Данное является частичным продолжением находящихся в процессе одновременного рассмотрения заявок на патент США №561024, поданной 13 декабря 1983 года; 582185, поданной 21 февраля 1984 года, и 655841, поданной 28 сентября 1984 года.
Предпосылки изобретения
Настоящее изобретение относится к манипуляции генетическими материалами и, в частности, к рекомбинантным методикам, позволяющим получить полипептиды, обладающие частично или полностью первичной структурной конформацией и/или одним или более из биологических свойств встречающегося в природе эритропоэтина.
А. Манипуляция генетическими материалами
Генетические материалы можно в широком смысле определить как те химические вещества, которые программируют и управляют производством компонентов клеток и вирусов и направляют ответы клеток и вирусов. Длинноцепочечное полимерное вещество, известное как дезоксирибонуклеиновая кислота (ДНК), содержит генетический материал всех живых клеток и вирусов, за исключением некоторых вирусов, программируемых рибонуклеиновой кислотой (РНК). Структурными звеньями в ДНК-полимерах являются четыре различных нуклеотида, каждый из которых состоит из пурина (аденин или гуанин) или из пиримидина (тимин или цитозин), связанных с дезоксирибозо-сахаром, к которому присоединена фосфатная группа. Присоединение нуклеотидов в форме линейного полимера происходит путем слияния 5'-фосфата одного нуклеотида с 3'-гидроксильной группой другого нуклеотида. Функциональная ДНК встречается в форме устойчивых двунитевых сообществ одиночных нитей нуклеотидов (известных как дезоксиолигонуклеотиды), чьи сообщества образуются путем водородной связи между пуриновыми и пиримидиновыми основаниями [т.е. "комплементарные" сообщества, существующие или между аденином (А) и тимином (Т), или между гуанином (G) и цитозином (С)]. По конвенции принято нуклеотиды называть по названиям составляющих их пуриновых или пиримидиновых оснований, а комплементарные сообщества нуклеотидов в двунитевой ДНК (т.е. А-Т и G-C) называть как "пары оснований". Рибонуклеиновая кислота представляет собой полинуклеотид, содержащий аденин, гуанин, цитозин и урацил (U), скорее, чем тимин, связанный с рибозой и фосфатной группой.
Короче говоря, программирующая функция ДНК осуществляется главным образом посредством процесса, в котором специфические последовательности нуклеотидов ДНК (гены) "транскрибируются" в относительно неустойчивые полимеры информационной РНК (мРНК). В свою очередь, мРНК служит в качестве матрицы для образования структурных, регуляторных и каталитических белков из аминокислот. Этот процесс "трансляции" мРНК включает функционирование мелких нитей РНК (тРНК), которые транспортируют и выравнивают индивидуальные аминокислоты вдоль нитей мРНК, разрешая образование нуклеотидов в надлежащих последовательностях аминокислот."Информация" мРНК, исходящая от ДНК и создающая основание для подачи тРНК и ориентации любой из двадцати аминокислот для "экспрессии" полипептида, находится в форме триплетных "кодонов" - последовательных группирований трех нуклеотидных оснований. В известном смысле, образование белка является окончательной формой "экспрессии" программированной генетической информации, обеспечиваемой нуклеотидной последовательностью гена.
"Промоторные" последовательности ДНК обычно "предшествуют" гену в ДНК-полимере и обеспечивают участок для инициирования транскрипции в РНК. "Регуляторные" последовательности ДНК обычно также "расположены выше по течению" (т.е. предшествуют) гена в данном ДНК-полимере, связывая белки, определяющие частоту (или скорость) транскрипционального инициирования. Совместно называемые как "промоторная/регуляторная" или "управляющая" последовательность ДНК, это такие последовательности, которые предшествуют отобранному гену (или ряду генов) в функциональном ДНК-полимере, способствуют определению того, произойдет ли транскрипция и, в конце концов, экспрессия гена. Последовательности ДНК, которые "сопровождают" ген в ДНК-полимере и обеспечивают подачу сигнала для окончания транскрипции в мРНК, называются как "терминаторные" последовательности транскрибирования.
В фокусе микробиологической технологии последнего десятилетия находится попытка индустриального производства и фармацевтически важных веществ с использованием микроорганизмов, которые или первоначально не имеют генетически кодированной информации, относящейся к желаемому продукту, включенному в их ДНК, или (в случае с клетками млекопитающих в культуре) не выражают обычным путем хромосомный ген на заметном уровне. Проще говоря, ген, устанавливающий структуру желаемого полипептидного продукта, или выделяют от "донорного" микроорганизма, или химически синтезируют и затем устойчиво интродуцируют в другой микроорганизм, предпочтительно, самореплицирующийся одноклеточный микроорганизм, такой как бактерии, дрожжи или клетки млекопитающих в культуре. Существующее машинное оборудование для экспрессии генов в "трансформированных" или "трансфектированных" микробных клетках-хозявах работает для создания желаемого продукта с использованием экзогенной ДНК в качестве матрицы для транскрипции мРНК, которая затем транслируется в непрерывную последовательность аминокислотных остатков.
Данная область техники представлена широко в патентных и литературных публикациях, относящихся к методологиям "рекомбинантной РНК" для выделения, синтеза, очистки и амплификации генетических материалов, используемых при трансформации отобранных микроорганизмов хозяина. Патент США №4237224, выданном Cohen, et al., например, относится к трансформации одноклеточных микроорганизмов хозяина с "гибридной" вирусной или кольцевой плазмидной ДНК, включающей отобранные последовательности экзогенной ДНК. Методики патента Cohen, et al. впервые включают получение вектора трансформации посредством ферментативно отщепляющейся вирусной или кольцевой плазмидной ДНК для образования нитей линейной ДНК. Отобранные инородные ("экзогенные" или "гетерологические") нити ДНК, обычно включающие последовательности, кодирующие желательный продукт, получают в линейной форме путем использования сходных ферментов. Линейную вирусную или плазмидную ДНК инкубируют с инородной ДНК в присутствии сшивающих ферментов, способных осуществлять процесс восстановления, а "гибридные" векторы образуются содержащими отобранный сегмент экзогенной ДНК, "сплетенный" в плазмиду вирусной или кольцевой ДНК.
Трансформация совместимых одноклеточных микроорганизмов хозяина с гибридным вектором заканчивается образованием многократных копий экзогенной ДНК в популяции клеток-хозяев. В некоторых случаях желаемым результатом является просто амплификация инородной ДНК, и собранный, выращенный в культуре "продукт" представляет собой ДНК. Более часто целью трансформации является экспрессия, осуществляемая при помощи клеток-хозяев экзогенной ДНК в форме крупномасштабного синтеза выделяемых количеств промышленно важных фрагментов белка или полипептида, кодируемых инородной ДНК. См. также, например, патенты США №№4264731 (выданный Shine), 4273875 (выданный Manis), 4293652 (выданный Cohen) и Европейскую патентную заявку №093619, опубликованную 9 ноября 1983 года.
Развитие специфических последовательностей ДНК для сплетения в векторы ДНК достигается путем применения целого ряда технических приемов, зависящих в значительной мере от степени "инородности" "донора" по отношению к проектируемому хозяину, а также от размера полипептида, выражаемого в хозяине. Опасаясь чрезмерного упрощения, можно утверждать, что существуют три основных альтернативных способа: (1) "выделение" двунитевой ДНК-последовательности от геномной ДНК донора; (2) химическое получение ДНК-последовательности, создающей код для представляющего интерес полипептида, и (3) синтез in vitro двунитевой ДНК-последовательности посредством ферментативной "обратной транскрипции" мРНК, выделенной из донорских клеток. Указанные выше способы, которые включают образование ДНК-"комплемента" мРНК, обычно называют как "кДНК" способы.
Получение последовательностей ДНК часто является методом выбора, когда известна полная последовательность аминокислотных остатков желаемого полипептида. Методики получения ДНК, описанные в одновременно рассматриваемой заявке на патент США №483451, Alton, et al. (поданной 15 апреля 1983 года, и соответствующей заявке РСТ US 83/00605, опубликованной 24 ноября 1983 года под номером WO 83/04053), например, создают средства для достижения таких чрезвычайно желаемых результатов, как: предусмотрение наличия чередующихся кодонов, часто встречающихся в микроорганизме хозяина, отобранном для экспрессии (например, предусмотрение "предпочтительных" кодонов дрожжей или Е. coli); избежание присутствия нетранслированных "интронных" последовательностей (обычно находящихся в геномных ДНК-последовательностях млекопитающих и их мРНК-матрицах), которые не без труда обрабатываются прокариотическими клетками-хозяевами; избежание нежелательных "ведущих" полипептидных последовательностей, обычно кодируемых геномными ДНК и кДНК-последовательностями, однако, часто не легко отщепляемых от представляющего интерес полипептида при помощи бактериальных или дрожжевых клеток-хозяев; предусмотрение легкой вставки ДНК в подходящие экспрессивные векторы в сообществах с желаемыми промоторными/регуляторными и терминаторными последовательностями; и предусмотрение легкого построения генов, кодирующих полипептидные фрагменты и аналоги желательных полипептидов.
Когда полная последовательность аминокислотных остатков желаемого полипептида не известна, непосредственное получение последовательностей ДНК невозможно и выделение последовательностей ДНК, кодирующих полипептиды при помощи кДНК-способа, становится методом отбора, несмотря на возможные недостатки в легкости сборки векторов экспрессии, способных обеспечить высокие уровни микробной экспрессивности, указанной выше. Среди стандартных методик для выделения кДНК-последовательностей находится получение плазмидонесуших кДНК- "библиотек", происходящих от обратной транскрипции мРНК, часто встречающейся в донорских клетках, отобранных в качестве ответственных за экспрессию генов высокого уровня (например, библиотеки кДНК, происходящие от гипофизарных клеток, которые выражают относительно большие количества продуктов гормонов роста). Где известны значительные части аминокислотной последовательности полипептидов, там меченые, зондовые последовательности однонитевой ДНК, дуплицирующей последовательность, предположительно находящуюся в "плановой" кДНК, можно использовать в методиках ДНК/ДНК-гибридизации, проводимых на клональных копиях кДНК, которая была денатурирована в однонитевую форму. [См., главным образом, раскрытие и обсуждения данной области техники, представленные в патенте США 4394443, выданном на имя Weissman, et al. и последние показы использования длинных зондов олигонуклеотидной гибридизации, сообщенные в работах Wallace, et al., Nuc. Acids Res., 6, pp.3543-3557 (1979), Reyes, et al., P.N.A.S. (U.S.A.), 79, pp.3270-3274 (1982) и Jaye, et al., Nuc. Acids Res., 11, pp.2325-2335 (1983). См. также патент США №4358535, выданный на имя Falkow, et al., касающийся методик ДНК/ДНК-гибридизации при проведении диагностики; опубликованные Европейские патентные заявки №0070685 и №0070687, относящиеся к светоиспускающим меткам на однонитевых полинуклеотидных зондах; работу Davis, et al., "А Manual for Genetic Engineering, Advanced Bacterial Genetics", Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1980) at pp.55-58 and 174-176, касающуюся методик гибридизации колоний и эпидемических заболеваний; и New England Nuclear (Boston, Mass.) брошюры по "Gene Screen" Hybridization Transfer Membrane materials, дающие руководства для переноса и гибридизации ДНК и РНК, Каталог №NEF-972].
Среди наиболее значительных последних достижений в методиках гибридизации для экранирования рекомбинантных клонов является использование меченых смешанных олигонуклеотидных зондов, каждый из которых потенциально является полным комплементом специфической последовательности ДНК в выборке гибридизации, включающей гетерогенную смесь однонитевых ДНК и РНК. Эти методики признаются особенно полезными при выявлении кДНК-клонов, получаемых из источников, которые создают крайне малые количества последовательностей мРНК для представляющего интерес полипептида. Короче говоря, использование строгих условий гибридизации, направленных на избежание неспецифического связывания, может позволить, например, авторадиографическую визуализацию специфического клона кДНК на исходе гибридизации плановой ДНК тому единственному зонду в смеси, который является ее полным комплементом. См., главным образом, Wallace, et al., Nuc. Acids Res., 9, pp.879-897 (1981); Suggs, et al., P.N.A.S. (U.S.A.), 78, pp.6613-6617 (1981); Choo, et al., Nature, 299, pp.178-180 (1982); Kurachi, et al., P.N.A.S. (U.S.A.), 79, pp.6461-6464 (1982); Ohkubo, et al., P.N.A.S. (U.S.A.), 80, pp.2196-2200 (1983); и Kornblihtt, et al., P.N.A.S. (U.S.A.), 80, pp.3218-3222 m(1983). Вообще, методики смешанных зондов в работе Wallace, et al. (1981) выше, были расширены различными разработчиками до момента, когда сообщалось о получении достоверных результатов в выделении кДНК-клонов с использованием 32-членного смешанного "пула" олигонуклеотидных зондов длиной в 16 оснований (16-mer) равномерно изменяющихся последовательностей ДНК вместе с одиночным 11-mer для выполнения двухпозиционного "положительного" подтверждения наличия кДНК, представляющей интерес. См. работу Singer-Sam, et al., P.N.A.S. (U.S.A.), 80, pp.802-806 (1983).
Использование изолятов геномной ДНК является наименее распространенным из трех указанных выше способов развития специфических последовательностей для использования в рекомбинантных методиках. Это особенно верно в области рекомбинантных методик, направленных на обеспечение микробной экспрессивности полипептидов млекопитающих, и обусловлено, главным образом, сложностью геномной ДНК млекопитающих. Таким образом, хотя и существуют надежные методики для развития фаг-несущих библиотек геномной ДНК человека и других видов млекопитающих [см., например, работу Lawn, et al., Cell, 15, pp.1157-1174 (1978), относящуюся к методикам для генерирования геномной библиотеки человека, обычно называемой как "Библиотека Maniatis"; работу Karn, et al., P.N.A.S. (U.S.A.), 77, pp.5172-5176 (1980), касающуюся геномной библиотеки человека на основе методики альтернативного ограничения эндонуклеазной фрагментации; работу Blattner, et al., Science, 196. pp.161-169 (1977), описывающую построение бычьей геномной библиотеки], предпринято несколько относительно успешных попыток использования методик гибридизации в выделении геномной ДНК при отсутствии далеко идущего предвидения аминокислотных или ДНК-последовательностей. В качестве одного примера можно привести работу Fiddes, et al., J. Mol. and App. Genetics, 1, pp.3-18 (1981), где сообщается об успешном выделении гена, кодирующего альфа-субъединицу гипофизарных гликопротеидных гормонов человека из Библиотеки Maniatis путем использования "предельно длинного" зонда, включающего полный фрагмент 621 пары азотистых оснований предварительно выделенной кДНК-последовательности для альфа-субъединицы. В качестве другого примера, в работе Das, et al., Р.N.А.S. (U.S.А.), 80, pp.1531-1535 (1983) сообщается о выделении геномных клонов человека для HLA-DR человека с использованием синтетического олигонуклеотида со 175 парами оснований. И, наконец, в работе Anderson, et al., P.N.A.S. (U.S.A.), 80, pp.6838-6842 (1983) сообщается о выделении геномного клона для бычьего панкреатического трипсинового ингибитора (BPTI) с использованием одиночного зонда, имеющего 86 пар оснований в длину и построенного в соответствии с известной аминокислотной последовательностью BPTI. Авторы отмечают определение скромных перспектив для выделения мРНК, пригодной для синтеза кДНК-библиотеки, ввиду очевидных низких уровней мРНК в первоначально нацеленных источниках околоушной железы и тканей легкого. Выражая затем надежды на успех в зондировании геномной библиотеки с использованием смеси меченых зондов, они констатируют: "Большей частью, олигодезоксинуклеотидные зонды смешанных последовательностей применялись для выделения белковых генов неизвестной последовательности из библиотек кДНК. Такие зонды обычно представляют собой смесь 8-32 олигонуклеотидов и имеют длину 14-17 нуклеотидов, являясь типичными представителями каждой возможной комбинации кодонов для небольшого растяжения (5-6 остатков) аминокислотной последовательности. В жестких условиях гибридизации, которые ставят в худшее положение зонды с неправильно спаренными азотистыми основаниями, эти смеси способны локализовывать специфические последовательности генов в клональных библиотеках малой сложности. Тем не менее, ввиду их незначительной длины и неоднородности, смешанные зонды часто лишены специфичности, необходимой для зондирования таких сложных последовательностей, как геном млекопитающих. Это делает такой способ непрактичным для выделения белковых генов млекопитающих, когда соответственные мРНК недоступны." (Ссылки опущены).
Таким образом, в данной области техники продолжает оставаться необходимость в совершенствовании способов выполнения быстрого и эффективного выделения кДНК-клонов в тех случаях, когда мало что известно об аминокислотной последовательности и когда "обогащенные" источники ткани мРНК не очень доступны, чтобы их можно было использовать в построении библиотек кДНК. Такие усовершенствованные способы были бы особенно полезны, если бы они применялись для выделения геномных клонов млекопитающих, где доступна неплотная информация относительно аминокислотных последовательностей полипептида, кодируемого искомым геном.
В. Эритропоэтин как представляющий интерес полипептид
Эритропоэз, образование красных кровяных телец, осуществляется непрерывно на протяжении всей жизни человека для компенсации разрушенных клеток. Эритропоэз представляет собой чрезвычайно точно регулируемый физиологический механизм, позволяющий вырабатываться в крови достаточному количеству красных кровяных телец для правильной оксигенации ткани, но не настолько большому, чтобы клетки воспрепятствовали кровообращению. Образование красных кровяных телец происходит в костном мозге под контролем гормона, эритропоэтина.
Эритропоэтин, кислотный гликопротеид с молекулярным весом приблизительно 34000 Дальтон, может встречаться в трех формах: α, β и асиало. Формы α и β отличаются незначительно в углеводных компонентах, однако, имеют одинаковую потенцию, биологическую активность и молекулярный вес. Форма асиало представляет собой α или β-форму с отщепленным концевым углеводом (сиаловая кислота). Эритропоэтин присутствует в очень низких концентрациях в плазме, когда тело находится в здоровом состоянии, при котором ткани получают достаточную оксигенацию от существующего количества эритроцитов. Эта нормальная низкая концентрация достаточна для стимулирования замены красных кровяных телец, которые обычно теряются в процессе старения.
Количество эритропоэтина в кровообращении возрастает в условиях гипоксии, когда снижается перенос кислорода кровяными клетками при кровообращении. Гипоксия может вызываться потерей большого количества крови при кровотечении, разрушением красных кровяных телец вследствие радиоактивного облучения, снижением потребления кислорода из-за больших высот или продолжительного бессознательного состояния, а также различными формами анемии. Под влиянием тканей, подверженных гипоксическому стрессу, эритропоэтин начинает увеличивать образование красных кровяных телец путем стимулирования конверсии первичных предшествующих клеток в костном мозге в проэритробласты, которые впоследствии доводят до созревания, синтезируют гемоглобин и реализуются в кровообращении в качестве красных кровяных телец. Когда число красных кровяных телец в кровообращении больше, чем необходимо для нормальной потребности тканей в кислороде, эритропоэтин в кровообращении снижается.
См., главным образом, работы Testa, et al., Exp. Hematol., 8(Supp. 8), 144-152 (1980); Tong, et al., J. Biol. Chem., 256(24), 12666-12672 (1981); Goldwasser, J. Cell. Physiol., 110(Supp. 1), 133-135 (1982); Finch, Blood, 60 (6), 1241-1246 (1982); Sytowski, et al., Expt. Hematol., 8 (Supp. 8), 52-64 (1980); Naughton, Ann. Clin. Lab. Sci., 13 (5), 432-438 (1983); Weiss, et al., Am. J. Vet. Res., 44 (10), 1832-1835 (1983); Lappin, et al., Exp. Hematol., 11 (7), 661-666 (1983); Baciu, et al., Ann. N.Y. Acad. Sci., 414, 66-72 (1983); Murphy, et al., Acta. Haematologica Japonica, 46 (7), 1380-1396 (1983); Dessypris, et al., Brit. J. Haematol., 56, 295-306 (1984); и Emmanouel, et al., Am. J. Physiol., 247 (1 Pt 2), F 168-76 (1984).
Так как эритропоэтин важен в процессе образования красных кровяных телец, гормон имеет потенциальное полезное применение как в диагностике, так и при лечении нарушений крови, характеризующихся низким или несовершенным образованием красных кровяных телец. См., главным образом, работы Pennathur-Das, et al., Blood, 63 (5), 1168-71 (1984) и Haddy, Am. Jour. Ped. Hematol./Oncol., 4, 191-196, (1982), касающиеся эритропоэтина в возможных терапиях серповидно-клеточных болезней, а также Eschbach, et al., J. Clin. Invest., 74 (2), pp.434-441, (1984), в которой описан терапевтический режим для уремической овцы на основании реакции in vivo на вливание плазмы, обогащенной эритропоэтином, и предложена дозировка 10 U ЕРО/кг в день в течение 15-40 дней как поправка анемии такого типа, который ассоциируется с хронической почечной недостаточностью. См. также работу Krane, Henry Ford Hosp. Med. J., 31 (3), 177-181 (1983).
Недавно было оценено, что наличие эритропоэтина в большом количестве позволит лечить каждый год анемии у 1600000 человек только в США. См., например, Morrison, "Bioprocessing in Space - an Overview", pp.557-571 in The World Biotech Report 1984, Volume 2:USA, (Online Publications, New York, N.Y. 1984). Последние исследования создали основание для предсказания эффективности эритропоэтиновой терапии в разнообразии состояний болезни, расстройств и состояний гематологического нарушения: Vedovato, et al., Acta. Haematol., 71, 211-213 (1984) (бета-талассемия); Vichinsky, et al., J. Pediatr., 105 (1), 15-21 (1984) (кистозный фиброз); Cotes, et al., Brit. J. Obstet. Gyneacol., 90 (4), 304-311. (1983) (беременность, расстройства менструаций); Haga, et al., Acta. Pediatr. Scand., 72, 827-831 (1983) (ранняя анемия преждевременности); Claus-Walker, et al., Arch. Phys. Med. Rehabil., 65, -370-374 (1984) (повреждение спинного мозга); Dunn, et al., Eur. J. Appl. Physiol., 52, 178-182 (1984) (космический полет); Miller, et al., Brit. J. Haematol., 52, 545-590 (1982) (острая потеря крови); Udupa, et al., J. Lab. Clin. Med., 103 (4), 574-580 и 581-588 (1984); и Lipschitz, et al., Blood, 63 (3), 502-509 (1983) (старение); и Dainiak, et al., Cancer, 51 (6), 1101-1106 (1983) и Schwartz, et al., Otolaryngol., 109, 269-272 (1983) (различные неопластические состояния болезни, сопровождаемые аномальным эритропоэзом).
Предыдущие попытки получить эритропоэтин с хорошим выходом выработки из плазмы крови или из мочи доказаны как относительно неудачные. Усложненная и утонченная лабораторная техника, неизбежно и, как правило, приводит к сбору очень малых количеств загрязненных и неустойчивых экстрактов, содержащих эритропоэтин.
В патенте США №3033753 описан способ частичной очистки эритропоэтина от плазмы крови овцы, что обеспечивает получение малых выходов общего беспримесного экстракта, содержащего эритропоэтин.
Первоначальные попытки выделить эритропоэтин из мочи привели к неустойчивым, биологически неактивным препаратам гормона. В патенте США №3865801 описан способ стабилизации биологической активности общего вещества, содержащего эритропоэтин, восстановленный из мочи. Получаемый препарат, содержащий эритропоэтин, сдерживает 90% активности эритропоэтина и является устойчивым.
Другой способ очистки эритропоэтина человека от мочи пациентов, страдающих апластической анемией, описан в работе Miyake, et al., J. Biol. Chem., Vol.252; №15 (August 10, 1977), pp.5558-5564. Эта семиэтапная методика включает ионообменную хроматографию, осаждение этанола, гель-фильтрацию, адсорбционную хроматографию и вырабатывает чистый эритропоэтин с потенцией 70400 единиц/мг белка при выходе 21%.
В патенте США №4397840, выданном Takezawa, et al., описаны способы получения "эритропоэтинового продукта" из образцов мочи здорового человека со слабо основными ионными изменениями и утверждается, что полученные продукты с низким молекулярным весом "не имеют ингибирующих действий против эритропоэтина".
В заявке на патент Великобритании №2085887, Sugimoto, et al., опубликованной 6 мая 1982 года, описан способ получения гибридных лимфобластоидных клеток человека и сообщается, что уровни продуцирования находятся в пределах от 3 до 420 единиц эритропоэтина на мл суспензии клеток (распределенных в культурах после размножения хозяином млекопитающего с содержанием до 107 клеток на мл). На самых высоких уровнях продуцирования, которые, как утверждается, должны быть достигнуты, скорость продуцирования эритропоэтина может быть рассчитана, чтобы составить от 40 до 4000 единиц/106 клеток/48 часов в культуре in vitro с последующим переносом клеток из систем размножения in vivo (См. также эквивалентный патент США №4377513). Были сделаны многочисленные предложения в отношении выделения эритропоэтина из тканевых источников, включающих неопластические клетки, однако выходы выработки были совершенно низкими. См., например, работы Jelkman, et al., Expt. Hematol., 11 (7), 581-588 (1983); Tambourin, et al., P.N.A.S. (U.S.A.), 80, 6269-6273 (1983); Katsuoka, et al., Gann, 74, 534-541 (1983); Hagiwara, et al., Blood, 63 (4), 828-835 (1984); и Choppin, et al., Blood, 64 (2), 341-347 (1984).
Другие технологии выделения, применяемые для получения очищенного эритропоэтина, включали в себя иммунологические методики. Поликлональное, являющееся производным сыворотки антитело, направленное против эритропоэтина, развивается путем инъецирования животного, предпочтительно, крысы или кролика, человеческим эритропоэтином. Инъецированный эритропоэтин человека распознается как инородное антигенное вещество иммунной системой животного и вызывает продуцирование антител против антигена. Различные клетки, реагирующие на стимулирование посредством антигенного вещества, продуцируют и высвобождают в кровообращении антитела, отличающиеся слегка от тех, которые продуцируются другими реагирующими клетками. Активность антител остается в сыворотке животного, когда его кровь экстрагирована. И хотя неочищенная сыворотка или препараты антител, очищенные как сывороточная фракция иммуноглобулина G, могут быть затем использованы в пробах для обнаружения комплексообразования с человеческим эритропоэтином, вещества испытывают главное неудобство. Это сывороточное антитело, состоящее из всех различных антител, продуцированных отдельными клетками, по природе является поликлональным и образует комплексы с компонентами в общих экстрактах иначе, чем одиночный эритропоэтин.
Интерес для предпосылок настоящего изобретения представляют последние достижения в области разработки целостных культур клеток, способных продуцировать одиночный вид антитела, который специфически иммунологически активен с одиночной антигенной детерминантой отобранного антигена. См., главным образом, работу Chisholm, High Technology, Vol.3, №1, 57-63 (1983). Предпринимались попытки употребить слияние клеток и методики гибридизации для развития "моноклональных" антител к эритропоэтину и применить эти антитела в выделении и количественном определении эритропоэтина человека. В качестве одного примера можно привести сообщение, появившееся в аннотированной форме в работе Lee-Huang, Abstract №1463 of Fed. Proc., 41, 520 (1982), в которой говорится об успешной разработке линий клеток гибридомы мышь-мышь, выделяющих моноклональные антитела к человеческому эритропоэтину. В качестве другого примера, подробное описание получения и использования моноклонального, противоэритропоэтинового антитела появилось в работе Weiss, et al., P.N.A.S. (U.S.A.), 79, 5465-5469 (1982). См. также работы Sasaki, Biomed. Biochim. Acta., 42 (11/12), 5202-5206 (1983); Yanagawa, et al., Blood, 64 (2), 357-364 (1984); Yanagawa, et al., J. Biol. Chem., 259 (5), 2707-2710 (1984); и патент США №4465624.
Для предпосылок изобретения также представляют интерес сообщения об иммунологической активности синтетических пептидов, которые по существу дуплицируют аминокислотную последовательность, сохранившуюся в встречающихся в природе белках, гликопротеидах и нуклепротеидах. Более конкретно, полипептиды с относительно низким молекулярным весом показаны участвующими в иммунных реакциях, сходных по продолжительности и протяженности с иммунными реакциями физиологически важных белков, таких как вирусные антигены, полипептидные гормоны и тому подобное. Среди иммунных реакций таких полипептидов включено и провоцирование образования специфических антител в иммунологически активных животных. См., например, работы Lerner, et al., Cell, 23, 309-310 (1981); Ross, et al., Nature, 294, 654-656 (1981); Walter, et al., P.N.A.S. (U.S.A.), 77, 5197-5200 (1980); Lerner, et al., P.N.A.S. (U.S.A.), 78, 3403-3407 (1981); Walter, et al., P.N.A.S. (U.S.A.), 78, 4882-4886 (1981); Wong, et al., P.N.A.S. (U.S.A.), 78, 7412-7416, (1981); Green, et al., Cell, 28, 477-487 (1982); Nigg, et al., P.N.A.S. (U.S.A.), 79, 5322-5326 (1982); Baron, et al., Cell, 28, 395-404 (1982); Dreesman, et al., Nature, 295, 158-160 (1982); и Lerner, Scientific American, 248, №2, 66-74 (1983). См. также работу Kaiser, et al., Science, 223, pp.249-255 (1984), касающуюся биологической и иммунологической активности синтетических пептидов, которые приблизительно обладают вторичными структурами пептидных гормонов, однако, могут не обладать их первичной структурной конформацией. Вышеуказанные исследования касаются, конечно, аминокислотных последовательностей белков, иных, нежели эритропоэтин, вещество, для которого не было опубликовано существенной информации об аминокислотной последовательности. В совместной заявке на патент США №463724, поданной 4 февраля 1983 года J. Egrie, опубликованной 22 августа 1984 года как европейская заявка на патент №0116446, описана линия клеток гибридомы мышь-мышь (А.Т.С.С. № НВ8209), которая продуцирует крайне специфическое моноклональное, противоэритропоэтиновое антитело, которое является также специфически иммунонеактивным с полипептидом, содержащим следующую последовательность аминокислот:
NH2-Ala-Pro-Pro-Arg-Leu-Ile-Cys-Asp-Ser-Arg-Val-Leu-Glu-Arg-Tyr-Leu-Leu-Glu-Ala-Lys-COOH.
Полипептидной последовательностью является та последовательность, которая закреплена за первыми двадцатью аминокислотными остатками созревшего эритропоэтина человека, изолированного в соответствии со способом, описанным в работе Miyake, et al., J. Biol. Chem., 252, 5558-5564 (1977), и над которой провели аминокислотный анализ посредством газофазового секвенсера (Applied Biosystems, Inc.) в соответствии с методикой Hewick, М., et al., J. Biol. Chem., 256, 7990-7997 (1981). См. также работу Sue, et al., Proc. Nat. Acad. Sci. (USA), 80, pp.3651-3655 (1983), касающуюся развития поликлональных антител против синтетического 26-mer на основе отличающейся аминокислотной последовательности, и работу Sytowski, et al., J. Immunol. Methods, 69, pp.181-186 (1984).
Хотя поликлональные или моноклональные антитела, как описано выше, располагают весьма полезными материалами для использования в иммуноиспытаниях для обнаружения и количественного определения эритропоэтина и могут быть пригодны в сродственной очистке эритропоэтина, кажется маловероятным, что эти материалы для крупномасштабного выделения могут обеспечить количества эритропоэтина млекопитающих источников, достаточные для проведения дальнейшего анализа, клинического обследования и возможного широконаправленного терапевтического использования вещества при лечении, например, хронической болезни почек, при которой пораженные ткани не в состоянии поддерживать продуцирование эритропоэтина. Поэтому предполагается, что наилучшие перспективы для полной характеристики эритропоэтина млекопитающих и создания больших количеств его для возможной диагностики и клинического применения содержит употребление рекомбинантных методик для осуществления крупномасштабного микробного синтеза соединения.
Несмотря на то, что в попытке выделить последовательности ДНК, кодирующие эритропоэтин человека или других видов млекопитающих, были предприняты значительные усилия, ни одно из них не было успешным. Это обусловлено, главным образом, дефицитом тканевых источников, особенно человеческих тканевых источников, обогащенных в мРНК так, чтобы позволить построение кДНК-библиотеки, из которой посредством применения известных методик можно выделить последовательность ДНК, кодирующую эритропоэтин. Кроме того, мало что известно о непрерывной последовательности аминокислотных остатков эритропоэтина, а поэтому невозможно построить, например, длинные полинуклеотидные зонды, подходящие для надежного использования в скрининге ДНК/ДНК-гибридизации кДНК и особенно геномных библиотек ДНК. Иллюстративно последовательность двадцати аминокислот, применяемая для генерирования вышеуказанного моноклонального антитела, продуцированного А.Т.С.С. №НВ8209, не допускает построения однозначного 60-основного олигонуклеотидного зонда способом, описанным Andersen, et al., выше. Найдено, что человеческий ген для эритропоэтина может проявляться как "ген одиночной копии" в геноме человека, и так или иначе, генетический материал, кодирующий эритропоэтин человека, вероятно, составляет менее 0,00005% общей геномной ДНК человека, которая должна присутствовать в геномной библиотеке.
На сегодняшний день наиболее успешные из известных попыток в рекомбинантных способах для создания последовательностей ДНК, пригодных при использовании в микробной экспрессии изолируемых количеств эритропоэтина млекопитающих, далеко не достигли цели. В качестве примера можно привести работу Farber, et al., Exp. Hematol., 11, Supp. 14, Abstract 101 (1983), в которой сообщается об экстракции мРНК из почечных тканей бабуинов, обработанных фенилгидразином, и инъецировании мРНК в ооциты Xenopus laevis с довольно временным результатом получения in vitro смеси "трансляционных продуктов", которые имели проявляющиеся биологические свойства эритропоэтина. Совсем недавно Farber, et al., Blood, 62, №5, Supp. №1, Abstract, 392, at page 122a (1983) доложили о трансляции in vitro мРНК почки человека лягушечьими ооцитами. Полученная смесь продукта трансляции была оценена как содержащая порядка 220 mU продукта трансляции, имеющего активность эритропоэтина на микрограмм введенной мРНК. И хотя такие уровни трансляции in vitro экзогенной мРНК, кодирующей эритропоэтин, были признаны совершенно низкими (по сравнению даже с ранее сообщенными уровнями трансляции мРНК бабуина в искомый продукт), посчитали, что результаты подтверждают существование почки человека в качестве участка экспрессивности эритропоэтина, позволяющей построение кДНК-библиотеки обогащенной почки человека, из которой можно выделить желаемый ген. [См. также Farber, Clin. Res., 31 (4), 769A (1983)].
С тех пор как были поданы заявки на патенты США №№561024 и 582185, появилось единственное сообщение о клонировании и экспрессии того, что признано кДНК эритропоэтина человека в Е.coli. Короче говоря, целый ряд клонов кДНК был внесен в плазмиды Е.coli, и продукты слияния β-лактамазы были отмечены как иммунореактивные с моноклональным антителом к неустановленному "эпитопу" эритропоэтина человека. См. Lee-Huang, Proc. Nat. Acad. Sci. (USA), 81, pp.2708-2712 (1984).
Краткое изложение
Предлагаются впервые новые очищенные и выделенные полипептидные продукты, имеющие частично или полностью первичную структурную конформацию (т.е. непрерывную последовательность аминокислотных остатков) и одно или более биологических свойств (например, иммунологические свойства и биологическая активность in vivo и in vitro) встречающегося в природе эритропоэтина, включая его аллельные варианты. Эти полипептиды также однозначно отличаются тем, что являются продуктом экспрессии прокариотического или эукариотического хозяина (например, бактериальными, дрожжевыми и клетками млекопитающих в культуре) последовательностей экзогенной ДНК, полученных путем геномного или кДНК-клонирования, или путем генного синтеза. Продукты микробной экспрессии в клетках позвоночных (например, млекопитающих и птиц) могут, кроме того, отличаться тем, что свободны от сообщества с белками человека или другими загрязнителями, которые могут ассоциироваться с эритропоэтином в его естественной для клеток млекопитающих окружающей среде или в экстрацеллюлярных текучих средах, таких как плазма крови или моча. Продукты типичных дрожжевых (например, Saccaromyces cerevisiae) или прокариотических (например, Е.coli) клеток-хозяев свободны от сообщества с любыми белками млекопитающих. В зависимости от применяемого хозяина предлагаемые полипептиды могут быть гликозилированы млекопитающими или другими эукариотическими углеводами, или могут быть негликозилированными. Предлагаемые полипептиды могут также включать аминокислотный остаток начального метионина (в положении - 1).
Предлагаемые новые гликопротеиновые продукты включают те, которые имеют первичную структурную конформацию, достаточно дуплицирующую конформацию встречающегося в природе (например, человеческого) эритропоэтина для того, чтобы дать возможность обладать одним или более из биологических свойств последнего, и те, которые имеют среднюю углеводную композицию, отличающуюся от композиции встречающегося в природе (например, человеческого) эритропоэтина.
Клетки позвоночных (например, COS-1 и СНО), предлагаемые в настоящем изобретении, содержат первые клетки, всегда доступные, которые могут размножаться непрерывно in vitro и которые во время роста в культуре способны продуцировать в среде их роста более 100U (предпочтительно, более 500U и наиболее предпочтительно, более 1000U-5000U) эритропоэтина на 106 клеток за 48 часов, как определено радиоиммуноисследованием.
Предлагаются также синтетические полипептиды, полностью или частично дуплицирующие непрерывные последовательности аминокислотных остатков эритропоэтина, которые здесь впервые разъяснены. Эти последовательности путем разделения первичных, вторичных или третичных структурных и конформационных характеристик со встречающимся в природе эритропоэтином могут обладать биологической активностью и/или иммунологическими свойствами совместно со встречающимся в природе продуктом так, что они могут применяться как биологически активные или иммунологические заместители эритропоэтина в терапевтических и иммунологических процессах. Соответственно, предлагаются моноклональные и поликлональные антитела, вырабатываемые стандартными способами, которые иммунореактивны с такими полипептидами и, предпочтительно, также иммунореактивны со встречающимся в природе эритропоэтином.
Настоящее изобретение иллюстрируют клонированные последовательности ДНК видовых начал обезьяны и человека, а также полипептидные последовательности, удобно выведенные оттуда и представляющие, соответственно, первичную структурную конформацию видовых начал обезьяны и человека.
Предлагаются также новые, биологически функциональные векторы вирусной и кольцевой плазмидной ДНК, охватывающие последовательности ДНК изобретения, и микробные (например, бактериальные, дрожжевые и клетки млекопитающих) организмы хозяина, устойчиво трансформируемые или трансфектируемые такими векторами. Соответственно, предлагаются новые способы получения полезных полипептидов, включающие культивируемое выращивание таких трансформируемых или трансфектируемых микробных хозяев в условиях, облегчающих крупномасштабную экспрессию экзогенных, вектор-несущих последовательностей ДНК и выделение желаемых полипептидов из питательной среды, клеточных лизатов или фракций клеточных мембран.
Выделение и очистку микробиально выраженных полипептидов, предлагаемых настоящим изобретением, можно осуществить при помощи традиционных способов, включающих, например, разделение препаративной хроматографией и иммунологическое разделение препаратов, содержащих моноклональные и/или поликлональные антитела.
Разъяснив последовательность аминокислотных остатков эритропоэтина, настоящее изобретение предлагает полное и/или частичное построение ДНК-последовательностей, кодирующих эритропоэтин и содержащих такие благоприятные характеристики, как внедрение кодонов, "предпочитаемых" для экспрессии отобранными хозяевами не млекопитающих, обеспечение участков для расщепления посредством ограничительных эндонуклеазных ферментов и обеспечение дополнительных начальных, конечных или промежуточных ДНК-последовательностей, которые способствуют построению легко выражаемых векторов. Соответственно, предлагается построение (и развитие посредством специфического мутагенеза кДНК и геномной ДНК) ДНК-последовательностей, кодирующих микробную экспрессию полипептидных аналогов или производных эритропоэтина, которые отличаются от встречающихся в природе форм с точки зрения идентичности и местоположения одного или более аминокислотных остатков (т.е. аналогов делеции, содержащих меньшее количество всех остатков, определенных для ЕРО, и/или аналогов замещения, в которых один или более установленных остатков заменены другими остатками, и/или дополнительных аналогов, в которых один или более аминокислотных остатков добавлены к конечной или средней части полипептида); и которые обладают некоторыми или всеми свойствами встречающихся в природе форм.
Предлагаемые новые ДНК-последовательности включают все последовательности, пригодные в обеспечении экспрессии в прокариотических или эукариотических клетках-хозяевах полипептидных продуктов, имеющих, по меньшей мере, часть первичной структурной конформации и одно или более из биологических свойств эритропоэтина, которые охватываются: (а) ДНК-последовательностями, приведенными в Таблицах V и VI, или их комплементарными нитями; (b) ДНК-последовательностями, которые гибридизируют (в условиях гибридизации, иллюстрируемых здесь, или более жестких условиях) к ДНК-последовательностям, которые определены в (а), или их фрагментам; и (с) ДНК-последовательностями, которые вследствие дегенерации генетического кода должны гибридизировать к ДНК-последовательностям, определенным в (а) и (b) выше. Особенно охвачены в части (b) геномные ДНК-последовательности, кодирующие формы аллельных вариантов обезьяньего и эритропоэтина человека и/или кодирующие эритропоэтин других видов млекопитающих. Особенно охвачены в части (с) построенные ДНК-последовательности, кодирующие ЕРО, ЕРО-фрагменты и ЕРО-аналоги, чьи ДНК-последовательности могут включать кодоны, способствующие трансляции информационной РНК в хозяевах беспозвоночных.
Охвачен предлагаемым изобретением тот класс полипептидов, который кодируется частями ДНК-комплемента в верхнюю часть последовательности геномной ДНК, приведенной в Таблице VI, т.е. "комплементарно инвертированные белки", как описано в работе Tramontano, et al., Nucleic Acids Research, 12, pp.5048-5059 (1984).
Также охвачены изобретением фармацевтические композиции, содержащие эффективные количества полипептидных продуктов настоящего изобретения вместе с пригодными разбавителями, стимуляторами и/или носителями, позволяющими создание эритропоэтиновой терапии, особенно при лечении анемических болезненых состояний и, более всего, таких анемических состояний, которые приводят к хронической почечной недостаточности.
Предлагаемые полипептидные продукты можно "метить" ковалентной ассоциацией с обнаруживаемым маркерным веществом (например, радиомеченные 125I) для обеспечения реагентами, пригодными при обнаружении и количественном определении эритропоэтина в твердой ткани и жидкостных образцах, таких как кровь или моча. ДНК-продукты можно также метить обнаруживаемыми маркерами (как, например, радиометки и неизотопные метки, такие как биотин) и использовать в процессах ДНК-гибридизации для локализации положения эритропоэтинового гена и/или положения любого родственного генного семейства в хромосомной карте человека, обезьяны и других видах млекопитающих. Их также можно использовать для идентификации нарушений эритропоэтинового гена на ДНК-уровне и использовать в качестве генных маркеров для идентификации соседних генов и их нарушений.
Как подробно описано ниже, настоящее изобретение, кроме того, обеспечивает значительные усовершенствования в способах обнаружения специфических однонитевых полинуклеотидов неизвестной последовательности в гетерогенном клеточном или вирусном образце, включающем кратные однонитевые полинуклеотиды, в котором:
(a) получают смесь меченых зондов однонитевых полинуклеотидов с равномерно изменяющимися последовательностями оснований, причем каждый из указанных зондов является потенциально специфически комплементарным к последовательности оснований, которая предположительно уникальна для обнаруживаемого полинуклеотида,
(b) фиксируют образец на плотном субстрате,
(c) обрабатывает субстрат, имеющий фиксированный на нем образец, для ослабления дальнейшего связывания с ним полинуклеотидов, за исключением пути гибридизации к полинуклеотидам в указанном образце,
(d) обработанный субстрат, имеющий фиксированный на нем образец, временно приводят в соприкосновение с указанной смесью меченых зондов в условиях, способствующих гибридизации только между полностью комплементарными полинуклеотидами, и
(e) специфический полинуклеотид обнаруживают мониторингом на наличие реакции гибридизации между ним и полностью комплементарным зондом внутри указанной смеси меченых зондов, как доказано наличием наивысшей плотности меченого материала на субстрате в локусе специфического полинуклеотида по сравнению с плотностью фона меченого материала, вытекающей из неспецифического связывания меченых зондов с субстратом.
Данные способы особенно эффективны в ситуациях, диктующих использование 64, 128, 256, 512, 1024 или более смешанных полинуклеотидных зондов, имеющих длину от 17 до 20 оснований в ДНК/ДНК, или РНК/РНК, или ДНК/РНК-гибридизациях.
Как описано ниже, указанные выше усовершенствованные способы наглядно позволили идентифицировать кДНК-клоны, кодирующие эритропоэтин обезьян в пределах библиотеки, полученной из мРНК почечных клеток анемических обезьян. Более конкретно, смесь 128 равномерно изменяющихся 20-mer зондов, основанных на информации аминокислотной последовательности, вытекающей из чередующихся фракций, эритропоэтина человека, использовали в методиках гибридизации колоний для идентификации семи "положительных" кДНК-клонов эритропоэтина в пределах совокупности 200000 колоний. Даже более того, практика применения усовершенствованных способов настоящего изобретения позволила быстро выделить три положительных клона из пределов экранирования 1500000 стерильных пятен, составляющих геномную библиотеку человека. Это было выполнено посредством использования указанной выше смеси 128 20-mer зондов вместе со вторым набором 128 17-mer зондов на основании аминокислотного анализа отличающейся непрерывной последовательности эритропоэтина человека.
Указанные выше иллюстративные способы составляют первый известный пример использования кратных смешанных олигонуклеотидных зондов в процессах ДНК/ДНК-гибридизации, направленных на выделение геномных клонов млекопитающих, и первый известный пример использования смеси более чем 32 олигонуклеотидных зондов в выделении кДНК-клонов.
Многочисленные аспекты и преимущества изобретения станут очевидными для специалистов в данной области при рассмотрении следующего подробного описания изобретения, создающего пример практического применения изобретения в предпочтительных вариантах его осуществления.
Подробное описание изобретения
В соответствии с настоящим изобретением были выделены и охарактеризованы ДНК-последовательности, кодирующие частично или полностью полипептидную последовательность вида эритропоэтина человека и обезьяны (в дальнейшем "ЕРО"). Кроме того, ДНК обезьяны и человека стала предметом эукариотической и прокариотической экспрессии, создающей выделяемые количества полипептидов, проявляющих биологические (например, иммунологические) свойства встречающегося в природе ЕРО, а также биологические активности in vivo и in vitro ЕРО.
Видовые ДНК обезьян сначала выделяли из кДНК-библиотеки, построенной мРНК, которая происходила от почечной ткани обезьяны в химически индуцированном анемическом состоянии и чья сыворотка, как было установлено иммунологическим путем, содержала более высокие уровни ЕРО по сравнению с сывороткой нормальной обезьяны. Выделение требуемых кДНК-клонов, содержащих ЕРО-кодирующую ДНК, осуществлялось путем использования гибридизации колоний ДНК/ДНК с применением пула 128 смешанных, радиомеченных, 20-mer олигонуклеотидных зондов, и включало быстрое экранирование 200000 колоний. Расчет олигонуклеотидных зондов был основан на информации аминокислотной последовательности, представленной ферментативной фрагментацией и определением последовательности малой пробы человеческого ЕРО.
ДНК человека была выделена из библиотеки геномной ДНК человека. Выделение клонов, содержащих ЕРО-кодирующую ДНК, осуществляли путем использования гибридизации бляшек ДНК/ДНК с применением указанных выше 128 смешанных 20-mer олигонуклеотидных зондов, а также второго пула 128 радиомеченных 17-mer зондов, чьи последовательности основывались на информации аминокислотной последовательности, полученной из отличающегося ферментативного фрагмента ЕРО человека.
Положительные колонии и бляшки были проверены при помощи дидеокси-программирования последовательности клональной ДНК с использованием набора 16 последовательностей в пределах пула 20-mer зондов, и отобранные клоны были подвергнуты анализу на нуклеотидную последовательность с окончательной дедукцией первичной структурной конформации ЕРО-полипептидов, кодируемых ею. Дедуцированные полипептидные последовательности проявляли высокую степень гомологии по отношению друг к другу и к частичной последовательности, генерированной аминокислотным анализом фрагментов ЕРО человека.
Отобранный положительный кДНК-клон обезьян и отобранный положительный геномный клон человека были внесены в "челночный" ДНК-вектор, который амплифицировали в Е.coli и применяли для трансфекции клеток млекопитающих в культуре. Культурный рост трансфектированных клеток-хозяев выражался в препаратах, плавающих на поверхности культуральной среды, содержащих по оценке до 3000 mU ЕРО на мл культуральной жидкости.
Следующие примеры представлены с целью иллюстрации изобретения и особенно направлены на способы, осуществленные ранее для идентификации ЕРО, кодирующего кДНК-клоны обезьяны и геномные клоны человека, на способы, являющиеся результатом этой идентификации, а также на последовательности, развитие систем экспрессии и иммунологическое подтверждение экспрессии ЕРО в таких системах.
Более конкретно. Пример 1 направлен на определение аминокислотной последовательности ЕРО-фрагментов человека и построение смесей радиомеченных зондов на основании результатов этого определения последовательностей. Пример 2 направлен, главным образом, на способы, включенные в идентификацию положительных кДНК-клонов обезьян, и таким образом обеспечивает получение информации, касающейся лечения животных и предварительного анализа радиоиммуноисследования (RIA) сывороток животных. Пример 3 направлен на получение кДНК-библиотеки, экранирование гибридизации колоний и проверку положительных клонов, определение последовательности ДНК положительного кДНК-клона, а также генерирование информации первичной структурной конформации (аминокислотной последовательности) полипептидов ЕРО обезьян. Пример 4 направлен на способы, включенные в идентифицирование положительных геномных клонов человека, и обеспечивает, таким образом, информацией, относящейся к источнику геномной библиотеки, способам гибридизации бляшек и проверке положительных клонов. Пример 5 направлен на определения последовательности ДНК положительного геномного клона и генерирование информации аминокислотной последовательности полипептидов ЕРО человека, включая сравнение с информацией последовательности ЕРО обезьяны. Пример 6 направлен на способы построения вектора, включающего ЕРО-кодирующую ДНК, произошедшую от положительного кДНК-клона обезьяны, использование вектора для трансфектирования клеток COS-1 и культурного роста трансфектированных клеток. Пример 7 направлен на способы построения вектора, включающего ЕРО- кодирующую ДНК, произошедшую от положительного геномного клона человека, использование вектора для трансфектирования клеток COS-1 и культурного роста трансфектированных клеток. Пример 8 направлен на способы иммуноисследований, проводимых на надосадочных жидкостях сред, полученных из культурного роста трансфектированных клеток в соответствии с Примерами 6 и 7. Пример 9 направлен на биологическую активность in vitro и in vivo микробиально выраженного ЕРО Примеров 6 и 7.
Пример 10 направлен на развитие систем экспрессии хозяина млекопитающих для кДНК ЕРО видов обезьян и геномной ДНК человека, включая клетки яичника китайского хомяка ("СНО"), и на иммунологическую и биологическую активности продуктов таких систем экспрессии, а также характеристику таких продуктов. Пример 11 направлен на получение генов, кодирующих ЕРО человека и аналоги ЕРО, гены которых включают ряд предпочтительных кодонов для экспрессии в Е.coli и дрожжевых клетках-хозяевах, а также на системы такой экспрессии. Пример 12 относится к профилям иммунологической и биологической активностей выраженных продуктов систем, приведенных в Примере 11.
Пример 1
А. Определение аминокислотной последовательности фрагмента ЕРО человека
ЕРО человека выделяют из мочи и подвергают триптическому расщеплению, приводящему к развитию и выделению 17 раздельных фрагментов в количествах, приближенно выраженных в 100-150 пикомолей.
Фрагментам произвольно приписывают номера и анализируют на аминокислотную последовательность микроанализом с использованием газофазового секвенсера (Applied Biosystems) для получения информации о последовательности, приведенной в Таблице I ниже, где применены буквенные коды и "X" обозначает остаток, однозначно не определенный.
В. Расчет и построение смесей олигонуклеотидных зондов
Аминокислотные последовательности, приведенные в Таблице 1, рассматриваются в смысле дегенерации генетического кода с целью выяснения того, можно ли применять методики смешанных зондов к приемам ДНК/ДНК-гибридизации на кДНК и/или библиотеках геномной ДНК. Этот анализ выявляет, что в пределах фрагмента №Т35 существует ряд из 7 аминокислотных остатков (Val-Asn-Phe-Tyr-Ala-Trp-Lys), который можно однозначно охарактеризовать как кодируемый одной из 128 возможных ДНК-последовательностей, охватывающей 20 пар оснований. Поэтому синтезируют первый набор из 128 20-mer олигонуклеотидов с использованием стандартных фосфоамидитных способов, см., например, работу Beaucage, et al., Tetrahedron Letters, 22, pp.1859-1862 (1981) на твердом носителе, в соответствии с последовательностью, приведенной ниже в Таблице II.
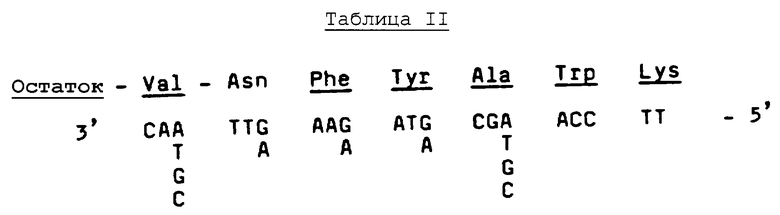
Дополнительный анализ выявляет, что в пределах фрагмента №Т38 существует ряд из 6 аминокислотных остатков (Gln-Pro-Trp-Glu-Pro-Leu), на основе которого можно получить пул из 128 смешанных олигонуклеотидных 17-mer зондов, как показано в Таблице III ниже.
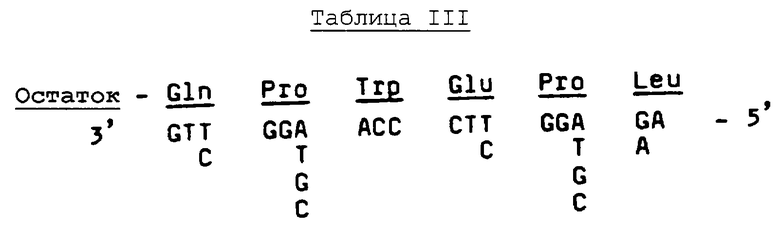
Олигонуклеотидные зонды метят в 5'-конце гамма-32Р-АТР, 7500-8000 Ci/ммоль (ICN), используя полинуклеотидную киназу Т4 (NEN).
Пример 2
А. Методики обработки обезьян и RIA-анализ
Самок Cynomolgus обезьян Масаса fasicularias (2,5-3 кг, 1,5-2 года) обрабатывают путем подкожного введения раствора гидрохлорида фенилгидразина (рН 7,0) в дозировке 12,5 мг/кг на 1, 3 и 5 день. Гематокрит контролируют перед каждым введением. На 7 день, или как только уровень гематокрита опускается ниже 25% от первоначального уровня, собирают сыворотку и почки после введения гидрохлорида кетамина в дозировке 25 мг/кг. Собранные материалы немедленно замораживают в жидком азоте и хранят при температуре -70°С.
В. RIA для ЕРО
Применяют радиоиммунологическое исследование для количественного определения ЕРО в образцах в соответствии со следующими методиками:
Стандарт эритропоэтина или неизвестный образец инкубируют вместе с антисывороткой в течение двух часов при температуре 37°С. После двухчасового инкубирования пробирки с образцами охлаждают на льду, добавляют эритропоэтин, меченный 125I, и пробирки инкубируют при температуре 0°С, по крайней мере, еще 15 часов. Каждая исследуемая пробирка содержит 500 мкл инкубацонной смеси, содержащей 50 мкл разведенной иммунной сыворотки, 10000 cpm 125I-эритропоэтина, 5 мкл трасилола и 0-250 мкл или стандарта ЕРО, или неизвестного образца, а также PBS, содержащий 0,1% BSA для восполнения оставшегося объема. Используемой сывороткой является спущенная во второй пробе кровь кролика, иммунизированного 1%-ным чистым препаратом мочевого эритропоэтина человека. Конечное разведение антисыворотки регулируют так, чтобы содержание 125I-эритропоэтина, связанного с антителом, не превышало 10-20% от общих входных количеств. В общем, это соответствует конечному разведению антисыворотки от 1:50000 до 1:100000.
125I-эритропоэтин, связанный с антителом, осаждают добавлением 150 мкл Staph А. После 40-минутного инкубирования образцы центрифугируют и осадки в пробирке промывают два раза 0,75 мл раствором 10 мМ Tris-HCl, имеющим рН=8,2, содержащим 0,15 М NaCl, 2 мМ EDTA и 0,05% Triton X-100. Определяют количество промытых остатков при помощи счетчика гамма-квантов для определения процента связи 125I-эритропоэтина. Количества, связанные пре-иммунными сыворотками, вычитают из всех конечных значений для корректировки неспецифического осаждения. Содержание эритропоэтина неизвестных образцов определяют путем сравнения со стандартной кривой.
Приведенную выше методику применяют на сыворотке обезьян, полученной выше, в части А, а также не сыворотке обработанных обезьян. Уровни нормальной сыворотки, как определено, составляют приблизительно 36 mU/мл, тогда как для сыворотки обработанных обезьян они составляют от 1000 до 1700 mU/мл.
Пример 3
А. Построение кДНК-библиотеки обезьян
Информационную ДНК выделяют из почек нормальных и анемических обезьян посредством методики с использованием тиоцианата гуанидиния, описанной в работе Chirgwin, et al., Biochemistry, 18, p.5294 (1979), и поли (А)+ мРНК очищают, пропуская дважды через хроматографическую колонку с олиго(dT)-целлюлозной, как описано на стр.197-198 в работе Maniatis, et al., "Molecular Cloning, A Laboratory Manual" (Cold Springs Harbor Laboratory, Cold Springs, Harbor, N.Y., 1982). Библиотеку кДНК строят в соответствии с видоизменением основных методик, изложенных в работе Okayama, et al., Mol. and Cell. Biol., 2, pp.161-170 (1982). Ключевыми моментами предпочтетаемых теперь методик являются следующие: (1) pUC8 используют в качестве подошвенного вектора, разрезают PstI и затем снабжают хвостом длиной в 60-80 оснований при помощи олиго dT; (2) расщепление HincII используют для отщепления олиго dT-хвоста от одного конца вектора; (3) синтез первой нити и присоединение хвоста при помощи олиго dG осуществляют в соответствии с опубликованной методикой; (4) расщепление BamHI применяют для отщепления олиго dG-хвоста от одного конца вектора; и (5) замену РНК-нити на ДНК производят в присутствии двух связующих звеньев (GATCTAAAGACCGTCCCCCCCCC и ACGGTCTTTA) при трехкратном молярном избытке по отношению к вектору с олиго dG-хвостом.
В. Методики гибридизации колоний для просеивания кДНК-библиотеки обезьян
Трансформированные Е.coli разбрасывают с плотностью 9000 колоний на 10×10 см питательных чашках, содержащих 50 мкг/мл ампициллина. GeneScreen фильтры (New England Nuclear Catalog №NEF-972) предварительно увлажняют на BHI-САМ чашке (37 г/л бактоэкстракта мозг/сердце, 2 г/л кислот Casamino и 15 г/л агара, содержащего 500 мкг/мл хлорамфеникола) и используют для поднятия колоний с чашки. Колонии выращивают в той же среде в течение 12 часов или более для амплифицирования численности плазмидной копии. Амплифицированные колонии (бока колоний) обрабатывают серийно, помещая фильтры над двумя кусками 3 MM ватмана, насыщенного следующими растворами:
(1) 50 мМ глюкоза - 25 мМ Tris-HCl (рН 8,0) - 10 мМ EDTA (рН 8,0) в течение 5 минут,
(2) 0,5 М NaOH в течение 10 минут, и
(3) 1,0 М Tris-HCl (рН 7,5) в течение трех минут.
Фильтры затем сушат на воздухе в вакууме в течение двух часов при температуре 80°С.
Затем фильтры подвергают расщеплению Протеиназой К через обработку раствором, содержащим 50 мкг/мл протеазного фермента в Буфере К [0,1 М Tris-HCl (рН 8,0) - 0,15 М NaCl - 10 мМ EDTA (рН 8,2) - 0,2% SDS]. Конкретно, добавляют к каждому фильтру 5 мл раствора и расщепление продолжают при температуре 55°С в течение 30 минут, после чего раствор удаляют.
Фильтры затем обрабатывают 4 мл буфера предварительной гибридизации (5 × SSPE - 0,5% SDS - 100 мкг/мл SS E.coli ДНК - 5 × BFP). Предварительную гибридизационную обработку осуществляют при температуре 55°С, как правило, в течение 4 часов или более, после чего раствор удаляют.
Процесс гибридизации проводят следующим образом. В каждый фильтр добавляют 3 мл гибридизационного буфера (5 × SSPE - 0,5% SDS - 100 мкг/мл дрожжевой тРНК), содержащего 0,025 пикомолей каждой из 128 последовательностей зондов, представленных в Таблице II (полная смесь обозначена как смесь EPV), и фильтры выдерживают при температуре 48°С в течение 20 часов. Эта температура на 2°С ниже, чем нижнее из значений температуры диссоциации (Td), определенной для любого из зондов.
По окончании гибридизации фильтры промывают три раза в течение 10 минут на качалке раствором 6 × SSC - 0,1% SDS при комнатной температуре и промывают от двух до трех раз раствором 6 × SSC - 1% SDS при температуре гибридизации (48°С).
Авторадиографией фильтров выявляют семь положительных клонов среди скринированных 200000 колоний.
Анализ начальной последовательности одного из мнимых кДНК-клонов обезьяны (обозначен как клон 83) проводят для проверки результатов путем видоизменения методики, описанной Wallace, et al., Gene, 16, pp.21-26 (1981). Короче, ДНК-плазмиду из кДНК-клона 83 обезьяны линеаризуют путем расщепления EcoRI и денатурируют посредством нагревания в ванне с кипящей водой. Нуклеотидную последовательность определяют дидеокси-способом, описанным Sanger, et al., P.N.A.S. (U.S.A.), 74, pp.5463-5467 (1977). Набор из EPV-смеси зондов, состоящих из 16 последовательностей, используют в качестве затравки для реакций последовательностей.
С. Определение кДНК-последовательности ЕРО обезьяны
Анализ нуклеотидных последовательностей клона 83 осуществляют посредством методик, описанных Messing, Methods in Enzymology, 101, pp.20-78 (1983). В таблице IV представлены результаты анализа карты предварительного ограничения для EcoRI/HindIII клонированного фрагмента клона 83 с приблизительно 1600 парами оснований. Приближенные местоположения участков распознавания ограничительных эндонуклеазных ферментов выражены числом оснований 3' и участку Е.coli в 5' конце фрагмента. Определение нуклеотидной последовательности осуществляют нахождением последовательности отдельных ограничительных фрагментов, с целью спаривания перекрывающихся фрагментов. Например, перекрывание информации последовательности, представленное анализом нуклеотидов в ограничительном фрагменте, обозначено С113 (Sаu3А при ˜111/SmaI при ˜324), а определение последовательности с обратным порядком обозначено С73 (AluI при ˜424/BstEII при ˜203).
Определение последовательности приблизительно 1342 пар оснований (в пределах области от участка 3' Sau3A до участка EcoRI и участка HindIII) и анализ всех возможных систем считывания способствуют развитию ДНК и информации аминокислотной последовательности, представленной в Таблице V. В таблице мнимый первоначальный аминокислотный остаток аминоконца созревшего ЕРО (как проверено путем корреляции ранее указанного анализа последовательности двадцати остатков аминоконцов) обозначен числом +1. Присутствие определяющего метионин АТС-кодона (обозначен -27), "направленного против течения" конечного аланинового остатка первоначального амина, как первого остатка, обозначенного для аминокислотной последовательности созревшего белка, является показателем правдоподобия того, что ЕРО первоначально выражен в цитоплазме в форме предшественника, включающей "ведущую" область из 27 аминокислотных остатков, которая вырезается до вхождения созревшего ЕРО в кровообращение. Участки потенциального гликозилирования в пределах полипептида обозначены звездочками. Вычисленный молекулярный вес области трансляции составляет 21117 дальтон, а молекулярный вес 165 остатков полипептида, составляющих созревший ЕРО обезьяны, равен 18236 дальтон.



Полипептидная последовательность в Таблице V может быть без труда подвергнута анализу на присутствие чрезвычайно гидрофильных областей и/или вторичных конформационных характеристик, являющихся показателем возможно очень иммуногенных областей, например, при помощи способов, предложенных Норр, et al., P.N.A.S. (U.S.A.), 78, pp.3824-3828 (1981), и Kyte, et al., J. Mol. Biol., 157, pp.105-132 (1982), и/или Chou, et al., Biochem., 13, pp.222-245 (1974) и Advances in Enzymology, 47, pp.45-47 (1978). Компьютеризация анализа в соответствии со способом Норр, et al., возможна благодаря программе, обозначенной PEP Reference Section 6.7, представленной Intelligenetics, Inc., 124 University Avenue, Palo Alto, California.
Пример 4
А. Геномная библиотека человека
Геномную библиотеку фетальной печени человека, несущей фаг Сh4А, полученную в соответствии с методиками Lawn, et al., Cell, 18, pp.533-543 (1979), сохраняют для использования в исследовании гибридизации бляшек.
В. Методики гибридизации бляшек для просеивания геномной библиотеки человека
Фаговые частицы подвергают лизису и ДНК фиксируют на фильтрах (50000 бляшек на фильтр) в соответствии с методиками, описанными Woo, Methods In Enzymology, 68, pp.389-395 (1979), за исключением того, что использовали GeneScreen Plus фильтры (New England Nuclear Catalog № NEF-976) и NZYAM чашки (NaCl, 5 г; MgCl2-6Н2О, 2 г; NZ-Амин А, 10 г; дрожжевой экстракт, 5 г; кислоты casamino, 2 г; мальтоза, 2 г; и агар, 15 г на литр).
Высушенные на воздухе фильтры обжигают при температуре 80°С в течение 1 часа и затем расщепляют протеиназой К, как описано в Примере 3, часть В. Предварительную гибридизацию осуществляют с использованием 1 М NaCl - 1% SDS-буфера при температуре 55°С в течение 4 часов или более, после чего буфер удаляют. Гибридизационные и постгибридизационные промывки осуществляют, как описано в Примере 3, часть В. Применяют как смесь 128 20-mer зондов, обозначенных EPV, так и смесь 128 17-mer зондов Таблицы III (обозначена как смесь EPQ). Гибридизацию осуществляют при температуре 48°С с использованием смеси зондов EPV. Гибридизацию с использованием смеси зондов EPQ осуществляют при температуре 46°С, т.е. на 4 градуса ниже наименьшей вычисленной Td для членов смеси. Удаление гибридизированных зондов для повторной гибридизации выполняют кипячением в 1 × SSC - 0,1% SDS в течение двух минут. Авторадиография фильтров выявляет три положительных клона (активных с обеими смесями зондов) среди скринированных 1500000 фаговых бляшек. Проверку положительных клонов на ЕРО-кодирование достигают посредством определения последовательности ДНК и электронной микрографической визуализации гетеродуплексной формации с кДНК обезьяны Примера 3. Эта методика также дает доказательство присутствия кратных интронов в геномной ДНК-последовательности.
Пример 5
Осуществляют анализ нуклеотидных последовательностей положительных клонов (обозначены λhEl), и результаты, полученные на сегодняшний день, приведены в Таблице VI.





В таблице VI первоначальная непрерывная ДНК-последовательность обозначает верхнюю нить 620 оснований в том, что, по-видимому, является нетранслированной последовательностью, немедленно предшествующей транслированной части гена ЕРО человека. Более конкретно, последовательность содержит 5'-конец гена, который ведет к транслированной области ДНК, кодирующей первые четыре аминокислоты (от -27 до -24) последовательности-лидера ("пре-последовательность"). Четыре пары оснований в последовательности, предшествующей последней, кодируют начало лидера, однако, еще не определены однозначно и поэтому обозначены "X". Затем следует интрон из приблизительно 639 пар оснований (из которых у 439 пар оснований определена последовательность, а оставшиеся 200 пар оснований обозначены "I.S."), немедленно предшествуя кодону для глутамина, который обозначен как остаток -23 транслированного полипептида. Следующая немедленно экзон-последовательность кодирует аминокислотные остатки через аланиновый остаток (обозначенный как остаток +1 аминокислотной последовательности созревшего ЕРО человека) до кодона, определяющего треонин в положении +26, после чего следует второй интрон, состоящий из 256 оснований, как специфически и обозначено. Следующая за этим интроном экзон-последовательность для аминокислотных остатков 27-55 продолжается третьим интроном, содержащим 612 пар оснований. Последующий экзон кодирует остатки 56-115 ЕРО человека, затем идет четвертый нитрон из 134 пар оснований, который продолжается экзоном, кодирующим остатки 116-166 и кодон "остановки" (TGA). Наконец, в Таблице VI идентифицируется последовательность 568 пар оснований в том, что составляет нетранслированную область 3' гена ЕРО человека, две пары оснований которой ("X") еще не определены однозначно на последовательность.
Таблица VI служит для идентификации первичной структурной конформации (аминокислотной последовательности) созревшего ЕРО человека, включающей 166 установленных аминокислотных остатков (с вычисленным молекулярным весом =18399). В Таблице также выявлена ДНК-последовательность, кодирующая последовательность-лидер из 27 остатков вместе с ДНК-последовательностью 5' и 3', которые могут быть важными для функций промотора/оператора, осуществляемых генным опероном. Участки для потенциального гликозилирования полипептида созревшего ЕРО человека обозначены в Таблице звездочками. Необходимо отметить, что специфическая аминокислотная последовательность в Таблице VI, вероятно, составляет последовательность встречающейся в природе аллельной формы эритропоэтина человека. Основание для такого положения найдено в результатах непрерывных попыток определения последовательностей мочевых изолятов эритропоэтина человека, которые позволили располагать информацией относительно того, что значительное количество молекул эритропоэтина имеют метионин в остатке 126 в противоположность серину, как показано в Таблице.
Таблица VII, ниже, иллюстрирует степень гомологии полипептидной последовательности между ЕРО человека и обезьяны. В верхней непрерывной линии Таблицы буквенные обозначения применены для представления дедуцированных транслированных последовательностей ЕРО человека, начиная с остатка -27, а нижняя непрерывная линия показывает дедуцированную полипептидную последовательность ЕРО обезьяны, начиная от числа -27, обозначающего остаток. Звездочки применяются для выделения гомологии последовательностей. Следует отметить, что дедуцированные последовательности ЕРО человека и обезьяны выявляют "дополнительный" остаток лизина (К) в положении 116 (для человека). Ссылка к Таблице VI означает, что этот остаток находится на краю мнимого мРНК-соединения в геномной последовательности. Присутствие остатка лизина в полипептидной последовательности человека проверяют путем определения последовательности клона кДНК человека, полученной из мРНК, выделенной из клеток COS-1, которые трансформированы с геномной ДНК человека в Примере 7, ниже.

Пример 6
При первоначальных попытках микробиологического синтеза изолируемых количеств эритропоэтина полипептидного материала, кодируемого кДНК обезьяны, получаемого по методике примера 3, выбранная система экспрессии представляет собой систему, содержащую клетки-хозяева млекопитающего (например, клетки COS-1, А.Т.С.С., №CRL-1650). Указанные клетки осуществляют трансфекцию посредством "челночного" вектора, обладающего способностью к спонтанной репликации в хозяине Е.coli (благодаря присутствию ДНК-производных от плазмид pBR322) и хозяевах млекопитающих (благодаря присутствию ДНК-производных от вирусов SV40).
Более конкретно, вектор экспрессии строят в соответствии со следующими методиками. Клон плазмид 83, полученный в примере 3, амплифицируют в бактерии Е.coli и при расщеплении EcoRI и HindIII выделяют приблизительно 1,4 т.п.н. ДНК обезьяны, кодирующую эритропоэтин. Из плазмиды pBR322 отдельно выделяют 4,0 т.п.н. фрагмента HindIII/SalI. Из ДНК, содержащей повторы M13mp10 RF ДНК (Р и L лаборатории), получают около 30 пар оснований фрагмента "линкера" EcoRI/SalI. Указанный "линкер" включает последовательно EcoRI-липкий конец, за которым следуют участки распознавания SstI, SmaI, BamHI и XbaI и липкий конец SalI. Указанные выше три фрагмента сшивают с получением примерно 5,4 т.п.н. промежуточной плазмиды ("pERS"), ЕРО-ДНК которой граничит с одной стороной при использовании "банк" полезных участков ограничения распознавания эндонуклеазы. После этого плазмиду pERS расщепляют HindIII и SalI с получением эритропоэтина ДНК и трансформацией EcoRI в "линкер" SalI (M13mp10). Указанный выше фрагмент ДНК длиной 1,4 т.п.н. сшивают с участком BamHI/SalI приблизительно длиной 4,0 т.п.н. плазмиды pBR322 и другим "линкером" фрагмента M13mp10 HindIII/BamHI RF, имеющий также приблизительно 30 пар оснований. Фрагмент "линкера" М13 представляет собой HindIII-липкий конец, после чего следуют участки узнавания PstI, SalI, XbaI и липкий конец BamHI. Продуктом сшивки вновь становится полезная промежуточная плазмида ("pBR-EPO"), содержащая ЕРО-ДНК, которая ограничена с обеих сторон "накопленными рядами" ограничительного участка.
Выбранный для экспрессии ЕРО-ДНК в клетках COS-1 ("pDSVL1") вектор строят заранее для возможности осуществления отбора и аутодупликации в Е.coli. Указанных характеристик достигают за счет начала репликации и порядка следований генов ДНК, стойких к действию ампициллина, которые присутствуют в области, осуществляющей стягивание нуклеотидов 2448 до 4362 плазмиды pBR322. Указанный порядок следования структурно модифицируют посредством введения "линкера", обеспечивающего распознавание HindIII сразу же при примыкании нуклеотида 2448 до включения в указанный вектор. Наряду с подбором векторов для экспрессии ЕРО-ДНК к другим полезным свойствам относятся способность к аутодупликации в клетках COS-1 и наличие последовательности, действующей как вирусный промотор в клетках млекопитающих. Указанных характеристик достигают посредством начала порядка следования ДНК-репликации и последовательности "позднего" гена ДНК, являющегося вирусным усилителем, который присутствует в цепи, содержащей 342 пары оснований, стягивающую число нуклеотидов от 5171 до 270 генома SV40. Уникальный ограничительный участок (BamHI) образуется на векторе и сразу же примыкает к ряду вирусного промотора благодаря использованию коммерчески доступной последовательности "линкера" (Collaborative Research). Дополнительно в вектор вводят последовательность 237 пар оснований (производная как нуклеотидное число от 2553 до 2770 вируса SV40), содержащий сигнал полиаденилирования "позднего гена" вирусной мРНК (обычно известный как терминатор транскрипции). Указанный фрагмент располагают в векторе в соответствующей ориентации относительно вирусного промотора "позднего гена" через уникальный участок BamHI. В указанном векторе также представлен другой ген млекопитающего, находящийся в положении не материальном относительно возможной транскрипции гена, который помещен на указанном уникальном участке между последовательностями вирусного промотора и терминатора. [Указанный ген млекопитающего содержал в себе приблизительно 2500 пар оснований дигидрофолатредуктазы (DHFR)-минигена мыши, выделенного из плазмиды pMG-1, Gasser, et al., P.N.A.S. (U.S.A.), 79, pp.6522-6526, (1982)]. И опять большинство действующих компонентов плазмиды pDSVL1 состоит из 2448-4362 нуклеотидов плазмиды pBR322 наряду с нуклеотидами 5171-270 (342 пары оснований) и 2553-2770 (237 пар оснований) вирусной ДНК, содержащей вирус SV40.
В соответствии с методиками, описанными ранее, например, Maniatis, et al., выше, эритропоэтин-кодирующую ДНК выделяют из плазмиды pBR-EPO в качестве фрагмента BamHI и вшивают в плазмиду pDSVL1, вырезанную BamHI. Для удостоверения введения ЕРО-гена в правильной ориентации в двух из полученных клонированных векторов (дуплицированные векторы Н и L) проводят анализ с использованием ограничительного фермента (см. фиг.2, иллюстрирующую плазмиду pDSVL1-MkE). Векторы с неправильной ориентацией ЕРО-генов оставляют в качестве регуляторов отрицательных величин в экспериментах трансфекции, предназначенных для определения уровней экспрессии ЕРО в хозяевах, которые трансформируются посредством векторов с правильной ориентацией ЕРО-ДНК.
Векторы Н, L, F, Х и G объединяют с носителем ДНК (ДНК печени и селезенки мыши). Для осуществления трансфекции используют дуплетные 60 мм пластинки, используя методы микроосаждения фосфатом кальция. Дупликатные 60 мм пластинки также трансфектируют ДНК-носителем в качестве "имитации" управления трансформацией негативных величин. Через 5 дней всю культуральную среду подвергают испытанию на присутствие полипептидов, которые обладают иммунологическими свойствами эритропоэтина природного происхождения.
Пример 7
А. Система экспрессии исходного ЕРО с использованием клеток COS-1
Выбранная на первоначальном этапе микробиологического синтеза подвергаемых выделению количеств полипептидного материала эритропоэтина человека, кодируемого посредством ЕРО-клона геномной ДНК человека, система также включает в себя экспрессию в клетках-хозяевах млекопитающих (например, клетки COS-1, А.Т.С.С. № CRL-1650). ЕРО-ген человека первоначально субклонируют в "челночный" вектор, который обладает способностью к автономной репликации в обоих хозяевах бактерии Е.coli (благодаря ДНК-производной от плазмиды pBR322) и в клетке линии COS-1 млекопитающего (благодаря наличию вируса SV-40 вирус-производных ДНК). Челночный вектор, содержащий ЕРО-ген, затем трансфектируют в клетки COS-1. ЕРО-полипептидный материал образуется в трансфектированных клетках и выделяется в клеточную культуральную среду.
Более конкретно, вектор экспрессии признаков строят в соответствии со следующими методиками. ДНК, выделенная из клона λhE1 барашка, содержащего геномный ЕРО-ген, расщепляют BamHI и HindIII ограничительных эндонуклеаз и выделяют 5,6 т.п.н. ДНК-фрагмента, как обнаружено, содержащего полный ЕРО-ген. Указанный фрагмент ДНК смешивают и сшивают с бактериальной плазмидой pUC8 (Bethesda Research Laboratories, Inc.), которую аналогичным образом расщепляют, создавая промежуточную плазмиду "pUCS-HuE", образуя, таким образом, подходящий источник ограничивающего фрагмента.
Выбранные векторы для экспрессии ЕРО-ДНК в клетках COS-1 строят заранее. Плазмида pSV4SEt, которая содержит ДНК- последовательности, дает возможность выбора и аутодупликации в Е.coli. Указанных характеристик достигают началом возникновения репликации и порядком следования гена ДНК, устойчивого к ампициллину, который присутствует в области, стягивающей нуклеотиды от 2448 до 4362 бактериальной плазмиды pBR322. Изменяют структуру указанной последовательности путем введения линкера, обеспечивая участок распознавания HindIII сразу же при прилегании к нуклеотиду 2448. Плазмида pSV4SEt также способна к аутодупликации в клетках COS-1. Указанной характеристики достигают посредством использования фрагмента 342 пар оснований, содержащего вирусное начало репликации с помощью вируса SV40 (количество нуклеотидов от 5171 до 270). Указанный фрагмент модифицируют при введении линкера, который обеспечивает участок распознавания EcoRI, прилегающий к нуклеотиду 270, и линкера, обеспечивающего участок распознавания SalI, примыкающий к нуклеотиду 5171. В данном векторе также присутствует фрагмент 1061 пары оснований вируса SV40 (число нуклеотидов от 1711 до 2772 с дополнительным линкером, который обеспечивает участок распознавания SalI, прилегая к следующему нуклеотиду 2772). В пределах указанного фрагмента имеется уникальная последовательность распознавания BamHI. Суммируя вышесказанное, плазмида pSV4SEt содержит в себе уникальные участки распознавания BamHI и HindIII, давая возможность введения гена ЕРО человека. Указанные последовательности дают возможность осуществления репликации и избирательного отбора в Е.coli и репликации в клетках COS-1.
Для введения ЕРО-гена в плазмиду pSV4SEt, плазмиду pUC8-HuE расщепляют BamHI и HindIII до ограничивающей эндонуклеазы и выделяют 5,6 т.п.н. ЕРО, кодирующего ДНК фрагмента. Плазмиду pSV4SEt также расщепляют BamHI и HindIII и основную часть фрагмента, содержащую 2513 пар оснований, выделяют (сохраняя при этом все необходимые функции). Указанные фрагменты смешивают и сшивают, образуя конечный вектор "pSVgHuEPO" (см. фиг.3). Этот вектор разводят в Е.coli, а вектор ДНК выделяют. Для подтверждения ввода ЕРО-гена проводят анализ ограничительного фермента.
ДНК плазмиды pSVgHuEPO используют для выражения признаков ЕРО-полипептидного материала в клетках COS-1. Детально излагая, ДНК плазмиды pSVgHuEPO объединяют с ДНК-носителем и трансфектируют в трипликатные 60 мм пластинки клеток COS-1. В качестве контроля ДНК носитель один трансфектируют в клетках COS-1. Клеточную культуральную среду отбирают спустя 5 и 7 дней и подвергают испытанию на присутствие полипептидов, которые обладают иммунологическими свойствами, характерными для встречающегося в природе ЕРО человека.
В. Вторая система экспрессии признаков ЕРО в клетках COS-1.
Еще одну систему экспрессии применяют для получения улучшенных продуктов человеческого эритропоэтин-полипептидного материала, кодируемого с помощью клона геномной ДНК человека, образующей ЕРО в клетках COS-1 (А.Т.С.С. № CRL-1650).
В предшествующей системе признаки ЕРО выражают в клетках COS-1 с использованием своего собственного промотора, который присутствует в пределах ограничительного фрагмента BamHI относительно HindIII, имеющего длину 5,6 т.п.н.. В следующем построении ЕРО-ген чередуют с получением экспрессии признаков, используя поздний промотор вируса SV40.
Более конкретно, ограничительный фрагмент геномного ЕРО человека, представляющий клонированный 5,6 т.п.н. BamHI относительно HindIII, модифицируют в соответствии со следующей методикой. Как описывается выше, плазмиду pUC8-HuE расщепляют BamHI и ограничительной эндонуклеазой BstEII. Ограничительная эндонуклеаза BstEII расщепляет в пределах 5,6 т.п.н. ЕРО-ген в положении, которое представляет 44 пары оснований 5' до инициирующих ATG, кодируя предшественник-пептид и примерно 680 пар оснований 3' до ограничительного участка HindIII. Затем выделяют фрагмент, включающий приблизительно 4900 пар оснований. Фрагмент ДНК с синтетическим линкером, содержащий Sall и BstEII липкие концы и внутренний участок распознавания BamHI, подвергают синтезу и очистке. Указанные два фрагмента смешивают и сшивают с плазмидой pBR322, которую вырезают с помощью Sall и BamHI для получения промежуточной плазмиды pBRgHE. Геномный ген ЕРО человека можно выделить из нее в виде фрагмента расщепления BamHI с 4900 пар оснований, несущего полную структуру гена с одиночной ATG 44 пары оснований 3' относительно участка BamHI, прилегающего к области окончания кодирования аминогрупп.
Полученный фрагмент выделяют и вводят как фрагмент BamHI в плазмиду pDSVL1 расщепленного вектора экспрессии BamHI (см. пример 6). Получающуюся в результате этого плазмиду pDSVLgHuEPO, как показано на фиг.4, используют для выражения признаков ЕРО полипептидного материала из клеток COS-1, как описано в примерах 6 и 7А.
Пример 8
Культуральную среду, выращенную из 6 трансфектированных культур клеток COS-1 примера 6, анализируют с помощью радиоиммунологического анализа согласно методике, изложенной в примере 2, часть В. Каждый образец анализируют при значениях аликвотной пробы 250, 125, 50 и 25 мкл. Надосадочные слои из роста клеток, "имитационно" трансфектированных или трансфектированных с помощью векторов с неправильной ориентацией генов ЕРО, имели отрицательные значения на иммунореактивность. Для каждого образца двух надосадочных слоев жидкости, полученных из роста клеток COS-1, трансфектированых векторами (Н и L) с правильной ориентацией ДНК ЕРО, % ингибирования 125I-EPO, связанного с антителом, колебался в пределах от 72 до 88%, все значения которых размещены в верхнем положении стандартной кривой. Поэтому нельзя точно рассчитать истинную концентрацию эритропоэтина в плавающих слоях культуры. Попредварительным расчетам она составляла 300 mU/мл, однако, выполненным при значениях самой большой аликвотной пробы (250 мкл).
Образец культуральной жидкости, полученной согласно примеру 6, и 5- и 7-дневные культуральные жидкости, описанные в примере 7А, подвергали радиоиммунологическому анализу для сравнения активности рекомбинантных ЕРО-материалов обезьяны и человека относительно встречающегося в природе ЕРО человека, отвечающего стандарту. Полученные результаты представлены в графической форме на Фиг.1. Короче говоря, представленные результаты неожиданно показали, что рекомбинантный ЕРО обезьяны существенно конкурентен для антитела анти-ЕРО человека, хотя не удалось полностью ингибировать связующее в условиях испытания. Максимальный % значений ингибирования для рекомбинантного эритропоэтина человека, тем не менее, почти приближается к стандартным показателям. Параллельный характер кривой доза-эффект предполагает иммунологическую идентичность последовательностей (эпитопов) в общем. Перед выполнением расчетов концентрации ЕРО обезьяны в культуральных жидкостях были проведены повторные оценки при указанных высоких значениях разведения и было обнаружено, что они колебались в интервале от 2,91 до 3,12 U/мл. Вычисленные уровни продуцирования ЕРО человека установлены соответственно при значениях 392 mU/мл для образца с 5-дневным ростом и 567 mU/мл для образца с семидневным ростом. Расчетные уровни продуцирования ЕРО обезьяны в примере 7 В системы экспрессии были аналогичными или лучшими.
Пример 9
Культуральные жидкости, полученные согласно примерам 6 и 7, подвергали анализу in vitro для определения активности ЕРО в соответствии с методикой Goldwasser, et al., Endocrinology, 97, 2, pp.315-323 (1975). Вычисленные значения ЕРО обезьяны для испытуемых культуральных жидкостей колебались в пределах от 3,2 до 4,3 U/мл. Культуральные жидкости ЕРО человека также проявляли активность в полученных расчетах при анализе in vitro и, кроме того, указанную активность можно нейтрализовать под действием антитела "анти-ЕРО". Культуральные жидкости рекомбинантного ЕРО обезьяны по примеру 6 также подвергали количественному определению in vivo биологической активности согласно общему методу Cotes, et al., Nature, 191, pp.1065-1067 (1961), и Hammond, et al., Ann. N.Y. Acad. Sci., 149, pp.516-527 (1968). Указанные уровни активности колебались от 0,94 до 1,24 U/мл.
Пример 10
В предыдущих примерах рекомбинантный ЕРО-материал человека или обезьяны получают от векторов, которые используются для трансфектирования клеток COS-1. Указанные векторы осуществляют репликацию в клетках COS-1 благодаря присутствию Т-антигена вируса SV40 внутри клетки начала возникновения репликации SV40 на указанных векторах. Хотя полученные векторы воспроизводят полезные качества ЕРО в клетках COS-1, их выраженность является только временной (в течение 7-14 дней) из-за возможных потерь вектора. Кроме того, только незначительный % воспроизведений COS-1 становится продуктивно трансфектированным указанными векторами. Настоящий пример описывает системы экспрессии признаков, использующих клетки DHFR- яичника китайского хомячка (СНО) и селектируемый маркер, DHFR. [Для обсуждения родственных систем экспрессии см. патент США №4399216 и заявки на выдачу Европейского патента №117058, 117059 и 117060, опубликованные 29 августа 1984 г.].
В клетках СНО DHFR- (Dux-Bll) CHO K1, Urlaub, et al., Proc. Nat. Acad. Sci. (U.S.A.), Vol. 77, 4461 (1980), не хватает фермент дигидрофолатредуктазы (DHFR) из-за мутаций, происходящих в структурных генах, и, следовательно, необходимо наличие глицина, гипоксантина и тимидина в культуральной среде. Плазмиды pDSVL-MkE (пример 6) или pDSVL-gHuEPO (пример 7 В) трансфектируют вместе с ДНК носителем в DHFR- клетки СНО, которые выращивают в среде, содержащей гипоксантин, тимидин и глицин в 60 мм культуральных пластинах. Плазмиду pSVgHuEPO (пример 7А) смешивают с плазмидой pMG2, содержащей ген мыши на основе дигидрофолатредуктазы, клонированный в вектор бактериальной плазмиды pBR322 (по Gasser, et al., выше). Смесь плазмид и ДНК носителя трансфектируют в клетки DHFR- СНО. (Клетки, которые захватывают одну плазмиду, обычно также захватят вторую плазмиду). Через 3 дня полученные клетки диспергируют посредством трипсинизирования на нескольких 100 мм культуральных пластинах в среде, в которой не хватает гипоксантина и тимидина. Только те клетки, которые трансформируются в устойчивое состояние, посредством гена DHFR, проявляют выживание в данной среде гена ЕРО. Через 7-21 день становятся видны колонии живущих клеток. Указанные колонии трансформантов после диспергирования путем трипсинизирования могут непрерывно размножаться в среде с недостаточностью гипоксантина и тимидина, создавая новые штаммы клеток (например, СНО pDSVL-MkEPO, CHO pSVgHuEPO, CHO pDSVL-gHuEPO).
Культуральные жидкости, полученные из вышеуказанных клеточных штаммов, подвергают радиоиммунологическому анализу на наличие рекомбинантного ЕРО обезьяны или человека. Среда для штаммов CHO pDSVL-MkEPO содержит ЕРО с такими же иммунологическими свойствами, как и ЕРО, полученный из клеток COS-1, трансфектированных плазмидой pDSVL-MkEPO. Образец культуральной жидкости с 65-часовой выдержкой содержит 0,60 U/мл ЕРО обезьяны.
Культуральные жидкости, полученные из CHO pSVgHuEPO и СНО pDSVL-gHuEPO, содержат рекомбинантный ЕРО человека, обладающий такими же иммунологическими свойствами, как и ЕРО, полученный из клеток COS-1, трансфектированных плазмидой pSVgHuEPO или pDSVL-gHuEPO. Образец культуральной жидкости с 3-дневной выдержкой, полученный из СНО pSVgHuEPO, содержит 2,99 U/мл ЕРО человека, а образец с 5,5-дневной выдержкой, полученный из СНО pDSVL-gHuEPO, содержит 18,2 U/мл ЕРО человека, как показывают результаты радиоиммунологического анализа.
Качество ЕРО, получаемого посредством вышеуказанных штаммов клеток, можно повысить за счет усиления генов, создавая новые штаммы клеток, обладающие большей эффективностью. Фермент дигидрофолатредуктазы (DHFR), который представляет собой продукт, кодируемый геном DHFR, может быть ингибирован с помощью метотрексата (МТХ). В частности, клетки, размножаемые в среде с недостаточностью гипоксантина и тимидина, ингибируются или убиваются метотрексатом. В соответствующих условиях (например, при минимальной концентрации МТХ) можно получить клетки, устойчивые и способные к росту в МТХ. Обнаружено, что указанные клетки становятся устойчивыми к МТХ благодаря усилению ряда их DHFR генов, в результате чего повышается продуцирование DHFR фермента. Живущие клетки можно, в свою очередь, обработать повышенными концентрациями метотрексата, в результате чего в штаммах клетки увеличивается количество генов DHFR. "Мимолетные гены" (например, ЕРО), переносимые на вектор экспрессии вместе с геном DHFR или трансформируемые с ним, как часто обнаруживается, повышают число своих воспроизведений.
В качестве примера практической реализации системы амплификации штамм клетки СНО pDSVL-MkE обрабатывают метотрексатом путем повышения его концентраций (0 нМ, 30 нМ и 100 нМ). Образцы, представляющие 65-часовую среду культивирования на каждом этапе амплификации, по данным радиоиммунологического анализа содержат 0,60, 2,45 и 6,10 U/мл эритропоэтина, соответственно. Штамм клеток СНО pDSVL-gHuEPO подвергают обработке метотрексатом (МТХ) с серийным повышением его концентраций: 30 нМ, 50 нМ, 100 нМ, 200 нМ, 1 мкМ и 5 мкМ. Образец, представляющий 3-дневную надосадочную жидкость, на этапе его обработки метотрексатом (МТХ) с концентрацией 100 нМ содержит по данным радиоиммунологического анализа 3089±129 U/мл эритропоэтина человека. Образцы, представляющие 48-часовую культуральную среду, обработанные метотрексатом с концентрацией 100 нМ и 1 мкМ, содержат по данным радиоиммунологического анализа 466 и 1352 U/мл эритропоэтина человека, соответственно (усредненное число, полученное в результате 3-разового испытания образцов). По указанной методике в 60 мм чашки Петри, содержащие 5 мл среды, высевают по 1×106 клеток. Спустя 24 часа среду удаляют, заменяя ее на 5 мл свежей среды, в которой отсутствует сыворотка (DMEM с высоким содержанием глюкозы, в которую дополнительно добавляют 0,1 нМ не-основных аминокислот и L-глутамин). Эритропоэтину дают возможность накапливаться в течение 48 часов в среде, не содержащей сыворотки. Затем среду собирают для испытания посредством радиоиммунологического анализа, а клетки трипсинизируют и затем подсчитывают. Средние значения образцов в результате радиоиммунологического анализа составляют порядка 467 и 1352 U/мл эритропоэтина для клеток, выращенных при обработке метотрексатом с концентрацией 100 нМ и 1 мкМ, соответственно, обеспечивают фактический выход эритропоэтина порядка 2335 и 6750 U на чашку. Среднее количество клеток на чашку составляет 1,94×106 и 3,12×106 клеток, соответственно. Таким образом, скорость эффективного продуцирования в указанных условиях культивирования имеет значения 1264 и 2167 U/106 клеток за 48 часов.
Описанные выше клеточные культуры представляют собой генетически разнородную популяцию. С целью выделении генетически неоднородных клонов, обладающих наивысшей способностью к продуцированию, используют стандартные методы отбора (см. раздел А, часть 2 "Points to Consider in the Characterization of Cell Lines Used to Produce Biologies", 1 июня 1984 г., Отдел по научной проверке биопрепаратов. Центр по лекарственным и биопрепаратам. Управление по пищевым и лекарственным продуктам США).
Описанную выше продуктивность линий клеток СНО, продуцирующих эритропоэтин, можно улучшить при использовании соответствующих методик культивирования клеток. Размножение клеток млекопитающих животных в культуральной среде обычно требует наличия сыворотки в ростовой среде. Способ получения эритропоэтина из клеток СНО в бессывороточной среде в значительной степени облегчает ректификацию эритропоэтина из культуральной среды. Описываемый ниже способ дает возможность экономного получения эритропоэтина в больших количествах в бессывороточной среде, достаточных для производства.
Клетки линии СНО pDSVL-gHuEPO, выращенные в стандартных условиях культивирования клеток, для посева используют флаконы с перевиваемой клеточной культурой. Указанные клетки размножаются в виде суспензионной клеточной линии во флаконах, содержащих перевиваемую клеточную культуру, в среде, которая состоит из 50:50 смеси DMEM с высоким содержанием глюкозы и F12 Ham, в которую дополнительно добавлено 5% фетальной бычьей сыворотки (FCS), L-глутамин, пенициллин и стрептомицин, 0,05 нМ не-основных аминокислот и метотрексат с соответствующей концентрацией. Клеточные культуры в виде суспензии дают возможность для быстрого размножения до больших объемов эритропоэтину, получающемуся из клеток СНО. Клетки СНО, выращиваемые в суспензии, используют для их посева во вращающиеся сосуды при первоначальной плотности засевания 1,5×107 жизнеспособных клеток на 850 см2 указанного сосуда, содержащего 200 мл среды. В течение 3-дневного периода указанным клеткам дают возможность для роста до образования монослоя относительно плотно прилегающей линии клеток. Применяемая в данной фазе роста клеток среда аналогична среде, используемой для культивирования клеток в суспензии. К концу 3-дневного периода роста клеток среду, содержащую сыворотку, отделяют и заменяют ее на 100 мл бессывороточной среды. Бессывороточная среда содержит 50:50 смеси DMEM с высоким содержанием глюкозы и F12 Ham, в которую дополнительно введено 0,05 мМ не-основных аминокислот и L-глутамин. Вращающиеся сосуды с приготовленной средой помещают на 2-3 часа в инкубатор и затем ее вновь удаляют, заменяя 100 мл свежей, не содержащей сыворотки среды. При 1-3-часовой инкубации в бессывороточной среде происходит снижение концентрации загрязнения белков сывороткой. Затем вращающиеся сосуды опять помещают на 7 дней в инкубатор, в течение которых происходит накопление эритропоэтина в бессывороточной культуральной среде. К концу 7-дневной фазы продуцирования эритропоэтина старую среду удаляют, заменяя ее свежей, не содержащей сыворотки средой, необходимой для проведения второго цикла продуцирования. В виде примера практической реализации указанной системы продуцирования представлен образец, находящийся в течение 7 дней в бессывороточной среде, который по данным радиоиммунологического анализа содержит 3892±409 U/мл эритропоэтина. Принимая во внимание данные расчета плотности клеток порядка 0,9-1,8×105 клеток на см2, в каждом вращающемся сосуде объемом 850 см2 содержится от 0,75 до 1,5×108 клеток, и исходя из этого, таким образом, скорость продуцирования эритропоэтина в течение 7 дней в 100 мл культуры достигает значений в пределах от 750 до 1470 U/106 клеток за 48 часов.
Надосадочные жидкости из клеточной линии СНО pDSVL-MkEPO, амплифицированные в метотрексате с концентрацией 10 нМ, подвергают радиоиммунологическому анализу для испытания активности эритропоэтина in vitro и in vivo. Образец, приготовленный в концентрированной среде при радиоиммунологическом анализе, содержит 41,2±1,4 U/мл; 41,2±0,064 U/мл Mk-эритропоэтина, как определено по данным анализа на биологическую активность in vitro, и 42,5±5 U/мл по данным анализа определения биологической активности in vivo. При определении последовательности аминокислотных остатков полипептидных продуктов обнаруживается наличие эритропоэтиновых продуктов, причем доминантный вид включает 3 остатка "лидирующей" последовательности, примыкающей к предполагаемому завершению аминогрупп аланином. В настоящее время не ясно, является ли это результатом неправильной обработки ядра полипептида в клетках СНО, или отражает различие в структуре конечного завершения аминогрупп эритропоэтина обезьяны относительно эритропоэтина человека.
Надосадочную жидкость, приготовленную из линии клеток СНО pDSVL-gHuEPO, подвергают трем испытаниям. По данным радиоиммунологического анализа 5,5-дневный образец содержит порядка 18,2 U/мл рекомбинантного эритропоэтина человека в указанной среде; при анализе in vitro 15,8±4,6 U/мл и при анализе in vivo 16,8±3,0 U/мл.
Надосадочную жидкость, приготовленную из линии клеток СНО pDSVL-gHuEPO с постепенным амплифицированием метотрексатом с концентрацией 100 нМ, подвергают трем испытаниям. По данным радиоиммунологического анализа, 3-дневный образец содержит 3089±129 U/мл рекомбинантного эритропоэтина человека; при испытании образца in vitro 2589±71,5 U/мл и при испытании образца in vivo 2040±160 U/мл. Последовательность аминокислот показывает завершение аминогрупп, соответствующее представленному в таблице VI.
Клеточную кондиционированную среду, приготовленную из клеток СНО, которая трансфектирована плазмидой pDSVL-MkE в метотрексате с концентрацией 10 нМ, объединяют и метотрексат подвергают диализу в течение нескольких дней, в результате чего образуется культура с активностью эритропоэтина порядка 221±5,1 U/мл (эритропоэтин-клеточная кондиционированная среда) (ЕРО-ССМ). Для определения действия ЕРО-ССМ на уровень гематокрита в линии Balb/C обычной мыши in vivo проводят следующий эксперимент. Клеточную кондиционированную среду, приготовленную из нетрансфектированных клеток СНО (ССМ) и ЕРО-ССМ, регулируют, используя забуференный фосфором раствор (PBS). Клеточную кондиционированную среду применяют на контрольной группе животных (3 мышы), а для экспериментальных групп (2 мышы на группу) используют 2 уровня дозировки ЕРО-ССМ: 4 единицы на инъекцию и 44 единицы на инъекцию. В течение 5-недельного курса 7 мышам вводят внутрибрюшинно инъекции по 3 раза в неделю. После 8-ой инъекции средние показатели гематокрита у контрольной группы, как установлено, составляют 50,4%, для группы при введении 4 единиц ЕРО-ССМ они составляют 55,1% и при введении 44 единиц ЕРО-ССМ - 67,9%.
Продукты экспрессии клеток млекопитающих животных можно без затруднения извлекать в довольно очищенном виде из культуральной среды с использованием ВЭЖХ (С4) с этанольным градиентом, предпочтительно при значении рН=7.
Была проведена предварительная попытка по определению свойств рекомбинантных гликопротеиновых продуктов, полученных из кондиционированной среды экспрессии клеток COS-1 и СНО гена эритропоэтина из мочи человека, при использовании как анализа Western blot (окрашивания), так и анализа SDS-PAGE. Указанные исследования показывают, что эритропоэтиновый материал, продуцируемый клетками СНО, имеет несколько больший молекулярный вес, чем продукты экспрессии клеток COS-1, который, в свою очередь, немного больше мочевого экстракта человека пулевого источника. Все продукты в некоторой степени гетерогенны. Обработка ферментом нейраминидазы для удаления сиаловой кислоты приводит к тому, что в результате рекомбинантные продукты клеток COS-1 и СНО имеют примерно одинаковый молекулярный вес, который, тем не менее, как один, так и другой, больше молекулярного веса получающегося мочевого экстракта человека, содержащего азиалогруппы. Обработка ферментом F-эндоглюкозидазы (ЕС 3.2.1) рекомбинантного продукта клеток СНО и продукта мочевого экстракта (для полного удаления углеводов из обоих продуктов) приводит в результате к получению почти гомогенных продуктов, которые, в основном, имеют одинаковый молекулярный вес.
Очищенный эритропоэтин, полученный из мочи человека, и рекомбинантный, продуцируемый из клеток СНО, эритропоэтин согласно настоящему изобретению подвергают анализу на определение углеводного состава по методике, описанной Ledeen, et al., Methods in Enzymology, 83 (Part D, 139-191 (1982), которая модифицирована при использовании метода гидролиза, описанного Nesser, et al., Anal. Biochem., 142, 58-67 (1984). Определенные экспериментальным путем показатели углеводного состава для мочевого изолята (выраженные в виде молярных отношений в указанных продуктах) имеют следующие значения: гексоза, 1,73; N-ацетилглюкозамин, 1; N-ацетилнейраминовая кислота, 0,93; фукоза, 0 и N-ацетилгалактозамин, 0. Соответствующие значения для рекомбинантных продуктов (производных от линии клеток СНО pDSVL-gHuEPO 3-дневной культуральной среды, усиленной метотрексатом при концентрации 100 нМ) имеет следующие показатели: гексоза, 15,09; N-ацетилглюкозамин, 1; N-ацетилнейраминовая кислота, 0,998; фукоза, 0 и N-ацетилгалактозамин, 0.
Полученные результаты совпадают с результатами описанных выше анализов Western blot и SDS-PAGE.
Гликопротеиновые продукты, получаемые согласно настоящему изобретению, таким образом, представляют собой достижимые продукты, первичная структурная конформация которых в достаточной степени воспроизводит конформацию эритропоэтинов естественного происхождения для придания им одного или нескольких биологических свойств, и количественный состав углеводов которых отличается от состава эритропоэтинов природного происхождения.
Пример 11
Предлагаемый пример относится ко всему циклу получения эритропоэтинов посредством сборки нуклеотидных оснований двух структурных генов, кодирующих последовательность эритропоэтина человека, представленных в таблице VI и содержащих, соответственно, "предпочтительные" кодоны для экспрессии признаков в Е.coli и дрожжевых клетках (S.cerevisiae).
В указанном примере дополнительно описана конструкция генов, кодирующих аналоги эритропоэтина. Короче говоря, используемые протоколы в общем представлены в указанном выше описании Alton, et al. (WO 83/04053). Указанные гены сконструированы для первоначальной сборки составляющих олигонуклеотидов в сложном дуплексе, которые, в свою очередь, собраны в три дискретные части. Указанные части построены для амплификации и при удалении из системы амплификации могут быть собраны последовательно или через лигатуру фрагмента со сложной структурой в подходящий вектор экспрессии.
В таблицах VIII-XIV показана схема и сборка полученного гена, кодирующего продукт трансляции эритропоэтина человека, в котором отсутствует какая-либо лидирующая или предшествующая последовательность, но содержится исходный остаток метионина в положении -1. Более того, ген, введенный в основную часть клетки Е.coli, отдает предпочтение кодонам и, таким образом, указанную конструкцию называют "ЕСЕРО" ген.





Более детально, Таблица VIII иллюстрирует применение олигонуклеотидов для генерирования Части 1 гена ЕСЕРО, кодирующего амино концевые остатки человеческого полипептида. Олигонуклеотиды собирают в дуплексы (1 и 2, 3 и 4, и т.д.), которые затем сшивают для получения Части 1 ЕСЕРО, как в Таблице IX. Следует отметить, что собранная часть включает соответственные липкие концы конечных EcoRI и BamHI, причем "по потоку" липкого конца EcoRI направлен участок распознавания ограничительного фермента XbaI, а "против потока" липкого конца BamHI направлен участок распознавания KpnI. Часть 1 можно без труда амплифицировать, используя вектор фага М13, применяемый для проверки последовательности данной части. Некоторые трудности неожиданно возникают при выделении части в качестве фрагмента XbaI/KpnI из ДНК RF, генерированной в Е.coli, вероятно, из-за метилирования оснований участка распознавания KpnI в пределах хозяина. Поэтому выделяют ДНК фага и воспроизводят в двунитевой форме in vitro посредством растяжения праймером, и затем выделяют требуемый двунитевый фрагмент.
Части 2 и 3 гена ЕСЕРО (Таблицы XI и XIII) строят подобным образом из олигонуклеотидов Таблиц Х и XII, соответственно. Каждую часть амплифицируют на векторе М13, применяемом для проверки последовательности, и выделяют из ДНК фага. Как явствует из Таблицы XI, Часть 2 ЕСЕРО строится с липкими концами EcoRI и BamHI и может быть выделена в качестве фрагмента KpnI/BglII. Точно так же Часть 3 ЕСЕРО получена с липкими концами BamHI и SalI и может быть выделена из ДНК RF как фрагмент BglII/SalI. Полученные таким образом три части можно без труда собрать в непрерывную последовательность ДНК (Таблица XIV), кодирующую полипептид полного ЕРО человека с метиониновым кодоном аминоконца (ATG) для трансляционного инициирования Е.coli. Следует отметить, что "против потока" первоначального ATG направлен ряд пар оснований, достаточно дуплицирующий последовательность участка связывания рибосомы сильно выраженного OMP-f гена Е.coli.
Для переноса ЕСЕРО можно использовать любой подходящий вектор экспрессии. Специфическим вектором, выбранным для экспрессии гена ЕСЕРО в качестве "температурно-чувствительной" плазмиды pCFM536, является производное плазмиды pCFM414 (А.Т.С.С.40076), как описано в одновременно рассматриваемой заявке на патент США №636727, поданной 6 августа 1984 г, Charles F. Morris. Более конкретно, pCFM536 расщепляют XbaI и HindIII; большой фрагмент выделяют и используют в двухстороннем сшивании с геном ЕСЕРО. Части 1 (XbaI/KpnI), 2 (KpnI/BgIII) и 3 (BglII/SalI) предварительно собирают в правильном порядке в М13 и выделяют оттуда ген ЕРО как одиночный фрагмент XbaI/HindIII. Этот фрагмент включает часть полилинкера из М13 mр9-фага, стягивающего в нем участки SalI и HindIII. Контроль за экспрессией в получаемой плазмиде экспрессии, р536, выполняют при помощи промотора лямбда РL, который сам может контролироваться репрессорным геном C1857 (таким как тот, который получен в K12ΔHtrp нити Е.coli).
Построенный ген ЕСЕРО может быть модифицирован различным образом для кодирования аналогов эритропоэтина, таких как [Asn2, des-Pro2 по Ile6] hEPO и [His7] hEPO, как описано ниже.
A. [Asn2, des-Pro2 по Ile6] hEPO
Плазмиду 536, несущую построенный ген ЕСЕРО Таблицы XIV как вставка XbaI в HindIII, расщепляют HindIII и XhoI. Эндонуклеазой последнего разрезают ген ЕСЕРО в уникальном участке распознавания, имеющем 6 пар оснований, стягивая последнее основание кодона, кодирующего Asp8 со вторым основанием кодона Arg10. Строят последовательность линкера XbaI/XhoI, имеющую следующую последовательность;
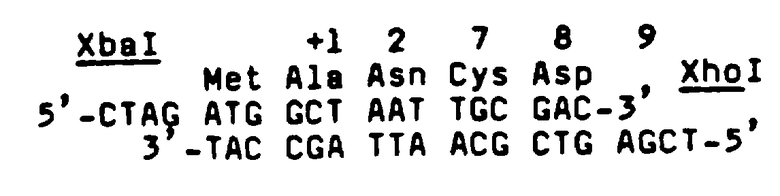
Линкер XbaI/XhoI и фрагмент последовательности гена ЕСЕРО XhoI/HindIII вставляют в большой фрагмент, получающийся в результате расщепления XbaI и HindIII плазмиды pCFM526 - производное плазмиды pCFM414 (А.Т.С.С. 40076) - как описано в одновременно рассматриваемой заявке на патент США №636727, поданной 6 августа 1984 года Charles F. Morris, для генерирования плазмида-несущей ДНК-последовательности, кодирующей экспрессию Е.coli формы Met-1 желаемого аналога.
В. [His7] hEPO
Плазмиду 536 расщепляют HindIII и XhoI, как в части А выше. Линкер XbaI/XhoI построен со следующей последовательностью:
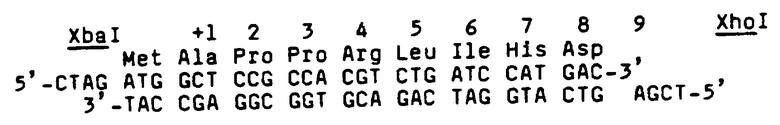
Линкер и фрагмент последовательности ЕСЕРО XhoI/HindIII затем вставляют в pCFM526 для генерирования плазмида-несущей ДНК-последовательности, кодирующей экспрессию Е.coli формы Met-1 желаемого аналога.
Построение изготовленного гена ("SCEPO"), включающего в себя дрожжевые предпочтительные кодоны, происходит, как описано в следующих Таблицах XV-XXI. Так же как и в случае с геном ЕСЕРО, полное построение содержит образование трех наборов олигонуклеотидов (Таблицы XV, XVII и XIX), которые сформированы в дуплексы и собраны в части (Таблицы XVI, XVIII и XX). Следует отметить, что синтез облегчается частично использованием некоторых субоптимальных кодонов в обоих SCEPO и ЕСЕРО построениях, т.е. олигонуклеотиды 7-12 части 1 обоих генов были идентичны, как были идентичны олигонуклеотиды 1-6 части 2 в каждом гене.

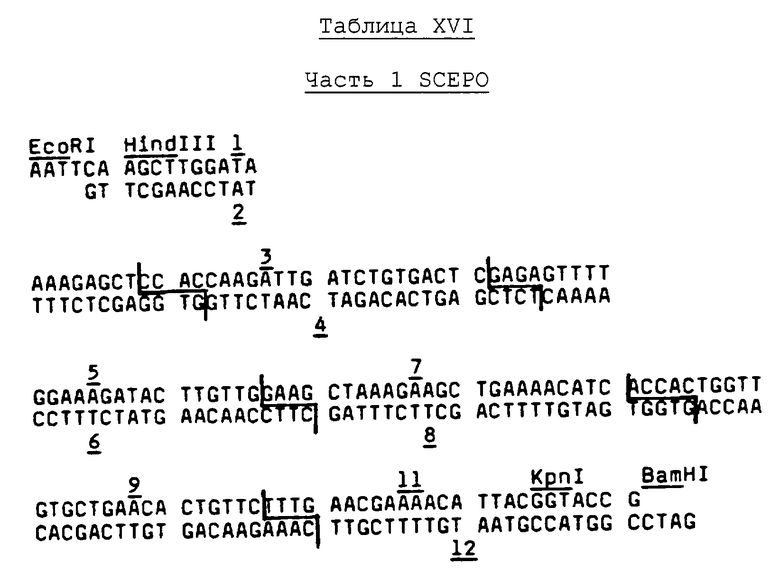

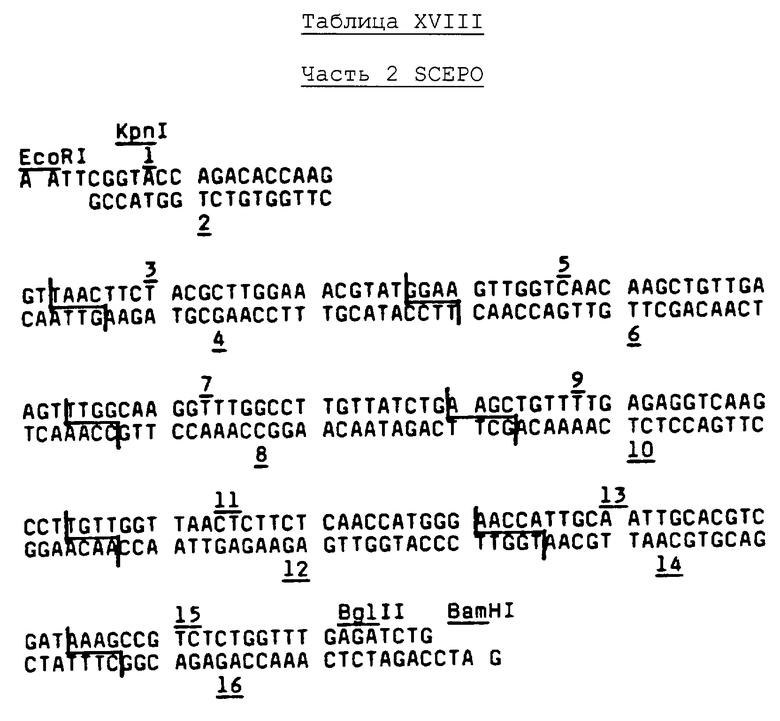

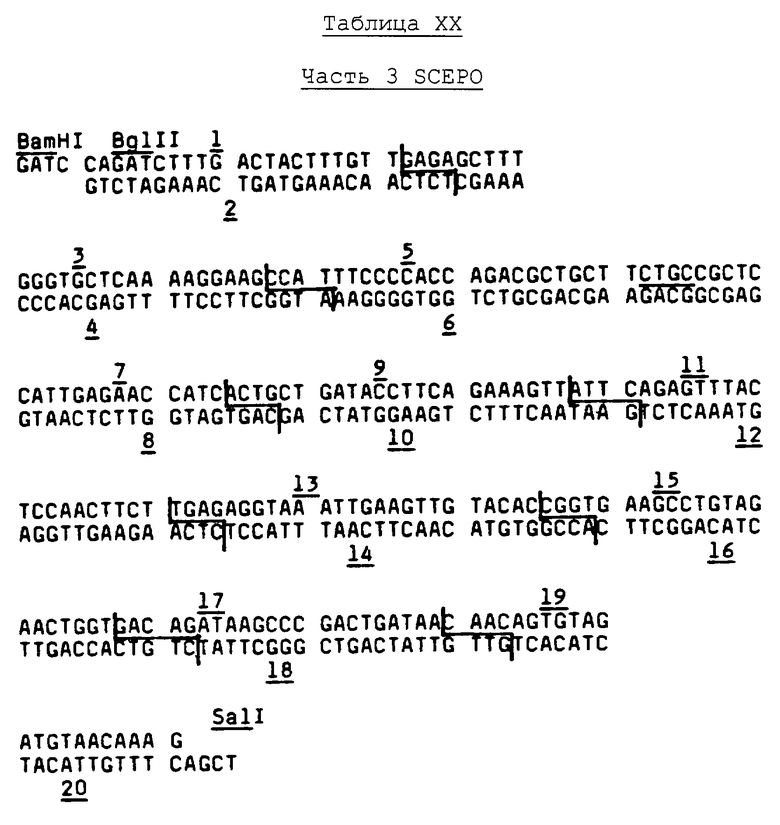

Собранные части SCEPO выстраивают в ряд в М13 и Части 1, 2 и 3 выделяют из фага в качестве фрагментов HindIII/KpnI, KpnI/BglII и BglII/SalI.
Предпочитаемой сейчас системой экспрессии для продуктов гена SCEPO является система секреции на основе секреции α-фактора S. cerevisiae, как описано в находящейся в процессе одновременного рассмотрения заявке на патент США №487753, поданной 22 апреля 1983 года Grant A. Bitter, опубликованной 31 октября 1984 года как заявка на европейский патент 0123294. Короче говоря, система включает конструкции, где ДНК, кодирующая ведущую последовательность гена α-фактора дрожжей, располагается сразу за 5' к кодирующей области выражаемого экзогенного гена. В результате чего транслируемый генный продукт включает ведущую или сигнальную последовательность, которая "обрабатывается" эндогенным ферментом дрожжей в направлении секреции оставшейся части продукта. Так как конструкция дает возможность использования кодона инициирования трансляции α-фактора (ATG), не было необходимости в создании такого кодона в положении -1 гена SCEPO. Как можно заключить из Таблицы XXI, аланин (+1), кодирующий последовательность, стоит за последовательностью линкера, что позволяет осуществить непосредственную вставку в плазмиду, включающую ДНК для первых 80 остатков проводника α-фактора, следующего за промотором α-фактора. Специфическое предпочитаемое построение для экспрессии гена SCEPO содержит четырехстороннее сшивание, включая указанные выше фрагменты части SCEPO и большой фрагмент расщепления HindIII/SalI плазмиды рαС3. Из получаемой плазмидной рαС3/SCEPO выделяют промоторную и лидирующую последовательности α-фактора, а также ген SCEPO путем расщепления BamHI и сшивания расщепленной BamHI в плазмиду pYE с образованием плазмидной pYE/SCEPO экспрессии.
Пример 12
Настоящий пример относится к экспрессии рекомбинантных продуктов построенных ЕСЕРО и SCEPO-генов в пределах системы экспрессии Примера 11.
При использовании системы экспрессии, рассчитанной для применения в клетках-хозяевах Е.coli, плазмиду р536 Примера 11 трансформируют в клетках Е.coli АМ7, ранее трансформированные с пригодной плазмидой, pMW1, принявшей к себе ген C1857. Культуры клеток в В бульоне (ампициллин 50 мкг/мл и канамицин 5 мкг/мл, предпочтительно с 10 мМ MgSO4) выдерживают при температуре 28°С и во время роста клеток в культуре до О.D.600=0,1, экспрессию ЕРО индуцируют, повышая температуру до 42°С. Клетки растут до приблизительно 40 O.D., обеспечивая получение ЕРО (оценено гелем) около 5 мг/OD л.
Клетки собирают, подвергают лизису, разбивают французским прессом (10000 psi) и обрабатывают лизоцимом и детергентом NP-40. Осадок, полученный в результате центрифугирования 24000хg, растворяют в HCl гуанидине и подвергают дальнейшей одноразовой очистке при помощи С4 (Vydac) ВЭЖХ с обращенной фазой (EtOH, 0-80%, 50 мМ NH4Ac, pH 4,5). Определение последовательности оснований белка выявляет, что продукт чист более чем на 95%, а полученные продукты выявляют два разных аминоконца, A-P-P-R... и P-P-R... при относительном количественном отношении, равном 3:1. Это последнее наблюдение hEPO и [des Ala1] hEPO-продуктов означает, что "обработка" аминоконцов в пределах клеток-хозяев служит для удаления конечного метионина и, в некоторых случаях, начального аланина. Активность, определенная радиоиммуноисследованием, для изолятов находится на уровне 150000-160000 U/мг; активность, выявленная in vitro, составляет порядка 30000-62000 U/мг; и активность, выявленная in vivo, варьируется в пределах от 120 до 720 U/мг. (Сравнено со стандартом человеческого мочевого изолята =70000 U/мг, в каждом испытании). Кривая доза-эффект для рекомбинантного продукта в испытании in vivo значительно отличается от кривой стандарта мочевого ЕРО человека.
Плазмиды ЕРО-аналога, образованные в частях А и В Примера 11, каждая трансформируется в pMW1-трансформированные клетки Е.coli AM7, и клетки культивируют, как описано выше. Очищенные изоляты испытывает как RIA, так и анализами in vitro. Значения, полученные при испытаниях RIA и in vitro, для продуктов экспрессии [Asn2, des-Pro2 по Ile6] hEPO, составляют приблизительно 11000 U/мг и 6000 U/мг белка, соответственно, тогда как значения анализа для [His7] hEPO составляют около 41000 U/мг и 14000 U/мг белка, соответственно. Это означает, что продукты аналогов являются на 1/4-1/10 "активными", так же как продукт экспрессии "родителя" в испытаниях.
В системе экспрессии, предназначенной для использования в клетках-хозяевах S. cerevisiae, плазмиду pYE/SCEPO трансформируют в два различных штамма, YSDP4 (генотип α рер 4-3 trp1) и RK81 (генотип αα рер 4-3 trp1). Трансформированных хозяев YSDP4 выращивают в среде 3D (Методы генетики дрожжей, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., p.62 (1983)), пополняемой кислотами casamino 0,5%, pH 6,5 при температуре 30°С. Среды, собранные, когда клетки вырастают до 36 O.D., содержат продукты ЕРО на уровне около 244 U/мл (97 мкг/OD л, что выявлено посредством RIA). Трансформированные клетки RK81, выращенные до 6,5 O.D. или 60 O.D., обеспечивают создание сред с концентрациями ЕРО около 80-90 U/мл (34 мкг/OD л, что выявлено RIA). Предварительные анализы выявляют значительную гетерогенность в продуктах, полученных посредством применения системы экспрессии, что, по-видимому, обусловлено изменениями в гликозилировании выраженных белков, а также относительно высоким содержанием маннозы присоединенного углевода.
Плазмиды РαС3 и pYE в клетках Е.coli HB101 были сданы для регистрации в соответствии с Правилами производства дел Патентного ведомства США от 27 сентября 1984 года американской коллекцией типовых культур, 12301 Parklawn Drive, Rockville, Maryland, под депозитными номерами А.Т.С.С. 39881 и А.Т.С.С. 39882 соответственно. Плазмиды pCFM526 в клетках АМ7, pCFM536 в клетках JM103 и pMW1 в клетках JM103 были зарегистрированы подобным образом 21 ноября 1984 года как А.Т.С.С. 33932, 33934 и 33933, соответственно. Штаммы YSPD4 и RK81 Saccharomyces cerevisiae были зарегистрированы 21 ноября 1984 года как А.Т.С.С. 20734 и 20733, соответственно.
Должно стать очевидным из рассмотрения представленных выше иллюстративных примеров, что настоящее изобретение в его многих аспектах предлагает исключительно ценные продукты и способы.
Предлагаемые полипептиды являются заметно полезными материалами, будь они микробиально выраженными продуктами или синтетическими продуктами, первичная, вторичная или третичная конформация которых стала впервые известна благодаря настоящему изобретению.
Как отмечалось выше, рекомбинантно продуцируемые и синтетические продукты изобретения обладают в различной степени биологической активностью in vitro изолятов ЕРО из натуральных источников и поэтому проектируются с тем, чтобы иметь пригодность в качестве заместителей изолятов ЕРО в культуральных средах, применяемых для роста клеток эритропоэтина в культуре. Также, поскольку полипептидные продукты настоящего изобретения обладают активностью in vivo натуральных изолятов ЕРО, они являются заметно пригодными для использования в методиках эритропоэтиновой терапии, применяемых к млекопитающим, включая человека, для разработки любого или всех эффектов, присущих ЕРО in vivo, например, стимулирование ответа ретикулоцитов, развитие феррокинетических эффектов (таких как эффекты кругооборота железа в плазме и эффекты времени перехода в мозговое вещество), массообмены эритроцита, стимулирование синтеза гемоглобина С (см. Eschbach, et al., выше) и, как отмечается в Примере 10, увеличение уровней гематокрита у млекопитающих. В группу людей, обрабатываемых предлагаемыми продуктами, включали пациентов, как правило, нуждающихся в переливании крови, включая жертв травм, хирургических больных, больных-почечников, включая диализных больных, и пациентов с разновидностью состава крови, вызывающего нарушения, такие как гемофилия, серповидно-клеточная болезнь, физиологический анемиаз и тому подобное. Минимизация необходимости в терапии с переливанием крови благодаря использованию терапии эритропоэтином, можно ожидать, приведет к пониженному переносу возбудителей инфекционного заболевания. Предлагаемые продукты благодаря их получению при помощи рекомбинантных способов, надо полагать, не содержат пирогены, натуральные угнетающие вещества и тому подобное, и поэтому несомненно должны обеспечивать повышенную общую отдачу в терапевтических методах по отношению к природным продуктам. Эритропоэтиновая терапия с использованием предлагаемых продуктов также должна стать полезной в повышении кислородопропускающей способности индивидуумов, сталкивающихся с гипоксическими условиями внешней среды и, возможно, в обеспечении благоприятных сердечно-сосудистых эффектов.
Предпочтительным способом введения предлагаемых полипептидных продуктов являются парентеральные (например, внутривенные, внутримышечные, подкожные или внутрибрюшинные) пути, и введенные композиции должны, как обычно, содержать терапевтически эффективные количества продукта в сочетании с приемлемыми разбавителями, носителями и/или стимуляторами. Предварительные фармакокинетические исследования отмечают более длительный период полураспада in vivo для продуктов ЕРО обезьяны при введении их внутримышечно, скорее, нежели внутривенно. Как предполагается, эффективные дозировки могут изменяться в значительной степени в зависимости от условий, однако, терапевтические дозы, как сейчас ожидают, должны варьироваться в пределах от 0,1 (˜7U) до 100 (˜7000U) мкг/кг веса тела активного материала. Стандартные разбавители, такие как сывороточный альбумин человека, предполагаются для использования в фармацевтических композициях настоящего изобретения наряду со стандартными носителями, такими как физиологический раствор.
Стимулирующие материалы, пригодные для использования в предлагаемых композициях, включают соединения, указанные самостоятельно для эритропоэтических стимулирующих эффектов, как например, тестостероны, стимуляторы предшествующих клеток, фактор роста инсулиноподобных, простагландины, серотонин, циклический АМФ, пролактин и триидотиронин, а также средства, обычно применяемые при лечении апластической анемии, такие как метенолен, станозол и нандролон [(см., например, Resegotti, et al., Panminerva Medica, 23, 243-248 (1981); McGonigle, et al., Kidney Int., 25(2), 437-444 (1984); Pavlovic-Kantera, et al., Expt. Hematol., 8 (Supp. 8), 283-291 (1980); и Kurtz, FEES Letters, 14a (1), 105-108 (1982)]. Также предполагается в качестве стимуляторов использовать вещества, увеличивающие действия или синергизирующие эритропоэтин или асиало-ЕРО, такие как адренергические агонисты, тироидные гормоны, андрогены и ВРА [см. Dunn, "Current Concepts in Erythropoiesis", John Wiley and Sons (Chichester, England, 1983); Weiland, et al., Blut, 44 (3), 173-175 (1982); Kalmanti, Kidney Int., 22, 383-391 (1982); Shahidi, New. Eng. J. Med., 289, 72-80 (1973), Fisher, et al., Steroids, 30 (6), 833-845 (1977); Urabe, et al., J. Exp. Med., 149, 1314-1325 (1979); и Billat, et al., Expt. Hematol., 10 (1), 133-140 (1982)], a также классы соединений, обозначающие "гепатические эритропоэтические факторы" [см. Naughton, et al., Acta. Haemat., 69, 171-179 (1983)] и "эритротропины" [как описано Congote, et al., in Abstract 364, Proceedings 7th Intrenational Congress of Endocrinology (Quebec City, Quebec, July 1-7, 1984]; Congote, Biochem. Biophys. Res. Comm., 115(2), 447-483 (1983) и Congote, Anal. Biochem., 140, 428-433 (1984)] и "эритрогенины" [как описано в работе Rothman, et al., J. Surg. Oncol., 20, 105-108 (1982)]. Предварительные скрининги, предназначенные для измерения эритропоэтических ответов экс-гипоксических полицитемических мышей, предварительно обработанных либо 5-α-дигидротестостероном, либо нандролоном, а затем предлагаемым эритропоэтином, проявляли сомнительные результаты.
Диагностические использования предлагаемых полипептидов также обширны и включают применение меченых и немеченых форм в разнообразии методик иммуноисследования, включая RIA, ELISA и тому подобное, а также в разнообразии анализов, проводимых для выявления активности in vitro и in vivo. См., например, работы Dunn, et al., Expt. Hematol., 11 (7), 590-600 (1983); Gibson, et al., Pathology, 16, 155-156 (1984); Krystal, Expt. Hematol., 11 (7), 649-660 (1983); Saito, et al., Jap. J. Med., 23 (1), 16-21 (1984); Nathan, et al., New. Eng. J. Med., 308 (9), 520-522 (1983); и различные ссылочные материалы, имеющие отношение к упоминаемым в этих работах исследованиям. Предлагаемые полипептиды, включающие синтетические пептиды, которые содержат последовательности остатков впервые выявленного ЕРО, также располагают чрезвычайно полезными чистыми материалами для производства поликлональных антител и "банков" моноклональных антител, специфических для отличающихся непрерывных и прерывистых эпитопов ЕРО. В качестве одного примера, предварительный анализ аминокислотных последовательностей Таблицы VI в смысле водолечимости в соответствии с Норр, et al., P.N.A.S. (U.S.A.), 78, pp.3824-3828 (1981) и вторичных структур в соответствии с Chou, et al., Ann. Rev. Biochem., 47, p.251 (1978) выявил, что синтетические пептиды, дуплицирующие непрерывные последовательности остатков, стягивающих позиции 41-57, включительно, 116-118, включительно, и 144-166, включительно, вероятно, должны продуцировать чрезвычайно антигенную реакцию и вырабатывать полезные моноклональные и поликлональные антитела, иммунореактивные как с синтетическим пептидом, так и целым белком. Такие материалы, как ожидается, должны быть полезны при обнаружении и сродственной очистке ЕРО и связанных с ним продуктов.
Иллюстративно были получены три синтетических пептида:
Предварительные исследования иммунизации с применением вышеуказанных полипептидов выявили относительно слабую положительную реакцию на hEPO 41-57, не поддающуюся оценке реакцию на hEPO 116-128 и сильную положительную реакцию на hEPO 144-166, как определено способностью сывороточных антител кролика иммуноосаждать изоляты мочевого ЕРО человека, меченного 125I. Предварительные исследования активности in vivo, проведенные на трех пептидах, не выявили значительной активности как каждого в отдельности, так и в комбинации.
Несмотря на то, что выведенные последовательности аминокислотных остатков ЕРО млекопитающих, представленные в иллюстративных примерах, по существу определяет первичную структурную конформацию созревшего ЕРО, следует понять, что специфическая последовательность 165 аминокислотных остатков ЕРО обезьяны в Таблице V и 166 остатков ЕРО человека в Таблице VI не ограничивает объема полезных полипептидов, предлагаемых изобретением. Охватываются настоящим изобретением те различные свободно встречающиеся аллельные формы ЕРО, которые в прошлом вошли в исследования биологически активных полипептидов млекопитающих, как например, γ интерферон человека (Срав., например, иммунный интерферон человека, имеющий остаток аргинина в положении №140 в ЕРО, о котором сообщалось в опубликованной заявке на патент 0077670, и вид, имеющий глутамин в положении №140, как сообщалось в работе Gray, et al., Nature, 295, pp.503-508 (1982). Оба вида отличаются тем, что составляют последовательности "созревшего" γ интерферона человека). Аллельные формы полипептидов созревшего ЕРО могут отличаться друг от друга и от последовательностей, представленных в Таблицах V и VI, в отношении длины и последовательности и/или в отношении делеций, замещений, вставок или добавлений аминокислот в последовательность наряду с логически вытекающими вариациями в способности для гликозилирования. Как отмечалось выше, одна предположительная аллельная форма ЕРО человека, как кажется, должна включать остаток метионина в положении 126. Предположительно, встречающиеся в природе аллельные формы последовательностей кДНК и ЕРО-кодирующей геномной ДНК, вероятно, также должны встретиться, причем кодирующими вышеуказанные типы аллельных полипептидов или просто использующими отличающиеся кодоны для обозначения таких же полипептидов, которые указаны.
В дополнение к встречающимся в природе аллельным формам созревшего ЕРО настоящее изобретение также включает в себя другие "продукты ЕРО", такие как полипептидные аналоги ЕРО и фрагменты "созревшего" ЕРО. Следуя методикам указанной выше опубликованной заявки на патент Alton, et al. (WO/83/04053), можно без труда рассчитать и изготовить гены, кодирующие микробиальную экспрессию полипептидов, имеющих первичную конформацию, которая отличается от конформации, указанной здесь для созревшего ЕРО, с точки зрения идентичности или местоположения одного или более остатков (например, замещения, конечные или промежуточные добавления и делеции). Альтернативно, модификации кДНК и генов геномного ЕРО могут быть легко осуществлены посредством хорошо известных методик сайт-направленного мутагенеза и использованы для генерирования аналогов и производных ЕРО. Такие продукты ЕРО должны обладать, по крайней мере, одним из биологических свойств ЕРО, но могут отличаться в других свойствах. Как примеры, проектируемые продукты ЕРО настоящего изобретения включают те, которые сокращены путем, например, делеции [Asn2, des-Pro2 по Ile6] hEPO, [des-Thr163 no Arg166] hEPO и "Δ27-55hЕРО", причем последняя имеет остатки, кодируемые удаленным целым экзоном; или которые более устойчивы к гидролизу (и, следовательно, могут иметь более выраженные или дольше продолжающиеся влияния, чем встречающийся в природе ЕРО); или которые были изменены для удаления одного или более потенциальных участков для гликозилирования (которое может привести к высокой активности для продуктов, продуцируемых дрожжами); или которые имеют один или более остатков цистеина, удаленных или замененных, например, остатками гистидина или серина (как например, аналог [His7] hEPO), и потенциально более легко выделяются в активной форме из микробиальных систем; или которые имеют один или более остатков тирозина, замененных фенилаланином (как например, аналоги [Phe15] hEPO, [Phe49] hEPO и [Phe145] hEPO) и могут связываться более или менее легко с рецепторами ЕРО на клетках-мишенях. Также охвачены полипептидные фрагменты, дуплицирующие только часть непрерывной аминокислотной последовательности или вторичных конформаций в пределах созревшего ЕРО, причем фрагменты могут обладать одной активностью ЕРО (например, рецепторным связыванием) и не другими (например, эритропоэтическая активность). Особенно важны в этом отношении те потенциальные фрагменты ЕРО, которые объяснены при рассмотрении последовательности геномной ДНК человека, представленной в Таблице VI, т.е. "фрагменты" полной непрерывной ЕРО-последовательности, которая очерчена интронными последовательностями и которая может составить резко выраженные "домены" биологической активности. Примечательно, что отсутствие активности in vivo для любого одного или более "продуктов ЕРО" настоящего изобретения в целом не препятствует терапевтической полезности (см. Weiland, et al., выше) или полезности в других контекстах, таких как ЕРО-испытания или ЕРО-антагонизм. Антагонисты эритропоэтина могут быть очень полезны при лечении полицитемиаза или в случаях перепроизводства ЕРО [см., например, Adamson, Hosp.Practice, 18 (12), 49-57 (1983) и Hellmann, et al., Clin. Lab. Haemat., 5, 335-342 (1983)].
Другой особенностью настоящего изобретения является то, что клонированные ДНК-последовательности, описанные здесь, которые кодируют полипептиды ЕРО человека и обезьяны, весьма ценны для информации, которую они предлагают в отношении аминокислотной последовательности эритропоэтина млекопитающих, который до сих пор был недоступен, несмотря на десятилетиями проводившуюся аналитическую обработку изолятов встречающихся в природе продуктов. ДНК-последовательности также весьма ценны как продукты, пригодные в осуществлении крупномасштабного микробиального синтеза эритропоэтина при помощи разнообразия рекомбинантных методик. Кроме того, предлагаемые ДНК-последовательности полезны в вырабатывании новых и пригодных векторов вирусной и кольцевой плазмидной ДНК, новых и полезных трансформированных и трансфектированных микробиальных прокариотических и эукариотических клеток-хозяев (включая бактериальные и дрожжевые клетки и клетки млекопитающих, выращенные в культуре), а также новые и пригодные способы культивируемого роста таких микробиальных клеток-хозяев, способных обеспечивать экспрессию ЕРО и продуктов ЕРО. ДНК-последовательности предлагаемого изобретения также являются весьма пригодными материалами для использования в качестве меченых зондов в выделении ЕРО и связанного с ним белка, кодирующего последовательности кДНК и геномной ДНК млекопитающих, других, нежели человек и обезьяна, иллюстрируемые здесь специфически. Степень возможного использования ДНК-последовательностей в различных альтернативных способах белкового синтеза (например, в клетках насекомых) или в генетической терапии людей и других млекопитающих, еще невозможно определить. Предлагаемые ДНК-последовательности, как ожидается, должны стать полезными в разработке трансгенных видов млекопитающих, которые могут служить в качестве эукариотных "хозяев" для получения эритропоэтина и продуктов эритропоэтина в большом количестве. См., главным образом, работу Palmiter, et al., Science, 222 (4625), 809-814 (1983).
В свете рассмотренного здесь, конкретное описание иллюстративных примеров, несомненно, не ограничивает объема настоящего изобретения, и специалистам в данной области будут очевидны многочисленные модификации и варианты его осуществления. В качестве одного примера, хотя ДНК-последовательности, предлагаемые иллюстративными примерами, включают последовательности кДНК и геномной ДНК, данная заявка обеспечивает информацию об аминокислотной последовательности, существенной для конструирования ДНК-последовательности, изобретение также предполагает такие изготовленные ДНК-последовательности, которые можно сконструировать на основании знания об аминокислотных последовательностях ЕРО. Они могут кодировать ЕРО (как в Примере 12), а также фрагменты ЕРО и полипептидные аналоги ЕРО (т.е. "продукты ЕРО"), которые могут обладать одним или более из биологических свойств встречающегося в природе ЕРО, однако, не обладать другими (или обладать другими в различной степени).
ДНК-последовательности, предлагаемые настоящим изобретением, таким образом, должны охватывать все ДНК-последовательности, пригодные для использования в обеспечении экспрессии в прокариотической или эукариотической клетке хозяина полипептидного продукта, имеющего, по крайней мере, часть первичной структурной конформации и одно или более из биологических свойств эритропоэтина, и отбираются из: (а) ДНК-последовательностей, представленных в Таблицах V и VI; (b) ДНК-последовательностей, которые гибридизируют к ДНК-последовательностям, определенным в (а), или их фрагментам; (с) ДНК-последовательностей, которые из-за дегенерации генетического кода должны гибридизировать к ДНК-последовательностям, определенным в (а) и (b). В этой связи заслуживает внимания, например, то, что существующие последовательности генов аллельного ЕРО человека и обезьяны и последовательности генов других видов млекопитающих, как ожидается, должны гибридизировать к последовательностям в Таблицах V и VI или их фрагментам. Кроме того, из-за дегенерации генетического кода гены SCEPO и ЕСЕРО и полученные или мутагенизированные последовательности кДНК или геномной ДНК, кодирующие различные фрагменты ЕРО и его аналоги, должны также гибридизировать к указанным выше ДНК-последовательностям. Такие гибридизации можно без труда осуществить в условиях гибридизации, описанных здесь в отношении первоначального выделения ДНК, кодирующей ЕРО человека и обезьяны, или более строгих условиях, если желательно снизить гибридизацию фона.
Подобным образом, хотя вышеуказанные примеры иллюстрируют изобретение в отношении микробиальной экспрессии продуктов ЕРО в контексте экспрессии ДНК клеток млекопитающих, вставленной в гибридный вектор бактериальных плазмидных и вирусных геномных видов, в объем изобретения входит большое разнообразие систем экспрессии. Особенно охвачены системы экспрессии, включающие векторы гомогенных видов, относящихся к разнообразию бактериальных, дрожжевых и клеток млекопитающих в культуре, а также к системам экспрессии, не включающим векторы (как например, трансфекция клеток фосфорнокислым кальцием). В этой связи станет понятно, что экспрессия, например, ДНК обезьяны в клетках-хозяев обезьяны в культуре и клетках-хозяев человека в культуре действительно составляет пример экспрессии "экзогенной" ДНК, так как ДНК ЕРО, чью экспрессию высокого уровня пытаются найти, не будет иметь своих источников в геноме хозяина. Системы экспрессии настоящего изобретения, кроме того, предполагают эти действия, вытекающие в цитоплазматическое образование продуктов ЕРО и накапливание гликозилированных и негликозилированных продуктов ЕРО в цитоплазме клетки-хозяина или мембранах (например, накапливание в бактериальных пространствах периплазмы), или в надосадочных жидкостях культуральных сред, как пояснено выше, или в довольно редких системах, как например, системах экспрессии Р. aeruginosa (описанных в работе Gray, et al., Biotechnology, 2, pp.161-165 (1984).
Усовершенствованные методологии гибридизации настоящего изобретения, хотя и были иллюстративно применены выше к скринингу ДНК/ДНК-гибридизации, в равной степени применимы к скринингу РНК/РНК и РНК/ДНК. Методики смешанных зондов, как проиллюстрировано здесь, главным образом составляют ряд усовершенствований в процессах гибридизации, позволяя осуществить более быстрое и надежное выделение полинуклеотидов. Эти многие отдельные усовершенствования обработки включают: улучшенный перенос колоний и методики сохранения; использование фильтров на основе нейлона, таких как GeneScreen и GeneScreen Plus, разрешающих повторное зондирование этими же фильтрами и повторное использование фильтра, применение новых методов лечения протеазой [срав., например, с Taub, et al., Anal. Biochem., 126, pp.222-230 (1982)]; использование очень низких отдельных концентраций (порядка 0,025 пикомоля) большого числа смешанных зондов (например, более чем 32); и проведение циклов гибридизации и пост-гибридизации в строгих температурных интервалах, близко приближающихся (т.е. в пределах 4°С и, предпочтительно, в пределах 2°С) к наименьшей рассчитанной температуре диссоциации любого из применяемых смешанных зондов. Эти усовершенствования объединяются с получением результатов, которых нельзя было ожидать. Это в полной мере поясняется тем, что методики смешанных зондов, включающие 4-разовое число зондов, которое, как прежде сообщалось, всегда с успехом использовалось даже в ситах кДНК на видах информационной ДНК относительно малой численности, были успешно применены к выделению гена уникальной последовательности в скрининге геномной библиотеки из 1500000 стерильных пятен. Этот легко было осуществлено, по существу, одновременно с публикацией мнения Andersen, et al., выше, что способы скрининга с использованием смешанных зондов были "...непрактичны для выделения белковых генов млекопитающих, когда соответственные ДНК недоступны".
Отличительные признаки изобретения, раскрытые в предшествующем описании, в следующей формуле изобретения и/или в прилагаемых чертежах, могут отдельно и в любой комбинации их стать материалом для осуществления изобретения в разнообразных его формах.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения человеческого эритропоэтина | 1987 |
|
SU1801118A3 |
| ХИМЕРНЫЙ БЕЛОК, СИНТЕТИЧЕСКАЯ ДНК, КОДИРУЮЩАЯ УКАЗАННЫЙ БЕЛОК, ЭКСПРЕССИОННЫЙ ВЕКТОР, ШТАММ-ПРОДУЦЕНТ СИНТЕТИЧЕСКОЙ ДНК И СПОСОБ ПОЛУЧЕНИЯ ПЛАЗМИДНОЙ ДНК | 2016 |
|
RU2736472C2 |
| РЕКОМБИНАНТНЫЙ ПОЛИПЕПТИД, СПОСОБ ИНГИБИРОВАНИЯ АГРЕГАЦИИ ТРОМБОЦИТОВ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК (ВАРИАНТЫ), РЕКОМБИНАНТНЫЙ ШТАММ E.COLI (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДА | 1991 |
|
RU2109816C1 |
| ГЕМОПОЭТИЧЕСКИЙ БЕЛОК (ВАРИАНТЫ), ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, СПОСОБ ПОЛУЧЕНИЯ ГЕМОПОЭТИЧЕСКОГО БЕЛКА, СПОСОБ СЕЛЕКТИВНОЙ ЭКСПАНСИИ КРОВЕТВОРНЫХ КЛЕТОК, СПОСОБ ПРОДУЦИРОВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СПОСОБ СТИМУЛЯЦИИ ПРОДУЦИРОВАНИЯ КРОВЕТВОРНЫХ КЛЕТОК У ПАЦИЕНТА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБ ЛЕЧЕНИЯ | 1997 |
|
RU2245887C2 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ ЛОВУШЕК GDF И АКТИВАТОРОВ РЕЦЕПТОРОВ ЭРИТРОПОЭТИНА ДЛЯ ПОВЫШЕНИЯ СОДЕРЖАНИЯ ЭРИТРОЦИТОВ | 2020 |
|
RU2814047C2 |
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ DEMEQUINA SEDIMINICOLA | 2019 |
|
RU2722933C1 |
| ПРИМЕНЕНИЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ PASTEURELLA PNEUMOTROPICA ДЛЯ МОДИФИКАЦИИ ГЕНОМНОЙ ДНК В КЛЕТКАХ | 2019 |
|
RU2724470C1 |
| ВАРИАНТЫ TREM2, УСТОЙЧИВЫЕ К ДЕЙСТВИЮ ШЕДДАЗЫ | 2018 |
|
RU2790661C2 |
| РЕКОМБИНАНТНЫЕ AAV ВЕКТОРЫ, ЭКСПРЕССИРУЮЩИЕ ОСТЕОПРОТЕКТИВНЫЕ ГЕНЫ, ВКЛЮЧАЯ HAS2 И ЛУБРИЦИН, ПРИГОДНЫЕ ПРИ ЛЕЧЕНИИ ОСТЕОАРТРИТА И СХОДНЫХ ЗАБОЛЕВАНИЙ СУСТАВОВ У МЛЕКОПИТАЮЩИХ | 2017 |
|
RU2771490C2 |
| Синергетическое действие SMN1 и miR-23a при лечении спинальной мышечной атрофии | 2021 |
|
RU2839898C2 |
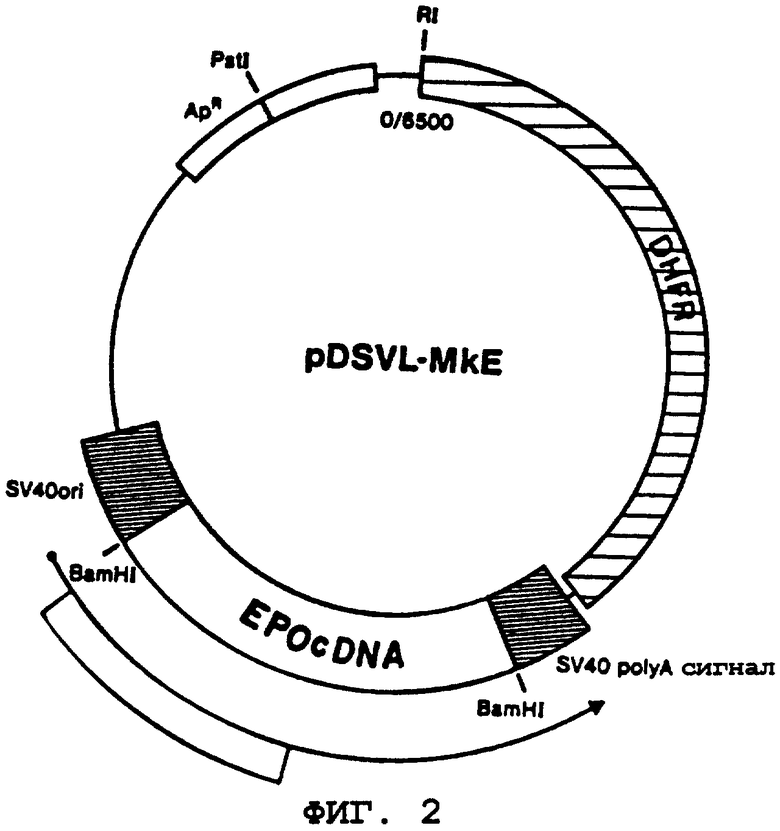
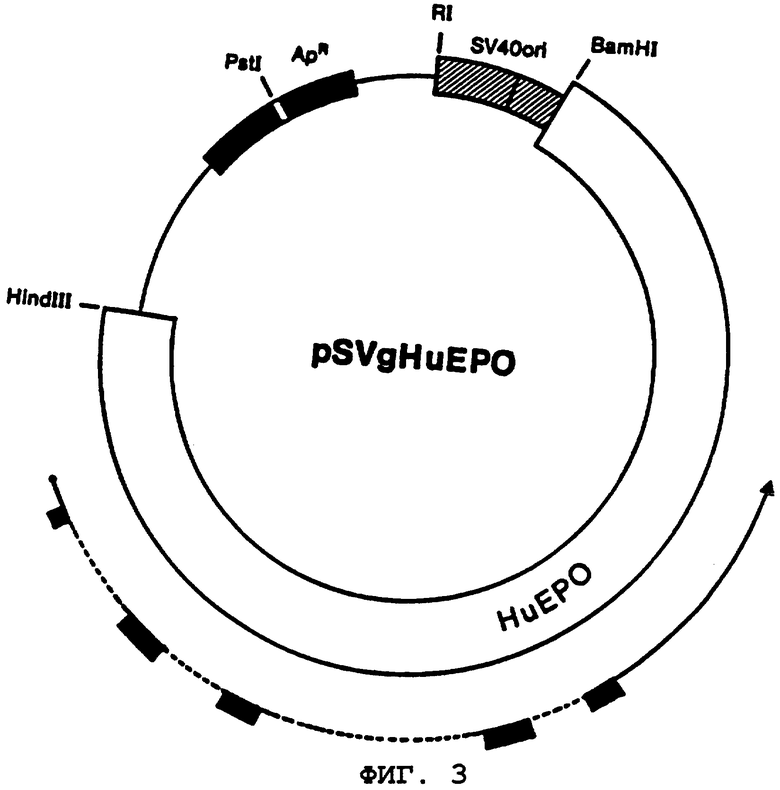
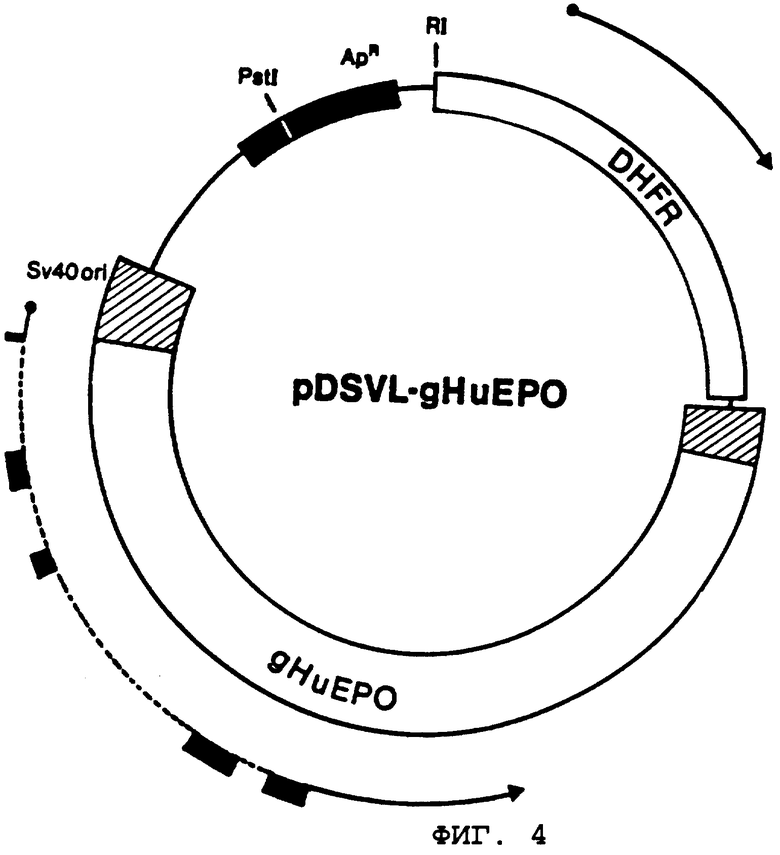
Изобретение относится к области генной инженерии и биотехнологии и может быть использовано в медицине. С помощью технологии рекомбинантных ДНК получен гликозилированный полипептид (гликопротеид) эритропоэтина человека, обладающий способностью увеличивать продукцию ретикулоцитов и эритроцитов, а также повышать уровень синтеза гемоглобина и потребление железа клетками костного мозга и характеризующийся более высокой, по сравнению с эритропоэтином, выделенным из мочи человека, молекулярной массой. Описаны варианты кодирующих его последовательностей ДНК, включающие эти последовательности векторные конструкции, получение трансформированных линий клеток млекопитающих, продуцирующих рекомбинантный эритропоэтин человека, а также способ его получения и очистки. Предложены фармацевтические композиции, включающие гликозилированный полипептид (гликопротеид) эритропоэтина по изобретению в качестве активного начала. Применение изобретения позволяет масштабировать процесс получения активного эритропоэтина человека, пригодного для использования в медицине. 13 н. и 9 з.п. ф-лы, 4 ил., 21 табл.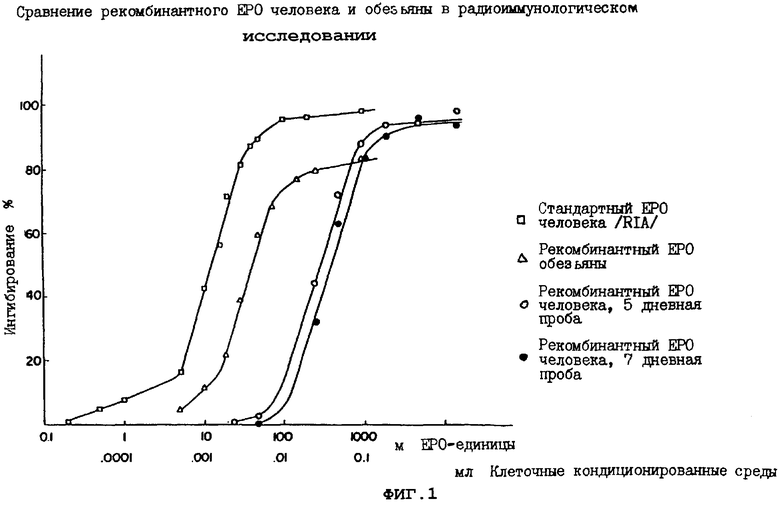
Ala Pro Pro Arg Leu Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Scr Leu Asn Glu Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Scr Ser Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile Thr Ala Asp Thr Phc Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp Arg.
Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp Arg.
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp Arg.
Ala Pro Pro Arg Leu Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp Arg,
более высокой по сравнению с эритропоэтином человека, выделенным из мочи, молекулярной массой, определяемой с помощью SDS-PAGE, и способностью увеличивать продукцию ретикулоцитов и эритроцитов и усиливать синтез гемоглобина и потребление железа клетками костного мозга, или часть указанного гликопротеида, сохраняющая активность полноразмерной формы.
Ala Pro Pro Arg Leu Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp Arg,
а указанный гликопротеид эритропоэтин характеризуется более высокой по сравнению с эритропоэтином человека, выделенным из мочи, молекулярной массой, определяемой с помощью SDS-PAGE, и способностью увеличивать продукцию ретикулоцитов и эритроцитов и усиливать синтез гемоглобина и потребление железа клетками костного мозга, или часть указанного гликопротеида, сохраняющая активность полноразмерной формы.
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| LEE-HUANG | |||
| Proc | |||
| Nat | |||
| Acad | |||
| Sci | |||
| Горный компас | 0 |
|
SU81A1 |

