Область техники, к которой относится изобретение
Настоящее изобретение касается производных сапогенина и их применения при лечении нарушений умственных способностей и подобных заболеваний, а также касается композиций, применяемых при таком лечении. Изобретение также касается лечения заболеваний, характеризующихся недостаточным числом или нарушением функции мембраносвязанных рецепторов. В дальнейшем при описании настоящего изобретения будет привлекаться главным образом лечение болезни Альцгеймера (AD) и старческой деменции по типу Альцгеймера (SDAT), при которых была показана недостаточность ряда типов рецепторов. Однако следует иметь в виду, что настоящее изобретение в общем касается лечения заболеваний, обусловленных собственным патологическим состоянием и/или воздействием неблагоприятных условий окружающей среды, причем именно таких заболеваний, которые характеризуются недостаточным числом или нарушением функции мембраносвязанных рецепторов либо нарушением синаптической передачи между нейронами или нейронами и эффекторными клетками.
Уровень техники
К заболеваниям указанного выше типа относятся болезнь Паркинсона, деменция, сопровождающаяся изменениями в тельцах Льюиса, постуральная (ортостатическая) гипотензия, аутизм, синдром хронического утомления, миастения, болезнь Ламберта-Итона (Lambert-Eaton), болезни и недомогания, связанные с синдромом войны в Персидском заливе, профессиональное воздействие фосфороорганических соединений и недомогания, связанные с возрастом.
Болезнь Альцгеймера (AD) и старческая деменция по типу Альцгеймера (SDAT) - это серьезные и все нарастающие проблемы во всех обществах, в которых, вследствие увеличения продолжительности жизни и сдерживания легких заболеваний, демографический профиль все более сдвигается в сторону стареющих популяций. Имеется настоятельная потребность в средствах, которые способны излечивать или способствовать лечению AD/SDAT.
Ухудшение памяти в связи с возрастом (AAMI) характерно для пожилых пациентов, которые, будучи психически и физически нормальными, жалуются на потерю памяти. Это мало изученный синдром, однако препараты, эффективные при лечении AD/SDAT, могут быть полезными также и для этих пациентов.
Исследования AD/SDAT проводятся традиционными и стандартными методами медицинских исследований и дисциплин. В традиционной медицине существует несколько подходов к лечению AD/SDAT. Известно, что биохимические процессы, лежащие в основе памяти в коре головного мозга, являются (по меньшей мере частично) холинергическими. Специалистам в этой области известно, что "холинергически опосредованные" механизмы непосредственно объясняются действием ацетилхолина на рецепторы, и эти эффекты - прямые. Другие клинически полезные эффекты могут быть вызваны модуляцией высвобождения ацетилхолина из пресинаптических нервных окончаний или ингибированием ферментов, разрушающих ацетилхолин. Эти модулирующие факторы могут действовать через нейроны с нехолинергическим медиатором, и такие эффекты относятся к косвенным. Некоторые попытки лечения были сосредоточены на роли других медиаторов, таких как 5-гидрокситриптамин, являющийся медиатором в других областях мозга, например ядрах среднего мозга. Однако поскольку волокна из этих областей выходят вперед к церебральной коре, где основным нейромедиатором является ацетилхолин, то на этом медиаторе было сосредоточено внимание при поиске подходящих лекарственных средств.
Холинергическая стратегия при лечении AD/SDAT была направлена на несколько точек биохимических путей образования, синаптического выброса и удаления высвобожденного ацетилхолина.
Один из подходов включает лечение высокими дозами лецитина и других предшественников ацетилхолина. Он имеет ограниченное применение в обеспечении продолжительного улучшения когнитивных функций.
Другой подход включает применение растительных препаратов типа экстракта корней Polygalae, который, как было показано, повышает активность холинацетилхолинтрансферазы (CAT) и секрецию фактора роста нервов (NGF) в мозге. Пероральное введение NGF не влияло на нейроны центральной нервной системы, так как это высокомолекулярный белок, который не проходит через гематоэнцефалический барьер. Однако для улучшения поведения, связанного с памятью, были предложены вещества, способные проходить через гематоэнцефалический барьер и оказывающие стимулирующий эффект на синтез NGF в центральной нервной системе.
Результаты третьего клинического подхода, в котором применяются ингибиторы холинэстеразы, такие как такрин гидрохлорид, были слегка более положительными, чем вышеуказанные. Препараты, полученные из растений, применяемых в китайской и западной медицине, к примеру гуперзин, галантамин и физостигмин, все оказывали некоторое, хотя и ограниченное, улучшение при лечении AD/SDAT в клинических испытаниях, а также на лабораторных моделях. Все эти вещества являются ингибиторами ацетилхолинэстеразы (AChE). У пациентов с AD/SDAT может происходить снижение синтеза ацетилхолина (ACh), уменьшение эффективности высвобождения ACh из пресинаптических терминалей и снижение числа или функции постсинаптических (M1) рецепторов. Также отмечалось снижение числа пресинаптических М2-рецепторов. Положительное действие ингибиторов AChE объясняется повышением содержания ACh в синапсах мозга вследствие замедленного разрушения высвобожденного медиатора.
Известно, что композиции, модулирующие холинергические функции, влияют на память и воспроизведение. Например, никотин стимулирует никотиновые рецепторы ацетилхолина и считается, что краткосрочное улучшение памяти при табакокурении обусловлено никотином. Антагонист ацетилхолина-скополамин вызывает амнезию и нарушение когнитивных функций, что в психомоторных тестах проявляется в виде замедления простых реакций, возможно, как следствие нарушения внимания, и он применяется с этой целью как сопутствующий анальгетик. Амнезирующее действие скополамина может антагонизироваться никотином.
Имеется два семейства субтипов никотиновых рецепторов (α и β), и каждое из них включает четыре подгруппы, различающиеся по специфичности к лиганду. Роль никотиновых рецепторов в ЦНС еще недостаточно изучена на молекулярном уровне. Возможно, что вещества, связывающиеся с никотиновыми рецепторами, модифицируют скорость метаболизма на сайтах мускариновых рецепторов мозга. Никотиновые рецепторы представляют собой лигандзависимые ионные каналы, активация которых вызывает кратковременное (миллисекундное) увеличение клеточной проводимости Na+ и Са++, деполяризацию и возбуждение.
Другой класс холинергических рецепторов стимулируется мускарином. Такие мускариновые (М) рецепторы относятся к рецепторам, связанным с G-белками. Ответы мускариновых рецепторов более медленные; они могут быть и возбуждающими, и тормозными. Они необязательно связаны с изменениями ионной проводимости. При клонировании холинергических рецепторов было обнаружено пять типов мускариновых рецепторов, которым присвоены обозначения M1-M5. Фармакологической активностью обладают четыре из клонированных рецепторов, обозначаемые как M1-M4 на основании их фармакологической специфичности.
Применение специфических рецепторных белков и моноклональных антител дало возможность установить локализацию в мозге мускариновых M1 (постсинаптические) и М2 (пресинаптические) рецепторов. Но в сердце М2-рецепторы являются постсинаптическими. Пресинаптические мускариновые рецепторы считаются тормозными, связывание ACh с этими рецепторами ослабляет дальнейшее высвобождение ACh, обеспечивая механизм отрицательной обратной связи при высвобождении ACh. Следовательно, избирательные антагонисты М2-рецепторов, преобладающих в мозге, могут быть полезными при лечении болезни Альцгеймера.
Известно, что при таких заболеваниях, как AD/SDAT, наблюдается общая гибель нейронов и недостаточность холинергической функции нервов. Высказывалось предположение, что при лечении таких заболеваний участки с высоким сродством к никотину на оставшихся холинергических нейронах могут превращаться в участки с низким сродством связывания, тем самым продлевая высвобождение медиатора. При снижении сродства участков связывания к никотину предотвращается процесс быстрой десенситизации.
Активация никотиновых рецепторов агонистами в мозге имеет быстрое начало и быстрый конец. Уменьшение сродства никотиновых рецепторов приводит к снижению процесса десенситизации. Schwartz R.D. et al. (J. Neuro Chem. 42 (1984), 1495-98) показали, что сайты связывания никотина локализованы пресинаптически на холинергических (а также 5-гидрокситриптаминергических и катехоламинергических) окончаниях аксонов. Изменения в участках с высоким сродством связывания при AD/SDAT могут повлечь за собой изменения модулирующего действия, которое никотиновые участки оказывают на другие медиаторные системы.
Пресинаптические холинергические механизмы также находятся под тормозным контролем ГАМК-ергических нейронов и полагают, что такое торможение может усиливаться при AD/SDAT. Снятие или ослабление торможения усиливает корковую пресинаптическую холинергическую активность и интенсифицирует когнитивную обработку информации.
Межнейронные взаимодействия волокон, имеющих никотиновую иннервацию (которая снижает сродство связывания), так же как и растормаживание ГАМК-ергических волокон, имеют пресинаптическую локализацию.
Эта модель передачи сигналов в ЦНС - упрощенная, но она обеспечивает основу для понимания попыток, предпринимавшихся с целью увеличения эффективной концентрации ацетилхолина в центральных синапсах. Кроме того, она иллюстрирует концепцию прямого и непрямого воздействия. Указанным выше трем традиционным подходам к лечению AD/SDAT: добавлению предшественников ACh, заместительной терапии агонистами и ингибированию АХЭ присущи и недостатки. Такое лечение может приводить к кратковременному увеличению количества ACh, который будет активировать механизмы обратной связи, приводящие к десенситизации постсинаптических рецепторов. Исходя из теории, не просматриваются долговременные положительные эффекты, а с прекращением лечения положительные эффекты на AD/SDAT и AAMI исчезнут и состояние может даже ухудшиться.
Было показано, что соединение, обладающее активностью M1-агониста и М2/М3-антагониста улучшает умственные способности пациентов с SDAT (Sramak et al., Life Sciences, vol 2, No. 3, 195-202 (1997)). Однако это соединение вызывает неприемлемые побочные холинергические эффекты, такие как утомление, понос и тошнота.
Более радикальный подход к AD/SDAT и AAMI заключается в увеличении числа постсинаптических (M1) рецепторов в мозге. Из китайского патента № CN1096031 A известно, что сарсасапогенин (SaG) способен увеличивать число холинергических M1-рецепторов.
Были патентные публикации, в которых заявлялось о полезности ряда стероидных сапогенинов, обладающих структурой спиростана, фуро-спиростана, спиросолана или соланидина, при лечении заболеваний, включая SDAT. Две патентные публикации имеют особое отношение к настоящему изобретению: китайская патентная публикация № CN 1096031 A о применении спиростанового сапогенина - сарсасапогенина при лечении SDAT. В этом документе, однако, дано лишь краткое описание. Другой документ, имеющий отношение к настоящему изобретению, - это патентная публикация DE 4303214 А1, в которой идет речь о применении очень большого числа сапонинов и сапогенинов при лечении целого ряда заболеваний, имеющих, по мнению авторов, вирусное происхождение. Раскрытие этого изобретения, однако, имеет сомнительную ценность, так как хорошо известно, что при очень большом числе заболеваний, характеризующихся недостаточностью синаптической передачи, отсутствует элемент инфекционности, поэтому основная предпосылка заявленного изобретения ошибочна. Кроме того, они не представили никаких данных, позволяющих специалисту в этой области выбрать предпочтительное соединение из большого числа заявленных.
Авторы настоящего изобретения обнаружили, что некоторые производные сапогенина проявляют способность к регуляции рецепторов. В частности, было обнаружено, что эти соединения увеличивают число М2-рецепторов в мозге. Поэтому один из аспектов изобретения предусматривает применение производных сапогенина общей формулы (I) или (II) в изготовлении лекарства для лечения заболеваний, характеризующихся недостаточным числом или нарушением функции мембраносвязанных рецепторов.
Специалистам в этой области известно о взаимоотношениях между сапонинами и их сапогенинами, как и то, что сапогенины преимущественно жирорастворимы, тогда как сапонины в основном водорастворимы. Сапогенины, таким образом, более способны к проникновению через гематоэнцефалический барьер. Специалистам также известно об эпимеризации ряда сапогенинов в условиях кислотного гидролиза.
Различия в фармакологических свойствах и фармакодинамическом действии разных видов сапогенинов подчеркивают необходимость отбора веществ, наиболее полезных для лечения AD/SDAT. Открытие новых фактов в отношении действия производных сапогенина дало возможность определить, какие именно вещества наиболее пригодны для лечения AD/SDAT и тому подобных заболеваний.
Авторы изобретения обнаружили, что описанные выше свойства проявляют те производные сапогенина, которым присуща cis-конформация кольца А/В в замкнутой кольцевой системе.
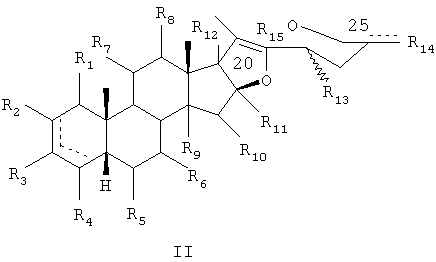
Соответственно, представляющие интерес производные сапогенина по настоящему изобретению имеют следующую общую формулу (I) или (II):
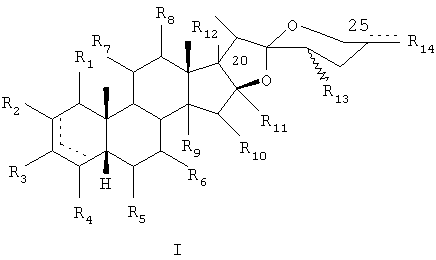

а также их стереоизомеры и рацемические смеси, их фармацевтически приемлемые пролекарства и соли.
В общей формуле (I):
- R1, R2, R3, R4, R5, R6, R7, R8, R10, независимо друг от друга, представляют собой Н, ОН, =O или OR, где R - необязательно замещенный алкил, необязательно замещенный ацил, необязательно замещенный карбамоил или алкоксикарбонил;
- R9, R11, R12, R13 представляют собой Н, ОН или OR, где R - необязательно замещенный алкил, необязательно замещенный ацил, необязательно замещенный карбамоил или алкоксикарбонил;
- R14 - необязательно замещенная алкильная группа,
 означает необязательно двойную связь,
означает необязательно двойную связь,
кроме тех случаев, когда одновременно:
- R1=R2=R4=R5=R6=R7=R8=R9=R10=R11=R12=R13=Н,
- R3=βОН,
- R14=СН3,
- метильная группа в С22 является α-группой,
- С20 является α-группой, a C25 обладает S-конфигурацией.
Предпочтительно в общей формуле (I):
- R4, R9, R12, R13=Н;
- R1, R2, R3, R5, R6, R7, R8, R10, независимо друг от друга, представляют собой Н, ОН, =O или OR, где R - необязательно замещенный алкил, необязательно замещенный ацил, необязательно замещенный карбамоил или алкоксикарбонил;
- R11=Н, ОН или OR, где R - необязательно замещенный алкил, необязательно замещенный ацил, необязательно замещенный карбамоил иди алкоксикарбонил;
- R14 - необязательно замещенная алкильная группа,
 означает необязательно двойную связь,
означает необязательно двойную связь,
кроме тех случаев, когда одновременно:
- R1=R2=R4=R5=R6=R7=R8=R9=R10=R11=R12=R13=Н,
- R3=βOH,
- R14=CH3,
- метильная группа в С22 является α-группой,
- С20 является α-группой, a C25 обладает S-конфигурацией.
Более предпочтительно в общей формуле (I):
- R1=R2=R4=R5=R6=R7=R8=R9=R10=R11=R12=R13=Н,
- R3=Н, -ОН, -ОМе, -ОСОСН3, =O, -O-CO-OEt или -O-СО-(СН2)2-CO2Н,
- R14=СН3,
кроме тех случаев, когда одновременно:
- R1=R2=R4=R5=R6=R7=R8=R9=R10=R11=R12=R13=Н,
- R3=βОН,
- R14=CH3,
- С25 обладает S-конфигурацией,
- С20 является α-группой и метильная группа в С22 является α-группой.
В общей формуле (II):
- R1, R2, R3, R4, R5, R6, R7, R8, R10, независимо друг от друга, представляют собой Н, ОН, =O или OR, где R - необязательно замещенный алкил, необязательно замещенный ацил, необязательно замещенный карбамоил или алкоксикарбонил;
- R9, R11, R12, R13 представляют собой Н, ОН или OR, где R - необязательно замещенный алкил, необязательно замещенный ацил, необязательно замещенный карбамоил или алкоксикарбонил;
- R14 = необязательно замещенная алкильная группа;
- R15 = Н, необязательно замещенный алкил, необязательно замещенный ацил или глюкозил;
 означает необязательно двойную связь.
означает необязательно двойную связь.
Предпочтительно в общей формуле (II):
- R4, R9, R12, R13=Н;
- R1, R2, R3, R5, R6, R7, R8, R10, независимо друг от друга, представляют собой Н, ОН, =O или OR, где R - необязательно замещенный алкил, необязательно замещенный ацил, необязательно замещенный карбамоил или алкоксикарбонил;
- R11=Н, ОН или OR, где R - необязательно замещенный алкил, необязательно замещенный ацил, необязательно замещенный карбамоил или алкоксикарбонил;
- R14 - необязательно замещенная алкильная группа;
- R15=Н, необязательно замещенный алкил, необязательно замещенный ацил или глюкозил;
 означает необязательно двойную связь.
означает необязательно двойную связь.
Особенно предпочтительны следующие соединения:


В настоящем изобретении применяются следующие обозначения:
"Ацил" означает группу Н-СО- или алкил-СО-, в которой алкильная группа соответствует описанию настоящего изобретения. Предпочтительно ацильные группы содержат низший алкил. К типичным ацильным группам относятся формил, ацетил, пропаноил, 2-метилпропаноил, бутаноил и пальмитоил.
"Алкил" означает алифатическую углеводородную группу, которая может быть линейной или разветвленной, с длиной цепи примерно от 1 до 20 углеродных атомов. Предпочтительно алкильные группы имеют длину цепи от 1 до 12 углеродных атомов. Разветвленная цепь означает, что к линейной цепи алкильной группы присоединена одна или несколько низших алкильных групп, таких как метил, этил или пропил. "Низший алкил" означает цепь примерно из 1 до 4 углеродных атомов, которая может быть линейной или разветвленной. К типичным алкильным группам относятся метил, этил, н-пропил, изопропил, н-бутил, трет-бутил, н-пентил, 3-пентил.
"Необязательно замещенный" означает, что указанная группа может содержать одну или несколько замещающих групп, которые могут быть одинаковыми или разными, к числу которых относятся гало, алкил, циклоалкил, гидрокси, алкокси, амино, ациламино, арил, ароиламино, карбокси, алкоксикарбонил, аралкоксикарбонил, гетероаралкоксикарбонил, необязательно замещенный карбамоил.
Термин "фармацевтическая композиция" обозначает композицию, содержащую соединение формулы I или II и, по меньшей мере, один компонент, выбранный из группы, включающей фармацевтически приемлемые носители, разбавители, адъюванты, инертные наполнители и растворители - консерванты, наполнители, дезинтегрирующие вещества, смачивающие вещества, диспергирующие вещества, суспендирующие вещества, подслащивающие вещества, ароматизирующие вещества, отдушки, антибактериальные вещества, противогрибковые вещества, смазывающие вещества и высвобождающие вещества, в зависимости от способа введения и дозовой формы.
"Фармацевтически приемлемый" означает, в рамках здравого медицинского рассуждения, годный для применения при контакте с клетками человека и низших животных без нежелательной токсичности, раздражения, аллергического ответа и т.п., и соразмерный с удовлетворительным соотношением польза/вред.
"Фармацевтически приемлемые дозовые формы" означает дозовые формы соединения по изобретению, к которым относятся, к примеру, таблетки, драже, порошки, эликсиры, сиропы, жидкие препараты, включая суспензии, аэрозоли, таблетки для ингаляции, леденцы, эмульсии, растворы, гранулы, капсулы и свечи, а также жидкие препараты для инъекций, включая препараты липосом. Основные методы и композиции можно найти в Remington, Pharmaceutical Sciences, Mack Publishing, Co., Easton, PA, последнее издание.
"Фармацевтически приемлемые пролекарства", в применении к настоящему изобретению, означает пролекарства из соединений, применяемых согласно настоящему изобретению, которые, в рамках здравого медицинского рассуждения, пригодны для применения при контакте с тканями человека и низших животных с нежелательной токсичностью, аллергическими реакциями и т.п., соразмерно с удовлетворительным соотношением польза/вред, и эффективны для своего назначения, а также цвиттерионные формы, если это возможно, соединений по изобретению. Термин "пролекарство" означает соединения, которые подвергаются быстрому превращению in vivo, образуя родительское соединение приведенной выше формулы, например, при гидролизе в крови. Функциональные группы, которые подвергаются быстрому превращению путем метаболического расщепления in vivo, составляют класс групп, реагирующих с карбоксильной группой соединений настоящего изобретения. Вследствие легкости, с которой метаболически расщепляемые группы в соединениях, применяемых согласно настоящему изобретению, расщепляются in vivo, соединения, несущие такие группы, действуют как пролекарства. Всестороннее обсуждение пролекарств представлено в следующих работах:
Design of Prodrugs, H. Bundgaard, ed., Elsevier, 1985; Methods in Enzymology, K. Widder et al., Ed., Academic Press, 42, p.309-396, 1985; A Textbook of Drug Design and Development, Krogsgaard-Larsen and H. Bundgaard, ed., Chapter 5; Design and Applications of Prodrugs, p.113-191, 1991; Advanced Drug Delivery Reviews, H. Bundgaard, 8, p.1-38, 1992; Journal of Pharmaceutical Sciences, 77, p.285, 1988; Chem. Pharm. Bull., N. Nakeya et al., 32, p.692, 1984; Pro-drugs as Novel Delivery Systems, T. Higuchi and V. Stella, Vol.14 of the A.C.S. Symposium Series, и Bioreversible Carriers in Drug Design, Edward B. Roche, ed., American Pharmaceutical Association and Pergamon Press, 1987, которые включены в настоящее изобретение в виде ссылок.
"Фармацевтически приемлемые соли" обозначает относительно нетоксичные, неорганические и органические соли соответствующих кислот и оснований из соединений настоящего изобретения. Эти соли могут быть получены in situ во время заключительного выделения и очистки соединений. В частности, соли соответствующих кислот можно получить путем проведения отдельной реакции между очищенным соединением в виде свободного основания и подходящей органической или неорганической кислотой и выделения образовавшейся при этом соли. Смотри, к примеру, работу S.M. Berge et al., Pharmaceutical Salts, J. Pharm. Sci., 66: p.1-19 (1977), которая включена в виде ссылки. Соли соответствующих оснований также можно получить путем проведения отдельной реакции между очищенным соединением в виде кислоты и подходящим органическим или неорганическим основанием и выделения образовавшейся при этом соли. К числу солей оснований относятся фармацевтически приемлемые соли металлов и аминов.
Некоторые производные сапогенина, представляющие интерес для настоящего изобретения, в природе могут встречаться в ряде растений, особенно из родов Smilax, Asparagus, Anemarrhena, Yucca и Agave. К видам, представляющим сейчас наибольший интерес, относятся Smilax regelii Kilip & Morton, известная как гондурасская сассапариль; Smilax aristolochiaefolia Miller, известная как мексиканская сассапариль; Smilax ornata Hooker, известная как ямайская сассапариль; Smilax aspera, известная как испанская сассапариль; Smilax glabra Roxburgh; Smilax febrifuga Kunth, известная как эквадорская или перуанская сассапариль; Anemarrhena asphodeloides Bunge; Yucca schidigera Roezl ex Ortgies и Yucca brevifolia Engelm. Представляющие интерес производные сапогенина могут происходить также из других родов, к примеру Dioscorea, Trillium, Solanum, Strophantus, Digitalis и Trigonella. Однако некоторые производные сапогенина из этих источников обладают нежелательными свойствами и поэтому не рекомендуются для применения в настоящем изобретении.
Производные сапогенина настоящего изобретения можно также приобрести коммерческим путем, а их поставщики хорошо известны в этой области и включают Sigma Aldrich, Research Plus Inc., Steraloids Inc. и др.
Следующий аспект изобретения предусматривает способ получения соединений настоящего изобретения.
Замещенные сапогенины настоящего изобретения можно получить синтетическим путем. Например, их можно получить из незамещенных производных сапогенина, встречающихся в природе или приобретаемых коммерческим путем, как указано выше.
Исходя из этих незамещенных сапогенинов, реакция может включать, по меньшей мере, одну стадию замещения, в которой функциональная группа производного сапогенина подвергается замещению. Обычно исходным материалом служит незамещенный сапогенин с требуемой стереохимией, а реакция включает замещение одной ОН-группы требуемым функциональным радикалом, при этом в качестве исходного материала предпочтительны смилагенин и эписмилагенин.
Соединения, применимые согласно настоящему изобретению, можно получить путем приложения или адаптации известных методов, что означает методы, применявшиеся до этого или описанные в литературе, например, описанные R.C. Larock in Comprehensive Organic Transformations, VCH publishers, 1989.
В описываемых далее реакциях может возникнуть необходимость в защите реакционноспособных функциональных групп, например гидроксильных или карбоксильных групп, если они должны присутствовать в конечном продукте, чтобы предотвратить нежелательное участие их в реакциях. Могут применяться общепринятые защитные группы в соответствии со стандартной практикой, примеры которых приведены T.W. Green and P.G.M. Wuts in "Protective Groups in Organic Chemistry" John Wiley and Sons, 1991; J.F.W. McOmie in "Protective Groups in Organic Chemistry" Plenum Press, 1973.
Полученное таким образом соединение можно выделить из реакционной смеси стандартными методами. Например, соединения можно выделить отгонкой растворителя из реакционной смеси или, при необходимости, после отгонки растворителя из реакционной смеси вылить остаток в воду и экстрагировать в несмешивающемся с водой органическом растворителе, а затем отогнать растворитель из экстракта. Кроме того, при желании продукт можно дополнительно очистить различными известными методами, такими как перекристаллизация, переосаждение или различные хроматографические методы, особенно методами колоночной хроматографии или препаративной тонкослойной хроматографии.
Следующий аспект настоящего изобретения предусматривает фармацевтическую композицию, обладающую свойством усиления когнитивных функций, которая содержит эффективное количество производного сапогенина по изобретению.
В следующем аспекте изобретения производные сапогенина по настоящему изобретению представляют собой стероиды, которые предпочтительно не обладают эстрогенным действием.
Другой аспект изобретения предусматривает фармацевтическую композицию, обладающую свойством усиления когнитивных функций, которая содержит эффективное количество производного сапогенина по изобретению в виде экстракта, происходящего из растений рода Smilax, Asparagus, Anemarrhena, Yucca или Agave.
Следует иметь в виду, что изобретение охватывает собой применение композиций, определенных выше. Таким образом, согласно пятому аспекту, настоящее изобретение предусматривает способ усиления когнитивных функций, включающий введение человеку или животному эффективной дозы композиции настоящего изобретения.
Изобретение также предусматривает способ усиления когнитивных функций у человека или животного, включающий введение эффективной дозы производные сапогенина по изобретению. Также оно касается применения производных сапогенина по изобретению в пищевых продуктах или напитках для усиления когнитивных функций.
В применении к настоящему изобретению термин "когнитивная функция" относится к таким функциям, как мышление, рассуждение, запоминание, воображение и обучение.
Следующий аспект изобретения также касается композиции, обладающей свойством усиления когнитивных функций, которая содержит, по меньшей мере, два и предпочтительно два производных сапогенина по изобретению.
При определении соединений, которые могут применяться при лечении SDAT и других заболеваний, характеризующихся уменьшением числа рецепторов или снижением синаптической передачи, авторы учитывали потребность в идентификации соединений, обладающих требуемым действием, но лишенных каких бы то ни было эстрогенных эффектов, которые были бы неприемлемыми, особенно для пациентов-мужчин. Ряд соединений, заявленных в качестве активных в патентной заявке DE 4303214 A1, обладают заметной эстрогенной активностью и поэтому неприемлемы. Предпочтительно, однако, производные сапогенина настоящего изобретения не проявляют эстрогенной активности. Кроме того, эти соединения тестировали на других стероидных рецепторах и они не проявляли активности на следующих рецепторах:
прогестерона, глюкокортикоидов, тестостерона.
Производные сапогенина по настоящему изобретению также тестировали на активность в ряде тестов in vitro. Следующие тесты/эксперименты считались наиболее важными в определении возможной активности повышения числа мембраносвязанных рецепторов: клетки яичников китайского хомячка (СНО), трансфецированные фрагментом ДНК, кодирующим мускариновый рецептор. В большинстве экспериментов использовали клеточную линию, экспрессирующую М2-рецептор.
Далее описываются методы и результаты этих экспериментов.
Эксперименты с клеточной линией СНО
Эффекты различных соединений на экспрессию М2-рецепторов изучали на клетках СНО, трансфецированных ДНК, кодирующей М2-рецептор. Число рецепторов определяли по связыванию меченного тритием QNB за вычетом неспецифического связывания. Соединения растворяли в DMSO, который также применяли в качестве контроля. Соединения тестировали при различных конечных концентрациях. Соединения также тестировали в присутствии и в отсутствие тамоксифена, чтобы отличить механизм, опосредованный эстрогеновыми рецепторами.
Соединения считались активными, если они повышали экспрессию рецептора, выраженную в % относительно контроля, более чем на 15%.
Результаты суммированы ниже в Таблице 1.
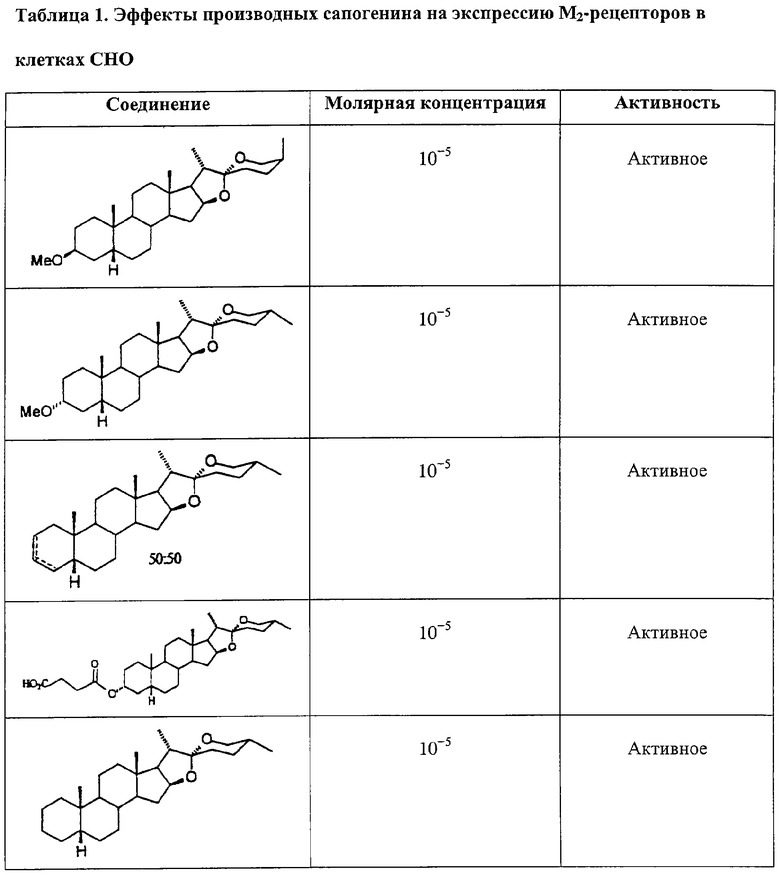


Таким образом, эти эксперименты показывают, что производные сапогенина по изобретению обладают способностью увеличивать число мускариновых рецепторов, экспрессированных на поверхности клеток СНО, культивированных in vitro. Тамоксифен не устранял этот эффект, значит, механизм не связан с эстрогеновыми рецепторами.
Проведенная экспериментальная работа показывает, что соединения настоящего изобретения способны нормализировать число мускариновых рецепторов, а именно они имеют тенденцию предотвращать снижение количества рецепторов со временем, а также восстанавливать число рецепторов до нормального уровня при добавлении к клеткам, в которых количество рецепторов снижено.
Мы предполагаем, что активные соединения, заявленные в настоящем изобретении, действуют через G-белки и что их эффекты на число рецепторов являются вторичными по отношению к эффекту на G-белки. При стимуляции связанных с G-белками мембранных рецепторов запускаются два основных процесса: эффекторный ответ и интернализация рецептора. Последующий процессинг рецептора до такого состояния, что он обретает способность вернуться на поверхность клетки или другой мембраны, где он может снова взаимодействовать с другим рецепторным лигандом, по-видимому, зависит от ряда факторов. Многие из этих факторов или механизмов, по-видимому, связаны с G-белками. Есть данные, что активация М3-рецепторов оказывает влияние на экспрессию G-белков или их уровень. Можно предположить, что эффекты соединений, описанных в настоящем патенте, обусловлены их влиянием на процессы регенерации рецепторов, соединения с G-белками или гомеостаза G-белков.
Альтернативная гипотеза состоит в том, что эти соединения усиливают синтез или высвобождение либо снижают скорость деградации нейротрофических факторов, таких как мозговой фактор роста и/или фактор роста нервов. Такие эффекты на факторы роста могут быть обусловлены влиянием соединения на цитозольный или ядерный рецептор либо связыванием соединения с промоторным участком и последующим действием его непосредственно на образование мРНК для фактора роста, или же быть следствием увеличения продукции другого материального фактора типа G-белка, наконец, эти эффекты могут быть вторичными по отношению к эффекту на процессинг рецептора или G-белка.
Повышенная экспрессия и/или аномальный процессинг предшественника амилоидного белка (АРР) связаны с образованием амилоидных бляшек и амилоидных отложений в сосудах мозга, которые являются основными морфологическими признаками болезни Альцгеймера. Особый интерес представляют процессы, регулирующие протеолитическое расщепление АРР на амилоидогенные и неамилоидогенные фрагменты. Расщепление АРР ферментом α-секретазой в β-амилоидной последовательности белка приводит к образованию неамилоидогенного С-концевого фрагмента и растворимого фрагмента APPsα; для последнего было показано, что он обладает нейротрофической и нейропротекторной активностью, а также улучшает память у мышей при введении в желудочки мозга (ICV). Наоборот, процессинг АРР β-секретазой обнажает N-конец β-амилоида, который удаляется при расщеплении γ-секретазой по вариабельному С-концу. При этом образуются β-амилоидные пептиды, содержащие по 39-43 аминокислоты, которые оказались нейротоксичными и накапливаются в бляшках, создавая помехи для межнейронных связей.
В ряде исследований было показано, что стимуляция связанных с протеинкиназой С (РКС) мускариновых M1- и М3-рецепторов приводит к повышению активности α-секретазы. В результате этого усиливается процессинг АРР до APPsα, обладающего нейропротекторным действием. Параллельно этому снижается процессинг АРРβ- и γ-секретазой, вследствие чего уменьшается образование β-амилоида. Другие медиаторы, такие как фактор роста нервов (NGF) и мозговой нейротрофический фактор (BDNF), а также брадикинин и вазопрессин, могут оказывать подобные эффекты, увеличивая процессинг АРР до APPsα. В эффектах NGF может также участвовать ряд других факторов, включая связывание фактора со связанным с тирозинкиназой (TrkA) рецептором и стимуляцию фосфолипазы Сγ с последующим фосфорилированием и активацией протеинкиназы С (РКС) и возрастанием активности α-секретазы.
Следует ожидать, что любое воздействие, приводящее к избирательной активации протеинкиназы С в мозге, будет полезным при лечении болезни Альцгеймера. До недавнего времени не было селективных агонистов M1-рецептора. А неселективные агонисты будут стимулировать пресинаптические М2-рецепторы, что должно привести к образованию отрицательной обратной связи и поэтому еще более ослабит мускариновую передачу. Сейчас уже появляются селективные агонисты M1-рецептора (талсаклидин) и их уже исследуют на предмет лечения болезни Альцгеймера (AD). Существует, однако, существенный риск, как и при хроническом введении любых рецепторных агонистов, что наблюдаемый положительный клинический эффект будет сильно ограничен в количественном отношении из-за уменьшения числа рецепторов или снижения чувствительности и в отношении побочных эффектов из-за отсутствия рецепторной специфичности. Таким образом, следует ожидать, что описанные в настоящем изобретении соединения, избирательно регулирующие число и функции мускариновых рецепторов, будут лишены тех проблем, которые наблюдаются с мускариновыми агонистами, и поэтому будут особенно полезными. Действительно, их преимущества могут проявляться по трем направлениям, перечисленным ниже.
1. Избирательное увеличение числа M1-рецепторов, ведущее к усилению синаптической передачи. Хроническое введение селективного агониста, по крайней мере, не будет оказывать отрицательного влияния на передачу.
2. Вторичное по отношению к увеличению числа рецепторов усиление стимуляции РКС с последующей активацией α-секретазы, что приводит к:
2.1) уменьшению образования β-амилоида с последующим снижением образования бляшек и гибели нейронов;
2.2) повышению APPsα с последующим улучшением корковых функций, о чем свидетельствует улучшение кратковременной и долговременной памяти.
Для дальнейшей иллюстрации изобретения не ограничивающими его примерами обратимся теперь к прилагаемым чертежам и последующим примерам.
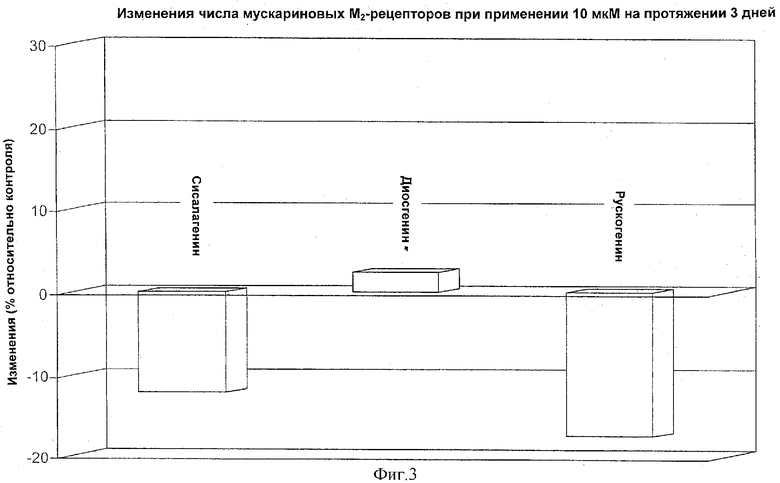
ФИГУРЫ 1, 2, 3 иллюстрируют результаты, полученные ниже в примере 1.
ФИГУРА 4 иллюстрирует гипотетический механизм действия производных сапогенина.
На Фиг.4 схематически представлено действие производных сапогенина по изобретению. Предполагается, что производные сапогенина действуют в основном на клеточные ядра, однако изобретение не ограничивается каким-либо определенным механизмом действия. Наблюдавшеейся увеличение числа мускариновых рецепторов после введения производных сапогенина объясняется усилением экспрессии белка мускаринового рецептора. На схеме показана возможная связь между секретазами и образованием β-амилоидного белка (обсуждавшаяся выше).
Следующие примеры приводятся для иллюстрации изобретения, не ограничиваясь ими.
ПРИМЕР 1
В линии клеток СНО, экспрессирующих рекомбинантные мускариновые рецепторы человека in vitro, число мускариновых рецепторов имеет тенденцию к снижению по времени. При инкубации с производными сапогенина по изобретению (1-10 мкМ) в течение 72 часов повышается плотность мускариновых рецепторов.
Методы
Действие производных сапогенина по изобретению на плотность мускариновых рецепторов в клетках СНО, экспрессирующих рекомбинантные мускариновые рецепторы человека.
Клетки яичников китайского хомячка (СНО), экспрессирующие высокий уровень рецептора (˜2,2 пмоль рецептора/мг белка), культивировали в колбах (150 мл) в течение 24 часов перед началом эксперимента. В среду добавляли растворитель (DMSO) и производные сапогенина (от 1 до 10 мкМ) на 48 часов. Культуральную среду удаляли, клетки снимали, отделяли от поверхности и ресуспендировали в растворе Хэнкса, центрифугировали и определяли уровень М-рецептора путем инкубации с [3H]-QNB в течение 30 минут с последующим измерением в жидком сцинтилляционном счетчике. Содержание белка определяли микрометодом Лоури.
Результаты
Результаты показаны на Фигурах 1-3. Обработка производными сапогенина по изобретению на период культивирования предотвращает снижение числа мускариновых рецепторов концентрационно-зависимым образом.
ПРИМЕР 2
3-O-Этоксикарбонил-5β, 20α, 22α, 25R-спиростан-3β-ол

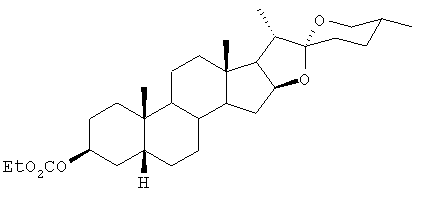
Этиловый эфир хлороформиата (1,40 г, 12,9 ммоль) добавляли по каплям с перемешиванием в раствор смилагенина (2,08 г, 5,0 ммоль) в безводном дихлорметане (15 мл) и безводном пиридине (1,02 г, 12,9 ммоль). Смесь перемешивали при комнатной температуре в течение 18 часов, обрабатывали водой (30 мл) с дихлорометаном и разделяли в делительной воронке. Водный слой дважды экстрагировали дихлорометаном и органические слои объединяли, промывали водой и высушивали над MgSO4 (безводн.). Растворитель выпаривали под вакуумом, получая масло (2,1 г), которое быстро закристаллизовывалось. Этот материал хроматографировали на силикагеле (около 70 г). Элюировали смесью этилацетата-гексана (1:9) и перекристаллизовывали из метанола, получая белые кристаллы 3-O-этоксикарбонил-5β, 20α, 22α, 25R-спиростан-3β-ола (1,08 г): т.пл. 154-156°С; m/z 488 (М+ для С30Н48О5); 1H-ЯМР (270 МГц, CDCl3) δ 0,76 (3Н, s, 18-СН3), 0,78 (3Н, s, 27-СН3), 0,95 (3Н, s, 21-СН3), 0,98 (3Н, s, 19-СН3), 1,0-2,05 (27Н, complex m, алифатические), 1,31 (3Н, t, J=7 Гц, CO2-С-СН3), 3,33-3,46 (2Н, m, 26-OCH2), 4,18 (2H, q, J=7 Гц, CO2СН2), 4,40 (1Н, m, 16-OCH), 4,95 (1Н, m, H-3) ppm; 13С-ЯМР (270 МГц, CDCl3) 14,3 (C-C-O2C), 14,5 16,5, 17,1, 20,9, 23,7, 25,0, 26,4, 28,8, 30,3, 30,6, 31,4, 31,8, 35,0, 35,3, 37,0, 40,0, 40,3, 40,7, 41,6, 56,4 (C-14), 62,3 (C-17), 63,3 (C-O2C), 66,9 (C-26), 74,8 (C-3), 80,9 (C-16), 109,2 (C-22), 154,8 (карбонил) ppm; Rf 0,65 (силикагель, этилацетат-гексан, 1:9).
ПРИМЕР 3
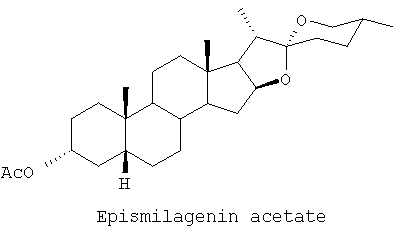
Эписмилагенин сукцинат

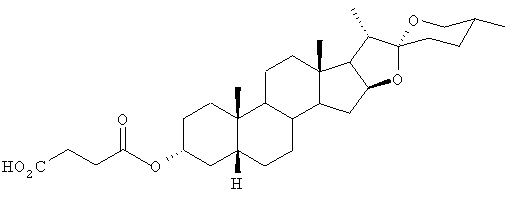
Раствор эписмилагенина (200 мг, 0,48 ммоль) и янтарного ангидрида (60 мг, 0,59 ммоль) в безводном пиридине перемешивали при комнатной температуре в атмосфере азота в течение ночи. Добавляли еще одну порцию янтарного ангидрида (120 мг, 1,18 ммоль) и реакционную смесь перемешивали в течение 24 часов. После добавления еще одной порции янтарного ангидрида (120 мг, 1,18 ммоль) реакционную смесь нагревали при 50°С с перемешиванием дальнейшие 24 часа. После охлаждения реакционной смеси добавляли воду (10 мл) и водный раствор экстрагировали диэтиловым эфиром (4×20 мл). Органические экстракты объединяли, промывали водой (3×20 мл), высушивали (MgSO4, безводн.) и фильтровали. Растворитель выпаривали под вакуумом, получая оранжевое масло (1,8 г), которое подвергали хроматографии на силикагеле и элюировали смесью этилацетата/петролейного эфира (1:4). После перекристаллизации продукта из ацетона получали белые кристаллы сукцината эписмилагенина (87 мг); т.пл. 180-182°С; 1H-ЯМР (CDCl3, 270 МГц): частичные результаты δ 4,75 (1Н, m), 4,6 (1H, m), 3,50 (1H, dd), 3,40 (1H, t), 2,6 (4H, br dd), 0,98 (3H, d), 0,95 (3H, s), 0,80 (3H, d), 0,75 (3H, s) ppm; спектр 13С-ЯМР (CDCl3, 68 МГц): δ 171,81, 109,27, 80,91, 74,90, 66,85, 62,25, 56,29, 41,84, 41,62, 40,65, 40,51, 40,18, 35,44, 35,01, 34,72, 32,17, 31,77, 31,38, 30,25, 29,33, 28,79, 26,93, 26,55, 23,58, 20,58, 17,11, 16,43, 14,48 ppm; Rf 0,11 (силикагель, этилацетат/петролейный эфир, 3:7).
СВОДКА РЕЗУЛЬТАТОВ
Модели и подробные результаты тестирования описаны в Приложении.
Результаты тестирования суммированы следующим образом: (X = нет или почти нет измеримой активности; пустая ячейка = не тестировали; √ = есть активность).
Как видно, по всем тестированным соединениям предсказания, исходящие из рецепторного скринирования (модель А), подтверждаются при тестировании на нейронах (модель В) и в тесте на усиление когнитивной способности in vivo (модель С), и таким образом подтверждается, что рецепторное скринирование является надежным прогнозирующим параметром усиления когнитивной способности и связанной с этим активности.
ПРИЛОЖЕНИЕ
Экспериментальные модели
Введение
Использовали три экспериментальные модели для оценки активности исследуемых соединений в отношении заболеваний, связанных с нарушением когнитивной способности.
При получении приведенных результатов использовали следующие модели.
А. Рецепторные модели. Исследовали влияние исследуемых соединений на зависимое от времени снижение плотности ацетилхолиновых мускариновых-2 рецепторов на клетках яичников китайского хомячка (СНО) и на культуре клеток сердечной мышцы (ССМ). На клетках СНО, котрансфецированных β2-адренергическим рецептором и ацетилхолиновым мускариновым-3 рецептором, показано, что (i) соединения не только предотвращают уменьшение ацетилхолиновых мускариновых-2 рецепторов, но (ii) они и не повышают плотность рецепторов, если не происходит зависимое от времени их уменьшение.
В. Нейронно-клеточные модели. Различные нейродегенеративные заболевания вызывают гибель клеток определенных типов нейронов. Используя разные типы нейронов, можно моделировать различные заболевания: корковые нейроны - болезнь Альцгеймера, мотонейроны - мотонейронная болезнь (амиотрофический боковой склероз), дофаминергические нейроны - болезнь Паркинсона. Кроме того, можно использовать соответствующие токсины: β-амилоид - болезнь Альцгеймера, глутамат - мотонейронная болезнь, МРР+ - болезнь Паркинсона. Нейронно-клеточные модели также применяются для исследования механизма действия соединений: предварительная или последующая обработка, апоптоз, нейротрофическое действие (разрастание нейритов).
С. Лабораторные модели in vivo. Старые крысы и обработанные нейротоксином (β-амилоид и иботеновая кислота) крысы проявляют нарушение когнитивных способностей, уменьшение плотности рецепторов и снижение уровня нейротрофинов, и их используют в качестве общей модели нейродегенеративных нарушений (старые крысы) или специфической модели болезни Альцгеймера (обработка крыс β-амилоидом и иботеновой кислотой). Можно измерять влияние исследуемых соединений на когнитивные способности, плотность рецепторов и уровень нейротрофинов.
А. Рецепторные модели
Дегенерацию и убыль рецепторов мозга можно моделировать in vitro на клетках, трансфецированных для экспрессии β2-рецепторов и/или м2- и/или м3-рецепторов.
Ацетилхолин является нейромедиатором в нервно-мышечных синапсах, во многих синапсах вегетативной нервной системы и в синапсах некоторых других частей нервной системы. Мускариновые ацетилхолиновые рецепторы - это рецепторы, на которые действует ацетилхолин. Мускариновые-2 ацетилхолиновые рецепторы являются пресинаптическими рецепторами и находятся в стволе головного мозга и в сердце. Активация мускариновых-2 ацетилхолиновых рецепторов обычно вызывает тормозящие эффекты, как правило, ингибирование аденилатциклазы. Мускариновые-3 ацетилхолиновые рецепторы находятся в гладких мышцах, коре головного мозга и эндокринных железах. Активация мускариновых-3 ацетилхолиновых рецепторов обычно вызывает возбуждение клеток.
Норадреналин является представителем другого семейства нейромедиаторов - катехоламинов, в которую входит и дофамин. Адренергические рецепторы - это рецепторы, на которые действует адреналин и норадреналин. Дофаминовые рецепторы - это рецепторы, на которые действует дофамин. β2-Адренергические рецепторы являются постсинаптическими рецепторами, которые играют важную роль в симпатической нервной системе, более чувствительны к адреналину, чем к норадреналину, и обладают сильным сродством к агонисту тербуталину.
Заявители использовали клетки яичников китайского хомячка (СНО), трансфецированные вектором для мускариновых-2 ацетилхолиновых рецепторов, и клетки СНО, котрансфецированные векторами для β2-адренергических рецепторов и для мускариновых-3 ацетилхолиновых рецепторов.
Заявители также использовали культуру клеток сердечной мышцы (ССМ), которые от природы экспрессируют мускариновые-2 ацетилхолиновые рецепторы.
Эти исследования описаны в Примерах 1 и 2, приведенных ниже.
Rimie et al. (Brain Res. 336, 19-25, 1985) и Selkoe (Sci. Am. 267, 134-142, 1992) сообщали, что связанное с возрастом ухудшение корковых функций у человека может коррелировать со снижением плотности ацетилхолиновых рецепторов (мускариновых и никотиновых) и/или снижением сопряженных с ними функций в корковых зонах. Narang (Mech. Ageing Dev. 78, 221-239, 1995) сообщал, что связывание с мускариновыми аце-тилхолиновыми рецепторами значительно уменьшается в гиппокампе у старых крыс, указывая на возможную корреляцию с возрастными нарушениями когнитивных способностей или повреждением нейронов. Biegon et al. (Neurobiol. Aging 10, 305-310, 1989) сообщали о подобном эффекте в стриатуме у старых крыс. В публикации Rinne et al. (см. выше) сообщается о подобном эффекте в коре у старых людей. Drakarch et al. (Expert. Opin. Investig. Drugs 10, 1855-1861, 2001) сообщали, что нейродегенеративные нарушения, такие как болезнь Паркинсона, сопровождаются уменьшением активности дофаминергических рецепторов.
Следовательно, можно полагать, что активные вещества, предупреждающие или предотвращающие убыль рецепторов на рецепторных моделях либо оказывающие влияние на экспрессию рецепторов на рецепторных моделях, будут по меньшей мере частично эффективными, к примеру, против болезни Альцгеймера, SDAT, деменции Lewi body и возрастных нарушений когнитивных способностей, когнитивных симптомов болезни Паркинсона, ортостатической гипотензии, аутизма, синдрома хронической усталости, миастении gravis, болезни Ламберта-Итона, синдрома войны в Персидском заливе и заболеваний, связанных с фосфорорганическими соединениями.
В. Нейронно-клеточные модели
Нейронную и нервно-мышечную дегенерацию и гибель клеток (апоптоз) можно моделировать in vitro на первичных культурах корковых нейронов крыс, дофаминергических нейронах среднего мозга крыс и мотонейронах спинного мозга крыс. Нейродегенерацию моделируют, подвергая первичные корковые нейроны крыс воздействию нейротоксина глутамата, а выживание клеток проверяют, измеряя активность лактатдегидрогеназы (LDH) в нейронах.
Нейродегенерацию в специфическом контексте болезни Альцгеймера моделируют, подвергая первичные корковые нейроны крыс воздействию β-амилоида. Нейродегенерацию в специфическом контексте болезни Паркинсона моделируют, подвергая дофаминергические нейроны среднего мозга крыс воздействию нейротоксина МРР+ в соответствии с публикацией Mytinlineou et al. (Science 225, 529-531, 1984). Нейродегенерацию в специфическом контексте мотонейронной болезни моделируют, подвергая мотонейроны спинного мозга крыс воздействию глутамата. Апоптоз определяют, измеряя активность каспазы-3 в первичных корковых нейронах крыс после воздействия глутамата или стауроспорина. Katzman et al. (Faseb J. 5, 278-286, 1991) сообщали о корреляции между индукцией отростков у первичных нейронов крыс и улучшением симптомов нейродегенеративных заболеваний. Рост отростков измеряют по средней длине отростков и числу нейронов с отростками.
Эти исследования описаны в Примерах 3-11, приведенных ниже. Микрофотографии, представленные на фиг.5, относятся к результатам Примера 8.
Вполне вероятно, что активные вещества, предупреждающие или предотвращающие повреждение нейронов и/или апоптоз на нейронно-клеточных моделях, будут в определенной степени эффективными, к примеру, против нейродегенеративных симптомов болезни Паркинсона, миастении gravis, болезни Ламберта-Итона, ортостатической гипотензии и синдрома аутизма.
С. Модели на крысах in vivo
Когнитивные способности можно тестировать in vivo, проводя анализ обучения и памяти на лабораторных крысах.
Заявители моделировали уменьшение нейротрофинов, убыль рецепторов и нарушение когнитивных способностей, используя старых или обработанных нейротоксином крыс. Измеряя плотность рецепторов мозга у забитых крыс, можно выявить корреляции между действием активных средств против нарушения когнитивных способностей и против дегенерации и убыли мускариновых ацетилхолиновых и/или дофаминовых рецепторов и/или против уменьшения нейротрофинов.
Эти исследования описаны в Примерах 12-14, приведенных ниже.
Такие корреляции можно использовать для подтверждения прогнозов вероятного действия активных средств против всех заболеваний, перечисленных выше в связи с методами тестирования А и В.
Результаты тестирования, включая методики и обсуждение
ПРИМЕР 4
Активность соединений: сарсасапогенина (для сравнения), анзурогенина-D и смилагенина тестировали следующими методами.
1. Эксперименты на клетках линии СНО
Исследовали эффекты определенных соединений на экспрессию мускариновых-2 ацетилхолиновых рецепторов в клетках СНО, трансфецированных с помощью ДНК для мускариновых-2 ацетилхолиновых рецепторов. Количество рецепторов определяли методами связывания радиолигандов. Соединения растворяли в ДМСО и ДМСО (0,5%) использовали в качестве контроля на растворитель. Соединения тестировали в серии конечных концентраций. Результаты приведены ниже в табл.3.
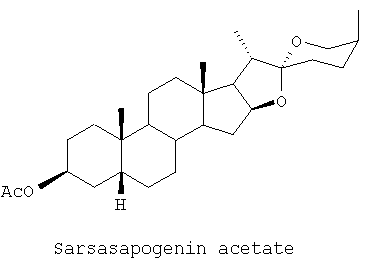
Некоторые соединения тестировали в концентрации 10-5 М на способность вызывать повышение экспрессии мускариновых-2 ацетилхолиновых рецепторов больше, чем на 15%, по сравнению с контролем на растворитель. При тестировании оказалось, что следующие соединения обладают этой способностью: сарсасапогенин, сарсасапогенин ацетат, смилагенин, смилагенин ацетат, смилагенин катилат, эписмилагенин, эписмилагенин ацетат и эписмилагенин сукцинат. При тестировании оказалось, что следующее соединение не обладает этой способностью: диосгенин.
2. Эксперименты на клетках сердечной мышцы
В данном исследовании также изучали эффекты вышеуказанных соединений.
Клетки сердечной мышцы выделяли из мышц желудочков новорожденных крыс Sprague-Dawley и культивировали in vitro. Плотность мускариновых ацетилхолиновых рецепторов, экспрессированных на поверхности клеток, измеряли в мембранных фрагментах после гомогенизации клеток, собранных в различные промежутки времени, с помощью методов связывания радиолигандов. Предварительные эксперименты показали, что плотность мускариновых ацетилхолиновых рецепторов снижалась после 10-дневного культивирования. Поэтому эксперименты ставили таким образом, чтобы исследовать эффекты данных соединений в ингибировании этого снижения плотности рецепторов. Результаты этих экспериментов приведены ниже в табл.4.
Результаты свидетельствуют, что данные соединения уменьшали зависимое от времени снижение плотности мускариновых ацетилхолиновых рецепторов in vitro.
ПРИМЕР 5
Исследовали эффекты сарсасапогенина, сарсасапогенина катилата, эписарсасапогенина катилата, эписарсасапогенина сукцината, эписмилагенина катилата и эписмилагенина ацетата на экспрессию мускариновых-2 ацетилхолиновых рецепторов в клетках СНО, трансфецированных вектором для мускариновых-2 ацетилхолиновых рецепторов.
Результаты приведены ниже в табл.5.
За время культивирования трансфецированные клетки СНО проявляют зависимое от времени снижение плотности мускариновых-2 ацетилхолиновых рецепторов. Обработка сарсасапогенином, сарсасапогенином катилатом, эписарсасапогенином катилатом, эписарсасапогенином сукцинатом и эписмилагенином катилатом предотвращает зависимое от времени снижение плотности мускариновых-2 ацетилхолиновых рецепторов.
В клетках СНО, котрансфецированных мускариновым-3 ацетилхолиновым рецептором и β2-адренергическим рецептором, отмечается зависимое от времени снижение плотности β2-адренергических рецепторов, но не наблюдается снижения плотности мускариновых-3 ацетилхолиновых рецепторов.
При инкубации клеток СНО, котрансфецированных мускариновым-3 ацетилхолиновым рецептором и β2-адренергическим рецептором, в присутствии эписмилагенина ацетата плотность мускариновых-3 ацетилхолиновых рецепторов изменяется незначительно, однако предотвращается зависимое от времени снижение плотности β2-адренергических рецепторов (см. табл.6).
Эти эксперименты свидетельствуют, что сарсасапогенин, сарсасапогенин катилат, эписарсасапогенин катилат, эписарсасапогенин сукцинат, эписмилагенин катилат и эписмилагенин ацетат повышают плотность мускариновых рецепторов, экспрессированных в клетках СНО, культивируемых in vitro. Кроме того, эписмилагенин ацетат предотвращает зависимое от времени снижение β2-адренергических рецепторов, не изменяя плотность мускариновых-3 ацетилхолиновых рецепторов, то есть рецепторов, не проявляющих зависимого от времени снижения плотности в клетках СНО.
ПРИМЕР 6
В этом исследовании изучали эффекты предварительной 24-часовой обработки сарсасапогенином, эписарсасапогенином, эписарсасапогенином катилатом, смилагенином, эписмилагенином и диосгенином на вызванное глутаматом повреждение нейронов у первичных корковых нейронов крыс.
Корковые нейроны крыс культивировали в течение 10 дней и на 10-й день среду заменяли на лишенную сыворотки среду определенного состава. На 12-й день культуры промывали свежей средой, содержащей исследуемое соединение или контрольный растворитель (ДМСО, 0,25%), и оставляли в ней. На 13-й день культуры подвергали воздействию глутамата (100 мкМ, 10 мин). После инкубационного периода культуры промывали свежей средой, содержащей исследуемое соединение или контрольный растворитель, и оставляли в ней еще на 24 часа.
Повреждение нейронов оценивали по измерению активности лактатдегидрогеназы (LDH), высвободившейся в среду за 24 часа обработки глутаматом, с помощью нерадиоактивного набора CytoTox 96 и выражали количественно по измерению оптического поглощения при 450 нм. Затем его пересчитывали на выживаемость нейронов.
Воздействие глутамата вызывало значительное повреждение нейронов. В культурах, предварительно обработанных сарсасапогенином, эписарсасапогенином катилатом или смилагенином в течение 24 часов, отмечалось значительное уменьшение вызванного глутаматом повреждения нейронов (табл.7). Напротив, диосгенин не вызывал значительного уменьшения вызванного глутаматом повреждения нейронов.
Исследование показало, что сарсасапогенин, эписарсасапогенин, эписарсасапогенин катилат, смилагенин и эписмилагенин, но не диосгенин обеспечивают защиту нейронов после воздействия глутамата у первичных корковых нейронов крыс in vitro.
ПРИМЕР 7
В этом исследовании изучали способность сарсасапогенина и смилагенина к предотвращению вызванного глутаматом повреждения нейронов у первичных корковых нейронов крыс.
Воздействие глутамата (100 мкМ, 10 мин) на первичные корковые нейроны крыс вызывало повышение активности лактатдегидрогеназы (LDH) при измерении через 24 часа, что указывает на значительное повреждение нейронов (табл.7).
После обработки глутаматом внесение сарсасапогенина и смилагенина на 24 часа значительно уменьшало активность LDH по сравнению с нейронами, обработанными одним глутаматом, что свидетельствует о существенной защите нейронов (табл.8).
Это исследование показало, что сарсасапогенин и смилагенин предотвращают вызванную глутаматом дегенерацию нейронов, что свидетельствует о терапевтическом потенциале при нейродегенеративных заболеваниях. Исследование также показывает, что диосгенин неэффективен.
ПРИМЕР 8
В этом исследовании изучали способность смилагенина к предотвращению вызванного глутаматом апоптоза в первичной культуре корковых нейронов крыс в течение 6 дней in vitro. Апоптоз оценивали по измерению активности каспазы-3. Относительную активность каспазы-3 измеряли колориметрически по расщеплению субстрата каспазы-3 по оптической плотности, приведенной к содержанию белка в пробе.
Воздействие глутамата (100 мкМ, 10 мин) на первичные культуры корковых нейронов вызывало повышение активности каспазы-3 при измерении через 24 часа, что указывает на значительный апоптоз (табл.8).
После обработки глутаматом внесение смилагенина на 6 часов уменьшало активность каспазы-3 по сравнению с нейронами, обработанными одним глутаматом, что свидетельствует о существенном антиапоптозном эффекте (табл.9).
Это исследование показало, что смилагенин предотвращает индуцированную глутаматом активность каспазы-3 в первичных корковых нейронах крыс in vitro.
ПРИМЕР 9
В этом исследовании изучали антиапоптозное действие сарсасапогенина и смилагенина на активность каспазы-3, маркера апоптоза, в первичных культурах корковых нейронов крыс при воздействии стауроспорина, сильного алкалоидного ингибитора киназ, вызывающего апоптозную смерть клеток у многих разных типов клеток.
Предварительная обработка корковых нейронов крыс сарсасапогенином вызывала значительное уменьшение активности каспазы-3, вызванной воздействием стауроспорина (табл.10).
Это исследование показало, что сарсасапогенин и смилагенин частично предотвращают индуцированную стауроспорином активность каспазы-3 в первичных корковых нейронах крыс in vitro.
ПРИМЕР 10
Нейродегенеративные заболевания характеризуются прогрессирующей гибелью нейронов и разрушением отростков нейронов (нейритов). Вещества, индуцирующие рост нейритов, могут способствовать образованию новых связей между нейронами и улучшить симптомы нейродегенеративных заболеваний (Katzman et al., Faseb J. 5, 278-286, 1991).
Обработка первичных корковых нейронов крыс сарсасапогенином и смилагенином вела к значительному увеличению длины существующих нейритов и процента нейронов, имеющих нейриты, по данным оптической микрометрии (табл.12 и 11).
Эти результаты показывают, что смилагенин и Сарсасапогенин обладают нейротрофическим действием in vitro.
ПРИМЕР 11
В этом исследовании изучали способность смилагенина, сарсасапогенина, эписарсасапогенина и эписмилагенина к предотвращению МРР+-индуцированного апоптоза в дофаминергических нейронах крыс.
Повреждения, вызываемые нейротоксином МРР+, метаболитом 1-метил-4-фенил-1,2,3,6-тетрагидропиридина (МРТР), напоминают дегенерацию дофаминергических нейронов нигростриатума при нейродегенеративных заболеваниях, таких как болезнь Паркинсона (Mytinlineou et al., Science 225, 529-531, 1984). Наиболее заметные биохимические изменения, вызываемые этим токсином, включают снижение уровня дофамина и его метаболитов в substantia nigra pars compacta и в хвостатом ядре (Burns et al., Proc. Nat. Acad. Sci. USA 80, 4546-4550, 1983) и уменьшение захвата дофамина в препаратах синаптосом из нигростриатума (Heikkila et al., J. Neurochem. 44, 310-313, 1985).
Предварительная обработка в течение 24 ч дофаминовых нейронов среднего мозга крыс сарсасапогенином, смилагенином, эписарсасапогенином и эписмилагенином значительно повышала выживаемость нейронов, подвергнутых воздействию одного МРР+, что свидетельствует о существенном нейрозащитном действии (табл.13).
Результаты этих опытов были получены в виде микрофотоснимков, а также проведен дополнительный опыт по проверке соединения эписмилагенина катилата (см. Фиг.5). Наблюдается впечатляющая сохранность роста нейритов при использовании исследованных соединений по сравнению с контролем и с опытами без обработки.
Это исследование показало, что сарсасапогенин, смилагенин, эписарсасапогенин и эписмилагенин предотвращают вызванное МРР+ повреждение нейронов у первичных дофаминовых нейронов среднего мозга, то есть на модели болезни Паркинсона in vitro.
ПРИМЕР 12
В этом исследовании изучали способность смилагенина и сарсасапогенина к предотвращению МРР+-индуцированного апоптоза в дофаминергических нейронах крыс.
Обработка нейронов среднего мозга крыс МРР+ (2 мкМ, 24 ч) вела к повреждению нейронов при измерении через 48 ч (табл.13).
После обработки МРР+ внесение сарсасапогенина и смилагенина на 48 ч вело к значительному уменьшению повреждений нейронов по сравнению с нейронами, обработанными одним МРР+, что свидетельствует о существенном нейрозащитном действии (табл.14).
Обработка МРР+ также значительно снижала процент нейронов, имеющих нейриты (табл.15). После воздействия МРР+ сарсасапогенин и смилагенин значительно уменьшали снижение числа нейронов, имеющих нейриты.
Из этих результатов следует ожидать, что данные соединения будут предотвращать повреждение нейронов.
ПРИМЕР 13
В этом исследовании изучали способность смилагенина к предотвращению вызванного глутаматом повреждения нейронов у первичных мотонейронов спинного мозга крыс, то есть на модели дегенерации мотонейронов in vitro.
Мотонейроны крыс получали в соответствии с описанным методом (Martinou et al., Neuron 8, 737-744, 1992). На 10-й день среду удаляли и культуры подвергали воздействию глутамата (4 мкМ) в течение 10 мин при 37°С в среде определенного состава. После обработки глутаматом культуры промывали модифицированной Дюльбекко средой Игла при 37°С, а затем помещали в свежую среду, содержащую исследуемые соединения. Через 48 ч определяли степень дегенерации мотонейронов по измерению количества лактатдегидрогеназы (LDH), высвобожденной в культуральную среду.
Результаты представлены ниже в табл.16.
Эти значения отличаются, потому что добавляли большее количество культуры (низкая токсичность глутамата в исходных данных).
Это исследование показало, что сарсасапогенин и смилагенин предотвращают вызванное глутаматом повреждение нейронов у мотонейронов спинного мозга крыс, то есть на модели мотонейронной болезни in vitro.
ПРИМЕР 14
В этом исследовании изучали способность смилагенина и сарсасапогенина к уменьшению вызванного β-амилоидом повреждения нейронов у первичных корковых нейронов крыс.
У первичных корковых нейронов крыс 24-часовая предобработка смилагенином и сарсасапогенином значительно уменьшала вызванное β-амилоидом (15 мкМ) выделение лактатдегидрогеназы (LDH) и повышала выживаемость нейронов по сравнению с нейронами, обработанными одним β-амилоидом (табл.17).
Это исследование показало, что сарсасапогенин и смилагенин предотвращают вызванное β-амилоидом повреждение нейронов у первичных корковых нейронов крыс, то есть на модели болезни Альцгеймера in vitro.
ПРИМЕР 15
Во второй половине жизни (у человека начиная с возраста 40 лет) плотность нейронов в головном мозге снижается (Selkoe D.J., Sci. Am., 267, 134-142, 1992). Изменения корковых функций могут быть вызваны уменьшением числа нейронов, их взаимосвязей, снижением уровня нейротрофинов, таких как мозговой нейротрофический фактор (BDNF; Bothwell M., Functional interactions of neurotrophins and neurotrophin receptors, Annu. Rev. Neurosci., 18, 223-253, 1995), снижением плотности ацетилхолиновых рецепторов (мускариновых и никотиновых) и/или уменьшением сопряженных с ними функций в корковых зонах (Rinne et al., Brain Res., 336, 19-25, 1985; Selkoe D.J., Sci. Am., 267, 134-142, 1992). Кроме того, при старении плотность мускариновых ацетилхолиновых рецепторов значительно снижается в гиппокампе (Narang, 1995) и стриатуме старых крыс (Biegon et al., Nerobiol. Aging, 10, 305-310, 1989) и человека (Rinne et al., Brain Res., 336, 19-25, 1985). Кроме того, при болезни Альцгеймера уменьшение холинергической активности связано с отложением β-амилоидных бляшек (von der Kammer et al., Biochem. Soc. Symp.131-140, 2001). При других нейродегенеративных заболеваниях, таких как болезнь Паркинсона, проявляется характерное снижение дофаминергической активности (Drukarch et al., Expert. Opin. Investig. Drugs 10, 1855-1861, 2001).
При приеме внутрь (предписанный клинический способ применения) сарсасапогенина, эписарсасапогенина катилата или смилагенина (18 мг/кг/день) на протяжении 2-3 месяцев у старых крыс Sprague-Dawley (SD, возраст 20 мес) предотвращались нарушения способности к обучению и памяти (измеряли с помощью Y-лабиринта, табл.18), снижение плотности мускариновых ацетилхолиновых рецепторов и частично уменьшалось падение уровня нейротрофина BDNF (табл.19).
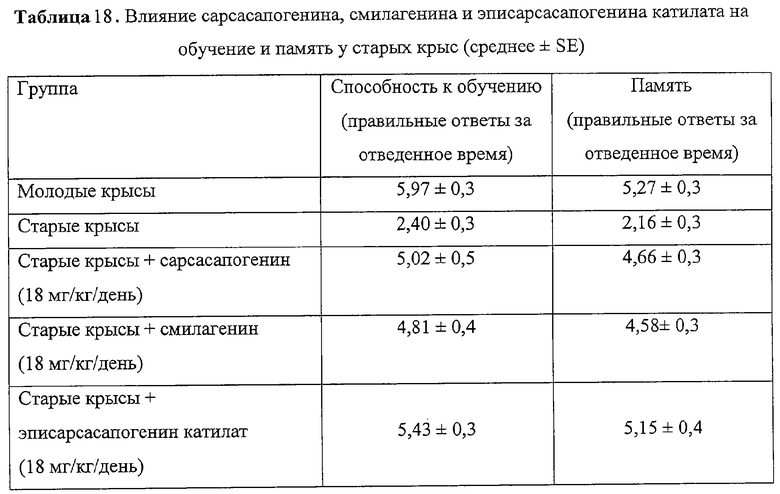
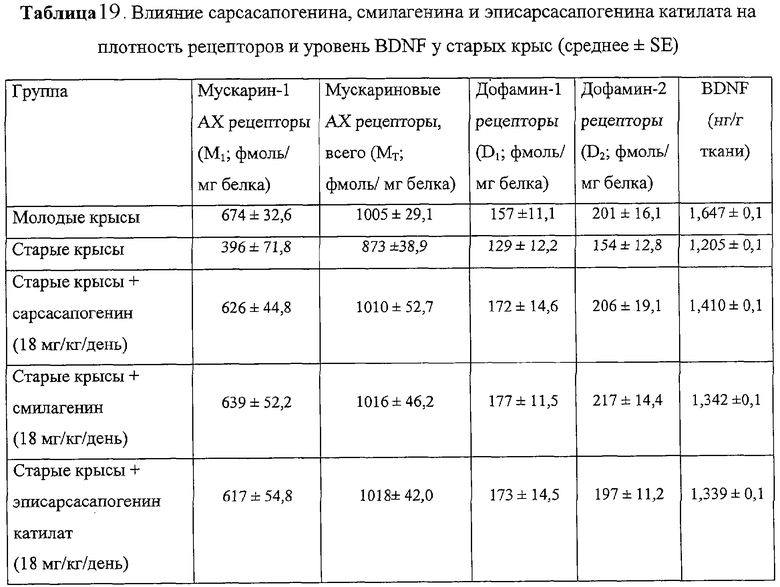
Это исследование показало, что сарсасапогенин, смилагенин и эписарсасапогенин катилат предотвращают нарушения когнитивных способностей, падение уровня BDNF и снижение плотности мускариновых ацетилхолиновых рецепторов и дофаминовых рецепторов у старых крыс.
ПРИМЕР 16
Введение β-амилоида и иботеновой кислоты в мозг крыс вызывает гибель нейронов, убыль рецепторов, нарушение когнитивных способностей и применяется в качестве модели болезни Альцгеймера in vivo. Предшествующие исследования показали, что местная инъекция β-амилоида в nucleus vasalis мозга крыс вызывает снижении функции ацетилхолиновых нейронов и нарушение поведенческих реакций вплоть до двух месяцев после операции (Giovanelli et al., 1995, Neuroscience, 66, 781-792). Кроме того, совместное введение β-амилоида и иботеновой кислоты в гиппокамп крыс синергично вызывает гибель нейронов с инфильтрацией глиальных клеток не только вблизи от, но и вдали от места инъекции (Morimoto et al., 1998, Neuroscience, 84, 479-487).
В данном эксперименте использовали метод Моримото (Morimoto et al., 1998, Neuroscience, 84, 479-487) с модификациями (односторонняя инъекция, 4 мкг β-амилоида и 1 мкг иботеновой кислоты). Использовали 3-месячных крыс Sprague-Dawley.
При приеме внутрь (предписанный клинический способ применения) сарсасапогенина, эписарсасапогенина катилата, эписарсасапогенина и эписмилагенина (18 мг/кг/день) на протяжении 2 месяцев предотвращались нарушения способности к обучению и памяти (измеряли с помощью тестов на перешагивание и спрыгивание, табл.19) и снижение плотности мускариновых ацетилхолиновых рецепторов (табл.19). Напротив, при приеме внутрь эписарсасапогенина этилсукцината и диосгенина (18 мг/кг/день) на протяжении 2 месяцев не наблюдалось предотвращения нарушения способности к обучению и памяти или снижения плотности мускариновых ацетилхолиновых рецепторов (табл.20).
Это исследование показало, что Сарсасапогенин, Эписарсасапогенин катилат, эписарсасапогенин и эписмилагенин предотвращают нарушение когнитивных способностей и снижение плотности мускариновых ацетилхолиновых рецепторов, происходящее в этой модели болезни Альцгеймера in vivo. Напротив, Эписарсасапогенин этилсукцинат и диосгенин не способны предотвратить нарушение когнитивных способностей и снижение плотности мускариновых ацетилхолиновых рецепторов, происходящие в этой модели болезни Альцгеймера in vivo.
ПРИМЕР 17
Использовали модель болезни Альцгеймера на животных, описанную в Примере 13, для оценки влияния смилагенина на нарушение когнитивных способностей, плотность мускариновых ацетилхолиновых рецепторов и уровень BDNF у крыс. Однако для оценки обучения и памяти использовали Y-лабиринт, как описано в Примере 12, вместо тестов на перешагивание и спрыгивание.
При приеме внутрь (предписанный клинический способ применения) смилагенина (3,6, 9 и 18 мг/кг/день) на протяжении 2-3 месяцев у 3-месячных крыс Sprague-Dawley предотвращались нарушения способности к обучению и памяти, снижение плотности мускариновых ацетилхолиновых рецепторов и частично уменьшалось падение уровня нейротрофина BDNF (табл.21).
Это исследование показало, что смилагенин предотвращает нарушения когнитивных способностей, снижение плотности мускариновых ацетилхолиновых рецепторов и частично уменьшает падение уровня нейротрофина BDNF, происходящие в этой модели болезни Альцгеймера in vivo.
| название | год | авторы | номер документа |
|---|---|---|---|
| СТЕРОИДНЫЕ САПОГЕНИНЫ И ИХ ПРОИЗВОДНЫЕ ДЛЯ ЛЕЧЕНИЯ | 2004 |
|
RU2297228C2 |
| ПРОИЗВОДНЫЕ САПОГЕНИНА, ИХ СИНТЕЗ, ПРИМЕНЕНИЕ И СПОСОБЫ, ОСНОВАННЫЕ НА ИХ ПРИМЕНЕНИИ | 2002 |
|
RU2311423C2 |
| СТЕРОИДНЫЕ САПОГЕНИНЫ И ИХ ПРОИЗВОДНЫЕ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ АЛЬЦГЕЙМЕРА | 1999 |
|
RU2242978C2 |
| ТЕРАПЕВТИЧЕСКИЕ СПОСОБЫ И ПРИМЕНЕНИЯ САПОГЕНИНОВ И ИХ ПРОИЗВОДНЫХ | 2003 |
|
RU2332999C2 |
| СПОСОБ СТЕРЕОСПЕЦИФИЧЕСКОГО ВОССТАНОВЛЕНИЯ САПОГЕН-3-ОНОВ | 2003 |
|
RU2326890C2 |
| КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2792562C2 |
| ОРГАНИЧЕСКИЕ СОЕДИНЕНИЯ | 2012 |
|
RU2610094C2 |
| КОМПОЗИЦИИ ИЗОИНДОЛИНА И СПОСОБЫ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНОГО ЗАБОЛЕВАНИЯ | 2015 |
|
RU2692258C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ НЕЙРОДЕГЕНЕРАТИВНОГО ЗАБОЛЕВАНИЯ | 2012 |
|
RU2618412C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ НЕЙРОДЕГЕНЕРАТИВНЫХ РАССТРОЙСТВ | 2012 |
|
RU2699038C2 |




В изобретении раскрыты производные 5β-сапогенина и псевдосапогенина общей формулы I или II, где в общей формуле I или II: R1, R2, R4, R5, R6, R7, R8, R9, R10, R11, R12, R13 и, если присутствует, R15 - H, независимо R1 и R13 - ОН или R3 - ОН, =O или OR, где R - необязательно замещенный алкил; низший ацил, необязательно замещенный карбамоилом или амино или низшим алкоксикарбонилом; низший алкоксикарбонил; R14 низший алкил,  необязательно двойная связь за исключением смилагенина и тех соединений формулы I, где одновременно R1=R2=R4=R5=R6=R7=R8=R9=R10=R11=R12=R13=H, R3=βOH, R14=CH3, и метил в С20 положении имеет α-ориентацию и С25 имеет S-конфигурацию. Соединения увеличивают, количество мускариновых рецепторов для усиления функции мускариотных рецепторов у человека или животного, а также полезны для усиления когнитивной функции у человека или животного. 3 н. и 8 з.п. ф-лы, 5 ил., 21 табл.
необязательно двойная связь за исключением смилагенина и тех соединений формулы I, где одновременно R1=R2=R4=R5=R6=R7=R8=R9=R10=R11=R12=R13=H, R3=βOH, R14=CH3, и метил в С20 положении имеет α-ориентацию и С25 имеет S-конфигурацию. Соединения увеличивают, количество мускариновых рецепторов для усиления функции мускариотных рецепторов у человека или животного, а также полезны для усиления когнитивной функции у человека или животного. 3 н. и 8 з.п. ф-лы, 5 ил., 21 табл.



или их рацемические смеси или их фармацевтически приемлемые пролекарства и соли,
где в общей формуле (I) или (II)
R1, R2, R4, R5, R6, R7, R8, R9, R10, R11, R12, R13 и, если присутствует, R15
представляют собой Н; или
независимо друг от друга R1 и R13, могут быть ОН, и R может быть ОН, =O или OR, где R - необязательно замещенный алкил; низший ацил, необязательно замещенный карбамоилом или амино или низшим алкоксикарбонилом; низший алкоксикарбонил;
R14 представляет собой низший алкил
 означает необязательно двойную связь;
означает необязательно двойную связь;
за исключением смилагенина и тех соединений формулы I, в которых одновременно R1=R2=R4=R5=R6=R7=R8=R9=R10=R11=R12=R13=H, R3=βOH, R14=CH3, и метильная группа в положении С20 является α-группой и С25 имеет S-конфигурацию,
для производства композиции, предназначенной для
(i) увеличения количества мускариотных рецепторов или усиления функции мускариотных рецепторов у человека или животного;
(ii) для усиления когнитивной функции у человека или животного;
(iii) для лечения состояний, характеризующихся присутствием нейрофибриллярных клубков или β-амилоидных бляшек.
R1, R2, R4, R5, R6, R7, R8, R9, R10, R11, R12, R13 представляют собой Н;
R3=H, -ОН, -ОМе, -ОСОСН3, =О, -O-CO2Et, -O-CO-(CH2)2-CO2H;
R14 - представляет собой СН3;








| WO 9916786 А, 08.04.1999 | |||
| Способ получения диосгенина | 1978 |
|
SU724521A1 |
| R.E.MARKER et al | |||
| J | |||
| of Amer | |||
| Chem | |||
| Soc | |||
| Способ приготовления пищевого продукта сливкообразной консистенции | 1917 |
|
SU69A1 |
| Электрический паяльник | 1924 |
|
SU2167A1 |

