Изобретение относится к способу получения человеческого активатора плазминогена тканевого типа, соответствующему тому, что находят в человеческой сыворотке и/или тканях, в терапевтически достаточных количествах.
Предпосылками изобретения являлось то, что были определены последовательность ДНК и укороченная аминокислотная последовательность человеческого активатора плазминогена. Это позволило получить человеческий активатор плазминогена с применением технологии рекомбинантной ДНК, что в свою очередь обеспечило возможность получения качественного материала в достаточном количестве и дало начало проведению испытаний на животных и клинических испытаний как предпосылку выпуска в продажу данного препарата.
Фибринолитическая система находится в динамическом равновесии с системой коагуляции, поддерживая интактный, открытый вазикулярный слой. Система коагуляции откладывает фибрин как матрицу, служащую для восстановления гемостатического условия. Фибринолитическая система удаляет фибриновую сетку после достижения гемостатического условия. Фибринолитический процесс вызывается протеолитическим ферментом плазмином, который генерирует из плазмы протеиновый предшественник плазминогена. Плазминоген превращается в плазмин при активации активатором.
В настоящее время коммерчески доступны два активатора: стрептокиназа и урокиназа. Оба используются для лечения острых сосудистых заболеваний, таких как инфаркт миокарда, паралич, легочная эмболия, глубокий тромбоз вен, периферическая артериальная окклюзия и другие тромбозы вен.
Этиологической основой этих заболеваний, как отмечено в частных или тяжелых случаях, является полная закупорка кровеносного сосуда сгустком крови тромбоз или тромбоэмболия. Традиционная антикоагулянтная терапия, например гепарином или кумарином, не приводит к увеличению растворимости тромба или тромбоэмболии. Тромболитические агенты, упоминавшиеся ранее, стрептокиназа и урокиназа, могут эффективно использоваться на практике. Однако имеются жесткие ограничения. Так как у них отсутствует высокое сродство к фибрину, то и активированный циркулирующий и связанный с фибрином плазминоген относительно неразличимы. Плазмин, образовавшийся в токе крови, довольно быстро нейтрализуется и не используется в тромболизе. Остаточный плазмин будет разлагать некоторые факторы свертывания протеинов, например фибриноген, фактор V и фактор VII, вызывая геморрагический потенциал. В дополнение к этому стрептокиназа является в значительной степени антигенной и пациенты с высоким содержанием антител плохо поддаются лечению и не могут подвергаться ему длительное время. Лечение урокиназой является дорогостоящим, потому что ее выделяют из человеческой мочи или культур ткани, поэтому ее обычно не применяют в клинической практике.
Так называемые активаторы плазминогена были выделены из различных тканей человека, например из маточной ткани, крови, сыворотки и из клеточной культуры. Активаторы плазминогена, происходящие из этих источников, были классифицированы на две основные группы: активаторы плазминогена типа урокиназы (у-АП) и активаторы плазминогена тканевого типа (т-АП), основываясь на различиях в их иммунологических свойствах. (Сокращение т-АП и у-АП были предложены на XXXIII Съезде Международного Комитета по тромбозу и гемостазу, Бергамо, Италия, 27 июля 1982 г.).
Недавно было установлено, что линия человеческой меланомы секретирует т-АП. Исследование этого меланомного активатора плазминогена показало, что он не отличим как иммунологически, так и по аминокислотному составу от активатора плазминогена, выделенного из обычной человеческой ткани.
Продукт был выделен в относительно чистом виде и охарактеризован. Было найдено, что он является высоко активным фибринолинотическим агентом. Несколько исследований, в которых был использован т-АП, выделенный от линии клеток меланомы, показали его высокое сродство к фибрину по сравнению с активаторами плазминогена урокиназного типа. Однако более интенсивное исследование человеческого т-АП в качестве потенциального тромболитического агента было затруднено его очень низким уровнем концентрации в крови, экстрактах тканей, перфузоре и в культурах клеток.
Понятно, что применение технологии рекомбинантной ДНК и связанных с ней технологий позволит получить большие количества высококачественного человеческого активатора плазминогена тканевого типа (ранее упоминавшегося как активатор человеческого плазминогена), свободного от других человеческих протеинов.
Молекулярные биологи способны с достаточной легкостью проводить рекомбинацию различных последовательностей ДНК, создавая по существу новые ДНК, способные продуцировать большие количества экзогенного протеинового продукта в трансформированных микробах и культурах клеток. Основные средства и методы работы заключаются в связывании in vitro различных тупоконечных или липких концевых фрагментов ДНК, получая мощные векторы экспрессии, используемые для трансформации конкретных организмов, чем обеспечивается эффективный синтез желаемого экзогенного продукта.
Для создания векторов экспрессии осуществляют рекомбинацию ДНК, а именно объединяют репликации, фрагмент, кодирующий генетический маркер, промотор экспрессии, гетерологический ген и вектор вне клетки-хозяина. Полученный рекомбинантный реплицирующийся вектор экспрессии или плазмиду вводят в клетки трансформацией и получают большие количества рекомбинантного вектора при росте трансформанта. Когда ген правильно вставлен относительно участков, которые управляют транскрипцией и трансляцией данного гена, полученный в результате вектор экспрессии полезен для нарабатывания полипептидной последовательности, которую кодирует встроенный ген. Целевой продукт можно получать лизированием, если необходимо, клеток реципиента в микробной системе и извлечением продукта при соответствующей очистке от других протеинов.
На практике при использовании технологии рекомбинантной ДНК можно экспрессировать гетерологичные полипептиды, так называемая прямая экспрессия, или можно экспрессировать гетерологичный полипептид, сплавленный с частью аминокислотной последовательности гомологичного полипептида.
Хорошо изучено культивирование клеток и тканей для изучения генетики и физиологии клеток с помощью средств и методов. Для этого сохраняют постоянные клеточные линии. Для использования в исследовании такую линию клеток выдерживают на твердом носителе в жидкой среде или выращивают в суспензии, содержащей источник питательных веществ.
Клетки, продуцирующие целевой протеин, также продуцируют сотни других протеинов, эндогенных продуктов клеточного метаболизма. Эти загрязняющие протеины, также как и другие соединения, если их не удалить из целевого протеина, могут оказаться токсичными при введении животным или людям во время терапевтического лечения. Следовательно, разрабатываются способы выделения и очистки целевого продукта, чем обеспечивается гомогенный продукт. Доказывается идентичность целевого продукта.
Ближайшим аналогом предлагаемых способа и штамма является способ получения активатора плазминогена тканевого типа, предусматривающий конструирование рекомбинантной плазмидной ДНК, кодирующей активатор плазминогена тканевого типа, трансформацию полученными ДНК штаммов, реципиентов, выращивание трансформантов, выделение и очистку целевого продукта [1]
Успешно использована технология рекомбинантной ДНК для получения активатора плазминогена человеческой ткани (т-АП) в количествах, достаточных для начала и проведения испытаний на животных и клинических испытаний как предпосылки для выпуска его в продажу. Продукт человеческой т-АП пригоден для использования во всех его формах для профилактического или терапевтического лечения людей с различными сердечно-сосудистыми расстройствами и заболеваниями.
Изобретение также относится к векторам экспрессии, содержащим фрагменты ДНК, кодирующие человеческий активатор плазминогена ткани и штаммам микроорганизмов или клеточных культур, трансформированных ими, способным продуцировать активатор плазминогена человеческой ткани.
Описание признаков штамма. АТСС GCL 60STRC (Роговица, кролик, Prycytolagus Cuniculus).
Общепринятая среда для размножения: минимальная поддерживающая среда Игла (МЕМ) с несущественными аминокислотами и сбалансированный солевой раствор Эрла (BSS) 90% плодовая коровья сыворотка 10%
Клеточная линия SIRC была получена М. Фолькертом (Государственный институт сыворотки, Дания, Копенгаген) из роговицы нормального кролика в 1957 г. Восприимчива к вирусу рубеллы. Пригодна для первичной изоляции вируса рубеллы. Ранее появление отчетливых цитопатических изменений делает эту клеточную линию особенно пригодной как для размножения, так и квантизации вируса рубеллы. Справочный семенной запас был заморожен в феврале 1966 г.
Описание депонированного справочного семенного запаса
Количество последовательных субкультур из исходной ткани: приблизительно 400 в лаборатории создателя, 19 в лаборатории представляющей организации, 11 в лаборатории, дающей отзыв.
Среда замораживания: культурная среда 90% диметилсульфоксид (DMSO) 10% без антибиотиков.
Жизнеспособность: приблизительно 85% (исключение красителя).
Культурная среда: сбалансированный солевой раствор Эрла (BSS) с уменьшенным количеством бикарбоната (0,84 г/л) плюс 10% инактивированной телячьей сыворотки; гидролизированный лактаальбумин 1,7 г/л; и дрожжевой экстракт, Дифко, 0,57 г/л; без антибиотиков.
Ростовая характеристика размороженных клеток: инокулят 4,0 ˙105жизнеспособных клеток на трехунцевый аптечный пузырек дает 2,5˙ 104клеток за 7 дней, когда среду меняют на 3-й день и рН поддерживают при 7,3.
Эффективность посева: менее чем 1% в вышеуказанной культуральной среде.
Морфология: фибробластоподобная.
Кариология: Хромосомное частотное распределение 50 клеток: 2 n 44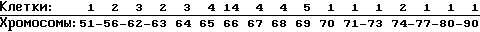
Модальное число (66) не позволяет определить пол донора цитологически. Большинство клеток содержит одну длинную неспаренную, субметацентрическую хромосому и 2-3 малых телоцентрических хромосомы с сателлитами. Полиплоидность в 5/200 метафазных клетках.
Стерильность: тесты на микоплазму, бактерии и грибки были отрицательны.
Вид: кролик.
Восприимчивость к вирусу: невосприимчив к полиовирусу 2, к вирусу Коксаки А-9 и В-5. Репликация вируса рубеллы с сопровождающими цитопатическими изменениями подтверждена Д.Джей. Мельником, Бейлорский университет, Хьюстон, Техас и Др.Джей. Биттлом, Питмэн-Мур Ко. Индианополис, Индиана.
Обратная траскриптаза: не обнаружены.
Представлена: Джей. Лирхой, Энтеровирусный факультет, Государственный институт сыворотки, Копенгаген, Дания.
Подготовлена и охарактеризована: Калифорнийский университет, Лаборатория морской биологии, Беркли, Калифорния и Американская коллекция типовых культур, Роквилл, Мэриленд.
АТСС CCL 61 CHO-KI (яичник, китайский хомячок), Cricetulus griseus) требующий пролина.
Общепринятая среда для размножения: среда F-12 Гэма 90% плодовая коровья сыворотка 10%
Клетки СНО-KI были получены в качестве субклона из родительской СНО-клеточной линии, начало было положено биопсией яичника взрослого китайского хомячка.
Оригинальная культуральная среда состояла из среды N 16 с физиологическим раствором 81% NCTC 109,4% плодовая коровья сыворотка 15% Из-за субклонов культуральную среду заменили на среду F-12 Гэма плюс плодовая коровья сыворотка. Клетки СНО-KI требуют пролина и имеют модальное хромосомное число 20. Очевидно, в клетках отсутствует активная форма гена, необходимая для синтеза пролина, и блок биосинтетической цепи лежит в стадии, превращающей глутаминовую кислоту в глутаминовый гамма-полуальдегид. Частота реверсии независимо от пролина равна приблизительно 10-6.
Используются для индукции и изоляции мутантов пищевой недостаточности и в исследованиях, относящихся к экспрессии гена посредством гибридизации. Клеточная линия была предоставлена в Американскую коллекцию типовых культур (АТСС) в июне 1970 г. после проведения субкультиваций от исходной ткани.
Описание депонированного справочного семенного запаса
Количество последовательных субкультур их исходной ткани: приблизительно 400; 7 в АТСС.
Среда замораживания: культуральная среда 92% глицерин 8% без антибиотиков.
Жизнеспособность: приблизительно 90% (исключение красителя).
Культуральная среда: среда F-12 Гэма 90% плодовая коровья сыворотка 10% без антибиотиков.
Ростовая характеристика размноженных клеток: инокулят 105жизнеспособных клеток/мл в вышеуказанной культуральной среде при 37оС увеличивался за 7 дней.
Эффективность посева: приблизительно 90% в вышеуказанной культуральной среде.
Морфология: эпителиальноподобная.
Кариология: Хромосомное частотное распределение 50 клеток: 2 n 22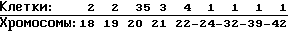
количество является гиподиплоидным.
Стерильность: тесты на микроплазмы, бактерии и грибки отрицательны.
Вид: китайский хомячок, подтвержден путем антителоцитотоксичной пробы с исключением красителя и изоэнзиматического анализа.
Восприимчивость к вирусу: восприимчив к везикулярному стоматиту (индийский штамм) и арбовирусам Гетана. Не восприимчив к полиовирусу, к арбовирусам Модок и Баттон Уиллоу.
Обратная транскриптаза: не обнаружена.
Характеристики: Справочные клетки требуют для своего роста пролин.
Представлен: Т. Т. Паком, Институт исследований рака им. Элеоноры Рузвельт, Медицинский центр Университета Колорадо, Денвер, Колорадо.
Подготовлена и охарактеризована: Американская коллекция типовых культур, Роквилл, Мэриленд.
АТСС CCL 62 FL (Амнион, человеческий, маркеры He/La)
Общепринятая среда для размножения: АТСС CRCM 30 90% телячья сыворотка 10%
Клеточная линия FL была получена из нормальной человеческой анионовой ткани в марке 1956 г Джей. Фогом и Р.Лундом. С помощью трипсинизации ткани суспендировали клетки в среде, содержащей 20% воловьей сыворотки и сбалансированный солевой раствор Эрла, содержащий 0,5% гидролизованного лактаальбумина, и затем высевали в чашки Петри. При первой пересадке использовали среду, содержащую 20% человечьей сыворотки, и пересадку осуществляли в пробирки. Рост улучшался на шестой пересадке, и линия затем выращивалась непрерывно в среде LY 80% сыворотка человека 20% Среда LY это обозначение, данное смеси, состоящей из 0,5% гидролизованного лактаальбумина, 0,1% дрожжевого экстракта и 0,4% декстрозы в балансированном солевом растворе Эрла. Клетки также способны к росту в различных средах, дополненных сывороткой из различных видов. Во время ранних пересадок клетки несколько увеличивались, но фибробластоподобная стадия не наблюдалась никогда. Клетки FL использовались и широко изучались в отношении различных свойств, включая вирусную восприимчивость, туморогенность, морфологию и кариологию.
Описание депонированного справочного семенного запаса
Количество последовательных субкультур из исходных ткани: неизвестно; по оценке 480.
Среда замораживания: базальная среда (Игл) 80% человечья сыворотка 15% глицерин 5% без антибиотиков.
Жизнеспособность: приблизительно 92% (исключение красителя).
Культуральная среда: среда LY 80% человечья сыворотка 20% без антибиотиков.
Ростовая характеристика размороженных клеток: инокулят 105жизнеспособных клеток/мл в вышеуказанной среде увеличивается приблизительно в 5-8 раз за 8-10 ней. Прикрепление клеток не максимально в первом пересеве. В последующих пересевах (0,5-0,5) ˙105 жизнеспособных клеток будут увеличиваться приблизительно в 15-20 раз за 7 дней.
Эффективность посева: приблизительно 20% в вышеназванной среде.
Морфология: эпителиальноподобная.
Кариология: Хpомосомное частотное распределение 100 клеток:
2 n 46 (как определено на справочных клетках Джей. Фогом)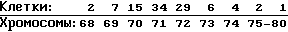
В 100% клеток присутствует крупная субметацентриковая маркерная хромосома.
He/La маркерные хромосомы: одна копия N 1, одна копия N 2, три копии N 3 и две копии N 4 как показывает G-характер исчерченности. Эта клеточная линия, возможно, была загрязнена клетками He/La.
Стерильность: тесты на микоплазму, бактерии и грибки были отрицательны.
На фиг. 1 изображена рестрикционная карта полной длины к-ДНК человеческого т-ПА; на фиг. 2 след пептида, переваренного трипсином человеческого т-АП при высокоэффективной жидкостной хроматографии; на фиг. 3 конструкция ДГФР (мутанта или дикого типа) плазмид, кодирующих т-АП, пригодных для трансформации культур клеток ткани млекопитающего; на фиг. 4 схематическая диаграмма человеческого активатора плазминогена ткани, полученного по способу, описанному в примере 1.
Нуклеотидная последовательность и уменьшенная аминокислотная последовательность полной длины к-ДНК человеческого т-АП дана в конце описания.
А. Определения
"Активатор плазминогена человеческой ткани", или "человеческий т-АП", или "т-АП" означает активатор плазминогена, присущего человеку (тканевого типа), полученный в микробной системе или в системе культуры клеток, активных формах, включающих протеазный участок. Предлагаемый активатор плазминогена человеческой ткани был выделен с помощью технологии рекомбинантной ДНК. Понятно, что существуют природные аллельные вариации генов и они встречаются от индивидуума к индивидууму. Эти вариации могут быть вызваны аминокислотными различиями в общей последовательности или делециями, замещениями, вставками, инверсиями или дополнениями аминокислоты (кислот) в указанную последовательность. Кроме того, местоположение и степень гликозилирования будут зависеть от клетки-реципиента.
При использовании технологии рекомбинантной ДНК для получения различных производных активатора плазминогена человеческой ткани осуществляют модификацию последовательности в результате единичных или множественных аминокислотных замещений, делеций, дополнений или замен аминокислот, например, с помощью сайт-специфического мутагенеза основной ДНК. Все такие аллельные вариации и модификации, приводящие в результате к производным активатора плазминогена человеческой ткани, включены в область предлагаемого изобретения, так же как и другие родственные активаторы плазминогена, присущие человеку (тканевого типа). Если основная биологическая активность активатора плазминогена человеческой ткани при этом сохраняется без изменения, активатор плазминогена человеческой ткани получают:
имеющим метионин в качестве первой аминокислоты (присутствует посредством вставки начального сигнального кодона АТГ перед структурным геном);
где метионин внутри- или внеклеточно отщеплен;
вместе или с его сигнальным полипептидом или с сопряженным протеином, иным по сравнению с традиционным сигнальным полипептидом, при этом сигнальный полипептид или конъюгат специфически отщепим во внутри- или внеклеточном окружении;
прямой экспрессией зрелой формы без необходимости отщепления любого периферического, излишнего полипептида.
Последнее особенно важно, когда данный хозяин не удаляет или удаляет неэффективно сигнальный полипептид. Вектор экспрессии предназначен для экспрессии активатора тканевого плазминогена вместе с его сигнальным полипептидом. В любом случае полученный таким образом человеческий т-АП в его различных формах извлекают и очищают до уровня, соответствующего его использованию при лечении различных сосудистых расстройств или заболеваний.
Кроме того, т-АП имеет формы, которые включают как одноцепочный (1-цепь) протеин, так 2-цепочечный протеин. Последний протеолитически происходит из 1-цепочечного протеина. Теоретически считается, что 2-цепочечный протеин ассоциирован с продуцированным фибрином и что протеолитическое превращение из 1-цепочечного в 2-цепочечный протеиновый материал происходит в месте превращения плазминогена в плазмин. Предлагаемое изобретение обеспечивает введение 1-цепочечного протеина для конверсии in vivo. Описано введение 2-цепочечного протеина, который также проявляет активность, 2-цепочечный протеин может быть получен in vitro протеолитическим превращением 1-цепочечного материала. Так называемый "крингл" участок расположен выше участка сериновой протеазы и, как полагают, играет важную роль при связывании активатора тканевого плазминогена с фибриновой матрицей. Активатор тканевого плазминогена получают с ферментативно активным участком.
Для краткости в предлагаемом изобретении человеческий т-АП способен катализировать превращение плазминогена в плазмин, связывать фибрин, и классифицируется как т-АП, исходя из иммунологических свойств.
"По существу чистая форма" используется для описании состояния человеческого т-АП, свободного от протеина или других материалов, обычно ассоциированных с человеческим т-АП, когда его продуцируют нерекомбинантные клетки.
"ДГФР-протеин" относится к протеину, который обладает дигидрофолат редуктазы (ДДГФР) активностью и для его продуцирования требуются клетки, которые способны выживать в среде, лишенной гипоксантина, глицина и тимидина (ГГТ-среда). Обычно клетки, дефектные по ДГФР, не способны к росту в этой среде. Клетки, которые содержат ДГФР-протеин, успешно растут на ней.
"Клетки, чувствительные к МТТ" относятся к клеткам, которые не способны расти на среде, которая содержит ингибитор-метотрексат (МТТ) ДГФР. Следовательно, "клетки, чувствительные к МТТ" являются клетками, которые не способны расти в обычных условиях и в среде, пригодной для типа клеток, когда концентрация МТТ составляет 0,2 мкг/мл или более. Некоторые клетки, такие как бактерии, не проявляют чувствительности к МТТ, так как они не пропускают МТТ через мембрану клетки, даже если они содержат ДГФР. Клетки, которые содержат дикого вида ДГФР, будут чувствительными к метотрексату, если они проницаемы или способны поглощать МТТ.
"ДГФР дикого типа" относится к дегидрофолатредуктазе такой, которая находится в конкретном рассматриваемом организме. ДГФР дикого типа обычно чувствительна in vitro к низким концентрациям метотрексата.
"ДГФР протеин с низким сродством к связыванию МТТ" этот ДГФР-протеин обеспечивает рост чувствительных к МТТ клеток в среде, содержащей 0,2 мкг/мл или более МТТ.
"Вектор экспрессии" включает векторы, которые способны экспрессировать последовательность ДНК, содержащиеся в нем, при этом такие последовательности действительно связаны с другими последовательностями, способными влиять на их экспрессию. Эти векторы экспрессии должны реплицироваться в организмах-реципиентах или как эписомы, или как интегральная часть хромосомной ДНК. Обычно векторы экспрессии, используемые в технологии рекомбинантной ДНК, часто находятся в форме плазмид, которые означают круглые, двунитевые ДНК, которые в их векторной форме не встроены в хромосому. В предлагаемом описании плазмида и вектор использованы взаимозаменяемо, так как чаще всего плазмида используется в форме вектора. Однако изобретение включает другие формы векторов экспрессии.
"Рекомбинантные клетки+реципиенты" означают клетки, которые трансформированы векторами, сконструированными с использованием техники рекомбинантной ДНК.
Культуры клеток-реципиентов и векторы (В)
Векторы и способы, описанные здесь, пригодны для использования в клетках-реципиентах широкого круга прокариотных и двукариотных организмов.
Разумеется, обычно прокариоты являются предпочтительными для клонирования последовательностей ДНК при конструировании векторов. Особенно полезен штамм 294 Е. coli K12 (АТСС N 31446). Другими микробными штаммами, которые могут быть использованы, являются штаммы E. coli B и E. coli Х1776 (АТСС N 31537). Разумеется, что эти примеры приведены для иллюстрации и не ограничивают изобретение.
Могут быть использованы E.coli W3110/F-λ- прототроф, (АТСС N 27325), бактерии, такие как Bacillus subtilis и другие энтеробактерии, такая как Salmonella typhimurium или Serratia marcescens и различные виды Psendomonas.
Обычно в сочетании с этими реципиентами используют плазмидные векторы, содержащие репликон и регуляторные последовательности, которые происходят из видов, совместимых с клетками-реципиентами. Обычно вектор несет участок репликации, а также маркерные последовательности для фенотипической селекции трансформированных клеток. Например, E. coli обычно трансформируют, используя pBP плазмиду рВР, 322. рВР, 322 содержит гены устойчивости к ампициллину и тетрациклину. рВР, 322 плазмида или другие микробные плазмиды могут содержать промоторы, которые могут быть использованы микробным организмом для экспрессии протеинов. Чаще всего промоторы, используемые в конструировании рекомбинантной ДНК, включают промоторные системы β-лактамазы.
Помимо прокариотов можно также использовать эукариоты, такие как культуры дрожжей. Широко используются среди эукариотных организмов Saccharomyces cerevisial или обычные пекарские дрожжи, хотя обычно доступны к другие штаммы. Для экспрессии в Saccharomyces обычно используют плазмиду YRp 7. Эта плазмида содержит trp 1 ген, который обеспечивает селективный отбор мугантного штамма дрожжей, потерявшего способность к росту в триптофане, например АТСС N 44076 или РЕР4-1.
Промоторные последовательности, используемые в дрожжевых клетках, включают промоторы для 3-фосфоглицераткиназы или других гликолитических ферментов, таких как энолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфат изомераза, 3-фосфоглицератмутаза, пируват киназа, триосефосфатизомераза, фосфоглюкозоизомераза и глюкокиназа. При конструировании плазмид экспрессии концевые последовательности, ассоциированные с вышеперечисленными генами, также встроены в вектор экспрессии, чтобы обеспечить полиаденилирование м-РНК и терминацию. Другими промоторами являются индуцибельные промотирующие области для алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, ферментов, ассоциированных с метаболизмом азота, и вышеупомянутых глицеральдегид-3-фосфатдегидрогеназы, и ферментов, ответственных за утилизацию мальтозы и галактозы. Пригодным является любой плазмидный вектор, содержащий дрожжесовместимый промотор, источник репликации и концевую последовательность. пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфат изомераза, 3-фосфоглицератмутаза, пируват киназа, триосефосфатизомераза, фосфоглюкозоизомераза и глюкокиназа. При конструировании плазмид экспрессии концевые последовательности, ассоциированные с вышеперечисленными генами, также встроены в вектор экспрессии, чтобы обеспечить полиаденилирование м-РНК и терминацию. Другими промоторами являются индуцибельные промотирующие области для алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, ферментов, ассоциированных с метаболизмом азота, и вышеупомянутых глицеральдегид-3- фосфатдегидрогеназы, и ферментов, ответственных за утилизацию мальтозы и галактозы. Пригодным является любой плазмидный вектор, содержащий дрожжесовместимый промотор, источник репликации и концевую последовательность.
Кроме того, в качестве реципиентов могут использоваться культуры эукариотических клеток. В принципе любая такая клеточная культура является работоспособной, независимо от того, является ли она культурой позвоночного или беспозвоночного. Наибольший интерес представляют клетки позвоночных, так как размножение клеток позвоночных в культуре тканей становится рутинной процедурой в последние несколько лет. Примерами таких полезных линий являются VERR и He/La клетки, линии клеток яичника китайского хомячка (СНО) и W138. ВНК, СOS 7 и ДМСК-линии клеток. Векторы экспрессии таких клеток обычно включают источник репликации, промотор, локализованный впереди гена, который должен быть экспрессирован, наряду с любыми участками, необходимыми для связывания рибосом, участком полиаденилирования, и транскрипционные последовательности.
Обычно используемые в культуре тканей промоторы происходят из полиомы. Аденовируса 2 и чаще Симиан Вирус 40 (SV40). Ранний и поздний промоторы SV40 вируса являются особенно полезными, потому что оба легко получаются из вируса в виде фрагмента, который также содержит источник репликации вируса SV40. Меньшие или большие фрагменты SV40 представляют собой последовательности размером 250 п.о. а именно Hind III Bgl I, фрагмент, локализованный в источнике репликации вируса. Желательно использовать промотирующие или регуляторные последовательности, ассоциированные с последовательностью экспрессируемого гена и совместимые с системами клеток-реципиентов.
Источник репликации может происходить из SV40 или другого вируса (например, Полиома, Адено, VSV, BPV и т.п.), или обеспечиваться механизмом хромосомной репликации клетки-хозяина, если вектор встроен в хромосому клетки-хозяина.
При отборе предпочтительной клетки-реципиента для трансфекции векторами изобретения, которые включают последовательности ДНК, кодирующие как т-АП, так и ДГФР-протеин, учитывают тип примененного ДГФР-протеина. При применении протеина ДГФР дикого типа выбирают клетку-реципиента, которая лишена ДГФР, в результате чего последовательность, кодирующая ДГФР, может использоваться в качестве маркера для отбора трансфектантов в среде, которая лишена гипоксантина, глицина и тимидина. Подходящей клеткой-реципиентом является линия клеток яичника китайского хомячка (СНО), лишенная ДГФР-активности.
В случае использования в качестве регуляторной последовательности ДГФР-протеина с низкой связывающей способностью к МТТ, нет необходимости использовать устойчивые к ДГФР клетки. При этом может быть использована среда, содержащая МТТ, в качестве средства для селекции. Большинство эукариотических клеток, которые способны адсорбировать МТТ, являются чувствительными к метотрексату. Одной линией клеток является линия СНО, СНО-ЕI АТСС N ССI 61.
Использованные методики
Если в качестве клеток-реципиентов используют клетки без труднопреодолимых мембранных барьеров клетки, трансфекцию проводят методом осаждения фосфатом кальция. Однако могут быть использованы другие методы для введения ДНК в клетку, такие как инъекция ядра или слияние протопластов.
Если используют клетки прокариота или клетки, которые содержат различные конструкции клеточной стенки, предпочтительным способом трансфекции является кальциевая обработка с использованием хлористого кальция.
При конструировании подходящих векторов, содержащих желаемые кодирующие и регуляторные последовательности, применяют стандартные методики связывания. Выделенные плазмиды или фрагменты ДНК обрабатывают рестриктазой и снова лигируют для образования требуемой плазмиды. Расцепление проводят в результате обработки рестрикционным ферментом (или ферментами) в подходящем буфере. Обычно используют около 1 мкг плазмиды или фрагментов ДНК и примерно 1 ед. фермента в 20 мкл буферного раствора. (Соответствующие буферы и количества субстрата для конкретных рестрикционных ферментов определены производителем). Пригодным является время инкубации около 1 ч при 37оС. После инкубаций протеин удаляют экстракцией фенолом и хлороформом, а нуклеиновую кислоту извлекают из водной фракции осаждением этанолом.
Если требуются тупые концы, фрагменты ДНК обрабатывают в течение 15 мин при 15оС 10 ед. полимеразы 1 (Кленова), экстрагируют фенолом-хлороформом и осаждают этанолом.
Проводят разделение по размерам полученных фрагментов, используя 6%-ный полиакриламидный гель.
Для связывания нужных фрагментов их обрабатывают примерно 10 ед. Т4 ДНК лигазы на 0,5 мкг ДНК. Для предотвращения повторного связывания расщепленного вектора полезна предварительная обработка бактериальной щелочной фосфатазой.
Сконструированные плазмиды используют для трансформации E. coli К12 штамма 294 (АТСС 31446) и отбирают трансформанты по устойчивости к ампициллину или тетрациклину. Из трансформантов получают плазмиду, анализируют рестрикционным и/или иными методами.
Проводят амплификацию последовательностей, кодирующих ДГФР-протеин,в результате выращивания культуры клеток-реципиентов в присутствии приблизительно 20-500000 нМ концентраций метотрексата, конкурентного ингибитора ДГФР-активности. При этом эффективный интервал концентраций зависит от природы ДГФР-гена, протеина и характеристик реципиента. Можно также использовать подходящие концентрации других аналогов фолиевой кислоты или других соединений, которые ингибируют ДГФР. Однако МТТ является удобным, легко доступным и эффективным.
Сущность изобретения состоит в том, что человеческий активатор плазминогена ткани получают следующим образом.
Культивируют клетки человеческой мелономы, продуцирующие активатор тканевого плазминогена, для слияния.
Из клеточных культур экстрагируют в присутствии ингибиторов рибонуклеазы цитоплазмическую ДНК.
На олиго-dT колонке изолируют всю мРНК в полиаденилированной форме. Эту м-РНК фракционируют по размерам с использованием электрофореза на агарозном геле.
Идентифицируют гель-фракцию, содержащую специфическую ДНК активатора тканевого плазминогена. Транслируют РНК от каждой гель-фракции в системе in vitro лизата ретикулоцитов кролика, дополненную микросомами поджелудочной железы собаки. Полученные в результате продукты трансляции затем иммуноосаждают специфическим IgG антителом активатора плазминогена человеческой ткани.
Соответствующую ДНК (21 по 24 S) превращают в соответствующую однонитевую комплементарную ДНК (кДНК), из которой получают двунитевую кДНК. После поли-dC подготовки ее встраивают в вектор, такой как плазмида, несущая один или более фенотипических маркеров.
Используют полученные таким образом векторы для трансформации бактериальных клеток, получают библиотеку клонированной кДНК. Готовят и используют для зондирования библиотеки колоний пул радиоактивно меченых синтетических дезоксиолигонуклеотидов, комплементарных к кодонам для известных аминокислотных последовательностей в т-АП, таких как, например, пул из 8-14-меров, 5'-dTC/GA/CA/GA/TA/CT/TCCCA-3/', компле- ментарный к последовательностям, кодирующим известную аминокислотную последовательность: триптофан глютаминовая кислота тирозинцистеин маспаргиновая кислота (W-E-Y-C-Д).
Выделяют из положительных клонов плазмиду, содержащую кДНК, и определяют последовательность.
Затем подготавливают ДНК, кодирующую t-РА, с определенной последовательностью для вставки in vitro в соответствующий вектор экспрессии, который используют для трансформации в подходящую клетку-реципиент, которую в свою очередь культивируют, и получают желаемый активатор плазминогена человеческой ткани.
Полученный таким образом активатор плазминогена человеческой ткани содержит примерно 251 аминокислоту в его серинпротеазном участке и участок tringle, с последовательностями, расположенными вверх от него, которые, как полагают в настоящее время, являются ответственными за связывание фибрина. Зрелый протеин плюс его сигнальная последовательность составляют в сумме 562 аминокислоты.
Описанная выше процедура позволяет получить чистый т-АП. Использование дополнительной кодирующей последовательности, чувствительной к метотрексату, позволяет получать в клетках-реципиентах при культивировании активный т-АП протеин в количествах более 0,1 п г на клетку в день. Применяя подходящие условия амплификации, можно получать количества более 20 пг на клетку в день. Иначе говоря, получают более 9˙ 10-6 ед. Плуга на клетку в день, а при подходящей амплификации более 18 ˙10-4 ед. Плуга на клетку в день активности т-АП.
Применение метотрексата, который, хотя обычно является губительным для клеток, в описанном способе способен поглощаться или обеспечивать рост клеток в присутствии контролируемых уровней МТТ при амплификации гена, кодирующего ДГФР.
Это демонстрирует тот аспект, что амплификация гена для ДГФР может вызвать амплификацию ассоциированных последовательностей, которые кодируют другие протеины.
Другие механизмы, обеспечивающие устойчивость к метотрексату, включают уменьшение связывающей способности ДГФР протеина, за счет чего он становится менее чувствительным к метотрексату.
Гены ДГФР дикого типа, и гены, кодирующие ДГФР, которые являются устойчивыми к МТТ благодаря его уменьшенной с ДГФР способности с ДГФР к связыванию, амплифицируются в присутствии МТТ. Следовательно, происходит воздействие амплификации последовательности кодирующей ДГФР на ассоциированные последовательности, кодирующие протеин, чем обеспечивается механизм контроля, позволяющий повысить уровень экспрессии т-АП последовательностей в присутствии МТТ или благодаря предварительной обработке трансформированных клеток МТТ.
Изобретение иллюстрируется следующими примерами. В примерах применяют в качестве культур клеток-реципиента культуру-хозяина Е. coli и линию СНО клеток, пригодную для последовательности, кодирующей ДГФР-протеин. Однако для использования в способе пригодны также другие эукариотные и прокариотные клетки.
На фиг. 1 представлена рестрикционная карта полной длины кДНК активатора плазминогена человеческой ткани. Число и размер фрагментов, полученных рестрикционным расщеплением, были оценены электрофорезом на 6%-ный акриламидный гель. Положение участков подтверждены последовательностью нуклеиновой кислоты (представлена на фиг. 5). Кодирующая область самого большого открытого читаемого каркаса оконтурена, заштрихованная область представляет собой предполагаемую сигнальную пептидную последовательность, тогда как пунктирная область представляет собой предположительную последовательность зрелого активатора плазминогена (527 аминокислот). 5' конец мРНК находится слева, тогда как 3' конец находится справа.
Нуклеотидная последовательность и укороченная аминокислотная последовательность полной длины кДНК активатора плазминогена человеческой ткани 35 аминокислот (-35 по -1), предшествующих зрелой последовательности, изображены в виде непрерывной последовательности. Полагают, что эта последовательность из 35 аминокислот представляет собой гидрофильную про-последовательность, предшествующую серину (+1) зрелого протеина, примерно 12-15 аминокислот, в свою очередь, предшествует "традиционному" гидрофобному сигналу (протяженностью от 5' до -35). Этот тип пре-про-структуры у секретируемых протеинов был описан ранее, например, у пре-про-альбумина. Следовательно, все молекулы секретированного активатора тканевого плазминогена будут начинаться с серина (+1) как аминоокончания. Однако гидрофильная последовательность может кодировать одну из функций активатора тканевого плазминогена. При этом пептид молекулярным весом 10000 Да может быть отщеплен от аминоконцевого участка нативного плазминогена (Glu-плазминоген, названный по аминоконцевому остатку), давая в результате меньшую молекулу с новым аминоокончанием, обозначенную Lys-плазминогеном. Lys-плазминоген еще легче активируется в плазмин, а также имеет большее сродство к фибрину, чем Glu-плазминоген. Плазмин был показан при катализе конверсии Glu- в Lys-плазминоген. Первые количества образовавшегося плазмина помимо разложения фибрина приводят в результате к генерации молекул плазминогена, которые легче активируются и также связываются плотнее с их субстратом, чем нативный плазминоген. Результатом является более быстрое разложение фибрина. Гидрофильный пептид активатора тканевого плазминогена может быть включен в подобный механизм, его отщепление приводит в результате к модифицированному связыванию фермента с фибрином. Однако в любом случае последовательность из 35 аминокислот рассматривается как предпоследовательность зрелого протеина.
На физической карте р Δ RIPA* плазмиды экспрессии активатора тканевого плазминогена исходную плазмиду рРА25Е10 сначала обрабатывают Pst 1 для выделения 376 bp фермента фрагмента размером 367 bp, который затем обрабатывают.
На фиг. 2 показана хромограмма полипептидов при высокоэффективной жидкостной хроматографии активатора тканевого плазминогена, переваренного тpипсином (поглощение при 210 нм). Стрелка показывает на пик, соответствующий пептиду, использованному для получения нуклеотидного зонда. Было найдено, что пептид, представленный этим пиком, имеет полную последовательность: L-T-W-E-Y-C-D--V-P-S-C-S-T-G-C-L. Была определена последовательность других подобных пиков и было найдено подтверждение правильной аминокислотной последовательности активатора плазминогена человеческой ткани. Каждая буква в коде пептида относится к аминокислоте, обозначенной следующим образом:
Asp D Аспарагиновая кислота Ile I Изолейцин
Thr T Треонин Leu L Лейцин
Ser S Серин Tyr Y Тирозин
Glu E Глютаминовая кислота Phe F Фенилаланин
Pro P Пролин His H Гистидин
Gly G Глицин Lys K Лизин
Ala A Аланин Arg R Аргинин
Gys C Цистеин Тrp W Триптофан
Val V Валин Glr Q Глютамин
Met M Метионин AsnN Аспарагин
Плазмида, кодирующая прямую экспрессию зрелого активатора плазминогена человеческой ткани в E. coli 50 мкг плазмида рРА17 обрабатывают с помощью Sau 3AI, Hin сП и Hha I и проводят электрофорез на 6%-ном полиакриламидном геле. Извлекают приблизительно 0,5 мкг фрагмента Sau 3AI-Hha I. Подобным размером 55 bp очищают приблизительно 3 мкг фрагмента HhaI-NarI из 80 мкг клона размером 263 bp рРА25Е10, выделяют фрагмент PstI-NarI размером 300 bp, затем обрабатывают фрагмент HhaI рестриктазой. Обработку рестриктазой проводят при 37оС в течение 1 ч, затем продукты реакции разделяют и подвергают электроэлюированию на 6% -ных полиакриламидных гелях. Два указанных дезоксиолигонуклеотида 5' d AATTCATGTCTTATCAAGT (I) и 5' GATCACTTGATAAGACATG (II) синтезируют твердофазным фосфотриэфирным методом, фосфорилируют 100 пмолей олигонуклеотида II в 30 мкл реакционной смеси, содержащей 60 мМ Трис (рН 8), 10 мМ MgCl, 15 мМ β-меркаптоэтанола и 50 мКи (γ 32Р/АТР/Амершам 5000 Мюри Ммоль-1), прибавляют 12 единиц Т4 полинуклеотидной киназы, реакционную смесь оставляют при 37оС на 15 мин. Затем прибавляют 1 мкл 10 мМ АТР и 12 ед Т4 киназы и проводят реакцию еще 30 мин. После экстракции фенолом/хлороформом объединяют фосфорилированный олигомер II и 5' гидроксильный олигомер I с 0,5 мкг элюированного Sau 3AI-HhaI фрагмента размером 55 bp и 2 мкг HhaI-NarI фрагмента размером 263 bp и осаждают этанолом. Эти фрагменты связывают при комнатной температуре в течение 4 ч в 60 мкл 20 мМ трис-HCl (рН 7,5), 10 мМ MgCl2, 10 мМ дитиотрейтола, 0,5 мМ АТР и 1000 ед. Т4 ДНК-лигазы. Смесь обрабатывают в течение 1 ч 48 ед. NarI, 20 ед. EcoRI и 40 ед. Bgl II и проводят электрофорез на 6%-ном геле. Электроэлюированием извлекают продукт 338 bp (приблизительно 0,1 мкг). Оставшиеся кодирующие последовательности т-АП) (аминокислоты 111528) изолируют на фрагменты размером 1645 bp в результате обработки плазмиды рРА25El0 Nar I и Bgl II. Плазмида рLeIFAtrp 103 является производной от плазмиды рLcIFA 25, в которой ECORI участок bLeI FA-гене был удален. Обрабатывают 3 мкг pLeIFAtrp 103 20 ед. ECORI и 20 ед. Bgl II в течение 90 мин при 37оС, проводят электрофорез на 6%-ном полиакриламидном геле и электроэлюированием извлекают большой фрагмент вектора (≈ 4 200 bp). Для окончательного конструирования связывают 80 нг EcoRI-Bgl II PLeIFA trp 103 со 100 нг фрагмента Nar I Bgl размером 1645 bp и 20 нг фрагмента EcoRI Nar I размером 338 bp в течение 10 ч при комнатной температуре. Эту связанную смесь используют для трансформации E.coli К-12 штамм 294. Получают плазмиду ДНК из 38 этих трансформантов и обрабатывают EcoRI. Десять из этих плазмид содержат целевые 600 bp и 472 bp EcoRI-фрагменты. Анализ последовательности ДНК подтверждает, что одна из этих плазмид (pt-PAtrp12) имеет желаемую нуклеотидную последовательность в результате правильного соединения между trp промотером, синтетической ДНК и кДНК.
П р и м е р 1. Используют клетки человеческой меланомы (Bower). Клетки меланомы культивируют до сливных монослоев в 100 мл минимальной основной среды Игла, дополненной бикарбонатом натрия (конечная концентрация 0,12%), 2 мМ глютамина и 10% инактивированной теплом сыворотки плода коровы. Для подтверждения того, что клетки меланомы активно продуцируют активатор плазмидогена человеческой ткани, культивируют клетки человеческой меланомы до слива в чашку микротитратора с 24 отверстиями. Независимо от наличия или отсутствия 0,33 мкМ ингибитора протеазы апротинина клетки промывают 1 раз фосфатным забуферированным раствором и прибавляют 0,3 мл среды, не содержащей сыворотки и метионина. Добавляют 75 мкКюри (35S)-метионина и клетки метят при 37оС в течение 3 ч. После 3 ч среду удаляют и обрабатывают или специфическим IgG активатора тканевого плазминогена или пре-иммунной сывороткой для иммуноосаждения. Продукты иммуноосаждения подвергают электрофорезу на 100%-ном SDS-акриламидном геле. Пластину геля фиксируют, сушат и подвергают флюорографии.
Экстрагируют всю РНК из культур клеток меланомы. Клетки таблетируют центрифугированием и повторно суспендируют в 10 мМ NaCl, 10 мМ трисHCl, рН 7,5, 1,5 мМ MgCl2. Клетки лизируют добавлением NP-40 (конечная концентрация 1% ), а ядра таблетируют центрифугированием. Надосадочный слой, содержащий всю РНК, затем очищают многократными экстракциями фенолом и хлороформом. Водную фазу делают 0,2 М по NaCl, а затем осаждают всю РНК добавлением двух объемов этанола. Используют хроматографию на олиго-dT целлюлозе для очистки иРНК от общей РНК. Типичные выходы из 10 культивированных клеток меланомы составляют 5-10 мг общей РНК и 50-200 мкг поли(А)плюс мРНК.
Проводят фракционирование поли А+мРНК (200 мкг) путем электрофореза на мочевиноагарозном геле. Пластина агарозного геля состоит из 1,75% агарозы, 0,025 М цитрата натрия, рН 3,8 и 6 М мочевины. Проводят электрофорез в течение 7 ч при 25 мА и 4оС. Затем гель фракционируют острым лезвием. Индивидуальные тонкие слои в виде ломтиков расплавляют при 70оС и экстрагируют дважды фенолом и один раз хлороформом. Затем фракции осаждают этанолом и последовательно испытывают in vitro трансляцией в систему лизата ретикулоситов, дополненную микросомами поджелудочной железы собаки.
Испытание проводят следующим образом.
Проводят трансляцию, используя 25 мкКюри (35S)-метионина и 500 нг из каждого тонкого слоя геля РНК с конечной объемной концентрацией 30 мкл, содержащей 25 мМ НЕРЕS, 48,3 мМ хлористого калия, 10 мМ креатин фосфата, 19 аминокислот по 50 мМ каждой, 1,1 мМ хлористого магния, 16,6 мМ ЭДТУК, 0,16 мМ дитиотрейтола, 8,3 мМ гомина, 16,6 мг/мл креатинкиназы, 0,33 мМ хлористого кальция, 0,66 мМ EGTa, 23,3 мМ хлористого натрия.
Проводят инкубирование при 30оС в течение 90 мин. Обрабатывают нуклеазой мембраны микросом поджелудочной железы собаки, полученной из красных микросом, с использованием ЭДТУК для удаления рибосом. Они находятся в трансляционной смеси при конечной концентрации 7А260 ед.мл. Продукты трансляции или иммуноосажденные продукты трансляции анализируют с помощью электрофореза на 10% -ных полиакриламидных гелях в додецилсульфате натрия. Фиксируют незапятнанные тонкие пластинки гелей, сушат и подвергают флюорографии.
Полученные в результате продукты трансляции из каждой гель-фракции иммуноосаждают специфическим анти-IgG активатора плазминогена человеческой ткани кролика. Наблюдается одна основная полоса иммуноосажденного полипептида в фракциях трансляции номера 7 и 8 (миграции от 21 до 24S), имеющая молекулярную массу приблизительно 63000 дальтон. Эта полоса не наблюдается при использовании пре-иммунного IgG для иммуноосаждения, которому подвергаются эти полипептиды, являющиеся специфическим активатором тканевого плазминогена.
Используют 5 мкг фракционированной на геле мРНК (тонкий ломтик геля 7 мРНК) для получения двунитевой кДНК с помощью стандартных процедур. Фракционируют по размеру кДНК на 6%-ном полиакриламидном геле. Электроэлюируют кДНК размером больше, чем 350 основных пар 3125 нг/ 30 нг кДНК/ достраивают дезокси /C/ остатками, с помощью концевой дезоксинуклеотидтрансферазы и лигируют с 300 нг плазмиды рBR322, которую предварительно достраивают дезокси/G/ остатками на PstI участке. Лигированную смесь затем трансформируют в Е. coli К12 штамм 294 (АТСС 31446). Получают приблизительно 4600 трансформантов.
Получают очищенный активатор плазминогена человеческой ткани.
Молекулу сканируют, чтобы локализовать лучше изученные области для маркировки синтетическими зондами, следующим образом.
Протеины восстанавливают и карбоксиметилируют. Сначала диализуют 2 мг образца активатора тканевого плазминогена против 0,01% Твина 80 в течение ночи при комнатной температуре. Затем растворяют лиофилизованный протеин в 12 мл 0,56 М Трис-HCl буфера (рН 8,6), 8 М мочевине и 5 мМ ЭДТУК. Восстанавливают дисульфидные связи добавлением 0,1 мл β-меркаптоэтанола. Эту реакцию проводят в азоте в течение 2 ч при 45оС. Восстановленные дисульфиды алкилируют карбоксиметилпроизводным при добавлении 1,0 мл 1,4 М иодуксусной кислоты в 1 н. NaOH. После 20 мин при комнатной температуре реакцию останавливают диализом против 0,01% Твина 80 в течение 18 ч при комнатной температуре и лиофилизуют. Полученный в результате лиофилизованный карбоксиметилированный протеин снова растворяют в 3 мл 0,1 М натрийфосфатного буфера (рН 7,5). Прибавляют трипсин (ТРСК) (соотношение 1:50) и выдерживают при 37оС. Отбирают аликвоты (0,1 мл) через 3, 6 и 12 ч. Второе добавление трипсина проводят через 12 ч.
Реакцию останавливают через 24 ч, замораживая образец до тех пор, пока его можно будет использовать для высокоэффективной жидкостной хроматографии. Степень обработки трипсином определяют с помощью SDS-гелей на аликвотах. Все гели были чистыми, за исключением четкой полосы на 3-часовом аликвоте. Это указывает, что после 24-часовой обработки не остается больших пептидов.
Помещают образец примерно 0,5 мл в колонку высокого разрешения Альтекс С-8 ультрасфера 5 мкм с двумя циклами. Устанавливают постепенный градиент ацетонитрила (от 1 до 5% за 5 мин, от 5 до 35% за 100 мин, 35-50% за 30 мин). В одном из двух препаративных циклов элюент контролируют по двум длинам волн (210 и 280 нм). Используют соотношение поглощения при двух длинах волн, чтобы указать пептиды, содержащие трептофан.
Пики пептидов, содержащие триптофан, были первыми направлены на определение последовательности. Это дает возможность определения последовательности вокруг большинства триптофанов. После определения последовательности у 25 лучших возможных пептидных пиков, все данные по последовательностям, которые могут быть выровнены в ряд, объединяют для получения предварительной модели первичной структуры активатора тканевого плазминогена. Из этих данных и модели локализуют несколько возможных зондов.
Индивидуально инокулируют колонии в отверстия пластин микротитратора, содержащие LB(93) +5 мкг/мл тетрациклина, и хранят при -20оС после добавления ДМСО до 7% Выращивают две копии пула колоний на нитроцеллюлозных фильтрах и фиксируют ДНК из каждой колонии на фильтре.
Готовят 32Р меченый TC/GA/CA/GA/TA/TC/TCCCA зонд (из синтетического олигомера (W-E-Y-C-D) 14-мерный пул, как описано выше). Фильтры, содержащие 4600 трансформантов, предварительно гибридизируют в течение 2 ч при комнатной температуре в 50 мМ натрий-фосфата рН 6,8 5X S,S 0,(80), 150 мкг/мл обработанной ультразвуком ДНК спермы лосося, 5% раствора Денхардта 10% формамида, а затем гидролизуют с 50 ˙106 счет/мин меченого зонда в том же растворе. После инкубации в течение ночи при комнатной температуре фильтры промывают 3 раза при комнатной температуре в 6Х S,SO, 0,1% SDS в течение 30 мин, один раз в 2Х SSC, а затем экспонируют на рентгеновской пленке Кодак ХР-5 с Дюрон Лайтинг Плюс интенсифицирующими экранами в течение 16 ч.
Выделяют плазмиду ДНК быстрым способом из всех колоний, показывающих положительную реакцию гибридизации. Затем определяют последовательности вставок к ДНК из этих клонов после субклонирования фрагментов в М13 векторе mp 7 и с помощью химической процедуры Максама Гилберта. Вставка к ДНК в клоне 25Е10 была продемонстрирована как имеющая ДНК, кодирующая активатор тканевого плазминогена путем сравнения его аминокислотной последовательности с пептидной последовательностью (см.выше), полученной из очищенного активатора тканевого плазминогена и с продуктом его экспрессии, полученным в Е.coli как детально описано ниже. Вставка кДНК клона 25Е10 (Плазмида рРА25Е10) имеет размер 2304 bp и наиболее длинную открыто читаемую структуру, кодирующую протеин из 508 аминокислот (молекулярная масса 56 756) и содержащую 3'-нетранслируемую область размером 772 bp. Этот кДНК клон потерял N-терминальные кодирующие последовательности.
Лигируют 50 мкг рРA25Е10, обработанной PstI и выделяют фрагмент размером 376 bp электрофорезом на 6%-ном полиакриламидном геле. Выделяют примерно 3 мкг этого фрагмента из гели электроэлюи- рованием, обрабатывают 30 ед. DdcI в течение 1 ч при 37оС, экстрагируют фенолом и хлороформом и осаждают этанолом. Полученные в результате DdcI липкие концы заполняют до тупых концов добавлением 5 ед. ДНК полимеразы I (фрагмент Кленова) и 0,1 мМ каждого из dATP. dCTP, dGTP, dTTР в реакционную смесь и инкубируют при 4оС в течение 3 ч. После экстракции фенолом и хлороформом ДНК обраба- cGTP, dTTP в реакционную смесь и инкубируют при 4оС в течение 3 ч. После экстракции фенолом и хлороформом ДНК обрабатывают 15 ед. NarI в течение 2 ч и подвергают реакционную смесь электрофорезу на 6%-ном полиакриламидном геле. Извлекают приблизительно 0,5 мкг желаемого фрагмента размером 125 bp с тупым концом NarI. Этот фрагмент кодирует аминокислоты с номера 69 по 110 зрелого протеина активатора тканевого плазминогена полной длины.
Для получения NarI-Bgl II фрагмента размером 1645 bp 30 мкг рРА52Е10 обрабатывают 30 ед. NarI и 35 ед. Bgl II в течение 2 ч при 37оС и реакционную смесь подвергают электрофорезу на 6%-ном полиакриламидном геле. Выделяют приблизительно 6 мкг NarI-Bgl II фрагмента размером 1645 bp.
Плазмида pdc ltaRISRC является производной плазмиды pSR Сeх 16, в которой EcoRI участки, проксимальные к trp промотору и дистальные к SRC-гену, были удалены репарацией ДНК полимеразой I и само-комплементарный олигонуклеотид AATTATGAATTCAT (синтезированный фосфотриэфирным методом) был вставлен в EcoRI участок в непосредственной близости к XbaI участку. Обрабатыват 20 мкг pdeltaRISRC эндопулмазой EсoRI, экстрагируют фенолом и хлороформом и осаждают этанолом. Затем обрабатывают плазмиду 100 ед. нуклеазы S I при 16оС в течение 30 мин в 25 мМ ацетат натрия (рН 4,6), 1 мМ ZnCl2 и 0,3 М NaCl для создания тупого конца с последовательностью ATG. После экстракции фенолом и хлороформом и осаждения этанолом обрабатывают ДНК BamHI, проводят электрофорез на 6%-ном полиакриламидном геле и электроэлюированием извлекают большой (4300 bp) фрагмент вектора.
Экспрессионную плазмиду собирают связыванием вместе 0,2 мкг вектора, 0,06 мкг фрагмента размером 125 bp,с тупым концом NarI и 0,6 мкг фрагмента NarI-Bgl II размером 1645 bp с 10 ед. Т4 ДНК лигазы в течение 7 ч при комнатной температуре и используют для трансформации E.coli штамм 294 (АТСС N 31446). Плазмиду ДНК получают из 26 колоний и обрабатывают XbaI и EcoRI, 12 из этих плазмид содержат желаемые фрагменты XbaI-EcoRI размером 4115 bp и EcoRI размером 472 bp. Анализ последовательности ДНК подтверждает, что некоторые из этих плазмид имеют АТС начальный кодон, правильно расположенный в начале аминокислоты номер 69 (серин). Одна из этих плазмид p ΔRIPAо была испытана и продуцирует желаемый активатор тканевого плазминогена.
П р и м е р 2. Используют 0,4 мкг синтетического олигонуклеотида 5' TTCCTGCAGA GGGGG3' для заполнения 7,5 мкг гель-фракции номер 8 мРНК (выше), чтобы получить двунитевую кДНК. кДНК фракционируют по размерам больше 300 основных пар (36 нг) и подвергают электроэлюированию. Достраивают 5 нг кДНК дезокси (С) остатками, используя терминальную дезоксицитидилтрансферазу и лигируют с 50 нг плазмиды рBR322, которая была подобным образом обработана дезокси (G) остатками на PstI участке. Отожженную смесь затем трансформируют в E.coli К12 штамм 294. Получают приблизительно 1500 трансформантов.
Так как реакция заполнения кДНК была осуществлена с использованием синтетического фрагмента, который гибридизируется с 13 основаниями N-терминального клона рРА25Е10, отсутствует удобный рестрикционный фрагмент в этой области (которая включает 16-мерную последовательность) для скрининга клонов кДНК. Следовательно, необходимо выделить геномный клон активатора плазминогена человеческой ткани, чтобы идентифицировать любые достроенные клоны кДНК, содержащие N-терминальные последовательности, кодирующие активатор тканевого плазминогена.
Первая стадия этого процесса включает установление факта, что только единственный гомологичный гек активатора тканевого плазминогена имеется в человечекой геномной ДНК. Для определения этого проводят гибридизацию Саузерна. 5 мкг ДНК человеческого лимфоцита высокой молекулярной массы обрабатывают различными рестрикционными эндонуклеазами, проводят электрофорез на 1,0% -ных агарозных гелях и помещают в виде пятна на нитроцеллюлозном фильтре. Готовят 32Р-меченую ДНК в виде зонда из фрагмента НраII-RsaI размером 230 bp плазмиды рРА25Е10 и гибридизуют на нитроцеллюлозном фильтре. 35 ˙106 счетов в минуту зонда гибридизируют в течение 40 ч, затем промывают. Два образца, обработанные эндонуклеазой, гибридизуются с зондом фрагмента Bgl II (5,7 Vsbp) и PVU II (4,2 Kbp). Два фрагмента ДНК гибридизуются с Нinc II/5,1 Kbp и 4,3 Kbp. Взятые вместе, эти данные подтверждают наличие только одного гена активатора тканевого плазминогена в человеческом геноме, а также то, что этот ген содержит по крайней мере одну внедрившуюся последовательность.
Накладывают 1 миллион рекомбинантного λ-фага на ДР 50 СарФ при плотности 10000 Рfn 15 см на пластины и готовят реплики на нитроцеллюлозных фильтрах для каждой пластины. Готовят зонд 32Р-меченой ДНК по стандартным процедурам из 230 пар оснований HpaII-RsaI, в котором 34 пары оснований локализованы от 5' конца плазмиды рРА25Е10. Каждый нитроцеллюлозный фильтр предварительно гибридизуют при 42оС в течение 2 ч в 50 мМ натрий-фосфате (рН 6,5), 5Х SSC (77), 0,05 мг/мл обработанной ультразвуком ДНК спермы лосося, раствора 5Х Денхардта, 58% формамида, а затем гибридизуют с 50x x106 счетами/мин меченого зонда в том же самом растворе, содержащем 10% декстрансульфата натрия. После инкубации в течение ночи при 42оС фильтры промывают 4 раза при 50оС в 0,2Х, SSC, 0,1% SDS в течение 30 мин, один раз в 2 х SSC при комнатной температуре, а затем экспонируют на рентгеновской пленке Кодак XR-5 с интенсифицирующими экранами Дюпон Кронекс в течение ночи. Всего было получено 19 клонов, гибридизующихся с зондом. Готовят фаг ДНК, как было описано ранее, из 6 рекомбинантов. Отбирают λ-клон С для получения фрагмента PVUII, предназначенного для скрининга колоний. Обрабатывают 30 мкг ДНК рестриктазой PVUII в течение 1 ч при 37оС и проводят электрофорез на 1,0% агарозных гелях. Электроэлюируют и очищают фрагмент 4,2 Kbp, предварительно было показано, что он содержит последовательности активатора тканевого плазминогена. Готовят 32Р-меченый зонд для гибридизации колоний.
Переносят колонии с пластинок и выращивают на нитроцеллюлозных фильтрах, затем фиксируют каждую колонию на фильтре по процедуре Гранштейна-Хогнесса. Готовят 32Р-меченый зонд PVUII фрагмент размером 4,2 Kbp из выделенного λ -геномного клона активатора тканевого плазминогена. Гибридизуют фильтры, содержащие 1500 трансформантов, с 112˙ 106 срм 32Р-геномного PVUII фрагмента. Гибридизацию проводят 16 ч. Фильтры экстенсивно промывают, а затем экспонируют на рентгеновской пленке Кодак Х-5R с интенсифицирующими экранами в течение 16-48 ч. Четко гибридизуется 18 колоний с геномным зондом. Выделяют плазмиду ДНК из каждой из этих колоний, наносят на нитроцеллюлозные фильтры и гибридизуют с 32Р-меченым синтетическим олигонуклеотидом (16-мер). Из 18 клонов с киназным 16 мером гибридизуется смесь. При анализе последовательности после субклонирования фрагментов в m13 вектор mp7один клон (рРА17) показал, что он содержит правильную 5' N-терминальную область активатора тканевого плазминогена, сигнальную лидерную последовательность и 84 bp 5' нетранслируемой области. Из двух клонов рРА2510 и РРА17 были определены полная нуклеотидная последовательность и рестрикционная карта полной длины клона активатора тканевого плазминогена.
Нативная молекула активатора тканевого плазминогена стабилизирована 17 дисульфидными мостиками, что основано на гомологии с другими серинпротеазами. Имеется четыре потенциальных участка II-гликозилирования: три локализованы в "крингл" областях при asn117, asn188, asn218 и один потенциальный участок в легкой области цепи при asn448. Варианты в структуре олигосахаридных лигандов могут быть ответственными за различные молекулярные формы (виды с молекулярной массой 65000 и 63000).
Из плазмиды рРА17 выделяют растрикционный фрагмент san 2AI-HhaI размером 55 bp, соответствующий аминокислотам 5-23. Рестрикционный участок San 3AI предположительно локализован на кодоне четыре зрелой кодирующей последовательности и использован для удаления сигнальной пептидной кодирующей области. Из плазмиды рРА25Е10 также был выделен фрагмент HhaI-NarI размером 263 bp (кодирующий аминокислоты 24-110). Конструируют два синтетических дезоксиолиго- нуклеотида, которые реконструируют кодоны для аминокислот 1-4, включают кодон инициирования трансляции ATG и создат ECoRI сайт. Эти три фрагмента затем связывают вместе с образованием фрагмента размером 338 bp, кодирующего аминокислоты 1-110. Этот фрагмент и фрагмент NarI-Bgl II размером 1645 bр из рРА25Е10 затем связывают между EcoRI и Bgl II участками плазмиды pLe IFAtr р 103 для получения плазмиды экспрессии pt-PAtrp 12. Клонированный ген-т-АП транскрибируется под контролем фрагмента размером 300 bp из trp оперона E. coli, который содержит trp промотор, оператор и последовательность Shine-Babgarno лидерного пептида, но теряет лидерный пептид с ATG инициирующего кодона.
Выращивают E. coli К12 штамм W 3110 (АТСС N 27325), содержащий плазмиду pt-PAtrp12, и готовят экстракты для проб на фибринолитическую активность. Измеряют количество образовавшегося плазмина измерением степени разрушения плазмином фибрина на агарозной пластине, содержащей плазминоген и фибрин. Плазмин дает четкую зону лизиса на фибриновой пластине и площадь этой зоны может быть скоррелирована с количеством активатора тканевого плазминогена в образце. При испытании экстрактов из клонов, содержащих pt-PAtrp12, на активность активатора тканевого плазминогена является очевидным использование пробы с фибриновой пластиной с образованием четкой зоны лизиса. Эта фибринолитическая активность ингибируется анти-т-АП Igb, но не ингибируется IgG или анти-урокиназа IgG, не показывает активности экстракт, полученный из клеток, содержащих в качестве контроля α-интерферонную плазмиду pL c IFtrp103. Используя стандартную кривую очищенного т-АП, можно оценить, что получают приблизительно 20 единиц экстрагированной активности на 109клеток (для очищенного т-АП, 90000, единиц Плуга 1 мг).
Анализ последовательности основан на разложении по Эдману. Образец вводят в чашку Бекмана 890В или 8900 спиннинговой чашки секвенатора. В качестве носителя в чашке используют полибрен ТМ (поли-N,N,N',N'-тетраметил-N-триметиленгексаметилендиаммоний диацетат). Секвенатор был модифицирован охлаждаемой ловушкой и введены некоторые изменения программы для снижения фоновых пиков. Реагентами были последовательность Бекмана типа 0,1 молярного Квадрольного буфера, фенилизотиоцианат и гептафтормасляная кислота.
Собранные циклы Эдмана превращают вручную в производные 2-анилино-5-тиазолина. 1-Хлорбутан сушат в азоте. Затем прибавляют 1,0 н.HCl в воде к 2-анилино-5-тиазолинону и нагревают до 70оС в течение 20 мин для превращения его в 3-фенил-2-тиогидантоин (ФТГ производное). Затем растворяют остаток ФТГ-аминокислоты в 50% ацетонитрила и воды и помещают в хроматограф для высокоэффективной жидкостной хроматографии с обратной фазой. Затем каждую ФТГ-аминокислоту идентифицируют сравнением времени и удерживания стандартной смеси ФТГ-аминокислот, которые обрабатывают таким же образом.
П р и м е р 3. Чувствительную пробу для активатора тканевого плазминогена можно получить путем контроля конверсии плазминогена, катализируемой активатором тканевого плазминогена, в плазмин. Плазмин является ферментом, для которого имеются пробы на хромогенный субстрат. Эти пробы основаны на протеолитическом отщеплении трипептида от хромофорной группы. Скорость отщепления непосредственно связана как со специфичностью, так и с концентрацией испытываемой протеазы. Основой пробы является определение количества плазмина, образовавшегося после инкубирования раствора активатора тканевого плазминогена с раствором плазминогена. Чем больше количество активатора, тем большее количество плазмина образуется. Определяют плазмин путем контроля его отщепления от хромогенного субстрата S2251 (получен от Кеби Групп, Инк. Гринвич, СТ).
Смешивают аликвот образца с 0,10 мл 0,7 мг/мл плазминогена (в 0,05 М Трис HCl, рН 7,4, содержащем 0,012 М NaCl) и устанавливают объем 0,15 мл. Смесь инкубируют при 37оС в течение 10 мин, прибавляют 0,35 мл S2251 (1,0 мМ раствор в приведенном выше буфере) и проводят реакцию в течение 30 мин при 37оС. Для завершения реакции прибавляют 25 мкл ледяной уксусной кислоты. Образцы центрифугируют и измеряют поглощение при 405 нм. Получают количественную оценку активности при сравнении со стандартным раствором урокиназы. Модифицируют условия опыта для делеции полной длины активатора тканевого плазминогена добавлением 0,2 мг фибриногена к раствору. Результатом добавления фибриногена при стимуляции активности активатора тканевого плазминогена является некоторое повышение уровня активности. Регистрируют активность в единицах Плуга, причем 90000 еиниц Плуга равны активности, проявляемой 1 мг очищенного активатора тканевого плазминогена.
Была разработана чувствительная проба на активность активатора тканевого плазминогена. Проба основана на определении образования плазмона при измерении степени разрушения фибрина плазмином на агаровой пластине, содержащей фибрин и плазминоген. Плазмин дает четкую зону лизиса на фибриновой пластине. Площадь этой зоны лизиса может быть скоррелирована с количеством активатора тканевого плазминогена в образце.
Следуя процедуре Гранелли-Пиперно и Рейха, инокулируют пластины 3,5 ч при 37оС и измеряют зоны лизиса. Количественную оценку получают при сравнении со стандартным раствором урокиназы.
Инокулируют колонию E. coli, содержащую плазмиду р Δ RIPAо в тестовую трубку, содержащую 5 мл среды роста LP, с 20 мкг/мл ампициллина. Клетки выращивают в течение ночи при 37оС. Аликвоту этой культуры разбавляют 1:100 в 300 мл М9 среды, содержащей 20 мкг/мл ампициллина. Клетки выращивают во стряхиваемой колбе при 37оС течение 4 ч, результирующее поглощение при 550 нм равно 0,419. Добавляют триптофановый аналог индолакриловой кислоты до концентрации 30 мкг/мл. Инкубируют клетки 90 мин с результирующим поглощением при 550 нм, равном 0,628. Собирают клетки центрифугированием и повторно суспендируют в 0,8 мл 0,01 М Трис, рН 8,0, содержащем 0,01 М. ЭДТУК. Полученную суспензию интенсивно перемешивают 18 ч при комнатной температуре. Образцы центрифугируют и проверяют надосадочный слой на активность активатора тканевого плазминогена.
В табл. 1 и 2 приведены результаты, полученные при активации плазминогена соответствующими экстрактами Е.coli. Генерируемая активность зависит от наличия плазминогена (табл.1). Эта активность не ингибируется предиммунной сывороткой кролика, но она четко ингибируется антисывороткой против активатора тканевого плазминогена из очищенных клеток меланомы (табл.1 и 2). Это показывает, что экстракты E.coli дают активность по активации плазминогена, которая ингибируется антителами против активатора тканевого плазминогена.
Добавляют стандартное количество урокиназы к центральному ряду при концентрациях слева направо 0,24, 0,14, 0,10, 0,05 и 0,02 единиц Плуга. Нижний ряд представляет собой образцы натурального активатора тканевого плазминогена с таким же количеством фермента, как и в каждом ряду. Источники содержат слева направо активатор тканевого плазминогена, анти-активатор плазминогена плюс предиммунная сыворотка и активатор тканевого плазминогена плюс антитела активатора тканевого плазминогена. Источники в каждом верхнем ряду содержат 8 мкл экстрактов Е.coli рекомбинантного активатора тканевого плазминогена. Первый источник является одним экстрактом, во второй источник добавили предиммунную сыворотку, а в третий источник добавили антителя активатора тканевого плазминогена. Очевидно, что предиммунная сыворотка не влияет на природный или рекомбинантный активатор тканевого плазминогена и что антитела активатора тканевого плазминогена ингибируют активность как природного продукта, так и экстрактов из Е.coli. Основываясь на урокиназном стандарте, экстракты содержат несколько меньшее, чем 2,5 единицы Плуга/мл. Это сравнение благоприятно для величины, полученной в табл. 1-3 единица Плуга/мл.
В табл. 1 и 2 представлены результаты опытов.
П р и м е р 4. Вставляют последовательность, кодирующую активатор тканевого плазминогена (т-АП), в экспрессионную плазмиду, содержащую мутантную ДГФР с низким связывающим сродством к МТТ.
Три фрагмента из плазмид рРА25Е10, и рРА17 и pt-pAtrp12 (выше) получают следующим образом.
Обрабатывают плазмиду рРА17DdI, наполняют с помощью ДНК полимеразы I Кленова и разрезают с помощью PstI; выделяют фрагмент размером приблизительно 200 bp, содержащий 5' терминальную последовательность т-АП, генерированную таким образом. Получают второй т-АП фрагмент обработкой pt-pAtrp 12 с помощью рестриктазы PstI и NarI, выделяют фрагмент размером приблизительно 310 bp. Третий т-АП фрагмент получают обработкой рРА25Е10 с помощью NarI и Bgl II и выделяют фрагмент размером приблизительно 1645 bp, который кроме того содержит большую часть кодирующей области т-АП, некоторое количество 3' нетранслируемых последовательностей.
Плазмиду рЕ342, которая экспрессирует поверхностный антиген HBV (также называемый рНВ S 348-Е), (фрагмент вируса SV40) выделяют обработкой ДНК SV40 с помощью Hind III и превращением Hind III концов в EcoRI концы при добавлении олигонуклеотида АСT АTTС. Эту ДНК обрабатывают PVU II и добавляют RI связки. Последующая обработка EcoRI приводит к получению фрагмента размером 348 пар оснований. Его клонируют в pBR322. Коструируют плазмиду экспресии рНВ Р348-Е путем клонирования фрагмента размером 1986 пар оснований, полученного в результате обработки HBV с помощью EcoRI и Bgl III (который охватывает ген, кодирующий HBgAg) в плазмиду pML на участках EcoRI и BamHI. (pML является производной от рВR322, которая имеет последовательности делеции, которые являются ингибиторами репликации плазмиды в клетках обезьяны). Затем линеаризуют полученную в результате плазмиду (pRI-Bgl) с EcoRI и вводят фрагмент размером 348 пар оснований, представляющий собой исходную область SV 40, а EcoRI участок pRI-Bgl. Исходный фрагмент может быть вставлен в любой ориентации. Так как этот фрагмент кодирует как ранний, так и поздний SV40 промоторы при добавлении к источнику репликации, HBV гены могут быть экспрессированы под контролем любого промотора в зависимости от его ориентации (р HOS 348-Е является результатом HBS экспрессии под контролем раннего промотора). Модифицируют рЕ342 частичным гидролизом EcoRI, заполняя отщепленный участок с использованием ДНК полимеразы I Кленова и связывая вместе полученные части плазмиды, удаляя таким образом EcoRI Сайм, предшествующий SV40 промотору в рЕ343. Полученную в результате плазмиду рЕ342 Δ РI обрабатывают EcoRI, заполняют с помощью ДНК полимеразы I Кленова и гидролизуют BamHI. После электрофореза на акриламидном геле электроэлюируют фрагмент размером приблизительно 3500 bp, экстрагируют фенолом и хлороформом, осаждают этанолом как описано выше.
Конструируют вектор р342 Е размером 3500 bp и связывают вместе описанные выше т-АП фрагменты размером приблизительно 2160 bp, используя стандартные методики. Выделяют плазмиду, содержащую три фрагмента кодирующих т-АП в четкой ориентации, характеризуют и обрабатывают рЕ342-АП. Эту плазмиду гидролизуют Sac II и обрабатывают бактериальной щелочной фосфатазой (BRL). Для встраивания ДГФР последовательности (наряду с контрольными последовательностями для ее экспрессии) выделяют фрагмент размером приблизительно 1700 bp обработкой pЕНЕR Sac II. (pEHER является плазмидой, экспрессирующей мутант ДГФР). Этот фрагмент встраивают в рЕ341-АП плазмиду для создания рЕТРАЕR400 плазмиды, которая является аналогичной рЕНЕR за исключением того, что HESAg кодирующая область заменена кДНК последовательностями т-АП.
Проводят трансфекцию рЕТРАЕ 400 (рЕТРЕR) как в d СНО-DUX BII клетки и DГФР+СНО-КI (ATCC CCL6I) клетки по методу Грэхэма и Ван дер Эба. Отбирают трансформированные dhfr клетки путем роста на среде,лишенной глицина, гипоксантина и тимидина. Отбирают трансформированные ДГФР+клетки выращиванием в ≥ 100 мМ МТТ. Колонии, которые возникают на соответствующей селекционной среде, выделяют, используя клонирующие циклы, и размножают на той же самой среде до нескольких генераций.
Для амилификации клетки из колоний расщепляют в среде, содержащей 5 ˙104, 105, 2,5 ˙105, 5 ˙105 и 106 нМ МТТ и пассируют несколько раз. Клетки помещают на пластины при очень низких плотностях (102-103клеток/пластину) в 10 см чашки и выделяют полученные в результате колонии.
Испытывают коамплификацию ДГФР и т-АП последовательностей путем выдедения ДНК из сливных монослоев амплифицированных колоний следующим образом. Промывают сливные монослои в 150 мм пластинах 50 мл стерильного pBS и лизируют добавлением 5 мл 0,1% SDS, 0,4 М СaCl2, 0,1 М ЗДАТУК, рН 8. Через 5-10 мин удаляют смесь, экстрагируют фенолом, экстрагируют хлороформом и осаждают этанолом. Повторно суспендируют ДНК в 1 мл (на 150 мм пластине) 10 мМ Трис-HCl рН 8, 1 мМ ЭДАТУК (ТЕ), добавляют RNase до 0,1 мг/мл и раствор инкубируют 30 мин при 37оС. Затем прибавляют SDS до 0,1% прибавляют проназу (сигма) до 0,5 мг/мл. После 3-16 ч инкубации при 37оС раствор снова экстрагируют фенолом, экстрагируют хлороформом и осаждают этанолом. Повторно суспендируют таблетку ДНК в 0,5 мл воды и обрабатывают рестриктазами. Приблизительно 5-10 мкг обработанной ДНК подвергают электрофорезу на агарозном геле 1% агарозы в Трис-ацетатном буфере (40 мМ Трис, 1 мМ ЭДАТУК, устанавливливают рН 8,2 уксусной кислотой).
После того как краситель бромфеноловый синий пройдет 2/3 пути по гелю, гель извлекают и окрашивают этидий-бромидом. После визуализации ДНК в ультрафиолетовом свете ДНК переносят из геля на нитроцеллюлозные фильтры. Затем фильтры гибридизуют с транслированным зондом, изготовленным из фрагмента 1700 bp Sac II pEHER или из Bgl II фрагмента размером приблизительно 1970 bp pETPER.
П р и м е р 5. По методике, аналогичной использованной при конструировании pETPER, конструируют плазмиду pETPFR, содержащую последовательность ДНК, кодирующую ДГФР дикого типа. Вместо плазмиды рЕНЕ в качестве источника ДГФР гена, кодирующего, используют плазмиду рЕ342.НВУ.Е400.Д22. Плазмида рЕ342. НВУ. Е400. Д22 является такой же, как и рЕРЕR с тем источником, что имеется единственное различие в основной паре оснований между диким типом и мутантом ДГФР. Следовательно, полученная в результате плазмида рЕТРFR является аналогичной рЕТРЕ, исключая последовательность ДНК, кодирующую дикий тип ДГФР, замененную на последовательность мутанта.
Используют pETPFR для трансфекции лишенных ДГФР клеток СНО, используя метод кальций-фосфатного осаждения. Испытывают 21 колонию, выросшую на селективной среде (-HGT), детекцией на образование плазмина по разрушению фибрина на агарозной пластине.
Затем количественно испытывают четыре из отобранных клона на образование плазмина на клеточную основу.
При таком количественном детектировании было обнаружено, что четыре испытуемых клона обнаруживают одинаковую или сравнимую т-АП секрецию в среде, определенную как единицы (клетку) день. Готовят субклоны, переносят инокулумы от двух клонов на отдельные пластины, содержащие HGT-среду. Для дальнейшего анализа используют два из полученных в результате субклона.
Указанные выше субклоны помещают в количестве 2˙ 105 клеток на пластины 100 мл в 50 нМ МТТ для индукции амплификации. Выжившие клетки во всех случаях дают примерно в 10 раз большую активность активатора тканевого плазминогена. Отбирают два из этих клонов для дальнейшего исследования и называют их 1-15 и 18В-9.
Субклон 1-15 снова амплифицируют, высевая 2 ˙105 клеток на пластины 100 мм, содержащие 500 нм МТТ. Биопробы с амплифицированными таким образом дают еще большее увеличение количества (примерно в 3 раза) т-АП, уровни находились в области 7 ˙10-4 единицы (клетку) день. Затем часть этих амплифицированных клеток переносят и выдерживают в присутствии 10000 нм МТТ. Затем испытывают субклоны 1-15 и 18В-9 после выдерживания примерно в течение 1-2 месяцев в условиях, приведенных в табл. 3.
Количественно испытывают т-АП в культуральной среде в радиоиммунном испытании следующим образом.
Серийно разбавляют очищенный т-АП и очищенный меченый иодом т-АП, полеченные из клеток меланомы, чтобы получить коцнентрации от 12,5 до 400 мг/мл в буфере, содержащем забуферированный фосфатом солевой раствор, рН 7,3, 0,5% сыворотки бытового альбумина, 0,01% Твина 30 и 0,02% NaN3. Образцы с соответствующими разбавлениями среды, подлежащие испытанию, прибавляют к радиоактивно меченым протеинам. Антигены инкубируют в течение ночи при комнатной температуре в присутствии 1:10000 разбавления IgG фракции анти-т-АП антисыворотки кролика. Осаждают комплекс антиген-антитело путем адсорбции коза-анти-кролик IgG иммунобусинок (Био Рад) в течение 2 ч при комнатной температуре. Шарики очищают добавлением разбавленного буфера с последующим центрифугированием в течение 10 мин при 2000 х g при 4оС. Надосадочный слой отбрасывают и измеряют радиоактивность осадков. Определяют коцнентрации при сравнении с внутренним стандартом.
Использовали следующие линии клеток. Линия клеток "1" представляет собой неамплифицированный клон из четырех детектированных клонов; "1-15500" представляет собой амплифицированный субклон линии клеток "1", который амплифицирован первоначально в 50 нМ МТТ, чтобы получить 1-15 и затем перенесен для дальнейшей амплификации в 500 нММТТ; 1-1510000является субклоном 1-15500, который был еще амплифицирован в присутствии 10000 нМ МТТ. Линия клеток 18В-9 представляет собой субклон одного из оригинальных четырех детектированных, который был амплифицирован на 40 нМ МТТ.
Все амплифицированные клетки показали повышенные уровни продуцирования т-АП по сравнению с теми, что показывают неамплифицированные культуры клеток. Если неамплифицированная культура продуцирует количества т-АП больше 0,5 пг (клетку) день, то амплификация в результате дает уровни, достигающие 50 пг (клетку) день.
П р и м е р 6. Активатор тканевого плазминогена объединяют в смесь с фармацевтически приемлемым носителем. Подходящие носители и другие человеческие протеины, например сывороточный человеческий альбумин, будут содержать эффективное количество протеина вместе с подходящим количеством носителя, чтобы получить фармацевтические приемлемые композиции, пригодные для эффективности введения хозяину.
Например, активатор плазминогена человеческой ткани может быть парентерально введен субъектам, страдающим ее сердечно-сосудистыми заболеваниями, расстройствами. Дозировка и частота введения дозы могут быть параллельными обычно применяемым при клинических исследованиях другим сердечно-сосудистых тромболитических агентом, например примерно 440 IV/кг веса тела, как внутривенная первичная доза с последующей постоянной внутривенной инфузией примерно 440 IV/кг/ч в течение 12 ч для пациентов, страдающих легочной эмболией.
В качестве примера может быть приведена рецептура, содежащая активатор тканевого плазминогена активностью 25000 IV 25 мг маннита и 45 мг NaCl, который может быть объединен с 5 мл стерильной воды для инъекции и смешан с подходящим объемом 0,9%-ного хлористого натрия для инъекций или 5%-ной декстрозой для инъекций для внутривенного введения.
Структура конкретного варианта человеческого т-АП, полученногоо в примерах, была изучена в некоторых деталях. Была определена последовательность нуклеотидов, кодирующая т-АТ, изучена биохимия протеинов.
Ранее было показано, что двуцепочечный человеческий т-АП образуется при протеолитическом отщеплении одноцепочечной молекулы в двух полипептидах, соединенных дисульфидными связями. Настоящая работа позволяет сделать вывод, что тяжелая цепь (30882 мол.масса) происходит из NH2-терминальной части, а легкая цепь (мол.масса 28126) включает СООН-терминальную область. Исследование N-терминального конца двуцепочечной молекулы позволяет сделать вывод, что двунитевая форма образуется при расщеплении единственной аргинин-изолейциновой связи (фиг.4, отмечено стрелкой).
Первичная структура части области тяжелой цепи человеческого т-АП (фиг. 4) подтверждает высокую степень гомологии последовательности с "кнингл"-областями плазминогена и протромбина. "Крингл"-область означает триплетную дисульфидную структуру, первоначально обнаруженную в "про/"фрагменте протромбина. В первичной последовательности т-АП имеется две так называемые "крингл"-области из 82 аминокислот, которые обладают высокой степенью гомологии с 5 "крингл"-областями плазминогена. Оставшиеся N-терминальные аминокислоты (91) обладают небольшой гомологией с традиционной "крингл"-областью. Можно предположить, что эта область содержит структуру с множественными дисульфидными связями, так как здесь было найдено 11 дополнительных цистеиновых остатков.
Каталитический участок легкой цепи человеческого т-АП, так называемая область сериновой протеазы, как в других серийновых ферментах образуется гистидин 322, аспаргин 371 и серин 478 остатками. Кроме того, аминокислотные последовательности, окружающие эти остатки, являются гомологичными к соответствующим частям других серин протеаз, таких как трипсин, промромбин и плазминоген.
GTTCTGAGCACAGGGCTGGAGAGAAAACCTCTGCGAGGAAAGGGAAGGAGCAAGCCGTGA
35 -30
met aspala met lys arg gly leu
ATTTAAGGGACGCTGTGAAGCAATGC ATG GAT GCA ATG AAG AGA GGG CTC
-20
cys cys val leu leu leu cys gly ala val phe val ser pro ser
TGC TGT GTG CTG CTG CTG TGT GGA GCA GTC TTC GTT TCG CCC AGC
-10 1
gln glu ile his ala arg phe arg arg gly ala arg SER TYR GLN
CAG CAA ATC CAT GCC CGA TTC AGA AGA GGA GCC AGA TCT TAC CAA
10
VAL ILA CYS ARG ASP GLU LYS THR GLN MET ILE TYR GLN GLN HIS
GTG ATC TGC AGA GAT GAA AAA ACG CAG ATG ATA TAC CAG CAA CAT
20 30
GLN SER TRP LEU ARG PRO VAL LEU ARG SER ASN ARG VAL GLU TYR
CAG TCA TGG CTG CGC CCT GTG CTC AGA AGC AAC CGG GTG GAA TAT
40
CYS TRP CYS ASN SER GLY ARG ALA GLN CYS HIS SER VAL PRO VAL
TGC TGG TGC AAC AGN GGC AGG GCA CAG TGC CAC TCA GG CCT GTC
50 60
LYS SER SYS SER GLU PRO ARG CYS PHE ASN GLY GLY THR CYS GLN
AAA AGT TGC AGC GAG CCA AGG TGT TTC AAC GGG GGC ACC TGC CAG
70
GLN ALA LEU TYR PHE SER ASP PHE VAL CYS GLN CES PRO GLU GLY
CAG GCC CTG TAC TTC TCA GAT TTC GTG TGC CAG TGC CCC GAA GGA
80 90
PHE ALA GCY LYS CYS CYS GLU ILE ASP THR ARC ALA THR CYS TYR
TTT GCT GGG AAG TGC TGT GAA ATA GAT ACC AGG GCC ACG TGC TAC
100
GLU ASP GLN GLY ILE SER TYE ARS GLY THR TRP SER THR ALA GLU
GAG GAC CAG GGC ATC AGC TAC AGG GGC ACG TGG AGC ACA GCG GAG
110 120
SER GLY ALA GLU CYS THR ASN TRP ASN SER SER ALA LEU ALA GLN
AGT GGC GCC GAG TGC ACC AAC TGG AAC AGC AGC GCG TTG GCC CAG
130
LYS PRO TYR SER GLY ARG ARG PRO ASP ALA ILE ARG LEU GLY LEU
AAG CCC TAC AGC GGG CGG AGG CCA GAC GCC ATC AGG CTG GGC CTG
140 150
GLY ASN HIS ASN TYR CYS ARG ASN PRO ASP ARG ASP SER LYS PRO
GGG AAC CAC AAC TAC TGC AGA AAC CCA GAT CGA GAC TCA AAG CCC
160
TRP CYS TYR VAL THE LYS ALA GLY LYS TYR SER SER GLU PHE CYS
TGG TGC TAC GTC TTT AAG GCG GGG AAG TAC AGC TCA GAG TTC TGC
170 180
SER THR PRO ALA CYS SER GLU GLY ASN SER ASP CYS TYR PHE GLY
AGC ACC CCT GCC TGC TCT GAG GGA AAC AGT GAC TGC TAC TTT GGG
190
ASN GLY SER ALA TYR ARG GLY THR HIS SER LEU THR GLU SER GLY
AAT GGG TCA GCC TAC CGT GGC ACG CAC AGC CTC ACC GAG TCG GGT
200 210
ALA SER CYS LEU PRO TRP ASN SER MET ILE LEU ILE GLY LYS VAL
GCC TCC TGC CTC CCG TGG AAT TCC ATG ATC CTG ATA GGC AAG GTT
220
TYR THR ALA GLN ASN PRO SER ALA GLN ALA LEU GLY LEU GLY LYS
TAC ACA GCA CAG AAC CCC AGT GCC CAG GCA CTG GGC CTG GGC AAA
230 240
HYS ASN TYR CYS ARG ASN PRO ASP GLY ASP ALA LYS PRO TRP CYS
CAT AAT TAC TGC CGG AAT CCT GAT GGG GAT GCC AAG CCC TGG TGC
250
HYS VAL LEU LYS ASN ARG ARG LEU THR TRP GLU TYR CYS ASP VAL
CAC GTG CTG AAG AAC CGC AGG CTG ACG TGG GAG TAC TGT GAT GTG
260 270
PRO SER CYS SER THR CYS GLY LEU ARG GLN TYR SER GLN PRO GLN
CCC TCC TGC TCC ACC TGC GGC CTG AGA CAG TAC AGC CAG CCT CAG
280
PHE ARG ILE LYS GLY GLY LEU PHE ALA ASP ILE ALA SER HIS PRO
TTT CGC ATC AAA GGA GGG CTC TTC GCC GAC ATC GCC TCC CAC CCC
280 300
TRP GLN ALA ALA ILE PHE ALA LYS HIS ARG ARG SER PRO GLY GLU
TGG CAG GCT GCC ATC TTT GCC AAG CAC AGG AGG TCG CCC GGA GAG
310
ARG PHE LEU CYS GLY GLY ILE LEU ILE SER SER CYS TRP ILE LEU
CGG TTC CTG TGC GGG GGC ATA CTC ATC AGC TCC TGC TGG ATT CTC
320 330
SER ALA ALA HIS CYS PHE GLN GLU ARG PHE PRO PRO HIS HIS LEU
TCT GCC GCC CAC TGC TTC CAG GAG AGG TTT CCG CCC CAC CAC CTG
340
THR VAL ILE LEU GLY ARG THR TYR ARG VAL VAL PRO GLY GLU GLU
ACG GTG ATC TTG GGC AGA ACA TAC CGG GTG GTC CCT GGC GAG GAG
350 360
GLU GLN LYS PHE GLU VAL GLU LYS TYR ILE VAL HIS LYS GLU PHE
GAG CAG AAA TTT GAA GTC GAA AAA TAC ATT GTC CAT AAG GAA TTC
370
ASP ASP ASP THR TYR ASP ASN ASP ILE ALA LEU LEU GLN LEU LYS
GAT GAT GAC ACT TAC GAC AAT GAC ATT GCG CTG CTG CAG CTG AAA
380 390
SER ASP SER SER ARG CYS ALA GLN GLU SER SER VAL VAL ARG THR
TCG GAT TCG TCC CGC TGT GCC CAG GAG AGC AGC GTG GTC CGC ACT
400
VAL CYS LEU PRO PRO ALA ASP LEU GLN LEU PRO ASP TRP THR GLU
GTG TGC CTT CCC CCG GCG GAC CTG CAG CTG CCG GAC TGG ACG GAG
410
CYS GLU LEU SER GLY TYR GLY LYS HIS GLU ALA LEU SER PRO PHE
TGT GAG CTC TCC GGC TAC GGC AAG CAT GAG GCC TTG TCT CCT TTC
430
TYR SER GLU ARG LEU LYS GLU ALA HIS VAL ARG LEU TYR PRO SER
TAT TCG GAG CGG CTG AAG GAG GCT CAT GTC AGA CTG TAC CCA TCC
440 450
SER ARG CYS THR SER GLN HIS LEU LEU ASN ARG THR VAL THR ASP
AGC CGC TGC ACA TCA CAA CAT TTA CTT AAC AGA ACA GTC ACC GAC
460
ASN MET LEU CYS ALA GLY ASP THR ARG SER GLY GLY PRO GLN ALA
AAC ATG CTG TGT GCT GGA GAC ACT CGG AGC GGC GGG CCC CAG GCA
470 480
ASN LEU HIS ASP ALA CYS GLN GLY ASP SER GLY GLY PRO LEU VAL
AAC TTG CAC GAC GCC TGC CAG GGG GAT TCG GGA GGC CCC CTG GTG
490
CYS LEU ASN ASP GLY ARG MET THR LEU VAL GLY ILE ILE SER TRP
TGT CTG AAC GAT GGC CGC ATG ACT TTG GTG GGC ATC ATC AGC TGG
500 510
GLY LEU GLY CYS GLY GLN LYS ASP VAL PRO GLY VAL TYR THR LYS
GGC CTG GGC TGT GGA CAG AAG GAT GTC CCG GGT GTG TAC ACA AAG
520 527
VAL THR ASN TYR LEU ASP TRP ILE ARG ASP ASN MET ARG PRO OP
GTT ACC AAC TAC CTA GAC TGG ATT CGT GAC AAC ATG CGA CCG TGA
CCAGGAACACCCGACTCCTCAAAAGCAAATGAGATCCCGCCTCTTCTTCTTCAGAAGACA
CTGCAAAGGCGCAGTGCTTCTCTACAGACTTCTCCAGACCCACCACACCGCAGAAGCGGG
ACGAGACCCTACAGGGAGAGGGAAGAGTGCATTTTCCCAGATACTTCCCATTTTGGAAGT
TTTCAGGACTTGGTCTGATTTCAGGATACTCTGTCAGATGGGAAGACATGAATGCACACT
AGCCTCTCCAGGAATGCCTCCTCCCTGGGCAGAAGTGGCCATGCCACCCTGTTTTCGCTA
AAGCCCAACCTCCTGACCTGTCACCGTGAGCAGCTTTGGAAACAGGACCACAAAAATGAA
AGCATGTCTCAATAGTAAAAGAAACAAGAGATCTTTCAGGAAAGACGGATTGCATTAGAA
ATAGACAGTATATTTATAGTCACAAGGGCCCAGCAGGGCTCAAAGTTGGGGCAGGCTGGC
TGGCCCGTCATGTTCCTCAAAAGCGCCCTTGACGTCAAGTCTCCTTCCCCTTTCCCCACT
CCCTGGCTCTCAGAAGGTATTCCTTTTGAGTACAGTGTGTAAAGTGTAAATCCTTTTTCT
TTATAAACTTTAGAGTAGCATGAGAGAATTGTATCATTTGAACAACTAGGCTTCAGCATA
TTTATAGCGATCCATCGTTAGTTTTTACTTTCCGTTGCCACAACCCTGTTTTATACCGTA
CTTAATAAATTCGGATATATTTTTTCACAGTTTTTTCCAAAAAAAAAAAAAA
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм бактерий ЕSснеRIснIа coLI - продуцент активатора плазминогена тканевого типа | 1983 |
|
SU1662352A3 |
| СПОСОБ ПОЛУЧЕНИЯ ЗИМОГЕННОЙ ФОРМЫ ЧЕЛОВЕЧЕСКОГО БЕЛКА С | 1989 |
|
RU2018535C1 |
| Способ конструирования рекомбинантной плазмидной ДНК, кодирующей зимоген С человека | 1988 |
|
SU1739854A3 |
| Способ получения человеческого эритропоэтина | 1987 |
|
SU1801118A3 |
| Способ получения рекомбинантной плазмидной ДНК и способ получения зимогенной формы белка С человека | 1991 |
|
SU1838411A3 |
| Способ получения водорастворимой части человеческого рецептора малого сродства F @ | 1987 |
|
SU1727533A3 |
| Способ получения рекомбинантного активированного белка С человека | 1988 |
|
SU1830081A3 |
| ИММУНОИНДУЦИРУЮЩИЙ АГЕНТ | 2017 |
|
RU2777992C2 |
| ИММУНОИНДУЦИРУЮЩЕЕ СРЕДСТВО | 2016 |
|
RU2744843C2 |
| АНТИТЕЛО ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2781148C2 |
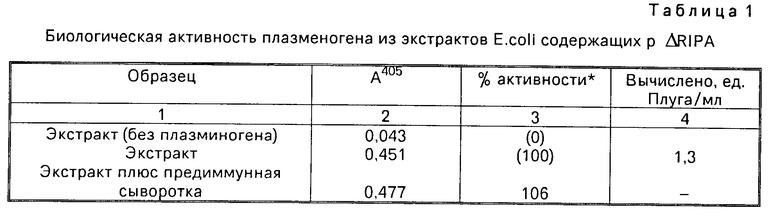

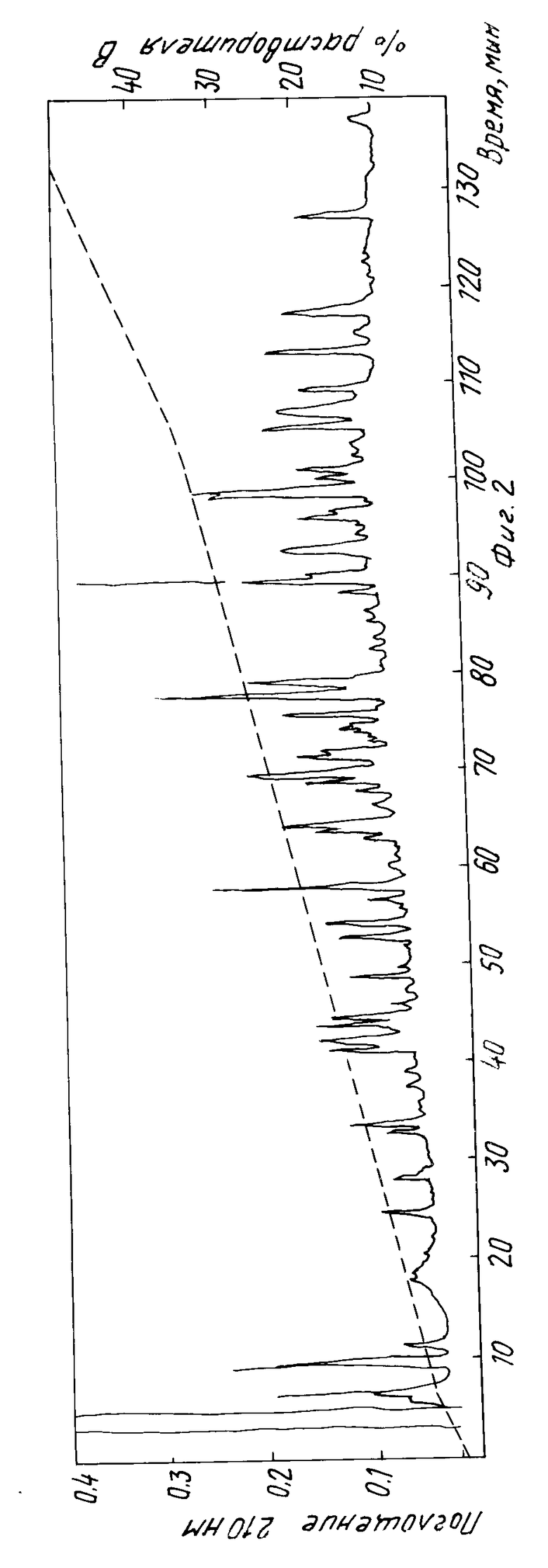
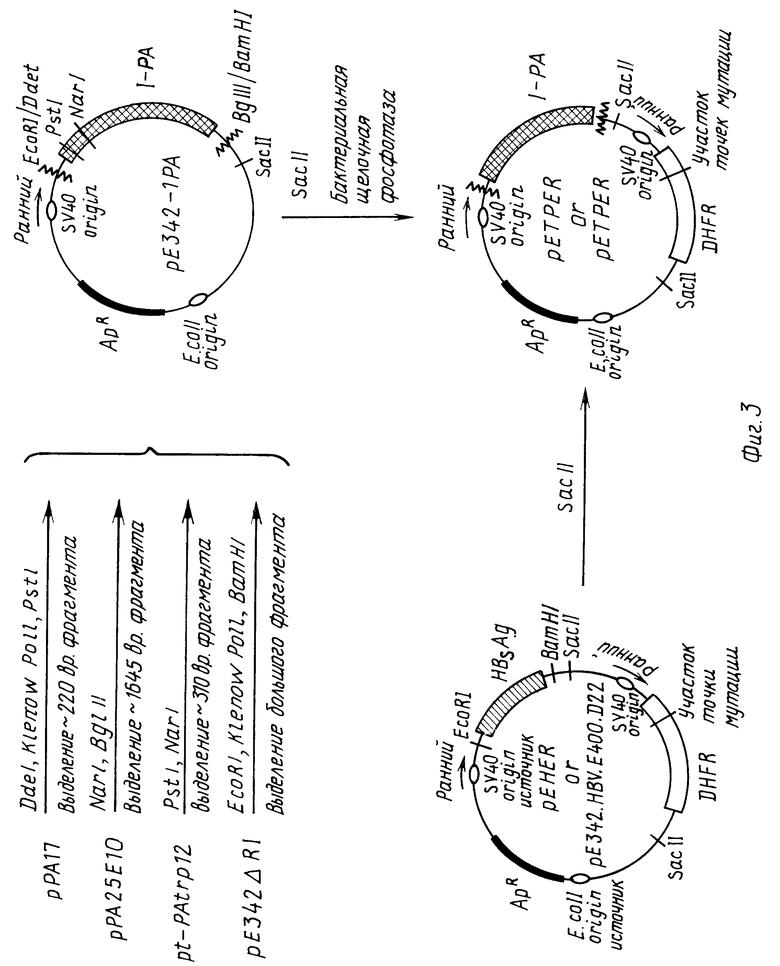

Использование: изобретение относится к генетической инженерии, в частности к способу получения активатора плазминогена тканевого типа с использованием штаммов культивируемых клеток животных. Сущность изобретения: способ получения активатора плазминогена тканевого типа предусматривает культивирование штамма CHO KI АТСС NCCL61, трансформированного рекомбинантной плазмидой ДНК PETPER, выращивание трансформантов в присутствии конкурентного ингибитора активности DHFR в концентрации 20-500 000 нм, выделение и очистку целевого продукта. 2 с. п. ф-лы, 4 ил. 3 табл.
| Устройство для сухой перегонки бересты | 1932 |
|
SU37687A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
