Настоящее изобретение относится к области медицины, в частности к способу определения тканевого активатора плазминогена (ТАП) в биологических образцах для диагностических целей.
ТАП и урокиназа являются эндогенными активаторами плазминогена. Они превращают плазминоген в плазмин, который растворяет фибрин сгустка. ТАП, обладающий сродством к фибрину, играет ключевую роль в фибринолизе in vivo. Нормальная плазма содержит 4-6 нг/мл ТАП. Уровень ТАП в крови человека понижен при тромботических и повышен при онкологических заболеваниях и кровотечениях. Кроме того, рекомбинантный ТАП вводится внутривенно пациентам при инфаркте миокарда и инсульте для растворения фибрина окклюдированного тромба. Таким образом, точное определение уровня ТАП в крови необходимо для диагноза тромбофилии и гемофилии, а также для контроля активности ТАП во время тромболитической терапии.
ТАП секретируется клетками эндотелия в циркуляцию и быстро удаляется печенью (τ 1/2=8 мин). В крови содержатся специфический ингибитор активаторов плазминогена ПАИ-1, α 2-антиплазмин и С1-ингибитор, которые образуют с ТАП неактивные комплексы ТАП/ингибитор. Типичный базальный уровень активного ТАП в крови равен 1 нг/мл (0.5 МЕ/мл) и неактивного комплекса ТАП/ингибитор - 5 нг/мл [Wiman В. et all, J. Lab. Clin. Med. 105, 265-270, 1985]. Обычно ТАП не определяется в цельной крови, большая часть активности ТАП найдена в плазме крови. Для предотвращения взаимодействия ТАП с плазменными ингибиторами пробу крови подкисляют до рН 4.0 и выделяют плазму [Chmielewska et all, Thromb. Res., 31, 427-436, 1983]. Эуглобулиновая фракция стабилизированной цитратом плазмы содержит активность ТАП и не содержит ингибиторов.
Существуют три различных способа определения ТАП: иммуноферментный метод, прямой и непрямой функциональные методы. Иммуноферментный метод [Bergsdorf N., Nilsson Т., Wallen P. Thromb. Haemost. 50, 740-744 (1993)] позволяет определять вплоть до 1-2 нг/мл антигена ТАП, но он не дает информации об активности фермента. К недостаткам этого метода относятся также малая доступность и высокая стоимость иммуноферментных реагентов. В прямом функциональном методе синтетический хромогенный или флуорогенный специфический субстрат ТАП инкубируют с образцом и измеряют скорость образования продукта гидролиза субстрата под действием ТАП [USA Patent N 4 278 762, (1981)]. Однако этот метод недостаточно чувствителен. В непрямом функциональном методе плазминоген активируют под действием ТАП в присутствии субстрата плазмина и активность ТАП определяют по гидролизу субстрата образующимся плазмином. К непрямым функциональным методам, в которых используют природный субстрат плазмина фибрин, относятся методы, основанные на измерении площади зоны лизиса фибриновой пластины [Astrup Т., Mullerts S. Arch. Biochem. Biophys. 40, 346-351 (1952)] или времени полного лизиса фибринового сгустка [Christensen L.R. Proc. Soc. Biol. Med. 46, 674-679 (1941)]. Недостатками этих методов являются неясность кинетики протекания реакций, вовлеченных в процесс растворения твердофазного субстрата, и трудность интерпретации полученных результатов.
Существенные недостатки трех вышеописанных способов исправляются наиболее близким по техническому решению способом (прототипом), описанным Verheijen J.P. et all [USA Patent №4563420 (1986)]. В этом способе образец, в котором измеряют активность ТАП, инкубируют с Лиз-плазминогеном и пептидным субстратом плазмина Н-D-Val-Leu-Lys-pNA (S-2251) в присутствии растворимых BrCN-фрагментов фибриногена в качестве стимулятора активации плазминогена ТАП. Плазмин (Пл), образующийся при активации плазминогена (Пг) ТАП, катализирует гидролиз субстрата (С), при этом образуется окрашенный продукт п-нитроанилин (НА), который измеряют спектрофотометрически при 405 им. Схему реакции можно представить уравнениями (1 и 2):
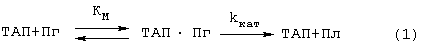
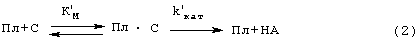
Преимуществами этого способа являются: 1) возможность количественного описания сопряженного процесса в растворе (кинетические параметры Км, kКАТ, К’М и k’КАТ могут быть определены из кинетики отдельных реакций) и 2) высокая чувствительность метода, так как фрагменты фибриногена повышают каталитическую эффективность (kКАТ/КМ) активации плазминогена ТАП на 2-3 порядка благодаря резкому снижению Км и некоторому увеличению kКАТ.
Недостатками способа являются использование: 1) высокотоксичного цианистого брома, необходимого для получения фрагментов фибриногена из фибриногена; 2) коммерческого дорогостоящего субстрата плазмина H-D-Val-Leu-Lys-pNA (S-2251), для гидролиза которого плазмином характерно высокое значение константы Михаэлиса (К’M=0.6 мМ) и недостаточно высокая каталитическая эффективность (kКАТ/KM=42 мМ-1с-1) [Kiss I., Biochem. Biophys. Res. Comm. 131, 928-934 (1985)]) и измерение активности ТАП проводится не при [С]0>>К’M (когда скорость максимальна), а при [C]0 ≈K’M, что снижает чувствительность метода и 3) Лиз-плазминогена, обычно содержащего примесь плазмина, который приводит к значительному фоновому гидролизу субстрата и повышает погрешность определения низких концентраций ТАП.
Задачей предлагаемого способа определения ТАП является повышение чувствительности и точности детекции ТАП за счет использования каталитически более эффективного субстрата плазмина и снижения примеси плазмина в препарате плазминогена, а также упрощение способа путем использования стимулятора активации плазминогена ТАП, полученного без применения цианистого брома.
Для решения поставленной задачи предлагается способ определения ТАП, который заключается в том, что образец ТАП инкубируют с раствором плазминогена в присутствии хромогенного специфического субстрата плазмина и стимулятора активации плазминогена ТАП с последующим измерением скорости накопления продукта гидролиза субстрата образующимся плазмином. Согласно изобретению в качестве субстрата ТАП используют Глу-плазминоген, практически не содержащий примеси плазмина, или Лиз-плазминоген желательно после ингибирования в нем примеси плазмина, в качестве субстрата плазмина используют новый отечественный специфический субстрат HCO-Ala-Phe-Lys-p-mtroanilide (APL-pNA) [Патент N 98100177/14 (1998), Россия] и в качестве стимулятора активации плазминогена ТАП используют растворимый Дез-АА-фибрин-мономер, полученный с помощью отечественного тромбиноподобного фермента анцистрона из яда змеи Agkistrodon halys.
Глу-плазминоген выделяют из свежезамороженной плазмы крови человека методом аффинной хроматографии на Lys-Sepharose 4B в присутствии панкреатического ингибитора трипсина (для исключения образования плазмина) по методике, описанной в работе [Castellino F.J et al., Methods Enzymol., 80, 365-378 (1981)]. Препарат электрофоретически гомогенен и практически не содержит примеси плазмина.
Лиз-плазминоген выделяют из плазмы человека аффинной хроматографией на Lys-sepharose 4B в отсутствии панкреатического ингибитора трипсина по методу, описанному в работе [Deutsch D.G. et al., Science, 170, 1095-1096 (1970)]. Полученный препарат Лиз-плазминогена, в котором обычно содержится 0.5-5% примеси активного плазмина, обрабатывают избытком панкреатического ингибитора и очищают путем ультрафильтрации.
Для выделения эуглобулиновой фракции образец плазмы (100 мкл), содержащей ТАП, разбавляют в 0.9 мл дистиллированной воды, затем раствор подкисляют добавлением 100 мкл 0.25% уксусной кислоты и инкубируют в течение 1 ч при 4° С. После центрифугирования в течение 5 минут при 2000 об/мин надосадочную жидкость декантируют. Полученный осадок растворяют в 100 мкл исследуемого буфера.
Для измерения большого числа образцов в рутинных экспериментах реакцию проводят в 96-луночных стриповых планшетах с использованием мультиканального спектрофотометра.
Реакцию проводят при 25° С или 37° С, но скорость реакции при 25° С в 3 раза ниже. Инкубацию реакционных растворов при 37° С проводят в закрытых ячейках во избежание испарения.
Реакцию проводят при рН 7.0-8.3 в присутствии 0.1% Тритона Х-100 для элиминирования сорбции ТАП к стенкам сосуда.
При насыщающих условиях (когда [Пг]0>>КM и [С]0>>К’M) и отсутствии спонтанного гидролиза субстрата образование продукта п-нитроанилина в ходе реакций (1 и 2), измеряемое спектрофотометрически при 405 нм, описывается уравнением
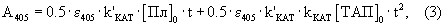
где ε 405 - коэффициент экстинкции п-нитроанилина.
Если плазминоген не содержит примеси плазмина ([Пл]0=0), то уравнение (3) преобразуется в уравнение
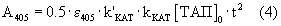
и скорость реакции пропорциональна концентрации ТАП.
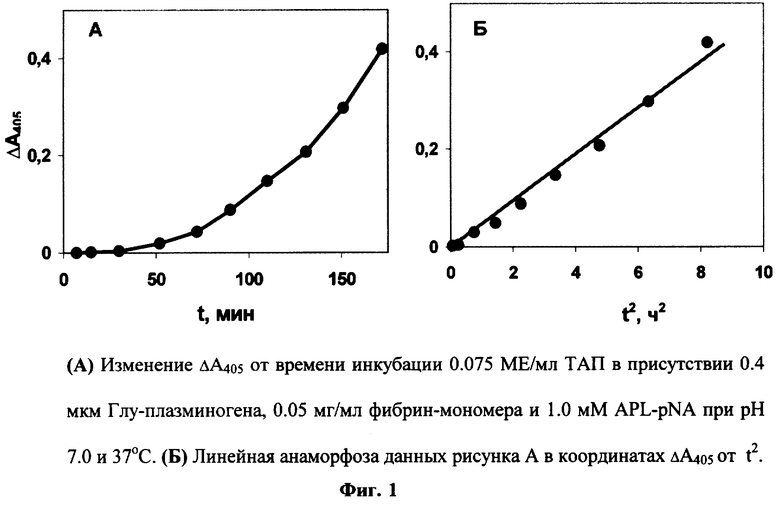
Пример 1
ТАП (0.075 МЕ/мл) инкубируют при 37° С в 0.2 мл Трис-ацетатного буфера (0.1 М, рН 7.0), содержащего 0.4 мкМ Глу-плазминогена, 0.05 мг/мл фибрин-мономера, 0.1% Тритона Х-100 и 1 мМ хромогенного субстрата плазмина - APL-pNA, в ячейке 96-луночной стриповой планшеты. Контрольная ячейка, не содержащая ТАП, служит для измерения фонового гидролиза. В ходе инкубации в течение 3 часов измеряют оптическую плотность образца (A405) и контроля (А405°) на планшетном фотометре Antos 2020. Скорость реакции прямо пропорциональна концентрации ТАП в образце, которую определяют либо из значения Δ А405(A405-А405°) через 2 часа реакции, либо из значения тангенса угла наклона зависимости Δ A405 от t2. Как видно, в присутствии ТАП зависимость Δ А405 от t имеет параболический вид (Фиг.1,А), а от t2 - линейный вид (Фиг.1,Б).
Пример 2
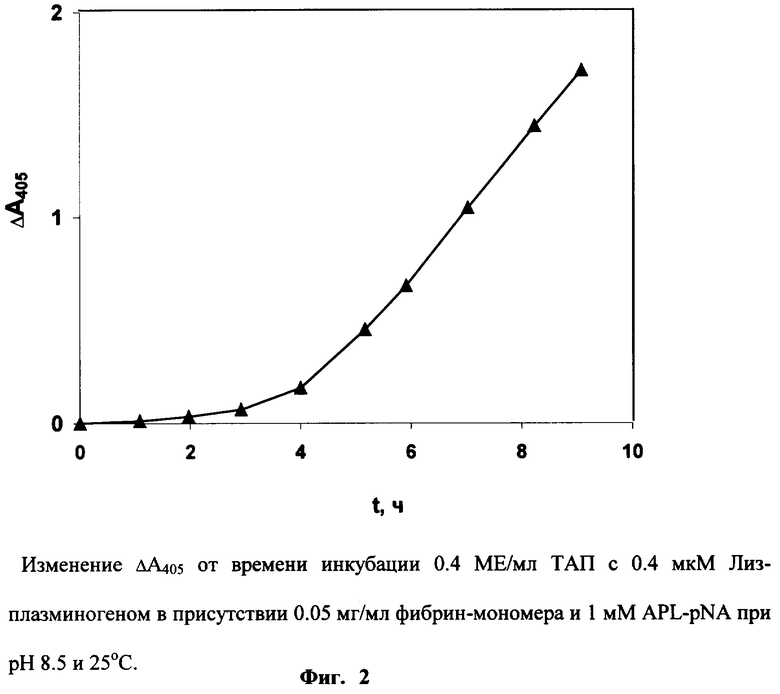
ТАП (0.4 МЕ/мл) инкубируют при 25° С в 0.2 мл Трис-ацетатного буфера (0.1 М, рН 8.5), содержащего 0.4 мкМ Лиз-плазминогена, 0.05 мг/мл фибрин-мономера, 0.1% Тритона Х-100 и 1 мМ APL-pNA. Изменение оптической плотности образца и контроля (без ТАП) регистрируют в течение 9 часов. Активность ТАП определяют по величине Δ А405, наблюдаемой через 7 часов (Фиг.2).
Пример 3
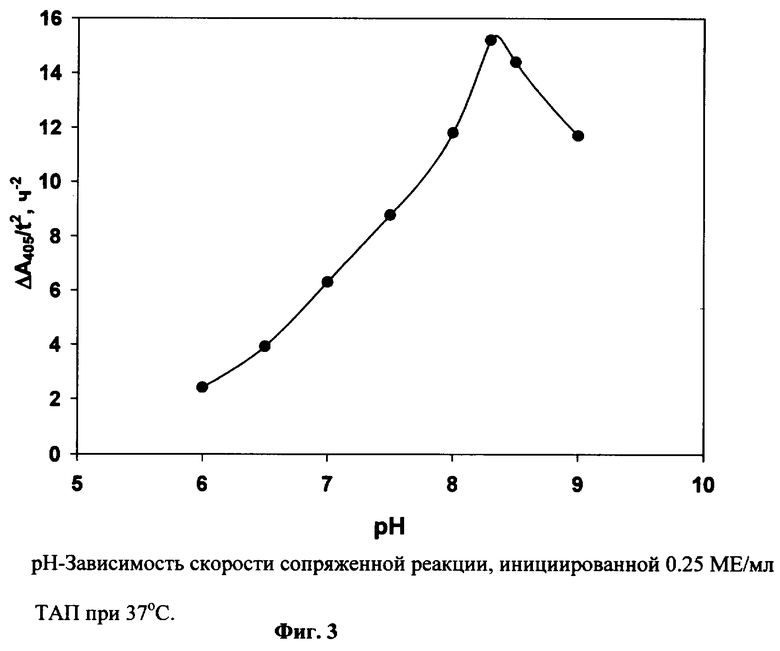
ТАП (0.25 МЕ/мл) инкубируют при 37° С в 0.2 мл 0.1 М Трис-ацетатного буфера (0.1% тритона Х-100) с различными значениями рН. Концентрации Глу-плазминогена, фибрин-мономера и APL-pNA такие же, как в примере 1. При каждом значении рН измеряют фоновый гидролиз субстрата в отсутствие ТАП (А405°). Фиг.3 показывает зависимость Δ A405/t2 от рН. Оптимальное значение рН сопряженной реакции 8.3.
Пример 4
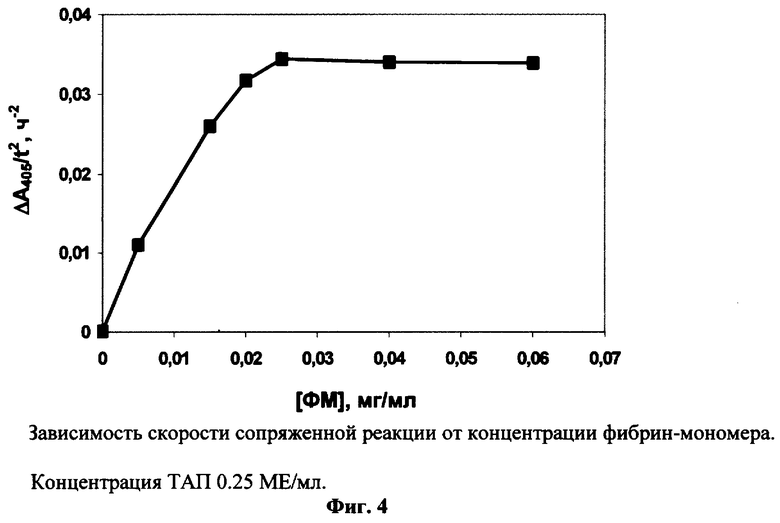
ТАЛ (0.25 МЕ/мл) инкубируют при 37° С в 0.2 мл 0.1 М Трис-ацетатного буфера рН 8.3, содержащего 0.4 мкМ Глу-плазминогена, 0.1% Тритона Х-100, 1 мМ APL-pNA и разные концентрации фибрин-мономера. Для каждой концентрации стимулятора измеряют фоновый гидролиз субстрата в отсутствие ТАП. Из зависимости Δ A405/t2 от концентрации фибрин-мономера видно, что плато достигается при концентрации стимулятора 0.025 мг/мл (Фиг.4).
Пример 5
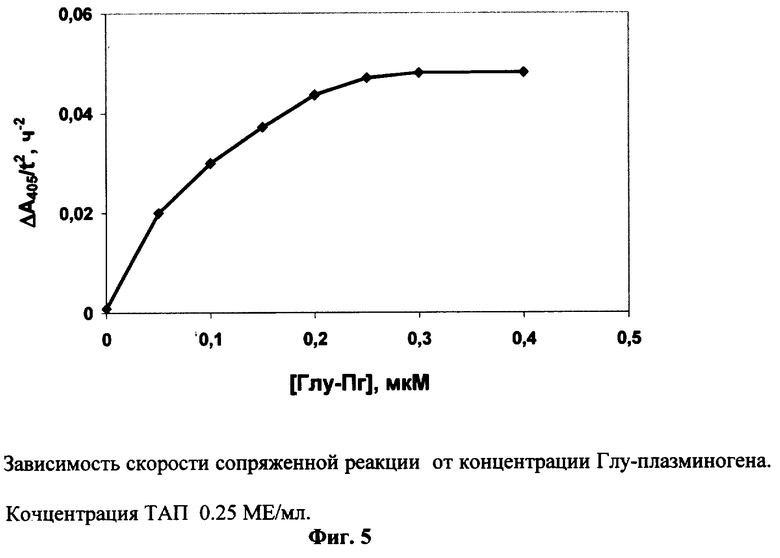
Эксперименты по примеру 4 проводят в присутствии 0.025 мг/мл фибрин-мономера и различных концентраций Глу-плазминогена. Для каждой концентрации Глу-плазминогена измеряют фоновый гидролиз субстрата в отсутствие ТАП. Скорость реакции (Δ A405/t2) достигает максимума при концентрации Глу-плазминогена 0.3 мкМ (Фиг.5).
Пример 6
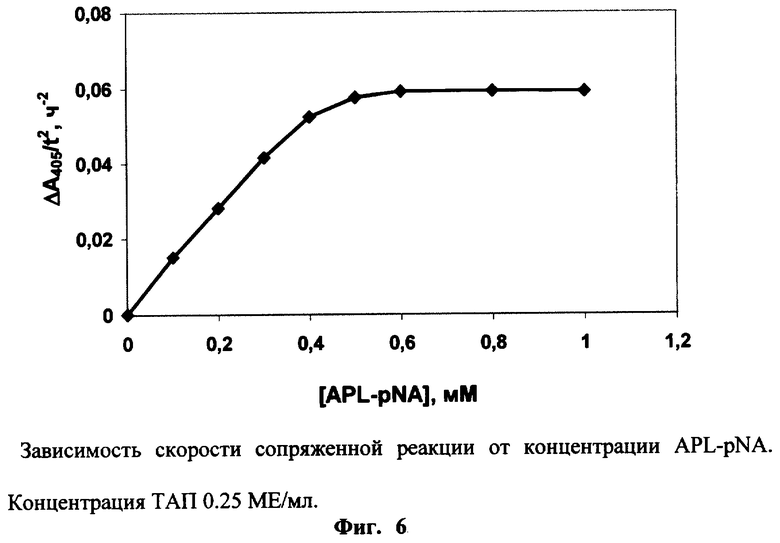
Эксперименты по примеру 4 проводят в присутствии 0.3 мкМ Глу-плазминогена, 0.025 мг/мл фибрин-мономера и различных концентраций хромогенного субстрата плазмина - APL-pNA. Измеряют фоновый гидролиз для каждой концентрации субстрата в отсутствие ТАП. Зависимость Δ A405/t2 от концентрации субстрата показывает, что плато достигается при 0.6 мМ APL-pNA (Фиг.6).
Пример 7
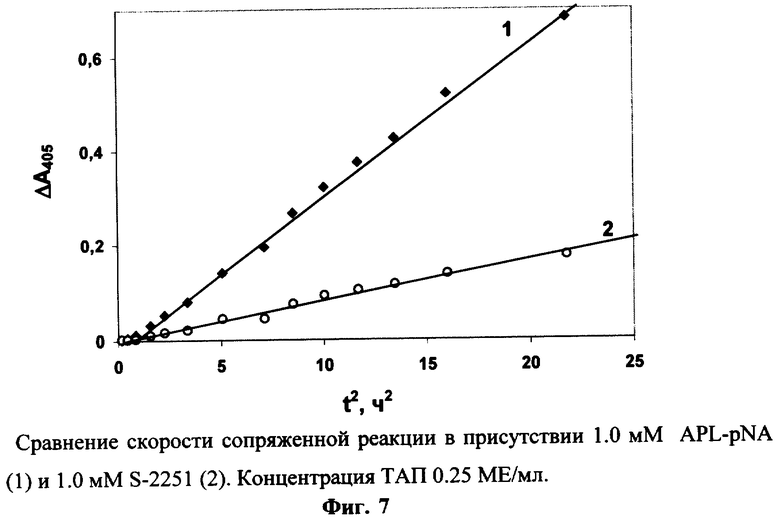
0.25 МЕ/мл ТАП инкубируют в присутствии 0.3 мкМ Глу-плазминогена, 0.025 мг/мл фибрин-мономера и 1 мМ APL-pNA или S-2251 (″ Sigma″ , США) при рН 8.3 и 37° С. Для каждого субстрата измеряют фоновый гидролиз в отсутствие ТАП. Фиг.7 показывает, что скорость сопряженной реакции под действием ТАЛ значительно выше в присутствии APL-pNA, используемого в предлагаемом способе, чем в присутствии S-2251, используемого в прототипе. Это связано с более высокой каталитической эффективностью гидролиза плазмином APL-pNA (kКАТ/Км=82 мМ-1с-1) в сравнении с гидролизом S-2251 (kКАТ/Км=42 мМ-1с-1 [Kiss I., Biochem. Biophys. Res. Comm. 131, 928-934 (1985)]). Кроме того, при используемой концентрации субстратов гидролиз плазмином APL-pNA (К’M=0.13 мМ) протекает в условиях, когда [С]0>>К’M и скорость гидролиза максимальна, в то время как S-2251 (К’M=0.6 мМ, [Kiss I., Biochem. Biophys. Res. Comm. 131, 928-034 (1985)]) гидролизуется плазмином в условиях [С]0 ≈К’M.
Пример 8
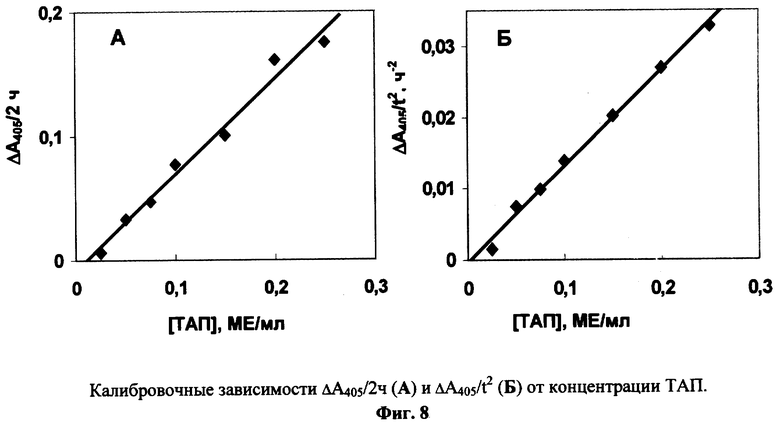
Эксперименты по примеру 4 проводят в присутствии 0.3 мкМ Глу-плазминогена, 0.025 мг/мл фибрин-мономера и различных концентраций стандартного рекомбинантного ТАЛ (3-й Международный стандарт N 98/714, National Institute of Biological Standards and Control, NIBSC, Великобритания). В контрольной ячейке ТАП отсутствует. Каждый тест повторяют трижды. Из кинетических кривых изменения оптической плотности от времени строят калибровочный график: либо зависимость Δ A405/2ч (изменение поглощения за 2 часа реакции) от концентрации ТАП (Фиг.8, А), либо зависимость Δ A405/t2 от концентрации ТАП (Фиг.8, Б). Эти линейные графики используют для определения ТАП в испытуемых образцах.
Пример 9
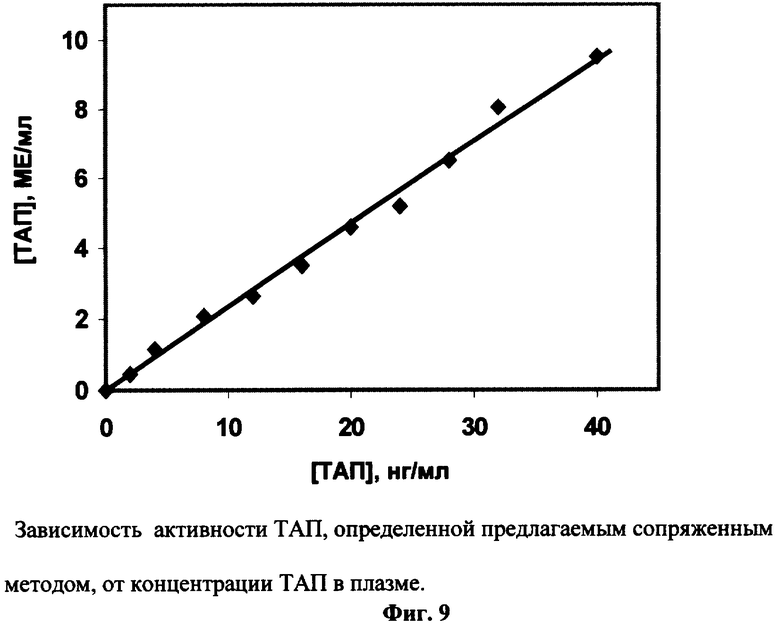
В стандартную плазму добавляют различные концентрации стандартного ТАП до конечной концентрации 1-40 нг/мл. Из образцов плазмы (100 мкл) с различным содержанием ТАП выделяют осадки эуглобулиновой фракции плазмы, как описано выше, и растворяют их в 100 мкл исследуемого буфера. В экспериментах по примеру 8 в качестве исследуемого образца используют предварительно разбавленные в 20 раз растворы эуглобулиновой фракции плазмы с различным содержанием ТАЛ. Активность ТАП в образцах определяют по величине Δ A405/t2, используя калибровочный график (Фиг.8, Б). Фиг.9 иллюстрирует линейную зависимость активности ТАП, измеренной по заявляемому способу, от концентрации ТАП в плазме человека.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РАСТВОРИМОГО ФИБРИН-МОНОМЕРА | 2003 |
|
RU2253474C1 |
| ПРОЛОНГИРОВАННОЕ ТРОМБОЛИТИЧЕСКОЕ СРЕДСТВО И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 1995 |
|
RU2121362C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ СОДЕРЖАНИЯ ПЛАЗМИНОГЕНА И ИНГИБИТОРОВ ПЛАЗМИНА В ПЛАЗМЕ КРОВИ | 1998 |
|
RU2121690C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВАЦИИ ПЛАЗМИНОГЕНА БАКТЕРИЯМИ В УСЛОВИЯХ IN VITRO | 2012 |
|
RU2514662C2 |
| ДИМЕР МОЛЕКУЛЯРНОГО ВАРИАНТА АПО-ЛИПОПРОТЕИНА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ | 1992 |
|
RU2134696C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ БИОЛОГИЧЕСКОЙ АКТИВНОСТИ ДЕФИБРОТИДА, ОСНОВАННЫЙ НА ПРИМЕНЕНИИ ЭУГЛОБУЛИНА | 2012 |
|
RU2627177C2 |
| ВАРИАНТЫ ПЛАЗМИНОГЕНА И ПЛАЗМИНА | 2010 |
|
RU2564131C2 |
| ВАРИАНТЫ ПЛАЗМИНОГЕНА И ПЛАЗМИНА | 2012 |
|
RU2604810C2 |
| ВАРИАНТЫ ПЛАЗМИНОГЕНА И ПЛАЗМИНА | 2012 |
|
RU2604807C2 |
| РЕКОМБИНАНТНЫЙ ПОЛИПЕПТИД СО СВОЙСТВАМИ ПЛАЗМИНОГЕНА ЧЕЛОВЕКА ПРЕВРАЩАТЬСЯ ПРИ АКТИВАЦИИ В ПЛАЗМИН, КОТОРЫЙ КАТАЛИЗИРУЕТ РАСЩЕПЛЕНИЕ ФИБРИНА, ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ПОЛИПЕПТИД, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК ДЛЯ ЭКСПРЕССИИ ПОЛИПЕПТИДА И ТРАНСФОРМИРОВАННАЯ КЛЕТКА Escherichia coli - ПРОДУЦЕНТ ПОЛИПЕПТИДА | 2009 |
|
RU2432396C2 |
Изобретение относится к медицине, а именно к диагностическим методам. Способ определения тканевого активатора плазминогена заключается в инкубации образца с плазминогеном, стимулятором превращения плазминогена в плазмин под действием тканевого активатора плазминогена и хромогенным синтетическим субстратом плазмина и определении активности тканевого активатора плазминогена по концентрации окрашенного продукта гидролиза субстрата плазмином. В качестве плазминогена используют Лиз-плазминоген, предварительно обработанный панкреатическим ингибитором трипсина, или Глу-плазминоген. В качестве субстрата плазмина используют специфический субстрат HCO-Ala-Phe-Lys-p-nitroanilide. В качестве стимулятора превращения плазминогена в плазмин под действием тканевого активатора плазминогена используют растворимый фибрин-мономер, полученный с помощью тромбино-подобного фермента анцистрона из яда змеи Agkistrodon halys. Инкубацию проводят при рН 7,0-8,5. Наиболее оптимальной концентрацией для Лиз- или Глу-плазминогена является
0,3 мкМ, для специфического субстрата HCO-Ala-Phe-Lys-p-nitroanilide – 0,6 мМ, для фибрин-мономера – 0,025 мг/мл. Способ обладает высокой чувствительностью и точностью. 9 ил.
| US 4563420 A1, 07.01.1986 | |||
| US 6297023 A1, 02.10.2001 | |||
| СПОСОБ ОПРЕДЕЛЕНИЯ СОДЕРЖАНИЯ ПЛАЗМИНОГЕНА И ИНГИБИТОРОВ ПЛАЗМИНА В ПЛАЗМЕ КРОВИ | 1998 |
|
RU2121690C1 |
| RANBY M | |||
| et al | |||
| In: Progress in Fibrinolysys | |||
| Vol.V, Editors: J.F.Davidson et al., Edinburg, 1981 | |||
| Способ определения агрегационной активности тромбоцитов | 1989 |
|
SU1691749A1 |
| СПОСОБ ПОЛУЧЕНИЯ АКТИВАТОРА ПЛАЗМИНОГЕНА ТКАНЕВОГО ТИПА, ШТАММ КУЛЬТИВИРУЕМЫХ КЛЕТОК ЖИВОТНЫХ СНО-ПРОДУЦЕНТ АКТИВАТОРА ПЛАЗМИНОГЕНА ТКАНЕВОГО ТИПА | 1987 |
|
RU2046827C1 |
| US 5472851 A1, 05.12.1995 | |||
| US 5175087 A1, 29.12.1992 | |||
| US 5096811 A1, 17.03.1992 | |||
| Гидравлический распределитель | 1959 |
|
SU142488A1 |

