Настоящее изобретение относится к растворимому CTLA4, являющемуся мутантным вариантом CTLA4 дикого типа, сохраняющим способность связывать CD80 и/или CD86.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Антиген-неспецифические межклеточные взаимодействия между Т-лимфоцитами и антигенпредставляющими клетками (АРС) вызывают Т- клеточные костимулирующие сигналы, вызывающие Т-клеточный ответ на антиген (Jenkins and Johnson (1993) Curr. Opin. Immunol. 5:361-3 67). Костимулирующие сигналы определяют величину Т-клеточного ответа на антиген и то, активирует или инактивирует этот ответ последующие реакции на антиген (Mueller et al. (1989) Annu. Rev. Immunol. 7:445-480).
Активация Т-клеток в отсутствие костимуляпии приводит к остановке Т-клеточного ответа или к анергическому Т-клеточному ответу (Schwartz,R.H. (1992) Cell 71:1065-1068). Один ключевой костимулирующий сигнал образуется при взаимодействии рецептора Т-клеточной поверхности CD28 с родственными В7 молекулами на антигенпрезентирующих клетках (например, так же известными как В7-1 и В7-2, или CD80 и CD86 соответственно) (P.Linsley and J.Ledbetter (1993) Annu.Rev. Immunol. 11:191-212). Вещество, известное в настоящее время как CD80 (В7-1), было первоначально описано как активационный антиген, связанный с человеческими В-клетками (Yokochi,T. et al. (1981) J. Immunol 128:823-827; Freeman, G.J. et al.(1989) J. Immunol. 143:2714-2722) и соответственно идентифицируемый как противорецептор в отношении родственных Т-клеточных молекул CD28 и CTLA4 (Linsley, Р., et al. (1990) Ргос. Natl. Acad. Sci. USA 87:5031-5035; Linsley, P.S. et al. (1991a) J. Exp. Med. 173:721-730; Linsley, P.S. et al. (1991b) J. Exp. Med. 174:561-570).
Позднее на антигенпредставляющих клетках был идентифицирован другой противорецептор для CTLA4 (Azuma.N. et al. (1993) Nature 366:76-79; Freeman (1993a) Science 262:909-911: Freeman. G.J. et. al. (1993b) J. Exp. Med. 178:2185-2192: Hathcock.K.L.S., et al. (1994) J. Exp. Med. 180:631-640: Lenschow, D.J. et al., (1993) Proc. Natl. Acad. Sci. USA 90:11054-11058; Ravi-Wolf. Z., 0993) Proc. Natl. Acad. Sci. USA 90:11182-11186: Wu, Yet al., (1993) J. Exp. Med. 178:1789-1793). Это вещество, известное в настоящее время как CD86 (Caux, С., et al. (1994) J. Exp. Med. 180:1841-1848), но также называемое В7-0 (Azuma et al. (1993, см. выше) или В7-2 (Freeman et al. (1993a), см. выше), имеет последовательность, примерно на 25% идентичную последовательности CD80 на ее внеклеточном участке (Azuma et al. (1993), см. выше; (Freeman et al. (1993a), см. выше, (1993b), см. выше). Клетки, трансфицируемые CD86, инициируют опосредуемые CD86 Т-клеточные реакции (Azuma et al. (1993), см. выше; Freeman et al. (1993a), (1993b), см. выше).
Сравнение экспрессии CD80 и CD86 являлось предметом нескольких исследований (Azuma et al. (1993), см. выше; Hathcock, et al. (1994), см. выше; Larsen, С.Р., et al. (1994) J. Immunol. 152:5208-5219; Stack, R.M., et al., (1994) J. Immunol 152:5723-5733). Современные данные показывают, что экспрессия CD80 и CD86 регулируется по-разному и наводят на мысль, что экспрессия CD86 имеет тенденцию предшествовать экспрессии CD80 в процессе иммунного ответа.
Растворимые формы CD28 и CTLA4 были созданы слиянием вариабельных (v)-подобных внеклеточных областей CD28 и CTLA4 с константными областями иммуноглобулина (Ig), что дает в результате CD28Ig и CTLA4Ig. CTLA4Ig связывает как CD80-позитивные, так и CD86-позитивные клетки более сильно, чем CD28Ig (Linsley, P., et al. (1994) Immunity 1:793-80). Многие из зависимых от Т- клеток иммунных реакций блокируются CTLA4Ig как in vitro, так и in vivo (Linsley, et al., (1991b), см. выше; Linsley, P.S. et. al. (1992а) Science 257:792-795: Linsley, P.S. etal. C1992bU.Exp.Med. 176:1595-1604: Lenschow, D.J. et al., (1992), Science 257:789-792; Tan, P. et al., (1992) J. Exp. Med. 177:165-173; Turka, L.A., Proc. Natl. Acad. Sci. USA 89:11102-11105).
Peach et al., (J. Exp. Med. (1994) 180:2049-2058) идентифицировал участки внеклеточной области CTLA4, которые являются важными для прочного связывания с CD80. Конкретно гексапептидный мотив (MYPPPY) в области, подобной гипервариабельному участку 3 (CDR3), был идентифицирован как полностью консервативная область у всех членов семейства CD28 и CTLA4. Мутагенез со сканированием аланина с помощью мотива MYPPPY в CTLA4 и в выбранных остатках CD28Ig ослабляет или прекращает связывание с CD80.
Были также созданы химерные молекулы, взаимодействующие с гомологичными областями CTLA4 и CD28. Молекулы HS4, HS4-A и HS4-B были конструированы "пересадкой" CDR3-подобных участков CTLA4, которые содержали также часть карбоксильного конца, продолженную так, чтобы включать определенные неконсервативные аминокислотные остатки на CD28Ig. Эти гомологичные мутанты проявили более высокую авидность связывания с CD80, чем CD28Ig.
В другой группе химерных гомологичных мутантов CDR1-подобную область CTLA4, которая не является консервативной в CD28 и, как прогнозировалось, пространственно прилегает к CDR3-подобному участку, "пересаживали" в HS4 и HS4-A. Эти химерные гомологичные мутантные молекулы (обозначенные HS7 и HS8) продемонстрировали даже более высокую авидность связывания с CD80, чем CD28Ig.
Химерные гомологичные мутантные молекулы были также получены пересадкой в HS7 и HS8 CDR2-подобного участка CTLA4, но эта комбинация дополнительно не улучшает авидность связывания с CD80. Следовательно, был определен мотив MYPPPY CTLA4 и CD28, но некоторые неконсервативные аминокислотные остатки CDR1- и CDR3-подобных участков также ответственны за повышенную авидность связывания CTLA4 с CD80.
Было показано, что CTLA4Ig эффективно блокирует обусловленную CD80 Т-клеточную дополнительную стимуляцию (костимуляцию), но не эффективен для блокады CD86-позитивных иммунных реакций. Были созданы молекулы растворимого мутантного CTLA4, в частности, имеющего более высокую авидность к CD86, чем CTLA4 дикого типа, как, возможно, более способные блокировать примирование антиген-специфических активированных клеток, чем CTLA4Ig.
Сохраняется необходимость в более совершенном CTLA4 для создания улучшенных фармацевтических композиций для супрессии иммунного ответа и для терапии рака, чем известные ранее растворимые формы CTLA4.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Соответственно изобретение предоставляет растворимый мутантный CTLA4, молекулы которого связываются с CD80 и/или CD86. Мутантные молекулы по изобретению включают такие молекулы, которые могут распознавать любой из CD80 и CD86 либо оба, и связываться с любым из CD80 и CD86 или с обоими. В некоторых вариантах изобретения мутантные молекулы по изобретению связывают CD80 и/или CD86 с более высокой авидностью, чем CTLA4.
Одним из примеров мутантной молекулы CTLA4 является L104EA29YIg (фиг.7), представленная в данном описании. Другим примером мутантной молекулы CTLA4 является L104EIg (фиг.8), представленная в данном описании. L104EA29YIg и L104EIg связываются с CD80 и CD86 с более высокой авидностью, чем CTLA4Ig.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фиг.1 показан анализ равновесного связывания L104EA29YIg, L104EIg и CTLA4Ig дикого типа с CD86Ig.
Фигуры 2А и 2В иллюстрирует данные анализа FACS, показывающего связывание L104EA29YIg, L104EIg и CTLA4Ig с человеческими CD80- и CD86-трансфицированными СНО клетками, как описано в примере 2, см. ниже.
На фигурах 3А и 3В показано ингибирование пролиферации CD80-позитивных и CD86-позитивных СНО-клеток, как описано ниже в примере 2.
На фигурах 4А и 4В показано, что L104EA29YIg более эффективно ингибирует пролиферацию первичных и вторичных аллостимулированных Т-клеток, чем CTLA4Ig, как описано ниже в примере 2.
Фигуры 5А-С иллюстрируют, что L104EA29YIg более эффективно, чем CTLA4Ig, ингибирует продуцирование цитокинов IL-2 (фиг.5А), IL-4 (фиг.5В) и γ-интерферона (фиг.5С) аллостимулированных человеческих Т-клеток, как описано ниже в примере 2.
На фигуре 6 показано, что L104EA29YIg более эффективно, чем CTLA4Ig ингибирует пролиферацию фитогемагглютинин(РНА)-стимулируемых Т-клеток обезьян, как описано ниже в примере 2.
На фигуре 7 изображена нуклеотидная и аминокислотная последовательность мутантной молекулы CTLA4 (L104EA29YIg), содержащая сигнальный пептид; мутантная внеклеточная область CTLA4, начиная с метионина в положении +1 и кончая аспарагиновой кислотой в положении +124, или начиная с аланина в положении -1 и кончая аспарагиновой кислотой в положении +124; и область Ig, как описано ниже в примере 1.
На фигуре 8 изображена нуклеотидная и аминокислотная последовательность мутантной молекулы CTLA4 (L104EIg), содержащая сигнальный пептид; мутантная внеклеточная область CTLA4, начиная с метионина в положении +1 и кончая аспарагиновой кислотой в положении +124, или начиная с аланина в положении -1 и кончая аспарагиновой кислотой в положении +124; и область Ig, как описано ниже в примере 1.
На фигуре 9 изображены нуклеотидная и аминокислотная последовательность CTLA4Ig, содержащая сигнальный пептид; аминокислотная последовательность внеклеточной области CTLA4 дикого типа, начиная с метионина в положении +1 до аспарагиновой кислоты в положении +124, или начиная с аланина в положении -1 до аспарагиновой кислоты в положении +124; и область Ig.
На фигурах 10А-С представлены результаты PAGE в SDS-геле (фиг.10А) для CTLA4Ig (дорожка 1), L104EIg (дорожка 2) и L104EA29YIg (дорожка 3А): и результаты эксклюзионной по размеру хроматографии для CTLA4Ig (фиг.10В) и L104EA29YIg (фиг.10С).
На фигурах 11А и 11В изображена ленточная диаграмма V-подобной складки внеклеточного Ig CTLA4, полученная решением структуры методом ЯМР-спектроскопии. На фиг.11В дан развернутый вид области S25-R33 и области MYPPPY, показывающий положение и ориентацию боковой цепи мутаций, повышающих авидность, L104 и А29.
На фигуре 12 дана схематическая диаграмма вектора, piLN-LEF29Y, с инсерцией L104EA29YIg.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ОПРЕДЕЛЕНИЯ
Следующие слова и фразы, применяемые в данном описании, имеют указанные ниже значения.
Применяемый в данном описании "CTLA4 дикого типа" имеет аминокислотную последовательность природного полноразмерного CTLA4 (Патенты США 5434131, 5844095, 5851795) или его внеклеточной области, которая связывает CD80 и/или CD86, и/или препятствует CD80 и/или CD86 связываться с их лигандами. В конкретных вариантах изобретения внеклеточная область CTLA4 дикого типа начинается с метионина в положении +1 и оканчивается аспарагиновой кислотой в положении +124, или внеклеточная область CTLA4 дикого типа начинается с аланина в положении -1 и заканчивается аспарагиновой кислотой в положении +124. CTLA4 дикого типа представляет собой белок клеточной поверхности, содержащий N-концевую внеклеточную область, трансмембранную область и С-концевую цитоплазматическую область. Внеклеточная область связывается с целевыми антигенами (мишенями), такими как CD80 и CD86. В природной клетке белок CTLA4 дикого типа транслируется в виде незрелого полипептида, который включает сигнальный пептид на N-конце. Незрелый белок претерпевает посттрансляционное, которое включает отщепление и удаление сигнального пептида с образованием продукта расщепления CTLA4, имеющего вновь образованный N-конец, который отличается от N-конца незрелой формы. Специалист в данной области техники понимает, что может происходить дополнительное посттрансляционное процессирование, при котором из вновь образованного N-конца продукта расщепления CTLA4 удаляется одна или более аминокислот. Молекула зрелой формы CTLA4 включает внеклеточную область CTLA4 или любую ее часть, связывающуюся с CD80 и/или CD86.
"CTLA4Ig" представляет собой растворимый слитый белок, содержащий внеклеточную область CTLA4 дикого типа или ее часть, которая связывает CD80 и/или CD86, соединенного с концом Ig. Особый вариант изобретения содержит внеклеточную область CTLA4 дикого типа, начиная с метионина в положении +1 и кончая аспарагиновой кислотой в положении +124; или начиная с аланина в положении -1 и заканчивая аспарагиновой кислотой в положении +124; участок соединения, аминокислотный остаток глутамин в положении +125; и иммуноглобулиновый участок, охватывающий область от глутаминовой кислоты в положении +126 до лизина в положении +357 (фигура 9).
Применяемое в данном описании понятие "слитый белок" определяется как одна или более аминокислотных последовательностей, соединенных друг с другом с помощью хорошо известных в технике методов и методов, описанных в патентах США 5434131 или 5637481. При этом соединенные аминокислотные последовательности образуют один слитый белок.
Применяемое в данном описании выражение "молекула мутантного CTLA4" ("мутантный CTLA4") может обозначать молекулу полноразмерного CTLA4 (полноразмерный CTLA4) или ее (его) части (производные или фрагменты), которые содержат мутацию или множественные мутации в CTLA4 (предпочтительно во внеклеточной области CTLA4), так, что она (он) становится аналогичной(-ым), но не идентичной(-ым) молекуле CTLA4 дикого типа (CTLA4 дикого типа). Молекулы мутантного CTLA4 могут включать в себя биологически или химически активную не-CTLA4 молекулу, либо эта молекула может быть присоединена к молекуле мутантного CTLA4. Молекулы мутантного соединения могут быть растворимыми (т.е. циркулирующими) или могут быть связаны с поверхностью. Молекулы мутантного CTLA4 могут включать всю внеклеточную область CTLA4 или ее части, например фрагменты или производные. Мутантный CTLA4 можно получать синтетически или методами рекомбинантнои ДНК.
Употребляемый в данном описании термин "мутация" обозначает изменение в нуклеотидной или аминокислотной последовательности полипептида дикого типа. В данном случае он обозначает изменение во внеклеточной области CTLA4 дикого типа. Изменение может представлять собой изменение аминокислоты, которое включает замены, делеции, добавления или усечения (сокращения). Мутантная молекула может содержать одну или несколько мутаций. Мутации в нуклеотидной последовательности могут вызывать или не вызывать мутации в аминокислотной последовательности, что очень понятно из уровня техники. А именно некоторые нуклеотидные кодоны кодируют одну и ту же аминокислоту. Примеры включают кодоны CGU, CGG, CGC и CGA, кодирующие аминокислоту аргинин (R); или кодоны GAU и GAC, кодирующие аминокислоту - аспарагиновую кислоту (D). Следовательно, белок может кодироваться одной или более нуклеиновой кислотой, которые отличаются специфической нуклеотидной последовательностью, но все же кодируют белки с идентичными последовательностями. Ниже даны последовательности, кодирующие аминокислоту:
Употребляемый в данном описании термин "внеклеточная область CTLA4" обозначает часть CTLA4, которая распознает CD80 и/или CD86 и связывается с ними. Например, внеклеточная область CTLA4 включает участок от метионина в положении +1 до аспарагиновой кислоты в положении +124. Или же внеклеточная область CTLA4 включает участок от аланина в положении -1 до аспарагиновой кислоты в положении +124 (фигура 9). Внеклеточная область включает фрагменты или производные CTLA4, которые связываются с CD80 и/или CD86.
Употребляемый в данном описании термин "не-CTLA4 белковая последовательность" или "не-CTLA4 молекула" обозначает любую молекулу, которая не связывается с CD80 и/или CD86 и не препятствует связыванию CTLA4 с ее целью. Пример включает, без ограничения, константную область иммуноглобулина (Ig) или ее часть. Предпочтительно константная область иммуноглобулина представляет собой константную область человеческого Ig или Ig обезьян, например человеческий С (гамма) 1, включая шарнир, области СН2 и СН3. Константная область Ig может мутировать, ослабляя его эффекторные функции (Патенты США 5637481 и 6132992).
Употребляемое в данном описании выражение "фрагмент мутантной молекулы CTLA4" обозначает часть мутантной молекулы CTLA4, предпочтительно внеклеточную область CTLA4 или ее участок, который распознает мишень, например CD80 и/или CD86, и связывается с нею.
Употребляемое в данном описании выражение "производное мутантного CTLA4" ("молекулы мутантного CTLA4") обозначает соединение, имеющее последовательность, по меньшей мере, на 70% сходную с последовательностью внеклеточной области CTLA4, и функции, аналогичные функциям этой области, т.е. оно распознает CD80 и/или CD86 и связывается с ними.
Употребляемый в данном описании "часть (участок) молекулы CTLA4" включает фрагменты и производные (молекулы) CTLA4, которые связываются с CD80 и/или CD86.
Для более полного понимания настоящего изобретения ниже представлено следующее описание.
КОМПОЗИЦИИ ПО ИЗОБРЕТЕНИЮ
Настоящее изобретение предоставляет растворимый мутантный CTLA4, который распознает CD80 и/или CD86 и связывается с ними. В некоторых вариантах изобретения растворимые мутантные CTLA4 проявляют более высокую авидность к CD80 и/или CD86, чем CTLA4Ig.
Примеры мутантого CTLA4 включают L104EA29YIg (Фигура 7). Аминокислотная последовательность L104EA29YIg может начинаться с аланина в положении -1 аминокислоты и оканчиваться лизином в положении +357 аминокислоты. Или же аминокислотная последовательность L104EA29YIg может начинаться метионином в положении +1 аминокислоты и заканчиваться лизином в положении +357 аминокислоты. CTLA4-участок L104EA29YIg охватывает метионин в положении аминокислоты +1 вплоть до аспарагиновой кислоты в положении +124 аминокислоты. L104EA29YIg содержит область соединения - аминокислотный остаток глутамин в положении +125 - и участок иммуноглобулина, охватывающий область от глутаминовой кислоты в положении +126 до лизина в положении +357 (фигура 7). Авидность L104EA29YIg при связывании с CD80, примерно, вдвое выше авидности CTLA4Ig дикого типа (далее называемого CTLA4Ig) и, примерно, в 4 раза выше при связывании с CD86. Такое более сильное связывание приводит к тому, что L104EA29YIg более эффективно блокирует иммунные реакции, чем CTLA4Ig.
Молекулы мутантного CTLA4 содержат, по меньшей мере, внеклеточную область CTLA4 или ее участок, который связывается с CD80 и/или CD86. Внеклеточная область молекулы мутантного CTLA4 содержит аминокислотную последовательность, начиная от метионина в положении +1 до аспарагиновой кислоты в положении +124 (фиг.7 или 8). Или же внеклеточная область CTLA4 может включать аминокислотную последовательность, начиная с аланина в положении -1 вплоть до аспарагиновой кислоты в положении +124 (фиг.7 или 8).
В одном варианте изобретения растворимый мутантный CTLA4 представляет собой слитый белок, включающий внеклеточную область CTLA4, имеющую одну или более мутаций в области аминокислотной последовательности, начиная от серина в положении +25 и кончая аргинином в положении +33 (S25-R33). Например, аланин в положении +29 CTLA4 дикого типа можно заменить на тирозин (кодоны: UAU, UAC). Или же аланин можно заменить на лейцин (кодоны: UUA, UUG, CUU, CUC, CUA, CUG), фенилаланин (кодоны: UUU, UUC), триптофан (кодон: UGG) или треонин (кодоны: ACU, АСС, АСА, ACG). Как легко поймут специалисты в данной области, урацил (U), нуклеотид последовательности РНК, соответствует тимину (Т), нуклеотиду последовательности ДНК.
В другом варианте изобретения растворимый мутантный CTLA4 представляет собой слитый белок, содержащий внеклеточную область CTLA4, имеющую одну или более мутаций в или близ области аминокислотной последовательности, начиная с метионина в положении +97 и кончая глицином в положении +107 (M97-G107). Например, лейцин в положении +104 CTLA4 дикого типа можно заменить на глутаминовую кислоту (кодоны: GAA, GAG). Мутантный CTLA4, содержащий такую замену, называется в данном описании L104EIg (фиг.8).
Еще в одном варианте изобретения растворимый мутантный CTLA4 представляет собой слитый белок, содержащий внеклеточную область CTLA4, содержащую одну или более мутаций в областях S25-R33 и M97-G107. Например, в одном варианте изобретения мутантный CTLA4 содержит в положении +29 тирозин вместо аланина; и в положении +104 глутаминовую кислоту вместо лейцина. Мутантный CTLA4, содержащий такую замену, называется в данном описании L104EA29YIg (фиг.7). Нуклеиновая кислота, которая кодирует L104EA29YIg, содержится в pD16 L104EA29YIg и депонирована 19 июня 2000 года Американской Коллекцией Культур Клеток (АТСС), 10801 University Blvd., Manasas, VA 20110-2209 (АТСС No. PTA-2104). Вектор pD16 L104EA29YIg является производным от вектора pcDNA3 (INVITROGEN).
Изобретение далее включает растворимый мутантный CTLA4, содержащий внеклеточную область CTLA4, как показано на фиг.7 или 8, или его часть (части), и фрагмент, который изменяет растворимость, аффинность и/или валентность мутантного CTLA4.
В соответствии с практикой применения изобретения фрагмент может представлять собой константную область иммуноглобулина или ее часть. Для применения in vivo предпочтительно, чтобы константная область иммуноглобулина не вызывала вредного иммунного ответа у субъекта. Например, в клинических протоколах может быть предпочтительным, чтобы молекулы мутанта включали константные области человеческого или обезьяньего иммуноглобулина. Одним из примеров подходящей области иммуноглобулина является человеческий С (гамма) 1, содержащий шарнирную, СН2 и СН3 области. Возможны другие изотипы. Кроме того, возможны другие константные области (предпочтительно другие слабо- или неиммуногенные константные области иммуноглобулина).
Другие фрагменты включают полипептидные метки. Примеры подходящих меток включают, без ограничения, молекулы р97, env gp120, Е7 и ova (Dash, В., etal. (1994) J.Gen. Virol. 75:1389-97: Ikeda, Т., et al. (1994) Gene 138:193-6; Falk, К., et al. (1993) Cell. Immunol 150:447-52: Fujisaka, K. et al. (1994) Virology 204:789-93). Возможно применять другие молекулы в качестве меток (Gerard, С. et al. (1994) Neuroscience 62:721-739; Byrn, R. et al. J. Virol. (1989) 63:4370-4375; Smith, D, et al. (1987) Science 238:1704-1707; Lasky, L., (1996) Science 233:209-212).
Изобретение далее включает растворимые мутантные слитые белки CTLA4Ig, предпочтительно более реактогенные в отношении антигена CD80 и/или CD86 по сравнению с CTLA4 дикого типа. Одним из примеров является L104EA29YIg, как показано на фиг.7.
В другом варианте изобретения молекула растворимого мутантного белка CTLA4 включает соединительный аминокислотный остаток, расположенный между участком CTLA4 и участком иммуноглобулина. Участок соединения аминокислот может быть любой аминокислотой, включающей глутамин. Соединительная аминокислота (участок соединения) может вводиться методами молекулярной генетики или синтетическими химическими методами, известными из уровня техники.
В другом варианте изобретения молекула растворимого мутантного белка CTLA4 включает участок иммуноглобулина (например, шарнирную СН2 и СН3 области), где любой остаток цистеина или все цистеиновые остатки в шарнирной области иммуноглобулинового участка заменены на серин, например цистеиновые остатки в положениях +130, +136 или +139 (фиг.7 или 8). Мутантные молекулы могут также включать пролин в положении +148, замененный на серин, как показано на фиг.7 или 8.
Молекула растворимого мутантного белка CTLA4 может включать последовательность сигнального пептида, связанную с N-концом внеклеточной области СТLА4-участка мутантной молекулы. Сигнальный пептид может представлять собой любую последовательность, которая позволяет мутантной молекуле секретировать, включая сигнальный пептид онкостатина М (Malick, et al. (1989) Molec. Cell. Biol. 9:2847-2853), или CD5 (Jones, N.H. et al., (1986) Nature 323:346-349), или сигнальный пептид любого внеклеточного белка.
Молекула мутантного соединения может включать сигнальный пептид онкостатина М, связанный с N-концом внеклеточной области CTLA4, и молекулу человеческого иммуноглобулина (например, шарнирную, СН2 и СН3 области), связанную с С-концом внеклеточной области CTLA4. Эта молекула включает сигнальный пептид онкостатина М, охватывающий аминокислотную последовательность, начиная с метионина в положении -26 и вплоть до аланина в положении -26, участок CTLA4, охватывающий аминокислотную последовательность от метионина в положении +1 и до аспарагиновой кислоты в положении +124, участок соединения аминокислот - глутаминовый остаток - в положении 125 и участок иммуноглобулина, охватывающий аминокислотную последовательность от глутаминовой кислоты в положении +126 до лизина в положении +357.
Растворимый мутантный белок CTLA4 можно получать методами рекомбинантной ДНК (молекулярными) или химическими синтетическими методами. Молекулярные методы включают следующие стадии: интродукция подходящей клетки-хозяина с молекулой нуклеиновой кислоты, которая экспрессирует и кодирует молекулу растворимого мутантного CTLA4; культивирование интродуцированной таким образом клетки-хозяина в условиях, которые позволяют клетке-хозяину экспрессировать мутантные молекулы; и выделение экспрессирующего мутантного белка. Участок сигнального пептида мутантной молекулы делает возможной экспрессию белка на поверхности клетки и секрецию его клеткой-хозяином. Транслируемые мутантные молекулы могут претерпевать посттрансляционную модификацию, включающую отщепление сигнального пептида с образованием зрелого белка, содержащего участки CTLA4 и иммуноглобулина. Отщепление может осуществляться после аланина в положении -1, что дает молекулу зрелого мутантного белка, содержащего метионин в положении +1 в качестве первой аминокислоты (фиг.7 или 8). Или же отщепление может проходить после метионина в положении -2, что дает молекулу зрелого мутантного белка, содержащего аланин в положении -1 в качестве первой аминокислоты.
Предпочтительным вариантом изобретения является молекула растворимого мутантного белка CTLA4, которая содержит внеклеточную область человеческого CTLA4, связанную с целой молекулой иммуноглобулина или ее частью (например, шарнирной, СН2 и СН3 областями). Молекула этого предпочтительного белка включает CTLA4-участок молекулы растворимого белка, охватывающий аминокислотную последовательность от метионина в положении +1 и до аспарагиновой кислоты в положении +124, участок соединения аминокислот - глутаминовый остаток - в положении +125 и участок иммуноглобулина, охватывающий аминокислотную последовательность от глутаминовой кислоты в положении +126 до лизина в положении +357. Участок, содержащий внеклеточную область CTLA4, мутирует так, что аланин в положении +29 заменяется на тирозин, а лейцин в положении +104 заменяется на глутаминовую кислоту. Иммуноглобулиновый участок мутантной молекулы может мутировать так, что цистеиновые остатки в положениях +130, +136 и +139 заменяются на серин, и пролин в положении +148 заменяется на серин. Этот мутантный белок (мутантная молекула) обозначена в данном описании L104EA29YIg (фиг.7).
Другим вариантом мутантной молекулы L104EA29YIg по изобретению является мутантная молекула, имеющая аминокислотную последовательность от аланина в положении -1 и до аспарагиновой кислоты в положении +124, участок соединения аминокислот - глутаминовый остаток - в положении +125 и участок иммуноглобулина, охватывающий глутаминовую кислоту в положении +126 (например, от +126 до лизина в положении +357). Участок, содержащий внеклеточную область CTLA4, мутирует так, что аланин в положении +29 заменяется на тирозин; а лейцин в положении +104 заменяется на глутаминовую кислоту. Иммуноглобулиновый участок мутантной молекулы мутирует так, что цистеиновые остатки в положениях +130, +136 и +139 заменяются на серин, и пролин в положении +148 заменяется на серин. Этот мутантный белок (мутантная молекула) обозначена в данном описании L104EA29YIg (фиг.7). После отщепления сигнальной последовательности L104EA29YIg может начинаться либо с метионина в положении +1, либо с аланина в положении -1.
Другой мутантной молекулой по изобретению является молекула растворимого мутантного белка CTLA4, имеющая внеклеточную область человеческого CTLA4, связанную с молекулой человеческого иммуноглобулина (например, шарнирной областью, СН2 и СН3). Эта молекула включает часть аминокислотной последовательности, кодирующей CTLA4, начиная с метионина в положении +1 и до аспарагиновой кислоты в положении +124, участок соединения аминокислот - глутаминовый остаток - в положении +125 и участок иммуноглобулина, охватывающий аминокислотную последовательность от глутаминовой кислоты в положении +126 до лизина в положении +357. Участок, содержащий внеклеточную область CTLA4, мутирует так, что лейцин в положении +104 заменяется на глутаминовую кислоту. Шарнирный участок молекулы мутирует так, что цистеиновые остатки в положениях +130, +136 и +139 заменяются на серин, и пролин в положении +148 заменяется на серин. Этот мутантный белок (мутантная молекула) обозначен в данном описании L104EIg (фиг.8).
Альтернативным вариантом L104EIg является молекула растворимого мутантного CTLA4, содержащая внеклеточную область CTLA4, связанную с молекулой человеческого иммуноглобулина (например, шарнирной областью, СН2 и СН3). Эта предпочтительная молекула включает часть аминокислотной последовательности от аланина в положении -1 и до аспарагиновой кислоты в положении +124, участок соединения аминокислот - глутаминовый остаток - в положении +125 и участок иммуноглобулина, охватывающий область от глутаминовой кислоты в положении +126 до лизина в положении +357. Участок, содержащий внеклеточную область CTLA4, мутирует так, что лейцин в положении +104 заменяется на глутаминовую кислоту. Шарнирный участок мутантной молекулы мутирует так, что цистеиновые остатки в положениях +130, +136 и +139 заменяются на серии, и пролин в положении +148 заменяется на серин. Этот мутантный белок (мутантная молекула) обозначен в данном описании L104EIg (фиг.8).
Кроме того, настоящее изобретение включает молекулу растворимого мутантного CTLA4, имеющую: (а) первую аминокислотную последовательность мембранного гликопротеина, например CD28, CD86, CD80, CD40 и gp39, которая блокирует пролиферацию Т-клеток, слитую со второй аминокислотной последовательностью; (b) вторую аминокислотную последовательность, являющуюся фрагментом внеклеточной области мутантного CTLA4, которая блокирует пролиферацию Т-клеток, такую как, например, молекула аминокислоты, включающая область от метионина в положении +1 до аспарагиновой кислоты в положении +124 (фиг.7 или 8); и (с) третью аминокислотную последовательность, которая действует как идентификационная метка или повышает растворимость молекулы. Например, третья аминокислотная последовательность может состоять исключительно из аминокислотных остатков шарнирной области, СН2- и СН3-областей молекулы неиммуногенного иммуноглобулина. Примеры подходящих молекул иммуноглобулина включают, без ограничения, человеческий или обезьяний иммуноглобулин, например С (гамма) 1. Также возможны другие изотипы.
Кроме того, изобретение включает нуклеотидные молекулы, имеющие нуклеотидные последовательности, кодирующие аминокислотные последовательности, соответствующие молекулам растворимого мутантного CTLA4 по изобретению. В одном варианте изобретения нуклеотидная молекула представляет собой ДНК (например, кДНК) или ее гибрид. Или же нуклеотидные молекулы представляют собой РНК или их гибриды.
Кроме того, данное изобретение включает вектор, который содержит нуклеотидные последовательности по изобретению. Также охватывается система вектора-хозяина. Система вектора-хозяина включает вектор по изобретению в подходящей клетке-хозяине. Примеры подходящих клеток-хозяев включают, без ограничения, клетки прокариот и эукариот.
Изобретение включает фармацевтические композиции для лечения заболеваний иммунной системы, содержащие фармацевтически эффективные количества растворимого мутантного CTLA4. В некоторых вариантах изобретения заболевания иммунной системы опосредуются взаимодействием CD28- и/или СТLА4-позитивных клеток с CD80- и/или СD86-позитивными клетками. Молекулы растворимого мутантного CTLA4 предпочтительно представляют собой молекулы CTLA4, содержащие одну или более мутаций во внеклеточной области CTLA4. Фармацевтическая композиция может включать растворимый мутантный белок CTLA4 и/или нуклеиновую кислоту и/или кодирующие их векторы. В предпочтительных вариантах изобретения растворимый мутантный CTLA4 имеет аминокислотную последовательность внеклеточной области CTLA4, как показано на одной из фигур, 7 или 8 (L104EA29Y или L104E соответственно). Еще более предпочтительно, когда растворимым мутантным CTLA4 является L104EA29YIg, представленный в данном описании. Композиции могут дополнительно содержать другие терапевтические агенты, включая, без ограничения, лекарственные токсины, ферменты, антитела или конъюгаты.
Фармацевтические композиции также предпочтительно включают подходящие носители или адъюванты, содержащие любой материал, который при соединении с соединением (молекулой) по изобретению (например, растворимым мутантным CTLA4, таким как L104EA29Y или L104E) сохраняет активность молекулы и не является реактивным в отношении иммунной системы субъекта. Примеры подходящих носителей и адъювантов включают, без ограничения, человеческий сывороточный альбумин; ионообменники; окись алюминия; лецитин; буферы, такие как фосфаты; глицин; сорбиновая кислота; сорбат калия; и соли или электролиты, такие как протаминсульфат. Другие примеры включают любые стандартные фармацевтические носители, такие как физиологический раствор с фосфатным буфером; вода; эмульсии, такие как эмульсия масла в воде; и различные увлажняющие средства. Другие носители могут также включать стерильные растворы; таблетки, включая таблетки, покрытые оболочкой, и капсулы. Обычно такие носители содержат эксципиенты, такие как крахмал, молоко, сахар, некоторые виды глин, желатин, стеариновую кислоту или ее соли, стеарат магния или кальция, тальк, растительные жиры или масла, смолы, гликоли или другие известные эксципиенты. Такие носители могут также включать вкусовые добавки и красители или другие ингредиенты. Композиции, содержащие такие носители, готовят хорошо известными обычными методами. Такие препараты можно также помещать в различные липидные композиции, такие как липосомы, а также в различные полимерные композиции, такие как полимерные микросферы.
Фармацевтические композиции по изобретению можно вводить обычными способами, включая, но без ограничения, внутривенный (вв), внутрибрюшинный (интраперитонеальный, вб), внутримышечный (вм), подкожный, пероральный способы введения, введение в виде суппозитория или при местном контакте, или имплантация устройства пролонгированного действия, такого как мини-осмотический насос, субъекту.
Фармацевтические композиции по изобретению могут быть в виде различных лекарственных форм, которые включают, без ограничения, жидкие растворы или суспензии, таблетки, пилюли, порошки, суппозитории, полимерные микрокапсулы или микровезикулы (микропузырьки), липосомы и растворы для инъекций или вливаний. Предпочтительная форма зависит от способа введения и терапевтического применения.
Наиболее эффективный способ применения и схема приема композиций по изобретению зависят от тяжести и течения заболевания, состояния здоровья больного и реакции на лечение и мнения лечащего врача. Соответственно дозировки композиций следует корректировать для каждого пациента.
Молекулы растворимого мутантного CTLA4 можно вводить субъекту в количестве и во времени (например, продолжительность и/или число раз), достаточном для того, чтобы блокировать связывание эндогенного В 7 (например, CD 80 и/или CD 86) с (их) соответствующими лигандами в организме субъекта. Тем самым блокада связывания эндогенный В7/лиганд ингибирует взаимодействие В7-позитивных клеток (например, CD80- и/или CD86-позитивных клеток) с CD28- и/или СТ1А4-позитивными клетками. Дозировка терапевтического агента зависит от многих факторов, включая, но без ограничения, характер ткани, в которую вводят агент, вид аутоиммунного (аутоаллергического) заболевания, подлежащего лечению, тяжесть заболевания, состояние здоровья субъекта и реакция субъекта на лечение с помощью агентов. Соответственно дозировка агентов может меняться в зависимости от субъекта и способа введения. Молекулы растворимого мутантного CTLA4 можно вводить в количестве 0,1-20,0 мг/кг веса больного в день, предпочтительно 0,5-10,0 мг/кг/день. Вводить фармацевтические композиции можно через различные промежутки времени. В одном варианте изобретения фармацевтическую композицию по изобретению можно вводить в течение одного часа или более. Кроме того, введение можно повторять в зависимости от тяжести заболевания, а также других факторов, как следует из уровня техники.
Изобретение далее включает способы получения белка, заключающиеся в выращивании системы хозяин-вектор по изобретению таким образом, чтобы продуцировать белок в хозяине, и в выделении полученного при этом белка.
Кроме того, изобретение включает способы регуляции функционального взаимодействия CTLA4- и/или CD28-позитивных Т-клеток с CD80- и/или CD86-позитивньми клетками. Способы включают контактирование CD80- и/или CD86-позитивных клеток с молекулой растворимого мутантного CTLA4 по изобретению таким образом, чтобы образовались комплексы мутантный CTLA4 / CD80 и/или мутантный CTLA4/ CD86, причем комплексы препятствуют реакции эндогенного антигена к CTLA4 с CD80 и/или CD86, и/или комплексы препятствуют реакции эндогенного антигена к CD28 с CD80 и/или CD86. В одном варианте изобретения молекула растворимого мутантного CTLA4 представляет собой слитый белок, который содержит, по меньшей мере, часть внеклеточной области мутантного CTLA4. В другом варианте изобретения молекула растворимого мутантного CTLA4 содержит: первую аминокислотную последовательность, включающую внеклеточную область CTLA4 от метионина в положении +1 до аспарагиновой кислоты в положении +124, содержащую, по меньшей мере, одну мутацию; и вторую аминокислотную последовательность, включающую шарнирную область, СН2- и СН3-области молекулы человеческого иммуноглобулина-гамма 1 (фиг.7 или 8).
В соответствии с практикой применения данного изобретения CD80- и/или CD86-позитивные клетки контактируют с фрагментами или производными молекулы растворимого мутантного CTLA4 по изобретению. Или же растворимый мутантный CTLA4 по изобретению представляет собой слитый белок CD28Ig/CTLA4Ig, имеющий первую аминокислотную последовательность, соответствующую части внеклеточной области рецептора CD28, слитую со второй аминокислотной последовательностью, соответствующей части внеклеточной области мутантного рецептора CTLA4, и третью аминокислотную последовательность, соответствующую шарнирной области, СН2- и СН3-области молекулы человеческого иммуноглобулина С-гамма-1 (фиг.7 или 8).
Предполагается, что растворимый мутантный CTLA4 проявляет ингибирующие свойства in vivo. В условиях, когда взаимодействие Т-клетка/АРС-клетка, например взаимодействие Т-клетка/В-клетка, происходит в результате контакта Т-клеток и АРС-клеток, связывание введенных с целью реакции с CD80- и/или CD86-позитивными клетками, например с В-клетками, молекул мутантного CTLA4 может препятствовать (т.е. ингибировать) взаимодействию Т-клетка/АРС-клетка, вызывающему регуляцию иммунных реакций.
Изобретение охватывает способы негативной регуляции (ингибирования) иммунных реакций. Негативная регуляция (модуляция) иммунного ответа молекулами растворимого мутантного CTLA4 может происходить путем ингибирования или блокады иммунного ответа уже в процессе развития или может предотвращать индукцию иммунного ответа. Молекулы растворимого CTLA4 по изобретению могут ингибировать функции активированных Т-клеток, такие как пролиферация Т-лимфоцитов и секреция цитокинов, подавляя Т-клеточные реакции или вызывая специфичную толерантность в Т-клетках, или тем и другим способом одновременно.
Кроме того, данное изобретение охватывает способы лечения заболеваний иммунной системы и индукции толерантности. В особенных вариантах изобретения заболевания иммунной системы опосредуются взаимодействием CD28- и/или СТLА4-позитивных клеток с CD80/CD86-позитивными клетками. В другом варианте изобретения Т-клеточное взаимодействие ингибируется. Заболевания иммунной системы включают, но без ограничения, аутоиммунные заболевания, иммунопролиферативные заболевания и заболевания, связанные с трансплантацией. Эти способы включают введение субъекту молекул растворимого мутантного CTLA4 по изобретению с целью регуляции Т-клеточного взаимодействия с CD80- и/или CD86-позитивными клетками. Или же можно вводить гибрид мутантного CTLA4, содержащий мембранный гликопротеин, соединенный с молекулой мутантного CTLA4. Примеры заболеваний, связанных с трансплантацией, включают вызванные реакцией "трансплантат против хозяина" (GVHD) (например, такую, которая может явиться результатом пересадки костного мозга или при индукции толерантности), расстройства иммунной системы, обусловленные отторжением трансплантата, хроническим отторжением, и алло- и ксенотрансплантатами тканей или клеток, включая плотные ткани (органы), островки, мышцы, гепатоциты, нейроны. Примеры иммунопролиферативных заболеваний включают, без ограничения, псориаз; Т-клеточную лимфому; Т-клеточный острый лимфобластный лейкоз; доброкачественный лимфоцитарный ангиит; и аутоиммунные заболевания, такие как волчанка (например, системная красная волчанка, lupus nephritis), тиреоидит Хасимото, первичная микседема, болезнь Грейвса, злокачественная анемия, аутоиммунный атрофический гастрит, болезнь Аддисона, диабет (например, инсулинзависимый сахарный диабет типа I), синдром Гудпасчера, тяжелая псевдопаралитическая миастения, пузырчатка, болезнь Крона, симпатическая офтальмия, симпатический увеит, рассеянный склероз, аутоиммунная гемолитическая анемия, идиопатическая тромбоцитопения, первичный билиарный цирроз, хронический активный гепатит, неспецифический язвенный колит, синдром Сегрена, ревматические заболевания (например, ревматоидный артрит), полимиозит, склеродерма и смешанная соединительнотканная болезнь.
Данное изобретение далее включает способ ингибирования отторжения субъектом трансплантатов однородных органов и/или плотных тканей, при этом субъект является реципиентом пересаженной ткани. Как правило, при трансплантации ткани отторжение трансплантата инициируется распознаванием его Т-клетками как чужеродного с последующим иммунным ответом, который разрушает трансплантат. Молекулы растворимого мутантного CTLA4 по данному изобретению, ингибируя пролиферацию Т-лимфоцитов и/или секрецию цитокинов, могут привести к уменьшению деструкции, а индукция антиген-специфичной иммунологической толерантности Т-клеток может привести к продолжительному приживлению трансплантата при отсутствии необходимости в общей иммуносупрессии. Кроме того, молекулы растворимого мутантного CTLA4 по изобретению можно применять вместе с другими фармацевтическими препаратами, такими как кортикостероиды, циклоспорин, рапамицин, микофенолят мофетил, азатиоприн, такролизмус, базиликсимаб и/или другие биологические соединения.
Настоящее изобретение также включает способы ингибирования гомологичной болезни у субъекта. Этот способ заключается во введении субъекту (молекул) растворимого мутантного CTLA4 по изобретению, одного или вместе с дополнительными лигандами, реактогенными с IL-2, IL-4 или γ-интерфероном. Например, растворимый мутантный CTLA4 по данному изобретению можно вводить реципиенту трансплантата костного мозга с целью ингибировать аллореактивность донорных Т-клеток. Или же донорные Т-клетки трансплантата костного мозга можно лишать иммуногенности в отношении аллоантигенов реципиента ex vivo перед трансплантацией.
Ингибирование Т-клеточного иммунного ответа растворимым мутантным CTLA4 может также быть полезным при лечении аутоиммунных нарушений. Многие аутоиммунные нарушения являются следствием неподходящей активации Т-клеток, реактивных в отношении аутоантигенов и промотирующих продуцирование цитокинов и аутоантител, "вовлеченных" в патологию заболевания. Введение (молекул) растворимого мутантного CTLA4 субъекту, страдающему аутоиммунным нарушением, может предупредить активацию аутореактивных Т-клеток и может уменьшить или снять симптомы заболевания. Этот метод может также заключаться во введении субъекту (молекул) растворимого мутантного CTLA4 по изобретению, одного или вместе с дополнительными лигандами, реактогенными с IL-2, IL-4 или γ-интерфероном.
Данное изобретение далее охватывает применение (молекул) растворимого мутантного CTLA4 вместе с другими иммуносупрессорами, например с циклоспорином (см. Mathiesen, в: "Prolonged Survival and Vascularization of Xenografted Glioblastoma Cells in the Central Nervous System of Cyclosporin A-treated Rats" (1989) Cancer Lett., 44:151-156); преднизоном, азатиоприном и метотрексатом (R. Handschumacher "Chapter 53: Drugs Used for Immunosuppression" pages 1264-1276). Возможно применение других иммуносупрессоров. Например, при лечении ревматоидного артрита (молекулы) растворимого мутантного CTLA4 можно вводить с фармацевтическими препаратами, включая, но без ограничения, кортикостероиды, нестероидные противовоспалительные лекарственные препараты/ингибиторы Сох-2, метотрексат, гидроксихлорохин, сульфасалазопирин, соли золота, этанерцепт, инфликсимаб, анакинру, азатиоприн и/или другие биологические препараты, такие как противо-ТГФ-препараты. При лечении системной красной волчанки молекулы растворимого мутантного CTLA4 можно вводить с фармацевтическими препаратами, включая, но без ограничения, кортикостероиды, цитоксан, азатиоприн, гидроксихлорохин, микофенолят мофетил и/или другие биологические препараты. Кроме того, при лечении рассеянного склероза молекулы растворимого мутантного CTLA4 можно вводить с фармацевтическими препаратами, включая, но без ограничения, кортикостероиды, интерферон бета-1 а, интерферон бета-1b, глатирамера ацетат, метоксантрона гидрохлорид и/или другие биологические препараты.
Молекулы растворимого мутантного CTLA4 (предпочтительно L104EA29YIg) можно также применять в комбинации с одним или более следующих агентов для регуляции иммунного ответа: растворимый gp 39 (также известный как лиганд CD40 (CD40 L), CD-154, Т-ВАМ, TRAP), растворимый CD40, растворимый CD80, растворимый CD86, растворимый CD28, растворимый CD56, растворимый Thy-1, растворимый CD3, растворимый TCR, растворимый VLA-4, растворимый VCAM-1, растворимый LECAM-1, растворимый ELAM-1, растворимый CD44, антитела, реактивные в отношении gp 39; антитела, реактивные в отношении CD40; антитела, реактивные в отношении В7; реактивные в отношении CD28; антитела, реактивные в отношении LFA-1; антитела, реактивные в отношении LFA-2; антитела, реактивные в отношении IL-2, антитела реактивные в отношении IL-12, антитела реактивные в отношении IFN-гамма; антитела, реактивные в отношении CD2; антитела, реактивные в отношении CD48; антитела, реактивные в отношении любых ICAM (например, ICAM-2); антитела, реактивные в отношении CTLA4; антитела, реактивные в отношении Thy-1; антитела, реактивные в отношении CD56; антитела, реактивные в отношении CD3; антитела, реактивные в отношении CD29; антитела, реактивные в отношении TCR; антитела, реактивные в отношении VLA-4; антитела, реактивные в отношении VCAM-1; антитела, реактивные в отношении LECAM-1; антитела, реактивные в отношении ELAM-1; антитела, реактивные в отношении CD44. В отдельных вариантах изобретения предпочтительными являются моноклональные антитела (mAbs). В других вариантах изобретения предпочтительными являются фрагменты антител. Все специалисты в данной области техники легко поймут, что комбинация может включать растворимый мутантный CTLA4 по изобретению и другой иммуносупрессор, растворимый мутантный CTLA4 с двумя другими иммуносупрессорами, растворимый мутантный CTLA4 с тремя другими иммуносупрессорами и т.д. Определение оптимальной комбинации и дозировок можно определить и оптимизировать хорошо известньми в технике методами.
Некоторые конкретные комбинации включают следующие: L104EA29YIg и CD80 mAbs; L104EA29YIg и CD86 mAbs; L104EA29YIg, CD80 mAbs и CD86 mAbs; L104EA29YIg и gp 39 mAbs; L104EA29YIg и CD40 mAbs; L104EA29YIg и CD28 mAbs; L104EA29YIg, CD80 и CD86 mAbs и gp 39 mAbs; L104EA29YIg, CD80 и CD86 mAbs и CD40 mAbs; и L104EA29YIg, анти-LFA1 mAb и анти-gp 39 mAb. Конкретным примером gp 39 mAb является MR1. Другие комбинации легко поймут и оценят по достоинству специалисты в данной области техники.
Растворимый мутантный CTLA4 (молекулы), например L104EA29Y, можно вводить как единственный (самостоятельный) активный ингредиент или вместе с другими лекарственными веществами для иммуномодуляции или для лечения или предупреждения острого или хронического отторжения алло- или ксенотрансплантата, или воспалительных или аутоиммунных заболеваний, или для индукции толерантности. Например, его можно использовать в комбинации с ингибитором кальциневрина, например циклоспорином А или FK506; с иммуносупрессорным макролидом, например рапамицином или его производным; например, 40-O-(2-гидрокси)этилрапамицином; с лимфоцитарным агентом хоминга, например, FTY720 или его аналогом; с кортикостероидами; с циклофосфамидом; с азатиоприном; с метотрексатом; лефлюномидом или его аналогом; с мизорибином; с микофенольной кислотой; с микофенолятом мофетила; с 15-дезоксипергуалином или его аналогом; с иммуносупрессорными моноклональными антителами, например моноклональными антителами к рецепторам лейкоцитов, например МНС, CD2, CD3, CD4, CD 11a / CD18, CD7, CD25, CD27, В7, CD40, CD45, CD58, CD 137, ICOS, CD150 (SLAM), 0Х40, 4-1BB или другие лиганды; или с другими иммуномодуляторными соединениями, например CTLA4/CD28-Ig, или молекулами других ингибиторов адгезии, например, mAbs или низкомолекулярными ингибиторами, включая антагонисты LFA-1, антагонисты Селектина и антагонисты VLA-4. Соединение в особенности применимо в комбинации с соединением, которое мешает CD40 и его лиганду, например антителам к CD40 и антителам к CD40-L, достичь вышеописанных показаний, например индукции толерантности.
Когда молекулы растворимого мутантного CTLA4 по изобретению вводят в сочетании с другими с иммуносупрессорными/иммуномодулирующими и противовоспалительными терапевтическими средствами, например, такими, как приведенные выше, дозировки совместно вводимого иммуносупрессора, иммуномодулятора или противовоспалительного средства, естественно, зависят от типа совместно вводимого лекарственного вещества, например, от того, является оно стероидом или циклоспорином, от конкретного применяемого лекарственного вещества, от состояния больного и т.п.
В соответствии с вышеописанным настоящее изобретение предоставляет в дополнительном аспекте указанные выше способы, заключающиеся в совместном введении, например одновременном (сопутствующем) или последовательном, терапевтически, эффективного количества (молекул) растворимого мутантного CTLA4 по изобретению, L104EA29YIg, в свободном виде или в виде фармацевтически приемлемой соли, и второго лекарственного вещества, при этом указанное второе лекарственное вещество является иммуносупрессором, иммуномодулятором или противовоспалительным лекарственным веществом, например, таким, как указанные выше. Кроме того охватываются терапевтические комбинации, например набор, например, для применения по любому из вышеуказанных методов, содержащий L104EA29YIg, в свободном виде или в виде фармацевтически приемлемой соли, вводимый одновременно или последовательно с, по меньшей мере, одной фармацевтической композицией, содержащей иммуносупрессор, иммуномодулятор или противовоспалительное лекарственное средство. Набор может содержать инструкции для их применения.
СПОСОБЫ ПОЛУЧЕНИЯ МОЛЕКУЛ ПО ИЗОБРЕТЕНИЮ
Экспрессию молекул мутантного CTLA4 можно осуществлять в клетках прокариот. Наиболее часто прокариоты представлены различными штаммами бактерий. Бактерии могут быть грамположительными и грамотрицательными. Как правило, предпочтительными являются грамотрицательные бактерии, такие как Е. coli. Также можно использовать другие штаммы микробов.
Последовательности, кодирующие молекулы мутантного CTLA4, можно встраивать в вектор, созданный для экспрессирования чужеродных последовательностей в клетках прокариот, таких как Е. coli. Эти векторы могут включать обычно используемые прокариотные регулярные последовательности, которые, как представлено в данном описании, включают промоторы инициации транскрипции, при необходимости, с оператором, наряду с сайтами связывания рибосом, включают такие обычно используемые промоторы, как промоторные системы бета-лактамазы (пенициллиназы) и лактозы (lac) (Chang, et al., (1977) Nature 198:1056), промоторная система триптофана (trp) (Goeddel, et al., (1980) Nucleic Acids Res. 8:4057) и промотор PL и сайт связывания рибосом N-гена фага лямбда (Shimatake, et al., (1981) Nature 292:128).
Такие векторы экспрессии также включают ориджины репликации и селективные маркеры, такие как ген бета-лактамазы или неомицин-фосфотрансферазы, придающие устойчивость к антибиотикам, так что векторы могут реплицировать в бактериях, и можно отбирать клетки, несущие плазмиды, выращенные в присутствии антибиотиков, таких как ампициллин или канамицин.
Плазмиду экспрессии можно вводить в клетки прокариот с помощью различных стандартных методов, включая, но без ограничения, CaCl2-шок (Cohen, (1972) Proc. Natl. Acad. Sci. USA 69:2110, и Sambrook et al. (eds.), "Molecular Cloning: A Laboratory Manual", 2nd Edition, Cold Spring Harbor Press, (1989) и электропорацию.
В соответствии с практикой применения данного изобретения клетки эукариот также пригодны в качестве клеток-хозяев. Примеры клеток эукариот включают любую животную клетку, первичную или иммортализованную, клетки дрожжей (например, Saccharomyces cerevisiae. Schizosaccharomyces pombe и Pichia pastoris) и растительные клетки. Клетки миеломы, COS и СНО являются примерами животных клеток, которые можно использовать в качестве клеток-хозяев. Конкретные СНО-клетки включают, но без ограничения, DG44 (Chasin, et la., 1986 Som. Cell. Molec. Genet. 12:555-556; Kolkekar 1997 Biochemistry 36:10901-10909), CHO-K1 (ATCCNo. CCL-61), CHO-K1 Tet-On клеточная линия (Clontech), СНО, обозначенные ЕСАСС 85050302 (CAMR, Salisbury, Wiltshire, UK), СНО клон 13 (GEIMG, Geneva, IT), СНО клон В (GEIMG, Geneva, IT), CHO-K1/SF, обозначенный ЕСАСС 93061607 (CAMR, Salisbury, Wiltshire, UK), и RR-CHOK1, обозначенный ЕСАСС 92052129 (CAMR, Salisbury, Wiltshire, UK). Примеры растительных клеток включают клетки табака (целое растение, культуру клеток или каллюс), пшеницы, сои культурной и риса. Также применимы семена пшеницы, сои культурной и риса.
Нуклеотидные последовательности, кодирующие молекулы мутантного CTLA4, можно также встраивать в вектор, предназначенный для экспрессирования чужеродных последовательностей в клетках эукариот. Регуляторные элементы вектора могут варьироваться в соответствии с конкретной эукариотной клеткой-хозяином.
Используемые обычно эукариотные регулярные последовательности для применения в векторах экспрессии включают промоторы и регулярные последовательности, совместимые с клетками млекопитающих, такие, например, как промотор CMV (вектор CDM8) и вирус саркомы птиц (ASV) (вектор πLN). Другие применяемые обычно промоторы включают ранние и поздние промоторы вируса зеленой мартышки (SV40) (Fiers, et al., Nature 273:113), или другие вирусные промоторы, такие как вирусные промоторы полиомы, Аденовируса 2 и вируса бычьей папилломы. Можно также использовать индуцибельный промотор, такой как hMTII (Karin, et al., (1982) Nature 299:797-802).
Векторы экспрессии (молекул) мутантного CTLA4 в клетках эукариот могут также нести последовательности, называемые энхансерами (энхансерная область). Они важны для оптимизации экспрессии гена и находятся выше (upstream) или ниже (downstream) промотора.
Примеры векторов экспрессии эукариотных клеток-хозяев включают, но без ограничения, векторы клеток-хозяев млекопитающих (например, BPV-1, pHyg, pRSV, hSV2, pTK2 (Maniatis); pIRES9 (Clontech); pRc/CMV2, pRc/RSV, pSFV1 (Life Technologies); векторы pVPakc, векторы pCMV, векторы pSG5 (Stratagene)), ретровирусные векторы (например, векторы pFB (Stratagene)), pCDNA-3 (Invitogene) или их модифицированные формы, аденовирусные векторы; векторы аденосвязанных вирусов, векторы бакуловирусов, векторы дрожжей (например, векторы pESC (Stratagene)).
Нуклеотидные последовательности, кодирующие молекулы мутантного CTLA4, можно интегрировать в геном эукариотной клетки-хозяина и реплицировать в виде реплик генома клетки-хозяина. Или же вектор, несущий молекулы мутантного CTLA4, может содержать ориджины репликации, делающие возможной внехромосомную репликацию.
Для экспрессии нуклеотидных последовательностей в Saccharomyces cerevisiae можно использовать ориджин репликации эндогенной дрожжевой плазмиды, виток (кольцо) 2μ. (Broach, (1983) Meth. Enz. 101:307). Или же можно использовать последовательности генома дрожжей, способные промотировать автономную репликацию (см., например, Stinchomb et al., (1979) Nature 282:39); Tschemper et al., (1980) Gene 10:157; и Clarke et al., Meth. Enz. 101:300).
Транскрипционный контроль последовательностей векторов дрожжей включает промоторы синтеза гликолитических ферментов (Hess et al., (1968) J Adv. Enzyme Reg. 7:149; Holland et al., (1978) Biochemistry 17:4900). Дополнительные промоторы, известные в технике, включают CMV-промотор, созданный в векторе CDM8 (Toyama and Okayama, (1990) FEBS 268:217-221); промотор 3-фосфоглицераткиназы (HitzemanetaL, (1980) J. Biol. Chem.255:2073) и промоторы других гликолитических ферментов.
Другие промоторы являются индуцибельными, так как они могут регулироваться раздражителями из окружающей среды или из питательной среды для выращивания клеток. Эти индуцибельные промоторы включают промоторы генов белков теплового шока, алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, ферментов, связанных с катаболизмом азота, и ферментов, ответственных за утилизацию мальтозы и галактозы.
Регуляторные последовательности можно помещать на 3'-конец кодирующих последовательностей. Эти последовательности могут стабилизировать матричную РНК. Такие терминаторы обнаружены на 3' нетранслируемом участке, следующем после кодирующих последовательностей нескольких генов дрожжей и млекопитающих.
Примеры векторов растений и растительных клеток включают, но без ограничения, плазмиды Тi агробактерий, вирус мозаики цветной капусты (CaMV) и вирус золотой мозаики томатов (TGMV).
Общие аспекты трансформации системы-хозяина клетки млекопитающего описаны Axel (Патент США 4399216, выдан 16 августа 1983 года). Клетки млекопитающих можно трансформировать методами, включая, но без ограничения, трансфекцию в присутствии фосфата кальция, микроинъекцию, электропорацию или трансдукцию при участии вирусных векторов.
Способы введения последовательности чужеродной ДНК в геномы растений или дрожжей включают (1) механические методы, такие как микроинъекция ДНК в одиночные клетки или протопласты, осуществляемая при интенсивном перемешивании клеток со стеклянными бусами в присутствии ДНК, или при "выстреливании" вольфрамовыми или золотыми шариками в клетки или протопласты; (2) введение ДНК путем превращения клеточных мембран в проницаемые для макромолекул обработкой их полиэтиленгликолем или с помощью электроимпульсов при высоком напряжении (электропорация); или (3) применение липосом (содержащих ДНК), которые сливаются с клеточными мембранами.
Экспрессию молекул мутантного CTLA4 можно обнаружить методами, известными в технике. Например, мутантные молекулы можно обнаружить по окрашиванию кумасси голубым гелей SDS-PAGE или иммуноблоттингом с применением антител, которые связываются с CTLA4. Выделение белка можно проводить, применяя стандартные способы очистки белка, например, аффинную хроматографию, ионообменную хроматографию, и получая практически чистый продукт (R. Scopes в: "Protein Purification. Principles and Practice". Third Edition, Springer-Verlag (1994)).
Данное изобретение далее охватывает молекулы растворимого мутантного CTLA4 (растворимый мутантный CTLA4), продуцируемые, как указано выше в данном описании.
МУТАГЕНЕЗ CTLA4Ig С ИСПОЛЬЗОВАНИЕМ КОДОНОВ
В одном варианте изобретения использованы сайт-направленный мутагенез и новая методика скрининга для идентификации нескольких мутаций во внеклеточной области CTLA4, которые повышают авидность связывания с CD86. В этом варианте изобретения получают мутации в остатках на участках внеклеточной области CTLA4 от серина 25 до аргинина 33, С' цепи (аланин 49 и треонин 51), F цепи (лизин 93, глутаминовая кислота 95 и лейцин 96) и на участке от метионина до тирозина 102, от тирозина 103 до глицина 107 и в G цепи в положениях глутамин 111, тирозин 114 и изолейцин 115. Эти сайты выбраны на основе исследований слитых белков химерный CD28/CTLA4 (Peach et al., J. Exp. Med., 1994, 180:2049-2058) и на основе модели, позволяющей предсказать, боковые цепи каких аминокислотных остатков подвержены действию растворителя, и отсутствие идентичности или гомологии аминокислотных остатков в некоторых положениях между CD28 и CTLA4. Далее любой остаток, который пространственно расположен в непосредственной близости (5-10 Å) к идентифицируемым остаткам, рассматривается как часть настоящего изобретения.
С целью синтеза и скрининга молекул растворимого мутантного CTLA4 с измененной аффинностью к CD80 и/или CD86 выбрана стратегия, включающая две стадии. Эксперименты повлекли за собой сначала создание библиотеки мутаций в специфичном кодоне внеклеточной области CTLA4, а затем ее скрининг методом анализа Biacore с целью идентифицировать мутанты с измененной реактивностью в отношении CD80 или CD86. Аналитическая система Biacore (Pharmacia, Piscataway, N.J.) использует детекторную систему резонанса поверхностного плазмона, которая в основном включает ковалентное связывание либо CD80Ig либо CD86Ig с покрытым декстраном сенсорным чипом, расположенном в детекторе. Затем испытуемое вещество (молекулы испытуемого вещества) можно инъецировать в камеру, содержащую сенсорный чип, и количество связывающегося комплементарного белка можно оценить на основе изменения молекулярной массы, которая физически связывается с покрытой декстраном стороной сенсорного чипа; изменение молекулярной массы можно измерять с помощью детекторной системы.
ПРЕИМУЩЕСТВА ДАННОГО ИЗОБРЕТЕНИЯ
Так как связывание CTLA4 с CD80 и CD86 характеризуется высокими скоростями протекания ("on", "к") и высокими скоростями диссоциации ("off, "от"), и так как комплексы CTLA4Ig- CD86 диссоциируют в 5-8 раз быстрее, чем комплексы CTLA4Ig-CD80, резонно предположить, что замедление скорости диссоциации CTLA4Ig в его комплексе с CD80 и/или с CD86 приводит к соединениям (молекулам) с более сильными иммуносупрессорными свойствами. Следовательно, ожидается, что молекулы растворимого мутантного CTLA4, обладающие более высокой авидностью в отношении CD80- или CD86-позитивных клеток по сравнению с CTLA4 дикого типа или не-мутантными формами CTLA4Ig, будут блокировать примирование антиген-специфичных активированных клеток с более высокой эффективностью, чем CTLA4 дикого типа или не-мутантные формы CTLA4Ig.
Кроме того, затраты на получение CTLA4Ig очень высоки. Мутантный CTLA4Ig (молекулы), обладающий более сильными иммуносупрессорными свойствами, может применяться в клинике в значительно меньших дозах, чем не-мутантный CTLA4Ig, давая аналогичные уровни иммуносупрессии. Следовательно, применение растворимого мутантного CTLA4, например L104EA29YIg, может быть очень эффективным с точки зрения стоимости.
Следующие примеры представлены для того, чтобы проиллюстрировать данное изобретение и помочь рядовым специалистам в его осуществлении и применении. Примеры никоим образом не претендуют на какое-либо ограничение объема изобретения.
ПРИМЕРЫ
ПРИМЕР 1
В данном примере представлено описание способов получения нуклеотидных последовательностей, кодирующих молекулы растворимого мутантного CTLA4 по изобретению. "Односайтовый" мутант (мутант по одному сайту) L104EIg получен и изучена кинетика его связывания с CD80 и/или с CD86. Нуклеотидная последовательность L104EIg использована в качестве матрицы для создания последовательности "двухсайтового" мутантного (мутантного по двум сайтам) CTLA4, L104EA29YIg, и изучена кинетика его связывания с CD80 и/или с CD86.
Мутагенез CTLA4 с использованием кодонов:
Для определения мутантных молекул CTLA4Ig с меньшими скоростями диссоциации (скорости "от" ("off')) в молекулах с CD80 и/или с CD86 выработана стратегия мутагенеза и скрининга. "Односайтовые" мутантные нуклеотидные последовательности получают, применяя CTLA4Ig (патенты США 5844095; 5851795; и 5885796; АТСС №68629) в качестве матрицы. Мутагенные нуклеотидные PCR праймеры созданы для случайного мутагенеза кодона специфичной кДНК, при этом в положениях 1 и 2 кодона допускается любое основание, но в положении 3 стоит только гуанин или тимин (XXG/T; так же известный как NNG/T). Таким манером можно осуществлять произвольную мутацию специфичного кодона, кодирующего аминокислоту, чтобы кодировать каждую из 20 аминокислот. При этом мутагенез ХХП/Т дает 32 потенциальных кодона, кодирующих каждую из 20 аминокислот. PCR-продукты, кодирующие мутации в непосредственной близости к -M97-G107 CTLA4Ig (см. фиг.7 или 8), отщепляют с помощью SacI/XbaI и субклонируют в аналогичным образом разрезанный вектор экспрессии CTLA4IgπLN. Этот метод используют, чтобы получить "односайтовую" СTLА4-мутантную молекулу, L104EIg (фиг.8). С целью мутагенеза в пространственной близости к S25-R33 CTLA4Ig молчащий сайт рестрикции Nhel сначала вводят 5' к этой петле с помощью PCR праймер-направленного мутагенеза. PCR-продукты расщепляют под действием NheI/XbaI и субклонируют в аналогичным образом разрезанные векторы экспрессии CTLA4Ig или L104EIg. Этот метод используют для получения молекулы "двухсайтового" мутантного CTLA4, L104EA29YIg (фиг.7). В частности, нуклеотидная молекула, кодирующая "односайтовую" мутантную молекулу CTLA4, L104EIg, использована в качестве матрицы для получения молекулы "двухсайтового" мутантного CTLA4, LI 04EA29YIg. Вектор piLN, содержащий L104EA29YIg, показан на фиг.12.
ПРИМЕР 2
Ниже дано описание методов скрининга, применяемых для идентификации "одно-" и "двухсайтовых" мутантных CTLA4-полипептидов, экспрессируемых с использованием конструкций, описанных в примере 1, которые проявляют более высокую авидность связывания с антигенами CD80 и CD86 по сравнению с молекулами немутантного CTLA4Ig.
Современные исследования in vitro и in vivo показывают, что CTLA4Ig сам по себе не способен полностью блокировать примирование антиген-специфичных активированных Т-клеток. Исследования in vitro по определению ингибирования пролиферации Т-клеток, проводимые с CTLA4Ig и любым антителом, связывающим CD80 или CD86, показывают, что моноклональное антитело, специфическое к CD80, не увеличивает ингибирования CTLA4Ig. Однако антитело к CD86 повышает ингибирование, это указывает, что CTLA4Ig неэффективно блокирует взаимодействие с CD86. Эти данные подтверждают более раннее открытие Linsley et al. (Immunity. (1994), 1:793-801), что для ингибирования опосредуемых CD80 клеточных реакций требуется, примерно, в 100 раз более низкая концентрация CTLA4Ig, чем для CD86-опосредуемых реакций. На основании этих открытий было сделано предположение, что молекулы растворимого мутантного CTLA4, обладающие более высокой авидностью в отношении CD86, чем CTLA4 дикого типа, способен лучше блокировать примирование антиген-специфичных активированных клеток, чем CTLA4Ig.
С этой целью молекулы растворимого мутантного CTLA4, описанного выше, в примере 1, подвергают скринингу, применяя новую методику, для того, чтобы идентифицировать некоторые мутации во внеклеточной области CTLA4, которые повышают авидность связывания с CD80 и CD86. Эта стратегия скрининга дает эффективный метод непосредственной идентификации мутантов со сравнительно более медленными "от" ("off")-скоростями, не требующий очистки белка или количественного определения, так как определение "от" ("off")-скорости не зависит от концентрации (O'Shannessy et al., (1993) Anal. Biochem., 212:457-468).
Клетки COS трансфицируют, используя мини-препарат очищенной плазмидной ДНК, и культивируют в течение нескольких дней. К BIAcore биосенсорным чипам (Pharmacia Biotech AB, Uppsala, Sweden), покрытым растворимыми CD80Ig или CD86Ig, добавляют трехдневные кондиционированные культуральные среды. Специфичное связывание и диссоциацию мутантных белков определяют с помощью резонанса поверхностного плазмона (O'Shannessy et al., (1993) Anal. Biochem., 212:457-468). Все эксперименты проводят на биосенсорах BIAcore™ и BIAcore™ 2000 при 25°С. Лиганды иммобилизуют на сенсорных чипах NCM5 для поиска соединений (Pharmacia), используя стандартное связывание с N-этил-N'-(диметиламинопропил)карбодиимид-N-гидроксисукцинимидом (Johnsson, В., et al. (1991) Anal. Biochem. 198:268-277; Khiiko, S.N., et al. (1993) J. Biol. Chem. 268:5425-15434 (?)).
Метод скрининга
Клетки COS, выращенные в 24-луночных планшетах для тканевых культур, транзиторно трансфицируют с использованием ДНК, кодирующей мутантный CTLA4Ig. Через 3 дня собирают культуральные среды, содержащие секретированный растворимый мутантный CTLA4Ig.
Ток кондиционированных COS-клеточных культуральных сред пропускают через биосенсорные чипы BIAcore, дериватизированные при использовании CD80Ig или CD86Ig (как описано в Greene et al., 1996 J.Biol. Chem. 271:26762-26771), или мутантные молекулы идентифицируют при "от" ("off")-скоростях, меньших, нежели "от"-скорости, наблюдаемые для CTLA4Ig дикого типа. кДНК, соответствующие образцам выбранных сред, секвенируют и получают ДНК для осуществления транзиторной трансфекции клеток COS в больших масштабах, с использованием которой получают мутантный белок CTLA4Ig после очистки от протеина А культуральной среды. Условия анализа BIAcore и методика анализа данных равновесного связывания такие, как описано в Greene et al., 1996 J. Biol. Chem. 271:26762-26771 и как представлено в данном описании.
Анализ данных BIAcore
Диаграмма чувствительности перед анализом нормализована по уровню нулевого отклика (RU). Для определения фоновой величины отклика (RU) на основании большой разницы значений коэффициентов рефракции растворов образцы пропускают через "mock-derivatized" проточные кюветы. Константы равновесной диссоциации (Кd) рассчитывают из графика зависимости Req от С, где Req обозначает отклик (ответ) в стационарном состоянии минус отклик на "mock-derivatized" чипе, а С обозначает молярную концентрацию аналита. Кривые связывания анализируют, используя выпускаемые промышленностью программы для вычерчивания нелинейных кривых (Prism, GraphPad Software).
Экспериментальные данные сначала аппроксимируют к связыванию одиночного лиганда с одиночным рецептором (1-сайтовая модель, т.е. простая ленгмюровская система, А+В↔АВ), и константы равновесной ассоциации (Кd=[А]·[В]\[АВ] вычисляют по уравнению R=Rmax·С/(Кd+С). Затем аппроксимируют данные к простейшей двухсайтовой модели связывания лиганда (т.е. к рецептору, имеющему два невзаимодействующих независимых сайта связывания, как описывается уравнением R=Rmax·С\(Кd1+С)+Rmах2·С\(Кd2+С)).
Соответствие (согласие, пригодность) этих двух моделей анализируют визуально, сравнивая с экспериментальными данными, и статистически с помощью F-критерия суммы квадратов. Простейшую односайтовую модель выбирают как наиболее соответствующую, если двухсайтовая модель не соответствует значительно лучше (р<0,1).
Анализ ассоциации и диссоциации проводят, используя программу BIA evaluation 2.1 Software (Pharmacia). Константы скорости ассоциации kon вычисляют двумя способами, принимая во внимание как гомогенное односайтовое взаимодействие, так и параллельное двухсайтовое взаимодействие. Для односайтового взаимодействия значения kon вычисляют по уравнению Rt=Req(1-exp-ks(t-t 0), где Rt обозначает ответ (отклик) в данное время, t; Req обозначает отклик (ответ) в стационарном состоянии; to обозначает время в начальный момент инъекции; и ks=dR/dt=kon·Ckoff и где С обозначает концентрацию аналита, вычисленную как функция мономерных сайтов связывания. Для двухсайтовых взаимодействий значения kon вычисляют по уравнению Rt=Req1(1-exp-ks1(t-t 0)+Req2(1-exp-ks2(t-t 0). Для каждой модели значения kon определяют по тангенсу угла наклона (примерно, при 70% максимальной ассоциации) графика зависимости ks от С.
Данные диссоциации анализируют в соответствии с односайтовой (АВ=А+В) и двухсайтовой (AiBj=Ai+Bj) моделями, а константы скорости (koff) вычисляют исходя из наиболее соответствующих кривых. Можно использовать модель сайта связывания, за исключением тех случаев, когда остатки превышают фон компьютера (2-10 RU, в зависимости от компьютера (ЭВМ)), в этом случае применяют модель сайта с двумя связываниями. Полупериод размещения рецептора вычисляют из соотношения t1/2=0,693/koff.
Проточная цитометрия:
Мышиные mAb L307.4 (анти-CD80) получены от Becton Dickinson (San Jose, California), a IT1.2 (анти-В7-0 [так же известные как CD86]) - от Pharmingen (San Diego, California). С целью иммунного окрашивания CD80-позитивные и/или CD86-позитивные СНО-клетки извлекают из сосудов с культурами, инкубируя в физиологическом растворе с фосфатньм буфером (PBS), содержащем 10 мМ ЭДТК (EDTA). Клетки СНО (1-10·105) сначала инкубируют с mAbs или со слитыми белками иммуноглобулина в DMEM, содержащем 10% эмбриональной бычьей сыворотки (FBS), затем отмывают и инкубируют с реагентами второй стадии конъюгированных с флуоресцеинизоцианатом антител козы к человеческому или мышиному иммуноглобулину (Tago, Burlingame, California). Клетки отмывают последний раз и анализируют на FACScan (Becton Dickinson).
SDS-PAGE и хроматография по размеру молекул
SDS-PAGE осуществляют Tris/глицин 4-20% акриламидных гелях (Novex, San Diego, СА). Аналитические гели окрашивают кумасси голубым, изображения влажных гелей получают цифровым сканированием. CTLA4Ig (25 мкг) и L104EA29YIg (25 мкг) анализируют хроматографией по размеру молекул на колонке TSK-GEL G300 SWXL (7,8×300 мм, Tosohaas, Montgomeryville, PA), уравновешенной забуференным фосфатом физиологическим раствором, содержащим 0,02% NAN3, при скорости тока 1,0 мл/мин.
CTLA4XC120S и L104EA29YXC120S.
Одноцепочечную CTLA4XC120S получают по описанной методике (Linsley et al., (1995) J. Biol. Chem.. 270:15417-15424). Короче говоря, в качестве матрицы используют плазмиду экспрессии онкостатина М CTLA 4 (OMCTLA4), "прямой" праймер, GAGGTGATAAAGCTTCACCAATGGGTGTACTGCTCACACAG выбирают, чтобы его последовательность совмещалась с последовательностями вектора; а "обратный" праймер,
GTGGTGTATTGGTCTAGATCAATCAGAATCTGGGCACGGTTC
соответствует последним семи аминокислотам (т.е. аминокислотам 118-124) внеклеточной области CTLA4 и содержит сайт рестрикции ферментом и терминирующий кодон (TGA). Обратный праймер определяет мутацию C120S (цистеин на серин в положении 120). В особенности нуклеотидная последовательность GCA (нуклеотиды 34-36) изображенного выше обратного праймера заменяется на одну из следующих нуклеотидных последовательностей: AGA, GGA, TGA, CGA, ACT или GCT. Как понимают специалисты в данной области техники, нуклеотидная последовательность GСФ является обратной комплементарной последовательностью к кодону TGC для цистеина. Аналогично нуклеотидные последовательности AGA, GGA, TGA, CGA, ACT или GCT являются обратными комплементарными последовательностями к кодонам для серина. Продукты полимеразной цепной реакции расщепляют с помощью HindIII/XbaI и направленно субклонируют в вектор экспрессии πLN (Bristol-Myers Squibb Company, Princeton, NJ). L104EA29YXC120S готовят аналогичным образом. Каждую конструкцию контролируют с помощью секвенирования ДНК.
Идентификация и биохимическая характеристика мутантов с высокой авидностью
Для мутагенеза выбирают двадцать четыре аминокислоты и образующиеся ˜2300 мутантных белков анализируют на связывание с CD86Ig методом поверхностного плазменного резонанса (SPR; как описано выше). Преобладающие эффекты мутагенеза на каждом сайте суммированы в таблице II. Случайный мутагенез аминокислот в S25-R33, по-видимому, не меняет связывания с лигандом. Мутагенез Е31 и R33 и остатков M97-Y102, по-видимому, к ослаблению связывания с лигандом. Мутагенез остатков S25, А29, и Т30, K93, L96, Y103, L104 и G105 приводит к белкам к замедленным "к" ("он") и/или замедленным "от" ("off"') скоростям. Эти результаты подтверждают предыдущие открытия, что остатки в области S25-R33 и остатки в или рядом с областью M97-Y102 влияют на связывание с лигандом (Peach et al., (1994) J. Exp. Med., 180:2049-2058.
Мутагенез сайтов S25, Т30, К93, L96, Y103 и G105 позволяет идентифицировать некоторые мутантные белки с замедленными скоростями "от" CD86Ig. Однако в этих примерах медленная скорость "от" компенсируются медленной скоростью "к", в результате получаются мутантные белки с общей авидностью к CD86Ig, которая, по-видимому, аналогична общей авидности, наблюдаемой у CTLA4Ig. Кроме того, мутагенез К93 приводит к значительной агрегации, которая может быть ответственна за наблюдаемые кинетические изменения.
Случайный мутагенез L104 с последующей COS-клеточной трансфекцией и скринингом методом SPR образцов культуральных сред над иммобилизованным CD86Ig дает шесть образцов сред, содержащих мутантные белки, имеющие, примерно, вдвое меньшие скорости "от", чем CTLA4Ig дикого типа. В результате секвенирования соответствующих кДНК этих мутантов найдено, что каждая кодирует мутацию лейцина в глутамин (L104E). По-видимому, замена лейцина на аспарагиновую кислоту (L104D) не влияет на связывание CD86Ig.
Затем мутагенез повторяют в каждом сайте, перечисленном в таблице II, на этот раз в качестве PCR-темплата (матрицы) вместо CTLA4Ig дикого типа используют L104E, как описано выше. SPR анализ опять с использованием иммобилизованного CD86Ig дает (определяет) шесть препаратов культуральной среды, полученных при мутагенезе аланина 29, с белками, имеющими скорость "от", примерно, в 4 раза меньшую, чем CTLA4Ig дикого типа. Два самых медленных представляют собой замены на тирозин (L104EA29Y), два являются заменами на лейцин (L104EA29L), один с заменой на триптофан (L104EA29W) и один с заменой на треонин (L104EA29T). По-видимому, никаких мутантов с медленной скоростью "от" не идентифицировано, когда случайно мутирует один аланин 29 в CTLA4Ig дикого типа.
Относительную молекулярную массу и состояние агрегации очищенного L104E и L104EA29YIg оценивают методом SDS-PAGE и хроматографией по размеру частиц. L104EA29YIg (˜1 мкг; дорожка 3) и L104EIg (˜1 мкг; дорожка 2), по-видимому, имеют такую же электрофоретическую подвижность, что и CTLA4Ig (˜1 мкг; дорожка 1) в присутствии восстановителя (˜50 кДа; +βМЕ; плюс 2-меркаптоэтанол) и без восстановления (˜100 кДа; -βМЕ) (фиг.10А). Хроматография по размеру частиц показывает, что L104EA29YIg (фиг.10С), по-видимому, имеет такую же подвижность, что и димерный CTLA4Ig (фиг.10В). Основные пики соответствуют димеру белка, тогда как быстрее элюирующий минорный пик на фиг.10В соответствует агрегатам с более высокой молекулярной массой. Примерно 5,0% CTLA4Ig представляет собой агрегаты с более высокой молекулярной массой, но нет доказательства присутствия агрегации L104EA29YIg или L104EIg. Следовательно, более прочное связывание с CD86IG, наблюдаемое для L104EIg и L104EA29YIg, нельзя отнести за счет агрегации, вызываемой мутагенезом.
Анализ равновесия и кинетики связывания
Анализ равновесия и кинетики связывания осуществляют на очищенном от протеина А CTLA4Ig, L104EIg и L104EA29YIg с помощью поверхностного плазменного резонанса (SPR). Результаты приведены в таблице I. Наблюдаемые константы равновесной диссоциации (Кd; таблица I) вычисляют на основе кривых связывания, полученных в интервале концентраций (5,0-200 нМ). L104EA29YIg сильнее связывается с CD86Ig, чем L104EIg и CTLA4Ig. Более низкое значение Кd для L104EA29YIg (3,21 нМ), чем для L104EIg (6,06 нМ) или CTLA4Ig (13,9 нМ), указывает на более высокую авидность связывания L104EA29YIg с CD86Ig. Более низкое значение Кd L104EA29YIg (3,66 нМ), чем L104EIg (4,47 нМ) или CTLA4Ig (6,51 нМ), указывает на более высокую авидность связывания L104EA29YIg с CD80Ig.
Кинетика связывания показывает, что сравнительные скорости "к" связывания CTLA4Ig, L104EIg и L104EA29YIg с CD80 аналогичны, как и скорости "к" для CD86Ig (таблица I). Однако скорости "от" ("off) для этих молекул не эквивалентны (таблица I). По сравнению с CTLA4Ig скорость L104EA29YIg "от" ("off) CD80Ig, примерно, в 2 раза медленнее, а скорость "от" CD86Ig, примерно в 4 раза медленнее. Так как введение этих мутаций незначительно влияет на скорости "к" ("on"), увеличение авидности к CD80Ig и к CD86Ig, наблюдаемое для L104EA29YIg, прежде всего, по-видимому, вызвано уменьшением скоростей "от" ("off).
Чтобы выяснить, вызвано увеличение авидности L104EA29YIg к CD80Ig и к CD86Ig мутациями, влияющими на то, как каждый мономер ассоциируется в димер, или же повышающими авидность структурными изменениями в каждом мономере, получают одноцепочечные конструкции внеклеточных областей CTLA4 и L104EA29YIg с последующим мутагенезом цистеина 120 на серин, как описано выше и в (Linsley et al., (1995) J. Biol. Chem., 270:15417-15424). Как было показано методом гель-фильтрации (Linsley et aL (1995). см. выше), очищенные белки CTLA4XС120S и L104EA29YXC120S являются мономерными до того, как их свойства связываться с лигандами анализируются методом SPR. Результаты показывают, что аффинность связывания обоих мономерных белков с CD86Ig, примерно, в 35-80 раз меньше, чем аффинность связывания, наблюдаемая для их соответствующих димеров (таблица I). Это подтверждает ранее опубликованные данные, устанавливающие, что для высокой авидности связывания с лигандом требуется димеризация CTLA4 (Greene et al., (1996) J. Biol. Chem., 271:26762-26771).
Аффинность связывания L104EA29YXC120S как с CD86Ig, так и с CD861g, примерно, в 2 раза выше, чем аффинность связывания с ними CTLA4XC120S, увеличение аффинности вызвано более чем в 3 раза замедленной скоростью диссоциации "от" обоих лигандов. Следовательно, вероятнее всего более сильное связывание L104EA29YIg с лигандом вызвано повышающими авидность структурными изменениями, которые вводятся в каждую мономерную цепь, нежели изменениями, вызывающими димеризацию молекулы.
Местоположение и структурный анализ мутаций, повышающих авидность
Структура внеклеточной IgV-подобной области CTLA4 недавно определена методом ЯМР-спектроскопии (Metzler et al., (1997) Nature Struct. Biol., 4:527-531. Это позволяет найти точное местоположение лейцина 104 и аланина 29 в трехмерной складке (фиг.11А-В). Лейцин 104 расположен рядом с высококонсервативной аминокислотной последовательностью MYPPPY. Аланин расположен близ С конца участка S25-R33,который пространственно близок к участку MYPPPY. Хотя между остатками этих двух участков существует значительное взаимодействие, по-видимому, нет непосредственного взаимодействия между L104 и А29, хотя оба содержат часть соседнего гидрофобного ядра в белке. Влияние двух мутантов, повышающих авидность, на структуру оценивают с помощью модели. Мутация A29Y легко располагается в кармане между участком S25-R33 и участком MYPPPY и может служить для стабилизации конформации участка MYPPPY. В CTLA4 дикого типа между L104 и L96 и V94 около участка MYPPPY существует сильное гидрофобное взаимодействие. По двум причинам маловероятно, что мутация глутаминовой кислоты приобретает конформацию, аналогичную конформации L104. Первая: без значительных "пертурбаций" на участке S25-R33 имеется слишком мало места для расположения более длинной боковой цепи глутаминовой кислоты. Вторая: велика энергия, затрачиваемая на то, чтобы "нести" отрицательный заряд боковой цепи глутаминовой кислоты в гидрофобной области. Напротив, исследования моделей позволяют предсказать, что боковая цепь глутаминовой кислоты выталкивается на поверхность, где ее заряд может стабилизироваться за счет сольватации. Такое конформационное изменение легко может разместиться около G105 при минимальном искажении других остатков на этих участках.
Связывание мутантов с высокой авидностью с клетками СНО, экспрессирующими CD80 или CD86.
Анализ FACS (фиг.2) связывания CTLA4Ig и мутантных молекул с устойчиво трансфицируемыми клетками CD80+ и CD86+CHO осуществляют так, как представлено в данном описании. CD80-позитивные клетки СНО инкубируют с увеличивающимися концентрациями CTLA4Ig, L104EA29YIg или L104EIg, а затем отмывают. Связанный слитый белок иммуноглобулина обнаруживают при использовании конъюгированных с флуоресцеинизоцианатом антител козы к человеческому иммуноглобулину.
Как показано на фиг.2, CD-позитивные или CD-позитивные СНО-клетки (1,5·105) инкубируют с указанными концентрациями CTLA4Ig (темные квадраты), L104EA29YIg (круги) или L104EIg (треугольники) в течение 2 ч при 23°С, отмывают и инкубируют с конъюгированными с флуоресцеином антителами козы к человеческому иммуноглобулину. Связывание в целом с 5000 жизнеспособных клеток анализируют (единственное определение) на FACScan и среднее значение интенсивности флуоресценции (MFI) определяют из данных гистограммы с помощью PC-LYSYS. Данные корректируют по фону флуоресценции, измеряемому на клетках, инкубированных только с реагентом второй стадии (MFI=7). Контрольное L6 mAb (80 мкг/мл) дает MFI<30. Эти результаты являются типичными для четырех независимых экспериментов.
Связывание L104EA29YIg, L104EIg и CTLA4Ig с человеческими CD80-трансфицированными СНО-клетками приблизительно эквивалентно. L104EA29YIg и L104EIg связываются более прочно с СНО-клетками, устойчиво трансфицируемыми с человеческими CD86, нежели CTLA4Ig (фиг.2В).
Функциональный анализ:
Человеческие CD4-позитивные Т-клетки выделяют иммуномагнитной негативной селекции (Linsley et al., (1992) J. Exp. Med. 176:1595-1604). Выделенные СD4-позитивные Т-клетки стимулируют форбаль-миристат-ацетатом (РМА) плюс CD80-позитивные или CD86-позитивные СНО-клетки в присутствии "титрующих" концентраций ингибитора. CD-позитивные Т-клетки (8-10·104/лунка) культивируют в присутствии 1 нМ РМА в присутствии или в отсутствие облученных стимуляторов СНО-клеток. Пролиферативный ответ измеряют, добавляя 1 мкКи/лунку [3Н]тимидина в течение последних 7 часов из 72 часов культивирования. Ингибирование Т-клеток, стимулированных с помощью РМА плюс CD80-позитивные CHO, или CD86-позитивные CHO, осуществляют при использовании L104EA29YIg и CTLA4Ig. Результаты показаны на фиг.3. L104EA29YIg ингибирует пролиферацию обработанных РМА СНО-клеток сильнее, чем CTLA4Ig (фиг.3А). L104EA29YIg также более эффективен, чем CTLA4Ig, при ингибировании пролиферации обработанных РМА С086-позитивных СНО-клеток (фиг.3В). Следовательно, L104EA29YIg представляет собой более мощный ингибитор опосредуемой как CD80-, так и CD-86 костимуляции Т-клеток.
На фиг.4 показано ингибирование с помощью L104EA29YIg и CTLA4Ig аллостимулированных человеческих Т-клеток, полученных как указано выше и дополнительно стимулированных при использовании линии человеческих В-лимфобластоидных клеток (LCL), называемой РМ, которая экспрессирует CD80 и CD86 (количество Т-клеток 3,0·104/лунка и РМ 8,0·103/лунка). Первичная аллостимуляция осуществляется в течение 6 дней, затем клетки выдерживают в пульсовом режиме с 3H-тимидином в течение 7 дней перед определением введенной радиоактивной метки.
Вторичную аллостимуляцию осуществляют следующим образом. Через семь дней первичные аллостимулированные Т-клетки собирают над средой для фракционирования лимфоцитов (LSM) (ICN, Aurora, ОН) и оставляют на 24 ч. Т-клетки снова стимулируют (вторично) в присутствии титрующихся количеств CTLA4Ig и L104EA29YIg при добавлении РМ в тех же соотношениях, какие указаны выше. Стимуляцию проводят в течение 3 дней, затем клетки выдерживают в пульсовом режиме с радиоактивной меткой и собирают как указано выше. Действие L104EA29YIg на первичные аллостимулированные клетки показано на фиг.4А. Действие L104EA29YIg на вторичные аллостимулированные клетки показано на Фиг.4В. L104EA29YIg ингибирует пролиферативный ответ как первичных, так и вторичных Т-клеток лучше, чем CTLA4Ig.
Для измерения продуцирования цитокинов (фиг.5) ставят дубликаты планшетов для вторичной аллостимуляции. Через 3 дня культуральные среды анализируют, используя наборы ELISA (Biosource, Camarillo, CA), в условиях, рекомендуемых производителем. Найдено, что L104EA29YIg более эффективно, чем CTLA4Ig, блокирует продуцирование Т-клетками IL-2, IL-4 и γ-IFN цитокинов после вторичного аллогенного раздражителя (фиг.5А-С).
Действие L104EA29YIg и CTLA4Ig на реакцию лимфоцитов обезьян в смешанной культуре (MLR) показано на фиг.6. Мононуклеарные клетки периферической крови (PBMC'S; 3,5·104 клеток/лунка от каждой обезьяны) от 2 обезьян очищают на среде для фракционирования лимфоцитов (LSM) и смешивают с2 мкг/мл фитогемагглютинина (РНА, ФГА). Клетки стимулируют в течение 3 дней, а затем в течение 16 ч выдерживают с радиоактивной меткой в пульсовом режиме перед сбором. L104EA29YIg ингибирует пролиферацию Т-клеток обезьяны лучше, чем CTLA4Ig.
Специалистам в области техники, к которой относится настоящее изобретение, ясно, что данное изобретение может быть воплощено в иных формах, нежели изложенные выше, не отступая при этом от сущности и основных характеристик изобретения. Следовательно, изложенные выше конкретные варианты изобретения следует рассматривать как иллюстрирующие, но не ограничивающие.






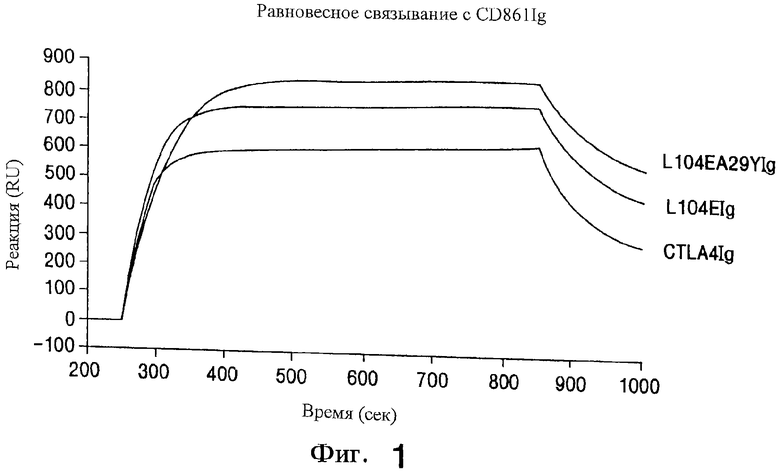

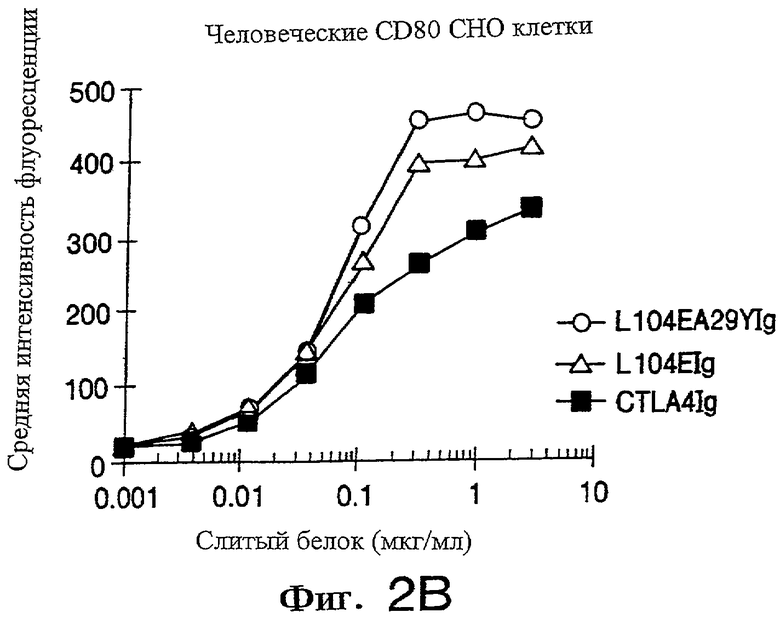


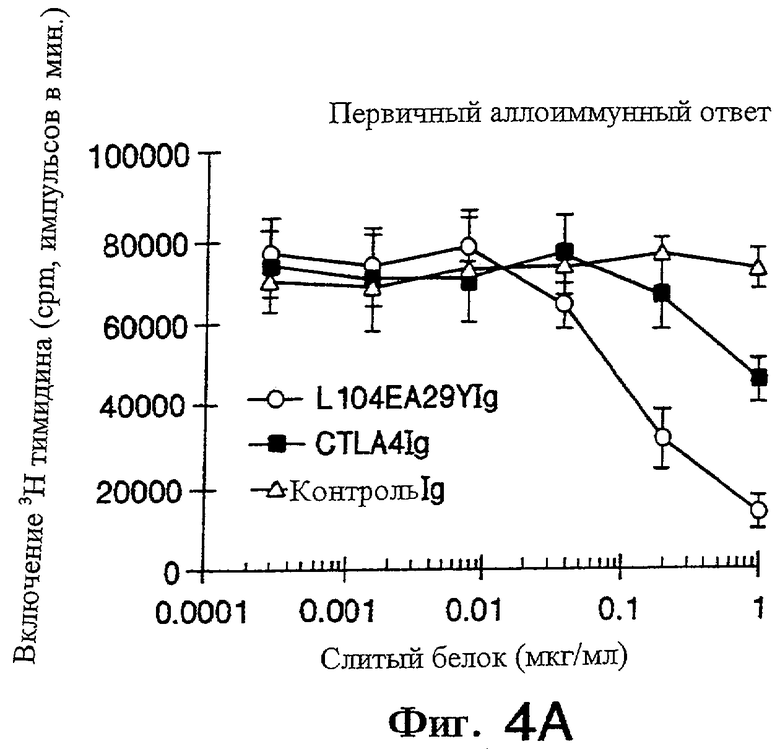




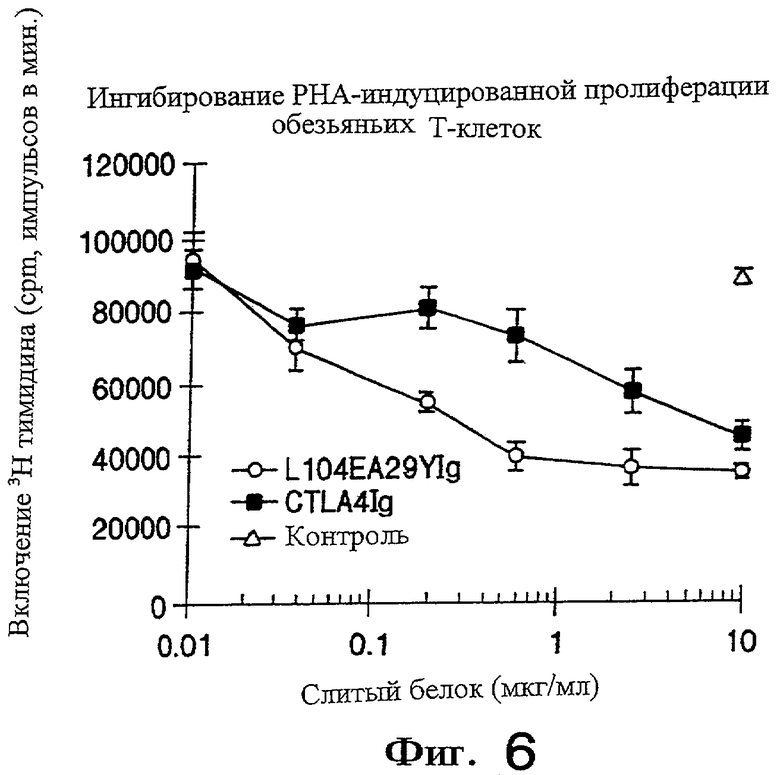



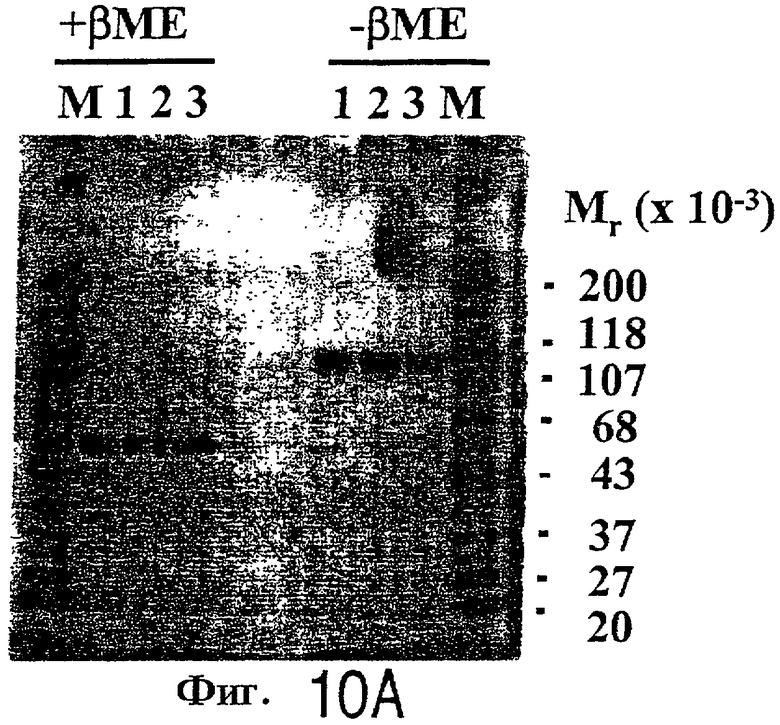




Изобретение относится к иммунобиотехнологии и касается растворимого CTLA4, являющегося мутантным вариантом CTLA4 дикого типа, сохраняющим способность связывать CD80 и/или CD86. Молекулы растворимого CTLA4 имеют первую аминокислотную последовательность, содержащую внеклеточную область CTLA4, где некоторые аминокислотные остатки в области S25-R33 и в области М97-G107 являются мутантными. Мутантные молекулы по изобретению могут также включать вторую аминокислотную последовательность, которая повышает растворимость мутантной молекулы. Описаны также молекулы нуклеиновой кислоты (НК), кодирующие указанные CTLA4 и включающие НК векторы. Изобретение раскрывает также способ получения мутантного CTLA4 и способы его использования: для регуляции взаимодействия Т-клетки с CD-80- и/или CD86-позитивной клеткой; для лечения заболеваний иммунной системы; для подавления реакции «трансплантат против хозяина». Растворимый мутантный CTLA4 по изобретению связывается с антигеном CD80 и/или CD86 с большей авидностью, чем CTLA4 дикого типа или немутированный CTLA41g. 33 н. и 32 з.п. ф-лы, 12 ил., 2 табл.
а) аминокислотную последовательность, показанную на фиг.7, начинающуюся с метионина в положении +1 и оканчивающуюся аспарагиновой кислотой в положении +124 или ее часть, которая связывается с CD80- и/или CD86, или
б) аминокислотную последовательность, показанную на фиг.7, начинающуюся с аланина в положении -1 и оканчивающуюся аспарагиновой кислотой в положении +124, или ее часть, которая связывается с CD80 и/или CD86, и
дополнительно содержащая аминокислотную последовательность, которая меняет растворимость, аффинность или валентность молекулы мутантного CTLA4.
Приоритет по пунктам:
| WO 9833513 A, 06.08.1998 г | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| СПОСОБ ИЗГОТОВЛЕНИЯ ИЗ ТОНКОЛИСТОВЫХ МАТЕРИАЛОВ ТОЛЩИНОЙ ОТ 0,2 мм И МЕНЕЕ СТУПЕНЧАТЫХ ОСЕСИММЕТРИЧНЫХ ОБОЛОЧЕК С ЦЕНТРАЛЬНЫМ ВЫСТУПОМ, ОТНОШЕНИЕ ВЫСОТЫ КОТОРОГО К ЕГО ДИАМЕТРУ РАВНО ИЛИ ПРЕВЫШАЕТ 0,5 | 2006 |
|
RU2327539C2 |
| WO 9949734 A, 07.10.1999 г | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Воздуховод | 1981 |
|
SU947582A1 |

