1. Область изобретения
Настоящее изобретение относится к глазным гипотензивным липидам. В частности, настоящее изобретение относится к 1-этаноламиду PGF2α (простагландина F2α) и родственным соединениям, и к содержащим эти соединения фармацевтическим композициям, а также к способам применения этих соединений для снижения внутриглазного давления у млекопитающих.
2. Краткое описание уровня техники
Из уровня техники хорошо известно о многочисленных глазных гипотензивных агентах, применяемых для лечения различных гипертензивных состояний глаза, включая первичную и вторичную глаукому, представляющую серьезную проблему для здоровья человека. Лекарственные препараты, использующиеся для лечения глазной гипертензии (глаукомы), включают в себя антагонисты β-адренорецепторов и различные простагландины. Имеется значительный объем научной и патентной литературы, касающейся простагландинов и их использования для лечения глазной гипертензии. Например, см. Bito L.Z. Biological Protection with Prostaglandins Cohen, M.M., ed., Boca Raton. Fla., CRC Press Inc., 1985, pp. 231-252; и Bito, L.Z., Applied Pharmacology in the Medical Treatment of Glaucomas Drance, S.M. and Neufeld, A.H. eds., New York, Grune & Stratton, 1984, pp. 477-505.
Патент US 5288754 содержит, кроме того, ссылки на уже известный уровень техники, направленные на простагландины и родственные соединения, активные в качестве средств для снижения внутриглазного давления у млекопитающих. Сам патент США описывает "Polar С-1 Esters of Prostaglandins" ("Полярные С-1-эфиры простагландинов"), включая С-1-амиды и С-1-замещенные амиды карбоновой кислоты соединения, известного как PGF2α. Дополнительные производные простагландина, родственные PGF2α, которые были выявлены из изучения уровня техники, проведенного в исследовательских лабораториях заявителя настоящей заявки, и которые проявляют высокую внутриглазную гипотензивную активность, показаны ниже формулой и идентифицированы произвольными номерами как "известное соединение 1 " и "известное соединение 2". Известное соединение 1 описано в патентах US 5352708, 5607978 и 5688819, а известное соединение 2 описано в патенте US 5545665.
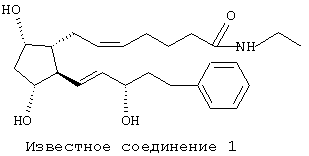
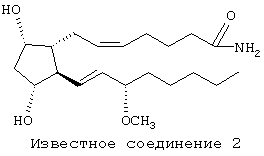
Значительное большинство глазных гипотензивных агентов, имеющих структуру простагландина или близко родственную ему структуру, действуют через известные "простагландиновые" рецепторы. В частности, известно, что соединение PGF2α проявляет свое глазное гипотензивное действие через FP-рецептор. Как раскрыто в работе Abramovitz с соавт. (J. Biol. Chem., 269:2632, 1994), включенной здесь в качестве ссылки, под термином "FP-рецептор" подразумевают рецептор к простагландину человека. Структура PGF2α, содержащая нумерацию, обычно используемую в номенклатуре простагландинов и родственных соединений, изображена ниже.
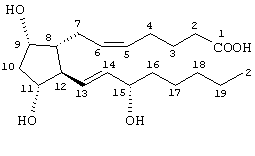
PGF2α
Настоящее изобретение направлено на класс соединений, родственных 1-этаноламиду PGF2, которые наряду с известными соединениями 1 и 2, были неожиданно обнаружены в качестве средств, обладающих сильной глазной гипотензивной активностью, но которые не проявляют своего глазного гипотензивного действия ни через FP-рецептор, ни через какой-либо другой известный к настоящему времени простагландиновый рецептор, например DP, ЕР, ЕP2, ЕР3, ЕP4, FP, IP и ТР.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
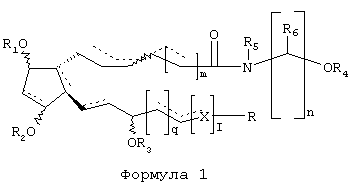
Настоящее изобретение направлено на соединения Формулы 1, где пунктирные линии означают отсутствие связи или связь при условии, что в формуле нет двух соседних двойных связей;
присоединения волнистых линий означают альфа(α, вниз)- или бета(β, вверх)-конфигурацию, там, где волнистые линии присоединены к двойной связи, они означают либо Z(цис)-, либо Е(транс)-конфигурацию;
штриховые линии обозначают альфа(α)-конфигурацию, а сплошные треугольники - бета(β)-конфигурацию;
m представляет собой целое число от 0 до 5;
n представляет собой целое число от 1 до 6 при условии, что соединение, представленное формулой, не является 1-этаноламидом PGF2α;
q и r независимо друг от друга являются целыми числами от 0 до 6;
X представляет собой СН2, O или S, при условии, что когда Х представляет собой O или S, пунктирная линия, примыкающая к X, обозначает отсутствие связи;
R означает СН3, фенил, фурил, тиенил, С3-8-циклоалкил, или фенил, фурил или тиенил, замещенные одним или двумя заместителями, выбранными из группы, состоящей из F, C1, Вr, С1-6-алкила, NO2, CN, СOOН и СOO-(С1-6-алкила);
R1, R2, R3 и R4 независимо друг от друга представляют собой Н или C1-6-алканоил с нормальной или разветвленной цепью, бензоил или низший С1-6-алкил;
R5 представляет собой Н или С1-6-алкил с нормальной или разветвленной цепью и
R6 представляет собой Н или С1-4-алкил с нормальной либо разветвленной цепью
или фармацевтически приемлемую соль данного соединения, причем указанные соединения снижают внутриглазное давление у млекопитающего, но не проявляют свою глазную гипотензивную активность ни через FP-простагландиновый рецептор, ни через какой-либо другой общеизвестный простагландиновый рецептор.
Настоящее изобретение также относится к выделенному в основном чистому 1-этаноламиду PGF2α, который, как было неожиданно установлено, присутствует в основных биологических системах в качестве природного соединения. Кроме того, настоящее изобретение направлено на фармацевтические композиции, содержащие в качестве активного компонента 1-этаноламид PGF2α и/или одно или более соединений в соответствии с Формулой 1, и на способ лечения млекопитающих, включая человека, при необходимости такого лечения, эффективным количеством фармацевтической композиции, содержащей в качестве активного компонента 1-этаноламид PGF2α и/или одно или более соединений в соответствии с Формулой 1, для снижения внутриглазного давления.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фиг.1 представляет собой график влияния возрастающих доз 1-этаноламида PGF2α на гладкую мышцу сфинктера радужной оболочки кошки. Точки обозначают средние значения ± SEM (среднеквадратичная ошибка, standard error of the mean); n=4.
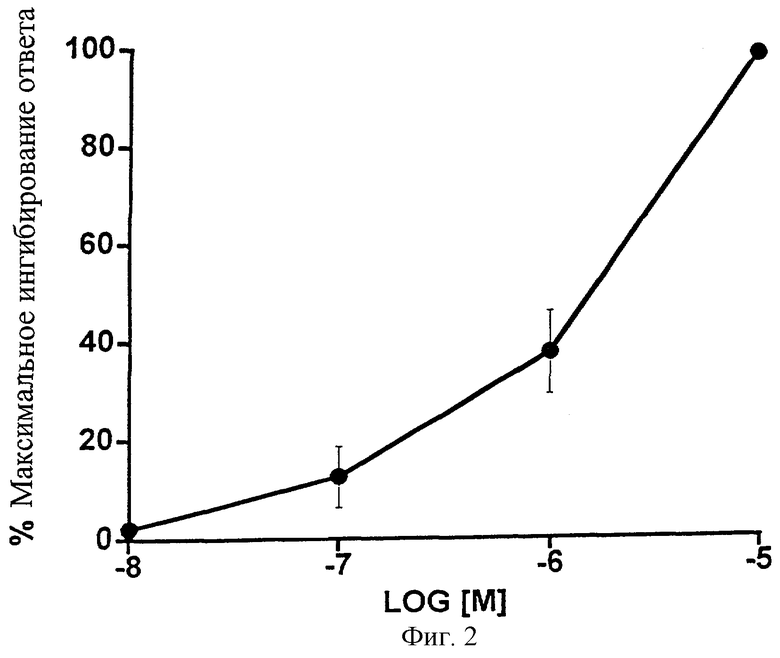
Фиг.2 представляет собой график, показывающий ингибирующее влияние возрастающих доз 1-этаноламида PGF2α на область стимулированного сокращения изолированного сосуда семенного протока морской свинки. Точки обозначают средние значения ± SEM; n=4.
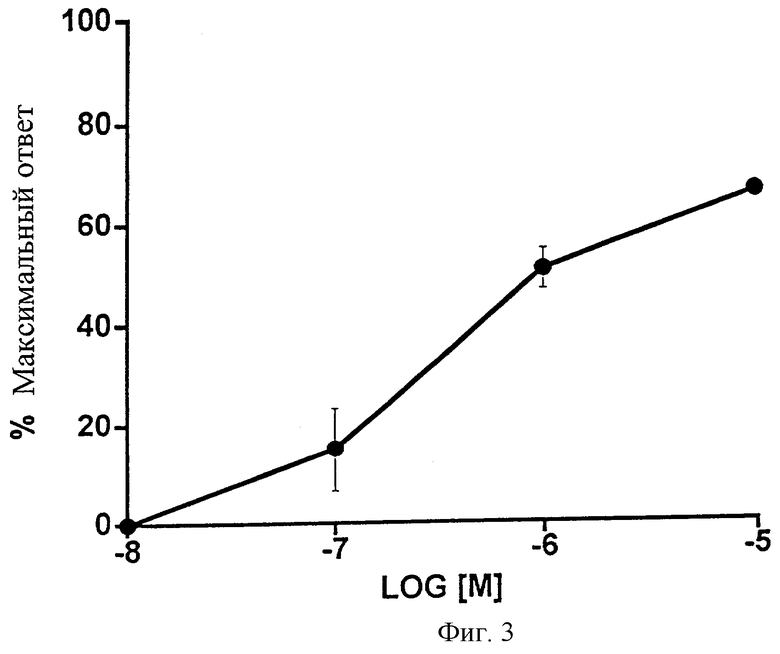
Фиг.3 представляет собой график, показывающий влияние возрастающих доз 1-этаноламида РGF2α на изолированную подвздошную кишку цыпленка. Точки обозначают средние значения ± SEM; n=4.
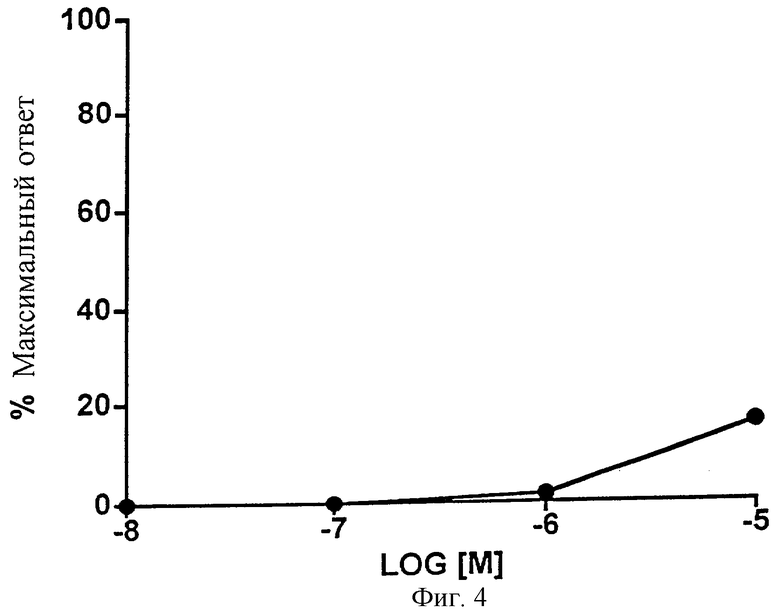
Фиг.4 представляет собой график, показывающий влияние возрастающих доз 1-этаноламида PGF2α на изолированную подвздошную кишку морской свинки. Точки обозначают средние значения ± SEM; n=4.
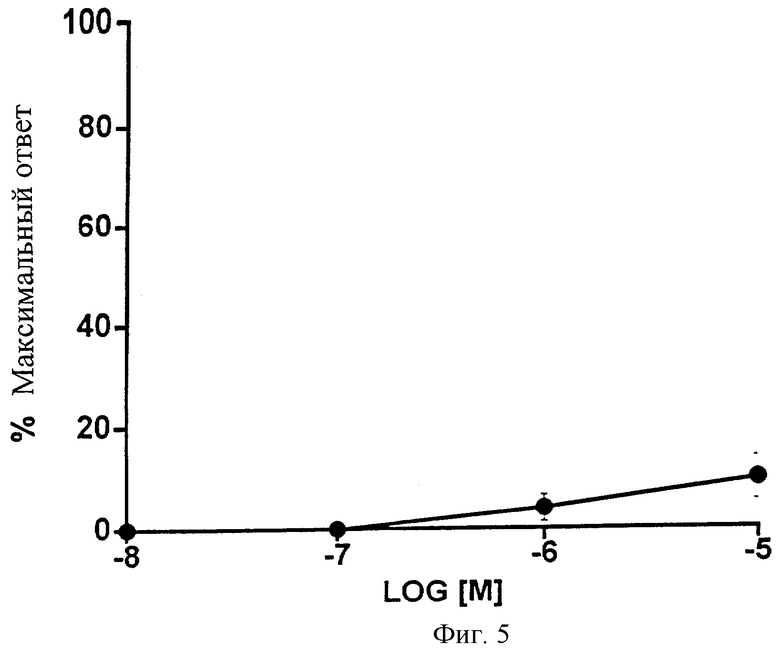
Фиг.5 представляет собой график, показывающий влияние возрастающих доз 1-этаноламида PGF2α на изолированную гладкую мышцу аорты крысы. Точки обозначают средние значения ± SEM; n=4.
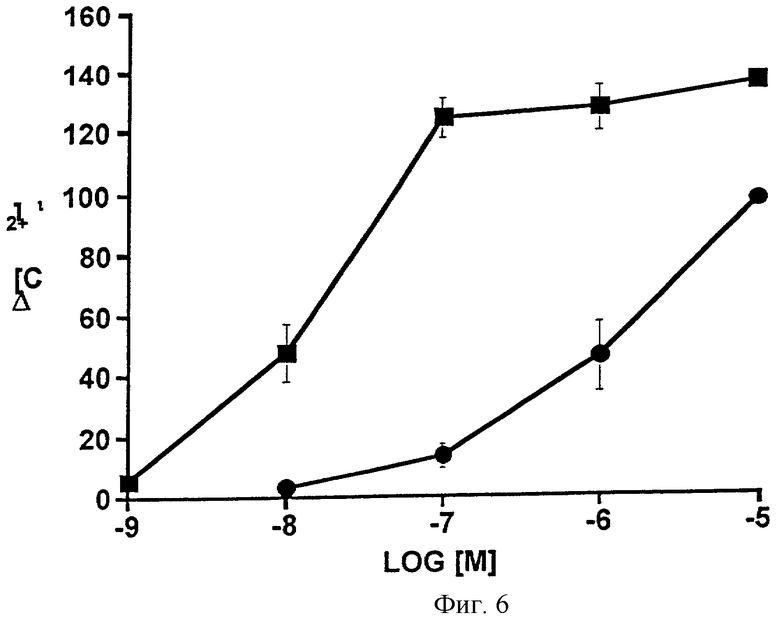
Фиг.6 представляет собой график, показывающий увеличение концентрации внутриклеточного Ca2+ [Ca2+];, вызванное возрастающими дозами PGF2α ( ) и 1-этаноламида PGF2α(•) в клетках CRL 1497. Точки являются средними значениями ± SEM; n=4.
) и 1-этаноламида PGF2α(•) в клетках CRL 1497. Точки являются средними значениями ± SEM; n=4.
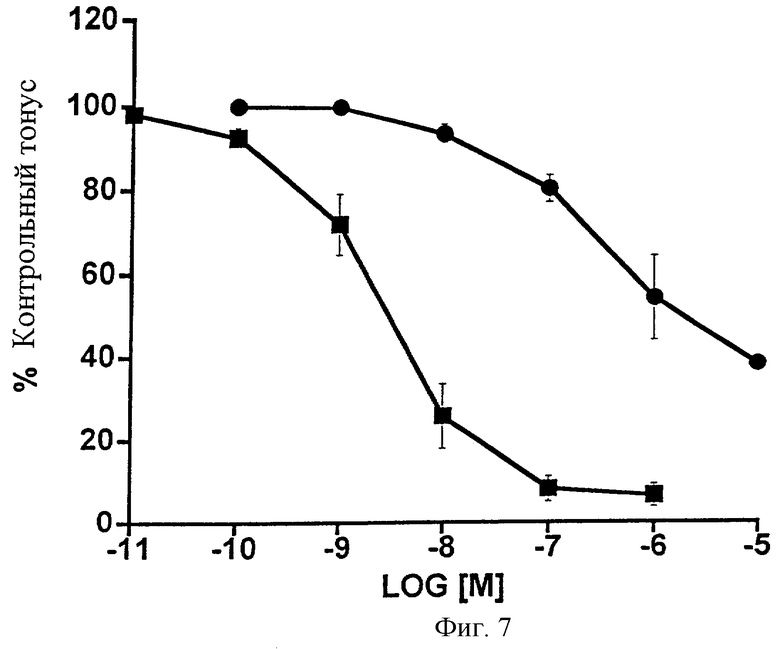
Фиг.7 представляет собой график, показывающий сосудорасширяющее действие 1-этаноламида простагландина F2α ( ) и простагландина F2α (•) на предварительно сокращенный гистамином сегмент яремной вены кролика. Точки являются средними значениями ± SEM; n=6 для 1-этаноламида PGF2α, n=7 для PGF2α.
) и простагландина F2α (•) на предварительно сокращенный гистамином сегмент яремной вены кролика. Точки являются средними значениями ± SEM; n=6 для 1-этаноламида PGF2α, n=7 для PGF2α.
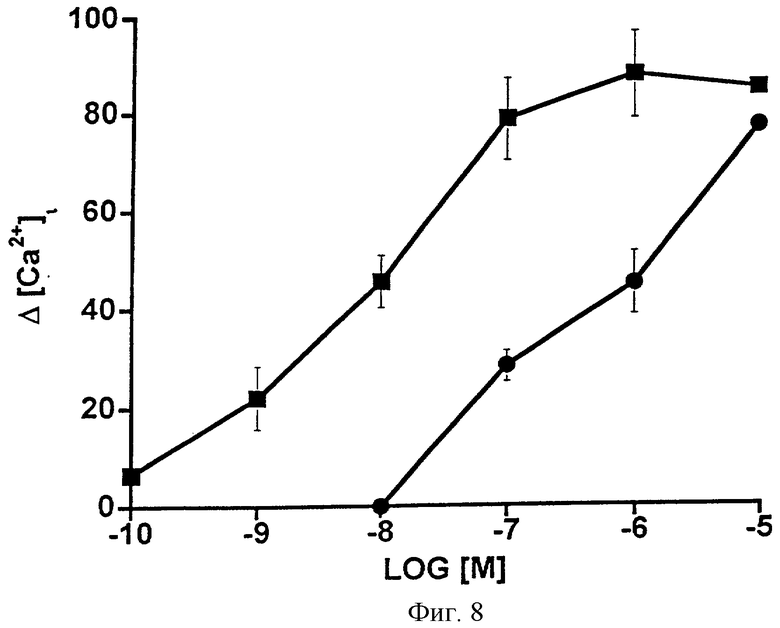
Фиг.8 представляет собой график, показывающий увеличение концентрации внутриклеточного Ca2+ [Ca2+]i, вызванное возрастающими дозами PGF2α ( ) и 1-этаноламида PGF2α (•) в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором кошки. Точки являются средними значениями ± SEM; n=3.
) и 1-этаноламида PGF2α (•) в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором кошки. Точки являются средними значениями ± SEM; n=3.
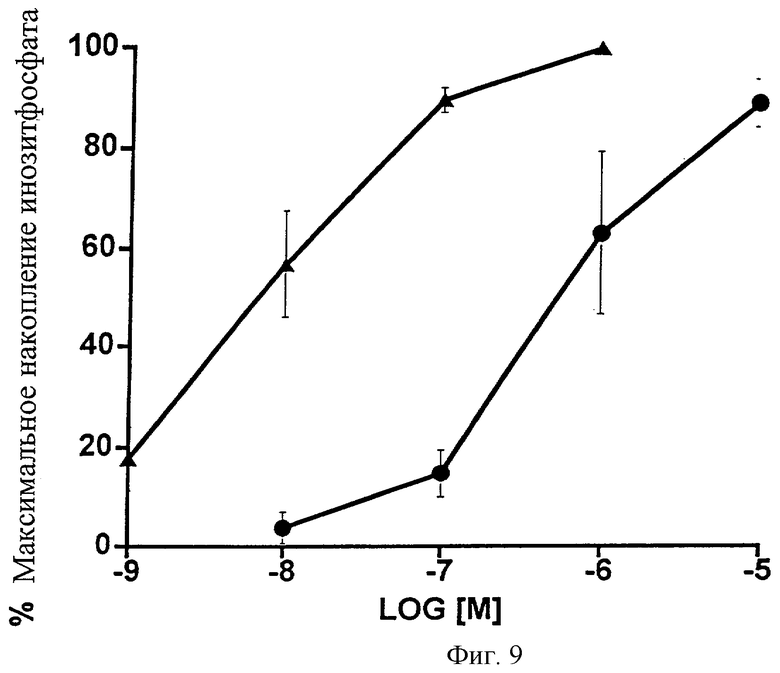
Фиг.9 представляет собой график, показывающий общее накопление инозитфосфата, вызванное возрастающими дозами 17-фенилпростагландина F2α ( ) и 1-этаноламида PGF2α (•) в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором кошки. Даны средние значения ± SEM; n=3.
) и 1-этаноламида PGF2α (•) в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором кошки. Даны средние значения ± SEM; n=3.
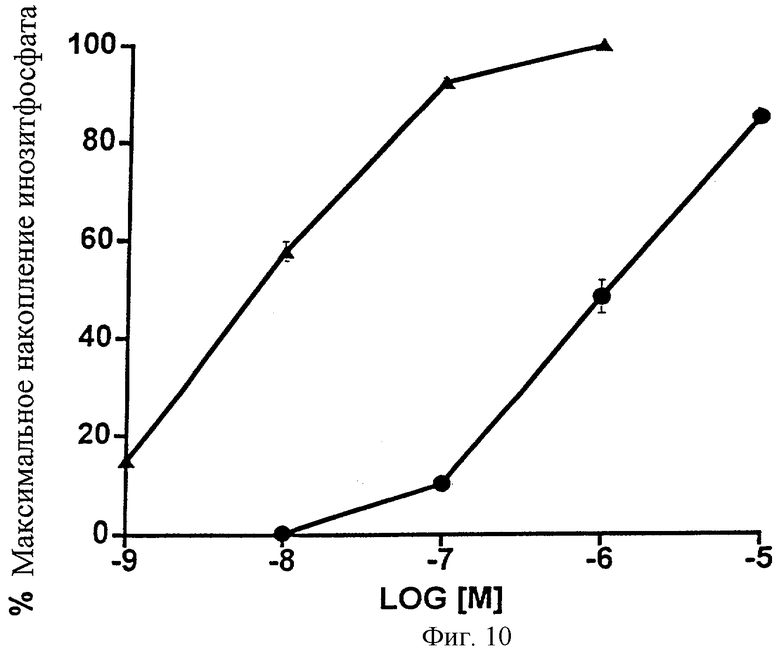
Фиг.10 представляет собой график, показывающий общее накопление инозитфосфата, вызванное возрастающими дозами 17-фенилпростагландина F2α ( ) и 1-этаноламида PGF2α (•) в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором человека. Даны средние значения ± SEM; n=3.
) и 1-этаноламида PGF2α (•) в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором человека. Даны средние значения ± SEM; n=3.
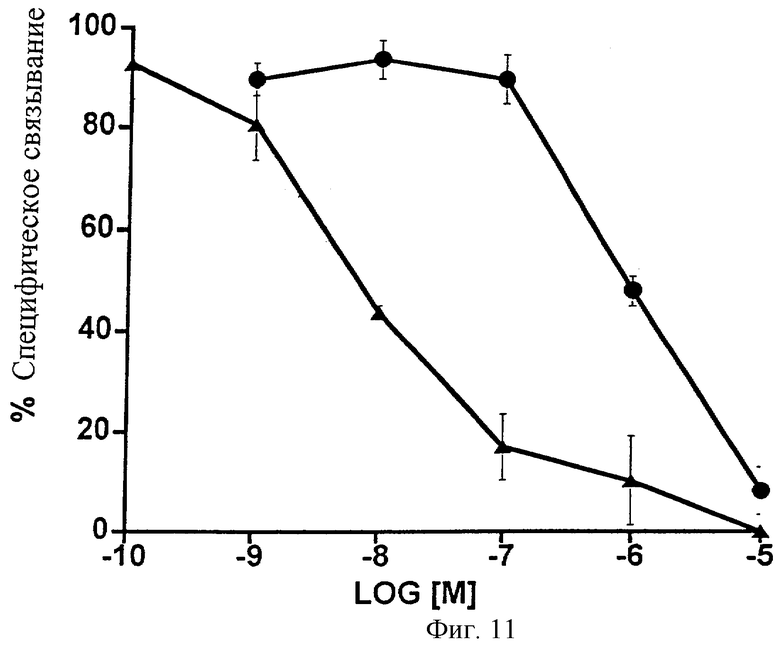
Фиг.11 представляет собой график, показывающий анализ конкуренции связывания меченого лиганда в сравнении с 5 нМ 3H-17-фенил-РGF2α с использованием клеток НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором кошки. Изображена конкуренция, осуществленная с помощью немеченого 1-этаноламида PGF2α (•) и немеченого 17-фенил-РGF2α ( ). Точки являются средними значениями ± SEM; n=3 экспериментам, выполненным с тремя повторностями.
). Точки являются средними значениями ± SEM; n=3 экспериментам, выполненным с тремя повторностями.
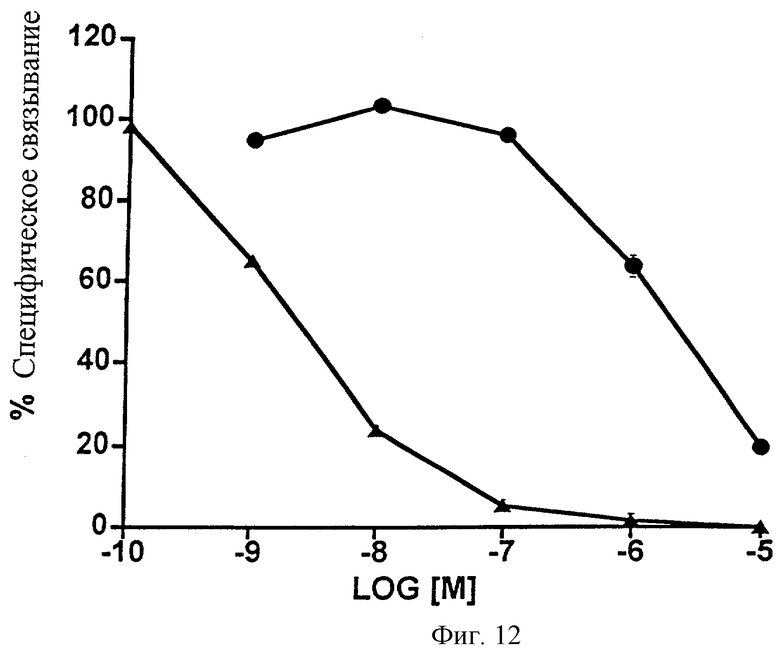
Фиг.12 представляет собой график, показывающий анализ конкуренции связывания меченого лиганда в сравнении с 5 нМ 3H-17-фенил-РGF2α с использованием клеток НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором человека. Изображена конкуренция, осуществленная с помощью немеченого 1-этаноламида PGF2α (•) и немеченого 17-фенил- PGF2α ( ). Точки являются средними значениями ± SEM; n=3 экспериментам, выполненным с тремя повторностями.
). Точки являются средними значениями ± SEM; n=3 экспериментам, выполненным с тремя повторностями.
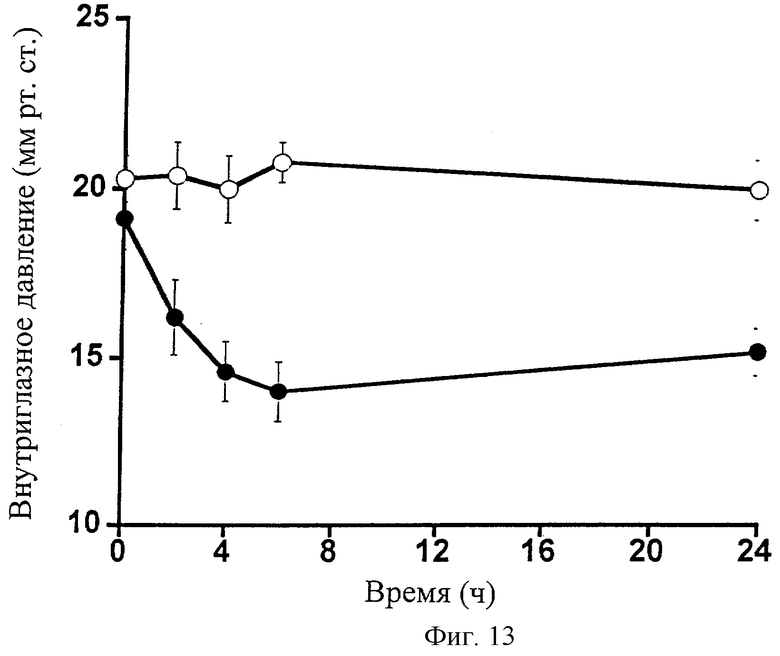
Фиг.13 представляет собой график, показывающий влияние 0,1% 1-этаноламида PGF2α (•) на внутриглазное давление собаки. Контралатеральные глаза для контроля обрабатывали наполнителем (о). Точки являются средними значениями ± SEM; n=6.
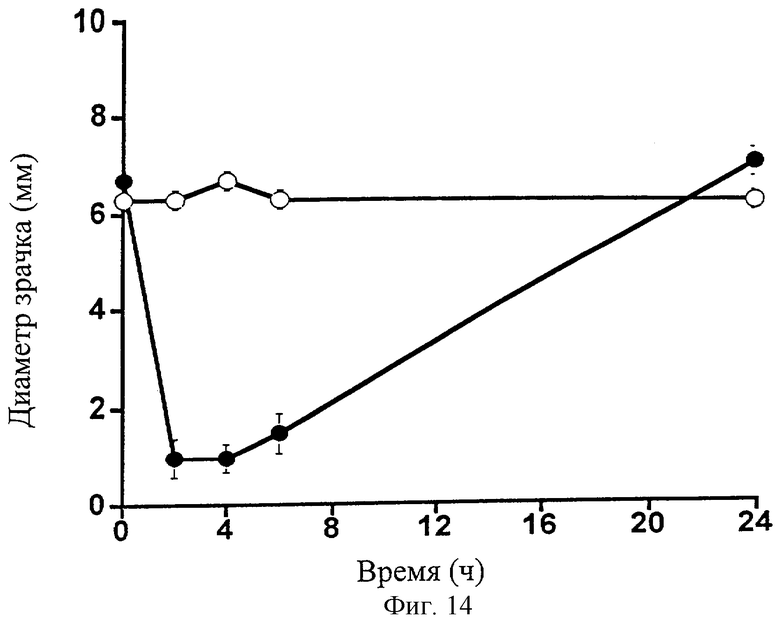
Фиг.14 представляет собой график, показывающий влияние 0,1% 1-этаноламида PGF2α (•) на диаметр зрачка собаки. Контралатеральные глаза для контроля обрабатывали наполнителем (о). Точки являются средними значениями ± SEM; n=6.
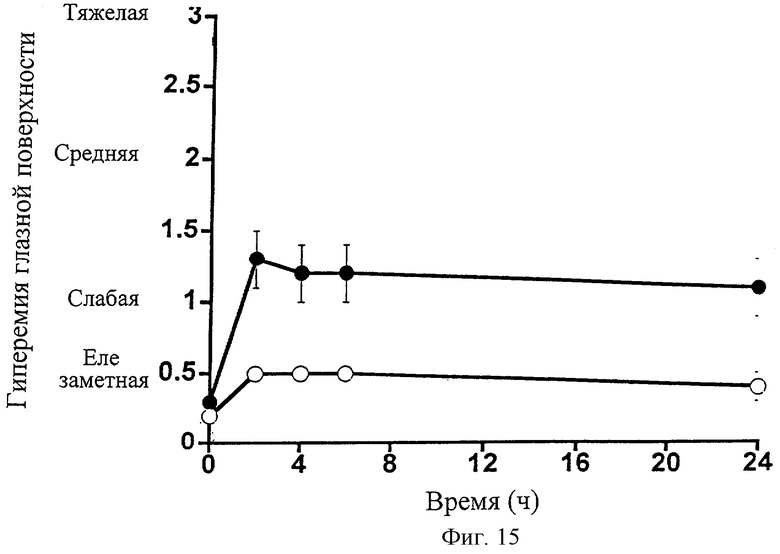
Фиг.15 представляет собой график, показывающий влияние 1-этаноламида PGF2α (•) на гиперемию поверхности глаза собаки. Контралатеральные глаза для контроля обрабатывали наполнителем (о). Точки являются средними значениями ± SEM; n=6.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В связи с настоящим изобретением было неожиданно обнаружено, что 1-этаноламид PGF2α является природным соединением и может быть образован из анандамида, природного каннабиномиметика, с помощью рекомбинантной СОХ-2 и PGF-синтазы. Известно, что СОХ-2 (циклооксигеназа-2) представляет собой эндопероксидсинтазу простагландина, которая является индуцибельной и не обязательно присутствует в тканях (см. Meade et al. (1993) J. Biol. Chem. 268: 6610-6614). Поскольку не ожидалось, что СОХ-2 будет присутствовать в нестимулированных тканях, полученные из настоящего изобретения данные о том, что значительные количества этаноламида PGF2α образовывались после введения белым мышам анандамида, являются неожиданными. Кроме того, этаноламид PGF2α также неожиданно был обнаружен у мышей, не получавших анандамида, что подтверждает тот факт, что 1-этаноламид PGF2α является конститутивным и может вести себя как эндогенно синтезируемый гормон.
В соответствии с изобретением, 1-этаноламид РGF2α был синтезирован с целью получения этого вещества в изолированном и химически чистом виде и для обеспечения возможности составления фармацевтических композиций, содержащих это вещество. Синтез этаноламида PGF2α также дает возможность синтезировать все соединения в пределах Формулы 1, но лишь с такими модификациями в процессе синтеза, которые легко доступны для практикующего химика-органика и, в частности, для химика, работающего в области химии простагландинов и родственных соединений. Таким образом, настоящее изобретение охватывает соединения Формулы 1.
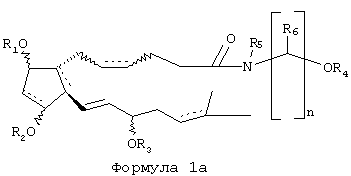
Предпочтительные соединения в объеме правовой охраны изобретения представлены Формулой 1а, и особенно предпочтительными являются соединения в пределах Формулы 1а, где пунктирные линии между атомами углерода 10 и 11, 8 и 12, 17 и 18 означают отсутствие связи, а между атомами углерода 5 и 6 означают связь. Кроме того, поскольку представляет интерес замещение гидроксильных функциональных групп соединений Формулы 1а или Формулы 1 (от R1-группы до R4), в настоящий момент предпочтительными являются соединения, в которых одна или более гидроксильных групп не этерифицированы и не алкилированы (один или более заместитель от R1 до R4 представляет собой Н), или, если эти группы этерифицированы, этерифицирующая группа представляет собой алканоил, имеющий от 1 до 6 углеродных атомов (формил, ацетил, пропаноил, бутаноил, пентаноил, гексаноил или какой-либо С3-6-алканоил с разветвленной цепью). Поскольку представляет интерес заместитель, обозначенный как R5 в Формуле 1а или Формуле 1, предпочтительными являются соединения, в которых R5 является Н или низшим С1-3-алкилом, наиболее предпочтителен Н. R6 предпочтительней представляет собой Н или низший С1-3-алкил, наиболее предпочтителен Н, и n предпочтительно равно 2.
Вновь возвращаясь к Формуле 1а и Формуле 1, конфигурация групп OR1, OR2 и OR3 предпочтительней представляет собой альфа(С-9, С-11 и С-15-α)-конфигурацию, а конфигурация двойной связи 5-6 - предпочтительней Z(цис)- конфигурацию.
Наиболее предпочтительными соединениями согласно настоящему изобретению являются 1-этаноламид PGF2α и его ацилированные или алкилированные производные, представленные Формулой 2, где заместители от R1 до R4 определены как для Формулы 1, но ацилированные соединения являются более предпочтительными, чем алкилированные.
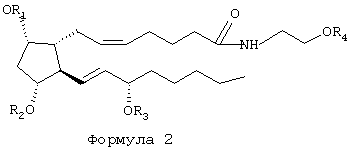
1-этаноламид PGF2α=R1=R2=R3=R4=H
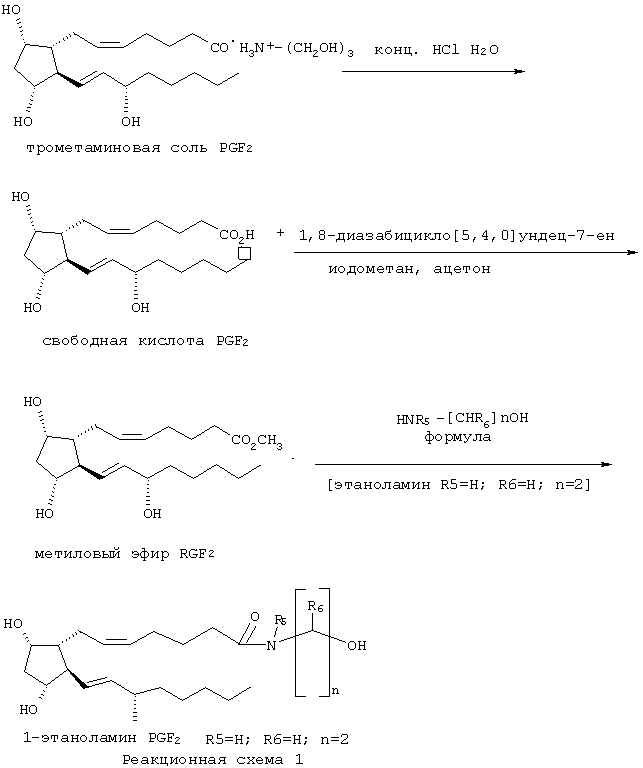
Синтез соединений изобретения проиллюстрирован с помощью синтеза 1-этаноламида PGF2α, представленного реакционной схемой 1, а также подробно раскрыт ниже. Обратимся теперь к схеме реакции. Коммерчески доступную триэтаноламмониевую соль (соль трометамина, трис(гидроксиметил)аминометана) карбоновой кислоты PGF2α обрабатывают кислотой для выделения свободной карбоновой кислоты. Свободную карбоновую кислоту PGF2α превращают в метиловый эфир обработкой метилирующим агентом (предпочтительно, йодистым метилом) в присутствии акцептора кислоты (предпочтительно, 1,8-диазабицикло[5.4.0]ундец-7-ена) с получением метилового эфира РGF2α. Затем метиловый эфир PGF2α обрабатывают этаноламином для получения 1-этаноламида PGF2α. Таким образом, вся "амидная" группа вводится в молекулу в ходе реакции между метиловым эфиром PGF2α и производным гидроксиалкиламина Формулы 3, где R5, R6 и n такие, как определены выше для Формулы 1, и где Формула 3 представляет собой этаноламин, в котором R5 и R6 означают Н, и n равно 2. Гидроксиалкиламины Формулы 3 доступны благодаря химической литературе и с успехом используются в работе практикующего химика-органика. Соединения, для которых в Формуле 1 одна или более пунктирных линий представляют собой связь, и/или где конфигурация гидроксильных групп или конфигурация заместителей около одной или более двойных связей иные, чем в 1-этаноламиде PGF2α, могут быть получены с помощью реакций, аналогичных изображенным на реакционной схеме 1, но при использовании в качестве исходного соединения производного С-1-карбоновой кислоты простагландина соответствующей структуры и стереохимии. Эти исходные соединения доступны благодаря химической литературе и используются в работе практикующего химика-органика. Соединения, для которых в Формуле 1 одна или более групп от R1 до R4 отличается от Н, то есть соединения, где один или более атомов углерода С-9, С-11, С-15 и гидроксигруппа гидроксиалкильного фрагмента в амидной части этерифицированы или алкилированы, могут быть получены реакциями этерификации или алкилирования из "свободных гидроксильных" соединений. Вообще говоря, модификации а- и ω-боковых цепей соединений Формулы 1, как определено для этой формулы, используются в работе практикующего химика-органика в соответствии с патентами США №5545665, 5834498 и 5352708 или по аналогии с ними. Описания этих патентов приводятся здесь в виде ссылок.
БИОЛОГИЧЕСКАЯ АКТИВНОСТЬ
Соединения изобретения являются уникальными, поскольку представляют собой сильнодействующие глазные гипотензивные агенты, которые, тем не менее, не действуют через FP-рецептор. Это показано с помощью нескольких аналитических методов, которые хорошо известны в данной области и используются для определения внутриглазной гипотензивной активности, а также для определения рецептора, через который действуют эти соединения. Внутриглазная гипотензивная активность 1-этаноламида PGF2α и его фактическая инертность в отношении FP- или других известных рецепторов простагландинов, продемонстрированная несколькими методами, являются особенно поразительными и неожиданными при сравнении биологического распределения этого соединения с "предшественником" PGF2α.
Описание биологических исследований и результаты исследований приведены ниже.
МЕТОДЫ ИССЛЕДОВАНИЯ ИЗОЛИРОВАННОЙ ТКАНИ
Напряжение гладкой мышцы изолированных тканей измеряли изометрически с помощью датчиков силового смещения (Grass FT-03) и записывали на Grass Polygraphs (модели 7G и 79Е). Ванночки для органов содержали раствор Кребса, выдержанный при 37°С и насыщенный смесью газов 95% O2/5% СO2 для получения рН 7,4. Раствор Кребса имел следующий состав (мМ): NaCl, 118,0; КСl, 4,7; КН2РO4 1,2; СаСl2, 1,9; MgSO4, 1,18; NaHCO3, 25,0; глюкоза, 11,7; индометацин, 0,001.
(а) РАДУЖНАЯ ОБОЛОЧКА КОШКИ
Взрослых домашних кошек умерщвляли введением внутривенно сверхдозы пентобарбитала натрия (Anthony, Arcadia, CA). Глаза сразу же энуклеировали и помещали в лед. Мышцу сфинктера радужной оболочки закрепляли вертикально с натяжением от 50 до 100 мг в ванночке для органов емкостью 10 мл, имеющей двойные стенки. Перед началом каждого эксперимента был предусмотрен период стабилизации длительностью 60 мин. Активность определяли по ответным реакциям сокращения. Соединения добавляли в ванночку кумулятивно и, по меньшей мере, 30 минут после полной отмывки отводили на восстановление и возвращение к исходному растяжению. В качестве контроля определяли ответную реакцию на 10-7 М PGF2α в начале и в конце каждого эксперимента и между кривыми зависимости "доза - ответ". Для каждой ткани было проанализировано не более двух соединений.
(б) АОРТА КРЫСЫ
Взрослых крыс Sprague Dawley весом 180-220 г подвергали наркозу с помощью ингаляции СО2, после чего декапитировали и обескровливали. Грудную аорту извлекали и очищали от всех прилегающих тканей. Для удаления крови просвет аорты промывали. Аорту разделяли на три небольших сегмента длиной 5-8 мм. Каждый сегмент закрепляли с натяжением 2 г в ванночке для органов емкостью 10 мл, имеющей двойные стенки, с помощью двух проволочных крючков, размещенных вдоль просвета сосуда. Такое размещение позволяет измерять силу сокращений, развиваемых круговым слоем гладкой мышцы. Ткани уравновешивали в течение 1 часа перед кумулятивным добавлением соединений в ванночки для органов. В качестве контроля в начале и в конце каждого эксперимента определяли ответную реакцию на 10-7 М U-46619 (тромбоксаномиметика). На восстановительный период после полной отмывки лекарственного препарата отводили 30-45 минут. Для каждой ткани было проанализировано только одно исследуемое соединение.
(в) ПОДВЗДОШНАЯ КИШКА ЦЫПЛЕНКА
Подвздошная кишка цыпленка представляет собой модель для изучения ЕР3-рецептора. Участки подвздошной кишки цыпленка длиной 1,5 см подвешивали с натяжением 1 г. После 1 часового периода уравновешивания получали стандартную кривую "доза-ответ" для PGF2α с 30 минутными отмывками между отдельными дозами. Затем некумулятивно добавляли анализируемое соединение. Под конец в качестве второго контроля добавляли максимальную дозу РGЕ2 (10-6 М). Затем для каждой концентрации вычисляли сократительную активность как % от максимального ответа на PGE2, достигнутого при концентрации 10-6 М.
(г) ПОДВЗДОШНАЯ КИШКА МОРСКОЙ СВИНКИ
Подвздошная кишка морской свинки представляет собой модель для изучения EP1-рецептора. Сегмент подвздошной кишки морской свинки размером приблизительно 1,5 см подвешивали с натяжением 1 г в ванночке для органов, имеющей двойные стенки. После 1-часового периода уравновешивания получали кривую зависимости "доза - ответ" для PGE2. Между дозами допускались 30-минутные периоды отмывки. Аналогичным образом получали кривую "доза - ответ" для 1-этаноламида РGF2α. В качестве контроля в конце вводили 10-6 М РGЕ2. Данные выражали как % от максимального ответа на РGЕ2.
(д) СОСУД СЕМЕННОГО ПРОТОКА МОРСКОЙ СВИНКИ
Активность в отношении подтипа простагландин-чувствительного ЕР3-рецептора определяли также по способности простаноидов ингибировать судорожную реакцию гладкой мышцы, вызванную действием электрического тока на изолированный сосуд семенного протока морской свинки. Участки сосуда семенного протока морской свинки размером 1,5 см подвешивали с начальным натяжением 1 г и уравновешивали в течение, по меньшей мере, 30 минут. Затем каждые 30 секунд ткани подвергали раздражению электрическим импульсом 20 вольт. После стабилизации судорожной реакции для PGE2 получали в кумулятивную стандартную кривую зависимости "доза - ответ". Затем с помощью кривой зависимости "кумулятивная доза - ответ" оценивали исследуемые соединения. Ткани промывали и уравновешивали в течение 1 часа между добавлением соединений. В качестве стандартного контроля в конце эксперимента снова оценивали PGE2. Активность вычисляли как процент ингибирования пика судорожной реакции мышцы.
(е) ЯРЕМНАЯ ВЕНА КРОЛИКА
Новозеландским кроликам-альбиносам обоего пола весом 2-4 кг вводили 1000 ед. гепарина в маргинальную глазную вену, после чего умерщвляли с помощью углекислого газа. Внешние яремные вены очищали от жира и прилегающей соединительной ткани и извлекали. Вены рассекали и каждое кольцо длиной 3-4 мм подвешивали между двумя металлическими крючками в ванночке для органов, имеющей двойные стенки. Ткани уравновешивали в течение 1 часа с натяжением 0,75 г, которое повторно регулировали по мере расслабления тканей. Для сокращения ткани и определения ответной реакции применяли однократные дозы гистамина: 10-5 М, затем 2-3×10-6 M, с отмывкой после каждой дозы. Затем на 5 минут прикладывали антагонист ТР-рецептора SQ29548 в концентрации 10-6 М (Ogletree et al. (1985) J. Pharmacol. Exp. Ther. 234 435-441), после чего добавляли 2-3×10-6 М гистамина для стимуляции сократительной реакции. Через 30 мин после предварительной обработки гистамином анализировали реакцию расслабления путем добавления кумулятивных доз исследуемых соединений с 10-8-10-7 М PGE2 в конце каждой кривой зависимости "доза - ответ" в качестве контроля. На восстановительный период после отмывки тканей отводили 30-50 минут. Релаксационную активность вычисляли как % от контрольного тонуса, вызванного гистамином.
(ж) ТРОМБОЦИТЫ ЧЕЛОВЕКА
Простаноидную активность по отношению к DP-, TP- и IP-рецепторам определяли по их способности вызывать агрегацию (ТР-рецепторная активность) или ингибировать АДФ-индуцированную агрегацию тромбоцитов человека in vitro (DP- и IP-рецепторная активность). Свежую цельную кровь получали от здоровых доноров-волонтеров и смешивали с цитратно-декстрозным раствором (ACD -(citric) acid, (sodium) citrate, dextrose). Кровь центрифугировали со скоростью 1000 об/мин для получения обогащенной тромбоцитами плазмы (PRP). К 400 мкл PRP добавляли 4,5 мкл простаноида или наполнителя и инкубировали 2 мин при 37°С в агрегометре Пэйтона, наблюдая за агрегационной активностью. Для полной агрегации добавляли АДФ до конечной концентрации 2×10-5 М. Ингибирование агрегации вычисляли как процентную разницу между агрегацией, вызванной 2×10-5 М АДФ в отсутствие и в присутствии лекарственного препарата. Агрегационную активность вычисляли как процент агрегации, вызванной простаноидом, относительно агрегации, вызванной 2×10-5 М АДФ. В начале и конце каждого эксперимента определяли контрольные агрегационные реакции лишь для 2×10-5 М АДФ.
(з) КЛЕТКИ CRL 1497: Са2+-СИГНАЛ
Клетки человека CRL 1497 сеяли в культуральные флаконы в модифицированной Дульбекко среде Игла (DMEM), содержащей 10% эмбриональной телячьей сыворотки, 2 мМ 1-глутамина и 0,05 мг/мл гентамицина (все получено от Gibco, Grand Island, NY). Клеточные культуры поддерживали во влажной атмосфере (95% воздуха, 5% СO2) и растили до получения монослоя.
Клетки извлекали из культуральных флаконов приблизительно через 1 мин после обработки смесью 0,05% трипсин/0,53 мМ ЭДТА в сбалансированном солевом растворе Хенкса (HBSS, Gibco, Grand Island, NY) при 37°С. Протеолитическую активность ингибировали добавлением 5 мл DMEM с 10% эмбриональной телячьей сыворотки. Клетки последовательно промывали раствором Хенкса и средой, содержащей 140 мМ NaCl, 50 мМ КСl, 1 мМ MgCl2, 1,5 мМ CaCl2, 10 мМ HEPES (N-2-гидроксиэтилпиперазин-N-2-этансульфоновая кислота): трис (тригидрооксиметиламинометан), 5 мМ глюкозы, 5 мМ пируватаМа, 0,1% бычьего сывороточного альбумина, при рН 7,4.
Центрифугирование между промывками осуществляли при комнатной температуре в течение 15 минут при 200 g. Клетки подсчитывали, ресуспендировали в упомянутой выше среде и инкубировали с 2×10-6 М смеси Fura 2 и ацетоксиметилового эфира на встряхиваемой водяной бане в течение 30 минут при 37°С. Далее клетки промывали в вышеуказанной среде и ресуспендировали до концентрации 2×106 клеток/мл. Затем аликвоты по 0,5 мл клеточной суспензии добавляли в микропробирки с автоматическими крышками для того, чтобы на каждое экспериментальное определение концентрации внутриклеточного свободного Са2+ ([Са2+]i) приходилось 106 клеток.
Флюоресценцию измеряли на флюоресцентном спектрофотометре Perkin-Elmer LS-5 в интервале длин волн возбуждения и излучения от 340 до 492 нм, соответственно, с обеими щелями по 10 нм. Для каждого экспериментального определения 106 клеток отмывали (при 200 g в течение 5 мин) и суспендировали в 2 мл кювете с буфером, содержащим 120 мМ NaCl, 6 мМ КСl, 1 мМ MgSO4, 1,5 мМ СаСl2, 20 мМ HEPES, 1 мг/мл глюкозы и 1 мг/мл пиру вата Na. Перемешивание осуществляли с помощью установленной наверху лопастной мешалки, при температуре 37°С. Для получения fmax клетки лизировали дигитонином (10 мкл из 100 мг/мл ДМСО). Затем для получения fmin последовательно добавляли EGTA (этиленгликольтетрауксусная кислота) (100 мМ) и 10 N NaOH для подведения рН раствора до 8,5.
(и) РЕКОМБИНАНТНЫЙ FP-РЕЦЕПТОР КОШКИ И ЧЕЛОВЕКА; СТАБИЛЬНЫЕ ТРАНСФЕКТАНТЫ
Клетки НЕК-293 выращивали в DMEM с 10% сыворотки плода коровы (FBS), 250 мкг/мл G418 (Life Technologies) и 200 мг/мл гентамицина или пенициллина/стрептомицина. Отбор стабильных трансфектантов проводили с помощью 200 мкг/мл гигромицина, причем оптимальную концентрацию определяли, исследуя кривые гибели клеток в зависимости от концентрации гигромицина.
Клетки для трансфекции выращивали на 10 см чашках до 50-60% монослоя. Плазмиду рСЕР4, содержащую вставки кДНК для FP-рецептора человека (20 мкг), добавляли к 500 мкл 250 мМ CaCl2. Затем по каплям прибавляли HBSS × 2 (280 мМ NaCl, 20 мМ HEPES кислоты, 1,5 мМ Na2HPO4, pH 7,05-7,12) до 500 мкл при постоянном перемешивании при комнатной температуре. Через 30 мин к смеси добавляли 9 мл DMEM. Затем к клеткам, предварительно отмытым 10 мл фосфдтно-солевого буфера (PBS), прибавляли смесь ДНК/ПМЕМ/фосфат кальция.
Клетки инкубировали в течение 5 часов при 37°С во влажной атмосфере (95% воздуха, 5% СO2). Затем раствор фосфата кальция удаляли и клетки в течение 2 мин обрабатывали 10% раствором глицерина в DMEM. Раствор глицерина меняли затем на среду DMEM, содержащую 10% сыворотки плода коровы (FBS). Клетки инкубировали в течение ночи, и среду меняли на DMEM/10% FBS, содержащую 250 мкг/мл G418 и пенициллин/стрептомицин. На следующий день добавляли гигромицин В до конечной концентрации 200 мг/мл.
Через десять дней после трансфекции отделяли гигромицин В-устойчивые клоны и помещали их в отдельную лунку на 24-лу ночной плате. После достижения монослоя каждый клон переносили в одну лунку 6-луночной платы, а затем в 10 см чашку. Клетки поддерживали в условиях селекции гигромицином до использования.
Стабильные трансфектанты, содержащие FP-рецептор кошки, получали аналогичным образом, используя липофектамин. Клетки также выращивали до 50-60% монослоя в 10 см чашках и с помощью липофектамина трансфицировали плазмидой рСЕР4, включающей вставки кДНК, кодирующей FP-рецептор кошки. Селекцию с помощью гигромицина В начинали через 48 часов после трансфекции. Через восемь дней после трансфекции устойчивые к гигромицину В клоны отделяли и переносили в 24-лу ночные платы.
Исследования Са2+-зависимой передачи сигналов на рекомбинантных FP-рецепторах кошки и человека проводили аналогично тому, как описано для клеток CRL 1497.
(к) ОБРАЗОВАНИЕ ИНОЗИТФОСФАТА
Рецептор-опосредованный гидролиз фосфоинозитида (PI) определяли измерением накопления суммарного инозитфосфата (IP) в клетках, предварительно инкубированных с 3H-миоинозитом. Постоянные клеточные линии НЕК 293, экспрессирующие FP-рецепторы человека или кошки, засевали в 10 см чашки (по 106 клеток на каждую чашку в DMEM с 10% FBS). На следующий день клетки инкубировали с 18 мкКи мио-[2 3H]инозита (Amersham; 10-20 мкКи/ммоль) в 6 мл DMEM с 10% FBS при 37°С в течение 24 часов. Затем клетки дважды промывали фосфатно-солевым буфером (PBS), инкубировали в течение 5 мин с 1 мл трипсин-ЭДТА и суспендировали в 10 мл DMEM, содержащей 25 мМ HEPES. Клетки осаждали центрифугированием при 1000 об/мин и ресуспендировали в DMEM/25 мМ HEPES с 10 MMLiCI в течение 10 мин. Аликвоты по 200 мкл клеточной суспензии инкубировали с 50 мкл лекарственного препарата в течение 30 мин при 37°С (двойное определение), после чего с помощью 750 мкл смеси хлороформ-метанол-4 н. НСl=100:200:2 останавливали стимуляцию агониста с последующим добавлением 250 мкл хлороформа и 0,5 N HCl для экстракции инозитфосфатов. 750 мкл водного слоя наносили на колонку, заполненную 0,5 мл анионобменной смолы Dowex AG 1-X8 (формиатная форма, 100-200 меш, BioRad) для отделения компонентов с радиоактивной меткой. Процедура элюирования состояла из трех 3 мл промывок с 5 мМ инозита, затем двух 750 мкл элюций 1,3 М формиатом аммония с 0,1 М муравьиной кислоты. Элюат смешивали с 10 мл сцинтилляционной жидкости Аквазоль (Packard Instrument Co.) и с помощью жидкостного сцинтилляционного счетчика определяли общую концентрацию 3Н] инозитфосфатов.
(л) СВЯЗЫВАНИЕ МЕЧЕНОГО ЛИГАНДА
Препараты плазматической мембраны клеток НЕК 293, стабильно трансфицированных FP-рецептором человека или кошки, получали следующим образом. Клетки, промытые ТМЕ-буфером, соскабливали со дна флакона и гомогенизировали в течение 30 с с помощью Brinkhman PT 10/35 политрона. При необходимости, для подведения объема до 40 мл в центрифужные пробирки добавляли ТМЕ-буфер, содержащий 100 мМ трис-основания, 20 мМ MgCl2, 2 мМ ЭДТА; физиологическое значение рН получали прибавлением 10 н. HCl.
Клеточный гомогенат центрифугировали со скоростью 19000 об/мин при 4-6°С в течение 20 мин на роторе Ti-60 Beckman. Осадок затем ресуспендировали в ТМЕ-буфере для получения конечной концентрации белка 1 мг/мл, как определено с помощью Biorad анализа. Количественные анализы связывания радиоактивных лигандов выполняли в объеме 200 мкл.
Связывание [3Н](N)17-фенил-РGF2α (удельная активность 85 Ки/ммоль) определяли в двух повторностях, и опыты повторяли три раза. Инкубации проводили в течение 60 мин при 25°С и останавливали добавлением 4 мл ледяного 50 мМ трис-НСl буфера с последующей быстрой фильтрацией через фильтры Whatman GF/B и тремя дополнительными промывками по 4 мл в клеточном коллекторе (Brandel). Проводили анализы конкурентного связывания, используя конечную концентрацию 5 нМ [3H](N)17-фенил-РGF2α, а неспецифическое связывание определяли с помощью 10-5 М немеченого 17-фенил-РGF2α.
(м) ВНУТРИГЛАЗНОЕ ДАВЛЕНИЕ (ВГД)
Исследования внутриглазного давления у собак с привлечением пневматонометрии проводили на находящихся в сознании собаках породы коротконогая гончая обоего пола (10-15 кг). Животные оставались в сознании на протяжении всего исследования и слегка удерживались рукой. Лекарственные препараты вводили местно в один глаз в виде капли объемом 25 мкл, другой глаз для контроля обрабатывали 25 мкл наполнителя (0,1% полисорбата 80-10 мМ трис). Для анестезии роговицы во время тонометрии использовали 0,1%-ный раствор пропаракаина. Внутриглазное давление определяли непосредственно перед введением препарата и через 2, 4 и 6 часов после введения каждый день на протяжении 5 дней исследования. Лекарство вводили сразу после регистрации первого значения ВГД.
(н) ДИАМЕТР ЗРАЧКА
Диаметр зрачка собаки измеряли с помощью оптического стержня (миллиметровой линейки, содержащей полукруглые отметки стандартной ширины (мм) в качестве контроля. Слегка придерживая собаку рукой, при нормальном комнатном освещении измеряли диаметр зрачка путем подбора полукруга к зрачку. Для собак с очень темными зрачками использовали специальный точечный фонарик, но только очень быстро во избежание сужения зрачка. Диаметр зрачка измеряли в то же время, что ВГД и гиперемию.
(о) ГИПЕРЕМИЯ ГЛАЗНОЙ ПОВЕРХНОСТИ
Гиперемию глазной поверхности определяли визуально и рассчитывали по системе, использующейся в обычной клинической практике.

Гиперемию глазной поверхности оценивали одновременно с измерением внутриглазного давления. Необходимо отметить, что необработанные глаза собак часто имели розовато-красный оттенок. Таким образом, следовая и даже слабая гиперемия не обязательно выходит за пределы нормального диапазона.
РЕЗУЛЬТАТЫ
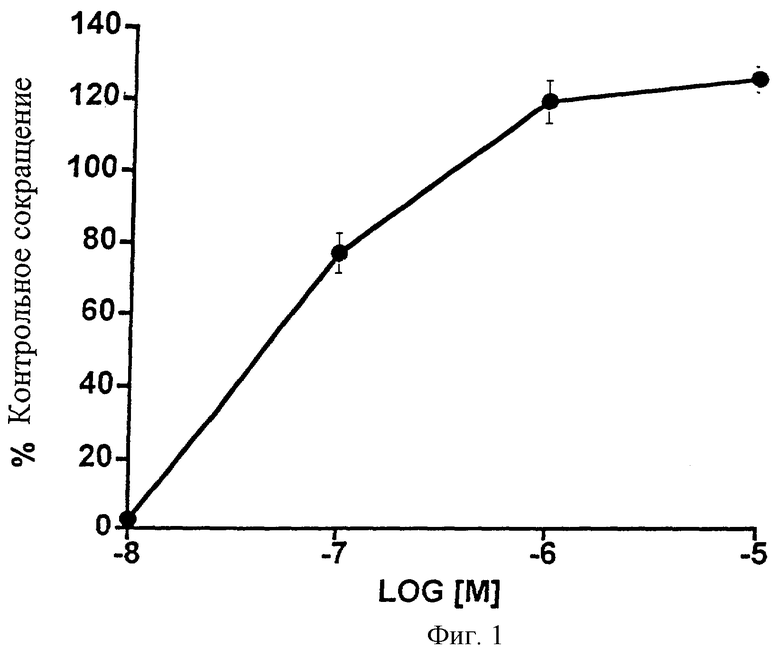
Эффекты 1-этаноламида простагландина F2α на сфинктер радужной оболочки кошки изображены на Фиг.1 и представлены в виде таблицы (Таблица 1). 1-Этаноламид простагландина F2α (1-этаноламид РGF2α) вызывал дозозависимое сокращение гладкой мышцы сфинктера радужной оболочки кошки. Была получена величина эффективной концентрации (ЭК)50, равная 58 нМ. Данные указывают на критическую концентрацию в области 10 нМ.
1-Этаноламид простагландина F2α был умеренно эффективен при ингибировании области раздражения, вызвавшего сокращение изолированного препарата сосуда семенного протока морской свинки. Данные показыны на Фиг.2 и представлены в виде таблицы (Таблица 2).
Фиг.3 показывает влияние 1-этаноламида РGF2α на изолированную подвздошную кишку цыпленка. Сравнение реакции с контрольным соединением (простагландином Е2, 10-6 М) подтверждает, что 1-этаноламид PGF2α может вести себя отчасти как слабый агонист. Данные представлены в Таблице 3. 1-Этаноламид РGF2α оказался менее эффективным для подвздошной кишки цыпленка (Фиг.4, Таблица 4), которая, как сообщалось, является моделью для изучения EP1-рецептора.
Эффекты возрастающих доз 1-этаноламида PGF2α на изолированную аорту крысы показаны на Фиг.5 и в Таблице 5. При высоких дозах, 10-6 и 10-5 М, наблюдалась остаточная активность. Поскольку аорта крысы является моделью для изучения ТР-рецептора, активность в отношении ТР-рецептора дополнительно оценивали во время агрегации тромбоцитов. Кроме того, определяли ингибирование АДФ, вызывающего агрегацию тромбоцитов человека, для того, чтобы обнаружить какое-либо взаимодействие с DP- или IP-рецепторами. Никакого влияния на тромбоциты не наблюдалось. Данные представлены в Таблице 6; график не прилагается, так как никакой реакции не было.
Са2+-сигнал, продуцируемый возрастающими дозами 1-этаноламида простагландина F2α в фибробластах кожи человека (клетки CRL 1497), сравнивают с сигналом, продуцируемым простагландином F2α, на Фиг.6. Данные также представлены в виде таблицы (Таблица 7). Полученные значения ЭК50 составляли 16 нМ для PGF2α и 1586 нМ для 1-этаноламида РGF2α, показывая почти стократное различие в активности по отношению к FP-рецептору человека.
Недавние исследования показали, что простагландин F2α может вызывать эндотелийзависимое, NO (окись азота)-опосредованное расширение сосудов (Chen et al. (1995) Br. J. Pharmacol. 116: 3035-3041). Таким образом, эффекты 1-этаноламида PGF2α сравнивали с эффектом PGF2α при продуцировании эндотелийзависимого расширения сосудов (вазорелаксации) предварительно сокращенного под действием гистамина изолированного сегмента яремной вены кролика. В то время как PGF2α продуцировал заметную вазорелаксацию, 1-этаноламид PGF2α проявлял относительно низкую активность (Фиг.7). Данные также представлены в виде таблицы (Таблица 8). Были получены следующие значения ЭК50 для предварительно сокращенного сегмента яремной вены кролика: 1-этаноламид PGF2α=2000 нМ, PGF2α=5,3.
Поскольку заметная активность наблюдалась для 1-этаноламида РGF2α в отношении препарата сфинктера радужной оболочки кошки, его активность сравнивали с активностью РGF2α в отношении рекомбинантного FP-рецептора кошки. Фиг.8 дает сравнение активности 1-этаноламида PGF2α и PGF2α при стимуляции Са2+-сигнала в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором кошки. Были получены значения ЭК50, равные 14 нМ для РGF2α и 1458 для 1-этаноламида PGF2α, которые указывают на почти стократное различие в активности. Данные также представлены в виде таблицы (Таблица 9).
Эффекты 1-этаноламида РGF2α и 17-фенил-РGF2α на общее накопление инозитфосфата в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором кошки, показаны на Фиг.9. Снова очевидно большое различие в активности. Значения ЭК50 составили 9 нМ для 17-фенил-PGF2α и 524 нМ для 1-этаноламида PGF2α. Данные также представлены в виде таблицы (Таблица 10).
Кроме того, были исследованы эффекты 1-этаноламида PGF2α и 17-фенил-PGF2α на общее накопление инозитфосфата в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором человека. Были получены значения ЭК50, равные 8 нМ для 17-фенил PGF2α и 1003 нМ. Данные представлены на Фиг.10 и в Таблице 11. Результаты были аналогичны результатам, полученным для рекомбинантного FP-рецептора кошки.
Согласно функциональным исследованиям, использующим стабильно трансфицированные FP-рецепторами препараты, 1-этаноламид PGF2α имел относительно умеренную аффинность (химическое сродство) к FP-рецептору. Таким образом, 1-этаноламид PGF2α (известное соединение 1) имел лишь слабое сродство к рецептору кошки, тогда как 17-фенил- PGF2α проявлял высокое сродство (Фиг.11; Таблица 12). 50%-ная ингибирующая концентрация (ИK50) для полученного 1-этаноламида PGF2α составила 882 нМ, по сравнению с величиной ИК50, равной 43,5, для 17-фенил-РGF2α. Аналогичные результаты были получены для рекомбинантного FP-рецептора человека: в случае 1-этаноламида РGF2α ИК50=2015 нМ, 17-фенил- РGF2α-ИK50=2,3 нМ. Эти данные по связыванию меченого лиганда изображены на Фиг.12 и представлены в виде таблицы (Таблица 13).
Глазное действие 1-этаноламида РGF2α изучали на собаках. 1-Этаноламид PGF2α в 0,1%-ной дозе вызывал заметное и очень значительное снижение внутриглазного давления (Фиг.13; Таблица 14). Действие сохранялось в течение всего 24-часового экспериментального периода. 1-Этаноламид PGF2α также вызывал заметное сужение зрачка (миоз) (Фиг.14; Таблица 15). В отличие от глазного гипотензивного действия, уменьшение диаметра зрачка устранялось к 24 ч временной точке. 1-Этаноламид PGF2α вызывал слабую гиперемию глазной поверхности, сравнимую со следовой для наполнителя, в течение всего 24-часового экспериментального периода (Фиг.15; Таблица 16).
В следующих Таблицах, если не указано особо, концентрация исследуемых или контрольных соединений показана в виде логарифмической экспоненты. В соответствии с этим и в качестве примера, "-8" в таблицах означает концентрацию 10-8 М исследуемого или контрольного соединения.
ТАБЛИЦА 1. Влияние возрастающих доз 1-этаноламида PGF2α на изолированную гладкую мышцу сфинктера радужной оболочки кошки. Контрольное сокращение создавали с помощью 10-7 М PGF2α. Даны средние значения ± SEM; n=4.
ТАБЛИЦА 2. Ингибирующее влияние возрастающих доз 1-этаноламида PGF2α на область стимулированного сокращения изолированного сосуда семенного протока морской свинки.
ТАБЛИЦА 3. Влияние возрастающих доз 1-этаноламида PGF2α на изолированную подвздошную кишку цыпленка. Контрольное сокращение создавали с помощью 10-6 М PGE2. Даны средние значения ± SEM; n=4.
ТАБЛИЦА 4. Влияние возрастающих доз 1-этаноламида PGF2α на изолированную подвздошную кишку морской свинки. Контрольное сокращение создавали с помощью 10-6 М PGE2. Даны средние значения ± SEM; n=4.
ТАБЛИЦА 5. Влияние возрастающих доз 1-этаноламида PGF2α на изолированную аорту крысы. Контрольное сокращение создавали с помощью 10-7 М U-46619. Даны средние значения ± SEM; n=4.
ТАБЛИЦА 6. Влияние возрастающих доз 1-этаноламида простагландина F2α на агрегацию тромбоцитов и ингибирование агрегации, индуцированной АДФ. Контрольный ответ вызывают с помощью 2×10-5 М АДФ. Даны средние значения ± SEM; n=4.
ТАБЛИЦА 7. Увеличение внутриклеточной концентрации свободного Ca2+ [Ca2+];, вызванное возрастающими дозами 1-этаноламида простагландина F2α и простагландина F2α в CRL 1497 кожных фибробластов человека.
ТАБЛИЦА 8. Вазорелаксация предварительно сокращенного сегмента яремной вены кролика под действием 1-этаноламида простагландина F2α и простагландина F2α. Контрольное расширение сосудов обеспечивают с помощью 10-7 М PGE2. Даны средние значения ± SEM; n=6 для 1-этаноламида PGF2α, n=7 для РGF2α.
ТАБЛИЦА 9. Увеличение внутриклеточной концентрации свободного Са2+ [Са2+]i, вызванное возрастающими дозами 1-этаноламида простагландина F2α и простагландина F2α в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором кошки. Даны средние значения ± SEM; n=3.
ТАБЛИЦА 10. Общее накопление инозитфосфата, вызванное возрастающими дозами 1-этаноламида простагландина F2α и 17-фенилпростагландина F2α, в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором кошки. Контрольное раздражение создают с помощью 10-6 М 17-фенил-РGF2α. Даны средние значения ± SEM. n=3 для 1-этаноламида РGF2α и 4 для 17-фенил-РGF2α в опытах, выполненных с двумя повторностями.
ТАБЛИЦА 11. Общее накопление инозитфосфата, вызванное возрастающими дозами 1-этаноламида простагландина F2α 17-фенилпростагландина F2α, в клетках НЕК-293, стабильно трансфицированных рекомбинантным FP-рецептором человека. Контрольное раздражение создают с помощью 10-6 М 17-фенил-РGF2α. Даны средние значения ± SEM. n=3 для 1-этаноламида PGF2α и 4 для 17-фенил-PGF2α в опытах, повторенных дважды.
ТАБЛИЦА 12. Изучение конкурентного связывания меченого лиганда (1-этаноламида РGF2α и 17-фенил-РGF2α против 5 нМ 3H-17-фенил-РGF2α) с рекомбинантным FP-рецептором кошки. Даны средние значения ± SEM; n=3 опытам, повторенным дважды.
ТАБЛИЦА 13. Изучение конкурентного связывания меченого лиганда (1-этаноламида PGF2α и 17-фенил- PGF2α против 5 нМ 3H-17-фeнил-PGF2α) с рекомбинантным FP-рецептором человека. Даны средние значения ± SEM; n=3 опытам, выполненным дважды.
1,1**
0,9**
0,9**
0,7**
ТАБЛИЦА 14. Влияние 0,1%-ного раствора 1-этаноламида простагландина F2α на внутриглазное давление собаки. Даны средние значения ± SEM; n=6. **р<0,01 существенное отклонение по сравнению с базовым внутриглазным давлением согласно t-распределению Стьюдента.
ТАБЛИЦА 15. Влияние 0,1%-ного раствора 1-этаноламида простагландина F2α на диаметр зрачка собаки. Даны средние значения ± SEM; n=6. **р<0,1 существенное отклонение по сравнению с базовым диаметром зрачка согласно t-распределению Стьюдента.
ТАБЛИЦА 16. Влияние 0,1%-ного раствора 1-этаноламида простагландина F2α на гиперемию глазной поверхности собаки. Даны средние значения ± SEM; n=6.
ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ, СПОСОБЫ ИСПОЛЬЗОВАНИЯ
Фармацевтические композиции могут быть приготовлены путем комбинирования терапевтически эффективного количества, по меньшей мере, одного соединения согласно настоящему изобретению или его фармацевтически приемлемой соли в качестве активного компонента со стандартными, приемлемыми в офтальмологии, фармацевтическими наполнителями и путем приготовления стандартных лекарственных форм, пригодных для местного глазного применения. Терапевтически эффективное количество обычно составляет от примерно 0,0001 до 5% (вес./об.), предпочтительно, от примерно 0,001 до 1,0% (вес./об.) для жидкой лекарственной формы.
Для офтальмологического применения предпочтительно готовят растворы, используя в качестве основного наполнителя физиологический солевой раствор. Предпочтительней поддерживать рН таких офтальмологических растворов в интервале между 6,5 и 7,2 с помощью подходящей буферной системы, наиболее предпочтительным является нейтральное значение рН. Лекарственные средства могут также содержать стандартные, фармацевтически приемлемые консерванты, стабилизаторы и поверхностно-активные вещества.
Предпочтительные консерванты, которые могут использоваться в фармацевтических композициях настоящего изобретения, включают, но не исчерпываются перечнем, хлорид бензалкония, хлорбутанол, тимеросал (натриевая соль этилртутьтиосалициловой кислоты), ртутьацетофенил и ртутьнитрофенил. Предпочтительным поверхностно-активным веществом является, например, Tween 80 (моноолеат полиоксиэтилен(20)сорбитана). Предпочтительней использовать в офтальмологических препаратах настоящего изобретения также различные наполнители. Эти наполнители включают, но не исчерпываются перечнем, поливиниловый спирт, повидон (поливинилпирролидон), гидроксипропилметилцеллюлозу, полоксамеры, карбоксиметилцеллюлозу, гидроксиэтилцеллюлозу и очищенную воду.
При необходимости или для удобства могут быть добавлены регуляторы тонуса. Они включают, но не исчерпываются перечнем, соли, в частности хлорид натрия, хлорид калия, маннит и глицерин или какой-либо другой подходящий офтальмологически приемлемый регулятор тонуса.
Могут использоваться различные буферы и средства для регуляции рН при условии, что полученный препарат будет офтальмологически приемлемым. Соответственно, буферные композиции включают ацетатные буферы, цитратные буферы, фосфатные и боратные буферы. Для регуляции рН таких лекарственных препаратов, при необходимости, могут быть использованы кислоты или основания.
Аналогично, офтальмологически приемлемый антиоксидант для использования в настоящем изобретении включает, но не исчерпывается перечнем, метабисульфит натрия, тиосульфат натрия, ацетилцистеин, бутилированный гидроксианизол и бутилированный гидрокситолуол.
Другими наполнителями, которые могут быть включены в офтальмологические препараты, являются хелатирующие агенты. Предпочтительным хелатирующим агентом является динатриевая соль этилендиаминтетрауксусной кислоты, хотя другие хелатирующие агенты также могут использоваться вместо нее или в сочетании с ней.
Компоненты обычно используют в следующих количествах:
Компонент, количество (% вес/об.)
активный компонент около 0,001-5
консервант 0-0,10
наполнитель 0-40
регулятор тонуса 0-10
буфер 0,01-10
регулятор рН, количество для подведения рН до
4,5-7,5
антиоксидант, при необходимости
поверхностно-активное вещество, при необходимости
очищенная вода, при необходимости для доведения объема до 100%
Фактическая доза активных соединений настоящего изобретения зависит от конкретного соединения и от условия введения; выбор соответствующей дозы с успехом осуществляется в пределах знаний квалифицированного специалиста.
Офтальмологические лекарственные средства настоящего изобретения удобно упаковывают в формы, подходящие для дозированного применения, например, в контейнеры, снабженные капельницей для облегчения введения в глаз. Контейнеры, подходящие для капельного применения, обычно изготовлены из подходящего инертного нетоксичного пластического материала и, как правило, вмещают от 0,5 до примерно 15 мл раствора.
Растворы, особенно не содержащие консервантов, часто готовят в емкостях для одноразового применения, содержащие примерно до десяти, предпочтительно, примерно до пяти стандартных доз, где обычная стандартная доза содержит от одной до примерно 8 капель, предпочтительно, от одной до примерно 3 капель. Объем одной капли обычно составляет около 20-35 мкл.
ПОДРОБНОЕ ОПИСАНИЕ СПОСОБОВ СИНТЕЗА
Пример 1.
РGF2α (свободная карбоновая кислота)
К перемешиваемому раствору 20 г трометаминовой соли PGF2α (трометамин -трис(гидроксиметил)аминометан) (51,9 ммоль) в 150 мл Н2О прибавляли концентрированную НСl так, что рН раствора доводили до 2. Раствор экстрагировали 3×200 мл хлороформа и концентрировали в вакууме. Свободная карбоновая кислота PGF2α появлялась, главным образом, в виде осадка, наслаивавшегося между водной и органической фазами. Осадок с этого слоя собирали и объединяли с осадком, полученным фильтрацией концентрированного хлороформенного раствора, после чего объединенные осадки промывали 50% смесью хлороформ/метанол.
Метиловый эфир PGF2α; метил-(5Z,8α,9α,11α,13E,15S)-9,11,15-тригидроксипроста-5,13-диен-1-оат
К перемешиваемому раствору свободной карбоновой кислоты PGF2α (2,528 г, 7,13 ммоль) в 50 мл ацетона прибавляли 3,2 мл 1,8-диазабицикло[5.4.0]ундец-7-ена (21,39 ммоль) и 1,33 мл йодистого метила (21,39 ммоль). Раствор перемешивали в течение ночи, разбавляли 150 мл этилацетата, промывали 2×50 мл 0,5 м LiOH, 1×50 мл солевого раствора и концентрировали в вакууме для получения указанного соединения, чистого метилового эфира (чистота проверена с помощью ЯМР 1Н).
1H ЯМР (СDCl3): 5,56-5,34 (м, 4Н), 4,01 (м, 1Н), 3,87 (м, 1H), 3,63 (с, 1Н), 3,18 (т, 2Н, J=5,8 Гц), 2,31-2,02 (м, 8Н), 1,69-1,25 (м, 12Н), 0,849 (т, 3Н, J=6,7 Гц).
1-Этаноламид PGF2α; (5Z,8α,9α,11α,13E,15S)-9,11,15-тригидрокси-N-(2-гидрокси-этил)проста-5,13-диен-1-амид
К перемешиваемому раствору метилового эфира PGF2α (235,4 мг, 0,639 ммоль) в 4,5 мл безводного метанола прибавляли 771 мкл этаноламина (12,78 ммоль). Трубку, содержащую смесь, герметизировали и в течение ночи нагревали при 50°С. Отбирали аликвоту, концентрировали в вакууме при нагревании для удаления избытка этаноламина и анализировали методом ЯМР 1H. Было показано, что реакция завершилась на 80%. Раствор для удаления остаточного этаноламина концентрировали в глубоком вакууме при 65°С в течение 1,5 часов, затем при 60°С в течение ночи с помощью перегонки Kugehlrohr'a. 1H ЯМР и ТСХ в системе 5% метанол/этилацетат показали удаление этаноламина (метиловый эфир: Rf=0,41, 1-этаноламид: Rf=0,04). Технический продукт был подвергнут флэш-хроматографии смесью 5% метанол/этилацетат для извлечения оставшегося непрореагировавшего исходного вещества, метилового эфира, в фракциях 4-8, и 50% смесью метанол/этилацетат для получения требуемого указанного соединения в фракциях 11-18. Дополнительная флэш-хроматография фракций 11-18 смесью 15% метанол/этилацетат дала указанное соединение, не содержащее исходного вещества, но все еще загрязненное эпимером С 15 в некоторых фракциях (фракции 7-10), и требуемый продукт в чистом виде (фракции 11-20). Спектры 1H ЯМР и 13С ЯМР указанного соединения и его С15-эпимера идентичны. Теоретический выход = 254,0 мг, фактически полученный выход 205,6 мг, 81%.
1Н ЯМР (CD3OD): 5,4-5,2 (м, 4Н), 3,98 (м, 1Н), 3,89 (м, 1Н), 3,72 (м, 1Н), 3,47 (т, 2Н, J=5,8 Гц), 3,18 (т, 2Н, J=5,8 Гц), 2,24-1,90 (м, 8Н), 1,50-1,21 (м, 12Н), 0,796 (т, 3Н, J=6,5 Гц).
| название | год | авторы | номер документа |
|---|---|---|---|
| Производные простагландина Fальфа для снижения внутриглазного давления | 2018 |
|
RU2718744C2 |
| ЛИШЕННЫЕ ПОБОЧНЫХ ЭФФЕКТОВ ПРОИЗВОДНЫЕ ПРОСТАГЛАНДИНОВ ДЛЯ ЛЕЧЕНИЯ ГЛАУКОМЫ | 1998 |
|
RU2207858C2 |
| ЗАЩИТНОЕ СРЕДСТВО ДЛЯ НЕЙРОНА СЕТЧАТКИ, СОДЕРЖАЩЕЕ В КАЧЕСТВЕ АКТИВНОГО КОМПОНЕНТА ПРОИЗВОДНОЕ ПРОСТАГЛАНДИНА F2α | 2006 |
|
RU2414904C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ТАФЛУПРОСТА | 2021 |
|
RU2761625C2 |
| ВНУТРИГЛАЗНЫЕ СИСТЕМЫ ДОСТАВКИ ЛЕКАРСТВЕННОГО СРЕДСТВА С ЗАМЕДЛЕННЫМ ВЫСВОБОЖДЕНИЕМ И СПОСОБЫ ЛЕЧЕНИЯ ГЛАЗНЫХ ЗАБОЛЕВАНИЙ | 2010 |
|
RU2532333C2 |
| СПОСОБ ЛЕЧЕНИЯ ПОВЫШЕННОГО ВНУТРИГЛАЗНОГО ДАВЛЕНИЯ С ПОМОЩЬЮ ВНУТРИГЛАЗНОЙ СИСТЕМЫ ДОСТАВКИ ЛЕКАРСТВЕННОГО СРЕДСТВА С ЗАМЕДЛЕННЫМ ВЫСВОБОЖДЕНИЕМ | 2014 |
|
RU2664686C2 |
| ВНУТРИКАМЕРНЫЕ ИМПЛАНТАТЫ С ПРОЛОНГИРОВАННЫМ ВЫСВОБОЖДЕНИЕМ ТЕРАПЕВТИЧЕСКОГО АГЕНТА | 2011 |
|
RU2565445C2 |
| СПОСОБ И КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ДЕГЕНЕРАЦИИ ЖЕЛТОГО ПЯТНА | 2010 |
|
RU2548762C2 |
| МОДУЛЯТОРЫ СВЯЗЫВАНИЯ PAI-1 ДЛЯ ЛЕЧЕНИЯ ГЛАЗНЫХ БОЛЕЗНЕЙ | 2007 |
|
RU2465898C2 |
| СОЕДИНЕНИЕ ГИДРОКСИТРИАЗИНА И ЕГО ПРИМЕНЕНИЕ В МЕДИЦИНСКИХ ЦЕЛЯХ | 2016 |
|
RU2772907C2 |
Изобретение описывает соединение: 1-этаноламид PGF2α, формулы I
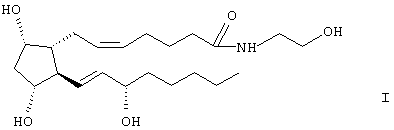
которое представляет собой эффективное соединение для снижения внутриглазного давления у млекопитающего, но не действует через FP-рецептор, через который действуют многие глазные гипотензивные простагландины. 2 н. и 2 з.п. ф-лы, 16 табл., 15 ил.
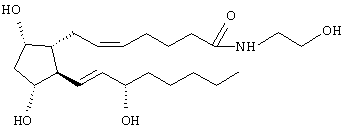
или его фармацевтически приемлемые соли, снижающие внутриглазное давление у млекопитающих, проявляющие глазную гипотензивную активность не через FP-простагландиновый рецептор.
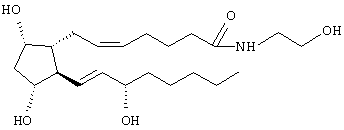
или его фармацевтически приемлемой соли, снижающих внутриглазное давление у млекопитающих, но проявляющих глазную гипотензивную активность не через FP-простагландиновый рецептор, и фармацевтически приемлемый наполнитель.
| WO 9925358 A, 27.05.1999 | |||
| WO 9636599 A, 21.11.1996 | |||
| Многоканальный спектроанализатор | 1987 |
|
SU1439511A2 |

