Область изобретения
Настоящее изобретение относится к области иммунотерапии, а также к получению и использованию генетически трансформированных клеток, обладающих способностью восстанавливать толерантность к собственным антигенам у больных, страдающих аутоиммунными заболеваниями. Более конкретно, изобретение относится к конструированию и созданию гена, кодирующего энцефалитогенный эпитоп протеолипидного белка (PLP), к способам экспрессии эпитопа PLP in vitro и in vivo, способам секреции эпитопа PLP in vivo, а также способам переноса части гена PLP хозяину для подавления прогрессии иммунного ответа на собственные антигены, образовавшиеся из белков миелина.
Уровень техники
Иммунная система может отвечать на антиген двумя способами. Положительный ответ приводит к дифференцировке Т- и В-клеток, продукции антител и развитию иммунологической памяти. Отрицательный ответ приводит к подавлению или инактивации специфических лимфоцитов и толерантности. Толерантность можно определить как неспособность организма генерировать иммунный ответ против специфического антигена. В норме организм толерантен к своим собственным антигенам.
Считается, что аутоиммунные заболевания являются результатом неконтролируемого иммунного ответа, направленного на собственные антигены. Например, имеются указания на то, что у больных рассеянным склерозом такого рода ответ направлен на белое вещество центральной нервной системы и, в частности, на белки белого вещества. В конце концов разрушается миелиновая оболочка, окружающая аксоны. Это может приводить к параличам, сенсорным нарушениям и проблемам со зрением. При рассеянном склерозе наблюдается инфильтрация мозга Т-клетками и макрофагами. У больных рассеянным склерозом были выделены аутореактивные миелин-специфические Т-клетки, хотя и у нормальных индивидуумов обнаруживаются Т-клетки с указанной специфичностью. J.M.LaSalle et al., J.Immunol. 147:774-780 (1991), J.M.LaSalle et al., J.Exp.Med. 176:177-186 (1992), J.Correale et al., Neurology. 45:1370-1378 (1995). В настоящее время среди белков миелина, являющихся мишенью иммунного ответа при рассеянном склерозе, выделяют основный белок миелина (МРВ), протеолипидный белок (PLP) и миелин-олигодендроцитный гликопротеин (MOG). Те индивидуумы, у которых не развивается аутоиммунный ответ на собственные белки, считаются “толерантными” к собственным антигенам. Таким образом, доказательства того, что рассеянный склероз вызывается патогенными Т-клетками, являются непрямыми, однако близкое сходство характеристик заболевания с таковыми, наблюдаемыми при изучении экспериментального аутоиммунного энцефаломиелита (ЕАЕ) на мышах, дают основания полагать, что рассеянный склероз вызывается аберрантным иммунным ответом, обусловленным Т-клетками.
Экспериментальный аутоиммунный энцефаломиелит (ЕАЕ), “мышиная” модель рассеянного склероза человека, является предметом интенсивных и плодотворных исследований в течение нескольких лет и характеризуется гистопатологическими и клиническими проявлениями, аналогичными с таковыми для возвратных форм рассеянного склероза. The T Lymphocyte in Experimental Allergic Encephalomyelitis. Ann.Rev.Immunol. 8:579-621 (1990). ЕАЕ можно вызвать у мышей SJL введением гомогената спинного мозга мыши (MSCH), МРВ, PLP, введением синтетических пептидов, чьи последовательности соотвествуют главным энцефалитогенным эпитопам основного белка миелина, МРВ 84-104, протеолипидного белка, PLP 139-151, или адоптивным переносом активированных клеток CD4' ТH1, но не ТH2 клеток, специфичных к энцефалитогенным эпитопам. При ЕАЕ главные энцефалитогенные эпитопы миелиновых последовательностей, такие как МРВ, также могут активировать человеческие Т-клетки нескольких различных гаплотипов, включая HLA-DR2. R. Martin, et al., J.Exp.Med. 173:19-24 (1992). Экспериментальное заболевание характеризуется прерывистым течением неврологической дисфункции, периваскулярной инфильтрацией мононуклеарами и демиелинизацией. Повреждения центральной нервной системы, по-видимому, обусловливаются воспалительными цитокинами, которые могут неспецифически активировать дополнительные моноциты и макрофаги. J.E.Blalock, The Immune System. Our Sixth Sense. The Immunologist, 2:8-15 (1994).
Притом, что первичная атака при ЕАЕ может быть индуцирована введением Т-клеток, специфичных как к МБР, так и к LPL, детальное изучение реактивностей Т-клеток при первичной и последующих вспышках заболевания показало наличие Т-клеток, взаимодействующих со специфичностью, отличающейся от специфичности индуцирующих эпитопов. Эта экспансия энцефалитогенных эпитопов была обозначена как “распространение детерминанты”. S.D.Miller and W.J.Karpus, Immunology Today 15:356-361 (1994), P.V.Lehman, T.Forsthuber, A.Miller and E.E.Sercarz, Nature 358:155-157 (1992), H.Jiang, S.-I.Zhang and B.Pernis, Sclence 256:1213-1215 (1992). Поэтому антиген-специфичное лечение было бы более эффективным при его возможно более раннем назначении до начала возрастания сложности эпитопов и развития возможного неспецифического воспаления.
Целью иммунотерапии является восстановление толерантности без подавления иммунной системы в целом, что могло бы привести к таким осложнениям, как инфекция, геморрагии и рак. Используемые в настоящее время препараты для лечения аутоиммунных заболеваний являются неспецифическими иммуносупрессорными агентами, такими как противовоспалительные агенты или препараты, блокирующие пролиферацию клеток или подавляющие провоспалительные цитокины. В целом данные агенты эффективны в течение ограниченного периода и приводят к тяжелым осложнениям.
Было бы желательно воздействовать на иммунную систему более специфическим образом, с тем чтобы контролировать ответ на собственные антигены и теоретически “лечить” заболевание без подавления всей иммунной системы в целом. В последние годы было предложено и испытано несколько схем специфической иммунотерапии, большинство из которых не нашли практического применения, поскольку оказались неэффективны в отношении людей. Например, могут быть синтезированы высокоаффинные пептиды, взаимодействующие с молекулами МНС класса II и предотвращающие связывание энцефалитогенных пептидов, и, тем самым, предотвращающие активацию патогенных Т-клеток. A.Franco et al., The Immunologist 2:97-102 (1994). Недостатком данного подхода является трудность достижения эффективных концентраций ингибиторных пептидов in vivo. G.Y.Ishioka et al., J Immunol. 152:4310-4319. При альтернативной стратегии пептиды, являющиеся аналогами энцефалитогенных последовательностей, антагонизируют с рецепторами антиген-специфических Т-клеток, делая их неактивными, хотя точный механизм пока остается неизвестным. S.C.Jameson et al., J.Exp.Med. 177:1541-1550 (1993), N.Karin et al., J.Exp.Med. 180:2227-2237 (1994), V.K.Kuchroo et al., J.Immunol. 153:3326-3336 (1994). Было изучено также оральное введение миелина, индуцирующее состояние иммунологической толератности, которое, по-видимому, обусловлено индукцией супрессорных Т-клеток или анергией. H.L.Weiner et al., Annu.Rev.Immunol. 12:809-837 (1994), C.C.Whitacre et al., J.Immunol. 147:2155-2163 (1991), SJ.Khoury et al., J.Exp.Med. 176:1355-1364 (1992). Показано, что подобное лечение эффективно для некоторых, но далеко не всех индивидуумов. H.L.Weiner et al., Science. 259:1321-1324 (1993). Таким образом, очевидна необходимость разработки новых способов лечения рассеянного склероза и других аутоиммунных заболеваний с использованием эффективных иммуноспецифических подходов.
Краткое описание изобретения
Настоящее изобретение направлено на устранение недостатков, известных из уровня техники. В целом изобретение основано на том открытии, что технологии рекомбинантных ДНК и переноса клеток могут быть использованы для восстановления толерантности к собственным тканям. Настоящее изобретение относится к способу лечения больного, страдающего рассеянным склерозом, который заключается во введении больному гистосовместимых фибробластов, притом, что данные гистосовместимые фибробласты были обработаны in vitro с введением в них фрагмента ДНК, кодирующего аминокислоты 101-157 протеолипидного белка, данный фрагмент ДНК был введен в указанные гистосовместимые фибробласты in vitro при помощи рекомбинантного ретровирусного вектора, данный фрагмент ДНК также содержит гидрофобную лидерную последовательность и данная гидрофобная лидерная последовательность обеспечивает возможность синтеза аминокислотной последовательности 101-157 протеолипидного белка в эндоплазматическом ретикулуме указанных гистосовместимых фибробластов с дальнейшей конститутивной секрецией, данный фрагмент ДНК также содержит один или несколько сайтов рестракции, что обеспечивает возможность встраивания дополнительных последовательностей генов и продукт гена или продукты генов данного фрагмента ДНК будут экспрессироваться в организме данного больного в терапевтически эффективных количествах с восстановлением Т-клеточной толерантности.
Объектом изобретения является также культура фибробластов, обработанная in vitro введением в нее фрагмента ДНК, кодирующего аминокислоты 101-157 протеолипидного белка, при этом фибробласты являются гистосовместимыми.
Фрагмент ДНК может быть введен с помощью рекомбинантного вектора, который может являться ретровирусным.
В предпочтительном варианте осуществление изобретения фрагмент ДНК также содержит бокс Козака, обеспечивающий эффективную трансляцию мРНК, транскрибируемой на данном фрагменте ДНК, данный фрагмент ДНК также содержит на 3'-конце кодон, соответствующий заряженной аминокислоте, в результате чего указанный белок не закрепляется на мембране.
При осуществлении предпочтительного варианта изобретения клетки продуцирующей линии РА317 трансдуцируют ретровирусным PLP-вектором и получают супернатант, содержащий рекомбинантный ретровирус. Продуцирующая линия РА317 была получена доктором A.Dusty Miller, тщательно охарактеризована и ее применение разрешено Администрацией по контролю пищевых продуктов и лекарственных препаратов США (FDA) для клинических испытаний способов лечения генетических заболеваний и рака. Miller and Baltimore, Mol.Cell.Biol. 6:2895-2902 (1986), W.F.Anderson, Science. 256:808-813.
Краткое описание чертежей
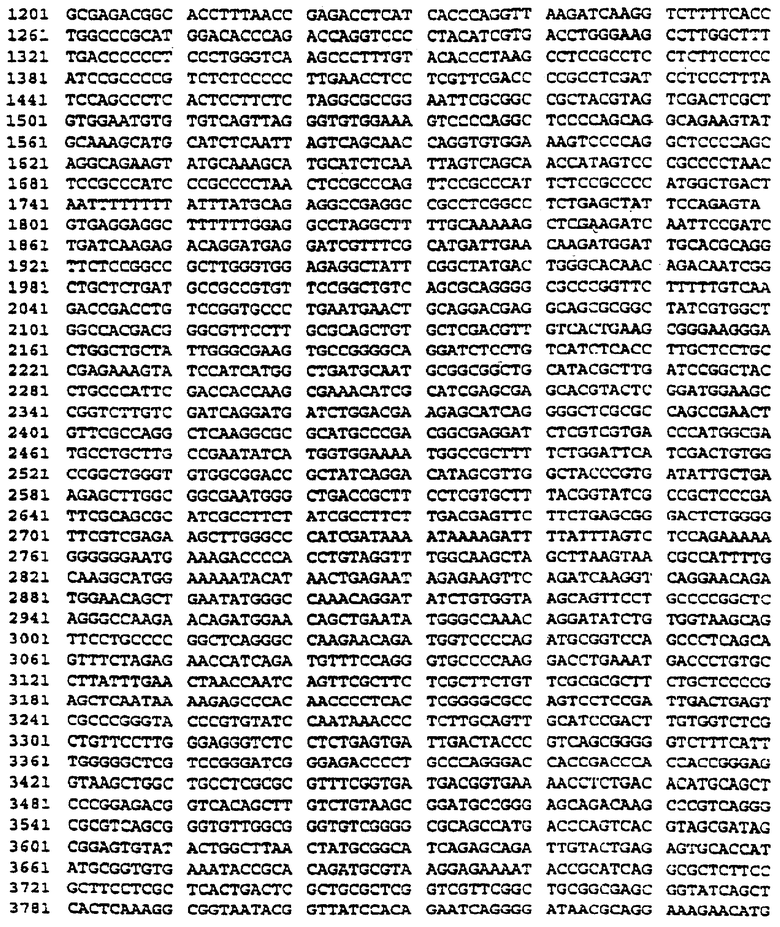
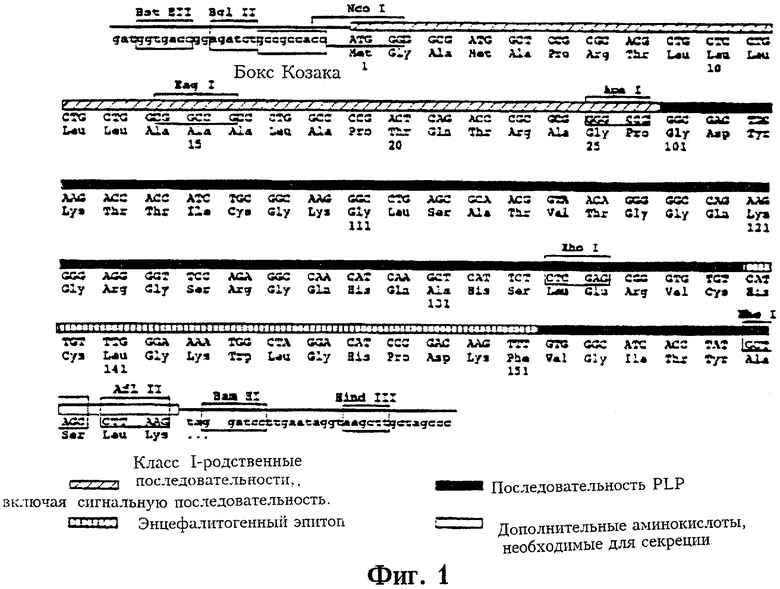
Фиг.1 представляет карту части гена PLP, на которой показаны сайты рестрикции и последовательность генного продукта.
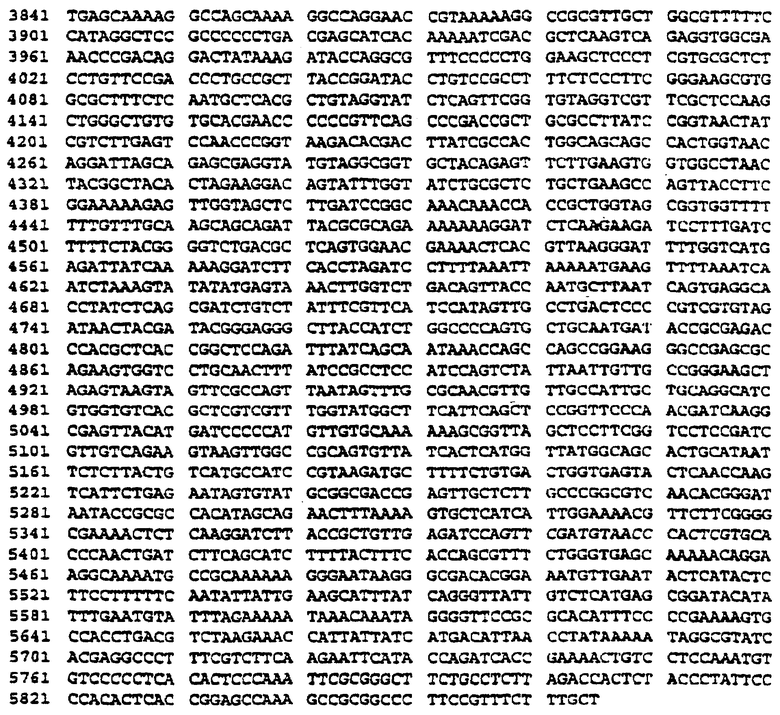
Фиг.2 представляет карту вектора G1XSvNa с сайтами рестрикции и функциональными особенностями. На Фиг.2b приведена полная последовательность ДНК вектора G1XSvNa.
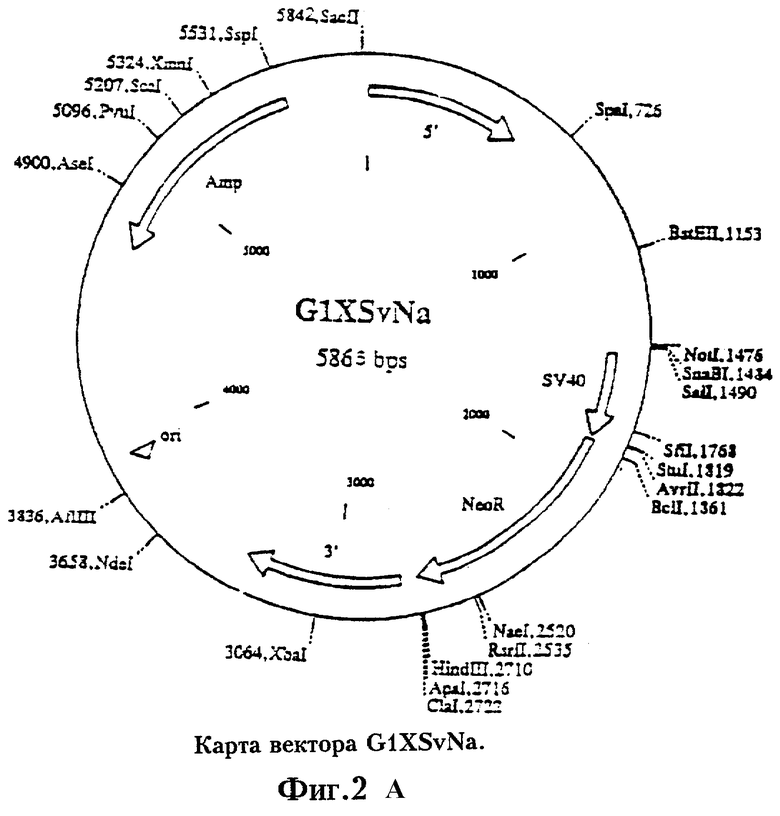
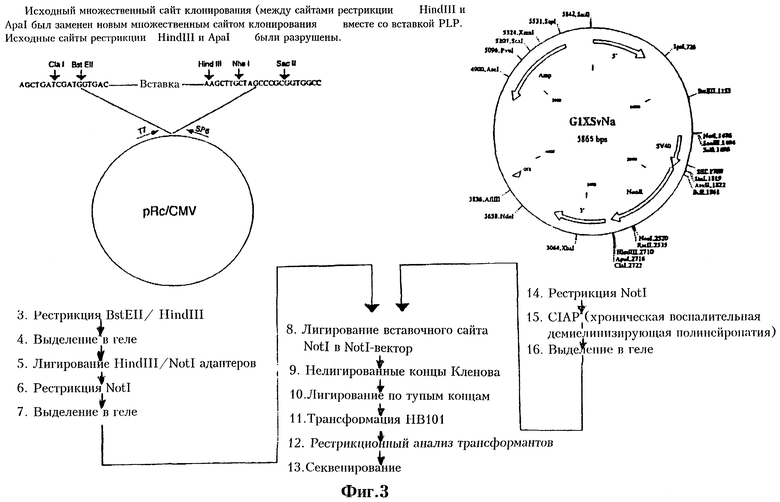
На Фиг.3 приведена схема конструирования вектора G1XSvNa PLP, содержащего вставку гена PLP.
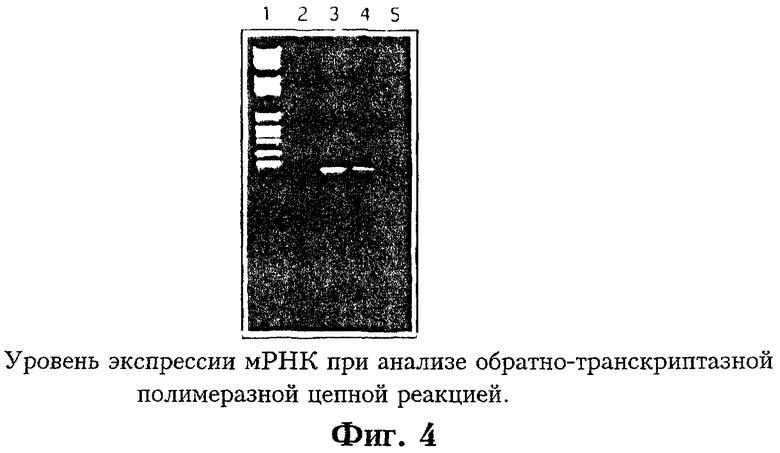
На Фиг.4 показан уровень мРНК, экспрессируемой в трансфицированных и трансдуцированных фибробластах при анализе обратно-транскриптазной полимеразной цепной реакцией. Линия 1 соответствует стандартам молекулярного веса, линия 2 - отрицательный контроль из мнимо трансфицированных фибробластов, линия 3 - положительный контроль с плазмидой, содержащей ген PLP, линия 4 - кДНК из PLP - трансфицированных фибробластов SJL, линия 5 - кДНК из PLP - трансдуцированных фибробластов SJL.
На Фиг.5 представлен уровень белка PLP в супернатантах трансдуцированных фибробластов при определении ELISA.
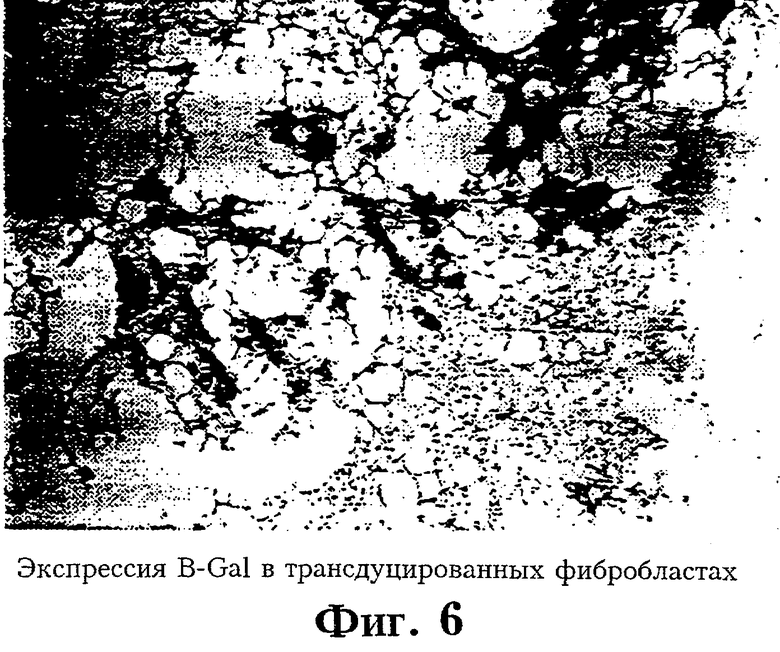
На Фиг.6 показан уровень экспрессии В-Gal в трансдуцированных фибробластах.
На Фиг.7 приведена система клинического учета хронического ЕАЕ.
На Фиг.8 приведена система гистологического учета хронической ЕАЕ.
Фиг.9 иллюстрирует клиническую оценку ЕАЕ у мышей, обработанных ретровирус-трансдуцированными фибробластами.
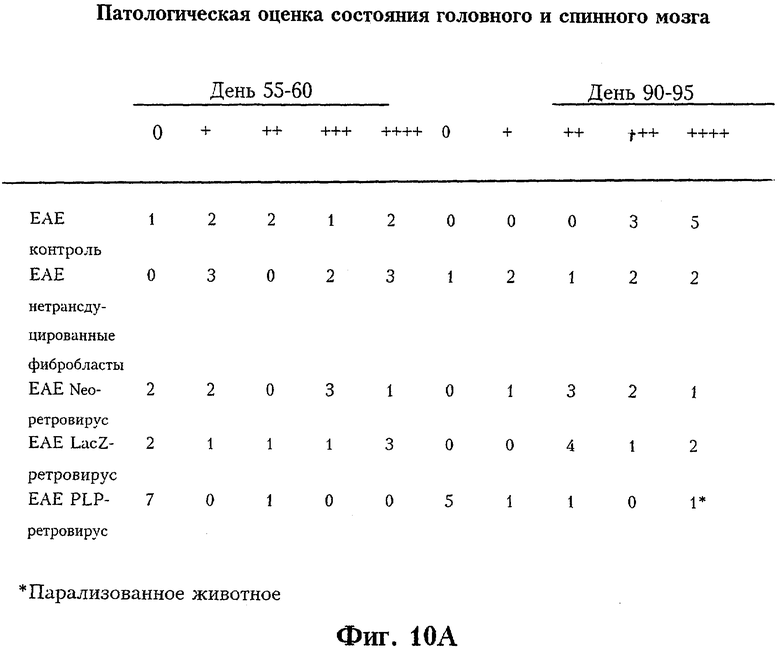
Фиг.10а отражает патологическую оценку спинного и головного мозга у мышей SJL, обработанных ретровирус-трансдуцированными фибробластами. На Фиг.10b суммирована патологическая оценка спинного и головного мозга от дня 55-60 до дня 90-95.
Фиг.11 демонстрирует гистологию мышей SJL, обработанных ретровирус-трансдуцированными фибробластами.
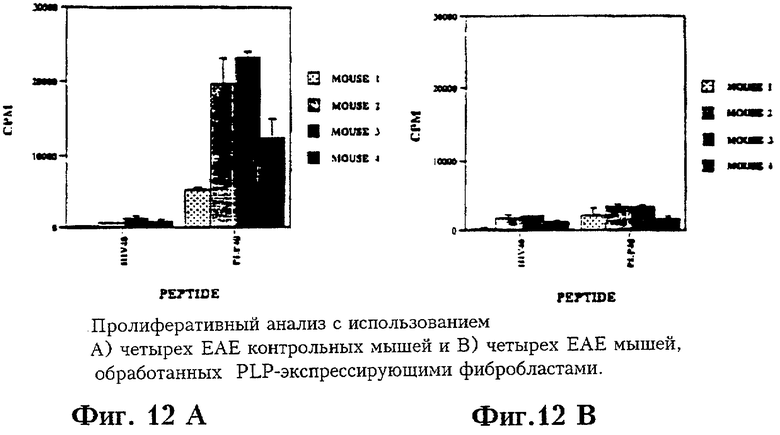
На Фиг.12 приведены результаты пролиферативных тестов на мышах с ЕАЕ, обработанных PLP- экспрессирующими фибробластами.
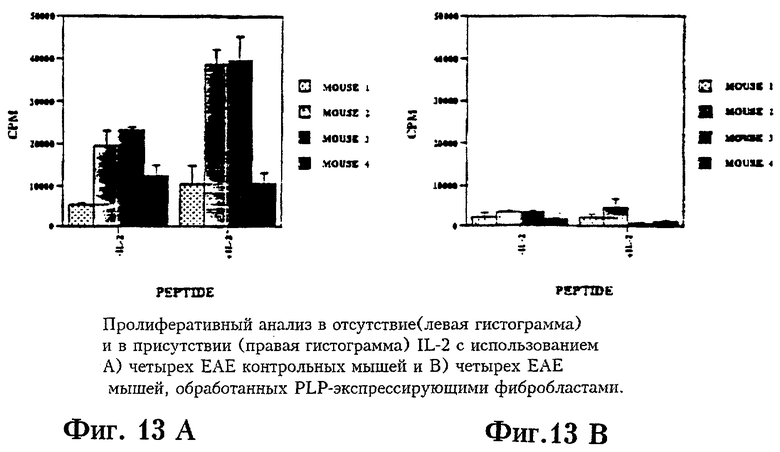
На Фиг.13 приведены результаты пролиферативных тестов с IL-2 и без IL-2 на мышах с ЕАЕ, обработанных PLP- экспрессирующими фибробластами.
Подробное описание изобретения
Как указывалось выше, настоящее изобретение относится к применению генетически трансформированных клеток для восстановления толератности к собственным антигенам у больных, страдающих аутоиммунным заболеванием. Генетически трансформированные клетки могут быть любыми клетками млекопитающих. Термин “генетически трансформированный” относится к клетке, в которую введены один или несколько рекомбинантных генов, таких как ген, кодирующий эпитоп собственного антигена.
Ген представляет собой дезоксирибонуклеотидную последовательность, кодирующую аминокислотную последовательность. Гены, вводимые рекомбинантными методами, могут быть в виде синтетического олигонуклеотида, кДНК (т.е. они не будут содержать интронов), в виде копии геномной последовательности или гибридного гена, слитого из двух или более последовательностей генов. Дополнительно, ген может быть связан с одной или более нуклеотидной последовательностью, способной направлять экспрессию продукта гена. Элементы последовательностей, способные влиять на экспрессию гена, включают, но не ограничиваются перечисленными, промоторы, энхансерные элементы, сигналы терминации транскрипции, сайты полиаденилирования, последовательности бокса Козака для повышения эффективности трансляции и лидерные последовательности. Дополнительно, последовательность гена может содержать рестрикционные сайты, позволяющие осуществлять вставки дополнительных последовательностей. Предпочтительно, ген должен содержать лидерную последовательность с тем, чтобы продукт гена синтезировался в цитоплазматическом ретикулуме с последующей конститутивной секрецией.
Рекомбинантные гены, предназначенные для введения в клетки с целью их генетической трансформации, могут кодировать один или несколько эпитопов, фрагментов, доменов или мини-белков белкового антигена. Примерами белков, из которых могут быть получены эпитопы, фрагменты, домены или мини-белки, могут служить (без ограничения перечисленными) белки миелина, рецептор ацетилхолина, рецептор TSH и коллаген.
Считается, что белковые аутоантигены, служащие мишенью аутоиммунного ответа, являются высококонсервативными как в пределах вида, так и между видами. Поэтому, несмотря на то, что настоящее изобретение прежде всего направлено на способ лечения людей, данный способ может быть применен и для лечения животных. Примерами аутоиммунных заболеваний, опосредованных Т-клетками, для лечения которых может применяться настоящее изобретение, служат (без ограничения перечисленными) рассеянный склероз, миастения гравис, системная красная волчанка, псориаз, ювенильный диабет, ревматоидный артрит, заболевания щитовидной железы и хроническая воспалительная демиелинизирующая полинейропатия (CDIP).
Векторы экспрессии обычно представляют собой дезоксирибонуклеотидные молекулы, сконструированные для контролируемой экспрессии одного или нескольких представляющих интерес генов. Векторы могут содержать одну или несколько оперативно связанных с геном нуклеотидных последовательностей, которые контролируют экспрессию нужного гена или генов. Имеется большое разнообразие доступных векторов экспрессии и квалифицированный специалист может легко выбрать соответствующий вектор. Кроме того, в обычных лабораторных руководствах описаны методы рекомбинантных ДНК и способы получения и использования векторов экспрессии. Дополнительно вектор может содержать селективный маркер, например, ген устойчивости к антибиотику.
Ген можно ввести в клетку млекопитающих при помощи любой методики переноса генов. Примерами таких методик, без ограничения перечисленными, служат перенос генов посредством вирусной РНК, т.е. ретровирусная трансдукция, перенос генов посредством вирусной ДНК, электропорация, кальций-фосфатная трансфекция, микроинъекция или перенос генов посредством липосом. Выбор процедуры, необходимой для получения генетически трансформированных клеток, секретирующих продукт нужного гена, будет зависеть от природы и свойств выбранных клеток. Конкретные технологии введения генов в клетки широко известны специалистам в данной области исследований.
Следующие примеры раскрывают конструирование части гена PLP, показывают экспрессию продукта гена PLP in vitro и in vivo и эффекты продукта гена PLP in vivo. Они приведены для иллюстрации изобретения и не ограничивают его объем.
ПРИМЕР 1
КОНСТРУИРОВАНИЕ ГЕНА PLP
У мышей SJL/J энцефалитогенный эпитоп PLP представлен аминокислотами 139-151. N.Takahashi et al., Cell, 42:139-148 (1985), K.Sakai et al., J.Neuroimmunol. 19:21-32 (1988), D.H.Kono et al., J.Exp.Med. 168:213-227. Описанный в настоящем изобретении вектор сконструирован таким образом, чтобы кодируемый им генный продукт конститутивно секретировался из фибробластов. Поскольку полный белок PLP является гидрофобным трансмембранным белком (H.-J.Diehl, M.Schaich, R.-M.Buzinski and W.Stoffel, PNAS U.S.A. 83:9807-9811 (1986)), а энцефалитогенный эпитоп является экстраклеточным, была сконструирована плазмида, кодирующая аминокислоты 101-157 и дополнительные аминокислоты, необходимые для секреции. Эта последовательность имеет гидрофильный характер.
1. Синтез олигонуклеотидов и конструирование вектора PLP pRc/CMV
Олигонуклеотиды могут быть синтезированы вручную, скажем, фосфотриэфирным методом, как описано, например, R.L.Letsinger et al., J.Am. Chem. Soc. 98:3655 (1967) (работа приводится в качестве ссылки). Из уровня техники хорошо известны и другие методы. См. также Maneucci and Caruthers, J.Am. Chem. Soc. 103:3185 (1981) (работа приводится в качестве ссылки).
Однако, предпочтительно получение последовательности нужного гена автоматическим синтезом индивидуальных олигонуклеотидов в концентрациях 2 мкМ. Для получения последовательности аминокислот PLP 101-157 синтез ДНК проводили на синтезаторе ДНК Perkin Elmer/Applied Biosystems Division Model 394 с использованием циатоэтил-защищенных фосфорамидатов. Диметокситритильную группу (DMT) не удаляли с 5'-гидроксильной группы для облегчения очистки. После обычного снятия олигонуклеотидов со смолы концентрированным гидроксидом аммония и депротекции при 55°С в течение 16 часов, олигонуклеотиды очищали с использованием патронов для очистки олигонуклеотидов (ОРС) согласно инструкциям производителя (Applied Biosystems Inc.). Были синтезированы пять олигонуклеотидов со следующими последовательностями:
O L G 1 5 ' -
CGGCGACTACAAGACCACCATCTGCGGCAAGGGCCTGAGCGCAACGGTA
АСА
GGGGGCCAGAAGGGGAGGGGTTCCAGAGGCCAACATCAAGCTCATTCTC
TCGAGC-3'
O L G 2 5 ' -
GAGCTTGATGTTGGCCTCTGGAACCCCTCCCCTTCCTGGCCCCCTGTTAC
CGTTGCG
CTCAGGCCCTTGCCGCAGATGGTGGTCTTGTAGTCGCCGGGCC-3'
O L G 3 5 ' -
GGGTGTGTCATTGTTTGGGAAAATGGCTAGGACATCCCGACAAGTTTGTG
GGCATCACCTATGCTAGCCTTAAGTAGGATCCTTGAATAGGTA-3'
O L G 4 5 ' -
AGCTTACCTATTCAAGGATCCTACTTAAGGCTAGCATAGGTGATGCCCA-3'
O L G 5 5 ' -
CAAACTTGTCGGGATGTCCTAGCCATTTTCCCAAACAATGACACACCCGCTCG
AGAGAAT-3'
Каждый очищенный олигонуклеотид высушивали в вакууме, промывали 1 мл стерильной дважды дистиллированной воды и затем опять упаривали досуха в вакууме (упариватель Speed vac, Savant Inc.). 80 пМ каждого олигомера подвергали кинированию при 37°С в течение 1 часа, ресуспендируя в 56,6 мкл 1X киназного буфера (полинуклеотидкиназный буфер, Boehringer Mannheim, Indianapolis, IN), содержащего 10 единиц полинуклеотидкиназы (Boehringer Mannheim, Indianapolis, IN) и 100 мкМ АТР. Для гибридизации каждый индивидуальный олигонуклеотид смешивали в 2xSSC (0.03 М цитрат натрия, рН 7.0 и 0.3 М NaCl) в пробирке для ПЦР с соответствующим комплементарным олигомером. Объем каждой гибридизационной смеси доводили до 200 мкл. Олигомер OLG1 гибридизовали с олигомером OLG2, а олигомеры OLG4 и OLG5 гибридизовали с олигомером OLG3. Гибридизацию проводили в термосайклере Perkin Elmer 9600 по следующей программе: 1) 99,9° 2 минуты и 2) от 99,9° до 4° 15 минут. При понижении температуры до 4°С, когда температура в термосайклере опускалась до 37°С, раствор, содержащий олигомерный дуплекс OLG1 и OLG2, смешивали с раствором, содержащим олигомеры OLG3, OLG4 и OLG5. Затем снижение температуры продолжали до 22°С. Затем добавляли 5 единиц (5 мкл) лигазы Т4 (Boehringer Mannheim, Indianapolis, IN) и 45 мкл фирменного 10Х Т4 ДНК-лигазного буфера (Boehringer Mannheim, Indianapolis, IN) и лигирование продолжали в течение ночи при 10°С.
Лигированную ДНК осаждали двумя объемами 100%-ного этанола и инкубировали при -70°С 1 час. Преципитат центрифугировали 30 минут при 17 000 × g и 4°С. Супернатант отбрасывали, а осадок промывали 1 мл 70%-ноги этанола и центрифугировали 10 минут при 17 000 × g и 4°С. Осадок ДНК высушивали в вакууме (упариватель Speed vac, Savant Inc.) и ресуспендировали в 45 мкл стерильной дважды дистиллированной воды.
ДНК соответствующего молекулярного веса выделяли при помощи электрофореза. К образцу добавляли 5 мкл 10Х буфера для нанесения (6.25 г Фиколла и 0.93 г динатриевой соли ЕДТА/ 25 мл 10% SDS, оранжевый G, ксилен цианол и бромфеноловый синий) и наносили на акриламидный гель с мочевиной размером 14.5 см × 16 см × 0.15 мм (7М мочевина/8% акриламид с 1.1% бисакриламида). В качестве буфера для геля и для электрофореза использовали ТВЕ (89 мМ Tris, 89 мМ борной кислоты и 2 мМ ЕДТА, рН 8.0). Образец подвергали электрофорезу при 35 мА до тех пор, пока полоса красителя оранжевого G не доходила до края геля на 1 см. Акриламидный гель дважды промывали водой по 5 минут. После последней промывки гель инкубировали 3 минуты в 500 мл раствора, содержащего 10 мкл раствора бромида этидия (10 мг/мл) и просматривали в УФ-свете. Полосу, соответствующую лигированной ДНК, вырезали из геля и фрагментировали на маленькие кусочки для электроэлюции в аппарате IBI (модель UEA: International Biotechnologies Inc., New Haven, CT).
Для электроэлюции солевую ловушку аппарата заполняли 125 мкл раствора 7М ацетата натрия с бромфеноловым синим. Камеру для буфера заполняли 1/2Х ТВЕ. Образец подвергали электроэлюции при 85 В в течение 1 часа. После удаления элюированной ДНК ячейку для образца промывали 1/2Х ТВЕ и смыв объединяли с первоначальным элюатом. Затем элюированную ДНК осаждали в течение ночи при -70°С двумя объемами 100%-ного этанола. Преципитат осаждали, промывали, как описано ранее, и ресуспендировали в 15 мкл стерильной дважды дистиллированной воды.
Перед лигированием элюированной части гена PLP в вектор pRc/CMV (Invitrogen, San Diego, CA), вектор pRc/CMV рестрицировали эндонуклеазами Ара I и Hind III согласно рекомендациям производителя (Boehringer Mannheim, Indianapolis, IN). Затем ресуспендированный конструкт гена PLP добавляли к 5 мкл смеси, содержащей 0.3 мкг разрезанного вектора pRc/CMV (2 мкл), 1 единицу лигазы Т4 (1 мкл) (Boehringer Mannheim, Indianapolis, IN) и 2 мкл 10Х T4 ДНК-лигазного буфера (Boehringer Mannheim, Indianapolis, IN). Затем лигированным вектором трансформировали компетентные клетки линии AG1.
Трансформацию осуществляли, смешивая смесь для лигирования с клетками AG1 и инкубируя на льду 20 минут. Затем смесь клетки/вектор инкубировали при 42°С 2 минуты и высевали на ночь на агар с бульоном Лурии (LB; Bio 101, Vista, CA), в состав которого входил ампициллин (80 мг/мл) (Sigma, St.Louis, МО). Колонии скринировали в отношении правильной последовательности вектора, выделяя плазмидную ДНК, с последующим ее секвенированием.
Для выделения плазмид использовали коммерческие наборы Wizard Minipreps (Promega, Madison, WI). Колонии перекалывали с чашек с LB/Amp агаром и подращивали 3,5 часа в 5 мл среды LB (Bio 101, Vista, CA) с добавлением 80 мг/мл ампициллина (Sigma, St. Louis, МО). Для осаждения клеток 3 мл среды центрифугировали при 17 000 × g при комнатной температуре в течение 1 минуты. Выделение плазмиды проводили согласно рекомендациям производителя. Для секвенирования использовали 1 мкг плазмиды.
Последовательности олигонуклеотидов могут быть проверены хорошо известными из уровня техники методами, такими как описанный Sanger et al., PNAS U.S.A. 70:1209 (1973) или методом Максама-Гилберта, Meth.Enzymology. 65:499 (1977) (оба метода упоминаются в настоящем описании в качестве ссылок). Предпочтительно секвенировать плазмиду при помощи автоматического секвенатора. В случае конструкта pRc/CMV PLP плазмида была секвенирована автоматическим флуоресцентным методом секвенирования ДНК (Perkin Elmer/Applied Biosystems Inc., Foster City, CA) с использованием следующих праймеров: GATTTAGGTGACACTATAG и TAATACGACTCACTATAGGG. Данные праймеры использовали в качестве затравки вектор, фланкированный боксом Козака и “стоп”-сайтом всего конструкта. На Фиг.1 приведена карта части гена PLP с указанием сайтов рестрикции и последовательности генного продукта. На 5'-конце конструкта авторами была ранее встроена гидрофобная лидерная последовательность гена МНС класса IId с тем, чтобы генный продукт синтезировался в эндоплазматическом ретикулуме (ER) для последующей конститутивной секреции. Linsk et al., J.Exp.Med. 164:794-813 (1996). Кроме того, на 3'-конце был добавлен лизиновый кодон, чтобы исключить прикрепление белка к мембране. В конструкт был включен бокс Козака, чтобы обеспечить эффективную трансляцию. В конструкт были также вставлены сайты рестрикции Afl II и Barn HI, с тем чтобы сделать возможной вставку других эпитопов.
ПРИМЕР 2
ЭКСПРЕССИЯ БЕЛКА PLP IN VITRO
Для того чтобы продемонстрировать, что PLP-вектор кодирует конститутивно секретируемый белок, были проведены следующие эксперименты. В частности, уровни мРНК PLP оценивали в фибробластах SJL, трансфицированных вектором pRc/CMV PLP, а уровни мРНК и белка PLP оценивали в фибробластах SJL, трансфицированных вектором pG1PLPSvNa.
1. Получение культур фибробластов
Сингенные фибробласты (полученные от мышей SJL), предоставленные доктором G.Dveskler (Uniformed Services University, Bethesda, MD), выращивали при 37°С в среде DMEM с добавлением 5% глутамина и 10% эмбриональной телячьей сыворотки. Клетки собирали и замораживали в концентрации 1×107 клеток на ампулу. Аликвоты замороженных клеток тестировали на присутствие микоплазмы, стерильность и жизнеспособность.
2. Ретровирусные конструкты
Был сконструирован рекомбинантный ретровирусный вектор, в который были встроены экзогенные гены. Стратегия клонирования была нацелена на получение вектора pG1XSvNa (W.French Anderson, University of Southern California), содержащего вставку PLP из вектора pRc/CMV-PLP. Вектор pG1XSvNa, как и большинство использованных в доклинических и клинических испытаниях ретровирусных векторов, был получен на основе ретровируса лейкоза мышей Молони (Mo-MuLV). Rosenberg et al., N.Eng.J.Med. 323:570-578 (1990), Culver et al., Science. 256:1550-1552 (1992). Вектор pGlXSvNa имеет размеры 5865 пар оснований. Его карта, функциональные особенности и полная последовательность ДНК приведены на Фиг.2а и 2b. На Фиг.3 приведена схема конструирования вектора pG1PLPSvNa. Как правило, вектор pRc/CMV-PLP расщепляли BstEII/HindIII и кодирующий PLP фрагмент выделяли электрофорезом в геле. После электроэлюции адаптеры HindIII/NotI (Stratagene, La Jolla, CA) лигировали в HindIII сайт элюированного фрагмента. Для получения Notl-концов проводили рестрикцию Notl. Вектор pGlXSvNa рестрицировали Notl и электроэлюцией выделяли фрагмент размером 5865 пар оснований, концы которого обрабатывают щелочной фосфатазой из кишечника теленка (ClAP). Вставку лигировали в Notl-сайт вектора. BstEII-концы вставки и Notl-сайта вектора достраивали с помощью фрагмента Кленова. Для замыкания вектора проводили лигирование по тупым концам. Клетки НВ101 трансформировали лигазной смесью, а затем проводили рестрикционный анализ для подверждения наличия вставки и определения ее ориентации. Рекомбинантные ретровирусы дефектны по репликации и неспособны образовывать инфекционный вирус.
3. Супернатант ретровирусного вектора
Для получения содержащего рекомбинантный PLP-вирус супернатанта PLP-трансдуцированные упаковочные клетки линии РАЗ17 выращивали в 4 мл соответствующей культуральной среды во флаконах Т25 (Corning, Cambridge, MA). Содержащий ретровирусный вектор супернатант получали, собирая культуральную среду по достижении клетками 80-90% монослоя, и хранили в 1-мл аликвотах при -70°С.
С клеточной линией-продуцентом и/или с вирусным супернатантом были проведены следующие исследования:
1. Титр вируса определяют на клетках ЗТЗ. Используют вирусные препараты с титром выше, чем 5×104 колониеобразующих единиц на мл.
2. Стерильность клеточной линии-продуцента и супернатанта подтверждают тестами на аэробные и анаэробные бактерии, грибы и микоплазму.
Препараты PLP-вектора, полученные из клеток РА317, должны быть тщательно исследованы, чтобы подтвердить отсутствие компетентного по репликации вируса. Данный аспект особенно важен при том варианте осуществления изобретения, когда оно применяется для лечения людей. Чтобы подтвердить отсутствие компетентного по репликации вируса, должны быть проведены тесты как вирусного супернатанта, так и трансдуцированных фибробластов. С клеточной линией-продуцентом и/или с вирусным супернатантом могут быть проведены следующие исследования:
1. Титр вируса определяют на клетках ЗТЗ. Используют вирусные препараты с титром выше, чем 5×104 колониеобразующих единиц на мл.
2. Клеточную линию-продуцент проверяют при помощи Саузерн-блоттинга на присутствие части гена PLP.
3. Определяют продукцию PLP клеточной линией-продуцентом, причем данная продукция должна значительно превышать исходные контрольные значения (по ELISA).
4. Стерильность клеточной линии-продуцента и супернатанта подтверждают тестами на аэробные и анаэробные бактерии, грибы и микоплазму.
5. К проводимым вирусологическим тестам относятся: МАР-тест, тест на вирус LCM, тест на тимический агент, S-L-тест на экотропный вирус, S+L-тест на ксенотропный вирус, S+L-тест на амфотропный вирус и ЗТЗ амплификацию.
6. Для того чтобы убедиться в отсутствии адвентиционных агентов, проводят электронную микроскопию.
После введения гена в фибробласты и до введения фибробластов больным проводят следующие исследования:
1. Жизнеспособность клеток должна составлять более 70%, о чем судят по окрашиванию трипановым синим.
2. Перед введением проводят цитологический анализ 200 клеток, чтобы исключить присутствие опухолевых клеток.
3. Подтверждают стерильность тестами на присутствие аэробных и анаэробных бактерий, грибов и микоплазмы.
4. S+L-тест должен быть отрицательным, включая амплификацию ЗТЗ.
5. ПЦР-анализ на присутствие гена поверхностного белка 4070А должен быть отрицательным.
6. Обратно-транскриптазный тест должен быть отрицательным.
7. Для подтверждения присутствия интактного провируса трансдуцированные фибробласты исследовали Саузерн-блоттингом.
8. Продукцию белка PLP подверждали тестом на белок PLP.
4. Трансфекция фибробластов
До трансфекции фибробластов SJL из трансформированных клеток AG1 выделяют высоко очищенный вектор PLP-pRc/CMV. Крупномасштабную очистку ДНК проводят с использованием коммерчески доступных наборов и градиентное центрифугирование в CsCl. Первоначальную очистку проводят с использованием набора Wizard Megaprep Kit (Promega, Madison, WI). Трансформированные клетки AG1, выращенные в 1000 мл ночной культуре в среде LB/Amp при 37°С, осаждают и выделяют плазмидную ДНК согласно инструкциям производителя. Затем выделенную ДНК, ресуспендированную в 3 мл буфера ТЕ (10 мМ Tris-HCl, pH 7.4 и 1 мМ динатриевой соли ЕДТА, рН 8.0) подвергают градиентному центрифугированию в CsCl. Модифицированную очистку ДНК градиентным центрифугированием в CsCl проводят по методике, опубликованной в “Current Protocols in Molecular Biology, Vol.1” (Greene Publishing Associates and Wiley-Interscience).
После того, как полосу ДНК извлекают из центрифужной пробирки, из образца удаляют этидий бромид, отмывая тремя объемами SSC-насыщенного изопропанола. Отмывки продолжают до тех пор, пока водная фаза не становится чистой. CsCl удаляют переосаждением. Затем к образцу добавляют 2 объема 0.2М NaCl/TE и 2 объема 100%-ного этанола (по отношению к общему объему раствора ДНК и 0.2М NaCl/TE), смешивают и помещают на 10 минут на лед. Приципитировавшую ДНК осаждают центрифугированием при 10 000 × g 10 минут при 4°С. Осадок промывают холодным 70%-ным этанолом, заново центрифугируют при 10 000 × g 10 минут при 4°С и высушивают в вакууме (упариватель Speed vac, Savant Inc.). Очищенную ДНК ресуспендируют в дважды дистиллированной стерильной воде и используют в экспериментах по трансфекции.
Опытные фибробласты SJL трансфицируют при помощи LipofectAMINE (Life Technologies Inc./Gibco BRL) согласно инструкциям производителя. Контрольные фибробласты SJL подвергают тем же процедурам, но без ДНК-конструкта. Для трансфекции используют 3 мкл очищенной в CsCl плазмиды PLP-pRc/CMV и 25 мкл Липофектамина. Накануне эксперимента приблизительно по 3×105 клеток засевают в 25 см2 культуральные флаконы (Corning Costar Corp., Cambridge, MA) и выращивают при 37°С в атмосфере 5% СO2 в 5 мл культуральной среды DMEM (среда Игла в модификации Дальбекко) (Irvine Scientific, Santa Ana, CA) с добавлением 5% глутамина, 10% эмбриональной телячьей сыворотки, 25 Ед/мл натриевой соли пенициллина G и 25 мг/мл стрептомицин сульфата. Затем клетки промывают 3 мл бессывороточной среды HL-1 (Hycor Biomedical Inc., Irvine, CA). После того, как клетки были проинкубированы с комплексом ДНК/Липофектамин 6 часов при 37°С в атмосфере 5% СO2, во флаконы добавляют по 1 мл среды DMEM. Через 36 часов после завершения процедуры трансфекции среду заменяют на 5 мл среды DMEM, содержащей 900 мкг G418 (Life Technologies Inc./Gibco BRL) на мл среды. Опытные клетки выращивают в присутствии 900 мкг G418 (на мл среды до того момента, пока контрольные клетки не погибали, а отдельном опытном флаконе более не наблюдалось гибели клеток). Затем концентрацию G418 снижают до 600 мкг/мл культуральной среды для продолжения культивирования клеток.
5. Трансдукция фибробластов
Для трансдукции фибробластов используют ретровирусные конструкты, содержащие селективный маркер neo, а также ген PLP или ген Lac-z. Трансдукцию ретровирусом проводят на жизнеспособных клетках (90%-ная жизнеспособность по окраске трипановым синим). В каждую лунку 24-луночного планшета (Falcon, Franklin Lakes, NJ) засевают 2×106 клеток в 0,5 мл среды DMEM-10 (среда DMEM с добавлением 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 50 Ед/мл пенициллина G, 50 мг/мл стрептомицина). Клетки переносят в инкубатор для прикрепления (37°, 5% СО2). После того, как клетки прикрепились к подложке, в каждую ячейку вносят 1 мл ретровирусного супернатанта и полибрен (Sigma, St.Louis, МО) (конечная концентрация 10 мкг/мл). Клетки инкубируют, как указано выше, в течение 2,5 часов без встряхивания. По истечении 2,5 часов клетки переносят во флаконы Т25 и добавляют среду DMEM-10 до конечного объема 8 мл. На третий день после трансдукции среду заменяют на селективную (среда DMEM-10 с добавлением 900 мкг/мл G418 (Gibco, Grand Island, NY). Затем концентрацию G418 снижают до 600 мкг/мл культуральной среды для продолжения культивирования клеток.
6. Анализ экспрессии мРНК
Выделение РНК проводили в асептических условиях с использованием РНКаза-свободных материалов и DEPC (диэтилпирокарбонат)-обработанных растворов. 4×106 экспериментальных и контрольных клеток дважды промывали ледяным забуференным фосфатным раствором, ресуспендировали в 200 мкл смеси для лизиса клеток (10 мМ Tris, рН 7.5, 0,15 мМ NaCl, 1.5 мМ MgCl2, 0.65% NP 40), перемешивали и центрифугировали при 17 000 × g и 4°С 5 минут. Супернатант переносили в пробирку, содержащую 200 мкл мочевинной смеси (7М мочевина, 1% SDS, 0,35 М NaCl, 10 мМ ЕДТА и 10 мМ Tris-HCl, рН 7.5) и 400 мкл смеси фенол:хлороформ:изоамиловый спирт (25:24:1). Раствор перемешивали и центрифугировали 1 минуту при 17 000 × g. Эту процедуру с водной фазой повторяли дважды, затем ее переносили в пробирку с 400 мкл фенола и отмывали, как описано ранее. Водную фазу опять переносили в новую пробирку и осаждали РНК 1 мл 100%-ного этанола при -20°С в течение ночи. Осажденную РНК один раз промывали 70%-ным этанолом. После удаления этанола осадок высушивали в вакууме. 1 мкг РНК использовали для анализа обратно-транскриптазной ПЦР (ОТ-ПЦР).
ОТ-ПЦР проводили с использованием коммерческого набора GenAmp RNA PCR Kit (Perkin Elmer/ABI) согласно инструкциям производителя. Для амплификации кДНК использовали следующие праймеры: 5'-GCGACTACAAGACCACCATCT-3' и 5'-TAAGGCTAGCATAGGTGATG-3'. Продукты ПЦР разделяли электрофорезом в 1,5%-ном геле агарозы (SeaKem GTG; FMC)/TAE с 1 мкл этидия бромида (10 мг/мл) на 1 мл раствора агарозы. Электрофорез проводили в буфере ТАЕ при постоянном токе в 40 мА. Электрофорез проводили в течение такого времени, которое было достаточно для четкого разделения маркеров молекулярного веса, что позволяло верифицировать приблизительный молекулярный вес продуктов ПЦР. Полосу представляющей интерес ДНК вырезали из геля и ДНК очищали с использованием коммерческого набора MERmaid (Bio 101, Vista, CA) согласно инструкциям производителя. Затем очищенную ДНК секвенировали автоматическим методом с флуоресцентной ДНК (Perkin Elmer/ABI, Foster City, CA).
На Фиг.4 показана фотография агарозного геля с PLP-специфическими продуктами ОТ-ПЦР. Результаты свидетельтвуют, что мРНК присутствует как в PLP-трансдуцированных, так и в PLP-трансфицированных клетках. Еще предстоит установить корреляцию между мРНК и секретируемым белком, поскольку концентрация пептида не обязательно соответствует уровню мРНК.
7. Анализ экспрессии белка
Количественную экспрессию in vitro кодируемых геном PLP белков оценивали иммунологически при помощи ELISA. Тестировали неразведенные супернатанты из культур фибробластов, трансдуцированных геном PLP. Супернатанты вносили в лунки 96-луночного планшета для микротитрования. Первичные анти-PLP 139-151 антитела 4Е10 139-151, полученные от доктора M.Lees (Harvard), специфичны к PLP 139-151. Они были добавлены в ячейки в виде неразведенного гибридомного супернатанта, а затем вносили конъюгат вторых антител (антитела козы к иммуноглобулинам мыши) с пероксидазой хрена в разведении 1:500. Планшет проявляли и анализировали поглощение при 490 нм на ELISA-ридере. На Фиг.5 приведены результаты ELISA супернатантов трансдуцированных фибробластов. Образцы 1 и 2 представляют собой PLP (аминокислоты 139-151) и пептиды gp120 HIV, использованные в концентрации 5 мкг/мл. Данный эксперимент доказывает, что PLP-трансдуцированные фибробласты действительно продуцируют и секретируют частичный белок PLP.
ПРИМЕР 3
ЭФФЕКТЫ БЕЛКА PLP IN VIVO
Критической для изложенного в настоящем примере варианте осуществления изобретения является возможность доставки генетически трансформированных фибробластов больному таким образом, чтобы клетки выживали в достаточном количестве и в течение достаточно длительного времени, обеспечивая больного постоянно секретируемым антигеном.
С тем, чтобы проследить судьбу трансплантированных фибробластов, фибробласты SJL, трансдуцированные несущим В-галактозидазу ретровирусом, вводили подкожно между лопатками мышам SJL. Все животные были самками линии SJL в возрасте 6-8 недель и были получены из Jackson Labs. Животных содержали согласно рекомендациям NIH (National Research Council, 1986). Подкожно введенные фибробласты обнаруживались в большом количестве спустя 60 дней после введения. Фибробласты, инъецированные в подушечку лапы или внутримышечно, не удавалось обнаружить уже на восьмой день.
1. Судьба In Vivo B-gal-трансдуцированных клеток
Активность маркера В-галактозидазы оценивали на двух группах нормальных мышей (каждая из 8 животных). Двум мышам инъекции Lac-Z-трансдуцированных клеток делали подкожно в спину, двум мышам - внутримышечно и двум мышам - в подушечку лапы. Одному животному вводили фибробласты, трансдуцированные только маркером neo, а последней мыши вводили не-трансдуцированные фибробласты. После сбора и промывки клетки различных линий суспендировали в концентрации 107 в 2 мл раствора Хэнкса и медленно вводили через иглу 25-го калибра в различные точки. Животных забивали на 10-й и 15-й дни после обработки и места инъекций подвергали гистохимическому исследованию. Кусочки тканей фиксировали в 4%-ном параформальдегиде 1 час, промывали три раза в PBS, а затем выдерживали ночь в 8,4%-ном растворе акриламида. На следующее утро образцы заключали в акриламид, который после отверждения разрезали и замораживали. Замороженные срезы толщиной 10 мкм, полученные с помощью криостата, окрашивали 1 мл 5-бром-4-хлор-3-индолил-В-d-галактопиранозид (X-Gal) в PBS. X-Gal растворяли в DMSO в концентрации 40 мг/мл и затем добавляли в реакционную смесь. Инкубацию проводили 14-18 часов при 37°С. На Фиг.6 показана экспрессия B-gal в трансдуцированных фибробластах через 60 дней in vivo. He имелось указаний на воспалительный ответ, что дает основание считать, что использованный для трансдукции сингенных фибробластов ретровирус не провоцирует иммунного ответа или процесса отторжения.
2. Эффект PLP на нормальных мышей SJL
Другим важным аспектом настоящего изобретения, отраженным в настоящем примере, является ответ на вопрос, могут ли трансдуцированные фибробласты, секретирующие PLP, вызывать ЕАЕ у нормальных животных. Для проверки этого предположения 12-ти нормальным мышам SJL вводили 107 PLP-секретирующих фибробластов SJL. Шести животным фибробласты вводили подкожно, а другим шести - интраперитонеально. На 16-й день животных забивали и обследовали на наличие воспаления или ЕАЕ. На Фиг.7 представлена система клинического учета хронического ЕАЕ. Lu et al., Mol.Immunol. 28:623-630 (1991), J.Williamson et al., J.Neuroimmunol. 32:199-207 (1991). На мышиной модели ЕАЕ для воспроизведения рассеянного склероза при использовании гомогенатов спинного мозга с адъювантом, воспаление в ЦНС можно обнаружить на 14-й день. В данном опыте у нормальных животных, получивших инъекцию PLP-секретирующих фибробластов, не обнаружено каких-либо клинических признаков заболевания даже на 60-й день обследования. Кроме того, у животных на 60-й день не обнаружено гистологических признаков воспаления в ЦНС. На Фиг.8 представлена система гистологического учета хронического ЕАЕ. J.Goverman et al., Cell. 72:551-560 (1993).
3. Клиническая и гистологическая оценка мышей с острым ЕАЕ после введения ретровирус-трансдуцированных фибробластов
Шестинедельным мышам SJL вводили гомогенат спинного мозга мышей (MSCH) в полном адъюванте Фрейнда (CFA) и через 7 дней - MSCH в неполном адъюванте Фрейнда (IFA). J.Immunol. 144:909-915 (1990). Первые признаки ЕАЕ наблюдались на 14-18-й день с полным выздоровлением к 21-му дню. У 95% животных наблюдались клинические признаки острого ЕАЕ. Этим животным на 21-й день вводили 107 PLP-секретирующих фибробластов SJL или контрольных фибробластов. Животных, у которых не развилось клинических признаков заболевания, исключали из эксперимента. На Фиг.9 представлена клиническая оценка мышей с ЕАЕ после введения ретровирус-трансдуцированных фибробластов. У животных, которым вводили PLP-секретирующие фибробласты, наблюдалось значительное снижение клинических признаков и резкое уменьшение числа клеток воспаления, особенно в мозге. На Фиг.10а представлена патологическая оценка головного и спинного мозга мышей SJL после введения PLP-секретирующих фибробластов. На Фиг.10b суммирована патологическая оценка головного и спинного мозга мышей на сроках 55-60 и 90-95 дней. Гистологическую оценку ЕАЕ проводили на гематоксилин-эозин окрашенных срезах головного и спинного мозга.
4. Клиническая и гистологическая оценка мышей с хроническим ЕАЕ после введения ретровирус-трансдуцированных фибробластов
150 мышам вводили MSCH в CFA. Вторая иммунизация была проведена через 7 дней. A.M.Brown and D.E.McFarlin, Laboratory Invest. 45:278-284 (1981). К дню +14-16 у 113-ти животных развились клинические признаки заболевания, продолжавшиеся 3-4 дня. Этих положительных животных отделяли для последующих экспериментов и к дню +55-60 у 100 мышей наблюдался первый рецидив. У 67 мышей наблюдался второй рецидив к дню +137. Через 8 дней после рецидива животным вводили 107 фибробластов и еще через 18-23 дня животных забивали. Использовали четыре различных типа фибробластов: трансдуцированные ретровирусом, кодирующим PLP ретровирусом, В-галактозидазу и пео-селективный маркер, а также нетрансдуцированные клетки. На Фиг.11 представлена гистология мышей с хроническим ЕАЕ, обработанных ретровирус-трансдуцированными фибробластами. Среди животных, которым вводили PLP-секретирующие фибробласты, не наблюдалось животных с воспалением степени 2+ и 3+.
1. Периферический иммунный статус обработанных мышей по сравнению с контрольными ЕАЕ мышами
Клетки селезенок четырех контрольных ЕАЕ мышей и четырех ЕАЕ мышей, которым были введены фибробласты, экспрессирующие белок PLP, были использованы в пролиферативных тестах, в ходе которых клетки селезенок инкубировали с 40 мкМ пептида PLP 140-151 или с 40 мкМ пептида 308-322 gp120 HIV в течение 4 дней, а затем помечены 3Н-тимидином в течение 24 часов.
Животных умерщвляли асфиксией СО2. Клетки селезенок диспергировали до состояния одноклеточной суспензии в среде RPMI1640, пропуская суспензию через сито #60. Клетки однократно промывали перед тем, как засеять в лунки 96-луночных планшетов (8×105 на ячейку) в 0,2 мл среды HL-1 (Нусог Biomedical, Irvine, CA) с добавлением 2 мМ глутамина, 100 Ед/мл пенициллина, 100 мкг стрептомицина. Клетки культивировали с добавлением или без добавления 40 мкМ пептида 4 дня при 37°С в атмосфере 5% СО2. Для антигенспецифической пролиферации использовали пептид PLP 140-151 и пептид МБР 89-101, а в качестве отрицательного контроля использовали пептид 308-322 gp120 HIV. Отдельные ячейки также содержали 100 Ед/мл рекомбинантного мышиного IL2 (Boehringer Mannheim, Indianapolis, IN). В последние 18-24 часов культивирования в каждую ячейку добавляли 1 мкКи3 Н-тимидина (ICN, Irvine, CA), клетки собирали на стекловолоконные фильтры “Xtal Scint” (Beckman, Fullerton, CA) и просчитывали на сцинтилляционном счетчике Beckman LS6000. Рассчитывали значения включения тимидина (число имульсов в эксперименте в минуту минус фоновые импульсы в минуту) и выводили средние значения для трех культур плюс - минус стандартное отклонение.
Результаты, приведенные на Фиг.12, дают основание полагать, что PLP-специфический пролиферативный ответ значительно снижается у мышей с ЕАЕ, которым были введены PLP-экспрессирующие фибробласты.
Фиг.13 иллюстрирует эксперимент, аналогичный приведенному на Фиг.12, с той разницей, что в течение 5-ти дней дополнительно вводили мышиный IL-2 (10 U/мл). Эти результаты показывают, что механизм, с помощью которого значительно снижается PLP-специфический пролиферативный ответ, может включать возможность делеции Т-клеток, а не анергию, поскольку данные лимфоциты не отвечают на 11-2.
Притом, что механизм, посредством которого заявленный согласно настоящему изобретению способ восстанавливает толерантость у индивидуумов, страдающих от обусловленного Т-клетками аутоиммунного заболевания, еще не полностью понят, очевидны преимущества заявленного способа лечения по сравнению с альтернативными подходами. Предложенный способ основан на генетическом подходе к иммуноспецифическому “выключению” патогенного Т-клеточного ответа и не подавляет иммунную систему в целом. В том случае, когда обусловленное Т-клетками аутоиммунное заболевание у данного индивидуума обусловлено Т-клетками со множественной специфичностью, заявленный в соответствии с настоящим изобретением способ может быть легко адаптирован применительно к данным специфичностям.
Объем настоящего изобретения не ограничивается описанными вариантами его осуществления, которые иллюстрируют отдельные аспекты изобретения. Клоны, ДНК или аминокислотные последовательности, которые являются функционально эквивалентными описанным, также включены в объем настоящего изобретения. Различные модификации изобретения, в дополнение к приведенным, будут очевидны для квалифицированного специалиста из предшествующего описания. Такие модификации также включаются в объем притязаний по данной заявке.
Различные публикации, процитированные в настоящем описании, включены в него исключительно в качестве ссылок.











Настоящее изобретение относится к применению генно-инженерных подходов для лечения аутоиммунного заболевания, в частности для лечения рассеянного склероза. Это достигается введением одного или нескольких рекомбинантных генов, кодирующих аутоантигены, которые являются мишенью аутоиммунного ответа. В частности, согласно настоящему изобретению заявлен способ конструирования гена, кодирующего энцефалитогенный эпитоп протеолипидного белка, а также экспрессия генного продукта in vivo при помощи рекомбинантного ретровирусного вектора. Экспрессия и секреция энцефалитогенного эпитопа улучшает гистопатологические и клинические показатели при экспериментальном аутоиммунном энцефаломиелите (ЕАЕ) у мышей, служащем моделью рассеянного склероза (MS). Преимущество изобретения заключается в разработке способа восстановления толерантности при лечении рассеянного склероза без подавления иммунной системы. 3 н. и 3 з.п. ф-лы, 13 ил.
| US 5468481 А, 21.11.1995 | |||
| US 5460945 A, 24.10.1995 | |||
| FYMAN et al., DNA Vaccines: Protective ummunization by parenteral, mucosal, and gene gun inoculations, Proc, Natl | |||
| Acad | |||
| Sci | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |

