Настоящее изобретение относится к системе для уничтожения растительных клеток и прежде всего к трансгенным растениям, которые несут в своем геноме химерный ген, при экспрессии которого образуется цитотоксический протеин.
Одним из известных селекционерам растений путей получения новых сортов является создание гибридов существующих сортов, несущих требуемые признаки. Гибриды, как правило, отличаются от любого из родителей улучшенными различными характеристиками, это явление называют гибридной силой (гетерозис). Такие гибридные скрещивания можно осуществлять с помощью перекрестного опыления вручную, что представляет собой утомительную и требующую много времени процедуру.
В процессе осуществления таких гибридных скрещиваний предупреждение самоопыления имеет решающее значение. Для предупреждения самоопыления материнское растение можно эмаскулировать вручную, например, при получении гибридной кукурузы путем удаления метелки. Однако крупномасштабная эмаскуляция видов с гермафродитными цветками невозможна с экономической точки зрения. Можно также получать материнские линии (с мужской стерильностью) с помощью генетической мужской стерильности, этот хорошо известный у многих растений признак, как правило, является рецессивным и моногенным. Связанная с использованием этого подхода проблема состоит в трудности получения для каждого скрещивания чистых родительских линий с мужской стерильностью. Наиболее широко применяемая для создания гибридов система получения мужской стерильности представляет собой систему цитоплазматической мужской стерильности (cms). В этом случае недоразвитие пыльцы обусловлено цитоплазматическими факторами. У тех культурных растений, у которых в зародышевой плазме выявлена cms, например у кукурузы и подсолнечника, этот признак широко используется для создания гибридов. Известно несколько недостатков указанной системы: цитоплазма с мужской стерильностью может нести другие нежелательные характеристики, например Т-цитоплазму у кукурузы и чувствительность к Helminthosporium maydis; при ее применении необходимо поддерживать линии с изогенной мужской фертильностью; и применение этой системы ограничено видами, у которых доступен цитоплазматический источник стерильности.
Другим преимуществом системы, основанной на использовании мужской стерильности, является то, что с ее помощью можно получать беспыльцевые растения. Это требуется для многих сортов декоративных цветковых растений и также может найти применение для борьбы с нежелательными генетическими признаками путем предупреждения ауткроссинга.
Еще одним ценным признаком основанной на использовании стерильности системы является возможность получать растения с женской стерильностью, у которых при развитии плодов не происходит образования семян. Бессеменные сорта плодов имеют преимущества при обработке, например, томатов и также являются предпочтительными для потребителя, например, дыни. В настоящее время бессеменные сорта известны и существуют соответствующие программы селекции, но создание бессеменных плодов ограничено отсутствием у многих видов растений пригодной зародышевой плазмы.
В тех случаях, когда генетический источник стерильности отсутствует или недоступен по другим причинам, можно применять метод генетической модификации, который обеспечивает стерильность с помощью системы для уничтожения клеток, при этом некроз происходит в специфических клетках репродуктивных тканей.
В WO 89/10396 описана система для уничтожения растительных клеток, при использовании которой в растение интродуцируют химерный ген, содержащий специфичный для пыльника промотор, связанный с протеином или полипептидом РНКазы, при экспрессии которого происходит нарушение клеточного метаболизма. Таким образом, экспрессия химерного гена приводит к некрозу клеток пыльника и, как следствие, к мужской стерильности растений.
Для создания женской стерильности у растений можно использовать аналогичную систему для уничтожения клеток, мишенью которой является семяпочка.
В WO 93/18170 и WO 92/04453 описаны системы для уничтожения растительных клеток, специфические для борьбы с заражением нематодами. Согласно описанным в WO 93/18170 и WO 92/04453 системам в определенные виды растений-хозяев интродуцируют ген, содержащий кодирующую последовательность, причем кодирующая последовательность кодирует продукт, снижающий вредоносное действие нематод. Ген содержит также промоторную область, которая контролирует экспрессию кодирующей последовательности таким образом, что экспрессия происходит при нападении нематод и осуществляется практически специфично в клетках областей, которыми питаются нематоды, или в соседних клетках. Для снижения вредоносного действия нематод продукт либо может оказывать неблагоприятное воздействие на растительные клетки, которые в результате дифференцировки превращаются в клетки областей, которыми питаются нематоды, либо на соседние с ними клетки, либо может оказывать неблагоприятное воздействие непосредственно на нематод.
Имеющие экономическое значение паразитирующие на растениях нематоды включают цистообразующие нематоды, такие как картофельные нематоды (Globodera rostochiensis и G.pallida), соевая нематода (Heterodera glycines), свекловичная нематода (Heterodera schachtii) и зерновая нематода (Heterodera avenae), и галловые нематоды, такие как Meloidogyne spp. Такие паразитирующие на растениях нематоды являются основными патогенами многих культурных растений во всем мире, например овощных, кормовых бобовых культур, томатов, арбузов, винограда, арахиса, табака и хлопчатника.
Наиболее широко распространенные современные средства борьбы с нематодами основаны на применении химических средств защиты растений, агротехнических приемов и устойчивых сортов растений, и их часто используют в интегрированной системе борьбы с паразитирующими на растениях нематодами. Имеется необходимость в совершенствовании системы борьбы с нематодами, поскольку указанные существующие в настоящее время подходы не обеспечивают надежную защиту посевов. Безопасность нематицидов для окружающей среды является проблематичной, и они не всегда обладают достаточно высокой эффективностью. Однако применение системы защиты культурных растений может быть убыточным для фермеров вследствие различных причин. Широкий круг хозяев галловых нематод ограничивает возможность применения экономически значимых культур, которые не являются мишенью для нематод. Эффективные устойчивые сорта часто не являются доступными, а те, которые фермер может применять, иногда при невысокой численности нематод оказываются хуже чувствительных сортов. Кроме того, устойчивость может снижаться при высоких температурах почвы, что характерно для тропической и субтропической зон.
Можно рассматривать и другие пути применения систем для уничтожения растительных клеток. Например, мишенью могут быть определенные части цветка, что обусловливает изменение морфологии цветка. В другом варианте мишенью могут быть боковые корни, шипы или жгучие волоски. Можно достигать отделения (абсцизии) листьев или плодов, когда мишенью является отделительный слой листа или плода. Можно достигать также облегчения сбрасывания семян в случае, когда мишенью является семяножка. При использовании в качестве мишеней других органов, таких как трихомы, которые, как правило, являются железистыми (гландулярными), производство химических субстанций в этих органах можно прекращать или предупреждать. Другим вариантом применения может быть индуцибельное отделение корней, листьев, цветков или плодов в конце вегетационного периода.
В NZ 260511 предложена система для уничтожения растительных клеток, отличающаяся повышенной тканеспецифичностью. Эта система позволяет осуществлять экспрессию цитотоксической молекулы (под контролем первого промотора, где первый промотор обеспечивает экспрессию в специфических клетках-мишенях и в одной или нескольких других областях растения) в сочетании с защитной молекулой (под контролем второго промотора, где второй промотор обеспечивает экспрессию во всех областях, в которых первый промотор проявляет активность, кроме специфических клеток-мишеней). Примерами приемлемых цитотоксической и защитной молекул являются протеазы и ингибиторы протеаз соответственно или нуклеазы и ингибиторы нуклеаз соответственно. В WO 93/10251 описано применение цитотоксической рибонуклеазы барназы в сочетании с защитной молекулой-ингибитором барстаром.
В WO 98/44138 описан метод повышения специфичности экспрессии гена путем направленного переноса специфического сайта экспрессии гена-мишени. Таким образом получают химерный ген, при этом химерный ген содержит промотор, который обеспечивает экспрессию более чем в одной области организма, подлежащего воздействию. Промотор связывают с агентом, оказывающим воздействие на функцию эндогенного гена в растении, который также экспрессируется более чем в одной области растения. Промотор и агент выбирают так, чтобы сайты их экспрессии перекрывались в одной или в нескольких требуемых областях. Наличие этого(их) перекрывающегося(ихся) сайта(ов) обеспечивает повышение специфичности и направленности экспрессии гена.
Инактивирующие рибосому протеины (RIP) представляют собой группу токсичных растительных протеинов, которые каталитически инактивируют рибосомы эукариотических организмов (Stirpe и Barbieri, 1986). RIP функционируют в качестве N-гликозидаз, осуществляя удаление определенного аденина в консервативной петле большой молекулы рРНК и тем самым предупреждает связывание фактора удлинения 2, блокируя в результате синтез клеточного протеина.
Описаны три формы RIP. RIP типа 1, такие как антивирусный протеин фитолакки американской (РАР) и ингибитор трансляции ячменя, каждый из которых имеет одну полипептидную цепь, величина относительной молекулярной массы (Mr) которой составляет примерно 30000. Все RIP типа 2, такие как рицин, абрин и модекцин, имеют по две полипептидные цепи. Один обладающий RIP-активностью полипептид (А-цепь) связывают с помощью дисульфидного мостика со связывающим галактозу лектином (Б-цепь; Stirpe и др., 1978). Величина Мr каждого RIP типа 2 составляет примерно 60000. RIP типа 3, такие как RIP кукурузы, имеют одну полипептидную цепь, которая затем подвергается протеолитическому расщеплению с высвобождением двух активных пептидных доменов.
Фитолакка американская (Phytolacca americana) продуцирует три различных антивирусных протеина, а именно PAP', PAPII и PAP-S, которые характерны для весенних листьев, летних листьев и семян соответственно. Установлено сходство аминокислотных последовательностей этих протеинов. В контексте настоящего описания "РАР" относится ко всем трем этим антивирусным протеинам. РАР' или PAPII находятся в матриксе клеточной оболочки мезофильных клеток листа и их можно выделять с помощью водной экстракции мацерированной листовой ткани (Ready и др., 1986). Было установлено, что при их экзогенном нанесении на поверхность листьев растений можно осуществлять защиту нескольких различных растений-хозяев от заражения широким спектром вирусов (Chen и др., 1991; Lodge и др., 1993).
В патенте US 6015940 описано создание клона кДНК РАР', полученного из весенних листьев фитолакки американской, и его применение под контролем конститутивного промотора (либо промотора 35S вируса мозаики цветной капусты, либо промотора 35S вируса мозаики норичника шишковатого) для получения трансгенных растений табака и картофеля, устойчивых к вирусам полиомы PVX и PVY. В качестве негативной характеристики устойчивого фенотипа следует отметить, что у растений, уровни экспрессии РАР' которых превышали 10 нг/мг протеина, обнаружена крапчатость листьев и задержка роста. Растения, накапливающие более высокие уровни РАР', были стерильными.
Трансгенные растения, содержащие характерные для летних листьев формы РАР, т.е. PAP-II, описаны в WO 99/60843. Большое количество полноразмерных и укороченных генных последовательностей PAP-II подвергали скринингу для выявления таких вариантов протеинов PAP-II, которые сохраняли антивирусную активность, но не обладали фитотоксичностью. Трансгенные растения обладали как антивирусной, так и фунгицидной активностью.
Ген РАР сначала экспрессируется в листьях in vivo с получением неактивного протеина Pro-РАР. Известно, что после трансляции происходит направленный перенос молекулы протеина Pro-РАР' в клеточную оболочку. На определенной стадии этого процесса N- и С-концевые удлиняющие сегменты молекулы Pro-РАР' отщепляются с образованием активированной молекулы РАР' (зрелый РАР'). Для PAP-S (который экспрессируется в семенах) известна клеточная локализация. Однако N-концевая процессированная область PAP-S, вероятно, имеет свойства, аналогичные свойствам сигнальных последовательностей, обеспечивающих направленный перенос.
Строение зрелого протеина PAP-S, т.е. протеина, у которого удалены N- и С-концевые удлиняющие сегменты, можно охарактеризовать наличием двух отдельных доменов, соответствующих RIP типа 3 или двум полипептидам RIP типа 2, т.е. домена связывания рибосомы и каталитического домена. Указанные в настоящем описании молекулы представляют собой рекомбинантные молекулы PAP-S, содержащие либо последовательность PAP-S α, либо последовательность PAP-S β. Исходя из структурных особенностей, можно предположить, что PAP-S α содержит области распознающего РНК мотива и домена связывания рибосомы, а PAP-S β содержит сайт имеющего решающее значение для катализа остатка. При создании настоящего изобретения неожиданно было установлено, что экспрессия только либо протеина PAP-S α, либо протеина PAP-S β приводит к значительному ингибированию активности рибосом.
Настоящее изобретение относится к способу индукции некротического действия в специфических клетках растения, согласно которому растение трансформируют химерным геном, где кодирующая последовательность гена кодирует зрелый антивирусный протеин фитолакки американской или его фрагмент, этот ген содержит промотор, который проявляет активность в ответ на специфическое раздражающее воздействие на это растение, в результате чего зрелый антивирусный протеин фитолакки американской или его фрагмент экспрессируются в специфических клетках растения.
Настоящее изобретение относится также к растению, трансформированному химерным геном, причем кодирующая последовательность гена кодирует зрелый антивирусный протеин фитолакки американской или его фрагмент, этот ген содержит промотор, который проявляет активность в ответ на специфическое раздражающее воздействие на это растение, в результате этот зрелый антивирусный протеин фитолакки американской или его фрагмент экспрессируются в специфических клетках растения.
Настоящее изобретение относится также к рекомбинантной растительной клетке, трансформированной химерным геном, где кодирующая последовательность гена кодирует зрелый антивирусный протеин фитолакки американской или его фрагмент, этот ген содержит промотор, который проявляет активность в ответ на специфическое раздражающее воздействие на это растение, в результате этот зрелый антивирусный протеин фитолакки американской или его фрагмент экспрессируются в специфических клетках растения.
Кроме того, настоящее изобретение к выделенной молекуле ДНК, содержащей химерный ген, где кодирующая последовательность гена кодирует зрелый антивирусный протеин фитолакки американской или его фрагмент, этот ген содержит промотор, который проявляет активность в ответ на специфическое раздражающее воздействие на это растение, в результате этот зрелый антивирусный протеин фитолакки американской или его фрагмент экспрессируются в специфических клетках растения.
Настоящее изобретение относится также к биологически функциональному экспрессионному вектору, содержащему химерный ген, причем кодирующая последовательность гена кодирует зрелый антивирусный протеин фитолакки американской или его фрагмент, этот ген содержит промотор, который проявляет активность в ответ на специфическое раздражающее воздействие на это растение, в результате этот зрелый антивирусный протеин фитолакки американской или его фрагмент экспрессируются в специфических клетках растения.
В контексте настоящего описания понятие "фрагмент" обозначает фрагмент гена, кодирующего антивирусный протеин фитолакки американской, причем этот фрагмент обладает активностью в отношении ингибирования синтеза протеинов.
Химерный ген предпочтительно кодирует зрелый протеин PAP-S, нуклеотидная последовательность которого представлена в SEQ. ID. No. 3, а аминокислотная последовательность представлена в SEQ. ID. No. 4, или PAP-S α, нуклеотидная последовательность которого представлена в SEQ. ID. No. 5, а аминокислотная последовательность представлена в SEQ. ID. No. 6, или PAP-S β, нуклеотидная последовательность которого представлена в SEQ. ID. No. 7, а аминокислотная последовательность представлена в SEQ. ID. No. 8, или зрелый протеин РАР' или его вариант, описанный в патенте США 6015940 (т.е. нуклеотиды 290-1076 SEQ. ID. No. 30 и 31), или зрелый протеин PAPII, описанный в международной заявке на патент WO 99/60843 (т.е. нуклеотиды 75-903 SEQ. ID. No. 32).
В зависимости от требуемой гомологии нуклеотидных последовательностей можно применять различные условия строгости гибридизации для скрининга аналогичных последовательностей. В качестве примера (но не ограничиваясь ими) для процесса гибридизации используют следующие строгие условия: гибридизация со связанной с фильтром ДНК в 0,5М NaHPO4, 7%-ном додецилсульфате натрия (ДСН), 1 мМ ЭДТК при 65°С и отмывка в 0,1×SSC/0,1% ДСН при 68°С (Ausubel F.M. и др., (ред.), 1989, Current Protocols in Molecular Biology, том I, изд-во Green Publishing Associates, Inc. и John Wiley и Sons, Inc., New York, c.2.10.3). Другие строгие условия, которые можно использовать, хорошо известны в данной области. Можно применять процессы гибридизации в следующих условиях умеренной строгости: гибридизация со связанной с фильтром ДНК в 0,5М NaHPO4, 7%-ном додецилсульфате натрия (ДСН), 1 мМ ЭДТК при 65°С и отмывка в 0,2×SSC/0,l% ДСН при 42°С (Ausubel и др., 1989, выше). Другие условия умеренной строгости, которые можно использовать, хорошо известны в данной области. Для процессов гибридизации можно применять другие растворы, такие как стандартный раствор хлорида и цитрата натрия (SSC) или хлорида натрия и фосфата ЭДТК (SSPE).
Приемлемые гомологичные последовательности представляют собой последовательности, которые по меньшей мере на 70%, предпочтительно на 80% и еще более предпочтительно на 90 или 95% гомологичны каждой из приведенных в настоящем описании последовательностей, причем такие гомологичные последовательности сохраняют требуемую ферментативную активность.
Приемлемые последовательности могут также представлять их варианты. Согласно настоящему изобретению вариант может представлять собой любую замену, вариацию, модификацию, замещение или делецию или добавление одной или нескольких нуклеотидов/аминокислот, позволяющую получать последовательность, которая экспрессирует или проявляет требуемую ферментативную активность. Согласно изобретению можно также использовать производное или мутацию. Производное имеет определенные модификации, как правило химические, по сравнению со встречающимся в естественных условиях полипептидом, экспрессируемым нуклеиновой кислотой.
Другим объектом настоящего изобретения является способ индукции некротического действия в специфических клетках растения, где растение трансформируют двумя химерными генами, причем кодирующая последовательность одного из указанных генов кодирует инактивированный антивирусный протеин фитолакки американской, а кодирующая последовательность другого из указанных генов кодирует вторую молекулу, которая представляет собой активатор инактивированного антивирусного протеина фитолакки американской, каждый из двух генов содержит промотор, причем эти промоторы действуют совместно в ответ на специфическое раздражающее воздействие на это растение, в результате этот инактивированный антивирусный протеин фитолакки американской активируется в специфических клетках растения.
Настоящее изобретение относится также к растению, трансформированному двумя химерными генами, причем кодирующая последовательность одного из указанных генов кодирует инактивированный антивирусный протеин фитолакки американской, а кодирующая последовательность другого из указанных генов кодирует вторую молекулу, которая представляет собой активатор инактивированного антивирусного протеина фитолакки американской, каждый из двух генов содержит промотор, причем эти промоторы действуют совместно в ответ на специфическое раздражающее воздействие на это растение, в результате этот инактивированный антивирусный протеин фитолакки американской активируется в специфических клетках растения.
Настоящее изобретение относится также к рекомбинантной растительной клетке, выделенной молекуле ДНК двух химерных генов и к биологически функциональному экспрессионному вектору, содержащему два химерных гена, где каждый химерный ген подпадает под второй объект настоящего изобретения.
В контексте настоящего описания понятие "инактивированный антивирусный протеин фитолакки американской" обозначает зрелый антивирусный протеин фитолакки американской, такой, например, как PAP-S, функционально связанный с гетерологичной или гомологичной N-или С-концевой блокирующей последовательностью.
Химерный ген предпочтительно кодирует зрелый протеин PAP-S, нуклеотидная последовательность которого представлена в SEQ. ID. No. 3, а аминокислотная последовательность представлена в SEQ. ID. No. 4, или PAP-S α, нуклеотидная последовательность которого представлена в SEQ. ID. No. 5, а аминокислотная последовательность которого представлена в SEQ. ID. No. 6, или PAP-S β, нуклеотидная последовательность которого представлена в SEQ. ID. No. 7, а аминокислотная последовательность которого представлена в SEQ. ID. No. 8, или зрелый PAP' или его вариант, описанный в патенте США 6015940 (т.е. нуклеотиды 290-1076 SEQ. ID. No. 30 и 31), или зрелый PAPII, описанный в международной заявке, опубликованной под No. WO 99/60843 (т.е. нуклеотиды 75-903 SEQ. ID. No. 32), каждый связанный с гетерологичной или гомологичной N- или С-концевой блокирующей последовательностью, как описано выше для получения инактивированного антивирусного протеина фитолакки американской.
Концевая блокирующая последовательность предпочтительно представляет собой последовательность, которая может оказывать дополнительное некротическое действие на клетку или патоген.
Предпочтительно концевая блокирующая последовательность представляет собой последовательность оризацистатина (Δ86 последовательность цистатина), представленную в настоящем описании как SEQ. ID. No. 17. Последовательность Δ86 цистатина блокирует действие встречающихся в естественных условиях протеаз, предотвращая тем самым встречающуюся в естественных условиях экспрессию кодирующей последовательности РАР. Цистатины представляют собой небольшие (˜100 аминокислот) ингибиторы протеинов цистеиновых протеаз, которые обнаружены во многих видах растений [Ryan, 1990; Richardson, 1991], и большого количества генов (>60), последовательность которых установлена. Фитоцистатины представляют собой класс, отличный от цистатинов типа I и II [Kondo и др., 1991]. Оризацистатин I (ОС-I) семян риса [Abe и др., 1987] представляет собой эффективный ингибитор, однако существенно менее эффективный, чем цистатины животных, такие как цистатин из белка куриных яиц (CEWC). Описаны рекомбинантные варианты ОС-1, полученные с помощью конструирования протеинов (Uwrin и др., 1995), и делеционный мутант Δ-Asp86 (ОС-1Δ86), обладающий более высокой ингибирующей активностью.
В другом варианте концевая блокирующая последовательность представляет собой только молекулу, инактивирующую молекулу зрелого РАР. Предпочтительно блокирующая концы последовательность представляет собой встречающуюся в естественных условиях концевую последовательность Pro-РАР, полученную из любой молекулы Pro-РАР.
Блокирующая последовательность может быть локализована на N-конце или на С-конце зрелой последовательности РАР.
Предпочтительно блокирующая последовательность представляет собой С-концевую блокирующую последовательность.
Для обеспечения реактивации блокированной последовательности зрелого РАР в инактивированном антивирусном протеине фитолакки американской также должен присутствовать специфический сайт расщепления, локализованный между последовательностью зрелого антивирусного протеина фитолакки американской и блокирующей последовательностью. Сайт расщепления предпочтительно локализован между зрелым антивирусным протеином фитолакки американской и блокирующей последовательностью.
Предпочтительно специфический сайт расщепления представляет собой сайт расщепления, специфический для протеазы NIa вируса табачной гравировки (TEV) (Carrington и Dougherty, 1998), представленный в настоящем описании как SEQ. ID. NO. 28 и SEQ. ID. No. 29. Распознаваемый указанной протеиназой сайт представляет собой гептапептид Glu-Xaa-Xaa-Tyr-Xaa-Gln-Gly или гептапептид Glu-Xaa-Xaa-Tyr-Xaa-Gln-Ser, причем расщепление происходит между остатками Gin и Gly (или Ser).
Кодирующая последовательность второй молекулы предпочтительно представляет собой протеазу, которая обладает способностью расщеплять специфический сайт расщепления. Когда специфический сайт расщепления представляет собой сайт расщепления протеазой (PCS) TEV, то протеаза может представлять собой NIa-протеазу TEV. Специалисты в данной области могут использовать другие специфические сайты расщепления протеазами. Например, сайтспецифическую протеазу энтерокиназу можно применять для расщепления соответствующего сайта расщепления, связанного со зрелым антивирусным протеином фитолакки американской.
Согласно одному из объектов изобретения указанное раздражающее воздействие на растение может быть вызвано нападением патогена. В другом варианте раздражающим воздействием может быть химическая индукция или индукция в результате естественного развития растения.
Как должно быть очевидно специалистам в данной области, первый и второй объекты изобретения могут найти широкое применение для всех представителей растительного царства с целью защиты их, например, от грибов, нематод, бактерий и вирусов, а также для других целей.
Следует понимать, что в контексте настоящего описания понятие "некротическое действие" подразумевает значительное нарушение метаболизма, чтобы при применении изобретения достигать требуемого воздействия, например обеспечивать защиту от болезней.
Предпочтительно согласно второму объекту настоящего изобретения промоторы (двухкомпонентная система) имеют перекрывающуюся (иеся) зону(ы) экспрессии. Характеристики перекрывающейся экспрессии двух промоторов и их соответствующие реакции на указанные воздействия являются такими, что они оказывают воздействие на направление экспрессии инактивированного антивирусного протеина фитолакки американской и его молекулы-активатора, приводящие к тому, что летальное или вредное воздействие затрагивает специфические клетки и только их.
В другом варианте промоторы могут иметь одни и те же сайты экспрессии или экспрессируемые протеины могут иным образом накапливаться в области-мишени.
Третьим объектом настоящего изобретения является способ индукции некротического действия в специфических клетках растения, согласно которому растение трансформируют химерным геном, при этом кодирующая последовательность гена кодирует предшественник молекулы РАР или ее С-концевого делеционного варианта, причем ген содержит промотор, который проявляет действие в ответ на специфическое раздражающее воздействие на растение, в результате экспрессируемый кодирующей последовательностью протеин экспрессируется в специфических клетках растения, причем указанный промотор следует выбирать таким образом, чтобы обеспечивать одно из следующих воздействий: снижение вредоносного действия нематод, стерильность, изменения в морфологии цветка, отделение (листьев или плодов), сбрасывание семян или развитие трихом.
Настоящее изобретение относится также к растению, трансформированному согласно методу, который является третьим объектом изобретения. Настоящее изобретение относится также к рекомбинантной растительной клетке, выделенной ДНК химерного гена и биологически функциональному экспрессионному вектору, где они все содержит химерный ген, описанный в третьем объекте изобретения.
Согласно третьему объекту изобретения кодирующая последовательность предпочтительно кодирует одну из следующих последовательностей: нуклеотидную последовательность Pro-PAP-S, представленную в SEQ. ID. No. 1, и аминокислотную последовательность, представленную в SEQ. ID. No. 2, или PAP' или его варианта, описанного в патенте США 6015940, который имеет последовательность, представленную в SEQ. ID. No. 30 и 31, или PAPII, последовательность которого описана в международной заявке на патент, опубликованной под No. WO 99/60843 и представленной в настоящем описании как SEQ. ID. No.32.
Выбор приемлемых промоторов должен зависеть от выбора специфических клеток. Если целью является защита от нападения нематод, то специфические клетки могут представлять собой клетки областей, которыми питаются нематоды. В этом случае согласно второму объекту изобретения приемлемые промоторы могут представлять собой промотор KNT1, действующий совместно с промотором TobRB7. Согласно первому и третьему объектам изобретения эти промоторы можно применять по отдельности. Метод выделения промотора KNT1 описан в новозеландском патенте 260511, а также дополнительно описан ниже, а метод выделения промотора TobRB7 описан в международной заявке на патент WO 94/17194. Объект изобретения указанной заявки, касающийся выделения промотора, включен в настоящее описание в качестве ссылки. Приемлемыми промоторами являются также промоторы Lemmi, выделение которых описано в международной заявке на патент, опубликованной под No. WO 92/21757.
Согласно второму объекту, например, кодирующая последовательность антивирусного протеина фитолакки американской может находиться под контролем промотора TobRB7, в то время как кодирующая последовательность молекулы-активатора под контролем промотора KNT1. Промотор KNT1 обеспечивает экспрессию в клетках областей, которыми питаются нематоды, кончиках корней и в меньшей степени в других видах меристемы, в то время как промотор TobRB7 обеспечивает экспрессию в корнях и гигантских клетках (Conkling и др., 1990). В указанных исследованиях продемонстрировано, что промотор TobRB7 может осуществлять экспрессию в массе корня, но не в кончиках корня. Таким образом, зоной перекрывающейся экспрессии промоторов KNT1 и TobRB7 являются гигантские растительные клетки (клетки областей, которыми питаются нематоды).
В другом варианте, когда необходимо обеспечивать мужскую стерильность, специфичные клетки могут представлять собой клетки пыльника растений. Примеры приемлемых промоторов описаны у Twell и др. (1991) и Mariani и др. (1990).
Согласно первому варианту осуществления настоящего изобретения область-мишень может представлять собой область, которой питаются нематоды. В случае, когда область-мишень представляет собой область, которой питаются нематоды, в качестве промотора выбирают промотор, который индуцируется в области, которой питаются нематоды, и/или в соседней области. Индукция такого промотора предпочтительно происходит при поражении растения нематодами. Примером приемлемого промотора является промотор KNT1. Другие приемлемые промоторы включают промотор TobRB7 и промоторы Lemmi.
Область, которой питаются нематоды, может включать, например, растительные клетки, находящиеся в локальной области поражения, которые впоследствии подвергаются редифференцировке, образуя синцитий (в случае цистообразующих нематод), или гигантские клетки и/или сопутствующие гипертрофические клетки (в случае галловых нематод), и/или одну или несколько клеток синцития, гигантских клеток и сопутствующих гипертрофических клеток.
При использовании в качестве мишени области, которой питаются нематоды, можно получать устойчивые к нематодам растения. Понятие "устойчивое к нематодам растение" относится к растению, которое при заражении паразитирующими на растении нематодами обладает способностью предупреждать, замедлять или иным образом оказывать отрицательное воздействие на рост и развитие нематод, поражающих растение, предупреждая тем самым достижения численности паразитирующих на растениях нематод экономически значимого уровня в период вегетации культуры. При этом подразумевается, что нематоды могут, например, погибать или жизненный цикл нематод может замедляться, что приводит к удлинению времени, необходимого для достижения половой зрелости и откладки яиц, или женские особи нематод могут иметь уменьшенный размер и вследствие этого пониженную способность к откладке яиц, которое происходит только после достижения женскими особями нематод минимального критического размера.
Настоящее изобретение применимо к следующим видам нематод (но не ограничиваясь ими): Globodera spp., Heterodera spp. и Meloidogyne spp.
Согласно второму варианту осуществления настоящего изобретения способ может быть направлен не на достижение устойчивости к нематодам, а на получение мужской стерильности у растений. Например, областью-мишенью может быть один или несколько из таких органов растения, как пыльца, пыльник или тапетум. Если областью-мишенью является, например, тапетум, то выбранный промотор представляет собой промотор, который индуцируется в тапетуме и/или в соседних с тапетумом областях. Примером приемлемого для тапетума промотора является промотор ТА29 табака, описанный у Mariani и др. (1990). Специфичные для пыльника промоторы описаны у Twell и др. (1991).
Согласно третьему варианту осуществления настоящего изобретения способ применяют для достижения женской стерильности растений. Например, область-мишень может представлять собой семяпочку растения. Это предусматривает, что выбранный промотор представляет собой промотор, который индуцируется в семяпочке и/или соседней области. Примером приемлемого промотора является промотор AGL15, описанный у Perry и др. (1996).
Согласно четвертому варианту осуществления настоящего изобретения оказывают воздействие на морфологическое строение цветка. Например, областью-мишенью являются определенные части цветка, при этом целью является, чтобы эти определенные части цветка не развивались и морфологическое строение цветка изменялось. В этом случае выбранный промотор представляет собой промотор, который индуцируется в чашелистике, плодолистике, лепестке и/или тычинке, или в соседних областях. Примерами приемлемых промоторов являются промоторы генов, обусловливающих бесполость (agamous), отсутствие лепестков (apetala3), шарообразность (globosa), пестичность (pistillata) и другие различные дефициты (Sieburth и Meyerowitz, 1997; Samach и др., 1997 и приведенных в этих публикациях ссылках).
Согласно пятому варианту осуществления настоящего изобретения способ по изобретению применяют для того, чтобы способствовать или усиливать абсцизию листьев и/или плодов у растений. Например, область-мишень может представлять собой отделительный слой листа и/или плода. Выбранный в этом случае промотор представляет собой промотор, который индуцируется в отделительном слое и/или в соседней области.
Согласно шестому варианту осуществления настоящего изобретения мишенью являются трихомы, при этом трихомы, как правило, являются гландулярными. Выбранный промотор представляет собой промотор, который индуцируется в трихомах и/или соседних областях. Вызывая некроз трихомов растения, можно прекращать или предупреждать производство трихомом химических субстанций.
Согласно седьмому варианту осуществления настоящего изобретения мишенью являются боковые корни, шипы или жгучие волоски.
Согласно восьмому варианту осуществления настоящего изобретения способ предназначен для борьбы с вирусной инфекцией. При вирусных инфекциях в значительной степени пораженных вирусом клетках специфично или практически специфично индуцируются многочисленные гены. Выбранный промотор представляет собой промотор, который индуцируется в пораженных вирусом клетках и/или в соседней области.
Согласно девятому варианту осуществления настоящего изобретения способ предназначен для облегчения сбрасывания семян с растения, когда мишенью являются семена. Известно большое количество генов, которые в процессе развития семян специфично или практически специфично индуцируются в определенных клетках/частях семени.
Согласно десятому варианту осуществления настоящего изобретения выбирают промотор, который индуцируется внешним стимулом и который индуцируется, например, в корнях растения. Такой промотор можно использовать для отделения корня в конце вегетационного периода. Для отделения листьев, цветков или плодов в конце вегетационного периода можно применять промоторы, которые индуцируются, например, в черешках листьев, цветоножках или плодоножках.
Каждый химерный ген может дополнительно содержать 3'-нетранслируемую последовательность терминатора. Последовательность терминатора можно получать из генов растений, бактерий или вирусов. Пригодные последовательности терминаторов представляют собой, например, последовательность терминатора rbcS гена Е9 гороха, последовательность терминатора Nos, выведенную из гена нопалинсинтазы Agrobacterium tumefaciens, и последовательность терминатора 35S вируса мозаики цветной капусты. Специалисту в данной области известны другие последовательности терминаторов. Химерный ген может также необязательно содержать последовательности энхансеров транскрипции или трансляции, например, описанные в международной заявке на патент, опубликованной под No. WO 97/20056, например внутриклеточные, обеспечивающие направленный перенос последовательности, и интроны, а также нуклеотидные последовательности, функцией которых является облегчение процесса трансформации и стабильной экспрессии химерного гена, такие как пограничные области Т-ДНК, области, обеспечивающие присоединение к гиалоплазме, и последовательности эксцизии/рекомбинации.
Методы трансформации растений хорошо известны в данной области и включают, например, опосредуемую Agrobacterium трансформацию. Как правило, при опосредуемой Agrobacterium трансформации бинарный вектор, несущий представляющую интерес чужеродную ДНК, т.е химерный ген, переносят из соответствующего штамма Agrobacterium в растение-мишень посредством совместного культивирования Agrobacterium с эксплантатами растения-мишени. Трансформированную ткань растения затем регенерируют на средах для селекции, причем среды для селекции содержат селектируемый маркер и гормоны роста растений.
Пригодные методы трансформации включают, например, также непосредственный перенос генов в протопласты с помощью полиэтиленгликоля или электропорации, бомбардировку частицами, микроинъекцию и применение волокон из карбида кремния.
Приемлемые виды растений, которые можно трансформировать согласно настоящему изобретению, включают (но не ограничиваясь ими) рис, пшеницу, кукурузу, канолу, картофель, табак, сахарную свеклу, сою, томаты, арахис, хлопчатник, виноград, арбуз, папайю, овощные культуры и кормовые бобовые растения.
Согласно второму объекту изобретения оба химерных гена интродуцируют в растение в одной кассете экспрессии. В другом варианте каждый химерный ген интродуцируют в растение в различных кассетах экспрессии путем последовательной или одновременной трансформации с помощью двух трансгенных конструкций. Такую двухкомпонентную систему можно также получать путем скрещивания двух растений, каждое из которых содержит один индивидуальный компонент.
Для облегчения понимания и осуществления на практике изобретение более подробно рассмотрено ниже со ссылкой на прилагаемые чертежи, на которых показано:
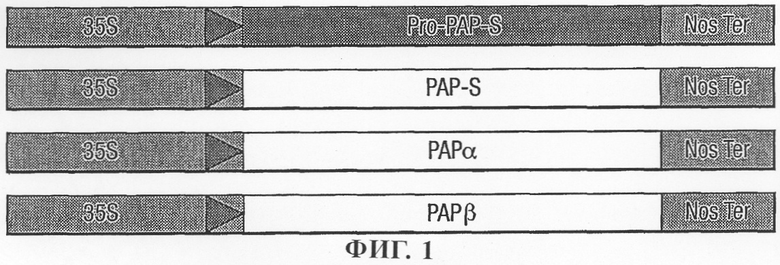
на фиг.1 - схематическое изображение промоторных конструкций для экспрессии РАР в протопластах;
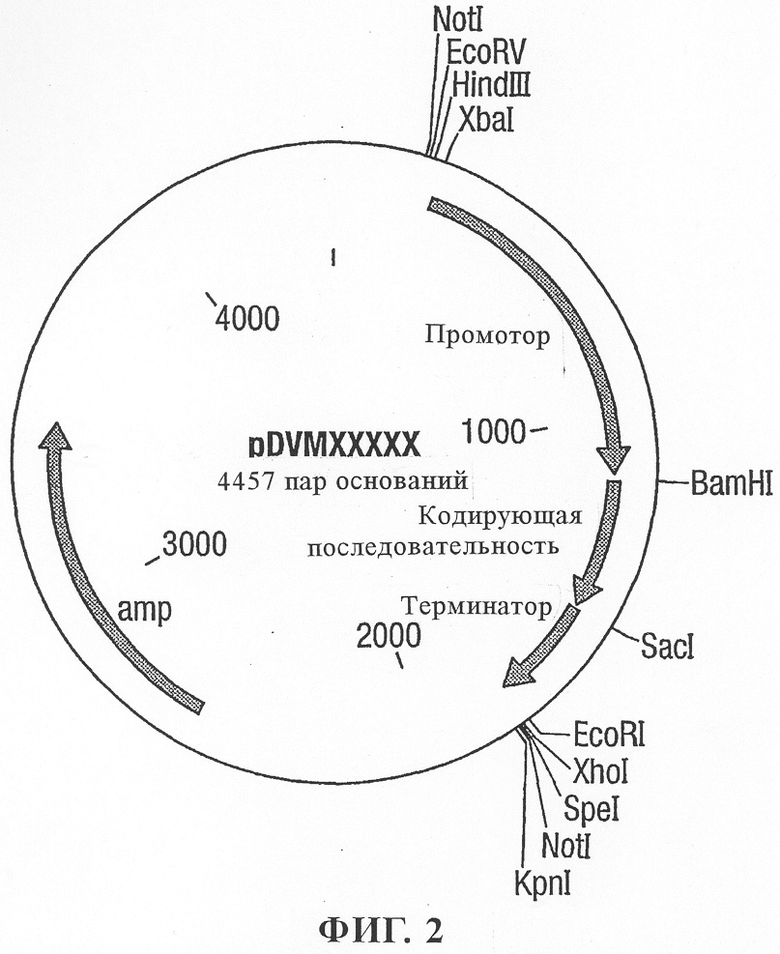
на фиг.2 - вектор pDVM;
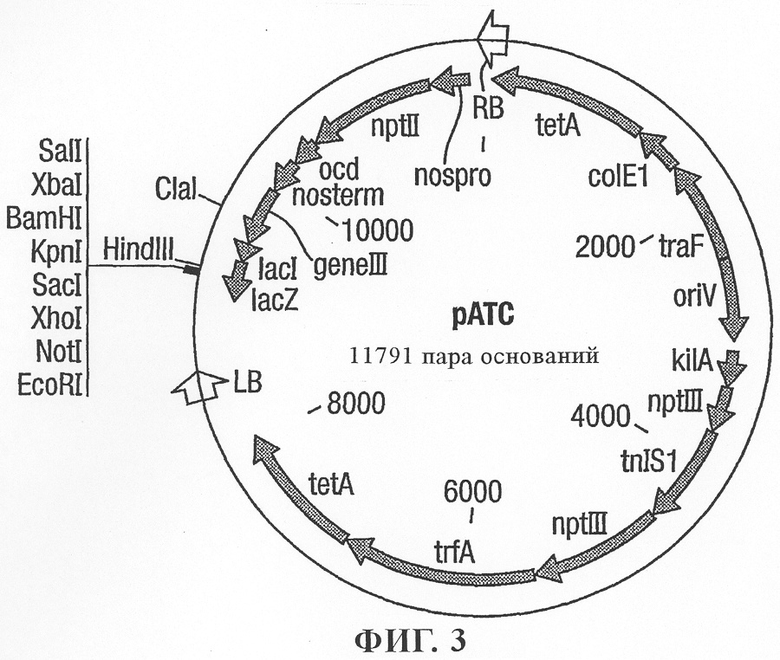
на фиг.3 - растительный трансформирующий вектор рАТС;
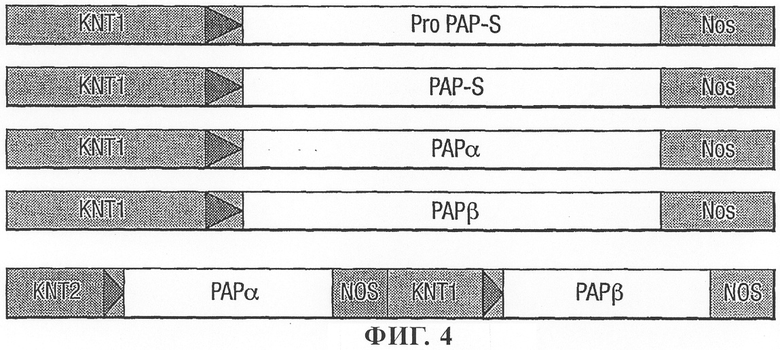
на фиг.4 - схематическое изображение индуцируемых нападением нематод промоторных конструкций для экспрессии РАР в трансгенных растениях;
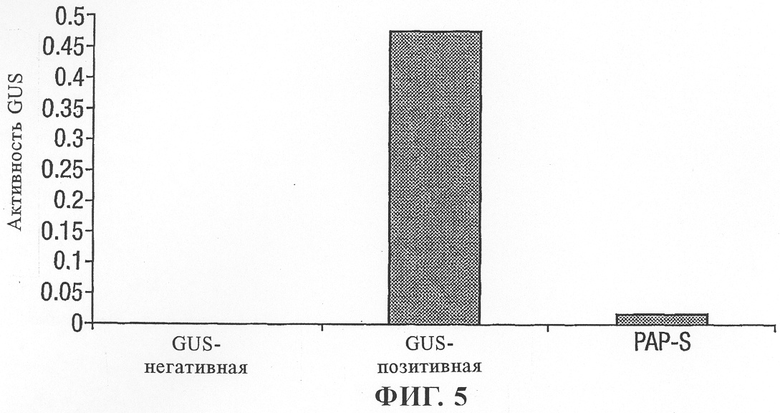
на фиг.5 - воздействие протеина PAP-S на рибосомы табака, оцененное по синтезу протеина GUS;
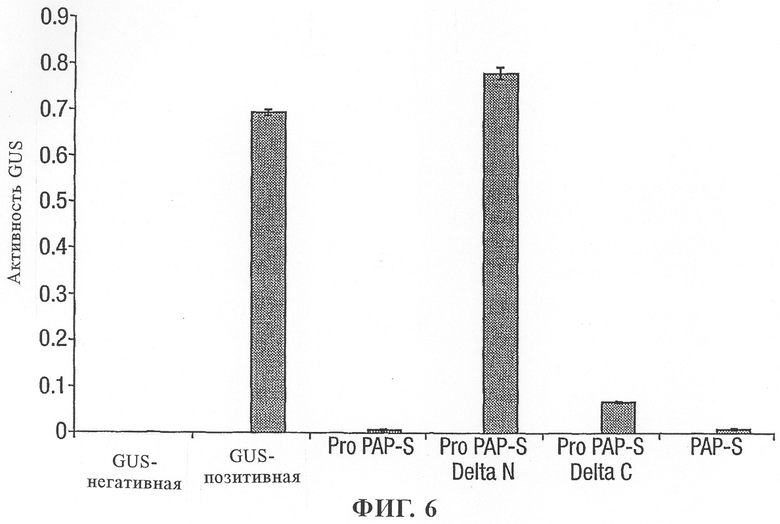
на фиг.6 - воздействие модифицированных протеинов PAP-S на рибосомы табака, оцененное по синтезу протеина GUS;
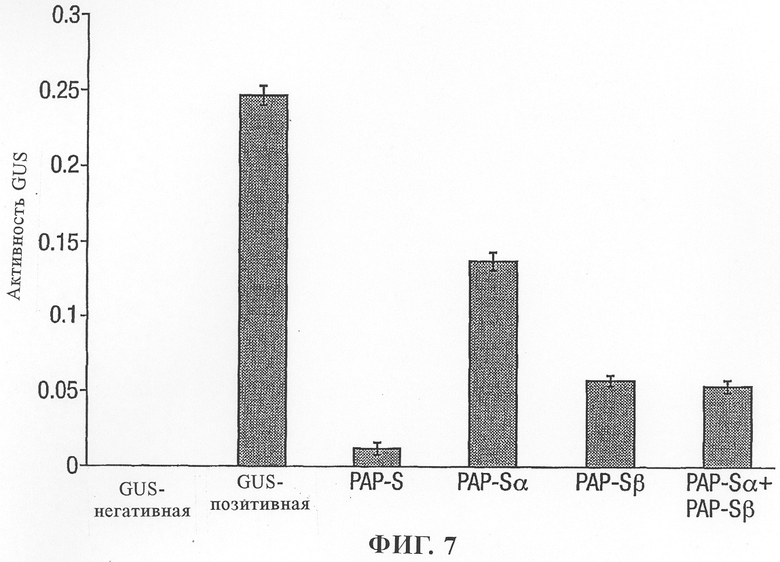
на фиг.7 - воздействие PAP-S α и PAP-S β на протопласты;
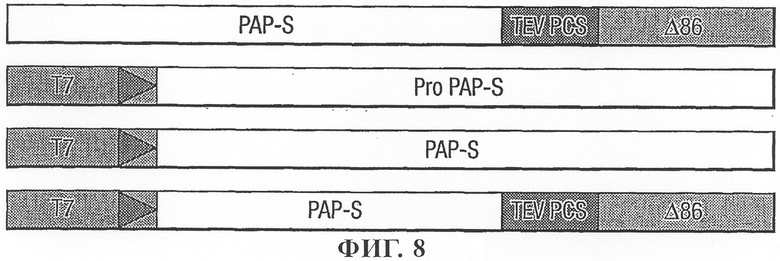
на фиг.8 - схематическое изображение слитой конструкции РАР-S/сайт расщепления протеазой (PCS)/цистатин ΔD86 и конструкций в векторах pBluescript, применяемых в экспериментах с использованием TnT;
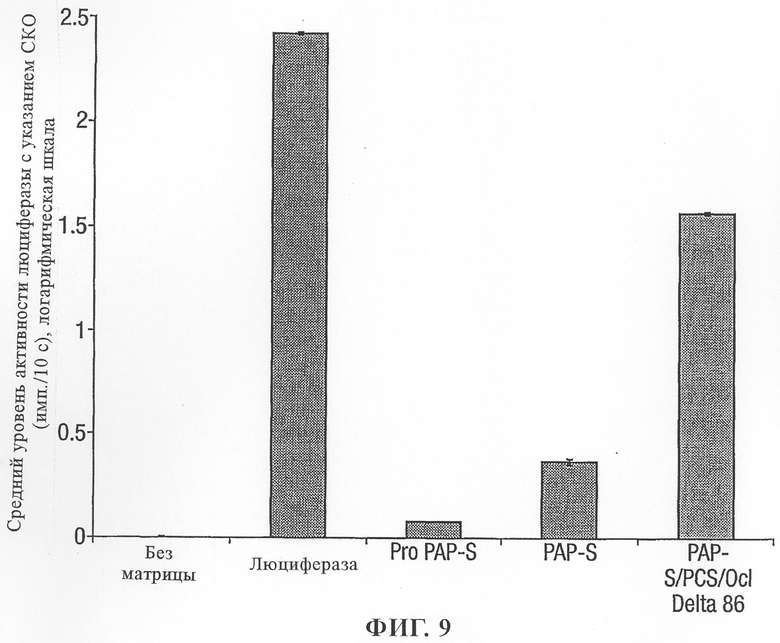
на фиг.9 - воздействие протеинов РАР на трансляцию протеина люциферазы в лизатах ретикулоцитов кроликов;
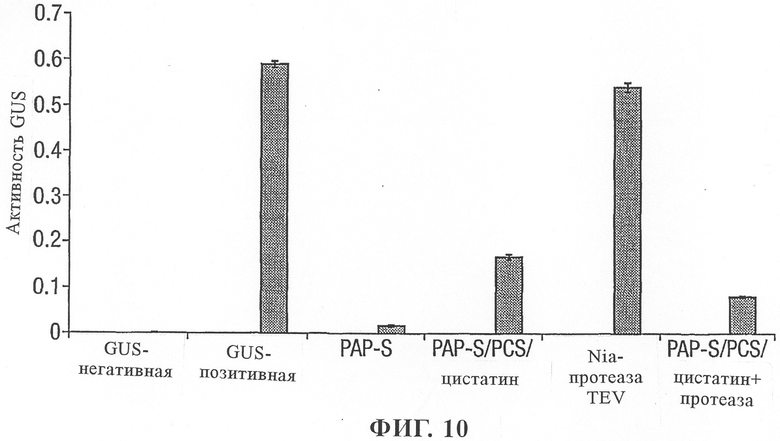
на фиг.10 - воздействие модифицированного протеина РАР-S/цистатин на рибосомы табака, оцененное по синтезу протеина GUS;
на фиг.11 - схематическое изображение конструкции индуцируемого нападением нематод промотора, предназначенного для регулирования экспрессии PAP-S-PCS-цистатин ΔD86 и NIa-протеазы TEV в трансгенных растениях;
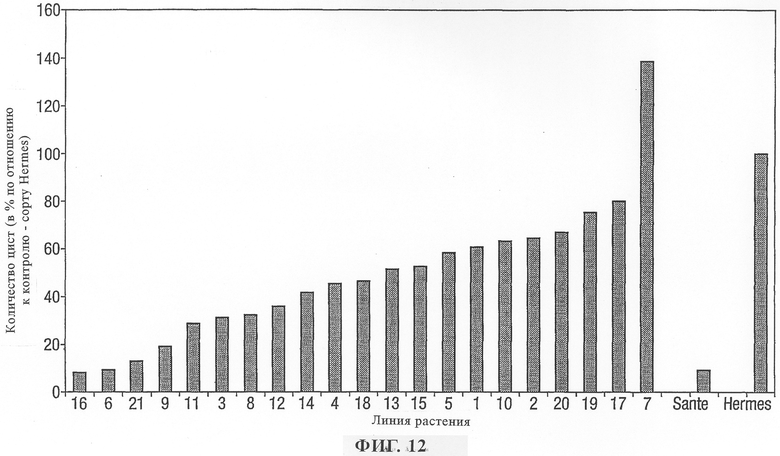
на фиг.12 - скрининг устойчивости растений картофеля, трансформированных Pro-PAP-S и зараженных картофельной (цистообразующей) нематодой линии 2/3, по сравнению с чувствительным контрольным (Hermes) и поступающим в продажу устойчивым сортом (Sante);
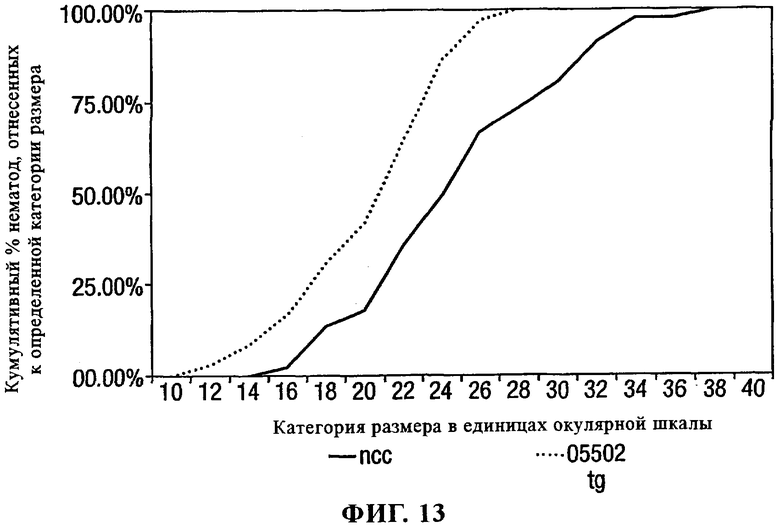
на фиг.13 - данные опыта по оценке устойчивости к яванской галловой нематоде растений табака, содержащих конструкцию Pro-PAP-S (рАТС05502) по сравнению с выращенными с помощью культуры ткани контрольными растениями (ncc). Размеры паразитирующих нематод выражены в единицах окулярной шкалы;
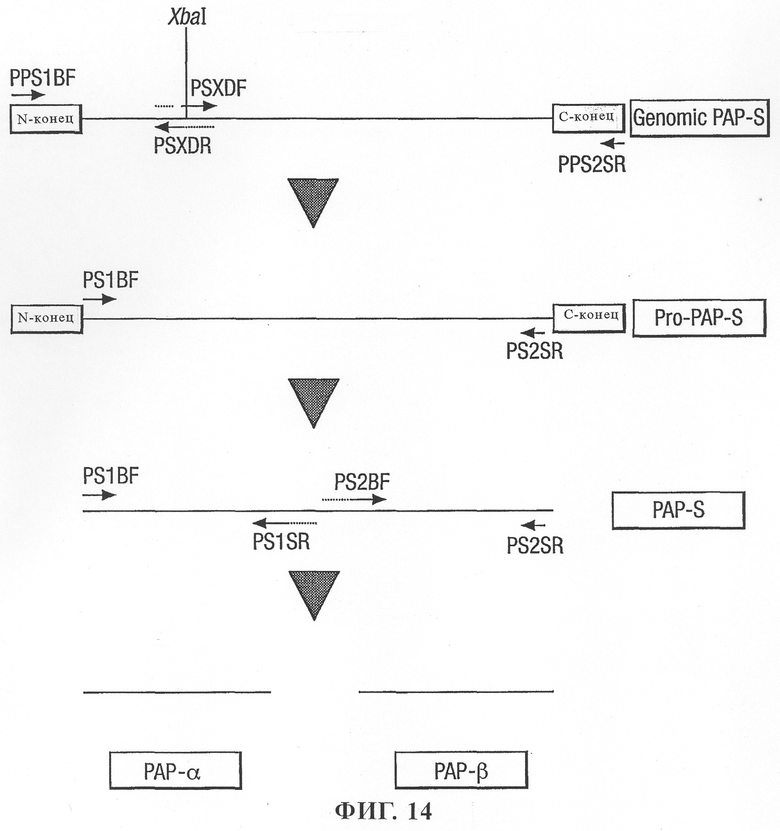
на фиг.14 - схематическая диаграмма получения вариантов PAP-S с помощью ПЦР;
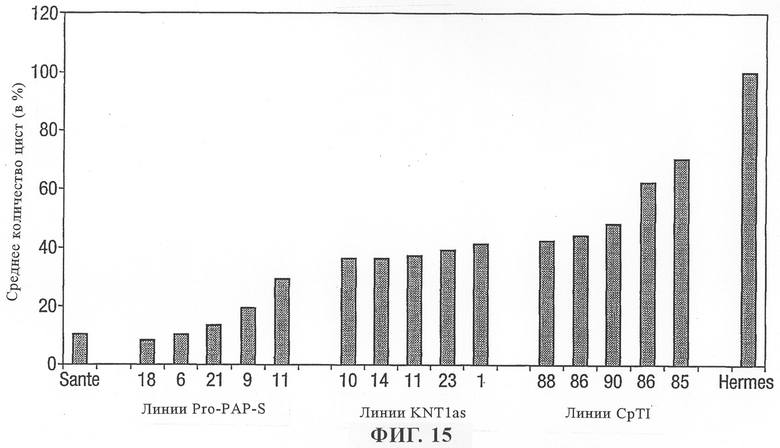
на фиг.15 - сравнение среднего количества цист на растениях картофеля, трансформированных генами, содержащими Pro PAP-S, антисмысловой промотор KNT1 или ингибитор трипсина коровьего гороха (CpTI).
Примеры
Клонирование и секвенирование антивирусного протеина фитолакки американской (PAP-S), выведенного из геномной ДНК фитолакки американской
Последовательность Pro-PAP-S выделяли из ДНК листьев фитолакки американской с помощью полимеразной цепной реакции (ПЦР) с использованием полимеразы Pfu. Последовательность выделяли в виде двух сегментов, которые затем объединяли, удаляя сайт XbaI, находящийся в центральной области молекулы. 5'-область молекулы амплифицировали с помощью праймеров PPS1BF и PSXDR (SEQ. ID. No. 9 и 16 соответственно), а 3'-область с помощью праймеров PSXDF и PPS2SR (SEQ. ID. No. 15 и 10 соответственно). Два ПЦР-продукта объединяли и перекрывающиеся фрагменты ДНК удлиняли с помощью полимеразы Pfu и повторно амплифицировали с использованием праймеров PPS1BF и PPS2SR (SEQ. ID. No. 9 и 10 соответственно). ПЦР-продукт расщепляли с помощью рестриктаз XbaI и SalI и клонировали в pBluescript, получая клон pBS/Pro-PAP-S (фиг.14).
Используя клон pBS/Pro-PAP-S в качестве матрицы последовательность PAP-S амплифицировали с помощью ПЦР с использованием праймеров PS1BF и PS2SR (SEQ. ID. No. 11 и 12 соответственно) для удаления N- и С-концевых областей. С помощью ПЦР-праймеров добавляли стартовый кодон метионина и стоп-кодон ТАА. Последовательность стартового и стоп-кодонов представлена в SEQ. ID. No. 3, но в SEQ. ID. No. 3 не представлены сайты клонирования фермента. Добавленный остаток метионина представлен в SEQ. ID. No. 4. ПЦР-продукт расщепляли рестриктазами XbaI и SalI и клонировали в pBluescript, получая клон pBS/PAP-S. Секвенирование клона PAP-S позволило выявить только одну нуклеотидную замену относительно полученной из базы данных последовательности. Приемлемые комбинации праймеров использовали для удаления либо N-, либо С-концевых областей независимо друг от друга.
После сравнения PAP-S и протеинов RIP кукурузы номинальные полипептидные домены PAP-S α и PAP-S β были определены относительно соответствующих областей RIP α, β и центрального домена кукурузы (эти данные представлены в находящейся в одновременном рассмотрении заявке этих же авторов, поданной одновременно с настоящей заявкой). Полипептидные домены PAP-S α и PAP-S β амплифицировали из плазмидной ДНК клона PAP-S с помощью пары праймеров PS1BF плюс PS1SR (SEQ. ID. No. 11 и 14 соответственно) и PS2BF плюс PS2SR соответственно (SEQ. ID. No. 13 и 12 соответственно). Стартовые кодоны метионина и стоп-кодоны ТАА добавляли с помощью ПЦР-праймеров. Последовательности стартового и стоп-кодонов представлены в SEQ. ID. No. 5 и 7, но сайты клонирования фермента не представлены в SEQ. ID. No. 5 и 7. Добавленный остаток метионина представлен в SEQ. ID. No. 6 и 8. Последовательности амплифицированных рекомбинантных PAP-S α и PAP-S β клонировали в различных векторах.
Выделение промотора, который индуцируется в области-мишени и/или соседних областях
Приведенный в настоящем описании метод выделения промотора дан только в качестве примера. Альтернативные методы выделения приемлемого промотора, предназначенного для применения согласно настоящему изобретению, хорошо известны специалисту в данной области, на некоторые из этих методов сделаны ссылки выше.
Метод выделения промотора KNT1 состоит в следующем.
Выращивание и заражение растений табака
Семена табака сорта С319 проращивали в компосте Fisons F1 в следующих условиях: интенсивность света 4500-5000 лк; 16 ч света/8 ч темноты; температура 20-25°С. Примерно через 3 недели проростки осторожно промывали водопроводной водой для удаления почвы и переносили в пакеты (фирма Northrup-King), по 2 растения на пакет, и выращивали в течение еще 1 недели в камере Conviron при 25°С и при указанном выше световом режиме. Корни с задней стороны пакета высвобождали и их кончики поддерживали с помощью бумаги из стекловолокна типа Whatman GF/A. Затем кончики корней заражали трехдневными нематодами (М.javanica), внося аликвотные количества по 10 мкл (50 нематод), и наверх помещали второй кусок бумаги типа GF/A, для того, чтобы полностью со всех сторон закрыть кончик корня. Через 24 ч после заражения бумагу GF/A удаляли для того, чтобы обеспечить синхронность заражения. Через 3 дня после заражения корневые узлы отсекали (оставляя здоровую часть корня и ткань кончика корня сзади) и немедленно замораживали в жидком азоте. С 80 инокулированных растений получали примерно 0,5-1,0 г зараженной ткани корня.
Окрашивание для визуализации нематод в зараженных корнях
Уровень заражения оценивали по количеству нематод, заразивших один кончик корня. Корни отделяли через 3 дня после заражения растений и погружали на 90 с при 95°С в лактофенол, содержащий 0,1% красителя Cotton Blue. После отмывки в течение 5 с в воде корни выдерживали в лактофеноле при комнатной температуре (КТ) в течение 3-4 дней до обесцвечивания. Окрашенные нематоды визуализировали с помощью светового микроскопа.
Выделение РНК из здоровой и зараженной корневой ткани
Корневую ткань измельчали с получением тонко измельченного порошка с помощью ступки и пестика при охлаждении в жидком азоте. Затем аликвотные количества примерно по 100 мг переносили в аналогичным образом охлажденные микроцентрифужные пробирки и добавляли 300 мкл горячего фенольного буфера для экстракции (50% фенола, 50% буфера для экстракции: 0,1М хлорид лития, 0,1М Трис-HCl рН 8,0 (КТ), 10 мМ ЭДТК, 1% ДСН) и инкубировали при 80°С в течение 5 мин. Затем добавляли равный объем хлороформа и гомогенат центрифугировали с помощью микроцентрифуги в течение 15 мин при 4°С. После этого водную фазу экстрагировали с помощью 600 мкл фенола/хлороформа и вновь центрифугировали с помощью микроцентрифуги, как описано выше. Водную фазу вновь отделяли и РНК осаждали с помощью равного объема хлорида лития при 4°С в течение ночи. Осадок пеллетировали с помощью микроцентрифуги в течение 15 мин при КТ и промывали в 70%-ном этаноле. Дебрис лиофилизировали, ресуспендировали в обработанной DEPC-водой и анализировали спектрофотометрически. Уровень РНК оценивали с помощью денатурирующего гель-электрофореза (адаптированный для целей исследования метод, описанный у Shirzadegan и др., 1991).
Субтрактивное клонирование инфекционных специфических кДНК
Поли(А)+ -РНК (мРНК) выделяли из 200 мг образцов общей РНК, полученных из здоровой и зараженной корневой ткани табака сорта С319 с помощью магнитных загруженных олиго-дТ гранул типа Dynabead согласно инструкциям производителя. Синтез первой цепи кДНК осуществляли in situ с использованием связанной с гранулами Dynabead поли(А)+ фракции, выделенной из здоровой ткани, получая драйвер-ДНК. Синтез первой и второй цепи осуществляли in situ с использованием связанной с гранулами Dynabead поли(A)+ фракции, выделенной из зараженной ткани, получая ДНК-мишень. Все реакции с использованием кДНК проводили с помощью набора для синтеза кДНК согласно инструкциям производителя (фирма Pharmacia). Три примененных олигонуклеотида, т.е. SUB21 (5' CTCTTGCTTGAATTCGGACTA 3'), обозначен как SEQ. ID. No. 30, SUB25 (5' TAGTCCGAATTCAAGCAAGAGCACA 3') (последовательности, описанные у Duguid и Dinauer, 1990 и обозначенные как SEQ. ID. No. 31) и LDT15 (5' GACAGAAGCGGATCCd(T)15 3') (O'Reilly, 1991), обозначена как SEQ. ID. No. 32, обрабатывали полинуклеотидкиназой фага Т4 согласно методу, описанному у Maniatis и др., (1982). Затем SUB21 и SUB25 "отжигали", получая линкер, который далее лигировали с ДНК-мишенью с помощью ДНК-лигазы фага Т4 согласно методу, описанному у King и Blakesley (1986). Затем гранулы, несущие ДНК-мишень, интенсивно промывали с помощью ТЕ и вторую цепь кДНК элюировали при 95°С в пятикратном (5х) SSC.
РНК, связанную с гранулами Dynabead, которые связаны с драйвер-ДНК, удаляли нагреванием и элюированную ДНК-мишень гибридизовали с драйвер-ДНК при 55°С в 5×SSC в течение 5 ч. Не вступившую в реакцию гибридизации ДНК-мишень отделяли от связанной с гранулами драйвер-ДНК при комнатной температуре (КТ) согласно инструкциям производителя, после чего вступившую в реакцию гибридизации ДНК-мишень также отделяли от связанной с гранулами драйвер-ДНК при 95°С. ДНК-мишень, полученную элюированием при комнатной температуре, вновь добавляли к драйвер-ДНК и гибридизацию повторяли. Этот процесс повторяли до тех пор, пока количество ДНК-мишени, вступившее в реакцию гибридизации с драйвер-ДНК, больше не превышало количество, не вступившее в реакцию гибридизации. Концентрации ДНК оценивали с помощью набора типа DNA Dipstick (фирма Invitrogen) согласно инструкциям производителя.
Аликвотные количества конечной фракции, полученной элюированием при комнатной температуре, использовали для ПЦР-амплификации (Eckert и Kunkel, 1990), получая двухцепочечную кДНК для клонирования в плазмидном векторе. Амплификацию ДНК-мишени осуществляли с помощью праймеров SUB21 и LDT15 в условиях, описанных у Frohman и др., 1988. Затем ПЦР-продукты встраивали путем лигирования в расщепленный с помощью Smal вектор pBluescript согласно методу, описанному у King и Blakesley (1986).
Скрининг субтрактивной библиотеки с помощью обратного Нозерн-блоттинга
Рекомбинанты идентифицировали с помощью колониальной ПЦР (Gussow и Clackson, 1989). Амплифицированные вставки анализировали с помощью Саузерн-блоттинга в трех повторностях на мембранах типа Pall Biodyne согласно рекомендациям производителя. Как предварительную гибридизацию, так и основную гибридизацию осуществляли при 42°С в 5×SSPE, 0,05% BLOTTO, 50% формамида. Мембраны гибридизовали по отдельности с кДНК-зондами (см. ниже), полученными из здоровой и зараженной ткани, и с зондом, содержащим амплифицированную ДНК-мишень из последней, полученной отделением (вычитанием) фракции. Клоны, проявившие сигнал гибридизации только с кДНК-зондом из зараженной ткани или проявившие сигнал гибридизации только с зондом из полученной отделением (вычитанием) фракции, отбирали для дальнейшего анализа.
Получение кДНК-зонда
Образцы, содержащие по 10 мкг общей РНК из здоровой и зараженной ткани, обрабатывали 2,5 ед ДНКазы1 при 37°С в течение 15 мин. Затем ДНКазу1 денатурировали при 95°С в течение 10 мин перед синтезом кДНК, который осуществляли с использованием протокола производителя (фирма Pharmacia). Затем РНК удаляли в присутствии 0,4 М гидроксида натрия в течение 10 мин при КТ и кДНК очищали с помощью вращающейся колонки Sephacryl 400HR. Выход и концентрацию определяли с помощью набора типа DNA Dipsticks (фирма Invitrogen). кДНК метили с помощью олигонуклеотидов из расчета примерно 35 нг/зонд с использованием стандартного протокола олигонуклеотидного мечения фирмы Pharmacia.
Нозерн-блоттинг
Для оценки профиля экспрессия клонов, выбранных с использованием анализа на основе обратного Нозерн-блоттинга, их применяли в качестве зондов для анализа с помощью Нозерн-блоттинга либо общей РНК, либо поли(А)+-РНК, полученной из здоровых и зараженных корней, стеблей, листьев и цветков. Блоты общей РНК содержали по 25 мкг РНК на полосу, в то время как блоты поли (А)+-РНК содержали по 0,5-1,0 мкг РНК на полосу. РНК подвергали электрофорезу на формальдегидных гелях и блоттировали на мембране типа Pall Biodyne В согласно методу, описанному у Fourney и др. (1988). Зонды метили и гибридизовали согласно описанным выше методикам.
Саузерн-блоттинг
Для определения того, имеют ли отобранные кДНК происхождение из растения или нематоды, ДНК табака сорта С319 и нематоды M.javanica получали согласно методу, описанному у Gawel и Jarret (1991). Готовили блоты для анализа методом Саузерн-блоттинга, которые содержали по 10 мкг расщепленной с помощью EcoRI и HindII ДНК на полосу. Блоты гибридизовали с меченными с помощью олигонуклеотидов зондами согласно описанному выше методу.
Гибридизация in situ
Для определения локализации экспрессии представляющих интерес кДНК в области, которой питаются нематоды, осуществляли гибридизацию in situ. Ткань зараженных и здоровых корней погружали в воск, делали срезы и гибридизовали с зондами согласно методу, описанному у Jackson (1991).
Выделение 5'-концов мРНК
Строение 5'-концов представляющих интерес РНК определяли с помощью 5'-RACE согласно методу, описанному у Frohman и др. (1988).
Выделение промоторных областей
Промоторные области представляющих интерес генов выделяли с помощью ПЦР с использованием лигированного вектора. 100 нг образцов расщепленной с помощью рестриктаз геномной ДНК табака сорта С319 лигировали в течение 4 ч при КТ (King и Blakesley, 1986) с 100 нг образцов вектора pBluescript (расщепленного с помощью рестриктаз с получением совместимых концов). Как правило, использовали такие ферменты, как EcoRI, BamHI, HindIII, BglII, XhoI, ClaI, SalI, KpnI, PstI и SstI. Затем осуществляли ПЦР с использованием в качестве матрицы продуктов лигирования с применением векторного праймера, такого как секвенирующий праймер-40 и праймер, комплементарный 5'-концу мРНК. Затем ПЦР-продукты клонировали и секвенировали. При необходимости процесс повторяли с использованием нового праймера, комплементарного 5'-концу промоторного фрагмента, для гарантии того, что выделены контролирующие последовательности промоторов.
С использованием описанных выше процедур был идентифицирован и выделен из растений табака ген KNT1. Промоторный фрагмент KNT1 длиной примерно 0,8 т.п.н., простирающийся от сайта инициации транскрипции, выделяли и встраивали в репортерный несущий ген GUS вектор pBI101 (Jefferson и др., 1987). Полученную в результате конструкцию pBIN05101 применяли для трансформации растений табака. При заражении М. javanica был обнаружен высокий уровень экспрессии гена GUS в областях, которыми питаются нематоды.
Установлено, что ген KNT1 гомологичен гену, присутствующему в других видах растений, отличных от табака. Эти растения включают (но не ограничиваясь ими) Solanum tuberosum (картофель), Lycopersicon esculentum (томаты) и Beta vulgaris (свекла). Ген KNT1 индуцировался как галловыми, так и цистообразующими видами нематод.
В соответствии с Будапештским договором о международном признании депонирования микроорганизмов для целей патентной процедуры конструкция pBIN05101 была депонирована фирмой Advanced Technologies (Кэмбридж) Limited, 210 Cambridge Science Park, Cambridge CB4 OWA, Великобритания в Национальных коллекциях промышленных, пищевых и морских бактерий (National Collections of Industrial, Food и Marine Bacteria, NCIMB), 23 St. Machar Street, Абердин, Шотландия, 20 марта 1997 г. под регистрационным номером NCIMB 40870. Вектор содержит левую и правую пограничные последовательности Т-ДНК штамма Agrobacterium tumefaciens C58. Между пограничными последовательностями находится множественный сайт клонирования и ген, обусловливающий устойчивость к канамицину, под контролем растительного промотора (Nos). Снаружи от пограничных последовательностей в векторе расположен бактериальный ген, обусловливающий устойчивость к канамицину. Вставка в вектор включает промотор KNT1 - кодирующую последовательность глюкуронидазы - терминатор Nos.
Получение конструкций PAP-S
Конструкции трансгенов PAP-S создавали в векторах, пригодных для применения в системе кратковременной экспрессии в протопласте (фиг.1). Конструкции создавали в векторе pDE4 с большим количеством копий (Denecke и др., 1990) с использованием конститутивного промотора 35S CaMV. Были созданы следующие конструкции:
1.pDE4/Pro-PAP-S
2. pDE4/PAP-S ΔN
3. pDE4/PAP-S ΔС
4. pDE4/PAP-S
5. pDE4/PAP-S α
6. pDE4/PAP-S β
Для экспериментов по трансформации растений также создавали содержащие эффекторные компоненты PAP-S-конструкции в бинарных векторах с использованием различных промоторов. Последовательности PAP-S сначала клонировали в векторах-донорах pDVM (как показано на фиг.2), содержащих индуцируемый нападением нематод промотор KNT1 и терминатор Nos. Затем кассеты промотор-ген-терминатор вырезали и клонировали в бинарном векторе рАТС (фиг.3) и интродуцировали путем электропорации в штамм Agrobacterium tumefaciens LBA4404.
Аналогичным образом получали конструкции, пригодные для применения согласно другим вариантам осуществления изобретения, с использованием методик, аналогичных описанным выше, и выбирая промоторы, соответствующие мишени.
Экспресс-анализ конструкций PAP-S в протопластах табака
В этом анализе опосредуемую PAP-S инактивацию рибосом определяли, оценивая синтез протеина GUS. Протопласты получали из листьев выращенных in vitro растений табака и подвергали электропорации с использованием конструкций PAP-S. Эффективность рибосом в отношении трансляции протеина в протопластах табака оценивали с помощью репортерного гена GUS в конструкции pDE4 под контролем промотора 35S CaMV. Несущую ген GUS конструкцию pDE4 подвергали совместной электропорации с каждой конструкцией PAP-S. Для получения данных об оптимальных уровнях синтеза протеина GUS в протопластах в условиях конкуренции со вторым протеином использовали GUS-позитивный контроль, для создания которого осуществляли совместную электропорацию протопластов табака с использованием несущей нетоксичный хаперонный протеин BiP конструкции pDE800 (Leborgna-Castel и др., 1999) и несущей GUS конструкции pDE4. Также применяли GUS-отрицательный контроль, для создания которого осуществляли совместную электропорацию протопластов табака с использованием незагруженного вектора pDE4 и несущей нетоксичный хаперонный протеин BiP конструкции.
(I) Зрелый PAP-S.
Протопласты табака трансформировали путем совместной электропорации конструкциями PAP-S и конструкцией, несущей GUS.
Воздействие активности PAP-S на рибосомы оценивали по уровням активности GUS после экспрессии в течение 24 ч (фиг.5). Результаты свидетельствовали о том, что протеин PAP-S эффективно инактивирует рибосомы табака. Поэтому при сравнении с GUS-положительным контролем были обнаружены лишь фоновые уровни GUS.
(II) Конструкции, содержащие модифицированный PAP-S
С использованием протопластов табака анализировали различные версии протеинов PAP-S (фиг.6).
Установлено, что интактный Pro-PAP-S обладал такой же эффективностью, что и зрелый протеин PAP-S в отношении ингибирования трансляции протеина, что поддерживает гипотезу о его активации эндогенными протеазами. Однако модификации последовательностей Pro-PAP-S препятствуют этой активации. Роль N- и С-концевых удлинений протеина PAP-S, вероятно является различной, так как предварительная делеция С-концевой области Pro-PAP-S оказывала небольшое воздействие на последующую инактивацию рибосом, а равноценная делеция N-концевой области Pro-PAP-S приводила к синтезу молекулы, которая не могла процессироваться с образованием активной формы.
Это позволяет создать двухкомпонентную систему активации, основанную на применении Δ-N Pro-PAP-S со специфическим сайтом расщепления, встроенным по соседству с С-концевой областью, или в альтернативном варианте или в дополнение к нему, основанную на применении Δ-С Pro-PAP-S с последовательностью, которая блокирует эндогенные протеазы, но действует в качестве специфического сайта расщепления, встроенного по соседству с N-концевой областью.
(III) Различные домены PAP-S
Полипептидные области PAP-S α и PAP-S β также экспрессировали либо индивидуально, либо в сочетании друг с другом в протопластах табака (фиг.7).
Неожиданно было установлено, что экспрессия одного любого протеина PAP-S α или PAP-S β приводила к значительному снижению активности GUS. С учетом предположений, основанных на структурных особенностях, PAP-S α содержит мотив распознавания РНК и области домена связывания рибосомы, в то время как PAP-S β содержит имеющий решающее значение остаток каталитического сайта. Протеин PAP-S α может предупреждать трансляцию протеина путем связывания с рибосомами и прекращения трансляции протеина. Результаты позволяют предположить, что PAP-S α адаптирован к правильной складчатой конформации и обладает способностью к специфическому распознаванию молекул. PAP-S β содержит остаток активного сайта, необходимый для депуринизации рибосом, но маловероятно, что он обладает способностью распознавать рибосому и взаимодействовать с ней. Однако эти результаты неожиданно продемонстрировали, что PAP-S β в действительности обладает способностью ингибировать синтез протеина. Это позволяет предположить, что при индивидуальном применении PAP-S β адаптирован к правильной складчатой конформации и обладает способностью к специфическому распознаванию молекул и каталитическому расщеплению. Инактивация рибосом, происходящая при индивидуальном применении PAP-S β, выше, чем при применении PAP-S α.
Одновременная экспрессия и PAP-S α и PAP-S β в протопластах не приводила к дополнительному снижению активности GUS по сравнению с экспрессией только протеина PAP-S β. Это отличается от ситуации, характерной для применения доменов RIP α и RIP β кукурузы, при совместной экспрессии которых происходило дополнительное снижение активности, что описано в находящейся в одновременном рассмотрении заявке этих же авторов, поданной одновременно с настоящей заявкой.
Конструкция, содержащая инактивированную с помощью структурной модификации форму PAP-S:
Ингибирование активности РАР путем предупреждения связывания рРНК может быть опосредовано образованием слитого протеина, простирающегося в активный сайт вблизи РАР.
Приемлемый компонент, выбранный для создания слитого протеина, представлял собой протеин цистатина риса (оризацистатин), и для оптимальной экспрессии в картофеле синтезировали синтетическую версию гена, содержащую наиболее часто встречающийся кодон (SEQ. ID. No. 17). Для гарантии того, что цистатин будет располагаться в положении в области активного сайта РАР, 7 аминокислот изымали путем делеции из С-конца РАР. Слияние с цистатином создавали, использую линкерную последовательность, кодирующую сайт расщепления протеазой NIa вируса табачной гравировки (TEV).
Описанные ниже праймеры создавали для конструирования слияния между PAP-S и цистатином риса ΔD86 с сайтом расщепления протеазой (PCS) TEV между ними в качестве линкера (фиг.8). Последовательность, кодирующую слитый протеин, амплифицировали из клонированной ДНК с использованием двух ПЦР-фрагментов; (I) PAP-S плюс PCS амплифицировали с использованием PS1BF и PCSPAPSR (SEQ. ID. No. 11 и 20), и (II) цистатин Δ86 с PCS амплифицировали с использованием праймеров PCSDelta86F и SYNPOTDelta86SR (SEQ. ID. No. 21 и 22). Затем два ПЦР-фрагмента применяли для амплификации полноразмерного слитого протеина (PAPS- PCS-цистатин Delta86) путем обеспечивающей перекрывающее удлинение ПЦР с использованием праймеров PS1BF и SYNPOTDelta86SR (SEQ. ID. No. 11 и 22).
Согласно этой процедуре 8 аминокислот из С-конца PAP-S удаляли и встраивали сайт расщепления протеазой NIa TEV (7 аминокислот) с помощью праймера PCSDelta86F (SEQ. ID. No. 21), обеспечивающего кодирование гептапептида: Glu Pro Val Tyr Phe Gln Gly. Затем добавляли цистатин в виде протеина, слитого с С- концом последовательности PAPS-PCS.
Слитый ПЦР-продукт клонировали в векторе pBluescript и его последовательность подтверждали путем анализа последовательности.
Функциональный анализ инактивированного протеина PAP-S в системе транскрипции/трансляции in vitro
Активность конструкций pBS/Pro PAP-S, pBS/PAP-S и pBS/PAP-S/PCS/цистатин ΔD86 оценивали с помощью системы лизата кроличьих ретикулоцитов. Применяли систему быстрой слитой транскрипции и трансляции на основе TnT фага Т7 (фирма Promega Corporation).
Содержащие TnT реакционные смеси инкубировали при 30°С всего в течение 60 мин с различными конструкциями РАР с использованием содержащей люциферазу конструкции в качестве положительного контроля. После начальной 30-минутной инкубации конструкций РАР с содержащими TnT реакционными смесями во все реакционные смеси добавляли равное количество матрицы-люциферазы и инкубацию продолжали в течение еще 30 мин. Аликвотные количества различных реакционных смесей смешивали с реагентом для анализа люциферазы (Luciferase Assay Reagent) и активность люциферазы оценивали с помощью люминометра в течение 10 с (фиг.9).
Результаты по оценке трансляции протеина Pro PAP-S и PAP-S свидетельствовали о том, что, как и ожидалось, протеины инактивировали трансляцию, осуществляемую с помощью кроличьих рибосом. Модифицированный слитый протеин РАР оказался относительно неактивным по сравнению с РАР. Трансляцию различных протеинов РАР подтверждали по включению биотинилированной тРНК лизила, которую использовали в качестве метки в процессе трансляции и которую выявляли с помощью стрептавидина, конъюгированного с щелочной фосфатазой. Не было возможно обнаружить транслированные протеины Pro PAP-S и PAP-S из-за быстрой инактивации рибосом, в то же время были выявлены относительно большие количества модифицированного слитого протеина РАР, что свидетельствовало об относительно слабой активности этого слитого протеина РАР в отношении кроличьих рибосом.
Клонирование NIa-протеазы TEV
Ген NIa-протеазы TEV амплифицировали из клона кДНК TEV. Для целей клонирования сайт рестрикции SpeI удаляли с помощью перекрывающихся обеспечивающих удаление праймеров. NIa-протеазу сначала амплифицировали в виде двух ПЦР-фрагментов с использованием праймеров NIAPROBF (SEQ. ID. No. 23) плюс NIAPPROSR (SEQ. ID. No. 26) и NIAPROSDF (SEQ. ID. NO. 25) плюс NIAPROSR (SEQ. ID. NO. 24) и затем полноразмерную протеазу амплифицировали с помощью обеспечивающей перекрывающееся удлинение ПЦР с использованием праймеров NIAPROBF (SEQ. ID. No. 23) плюс NIAPROSR (SEQ. ID. No. 24).
Экспресс-анализ в протопласте слитого протеина PAP-S
Воздействие модифицированного PAP-S/цистатина на рибосомы табака анализировали в протопластах, и было установлено, что протеин обладал значительно более низкой активностью по сравнению с немодифицированным протеином (фиг.10). Однако при совместной экспрессии NIa-протеазы TEV в сочетании с протеином модифицированный PAP-S/цистатин обнаружено повышение инактивации рибосом. Раздельная экспрессия NIa-протеазы TEV не оказывала воздействия на синтез протеина GUS.
Двухкомпонентная система для индуцируемой нематодами экспрессии модифицированного PAP-S
Для оценки эффективности модифицированного протеина PAP-S и его активации с помощью NIa-протеазы TEV создавали двойную конструкцию с использованием двух индуцируемых нематодами промоторов KNT1 (описан выше) и KNT2 (SEQ. ID. No. 27). KNT2 обеспечивал экспрессию в массе корня (но не в кончиках корней) и в гигантских клетках пораженного нематодами растения.
Ген NIa-протеазы TEV амплифицировали с помощью ДНК-полимеразы Pfu и ПЦР-продукт клонировали в виде XbaI- и SalI-фрагмента в векторах pBluescript. Рекомбинантный клон секвенировали с целью подтверждения его идентичности. Гены слияния PAP-S-PCS-цистатин ΔD86 NIa-протеазы TEV клонировали по отдельности в векторах-донорах pDVM в виде отдельных конструкций с помощью промоторов KNT2 и KNT1 соответственно. Двойную конструкцию (фиг.11) создавали в промежуточном векторе-доноре pDVM и повторно клонировали в бинарном растительном трансформирующем векторе рАТС для получения трансгенных растений и признака устойчивости к нематодам.
Получение трансгенных растений
Две конструкции, а именно рАТС05501, содержащую полностью процессированную зрелую версию PAP-S, и рАТС05502, содержащую предшественник Pro-PAP-S (фиг.4), интродуцировали с помощью штамма Agrobacterium tumefaciens LBA4404 в растения табака и картофеля с использованием метода совместного культивирования листовых дисков, описанного у Horsch и др. (1985).
Попытки трансформировать растения табака с помощью рАТС05501 (зрелый PAP-S) не привели к получению каллюса или проростков, несмотря на повторные эксперименты. При использовании контрольных конструкций, как и ожидалось, были получены трансгенные проростки, что свидетельствовало о зависимости указанного явления от зрелой последовательности PAP-S.
В противоположность этому при использовании рАТС05502 (Pro- PAP-S) были получены трансгенные растения табака.
При трансформации растений картофеля не возникало трудностей с получением трансгенных растений ни при использовании конструкций, содержащих зрелый PAP-S, ни при использовании конструкций, содержащих предшественник Pro-PAP-S.
Скрининг в отношении устойчивости к нематодам растений табака, трансформированных конструкциями PAP-S
Трансгенные несущие Pro-PAP-S растения табака использовали в стандартных тестах по определению устойчивости в отношении галловых нематод Meloidogyne javanica. Трансгенные и контрольные проростки табака высаживали случайным образом вслепую в устройства, позволяющие увлажнять корни [(рутрайнеры (rootrainer)], заполненные не содержащим удобрений перегнойным компостом высокого качества, и наиболее крупные листья подрезали наполовину. Растения накрывали полиэтиленом для поддержания высокой влажности до тех пор, пока не произойдет приживление. Перед завершением приживления в полиэтилене постепенно делали отверстия для снижения влажности. Через неделю после приживления небольшие растения заражали 200 вылупившимися нематодами второго возраста (J2) Meloidogyne javanica. Полив жидкой подкормкой осуществляли только после того, как почва становилась достаточно сухой для начала увядания листьев. Рутрайнеры помещали в ящики на нагреваемое утепляющее покрытие для поддержания температуры почвы на уровне 25-30°С. Листья повторно обрезали один раз в неделю для прекращения их роста и предотвращения покрытия точек роста более крупными листьями из-за плотности посадки. Растения собирали для их анализа примерно через 4 недели после заражения и корни отмывали от почвы и отбеливали в течение 4 мин в 1%-ном растворе гипохлорита натрия. Отбеливатель удаляли промывкой водой и затем корни замачивали в большом объеме воды в течение 15 мин при периодическом перемешивании. Затем корни помещали в 10-15 мл разбавленного в соотношении 1:500 маточного раствора кислого фуксина в 5% уксусной кислоте (маточный раствор кислого фуксина получали согласно методу, описанному в "Introduction to Plant Nematology" у V.Н.Dropkin, ISBN 0-471-85268-6. Для его приготовления 0,35 г кислого фуксина растворяли в 100 мл смеси 1:3 ледяной уксусной кислоты и дистиллированной воды). Для окраски образцы помещали в кипящую водяную баню на 4 мин и затем переносили в баню с температурой 37°С на 4 ч. Краситель декантировали и образцы осветляли, добавляя подкисленный глицерин, и инкубировали при 37°С в течение ночи.
Затем осветленные корни с окрашенными нематодами помещали в чашки Петри (образец помещали на внутреннюю сторону крышки чашки Петри и основание чашки Петри использовали для распределения и придавливания образца с целью облегчения его анализа с помощью микроскопа). Образцы анализировали при увеличении в 20-100 раз и популяцию нематод анализировали несколькими путями. Галловые нематоды разделяли на 3 группы: а) паразитиформные нематоды, б) инкапсулированные нематоды, у которых не образуются яйца, и в) инкапсулированные нематоды, у которых образуются яйца. Диаметры практически полностью инкапсулированных нематод оценивали визуально с помощью окулярной шкалы.
Размеры паразитирующих на растении нематод оказались существенно сниженными на трансгенных растениях табака линии Pro-PAP-S по сравнению с размером нематод на контрольных зараженных растениях (фиг.13). Это свидетельствует о том, что растения, экспрессирующие Pro-PAP-S, оказывают ингибирующее действие на развитие нематод.
Скрининг в отношении устойчивости к нематодам растений табака, трансформированных конструкциями PAP-S
Для опыта по оценке устойчивости в отношении картофельных (цистообразующих) нематод Globodera pallida, патогенный вариант 2/3 (КЦН), использовали 21 трансгенную линию, несущую конструкцию Pro-PAP-S. Для опыта использовали контрольный чувствительный (Hermes) и частично устойчивый (Sante) сорта.
Первичную оценку устойчивости трансгенных растений осуществляли с помощью случайных опытов вслепую с использованием групп, включающих 20-25 трансгенных линий, и с использованием по меньшей мере 10 повторностей для двух или большего количества контрольных линий. Рутрайнеры или контейнеры заполняли смесью 50:50 суглинка и песка. 12 л смеси суглинка и песка увлажняли 1250 мл воды, получая смесь с концентрацией воды 40%. По 3 цисты помещали на корни каждого проростка, который затем вносили в отверстие в компосте и компостом осторожно закрывали корни. Растения приживлялись и после этого их поливали только 1 раз в неделю жидкой подкормкой или только тогда, когда почва становилась достаточно сухой. После достижения растениями высоты примерно 10 см их верхушки отрезали, препятствуя дальнейшему росту. Растения выращивали в течение примерно 3 месяцев, позволяя цистам созреть. Затем растениям давали засохнуть в течение 1 месяца. Цисты выделяли из растений, интенсивно промывая почву и корни в химическом стакане с 250 мл воды. Почве давали осадиться в течение нескольких минут и супернатант сливали в большую фильтровальную воронку с помещенным в нее диском из фильтровальной бумаги типа Whatman No.1 диаметром 32 см. Супернатанту давали выстояться в воронке в течение 1 мин и затем в центральную область поверхности раствора вносили каплю детергента Hederol для смещения материала поверхности мениска раствора к боковой стороне фильтра. Затем основание фильтра прокалывали для удаления остатка раствора. Диск из фильтровальной бумаги удаляли и подсчитывали количество прикрепившихся цист.
Растения рассматривались как обладающие признаками устойчивости, если они были заражены меньшим количеством цист, чем чувствительные контрольные линии (фиг.12). Популяция трансгенных растений в целом была в значительной степени меньше поражена КЦН. По меньшей мере у двух трансгенных линий (а именно рАТС05502 - 6, 8 16) обнаружен уровень устойчивости такой же, как и у имеющегося в продаже устойчивого сорта Sante.
Сравнение растений картофеля, трансформированных Pro PAP-S, антисмысловым KNT1 или ингибитором трипсина коровьего гороха (CpTI) под контролем промотора KNT1, позволило установить, что Pro PAP-S придает более высокий уровень устойчивости по сравнению с уровнем устойчивости трансгенных линий, содержащих либо антисмысловой KNT1, либо CpTI (фиг.15).
KNT1 представляет собой ген, который практически специфично индуцируется в гигантских клетках при заражении нематодами. Вероятно, KNT1 играет важную роль в процессе заражения и развития гигантских клеток. Внесение антисмысловой последовательности KNT1, вероятно, ингибирует процесс заражения нематодами.
Ингибитор трипсина коровьего гороха (CpTI) представляет собой протеин, который ингибирует у нематод участвующие в расщеплении ферменты, и его экспрессия должна препятствовать питанию и развитию нематод. Это не является системой, вызывающей гибель клеток per se, но оказывает непосредственное воздействие на нематод.
Сравнительные данные позволяют предположить, что трансформация растений конструкцией, содержащей Pro PAP-S, оказывает более заметное воздействие на снижение заражения нематодами, чем действие линий, содержащих либо антисмысловой KNT1, либо CpTI.















Изобретение относится к биотехнологии, в частности к системе для уничтожения растительных клеток. Проводят трансформацию растения химерным геном, где кодирующая последовательность гена кодирует РАР-S α протеин или PAP-S β протеин, или его фрагмент, этот ген содержит промотор, который проявляет активность в ответ на специфическое раздражающее воздействие на растение. Растение также может быть трансформировано либо одной конструкцией, включающей два химерных гена, один из которых кодирует инактивированный антивирусный протеин фитоллаки американской, а другой кодирует активатор инактивированного антивирусного протеина фитоллаки американской, либо набором конструкций, где каждый упомянутый выше химерный ген находится в отдельной конструкции. Изобретение позволяет эффективно удалять заданные клетки в организме растения. 5 н. и 32 з.п. ф-лы, 15 ил.
| WO 9318170, 16.09.1993 | |||
| US 5332808, 26.07.1994 | |||
| Способ получения сложных эпоксиэфиров ненасыщенных кислот | 1973 |
|
SU466222A1 |
| Испаритель в приборах для нефтяного отопления | 1930 |
|
SU21293A1 |

