Предпосылки изобретения
Настоящее изобретение относится к применению ксенона в качестве нейропротектора и/или при ингибировании синаптической пластичности. В частности, настоящее изобретение относится к применению ксенона в качестве антагониста NMDA, и к его применению при лечении состояний, связанных с активностью рецепторов NMDA.
Рецепторы NMDA (N-метил-D-аспартат) представляют собой большой подкласс глютаматных рецепторов глютаматов, а глютамат, как предполагается, является наиболее важным нейротрансмиттером возбуждения в центральной нервной системе млекопитающих. Что важно, активация рецептора NMDA, как показано, является центральным событием, которое ведет к эксцитотоксичности и гибели нервных клеток при многих болезненных состояниях, а также в результате гипоксии и ишемии как следствия черепно-мозговой травмы, инсульта и остановки сердца.
В данной области известно, что рецептор NMDA играет главную роль в синаптической пластичности, которая лежит в основе многих высших познавательных функций, таких как память и обучаемость, а также в некоторых ноцицептивных путях и при восприятии боли (Collingridge et al. The NMDA Receptor, Oxford University Press, 1994). Кроме того, некоторые свойства рецепторов NMDA говорят о том, что они могут быть вовлечены в обработку информации в мозге, которая лежит в основе сознания как такового.
Антагонисты рецепторов NMDA обладают терапевтической ценностью по ряду причин, таких как следующие далее три конкретных причины. Во-первых, антагонисты рецепторов NMDA соотносятся с выраженной глубокой аналгезией, в высшей степени желательным компонентом общей анестезии и седативного воздействия. Во-вторых, антагонисты рецепторов NMDA являются нейропротекторами во многих клинически важных обстоятельствах (включая ишемию, черепно-мозговую травму, невропатологические болезненные состояния и определенные типы конвульсий). В-третьих, антагонисты рецепторов NMDA связывают со значительной степенью амнезии.
Однако из литературных данных ясно, что существует ряд недостатков, связываемых с современными антагонистами рецепторов NMDA. Они включают совершение непроизвольных движений, стимуляцию симпатической нервной системы, индуцирование нейротоксичности при высоких дозах (что является важным, поскольку антагонисты рецепторов NMDA имеют слабое воздействие в качестве общих анестетиков), подавление активности миокарда и проконвульсии в некоторых эпилептогенных парадигмах, например, "разжигание" (Wlaz P. et al, Eur. J.Neurosci. 1994; 6:1710-1719). В частности, существуют значительные сложности при разработке новых антагонистов рецепторов NMDA, которые способны преодолевать гематоэнцефалический барьер. Этот фактор также ограничивает терапевтические применения многих известных антагонистов NMDA.
Настоящее изобретение, таким образом, относится к усовершенствованному антагонисту рецепторов NMDA для общего фармацевтического использования, который может легко диффундировать через гематоэнцефалический барьер.
Сущность изобретения
Аспекты настоящего изобретения представлены в прилагаемой формуле изобретения и в следующем далее описании.
Таким образом, настоящее изобретение относится к применению ксенона в качестве нейропротектора и/или в качестве ингибитора синаптической пластичности. В предпочтительном аспекте, ксенон действует в качестве антагониста рецептора NMDA.
Во втором аспекте, настоящее изобретение относится к способу лечения, включающему модуляцию активности рецепторов NMDA у млекопитающих, способ включает модуляцию активности рецептора NMDA путем введения млекопитающему терапевтически эффективного количества ксенона.
В предпочтительном аспекте настоящего изобретения, ксенон вводится в сочетании с фармацевтически приемлемым носителем, разбавителем или наполнителем.
Предпочтительно, способ лечения может быть использован для лечения млекопитающего, страдающего от состояния, связанного с активностью рецепторов NMDA. В более предпочтительном аспекте, млекопитающее лечится от состояния, которое связывается с активацией рецепторов NMDA.
Еще более предпочтительно, настоящее изобретение относится к способу лечения, где ксенон понижает уровень активации рецепторов NMDA.
Другое воплощение настоящего изобретения относится к способу получения фармацевтической композиции, пригодной для использования при модуляции активности рецептора NMDA, этот способ включает добавление антагониста NMDA к фармацевтически приемлемому носителю, наполнителю или разбавителю, где усовершенствование включает использование ксенона в качестве антагониста NMDA.
Следующее воплощение настоящего изобретения относится к фармацевтической композиции для модуляции активности NMDA, которая содержит антагонист NMDA и фармацевтически приемлемый носитель, наполнитель или разбавитель, где усовершенствование включает использование ксенона в качестве антагониста NMDA.
Еще одно воплощение настоящего изобретения относится к использованию ксенона при получении фармацевтического средства для использования при модуляции активности рецепторов NMDA у млекопитающих.
В предпочтительном аспекте, ксенон в фармацевтическом средстве используется в сочетании с фармацевтически приемлемым носителем, разбавителем или наполнителем.
Предпочтительно, фармацевтическое средство предназначается для использования при лечении состояния, связанного с активностью рецепторов NMDA. В более предпочтительном аспекте, фармацевтическое средство предназначено для использования при лечении состояния, связанного с активацией рецепторов NMDA. Еще более предпочтительно, фармацевтическое средство предназначено для использования при понижении уровня активации рецептора NMDA.
Подробное описание изобретения
В широком аспекте, настоящее изобретение относится к применению ксенона в качестве нейропротектора и/или в качестве ингибитора синаптической пластичности. В предпочтительном аспекте, ксенон действует в качестве антагониста рецепторов NMDA.
Ксенон представляет собой химически инертный газ, анестетические свойства которого известны в течение более 50 лет (Lawrence JH et al, J.Physiol. 1946; 105:197-204). Co времени его первого использования в хирургии (Cullen SC et al, Science 1951; 113:580-582), несколько исследовательских групп показали, что он имеет превосходные фармакологические свойства, включая отсутствие побочных продуктов метаболизма, глубокую аналгезию, быстрое начало воздействия и его окончание и минимальное воздействие на сердечно-сосудистую систему (Lachmann in et al, Lancet 1990; 335:1413-1415; Kennedy RR et al, Anaesth. Intens. Care 1992; 20:66-70; Luttropp HH et al, Acta Anaesthesiol. Scand. 1994; 38:121-125; Goto Т. et al, Anesthesiology 1997; 86:1273-1278; Marx Т. et al, Br. J. Anaesth. 1997; 78:326-327). Однако, до сих пор, молекулярные механизмы, лежащие в основе клинической активности ксенона, остаются известными только приблизительно.
Авторы описали применение ксенона при фармацевтическом применении в заявке на патент Соединенного Королевства номер GB 9913677.2 (подано 11 июня 1999 года), содержание которой включается сюда в качестве ссылки.
Термин "антагонист" используется в его обычном смысле, используемом в данной области, то есть как химическое соединение, которое предотвращает функциональную активацию рецептора его природным агонистом (глютаматом, в данном случае).
Является широко известным, что другие анестетики усиливают активность рецепторов ингибиторной ГАМК (у-аминомасляная кислота типа A) (Franks NP et al Nature 1994; 367: 607-614; Mihic SJ et al, Nature 1997; 389: 385-389). Однако воздействие ксенона на эти рецепторы, как предполагается, является очень слабым. Вместо этого, ксенон сильно ингибирует каналы рецепторов NMDA, обуславливающих возбуждение. Это воздействие участвует во многих важных особенностях его фармакологических свойств и, видимо, является инструментом анестетического и аналгетического воздействия этого инертного газа.
Необходимо заметить, что в литературе нигде не описывается и не предполагается использование ксенона в качестве нейропротектора и/или ингибитора синаптической пластичности. В литературе также не описывается и не предполагается использование ксенона в качестве антагониста рецепторов NMDA.
В отличие от многих других антагонистов NMDA, ксенон способен быстро достигать равновесия в мозге путем диффузии через гематоэнцефалический барьер. Дополнительным преимуществом использования ксенона в качестве антагониста NMDA является то, что молекула представляет собой инертный, летучий газ, который легко удаляется при выдохе.
Таким образом, настоящее изобретение относится к использованию ксенона для достижения одной или нескольких из следующих целей:
a) обеспечение нейропротекции;
b) ингибирование синаптической пластичности, например ингибирование развития толерантности к опиатам.
Термин "нейропротекция" означает защиту объекта нервной системы, такого как нейрон, в месте повреждения, например ишемического повреждения или травматического повреждения.
В следующем воплощении, настоящее изобретение предусматривает способ осуществления нейропротекции и/или ингибирования синаптической пластичности у млекопитающих, который включает введение млекопитающему терапевтически эффективного количества ксенона.
Предпочтительно, ксенон вводится в сочетании с фармацевтически приемлемым носителем, разбавителем или наполнителем. В качестве примера, в фармацевтических композициях по настоящему изобретению, ксенон может быть смешан с любым соответствующим связующим (связующими, любрикантом (любрикантами), суспендирующим агентом (агентами), агентом (агентами) для покрытий, солюбилизирующим агентом (агентами), выбираемыми в соответствии с предлагаемым способом введения и обычной фармацевтической практикой.
Ксенон может также быть введен в сочетании с другим фармацевтически активным агентом. Этот агент может быть любым соответствующим фармацевтически активным агентом, включая анестетические или седативные агенты, которые способствуют ГАМКэргической активности. Примеры таких ГАМКэргических агентов включают изофлуран, пропофол и бензодиазапины.
В другом воплощении, настоящее изобретение относится к способу понижения уровня активации рецепторов NMDA у млекопитающих, который включает модуляцию активности рецептора NMDA путем введения млекопитающему терапевтически эффективного количества ксенона, где указанное понижение достигает нейропротекции и/или ингибирования синаптической пластичности.
Дальнейшее воплощение настоящего изобретения относится к способу получения фармацевтической композиции, пригодной для использования при нейропротекции и/или ингибировании синаптической пластичности, который включает добавление ксенона к фармацевтически приемлемому носителю, наполнителю или разбавителю, где усовершенствование включает использование ксенона в качестве нейропротектора и/или ингибитора синаптической пластичности.
В другом воплощении, настоящее изобретение относится к фармацевтической композиции для нейропротекции и/или ингибирования синаптической пластичности, которая содержит ксенон и фармацевтически приемлемый носитель, наполнитель или разбавитель, где усовершенствование включает использование ксенона в качестве нейропротектора и/или ингибитора синаптической пластичности.
Как правило, фармацевтические композиции по настоящему изобретению могут доставляться внутривенно (либо путем приема пилюль, либо путем вливания), в осевую часть центральной нервной системы (либо субдурально, либо субарахноидально), трансдермально, либо путем ингаляции.
В качестве примера, настоящее изобретение также относится к применению ксенона для получения лекарственного средства для одной или нескольких из следующих целей:
a) для обеспечения нейропротекции;
b) для ингибирования синаптической пластичности, например, для ингибирования развития толерантности к опиатам.
Концентрация ксенона, используемая в композиции, может представлять собой минимальную концентрацию, требуемую для достижения желаемого клинического воздействия. Для врача является обычным определение реальной дозы, которая была бы наиболее соответствующей для конкретного пациента, и эта доза будет изменяться с возрастом, массой и восприимчивостью конкретного пациента. Могут, разумеется, существовать индивидуальные случаи, где предполагаются диапазоны более высоких или более низких доз, и они находятся в рамках настоящего изобретения.
Фармацевтическая композиция по настоящему изобретению может предназначаться для введения людям или для введения животным.
Таким образом, композиция по настоящему изобретению может также использоваться в качестве лекарственного средства для животных. В этом отношении, настоящее изобретение дополнительно относится к ветеринарной композиции, содержащей композицию по настоящему изобретению и ветеринарно приемлемый разбавитель, наполнитель или носитель.
Для ветеринарного использования композиция по настоящему изобретению или его ветеринарно-приемлемая композиция, как правило, вводится в соответствии с обычной ветеринарной практикой, и ветеринар будет определять режим дозировки и способ введения, которые будут наиболее соответствовать конкретному животному.
Далее настоящее изобретение описывается со ссылкой на прилагаемые чертежи, где:
Фиг.1 демонстрирует контрольные постсинаптические отклики возбуждения и рефрактерности.
Фиг.2 демонстрирует репрезентативные графики, иллюстрирующие воздействие ксенона на постсинаптические отклики возбуждения и рефрактерности.
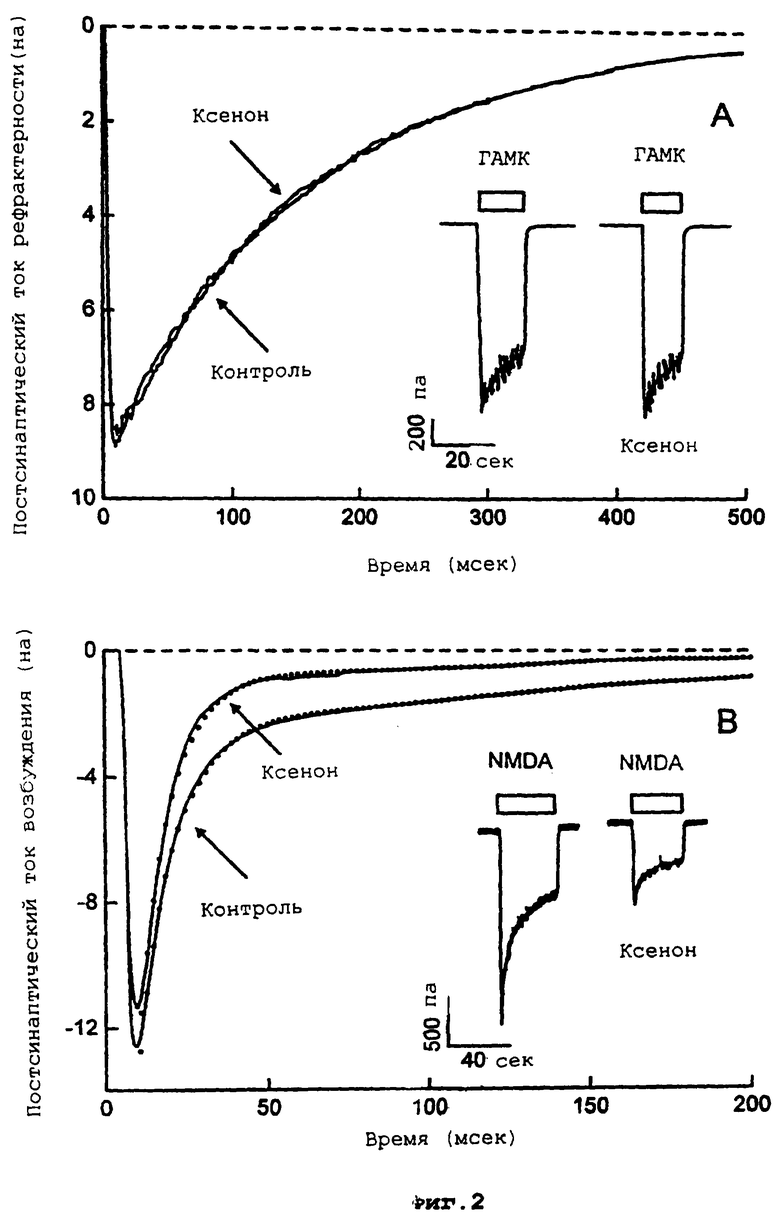
Фиг.3 демонстрирует процент от максимального тока в контроле как функцию концентрации NMDA, в присутствии и в отсутствие ксенона.
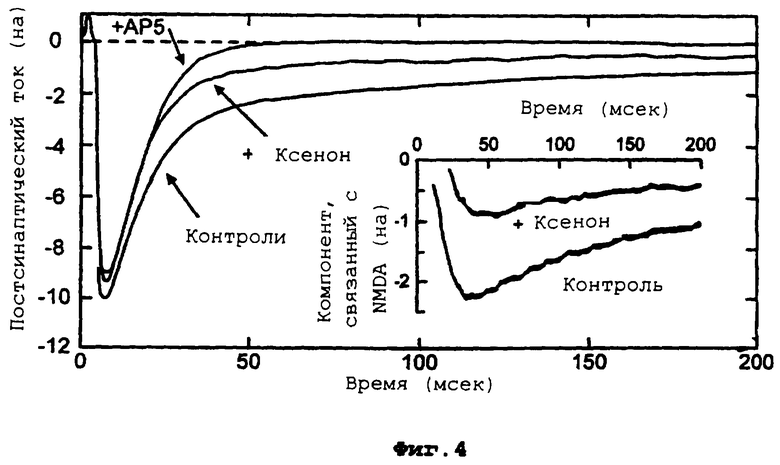
Фиг.4 демонстрирует воздействие АР5 и ксенона на постсинаптические отклики возбуждения, и на врезке компонент, медиируемый рецептором NMDA.
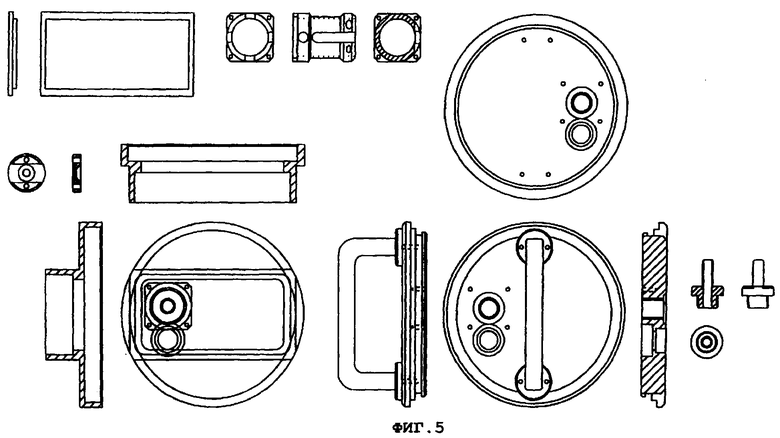
Фиг.5 демонстрирует конструкцию и рабочие характеристики устройства, используемого для экспонирования клеток в культуре.
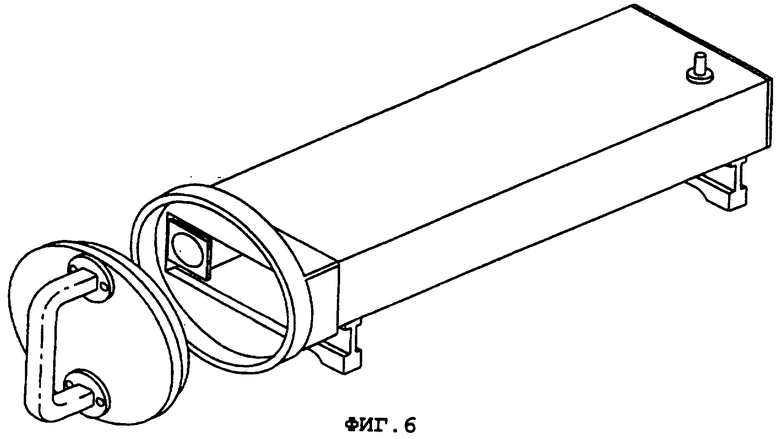
Фиг.6 демонстрирует общий вид всей камеры, используемой для экспонирования клеток в культуре.
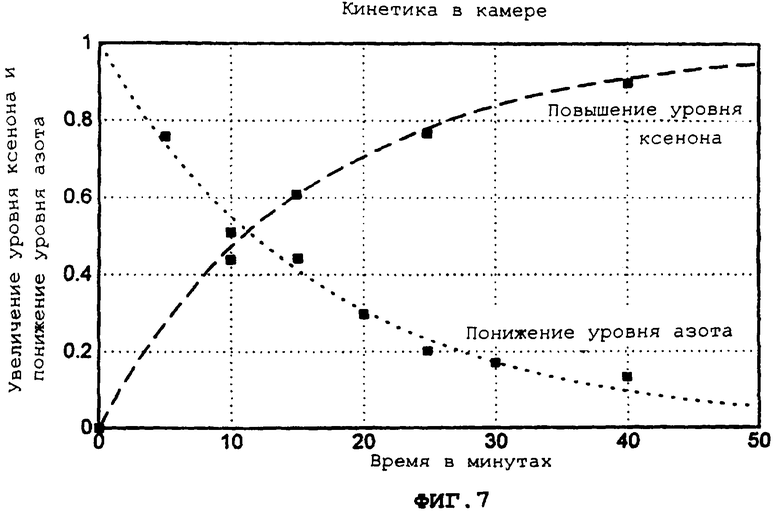
Фиг.7 демонстрирует кинетику процессов в камере для типичного эксперимента, в котором ксенон вводится в камеру, демонстрируя увеличение уровня ксенона и увеличение уровня азота со временем (в минутах).
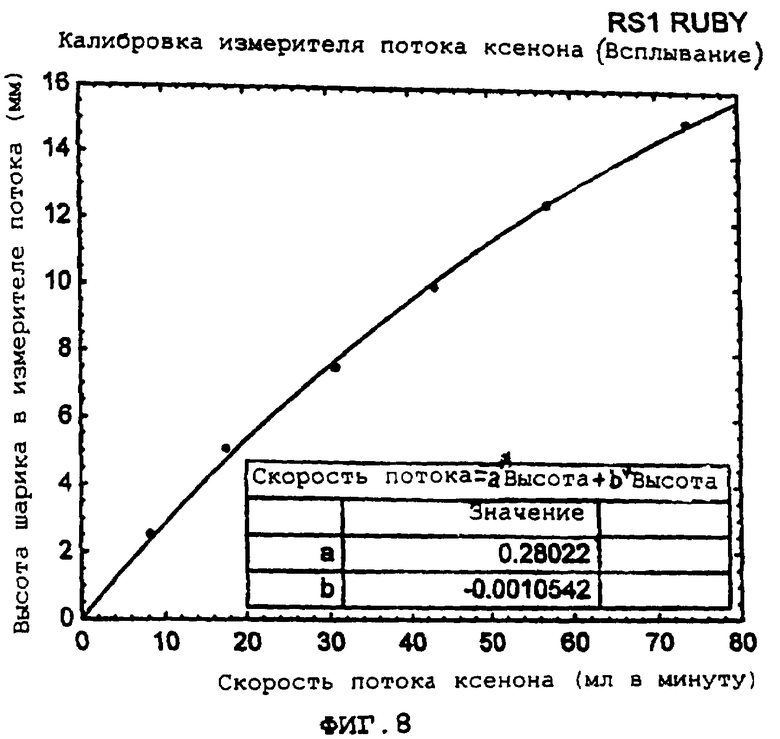
Фиг.8 демонстрирует калибровочный график для ксенона.
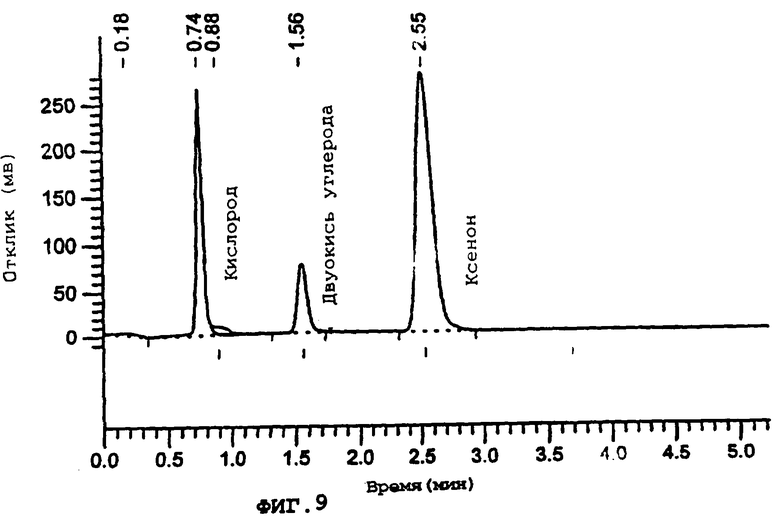
Фиг.9 демонстрирует типичный пример анализа газов из камеры.
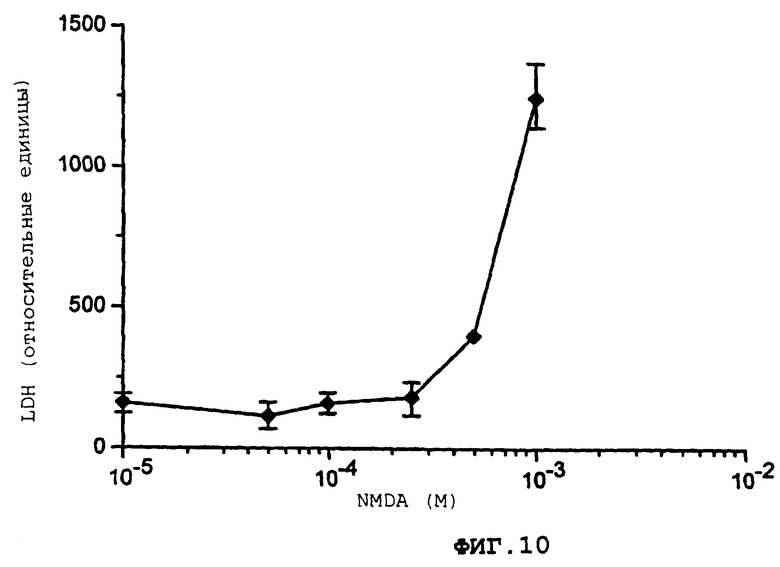
Фиг.10 демонстрирует кривую дозовой зависимости для вызванного NMDA повреждения нейрона.
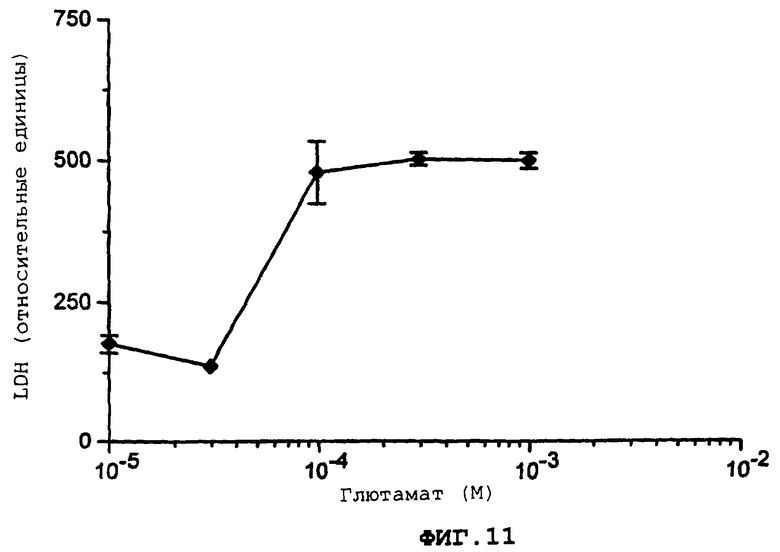
Фиг.11 демонстрирует кривую дозовой зависимости для вызванного глютаматом повреждения нейрона.
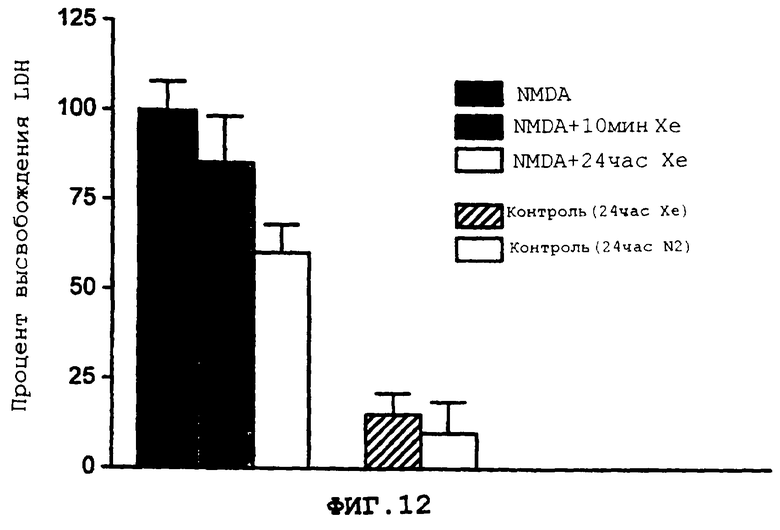
Фиг.12 демонстрирует воздействие 75% ксенона во время вызванного NMDA повреждения нейрона и после него.
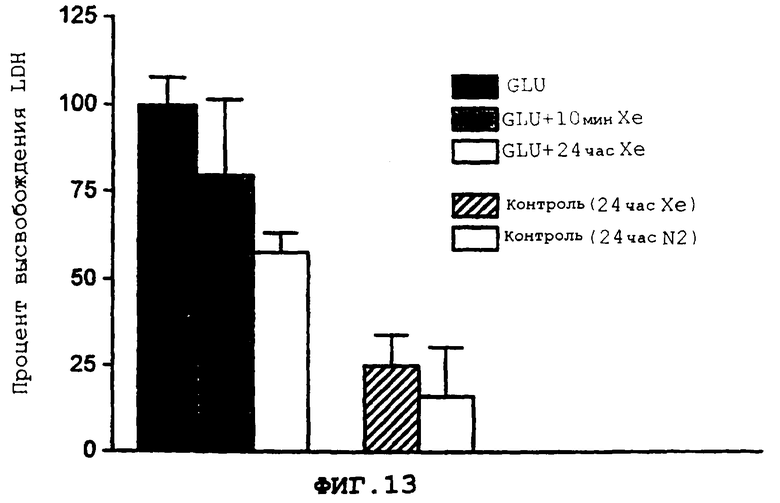
Фиг.13 демонстрирует воздействие 75% ксенона во время вызванного глютаматом повреждения нейрона и после него.
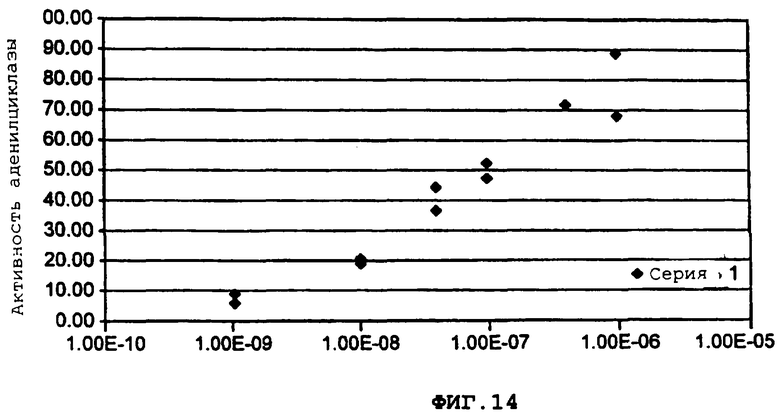
Фиг.14 демонстрирует воздействие увеличения дозы простагландина E1 (PGE1) на аденилилциклазу (AC).
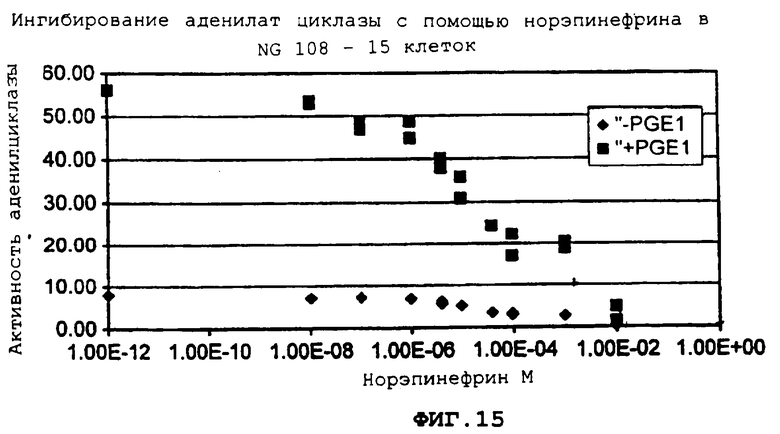
Фиг.15 демонстрирует воздействие совместного введения норэпинефрина на активность аденилилциклазы (AC).
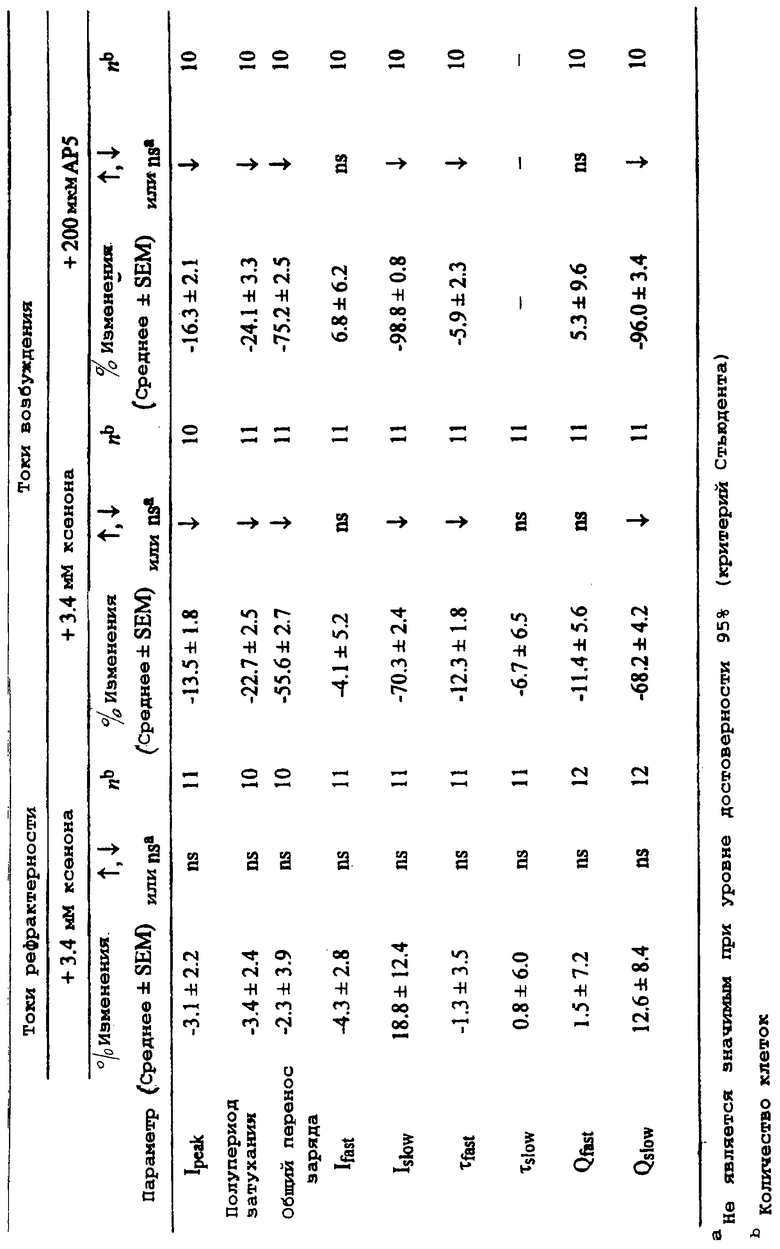
Более подробно, фигура 1 демонстрирует А) пример постсинаптического тока возбуждения (EPSC) и его чувствительность к 1 мМ кинуреновой кислоты, и нечувствительность к 10 мкМ бикукуллина. Врезка демонстрирует линейную зависимость тока от напряжения для пика EPSC, нормированную на значение тока, измерянное при -60 мВ; точки данных представляют собой средние значения (для усреднения по 5 клеткам). В) Пример постсинаптического тока рефрактерности (IPSC) и его чувствительность к 10 мкм бикукуллина и нечувствительность к 1 мМ кинуреновой кислоты. Врезка демонстрирует линейную зависимость тока от напряжения для пика IPSC, нормированную на значение тока, измеренное при -60 мВ; точки данных представляют собой средние значения (для усреднения по 6 клеткам). На врезках значки ошибок представляют собой SEM (среднеквадратичное отклонение), но они не показаны там, где они меньше, чем размер символа.
Более подробно, фиг.2 демонстрирует А) ГАМКэргические постсинаптические токи в присутствии и в отсутствие 3,4 мМ ксенона. Ксенон не оказывает какого-либо значительного воздействия на IPSC. Врезка демонстрирует репрезентативные графики, иллюстрирующие отсутствие какого-либо воздействия 4,3 мМ ксенона на ток, вызванный 3 мкМ ГАМК. В) Глютаматэргические постсинаптические токи в присутствии и отсутствие 3,4 мМ ксенона. Принципиальное воздействие представляет собой уменьшение в медленном компоненте тока (в этом примере, примерно на 70%). Точки представляют собой результаты двухэспоненциальной подгонки к измерянным токам. Врезка демонстрирует примеры, иллюстрирующие воздействие 3,4 мМ ксенона на ток, вызванный 100 мкм NMDA.
Более подробно, фиг.3 демонстрирует, что NMDA активирует внутренний ток (в нейронах, поддерживаемых при фиксированном напряжении -60 мВ) при EC50 24±2 мкМ NMDA и коэффициенте Хилла 1,2±0,1. Ксенон ингибирует ток приблизительно на 60%, но не вызывает значительных изменений ни EC50, ни коэффициента Хилла. Каждая точка данных представляет средний пиковый ток, по меньшей мере, от 6 клеток.
Более подробно, фиг.4 демонстрирует, что ксенон селективно ингибирует медиируемый рецептором NMDA компонент глютаматэргических постсинаптических токов возбуждения (EPCS). Нейроны поддерживаются при фиксированном напряжении -60 мВ; синаптические отклики стимулируются с помощью 2 мс деполяризующего импульса до +20 мВ. Контрольные глютаматэргические EPSC проявляют характерное двухфазное затухание. Медленный компонент полностью блокируется с помощью 200 мкм АР5, оставляя быстрый компонент почти без изменения. Врезка представляет медиируемый рецептором NMDA компонент (разницу между контрольным EPCS и его значением в присутствии АР5) и его значение в присутствии ксенона (вычисляется как разница между EPSC в присутствии ксенона и его значением в присутствии АР5). Контрольные растворы уравновешиваются при комнатной температуре с 80% N2 и 20% O2, а исследуемые растворы с 80% ксенона и 20% O2,
Более подробно, фиг.5 демонстрирует детали лицевой панели и узла вентилятора, разводку газа и электричества. Камера изготавливается из нержавеющей стали или анодированного алюминия и легко может быть стерилизована. Газы входят через входной узел, соединенный с высокоскоростным вентилятором, который обеспечивает эффективное перемешивание для того, чтобы обеспечить то, что клетки экспонируются для точно определенных концентраций ксенона. Если ксенон вводится в камеру со скоростью ν мл/мин и камера имеет объем V, и концентрация ксенона, входящего в камеру, равна cin, тогда концентрация выходящего газа cout, дается уравнением:
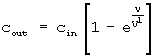
Более подробно, фиг.7 демонстрирует экспериментальные испытания, иллюстрирующие, что ксенон заменяет азот точно с теоретической скоростью. В присутствии одной чашки с культурой клеток, объем газа равен 570 мл. Таким образом, для скорости 40 мл ксенона/мин и для одной чашки с культурой клеток, газ в камере теоретически должен достичь 95%-ной конечной концентрации, когда t=43 мин, если газы однородно смешиваются.
Более подробно, фигура 8 представляет собой пример калибровочного графика для ксенона, демонстрирующего высоту шарика в измерителе потока (мм) в зависимости от скорости потока ксенона (мл/мин). Композиция газообразной окружающей среды, для которой экспонируются клетки, определяется с помощью прецизионных проточных ротаметров, которые калибруются для ксенона (или альтернативно, для азота, кислорода или двуокиси углерода).
Более подробно, фигура 9 демонстрирует анализ газов, когда заданная смесь газов (ксенон, кислород, азот и двуокись углерода) проходит через камеру, где смешиваются газы, обеспечивая стабильную и прецизионно контролируемую окружающую среду. Состав газов подтверждается с использованием газовой хроматографии. Газы отбираются на выходе и герметизуются в стеклянном флаконе. Флакон переносится в автоматический самплер Perkin Elmer HS 40 XL, который подготавливает газы во флаконе для анализа (объем пробы 40 мкл), а затем они вводятся в газовый хроматограф Perkin Elmer XL, соединенный с колонкой 2 м×1/8ss Chromosorb 102, 80-100 меш, работающую при 60°С, с гелием в качестве газа-носителя (2 мл/мин). Газы детектируются с использованием детектора на основе теплопроводности при 110°С, при температуре инжектора 150°С.
Более подробно, фиг.10 демонстрирует кривую дозовой зависимости вызванного NMDA повреждения нейрона в совместной культуре возрастом 15 дней. Контрольные эксперименты демонстрируют, что лежащий внизу глиальный монослой не подвергается действию NMDA при максимальной используемой дозе (Обработка: 10 мин, выход LDH через 24 ч).
Более подробно, фиг.11 демонстрирует кривую дозовой зависимости вызванного глютаматом повреждения нейрона в совместной культуре возрастом 15 дней. Контрольные эксперименты демонстрируют, что лежащий внизу глиальный монослой не подвергается воздействию глютамата при максимальной используемой дозе (Обработка: 30 мин, выход LDH через 6 ч).
Более подробно, фиг.12 демонстрирует воздействие 75% ксенона во время вызванного NMDA повреждения нейрона, при котором сестринские культуры (n=3) экспонируются для NMDA (750 мкмоль в течение 10 мин), и после него, и последующее повреждение нейронов количественно определяется с помощью появления LDH через 24 ч после экспонирования.
Более подробно, фиг.13 демонстрирует воздействие 75% ксенона во время вызванного глютаматом повреждения нейрона, при котором сестринские культуры (n=3) экспонируются для глютамата (75 мкМ в течение 10 мин), и после него, и последующее повреждение нейронов количественно определяется с помощью появления LDH через 24 ч после экспонирования.
Более подробно, фиг.14 демонстрирует, что увеличение доз PGE1 зависимым от дозы образом стимулирует аденилилциклазу (AC).
Более подробно, фиг.15 демонстрирует, что когда агонист альфа-2, норэпинефрин дополнительно вводится при условиях, в которых активность АС стимулируется PGE1, активность АС зависимым от дозы образом ингибируется норэпинефрином. Даже при базовых условиях ("-PGE1") эффект ингибирования является очевидным.
Исследования, проведенные авторами заявки, показывают, что ксенон не оказывает явного воздействия на рецепторы ГАМКА. Токи, активируемые с помощью 3 мкм ГАМК, как у поддерживаемых при фиксированном напряжении культивируемых гиппокампальных нейронов крыс, так и в поддерживаемых при фиксированном напряжении клетках РАЗ (Hadingham KL et al, Proc. Natl. Acad. Sci. USA, 1992; 89, 6378-6382), которые стабильно экспрессируют определенные субъединицы ГАМК, не подвергаются значительному воздействию даже с помощью 100% ксенона. Значение этого наблюдения становится ясным, если иметь в виду, что при функционировании в качестве анестетика для людей, половина максимальной эффективной концентрации EC50 составляет 71% объем/объем (Cullen et al, Anesthsiology 1969, 305-309). Ксенон также, как показано, оказывает небольшое воздействие на функциональные синапсы, высвобождающие ГАМК, в гиппокампальных нейронах, при уменьшении 80% ксеноном пиковых постсинаптических токов рефрактерности только на 8±2%, указывая на то, что пресинаптическе воздействия ксенона должны быть очень умеренными.
В противоположность этому, ксенон, как показано, обладает значительным воздействием на рецепторы NMDA. Механистические исследования на культуре гиппокампальных нейронов показывают, что 80% ксенона (который будет поддерживать хирургическую анестезию) уменьшает активируемые NMDA токи вплоть до 60%, без какого-либо значительного изменения значения EC50 или коэффициента Хилла для NMDA. Это неконкурентное ингибирование указывает на то, что ксенон должен сильно ингибировать передачу импульса в нейроне, несмотря на высокие концентрации глютамата в синаптических щелях.
Дополнительные исследования осуществляются с использованием микроостровковых культур гиппокампальных нейронов, которые образуют синапсы сами с собой (аутапсы) (Bekkers JM et al, Proc. Natl. Acad. Sci. USA, 1991; 88, 7834-7838). Типичный глютаматэргический постсинаптический ток, полученный от гиппокампального нейрона, представлен на фигуре 4. Контрольная запись показывает характерную двухфазную зависимость от времени, при этом быстрый компонент медиируется рецепторами вещества иного, чем NMDA, а гораздо более медленный компонент медиируется рецепторами NMDA. Компонент, медиируемый рецепторами NMDA, легко идентифицируется, поскольку он блокируется с помощью высокоселективного конкурентного антагониста АР5, DL-2-амино-5-фосфононпентаноата (Watkins JC et al, Annu. Rev. Pharmacol. Toxicol. 1981; 21,165-204).
Добавление 200 мкм АР5, как показано, почти полностью блокирует медленный компонент, оставляя только быстрый компонент с зависимостью от времени в виде одной экспоненты, очень похожей на зависимость быстрого компонента для контроля. Воздействие ксенона на глютаматэргический постсинаптический ток напоминает воздействие АР5 (фиг.4). Медленный компонент, медиируемый рецепторами NMDA, уменьшается более чем на 70%, в то время как быстрый компонент едва изменяется. Таким образом, ксенон не только ингибирует синаптические рецепторы NMDA, он имеет небольшое видимое воздействие на рецепторы веществ, иных, чем NMDA.
С точки зрения воздействия на синаптические токи, селективность воздействия, наблюдаемая для ксенона, является неожиданной. Ввиду того факта, что почти все общие анестетики усиливают воздействие ГАМК на рецепторы ГАМКА (Tanelian DL et al, Anesthesiology 1993; 78: 757-776; Franks NP et al. Nature 1994; 367:607-614), ожидается, что и ксенон не будет исключением. Однако полное отсутствие воздействия ксенона на рецепторы ГАМКА помещает их в тот же класс агентов, что и кетамин, так называемый "диссоциативный" анестетик, который также не действует на рецепторы ГАМКА (Brockmeyer DM et al, Br. J.Anesesth. 1995; 74:79-84) и, как полагают, действует в основном на рецепторы NMDA. Подобным же образом, результаты показывают, что ксенон селективно блокирует рецепторы NMDA при небольшом воздействии на АМРА/КА. Последний результат является сильным аргументом в пользу того, что воздействие ксенона является постсинаптическим по природе. Отсутствие воздействия ксенона на время затухания компонента, связанного с рецепторами NMDA, однако, говорит о простом механизме ингибирования, блокировании открытых каналов, что следует из наблюдения того, что концентрация EC50 для токов, вызываемых NMDA, не изменяется в присутствии ксенона. Чтобы ни было конкретной молекулярной основой для неожиданной селективности ксенона по отношению к рецепторам NMDA, оно скорее связано с множеством особенностей его необычных фармакологических свойств, включая способность вызывать глубокую аналгезию и психотомиметические воздействия.
Таким образом, является ясным, что ксенон является неожиданно селективным в своем действии, оказывая очень разное воздействие на синаптическую передачу возбуждения и рефрактерности. Воздействие ксенона может быть описано только с точки зрения воздействия на глютаматэргические синапсы, хотя и другие мишени также могут быть определены в будущем. Тем не менее, нечувствительность ГАМКэргических синапсов к ксенону указывает на то, что механизмы его воздействия являются явно отличными от механизмов для различных анестетиков. На механистическом уровне является ясным, что для ксенона постсинаптические рецепторы являются наиболее важными молекулярными мишенями.
Далее, настоящее изобретение описывается с помощью примеров.
Примеры
Культивирование гиппокампальных нейронов
Гиппокампальные нейроны выращивают в культуре с использованием способов, описанных ранее (Segal MM et al, J. Neurophysiol. 1990, 64:1390-1399; Bekkers JM et al, Proc. Natl. Acad. Sci. U.S.A. 1991, 88:7834-7838; Mennerick S. et al, J. Neurophysiol. 1995, 73:320-332). Вкратце, гиппокампы крыс линии Sprague Dawley (день после рождения 1-3) иссекаются, крупно нарезаются и перемешиваются в содержащем папаин растворе (20 единиц/мл) в течение 30 минут при 37°С. После промывки не содержащим фермента раствором, ткань осторожно перемешивается с помощью обработанной на горелке пипетки Пастера, и клетки наносят на субстрат с плотностью 8-10×104 клеток мл-1 и культивируются (95% воздух/5% CO2) при 37°С. Покровные стекла, используемые для культивирования клеток, сначала покрывают агарозой (0,15% масс/объем), а затем на них наносят распылением мелкодисперсный аэрозоль из поли-D-лизина (PDL) и коллагена (0,1 мг/мл PDL и 0,5 мг/мл коллагена из хвостов крыс) из стеклянного микрораспылителя, и стерилизуют путем экспонирования для УФ-света. При этом получаются микроостровки пермиссивного субстрата с диаметрами в пределах между 100 и 1000 мкм. На 3-4 дни после нанесения на субстрат, когда слой глиальных клеток становится сплошным примерно на 80%, добавляют противомитотический агент (цитозин β-D-арабинофуранозид, 5 мкМ) для приостановки пролиферации глиальных клеток. Затем культурам нейронов дают возможность созреть в течение дополнительных 4-9 дней. Авторы используют микроостровки, которые содержат единичные изолированные нейроны, у которых процессы в аксонах и дендритные разветвления формируют множество аутосинапсов (аутапсов). Этот метод позволяет получить большую популяцию фенотипически идентичных моносинаптических соединений.
Электрофизиология
Нейроны поддерживаются при фиксированном напряжении с использованием методики регистрации для целых клеток (усилитель Axopatch 200, Axon Instruments, Foster City, CA). Электроды изготавливаются из боросиликатного стекла и, как правило, имеют сопротивление в пределах между 3-5 МОм. Сопротивление цепи компенсируется на 75-90%. Нейроны поддерживаются при фиксированном напряжении -60 мВ, и синаптические отклики стимулируются с помощью 2 мс деполяризующего импульса до +20 мВ. Вскоре после восстановления потенциала мембраны до -60 мВ наблюдается и регистрируется большой (1-20 на) постсинаптический ток. Для синаптических измерений отбираются данные на 50 кГц, фильтруются на 20 кГц (-3 дб, 8-полюсный фильтр Бесселя) и сохраняются в компьютере. Внеклеточный регистрационный раствор представляет собой (мМ) 137 NaCl, 5 KCl, 3 CaCl2, 5 HEPES, 10 глюкозы, 0,001 глицина, 0,0001 стрихнина-HCl, титруют до рН 7,3 с помощью NaOH; и внутриклеточный (пипеточный) раствор представляет собой (мМ) 140 KCl, 4 NaCl, 0,5 EGTA, 2 MgATF, 10 HEPES, титруют до рН 7,25 с помощью КОН.
Для экспериментов, где ГАМК, NMDA или глютамат добавляют экзогенно, нейроны выращивают в массовой культуре и используют 3-11 дней после нанесения на субстрат. Данные получают на 200 Гц и фильтруют на 100 Гц (-3 дб, 8-полюсный фильтр Бесселя). Внеклеточный регистрационный раствор для откликов, вызванных NMDA - и глютаматом, представляет собой (мМ) 150 NaCl, 4 KCl, 2 CaCl2, 10 HEPES, 10 глюкозы, 0,0002 тетродотоксин цитрата (Tocris Cookson, Bristol, UK), 0,1 пикротоксина, 0,0001 стрихнина-HCl, 0,001 глицина, титруют до рН 7,40 с помощью NaOH; внеклеточный регистрационный раствор для откликов, вызванных ГАМК, представляет собой (мМ) 150 NaCl, 4 KCl, 1 CaCl2,1 MgCl2, 10 HEPES, 10 глюкозы, 0,0002 тетродотоксин цитрата, титруют до рН 7,40 с помощью NaOH; и внутриклеточный (пипеточный) раствор для откликов, вызванных ГАМК-, NMDA- и глютаматом, представляет собой (мМ) 140 CsCl, 3 NaCl, 11 EGTA, 2 MgATF, 10 HEPES, титруют до рН 7,20 с помощью CsOH. Если не утверждается иного, все химикалии получают от Sigma Chemical Co. (Poole, Dorset, UK). Растворы для исследований доставляются в клетки с использованием системы перфузии с быстрым переключением (Downie DL et al, Br. J.Pharmacol. 1996, 118:493-502). Все электрофизиологические измерения проводятся при комнатной температуре (20-23°С).
Приготовление анестетических растворов
Растворы ксенона готовят путем, сначала, барботирования чистых газов (кислорода, азота или ксенона) через барботеры из мелкопористого спеченного стекла в 250- или 500-мл колбах Дрехселя, заполненных внеклеточным солевым раствором для регистрации. Растворы барботируются в течение 1,5-2 ч, хотя равновесие, как обнаружено, происходит в пределах 45 мин (Для сведения окисления к минимуму, нейротоксины и нейротрансмиттеры исключаются из полностью оксигенированного солевого раствора). Во время барботирования растворы непрерывно перемешиваются при комнатной температуре. Затем эти растворы смешиваются для достижения желаемых конечных концентраций газов. У авторов, контрольные растворы обычно содержат 80% раствора азота и 20%-ного раствора кислорода, в то время как растворы для исследований у авторов обычно содержат 80% раствора ксенона и 20%-ного раствора кислорода. Используя Бунзеновский коэффициент распределения вода/газ 0,0965 (Smith RA et al, Biochim. Biophys. Acta 1981, 645:327-338), авторы вычислили, что стандартный раствор для исследований у авторов содержит 3,4 мМ ксенона. Ксенон (чистота для исследований, степень чистоты 99,993%) поставляется ВОС gases, Guildford, Surrey, UK. Во всех случаях ксенон предварительно подводится к нейронам в течение, по меньшей мере, 30 секунд перед инициированием синаптических токов.
Интегрирование синаптических откликов
В порядке получения оценки для общего переноса заряда значения EPSC или IPSC численно интегрируются. Однако, поскольку в некоторых случаях токи не затухают до базовой линии к концу периода записи, вводится коррекция (которая неизменно является меньшей, чем 5% от общего переноса заряда) путем экстраполяции наблюдаемого тока до базовой линии с использованием двухэкспоненциальной подгонки для фазы затухания отклика.
Контрольные синаптические токи
Контрольные синаптические токи характеризуются и почти неизменно попадают в одну из двух популяций в соответствии с данными предыдущих исследований (Bekkers JM et al, Proc. Natl. Acad. Sci. U.S.A. 1991, 88:7834-7838; Mennerick S. et al, J. Neurophysiol. 1995, 73:320-332). Примерно половина клеток демонстрирует постсинаптические токи, которые затухают относительно быстро (полупериод жизни ˜8 мс), в то время как другая половина демонстрирует токи, которые являются значительно более медленными (полупериод жизни ˜40 мс) (см. фиг.1 и таблицу 1). Более быстрые отклики регистрируются от клеток округлого вида и со сложными дендритными разветвлениями. Их чувствительность к кинуреновой кислоте (80±3% ингибирование пикового тока с помощью 1 мМ кинуреновой кислоты; n=7 клеток) и нечувствительность к бикукуллину (0,4±1,3% ингибирование пикового тока с помощью 10 мкМ бикукуллина; n=13 клеток) идентифицирует эти клетки как глютаматэргические нейроны с передачей возбуждения. В противоположность этому, более медленные синаптические токи почти полностью блокируются с помощью бикукуллина (94±1% ингибирование пикового тока с помощью 10 мкМ бикукуллина; n=8 клеток) и не подвергаются воздействию кинуреновой кислоты (6±4% ингибирование пикового тока с помощью 1 мМ кинуреновой кислоты; n=5 клеток), что идентифицирует эти отклики как ГАМКэргические. Эти ингибиторные нейроны имеют тенденцию быть плоскими с более простыми дендритными разветвлениями. Благодаря недавнему открытию, что ГАМК и глицин могут совместно высвобождаться интернейронами спинного мозга (Jonas P. et al. Science 1998,18 281:419-424), авторы рассматривают возможность, что отклики рефрактерности, которые они регистрируют, медиируются, частично, рецепторами глицина. Однако контрольный ток очень слабо изменяется (4±2% ингибирование; n=6 клеток) под действием 100 нМ стрихнина, подтверждая, что токи рефрактерности являются полностью ГАМКэргическими.
Токи возбуждения и рефрактерности имеют по существу идентичные времена нарастания (см. таблицу 1), и пиковые значения токов изменяются линейно с прилагаемым потенциалом (см. врезки на фиг.1). Фаза затухания синаптического тока I(t), где t - это время, измерянное для пика тока, подгоняется с помощью двухэкспоненциального уравнения формы
I(t)=Ifaste-t/τ fast+Islowe-t/τ slow,
где Ifgst и Isiow представляют собой амплитуды, a τfast и τslow являются временными константами быстрого и медленного компонентов соответственно. Значения этих временных констант затухания, измеренных по контрольным откликам возбуждения и рефрактерности, приведены в таблице 1. В обоих случаях, примерно две-три общего переноса заряда осуществляется с помощью медленного компонента. Для глютаматэрги-ческих откликов возбуждения этот медленный компонент может быть легко идентифицирован как медиируемый с помощью рецепторов NMDA, поскольку он полностью (99±1%; n=10 клеток) блокируется с помощью 200 мкм АР5 (DL-2-амино-5-фосфонопентаноата), высоко селективного антагониста рецепторов NMDA (Davies J. et al, Neurosci. Letts 1981, 21:77-81). В присутствии этой концентрации АР5, фаза затухания синаптического тока может хорошо быть описана с помощью подгонки одной экспонентой с амплитудой и временной зависимостью, слегка отличной от зависимости для быстрого компонента контроля. Этот быстрый компонент, который в большой степени определяет амплитуду пикового тока возбуждения (Ifast/Itotal=92±1%; n=13 клеток), может быть приписан токам, медиируемым с помощью рецепторов АМРА/КА (Bekkers JM et при, Proc. Natl. Acad. Sci. U.S.A. 1991, 88:7834-7838; Mennerick S. et al, J.Neurophysiol. 1995, 73:320-332).
Воздействие ксенона на синаптические токи
Концентрация ксенона в газовой фазе, которая предотвращает отклик на болезненный стимул (то есть МКК), видимо, меняется в зависимости от вида, составляя 71% атм у людей (Cullen SC et al, Anesthesiology 1969, 31:305-309), 98% атм у макак резус (Whitehurst SL et al, J. Neurosurg. Anesthesiol. 1994, 6:275-279) и 161% атм у крыс (Koblin DD et al, Anesth. Analg. 1998, 87:419-424). Когда эти значения преобразуются в концентрации свободного газа в воде при 37°С (Franks NP et al, Br. J.Anaesth. 1993, 71:65-76; Franks NP et al, Anesthesiology, 1996, 84:716-720) с использованием коэффициента распределения Оствальда вода/газ 0,0887 (Weathersby PK et al. Undersea Biomed. Res. 1980, 7:277-296), полученные значения составляют 2,5 мМ, 3,4 мМ и 5,6 мМ для людей, обезьян и крыс соответственно, при этом среднее значение составляет 3,8 мМ. Для экспериментов, описываемых здесь, осуществляемых при комнатной температуре, концентрация ксенона, присутствующего в стандартном растворе для исследований, составляет 3,4 мМ. При этой концентрации ксенон имеет пренебрежимо малое воздействие на синаптические токи рефрактерности, но сильно подавляет токи возбуждения. Это иллюстрируется с помощью соответствующих графиков на фиг.2.
Для ГАМКэргических синаптических токов 3,4 мМ ксенона не воздействует ни на пиковое значение, ни на временную зависимость постсинаптических токов. Процентные изменения различных параметров рефрактерности синапсов перечислены в таблице 2, где, как можно заметить, ни один из них значительно не изменяется. Это воздействие ксенона на токи, вызываемые с помощью низкой (3 мкМ) концентрации экзогенно вводимой ГАМК, также исследуется. Здесь 4,3 мМ ксенона не оказывают никакого значительного воздействия на ток, индуцируемый ГАМК (увеличение 2±3%, n=4 клетки). Репрезентативная пара графиков представлена на врезке, на фиг.2А. В противоположность этому, 3,4 мМ ксенона сильно подавляют глютаматэргический синаптический ток, при этом воздействие ограничивается почти исключительно медленным компонентом тока, медиируемым рецепторами NMDA (фиг.2В). Это очевидно по процентным изменениям различных синаптических параметров возбуждения, которые перечислены в таблице 2. Здесь можно увидеть, что качественное воздействие ксенона с замечательной точностью воспроизводится при воздействии АР5. При заданной концентрации АР5 (200 мкМ), компонент, связанный с рецепторами NMDA, как ожидается, должен быть почти полностью блокирован (Davies J. et al, Neurosci. Letts 1981,21:77-81), и это согласуется с 99% блокировкой Islow (см. таблицу 2). Это сопровождается 75% уменьшением общего переноса заряда, что близко к 61% общего заряда, который, по оценкам авторов, должен переноситься с помощью медленного компонента, медиируемого рецепторами NMDA (см. таблицу 1). Это различие может быть связано с небольшим, но значимым уменьшением быстрой временной константы τfast с помощью АР5 (таблица 2). Ксенон, подобным же образом, вызывает большое ингибирование (70%) Islow и большое ингибирование (56%) общего переноса заряда, оказывая при этом только небольшое воздействие на быстрый компонент, медиируемый рецепторами АМРА/КА.
Нейропротекторное воздействие
Совместное культивирование неонатальных глиальных и фетальных нейрональных клеток
Разбавленные культуры мышиных кортикальных клеток готовят с использованием целых новых клеток коры головного мозга зародышей мышей (14-17 дней беременности). Культуры кортикальных глиальных клеток приготавливают из недавно рожденных мышей. После осуществления отделения ненейрональных клеток с помощью цитозин арабинозида, выжившие кортикальные нейрональные клетки наносят на подложку из глиальных клеток и культивируют совместно.
Повреждение совместной нейрональной культуры с помощью NMDA
Через 15 дней после исходного сбора фетальных кортикальных клеток чашки с совместной культурой экспонируют при комнатной температуре для N-метил, D-аспартата (NMDA), 10-1000 мкМ, в контрольном солевом растворе (CCS) в течение 10 мин. После осторожной промывки с помощью CCS клетки инкубируются в течение 6-24 ч в минимальной основной среде Игла, не содержащей глютамата или лактат дегидрогеназы (LDH), но содержащей глюкозу, 25 мМ и NaHCO3, 38 мМ, при 37°С в увлажненной атмосфере СО2 (5%). Степень повреждения определяется путем измерения количества LDH, высвобожденного в среду. Кривая дозовой зависимости для повреждения, вызванного NMDA, представлена на фиг.10, как описывается перед этим.
Повреждение совместной нейрональной культуры с помощью глютамата
Через 15 дней после исходного сбора кортикальных клеток чашки с совместной культурой экспонируют при комнатной температуре для глютамата, 10-1000 мкМ, в контрольном солевом растворе (CCS) в течение 10-30 мин. После осторожной промывки с помощью CCS, клетки инкубируются в течение 6-24 ч в минимальной основной среде Игла, не содержащей глютамат или лактат дегидрогеназу (LDH), но содержащей глюкозу, 25 мМ и NaHCO3, 38 мМ, при 37°С в увлажненной атмосфере СОз (5%). Степень повреждения определяется путем измерения количества LDH, высвобожденного в среду. Кривая дозовой зависимости для повреждения, вызванного глютаматом, представлена на фиг.11, как описывается перед этим.
Защита совместной культуры нейрональных-глиальных клеток, поврежденных NMDA, с помощью ксенона
75% от одной атмосферы ксенона доставляют к клеткам во время экспонирования, и после него, для 750 мкМ NMDA, концентрации, которая вызывает повреждение, близкое к максимальному. Степень повреждения определяется с помощью измерения уровня LDH, выделяемого в среду (после коррекции на "фоновое" высвобождение LDH). Ксенон уменьшает повреждение, вызванное NMDA, более чем на 30%; когда поступление ксенона продолжается после экспонирования для NMDA, повреждение дополнительно уменьшается более чем на 55%. Результаты этого защитного воздействия ксенона иллюстрируется на фиг.12, как описывается выше.
Защита совместной культуры нейрональных-глиальных клеток, поврежденных глютаматом, с помощью ксенона 75% от одной атмосферы ксенона доставляют к клеткам во время экспонирования, и после него, для 75 мкМ глютамата, концентрации, которая вызывает повреждение, близкое к максимальному. Степень повреждения определяется с помощью измерения уровня LDH, выделяемого в среду (после коррекции на "фоновое" высвобождение LDH). Ксенон уменьшает повреждение, вызванное глютаматом, более чем на 35%; когда поступление ксенона продолжается после экспонирования для глютамата, повреждение дополнительно уменьшается более чем на 60%. Результаты этого защитного воздействия ксенона иллюстрируются на фиг.13, как описывается выше.
Ингибирование синаптической пластичности
Нарушение функций центральной нервной системы может быть вызвано изменением прочности соединений между нейронами, которое упоминается как синаптическая пластичность. Авторы настоящего изобретения используют линию гибридных клеток нейробластома Х глиома для исследования воздействия ксенона на синаптическую пластичность. В этой парадигме, адренэргические рецепторы альфа-2, которые, в основном, включены в синаптическую передачу, ингибируют аденилил циклазу (AC).
Для исследования ингибирующего воздействия адренэргических рецепторов альфа-2 на АС, сначала, необходимо стимулировать эндогенную АС с помощью простагландина E1 (PGE1). Увеличение доз PGE1 зависимым от дозы образом стимулирует АС (фиг.14). Когда агонист альфа-2, норэпинефрин вводится совместно при условиях, при которых активность АС стимулируется с помощью PGE1, эта активность ингибируется зависимым от дозы образом с помощью норэпинефрина (фиг.15).
Для индуцирования "синаптической пластичности" клетки предварительно обрабатываются в течение 8 часов с помощью норэпинефрина; после приготовления гомогенатов из тщательно отмытых клеток, вводимый повторно норэпинефрин теряет свое ингибирующее воздействие на активность АС, стимулируемую PGE1. Если эти клетки одновременно экспонируются для ксенона, во время фазы предварительной обработки, ингибиторное воздействие норэпинефрина сохраняется.
Модификации настоящего изобретения будут очевидны специалисту в данной области. Литературные источники, рассмотренные здесь, включаются сюда в качестве ссылок.
b Itotal=Ifast+Islow
Qtotal=Qfast+Qslow
Воздействия 3,4 мМ (примерно 1 MAC) ксенона на синаптические токи и 200 мкМ АР5 на синаптические токи возбуждения
| название | год | авторы | номер документа |
|---|---|---|---|
| АНЕСТЕТИЧЕСКАЯ КОМПОЗИЦИЯ, ВКЛЮЧАЮЩАЯ АНТАГОНИСТ NMDA И АЛЬФА-2 АДРЕНЕРГИЧЕСКИЙ АГОНИСТ | 2000 |
|
RU2244558C2 |
| СОЕДИНЕНИЯ, АКТИВНЫЕ ПРИ НОВОМ САЙТЕ НА ОПЕРИРУЕМЫХ РЕЦЕПТОРОМ КАЛЬЦИЕВЫХ КАНАЛАХ, ПРИМЕНИМЫЕ ДЛЯ ЛЕЧЕНИЯ НЕВРОЛОГИЧЕСКИХ НАРУШЕНИЙ | 1996 |
|
RU2246300C2 |
| КОМБИНИРОВАННОЕ ЛЕКАРСТВЕННОЕ СРЕДСТВО ДЛЯ ПЕРВИЧНОЙ НЕЙРОПРОТЕКЦИИ | 2017 |
|
RU2636616C1 |
| МОДУЛЯТОР NMDA-РЕЦЕПТОРА СО СТАБИЛИЗИРОВАННОЙ ВТОРИЧНОЙ СТРУКТУРОЙ И ЕГО ПРИМЕНЕНИЕ | 2011 |
|
RU2566821C2 |
| ПРОИЗВОДНЫЕ 5БЕТА-САПОГЕНИНА И ПСЕВДОСАПОГЕНИНА И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ ДЕМЕНЦИИ | 2000 |
|
RU2325396C2 |
| КОМБИНАЦИИ СОЕДИНЕНИЙ, МОДУЛИРУЮЩИХ NMDA-РЕЦЕПТОР | 2015 |
|
RU2721948C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ НЕЙРОДЕГЕНЕРАТИВНОГО ЗАБОЛЕВАНИЯ | 2012 |
|
RU2618412C2 |
| ТЕРАПЕВТИЧЕСКОЕ ЛЕЧЕНИЕ МЕТАБОЛИЧЕСКОГО СИНДРОМА, ДИАБЕТА 2 ТИПА, ОЖИРЕНИЯ ИЛИ ПРЕДИАБЕТА | 2008 |
|
RU2467743C2 |
| ЦИКЛОПРОПИЛПИПЕРИДИНОВЫЕ ИНГИБИТОРЫ ТРАНСПОРТЕРА ГЛИЦИНА | 2005 |
|
RU2387644C2 |
| АНТАГОНИСТЫ РЕЦЕПТОРА СВ1 | 2012 |
|
RU2593751C2 |
Предложено: применение ксенона как антагониста NMDA в качестве нейропротектора и/или ингибитора синаптической пластичности; способ обеспечения нейропротекции и/или ингибирования синаптической пластичности; способ получения фармацевтической композиции и фармацевтическая композиция с указанным назначением; применение ксенона для получения фармацевтического средства для нейропротекции и/или ингибирования синаптической пластичности. Изобретение отличается тем, что выявлена возможность известного антагониста NMDA ксенона уменьшать более чем на 30% повреждение нейронов и глиальных клеток, вызванное глютаматом, и если поступление ксенона продолжается действия NMDA, указанное повреждение дополнительно уменьшается более чем на 55%. 5 н. и 9 з.п. ф-лы, 2 табл., 15 ил.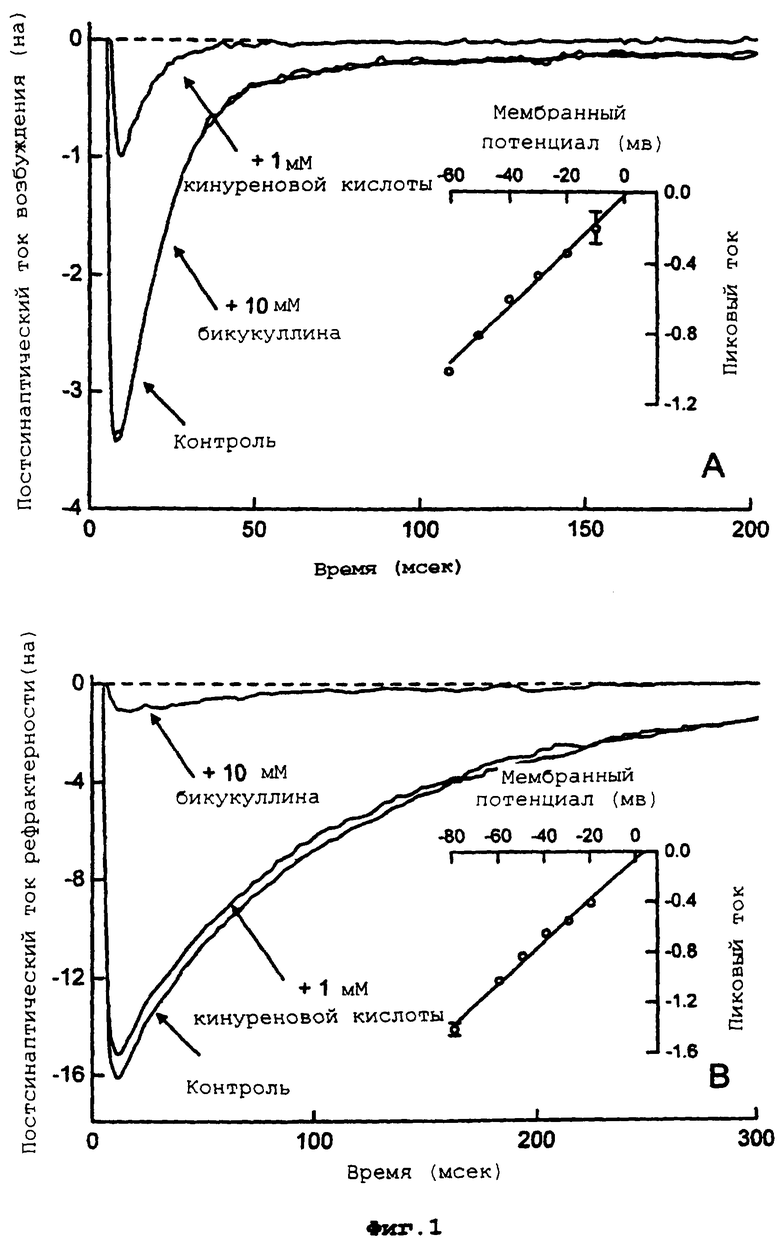
| реферат из АБД Medline: Sagratella S | |||
| NMDA antagonists: antiepileptic-neuro-protective drugs with diversified neuropharmacological profiles.Pharmacol | |||
| Res | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| Smith B.W | |||
| et al | |||
| How does xenon produce anaestesia | |||
| Nature | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| Устройство для моделирования нейрона | 1982 |
|
SU1075632A1 |
| Способ приготовления искусственно-радиоактивных лечебных ванн | 1949 |
|
SU120307A1 |
| Устройство для прижима ленточного носителя информации | 1979 |
|
SU864329A1 |

