ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к системе доставки для антигена в виде Т-клетки, применению такой Т-клетки для мониторинга иммунного ответа и к способу получения данной Т-клетки.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Перенос генов в человеческие клетки исследуется в плане лечения разнообразных генетических и приобретенных человеческих заболеваний. Генетические заболевания, развивающиеся вследствие дефекта единичного гена, были первыми предложены в качестве первичных целей генной терапии. Однако большинство испытаний по генной терапии человека, одобренных к настоящему времени, относятся к лечению приобретенных заболеваний, таких как злокачественные опухоли и СПИД. Один из подходов к генной терапии опухолей основывается на стимуляции иммунной системы хозяина против опухолевых антигенов путем вакцинации пациента генетически модифицированными опухолевыми клетками, в которых трансген (цитокин, алло-HLA и т.д.) направлен на повышение их иммуногенности. Альтернативный подход основан на адаптивном переносе полученных ex vivo опухолеспецифических рецепторов. Вводимые путем инфузии эффекторы обычно представляют собой аутологичные или совместимые по HLA лимфоциты, пролиферация которых была стимулирована in vitro специфичными, а также неспецифическими стимулами. Амплификация эффекторной функции введенных путем инфузии клеток достигалась путем их трансдукции кодирующих цитокины генов.
В EP 0904786 описан способ опухолевой вакцинации путем, по меньшей мере, двух последовательных введений композиций, состоящих по существу из аутологичных или родственных по HLA клеток, которые способны представлять антиген в пациенте, причем данные клетки представляют собой опухолевые клетки, трансдуцированные чужеродным геном, или антигенпрезентирующие клетки, трансдуцированные опухолевым антигеном или чужеродным антигеном.
В WO 97/19169 описана противоопухолевая вакцина, содержащая опухолевые клетки, по меньшей мере, часть которых содержит, по меньшей мере, один гаплотип MHC-I пациента на клеточной поверхности и которые загружены антигеном.
Таким образом, предшествующий уровень техники использует подход применения опухолевых клеток в качестве системы доставки антигена, или антигенпрезентирующей клетки, которая считается представляющей антиген непосредственно Т-клетке.
Однако понятно, что имеет место постоянная потребность в предоставлении вакцин и подходов к вакцинации. Настоящее изобретение относится к такому подходу.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к подвергнутым манипуляции Т-клеткам, которые при введении пациенту способны приводить в контакт с профессиональными антигенпрезентирующими клетками (APC) антигены, такие как опухолевые, бактериальные или вирусные антигены. Более конкретно, изобретение относится к трансдуцированной Т-клетке, содержащей экзогенный (или чужеродный) ген, кодирующий антиген, для стимуляции ответа против данного антигена, и Т-клетке, нагруженной антигенным полипептидом для стимуляции ответа. Т-клетка также может включать в себя другой антиген, такой как HSV-Tk, который может использоваться для мониторинга эффективности вакцинации. Данные Т-клетки удобно получают in vitro, и затем их вводят инъекцией или иным способом in vivo, так что они несут продукты трансгена к профессиональным антигенпрезентирующим клеткам, таким образом стимулируя иммунный ответ против продуктов трансгена. Изобретение также относится к методологии, в соответствии с которой можно проводить мониторинг иммунологического статуса пациента. Настоящее изобретение, кроме того, относится к способу культивирования Т-клеток.
Таким образом, авторы изобретения в настоящее время обнаружили, что Т-клетки, содержащие антиген, действуют, доставляя антиген к антигенпрезентирующим клеткам (APC), так что APC впоследствии представляют антиген с формированием иммунного ответа. Поэтому настоящее изобретение характеризуется иной стратегией по сравнению с предшествующим уровнем техники.
ИЗЛОЖЕНИЕ ИЗОБРЕТЕНИЯ
Согласно одному из аспектов настоящего изобретения предоставляется система доставки, включающая в себя Т-клетку, содержащую, по меньшей мере, один антиген и способную доставлять антиген в лимфатический узел, или другую часть лимфатической системы.
Иными словами, настоящее изобретение относится к системе доставки, включающей в себя Т-клетку, содержащую, по меньшей мере, один антиген, способную нагружать антигенпрезентирующие клетки (APC) данным антигеном.
Антиген в основном чужероден в отношении Т-клетки. В определение «чужеродный» авторы изобретения включают антигены, которые в норме экзогенны относительно Т-клетки.
Антиген может вводиться в Т-клетку в качестве полинуклеотида, кодирующего антиген, т.е. Т-клетка может трансфицироваться или трандуцироваться данным полинуклеотидом или Т-клетка может быть нагружена антигеном в виде полипептида, который с тем же успехом может быть белком. Для простоты употребления, термины «полипептид» и «белок», в общем, используются в данном описании взаимозаменяемо. Таким образом, под Т-клеткой, содержащей антиген, авторы изобретения подразумевают, что антиген ассоциирован с Т-клеткой, так что Т-клетка способна доставлять антиген в лимфатический узел и/или APC.
Согласно другому аспекту настоящего изобретения предоставляется система доставки, включающая в себя Т-клетку, содержащую, по меньшей мере, один антиген, и способную доставлять антиген к APC для последующего представления на APC.
В одном из осуществлений чужеродный антиген представляет собой бактериальный или вирусный антиген. Предпочтительно, чужеродный антиген представляет собой опухолевый антиген.
Предпочтительно Т-клетка далее включает в себя, по меньшей мере, одну другую чужеродную полипептидную или полинуклеотидную последовательность.
В одном из осуществлений еще один чужеродный ген представляет собой маркер, т.е. маркерный ген или ген селекции.
Предпочтительно маркер представляет собой ген устойчивости бактерий, например ген устойчивости бактерий, который обеспечивает устойчивость к неомицину.
В другом осуществлении маркер представляет собой еще один антиген, предпочтительно HSV-Tk или CD20. Данные антигены особенно предпочтительны, поскольку они могут использоваться в так называемых суицидных системах.
В дальнейшем осуществлении система доставки, кроме того, включает в себя еще один антиген и маркерный ген.
Предпочтительно Т-клетка экспрессирует, по меньшей мере, один из следующих маркеров: HLA-I, HLA-II, CD80, CD86, CD27, CD40L, CD62L, CCR7, CD54 и CD25.
Таким образом, в другом аспекте настоящего изобретения предоставляется Т-клетка, которая экспрессирует, по меньшей мере, один из следующих маркеров: HLA-I, HLA-II, CD80, CD86, CD27, CD40L, CD62L, CCR7, CD54 и CD25.
По другому аспекту настоящего изобретения предоставляется применение Т-клетки, содержащей, по меньшей мере, два антигена, способных вызывать иммунный ответ, в котором ответ против одного из антигенов используется для мониторинга иммунного ответа против другого антигена.
По другому аспекту настоящего изобретения предоставляется способ мониторинга иммунного ответа против Т-клетки, содержащей первый антиген, способный вызывать иммунный ответ, причем способ охватывает введение в Т-клетку второго антигена.
По другому аспекту настоящего изобретения предоставляется способ загрузки APC in vivo, охватывающий воздействие на APC Т-клетки, содержащей антиген.
Еще по одному аспекту настоящего изобретения предоставляется способ получения Т-клетки для применения по заявленному способу, охватывающий:
выделение Т-клетки;
активацию Т-клетки;
культивирование Т-клетки;
введение в данную Т-клетку антигена.
Предпочтительно Т-клетка подлежит трансдукции или трансфекции полинуклеотидной последовательностью, кодирующей антиген.
Предпочтительно Т-клетка активируется фитогемагглютинином, гранулами, покрытыми моноклональным антителом против CD3 или моноклональным антителом против CD3/CD28.
Предпочтительно Т-клетка культивируется в присутствии факторов роста, таких как hu-r-IL-2.
Предпочтительно Т-клетка культивируется в культуральной среде, которая содержит 5% аутологичной сыворотки.
Предпочтительно Т-клетка культивируется в количестве 1×106 клеток/мл.
По дальнейшему аспекту настоящего изобретения предоставляется Т-клетка, которую можно получить способом по настоящему изобретению и которую можно использовать в качестве системы доставки по настоящему изобретению.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Различные предпочтительные характеристики и осуществления настоящего изобретения теперь будут описаны посредством неограничивающих примеров.
Хотя, в общем, указанные здесь способы хорошо известны в данной области, может быть сделана конкретная ссылка на Sambrook et al., Molecular Cloning, A Laboratory Manual (1989) и Ausubel et al., Short Protocols in Molecular Biology (1999) 4th Ed, John Wiley & Sons, Inc (а также на полную версию Current Protocols in Molecular Biology).
В общем плане настоящее изобретение относится к Т-лимфоцитам (Т-клеткам), подвергаемым манипуляциям in vitro, которые при инъекции in vivo способны приводить антигены (например, опухолевые, бактериальные или вирусные антигены) в контакт с профессиональными антигенпрезентирующими клетками. Однако настоящее изобретение также относится к манипуляциям Т-клетками in vivo. Более конкретно, изобретение относится к: а) трансдуцированным/трансфицированным in vitro Т-лимфоцитам, содержащим экзогенные гены, кодирующие опухолевый антиген, для стимуляции иммунного ответа против опухоли; и b) Т-лимфоцитам, нагруженным in vitro пептидами, кодирующими эпитопы опухолевых антигенов. Т-клетки могут также содержать сильный антиген, например HSV-Tk, для мониторинга эффективности вакцинации.
Имелись сообщения о том, что Т-клетки являются подходящими мишенями для переноса генов, например, посредством ретровирусных векторов. В частности, описано применение Т-клеток, экспрессирующих опухолевые антигены, для противоопухолевой вакцинации. Однако также сообщалось, например в EP 0905786, о том, что некий эффект имеет место посредством Т-клеток, действующих в качестве антигенпрезентирующих клеток (APC). В отличие от данной точки зрения, авторами изобретения в настоящее время показано, что Т-клетки действуют путем воздействия на APC трансгена, который затем представляется APC. Иными словами, Т-клетки воздействуют на APC трансгенным продуктом для процессинга и представления антигена. В термин «воздействие» авторы изобретения включают приведение трансгена в контакт с APC, так что происходит связывание и/или захват трансгенного продукта. APC интернализует антиген и процессирует его для представления.
В термин «процессинг антигена» авторы изобретения включают деградацию антигена с образованием более коротких пептидных последовательностей и ассоциацию пептида с молекулами МНС. Два различных класса молекул МНС, МНС класса I и МНС класса II, регулируют представление антигенов CD8+ или CD4+ клеткам соответственно. Представление антигена, строго говоря, представляет собой активацию T-клеток через Т-клеточные рецепторы, которые специфично распознают антигенный пептид в ассоциации с молекулами MHC класса I или класса II на поверхности APC; однако B-клетки также способны распознавать и связывать некоторые антигены.
Авторами изобретения также обнаружено, что настоящая система предоставляет эффективный способ доставки антигена в лимфатическую систему и, особенно, в лимфатические узлы. T-клетки могут вводиться местно, но авторы изобретения также полагают, что T-клетки способны эффективно мигрировать в лимфатические узлы, даже если их вводят в удаленном участке. Лимфатические узлы функционируют как иммунный фильтр для жидкости организма, известной как лимфа. Лимфатические узлы расположены по всему организму. При том, что они состоят по большей части из В-клеток, дендритных клеток и макрофагов, узлы дренируют жидкость из большей части тканей человека. Антигены фильтруются из лимфы в лимфатическом узле до возвращения лимфы в циркуляцию. Сходным образом в селезенке макрофаги и дендритные клетки, которые захватывают антигены, представляют данные чужеродные вещества T- и B-клетки, инициирующие впоследствии иммунный ответ.
Изобретение далее относится к клеточно-зависимой системе доставки антигена. В данной системе антиген, или полинуклеотид, кодирующий антиген, вводят в одну или несколько клеток ex vivo и затем вводят пациенту.
T-клетки по настоящему изобретению могут вводиться отдельно, но в общем их вводят в виде фармацевтической композиции.
APC и иммунный ответ
Как указывалось выше, настоящее изобретение относится к доставке антигена антигенпрезентирующей клетке (APC). APC включают в себя макрофаги, дендритные клетки, В-клетки и любые другие клеточные типы, экспрессирующие молекулу MHC.
Макрофаги и клетки-фагоциты моноцитарного ростка постоянно находятся в тканях и особо хорошо приспособлены для эффективного представления антигена. Они в основном экспрессируют молекулы MHC класса II и со своими фагоцитарными характеристиками исключительно эффективны в плане поглощения макромолекулярного материала или материала в виде частиц, его расщепления, его процессинга в интенсивной лизосомальной системе с образованием формы антигенных пептидов и экспрессии пептидов на поверхности клетки для распознавания T-лимфоцитами.
Дендритные клетки, названные так из-за своего разветвленного строения, обнаруженные во многих органах по всему организму, имеют костномозговое происхождение и обычно экспрессируют большие количества антигена MHC класса II. Дендритные клетки активно двигаются и могут рециркулировать между кровяным руслом и тканями. Дендритные клетки являются антигенпрезентирующими клетками с уникальной способностью индуцировать первичные иммунные реакции, обеспечивая таким образом формирование иммунологической памяти. Дендритные клетки в незрелом состоянии имеют высокую фагоцитарную способность. После захвата антигена незрелые дендритные клетки мигрируют в лимфатические узлы, где после созревания активируют циркулирующие антигенспецифические лимфоциты. Таким образом, они тоже считаются важными APC. Клетки Лангерганса являются примером дендритных клеток, которые локализованы в коже.
B-лимфоциты, хотя и не активные в плане фагоцитоза, являются позитивными по классу II и обладают антигенспецифическими рецепторами клеточной поверхности, иммуноглобулинами или молекулами антител. Вследствие своего потенциала к связыванию антигена с высоким сродством, B-клетки наделены уникальной способностью концентрировать на своей поверхности малые количества антигена, эндоцитировать его, процессировать его и представлять его в виде антигенного пептида в ассоциации с антигеном MHC на своей поверхности. За счет этого B-клетки становятся исключительно эффективными APC.
Таким образом, иммунный ответ на чужеродный антиген требует присутствия антигенпрезентирующей клетки (APC) (обычно макрофага или дендритной клетки) в комбинации с B-клеткой или Т-клеткой. Когда APC представляет антиген на своей клеточной поверхности B-клетке, В-клетка получает сигнал к пролиферации и продукции антител, которые специфично связывают тот самый антиген. Если антитело связывается с антигенами на бактериях или паразитах, оно действует как сигнал для полиморфно-ядерных лейкоцитов или макрофагов к их поглощению (фагоцитозу) и уничтожению. Другой важной функцией антител является инициация «разрушающего каскада комплемента». При связывании антител с клетками или бактериями белки сыворотки, называемые комплементом, связываются с иммобилизованными антителами и разрушают бактерии путем образования в них отверстий. Антитела также могут служить сигналом для клеток-естественных киллеров и макрофагов к уничтожению клеток, инфицированных вирусами или бактериями.
Если APC представляет антиген Т-клеткам, T-клетки становятся активированными. Активированные T-клетки пролиферируют и становятся секреторными в случае CD4+ T-клеток, или, если они являются CD8+ T-клетками, они становятся активированными в плане уничтожения клеток-мишеней, которые специфично экспрессируют антиген, представленный APC. Продукция антител и активность CD8+ киллерных T-клеток преимущественно регулируется субпопуляцией CD4+ хелперных Т-клеток. CD4+ T-клетки обеспечивают данные клетки факторами роста или сигналами, которые дают им сигнал к пролиферации и более эффективному функционированию.
Т-клетки
T-клетки или Т-лимфоциты обычно подразделяют на две основных субпопуляции, которые являются функционально и фенотипически (что можно идентифицировать) различными. Субпопуляция T-хелперов, также называемых CD4+ T-клетками, является подходящим координатором иммунной регуляции. Основной функцией Т-хелперной клетки является усиление или потенцирование иммунных реакций путем секреции специализированных факторов, которые активируют другие лейкоциты для борьбы с инфекцией.
Другой важный тип T-клетки называется Т-киллерной субпопуляцией или CD8+ Т-клеткой. Данные клетки имеют значение при непосредственном уничтожении некоторых опухолевых клеток, инфицированных вирусом клеток и иногда паразитов. Они часто зависят от вторичных лимфоидных органов (лимфатических узлов и селезенки), как от участков, где происходит активация, но также их находят в других тканях организма, более очевидно, в печени, легких, крови и кишечном и репродуктивном трактах.
При использовании Т-клеток в способах ex vivo по изобретению, данные T-клетки обычно представляют собой Т-лимфоциты, выделенные из крови пациента или донора. T-клетки получают любым подходящим способом (например, описанным в US-A-4663058) и могут быть обогащены и/или очищены стандартными способами, включая опосредованное антителами разделение. Т-клетки могут использоваться в комбинации с другими иммунными клетками, полученными из того же или другого субъекта. Альтернативно в качестве источника Т-клеток и других клеточных типов может использоваться цельная кровь или обогащенная лейкоцитами кровь или очищенные лейкоциты. Особенно предпочтительно применение хелперных T-клеток (CD4+). Альтернативно могут использоваться другие Т-клетки, такие как CD8+ клетки. Также может быть удобным применять клеточные линии, такие как Т-клеточные гибридомы, незрелые Т-клетки периферического или тимусного происхождения и NK-T-клетки. В предпочтительном осуществлении Т-клетки, используемые по настоящему изобретению, представляют собой Т-клетки, которые могут переносить антигенспецифичную супрессию на другие T-клетки.
Введение полипептидов и последовательностей нуклеиновой кислоты в Т-клетки
Антигенные полипептидные вещества могут вводиться в Т-клетки в виде полипептидов как таковых или путем введения конструкций нуклеиновой кислоты/вирусных векторов, кодирующих полипептид, в клетки в условиях, которые обеспечивают экспрессию полипептида в Т-клетке.
Предпочтительно полинуклеотид для применения по изобретению в векторе функционально связан с последовательностью контроля, которая способна обеспечивать экспрессию кодирующей последовательности клеткой хозяина, то есть вектор является экспрессирующим вектором. Термин «функционально связанный» означает, что описанные компоненты находятся во взаимоотношениях, позволяющих им функционировать в присущей им манере. Регуляторная последовательность, «функционально связанная» с кодирующей последовательностью, лигирована таким путем, что экспрессия кодирующей последовательности достигается в условиях, совместимых с такими контрольными последовательностями.
Контрольные последовательности могут модифицироваться, например, путем добавления дальнейших элементов регуляции транскрипции, чтобы уровень транскрипции, направляемой последовательностями контроля, лучше реагировал на модуляторы транскрипции.
Векторы по изобретению могут вводиться в Т-клетки путем трансформации или трансфекции, как описано ниже для обеспечения экспрессии антигена.
Настоящее изобретение также относится к Т-клеткам, в которые введены, то есть загружены антигены, например, в виде полипептидов.
Для простоты ссылки антигенные полипептиды и кодирующие их полинуклеотиды будут указываться как «антиген».
Подходящие неограничивающие способы введения обоих генов и полипептидов в Т-клетки обсуждаются ниже.
Любой подходящий способ трансформации Т-клетки может использоваться. Неограничивающими примерами доступных в настоящее время механизмов доставки являются электропорация, трансформация с применением фосфата кальция и бомбардировка частицами. Однако перенос конструкции может проводиться любым из указанных способов, которые физически или химически повышают проницаемость клеточной мембраны. Подходящие способы описаны более подробно ниже.
1. Электропорация
В некоторых предпочтительных осуществлениях настоящего изобретения антиген вводят в клетки путем электропорации. В электропорации задействуется воздействие на суспензию клеток и ДНК высоковольтного электрического разряда.
Считается, что условия электропорации могут оптимизироваться для Т-клеток. Можно, в частности, оптимизировать такие параметры, как вольтаж, емкость, время и композиция среды для электропорации. Выполнение других рутинных настроек известно специалистам в данной области.
2. Бомбардировка частицами
В одном из способов переноса голой ДНК-конструкции в клетки участвует бомбардировка частицами. Данный способ зависит от способности ускорять покрытые ДНК микроснаряды до высоких скоростей, что позволяет им пронизывать клеточные мембраны и проникать в клетки без их уничтожения. Используемые микроснаряды состоят из биологически инертных веществ, таких как вольфрамовые, платиновые и золотые гранулы.
Считается, что в некоторых случаях для доставки ДНК в клетку-реципиент нет необходимости в осаждении ДНК на металлических частицах с использованием бомбардировки частиц. Считается, что частицы могут содержать ДНК, а не быть покрытыми ДНК. Следовательно, предполагается, что покрытые ДНК частицы могут повышать уровень доставки ДНК посредством бомбардировки частицами, но само по себе это не обязательно.
Разработаны некоторые устройства для ускорения малых частиц. Одно из таких устройств основывается на разряде высокого напряжения, генерирующем электрический ток, который, в свою очередь, обеспечивает движущую силу. Другой способ задействует применение системы доставки Biolistic Particle Delivery System, которая может использоваться для пролета частиц, покрытых ДНК, через экран, такой как экран из нержавеющей стали или материала Nytex, на поверхность фильтра, покрытую клетками из суспензии. Данный экран диспергирует частицы так, что они не доставляются в клетки-реципиенты в виде больших агрегатов. Полагают, что встраивание экрана между устройством бомбардировки и бомбардируемыми клетками снижает размер бомбардируемых агрегатов и может вносить вклад в более высокую частоту трансформации за счет снижения повреждения, наносимого клеткам-реципиентам снарядами, которые слишком велики.
Для бомбардировки клетки в суспензии предпочтительно концентрируют на фильтрах или, альтернативно, на твердой культуральной среде. Клетки, подлежащие бомбардировке, располагают на подходящем расстоянии ниже планшета, останавливающего макроскопические снаряды. Если это требуется, один или несколько экранов также помещают между устройством ускорения и бомбардируемыми клетками.
Считается, что может возникнуть намерение настроить различные параметры бомбардировки в мелкомасштабных исследованиях с целью полной оптимизации условий. В частности, может возникнуть намерение настроить физические параметры, такие как расстояние пропуска, расстояние полета, расстояние до ткани и давление гелия. Также можно оптимизировать факторы снижения травмы путем модификации условий, которые влияют на физиологическое состояние клеток реципиента и которые поэтому могут влиять на эффективность трансформации и интеграции. Например, осмотическое состояние, гидратация ткани и стадия субкультивирования или клеточный цикл Т-клетки-реципиента могут доводиться до оптимальной трансформации. Выполнение других рутинных настроек известно специалистам в данной области.
3. Трансформация вирусами
a. Аденовирусная инфекция
В одном из способов доставки конструкций нуклеиновой кислоты используется аденовирусный экспрессирующий вектор. Хотя аденовирусные векторы, как известно, обладают низкой способностью к интеграции в геномную ДНК, данная характеристика уравновешивается высокой эффективностью переноса генов, предоставляемой данными векторами. «Аденовирусный экспрессирующий вектор», как подразумевается, включает в себя те конструкции, содержащие последовательности аденовируса, которых достаточно для (a) поддержки упаковки конструкции и (b) конечной экспрессии конструкции, которая была клонирована.
Вектор включает в себя генноинженерную форму аденовируса. Знание генной организации аденовируса, вируса с линейной, двухцепочечной ДНК размером 36 т.п.н., позволяет осуществлять замену больших фрагментов аденовирусной ДНК чужеродными последовательностями размером до 7 т.п.н. В отличие от ретровируса, аденовирусная инфекция клетки хозяина не приводит к хромосомной интеграции, поскольку аденовирусная ДНК может реплицироваться по типу эписомы без потенциальной генотоксичности. Также аденовирусы структурно стабильны, и после интенсивной амплификации не отмечали никаких перестановок в геноме.
Аденовирус особенно подходит для применения в качестве вектора переноса генов из-за его генома средних размеров, простоты манипуляций с ним, высокого титра, широкого интервала клеток-мишеней и высокой инфекционности. Оба конца вирусного генома содержат инвертированные повторы (ITR) длиной 100-200 пар оснований, где находятся цис-элементы, необходимые для репликации и упаковки вирусной ДНК. Ранние (E) и поздние (L) области генома содержат различные транскрипционные единицы, которые подразделены по началу репликации вирусной ДНК. Область E1 (E1A и E1B) кодирует белки, ответственные за регуляцию транскрипции вирусного генома и небольшого количества клеточных генов. Экспрессия области E2 (E2A и E2B) приводит к синтезу белков для репликации вирусной ДНК. Данные белки вовлечены в репликацию ДНК, экспрессию поздних генов и запирание клетки хозяина. Продукты поздних генов, включающие большинство белков вирусного капсида, экспрессируются только после значительного процессинга единичного первичного транскрипта, выходящего с главного позднего промотора (MLP). MLP (расположенный на 16,8 m.u.) является особенно эффективным во время поздней фазы инфекции, и все мРНК, выходящие с данного промотора, обладают 5'-концевой тройственной лидерной (TPL) последовательностью, которая обеспечивает этим мРНК предпочтение для трансляции.
В настоящей системе рекомбинантный аденовирус получают путем гомологичной рекомбинации между вектором-челноком и провирусным вектором. Вследствие возможной рекомбинации между двумя провирусными векторами в данном процессе может генерироваться аденовирус дикого типа. Поэтому критичным является выделение единственного клона вируса из индивидуальной бляшки и оценка структуры его генома.
Получение и размножение настоящих аденовирусных векторов, которые характеризуются дефицитом репликации, зависит от уникальной вспомогательной клеточной линии, обозначенной 293, которая трансформирована из клеток эмбриональной почки человека введением ДНК-фрагментов Ad5, и полученная линия конститутивно экспрессирует белки E1. Поскольку область E3 из генома аденовируса несущественна, настоящие аденовирусные векторы, функционирующие с помощью клеток 293, несут чужеродную ДНК в E1, в D3 или в обеих областях. В природе аденовирус может упаковывать примерно 105% генома дикого типа, обеспечивая возможность добавления примерно 2 дополнительных т.п.н. ДНК. В комбинации примерно с 5,5 т.п.н. ДНК, которой замещены E1- и E3-области, максимальная емкость настоящих аденовирусных векторов составляет до 7,5 т.п.н., или примерно 15% от общей длины вектора. Более 80% аденовирусного генома остается в виде каркаса вектора.
Вспомогательные клеточные линии могут происходить от человеческих клеток, таких как клетки эмбриональной почки человека, мышечные клетки, кроветворные клетки или другие человеческие эмбриональные мезенхимальные или эпителиальные клетки. Альтернативно вспомогательные клетки могут происходить от клеток других видов млекопитающих, которые переносят аденовирус человека. Такие клетки включают в себя, например, клетки Vero, или другие эмбриональные мезенхимальные или эпителиальные клетки обезьян. Как указывалось выше, предпочтительной вспомогательной клеточной линией является 293.
Кроме того требования, что аденовирусный вектор должен быть дефективным по репликации, или, по меньшей мере, дефективным при определенных условиях, природа аденовирусного вектора не считается значимой для успешного воплощения изобретения. Аденовирус может относиться к любому из 42 различных известных серотипов из подгрупп A-F. Аденовирус типа 5 подгруппы C является предпочтительным исходным материалом для получения дефективного по репликации при определенных условиях аденовирусного вектора для применения по настоящему изобретению. Такое условие связано с тем, что аденовирус типа 5 является человеческим аденовирусом, о котором накоплено большое количество биохимической и генетической информации, и он исторически использовался в большинстве конструкций, использующих аденовирус в качестве вектора.
Как утверждалось выше, типичный вектор по настоящему изобретению дефективен по репликации и не содержит области аденовируса E1. Таким образом, наиболее удобным является введение трансформирующей конструкции в положении, из которого удалены кодирующие E1 последовательности. Однако положение вставки конструкции в аденовирусные последовательности не являются критичными в контексте изобретения. Полинуклеотид, кодирующий интересующий ген, может также встраиваться в место удаленной области E3 в векторах замещения E3 или в область E4, где вспомогательная клеточная линия или вспомогательный вирус компенсируют дефект E4.
Рост и манипуляции с аденовирусом известны специалистам в данной области, и данный вирус характеризуется большим разнообразием хозяев in vitro и in vivo. Данная группа вирусов может быть получена с высоким титром, например 109-1011 бляшкообразующих единиц на мл, и они высокоинфекционны. Жизненный цикл аденовируса не требует интеграции в геном клетки хозяина. Чужеродные гены, доставляемые аденовирусными векторами, являются эписомными и поэтому характеризуются низкой генотоксичностью в отношении клеток хозяина. Не сообщалось о побочных эффектах в исследованиях по вакцинации аденовирусами дикого типа, что демонстрирует их безопасность и терапевтический потенциал также в качестве векторов для переноса генов in vivo.
b. Инфекция AAV
Адено-ассоциированный вирус (AAV) является привлекательной векторной системой для применения по настоящему изобретению, поскольку он характеризуется высокой частотой интеграции, и он может инфицировать неделящиеся клетки, что, таким образом, делает его удобным для доставки генов в клетки млекопитающих в культуре ткани. AAV характеризуется широким интервалом инфекционности. Подробности, касающиеся получения и применения rAAV-векторов, описаны в патенте США № 5139941 и в патенте США № 4797368, каждый из которых включен сюда в качестве ссылки.
Рекомбинантные AAV-векторы также успешно используют для трансдукции in vivo и in vitro маркерных генов и генов, вовлеченных в заболевания человека. Недавно AAV-вектор был одобрен для испытаний фазы I на людях для лечения кистозного фиброза.
AAV представляет собой зависимый парвовирус, который требует совместной инфекции другим вирусом (аденовирусом или представителем семейства герпесвирусов) для прохождения продуктивной инфекции культивируемых клеток. В отсутствие совместной инфекции вспомогательным вирусом геном AAV дикого типа интегрируется посредством его концов в человеческую хромосому 19, где он остается в латентном состоянии как провирус. Однако rAAV не ограничен в плане интеграции хромосомой 19, если также не экспрессируется белок AAV Rep. Когда клетка, несущая провирус AAV, подвергается суперинфекции вспомогательным вирусом, геном AAV «спасается» из хромосомы или из рекомбинантной плазмиды, и устанавливается нормальная продуктивная инфекция.
Обычно рекомбинантный вирус AAV (rAAV) получают путем совместной трансфекции плазмидой, содержащей интересующий ген, фланкированный двумя терминальными повторами AAV, и экспрессирующей плазмидой, содержащей кодирующие последовательности AAV дикого типа без терминальных повторов, например pIM45. Клетки также инфицировали или трансфицировали аденовирусом или плазмидами, несущими аденовирусные гены, требуемые для вспомогательной функции для AAV. Запасы вируса rAAV, полученные таким способом, контаминируют аденовирусом, которые должны быть физически отделены от частиц rAAV (например, центрифугированием в градиенте плотности хлорида цезия). Альтернативно могут использоваться аденовирусные векторы, содержащие кодирующие области AAV, или клеточные линии, содержащие кодирующие области AAV и некоторые или все аденовирусные гены. Также могут использоваться клеточные линии, несущие ДНК rAAV в качестве интегрированного провируса.
c. Ретровирусная инфекция
В одном из предпочтительных способов по настоящему изобретению применяются ретровирусы. Ретровирусный вектор по настоящему изобретению может происходить или быть получаемым из любого подходящего ретровируса. Идентифицировано большое количество различных ретровирусов. Примеры включают в себя: вирус лейкоза мыши (MLV), вирус иммунодефицита человека (ВИЧ), вирус иммунодефицита обезьян, вирус Т-клеточного лейкоза человека (HTLV), вирус инфекционной анемии лошади (EIAV), вирус опухоли молочной железы мыши (MMTV), вирус саркомы Рауса (RSV), вирус саркомы Фуджинами (FuSV), вирус мышиного лейкоза Молони (Mo-MLV), вирус мышиной остеосаркомы FBR (FBR MSV), вирус мышиной саркомы Молони (Mo-MSV), вирус мышиного лейкоза Абельсона (A-MLV), вирус птичьего миелоцитоматоза 29 (MC29) и вирус птичьего эритробластоза (AEV). Подробный список ретровирусов может быть найден в Coffin et al., 1997, "retroviruses", Cold Spring Harbour Laboratory Press Eds: JM Coffin, SM Hughes, HE Varmus p. 758-763.
Подробности структуры генома некоторых ретровирусов могут быть найдены в данной области. Для примера, подробности по ВИЧ и Mo-MLV могут быть найдены в NCBI Genbank (инвентарные номера геномов № AF033819 и AF033811 соответственно).
Ретровирусы в общих чертах могут быть разделены на две категории: а именно «простые» и «комплексные». Ретровирусы могут далее подразделяться на семь групп. Пять из этих групп представляют собой ретровирусы с онкогенным потенциалом. Оставшиеся две группы представляют собой лентивирусы и спумавирусы. Обзор данных ретровирусов представлен в Coffin et al., 1997 (ibid).
Группа лентивирусов может даже далее подразделяться на вирусы «приматов» и «организмов, не относящихся к приматам». Примеры лентивирусов приматов включают в себя вирус иммунодефицита человека (ВИЧ), агент, вызывающий синдром приобретенного иммунодефицита человека (СПИД), и вирус иммунодефицита обезьян (SIV). Группа лентивирусов организмов, не относящихся к приматам, включает в себя прототип «медленный вирус» visna/maedi (VMV), а также родственный ему вирус артрита-энцефалита козы (CAEV), вирус инфекционной анемии лошади (EIAV) и описанные позднее вирус иммунодефицита кошек (FIV) и вирус иммунодефицита крупного рогатого скота (BIV).
Различие между семейством лентивирусов и другими типами ретровирусов состоит в том, что лентивирусы обладают способностью инфицировать делящиеся и неделящиеся клетки. Другие лентивирусы, такие как MLV, наоборот, не способны инфицировать неделящиеся клетки, такие как те, что составляют ткань мышцы, головного мозга, легкого и печени.
Каждый ретровирусный геном содержит гены, называемые gag, pol и env, которые кодируют белки вириона и ферменты. В провирусе данные гены фланкированы с обоих концов областями, называемыми длинными концевыми повторами (LTR). LTR ответственны за интеграцию и транскрипцию провируса. LTR также служат как энхансерные-промоторные последовательности и могут контролировать экспрессию вирусных генов. Инкапсулирование ретровирусных РНК происходит за счет последовательности psi, находящейся на 5'-конце вирусного генома.
LTR сами по себе являются идентичными последовательностями, которые могут подразделяться на три элемента, которые называют U3, R и U5. U3 происходит от последовательности, уникальной для 3'-конца РНК. R происходит от последовательности, происходящей из последовательности, повторяемой на обоих концах РНК, а U5 происходит от последовательности, уникальной для 5'-конца РНК. Размеры трех элементов могут значительно варьироваться в разных ретровирусах.
Основная молекулярная организация РНК-генома всех инфекционных ретровирусов представляет собой (5') R-US-gag, pol, env-U3-R (3'). В дефектном ретровирусном векторном геноме gag, pol и env могут отсутствовать или быть нефункциональными. R-области с обоих концов РНК являются повторяемыми последовательностями. U5 и U3 представляют собой уникальные последовательности на 5'- и 3'-концах РНК-генома соответственно.
Для конструирования ретровирусного вектора нуклеиновую кислоту, кодирующую интересующий ген, встраивают в вирусный геном вместо некоторых вирусных последовательностей с получением вируса, дефектного по репликации. Для продукции вирионов конструируют упаковывающую клеточную линию, содержащую гены gag, pol и env, но без LTR и компонентов упаковки (Mann et al., 1983). Когда рекомбинантную плазмиду, содержащую кДНК, вместе с ретровирусными LTR и последовательностями упаковки вводят в данную клеточную линию (например, путем осаждения фосфатом кальция), последовательность упаковки позволяет РНК-транскрипту рекомбинантной плазмиды упаковываться в вирусные частицы, которые затем секретируются в культуральную среду. Затем среду, содержащую рекомбинантные ретровирусы, собирают, необязательно концентрируют и применяют для переноса генов. Ретровирусные векторы способны инфицировать много различных клеточных типов. Однако для интеграции и стабильной экспрессии требуется разделение клеток хозяина.
Проблема, связанная с применением дефектных ретровирусных векторов, представляет собой потенциальное возникновение в упаковывающих клетках способного к репликации вируса дикого типа. Это может происходить из-за событий рекомбинации, при которой интактная последовательность из рекомбинантного вируса встраивается выше последовательности gag, pol, env, интегрированной в геном клетки хозяина. Однако в настоящее время доступны новые упаковывающие клеточные линии, которые должны резко снижать вероятность рекомбинации.
d. Другие вирусные векторы
Другие вирусные векторы могут использоваться в качестве конструкций в описанных здесь способах и композициях. Могут использоваться векторы, происходящие от таких вирусов, как вирусы коровьей оспы и герпесвирусы.
4. Соосаждение фосфатом кальция или обработка DEAE-декстраном
В других предпочтительных осуществлениях антиген вводят в клетки с использованием соосаждения фосфатом кальция. В другом осуществлении экспрессирующая конструкция доставляется в клетки с использованием DEAE-декстрана с последующей обработкой полиэтиленгликолем.
5. Прямая микроинъекция или загрузка обработкой ультразвуком
Дальнейшие осуществления включают в себя введение антигена путем прямой микроинъекции или загрузки обработкой ультразвуком.
6. Опосредованная липосомами трансформация
В дальнейшем осуществлении антиген может быть захвачен липосомой. Липосомы представляют собой везикулярные структуры, характеризующиеся фосфолипидной бислойной мембраной и внутренней водной средой. Мультиламеллярные липосомы характеризуются множественными липидными слоями, разделенными водной средой. Они образуются спонтанно, когда фосфолипиды суспендируют в избытке водного раствора. Липидные компоненты претерпевают самопроизвольную перестройку перед образованием закрытых структур и захватывают воду и растворенные вещества между липидных бислоев. Также рассматриваются конструкции нуклеиновой кислоты в комплексе с липофектамином (Gibco BRL).
В некоторых осуществлениях изобретения липосома может образовывать комплекс с гемагглютинирующим вирусом (HVJ). Как было показано, это ослабляет слияние с клеточной мембраной и способствует проникновению в клетку инкапсулированной в липосомы ДНК. В других осуществлениях липосома может образовывать комплекс или использоваться в сочетании с ядерными негистоновыми хромосомными белками (HMG-1). Еще в одном осуществлении липосома может образовывать комплекс или использоваться в сочетании с HVJ и HMG-1.
7. Трансфекция посредством аденовирусов
В некоторых осуществлениях конструкцию нуклеиновой кислоты вводят в клетку с использованием трансфекции посредством аденовируса. О повышенной эффективности трансфекции сообщалось для клеточных систем, использующих сочетанные с аденовирусом системы, и авторы изобретения рассматривают применение того же способа для повышения эффективности трансфекции.
Культивирование
Настоящее изобретение также относится к способу культивирования T-клеток для применения по настоящему изобретению.
В частности, мононуклеарные клетки периферической крови (PBMC), выделенные из периферической крови, могут активироваться in vitro для пролиферации. Митогенная стимуляция может быть достигнута с применением таких соединений, как фитогемагглютинин (PHA, 2 мкг/мл) или mAb против CD3 (OKT3; 30 нг/мл).
Активированные Т-клетки могут культивироваться в концентрации 1×106 клеток/мл, в среде, дополненной 5% аутологичной сывороткой в присутствии факторов роста, таких как hu-r-IL-2 (например, 100 ед./мл).
В одном предпочтительном осуществлении Т-клетки культивируются в присутствии магнитных гранул, покрытых антителами к CD3 и CD28, предпочтительно в присутствии hu-r-IL2 (например, 100 ед./мл). Обычно гранулы используются в соотношении три гранулы/T-клетка.
Затем Т-клетки могут подвергаться трансдукции/трансфекции путем воздействия на активированные T-клетки, например, с использованием вирусного(ых) вектора(ов), кодирующего(их) антиген(ы). Экспрессия трансгена(ов) может использоваться для оценки эффективности трансдукции. Гены и маркеры селекции обсуждаются более подробно ниже. Альтернативно лимфоциты могут культивироваться, как описано выше, но не подвергаться трансдукции/трансфекции. Вместо этого до введения пациенту активированные Т-клетки загружают антигеном, например, в виде синтетических пептидов, кодирующих, например, хорошо известные опухолевые, бактериальные и вирусные антигены.
Весь процесс манипуляции длится, в общем, в течение 5-15 суток.
Т-клетки, подвергаемые манипуляциям, могут замораживаться для целей хранения.
В конце культивирования подвергаемые манипуляциям T-клетки должны экспрессировать, по меньшей мере, один из следующих маркеров: HLA-I, HLA-II, CD80, CD86, CD27, CD40L, CD62L, CCR7, CD54 и CD25. Таким образом, авторами изобретения обнаружено, что значимым является фенотип T-клетки.
Вакцина
Изобретение относится к способу индукции иммунного ответа у субъекта, особенно у млекопитающего, предпочтительно у человека, который охватывает инокуляцию субъекта содержащей антиген Т-клеткой по настоящему изобретению, так что антиген продуцирует гуморальный и/или Т-клеточный иммунный ответ для защиты данного субъекта, например, от опухоли или инфекции, такой как бактериальная или вирусная инфекция. Также предоставляются способы, посредством которых такой иммунный ответ замедляет рост опухоли или репликации бактерии или вируса.
Таким образом, изобретение относится к иммунологической композиции, которая при введении субъекту, предпочтительно человеку, способному к индукции в его организме иммунного ответа, индуцирует иммунный ответ у такого субъекта. Иммунный ответ может использоваться терапевтически или профилактически и может принимать формы гуморального иммунитета и/или клеточного иммунитета, такого как клеточный иммунитет, возникающий от CTL или CD4+ T-клеток.
Получение вакцин, которые содержат иммуногенный(ые) полипептид(ы) в качестве активного(ых) ингредиента(-ов), известно специалисту в данной области. Обычно такие вакцины получают в виде средств для инъекции или жидких растворов или суспензий; твердые формы, подходящие для растворения или суспендирования в жидкости перед инъекцией, могут также быть получены. Препараты также могут быть эмульгированы, или белки в них могут быть инкапсулированы в липосомах. Активные иммуногенные ингредиенты часто смешивают с наполнителями, которые являются фармацевтически приемлемыми и совместимы с активными ингредиентами. Подходящие наполнители представляют собой, например, воду, солевой раствор, декстрозу, глицерин, этанол, или тому подобное и их комбинации.
Кроме того, если требуется, вакцина может содержать малые количества добавочных веществ, таких как увлажняющие или эмульгирующие средства, средства поддержания pH, и/или адъюванты, которые усиливают эффективность вакцины.
Препарат вакцины по изобретению относится и/или содержит адъювантную систему для усиления иммуногенности препарата. Предпочтительно, адъювантная система стимулирует предпочтительно ответ типа TH1.
Иммунный ответ может, в общем, подразделяться на две противоположные категории, представляющие собой гуморальный или клеточно-опосредованный иммунный ответ (традиционно характеризующийся антительными и клеточными эффекторными механизмами защиты соответственно). Данные категории ответа называются реакциями TH1-типа (клеточно-опосредованными ответами) и иммунными реакциями TH2-типа (гуморальными ответами).
Особые иммунные реакции TH1-типа могут характеризоваться образованием антигенспецифических, ограниченных по гаплотипу цитотоксических T-лимфоцитов и реакциями естественных киллеров. Реакции TH1-типа мыши часто характеризуются образованием антител субтипа IgG2a, тогда как у людей они соответствуют антителам типа IgG1. Иммунные реакции TH2-типа характеризуются образованием большого разнообразия иммуноглобулиновых изотипов, включая IgG1, IgA и IgM у мышей.
Можно считать, что движущей силой развития данных двух типов иммунного ответа являются цитокины. Высокие уровни цитокинов TH1-типа обеспечивают тенденцию к предпочтительной индукции опосредованных клетками иммунных реакций на данный антиген, тогда как высокие уровни цитокинов TH2-типа обеспечивают тенденцию к предпочтительной индукции гуморальных иммунных реакций на антиген.
Иммуномодуляторы, такие как вакцины, могут быть получены из Т-клеток, включающих в себя один или несколько полипептидов, или даже нуклеотидные последовательности по настоящему изобретению. Получение иммуномодуляторов, которые содержат иммуногенный(е) пептид(ы) в качестве активного(ых) ингредиента(ов), известно специалистам в данной области. Обычно такие иммуномодуляторы получают в виде средств для инъекции, или жидких растворов или суспензий; твердые формы, подходящие для растворения или суспендирования в жидкости перед инъекцией, могут также быть получены. Препараты также могут быть эмульгированы, или белки в них могут быть инкапсулированы в липосомах. Активные иммуногенные ингредиенты часто смешивают с наполнителями, которые являются фармацевтически приемлемыми и совместимы с активными ингредиентами. Подходящие наполнители представляют собой, например, воду, солевой раствор, декстрозу, глицерин, этанол или тому подобное и их комбинации.
Кроме того, если требуется, иммуномодуляторы могут содержать малые количества добавочных веществ, таких как увлажняющие или эмульгирующие средства, средства поддержания pH, и/или адъюванты, которые усиливают эффективность иммуномодуляторов. Неограничивающие примеры адъювантов, которые могут быть эффективными, включают в себя гидроксид алюминия, N-ацетилмурамил-L-треонил-D-изоглутамин (thr-MDP), N-ацетил-нормурамил-L-аланил-D-изоглутамин (CGP 11637, обозначаемый как nor-MDP), N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2-(1'-2'-дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)этиламин (CGP 19835A, обозначаемый как MTP-PE), и RIBI, который содержит три компонента, выделенные из бактерий, монофосфориллипид A, димиколат трегалозы и скелет клеточной стенки (MPL+TDM+CWS) в 2%-ной эмульсии в смеси сквален/Tween 80.
Дальнейшие примеры адъювантов и других средств включают гидроксид алюминия, фосфат алюминия, сульфат алюминия-калия (квасцы), сульфат бериллия, силикагель, каолин, уголь, эмульсии «вода в масле», эмульсии «масло в воде», мурамилдипептид, бактериальный эндотоксин, липид X, Corynebacterium parvum (Propionobacterium acnes), Bordetella pertussis, полирибонуклеотиды, альгинат натрия, ланолин, лизолецитин, витамин A, сапонин, липосомы, левамизол, DEAE-декстран, блок-сополимеры или другие синтетические адъюванты. Такие адъюванты доступны коммерчески из различных источников, например Merck Adjuvant 65 (Merck and Company, Inc., Rahway, Нью-Джерси) или неполный адъювант и полный адъювант Фрейнда (Difco Laboratories, Детройт, Мичиган).
Обычно используют такие адъюванты, как Amphigen (масло в воде), Alhydrogel (гидроксид алюминия), или используют смесь Amphigen и Alhydrogel. Для использования у человека одобрен только гидроксид алюминия.
Отношение иммуногена и адъюванта может варьироваться в широком интервале, при том что оба должны присутствовать в эффективных количествах. Например, гидроксид алюминия может присутствовать в количестве, равном примерно 0,5%, в смеси иммуномодуляторов (основа Al2O3). Обычно иммуномодуляторы включают в состав препарата так, что он содержит конечную концентрацию иммуногена в интервале от 0,2 до 200 мкг/мл, предпочтительно от 5 до 50 мкг/мл, наиболее предпочтительно 15 мкг/мл.
После получения препарата иммуномодуляторы могут вводиться в стерильный контейнер, который затем закупоривают и хранят при низкой температуре, например при 4°C, или препарат может высушиваться вымораживанием. Лиофилизация обеспечивает долгосрочное хранение в стабилизированной форме.
Иммуномодуляторы обычно вводят парентерально, путем инъекции, например, или подкожно или внутримышечно. Дополнительные препараты, которые подходят для других путей введения, включают в себя суппозитории и, в некоторых случаях, пероральные препараты. Традиционные связующие вещества и носители для суппозиториев могут включать в себя, например, полиалкиленгликоли или триглицериды; такие суппозитории могут быть образованы из смесей, содержащих активный ингредиент в интервале от 0,5% до 10%, предпочтительно от 1% до 2%. Пероральные препараты включают в себя такие обычно используемые наполнители, как, например, фармацевтические марки маннита, лактозы, крахмала, стеарата магния, сахарина натрия, целлюлозы, карбоната магния, и тому подобного. Данные композиции принимают форму растворов, суспензий, таблеток, пилюль, капсул, препаратов замедленного высвобождения или порошков и содержат от 10% до 95% активного ингредиента, предпочтительно от 25% до 70%. Там, где композиция иммуномодуляторов лиофилизована, лиофилизованный материал может восстанавливаться перед введением, например, в виде суспензии. Восстановление предпочтительно осуществляется в буфере.
Полипептиды по изобретению могут вводиться в состав препарата иммуномодуляторов в нейтральном виде или в виде соли. Фармацевтически приемлемые соли включают в себя кислотно-аддитивные соли (образованные свободными аминогруппами пептида) и соли, которые образованы с неорганическими кислотами, такими как, например, соляная или фосфорная кислота, или с такими органическими кислотами, как уксусная, щавелевая, винная или малеиновая. Соли, образованные свободными карбоксильными группами, могут также быть основаны на неорганических основаниях, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа, и таких органических основаниях, как изопропиламин, триметиламин, 2-этиламиноэтанол, гистидин и прокаин.
Антиген
В общих представлениях антиген является любым соединением, которое способно взаимодействовать с антигенным рецептором. Антиген связывается со специфическим антителом для обеспечения механизма, по которому антиген распознается и инактивируется, таким образом, антиген образует комплекс со специфическим антителом, так что комплекс может присоединяться к специализированным иммунным клеткам, которые либо интернализуют его с целью разрушения, либо высвобождают биологические медиаторы, такие как гистамин для продукции аллергического/воспалительного ответа. Иммуноген представляет собой антиген, который активирует иммунные клетки с целью развития против него иммунного ответа. Таким образом, иммуноген представляет собой антиген, но антиген не обязательно является иммуногеном. Однако для простоты ссылки здесь термины «антиген» и «иммуноген» используются взаимозаменяемо.
Настоящее изобретение, в общем, относится к новому способу представления антигена иммунной системы. Некоторые из факторов, которые определяют иммуногенный потенциал, и, следовательно, его способность генерировать иммунный ответ, включают в себя доступность антигенного эпитопа для распознавания иммунной системой и путь введения антигена. Настоящее изобретение относится к системе улучшения данных факторов, так что иммуногенность антигена оптимизируется. В результате субъект может получать две или три дозы лишь за одно введение. Таким образом, улучшается не только эффективность вакцины, но также приверженность пациента.
Настоящее изобретение применимо к любому антигену, который может быть введен в T-клетку, так что антиген представляется иммунной системе и более конкретно APC. В особенно предпочтительном осуществлении антиген вводят в Т-клетку в виде полинуклеотида, который способен экспрессироваться в Т-клетке.
Известен широкий спектр подходящих антигенов. Их последовательности могут быть выбраны, например, на основе полипептидных последовательностей, известных в литературе. Для полипептидов, которые характеризуются частичной последовательностью белка с иммуногенной активностью, возможно установить, какие пептиды являются подходящими кандидатами посредством сравнения последовательностей. Предпочтительными кандидатами являются полипептиды, иммуногенность которых уже была продемонстрирована, то есть полипептиды, происходящие из известных иммуногенов, таких как вирусные или бактериальные белки. Полипептиды данного типа предпочтительно характеризуются сильным взаимодействием в тесте MLC, характеризующем их иммуногенность. Термин «белок» включает в себя одноцепочечные полипептидные молекулы, а также комплексы нескольких полипептидов, где индивидуальные составляющие полипептиды ковалентно или нековалентно связаны. Термин «полипептид» включает в себя пептиды из двух или более аминокислот длиной, обычно имеющие более 5, 10 или 20 аминокислот.
Будет понятно, что полипептидные последовательности для применения не ограничены конкретными последовательностями или их фрагментами или последовательностями, полученными из конкретного белка, но также включают в себя гомологичные последовательности, полученные из любого источника, например родственные вирусные/бактериальные антитела, клеточные гомологи и синтетические пептиды, а также их варианты и производные. Полипептидные последовательности по настоящему изобретению также включают в себя полипептиды, кодируемые полинуклеотидами по настоящему изобретению.
Таким образом, настоящее изобретение охватывает варианты, гомологи или производные аминокислотных последовательностей антигенов, а также варианты, гомологи или производные аминокислотных последовательностей, кодируемые нуклеотидными последовательностями по настоящему изобретению.
В контексте настоящего изобретения используется гомологичная последовательность, которая включает в себя аминокислотную последовательность, которая идентична, по меньшей мере, на 60, 70, 80 или 90%, предпочтительно идентична, по меньшей мере, на 95 или 98% на аминокислотном уровне, в отношении конкретной последовательности. В частности, гомология обычно должна рассматриваться в отношении тех областей последовательности, которые, как известно, существенны для антигенности, а не соседних несущественных областей. Наличие областей, которые являются консервативными среди представителей семейства, должно требоваться для получения относительно высоких коэффициентов гомологии: это помогает при определении функциональных молекул. Области, которые являются уникальными и малоконсервативными среди представителей семейства, должны/могут характеризоваться более низким коэффициентом по принципу поэтапного рассмотрения случаев: это помогает установить новизну. Данный поэтапный подход, в общем, более полезен, чем общий коэффициент гомологии. Хотя гомология также может рассматриваться в плане сходства (т.е. аминокислотных остатков, характеризующих сходные химические свойства/функции), в контексте настоящего изобретения предпочтительно выражать гомологию в плане идентичности последовательности.
Сравнения гомологии могут проводиться на глаз или, что более распространено, с помощью легко доступных программ сравнения последовательности. Данные коммерчески доступные компьютерные программы могут рассчитывать % гомологии между двумя или более последовательностями.
Термины «вариант» или «производное» в отношении аминокислотных последовательностей по настоящему изобретению включает в себя любую замену, вариант, модификацию, замену, делецию или добавление одной (или нескольких) аминокислоты по отношению к последовательности, что обеспечивает полученную в результате аминокислотную последовательность, которая характеризуется антигенной активностью, предпочтительной, составляющей, по меньшей мере, от 25 до 50% от активности полипептидов, представленных в списке последовательностей, более предпочтительно, по меньшей мере, с той же активностью.
Таким образом, конкретные последовательности по изобретению могут модифицироваться для применения по настоящему изобретению. Обычно делают модификации, которые поддерживают антигенность последовательности. Таким образом, в одном из осуществлений могут быть сделаны аминокислотные замены, например, от 1, 2 или 3 до 10, 20 или 30 замен, при обеспечении того, что модифицированная последовательность сохраняет, по меньшей мере, примерно от 25 до 50% активности, или имеет, по существу, прежнюю активность. Однако в качестве альтернативного осуществления модификации аминокислотных последовательностей полипептида по изобретению могут быть сделаны специально для снижения биологической активности полипептида. Например, укороченные полипептиды, которые сохраняют способность связываться с целевой молекулой, но лишены функционального эффекторного домена, могут использоваться в качестве ингибиторов биологической активности полноразмерной молекулы.
В общем, предпочтительно менее 20%, 10% или 5% аминокислотных остатков варианта или производного изменяют по сравнению с соответствующей областью, обозначенной в списке последовательностей.
Аминокислотные замены могут исключать применение не встречающихся в природе аналогов, например, для повышения периода полужизни в плазме крови терапевтически введенного полипептида (смотри ниже для дальнейших подробностей по продукции пептидных производных для применения в терапии).
Консервативные замены могут быть сделаны, например, по таблице ниже. Аминокислоты в пределах одной ячейки во второй колонке и предпочтительно в одной строке в третьей колонке могут замещаться одна на другую:
Полипептиды по изобретению также включают в себя фрагменты полноразмерных полипептидов и их варианты. Предпочтительные фрагменты включают те, что содержат эпитоп. Подходящие фрагменты составляют, по меньшей мере, около 5, например 10, 12, 15 или 20 аминокислот в длину. Они могут также составлять менее 200, 100 или 50 аминокислот в длину. Полипептидные фрагменты белков и их аллельные и видовые варианты могут содержать один или несколько (например, 2, 3, 5 или 10) замен, делеций, или вставок, включая консервативные замены. Там, где были сделаны замены, делеции и/или вставки, например, посредством рекомбинантной технологии, изменены, предпочтительно, менее 20%, 10% или 5% аминокислотных остатков, обозначенных в списке последовательностей.
Особенно предпочтительные фрагменты включают в себя те фрагменты, что содержат антигенные домены.
Полинуклеотиды для применения по изобретению включают в себя последовательности нуклеиновой кислоты, кодирующие последовательности по изобретению. Специалисту в данной области будет понятно, что многочисленные различные полинуклеотиды могут кодировать один и тот же полипептид в результате вырожденности генетического кода. Кроме того, следует понимать, что специалисты в данной области могут с использованием рутинных способов проводить нуклеотидные замены, которые не влияют на полипептидную последовательность, кодируемую полинуклеотидами по изобретению, для отражения использования кодонов каким-либо конкретным организмом хозяина, в котором надлежит экспрессировать полипептиды по изобретению.
Полинуклеотиды по изобретению могут включать в себя ДНК или РНК. Они могут быть одноцепочечными или двухцепочечными. Они также могут являться полинуклеотидами, которые включают в себя синтетические или модифицированные нуклеотиды. Некоторое число различных типов модификации олигонуклеотидов известно в данной области. Они включают в себя метилфосфонатные и фосфоротиоатные остовы, добавление акридиновых или полилизиновых цепей с 3' и/или 5' концов молекулы. Для целей настоящего изобретения следует понимать, что описанные здесь полинуклеотиды могут модифицироваться любым способом, доступным в данной области. Такие модификации могут проводиться с целью усиления активности in vivo или периода жизни полинуклеотидов по изобретению.
Термины «вариант», «гомолог» или «производное» в отношении нуклеотидной последовательности включают в себя любую замену, вариацию, модификацию, перемещение, делецию или добавление одной (или нескольких) нуклеиновой кислоты в отношении последовательности, с предоставлением того, что полученная в результате нуклеотидная последовательность кодирует полипептид, характеризующийся антигенной активностью.
Как указано выше в отношении гомологии последовательности, предпочтительно имеет место гомология, составляющая, по меньшей мере, 75%, более предпочтительно, по меньшей мере, 85%, более предпочтительно, по меньшей мере, 90% гомологии в отношении последовательностей, показанных здесь в списке последовательностей. Более предпочтительно, имеет место гомология, составляющая, по меньшей мере, 95%, более предпочтительно, по меньшей мере, 98%. Сравнение гомологии по нуклеотидам может проводиться, как описано выше. Предпочтительной программой для сравнения является описанная выше программа GCG Wisconsin Bestfit. Матрица коэффициентов, принятая по умолчанию, имеет значение совпадения, равное 10 для каждого идентичного нуклеотида, и -9 для каждого несовпадения. Принятый по умолчанию штраф за открытие вставки составляет -50, а принятый по умолчанию штраф за продление вставки составляет -3 для каждого нуклеотида.
Настоящее изобретение также относится к нуклеотидным последовательностям, которые способны селективно гибридизоваться с антигенными последовательностями, или с любым их вариантом, фрагментом или производным, или с последовательностью, комплементарной любой из указанных выше. Нуклеотидные последовательности предпочтительно составляют, по меньшей мере, 15 нуклеотидов в длину, более предпочтительно, по меньшей мере, 20, 30, 40 или 50 нуклеотидов в длину.
Используемый здесь термин «гибридизация» включает в себя «процесс, в котором цепь нуклеиновой кислоты соединяется с комплементарной цепью за счет спаривания оснований», а также процесс амплификации, проводимый по технологиям полимеразной цепной реакции.
Полинуклеотиды по изобретению, способные к селективной гибридизации с представленными здесь нуклеотидными последовательностями, или с комплементарными им последовательностями, в общем, по меньшей мере, на 70%, предпочтительно, по меньшей мере, на 80 или 90% и, более предпочтительно, по меньшей мере, на 95% или 98% гомологичны соответствующим нуклеотидным последовательностям по области, равной, по меньшей мере, 20, предпочтительно, по меньшей мере, от 25 до 30, например, по меньшей мере, 40, 60 или 100 или большему количеству следующих друг за другом нуклеотидов.
Термин «селективно гибридизуемый» означает, что полинуклеотид, использованный в качестве зонда, применяется в условиях, где целевой полинуклеотид по изобретению, как это обнаружено, гибридизуется с зондом на уровне, значительно превышающем фоновый. Фоновая гибридизация может происходить по причине наличия других полинуклеотидов, например при скрининге библиотеки кДНК или геномной ДНК. В таком случае фон подразумевает уровень сигнала, генерируемый путем взаимодействия между зондом и неспецифическим представителем библиотеки ДНК, который в 10 раз менее интенсивен, предпочтительно в 100 раз менее интенсивен, чем специфичное взаимодействие, наблюдаемое с целевой ДНК. Интенсивность взаимодействия может быть измерена, например, путем радиоактивного мечения зонда, например, 32P.
Условия гибридизации основаны на температуре плавления (Tm) комплекса связывания нуклеиновой кислоты, как указано в Berger and Kimmel (1987, Guide to Molecular Cloning Techniques, Methods in Enzymology, Vol 152, Academic Press, San Diego CA), и дают определенную «жесткость», как объяснено ниже.
Условия максимальной жесткости обычно имеют место при температуре, примерно равной Tm -5 °C (5°C ниже Tm зонда); высокой жесткости - примерно при 5°C-10°C ниже Tm; умеренной жесткости - примерно при 10°C-20°C ниже Tm; и низкой жесткости примерно при 20°C-25°C ниже Tm. Как станет ясно специалистам в данной области, максимальная жесткость гибридизации может использоваться для идентификации или выявления идентичных полинуклеотидных последовательностей, в то время как гибридизация при умеренной (или низкой) жесткости может использоваться для идентификации или выявления сходных или родственных полинуклеотидных последовательностей.
В предпочтительном аспекте настоящее изобретение относится к нуклеотидным последовательностям, которые могут гибридизоваться с антигенной нуклеотидной последовательностью в жестких условиях (например, 65°C и 0,1×SSC {1×SSC=0,15 M NaCl, 0,015 M Na3-цитрат pH 7,0}).
Там, где полинуклеотид по изобретению является двухцепочечным, обе цепи дуплекса, отдельно или в сочетании, относятся к настоящему изобретению. Там, где полинуклеотид по изобретению является одноцепочечным, следует понимать, что комплементарная этому полинуклеотиду последовательность также входит в объем настоящего изобретения.
Полинуклеотиды, которые не на 100% гомологичны последовательностям по настоящему изобретению, но относятся к объему изобретения, могут быть получены некоторыми путями. Другие варианты описанных здесь последовательностей могут быть получены, например, путем зондирования библиотек ДНК, полученных из некоторого числа субъектов, например субъектов из различных популяций. Кроме того, могут быть получены другие вирусные/бактериальные или клеточные гомологи, в особенности, клеточные гомологи, найденные в клетках млекопитающих (например, в клетках крыс, мышей, крупного рогатого скота и приматов), и такие гомологи и их фрагменты, в общем, способны селективно гибридизоваться с последовательностями, показанными в приведенном ниже списке последовательностей. Такие последовательности могут быть получены путем зондирования библиотек ДНК, полученных из геномных библиотек ДНК других видов животных, и путем зондирования таких библиотек зондами, включающими в себя все известные последовательности или их часть, в условиях от средней до высокой жесткости. Сходные соображения переносят на получение видовых гомологов и аллельных вариантов.
Варианты и видовые/линейные гомологи также могут быть получены с использованием вырожденной PCR, в которой применяются праймеры, сконструированные для направленного действия на целевые последовательности в пределах вариантов и гомологов, кодирующих консервативные аминокислотные последовательности внутри последовательностей по настоящему изобретению. Консервативные последовательности могут предсказываться, например, путем выравнивания аминокислотных последовательностей из нескольких вариантов/гомологов. Выравнивания последовательности могут проводиться с использованием известного в данной области компьютерного программного обеспечения. Например, широко применяется GCG Wisconsin PileUp.
Праймеры, использованные для вырожденной PCR, содержат одну или несколько вырожденных позиций и будут использоваться при условиях жесткости, более низкой, чем та, что используется для клонирования последовательностей с праймерами для единственной последовательности против известных последовательностей.
Альтернативно такие полинуклеотиды могут быть получены путем сайтспецифического мутагенеза характеризованных последовательностей. Это может использоваться, например, для молчащих изменений кодона, которые требуются для последовательностей с целью оптимизации кодонных предпочтений для конкретной клетки-хозяина, в которой экспрессированы полинуклеотидные последовательности. Другие изменения последовательности могут потребоваться для введения участков распознавания ферментов рестрикции или для изменения характеристики или функции полипептидов, кодируемых полинуклеотидами.
Полинуклеотиды, такие как ДНК-полинуклеотиды и зонды по изобретению, могут продуцироваться рекомбинантно, синтетически или любыми другими средствами, доступными специалистам в данной области. Их также можно клонировать стандартными способами.
Когда используют белки или белковые фрагменты, соответствие процессированного конечного продукта может быть продемонстрировано путем химического анализа или путем биологических анализов (способность APC стимулировать T-клетки, которые специфичны в отношении процессированных фрагментов).
В принципе, пептидные кандидаты выбирают по их совместимости в качестве чужеродных пептидов на различных стадиях; в общем, кандидатов вначале тестируют в тесте по связыванию пептидов на предмет их способности к связыванию с молекулой MHC-I, предпочтительно в серии тестов.
Одним из подходящих способов исследования является, например, FACS-анализ, основанный на проточной цитометрии. Пептид метят флуоресцентной меткой, например, FITC (флуоресцеинизотиоцианат) и воздействуют им на опухолевые клетки, которые экспрессируют молекулу MHC-1. В потоке отдельные клетки облучают лазером на конкретной длине волны; измеряют испускаемую флуоресценцию, которая зависит от количества пептида, связанного с клеткой.
Другим способом определения количества связанного пептида является график Скетчарда. Для этого применяют пептиды, меченные I125 или ионами редкоземельных металлов (например, европием). Клетки при 4°C заряжают различными определенными концентрациями пептида в течение периода от 30 до 240 минут. Для определения неспецифического взаимодействия пептида с клетками избыток немеченого пептида добавляют к некоторым из образцов, что предотвращает специфическое взаимодействие меченого пептида. Затем клетки отмывают для удаления любого неспецифического ассоциированного с клетками материала. Затем определяют количество связанного с клеткой пептида в сцинтилляционном счетчике с использованием испускаемой радиоактивности или в фотометре, который подходит для измерения долгоживущей флуоресценции. Полученные таким образом данные оценивают с использованием стандартных способов.
На второй стадии кандидаты с хорошим качеством связывания тестируют на предмет их иммуногенности.
Иммуногенность ксенопептидов, происходящих из белков, иммуногенная активность которых неизвестна, может тестироваться, например, в тесте MLC. Пептиды, которые провоцируют особо сильную реакцию в данном тесте, который предпочтительно также тестируют в серии с различными пептидами, с использованием в качестве стандарта пептида с известной иммуногенной активностью, подходят для целей настоящего изобретения.
Другим возможным путем тестирования MHC-I-связывающих пептидных кандидатов на их иммуногенность является исследование связывания данных пептидов с клетками T2. Один из таких тестов основан на характерной природе клеток T2 или клеток RMA-S, состоящей в том, что они дефективны по механизму транспорта пептидов TAP и представляют только стабильные молекулы MHC-I, которыми они воздействуют на пептиды, представляемые в контексте MHC-I.
Клетки T2 или клетки RMA-S, стабильно трансфицированные геном HLA, например генами HLA-A1 и/или HLA-A2, используют в данном тесте. Если клетки действуют посредством пептидов, которые являются подходящими лигандами MHC-I, и представляются в контексте MHC-I таким образом, что они распознаются иммунной системой как чужеродные, данные пептиды вызывают появление молекул HLA на клеточной поверхности в значительных количествах. Детекция HLA на клеточной поверхности, например, посредством моноклональных антител дает возможность для идентификации подходящих пептидов. Здесь опять подходящим образом используют стандартный пептид, о котором известно, что он обладает хорошей способностью связывать HLA или MHC.
Антиген может происходить из грибов, паразитов, бактерий и вирусов. Таким образом, одно из осуществлений изобретения относится к предотвращению или лечению инфекционных заболеваний. Примеры инфекционных заболеваний, к которым может применяться настоящее изобретение, включают в себя: аденовирусную инфекцию, сибирскую язву, холеру, дифтерию, столбняк, коклюш, малярию, грипп, включая Haemophilus influenzae типа B, гепатит A, гепатит B, энцефалит, корь, краснуху, менингит, например менингококк типов A, C, Y и W-135, чуму, пневмококк, бешенство, оспу, сальмонеллез, брюшной тиф, ветряную оспу, желтую лихорадку, лихорадку Рифт-Валли. Однако настоящее изобретение может также относиться к применению любого нового иммуногена, который становится доступным. В действительности, поскольку настоящее изобретение улучшает представление антигена иммунной системе, оно может быть особо применимым в отношении заболеваний и инфекций, которые, в общем, не поддаются лечению вакцинным подходом, например ВИЧ.
В особенно предпочтительном осуществлении настоящее изобретение относится к противораковой вакцине. Таким образом, настоящее изобретение может использоваться при лечении опухолей, таких как меланома, рак предстательной железы, легких, молочной железы и толстого кишечника, и лимфомы.
Одним из недостатков общепринятого лечения злокачественных опухолей, такого как химиотерапия и облучение, является то, что при разрушении раковых клеток они также повреждают некоторую часть здоровой ткани. Противораковые вакцины призваны уменьшить значение данных проблем. Настоящее изобретение может, в частности, использоваться в отношении опухолево-ассоциированных антигенов. Даже несмотря на то, что иммунная система не распознает раковые клетки как чужеродные, было обнаружено, что раковые клетки несут на своей поверхности конкретные антигены. Они могут быть уникальными для отдельных опухолей, распространяться на несколько типов, или экспрессироваться нормальной тканью, из которой возникает опухоль. Настоящее изобретение также может применяться в отношении антигенов, ассоциированных с продуктами генов, вовлеченных в образование опухоли, таких как онкогены и гены супрессии опухоли. Настоящее изобретение далее относится к вирусам, которые ассоциированы со злокачественной опухолью, таким как вирус гепатита B, который, как было давно установлено, имеет отношение к развитию рака печени и EBV.
Настоящее изобретение может использоваться после диагностики заболевания или после начальной терапии, такой как хирургия или химотерапия.
Неограничивающие примеры опухолевых антигенов, которые могут использоваться по настоящему изобретению, будут обсуждаться ниже.
Поскольку клонирование MAGE-1, первого гена, о котором сообщается, что он кодирует опухолевый антиген человека, распознаваемый Т-клетками, для меланомы идентификация и характеристика опухолевых антигенов была достигнута. Основной причиной этого является сложность получения клеточных линий других типов злокачественных опухолей in vitro, причем такие линии необходимы для получения опухолеспецифических линий или клонов CTL, подлежащих использованию в генетическом или биохимическом подходе, направленном на молекулярную идентификацию новых раковых антигенов. Недавно, однако, новые подходы позволили открыть новые антигены, распознаваемые Т-клетками, даже в опухолях, отличных от меланомы. Примеры различных категорий антигенов приведены ниже. Однако, как обсуждалось выше, аналоги или искусственно модифицированные эпитопы также могут использоваться по настоящему изобретению. Другие антигены, идентифицированные посредством антител, также могут использоваться, и их большая коллекция, выявленная по технологии Serex, может быть обнаружена в базе Institute for Cancer Research (www.licr.org/SEREX.htm). Следует заметить, что многие опухолевые антигены (например, MAGE, NY-ESO-1a), как известно в настоящее время, распознаются обоими типами T-клеток и антителами у одних и тех же пациентов со злокачественной опухолью. Кроме того, с использованием современных технологий (например, вычитающей гибридизации, анализа репрезентативных различий, микрополей) выявляются тысячи генов, которые преимущественно экспрессируются или гиперэкспрессируются в клетках неоплазий по сравнению с нормальными клетками, или же они экспрессируются в метастазах, но не в первичных, ранних очагах (например, меланома, рак молочной железы, лимфома и т.д.). С использованием подходящего компьютерного алгоритма идентифицируется некоторое количество новых эпитопов, которые могут связывать MHC. С применением таких подходов большой массив генных продуктов может подлежать скринингу на предмет их потенциальной антигенной функции. Иммуногенные эпитопы могут выбираться посредством подходящих функциональных анализов. Все могут также использоваться по настоящему изобретению.
Классификация опухолевых антигенов
Группа 1. Рестрицированные по классу I HLA раковые антигены/антигены яичек
Вехой в опухолевой иммунологии, конечно, являлось клонирование MAGE-1, и последовавшая через год характеристика первого определенного T-клетками антигенного эпитопа. За данными открытиями быстро последовала идентификация новых представителей данной группы. Были образованы генные семейства MAGE, BAGE и GAGE. Антигены, принадлежащие к данной группе, в настоящее время включающей в себя также NY-ESO-1, названы раковые антигены/антигены яичек (CT) по причине их экспрессии в гистологически различных опухолях человека и, среди нормальных тканей, в сперматоцитах/сперматогониях яичек и изредка в плаценте. Данные антигены в настоящее время представляют один из основных компонентов для разработки противоопухолевых вакцин. Образование антигенов CT может быть результатом реактивации генов, обычно молчащих в тканях взрослых, но их транскрипция активируется в некоторых опухолях. Гены CT, возможно, являются наиболее охарактеризованными. Новые гены в группе антигенов CT, которые были клонированы, включают в себя CT9, CT10, LAGE, MAGE-B5, -B6, -C2, -C3 и -D, HAGE, SAGE.
Группа 2. Рестрицированные по классу I HLA антигены дифференцировки
Данные антигены распределены между опухолью и нормальной тканью, из которой возникает опухоль; большая часть обнаружена в меланомах и нормальных меланоцитах. Многие из белков, родственных меланоцитарным белкам, вовлечены в биосинтез меланина. Из белков меланосомы могут происходить эпитопы, распознаваемые как CD8+, так CD4+ T-клетками.
Группа 3. Рестрицированные по классу I HLA широко экспрессированные антигены
Гены, кодирующие широко экспрессированные опухолевые антигены, были выявлены во многих нормальных тканях, а также в гистологически различных типах опухолей без предпочтительной экспрессии в конкретном виде рака. Возможно, что многие эпитопы, экспрессированные в нормальных тканях, находятся ниже барьерного уровня распознавания Т-клетками, в то время как их гиперэкспрессия в опухолевых клетках может переключать противораковый ответ путем поломки установившейся ранее устойчивости. Данные широко экспрессированные генные продукты характеризуются широким спектром механизмов, которые вовлечены в образование определенных Т-клетками эпитопов, за счет изменений в транскрипции и трансляции генов. В качестве примера следует выделить эпитоп CEA, который происходит из не определенной AUG альтернативной ORF, в то время как ген RU2 создает свой эпитоп путем транскрипции обратной цепи.
Группа 4. Рестрицированные по классу I HLA опухолеспецифичные антигены
Уникальные опухолевые антигены возникают вследствие точечных мутаций нормальных генов (таких как катенин, CDK4), молекулярные изменения в которых часто сопутствуют неопластической трансформации или прогрессии. Данные антигены, таким образом, экспрессируются только в индивидуальной опухоли, где их идентифицируют, поскольку низка вероятность того, что одна и та же мутация может возникнуть в двух различных неоплазиях, если только она не затрагивает гены (например, RAS), изменение которых представляет собой обязательную стадию неопластической трансформации. В мышиных моделях уникальные антигены, как показано, являются более иммуногенными, чем другие группы распространенных антигенов, поскольку уникальные антигены отвечают за отторжение опухолевых трансплантатов у мышей, и их определяют как опухолеспецифические антигены трансплантации (TSTA).
Описаны другие опухолеспецифические, но распространенные антигены, которые генерированы путем изменения механизмов сплайсинга и которые имеют место в опухолях, но не в нормальных клетках, как в случае TRP-2/INT2.
Группа 5. Рестрицированные по классу II HLA антигены
Стимуляция CD4+ T-хелперных клеток опухолевыми антигенами, как считается, ослаблена или отсутствует у пациентов со злокачественными опухолями, и это может являться причиной недостаточного иммунного ответа на опухоли. Поэтому идентификация опухолевых антигенных эпитопов, распознаваемых такими лимфоцитами, является ключевой стадией давно требуемого улучшения противоопухолевого иммунного ответа, который может приводить к клинической эффективности. Первый эпитоп, представленный HLA II класса и способный провоцировать ответ CD4+ T-клеток был идентифицирован в 1994 г. в тирозиназе меланомы. Затем последовал пропуск длиной 4 года, во время которого был охарактеризован только один дополнительный эпитоп, до того как были открыты другие гены, кодирующие рестрицированные по классу II пептиды. Однако, как только стали доступны технические и методологические подходы к идентификации CD4+ T-клеточных эпитопов опухолевых антигенов, стал наблюдаться экспоненциальный рост числа сообщений о таких эпитопах. Фактически, с 1998 г. была проведена молекулярная идентификация 27 новых рестрицированных по классу II HLA эпитопов из 14 антигенов с использованием, среди прочих подходов, библиотек Ii-кДНК слияния, иммунизированных трансгенных мышей и биохимических подходов. Следует заметить, что даже рестрикцированные по классу II антигены включают в себя подгруппу мутированных белков, которые, следовательно, представляют собой истинные опухолеспецифические антигены.
Группа 6. Белки слияния
При некоторых злокачественных опухолях, в частности при некоторых формах лейкозов, молекулярный механизм канцерогенеза включает в себя транслокацию хромосом, что приводит к слиянию удаленных генов. Это часто приводит к синтезу белков слияния, которые характеризуют каждый тип заболевания (например, bcr-abl, pml-RAR при CML и APL соответственно) и генерируют новые эпитопы, которые могут распознаваться T-клетками, CD8+ или CD4+ при рестрикции по классу I или классу II HLA соответственно. Хотя оказывается, что данные эпитопы являются у пациентов с лейкозами слабо иммуногенными, некоторые из данных пептидов или белков могут, тем не менее, использоваться для стимуляции дендритных клеток при вакцинации.
Неограничивающие примеры опухолевых антигенов, которые особенно подходят для настоящего изобретения, включают в себя: MAGE, BAGE и GAGE, распознающие CTL эпитопы из тирозиназы, MelanAMart1, gp100mel7 и gp75TRP1.
Ген селекции или маркер
Т-клетка по настоящему изобретению предпочтительно содержит один или несколько чужеродных генов, который представляет собой ген селекции или маркерный ген. Как хорошо известно, гены селекции могут использоваться для отбора трансформированных клеток из нетрансформированных клеток. Многие различные селектируемые маркеры успешно использовались в качестве векторов, и они в равной степени хорошо могут использоваться по настоящему изобретению. Используют подходящие бактериальные или животные антигены. В предпочтительном осуществлении применяют, например, бактериальные антигены (например, β-галактозидазу), бактериальные гены или другие гены, несущие устойчивость к антибиотикам (например, neo, amp, Kan и tet) или суицидный ген (например, HSV-Tk).
В предпочтительном осуществлении Т-клетка включает в себя, по меньшей мере, один суицидный ген в качестве еще одного чужеродного антигена. Преимущество, предоставляемое применением такого суицидного гена, состоит в том, что активация данного гена может контролироваться системным введением дальнейшего соединения, такого как циклоспорин или 5'-фторцитозин. Пример суицидного гена включает в себя ген тимидинкиназы вируса простого герпеса (HSV Tk), который может уничтожать инфицированные и окружающие клетки после обработки ганцикловиром.
Другие суицидные гены включают в себя, например, ген цитозиндезаминазы, который дает летальную чувствительность к 5-фторцитозину, и человеческую молекулу клеточной поверхности CD20, которая обеспечивает уничтожение трансдуцированных клеток путем введения ритуксимаба (Introna et al. Hum Gene Ther 11:611, 2000). Преимуществом CD20 является то, что, будучи человеческой молекулой, она не иммуногенна.
Особым преимуществом применения суицидного гена является его безопасность в том плане, что он может использоваться для уничтожения Т-клетки в случае возникновения нежелательных побочных эффектов.
По особо предпочтительному осуществлению настоящего изобретения ген селекции или маркерный ген сам является иммуногенным или экспрессирует белок, который является иммуногенным. Преимущество такой системы состоит в том, что она предоставляет мониторинг иммунологического статуса пациента и поэтому эффективности вакцинного лечения. Примеры таких подходящих дополнительных чужеродных антигенов включают HSV-Tk и Neo, причем оба они генерируют иммунный ответ у иммунокомпетентного пациента. Например, мониторинг иммунизации пациента первым антигеном может проводиться путем выявления цитолитической активности против HSV-Tk, как это более подробно описано ниже.
По соображениям безопасности может также осуществляться прекращение роста T-клеток по настоящему изобретению, например, путем облучения или обработки митомицина. Такое облучение может проводить при комнатной температуре в дозе 100 рад/минуту. Клетки, подвергнутые такому облучению, больше не пролиферируют, но за счет их мРНК, которая пока присутствует в клетке, способны экспрессировать белки.
Лечение
Оно включает в себя любое терапевтическое применение, которое может принести пользу человеку или животному, не относящемуся к человеку. Лечение млекопитающих является особенно предпочтительным. В объем настоящего изобретения входят как способы лечения человека, так и ветеринарные способы лечения.
Лечение может проводиться в отношении существующего состояния, или оно может быть профилактическим. Оно может осуществляться в отношении взрослого субъекта, подростка, ребенка, плода или любой части указанных субъектов (например, органа, ткани, клетки или молекулы нуклеиновой кислоты).
Т-клетки, полученные способом по изобретению, могут вводиться пациенту, страдающему злокачественной опухолью.
В общем, в подходе ex vivo пациент является тем же пациентом, из которого происходят обработанные Т-клетки. Примеры злокачественных опухолей, которые могут подлежать лечению, включают в себя злокачественные опухоли молочной железы, шейки матки, толстого кишечника, прямой кишки, эндометрия, почки, легких, яичника, поджелудочной железы, предстательной железы, желез, кожи, желудка, мочевого пузыря, ЦНС, пищевода, головы и шеи, печени, яичек, тимуса и щитовидной железы. Также могут подлежать лечению злокачественные опухоли клеток крови, клеток костного мозга, B-лимфоцитов, T-лимфоцитов, лимфоцитарных предшественников или предшественников миелоидных клеток.
Опухоль может представлять собой солидную опухоль и несолидную опухоль и может представлять собой первичную опухоль или диссеминированную метастазированную (вторичную) опухоль. Несолидные опухоли включают в себя миелому; лейкоз (острый или хронический, лимфоцитарный или миелоцитарный), такой как острый миелобластный, острый промиелоцитарный, острый миеломоноцитарный, острый моноцитарный, эритролейкоз; и лимфомы, такие как ходжкинская, неходжкинская и лимфома Беркитта. Солидные опухоли включают в себя карциному, карциному толстого кишечника, мелкоклеточную карциному легких, немелкоклеточную карциному легких, карциному почек, аденокарциному, меланому, карциному базальных клеток или плоскоклеточную карциному, мезотелиому, нейробластому, глиому, астроцитому, медуллобластому, ретинобластому, саркому, остеосаркому, рабдомиосаркому, фибросаркому, остеогенную саркому, гепатому и семиному.
Обычно композиции по настоящему изобретению могут вводиться с опухолеспецифическим антигеном, таким как антигены, которые гиперэкспрессированы на поверхности опухолевых клеток.
Т-клетки могут использоваться для лечения имеющегося иммунного ответа (такого как аллергическое состояние или аутоиммунное заболевание) или могут использоваться для создания у пациента толерантности. Таким образом, клетки по настоящему изобретению могут использоваться в терапевтических способах для лечения и профилактики заболеваний, характеризующихся неподходящей активности лимфоцитов у животных и человека. T-клетки могут использоваться для придания толерантности в отношении единственного или множественных антигенов.
Обычно Т-клетки получают от пациента или донора и стимулируют, как описано выше, перед возвращением пациенту (терапия ex vivo).
Фармацевтические композиции
Фармацевтическая композиция представляет собой композицию, которая включает в себя терапевтически эффективное количество фармацевтически активного средства или состоит из него. Она предпочтительно включает в себя фармацевтически приемлемый носитель, растворитель или наполнители (включая их комбинации). Приемлемые носители или растворители для терапевтического применения хорошо известны в области фармации и описаны, например, в Remington's Pharmaceutical Sciences, Mack Publishing Co. (A. R. Gennaro edit. 1985). Выбор фармацевтического носителя, наполнителя или растворителя может быть сделан в контексте предполагаемого пути введения и стандартной фармацевтической практики. Фармацевтические композиции могут включать в себя в качестве носителя, наполнителя или растворителя или в дополнение к ним любое(ые) подходящее(ие) связующее(ие) средство(а), смазку(и), суспендирующее(ие) средство(а), покрывающее(ие) средство(а), солюбилизирующее(ие) средство(а).
«Терапевтически эффективное количество» относится к количеству терапевтического средства, которое эффективно для достижения предназначенной для него цели. В то время как потребности конкретного пациента могут изменяться, определение оптимальных интервалов для эффективных количеств каждого аддукта окиси азота доступно специалисту в данной области. В общем, режим дозировки для лечения состояния соединением и/или композицией по данному изобретению выбирают согласно различным факторам, включая тип, возраст, вес, пол, диету и медицинское состояние пациента, тяжесть дисфункции, путь введения, фармакологические соображения, такие как активность, эффективность, профили фармакокинетики и токсикологии конкретного применяемого соединения, то, используется ли система доставки лекарственного средства, и то, вводится ли соединение как часть лекарственной комбинации, и он может регулироваться специалистом в данной области. Таким образом, используемый в действительности режим дозировки может широко варьироваться и поэтому может отклоняться от предпочтительного режима дозировки, установленного здесь.
Примеры фармацевтически приемлемых носителей включают в себя, например, воду, растворы солей, алкоголь, силикон, воски, вазелин, растительные масла, полиэтиленгликоли, пропиленгликоль, липосомы, сахара, желатин, лактозу, амилозу, стеарат магния, тальк, поверхностно-активные вещества, кремниевую кислоту, вязкий парафин, парфюмерное масло, моноглицериды и диглицериды жирных кислот, сложные эфиры petroethral жирных кислот, гидроксиметилцеллюлозу, поливинилпирролидон и тому подобное.
Требуемый интервал дозировки зависит от выбора пептида, пути введения, природы препарата, природы состояния субъекта и суждения лечащего врача. Подходящие дозировки, однако, лежат в интервале 0,1-100 мкг/кг субъекта.
Вакцинная композиция обычно находится в инъецируемой форме. Для усиления иммунного ответа могут использоваться общепринятые адъюванты. Подходящая единичная доза для вакцинации составляет 0,5-5 микрограмм/кг антигена, и такую дозу предпочтительно вводят 1-3 раза и с интервалом 1-3 недель. При указанном интервале дозировки не будет наблюдаться неблагоприятных токсикологических эффектов соединений по изобретению, которые будут препятствовать их введению подходящим субъектам.
Больших вариаций требуемых дозировок, однако, следует ожидать в свете разнообразия доступных соединений и различной эффективности различных путей введения. Например, пероральное введение, как можно ожидать, требует более высоких дозировок, чем введение путем внутривенной инфекции. Вариации в данных уровнях дозировки могут регулироваться с использованием стандартных эмпирических рутинных способов оптимизации, хорошо понятных специалистам в данной области.
Предпочтительные характеристики и осуществление настоящего изобретения будут описаны далее со ссылкой на следующие примеры и фигуры.
Описание чертежей
На фигуре 1 представлены результаты экспериментов по клеточному лизису, в которых используются эффекторные клетки, выделенные из пациентов, подвергнутых инфузии, в отношении стимуляторных клеток (закрашенные символы) и в отношении не относящихся к делу мишеней (пустые символы). Эффекторные клетки генерированы путем стимуляции мононуклеарных клеток периферической крови (PBMC), выделенных из 7 подвергшихся лечению пациентов, аутологичными трансдуцированными T-клетками (кружки) и аллогенными T-клетками (квадрат). Трансдукцию проводили векторами, кодирующими белок HSV-TK и маркер клеточной поверхности ΔLNGFr (человеческий низкоаффинный рецептор фактора роста нервов с делецией).
На фигуре 2 представлены результаты экспериментов по клеточному лизису, где используются эффекторные клетки, выделенные из подвергнутого лечению пациента, в отношении немодифицированных клеток и клеток, экспрессирующих HSV-TK или ΔLNGFr.
На фигуре 3 показано отношение между временем инфузии, числом введенных путем инъекции инженерных Т-клеток и развитием специфичного в отношении трансгена иммунного ответа. Точки данных представляют инфузию клеток, ответственных (закрашенные символы) или не ответственных (пустые символы) за индукцию иммунного ответа.
На фигуре 4 представлены результаты экспериментов по клеточному лизису, в которых используются эффекторные клетки, выделенные из двух здоровых доноров, в отношении стимуляторных клеток (закрашенные символы) и в отношении не относящихся к делу мишеней (пустые символы). Эффекторные клетки генерированы путем стимуляции мононуклеарных клеток периферической крови (PBMC), выделенных из 2 здоровых доноров, аутологичными трансдуцированными T-клетками (кружки) и аллогенными T-клетками (квадраты).
На фигуре 5 показано схематичное представление клинической истории и ведения пациента UPN-15. Мониторинг наличия (закрашенный кружок) или отсутствия (пустой кружок) циркулирующих трансдуцированных T-клеток проводили в указанные моменты времени посредством PCR с HSV-TK-специфичными праймерами. Мониторинг развития иммунного ответа проводили путем стимуляции ex vivo циркулирующих лимфоцитов аутологичными трансдуцированными и аллогенными Т-клетками. Пациента считали не иммунизированным (пустые звездочки) или иммунизированным (закрашенные звездочки) против трансгена в зависимости выявленной цитолитической активности против HSV-TK.
На фигуре 6 показаны результаты экспериментов ELISPOT, в которых использованы свежие лимфоциты, выделенные из подвергавшихся лечению пациентов с меланомой, в отношении аутологичных клеток, экспрессирующих MAGE-3 или HSV-Tk, и в отношении аутологичных клеточных линий EBV и меланомы.
На фигуре 7 показаны результаты экспериментов по высвобождению IFNγ, в которых использованы эффекторные клетки, выделенные in vitro из подвергнутого лечению пациента с меланомой, в отношении трансдуцированных MAGE-3 и нетрансдуцированных T-клеток (т.е. PHA-M3 и PHA), соответственно, и аутологичных и неродственных клеточных линий меланомы. Эффекторные клетки получали путем стимуляции PBMC, выделенных из подвергнутого лечению пациента аутологичными, трансдуцированными MAGE-3 T-клетками.
Примеры
Пример 1
Инфузия донорских лимфоцитов в контексте аллогенной трансплантации костного мозга (алло-BMT) по случаю лейкоза является установленной стратегией для контроля за рецидивом опухоли и вирусными инфекциями. Для предотвращения обычного риска болезни «трансплантат против хозяина» (GVHD) BM-донорные лимфоциты могут подвергаться трансдукции суицидным геном, несущим чувствительность к лекарственному средству, что может обеспечить селективную элиминацию трансдуцированных клеток для терапевтических целей и целей безопасности.
Мононуклеарные клетки периферической крови (PBMC) выделяли посредством центрифугирования в градиенте плотности Lymphoprep (Nycomed, Осло, Норвегия) из всех исследованных BM-доноров и здоровых контрольных субъектов.
Активация. Различные сигналы активации использовали для активации Т-клеток (здесь и далее обозначенные как Т-бласты): фитогемагглютинин (PHA) 2 мкг/мл (Boehringer Mannheim); mAb против CD3 (OKT3) 30 нг/мл (Orthoclone, Милан, Италия); OКT3 (30 нг/мл). Активированные Т-клетки культивировали в количестве 1×106 клеток/мл, в RPMI 1640, дополненной 5% аутологичной сывороткой в присутствии hu-r-IL-2 100 ед./мл (EuroCetus Italia S.r.l., Милан, Италия). Среду культивирования меняли каждые 3-4 суток.
Трансдукция. Использовали два способа трансдукции лимфоцитов. 1 - совместное культивирование: активацию лимфоцитов проводили на полусомкнутом монослое летально облученных упаковывающих клеток (100 Грэй; GIL RAD, Gilardoni, Mandello, Италия), во флаконах для культуры ткани в присутствии полибрена (8 мкг/мл). Совместное культивирование проводили в течение 72 часов. 2 - спинокуляция: через 3-4 суток активации лимфоциты получали и подвергали трансдукции за 2 цикла 2-часового центрифугирования в присутствии образцов бесклеточной ретровирусной надосадочной жидкости и полибрена (8 мкг/мл).
После процедур трансдукции лимфоциты собирали и высевали в свежей среде (106/мл). Трансдуцированные ретровирусом клетки анализировали на предмет экспрессии ΔLNGFr на 5 сутки посредством FACS с использованием мышиных mAb против человеческого LNGFR 20.4 (ATCC, Rockville, Мэриленд).
Отбор. Чистую популяцию трансдуцированных клеток получали двумя способами отбора. Лимфоциты, трансдуцированные вектором SFCMM2, кодирующие белок слияния HSV-TK/neo (TN), бифункциональный белок, несущий устойчивость к неомицину и активность HSV-TK, отбирали путем добавления неомицина (0,8 мкг/мл) в культуральную среду. Свежий неомицин добавляли на 3, 7 и 10 сутки культивирования.
Лимфоциты, трансдуцированные вектором SFCMM3, кодирующим HSV-TK дикого типа и ΔLNGFr, подвергали иммунной селекции путем магнитной сортировки с использованием mAb 20.4 и гранул, покрытых крысиными антителами против мышиных IgG1 (Dynabeads M-450, Dynal A. S. N0212, Осло, Норвегия).
Чистые популяции трансдуцированных клеток замораживали и затем вводили пациенту или использовали ex vivo в качестве стимулятора для мониторинга антигенспецифичных иммунных реакций.
Образцы PBMC получали из 7 обработанных пациентов, и обеспечивали им контакт с облученными трансдуцированными Т-клетками соответствующего BM-донора, или с облученными аллогенными T-бластами. Через 2 дня культивирования добавляли 10 ед./мл IL2. Смеси наблюдали на предмет лизиса стимуляторных клеток, что указывало на то, что в образцах присутствуют цитотоксические Т-лимфоциты (CTL), специфичные в отношении трансгенного продукта, экспрессируемого трансдуцированными Т-клетками, или в отношении аллоантигена. Использованный анализ лизиса представлял собой анализ высвобождения хрома. Результаты, представленные на фигуре 1, показывают, что от всех пациентов получали цитотоксические Т-клетки со специфичностью в отношении трансдуцированных мишеней. Для оценки иммунокомпетентности тестировали специфичный иммунный ответ против аллогенных мишеней и детектировали его во всех отвечающих пациентах. Это указывает на то, что инфузия Т-бластов, трансдуцированных в описанных выше условиях, может вызывать иммунный ответ против трансдуцированных клеток.
Пример 2
Дальнейшие исследования проводили для идентификации целевых для иммунного ответа антигенов. Эффекторные клетки, выделенные из одного пациента, тестировали против аутологичных T-бластов, экспрессирующих единственные компоненты вектора. T-бласты, экспрессирующие HSV-TK, подвергались лизису, в то время как экспрессирующие ΔLNGFr не распознавали вовсе (фиг. 2). Аналогичные результаты получали с эффекторными клетками из другого пациента.
Пример 3
Результаты из примера 1 (т.е. генерирование HSV-TK-специфичного иммунитета) анализировали в отношении времени инфузий и числа введенных путем инфузии инженерных Т-клеток. Представлено время инфузий, измеренное в месяцах от трансплантации костного мозга (абсцисса) и числа инъецированных инженерных Т-клеток (ордината). Развитие иммунного ответа против трансдуцированных клеток оценивали путем стимуляции ex vivo PBL против аутологичных трансдуцированных и аллогенных T-клеток, как описано в примере 1. Если присутствовал позитивный аутоиммунный ответ, пациента считали неиммунизированным (пустые символы) или иммунизированным (закрашенные символы) трансгеном, в зависимости от цитолитической активности в отношении трансдуцированных клеток. Точки данных представляют инфузию клеток, ответственных (закрашенные символы) или не ответственных (пустые символы) за индукцию иммунного ответа. Для пациента UPN-15 представлены два различных момента времени. Данные результаты указывают на то, что индукция иммунного ответа клетками, с которыми манипулировали in vitro, была независимой от числа введенных путем инфузии клеток и была точно связана с наличием функциональной иммунной системы во время инфузии.
Пример 4
Дальнейшие исследования проводили для идентификации механизма, используемого трансдуцированными Т-бластами, для генерирования специфичного в отношении трансгена иммунного ответа in vivo. Образцы PBMC получали от 2 здоровых доноров и стимулировали по примеру 1 облученными трансдуцированными T-бластами, или облученными аллогенными T-бластами. Через 2 суток культивирования добавляли 10 ед./мл IL2. Смеси наблюдали на предмет лизиса стимуляторных клеток, что указывало на то, что в образцах присутствуют CTL, специфичные в отношении трансгенного продукта, экспрессируемого трансдуцированными Т-бластами, или в отношении аллоантигена. Результаты, представленные на фигуре 4, показывают, что цитотоксические Т-клетки со специфичностью в отношении трансдуцированных мишеней не получены от здоровых контрольных субъектов. Это указывает на то, что в экспериментальных установках авторов изобретения трансдуцированные Т-бласты не могут действовать в качестве профессиональных антигенпрезентирующих клеток, сами по себе активируя наивные Т-клетки-предшественники. Путь перекрестного представления in vivo, опосредованный резидентными APC, может быть ответственным за индукцию специфичного в отношении трансгена иммунного ответа in vivo у пациентов, подвергнутых инфузии.
Пример 5
Дальнейшие эксперименты проводили для определения клеточного механизма, ответственного за специфичный в отношении трансгена иммунный ответ. Способность генерировать эффективные иммунные реакции и присутствие циркулирующих трансдуцированных T-бластов оценивали по времени ведения пациента UPN-15 (фиг. 5). Присутствие циркулирующих индуцированных Т-бластов анализировали путем PCR с HSV-TK-специфичными праймерами, на свежих образцах крови. Восстановление иммунной системы и HSV-TK-специфичную иммунизацию оценивали, как описано в примере 1. Пациенту UPN-15 вводили клетки путем инъекции сразу после трансплантации в отсутствие функциональной иммунной системы. Введенные путем инфузии клетки выявляли в циркуляции путем PCR в течение длительного времени (фиг. 5, закрашенные кружки), даже при полном восстановлении иммунной системы. Восстановление иммунной системы документировали по присутствию активного иммунного репертуара против аллоантигенов в отсутствие HSV-TK-специфичного иммунного ответа (фиг. 5, пустые звездочки). Поэтому во время инъекции иммунизация не происходила, поскольку способность пациента генерировать адекватный Т-клеточный иммунный ответ сильно подорвана в плане способности представления антигена и ответа на антиген. Впоследствии, когда иммунная система полностью восстанавливалась, циркулирующие трансдуцированные Т-бласты не были способны обеспечивать сигналы активации, они не действовали как APC, и поэтому не стимулировались цитотоксические эффекторные клетки, специфичные в отношении трансдуцированных клеток.
Вторая инъекция HSV-TK-инженерных Т-бластов, проведенная при полном восстановлении иммунной системы UPN-15, приводила к индукции HSV-TK-специфического иммунитета (фиг. 5, закрашенные звездочки), вызывающего деструкцию инженерных клеток (фиг. 5, пустые кружки).
Поэтому иммунизация происходила, когда T-клетки, с которыми проводили манипуляции, как описано в примере 1, доставляют в иммунокомпетентную среду. В частности, Т-клетки действуют in vivo в качестве носителя, несущего антиген в правильное место с точки зрения индукции иммунного ответа. Здесь частичная клеточная гибель инъецированных клеток, опосредованных депривацией фактора роста (то есть IL2), приводит к высвобождению несомых антигенов; за этим следует представление высвобожденного антигенного материала APC хозяина и активация наивных HSV-TK-специфичных Т-клеток-предшественников.
В предшествующих экспериментах описана новая методология, посредством которой Т-лимфоциты могут модифицироваться in vitro так, что они становятся носителями для доставки антигена-мишени in vivo.
Пример 6
T-лимфоциты могут модифицироваться in vitro так, что они становятся носителями для доставки антигена-мишени in vivo. Аутологичные лимфоциты от пациента, пораженного MAGE-3-позитивной меланомой, трансдуцировали ретровирусным вектором, кодирующим опухолевый антиген MAGE-3 и белок слияния HSV-TK/neo (TN), бифункциональный белок, несущий устойчивость к неомицину и активность HSV-TK.
Использованные здесь процедуры активированной трансдукции и отбора были теми же, что сообщались в примере 1. В конце процесса манипуляции чистые популяции трансдуцированных клеток замораживали и затем вводили пациенту по протоколу вакцинации с повышением дозы. Трансдуцированные клетки также использовали ex vivo в качестве стимуляторов для мониторинга антигенспецифических иммунных реакций.
Образцы PBMC получали от пациента с меланомой перед лечением и после пятой вакцинации. Собранные клетки немедленно приводили в контакт с аутологичными клетками меланомы EBV и с EBV, трансдуцированными MAGE-3 или HSV-TK. Тест проводили на планшетах ELISPOT, покрытых mAb против IFNγ. Через 24 ч культивирования клетки удаляли, и планшеты обрабатывали для оценки присутствия IFNγ. Результаты, показанные на фигуре 6, показывают, что высвобождающие IFNγ T-клетки со специфичностью в отношении трансгенов (т.е. MAGE-3 и HSV-TK) получали после вакцинации. Также выявляли значимое увеличение частоты опухолеспецифических Т-клеток. Это указывало на то, что инфузия T-бластов, трансдуцированных в описанных выше условиях, могла вызывать иммунные реакции против трансдуцированных клеток.
Пример 7
PBMC получали от пациента с меланомой после вакцинаций и приводили в контакт с облученными, трансдуцированными MAGE-3 Т-клетками. Через 2 суток культивирования добавляли 10 ед./мл IL2. Смеси наблюдали на предмет высвобождения IFNγ в присутствии стимуляторных клеток, что указывало на присутствие в образцах эффекторных клеток, специфичных в отношении трансгенного продукта MAGE-3, экспрессируемого трандуцированными Т-клетками. Результаты, представленные на фигуре 7, показывают, что получали эффекторные Т-клетки со специфичностью в отношении MAGE-3 и аутологичных опухолей. Это далее подтверждает, что инфузия Т-бластов, трансдуцированных в описанных выше условиях, может вызывать иммунные реакции против трансгенных продуктов.
Пример 8
Для исследования кинетики получения специфичных в отношении трансгенов иммунных эффекторов, в отношении дозы инфузии трансдуцированных клеток, PBMC получали от пациента с меланомой до лечения и в различные моменты времени протокола вакцинации (т.е., после 3, 5 и 15 вакцинации). Собранные клетки приводили в контакт с облученными, трансдуцированными MAGE-3 или трансдуцированными HSV-TK Т-клетками. Через 2 суток культивирования добавляли 10 ед./мл IL2. Для каждого момента времени получали несколько независимых микрокультур (в интервале от 30 до 48). После нескольких раундов стимуляции микрокультуры наблюдали на предмет высвобождения IFNγ в присутствии стимуляторных клеток, что указывает на присутствие в образцах эффекторных клеток, специфичных в отношении MAGE-3 или HSV-TK. Результаты, суммированные в следующей таблице, показывают существование корреляции между инъекцией трансдуцированных Т-клеток и развитием иммунного ответа против трансгенов. Непрерывное повышение количества реакционноспособных в отношении MAGE-3 Т-клеток-предшественников, представленное числом позитивных микрокультур, наблюдаемое при инфузии in vivo экспрессирующих MAGE-3 Т-клеток, ответственно за индукцию специфичных в отношении MAGE-3 иммунных реакций.
Все указанные в приведенной выше спецификации публикации включены сюда в качестве ссылки. Различные модификации и вариации описанных способов и системы по изобретению будут понятны специалисту в данной области без уклонения от объема и сущности изобретения. Хотя изобретение описано в связи с конкретными предпочтительными осуществлениями, следует понимать, что заявленное изобретение не должно чрезмерно ограничиваться такими конкретными осуществлениями. В действительности, различные модификации описанных способов осуществления изобретения, понятные специалистам в области молекулярной биологии или связанных областях, как подразумевается, лежат в объеме следующей формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ EX VIVO ЭКСПАНСИИ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2814083C2 |
| СПОСОБЫ ЭКСПАНДИРОВАНИЯ АНТИГЕНСПЕЦИФИЧЕСКИХ CAR-T-КЛЕТОК, КОМПОЗИЦИИ И ПРИМЕНЕНИЯ, СВЯЗАННЫЕ С НИМИ | 2019 |
|
RU2800920C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ С MND-ПРОМОТОРОМ | 2015 |
|
RU2708311C2 |
| ТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ РАКА | 2016 |
|
RU2720984C2 |
| АНТИГЕННЫЕ РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2792042C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ С MND-ПРОМОТОРОМ | 2015 |
|
RU2799573C2 |
| АНТИГЕННЫЕ РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2794945C2 |
| МНС-ДИСПЛЕЙ ПЕПТИДОВ МЛЕКОПИТАЮЩЕГО КАК ИНСТРУМЕНТ ДЛЯ ВЫБОРА АНТИГЕННОЙ ДЕТЕРМИНАНТЫ ПРИ СОЗДАНИИ ВАКЦИН | 2019 |
|
RU2795983C2 |
| МОДИФИЦИРОВАННЫЕ МОНОЦИТЫ/МАКРОФАГИ, ЭКСПРЕССИРУЮЩИЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2766690C2 |
| ВАКЦИНА НА ОСНОВЕ В-КЛЕТОК, НАГРУЖЕННЫХ ЛИГАНДОМ Т-КЛЕТОК-ПРИРОДНЫХ КИЛЛЕРОВ И АНТИГЕНОМ | 2006 |
|
RU2420311C2 |
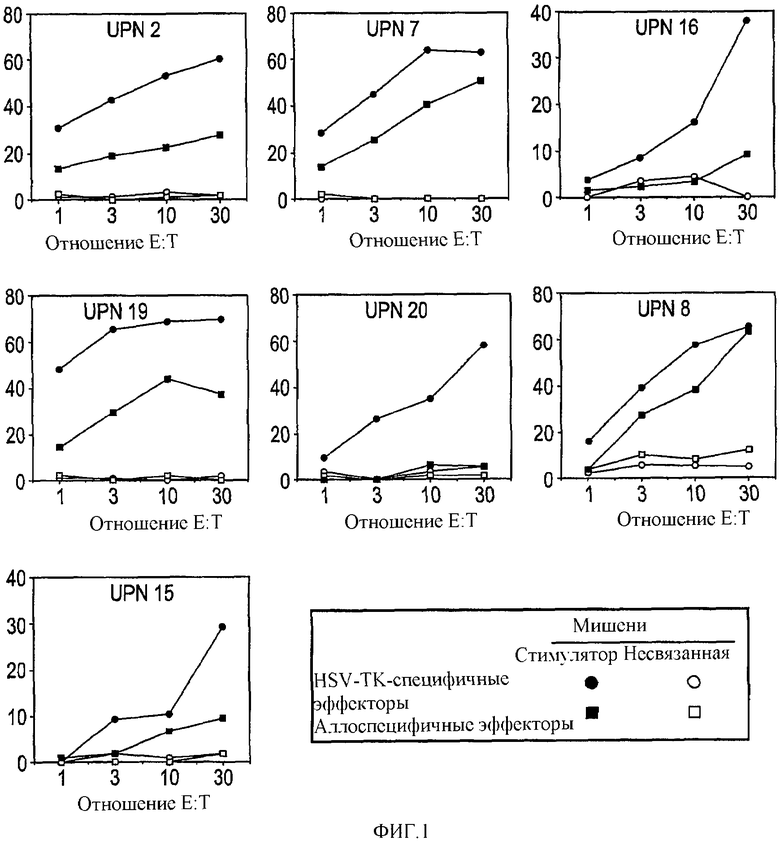
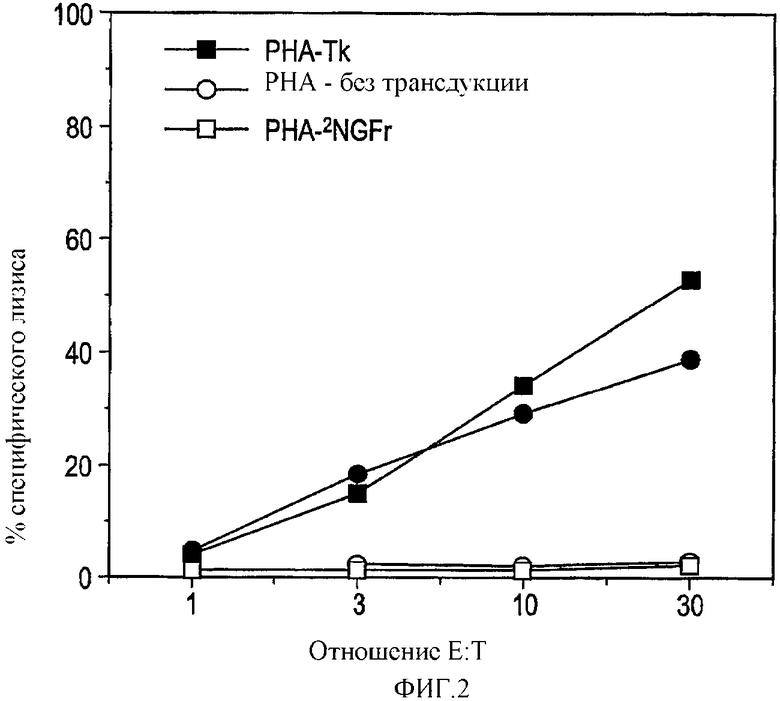

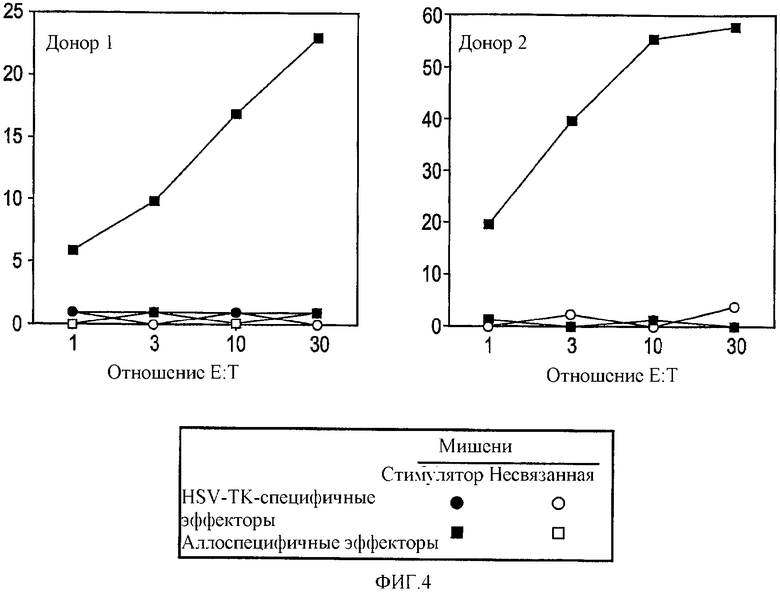
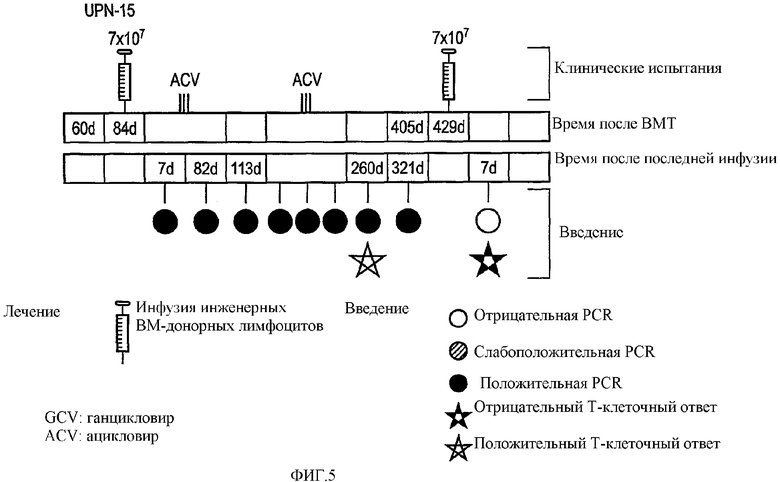
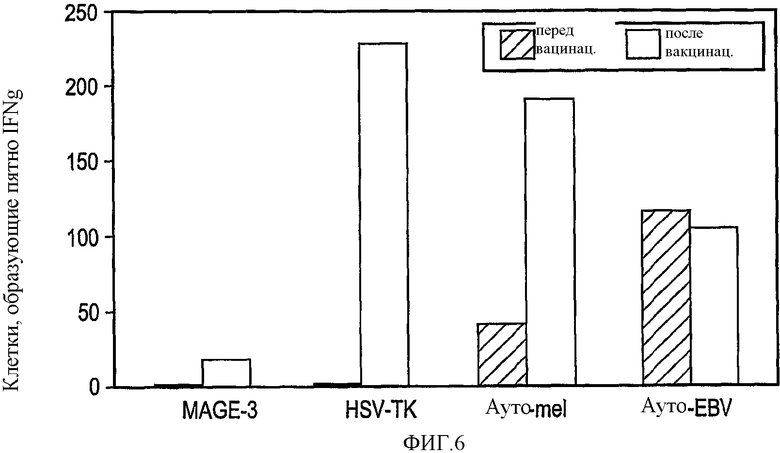
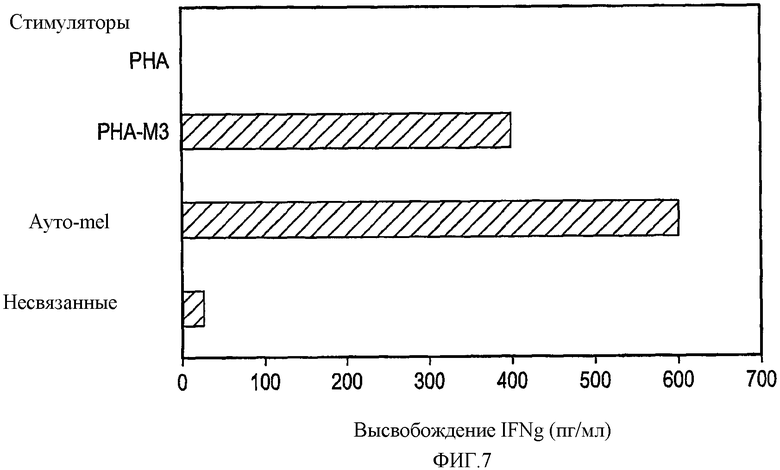
Группа изобретений относится к системе доставки для антигена в виде Т-клетки, содержащую, по меньшей мере, один антиген, способную загружать данным антигеном антигенпрезентирующие клетки. Предложено применение Т-клетки, содержащей по меньшей мере один антиген, в приготовлении лекарственного средства для доставки антигена в лимфатический узел. Предложено применение антигена, ассоциированного с Т-клеткой, содержащей по меньшей мере один другой антиген, способный вызывать иммунный ответ, в приготовлении лекарственного средства для мониторинга иммунного ответа против по меньшей мере одного другого антигена. Предложено применение Т-клетки, содержащей по меньшей мере один антиген, для приготовления лекарственного средства для загрузки антигенпрезентирующих клеток (АРС) антигеном. Предложен способ получения Т-клетки для применения по любому предшествующему пункту, включающий выделение Т-клетки, активацию Т-клетки, культивирование Т-клетки и введение антигена в Т-клетку, где Т-клетка трансдуцирована антигеном. 4 н. и 20 з.п. ф-лы, 1 табл., 7 ил.
выделение Т-клетки;
активацию Т-клетки;
культивирование Т-клетки; и
введение антигена в Т-клетку, где Т-клетка трансдуцирована антигеном.
| Устройство для электрообработки пульп | 1979 |
|
SU904786A1 |
| WO 9601126 А1, 18.01.1996 | |||
| WO 9719169 А1, 29.05.1997 | |||
| МЕТОД УНИЧТОЖЕНИЯ НЕЖЕЛАТЕЛЬНЫХ КЛЕТОК-МИШЕНЕЙ | 1997 |
|
RU2182493C2 |

