ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка заявляет приоритет предварительной заявки на патент США № 62/410543, поданной 20 октября 2016 г.; предварительной заявки на патент США № 62/444093, поданной 9 января 2017 г.; предварительной заявки на патент США № 62/458324, поданной 13 февраля 2017; предварительной заявки на патент США № 62/502058, поданной 5 мая 2017; предварительной заявки на патент США № 62/516373, поданной 7 июня 2017; и предварительной заявки на патент США № 62/552792, поданной 31 августа 2017, каждая из которых в полном объеме включена в данный документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0002] Настоящее описание относится к области профилактики и/или лечения болезни Фабри и генной терапии.
УРОВЕНЬ ТЕХНИКИ
[0003] Генная терапия имеет огромный потенциал для новой эры терапии человека. Эти методики позволят лечить состояния, которые ранее были неразрешимыми в рамках стандартной медицинской практики. Одной из областей, которая является особенно многообещающей, является способность добавлять в клетку трансген, чтобы заставить эту клетку экспрессировать продукт, который ранее не продуцировался в этой клетке или продуцировался недостаточно оптимально. Примеры применений этой технологии включают вставку гена, который кодирует терапевтический белок, вставку кодирующей последовательности, которая кодирует белок, который так или иначе отсутствует в клетке или у субъекта, и вставку последовательности, которая кодирует структурную нуклеиновую кислоту, такую как микроРНК.
[0004] Трансгены могут быть доставлены в клетку множеством путей таким образом, что трансген становится интегрированным в собственный геном клетки и содержится там. В последние годы была разработана стратегия интеграции трансгена, которая использует расщепление сайт-специфическими нуклеазами для направленной вставки в выбранный геномный локус (см., например, совместный патент США 7888121). Нуклеазы, такие как цинк-пальцевые нуклеазы (ZFN), эффекторные нуклеазы, подобные активаторам транскрипции, (TALEN) или нуклеазные системы, такие как РНК-управляемая система CRISPR/Cas (которая использует сконструированную направляющую РНК), являются специфическими по отношению к целевым генам и могут быть использованы таким образом, что конструкция трансгена вставляется путем либо направляемой гомологией репарации (HDR), либо с помощью захвата конца во время процессов, управляемых негомологичным соединением концов (NHEJ). Смотри, например, патенты США №№ 99,394,545; 9,255,250; 9,200,266; 9,045,763; 9,005,973; 9,150,847; 8,956,828; 8,945,868; 8,703,489; 8,586,526; 6,534,261; 6,599,692; 6,503,717; 6,689,558; 7,067,317; 7,262,054; 7,888,121; 7,972,854; 7,914,796; 7,951,925; 8,110,379; 8,409,861; публикации патентов США 20030232410; 20050208489; 20050026157; 20050064474; 20060063231; 20080159996; 201000218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983; 20130196373; 20140120622; 20150056705; 20150335708; 20160030477 и 20160024474, описания которых включено в качестве ссылки во всей их полноте.
[0005] Трансгены могут быть введены или содержаться в клетках разными способами. Следуя подходу на основе «кДНК», трансген вводят в клетку таким образом, что трансген содержится внешнехромосомно, а не посредством интеграции в хроматин клетки. Tрансген может содержаться на круговом векторе (например, плазмиде или неинтегрирующем вирусном векторе, таком как AAV или Lentivirus), где вектор может содержать транскрипционные регуляторные последовательности, такие как промоторы, энхансеры, сигнальные polyA поcледовательности, интроны и сигналы сплайсинга (публикация патента США № 20170119906). Альтернативный подход включает вставку трансгена в экспрессируемой на высоком уровне локализации «safe harbor», например, ген альбумина (смотри, патент США № 9394545). Этот подход был назван In Vivo Protein Replacement Platform® или IVPRP. Следуя этому подходу, трансген вставляют в «safe harbor» ген (например, альбумин) посредством опосредованной нуклеазой направленной вставки, где экспрессия трансгена управляется промотором альбумина. Tрансген сконструирован таким образом, чтобы включать сигнальную последовательность, которая способствует секреции/экскреции белка, кодируемого трансгеном.
[0006] Локусы «safe harbor» включают такие локусы как гены AAVS1, HPRT, Albumin и CCR5 в клетках человека и Rosa26 в клетках мыши. См., например, патенты США №№ 7,888,121; 7,972,854; 7,914,796; 7,951,925; 8,110,379; 8,409,861; 8,586,526; публикации патентов США 20030232410; 20050208489; 20050026157; 20060063231; 20080159996; 201000218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983; 20130177960 и 20140017212. Опосредованная нуклеазой интеграция предлагает перспективу улучшения экспрессии трансгена, повышенную безопасность и продолжительность экспрессии, по сравнению с классическими подходами к интеграции, которые основаны на случайной интеграции трансгена, поскольку она делает возможным точное позиционирование трансгена для минимального риска сайленсинга гена или активации соседних онкогенов.
[0007] Хотя доставка трансгена в клетку-мишень является одним препятствием, которое необходимо преодолеть, чтобы полностью внедрить эту технологию, еще одна проблема, которую необходимо преодолеть, заключается в том, чтобы гарантировать, что после того, как трансген введен в клетку и экспрессируется, генный продукт, кодируемый таким образом, должен быть доставлен в необходимое местоположение в организме и быть создан в достаточных локальных концентрациях, чтобы быть эффективным. Для болезней, характеризующихся отсутствием белка или наличием аберрантного нефункционального белка, доставка трансгена, кодирующего белок дикого типа, может быть чрезвычайно полезной.
[0008] Лизосомные болезни накопления (LSD - англ.: lysosomal storage diseases) представляют собой группу редких метаболических моногенных заболеваний, характеризующихся отсутствием функциональных индивидуальных лизосомальных белков, обычно участвующих в расщеплении отработанных липидов, гликопротеинов и мукополисахаридов. Эти болезни характеризуются накоплением этих соединений в клетке, поскольку они не могут перерабатывать их в метаболическом цикле из-за неправильного функционирования специфического фермента. Наиболее распространенными примерами являются болезни Гоше (дефицит глюкоцереброзидазы - название гена: GBA), Фабри (дефицит α галактозидазы A - GLA), Хантера (дефицит идуронат-2-сульфатазы - IDS), Харлера (дефицит альфа-L идуронидазы - IDUA), Помпе (альфа-глюкозидазы (GAA)) и Ниманна-Пика (дефицит сфингомиелинфосфодиэстеразы 1 - SMPD1). При объединении всех вместе, LSD имеют частоту в популяции около 1 на 7000 родов. См., также, публикацию патента США №№ 20140017212; 2014-0112896; и 20160060656.
[0009] Например, болезнь Фабри представляет собой Х-сцепленное расстройство метаболизма гликосфинголипидов, вызванное дефицитом фермента α-галактозидазы A (α-GalA). Это связано с прогрессирующим отложением гликосфинголипидов, в том числе глоботриаозилцерамида (также известного как GL-3 и Gb3) и глоботриаозилсфингозина (lyso-Gb3), галабиоазилцерамида и вещества группы В. Симптомы болезни разнообразны и могут включать жжение, покалывающую боль (акропарестезию) или эпизоды интенсивной боли, которые называют «кризисами Фабри», которые могут длиться от нескольких минут до нескольких дней. Другие симптомы включают нарушение потоотделения, низкую толерантность к физической нагрузке, красновато-пурпурную сыпь, называемую ангиокератомой, нарушения зрения, желудочно-кишечные проблемы, проблемы с сердцем, такие как увеличение сердца и инфаркт миокарда, проблемы с почками, которые могут привести к почечной недостаточности и проблемам с ЦНС и, в общем, средняя продолжительность жизни у пациентов с болезнью Фабри существенно сокращается.
[0010] Современное лечение болезни Фабри может включать ферментозаместительную терапию (ФЗТ) двумя различными препаратами α-GalA человека, агалзидазы бета или агалзидазы альфа, что требует дорогостоящих и длительных инфузий (обычно от около 0,2 до 1 мг/кг) пациенту каждые две недели. Такое лечение предназначено только для лечения симптомов и не оказывает лечебного воздействия, поэтому пациенту необходимо давать повторную дозировку этих белков до конца их жизни, и к вводимому белку потенциально могут вырабатываться нейтрализующие антитела.
[0011] Кроме того, побочные реакции связаны с ФЗТ, в том числе с такими иммунными реакциями, как развитие анти- α-GalA антител у субъектов, которых лечили препаратами α-GalA. На самом деле, у 50% мужчин, которых лечили агалзидазой альфа, а у 88% мужчин, которых лечили агалзидазой бета, развивались антитела α-GalA. Важно отметить, что значительная часть этих антител представляет собой нейтрализующие антитела и, соответственно, снижают терапевтическое воздействие терапии (Meghdari et al (2015) PLoS One 10(2):e0118341. Doi:10.1371/journal.pone.0118341). Кроме того, ФЗТ не останавливает прогрессирование болезни у всех пациентов.
[0012] Таким образом, остается потребность в способах и композициях, не относящихся к ФЗТ, которые могут быть применены для лечения болезни Фабри, в том числе лечения посредством редактирования генома, например, для доставки экспрессированного трансгена, кодируемого генным продуктом, на терапевтически значимом уровне.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0013] Описанное в данном документе представляет собой способы и композиции для лечения и/или профилактики болезни Фабри. Изобретение описывает способы вставки последовательность трансгена в подходящую клетку-мишень (например, субъекту с болезнью Фабри), где трансген кодирует по меньшей мере один белок (например, по меньшей мере один белок α-GalA), который лечит болезнь. Способы могут быть in vivo (доставку последовательности трансгена в клетку у живого субъекта) или ex vivo (доставку модифицированных клеток живому субъекту). Изобретение также описывает способы трансфекции и/или трансдукции подходящей клетки-мишени с экспрессирующей системой, таким образом, что трансген, который кодирует α-GalA, экспрессирует белок, который лечит (например, частично снимает один или более симптомов, связанных с) болезнь. Белок α-GalA может быть экскретирован (секретирован) из клетки-мишени таким образом, что он может воздействовать или быть поглощенным другими клетками, которые не несут трансген (перекрестная коррекция). Изобретение также обеспечивает способы получения клетки (например, зрелой или недифференцированной клетки), в которых получают высокие уровни α-GalA, где введение популяции этих измененных клеток пациенту обеспечит этот необходимый для лечения болезни или состояния белок. Кроме того, изобретение обеспечивает способы получения клетки (например, зрелой или недифференцированной клетки), в которых получают высокоактивную форму (терапевтическую) α-GalA, где введение, или создание, популяции этих измененных клеток пациенту обеспечит активность белка, необходимую для лечения (например, для снижения или устранения одного или более симптомов) болезни Фабри активность белка. Высокоактивная форма α-GalA, продуцируемая таким образом, как описано в данном документе, также может быть выделена из клеток, как описано в данном документе, и введенная нуждающемуся в этом пациенту, с применением стандартных ферментозаместительных процедур, известных специалисту в данной области техники.
[0014] Описанное в данном документе представляет собой способы и композиции для экспрессии по меньшей мере одного белка α галактозидазы A (α-Gal A). Композиции и способы могут быть для применения in vitro, in vivo или ex vivo и включают введение трансгена GLA (например, кДНК с последовательностями GLA дикого типа или кодон-оптимизированными последовательностями GLA), который кодирует по меньшей мере один белок α-Gal A в клетку таким образом, что в клетке экспрессируется белок α-Gal A. В некоторых вариантах осуществления настоящего изобретения клетка находится у субъекта с болезнью Фабри. В любом из способов, описанных в данном документе, трансген может быть введен в печень субъекта. Необязательно, способы дополнительно включают введение одной или более нуклеаз, которые расщепляют эндогенный ген альбумина в клетке печени у субъекта таким образом, что трансген интегрируется в и экспрессируется из гена альбумина. В любом из способов, описанные в данном документе, белок α-Gal A, экспрессированный из трансгена, может уменьшать количество гликосфинголипидов у субъекта по меньшей мере в 2 раза. Трансген GLA может дополнительно включать дополнительные элементы, в том числе, например, сигнальный пептид и/или один или более контрольных элементов. Также обеспечивают генетически модифицированные клетки (например, стволовые клетки, клетки-предшественники, клетки печени, мышечные клетки и т.д.), содержащие экзогенный трансген GLA (интегральный или экстрахромосомный), в том числе клетки, созданные с помощью способов, описанных в данном документе. Эти клетки могут быть применены для обеспечения белка α-Gal A субъекту с болезнью Фабри, например, путем введения клетки(ок) нуждающемуся в этом субъекту или, в качестве альтернативы, путем выделения белка α-Gal A, продуцируемого клеткой, и введения белка нуждающемуся в этом субъекту (ферментозаместительные терапии). Также предлагаются векторы (например, вирусные векторы, такие как AAV или Ad или липидные наночастицы), содержащие трансген GLA, для применения в любом из способов, описанных в данном документе, в том числе для применения при лечении болезни Фабри.
[0015] В одном аспекте изобретение описывает способ экспрессии трансгена, который кодирует один или более корректирующих трансгенов GLA в клетке субъекта. Трансген может быть вставлен в геном подходящей клетки-мишени (например, клетки крови, клетки печени, клетки мозга, стволовой клетки, клетки-предшественника и т.д.) таким образом, что продукт α-GalA, кодируемый с помощью этого корректирующего трансгена, стабильно интегрируется в геном клетки (также называемый подходом на основе «IVPRP®») или, в качестве альтернативы, трансген может содержаться в клетке внешнехромосомно (также называемый подходом на основе кДНК). В одном варианте осуществления настоящего изобретения, корректирующий трансген GLA вводят (стабильно или внешнехромосомно) в клетки клеточной линии для in vitro получения замещающего белка, который (необязательно очищенный и/или выделена) может затем быть введен субъекту для лечения субъекта с болезнью Фабри (например, путем снижения и/или устранения одного или более симптомов, связанных с болезнью Фабри). В некоторых вариантах осуществления настоящего изобретения, продукт α-GalA, кодируемый с помощью этого корректирующего трансгена, увеличивает активность α-GalA в ткани субъекта на любую величину, по сравнению с субъектами, которые не подвергались лечению, например, в от 2 до более 1000 раз (или любая величина между ними), в том числе, но без ограничения этим, от 2 до 100 раз (или любое значение между ними, в том числе в 10, 20, 30, 40, 50, 60, 70, 80, 90, 100 раз), в от 100 до 500 раз (или любое значение между ними) или в от 500 до 1000 раз или более.
[0016] В другом аспекте, описанное в данном документе представляет собой ex vivo или in vivo способы лечения субъекта с болезнью Фабри (например, путем снижения и/или устранения одного или более симптомов, связанных с болезнью Фабри), способы, включающие вставку трансгена GLA в клетку, как описано в данном документе (подходы на основе кДНК и/или IVPRP), таким образом, что белок получают от субъекта с болезнью Фабри. В некоторых вариантах осуществления настоящего изобретения, выделенные клетки, содержащие трансген GLA, могут быть применены для лечения нуждающегося в этом пациента, например, путем введения клеток субъекту с болезнью Фабри. В других вариантах осуществления настоящего изобретения, корректирующий трансген GLA вставляют в ткань-мишень в теле таким образом, что замещающий белок получают in vivo. В некоторых предпочтительных вариантах осуществления настоящего изобретения, корректирующий трансген вставляют в геном клеток в ткани-мишени, в то время как в других предпочтительных вариантах осуществления настоящего изобретения, корректирующий трансген является вставленным в клетки ткани-мишени и содержится в клетках внешнехромосомно. В любом из способов, описанном в данном документе, экспрессированный белок α-GalA может быть экскретирован из клетки, чтобы действовать или быть захваченным второстепенными мишенями, в том числе другими клетками в других тканях (например, посредством экспортация в кровь), в которых отсутствует трансген GLA (перекрестная коррекция). В некоторых случаях, первичная и/или вторичная ткань-мишень представляет собой печень. В других случаях первичная и/или вторичная ткань-мишень представляет собой мозг. В других случаях первичная и/или вторичная мишень представляет собой кровь (например, сосудистую сеть). В других случаях первичная и/или вторичная мишень представляет собой скелетную мышцу.
[0017] В некоторых вариантах осуществления настоящего изобретения, способы и композиции, описанные в данном документе, применяют для уменьшения количества гликосфинголипидов, в том числе глоботриаозилцерамида (также известного как GL-3 и Gb3) и глоботриаозилсфингозина (lyso-Gb3), галабиоазилцерамида, откладывающихся в тканях субъекта, страдающего от болезни Фабри. В некоторых вариантах осуществления настоящего изобретения, продукт α-GalA, кодируемый с помощью этого корректирующего трансгена, уменьшает гликосфинголипиды в ткани субъекта на любую величину, по сравнению с субъектами, которые не подвергались лечению, например, в от 2 до более 100 (или любая величина между ними) раз, в том числе, но без ограничения этим, в от 2 до 100 раз (или любое значение между ними, в том числе в 10, 20, 30, 40, 50, 60, 70, 80, 90, 100 раз).
[0018] В любом из способов, описанных в данном документе, корректирующий трансген GLA содержит последовательность дикого типа функционирующего гена GLA, тогда как в других вариантах осуществления настоящего изобретения, последовательность корректирующего трансгена GLA каким-то образом изменяют, чтобы добиться усиленной биологической активности (например, оптимизированных кодонов, чтобы повысить биологическую активность и/или изменения транскрипционных и трансляционных регуляторных последовательностей для улучшения экспрессии гена). В некоторых вариантах осуществления настоящего изобретения ген GLA модифицируют для улучшения характеристик экспрессии. Подобные модификации могут включать, но без ограничения этим, вставку сайта начала трансляции (например, метионин), добавление оптимизированной последовательности Kozak, вставку сигнального пептида и/или оптимизацию кодона. В некоторых вариантах осуществления настоящего изобретения сигнальный пептид может быть выбран из сигнального пептида альбумина, сигнального пептида F.IX, сигнального пептида IDS и/или сигнального пептида α-GalA. В любом из вариантов осуществления настоящего изобретения, описанном в данном документе, донор GLA может содержать донор, как показано на любой из Фигур 1B, 1C, 10 и/или 13.
[0019] В любом из способов, описанных в данном документе, трансген GLA может быть вставлен в геном клетки-мишени с применением нуклеазы. Неограничивающие примеры подходящих нуклеаз включают цинк-пальцевые нуклеазы (ZFN), TALEN (нуклеазы подобного активаторам транскрипции белка) и/или нуклеазные системы CRISPR/Cas, которые включают ДНК-связывающую молекулу, которая связывается с сайтом-мишенью в представляющей интерес области (например, связанный с болезнью ген, экспрессируемый на высоком уровне ген, ген альбумина или другой или ген «safe harbor») в геноме клетки и одном или более доменах нуклеазы (например, домен расщепления и/или расщепляющий полудомен). Расщепляющие домены и расщепляющие полудомены могут быть получены, например, из различных рестрикционных эндонуклеаз, белков Cas и/или хоуминг-эндонуклеаз. В некоторых вариантах осуществления настоящего изобретения цинк-пальцевый домен распознает сайт-мишень в гене альбумина или гене глобина в клетках-предшественниках красной крови (RBC). Смотри, например, публикацию патента США № 2014001721, включенную в данный документ посредством ссылки во всей своей полноте. В других вариантах осуществления настоящего изобретения нуклеаза (например, система ZFN, TALEN, и/или CRISPR/Cas) связывается с и/или расщепляет ген «safe-harbor», например, ген CCR5, ген PPP1R12C (также известного как AAVS1), ген альбумина, HPRT или Rosa. См., например, патенты США №№ 7,888,121; 7,972,854; 7,914,796; 7,951,925; 8,110,379; 8,409,861; 8,586,526; публикации патентов США 20030232410; 20050208489; 20050026157; 20060063231; 20080159996; 201000218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983; 20130177960 и 20140017212. Нуклеазы (или их компоненты) могут быть предложены в качестве полинуклеотида, который кодирует одну или более нуклеаз (например, система ZFN, TALEN и/или CRISPR/Cas), описанных в данном документе. Полинуклеотид может представлять собой, например, мРНК. В некоторых аспектах мРНК может быть химически модифицированной (Смотри, например, Kormann et al, (2011) Nature Biotechnology 29(2):154-157). В других аспектах мРНК может содержать кэп ARCA (смотри патенты США 7074596 и 8153773). В дополнительных вариантах осуществления настоящего изобретения мРНК может содержать смесь немодифицированных и модифицированных нуклеотидов (смотри публикацию патента США 20120195936). В еще одних дополнительных вариантах осуществления настоящего изобретения, мРНК может содержать элемент WPRE (смотри публикацию патента США № 20160326548).
[0020] В другом аспекте изобретение включает генетически модифицированные клетки (например, стволовые клетки, клетки-предшественники, клетки печени, мышечные клетки и т.д.) с желаемым трансгеном GLA (необязательно интегрированным с применением нуклеазы). В некоторых аспектах, отредактированные стволовые клетки или клетки-предшественники затем размножают и могут быть индуцированы для дифференцирования в зрелые отредактированные клетки ex vivo, а затем клетки дают пациенту. Таким образом, для применения в этом изобретении могут быть выбраны клетки, происходящие от генетически отредактированных (модифицированных) GLA-продуцирующих стволовых клеток или клеток-предшественников, как описано в данном документе. В других аспектах, отредактированные прекурсоры (например, CD34+ стволовые клетки) дают в трансплантате костного мозга, который, после удачной имплантации, пролиферирует продуцирование отредактированных клеток, которые затем дифференцируют и зреют in vivo и содержат биологический, экспрессированный из трансгена GLA. В некоторых вариантах осуществления настоящего изобретения, отредактированные CD34+ стволовые клетки дают пациенту внутривенно таким образом, что отредактированные клетки мигрируют в костный мозг, дифференцируются и зреют, продуцируя белок α-Gal A. В других аспектах, отредактированные стволовые клетки представляют собой мышечные стволовые клетки, которые затем вводят в мышечную ткань. В некоторых аспектах, сконструированная нуклеаза представляет собой цинк-пальцевую нуклеазу (ZFN) (термин «ZFN» включает пару ZFN) и в других аспектах нуклеаза представляет собой нуклеазу TALE (TALEN) (термин «TALEN» включает пару TALEN), а в других аспектах применяют систему CRISPR/Cas. Нуклеазы могут быть сконструированы чтобы иметь специфичность по отношению к локусу «safe harbor», гену, связанному с болезнью, или по отношению к гену, который экспрессируется на высоком уровне в клетках. Только в качестве не ограничивающего примера, локус «safe harbor» может представлять собой сайт AAVS1, ген CCR5, ген альбумина или ген HPRT, тогда как связанный с болезнью ген может представлять собой ген GLA, кодирующий α-галактозидазу A.
[0021] В другом аспекте, описанное в данном документе представляет собой вектор экспрессии нуклеазы (например, ZFN, пару ZFN, TALEN, пару TALEN и/или систему CRISPR/Cas), содержащий полинуклеотид, который кодирует одну или более нуклеаз, как описано в данном документе, функционально связанных с промотором. В одном варианте осуществления настоящего изобретения, вектор экспрессии представляет собой вирусный вектор. В дополнительном аспекте, описанное в данном документе представляет собой вектор экспрессии GLA, содержащий полинуклеотид, который кодирует α-GalA, как описано в данном документе, функционально связанный с промотором. В одном варианте осуществления настоящего изобретения, вектор экспрессии представляет собой вирусный вектор.
[0022] В другом аспекте, описанное в данном документе представляет собой клетку-хозяина, содержащую одну или более векторов экспрессии нуклеаз (например, ZFN, пару ZFN, TALEN, пару TALEN и/или систему CRISPR/Cas) и/или вектор экспрессии α-GalA, как описано в данном документе. Клетка-хозяин может быть стабильно трансформированной или временно трансфицированной или их комбинацией с одним или более векторами экспрессии нуклеазы. В некоторых вариантах осуществления настоящего изобретения клетка-хозяин представляет собой клетку печени.
[0023] В других вариантах осуществления настоящего изобретения, способы обеспечивают замещение геномной последовательности в любом гене-мишени с терапевтическим трансгеном GLA, как описано в данном документе, например, с применением нуклеазы (например, ZFN, пары ZFN, TALEN, пары TALEN и/или системы CRISPR/Cas) (или одного или более векторов, которые кодируют указанную нуклеазу), как описано в данном документе, и «донорной» последовательности или трансгена GLA, который вставляют в ген после целевого расщепления с помощью нуклеазы. Донорная последовательность GLA может присутствовать в векторе, несущем нуклеазу (или ее компонент), присутствующей в отдельном векторе (например, векторе Ad, AAV или LV или мРНК) или, в качестве альтернативы, может быть введена в клетку с применением другого механизма доставки нуклеиновых кислот. Подобная вставка донорной нуклеотидной последовательности в локус-мишень (например, экспрессируемый на высоком уровне ген, связанный с болезнью ген, другой ген «safe harbor» и т. д.) приводит к экспрессии трансгена GLA под контролем эндогенных элементов генетического контроля локусов-мишеней (например, альбумин, глобин и т. д.). В некоторых аспектах вставка трансгена GLA, например, в ген-мишень (например, альбумин), приводит к экспрессии интактной белковой последовательности α-GalA и отсутствию каких-либо аминокислот, кодируемых мишенью (например, альбумином). В других аспектах экспрессированный экзогенный белок α-GalA представляет собой слитый белок и содержит аминокислоты, кодируемые трансгеном GLA и эндогенным локусом, в который вставляют трансген GLA (например, из эндогенного локуса-мишени или, в качестве альтернативы, из последовательностей на трансгене, которые кодируют последовательности локуса-мишени). Мишень может представлять собой любой ген, например, ген «safe harbor», такой как ген альбумина, ген AAVS1, ген HPRT; ген CCR5; или экспрессируемый на высоком уровне ген, такой как ген глобина в клетке-предшественнике RBC (например, бета-глобина или гамма-глобина). В некоторых случаях, эндогенные последовательности будут присутствовать на амино-(N)-концевой части экзогенного белка α-GalA, тогда как в других случаях, эндогенные последовательности будут присутствовать на карбокси-(C)-концевой части экзогенного белка α-GalA. В других случаях эндогенные последовательности будут присутствовать на обоих, и N-, и C-терминальных частях экзогенного белка α-GalA. В некоторых вариантах осуществления настоящего изобретения эндогенные последовательности кодируют сигнальный пептид секреции, который удаляют во время процесса секреции белка α-GalA из клетки. Эндогенные последовательности могут содержать полноразмерные последовательности дикого типа или мутантные эндогенные последовательности или, в качестве альтернативы, могут содержать частичные эндогенные аминокислотные последовательности. В некоторых вариантах осуществления настоящего изобретения эндогенное слияние ген-трансген расположено на эндогенном локусе внутри клетки, тогда как в других вариантах осуществления настоящего изобретения, эндогенную последовательность-кодирующую трансген последовательность вставляют в другой локус в пределах генома (например, последовательность GLA-трансгена, вставленного в локус альбумина, HPRT или CCR5). В некоторых вариантах осуществления настоящего изобретения трансген GLA экспрессируется таким образом, что терапевтический белковый продукт α-GalA сохраняют внутри клетки (например, прекурсор или зрелая клетка). В других вариантах осуществления настоящего изобретения трансген GLA гибридизуют с внеклеточным доменом белка мембраны таким образом, что после экспрессии слияние трансгена α-GalA будет приводить к локализации терапевтического белка на поверхности. В некоторых аспектах внеклеточный домен выбирают из тех белков, которые приведены в Таблице 1. В некоторых аспектах, отредактированные клетки дополнительно включают трансмембранный белок для направления миграции клеток к определенному типу ткани. В одном аспекте трансмембранный белок содержит антитело, тогда как в других, трансмембранный белок содержит рецептор. В некоторых вариантах осуществления настоящего изобретения клетка представляет собой прекурсор (например, CD34+ или гемопоэтическую стволовую клетку) или зрелую RBC (происходящую из генетически модифицированной GAL-продуцирующей клетки, как описано в данном документе). В некоторых аспектах терапевтический белковый продукт α-GalA, кодируемый на трансгене, экспортируют наружу из клетки, чтобы влиять на или поглощаться клетками, в которых отсутствует трансген. В некоторых вариантах осуществления настоящего изобретения клетка представляет собой клетку печени, которая высвобождает терапевтический белок α-GalA в ток крови, чтобы действовать на дистальные ткани (например, почки, селезенки, сердца, мозга и т.д.).
[0024] Изобретение также предоставляет способы и композиции получения клетки (например, RBC), которая несет терапевтический белок α-GalA для лечения болезни Фабри, который может быть применен универсально для всех пациентов в качестве аллогенного продукта. Это делает возможным развитие отдельного продукта для лечения пациентов с болезнью Фабри, например. Эти носители могут содержать трансмембранные белки для принятия участия в направленной миграции клетки. В одном аспекте трансмембранный белок содержит антитело, тогда как в других, трансмембранный белок содержит рецептор.
[0025] В одном варианте осуществления настоящего изобретения, трансген GLA экспрессируется из промотора альбумина с последующей вставкой в локус альбумина. Биологическое вещество, кодируемое трансгеном GLA, затем может быть высвобождено в ток крови, при условии, что трансген вставляют в гепатоцит in vivo. В некоторых аспектах трансген GLA доставляют в печень in vivo в вирусном векторе через внутривенное введение. В некоторых вариантах осуществления настоящего изобретения донорный трансген GLA содержит консенсусную последовательность Kozak перед кодирующей последовательностью α-GalA (Kozak (1987) Nucl Acid Res 15(20):8125-48), таким образом, что в экспрессированном продукте отсутствует сигнальный пептид альбумина. В некоторых вариантах осуществления настоящего изобретения донорный трансген α-GalA содержит альтернативный сигнальный пептид, такой как те, которые от генов Albumin, IDS или F9, вместо нативной сигнальной последовательности GLA. Таким образом, донор может содержать сигнальный пептид, как показано в любой из SEQ ID NO:1-5 или последовательности, проявляющей гомологичность по отношению к этим последовательностям, которые действуют в качестве сигнального пептида (смотри, например, Фигуры 1B, 10, 13 и 25).
[0026] В некоторых вариантах осуществления настоящего изобретения донор трансгена GLA трансфицируют или трансдуцируют в клетку для эписомальной или экстрахромосомной стабилизации трансгена. В некоторых аспектах донор трансгена GLA содержится в векторе, содержащем регуляторные домены для регулирования экспрессии донора трансгена. В некоторых случаях, регуляторные домены для регулирования экспрессии трансгена являются эндогенными доменами по отношению к трансгену, который экспрессируется, тогда как в других случаях, регуляторные домены являются гетерологичными по отношению к трансгену. В некоторых вариантах осуществления настоящего изобретения трансген GLA содержится в вирусном векторе, тогда как в других, содержится в плазмиде или миникольце. В некоторых вариантах осуществления настоящего изобретения вирусный вектор представляет собой AAV, Ad или LV. В дополнительных аспектах, вектор, содержащий донор трансгена, доставляют в подходящую клетку-мишень in vivo, таким образом, что терапевтический белок α-GalA, кодируемый донором трансгена, высвобождают в ток крови, когда вектор донора трансгена доставляют в гепатоцит.
[0027] В другом варианте осуществления настоящего изобретения изобретение описывает клетки-предшественники или CD34+ гемопоэтическую стволовую клетку (HSPC) клетки), в которых трансген GLA вставляют таким образом, что зрелые клетки, полученные из этих прекурсоров содержат высокие уровни кодируемого трансгеном продукта α-GalA. В некоторых вариантах осуществления настоящего изобретения эти прекурсоры индуцируют плюрипотентные стволовые клетки (iPSC).
[0028] В некоторых вариантах осуществления настоящего изобретения способы изобретению могут быть применены in vivo в трансгенных животных системах. В некоторых аспектах, трансгенное животное может быть применено в разработке модели, где трансген кодирует белок α-GalA человека. В некоторых случаях, трансгенное животное может быть инактивировано по соответствующему эндогенному локусу, делая возможным разработку in vivo систем, где белок человека может быть исследован в изолированной среде. Подобные трансгенные модели могут быть применены в целях скрининга для выявления малых молекул, или больших биомолекул или других объектов, которые могут взаимодействовать с или модифицировать представляющий интерес белок человека. В некоторых аспектах, трансген GLA интегрируют в выбранный локус (например, экспрессируемый на высоком уровне или «safe harbor») в стволовую клетку (например, эмбриональную стволовую клетку, индуцированную плюрипотентную стволовую клетку, стволовую клетку печени, нервную стволовую клетку и т.д.) или получают эмбрион отличного от человека животного с помощью любых способов, описанных в данном документе, и тех, которые являются стандартными в данной области техники, а затем эмбрион имплантируют таким образом, что рождается живое животное. Затем животное растят до половой зрелости и позволяют производить потомство, где по меньшей мере некоторые из потомков имеют интегральный трансген GLA.
[0029] В еще одном дополнительном аспекте, предлагаемое в данном документе представляет собой способ сайт-специфической интеграции нуклеотидной последовательности в эндогенный локус (например, связанный с болезнью, экспрессируемый на высоком уровне, такой как локус альбумина в клетке печени или локус глобина в клетках-предшественниках RBC хромосомы, например, в хромосоме эмбриона отличного от человека животного. В некоторых вариантах осуществления настоящего изобретения, способ включает: (a) инъектирование эмбриона отличного от человека животного с (i) по меньшей мере одним вектором ДНК, где вектор ДНК содержит последовательность против хода транскрипции и последовательность по ходу транскрипции, фланкирующие α-GalA, который кодирует нуклеотидную последовательность, для интеграции и (ii) по меньшей мере одна молекула полинуклеотида, которая кодирует по меньшей мере одну нуклеазу (цинк-пальцевую, пару ZFN, нуклеазу TALE, пару TALEN или систему CRISPR/Cas), которая распознает сайт интеграции в локусе-мишени и (b) культивирование эмбриона, чтобы сделать возможной экспрессию нуклеазы (системы ZFN, TALEN и/или CRISPR/Cas, где двухцепочечный разрыв, введенный в сайт интеграции с помощью нуклеазы, репарируют посредством гомологичной рекомбинации с вектором ДНК, таким образом, чтобы интегрировать нуклеотидную последовательность в хромосому. В некоторых вариантах осуществления настоящего изобретения, полинуклеотид, который кодирует нуклеазу, представляет собой РНК.
[0030] В любом из предыдущих вариантов осуществления настоящего изобретения, способы и соединения по изобретению могут быть объединены с другими терапевтическими средствами для лечения субъектов с болезнью Фабри. В некоторых вариантах осуществления настоящего изобретения, способы и композиции включают применение молекулярного шаперона (Hartl et al (2011) Nature 465: 324-332), чтобы обеспечить правильное сворачивание белка Фабри. В некоторых аспектах шаперон может быть выбран из хорошо известных белков-телохранителей, таких как AT1001 (Benjamin et al (2012) Mol Ther 20(4):717-726), AT2220 (Khanna et al (2014) PLoS ONE 9(7): e102092, doi:10.1371) и Migalastat (Benjamin et al (2016) Genet Med doi: 10.1038/gim.2016.122). В некоторых аспектах, способы и композиции применяют в комбинации со способами и композициями, которые дают возможность проходить через гематоэнцефалический барьер. В других аспектах, способы и композиции применяют в комбинации с соединением, известным для подавления иммунного ответа субъекта.
[0031] Также обеспечивают набор, содержащий нуклеазную систему и/или донор GLA, как описано в данном документе. Набор может содержать нуклеиновые кислоты, которые кодируют одну или более нуклеаз (ZFN, пару ZFN, TALEN, пару TALEN и/или систему CRISPR/Cas), (например, молекулы РНК или система ZFN, TALEN и/или CRISPR/Cas, которые кодируют гены, содержащиеся в подходящем векторе экспрессии), молекулы-доноры, векторы экспрессии, которые кодируют одиночную направляющую РНК подходящих линий клеток-хозяев, инструкции по выполнению способов по изобретению и тому подобное.
[0032] Эти и другие аспекты будут очевидны для специалиста в данной области техники в свете раскрытия в целом.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0033] Фигуры 1A-1C иллюстрируют ферментативную реакцию, осуществляемую с помощью фермента α-GalA дикого типа и исходного донора и кассеты экспрессии трансгена. Фигура 1A иллюстрирует реакцию, осуществляемую с помощью α-GalA, где у млекопитающих дикого типа поврежден субстрат Gb3. У организмов с болезнью Фабри субстрат Gb3 накапливается до токсичных уровней. Фигура 1B иллюстрирует исходный вирусный вектор, применяемый для экспрессии α-GalA из кДНК, тогда как Фигура 1C иллюстрирует первичный вирусный вектор, применяемый для экспрессии α-GalA, со следующей опосредованной нуклеазой вставкой в ген альбумина.
[0034] Фигура 2 представляет собой график, иллюстрирующий активность α-GalA, обнаруженной в клеточной среде HepG2/C3A за период времени семь дней, когда клетки трансдуцируют с альбумин-специфическими нуклеазами (ZFN) и донором, изображенным на Фигуре 1C (проиллюстрированным на правой панели с пометкой «IVPRP», которая является аббревиатурой от «In Vivo Protein Replacement Platform®»). Уровни активности в среде из клеток, которые претерпели макетную процедуру трансдукции, иллюстрируют в левой панели. Столбики слева направо иллюстрируют активность в день 3, день 5, день 7 и клетки отдельно.
[0035] Фигуры 3A и 3B представляют собой графики, иллюстрирующие уровни активности α-GalA, выявленной с применением подхода на основе кДНК. Фигура 3A иллюстрируют активность в клеточной среде HepG2/C3A, выявленную за период времени 6 дней при различных дозах вируса AAV, содержащих кассету экспрессии кДНК, проиллюстрированной на Фигуре 1B (столбики слева направо иллюстрируют макетные трансфекции, 10K, 30K, 100K, 300K, 1000K, 3000K и 9000K). Фигура 3B представляет собой график, иллюстрирующий активность, выявленную в клеточных осадках клеток с Фигуры 3A в последний момент эксперимента.
[0036] Фигуры 4A и 4B представляют собой графики, отображающие in vivo активность у GLAKO мышей, которых лечили с кДНК, содержащей AAV. Фигура 4A иллюстрирует результаты для каждой отдельной мыши, которую лечили с помощью 2,0e12 геномов вектора на килограмм массы тела (гв/кг) AAV2/6, содержащего конструкцию кДНК, тогда как Фигура 4B иллюстрирует результаты для каждой мыши, которую лечили с помощью 2,0e13 гв/кг AAV2/6-кДНК. На Фигуре 4A, одну мышь дополнительно лечили с помощью молекулярного шаперона DGJ в указанный день. Также проиллюстрированное пунктиром на обеих фигурах представляет собой уровни активности α-GalA, обнаруженные у мышей дикого типа. Как проиллюстрировано, мыши, получавшие лечение, иллюстрируют уровни выше дикого типа, что указывает на терапевтически эффективные уровни.
[0037] Фигуры 5A-5F представляют собой графики, отображающие уровни липидного субстрата Gb3 у GLAKO мышей и у мышей, получавших лечение с помощью содержащей AAV2/6 конструкции кДНК. Фигура 5A иллюстрирует уровни субстрата, обнаруженные в плазме крови, а Фигура 5B иллюстрирует субстрат в ткани сердца. Фигура 5C иллюстрирует субстрат, обнаруженный в печени, и Фигура 5D иллюстрирует субстрат, обнаруженный в тканях почки. Во всех проиллюстрированных тканях, уровни Gb3 являются более низкими, чем у не получавших лечение мышей GLAKO. Также указанное на Фигуре 5D представляет собой нижний предел количественного определения (LLOQ) для этого анализа. Уровни Gb3 и lyso-Gb3 у мышей, получавших лечение, также экспрессировали в единицах количества обнаруженного субстрата, по сравнению с не получавшими лечение мышами. Фигура 5E иллюстрирует процент Gb3, остающийся в специфических тканях, по сравнению с не получавшими лечение мышами GLAKO и Фигура 5F иллюстрирует процент lyso-Gb3, остающийся в специфических тканях, по сравнению с не получавшими лечение мышами GLAKO. Наборы данных тканей на 5E и 5F иллюстрируют для каждой группе лечения (не получавшие лечение GLAKO), получавшие лечение низкой и высокой дозой GLAKO и мыши дикого типа), где столбики представляют данные от (слева направо) плазмы, печени, сердца и почки.
[0038] Фигуры 6A-6E изображает результаты для подхода на основе IVPRP, исследованного in vivo. Фигура 6A изображает активность α-Gal A, обнаруженного в плазме мышей GLAKO, получавших лечение, с вирусом AAV2/8, содержащим донор трансгена, проиллюстрированный на Фигуре 1C с течением времени, где некоторые мыши получали иммуносупрессию (смотри Пример 4). Также проиллюстрированное представляет собой уровень, обнаруженный у мышей дикого типа. Фигура 6B представляет собой график, иллюстрирующий уровень инсерционно-делеционных мутаций, обнаруженных в печени получавших лечение животных в день 90. Инсерционно-делеционные мутации (вставки и/или делеции) являются индикацией активности нуклеазы. Фигуры 6C, 6D и 6E представляют собой временную динамику активности, обнаруженной в плазме мышей, получавших лечение, за период времени близко 30 дней. Фигура 6C иллюстрирует активность у животных, которые дополнительно лечат с помощью низкой иммуносупрессии, тогда как Фигура 6D иллюстрирует активность у получавших лечение животных с умеренной иммуносупрессией и Фигура 6E иллюстрирует получавших лечение животных с высокими уровнями иммуносупрессии. Также проиллюстрированное на Фигурах 6C, 6D и 6E представляет собой уровни, обнаруженные у мышей дикого типа, для сравнения (пунктирная линия).
[0039] Фигуры 7A-7C представляют собой графики, отображающие активность α-Gal A, выявленные с течением времени у получивших лечение животных, и с иммуносупрессией («IS»), и с шапероном DGJ. Фигура 7A иллюстрирует результаты для получавших лечение животных с низкими уровнями иммуносупрессии, где стрелки изображают хронометрирование дозы шаперона и получавших лечение мышей. На Фигуре 7A, всех мышей лечили с помощью шаперона и результаты демонстрируют, что активность повышена. Фигура 7B иллюстрирует результаты для животных под умеренной иммуносупрессией, где двух мышей лечили с помощью DGJ. У этих двух мышей видели повышение в активности α-Gal A в их плазме. Фигура 7C изображает результаты для мышей с высоким уровнем иммуносупрессии, и снова указывает, когда трех мышей лечили с помощью DGJ. Эти результаты демонстрируют, что шаперон повышает уровень выявленной активности. Пунктирная линия указывает уровни активности, обнаруженные у мышей дикого типа для сравнения.
[0040] Фигура 8 представляет собой график, иллюстрирующий сравнение активности α-Gal A в тканях получавших лечение мышей посредством подхода на основе либо кДНК, либо IVPRP. Также проиллюстрированное для сравнения представляет собой уровни у мышей дикого типа и у не получавших лечение мышей GLAKO. Проиллюстрированные ткани представляют собой печень, плазму, селезенку, сердце и почку. Обратите внимание, что ось Y разделена, указывая на то, что подход на основе кДНК при дозе 2,0e13 гв/кг продуцирует активность α-GalA почти в 100 раз выше уровня дикого типа и эта активность обнаруживается во всех исследуемых тканях.
[0041] Фигуры 9A-9C изображают уровни активности α-GalA и липидного субстрата Gb3, обнаруживаемые в качестве результата как подходов на основе кДНК, так и In Vivo Protein Replacement Platform® (IVPRP). Фигура 9A иллюстрирует средние уровни активности, выявленные у разных групп лечения. Фигура 9B иллюстрирует количество Gb3, обнаруженного в тканях плазме, печени и сердца для различных групп, и демонстрирует, что подход на основе кДНК приводит к уменьшению Gb3, приближающемуся к мышам дикого типа, указывая на то, что белок, экспрессированный из трансгена, является эффективным в воздействии на его целевой субстрат. Фигура 9C представляет собой график, иллюстрирующий уровень активности α-GalA у отдельных мышей из таблицы в 9A (группа ZFN+Донор+DGJ не показана). Мыши, получавшие высокую дозу кДНК (2,0e13 гв/кг донора вектора кДНК), иллюстрируют с помощью черных кругов на черной линии. Мыши, получавшие низкую дозу кДНК (2e12 гв/кг донора вектора кДНК), иллюстрируют с помощью заштрихованных треугольников на пунктирной линии. Мышей дикого типа иллюстрируют с помощью черных открытых кругов на серой линии, а мышей GLAKO иллюстрируют с помощью черных квадратов на черной линии. У трех из четырех мышей, получавших высокую дозу кДНК, имеют уровни в 100 раз выше, чем у мышей дикого типа.
[0042] Фигура 10 представляет собой схему, иллюстрирующую различные приводимые в качестве примера конструкции донора (Варианты №A-№L, также соответствуют Вариантам A-L), применяемые для подхода на основе IVPRP®. Аббревиатуры на схемах являются следующими: «ITR» представляет собой инвертированную AAV область терминальных повторов. «HA-R» и «HA-L» представляют собой правое (R) и левое (L) плечи гомологичности, которые имеют гомологичность по отношению к последовательности альбумина, фланкирующей сайт расщепления ZFN. «SA» представляет собой сайт акцептора сплайсинга из гена F9, тогда как «HBB-IGG» представляет собой последовательность интрона, «GLAco» представляет собой кодон-оптимизированную кодирующую последовательность α-GalA, тогда как «GLAco v.2» представляет собой альтернативную оптимизацию кодона кодирующей последовательность α-GalA, «bGHpA» представляет собой poly A последовательность из бычьего гормона роста, «GLA Signal pept» представляет собой сигнальный пептид из гена GLA, «слияние» соответствует конструкции с 2-5 дополнительными аминокислотами, вставленными между сайтом акцептора сплайсинга и трансгеном GLA, «T2A» и «F2A» представляют собой саморасщепляющие последовательности из T. assigna и вируса ящера, соответственно. «IDS Signal pept» представляет собой сигнальный пептид в случае гена IDS, тогда как «FIX Signal pept» представляет собой сигнальный пептид из гена FIX. «TI» представляет собой 5' последовательность связывания праймера NGS, добавленную на 3' конце трансгена, за которой следует целевая интеграция (TI)-специфической последовательности с тем же самым нуклеотидным составом, что и локус дикого типа, что делает возможным секвенирование следующего поколения для измерения инсерционно-делеционных мутаций и HDR-опосредованной интеграции трансгена, одновременно. Смотрите Примеры для более подробной информации.
[0043] Фигуры 11A и 11B представляют собой графики, отображающие активность α-GalA in vitro в клетках HepG2/C3A. Проиллюстрированное на Фигуре 11A представляет собой активность, выявленную в клетках и в клеточном супернатанте, с применением исходного донора и варианта доноров №A, №B и №E, как проиллюстрировано на Фигуре 10. «Z+D» соответствует введению ZFN и донора. Данные показывают, что Варианты №A и №B имеют большую активность, чем исходный донор. Фигура 11B представляет собой график, иллюстрирующий активность α-GalA по сравнению с Вариантами №A, №K, №J, №H и №I (Варианты A, K, J, H и I) при либо низкой (300000/600000 гв/клетку ZFN/донор), либо высокой (600000/1200000 гв/клетку ZFN/донор) дозе ZFN и доноров GLA. Набор данных «Только донор» представляет клетки, которые получали лечение только с помощью конструкции донора, без какого-либо из ZFN. Столбики представляют средние значения по группе со стандартными отклонениями, указанными с помощью планок погрешностей. Данные указывают на то, что Вариант №K приводит к самой высокой активности в этом наборе.
[0044] Фигура 12 представляет собой график, иллюстрирующий активность вариантов №A, №B и №E in vivo. Применяют мышей GLAKO и отбирают образцы плазмы один раз в неделю. Фигура 12 иллюстрирует данные для каждой группы в день 56 после инъекции, и также иллюстрирует данные подхода на основе кДНК, для сравнения. В день 28, мыши, получавшие лечение с донорами «новых» вариантов имели значительно большую активность α-GalA, чем исходный донор. «Первичный» донор соответствует донору, применяемому перед оптимизацией, смотри Фигуру 10 и проиллюстрированное на Фигуре 12 в виде черного столбика слева от каждой группировки. Результаты кДНК представлены только для дня 56 в правом нижнем углу графика. Пунктирная линия указывает 50-кратный уровень активность у мышей дикого типа, что указывает на то, что все образцы проявляли активность по меньшей мере в 40 раз больше, чем у дикого типа в 28 день.
[0045] Фигуры 13A и 13B представляют собой схемы приводимых в качестве примера кассет экспрессии кДНК. Фигура 13A иллюстрирует планирование экспрессирующей кДНК системы, описанной ранее (смотри публикацию патента США № 20170119906), где кодирующую GLA последовательность вставляют с применением различных протоколов оптимизации кодона (ДНК 2.0 v1 в сравнении с GeneArt v2, «GLAco v.2»). Фигура 13B иллюстрирует кассету экспрессии кДНК, применяемую в этой работе, с альтернативным протоколом оптимизации кодона и иллюстрирует Варианты №№1-6 (также называемые Варианты 1-6) с применением сигнальных пептидов из генов IDS, FIX или ALB в комбинации с кодирующими GLA последовательностями, оптимизированными с применением двух различных протоколов.
[0046] Фигуры 14A и 14B представляют собой графики, иллюстрирующие экспрессию активности α-GalA с применением подхода на основе кДНК. На фигуре клетки HepG2/C3A трансдуцируют с помощью AAV, содержащего указанную конструкцию кДНК, где исследовали влияние различных сигнальных пептидов, как проиллюстрировано на Фигуре 13B. Активность α-Gal A измеряли в клеточном супернатанте в день 3 и день 5, а результаты указывают, что лидерная последовательность IDS и FIX (F9) приводит к более высоким уровням активности, чем лидерные последовательности либо GLA, либо альбумина (ALB). Фигура 14B иллюстрирует активность α-Gal A в день 5 для Вариантов №1, №2, №4, №5 и №6. Для этих исследований клетки получают 3,0 e5 гв/клетку векторов кДНК GLA AAV2/6. Столбики представляют средние значения по группе, а планки погрешностей иллюстрируют стандартные отклонения.
[0047] Фигуры 15A-15C представляют собой графики, отображающие активность α-Gal A в либо плазме (Фигура 15A), либо в выбранных тканях (Фигура 15B). Мышам GLAKO инъецируют 3e11 гв ZFN, предназначенного для создания двухцепочечного разрыва в интроне Albumin 1, и 1,2e12 гв исходной донорной конструкции GLA или вариантов A, B, E или J (общая доза AAV/мышь=6e13 гв/кг). Фигура 15A изображает активность плазмиды α-Gal A у мышей, которые сопровождали оценкой в течение 2 месяцев каждую неделю или каждые две недели. На левой панели проиллюстрированы результаты для животных, получающих исходный донор, вариант A, вариант E или вариант B. На правой панели проиллюстрированы результаты для животных дикого типа или животных, получающих вариант E или J. Фигура 15B иллюстрирует активность α-Gal A, которую измеряли в печени, сердце, почке и селезенке, что анализировали после того, как животных, проиллюстрированных на Фигуре 15A, умерщвляли. График слева на Фигуре 15B иллюстрирует данные спустя 2 месяца после лечения с помощью исходной донорной конструкции GLA («Первичная», проиллюстрированная в самых левых столбиках каждой группы), после лечения с помощью варианта A (вторые столбики слева в каждой группе), Вариант B (средние столбики для каждой группы), Вариант E (вторые столбики справа в каждой группе) и у животных дикого типа («Дикого типа», проиллюстрированная в самых правых столбиках каждой группы). График справа на Фигуре 15B изображает активность Вариантов E и J, где в каждом наборе данных активность у не получавших лечение мышей GLAKO иллюстрируют в самом левом столбике; у мышей дикого типа, вторые столбики слева в каждой группе; активность у GLAKO мышей, получавших лечение с помощью Варианта №E, проиллюстрирована в третьих столбиках слева, тогда как активность в случае Варианта J иллюстрируют в самом правом столбике. α-Gal A был многократно выше дикого типа в плазме и всех измеренных тканей для вариантов донора GLA A, B, E и J. Фигура 15C изображает уровень активности α-Gal A в плазме, где проиллюстрированы данные для каждой мыши, которая получала лечение с помощью пары ZFN и Варианта A донора. Следует обратить внимание, что это тот же эксперимент, что и проиллюстрированный на Фигуре 15A, с пометкой Вариант A, за исключением того, что на Фигуре 15A проиллюстрированы данные для мышей как группы, тогда как на Фигуре 15C проиллюстрированы данные для каждой мыши, которая получала лечение.
[0048] Фигуры 16A и 16B представляют собой графики, отображающие количество гликолипидного субстрата α-Gal A (Gb3 и lyso-Gb3), остающегося после лечения с помощью ZFN + различных вариантов донора. Содержание Gb3 (Фигура 16A) и lyso-Gb3 (Фигура 16B) измеряют в плазме, сердце, печени, почке и селезенке (данные селезенки не проиллюстрированы) посредством масс-спектрофотометрии. Каждый набор данных иллюстрируют в группах по 4, отображающих уровни (слева направо в каждой группе) в плазме, печени, сердце и почке. Количество субстрата выражают в виде остающейся фракции, по сравнению с не получавшими лечение мышами GLAKO. Количество и Gb3, и lyso-Gb3 было значительно снижено в тканях мышей, которые получали лечение с помощью вариантов A, B или E донора GLA.
[0049] Фигуры 17A-17C иллюстрируют эффект лечения с помощью белка α-Gal A с ферментом дегликозилирования PNGaseF или Endo H. Фигура 17A иллюстрирует результаты вестерн-блоттинг анализа, полученные из гомогената, полученного из печени мыши получавших лечение животных путем подхода на основе IVPRP. На верхней панели показаны образцы трех мышей (с пометкой «Вариант A донора GLA»), а также образец мыши дикого типа («WT»), не получавшей лечение мыши GLAKO («GLAKO») и образец рекомбинантного Gal A человека («rec. hGal A»). На нижней панели, с пометкой «Вариант J донора GLA», образцы от двух мышей иллюстрируют вместе с образцом мыши дикого типа и образцом не получавшей лечение GLAKO мыши, а также образец рекомбинантного Gal A человека. (+) и (-) на обоих пятнах указывают на лечение с помощью PNGase F или Endo H. Фигура 17B иллюстрирует результаты вестерн-блоттинг анализа, полученные таким образом, как описано на Фигуре 17A, за исключением того, что мышей лечили с применением подхода на основе кДНК («первичная» конструкция). Фигура 17C представляет собой схему, отображающее расщепление сложных структур гликозилирования с помощью PNGaseF. Данные демонстрируют, что фермент Gal A, экспрессируемый у получавших лечение животных GLAKO, после подхода на основе либо IVPRP®, либо кДНК, иллюстрирует подобное дегликозилирование, как и дегликозилированный рекомбинантный белок человека после лечения с помощью PNGaseF.
[0050] Фигуры 18A-18C представляют собой графики, отображающие активности, измеренные с применением исходной конструкции кДНК, по сравнению с Вариантом №4 (проиллюстрированным на 13B выше). Фигура 18A изображает активность α-GalA в плазме у GLAKO мышей, получавших лечение с помощью 2e12 гв/кг GLA кДНК, содержащей AAV2/6, как указано. Активность измеряли в течение до 60 дней после инъекции. На Фигуре 18B указана активность α-GalA в тканях, как показано на мышах на Фигуре 18A. Наборы данных, слева направо, иллюстрируют активность α-GalA у не получавших лечение мышей GLAKO (самый левый столбик); мышей дикого типа (второй слева столбик); мышей GLAKO, получавших лечение с помощью исходного варианта кДНК (третий слева столбик); и мышей GLAKO, получавших лечение с помощью варианта D кДНК. Горизонтальные пунктирные линии указывают активность, соответствующую 10-кратному уровню дикого типа, для сравнения. Фигура 18C изображает результаты вестерн-блоттинг анализа, выявляющего α-GalA человека в печени 3 мышей GLAKO, получавших лечение с помощью Варианта №4 кДНК. Для сравнения иллюстрируют активность у мыши дикого типа («WT») и не получавшей лечение мыши GLAKO. Для целей сравнения, проиллюстрированное также представляет собой рекомбинантный hGalA. Образцы обрабатывали с помощью PNGasdF или EndoH, как описано на Фигуре 17.
[0051] Фигура 19 представляет собой график, отображающий уровень активности α-Gal A в плазме мышей, которые получали лечение с помощью исходной конструкции кДНК (проиллюстрированной на Фигуре 13). Каждую группу подвергали лечению с помощью AAV, содержащим конструкцию в указанных дозах, от 1,25e11 до 5,0e12 гв/кг (сплошные линии, средние значения по группе, указанные с помощью планки погрешностей). Также были включены мыши дикого типа и не получавшие лечение GLAKO мыши и указанные на фигуре.
[0052] Фигуры 20A и 20B представляют собой графики, отображающие активность α-Gal A, выявленную после in vivo экспрессии Вариантов E и J. Фигура 20A иллюстрирует активность α-Gal A, обнаруженную в плазме после лечения мышей GLAKO с помощью ZFN, специфического по отношению к альбумину и донорам либо Варианта E, либо Варианта J (смотри Фигуру 10). Фигура 20B иллюстрирует активность α-Gal A, обнаруженную в различных тканях, представляющих интерес (печени, сердце, почке и селезенке). В каждом наборе данных на Фигуре 20B, слева направо, столбики иллюстрируют результаты для мышей GLAKO, мышей дикого типа (WT), Варианта E донора или Варианта J донора.
[0053] Фигуры 21A и 21B представляют собой графики, отображающие количество субстрата α-Gal A, обнаруженного в различных представляющих интерес тканях (плазме, печени, сердце и почке). Фигура 21A изображает количество GB3, выявленное в процентах от того, что обнаружено у мышей GLAKO (устанавливают в качестве 100%). Фигура 21B изображает количество lyso-GB3, выявленное в процентах от того, что обнаружено у мышей GLAKO (устанавливают в качестве 100%). А на Фигурах 21A и 21B каждый набор данных, слева направо, иллюстрирует результаты, обнаруженные в плазме, печени, сердце и почке.
[0054] Фигура 22 представляет собой график, отображающий длительную модификацию гепатоцитов в модели болезни Фабри у мышей GLAKO, после опосредованной нуклеазой целевой интеграции трансгена GLA, и иллюстрирует процентное содержание инсерционно-делеционных мутаций в клетках печени, получавших лечение при указанных состояниях.
[0055] Фигуры 23A и 23B представляют собой графики, отображающие α-Gal A, экспрессированный из интегрального трансгена, секретированного в ток крови и поглощенного вторичными тканями. Мышей GLAKO подвергали лечению с помощью ZFN и одним из двух конструкций донора hGLA. Фигура 23A изображает активность GalA в плазме от получавших лечение животных с указанными конструкциями или не получавших лечение животных. Фигура 23B иллюстрирует активность GalA в указанных тканях (печени, селезенке, сердце и почке) при указанных состояниях. Самый левый столбик иллюстрирует активность у не получавших лечение животных; второй слева столбик иллюстрирует активность у получивших лечение только с помощью Варианта E донора животных; средний столбик иллюстрирует активность у животных дикого типа; второй справа столбик иллюстрирует активность у получивших лечение с помощью ZFN и Варианта A донора животных; и крайний справа столбик иллюстрирует активность у получивших лечение с помощью ZFN и Варианта E донора животных. Не получавшие лечение мыши GLAKO, не получавшие лечение мыши дикого типа и мыши GLAKO, получавшие лечение с помощью донор, но не ZFN, включают в качестве контроля. Стабильная активность в плазме достигала до 80-кратного уровня дикого типа. Графики отображают активность α-Gal A в плазме с течением времени и активность в ткани по окончанию исследования (День 56).
[0056] Фигуры 24A и 24B представляют собой графики, отображающие содержание субстрата Фабри в указанных тканях. Фигура 24A иллюстрирует содержание Gb3 и Фигура 24B иллюстрирует содержание lyso-Gb3 в виде % снижения от не получавших лечение мышей GLAKO в указанной состоянии. Столбики под каждым состоянием иллюстрируют уровни в плазме, печени, сердце и почке, слева направо. Мыши, получавшие лечение с помощью ZFN и любого варианта донора hGLA, имели значительно сниженное содержание субстрата.
[0057] Фигуры 25A и 25B иллюстрируют схемы Варианта L и Варианта M и целевой интеграции в локус альбумина дикого типа. Фигура 25A изображает варианты L и M и иллюстрирует то, что Вариант M отличается от Варианта L тем, что он содержит сигнальный пептид IDS, а не сигнальный пептид GLA. Аббревиатуры являются такими же, как и на Фигуре 10. Фигура 25B иллюстрирует интеграцию трансгена GLA в локус Albumin. «TI» представляет собой 5' последовательность связывания праймера секвенирования следующего поколения (NGS), добавленную на 3' конце трансгена, за чем следует целевая интеграция (TI)-специфической последовательности с тем же самым нуклеотидным составом, что и локус дикого типа, что делает возможным секвенирование следующего поколения для измерения инсерционно-делеционных мутаций и HDR-опосредованной интеграции трансгена, одновременно.
[0058] Фигуры 26A и 26B представляют собой графики, отображающие модификацию (процент инсерционно-делеционных мутаций или процент TI) с применением указанных доноров в HepG2 клеточной линии гематокарциномы человека при указанных дозировках. Фигура 26A иллюстрирует результаты с применением Варианта L донора и Фигура 26B иллюстрирует результаты с применением Варианта M донора.
[0059] Фигуры 27A и 27B представляют собой графики, отображающие каким образом продуцируемый в печени α-Gal A секретируется в ток крови и поглощается вторичными тканями. Конструкцию донора GLA, содержащую сигнальный пептид IDS и 3' последовательность для анализа целевой интеграции (TI), применяют для лечения мышей GLAKO. Фигура 27A изображает активность GalA в плазме от получавших лечение животных с указанными конструкциями или не получавших лечение животных. Фигура 27B иллюстрирует активность GalA в указанных тканях (печени, селезенке, сердце и почке) при указанных состояниях. Самый левый столбик иллюстрируют активность у не получавших лечение животных; второй слева столбик иллюстрирует активность у получивших лечение только с помощью Варианта М донора животных; средний столбик иллюстрирует активность у животных дикого типа; второй справа столбик иллюстрирует активность у получивших лечение с помощью ZFN и низкой дозы Варианта М донора животных; и крайний справа столбик иллюстрирует активность у получивших лечение с помощью ZFN и высокой дозы Варианта M донора животных. Как проиллюстрировано, наблюдалась 250-кратная стабильная активность в плазме относительно дикого типа, а активность α-Gal A в сердце и почке была более чем 20-кратной относительно дикого типа и 4-кратной относительно дикого типа, соответственно.
[0060] Фигуры 28A и 28B представляют собой графики, отображающие активность α-GAL A в клетках, получавших лечение с помощью печеночноспецифических конструкций, содержащих конструкцию GLA. Фигура 28A иллюстрирует активность клеток HepG2 в супернатанте, а Фигура 28B иллюстрирует активность K562 в клеточных осадках, культивированного в присутствие супернатанта от получавших лечение или не получавших лечение клеток HepG2, как проиллюстрировано на Фигуре 28A.
[0061] Фигура 29 представляет собой график, отображающий активность α-GAL A в плазме мышей GLAXO, дозированный с от 1,25e11 до 5,0e12 гв/кг исходной конструкции кДНК (сплошные линии, средние значения по группе, n = от 4 до 7 на группу) и наблюдали в течение 6 месяцев. Также проиллюстрированы мыши GLAKO дикого типа (серая пунктирная линия, указанная стрелкой) и не получавшие лечение (черная пунктирная линия, указанную стрелкой).
[0062] Фигура 30 иллюстрирует графики, отображающие активность α-Gal A в указанных тканях (печени, селезенке, сердце и почке) через 6 месяцев после лечения с указанными дозировками. Также проиллюстрированы животные дикого типа и не получавшие лечение.
[0063] Фигура 31 иллюстрирует графики, отображающие дозозависимое снижение содержания субстрат Gb3 Фабри в указанных тканях (печени, селезенке, сердце и почке) у GLAKO мышей с от 1,25e11 до 5,0e12 гв/кг исходной конструкции кДНК в виде % снижения относительно не получавших лечение мышей GLAKO (средние значения по группе, n = от 4 до 7 на группу). Мыши демонстрируют измеренное дозозависимое снижение содержания Gb3 во всех тканях.
[0064] Фигуры 32A и 32B иллюстрируют графики, отображающие процент субстрата Gb3, остающегося в различных тканях, представляющих интерес (плазме, печени, сердце и почке), после указанного протокола лечения (смотри также Фигуру 18). Фигура 32A изображает количество GB3, выявленное в процентах от того, что обнаружено у не получавших лечение мышей GLAKO (устанавливают в качестве 100%). Фигура 32B изображает количество lyso-Gb3, выявленное в виде процента от того, что обнаруживают у не получавших лечение мышей GLAKO (устанавливают в качестве 100%). А на Фигурах 32A и 32B каждый набор данных, слева направо, иллюстрирует результаты, обнаруженные в плазме, печени, сердце и почке.
[0065] Фигуры 33A и 33B представляют собой графики, отображающие процент субстрата Gb3, остающийся в различных тканях, представляющих интерес (плазме, печени, сердце и почке), после указанного протокола лечения (смотри также Фигуру 27). Фигура 33A изображает количество GB3, выявленное в процентах от того, что обнаружено у не получавших лечение мышей GLAKO (устанавливают в качестве 100%). Фигура 33B изображает количество lyso-Gb3, выявленное в виде процента от того, что обнаруживают у не получавших лечение мышей GLAKO (устанавливают в качестве 100%). А на Фигурах 33A и 33B каждый набор данных, слева направо, иллюстрирует результаты, обнаруженные в плазме, печени, сердце и почке.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0066] Описанное в данном документе представляет собой способы и композиции для лечения или профилактики болезни Фабри. Изобретение обеспечивает способы и композиции вставки трансгена GLA кодирующего белок, который отсутствует или недостаточно экспрессируется у субъекта с болезнью Фабри, таким образом, что в печени экспрессируется ген и экспрессируется терапевтический (замещающий) белок. Изобретение также описывает изменение клетки (например, прекурсора или зрелой RBC, iPSC или клетки печени) таким образом, что оно продуцирует высокие уровни терапевтического средства, а введение популяции этих измененных клеток пациенту обеспечит этот необходимый белок. Трансген может кодировать желаемый белок или структурную РНК, которая является эффективной терапевтически у нуждающегося в этом пациента.
[0067] Таким образом, способы и композиции по изобретению могут быть применены для экспрессии, из трансгена, одного или более терапевтически эффективных белков α-GalA из любого локуса (например, экспрессируемого на высоком уровне локуса альбумина) для замены фермента, который является дефективным и/или отсутствующим при болезни Фабри. Дополнительно, изобретение обеспечивает способы и композиции для лечения (в том числе частичного снятия одного или более симптомов) болезни Фабри путем вставки последовательностей трансгена в экспрессируемые на высоком уровне локусы в клетках, таких как клетки печени. Раскрытое в изобретении представляет собой способы и композиции для доставки α-GalA, которая кодирует трансген посредством вирусного вектора, в печень нуждающемуся в этом субъекта, где вирус может быть введен посредством инъекции в периферическую венозную систему или путем прямой инъекции в идущий к печени кровеносный сосуд (например, воротную вену). Способы и композиции могут быть применены для индуцирования вставки трансгена в локус «safe harbor» (например, альбумин) или могут быть применены для вызывания экстрахромосомной стабилизации виральной конструкции кДНК в клетке печени. В любом случае, трансген экспрессируется на высоком уровне и обеспечивает терапевтический эффект нуждающемуся в этом пациенту с болезнью Фабри.
[0068] Кроме того, трансген может быть введен в клетки, полученные от пациента, например, полученные от пациента индуцированные плюрипотентные стволовые клетки (iPSC) или другие типы стволовых клеток (эмбриональных или гематопоэтических) для применения при возможной имплантации. Особенно полезным является вставка терапевтического трансгена в гемопоэтическую стволовую клетку для имплантации нуждающемуся в этом пациенту. Поскольку стволовые клетки дифференцируются в зрелые клетки, они будут содержать высокие уровни терапевтического белка для доставки в ткани.
Общая информация
[0069] Практика способов, а также получение и применение композиций, описанных в данном документе, использует, если не указано иное, общепринятые техники в области молекулярной биологии, биохимии, структуры и анализа хроматина, вычислительной химии, культивирования клеток, рекомбинантной ДНК и смежных областей. Эти методики полностью раскрыты в литературе. Смотри, например, Sambrook et al. MOLECULAR CLONING: A LABORATORY MANUAL, Second edition, Cold Spring Harbor Laboratory Press, 1989 and Third edition, 2001; Ausubel et al., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, New York, 1987 and periodic updates; the series METHODS IN ENZYMOLOGY, Academic Press, San Diego; Wolffe, CHROMATIN STRUCTURE AND FUNCTION, Third edition, Academic Press, San Diego, 1998; METHODS IN ENZYMOLOGY, Vol. 304, ʺChromatinʺ (P.M. Wassarman and A. P. Wolffe, eds.), Academic Press, San Diego, 1999; and METHODS IN MOLECULAR BIOLOGY, Vol. 119, ʺChromatin Protocolsʺ (P.B. Becker, ed.) Humana Press, Totowa, 1999.
Определения
[0070] Термины «нуклеиновая кислота», «полинуклеотид» и «олигонуклеотид» применяются взаимозаменяемо и соответствуют дезоксирибонуклеотидному или рибонуклеотидному полимеру, в линейной или круговой конформации и в либо одно-, либо двухцепочечной форме. Для целей настоящего раскрытия эти термины не следует истолковывать как ограничивающие в отношении длины полимера. Термины могут включать известные аналоги природных нуклеотидов, а также нуклеотиды, которые являются модифицированными по функциональным группам основания, сахара и/или фосфата (например, фосфоротиоатные остовы). В общем, аналог конкретного нуклеотида имеет аналогичную специфичность спаривания оснований; т. е., аналог A будет комплементарной парой для основания T.
[0071] Термины «полипептид», «пептид» и «белок» используют взаимозаменяемо для обозначения полимера из аминокислотных остатков. Термин также применяют по отношению к аминокислотным полимерам, в которых одна или более аминокислот являются химическими аналогами или модифицированными производными соответствующих природных аминокислот.
[0072] «Связывание» относится к специфичному по отношению к последовательности нековалентному взаимодействию между макромолекулами (например, между белком и нуклеиновой кислотой). Не все компоненты связывающего взаимодействия должны быть последовательность-специфическими (например, контакты с фосфатными остатками в остове ДНК), до тех пор, пока взаимодействие в целом зависит от последовательности. Подобные взаимодействия в общем характеризуют с помощью константы диссоциации (Kd) 10-6 M-1 или ниже. «Аффинность» соответствует силе связывания: увеличенная аффинность связывания коррелируется с более низким Kd.
[0073] «Связывающий домен»представляет собой молекулу, которая способна нековалентно связываться с другой молекулой. Связывающая молекула может связываться с, например, молекулой ДНК (ДНК-связывающим белком, таким как белок цинкового пальца, или белком TAL-эффекторного домена или одиночной направляющей РНК), молекулой РНК (РНК-связывающим белком) и/или молекулой белка (белок-связывающим белком). В случае с белок-связывающей молекулой, она может связываться сама с собой (чтобы образовать гомодимеры, гомотримеры, и т. д.) и/или она может связываться с одной или более молекулами другого белка или белков. Связывающая молекула может иметь более одного типа связывающей активности. Например, цинк-пальцевые белки имеют ДНК-связывающую, РНК-связывающую и белок-связывающую активность. Таким образом, ДНК-связывающие молекулы, в том числе ДНК-связывающие компоненты искусственных нуклеаз и факторы транскрипции, включают, но без ограничения этим, ZFP, TALE и sgРНК.
[0074] «Цинк-пальцевый ДНК-связывающий белок» (или связывающий домен) представляет собой белок или домен в пределах большего белка, который связывает ДНК специфичным по отношению к последовательности образом одним или несколькими цинковыми пальцами, которые являются областями аминокислотной последовательности в пределах связывающего домена, чью структуру стабилизируют благодаря координации иона цинка. Термин цинк-пальцевый ДНК-связывающий белок часто сокращается как цинк-пальцевый белок или ZFP. Искусственные нуклеазы и факторы транскрипции могут включать ДНК-связывающий домен ZFP и функциональный домен (домен нуклеазы в случае ZFN или транскрипционный регуляторный домен в случае ZFP-TF). Термин «цинк-пальцевая нуклеаза» включает один ZFN, а также пару ZFN, который димеризует для расщепления гена-мишени.
[0075] «ДНК-связывающий домен TALE» или «TALE» представляет собой полипептид, содержащий один или более повторяющихся доменов/единиц TALE. Повторяющиеся домены берут участие в связывании TALE с его родственной целевой последовательностью ДНК. Отдельная «повторяющаяся единица» (также упоминаются как «повторяющаяся») обычно имеет 33-35 аминокислот в длину и проявляет по меньшей мере некоторую гомологичность последовательности с другими повторяющимися последовательностями TALE в пределах встречающихся в природе белок TALE. Смотри, например, патент США № 8586526. Искусственные нуклеазы и факторы транскрипции могут включать ДНК-связывающий домен TALE и функциональный домен (домен нуклеазы в случае TALEN или транскрипционный регуляторный домен в случае TALEN-TF). Термин «TALEN» включает один TALEN, а также пару TALEN, которые димеризуются для расщепления гена-мишени.
[0076] Цинк-пальцевые и TALE-связывающие домены могут быть «сконструированы» для связывания с предетерминированной нуклеотидной последовательностью, например, посредством конструирования (изменение одной или более аминокислот) области спирали распознавания встречающегося в природе цинк-пальцевого белка или белка TALE. Следовательно, сконструированные ДНК-связывающие белки (цинк-пальцевые или TALE) представляют собой белки, которые не встречаются в природе. Не ограничивающими примерами способов конструирования ДНК-связывающих белков являются дизайн и селекция. Созданный ДНК-связывающий белок представляет собой не встречающийся в природе белок, дизайн/композиция которого главным образом вытекает из рационального критерия. Рациональный критерий дизайн включает применение правил замещения и компьютеризированных алгоритмов для обработки информации в базе данных, хранящей информацию о существующих конструкциях ZFP и/или дизайнах TALE и данных связывания. Смотри, например, патенты США №№ 8568526; 6140081; 6453242; и 6534261; смотри также WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[0077] «Выбранный» цинк-пальцевый белок или TALE представляет собой не обнаруженный в природе белок, продуцирование которого происходит в результате эмпирического процесса, такого как фаговое отображение, ловушка белок-белковых взаимодействий или гибридная селекция. Смотри, например, патенты №№ 8586526; 5789538; US 5925523; US 6007988; US 6013453; US 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970; WO 01/88197; WO 02/099084.
[0078] «Рекомбинация» соответствует способу обмена генетической информацией между двумя полинуклеотидами. Для целей настоящего раскрытия, «гомологическая рекомбинация (HR)» соответствует специализированной форме подобного обмена, который имеет место, например, во время репарации двухцепочечных разрывов в клетках посредством механизмов репарации, направляемой гомологией. Этот процесс требует нуклеотидной гомологии последовательности, использует «донорные» молекулы для шаблонного репарирования молекулы-«мишени» (т.е. той, которая претерпевает двухцепочечный разрыв) и по-другому известен как «не-перекрестное преобразование гена» или «конверсия гена короткого тракта», потому что он приводит к передаче генетической информации от донора к мишени. Не желая быть связанными какой-либо конкретной теорией, подобная передача может затрагивать исправление ошибок спаривания оснований гетеродуплексной ДНК, которая образуется между нарушенной мишенью и донором, и/или «зависимую от синтеза ренатурацию цепи», в которой донор применяют для повторного синтезирования генетической информации, которая может стать частью мишени и/или связанных процессов. Такая специализированная HR часто приводит к изменению последовательности молекулы-мишени, таким образом, что часть или вся последовательность донорного полинуклеотида включают в полинуклеотид-мишень.
[0079] В способах раскрытия одна или более целевых нуклеаз, как описано в данном документе, создают двухцепочечный разрыв в последовательности-мишени (например, клеточном хроматине) на предетерминированном сайте, а в клетку может быть введен «донорный» полинуклеотид, имеющий гомологичность по отношению к нуклеотидной последовательности в области разрыва. Присутствие двухцепочечного разрыва было проиллюстрировано для облегчения интеграции донорной последовательности. Донорная последовательность может быть физически интегральной или, в качестве альтернативы, донорный полинуклеотид применяют в качестве шаблона для репарации разрыва посредством гомологической рекомбинации, что приводит к введению всей или части нуклеотидной последовательности, как в доноре, в клеточный хроматин. Таким образом, первая последовательность в клеточном хроматине может быть изменена и, в некоторых вариантах осуществления настоящего изобретения, может быть преобразована в последовательность, присутствующую в донорном полинуклеотиде. Таким образом, применение терминов «замена» или «замещение» могут пониматься как замена одной нуклеотидной последовательности на другую, (т. e., замена последовательности в информационном смысле), и не обязательно требуют физического или химического замещения одного полинуклеотида на другой.
[0080] В любом из способов, описанных в данном документе, дополнительные пары цинк-пальцевых белков или белков TALEN могут быть применены для дополнительного двухцепочечного расщепления дополнительных сайтов-мишеней внутри клетки.
[0081] В некоторых вариантах осуществления способов целевой рекомбинации и/или замещения и/или изменения последовательности в представляющей интерес области в клеточном хроматина, хромосомную последовательность изменяют путем гомологической рекомбинации с экзогенной «донорной» нуклеотидной последовательностью. Подобную гомологическую рекомбинацию стимулируют присутствием двухцепочечного разрыва в клеточном хроматине, если присутствуют последовательности, гомологичные области разрыва.
[0082] В любом из способов, описанных в данном документе, первая нуклеотидная последовательность («донорная последовательность») может содержать последовательности, которые являются гомологичными, но не идентичными, геномным последовательностям в представляющей интерес области, тем самым стимулируя гомологическую рекомбинацию для вставки неидентичной последовательности в представляющей интерес области. Таким образом, в некоторых вариантах осуществления настоящего изобретения, части донорной последовательности, которые являются гомологичными последовательностям в представляющей интерес области, проявляет от около 80 до 99% (или любое целое число между ними) идентичности последовательности по отношению к геномной последовательности, которую замещают. В других вариантах осуществления настоящего изобретения гомологичность между донором и геномной последовательность выше 99%, например, если между донором и геномными последовательностями из более 100 смежных пар оснований отличается только 1 нуклеотид. В некоторых случаях, негомологичная часть донорной последовательности может содержать последовательности, не присутствующие в представляющей интерес области, таким образом, что новые последовательности вводят в представляющую интерес область. В этих случаях, негомологичную последовательность в общем фланкируют последовательностями из 50-1000 пар оснований (или любое целое число между ними) или любого количества пар оснований больше 1000, которые являются гомологичными или идентичными последовательностям в представляющей интерес области. В других вариантах осуществления настоящего изобретения донорная последовательность является негомологичной по отношению к первой последовательности и вставленной в геном путем механизмов негомологичной рекомбинации.
[0083] Любые способы, описанные в данном документе, могут быть применены для частичной или полной идентификации одной или более последовательностей-мишеней в клетке путем целевой интеграции донорной последовательности, которая нарушает экспрессию представляющего(их) интерес гена(ов). Также обеспечивают клеточные линии с частично или полностью инактивированными генами.
[0084] Более того, способы целевой интеграции, как описано в данном документе, также могут быть применены для интегрирования одной или более экзогенных последовательностей. Экзогенная нуклеотидная последовательность может содержать, например, один или более генов или молекул кДНК, или любой тип кодирующей или некодирующей последовательности, а также одного или более контрольных элементов (например, промоторов). Кроме того, экзогенная нуклеотидная последовательность может продуцировать одну или более молекул РНК (например, коротких шпилечных РНК (кшРНК), РНК ингибиторов (РНКи), микроРНК (миРНК) и т. д.).
[0085] «Расщепление» соответствует разрыванию ковалентного остова молекулы ДНК. Расщепление может быть инициировано различными способами, в том числе, но без ограничения ими, ферментативным или химическим гидролизом фосфодиэфирной связи. Оба, и одноцепочечное расщепление, и двухцепочечное расщепление, являются возможными, а двухцепочечное расщепление может происходить в результате двух отдельных случаев одноцепочечного расщепления. Расщепление ДНК может привести к получению либо тупых концов, либо ступенчатых концов. В некоторых вариантах осуществления настоящего изобретения, слитые полипептиды применяют для целевого расщепления двухцепочечной ДНК.
[0086] «Расщепляющий полудомен» представляет собой полипептидную последовательность, которая в сочетании со вторым полипептидом (либо идентичным, либо отличным) образует комплекс, имеющий расщепляющую активность (предпочтительно, двухцепочечную расщепляющую активность). Термины «первый и второй расщепляющие полудомены» «+ и - расщепляющие полудомены» и «право- и лево- расщепляющие полудомены» применяют взаимозаменяемо для обозначения пары расщепляющих полудоменов, которые димеризуются.
[0087] «Сконструированный расщепляющий полудомен» представляет собой расщепляющий полудомен, который может быть модифицирован таким образом, чтобы образовывать облигатные гетеродимеры с другим расщепляющим полудоменом (например, другим сконструированным расщепляющим полудоменом). Смотри, патенты США №№ 7888121; 7914796; 8034598 и 8823618, которые включены в настоящий документ в виде ссылки
[0088] Термин «последовательность» соответствует нуклеотидной последовательности любой длины, которая может представлять собой ДНК или РНК; может быть линейной, циклической или разветвленной и может быть как одноцепочечной, так и двухцепочечной. Термин «донорная последовательность» соответствует нуклеотидной последовательности, которую вставляют в геном. Донорная последовательность может быть любой длины, например, от 2 до 10000 нуклеотидов в длину (или любое целое число между или выше), предпочтительно, от около 100 до 1000 нуклеотидов в длину (или любое целое число между ними), более предпочтительно, от около 200 до 500 нуклеотидов в длину.
[0089] «Связанный с болезнью ген» представляет собой ген, который каким-то образом является дефектным при моногенной болезни. Неограничивающие примеры моногенных болезней включают тяжелый комбинированный иммунодефицит, муковисцидоз, гемофилии, лизосомные болезни накопления (например, болезни Гоше, Херлера, Хантера, Фабри, Неймана-Пика, Тея-Саха и т. д.), серповидноклеточную анемию и талассемию.
[0090] «Хроматин» представляет собой структуру нуклеопротеина, содержащую клеточный геном. Клеточный хроматин содержит нуклеиновую кислоту, преимущественно, ДНК и белок, в том числе гистоны и негистоновые хромосомные белки. Большая часть эукариотического клеточного хроматина существует в форме нуклеосом, где нуклеосомное ядро содержит приблизительно 150 пар оснований, связанных с ДНК, с октамером, содержащим два гистона из H2A, H2B, H3 и H4; а линкерная ДНК (различной длины, в зависимости от организма) распространяется между нуклеосомными ядрами. Молекула гистона H1, в общем, связана с линкерной ДНК. Для целей настоящего раскрытия термин «хроматин» предназначен для охватывания всех типов клеточного нуклеопротеина, и прокариотического, и эукариотического. Клеточный хроматин включает и хромосомный, и эписомальный хроматин.
[0091] «Хромосом» представляет собой комплекс хроматина, содержащий весь или часть генома клетки. Геном клетки часто характеризуется по своему кариотипу, который представляет собой совокупность всех хромосом, которые составляют геном клетки. Геном клетки может содержать одну или более хромосом.
[0092] «Эписома» представляет собой реплицирующуюся нуклеиновую кислоту, нуклеопротеиновый комплекс или другую структуру, содержащую нуклеиновую кислоту, которая не является частью хромосомного кариотипа клетки. Примеры эписом включают плазмиды и определенные вирусные геномы.
[0093] «Сайт-мишень» или «последовательность-мишень» представляет собой нуклеотидную последовательность, которая определяет часть нуклеиновой кислоты, с которой связывающая молекула будет связываться, при условии наличия достаточных условий для связывания.
[0094] «Экзогенная» молекула представляет собой молекулу, которая обычно не присутствует в клетке, но может быть введена в клетку с помощью одного или более генетических, биохимических или других способов. «Нормальное присутствие в клетке» определяют с учетом конкретной стадии развития и условий окружающей среды клетки. Таким образом, например, молекула, которая присутствует только во время эмбрионального развития мышц, является экзогенной молекулой по отношению к мышечной клетке взрослого человека. Точно так же молекула, индуцированная тепловым шоком, является экзогенной молекулой по отношению к клетке, не получавшей тепловой шок. Экзогенная молекула может содержать, например, функционирующую версию неправильно функционирующей эндогенной молекулы или неправильно функционирующую версию нормально функционирующей эндогенной молекулы.
[0095] Экзогенная молекула может представлять собой, среди прочего, малую молекулу, такую как полученные способом комбинаторной химии, или макромолекулу, такую как белок, нуклеиновая кислота, углевод, липид, гликопротеин, липопротеин, полисахарид, любое модифицированное производное вышеуказанных молекул или любой комплекс, включающий одну или несколько вышеуказанных молекул. Нуклеиновые кислоты включают ДНК и РНК, могут быть одно- или двухцепочечными; могут быть линейными, разветвленными или циклическими; и могут быть любой длины. Нуклеиновые кислоты включают те, которые способны образовывать дуплексы, а также триплексообразующие нуклеиновые кислоты. Смотри, например, патенты США №№ 5176996 и 5422251. Белки включают, но без ограничения этим, ДНК-связывающие белки, факторы транскрипции, факторы реконструкция хроматина, метилированные ДНК-связывающие белки, полимеразы, метилазы, деметилазы, ацетилазы, деацетилазы, киназы, фосфатазы, интегразы, рекомбиназы, лигазы, томоизомеразы, гиразы и геликазы.
[0096] Экзогенная молекула может быть молекулой такого же типа, что и эндогенная молекула, например, экзогенный белок или нуклеиновая кислота. Например, экзогенная нуклеиновая кислота может содержать заражающий вирусный геном, плазмиду или эписому, введенные в клетку, или хромосому, которая в обычном состоянии не присутствует в клетке. Способы введения экзогенных молекул в клетки известны специалистам в данной области техники и включают, но без ограничения этим, липид-опосредованную передачу (т.е., липосомы, в том числе нейтральные и катионные липиды), электропорацию, прямое инъектирование, слияние клеток, бомбардировку частицами, способ соосаждения фосфата кальция, DEAE-декстран-опосредованную передачу и вирусную вектор-опосредованную передачу. Экзогенная молекула также может представлять собой молекулу того же типа, что и эндогенная молекула, но полученная из отличного от того, из которого получена клетка, вида. Например, нуклеотидная последовательность человека может быть введена в клеточную линию, которую обычно получают от мыши или хомяка.
[0097] В противоположность этому, «эндогенная» молекула, которая обычно присутствует в конкретной клетке на конкретной стадии развития под конкретными условиями окружающей среды. Например, эндогенная нуклеиновая кислота может содержать хромосому, геном митохондрии, хлоропласт или другую органеллу или встречающуюся в природе эписомальную нуклеиновую кислоту. Дополнительные эндогенные молекулы могут включать белки, например, факторы транскрипции и ферменты.
[0098] «Слитая» молекула представляет собой молекулу, в которой две или более молекулы субъединиц связаны, предпочтительно, ковалентно. Молекулы субъединиц могут быть молекулами одного и того же химического типа или могут быть молекулами различных химических типов. Примеры первого типа слитой молекулы включают, но без ограничения этим, слитые белки (например, слияние между ZFP или ДНК-связывающим доменом TALE и одним или более доменами активации) и слитые нуклеиновые кислоты (например, нуклеиновую кислоту, которая кодирует слитый белок, описанный supra). Примеры второго типа слитой молекулы включают, но без ограничения этим, слияние между триплексообразующей нуклеиновой кислотой и полипептидом и слияние между связующим с малой бороздой веществом и нуклеиновой кислотой.
[0099] Экспрессия слитого белка в клетке может быть результатом доставки слитого белка в клетку или путем доставки полинуклеотида, который кодирует слитый белок, в клетку, где полинуклеотид транскрибируют, а транскрипт транслируют для получения слитого белка. Транс-сплайсинг, расщепление полипептида и лигирование полипептида также могут быть вовлечены в экспрессию белка в клетке. Способы доставки полинуклеотида и полипептида в клетки представлены в другом месте данного раскрытия.
[0100] «Ген», для целей настоящего раскрытия, содержит область ДНК, которая кодирует генный продукт (смотри infra), также как области ДНК, которые регулируют продуцирование генного продукта, являются ли такие регуляторные последовательности смежными с кодирующими и/или транскрибируемыми последовательностями или нет. Соответственно, ген включает, но не обязательно ограничивается этим, промотор последовательности, терминаторы, трансляционные регуляторные последовательности, такие как сайты связывания рибосом и внутренние участки посадки рибосомы, энхансеры, сайленсеры, инсуляторы, граничные элементы, точки начала репликации, сайты интеграции матрицы и контрольные участки локуса.
[0101] «Экспрессия гена» соответствует конверсии информации, содержащейся в гене, в генный продукт. Генный продукт может быть прямым транскрипционным продуктом гена (например, мРНК, тРНК, рРНК, антисмысловой РНК, рибозимом, структурной РНК или любым другим сайтом РНК) или белком, получаемым трансляцией мРНК. Генные продукты также включают РНК, которые модифицируют с помощью таких процессов, как кэпирование, полиаденилирование, метилирование и редактирование, и белки, которые модифицируют с помощью, например, метилирования, ацетилирования, фосфорилирования, убиквитинирования, ADP-рибозилирования, миристилирования и гликозилирования.
[0102] «Модуляция» экспрессии гена соответствует изменению в активности гена. Модуляция экспрессии может включать, но без ограничения этим, ген активации и ген репрессии. Редактирование генома (например, расщепление, изменение, идентификация, случайная мутация) может быть применено для модуляции экспрессии. Идентификация гена соответствует любому снижению экспрессии гена по сравнению с клеткой, которая не содержит систему ZFP, TALE или систему CRISPR/Cas, как описано в данном документе. Таким образом, идентификация гена может быть частичной или полной.
[0103] «Представляющая интерес область» представляет собой любую область клеточного хроматина, такие как, например, ген или некодирующая последовательность в пределах или примыкающая к гену, в котором желательно связать экзогенную молекулу. Связывание может быть для целей целевого расщепления ДНК и/или целевой рекомбинации. Представляющая интерес область может присутствовать в хромосоме, эписоме, геноме органеллы (например, митохондриальном, хлоропласта) или заражающем вирусном геноме, например. Представляющая интерес область может находиться в пределах кодирующей области гена, в пределах таких транскрибирующихся некодирующих областей как, например, лидерные последовательности, трейлерные последовательности или интроны, или в пределах нетранскрибирующихся областей, либо против хода транскрипции, либо по ходу транскрипции кодирующей области. Представляющая интерес область может как одной нуклеотидной парой, так и иметь до 2000 нуклеотидных пар в длину или любое целочисленное значение нуклеотидных пар.
[0104] «Эукариотические» клетки включают, но без ограничения этим, грибковые клетки (такие как дрожжи), клетки растения, клетки животного, клетки млекопитающего и клетки человека (например, клетки печени, мышечные клетки, RBC, T-клетки и т. д.), в том числе стволовые клетки (плюрипотентные и мультипотентные).
[0105] «Красные кровяные тельца» (RBC) или эритроциты являются терминально дифференцированными клетками, происходящими из гемопоэтических стволовых клеток. В них отсутствуют нуклеаза и большинство клеточных органелл. RBC содержат гемоглобин для переноса кислорода из легких в периферические ткани. Фактически, 33% отдельного RBC представляет собой гемоглобин. Они также переносят CO2, вырабатываемый клетками во время метаболизма, из тканей и обратно в легкие для выброса во время выдоха. RBC продуцируются в костном мозге в ответ на гипоксию крови, которая опосредует высвобождение эритропоэтина (EPO) почкой. EPO вызывает повышение количества проэритробластов и сокращает время, необходимое для полного созревания RBC. Спустя приблизительно 120 дней, поскольку RBC не содержат ядра или каких-либо другие регенеративные способности, клетки удаляют из циркуляции либо фагоцитарной активностью макрофагов в печени, селезенке и лимфатических узлах (~90%), либо гемолизом в плазме (~10%). После поглощения макрофагами, химические компоненты RBC расщепляются в пределах вакуоли макрофагов в соответствиями с действием лизосомальных ферментов. RBC, in vitro или in vivo, могут происходить от генетически модифицированной стволовой клетке или клеток-предшественников RBC, как описано в данном документе.
[0106] «Секреторные ткани» представляют собой те ткани у животного, которые секретируют продукты из отдельной клетки, которые обычно получают из эпителия в просвет некоторого типа. Примеры секреторных тканей, которые локализуются в желудочно-кишечном тракте, включают клетки, которые выстилают кишечник, поджелудочную железу и желчный пузырь. Другие секреторные ткани включают печень, ткани, связанные с глазом и слизистыми оболочками, такими как слюнные железы, молочные железы, предстательная железа, гипофиз и другие члены эндокринной системы. Дополнительно, секреторные ткани включают отдельные клетки ткани такого типа, которые способны к секреции.
[0107] Термины «функциональная связь» и «функциональным образом связанный» (или «функционально связанный») применяют взаимозаменяемо со ссылкой на сопоставление двух или более компонентов (такие как элементы последовательности), в которых компоненты расположены таким образом, что оба компонента функционируют нормально и допускают возможность того, что по меньшей мере один из компонентов может выполнять функцию, которая действует по меньшей мере на один из других компонентов. В качестве иллюстрации, транскрипционная регуляторная последовательность, такая как промотор, функционально связана с кодирующей последовательностью, если транскрипционная регуляторная последовательность контролируют уровень транскрипции кодирующей последовательности в ответ на наличие или отсутствие одного или более транскрипционных регуляторных факторов. Транскрипционная регуляторная последовательность, в общем, функционально связана в cis положении с кодирующей последовательностью, но не обязательно должны быть в непосредственной близости к ней. Например, энхансер представляет собой транскрипционную регуляторную последовательность, которая функционально связана с кодирующей последовательностью, даже если они не являются смежными.
[0108] Что касается слитых полипептидов, термин «функционально связанный» может означать, что каждый из компонентов выполняет ту же функцию в связи с другим компонентом, как это было бы, если бы они не были связаны между собой таким образом. Например, по отношению к слитому полипептиду, в котором ДНК-связывающий домен ZFP, TALE или Cas гибридизуют в домен активации, ZFP или ДНК-связывающий домен TALE и домен активации являются функционально связанными, если, в слитом полипептиде, часть ДНК-связывающего домена ZFP или TALE способна связывать свой сайт-мишень и/или свой связывающий сайт, тогда как домен активации способен активировать экспрессию гена. Когда слитый полипептид, в котором ДНК-связывающий домен ZFP или TALE гибридизуют в домен расщепления, ДНК-связывающий домен ZFP или TALE и домен расщепления являются функционально связанными, если, в слитом полипептиде, часть ДНК-связывающего домена ZFP или TALE способна связывать свой сайт-мишень и/или свой связывающий сайт, тогда как домен расщепления способен расщеплять ДНК в непосредственной близости к сайту-мишени.
[0109] «Функциональный фрагмент» белка, полипептида или нуклеиновой кислоты представляет собой белок, полипептид или нуклеиновую кислоту, чья последовательность является не идентичной полноразмерному белку, полипептиду или нуклеиновой кислоте, сохраняет ту же функцию, что и полноразмерный белок, полипептид или нуклеиновая кислота. Функциональный фрагмент может содержать больше, меньше или такое же количество остатков, что и соответствующая нативная молекула, и/или может содержать одну или более аминокислотных или нуклеотидных замен. Способы определения функции нуклеиновой кислоты (например, кодирующая функция, способность гибридизоваться с другой нуклеиновой кислотой) хорошо известна в данной области техники. Подобным образом, способы определения функции белка являются хорошо известными. Например, ДНК-связывающая функция полипептида может быть определена, например, с помощью анализа связывания с фильтрами, электрофоретической подвижности или иммунопреципитации. Расщепление ДНК может быть проанализировано с помощью гель-электрофореза. Смотри Ausubel et al., supra. Способность белка взаимодействовать с другим белком может быть определена, например, путем совместной иммунопреципитации, двухгибридных анализов или комплементации, как генетической, так и биохимической. Смотри, например, Fields et al. (1989) Nature 340:245-246; патент США № 5585245 и PCT WO 98/44350.
[0110] «Вектор» способен переносить генные последовательности в клетки-мишени. Обычно, «конструкция вектора», «вектор экспрессии» и «вектор для переноса генов» означает любую конструкцию нуклеиновой кислоты, способную направлять экспрессию представляющего интерес гена и которая может передавать клеткам-мишеням последовательности гена. Таким образом, термин включает средства клонирования и экспрессии, а также интегрирующие векторы.
[0111] «Репортерный ген» или «репортерная последовательность» соответствует любой последовательности, которая продуцирует белковый продукт, который легко определен, предпочтительно, хотя и не обязательно, обычным анализом. Подходящие репортерные гены включают, но без ограничения этим, последовательности, которые кодируют белки, которые опосредуют устойчивость к антибиотикам (например, устойчивость к ампициллину, устойчивость к неомицину, устойчивость к G418, устойчивость к пуромицину), последовательности, которые кодируют цветные или флуоресцентные или люминесцентные белки (например, зеленый флуоресцентный белок, усиленный зеленый флуоресцентный белок, красный флуоресцентный белок, люцифераза), и белки, которые опосредуют усиленный клеточный рост и/или амплификацию гена (например, дигидрофолатредуктаза). Метки эпитопа включают, например, одну или более копий FLAG, His, myc, Tap, HA или любую обнаруживаемую аминокислотную последовательность. «Метки экспрессии» включают последовательности, которые кодируют репортеры, которые могут быть функционально связаны с желаемой генной последовательностью, для того, чтобы контролировать экспрессию представляющего интерес гена.
[0112] Термины «субъект» и «пациент» применяют взаимозаменяемо и относятся к млекопитающим, таким как пациенты-люди и отличные от человека приматы, а также экспериментальным животным, таким как кролики, собаки, кошки, крысы, мыши и другие животные. Соответственно, термин «субъект» или «пациент», как применяют в данном документе, означает любого пациента млекопитающего или субъекта, которому могут быть введены измененные по изобретению клетки и/или белки, полученные с помощью измененных по изобретению клеток. Субъекты по настоящему изобретению включают тех, кто имеет LSD.
Нуклеазы
[0113] В практике настоящего изобретения может быть применена любая нуклеаза, в том числе, но без ограничения этим, по меньшей мере одна ZFN, TALEN, хоуминг-эндонуклеазы и системы, содержащие направляющие РНК CRISPR/Cas и/или Ttago, которые являются полезными для in vivo расщепления молекулы-донора, несущей трансген, и нуклеазы для расщепления генома клетки таким образом, что трансген целенаправленно интегрируют в геном. Таким образом, описанное в данном документе представляет собой композиции, содержащие одну или более нуклеаз, которые расщепляют выбранный ген, чье расщепление приводит к геномной модификации гена (например, вставкам и/или делециям в расщепленный ген). В некоторых вариантах осуществления настоящего изобретения одна или более нуклеаз являются встречающимися в природе. В других вариантах осуществления настоящего изобретения одна или более нуклеаз являются встречающимися в природе, т.е., сконструированными в ДНК-связывающей молекуле (также упоминаются как ДНК-связывающий домен) и/или домене расщепления. Например, ДНК-связывающий домен встречающейся в природе нуклеазы может быть изменен для связывания с выбранным сайтом-мишенью (например, ZFP, TALE и/или онРНК CRISPR/Cas, которая сконструирована для связывания с выбранным сайтом-мишенью). В других вариантах осуществления настоящего изобретения нуклеаза содержит гетерологичные ДНК-связывающие домены и домены расщепления (например, цинк-пальцевые нуклеазы; ДНК-связывающие белки TAL-эффекторного домена; ДНК-связывающие домены мегануклеазы с гетерологичными доменами расщепления). В других вариантах осуществления настоящего изобретения нуклеаза содержит такую систему как CRISPR/Cas системы Ttago.
A. ДНК-связывающие домены
[0114] В некоторых вариантах осуществления настоящего изобретения, композиция и способы, описанные в данном документе, используют ДНК-связывающий домен мегануклеазы (хоуминг-эндонуклеазы) для связывания с молекулой-донором и/или представляющей интерес связывающей области в геноме клетки. Встречающиеся в природе мегануклеазы распознают 15-40 сайтов расщепления пар оснований и обычно объединяют в четыре семейства: семейство LAGLIDADG, семейство GIY-YIG, семейство His-Cyst box и семейство HNH. Приводимые в качестве примера хоуминг-эндонуклеазы включают I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. Известны их последовательности распознавания. Смотри также патент США № 5420032; патент США № 6833252; Belfort et al. (1997) Nucleic AcidsRes,25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet.12:224-228; Gimble et al. (1996) J. Mol. Biol. 263:163-180; Argast et al. (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs. Кроме того, ДНК-связывающая специфичность хоуминг-эндонуклеаз и мегануклеаз может быть сконструирована для связывания не встречающихся в природе сайтов-мишеней. Смотри, например, Chevalier et al. (2002) Molec. Cell 10:895-905; Epinat et al. (2003) Nucleic Acids Res. 31:2952-2962; Ashworth et al. (2006) Nature 441:656-659; Paques et al. (2007) Current Gene Therapy7:49-66; публикация патента США № 20070117128. ДНК-связывающие домены хоуминг-эндонуклеаз и мегануклеаз могут быть изменены в контексте нуклеазы вцелом (т. е., таким образом, что нуклеаза включает родственный домен расщепления) или могут быть слиты с гетерологичным доменом расщепления.
[0115] В других вариантах осуществления настоящего изобретения ДНК-связывающий домен одной или более нуклеаз, применяемых в способах и композициях, описанных в данном документе, содержат встречающийся в природе или сконструированный (не встречающихся в природе) ДНК-связывающий домен TAL эффектора. Смотри, например, патент США № 8586526, включенный в данный документ посредством ссылки во всей своей полноте. Известно, что патогенные для растений бактерии рода Xanthomonas вызывают многие заболевания у важных сельскохозяйственных культур. Патогенность Xanthomonas зависит от консервативной системы секреции типа III (T3S), которая вводит более 25 различных эффекторных белков в клетку растения. Среди этих инъектированных белков присутствуют подобные активаторам транскрипции (TAL) эффекторы, которые имитируют транскрипционные активаторы растения и манипулируют растительным транскриптомом (смотри, Kay et al (2007) Science 318:648-651). Эти белки содержат ДНК-связывающий домен и транскрипционный домен активации. Одним из наиболее хорошо исследованных TAL-эффекторов является AvrBs3 из Xanthomonas campestgris pv. Vesicatoria (смотри Bonas et al (1989) Mol Gen Genet 218: 127-136 и WO 2010079430). TAL-эффекторы содержат центральный домен тандемных повторов, каждый из которых содержит приблизительно 34 аминокислоты, которые являются ключом к ДНК-связывающей специфичности этих белков. Кроме того, они содержат последовательность ядерной локализации и кислый транскрипционный домен активации (для обзора смотри SchoРНКck S, et al (2006) J Plant Physiol 163(3): 256-272). Кроме того, в фитопатогенных бактериях Ralstonia solanacearum были обнаружены два гена, обозначенные brg11 и hpx17, которые являются гомологичными AvrBs3 семейству Xanthomonas в R. solanacearumbiovar 1 штамма GMI1000 и в biovar 4 штамме RS1000 (Смотря Heuer et al (2007) Appl and Envir Micro 73(13): 4379-4384). Эти гены являются на 98,9% идентичными друг другу в нуклеотидной последовательности, но отличаются друг от друга делецией 1575 п.о. в повторяющемся домене hpx17. Однако оба генных продукта имеют менее 40% идентичности последовательности с семейством белков AvrBs3 Xanthomonas. Смотри, например, патент США № 8586526, включенный в данный документ посредством ссылки во всей своей полноте.
[0116] Специфичность этих TAL эффекторов зависит от последовательностей, обнаруженных в тандемных повторах. Повторяющаяся последовательность содержит приблизительно 102 п.о. и повторяющиеся последовательности обычно на 91-100% гомологичны одна другой (Bonas et al, ibid). Полиморфизм повторяющихся последовательностей обычно находится в позициях 12 и 13, и, как представляется, существует взаимно-однозначное соответствие между идентичностью гиперварибельных диостатков (RVD) в положениях 12 и 13 с идентичностью смежных нуклеотидов в последовательности-мишени TAL-эффекторов (смотри Moscou and Bogdanove, (2009) Science 326:1501 и Boch et al (2009) Science 326:1509-1512). Экспериментально природный код для распознавания ДНК этих TAL-эффекторов определяют таким образом, что последовательность HD в положениях 12 и 13 приводит к связыванию с цитозином (C), NG связывается с T, NI с A, C, G или T, NN связывается с A или G и ING связывается с T. Эти ДНК-связывающие повторы могут быть собраны в белки с новыми комбинациями и количествами повторов, чтобы создать искусственные факторы транскрипции, которые способны взаимодействовать с новыми последовательностями и активировать экспрессию не-эндогенного репортерного гена в клетках растения (Boch et al, ibid). Сконструированные TAL белки были связаны с расщепляющим полудоменом FokI для получения слияния нуклеазы с эффекторным доменом TAL (TALEN), проявляющим активность в анализе по гену-репортеру дрожжей (мишени на основе плазмиды). Смотри, например, патент США № 8586526; Christian et al ((2010) Genetics epub 10.1534/genetics.110.120717).
[0117] В некоторых вариантах осуществления настоящего изобретения ДНК-связывающий домен одной или более нуклеаз, применяемых для in vivo расщепления и/или целевого расщепления генома клетки, содержат цинк-пальцевый белок. Предпочтительно, цинк-пальцевый белок является не встречающимся в природе в том, что его конструируют для связывания с выбранным сайтом-мишенью. Смотри, например, Смотри, например, Beerli et al. (2002) Nature Biotechnol.20:135-141; Pabo et al. (2001) Ann. Rev. Biochem.70:313-340; Isalan et al. (2001) Nature Biotechnol.19:656-660; Segal et al. (2001) Curr. Opin. Biotechnol.12:632-637; Choo et al. (2000) Curr. Opin. Struct. Biol,10:411-416; патенты США №№ 6453242; 6534261; 6599692; 6503717; 6689558; 7030215; 6794136; 7067317; 7262054; 7070934; 7361635; 7253273; и публикацию патента США №№ 2005/0064474; 2007/0218528; 2005/0267061, все из которых включены в данный документ в виде ссылки во всей их полноте.
[0118] Сконструированный цинк-пальцевый связывающий домен может иметь новую связывающую специфичность, по сравнению с встречающимся в природе цинк-пальцевым белком. Способы разработки включают, но без ограничения этим, рациональный дизайн и различные типы селекции. Рациональный дизайн включает, например, применение баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и отдельные цинк-пальцевые аминокислотные последовательности, в которых каждая триплетная или четверная нуклеотидная последовательность связана с одной или несколькими аминокислотными последовательностями цинковых пальцев, которые связывают конкретную триплетную или квадруплетную последовательность. Смотри, например, совместные патенты США 6453242 и 6534261, включенные в данный документ посредством ссылки во всей своей полноте.
[0119] Приводимые в качестве примера селекция способы, в том числе фаговое отображение и системы двойных гибридов, раскрыты в патентах США 5789538; 5925523; 6007988; 6013453; 6410248; 6140466; 6200759; и 6242568; а также WO 98/37186; WO 98/53057; WO 00/27878; и WO 01/88197. Кроме того, усиление связывающей специфичности цинк-пальцевых связывающих доменов было описано, например, в совместном патенте WO 02/077227.
[0120] Кроме того, как раскрыто в этих и других ссылках, цинк-пальцевые домены и/или мультипальцевые цинк-пальцевые белки могут быть связаны вместе с применением любых подходящих линкерных последовательностей, в том числе, например, линкеров в 5 или более аминокислот в длину. Смотри, также, патенты США №№ 8772453; 6479626; 6903185; и 7153949 для приводимых в качестве примера линкерных последовательностей. Описанные в данном документе белки могут содержать любую комбинацию подходящих линкеров между отдельными цинк-пальцевыми белками.
[0121] Выбор сайтов-мишеней; ZFP и способов для дизайна и конструирования слитых белков (и полинуклеотидов, которые кодируют то же самое) известны специалистам в данной области техники и описаны подробно в патентах США №№ 6140081; 5789538; 6453242; 6534261; 5925523; 6007988; 6013453; 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970; WO 01/88197; WO 02/099084; WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[0122] Кроме того, как раскрыто в этих и других ссылках, цинк-пальцевые домены и/или мультипальцевые цинк-пальцевые белки могут быть связаны вместе с применением любых подходящих линкерных последовательностей, в том числе, например, линкеров в 5 или более аминокислот в длину. Смотри, также, патенты США №№ 6479626; 6903185; и 7153949 для приводимых в качестве примера линкерных последовательностей из 6 или более аминокислот в длину. Описанные в данном документе белки могут содержать любую комбинацию подходящих линкеров между отдельными цинк-пальцевыми белками.
[0123] В некоторых вариантах осуществления настоящего изобретения ДНК-связывающий домен является частью нуклеазной системы CRISPR/Cas, в том числе, например, одиночной направляющей РНК (онРНК). Смотри, например, патент США № 8697359 и публикацию патента США № 20150056705. Локус CRISPR (кластерные короткие палиндромные повторы, разделенные регулярными промежутками), который кодирует компоненты РНК системы и локус (CRISPR-связанный) Cas, который кодирует белки (Jansen et al., 2002. Mol. Microbiol. 43: 1565-1575; Makarova et al., 2002. Nucleic Acids Res. 30: 482-496; Makarova et al., 2006. Biol. Direct 1: 7; Haft et al., 2005. PLoS Comput. Biol. 1: e60), составляют ген последовательности нуклеазной системы CRISPR/Cas. Локусы CRISPR у микробных хозяев содержат комбинацию CRISPR-связанных (Cas) генов, а также некодирующих элементов РНК, способных к программированию специфичности расщепления CRISPR-опосредованной нуклеиновой кислоты.
[0124] CRISPR типа II является одной из наиболее хорошо исследованных систем и осуществляет целевой разрыв двухцепочечной ДНК в четыре последовательных этапа. Сначала две некодирующие РНК, tracrРНК и массив пре-crРНК, транскрибируют из локуса CRISPR. После tracrРНК гибридизуется в повторяющиеся области пре-crРНК и опосредует обработку пре-crРНК в зрелые crРНК, содержащие отдельные спейсерные последовательности. После этого комплекс зрелая crРНК:tracrРНК направляет Cas9 к целевой ДНК посредством спаривания оснований по Уотсону-Крику между спейсером на crРНК и протоспейсером на целевой ДНК рядом с соседним к протоспейсеру мотивом (PAM), дополнительное требование для целевого распознавания. И в конце Cas9 опосредует расщепление целевой ДНК для создания двухцепочечного разрыва в пределах протоспейсера. Активность системы CRISPR/Cas содержит три этапа: (i) вставки последовательностей чужеродной ДНК в массив CRISPR для предотвращения будущих атак, в процессе, называемом «адаптация», (ii) экспрессии соответствующих белков, а также экспрессию и обработку массива, с последующей (iii) РНК-опосредованной интерференции с чужеродной нуклеиновой кислотой. Таким образом, в бактериальной клетке, несколько из так называемых белков «Cas» связаны с естественной функцией системы CRISPR/Cas и выполняют роли в таких функциях, как вставка чужеродной ДНК и т. д.
[0125] В некоторых вариантах осуществления настоящего изобретения белок Cas может быть «функциональным производным» встречающегося в природе Cas белка. «Функциональное производное» полипептида нативной последовательности представляет собой соединение, имеющее общее качественное биологическое свойство с полипептидом нативной последовательности. «Функциональные производные» включают, но без ограничения этим, фрагменты нативной последовательности и производные полипептида нативной последовательности и их фрагменты, предлагая, что у них есть общая с полипептидом соответствующей нативной последовательности биологическая активность. Биологическая активность, рассматриваемая в данном документе, представляет собой способность функционального производного гидролизовать субстрат ДНК на фрагменты. Термин «производное» охватывает и варианты аминокислотной последовательности полипептида, ковалентные модификации и их слияния. Подходящие производные полипептида Cas или их фрагмент включают, но без ограничения этим, мутанты, слияния, ковалентные модификации белка Cas или их фрагмент. Белок Cas, который включает белок Cas или его фрагмент, а также производные белка Cas или их фрагмент, может быть получен из клетки или синтезирован химически или путем комбинации этих двух процедур. Клетка может представлять собой клетку, которая естественным образом продуцирует белок Cas, или клетку, которая естественным образом продуцирует белок Cas и является генетически сконструированной для продуцирования эндогенного белка Cas на более высоком уровне экспрессии или для продуцирования белка Cas из экзогенно введенной нуклеиновой кислоты, причем эта нуклеиновая кислота кодирует Cas, который является таким же или отличным от эндогенного Cas. В некоторых случаях, клетка естественным образом не продуцирует белок Cas и генетически сконструирована для продуцирования белка Cas. Дополнительные не ограничивающие примеры РНК-управляемых нуклеаз, которые могут быть применены для добавления к и/или для замены белков Cas, включают белки CRISPR Class 2, такие как Cpf1. Смотри, например, Zetsche et al. (2015) Cell 163:1-13.
[0126] Система CRISPR-Cpf1, определенная в Francisella spp, представляет собой систему CRISPR-Cas класса 2, которая опосредует усиленную интерференцию ДНК в клетках человека. Являясь функционально консервативными, Cpf1 и Cas9 тем не менее различаются во многих аспектах в том, что касается направляющих РНК и специфичности субстрата (смотри Fagerlund et al, (2015) Genom Bio 16:251). Основное различие между белками Cas9 и Cpf1 заключается в том, что Cpf1 не использует tracrРНК и, следовательно, требует только crРНК. crРНК FnCpf1 имеют 42-44 нуклеотидов в длину (19-нуклеотидный повтор и 23-25-нуклеотидный спейсер) и содержат одну шпильку, которая допускает изменения последовательности, которые сохраняют вторичную структуру. Кроме того, crРНК Cpf1 значительно короче, чем ~100-нуклеотидные сконструированные sgРНК, требуемые Cas9, и требования PAM для FnCpfl представляют собой 5'-TTN-3' и 5'-CTA-3' на смещенной цепи. Хотя и Cas9, и Cpf1 делают двухцепочечные разрывы в целевой ДНК, Cas9 использует свои RuvC- и HNH-подобные домены для осуществления тупоконечных срезов в пределах затравочной последовательности, направляющей РНК, тогда как Cpf1 использует RuvC-подобный домен для продуцирования ступенчатых разрывов вне затравочной последовательности. Поскольку Cpf1 делает ступенчатые срезы вдали от критической затравочной области, NHEJ не будет нарушать сайт-мишень, следовательно, гарантируя, что Cpf1 может продолжать вырезать тот же сайт, пока не произойдет желаемой HDR рекомбинации. Таким образом, в способах и композициях, описанных в данном документе, следует понимать, что термин «Cas» включает и Cas9, и Cfp1 белки. Таким образом, как применяют в данном документе, «система CRISPR/Cas» соответствует и CRISPR/Cas, и/или CRISPR/Cfp1 системам, в том числе как нуклеазным, так и/или системам фактора транскрипции.
[0127] В некоторых вариантах осуществления настоящего изобретения ДНК-связывающий домен представляет собой часть системы TtAgo (смотри Swarts et al, ibid; Sheng et al, ibid). В эукариотах сайленсинг гена опосредуют с помощью семейства белков Argonaute (Ago). В этой парадигме Ago связан с небольшими (19-31 нт) РНК. Этот комплекс сайленсинга белок-РНК распознает целевые РНК посредством спаривания оснований по Уотсону-Крику между малой РНК и мишенью и эндонуклеолитически расщепляет целевую РНК (Vogel (2014) Science 344:972-973). Напротив, прокариотические белки Ago связываются с небольшими одноцепочечными фрагментами ДНК и, вероятно, функционируют для обнаружения и удаления чужеродной (часто вирусной) ДНК (Yuan et al., (2005) Mol. Cell 19, 405; Olovnikov, et al. (2013) Mol. Cell 51, 594; Swarts et al., ibid). Приводимые в качестве примера прокариотические белки Ago включают белки из Aquifex aeolicus, Rhodobacter sphaeroides и Thermus thermophilus.
[0128] Один из наиболее хорошо исследованных прокариотических белков Ago является одним из T. thermophilus (TtAgo; Swarts et al. ibid). TtAgo связаны с либо 15 нт, либо 13-25 нт одноцепочечными фрагментами ДНК с 5' фосфатными группами. Эта «направляющая ДНК», связанная с TtAgo, служит для направления комплекса белок-ДНК с целью связывания по Уотсону-Крику комплементарно последовательности ДНК в молекуле ДНК третьей стороны. Как только информация о последовательности в этих направляющих ДНК позволяет идентифицировать целевую ДНК, комплекс TtAgo-направляющая ДНК расщепляет целевую ДНК. Подобный механизм также поддерживается с помощью структуры комплекса TtAgo-направляющая ДНК, пока он связан со своей целевой ДНК (G. Sheng et al., ibid). Ago из Rhodobacter sphaeroides (RsAgo) имеют подобные свойства (Olivnikov et al. ibid).
[0129] Экзогенные направляющие ДНК произвольной последовательности ДНК могут быть загружены на белок TtAgo (Swarts et al. ibid.). Поскольку специфичность расщепления TtAgo направляют с помощью направляющей ДНК, комплекс TtAgo-ДНК, образованный с экзогенной, указанной исследователем направляющей ДНК, будет, следовательно, направлять расщепление целевой ДНК TtAgo к комплементарной, определенной исследователем целевой ДНК. Таким образом, можно создать целевой двухцепочечный разрыв в ДНК. Применение системы TtAgo-направляющая ДНК (или ортологических систем Ago-направляющая ДНК от других организмов) делает возможным целевое расщепление геномной ДНК в клетках. Подобное расщепление может быть либо одно-, либо двухцепочечным. Для расщепления геномной ДНК млекопитающего было бы предпочтительнее использовать версию кодон-оптимизированного TtAgo для экспрессии в клетках млекопитающего. Кроме того, это может быть предпочтительным для лечения клетки с комплексом TtAgo-ДНК, образованным in vitro, где белок TtAgo гибридизуют с проникающим в клетку пептидом. Кроме того, может быть предпочтительнее использовать версию белка TtAgo, которая была изменена посредством мутагенеза, чтобы иметь улучшенную активность при 37 градусах Цельсия. TtAgo-РНК-опосредованное расщепление ДНК может быть использовано для того, чтобы влиять на совокупность результатов, в том числе нокаут гена, добавление целевого гена, коррекцию гена, делецию целевого гена с использованием способов, известных в данной области техники, для эксплуатирования разрывов ДНК.
[0130] Таким образом, нуклеаза содержит ДНК-связывающий домен, который специфически связывается с сайтом-мишенью в любом гене, в который является желательным вставить донора (трансген).
B. Домены расщепления
[0131] С целью образования нуклеазы, любой подходящий домен расщепления может быть функционально связан с ДНК-связывающим доменом. Например, ДНК-связывающие домены ZFP могут быть слитыми с доменами нуклеазы для создания ZFN - функционального объекта, который способен распознавать предполагаемую мишень для нуклеиновой кислоты через свой сконструированный (ZFP) ДНК-связывающий домен и вызывает разрезание ДНК возле ZFP связывающего сайта посредством активности нуклеазы. Смотри, например, Kim et al. (1996) Proc Natl Acad Sci USA 93(3):1156-1160. Термин ʺZFNʺ включает пару ZFN, которые димеризуются для расщепления ген-мишень. В последнее время ZFN были использованы для модификации генома в различных организмах. Смотри, например, публикации патентов США 20030232410; 20050208489; 20050026157; 20050064474; 20060188987; 20060063231; и международную публикацию WO 07/014275. Также, ДНК-связывающие домены TALE могут быть слитыми с доменами нуклеазы для создания TALEN. Смотри, например, патент США № 8586526. Также были описаны нуклеазные системы CRISPR/Cas, содержащие одиночные направляющие РНК (sgРНК), которые связываются с ДНК и ассоциируются с доменами расщепления (например, домены Cas) для индуцирования целевого расщепления. Смотри, например, патенты США №№ 8697359 и 8932814 и публикацию патента США № 20150056705.
[0132] Как отмечено выше, домен расщепления может быть гетерологичным по отношению к ДНК-связывающему домену, например, цинк-пальцевому ДНК-связывающему домену и домену расщепления из нуклеазы или ДНК-связывающему домену TALEN и домену расщепления из нуклеазы; ДНК-связывающему домену sgРНК и домену расщепления из нуклеазы (CRISPR/Cas); и/или ДНК-связывающему домену мегануклеазы и домену расщепления из различных нуклеаз. Гетерологичные домены расщепления могут быть получены из любой эндонуклеазы или экзонуклеазы. Приводимые в качестве примера эндонуклеазы, из которых может быть получен домен расщепления, включают, но без ограничения этим, рестрикционные эндонуклеазы и хоуминг-эндонуклеазы. Смотри, например, 2002-2003 Catalogue, New England Biolabs, Beverly, MA; и Belfort et al. (1997) Nucleic Acids Res.25:3379-3388. Известны дополнительные ферменты, которые расщепляют ДНК (например, нуклеаза S1; нуклеаза золотистой фасоли; панкреатическая ДНКаза I; микрококковая нуклеаза; эндонуклеаза HO дрожжей; смотри также Linn et al. (eds.) Nucleases, Cold Spring Harbor Laboratory Press,1993). Один или более этих ферментов (или их функциональные фрагменты) могут быть применены в качестве источника доменов расщепления и расщепляющих полудоменов.
[0133] Подобным образом, расщепляющий полудомен может быть получен из любой нуклеазы или ее части, как изложено выше, это требует димеризации для расщепляющей активности. В общем, для расщепления требуются два слитых белка, если слитые белки содержат расщепляющие полудомены. В качестве альтернативы могут быть применены отдельный белок, содержащий два расщепляющих полудомена. Два расщепляющие полудомена могут быть получены из той же самой эндонуклеазы (или ее функциональных фрагментов) или каждый расщепляющий полудомен может быть получен из различных эндонуклеаз (или их функциональных фрагментов). Кроме того, сайты-мишени для двух слитых белков предпочтительно расположены по отношению друг к другу таким образом, что связывание двух слитых белков с их соответствующими сайтами-мишенями размещает полудомены расщепления в пространственной ориентации друг к другу, что позволяет полудоменам расщепления образовывать функциональный домен расщепления, например, путем димеризации. Таким образом, в некоторых вариантах осуществления настоящего изобретения, ближние края сайтов-мишеней разделены 5-8 нуклеотидами или 15-18 нуклеотидами. Тем не менее, между двумя сайтами-мишенями может находиться любое целое число нуклеотидов или нуклеотидных пар (например, от 2 до 50 нуклеотидных пар или более). В общем, сайт расщепления лежит между сайтами-мишенями.
[0134] Эндонуклеазы рестрикции (ферменты рестрикции) присутствуют во многих видах и способны специфически связываться с последовательностью ДНК (на сайте распознавания) и расщепляя ДНК на или рядом с сайтом связывания. Определенные ферменты рестрикции (например, Type IIS) расщепляют ДНК на сайтах, удаленных из сайта распознавания и имеют разделяемые домены связывания и расщепления. Например, фермент Fok I Type IIS катализирует двухцепочечное расщепление ДНК, на 9 нуклеотидах от ее сайта распознавания на одной цепи и 13 нуклеотидах от ее сайта распознавания на другой. Смотри, например, патенты США 5356802; 5436150 и 5487994; а также Li et al. (1992) Proc. Natl. Acad. Sci. USA 89:4275-4279; Li et al. (1993) Proc. Natl. Acad. Sci. USA 90:2764-2768; Kim et al. (1994a) Proc. Natl. Acad. Sci. USA 91:883-887; Kim et al. (1994b) J. Biol. Chem. 269:31978-31982. Таким образом, в одном варианте осуществления настоящего изобретения, слитые белки содержат домен расщепления (или расщепляющий полудомен) от по меньшей мере одного фермента рестрикции Type IIS и одного или более цинк-пальцевых связывающих доменов, который может быть или может быть не быть сконструированным.
[0135] Приводимый в качестве примера фермент рестрикции Type IIS, чей домен расщепления является отделимым от связывающего домена, представляет собой Fok I. Этот конкретный фермент является активным в виде димера. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10570-10575. Соответственно, для целей настоящего раскрытия, часть фермента Fok I, применяемая в раскрытых слитых белках, считают расщепляющим полудоменом. Таким образом, целевое двухцепочечное расщепление и/или целевое замещение клеточных последовательностей с применением цинк-пальцевых-Fok I слияний, двух слитых белков, каждый из которых содержит FokI расщепляющий полудомен, может быть применен для восстановления каталитически активного домена расщепления. В качестве альтернативы молекула одиночного полипептида содержит цинк-пальцевый связывающий домен и также могут быть применены два Fok I расщепляющих полудомена. Параметры целевого расщепления и изменения целевой последовательности с применением слияний цинковый палец-Fok I обеспечивают в других местах в данном описании.
[0136] Домен расщепления или расщепляющий полудомен могут представлять собой любую часть белка, которая сохраняет активность расщепления или которая сохраняет способность к мультимеризации (например, димеризации) для образования функционального домена расщепления.
[0137] Приводимые в качестве примера ферменты рестрикции Type IIS описывают в патенте США 7888121, включенном в данное описание во всей своей полноте. Дополнительные ферменты рестрикции также содержат разделяемые связывающие домены и домены расщепления, и они предусмотрены настоящим раскрытием. Смотри, например, Roberts et al. (2003) Nucleic Acids Res.31:418-420.
[0138] В некоторых вариантах осуществления настоящего изобретения, домен расщепления содержит один или более сконструированных расщепляющих полудоменов (также упоминаются как мутанты домена димеризации), которые минимизируют или предотвращают гомодимеризацию, как описано, например, в патентах США №№ 8772453; 8623618; 8409861; 8034598; 7914796; и 7888121, раскрытия всех из которых включены посредством ссылки во всей их полноте в данном документе. Аминокислотные остатки в положениях 446, 447, 479, 483, 484, 486, 487, 490, 491, 496, 498, 499, 500, 531, 534, 537 и 538 FokI являются мишенями для воздействия на димеризацию расщепляющих полудоменов FokI.
[0139] Приводимые в качестве примера сконструированные расщепляющие полудомены FokI, которые образуют облигатные гетеродимеры, включают пару, в которой первый расщепляющий полудомен включает мутации по аминокислотным остаткам в положениях 490 и 538 FokI, а второй расщепляющий полудомен включает мутации по аминокислотным остаткам 486 и 499.
[0140] Таким образом, в одном варианте осуществления настоящего изобретения, мутация по 490 замещает Glu (E) на Lys (K); мутация по 538 замещает Iso (I) на Lys (K); мутация по 486 замещает Gln (Q) на Glu (E); а мутация в положении 499 замещает Iso (I) на Lys (K). В частности, сконструированные расщепляющие полудомены, описанные в данном документе, готовят путем мутаций по позициям 490 (E→K) и 538 (I→K) в одном расщепляющем полудомене для продуцирования сконструированного расщепляющего полудомена, обозначенного «E490K:I538K» («KK») и с помощью мутаций по позициям 486 (Q→E) и 499 (I→L) в другом расщепляющем полудомене для продуцирования сконструированного расщепляющего полудомена, обозначенного «Q486E:I499L» («EL»). Сконструированные расщепляющие полудомены, описанные в данном документе, представляют собой облигатные мутанты гетеродимеров, в которых аберрантное расщепление минимизировано или прекращается. Патенты США №№ 7914796 и 8034598, раскрытия которых включены в данный документ посредством ссылки в полном объеме. В некоторых вариантах осуществления настоящего изобретения, сконструированный расщепляющий полудомен содержит мутации в положениях 486, 499 и 496 (пронумерованы по сравнению с FokI дикого типа), к примеру, мутации, которые заменяют остаток Gln (Q) дикого типа в положении 486 на остаток Glu(E), остаток Iso (I) дикого типа в положении 499 на остаток Leu (L) и остаток Asn (N) дикого типа в положении 496 на остаток Asp (D) или Glu (E) (также упоминаются как домены «ELD» и «ELE», соответственно). В других вариантах осуществления настоящего изобретения, сконструированный расщепляющий полудомен содержит мутации в положениях 490, 538 и 537 (пронумерованы по сравнению с дикого типа FokI), к примеру, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 на остаток Lys (K), остаток Iso (I) дикого типа в положении 538 на Lys (K) остаток и остаток His (H) дикого типа в положении 537 на остаток Lys (K) или остаток Arg (R) (также упоминаются как домены «KKK» и «KKR», соответственно). В других вариантах осуществления настоящего изобретения, сконструированный расщепляющий полудомен содержит мутации в положениях 490 и 537 (пронумерованы по сравнению с дикого типа FokI), к примеру, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 на Lys (K) остаток и остаток His (H) дикого типа в положении 537 на остаток Lys (K) или остаток Arg (R) (также упоминаются как домены «KIK» и «KIR», соответственно). Смотри, например, патент США № 8772453. В других вариантах осуществления настоящего изобретения, сконструированный расщепляющий полудомен содержит мутации «Sharkey» (смотри Guo et al, (2010) J. Mol. Biol. 400(1):96-107).
[0141] Сконструированные расщепляющие полудомены, описанные в данном документе, могут быть получены с применением любого подходящего способа, например, с помощью сайт-направленного мутагенеза расщепляющих полудоменов дикого типа (Fok I), как описано в патентах США №№ 8623618; 8409861; 8034598; 7914796; и 7888121.
[0142] Способы и композиции также применяют, чтобы повысить специфичность пары нуклеаз к его предполагаемой цели по сравнению с другими непреднамеренными сайтами расщепления, известными как побочные сайты-мишени (смотри публикацию патента США № US-2017-0218349-A1). Таким образом, нуклеазы, описанные в данном документе, могут содержать мутации в одной или более из их ДНК-связывающих остов домена областей и/или одну или более мутаций в их доменах расщепления нуклеазы. Эти нуклеазы могут включать мутации в аминокислоте в пределах ДНК-связывающего домена ZFP («остов ZFP»), который может неспецифически взаимодействовать с фосфатами на остове ДНК, но они не содержат изменений в спиралях распознавания ДНК. Таким образом, изобретение включает мутации катионных аминокислотных остатков в остове ZFP, которые не требуются для целевой специфичности нуклеотида. В некоторых вариантах осуществления настоящего изобретения эти мутации в остове ZFP включают мутирование катионного аминокислотного остатка в нейтральный или анионный аминокислотный остаток. В некоторых вариантах осуществления настоящего изобретения эти мутации в остове ZFP включают мутирование полярного аминокислотного остатка в нейтральный или неполярный аминокислотный остаток. В предпочтительных вариантах осуществления настоящего изобретения мутации в положении (-5), (-9) и/или положении (-14) по сравнению с ДНК-связывающей спирали. В некоторых вариантах осуществления настоящего изобретения цинковый палец может содержать одну или более мутаций в (-5), (-9) и/или (-14). В дополнительных вариантах осуществления настоящего изобретения один или более цинковых пальцев в мультипальцевом цинк-пальцевом белке могут содержать мутации в (-5), (-9) и/или (-14). В некоторых вариантах осуществления настоящего изобретения аминокислоты в (-5), (-9) и/или (-14) (например, аргинин (R) или лизин (K)) мутируют в аланин (A), лейцин (L), Ser (S), Asp (N), Glu (E), Tyr (Y) и/или глутамин (Q).
[0143] В некоторых вариантах осуществления настоящего изобретения, сконструированные расщепляющие полудомены получают из домена нуклеазы FokI и содержат мутацию в одном или более аминокислотных остатков 416, 422, 447, 448 и/или 525, пронумерованы по сравнению с полноразмерным FokI дикого типа. В некоторых вариантах осуществления настоящего изобретения мутации в аминокислотных остатках 416, 422, 447, 448 и/или 525 вводят в FokI «ELD», «ELE», «KKK», «KKR», «KK», «EL», «KIK», «KIR» и/или Sharkey, как описано выше.
[0144] Дополнительно, описанное в данном документе представляет собой способы для повышения специфичности расщепляющей активности посредством независимого титрования сконструированных партнеров полудоменов расщепления нуклеазного комплекса. В некоторых вариантах осуществления настоящего изобретения соотношение двух партнеров (домены полурасщепления) составляет соотношение 1:2, 1:3, 1:4, 1:5, 1:6, 1:8, 1:9, 1:10 или 1:20 или любая величина между ними. В других вариантах осуществления настоящего изобретения соотношение двух партнеров составляет более 1:30. В других вариантах осуществления настоящего изобретения два партнера развернуты в соотношении, которое выбрано, чтобы отличаться от 1:1. При использовании по отдельности или в комбинации, способы и композиции обеспечивают удивительное и неожиданное увеличение специфичности нацеливания посредством снижения побочной расщепляющей активности. Нуклеазы, применяемые в этих вариантах осуществления настоящего изобретения, могут содержать ZFN, пару ZFN, TALEN, пару TALEN, CRISPR/Cas, CRISPR/dCas и TtAgo или любую их комбинацию.
[0145] В качестве альтернативы нуклеазы могут быть выбраны in vivo на сайте-мишени нуклеиновой кислоты с применением так называемой технологии «расщепленных ферментов» (смотри, например, публикацию патента США № 20090068164). Компоненты подобных расщепленных ферментов могут быть экспрессированы либо на разделенных конструкциях экспрессии, либо могут быть связаны в одной открытой рамке считывания, где отдельные компоненты разделяют, например, с помощью саморасщепления пептида 2A или последовательности IRES. Компоненты могут быть отдельными цинк-пальцевыми связывающими доменами или доменами домена, связывающего нуклеиновую кислоту мегануклеазы.
[0146] Нуклеазы могут быть проверены на активность перед использованием, например, в хромосомной системе на основе дрожжей, как описано в патенте США № 8563314. Экспрессия нуклеазы может находиться под контролем конститутивного промотора или индуцируемого промотора, например, промотора галактокиназы, который активируют (дерепрессируют) в присутствие раффинозы и/или галактозы и репрессируют в присутствие глюкозы.
[0147] Связанная с Cas9 система CRISPR/Cas содержит два не кодирующих РНК компонента: массив tracrРНК и пре-crРНК, содержащий направляющие последовательности (спейсеры) нуклеазы, разделенные идентичными прямыми повторами (DR). Для применения системы CRISPR/Cas для осуществления геномной инженерии, должны присутствовать обе функции этих РНК (смотри Cong et al, (2013) Sciencexpress 1/10.1126/science 1231143). В некоторых вариантах осуществления настоящего изобретения, tracrРНК и пре-crРНК обеспечивают посредством разделенных конструкций экспрессий или разделенных РНК. В других вариантах осуществления настоящего изобретения, конструируют химерную РНК, где сконструированную зрелую crРНК (обеспечивая целевую специфичность) гибридизуют в tracrРНК (обеспечивая взаимодействие с Cas9) для создания химерного гибрида cr-РНК-tracrРНК (также называемая одиночной направляющей РНК). (смотри Jinek ibid и Cong, ibid).
Целевые сайты
[0148] Как описано подробно выше, домены ДНК могут быть сконструированы для связывания с любой выбранной последовательностью в локусе, например, альбумина или другого гена «safe harbor». Сконструированный ДНК-связывающий домен может иметь новую связывающую специфичность, по сравнению со встречающимся в природе ДНК-связывающим доменом. Способы разработки включают, но без ограничения этим, рациональный дизайн и различные типы селекции. Рациональный дизайн включает, например, применение баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и отдельные (например, цинк-пальцевые) аминокислотные последовательности, в которых каждая триплетная или четверная нуклеотидная последовательность связана с одной или несколькими аминокислотными последовательностями ДНК-связывающего домена, которые связывают конкретную триплетную или квадруплетную последовательность. Смотри, например, совместные патенты США 6453242 и 6534261, включенные в данный документ посредством ссылки во всей своей полноте. Также может быть осуществлен рациональный дизайн TAL-эффекторных доменов. Смотри, например, публикацию патента США № 20110301073.
[0149] Приводимые в качестве примера селекция способы, применимые по отношению к ДНК-связывающих доменов, в том числе фаговое отображение и системы двойных гибридов, раскрыты в патентах США 5789538; 5925523; 6007988; 6013453; 6410248; 6140466; 6200759; и 6242568; а также WO 98/37186; WO 98/53057; WO 00/27878; WO 01/88197 и GB 2338237.
[0150] Выбор сайтов-мишеней; нуклеаз и способов дизайна и конструирования слитых белков (и полинуклеотидов, которые кодируют тоже самое) известен специалистам в данной области техники и описаны подробно в патентах США №№ 7888121 и 8409891, включенные в данный документ посредством ссылки во всей своей полноте.
[0151] Кроме того, как раскрыто в этих и других ссылках, ДНК-связывающие домены (например, мультипальцевые цинк-пальцевые белки) могут быть связаны вместе с применением любого подходящего линкера последовательности, в том числе например, линкеров из 5 или более аминокислот. Смотри, например, патенты США №№ 6479626; 6903185; и 7153949 для приводимых в качестве примера линкерных последовательностей из 6 или более аминокислот в длину. Описанные в данном документе белки могут содержать любую комбинацию подходящих линкеров между отдельными ДНК-связывающими доменами белка. Смотри, также, публикацию патента США № 20110301073.
Доноры
[0152] Как указано выше, обеспечивают вставку экзогенной последовательности (также называемой «донорной последовательностью» или «донором»), например, для коррекции мутантного гена или для повышенной экспрессии гена, кодирующего белок, отсутствующий или дефицитный при болезни Фабри (например, α-GalA). Очевидно, что донорная последовательность обычно не является идентичной по отношению к геномной последовательности, в которой она находится. Донорная последовательность может содержать негомологичную последовательность, фланкированную двумя областями гомологичности («плечи гомологичности»), для обеспечения эффективной HDR в представляющей интерес локализации. Дополнительно, донор последовательности может содержать векторную молекулу, содержащую последовательности, которые являются гомологичными по отношению к представляющей интерес области в клеточном хроматине. Молекула-донор может содержать несколько прерывистых областей гомологичности с клеточным хроматином. Например, для направленной вставки последовательностей, которые в обычных условиях отсутствуют в представляющей интерес области, указанные последовательности могут присутствовать в донорной молекуле нуклеиновой кислоты и быть фланкированными областями гомологичности к последовательности в представляющей интерес области.
[0153] Описанное в данном документе представляет собой способы направленной вставки трансгена, которые кодируют белок α-GalA для вставки в выбранную локализацию. Трансген GLA может кодировать полноразмерный белок α-GalA или может кодировать усеченный белок α-GalA. Полинуклеотиды для вставки могут также называться «экзогенными» полинуклеотидами, «донорными» полинуклеотидами или молекулами или «трансгенами». Не ограничивающие приводимые в качестве примера доноры GLA проиллюстрированы на Фигурах 1B, 1C, 10, 13 и 25.
[0154] Донорный полинуклеотид может представлять собой ДНК или РНК, одноцепочечную и/или двухцепочечную, и может быть введен в клетку в линейной или циклической форме. Смотри, например, патенты США №№ 8703489 и 9255259. Донорная(ые) последовательность(и) может(могут) также содержаться в пределах MC ДНК, которая(ые) может(могут) быть введен(ы) в клетку в циклической или линейной форме. Смотри, например, публикацию патента США № 20140335063. Если вводят в линейной форме, концы донорной последовательности могут быть защищены (например, от экзонуклеолитической деградации) способами, известными специалистам в данной области техники. Например, один или более дидезоксинуклеотидных остатков добавляют на 3' конец линейной молекулы и/или самокомплементарные олигонуклеотиды лигируют с одного или обоих концов. Смотри, например, Chang et al. (1987) Proc. Natl. Acad. Sci. USA 84:4959-4963; Nehls et al. (1996) Science 272:886-889. Дополнительные способы защиты экзогенных полинуклеотидов от деградации, содержат, но без ограничения ими, добавление концевой аминогруппы(аминогрупп) и использование модифицированных межнуклеотидных связей, таких как, например, фосфоротиоаты, фосфорамидаты и остатки O-метилрибозы или дезоксирибозы.
[0155] Полинуклеотид может быть введен в клетку в качестве части вирусной или невирусной векторной молекулы, которая имеет дополнительные последовательности, такие как, например, начала репликации, промоторы и гены, кодирующие устойчивость к антибиотикам. Более того, донорные полинуклеотиды могут быть введены в качестве открытой нуклеиновой кислоты, в качестве нуклеиновой кислоты, образующей комплекс с таким средством, как липосома или полоксамер, или могут быть доставлены с помощью вирусов (например, аденовируса, AAV, герпесвируса, ретровируса, лентивируса и дефектного интегразой лентивируса (IDLV)).
[0156] Донор, как правило, вставляют таким образом, что его экспрессия управляется эндогенным промотором в сайте интеграции, а именно промотором, который управляет экспрессией эндогенного гена, в который вставлен донор (например, экспрессируемый на высоком уровне, альбумин, AAVS1, HPRT и т. д.). Однако будет очевидно, что донор может содержать промотор и/или энхансер, например, конститутивный промотор или индуцируемый или тканеспецифичный промотор. В некоторых вариантах осуществления настоящего изобретения донор содержится в клетке в плазмиде экспрессии таким образом, что ген экспрессируется внешнехромосомно.
[0157] Молекула-донор может быть вставлена в эндогенный ген таким образом, что весь эндогенный ген не экспрессируется полностью или частично. Например, трансген, как описано в данном документе, может быть вставлен в альбумин или другой локус таким образом, что экспрессируют некоторые (N-терминальные и/или C-терминальные по отношению к трансгену, который кодирует лизосомальный фермент) или ни одну из эндогенных последовательностей альбумина, например, в качестве слияния с трансгеном, который кодирует белок(белки) α-GalA. В других вариантах осуществления настоящего изобретения трансген (например, с или без дополнительных кодирующих последовательностей, таких как в случае альбумина) интегрируют в любой эндогенный локус, например, локус «safe-harbor».
[0158] Когда эндогенные последовательности (эндогенные или часть трансгена) экспрессируют с трансгеном, эндогенные последовательности (например, альбумина и т. д.) могут быть полноразмерными последовательностями (дикого типа или мутантными) или частичными последовательностями. Предпочтительно эндогенные последовательности являются функциональными. Неограничивающие примеры функции этих полноразмерных или частичных последовательностей (например, альбумина) включают увеличение периода полувыведения полипептида в сыворотке, экспрессированного трансгеном (например, терапевтическим геном), и/или действия в качестве носителя.
[0159] Более того, хотя не требуются для экспрессии, экзогенные последовательности могут также включать транскрипционные или трансляционные регуляторные последовательности, например, промоторы, энхансеры, инсуляторы, внутренние участки посадки рибосомы, последовательности, которые кодируют пептиды 2A и/или сигналы полиаденилирования.
[0160] Экзогенные последовательности, связанные с трансгеном, могут также включать сигнальные пептиды для принятия участия в обработке и/или секреции кодируемого белка. Неограничивающие примеры этих сигнальных пептидов включают пептиды из Albumin, IDS и Factor IX (смотри, например, Фигуру 13).
[0161] В некоторых вариантах осуществления настоящего изобретения экзогенная последовательность (донор) содержит слияние представляющего интерес белка и, в качестве его партнера по слиянию, внеклеточный домен белка мембраны, в результате чего слитый белок находится на поверхности клетки. Это позволяет белку, кодируемому трансгеном, потенциально действовать в сыворотке. В случае болезни Фабри фермент α-GalA, кодируемый слиянием трансгена, воздействует на метаболические продукты, которые накапливаются в сыворотке из ее локализации на поверхности клетки (например, RBC). Кроме того, если RBC поглощается макрофагом селезенки, что является нормальным ходом деградации, лизосома, образованная, когда макрофаг охватывает клетку, подвергает мембранно-связанный слитый белок воздействию высоких концентраций продуктов метаболизма в лизосоме при рН, более благоприятных для этого фермента в природных условиях. Неограничивающие примеры потенциальных партнеров слияния иллюстрируют ниже в Таблице 1.
Таблица 1: Примеры потенциальных партнеров слияния
[0162] В некоторых случаях, донор может быть эндогенным геном (GLA), который может быть модифицирован. В качестве примера, для продуцирования донора оптимизация кодона может быть выполнена на эндогенном гене. Более того, хотя ответ антитела на ферментозаместительную терапию варьируется по отношению к специфическому терапевтическому ферменту в вопросе и с отдельным пациентом, значительный иммунный ответ был замечен у многих пациентов с болезнью Фабри, подвергавшихся лечению с помощью замещения фермента на α-GalA дикого типа. Считается, что трансген обеспечивает терапевтический белок, когда он увеличивает количество белка (и/или его активность) по сравнению с субъектами без трансгена. Кроме того, релевантность этих антител к эффективности лечения также является переменной (смотри Katherine Ponder, (2008) J Clin Invest 118(8):2686). Таким образом, способы и композиции по настоящему изобретению могут содержать поколение донора с модифицированными последовательностями по сравнению с GLA дикого типа, в том числе, но без ограничения этим, модификации, которые продуцируют изменения функционально молчащей аминокислоты на сайтах, о которых известно, что они являются инициирующими эпитопами для эндогенных иммунных ответов, и/или усечения таким образом, что полипептид, полученный с помощью подобного донора, является менее иммуногенным.
[0163] Пациенты с болезнью Фабри часто имеют неврологические осложнения из-за отсутствия недостающего фермента α-GalA в мозге. К сожалению, зачастую трудно доставить терапевтические средства в мозг посредством крови в соответствии с непроницаемостью гематоэнцефалического барьера. Таким образом, способы и композиции изобретению могут быть применены в сочетании со способами повышения доставки терапевтического средства в мозг, в том числе, но без ограничения этим, способами, которые вызывают временное открытие плотных контактов между клетками капилляров мозга, такое как преходящее осмотическое нарушение, путем использования внутрикаротидного введения гипертонического раствора маннита, использования сфокусированного ультразвука и введения аналога брадикинина (Matsukado et al (1996) Neurosurgery 39:125). В качестве альтернативы терапевтические средства могут быть созданы для использования рецепторов или механизмов транспорта для специфического транспорта в мозг. Примеры специфических рецепторов, которые могут быть применены, включают трансферриновый рецептор, инсулиновый рецептор или связанные с рецепторами липопротеинов низкой плотности белки 1 и 2 (LRP-1 и LRP-2). Известно, что LRP взаимодействует с рядом секретируемых белков, таких как apoE, tPA, PAI-1 и т.д., и, таким образом, гибридизует последовательность распознавания одного из этих белков для LRP может облегчать транспорт фермента в мозг, после экспрессии в печени терапевтического белка и секреции в ток крови (смотри Gabathuler, (2010) ibid).
Клетки
[0164] Предлагаемое в данном документе также представляет собой генетически модифицированные клетки, например, клетки печени или стволовые клетки, содержащие трансген, который кодирует белок α-GalA, в том числе клетки получают способами, описанными в данном документе. Трансген GLA может быть полноразмерным или модифицированным и может быть экспрессированным внешнехромосомно или может интегрироваться целевым образом в геном клетки с использованием одной или более нуклеаз. В отличие от случайной интеграции, нуклеазо-опосредованная целевая интеграция гарантирует, что трансген интегрируется в определенный ген. Трансген может быть интегрирован в любое место гена-мишени. В некоторых вариантах осуществления настоящего изобретения, трансген интегрируют в или около сайта связывания и/или расщепления нуклеазы, например, в пределах 1-300 (или любого количества пар оснований между ними) пар оснований против хода транскрипции или по ходу транскрипции сайта расщепления и/или связывающего сайта, более предпочтительно, в пределах 1-100 пар оснований (или любого количества пар оснований между ними) с каждой стороны сайта расщепления и/или связывающего сайта, еще более предпочтительно, в пределах 1-50 пар оснований (или любого количества пар оснований между ними) с каждой стороны сайта расщепления и/или связывающего сайта. В некоторых вариантах осуществления настоящего изобретения, интегрированная последовательность не включает какие-либо векторные последовательности (например, вирусную векторную последовательность).
[0165] Любой тип клеток может быть генетически модифицирован, как описано в настоящем документе, чтобы включать трансген, в том числе, но без ограничения этим, клетки или клеточные линии. Другие не ограничивающие примеры генетически модифицированных клеток, как описано в данном документе, включают T-клетки (например, CD4+, CD3+, CD8+ и т. д.); дендритные клетки; B-клетки; аутологичные (например, получаемые от пациента), мышечные клетки, клетки мозга и подобное. В некоторых вариантах осуществления настоящего изобретения клетки представляют собой клетки печени и являются модифицированными in vivo. В некоторых вариантах осуществления настоящего изобретения клетки представляют собой стволовые клетки, в том числе гетерологичные плюрипотентные, тотипотентные или мультипотентные стволовые клетки (например, клетки CD34+, индуцированные плюрипотентные стволовые клетки (iPSC), эмбриональные стволовые клетки или подобное). В некоторых вариантах осуществления настоящего изобретения, клетки, как описано в данном документе, представляют собой стволовые клетки, полученные от пациента.
[0166] Клетки, как описано в данном документе, являются полезными при лечении и/или профилактике болезни Фабри у субъекта с расстройством, например, с помощью in vivo терапий. Также обеспечивают еx vivo терапии, например, когда нуклеаза-модифицированные клетки могут быть размножены, а затем заново введены пациенту с применением стандартных методик. Смотри, например, Tebas et al (2014) New Eng J Med 370(10):901. В случае стволовых клеток, после инфузии субъекту, также наблюдается дифференцирование этих прекурсоров in vivo в клетки, экспрессирующие функциональный белок (из вставленного донора).
[0167] Также обеспечивают фармацевтические композиции, содержащие клетки, как описано в данном документе. Кроме того, перед введением пациенту клетки могут быть криоконсервированы.
Доставка
[0168] Нуклеазы, полинуклеотиды, которые кодируют эти нуклеазы, донорные полинуклеотиды и/или композиции (например, клетки, белки, полинуклеотиды и т. д.), описанные в данном документе, могут быть доставлены in vivo или ex vivo любыми подходящими способами.
[0169] Способы доставки нуклеазы, как описано в данном документе, описывают, например, в патентах США №№ 6453242; 6503717; 6534261; 6599692; 6607882; 6689558; 6824978; 6933113; 6979539; 7013219; и 7163824, раскрытия всех из которых включены посредством ссылки во всей их полноте в данном документе.
[0170] Нуклеазы и/или донорные конструкции, как описано в данном документе, могут также быть доставлены с применением векторов, которые содержат последовательности, кодирующие один или более цинк-пальцевых, TALEN и/или Cas белков. Любые векторные системы могут быть применены, в том числе, но без ограничения этим, плазмидные векторы, ретровирусные векторы, лентивирусные векторы, аденовирусные векторы, поксвирусные векторы; герпесвирусные векторы и векторы аденоассоциированного вируса и т.д. Смотри, также, патенты США №№ 6534261; 6607882; 6824978; 6933113; 6979539; 7013219; и 7163824, включенные в данный документ посредством ссылки во всей своей полноте. Более того, будет очевидно, что любой из этих векторов может содержать одну или более последовательностей, необходимых для лечения. Таким образом, когда одна или более нуклеаз или донорную конструкцию вводят в клетку, нуклеазы и/или донорный полинуклеотид могут переноситься на одном и том же векторе или на различных векторах. Когда применяют множественные векторы, каждый вектор может содержать последовательность, которая кодирует одну или множество нуклеаз и/или донорных конструкций.
[0171] Обычные вирусные и не вирусные способы передачи гена могут быть применены для введения нуклеиновых кислот, которые кодируют нуклеазы и донорные конструкции в клетках (например, млекопитающего клетки) и тканых-мишенях. Системы доставки невирусного вектора включают плазмиды ДНК, открытую нуклеиновую кислоту и нуклеиновую кислоту, образующую комплекс со средством доставки, таким как липосома или полоксамер. Системы доставки вирусного вектора включают вирусы ДНК и РНК, которые имеют либо эписомальные, либо интегральные геномы после доставки в клетку. Для обзора процедур генной терапии, смотри Anderson, Science 256:808-813 (1992); Nabel & Felgner, TIBTECH 11:211-217 (1993); Mitani & Caskey, TIBTECH 11:162-166 (1993); Dillon, TIBTECH 11:167-175 (1993); Miller, Nature 357:455-460 (1992); Van Brunt, Biotechnology 6(10):1149-1154 (1988); Vigne, Restorative Neurology and Neuroscience 8:35-36 (1995); Kremer & Perricaudet, British Medical Bulletin 51(1):31-44 (1995); Haddada et al., in Current Topics in Microbiology and Immunology Doerfler and Böhm (eds.) (1995); и Yu et al., Gene Therapy 1:13-26 (1994).
[0172] Способы невирусной доставки нуклеиновых кислот включают электропорацию, липофекцию, микроинъекцию, биолистику, виросомы, липосомы, иммунолипосомы, поликатион или конъюгаты липид:нуклеиновая кислота, открытую ДНК, искусственные вирионы и усиленное средством поглощение ДНК. Для доставки нуклеиновых кислот также может быть использовано применение сонопорации с применением, например, системы Sonitron 2000 (Rich-Mar).
[0173] Дополнительные приводимые в качестве примера системы доставки нуклеиновой кислоты включают те, которые предлагают Amaxa Biosystems (Кельн, Германия), Maxcyte, Inc. (Роквилл, Мэриленд), BTX Molecular Delivery Systems (Холлистон, Массачусетс) и Copernicus Therapeutics Inc. (смотри, например, US6008336). Липофекцию описывают в, например, патентах США №№ 5049386; 4946787; и 4897355), а реагенты липофекции доступны коммерчески (например, Transfectam™ и Lipofectin™). Катионные и нейтральные липиды, которые подходят для эффективного распознавания рецепторов липофекции полинуклеотидов включают таковые от Felgner, WO 91/17424, WO 91/16024.
[0174] Получение комплексов липид:нуклеиновая кислота, в том числе целевых липосом, таких как иммунолипидные комплексы, хорошо известны специалисту в данной области техники (смотри, например, Crystal, Science 270:404-410 (1995); Blaese et al., Cancer Gene Ther. 2:291-297 (1995); Behr et al., Bioconjugate Chem. 5:382-389 (1994); Remy et al., Bioconjugate Chem. 5:647-654 (1994); Gao et al., Gene Therapy 2:710-722 (1995); Ahmad et al., Cancer Res. 52:4817-4820 (1992); патенты США №№ 4186183, 4217344, 4235871, 4261975, 4485054, 4501728, 4774085, 4837028 и 4946787).
[0175] Композиции, описанные в данном документе (кДНК и/или нуклеазы), могут также быть доставлены с применением наночастиц, например, липидных наночастиц (LNP). Смотри, например, Lee et al (2016) Am J Cancer Res 6(5):1118-1134; публикацию патента США № 20170119904; и предварительную заявку на патент США 62/559186.
[0176] Дополнительные способы доставки включают применение упаковки нуклеиновых кислот для доставки в средства доставки EnGeneIC (EDV). Эти EDV являются специфически доставленными в ткани-мишени с применением биспецифических антител, где одно плечо антитела имеет специфичность по отношению к ткани-мишени, а другое имеет специфичность по отношению к EDV. Антитело связывает EDV с поверхностью клетки-мишени, а затем EDV заносят в клетку с помощью эндоцитоза. Оказавшись в ячейке, содержимое высвобождается (смотри MacDiarmid et al (2009) Nature Biotechnology 27(7):643).
[0177] Применение основанных на вирусных РНК или ДНК систем для доставки нуклеиновых кислот, которые кодируют сконструированные ZFP, используют преимущества высокоразвитых процессов для нацеливания вируса на специфические клетки в теле и направления миграции нагрузки вируса в ядро. Вирусные векторы могут быть введены непосредственно субъектам (in vivo) или они могут быть применены для лечения клетки in vitro, а субъектам вводят модифицированные клетки (ex vivo). Обычные вирусные системы для доставки ZFP включают, но без ограничения этим, ретровирусные, лентивирусные, аденовирусные, адено-связанные, векторы вируса коровьей оспы и простого герпеса для переноса генов. Интеграция в геном-хозяин возможна с помощью методов переноса генов ретровируса, лентивируса и аденоассоциированного вируса, что часто приводит к длительной экспрессии вставленного трансгена. Кроме того, высокая эффективность трансдукции наблюдалась во многих различных типах клеток и тканей-мишеней.
[0178] Тропизм ретровируса может быть изменен путем включения чужеродных белков оболочки, размножая потенциальную целевую популяцию клеток-мишеней. Лентивирусные векторы представляют собой ретровирусные векторы, которые способны трансдуцировать или инфицировать не делящиеся клетки и обычно продуцируют высокие вирусные титры. Выбор ретровирусной системы переноса генов зависит от ткани-мишени. Ретрoвирусные векторы состоят из длинных концевых повторов cis-действия с упаковочной способностью до 6-10 т.п.н. чужеродной последовательности. Минимальные cis-действующие LTR являются достаточными для репликации и упаковки векторов, которые затем применяют для интегрирования терапевтического гена в клетку-мишень с целью обеспечения перманентной экспрессии трансгена. Широкоприменяемые ретровирусные векторы включают векторы, основанные на вирусе мышиного лейкоза (MuLV), вирусе лейкемии гиббоновых обезьян (GaLV), вирусе Simian Immunodeficiency (SIV), вирусе иммунодефицита человека (HIV) и их комбинациях (смотри, например, Buchscher et al., J. Virol. 66:2731-2739 (1992); Johann et al., J. Virol. 66:1635-1640 (1992); Sommerfelt et al., Virol. 176:58-59 (1990); Wilson et al., J. Virol. 63:2374-2378 (1989); Miller et al., J. Virol. 65:2220-2224 (1991)).
[0179] В применениях, в которых предпочтительна временная экспрессия, могут быть применены системы на основе аденовируса. Векторы на основе аденовируса способны к очень высокой эффективности трансдукции во многих типах клеток и не требуют деления клеток. С такими векторами были получены высокие титры и высокие уровни экспрессии. Этот вектор может быть получен в больших количествах в относительно простой системе. Векторы аденоассоциированного вируса («AAV») также применяют для трансдуцирования клеток целевыми нуклеиновыми кислотами, например, в in vitro продуцировании нуклеиновых кислот и пептидов, и для in vivo и ex vivo процедур генной терапии (смотри, например, West et al., Virology 160:38-47 (1987); патент США № 4797368; WO 93/24641; Kotin, Human Gene Therapy 5:793-801 (1994); Muzyczka, J. Clin. Invest. 94:1351 (1994). Конструкцию рекомбинантных векторов AAV описывают в ряде публикаций, в том числе патенте США № 5173414; Tratschin et al., Mol. Cell. Biol. 5:3251-3260 (1985); Tratschin, et al., Mol. Cell. Biol. 4:2072-2081 (1984); Hermonat & Muzyczka, PNAS 81:6466-6470 (1984); и Samulski et al., J. Virol. 63:03822-3828 (1989).
[0180] По меньшей мере, шесть подходов к вирусным векторам в настоящее время доступны для переноса генов в клинических испытаниях, в которых применяют подходы, которые включают комплементацию дефективных векторов генами, вставленными в линии клеток-хелперов, для получения трансдуцирующего средства.
[0181] pLASN и MFG-S являются примерами ретровирусных векторов, которые были использованы в клинических испытаниях (Dunbar et al., Blood 85:3048-305 (1995); Kohn et al., Nat. Med. 1:1017-102 (1995); Malech et al., PNAS 94:22 12133-12138 (1997)). PA317/pLASN был первым терапевтическим вектором, применяемым в испытании генной терапии. (Blaese et al., Science 270:475-480 (1995)). Эффективность трансдукции в 50% или более наблюдали в случае упакованных векторов MFG-S. (Ellem et al., Immunol Immunother. 44(1):10-20 (1997); Dranoff et al., Hum. Gene Ther. 1:111-2 (1997).
[0182] Рекомбинантные аденоассоциированные вирусные векторы (rAAV) представляют собой многообещающие альтернативные системы доставки генов, основанные на дефектном и непатогенном парвовирусном аденоассоциированном вирусе типа 2. Все векторы получены из плазмиды, которая сохраняет только инвертированные концевые повторы AAV 145 п.н., фланкирующие кассету экспрессии трансгена. Эффективная передача гена и стабильная доставка трансгена в соответствии с интеграцией в геномы трансдуцирующей клетки являются ключевыми особенностями для этой векторной системы. (Wagner et al., Lancet 351:9117 1702-3 (1998), Kearns et al., Gene Ther. 9:748-55 (1996)). В соответствии с настоящим изобретением также могут быть применены другие серотипы AAV, в том числе не ограничивающий пример, rh10 AAV1, AAV3, AAV4, AAV5, AAV6, AAV8, AAV 8.2, AAV9 и AAV и псевдотипированный AAV, такой как AAV2/8, AAV2/5 и AAV2/6.
[0183] Дефицитные по репликации рекомбинантные аденовирусные векторы (Ad) могут быть получены при высоком титре и легко инфицируют ряд различных типов клеток. Большинство аденовирусных векторов сконструированы таким образом, что трансген замещает гены Ad E1a, E1b и/или E3; впоследствии, вектор с дефектом репликации размножают в 293 клетках человека, которые обеспечили функцию удаленного гена в векторах trans. Векторы Ad могут трансдуцировать различные типы тканей in vivo, в том числе неделящиеся, дифференцированные клетки, такие как обнаруженные в печени, почке или мышце. Обычные Ad векторы имеют большой переносимый объем. Пример применения векторa Ad в клинических испытаниях включал полинуклеотидную терапию для противоопухолевой иммунизации с внутримышечным введением (Sterman et al., Hum. Gene Ther. 7:1083-9 (1998)). Дополнительные примеры применения аденовирусных векторов для передачи гена в клинических испытаниях включают Rosenecker et al., Infection 24:1 5-10 (1996); Sterman et al., Hum. Gene Ther. 9:7 1083-1089 (1998); Welsh et al., Hum. Gene Ther. 2:205-18 (1995); Alvarez et al., Hum. Gene Ther. 5:597-613 (1997); Topf et al., Gene Ther. 5:507-513 (1998); Sterman et al., Hum. Gene Ther. 7:1083-1089 (1998).
[0184] Упаковывающие клетки применяют для образования вирусных частиц, которые способны заражать клетку-хозяина. Подобные клетки включают 293 клетки, которые упаковывают аденовирус, и клетки cells2 или клетки PA317, которые упаковывают ретровирус. Вирусные векторы, применяемые в генной терапии, обычно генерируют с помощью клеточной линии-продуцента, которая упаковывает вектор нуклеиновой кислоты в вирусную частицу. Векторы обычно содержат минимальные вирусные последовательности, требуемые для упаковки и последующей интеграции в хозяина (если это применимо), другие вирусные последовательности заменяются экспрессией кассеты, которая кодирует белок, подлежащий экспрессированию. Недостающие вирусные функции поставляют в trans с помощью упаковки клеточной линии. Например, векторы AAV, применяемые в генной терапии, обычно имеют только инвертированные концевые повторяющиеся (ITR) последовательности из генома AAV, который требуют для упаковки и интеграции в геном-хозяин. Вирусную ДНК упаковывают в клеточную линию, которая содержит плазмиду-хелпер, который кодирует другие гены AAV, называемые rep и cap, но отсутствуют последовательности ITR. Клеточную линию также инфектируют с помощью аденовируса в виде хелпера. Вирус-хелпер промотирует репликацию вектора AAV и экспрессию генов AAV из плазмиды-хелпера. Плазмида-хелпер не упакована в значительных количествах из-за отсутствия последовательностей ITR. Загрязнение аденовирусом может быть уменьшено, например, тепловой обработкой, к которой аденовирус более чувствителен, чем AAV.
[0185] Во многих применениях генной терапии желательно, чтобы генотерапевтический вектор доставлялся с высокой степенью специфичности к конкретному типу ткани. Соответственно, вирусный вектор может быть модифицированным, чтобы иметь специфичность по отношению к данному типу клеток путем экспрессии лиганда в виде слитого белка с вирусным белком оболочки на внешней поверхности вируса. Выбранный лиганд обладает аффинностью к рецептору, о котором известно, что он присутствует в интересующем типе клеток. Например, Han et al., Proc. Natl. Acad. Sci. USA 92:9747-9751 (1995), сообщили, что вирус мышиного лейкоза Молони может быть модифицирован для экспрессии герегулина человека, слитого с gp70, а рекомбинантный вирус инфицирует некоторые клетки рака молочной железы человека, экспрессируя рецептор эпидермального фактора роста человека. Этот принцип может быть распространен на другие пары вирус - клетка-мишень, в которых клетка-мишень экспрессирует рецептор, а вирус экспрессирует слитый белок, содержащий лиганд для рецептора клеточной поверхности. Например, нитчатый фаг может быть сконструирован для того, чтобы отображать фрагменты антитела (например, FAB или Fv), имеющие специфическую связывающую аффинность практически для любого выбранного клеточного рецептора. Хотя вышеприведенное описание применяют преимущественно по отношению к вирусным векторам, эти же принципы могут применяться и к невирусным атакам. Подобные векторы могут быть сконструированы для того, чтобы содержать специфические последовательности поглощения, которые способствуют поглощению конкретными клетками-мишенями.
[0186] Генотерапевтические векторы могут быть доставлены in vivo путем введения отдельному пациенту, обычно с помощью систематического введения (например, внутривенной, внутрибрюшинной, внутримышечной, подкожной или внутричерепной инфузии) или местного применение, как описано ниже. В качестве альтернативы векторы могут быть доставлены в клетки ex vivo, такие как эксплантированные от отдельного пациента клетки (например, лимфоциты, пунктаты костного мозга, биопсия ткани) или гематопоэтические стволовые клетки универсального донора, после реимплантации клеток пациенту, обычно после селекции клеток, которые имеют встроенный вектор.
[0187] Векторы (например, ретровирусы, аденовирусы, липосомы и т. д.), содержащие нуклеазы и/или донорные конструкции, также могут быть введены в организм направленно для трансдукции клеток in vivo. В качестве альтернативы может быть введена открытая ДНК. Введение осуществляют любым из путей, обычно применяемых для введения молекулы в максимальный контакт с клетками крови или ткани, в том числе, но без ограничения этим, инъекцией, инфузией, местным применением и электропорацией. Подходящие способы введения подходящих нуклеиновых кислот являются доступными и хорошо известно специалистам в данной области техники, и хотя для осуществления введения конкретной композиции может быть применен более чем один путь, конкретный путь может часто обеспечивать более оперативную и более эффективную реакцию, чем другой путь.
[0188] Векторы, подходящие для введения полинуклеотидов, описанных в данном документе, включают неинтегрирующие лентивирус векторы (IDLV). Смотри, например, Ory et al. (1996) Proc. Natl. Acad. Sci. USA 93:11382-11388; Dull et al. (1998) J. Virol. 72:8463-8471; Zuffery et al. (1998) J. Virol. 72:9873-9880; Follenzi et al. (2000) Nature Genetics 25:217-222; публикацию патента США № 2009/054985.
[0189] Фармацевтически приемлемые носители частично определяют с помощью конкретной вводимой композиции, а также с помощью конкретного способа, применяемого для осуществления введения композиции. Соответственно, существует большое разнообразие подходящих составов доступных фармацевтических композиций, как описано ниже (смотри, например, Remington's Pharmaceutical Sciences, 17th ed., 1989).
[0190] Будет очевидно, что кодирующие нуклеазы последовательности и донорные конструкции могут быть доставлены с применением одних тех же или отличных систем. Например, донорный полинуклеотид может быть перенесен с помощью плазмиды, тогда как одна или более нуклеаз могут быть перенесены с помощью вектора AAV. Более того, с помощью того же или отличных путей могут быть введены различные векторы (внутримышечной инъекции, инъекции в хвостовую вену, другой внутривенной инъекции, внутрибрюшинного введения и/или внутримышечной инъекции. Векторы могут быть доставлены одновременно или в любом последовательном порядке.
[0191] Составы для и ex vivo, и in vivo введений включают суспензии в жидкости или эмульгированных жидкостях. Активные ингредиенты часто смешивают с эксципиентом, которые являются фармацевтически приемлемыми и совместимыми с активным ингредиентом. Подходящие эксципиенты включают, например, воду, физиологический раствор, декстрозу, глицерин, этанол или подобное и их комбинации. Кроме того, композиция может содержать минорные количества вспомогательных веществ, таких как смачивающие или эмульгирующие средства, рН-буферные средства, стабилизирующие средства или другие средства, которые усиливают эффективность фармацевтической композиции.
Применения
[0192] Способы по данному изобретению рассматривают лечение и/или профилактику болезни Фабри (например, лизосомной болезни накопления). Лечение может включать вставку корректирующего связанного с болезнью трансгена GLA в локус «safe harbor» (например, альбумин) в клетке для экспрессии требуемого фермента и высвобождения в ток крови. Корректирующая α-GalA, которая кодирует трансген, может кодировать белок дикого типа или модифицированный белок; и/или может содержать кодон-оптимизированный трансген GLA; и/или трансген, в котором эпитопы могут быть удалены без функционально измененного белка. В некоторых случаях, способы включают вставку эписомы, экспрессирующей α-GalA, которая кодирует трансген в клетку для экспрессии требуемого фермента и высвобождения в ток крови. Вставка в секреторную клетку, такую как клетка печени, для высвобождения продукта в ток крови, является особенно полезным. Способы и композиции по изобретению также могут быть применены в любых обстоятельствах, когда это является желательным для обеспечения трансгена GLA, который кодирует одно или более терапевтических средств в гемопоэтической стволовой клетке таким образом, что зрелые клетки (например, RBC), получаемые из (происходят из) этих клеток, содержат терапевтический белок α-GalA. Эти стволовые клетки могут быть дифференцированы in vitro или in vivo и могут быть получены от универсального донорского типа клеток, которые могут быть применены для всех пациентов. Дополнительно, для направления миграции клеток в теле клетки могут содержать трансмембранный белок. Лечение может также содержать применение клеток пациента, содержащих терапевтический трансген, где клетки размножают ex vivo и затем вводят обратно пациенту. Например, HSC, содержащая подходящую α-GalA, которая кодирует трансген, может быть вставлена пациенту посредством трансплантата костного мозга. В качестве альтернативы, стволовые клетки, такие как стволовые клетки мышц или iPSC, которые были отредактированы с применением α-GalA, которая кодирует трансген, также могут быть инъектированы в мышечную ткань.
[0193] Таким образом, эта технология может быть применена при состоянии, когда у пациента присутствует дефицит какого-либо белка, вызванный проблемами (например, проблемами в уровне экспрессии или проблемами с белком, экспрессированным в виде недо- или не-функционирования). Особо полезным по данному изобретению является экспрессия трансгенов для корректировки или восстановления функциональности у субъектов с болезнью Фабри.
[0194] В качестве не ограничивающих примеров различные способы продуцирования функционального белка α-Gal A для замены дефективного или недостающего белка α-Gal A совершенствуют и применяют для лечения болезнь Фабри. Доноры нуклеиновой кислоты, которые кодируют белки, могут быть вставлены в локус «safe harbor» (например, альбумин или HPRT) и экспрессированы либо с применением экзогенного промотора или с применением промотора, присутствующего в «safe harbor». Особенно полезным является вставка трансгена GLA в локусе альбумина в клетке печени, где трансген GLA дополнительно включает последовательности, кодирующие сигнальный пептид, который опосредует секрецию экспрессированного белка α-Gal A из клетки печени в ток крови. В качестве альтернативы доноры могут быть применены для корректирования дефективного гена in situ. Желаемая α-GalA, которая кодирует трансген, может быть вставлена в CD34+ стволовую клетку и возвращена пациенту во время трансплантации костного мозга. В заключение донор нуклеиновой кислоты может быть вставлен в CD34+ стволовую клетку в бета-локусе глобина таким образом, что зрелая красная кровяная клетка, получаемая из этой клетки, имеет высокую концентрация биологически кодируемого нуклеиновой кислоты донора. Содержащая биологические препараты RBC может затем быть нацелена на правильную ткань посредством трансмембранных белков (например, рецептора или антитела). Дополнительно, RBC могут быть сенсибилизированы ex vivo посредством электросенсибилизации, чтобы сделать их более восприимчивыми к разрушению после воздействия источника энергии (смотри WO2002007752).
[0195] В некоторых применениях эндогенный ген может быть инактивирован путем применения способов и композиций по изобретению. Примеры этого аспекта включают инактивирование регулятора аберрантного гена или аберрантного связанного с болезнью гена. В некоторых применениях аберрантный эндогенный ген может быть замещен, либо функционально, либо in situ, с версией гена дикого типа. Вставленный ген может также быть изменен для улучшения экспрессии терапевтического белка α-GalA или для снижения его иммуногенности. В некоторых применениях, вставленная α-GalA, которая кодирует трансген, представляет собой слитый белок для повышения его транспорта в выбранную ткань, такую как мозг.
[0196] Следующие Примеры относятся к приводимым в качестве примера вариантам осуществления настоящего изобретения, в которых нуклеаза содержит цинк-пальцевую нуклеазу (ZFN) (или пару ZFN) или TALEN (или пару TALEN). Следует принять во внимание, что это только для целей иллюстративного примера и что могут быть применены другие нуклеазы или нуклеазные системы, в качестве примера, хоуминг-эндонуклеазы (мегануклеазы) со сконструированными ДНК-связывающими доменами и/или слияния встречающихся в природе ДНК-связывающих доменов сконструированных хоуминг-эндонуклеаз (мегануклеаз) и гетерологичных доменов расщепления и/или система CRISPR/Cas, содержащая сконструированную одиночную направляющую РНК. Подобным образом, следует принимать во внимание, что подходящие доноры GLA не ограничиваются приведенными ниже примерами, а включают любой трансген GLA.
ПРИМЕРЫ
Пример 1. Дизайн и конструирование α-GalA, которая кодирует трансгены
[0197] Для экспрессии трансгенов GLA принимали два подхода. Один подход, называемый In Vivo Protein Replacement Platform® («IVPRP»), применяет сконструированные нуклеазы для вставки трансгена в локус альбумина таким образом, что экспрессией управляли с помощью промотора альбумин (смотри, патенты США №№ 9394545 и 9150847). Второй подход включает трансдукцию клетки с AAV, содержащим копию кДНК трансгена, где кДНК дополнительно включает промотор и другие регуляторные последовательности. Дизайны кассета экспрессии трансгена GLA для этих двух подходов проиллюстрированы на Фигуре 1.
Пример 2. Способы
Трансдукция клетки HepG2/C3a и K562
[0198] Клетки HepG2 трансдуцируют с применением стандартных методик и в кДНК, и IVPRP® системах.
A. кДНК
[0199] Подход на основе кДНК может включать применение доставляемой с помощью AAV экспрессирующей конструкции, содержащей энхансер APOE, связанный с промотором hAAT (Okuyama et al (1996) Hum Gene Ther 7(5):637-45), интрон HBB-IGG (химерный интрон, состоящий из 5´-донорного сайта из первого интрона бета-гена глобина человека и ветвь и 3´-акцепторный сайт из интрона вариабельной области тяжелой цепи гена иммуноглобулина), сигнальный пептид, кодирующую последовательность (причем кодирующая последовательность является необязательно кодон-оптимизированной) и poly A сигнальную последовательность гормона роста крупного рогатого скота (bGH).
[0200] Для кДНК систем клетки HepG2 трансдуцировали с помощью векторов кДНК GLA AAV, как описано в данном документе, а супернатант собирали и исследовали на активность α-Gal A. Кроме того, клетки K562 культивировали в супернатанте, собранном из трансдуцированных клеток HepG2, в отсутствие и присутствие избытка манноза-6 фосфата (M6P, 5 мM), который насыщает рецепторы M6P на поверхности клетки и блокирует поглощение α-Gal A. Клеточные осадки собирали и исследовали на активность α-Gal A.
B. IVPRP®
[0201] Существуют три компонента для IVPRP® при болезни Фабри: два вектора rAAV2/6, которые кодируют SBS 47171 и SBS 47898 ZFN, созданные расщеплять специфический локус в интроне 1 альбумина человека, и один вектор rAAV2/6, который кодирует донорный шаблон hGLA. Донорный шаблон hGLA представляет собой кодон-оптимизированную версию кДНК hGLA, фланкированную плечами гомологичности для облегчения интеграции репарации, направляемой гомологией, (HDR) донора в альбумин человека.
[0202] Клетки HepG2/C3A (также упоминаются как клетки «HepG2») (ATCC, CRL 10741) содержали в минимальной поддерживающей среде (MEM) с солями Эрла и L-глутамином (Corning) с 10% фетальной бычьей сывороткой (FBS) (Life Technologies) и 1X пенициллином/стрептомицином/глутамином (Life Technologies) и инкубировали при 37°C и 5% CO2. Клетки перевивали каждые 3 4 дня.
[0203] Для трансдукции IVPRP®, клетки промывали и трипсинизировали с помощью 0,25% трипсина/2,21 мМ EDTA (Corning) и ресуспендировали в среде для выращивания клеток. Малые аликвоты перемешивали 1:1 с раствором трипанового синего 0,4% (масс./об.) в забуференном фосфатом физиологическим раствором (PBS; Corning) и проводили подсчет на Automated Cell Counter TC20 (Bio Rad). Клетки ресуспендировали при плотности 2e5 на мл в среде для выращивания клеток и высевали в 24-луночный планшет (Corning) при 1e5 в 0,5 мл среды на лунку. Рекомбинантные частицы AAV2/6 перемешивали при соответствующей множественности заражения (MOI) со средой для выращивания клеток и добавляли в клетки.
[0204] Клетки HepG2 трансдуцировали с помощью либо только донора hGLA (в двух экземплярах; контроль), либо с двумя SB 47171 и SB 47931 ZFN hALB плюс донор IDS SB (в трех экземплярах). MOI для донора только трансдукции составляла 6e5 геномов вектора (гв)/клетку. MOI для трансдукции ZFN+донор составляла 3e5 гв/клетку для каждой ZFN и 6e5 гв/клетку для донора hGLA. Это представляет соотношение ZFN1:ZFN2:донор 1:1:2, которое было ранее определено в качестве оптимального соотношения для in vitro экспериментов. Донор hGLA добавляли через 24 часа после векторов ZFN, чтобы максимизировать эффективность трансдукции in vitro.
[0205] После трансдукции клетки возвращали в культуру на 6-10 дней. Супернатант собирали в Дни 3, 5, 7 и 10 (где это применимо) и заменяли на свежую среду. После заключительного этапа сбора супернатанта, клетки трипсинизировали и ресуспендировали, как описано выше, затем центрифугировали для создания осадок клеток, промывали с помощью PBS и хранили при -80C.
[0206] Подобный способ применяли для трансдуцирования клеток HepG2 с конструкциями кДНК GLA. MOI для конструкции кДНК GLA составляла либо 3e4, 1e5, 3e5, либо 1e6 гв/клетку.
Анализ активности α-GalA
[0207] Активность α-GalA анализировали во флуориметрическом анализе с применением синтетического субстрата 4-метилумбеллиферил-α-D-галактопиранозида (4MU-α-Gal, Sigma).
[0208] Вкратце, 10 микролитров супернатанта клеточной культуры HepG2 перемешивали с 40 мкл 5 мМ 4MU-α-Gal, растворенного в фосфатном буфере (0,1 M цитрата/0,2 M фосфатного буфера, pH 4,6, 1% Triton X-100). Реакции инкубировали при 37°C и останавливали путем добавления 100 мкл 0,5 M глицинового буфера, pH 10,3. Высвобождение 4-метилумбеллиферона (4MU) определяли путем измерения флуоресценции (Ex365/Em450) с применением флуоресцентного ридера SpectraMax Gemini XS (Molecular Devices, Саннивейл, Калифорния).
[0209] Стандартную кривую получали с использованием серийных 2-кратных разведений 4MU. Получаемые данные подгоняли под кривую в двойном логарифмическом масштабе; концентрация 4MU в исследуемых образцах рассчитывали с применением наиболее лучшим образом подогнанной модели. Ферментативную активность выражали в виде нмоль 4MU, высвобождаемых за час времени инкубации для анализа, на мл супернатанта клеточной культуры (нмоль/ч/мл).
Определение Gb3
Количественное определение и анализ Gb3 и субстрата Lyso-Gb3:
[0210] Субстрат Фабри глоботриаозилцерамида (Gb3) измеряли в выбранных клетках плазмы и тканях посредством масс-спектрометрии. Вкратце, ткани взвешивали и механически разрушали в жидкости для разрушения тканей (5% MeOH, 95% воды и 0,1% уксусной кислоты) при соотношении 5 мл жидкости на мг ткани. 10 мкл плазмы или суспензии ткани затем добавляли к 90 мкл раствора для преципитации (MeOH с внутренним стандартом N-трикозаноилцерамидтригексозида (C23:0, Matreya) вносили в раствор) в силиконизированной пробирке, перемешивали вихревым способом и помещали на качающуюся тарелку при комнатной температуре в течение 30 минут. Образец затем центрифугировали и добавляли 10 мкл образца к 90 мкл одиночного холостого образца (DMSO/MeOH 1:1+0,1% FA) в стеклянной пробирке для ЖХ-МС. Образец анализировали на длину цепи Gb3 24:0, предоминантные виды Gb3, присутствующие у мышей GLAKO, и измеряли против стандартной кривой, состоящей из тригексозида керамида (Gb3, Matreya).
[0211] Глоботриаозилсфингозин (lyso-Gb3) измеряли подобным образом с применением глюкозилсфингозина (Matreya) в качестве внутреннего стандарта и лизо-керамидтригексозида (lyso-Gb3, Matreya) для создания стандартной кривой.
Оценка генной модификации (% инсерционно-делеционных мутаций)
[0212] Сайт-мишень ZFN подвергали анализу последовательности с применением системы MiSeq (Illumina, Сан-Диего, Калифорния) Пару олигонуклеотидных праймеров создавали для амплификации фрагмента из 194 п.о., перекрывая сайт-мишень ZFN в локусе альбумина человека или локусе альбумина мыши, и для введения связывающих последовательностей для второго раунда амплификации. Продукты этой PCR амплификации очищали и подвергали второму раунду PCR с олигонуклеотидами, созданными для введения ампликон-специфической последовательности идентификатора («штрих-код»), а также концевых областей, предназначенных для связывания секвенирующих олигонуклеотидных праймеров. Затем смешанную популяцию ампликонов со штрих-кодом подвергали анализу MiSeq, процедуре твердофазного секвенирования, которая позволяет проводить параллельный анализ тысяч образцов на одном аналитическом чипе.
In vivo исследование IVPRP® с болезнью Фабри и векторов кДНК в модели мышей GLAKO
[0213] Для демонстрирования эффективности этих терапевтических средств в животной модели болезни Фабри, мышей GLAKO трансдуцировали с помощью такой же конструкции кДНК GLA AAV2/6, применяемой в клетках HepG2. Других мышей GLAKO трансдуцировали с помощью мышиной версии IVPRP при болезни Фабри, которая состоит из двух векторов rAAV2/8, которые кодируют SB-48641 и SB-31523 ZFN, созданные для расщепления альбумина мыши, и один вектор rAAV2/8, который кодирует донорный шаблон кДНК hGLA с плечами гомологичности мыши. В качестве контролей, дополнительным мышам GLAKO и мышам дикого типа инъектировали с помощью буфера для состава с вектором AAV (PBS, 35 нM NaCl, 1% сахарозы, 0,05% плюрона) F-68, pH 7,1), не содержащим частицы вектора. Животные получали 50 мг/кг циклофосфамида каждые две недели, начиная со дня перед инъекцией AAV. Все мыши были в возрасте 4-12 недель во время инъекции. Мышей держали под контролем в течение 2-3 месяцев, с плазмой, которую брали еженедельно или раз в две недели посредством подчелюстной пункции для измерения активности α-GalA в плазме. Мышей подвергали эвтаназии в конце эксперимента и измеряли активность α-GalA в плазме, печени, почке, сердце и селезенке, как описано выше. Уровни субстрат Gb3 и lyso-Gb3 измеряли в плазме, печени, почке, сердце и селезенке посредством масс-спектрометрии. У мышей, которые получали лечение с помощью IVPRP при болезни Фабри, инсерционно-делеционные мутации в ткани печени измеряли посредством MiSeq, как описано выше.
Процедуры вестерн-блоттинга и дегликозилирования:
[0214] Печени мышей гомогенизировали в 0,1 M цитрата/0,2 M фосфатного буфера, pH 4,6. Гомогенаты печени кипятили в течение 10 минут, затем аликвоты каждого образца дегликозилировали путем обработки с помощью PNGase F (New England Biolabs, NEB) в течение 1 часа, в соответствии с протоколом NEB.
[0215] 1 мкг общего белка загружали на 4-12% гель Bis-Tris Midi Gel NuPage (Invitrogen). 0,5 нг рекомбинантного GLA человека, который загружали (R&D Systems) перед и после лечения с помощью PNGase F, включали в качестве ссылки на размер.
[0216] Антитела, которые применяли для вестерн-блоттинг, представляли собой: Первичное антитело: α-GLA, моноклональное антитело кролика Sino Biological, 1:1000; Вторичное антитело: α-кроличий IgG-HRP козла, Thermo Fisher, 1:10000.
Пример 3. Экспрессия трансгена GLA in vitro
[0217] Подход на основе IVPRP®: Способы описаны выше в Примере 2. Вкратце, клетки HepG2/C3a трансдуцировали с помощью ZFN AAV2/6 и донорных векторов hGLA в дозе 100 тыс. гв/клетку для каждого ZFN и 200 тыс. гв/клетку для донора GLA или дозы 300 тыс./600 тыс. для ZFN и донорных, соответственно.
[0218] Как проиллюстрировано на Фигуре 2, трансдуцированные клетки имели повышенную активность α-GalA в супернатанте и клеточных осадках, и активность достигала 3-кратных уровней трансдуцированных макетом HepG2 в группах ZFN+донора. Инсерционно-делеционные мутации в локусе альбумина, измерение активности ZFN, измеряли при каждой дозе вектора для конструкций A и B донора GLA. Инсерционно-делеционные мутации в донорах A и B составляли 43,46% и 39,81% для дозы вектора 300/600 и 8,81% и 9,69% для дозы вектора 100/200.
[0219] Подход на основе кДНК: конструкцию кДНК, проиллюстрированную на Фигуре 1B, также исследовали в клетках HepG2/G3, как описано выше. Как проиллюстрировано на Фигуре 3, клетки HepG2/C3a, трансдуцированные с помощью векторов кДНК GLA AAV2/6, имели дозозависимую повышенную активность α-GalA в супернатанте и клеточных осадках. Каждую дозу отмечали на Фигуре 3 и указывали тысячи (тыс.) копий вирусного вектора на клетку. Активность супернатанта α-GalA достигала 200-кратных макетных уровней при высоких дозах кДНК.
[0220] Белки могут быть выделены и введены субъекту с помощью ферментозаместительных терапий.
Пример 4. In vivo исследование двух подходов
[0221] Далее два типа подходов (на основе кДНК и на основе IVPRP®) исследовали in vivo. Конструкции упаковывали в AAV 2/6 или AAV 2/8 и затем инъектировали внутривенно нокаутным по GLA (GLAKO) мышам. Это мышиная модель болезни Фабри (Bangari et al (2015) Am J Pathol. 185(3):651-65). Исследуемые препараты проиллюстрированы ниже (Таблица 2) вместе с режимами дозирования (Таблица 3).
Таблица 2: Исследуемые препараты для подходов на основе IVPRP® и на основе кДНК
Таблица 3: Режимы дозирования для in vivo исследования подходов IVPRP® и кДНК
*Животные, получавшие дозу на основе гв/мышь. При том условии, что навешивали 0,020 кг тела для всех мышей, уровень дозы общего AAV составляет 7,5e13 гв/кг для животных, получающих ZFN+донор
Подход на основе кДНК:
[0222] Как проиллюстрировано на Фигуре 4, мыши GLAKO из групп, получавших лечение с помощью кДНК, показывали супрафизиологическую активность α-GalA в плазме уже на 7 день после введения AAV. Показанное на фигурах представляет собой результаты от отдельных мышей. Активность α-GalA в плазме измеряли еженедельно и высокие, дозозависимые уровни активности поддерживали на протяжении всей продолжительности исследования. Активность в плазме достигала 6-кратного уровня дикого типа в группе низкой дозе (2,0e12 гв/кг) и 280-кратного уровня дикого типа в группе высокой дозы (2,0e13 гв/кг). Мышей подвергали эвтаназии спустя два месяца и анализировали на активность α-GalA и накопление Gb3 в печени и вторичных, дистальных тканях.
[0223] Как проиллюстрировано на Фигуре 5, дозозависимое повышение активности α-GalA обнаруживали в печени, сердце и почках, наряду с соответствующим снижением содержания субстрата Gb3. Gb3 не обнаруживался в тканях некоторых мышей GLAKO, которым вводили высокую дозу кДНК AAV2/6. Данные также анализировали с точки зрения объема выведения субстратов по сравнению с не получавшими лечение мышами GLAKO (Фигура 5E и Фигура 5F) и продемонстрировали, что мыши, получавшие лечение с помощью высокой дозы кДНК, имели в среднем менее 10% субстрата, который обнаруживали у не получавших лечение мышей GLAKO.
Подход на основе IVPRP®:
[0224] Уровни в плазме брали для мышей GALKO, которые получали дозу в соответствии с подходом на основе IVPRP® за период времени в 90 дней. Данные (Фигура 6A) показывают, что активность белка α-Glа обнаруживали в сыворотке на уровне приблизительно в 25-30% от того, что видели у мышей дикого типа. В этом эксперименте одна группа клеток назначали умеренный режим иммуносупрессии (50 мг/кг циклофосфамида каждые 2 недели). Измерение активности ZFN в печени (инсерционно-делеционные мутации) обнаруживало, что получавшие лечение животные с умеренным режимом иммуносупрессии имели немного более высокий уровень инсерционно-делеционных мутаций (Фигура 6B), но обе группы имели ожидаемый диапазон инсерционно-делеционных мутаций.
[0225] Второй эксперимент осуществляли с применением значительно более жесткой иммуносупрессии (дозирование показано ниже в Таблице 4) и данные (Фигуры 6C, 6D, и 6E) продемонстрировали, что иммуносупрессия значительно не увеличивала активность белка α-Gal A.
Таблица 4: IVPRP® In vivo исследование №2, титрование иммуносупрессии
каждые 2 недели
каждые 2 недели
каждые 2 недели
[0226] Считается, что α-Gal A подвержен инактивации из-за неправильного сворачивания, поскольку некоторые мутации, которые являются дистальными по отношению к активному участку белка, приводят к болезни Фабри (Garman and Garboczi (2004) J Mol Biol 337(2):319-35), и что применение молекулярных шаперонов, в том числе дезоксигалактоноиримицина (DGJ), было предложено для применения с некоторыми мутантами GLA (Moise et al (2016) J. Am. Soc. Mass Spectrom 27(6): 1071-8). Таким образом, в описанном выше исследование, добавляли DGJ приблизительно в день 30-35. В частности, 3мг/кг, разведенные в 200 мкл воды, давали через желудочный зонд ежедневно. Быстрый рост активности α-Gal A обнаруживали у получавших лечение с помощью DGJ животных (Фигура 7).
[0227] Ткани от животных в этом исследовании также проверяли на активность α-GLA, как описано выше. Результаты (Фигура 8) демонстрировали, что активность может быть обнаружена в тканях, в частности, в печени и селезенке. Во всех тканях, активность, обнаруженная в случае подхода на основе кДНК с высокой дозой, была выше, чем для мышей дикого типа.
[0228] Уровни первичного субстрата α-Gal A также измеряли в плазме, ткани печени и сердца, как описано выше. Данные (Фигура 9) показывали уменьшение обнаруживаемого в плазме Gb3 для образцов IVPRP®, и не обнаруживаемого Gb3 для образцов кДНК (эквивалентно мышам дикого типа). Для ткани печени и сердца, образцы IVPRP® также показывали уменьшение обнаруживаемого Gb3, что также справедливо для образцов с низкой дозой кДНК. Для образцов с высокой дозой кДНК, уровни были почти такими же, как у образцов дикого типа.
[0229] Эти результаты иллюстрируют то, что внедрение трансгена GLA с помощью подхода либо на основе кДНК, либо на основе IVPRP®, как описано в данном документе, обеспечивает терапевтические эффекты in vivo.
Пример 5. Оптимизация дизайна донора IVPRP®
[0230] Дизайн донора также исследовали для подхода на основе IVPRP® с целью оптимизации дизайна кодирующей GLA области и с целью оптимизации сигнального пептида. Для начала, дизайн донора изменяли для введения сигнального пептида α-Gal A (GLA) (последовательность: MQLRNPELHLGCALALRFLALVSWDIPGARA, SEQ ID NO:1) перед кодирующей GLA последовательностью и вставляли последовательность Kozak (последовательность: GCCACCATG, SEQ ID NO:2) перед сигнальным пептидом α-Gal A для провоцирования нового события трансляции, отдельно от сигнального пептида альбумина (смотри Фигуру 10, примеры представляют собой Вариант №A, Вариант №B). Кроме того, анализировали применение альтернативного сигнального пептида IDS (последовательность: MPPPRTGRGLLWLGLVLSSVCVALG, SEQ ID NO:3) (Фигура 10, Вариант №H), в том числе и применение 2A-подобной последовательности от T. asigna («T2A») (Luke et al (2008) J Gen Virol. 89(Pt 4): 1036-1042) для удаления последовательности 5' сигнального пептида во время трансляции. Новые конструкции исследовали в клетках HepG2/C3A, как описано ранее.
[0231] Результаты показывали, что Вариант №A и Вариант №B имели намного более высокие уровни активности α-GalA, чем исходного донора (Фигура 11A) in vitro. Кроме того, Вариант K демонстрировал даже более высокие уровни активности α-GalA по сравнению с Вариантом A или исходным донором (Фигура 11B).
[0232] Конструкции затем исследовали in vivo у мышей GLAKO с применением протокола дозирования, приведенного ниже в Таблице 5.
Таблица 5: In vivo исследование дизайна доноров IVPRP® у мышей GLAKO
+GLAsp
+Kozak, +GLAsp
+T2A, +GLAsp
[0233] Плазму брали один раз в неделю для измерения активности α-Gal A, как описано выше. Активность обнаруживали во всех образцах у каждой мыши, с новыми дизайнами, демонстрирующими улучшение по сравнению с исходным донором (Фигура 12), и уровни были по меньшей мере в 40 раз выше, чем у дикого типа на 28 день (указано пунктирной линией). Образцы с течением времени показали увеличение, где активность измеряли при 80-кратном уровне дикого типа для Группы 2 (Донор №A) и 50-кратном уровне дикого типа для Группы 4 (Донор №E). Образцы ткани брали у мышей и измеряли уровни Gb3 и обнаруживали, что они снижены по сравнению с не получавшими лечение мышами GLAKO.
[0234] Эксперимент, описанный выше, осуществляли в течение 56 дней, после чего животных умерщвляли и анализировали на активность α-Gal A в печени, сердце, почке и селезенке. Детальные данные (Фигура 15) демонстрируют, что этот подход приводит к увеличению активности α-Gal A в исследованных тканях, в том числе 100-кратное повышение активности α-Gal A в плазме получавших лечение животных по сравнению с плазмой животных дикого типа, 9-кратное повышение активности α-Gal A в сердце получавших лечение животных по сравнению с сердцами животных дикого типа (не получавших лечение) и 80% повышение активности α-Gal A в почках получавших лечение животных по сравнению с не получавшими лечение (дикого типа) животными.
[0235] Анализ ткани затем проводили для определения уровней гликолипидного субстрата α-Gal As (Gb3 (Фигура 16A) и lyso-Gb3 (Фигура 16B)) в различных тканях (плазме, печени, сердце и почке) после лечения. Как проиллюстрировано на Фигуре 16, лечение, как описано в данном документе, приводили к сниженным уровням обоих субстратов (Gb3 и lyso-Gb3) во всех исследованных тканях (плазме, печени, сердце и почке) у получавших лечение животных с вариантами A, B или E по сравнению с таковыми перед лечением (первичным) и у не получавшие лечение животных (дикого типа), демонстрируя то, что композиции и способы, описанные в данном документе, обеспечивают терапевтически эффективные уровни белка in vivo.
[0236] Эксперименты повторяли таким образом, как описано выше, для анализа активности α-Gal A в плазме и в различных тканях (печень, ухе, почке и селезенке) после введения Варианта E и Варианта J (смотри Фигуру 10) с альбумин-нацеленными ZFN. Как проиллюстрировано на Фигурах 20 и 21, активность α-Gal A в плазме (Фигура 20A) и в печени, сердце, почке и селезенке (Фигура 20B) животных, получающих донор Варианта J, продуцировали активность α-Gal A в плазме около 300-кратного уровня дикого типа и 10-100-кратную или более активность α-Gal A в ткани относительно уровня в печени, сердце и селезенке дикого типа.
[0237] Концентрации гликолипидных субстратов α-Gal A (Gb3 (Фигура 21A) и lyso-Gb3 (Фигура 21B)) в различных тканях (плазме, печени, сердце и почке) после лечения измеряли таким образом, как описано в данном документе. Как проиллюстрировано на Фигуре 21, экспрессия Варианта J значительно снижала уровни субстрата.
Пример 6. Оптимизация дизайна кассеты трансгена GLA для подхода на основе кДНК
[0238] Кассету трансгена GLA для подхода на основе кДНК также оптимизировали. Трансген связывали с последовательностями, которые кодируют различные сигнальные пептиды, в том числе пептид α-Gal A, сигнальный пептид для гена IDS (идуронат-2-сульфатазы), ген FIX (Фактор IX, (последовательность: MQRVNMIMAESPGLITICLLGYLLSAEC, SEQ ID NO:4)) и сигнальные пептиды (Фигура 13B) альбумина (последовательность: MKWVTFISLLFLFSSAYS, SEQ ID NO:5)
. Кроме того, трансген GLA вставляли в альтернативный оптимизированный вектор экспрессии кДНК (Фигура 13A, также публикация патента США № 20170119906). Все конструкции исследовали, как описано выше, в клетках HepG2/C3A in vitro в дозах от 30 до 600 тысяч (тыс.) копий вирусного вектора на клетку и указывали, что лидерные последовательности IDS и FIX (F9) приводят к более высокой активности α-GalA, чем применение лидерных последовательностей GLA или ALB (альбумин) (Фигура 14A). Данные для вариантов №4, №5 и №6 кДНК (Фигура 13) иллюстрировали на Фигуре 14B.
[0239] Конструкции также исследовали у GLAKO мышей таким образом, как описано выше, и они являются активными in vivo.
Пример 7. Анализ белка α-Gal A с помощью вестерн-блоттинга и дегликозилирования
[0240] Плазму от мышей, получавших лечение с помощью либо подхода на основе IVPRP®, либо подхода кДНК, анализировали на присутствие белка α-GalA, как описано в Примере 2. Дополнительно, образцы также обрабатывали с помощью PNGaseF для вызывания дегликозилирования.
[0241] Как проиллюстрировано на Фигуре 17, белок α-GalA получали in vivo у мышей GLAKO после либо IVPRP® (Фигура 17A, отображающая результаты для Варианта A и Варианта J), либо лечения с помощью исходной конструкции кДНК (конструкции, изображенной на Фигуре 13B, данные проиллюстрированный на Фигуре 17B) ведет себя аналогично рекомбинантному белку hGalA, указывая на то, что композиция и способы, описанные в настоящем документе, обеспечивают белки на клинически релевантных уровнях, а именно терапевтических уровнях, подобных таковым для рекомбинантных терапевтических белков, которые применяют в настоящее время в ферментативных заместительнях терапиях.
Пример 8. In vitro продуцирование белка после введения кДНК
[0242] Клетки Hep2G трансдуцировали с помощью Вариант №4 кДНК GLA AAV и собирали супернатант спустя 5 дней и исследовали на активность α-Gal A, а супернатант применяли на культуре клеток K562, как описано в Примере 2.
[0243] Как проиллюстрировано на Фигуре 28A, супернатант собирали через 5 дней после трансдукции клеток HepG2 с Вариантом №4 кДНК GLA AAV, демонстрирующим высокие количества активности α-Gal A. Фигура 28B проиллюстрировано, что α-Gal A от супернатанта HepG2 поглощали с помощью K562 в пределах 24 часов и что поглощение блокировали с помощью M6P.
[0244] Следовательно, клетки, как описано в данном документе, продуцируют и секретируют α-Gal A в высоких количествах, которые секретируют α-Gal A, которая затем поглощается другими клетками. Соответственно, системы, описанные в данном документе, могут быть применены для продуцирования α-Gal A для введения нуждающимся в этом субъектам, например, в ферментозаместительных терапиях.
Пример 9. In vivo активность мышей, которые получали лечение с помощью Варианта №4 кДНК
[0245] Мышей GLAKO лечили внутривенно с помощью 200 мкл буфера для состава, содержащего 5,0e10 гв (2,0e12 гв/кг) AAV, содержащего вариант №4 кДНК (смотри, Фигура 13) или исходную конструкцию кДНК (Фигура 13B), а активность α-GalA в плазме анализировали в течение периода времени 2 месяца. Активность α-GalA в плазме мышей GLAKO, получавших лечение с помощью Варианта №4, была 10-кратной по сравнению с наблюдаемой в случае исходной конструкции кДНК (Фигура 18A). Как описано выше, активность также измеряли в печени, сердце, почке и селезенке для двух групп лечения и иллюстрировали на Фигуре 18B. Дополнительно, белок α-GalA анализировали в печени мышей, получавших лечение, и наблюдали изменения в молекулярной массе после лечения с помощью PNGase F или Endo H, как обсуждалось выше (Фигура 18C). Дополнительно, как проиллюстрировано на Фигуре 32, мыши GLAKO, получавшие лечение с помощью как исходной, так и Варианта №4 кДНК, проявляли сниженную концентрацию субстрата Фабри во всех исследованных тканях.
[0246] Эти данные демонстрируют. что подход на основе кДНК является также робастной платформой для продуцирования белка α-GalA на терапевтически эффективных уровнях in vivo.
Пример 10. Титрование дозы in vivo у мышей, получавших лечение с помощью исходной конструкции кДНК
[0247] Мышей GLAKO лечили внутривенно с помощью дозы AAV, содержащей исходную конструкцию кДНК (Фигура 13B), изменяющейся в диапазоне от 1,25e11 гв/кг до 5,0e12 гв/кг и анализировали активность α-GalA в плазме в течение периода времени 6 месяцев, как описано в Примерах 4 и 5.
[0248] Мыши GLAKO, получавшие лечение с помощью исходной кДНК, имели дозозависимую активность α-GalA в плазме изменяющуюся в диапазоне от 1-кратной дикого типа до 40-кратной дикого типа (Фигура 19). Кроме того, как проиллюстрировано на Фигуре 29, активность α-Gal A оставалась на терапевтических уровнях (в зависимости от дозы) в течение 6 месяцев после трансдукции, что свидетельствует о долгосрочной терапевтической пользе. Фигура 30 иллюстрирует активность α-Gal A в печени, селезенке, сердце и почке в день 180 (6 месяцев после лечения) и также иллюстрирует терапевтические уровни в этих тканях. Дозозависимое повышение активности α-Gal A также соответствовало снижению содержания субстрата Gb3. Смотри, Фигуры 31 и 32, иллюстрирующие дозозависимое снижение содержание Gb3 во всех оцениваемых тканях.
[0249] Данные демонстрируют, что терапевтические уровни белка α-Gal A получали у субъектов, получавших лечение с помощью подхода на основе кДНК, описанного в данном документе.
Пример 11. Дополнительные in vivo исследования IVPRP®
[0250] Мышей GLAKO лечили с помощью ZFN и различных приводимых в качестве примера донорных конструкций hGLA и оценивали таким образом, как описано выше, на геномную модификацию, активность GLA in vivo и снижение субстратов Фабри in vivo. Смотри Пример 5.
[0251] Как проиллюстрировано на Фигуре 22, опосредованная нуклеазой интеграция доноров GLA приводила к длительной модификации гепатоцитов в моделях мышей GLAKO болезни Фабри. Фигура 23 иллюстрируют активность α-Gal A в указанных тканях с течением времени (Фигура 23A) и через два месяца (Фигура 23B) после введения нуклеазы и доноров GLA животным. Как проиллюстрировано, продуцируемая в печени α-Gal A секретируется в ток крови и поглощается вторичными тканями, в том числе то, что ее стабильная активность в плазме достигала 80-кратного уровня дикого типа. Фигура 24 показывает животных, получавших лечение с помощью нуклеазы и доноров, проявлявших большее снижение концентрации субстрата во всех тканях, исследовали по сравнению с не получавшими лечение животными, животными дикого типа и животными, получавшими лечение только с помощью доноров.
[0252] Эксперименты на мышах HepG2 и GLAKO проводили с применением Варианта L и модифицированного донора, обозначенного как Вариант M (Фигура 25), который включает сигнальный пептид IDS вместо сигнального пептида GLA Варианта L.
[0253] Для детектирования интеграции доноров L и M, применяли подход на основе NGS, основанный на неискаженной схеме PCR, которая генерирует различные продукты, когда донор интегрируется по сравнению с продуктом, сгенерированным для гена дикого типа, лишенного события интеграции. Вкратце, последовательность праймера 5- NGS (идентичного Праймеру 1 на Фигуре 25B) добавляли на 3' конце трансгена (смотри Фигуру 25A). Незамедлительно по ходу транскрипции последовательности праймера NGS добавляли последовательность целевой интеграции (TI). Последовательность TI имеет тот же самый нуклеотидный состав и длину, что и соответствующая последовательность в локусе альбумина, но основную последовательность рандомизировали таким образом, что для амплификации продукта PCR, связанного с локусом дикого типа без интеграции, не вводят сдвиги PCR, по сравнению с локусом, содержащим интеграцию трансгена. Два продукта PCR, таким образом, используют идентичные праймеры и продуцируют продукты PCR одинакового размера и композиции, но имеют отличающиеся последовательности, делая возможным легкую идентификацию событий TI с помощью NGS, а также одновременный анализ инсерционно-делеционных мутаций и событий TI с помощью NGS.
[0254] Для анализа событий интеграции в клетках человека, применяли следующие праймеры:
Праймер 1: 5' GCACTAAGGAAAGTGCAAAG (SEQ ID NO:6)
Праймер 2: 5' TAATACTCTTTTAGTGTCTA (SEQ ID NO:7)
[0255] Последовательность TI, применяемую в клетках человека, иллюстрировали ниже, где рандомизированная последовательность выделена курсивом, а локализация связывающего Праймер 1 сайта выделена подчеркиванием:
5'GCACTAAGGAAAGTGCAAAGTAAGATTGACCAGACCAGATAGAAGAATGTAACTGTAGTTCTAATAGGACTTATTATCCCAAAGAC (SEQ ID NO:8).
[0256] Амплификация с применением двух праймеров продуцирует ампликон из 222 п.о., как проиллюстрировано ниже:
Ампликон дикого типа (без вставки): 5'GCACTAAGGAAAGTGCAAAGTAACTTAGAGTGACTGAAACTTCACAGAATAGGGTTGAAGATTGAATTCATAACTATCCCAAAGACCTATCCATTGCACTATGCTTTATTTAAAAACCACAAAACCTGTGCTGTTGATCTCATAAATAGAACTTGTATTTATATTTATTTTCATTTTAGTCTGTCTTCTTGGTTGCTGTTGATAGACACTAAAAGAGTATTA (SEQ ID NO:9).
Ампликон TI (курсив указывает на рандомизированную последовательность): 5'GCACTAAGGAAAGTGCAAAGTAAGATTGACCAGACCAGATAGAAGAATGTAACTGTAGTTCTAATAGGACTTATTATCCCAAAGACCTATCCATTGCACTATGCTTTATTTAAAAACCACAAAACCTGTGCTGTTGATCTCATAAATAGAACTTGTATTTATATTTATTTTCATTTTAGTCTGTCTTCTTGGTTGCTGTTGATAGACACTAAAAGAGTATTA (SEQ ID NO:10).
[0257] Подобным образом, последовательность TI и праймеры, применяемые для клеток мыши, иллюстрировали ниже. Для анализа событий интеграции применяли следующие праймеры:
Праймер 1: 5' TTGAGTTTGAATGCACAGAT (SEQ ID NO:11)
Праймер 2: 5' GAAACAGGGAGAGAAAAACC (SEQ ID NO:12).
[0258] Последовательность TI, применяемую в клетках мыши, иллюстрировали ниже, где рандомизированная последовательность выделена курсивом, а локализация связывающего Праймер 1 сайта выделена подчеркиванием:
5'AAATCTTGAGTTTGAATGCACAGATCAATTGTAAACTAAAGAAATAGTAATATAGAGTTTAAATATAGATAGCTATGACTGCACTTGATAGAAGGTAACGGTGCCACCTTCAGATTT (SEQ ID NO:13)
[0259] Амплификация с применением двух праймеров продуцирует ампликон из 247 п.о., как проиллюстрировано ниже:
Ампликон дикого типа (без вставки): 5'TTGAGTTTGAATGCACAGATATAAACACTTAACGGGTTTTAAAAATAATAATGTTGGTGAAAAAATATAACTTTGAGTGTAGCAGAGAGGAACCATTGCCACCTTCAGATTTTCCTGTAACGATCGGGAACTGGCATCTTCAGGGAGTAGCTTAGGTCAGTGAAGAGAAGAACAAAAAGCAGCATATTACAGTTAGTTGTCTTCATCAATCTTTAAATATGTTGTGTGGTTTTTCTCTCCCTGTTTC (SEQ ID NO:14)
Ампликон TI (курсив указывает на рандомизированную последовательность): 5'TTGAGTTTGAATGCACAGATCAATTGTAAACTAAAGAAATAGTAATATAGAGTTTAAATATAGATAGCTATGACTGCACTTGATAGAAGGTAACGGTGCCACCTTCAGATTTTCCTGTAACGATCGGGAACTGGCATCTTCAGGGAGTAGCTTAGGTCAGTGAAGAGAAGAACAAAAAGCAGCATATTACAGTTAGTTGTCTTCATCAATCTTTAAATATGTTGTGTGGTTTTTCTCTCCCTGTTTC (SEQ ID NO:15)
[0260] Дополнительно, эта методика может быть применена по отношению к отличным от человека приматов (макака-резус, NHP), применяя праймеры и вставленную последовательность TI, проиллюстрированную ниже:
Праймер 1: 5' CCACTAAGGAAAGTGCAAAG (SEQ ID NO:16)
Праймер 2: 5' TGAAAGTAAATATAAATACAAGTTC (SEQ ID NO:17)
[0261] Последовательность TI, применяемую в клетках NHP, иллюстрировали ниже, где рандомизированная последовательность выделена курсивом, а локализация связывающего Праймер 1 сайта выделена подчеркиванием:
5'CCACTAAGGAAAGTGCAAAGGAGCGCTAACTGGAACATACTCGCTATTTAAGAACATTATAAGATACTAATTCAGTATTCGAAGAC (SEQ ID NO:18).
[0262] Амплификация с применением двух праймеров продуцирует ампликон из 173 п.о., как проиллюстрировано ниже:
Ампликон дикого типа (без вставки): 5'CCACTAAGGAAAGTGCAAAG TAACTTAGAGTGACTTAAACTTCACAGAACAGAGTTGAAGATTGAATTCATAACTGTCCCTAAGACCTATCCATTGCACTATGCTTTATTTAAAAGCCACAAAACCTGTGCTGTTGATCTCATAAATAGAACTTGTATTTATATTTACTTTCA (SEQ ID NO:19)
Ампликон TI (курсив указывает на рандомизированную последовательность): 5'CCACTAAGGAAAGTGCAAAGGAGCGCTAACTGGAACATACTCGCTATTTAAGAACATTATAAGATACTAATTCAGTATTCGAAGACCTATCCATTGCACTATGCTTTATTTAAAAGCCACAAAACCTGTGCTGTTGATCTCATAAATAGAACTTGTATTTATATTTACTTTCA (SEQ ID NO:20).
[0263] Таким образом, клетки человека из HepG2 клеточной линии гепатокарциномы лечили с помощью ZFN и варианта №L донора GLA, содержат последовательность TI для анализа HDR. DNA очищали от трансдуцированных клеток спустя 7 дней после трансдукции и анализировали посредством NGS.
[0264] Как проиллюстрировано на Фигуре 26, in vitro инсерционно-делеционные мутации и TI (HDR) показали дозозависимый ответ на фиксированное соотношение ZFN и донор TI. Более того, как проиллюстрировано у GLAKO мышей, опосредованная нуклеазой целевая интеграция (TI) Варианта M давала 250-кратную стабильную активность в плазме относительно дикого типа, а активность α-Gal A в сердце и почке была более чем 20-кратной относительно дикого типа и 4-кратной относительно дикого типа, соответственно.
[0265] Также были проведены анализы для дальнейшей оценки того, поглощается ли α-Gal A вторичными тканями после опосредованной нуклеазой TI конструкции донора GLA. Вкратце, как описано выше, конструкцию донора GLA, содержащую сигнальный пептид IDS и 3' последовательность для анализа целевой интеграции (TI) (Вариант M донора), применяли для лечения мыши GLAKO и анализировали образцы плазмы и ткани (например, печени, сердца, селезенки, почки, мозга и т. д.) и на активность α-Gal A, и на концентрацию субстрата.
[0266] Как проиллюстрировано на Фигуре 27, стабильная активность α-Gal A в плазме достигала 250-кратной стабильной активности в плазме относительно дикого типа, а активность α-Gal A в сердце и почке была более чем 20-кратной относительно дикого типа и 4-кратной относительно дикого типа, соответственно.
[0267] Данные демонстрируют, что терапевтические уровни белка α-Gal A получали у субъектов (в том числе из вторичной ткани), получавших лечение с помощью подхода на основе IVPRP, описанного в данном документе.
[0268] Все публикации, патенты и патентные заявки, упоминаемые в данном документе, включены в настоящее описание посредством ссылки во всей своей полноте.
[0269] Хотя раскрытие было предоставлено в некоторых деталях в качестве иллюстрации и примера для целей ясности понимания, для специалистов в данной области техники будет очевидно, что различные изменения и модификации могут быть осуществлены на практике, не отступая от сущности или объема. Соответственно, приведенные выше описания и примеры не должны рассматриваться как ограничивающие.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SANGAMO THERAPEUTICS, INC.
<120> СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ
<130> 8325-0142.40
<140> PCT/US2017/057328
<141> 2017-10-19
<150> 62/552792
<151> 31.08.2017
<150> 62/516 373
<151> 07.06.2017
<150> 62/502 058
<151> 05.05.2017
<150> 62/458 324
<151> 13.02.2017
<150> 62/444 093
<151> 09.01.2017
<150> 62/410 543
<151> 20.10.2016
<160> 21
<170> PatentIn версия 3.5
<210> 1
<211> 31
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 1
Met Gln Leu Arg Asn Pro Glu Leu His Leu Gly Cys Ala Leu Ala Leu
1 5 10 15
Arg Phe Leu Ala Leu Val Ser Trp Asp Ile Pro Gly Ala Arg Ala
20 25 30
<210> 2
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 2
gccaccatg 9
<210> 3
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 3
Met Pro Pro Pro Arg Thr Gly Arg Gly Leu Leu Trp Leu Gly Leu Val
1 5 10 15
Leu Ser Ser Val Cys Val Ala Leu Gly
20 25
<210> 4
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 4
Met Gln Arg Val Asn Met Ile Met Ala Glu Ser Pro Gly Leu Ile Thr
1 5 10 15
Ile Cys Leu Leu Gly Tyr Leu Leu Ser Ala Glu Cys
20 25
<210> 5
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 5
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser
<210> 6
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 6
gcactaagga aagtgcaaag 20
<210> 7
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 7
taatactctt ttagtgtcta 20
<210> 8
<211> 86
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 8
gcactaagga aagtgcaaag taagattgac cagaccagat agaagaatgt aactgtagtt 60
ctaataggac ttattatccc aaagac 86
<210> 9
<211> 222
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 9
gcactaagga aagtgcaaag taacttagag tgactgaaac ttcacagaat agggttgaag 60
attgaattca taactatccc aaagacctat ccattgcact atgctttatt taaaaaccac 120
aaaacctgtg ctgttgatct cataaataga acttgtattt atatttattt tcattttagt 180
ctgtcttctt ggttgctgtt gatagacact aaaagagtat ta 222
<210> 10
<211> 222
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 10
gcactaagga aagtgcaaag taagattgac cagaccagat agaagaatgt aactgtagtt 60
ctaataggac ttattatccc aaagacctat ccattgcact atgctttatt taaaaaccac 120
aaaacctgtg ctgttgatct cataaataga acttgtattt atatttattt tcattttagt 180
ctgtcttctt ggttgctgtt gatagacact aaaagagtat ta 222
<210> 11
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 11
ttgagtttga atgcacagat 20
<210> 12
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 12
gaaacaggga gagaaaaacc 20
<210> 13
<211> 117
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 13
aaatcttgag tttgaatgca cagatcaatt gtaaactaaa gaaatagtaa tatagagttt 60
aaatatagat agctatgact gcacttgata gaaggtaacg gtgccacctt cagattt 117
<210> 14
<211> 247
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 14
ttgagtttga atgcacagat ataaacactt aacgggtttt aaaaataata atgttggtga 60
aaaaatataa ctttgagtgt agcagagagg aaccattgcc accttcagat tttcctgtaa 120
cgatcgggaa ctggcatctt cagggagtag cttaggtcag tgaagagaag aacaaaaagc 180
agcatattac agttagttgt cttcatcaat ctttaaatat gttgtgtggt ttttctctcc 240
ctgtttc 247
<210> 15
<211> 247
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 15
ttgagtttga atgcacagat caattgtaaa ctaaagaaat agtaatatag agtttaaata 60
tagatagcta tgactgcact tgatagaagg taacggtgcc accttcagat tttcctgtaa 120
cgatcgggaa ctggcatctt cagggagtag cttaggtcag tgaagagaag aacaaaaagc 180
agcatattac agttagttgt cttcatcaat ctttaaatat gttgtgtggt ttttctctcc 240
ctgtttc 247
<210> 16
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 16
ccactaagga aagtgcaaag 20
<210> 17
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 17
tgaaagtaaa tataaataca agttc 25
<210> 18
<211> 86
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 18
ccactaagga aagtgcaaag gagcgctaac tggaacatac tcgctattta agaacattat 60
aagatactaa ttcagtattc gaagac 86
<210> 19
<211> 173
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 19
ccactaagga aagtgcaaag taacttagag tgacttaaac ttcacagaac agagttgaag 60
attgaattca taactgtccc taagacctat ccattgcact atgctttatt taaaagccac 120
aaaacctgtg ctgttgatct cataaataga acttgtattt atatttactt tca 173
<210> 20
<211> 173
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 20
ccactaagga aagtgcaaag gagcgctaac tggaacatac tcgctattta agaacattat 60
aagatactaa ttcagtattc gaagacctat ccattgcact atgctttatt taaaagccac 120
aaaacctgtg ctgttgatct cataaataga acttgtattt atatttactt tca 173
<210> 21
<211> 9
<212> БЕЛОК
<213> Неизвестно
<220>
<223> Описание неизвестного:
Последовательность хоуминг-эндонуклеазы «LAGLIDADG»
<400> 21
Leu Ala Gly Leu Ile Asp Ala Asp Gly
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2020 |
|
RU2822369C2 |
| МОДИФИКАЦИЯ B-КЛЕТОК | 2018 |
|
RU2783116C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ГЕМОФИЛИИ В | 2011 |
|
RU2608643C2 |
| ПЕЧЕНЬ-СПЕЦИФИЧНЫЕ КОНСТРУКЦИИ, ЭКСПРЕССИОННЫЕ КАССЕТЫ ФАКТОРА VIII И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2774874C2 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОПОСРЕДОВАННОЙ НУКЛЕАЗОЙ НАПРАВЛЕННОЙ ИНТЕГРАЦИИ ТРАНСГЕНОВ | 2013 |
|
RU2650819C2 |
| СПОСОБЫ ОБНАРУЖЕНИЯ ДНК ДЛЯ САЙТ-СПЕЦИФИЧЕСКОЙ НУКЛЕАЗНОЙ АКТИВНОСТИ | 2013 |
|
RU2678001C2 |
| СПОСОБЫ И СОСТАВЫ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2013 |
|
RU2639277C2 |
| БЫСТРЫЙ НАПРАВЛЕННЫЙ АНАЛИЗ СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР ДЛЯ ОПРЕДЕЛЕНИЯ ДОНОРНОЙ ВСТАВКИ | 2014 |
|
RU2668819C2 |
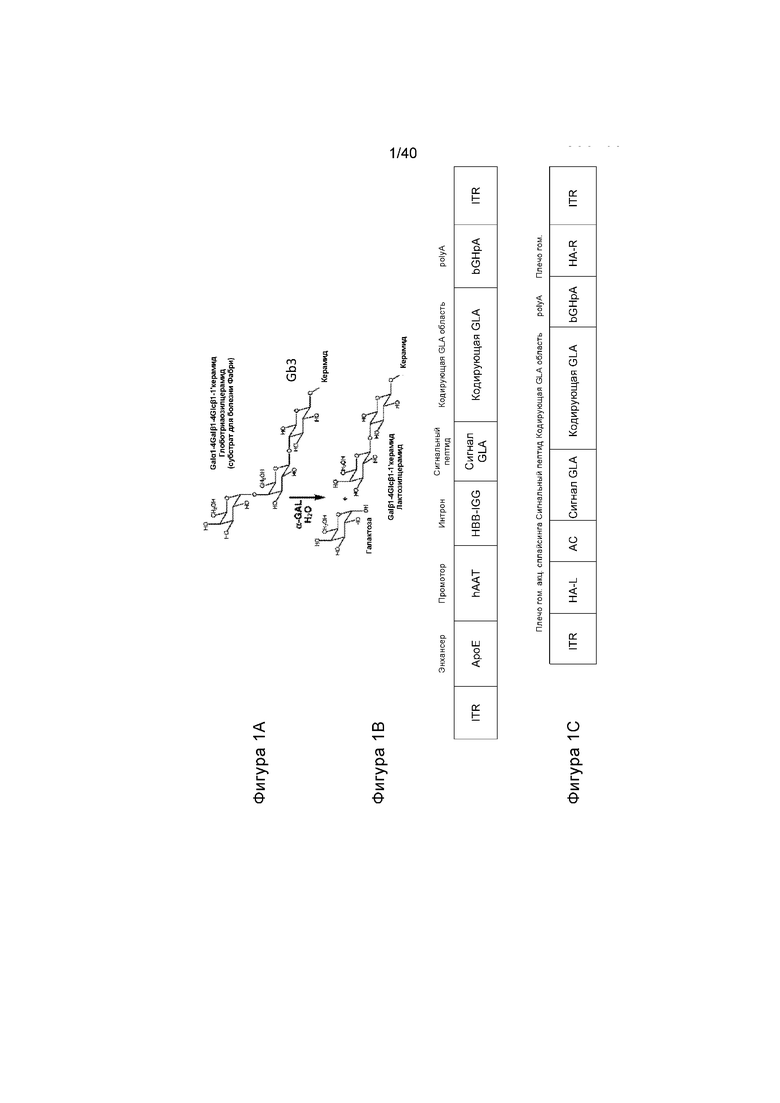
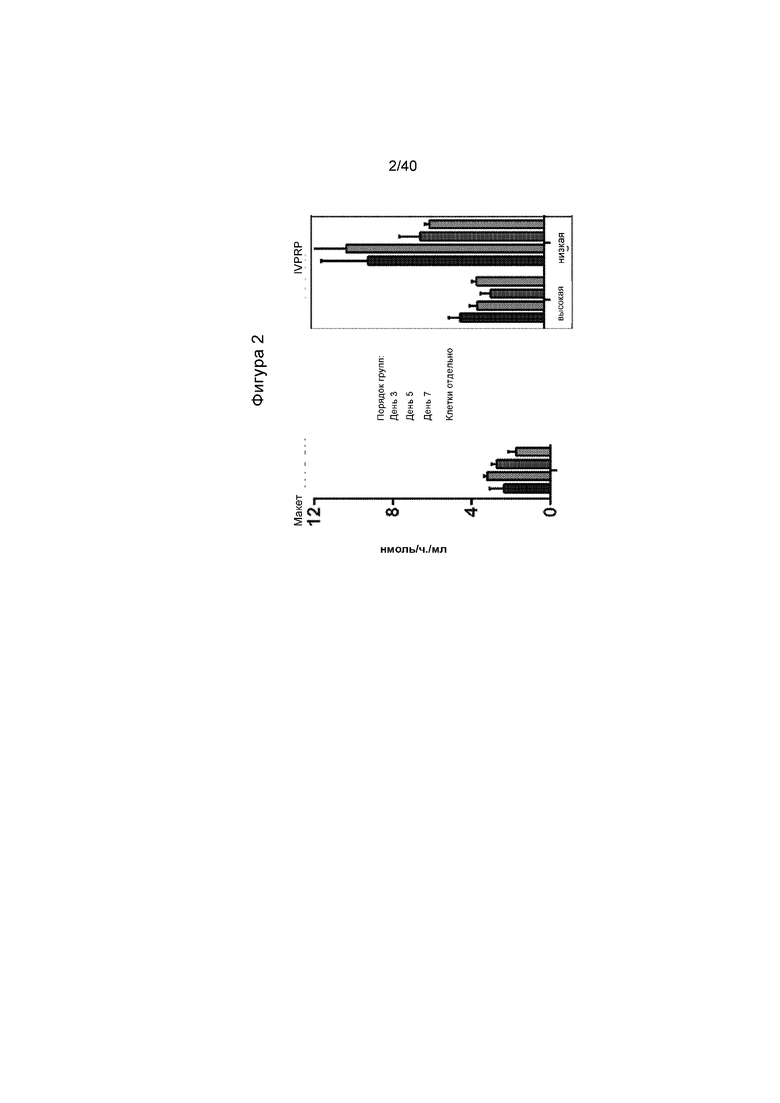
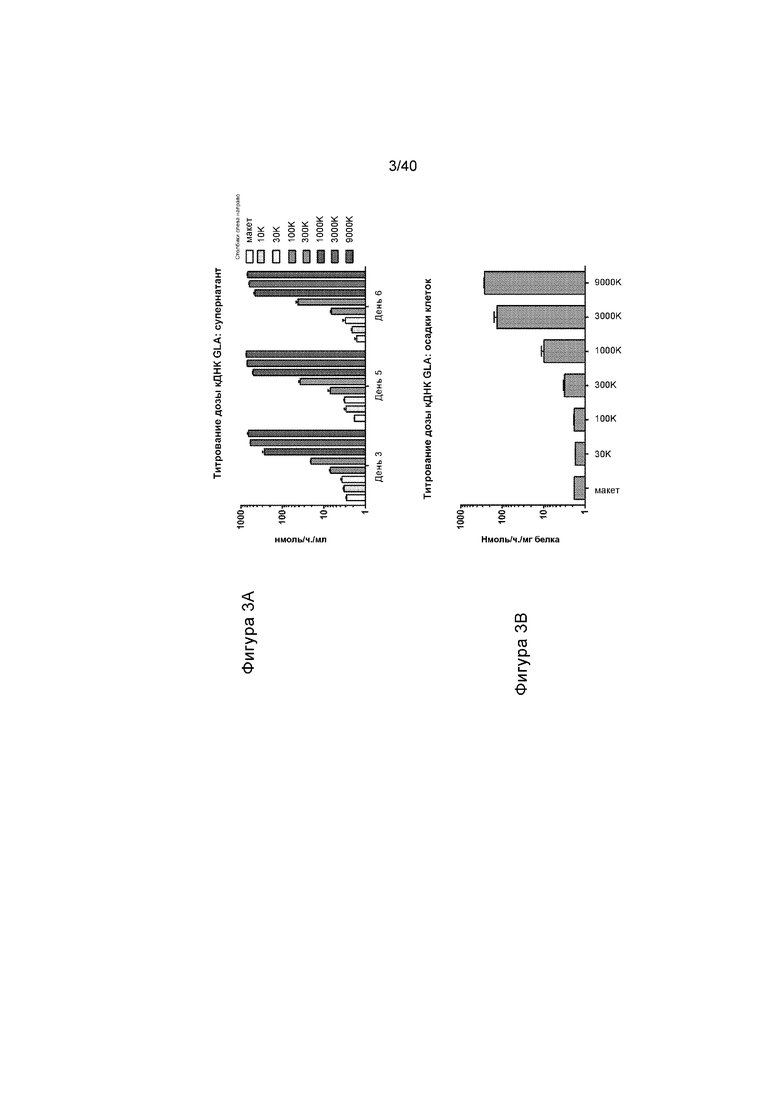
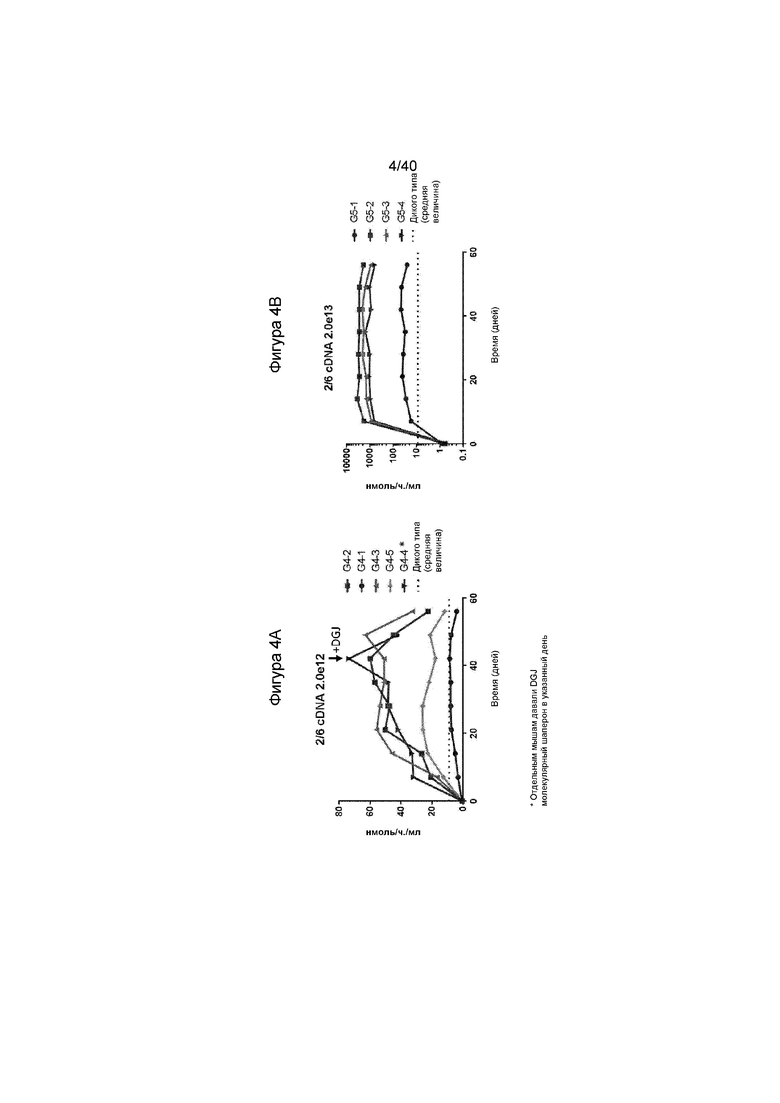
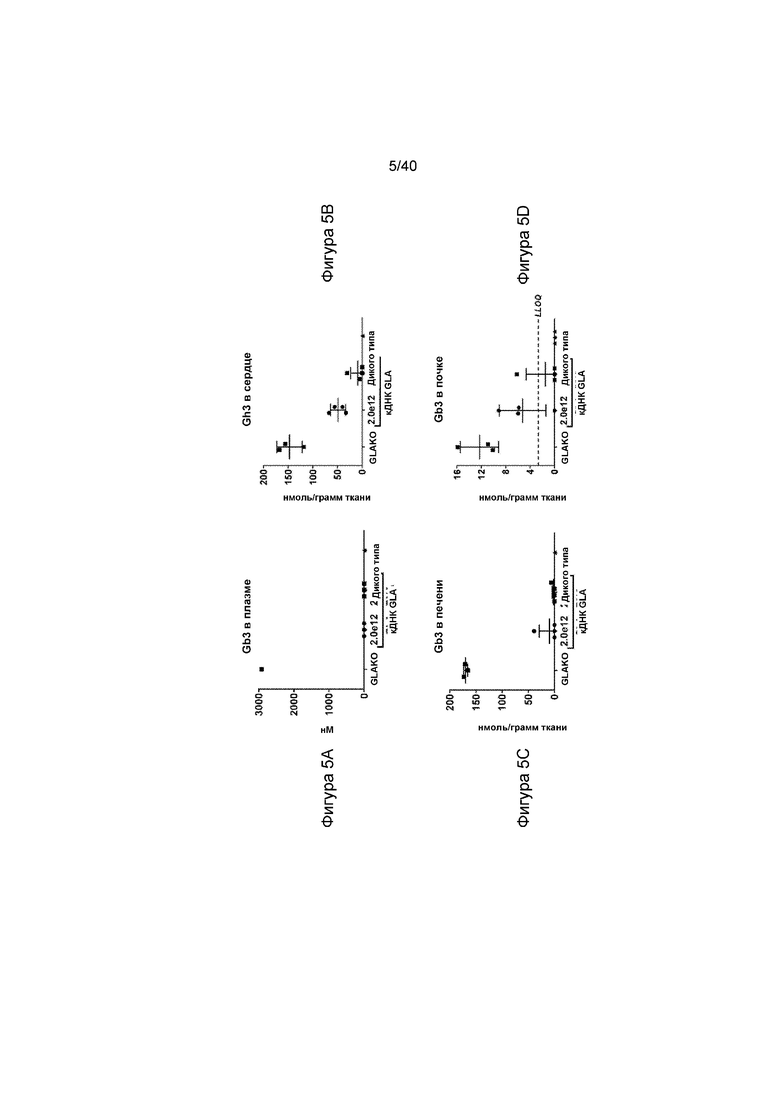

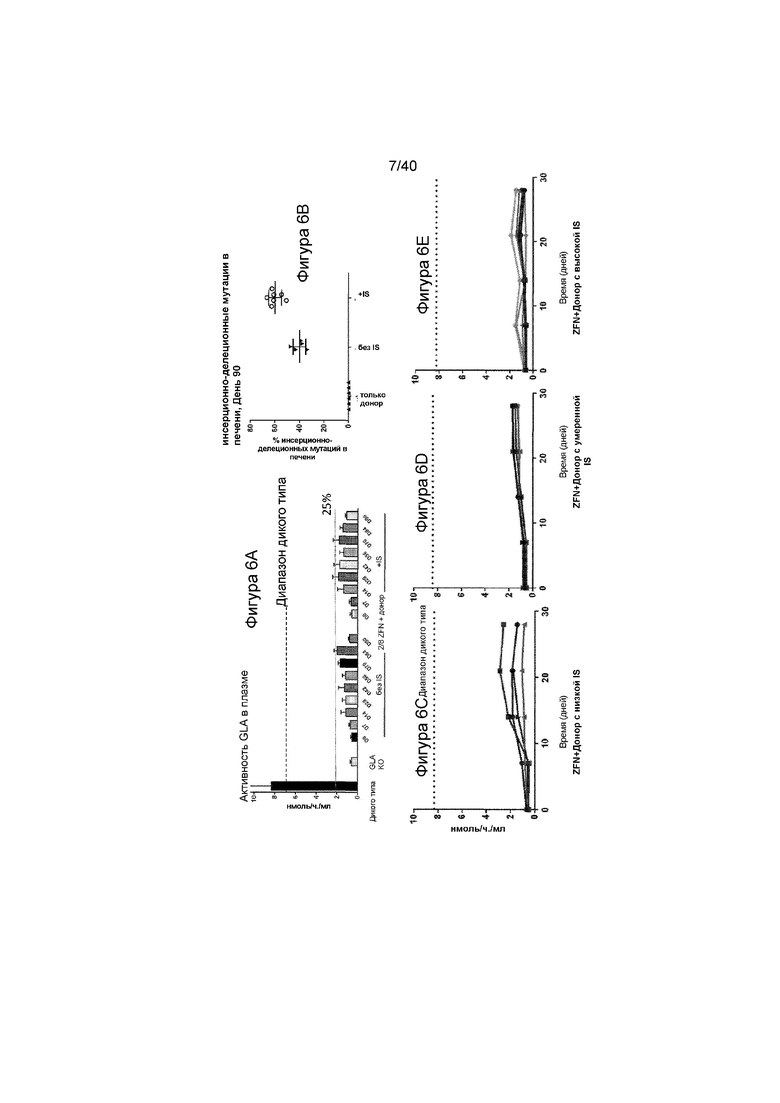
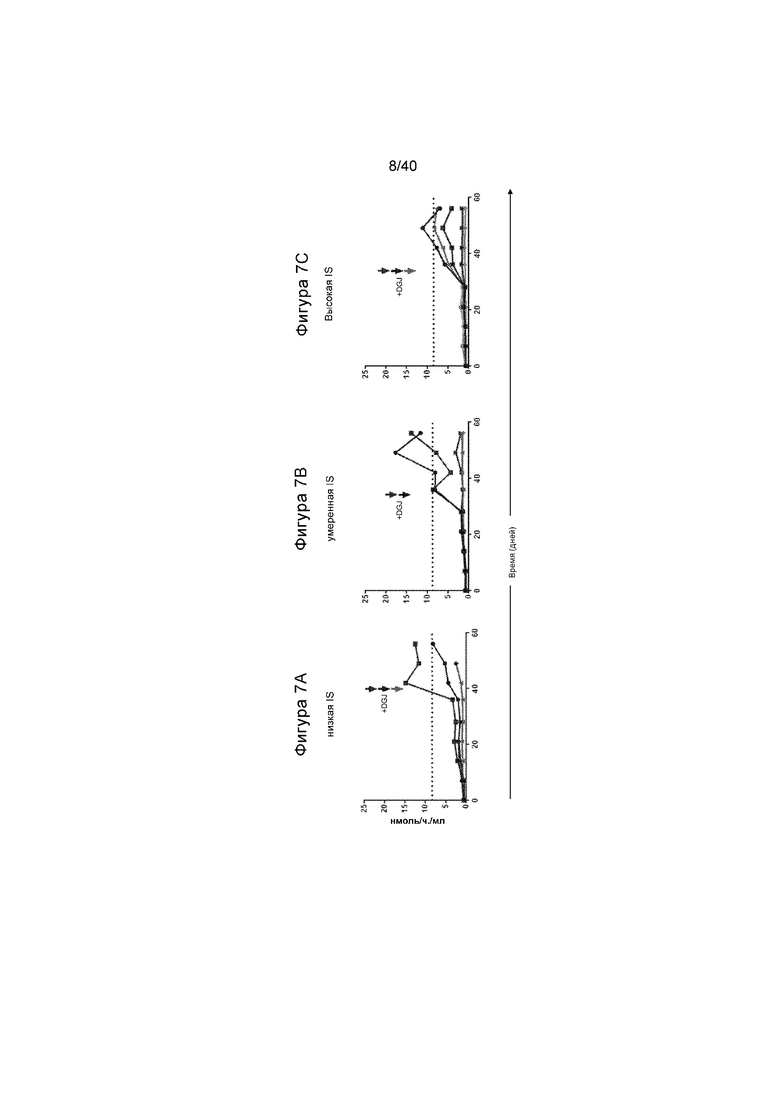
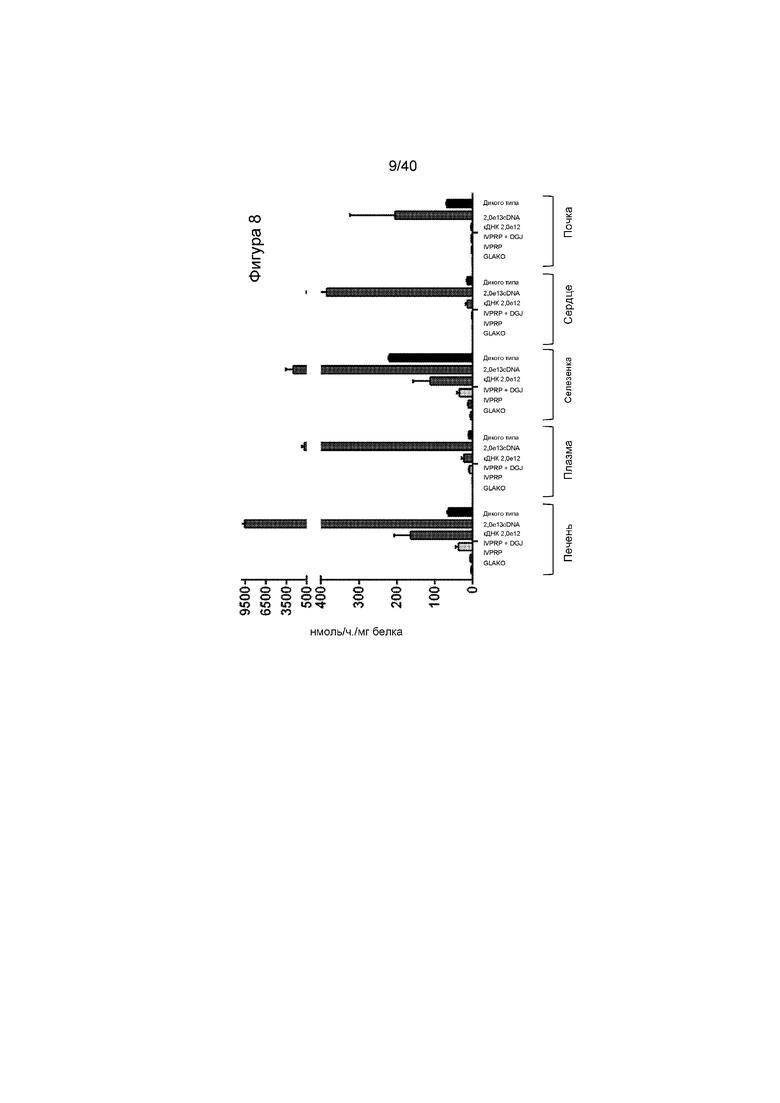


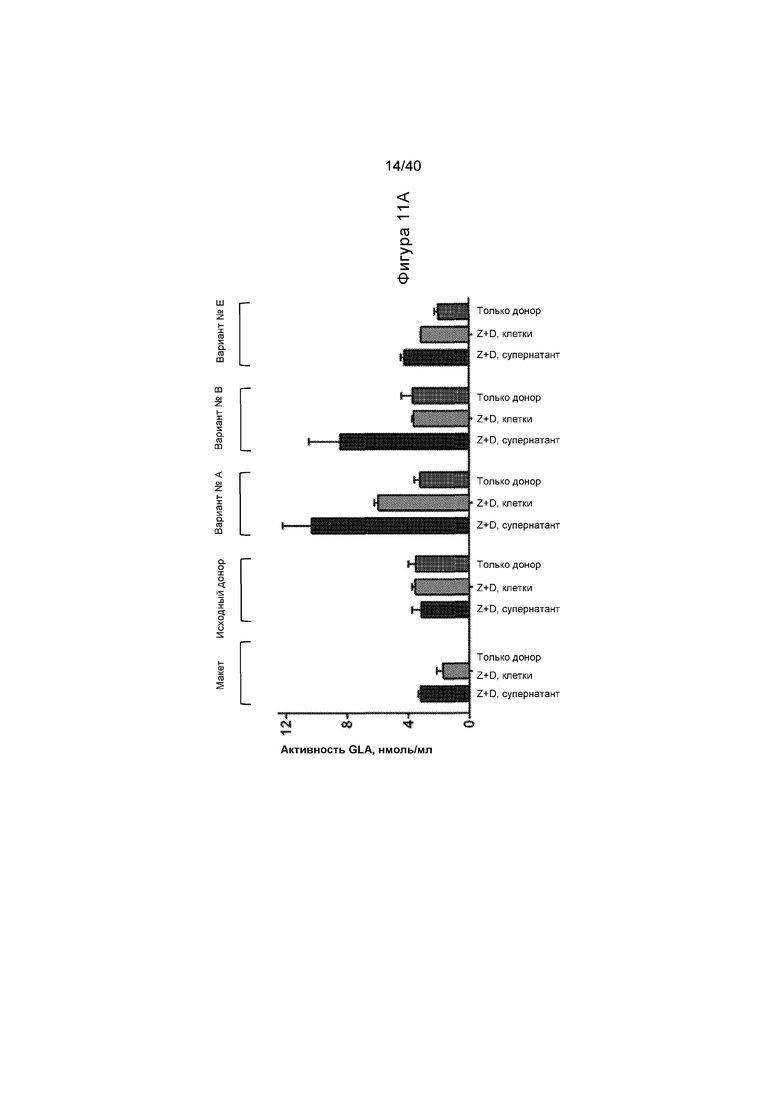
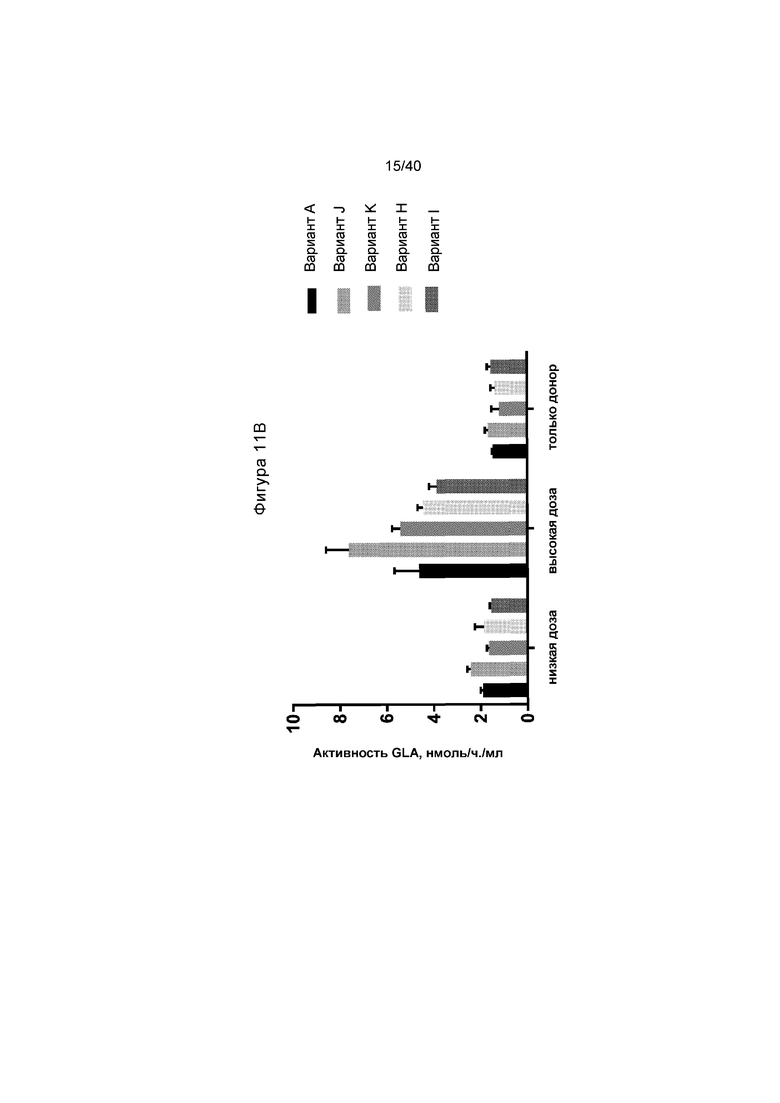
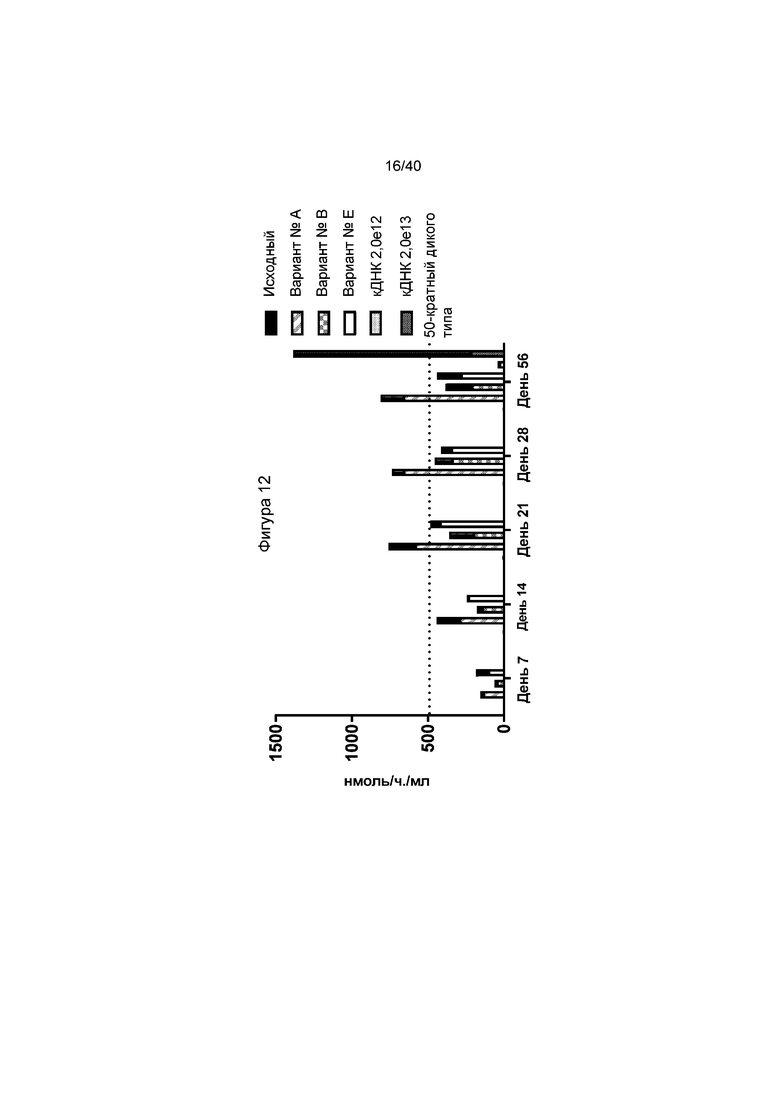
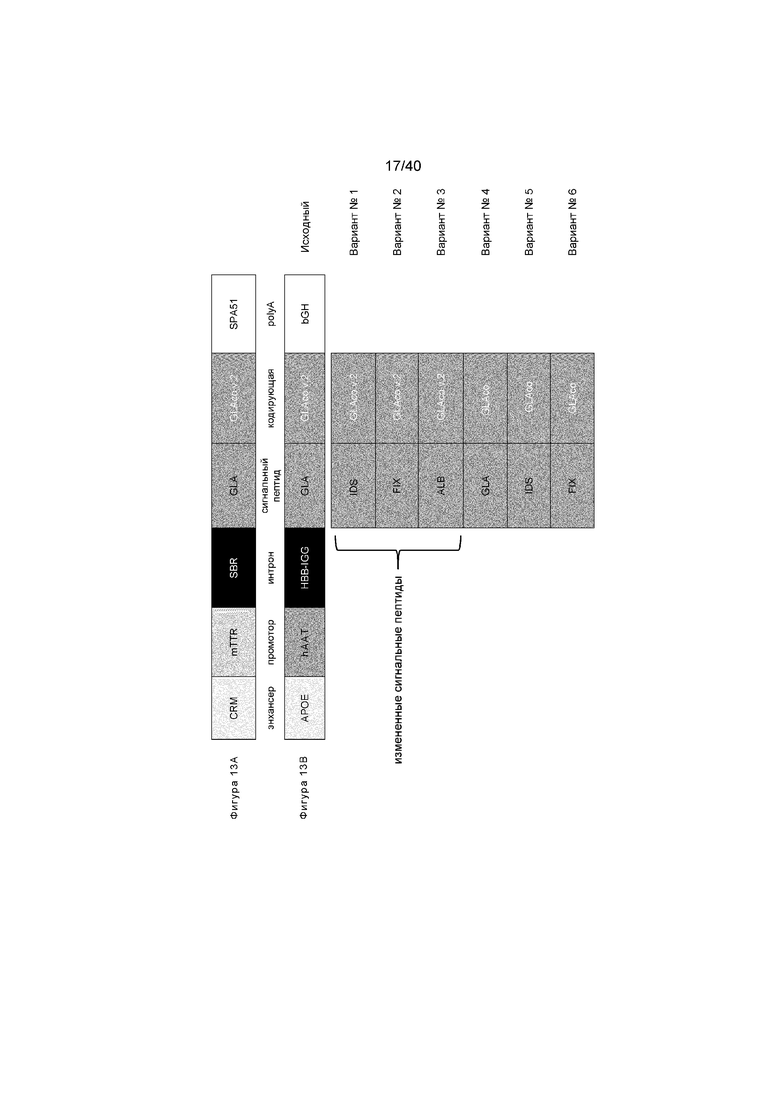

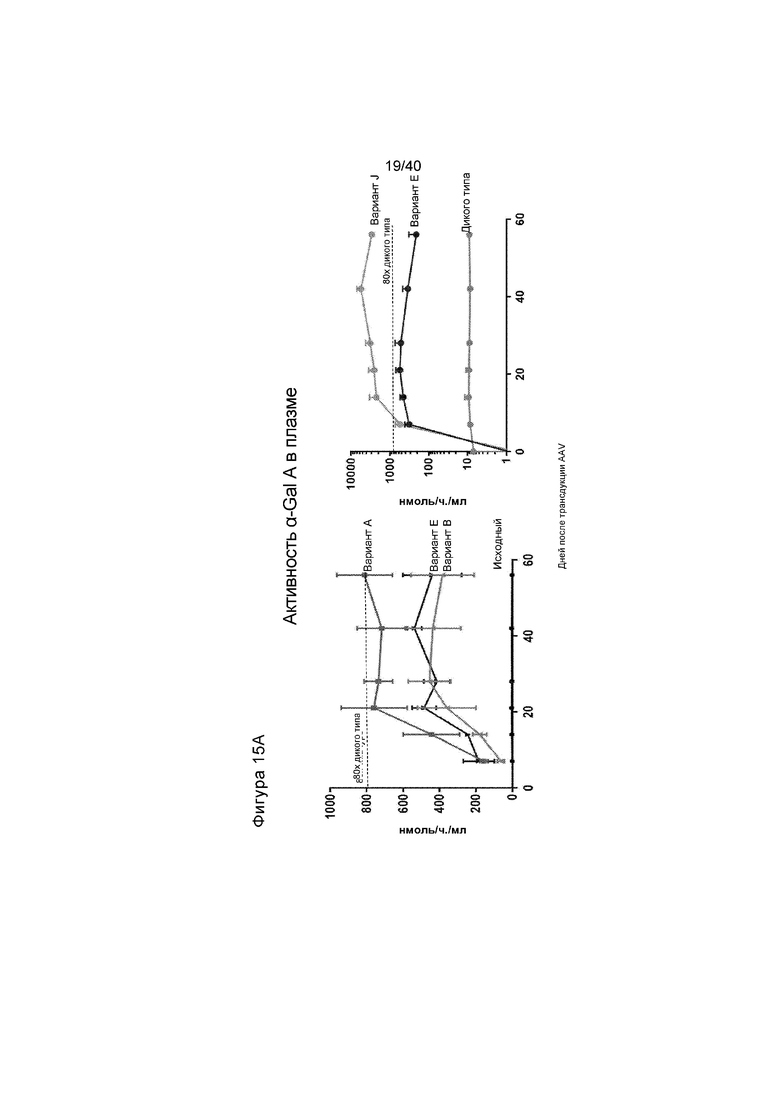
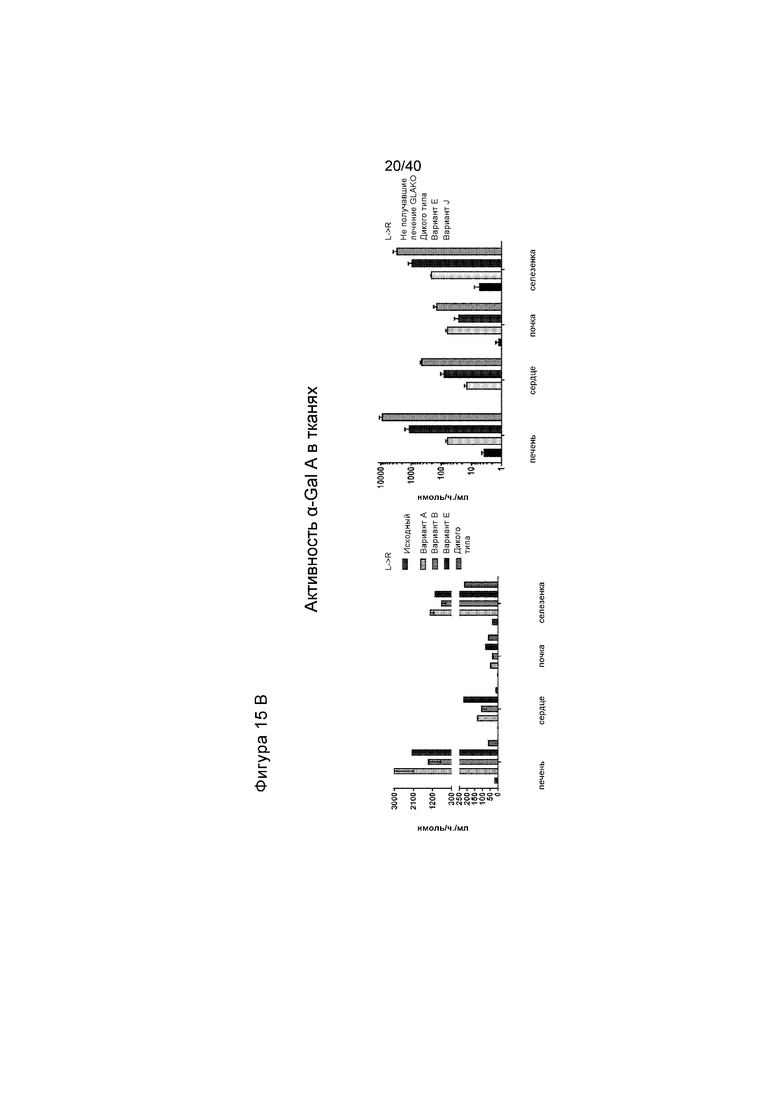
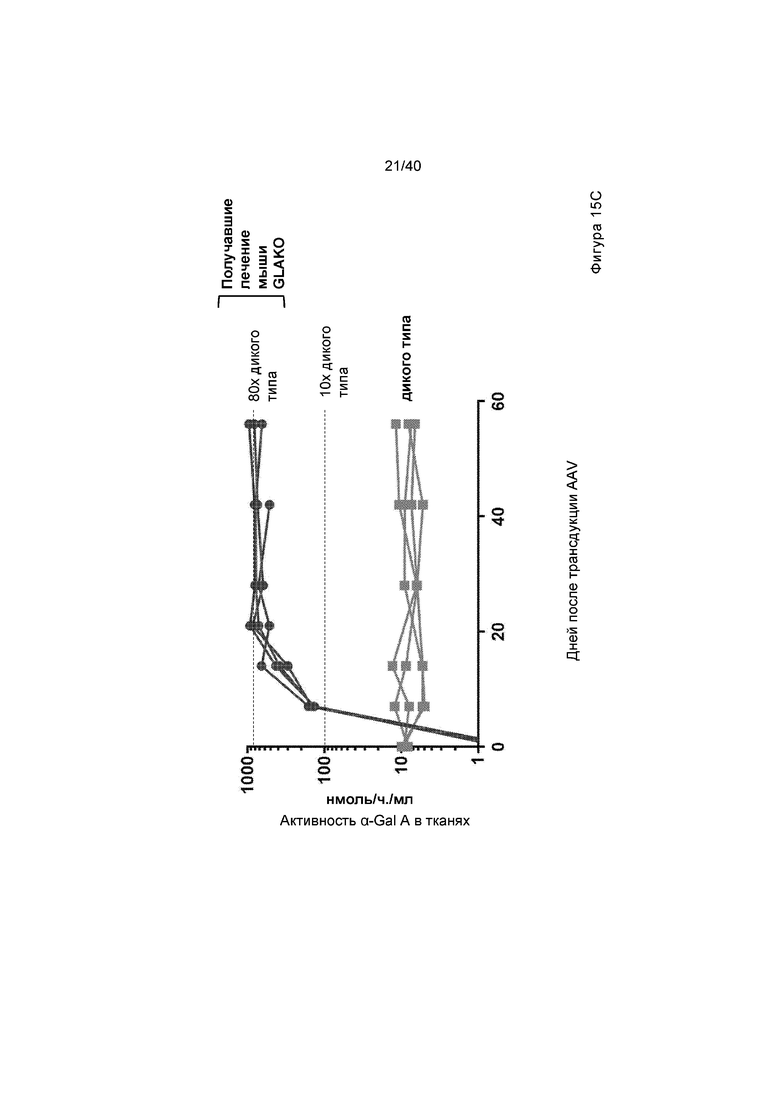
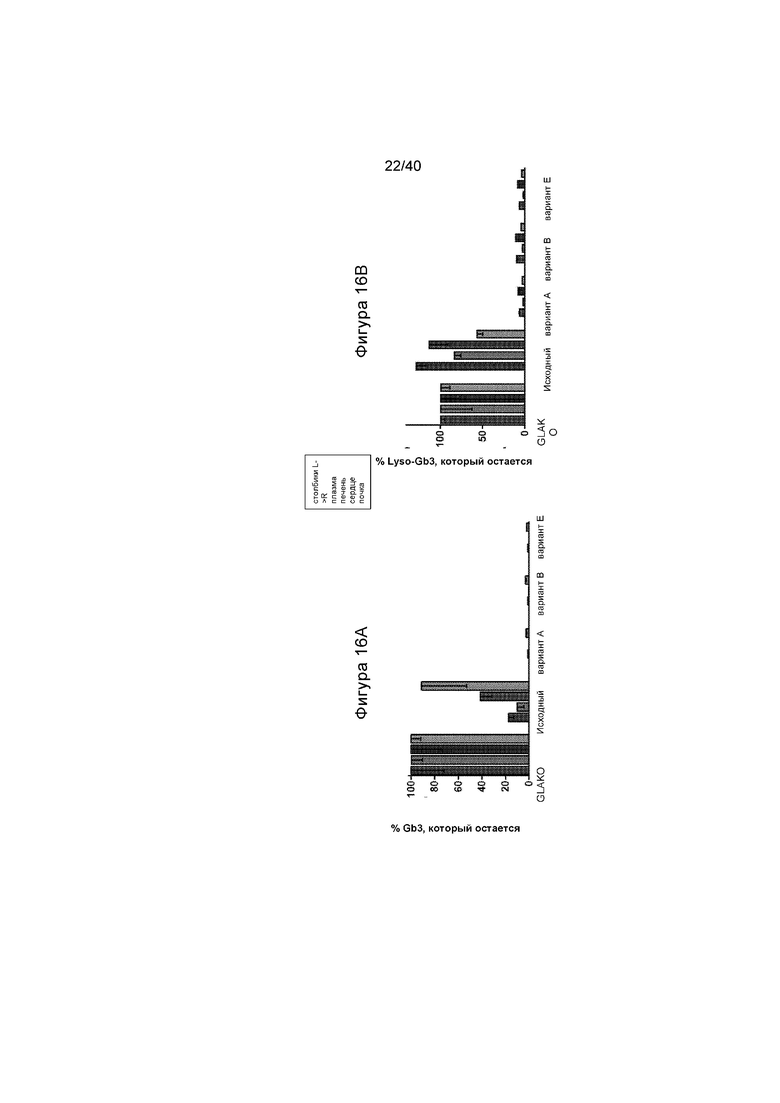

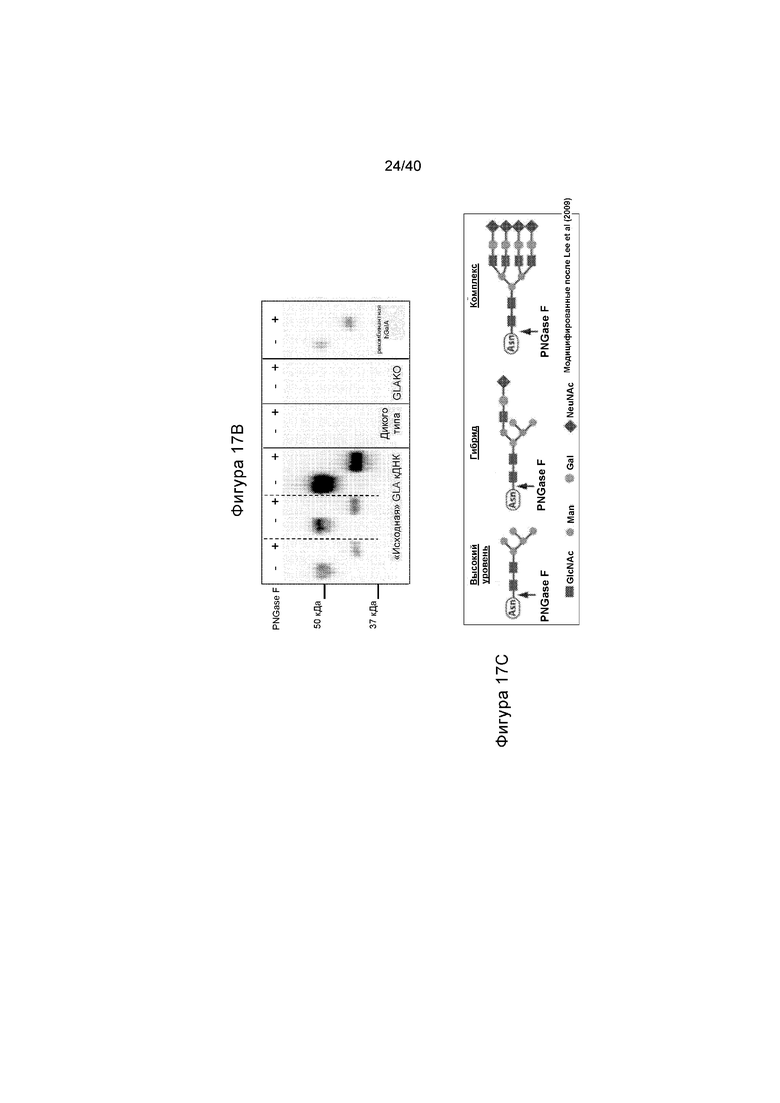
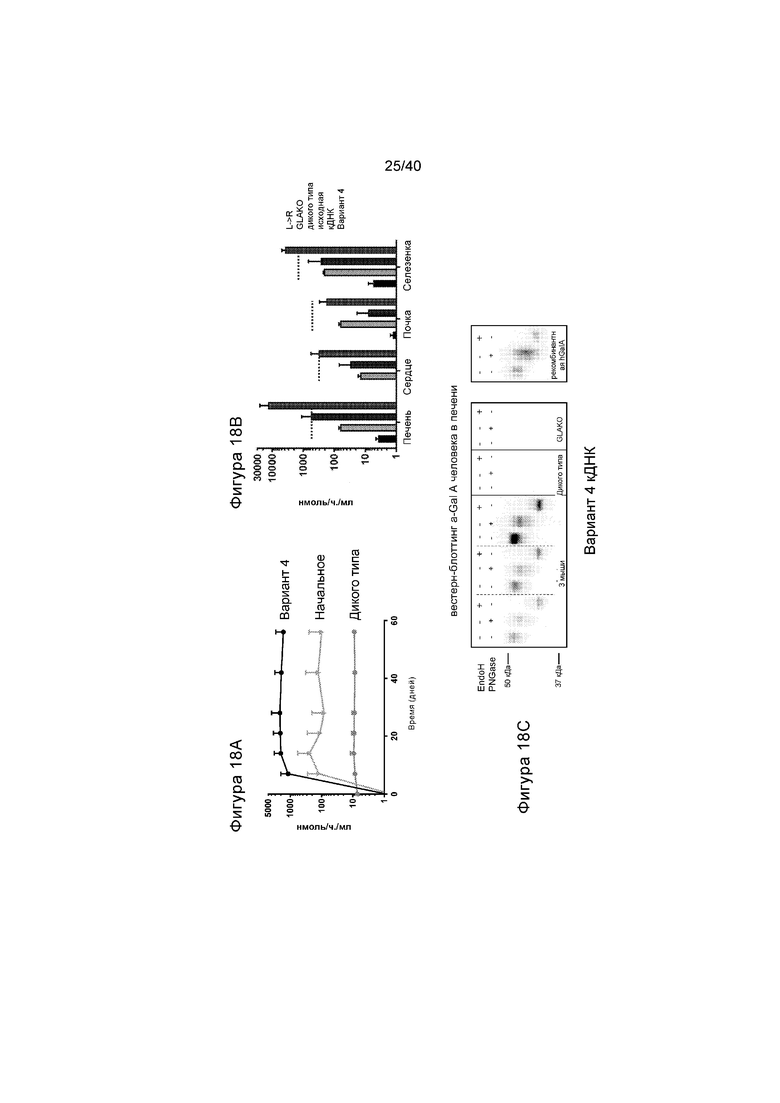

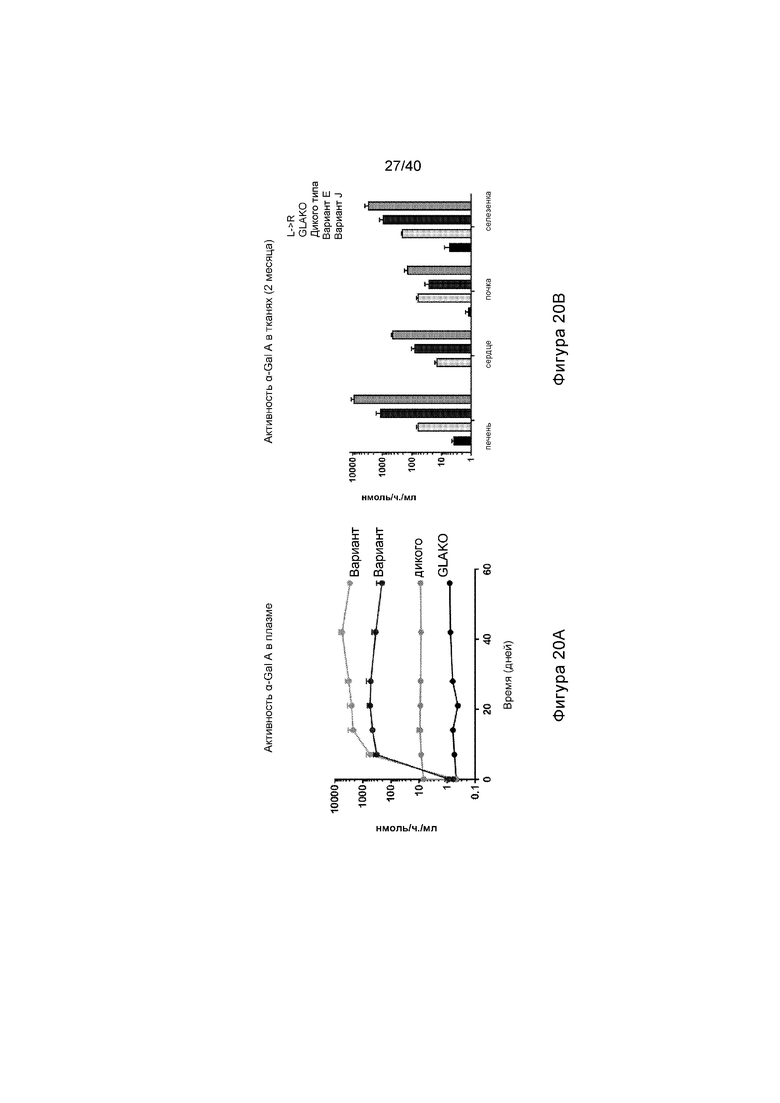
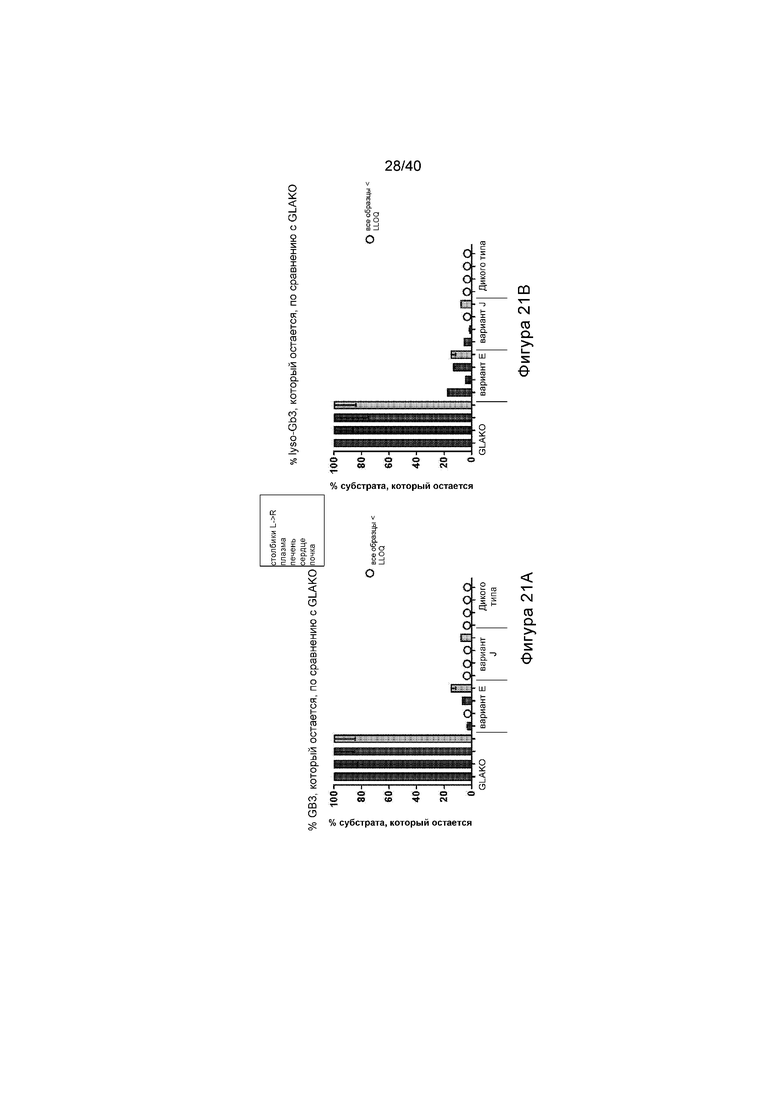
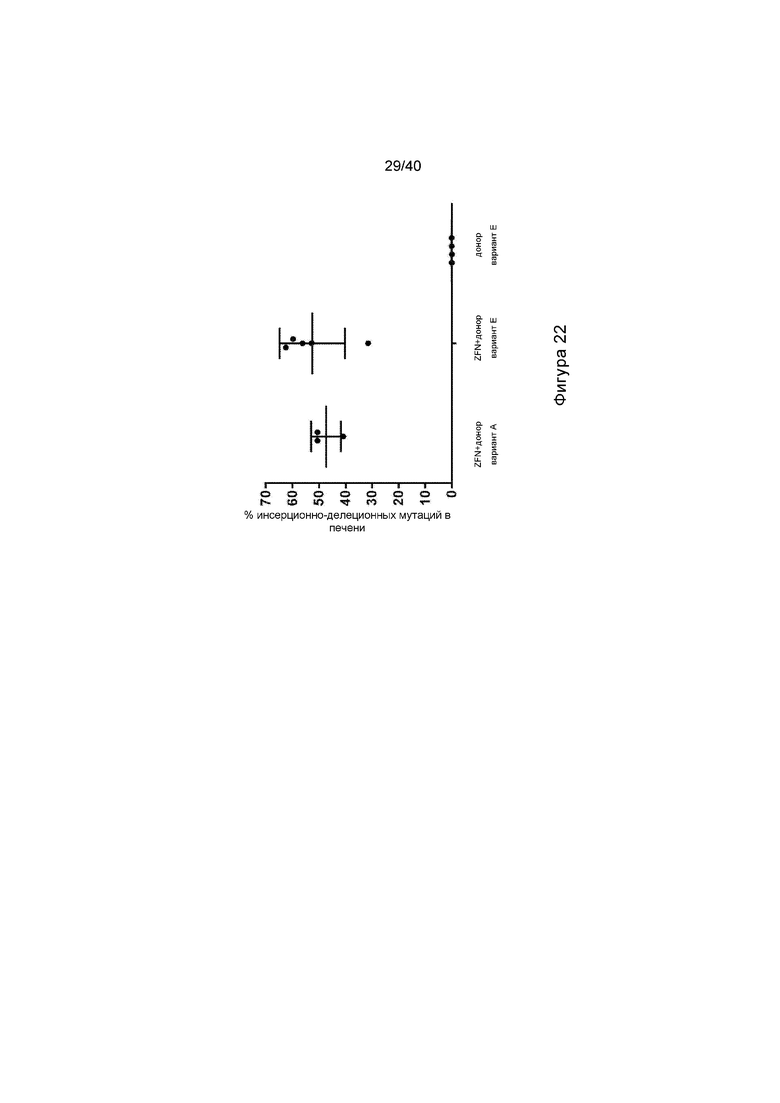
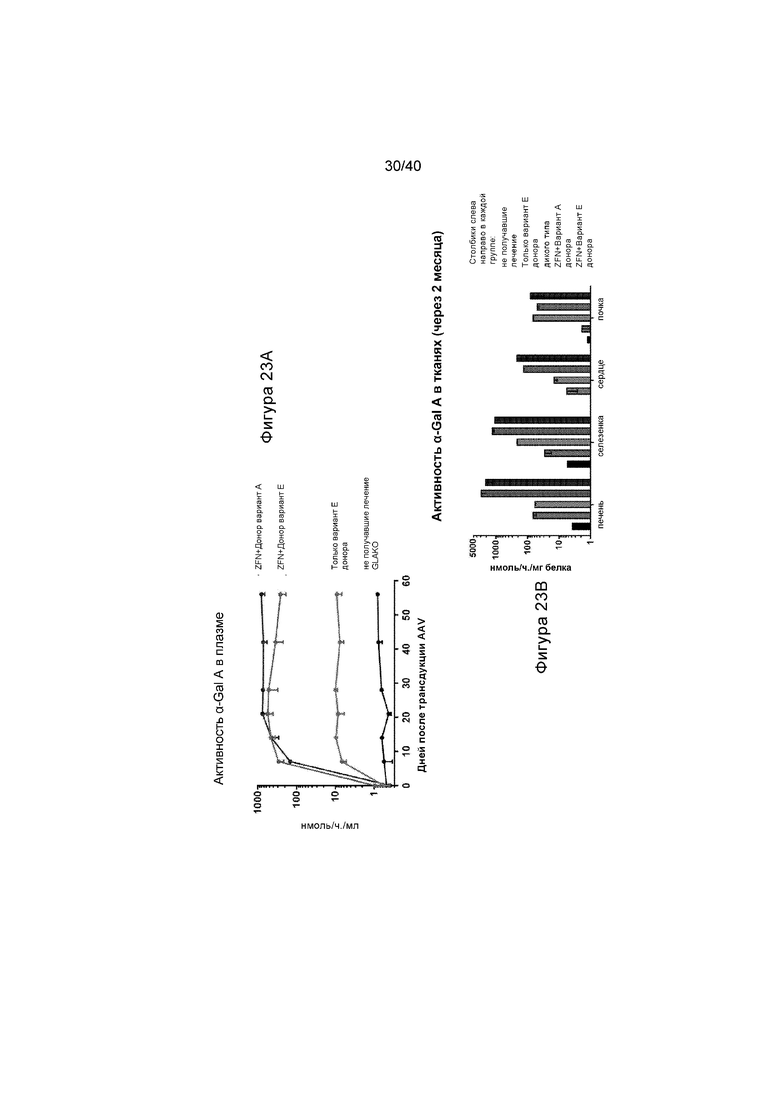
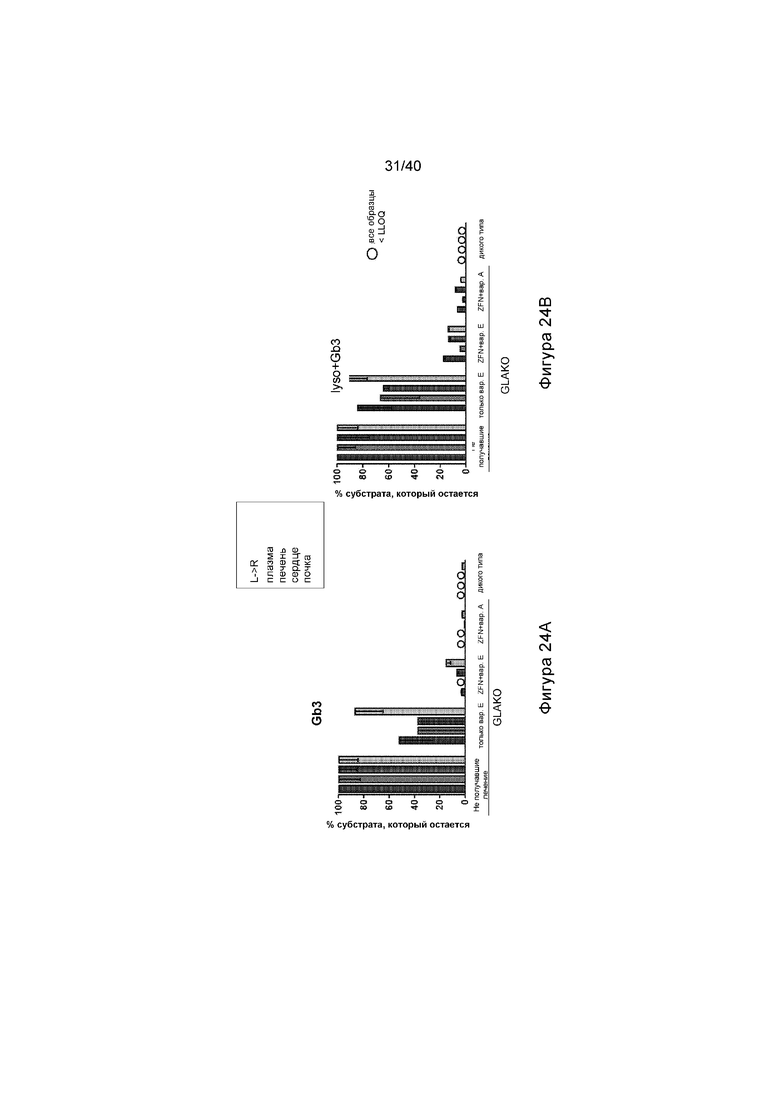
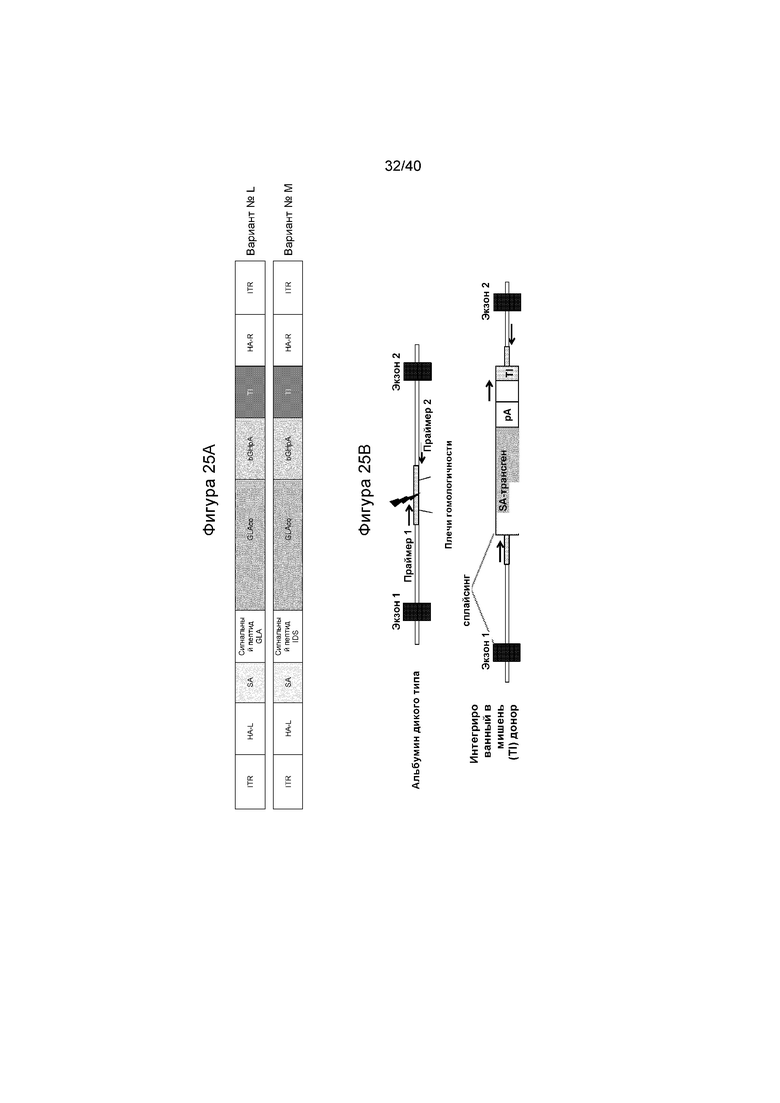
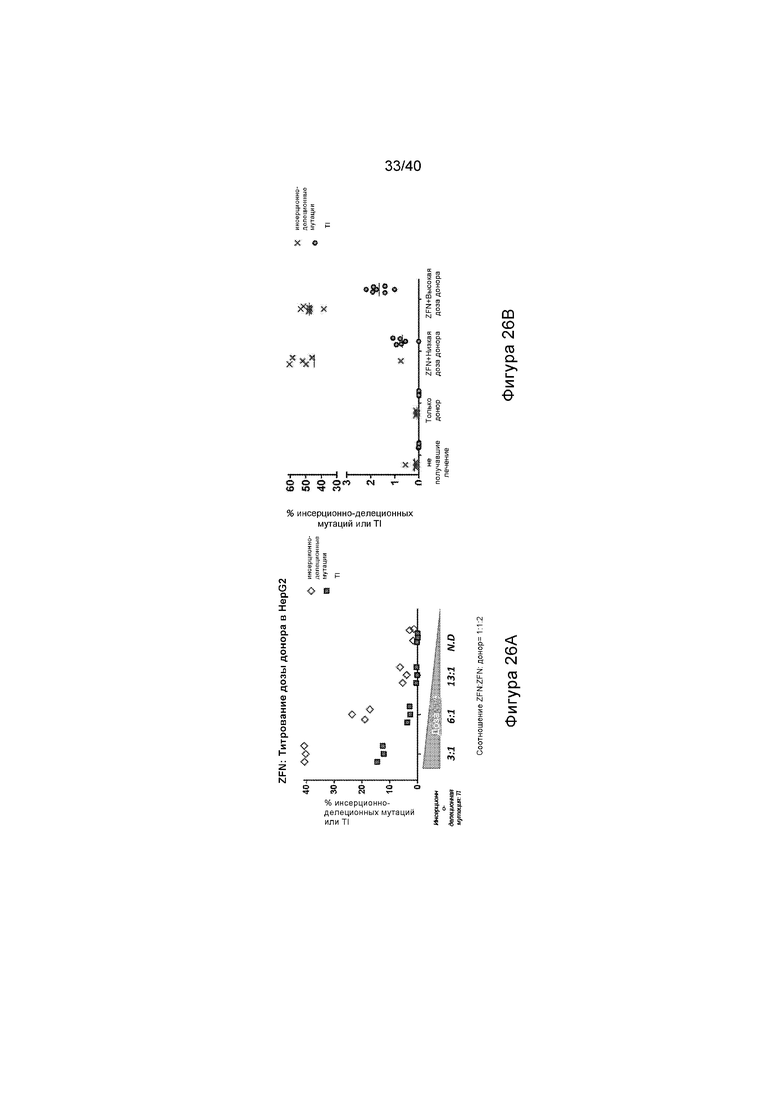
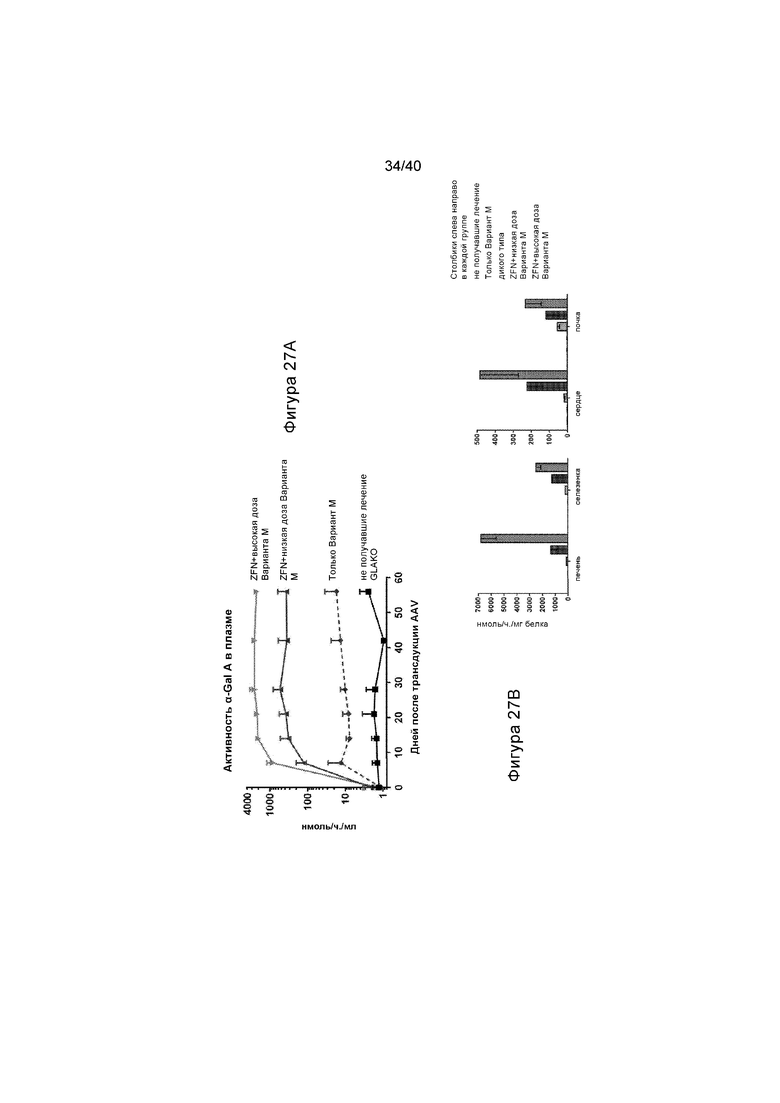


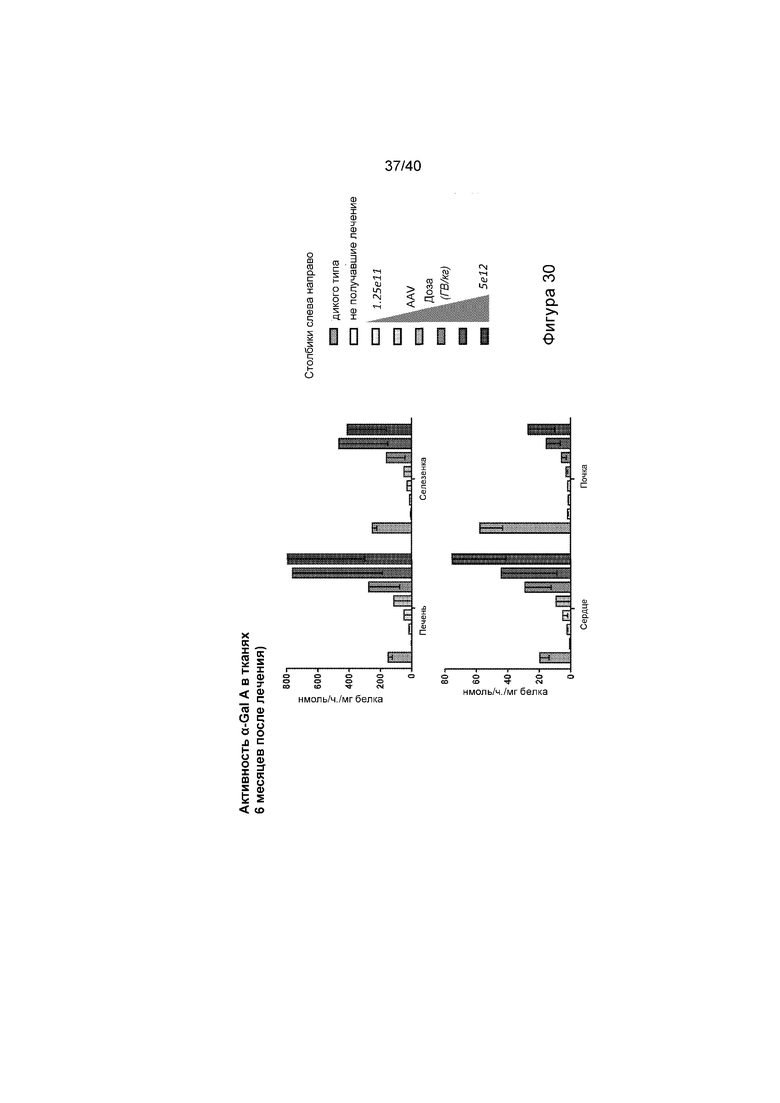

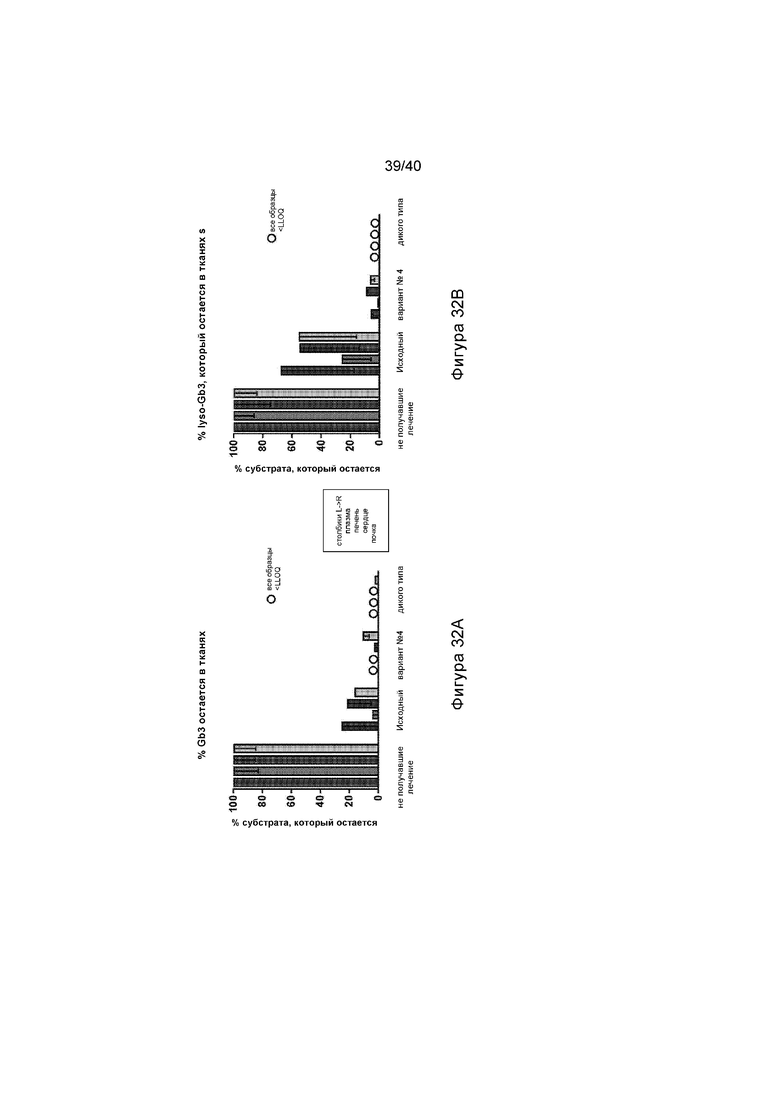
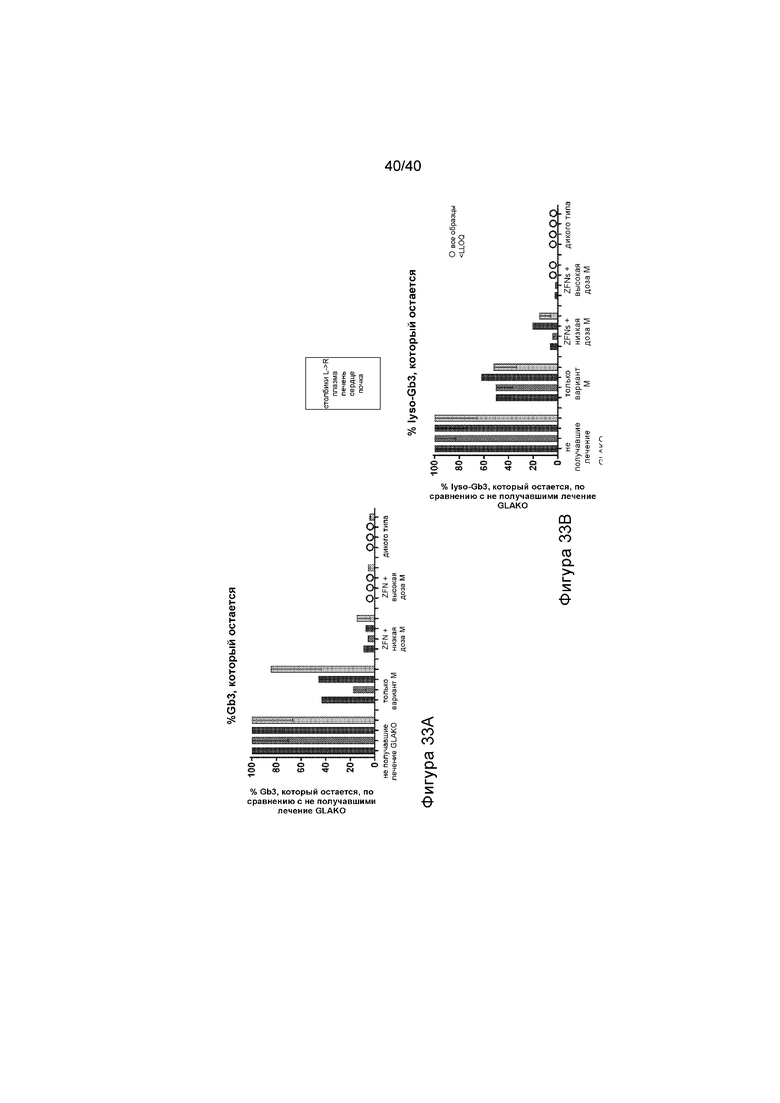
Изобретение относится к биотехнологии и представляет собой способ экспрессии по меньшей мере одного белка α галактозидазы A (α-Gal A) у нуждающегося в этом субъекта, включающий введение субъекту экспрессирующей аденоассоциированный вирус (AVV) конструкции, содержащей энхансер аполипротеина Е (APOE), связанный с промотором альфа-1-антитрипсина (hAAT), интрон гемоглобина бета человека (HBB)-IGG, сигнальный пептид, трансген кДНК, кодирующий по меньшей мере один белок альфа-галактозидазу (α-Gal A), и poly A сигнальную последовательность гормона роста крупного рогатого скота. Изобретение касается также фармацевтически приемлемой композиции, содержащей AVV экспрессирующую конструкцию, для лечения болезни Фабри. Изобретение позволяет эффективно лечить болезнь Фабри. 6 н. и 8 з.п. ф-лы, 33 ил., 5 табл., 11 пр.
1. Способ экспрессии по меньшей мере одного белка α галактозидазы A (α-Gal A) у нуждающегося в этом субъекта, включающий введение субъекту экспрессирующей аденоассоциированный вирус (AVV) конструкции, содержащей энхансер аполипротеина Е (APOE), связанный с промотором альфа-1-антитрипсина (hAAT), интрон гемоглобина бета человека (HBB)-IGG, последовательность, кодирующую сигнальный пептид, трансген кДНК, кодирующий по меньшей мере один белок альфа-галактозидазу (α-Gal A), и poly A сигнальную последовательность гормона роста крупного рогатого скота.
2. Способ по п. 1, отличающийся тем, что субъект страдает болезнью Фабри.
3. Способ по любому из пп. 1-2, отличающийся тем, что белок α-Gal A, экспрессированный из трансгена, снижает количество гликосфинголипидов у субъекта по меньшей мере в 2 раза.
4. Способ по п. 1, отличающийся тем, что трансген содержит последовательность α-GalA дикого типа или кодон-оптимизированную последовательность α-GalA.
5. Способ по любому из пп. 1-4, отличающийся тем, что сигнальный пептид представляет собой сигнальный пептид α-Gal A.
6. Способ по любому из пп. 1-5, где AVV экспрессирующую конструкцию вводят парентерально.
7. Способ по любому из пп. 1-6, где AVV экспрессирующую конструкцию вводят внутривенно.
8. Генетически модифицированная клетка для экспрессии белка α-Gal A, содержащая экспрессирующую аденоассоциированный вирус (AVV) конструкцию, содержащую энхансер аполипротеина Е (APOE), связанный с промотором альфа-1-антитрипсина (hAAT), интрон гемоглобина бета человека (HBB)-IGG, последовательность, кодирующую сигнальный пептид, трансген кДНК, кодирующий по меньшей мере один белок альфа-галактозидазу (α-Gal A), и poly A сигнальную последовательность гормона роста крупного рогатого скота.
9. Генетически модифицированная клетка по п. 8, отличающаяся тем, что клетка представляет собой стволовую клетку или клетку-предшественника.
10. Генетически модифицированная клетка по п. 8 или 10, отличающаяся тем, что клетка представляет собой клетку печени или мышц.
11. Применение AVV экспрессирующей конструкции для лечения болезни Фабри, где указанная AVV экспрессирующая конструкция содержит энхансер APOE, связанный с промотором hAAT, интрон HBB-IGG, последовательность, кодирующую сигнальный пептид, трансген кДНК, кодирующий по меньшей мере один белок альфа-галактозидазу (α-Gal A), и poly A сигнальную последовательность гормона роста крупного рогатого скота.
12. Фармацевтически приемлемая композиция, содержащая AVV экспрессирующую конструкцию, для лечения болезни Фабри, где указанная AVV экспрессирующая конструкция содержит энхансер APOE, связанный с промотором hAAT, интрон HBB-IGG, последовательность, кодирующую сигнальный пептид, трансген кДНК, кодирующий по меньшей мере один белок альфа-галактозидазу (α-Gal A), и poly A сигнальную последовательность гормона роста крупного рогатого скота, и фармацевтически приемлемый носитель или эксципиент.
13. Способ продуцирования белка α-Gal A для лечения болезни Фабри, при этом способ содержит экспрессирование белка α-Gal A в выделенной клетке в соответствии со способом по любому из пп. 1-8 и выделение белка α-Gal A, продуцируемого клеткой.
14. AVV экспрессирующая конструкция, содержащая энхансер APOE, связанный с промотором hAAT, интрон HBB-IGG, последовательность, кодирующую сигнальный пептид, трансген кДНК, кодирующий по меньшей мере один белок альфа-галактозидазу (α-Gal A), и poly A сигнальную последовательность гормона роста крупного рогатого скота, для применения в способе по любому из пп. 1-8.
| US 20060189561 A1, 24.08.2006 | |||
| US 20160208284 A1, 21.07.2016 | |||
| US 20140112896 A1, 24.04.2014 | |||
| Водогрейный прибор | 1928 |
|
SU11389A1 |
| ЛЕЧЕНИЕ α-ГАЛАКТОЗИДАЗНОЙ А НЕДОСТАТОЧНОСТИ | 2000 |
|
RU2248213C2 |

