAN™ (StemCell Technologies, Vancouver, British Columbia), дополненной rhIL-3 (10 нг/мл), rhIL-6 (50 нг/мл), rhTPO (50 нг/мл), rhFlt-3L (50 нг/мл), rhSCF (50 нг/мл).
[203] СИН лентивирусный вектор, который управляет коэкспрессией CD19-специфического химерного антигенного рецептора scFvFc:CD28:ζ и селекционной суицидной конструкции huEGFRt, трансдуцировали в Notch-размноженные стволовые клетки ПК на 3 или 4 день путем центрифугирования при 800 × g на протяжении 45 минут при 32 °C с лентивирусным супернатантом (MOI 3) и 4 мкг/мл протамина сульфата. В альтернативном варианте СИН лентивирусный вектор кодировал костимуляцию 4-1BB (смотрите Краткое описание графических материалов). Ввиду экспрессии ХАР на HSPC с высокой сигнальной способностью, на 7 день культивирования добавляли облученные ЛКЛ в соотношении 1:1, чтобы обеспечить антигенную стимуляцию.
[204] В конце культивирования с целью размножения NK-клетки и нейтрофилы все еще являются незрелыми. Чтобы полностью оценить лизирующую способность, были разработаны способы культивирования для повышения степени созревания. В случае NK-клеток культуру пересевали в среду RPMI, дополненную человеческой сывороткой, IL-2 в концентрации 50 Е/мл и IL-15 в концентрации 500 нг/мл, или среду RPMI, дополненную человеческой сывороткой, L-глутамином, IL-2 в концентрации 50 Е/мл и IL-15 в концентрации 500 нг/мл, на протяжении дополнительной недели культивирования.
[205] Чтобы оценить приживление размноженных клеток ПК использовали модель нулевых мышей (NOG) NOD/SCID IL2R. После прохождения сублетального облучения в организме мышей возможно надежное приживление размноженных клеток ПК. Чтобы посмотреть на трансплантат с трансдуцированными размноженными клетками ПК, мышей NOG облучали дозой 325cGy при помощи линейного ускорителя и инфузировали им путем инъекции в хвостовую вену потомство, полученное от 10000-30000 клеток CD34+ ПК, культивируемых на Delta-1ext-IgG.
[206] Результаты. Эффективность трансдукции соответствовала диапазону от 10 до > 50% и в общем случае наблюдали одинаковую трансдукцию для клеток CD34+ и CD34-. Анализ числа копий продемонстрировал наличие 1-4 копий/клетку согласно данным валидированного анализа методом количественной ПЦР в режиме реального времени, что соответствует требованиям FDA к клеточным продуктам для клинической генной терапии.
[207] Клетки CD34+ ПК, культивируемые с лигандом Notch, содержат различные типы клеток, которые можно определить на основе иммунофенотипирования. Культуры, трансдуцированные лентивирусом CD19-ХАР, сравнивали с нетрансдуцированной культурой, полученной из той же самой единицы пуповинной крови, при этом в отношении конечного иммунофенотипирования во время сбора или общего роста клеток в культуре, содержащей кратно размноженные CD34 и кратно размноженные TNC, не наблюдали существенной разницы.
[208] Экспрессия трансгена не влияла на конечный фенотип культуры через 14 дней, при этом экспрессия трансгена наблюдалась во всех подгруппах клеток и оказалась относительно стабильной на протяжении периода культивирования.
[209] Проводили дополнительные эксперименты, подвергая культуры клеток воздействию CD19+ ЛКЛ, чтобы определить, оказывает ли воздействие антигена неблагоприятный эффект на культуру. Добавление в культуру облученных ЛКЛ на 7 день в соотношении 1:1 не оказывало неблагоприятный эффект и де-факто повышало рост и жизнеспособность как трансдуцированных, так и нетрансдуцированных клеткок. ЛКЛ не увеличивали популяцию ХАР+, что дает возможность предположить, что антиген не повышает пролиферацию ХАР, экспрессирующих незрелые клетки. Кроме того, во всех фенотипических подгруппах клеток конечного продукта трансген был выявлен на одинаковом уровне. Графическое изображение результатов приведено на ФИГ.. 30A, 30B, 31, 32 и 33.
[210] Перенос эффекторной функции после взаимодействия с CD19 посредством экспрессии CD19-ХАР является важным для окончательной противораковой (например, противолейкозной) активности модифицированных клеток HSPC ПК. Дифференцировочные культуральные условия приводили к увеличению количества NK-клеток (ФИГ. 34). Проводили сортировку фракции клеток CD56+, которые использовали в ХАР с клетками-мишенями K562 и ЛКЛ. Как и ожидалось, как нетрансдуцированные, так и трансдуцированные клетки были способны уничтожать K562 и, хотя оба вида клеток также уничтожали ЛКЛ, лизис ЛКЛ был значительно усилен посредством экспрессии ХАР. В частности, экспрессирующие CD19-ХАР NK-клетки имели повышенную цитотоксическую активность по сравнению с нетрансдуцированными NK-клетками (50 против 30%), при этом оба вида клеток одинаково уничтожали K562-мишени (75 против 80%). Смотрите ФИГ. 35.
[211] Трансплантация размноженных клеток ПК модели NOG приводила к росту большой популяции клеток CD19+, начиная приблизительно с 4-5 недели после трансплантации. Никакого влияния на ранее приживление трансдуцированных клеток не наблюдали, при этом наблюдали значительное снижение приживления CD19 у мышей, которым трансплантировали экспрессирующие CD19-ХАР клетки, по сравнению с нетрансдуцированными клетками, при котором популяция CD19 составляла >20% приживленных клеток, что свидетельствует о анти-CD19 активности. Популяции NK-клеток увеличивали, используя клетки, секретирующие NS0-IL15, облученные и инъецируемые подкожно три раза в неделю, начиная с 3 недели, чтобы обеспечить усиленную эффекторную функцию. Этот эффект увеличивает количество клеток CD56+ in vivo. Смотрите ФИГ. 36 и 37.
[212] Данные показывают, что трансдукция размноженных клеток ПК во время культивирования в присутствии иммобилизованного Delta1ext-IgG для экспрессии CD19-специфического ХАР не оказывала заметного влияния на качественные и количественные показатели размножения, а также на его репопулирующую способность в мышиной модели. Эти результаты являются перспективными как способ обеспечения эффекта «трансплантат против рака (например, лейкемии)» для трансплантата пуповинной крови. Кроме того, трансдукция CD19-ХАР в размноженные при помощи универсального донора HSPC ПК позволяет незамедлительно (например, когда перед введением не нужна проверка иммунологической совместимости) проводить инфузию анти-CD19 клеточного продукта после выявления субъекта с клиническими показаниями к терапии, например, субъекта с рецидивом или персистентным МОЗ. Была продемонстрирована надежная трансдукция клеток CD34+ пуповинной крови, размноженных с лигандом Notch, не влияющая ни на культуру в целом, ни на in vivo способность приживления, и в то же время обеспечивающая анти-CD19 активность. Так как размноженные клетки пуповинной крови уже применяли в клинических условиях как готовую клеточную терапию без HLA-совместимости, описанные примеры демонстрируют дополнительное применение в качестве готовой клеточной терапии, которая дает возможность пациентам проходить иммунотерапию даже в случае невозможности получить и сконструировать аутологичный Т-клеточный продукт.
[213] Как указано, при практической реализации настоящего изобретения можно применять, если не указано иное, традиционные методы вирусологии, микробиологии, молекулярной биологии и технологии рекомбинантных ДНК, известные в данной области техники. Такие методы полностью описаны в литературе; смотрите, например, Sambrook, et al. Molecular Cloning: A Laboratory Manual (Current Edition); DNA Cloning: A Practical Approach, vol. I & II (D. Glover, ed.); Oligonucleotide Synthesis (N. Gait, ed., Current Edition); Nucleic Acid Hybridization (B. Hames & S. Higgins, eds., Current Edition); Transcription and Translation (B. Hames & S. Higgins, eds., Current Edition); CRC Handbook of Parvoviruses, vol. I & II (P. Tijessen, ed.); Fundamental Virology, 2nd Edition, vol. I & II (B. N. Fields and D. M. Knipe, eds.), каждая из которых включена в данный документ посредством ссылки в отношении приведенного в них материала.
[214] Как понятно специалисту в данной области техники, каждый раскрытый в данном документе вариант реализации изобретения может включать, преимущественно состоять или состоять из конкретных указанных элементов, этапов, ингридиентов или компонентов. «Включает» или «включающий» означает «содержит, состоит преимущественно или состоит из». Переходный термин «содержат» или «содержит» включает, но не ограничивается этим, и предполагает возможность включения неуказанных элементов, этапов, ингредиентов или компонентов даже в больших количествах. Переходная фраза «состоящий из» исключает любой не указанный элемент, этап, ингредиент или компонент. Переходная фраза «состоящий преимущественно из» ограничивает объем варианта реализации изобретения указанными элементами, этапами, ингредиентами или компонентами и теми, которые не оказывают материального влияния на вариант реализации изобретения. Материальное влияние приводит к (i) статистически значимому снижению эффективности введения клеток для создания противоракового эффекта в организме субъекта и/или (ii) статистически значимому снижению эффективности введения клеток для восстановления популяции иммунной системы субъекта.
[215] Если не указано иное, все числовые значения, выражающие количества ингредиентов, свойства, такие как молекулярная масса, реакционные условия и так далее, применяемые в описании и формуле изобретения, следует понимать как такие, которые во всех возможных случаях могут изменяться при помощи термина «около». Соответственно, если не указано противоположное, приведенные в описании и прилагаемой формуле изобретения числовые параметры, являются приблизительными величинами, которые могут варьироваться в зависимости от необходимых свойств, которые требуется получить посредством настоящего изобретения. Без ограничения доктрины эквивалентов объемом формулы изобретения, каждый числовой параметр должен быть представлен по меньшей мере в соответствии с указанным количеством значащих цифр с применением обычного способа округления. Там, где необходимо дополнительное уточнение, термин «около» имеет значение, которое обычно подразумевается специалистом в данной области техники при применении в сочетании с указанным числовым значением или диапазоном, т.е. обозначающее несколько больше или несколько меньше указанного значения или диапазона до диапазона, составляющего ±20% от указанного значения; ±19% от указанного значения; ±18% от указанного значения; ±17% от указанного значения; ±16% от указанного значения; ±15% от указанного значения; ±14% от указанного значения; ±13% от указанного значения; ±12% от указанного значения; ±11% от указанного значения; ±10% от указанного значения; ±9% от указанного значения; ±8% от указанного значения; ±7% от указанного значения; ±6% от указанного значения; ±5% от указанного значения; ±4% от указанного значения; ±3% от указанного значения; ±2% от указанного значения; или ±1% от указанного значения.
[216] Несмотря на то, что числовые диапазоны и параметры, составляющие полный объем изобретения, являются приблизительными, числовые значения, приведенные в конкретных примерах, поданы настолько точно, насколько это возможно. При этом любые числовые значения по определению содержат некоторые погрешности как результат среднеквадратического отклонения, возникающего при проведении соответствующих измерений.
[217] Следует учитывать, что форма единственного числа, применяемая в контексте описания изобретения (в особенности в контексте нижеприведенной формулы изобретения), включает как форму единственного, так и множественного числа, если в данном документе не указано иное или если это не противоречит контексту. Приводимое в данном документе перечисление диапазонов значений служит просто сокращенным способом ссылки на каждое отдельное значение, попадающее в этот диапазон. Если в данном документе не указано иное, каждое отдельное значение включено в описание изобретения, как если бы оно было приведено в данном документе отдельно. Все описанные в данном документе способы можно осуществлять в любом подходящем порядке, если в данном документе не указано иное или если это не противоречит контексту. Предложенное в данном документе применение любого или всех примеров или иллюстративные выражения (например, «такой как») предназначены исключительно для лучшего освещения изобретения и не ограничивают объем заявляемого изобретения. Никакие выражения в описании изобретения не следует воспринимать как такие, которые указывают на незаявленный элемент, существенный для практической реализации изобретения.
[218] Группы альтернативных элементов или вариантов реализации изобретения, раскрытые в данном документе, не следует воспринимать как ограничения. Каждый представитель группы может быть указан или заявлен отдельно или в комбинации с другими представителями группы или другими элементами, приведенными в данном документе. Предусматривается, что один или более представителей группы могут быть включены в группу или удалены из группы в целях удобства и/или патентоспособности. В случае если происходит такое включение или удаление, считается, что описание изобретения содержит группу с учетом изменений, таким образом, удовлетворяя описанию всех групп Маркуша, применяемых в прилагаемой формуле изобретения.
[219] В данном документе описаны конкретные варианты реализации изобретения, включая наилучший способ осуществления изобретения, известный авторам изобретения. Конечно, после прочтения вышеприведенного описания специалистам в данной области техники станут очевидны вариации этих описанных вариантов реализации изобретения. Авторы изобретения рассчитывают, что специалисты будут применять такие вариации надлежащим образом, и полагают, что практическая реализация данного изобретения может быть осуществлена способом, отличным от того, который конкретно описан в данном документе. Соответственно, данное изобретение включает все предусмотренные действующим законом изменения и эквиваленты объекта изобретения, приведенного в прилагаемой формуле изобретения. Кроме того, любая комбинация вышеописанных элементов во всех возможных вариациях включена в изобретение, если в данном документе не указано иное или если это не противоречит контексту.
[220] Кроме того, в тексте данного описания приведены многочисленные ссылки на книги, журнальные статьи, трактаты, патенты, печатные публикации и т.д. (вместе называемые «ссылками»). Каждая из вышеперечисленных ссылок отдельно включена посредством ссылки в отношении изложенного в них материала.
[221] В завершении, следует понимать, что раскрытые в данном документе варианты реализации изобретения иллюстрируют принципы настоящего изобретения. Другие модификации, которые можно применять, входят в объем данного изобретения. Таким образом, в качестве примера, но не ограничения, в соответствии с изложенным в данном документе материалом можно применять альтернативные формы настоящего изобретения. Соответственно, настоящее изобретение не ограничено тем, что конкретно показано и описано.
[222] Показанные в данном документе конкретные варианты приведены исключительно в качестве примера и в целях иллюстративного обсуждения предпочтительных вариантов реализации настоящего изобретения и представлены, чтобы предоставить материал, который, как считается, является наиболее необходимым и понятным описанием принципов и концептуальных аспектов различных вариантов реализации изобретения. В связи с этим не было предпринято попыток привести структурные детали изобретения более подробно, чем это необходимо для понимания принципов изобретения, описания вместе с графическими материалами и/или примеров, которых специалистам в данной области техники достаточно для понимания того, как осуществить на практике некоторые формы изобретения.
[223] Подразумевается, что используемые в данном документе определения и объяснения имеют преимущественную силу в любом будущем толковании, если они четко и однозначно не изменены в следующих примерах или если используемое значение не делает какое-либо толкование бессмысленным или практически бессмысленным. В случаях, когда толкование термина делает его бессмысленным или практически бессмысленным, следует пользоваться определением, взятым из Webster's Dictionary, 3rd Edition или другого известного специалистам в данной области техники словаря, такого как Oxford Dictionary of Biochemistry and Molecular Biology (Ed. Anthony Smith, Oxford University Press, Oxford, 2004).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Fred Hutchinson Cancer Research Center
Seattle Children's Hospital, D/B/A Seattle Children's
Research Institute
<120> МОДИФИЦИРОВАННЫЕ ГЕМОПОЭТИЧЕСКИЕ СТВОЛОВЫЕ КЛЕТКИ/КЛЕТКИ-ПРЕДШЕСТВЕННИКИ И НЕ-Т ЭФФЕКТОРНЫЕ КЛЕТКИ И ИХ ПРИМЕНЕНИЕ
<130> F053-0012PCT
<150> 61/898387
<151> 2013-10-31
<160> 117
<170> PatentIn версия 3.5
<210> 1
<211> 126
<212> ДНК
<213> Homo sapiens
<400> 1
aaacggggca gaaagaaact cctgtatata ttcaaacaac catttatgag accagtacaa 60
actactcaag aggaagatgg ctgtagctgc cgatttccag aagaagaaga aggaggatgt 120
gaactg 126
<210> 2
<211> 135
<212> ДНК
<213> Homo sapiens
<400> 2
aaacggggca gaaagaaact cctgtatata ttcaaacaac catttatgag accagtacaa 60
actactcaag aggaagatgg ctgtagctgc cgatttccag aagaagaaga aggaggatgt 120
gaactgcggg tgaag 135
<210> 3
<211> 22
<212> Белок
<213> Homo sapiens
<400> 3
Met Ala Leu Ile Val Leu Gly Gly Val Ala Gly Leu Leu Leu Phe Ile
1 5 10 15
Gly Leu Gly Ile Phe Phe
20
<210> 4
<211> 45
<212> Белок
<213> Homo sapiens
<400> 4
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys
35 40 45
<210> 5
<211> 210
<212> ДНК
<213> Homo sapiens
<400> 5
atgttctggg tgctggtggt ggtgggcggg gtgctggcct gctacagcct gctggtgaca 60
gtggccttca tcatcttttg ggtgaaacgg ggcagaaaga aactcctgta tatattcaaa 120
caaccattta tgagaccagt acaaactact caagaggaag atggctgtag ctgccgattt 180
ccagaagaag aagaaggagg atgtgaactg 210
<210> 6
<211> 42
<212> Белок
<213> Homo sapiens
<400> 6
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 7
<211> 556
<212> Белок
<213> Homo sapiens
<400> 7
Met Pro Pro Pro Arg Leu Leu Phe Phe Leu Leu Phe Leu Thr Pro Met
1 5 10 15
Glu Val Arg Pro Glu Glu Pro Leu Val Val Lys Val Glu Glu Gly Asp
20 25 30
Asn Ala Val Leu Gln Cys Leu Lys Gly Thr Ser Asp Gly Pro Thr Gln
35 40 45
Gln Leu Thr Trp Ser Arg Glu Ser Pro Leu Lys Pro Phe Leu Lys Leu
50 55 60
Ser Leu Gly Leu Pro Gly Leu Gly Ile His Met Arg Pro Leu Ala Ser
65 70 75 80
Trp Leu Phe Ile Phe Asn Val Ser Gln Gln Met Gly Gly Phe Tyr Leu
85 90 95
Cys Gln Pro Gly Pro Pro Ser Glu Lys Ala Trp Gln Pro Gly Trp Thr
100 105 110
Val Asn Val Glu Gly Ser Gly Glu Leu Phe Arg Trp Asn Val Ser Asp
115 120 125
Leu Gly Gly Leu Gly Cys Gly Leu Lys Asn Arg Ser Ser Glu Gly Pro
130 135 140
Ser Ser Pro Ser Gly Lys Leu Met Ser Pro Lys Leu Tyr Val Trp Ala
145 150 155 160
Lys Asp Arg Pro Glu Ile Trp Glu Gly Glu Pro Pro Cys Val Pro Pro
165 170 175
Arg Asp Ser Leu Asn Gln Ser Leu Ser Gln Asp Leu Thr Met Ala Pro
180 185 190
Gly Ser Thr Leu Trp Leu Ser Cys Gly Val Pro Pro Asp Ser Val Ser
195 200 205
Arg Gly Pro Leu Ser Trp Thr His Val His Pro Lys Gly Pro Lys Ser
210 215 220
Leu Leu Ser Leu Glu Leu Lys Asp Asp Arg Pro Ala Arg Asp Met Trp
225 230 235 240
Val Met Glu Thr Gly Leu Leu Leu Pro Arg Ala Thr Ala Gln Asp Ala
245 250 255
Gly Lys Tyr Tyr Cys His Arg Gly Asn Leu Thr Met Ser Phe His Leu
260 265 270
Glu Ile Thr Ala Arg Pro Val Leu Trp His Trp Leu Leu Arg Thr Gly
275 280 285
Gly Trp Lys Val Ser Ala Val Thr Leu Ala Tyr Leu Ile Phe Cys Leu
290 295 300
Cys Ser Leu Val Gly Ile Leu His Leu Gln Arg Ala Leu Val Leu Arg
305 310 315 320
Arg Lys Arg Lys Arg Met Thr Asp Pro Thr Arg Arg Phe Phe Lys Val
325 330 335
Thr Pro Pro Pro Gly Ser Gly Pro Gln Asn Gln Tyr Gly Asn Val Leu
340 345 350
Ser Leu Pro Thr Pro Thr Ser Gly Leu Gly Arg Ala Gln Arg Trp Ala
355 360 365
Ala Gly Leu Gly Gly Thr Ala Pro Ser Tyr Gly Asn Pro Ser Ser Asp
370 375 380
Val Gln Ala Asp Gly Ala Leu Gly Ser Arg Ser Pro Pro Gly Val Gly
385 390 395 400
Pro Glu Glu Glu Glu Gly Glu Gly Tyr Glu Glu Pro Asp Ser Glu Glu
405 410 415
Asp Ser Glu Phe Tyr Glu Asn Asp Ser Asn Leu Gly Gln Asp Gln Leu
420 425 430
Ser Gln Asp Gly Ser Gly Tyr Glu Asn Pro Glu Asp Glu Pro Leu Gly
435 440 445
Pro Glu Asp Glu Asp Ser Phe Ser Asn Ala Glu Ser Tyr Glu Asn Glu
450 455 460
Asp Glu Glu Leu Thr Gln Pro Val Ala Arg Thr Met Asp Phe Leu Ser
465 470 475 480
Pro His Gly Ser Ala Trp Asp Pro Ser Arg Glu Ala Thr Ser Leu Gly
485 490 495
Ser Gln Ser Tyr Glu Asp Met Arg Gly Ile Leu Tyr Ala Ala Pro Gln
500 505 510
Leu Arg Ser Ile Arg Gly Gln Pro Gly Pro Asn His Glu Glu Asp Ala
515 520 525
Asp Ser Tyr Glu Asn Met Asp Asn Pro Asp Gly Pro Asp Pro Ala Trp
530 535 540
Gly Gly Gly Gly Arg Met Gly Thr Trp Ser Thr Arg
545 550 555
<210> 8
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер CD19Rop
<400> 8
aggaagatat cgccacctac t 21
<210> 9
<211> 245
<212> Белок
<213> Homo sapiens
<400> 9
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Lys Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr His Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr Gly Ser Thr Ser Gly
100 105 110
Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Lys
115 120 125
Leu Gln Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Gln Ser Leu Ser
130 135 140
Val Thr Cys Thr Val Ser Gly Val Ser Leu Pro Asp Tyr Gly Val Ser
145 150 155 160
Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu Glu Trp Leu Gly Val Ile
165 170 175
Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser Ala Leu Lys Ser Arg Leu
180 185 190
Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln Val Phe Leu Lys Met Asn
195 200 205
Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr Tyr Cys Ala Lys His Tyr
210 215 220
Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser
225 230 235 240
Val Thr Val Ser Ser
245
<210> 10
<211> 735
<212> ДНК
<213> Homo sapiens
<400> 10
gacatccaga tgacccagac cacctccagc ctgagcgcca gcctgggcga ccgggtgacc 60
atcagctgcc gggccagcca ggacatcagc aagtacctga actggtatca gcagaagccc 120
gacggcaccg tcaagctgct gatctaccac accagccggc tgcacagcgg cgtgcccagc 180
cggtttagcg gcagcggctc cggcaccgac tacagcctga ccatctccaa cctggaacag 240
gaagatatcg ccacctactt ttgccagcag ggcaacacac tgccctacac ctttggcggc 300
ggaacaaagc tggaaatcac cggcagcacc tccggcagcg gcaagcctgg cagcggcgag 360
ggcagcacca agggcgaggt gaagctgcag gaaagcggcc ctggcctggt ggcccccagc 420
cagagcctga gcgtgacctg caccgtgagc ggcgtgagcc tgcccgacta cggcgtgagc 480
tggatccggc agccccccag gaagggcctg gaatggctgg gcgtgatctg gggcagcgag 540
accacctact acaacagcgc cctgaagagc cggctgacca tcatcaagga caacagcaag 600
agccaggtgt tcctgaagat gaacagcctg cagaccgacg acaccgccat ctactactgc 660
gccaagcact actactacgg cggcagctac gccatggact actggggcca gggcaccagc 720
gtgaccgtga gcagc 735
<210> 11
<211> 297
<212> Белок
<213> Homo sapiens
<400> 11
Met Thr Thr Pro Arg Asn Ser Val Asn Gly Thr Phe Pro Ala Glu Pro
1 5 10 15
Met Lys Gly Pro Ile Ala Met Gln Ser Gly Pro Lys Pro Leu Phe Arg
20 25 30
Arg Met Ser Ser Leu Val Gly Pro Thr Gln Ser Phe Phe Met Arg Glu
35 40 45
Ser Lys Thr Leu Gly Ala Val Gln Ile Met Asn Gly Leu Phe His Ile
50 55 60
Ala Leu Gly Gly Leu Leu Met Ile Pro Ala Gly Ile Tyr Ala Pro Ile
65 70 75 80
Cys Val Thr Val Trp Tyr Pro Leu Trp Gly Gly Ile Met Tyr Ile Ile
85 90 95
Ser Gly Ser Leu Leu Ala Ala Thr Glu Lys Asn Ser Arg Lys Cys Leu
100 105 110
Val Lys Gly Lys Met Ile Met Asn Ser Leu Ser Leu Phe Ala Ala Ile
115 120 125
Ser Gly Met Ile Leu Ser Ile Met Asp Ile Leu Asn Ile Lys Ile Ser
130 135 140
His Phe Leu Lys Met Glu Ser Leu Asn Phe Ile Arg Ala His Thr Pro
145 150 155 160
Tyr Ile Asn Ile Tyr Asn Cys Glu Pro Ala Asn Pro Ser Glu Lys Asn
165 170 175
Ser Pro Ser Thr Gln Tyr Cys Tyr Ser Ile Gln Ser Leu Phe Leu Gly
180 185 190
Ile Leu Ser Val Met Leu Ile Phe Ala Phe Phe Gln Glu Leu Val Ile
195 200 205
Ala Gly Ile Val Glu Asn Glu Trp Lys Arg Thr Cys Ser Arg Pro Lys
210 215 220
Ser Asn Ile Val Leu Leu Ser Ala Glu Glu Lys Lys Glu Gln Thr Ile
225 230 235 240
Glu Ile Lys Glu Glu Val Val Gly Leu Thr Glu Thr Ser Ser Gln Pro
245 250 255
Lys Asn Glu Glu Asp Ile Glu Ile Ile Pro Ile Gln Glu Glu Glu Glu
260 265 270
Glu Glu Thr Glu Thr Asn Phe Pro Glu Pro Pro Gln Asp Gln Glu Ser
275 280 285
Ser Pro Ile Glu Asn Asp Ser Ser Pro
290 295
<210> 12
<211> 84
<212> ДНК
<213> Homo sapiens
<400> 12
atgttctggg tgctggtggt ggtcggaggc gtgctggcct gctacagcct gctggtcacc 60
gtggccttca tcatcttttg ggtg 84
<210> 13
<211> 28
<212> Белок
<213> Homo sapiens
<400> 13
Met Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser
1 5 10 15
Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
20 25
<210> 14
<211> 84
<212> ДНК
<213> Homo sapiens
<400> 14
atgttctggg tgctggtggt ggtgggcggg gtgctggcct gctacagcct gctggtgaca 60
gtggccttca tcatcttttg ggtg 84
<210> 15
<211> 112
<212> Белок
<213> Homo sapiens
<400> 15
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 16
<211> 336
<212> ДНК
<213> Homo sapiens
<400> 16
cgggtgaagt tcagcagaag cgccgacgcc cctgcctacc agcagggcca gaatcagctg 60
tacaacgagc tgaacctggg cagaagggaa gagtacgacg tcctggataa gcggagaggc 120
cgggaccctg agatgggcgg caagcctcgg cggaagaacc cccaggaagg cctgtataac 180
gaactgcaga aagacaagat ggccgaggcc tacagcgaga tcggcatgaa gggcgagcgg 240
aggcggggca agggccacga cggcctgtat cagggcctgt ccaccgccac caaggatacc 300
tacgacgccc tgcacatgca ggccctgccc ccaagg 336
<210> 17
<211> 109
<212> Белок
<213> Homo sapiens
<400> 17
Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln
1 5 10 15
Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu
20 25 30
Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg
35 40 45
Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met
50 55 60
Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly
65 70 75 80
Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp
85 90 95
Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105
<210> 18
<211> 327
<212> ДНК
<213> Homo sapiens
<400> 18
ttcagcagaa gcgccgacgc ccctgcctac cagcagggcc agaatcagct gtacaacgag 60
ctgaacctgg gcagaaggga agagtacgac gtcctggata agcggagagg ccgggaccct 120
gagatgggcg gcaagcctcg gcggaagaac ccccaggaag gcctgtataa cgaactgcag 180
aaagacaaga tggccgaggc ctacagcgag atcggcatga agggcgagcg gaggcggggc 240
aagggccacg acggcctgta tcagggcctg tccaccgcca ccaaggatac ctacgacgcc 300
ctgcacatgc aggccctgcc cccaagg 327
<210> 19
<211> 110
<212> Белок
<213> Homo sapiens
<400> 19
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 20
<211> 330
<212> ДНК
<213> Homo sapiens
<400> 20
gcccccgagt tcctgggcgg acccagcgtg ttcctgttcc cccccaagcc caaggacacc 60
ctgatgatca gccggacccc cgaggtgacc tgcgtggtgg tggacgtgag ccaggaagat 120
cccgaggtcc agttcaattg gtacgtggac ggcgtggaag tgcacaacgc caagaccaag 180
cccagagagg aacagttcaa cagcacctac cgggtggtgt ctgtgctgac cgtgctgcac 240
caggactggc tgaacggcaa agaatacaag tgcaaggtgt ccaacaaggg cctgcccagc 300
agcatcgaaa agaccatcag caaggccaag 330
<210> 21
<211> 321
<212> ДНК
<213> Homo sapiens
<400> 21
ggccagcctc gcgagcccca ggtgtacacc ctgcctccct cccaggaaga gatgaccaag 60
aaccaggtgt ccctgacctg cctggtgaag ggcttctacc ccagcgacat cgccgtggag 120
tgggagagca acggccagcc tgagaacaac tacaagacca cccctcccgt gctggacagc 180
gacggcagct tcttcctgta cagccggctg accgtggaca agagccggtg gcaggaaggc 240
aacgtcttta gctgcagcgt gatgcacgag gccctgcaca accactacac ccagaagagc 300
ctgagcctgt ccctgggcaa g 321
<210> 22
<211> 107
<212> Белок
<213> Homo sapiens
<400> 22
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
100 105
<210> 23
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ЦМВ-праймер
<400> 23
tagcggtttg actcacgg 18
<210> 24
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер CoE1 ori
<400> 24
caggtatccg gtaagcgg 18
<210> 25
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер delU3
<400> 25
ccgtaccttt aagaccaatg acttac 26
<210> 26
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер EF1p
<400> 26
tcgcaacggg tttgcc 16
<210> 27
<211> 1074
<212> ДНК
<213> Homo sapiens
<400> 27
atgcttctcc tggtgacaag ccttctgctc tgtgagttac cacacccagc attcctcctg 60
atcccacgca aagtgtgtaa cggaataggt attggtgaat ttaaagactc actctccata 120
aatgctacga atattaaaca cttcaaaaac tgcacctcca tcagtggcga tctccacatc 180
ctgccggtgg catttagggg tgactccttc acacatactc ctcctctgga tccacaggaa 240
ctggatattc tgaaaaccgt aaaggaaatc acagggtttt tgctgattca ggcttggcct 300
gaaaacagga cggacctcca tgcctttgag aacctagaaa tcatacgcgg caggaccaag 360
caacatggtc agttttctct tgcagtcgtc agcctgaaca taacatcctt gggattacgc 420
tccctcaagg agataagtga tggagatgtg ataatttcag gaaacaaaaa tttgtgctat 480
gcaaatacaa taaactggaa aaaactgttt gggacctccg gtcagaaaac caaaattata 540
agcaacagag gtgaaaacag ctgcaaggcc acaggccagg tctgccatgc cttgtgctcc 600
cccgagggct gctggggccc ggagcccagg gactgcgtct cttgccggaa tgtcagccga 660
ggcagggaat gcgtggacaa gtgcaacctt ctggagggtg agccaaggga gtttgtggag 720
aactctgagt gcatacagtg ccacccagag tgcctgcctc aggccatgaa catcacctgc 780
acaggacggg gaccagacaa ctgtatccag tgtgcccact acattgacgg cccccactgc 840
gtcaagacct gcccggcagg agtcatggga gaaaacaaca ccctggtctg gaagtacgca 900
gacgccggcc atgtgtgcca cctgtgccat ccaaactgca cctacggatg cactgggcca 960
ggtcttgaag gctgtccaac gaatgggcct aagatcccgt ccatcgccac tgggatggtg 1020
ggggccctcc tcttgctgct ggtggtggcc ctggggatcg gcctcttcat gtga 1074
<210> 28
<211> 357
<212> Белок
<213> Homo sapiens
<400> 28
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Arg Lys Val Cys Asn Gly Ile Gly Ile Gly
20 25 30
Glu Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr Asn Ile Lys His Phe
35 40 45
Lys Asn Cys Thr Ser Ile Ser Gly Asp Leu His Ile Leu Pro Val Ala
50 55 60
Phe Arg Gly Asp Ser Phe Thr His Thr Pro Pro Leu Asp Pro Gln Glu
65 70 75 80
Leu Asp Ile Leu Lys Thr Val Lys Glu Ile Thr Gly Phe Leu Leu Ile
85 90 95
Gln Ala Trp Pro Glu Asn Arg Thr Asp Leu His Ala Phe Glu Asn Leu
100 105 110
Glu Ile Ile Arg Gly Arg Thr Lys Gln His Gly Gln Phe Ser Leu Ala
115 120 125
Val Val Ser Leu Asn Ile Thr Ser Leu Gly Leu Arg Ser Leu Lys Glu
130 135 140
Ile Ser Asp Gly Asp Val Ile Ile Ser Gly Asn Lys Asn Leu Cys Tyr
145 150 155 160
Ala Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly Thr Ser Gly Gln Lys
165 170 175
Thr Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser Cys Lys Ala Thr Gly
180 185 190
Gln Val Cys His Ala Leu Cys Ser Pro Glu Gly Cys Trp Gly Pro Glu
195 200 205
Pro Arg Asp Cys Val Ser Cys Arg Asn Val Ser Arg Gly Arg Glu Cys
210 215 220
Val Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro Arg Glu Phe Val Glu
225 230 235 240
Asn Ser Glu Cys Ile Gln Cys His Pro Glu Cys Leu Pro Gln Ala Met
245 250 255
Asn Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn Cys Ile Gln Cys Ala
260 265 270
His Tyr Ile Asp Gly Pro His Cys Val Lys Thr Cys Pro Ala Gly Val
275 280 285
Met Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr Ala Asp Ala Gly His
290 295 300
Val Cys His Leu Cys His Pro Asn Cys Thr Tyr Gly Cys Thr Gly Pro
305 310 315 320
Gly Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys Ile Pro Ser Ile Ala
325 330 335
Thr Gly Met Val Gly Ala Leu Leu Leu Leu Leu Val Val Ala Leu Gly
340 345 350
Ile Gly Leu Phe Met
355
<210> 29
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер EGFRt
<400> 29
atgcttctcc tggtgacaag 20
<210> 30
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 30
Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser Thr
1 5 10 15
Lys Gly
<210> 31
<211> 66
<212> ДНК
<213> Homo sapiens
<400> 31
atgctgctgc tggtgaccag cctgctgctg tgcgagctgc cccaccccgc ctttctgctg 60
atcccc 66
<210> 32
<211> 22
<212> Белок
<213> Искусственная последовательность
<220>
<223> GMCSFRss
<400> 32
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro
20
<210> 33
<211> 2529
<212> ДНК
<213> Искусственная последовательность
<220>
<223> GMCSFRss-CD19scFv-IgG4hinge-CD28tm-41BB-Zeta-T2A-EGFRt
<400> 33
atgctgctgc tggtgaccag cctgctgctg tgcgagctgc cccaccccgc ctttctgctg 60
atccccgaca tccagatgac ccagaccacc tccagcctga gcgccagcct gggcgaccgg 120
gtgaccatca gctgccgggc cagccaggac atcagcaagt acctgaactg gtatcagcag 180
aagcccgacg gcaccgtcaa gctgctgatc taccacacca gccggctgca cagcggcgtg 240
cccagccggt ttagcggcag cggctccggc accgactaca gcctgaccat ctccaacctg 300
gaacaggaag atatcgccac ctacttttgc cagcagggca acacactgcc ctacaccttt 360
ggcggcggaa caaagctgga aatcaccggc agcacctccg gcagcggcaa gcctggcagc 420
ggcgagggca gcaccaaggg cgaggtgaag ctgcaggaaa gcggccctgg cctggtggcc 480
cccagccaga gcctgagcgt gacctgcacc gtgagcggcg tgagcctgcc cgactacggc 540
gtgagctgga tccggcagcc ccccaggaag ggcctggaat ggctgggcgt gatctggggc 600
agcgagacca cctactacaa cagcgccctg aagagccggc tgaccatcat caaggacaac 660
agcaagagcc aggtgttcct gaagatgaac agcctgcaga ccgacgacac cgccatctac 720
tactgcgcca agcactacta ctacggcggc agctacgcca tggactactg gggccagggc 780
accagcgtga ccgtgagcag cgagagcaag tacggaccgc cctgcccccc ttgccctatg 840
ttctgggtgc tggtggtggt cggaggcgtg ctggcctgct acagcctgct ggtcaccgtg 900
gccttcatca tcttttgggt gaaacggggc agaaagaaac tcctgtatat attcaaacaa 960
ccatttatga gaccagtaca aactactcaa gaggaagatg gctgtagctg ccgatttcca 1020
gaagaagaag aaggaggatg tgaactgcgg gtgaagttca gcagaagcgc cgacgcccct 1080
gcctaccagc agggccagaa tcagctgtac aacgagctga acctgggcag aagggaagag 1140
tacgacgtcc tggataagcg gagaggccgg gaccctgaga tgggcggcaa gcctcggcgg 1200
aagaaccccc aggaaggcct gtataacgaa ctgcagaaag acaagatggc cgaggcctac 1260
agcgagatcg gcatgaaggg cgagcggagg cggggcaagg gccacgacgg cctgtatcag 1320
ggcctgtcca ccgccaccaa ggatacctac gacgccctgc acatgcaggc cctgccccca 1380
aggctcgagg gcggcggaga gggcagagga agtcttctaa catgcggtga cgtggaggag 1440
aatcccggcc ctaggatgct tctcctggtg acaagccttc tgctctgtga gttaccacac 1500
ccagcattcc tcctgatccc acgcaaagtg tgtaacggaa taggtattgg tgaatttaaa 1560
gactcactct ccataaatgc tacgaatatt aaacacttca aaaactgcac ctccatcagt 1620
ggcgatctcc acatcctgcc ggtggcattt aggggtgact ccttcacaca tactcctcct 1680
ctggatccac aggaactgga tattctgaaa accgtaaagg aaatcacagg gtttttgctg 1740
attcaggctt ggcctgaaaa caggacggac ctccatgcct ttgagaacct agaaatcata 1800
cgcggcagga ccaagcaaca tggtcagttt tctcttgcag tcgtcagcct gaacataaca 1860
tccttgggat tacgctccct caaggagata agtgatggag atgtgataat ttcaggaaac 1920
aaaaatttgt gctatgcaaa tacaataaac tggaaaaaac tgtttgggac ctccggtcag 1980
aaaaccaaaa ttataagcaa cagaggtgaa aacagctgca aggccacagg ccaggtctgc 2040
catgccttgt gctcccccga gggctgctgg ggcccggagc ccagggactg cgtctcttgc 2100
cggaatgtca gccgaggcag ggaatgcgtg gacaagtgca accttctgga gggtgagcca 2160
agggagtttg tggagaactc tgagtgcata cagtgccacc cagagtgcct gcctcaggcc 2220
atgaacatca cctgcacagg acggggacca gacaactgta tccagtgtgc ccactacatt 2280
gacggccccc actgcgtcaa gacctgcccg gcaggagtca tgggagaaaa caacaccctg 2340
gtctggaagt acgcagacgc cggccatgtg tgccacctgt gccatccaaa ctgcacctac 2400
ggatgcactg ggccaggtct tgaaggctgt ccaacgaatg ggcctaagat cccgtccatc 2460
gccactggga tggtgggggc cctcctcttg ctgctggtgg tggccctggg gatcggcctc 2520
ttcatgtga 2529
<210> 34
<211> 842
<212> Белок
<213> Искусственная последовательность
<220>
<223> GMCSFRss-CD19scFv-IgG4hinge-CD28tm-41BB-Zeta-T2A-EGFRt
<400> 34
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Glu Ser Lys Tyr Gly
260 265 270
Pro Pro Cys Pro Pro Cys Pro Met Phe Trp Val Leu Val Val Val Gly
275 280 285
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
290 295 300
Phe Trp Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln
305 310 315 320
Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser
325 330 335
Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys
340 345 350
Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln
355 360 365
Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu
370 375 380
Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg
385 390 395 400
Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met
405 410 415
Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly
420 425 430
Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp
435 440 445
Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Leu Glu Gly
450 455 460
Gly Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu
465 470 475 480
Asn Pro Gly Pro Arg Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys
485 490 495
Glu Leu Pro His Pro Ala Phe Leu Leu Ile Pro Arg Lys Val Cys Asn
500 505 510
Gly Ile Gly Ile Gly Glu Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr
515 520 525
Asn Ile Lys His Phe Lys Asn Cys Thr Ser Ile Ser Gly Asp Leu His
530 535 540
Ile Leu Pro Val Ala Phe Arg Gly Asp Ser Phe Thr His Thr Pro Pro
545 550 555 560
Leu Asp Pro Gln Glu Leu Asp Ile Leu Lys Thr Val Lys Glu Ile Thr
565 570 575
Gly Phe Leu Leu Ile Gln Ala Trp Pro Glu Asn Arg Thr Asp Leu His
580 585 590
Ala Phe Glu Asn Leu Glu Ile Ile Arg Gly Arg Thr Lys Gln His Gly
595 600 605
Gln Phe Ser Leu Ala Val Val Ser Leu Asn Ile Thr Ser Leu Gly Leu
610 615 620
Arg Ser Leu Lys Glu Ile Ser Asp Gly Asp Val Ile Ile Ser Gly Asn
625 630 635 640
Lys Asn Leu Cys Tyr Ala Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly
645 650 655
Thr Ser Gly Gln Lys Thr Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser
660 665 670
Cys Lys Ala Thr Gly Gln Val Cys His Ala Leu Cys Ser Pro Glu Gly
675 680 685
Cys Trp Gly Pro Glu Pro Arg Asp Cys Val Ser Cys Arg Asn Val Ser
690 695 700
Arg Gly Arg Glu Cys Val Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro
705 710 715 720
Arg Glu Phe Val Glu Asn Ser Glu Cys Ile Gln Cys His Pro Glu Cys
725 730 735
Leu Pro Gln Ala Met Asn Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn
740 745 750
Cys Ile Gln Cys Ala His Tyr Ile Asp Gly Pro His Cys Val Lys Thr
755 760 765
Cys Pro Ala Gly Val Met Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr
770 775 780
Ala Asp Ala Gly His Val Cys His Leu Cys His Pro Asn Cys Thr Tyr
785 790 795 800
Gly Cys Thr Gly Pro Gly Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys
805 810 815
Ile Pro Ser Ile Ala Thr Gly Met Val Gly Ala Leu Leu Leu Leu Leu
820 825 830
Val Val Ala Leu Gly Ile Gly Leu Phe Met
835 840
<210> 35
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Лидерная последовательность GMCSFss
<400> 35
atgcttctcc tggtgacaag ccttctgctc tgtgagttac cacacccagc attcctcctg 60
atccca 66
<210> 36
<211> 2529
<212> ДНК
<213> Искусственная последовательность
<220>
<223> GMCSFss-Her2scFv-IgG4hinge-CD28tm-41BB-Zeta-T2A-EGFRt
<400> 36
atgcttctcc tggtgacaag ccttctgctc tgtgagttac cacacccagc attcctcctg 60
atcccagata tccagatgac ccagtccccg agctccctgt ccgcctctgt gggcgatagg 120
gtcaccatca cctgccgtgc cagtcaggat gtgaatactg ctgtagcctg gtatcaacag 180
aaaccaggaa aagctccgaa actactgatt tactcggcat ccttcctcta ctctggagtc 240
ccttctcgct tctctggttc cagatctggg acggatttca ctctgaccat cagcagtctg 300
cagccggaag acttcgcaac ttattactgt cagcaacatt atactactcc tcccacgttc 360
ggacagggta ccaaggtgga gatcaaaggc agtactagcg gcggtggctc cgggggcgga 420
tccggtgggg gcggcagcag cgaggttcag ctggtggagt ctggcggtgg cctggtgcag 480
ccagggggct cactccgttt gtcctgtgca gcttctggct tcaacattaa agacacctat 540
atacactggg tgcgtcaggc cccgggtaag ggcctggaat gggttgcaag gatttatcct 600
acgaatggtt atactagata tgccgatagc gtcaagggcc gtttcactat aagcgcagac 660
acatccaaaa acacagccta cctgcagatg aacagcctgc gtgctgagga cactgccgtc 720
tattattgtt ctagatgggg aggggacggc ttctatgcta tggactactg gggtcaagga 780
accctggtca ccgtctcgag tgagagcaag tacggaccgc cctgcccccc ttgccctatg 840
ttctgggtgc tggtggtggt cggaggcgtg ctggcctgct acagcctgct ggtcaccgtg 900
gccttcatca tcttttgggt gaaacggggc agaaagaaac tcctgtatat attcaaacaa 960
ccatttatga gaccagtaca aactactcaa gaggaagatg gctgtagctg ccgatttcca 1020
gaagaagaag aaggaggatg tgaactgcgg gtgaagttca gcagaagcgc cgacgcccct 1080
gcctaccagc agggccagaa tcagctgtac aacgagctga acctgggcag aagggaagag 1140
tacgacgtcc tggataagcg gagaggccgg gaccctgaga tgggcggcaa gcctcggcgg 1200
aagaaccccc aggaaggcct gtataacgaa ctgcagaaag acaagatggc cgaggcctac 1260
agcgagatcg gcatgaaggg cgagcggagg cggggcaagg gccacgacgg cctgtatcag 1320
ggcctgtcca ccgccaccaa ggatacctac gacgccctgc acatgcaggc cctgccccca 1380
aggctcgagg gcggcggaga gggcagagga agtcttctaa catgcggtga cgtggaggag 1440
aatcccggcc ctaggatgct tctcctggtg acaagccttc tgctctgtga gttaccacac 1500
ccagcattcc tcctgatccc acgcaaagtg tgtaacggaa taggtattgg tgaatttaaa 1560
gactcactct ccataaatgc tacgaatatt aaacacttca aaaactgcac ctccatcagt 1620
ggcgatctcc acatcctgcc ggtggcattt aggggtgact ccttcacaca tactcctcct 1680
ctggatccac aggaactgga tattctgaaa accgtaaagg aaatcacagg gtttttgctg 1740
attcaggctt ggcctgaaaa caggacggac ctccatgcct ttgagaacct agaaatcata 1800
cgcggcagga ccaagcaaca tggtcagttt tctcttgcag tcgtcagcct gaacataaca 1860
tccttgggat tacgctccct caaggagata agtgatggag atgtgataat ttcaggaaac 1920
aaaaatttgt gctatgcaaa tacaataaac tggaaaaaac tgtttgggac ctccggtcag 1980
aaaaccaaaa ttataagcaa cagaggtgaa aacagctgca aggccacagg ccaggtctgc 2040
catgccttgt gctcccccga gggctgctgg ggcccggagc ccagggactg cgtctcttgc 2100
cggaatgtca gccgaggcag ggaatgcgtg gacaagtgca accttctgga gggtgagcca 2160
agggagtttg tggagaactc tgagtgcata cagtgccacc cagagtgcct gcctcaggcc 2220
atgaacatca cctgcacagg acggggacca gacaactgta tccagtgtgc ccactacatt 2280
gacggccccc actgcgtcaa gacctgcccg gcaggagtca tgggagaaaa caacaccctg 2340
gtctggaagt acgcagacgc cggccatgtg tgccacctgt gccatccaaa ctgcacctac 2400
ggatgcactg ggccaggtct tgaaggctgt ccaacgaatg ggcctaagat cccgtccatc 2460
gccactggga tggtgggggc cctcctcttg ctgctggtgg tggccctggg gatcggcctc 2520
ttcatgtga 2529
<210> 37
<211> 2850
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Конструкция Her 2 со спейсером средней длины
<400> 37
atgcttctcc tggtgacaag ccttctgctc tgtgagttac cacacccagc attcctcctg 60
atcccagata tccagatgac ccagtccccg agctccctgt ccgcctctgt gggcgatagg 120
gtcaccatca cctgccgtgc cagtcaggat gtgaatactg ctgtagcctg gtatcaacag 180
aaaccaggaa aagctccgaa actactgatt tactcggcat ccttcctcta ctctggagtc 240
ccttctcgct tctctggttc cagatctggg acggatttca ctctgaccat cagcagtctg 300
cagccggaag acttcgcaac ttattactgt cagcaacatt atactactcc tcccacgttc 360
ggacagggta ccaaggtgga gatcaaaggc agtactagcg gcggtggctc cgggggcgga 420
tccggtgggg gcggcagcag cgaggttcag ctggtggagt ctggcggtgg cctggtgcag 480
ccagggggct cactccgttt gtcctgtgca gcttctggct tcaacattaa agacacctat 540
atacactggg tgcgtcaggc cccgggtaag ggcctggaat gggttgcaag gatttatcct 600
acgaatggtt atactagata tgccgatagc gtcaagggcc gtttcactat aagcgcagac 660
acatccaaaa acacagccta cctgcagatg aacagcctgc gtgctgagga cactgccgtc 720
tattattgtt ctagatgggg aggggacggc ttctatgcta tggactactg gggtcaagga 780
accctggtca ccgtctcgag tgagagcaag tacggaccgc cctgcccccc ttgccctggc 840
cagcctagag aaccccaggt gtacaccctg cctcccagcc aggaagagat gaccaagaac 900
caggtgtccc tgacctgcct ggtcaaaggc ttctacccca gcgatatcgc cgtggaatgg 960
gagagcaacg gccagcccga gaacaactac aagaccaccc cccctgtgct ggacagcgac 1020
ggcagcttct tcctgtactc ccggctgacc gtggacaaga gccggtggca ggaaggcaac 1080
gtcttcagct gcagcgtgat gcacgaggcc ctgcacaacc actacaccca gaagtccctg 1140
agcctgagcc tgggcaagat gttctgggtg ctggtggtgg tcggaggcgt gctggcctgc 1200
tacagcctgc tggtcaccgt ggccttcatc atcttttggg tgaaacgggg cagaaagaaa 1260
ctcctgtata tattcaaaca accatttatg agaccagtac aaactactca agaggaagat 1320
ggctgtagct gccgatttcc agaagaagaa gaaggaggat gtgaactgcg ggtgaagttc 1380
agcagaagcg ccgacgcccc tgcctaccag cagggccaga atcagctgta caacgagctg 1440
aacctgggca gaagggaaga gtacgacgtc ctggataagc ggagaggccg ggaccctgag 1500
atgggcggca agcctcggcg gaagaacccc caggaaggcc tgtataacga actgcagaaa 1560
gacaagatgg ccgaggccta cagcgagatc ggcatgaagg gcgagcggag gcggggcaag 1620
ggccacgacg gcctgtatca gggcctgtcc accgccacca aggataccta cgacgccctg 1680
cacatgcagg ccctgccccc aaggctcgag ggcggcggag agggcagagg aagtcttcta 1740
acatgcggtg acgtggagga gaatcccggc cctaggatgc ttctcctggt gacaagcctt 1800
ctgctctgtg agttaccaca cccagcattc ctcctgatcc cacgcaaagt gtgtaacgga 1860
ataggtattg gtgaatttaa agactcactc tccataaatg ctacgaatat taaacacttc 1920
aaaaactgca cctccatcag tggcgatctc cacatcctgc cggtggcatt taggggtgac 1980
tccttcacac atactcctcc tctggatcca caggaactgg atattctgaa aaccgtaaag 2040
gaaatcacag ggtttttgct gattcaggct tggcctgaaa acaggacgga cctccatgcc 2100
tttgagaacc tagaaatcat acgcggcagg accaagcaac atggtcagtt ttctcttgca 2160
gtcgtcagcc tgaacataac atccttggga ttacgctccc tcaaggagat aagtgatgga 2220
gatgtgataa tttcaggaaa caaaaatttg tgctatgcaa atacaataaa ctggaaaaaa 2280
ctgtttggga cctccggtca gaaaaccaaa attataagca acagaggtga aaacagctgc 2340
aaggccacag gccaggtctg ccatgccttg tgctcccccg agggctgctg gggcccggag 2400
cccagggact gcgtctcttg ccggaatgtc agccgaggca gggaatgcgt ggacaagtgc 2460
aaccttctgg agggtgagcc aagggagttt gtggagaact ctgagtgcat acagtgccac 2520
ccagagtgcc tgcctcaggc catgaacatc acctgcacag gacggggacc agacaactgt 2580
atccagtgtg cccactacat tgacggcccc cactgcgtca agacctgccc ggcaggagtc 2640
atgggagaaa acaacaccct ggtctggaag tacgcagacg ccggccatgt gtgccacctg 2700
tgccatccaa actgcaccta cggatgcact gggccaggtc ttgaaggctg tccaacgaat 2760
gggcctaaga tcccgtccat cgccactggg atggtggggg ccctcctctt gctgctggtg 2820
gtggccctgg ggatcggcct cttcatgtga 2850
<210> 38
<211> 3180
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Конструкция Her 2 с длинным спейсером
<400> 38
atgcttctcc tggtgacaag ccttctgctc tgtgagttac cacacccagc attcctcctg 60
atcccagata tccagatgac ccagtccccg agctccctgt ccgcctctgt gggcgatagg 120
gtcaccatca cctgccgtgc cagtcaggat gtgaatactg ctgtagcctg gtatcaacag 180
aaaccaggaa aagctccgaa actactgatt tactcggcat ccttcctcta ctctggagtc 240
ccttctcgct tctctggttc cagatctggg acggatttca ctctgaccat cagcagtctg 300
cagccggaag acttcgcaac ttattactgt cagcaacatt atactactcc tcccacgttc 360
ggacagggta ccaaggtgga gatcaaaggc agtactagcg gcggtggctc cgggggcgga 420
tccggtgggg gcggcagcag cgaggttcag ctggtggagt ctggcggtgg cctggtgcag 480
ccagggggct cactccgttt gtcctgtgca gcttctggct tcaacattaa agacacctat 540
atacactggg tgcgtcaggc cccgggtaag ggcctggaat gggttgcaag gatttatcct 600
acgaatggtt atactagata tgccgatagc gtcaagggcc gtttcactat aagcgcagac 660
acatccaaaa acacagccta cctgcagatg aacagcctgc gtgctgagga cactgccgtc 720
tattattgtt ctagatgggg aggggacggc ttctatgcta tggactactg gggtcaagga 780
accctggtca ccgtctcgag tgagagcaag tacggaccgc cctgcccccc ttgccctgcc 840
cccgagttcc tgggcggacc cagcgtgttc ctgttccccc ccaagcccaa ggacaccctg 900
atgatcagcc ggacccccga ggtgacctgc gtggtggtgg acgtgagcca ggaagatccc 960
gaggtccagt tcaattggta cgtggacggc gtggaagtgc acaacgccaa gaccaagccc 1020
agagaggaac agttcaacag cacctaccgg gtggtgtctg tgctgaccgt gctgcaccag 1080
gactggctga acggcaaaga atacaagtgc aaggtgtcca acaagggcct gcccagcagc 1140
atcgaaaaga ccatcagcaa ggccaagggc cagcctcgcg agccccaggt gtacaccctg 1200
cctccctccc aggaagagat gaccaagaac caggtgtccc tgacctgcct ggtgaagggc 1260
ttctacccca gcgacatcgc cgtggagtgg gagagcaacg gccagcctga gaacaactac 1320
aagaccaccc ctcccgtgct ggacagcgac ggcagcttct tcctgtacag ccggctgacc 1380
gtggacaaga gccggtggca ggaaggcaac gtctttagct gcagcgtgat gcacgaggcc 1440
ctgcacaacc actacaccca gaagagcctg agcctgtccc tgggcaagat gttctgggtg 1500
ctggtggtgg tgggcggggt gctggcctgc tacagcctgc tggtgacagt ggccttcatc 1560
atcttttggg tgaaacgggg cagaaagaaa ctcctgtata tattcaaaca accatttatg 1620
agaccagtac aaactactca agaggaagat ggctgtagct gccgatttcc agaagaagaa 1680
gaaggaggat gtgaactgcg ggtgaagttc agcagaagcg ccgacgcccc tgcctaccag 1740
cagggccaga atcagctgta caacgagctg aacctgggca gaagggaaga gtacgacgtc 1800
ctggataagc ggagaggccg ggaccctgag atgggcggca agcctcggcg gaagaacccc 1860
caggaaggcc tgtataacga actgcagaaa gacaagatgg ccgaggccta cagcgagatc 1920
ggcatgaagg gcgagcggag gcggggcaag ggccacgacg gcctgtatca gggcctgtcc 1980
accgccacca aggataccta cgacgccctg cacatgcagg ccctgccccc aaggctcgag 2040
ggcggcggag agggcagagg aagtcttcta acatgcggtg acgtggagga gaatcccggc 2100
cctaggatgc ttctcctggt gacaagcctt ctgctctgtg agttaccaca cccagcattc 2160
ctcctgatcc cacgcaaagt gtgtaacgga ataggtattg gtgaatttaa agactcactc 2220
tccataaatg ctacgaatat taaacacttc aaaaactgca cctccatcag tggcgatctc 2280
cacatcctgc cggtggcatt taggggtgac tccttcacac atactcctcc tctggatcca 2340
caggaactgg atattctgaa aaccgtaaag gaaatcacag ggtttttgct gattcaggct 2400
tggcctgaaa acaggacgga cctccatgcc tttgagaacc tagaaatcat acgcggcagg 2460
accaagcaac atggtcagtt ttctcttgca gtcgtcagcc tgaacataac atccttggga 2520
ttacgctccc tcaaggagat aagtgatgga gatgtgataa tttcaggaaa caaaaatttg 2580
tgctatgcaa atacaataaa ctggaaaaaa ctgtttggga cctccggtca gaaaaccaaa 2640
attataagca acagaggtga aaacagctgc aaggccacag gccaggtctg ccatgccttg 2700
tgctcccccg agggctgctg gggcccggag cccagggact gcgtctcttg ccggaatgtc 2760
agccgaggca gggaatgcgt ggacaagtgc aaccttctgg agggtgagcc aagggagttt 2820
gtggagaact ctgagtgcat acagtgccac ccagagtgcc tgcctcaggc catgaacatc 2880
acctgcacag gacggggacc agacaactgt atccagtgtg cccactacat tgacggcccc 2940
cactgcgtca agacctgccc ggcaggagtc atgggagaaa acaacaccct ggtctggaag 3000
tacgcagacg ccggccatgt gtgccacctg tgccatccaa actgcaccta cggatgcact 3060
gggccaggtc ttgaaggctg tccaacgaat gggcctaaga tcccgtccat cgccactggg 3120
atggtggggg ccctcctctt gctgctggtg gtggccctgg ggatcggcct cttcatgtga 3180
<210> 39
<211> 735
<212> ДНК
<213> Homo sapiens
<400> 39
gatatccaga tgacccagtc cccgagctcc ctgtccgcct ctgtgggcga tagggtcacc 60
atcacctgcc gtgccagtca ggatgtgaat actgctgtag cctggtatca acagaaacca 120
ggaaaagctc cgaaactact gatttactcg gcatccttcc tctactctgg agtcccttct 180
cgcttctctg gttccagatc tgggacggat ttcactctga ccatcagcag tctgcagccg 240
gaagacttcg caacttatta ctgtcagcaa cattatacta ctcctcccac gttcggacag 300
ggtaccaagg tggagatcaa aggcagtact agcggcggtg gctccggggg cggatccggt 360
gggggcggca gcagcgaggt tcagctggtg gagtctggcg gtggcctggt gcagccaggg 420
ggctcactcc gtttgtcctg tgcagcttct ggcttcaaca ttaaagacac ctatatacac 480
tgggtgcgtc aggccccggg taagggcctg gaatgggttg caaggattta tcctacgaat 540
ggttatacta gatatgccga tagcgtcaag ggccgtttca ctataagcgc agacacatcc 600
aaaaacacag cctacctgca gatgaacagc ctgcgtgctg aggacactgc cgtctattat 660
tgttctagat ggggagggga cggcttctat gctatggact actggggtca aggaaccctg 720
gtcaccgtct cgagt 735
<210> 40
<211> 753
<212> ДНК
<213> Homo sapiens
<400> 40
gcattcctcc tgatcccaga tatccagatg acccagtccc cgagctccct gtccgcctct 60
gtgggcgata gggtcaccat cacctgccgt gccagtcagg atgtgaatac tgctgtagcc 120
tggtatcaac agaaaccagg aaaagctccg aaactactga tttactcggc atccttcctc 180
tactctggag tcccttctcg cttctctggt tccagatctg ggacggattt cactctgacc 240
atcagcagtc tgcagccgga agacttcgca acttattact gtcagcaaca ttatactact 300
cctcccacgt tcggacaggg taccaaggtg gagatcaaag gcagtactag cggcggtggc 360
tccgggggcg gatccggtgg gggcggcagc agcgaggttc agctggtgga gtctggcggt 420
ggcctggtgc agccaggggg ctcactccgt ttgtcctgtg cagcttctgg cttcaacatt 480
aaagacacct atatacactg ggtgcgtcag gccccgggta agggcctgga atgggttgca 540
aggatttatc ctacgaatgg ttatactaga tatgccgata gcgtcaaggg ccgtttcact 600
ataagcgcag acacatccaa aaacacagcc tacctgcaga tgaacagcct gcgtgctgag 660
gacactgccg tctattattg ttctagatgg ggaggggacg gcttctatgc tatggactac 720
tggggtcaag gaaccctggt caccgtctcg agt 753
<210> 41
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Шарнирный спейсер
<400> 41
gagagcaagt acggaccgcc ctgcccccct tgccctggcc agcctagaga accccaggtg 60
tacaccctgc ctcccagcca ggaagagatg accaagaacc aggtgtccct gacctgcctg 120
gtcaaaggct tctaccccag cgatatcgcc gtggaatggg agagcaacgg ccagcccgag 180
aacaactaca agaccacccc ccctgtgctg gacagcgacg gcagcttctt cctgtactcc 240
cggctgaccg tggacaagag ccggtggcag gaaggcaacg tcttcagctg cagcgtgatg 300
cacgaggccc tgcacaacca ctacacccag aagtccctga gcctgagcct gggcaag 357
<210> 42
<211> 356
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Шарнирная область/спейсер
<400> 42
taggaccgcc ctgcccccct tgccctgccc ccgagttcct gggcggaccc agcgtgttcc 60
tgttcccccc caagcccaag gacaccctga tgatcagccg gacccccgag gtgacctgcg 120
tggtggtgga cgtgagccag gaagatcccg aggtccagtt caattggtac gtggacggcg 180
tggaagtgca caacgccaag accaagccca gagaggaaca gttcaacagc acctaccggg 240
tggtgtctgt gctgaccgtg ctgcaccagg actggctgaa cggcaaagaa tacaagtgca 300
aggtgtccaa caagggcctg cccagcagca tcgaaaagac catcagcaag gccaag 356
<210> 43
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Шарнирная область/спейсер
<400> 43
tacggaccgc cctgcccccc ttgccctggc cagcctcgcg agccccaggt gtacaccctg 60
cctccctccc aggaagagat gaccaagaac caggtgtccc tgacctgcct ggtgaagggc 120
ttctacccca gcgacatcgc cgtggagtgg gagagcaacg gccagcctga gaacaactac 180
aagaccaccc ctcccgtgct ggacagcgac ggcagcttct tcctgtacag ccggctgacc 240
gtggacaaga gccggtggca ggaaggcaac gtctttagct gcagcgtgat gcacgaggcc 300
ctgcacaacc actacaccca gaagagcctg agcctgtccc tgggcaag 348
<210> 44
<211> 15
<212> Белок
<213> Homo sapiens
<400> 44
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 45
<211> 16
<212> Белок
<213> Homo sapiens
<400> 45
Glu Leu Lys Thr Pro Leu Gly Asp Thr His Thr Cys Pro Arg Cys Pro
1 5 10 15
<210> 46
<211> 15
<212> Белок
<213> Homo sapiens
<400> 46
Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
1 5 10 15
<210> 47
<211> 12
<212> Белок
<213> Homo sapiens
<400> 47
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro
1 5 10
<210> 48
<211> 12
<212> Белок
<213> Homo sapiens
<400> 48
Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro
1 5 10
<210> 49
<211> 27
<212> ДНК
<213> Homo sapiens
<400> 49
tacggaccgc cctgcccccc ttgccct 27
<210> 50
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 50
gaatctaagt acggaccgcc ctgcccccct tgccct 36
<210> 51
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 51
gagagcaagt acggaccgcc ctgcccccct tgccct 36
<210> 52
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер средней длины
<400> 52
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Gly Gln Pro Arg
1 5 10 15
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
20 25 30
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
35 40 45
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
50 55 60
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
65 70 75 80
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
85 90 95
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
100 105 110
Leu Ser Leu Ser Leu Gly Lys
115
<210> 53
<211> 838
<212> Белок
<213> Искусственная последовательность
<220>
<223> Leader _R11- Hinge- CD28tm/41BB-Z-T2A-tEGFR
<400> 53
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Gln Ser Val Lys Glu Ser Glu Gly Asp Leu
20 25 30
Val Thr Pro Ala Gly Asn Leu Thr Leu Thr Cys Thr Ala Ser Gly Ser
35 40 45
Asp Ile Asn Asp Tyr Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Ile Gly Phe Ile Asn Ser Gly Gly Ser Thr Trp Tyr
65 70 75 80
Ala Ser Trp Val Lys Gly Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr
85 90 95
Val Asp Leu Lys Met Thr Ser Leu Thr Thr Asp Asp Thr Ala Thr Tyr
100 105 110
Phe Cys Ala Arg Gly Tyr Ser Thr Tyr Tyr Gly Asp Phe Asn Ile Trp
115 120 125
Gly Pro Gly Thr Leu Val Thr Ile Ser Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu Val Met Thr Gln Thr
145 150 155 160
Pro Ser Ser Thr Ser Gly Ala Val Gly Gly Thr Val Thr Ile Asn Cys
165 170 175
Gln Ala Ser Gln Ser Ile Asp Ser Asn Leu Ala Trp Phe Gln Gln Lys
180 185 190
Pro Gly Gln Pro Pro Thr Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala
195 200 205
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Arg Ser Gly Thr Glu Tyr
210 215 220
Thr Leu Thr Ile Ser Gly Val Gln Arg Glu Asp Ala Ala Thr Tyr Tyr
225 230 235 240
Cys Leu Gly Gly Val Gly Asn Val Ser Tyr Arg Thr Ser Phe Gly Gly
245 250 255
Gly Thr Glu Val Val Val Lys Glu Ser Lys Tyr Gly Pro Pro Cys Pro
260 265 270
Pro Cys Pro Met Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala
275 280 285
Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Lys
290 295 300
Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg
305 310 315 320
Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro
325 330 335
Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg Ser
340 345 350
Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu
355 360 365
Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg
370 375 380
Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln
385 390 395 400
Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr
405 410 415
Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp
420 425 430
Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala
435 440 445
Leu His Met Gln Ala Leu Pro Pro Arg Leu Glu Gly Gly Gly Glu Gly
450 455 460
Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro
465 470 475 480
Arg Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His
485 490 495
Pro Ala Phe Leu Leu Ile Pro Arg Lys Val Cys Asn Gly Ile Gly Ile
500 505 510
Gly Glu Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr Asn Ile Lys His
515 520 525
Phe Lys Asn Cys Thr Ser Ile Ser Gly Asp Leu His Ile Leu Pro Val
530 535 540
Ala Phe Arg Gly Asp Ser Phe Thr His Thr Pro Pro Leu Asp Pro Gln
545 550 555 560
Glu Leu Asp Ile Leu Lys Thr Val Lys Glu Ile Thr Gly Phe Leu Leu
565 570 575
Ile Gln Ala Trp Pro Glu Asn Arg Thr Asp Leu His Ala Phe Glu Asn
580 585 590
Leu Glu Ile Ile Arg Gly Arg Thr Lys Gln His Gly Gln Phe Ser Leu
595 600 605
Ala Val Val Ser Leu Asn Ile Thr Ser Leu Gly Leu Arg Ser Leu Lys
610 615 620
Glu Ile Ser Asp Gly Asp Val Ile Ile Ser Gly Asn Lys Asn Leu Cys
625 630 635 640
Tyr Ala Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly Thr Ser Gly Gln
645 650 655
Lys Thr Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser Cys Lys Ala Thr
660 665 670
Gly Gln Val Cys His Ala Leu Cys Ser Pro Glu Gly Cys Trp Gly Pro
675 680 685
Glu Pro Arg Asp Cys Val Ser Cys Arg Asn Val Ser Arg Gly Arg Glu
690 695 700
Cys Val Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro Arg Glu Phe Val
705 710 715 720
Glu Asn Ser Glu Cys Ile Gln Cys His Pro Glu Cys Leu Pro Gln Ala
725 730 735
Met Asn Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn Cys Ile Gln Cys
740 745 750
Ala His Tyr Ile Asp Gly Pro His Cys Val Lys Thr Cys Pro Ala Gly
755 760 765
Val Met Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr Ala Asp Ala Gly
770 775 780
His Val Cys His Leu Cys His Pro Asn Cys Thr Tyr Gly Cys Thr Gly
785 790 795 800
Pro Gly Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys Ile Pro Ser Ile
805 810 815
Ala Thr Gly Met Val Gly Ala Leu Leu Leu Leu Leu Val Val Ala Leu
820 825 830
Gly Ile Gly Leu Phe Met
835
<210> 54
<211> 1055
<212> Белок
<213> Искусственная последовательность
<220>
<223> Leader _R11- Hinge-CH2-CH3- CD28tm/41BB-Z-T2A-tEGFR
<400> 54
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Gln Ser Val Lys Glu Ser Glu Gly Asp Leu
20 25 30
Val Thr Pro Ala Gly Asn Leu Thr Leu Thr Cys Thr Ala Ser Gly Ser
35 40 45
Asp Ile Asn Asp Tyr Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Ile Gly Phe Ile Asn Ser Gly Gly Ser Thr Trp Tyr
65 70 75 80
Ala Ser Trp Val Lys Gly Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr
85 90 95
Val Asp Leu Lys Met Thr Ser Leu Thr Thr Asp Asp Thr Ala Thr Tyr
100 105 110
Phe Cys Ala Arg Gly Tyr Ser Thr Tyr Tyr Gly Asp Phe Asn Ile Trp
115 120 125
Gly Pro Gly Thr Leu Val Thr Ile Ser Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu Val Met Thr Gln Thr
145 150 155 160
Pro Ser Ser Thr Ser Gly Ala Val Gly Gly Thr Val Thr Ile Asn Cys
165 170 175
Gln Ala Ser Gln Ser Ile Asp Ser Asn Leu Ala Trp Phe Gln Gln Lys
180 185 190
Pro Gly Gln Pro Pro Thr Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala
195 200 205
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Arg Ser Gly Thr Glu Tyr
210 215 220
Thr Leu Thr Ile Ser Gly Val Gln Arg Glu Asp Ala Ala Thr Tyr Tyr
225 230 235 240
Cys Leu Gly Gly Val Gly Asn Val Ser Tyr Arg Thr Ser Phe Gly Gly
245 250 255
Gly Thr Glu Val Val Val Lys Glu Ser Lys Tyr Gly Pro Pro Cys Pro
260 265 270
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
275 280 285
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
290 295 300
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
305 310 315 320
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
325 330 335
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
340 345 350
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
355 360 365
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
370 375 380
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
385 390 395 400
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
405 410 415
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
420 425 430
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
435 440 445
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
450 455 460
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
465 470 475 480
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Met Phe Trp Val
485 490 495
Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr
500 505 510
Val Ala Phe Ile Ile Phe Trp Val Lys Arg Gly Arg Lys Lys Leu Leu
515 520 525
Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu
530 535 540
Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys
545 550 555 560
Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln
565 570 575
Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu
580 585 590
Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly
595 600 605
Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu
610 615 620
Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly
625 630 635 640
Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser
645 650 655
Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro
660 665 670
Pro Arg Leu Glu Gly Gly Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys
675 680 685
Gly Asp Val Glu Glu Asn Pro Gly Pro Arg Met Leu Leu Leu Val Thr
690 695 700
Ser Leu Leu Leu Cys Glu Leu Pro His Pro Ala Phe Leu Leu Ile Pro
705 710 715 720
Arg Lys Val Cys Asn Gly Ile Gly Ile Gly Glu Phe Lys Asp Ser Leu
725 730 735
Ser Ile Asn Ala Thr Asn Ile Lys His Phe Lys Asn Cys Thr Ser Ile
740 745 750
Ser Gly Asp Leu His Ile Leu Pro Val Ala Phe Arg Gly Asp Ser Phe
755 760 765
Thr His Thr Pro Pro Leu Asp Pro Gln Glu Leu Asp Ile Leu Lys Thr
770 775 780
Val Lys Glu Ile Thr Gly Phe Leu Leu Ile Gln Ala Trp Pro Glu Asn
785 790 795 800
Arg Thr Asp Leu His Ala Phe Glu Asn Leu Glu Ile Ile Arg Gly Arg
805 810 815
Thr Lys Gln His Gly Gln Phe Ser Leu Ala Val Val Ser Leu Asn Ile
820 825 830
Thr Ser Leu Gly Leu Arg Ser Leu Lys Glu Ile Ser Asp Gly Asp Val
835 840 845
Ile Ile Ser Gly Asn Lys Asn Leu Cys Tyr Ala Asn Thr Ile Asn Trp
850 855 860
Lys Lys Leu Phe Gly Thr Ser Gly Gln Lys Thr Lys Ile Ile Ser Asn
865 870 875 880
Arg Gly Glu Asn Ser Cys Lys Ala Thr Gly Gln Val Cys His Ala Leu
885 890 895
Cys Ser Pro Glu Gly Cys Trp Gly Pro Glu Pro Arg Asp Cys Val Ser
900 905 910
Cys Arg Asn Val Ser Arg Gly Arg Glu Cys Val Asp Lys Cys Asn Leu
915 920 925
Leu Glu Gly Glu Pro Arg Glu Phe Val Glu Asn Ser Glu Cys Ile Gln
930 935 940
Cys His Pro Glu Cys Leu Pro Gln Ala Met Asn Ile Thr Cys Thr Gly
945 950 955 960
Arg Gly Pro Asp Asn Cys Ile Gln Cys Ala His Tyr Ile Asp Gly Pro
965 970 975
His Cys Val Lys Thr Cys Pro Ala Gly Val Met Gly Glu Asn Asn Thr
980 985 990
Leu Val Trp Lys Tyr Ala Asp Ala Gly His Val Cys His Leu Cys His
995 1000 1005
Pro Asn Cys Thr Tyr Gly Cys Thr Gly Pro Gly Leu Glu Gly Cys
1010 1015 1020
Pro Thr Asn Gly Pro Lys Ile Pro Ser Ile Ala Thr Gly Met Val
1025 1030 1035
Gly Ala Leu Leu Leu Leu Leu Val Val Ala Leu Gly Ile Gly Leu
1040 1045 1050
Phe Met
1055
<210> 55
<211> 945
<212> Белок
<213> Искусственная последовательность
<220>
<223> Leader _R11- Hinge-CH3- CD28tm/41BB-Z-T2A-tEGFR
<400> 55
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Gln Ser Val Lys Glu Ser Glu Gly Asp Leu
20 25 30
Val Thr Pro Ala Gly Asn Leu Thr Leu Thr Cys Thr Ala Ser Gly Ser
35 40 45
Asp Ile Asn Asp Tyr Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Ile Gly Phe Ile Asn Ser Gly Gly Ser Thr Trp Tyr
65 70 75 80
Ala Ser Trp Val Lys Gly Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr
85 90 95
Val Asp Leu Lys Met Thr Ser Leu Thr Thr Asp Asp Thr Ala Thr Tyr
100 105 110
Phe Cys Ala Arg Gly Tyr Ser Thr Tyr Tyr Gly Asp Phe Asn Ile Trp
115 120 125
Gly Pro Gly Thr Leu Val Thr Ile Ser Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu Val Met Thr Gln Thr
145 150 155 160
Pro Ser Ser Thr Ser Gly Ala Val Gly Gly Thr Val Thr Ile Asn Cys
165 170 175
Gln Ala Ser Gln Ser Ile Asp Ser Asn Leu Ala Trp Phe Gln Gln Lys
180 185 190
Pro Gly Gln Pro Pro Thr Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala
195 200 205
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Arg Ser Gly Thr Glu Tyr
210 215 220
Thr Leu Thr Ile Ser Gly Val Gln Arg Glu Asp Ala Ala Thr Tyr Tyr
225 230 235 240
Cys Leu Gly Gly Val Gly Asn Val Ser Tyr Arg Thr Ser Phe Gly Gly
245 250 255
Gly Thr Glu Val Val Val Lys Glu Ser Lys Tyr Gly Pro Pro Cys Pro
260 265 270
Pro Cys Pro Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
275 280 285
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
290 295 300
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
305 310 315 320
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
325 330 335
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
340 345 350
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
355 360 365
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Met Phe
370 375 380
Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu
385 390 395 400
Val Thr Val Ala Phe Ile Ile Phe Trp Val Lys Arg Gly Arg Lys Lys
405 410 415
Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr
420 425 430
Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly
435 440 445
Gly Cys Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala
450 455 460
Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg
465 470 475 480
Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu
485 490 495
Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
500 505 510
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
515 520 525
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
530 535 540
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
545 550 555 560
Leu Pro Pro Arg Leu Glu Gly Gly Gly Glu Gly Arg Gly Ser Leu Leu
565 570 575
Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Arg Met Leu Leu Leu
580 585 590
Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro Ala Phe Leu Leu
595 600 605
Ile Pro Arg Lys Val Cys Asn Gly Ile Gly Ile Gly Glu Phe Lys Asp
610 615 620
Ser Leu Ser Ile Asn Ala Thr Asn Ile Lys His Phe Lys Asn Cys Thr
625 630 635 640
Ser Ile Ser Gly Asp Leu His Ile Leu Pro Val Ala Phe Arg Gly Asp
645 650 655
Ser Phe Thr His Thr Pro Pro Leu Asp Pro Gln Glu Leu Asp Ile Leu
660 665 670
Lys Thr Val Lys Glu Ile Thr Gly Phe Leu Leu Ile Gln Ala Trp Pro
675 680 685
Glu Asn Arg Thr Asp Leu His Ala Phe Glu Asn Leu Glu Ile Ile Arg
690 695 700
Gly Arg Thr Lys Gln His Gly Gln Phe Ser Leu Ala Val Val Ser Leu
705 710 715 720
Asn Ile Thr Ser Leu Gly Leu Arg Ser Leu Lys Glu Ile Ser Asp Gly
725 730 735
Asp Val Ile Ile Ser Gly Asn Lys Asn Leu Cys Tyr Ala Asn Thr Ile
740 745 750
Asn Trp Lys Lys Leu Phe Gly Thr Ser Gly Gln Lys Thr Lys Ile Ile
755 760 765
Ser Asn Arg Gly Glu Asn Ser Cys Lys Ala Thr Gly Gln Val Cys His
770 775 780
Ala Leu Cys Ser Pro Glu Gly Cys Trp Gly Pro Glu Pro Arg Asp Cys
785 790 795 800
Val Ser Cys Arg Asn Val Ser Arg Gly Arg Glu Cys Val Asp Lys Cys
805 810 815
Asn Leu Leu Glu Gly Glu Pro Arg Glu Phe Val Glu Asn Ser Glu Cys
820 825 830
Ile Gln Cys His Pro Glu Cys Leu Pro Gln Ala Met Asn Ile Thr Cys
835 840 845
Thr Gly Arg Gly Pro Asp Asn Cys Ile Gln Cys Ala His Tyr Ile Asp
850 855 860
Gly Pro His Cys Val Lys Thr Cys Pro Ala Gly Val Met Gly Glu Asn
865 870 875 880
Asn Thr Leu Val Trp Lys Tyr Ala Asp Ala Gly His Val Cys His Leu
885 890 895
Cys His Pro Asn Cys Thr Tyr Gly Cys Thr Gly Pro Gly Leu Glu Gly
900 905 910
Cys Pro Thr Asn Gly Pro Lys Ile Pro Ser Ile Ala Thr Gly Met Val
915 920 925
Gly Ala Leu Leu Leu Leu Leu Val Val Ala Leu Gly Ile Gly Leu Phe
930 935 940
Met
945
<210> 56
<211> 845
<212> Белок
<213> Искусственная последовательность
<220>
<223> Leader _R12 - CD28tm/41BB-Z-T2A-tEGFR
<400> 56
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Gln Glu Gln Leu Val Glu Ser Gly Gly Arg
20 25 30
Leu Val Thr Pro Gly Gly Ser Leu Thr Leu Ser Cys Lys Ala Ser Gly
35 40 45
Phe Asp Phe Ser Ala Tyr Tyr Met Ser Trp Val Arg Gln Ala Pro Gly
50 55 60
Lys Gly Leu Glu Trp Ile Ala Thr Ile Tyr Pro Ser Ser Gly Lys Thr
65 70 75 80
Tyr Tyr Ala Thr Trp Val Asn Gly Arg Phe Thr Ile Ser Ser Asp Asn
85 90 95
Ala Gln Asn Thr Val Asp Leu Gln Met Asn Ser Leu Thr Ala Ala Asp
100 105 110
Arg Ala Thr Tyr Phe Cys Ala Arg Asp Ser Tyr Ala Asp Asp Gly Ala
115 120 125
Leu Phe Asn Ile Trp Gly Pro Gly Thr Leu Val Thr Ile Ser Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu
145 150 155 160
Val Leu Thr Gln Ser Pro Ser Val Ser Ala Ala Leu Gly Ser Pro Ala
165 170 175
Lys Ile Thr Cys Thr Leu Ser Ser Ala His Lys Thr Asp Thr Ile Asp
180 185 190
Trp Tyr Gln Gln Leu Gln Gly Glu Ala Pro Arg Tyr Leu Met Gln Val
195 200 205
Gln Ser Asp Gly Ser Tyr Thr Lys Arg Pro Gly Val Pro Asp Arg Phe
210 215 220
Ser Gly Ser Ser Ser Gly Ala Asp Arg Tyr Leu Ile Ile Pro Ser Val
225 230 235 240
Gln Ala Asp Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Asp Tyr Ile Gly
245 250 255
Gly Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly Glu Ser
260 265 270
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Met Phe Trp Val Leu Val
275 280 285
Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala
290 295 300
Phe Ile Ile Phe Trp Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile
305 310 315 320
Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp
325 330 335
Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
340 345 350
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
355 360 365
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
370 375 380
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
385 390 395 400
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
405 410 415
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
420 425 430
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
435 440 445
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
450 455 460
Leu Glu Gly Gly Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp
465 470 475 480
Val Glu Glu Asn Pro Gly Pro Arg Met Leu Leu Leu Val Thr Ser Leu
485 490 495
Leu Leu Cys Glu Leu Pro His Pro Ala Phe Leu Leu Ile Pro Arg Lys
500 505 510
Val Cys Asn Gly Ile Gly Ile Gly Glu Phe Lys Asp Ser Leu Ser Ile
515 520 525
Asn Ala Thr Asn Ile Lys His Phe Lys Asn Cys Thr Ser Ile Ser Gly
530 535 540
Asp Leu His Ile Leu Pro Val Ala Phe Arg Gly Asp Ser Phe Thr His
545 550 555 560
Thr Pro Pro Leu Asp Pro Gln Glu Leu Asp Ile Leu Lys Thr Val Lys
565 570 575
Glu Ile Thr Gly Phe Leu Leu Ile Gln Ala Trp Pro Glu Asn Arg Thr
580 585 590
Asp Leu His Ala Phe Glu Asn Leu Glu Ile Ile Arg Gly Arg Thr Lys
595 600 605
Gln His Gly Gln Phe Ser Leu Ala Val Val Ser Leu Asn Ile Thr Ser
610 615 620
Leu Gly Leu Arg Ser Leu Lys Glu Ile Ser Asp Gly Asp Val Ile Ile
625 630 635 640
Ser Gly Asn Lys Asn Leu Cys Tyr Ala Asn Thr Ile Asn Trp Lys Lys
645 650 655
Leu Phe Gly Thr Ser Gly Gln Lys Thr Lys Ile Ile Ser Asn Arg Gly
660 665 670
Glu Asn Ser Cys Lys Ala Thr Gly Gln Val Cys His Ala Leu Cys Ser
675 680 685
Pro Glu Gly Cys Trp Gly Pro Glu Pro Arg Asp Cys Val Ser Cys Arg
690 695 700
Asn Val Ser Arg Gly Arg Glu Cys Val Asp Lys Cys Asn Leu Leu Glu
705 710 715 720
Gly Glu Pro Arg Glu Phe Val Glu Asn Ser Glu Cys Ile Gln Cys His
725 730 735
Pro Glu Cys Leu Pro Gln Ala Met Asn Ile Thr Cys Thr Gly Arg Gly
740 745 750
Pro Asp Asn Cys Ile Gln Cys Ala His Tyr Ile Asp Gly Pro His Cys
755 760 765
Val Lys Thr Cys Pro Ala Gly Val Met Gly Glu Asn Asn Thr Leu Val
770 775 780
Trp Lys Tyr Ala Asp Ala Gly His Val Cys His Leu Cys His Pro Asn
785 790 795 800
Cys Thr Tyr Gly Cys Thr Gly Pro Gly Leu Glu Gly Cys Pro Thr Asn
805 810 815
Gly Pro Lys Ile Pro Ser Ile Ala Thr Gly Met Val Gly Ala Leu Leu
820 825 830
Leu Leu Leu Val Val Ala Leu Gly Ile Gly Leu Phe Met
835 840 845
<210> 57
<211> 952
<212> Белок
<213> Искусственная последовательность
<220>
<223> Leader _R12- Hinge- CH3- CD28tm/41BB-Z-T2A-tEGFR
<400> 57
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Gln Glu Gln Leu Val Glu Ser Gly Gly Arg
20 25 30
Leu Val Thr Pro Gly Gly Ser Leu Thr Leu Ser Cys Lys Ala Ser Gly
35 40 45
Phe Asp Phe Ser Ala Tyr Tyr Met Ser Trp Val Arg Gln Ala Pro Gly
50 55 60
Lys Gly Leu Glu Trp Ile Ala Thr Ile Tyr Pro Ser Ser Gly Lys Thr
65 70 75 80
Tyr Tyr Ala Thr Trp Val Asn Gly Arg Phe Thr Ile Ser Ser Asp Asn
85 90 95
Ala Gln Asn Thr Val Asp Leu Gln Met Asn Ser Leu Thr Ala Ala Asp
100 105 110
Arg Ala Thr Tyr Phe Cys Ala Arg Asp Ser Tyr Ala Asp Asp Gly Ala
115 120 125
Leu Phe Asn Ile Trp Gly Pro Gly Thr Leu Val Thr Ile Ser Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu
145 150 155 160
Val Leu Thr Gln Ser Pro Ser Val Ser Ala Ala Leu Gly Ser Pro Ala
165 170 175
Lys Ile Thr Cys Thr Leu Ser Ser Ala His Lys Thr Asp Thr Ile Asp
180 185 190
Trp Tyr Gln Gln Leu Gln Gly Glu Ala Pro Arg Tyr Leu Met Gln Val
195 200 205
Gln Ser Asp Gly Ser Tyr Thr Lys Arg Pro Gly Val Pro Asp Arg Phe
210 215 220
Ser Gly Ser Ser Ser Gly Ala Asp Arg Tyr Leu Ile Ile Pro Ser Val
225 230 235 240
Gln Ala Asp Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Asp Tyr Ile Gly
245 250 255
Gly Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly Glu Ser
260 265 270
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Gly Gln Pro Arg Glu Pro
275 280 285
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
290 295 300
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
305 310 315 320
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
325 330 335
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
340 345 350
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
355 360 365
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
370 375 380
Leu Ser Leu Gly Lys Met Phe Trp Val Leu Val Val Val Gly Gly Val
385 390 395 400
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
405 410 415
Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe
420 425 430
Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg
435 440 445
Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser
450 455 460
Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr
465 470 475 480
Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys
485 490 495
Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn
500 505 510
Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu
515 520 525
Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly
530 535 540
His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr
545 550 555 560
Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Leu Glu Gly Gly Gly
565 570 575
Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro
580 585 590
Gly Pro Arg Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu
595 600 605
Pro His Pro Ala Phe Leu Leu Ile Pro Arg Lys Val Cys Asn Gly Ile
610 615 620
Gly Ile Gly Glu Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr Asn Ile
625 630 635 640
Lys His Phe Lys Asn Cys Thr Ser Ile Ser Gly Asp Leu His Ile Leu
645 650 655
Pro Val Ala Phe Arg Gly Asp Ser Phe Thr His Thr Pro Pro Leu Asp
660 665 670
Pro Gln Glu Leu Asp Ile Leu Lys Thr Val Lys Glu Ile Thr Gly Phe
675 680 685
Leu Leu Ile Gln Ala Trp Pro Glu Asn Arg Thr Asp Leu His Ala Phe
690 695 700
Glu Asn Leu Glu Ile Ile Arg Gly Arg Thr Lys Gln His Gly Gln Phe
705 710 715 720
Ser Leu Ala Val Val Ser Leu Asn Ile Thr Ser Leu Gly Leu Arg Ser
725 730 735
Leu Lys Glu Ile Ser Asp Gly Asp Val Ile Ile Ser Gly Asn Lys Asn
740 745 750
Leu Cys Tyr Ala Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly Thr Ser
755 760 765
Gly Gln Lys Thr Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser Cys Lys
770 775 780
Ala Thr Gly Gln Val Cys His Ala Leu Cys Ser Pro Glu Gly Cys Trp
785 790 795 800
Gly Pro Glu Pro Arg Asp Cys Val Ser Cys Arg Asn Val Ser Arg Gly
805 810 815
Arg Glu Cys Val Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro Arg Glu
820 825 830
Phe Val Glu Asn Ser Glu Cys Ile Gln Cys His Pro Glu Cys Leu Pro
835 840 845
Gln Ala Met Asn Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn Cys Ile
850 855 860
Gln Cys Ala His Tyr Ile Asp Gly Pro His Cys Val Lys Thr Cys Pro
865 870 875 880
Ala Gly Val Met Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr Ala Asp
885 890 895
Ala Gly His Val Cys His Leu Cys His Pro Asn Cys Thr Tyr Gly Cys
900 905 910
Thr Gly Pro Gly Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys Ile Pro
915 920 925
Ser Ile Ala Thr Gly Met Val Gly Ala Leu Leu Leu Leu Leu Val Val
930 935 940
Ala Leu Gly Ile Gly Leu Phe Met
945 950
<210> 58
<211> 1062
<212> Белок
<213> Искусственная последовательность
<220>
<223> Leader _R12- Hinge-CH2-CH3- CD28tm/41BB-Z-T2A-tEGFR
<400> 58
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Gln Glu Gln Leu Val Glu Ser Gly Gly Arg
20 25 30
Leu Val Thr Pro Gly Gly Ser Leu Thr Leu Ser Cys Lys Ala Ser Gly
35 40 45
Phe Asp Phe Ser Ala Tyr Tyr Met Ser Trp Val Arg Gln Ala Pro Gly
50 55 60
Lys Gly Leu Glu Trp Ile Ala Thr Ile Tyr Pro Ser Ser Gly Lys Thr
65 70 75 80
Tyr Tyr Ala Thr Trp Val Asn Gly Arg Phe Thr Ile Ser Ser Asp Asn
85 90 95
Ala Gln Asn Thr Val Asp Leu Gln Met Asn Ser Leu Thr Ala Ala Asp
100 105 110
Arg Ala Thr Tyr Phe Cys Ala Arg Asp Ser Tyr Ala Asp Asp Gly Ala
115 120 125
Leu Phe Asn Ile Trp Gly Pro Gly Thr Leu Val Thr Ile Ser Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu
145 150 155 160
Val Leu Thr Gln Ser Pro Ser Val Ser Ala Ala Leu Gly Ser Pro Ala
165 170 175
Lys Ile Thr Cys Thr Leu Ser Ser Ala His Lys Thr Asp Thr Ile Asp
180 185 190
Trp Tyr Gln Gln Leu Gln Gly Glu Ala Pro Arg Tyr Leu Met Gln Val
195 200 205
Gln Ser Asp Gly Ser Tyr Thr Lys Arg Pro Gly Val Pro Asp Arg Phe
210 215 220
Ser Gly Ser Ser Ser Gly Ala Asp Arg Tyr Leu Ile Ile Pro Ser Val
225 230 235 240
Gln Ala Asp Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Asp Tyr Ile Gly
245 250 255
Gly Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly Glu Ser
260 265 270
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly
275 280 285
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
290 295 300
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
305 310 315 320
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
325 330 335
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
340 345 350
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
355 360 365
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
370 375 380
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
385 390 395 400
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
405 410 415
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
420 425 430
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
435 440 445
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
450 455 460
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
465 470 475 480
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
485 490 495
Leu Gly Lys Met Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala
500 505 510
Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Lys
515 520 525
Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg
530 535 540
Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro
545 550 555 560
Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg Ser
565 570 575
Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu
580 585 590
Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg
595 600 605
Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln
610 615 620
Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr
625 630 635 640
Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp
645 650 655
Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala
660 665 670
Leu His Met Gln Ala Leu Pro Pro Arg Leu Glu Gly Gly Gly Glu Gly
675 680 685
Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro
690 695 700
Arg Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His
705 710 715 720
Pro Ala Phe Leu Leu Ile Pro Arg Lys Val Cys Asn Gly Ile Gly Ile
725 730 735
Gly Glu Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr Asn Ile Lys His
740 745 750
Phe Lys Asn Cys Thr Ser Ile Ser Gly Asp Leu His Ile Leu Pro Val
755 760 765
Ala Phe Arg Gly Asp Ser Phe Thr His Thr Pro Pro Leu Asp Pro Gln
770 775 780
Glu Leu Asp Ile Leu Lys Thr Val Lys Glu Ile Thr Gly Phe Leu Leu
785 790 795 800
Ile Gln Ala Trp Pro Glu Asn Arg Thr Asp Leu His Ala Phe Glu Asn
805 810 815
Leu Glu Ile Ile Arg Gly Arg Thr Lys Gln His Gly Gln Phe Ser Leu
820 825 830
Ala Val Val Ser Leu Asn Ile Thr Ser Leu Gly Leu Arg Ser Leu Lys
835 840 845
Glu Ile Ser Asp Gly Asp Val Ile Ile Ser Gly Asn Lys Asn Leu Cys
850 855 860
Tyr Ala Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly Thr Ser Gly Gln
865 870 875 880
Lys Thr Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser Cys Lys Ala Thr
885 890 895
Gly Gln Val Cys His Ala Leu Cys Ser Pro Glu Gly Cys Trp Gly Pro
900 905 910
Glu Pro Arg Asp Cys Val Ser Cys Arg Asn Val Ser Arg Gly Arg Glu
915 920 925
Cys Val Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro Arg Glu Phe Val
930 935 940
Glu Asn Ser Glu Cys Ile Gln Cys His Pro Glu Cys Leu Pro Gln Ala
945 950 955 960
Met Asn Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn Cys Ile Gln Cys
965 970 975
Ala His Tyr Ile Asp Gly Pro His Cys Val Lys Thr Cys Pro Ala Gly
980 985 990
Val Met Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr Ala Asp Ala Gly
995 1000 1005
His Val Cys His Leu Cys His Pro Asn Cys Thr Tyr Gly Cys Thr
1010 1015 1020
Gly Pro Gly Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys Ile Pro
1025 1030 1035
Ser Ile Ala Thr Gly Met Val Gly Ala Leu Leu Leu Leu Leu Val
1040 1045 1050
Val Ala Leu Gly Ile Gly Leu Phe Met
1055 1060
<210> 59
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Лидерная последовательность
<400> 59
atgcttctcc tggtgacaag ccttctgctc tgtgagttac cacaccca 48
<210> 60
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкерный пептид
<400> 60
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 61
<211> 229
<212> Белок
<213> Искусственная последовательность
<220>
<223> Длинный спейсер
<400> 61
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 62
<211> 687
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Длинный спейсер
<400> 62
gagagcaagt acggaccgcc ctgcccccct tgccctgccc ccgagttcct gggcggaccc 60
agcgtgttcc tgttcccccc caagcccaag gacaccctga tgatcagccg gacccccgag 120
gtgacctgcg tggtggtgga cgtgagccag gaagatcccg aggtccagtt caattggtac 180
gtggacggcg tggaagtgca caacgccaag accaagccca gagaggaaca gttcaacagc 240
acctaccggg tggtgtctgt gctgaccgtg ctgcaccagg actggctgaa cggcaaagaa 300
tacaagtgca aggtgtccaa caagggcctg cccagcagca tcgaaaagac catcagcaag 360
gccaagggcc agcctcgcga gccccaggtg tacaccctgc ctccctccca ggaagagatg 420
accaagaacc aggtgtccct gacctgcctg gtgaagggct tctaccccag cgacatcgcc 480
gtggagtggg agagcaacgg ccagcctgag aacaactaca agaccacccc tcccgtgctg 540
gacagcgacg gcagcttctt cctgtacagc cggctgaccg tggacaagag ccggtggcag 600
gaaggcaacg tctttagctg cagcgtgatg cacgaggccc tgcacaacca ctacacccag 660
aagagcctga gcctgtccct gggcaag 687
<210> 63
<211> 622
<212> Белок
<213> Homo sapiens
<400> 63
Met Ala Leu Pro Thr Ala Arg Pro Leu Leu Gly Ser Cys Gly Thr Pro
1 5 10 15
Ala Leu Gly Ser Leu Leu Phe Leu Leu Phe Ser Leu Gly Trp Val Gln
20 25 30
Pro Ser Arg Thr Leu Ala Gly Glu Thr Gly Gln Glu Ala Ala Pro Leu
35 40 45
Asp Gly Val Leu Ala Asn Pro Pro Asn Ile Ser Ser Leu Ser Pro Arg
50 55 60
Gln Leu Leu Gly Phe Pro Cys Ala Glu Val Ser Gly Leu Ser Thr Glu
65 70 75 80
Arg Val Arg Glu Leu Ala Val Ala Leu Ala Gln Lys Asn Val Lys Leu
85 90 95
Ser Thr Glu Gln Leu Arg Cys Leu Ala His Arg Leu Ser Glu Pro Pro
100 105 110
Glu Asp Leu Asp Ala Leu Pro Leu Asp Leu Leu Leu Phe Leu Asn Pro
115 120 125
Asp Ala Phe Ser Gly Pro Gln Ala Cys Thr His Phe Phe Ser Arg Ile
130 135 140
Thr Lys Ala Asn Val Asp Leu Leu Pro Arg Gly Ala Pro Glu Arg Gln
145 150 155 160
Arg Leu Leu Pro Ala Ala Leu Ala Cys Trp Gly Val Arg Gly Ser Leu
165 170 175
Leu Ser Glu Ala Asp Val Arg Ala Leu Gly Gly Leu Ala Cys Asp Leu
180 185 190
Pro Gly Arg Phe Val Ala Glu Ser Ala Glu Val Leu Leu Pro Arg Leu
195 200 205
Val Ser Cys Pro Gly Pro Leu Asp Gln Asp Gln Gln Glu Ala Ala Arg
210 215 220
Ala Ala Leu Gln Gly Gly Gly Pro Pro Tyr Gly Pro Pro Ser Thr Trp
225 230 235 240
Ser Val Ser Thr Met Asp Ala Leu Arg Gly Leu Leu Pro Val Leu Gly
245 250 255
Gln Pro Ile Ile Arg Ser Ile Pro Gln Gly Ile Val Ala Ala Trp Arg
260 265 270
Gln Arg Ser Ser Arg Asp Pro Ser Trp Arg Gln Pro Glu Arg Thr Ile
275 280 285
Leu Arg Pro Arg Phe Arg Arg Glu Val Glu Lys Thr Ala Cys Pro Ser
290 295 300
Gly Lys Lys Ala Arg Glu Ile Asp Glu Ser Leu Ile Phe Tyr Lys Lys
305 310 315 320
Trp Glu Leu Glu Ala Cys Val Asp Ala Ala Leu Leu Ala Thr Gln Met
325 330 335
Asp Arg Val Asn Ala Ile Pro Phe Thr Tyr Glu Gln Leu Asp Val Leu
340 345 350
Lys His Lys Leu Asp Glu Leu Tyr Pro Gln Gly Tyr Pro Glu Ser Val
355 360 365
Ile Gln His Leu Gly Tyr Leu Phe Leu Lys Met Ser Pro Glu Asp Ile
370 375 380
Arg Lys Trp Asn Val Thr Ser Leu Glu Thr Leu Lys Ala Leu Leu Glu
385 390 395 400
Val Asn Lys Gly His Glu Met Ser Pro Gln Val Ala Thr Leu Ile Asp
405 410 415
Arg Phe Val Lys Gly Arg Gly Gln Leu Asp Lys Asp Thr Leu Asp Thr
420 425 430
Leu Thr Ala Phe Tyr Pro Gly Tyr Leu Cys Ser Leu Ser Pro Glu Glu
435 440 445
Leu Ser Ser Val Pro Pro Ser Ser Ile Trp Ala Val Arg Pro Gln Asp
450 455 460
Leu Asp Thr Cys Asp Pro Arg Gln Leu Asp Val Leu Tyr Pro Lys Ala
465 470 475 480
Arg Leu Ala Phe Gln Asn Met Asn Gly Ser Glu Tyr Phe Val Lys Ile
485 490 495
Gln Ser Phe Leu Gly Gly Ala Pro Thr Glu Asp Leu Lys Ala Leu Ser
500 505 510
Gln Gln Asn Val Ser Met Asp Leu Ala Thr Phe Met Lys Leu Arg Thr
515 520 525
Asp Ala Val Leu Pro Leu Thr Val Ala Glu Val Gln Lys Leu Leu Gly
530 535 540
Pro His Val Glu Gly Leu Lys Ala Glu Glu Arg His Arg Pro Val Arg
545 550 555 560
Asp Trp Ile Leu Arg Gln Arg Gln Asp Asp Leu Asp Thr Leu Gly Leu
565 570 575
Gly Leu Gln Gly Gly Ile Pro Asn Gly Tyr Leu Val Leu Asp Leu Ser
580 585 590
Val Gln Glu Ala Leu Ser Gly Thr Pro Cys Leu Leu Gly Pro Gly Pro
595 600 605
Val Leu Thr Val Leu Ala Leu Leu Leu Ala Ser Thr Leu Ala
610 615 620
<210> 64
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер mid-Ampr
<400> 64
ttgagagttt tcgccccg 18
<210> 65
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Модифицированный человеческий IgG4
<400> 65
Glu Val Val Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 66
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Модифицированный человеческий IgG4
<400> 66
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 67
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Модифицированный человеческий IgG4
<400> 67
Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5
<210> 68
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Модифицированный человеческий IgG4
<400> 68
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 69
<211> 750
<212> Белок
<213> Homo sapiens
<400> 69
Met Trp Asn Leu Leu His Glu Thr Asp Ser Ala Val Ala Thr Ala Arg
1 5 10 15
Arg Pro Arg Trp Leu Cys Ala Gly Ala Leu Val Leu Ala Gly Gly Phe
20 25 30
Phe Leu Leu Gly Phe Leu Phe Gly Trp Phe Ile Lys Ser Ser Asn Glu
35 40 45
Ala Thr Asn Ile Thr Pro Lys His Asn Met Lys Ala Phe Leu Asp Glu
50 55 60
Leu Lys Ala Glu Asn Ile Lys Lys Phe Leu Tyr Asn Phe Thr Gln Ile
65 70 75 80
Pro His Leu Ala Gly Thr Glu Gln Asn Phe Gln Leu Ala Lys Gln Ile
85 90 95
Gln Ser Gln Trp Lys Glu Phe Gly Leu Asp Ser Val Glu Leu Ala His
100 105 110
Tyr Asp Val Leu Leu Ser Tyr Pro Asn Lys Thr His Pro Asn Tyr Ile
115 120 125
Ser Ile Ile Asn Glu Asp Gly Asn Glu Ile Phe Asn Thr Ser Leu Phe
130 135 140
Glu Pro Pro Pro Pro Gly Tyr Glu Asn Val Ser Asp Ile Val Pro Pro
145 150 155 160
Phe Ser Ala Phe Ser Pro Gln Gly Met Pro Glu Gly Asp Leu Val Tyr
165 170 175
Val Asn Tyr Ala Arg Thr Glu Asp Phe Phe Lys Leu Glu Arg Asp Met
180 185 190
Lys Ile Asn Cys Ser Gly Lys Ile Val Ile Ala Arg Tyr Gly Lys Val
195 200 205
Phe Arg Gly Asn Lys Val Lys Asn Ala Gln Leu Ala Gly Ala Lys Gly
210 215 220
Val Ile Leu Tyr Ser Asp Pro Ala Asp Tyr Phe Ala Pro Gly Val Lys
225 230 235 240
Ser Tyr Pro Asp Gly Trp Asn Leu Pro Gly Gly Gly Val Gln Arg Gly
245 250 255
Asn Ile Leu Asn Leu Asn Gly Ala Gly Asp Pro Leu Thr Pro Gly Tyr
260 265 270
Pro Ala Asn Glu Tyr Ala Tyr Arg Arg Gly Ile Ala Glu Ala Val Gly
275 280 285
Leu Pro Ser Ile Pro Val His Pro Ile Gly Tyr Tyr Asp Ala Gln Lys
290 295 300
Leu Leu Glu Lys Met Gly Gly Ser Ala Pro Pro Asp Ser Ser Trp Arg
305 310 315 320
Gly Ser Leu Lys Val Pro Tyr Asn Val Gly Pro Gly Phe Thr Gly Asn
325 330 335
Phe Ser Thr Gln Lys Val Lys Met His Ile His Ser Thr Asn Glu Val
340 345 350
Thr Arg Ile Tyr Asn Val Ile Gly Thr Leu Arg Gly Ala Val Glu Pro
355 360 365
Asp Arg Tyr Val Ile Leu Gly Gly His Arg Asp Ser Trp Val Phe Gly
370 375 380
Gly Ile Asp Pro Gln Ser Gly Ala Ala Val Val His Glu Ile Val Arg
385 390 395 400
Ser Phe Gly Thr Leu Lys Lys Glu Gly Trp Arg Pro Arg Arg Thr Ile
405 410 415
Leu Phe Ala Ser Trp Asp Ala Glu Glu Phe Gly Leu Leu Gly Ser Thr
420 425 430
Glu Trp Ala Glu Glu Asn Ser Arg Leu Leu Gln Glu Arg Gly Val Ala
435 440 445
Tyr Ile Asn Ala Asp Ser Ser Ile Glu Gly Asn Tyr Thr Leu Arg Val
450 455 460
Asp Cys Thr Pro Leu Met Tyr Ser Leu Val His Asn Leu Thr Lys Glu
465 470 475 480
Leu Lys Ser Pro Asp Glu Gly Phe Glu Gly Lys Ser Leu Tyr Glu Ser
485 490 495
Trp Thr Lys Lys Ser Pro Ser Pro Glu Phe Ser Gly Met Pro Arg Ile
500 505 510
Ser Lys Leu Gly Ser Gly Asn Asp Phe Glu Val Phe Phe Gln Arg Leu
515 520 525
Gly Ile Ala Ser Gly Arg Ala Arg Tyr Thr Lys Asn Trp Glu Thr Asn
530 535 540
Lys Phe Ser Gly Tyr Pro Leu Tyr His Ser Val Tyr Glu Thr Tyr Glu
545 550 555 560
Leu Val Glu Lys Phe Tyr Asp Pro Met Phe Lys Tyr His Leu Thr Val
565 570 575
Ala Gln Val Arg Gly Gly Met Val Phe Glu Leu Ala Asn Ser Ile Val
580 585 590
Leu Pro Phe Asp Cys Arg Asp Tyr Ala Val Val Leu Arg Lys Tyr Ala
595 600 605
Asp Lys Ile Tyr Ser Ile Ser Met Lys His Pro Gln Glu Met Lys Thr
610 615 620
Tyr Ser Val Ser Phe Asp Ser Leu Phe Ser Ala Val Lys Asn Phe Thr
625 630 635 640
Glu Ile Ala Ser Lys Phe Ser Glu Arg Leu Gln Asp Phe Asp Lys Ser
645 650 655
Asn Pro Ile Val Leu Arg Met Met Asn Asp Gln Leu Met Phe Leu Glu
660 665 670
Arg Ala Phe Ile Asp Pro Leu Gly Leu Pro Asp Arg Pro Phe Tyr Arg
675 680 685
His Val Ile Tyr Ala Pro Ser Ser His Asn Lys Tyr Ala Gly Glu Ser
690 695 700
Phe Pro Gly Ile Tyr Asp Ala Leu Phe Asp Ile Glu Ser Lys Val Asp
705 710 715 720
Pro Ser Lys Ala Trp Gly Glu Val Lys Arg Gln Ile Tyr Val Ala Ala
725 730 735
Phe Thr Val Gln Ala Ala Ala Glu Thr Leu Ser Glu Val Ala
740 745 750
<210> 70
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер post-Ampr
<400> 70
aatagacaga tcgctgagat aggt 24
<210> 71
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер pre-U5
<400> 71
atcaaaagaa tagaccgaga tagggt 26
<210> 72
<211> 123
<212> Белок
<213> Homo sapiens
<400> 72
Met Lys Ala Val Leu Leu Ala Leu Leu Met Ala Gly Leu Ala Leu Gln
1 5 10 15
Pro Gly Thr Ala Leu Leu Cys Tyr Ser Cys Lys Ala Gln Val Ser Asn
20 25 30
Glu Asp Cys Leu Gln Val Glu Asn Cys Thr Gln Leu Gly Glu Gln Cys
35 40 45
Trp Thr Ala Arg Ile Arg Ala Val Gly Leu Leu Thr Val Ile Ser Lys
50 55 60
Gly Cys Ser Leu Asn Cys Val Asp Asp Ser Gln Asp Tyr Tyr Val Gly
65 70 75 80
Lys Lys Asn Ile Thr Cys Cys Asp Thr Asp Leu Cys Asn Ala Ser Gly
85 90 95
Ala His Ala Leu Gln Pro Ala Ala Ala Ile Leu Ala Leu Leu Pro Ala
100 105 110
Leu Gly Leu Leu Leu Trp Gly Pro Gly Gln Leu
115 120
<210> 73
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер psi
<400> 73
gcagggagct agaacgattc 20
<210> 74
<211> 10014
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ХАР R11 со спейсером средней длины: PJ_R11-CH3-41BB-Z-T2A-tEGFR
<400> 74
gttagaccag atctgagcct gggagctctc tggctaacta gggaacccac tgcttaagcc 60
tcaataaagc ttgccttgag tgcttcaagt agtgtgtgcc cgtctgttgt gtgactctgg 120
taactagaga tccctcagac ccttttagtc agtgtggaaa atctctagca gtggcgcccg 180
aacagggact tgaaagcgaa agggaaacca gaggagctct ctcgacgcag gactcggctt 240
gctgaagcgc gcacggcaag aggcgagggg cggcgactgg tgagtacgcc aaaaattttg 300
actagcggag gctagaagga gagagatggg tgcgagagcg tcagtattaa gcgggggaga 360
attagatcga tgggaaaaaa ttcggttaag gccaggggga aagaaaaaat ataaattaaa 420
acatatagta tgggcaagca gggagctaga acgattcgca gttaatcctg gcctgttaga 480
aacatcagaa ggctgtagac aaatactggg acagctacaa ccatcccttc agacaggatc 540
agaagaactt agatcattat ataatacagt agcaaccctc tattgtgtgc atcaaaggat 600
agagataaaa gacaccaagg aagctttaga caagatagag gaagagcaaa acaaaagtaa 660
gaaaaaagca cagcaagcag cagctgacac aggacacagc aatcaggtca gccaaaatta 720
ccctatagtg cagaacatcc aggggcaaat ggtacatcag gccatatcac ctagaacttt 780
aaatgcatgg gtaaaagtag tagaagagaa ggctttcagc ccagaagtga tacccatgtt 840
ttcagcatta tcagaaggag ccaccccaca agatttaaac accatgctaa acacagtggg 900
gggacatcaa gcagccatgc aaatgttaaa agagaccatc aatgaggaag ctgcaggcaa 960
agagaagagt ggtgcagaga gaaaaaagag cagtgggaat aggagctttg ttccttgggt 1020
tcttgggagc agcaggaagc actatgggcg cagcgtcaat gacgctgacg gtacaggcca 1080
gacaattatt gtctggtata gtgcagcagc agaacaattt gctgagggct attgaggcgc 1140
aacagcatct gttgcaactc acagtctggg gcatcaagca gctccaggca agaatcctgg 1200
ctgtggaaag atacctaaag gatcaacagc tcctggggat ttggggttgc tctggaaaac 1260
tcatttgcac cactgctgtg ccttggatct acaaatggca gtattcatcc acaattttaa 1320
aagaaaaggg gggattgggg ggtacagtgc aggggaaaga atagtagaca taatagcaac 1380
agacatacaa actaaagaat tacaaaaaca aattacaaaa attcaaaatt ttcgggttta 1440
ttacagggac agcagagatc cagtttgggg atcaattgca tgaagaatct gcttagggtt 1500
aggcgttttg cgctgcttcg cgaggatctg cgatcgctcc ggtgcccgtc agtgggcaga 1560
gcgcacatcg cccacagtcc ccgagaagtt ggggggaggg gtcggcaatt gaaccggtgc 1620
ctagagaagg tggcgcgggg taaactggga aagtgatgtc gtgtactggc tccgcctttt 1680
tcccgagggt gggggagaac cgtatataag tgcagtagtc gccgtgaacg ttctttttcg 1740
caacgggttt gccgccagaa cacagctgaa gcttcgaggg gctcgcatct ctccttcacg 1800
cgcccgccgc cctacctgag gccgccatcc acgccggttg agtcgcgttc tgccgcctcc 1860
cgcctgtggt gcctcctgaa ctgcgtccgc cgtctaggta agtttaaagc tcaggtcgag 1920
accgggcctt tgtccggcgc tcccttggag cctacctaga ctcagccggc tctccacgct 1980
ttgcctgacc ctgcttgctc aactctacgt ctttgtttcg ttttctgttc tgcgccgtta 2040
cagatccaag ctgtgaccgg cgcctacggc tagcgaattc gccaccatgc tgctgctggt 2100
gacaagcctg ctgctgtgcg agctgcccca ccccgccttt ctgctgatcc cccagagcgt 2160
gaaagagtcc gagggcgacc tggtcacacc agccggcaac ctgaccctga cctgtaccgc 2220
cagcggcagc gacatcaacg actaccccat ctcttgggtc cgccaggctc ctggcaaggg 2280
actggaatgg atcggcttca tcaacagcgg cggcagcact tggtacgcca gctgggtcaa 2340
aggccggttc accatcagcc ggaccagcac caccgtggac ctgaagatga caagcctgac 2400
caccgacgac accgccacct acttttgcgc cagaggctac agcacctact acggcgactt 2460
caacatctgg ggccctggca ccctggtcac aatctctagc ggcggaggcg gcagcggagg 2520
tggaggaagt ggcggcggag gatccgagct ggtcatgacc cagaccccca gcagcacatc 2580
tggcgccgtg ggcggcaccg tgaccatcaa ttgccaggcc agccagagca tcgacagcaa 2640
cctggcctgg ttccagcaga agcccggcca gccccccacc ctgctgatct acagagcctc 2700
caacctggcc agcggcgtgc caagcagatt cagcggcagc agatctggca ccgagtacac 2760
cctgaccatc tccggcgtgc agagagagga cgccgctacc tattactgcc tgggcggcgt 2820
gggcaacgtg tcctacagaa ccagcttcgg cggaggtact gaggtggtcg tcaaatagga 2880
ccgccctgcc ccccttgccc tgcccccgag ttcctgggcg gacccagcgt gttcctgttc 2940
ccccccaagc ccaaggacac cctgatgatc agccggaccc ccgaggtgac ctgcgtggtg 3000
gtggacgtga gccaggaaga tcccgaggtc cagttcaatt ggtacgtgga cggcgtggaa 3060
gtgcacaacg ccaagaccaa gcccagagag gaacagttca acagcaccta ccgggtggtg 3120
tctgtgctga ccgtgctgca ccaggactgg ctgaacggca aagaatacaa gtgcaaggtg 3180
tccaacaagg gcctgcccag cagcatcgaa aagaccatca gcaaggccaa gggccagcct 3240
cgcgagcccc aggtgtacac cctgcctccc tcccaggaag agatgaccaa gaaccaggtg 3300
tccctgacct gcctggtgaa gggcttctac cccagcgaca tcgccgtgga gtgggagagc 3360
aacggccagc ctgagaacaa ctacaagacc acccctcccg tgctggacag cgacggcagc 3420
ttcttcctgt acagccggct gaccgtggac aagagccggt ggcaggaagg caacgtcttt 3480
agctgcagcg tgatgcacga ggccctgcac aaccactaca cccagaagag cctgagcctg 3540
tccctgggca agatgttctg ggtgctggtg gtggtgggcg gggtgctggc ctgctacagc 3600
ctgctggtga cagtggcctt catcatcttt tgggtgaaac ggggcagaaa gaaactcctg 3660
tatatattca aacaaccatt tatgagacca gtacaaacta ctcaagagga agatggctgt 3720
agctgccgat ttccagaaga agaagaagga ggatgtgaac tgcgggtgaa gttcagcaga 3780
agcgccgacg cccctgccta ccagcagggc cagaatcagc tgtacaacga gctgaacctg 3840
ggcagaaggg aagagtacga cgtcctggat aagcggagag gccgggaccc tgagatgggc 3900
ggcaagcctc ggcggaagaa cccccaggaa ggcctgtata acgaactgca gaaagacaag 3960
atggccgagg cctacagcga gatcggcatg aagggcgagc ggaggcgggg caagggccac 4020
gacggcctgt atcagggcct gtccaccgcc accaaggata cctacgacgc cctgcacatg 4080
caggccctgc ccccaaggct cgagggcggc ggagagggca gaggaagtct tctaacatgc 4140
ggtgacgtgg aggagaatcc cggccctagg atgcttctcc tggtgacaag ccttctgctc 4200
tgtgagttac cacacccagc attcctcctg atcccacgca aagtgtgtaa cggaataggt 4260
attggtgaat ttaaagactc actctccata aatgctacga atattaaaca cttcaaaaac 4320
tgcacctcca tcagtggcga tctccacatc ctgccggtgg catttagggg tgactccttc 4380
acacatactc ctcctctgga tccacaggaa ctggatattc tgaaaaccgt aaaggaaatc 4440
acagggtttt tgctgattca ggcttggcct gaaaacagga cggacctcca tgcctttgag 4500
aacctagaaa tcatacgcgg caggaccaag caacatggtc agttttctct tgcagtcgtc 4560
agcctgaaca taacatcctt gggattacgc tccctcaagg agataagtga tggagatgtg 4620
ataatttcag gaaacaaaaa tttgtgctat gcaaatacaa taaactggaa aaaactgttt 4680
gggacctccg gtcagaaaac caaaattata agcaacagag gtgaaaacag ctgcaaggcc 4740
acaggccagg tctgccatgc cttgtgctcc cccgagggct gctggggccc ggagcccagg 4800
gactgcgtct cttgccggaa tgtcagccga ggcagggaat gcgtggacaa gtgcaacctt 4860
ctggagggtg agccaaggga gtttgtggag aactctgagt gcatacagtg ccacccagag 4920
tgcctgcctc aggccatgaa catcacctgc acaggacggg gaccagacaa ctgtatccag 4980
tgtgcccact acattgacgg cccccactgc gtcaagacct gcccggcagg agtcatggga 5040
gaaaacaaca ccctggtctg gaagtacgca gacgccggcc atgtgtgcca cctgtgccat 5100
ccaaactgca cctacggatg cactgggcca ggtcttgaag gctgtccaac gaatgggcct 5160
aagatcccgt ccatcgccac tgggatggtg ggggccctcc tcttgctgct ggtggtggcc 5220
ctggggatcg gcctcttcat gtgagcggcc gctctagacc cgggctgcag gaattcgata 5280
tcaagcttat cgataatcaa cctctggatt acaaaatttg tgaaagattg actggtattc 5340
ttaactatgt tgctcctttt acgctatgtg gatacgctgc tttaatgcct ttgtatcatg 5400
ctattgcttc ccgtatggct ttcattttct cctccttgta taaatcctgg ttgctgtctc 5460
tttatgagga gttgtggccc gttgtcaggc aacgtggcgt ggtgtgcact gtgtttgctg 5520
acgcaacccc cactggttgg ggcattgcca ccacctgtca gctcctttcc gggactttcg 5580
ctttccccct ccctattgcc acggcggaac tcatcgccgc ctgccttgcc cgctgctgga 5640
caggggctcg gctgttgggc actgacaatt ccgtggtgtt gtcggggaaa tcatcgtcct 5700
ttccttggct gctcgcctgt gttgccacct ggattctgcg cgggacgtcc ttctgctacg 5760
tcccttcggc cctcaatcca gcggaccttc cttcccgcgg cctgctgccg gctctgcggc 5820
ctcttccgcg tcttcgcctt cgccctcaga cgagtcggat ctccctttgg gccgcctccc 5880
cgcatcgata ccgtcgacta gccgtacctt taagaccaat gacttacaag gcagctgtag 5940
atcttagcca ctttttaaaa gaaaaggggg gactggaagg gctaattcac tcccaaagaa 6000
gacaagatct gctttttgcc tgtactgggt ctctctggtt agaccagatc tgagcctggg 6060
agctctctgg ctaactaggg aacccactgc ttaagcctca ataaagcttg ccttgagtgc 6120
ttcaagtagt gtgtgcccgt ctgttgtgtg actctggtaa ctagagatcc ctcagaccct 6180
tttagtcagt gtggaaaatc tctagcagaa ttcgatatca agcttatcga taccgtcgac 6240
ctcgaggggg ggcccggtac ccaattcgcc ctatagtgag tcgtattaca attcactggc 6300
cgtcgtttta caacgtcgtg actgggaaaa ccctggcgtt acccaactta atcgccttgc 6360
agcacatccc cctttcgcca gctggcgtaa tagcgaagag gcccgcaccg atcgcccttc 6420
ccaacagttg cgcagcctga atggcgaatg gaaattgtaa gcgttaatat tttgttaaaa 6480
ttcgcgttaa atttttgtta aatcagctca ttttttaacc aataggccga aatcggcaaa 6540
atcccttata aatcaaaaga atagaccgag atagggttga gtgttgttcc agtttggaac 6600
aagagtccac tattaaagaa cgtggactcc aacgtcaaag ggcgaaaaac cgtctatcag 6660
ggcgatggcc cactacgtga accatcaccc taatcaagtt ttttggggtc gaggtgccgt 6720
aaagcactaa atcggaaccc taaagggagc ccccgattta gagcttgacg gggaaagccg 6780
gcgaacgtgg cgagaaagga agggaagaaa gcgaaaggag cgggcgctag ggcgctggca 6840
agtgtagcgg tcacgctgcg cgtaaccacc acacccgccg cgcttaatgc gccgctacag 6900
ggcgcgtcag gtggcacttt tcggggaaat gtgcgcggaa cccctatttg tttatttttc 6960
taaatacatt caaatatgta tccgctcatg agacaataac cctgataaat gcttcaataa 7020
tattgaaaaa ggaagagtat gagtattcaa catttccgtg tcgcccttat tccctttttt 7080
gcggcatttt gccttcctgt ttttgctcac ccagaaacgc tggtgaaagt aaaagatgct 7140
gaagatcagt tgggtgcacg agtgggttac atcgaactgg atctcaacag cggtaagatc 7200
cttgagagtt ttcgccccga agaacgtttt ccaatgatga gcacttttaa agttctgcta 7260
tgtggcgcgg tattatcccg tattgacgcc gggcaagagc aactcggtcg ccgcatacac 7320
tattctcaga atgacttggt tgagtactca ccagtcacag aaaagcatct tacggatggc 7380
atgacagtaa gagaattatg cagtgctgcc ataaccatga gtgataacac tgcggccaac 7440
ttacttctga caacgatcgg aggaccgaag gagctaaccg cttttttgca caacatgggg 7500
gatcatgtaa ctcgccttga tcgttgggaa ccggagctga atgaagccat accaaacgac 7560
gagcgtgaca ccacgatgcc tgtagcaatg gcaacaacgt tgcgcaaact attaactggc 7620
gaactactta ctctagcttc ccggcaacaa ttaatagact ggatggaggc ggataaagtt 7680
gcaggaccac ttctgcgctc ggcccttccg gctggctggt ttattgctga taaatctgga 7740
gccggtgagc gtgggtctcg cggtatcatt gcagcactgg ggccagatgg taagccctcc 7800
cgtatcgtag ttatctacac gacggggagt caggcaacta tggatgaacg aaatagacag 7860
atcgctgaga taggtgcctc actgattaag cattggtaac tgtcagacca agtttactca 7920
tatatacttt agattgattt aaaacttcat ttttaattta aaaggatcta ggtgaagatc 7980
ctttttgata atctcatgac caaaatccct taacgtgagt tttcgttcca ctgagcgtca 8040
gaccccgtag aaaagatcaa aggatcttct tgagatcctt tttttctgcg cgtaatctgc 8100
tgcttgcaaa caaaaaaacc accgctacca gcggtggttt gtttgccgga tcaagagcta 8160
ccaactcttt ttccgaaggt aactggcttc agcagagcgc agataccaaa tactgttctt 8220
ctagtgtagc cgtagttagg ccaccacttc aagaactctg tagcaccgcc tacatacctc 8280
gctctgctaa tcctgttacc agtggctgct gccagtggcg ataagtcgtg tcttaccggg 8340
ttggactcaa gacgatagtt accggataag gcgcagcggt cgggctgaac ggggggttcg 8400
tgcacacagc ccagcttgga gcgaacgacc tacaccgaac tgagatacct acagcgtgag 8460
ctatgagaaa gcgccacgct tcccgaaggg agaaaggcgg acaggtatcc ggtaagcggc 8520
agggtcggaa caggagagcg cacgagggag cttccagggg gaaacgcctg gtatctttat 8580
agtcctgtcg ggtttcgcca cctctgactt gagcgtcgat ttttgtgatg ctcgtcaggg 8640
gggcggagcc tatggaaaaa cgccagcaac gcggcctttt tacggttcct ggccttttgc 8700
tggccttttg ctcacatgtt ctttcctgcg ttatcccctg attctgtgga taaccgtatt 8760
accgcctttg agtgagctga taccgctcgc cgcagccgaa cgaccgagcg cagcgagtca 8820
gtgagcgagg aagcggaaga gcgcccaata cgcaaaccgc ctctccccgc gcgttggccg 8880
attcattaat gcagctggca cgacaggttt cccgactgga aagcgggcag tgagcgcaac 8940
gcaattaatg tgagttagct cactcattag gcaccccagg ctttacactt tatgcttccg 9000
gctcgtatgt tgtgtggaat tgtgagcgga taacaatttc acacaggaaa cagctatgac 9060
catgattacg ccaagctcga aattaaccct cactaaaggg aacaaaagct ggagctccac 9120
cgcggtggcg gcctcgaggt cgagatccgg tcgaccagca accatagtcc cgcccctaac 9180
tccgcccatc ccgcccctaa ctccgcccag ttccgcccat tctccgcccc atggctgact 9240
aatttttttt atttatgcag aggccgaggc cgcctcggcc tctgagctat tccagaagta 9300
gtgaggaggc ttttttggag gcctaggctt ttgcaaaaag cttcgacggt atcgattggc 9360
tcatgtccaa cattaccgcc atgttgacat tgattattga ctagttatta atagtaatca 9420
attacggggt cattagttca tagcccatat atggagttcc gcgttacata acttacggta 9480
aatggcccgc ctggctgacc gcccaacgac ccccgcccat tgacgtcaat aatgacgtat 9540
gttcccatag taacgccaat agggactttc cattgacgtc aatgggtgga gtatttacgg 9600
taaactgccc acttggcagt acatcaagtg tatcatatgc caagtacgcc ccctattgac 9660
gtcaatgacg gtaaatggcc cgcctggcat tatgcccagt acatgacctt atgggacttt 9720
cctacttggc agtacatcta cgtattagtc atcgctatta ccatggtgat gcggttttgg 9780
cagtacatca atgggcgtgg atagcggttt gactcacggg gatttccaag tctccacccc 9840
attgacgtca atgggagttt gttttggcac caaaatcaac gggactttcc aaaatgtcgt 9900
aacaactccg ccccattgac gcaaatgggc ggtaggcgtg tacggaattc ggagtggcga 9960
gccctcagat cctgcatata agcagctgct ttttgcctgt actgggtctc tctg 10014
<210> 75
<211> 10015
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ХАР R11 с длинным спейсером: PJ_R11-CH2-CH3-41BB-Z-T2A-tEGFR
<400> 75
gttagaccag atctgagcct gggagctctc tggctaacta gggaacccac tgcttaagcc 60
tcaataaagc ttgccttgag tgcttcaagt agtgtgtgcc cgtctgttgt gtgactctgg 120
taactagaga tccctcagac ccttttagtc agtgtggaaa atctctagca gtggcgcccg 180
aacagggact tgaaagcgaa agggaaacca gaggagctct ctcgacgcag gactcggctt 240
gctgaagcgc gcacggcaag aggcgagggg cggcgactgg tgagtacgcc aaaaattttg 300
actagcggag gctagaagga gagagatggg tgcgagagcg tcagtattaa gcgggggaga 360
attagatcga tgggaaaaaa ttcggttaag gccaggggga aagaaaaaat ataaattaaa 420
acatatagta tgggcaagca gggagctaga acgattcgca gttaatcctg gcctgttaga 480
aacatcagaa ggctgtagac aaatactggg acagctacaa ccatcccttc agacaggatc 540
agaagaactt agatcattat ataatacagt agcaaccctc tattgtgtgc atcaaaggat 600
agagataaaa gacaccaagg aagctttaga caagatagag gaagagcaaa acaaaagtaa 660
gaaaaaagca cagcaagcag cagctgacac aggacacagc aatcaggtca gccaaaatta 720
ccctatagtg cagaacatcc aggggcaaat ggtacatcag gccatatcac ctagaacttt 780
aaatgcatgg gtaaaagtag tagaagagaa ggctttcagc ccagaagtga tacccatgtt 840
ttcagcatta tcagaaggag ccaccccaca agatttaaac accatgctaa acacagtggg 900
gggacatcaa gcagccatgc aaatgttaaa agagaccatc aatgaggaag ctgcaggcaa 960
agagaagagt ggtgcagaga gaaaaaagag cagtgggaat aggagctttg ttccttgggt 1020
tcttgggagc agcaggaagc actatgggcg cagcgtcaat gacgctgacg gtacaggcca 1080
gacaattatt gtctggtata gtgcagcagc agaacaattt gctgagggct attgaggcgc 1140
aacagcatct gttgcaactc acagtctggg gcatcaagca gctccaggca agaatcctgg 1200
ctgtggaaag atacctaaag gatcaacagc tcctggggat ttggggttgc tctggaaaac 1260
tcatttgcac cactgctgtg ccttggatct acaaatggca gtattcatcc acaattttaa 1320
aagaaaaggg gggattgggg ggtacagtgc aggggaaaga atagtagaca taatagcaac 1380
agacatacaa actaaagaat tacaaaaaca aattacaaaa attcaaaatt ttcgggttta 1440
ttacagggac agcagagatc cagtttgggg atcaattgca tgaagaatct gcttagggtt 1500
aggcgttttg cgctgcttcg cgaggatctg cgatcgctcc ggtgcccgtc agtgggcaga 1560
gcgcacatcg cccacagtcc ccgagaagtt ggggggaggg gtcggcaatt gaaccggtgc 1620
ctagagaagg tggcgcgggg taaactggga aagtgatgtc gtgtactggc tccgcctttt 1680
tcccgagggt gggggagaac cgtatataag tgcagtagtc gccgtgaacg ttctttttcg 1740
caacgggttt gccgccagaa cacagctgaa gcttcgaggg gctcgcatct ctccttcacg 1800
cgcccgccgc cctacctgag gccgccatcc acgccggttg agtcgcgttc tgccgcctcc 1860
cgcctgtggt gcctcctgaa ctgcgtccgc cgtctaggta agtttaaagc tcaggtcgag 1920
accgggcctt tgtccggcgc tcccttggag cctacctaga ctcagccggc tctccacgct 1980
ttgcctgacc ctgcttgctc aactctacgt ctttgtttcg ttttctgttc tgcgccgtta 2040
cagatccaag ctgtgaccgg cgcctacggc tagcgaattc gccaccatgc tgctgctggt 2100
gacaagcctg ctgctgtgcg agctgcccca ccccgccttt ctgctgatcc cccagagcgt 2160
gaaagagtcc gagggcgacc tggtcacacc agccggcaac ctgaccctga cctgtaccgc 2220
cagcggcagc gacatcaacg actaccccat ctcttgggtc cgccaggctc ctggcaaggg 2280
actggaatgg atcggcttca tcaacagcgg cggcagcact tggtacgcca gctgggtcaa 2340
aggccggttc accatcagcc ggaccagcac caccgtggac ctgaagatga caagcctgac 2400
caccgacgac accgccacct acttttgcgc cagaggctac agcacctact acggcgactt 2460
caacatctgg ggccctggca ccctggtcac aatctctagc ggcggaggcg gcagcggagg 2520
tggaggaagt ggcggcggag gatccgagct ggtcatgacc cagaccccca gcagcacatc 2580
tggcgccgtg ggcggcaccg tgaccatcaa ttgccaggcc agccagagca tcgacagcaa 2640
cctggcctgg ttccagcaga agcccggcca gccccccacc ctgctgatct acagagcctc 2700
caacctggcc agcggcgtgc caagcagatt cagcggcagc agatctggca ccgagtacac 2760
cctgaccatc tccggcgtgc agagagagga cgccgctacc tattactgcc tgggcggcgt 2820
gggcaacgtg tcctacagaa ccagcttcgg cggaggtact gaggtggtcg tcaaatacgg 2880
accgccctgc cccccttgcc ctgcccccga gttcctgggc ggacccagcg tgttcctgtt 2940
cccccccaag cccaaggaca ccctgatgat cagccggacc cccgaggtga cctgcgtggt 3000
ggtggacgtg agccaggaag atcccgaggt ccagttcaat tggtacgtgg acggcgtgga 3060
agtgcacaac gccaagacca agcccagaga ggaacagttc aacagcacct accgggtggt 3120
gtctgtgctg accgtgctgc accaggactg gctgaacggc aaagaataca agtgcaaggt 3180
gtccaacaag ggcctgccca gcagcatcga aaagaccatc agcaaggcca agggccagcc 3240
tcgcgagccc caggtgtaca ccctgcctcc ctcccaggaa gagatgacca agaaccaggt 3300
gtccctgacc tgcctggtga agggcttcta ccccagcgac atcgccgtgg agtgggagag 3360
caacggccag cctgagaaca actacaagac cacccctccc gtgctggaca gcgacggcag 3420
cttcttcctg tacagccggc tgaccgtgga caagagccgg tggcaggaag gcaacgtctt 3480
tagctgcagc gtgatgcacg aggccctgca caaccactac acccagaaga gcctgagcct 3540
gtccctgggc aagatgttct gggtgctggt ggtggtgggc ggggtgctgg cctgctacag 3600
cctgctggtg acagtggcct tcatcatctt ttgggtgaaa cggggcagaa agaaactcct 3660
gtatatattc aaacaaccat ttatgagacc agtacaaact actcaagagg aagatggctg 3720
tagctgccga tttccagaag aagaagaagg aggatgtgaa ctgcgggtga agttcagcag 3780
aagcgccgac gcccctgcct accagcaggg ccagaatcag ctgtacaacg agctgaacct 3840
gggcagaagg gaagagtacg acgtcctgga taagcggaga ggccgggacc ctgagatggg 3900
cggcaagcct cggcggaaga acccccagga aggcctgtat aacgaactgc agaaagacaa 3960
gatggccgag gcctacagcg agatcggcat gaagggcgag cggaggcggg gcaagggcca 4020
cgacggcctg tatcagggcc tgtccaccgc caccaaggat acctacgacg ccctgcacat 4080
gcaggccctg cccccaaggc tcgagggcgg cggagagggc agaggaagtc ttctaacatg 4140
cggtgacgtg gaggagaatc ccggccctag gatgcttctc ctggtgacaa gccttctgct 4200
ctgtgagtta ccacacccag cattcctcct gatcccacgc aaagtgtgta acggaatagg 4260
tattggtgaa tttaaagact cactctccat aaatgctacg aatattaaac acttcaaaaa 4320
ctgcacctcc atcagtggcg atctccacat cctgccggtg gcatttaggg gtgactcctt 4380
cacacatact cctcctctgg atccacagga actggatatt ctgaaaaccg taaaggaaat 4440
cacagggttt ttgctgattc aggcttggcc tgaaaacagg acggacctcc atgcctttga 4500
gaacctagaa atcatacgcg gcaggaccaa gcaacatggt cagttttctc ttgcagtcgt 4560
cagcctgaac ataacatcct tgggattacg ctccctcaag gagataagtg atggagatgt 4620
gataatttca ggaaacaaaa atttgtgcta tgcaaataca ataaactgga aaaaactgtt 4680
tgggacctcc ggtcagaaaa ccaaaattat aagcaacaga ggtgaaaaca gctgcaaggc 4740
cacaggccag gtctgccatg ccttgtgctc ccccgagggc tgctggggcc cggagcccag 4800
ggactgcgtc tcttgccgga atgtcagccg aggcagggaa tgcgtggaca agtgcaacct 4860
tctggagggt gagccaaggg agtttgtgga gaactctgag tgcatacagt gccacccaga 4920
gtgcctgcct caggccatga acatcacctg cacaggacgg ggaccagaca actgtatcca 4980
gtgtgcccac tacattgacg gcccccactg cgtcaagacc tgcccggcag gagtcatggg 5040
agaaaacaac accctggtct ggaagtacgc agacgccggc catgtgtgcc acctgtgcca 5100
tccaaactgc acctacggat gcactgggcc aggtcttgaa ggctgtccaa cgaatgggcc 5160
taagatcccg tccatcgcca ctgggatggt gggggccctc ctcttgctgc tggtggtggc 5220
cctggggatc ggcctcttca tgtgagcggc cgctctagac ccgggctgca ggaattcgat 5280
atcaagctta tcgataatca acctctggat tacaaaattt gtgaaagatt gactggtatt 5340
cttaactatg ttgctccttt tacgctatgt ggatacgctg ctttaatgcc tttgtatcat 5400
gctattgctt cccgtatggc tttcattttc tcctccttgt ataaatcctg gttgctgtct 5460
ctttatgagg agttgtggcc cgttgtcagg caacgtggcg tggtgtgcac tgtgtttgct 5520
gacgcaaccc ccactggttg gggcattgcc accacctgtc agctcctttc cgggactttc 5580
gctttccccc tccctattgc cacggcggaa ctcatcgccg cctgccttgc ccgctgctgg 5640
acaggggctc ggctgttggg cactgacaat tccgtggtgt tgtcggggaa atcatcgtcc 5700
tttccttggc tgctcgcctg tgttgccacc tggattctgc gcgggacgtc cttctgctac 5760
gtcccttcgg ccctcaatcc agcggacctt ccttcccgcg gcctgctgcc ggctctgcgg 5820
cctcttccgc gtcttcgcct tcgccctcag acgagtcgga tctccctttg ggccgcctcc 5880
ccgcatcgat accgtcgact agccgtacct ttaagaccaa tgacttacaa ggcagctgta 5940
gatcttagcc actttttaaa agaaaagggg ggactggaag ggctaattca ctcccaaaga 6000
agacaagatc tgctttttgc ctgtactggg tctctctggt tagaccagat ctgagcctgg 6060
gagctctctg gctaactagg gaacccactg cttaagcctc aataaagctt gccttgagtg 6120
cttcaagtag tgtgtgcccg tctgttgtgt gactctggta actagagatc cctcagaccc 6180
ttttagtcag tgtggaaaat ctctagcaga attcgatatc aagcttatcg ataccgtcga 6240
cctcgagggg gggcccggta cccaattcgc cctatagtga gtcgtattac aattcactgg 6300
ccgtcgtttt acaacgtcgt gactgggaaa accctggcgt tacccaactt aatcgccttg 6360
cagcacatcc ccctttcgcc agctggcgta atagcgaaga ggcccgcacc gatcgccctt 6420
cccaacagtt gcgcagcctg aatggcgaat ggaaattgta agcgttaata ttttgttaaa 6480
attcgcgtta aatttttgtt aaatcagctc attttttaac caataggccg aaatcggcaa 6540
aatcccttat aaatcaaaag aatagaccga gatagggttg agtgttgttc cagtttggaa 6600
caagagtcca ctattaaaga acgtggactc caacgtcaaa gggcgaaaaa ccgtctatca 6660
gggcgatggc ccactacgtg aaccatcacc ctaatcaagt tttttggggt cgaggtgccg 6720
taaagcacta aatcggaacc ctaaagggag cccccgattt agagcttgac ggggaaagcc 6780
ggcgaacgtg gcgagaaagg aagggaagaa agcgaaagga gcgggcgcta gggcgctggc 6840
aagtgtagcg gtcacgctgc gcgtaaccac cacacccgcc gcgcttaatg cgccgctaca 6900
gggcgcgtca ggtggcactt ttcggggaaa tgtgcgcgga acccctattt gtttattttt 6960
ctaaatacat tcaaatatgt atccgctcat gagacaataa ccctgataaa tgcttcaata 7020
atattgaaaa aggaagagta tgagtattca acatttccgt gtcgccctta ttcccttttt 7080
tgcggcattt tgccttcctg tttttgctca cccagaaacg ctggtgaaag taaaagatgc 7140
tgaagatcag ttgggtgcac gagtgggtta catcgaactg gatctcaaca gcggtaagat 7200
ccttgagagt tttcgccccg aagaacgttt tccaatgatg agcactttta aagttctgct 7260
atgtggcgcg gtattatccc gtattgacgc cgggcaagag caactcggtc gccgcataca 7320
ctattctcag aatgacttgg ttgagtactc accagtcaca gaaaagcatc ttacggatgg 7380
catgacagta agagaattat gcagtgctgc cataaccatg agtgataaca ctgcggccaa 7440
cttacttctg acaacgatcg gaggaccgaa ggagctaacc gcttttttgc acaacatggg 7500
ggatcatgta actcgccttg atcgttggga accggagctg aatgaagcca taccaaacga 7560
cgagcgtgac accacgatgc ctgtagcaat ggcaacaacg ttgcgcaaac tattaactgg 7620
cgaactactt actctagctt cccggcaaca attaatagac tggatggagg cggataaagt 7680
tgcaggacca cttctgcgct cggcccttcc ggctggctgg tttattgctg ataaatctgg 7740
agccggtgag cgtgggtctc gcggtatcat tgcagcactg gggccagatg gtaagccctc 7800
ccgtatcgta gttatctaca cgacggggag tcaggcaact atggatgaac gaaatagaca 7860
gatcgctgag ataggtgcct cactgattaa gcattggtaa ctgtcagacc aagtttactc 7920
atatatactt tagattgatt taaaacttca tttttaattt aaaaggatct aggtgaagat 7980
cctttttgat aatctcatga ccaaaatccc ttaacgtgag ttttcgttcc actgagcgtc 8040
agaccccgta gaaaagatca aaggatcttc ttgagatcct ttttttctgc gcgtaatctg 8100
ctgcttgcaa acaaaaaaac caccgctacc agcggtggtt tgtttgccgg atcaagagct 8160
accaactctt tttccgaagg taactggctt cagcagagcg cagataccaa atactgttct 8220
tctagtgtag ccgtagttag gccaccactt caagaactct gtagcaccgc ctacatacct 8280
cgctctgcta atcctgttac cagtggctgc tgccagtggc gataagtcgt gtcttaccgg 8340
gttggactca agacgatagt taccggataa ggcgcagcgg tcgggctgaa cggggggttc 8400
gtgcacacag cccagcttgg agcgaacgac ctacaccgaa ctgagatacc tacagcgtga 8460
gctatgagaa agcgccacgc ttcccgaagg gagaaaggcg gacaggtatc cggtaagcgg 8520
cagggtcgga acaggagagc gcacgaggga gcttccaggg ggaaacgcct ggtatcttta 8580
tagtcctgtc gggtttcgcc acctctgact tgagcgtcga tttttgtgat gctcgtcagg 8640
ggggcggagc ctatggaaaa acgccagcaa cgcggccttt ttacggttcc tggccttttg 8700
ctggcctttt gctcacatgt tctttcctgc gttatcccct gattctgtgg ataaccgtat 8760
taccgccttt gagtgagctg ataccgctcg ccgcagccga acgaccgagc gcagcgagtc 8820
agtgagcgag gaagcggaag agcgcccaat acgcaaaccg cctctccccg cgcgttggcc 8880
gattcattaa tgcagctggc acgacaggtt tcccgactgg aaagcgggca gtgagcgcaa 8940
cgcaattaat gtgagttagc tcactcatta ggcaccccag gctttacact ttatgcttcc 9000
ggctcgtatg ttgtgtggaa ttgtgagcgg ataacaattt cacacaggaa acagctatga 9060
ccatgattac gccaagctcg aaattaaccc tcactaaagg gaacaaaagc tggagctcca 9120
ccgcggtggc ggcctcgagg tcgagatccg gtcgaccagc aaccatagtc ccgcccctaa 9180
ctccgcccat cccgccccta actccgccca gttccgccca ttctccgccc catggctgac 9240
taattttttt tatttatgca gaggccgagg ccgcctcggc ctctgagcta ttccagaagt 9300
agtgaggagg cttttttgga ggcctaggct tttgcaaaaa gcttcgacgg tatcgattgg 9360
ctcatgtcca acattaccgc catgttgaca ttgattattg actagttatt aatagtaatc 9420
aattacgggg tcattagttc atagcccata tatggagttc cgcgttacat aacttacggt 9480
aaatggcccg cctggctgac cgcccaacga cccccgccca ttgacgtcaa taatgacgta 9540
tgttcccata gtaacgccaa tagggacttt ccattgacgt caatgggtgg agtatttacg 9600
gtaaactgcc cacttggcag tacatcaagt gtatcatatg ccaagtacgc cccctattga 9660
cgtcaatgac ggtaaatggc ccgcctggca ttatgcccag tacatgacct tatgggactt 9720
tcctacttgg cagtacatct acgtattagt catcgctatt accatggtga tgcggttttg 9780
gcagtacatc aatgggcgtg gatagcggtt tgactcacgg ggatttccaa gtctccaccc 9840
cattgacgtc aatgggagtt tgttttggca ccaaaatcaa cgggactttc caaaatgtcg 9900
taacaactcc gccccattga cgcaaatggg cggtaggcgt gtacggaatt cggagtggcg 9960
agccctcaga tcctgcatat aagcagctgc tttttgcctg tactgggtct ctctg 10015
<210> 76
<211> 241
<212> Белок
<213> Homo sapiens
<400> 76
Gln Ser Val Lys Glu Ser Glu Gly Asp Leu Val Thr Pro Ala Gly Asn
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Ser Asp Ile Asn Asp Tyr Pro
20 25 30
Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Phe Ile Asn Ser Gly Gly Ser Thr Trp Tyr Ala Ser Trp Val Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Ser Leu Thr Thr Asp Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Tyr
85 90 95
Ser Thr Tyr Tyr Gly Asp Phe Asn Ile Trp Gly Pro Gly Thr Leu Val
100 105 110
Thr Ile Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Glu Leu Val Met Thr Gln Thr Pro Ser Ser Thr Ser Gly
130 135 140
Ala Val Gly Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Ile
145 150 155 160
Asp Ser Asn Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Thr
165 170 175
Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser Arg
180 185 190
Phe Ser Gly Ser Arg Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Gly
195 200 205
Val Gln Arg Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly Val Gly
210 215 220
Asn Val Ser Tyr Arg Thr Ser Phe Gly Gly Gly Thr Glu Val Val Val
225 230 235 240
Lys
<210> 77
<211> 801
<212> ДНК
<213> Homo sapiens
<400> 77
gaattcgcca ccatgctgct gctggtgaca agcctgctgc tgtgcgagct gccccacccc 60
gcctttctgc tgatccccca gagcgtgaaa gagtccgagg gcgacctggt cacaccagcc 120
ggcaacctga ccctgacctg taccgccagc ggcagcgaca tcaacgacta ccccatctct 180
tgggtccgcc aggctcctgg caagggactg gaatggatcg gcttcatcaa cagcggcggc 240
agcacttggt acgccagctg ggtcaaaggc cggttcacca tcagccggac cagcaccacc 300
gtggacctga agatgacaag cctgaccacc gacgacaccg ccacctactt ttgcgccaga 360
ggctacagca cctactacgg cgacttcaac atctggggcc ctggcaccct ggtcacaatc 420
tctagcggcg gaggcggcag cggaggtgga ggaagtggcg gcggaggatc cgagctggtc 480
atgacccaga cccccagcag cacatctggc gccgtgggcg gcaccgtgac catcaattgc 540
caggccagcc agagcatcga cagcaacctg gcctggttcc agcagaagcc cggccagccc 600
cccaccctgc tgatctacag agcctccaac ctggccagcg gcgtgccaag cagattcagc 660
ggcagcagat ctggcaccga gtacaccctg accatctccg gcgtgcagag agaggacgcc 720
gctacctatt actgcctggg cggcgtgggc aacgtgtcct acagaaccag cttcggcgga 780
ggtactgagg tggtcgtcaa a 801
<210> 78
<211> 9685
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ХАР R11 с коротким спейсером: PJ_R11- 41BB-Z-T2A-tEGFR
<400> 78
gttagaccag atctgagcct gggagctctc tggctaacta gggaacccac tgcttaagcc 60
tcaataaagc ttgccttgag tgcttcaagt agtgtgtgcc cgtctgttgt gtgactctgg 120
taactagaga tccctcagac ccttttagtc agtgtggaaa atctctagca gtggcgcccg 180
aacagggact tgaaagcgaa agggaaacca gaggagctct ctcgacgcag gactcggctt 240
gctgaagcgc gcacggcaag aggcgagggg cggcgactgg tgagtacgcc aaaaattttg 300
actagcggag gctagaagga gagagatggg tgcgagagcg tcagtattaa gcgggggaga 360
attagatcga tgggaaaaaa ttcggttaag gccaggggga aagaaaaaat ataaattaaa 420
acatatagta tgggcaagca gggagctaga acgattcgca gttaatcctg gcctgttaga 480
aacatcagaa ggctgtagac aaatactggg acagctacaa ccatcccttc agacaggatc 540
agaagaactt agatcattat ataatacagt agcaaccctc tattgtgtgc atcaaaggat 600
agagataaaa gacaccaagg aagctttaga caagatagag gaagagcaaa acaaaagtaa 660
gaaaaaagca cagcaagcag cagctgacac aggacacagc aatcaggtca gccaaaatta 720
ccctatagtg cagaacatcc aggggcaaat ggtacatcag gccatatcac ctagaacttt 780
aaatgcatgg gtaaaagtag tagaagagaa ggctttcagc ccagaagtga tacccatgtt 840
ttcagcatta tcagaaggag ccaccccaca agatttaaac accatgctaa acacagtggg 900
gggacatcaa gcagccatgc aaatgttaaa agagaccatc aatgaggaag ctgcaggcaa 960
agagaagagt ggtgcagaga gaaaaaagag cagtgggaat aggagctttg ttccttgggt 1020
tcttgggagc agcaggaagc actatgggcg cagcgtcaat gacgctgacg gtacaggcca 1080
gacaattatt gtctggtata gtgcagcagc agaacaattt gctgagggct attgaggcgc 1140
aacagcatct gttgcaactc acagtctggg gcatcaagca gctccaggca agaatcctgg 1200
ctgtggaaag atacctaaag gatcaacagc tcctggggat ttggggttgc tctggaaaac 1260
tcatttgcac cactgctgtg ccttggatct acaaatggca gtattcatcc acaattttaa 1320
aagaaaaggg gggattgggg ggtacagtgc aggggaaaga atagtagaca taatagcaac 1380
agacatacaa actaaagaat tacaaaaaca aattacaaaa attcaaaatt ttcgggttta 1440
ttacagggac agcagagatc cagtttgggg atcaattgca tgaagaatct gcttagggtt 1500
aggcgttttg cgctgcttcg cgaggatctg cgatcgctcc ggtgcccgtc agtgggcaga 1560
gcgcacatcg cccacagtcc ccgagaagtt ggggggaggg gtcggcaatt gaaccggtgc 1620
ctagagaagg tggcgcgggg taaactggga aagtgatgtc gtgtactggc tccgcctttt 1680
tcccgagggt gggggagaac cgtatataag tgcagtagtc gccgtgaacg ttctttttcg 1740
caacgggttt gccgccagaa cacagctgaa gcttcgaggg gctcgcatct ctccttcacg 1800
cgcccgccgc cctacctgag gccgccatcc acgccggttg agtcgcgttc tgccgcctcc 1860
cgcctgtggt gcctcctgaa ctgcgtccgc cgtctaggta agtttaaagc tcaggtcgag 1920
accgggcctt tgtccggcgc tcccttggag cctacctaga ctcagccggc tctccacgct 1980
ttgcctgacc ctgcttgctc aactctacgt ctttgtttcg ttttctgttc tgcgccgtta 2040
cagatccaag ctgtgaccgg cgcctacggc tagcgaattc gccaccatgc tgctgctggt 2100
gacaagcctg ctgctgtgcg agctgcccca ccccgccttt ctgctgatcc cccagagcgt 2160
gaaagagtcc gagggcgacc tggtcacacc agccggcaac ctgaccctga cctgtaccgc 2220
cagcggcagc gacatcaacg actaccccat ctcttgggtc cgccaggctc ctggcaaggg 2280
actggaatgg atcggcttca tcaacagcgg cggcagcact tggtacgcca gctgggtcaa 2340
aggccggttc accatcagcc ggaccagcac caccgtggac ctgaagatga caagcctgac 2400
caccgacgac accgccacct acttttgcgc cagaggctac agcacctact acggcgactt 2460
caacatctgg ggccctggca ccctggtcac aatctctagc ggcggaggcg gcagcggagg 2520
tggaggaagt ggcggcggag gatccgagct ggtcatgacc cagaccccca gcagcacatc 2580
tggcgccgtg ggcggcaccg tgaccatcaa ttgccaggcc agccagagca tcgacagcaa 2640
cctggcctgg ttccagcaga agcccggcca gccccccacc ctgctgatct acagagcctc 2700
caacctggcc agcggcgtgc caagcagatt cagcggcagc agatctggca ccgagtacac 2760
cctgaccatc tccggcgtgc agagagagga cgccgctacc tattactgcc tgggcggcgt 2820
gggcaacgtg tcctacagaa ccagcttcgg cggaggtact gaggtggtcg tcaaatacgg 2880
accgccctgc cccccttgcc ctggccagcc tcgcgagccc caggtgtaca ccctgcctcc 2940
ctcccaggaa gagatgacca agaaccaggt gtccctgacc tgcctggtga agggcttcta 3000
ccccagcgac atcgccgtgg agtgggagag caacggccag cctgagaaca actacaagac 3060
cacccctccc gtgctggaca gcgacggcag cttcttcctg tacagccggc tgaccgtgga 3120
caagagccgg tggcaggaag gcaacgtctt tagctgcagc gtgatgcacg aggccctgca 3180
caaccactac acccagaaga gcctgagcct gtccctgggc aagatgttct gggtgctggt 3240
ggtggtgggc ggggtgctgg cctgctacag cctgctggtg acagtggcct tcatcatctt 3300
ttgggtgaaa cggggcagaa agaaactcct gtatatattc aaacaaccat ttatgagacc 3360
agtacaaact actcaagagg aagatggctg tagctgccga tttccagaag aagaagaagg 3420
aggatgtgaa ctgcgggtga agttcagcag aagcgccgac gcccctgcct accagcaggg 3480
ccagaatcag ctgtacaacg agctgaacct gggcagaagg gaagagtacg acgtcctgga 3540
taagcggaga ggccgggacc ctgagatggg cggcaagcct cggcggaaga acccccagga 3600
aggcctgtat aacgaactgc agaaagacaa gatggccgag gcctacagcg agatcggcat 3660
gaagggcgag cggaggcggg gcaagggcca cgacggcctg tatcagggcc tgtccaccgc 3720
caccaaggat acctacgacg ccctgcacat gcaggccctg cccccaaggc tcgagggcgg 3780
cggagagggc agaggaagtc ttctaacatg cggtgacgtg gaggagaatc ccggccctag 3840
gatgcttctc ctggtgacaa gccttctgct ctgtgagtta ccacacccag cattcctcct 3900
gatcccacgc aaagtgtgta acggaatagg tattggtgaa tttaaagact cactctccat 3960
aaatgctacg aatattaaac acttcaaaaa ctgcacctcc atcagtggcg atctccacat 4020
cctgccggtg gcatttaggg gtgactcctt cacacatact cctcctctgg atccacagga 4080
actggatatt ctgaaaaccg taaaggaaat cacagggttt ttgctgattc aggcttggcc 4140
tgaaaacagg acggacctcc atgcctttga gaacctagaa atcatacgcg gcaggaccaa 4200
gcaacatggt cagttttctc ttgcagtcgt cagcctgaac ataacatcct tgggattacg 4260
ctccctcaag gagataagtg atggagatgt gataatttca ggaaacaaaa atttgtgcta 4320
tgcaaataca ataaactgga aaaaactgtt tgggacctcc ggtcagaaaa ccaaaattat 4380
aagcaacaga ggtgaaaaca gctgcaaggc cacaggccag gtctgccatg ccttgtgctc 4440
ccccgagggc tgctggggcc cggagcccag ggactgcgtc tcttgccgga atgtcagccg 4500
aggcagggaa tgcgtggaca agtgcaacct tctggagggt gagccaaggg agtttgtgga 4560
gaactctgag tgcatacagt gccacccaga gtgcctgcct caggccatga acatcacctg 4620
cacaggacgg ggaccagaca actgtatcca gtgtgcccac tacattgacg gcccccactg 4680
cgtcaagacc tgcccggcag gagtcatggg agaaaacaac accctggtct ggaagtacgc 4740
agacgccggc catgtgtgcc acctgtgcca tccaaactgc acctacggat gcactgggcc 4800
aggtcttgaa ggctgtccaa cgaatgggcc taagatcccg tccatcgcca ctgggatggt 4860
gggggccctc ctcttgctgc tggtggtggc cctggggatc ggcctcttca tgtgagcggc 4920
cgctctagac ccgggctgca ggaattcgat atcaagctta tcgataatca acctctggat 4980
tacaaaattt gtgaaagatt gactggtatt cttaactatg ttgctccttt tacgctatgt 5040
ggatacgctg ctttaatgcc tttgtatcat gctattgctt cccgtatggc tttcattttc 5100
tcctccttgt ataaatcctg gttgctgtct ctttatgagg agttgtggcc cgttgtcagg 5160
caacgtggcg tggtgtgcac tgtgtttgct gacgcaaccc ccactggttg gggcattgcc 5220
accacctgtc agctcctttc cgggactttc gctttccccc tccctattgc cacggcggaa 5280
ctcatcgccg cctgccttgc ccgctgctgg acaggggctc ggctgttggg cactgacaat 5340
tccgtggtgt tgtcggggaa atcatcgtcc tttccttggc tgctcgcctg tgttgccacc 5400
tggattctgc gcgggacgtc cttctgctac gtcccttcgg ccctcaatcc agcggacctt 5460
ccttcccgcg gcctgctgcc ggctctgcgg cctcttccgc gtcttcgcct tcgccctcag 5520
acgagtcgga tctccctttg ggccgcctcc ccgcatcgat accgtcgact agccgtacct 5580
ttaagaccaa tgacttacaa ggcagctgta gatcttagcc actttttaaa agaaaagggg 5640
ggactggaag ggctaattca ctcccaaaga agacaagatc tgctttttgc ctgtactggg 5700
tctctctggt tagaccagat ctgagcctgg gagctctctg gctaactagg gaacccactg 5760
cttaagcctc aataaagctt gccttgagtg cttcaagtag tgtgtgcccg tctgttgtgt 5820
gactctggta actagagatc cctcagaccc ttttagtcag tgtggaaaat ctctagcaga 5880
attcgatatc aagcttatcg ataccgtcga cctcgagggg gggcccggta cccaattcgc 5940
cctatagtga gtcgtattac aattcactgg ccgtcgtttt acaacgtcgt gactgggaaa 6000
accctggcgt tacccaactt aatcgccttg cagcacatcc ccctttcgcc agctggcgta 6060
atagcgaaga ggcccgcacc gatcgccctt cccaacagtt gcgcagcctg aatggcgaat 6120
ggaaattgta agcgttaata ttttgttaaa attcgcgtta aatttttgtt aaatcagctc 6180
attttttaac caataggccg aaatcggcaa aatcccttat aaatcaaaag aatagaccga 6240
gatagggttg agtgttgttc cagtttggaa caagagtcca ctattaaaga acgtggactc 6300
caacgtcaaa gggcgaaaaa ccgtctatca gggcgatggc ccactacgtg aaccatcacc 6360
ctaatcaagt tttttggggt cgaggtgccg taaagcacta aatcggaacc ctaaagggag 6420
cccccgattt agagcttgac ggggaaagcc ggcgaacgtg gcgagaaagg aagggaagaa 6480
agcgaaagga gcgggcgcta gggcgctggc aagtgtagcg gtcacgctgc gcgtaaccac 6540
cacacccgcc gcgcttaatg cgccgctaca gggcgcgtca ggtggcactt ttcggggaaa 6600
tgtgcgcgga acccctattt gtttattttt ctaaatacat tcaaatatgt atccgctcat 6660
gagacaataa ccctgataaa tgcttcaata atattgaaaa aggaagagta tgagtattca 6720
acatttccgt gtcgccctta ttcccttttt tgcggcattt tgccttcctg tttttgctca 6780
cccagaaacg ctggtgaaag taaaagatgc tgaagatcag ttgggtgcac gagtgggtta 6840
catcgaactg gatctcaaca gcggtaagat ccttgagagt tttcgccccg aagaacgttt 6900
tccaatgatg agcactttta aagttctgct atgtggcgcg gtattatccc gtattgacgc 6960
cgggcaagag caactcggtc gccgcataca ctattctcag aatgacttgg ttgagtactc 7020
accagtcaca gaaaagcatc ttacggatgg catgacagta agagaattat gcagtgctgc 7080
cataaccatg agtgataaca ctgcggccaa cttacttctg acaacgatcg gaggaccgaa 7140
ggagctaacc gcttttttgc acaacatggg ggatcatgta actcgccttg atcgttggga 7200
accggagctg aatgaagcca taccaaacga cgagcgtgac accacgatgc ctgtagcaat 7260
ggcaacaacg ttgcgcaaac tattaactgg cgaactactt actctagctt cccggcaaca 7320
attaatagac tggatggagg cggataaagt tgcaggacca cttctgcgct cggcccttcc 7380
ggctggctgg tttattgctg ataaatctgg agccggtgag cgtgggtctc gcggtatcat 7440
tgcagcactg gggccagatg gtaagccctc ccgtatcgta gttatctaca cgacggggag 7500
tcaggcaact atggatgaac gaaatagaca gatcgctgag ataggtgcct cactgattaa 7560
gcattggtaa ctgtcagacc aagtttactc atatatactt tagattgatt taaaacttca 7620
tttttaattt aaaaggatct aggtgaagat cctttttgat aatctcatga ccaaaatccc 7680
ttaacgtgag ttttcgttcc actgagcgtc agaccccgta gaaaagatca aaggatcttc 7740
ttgagatcct ttttttctgc gcgtaatctg ctgcttgcaa acaaaaaaac caccgctacc 7800
agcggtggtt tgtttgccgg atcaagagct accaactctt tttccgaagg taactggctt 7860
cagcagagcg cagataccaa atactgttct tctagtgtag ccgtagttag gccaccactt 7920
caagaactct gtagcaccgc ctacatacct cgctctgcta atcctgttac cagtggctgc 7980
tgccagtggc gataagtcgt gtcttaccgg gttggactca agacgatagt taccggataa 8040
ggcgcagcgg tcgggctgaa cggggggttc gtgcacacag cccagcttgg agcgaacgac 8100
ctacaccgaa ctgagatacc tacagcgtga gctatgagaa agcgccacgc ttcccgaagg 8160
gagaaaggcg gacaggtatc cggtaagcgg cagggtcgga acaggagagc gcacgaggga 8220
gcttccaggg ggaaacgcct ggtatcttta tagtcctgtc gggtttcgcc acctctgact 8280
tgagcgtcga tttttgtgat gctcgtcagg ggggcggagc ctatggaaaa acgccagcaa 8340
cgcggccttt ttacggttcc tggccttttg ctggcctttt gctcacatgt tctttcctgc 8400
gttatcccct gattctgtgg ataaccgtat taccgccttt gagtgagctg ataccgctcg 8460
ccgcagccga acgaccgagc gcagcgagtc agtgagcgag gaagcggaag agcgcccaat 8520
acgcaaaccg cctctccccg cgcgttggcc gattcattaa tgcagctggc acgacaggtt 8580
tcccgactgg aaagcgggca gtgagcgcaa cgcaattaat gtgagttagc tcactcatta 8640
ggcaccccag gctttacact ttatgcttcc ggctcgtatg ttgtgtggaa ttgtgagcgg 8700
ataacaattt cacacaggaa acagctatga ccatgattac gccaagctcg aaattaaccc 8760
tcactaaagg gaacaaaagc tggagctcca ccgcggtggc ggcctcgagg tcgagatccg 8820
gtcgaccagc aaccatagtc ccgcccctaa ctccgcccat cccgccccta actccgccca 8880
gttccgccca ttctccgccc catggctgac taattttttt tatttatgca gaggccgagg 8940
ccgcctcggc ctctgagcta ttccagaagt agtgaggagg cttttttgga ggcctaggct 9000
tttgcaaaaa gcttcgacgg tatcgattgg ctcatgtcca acattaccgc catgttgaca 9060
ttgattattg actagttatt aatagtaatc aattacgggg tcattagttc atagcccata 9120
tatggagttc cgcgttacat aacttacggt aaatggcccg cctggctgac cgcccaacga 9180
cccccgccca ttgacgtcaa taatgacgta tgttcccata gtaacgccaa tagggacttt 9240
ccattgacgt caatgggtgg agtatttacg gtaaactgcc cacttggcag tacatcaagt 9300
gtatcatatg ccaagtacgc cccctattga cgtcaatgac ggtaaatggc ccgcctggca 9360
ttatgcccag tacatgacct tatgggactt tcctacttgg cagtacatct acgtattagt 9420
catcgctatt accatggtga tgcggttttg gcagtacatc aatgggcgtg gatagcggtt 9480
tgactcacgg ggatttccaa gtctccaccc cattgacgtc aatgggagtt tgttttggca 9540
ccaaaatcaa cgggactttc caaaatgtcg taacaactcc gccccattga cgcaaatggg 9600
cggtaggcgt gtacggaatt cggagtggcg agccctcaga tcctgcatat aagcagctgc 9660
tttttgcctg tactgggtct ctctg 9685
<210> 79
<211> 9721
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ХАР R12 со спейсером средней длины: PJ_R12-CH3-41BB-Z-T2A-tEGFR
<400> 79
gttagaccag atctgagcct gggagctctc tggctaacta gggaacccac tgcttaagcc 60
tcaataaagc ttgccttgag tgcttcaagt agtgtgtgcc cgtctgttgt gtgactctgg 120
taactagaga tccctcagac ccttttagtc agtgtggaaa atctctagca gtggcgcccg 180
aacagggact tgaaagcgaa agggaaacca gaggagctct ctcgacgcag gactcggctt 240
gctgaagcgc gcacggcaag aggcgagggg cggcgactgg tgagtacgcc aaaaattttg 300
actagcggag gctagaagga gagagatggg tgcgagagcg tcagtattaa gcgggggaga 360
attagatcga tgggaaaaaa ttcggttaag gccaggggga aagaaaaaat ataaattaaa 420
acatatagta tgggcaagca gggagctaga acgattcgca gttaatcctg gcctgttaga 480
aacatcagaa ggctgtagac aaatactggg acagctacaa ccatcccttc agacaggatc 540
agaagaactt agatcattat ataatacagt agcaaccctc tattgtgtgc atcaaaggat 600
agagataaaa gacaccaagg aagctttaga caagatagag gaagagcaaa acaaaagtaa 660
gaaaaaagca cagcaagcag cagctgacac aggacacagc aatcaggtca gccaaaatta 720
ccctatagtg cagaacatcc aggggcaaat ggtacatcag gccatatcac ctagaacttt 780
aaatgcatgg gtaaaagtag tagaagagaa ggctttcagc ccagaagtga tacccatgtt 840
ttcagcatta tcagaaggag ccaccccaca agatttaaac accatgctaa acacagtggg 900
gggacatcaa gcagccatgc aaatgttaaa agagaccatc aatgaggaag ctgcaggcaa 960
agagaagagt ggtgcagaga gaaaaaagag cagtgggaat aggagctttg ttccttgggt 1020
tcttgggagc agcaggaagc actatgggcg cagcgtcaat gacgctgacg gtacaggcca 1080
gacaattatt gtctggtata gtgcagcagc agaacaattt gctgagggct attgaggcgc 1140
aacagcatct gttgcaactc acagtctggg gcatcaagca gctccaggca agaatcctgg 1200
ctgtggaaag atacctaaag gatcaacagc tcctggggat ttggggttgc tctggaaaac 1260
tcatttgcac cactgctgtg ccttggatct acaaatggca gtattcatcc acaattttaa 1320
aagaaaaggg gggattgggg ggtacagtgc aggggaaaga atagtagaca taatagcaac 1380
agacatacaa actaaagaat tacaaaaaca aattacaaaa attcaaaatt ttcgggttta 1440
ttacagggac agcagagatc cagtttgggg atcaattgca tgaagaatct gcttagggtt 1500
aggcgttttg cgctgcttcg cgaggatctg cgatcgctcc ggtgcccgtc agtgggcaga 1560
gcgcacatcg cccacagtcc ccgagaagtt ggggggaggg gtcggcaatt gaaccggtgc 1620
ctagagaagg tggcgcgggg taaactggga aagtgatgtc gtgtactggc tccgcctttt 1680
tcccgagggt gggggagaac cgtatataag tgcagtagtc gccgtgaacg ttctttttcg 1740
caacgggttt gccgccagaa cacagctgaa gcttcgaggg gctcgcatct ctccttcacg 1800
cgcccgccgc cctacctgag gccgccatcc acgccggttg agtcgcgttc tgccgcctcc 1860
cgcctgtggt gcctcctgaa ctgcgtccgc cgtctaggta agtttaaagc tcaggtcgag 1920
accgggcctt tgtccggcgc tcccttggag cctacctaga ctcagccggc tctccacgct 1980
ttgcctgacc ctgcttgctc aactctacgt ctttgtttcg ttttctgttc tgcgccgtta 2040
cagatccaag ctgtgaccgg cgcctacggc tagcgaattc ctcgaggcca ccatgctgct 2100
gctggtgaca agcctgctgc tgtgcgagct gccccacccc gcctttctgc tgatccccca 2160
ggaacagctc gtcgaaagcg gcggcagact ggtgacacct ggcggcagcc tgaccctgag 2220
ctgcaaggcc agcggcttcg acttcagcgc ctactacatg agctgggtcc gccaggcccc 2280
tggcaaggga ctggaatgga tcgccaccat ctaccccagc agcggcaaga cctactacgc 2340
cacctgggtg aacggacggt tcaccatctc cagcgacaac gcccagaaca ccgtggacct 2400
gcagatgaac agcctgacag ccgccgaccg ggccacctac ttttgcgcca gagacagcta 2460
cgccgacgac ggcgccctgt tcaacatctg gggccctggc accctggtga caatctctag 2520
cggcggaggc ggatctggtg gcggaggaag tggcggcgga ggatctgagc tggtgctgac 2580
ccagagcccc tctgtgtctg ctgccctggg aagccctgcc aagatcacct gtaccctgag 2640
cagcgcccac aagaccgaca ccatcgactg gtatcagcag ctgcagggcg aggcccccag 2700
atacctgatg caggtgcaga gcgacggcag ctacaccaag aggccaggcg tgcccgaccg 2760
gttcagcgga tctagctctg gcgccgaccg ctacctgatc atccccagcg tgcaggccga 2820
tgacgaggcc gattactact gtggcgccga ctacatcggc ggctacgtgt tcggcggagg 2880
cacccagctg accgtgaccg gcgagtctaa gtacggaccg ccctgccccc cttgccctgg 2940
ccagcctcgc gagccccagg tgtacaccct gcctccctcc caggaagaga tgaccaagaa 3000
ccaggtgtcc ctgacctgcc tggtgaaggg cttctacccc agcgacatcg ccgtggagtg 3060
ggagagcaac ggccagcctg agaacaacta caagaccacc cctcccgtgc tggacagcga 3120
cggcagcttc ttcctgtaca gccggctgac cgtggacaag agccggtggc aggaaggcaa 3180
cgtctttagc tgcagcgtga tgcacgaggc cctgcacaac cactacaccc agaagagcct 3240
gagcctgtcc ctgggcaaga tgttctgggt gctggtggtg gtgggcgggg tgctggcctg 3300
ctacagcctg ctggtgacag tggccttcat catcttttgg gtgaaacggg gcagaaagaa 3360
actcctgtat atattcaaac aaccatttat gagaccagta caaactactc aagaggaaga 3420
tggctgtagc tgccgatttc cagaagaaga agaaggagga tgtgaactgc gggtgaagtt 3480
cagcagaagc gccgacgccc ctgcctacca gcagggccag aatcagctgt acaacgagct 3540
gaacctgggc agaagggaag agtacgacgt cctggataag cggagaggcc gggaccctga 3600
gatgggcggc aagcctcggc ggaagaaccc ccaggaaggc ctgtataacg aactgcagaa 3660
agacaagatg gccgaggcct acagcgagat cggcatgaag ggcgagcgga ggcggggcaa 3720
gggccacgac ggcctgtatc agggcctgtc caccgccacc aaggatacct acgacgccct 3780
gcacatgcag gccctgcccc caaggctcga gggcggcgga gagggcagag gaagtcttct 3840
aacatgcggt gacgtggagg agaatcccgg ccctaggatg cttctcctgg tgacaagcct 3900
tctgctctgt gagttaccac acccagcatt cctcctgatc ccacgcaaag tgtgtaacgg 3960
aataggtatt ggtgaattta aagactcact ctccataaat gctacgaata ttaaacactt 4020
caaaaactgc acctccatca gtggcgatct ccacatcctg ccggtggcat ttaggggtga 4080
ctccttcaca catactcctc ctctggatcc acaggaactg gatattctga aaaccgtaaa 4140
ggaaatcaca gggtttttgc tgattcaggc ttggcctgaa aacaggacgg acctccatgc 4200
ctttgagaac ctagaaatca tacgcggcag gaccaagcaa catggtcagt tttctcttgc 4260
agtcgtcagc ctgaacataa catccttggg attacgctcc ctcaaggaga taagtgatgg 4320
agatgtgata atttcaggaa acaaaaattt gtgctatgca aatacaataa actggaaaaa 4380
actgtttggg acctccggtc agaaaaccaa aattataagc aacagaggtg aaaacagctg 4440
caaggccaca ggccaggtct gccatgcctt gtgctccccc gagggctgct ggggcccgga 4500
gcccagggac tgcgtctctt gccggaatgt cagccgaggc agggaatgcg tggacaagtg 4560
caaccttctg gagggtgagc caagggagtt tgtggagaac tctgagtgca tacagtgcca 4620
cccagagtgc ctgcctcagg ccatgaacat cacctgcaca ggacggggac cagacaactg 4680
tatccagtgt gcccactaca ttgacggccc ccactgcgtc aagacctgcc cggcaggagt 4740
catgggagaa aacaacaccc tggtctggaa gtacgcagac gccggccatg tgtgccacct 4800
gtgccatcca aactgcacct acggatgcac tgggccaggt cttgaaggct gtccaacgaa 4860
tgggcctaag atcccgtcca tcgccactgg gatggtgggg gccctcctct tgctgctggt 4920
ggtggccctg gggatcggcc tcttcatgtg agcggccgct ctagacccgg gctgcaggaa 4980
ttcgatatca agcttatcga taatcaacct ctggattaca aaatttgtga aagattgact 5040
ggtattctta actatgttgc tccttttacg ctatgtggat acgctgcttt aatgcctttg 5100
tatcatgcta ttgcttcccg tatggctttc attttctcct ccttgtataa atcctggttg 5160
ctgtctcttt atgaggagtt gtggcccgtt gtcaggcaac gtggcgtggt gtgcactgtg 5220
tttgctgacg caacccccac tggttggggc attgccacca cctgtcagct cctttccggg 5280
actttcgctt tccccctccc tattgccacg gcggaactca tcgccgcctg ccttgcccgc 5340
tgctggacag gggctcggct gttgggcact gacaattccg tggtgttgtc ggggaaatca 5400
tcgtcctttc cttggctgct cgcctgtgtt gccacctgga ttctgcgcgg gacgtccttc 5460
tgctacgtcc cttcggccct caatccagcg gaccttcctt cccgcggcct gctgccggct 5520
ctgcggcctc ttccgcgtct tcgccttcgc cctcagacga gtcggatctc cctttgggcc 5580
gcctccccgc atcgataccg tcgactagcc gtacctttaa gaccaatgac ttacaaggca 5640
gctgtagatc ttagccactt tttaaaagaa aaggggggac tggaagggct aattcactcc 5700
caaagaagac aagatctgct ttttgcctgt actgggtctc tctggttaga ccagatctga 5760
gcctgggagc tctctggcta actagggaac ccactgctta agcctcaata aagcttgcct 5820
tgagtgcttc aagtagtgtg tgcccgtctg ttgtgtgact ctggtaacta gagatccctc 5880
agaccctttt agtcagtgtg gaaaatctct agcagaattc gatatcaagc ttatcgatac 5940
cgtcgacctc gagggggggc ccggtaccca attcgcccta tagtgagtcg tattacaatt 6000
cactggccgt cgttttacaa cgtcgtgact gggaaaaccc tggcgttacc caacttaatc 6060
gccttgcagc acatccccct ttcgccagct ggcgtaatag cgaagaggcc cgcaccgatc 6120
gcccttccca acagttgcgc agcctgaatg gcgaatggaa attgtaagcg ttaatatttt 6180
gttaaaattc gcgttaaatt tttgttaaat cagctcattt tttaaccaat aggccgaaat 6240
cggcaaaatc ccttataaat caaaagaata gaccgagata gggttgagtg ttgttccagt 6300
ttggaacaag agtccactat taaagaacgt ggactccaac gtcaaagggc gaaaaaccgt 6360
ctatcagggc gatggcccac tacgtgaacc atcaccctaa tcaagttttt tggggtcgag 6420
gtgccgtaaa gcactaaatc ggaaccctaa agggagcccc cgatttagag cttgacgggg 6480
aaagccggcg aacgtggcga gaaaggaagg gaagaaagcg aaaggagcgg gcgctagggc 6540
gctggcaagt gtagcggtca cgctgcgcgt aaccaccaca cccgccgcgc ttaatgcgcc 6600
gctacagggc gcgtcaggtg gcacttttcg gggaaatgtg cgcggaaccc ctatttgttt 6660
atttttctaa atacattcaa atatgtatcc gctcatgaga caataaccct gataaatgct 6720
tcaataatat tgaaaaagga agagtatgag tattcaacat ttccgtgtcg cccttattcc 6780
cttttttgcg gcattttgcc ttcctgtttt tgctcaccca gaaacgctgg tgaaagtaaa 6840
agatgctgaa gatcagttgg gtgcacgagt gggttacatc gaactggatc tcaacagcgg 6900
taagatcctt gagagttttc gccccgaaga acgttttcca atgatgagca cttttaaagt 6960
tctgctatgt ggcgcggtat tatcccgtat tgacgccggg caagagcaac tcggtcgccg 7020
catacactat tctcagaatg acttggttga gtactcacca gtcacagaaa agcatcttac 7080
ggatggcatg acagtaagag aattatgcag tgctgccata accatgagtg ataacactgc 7140
ggccaactta cttctgacaa cgatcggagg accgaaggag ctaaccgctt ttttgcacaa 7200
catgggggat catgtaactc gccttgatcg ttgggaaccg gagctgaatg aagccatacc 7260
aaacgacgag cgtgacacca cgatgcctgt agcaatggca acaacgttgc gcaaactatt 7320
aactggcgaa ctacttactc tagcttcccg gcaacaatta atagactgga tggaggcgga 7380
taaagttgca ggaccacttc tgcgctcggc ccttccggct ggctggttta ttgctgataa 7440
atctggagcc ggtgagcgtg ggtctcgcgg tatcattgca gcactggggc cagatggtaa 7500
gccctcccgt atcgtagtta tctacacgac ggggagtcag gcaactatgg atgaacgaaa 7560
tagacagatc gctgagatag gtgcctcact gattaagcat tggtaactgt cagaccaagt 7620
ttactcatat atactttaga ttgatttaaa acttcatttt taatttaaaa ggatctaggt 7680
gaagatcctt tttgataatc tcatgaccaa aatcccttaa cgtgagtttt cgttccactg 7740
agcgtcagac cccgtagaaa agatcaaagg atcttcttga gatccttttt ttctgcgcgt 7800
aatctgctgc ttgcaaacaa aaaaaccacc gctaccagcg gtggtttgtt tgccggatca 7860
agagctacca actctttttc cgaaggtaac tggcttcagc agagcgcaga taccaaatac 7920
tgttcttcta gtgtagccgt agttaggcca ccacttcaag aactctgtag caccgcctac 7980
atacctcgct ctgctaatcc tgttaccagt ggctgctgcc agtggcgata agtcgtgtct 8040
taccgggttg gactcaagac gatagttacc ggataaggcg cagcggtcgg gctgaacggg 8100
gggttcgtgc acacagccca gcttggagcg aacgacctac accgaactga gatacctaca 8160
gcgtgagcta tgagaaagcg ccacgcttcc cgaagggaga aaggcggaca ggtatccggt 8220
aagcggcagg gtcggaacag gagagcgcac gagggagctt ccagggggaa acgcctggta 8280
tctttatagt cctgtcgggt ttcgccacct ctgacttgag cgtcgatttt tgtgatgctc 8340
gtcagggggg cggagcctat ggaaaaacgc cagcaacgcg gcctttttac ggttcctggc 8400
cttttgctgg ccttttgctc acatgttctt tcctgcgtta tcccctgatt ctgtggataa 8460
ccgtattacc gcctttgagt gagctgatac cgctcgccgc agccgaacga ccgagcgcag 8520
cgagtcagtg agcgaggaag cggaagagcg cccaatacgc aaaccgcctc tccccgcgcg 8580
ttggccgatt cattaatgca gctggcacga caggtttccc gactggaaag cgggcagtga 8640
gcgcaacgca attaatgtga gttagctcac tcattaggca ccccaggctt tacactttat 8700
gcttccggct cgtatgttgt gtggaattgt gagcggataa caatttcaca caggaaacag 8760
ctatgaccat gattacgcca agctcgaaat taaccctcac taaagggaac aaaagctgga 8820
gctccaccgc ggtggcggcc tcgaggtcga gatccggtcg accagcaacc atagtcccgc 8880
ccctaactcc gcccatcccg cccctaactc cgcccagttc cgcccattct ccgccccatg 8940
gctgactaat tttttttatt tatgcagagg ccgaggccgc ctcggcctct gagctattcc 9000
agaagtagtg aggaggcttt tttggaggcc taggcttttg caaaaagctt cgacggtatc 9060
gattggctca tgtccaacat taccgccatg ttgacattga ttattgacta gttattaata 9120
gtaatcaatt acggggtcat tagttcatag cccatatatg gagttccgcg ttacataact 9180
tacggtaaat ggcccgcctg gctgaccgcc caacgacccc cgcccattga cgtcaataat 9240
gacgtatgtt cccatagtaa cgccaatagg gactttccat tgacgtcaat gggtggagta 9300
tttacggtaa actgcccact tggcagtaca tcaagtgtat catatgccaa gtacgccccc 9360
tattgacgtc aatgacggta aatggcccgc ctggcattat gcccagtaca tgaccttatg 9420
ggactttcct acttggcagt acatctacgt attagtcatc gctattacca tggtgatgcg 9480
gttttggcag tacatcaatg ggcgtggata gcggtttgac tcacggggat ttccaagtct 9540
ccaccccatt gacgtcaatg ggagtttgtt ttggcaccaa aatcaacggg actttccaaa 9600
atgtcgtaac aactccgccc cattgacgca aatgggcggt aggcgtgtac ggaattcgga 9660
gtggcgagcc ctcagatcct gcatataagc agctgctttt tgcctgtact gggtctctct 9720
g 9721
<210> 80
<211> 10051
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ХАР R12 с длинным спейсером: PJ_R12-CH2-CH3-41BB-Z-T2A-tEGFR
<400> 80
gttagaccag atctgagcct gggagctctc tggctaacta gggaacccac tgcttaagcc 60
tcaataaagc ttgccttgag tgcttcaagt agtgtgtgcc cgtctgttgt gtgactctgg 120
taactagaga tccctcagac ccttttagtc agtgtggaaa atctctagca gtggcgcccg 180
aacagggact tgaaagcgaa agggaaacca gaggagctct ctcgacgcag gactcggctt 240
gctgaagcgc gcacggcaag aggcgagggg cggcgactgg tgagtacgcc aaaaattttg 300
actagcggag gctagaagga gagagatggg tgcgagagcg tcagtattaa gcgggggaga 360
attagatcga tgggaaaaaa ttcggttaag gccaggggga aagaaaaaat ataaattaaa 420
acatatagta tgggcaagca gggagctaga acgattcgca gttaatcctg gcctgttaga 480
aacatcagaa ggctgtagac aaatactggg acagctacaa ccatcccttc agacaggatc 540
agaagaactt agatcattat ataatacagt agcaaccctc tattgtgtgc atcaaaggat 600
agagataaaa gacaccaagg aagctttaga caagatagag gaagagcaaa acaaaagtaa 660
gaaaaaagca cagcaagcag cagctgacac aggacacagc aatcaggtca gccaaaatta 720
ccctatagtg cagaacatcc aggggcaaat ggtacatcag gccatatcac ctagaacttt 780
aaatgcatgg gtaaaagtag tagaagagaa ggctttcagc ccagaagtga tacccatgtt 840
ttcagcatta tcagaaggag ccaccccaca agatttaaac accatgctaa acacagtggg 900
gggacatcaa gcagccatgc aaatgttaaa agagaccatc aatgaggaag ctgcaggcaa 960
agagaagagt ggtgcagaga gaaaaaagag cagtgggaat aggagctttg ttccttgggt 1020
tcttgggagc agcaggaagc actatgggcg cagcgtcaat gacgctgacg gtacaggcca 1080
gacaattatt gtctggtata gtgcagcagc agaacaattt gctgagggct attgaggcgc 1140
aacagcatct gttgcaactc acagtctggg gcatcaagca gctccaggca agaatcctgg 1200
ctgtggaaag atacctaaag gatcaacagc tcctggggat ttggggttgc tctggaaaac 1260
tcatttgcac cactgctgtg ccttggatct acaaatggca gtattcatcc acaattttaa 1320
aagaaaaggg gggattgggg ggtacagtgc aggggaaaga atagtagaca taatagcaac 1380
agacatacaa actaaagaat tacaaaaaca aattacaaaa attcaaaatt ttcgggttta 1440
ttacagggac agcagagatc cagtttgggg atcaattgca tgaagaatct gcttagggtt 1500
aggcgttttg cgctgcttcg cgaggatctg cgatcgctcc ggtgcccgtc agtgggcaga 1560
gcgcacatcg cccacagtcc ccgagaagtt ggggggaggg gtcggcaatt gaaccggtgc 1620
ctagagaagg tggcgcgggg taaactggga aagtgatgtc gtgtactggc tccgcctttt 1680
tcccgagggt gggggagaac cgtatataag tgcagtagtc gccgtgaacg ttctttttcg 1740
caacgggttt gccgccagaa cacagctgaa gcttcgaggg gctcgcatct ctccttcacg 1800
cgcccgccgc cctacctgag gccgccatcc acgccggttg agtcgcgttc tgccgcctcc 1860
cgcctgtggt gcctcctgaa ctgcgtccgc cgtctaggta agtttaaagc tcaggtcgag 1920
accgggcctt tgtccggcgc tcccttggag cctacctaga ctcagccggc tctccacgct 1980
ttgcctgacc ctgcttgctc aactctacgt ctttgtttcg ttttctgttc tgcgccgtta 2040
cagatccaag ctgtgaccgg cgcctacggc tagcgaattc ctcgaggcca ccatgctgct 2100
gctggtgaca agcctgctgc tgtgcgagct gccccacccc gcctttctgc tgatccccca 2160
ggaacagctc gtcgaaagcg gcggcagact ggtgacacct ggcggcagcc tgaccctgag 2220
ctgcaaggcc agcggcttcg acttcagcgc ctactacatg agctgggtcc gccaggcccc 2280
tggcaaggga ctggaatgga tcgccaccat ctaccccagc agcggcaaga cctactacgc 2340
cacctgggtg aacggacggt tcaccatctc cagcgacaac gcccagaaca ccgtggacct 2400
gcagatgaac agcctgacag ccgccgaccg ggccacctac ttttgcgcca gagacagcta 2460
cgccgacgac ggcgccctgt tcaacatctg gggccctggc accctggtga caatctctag 2520
cggcggaggc ggatctggtg gcggaggaag tggcggcgga ggatctgagc tggtgctgac 2580
ccagagcccc tctgtgtctg ctgccctggg aagccctgcc aagatcacct gtaccctgag 2640
cagcgcccac aagaccgaca ccatcgactg gtatcagcag ctgcagggcg aggcccccag 2700
atacctgatg caggtgcaga gcgacggcag ctacaccaag aggccaggcg tgcccgaccg 2760
gttcagcgga tctagctctg gcgccgaccg ctacctgatc atccccagcg tgcaggccga 2820
tgacgaggcc gattactact gtggcgccga ctacatcggc ggctacgtgt tcggcggagg 2880
cacccagctg accgtgaccg gcgagtctaa gtacggaccg ccctgccccc cttgccctgc 2940
ccccgagttc ctgggcggac ccagcgtgtt cctgttcccc cccaagccca aggacaccct 3000
gatgatcagc cggacccccg aggtgacctg cgtggtggtg gacgtgagcc aggaagatcc 3060
cgaggtccag ttcaattggt acgtggacgg cgtggaagtg cacaacgcca agaccaagcc 3120
cagagaggaa cagttcaaca gcacctaccg ggtggtgtct gtgctgaccg tgctgcacca 3180
ggactggctg aacggcaaag aatacaagtg caaggtgtcc aacaagggcc tgcccagcag 3240
catcgaaaag accatcagca aggccaaggg ccagcctcgc gagccccagg tgtacaccct 3300
gcctccctcc caggaagaga tgaccaagaa ccaggtgtcc ctgacctgcc tggtgaaggg 3360
cttctacccc agcgacatcg ccgtggagtg ggagagcaac ggccagcctg agaacaacta 3420
caagaccacc cctcccgtgc tggacagcga cggcagcttc ttcctgtaca gccggctgac 3480
cgtggacaag agccggtggc aggaaggcaa cgtctttagc tgcagcgtga tgcacgaggc 3540
cctgcacaac cactacaccc agaagagcct gagcctgtcc ctgggcaaga tgttctgggt 3600
gctggtggtg gtgggcgggg tgctggcctg ctacagcctg ctggtgacag tggccttcat 3660
catcttttgg gtgaaacggg gcagaaagaa actcctgtat atattcaaac aaccatttat 3720
gagaccagta caaactactc aagaggaaga tggctgtagc tgccgatttc cagaagaaga 3780
agaaggagga tgtgaactgc gggtgaagtt cagcagaagc gccgacgccc ctgcctacca 3840
gcagggccag aatcagctgt acaacgagct gaacctgggc agaagggaag agtacgacgt 3900
cctggataag cggagaggcc gggaccctga gatgggcggc aagcctcggc ggaagaaccc 3960
ccaggaaggc ctgtataacg aactgcagaa agacaagatg gccgaggcct acagcgagat 4020
cggcatgaag ggcgagcgga ggcggggcaa gggccacgac ggcctgtatc agggcctgtc 4080
caccgccacc aaggatacct acgacgccct gcacatgcag gccctgcccc caaggctcga 4140
gggcggcgga gagggcagag gaagtcttct aacatgcggt gacgtggagg agaatcccgg 4200
ccctaggatg cttctcctgg tgacaagcct tctgctctgt gagttaccac acccagcatt 4260
cctcctgatc ccacgcaaag tgtgtaacgg aataggtatt ggtgaattta aagactcact 4320
ctccataaat gctacgaata ttaaacactt caaaaactgc acctccatca gtggcgatct 4380
ccacatcctg ccggtggcat ttaggggtga ctccttcaca catactcctc ctctggatcc 4440
acaggaactg gatattctga aaaccgtaaa ggaaatcaca gggtttttgc tgattcaggc 4500
ttggcctgaa aacaggacgg acctccatgc ctttgagaac ctagaaatca tacgcggcag 4560
gaccaagcaa catggtcagt tttctcttgc agtcgtcagc ctgaacataa catccttggg 4620
attacgctcc ctcaaggaga taagtgatgg agatgtgata atttcaggaa acaaaaattt 4680
gtgctatgca aatacaataa actggaaaaa actgtttggg acctccggtc agaaaaccaa 4740
aattataagc aacagaggtg aaaacagctg caaggccaca ggccaggtct gccatgcctt 4800
gtgctccccc gagggctgct ggggcccgga gcccagggac tgcgtctctt gccggaatgt 4860
cagccgaggc agggaatgcg tggacaagtg caaccttctg gagggtgagc caagggagtt 4920
tgtggagaac tctgagtgca tacagtgcca cccagagtgc ctgcctcagg ccatgaacat 4980
cacctgcaca ggacggggac cagacaactg tatccagtgt gcccactaca ttgacggccc 5040
ccactgcgtc aagacctgcc cggcaggagt catgggagaa aacaacaccc tggtctggaa 5100
gtacgcagac gccggccatg tgtgccacct gtgccatcca aactgcacct acggatgcac 5160
tgggccaggt cttgaaggct gtccaacgaa tgggcctaag atcccgtcca tcgccactgg 5220
gatggtgggg gccctcctct tgctgctggt ggtggccctg gggatcggcc tcttcatgtg 5280
agcggccgct ctagacccgg gctgcaggaa ttcgatatca agcttatcga taatcaacct 5340
ctggattaca aaatttgtga aagattgact ggtattctta actatgttgc tccttttacg 5400
ctatgtggat acgctgcttt aatgcctttg tatcatgcta ttgcttcccg tatggctttc 5460
attttctcct ccttgtataa atcctggttg ctgtctcttt atgaggagtt gtggcccgtt 5520
gtcaggcaac gtggcgtggt gtgcactgtg tttgctgacg caacccccac tggttggggc 5580
attgccacca cctgtcagct cctttccggg actttcgctt tccccctccc tattgccacg 5640
gcggaactca tcgccgcctg ccttgcccgc tgctggacag gggctcggct gttgggcact 5700
gacaattccg tggtgttgtc ggggaaatca tcgtcctttc cttggctgct cgcctgtgtt 5760
gccacctgga ttctgcgcgg gacgtccttc tgctacgtcc cttcggccct caatccagcg 5820
gaccttcctt cccgcggcct gctgccggct ctgcggcctc ttccgcgtct tcgccttcgc 5880
cctcagacga gtcggatctc cctttgggcc gcctccccgc atcgataccg tcgactagcc 5940
gtacctttaa gaccaatgac ttacaaggca gctgtagatc ttagccactt tttaaaagaa 6000
aaggggggac tggaagggct aattcactcc caaagaagac aagatctgct ttttgcctgt 6060
actgggtctc tctggttaga ccagatctga gcctgggagc tctctggcta actagggaac 6120
ccactgctta agcctcaata aagcttgcct tgagtgcttc aagtagtgtg tgcccgtctg 6180
ttgtgtgact ctggtaacta gagatccctc agaccctttt agtcagtgtg gaaaatctct 6240
agcagaattc gatatcaagc ttatcgatac cgtcgacctc gagggggggc ccggtaccca 6300
attcgcccta tagtgagtcg tattacaatt cactggccgt cgttttacaa cgtcgtgact 6360
gggaaaaccc tggcgttacc caacttaatc gccttgcagc acatccccct ttcgccagct 6420
ggcgtaatag cgaagaggcc cgcaccgatc gcccttccca acagttgcgc agcctgaatg 6480
gcgaatggaa attgtaagcg ttaatatttt gttaaaattc gcgttaaatt tttgttaaat 6540
cagctcattt tttaaccaat aggccgaaat cggcaaaatc ccttataaat caaaagaata 6600
gaccgagata gggttgagtg ttgttccagt ttggaacaag agtccactat taaagaacgt 6660
ggactccaac gtcaaagggc gaaaaaccgt ctatcagggc gatggcccac tacgtgaacc 6720
atcaccctaa tcaagttttt tggggtcgag gtgccgtaaa gcactaaatc ggaaccctaa 6780
agggagcccc cgatttagag cttgacgggg aaagccggcg aacgtggcga gaaaggaagg 6840
gaagaaagcg aaaggagcgg gcgctagggc gctggcaagt gtagcggtca cgctgcgcgt 6900
aaccaccaca cccgccgcgc ttaatgcgcc gctacagggc gcgtcaggtg gcacttttcg 6960
gggaaatgtg cgcggaaccc ctatttgttt atttttctaa atacattcaa atatgtatcc 7020
gctcatgaga caataaccct gataaatgct tcaataatat tgaaaaagga agagtatgag 7080
tattcaacat ttccgtgtcg cccttattcc cttttttgcg gcattttgcc ttcctgtttt 7140
tgctcaccca gaaacgctgg tgaaagtaaa agatgctgaa gatcagttgg gtgcacgagt 7200
gggttacatc gaactggatc tcaacagcgg taagatcctt gagagttttc gccccgaaga 7260
acgttttcca atgatgagca cttttaaagt tctgctatgt ggcgcggtat tatcccgtat 7320
tgacgccggg caagagcaac tcggtcgccg catacactat tctcagaatg acttggttga 7380
gtactcacca gtcacagaaa agcatcttac ggatggcatg acagtaagag aattatgcag 7440
tgctgccata accatgagtg ataacactgc ggccaactta cttctgacaa cgatcggagg 7500
accgaaggag ctaaccgctt ttttgcacaa catgggggat catgtaactc gccttgatcg 7560
ttgggaaccg gagctgaatg aagccatacc aaacgacgag cgtgacacca cgatgcctgt 7620
agcaatggca acaacgttgc gcaaactatt aactggcgaa ctacttactc tagcttcccg 7680
gcaacaatta atagactgga tggaggcgga taaagttgca ggaccacttc tgcgctcggc 7740
ccttccggct ggctggttta ttgctgataa atctggagcc ggtgagcgtg ggtctcgcgg 7800
tatcattgca gcactggggc cagatggtaa gccctcccgt atcgtagtta tctacacgac 7860
ggggagtcag gcaactatgg atgaacgaaa tagacagatc gctgagatag gtgcctcact 7920
gattaagcat tggtaactgt cagaccaagt ttactcatat atactttaga ttgatttaaa 7980
acttcatttt taatttaaaa ggatctaggt gaagatcctt tttgataatc tcatgaccaa 8040
aatcccttaa cgtgagtttt cgttccactg agcgtcagac cccgtagaaa agatcaaagg 8100
atcttcttga gatccttttt ttctgcgcgt aatctgctgc ttgcaaacaa aaaaaccacc 8160
gctaccagcg gtggtttgtt tgccggatca agagctacca actctttttc cgaaggtaac 8220
tggcttcagc agagcgcaga taccaaatac tgttcttcta gtgtagccgt agttaggcca 8280
ccacttcaag aactctgtag caccgcctac atacctcgct ctgctaatcc tgttaccagt 8340
ggctgctgcc agtggcgata agtcgtgtct taccgggttg gactcaagac gatagttacc 8400
ggataaggcg cagcggtcgg gctgaacggg gggttcgtgc acacagccca gcttggagcg 8460
aacgacctac accgaactga gatacctaca gcgtgagcta tgagaaagcg ccacgcttcc 8520
cgaagggaga aaggcggaca ggtatccggt aagcggcagg gtcggaacag gagagcgcac 8580
gagggagctt ccagggggaa acgcctggta tctttatagt cctgtcgggt ttcgccacct 8640
ctgacttgag cgtcgatttt tgtgatgctc gtcagggggg cggagcctat ggaaaaacgc 8700
cagcaacgcg gcctttttac ggttcctggc cttttgctgg ccttttgctc acatgttctt 8760
tcctgcgtta tcccctgatt ctgtggataa ccgtattacc gcctttgagt gagctgatac 8820
cgctcgccgc agccgaacga ccgagcgcag cgagtcagtg agcgaggaag cggaagagcg 8880
cccaatacgc aaaccgcctc tccccgcgcg ttggccgatt cattaatgca gctggcacga 8940
caggtttccc gactggaaag cgggcagtga gcgcaacgca attaatgtga gttagctcac 9000
tcattaggca ccccaggctt tacactttat gcttccggct cgtatgttgt gtggaattgt 9060
gagcggataa caatttcaca caggaaacag ctatgaccat gattacgcca agctcgaaat 9120
taaccctcac taaagggaac aaaagctgga gctccaccgc ggtggcggcc tcgaggtcga 9180
gatccggtcg accagcaacc atagtcccgc ccctaactcc gcccatcccg cccctaactc 9240
cgcccagttc cgcccattct ccgccccatg gctgactaat tttttttatt tatgcagagg 9300
ccgaggccgc ctcggcctct gagctattcc agaagtagtg aggaggcttt tttggaggcc 9360
taggcttttg caaaaagctt cgacggtatc gattggctca tgtccaacat taccgccatg 9420
ttgacattga ttattgacta gttattaata gtaatcaatt acggggtcat tagttcatag 9480
cccatatatg gagttccgcg ttacataact tacggtaaat ggcccgcctg gctgaccgcc 9540
caacgacccc cgcccattga cgtcaataat gacgtatgtt cccatagtaa cgccaatagg 9600
gactttccat tgacgtcaat gggtggagta tttacggtaa actgcccact tggcagtaca 9660
tcaagtgtat catatgccaa gtacgccccc tattgacgtc aatgacggta aatggcccgc 9720
ctggcattat gcccagtaca tgaccttatg ggactttcct acttggcagt acatctacgt 9780
attagtcatc gctattacca tggtgatgcg gttttggcag tacatcaatg ggcgtggata 9840
gcggtttgac tcacggggat ttccaagtct ccaccccatt gacgtcaatg ggagtttgtt 9900
ttggcaccaa aatcaacggg actttccaaa atgtcgtaac aactccgccc cattgacgca 9960
aatgggcggt aggcgtgtac ggaattcgga gtggcgagcc ctcagatcct gcatataagc 10020
agctgctttt tgcctgtact gggtctctct g 10051
<210> 81
<211> 822
<212> ДНК
<213> Homo sapiens
<400> 81
accatgctgc tgctggtgac aagcctgctg ctgtgcgagc tgccccaccc cgcctttctg 60
ctgatccccc aggaacagct cgtcgaaagc ggcggcagac tggtgacacc tggcggcagc 120
ctgaccctga gctgcaaggc cagcggcttc gacttcagcg cctactacat gagctgggtc 180
cgccaggccc ctggcaaggg actggaatgg atcgccacca tctaccccag cagcggcaag 240
acctactacg ccacctgggt gaacggacgg ttcaccatct ccagcgacaa cgcccagaac 300
accgtggacc tgcagatgaa cagcctgaca gccgccgacc gggccaccta cttttgcgcc 360
agagacagct acgccgacga cggcgccctg ttcaacatct ggggccctgg caccctggtg 420
acaatctcta gcggcggagg cggatctggt ggcggaggaa gtggcggcgg aggatctgag 480
ctggtgctga cccagagccc ctctgtgtct gctgccctgg gaagccctgc caagatcacc 540
tgtaccctga gcagcgccca caagaccgac accatcgact ggtatcagca gctgcagggc 600
gaggccccca gatacctgat gcaggtgcag agcgacggca gctacaccaa gaggccaggc 660
gtgcccgacc ggttcagcgg atctagctct ggcgccgacc gctacctgat catccccagc 720
gtgcaggccg atgacgaggc cgattactac tgtggcgccg actacatcgg cggctacgtg 780
ttcggcggag gcacccagct gaccgtgacc ggcgagtcta ag 822
<210> 82
<211> 248
<212> Белок
<213> Homo sapiens
<400> 82
Gln Glu Gln Leu Val Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Gly
1 5 10 15
Ser Leu Thr Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Ser Ala Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Thr Ile Tyr Pro Ser Ser Gly Lys Thr Tyr Tyr Ala Thr Trp Val
50 55 60
Asn Gly Arg Phe Thr Ile Ser Ser Asp Asn Ala Gln Asn Thr Val Asp
65 70 75 80
Leu Gln Met Asn Ser Leu Thr Ala Ala Asp Arg Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Asp Ser Tyr Ala Asp Asp Gly Ala Leu Phe Asn Ile Trp Gly
100 105 110
Pro Gly Thr Leu Val Thr Ile Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu Val Leu Thr Gln Ser Pro
130 135 140
Ser Val Ser Ala Ala Leu Gly Ser Pro Ala Lys Ile Thr Cys Thr Leu
145 150 155 160
Ser Ser Ala His Lys Thr Asp Thr Ile Asp Trp Tyr Gln Gln Leu Gln
165 170 175
Gly Glu Ala Pro Arg Tyr Leu Met Gln Val Gln Ser Asp Gly Ser Tyr
180 185 190
Thr Lys Arg Pro Gly Val Pro Asp Arg Phe Ser Gly Ser Ser Ser Gly
195 200 205
Ala Asp Arg Tyr Leu Ile Ile Pro Ser Val Gln Ala Asp Asp Glu Ala
210 215 220
Asp Tyr Tyr Cys Gly Ala Asp Tyr Ile Gly Gly Tyr Val Phe Gly Gly
225 230 235 240
Gly Thr Gln Leu Thr Val Thr Gly
245
<210> 83
<211> 9384
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ХАР R12 с коротким спейсером: PJ_R12-Hinge-41BB-Z-T2A-tEGFR
<400> 83
gttagaccag atctgagcct gggagctctc tggctaacta gggaacccac tgcttaagcc 60
tcaataaagc ttgccttgag tgcttcaagt agtgtgtgcc cgtctgttgt gtgactctgg 120
taactagaga tccctcagac ccttttagtc agtgtggaaa atctctagca gtggcgcccg 180
aacagggact tgaaagcgaa agggaaacca gaggagctct ctcgacgcag gactcggctt 240
gctgaagcgc gcacggcaag aggcgagggg cggcgactgg tgagtacgcc aaaaattttg 300
actagcggag gctagaagga gagagatggg tgcgagagcg tcagtattaa gcgggggaga 360
attagatcga tgggaaaaaa ttcggttaag gccaggggga aagaaaaaat ataaattaaa 420
acatatagta tgggcaagca gggagctaga acgattcgca gttaatcctg gcctgttaga 480
aacatcagaa ggctgtagac aaatactggg acagctacaa ccatcccttc agacaggatc 540
agaagaactt agatcattat ataatacagt agcaaccctc tattgtgtgc atcaaaggat 600
agagataaaa gacaccaagg aagctttaga caagatagag gaagagcaaa acaaaagtaa 660
gaaaaaagca cagcaagcag cagctgacac aggacacagc aatcaggtca gccaaaatta 720
ccctatagtg cagaacatcc aggggcaaat ggtacatcag gccatatcac ctagaacttt 780
aaatgcatgg gtaaaagtag tagaagagaa ggctttcagc ccagaagtga tacccatgtt 840
ttcagcatta tcagaaggag ccaccccaca agatttaaac accatgctaa acacagtggg 900
gggacatcaa gcagccatgc aaatgttaaa agagaccatc aatgaggaag ctgcaggcaa 960
agagaagagt ggtgcagaga gaaaaaagag cagtgggaat aggagctttg ttccttgggt 1020
tcttgggagc agcaggaagc actatgggcg cagcgtcaat gacgctgacg gtacaggcca 1080
gacaattatt gtctggtata gtgcagcagc agaacaattt gctgagggct attgaggcgc 1140
aacagcatct gttgcaactc acagtctggg gcatcaagca gctccaggca agaatcctgg 1200
ctgtggaaag atacctaaag gatcaacagc tcctggggat ttggggttgc tctggaaaac 1260
tcatttgcac cactgctgtg ccttggatct acaaatggca gtattcatcc acaattttaa 1320
aagaaaaggg gggattgggg ggtacagtgc aggggaaaga atagtagaca taatagcaac 1380
agacatacaa actaaagaat tacaaaaaca aattacaaaa attcaaaatt ttcgggttta 1440
ttacagggac agcagagatc cagtttgggg atcaattgca tgaagaatct gcttagggtt 1500
aggcgttttg cgctgcttcg cgaggatctg cgatcgctcc ggtgcccgtc agtgggcaga 1560
gcgcacatcg cccacagtcc ccgagaagtt ggggggaggg gtcggcaatt gaaccggtgc 1620
ctagagaagg tggcgcgggg taaactggga aagtgatgtc gtgtactggc tccgcctttt 1680
tcccgagggt gggggagaac cgtatataag tgcagtagtc gccgtgaacg ttctttttcg 1740
caacgggttt gccgccagaa cacagctgaa gcttcgaggg gctcgcatct ctccttcacg 1800
cgcccgccgc cctacctgag gccgccatcc acgccggttg agtcgcgttc tgccgcctcc 1860
cgcctgtggt gcctcctgaa ctgcgtccgc cgtctaggta agtttaaagc tcaggtcgag 1920
accgggcctt tgtccggcgc tcccttggag cctacctaga ctcagccggc tctccacgct 1980
ttgcctgacc ctgcttgctc aactctacgt ctttgtttcg ttttctgttc tgcgccgtta 2040
cagatccaag ctgtgaccgg cgcctacggc tagaccatgc tgctgctggt gacaagcctg 2100
ctgctgtgcg agctgcccca ccccgccttt ctgctgatcc cccaggaaca gctcgtcgaa 2160
agcggcggca gactggtgac acctggcggc agcctgaccc tgagctgcaa ggccagcggc 2220
ttcgacttca gcgcctacta catgagctgg gtccgccagg cccctggcaa gggactggaa 2280
tggatcgcca ccatctaccc cagcagcggc aagacctact acgccacctg ggtgaacgga 2340
cggttcacca tctccagcga caacgcccag aacaccgtgg acctgcagat gaacagcctg 2400
acagccgccg accgggccac ctacttttgc gccagagaca gctacgccga cgacggcgcc 2460
ctgttcaaca tctggggccc tggcaccctg gtgacaatct ctagcggcgg aggcggatct 2520
ggtggcggag gaagtggcgg cggaggatct gagctggtgc tgacccagag cccctctgtg 2580
tctgctgccc tgggaagccc tgccaagatc acctgtaccc tgagcagcgc ccacaagacc 2640
gacaccatcg actggtatca gcagctgcag ggcgaggccc ccagatacct gatgcaggtg 2700
cagagcgacg gcagctacac caagaggcca ggcgtgcccg accggttcag cggatctagc 2760
tctggcgccg accgctacct gatcatcccc agcgtgcagg ccgatgacga ggccgattac 2820
tactgtggcg ccgactacat cggcggctac gtgttcggcg gaggcaccca gctgaccgtg 2880
accggcgagt ctaagtacgg accgccctgc cccccttgcc ctatgttctg ggtgctggtg 2940
gtggtgggcg gggtgctggc ctgctacagc ctgctggtga cagtggcctt catcatcttt 3000
tgggtgaaac ggggcagaaa gaaactcctg tatatattca aacaaccatt tatgagacca 3060
gtacaaacta ctcaagagga agatggctgt agctgccgat ttccagaaga agaagaagga 3120
ggatgtgaac tgcgggtgaa gttcagcaga agcgccgacg cccctgccta ccagcagggc 3180
cagaatcagc tgtacaacga gctgaacctg ggcagaaggg aagagtacga cgtcctggat 3240
aagcggagag gccgggaccc tgagatgggc ggcaagcctc ggcggaagaa cccccaggaa 3300
ggcctgtata acgaactgca gaaagacaag atggccgagg cctacagcga gatcggcatg 3360
aagggcgagc ggaggcgggg caagggccac gacggcctgt atcagggcct gtccaccgcc 3420
accaaggata cctacgacgc cctgcacatg caggccctgc ccccaaggct cgagggcggc 3480
ggagagggca gaggaagtct tctaacatgc ggtgacgtgg aggagaatcc cggccctagg 3540
atgcttctcc tggtgacaag ccttctgctc tgtgagttac cacacccagc attcctcctg 3600
atcccacgca aagtgtgtaa cggaataggt attggtgaat ttaaagactc actctccata 3660
aatgctacga atattaaaca cttcaaaaac tgcacctcca tcagtggcga tctccacatc 3720
ctgccggtgg catttagggg tgactccttc acacatactc ctcctctgga tccacaggaa 3780
ctggatattc tgaaaaccgt aaaggaaatc acagggtttt tgctgattca ggcttggcct 3840
gaaaacagga cggacctcca tgcctttgag aacctagaaa tcatacgcgg caggaccaag 3900
caacatggtc agttttctct tgcagtcgtc agcctgaaca taacatcctt gggattacgc 3960
tccctcaagg agataagtga tggagatgtg ataatttcag gaaacaaaaa tttgtgctat 4020
gcaaatacaa taaactggaa aaaactgttt gggacctccg gtcagaaaac caaaattata 4080
agcaacagag gtgaaaacag ctgcaaggcc acaggccagg tctgccatgc cttgtgctcc 4140
cccgagggct gctggggccc ggagcccagg gactgcgtct cttgccggaa tgtcagccga 4200
ggcagggaat gcgtggacaa gtgcaacctt ctggagggtg agccaaggga gtttgtggag 4260
aactctgagt gcatacagtg ccacccagag tgcctgcctc aggccatgaa catcacctgc 4320
acaggacggg gaccagacaa ctgtatccag tgtgcccact acattgacgg cccccactgc 4380
gtcaagacct gcccggcagg agtcatggga gaaaacaaca ccctggtctg gaagtacgca 4440
gacgccggcc atgtgtgcca cctgtgccat ccaaactgca cctacggatg cactgggcca 4500
ggtcttgaag gctgtccaac gaatgggcct aagatcccgt ccatcgccac tgggatggtg 4560
ggggccctcc tcttgctgct ggtggtggcc ctggggatcg gcctcttcat gtgagcggcc 4620
gctctagacc cgggctgcag gaattcgata tcaagcttat cgataatcaa cctctggatt 4680
acaaaatttg tgaaagattg actggtattc ttaactatgt tgctcctttt acgctatgtg 4740
gatacgctgc tttaatgcct ttgtatcatg ctattgcttc ccgtatggct ttcattttct 4800
cctccttgta taaatcctgg ttgctgtctc tttatgagga gttgtggccc gttgtcaggc 4860
aacgtggcgt ggtgtgcact gtgtttgctg acgcaacccc cactggttgg ggcattgcca 4920
ccacctgtca gctcctttcc gggactttcg ctttccccct ccctattgcc acggcggaac 4980
tcatcgccgc ctgccttgcc cgctgctgga caggggctcg gctgttgggc actgacaatt 5040
ccgtggtgtt gtcggggaaa tcatcgtcct ttccttggct gctcgcctgt gttgccacct 5100
ggattctgcg cgggacgtcc ttctgctacg tcccttcggc cctcaatcca gcggaccttc 5160
cttcccgcgg cctgctgccg gctctgcggc ctcttccgcg tcttcgcctt cgccctcaga 5220
cgagtcggat ctccctttgg gccgcctccc cgcatcgata ccgtcgacta gccgtacctt 5280
taagaccaat gacttacaag gcagctgtag atcttagcca ctttttaaaa gaaaaggggg 5340
gactggaagg gctaattcac tcccaaagaa gacaagatct gctttttgcc tgtactgggt 5400
ctctctggtt agaccagatc tgagcctggg agctctctgg ctaactaggg aacccactgc 5460
ttaagcctca ataaagcttg ccttgagtgc ttcaagtagt gtgtgcccgt ctgttgtgtg 5520
actctggtaa ctagagatcc ctcagaccct tttagtcagt gtggaaaatc tctagcagaa 5580
ttcgatatca agcttatcga taccgtcgac ctcgaggggg ggcccggtac ccaattcgcc 5640
ctatagtgag tcgtattaca attcactggc cgtcgtttta caacgtcgtg actgggaaaa 5700
ccctggcgtt acccaactta atcgccttgc agcacatccc cctttcgcca gctggcgtaa 5760
tagcgaagag gcccgcaccg atcgcccttc ccaacagttg cgcagcctga atggcgaatg 5820
gaaattgtaa gcgttaatat tttgttaaaa ttcgcgttaa atttttgtta aatcagctca 5880
ttttttaacc aataggccga aatcggcaaa atcccttata aatcaaaaga atagaccgag 5940
atagggttga gtgttgttcc agtttggaac aagagtccac tattaaagaa cgtggactcc 6000
aacgtcaaag ggcgaaaaac cgtctatcag ggcgatggcc cactacgtga accatcaccc 6060
taatcaagtt ttttggggtc gaggtgccgt aaagcactaa atcggaaccc taaagggagc 6120
ccccgattta gagcttgacg gggaaagccg gcgaacgtgg cgagaaagga agggaagaaa 6180
gcgaaaggag cgggcgctag ggcgctggca agtgtagcgg tcacgctgcg cgtaaccacc 6240
acacccgccg cgcttaatgc gccgctacag ggcgcgtcag gtggcacttt tcggggaaat 6300
gtgcgcggaa cccctatttg tttatttttc taaatacatt caaatatgta tccgctcatg 6360
agacaataac cctgataaat gcttcaataa tattgaaaaa ggaagagtat gagtattcaa 6420
catttccgtg tcgcccttat tccctttttt gcggcatttt gccttcctgt ttttgctcac 6480
ccagaaacgc tggtgaaagt aaaagatgct gaagatcagt tgggtgcacg agtgggttac 6540
atcgaactgg atctcaacag cggtaagatc cttgagagtt ttcgccccga agaacgtttt 6600
ccaatgatga gcacttttaa agttctgcta tgtggcgcgg tattatcccg tattgacgcc 6660
gggcaagagc aactcggtcg ccgcatacac tattctcaga atgacttggt tgagtactca 6720
ccagtcacag aaaagcatct tacggatggc atgacagtaa gagaattatg cagtgctgcc 6780
ataaccatga gtgataacac tgcggccaac ttacttctga caacgatcgg aggaccgaag 6840
gagctaaccg cttttttgca caacatgggg gatcatgtaa ctcgccttga tcgttgggaa 6900
ccggagctga atgaagccat accaaacgac gagcgtgaca ccacgatgcc tgtagcaatg 6960
gcaacaacgt tgcgcaaact attaactggc gaactactta ctctagcttc ccggcaacaa 7020
ttaatagact ggatggaggc ggataaagtt gcaggaccac ttctgcgctc ggcccttccg 7080
gctggctggt ttattgctga taaatctgga gccggtgagc gtgggtctcg cggtatcatt 7140
gcagcactgg ggccagatgg taagccctcc cgtatcgtag ttatctacac gacggggagt 7200
caggcaacta tggatgaacg aaatagacag atcgctgaga taggtgcctc actgattaag 7260
cattggtaac tgtcagacca agtttactca tatatacttt agattgattt aaaacttcat 7320
ttttaattta aaaggatcta ggtgaagatc ctttttgata atctcatgac caaaatccct 7380
taacgtgagt tttcgttcca ctgagcgtca gaccccgtag aaaagatcaa aggatcttct 7440
tgagatcctt tttttctgcg cgtaatctgc tgcttgcaaa caaaaaaacc accgctacca 7500
gcggtggttt gtttgccgga tcaagagcta ccaactcttt ttccgaaggt aactggcttc 7560
agcagagcgc agataccaaa tactgttctt ctagtgtagc cgtagttagg ccaccacttc 7620
aagaactctg tagcaccgcc tacatacctc gctctgctaa tcctgttacc agtggctgct 7680
gccagtggcg ataagtcgtg tcttaccggg ttggactcaa gacgatagtt accggataag 7740
gcgcagcggt cgggctgaac ggggggttcg tgcacacagc ccagcttgga gcgaacgacc 7800
tacaccgaac tgagatacct acagcgtgag ctatgagaaa gcgccacgct tcccgaaggg 7860
agaaaggcgg acaggtatcc ggtaagcggc agggtcggaa caggagagcg cacgagggag 7920
cttccagggg gaaacgcctg gtatctttat agtcctgtcg ggtttcgcca cctctgactt 7980
gagcgtcgat ttttgtgatg ctcgtcaggg gggcggagcc tatggaaaaa cgccagcaac 8040
gcggcctttt tacggttcct ggccttttgc tggccttttg ctcacatgtt ctttcctgcg 8100
ttatcccctg attctgtgga taaccgtatt accgcctttg agtgagctga taccgctcgc 8160
cgcagccgaa cgaccgagcg cagcgagtca gtgagcgagg aagcggaaga gcgcccaata 8220
cgcaaaccgc ctctccccgc gcgttggccg attcattaat gcagctggca cgacaggttt 8280
cccgactgga aagcgggcag tgagcgcaac gcaattaatg tgagttagct cactcattag 8340
gcaccccagg ctttacactt tatgcttccg gctcgtatgt tgtgtggaat tgtgagcgga 8400
taacaatttc acacaggaaa cagctatgac catgattacg ccaagctcga aattaaccct 8460
cactaaaggg aacaaaagct ggagctccac cgcggtggcg gcctcgaggt cgagatccgg 8520
tcgaccagca accatagtcc cgcccctaac tccgcccatc ccgcccctaa ctccgcccag 8580
ttccgcccat tctccgcccc atggctgact aatttttttt atttatgcag aggccgaggc 8640
cgcctcggcc tctgagctat tccagaagta gtgaggaggc ttttttggag gcctaggctt 8700
ttgcaaaaag cttcgacggt atcgattggc tcatgtccaa cattaccgcc atgttgacat 8760
tgattattga ctagttatta atagtaatca attacggggt cattagttca tagcccatat 8820
atggagttcc gcgttacata acttacggta aatggcccgc ctggctgacc gcccaacgac 8880
ccccgcccat tgacgtcaat aatgacgtat gttcccatag taacgccaat agggactttc 8940
cattgacgtc aatgggtgga gtatttacgg taaactgccc acttggcagt acatcaagtg 9000
tatcatatgc caagtacgcc ccctattgac gtcaatgacg gtaaatggcc cgcctggcat 9060
tatgcccagt acatgacctt atgggacttt cctacttggc agtacatcta cgtattagtc 9120
atcgctatta ccatggtgat gcggttttgg cagtacatca atgggcgtgg atagcggttt 9180
gactcacggg gatttccaag tctccacccc attgacgtca atgggagttt gttttggcac 9240
caaaatcaac gggactttcc aaaatgtcgt aacaactccg ccccattgac gcaaatgggc 9300
ggtaggcgtg tacggaattc ggagtggcga gccctcagat cctgcatata agcagctgct 9360
ttttgcctgt actgggtctc tctg 9384
<210> 84
<211> 937
<212> Белок
<213> Homo sapiens
<400> 84
Met His Arg Pro Arg Arg Arg Gly Thr Arg Pro Pro Leu Leu Ala Leu
1 5 10 15
Leu Ala Ala Leu Leu Leu Ala Ala Arg Gly Ala Ala Ala Gln Glu Thr
20 25 30
Glu Leu Ser Val Ser Ala Glu Leu Val Pro Thr Ser Ser Trp Asn Ile
35 40 45
Ser Ser Glu Leu Asn Lys Asp Ser Tyr Leu Thr Leu Asp Glu Pro Met
50 55 60
Asn Asn Ile Thr Thr Ser Leu Gly Gln Thr Ala Glu Leu His Cys Lys
65 70 75 80
Val Ser Gly Asn Pro Pro Pro Thr Ile Arg Trp Phe Lys Asn Asp Ala
85 90 95
Pro Val Val Gln Glu Pro Arg Arg Leu Ser Phe Arg Ser Thr Ile Tyr
100 105 110
Gly Ser Arg Leu Arg Ile Arg Asn Leu Asp Thr Thr Asp Thr Gly Tyr
115 120 125
Phe Gln Cys Val Ala Thr Asn Gly Lys Glu Val Val Ser Ser Thr Gly
130 135 140
Val Leu Phe Val Lys Phe Gly Pro Pro Pro Thr Ala Ser Pro Gly Tyr
145 150 155 160
Ser Asp Glu Tyr Glu Glu Asp Gly Phe Cys Gln Pro Tyr Arg Gly Ile
165 170 175
Ala Cys Ala Arg Phe Ile Gly Asn Arg Thr Val Tyr Met Glu Ser Leu
180 185 190
His Met Gln Gly Glu Ile Glu Asn Gln Ile Thr Ala Ala Phe Thr Met
195 200 205
Ile Gly Thr Ser Ser His Leu Ser Asp Lys Cys Ser Gln Phe Ala Ile
210 215 220
Pro Ser Leu Cys His Tyr Ala Phe Pro Tyr Cys Asp Glu Thr Ser Ser
225 230 235 240
Val Pro Lys Pro Arg Asp Leu Cys Arg Asp Glu Cys Glu Ile Leu Glu
245 250 255
Asn Val Leu Cys Gln Thr Glu Tyr Ile Phe Ala Arg Ser Asn Pro Met
260 265 270
Ile Leu Met Arg Leu Lys Leu Pro Asn Cys Glu Asp Leu Pro Gln Pro
275 280 285
Glu Ser Pro Glu Ala Ala Asn Cys Ile Arg Ile Gly Ile Pro Met Ala
290 295 300
Asp Pro Ile Asn Lys Asn His Lys Cys Tyr Asn Ser Thr Gly Val Asp
305 310 315 320
Tyr Arg Gly Thr Val Ser Val Thr Lys Ser Gly Arg Gln Cys Gln Pro
325 330 335
Trp Asn Ser Gln Tyr Pro His Thr His Thr Phe Thr Ala Leu Arg Phe
340 345 350
Pro Glu Leu Asn Gly Gly His Ser Tyr Cys Arg Asn Pro Gly Asn Gln
355 360 365
Lys Glu Ala Pro Trp Cys Phe Thr Leu Asp Glu Asn Phe Lys Ser Asp
370 375 380
Leu Cys Asp Ile Pro Ala Cys Asp Ser Lys Asp Ser Lys Glu Lys Asn
385 390 395 400
Lys Met Glu Ile Leu Tyr Ile Leu Val Pro Ser Val Ala Ile Pro Leu
405 410 415
Ala Ile Ala Leu Leu Phe Phe Phe Ile Cys Val Cys Arg Asn Asn Gln
420 425 430
Lys Ser Ser Ser Ala Pro Val Gln Arg Gln Pro Lys His Val Arg Gly
435 440 445
Gln Asn Val Glu Met Ser Met Leu Asn Ala Tyr Lys Pro Lys Ser Lys
450 455 460
Ala Lys Glu Leu Pro Leu Ser Ala Val Arg Phe Met Glu Glu Leu Gly
465 470 475 480
Glu Cys Ala Phe Gly Lys Ile Tyr Lys Gly His Leu Tyr Leu Pro Gly
485 490 495
Met Asp His Ala Gln Leu Val Ala Ile Lys Thr Leu Lys Asp Tyr Asn
500 505 510
Asn Pro Gln Gln Trp Thr Glu Phe Gln Gln Glu Ala Ser Leu Met Ala
515 520 525
Glu Leu His His Pro Asn Ile Val Cys Leu Leu Gly Ala Val Thr Gln
530 535 540
Glu Gln Pro Val Cys Met Leu Phe Glu Tyr Ile Asn Gln Gly Asp Leu
545 550 555 560
His Glu Phe Leu Ile Met Arg Ser Pro His Ser Asp Val Gly Cys Ser
565 570 575
Ser Asp Glu Asp Gly Thr Val Lys Ser Ser Leu Asp His Gly Asp Phe
580 585 590
Leu His Ile Ala Ile Gln Ile Ala Ala Gly Met Glu Tyr Leu Ser Ser
595 600 605
His Phe Phe Val His Lys Asp Leu Ala Ala Arg Asn Ile Leu Ile Gly
610 615 620
Glu Gln Leu His Val Lys Ile Ser Asp Leu Gly Leu Ser Arg Glu Ile
625 630 635 640
Tyr Ser Ala Asp Tyr Tyr Arg Val Gln Ser Lys Ser Leu Leu Pro Ile
645 650 655
Arg Trp Met Pro Pro Glu Ala Ile Met Tyr Gly Lys Phe Ser Ser Asp
660 665 670
Ser Asp Ile Trp Ser Phe Gly Val Val Leu Trp Glu Ile Phe Ser Phe
675 680 685
Gly Leu Gln Pro Tyr Tyr Gly Phe Ser Asn Gln Glu Val Ile Glu Met
690 695 700
Val Arg Lys Arg Gln Leu Leu Pro Cys Ser Glu Asp Cys Pro Pro Arg
705 710 715 720
Met Tyr Ser Leu Met Thr Glu Cys Trp Asn Glu Ile Pro Ser Arg Arg
725 730 735
Pro Arg Phe Lys Asp Ile His Val Arg Leu Arg Ser Trp Glu Gly Leu
740 745 750
Ser Ser His Thr Ser Ser Thr Thr Pro Ser Gly Gly Asn Ala Thr Thr
755 760 765
Gln Thr Thr Ser Leu Ser Ala Ser Pro Val Ser Asn Leu Ser Asn Pro
770 775 780
Arg Tyr Pro Asn Tyr Met Phe Pro Ser Gln Gly Ile Thr Pro Gln Gly
785 790 795 800
Gln Ile Ala Gly Phe Ile Gly Pro Pro Ile Pro Gln Asn Gln Arg Phe
805 810 815
Ile Pro Ile Asn Gly Tyr Pro Ile Pro Pro Gly Tyr Ala Ala Phe Pro
820 825 830
Ala Ala His Tyr Gln Pro Thr Gly Pro Pro Arg Val Ile Gln His Cys
835 840 845
Pro Pro Pro Lys Ser Arg Ser Pro Ser Ser Ala Ser Gly Ser Thr Ser
850 855 860
Thr Gly His Val Thr Ser Leu Pro Ser Ser Gly Ser Asn Gln Glu Ala
865 870 875 880
Asn Ile Pro Leu Leu Pro His Met Ser Ile Pro Asn His Pro Gly Gly
885 890 895
Met Gly Ile Thr Val Phe Gly Asn Lys Ser Gln Lys Pro Tyr Lys Ile
900 905 910
Asp Ser Lys Gln Ala Ser Leu Leu Gly Asp Ala Asn Ile His Gly His
915 920 925
Thr Glu Ser Met Ile Ser Ala Glu Leu
930 935
<210> 85
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер RRE
<400> 85
attgtctggt atagtgcagc ag 22
<210> 86
<211> 801
<212> ДНК
<213> Homo sapiens
<400> 86
gaattcgcca ccatgctgct gctggtgaca agcctgctgc tgtgcgagct gccccacccc 60
gcctttctgc tgatccccca gagcgtgaaa gagtccgagg gcgacctggt cacaccagcc 120
ggcaacctga ccctgacctg taccgccagc ggcagcgaca tcaacgacta ccccatctct 180
tgggtccgcc aggctcctgg caagggactg gaatggatcg gcttcatcaa cagcggcggc 240
agcacttggt acgccagctg ggtcaaaggc cggttcacca tcagccggac cagcaccacc 300
gtggacctga agatgacaag cctgaccacc gacgacaccg ccacctactt ttgcgccaga 360
ggctacagca cctactacgg cgacttcaac atctggggcc ctggcaccct ggtcacaatc 420
tctagcggcg gaggcggcag cggaggtgga ggaagtggcg gcggaggatc cgagctggtc 480
atgacccaga cccccagcag cacatctggc gccgtgggcg gcaccgtgac catcaattgc 540
caggccagcc agagcatcga cagcaacctg gcctggttcc agcagaagcc cggccagccc 600
cccaccctgc tgatctacag agcctccaac ctggccagcg gcgtgccaag cagattcagc 660
ggcagcagat ctggcaccga gtacaccctg accatctccg gcgtgcagag agaggacgcc 720
gctacctatt actgcctggg cggcgtgggc aacgtgtcct acagaaccag cttcggcgga 780
ggtactgagg tggtcgtcaa a 801
<210> 87
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер SV40
<400> 87
cgaccagcaa ccatagtcc 19
<210> 88
<211> 72
<212> ДНК
<213> Homo sapiens
<400> 88
ctcgagggcg gcggagaggg cagaggaagt cttctaacat gcggtgacgt ggaggagaat 60
cccggcccta gg 72
<210> 89
<211> 24
<212> Белок
<213> Homo sapiens
<400> 89
Leu Glu Gly Gly Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp
1 5 10 15
Val Glu Glu Asn Pro Gly Pro Arg
20
<210> 90
<211> 5844
<212> ДНК
<213> Homo sapiens
<400> 90
atgcttctcc tggtgacaag ccttctgctc tgtgagttac cacacccagc attcctcctg 60
atcccacgca aagtgtgtaa cggaataggt attggtgaat ttaaagactc actctccata 120
aatgctacga atattaaaca cttcaaaaac tgcacctcca tcagtggcga tctccacatc 180
ctgccggtgg catttagggg tgactccttc acacatactc ctcctctgga tccacaggaa 240
ctggatattc tgaaaaccgt aaaggaaatc acagggtttt tgctgattca ggcttggcct 300
gaaaacagga cggacctcca tgcctttgag aacctagaaa tcatacgcgg caggaccaag 360
caacatggtc agttttctct tgcagtcgtc agcctgaaca taacatcctt gggattacgc 420
tccctcaagg agataagtga tggagatgtg ataatttcag gaaacaaaaa tttgtgctat 480
gcaaatacaa taaactggaa aaaactgttt gggacctccg gtcagaaaac caaaattata 540
agcaacagag gtgaaaacag ctgcaaggcc acaggccagg tctgccatgc cttgtgctcc 600
cccgagggct gctggggccc ggagcccagg gactgcgtct cttgccggaa tgtcagccga 660
ggcagggaat gcgtggacaa gtgcaacctt ctggagggtg agccaaggga gtttgtggag 720
aactctgagt gcatacagtg ccacccagag tgcctgcctc aggccatgaa catcacctgc 780
acaggacggg gaccagacaa ctgtatccag tgtgcccact acattgacgg cccccactgc 840
gtcaagacct gcccggcagg agtcatggga gaaaacaaca ccctggtctg gaagtacgca 900
gacgccggcc atgtgtgcca cctgtgccat ccaaactgca cctacggatg cactgggcca 960
ggtcttgaag gctgtccaac gaatgggcct aagatcccgt ccatcgccac tgggatggtg 1020
ggggccctcc tcttgctgct ggtggtggcc ctggggatcg gcctcttcat gtgagcggcc 1080
gctctagacc cgggctgcag gaattcgata tcaagcttat cgataatcaa cctctggatt 1140
acaaaatttg tgaaagattg actggtattc ttaactatgt tgctcctttt acgctatgtg 1200
gatacgctgc tttaatgcct ttgtatcatg ctattgcttc ccgtatggct ttcattttct 1260
cctccttgta taaatcctgg ttgctgtctc tttatgagga gttgtggccc gttgtcaggc 1320
aacgtggcgt ggtgtgcact gtgtttgctg acgcaacccc cactggttgg ggcattgcca 1380
ccacctgtca gctcctttcc gggactttcg ctttccccct ccctattgcc acggcggaac 1440
tcatcgccgc ctgccttgcc cgctgctgga caggggctcg gctgttgggc actgacaatt 1500
ccgtggtgtt gtcggggaaa tcatcgtcct ttccttggct gctcgcctgt gttgccacct 1560
ggattctgcg cgggacgtcc ttctgctacg tcccttcggc cctcaatcca gcggaccttc 1620
cttcccgcgg cctgctgccg gctctgcggc ctcttccgcg tcttcgcctt cgccctcaga 1680
cgagtcggat ctccctttgg gccgcctccc cgcatcgata ccgtcgacta gccgtacctt 1740
taagaccaat gacttacaag gcagctgtag atcttagcca ctttttaaaa gaaaaggggg 1800
gactggaagg gctaattcac tcccaaagaa gacaagatct gctttttgcc tgtactgggt 1860
ctctctggtt agaccagatc tgagcctggg agctctctgg ctaactaggg aacccactgc 1920
ttaagcctca ataaagcttg ccttgagtgc ttcaagtagt gtgtgcccgt ctgttgtgtg 1980
actctggtaa ctagagatcc ctcagaccct tttagtcagt gtggaaaatc tctagcagaa 2040
ttcgatatca agcttatcga taccgtcgac ctcgaggggg ggcccggtac ccaattcgcc 2100
ctatagtgag tcgtattaca attcactggc cgtcgtttta caacgtcgtg actgggaaaa 2160
ccctggcgtt acccaactta atcgccttgc agcacatccc cctttcgcca gctggcgtaa 2220
tagcgaagag gcccgcaccg atcgcccttc ccaacagttg cgcagcctga atggcgaatg 2280
gaaattgtaa gcgttaatat tttgttaaaa ttcgcgttaa atttttgtta aatcagctca 2340
ttttttaacc aataggccga aatcggcaaa atcccttata aatcaaaaga atagaccgag 2400
atagggttga gtgttgttcc agtttggaac aagagtccac tattaaagaa cgtggactcc 2460
aacgtcaaag ggcgaaaaac cgtctatcag ggcgatggcc cactacgtga accatcaccc 2520
taatcaagtt ttttggggtc gaggtgccgt aaagcactaa atcggaaccc taaagggagc 2580
ccccgattta gagcttgacg gggaaagccg gcgaacgtgg cgagaaagga agggaagaaa 2640
gcgaaaggag cgggcgctag ggcgctggca agtgtagcgg tcacgctgcg cgtaaccacc 2700
acacccgccg cgcttaatgc gccgctacag ggcgcgtcag gtggcacttt tcggggaaat 2760
gtgcgcggaa cccctatttg tttatttttc taaatacatt caaatatgta tccgctcatg 2820
agacaataac cctgataaat gcttcaataa tattgaaaaa ggaagagtat gagtattcaa 2880
catttccgtg tcgcccttat tccctttttt gcggcatttt gccttcctgt ttttgctcac 2940
ccagaaacgc tggtgaaagt aaaagatgct gaagatcagt tgggtgcacg agtgggttac 3000
atcgaactgg atctcaacag cggtaagatc cttgagagtt ttcgccccga agaacgtttt 3060
ccaatgatga gcacttttaa agttctgcta tgtggcgcgg tattatcccg tattgacgcc 3120
gggcaagagc aactcggtcg ccgcatacac tattctcaga atgacttggt tgagtactca 3180
ccagtcacag aaaagcatct tacggatggc atgacagtaa gagaattatg cagtgctgcc 3240
ataaccatga gtgataacac tgcggccaac ttacttctga caacgatcgg aggaccgaag 3300
gagctaaccg cttttttgca caacatgggg gatcatgtaa ctcgccttga tcgttgggaa 3360
ccggagctga atgaagccat accaaacgac gagcgtgaca ccacgatgcc tgtagcaatg 3420
gcaacaacgt tgcgcaaact attaactggc gaactactta ctctagcttc ccggcaacaa 3480
ttaatagact ggatggaggc ggataaagtt gcaggaccac ttctgcgctc ggcccttccg 3540
gctggctggt ttattgctga taaatctgga gccggtgagc gtgggtctcg cggtatcatt 3600
gcagcactgg ggccagatgg taagccctcc cgtatcgtag ttatctacac gacggggagt 3660
caggcaacta tggatgaacg aaatagacag atcgctgaga taggtgcctc actgattaag 3720
cattggtaac tgtcagacca agtttactca tatatacttt agattgattt aaaacttcat 3780
ttttaattta aaaggatcta ggtgaagatc ctttttgata atctcatgac caaaatccct 3840
taacgtgagt tttcgttcca ctgagcgtca gaccccgtag aaaagatcaa aggatcttct 3900
tgagatcctt tttttctgcg cgtaatctgc tgcttgcaaa caaaaaaacc accgctacca 3960
gcggtggttt gtttgccgga tcaagagcta ccaactcttt ttccgaaggt aactggcttc 4020
agcagagcgc agataccaaa tactgttctt ctagtgtagc cgtagttagg ccaccacttc 4080
aagaactctg tagcaccgcc tacatacctc gctctgctaa tcctgttacc agtggctgct 4140
gccagtggcg ataagtcgtg tcttaccggg ttggactcaa gacgatagtt accggataag 4200
gcgcagcggt cgggctgaac ggggggttcg tgcacacagc ccagcttgga gcgaacgacc 4260
tacaccgaac tgagatacct acagcgtgag ctatgagaaa gcgccacgct tcccgaaggg 4320
agaaaggcgg acaggtatcc ggtaagcggc agggtcggaa caggagagcg cacgagggag 4380
cttccagggg gaaacgcctg gtatctttat agtcctgtcg ggtttcgcca cctctgactt 4440
gagcgtcgat ttttgtgatg ctcgtcaggg gggcggagcc tatggaaaaa cgccagcaac 4500
gcggcctttt tacggttcct ggccttttgc tggccttttg ctcacatgtt ctttcctgcg 4560
ttatcccctg attctgtgga taaccgtatt accgcctttg agtgagctga taccgctcgc 4620
cgcagccgaa cgaccgagcg cagcgagtca gtgagcgagg aagcggaaga gcgcccaata 4680
cgcaaaccgc ctctccccgc gcgttggccg attcattaat gcagctggca cgacaggttt 4740
cccgactgga aagcgggcag tgagcgcaac gcaattaatg tgagttagct cactcattag 4800
gcaccccagg ctttacactt tatgcttccg gctcgtatgt tgtgtggaat tgtgagcgga 4860
taacaatttc acacaggaaa cagctatgac catgattacg ccaagctcga aattaaccct 4920
cactaaaggg aacaaaagct ggagctccac cgcggtggcg gcctcgaggt cgagatccgg 4980
tcgaccagca accatagtcc cgcccctaac tccgcccatc ccgcccctaa ctccgcccag 5040
ttccgcccat tctccgcccc atggctgact aatttttttt atttatgcag aggccgaggc 5100
cgcctcggcc tctgagctat tccagaagta gtgaggaggc ttttttggag gcctaggctt 5160
ttgcaaaaag cttcgacggt atcgattggc tcatgtccaa cattaccgcc atgttgacat 5220
tgattattga ctagttatta atagtaatca attacggggt cattagttca tagcccatat 5280
atggagttcc gcgttacata acttacggta aatggcccgc ctggctgacc gcccaacgac 5340
ccccgcccat tgacgtcaat aatgacgtat gttcccatag taacgccaat agggactttc 5400
cattgacgtc aatgggtgga gtatttacgg taaactgccc acttggcagt acatcaagtg 5460
tatcatatgc caagtacgcc ccctattgac gtcaatgacg gtaaatggcc cgcctggcat 5520
tatgcccagt acatgacctt atgggacttt cctacttggc agtacatcta cgtattagtc 5580
atcgctatta ccatggtgat gcggttttgg cagtacatca atgggcgtgg atagcggttt 5640
gactcacggg gatttccaag tctccacccc attgacgtca atgggagttt gttttggcac 5700
caaaatcaac gggactttcc aaaatgtcgt aacaactccg ccccattgac gcaaatgggc 5760
ggtaggcgtg tacggaattc ggagtggcga gccctcagat cctgcatata agcagctgct 5820
ttttgcctgt actgggtctc tctg 5844
<210> 91
<211> 356
<212> Белок
<213> Homo sapiens
<400> 91
Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro Ala
1 5 10 15
Phe Leu Leu Ile Pro Arg Lys Val Cys Asn Gly Ile Gly Ile Gly Glu
20 25 30
Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr Asn Ile Lys His Phe Lys
35 40 45
Asn Cys Thr Ser Ile Ser Gly Asp Leu His Ile Leu Pro Val Ala Phe
50 55 60
Arg Gly Asp Ser Phe Thr His Thr Pro Pro Leu Asp Pro Gln Glu Leu
65 70 75 80
Asp Ile Leu Lys Thr Val Lys Glu Ile Thr Gly Phe Leu Leu Ile Gln
85 90 95
Ala Trp Pro Glu Asn Arg Thr Asp Leu His Ala Phe Glu Asn Leu Glu
100 105 110
Ile Ile Arg Gly Arg Thr Lys Gln His Gly Gln Phe Ser Leu Ala Val
115 120 125
Val Ser Leu Asn Ile Thr Ser Leu Gly Leu Arg Ser Leu Lys Glu Ile
130 135 140
Ser Asp Gly Asp Val Ile Ile Ser Gly Asn Lys Asn Leu Cys Tyr Ala
145 150 155 160
Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly Thr Ser Gly Gln Lys Thr
165 170 175
Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser Cys Lys Ala Thr Gly Gln
180 185 190
Val Cys His Ala Leu Cys Ser Pro Glu Gly Cys Trp Gly Pro Glu Pro
195 200 205
Arg Asp Cys Val Ser Cys Arg Asn Val Ser Arg Gly Arg Glu Cys Val
210 215 220
Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro Arg Glu Phe Val Glu Asn
225 230 235 240
Ser Glu Cys Ile Gln Cys His Pro Glu Cys Leu Pro Gln Ala Met Asn
245 250 255
Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn Cys Ile Gln Cys Ala His
260 265 270
Tyr Ile Asp Gly Pro His Cys Val Lys Thr Cys Pro Ala Gly Val Met
275 280 285
Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr Ala Asp Ala Gly His Val
290 295 300
Cys His Leu Cys His Pro Asn Cys Thr Tyr Gly Cys Thr Gly Pro Gly
305 310 315 320
Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys Ile Pro Ser Ile Ala Thr
325 330 335
Gly Met Val Gly Ala Leu Leu Leu Leu Leu Val Val Ala Leu Gly Ile
340 345 350
Gly Leu Phe Met
355
<210> 92
<211> 327
<212> Белок
<213> Homo sapiens
<400> 92
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 93
<211> 220
<212> Белок
<213> Homo sapiens
<400> 93
Met Leu Arg Leu Leu Leu Ala Leu Asn Leu Phe Pro Ser Ile Gln Val
1 5 10 15
Thr Gly Asn Lys Ile Leu Val Lys Gln Ser Pro Met Leu Val Ala Tyr
20 25 30
Asp Asn Ala Val Asn Leu Ser Cys Lys Tyr Ser Tyr Asn Leu Phe Ser
35 40 45
Arg Glu Phe Arg Ala Ser Leu His Lys Gly Leu Asp Ser Ala Val Glu
50 55 60
Val Cys Val Val Tyr Gly Asn Tyr Ser Gln Gln Leu Gln Val Tyr Ser
65 70 75 80
Lys Thr Gly Phe Asn Cys Asp Gly Lys Leu Gly Asn Glu Ser Val Thr
85 90 95
Phe Tyr Leu Gln Asn Leu Tyr Val Asn Gln Thr Asp Ile Tyr Phe Cys
100 105 110
Lys Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu Lys Ser
115 120 125
Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro Ser Pro
130 135 140
Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val Val Gly
145 150 155 160
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
165 170 175
Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met
180 185 190
Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro
195 200 205
Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser
210 215 220
<210> 94
<211> 164
<212> Белок
<213> Homo sapiens
<400> 94
Met Lys Trp Lys Ala Leu Phe Thr Ala Ala Ile Leu Gln Ala Gln Leu
1 5 10 15
Pro Ile Thr Glu Ala Gln Ser Phe Gly Leu Leu Asp Pro Lys Leu Cys
20 25 30
Tyr Leu Leu Asp Gly Ile Leu Phe Ile Tyr Gly Val Ile Leu Thr Ala
35 40 45
Leu Phe Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr
50 55 60
Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg
65 70 75 80
Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met
85 90 95
Gly Gly Lys Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
100 105 110
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
115 120 125
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
130 135 140
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
145 150 155 160
Leu Pro Pro Arg
<210> 95
<211> 240
<212> Белок
<213> Homo sapiens
<400> 95
Met Gly Asn Ser Cys Tyr Asn Ile Val Ala Thr Leu Leu Leu Val Leu
1 5 10 15
Asn Phe Glu Arg Thr Arg Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro
20 25 30
Ala Gly Thr Phe Cys Asp Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys
35 40 45
Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile
50 55 60
Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu Cys Ser Ser
65 70 75 80
Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His Cys Leu Gly
85 90 95
Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu
100 105 110
Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln
115 120 125
Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys
130 135 140
Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro
145 150 155 160
Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Val Thr Pro Pro Ala
165 170 175
Pro Ala Arg Glu Pro Gly His Ser Pro Gln Ile Ile Ser Phe Phe Leu
180 185 190
Ala Leu Thr Ser Thr Ala Leu Leu Phe Leu Leu Phe Phe Leu Thr Leu
195 200 205
Arg Phe Ser Val Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe
210 215 220
Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly
225 230 235 240
<210> 96
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер WPRE
<400> 96
actgtgtttg ctgacgcaac 20
<210> 97
<211> 168
<212> Белок
<213> Homo sapiens
<400> 97
Met Gly His His His His His His His His His His Ser Ser Gly His
1 5 10 15
Ile Glu Gly Arg His Met Arg Arg Val Pro Gly Val Ala Pro Thr Leu
20 25 30
Val Arg Ser Ala Ser Glu Thr Ser Glu Lys Arg Pro Phe Met Cys Ala
35 40 45
Tyr Pro Gly Cys Asn Lys Arg Tyr Phe Lys Leu Ser His Leu Gln Met
50 55 60
His Ser Arg Lys His Thr Gly Glu Lys Pro Tyr Gln Cys Asp Phe Lys
65 70 75 80
Asp Cys Glu Arg Arg Phe Phe Arg Ser Asp Gln Leu Lys Arg His Gln
85 90 95
Arg Arg His Thr Gly Val Lys Pro Phe Gln Cys Lys Thr Cys Gln Arg
100 105 110
Lys Phe Ser Arg Ser Asp His Leu Lys Thr His Thr Arg Thr His Thr
115 120 125
Gly Glu Lys Pro Phe Ser Cys Arg Trp Pro Ser Cys Gln Lys Lys Phe
130 135 140
Ala Arg Ser Asp Glu Leu Val Arg His His Asn Met His Gln Arg Asn
145 150 155 160
Met Thr Lys Leu Gln Leu Ala Leu
165
<210> 98
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсерная область
<220>
<221> ВАРИАНТ
<222> (1)..(1)
<223> Xaa представляет собой цистеин, глицин или аргинин
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> Xaa представляет собой цистеин или треонин
<400> 98
Xaa Pro Pro Xaa Pro
1 5
<210> 99
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Zeta
<400> 99
cgggtgaagt tcagcagaag 20
<210> 100
<211> 9
<212> Белок
<213> Homo sapiens
<400> 100
Ala Asp Arg Ala Thr Tyr Phe Cys Ala
1 5
<210> 101
<211> 11
<212> Белок
<213> Homo sapiens
<400> 101
Ala Ser Gly Phe Asp Phe Ser Ala Tyr Tyr Met
1 5 10
<210> 102
<211> 6
<212> Белок
<213> Homo sapiens
<400> 102
Asp Thr Ile Asp Trp Tyr
1 5
<210> 103
<211> 5
<212> Белок
<213> Homo sapiens
<400> 103
Asp Tyr Gly Val Ser
1 5
<210> 104
<211> 9
<212> Белок
<213> Homo sapiens
<400> 104
Gly Asn Thr Leu Pro Tyr Thr Phe Gly
1 5
<210> 105
<211> 7
<212> Белок
<213> Homo sapiens
<400> 105
Ile Asn Ser Gly Gly Ser Thr
1 5
<210> 106
<211> 8
<212> Белок
<213> Homo sapiens
<400> 106
Asn Val Ser Tyr Arg Thr Ser Phe
1 5
<210> 107
<211> 16
<212> Белок
<213> Homo sapiens
<400> 107
Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
1 5 10 15
<210> 108
<211> 11
<212> Белок
<213> Homo sapiens
<400> 108
Arg Ala Ser Gln Asp Ile Ser Lys Tyr Leu Asn
1 5 10
<210> 109
<211> 11
<212> Белок
<213> Homo sapiens
<400> 109
Ser Gly Ser Asp Ile Asn Asp Tyr Pro Ile Ser
1 5 10
<210> 110
<211> 5
<212> Белок
<213> Homo sapiens
<400> 110
Ser Asn Leu Ala Trp
1 5
<210> 111
<211> 7
<212> Белок
<213> Homo sapiens
<400> 111
Ser Arg Leu His Ser Gly Val
1 5
<210> 112
<211> 7
<212> Белок
<213> Homo sapiens
<400> 112
Thr Ile Tyr Pro Ser Ser Gly
1 5
<210> 113
<211> 16
<212> Белок
<213> Homo sapiens
<400> 113
Val Gln Ser Asp Gly Ser Tyr Thr Lys Arg Pro Gly Val Pro Asp Arg
1 5 10 15
<210> 114
<211> 16
<212> Белок
<213> Homo sapiens
<400> 114
Val Thr Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser Ala Leu Lys Ser
1 5 10 15
<210> 115
<211> 7
<212> Белок
<213> Homo sapiens
<400> 115
Tyr Ala Met Asp Tyr Trp Gly
1 5
<210> 116
<211> 8
<212> Белок
<213> Homo sapiens
<400> 116
Tyr Phe Cys Ala Arg Gly Tyr Ser
1 5
<210> 117
<211> 8
<212> Белок
<213> Homo sapiens
<400> 117
Tyr Ile Gly Gly Tyr Val Phe Gly
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КЛЕТКА | 2014 |
|
RU2717984C2 |
| КЛЕТКА | 2014 |
|
RU2732236C2 |
| КЛЕТКА | 2015 |
|
RU2768019C2 |
| ТРАНСГЕННЫЕ ГЕНЕТИЧЕСКИЕ МЕТКИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2822461C1 |
| ТРАНСГЕННЫЕ ГЕНЕТИЧЕСКИЕ МЕТКИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2729112C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2020 |
|
RU2833880C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2018 |
|
RU2809244C2 |
| КЛЕТКИ ДЛЯ ИММУНОТЕРАПИИ, СКОНСТРУИРОВАННЫЕ ДЛЯ НАЦЕЛИВАНИЯ НА АНТИГЕН, ПРИСУТСТВУЮЩИЙ ОДНОВРЕМЕННО НА ИММУННЫХ КЛЕТКАХ И НА ПАТОЛОГИЧЕСКИХ КЛЕТКАХ | 2015 |
|
RU2714258C2 |
| Т-КЛЕТКИ, МОДИФИЦИРОВАННЫЕ ХИМЕРНЫМ РЕЦЕПТОРОМ АНТИГЕНА, НАЦЕЛЕННЫМ НА CS1 | 2015 |
|
RU2727451C2 |
| Т-КЛЕТКИ С ДВУМЯ ХИМЕРНЫМИ АНТИГЕННЫМИ РЕЦЕПТОРАМИ CAR | 2021 |
|
RU2839662C1 |














































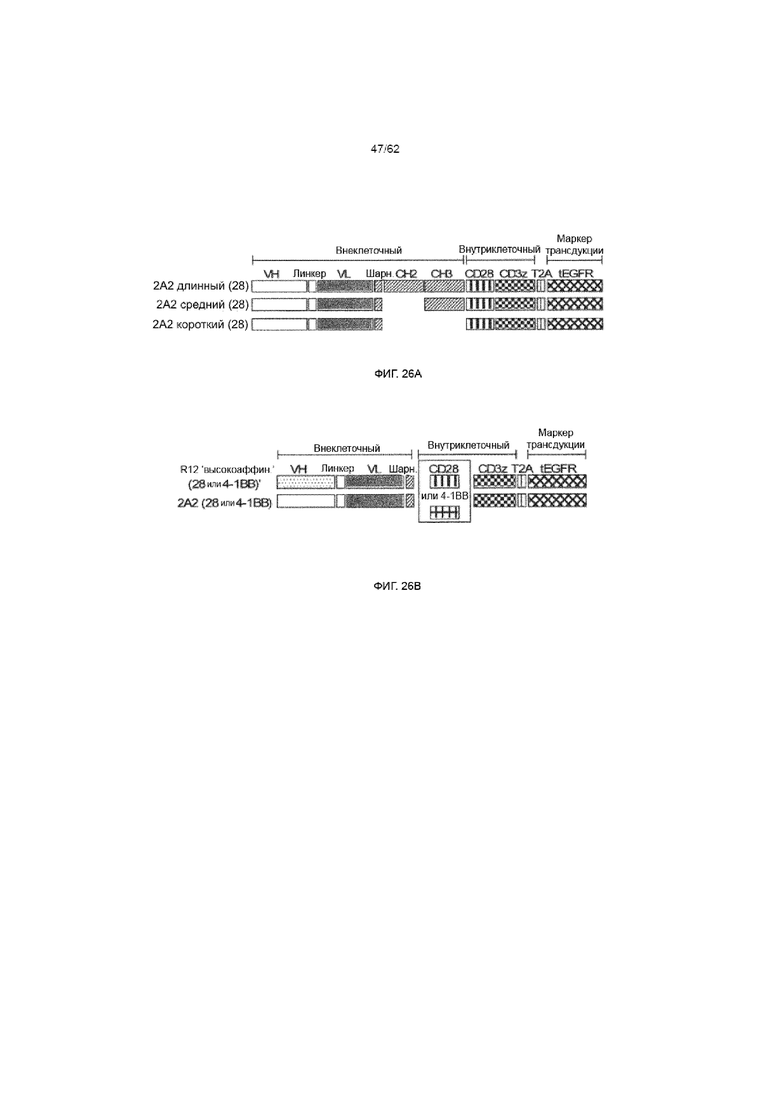
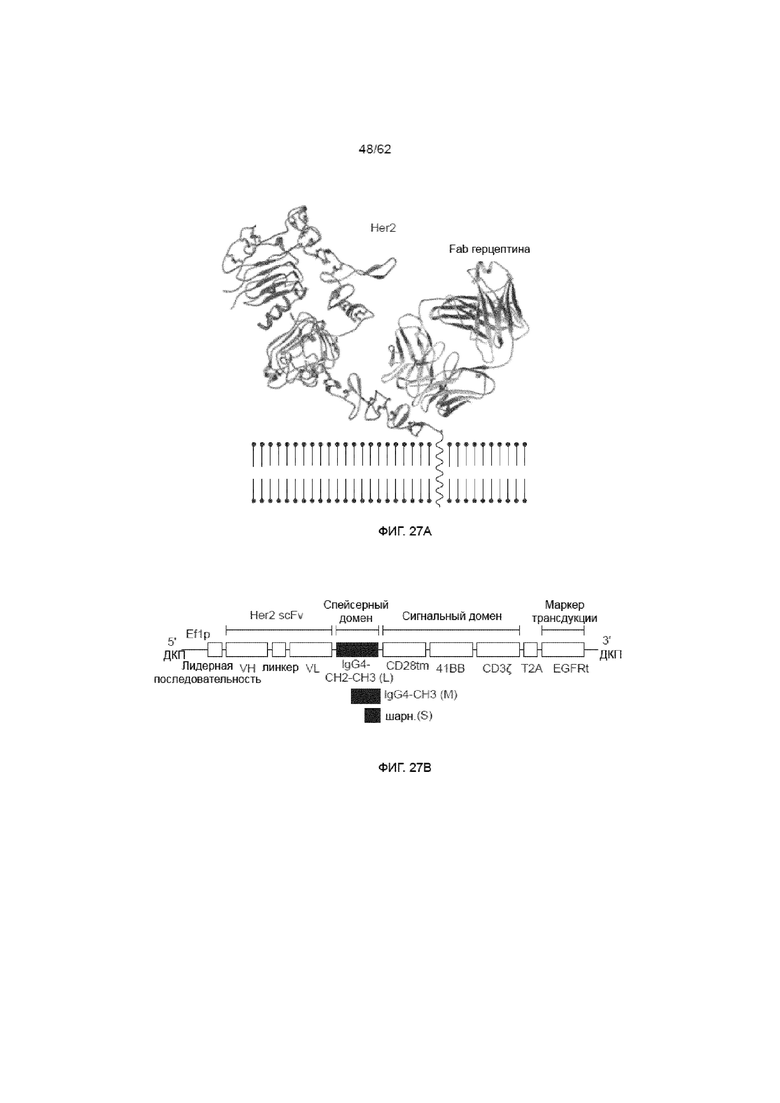

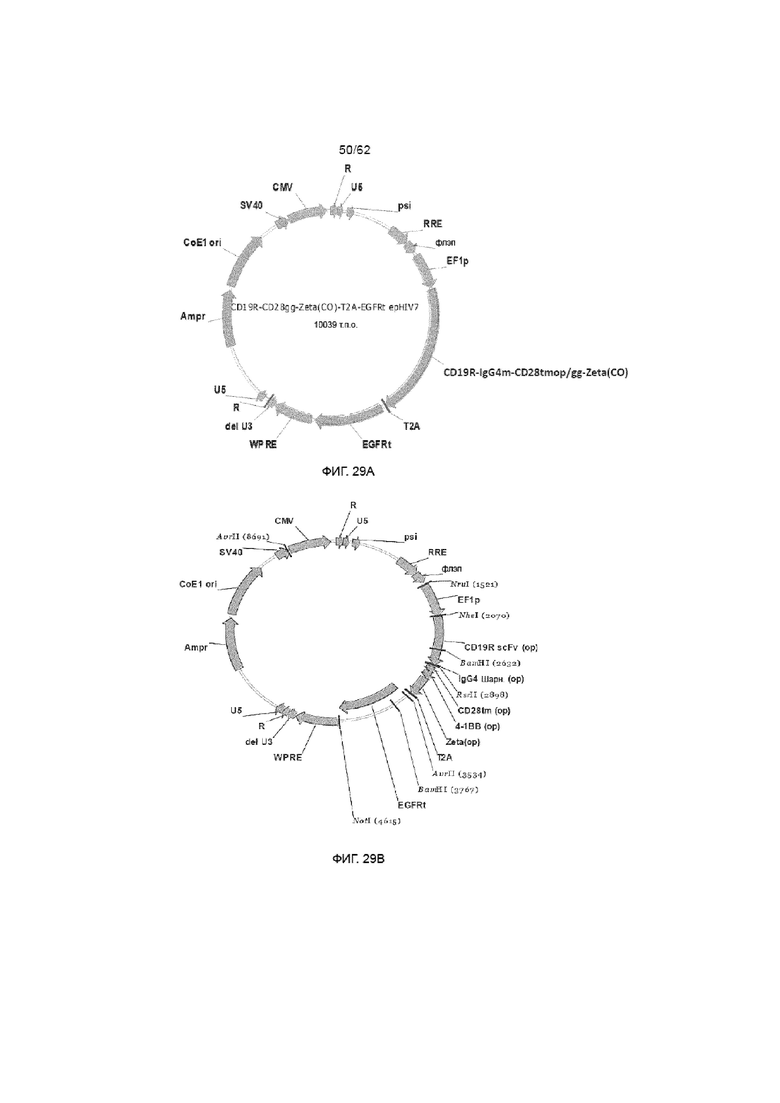
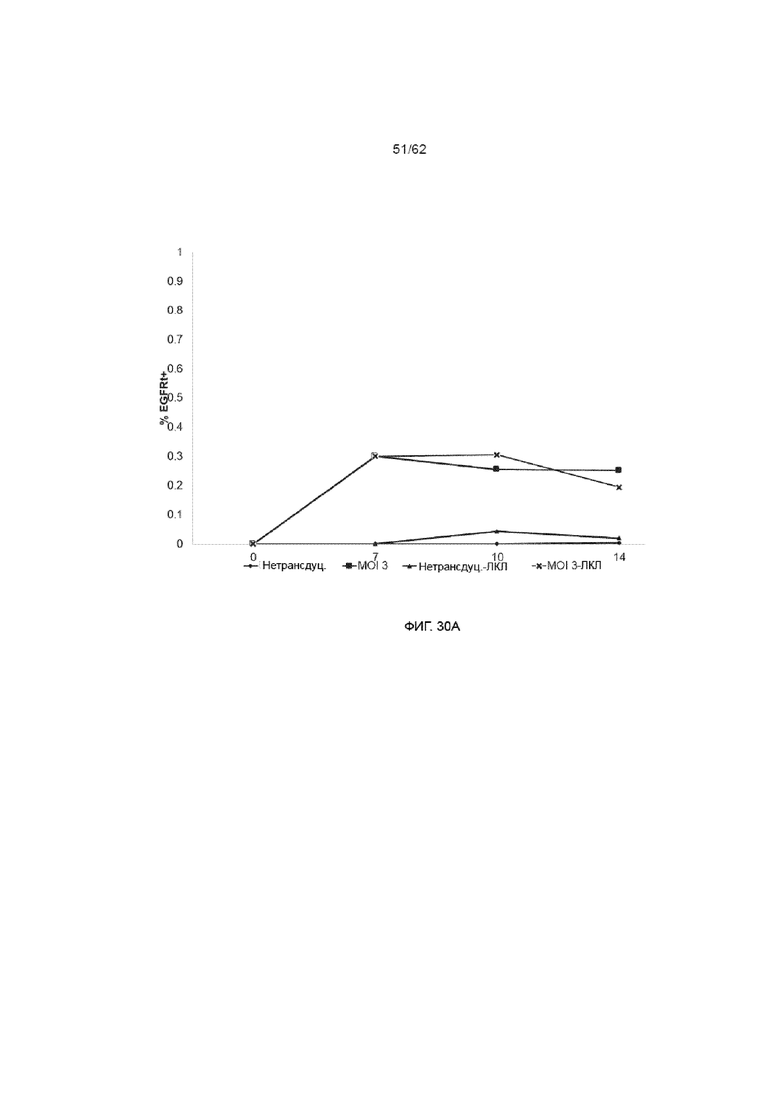
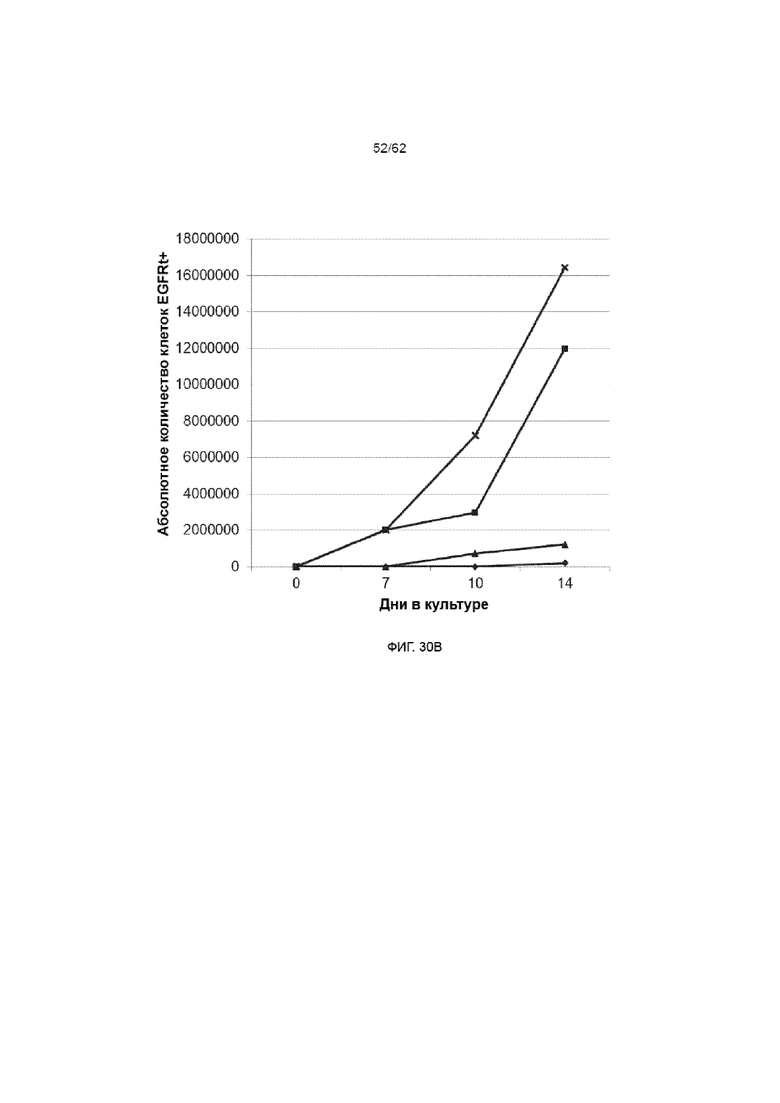
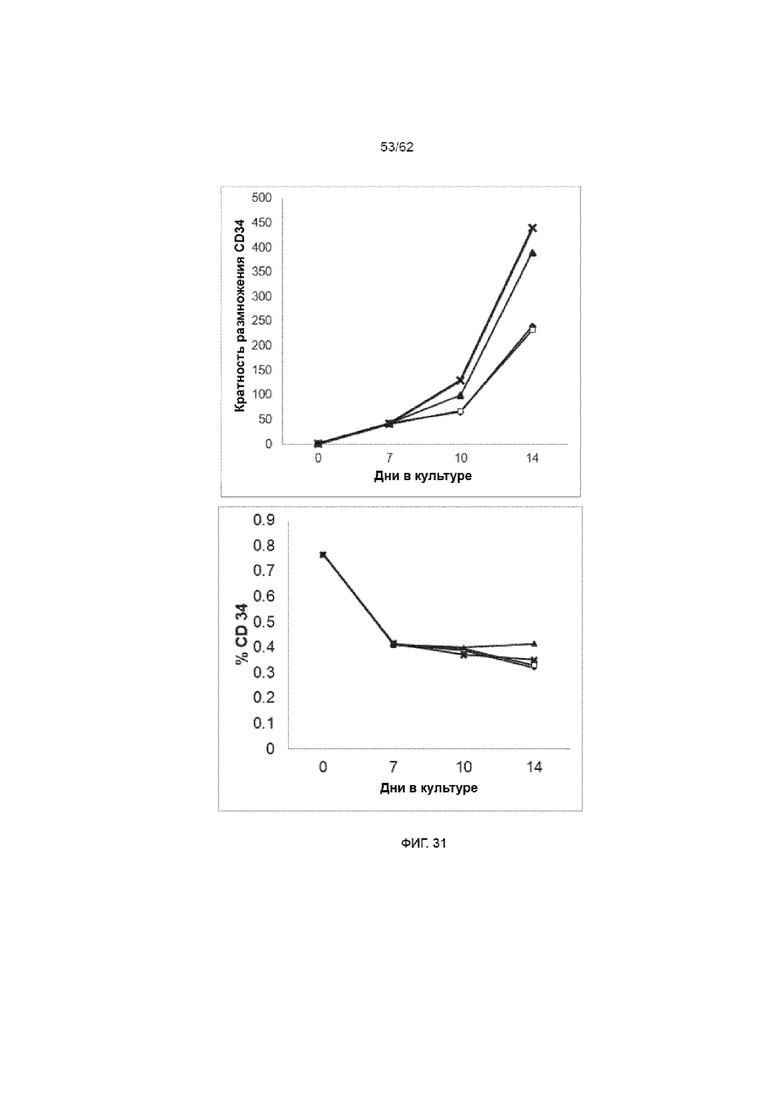
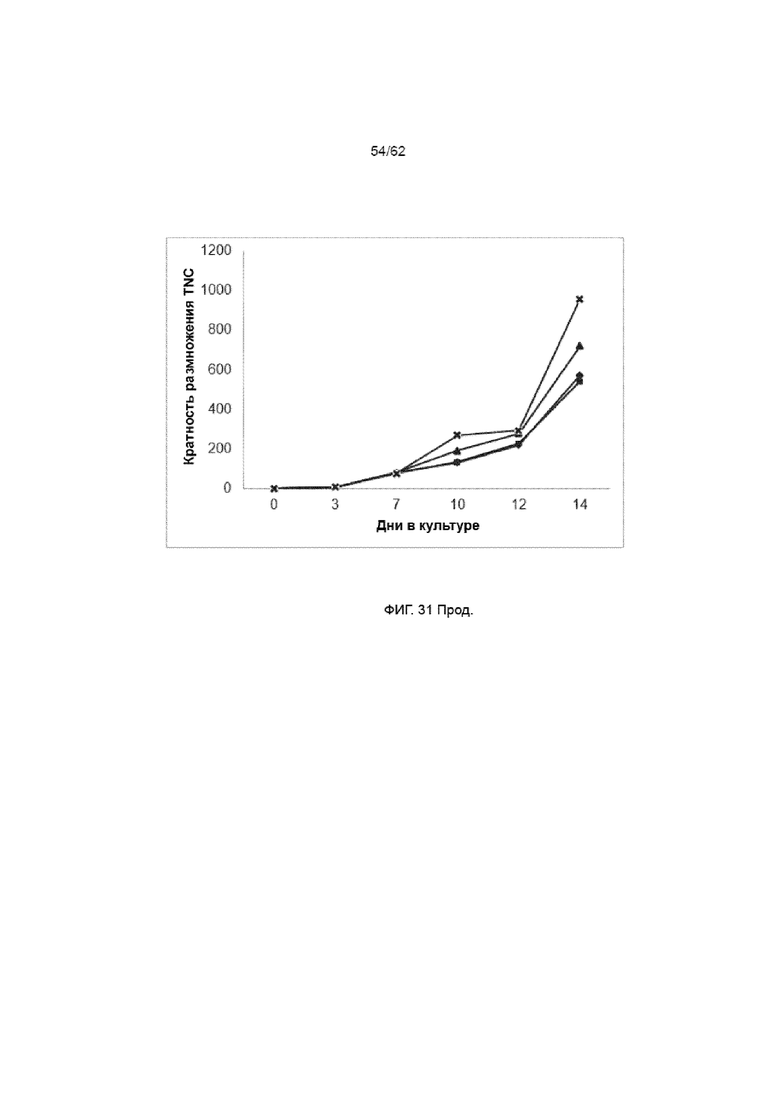
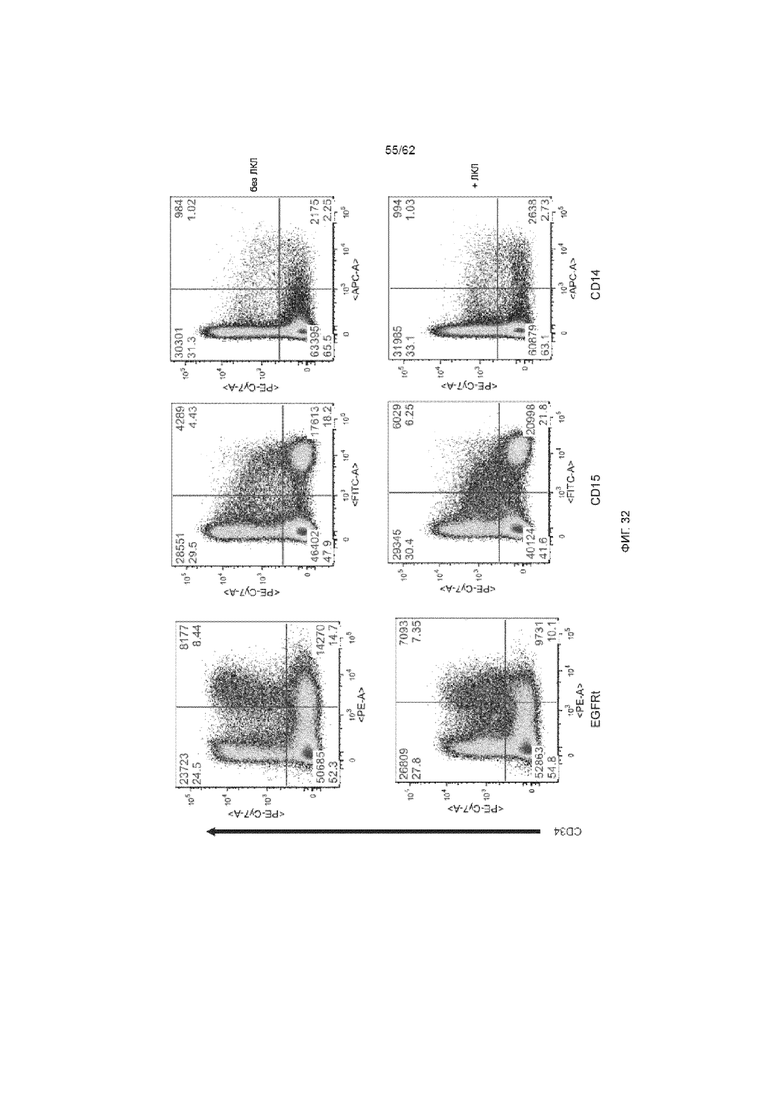
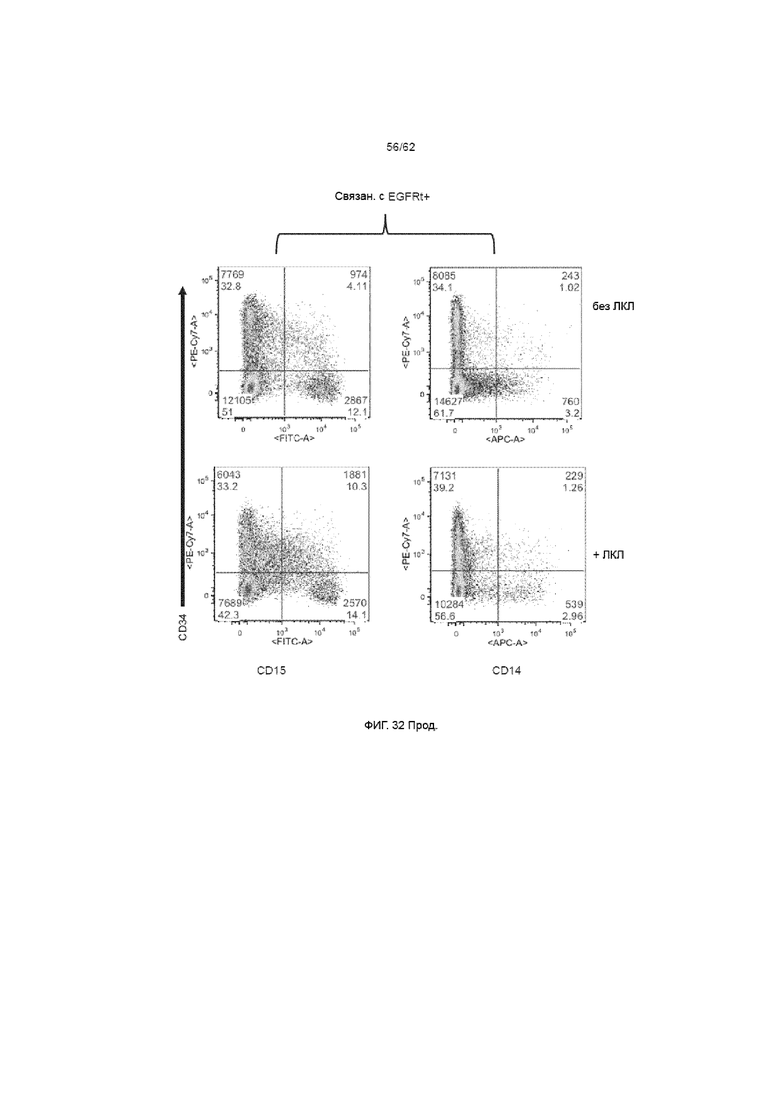
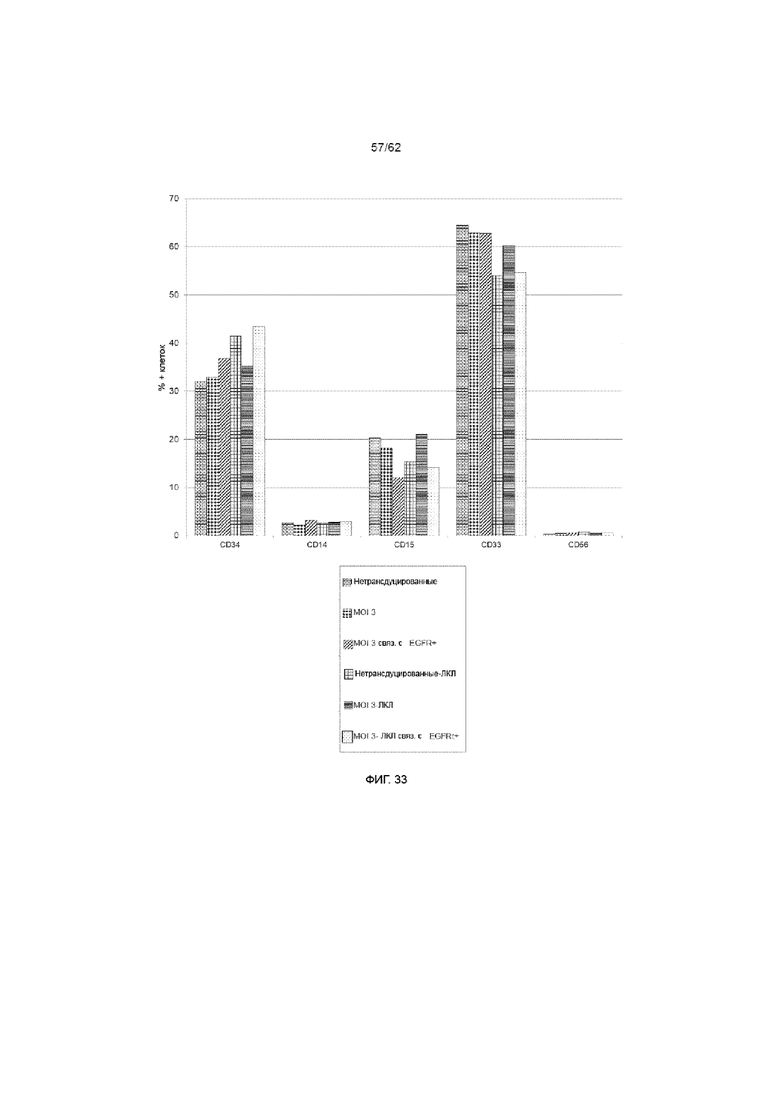
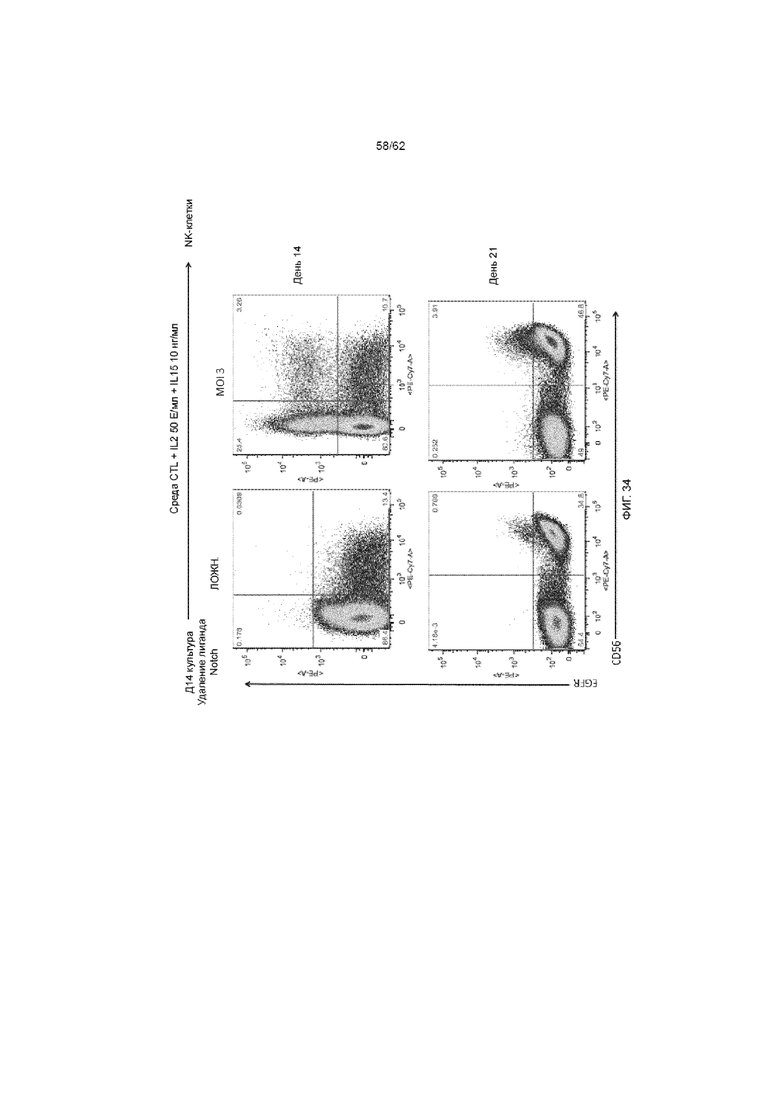
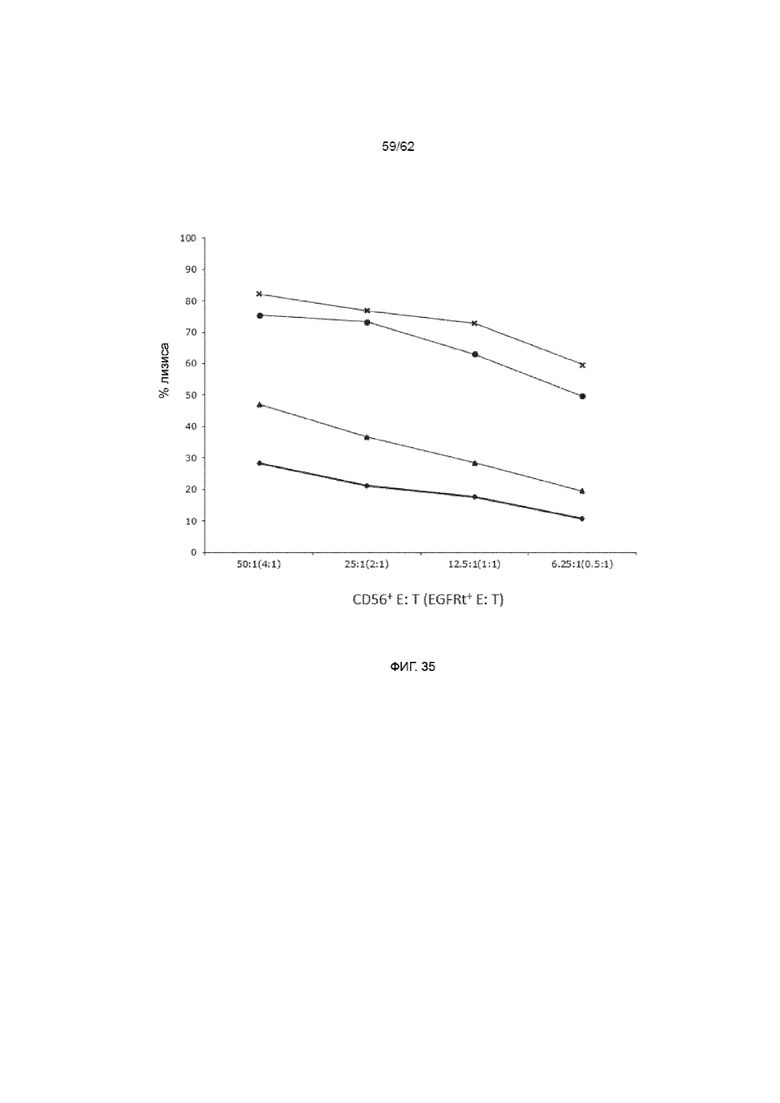
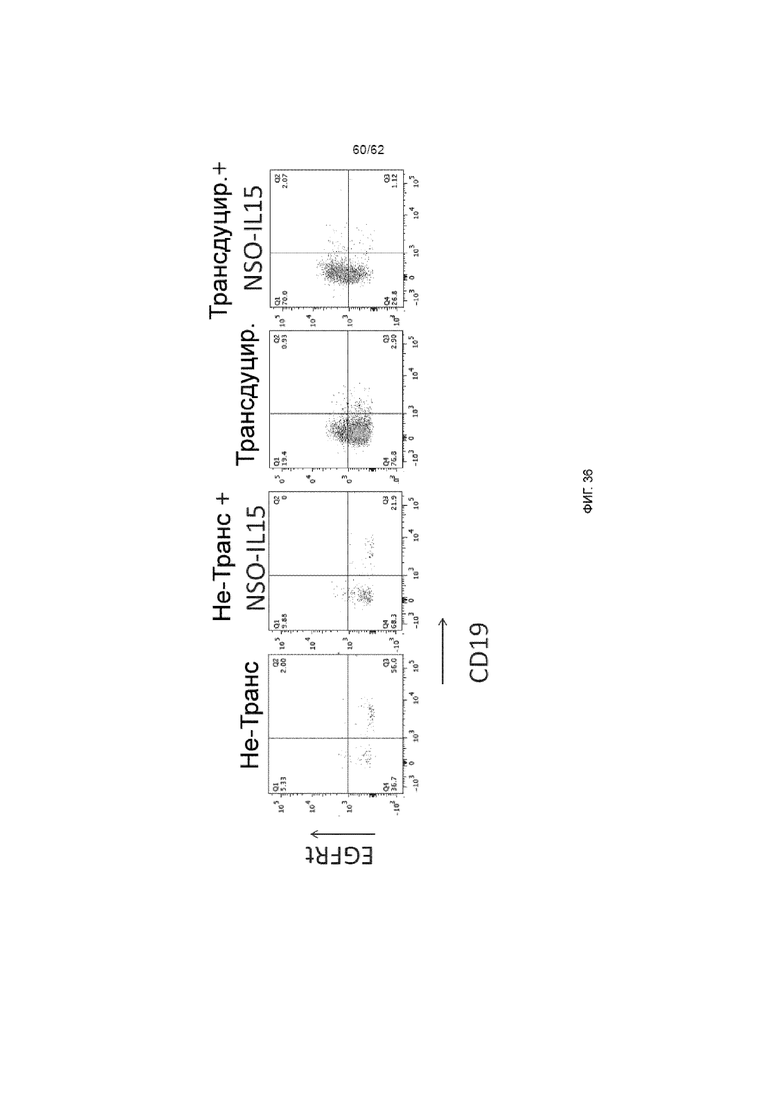
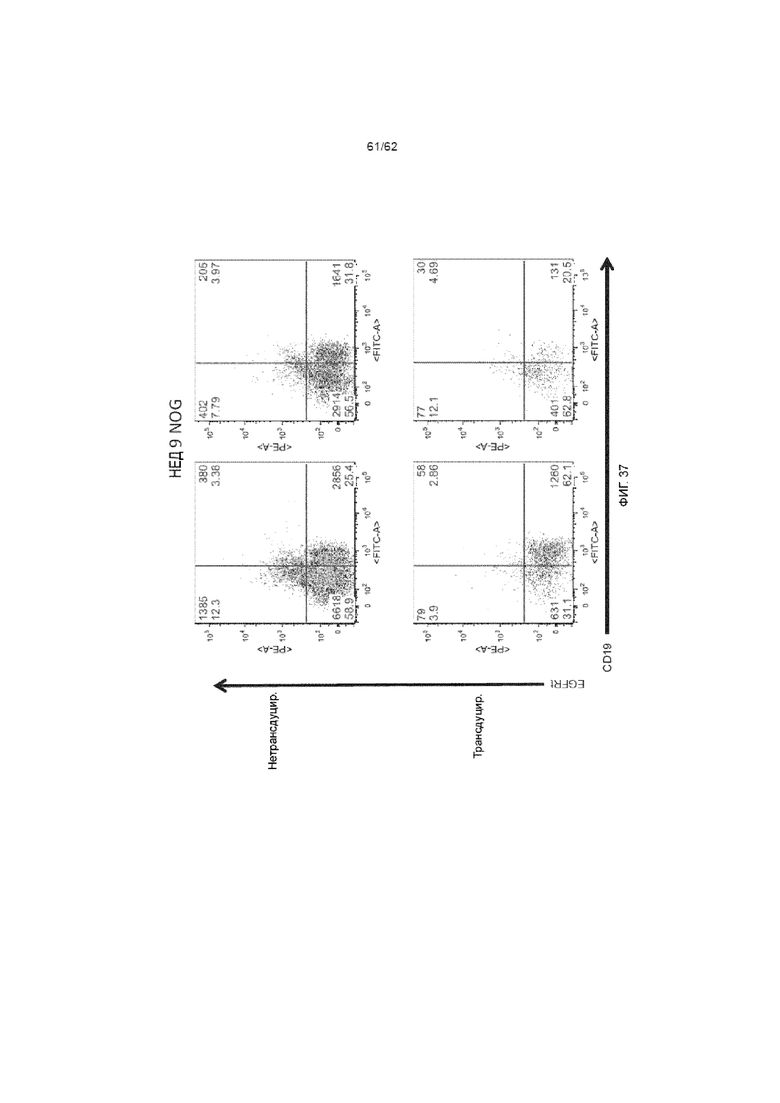
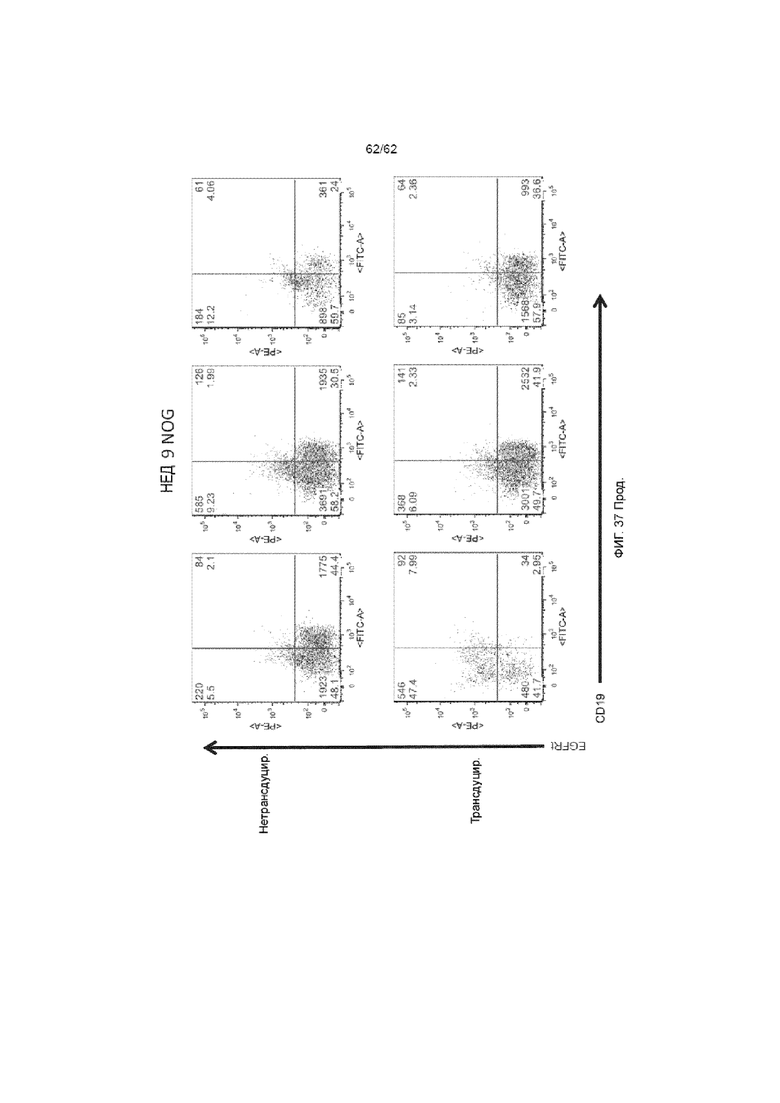
Изобретение относится к области биотехнологии. Предложен способ обеспечения анти-CD19 активности против клеток, экспрессирующих CD19, у субъекта. Способ включает введение субъекту терапевтически эффективного количества размноженных с агонистом Notch генетически модифицированных гемопоэтических стволовых клеток/клеток-предшественников (HSPC) CD34+ и/или терапевтически эффективного количества генетически модифицированных естественных клеток-киллеров, дифференцированных из размноженных с агонистом Notch HSPC CD34+. Генетическая модификация приводит к экспрессии молекул, содержащих внеклеточный компонент, содержащий CD19-лигандсвязывающий домен, связанный с внутриклеточным компонентом через трансмембранный домен. Также изобретение относится к набору и составу для обеспечения анти-CD19 активности против экспрессирующих CD19 клеток, содержащим терапевтически эффективное количество указанных генетически модифицированных клеток. Изобретение обеспечивает возможность введения модифицированных клеток пациентам для нацеливания на раковые клетки без необходимости проверки иммунологической совместимости перед введением. 4 н. и 45 з.п. ф-лы, 37 ил., 2 пр.
1. Способ обеспечения анти-CD19 активности против клеток, экспрессирующих CD19, у субъекта, включающий введение указанному субъекту терапевтически эффективного количества размноженных с агонистом Notch генетически модифицированных гемопоэтических стволовых клеток/клеток-предшественников (HSPC) CD34+ и/или терапевтически эффективного количества генетически модифицированных естественных клеток-киллеров, дифференцированных из размноженных с агонистом Notch HSPC CD34+,
где вводимые генетически модифицированные клетки не требуют проверки иммунологической совместимости с субъектом и где указанная генетическая модификация приводит к экспрессии молекул, содержащих внеклеточный компонент, связанный с внутриклеточным компонентом через трансмембранный домен,
где внеклеточный компонент содержит CD19-лигандсвязывающий домен и внутриклеточный компонент содержит эффекторный домен, содержащий (i) весь или часть внутриклеточного сигнального домена CD3ζ, (ii) весь или часть внутриклеточного сигнального домена CD28 и/или (iii) весь или часть внутриклеточного сигнального домена 4-1BB,
посредством чего обеспечивается анти-CD19 активность против клеток, экспрессирующих CD19, у субъекта.
2. Способ по п. 1, где субъект является педиатрическим пациентом с рецидивом острой лимфобластической лейкемии.
3. Способ по п. 1, где субъект имеет лейкемию или лимфому.
4. Способ по п. 1, где CD19-лигандсвязывающий домен содержит фрагмент антитела.
5. Способ по п. 1, где CD19-лигандсвязывающий домен содержит одноцепочечный вариабельный фрагмент (scFv) антитела.
6. Способ по п. 1, где лигандсвязывающий домен содержит одноцепочечный фрагмент Fv (scFv), содержащий последовательность CDRL1 SEQ ID NO. 108, последовательность CDRL2 SEQ ID NO. 111, последовательность CDRL3 SEQ ID NO. 104, последовательность CDRH1 SEQ ID NO. 103, последовательность CDRH2 SEQ ID NO. 114 и последовательность CDRH3 SEQ ID NO. 115, где остатки CDR пронумерованы согласно Кабату.
7. Способ по п. 1, где эффекторный домен содержит вариант внутриклеточного сигнального домена CD3ζ и/или часть внутриклеточного сигнального домена 4-1BB.
8. Способ по п. 1, где экспрессированные молекулы дополнительно содержат спейсерную область.
9. Способ по п. 8, где спейсерная область расположена между CD19-лигандсвязывающим доменом и внутриклеточным компонентом.
10. Способ по п. 8, где спейсерная область содержит часть шарнирной области человеческого антитела.
11. Способ по п. 8, где спейсерная область содержит шарнирную область и по меньшей мере одну другую часть Fc-домена человеческого антитела, выбранную из CH1, CH2, CH3 или их комбинаций.
12. Способ по п. 8, где спейсерная область содержит Fc-домен и шарнирную область тяжелой цепи человеческого IgG4.
13. Способ по п. 8, где спейсерная область имеет длину, выбранную из 12 аминокислот или менее, 119 аминокислот или менее или 229 аминокислот или менее.
14. Способ по п. 8, где спейсерная область имеет последовательность, приведенную в SEQ ID NO. 47, SEQ ID NO. 52 или SEQ ID NO. 61.
15. Способ по п. 1, где трансмембранный домен содержит трансмембранный домен CD28.
16. Способ по п. 15, где молекулы содержат спейсерную область между CD19-лигандсвязывающим доменом и трансмембранным доменом.
17. Способ по п. 1, где трансмембранный домен CD28 содержит трансмембранный домен CD4.
18. Способ по п. 1, где генетически модифицированные клетки дополнительно экспрессируют селектируемый маркер.
19. Способ по п. 18, где селектируемый маркер совместно экспрессируется с указанными молекулами с использованием рибосомного элемента пропуска.
20. Способ по п. 18, где селектируемый маркер представляет собой рецептор эпидермального фактора роста (EGFR), в котором отсутствует внутриклеточный сигнальный домен.
21. Способ по п. 1, где эффекторный домен содержит внутриклеточный сигнальный домен CD28 или 4-1BB и где внеклеточный компонент связан с внутриклеточным компонентом посредством спейсерной области, содержащей: (i) шарнирную область человеческого IgG4 и (ii) человеческий трансмембранный домен CD4 или CD28.
22. Способ по п. 1, где экспрессируемые молекулы имеют последовательность, приведенную в SEQ ID NO. 34.
23. Набор для обеспечения анти-CD19 активности против клеток, экспрессирующих CD19, содержащий состав, содержащий терапевтически эффективное количество размноженных с агонистом Notch генетически модифицированных гемопоэтических стволовых клеток/клеток-предшественников (HSPC) CD34+ и/или терапевтически эффективное количество генетически модифицированных естественных клеток-киллеров, дифференцированных из размноженных с агонистом Notch HSPC CD34+,
где указанная генетическая модификация приводит к экспрессии молекул, содержащих внеклеточный компонент, связанный с внутриклеточным компонентом через трансмембранный домен, где внеклеточный компонент содержит CD19-лигандсвязывающий домен и внутриклеточный компонент содержит эффекторный домен, содержащий (i) весь или часть внутриклеточного сигнального домена CD3ζ, (ii) весь или часть внутриклеточного сигнального домена CD28 и/или (iii) весь или часть внутриклеточного сигнального домена 4-1BB; и
инструкции по применению, указывающие, что все клетки в наборе можно вводить неаутологичному субъекту без проверки иммунологической совместимости.
24. Набор по п. 23, где CD19-лигандсвязывающий домен содержит одноцепочечный вариабельный фрагмент (ScFv) антитела.
25. Набор по п. 23, где CD19-лигандсвязывающий домен содержит одноцепочечный фрагмент Fv (scFv), содержащий последовательность CDRL1 SEQ ID NO. 108, последовательность CDRL2 SEQ ID NO. 111, последовательность CDRL3 SEQ ID NO. 104, последовательность CDRH1 SEQ ID NO. 103, последовательность CDRH2 SEQ ID NO. 114 и последовательность CDRH3 SEQ ID NO. 115, где остатки CDR пронумерованы согласно Кабату.
26. Набор по п. 23, где экспрессируемые молекулы дополнительно содержат спейсерную область между CD19-лигандсвязывающим доменом и внутриклеточным компонентом.
27. Набор по п. 26, где спейсерная область содержит Fc-домен и шарнирную область тяжелой цепи человеческого IgG4.
28. Набор по п. 26, где спейсерная область имеет последовательность, приведенную в SEQ ID NO. 47, SEQ ID NO. 52 или SEQ ID NO. 61.
29. Набор по п. 23, где трансмембранный домен содержит трансмембранный домен CD28.
30. Набор по п. 23, где трансмембранный домен содержит трансмембранный домен CD4.
31. Набор по п. 23, где генетически модифицированные клетки дополнительно экспрессируют селектируемый маркер.
32. Набор по п. 31, где селектируемый маркер представляет собой рецептор эпидермального фактора роста (EGFR), в котором отсутствует внутриклеточный сигнальный домен.
33. Набор по п. 23, где эффекторный домен содержит внутриклеточный сигнальный домен CD28 или 4-1BB и где внеклеточный компонент связан с внутриклеточным компонентом посредством спейсерной области, содержащей: (i) шарнирную область человеческого IgG4 и (ii) человеческий трансмембранный домен CD4 или CD28.
34. Набор по п. 23, где экспрессируемые молекулы имеют последовательность, приведенную в SEQ ID NO. 34, 53, 54, 55, 56, 57 или 58.
35. Применение набора по п. 23 для обеспечения анти-CD19 активности против клеток, экспрессирующих CD19.
36. Применение по п. 35, где субъект является педиатрическим пациентом с рецидивом острой лимфобластической лейкемии.
37. Состав для обеспечения анти-CD19 активности против клеток, экспрессирующих CD19, у субъекта, содержащий терапевтически эффективное количество размноженных с агонистом Notch генетически модифицированных гемопоэтических стволовых клеток/клеток-предшественников (HSPC) CD34+ и/или терапевтически эффективное количество генетически модифицированных естественных клеток-киллеров, дифференцированных из размноженных с агонистом Notch HSPC CD34+,
где указанные генетически модифицированные клетки не требуют проверки иммунологической совместимости с субъектом во время введения и где указанная генетическая модификация приводит к экспрессии молекул, содержащих внеклеточный компонент, связанный с внутриклеточным компонентом через трансмембранный домен, где внеклеточный компонент содержит лигандсвязывающий домен, который связывает CD19; и где внутриклеточный компонент содержит эффекторный домен, содержащий (i) весь или часть внутриклеточного сигнального домена CD3ζ, (ii) весь или часть внутриклеточного сигнального домена CD28 и/или (iii) весь или часть внутриклеточного сигнального домена 4-1BB.
38. Состав по п. 37, где CD19-лигандсвязывающий домен содержит одноцепочечный вариабельный фрагмент (scFv) антитела.
39. Состав по п. 37, где CD19-лигандсвязывающий домен содержит одноцепочечный фрагмент Fv (scFv), содержащий последовательность CDRL1 SEQ ID NO. 108, последовательность CDRL2 SEQ ID NO. 111, последовательность CDRL3 SEQ ID NO. 104, последовательность CDRH1 SEQ ID NO. 103, последовательность CDRH2 SEQ ID NO. 114 и последовательность CDRH3 SEQ ID NO. 115, где остатки CDR пронумерованы согласно Кабату.
40. Состав по п. 37, где экспрессируемые молекулы дополнительно содержат спейсерную область между CD19-лигандсвязывающим доменом и внутриклеточным компонентом.
41. Состав по п. 40, где спейсерная область содержит Fc-домен и шарнирную область тяжелой цепи человеческого IgG4.
42. Состав по п. 40, где спейсерная область имеет последовательность, приведенную в SEQ ID NO. 47, SEQ ID NO. 52 или SEQ ID NO. 61.
43. Состав по п. 37, где трансмембранный домен содержит трансмембранный домен CD28.
44. Состав по п. 37, где трансмембранный домен содержит трансмембранный домен CD4.
45. Состав по п. 37, где генетически модифицированные клетки дополнительно экспрессируют селектируемый маркер.
46. Состав по п. 45, где селектируемый маркер представляет собой рецептор эпидермального фактора роста (EGFR), в котором отсутствует внутриклеточный сигнальный домен.
47. Состав по п. 37, где эффекторный домен содержит внутриклеточный сигнальный домен CD28 или 4-1BB и где внеклеточный компонент связан с внутриклеточным компонентом посредством спейсерной области, содержащей: (i) шарнирную область человеческого IgG4 и (ii) человеческий трансмембранный домен CD4 или CD28.
48. Состав по п. 37, где экспрессируемые молекулы имеют последовательность, приведенную в SEQ ID NO. 34, 53, 54, 55, 56, 57 или 58.
49. Состав по п. 37, полученный для инфузии или инъекции.
| WO 2013123061 A1, 22.08.2013 | |||
| WO 2009091826 A2, 23.07.2009 | |||
| Andrade P.Z | |||
| et al | |||
| Нивелир для отсчетов без перемещения наблюдателя при нивелировании из средины | 1921 |
|
SU34A1 |
| Journal of cellular biochemistry, 2011, V.112, No.7, pp.1822-1831 | |||
| АНТИТЕЛА, МОДИФИЦИРУЮЩИЕ РАКОВЫЕ ЗАБОЛЕВАНИЯ | 2008 |
|
RU2468036C2 |
| COOPER L.J.N | |||
| et al., T-cell clones can be rendered | |||

