ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Пролиферация и дифференцировка клеток многоклеточных организмов регулируются гормонами и полипептидными факторами роста. Эти диффундируемые молекулы позволяют клеткам осуществлять коммуникацию друг с другом и действовать совместно для образования клеток, тканей и органов и репарации поврежденной ткани. Примеры гормонов и факторов роста включают в себя стероидные гормоны (например, эстроген, тестостерон), паратиреоидный гормон, фолликулостимулирующий гормон, интерлейкины, тромбоцитарный фактор роста (PDGF), эпидермальный фактор роста (EGF), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), эритропоэтин (ЕРО) и кальцитонин.
Гормоны и факторы роста влияют на клеточный метаболизм путем связывания с рецепторами. Рецепторы могут быть интегральными мембранными белками, которые связаны с путями передачи сигналов в клетке, такими как системы вторичных мессенджеров. Другие классы рецепторов являются растворимыми молекулами, такими как факторы транскрипции.
Цитокины обычно стимулируют пролиферацию и/или дифференцировку клеток гемопоэтической линии дифференцировки или участвуют в механизмах иммунных и воспалительных реакций организма. Примерами цитокинов, которые влияют на гемопоэз, являются эритропоэтин (ЕРО), который стимулирует развитие эритроцитов; тромбопоэтин (ТРО), который стимулирует развитие клеток мегакариоцитарной линии дифференцировки; и гранулоцитарный колониестимулирующий фактор (G-CSF), который стимулирует развитие нейтрофилов. Эти цитокины применимы в восстановлении нормальных уровней клеток крови в пациентах, страдающих от анемии, тромбоцитопении и нейтропении или получающих химиотерапию по поводу рака.
Интерлейкины представляют собой семейство цитокинов, которые опосредуют иммунологические реакции, в том числе воспаление. Интерлейкины опосредуют различные воспалительные патологии. Центральной для иммунной реакции является Т-клетка, которая продуцирует многие цитокины и искусственный (приобретенный) иммунитет к антигенам. Цитокины, продуцируемые Т-клеткой, были классифицированы как цитокины типа 1 и типа 2 (Kelso, A. Immun. Cell Biol. 76:300-317, 1998). Цитокины типа 1 включают в себя IL-2, IFN-γ, LT-α, и участвуют в воспалительных реакциях, противовирусном иммунитете, иммунитете против внутриклеточных паразитов и отторжении трансплантата. Цитокины типа 2 включают в себя IL-4, IL-5, IL-6, IL-10 и IL-13 и участвуют в гуморальных ответах, противогельминтном иммунитете и аллергических реакциях. Общие цитокины между типом 1 и типом 2 включают в себя IL-3, GM-CSF и TNF-α. Существуют доказательства в пользу предположения, что популяции Т-клеток, продуцирующие цитокины типа 1 и типа 2, преимущественно мигрируют в различные типы воспаленной ткани.
Зрелые Т-клетки могут быть активированы, а именно антигеном или другим стимулом, для продуцирования, например, цитокинов, молекул биохимической передачи сигналов или рецепторов, которые дополнительно влияют на судьбу популяции Т-клеток.
В-клетки могут быть активированы через рецепторы на их клеточной поверхности, в том числе рецептор В-клеток, и другие вспомогательные молекулы для выполнения вспомогательных клеточных функций, таких как продуцирование цитокинов.
Природные клетки-киллеры (МК-клетки) имеют общую клетку-предшественник с Т-клетами и В-клетками и играют роль в иммунологическом контроле. NK-клетки, которые содержат до 15% лимфоцитов крови, не экспрессируют рецепторы антигенов и, следовательно, не используют МНС-узнавание в качестве требования для связывания с клеткой-мишенью. NK-клетки участвуют в узнавании и убивании некоторых опухолевых клеток и инфицированных вирусом клеток. Считается что, in vivo МК-клетки требуют активации, однако было показано, что in vitro МК-клетки убивают некоторые типы опухолевых клеток без активации.
Продемонстрированные in vivo активности семейства цитокинов иллюстрируют огромный клинический потенциал и потребность в отношении других цитокинов, агонистов цитокинов и антагонистов цитокинов. Данное изобретение направлено на удовлетворение этих потребностей путем обеспечения нового цитокина, который стимулирует клетки гемопоэтической линии дифференцировки, а также относящихся к нему композиций и способов.
Данное изобретение обеспечивает такие полипептиды для этих и других применений, которые должны быть очевидны лицам с обычной квалификацией в данной области из приведенных здесь описаний.
Чертеж является иллюстрацией множественного сопоставления IL-2 человека, IL-15 человека, zalpha11-лиганда (SEQ ID NO:2), IL-4 человека, IL-4-мыши, GM-CSF человека и GM-CSF мыши.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Перед подробным изложением данного изобретения может быть полезным для его понимания определение следующих терминов:
Термин "аффинная метка" используется здесь для обозначения полипептидного сегмента, который может быть присоединен ко второму полипептиду для обеспечения очистки или детектирования второго полипептида или обеспечения сайтов для присоединения второго полипептида к субстрату. В принципе, любой пептид или белок, для которого доступны антитело или другой агент специфического связывания, может быть использован в качестве аффинной метки. Афинные метки включают в себя полигистидиновый участок (тракт), белок А (Nilsson et al., EMBO J. 4:1075, 1985; Nilsson et al., Methods Enzvmol. 198:3, 1991), глутатион S-трансферазу (Smith and Johnson, Gene 67:31, 1988), аффинную метку Glu-Glu (Grussenmeyer et al., Proc. Natl. Acad. Sci. USA 82:7952-4, 1985), вещество Р, пептид Flag™ (Hopp et al., Biotechnology 6:1204-10, 1988), стрептавидинсвязывающий пептид или другой антигенный эпитоп или связывающий домен. См., в общем, Ford et al., Protein Expression and Purification 2:95-107, 1991. ДНК, кодирующие аффинные метки, доступны от коммерческих поставщиков (например, Pharmacia Biotech, Piscataway, NJ).
Термин "аллельный вариант" используется здесь для обозначения любой из двух или нескольких альтернативных форм гена, занимающих один и тот же хромосомный локус. Аллельная вариация возникает природно через мутацию и может приводить к фенотипическому полиморфизму в популяциях. Мутации генов могут быть молчащими (без изменения в кодируемом полипептиде) или могут кодировать полипептиды, имеющие измененную аминокислотную последовательность. Термин аллельный вариант используется здесь также для обозначения белка, кодируемого аллельным вариантом гена.
Термины "амино-концевой" и "карбоксил-концевой" используются здесь для обозначения положений внутри полипептидов. Там, где позволяет контекст, эти термины используются со ссылкой на конкретную последовательность или часть полипептида для обозначения близости или относительного положения. Например, некоторая последовательность, расположенная карбоксил-терминально относительно ссылочной последовательности в полипептиде, расположена проксимально относительно карбоксил-конца ссылочной последовательности, но не обязательно находится при карбоксил-конце полного полипептида.
Термин "пара комплемент/антикомплемент" обозначает неидентичные части молекулы, которые образуют нековалентно связанную, стабильную пару при подходящих условиях. Например, биотин и авидин (или стрептавидин) являются членами-прототипами пары комплемент/антикомплемент. Другие примеры пар комплемент/антикомплемент включают в себя пары рецептор/лиганд, пары антитело/антиген (или гаптен, или зпитоп), пары смысловой/антисмысловой полинуклеотид и т.п. В случае, когда желательна последующая диссоциация пары комплемент/антикомплемент, пара комплемент/антикомплемент предпочтительно имеет аффинность связывания <109 М-1.
Термин "комплементы полинуклеотидной молекулы" обозначает полинуклеотидную молекулу, имеющую комплементарную последовательность оснований и обращенную ориентацию в сравнении со ссылочной последовательностью. Например, последовательность 5'-ATGCACGGG-3' комплементарна 5'-CCCGTGCAT-3'.
Термин "вырожденная нуклеотидная последовательность" обозначает последовательность нуклеотидов, которая включает в себя один или более вырожденных кодонов (в сравнении со ссылочной полинуклеотидной молекулой, которая кодирует полипептид). Вырожденные кодоны содержат различные триплеты нуклеотидов, но кодируют один и тот же аминокислотный остаток (т.е. триплеты GAU и GAC, каждый, кодируют Asp).
Термин "экспрессирующий вектор" используется для обозначения молекулы ДНК, линейной или кольцевой, которая содержит сегмент, кодирующий представляющий интерес полипептид, функционально связанный с дополнительными сегментами, которые обеспечивают его транскрипцию. Такие дополнительные элементы включают в себя промоторную и терминаторную последовательности и могут также включать в себя одну или несколько точек начала репликации, один или несколько селектируемых маркеров, энхансер, сигнал полиаденилирования и т.д. Экспрессирующие векторы обычно произведены из плазмидной или вирусной ДНК или могут содержать элементы обеих.
Термин "выделенный" в применении к полинуклеотиду обозначает, что этот полинуклеотид был удален из его природной генетической среды и, следовательно, не содержит других посторонних или нежелательных кодирующих последовательностей и находится в форме, подходящей для применения в генетически сконструированных системах продуцирования белков. Такие выделенные молекулы являются молекулами, которые выделены из их природного окружения и включают в себя кДНК-клоны и геномные клоны. Выделенные молекулы ДНК данного изобретения не содержат других генов, с которыми они обычно связаны, но могут включать в себя природно встречающиеся 5'- и 3'-нетранслируемые районы, такие как промоторы и терминаторы. Идентификация связанных районов будет очевидной лицу с обычной квалификацией в данной области (см., например, Dynan and Tijan, Nature 316:774-78, 1985).
"Выделенным" полипептидом или белком является полипептид или белок, который обнаруживается в условиях, иных, чем его природное окружение, например, отдельно от крови и ткани животного. В предпочтительной форме выделенный полипептид является по существу не содержащим других полипептидов, в частности других полипептидов животного происхождения. Предпочтительно обеспечение полипептидов в высокоочищенной форме, т.е. имеющих чистоту более 95%, более предпочтительно чистоту более 99%. При использовании в этом контексте термин "выделенный" не исключает присутствия того же самого полипептида в альтернативных физических формах, таких как димеры или альтернативно гликозилированные или дериватизованные формы.
Термин "неопластический" при применении к клеткам указывает клетки, подвергающиеся новой и отклоняющейся от нормы пролиферации, в частности, в ткани, где эта пролиферация является неконтролируемой и прогрессирующей, приводящей к неоплазме (новообразованию, т.е. опухоли). Неопластические (относящиеся к опухоли) клетки могут быть либо злокачественными, т.е. инвазивными и метастатическими, либо доброкачественными.
Термин "функционально (операбельно) связанные", при ссылке на ДНК-сегменты, указывает, что эти сегменты расположены таким образом, что они функционируют совместно для их предполагаемых целей, например, транскрипция инициируется в промоторе и протекает через кодирующий сегмент до терминатора.
Термин "ортолог" обозначает полипептид или белок, полученный из одного вида, который является функциональной копией полипептида или белка из другого вида. Различия последовательностей среди ортологов являются результатом видообразования.
"Паралоги" являются отличающимися, но структурно родственными белками, производимыми организмом. Считается, что паралоги возникают в результате дупликации генов. Например, α-глобин, β-глобин и миоглобин являются паралогами друг друга.
Термин "полинуклеотид" обозначает одно- или двухцепочечный полимер дезоксирибонуклеотидных или рибонуклеотидных оснований, считываемых от 5'-конца к 3'-концу. Полинуклеотиды включают в себя РНК и ДНК и могут быть изолированы из природных источников, синтезированы in vitro или получены из комбинации природных и синтетических молекул. Размеры полинуклеотидов выражаются как пары нуклеотидов (сокращенно "п.н."), нуклеотиды ("нт") или тысячи пар нуклеотидов ("т.п.н."). Там, где позволяет контекст, два последние термина могут описывать полинуклеотиды, которые являются одноцепочечными или двухцепочечными. При применении этого термина к двухцепочечным молекулам его используют для обозначения общей длины, и должно быть понятно, что он является эквивалентным термину "пары нуклеотидов". Специалистам в данной области будет понятно, что две цепи двухцепочечного полинуклеотида могут слегка отличаться по длине и что их концы могут быть расположены зигзагами в результате ферментативного расщепления; таким образом, не все нуклеотиды в двухцепочечной полинуклеотидной молекуле могут быть спаренными.
"Полипептид" является полимером аминокислотных остатков, соединенных пептидными связями, получен ли он природным путем или синтетическим путем. Полипептиды, имеющие менее приблизительно 10 аминокислотных остатков, обычно называют "пептидами".
Термин "промотор" используется здесь в его признанном в данной области значении для обозначения части гена, содержащей последовательности ДНК, которые обеспечивают связывание РНК-полимеразы и инициацию транскрипции. Промоторные последовательности обнаруживаются обычно, но не всегда, в 5'-некодирующих районах генов.
"Белок" обозначает макромолекулу, содержащую одну или несколько полипептидных цепей. Белок может включать в себя также непептидные компоненты, такие как углеводные группы. Углеводы и другие непептидные компоненты могут быть присоединены к белку клеткой, в которой продуцируется данный белок, и будут варьироваться в зависимости от типа клетки. Белки определены здесь в терминах их аминокислотных каркасных структур; заместители, такие как углеводные группы, обычно не указаны, но тем не менее они могут присутствовать.
Термин "рецептор" используется здесь для обозначения связанного клеткой белка, который связывается с биоактивной молекулой ("лигандом") и медиирует действие этого лиганда на клетку. Мембраносвязанные рецепторы характеризуются мультипептидной структурой, содержащей внеклеточный лиганд связывающий домен и внутриклеточный домен, который обычно участвует в передаче сигнала. Связывание лиганда с рецептором приводит к конформационному изменению в рецепторе, которое вызывает взаимодействия между эффекторным доменом (доменами) и другой молекулой (молекулами) в клетке. Эти взаимодействия, в свою очередь, приводят к изменениям в метаболизме клетки. Метаболические события, связанные с взаимодействиями рецептор-лиганд, включают в себя транскрипцию генов, фосфорилирование, дефосфорилирование, увеличения продуцирования циклического АМФ, мобилизацию клеточного кальция, мобилизацию мембранных липидов, клеточную адгезию, гидролиз инозитлипидов и гидролиз фосфолипидов. Обычно рецепторы могут быть мембраносвязанными, цитозольными или ядерными; мономерными (например, рецептор тиреоидстимулирующего гормона, рецептор β-адренергического гормона) или мультимерными (например, PDGF-рецептор, рецептор гормона роста, IL-3-рецептор, GM-CSF-рецептор, G-CSF-рецептор, рецептор эритропоэтина и IL-6-рецептор).
Термин "секреторная сигнальная последовательность" обозначает здесь последовательность ДНК, которая кодирует полипептид ("секреторный полипептид"), который как компонент большего полипептида направляет этот больший полипептид через секреторный путь клетки, в которой он синтезируется. Этот больший полипептид обычно расщепляется с удалением секреторного пептида во время прохождения через этот секреторный путь.
Термин "сплайсинговый вариант" используется здесь для обозначения альтернативных форм РНК, транскрибированных из гена. Сплайсинговая вариация возникает природно посредством использования альтернативных сайтов сплайсинга в транскрибируемой молекуле РНК или менее обычно между раздельно транскрибируемыми молекулами РНК и может приводить к нескольким мРНК, транскрибируемым из одного и того же гена. Сплайсинговые варианты могут кодировать полипептиды, имеющие измененную аминокислотную последовательность. Термин сплайсинговый вариант используют здесь также для обозначения белка, кодируемого сплайсинговым вариантом мРНК, транскрибируемым из гена.
Должно быть понятно, что молекулярные массы и длины полимеров, определяемые неточными аналитическими методами (например, гель-электрофорезом), являются приблизительными величинами. Когда такая величина выражается как "около" Х или "приблизительно" X, указанная величина Х должна пониматься как величина, определенная в точностью до ±10%.
Все цитируемые здесь ссылки включены в качестве ссылки в полном виде.
Данное изобретение основано частично на открытии новой последовательности ДНК, которая кодирует белок, имеющий структуру имеющего четырехспиральный пучок цитокина. Посредством способов клонирования, анализов пролиферации и исследований связывания, описанных подробно здесь, была идентифицирована полинуклеотидная последовательность, кодирующая новый лигандный полипептид, т.е. лиганд с высокой специфичностью в отношении ранее описанного orphan ("сирота")-рецептора zalpha11. Этот полипептидный лиганд, названный zalpha11-лигандом, был выделен из библиотеки кДНК, генерированной из активированных клеток периферической крови человека (hPBCs), которые были отобраны на CD3. CD3 является маркером клеточной поверхности, уникальным для клеток лимфоидного происхождения, в частности, Т-клеток.
В приведенных ниже примерах, линию клеток, которая является зависимой от связанного с orphan-рецептором zalpha11 пути для выживания и роста клеток в отсутствие других факторов роста, использовали для скрининга на источник кДНК, кодирующей zalpha11-лиганд. Предпочтительной зависимой от факторов роста клеточной линией, которую использовали для трансфекции и экспрессии рецептора zalpha11, была линия BaF3 (Palados and Steinmetz, Cell 41:727-734, 1985; Mathey-Prevot et al., Mol. Cell. Biol. 6:4133-4135, 1986). Однако другие зависимые от факторов роста клеточные линии, такие как FDC-P1 (Hapel et al., Blood 64:786-790, 1984) и M07e (Kiss et al., Leukemia 7:235-240. 1993), являются пригодными для этой цели.
Аминокислотная последовательность для zalpha11-рецептора показала, что этот кодируемый рецептор принадлежит к подсемейству рецепторов цитокинов Класса I, которое включает в себя, но не ограничивайся ими, рецепторы для IL-2, IL-4, IL-7, IL-15, ЕРО, ТРО, GM-CSF и G-CSF (Обзор см. в Cosman, "The Hematopoietin Receptor Superfamily" in Cytokine 5(2):95-106, 1993). Zalpha11-рецептор полно описан в патентной заявке РСТ совместного владения с номером US99/22149. Анализ тканевого распределения мРНК zalpha11-рецептора показал экспрессию в лимфатическом узле, лейкоцитах периферической крови (PBL), селезенке, костном мозгу и вилочковой железе (тимусе). Кроме того, эта мРНК была обильной в линии клеток Raji (ATCC No. CCL-86), полученной из лимфомы Беркитта. Тканевое распределение этого рецептора предполагает, что мишенью для предсказанного zalpha11-лиганда являются клетки гемопоэтического происхождения, в частности лимфоидные клетки-предшественники и лимфоидные клетки. Другие имеющие структуру четырехспирального пучка цитокины, которые действуют на лимфоидные клетки, включают в себя IL-2, IL-4, IL-7 и IL-15. В отношении обзора имеющих структуру четырехспирального пучка цитокинов см. Nicola et al., Advances in Protein Chemistry 52:1-65, 1999 и Kelso, A., Immunol. Cell BjoL76:300-317, 1998.
Кондиционированные среды (КС) отобранных на CD3+, ФМА/Иономицин-стимулированных клеток периферической крови человека поддерживали рост клеток BaF3, которые экспрессировали zalpha11-рецептор и были в ином случае зависимыми от IL-3. Кондиционированные среды из клеток, которые не были 1) ФМА/Иономицин-стимулированными; или не были 2) CD3 отобранными (с ФМА/Иономицин-стимуляцией или без нее), не поддерживали рост клеток BaF3/zalpha11-рецептор. Контрольные эксперименты показали, что эта пролиферативная активность не была связана с другими известными факторами роста и что способность таких кондиционированных сред стимулировать пролиферацию экспрессирующих zalpha11-рецептор клеток могла быть нейтрализована растворимой формой этого рецептора.
Пролиферацию zalpha11-рецептор-экспрессирующих клеток BaF3, подвергнутых воздействию КС (кондиционированной среды) от CD3+ отобранных ФМА/Иономицин-стимулированных клеток периферической крови человека, идентифицировали визуальным обследованием культур и/или анализом пролиферации. Многочисленные подходящие анализы пролиферации известны в данной области и они включают в себя анализы на уменьшение красителя, такого как alamarBlue™ (AccuMed International, Inc. Westlake, Ohio), 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолийбромид (Mosman, J. Immunol. Meth. 65:55-63. 1983); 3- (4,5-диметилтиазол-2-ил)-5-3-карбоксиметоксифенил-2Н-тетразолий; 2,3-бис(2-метокси-4-нитро-5-сульфофенил)-5-[(фениламино)карбонил]-2Н-тетразолийгидроксид и цианодитолилтетразолийхлорид (которые являются коммерчески доступными из Polysciences, Inc., Warrington, PA); анализы митогенеза, такие как, например, измерение включения 3H-тимидина; анализы вытеснения красителя, использующие, например, красители нафталиновый черный или трипановый синий; поглощение красителя с использованием диацетилфлуоресцеина; и высвобождение хрома. См. в общем, Freshney, Culture of Animal Cells: A Manual of Basic Technique. 3rd ed., Wiley-Liss, 1994, включ. здесь в качестве ссылки.
Библиотеку кДНК получали из CD3+ отобранных ФМА/Иономицин-стимулированных клеток периферической крови человека. Библиотеку кДНК CD3+ отобранных ФМА/Иономицин-стимулированных клеток периферической крови человека делили на пулы, содержащие множественные молекулы кДНК, и трансфицировали в линию клеток-хозяев, например, в клетки ВНК 570 (АТСС accession no. 10314). Трансфицированные клетки-хозяева культивировали в среде, которая не содержала экзогенных факторов роста, и кондиционированную среду собирали. Кондиционированные среды анализировали на способность стимулировать пролиферацию клеток BaF3, трансфицированных zalpha11-рецептором. Были идентифицированы пулы кДНК, продуцирующие кондиционированную среду, которая стимулировала клетки BaF3/zalpha11-рецептор. Эту объединенную плазмидную кДНК электропорировали в Е.coli. кДНК выделяли из одиночных колоний и трансфицировали индивидуально в клетки ВНК 570. Положительные клоны идентифицировали по положительному результату в анализе пролиферации клеток ВаР3/zаlрhа11-рецептор и специфичность испытывали нейтрализацией пролиферации с использованием растворимого zalpha11-рецептора.
Положительные клоны выделяли, и анализ секвенирования выявил, что полинуклеотидная последовательность, содержащаяся в этой плазмидной ДНК, была новой. Секреторная сигнальная последовательность состоит из аминокислотных остатков 1 (Met) - 31 (Gly), а зрелый полипептид состоит из аминокислотных остатков 32 (Gln) - 162 (Ser) (как показано в SEQ ID NO:2).
Обычно прогнозируется, что цитокины имеют структуру, состоящую из четырех альфа-спиралей, причем спирали А, С и D являются наиболее важными во взаимодействиях лиганд-рецептор и являются более высококонсервативными среди членов этого семейства. Со ссылкой на аминокислотную последовательность zalpha11-лиганда человека, показанную в SEQ ID NO:2, сопоставление аминокислотных последовательностей zalpha11-лиганда человека, IL-15 человека, IL-4 человека и GM-CSF человека, прогнозируется, что спираль А zalpha11-лиганда определяется аминокислотными остатками 41-56; спираль В аминокислотными остатками 69-84; спираль С аминокислотными остатками 92-105 и спираль D аминокислотными остатками 135-148; показанными в SEQ ID NO:2. Структурный анализ предполагает, что А/В-петля является длинной, В/С-петля является короткой и C/D-петля является параллельно длинной. Эта структура петель приводит к спиральной организации вверх-вверх-вниз-вниз. Остатки цистеина являются абсолютно консервативными между zalpha11-лигандом и IL-15, как показано на чертеже. Остатки цистеина, которые являются консервативными между IL-15 и zalpha11-лигандом, соответствуют аминокислотным остаткам 71, 78, 122 и 125 SEQ ID NO:2. Сохранение некоторых остатков цистеина обнаружено также в IL-2, IL-4, GM-CSF и zalpha11-лиганде, соответствующих аминокислотным остаткам 78-125 SEQ ID NO:2, как показано на чертеже. Совпадающее расположение цистеинов является дополнительным подтверждением структуры четырехспирального пучка. Также высококонсервативной в этом семействе, содержащем IL-15, IL-2, IL-4, GM-CSF и zalpha11-лиганд, является последовательность Glu-Phe-Leu, показанная в SEQ ID NO:2 в остатках 136-138 на чертеже.
Последующий анализ zalpha11-лиганда на основе множественных сопоставлений (показанных на чертеже) прогнозирует, что аминокислотные остатки 44, 47 и 135 (показанные в SEQ ID NO:2) играют важную роль в связывании zalpha11-лиганда с его родственным рецептором. Кроме того, предсказанная аминокислотная последовательность мышиного zalpha11-лиганда обнаруживает 57% идентичность с предсказанным белком человека. На основе сравнения последовательностей zalpha11-лиганда человека и мыши хорошо сохраненные остатки были обнаружены в районах, предсказанных для кодирования альфа-спиралей А и D. Соответствующие полинуклеотиды, кодирующие районы, домены, мотивы, остатки и последовательности zalpha11-лиганда, описанные здесь, показаны в SEQ ID NO:1.
Подробный мутационный анализ проводили для IL-4 и IL-2, оба из которых являются высокородственными zalpha11-лиганду. Анализ мышиного IL-2 (Zurawski et al., EMBO J. 12:5113-5119, 1993) показывает, что остатки в спиралях А и С являются важными для связывания с IL-2Rβ; критическими остатками являются Asp34, Asn99 и Asn103. Многочисленные остатки в петле А/В мышиного IL-2 и спираль В являются важными для связывания IL-2Rα, тогда как только единственный остаток, Gln141 в спирали D, является очень важным для связывания с IL-2Rα. Подобным образом, спирали А и С являются сайтами взаимодействия между IL-4 и IL-4Rα (структурно сходным с IL-2Rα), а остатки в спирали D являются существенными для взаимодействия IL-2Rα (Wang et al., Proc. Natl. Acad. Sci. USA 94:1657-1662, 1997; Kruse et al., EMBQ.J. 11:3237-3244, 1992). В частности, мутация Tyr124 в Asp в IL-4 человека создает антагонист, который связывается с IL-4Rα, но не с IL-2Rα и, следовательно, не может передавать сигнал (Kruse et al. ibid. 1992).
В то время как спираль А является относительно хорошо сохраненной между человеческим и мышиным zalpha11-лигандом, спираль С является более расходящейся. Хотя оба вида имеют преобладающие кислые аминокислоты в этом районе, эти различия могут быть ответственными за видовую специфичность взаимодействия между zalpha11-лигандом и его рецептором "β"-типа, zalpha11. Петля А/В и спираль В zalpha11-лиганда являются вполне консервативными между видами; хотя субъединица рецептора, соответствующая IL-2Rα, еще не была идентифицирована, консервативность в этом районе предполагает, что она является функционально значимой. D-спирали человеческого и мышиного zalpha11-лиганда являются также высококонсервативными. Антагонисты zalpha11-рецептора могут быть сконструированы через мутации в спирали D zalpha11-лиганда. Они могут включать в себя укорочение этого белка от остатка Gln145 (SEQ ID NO:2) или мутации Gln145 или lle148 (SEQ ID NO:2; соответствующего Tyr124 в IL-4 человека) в остатки, такие как Ala или Asp. Любая мутация, которая разрушает спиральную структуру zalpha11-лиганда, может устранять связывание его рецептора и, следовательно, ингибировать передачу сигнала.
Цитокины со структурой четырехспирального пучка группируют также по длине их компонентов-спиралей. Цитокины "длинноспиральной" формы обычно состоят из спиралей из 24-30 остатков и включают в себя IL-6, цилиарный нейротрофический фактор (CNTF), ингибирующий лейкоз фактор (LIF) и гормон роста человека (hGH). Цитокины "короткоспиральной" формы обычно состоят из спиралей из 18-21 остатков и включают в себя IL-2, IL-4 и GM-CSF. Авторы считают, что zalpha11-лиганд является новым членом группы цитокинов короткоспиральной формы. Исследования с использованием CNTF и IL-6 показали, что спираль CNTF может заменять эквивалентную спираль в IL-6, придавая этой химере свойства связывания CNTF. Таким образом, функциональные домены четырехспиральных цитокинов, по-видимому, определяются на основе структурной гомологии независимо от идентичности последовательности и могут сохранять функциональную целостность в химере (Kallen et al., J. Biol. Chem. 274:11859-11867, 1999). Таким образом, спиральные домены zalpha11-лиганда будут применимы для получения химерных слитых молекул, в частности, с другими цитокинами короткоспиральной формы, для определения и модулирования специфичности связывания рецептора. Особый интерес представляют слитые (гибридные) белки, сконструированные со спиралью А и/или спиралью D, или слитые белки, которые объединяют спиральный и петлевой домены из других цитокинов короткоспиральной формы, таких как IL-2, IL-4, IL-15 и GM-CSF. Аминокислотные остатки, составляющие спирали А, В, С и D и петли А/В, В/С и C/D, для zalpha11-лиганда, IL-2, IL-4, IL-15 и GM-CSF показаны в таблице 1.
Данное изобретение обеспечивает полинуклеотидные молекулы, в том числе молекулы ДНК и РНК, которые кодируют описанные здесь полипептиды zalpha11-лиганда. Специалистам в данной области будет понятно, что в связи с вырожденностью генетического кода возможна значительная вариация среди этих полинуклеотидных молекул. SEQ ID NO:3 является вырожденной последовательностью ДНК, которая включает в себя все ДНК, которые кодируют полипептид zalpha11-лиганда SEQ ID NO:2. Специалистам в данной области будет понятно, что вырожденная последовательность SEQ ID NO:3 обеспечивает также все последовательности РНК, кодирующие SEQ ID NO:2, путем замены U (урацилом) Т (тимина). Таким образом, кодирующие полипептид zalpha11-лиганда полинуклеотиды, содержащие район от нуклеотида 1 или 97 до нуклеотида 486 SEQ ID NO:3, и их РНК-эквиваленты рассматриваются данным изобретением. Таблица 2 дает однобуквенные коды, используемые в SEQ ID NO:3 для обозначения положений вырожденных нуклеотидов. "Разрешения" представляют собой нуклеотиды, обозначенные кодовой буквой. "Комплемент" указывает код для комплементарного нуклеотида (комплементарных нуклеотидов). Например, код Y обозначает либо С, либо Т, а его комплемент R обозначает А или G, причем А является комплементарным Т, а G является комплементарным С.
Вырожденные кодоны, используемые в SEQ ID NO:3, включающие в себя все возможные кодоны для конкретной аминокислоты, представлены в таблице 3.
Специалисту с обычной квалификацией в данной области будет понятно, что некоторая двусмысленность вводится в определение вырожденного кодона, представляющего все возможные кодоны, кодирующие каждую аминокислоту. Например, вырожденный кодон для серина (WSN) может в некоторых обстоятельствах кодировать аргинин (AGR), а вырожденный кодон для аргинина (MGN) может в некоторых обстоятельствах кодировать серин (AGY). Сходная взаимосвязь существует между кодонами, кодирующими фенилаланин и лейцин. Таким образом, некоторые полинуклеотиды, охватываемые вырожденной последовательностью, могут кодировать вариантные аминокислотные последовательности, но специалист с обычной квалификацией в данной области может легко идентифицировать такие вариантные последовательности сравнением с аминокислотной последовательностью SEQ ID NO:2. Вариантные последовательности могут быть легко тестированы на функциональность, как описано здесь. Так, предложена полинуклеотидная молекула, где указанные нуклеотиды являются такими, как показано в SEQ ID NO:3 от нуклеотида 121 до нуклеотида 444 или от нуклеотида 94 до нуклеотида 486.
Специалисту с обычной квалификацией в данной области будет также понятно, что различные виды могут проявлять "предпочтительное (преферентивное) использование кодонов". В общем, см., Grantham, et al., Nuc. Acids Res. 8:1893-912, 1980; Haas, et al., Curr. Biol. 6:315-24, 1996; Wain-Hobson, et al., Gene 13: 355-64, 1981; Grosjean and Fiers, Gene 18:199-209, 1982; Holm. Nuc. Adds Res. 14:3075-87, 1986; Ikemura, J. Mol. Biol. 158:573-97, 1982. В применении здесь термин "предпочтительное (преферентивное) использование кодонов" или "предпочтительные (преферентивные) кодоны" является термином данной области, относящимся к кодонам трансляции белка, которые наиболее часто используются в клетках определенных видов, отдавая, следовательно, предпочтение одному или немногим представителям возможных кодонов, кодирующих каждую аминокислоту (См. таблицу 3). Например, аминокислота треонин (Thr) может кодироваться АСА, АСС, ACG или ACT, но в клетках млекопитающих наиболее обычно используемым кодоном является АСС; в других видах, например, в клетках насекомых, дрожжей, вирусов или бактерий, могут быть предпочтительными другие кодоны Thr. Предпочтительные кодоны для конкретных видов могут быть введены в полинуклеотиды данного изобретения различными способами, известными в данной области. Введение предпочтительных последовательностей кодонов в рекомбинантную ДНК может, например, усиливать продуцирование этого белка, делая трансляцию белка более эффективной в конкретном типе клеток или виде. Таким образом, вырожденная последовательность кодонов, описанная в SEQ ID NO:3, служит в качестве матрицы для оптимизации экспрессии полинуклеотидов в различных типах клеток и видах, обычно используемых в данной области и описанных здесь. Последовательности, содержащие предпочтительные кодоны, могут быть тестированы и оптимизированы для экспрессии в различных видах и тестированы на функциональность, как описано здесь.
Как отмечалось ранее, выделенные полинуклеотиды данного изобретения включают в себя ДНК и РНК. Способы выделения ДНК и РНК хорошо известны в данной области. Обычно РНК выделяют из ткани или клетки, которая продуцирует большие количества РНК zalpha11-лиганда. Такие ткани и клетки идентифицируют Нозерн-блоттингом (Thomas, Proc. Natl. Acad. Sci. USA 77:5201, 1980) или скринингом кондиционированной среды от различных типов клеток на активность на клетках- или тканях-мишенях. Как только эта активность или продуцирующая РНК клетка или ткань идентифицирована, тотальная РНК может быть получена с использованием экстракции изотиоцианатом гуанидиния с последующим выделением центрифугированием в градиенте CsCl (Chirgwin et al., Biochemistry 18:52-94, 1979). Поли(А)+ РНК получают из тотальной РНК с использованием способа Aviv and Leder, Proc. Natl. Acad. Sci. USA 69:1408-1412 (1972). Комплементарную ДНК (кДНК) получают из пол(А)+ РНК при помощи известных способов. Альтернативно, может быть выделена геномная ДНК. Затем полинуклеотиды, кодирующие полипептиды zalpha11-лиганда, идентифицируют и выделяют, например, с использованием гибридизации или полимеразной цепной реакции (ПЦР).
Полноразмерный клон, кодирующий zalpha11-лиганд, может быть получен общепринятыми процедурами клонирования. Клоны комплементарной ДНК (кДНК) являются предпочтительными, хотя для некоторых применений (например, экспрессиии в трансгенных животных) могут быть предпочтительными использование геномного клона или модификация клона кДНК с целью включения по меньшей мере одного геномного интрона. Способы получения клонов кДНК и геномных клонов хорошо известны и находятся в пределах обычной квалификации в данной области и включают в себя использование описанной здесь последовательности или ее частей для зондирования или праймирования библиотеки. Экспрессионные библиотеки могут быть зондированы антителами к фрагментам zalpha 11-рецептора или другим партнерам специфического связывания.
Полинуклеотидные последовательности zalpha11-лиганда, описанные здесь, могут быть также использованы в качестве зондов или праймеров для клонирования 5'-некодирующих районов гена zalpha11-лиганда. В связи с тканеспецифической экспрессией, наблюдаемой для zalpha11-лиганда, ожидается, что этот район гена обеспечивает гемопоэз-специфическую и лимфоид-специфическую экспрессию. Таким образом, промоторные элементы из гена zalpha11-лиганда могли бы направлять тканеспецифическую экспрессию гетерологичных генов, например, в трансгенных животных или пациентах, подвергаемых генной терапии. Клонирование 5'-фланкирующих последовательностей также облегчает получение белков zalpha11-лиганда "активацией генов", как описано в патенте США US Patent No. 5641670. Вкратце, экспрессию эндогенного гена zalpha11-лиганда в клетке изменяют введением в локус zalpha11-лиганда конструкции ДНК, содержащей по меньшей мере нацеливающую последовательность, регуляторную последовательность, экзон и неспаренный донорный сайт сплайсинга. Нацеливающей последовательностью является 5'-некодирующая последовательность zalpha11-лиганда, которая делает возможной гомологичную рекомбинацию этой конструкции с эндогенным локусом zalpha11-лиганда, посредством чего последовательности внутри конструкции становятся функционально связанными с эндогенной кодирующей zalpha11-лиганд последовательностью. Таким путем эндогенный промотор zalpha11-лиганда может быть заменен или дополнен другими регуляторными последовательностями для обеспечения усиленной тканеспецифической или иным образом регулируемой экспрессии.
Далее, данное изобретение обеспечивает копии полипептидов и полинуклеотидов из других видов (ортологи). Эти виды включают в себя, но не ограничиваются ими, виды млекопитающих, птиц, земноводных, пресмыкающихся, рыб, насекомых и другие виды позвоночных и беспозвоночных. Особый интерес представляют собой полипептиды zalpha11-лиганда из других видов млекопитающих, в том числе полипептиды мышей, свиньи, овцы, коровы, псовых, кошачьих, лошадиных видов и полипептиды других приматов. Ортологи zalpha11-лиганда человека могут быть клонированы с использованием информации и композиций, обеспеченных данным изобретением, в сочетании с общепринятыми способами клонирования. Например, кДНК может быть клонирована с использованием мРНК, полученной из типа ткани или клеток, который экспрессирует zalpha11-лиганд, как описано здесь. Подходящие источники мРНК могут быть идентифицированы зондированием Нозерн-блотов зондами, сконструированными из описанных здесь последовательностей. Затем готовят библиотеку из мРНК позитивной ткани или линии клеток. Затем кодирующая лиганд zalpha11 кДНК может быть выделена различными способами, такими как зондирование полной или частичной кДНК человека или одним или несколькими наборами вырожденных зондов, основанных на описанных последовательностях. Эта кДНК может быть также клонирована с использованием полимеразной цепной реакции, или ПЦР (Mullis, US Patent No. 4683202), с применением праймеров, сконструированных из описанной здесь характерной последовательности zalpha11-лиганда человека. В дополнительном способе эта библиотека кДНК может быть использована для трансформации или трансфекции клеток-хозяев, и экспрессия представляющей интерес кДНК может быть детектирована антителом к полипептиду zalpha11-лиганда, исследованиями связывания или анализами активности. Подобные способы могут быть применены также для выделения геномных клонов.
Полинуклеотидная последовательность для мышиного ортолога zalpha11-лиганда была идентифицирована и показана в SEQ ID NO:55, а соответствующая аминокислотная последовательность показана в SEQ ID NO:56. Существует 62% идентичность между мышиной и человеческой последовательностями на протяжении района из 124 аминокислот, который соответствует остаткам 30-153 в SEQ ID NO:2 и остаткам 23-146 SEQ ID NO:56 zalpha11-лиганда. Зрелая последовательность для мышиного zalpha11-лиганда предположительно начинается при His18 (как показано в SEQ ID NO:56), который соответствует His25 (как показано в SEQ ID NO:2) в человеческой последовательности. Поскольку укороченная форма человеческого полипептида является активной, кажется вероятным, что эквивалентный полипептид мышиного zalpha11-лиганда (т.е. без остатков His18-Pro22 SEQ ID NO:56) является также активным. Тканевый анализ выявил, что экспрессия мышиного zalphall-лиганда обнаруживается в яичке, селезенке и вилочковой железе (тимусе).
Специалистам в данной области будет понятно, что последовательность, описанная в SEQ ID NO:1, представляет единственный аллель zalpha11-лиганда человека и что ожидается существование аллельной вариации и альтернативного сплайсинга. Аллельные варианты этой последовательности могут быть клонированы зондированием библиотек кДНК или геномных библиотек из различных индивидуумов в соответствии со стандартными процедурами. Аллельные варианты последовательности ДНК, показанной в SEQ ID NO:1, в том числе варианты, содержащие молчащие мутации, и варианты, в которых мутации приводят к изменениям в последовательности аминокислот, находятся в объеме данного изобретения, так же как и белки, которые являются аллельными вариантами SEQ ID NO:2. кДНК, генерируемые из образованных альтернативным сплайсингом мРНК, которые сохраняют свойства полипептида zalpha11-лиганда, включены в объем данного изобретения, так же как полипептиды, кодируемые такими кДНК и мРНК. Аллельные варианты и сплайсинговые варианты этих последовательностей могут быть клонированы зондированием библиотек кДНК или геномных библиотек из различных индивидуумов или тканей в соответствии со стандартными процедурами, известными в данной области. Так, предложена полинуклеотидная молекула, где указанные нуклеотиды являются такими, как показано в SEQ ID NO:1 от нуклеотида 167 до нуклеотида 490 или от нуклеотида 140 до нуклеотида 532.
Ген zalpha11-лиганда был картирован относительно маркера SHGC-12342 каркаса IL-2 с помещением zalpha11-лиганда на расстоянии 180 т.п.н. от этого маркера IL-2. Применение окружающих маркеров помещает ген zalpha11-лиганда в район 4q27 на карте интегрированной LDB хромосоме 4 (The Genetic Location Database, University of Southhampton). Данное изобретение обеспечивает также реагенты, которые найдут применение в диагностических приложениях. Например, ген zalpha11-лиганда, зонд, содержащий ДНК или РНК zalpha11-лиганда или их субпоследовательность, могут быть использованы для определения, присутствует ли ген zalpha11-лиганда на хромосоме человека, такой как хромосома 4, или имела ли место мутация гена. На основе аннотации фрагмента геномной ДНК человека, содержащей часть геномной ДНК zalpha11-лиганда (Genbank Accession No. (номер доступа Банка генов) АС007458), zalpha11-лиганд локализован в районе 4q27 хромосомы 4. Детектируемые хромосомные аберрации в генном локусе zalpha11-лиганда включают в себя, но не ограничиваются ими, анеуплоидию, изменения копийности гена, потерю гетерогенности (LOH), транслокации, инсерции, делеции, изменения сайтов рестрикции и реаранжировки. Такие аберрации могут быть детектированы с использованием полинуклеотидов данного изобретения путем применения молекулярно-генетических способов, таких как анализ полиморфизма длины фрагментов рестрикции (RFLP), анализ коротких тандемных повторов (STR) с использованием ПЦР-способов, и других способов анализа генетического сцепления, известных в данной области (Sambrook et al., ibid; Ausubel et al., ibid; Marian, Chest 108:255-65, 1995).
Точное знание положения гена может быть полезным для ряда целей, в том числе: 1) определения, является ли последовательность частью существующего контига, и получения дополнительных окружающих генетических последовательностей в различных формах, таких как YAC, ВАС или кДНК-клоны; 2) обеспечения возможного гена-кандидата для наследственного заболевания, который обнаруживает сцепление с тем же самым хромосомным районом; и 3) перекрестного сравнения с модельными организмами, такими как мышь, которое может способствовать определению, какую функцию может иметь конкретный ген.
Как утверждалось выше, ген zalpha11-лиганда человека локализован вблизи гена IL-2, который находится в районе хромосомы 4q, который, как было показано, имеет сцепление с восприимчивостью к воспалительному заболеванию пищеварительного тракта (IBD) (в том числе болезни Крона (CD) и язвенному колиту) в некоторых семьях (Hampe et al., Am. J. Hum. Genet. 64:808-816, 1999; Cho et al., Proc. Natl. Acad. Sci. 95:7502-7507, 1998). Кроме того, ген zalpha11-рецептора картирован относительно 16р11, другого геномного района, который ассоциирован с восприимчивостью к болезни Крона (CD) (Hugot et al., Nature 379:821-823, 1996; Ohmen et al., Hum. Mol. Genet. 5:1679-1683. 1996). Болезнь Крона (CD) представляет собой хроническое воспаление пищеварительного тракта с частым системным участием; хотя точная этиология неизвестна, иммунорегуляторная дисфункция, включающая в себя отсутствие толерантности к обычным антигенам пищеварительного тракта, является основным компонентом (в отношении обзора см. Braegger et al., Annals Allergy 72:135-141, 1994; Sartor, Am. J. Gastroenterol. 92:58-1 IS, 1997)). Несколько исследований обнаружили отклоняющуюся от нормы активность МК-клеток в пациентах с болезнью Крона (CD) (см., например, Egawa et al., J. Clin. Lab. Immunol. 20:187-192. 1986; Aparicio-Pages et al., J. Clin. Lab. Immunol. 29:119-124, 1989; Tol et al., Scand. J. Gastroenterol. 27:999-1005, 1992)), и было также документировано образование В-клеток с дефектной памятью (Brogan et al., J. Clin. Lab. Immunol. 24:69-74. 1987). Поскольку zalpha11-лиганд играет роль в иммунорегуляции и поскольку гены как рецептора, так и лиганда находятся в районах восприимчивости к CD (болезни Крона), как рецептор, так и лиганд являются генами-кандидатами для генетического предрасположения к болезни Крона.
Определение участия zalpha 11-рецептора и/или zalpha11-лиганда в патологии IBD (воспалительного заболевания пищеварительного тракта) может быть выполнено несколькими способами. Секвенирование экзонов из геномной ДНК может выявить кодирующие мутации (в том числе миссенс-мутации, нонсенс-мутации и мутации со сдвигом рамки (знаковые мутации)), как и секвенирование кДНК. Дополнительное преимущество секвенирования из геномной ДНК заключается в том, что границы сплайсинга также содержатся в секвенируемых фрагментах и могут выявить отклонения сплайсинга, которые могут не проявляться в пробах кДНК, если, например, неправильно сплайсированные РНК быстро деградировались. Была определена геномная структура zalpha11-лиганда. Другие способы анализа zalpha 11-лиганд а и zalpha 11-рецептора в пациентах с IBD включают в себя: (1) оценку продуцирования лиганда из активированных Т-клеток из пациентов в сравнении с нормальными контролями (т.е. при помощи биоанализа); (2) in situ гибридизацию РНК zalpha 11-рецептора или zalpha11-лиганда со срезами воспаленного кишечника из пациентов IBD в сравнении с подобными срезами из нормальных контролей; (3) иммунохимию на срезах из пациентов IBD в сравнении с нормальными контролями; и (4) оценку отвечаемости периферических В-клеток на zalpha11-лиганд, измеряемой при помощи анализов митогенеза.
Диагностический агент мог бы помогать врачам в определении типа заболевания и подходящей ассоциированной терапии или мог бы помогать в генетическом консультировании. Сами по себе антитела против zalpha11-лиганда, полинуклеотиды и полипептиды zalpha11-лиганда данного изобретения могут быть использованы для обнаружения полипептида, мРНК zalpha11-лиганда или антител против zalpha11-лиганда, служа, таким образом, в качестве маркеров, и могут быть непосредственно использованы для обнаружения генетических заболеваний или раков, как описано здесь, с использованием способов, известных в данной области и описанных здесь. Далее, полинукпеотидные зонды zalpha11-лиганда могут быть использованы для обнаружения отклонений от нормы, включающих в себя хромосому 4q27, как описано здесь. Эти отклонения от нормы могут быть связаны с заболеваниями человека или онкогенезом, самопроизвольным выкидышем или другими генетическими нарушениями. Таким образом, полинуклеотидные зонды zalpha11-лиганда могут быть использованы для обнаружения отклонений от нормы или генотипов, ассоциированных с этими дефектами.
Как обсуждалось выше, дефекты в гене zalpha11-лиганда сами могут приводить к наследственным патологическим состояниям человека. Молекулы данного изобретения, такие как полипептиды, антагонисты, агонисты, полинуклеотиды и антитела данного изобретения могут способствовать детектированию, диагностике, предупреждению и лечению заболеваниий, связанных с генетическим дефектом zalpha11-лиганда. Кроме того, полинукпеотидные зонды zalpha11-лиганда могут быть использованы для обнаружения аллельных различий между поражанными заболеванием и не пораженными заболеванием индивидуумами в хромосомном локусе zalpha11-лиганда. Сами по себе последовательности zalpha11-лиганда могут быть использованы в качестве диагностических агентов в профилировании ДНК в судебной медицине.
В общем, диагностические способы, используемые в анализе генетического сцепления для обнаружения генетического отклонения от нормы или аберрации в пациенте, известны в данной области. Большинство диагностических способов предусматривают стадии (i) получения генетической пробы из потенциально заболевшего пациента, заболевшего пациента или потенциального не заболевшего носителя рецессивного аллеля заболевания; (ii) получения продукта первой реакции инкубированием этой генетической пробы с полинуклеотидным зондом zalpha11-лиганда, где этот полинуклеотид будет гибридизоваться с комплементарной полинуклеотидной последовательностью, например, в RFLP-анализе, или инкубированием этой генетической пробы со смысловым и антисмысловым праймером в ПЦР-реакции при подходящих условиях ПЦР-реакции; (iii) визуализации продукта первой реакции гель-электрофорезом и/или другим известным способом, таким как визуализация продукта первой реакции полинуклеотидным зондом zalpha11-лиганда, где этот полинуклеотид будет гибридизоваться с комплементарной полинуклеотидной последовательностью первой реакции; и (iv) сравнения визуализированного первого продукта реакции с продуктом второй контрольной реакции генетической пробы из нормального или контрольного индивидуума. Различие между продуктом первой реакции и продуктом контрольной реакции является признаком генетического отклонения от нормы в заболевшем или потенциально заболевшем пациенте или признаком присутствия фенотипа гетерозиготного рецессивного носителя для незаболевшего пациента или присутствия генетического дефекта в опухоли из заболевшего пациента или присутствия генетической аномалии в плоде или предимплантационном эмбрионе. Например, различие в распределении рестрикционных фрагментов, длине ПЦР-продуктов, длине повторяющихся последовательностей в генетическом локусе zalpha11-лиганда и т.п. является признаком генетического отклонения от нормы, генетической аберрации или аллельного различия в сравнении с нормальным контролем. Контроли могут быть из не пораженных болезнью членов семьи или неродственных индивидуумов в зависимости от теста и доступности проб. Генетические пробы для применения в данном изобретении включают в себя геномную ДНК, мРНК и кДНК, выделенные из любой ткани или другой биологической пробы пациента, такой как, но не только, кровь, слюна, сперма, клетки эмбриона, амниотическая жидкость и т.п. Полинуклеотидный зонд или праймер может быть РНК или ДНК и будет содержать часть SEQ ID NO:1, комплемент SEQ ID NO:1 или их РНК-эквивалент. Такие способы, показывающие анализ генетического сцепления в отношении фенотипов заболевания человека, хорошо известны в данной области. В отношении способов на основе ПЦР в диагностике, см., в общем, Mathew (ed.), Protocols in Human Molecular Genetics (Humana Press, Inc. 1991), White (ed.), PCR Protocols: Current Methods and Applications (Humana Press, Inc. 1993), Cotter (ed.), Molecular Diagnosis of Cancer (Humana Press, Inc. 1996), Hanausek and Walaszek (eds.), Tumor Marker Protocols (Humana Press, Inc. 1998), Lo (ed.), Clinical Applications of PCR (Humana Press, Inc. 1998) и Meltzer (ed.), PCR in Bioanalysis (Humana Press, Inc. 1998)).
Мутации, ассоциированные с локусом zalpha11-лиганда, могут быть обнаружены с использованием молекул нуклеиновых кислот данного изобретения с примененим стандартных способов для прямого мутационного анализа, таких как анализ полиморфизма длин рестрикционных фрагментов, анализ коротких тандемных повторов с использованием ПЦР-способов, анализ системы амплификации-рефрактерной (резистентной) мутации, детектирование полиморфизма одноцепочечной конформации, способы расщепления при помощи РНКазы, денатурирующий градиентный гель-электрофорез, анализ ошибочного спаривания с использованием флуоресценции и другие генетические способы анализа, известные в данной области (см., например, Mathew (ed.), Protocols in Human Molecular Genetics (Humana Press, Inc. 1991), Marian, Chest 108:255 (1995), Coleman and Tsongalis, Molecular Diagnostics (Humana Press, Inc. 1996), Elles (ed.) Molecular Diagnosis of Genetic Diseases (Humana Press, Inc. 1996), Landegren (ed.), Laboratory Protocols for Mutation Detection (Oxford University Press 1996), Birren et al., (eds.), Genome Analysis, Vol. 2: Detecting Genes (Cold Spring Harbor Laboratory Press 1998), Dracopoli et al., (eds.), Current Protocols in Human Genetics, (John Wiley & Sons 1998) и Richards and Ward, "Molecular Diagnostic Testing", in Principles of Molecular Medicine, pages 83-88 (Humana Press, Inc. 1998). Прямой анализ гена zalpha11-лиганда на мутацию может проводиться с использованием геномной ДНК человека. Способы амплификации геномной ДНК, полученной, например, из лимфоцитов периферической крови, хорошо известны специалистам в данной области (см., например, Dracopoli et al., (eds.), Current Protocols in Human Genetics, на страницах 7.1.6-7.1.7 (John Wiley & Sons 1998)).
Положения интронов в гене zalpha11-лиганда определяли идентификацией геномных клонов с последующим секвенированием границ интрон/экзон. Первый интрон лежит между аминокислотным остатком 56 (Leu) и остатком 57 (Val) в SEQ ID NO:2 и имеет длину 115 п.н. Второй интрон имеет максимальную длину 4,4 т.п.н. и лежит между аминокислотным остатком 68 (Glu) и остатком 69 (Thr) в SEQ ID NO:2. Третий интрон имеет длину 2,6 т.п.н. и лежит между аминокислотным остатком 120 (Leu) и остатком 121 (Thr) в SEQ ID NO:2. Конечный интрон, 89 п.н., лежит между аминокислотным остатком 146 (Lys) и остатком 147 (Met) в SEQ ID NO:2. Полный ген простирается на приблизительно 8 т.п.н.
Структура гена zalpha11-лиганда является сходной со структурой гена IL-2 (Fujita et al., Proc. Natl. Acad. Sci. 80:7437-7441, 1983), хотя ген zalpha11-лиганда содержит один дополнительный интрон (интрон 4). Характер короткого первого интрона и длинных второго и третьего интронов является консервативным между этими двумя генами, хотя ген IL-2 является в целом несколько меньшим (около 6 т.п.н.). Ген IL-15, с другой стороны, состоит из 8 экзонов и простирается по меньшей мере на 34 т.п.н. (Anderson et al. Genomics 25:701-706, 1995). Таким образом, ген zalpha11-лиганда более сходен по структуре с геном IL-2, чем с геном IL-15.
В предпочтительных вариантах данного изобретения выделенные кодирующие zalpha11-лиганд молекулы нуклеиновых кислот будут гибридизоваться при строгих условиях с молекулами нуклеиновых кислот, имеющими нуклеотидную последовательность SEQ ID NO:1, с молекулами нуклеиновых кислот, имеющими нуклеотидную последовательность нуклеотидов 47-532 SEQ ID NO:1, или молекулами нуклеиновых кисллот, комплементарными SEQ ID NO:1. Обычно строгие условия выбирают таким образом, чтобы температура была на приблизительно 5°С ниже, чем точка термического плавления (Тm) для этой специфической последовательности при определенных ионной силе и рН. Тm является температурой (при определенных ионной силе и рН), при которой 50% последовательности-мишени гибридизуются с точно совместимым зондом.
Пара молекул нуклеиновых кислот, такая как ДНК-ДНК, РНК-РНК и ДНК-РНК, может гибридизоваться, если нуклеотидные последовательности имеют некоторую степень комплементарности. Гибриды могут допускать неправильно спаренные основания в двойной спирали, но на стабильность гибрида влияет степень ошибочного спаривания. Тm ошибочно спаренного гибрида снижается на 1°С для каждого 1-1,5% ошибочного спаривания. Варьирование строгости условий гибридизации позволяет контролировать степень ошибочного спаривания, которая будет присутствовать в данном гибриде. Степень строгости увеличивается по мере увеличении температуры гибридизации и уменьшении ионной силы гибридизационного буфера.
Приспособление этих условий для применения с конкретным полинуклеотидным гибридом находится вполне в рамках квалификации специалиста в данной области. Тm для конкретной последовательности мишени является температурой (при определенных условиях), при которой 50% последовательности-мишени будут гибридизоваться с точно совместимой последовательностью зонда. Условия, которые влияют на Тm, включают в себя размер и содержание пар оснований данного полинуклеотидного зонда, ионную силу гибридизационного раствора и присутствие дестабилизирующих агентов в гибридизационном растворе. Многочисленные уравнения для расчета Тm известны в данной области и являются специфическими для ДНК, РНК и гибридов ДНК-РНК и полинуклеотидных последовательностей-зондов варьирующей длины (см., например, Sambrook et al., (Molecular Cloning. A Laboratory Manual, Second Edition Cold Spring Harbor Laboratory Press 1989); Ausubel et al., (eds.), Current Protocols in Molecular Biology (John Wiley and Sons, Inc., 1987); Berger and Kimmel (eds.), Guide to Molecular Cloning Techniques, (Academic Press, Inc. 1987); и Wetmur, Crit. Rev. Biochim. Mol. Biol. 26:227 (1990)). Программное обеспечение для анализа последовательности, такое как OLIGO 6.0 (LSR; Long Lake, MN) и Primer Premier 4.0 (Premier Biosoft International; Palo Alto, CA), а также сайты в Интернете, являются доступными инструментами для анализа конкретной последовательности и расчета Тm на основе определенных пользователем критериев. Такие программы могут также анализировать конкретную последовательность при определенных условиях и идентифицировать подходящие последовательности зондов. Обычно, гибридизацию более длинных полинуклеотидных последовательностей, >50 пар оснований, выполняют при температуре на приблизительно 20-25°С ниже рассчитанной Тm. Для меньших зондов, <50 пар оснований, гибридизацию обычно проводят при Тm или на 5-10°С ниже. Это позволяет достичь максимальной скорости гибридизации для гибридов ДНК-ДНК и ДНК-РНК.
После гибридизации молекулы нуклеиновых кислот могут быть промыты для удаления не гибридизовавшихся при строгих условиях или при условиях высокой строгости молекул нуклеиновых кислот. Типичные строгие условия промывки включают в себя промывание в растворе 0,5х-2х SSC с 0,1% додецилсульфатом натрия (ДСН) при 55-65°С. То есть молекулы нуклеиновых кислот, кодирующие вариантный полипептид zalpha11-лиганда, гибридизуются с молекулой нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент), при строгих условиях промывки, в которых строгость промывки эквивалентна 0,5х-2х SSC с 0,1% ДСН при 55-65°С, в том числе 0,5х SSC с 0,1% ДСН при 55°С или 2х SSC с 0,1% ДСН при 65°С. Специалист в данной области легко придумает эквивалентные условия, например, с использованием замены SSC на SSPE в промывочном растворе.
Типичные условия высокой строгости промывки включают в себя промывание в растворе 0,1х-0,2х SSC с 0,1% додецилсульфатом натрия (ДСН) при 50-65°С. Другими словами, молекулы нуклеиновых кислот, кодирующие вариантный полипептид zalpha11-лиганда, гибридизуются с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент), при условиях высокой строгости промывки, в которых строгость промывки эквивалентна 0,1х-0,2х SSC с 0,1% ДСН при 50-65°С, в том числе 0,1х SSC с 0,1% ДСН при 50°С или 0,2х SSC с 0,1% ДСН при 65°С.
Данное изобретение обеспечивает также выделенные полипептиды zalpha11-лиганда, которые имеют по существу одинаковую идентичность последовательности с полипептидами SEQ ID NO:2 или их ортологами. Термин "по существу одинаковая идентичность последовательности" используется здесь для обозначения полипептидов, имеющих по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или более высокую, чем 95% идентичность последовательностям, показанным в SEQ ID NO:2, или их ортологам. Данное изобретение включает в себя также полипептиды, которые содержат аминокислотную последовательность, имеющую по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или более высокую, чем 95% идентичность последовательности аминокислотных остатков 1-162 или 33-162 SEQ ID NO:2. Данное изобретение включает в себя также молекулы нуклеиновых кислот, которые кодируют такие полипептиды. Способы определения процентной идентичности описаны ниже.
Данное изобретение рассматривает также вариантные молекулы нуклеиновых кислот zalpha11-лиганда, которые могут быть идентифицированы с использованием двух критериев: определения сходства между кодируемым полипептидом с аминокислотной последовательностью SEQ ID NO:2 и/или гибридизационного анализа, описанного выше. Такие варианты zalpha11-лиганда включают в себя молекулы нуклеиновых кислот: (1) которые гибридизуются с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент) при строгих условиях промывки, в которых строгость промывки эквивалентна 0,5х-2х SSC с 0,1% ДСН при 55-65°С; или (2) которые кодируют полипептид, имеющий по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или более высокую, чем 95% идентичность аминокислотной последовательности SEQ ID NO:2. Альтернативно, варианты zalpha11-лиганда могут быть охарактеризованы молекулами нуклеиновых кислот: (1) которые гибридизуются с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент) при условиях промывки высокой строгости, в которых строгость промывки эквивалентна 0,1х-0,2х SSC с 0,1% ДСН при 50-65°С; и (2) которые кодируют полипептид, имеющий по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или более высокую, чем 95% идентичность аминокислотной последовательности SEQ ID NO:2.
Процентную идентичность последовательности определяют общепринятыми способами. См., например, Altschul et al., Bull. Math. Bio. 48:603 (1986) и Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA 89:10915 (1992). Вкратце, две последовательности аминокислот сопоставляют выстраиванием для оптимизации оценок сопоставления с использованием penalty открывания гэпа 10 и penalty удлинения гэпа 1 и оценочной матрицы "BLOSUM62" Henikoff and Henikoff (ibid.), показаной в таблице 4 (аминокислоты представлены с использованием стандартных однобуквенных кодов). Затем процентную идентичность рассчитывают как
Общее число идентичных совпадений ×100
[длина более длинной последовательности плюс число гэпов, введенных в более длинную последовательность для сопоставления этих двух последовательностей]
Специалистам в данной области должно быть понятно, что имеется много установленных алгоритмов для сопоставления двух аминокислотных последовательностей. Алгоритм поиска сходства "FASTA" Pearson и Lipman представляет собой подходящий способ сопоставления белков для исследования уровня идентичности, общей для описанной здесь аминокислотной последовательности и аминокислотной последовательности предположительного варианта zalpha11-лиганда. Алгоритм FASTA описан Pearson and Lipman, Proc. Natl. Acad. Sci. USA 85:2444 (1988) и Pearson, Meth. Enzvmol. 183:63 (1990).
Вкратце, FASTA сначала характеризует сходство последовательностей путем идентификации районов, общих у запрашиваемой последовательности (например, SEQ ID NO:2) и тест-последовательности, которые имеют либо наивысшую плотность идентичностей (если переменная ktup равна 1), либо пар идентичностей (если ktup=2), без учета консервативных аминокислотных замен, инсерций или делеций. Затем десять районов с наивысшей плотностью идентичностей повторно оценивают сравнением сходства всех спаренных аминокислот с использованием матрицы аминокислотных замен, а концы районов "подравнивают" для включения только остатков, которые способствуют наивысшей оценке. Если имеется несколько районов с оценками, большими, чем величина "отсечения" (рассчитанная по предварительно определенной формуле на основе длины данной последовательности и ее величины ktup), то приведенные в порядок подравниванием первоначальные районы исследуют для определения, могут ли эти районы быть соединены с образованием приближенного сопоставления с гэпами. Наконец, районы с наивысшей оценкой двух аминокислотных последовательностей сопоставляют с использованием модификации алгоритма Needleman-Wunsch-Sellers (Needleman and Wunsch, J. Mol. Biol. 48:444 (1970); Sellers, SIAM J. Appl. Math. 26:787 (1974)), который делает возможными инсерций и делеций аминокислот. Предпочтительными параметрами для анализа FASTA являются: ktup=1, penalty открывания гэпа=10, penalty удлинения гэпа=1 и матрица замен=ВLOSUM62. Эти параметры могут быть введены в программу FASTA модификацией файла оценочной матрицы ("SMATRIX"), как объясняется в Приложении 2 Pearson, Meth. Enzymol. 183:63 (1990).
Программа FASTA может быть также использована для определения идентичности молекул нуклеиновых кислот с использованием приведенного выше соотношения. Для сравнений нуклеотидных последовательностей величина ktup может быть в диапазоне между 1 и 6, предпочтительно от 3 до 6, наиболее предпочтительно 3, с другими параметрами, установленными по умолчанию.
Вариантные полипептиды zalpha11-лиганда с по существу одинаковой идентичностью последовательности характеризуются тем, что они имеют одну или несколько аминокислотных замен, делеций или присоединений. Эти изменения предпочтительно имеют минорный характер, то есть замены консервативных аминокислот (см. таблицу 5) и другие замены, которые не влияют значимо на укладку или активность полипептида; небольшие делеций, обычно от одной до примерно 30 аминокислот; и небольшие амино- или карбоксил-концевые удлинения, такие как аминоконцевой остаток метионина, небольшой линкерный пептид до приблизительно 20-25 остатков или аффинная метка. Таким образом, данное изобретение включает в себя полипептиды из приблизительно от 108 до приблизительно 216 аминокислотных остатков, которые включают в себя последовательность, которая по меньшей мере на 70%, предпочтительно на 90% и более предпочтительно на 95% или более идентична соответствующему району SEQ ID NO:2. Полипептиды, содержащие аффинные метки, могут дополнительно содержать сайт протеолитического расщепления между полипептидом zalpha11-лиганда и этой аффинной меткой. Подходящие сайты включают в себя сайты расщепления тромбином и сайты расщепления фактором Ха.
Могут быть выполнены определения аминокислотных остатков, которые содержат районы или домены, которые являются критическими для сохранения структурной целостности. В этих районах можно определить специфические остатки, которые будут более или менее устойчивыми к изменению или сохранять общую третичную структуру этой молекулы. Способы анализа структуры последовательности включают в себя, но не ограничиваются ими, сопоставление множественных последовательностей с высокой идентичностью аминокислот или нуклеотидов, склонности к образованию вторичной структуры, бинарные распределения, комплементарную упаковку и скрытые полярные взаимодействия (Barton, Current Qpin. Struct. Biol. 5:372-376, 1995 и Cordes et al., Current Opin. Struct. Biol. 6:3-10, 1996). Обычно при конструировании модификаций для молекул или идентификации специфических фрагментов определение структуры будет сопровождаться оценкой активности модифицированных молекул.
Изменения аминокислотной последовательности выполняют в полипептидах zalpha11-лиганда таким образом, чтобы минимизировать разрушение структуры более высокого порядка, существенной для биологической активности. Например, когда полипептид zalpha11-лиганда содержит одну или несколько спиралей, изменения в аминокислотных остатках будут выполняться таким образом, чтобы не нарушить геометрию спиралей и других компонентов молекулы, где изменения в конформации уменьшают некоторую критическую функцию, например, связывание данной молекулы с ее партнерами связывания, например, спиралей А и D, остатков 44, 47 и 135 SEQ ID NO:2. Эффекты изменений аминокислотной последовательности могут быть прогнозированы, например, компьютерным моделированием, как описано выше, или определены анализом кристаллической структуры (см., например, Lapthorn et al., Nat. Struct. Biol. 2:266-268, 1995). Другие способы, которые хорошо известны в данной области, сравнивают укладку вариантного белка со стандартной молекулой (например, нативным белком). Например, может быть сделано сравнение распределения цистеина в вариантной и стандартной молекулах. Масс-спектрометрия и химическая модификация с использованием восстановления и алкилирования обеспечивают способы для определения остатков цистеина, которые ассоциированы с дисульфидными связями или свободны от таких ассоциаций (Bean et al., Anal. Biochem. 201:216-226, 1992; Gray, Protein Sci. 2:1732-1748, 1993 и Patterson et al., Anal. Chem. 66:3727-3732, 1994). Обычно считается, что, если модифицированная молекула не имеет такого же распределения цистеина, что и стандартная молекула, укладка может быть нарушенной. Другим хорошо известным и общепринятым способом измерения укладки является круговой дихроизм (КД). Измерение и сравнение КД-спектров, генерируемых модифицированной молекулой и стандартной молекулой, является рутиной (Johnson, Proteins 7:205-214. 1990). Кристаллография представляет собой другой хорошо известный способ для анализа укладки и структуры. Ядерный магнитный резонанс (ЯМР), ферментативное пептидное картирование и картирование эпитопов также являются известными способами для анализа сходства укладки и структуры между белками и полипептидами (Schaanan et al., Science 257:961-964, 1992).
Может быть получен профиль гидрофильности Hopp/Woods белковой последовательности zalpha11-лиганда, показанной в SEQ ID NO:2 (Норр et al., Ргос. Natl. Acad. Sci. 78:3824-3828, 1981; Норр, J. Immun. Meth. 88:1-18, 1986 и Triquier et al., Protein Engineering 11:153-169, 1998). Этот профиль основан на скольжении окна для шести остатков. Скрытые остатки G, S и Т и экспонированные (открытые) остатки Н, Y и W не принимаются во внимание. Например, в zalpha11-лиганде гидрофильные районы включают в себя аминокислотные остатки 114-119 SEQ ID NO:2, аминокислотные остатки 101-105 SEQ ID NO:2, аминокислотные остатки 126-131 SEQ ID NO:2, аминокислотные остатки 113-118 SEQ ID NO:2 и аминокислотные остатки 158-162 SEQ ID NO:2.
Специалистам в данной области будет понятно, что гидрофильность или гидрофобность следует учитывать при конструировании модификаций в аминокислотной последовательности полипептида zalpha11-лиганда, так чтобы не нарушить общий структурный и биологический профиль. Особый интерес для замены представляют гидрофобные остатки, выбранные из группы, состоящей из Val, Leu и Не, или из группы, состоящей из Met, Gly, Ser, Ala, Tyr и Trp. Например, остатки, устойчивые к замене, могли бы включать в себя остатки 100 и 103, показаные в SEQ ID NO:2. Остатки цистеина в положениях 71, 78, 122 и 125 SEQ ID NO:2 будут относительно неустойчивыми к замене.
Идентичности незаменимых аминокислот могут быть также выведены из анализа сходства последовательности между IL-15, IL-2, IL-4 и GM-CSF с zalpha11-лигандом. С использованием способов, таких как анализ "FASTA", описанный выше, районы высокого сходства идентифицируют в семействе белков и используют для анализа аминокислотной последовательности для консервативных районов. Альтернативным подходом к идентификации вариантного полинуклеотида zalpha11-лиганда на основе структуры является определение, может ли молекула нуклеиновой кислоты, кодирующая ген потенциального вариантного zalpha11-лиганда, гибридизоваться с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1, как обсуждалось выше.
Другими способами идентификации незаменимых аминокислот в полипептидах данного изобретения являются процедуры, известные в данной области, такие как сайт-направленный мутагенез или аланинсканирующий мутагенез (Cunningham and Wells, Science 244:1081 (1989); Bass et al., Proc. Natl. Acad. Sci. USA 88:4498 (1991), Coombs and Corey, "Site-Directed Mutagenesis and Protein Engineering" in Proteins: Analysis and Design. Angeletti (ed.), pages 259-311 (Academic Press, Inc. 1998)). В последнем способе отдельные мутации аланина вводят при каждом остатке в молекуле и полученные в результате мутантные молекулы тестируют на биологическую или биохимическую активность, как описано ниже, для идентификации аминокислотных остатков, которые являются критическими для активности данной молекулы. См. также Hilton et al., J. Biol. Chem. 271:4699 (1996).
Данное изобретение относится также к функциональным фрагментам полипептидов zalpha11-лиганда и молекулам нуклеиновых кислот, кодирующим такие функциональные фрагменты. "Функциональный" zalpha11-лиганд или его фрагмент, определенный здесь, отличается его пролиферативной или дифференцирующей активностью, его способностью индуцировать или ингибировать специализированные клеточные функции или его способностью специфически связываться с антителом против zalpha11-лиганда или рецептором zalpha11-лиганда (растворимым или иммобилизованным). Как описано ранее, zalpha11-лиганд характеризуется структурой четырехспирального пучка, содержащего спираль А (аминокислотные остатки 41-56), спираль В (аминокислотные остатки 69-84), спираль С (аминокислотные остатки 92-105) и спираль D (аминокислотные остатки 135-148), как показано в SEQ ID NO:2. Таким образом, данное изобретение дополнительно обеспечивает слитые (гибридные) белки, включающие в себя: (а) полипептидные молекулы, содержащие одну или более спиралей, описанных здесь; и (b) функциональные фрагменты, содержащие одну или несколько этих спиралей. Другая полипептидная часть слитого белка может состоять из другого состоящего из четырехспирального пучка цитокина, такого как IL-15, IL-2, IL-4 и GM-CSF, или неприродного и/или неродственного секреторного сигнального пептида, который облегчает секрецию слитого белка.
Таким образом, данное изобретение обеспечивает слитые белки, содержащие по меньшей мере четыре полипептида, где порядок полипептидов от N-конца к С-концу является следующим: первый полипептид содержит аминокислоты, выбранные из группы, состоящей из: (а) аминокислотных остатков 36-46 SEQ ID NO:111 спирали A IL-2; (b) аминокислотных остатков 29-43 SEQ ID NO:112 спирали А IL-15; (с) аминокислотных остатков 45-68 SEQ ID NO:113 спирали A IL-4; (d) аминокислотных остатков 30-44 SEQ ID NO:114 спирали А GM-CSF и (е) аминокислотных остатков 41-56 SEQ ID NO:2; первый спейсер из 6-27 аминокислот; и второй полипептид, который содержит аминокислотные остатки, выбранные из группы, состоящей из: (а) аминокислотных остатков 53-75 SEQ ID NO:111 спирали В IL-2; (b) аминокислотных остатков 65-83 SEQ ID NO:112 спирали В IL-4; (с) аминокислотных остатков 84-101 SEQ ID NO:113 спирали В IL-15; (d) аминокислотных остатков 72-81 SEQ ID NO:114 спирали В GM-CSF и (е) аминокислотных остатков 69-84 SEQ ID NO:2; второй спейсер из 5-11 аминокислотных остатков; третий полипептид, который содержит аминокислотные остатки, выбранные из группы, состоящей из (а) аминокислотных остатков 87-99 SEQ ID NO:111 спирали С IL-2; (b) аминокислотных остатков 95-118 SEQ ID NO:112 спирали С IL-4; (с) аминокислотных остатков 107-119 SEQ ID NO:113 спирали С IL-15; (d) аминокислотных остатков 91-102 SEQ ID NO:114 спирали С GM-CSF и (е) аминокислотных остатков 92-105 SEQ ID NO:2; третий спейсер из 3-29 аминокислотных остатков; и четвертый полипептид, который содержит аминокислотные остатки, выбранные из группы, состоящей из (а) аминокислотных остатков 103-121 SEQ ID NO:111 спирали D IL-2; (b) аминокислотных остатков 134-157 SEQ ID NO:112 спирали D IL-15; (с) аминокислотных остатков 134-160 SEQ ID NO:113 спирали D IL-4; (d) аминокислотных остатков 120-131 SEQ ID NO:114 спирали D GM-CSF и (е) аминокислотных остатков 135-148 SEQ ID NO:2; где по меньшей мере один из четырех полипептидов представляет собой zalpha1 1-лиганд. В других вариантах спейсерные пептиды будут выбраны из петель А/В и C/D zalpha11-лиганда, IL-2, IL-4, IL-15 или GM-CSF, как показано в таблице 1.
Рутинные делеционные анализы молекул нуклеиновых кислот могут быть проведены для получения функциональных фрагментов молекулы нуклеиновой кислоты, кодирующей полипептид zalpha11-лиганда. В качестве иллюстрации, молекулы ДНК, имеющие нуклеотидную последовательность SEQ ID NO:1, или их фрагменты, могут быть расщеплены нуклеазой Ваl31 с получением ряда "вмонтированных" делеций. Затем эти фрагменты ДНК инсертируют в экспрессирующие векторы в правильной рамке считывания и экспрессированные полипептиды выделяют и тестируют на активность zalpha11-лиганда, или на способность связываться с антителами против zalpha11-лиганда или рецептором zalpha1 1-лиганда. Одной из альтернатив расщеплению экзонуклеазами является использование олигонуклеотид-направленного мутагенеза для введения делеций или стоп-кодонов для указания продуцирования желательного фрагмента zalpha11-лиганда. Альтернативно, определенные фрагменты гена zalpha11-лиганда могут быть синтезированы с использованием полимеразной цепной реакции.
Стандартные способы идентификации функциональных доменов хорошо известны лицам с обычной квалификацией в данной области. Например, исследования на одном или на обоих концах укороченных интерферонов суммированы Horisberger and Di Marco, Pharmac. Ther. 66:507 (1995). Кроме того, стандартные способы для функционального анализа белков описаны, например, Treuter et al., Molec. Gen. Genet. 240:113 (1993); Content et al., "Expression and preliminary deletion analysis of the 42 kDa 2-5A synthetase induced by human interferon," in Biological Interferon Systems. Proceedings of ISIR-TNO Meeting on Interferon Systems. Cantell (ed.), pages 65-72 (Nijhoff 1987); Herschman, "The EGF Receptor," in Control of Animal Cell Proliferation 1, Boynton et al., (eds.) pages 169-199 (Academic Press 1985); Coumailleau et al., J. Biol. Chem. 270:29270 (1995); Fukunaga et al., J. Biol. Chem. 270:25291 (1995); Yamaguchi et al., Biochem. Pharmacol. 50:1295 (1995) и Meisel et al., Plant Molec. Biol. 30:1 (1996).
Многочисленные аминокислотные замены могут быть произведены и тестированы с использованием известных способов мутагенеза и скрининга, таких как описанные Reidhaar-Olson and Sauer (Science 241:53. 1988) или Bowie and Sauer (Proc. Natl. Acad. Sci. USA 86:2152, 1989). Вкратце, эти авторы описывают способы одновременной рандомизации двух или более положений в полипептиде, отбора на функциональный полипептид и затем секвенирования мутагенизированных полипептидов для определения спектра допустимых замен в каждом положении. Другие способы, которые могут быть использованы, включают в себя способ фагового представления, например, Lowman et al., Biochem. 30:10832, 1991; Ladner et al., U.S. Patent No. 5223409; Huse, WIPO Publication WO 92/06204) и район-направленный мутагенез (Derbyshire et al., Gene 46:145, 1986; Ner et al., DNA7:127, 1988).
Варианты описанных нуклеотидной и полипептидной последовательностей zalpha11-лиганда могут быть образованы перетасовкой ДНК, описанной Stemmer, Nature 370:389, 1994, Stemmer, Proc. Natl. Acad. Sci. USA 91:10747, 1994 и в международной публикации WO 97/20078. Вкратце, вариантные молекулы ДНК генерируют посредством гомологичной рекомбинации in vitro случайной фрагментацией исходной ДНК с последующей повторной сборкой при помощи ПЦР, приводящей к случайно введенным точковым мутациям. Этот способ может быть модифицирован с использованием семейства исходных молекул ДНК, таких как аллельные варианты или молекулы ДНК из различных видов, для введения дополнительной вариабельности в этот процесс. Отбор или скрининг на желательную активность с последующими дополнительными повторениями мутагенеза и анализа обеспечивает быструю "эволюцию" последовательностей посредством отбора на желательные мутации с одновременным отбором против вредных изменений.
Способы мутагенеза, описанные выше, могут комбинироваться с высокопроизводительными автоматизированными способами скрининга для обнаружения активности клонированных мутагенизированных полипептидов в клетках-хозяевах. Мутагенизированные молекулы ДНК, кодирующие биологически активные полипептиды или полипептиды, которые могут связываться с антителами против zalpha11-лиганда или с растворимым zalpha11-рецептором, могут быть извлечены из клеток-хозяев и быстро секвенированы с использованием современного оборудования. Эти способы делают возможным быстрое определение важности индивидуальных аминокислотных остатков в представляющем интерес полипептиде и могут применяться к полипептидам неизвестной структуры.
Кроме того, белки данного изобретения (или их полипептидные фрагменты) могут быть соединены с другими биоактивными молекулами, в частности, другими цитокинами, для обеспечения полифункциональных молекул. Например, одна или несколько спиралей из zalpha11-лиганда могут быть соединены с другими цитокинами для усиления из биологических свойств или эффективности получения.
Таким образом, данное изобретение обеспечивает ряд новых гибридных молекул, в которых сегмент, содержащий одну или несколько спиралей zalpha11-лиганда, слит с другим полипептидом. Слияние предпочтительно производят сплайсингом на уровне ДНК, чтобы сделать возможной экспрессию химерных молекул в рекомбинантных системах получения. Полученные молекулы анализируют затем на такие свойства, как улучшенная растворимость, улучшенная стабильность, пролонгированный полупериод клиренса, улучшенные уровни экспрессии и секреции и фармакодинамику. Такие гибридные молекулы могут, кроме того, содержать дополнительные аминокислотные остатки (например, полипептидный линкер) между компонентами-белками или компонентами-полипептидами.
Не встречающиеся природно аминокислоты включают в себя, без ограничения, транс-3-метилпролин, 2,4-метанопролин, цис-4-гидроксипролин, транс-4-гидроксипролин, N-метилглицин, алло-треонин, метилтреонин, гидроксиэтилцистеин, гидроксиэтилгомоцистеин, нитроглутамин, гомоглутамин, пипеколиновую кислоту, тиазолидинкарбоновую кислоту, дегидропролин, 3- и 4-метилпролин, 3,3-диметилпролин, трет-лейцин, норвалин, 2-азафенилаланин, 3-азафенилаланин, 4-азафенилаланин и 4-фторфенилаланин. В данной области известны несколько способов для включения природно не встречающихся аминокислотных остатков в белки. Например, может быть использована система in vitro, в которой нонсенс-мутации подавляются с использованием химически аминоацилированных супрессорных тРНК. Способы синтеза аминокислот и аминоацилирования тРНК известны в данной области. Транскрипцию и трансляцию плазмид, содержащих нонсенс-мутации, проводят обычно в бесклеточной системе, содержащей экстракт Е.coli S30 и коммерчески доступные ферменты и другие реагенты. Белки очищают хроматографией. См., например, Robertson et al., J. Am. Chem. Soc. 113:2722. 1991; Ellman et al., Methods Enzvmol. 202:301, 1991; Chung et al.. Science 259:806. 1993 и Chung et al.. Proc. Natl. Acad. Sci. USA 90:10145. 1993).
Во втором способе трансляцию проводят в ооцитах Xenopus микроинъекцией мутированной мРНК и химически аминоацилированных супрессорных тРНК (Turcatti et al., J. Biol. Chem. 271:19991, 1996). В третьем способе клетки Е. coli культивируют в отсутствие природной аминокислоты, которую предстоит заменить (например, фенилаланина), и в присутствии желательной природно не встречающейся аминокислоты (аминокислот) (например, 2-азафенилаланина, 3-азафенилаланина, 4-азафенилаланина или 4-фторфенилаланина). Природно не встречающаяся аминокислота включается в этот белок вместо ее природной копии. См. Koide et al., Biochem. 33:7470, 1994. Природно встречающиеся аминокислотные остатки могут быть превращены в природно не встречающиеся разновидности химической модификацией in vitro. Химическая модификация может комбинироваться с сайт-направленным мутагенезом для дополнительного расширения диапазона замен (Wynn and Richards, Protein Sci. 2:395. 1993).
Может быть полезным стабилизировать zalpha11-лиганд для продления полупериода существования этой молекулы, в частности, для продления метаболической персистенции в активном состоянии. Для достижения продленного полупериода существования молекулы zalpha11-лиганда могут быть химически модифицированы с использованием описанных здесь способов. ПЭГилирование является одним из способов, обычно применяемых, который показал увеличение полупериода существования в плазме, повышенную растворимость и пониженные антигенность и иммуногенность (Nucci et al., Advanced Drug Delivery Reviews 6:133-155, 1991 и Lu et al. Int. J. Peptide Protein Res. 43:127-138, 1994).
Аминокислотные остатки zalpha11-лиганда могут быть заменены ограниченным числом неконсервативных аминокислот, аминокислот, которые не кодируются генетическим кодом, не встречающихся природно аминокислот и неприродных аминокислот.
Данное изобретение обеспечивает также полипептидные фрагменты или пептиды, содержащие эпитоп-несущую часть полипептида zalpha11-лиганда, описанного здесь. Такие фрагменты или пептиды могут содержать "иммуногенный эпитоп", который является частью белка, которая вызывает гуморальный ответ (антителогенез) при использовании всего белка в качестве иммуногена. Иммуногенные эпитоп-несущие пептиды могут быть идентифицированы с использованием стандартных способов (см., например, Geysen et al., Proc. Natl. Acad. Sci. USA 81:3998 (1998)).
В противоположность этому полипептидные фрагменты или пептиды могут содержать "антигенный эпитоп", который является районом белковой молекулы, с которой может специфически связываться антитело. Определенные эпитопы состоят из линейного или непрерывного отрезка аминокислот, и антигенность такого эпитопа не разрушается денатурирующими агентами. В данной области известно, что относительно короткие синтетические пептиды, которые могут имитировать эпитопы белка, могут быть использованы для стимуляции образования антител против этого белка (см., например, Sutcliffe et al., Science 219:660 (1983)). Таким образом, антигенные эпитоп-несущие пептиды и полипептиды данного изобретения применимы для индукции антител, которые связываются с описанными здесь полипептидами. Профили гидрофильности Норр-Woods могут быть использованы для определения районов, которые имеют наиболее активный антигенный потенциал (Норр et al., 1981, ibid и Норр, 1986, ibid.). В zalpha11-лиганде эти районы включают в себя аминокислотные остатки 114-119, 101-105, 126-131, 113-118 и 158-162 SEQ ID NO:2.
Антигенные эпитоп-несущие пептиды и полипептиды предпочтительно содержат по меньшей мере четыре-десять аминокислот, по меньшей мере десять-четырнадцать аминокислот или около четырнадцати - около тридцати аминокислот SEQ ID NO:2 или SEQ ID NO:56. Такие эпитоп-несущие пептиды и полипептиды могут быть получены фрагментированием полипептида zalpha11-лиганда или химическим синтезом пептидов, как описано здесь. Кроме того, эпитопы могут быть выбраны с использованием фагового представления библиотек случайных пептидов (см., например, Lane and Stephen, Curr. Opin. Immunol. 5:268 (1993) и Cortese et al., Curr. Qpin. Biotechnol. 7:616 (1996)). Стандартные способы идентификации эпитопов и получения антител из малых пептидов, которые содержат эпитоп, описаны, например, Mole, "Epitope Mapping", in Methods in Molecular Biology. Vol. 10, Manson (ed.), pages 105-116 (The Humana Press, Inc. 1992); Price, "Production and Characterization of Synthetic Peptide-Derived Antibodies," in Monoclonal Antibodies: Production. Engineering and Clinical Application, Ritter and Ladyman (eds.), Pages 60-84 (Cambridge University Press 1995) и Coligan et al., (eds.), Current Protocols in Immunology, pages 9.3.1-9.3.5 and pages 9.4.1-9.4.11 (John Wiley & Sons 1997).
Независимо от конкретной нуклеотидной последовательности вариантного полинуклеотида zalpha11-лиганда этот полинуклеотид кодирует полипептид, который характеризуется его пролиферативной или дифференцирующей активностью, его способностью индуцировать или ингибировать специализированную клеточную функцию или способностью специфически связываться с антителом против zalpha11-лиганда или zalpha11-рецептором. Более конкретно, вариантные полинуклеотиды zalpha11-лиганда будут кодировать полипептиды, которые проявляют по меньшей мере 50% и предпочтительно больше, чем 70, 80 или 90% активности полипептида, показанного в SEQ ID NO:2.
Для любого полипептида zalpha11-лиганда, в том числе вариантов и слитых белков, специалист в данной области может легко генерировать полностью вырожденную полинуклеотидную последовательность, кодирующую этот вариант, с использованием информации, представленной в таблицах 1 и 2 выше.
Далее, данное изобретение обеспечивает различные другие полипептидные слияния (и родственные мультимерные белки, содержащие один или несколько слитых полипептидов). Например, полипептид zalpha11-лиганда может быть получен в виде слияния с димеризующим белком, как описано в патентах США с номерами 5155027 и 5567584. Предпочтительные димеризующие белки в этой связи включают в себя домены константных областей иммуноглобулинов. Полипептидные слияния иммуноглобулин-zalрhа11-лиганд могут быть экспрессированы в генетически сконструированных клетках (с получением различных мультимерных аналогов zalpha11-лиганда). Вспомогательные домены могут быть слиты с полипептидами zalpha11-лиганда для нацеливания их на специфические клетки, ткани или макромолекулы. Например, полипептид или белок zalpha11-лиганда мог бы быть нацелен на заданный тип клеток слиянием полипептида zalpha11-лиганда с лигандом, который специфически связывается с рецептором на поверхности этой клетки-мишени. Таким путем, полипептиды и белки могут быть нацелены для терапевтических или диагностических целей. Полипептид zalpha11-лиганда может быть слит с двумя или более частями молекулы, такими как аффинная метка для очистки и нацеливающий домен. Полипептидные слияния могут также содержать один или несколько сайтов расщепления, в частности, между доменами. См. Tuan et al., Connective Tissue Research 34:1-9, 1996.
С использованием обсуждаемых здесь способов специалист с квалификацией в данной области может идентифицировать и/или получить различные полипептиды, которые имеют по существу одинаковую идентичность последовательности с остатками 1-162 или 33-162 SEQ ID NO:2, или их функциональные фрагменты и слияния, где такие полипептиды или фрагменты сохраняют свойства белка дикого типа, такие как способность стимулировать пролиферацию, дифференцировку, индуцировать специализированную клеточную функцию или связывать zalpha11-рецептор или антитела против zalpha11-лиганда.
Полипептиды zalpha11-лиганда данного изобретения, в том числе полноразмерные полипептиды, функциональные фрагменты и гибридные полипептиды, могут продуцироваться в генетически сконструированных клетках-хозяевах в соответствии с общепринятыми способами. Подходящими клетками-хозяевами являются те типы клеток, которые могут быть трансформированы или трансфицированы экзогенной ДНК и выращены в культуре, и они включают в себя бактерии, грибковые клетки и культивируемые высшие эукариотические клетки. Эукариотические клетки, в частности, культивируемые клетки многоклеточных организмов, являются предпочтительными. Способы манипулирования клонированными молекулами ДНК и введения экзогенной ДНК в различные клетки-хозяева описаны в Sambrook et al., Molecular Cloning: A Laboratory Manual. (2nd ed.) (Cold Sprinq Harbor Laboratory Press, Cold Spring Harbor, NY, 1989 и Ausubel et al. (eds.) Current Protocols in Molecular Biology. John Wiley and Sons, Inc., NY, 1987.
В общих чертах, последовательность ДНК, кодирующую полипептид zalpha11-лиганда, функционально связывают с другими генетическими элементами, требующимися для ее экспрессии, обычно включающими в себя промотор и терминатор транскрипции, в экспрессирующем векторе. Этот вектор обычно будет также содержать один или несколько селектируемых маркеров и одну или несколько точек инициации репликации, хотя специалистам в данной области будет понятно, что в некоторых системах селектируемые маркеры могут быть обеспечены на отдельных векторах и репликация экзогенной ДНК может быть обеспечена интеграцией в геном клетки-хозяина. Выбор промоторов, терминаторов, селектируемых маркеров, векторов и других элементов является предметом рутинного конструирования в пределах обычной квалификации в данной области. Многие такие элементы описаны в литературе и являются доступными через коммерческих поставщиков.
Для направления полипептида zalpha11-лиганда в секреторный путь клетки-хозяина в экспрессирующем векторе обеспечивают секреторную сигнальную последовательность (также известную как лидерная последовательность, пре-пропоследовательность или пре-последовательность). Секреторная сигнальная последовательность может быть сигнальной последовательностью zalpha11-лиганда или может быть произведена из другого секретируемого белка (например, t-PA (тканевого активатора плазминогена)) или синтезирована de novo. Секреторная сигнальная последовательность функционально соединена с последовательностью ДНК zalpha11-лиганда, т.е. эти две последовательности соединены в правильной рамке считывания и правильно помещены для направления вновь синтезированного полипептида в секреторный путь клетки-хозяина. Секреторные сигнальные последовательности обычно расположены 5' (слева) от последовательности ДНК, кодирующей представляющий интерес полипептид, хотя некоторые сигнальные последовательности могут быть расположены в другом месте в представляющей интерес последовательности ДНК (см., например, Welch et al., U.S. Patent No. 5037743; Holland et al., U.S. Patent No. 5143830).
Альтернативно, секреторная сигнальная последовательность, содержащаяся в полипептидах данного изобретения, используется для направления других полипептидов в секреторный путь. Данное изобретение обеспечивает такие гибридные (слитые) полипептиды. Может быть изготовлен сигнальный гибридный полипептид, в котором секреторная сигнальная последовательность, полученная из аминокислотных остатков 1-31 SEQ ID NO:2, функционально связана с последовательностью ДНК, кодирующей другой полипептид, с использованием способов, известных в этой области и описанных здесь. Секреторная сигнальная последовательность, содержащаяся в гибридных полипептидах данного изобретения, предпочтительно слита на амино-конце с дополнительным пептидом для направления этого дополнительного пептида в секреторный путь. Такие конструкции имеют многочисленные применения, известные в данной области. Например, эти новые гибридные конструкции с секреторной сигнальной последовательностью могут направлять секрецию активного компонента обычно не секретируемого белка. Такие гибридные белки могут быть использованы in vivo и in vitro для направления пептидов через секреторный путь.
Культивируемые клетки млекопитающих являются подходящими хозяевами в данном изобретении. Способы введения экзогенной ДНК в клетки-хозяева млекопитающих включают в себя опосредованную фосфатом кальция трансфекцию (Wigler et al., Cell.14:725, 1978; Corsaro and Pearson, Somatic Cell Genetics 7:603, 1981; Graham and Van der Eb, Virology 52:456, 1973), электропорацию (Neumann et al., EMBO J. 1:841-845, 1982), опосредованную ДЭАЭ-декстраном трансфекцию (Ausubel et at., ibid.) и опосредованную липосомами трансфекцию (Hawley-Nelson et al., Focus 15:73, 1993; Ciccarone et al., Focus 15:80, 1993) и вирусные векторы (Miller and Rosman, BioTechnigues 7:980-90, 1989; Wang and Finer, Nature Med. 2:714-6, 1996). Получение рекомбинантных полипептидов в культивируемых клетках млекопитающих описано, например, Levinson et al., U.S. Patent No. 4713339; Hagen et al., U.S. Patent No. 4784950; Palmiter et al., U.S. Patent No. 4579821; и Ringold, U.S. Patent No. 4656134. Подходящие культвируемые клетки млекопитающих включают в себя клеточные линии COS-1 (АТСС No. CRL 1650), COS-7 (АТСС No. CRL 1651), ВНК (АТСС No. CRL 1632), ВНК 570 (АТСС No. CRL 10314), 293 (АТСС No. CRL 1573; Graham et al., J. Gen. Virol. 36:59-72 (1977) и линии клеток яичника китайского хомячка (например, СНО-К1; АТСС No. CCL 61). Дополнительные подходящие клеточные линии известны в данной области и доступны из общественных депозитариев, таких как Американская коллекция типовых культур (American Type Culture Collection, Manassas, VA). В общем, предпочтительны сильные промоторы транскрипции, такие как промоторы из SV-40 или цитомегаловируса. См., например, U.S. Patent No. 4956288. Другие подходящие промоторы включают в себя промоторы из генов металлотионеина (U.S. Patent No. 4579821 и U.S. Patent No. 4601978) и основной поздний промотор аденовируса.
Отбор с лекарственным средством обычно применяют для отбора на культивируемые клетки млекопитающих, в которые была встроена чужеродная ДНК. Такие клетки обычно называют "трансфектантами". Клетки, которые культивировались в присутствии селективного агента и способны передавать представляющий интерес ген их потомству, называют "стабильными трансфектантами". Предпочтительным селектируемым маркером является ген, кодирующий устойчивость к антибиотику неомицину. Отбор проводят в присутствии лекарственного средства типа неомицина, такого как G-418, или т.п. Системы отбора могут быть также использованы для увеличения уровня экспрессии представляющего интерес гена, способа, называемого "амплификацией". Амплификацию проводят культивированием трансфектантов в присутствии низкого уровня селективного агента и затем увеличением количества селективного агента для отбора клеток, которые продуцируют высокие уровни продуктов введенных генов. Предпочтительным амплифицируемым селектируемым маркером является дигидрофолатредуктаза, которая сообщает клеткам устойчивость к метотрексату. Другие гены устойчивости к лекарственным средствам (например, устойчивости к гигромицину, множественной устойчивости к лекарственным средствам, пуромицинацетилтрансферазе) также могут быть использованы. Альтернативные маркеры, которые вводят измененный фенотип, такие как зеленый флуоресцентный белок или белки клеточной поверхности, такие как CD4, CD8, МНС Класса I, щелочная фосфатаза плаценты, могут быть использованы для сортинга трансфицированных клеток от нетрансфицированных клеток такими способами, как FACS-сортинг (сортинг клеток с возбуждением флуоресценции) или способ разделения при помощи магнитных гранул.
Другие высшие эукариотические клетки могут быть также использованы в качестве хозяев, в том числе клетки растений, клетки насекомых и клетки птиц. Применение Agrobacterium rhizogenes в качестве вектора для экспрессии генов в клетках растений рассмотрено Sinkar et al., J. Biosci. (Bangalore) 11:47-58, 1987. Трансформация клеток насекомых и получение в них чужеродных полипептидов описаны Guarino et al., U.S. Patent No. 5162222 и WIPO publication WO 94/06463. Клетки насекомых могут быть инфицированы рекомбинантным бакуловирусом, обычно произведенным из вируса ядерного полиэдроза Autographa califomica (AcNPV). См. King, L.A. and Possee R.D., The Baculovirus Expression System: A Laboratory Guide. London, Chapman and Hall; O'Reilly D.R. et al., Baculovirus Expression Vectors: A Laboratory Manual. New York, Oxford University Press., 1994; и Richardson, C.D., Ed., Baculovirus Expression Protocols. Methods in Molecular Bioloav. Totowa, NJ, Humana Press, 1995. Второй способ получения рекомбинантного бакуловируса использует систему на основе транспозона, описанную Luckow (Luckow et al., J. Virol. 67:4566-79, 1993). Эта система продается в наборе Bac-to-Вас™ (Life Technologies, Rockville, MD). Эта система использует вектор-переносчик, pFastBac1™ (Life Technologies), содержащий транспозон Тn7, для перемещения ДНК, кодирующей полипептид zalpha11-лиганда, в геном бакуловируса, поддерживаемый в Е. coli, в виде большой плазмиды, названной "бакмидой". Вектор-переносчик pFastBac1™ использует промотор полиэдрина AcNPV для запуска экспрессии представляющего интерес гена, в данном случае zalpha11-лиганда. Однако pFastBac1™ может быть модифицирован до значительной степени. Промотор полиэдрина может быть удален и заменен промотором основного белка бакуловируса (также известного как Рсог, р6.9 или промотор МР), который экспрессируется раньше в инфекции бакуловирусом, и было показано, что он является предпочтительным для экспрессии секретируемых белков. См. Hill-Perkins, M.S. and Possee, R.D. J. Gen. Virol. 71:971-6. 1990; Bonning, B.C. et al., J, Gen. Virol. 75:1551-6, 1994 и Chazenbalk, G.D. and Rapoport, B. J. Biol. Chem. 270:1543-9, 1995. В таких конструкциях векторов-переносчиков может быть использована короткая или длинная версия промотора основного белка. Кроме того, могут быть сконструированы векторы-переносчики, которые заменяют секреторные сигнальные последовательности нативного zalpha11-лиганда секреторными сигнальными последовательностями, полученными из белков насекомых. Например, секреторная сигнальная последовательность из экдистероидглюкозилтрансферазы (EGT), Меллитина медоносной пчелы (Invitrogen, Carlsbad, СА) или gр67 бакуловируса (PharMingen, San Diego, CA) могут быть использованы в конструкциях для замены секреторной сигнальной последовательности нативного zalpha11-лиганда. Кроме того, векторы-переносчики могут содержать слияния в рамке считывания с ДНК, кодирующей метку эпитопа, на С- или N-конце экспрессируемого полипептида zalpha11-лиганда, например метку эпитопа Glu-Glu (Grussenmeyer, Т. et al., Proc. Natl. Acad. Sci. 82:7952-4, 1985). При помощи способа, известного в данной области, вектор-переносчик, содержащий zalpha11-лиганд, трансформируют в Е. coli и подвергают скринингу на бакмиды, которые содержат прерванный ген lacZ, свидетельствующий о рекомбинантном бакуловирусе. Бакмидную ДНК, содержащую рекомбинантный геном бакуловируса, выделяют при помощи известных способов и используют для трансфекции клеток Spodoptera frugiperda, например, клеток Sf9. Затем получают рекомбинантный вирус, экспрессирующий zalpha11-лиганд. Исходные материалы рекомбинантного вируса готовят способами, обычно используемыми в данной области.
Этот рекомбинантный вирус используют для инфицирования клеток-хозяев, обычно клеточной линии, полученной из осенних "походных (ратных) червей", Spodoptera frugiperda. См., в общих чертах, Glick and Pasternak, Molecular Biotechnology: Principles and Applications of Recombinant DNA. ASM Press, Washington, D.C., 1994. Другой подходящей клеточной линией является клеточная линия High FiveO™ (Invitrogen), произведенная из Trichoplusia ni (Патент США №5300435).
Грибковые клетки, в том числе дрожжевые клетки, могут быть также использованы в данном изобретении. Виды дрожжей, представляющие особый интерес в этом отношении, включают в себя Saccharomyces cerevisiae, Pichia pastohs и Pichia methanolica. Способы трансформации клеток S. cerevisiae экзогенной ДНК и получения из них рекомбинантных полипептидов описаны, например, Kawasaki, U.S. Patent No. 4599311; Kawasaki etal., U.S. Patent No. 4931373; Brake, U.S. Patent No. 4870008; Welch etal., U.S. Patent No. 5037743 и Murray et al., U.S. Patent No. 4845075. Трансформированные клетки отбирают по фенотипу, определяемому селектируемым маркером, обычно по устойчивости к лекарственному средству или по способности расти в отсутствие определенного питательного вещества (например, лейцина). Предпочтительной векторной системой для применения в Saccharomyces cerevisiae является векторная система РОТ1, описанная Kawasaki et al. (U.S. Patent No. 4931373), которая позволяет отбирать трансформированные клетки по росту в содержащей глюкозу среде. Подходящие промоторы и терминаторы для применения в дрожжах включают в себя промоторы и терминаторы из генов гликолитических ферментов (см., например, Kawasaki, U.S. Patent No. 4599311; Kingsman et al., U.S. Patent No. 4615974 и Bitter, U.S. Patent No. 4977092) и генов алкогольдегидрогеназы. См. также Патенты США №№4990446, 5063154, 5139936 и 4661454. Системы трансформации для других дрожжей, в том числе Hansenula polymorpha, Schizosaccharomyces pombe, Kluyveromyces lactis, Kluyveromyces fragilis, Ustilago maydis, Pichia pastoris, Pichia methanolica, Pichia guillennondii и Candida maltosa, известны в данной области. См., например, Gleeson et al., J. Gen. Microbiol. 132:3459-65, 1986 и Cregg, U.S. Patent No. 4882279. Клетки Aspergitlus могут быть использованы в соответствии со способами McKnight et al., U.S. Patent No. 4935349. Способы трансформации Acremonium chrysogenum описаны Sumino et at., U.S. Patent No. 5162228. Способы трансформации Neurospora описаны Lambowitz, U.S. Patent No. 4486533.
Применение Pichia methanolica в качестве хозяина для получения рекомбинантных белков описано в WIPO Publications WO 97/17450, WO 97/17451, WO 98/02536 и WO 98/02565. Молекулы ДНК для применения в трансформации Р. methanolica обычно получают в виде двухцепочечных кольцевых плазмид, которые предпочтительно линеаризуют перед трансформацией. Для получения полипептидов в Р. methanolica предпочтительно, чтобы промотор и терминатор в этой плазмиде были промотором и терминатором гена Р. methanolica, такого как ген утилизации спирта Р. methanolica (AUG1 или AUG2). Другие применимые промоторы включают в себя промоторы генов дигидроксиацетонсинтазы (DHAS), формиатдегидрогеназы (FMD) и каталазы (CAT). Для облегчения интеграции этой ДНК в хромосому хозяина предпочтительно иметь весь сегмент экспрессии этой плазмиды, фланкированный на обоих концах последовательностями ДНК хозяина. Предпочтительным селектируемым маркером для применения в Pichia methanolica является ген ADE2 Р. methanolica, который кодирует фосфорибозил-5-аминоимидазолкарбоксилазу (AIRC; ЕС 4.1.1.21), которая позволяет клеткам-хозяевам ade2 расти в отсутствие аденина. Для крупномасштабных промышленных процессов, где желательно минимизировать применение метанола, предпочтительно использовать клетки-хозяева, в которых делегированы оба гена утилизации метанола (AUG1 и AUG2). Для получения секретируемых белков предпочтительны клетки-хозяева, дефектные по генам вакуолярных протеаз (РЕР4 и PRB1). Электропорацию используют для облегчения введения плазмиды, содержащей ДНК, кодирующую представляющий интерес полипептид, в клетки Р. methanolica. Предпочтительно трансформировать клетки Р. methanolica электропорацией с использованием экспоненциально затухающего пульсирующего электрического поля, имеющего напряженность поля от 2,5 до 4,5 кВ/см, предпочтительно примерно 3,75 кВ/см, и константу времени ( Ω) от 1 до 40 миллисекунд, наиболее предпочтительно примерно 20 миллисекунд.
Прокариотические клетки-хозяева, в том числе штаммы бактерий Escheiichia coli, Bacillus и других родов, также являются применимыми клетками-хозяевами в данном изобретении. Способы трансформации этих хозяев и экспрессии клонированных в них чужеродных ДНК хорошо известны в данной области (см, например, Sambrook et al., ibid.). При экспрессии полипептида zalpha11-лиганда в бактериях, таких как Е. coli, этот полипептид может сохраняться в цитоплазме, обычно в виде нерастворимых гранул, или может быть направлен в периплазматическое пространство бактериальной секреторной последовательностью. В первом случае эти клетки лизируют и гранулы извлекают и денатурируют при помощи, например, гуанидинизотиоцианата или мочевины. Затем денатурированный полипептид может быть повторно уложен и димеризован путем разведения продукта денатурации, например, диализом против раствора мочевины и комбинацией восстановленного и окисленного глутатиона, с последующим диализом против забуференного солевого раствора. В последнем случае этот полипептид может быть извлечен из периплазматического пространства в растворимой и функциональной форме путем разрушения этих клеток (например, обработкой ультразвуком или осмотическим шоком) для высвобождения содержимого периплазматического пространства и извлечения этого белка, посредством чего устраняется необходимость денатурации и повторной укладки.
Трансформированные и трансфицированные клетки-хозяева культивируют в соответствии с общепринятыми процедурами в культуральной среде, содержащей питательные вещества и другие компоненты, необходимые для роста выбранных клеток-хозяев. Различные подходящие среды, в том числе среды с определенным составом среды и комплексные среды, известны в данной области и обычно включают в себя источник углерода, источник азота, незаменимые аминокислоты, витамины и минеральные соединения. Среды могут также содержать такие компоненты, как факторы роста или сыворотка, если требуется. Среда для выращивания будет обычно производить отбор на клетки, содержащие экзогенно добавленную ДНК, посредством, например, отбора с использованием лекарственного средства или недостаточности незаменимого питательного компонента, который дополняется селектируемым маркером, имеющимся на экспрессирующем векторе или котрансфицируемым в клетку-хозяина. Клетки Р. methanolica культивируют в среде, содержащей адекватные источники углерода, азота и микроэлементов, при температуре приблизительно 25-35°С. Жидкие культуры обеспечивают достаточной аэрацией с использованием общепринятых средств, таких как встряхивание небольших колб или барботирование ферментеров. Предпочтительной культуральной средой для Р. methanolica является YEPD (2% D-глюкоза, 2% Бакто™ Пептон (Difco Laboratories, Detroit, Ml), 1% Бакто™ дрожжевой экстракт (Difco Laboratories), 0,004% аденин и 0,006% L-лейцин).
Предпочтительно очищать полипептиды данного изобретения до ≥80% чистоты, более предпочтительно до ≥90% чистоты, еще более предпочтительно до ≥95% чистоты и особенно предпочтительно до фармакологически чистого состояния, т.е. до чистоты более 99,9%, в отношении загрязняющих макромолекул, в частности других белков и нуклеиновых кислот, и инфекционных и пирогенных агентов. Предпочтительно очищенный полипептид является по существу не содержащим других полипептидов, в частности других полипептидов животного происхождения.
Экспрессируемые рекомбинантные полипептиды zalpha11-лиганда (или химерные полипептиды zalpha11-лиганда) могут быть очищены с использованием фракционирования и/или общепринятых способов и сред очистки. Для фракционирования проб могут быть использованы осаждение сульфатом аммония и кислотная или хаотропная экстракция. Примеры стадий очистки могут включать в себя применение гидроксиапатита, гель-фильтрацию, жидкостную экспресс-хроматографию белков (FPLC) и высокоэффективную жидкостную хроматографию с обращенной фазой. Подходящие хроматографические среды включают в себя производные декстрана, агарозу, целлюлозу, полиакриламид, специальные диоксиды кремния и т.п. Предпочтительными являются производные PEI, DEAE, QAE и Q. Примеры хроматографических сред включают в себя среды, дериватизованные фенильной, бутильной или октильной группами, такие как Phenyl-Sepharose FF (Pharmacia), Toyopearl butyl 650 (Toso Haas, Montgomeryville, PA), Octyl-Sepharose (Pharmacia) и т.п.; или полиакриловые смолы, такие как Amberchrom CG 71 (Toso Haas) и т.п. Подходящие твердые носители включают в себя стеклянные гранулы, смолы на основе диоксида кремния, целлюлозные смолы, агарозные гранулы, сшитые агарозные гранулы, полистироловые гранулы, сшитые полиакриламидные смолы и т.п., которые являются нерастворимыми в условиях, в которых они должны использоваться. Эти носители могут быть модифицированы реакционноспособными группами, позволяющими присоединение белков посредством аминогрупп, карбоксильных групп, сульфгидрильных групп, гидроксильных групп и/или углеводных частей молекул. Примеры химических способов связывания включают в себя активацию цианогенбромидом, активацию N-гидроксисукцинимидом, активацию эпоксидом, активацию сульфгидрильными группами, активацию гидразидом и карбоксил- и аминопроизводные для способов сочетания с использованием карбодиимида. Эти и другие твердые среды являются хорошо известными и широко используемыми в данной области и доступны из коммерческих источников. Способы связывания рецепторных полипептидов со средами-носителями хорошо известны в данной области. Выбор конкретного способа является рутинным и определяется отчасти свойствами выбранного носителя. См., например, Affinity Chromatographv: Principles & Methods. Pharmacia LKB Biotechnology, Uppsala, Sweden, 1988.
Полипептиды данного изобретения могут быть выделены путем использования их физических или биохимических свойств. Например, адсорбционная хроматография с иммобилизованным ионом металла (IMAC) может быть использована для очистки богатых гистидином белков или белков, имеющих полигистидиновые метки. Вкратце, гель сначала загружают ионами двухвалентного металла с образованием хелата (Sulkowski, Trends in Biochem. 3:1-7, 1985). Богатые гистидином белки будут адсорбироваться на этом матриксе с различными аффинностями в зависимости от используемого иона металла и будут элюироваться конкурентной элюцией, понижением рН или применением сильных хелатообразователей. Другие способы очистки включают в себя очистку гликозилированных белков аффинной хроматографией с иммобилизованным пектином и ионообменной хроматографией (Methods in Enzvmol., Vol. 182:529-39, "Guide to Protein Purification", M. Deutscher, (ed.), (Acad. Press, San Diego, 1990, pp.) и использование растворимого zalpha11-рецептора. В дополнительных вариантах данного изобретения может быть сконструирован гибрид представляющего интерес полипептида и аффинной метки (например, мальтозусвязывающего белка, домена иммуноглобулина) для облегчения очистки.
Кроме того, при помощи способов, описанных в данной области, слитые полипептиды или гибридные белки zalpha11-лиганда конструируют с использованием районов или доменов zalpha11-лиганда данного изобретения в комбинации с районами или доменами других белков семейства цитокинов человека (таких как интерлейкины или GM-CSF) или гетерологичных белков (Sambrook et al., ibid., Altschul et al., ibid., Picard, Cur. Opin. Biology, 5:511-5, 1994 и приведенные в них ссылки). Эти способы позволяют определить биологическую важность более крупных доменов или районов в представляющем интерес полипептиде. Такие гибриды могут изменять кинетику реакции, связывание, сужать или расширять субстратную специфичность или изменять тканевую и клеточную локализацию полипептида и могут быть применены к полипептидам неизвестной структуры.
Слитые белки могут быть получены способами, известными специалистам в данной области, посредством получения каждого компонента слитого белка и химического конъюгирования его. Альтернативно, полинуклеотид, кодирующий один или несколько компонентов слитого белка в правильной рамке считывания, может быть получен с использованием известных способов и экспрессирован при помощи описанных здесь способов. Например, часть спирали или вся спираль, придающая биологическую функцию, может быть подвергнута обмену между zalpha11-лигандом данного изобретения и функционально эквивалентными спиралями из другого члена семейства, такого как IL-15, IL-2, IL-4 или GM-CSF. Такие компоненты включают в себя, но не ограничиваются ими, секреторную сигнальную последовательность, спирали А, В, С и D; петли А/В, В/С, C/D; представляющих четырехспиральный пучок цитокинов. Можно было бы ожидать, что такие гибридные белки имеют биологические функциональные особенности, которые являются одинаковыми с полипептидами данного изобретения или подобными особенностям полипептидов данного изобретения или других известных представляющих собой четырехспиральный пучок белков семейства цитокинов семейств в зависимости от сконструированного слитого белка. Кроме того, такие слитые белки могут проявлять другие свойства, как описано здесь.
Стандартные способы молекулярной биологии и клонирования могут быть использованы для обмена эквивалентных доменов между полипептидом zalpha11-лиганда и полипептидами, с которыми их сливают. Обычно сегмент ДНК, который кодирует представляющий интерес домен, например описанные здесь спирали А-D zalpha11-лиганда или другой домен, функционально (операбельно) связывают в рамке считывания с по меньшей мере одним другим сегментом ДНК, кодирующим дополнительный полипептид (например, домен или район из другого цитокина, такого как IL-2 или т.п.), и встраивают в подходящий экспрессирующий вектор, как описано здесь. Обычно конструкции ДНК конструируют таким образом, что несколько сегментов ДНК, кодирующих соответствующие районы полипептида, функционально связывают в рамке считывания для получения единой конструкции, кодирующей весь слитый белок или его функциональную часть. Например, конструкция ДНК кодировала бы от М-конца до С-конца слитый белок, содержащий сигнальный полипептид, за которым следует слитый белок зрелого содержащего четырехспиральный пучок цитокина, содержащий спираль А, затем спираль В, затем спираль С, затем спираль D. Такие слитые белки могут быть экспрессированы, выделены и тестированы на активность, как описано здесь.
Полипептиды zalpha11-лиганда или их фрагменты могут быть также получены при помощи химического синтеза. Полипептиды zalpha11-лиганда могут быть мономерами или мультимерами; гликозилированными или негликозилированными; пэгилированными или непэгилированными и могут содержать или могут не содержать инициирующий аминокислотный остаток метионин. Например, полипептиды могут быть получены твердофазным пептидным синтезом, например,, описанным Merrifield, J. Am. Chem. Soc. 85:2149, 1963.
Активность молекул данного изобретения может быть измерена с использованием различных способов, которые измеряют пролиферацию и/или связывание с клетками, экспрессирующими zalpha11-рецептор. Особый интерес представляют изменения zalpha11-лиганд-зависимых клеток. Подходящие клеточные линии, которые могут быть сконструированы, включают в себя IL-3-зависимую клеточную линию BaF3 (Pakacios and Steinmetz, Cell 41:727-734, 1985; Mathey-Prevot et al, Mol. Cell. Biol. 6:4133-4135, 1986), FDC-P1 (Hapel et al., Blood 64:786-790, 1984) и MО7e (Kiss et al., Leukemia 7:235-240, 1993). Зависимые от фактора роста клеточные линии могут быть установлены в соответствии с опубликованными способами (например, Greenberger et al., Leukemia Res. 8:363-375, 1984; Dexter et al., in Baum et al., Eds., Experimental Hematology Today. 8th Ann. Mtg. Int. Soc. Esp. Hematol. 1979, 145-156, 1980).
Белки данного изобретения применимы для стимуляции пролиферации, активации, дифференцировки и/или индукции или ингибирования специализированной клеточной функции клеток, участвующих в гомеостазе гемопоэза и иммунной функции. В частности, полипептиды zalpha11-лиганда применимы для стимуляции пролиферации, активации, дифференцировки, индукции или ингибирования специализированных клеточных функций клеток гемопоэтических линий, в том числе, но не только, Т-клеток, В-клеток, МК-клеток, дендритных клеток, моноцитов и макрофагов, а также эпителиальных клеток. Пролиферация и/или дифференцировка гемапоэтических клеток может быть измерена in vitro с использованием культивируемых клеток или in vivo введением молекул данного изобретения в подходящую модель животного. Анализы измерения пролиферации или дифференцировки клеток хорошо известны в данной области. Например, анализы измерения пролиферации включают в себя такие анализы, как чувствительность к химическим раздражителям, например, к красителю нейтральному красному (Cavanaugh et al., Investigational New Drugs 8:347-354, 1990, включ. здесь в качестве ссылки), включение радиоактивно меченных нуклеотидов (Cook et al., Analytical Biochem. 179:1-7, 1989, включ. здесь в качестве ссылки), включение 5-бром-2'-дезоксиуридина (BrdU) в ДНК пролиферирующихся клеток (Porstmann et al., J. Immunol. Methods 82:169-179, 1985, включ. здесь в качестве ссылки) и применение солей тетразолия (Mosmann, J. Immunol. Methods 65:55-63. 1983; Alley et al., Cancer Res. 48:589-601. 1988; Marshall et al., Growth Reg. 5:69-84, 1995; и Scudiero et al., Cancer Res. 48:4827-4833, 1988; все включены здесь в качестве ссылки). Анализы измерения дифференцировки включают в себя, например, измерение маркеров клеточной поверхности, ассоциированных со стадиеспецифической экспрессией ткани, ферментативной активности, функциональной активности или морфологических изменений (Watt, FASEB. 5:281-284, 1991; Francis, Differentiation 57:63-75, 1994; Raes, Adv. Anim. Cell Biol. Technol. Bioprocesses. 161-171, 1989; все включены здесь в качестве ссылки).
Молекулы данного изобретения могут быть анализированы in vivo с использованием вирусных систем доставки. Примеры вирусов для этой цели включают в себя аденовирус, герпесвирус, ретровирусы, вирус коровьей оспы и аденоассоциированный вирус (AAV). Аденовирус, двухцепочечный ДНК-вирус, является в настоящее время наиболее исследованным вектором переноса генов для доставки гетерологичной нуклеиновой кислоты (в отношении обзора см. Т.С. Becker et al., Meth. Cell Biol. 43:161-89, 1994 и J.T. Douglas and D.T. Curiel, Science & Medicine 4:44-53, 1997).
Активность полипептида zalpha11-лиганда в качестве лиганда может быть измерена биосенсорным микрофизиометром на основе кремния, который измеряет скорость внеклеточного подкисления или экскрецию протонов, связанную со связыванием рецептора и последующими физиологическими клеточными ответами. Примером устройства является микрофизиометр Cytosensor™, изготовляемый Molecular Devices, Sunnyvale, CA. Разнообразные клеточные ответные реакции, такие как пролиферация клеток, ионный транспорт, продуцирование энергии, воспалительный ответ, регуляторная и рецепторная активация и т.п., могут быть измерены этим способом. См., например, McConnel, Н.М. et al., Science 257:1906-1912, 1992; Pitchford, S. et al., Meth. Enzvmol. 228:84-108, 1997; Arimilli, S. et al., J. Immunol. Meth. 212:49-59, 1998; Van Uefde, I. et al., Eur. J. Pharmacol. 346:87-95, 1998.
Кроме того, zalpha11-лиганд может быть использован для идентификации клеток, тканей или клеточных линий, которые отвечают на стимулируемый zalpha11-лигандом путь. Микрофизиометр, описанный выше, может быть использован для быстрой идентификации чувствительных к лиганду клеток, таких как клетки, чувствительные к zalpha11-лиганду данного изобретения. Клетки могут культивироваться в присутствии или в отсутствие полипептида zalpha11-лиганда. Клетки, которые вызывают измеримое изменение во внеклеточном подкислении в присутствии zalpha11-лиганда, являются чувствительными к zalpha11-лиганду. Такие клетки или клеточные линии могут быть использованы для идентификации антагонистов или агонистов полипептида zalpha11-лиганда, как описано выше.
Ввиду тканевого распределения, наблюдаемого для zalpha11-рецептора, агонисты (в том числе природный zalpha11-лиганд/субстрат/кофактор и т.д.) и/или антагонисты имеют огромный потенциал в применениях как in vitro, так и in vivo. Соединения, идентифицированные в качестве агонистов zalpha11-лиганда, применимы для размножения, пролиферации, активации, дифференцировки и/или индукции или ингибирования специализированных клеточных функций клеток, участвующих в гомеостазе гемопоэза и иммунной функции. Например, zalpha11-лиганд и соединения-агонисты применимы в качестве компонентов определенных сред для культуры клеток и могут использоваться отдельно или в комбинации с другими цитокинами и гормонами для замены сыворотки, которую обычно применяют в клеточной культуре. Таким образом, агонисты применимы в специфической стимуляции роста и/или развития Т-клеток, В-клеток, NK-клеток, цитотоксических лимфоцитов и других клеток лимфоидной и миелоидной линий дифференцировки в культуре.
Антагонисты применимы также в качестве реагентов исследования для характеристики сайтов взаимодействия лиганд-рецептор. Антагонисты применимы для ингибирования размножения, пролиферации, активации и/или дифференцировки клеток, участвующих в регуляции гемопоэза. Ингибиторы активности zalpha11-лиганда (антагонисты zalpha11-лиганда) включают в себя антитела против zalpha11-лиганда и растворимые рецепторы zalpha11-лиганда, а также другие пептидные и непептидные агенты (в том числе рибозимы).
Zalpha11-лиганд может быть использован также для идентификации ингибиторов (антагонистов) его активности. Тест-соединения добавляют к описанным здесь тестам для идентификации соединений, ингибирующих активность zalpha11-лиганда. Кроме этих описанных здесь тестов пробы могут быть тестированы на ингибирование активности zalpha11-лиганда в многочисленных анализах, предназначенных для измерения связывания рецептора, стимуляции/ингибирования zalpha11-лиганд-зависимых клеточных ответных реакций или пролиферации экспрессирующих zаlрhа11-рецептор клеток.
Полипептид zalpha11-лиганда может экспрессироваться в виде слитого белка с константной областью тяжелой цепи иммуноглобулина, обычно Fc-фрагментом, который содержит два домена константной области и не содержит вариабельной области. Способы получения таких слияний описаны в патентах США №№5155027 и 5567584. Такие слияния обычно секретируются в виде мультимерных молекул, где Fc-части связаны друг с другом дисульфидной связью, а два не-lg-полипептида расположены в тесной близости друг с другом. Слияния этого типа могут быть использованы, например, для димеризации, увеличения стабильности и полупериода существования in vivo, для аффинной очистки лиганда, в качестве инструмента анализа in vitro или антагониста. Для применения в анализах эти химеры связывают с носителем через Fc-район и используют в формате ELISA.
Zalpha11-лиганд-связывающий полипептид может быть также использован для очистки лиганда. Этот полипептид иммобилизуют на твердом носителе, таком как гранулы из агарозы, сшитой агарозы, стекла, целлюлозных смол, смол на основе диоксида кремния, полистирола, сшитого полиакриламида, или подобных материалах, которые являются стабильными в условиях использования. Способы связывания полипептидов с твердыми носителями известны в данной области и включают в себя аминную химию, активацию цианогенбромидом, активацию N-гидроксисукцинимидом, активацию эпоксидом, сульфгидрильную активацию и гидразидную активацию. Полученную среду обычно готовят в форме колонки и жидкости, содержащие лиганд, пропускают через эту колонку один раз или несколько раз, чтобы позволить лиганду связаться с рецепторным полипептидом. Затем этот лиганд элюируют при помощи изменений концентрации соли, хаотропных агентов (гуанидина-HCl) или рН для разрушения связывания лиганд-рецептор.
Тест-система, которая использует лигандсвязывающий рецептор (или антитело, один член пары комплемент/антикомплемент) или его связывающий фрагмент, и коммерчески доступный биосенсорный прибор (BIAcore, Pharmacia Biosensor, Piscataway, NJ) могут быть выгодно использованы. Такой рецептор, антитело, член пары комплемент/антикомплемент или фрагмент иммобилизуют на поверхности рецепторного чипа. Применение этого прибора описано Karlsson, J. Immunol. Methods 145:229-40, 1991 и Cunningham and Wells, J. Mol. Biol. 234:554-63, 1993. Такой рецептор, антитело, член пары или фрагмент присоединяют ковалентно при помощи аминной или сульфгидрильной химии к волокнам декстрана, которые присоединены к пленке золота в проточной ячейке. Тест-пробу пропускают через эту ячейку. Если лиганд, эпитоп или противоположный член пары комплемент/антикомплемент присутствует в этой пробе, он будет связываться с иммобилизованным рецептором, антителом или членом пары соответственно, вызывая изменение в показателе преломления среды, которое детектируется как изменение в резонансе поверхностных плазмонов золотой пленки. Эта система позволяет определить скорости включения и выключения (on- и off-скорости), из которых может быть рассчитана аффинность связывания и сделана оценка стехиометрии связывания. Альтернативно, связывание лиганд-рецептор может анализироваться с использованием способа SELDI(TM) (Ciphergen, Inc., Palo Alto, CA).
Лигандсвязывающие рецепторные полипептиды могут быть также использованы в других системах анализа, известных в данной области. Такие системы включают в себя анализ Скетчарда для определения аффинности связывания (см. Scatchard, Ann. NY Acad. Sci. 51:660-72, 1949) и калориметрические анализы (Cunningham et al., Science 253:545-48, 1991; Cunningham et al., Science 245:821-25, 1991).
Полипептиды zalpha11-лиганда могут быть также использованы для получения антител, которые связываются с эпитопами, пептидами или полипептидами zalpha11-лиганда. Полипептид zalpha11-лиганд или его фрагмент служит в качестве антигена (иммуногена) для инокуляции животного и индукции иммунного ответа. Специалисту с квалификацией в данной области будет ясно, что антигены, эпитоп-несущие полипептиды содержат последовательность из по меньшей мере 6, предпочтительно по меньшей мере 9 и более предпочтительно по меньшей мере примерно 15-30 непрерывных аминокислотных остатков полипептида zalpha11-лиганда (например, SEQ ID NO:2). Включены полипептиды, содержащие большую часть полипептида zalpha11-лиганда, т.е. от 30-100 остатков до полной длины этой аминокислотной последовательности. Антигены или иммуногенные эпитопы могут также включать в себя присоединенные метки, адъюванты и носители, описанные здесь. Подходящие антигены включают в себя полипептид zalpha11-лиганда, кодируемый SEQ ID NO:2 от номера аминокислоты 32 до номера аминокислоты 162 или его фрагмент из непрерывных аминокислот 9-131. Другие подходящие антигены включают в себя полноразмерный и зрелый zalpha11-лиганд, спирали A-D и индивидуальные или множественные спирали А, В, С и D структуры четырехспирального пучка zalpha11-лиганда, описанной здесь. Предпочтительными пептидами для применения в качестве антигенов являются гидрофильные пептиды, такие как предсказанные специалистом в данной области из графика гидрофобности, как описано здесь, например, аминокислотные остатки 114-119, 101-105, 126-131, 113-118 и 158-162 SEQ ID NO:2.
Антитела из иммунной реакции, генерированной инокуляцией животного этими антигенами, могут быть выделены и очищены, как описано выше. Способы получения и выделения поликлональных и моноклональных антител хорошо известны в данной области (см., например, Current Protocols in Immunology. Cooligan, et al., (eds.), National Institutes of Health, John Wiley and Sons, Inc., 1995; Sambrook et al., Molecular Cloning: A Laboratory Manual. Second Edition. Cold Spring Harbor, NY, 1989 и Hurrell, J.G.R., Ed., Monoclonal Hvbridoma Antibodies: Techniques and Applications. CRC Press, Inc., Boca Raton, FL, 1982).
Как должно быть очевидно специалисту с обычной квалификацией в данной области, поликлональные антитела могут быть получены инокуляцией различных теплокровных животных, таких как лошади, коровы, козы, овцы, собаки, куры, кролики, мыши и крысы, полипептидом zalpha11-лиганда или его фрагментом. Иммуногенность полипептида zalpha11-лиганда может быть увеличена путем применения адъюванта, такого как квасцы (гидроксид алюминия) или полный или неполный адъювант Фрейнда. Полипептиды, применимые для иммунизации, включают в себя также слитые (гибридные) полипептиды, такие как гибриды zalpha11-лиганда или его части с полипептидом иммуноглобулина или с мальтозусвязывающим белком. Полипептидный иммуноген может быть полноразмерной молекулой или ее частью. Если эта часть полипептида является "гаптен-подобной", такая часть может быть предпочтительно соединена или связана с макромолекулярным носителем (таким как гемоцианин фиссуреллы (KLH), бычий сывороточный альбумин (БСА) или столбнячный токсоид) для иммунизации.
В применении здесь, термин "антитела" включает в себя поликлональные антитела, аффинно-очищенные поликлональные антитела, моноклональные антитела, и антигенсвязывающие фрагменты, такие как протеолитические F(ab')2- и Fab-фрагменты. Также включены генетически сконструированные интактные антитела или фрагменты, такие как химерные антитела, Fv-фрагменты, одноцепочечные антитела и т.п., а также синтетические антигенсвязывающие пептиды и полипептиды. Антитела не из человека могут быть "очеловечены" (гуманизированы) прививкой CDR не человека на каркасные и константные области иммуноглобулина человека или включением полных вариабельных доменов не человека (необязательно "окутыванием" их подобной человеческой поверхностью путем замены экспонированных остатков, результатом чего является "облицованное" антитело). В некоторых случаях "гуманизированные" антитела могут сохранять остатки иммуноглобулина не человека в каркасных доменах вариабельной области человека для усиления характеристик правильного связывания. Посредством "гуманизирования" антител может быть увеличен биологический полупериод существования в организме, и потенциал неблагоприятных иммунных реакций при введении людям уменьшается. Кроме того, антитела человека могут продуцироваться в трансгенных животных, не являющихся человеком, которые были сконструированы таким образом, чтобы содержать гены иммуноглобулина человека, как описано в WIPO Publication WO 98/24893. Предпочтительно, чтобы эндогенные гены иммуноглобулина в этих животных были инактивированы или элиминированы, например, гомологичной рекомбинацией.
Антитела рассматриваются как специфически связывающие, если: 1) они проявляют пороговый уровень связывающей активности и 2) они не имеют значимой перекрестной реактивности с родственными полипептидными молекулами. Пороговый уровень связывания определяется, если антитела против zalpha11-лиганда, описанного здесь, связываются с полипептидом, пептидом или эпитопом zalpha11-лиганда с аффинностью, по меньшей мере в 10 раз большей, чем аффинность связывания с контрольным (не zalpha11-лигандом) полипептидом. Предпочтительно, чтобы эти антитела проявляли аффинность связывания (Kа) 106 М-1 или большую, предпочтительно 107 М-1 или большую, более предпочтительно 108 М-1 или большую, и наиболее предпочтительно 109 М-1 или большую. Аффинность связывания антитела может быть легко определена специалистом с обычной квалификацией в этой области, например, при помощи анализа Скетчарда (Scatchard. Ann. NYAcad. Sci. 51:660-672, 1949).
То, что антитела против zalpha11-лиганда не имеют значимой перекрестной реактивности с родственными полипептидами, показано, например, детектированием этим антителом полипептида zalpha11-лиганда, но не известных родственных полипептидов, с использованием стандартного Вестерн-блот-анализа (Ausubel et al., ibid.). Примеры известных родственных полипептидов включают в себя ортологи и паралоги и сходные известные члены этого семейства белков. Скрининг может быть также выполнен с использованием zalpha11-лиганда не человека и мутантных полипептидов zalpha11-лиганда. Кроме того, антитела могут быть подвергнуты "скринингу против" известных родственных полипептидов для выделения популяции, которая специфически связывается с полипептидами zalpha11-лиганда. Например, антитела, индуцированные к zalpha11-лиганду, адсорбируют родственными полипептидами, прикрепленными к нерастворимому матриксу; антитела, специфические для zalpha11-лиганда будут протекать через этот матрикс при подходящих буферных условиях. Скрининг позволяет выделить поликлональные и моноклональные антитела, не имеющие перекрестной реактивности с известными близкородственными полипептидами (Antibodies: A Laboratory Manual. Hariow and Lane (eds.), Cold Spring Harbor Laboratory Press, 1988; Current Protocols in Immunology. Cooligan, et al. (eds.), National Institutes of Health, John Wiley and Sons, Inc., 1995). Скрининг и выделение специфических антител хорошо известны в этой области (см. Fundamental Immunology. Paul (eds.), Raven Press, 1993; Getzoff et al., Adv. in Immunol. 43:1-98, 1988; Monoclonal Antibodies: Principles and Practice. Coding, J.W. (eds.), Academic Press Ltd.. 1996; Benjamin et al., Ann. Rev. Immunol. 2: 67-101, 1984). Специфическое связывание антител против zalpha11-лиганда может быть определено многими способами, известными в этой области и описанными ниже.
Различные анализы, известные лицам с обычной квалификацией в данной области, могут быть использованы для детектирования антител, которые специфически связывают белки и полипептиды zalpha11-лиганда. Примеры анализов описаны подробно в Antibodies: A Laboratory Manual, Harlow and Lane (Eds.), Cold Spring Harbor Laboratory Press, 1988. Характерные примеры таких анализов включают в себя: конкурентный иммуноэлектрофорез, радиоиммуноанализ, радиоиммунопреципитацию, твердофазный иммуноферментный анализ (ELISA), дот-блот- или Вестерн-блот-анализ, ингибиторный или конкурентный анализ и сэндвич-анализ. Кроме того, антитела могут быть подвергнуты скринингу на связывание с белком или полипептидом дикого типа в сравнении с мутантным белком или полипептидом zalpha11-лиганда.
Антитела к zalpha11 могут быть использованы для мечения клеток, экспрессирующих zalpha11-лиганд; для выделения zalpha11-лиганда аффинной очисткой; для диагностических тестов для определения уровней в кровотоке полипептидов zalpha11-лиганда; для обнаружения или количественного определения растворимого zalpha11-лиганда как маркера скрытых патологии или заболевания; в аналитических способах с использованием FACS; для скрининга экспрессионных библиотек; для генерирования антиидиотипических антител и в качестве нейтрализующих антител или в качестве антагонистов для блокирования активности zalpha11-лиганда in vitro и in vivo. Подходящие прямые тэги (маркеры) или метки включают в себя радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные маркеры, хемилюминесцентные маркеры, магнитные частицы и т.п.; непрямые тэги (маркеры) или метки могут выводить на первое место применение пары биотин-авидин или других пар комплемент/антикомплемент в качестве промежуточных продуктов. Описанные здесь антитела могут быть также прямо или опосредованно конъюгированы с лекарственными средствами, токсинами, радионуклидами и т.п., и эти конъюгаты могут быть использованы для диагностических или терапевтических применений in vivo. Кроме того, антитела к zalpha11-лиганду или его фрагментам могут быть использованы in vitro для обнаружения денатурированного zalpha11-лиганда или его фрагментов в анализах, например, Вестерн-блотах или других анализах, известных в этой области.
Подходящие детектируемые молекулы могут быть прямо или опосредованно присоединены к полипептиду или антителу и включают в себя радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные маркеры, хемилюминесцентные маркеры, магнитные частицы и т.п. Подходящие цитотоксические молекулы могут быть прямо или опосредованно присоединены к этому полипептиду или антителу и включают в себя бактериальные или растительные токсины (например, дифтерийный токсин, сапорин, экзотоксин Pseudomonas, рицин, абрин и т.п.), а также терапевтические радионуклиды, такие как иод-131, рений-188 или иттрий-90 (либо непосредственно присоединенные к полипептиду или антителу, либо опосредованно присоединенные, например, через хелатирующую часть молекулы). Полипептиды или антитела могут быть также конъюгированы с цитотоксическими лекарственными средствами, такими как адриамицин. Для непрямого присоединения детектируемой или цитотоксической молекулы эта детектируемая или цитотоксическая молекула может быть конъюгирована с членом пары комплемент/антикомплемент, где другой член связан с этим полипептидом или частью антитела. Для этих целей примером комплементарной/антикомплементарной пары является биотин/стрептавидин.
Связывающие полипептиды могут также действовать как "антагонисты" zalpha11-лиганда для блокирования связывания zalpha11-лиганда и трансдукции сигнала in vitro и in vivo. Эти полипептиды, действующие против связывания zalpha11-лиганда, были бы применимы для ингибирования активности zalpha11-лиганда или связывания белка.
Гибридные белки полипептид-токсин или гибридные белки антитело-токсин могут быть использованы для нацеленного ингибирования или разрушения клетки или ткани (например, для обработки раковых клеток или тканей). Альтернативно, если данный полипептид имеет множественные функциональные домены (т.е. активирующий домен или рецепторсвязывающий домен, плюс нацеливающий домен), гибридный белок, включающий в себя только нацеливающий домен, может быть пригодным для направления детектируемой молекулы, цитотоксической молекулы или комплементарной молекулы в представляющий интерес тип клеток или тканей. В тех случаях когда гибридный белок только с единственным доменом включает в себя комплементарную молекулу, антикомплементарная молекула может быть конъюгирована с детектируемой или цитотоксической молекулой. Таким образом, такие гибридные белки, содержащие домен-комплементарную молекулу, представляют характерный для определенного типа клеток нацеливающий носитель для клетко/тканеспецифической доставки характерных для этого типа клеток конъюгатов антикомплементарно-детектируемых/цитотоксических молекул.
Гибридные белки zalpha11-лиганд-цитокин или гибридные белки антитело-цитокин могут быть использованы для усиления in vivo убивания тканей-мишеней (например, раков крови и костного мозга), если полипептид zalpha11-лиганда или антитело против zalpha11-лиганда поражает гиперпролиферативную клетку крови или костного мозга (см., в общем, Hornick et al., Blood 89:4437-47, 1997). Описанные слитые белки позволяют нацеливать цитокин в желаемый участок действия, обеспечивая посредством этого повышенную локальную концентрацию цитокина. Подходящие полипептиды zalpha11-лиганда или антитела против zalpha11-лиганда поражают нежелательную клетку или ткань (т.е. опухоль или лейкоз) и слитый цитокин медиирует улучшенный лизис клетки-мишени эффекторными клетками. Подходящие цитокины для этой цели включают в себя, например, интерлейкин 2 и гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF).
Дифференцировка представляет собой прогрессирующий и динамичный процесс, начинающийся плюрипотентными стволовыми клетками и заканчивающийся окончательно дифференцированными клетками. Плюрипотентные стволовые клетки, которые могут регенерироваться без коммитирования до линии дифференцировки, экспрессируют набор маркеров дифференцировки, которые утрачиваются когда выполняется коммитирование в отношении последовательности клеточных поколений. Клетки-предшественники экспрессируют набор маркеров дифференцировки, которые могут продолжать экспрессироваться или могут не продолжать экспрессироваться по мере прогрессирования этих клеток по пути направления дифференцировки к достижению зрелости. Маркеры дифференцировки, которые экспрессируются исключительно зрелыми клетками, являются обычно функциональными свойствами, такими как клеточные продукты, ферменты для получения клеточных продуктов и рецепторы. Стадию дифференцировки клеточной популяции подвергают мониторингу путем идентификации маркеров, присутствующих в данной клеточной популяции.
Есть доказательство, позволяющее предполагать, что факторы, которые стимулируют специфические типы клеток в направлении пути к конечной дифференцировке или дедифференцировке, воздействуют на всю клеточную популяцию, происходящую из общего предшественника или общей стволовой клетки. Таким образом, данное изобретение включает в себя стимуляцию или ингибирование пролиферации лимфоидных клеток, гемопоэтических клеток и эпителиальных клеток.
Zalpha11-лиганд выделяли из ткани, о которой известно, что она имеет важную иммунологическую функцию и которая содержит клетки, которые играют роль в иммунной системе. Zalpha11-лиганд экспрессируется в CD3+ отобранных активированных клетках периферической крови, и было показано, что экспрессия zalpha11-лиганда увеличивается после активации Т-клеток. Кроме того, результаты экспериментов, описанных в разделе примеров, демонстрируют, что полипептиды данного изобретения обладают действием на рост/размножение и/или дифференцированное состояние NK-клеток или предшественников NK-клеток. Дополнительное доказательство демонстрирует, что zalpha11-лиганд влияет на пролиферацию и/или дифференцировку Т-клеток и В-клеток in vivo. Факторы, которые как стимулируют пролиферацию гемапоэтических клеток-предшественников, так и активируют зрелые клетки, в общем известны. NK-клетки являются чувствительными только к IL-2, но пролиферация и активация обычно требует дополнительных факторов роста. Например, было показано, что IL-7 и Steel-фактор (c-kit-лиганд) требовались для образования колоний NK-клеток-предшественников. IL-15+IL-2 в комбинации с IL-7 и Steel-фактором были более эффективными (Mrozek et al., Blood 87:2632-2640, 1996). Однако неидентифицированные цитокины могут быть необходимы для пролиферации специфических субпопуляций NK-клеток и/или NK-клеток-предшественников (Robertson et al., Blood 76:2451-2438, 1990). Композиция, содержащая zalpha11-лиганд и IL-15, стимулирует NK-клетки-предшественники и NK-клетки, показывая, что эта композиция является более сильнодействующей, чем описанные ранее факторы и комбинации факторов.
Анализы для измерения дифференцировки включают в себя, например, измерение клеточных маркеров, ассоциированных со стадиеспецифической экспрессией ткани, ферментативной активности, функциональной активности или морфологических изменений (Watt, FASEB 5:281-284, 1991; Francis, Differentiation 57:63-75, 1994; Raes, Adv. Anim. Cell Biol. Technol. Bioprocesses. 161-171, 1989; включенные здесь в качестве ссылки). Альтернативно, полипептид zalpha11-лиганд сам может служить в качестве дополнительного маркера клеточной поверхности или секретируемого маркера, ассоциированного со стадиеспецифической экспрессией ткани. Само по себе прямое измерение полипептида zalpha11-лиганда или утраты его экспрессии в ткани по мере дифференцировки может служить в качестве маркера дифференцировки тканей.
Подобным образом, прямое измерение полипептида zalpha11-лиганда или утраты его экспрессии в ткани может быть выполнено в ткани или в клетках, когда они подвергаются прогрессированию опухоли. Увеличения инвазивности и подвижности клеток или увеличение или утрата экспрессии zalpha11-лиганда в предраковом или раковом состоянии в сравнении с нормальной тканью может служить в качестве диагностического средства для трансформации, инвазии и метастазирования в прогрессировании опухоли. Само по себе знание состояния прогрессирования опухоли или метастазирования поможет врачу выбрать наиболее правильную терапию и агрессивность лечения для конкретного индивидуального ракового пациента. Способы измерения увеличения или утраты экспрессии (либо мРНК, либо белка) хорошо известны в данной области и описаны здесь и могут быть использованы для экспрессии zalpha11-лиганда. Например, появление или исчезновение полипептидов, регулирующих клеточную подвижность, может быть использовано для помощи в диагностике и прогнозировании рака предстательной железы (Banyard, J. and Zetter, B.R., Cancer and Metast. Rev. 17:449-458, 1999). В качестве эффектора клеточной подвижности, увеличение или утрата экспрессии zalpha11-лиганда может служить в качестве диагностического инструмента для лимфоидных, В-клеточных, эпителиальных, гемопозтических и других раков.
Кроме того, активность и действие zalpha11-лиганда на развитие и метастазирование опухоли могут быть измерены in vivo. Несколько моделей сингенных мышей были разработаны для исследования влияния полипептидов, соединений или других обработок на развитие опухоли. В этих моделях опухолевые клетки, пассируемые в культуре, имплантируют в мышей того же самого штамма, что и донор опухоли. Эти клетки будут развиваться в опухоли, имеющие сходные характеристики, в реципиентных мышах, и метастазирование также будет иметь место в некоторых из этих моделей. Подходящие опухолевые модели для исследований авторов включают в себя карциному легких Льюиса (АТСС No. CRL-1642) и меланому В16 (АТСС No. CRL-6323) среди прочих. Обе они являются обычно используемыми опухолевыми линиями, сингенными относительно мышей C57BL6/J, которые легко поддаются культивированию и манипулированию in vitro. Опухоли, полученные в результате имплантации любой из этих клеточных линий, способны метастазировать в легкое в мышах C57BL6/J. Модель карциномы легкого Льюиса была недавно использована в мышах для идентификации ингибитора ангиогенеза (O'Reilly MS, et al., Cell 79:315-328, 1994). Мышей C57BL6/J обрабатывают экспериментальным агентом либо инъекцией один раз в день рекомбинантным белком, агонистом или антагонистом, либо однократной инъекцией рекомбинантного аденовируса. Спустя три дня после этой обработки 105-106 клеток имплантируют под кожу спинки. Альтернативно, сами эти клетки могут быть инфицированы рекомбинантным аденовирусом, например экспрессирующим zalpha11-лиганд, перед имплантацией, так что этот белок синтезируется в участке опухоли или внутриклеточно, а не системно. Мыши обычно развивают видимые опухоли в пределах 5 дней. Опухолям дают расти в течение периода до 3 недель, и во время этого периода они могут достигать размера 1500-1800 мм3 в контрольной обработанной группе. Размер опухоли и вес тела подвергают тщательному мониторингу на протяжении этого эксперимента. Во время умерщвления опухоль удаляют и взвешивают вместе с легкими и печенью. Было показано, что вес легкого коррелирует хорошо с метастатической нагрузкой опухоли. В качестве дополнительного измерения считали метастазы поверхности легких. Иссеченные опухоль, легкие и печень готовят для гистопатологического исследования, иммуногистохимии и in situ гибридизации с использованием способов, известных в данной области и описанных здесь. Таким образом может быть оценено влияние рассматриваемого экспрессируемого полипептида, например, zalpha11-лиганда, на способность опухоли рекрутировать сосудистую сеть и подвергаться метастазированию. Кроме того, наряду с использованием аденовируса, имплантированные клетки могут быть временно трансфицированы zalpha11-лигандом. Применение стабильных zalphall-лиганд-транфектантов, а также применение индуцируемых промоторов для активации экспрессии zalpha11-лиганда in vivo является известным в данной области и может быть использовано в этой системе для оценки индукции метастазирования zalpha11-лигандом. Кроме того, очищенный zalpha11-лиганд или кондиционированные zalphall-лигандом среды могут быть непосредственно инъецированы в эту мышиную модель и, следовательно, использованы в данной системе. В отношении общей ссылки см. O'Reilly MS, et al., Cell 79:315-328, 1994 и Rusciano D., et al., Murine Models of Liver Metastasis. Invasion Metastasis 14:349-361, 1995.
Zalpha11-лиганд будет применим в лечении онкогенеза и, следовательно, мог бы быть применим в лечении рака. Zalpha11-лиганд ингибирует стимулируемую IL-4 пролиферацию стимулируемых анти-lgM нормальных В-клеток, и сходный эффект наблюдается в линиях В-клеточных опухолей, что предполагает, что может быть терапевтическая польза в лечении пациентов zalpha11-лигандом для индукции клеток В-клеточных опухолей в менее пролиферативное состояние. Этот лиганд мог бы вводиться в комбинации с другими уже использующимися агентами, в том числе как с общепринятыми химиотерапевтическими агентами, так и иммуномодуляторами, такими как интерферон альфа. Было показано, что альфа/бета-интерфероны являются эффективными в лечении некоторых лейкозов и моделях болезней животных и ингибирующие рост эффекты интерферона-альфа и zalpha11-лиганд а являются аддитивными в отношении по меньшей мере одной полученной из В-клеточной опухоли клеточной линии.
Данное изобретение обеспечивает способ снижения пролиферации неопластических (опухолевых) В- и Т-клеток, предусматривающий введение млекопитающему с В- или Т-клеточной неоплазмой количества композиции zalpha11-лиганда, достаточного для снижения пролиферации неопластических В- или Т-клеток. В других вариантах, эта композиция может содержать по меньшей мере один другой цитокин, выбранный из группы, состоящей из IL-2, IL-15, IL-4, GM-CSF, Flt3-лиганда или фактора стволовых клеток.
В другом аспекте данное изобретение обеспечивает способ снижения пролиферации неопластических В- или Т-клеток, предусматривающий введение млекопитающему с В- или Т-клеточной неоплазмой количества антагониста zalpha11-лиганда, достаточного для снижения пролиферации неопластических В- или Т-клеток. В других вариантах эта композиция может содержать по меньшей мере один другой цитокин, выбранный из группы, состоящей из IL-2, IL-15, IL-4, GM-CSF, Flt3-лиганда или фактора стволовых клеток. Кроме того, антагонист zalpha11-лиганда может быть слитым белком лиганд/токсин.
Слитый токсин zalpha11-лиганд-сапорин может использоваться против сходной серии лейкозов и лимфом, расширяя диапазон лейкозов, которые могут лечиться zalpha11-лигандом. Опосредуемая слитым токсином активация zalpha11-рецептора обеспечивает два независимых средства для ингибирования роста клеток-мишеней, причем первое средство является идентичным эффектам, наблюдаемым с использованием одного лиганда, а второе обусловлено доставкой токсина через интернализацию рецептора. Ограничивающаяся лимфоидами картина экспрессии zalpha11-рецептора предполагает, что конъюгат лиганд-сапорин может быть переносимым пациентами.
Когда лечение злокачественных заболеваниий включает в себя трансплантацию костного мозга или стволовых клеток, zalpha11-лиганд может быть полезным в усилении реакции трансплантат против опухоли. Zalpha11-лиганд стимулирует генерирование литических МК-клеток из предшественников костного мозга и стимулирует пролиферацию Т-клеток после активации рецепторов антигена. Таким образом, когда пациенты получают аллогенные трансплантаты костного мозга, zalpha11-лиганд будет усиливать генерирование противоопухолевых реакций, с инфузией или без инфузии лимфоцитов донора.
Тканевое распределение рецептора для конкретного цитокина дает сильное указание на потенциальные участки действия данного цитокина. Нозерн-анализ zalpha11-рецептора выявил транскрипты в селезенке, тимусе, лимфатическом узле, костном мозгу и лейкоцитах периферической крови человека. Конкретные типы клеток были идентифицированы в качестве клеток, экспрессирующих zalpha11-рецепторы, и сильные сигналы наблюдали в реакции смешанной культуры лимфоцитов (СКЛ) и в клетках Raji лимфомы Беркита. Две линии моноцитов, ТНР-1 (Tsuchiya et al., Int. J. Cancer 26:171-176. 1980) и U937 (Sundstrom et al., Int. J. Cancer 17:565-577, 1976) были отрицательными.
Zalpha11-рецептор экспрессируется при относительно высоких уровнях в СКЛ, в которой смешаны мононуклеарные клетки периферической крови (PBMNC) из двух индивидуумов, что приводит к взаимной активации. Детектирование высоких уровней транскрипта в этой СКЛ, но не в покоящихся Т- или В-клеточных популяциях, предполагает, что экспрессия zalpha11-рецептора может быть индуцирована в одном или более типах клеток во время активации. Активация изолированных популяций Т- и В-клеток может быть искусственно достигнута стимуляцией клеток при помощи ФМА и иономицина. При подвергании прошедших сортинг клеток этим условиям активации уровни транскрипта zalpha11-рецептора увеличивались в обоих типах клеток, подтверждая роль для этого рецептора и zalpha11-лиганда в иммунных реакциях, в частности в аутокринном и паракринном размножениях Т-клеток и В-клеток во время активации. Zalpha11-лиганд может также играть роль в размножении более примитивных предшественников, участвующих в лимфопоэзе.
Было обнаружено, что zalpha11-рецептор присутствовал при низких уровнях в покоящихся Т- и В-клетках и положительно регулировался во время активации в обоих клеточных типах. Интересно, что В-клетки также отрицательно регулировались этим посредником более быстро, чем Т-клетки, что предполагает, что амплитуда сигнала и тайминг гашения сигнала являются важными для подходящей регуляции В-клеток.
Кроме того, большая доля клеток собственного слоя слизистой оболочки кишечника (lamina propria) обнаруживает положительные сигналы гибридизации с zalpha11-рецептором. Эта ткань состоит из смешанной популяции лимфоидных клеток, в том числе активированных CD4+ Т-клеток и активированных В-клеток. Иммунная дисфункция, в частности хроническая активация иммунной реакции слизистой оболочки, играет роль в этиологии болезни Крона; отклоняющаяся от нормы реакция на провоспалительные цитокины и/или на продуцирование провоспалительных цитокинов является также предположительным фактором (Braegger et al., Annals Allergy 72:135-141, 1994; Sartor RB Am. J. Gastroenterol. 92:53-11S, 1997). Zalpha11-лиганд вместе с IL-15 размножает NK-клетки из клеток-предшественников костного мозга и усиливает эффекторную функцию NK-клеток. Zalpha11-лиганд также костимулирует зрелые В-клетки, стимулированные антителами к CD40, но ингибирует пролиферацию В-клеток на сигналы, передаваемые через IgM. Zalpha11-лиганд усиливает пролиферацию Т-клеток вместе с сигналом через рецептор Т-клеток, и сверхэкспрессия в трансгенных мышах приводит к лимфопении и размножению моноцитов и гранулоцитов. Эти плейотропные эффекты zalpha11-лиганда предполагают, что он может обеспечивать терапевтическую примененимость для широкого диапазона заболеваний, возникающих из дефектов в иммунной системе, в том числе (но не ограничивающихся ими) системной красной волчанки (SLE), ревматоидного артрита (RA), множественного склероза (MS), тяжелой псевдопаралитической миастении и диабета. Важно отметить, что эти заболевания являются результатом комплексной сети иммунной дисфункции (например, SLE является манифестацией дефектов как в Т-, так и в В-клетках) и что иммунные клетки зависят от взаимодействия друг с другом для индукции сильной иммунной реакции. Таким образом, zalpha11-лиганд (или антагонист этого лиганда), который может быть использован для манипулирования более чем одним типом иммунной клетки, является привлекательным терапевтическим кандидатом для вмешательства в многочисленных стадиях заболевания.
Полипептиды и белки данного изобретения могут быть также использованы ex vivo, например, в аутологичной культуре костного мозга. Вкратце, костный мозг удаляют из пациента перед химиотерапией или трансплантацией органа и обрабатывают zalpha11-лигандом, необязательно в комбинации с одним или несколькими другими цитокинами. Обработанный костный мозг затем возвращают пациенту после химиотерапии для ускорения восстановления костного мозга или после трансплантации для супрессии реакции трансплантат против хозяина. Кроме того, белки данного изобретения могут быть также использованы для размножения ех vivo костного мозга или клеток-предшественников периферической крови (РВРС). Перед обработкой костный мозг может быть стимулирован фактором стволовых клеток (SCF) для высвобождения ранних клеток-предшественников в кровоток периферической крови. Эти клетки-предшественники могут быть собраны и сконцентрированы из периферической крови и затем обработаны в культуре zalpha11-лигандом, необязательно в комбинации с одним или несколькими другими цитокинами, в том числе, но не только, SCF, IL-2, IL-4, IL-7 или IL-15, для дифференцировки и пролиферации в лимфоидные культуры высокой плотности, которые затем возвращают пациенту после химиотерапии или трансплантации.
Данное изобретение обеспечивает способ размножения гемопоэтических клеток и предшественников гемопоэтических клеток, предусматривающий культивирование клеток костного мозга или клеток периферической крови с композицией, содержащей количество zalpha11-лиганда, достаточное для получения увеличения числа лимфоидных клеток в клетках костного мозга или клетках периферической крови в сравнении с клетками костного мозга или периферической крови, культивируемых в отсутствие zalpha11-лиганда. В других вариантах гемопоэтическими клетками и гемопоэтическими клетками-предшественниками являются лимфоидные клетки. В другом варианте лимфоидными клетками являются МК-клетки или цитотоксичные Т-клетки. Кроме того, композиция может также содержать по меньшей мере один другой цитокин, выбранный из группы, состоящей из IL-2, IL-15, IL-4, GM-CSF, Flt3 и фактора стволовых клеток.
Альтернативно zalpha11-лиганд может активировать иммунную систему, что было бы важным в бустинге иммунитета к инфекционным заболеваниям, лечении пациентов с ослабленным иммунитетом, таких как ВИЧ+ пациенты, или в улучшении вакцин. В частности, стимуляция zalpha11-лигандом или размножение NK-клеток или их предшественников могло бы обеспечить терапевтическую пользу в лечении вирусной инфекции и в качестве противоопухолевого фактора. Считается, что NK-клетки играют основную роль в элиминации метастатических опухолевых клеток, и пациенты как с метастазами, так и с твердыми опухолями, имеют уменьшенные уровни активности NK-клеток (Whiteside et al., Curr. Top. Microbiol. Immunol. 230:221-224, 1998). Подобным образом, стимуляция zalpha11-лигандом иммунной реакции против вирусных и невирусных патогенных агентов (в том числе бактерий, простейших и грибков) дало бы терапевтическую пользу в лечении таких инфекций путем ингибирования роста таких инфекционных агентов. Определение прямо или опосредованно уровней патогена или антигена, такого как опухолевая клетка, присутствующих в организме, может быть достигнуто рядом способов, известных в данной области и описанных здесь.
Данное изобретение включает в себя способ стимуляции иммунной реакции в млекопитающем, подвергнутом воздействию антигена или патогена, предусматривающий стадии: (1) определения прямо или опосредованно уровня антигена или патогена, присутствующего в указанном млекопитающем; (2) введения композиции, содержащей полипептид zalpha11-лиганда в фармацевтически приемлемом носителе; (3) определения прямо или опосредованно уровня антигена или патогена, присутствующего в указанном млекопитающем; и (4) сравнения уровня антигена или патогена в стадии 1 с уровнем антигена или патогена в стадии 3, причем изменение в этом уровне является признаком стимуляции иммунной реакции. В другом варианте композицию zalpha11-лиганда вводят повторно. В других вариантах антиген является В-клеточной опухолью; вирусом; паразитом или бактерией.
В другом аспекте данное изобретение обеспечивает способ стимуляции иммунной реакции в млекопитающем, подвергнутом воздействию антигена или патогена, предусматривающий стадии: (1) определения уровня антиген- или патоген-специфического антитела; (2) введения композиции, содержащей полипептид zalpha11-лиганда в фармацевтически приемлемом носителе; (3) определения уровня антиген- или патоген-специфического антитела после введения; (4) сравнения уровня антитела в стадии (1) с уровнем антитела в стадии (3), причем увеличение уровня антитела является признаком стимуляции иммунной реакции.
Полинуклеотиды, кодирующие полипептиды zalpha11-лиганда, применимы в способах генной терапии, где желательно увеличить или ингибировать активность zalpha11-лиганда. Если млекопитающее имеет мутированный или отсутствующий ген zalpha11-лиганда, этот ген zalpha11-лиганда может быть введен в клетки этого млекопитающего. В одном варианте ген, кодирующий полипептид zalpha11-лиганда, вводят in vivo в вирусном векторе. Такие векторы включают в себя аттенуированный или дефектный ДНК-вирус, такой как, но не ограничиваемый ими, вирус простого герпеса (HSV), вирус папилломы, вирус Эпстайна-Барр (EBV), аденовирус, аденоассоциированный вирус (AAV), и т.п. Предпочтительными являются дефектные вирусы, которые полностью или почти полностью лишены вирусных генов. Дефектный вирус не является инфекционным после введения в клетку. Применение дефектных вирусных векторов делает возможным введение в клетки в специфической, локализованной зоне, без опасения по поводу того, что этот вектор может инфицировать другие клетки. Примеры конкретных векторов включают в себя, но не ограничиваются ими, вектор дефектного вируса простого герпеса 1 (HSV1) (Kaplitt et al., Molec. Cell. Neurosci., 2:320-30, 1991), аттенуированный аденовирусный вектор, такой как вектор, описанный Stratford-Perricaudet et al., J. Clin. Invest. 90:626-30, 1992), и дефектный аденоассоциированный вирусный вектор (Samulski et al., J. Virol., 61: 3096-101, 1987; Samulski et al., J. Virol., 63:3822-8, 1989).
В другом варианте ген zalpha11-лиганда может быть введен в ретровирусном векторе, например, как описано в Anderson et al., U.S. Patent No. 5399346; Mann et al., Cell. 33:153, 1983; Temin et al., U.S. Patent No. 4650764; Temin et al., U.S. Patent No. 4980289; Markowitz et al., J. Virol. 62:1120, 1988; Temin et al., U.S. Patent No. 5124263; International Patent Publication No. WO 95/07358, published March 16, 1995 by Dougherty et al.; и Kuo et al., Blood, 82:845, 1993. Альтернативно, этот вектор может быть введен липофекцией in vivo с использованием липосом. Синтетические катионные липиды могут быть использованы для получения липосом для трансфекции in vivo гена, кодирующего маркер (Felgner et al., Proc. Natl. Acad. Sci. USA. 84, 7413-7, 1987; Mackey et al., Proc. Natl. Acad. Sci. USA 85:8027-31, 1988). Использование липофекции для введения экзогенных генов в специфические органы in vivo имеет определенные практические преимущества. Молекулярное нацеливание липосом в специфические клетки является одной областью преимущества. Более конкретно, направление трансфекции в конкретные клетки представляет одну область преимущества. Например, направление трансфекции в конкретные типы клеток было бы особенно выгодным в ткани с клеточной гетерогенностью, такой как иммунная система, поджелудочная железа, печень, почка и мозг. Липиды могут быть химически связаны с другими молекулами для нацеливания. Нацеленные пептиды (например, гормоны или нейротрансмиттеры), белки, такие как антитела, или непептидные молекулы могут быть связаны с липосомами химически.
Можно удалить клетки-мишени из тела; ввести этот вектор в виде "голой" ДНК-плазмиды и затем повторно имплантировать эти трансформированные клетки в тело. "Голые" ДНК-векторы для генной терапии могут быть введены в желательные клетки-хозяева способами, известными в данной области, например, трансфекцией, электропорацией, микроинъекцией, трансдукцией, клеточным слиянием, ДЭАЭ-декстрановым способом, кальций-фосфатным осаждением с использованием генного "дробовика" (шотган-способа) или с использованием вектора-переносчика ДНК. См., например, Wu et al., J. Biol. Chem. 267:963-7, 1992; Wu et al., J. Biol. Chem. 263:14621-4, 1988.
Антисмысловая методология может быть использована для ингибирования транскрипции гена zalpha11-лиганда, например, для ингибирования клеточной пролиферации in vivo. Конструируются полинуклеотиды, которые комплементарны сегменту кодирующего zalpha11-лиганд полинуклеотида (например, полинуклеотида, представленного в SEQ ID NO:1) для связывания с кодирующей zalphall-лиганд мРНК и ингибирования трансляции такой мРНК. Такие антисмысловые полинуклеотиды применимы для ингибирования экспрессии кодирующих полипептиды zalpha11-лиганда генов в клеточной культуре или в субъекте.
Мыши, "сконструированные" для экспрессии гена zalpha11-лиганда, называемые "трансгенными мышами", и мыши, которые обнаруживают полное отсутствие функции гена zalpha11-лиганда, называемые "мышами с нокаутом (гена)", также могут быть получены (Snouwaert et al., Science 257: 1083, 1992; Lowell et al., Nature 366: 740-42, 1993; Capecchi, M.R., Science 244:1288-1292, 1989; Palmiter, R.D. et al., Annu Rev Genet. 20:465-499, 1986). Например, трансгенные мыши, которые сверхэкспрессируют zalpha11-лиганд, либо повсеместно, либо под контролем тканеспецифического промотора или рестриктированного в отношении ткани промотора, могут быть использованы для выяснения, обусловливает ли сверхэкспрессия определенный фенотип. Например, сверхэкспрессия полипептида zalpha11-лиганда дикого типа, фрагмента полипептида или его мутанта может изменять нормальные клеточные процессы, приводя к фенотипу, который идентифицирует ткань, в которой экспрессия zalpha11-лиганда является функционально релевантной, и может указывать на терапевтическую мишень для zalpha11-лиганда, его агонистов или антагонистов. Например, предпочтительной трансгенной мышью для "конструирования" является мышь, которая сверхэкспрессирует zalpha11-лиганд (аминокислотные остатки 32-162 SEQ ID NO:2). Кроме того, такая сверхэкспрессия может приводить к фенотипу, который обнаруживает сходство с заболеваниями человека. Подобным образом, мыши с нокаутом zalpha11-лиганда могут быть использованы для определения, где zalpha11-лиганд является абсолютно необходимым in vivo. Фенотип мышей с нокаутом предсказывает эффекты in vivo, которые может иметь антагонист zalpha11-лиганда, такой как описанные здесь антагонисты. кДНК мышиного или человеческого zalpha11 может быть использована для получения мышей с нокаутом гена. Эти мыши могут быть использованы для исследования гена zalpha11-лиганда и белка, кодируемого им, в системе in vivo и могут быть использованы в качестве моделей in vivo для соответствующих заболеваний человека. Кроме того, экспрессия трансгенными мышами антисмысловых полинуклеотидов zalpha11-лиганда или направленных против zalpha11-лиганда рибозимов, описанных здесь, может быть использована аналогично экспрессии трансгенных мышей, описанной выше. Исследования могут также проводится посредством введения очищенного белка zalpha11-лиганда.
Для фармацевтического применения белки данного изобретения готовят для парентеральной, в частности, внутривенной или подкожной, доставки в соответствии с общепринятыми способами. Биологически активный полипептид или конъюгаты антител, описанные здесь, могут быть доставлены внутривенно, внутриартериально или внутрипротоково или могут быть введены локально в предполагаемом месте действия. Внутривенное введение может быть болюсной инъекцией или инфузией на протяжении обычного периода времени одного-нескольких часов. Обычно фармацевтические композиции будут включать в себя белок zalpha11-лиганда в комбинации с фармацевтически приемлемым носителем, таким как солевой раствор, забуференный солевой раствор, 5% декстроза в воде или т.п. Композиции могут дополнительно содержать один или несколько наполнителей, консервантов, солюбилизаторов, буферящих агентов, альбумин для предупреждения потери белка на поверхностях флаконов и т.д. Способы приготовления композиций хорошо известны в данной области и описаны, например, в Remington: The Science and Practice of Pharmacy. Gennaro, ed., Mack Publishing Co., Easton, PA, 19th ed., 1995. Терапевтические дозы обычно будут находиться в диапазоне от 0,1 до 100 мкг/кг массы тела в сутки, предпочтительно 0,5-20 мкг/кг в сутки, причем точная доза должна определяться врачом в соответствии с принятыми стандартами с учетом природы и тяжести подлежащего лечению состояния, особенностей пациента и т.д. Определение дозы находится в пределах обычной квалификации специалиста в данной области. Белки могут вводиться для остро требующегося лечения, на протяжении одной недели или менее, часто на протяжении периода одного-трех дней или могут вводиться в продолжительном лечении на протяжении нескольких месяцев или лет. Обычно терапевтически эффективное количество растворимого рецепторного полипептида zalpha11-лиганда составляет количество, достаточное для получения клинически значимого эффекта.
Далее данное изобретение иллюстрируется следующими ниже неограничительными примерами.
ПРИМЕРЫ
Пример 1
Конструирование химеры MPL-полипептид zalpha11: внеклеточный и ТМ-домен MPL, слитые с внутриклеточным доменом передачи сигнала zalpha11
Внеклеточный и трансмембранный домены мышиного MPL-рецептора выделяли из плазмиды, содержащей мышиный MPL-рецептор (плазмиды PHZ1/MPL) с использованием ПЦР с праймерами ZC17212 (SEQ ID NO:5) и ZC19914 (SEQ ID NO:6). Условия реакции были следующими: 95°С в течение 1 минуты; 35 циклов при 95°С в течение 1 минуты, 45°С в течение 1 минуты, 72°С в течение 2 минут; затем 72°С в течение 10 минут; затем пропитывание при 10°С. ПЦР-продукт подвергали электрофорезу на 1% агарозе с низкой точкой плавления (Boehringer Mannheim, Indianapolis, IN) и фрагмент MPL-рецептора приблизительно 1,5 т.п.н. выделяли с использованием набора для экстракции гелей Qiaquick™ Gel Extraction Kit (Qiagen) согласно инструкциям изготовителя.
Внутриклеточные домены zalpha11 человека выделяли из плазмиды, содержащей кДНК zalpha11-рецептора с использованием ПЦР с праймерами ZC19913 (SEQ ID NO:8) и ZC20097 (SEQ ID NO:9). Полинуклеотидная последовательность, соответствующая кодирующей zalpha11-рецептор последовательности, показана в SEQ ID NO:7, а соответствующая аминокислотная последовательность показана в SEQ ID NO:115. Условия реакции были такие же, что и описанные выше. ПЦР-продукт подвергали электрофорезу на 1% агарозе с низкой точкой плавления (Boehringer Mannheim) и фрагмент zalpha11 приблизительно 900 т.п.н. выделяли с использованием набора для экстракции гелей Qiaquick согласно инструкциям изготовителя.
Каждый из выделенных фрагментов, описанных выше, смешивали при отношении объемов 1:1 и использовали в ПЦР-реакции с применением ZC17212 (SEQ ID NO:5) и ZC20097 (SEQ ID NO:9) для создания химеры MPL-zalpha11. Условия реакции были следующими; 95°С в течение 1 минуты; 35 циклов при 95°С в течение 1 минуты; 55°С в течение 1 минуты, 72°С в течение 2 минут; затем 72°С в течение 10 минут; затем пропитывание при 10°С. Полный ПЦР-продукт подвергали электрофорезу на 1% агарозе с низкой точкой плавления (Boehringer Mannheim) и химерный фрагмент MPL-zalpha11 приблизительно 2,4 т.п.н. выделяли с использованием набора для экстракции гелей Qiaquick (Qiagen) согласно инструкциям изготовителя. Химерный фрагмент MPL-zalpha11 расщепляли EcoRI (BRL) и Xbal (Boehringer Mannheim) согласно инструкциям изготовителя. Весь продукт расщепления подвергали электрофорезу на 1% агарозе с низкой точкой плавления (Boehringer Mannheim) и расщепленную химеру MPL-zalpha11 выделяли с использованием набора для экстракции гелей Qiaquick (Qiagen) согласно инструкциям изготовителя. Полученную расщепленную химеру MPL-zalpha11 встраивали в экспрессирующий вектор, как описано ниже.
Реципиентный экспрессирующий вектор pZP-5N расщепляли EcoRI (BRL) и Hindlll (BRL) согласно инструкциям изготовителя и очищали на геле, как описано выше. Этот векторный фрагмент объединяли с расщепленной EcoRI и Xbal химерой MPL-zalpha11, выделенной, как описано выше, и Xbal/Hindlll-линкерным фрагментом в реакции лигирования. Лигирование проводили с лигазой Т4 (BRL) при 15°С в течение ночи. Пробу лигирования электропорировали в электрокомпетентные относительно DH10B ElectroMAX™ клетки Е. coli (25 мкф, 200 Ом, 2,3 В). Трансформанты высевали на чашки с LB+ампициллин и отдельные колонии подвергали скринингу при помощи ПЦР для подтверждения химеры MPL-zalpha11 с использованием ZC17212 (SEQ ID NO:5) и ZC20097 (SEQ ID NO:9) с применением условий ПЦР, описанных выше.
Подтверждение последовательности химеры MPL-zalpha11 выполняли при помощи анализов последовательности с использованием следующих праймеров: ZC12700 (SEQ ID NO:10), ZC5020 (SEQ ID NO:11), ZC6675 (SEQ ID NO:12), ZC7727 (SEQ ID NO:13), ZC8290 (SEQ ID NO:14), ZC19572 (SEQ ID NO:15), ZC6622 (SEQ ID NO:16), ZC7736 (SEQ ID NO:17) и ZC9273 (SEQ ID NO:18). Вставка (инсерт) была приблизительно 2,4 т.п.н. и была полноразмерной.
Пример 2
Пролиферация на основе химеры MPL-zalpha11 в ВАF3-тесте с использованием Alamar Blue
А. Конструирование клеток BaF3, экспрессирующих химеру MPL-zalpha11
BaF3, интерлейкин-3- (IL-3)-зависимую прелимфоидную клеточную линию, полученную из мышиного костного мозга (Palacios and Steinmetz, Cell 41:727-734, 1985; Mathey-Prevot et al., Mol. Cell. Biol. 6:4133-4135, 1986), поддерживали в полной среде (среде RPMI (JRH Bioscience Inc., Lenexa, KS), дополненной 10% инактивированной нагреванием фетальной телячьей сывороткой, 2 нг/мл мышиного IL-3 (mlL-3) (R & D, Minneapolis, MN), 2 мМ L-glutaMax-1™ (Gibco BRL), 1 мМ пируватом натрия (Gibco BRL) и PSN-антибиотиками (Gibco BRL)). Перед электропорацией ДНК плазмиды pZP-5N/MPL-zalpha11 (пример 1) получали и очищали с использованием набора Qiagen Maxi Prep kit (Qiagen) согласно инструкциям изготовителя. Клетки BaF3 для электропорации промывали один раз в среде RPMI и затем ресуспендировали в среде RPMI при плотности клеток 107 клеток/мл. Один мл ресуспендированных клеток BaF3 смешивали с 30 мкг ДНК плазмиды pZP-5N/MPL-zalpha11 и переносили в отдельные одноразовые камеры для электропорации (Gibco BRL). После 15-минутного инкубирования при комнатной температуре клетки подвергали двум последовательным электроимпульсам (800 МФ/300 В; 1180 МФ/300 В), подаваемым прибором для электропорации (CELL-PORATOR™; Gibco BRL). После 5 минут восстановления до нормы электропорированные клетки переносили в 50 мл полной среды и помещали в термостат на 15-24 ч (37°С, 5% CO2). Затем эти клетки откручивали и ресуспендировали в 50 мл полной среды, содержащей Geneticin™ (Gibco) для отбора (500 мкг/мл G418), в колбе Т-162 для выделения С418-резистентного пула. Пулы трансфицированных клеток BaF3, далее называемых клетками BaF3/MPL-zalpha11, анализировали на способность передачи сигнала, как описано ниже.
В. Тестирование способности передачи сигнала клеток BaF3/MPL-zalpha11 с использованием теста пролиферации с Alamar Blue
Клетки BaF3/MPL-zalpha11 откручивали и промывали в полной среде, как описано выше, но без mlL-3 (далее называемой "не содержащей mlL-3 средой"). Клетки откручивали и промывали 3 раза для гарантии удаления mlL-3. Затем клетки считали в гемоцитометре. Клетки высевали в 96-луночный планшет при 5000 клеток на лунку в объеме 100 мкл на лунку с использованием не содержащей mlL-3 среды.
Пролиферацию клеток BaF3/MPL-zalpha11 оценивали с использованием мышиного тромбопоэтина (mTPO), разведенного не содержащей mlL-3 средой до концентраций 500 нг/мл, 250 нг/мл, 125 нг/мл, 62 нг/мл, 30 нг/мл, 15 нг/мл, 7,5 нг/мл, 3,75 нг/мл, 1,8 нг/мл, 0,9 нг/мл, 0,5 нг/мл и 0,25 нг/мл. 100 мкл разведенного тТРО добавляли к клеткам BaF3/MPL-zalpha11. Общий объем теста равен 200 мкл. Негативные контроли оценивали параллельно с использованием только не содержащей mlL-3 среды, без добавления mTPO. Тест-планшеты инкубировали при 37°С, 5% СО2 в течение 3 дней, после чего добавляли Alamar Blue (Accumed, Chicago, IL) при 20 мкл на лунку. Alamar Blue дает флуорометрические показания на основе числа живых клеток и является, следовательно, прямым измерением пролиферации клеток в сравнении с негативным контролем. Планшеты опять инкубировали при 37°С, 5% СО2 в течение 24 часов. Планшеты считывали на планшет-ридере Fmax™ (Molecular Devices Sunnyvale, CA) с использованием программы SoftMax™ Pro при длинах волн 544 (возбуждение) и 590 (эмиссия).
Результаты подтвердили способность передачи сигнала внутриклеточной части zalpha11-рецептора, так как тромбопоэтин индуцировал пролиферацию приблизительно в 10 раз более высокую, чем уровень фона, при концентрациях mTPO 62 нг/мл и выше.
Пример 3
Конструирование экспрессирующего вектора, экспрессирующего полноразмерный zalpha11
Полный zalpha11-рецептор выделяли из плазмиды, содержащей кДНК zalpha11-рецептора (SEQ ID NO:7), при помощи ПЦР с использованием праймеров ZC19905 (SEQ ID NO:19) и ZC19906 (SEQ ID NO:20). Условия реакции были следующими; 95°С в течение 1 минуты; 35 циклов при 95°С в течение 1 минуты; 55°С в течение 1 минуты, 72°С в течение 2 минут; затем 72°С в течение 10 минут; затем пропитывание при 10°С. ПЦР-продукт подвергали электрофорезу на 1% агарозе с низкой точкой плавления (Boehringer Mannheim) и кДНК zalpha11 приблизительно 1,5 т.п.н. выделяли с использованием набора для экстракции гелей Qiaquick™ (Qiagen) согласно инструкциям изготовителя.
Очищенную кДНК zalpha11 расщепляли BamHI (Boehringer Mannheim) и EcoRI (BRL) согласно инструкциям изготовителя. Весь продукт расщепления подвергали электрофорезу на 1% агарозе с низкой точкой плавления (Boehringer Mannheim) и очищали отщепленный фрагмент zalpha11 с использованием набора для экстракции гелей Qiaquick согласно инструкциям изготовителя. Полученный отщепленный zalpha11-фрагмент встраивали в экспрессирующий вектор, как описано ниже.
Реципиентный экспрессирующий вектор pZP-5N расщепляли BamHI (Boehringer Mannheim) и EcoRI (BRL) согласно инструкциям изготовителя и очищали на геле, как описано выше. Этот векторный фрагмент объединяли с растщепленным BamHI и EcoRI zalpha11-фрагментом, выделенным, как описано выше, в реакции лигирования с использованием лигазы Т4 (BRL). Лигирование проводили инкубированием при 15°С в течение ночи. Пробу лигирования подвергали электропорации в электрокомпетентные относительно DH10B ElectroMAX™ клетки Е. coli (25 мкФ, 200 Ом, 2,3 В). Трансформанты высевали на чашки с LB+ампициллин и отдельные колонии подвергали скринингу при помощи ПЦР для подтверждения последовательности zalpha11 с использованием ZC19905 (SEQ ID NO:19) и ZC19906 (SEQ ID NO:20) с применением условий ПЦР, описанных выше.
Подтверждение последовательности MPL-zalpha11 выполняли при помощи анализов последовательности с использованием следующих праймеров: ZC12700 (SEQ ID NO:10), ZC5020 (SEQ ID NO:11), ZC20114 (SEQ ID NO:21), ZC19459 (SEQ ID NO:22), ZC19954 (SEQ ID NO:23) и ZC20116 (SEQ ID NO:24). Вставка была приблизительно 1,6 т.п.н. и была полноразмерной.
Пример 4
Пролиферация на основе zalpha11 в ВАF3-тесте с использованием Alamar Blue А. Конструирование клеток BaF3, экспрессирующих zalpha11-рецептор
Клетки BaF3, экспрессирующие полноразмерный zalpha11-рецептор, конструировали, как в примере 2А выше, с использованием 30 мкг экспрессирующего вектора zalpha11, описанного в примере 3 выше. Клетки BaF3, экспрессирующие мРНК zalpha11-рецептора, были названы BaF3/zalpha11. Эти клетки использовали для скрининга на активность zalpha11, как описано ниже в примерах 5 и 6.
Пример 5
Скрининг на zalpha11-лиганд с использованием клеток BaF3/zalpha11 при помощи теста пролиферации с Alamar Blue
А. Активация первичных спленоцитов обезьян для тестирования на присутствие zalpha11-лиганда
Спеленоциты обезьяны стимулировали in vitro с получением кондиционированной среды для тестирования на присутствие активности zalpha11-лиганда, как описано ниже. Селезенки обезьян получали из 8-летних самок обезьян М. nesestrian. Селезенки разрывали на части для получения суспензии отдельных клеток. Мононуклеарные клетки выделяли при помощи градиента плотности Ficoll-Paque® PLUS (Pharmacia Biotech, Uppsala, Sweden). Мононуклеарные клетки высевали при 2×106 клеток/мл в среду RPMI-1640, дополненную 10% ФТС, и активировали 5 нг/мл форбол-12-миристат-13-ацетата (ФМА) (Calbiochem, San Diego, СА) и 0,5 мг/мл Иономицина™ (Calbiochem) в течение 48 часов. Супернатант от стимулированных клеток селезенки обезьяны использовали для анализа пролиферации клеток BaF3/zalpha11, как описано ниже.
В. Скрининг на zalpha11-лиганд с использованием клеток BaF3/zalpha11 при помощи теста пролиферации с Alamar Blue
Клетки BaF3/zalpha11 откручивали и промывали в не содержащей mlL-3 среде. Клетки откручивали и промывали 3 раза для гарантии удаления mlL-3. Затем клетки считали в гемоцитометре. Клетки высевали в 96-луночный планшет при 5000 клеток на лунку в объеме 100 мкл на лунку с использованием не содержащей mlL-3 среды.
Пролиферацию клеток BaF3/zalpha11 оценивали с использованием кондиционированной среды от активированных клеток селезенки обезьяны (см. пример 5А выше). Кондиционированную среду разводили не содержащей mlL-3 средой до концентраций 50%, 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75% и 0,375%. 100 мкл разведенной кондиционированной среды добавляли к клеткам BaF3/zalpha11. Общий объем теста равен 200 мкл. Тест-планшеты инкубировали при 37°С, 5% CO2 в течение 3 дней, после чего добавляли Alamar Blue (Accumed, Chicago, IL) при 20 мкл на лунку. Планшеты опять инкубировали при 37°С, 5% СО2 в течение 24 часов. Планшеты считывали на планшет-ридере Fmax™ (Molecular Devices), как описано выше (пример 2).
Результаты подтвердили пролиферативный ответ клеток BaF3/zalpha11 на фактор, присутствующий в активированных кондиционированных средах клеток селезенки обезьяны. Этот ответ, согласно измерениям, был приблизительно в 4 раза выше фона при концентрации 50%. Нетрансфицированные клетки BaF3 не пролиферировали в ответ на этот фактор, что указывает на то, что этот фактор является специфическим для Zalpha11-рецептора.
С. Источник первичных клеток человека, используемый для выделения Zalpha11-лиганда
100 мл крови брали из каждого из шести доноров. Кровь извлекали с использованием 10Х 10 мл вакуумных пробирок (vacutainer), содержащих гепарин. Кровь объединяли из шести доноров (600 мл), разводили 1:1 в ЗФР и разделяли с использованием Ficoll-Paque® PLUS (Pharmacia Biotech). Выход выделенных первичных клеток человека после разделения на градиенте фиколла был равен 1,2×109 клеток.
Клетки суспендировали в 9,6 мл буфера MACS (ЗФР, 0,5% ЭДТА, 2 мМ ЭДТА). 1,6 мл клеточной суспензии извлекали и добавляли 0,4 мл микрогранул CD3 (Miltenyi Biotec, Auburn, СА). Смесь инкубировали в течение 15 минут при 4°С. Клетки, меченые CD3 гранулами, промывали 30 мл буфера MACS и затем ресуспендировали в 2 мл буфера MACS.
Колонку VS+ (Miltenyi) готовили согласно инструкциям изготовителя. Затем колонку VS+ помещали в магнитное поле VarioMACS™ (Miltenyi). Колонку уравновешивали 5 мл буфера MACS. Затем выделенные первичные клетки человека наносили на эту колонку. СD3-негативным клеткам давали пройти через колонку. Колонку промывали 9 мл (3×3мл) буфера MACS. Затем колонку удаляли из магнитного поля и помещали над пробиркой falcon на 15 мл. CD3+ клетки элюировали добавлением 5 мл буфера MACS на колонку и связавшиеся клетки вымывали с использованием поршня (плунжера), обеспечиваемого изготовителем. Инкубирование этих клеток с СD3-магнитными гранулами, промывки и стадии колонки VS+ (инкубирование-элюцию), описанные выше, повторяли еще пять раз. Полученные CD3+ фракции из шести разделений на колонке объединяли. Общий выход CD3+ отобранных Т-клеток человека был равен 3×108 клеток.
Пробу объединенных CD3+ отобранных Т-клеток человека брали для окрашивания и сортинга на клеточном сортере с возбуждением флуоресценции (FACS) для оценки их чистоты. CD3+ отобранные Т-клетки человека содержали 91% CD3+ клеток.
CD3+ отобранные Т-клетки человека активировали инкубированием в RPMI + 5% ФТС + ФМА 10 нг/мл и иномицине 0,5 мкг/мл (Calbiochem) в течение 13 часов при 37°С. Супернатант от этих активированных CD3+ отобранных Т-клеток человека тестировали на активность zalpha11-лиганда, как описано ниже. Кроме того, активированные CD+ отобранные клетки человека использовали для получения библиотеки кДНК, как описано в примере 6 ниже.
D. Тестирование супернатанта от активированных CD3+ отобранных Т-клеток человека на zalpha11-лиганд с использованием клеток BaF3/zalpha11 и теста пролиферации с использованием Alamar Blue
Клетки BaF3/zalpha11 откручивали и промывали в не содержащей mlL-3 среде. Клетки откручивали и промывали 3 раза для гарантии удаления mlL-3. Затем клетки считали в гемоцитометре. Клетки высевали в 96-луночный планшет при 5000 клеток на лунку в объеме 100 мкл на лунку с использованием не содержащей mlL-3 среды.
Пролиферацию клеток BaF3/zalpha11 оценивали с использованием кондиционированной среды от активированных CD3+ отобранных Т-клеток человека (см. пример 5С) выше), разведенной не содержащей mlL-3 средой до концентраций 50%, 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75% и 0,375%. 100 мкл разведенной кондиционированной среды добавляли к клеткам BaF3/zalpha11. Общий объем теста равен 200 мкл. Тест-планшеты инкубировали и анализировали, как описано в примере 5В.
Результаты подтвердили пролиферативный ответ клеток BaF3/zalpha11 на фактор, присутствующий в активированной кондиционированной среде CD3+ отобранных Т-клеток человека. Этот ответ, согласно измерениям, был приблизительно в 10 раз выше фона при концентрации 50%. Нетрансфицированные клетки BaF3 не пролиферировали в ответ на этот фактор, что указывает на то, что этот фактор является специфическим для Zalpha11-рецептора. Кроме того, растворимый zalpha11-рецептор блокировал эту пролиферативную активность в клетках BaF3/zalpha11 (см. пример 11).
Пример 6
Клонирование человеческого zalpha11-лиганда из библиотеки CD3+ отобранных клеток человека
Скрининг библиотеки кДНК первичных CD3+ отобранных активированных клеток человека выявил выделенную кДНК, которая является новым членом семейства цитокинов со структурой четырехспирального пучка. Эта кДНК кодировала zalpha11-лиганд. Эту кДНК идентифицировали скринингом на активность zalpha11-лиганда с использованием zalpha11-рецептора.
А. Вектор для конструирования CD3+ отобранной библиотеки
Вектором для конструирования CD3+ отобранной библиотеки была плазмида pZP7NX. Вектор pZP7NX конструировали следующим образом. Кодирующий район для селективного маркера DHFR в векторе pZP7 удаляли расщеплением ДНК рестрикционными ферментами (рестриктазами) Ncol и PstI (Boehringer Mannheim). Расщепленную ДНК подвергали электрофорезу на 1% агарозном геле, вырезали и очищали от геля с использованием набора для экстракции гелей Qiaquick™ Gel Extraction Kit (Qiagen) согласно инструкциям изготовителя. ДНК-фрагмент, представляющий кодирующий район селективного маркера Zeocin, амплифицировали с использованием ПЦР с праймерами ZC13946 (SEQ ID NO:25) и ZC13945 (SEQ ID NO:26) и pZeoSV2(+) в качестве матрицы. В праймере ZC13946 (SEQ ID NO:25) имелись дополнительные сайты рестрикции PstI и Bell, и дополнительные сайты Ncol и Sful имелись в праймере ZC13945 (SEQ ID NO:26). ПЦР-фрагмент вырезали рестриктазами PstI и Ncol и клонировали в вектор pZP7, полученный расщеплением теми же самыми двумя ферментами с последующей очисткой от геля. Этот вектор был назван pZP7Z. Затем кодирующий район Zeocin удаляли расщеплением ДНК вектора pZP7Z рестриктазами Bell и Sful. Расщепленную ДНК подвергали электрофорезу на 1% агарозном геле, вырезали и очищали от геля и затем лигировали с ДНК-фрагментом кодирующего неомицин района, вырезанным из вектора pZem228 (депонированного в Американской коллекции типовых культур (АТСС), Manassas, VA; ATCC Deposit No. 69446) с использованием тех же самых рестриктаз (Bell и Sful).
Этот новый вектор был назван pZP7N, в котором кодирующий район для селективного маркера DHFR был заменен кодирующим районом для селективного маркера неомицина из вектора pZem228. Фрагмент-наполнитель, содержащий сайт Xhol, добавляли к pZP7N для создания вектора, пригодного для высокоэффективного направленного клонирования кДНК; этот новый вектор был назван pZP7NX. Для подготовки этого вектора для кДНК, 20 мкг pZP7NX расщепляли 20 единицами EcoRI (Life Technologies, Gaithersburg, MD) и 20 единицами Xhol (Boehringer Mannheim, Indianapolis, IN) в течение 5 часов при 37°С, затем 68°С в течение 15 минут. Продукт расщепления затем подвергали электрофорезу на 0,8% низкоплавком агарозном геле 1ХТАЕ для отделения наполнителя от вектора. Полосу вектора вырезали и расщепляли "бета-агаразой" (New England Biolabs, Beverly, MA) согласно рекомендациям изготовителя. После осаждения этанолом расщепленный вектор ресуспендировали в воде до 45 нг/мл в подготовке для лигирования CD3+ отобранной библиотеки кДНК, описанной ниже.
В. Получение библиотеки кДНК первичных CD3+ отобранных активированных клеток человека
Приблизительно 1,5×108 первичных CD3+ отобранных клеток человека, стимулированных иономицином/ФМА, выделяли центрифугированием после культивирования при 37°С в течение 13 часов (пример 5С). Тотальную РНК выделяли из осадка клеток с использованием набора "RNeasy Midi" из Qiagen, Inc. (Valencia, CA). мРНК выделяли из 225 мкг тотальной РНК с использованием набора для очистки мРНК "MPG mRNA purification kit" из CPG Inc. (Lincoln Park, NJ). 3,4 микрограммов мРНК выделяли и превращали в двухцепочечную кДНК с использованием следующей процедуры.
кДНК первой цепи из стимулированных CD3+ отобранных клеток человека синтезировали следующим образом. Девять мкл олиго (d(T)-отобранной поли(А) CD3+ РНК в концентрации 0,34 мкг/мкл и 1,0 мкл 1 мкг/мкл праймера первой цепи ZC18698 (SEQ ID NO:27), содержащего сайт рестрикции Xhol, смешивали и нагревали при 65°С в течение 4 минут и охлаждали на льду. Синтез кДНК первой цепи инициировали добавлением 9 мкл буфера для первой цепи (5х SUPERSCRIPT ®-буфер; (Life Technologies), 4 мкл 100 мМ дитиотреитола и 2 мкл раствора дезоксинуклеотидтрифосфатов, содержащего 10 мМ каждого из dATP, dGTP, dTTP и 5-метил-аСТР (Pharmacia Biotech, Inc.), к смеси РНК-праймер. Реакционную смесь инкубировали при 45°С в течение 4 минут с последующим добавлением 8 мкл 200 Е/мкл SUPERSCRIPT®, РНКаза Н- обратной транскриптазы (Life Technologies). Реакцию инкубировали при 45°С в течение 45 минут с последующим снижением температуры инкубации на 1°С каждые 2 минуты до 50°С и реакцию поддерживали при 50° в течение 10 минут. Для денатурации любой вторичной структуры и позволения дополнительного удлинения этой кДНК реакцию затем нагревали до 70°С в течение 2 минут, затем снижали температуру до 55°С в течение 4 минут, после чего добавляли 2 мкл ОТ (обратной транскриптазы) SUPERSCRIPT® и инкубировали дополнительно в течение 15 минут с последующим снижением температуры до 70°С по 1 минуте/1°С. Невключенные нуклеотиды удаляли из кДНК осаждением дважды в присутствии 2 мкг гликогенового носителя, 2,0 М ацетата аммония и 2,5 объема этанола с последующей промывкой 100 мкл 70% этанола. Эту кДНК ресуспендировали в 98 мкл воды для использования в синтезе второй цепи.
Синтез второй цепи проводили на кДНК первой цепи в условиях, которые промотировали праймирование первой цепью синтеза второй цепи, что приводило к образованию ДНК-шпильки. Реакция второй цепи содержала 98 мкл кДНК первой цепи, 30 мкл 5х буфера для полимеразы I (100 мМ Трис-HCI, рН 7,5, 500 мМ KCI, 25 мМ MgCl2, 50 мМ (NH4)2SO4, 2 мкп 100 мМ дитиотреитола, 6 мкл раствора, содержащего 10 мМ каждого из дезоксинуклеотидтрифосфатов, 5 мкл 5 мМ b-NAD, 1 мкл 3 Е/мкл ДНК-лигазы Е. coli (New England Biolabs Inc.) и 4 мкл 10 Е/мкл ДНК-полимеразы 1 Е. coli (New England Biolabs Inc.). Реакцию собирали при комнатной температуре и инкубировали при комнатной температуре в течение 2 минут с последующим добавлением 4 мкл 3,8 Е/мкл РНКазы Н (Life Technologies). Реакцию инкубировали при 15°С в течение двух часов с последующим 15-минутным инкубированием при комнатной температуре. К реакции добавляли 10 мкл 1 М ТРИС рН 7,4 и экстрагировали дважды смесью фенол-хлороформ и один раз хлороформом, затем органические фазы обратно экстрагировали 50 мкл ТЭ (10 мМ ТРИС рН 7,4, 1 мМ ЭДТА), объединяли с другими водными фазами и осаждали этанолом в присутствии 0,3 М ацетата натрия. Осадок промывали 100 мкл 70% этанола, сушили на воздухе и ресуспендировали в 40 мкл воды.
Одноцепочечную ДНК шпилечной структуры расщепляли нуклеазой фасоли золотистой. Реакционная смесь содержала 40 мкл второй цепи кДНК, 5 мкл 10х буфера для нуклеазы фасоли золотистой (Life Technologies), 5 мкл нуклеазы фасоли золотистой (Pharmacia Biotech, Corp.), разбавленной до 1 Е/мкл в 1х буфере для нуклеазы фасоли золотистой. Реакцию инкубировали при 37°С в течение 45 минут. Реакцию останавливали добавлением 10 II 1 М Трис-HCl, рН 7,4, с последующими последовательными экстракциями смесью фенол/хлороформ и хлороформом, как описано выше. После экстракций кДНК осаждали этанолом в присутствии 0,3 М ацетата натрия. Осадок промывали 100 мкл 70% этанола, сушили на воздухе и ресуспендировали в 38 мкл воды.
Ресуспендированную кДНК затупляли на концах ДНК-полимеразой Т4. кДНК, которую ресуспендировали в 38 мкл воды, смешивали с 12 мкл 5х буфера для ДНК-полимеразы Т4 (250 мМ Трис-HCl, рН 8,0, 250 мМ KCL, 25 мМ MgCl2), 2 мкл 0,1 М дитиотреитола, 6 мкл раствора, содержащего 10 мМ каждого из дезоксинуклеотидтрифосфатов, и 2 мкл 1 Е/мкл ДНК-полимеразы Т4 (Boehringer Mannheim, Corp.). После инкубации в течение 45 минут при 15°С реакцию останавливали добавлением 30 мкл ТЭ с последующими последовательными экстракциями смесью фенол/хлороформ и хлороформом и обратно экстрагировали 20 мкл ТЭ, как описано выше. Эту ДНК осаждали этанолом в присутствии 2 мкл носителя Pellet Paint™ (краски для осадка) (Novagen) и 0,3 М ацетата натрия и ресуспендировали в 11 мкл воды.
Адаптеры EcoRI лигировали на 5'-концах кДНК, описанной выше, для облегчения клонирования в экспрессирующий вектор. 11 мкл кДНК и 4 мкл 65 пмоль/мкл полуфосфорилированного адаптора Eco R1 (Pharmacia Biotech, Corp.) смешивали с 5 мкл 5х лигазного буфера (Life Technologies), 2 мкл 10 мМ АТФ и 3 мкл 1 Е/мкл ДНК-лигазы Т4 (Life Technologies), 1 мкл 10х буфера для лигирования (Promega, Corp.), 9 мкл воды. Еще одно разбавление 1х буфером производили для предотвращения осаждения краски Pellet Paint. Реакцию инкубировали 9 часов на водяной бане с изменением температуры от 10 до 22°С на протяжении 9 часов, затем выдерживали в течение 45 минут при 25°С. Реакцию останавливали инкубированием при 68°С в течение 15 минут.
Для облегчения направленного клонирования кДНК в экспрессирующий вектор кДНК расщепляли Xhol с получением кДНК, имеющей 5'-EcoRl-липкий конец и 3'-Xhol-липкий конец. Сайт рестрикции Xhol на 3'-конце этой "ДНК был предварительно введен с использованием праймера ZC18698 (SEQ ID NO:27). Расщепление рестриктазами проводили в реакционной смеси, содержащей 35 мкл смеси для лигирования, описанной выше, 6 мкл 10х Н-буфера (Boehringer Mannheim Corp.), 1 мкл 2 мг/мл БСА (Biolabs Corp.), 17 мкл воды и 1,0 мкл 40 Е/мкл Xhol (Boehringer Mannheim). Расщепление проводили при 37°С в течение 1 часа. Реакцию останавливали инкубированием при 68°С в течение 15 минут с последующим осаждением этанолом, промыванием и сушкой, как описано выше, и ресуспендированием в 30 мкл воды.
Ресуспендированную кДНК нагревали до 65°С в течение 5 минут и охлаждали на льду, добавляли 4 мкл 5х красителя для нанесения на гель (Research Genetics Corp.) КДНК наносили на 1Х ТАЕ 0,8% низкоплавкой агарозы (SEA PLAQUE GTG™ low melt agarose; FMC Corp.) и подвергали электрофорезу. Примесные адаптеры и кДНК длиной ниже 0,6 т.п.н. вырезали из геля. Электроды меняли местами, расплавленную агарозу добавлялии для заполнения колодцев, буфер заменяли и кДНК подвергали электрофорезу до ее концентрирования вблизи начала дорожки. Зону геля, содержащую концентрированную кДНК, вырезали и помещали в микрофужную пробирку и агарозу расплавляли нагреванием до 65°С в течение 15 минут. После уравновешивания пробы до 45°С добавляли 2 мкл 1 Е/мкл бета-агарозы I (Biolabs Inc.) и смесь инкубировали в течение 90 минут при 45°С для расщепления агарозы. После инкубирования 1/10 объема 3 М Na-ацетатного буфера добавляли к пробе и смесь инкубировали на льду в течение 15 минут. Пробу центрифугировали при 14000 × g в течение 15 минут при комнатной температуре для удаления нерасщепленной агарозы, кДНК осаждали этанолом, промывали в 70% этаноле, сушили на воздухе и ресуспендировали в 40 мкл воды.
Для определения оптимального отношения кДНК к вектору несколько лигирований собирали и электропорировали. Вкратце, 2 мкл 5х буфера для лигазы Т4 (Life Technologies), 1 мкл 10 мМ АТФ. 1 мкл pZP7NX, расщепленной EcoRl-Xhol, 1 II ДНК-лигазы Т4 разбавляли до 0,25 Е/мкл (Life Technologies) воды до 10 мкл и 0,5, 1,2 или 3 мкл кДНК смешивали в 4 отдельных лигированиях, инкубировали при 22°С в течение 4 часов, 68°С в течение 20 минут, осаждали смесью ацетат натрия-этанол, промывали, сушили и ресуспендировали в 10 II. Один микролитр каждого лигирования электропорировали в 40 мкл DH10b ElectroMAX™-электрокомпетентных бактерий (Life Technologies) с использованием 0,1 см-кюветы (Biorad) и Genepulser, контроллера импульсов™ (Biorad), установленного на 2,5 кВ, 251 Ф, 200 Ом. Эти клетки немедленно ресуспендировали в 1 мл бульона SOC (Manniatis et al., supra) с последующим добавлением 500 Ч смеси 50% глицерин-SOC в качестве консерванта. Эти "исходные глицериновые растворы" замораживали в виде нескольких аликвот при -70°С. Аликвоту каждого оттаивали и высевали серийно на чашки с LB-агаром, дополненным ампициллином при 100 мкг/мл. Числа колоний показали, что оптимальное отношение CD3+ кДНК к вектору pZP7NX было 1 мкл к 45 нг; такое лигирование давало 4,5 млн первичных клонов.
Для цели скрининга этой библиотеки с использованием теста пролиферации на основе BaF3-zalpha11 (пример 5) глицериновые исходные растворы, описанные выше, разбавляли в жидкие культуры 100 или 250 клонов на пул в микротитрационных планшетах с глубокими лунками, выращивали 24 часа при 37°С со встряхиванием и плазмиду выделяли с использованием набора Qiagen в соответствии с инструкциями изготовителя. Такую ДНК затем трансфицировали в клетки ВНК, мреду кондиционировали 72 часа и помещали на клетки 5К BaF3-zalpha11 и выдерживали в течение 72 часов, после чего пролиферацию оценивали с использованием флуоресцентного анализа с Alamar blue (пример 5В и пример 2В).
Для цели скрининга этой библиотеки клонированием с секреторной ловушкой комплексная амплифицированная форма библиотеки требовалась для трансфекции клеток COS-7. Около 4,8 миллионов клонов высевали на 110 чашек 15 см с LB-агаром, дополненным 100 мкг/мл ампициллина, 10 мкг/мл метициллина. После выращивания чашек в течение ночи при 37°С бактерии собирали выскребанием и осаждали. Плазмидную ДНК экстрагировали из осажденных бактерий с использованием Nucleobond-giga™ (Clontech) в соответствии с инструкциями изготовителя. Затем эту плазмиду использовали для трансфекции клеток COS-7 (ATCC No. CRL 1651) на предметных стеклах и подвергали скринингу с использованием способа секреторной ловушки, описанного ниже (пример 12).
Пример 7
Экспрессионное клонирование zalpha11-лиганда человека
Глицериновые исходные растворы из библиотеки активированных CD3+ отобранных клеток человека (пример 6) добавляли к SuperBroth II™ (Becton Dickinson, Cockeysville, MD) +0,1 мг/мл ампициллина (амп) в концентрации 250 клеток на 800 микролитров. Е. coli давали уравновешиваться в течение 24 часов при комнатной температуре. Во время инокуляции 400 микролитров высевали на чашки с LB + амп для определения фактического титра инокуляции. Спустя 24 часа чашки считали и затем конечную концентрацию SuperBroth II™ + Е. coli доводили таким образом, что конечная концентрация была 250 клеток на 1,2 мл. Инокулировали 3 раза по 2 л с получением в целом 6 литров. Затем эту среду высевали в 96-луночные блоки с глубокими лунками (Qiagen). Засев производили на 8-канальном разливающем устройстве Q-Fill2™ (Genetix, Christchurch, Dorset, UK). E. coli выращивали в течение ночи при 37°С с встряхиванием при 250 об/мин на многоярусном шейкере New Brunswick Scientific Innova 4900 multi-tier environment shaker. E. coli откручивали из раствора при 3000 об/мин с использованием центрифуги Бекман GS-6KR. Осадки E. coli замораживали при -20°С или использовали в свежем виде перед получением минипрепаратов плазмидной ДНК. Каждый осадок содержит приблизительно 250 кДНК-клонов из библиотеки CD3+ отобранных клеток человека.
Затем эти пулы 250 кДНК-клонов готовили с использованием набора QIAprep ™ 96 Turbo Miniprep (Qiagen). Плазмидную ДНК элюировали с использованием 125 мкл ТЭ (10 мМ Трис рН 8, 1 мМ ЭДТА). Затем эту плазмидную ДНК использовали для трансфекции клеток ВНК.
Трансфекция ВНК
Клетки ВНК высевали в 96-луночные планшеты для культуры ткани при плотности 12000 клеток на лунку в объеме 100 мкл на лунку. Культуральной средой была DMEM (Gibco BRL), 5% инактивированная нагреванием фетальная телячья сыворотка, 2 мМ L-глутамин (Gibco BRL), 1x PSN (Gibco BRL), 1 мМ натрий-пируват (Gibco BRL).
На следующий день клетки ВНК промывали один раз 100 мкл SFA. SFA представляет собой бессывороточную среду, которая является DMEM/F12 (Gibco BRL), 2 мМ GlutaMax™ (Gibco BRL), 1 мМ натрий-пируват, 10 мкг/мл трансферрина, 5 мкг/мл инсулина, 10 мкг/мл фетуина, 2 мкг/мл селена, 25 мМ HEPES (Gibco BRL), 100 мкМ не-незаменимые аминокислоты (Gibco BRL).
Смесь ДНК/Липофектамин™ готовят следующим образом: 2,2 мкл реагента Липофектамин™ (Gibco BRL) объединяют с 102,8 мкл SFA при комнатной температуре; затем добавляют приблизительно 5 мкл плазмидной ДНК (200 нг/мкл) к Липофектамин™/8РА с образованием смеси ДНК/Липофектамин™, которую инкубируют при комнатной температуре в течение 30 минут. SFA удаляли из клеток ВНК и клетки инкубировали с 50 мкл смеси ДНК/Липофектамин™ в течение 5 часов при 37°С с 5% CO2. Пятьдесят мкл смеси ДНК/Липофектамин™ добавляли к каждой из двух лунок клеток ВНК, так что трансфекции проводили в двух повторностях.
После инкубирования клеток ВНК со смесью ДНК/Липофектамин™ в течение 5 часов смесь ДНК/Липофектамин™ удаляли и добавляли 100 мкл культуральной среды. Клетки инкубировали в течение ночи, среду удаляли и заменяли 100 мкл культуральной среды. После культивирования клеток в течение 72 часов кондиционированную среду удаляли, замораживали при -80°С в течение минимально 20 минут, оттаивали и затем 50 мкл анализировали в тесте пролиферации zalpha11/BaF3, описанном в примере 2В и примере 5, для идентификации пулов 250 клонов с активностью лиганда.
Тридцать пять 96-луночных планшетов подвергали скринингу в одном анализе. Это представляло приблизительно 250 кДНК на лунку или всего 840000 кДНК. Из них кондиционированную среду из 54 лунок (представляющую 250 кДНК на лунку) тестировали как положительную в тесте пролиферации. Кондиционированную среду из этих положительных пулов повторно тестировали во втором тесте (секреторной ловушки) с растворимым рецептором или без него (см. пример 12). Растворимый zalpha11CEE-рецептор (пример 10А) использовали в конечной концентрации около 1 мкг/мл). Для всех 54 позитивных клонов по существу вся активность нейтрализовалась добавлением растворимого zalpha11-рецептора, что свидетельствует о том, что эти пулы содержали кДНК zalpha11-лиганда. Четыре положительных клона выбирали для разрушения колоний и выделения отдельной кДНК, которая кодирует zalpha11-лиганд. Этими клонами были 45С5, 46G11, 40Н12 и 60А1.
Для каждого из этих 4 пулов 1 мкл ДНК использовали для трансформации ElectroMax™ клеток DH10B (Gibco BRL) злектропорацией. Трансформанты высевали на LB + амп (100 мкг/мл) + метициллин (10 мкг/мл) с получением отдельных колоний. Для каждого электропорированного пула 960 индивидуальных колоний выскребали зубочисткой в десять 96-луночных планшетов, содержащих 1,2 мл SuperBroth II™ на лунку. Эти планшеты были пронумерованы №1-10 для каждого из разбитых пулов (45С5, 46G11, 401-112 и 60А1). Их культивировали в течение ночи и плазмидную ДНК получали в виде минипрепарата, как описано выше. Для 46G11, 40Н12 и 60А1 плазмидную ДНК из чашек с разрушенными колониями трансфицировали в клетки ВНК, как описано выше.
Для 45С5 использовали протокол "быстрого прослеживания" для ускорения идентификации кДНК zalpha11-лиганда. Клетки ВНК трансфицировали плазмидной ДНК из чашек с разрушенными колониями, как описано выше, смесь ДНК/Липофектамин™ удаляли после 5 часов инкубации и добавляли культуральную среду. Поскольку трансфекции производили в двух повторностях, культуральную среду собирали на следующий день после 24 часов из одной из трансфицированных чашек и собирали на следующий день после 48 часов из другой трансфицированной чашки. Кондиционированную среду 24 часов анализировали, как описано выше, на активность zalpha11-лиганда с использованием теста пролиферации, как описано выше.
Плазмидную ДНК объединяли из чашек с разрушенными колониями 45С5 №1-4 и анализировали на связывание растворимого zalpha11-рецептора с его лигандом по протоколу "секреторной ловушки" (см. пример 12, ниже). Восемь положительных клонов идентифицировали из 384 последовательностей в целом. Результаты из теста пролиферации подтвердили активность zalpha11-лиганда и они коррелировали с результатами теста секреторной ловушки (см. пример 12). Параллельно плазмидную ДНК, полученную в виде минипрепарата из чашек №1-4 разрушенного пула 45С5, секвенировали для определения последовательности ДНК каждого из 384 клонов.
Несколько клонов, которые были идентифицированы как положительные в тестах пролиферации и секреторной ловушки, также секвенировали с использованием следующих праймеров: ZC14063 (SEQ ID NO:28), ZC7764a (SEQ ID NO:38), ZC7764b (SEQ ID NO:39), ZC22034 (SEQ ID NO:40) и ZC22035 (SEQ ID NO:41). Полинуклеотидная последовательность zalpha11-лиганда была полноразмерной (SEQ ID NO:1), и ее соответствующая аминокислотная последовательность показана в (SEQ ID NO:2).
Пример 8
Конструирование экспрессирующих векторов млекопитающих, которые экспрессируют растворимые рецепторы zalpha11: zalpha11CEE, zalpha11CFLG, zalpha11CHIS и zalpha11-Fc4
А. Конструирование экспрессирующего вектора zalpha11 млекопитающих, содержащего zalpha11CEE. ZaIphaUCFLG, zalpha11CHIS
Получали экспрессирующий вектор для экспрессии растворимого внеклеточного домена полипептида zalpha11, pC4zalpha11CEE, причем эта конструкция предназначена для экспрессии полипептида zalpha11, содержащего предсказанный инициирующий метионин и укороченного вблизи предсказанного трансмембранного домена, имеющего С-концевую Glu-Glu-метку (SEQ ID NO:29).
ПЦР-генерируемый фрагмент ДНК zalpha11 700 п.н. создавали с использованием ZC19931 (SEQ ID NO:30) и ZC19932 (SEQ ID NO:31) в качестве ПЦР-праймеров для добавления сайтов рестрикции Asp718 и BamHI. Плазмиду, содержащую кДНК zalpha11-рецептора (SEQ ID NO:7), использовали в качестве матрицы. ПЦР-амплификацию фрагмента zalpha11 проводили следующим образом: двадцать пять циклов при 94°С в течение 0,5 минут; пять циклов при 94°С в течение 10 секунд, 50°С в течение 30 секунд, 68°С в течение 45 секунд; затем выдерживание при 4°С. Реакцию очищали экстракцией смесью хлороформ/фенол и осаждением изопропанолом и расщепляли Asp718 и BamHI (Gibco BRL) согласно инструкциям изготовителя. Полосу предсказанного размера, 700 п.н., визуализировали при помощи электрофореза на 1% агарозном геле, вырезали и эту ДНК очищали с использованием системы очистки Qiaexll™ (Qiagen) согласно инструкциям изготовителя.
Вырезанную ДНК субклонировали в плазмиду рС4ЕЕ, которая была разрезана BamHI и Asp718. Экспрессирующий вектор pC4zalpha11CEE использует сигнальный пептид нативного zalpha11 и присоединяет Glu-Glu-метку (SEQ ID NO:29) к С-концу полинуклеотидной последовательности, кодирующей полипептид zalpha11. Плазмида рС4ЕЕ является экспрессирующим вектором млекопитающих, содержащим экспрессионную кассету, имеющую мышиный промотор металлотионеина-1, множественные сайты рестрикции для встраивания кодирующих последовательностей, стоп-кодон и терминатор гормона роста человека. Эта плазмида имеет также начало репликации Е. coli, экспрессионную единицу селектируемых маркеров млекопитающих, имеющую промотор, энхансер и начало репликации SV40, ген DHFR и терминатор SV40.
Приблизительно 30 нг рестрикционно отщепленной zalpha11-вставки и приблизительно 12 нг расщепленного вектора лигировали в течение ночи при 16°С. Один микролитр каждой реакции лигирования независимо электропорировали в DH10B-компетентные клетки (Gibco BRL, Gaithersburg, MD) согласно инструкциям изготовителя и высевали на LB-планшеты, содержащие 50 мг/мл ампициллина, и инкубировали в течение ночи. Колонии подвергали скринингу при помощи рестрикционного анализа ДНК, полученных из 2 мл жидких культур индивидуальных колоний. Последовательность вставки положительных клонов подтверждали анализом последовательности. Крупномасштабное получение плазмиды проводили с использованием набора QIAGEN® Maxi prep kit (Qiagen) согласно инструкциям изготовителя.
Тот же самый процесс использовали для получения растворимых рецепторов zalpha11 с С-концевой His-меткой, состоящей из 6 остатков His подряд, и С-концевой FLAG-меткой (SEQ ID NO:37), zalpha11CFLAG. Для конструирования этих конструкций вышеуказанный вектор имеет либо HIS, либо Р1-АО®-метку вместо glu-glu-метки (SEQ ID NO:29).
В. Экспрессионная конструкция млекопитающих растворимого zalpha11-рецептора. zalpha11-Fc4
Экспрессионную плазмиду, содержащую весь полинуклеотид или его часть, кодирующую zalpha11, конструировали гомологичной рекомбинацией. Внеклеточный домен zalpha11-рецептора сливали с Fc-районом, полученным из IgG человека, он был назван "Fc4" (SEQ ID NO:33) и содержит мутацию, так что он уже не связывается с Fc-рецептором. При помощи ПЦР выделяли фрагмент кДНК zalpha11, который включает в себя полинуклеотидную последовательность из внеклеточного домена zalpha11-рецептора. Два праймера использовали в получении этого фрагмента zalpha11: (1) каждый из праймеров для ПЦР включает в себя от 5' к 3' концу: 40 п.н. фланкирующей последовательности вектора (5' от вставки) и 17 п.н., соответствующих 5'-концу внеклеточного домена zalpha11 (SEQ ID NO:32); и (2) 40 п.н. 5'-конца полинуклеотидной последовательности Fc4 (SEQ ID NO:33) и 17 п.н., соответствующих 3'-концу внеклеточного домена zalpha11 (SEQ ID NO:34). Фрагмент Fc4 для слияния с zalpha11 генерировали при помощи ПЦР сходным образом. Двумя праймерами в получении фрагмента Fc4 были: (1) 5'-праймер, состоящий из 40 п.н. последовательности из 3'-конца внеклеточного домена zalpha11 и 17 п.н. 5'-конца Fc4 (SEQ ID NO:35) и (2) 3'-праймер, состоящий из 40 п.н. векторной последовательности (3' от вставки) и 17 п.н. 3'-конца Fc4 (SEQ ID NO:36).
ПЦР-амплификацию каждой из реакций, описанных выше, проводили следующим образом: один цикл при 94°С в течение 2 минут; двадцать пять циклов при 94°С в течение 30 секунд, 60°С в течение 30 секунд, 72°С в течение 1 минуты; один цикл при 72°С в течение 5 минут; с последующим выдерживанием при 4°С. Десять мкл из 100 мкл ПЦР-реакции подвергали электрофорезу на 0,8% LMP-агарозном геле (Seaplaque GTG) с 1 × ТВЕ-буфером для анализа. Остальные 90 мкл ПЦР-реакции осаждали добавлением 5 мкл 1 М NaCl и 250 мкл абсолютного этанола. Используемый экспрессирующий вектор получали из плазмиды pCZR199, полученной из pZP9 (ATCC депозит №98668) и разрезали Smal (BRL). Экспрессирующий вектор был произведен из плазмиды pCZR199 и представляет собой экспрессирующий вектор млекопитающих, содержащий экспрессионную кассету, имеющую немедленно ранний промотор CMV, консенсусный интрон из вариабельной области локуса тяжелой цепи мышиного иммуноглобулина, множественные сайты рестрикции для встраивания кодирующих последовательностей, стоп-кодон и терминатор гормона роста человека. Этот экспрессирующий вектор имеет также начало репликации Е. coli, экспрессионную единицу селектируемых маркеров млекопитающих, имеющую промотор, энхансер и начало репликации SV40, ген DHFR и терминатор SV40. Этот используемый экспрессирующий вектор конструировали из pCZR199 заменой промотора металлотионеина немедленно ранним промотором CMV.
100 мкл компетентных дрожжевых клеток (S. cerevisiae) объединяли с 10 мкл, содержащими приблизительно 1 мкг каждой из вставок zalpha11 и Fc4 и 100 нг расщепленного Smal (BRL) экспрессирующего вектора, и переносили в кювету для электропорации 0,2 см. Смеси дрожжи/ДНК подвергали электрошокам при 0,75 кВ (5 кВ/см), "неопределенном" числе Ом, 25 мкФ. В каждую кювету добавляли 600 мкл 1,2 М сорбита и дрожжи высевали в двух аликвотах по 300 мкл на две чашки URA-D и инкубировали при 30°С.
Спустя приблизительно 48 часов Ura+ дрожжевые трансформанты из отдельной чашки ресуспендировали в 1 мл H2O и откручивали быстро для осаждения дрожжевых клеток. Клеточный осадок ресуспендировали в 1 мл лизисного буфера (2% Тритон Х-100, 1% ДСН, 100 мМ NaCl, 10 мМ Трис, рН 8,0, 1 мМ ЭДТА). 500 мкл лизисной смеси добавляли в пробирку Эппендорфа, содержащую 300 мкл промытых кислотой стеклянных гранул и 200 мкл смеси фенол/хлороформ, встряхивали на вортексе два или три раза с интервалами 1 мин с последующим 5-минутным откручиванием в центрифуге Эппендорфа при максимальной скорости. Триста микролитров водной фазы переносили в свежую пробирку и ДНК осаждали 600 мкл этанола (EtOH) с последующим центрифугированием в течение 10 минут при 4°С. Осадок ДНК ресуспендировали в 100 мкл Н2O.
Трансформацию электрокомпетентных клеток Е. coli (DH10B, Gibco BRL) проводят с 0,5-2 мл препарата дрожжевой ДНК и 40 мкл клеток DH10B. Клетки подвергали электроимпульсам при 2,0 кВ, 25 мФ и 400 Ом. После электропорации 1 мл SOC (2% Бакто™ триптон (Difco, Detroit, Ml), 0,5% дрожжевой экстракт (Difco), 10 мМ NaCl, 2,5 мМ KCI, 10 мМ MgCb, 10 мМ MgSO4, 20 мМ глюкоза) высевали в аликвотах 250 мкл на четыре LB АМР чашки (LB-бульон (Lennox), 1,8% Бактоагар (Difco), 100 мг/л ампициллина).
Индивидуальные клоны, несущие правильную экспрессионную конструкцию для zalpha11-Fc4, идентифицировали рестрикционным расщеплением для подтверждения присутствия вставки zalpha11-Fc4 и для подтверждения, что различные последовательности ДНК были правильно соединены друг с другом. Вставку положительных клонов подвергали анализу последовательности. При крупномасштабном получении плазмидную ДНК выделяют с использованием набора QIAGEN Maxi kit (Qiagen) согласно инструкциям изготовителя.
Пример 9
Трансфекция и экспрессия полипептидов растворимого zalpha11-рецептора
А. Экспрессия в млекопитающих растворимого zalpha11-рецептора zalpha11CEE. Zalpha11CFLG и zalpha11CHIS
Клетки ВНК 570 (АТСС No. CRL-10314), пассаж 27, высевали в чашки при 1,2×106 клеток на лунку (6-луночная чашка) в 800 мкл бессывороточной (БС) среды DMEM (DMEM, Gibco/BRL High Glucose), (GIBCO BRL, Gaithersburg, MD). Эти клетки трансфицировали экспрессионными плазмидами, содержащими zalpha11CEE, zalpha11CFLG или zalpha11CHIS, описанными выше (см. пример 8), с использованием Липофектина™ (Gibco BRL) в бессывороточной (БС) среде DMEM. Три микрограмма zalpha11CEE, zalpha11CFLG или zalpha11CHIS отдельно разводили в пробирки на 1,5 мл до общего объема 100 мкл БС-DMEM. В отдельных пробирках 15 мкл Липофектина™ (Gibco BRL) смешивали с 100 мкл БС-DMEM. Смесь с Липофектином™ инкубировали при комнатной температуре в течение 30-45 минут, затем добавляли смесь ДНК и давали инкубироваться приблизительно 10-15 минут при комнатной температуре.
Всю смесь ДНК:Липофектин™ добавляли к посеянным клеткам и равномерно распределяли на них. Чашки инкубировали при 37°С в течение приблизительно пяти часов, затем переносили в отдельные чашки MAXI 150 мм в конечном объеме 30 мл DMEM/15% фетальная телячья сыворотка (ФТС) (Hyclone, Logan, UT). Чашки инкубировали при 37°С, 5% CO2 в течение ночи и смесь ДНК:Липофектин™ заменяли свежей селективной средой (5% ФТС/DMEM с 1 мкМ метотрексата (МТХ) на следующий день.
Приблизительно при 10-12 днях после трансфекции чашки промывали 10 мл БС-DMEM. Промывочную среду отсасывали и заменяли 7,25 мл бессывороточной DMEM. Затем стерильные тефлоновые сетки (Spectrum Medical Industries, Los Angeles, CA), предварительно пропитанные в БС-DMEM, помещали на колонии клональных клеток. Затем на эту сетку помещали стерильный нитроцеллюлозный фильтр, предварительно пропитанный в БС-DMEM. Метки для ориентации на нитроцеллюлозе переносили на чашку с культурой. Затем эти чашки инкубировали в течение 5-6 часов в термостате при 37°С, 5% CO2.
После инкубирования фильтры/сетки удаляли и среду отсасывали и заменяли средой 5% ФТС/DMEM с 1 мкМ МТХ. Затем фильтры блокировали в 10% нежирном сухом молоке/буфере Вестерн А (Вестерн А: 50 мМ Трис, рН 7,4, 5 мМ ЭДТА, 0,05% NP-40, 150 мМ NaCl и 0,25% желатин) в течение 15 минут при комнатной температуре на ротационном шейкере. Затем фильтры инкубировали с антителом против Glu-Glu, антителом против FLAG® или конъюгатами анти-His-HRP (пероксидаза хрена) соответственно в 2,5% нежирном сухом молоке/буфере Вестерн А в течение одного часа при комнатной температуре на ротационном шейкере. Затем фильтры промывали три раза при комнатной температуре буфером Вестерн А в течение 5-10 минут на одну промывку. Фильтры проявляли реагентом ultra ECL (Amersham Corp., Arlington Heights, IL) в соответствии с указаниями изготовителя и визуализировали на приборе для получения изображений Lumi-lmager (Roche Corp.).
Положительные экспрессирующие клональные колонии механически выскребали в 12-луночные чашки в одном мл 5% ФТС/DMEM с 5 мкМ МТХ, затем выращивали до конфлюэнтности. Затем пробы кондиционированных сред тестировали на уровни экспрессии при помощи электрофореза в ДСН-ПААГ и Вестерн-анализа. Три наиболее сильно экспрессирующих клона для каждой конструкции выскребали; два из трех замораживали в качестве резерва, а один размножали для тестирования на микоплазму и крупномасштабного посева в клеточной фабрике.
В. Экспрессия растворимого zalpha11-рецептора zalpha11-Fc4 в клетках млекопитающих
Клетки ВНК 570 (АТСС No. CRL-10314) высевали в чашки 10 см для культуры ткани и давали им расти до приблизительно 50-70% конфлюентности в течение ночи при 37°С, 5% CO2, в среде DMEM/ФТС (DMEM, Gibco/BRL High Glucose, (GIBCO BRL, Gaithersburg, MD), 5% фетальной телячьей сыворотке (Hyclone, Logan, UT), 1 мМ L-глутамине (JRH Biosciences, Lenexa, KS), 1 мМ пирувате натрия (Gibco BRL). Затем клетки трансфицировали плазмидой, содержащей zalpha11-Fc4 (см. пример 8), с использованием Липофектамина™ (Gibco BRL) в бессывороточной (БС) среде (DMEM, 10 мг/мл трансферрина, 5 мг/мл инсулина, 2 мг/мл фетуина, 1% L-глутамина и 1% пирувата натрия). Плазмиду, содержащую zalpha11-Fc4, разводили в пробирках на 15 мл до общего конечного объема 640 мкл БС-средой. 35 мл Липофектамина™ (Gibco BRL) смешивали с 605 мкл БС-среды. Эту смесь с Липофектамином™ добавляли к смеси ДНК и давали инкубироваться в течение приблизительно 30 минут при комнатной температуре. Пять миллилитров БС-среды добавляли к смеси ДНК: Липофектамин™. Клетки промывали один раз 5 мл БС-среды, отсасывали и добавляли смесь ДНК: Липофектамин™. Клетки инкубировали при 37°С в течение пяти часов, затем в каждую чашку добавляли 6,4 мл среды DMEM/10% ФТС, 1% PSN. Чашки инкубировали при 37°С в течение ночи и смесь ДНК: Липофектамин™ заменяли свежей средой 5% ФТС/DMEM на следующий день. В день 2 после трансфекции клетки разделяли в селективную среду (среду DMEM/ФТС, описанную выше, с добавлением 1 мМ метотрексата (Sigma Chemical Co., St. Louis, МО)) в чашках 150 мм при разведении 1:1, 1:20 и 1:50. Среду на этих клетках заменяли свежей селективной средой в день 5 после трансфекции. Приблизительно в день 10 после трансфекции две культуральные чашки 150 мм с устойчивыми к метотрексату колониями из каждой трансфекции трипсинизировали и клетки объединяли и высевали в колбу Т-162 и переносили в крупномасштабную культуру.
Пример 10
Очистка растворимых zalpha11-рецепторов из клеток ВНК 570
А. Очистка полипептида zalpha11CEE из ВНК 570
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки полипептида zalpha11, содержащего С-концевые метки Glu-Glu (ЕЕ). Тридцать литров кондиционированной среды из клеточной фабрики концентрировали до 1,6 литров при помощи спирального картриджа Amicon S10Y3 на ProFlux A30. Раствор протеазного ингибитора добавляли к концентрированным 1,6 л кондиционированной среды клеточной фабрики от трансфицированных клеток ВНК 570 (см. пример 9) до конечных концентраций 2,5 мМ этилендиаминотетрауксусной кислоты (ЭДТА, Sigma Chemical Со. St. Louis, МО), 0,003 мМ лейпептина (Boehringer Mannheim, Indianapolis, IN), 0,001 мМ пепстатина (Boehringer Mannheim) и 0,4 мМ Pefabloc (Boehringer Mannheim). Пробы извлекали для анализа и общий объем замораживали при -80°С до начала очистки. Концентрации общего целевого белка концентрированной кондиционированной среды клеточной фабрики определяли при помощи электрофореза в ДСН-ПААГ и Вестерн-блот-анализа с антителами против ЕЕ, конъюгированными с HRP (пероксидазой хрена).
100 мл субстрата для колонки анти-EE-G-Sepharose (приготовленного, как описано ниже) выливали в стеклянную колонку Waters АР-5, 5 см × 10 см. Колонку упаковывали проточно и уравновешивали на BioCad Sprint (PerSeptive BioSystems, Framingham, MA) забуференным фосфатом солевым раствором (ЗФР) рН 7,4. Концентрированную кондиционированную среду клеточной фабрики оттаивали, стерильно фильтровали с фильтром 0,2 микрон, рН доводили до 7,4, затем наносили на колонку в течение ночи со скоростью тока 1 мл/мин. Колонку промывали 10 колоночными объемами (КО) зафуференного фосфатом солевого раствора (ЗФР, рН 7,4), затем элюировали с подачей элюента через пробку 200 мл ЗФР (рН 6,0), содержащего 0,5 мг/мл ЕЕ-пептида (Anaspec, San Jose, CA), при скорости тока 5 мл/мин. Используемый ЕЕ-пептид имеет последовательность EYMPME (SEQ ID NO:29). Колонку промывали 10 колоночными объемами (КО) ЗФР, затем элюировали 5 колоночными объемами 0,2 М глицина, рН 3,0. рН элюируемой глицином колонки доводили до 7,0 2 колоночными объемами 5Х ЗФР, затем уравновешиали в ЗФР (рН 7,4). Фракции по 5 мл собирали на протяжении всей элюционной хроматографии и поглощение при 280 и 215 нм подвергали мониторингу; проходящие через колонку и промывочные пулы также сохраняли и анализировали. Фракции пика элюции ЕЕ-полипептида анализировали на целевой белок электрофорезом в ДСН-ПААГ с окрашиванием серебром и Вестерн-блоттингом с конъюгированным антителом анти-EE-HRP. Представляющие интерес фракции элюции полипептида объединяли и концентрировали с 60 до 5,0 мл с использованием центрифужного концентратора с мембраной, отсекающей молекулярную массу 10000 Дальтон (Millipore, Bedford, MA), согласно инструкциям изготовителя.
Для отделения zalpha11CEE от других соочищающихся белков концентрированные слитые фракции элюции полипептида подвергали очистке на POROS HQ-50 (сильной анионообменной колонке из PerSeptive BioSystems, Framingham, MA) при рН 8,0. Колонку 1,0×6,0 см заливали и проточно упаковывали на BioCad Sprint. Колонку загружали противоионом и затем уравновешивали в 20 мМ Трис рН 8,0 (Трис(гидроксиметиламинометан)). Пробу разводили 1:13 (для уменьшения ионной силы ЗФР), затем наносили на колонку POROS HQ при скорости тока 5 мл/мин. Колонку промывали 10 колоночными объемами (КО) 20 мМ Трис рН 8,0, затем элюировали 40 колоночными объемами градиента 20 мМ Трис/1 М хлорид натрия (NaCl) при скорости тока 10 мл/мин. Фракции по 1,5 мл собирали на протяжении всей хроматографии и поглощение при 280 и 215 нм подвергали мониторингу. Фракции пика элюции анализировали электрофорезом в ДСН-ПААГ с окрашиванием серебром. Представляющие интерес фракции объединяли и концентрировали до 1,5-2 мл с использованием центрифужного концентратора с мембраной, отсекающей молекулярную массу 10000 Дальтон (Millipore, Bedford, MA), согласно инструкциям изготовителя.
Для отделения полипептида zalpha11CEE от свободного пептида ЕЕ и любых примесных соочищающихся белков, объединенные концентрированные фракции подвергали хроматографии на колонке 1,5×90 см Sephadex S200 (Pharmacia, Piscataway, NJ), уравновешенной и загруженной в ЗФР при скорости тока 1,0 мл/мин, при помощи BioCad Sprint. Фракции по 1,5 мл собирали на протяжении всей хроматографии и поглощение при 280 и 215 нм подвергали мониторингу. Фракции пика элюции характеризовали при помощи электрофореза в ДСН-ПААГ с окрашиванием серебром и объединяли только наиболее чистые фракции. Этот материал представлял очищенный полипептид zalpha11CEE.
Этот очищенный материал подвергали наконец очистке на колонке 4 мл ActiClean Etox (Sterogene) для удаления любых оставшихся эндотоксинов. Пробу пропускали через уравновешенную ЗФР колонку под действием силы тяжести четыре раза, затем колонку промывали один раз 3 мл ЗФР и эту промывку объединяли с "очищенной" пробой. Затем этот материал стерильно фильтровали (0,2 микрон) и хранили при -80°С до приготовления аликвот.
На Вестерн-блотированных окрашенных Кумасси синим и серебром гелях электрофореза в ДСН-ПААГ полипептид zalpha11CEE был одной основной полосой со средней молекулярной массой приблизительно 50000 Дальтон. Подвижность этой полосы была одинаковой на восстанавливающем и на невосстанавливающем гелях.
Концентрацию белка очищенного материала определяли согласно анализу с использованием БСА (Pierce, Rockford, IL) и белок делили на аликвоты и хранили при -80°С в соответствии с обычными процедурами авторов изобретения. На IEF-гелях (изоэлектрическое фокусирование) белок перемещался с PI менее 4,5. Концентрация полипептида zalpha11CEE была 1,0 мг/мл.
Для получения анти-ЕЕ-Сефарозы объем слоя 100 мл протеин G-Сефарозы (Pharmacia, Piscataway, NJ) промывали 3 раза 100 мл ЗФР, содержащего 0,02% азида натрия с использованием фильтрующего элемента 0,45 микрон Nalgene на 500 мл. Гель промывали 6,0 объемами 200 мМ триэтаноламина, рН 8,2 (ТЭА, Sigma, St. Louis, МО) и добавляли равный объем раствора антител против ЕЕ, содержащего 900 мг антитела. После инкубирования в течение ночи при 4°С несвязавшееся антитело удаляли промыванием смолы 5 объемами 200 мМ ТЭА, как описано выше. Смолу ресуспендировали в 2 объемах ТЭА, переносили в подходящий резервуар и добавляли диметилпимилимидат-2НСl (Pierce, Rockford, IL), растворенный в ТЭА, до конечной концентрации 36 мг/мл G-Sepharose-геля. Гель качали при комнатной температуре в течение 45 минут и жидкость удаляли при помощи фильтрующего элемента, как описано выше. Затем неспецифические сайты на этом геле блокировали инкубированием в течение 10 минут при комнатной температуре с 5 объемами 20 мМ этаноламина в 200 мМ ТЭА. Затем гель промывали 5 объемами ЗФР, содержащего 0,02% азида натрия, и хранили в этом растворе при 4°С.
В. Очистка полипептида zalpha11 CFLAG из ВНК 570
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки полипептида zalpha11, содержащего С-концевые метки FLAG® (FLG) (Sigma-Aldrich Co.), Тридцать литров кондиционированной среды из клеточной фабрики концентрировали до 1,7 литров при помощи спирального картриджа Amicon S10Y3 на ProFlux A30. Раствор протеазного ингибитора добавляли к концентрированным 1,7 л кондиционированной среды клеточной фабрики от трансфицированных клеток ВНК 570 (см. пример 9) до конечных концентраций 2,5 мМ этилендиаминотетрауксусной кислоты (ЭДТА, Sigma Chemical Co. St. Louis, МО), 0,003 мМ лейпептина (Boehringer Mannheim, Indianapolis, IN), 0,001 мМ пепстатина (Boehringer Mannheim) и 0,4 мМ Pefabloc (Boehringer Mannheim). Пробы брали для анализа и общий объем замораживали при -80°С до начала очистки. Концентрации общего целевого белка кондиционированной среды клеточной фабрики определяли при помощи электрофореза в ДСН-ПААГ и Вестерн-блот-анализа с антителами против FLAG® (Kodak), конъюгированными с HRP (пероксидазой хрена). Колонку 125 мл анти-FLAG® М2-агарозного аффинного геля (SIGMA-Aldrich Co.) выливали в стеклянную колонку Waters АР-5, 5 см × 10 см. Колонку упаковывали проточно и уравновешивали на BioCad Sprint (PerSeptive BioSystems, Framingham, MA) забуференным фосфатом солевым раствором (ЗФР) рН 7,4. Концентрированную кондиционированную среду клеточной фабрики оттаивали, стерильно фильтровали с фильтром 0,2 микрон, рН доводили до 7,4, затем наносили на колонку в течение ночи со скоростью тока 1 мл/мин. Колонку промывали 10 колоночными объемами (КО) зафуференного фосфатом солевого раствора (ЗФР, рН 7,4), затем элюировали с подачей элюента через пробку 250 мл ЗФР (рН 6,0), содержащего 0,5 мг/мл FLAG® (Sigma-Aldrich Co.) пептида, при скорости тока 5 мл/мин. Используемый FLAG®-пептид имеет последовательность DYKDDDDK (SEQ ID NO:37). Колонку промывали 10 колоночными объемами (КО) ЗФР, затем элюировали 5 колоночными объемами 0,2 М глицина, рН 3,0. рН элюируемой глицином колонки доводили до 7,0 2 колоночными объемами 5Х ЗФР, затем уравновешивали в ЗФР (рН 7,4). Фракции по 5 мл собирали на протяжении всей элюционной хроматографии и поглощение при 280 и 215 нм подвергали мониторингу; проходящие через колонку и промывочные пулы также сохраняли и анализировали. Фракции пика элюции FLAG®-полипептида анализировали на целевой белок электрофорезом в ДСН-ПААГ с окрашиванием серебром и Вестерн-блоттингом с конъюгированным антителом анти-FLAG®-HRP. Представляющие интерес фракции элюции полипептида объединяли и концентрировали с 80 до 12 мл с использованием центрифужного концентратора с мембраной, отсекающей молекулярную массу 10000 Дальтон (Millipore, Bedford, MA), согласно инструкциям изготовителя.
Для отделения zalpha11CFLG от других соочищающихся белков, слитые фракции элюции полипептида подвергали очистке на POROS HQ-50 (сильной анионообменной колонке из PerSeptive BioSystems, Framingham, MA) при рН 8,0. Колонку 1,0×6,0 см заливали и проточно упаковывали на BioCad Sprint. Колонку загружали противоионом и затем уравновешивали в 20 мМ Трис рН 8,0 (Трис(гидроксиметиламинометан)). Пробу разводили 1:13 (для уменьшения ионной силы ЗФР), затем наносили на колонку POROS HQ-50 при скорости тока 5 мл/мин. Колонку промывали 10 колоночными объемами (КО) 20 мМ Трис рН 8,0, затем элюировали 40 колоночными объемами градиента 20 мМ Трис/1 М хлорид натрия (NaCl) при скорости тока 10 мл/мин. Фракции по 1,5 мл собирали на протяжении всей хроматографии и поглощение при 280 и 215 нм подвергали мониторингу, Фракции пика элюции анализировали электрофорезом в ДСН-ПААГ с окрашиванием серебром. Представляющие интерес фракции объединяли и концентрировали до 1,5-2 мл с использованием центрифужного концентратора с мембраной, отсекающей молекулярную массу 10000 Дальтон (Millipore, Bedford, МА), согласно инструкциям изготовителя.
Для отделения полипептида zalpha11CFLG от свободного пептида FLAG® и любых примесных соочищающихся белков объединенные концентрированные фракции подвергали хроматографии на колонке 1,5×90 см Sephacryl S200 (Pharmacia, Piscataway, NJ), уравновешенной и загруженной в ЗФР при скорости тока 1,0 мл/мин, при помощи BioCad Sprint. Фракции по 1,5 мл собирали на протяжении всей хроматографии и поглощение при 280 и 215 нм подвергали мониторингу. Фракции пика элюции характеризовали при помощи электрофореза в ДСН-ПААГ с окрашиванием серебром и объединяли только наиболее чистые фракции. Этот материал представлял очищенный полипептид zalpha11CFLG.
Этот очищенный материал подвергали наконец очистке на колонке 4 мл ActiClean Etox (Sterogene) для удаления любых оставшихся эндотоксинов. Пробу пропускали через уравновешенную ЗФР колонку под действием силы тяжести четыре раза, затем колонку промывали один раз 3 мл ЗФР и эту промывку объединяли с "очищенной" пробой. Затем этот материал стерильно фильтровали (0,2 микрон) и хранили при -80°С до приготовления аликвот.
На Вестерн-блотированных окрашенных Кумасси синим и серебром гелях электрофореза в ДСН-ПААГ полипептид zalpha11CFLG был одной основной полосой со средней молекулярной массой приблизительно 50000 Дальтон. Подвижность этой полосы была одинаковой на восстанавливающем и на невосстанавливающем гелях.
Концентрацию белка очищенного материала определяли согласно анализу с использованием БСА (Pierce, Rockford, IL) и белок делили на аликвоты и хранили при -80°С в соответствии с обычными процедурами авторов изобретения. На IEF-гелях (изоэлектрическое фокусирование) белок перемещался с PI менее 4,5. Концентрация полипептида zalpha11CFLG была 1,2 мг/мл.
С. Очистка полипептида zalpha11-Fc4 из трансфицированных клеток ВНК 570
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки полипептида zalpha11, содержащего С-концевое слияние с IgG/Fc человека (zalpha11-Fc4; примеры 8 и 9). 12000 мл кондиционированной среды от клеток ВНК 570, трансфицированных zalpha11-Fc4 (пример 9), фильтровали через стерилизующий фильтр 0,2 мм и затем дополняли раствором протеазных ингибиторов до конечных концентраций 0,001 мМ лейпептина (Boehringer Mannheim, Indianapolis, IN), 0,001 мМ пепстатина (Boehringer Mannheim) и 0,4 мМ Pefabloc (Boehringer Mannheim). Протеин G-Сефарозу (объем слоя 6 мл, Pharmacia Biotech) упаковывали и промывали 500 мл ЗФР (Gibco/BRL). Дополненную кондиционированную среду пропускали через колонку при скорости тока 10 мл/мин с последующим промыванием 1000 мл ЗФР (BRL/Gibco). Полипептид zalpha11-Fc4 элюировали из колонки 0,1 М глицином рН 3,5 и фракции по 2 мл собирали непосредственно в 0,2 мл 2 М триса рН 8,0 для доведения конечного рН до 7,0 в этих фракциях.
Элюированные фракции характеризовали при помощи электрофореза в ДСН-ПААГ и Вестерн-блоттинга с антителами против Fc человека (Amersham). Вестерн-блот-анализ гелей восстанавливающего электрофореза в ДСН-ПААГ выявил иммунореактивный белок приблизительно 80000 кДа во фракциях 2-10. Окрашенные серебром гели электрофореза в ДСН-ПААГ также выявили полипептид zalpha11-Fc4 80000 "Да во фракциях 2-10. Фракции 2-10 объединяли.
Концентрацию белка объединенных фракций определяли согласно анализу с использованием БСА (Pierce, Rockford, IL) и материал делили на аликвоты и хранили при -80°С в соответствии с обычными процедурами заявителей. Концентрация белка объединенных фракций была 0,26 мг/мл.
Пример 11
Тест с использованием растворимого zalpha11-рецептора zalpha11CEE, растворимых рецепторов zalpha11CFLG и zalpha11-Fc4 (мутанта) в конкурентном ингибиторном анализе
Клетки BaF3/-zalpha11 откручивали и промывали в не содержащей mlL-3 среде. Клетки откручивали и промывали 3 раза для гарантии удаления mlL-3. Затем клетки считали в гемоцитометре. Клетки высевали в 96-луночный планшет при 5000 клеток на лунку в объеме 100 мкл на лунку с использованием не содержащей mlL-3 среды.
Как кондиционированную среду от активации клеток селезенки обезьяны, так и CD3+ отобранные клетки человека, описанные в примере 5 выше, добавляли в отдельных экспериментах при концентрациях 50%, 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75% и 0,375%, с растворимыми zalpha11-рецепторами или без них (конструкциями СЕЕ, C-FLAG и Fc4; см. пример 9 и 10) при 10 мкг/мл. Общий объем теста был 200 мл.
Тест-планшеты инкубировали при 37°С, 5% CO2 в течение 3 дней, после чего добавляли Alamar Blue (Accumed) при 20 мкл на лунку. Планшеты опять инкубировали при 37°С, 5% СО2 в течение 24 часов. Планшеты считывали на планшет-ридере Fmax™ (Molecular Devices), как описано в примере 2. Результаты продемонстрировали полное ингибирование клеточного роста каждой из различных конструкций растворимых zalpha11-рецепторов при 10 мкг/мл, подтверждая, что этот фактор в каждой пробе был специфическим для zalpha11-рецептора.
Кривые титрования с разведениями этих растворимых рецепторов также определяли с использованием описанного выше теста. Как zalpha11CEE, так и zalpha11CFLG растворимые рецепторы zalpha11 были способны полностью ингибировать рост при такой низкой концентрации, как 20 нг/мл. Мутантный растворимый рецептор zalpha11-Fc4 был эффективным только при 1,5 мкг/мл.
Пример 12
Тест секреторной ловушки
Тест секреторной ловушки использовали для идентификации кДНК для zalpha11-лиганда. Положительные ДНК-пулы получали из экспрессионного клонирования, описанного в примере 7. Положительные ДНК-пулы 250 клонов трансфицировали в клетки ВНК в формате 96-луночного планщета и кондиционированную среду использовали в тесте пролиферации с использованием клеток BaF3/zalpha11, описанных в примерах 4 и 5. Несколько ДНК-пулов давали положительные активности, которые повторяли и нейтрализовали растворимыми zalpha11-рецепторами (см. пример 11).
Один из положительных ДНК-пулов, 45С5, трансфицировали в клетки COS-7 в 12-луночном формате с использованием способа с Липофектамином™, описанным ниже. Затем тест секреторной ловушки выполняли с использованием растворимых zalpha11-рецепторов (меченных на С-конце меткой Glu-Glu с биотинилированием или без биотинилирования; меченных на С-конце FLAG; или слияний Рс4-растворимый zalpha11-рецептор) (пример 9) для тестирования прямого связывания между потенциальным лигандом zalpha11-рецептора в пуле 45С5 и zalpha11-рецептором (см. ниже). Результат был положительным. Таким образом, ДНК пула 45С5 электропорировали в Е. coli и отдельные колонии выскребали в десять 96-луночных планшетов. Планшеты встряхивали при 37°С в течение 24 часов и затем минипрепараты ДНК (QiaPrep™ 96 Turbo Miniprep Kit; Qiagen) готовили в 96-луночном формате с использованием TomTech Quadra 9600. Затем плазмидную ДНК объединяли в формате рядов и столбцов, трансфицировали в клетки COS и затем определяли положительные пулы при помощи секреторной ловушки, как описано ниже.
Трансфекция клеток COS
Трансфекцию клеток COS выполняли следующим образом: Смесь 3 мкл объединенной ДНК и 5 мкл Липофектамина™ в 92 мкл бессывороточной среды DMEM (55 мг пирувата натрия, 146 мг L-глутамина, 5 мг трансферрина, 2,5 мг инсулина, 1 мкг селена и 5 мг фетуина в 500 мл DMEM), инкубировали при комнатной температуре в течение 30 минут и затем добавляли 400 мкл бессыворточной среды DMEM. Эту смесь 500 мкл добавляли на 1,5×105 клеток COS на лунку, высеянных на 12-луночном планшете для культуры ткани, и инкубировали в течение 5 часов при 37°С. 500 мкл 20% среды ФТС-DMEM (100 мл ФТС, 55 мг пирувата натрия и 146 мг L-глутамина в 500 мл DMEM) добавляли и инкубировали в течение ночи.
Тест секреторной ловушки
Тест секреторной ловушки выполняли следующим образом: среду отмывали от клеток при помощи ЗФР и затем фиксировали в течение 15 минут 1,8% формальдегидом в ЗФР. Затем клетки промывали TNT (0,1 М Трис-HCl, 0,15 М NaCl и 0,05% Твин-20 в H2O) и пропитывали 0,1% Тритоном-Х в ЗФР в течение 15 минут и опять промывали TNT. Клетки блокировали в течение 1 часа при помощи TNB (0,1 М Трис-HCl, 0,15 М NaCl и 0,5% блокирующзий реагент (NEN Renaissance TSA-Direct Kit) в H2O) и опять промывали TNT. При использовании биотинилированного белка клетки блокировали в течение 15-минутных инкубации с авидином и затем биотином (Vector Labs), с промыванием между ними TNT. В зависимости от того, какой растворимый рецептор использовали, клетки инкубировали в течение 1 часа с: (А) 1-3 мкг/мл слитого белка zalpha11 растворимый zаlрhа11-рецептор-Рс4 (пример 10); (В) 3 мкг/мл растворимого zalpha11-рецептора, меченного на С-конце FLAG, zalpha11CFLG (пример 10); (С) 3 мкг/мл растворимого zalpha11-рецептора, меченного на конце Glu-Glu, zalpha11CEE (пример 10); или (D) 3 мкг/мл биотинилированного растворимого zalpha11-рецептора zalpha11CEE в TNB. Затем клетки промывали TNT. В зависимости от того, какой растворимый рецептор использовали, клетки инкубировали в течение еще одного часа с: (А) разведенными 1:200 козьими антителами против человеческого Ig-HRP (Fc-специфическими); (В) разведенным 1:1000 M2-HRP; (С) разведенными 1:1000 антителами против Glu-Glu-HRP; или (D) разведенным 1:300 комплексом стрептавидин-HRP (набор NEN) в TNB. Опять клетки промывали TNT.
Положительное связывание детектировали флуоресцеин-тирамидным реагентом, разбавленным 1:50 в буфере для разведения (набор NEN) и инкубировали в течение 4-6 минут и промывали TNT. Клетки консервировали средой для монтировки препаратов Vectashield Mounting Media (Vector Labs Burlingame, CA), разбавленной 1:5 в TNT. Клетки визуализировали с использованием ПТС-фильтра на флуоресцентном микроскопе.
Пример 13
Хромосомное отнесение и определение места в хромосоме для гена zalpha11-лиганда
Ген для zalpha11-лиганда картировали на хромосоме 4 с использованием коммерчески доступной версии панели "Stanford G3 Radiation Hybrid Mapping Panel" (Research Genetics, Inc., Huntsville, AL). "Stanford G3 R.H. Panel" содержит ДНК из каждого из 83 радиационных (облученных) гибридных клонов целого генома человека, плюс две контрольные ДНК (RM-донор и А3-реципиент). Общественно доступный сервер WWW (http://shgc-www.stanford.edu) позволяет определить хромосомную локализацию маркеров.
Для картирования гена zalpha11-лиганда с панелью "Stanford G3 RH Panel" реакции по 20 мкл устанавливали в 96-луночном микротитрационном планшете (Stratagene, La Jolla, CA) и использовали в термоциклере "RoboCycler Gradient 96" (Stratagene). Каждая из этих 85 ПЦР-реакций состояла из 2 мкл 10Х KlenTaq PCR реакционного буфера (Clontech Laboratories, Inc., Palo Alto, CA), 1,6 мкл смеси dNTP (2,5 мМ каждый, PERKIN-ELMER, Foster City, CA), 1 мкл смыслового праймера ZC 22050 (SEQ ID NO:42), 1 мкл антисмыслового праймера ZC22051 (SEQ ID NO:43), 2 мкл "RediLoad" (Research Genetics, Inc., Huntsville, AL), 0,4 мкл 50Х смеси Advantage KlenTaq Polymerase (Clontech Laboratories, Inc.), 25 нг ДНК из индивидуального гибридного клона или контроля и ddH2O для общего объема 20 мкл. На реакции наслаивали равное количество минерального масла и их герметизировали. Условия ПЦР-циклера были следующими: 1 начальный цикл 4 минут денатурации при 94°С, 35 циклов 45 секунд денатурации при 94°С, 45 секунд отжига при 60°С и 1 минута и 15 секунд удлинения при 72°С; с конечным 1 циклом удлинения 7 минут при 72°С. Реакции разделяли электрофорезом на 2% агарозном геле (Life Technologies, Gaithersburg, MD).
Результаты показали сцепление гена zalpha11-лиганда относительно каркасного маркера SHGC-12342 IL-2 с LOD-баллом >12 на расстоянии 6 cR 10000 (приблизительно 180 т.п.н.) от маркера. Применение окружающих маркеров помещает ген zalpha11-лиганда в район 4q27 на интегрированной карте LDB хромосомы 4 (The Genetic Location Database, University of Southhampton, сервер WWW: http://cedar.geneties.soton.ac.uk/public html/.
Пример 14
Идентификация и клонирование мышиного zalpha11-лиганда
Использование EST-последовательности для получения полноразмерного мышиного zalpha11-лиганда
А. EST-последовательность мышиного zalpha11-лиганда
Поиском в базе данных с "ДНК-последовательностью человеческого zalpha11-лиганда (SEQ ID NO:1) в качестве запроса, мышиная EST (маркерная экспрессирующаяся последовательность) (EST1483966) была идентифицирована в качестве потенциальной частичной последовательности для мышиного zalpha11-лиганда. EST1483966 представляет мышиный геномный фрагмент, в котором пептидная последовательность, полученная из двух потенциальных экзонов, имеет общую высокую идентичность последовательности с пептидным сегментом человеческого zalpha11-лиганда (аминокислота 13 (lie) - аминокислота 80 (Gin) SEQ ID NO:2).
В. ПЦР-скрининг мышиной marathon-кДНК-панели
Одиннадцать мышиных проб Marathon-кДНК (Clontech) подвргали скринингу при помощи ПЦР, описанному ниже. Мышиные пробы marathon кДНК получали из тканей мозга, поджелудочной железы, почки, плаценты, слюнных желез, кожи, яичка, матки, эмбриона и селезенки. Авторы готовили их у себя в лаборатории с использованием набора для амплификации Marathon™-кДНК (Clontech) в соответствии с инструкциями изготовителя. На основе EST-последовательности два ПЦР-праймера, ZC22056 (SEQ ID NO:44) и ZC22057 (SEQ ID NO:45) использовали для идентификации источника мышиного zalpha11-лиганда при помощи ПЦР, Условия ПЦР-реакции были следующими: 94°С в течение 2 минут; 35 циклов при 94°С в течение 30 секунд, 68°С в течение 2 минут; затем 68°С в течение 4 минут; затем пропитывание при 10°С. ПЦР-продукты подвергали электрофорезу на 1% агарозном геле. Сильная полоса 150 п.н., представляющая амплифицированный "ДНК-фрагмент, была визуализирована. Это указывает на то, что marathon-кДНК селезенки является источником для кДНК-клонирования zalpha11-лиганда. Мышиная marathon-кДНК селезенки содержала положительную "ДНК, которую затем идентифицировали анализом последовательности как частичную кДНКдля мышиного zalpha11-лиганда.
С. Композиционная последовательность для полноразмерной кДНК была генерирована при помощи 5'- и 3'-RACE (быстрой амплификации концов кДНК)
5' и 3' фланкирующие последовательности частичной кДНК-последовательности мышиного zalpha11-лиганда получали при помощи амплификации 5'- и 3'-RACE. Два раунда ПЦР-амплификации с вмонтированными праймерами выполняли с дополнительными ген-специфическими олиго-праймерами ZC22205 (SEQ ID NO:46) и ZC22206 (SEQ ID NO:47), ZC22056 (SEQ ID NO:44) и ZC22057 (SEQ ID NO:45) и двумя адапторными олиго-праймерами ZC9739 (SEQ ID NO:48) и ZC9719 (SEQ ID NO:49). ПЦР-реакции выполняли следующим образом: 94°С в течение 2 минут; 35 циклов при 94°С в течение 30 секунд, 68°С в течение 2 минут; затем 68°С в течение 4 минут; затем пропитывание при 10°С. ПЦР-продукты подвергали электрофорезу на 1% агарозном геле, и был идентифицирован 5'-КАСЕ-продукт приблизительно 300 п.н. и 3'-РАСЕ-продукт приблизительно 800 п.н. Эти фрагменты выделяли с использованием набора для экстракции гелей Qiaquick™ (Qiagen).
Очищенные ПЦР-продукты секвенировали с использованием следующих праймеров: ZC9719 (SEQ ID NO:49), ZC22205 (SEQ ID NO:46) и ZC22206 (SEQ ID NO:47). Предварительная композиционная полноразмерная последовательность мышиного zalpha11-лиганда была идентифицирована комбинированием 5'- и 3'-RACE-фрагментов. Полноразмерный мышиный клон выделяли, как описано в примере 15 ниже.
Пример 15
Выделение кДНК-клона мышиного zalpha11 из активированной библиотеки мышиной селезенки
А. Мышиный первичный источник, используемый для выделения мышиного zalpha11-лиганда
Мышиные селезенки из мышей BALB/c собирали и раздавливали между предметными стеклами с матированными краями с получением клеточной суспензии. Выход выделенных первичных мышиных клеток был 6,4х108 клеток перед отбором, описанным ниже.
Клетки селезенки суспендировали в 9,6 мл MACS-буфера (ЗФР, 0,5% ЭДТА, 2 мМ ЭДТА). 1,6 мл клеточной суспензии удаляли и добавляли 0,4 мл CD90 (Тhу1.2)-микрогранул (Miltenyi Biotec). Смесь инкубировали в течение 15 минут при 4°С. Эти клетки, меченные гранулами CD90, промывали 30 мл MACS-буфера и затем ресуспендировали в 2 мл MACS-буфера.
VS+ колонку (Miltenyi) готовили в соответствии с инструкциями изготовителя. Затем VS+ колонку помещали в магнитное поле VarioMACS™ (Miltenyi). Колонку уравновешивали 5 мл MACS-буфера. Выделенные первичные мышиные клетки наносили затем на эту колонку. CD3-отрицательным клеткам давали пройти через колонку. Колонку промывали 9 мл (3×3 мл) MACS-буфера. Затем колонку удаляли из магнита и помещали на пробирку falcon на 15 мл. CD90+ клетки элюировали добавлением 5 мл MACS-буфера на колонку и связанные клетки смывали с использованием поршня (плунжера), обеспечиваемого изготовителем. Инкубирование этих клеток с CD90-магнитными гранулами, промывки и стадии VS+ колонки (инкубирование-элюция), описанные выше, повторяли еще раз. Полученные CD90+ фракции из 2 колоночных разделений объединяли. Общий выход CD90+ отобранных клеток мышиной селезенки был 1×108 клеток.
Пробу объединенных CD90+ отобранных мышиных клеток удаляли для окрашивания и сортинга на клеточном сортере с флуоресцентными антителами (FACS) для оценки их чистоты. РЕ-конъюгированное антитело против мышиного CD3ε хомячка (PharMingen) использовали для окрашивания и сортинга CD90+ отобранных клеток. Мышиные CD90+ отобранные клетки были на 93% CD3+ клетками, что предполагает, что эти клетки были на 93% Т-клетками.
Мышиные CD90+ отобранные клетки активировали инкубированием 3×106 клеток/мл в RPMI + 5% ФТС + ФМА 10 нг/мл и Иономицин 0,5 мкг/мл (Calbiochem) в течение ночи при 37°С. Супернатант от этих активированных CD90+ отобранных мышиных клеток тестировали на активность zalpha11-лиганда, как описано ниже. Кроме того, активированные CD90+ отобранные мышиные клетки использовали для получения библиотеки кДНК, как описано в примере 16 ниже.
Пример 16
Клонирование мышиного zalpha11-лиганда из библиотеки мышиных CD90+ отобранных клеток
Скрининг библиотеки "ДНК первичных мышиных активированных CD90+ отобранных клеток выявил выделенную "ДНК, которая является новым членом семейства цитокинов со структурой четырехспирального пучка. Эта кДНК кодировала мышиный ортолог zalpha11-лиганда человека. Эту кДНК идентифицировали гибридизационным скринингом.
А. Вектор для конструирования CD90+ отобранной библиотеки
Вектор pZP7N использовали для конструирования CD90+ отобранной библиотеки (см. пример 6А).
В. Получение кДНК-библиотеки первичных мышиных активированных CD90+ отобранных клеток
Приблизительно 1,5×108 первичных мышиных CD90+ отобранных клеток, стимулированных в смеси иономицин/ФМА (пример 15), выделяли центрифугированием. Тотальную РНК выделяли из осадка клеток и превращали в двухцепочечную кДНК, как описано в примере 6В. Затем эту ДНК трансфицировали в клетки ВНК, как описано в примере 6В, и пролиферацию оценивали с использованием флуоресцентного анализа с "Alamar blue" (пример 2В).
Для цели скрининга библиотеки клонированием с секреторной ловушкой была необходима комплексная амплифицированная форма библиотеки для трансфекции клеток COS-7. 4,8 миллионов клонов высевали на 110 чашек 15 см с LB-агаром, дополненным 100 мкг/мл ампициллина, 10 мкг/мл метициллина. После выращивания на чашках в течение ночи при 37°С бактерии собирали выскребанием и осаждали. Плазмидную ДНК экстрагировали из осажденных бактерий с использованием Nucleobond-giga™ (Clontech) в соответствии с инструкциями изготовителя. Затем эту плазмиду использовали для трансфекции клеток COS-7 на предметных стеклах и подвергали скринингу с использованием способа секреторной ловушки, описанного ниже (пример 17).
С. Скрининг активированной мышиной библиотеки кДНК
Приблизительно 5×1010 клонов высевали на 10 LB/Amp Maxi-чашек. Колонии снимали, денатурировали, нейтрализовали и сшивали с использованием стандартной процедуры (Sambrook, J. et al., supra). Пятьдесят нанограммов 5'RACE ПЦР-фрагмента 300 п.н. (пример 14) метили 32Р с использованием набора мечения случайных праймеров Prime-ltr RmT (Stratagene). 10 фильтров гибридизовали с этим меченым зондом при 65°С в течение ночи с использованием раствора для гибридизации ExpressHyb™ (Clontech). Затем эти фильтры промывали последовательно при 60°С в течение 1 часа три раза 0,1×SSC (30 мМ NaCl, 3 мМ цитрат натрий, рН 7,0), 0,1% ДСН; и затем при 65°С в течение 1 часа. Фильтры экспонировали при -80°С в течение ночи и рентгеновскую пленку проявляли. Агаровые пробки, содержащие положительные колонии, извлекали и эти клоны высевали на LB/Amp-чашки 10 см. Затем колонии снимали фильтром и опять гибридизовали согласно процедуре, описанной выше.
Один ДНК-клон, названный M11L/pZP7, выделяли и секвенировали с использованием следующих праймеров: ZC14063 (SEQ ID NO:50), ZC5020 (SEQ ID NO:51), ZC22421 (SEQ ID NO:52), ZC22604 (SEQ ID NO:53) и ZC22641 (SEQ ID NO:54), Полинуклеотидная последовательность этого клона является полноразмерным мышиным zalpha11-лигандом (SEQ ID NO:55) и согласуется с композиционной последовательностью, полученной из 5'- и 3'-РАСЕ-продуктов. Соответствующая аминокислотная последовательность для мышиного zalpha11-лиганда показана в SEQ ID NO:56.
Пример 17
Мышиный zalpha11-лиганд связывается с растворимым zalpha11-рецептором человека в тесте секреторной ловушки
ДНК для мышиного клона M11L/pZP7 трансфицировали в клетки COS и связывание растворимого zalpha11-рецептора человека слитого белка zalpha11-Fc4 (пример 10С) с трансфицированными клетками COS тестировали при помощи теста секреторной ловушки (пример 12). Этот тест подтвердил, что мышиный zalpha11-лиганд связывается с растворимым zalpha11-рецептором человека.
Трансфекцию клеток COS проводили, как в примере 12, с использованием 0,7 мкг ДНК М11L/pZP7 (пример 16) в 3 мкл.
Тест секреторной ловушки выполняли, как в примере 12, с использованием 1 мкг/мл слитого белка растворимый zalpha11-рецептор человека-Рс4 (пример 10С) в TNB и 1:200 разведенного козьего антитела против Ig человека-HRP (Fc-специфического) в TNB для детектируемого антитела. Положительное связывание растворимого zalpha11-рецептора человека с приготовленными фиксированными клетками детектировали флуоресцеин-тирамидным реагентом, как в примере 12. Клетки консервировали и визуализировали согласно примеру 12.
Положительный результат указывал на то, что мышиный zalpha11-лиганд связывается с растворимым zalpha11-рецептором человека.
Пример 18
Экспрессия мышиного zalpha11-лиганда в клетках млекопитающих
А. Конструирование экспрессирующего вектора M11L/pZP9
Готовили экспрессирующий вектор для экспрессии мышиного zalpha11-лиганда в клетках млекопитающих. Генерированный ПЦР ДНК-фрагмент zalpha11-лиганда 500 п.н. создавали с использованием ZC22283 (SEQ ID NO:57) и ZC22284 (SEQ ID NO:58) в качестве ПЦР-праймеров для амплификации кодирующего района мышиного zalpha11-лиганда и сайтов рестрикции Xhol и Xbal. Клон M11L/pZP7 мышиного zalpha11-лиганда (пример 16) использовали в качестве матрицы. Условия ПЦР-реакции были следующими: 94°С в течение 2 минут; 25 циклов при 94°С в течение 30 секунд, 68°С в течение 2 минут; затем 68°С в течение 4 минут; затем пропитывание при 10°С. Полосу предсказанного размера, около 500 п.н., визуализировали гель-электрофорезом на 1% агарозе, вырезали и ДНК очищали с использованием системы очистки Qiaexll™ (Qiagen) в соответствии с инструкциями изготовителя. Очищенную ДНК расщепляли Xhol и Xbal (Boehringer Mannheim) при 37°С в течение 2 часов. Затем эту ДНК выделяли из геля и очищали согласно указанной выше процедуре.
Вырезанную ДНК субклонировали в плазмиду pZP9, которая была разрезана Xhol и Xbal (Boehringer Mannheim). Плазмида pZP9 является экспрессирующим вектором млекопитающих, содержащим экспрессионную кассету, имеющую промотор мышиного металлотионеина-1 (МТ-1), множественные сайты рестрикции для встраивания кодирующих последовательностей и терминатор гормона роста человека. Эта плазмида имеет также начало репликации Е. coli, экспрессионную единицу селективного маркера экспрессии млекопитающих, имеющую промотор, энхансер и начало репликации SV40, ген DHFR и терминатор SV40.
Приблизительно 30 нг расщепленного рестриктазами фрагмента мышиного zalpha11-лиганда и около 10 нг расщепленного вектора pZP9 лигировали при комнатной температуре в течение 2 часов. Два мкг реакции лигирования трансформировали в INVaF'-компетентные клетки (Invitrogen) в соответствии с протоколом изготовителя и высевали на LB-чашки, содержащие 50 мкг/мл ампициллина, и инкубировали при 37°С в течение ночи. Колонии подвергали скринингу рестрикционным анализом с использованием Xhol и Xbal (Boehringer Mannheim) ДНК, полученной из жидких культур отдельных колоний. Анализ секвенирования подтвердил, что последовательность инсерта положительных клонов является последовательностью мышиного zalpha11-лиганда. Крупномасштабное получение плазмиды выполняли с использованием набора Qiagen® Maxi prep kit (Qiagen) в соответствии с инструкциями изготовителя. Экспрессирующий вектор, который содержит мышиный zalpha11-лиганд, был назван M11L/pZP9.
В. Экспрессия мышиного zalpha11-лиганда в млекопитающих
Клетки ВНК 570 (АТСС No. CRL-10314) высевали в чашки для культуры ткани 10 см и давали им расти до приблизительно 20% конфлюентности в течение ночи при 37°С, 5% CO2, в среде DMEM/ФТС (DMEM, Gibco/BRL High glucose media; Gibco BRL, Gaithersburg, MD), 5% фетальная сыворотка (Hyclone, Logan, UT), 1 мМ L-глутамин (JRH Biosciences, Lenexa, KS), 1 мМ пируват натрия (Gibco BRL)). Затем клетки трансфицировали плазмидой M11L/pZP9 (пример 18А) с использованием набора для стабильной СаРО4-трансфекции млекопитающих (Stratagene) в соответствии с инструкциями изготовителя.
Спустя один день после трансфекции клетки расщепляли 1:10 и 1:20 в селективные среды (среду DMEM/ФТС с добавлением 1 мкМ метотрексата (Sigma Chemical Co., St. Louis, МО)) в чашках 150 мм. Среду на этих клетках заменяли свежей селективной средой в день 5 после трансфекции. Спустя приблизительно 10 дней после трансфекции устойчивые к метотрексату колонии трипсинизировали и клетки объединяли и высевали в культуральные колбы для крупномасштабного получения. Как только эти клетки размножались до приблизительно 90% конфлюентности, их промывали ЗФР три раза и культивировали с бессывороточной средой ESTEP2 (DMEM (Gibco BRL), 0,11 г/л пирувата натрия, 3,7 г/л NaHCO3, 2,5 мг/л инсулина, 5 мг/л трансферрина, рН 7,0) с получением кондиционированной среды. Кондиционированные среды собирали спустя три дня и помещали в тест пролиферации BaF3 с использованием Alamar Blue, описанный в примере 19 ниже.
Пример 19
Мышиный zalpha11-лиганд активирует zalpha11-рецептор человека в BaFS-тесте с использованием Alamar Blue
Пролиферацию клеток BaF3/zalpha11 (пример 4 и 5В) оценивали с использованием бессывороточной кондиционированной среды от ВНК-клеток, экспрессирующих мышиный zalpha11-лиганд (пример 18).
Клетки BaF3/zalpha11 откручивали, промывали и высевали в не содержащую mlL-3 среду, как описано в примере 5В.
Пролиферацию клеток BaF3/zalpha11 оценивали с использованием бессывороточной кондиционированной среды от клеток ВНК, экспрессирующих мышиный zalpha11-лиганд (пример 18). Кондиционированную среду разводили не содержащей mlL-3 средой до концентраций: 50%, 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75% и 0,375%. Анализ пролиферации проводили, как в примере 5В.
Результаты подтвердили пролиферативную реакцию клеток BaF3/zalpha11 на мышиный zalpha11-лиганд. Эта реакция, как измерено, была приблизительно в 5 раз выше фона при концентрации 50%.
Пример 20
Zalpha11-лиганд активирует zalphall-рецептор человека в люциферазном тесте
А. Конструирование клеточной линии BaF3/KZ134/zalpha11
Плазмиду KZ134 конструировали с комплементарными олигонуклеотидами ZC12749 (SEQ ID NO:59) и ZC12748 (SEQ ID NO:60), которые содержат элементы связывания фактора транскрипции STAT из 4 генов: модифицированный индуцируемый элемент c-fos Sis (m67SIE или hSIE) (Sadowski, H. et al., Science 261:1739-1744, 1993), p21 SIE1 из гена р21 WAFI (Chin, Y. et al., Science 272:719-722, 1996), чувствительный элемент гена β-казеина молочной железы (Schmitt-Ney, М. et al., Mol. Cell. Biol. 11:3745-3755, 1991) и индуцируемый элемент STAT гена Fcg Rl (Seidel, H. et al., Proc. Natl. Acad. Sd. 92:3041-3045, 1995). Эти олигонуклеотиды содержат Asp718-Xhol-совместимые концы и их лигировали с использованием стандартных способов в реципиентный репортерный вектор светляка с c-fos-промотором (Poulsen, L.K. et al., J. Biol. Chem. 273:6229-6232, 1998), расщепленный теми же самыми ферментами и содержащий селектируемый маркер неомицина. Плазмиду KZ134 использовали для стабильной трансфекции клеток BaF3 с использованием стандартных способов трансфекции и отбора с получением клеточной линии BaF3/KZ134.
Стабильную индикаторную клеточную линию BaF3/KZ134, экспрессирующую полноразмерный zalpha11-рецептор, конструировали, как в примере 2А, с использованием около 30 мкг экспрессирующего вектора zalpha11, описанного в примере 3. Клоны разбавляли, высевали и отбирали с использованием стандартных способов. Клоны подвергали скринингу при помощи люциферазного теста (см. пример 20В, ниже) с использованием кондиционированной среды zalpha11-лиганда человека в качестве индуктора. Отбирали клоны с наивысшей люциферазной ответной реакцией (через люциферазу STAT) и с наименьшим фоном. Была отобрана линия стабильных трансфектантов. Эта линия клеток была названа BaF3/KZ134/zalpha11.
В. Человеческий и мышиный zalpha11-лиганды активируют zalpha11-рецептор в люциферазном тесте BaF3/KZ134/zalpha11
Клетки BaF3/KZ134/zalpha11 откручивали и промывали в не содержащей mlL-3 среде. Клетки откручивали и промывали 3 раза для гарантии удаления mlL-3. Затем клетки считали в гемоцитометре. Клетки высевали в 96-луночном формате при приблизительно 30000 клеток на лунку в объеме 100 мкл на лунку с использованием не содержащей mlL-3 среды. Ту же самую процедуру использовали для нетрансфицированных клеток BaF3/KZ134 для применения в качестве контроля в последующем анализе.
STAT-активацию клеток BaF3/KZ134/zalpha11 оценивали с использованием кондиционированной среды от (1) клеток ВНК570, трансфицированных zalpha11-лигандом человека (пример 7), или (2) клеток ВНК570, трансфицированных мышиным zalpha11-лигандом (пример 18), или (4) не содержащей mlL-3 средой для измерения реакции контроля "только среда". Кондиционированную среду разбавляли не содержащей mlL-3 средой RPMI до концентраций 50%, 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75% и 0,375%. 100 мкл разведенной кондиционированной среды добавляли к клеткам BaF3/KZ134/zalpha11. Анализ с использованием кондиционированной среды выполняли параллельно на клетках BaF3/KZ134 в качестве контроля. Общий объем реакционной смеси был 200 мкл. Тест-чашки инкубировали при 37°С, 5% CO2 в течение 24 часов, после чего эти клетки осаждали центрифугированием при 2000 об/мин в течение 10 минут и среду отсасывали и добавляли 25 мкл лизисного буфера (Promega). После 10 минут при комнатной температуре чашки измеряли на активацию STAT-репортерной конструкции считыванием их на люменметре (Labsystems Luminoskan, model RS), который добавлял 40 мкл субстрата люциферазного теста (Promega) при пяти секундах интеграции.
Результаты подтвердили STAT-репортерный ответ клеток BaF3/KZ134/zalpha11 на человеческий zalpha11-лиганд. Этот ответ, согласно измерению, был приблизительно 50-кратным в сравнении с контролем только-среда при концентрации 50%. STAT-активация в ответ на zalpha11-лиганд человека отсутствовала в необработанных контрольных клетках BaF3/KZ134, что указывает на то, что этот ответ является опосредованным zalpha11-рецептором.
Результаты подтвердили также STAT-репортерный ответ клеток BaF3/KZ134/zalpha11 на мышиный zalpha11-лиганд. Этот ответ, согласно измерениям, был приблизительно 40-кратным над контролем только-среда при концентрации 50%. Кроме того, STAT-активация в ответ на мышиный zalpha11-лиганд была очевидной (приблизительно 5-кратной) на нетрансфицированных контрольных клетках BaF3/KZ134, что предполагает, что мышиные клетки BaF3 могут иметь эндогенный мышиный рецептор.
Пример 21
Мышиный zalpha11-лиганд является активным в анализе с мышиным костным мозгом
А. Выделение неприкрепленных клеток костного мозга низкой плотности
Свежий аспират мышиной бедренной кости (костный мозг) получали из 6-10-недельных самцов мышей BALB/C или C57BL/6. Затем костный мозг промывали RPMI+10% ФТС (JRH, Lenexa KS; Hyclone, Logan UT) и суспендировали в RPMI+10% ФТС в виде суспензии целых клеток костного мозга. Затем суспензию целых клеток костного мозга подвергали градиенту плотности (Nycoprep, 1.077, Animal; Gibco BRL) для обогащения в отношении имеющих низкую плотность в основном мононуклеарных клеток следующим образом: суспензию целых клеток костного мозга (около 8 мл) осторожно пипетировали на верхнюю поверхность приблизительно 5 мл градиентного раствора Nycoprep в конической пробирке на 15 мл и затем центрифугировали при 600xg в течение 20 минут. Затем межфазный слой, содержащий мононуклеарные клетки низкой плотности, удаляли, промывали избытком RPMI+10% ФТС и осаждали центрифугированием при 400х g в течение 5-10 минут. Этот осадок ресуспендировали в RPMI+10% ФТС и высевали в колбу Т-75 при приблизительно 106 клеток/мл и инкубировали при 37°С, 5% CO2 в течение приблизительно 2 часов. Полученные клетки в суспензии были неприкрепленными клетками костного мозга низкой плотности (NA LD).
В. 96-луночный тест
Клетки мышиного костного мозга NA LD высевали при 25000-45000 клетках на лунку в 96-луночных планшетах для культуры ткани в RPMI+10% ФТС + 1 нг/мл фактора мышиных стволовых клеток (mSCF) (R&D Systems, Minneapolis, MN) плюс 5% кондиционированная среда от одного из следующих вариантов клеток: (1) клетки ВНК 570, экспрессирующие мышиный zalpha11-лиганд (пример 18), (2) клетки ВНК 570, экспрессирующие человеческий zalpha11-лиганд (пример 7), или (3) контрольные клетки ВНК 570, содержащие вектор и не экспрессирующие никакой лиганд. Эти клетки затем подвергали различным обработкам цитокинами для анализа размножения или дифференцировки гемопоэтических клеток из костного мозга. Для этого теста посеянные мышиные клетки костного мозга NA LD подвергали действию IL-15 человека (hlL-15) (R&D Systems) или одного из панели других цитокинов (R&D Systems). Серийное разведение hlL-15 или других цитокинов тестировали, с 2-кратным серийным разведением от концентрации приблизительно 50 нг/мл до концентрации приблизительно 6025 нг/мл. После 8-12 дней 96-луночные тесты оценивали в баллах на пролиферацию клеток при помощи теста с красителем Alamar blue, как описано в примере 5В.
С. Результаты из 96-луночного теста мышиных клеток костного мозга NA LD
Кондиционированная среда от клеток ВНК, экспрессирующая как мышиный, так и человеческий zalpha11-лиганд, действовала синергически с hlL-15 в промотировании размножения популяции гемопоэтических клеток в NA LD-клетках мышиного костного мозга. Это размножение гемопоэтических клеток не было обнаружено с кондиционированной контрольными клетками ВНК средой плюс IL-15. Популяцию гемопоэтических клеток, размноженную мышиным zalphall-лигандом с hlL-15, и популяцию гемопоэтических клеток, размноженную человеческим zalpha11-лигандом с hlL-15, дополнительно размножали в культуре клеток. Эти гемопоэтические клетки окрашивали фикоэритрин-меченым антителом против Pan NK-клеток (Pharmingen) и подвергали анализу проточной цитометрией, который показал, что размноженные клетки окрашивались положительно на этот маркер природных клеток-убийц (NK-клеток).
Тот же самый 96-луночный тест проводили с использованием свежих клеток костного мозга человека, приобретенных из Poietic Technologies, Gaithersburg, MD. Опять, в сочетании с IL-15, мышиный и человеческий zalpha11-лиганд размножал популяцию гемопоэтических клеток, которая окрашивалась положительно на маркер NK-клеток с использованием описанных выше антител.
Пример 22
Конструкции для получения содержащих zalpha11-лиганд человека трансгенных мышей
А. Конструкция для экспрессии zalpha11-лиганда человека от печень-специфического промотора МТ-1
Были сконструированы олигонуклеотиды для генерирования ПЦР-фрагмента, содержащего консенсусную последовательность Козака и кодирующий район zalpha11-лиганда человека. Эти олигонуклеотиды были сконструированы с сайтом Fsel на 5'-конце и сайтом Asel на 3'-конце для облегчения клонирования в (а) рМТ12-8, стандартный трансгенный вектор авторов, или (b) pKF051, лимфоид-специфический трансгенный вектор (пример 22В).
ПЦР-реакции проводили с 200 нг zalpha11-лиганда человека в качестве матрицы (пример 7) и олигонуклеотидами ZC22143 (SEQ ID NO:61) и ZC22144 (SEQ ID NO:62). Условия ПЦР-реакции были следующими: 95°С в течение 5 минут, где добавляли кДНК-полимеразу Advantage™ (Clontech); 15 циклов 95°С в течение 60 секунд, 60°С в течение 60 секунд и 72°С в течение 90 секунд; и 72°С в течение 7 минут. ПЦР-продукты разделяли электрофорезом в агарозном геле и очищали с использованием набора для экстракции гелей QiaQuick™ (Qiagen). Выделенный ДНК-фрагмент 488 п.н. расщепляли Fsel и Asel (Boehringer Mannheim), осаждали этанолом и лигировали в рМТ12-8, предварительно расщепленную Fsel и Asel. Плазмида рМТ12-8, сконструированная для экспрессии представляющего интерес гена в печени и других тканях трансгенных мышей, содержит экспрессионную кассету, фланкированную 10 т.п.н. 5'-ДНК МТ-1 и 7 т.п.н. 3'-ДНК МТ-1. Эта экспрессионная кассета содержит промотор МТ-1, интрон инсулина II мыши, полилинкер для встраивания желательного клона и поли А-последовательность гормона роста человека (hGH).
Около одного микролитра каждой реакции лигирования электропорировали в электрокомпетентные относительно DH10B ElectroMAX™ клетки (GIBCO BRL, Gaithesburg, MD) в соответствии с инструкциями изготовителя и высевали на LB-чашки, содержащией 100 мкг/мл ампициллина, и инкубировали в течение ночи. Колонии выскребали и выращивали в LB-среде, содержащей 100 мкг/мл ампициллина. Минипрепарат ДНК получали из извлеченных клонов и подвергали скринингу на инсерт zalpha11-лиганда человека рестрикционным расщеплением только EcoRl или комбинацией Fsel и Asel и затем гель-электрофорезу. Получали максипрепараты точного pMT-zalpha11-лиганда человека. Был получен Sall-фрагмент, содержащий 5'- и 3'-фланкирующие последовательности, промотор МТ-1, интрон инсулина II крысы, кДНК zalpha11-лиганда человека и поли А-последовательность hGH для применения для микроинъекции в оплодотворенные мышиные ооциты. Микроинъекцию и получение трансгенных мышей выполняли, как описано в Hogan, В. et al., Manipulating the Mouse Embryo. 2nd ed., Cold Spring Harbor Laboratory Press, NY, 1994.
В. Конструкция для экспрессии zalpha11-лиганда человека от лимфоид-специфического промотора EμLCK
Конструировали олигонуклеотиды для генерирования ПЦР-фрагмента, содержащего консенсусную последовательность Козака и кодирующий район zalpha11-лиганда человека. Эти олигонуклеотиды были сконструированы с сайтом Fsel на 5'-конце и сайтом Asel на 3'-конце для облегчения клонирования в pKF051, лимфоид-специфический трансгеный вектор.
ПЦР-реакции проводили с 200 нг zalpha11-лиганда человека в качестве матрицы (пример 7) и олигонуклеотидами ZC22143 (SEQ ID NO:61) и ZC22144 (SEQ ID NO:62). ПЦР-реакцию выполняли с использованием кДНК-полимеразы Advantage ™ (Clontech) при следующих условиях: 95°С в течение 5 минут, 15 циклов 95°С в течение 60 секунд, 60°С в течение 60 секунд и 72°С в течение 90 секунд; и 72°С в течение 7 минут. ПЦР-продукты очищали, как описано выше. Выделенный ДНК-фрагмент 488 п.н. расщепляли Fsel и Asel (Boehringer Mannheim), осаждали зтанолом и лигировали в pKF051, предварительно расщепленную Fsel и Asel. Трансгенный вектор pKF051 получен из р1026Х (Iritani, B.M., et al., EMBO J. 16:7019-31, 1997) и содержит Т-клетко-специфический проксимальный промотор Ick В/Т-специфический энхансер тяжелой цепи иммуноглобулина μ, полилинкер для встраивания желательного клона и мутированный ген hGH, который кодирует белок неактивного фактора роста (обеспечивающий 3'-интроны и сигнал полиаденилирования).
Около одного микролитра каждой реакции лигирования электропорировали, высевали, клоны выскребали и подвергали скринингу на инсерт zalpha11-лиганда человека рестрикционным расщеплением, как описано выше. Точный клон pKF051-zalpha11-лиганд подтверждали секвенированием и проводили получение максипрепарата этого клона. Notl-фрагмент, содержащий проксимальный промотор Ick и энхансер иммуноглобулина μ (EμLCK), кДНК zalpha11-лиганда и мутированный ген hGH, получали для применения для микроинъекции в оплодотворенные мышиные ооциты.
Пример 23
Тканевое распределение мышиного zalpha11-лиганда
Нозерн-блоты множественных мышиных тканей (Mouse, Mouse Embrio, Clontech; MB1010, MB1012 Origene) зондировали для определения тканевого распределения экспрессии мышиного zalpha11-лиганда. ПЦР-полученный зонд приблизительно 484 п.н. амплифицировали с использованием плазмиды M11L/pZP7 (пример 16) в качестве матрицы и олигонуклеотидов ZC22283 (SEQ ID NO:57) и ZC22284 (SEQ ID NO:58) в качестве праймеров. ПЦР-амплификацию проводили следующим образом: 1 цикл при 94°С в течение 1,0 минуты; 35 циклов 94°С в течение 30 секунд, 50°С в течение 30 секунд и 72°С в течение 30 секунд; затем 1 цикл при 72°С в течение 10 минут. ПЦР-продукты визуализировали электрофорезом на агарозном геле и ПЦР-продукт приблизительно 484 п.н. очищали с использованием набора для экстракции гелей (Qiagen) в соответствии с инструкциями изготовителя. Зонд радиоактивно метили с использованием набора для мечения REDIPRIME™ (Amersham) в соответствии с инструкциями изготовителя. Зонд очищали с использованием колонки NUCTRAP™ push Column (Stratagene). Раствор EXPRESSHYB™ (Clontech) использовали для предгибридизации и в качестве раствора для гибридизации для Нозерн-блотов. Гибридизация имела место в течение ночи при 65°С с применением 106 имп/мин/мл меченого зонда. Затем блоты промывали три раза в 2х SSC и 0,1% ДСН при комнатной температуре с последующими 2 промывками в 0,1х SSC и 0,1% ДСН при 55°С. Два транскрипта приблизительно 1,2 и 3,5 т.п.н. наблюдали в яичке. Более высоко идущий транскрипт наблюдали только в вилочковой железе.
Master Dot Blot мышиной РНК (Clontech), который содержал РНК из различных тканей, которые были нормализованы относительно 8 генов "домашнего хозяйства", также зондировали и гибридизовали, как описано выше. Экспрессию наблюдали в яичке.
Пример 24
Очистка немеченых человеческого и мышиного zalpha11-лиганда из ВНК 570
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки человеческого и мышиного zalpha11-лиганда из кондиционированных сред от клеток ВНК 570, трансфицированных конструкцией, экспрессирующей либо zalpha11-лиганд человека (пример 25), либо мышиный zalpha11-лиганд (M11L/pZP9) (пример 18). Кондиционированную среду концентрировали стандартными способами. Концентрированную кондиционированную среду (КМ) стерильно фильтровали через фильтры 0,45 и 0,22 микрон. Затем эту среду разбавляли до низкой ионной силы (≤2 mS) в 0,01 М HEPES (JRH Bioscience, Lenexa, KS) при рН 7,0. Затем разбавленную до низкой ионной силы KM наносили на колонку 10×66 мм (6 мл) Poros HS 50 (PerSeptive BioSystems, Framingham, MA) в течение ночи при 4 мл/мин с использованием BioCAD SPRINT (Perceptive BioSystems). Эту колонку промывали 10-20 колоночными объемами (КО) 0,01 М HEPES рН 7,0. Затем связанные белки ступенчато элюировали 1 М NaCl (Mallinckrodt, Paris, KY) в 0,01 M HEPES pH 7,0 при 5 мл/мин; фракции по два мл собирали на протяжении всей хроматографии и поглощение при 280 и 215 нм подвергали мониторингу. Фракции пиков анализировали при помощи биоанализа и при помощи гель-электрофореза в ПААГ-ДСН с окрашиванием серебром (Geno Technology, St. Louis, МО) и Кумасси (Signna, St. Louis, МО). Фракции пиков объединяли, стерильно фильтровали и разбавляли до ≤19 mS забуференным сульфатом солевым раствором (PBS, Gibco BRL) рН 7,2.
Затем разбавленную пробу наносили при 2 мл/мин с использованием BioCad SPRINT на колонку 0,8 мл Poros AL, которая имела либо растворимый рецептор zalpha11CFLAG (пример 10В), либо слитый растворимый рецептор zalpha11-Fc4 (пример 10С), иммобилизованные на смоле (см. ниже). Затем эту колонку промывали по меньшей мере 20 КО ЗФР при 10 мл/мин. Затем колонку быстро элюировали инжекцией 600 мкл 0,1 М глицина (Aminoacetic Acid; Glycocol, Spectrum, Gardena, CA) pH 2,5 при скорости тока 10 мл/мин с ЗФР на BioCAD 700E. Фракции по 1 мл собирали в течение 6 секунд каждую и сразу же нейтрализовали рН 55 мкл 2 М Триса (Трис(гидроксиметил)аминометана, ЕМ Science, Gibbstown, NJ) pH 8,8. Поглощение при 280 и 215 нм подвергали мониторингу на протяжении всей хроматографии.
Фракции пиков анализировали при помощи биоанализа и электрофореза в ПААГ-ДСН с окрашиванием серебром (Geno Technology) и Кумасси (Sigma). Две полосы, приблизительно 24 кД и 18 кД. наблюдали как на окрашенных серебром, так и на окрашенных Кумасси гелях для мышиного zalpha11-лиганда. Единственную полосу при приблизительно 18 кД можно было видеть на окрашенных как серебром, так и Кумасси гелях для zalpha11-лиганда человека.
Иммобилизация полипептидов растворимого zalpha11 рецептора человека на среде POROS AL
Готовили колонки Poros AL, имеющие иммобилизованный растворимый рецептор zalpha11CFLAG (пример 10В) или слитый растворимый рецептор zalpha11-Fc4. Использовали приблизительно 3 мг растворимого рецептора zalpha11CFLAG и приблизительно 10 мг слитого рецептора zalpha11-Fc4. Все операции проводили при комнатной температуре на BioCAD 700E. Колонку 4,5×50 мм со средой POROS AL упаковывали проточно в 2 М NaCl в соответствии с инструкциями изготовителя. Затем колонку уравновешивали в смеси 1,1 М Na2SO4/500 мМ Na-фосфат рН 7,2. Рецептор концентрировали до 4 мг/мл с использованием центрифужного концентратора Millipore 30 MWKO, затем разбавляли 1:1 в смеси 1,1 М Na2SO4/50 мМ Na-фосфат рН 7,2. Через колонку пропускали при 2 мл/мин смесь 1,1 М Na2SO4/50 мМ Na-фосфат рН 7,2 и инжекции по 100 мкл разбавленного лиганда производили после каждых 9 КО, пока не достигали стационарного состояния насыщения или прерывания через колонку. Затем пропускали 62 КО градиента от 1,1 М Na2SO4/50 мМ Na-фосфат рН 7,2 до 550 мМ Na2SO4/50 мМ Na-фосфат рН 7,2/5 мг/мл цианоборгидрида натрия. Затем колонку выдерживали в течение около 2 часов для завершения иммобилизации. Затем колонку уравновешивали в 0,2 М ТРИС рН 7,2/5 мг/мл цианоборгидрида натрия и давали стоять в течение приблизительно 1 часа для закрывания колонки, Наконец колонку уравновешивали в ЗФР/0,02% азид натрия и хранили при 4°С до использования. Перед использованием колонку предварительно элюировали 0,1 М глицином для гарантии, что были удалены неспецифические белки и что эта колонка не выщелачивала иммобилизованный растворимый zalpha11-рецептор человека.
Пример 25
Экспрессия zalpha11-лиганда человека в клетках млекопитающих
А. Конструирование экспрессирующего вектора PZMP11/zalpha11Lig
Экспрессионную плазмиду, содержащую весь полинуклеотид или часть полинуклеотида, кодирующего zalpha11-лиганд человека, конструировали посредством гомологичной рекомбинации. Фрагмент "ДНК zalpha11-лиганда человека (SEQ ID NO:63) выделяли с использованием ПЦР. Два праймера использовали в получении этого фрагмента zalpha11-лиганда человека в ПЦР-реакции: (1) праймер ZC22052 (SEQ ID NO:64), содержащий 40 п.н. фланкирующей последовательности вектора и 17 п.н., соответствующих амино-концу zalpha11-лиганда человека, и (2) праймер ZC22053 (SEQ ID NO:65), содержащий 40 п.н. 3'-конца, соответствующих фланкирующей последовательности вектора, и 17 п.н., соответствующих карбоксил-концу zalpha11-лиганда человека. Условия ПЦР-реакции были следующими: 1 цикл при 94°С в течение 2,0 минут; 25 циклов 94°С в течение 30 секунд, 60°С в течение 30 секунд и 72°С в течение 30 секунд; затем 1 цикл при 72°С в течение 5 минут, пропитывание 4°С. Десять мкл из 100 мкл ПЦР-реакции подвергали электрофорезу на 0,8% LMP-агарозном геле (Seplaque GTG) с 1 х ТВЕ-буфером для анализа, и наблюдали ожидаемый фрагмент приблизительно 560 п.н. Остальные 90 мкл ПЦР-реакции осаждали добавлением 5 мкл 1 М NaCl и 250 мкл абсолютного этанола для использования для рекомбинации на реципиентный вектор pZMPH, как описано ниже. Реципиентную плазмиду pZMPH предварительно разрезали Smal.
Плазмида pZMPH была получена из плазмиды pCZR199 (описанной здесь, например, в примере 8). Плазмида pCZR199 является экспрессирующим вектором млекопитающих, содержащим экспрессионную кассету, имеющую непосредственно ранний промотор CMV, консенсусный интрон из вариабельной области локуса тяжелой цепи мышиного иммуноглобулина, множественные сайты рестрикции для встраивания кодирующих последовательностей, стоп-кодон и терминатор гормона роста человека. Эта плазмида имеет также начало репликации Е. coli, экспрессионную единицу селектируемого маркера млекопитающих, имеющую промотор, энхансер и начало репликации SV40, ген DHFR и терминатор SV40. Вектор pZMPH конструировали из pCZR199 и он включает в себя замену промотора металлотионеина немедленно ранним промотором CMV и последовательности Козака на 5'-конце открытой рамки считывания.
Сто микролитров компетентных клеток дрожжей (S. cerevisiae) объединяли с 10 мкл смеси, содержащей приблизительно 1 мкг инсерта zalpha11-лиганда человека, и 100 нг расщепленного Smal вектора pZMPH, и переносили в кювету для электропорации 0,2 см. Смеси дрожжи/ДНК подвергали электроимпульсам при 0,75 кВ (5 кВ/см), неопределенному числу омов, 25 мкФ. В каждую кювету добавляли 600 мкл 1,2 М сорбита и дрожжи высевали в двух аликвотах по 300 мкл на две чашки URA-D и инкубировали при 30°С.
Спустя приблизительно 48 часов, Ura+ дрожжевые трансформанты из отдельной чашки ресуспендировали в 1 мл Н2О и откручивали быстро для осаждения дрожжевых клеток. Клеточный осадок ресуспендировали в 1 мл лизисного буфера (2% Тритон Х-100, 1% ДСН, 100 мМ NaCl, 10 мМ Трис, рН 8,0, 1 мМ ЭДТА). 500 мкл лизисной смеси добавляли в пробирку Эппендорфа, содержащую 300 мкл промытых кислотой стеклянных гранул и 200 мкл смеси фенол/хлороформ, встряхивали на вортексе два или три раза с интервалами 1 мин с последующим 5-минутным откручиванием в центрифуге Эппендорфа при максимальной скорости. Триста микролитров водной фазы переносили в свежую пробирку и ДНК осаждали 600 мкл этанола (EtOH) с последующим центрифугированием в течение 10 минут при 4°С. Осадок ДНК ресуспендировали в 100 мкл Н2О.
Трансформацию электрокомпетентных клеток Е. coli (DH10B, Gibco BRL) проводили с 0,5-2 мл препарата дрожжевой ДНК и 40 мкл клеток DH10B. Клетки подвергали электроимпульсам при 2,0 кВ, 25 мФ и 400 Ом. После электропорации 1 мл SOC (2% Бакто™ триптон (Difco, Detroit, Ml), 0,5% дрожжевой экстракт (Difco), 10 мМ NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза) высевали в аликвотах 250 мкл на четыре LB АМР чашки (LB-бульон (Lennox), 1,8% Бактоагар (Difco), 100 мг/л ампициллина).
Индивидуальные клоны, несущие правильную экспрессионную конструкцию для zalpha11-лиганда человека, идентифицировали рестрикционным расщеплением для подтверждения присутствия инсерта и для подтверждения, что различные последовательности ДНК были правильно соединены друг с другом. Инсерт положительных клонов подвергали анализу последовательности. При крупномасштабном получении плазмидную ДНК выделяли с использованием набора QIAGEN Maxi kit (Qiagen) согласно инструкциям изготовителя.
В. Экспрессия zalpha11-лиганда человека в млекопитающих
Клетки ВНК 570 (АТСС No. CRL-10314) высевали в чашки 10 см для культуры ткани и давали им расти до приблизительно 50-70% конфлюэнтности в течение ночи при 37°С, 5% CO2, в среде DMEM/ФТС (DMEM, Gibco/BRL High Glucose, (GIBCO BRL, Gaithersburg, MD), 5% фетальной телячьей сыворотке (Hyclone, Logan, UT), 1 мМ L-глутамине (JRH Biosciences, Lenexa, KS), 1 мМ пирувате натрия (Gibco BRL). Затем клетки трансфицировали плазмидой PZMPH/zalpha11Lig (пример 25А) с использованием Липофектамина™ (Gibco BRL) в бессывороточной (БС) среде (DMEM, 10 мг/мл трансферрина, 5 мг/мл инсулина, 2 мг/мл фетуина, 1% L-глутамина и 1% пирувата натрия). Плазмиду zalpha11-Fc4/pZMP6 (пример 8В) разводили в пробирках на 15 мл до общего конечного объема 640 мкл БС-средой. 35 мл Липофектамина™ (Gibco BRL) смешивали с 605 мкл БС-среды. Эту смесь Липофектамина™ добавляли к смеси ДНК и давали инкубироваться в течение приблизительно 30 минут при комнатной температуре. Пять миллилитров БС-среды добавляли к смеси ДНК: Липофектамин™. Клетки промывали один раз 5 мл БС-среды, отсасывали и добавляли смесь ДНК: Липофектамин™. Клетки инкубировали при 37°С в течение пяти часов, затем в каждую чашку добавляли 6,4 мл среды DMEM/10% ФТС, 1% PSN. Чашки инкубировали при 37°С в течение ночи и смесь ДНК: Липофектамин™ заменяли свежей средой 5% ФТС/DMEM на следующий день. В день 5 после трансфекции клетки разрушали в колбу Т-162 в селективную среду (среду DMEM/5% ФТС, 1% L-GLU, 1% Na-Pyr). Приблизительно спустя 10 дней после трансфекции две культуральные чашки 150 мм с устойчивыми к метотрексату колониями из каждой трансфекции трипсинизировали и клетки объединяли и высевали в колбу Т-162 и переносили в крупномасштабную культуру. Кондиционированную среду из крупномасштабной культуры использовали для очистки полипептида zalpha11-лиганда человека, как описано в примере 24.
Пример 26
Конструкция для получения содержащих мышиный zalpha11-лиганд трансгенных мышей
А. Конструкция для экспрессии мышиного zalpha11-лиганда от лимфоид-специфического промотора EμLCK
Были сконструированы олигонуклеотиды для генерирования ПЦР-фрагмента, содержащего консенсусную последовательность Козака и кодирующий район zalpha11-лиганда мыши. Эти олигонуклеотиды были сконструированы с сайтом Fsel на 5'-конце и сайтом Asel на 3'-конце для облегчения клонирования в (a) pKFO51, лимфоид-специфический трансгенный вектор, или (b) pTG12-8, стандартный трансгенный вектор авторов.
ПЦР-реакции проводили с 200 нг zalpha11-лиганда мыши в качестве матрицы (SEQ ID NO:55; пример 16) и олигонуклеотидами ZC23115 (SEQ ID NO:66) и ZC23116 (SEQ ID NO:67). ПЦР-реакцию проводили с использованием кДНК-полимеразы Advantage™ (Clontech) в условиях ПЦР, описанных в примере 22В. ПЦР-продукт выделяли, как описано в примере 22В. Выделенный ДНК-фрагмент 440 п.н. расщепляли и лигировали в pKFО51, предварительно расщепленный Fsel и Asel, как описано в примере 22 В.
Около одного микролитра каждой реакции лигирования электропорировали, высевали, клоны выскребали и подвергали скринингу на инсерт zalpha11-лиганд человека, как описано в примере 22. Правильный клон рКFO51-zаlрhа11-лиганд подтверждали секвенированием и получали максипрепарат этого клона. Notl-фрагмент, содержащий проксимальный промотор Ick, энхансер иммуноглобулина μ, кДНК zalpha11-лиганда и мутированный ген hGH, получали для использования для микроинъекции в оплодотворенные мышиные ооциты.
В. Конструкция для экспрессии мышиного zalpha11-лиганда от печень-специфического промотора МТ-1
Тот же самый инсерт мышиный zalpha11-лиганд из примера 26А субклонировали в вектор pTG12-8, как описано в примере 22А. Для этой конструкции около 10 мг максипрепарата ДНК рКFO51-zаlрhа11-пиганд расщепляли объединенными Fsel и Asel, осаждали этанолом и фрагмент мышиного zalpha11-лиганда очищали, как описано в примере 22. Затем этот фрагмент лигировали в вектор pTG12-8, который был предварительно расщеплен Fsel и Asel, как описано в примере 22А. Электропорацию, скрининг клонов и получение максипрепарата выполняли, как описано в примере 22. Sall-фрагмент, содержащий 5'- и 3'-фланкирующие последовательности, промотор МТ-1, интрон инсулина II крысы, кДНК мышиного zalpha11-лиганда и поли А-последовательность hGH, получали для использования для микроинъекции в оплодотворенные мышиные ооциты.
Пример 27
Поликлональные антитела к мышиному zalpha11-лиганду
Поликлональные антитела получали иммунизацией двух самок Новозеландских белых кроликов очищенным полипептидом muzalpha11/MBP-6H (пример 32). Каждый кролик получал исходную внутрибрюшинную инъекцию (ip) 200 мг очищенного белка в полном адъюванте Фрейнда с последующими бустерными инъекциями ip 100 мг пептида в неполном адъюванте Фрейнда каждые три недели. Спустя семь-десять дней после введения второй бустерной инъекции (всего 3 инъекции) производили кровоизвлечение у животных и собирали сыворотку. Затем кроликам продолжали вводить бустер-инъекции и брали кровь каждые три недели.
Muzalpha11L/MBP-6Н-специфическую кроличью сыворотку предварительно адсорбировали от антител анти-МВР с использованием колонки CNBr-SEPHAROSE 4В protein (Pharmacia LKB), которую готовили с использованием 10 мг очищенного рекомбинантного мальтозусвязывающего белка (МБР) на грамм CNBr-SEPHAROSE. Рекомбинантный МБР получали и очищали на амилозной колонке в лаборатории авторов с использованием способов, хорошо известных в данной области, Поликлональные muzalpha11-лиганд-специфические антитела очищали аффинно из кроличьей сыворотки с использованием колонки CNBr-SEPHAROSE 4В protein (Pharmacia LKB), которую готовили с использованием 10 мг очищенного рекомбинантного белка специфического антигена muzalpha11L/MBP-6H (пример 32) с последующим 20Х диализом в ЗФР в течение ночи. Muzalpha11-лиганд-специфические антитела характеризовали при помощи ELISA с применением 1 мкг/мл очищенного рекомбинантного белка muzalpha11L-MBP-6H (пример 32) или huzalpha11L-MBP-6H (пример 32) в качестве мишеней антител. Нижним порогом детектирования (LLD) кроличьего аффинно очищенного антитела анти-muzalpha11L/MBP-6H является разведение 100 пг/мл на его специфическом очищенном рекомбинантном антигене muzalpha11L/MBP-6H и 500 пг/мл на очищенном рекомбинантном huzalpha11L-MBP-6H.
Пример 28
Конструирование зкспрессирующего вектора млекопитающих и крупномасштабная экспрессия zalpha11-лиганда человека в клетках СНО DG44
Экспрессирующий вектор млекопитающих для zalpha11-лиганда человека (SEQ ID NO:1), предназначенный для добавления сайта Sall на 5'-конце и сайта Pmel к 3'-концу кДНК, конструировали посредством амплификации при помощи ПЦР из плазмиды, содержащей zalpha11-лиганд человека (пример 7) с олигонуклеотидными праймерами ZC22054 (SEQ ID NO:70) и ZC22055 (SEQ ID NO:71). Условия ПЦР-реакции были следующими: 94°С в течение 4 минут; 25 циклов 94°С в течение 45 секунд, 50°С в течение 45 секунд и 72°С в течение 3 минут; и 72°С в течение 10 минут. ПЦР-продукт выделяли, как описано здесь, и разрезали Sall и Pmel, затем лигировали в плазмиду pDC312, предварительно разрезанную в подходящих сайтах рестрикции в полилинкере, с использованием описанных здесь стандартных способов. Плазмида pDC312 описана в Morris, A. et al., "Expression Augmenting Sequence Element (EASE) isolated from Chinese Hamster Ovary Cells", in Animal Cell Technology. Carrondo, MJT et al., (eds.) (1997) Kluwer Academic Publishers, The Netherlands, p.529-534.
Лигированный вектор трансфицировали в адаптированные к суспензии клетки СНО DG44 (у себя, в Novo Nordisk, Denmark) с использованием липофекции при помощи реагента ЛипофектаминПлюс™ (Gibco/BRL) в соответствии с инструкциями изготовителя. Трансфектанты отбирали на среде PFCHO (JRH, Lenexa, Kansas), не содержащей тимидина и гипоксантина, с последующим отбором на 200 нМ метотрексате (Sigma, St. Louis, МО). Устойчивые к метотрексату клетки клонировали разведением и тестированием на продуцирование zalpha11-лиганда при помощи анализа активности BaF3 (пример 5В).
Продуктивный клон размножали и выращивали в 7-20-литровом биореакторе (Applicon Bioreactors, Schiedam, The Netherlands) в среде PFCHO с получением материала для очистки (пример 29).
Пример 29
Крупномасштабная очистка немеченого zalpha11-лиганда человека и мыши из экспрессионных клеточных линий млекопитающих ВНК и СНО
А. Экспрессируемый СНО zalpha11-лиганд человека
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки zalpha11-лиганда человека из по меньшей мере 30 литров СНО-кондиционированной среды (см. пример 28). Концентрированную или неконцентрированную кондиционированную среду (КС) стерильно фильтровали через фильтры 0,45 и 0,22 микрон. Затем концентрированную среду забуферивали 0,01 М MES (Fluka BioChemica, Switzerland)) и рН доводили до 6,0 и затем наносили на колонку 50×100 мм (196 мл) Poros 50 HS (сильный катионообменник из PerSeptive BioSystems, Framingham, МА) в течение ночи при 4-10 мл/мин с использованием BioCAD SPRINT (Perceptive BioSystems). Эту колонку промывали 10-20 колоночными объемами (КО) 0,01 М MES/0,130 М NaCl (Mallinckrodt, Paris, KY) рН 6,0 Затем связанные белки элюировали 10 КО градиента 0,130 М - 1 М NaCl в 0,01 М MES рН 6,0 при 30 мл/мин; фракции по 25 мл собирали на протяжении всей хроматографии и поглощение при 280 и 215 нм подвергали мониторингу. Фракции пиков анализировали при помощи гель-электрофореза в ПААГ-ДСН с окрашиванием серебром (Geno Technology, St. Louis, МО), Кумасси (Sigma, St. Louis, МО) и Вестерн-иммуноблоттинга с использованием антител против zalpha11-лиганда человека (пример 33 и пример 34).
Фракции пиков объединяли, затем концентрировали в концентраторе с перемешиваемой ячейкой на мембране YM10 (Millipore/Amicon, Bedford, MA) до обычного объема (1-10 мл). Затем эту пробу наносили на подходящую гель-фильтрационную колонку быстрого разрешения (52-600 мл) Sephacryl S-200 (Pharmacia, Uppsala Sweden), уравновешенную в ЗФР (Gibco BRL), при скоростях тока 1-2 мл/мин; фракции 1-2 мл собирали на протяжении всей хроматографии и поглощение при 280 и 215 нм подвергали мониторингу. Фракции пиков анализировали гель-электрофорезом на ПААГ-ДСН с окрашиванием серебром (Geno Technology, St. Louis, МО) и Кумасси (Sigma, St. Louis, МО).
Представляющие интерес фракции объединяли и концентрировали с использованием центрифужных концентраторов Millipore 5 kD MWKO до минимального объема. Затем конечный продукт анализировали при помощи гель-электрофореза на ПААГ-ДСН с окрашиванием Кумасси (Sigma, St. Louis, МО), Вестерн-иммуноблоттинга, N-концевого секвенирования, аминокислотного анализа и БСА-анализа (Pierce, Rockford, Illinois) для определения чистоты и концентрации белка.
В. Экспрессируемый ВНК 570 мышиный zalpha11-лиганд
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки zalpha11-лиганда мыши из ВНК-кондиционированной среды (пример 18). Концентрированную или неконцентрированную кондиционированную среду (КС) стерильно фильтровали через фильтры 0,45 и 0,22 микрон. Затем среду забуферивали 0,01 М MES (Fluka BioChemica, Switzerland)) и рН доводили до 6,0. Эту КС анализировали, наносили на колонку AS и элюировали с колонки, как описано в примере 29А.
Представляющие интерес фракции объединяли, затем концентрировали в концентраторе с перемешиваемой ячейкой, как в примере 29А, до объема 20-30 мл. рН доводили до 7,0, затем эту пробу наносили либо на колонку Poros AL 0,8 мл, которая имела около 3 мг zalpha11CFLAG-меченого растворимого рецептора (пример 10В), либо на колонку с приблизительно 10 мг слитого zalpha11-Fc4 рецептора (пример 10С), иммобилизованного на этой смоле (см. способ ниже), при 1 мл/мин на BioCAD SPRINT. Затем колонку промывали по меньшей мере 20 КО 0,3 М NaCl/3ФР (Gibco BRL)/0,01 M MES при 10 мл/мин. Затем колонку быстро элюировали инжекцией 600 мкл 0,1 М глицина (Aminoacetic Add; Glycocol, Spectrum, Gardena, CA) pH 2,5 при скорости тока 10 мл/мин с 3ФР на BioCAD SPRINT. Фракции по 1 мл собирали в течение 6 секунд каждую и сразу же нейтрализовали рН 55 мкл 2 М Триса рН 8,8 (Трис(гидроксиметил)аминометана, ЕМ Science, Gibbstown, NJ). Поглощение при 280 и 215 нм подвергали мониторингу на протяжении всей хроматографии. Фракции пиков анализировали при помощи гель-электрофореза на ПААГ-ДСН с окрашиванием серебром (Geno Technology, St. Louis, МО), Кумасси (Sigma, St. Louis, МО) и Вестерн-иммуноблоттингом, как описано выше.
Фракции пиков объединяли, затем концентрировали в концентраторе с перемешиваемой ячейкой, как в примере 29А до минимального объема (1-10 мл). Затем пробу наносили, уравновешивали и анализировали, как в примере 29А, на подходящей гель-фильтрационной колонке быстрого разрешения Sephacryl S-200 (Pharmacia). Фракции пиков анализировали гель-электрофорезом на ПААГ-ДСН с окрашиванием серебром (Geno Technology, St. Louis, МО) и Кумасси (Sigma, St. Louis, МО). Представляющие интерес фракции объединяли и концентрировали и анализировали, как в примере 29А.
С. Иммобилизация белков на носителе POROS AL
Все операции проводили при комнатной температуре на BioCAD 700E. Колонку 4,5×50 мм проточно упаковывали носителем POROS AL в 2 М NaCl в соответствии с описаниями изготовителя. Затем колонку уравновешивали в 1,1 М Na2SO4 и 50 мМ фосфате натрия рН 7,2. Рецептор концентрировали до 4 мг/мл с использованием центрифужного концентратора Millipore 30 kD MWCO, затем разбавляли 1:1 в 1,1 М Na2SO4 и 50 мМ фосфате натрия рН 7,2. Через колонку пропускали при 2 мл/мин 1,1 М Na2SO4 и 50 мМ фосфат натрия рН 7,2 и производили инжекции 100 мкл разведенного лиганда каждые 9 КО, пока не достигали стационарного состояния насыщения или прерывания через колонку. Затем пропускали 62 КО градиента от 1,1 М Na2SO4 и 50 мМ Na-фосфата рН 7,2 до 550 мМ Na2SO4 и 50 мМ Na-фосфата рН 7,2 с 5 мг/мл цианоборгидрида натрия. Колонку выдерживали в течение 2 часов для завершения иммобилизации. Затем колонку уравновешивали в 0,2 М ТРИС рН 7,2 с 5 мг/мл цианоборгидрида натрия и давали стоять в течение приблизительно 1 часа. Наконец колонку уравновешивали в ЗФР с 0,02% азидом натрия и хранили при 4°С до использования. Перед использованием колонку предварительно элюировали 0,1 М глицином для гарантии, что были удалены неспецифические белки и что эта колонка не выщелачивала иммобилизованный рецептор.
Пример 30
Конструирование экспрессирующего вектора, экспрессия и очистка немеченого zalpha11-лиганда человека и мыши из бакуловируса
А. Конструкция для экспрессии zalpha11-лиганд а человека в бакуловирусе
Экспрессирующий вектор, pzalpha11L, готовили для экспрессии полипептидов zalpha11-лиганда человека в клетках насекомых. Фрагмент 517 п.н., содержащий последовательность zalpha11-лиганда человека и кодируемые сайты рестрикции BamHI и Xhol на 5'- и 3'-концах соответственно, генерировали ПЦР-амплификацией из плазмиды, содержащей кДНК zalpha11-лиганда человека (пример 7) с использованием праймеров ZC23444 (SEQ ID NO:74) и ZC23445 (SEQ ID NO:75). Условия ПЦР-реакции были следующими: 1 цикл 94°С в течение 4 минут; затем 25 циклов при 94°С в течение 45 секунд, 50°С в течение 45 секунд и 72°С в течение 2 минут; затем 1 цикл при 72°С в течение 10 минут; затем пропитывание при 4°С. Фрагмент визуализировали при помощи гель-электрофореза (1% SeaPlaque/1% NuSieve). Полосу вырезали, разбавляли до 0,5% агарозы 2 мМ MgCl2, расплавляли при 65°С, расщепляли BamHI и Xhol (Boehringer Mannheim) и лигировали в расщепленный BamHI/Xhol экспрессирующий бакуловирусный вектор, pZBV3L. Вектор pZBV3L является модификацией экспрессирующего вектора pFastBac1™ (Life Technologies), где промотор полиэдрона был удален и заменен поздним промотором активирующего основного белка. Около 14 нанограммов расщепленного рестриктазами инсерта zalpha11-лиганда и около 40 нг соответствующего вектора лигировали в течение ночи при 16°С.
Смесь для лигирования разбавляли в 3 раза в ТЭ (10 мМ Трис-HCl, рН 7,5 и 1 мМ ЭДТА) и около 4 фмоль разбавленной смеси для лигирования трансформировали в Library Efficiency-компетентные клетки DH5α (Life Technologies) в соответствии с инструкциями изготовителя посредством теплового шока в течение 45 секунд на водяной бане при 42°С. Трансформированную ДНК и клетки разбавляли в 450 мкл среды SOC (2% Бакто™ Триптон, 0,5% Бакто-Дрожжевой экстракт, 10 мл 1 М NaCl, 1,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4 и 20 мМ глюкоза) и высевали на LB-чашки, содержащие 100 мкг/мл ампициллина. Клоны анализировали с использованием продуктов расщепления рестриктазами и 1 мкл положительного клона трансформировали в 20 мкл DН10Вас Мах Efficiency-компетентных клеток (GIBCO-BRL, Gaithesburg, MD) в соответствии с инструкцией изготовителя посредством теплового шока, как описано выше. Затем трансформированные клетки разбавляли в 980 мкл среды SOC (2% Бакто™ Триптон, 0,5% Бакто-Дрожжевой экстракт, 10 мл 1 М NaCl, 1,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4 и 20 мМ глюкоза), выращивали во встряхиваемом термостате при 37°С в течение 4 часов и высевали на чашки с агаром Luria, содержащим 50 мкг/мл канамицина, 7 мкг/мл гентамицина (Life Technologies), 10 мкг/мл тетрациклина, IPTG (Pharmacia Biotech) и Bluo-Gal (Life Technologies). Посеянные клетки инкубировали в течение 48 часов при 37°С. Отбор по цвету использовали для идентификации клеток, имеющих донорный инсерт, кодирующий zalpha11-лиганд человека, который был включен в плазмиду (называемую "бакмидой"). Колонии, которые имели белую окраску, выскребали для анализа. ДНК бакмиды zalpha11-лиганда человека выделяли из положительных колоний с использованием системы QiaVac Miniprep8 (Qiagen) в соответствии с инструкциями изготовителя. Клоны подвергали скринингу на правильный инсерт амплификацией ДНК с использованием праймеров к транспозируемому элементу в бакмиде путем применения ПЦР-праймеров ZC447 (SEQ ID NO:76) и ZC976 (SEQ ID NO:77). Условия ПЦР-реакции были следующими: 35 циклов 94°С в течение 45 секунд, 50°С в течение 45 секунд и 72°С в течение 5 минут; 1 цикл при 72°С в течение 10 минут; затем пропитывание при 4°С. ПЦР-продукт подвергали электрофорезу на 1% агарозе для проверки размера инсерта. Клоны, имеющие правильный инсерт, использовали для трансфекции клеток Spodoptera frugiperda (Sf9).
В. Экспрессия и генерирование материала для очистки zalpha11-лиганда человека из бакуловируса
Клетки Sf9 высевали при 5×106 клеток на чашку 35 мм и давали прикрепляться в течение 1 часа при 27°С. Пять микролитров бакмидной ДНК zalpha11-лиганда человека (описанной выше) разбавляли 100 мкл среды Sf9-900 II SFM (Life Technologies). Шесть мкл реагента CellFECTIN (Life Technologies) разбавляли 100 мкл Sf-900 II SFM. Бакмидную ДНК и растворы липидов осторожно смешивали и инкубировали 30-45 минут при комнатной температуре. Среду из одной чашки отсасывали, клетки промывали 1×2 мл свежей среды Sf-900 II SFM. 800 микролитров Sf-900 II SFM добавляли к смеси липид-ДНК. Промывочную среду удаляли аспирацией и смесь ДНК-липид добавляли к клеткам. Клетки инкубировали при 27°С в течение 4-5 часов. Смесь ДНК-липид удаляли аспирацией и 2 мл среды Sf-900 II добавляли в каждую чашку. Чашки инкубировали при 27°С, 90% влажности в течение 96 часов, после чего вирус собирали.
Для первичной амплификации клетки Sf9 выращивали в 50 мл среды Sf-900 II SFM во встряхиваемой колбе на 125 мл до приблизительной плотности 0,41-0,52 ×105 клеток/мл. Затем клетки инфицировали 150 мкл исходного вирусного раствора, описанного выше, и инкубировали при 27°С в течение 3 дней, после чего вирус собирали в соответствии со стандартными способами, известными в данной области. Пробу 500 мкл использовали для определения активности в ВаF3-тесте (пример 5) для подтверждения, что она была биологически активной.
Для вторичной амплификации клетки Sf9 выращивали в 1 л среды Sf-900 II SFM во встряхиваемой колбе на 2800 мл до приблизительной плотности 0,5×105 клеток/мл. Ее инфицировали 500 мкл первичного исходного вирусного раствора, описанного выше, и инкубировали при 27°С в течение 4 дней, после чего вирус собирали в соответствии со стандартными способами, известными в данной области. Вирус титровали и выращивали до большого масштаба для очистки продуцируемого бакуловирусом zalpha11-лиганда человека (huzalpha11L-Bv), как описано в примере 30С и примере 30D ниже.
С. Крупномасштабная очистка экспрессируемого бакуловирусом zalpha11-лиганда человека/мыши
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки zalpha11-лиганда человека (huzalpha11L-Bv) из BV-кондиционированной среды (пример 30В). Кондиционированную среду (КС) стерильно фильтровали через фильтры 0,45 и 0,22 микрон, затем забуферивали 0,01 М MES (Fluka BioChemica, Switzerland)) и рН доводили до 6,0. Затем КС наносили на колонку POROS 50 HS и подвергали хроматографии, фракции собирали, анализировали, как описано в примере 29А.
Полученные фракции пиков объединяли, концентрировали, подвергали хроматографии на гель-фильтрационной колонке высокого разрешения и анализировали, как описано в примере 29А.
Представляющие интерес фракции из гель-фильтрационной колонки объединяли и концентрировали центрифужными концентраторами 5 кД MWCO Millipore до минимального объема. Затем конечный продукт анализировали при помощи гель-электрофореза на ПААГ-ДСН с окрашиванием Кумасси (Sigma, St. Louis, МО), Вестерн-иммуноблоттинга, N-концевого секвенирования, аминокислотного анализа и БСА-анализа (Pierce, Rockford, Illinois) для определения чистоты и концентрации белка, как описано в примере 29А. Весь белок хранили при -80°С.
Р. Очистка в небольшом масштабе (<2 мг) экспрессируемого бакуловирусом zalpha11-лиганда человека/мыши
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки <2 мг zalpha11-лиганда человека или мыши из BV-кондиционированной среды. Кондиционированную среду (КС) фильтровали, забуферивали и рН доводили, как в примере 30. Затем КС наносили на колонку, элюировали и хроматографию POROS 50 HS анализировали, как описано в примере 30С.
Фракции объединяли, затем концентрировали диафильтрованием в концентраторе с перемешиваемой ячейкой на мембране YM10 (10 кД MWCO (отсечение мол массы) (Millipore/Amicon, Bedford, MA) до незначительного объема (20-30 мл). рН доводили до 7,0, затем пробу наносили либо на колонку 0,8 мл Poros AL, которая имела около 3 мг zalpha11CFLAG-меченого растворимого рецептора (пример 10В), либо на колонку с приблизительно 10 мг слитого zalpha11-Fc4 растворимого рецептора (пример 10С), иммобилизованного на этой смоле (см. способ в примере 29С), при 1 мл/мин на BioCAD SPRINT. Затем колонку промывали по меньшей мере 20 КО 0,3 М NaCl/3ФР (Gibco BRL)/0,01 M MES при 10 мл/мин. Затем колонку быстро элюировали инжекцией 600 мкл 0,1 М глицина (Aminoacetic Acid; Glycocol, Spectrum, Gardena, CA) pH 2,5 при скорости тока 10 мл/мин с 3ФР на BioCAD SPRINT. Фракции по 1 мл собирали в течение 6 секунд каждую и сразу же нейтрализовали pH 55 мкл 2 М Триса pH 8,8 (Трис(гидроксиметил)аминометана, ЕМ Science, Gibbstown, NJ). Поглощение при 280 и 215 нм подвергали мониторингу на протяжении всей хроматографии. Фракции анализировали, как описано выше.
Фракции пиков объединяли, затем концентрировали диафильтрованием в концентраторе с перемешиваемой ячейкой на мембране YM10 (10 кД MWCO (отсечение мол массы) (Millipore/Amicon, Bedford, MA) до 1-2 мл. Затем пробу наносили на подходящую гель-фильтрационную колонку быстрого разрешения Sephacryl S-200 (Pharmacia, Uppsala, Sweden), уравновешенную в 3ФР (Gibco BRL), при оптимальной скорости тока; фракции собирали на протяжении всей хроматографии и поглощение при 280 и 215 нм подвергали мониторингу. Фракции анализировали, как описано выше.
Представляющие интерес фракции объединяли, затем концентрировали центрифужными концентраторами 5 кД MWCO Millipore до незначительного объема. Затем конечный продукт анализировали при помощи гель-электрофореза на ПААГ-ДСН с окрашиванием Кумасси (Sigma, St. Louis, МО), Вестерн-иммуноблоттинга, N-концевого секвенирования, аминокислотного анализа и БСА-анализа (Pierce, Rockford, Illinois) для определения чистоты и концентрации белка. Весь белок хранили как описано выше.
Е. Конструкция для экспрессии мышиного zalpha11-лиганда в бакуловирусе: pzalpha11lig.M
Готовили экспрессирующий вектор, pzalpha11LM, для экспрессии полипептидов мышиного zalpha11-лиганда в клетках насекомых. Фрагмент 413 п.к, содержащий последовательность для мышиного zalpha11-лиганда и кодируемые сайты рестрикции BspEl и Xbal на 5'- и 3'-концах соответственно, генерировали ПЦР-амплификацией из плазмиды, содержащей кДНК мышиного zalpha11-лиганда (пример 16), с использованием праймеров ZC25970 (SEQ ID NO:109) и ZC25969 (SEQ ID NO:110) с применением ПЦР-системы высокой точности (Expand High Fidelity PCR System, Boehringer Mannheim) в соответствии с инструкциями изготовителя. Условия ПЦР были следующими: 1 цикл 94°С в течение 2 минут с последующими 35 циклами 94°С в течение 15 секунд, 50°С в течение 30 секунд и 72°С в течение 2 минут; 1 цикл при 72°С в течение 10 минут, с последующим пропитыванием при 4°С. Небольшую часть этого ПЦР-продукта визуализировали гель-электрофорезом (1% NuSieve агароза). Остальную часть этого фрагмента очищали с использованием набора для очистки ПЦР-продукта Qiagen в соответствии с инструкциями изготовителя и элюировали в 30 мкл H2O. Затем этот фрагмент расщепляли рестриктазами Bspel и Xbal (Boehringer Mannheim) при 37°С в течение приблизительно 2 часов, затем подвергали электрофорезу на агарозе, как описано выше. Полосу вырезали, очищали и элюировали с использованием набора для экстракции гелей Qiagen в соответствии с инструкциями изготовителя. Очищенный фрагмент лигировали в расщепленный Bspel/Xbal бакуловирусный экспрессирующий вектор, pZBV37L. Вектор pZBV37L является модификацией экспрессирующего вектора pFastBac1™ (Life Technologies), где промотор полиэдрона был удален и заменен поздним промотором активирующего основного белка с последующей секреторной сигнальной последовательностью из экдистероид-УДФ-глюкозилтрансферазы (EGT). Около 5 мкл расщепленного рестриктазами инсерта мышиного zalpha11-лиганда и около 100 нг соответствующим образом разрезанного вектора лигировали в течение ночи при 16°С в приблизительно 20 мкл. Пять мкл смеси для лигирования электропорировали в 50 мкл DН12S-элекгрокомпетентных бактериальных клеток Life Technologies, с использованием кюветы 2 мм с установкой 2 кВ, 25 мкФ и 400 Ом. Электропорированные клетки помещали в 1 мл среды SOC (2% Бакто™ Триптон, 0,5% Бакто-Дрожжевой экстракт, 10 мл 1 М NaCl, 1,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4 и 20 мМ глюкоза (Gibco BRL)), выращивали при 37°С в течение приблизительно 1 часа и высевали на LB-чашки, содержащие 100 мкг/мл ампициллина. ДНК из этих клонов выделяли и анализировали с использованием продуктов рестрикционного расщепления для идентификации положительных клонов. Приблизительно 5 нг ДНК из положительного клона трансформировали в 20 мкл DH10Bac Мах Efficiency-компетентных клеток (GIBCO-BRL, Gaithesburg, MD) в соответствии с инструкцией изготовителя, посредством теплового шока в течение 45 секунд на водяной бане при 42°С. Затем трансформированные клетки разбавляли и выращивали, как описано в примере ЗОЛ. Бакмиды, содержащие инсерт мышиного zalpha11-лиганда, идентифицировали и выделяли, как описано в примере 30А. Клоны подвергали скринингу на правильный инсерт амплификацией ДНК с использованием праймеров к транспозируемому элементу в этой бакмиде посредством ПЦР с использованием праймеров ZC447 (SEQ ID NO:76) и ZC976 (SEQ ID NO:77). Условия ПЦР-реакции были следующими: 35 циклов 94°С в течение 45 секунд, 50°С в течение 45 секунд и 72°С в течение 5 минут; 1 цикл при 72°С в течение 10 минут; затем пропитывание при 4°С. ПЦР-продукт подвергали электрофорезу на 1% агарозе для проверки размера инсерта. Клоны, имеющие правильный инсерт, использовали для трансфекции клеток Spodoptera frugiperda (Sf9).
F. Экспрессия и генерирование материала для очистки мышиного zalpha11 из бакуловируса
Клетки Sf9 высевали при 1 миллионе клеток на чашку 35 мм и давали прикрепляться в течение 1 часа при 27°С. Бакмидную ДНК zalpha11-лиганда мыши трансфицировали, как описано в примере 30В, и вирус собирали.
Для первичной амплификации клетки Sf9 высевали, как описано выше, и 500 мкл супернатанта спустя 72 часа после трансфекции добавляли и культурам давали продолжаться в течение 96 часов, после чего вирус собирали в соответствии со стандартными способами.
Для вторичной амплификации клетки Sf9 высевали, как описано выше, и добавляли 200 мкл первичного исходного вирусного раствора. Культуры инкубировали при 27°С в течение 72 часов, после чего вирус собирали в соответствии со стандартными способами.
Для третичной амплификации 10 мкл вторичного амплифицированного исходного вирусного раствора помещали на клетки Sf9 при 500000 клеток на лунку в 50 мл среды SF900II во встряхиваемой колбе на 250 мл в течение 6 дней и вирус собирали, как описано выше. Вирус титровали и выращивали до большого масштаба для очистки продуцируемого бакуловирусом мышиного zalpha11-лиганда (muzalpha11L-Bv), как описано в примере 30С и примере 30D.
Присутствие белка с предсказанной молекулярной массой в супернатанте определяли Вестерн-анализом с использованием поликлонального антитела против muzalpha11L/MBP-6H (пример 27). Анализ теста пролиферации на основе ВаРЗ (пример 5) также показал, что секретируемый лигад был активным.
Пример 31
Экспрессия zalpha11-лиганда человека и мыши в Е. coli
А. Конструирование вектора рТАР98/га1рПа11-лиганд, экспрессирующего слияние zalpha11-лиганд человека-MBP
Экспрессионную плазмиду, содержащую полинуклеотид, кодирующий zalpha11-лиганд человека, слитый на N-конце с мальтозусвязывающим белком (МВР), конструировали гомологичной рекомбинацией. Фрагмент кДНК zalpha11-лиганда человека (SEQ ID NO:1) выделяли с использованием ПЦР. Два праймера использовали в получении фрагмента zalpha11-лиганда человека в ПЦР-реакции: (1) Праймер ZC22128 (SEQ ID NO:78), содержащий 40 п.н. векторной фланкирующей последовательности и 26 п.н., соответствующих амино-концу zalpha11-лиганда человека, и (2) праймер ZC22127 (SEQ ID NO:79), содержащий 40 п.н. 3'-конца, соответствующих фланкирующей векторной последовательности, и 28 п.н., соответствующих карбоксил-концу zalpha11-лиганда человека. Условия ПЦР-реакции были следующими: 25 циклов 94°С в течение 30 секунд, 50°С в течение 30 секунд и 72°С в течение 1 минуты, с последующим пропитыванием при 4°С, реакции проводили в двух повторностях. Два мкл из 100 мкл ПЦР-реакции подвергали электрофорезу на 1,0% агарозном геле с 1 х ТВЕ-буфером для анализа и наблюдали ожидаемую полосу приблизительно 472 п.н. Оставшиеся 90 мкл ПЦР-реакции объединяли со второй ПЦР-пробиркой, осаждали 400 мкл абсолютного этанола для использования для рекомбинации в разрезанный Smal реципиентный вектор рТАР98 для получения конструкции, кодирующей слитый полипептид МВР-zalpha11-лиганд, как описано ниже.
Плазмиду рТАР98 получали из плазмид pRS316 и pMAL-c2. Плазмида pRS316 является челночным вектором Saccharomyces cerevisiae (Hieter P. and Sikorski, R., Genetics 122:19-27, 1989). pMAL-C2 (NEB) является экспрессионной плазмидой Е. coli. Она несет промотор tac, запускающий MalE (ген, кодирующий МВР), за которым следуют His-метка, сайт расщепления тромбина, клонирующий сайт и терминатор rrnB. Вектор рТАР98 конструировали с использованием гомологичной рекомбинации дрожжей. 100 нг разрезанной EcoRI pMAL-c2 рекомбинировали с 1 мкг разрезанной Pvul pRS316, 1 мкг линкера и 1 мкг разрезанной Scat/EcoRI pRS316. Линкер состоял из олигонуклеотидов ZC19372 (SEQ ID NO:80) (100 пмоль): ZC19351 (SEQ ID NO:81) (1 пмоль): ZC19352 (SEQ ID NO:82) (1 пмоль) и ZC19371 (SEQ ID NO:83) (100 пмоль), объединенных в ПЦР-реакции. Условия ПЦР-реакции были следующими: 10 циклов 94°С в течение 30 секунд, 50°С в течение 30 секунд и 72°С в течение 30 секунд; затем пропитывание при 4°С. ПЦР-продукты концентрировали осаждением 100% этанолом.
100 мкл компетентных дрожжевых клеток (S. cerevisiae) объединяли с 10 мкл, содержащей приблизительно 1 мкг ПЦР-продукта zalpha11-лиганда человека и 100 нг расщепленного Smal вектора рТАР98, и переносили в кювету для электропорации 0,2 см. Смесь дрожжи/ДНК подвергали электрошокам при 0,75 кВ (5 кВ/см), "неопределенном" числе Ом, 25 мкФ. В каждую кювету добавляли 600 мкл 1,2 М сорбита и дрожжи высевали в двух аликвотах по 300 мкл на две чашки URA-D и инкубировали при 30°С.
Спустя приблизительно 48 часов Ura+ дрожжевые трансформанты из отдельной чашки ресуспендировали в 1 мл H2O и откручивали быстро для осаждения дрожжевых клеток. Клеточный осадок ресуспендировали в 1 мл лизисного буфера (2% Тритон Х-100, 1% ДСН, 100 мМ NaCl, 10 мМ Трис, рН 8,0, 1 мМ ЭДТА). 500 мкл лизисной смеси добавляли в пробирку Эппендорфа, содержащую 300 мкл промытых кислотой стеклянных гранул и 200 мкл смеси фенол/хлороформ, встряхивали на вортексе два или три раза с интервалами 1 мин, с последующим 5-минутным откручиванием в центрифуге Эппендорфа при максимальной скорости. Триста микролитров водной фазы переносили в свежую пробирку и ДНК осаждали 600 мкл этанола (EtOH) с последующим центрифугированием в течение 10 минут при 4°С. Осадок ДНК ресуспендировали в 100 мкл Н2O.
Трансформацию электрокомпетентных клеток Е. coli (MC1061, Casadaban et al., J. Mol. Biol. 138, 179-207) проводили с 1 мкл препарата дрожжевой ДНК и 40 мкл клеток MC1061. Клетки подвергали электроимпульсам при 2,0 кВ, 25 мФ и 400 Ом. После электропорации 0,6 мл SOC (2% Бакто™ Триптон (Difco, Detroit, Ml), 0,5% дрожжевой экстракт (Difco), 10 мМ NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза) высевали в виде одной аликвоты на чашки с LB АМР (LB-бульон (Lennox), 1,8% Бакто™ агар (Difco), 100 мг/л ампициллина).
Индивидуальные клоны, несущие правильную экспрессионную конструкцию для zalpha11-лиганда человека, идентифицировали по экспрессии. Клетки выращивали в Superbroth II (Becton Dickinson) с 100 мкг/мл ампициллина в течение ночи. 50 мкл ночной культуры использовали для инокуляции 2 мл свежей Superbroth II + 100 мкг/мл ампициллина. Культуры выращивали при 37°С со встряхиванием в течение 2 часов. 1 мл этой культуры индуцировали 1 мМ IPTG. Спустя 2-4 часа 250 мкл каждой культуры смешивали с 250 мкл промытых кислотой стеклянных гранул и 250 мкл буфера Торнера с 5% β-МЭ и красителем (8 М мочевина, 100 мМ Трис рН 7,0, 10% глицерин, 2 мМ ЭДТА, 5% ДСН). Пробы встряхивали на вортексе в течение одной минуты и нагревали до 65°С в течение 5-10 минут. 20 мкл наносили на дорожку на 4-12% ПААГ для электрофореза (NOVEX). Разделение на гелях происходило в буфере 1xMES. Положительные клоны были названы рТАР126 и их подвергали анализу секвенированием. Полинуклеотидная последовательность слитого белка MBP-zalpha11-лиганд человека показана в SEQ ID NO:84, а соответствующий полипептид в SEQ ID NO:85.
В. Бактериальная экспрессия zalpha11-лиганда человека
Один мкл ДНК секвенирования использовали для трансформации штамма W3110 (АТСС). Клетки подвергали электроимпульсам при 2,0 кВ, 25 мкФ и 400 Ом. После электропорации добавляли 0,6 мл SOC (2% Бакто™ Триптон (Difco, Detroit, Ml), 0,5% Дрожжевой экстракт (Difco), 10 мл NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4 и 20 мМ глюкоза) высевали в виде одной аликвоты на чашки LB АМР (LB-бульон (Lennox), 1,8% Бакто™ агар (Difco), 100 мг/л ампициллина).
Индивидуальные клоны экспрессировали. Клетки выращивали в Superbroth II (Becton Dickinson) с 100 мкг/мл ампициллина в течение ночи. 50 мкл ночной культуры использовали для инокуляции 2 мл свежей Superbroth II + 100 мкг/мл ампициллина. Культуры выращивали при 37°С со встряхиванием в течение 2 часов. 1 мл этой культуры индуцировали 1 мМ IPTG. Спустя 2-4 часа 250 мкл каждой культуры смешивали с 250 мкл промытых кислотой стеклянных гранул и 250 мкл буфера Торнера с 5% β-МЭ и красителем (8 М мочевина, 100 мМ Трис рН 7,0, 10% глицерин, 2 мМ ЭДТА, 5% ДСН). Пробы встряхивали на вортексе в течение одной минуты и нагревали до 65°С в течение 10 минут. 20 мкл наносили на дорожку на 4-12% ПААГ гель для электрофореза (NOVEX). Разделение на гелях происходило в буфере 1xMES. Положительные клоны использовали для выращивания для очистки слитого белка huzalpha11L/MBP-6H (пример 32 ниже).
С. Конструирование вектора рТАР98/zаlрhа11-пиганд мыши, экспрессирующего слияние zalpha11-лиганд мыши/МБР
Экспрессионную плазмиду, содержащую полинуклеотид, кодирующий часть zalpha11-лиганда мыши, слитый на М-конце с мальтозусвязывающим белком (МВР), конструировали гомологичной рекомбинацией как описано в примере 31А. Фрагмент кДНК zalpha11-лиганда мыши (SEQ ID NO:55) выделяли с использованием ПЦР. Два праймера использовали в получении фрагмента zalpha11-лиганда мыши в ПЦР-реакции: (1) Праймер ZC22849 (SEQ ID NO:86), содержащий 40 п.н. векторной фланкирующей последовательности и 24 п.н., соответствующих амино-концу zalpha11-лиганда мыши, и (2) праймер ZC22850 (SEQ ID NO:87), содержащий 40 п.н. 3'-конца, соответствующих фланкирующей векторной последовательности, и 21 п.н., соответствующих карбоксил-концу zalpha11-лиганда мыши. Условия ПЦР-реакции были такими же, как описанные выше. Фрагмент приблизительно 450 п.н. клонировали в рТАР98, как описано выше. Клоны трансформировали, идентифицировали и выращивали, как описано выше. Положительные клоны были названы рТАР134 и их подвергали анализу секвенированием. Полинуклеотидная последовательность слитого белка МБР-мышиный zalpha11-лиганд в рТАР134 показана в SEQ ID NO:88, а соответствующая полипептидная последовательность показана в SEQ ID NO:89. Положительные клоны использовали для выращивания в Е. coli, как описано выше, для очистки слитого белка muzalpha11L/MBP-6H (пример 32).
Пример 32
Очистка zalpha11-MBP-лиганда или zalphall-MBP-рецептора
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки слияний zalpha11-MBP-лиганд для человеческого zalpha11-MBP-лиганда (huzalpha11L/MBP-6H) или мышиного zalpha11-MBP-лиганда (muzalpha11L/MBP-6H) из Е. coli. Слитые белки человеческий или мышиный zalpha11-MBP-рецептор получали с использованием одного и того же способа. Предварительно открученную замороженную пасту Е. coli оттаивали и разбавляли в 2 литра буфера В (0,02 М ТРИС (ЕМ Science); 0,2 М NaCl (Mallincrodt); 0,01 М 2-меркаптоэтанол (ЕМ Science); pH 8,0; с 5 мг/л пепстатина А (Boehringer Mannheim); 5 мг/л апротинина (Boehringer Mannheim) и 1 мг/л ПМСФ (Fluka)) плюс 1-2 мл противовспенивающего реагента AF289 (Sigma). Смесь обрабатывали в предварительно охлажденном разрушителе клеток French Press (Constant Systems LTD) с 20-30 кф/кв. дюйм.
Затем лизат центрифугировали при 18000 × g в течение 45 минут при 4°С; супернатант сохраняли. 200 мл суспензии смолы Amylose (New England BioLabs), предварительно уравновешенной в буфере А (0,02 М ТРИС (ЕМ Science); 0,2 М NaCl (Mallincrodt); 0,01 М 2-меркаптоэтанол (ЕМ Science); pH 8,0), добавляли к супернатанту-лизату и инкубировали в течение ночи в роллерных флаконах на 2 л для максимальной абсорбции слитого МВР-белка в загрузочной смеси. Смолу промывали в формате загрузочной колонки S 5 колоночными объемами буфера А, затем загрузку элюировали буфером С (буфер А с 0,02 мальтозой (Sigma)). Неочищенные фракции собирали и подвергали мониторингу по поглощению при 280 нм.
Элюированный белок анализировали при помощи электрофореза SDS NuPAGE (NOVEX) с окрашиванием Кумасси (Sigma). Пробу и весь объем белка хранили при -80°С.
Пример 33
Поликлональные антитела к zalpha11-лиганду человека
Поликлональные антитела к zalpha11-лиганду человека получали иммунизацией двух самок Ново-Зеландских белых кроликов очищенным полипептидом huzalpha11L/MBP-6H (пример 32) или очищенным СНО рекомбинантным белком huzalpha11L-CHO (пример 29). Каждый кролик получал исходную внутрибрюшинную инъекцию (ip) 200 мг очищенного белка в полном адъюванте Фрейнда с последующими бустерными инъекциями ip 100 мг очищенного белка в неполном адъюванте Фрейнда каждые три недели. Спустя семь-десять дней после введения второй бустерной инъекции (всего три инъекции) производили кровоизвлечение у животных и собирали сыворотку. Затем кроликам продолжали вводить бустер-инъекции и брали кровь каждые три недели.
Кроличью сыворотку, индуцированную к huzalpha11L/MBP-6H, предварительно адсорбировали антителами против МБР с использованием колонки CNBr-SEPHAROSE 4B protein (Pharmacia LKB), которую готовили с использованием 10 мг очищенного рекомбинантного МВР на грамм CNBr-SEPHAROSE (Pharmacia). Рекомбинантный МВР готовили и очищали на амилозной колонке в лаборатории с использованием способов, хорошо известных в данной области. Huzalpha11-лиганд-специфические поликлональные антитела очищали аффинно из сыворотки кроликов с использованием колонки CNBr-SEPHAROSE 4B protein (Pharmacia LKB), которую готовили с использованием 10 мг специфического антигена очищенного рекомбинантного белка huzalpha11L/MBP-6H или 10 мг очищенного СНО рекомбинантного белка huzalpha11L-CHO на грамм CNBr-SEPHAROSE с последующим 20Х диализом в 3ФР в течение ночи. Huzalpha11-лиганд-специфические антитела характеризовали при помощи ELISA с применением 1 мкг/мл очищенных рекомбинантных белков huzalpha11L/MBP-6H (пример 32), zalpha11-лиганда человека (huzalpha11L-CHO) (пример 29) или muzalpha11L-MBP/6H (пример 32) в качестве мишеней антител.
Нижним порогом детектирования (LLD) кроличьего аффинно очищенного антитела анти-huzаlрhа11/МВР-6Н было разведение 10 пг/мл на его специфическом очищенном рекомбинантном антигене huzalpha11L/MBP-6H, 500 пг/мл на очищенном рекомбинантном huzalpha11L-CHO и 100 пг/мл на очищенном рекомбикантном muzalpha11L-MBP/6H (пример 32). LLD кроличьего аффинно очищенного антитела анти-huzalpha11L-CHO было разведение 20 пг/мл на его специфическом очищенном рекомбинантном антигене huzalpha11L-CHO, 500 пг/мл на очищенном рекомбинантном huzalpha11L/MBP-6H и 50 нг/мл на очищенном рекомбинантном muzalphal 1L/MBP-6H.
Пример 34
Поликлональные антитела к пептиду zalpha11-лиганда человека
Поликлональные антитела к пептиду zalpha11-лиганда человека получали иммунизацией двух самок Ново-Зеландских белых кроликов пептидом zalpha11-лиганда человека, huzalpha11L-1 (SEQ ID NO:72) или huzalpha11L-3 (SEQ ID NO:73). Эти пептиды синтезировали с использованием синтезатора пептидов Applied Biosystems Model 431A (Applied Biosystems, Inc., Foster City, CA) в соответствии с инструкциями изготовителя. Затем пептиды конъюгировали с белком-носителем гемоцианином фиссуреллы (KLH) с активацией малеимидом. Каждый кролик получал исходную внутрибрюшинную инъекцию (ip) 200 мг очищенного белка в полном адъюванте Фрейнда с последующими бустерными инъекциями ip 100 мг очищенного белка в неполном адъюванте Фрейнда каждые три недели. Спустя семь-десять дней после введения второй бустерной инъекции (всего три инъекции) производили кровоизвлечение у животных и собирали сыворотку. Затем кроликам продолжали вводить бустер-инъекции и брали кровь каждые три недели.
Сыворотки кроликов, индуцированные к пептидам zalpha11-лиганда человека, характеризовали проверкой титра ELISA с использованием 1 мкг/мл соответствующего пептида, использованного для получения антитела (SEQ ID NO:72 или SEQ ID NO:73) в качестве мишени антитела. Две кроличьи сыворотки к пептиду huzalpha11L-1 имели титр в отношении их специфического пептида при разведении 1:5000000 (1:5×106). Две кроличьи сыворотки к пептиду huzalpha11L-3 имели титр в отношении их специфического пептида при разведении 1:5×106.
Поликлональные антитела, специфические для пептида zalpha11-лиганда человека, очищали аффинно из кроличьей сыворотки с использованием колонки CNBr-SEPHAROSE 4B protein (Pharmacia LKB), которую готовили с использованием 10 мг соответствующего специфического пептида (SEQ ID NO:72 или SEQ ID NO:73) на грамм CNBr-SEPHAROSE с последующим 20Х диализом в 3ФР в течение ночи. Huzalpha11-лиганд-специфические антитела характеризовали проверкой титра при помощи ELISA с применением 1 мкг/мл соответствующего очищенного пептидного антигена или очищенных рекомбинантных полноразмерных белков в качестве мишеней антител.
Нижним порогом детектирования (LLD) кроличьего аффинно очищенного антитела анти-huzalpha11L-l является разведение 500 пг/мл на его специфическом пептидном антигене (huzalpha11L-1; SEQ ID NO:72), 500 пг/мл на очищенном рекомбинантном huzalpha11L/MBP-6H (пример 32) и 500 пг/мл на очищенном СНО-рекомбинантном huzalpha11L-CHO (пример 29). Не наблюдали перекрестной реактивности относительно очищенного рекомбинантного muzalpha11L/MBP-6H (пример 32). LLD кроличьего аффинно очищенного антитела анти-Huzаlрhа11-3 равен 50 пг/мл на его специфическом пептидном антигене (huzalpha11L-3; SEQ ID NO:73), 50 пг/мл на очищенном СНО-рекомбинантном huzalpha11L-CHO (пример 29) и 100 пг/мл на очищенном бакуловирусном рекомбинантном huzalpha11L-Bv (пример 30). Наблюдали перекрестную реактивность в отношении очищенного muzalpha11L/MBP-6H (пример 32) с LLD 5 нг/мл.
Пример 35
Моноклональные антитела к zalpha11-рецептору человека
Моноклональные антитела к zalpha11-рецептору получали иммунизацией 5 мышей BALB/c (Harlan Sprague-Dawley, Indianapolis, IN) рекомбинантным очищенным белком растворимого рецептора, zalpha11CEE (huzalpha11-CEE-BHK) (пример 10А). Каждая мышь получала исходную внутрибрюшинную (ip) инъекцию 20 мг очищенного белка в полном адъюванте Фрейнда (Pierce, Rockford, IL) с последующими бустерными инъекциями ip 10 мг очищенного белка в неполном адъюванте Фрейнда каждые две недели. Спустя семь-десять дней после введения третьей бустерной инъекции производили кровоизвлечение у животных и собирали сыворотку.
Пробы сывороток мышей, индуцированных к huzalpha11-CEE-BHK, характеризовали проверкой титра при помощи ELISA с применением очищенного рекомбинантного СНО huzalpha11-Fc-белка (пример 10С) в качестве мишени антител. Проба одной мышиной сыворотки имела титр в отношении специфической мишени антитела при разведении 1:1000000 (1:1×106). Пробы четырех мышиных сывороток имели титр в отношении специфической мишени антитела при разведении 1:100000 (1:1×106).
Спленоциты собирали из 4 мышей с высоким титром и сливали с клетками мышиной миеломы SP2/0 с использованием ПЭГ 1500 (Boehringer Mannheim, UK) в двух отдельных процедурах слияния с применением отношения слияния 4:1 спленоцитов к миеломным клеткам (Antibodies: A Laboratory Manual. E. Harlow and D. Lane, Cold Spring Harbor Press). После 10 дней роста после слияния продуцирующие специфические антитела гибридомы идентифицировали при помощи ELISA с использованием очищенного рекомбинантного ВНК белка zalpha11 человека-Рс4 (пример 10С) в качестве мишени антитела и при помощи FACS с использованием клеток BaFS, экспрессирующих последовательность huzalpha11 (пример 4 и пример 2), в качестве мишени антител. Полученные 4 гибридомы, положительные согласно обоим способам, клонировали три раза лимитирующим разведением. Антитела были обозначены: 249.28.2.1.2.2; 247.10.2.15.4.6; 249.19.2.2.3.5 и 249.15.2.4.2.7.
Пример 36
Zalpha11-лиганд-трансгенные мыши
А. Получение трансгенных мышей, экспрессирующих zalpha11-лиганд человека и мыши
ДНК-фрагменты из трансгенных векторов (пример 22 и пример 26), содержащие 5' и 3' фланкирующие последовательности соответствующего промотора (печень-специфического промотора МТ-1 (zalpha11-лиганд мыши (пример 26В) или лимфоид-специфического промотора LCK (zalpha11-лиганд мыши и человека (примеры 26А и 22В), интрон инсулина II крысы, кДНК zalpha11-лиганда и поли А-последовательность гормона роста человека, получали и использовали для микроинъекции в оплодотворенные мышиные ооциты B6C3f1 (Taconic, Germantown, NY) с использованием стандартного протокола микроинъекции. См. Нодап, В. et al., Manipulating the Mouse Embyio. A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1994.
Восемь трансгенных мышей, экспрессирующих zalpha11-лиганд человека от лимфоид-специфического промотора EμLCK, идентифицировали среди 44 детенышей. Четыре из этих детенышей умерли, а 4 выросли до взрослого возраста. Уровни экспрессии были явно низкими в этих животных. Двадцать трансгенных мышей, экспрессирующих мышиный zalpha11-лиганд от лимфоид-специфического промотора EμLCK, идентифицировали среди 77 детенышей. Все 20 росли до взрослого состояния. Уровни экспрессии были явно низкими в этих животных. Три трансгенные мыши, экспрессирующие мышиный zalpha11-лиганд от печень-специфического промотора МТ-1, идентифицировали среди 60 детенышей. Два из этих детенышей умерли, а 1 вырос до взрослого состояния. Уровни экспрессии были явно низкими в этих животных. Ткани препарировали и гистологически исследовали, как описано ниже.
В. Микроскопическая оценка тканей трансгенных мышей
Селезенку, вилочковую железу и мезентериальные лимфатические узлы собирали и препарировали для гистологического исследования из трансгенных животных, экспрессирующих zalpha11-лиганд мыши и человека (пример 36А). Другие ткани, которые собирали рутинно, включают в себя следующие ткани: печень, сердце, легкое, почку, кожу, молочную железу, поджелудочную железу, желудок, тонкую и толстую кишку, мозг, слюнные железы, трахею, пищевод, надпочечник, гипофиз, половые пути, добавочные мужские половые железы, скелетную мышцу, включающую в себя периферический нерв, и бедренную кость с костным мозгом. Эти ткани собирали из неонатального детеныша, который умирал неожиданно, и нескольких взрослых трансгенных мышей, как описано ниже. Пробы фиксировали в 10% забуференном формалине, обрабатывали рутинным образом, заливали в парафин, делали срезы при 5 микронах и окрашивали их гематоксилином и эозином. Предметные стекла исследовал и оценивал в баллах в отношении тяжести изменений тканей (0=нет изменений, 1=легкое изменение, 2=умеренное, 3=тяжелое) ветеринар-патолог, имеющий сертификат министерства, не осведомленный о той или иной обработке.
Детеныш и 2 самки взрослых мышей, экспрессирующие zalpha11-лиганд человека, и 3 из 6 самцов взрослых мышей, экспрессирующие zalpha11-лиганд мыши, показали воспалительные инфильтраты во многих испытанных тканях. Пораженные органы варьировались в некоторой степени от мыши к мыши. Воспалительный инфильтрат состоял в основном из нейтрофилов и макрофагов в варьирующих количествах и соотношениях и имел слабую - умеренную степень по тяжести. Кроме того, эти животные обнаружили изменения в лимфоидных органах, в том числе от умеренной до тяжелой лимфопении в селезенке и вилочковой железе (с трансгенами zalpha11-лиганда человека и мыши); и тяжелую лимфопению (с трансгенами zalpha11-лиганда человека) или от слабого до тяжелого гнойного - пиогрануломатозного лимфаденита (с трансгеном zalpha11-лиганда мыши) в лимфатических узлах. Кроме того, повышенный внемедуллярный гемопоэз был очевидным в селезенках. Эти изменения не наблюдали в контрольных мышах такого же возраста.
С. Проточный цитометрический анализ тканей из трансгенных мышей, сверхэкспрессирующих zalpha11-лиганд
Трансгенных животных, сверхэкспрессирующих zalpha11-лиганд человека или мыши (пример 36А), умерщвляли для проточного цитометрического анализа периферической крови, вилочковой железы (тимуса), лимфатического узла, костного мозга и селезенки.
Клеточные суспензии готовили из селезенки, тимуса и лимфатических узлов разрыванием органа пинцетом в охлажденной на льду культуральной среде (500 мл среды RPMI 1640 (JRH Biosciences, Lenexa, KS); 5 мл 100х L-глутамина (Gibco BRL, Grand Island, NY); 5 мл 100х Na-пирувата (Gibco BRL); 5 мл 100х пенициллина, стрептомицина, неомицина (PSN) (Gibco BRL) и затем осторожно раздавливали эти клетки через клеточный деформатор (Falcon, VWR Seattle, WA). Периферическую кровь (200 мл) собирали в гепаринизированные пробирки и разбавляли до 10 мл HBSS, содержащим 10 Е гепарина/мл. Эритроциты удаляли из препаратов селезенки и периферической крови гипотоническим лизисом. Суспензии костного мозга готовили вымыванием костного мозга из бедренных костей охлажденной на льду культуральной средой. Клетки считали и испытывали на жизнеспособность с использованием Трипанового синего (GIBCO BRL, Gaithesburg, MD). Клетки ресуспендировали в охлажденной на льду окрашивающей среде (HBSS, 1% фетальная телячья сыворотка, 0,1% азид натрия) при концентрации десять миллионов на миллилитр. Блокирование Fc-рецептора и неспецифического связывания антител с клетками достигалось добавлением 10% нормальных козьих сывороток и Fc-блока (Pharmingen, La Jolla, CA) к суспензии клеток.
Суспензии клеток смешивали с равными объемами меченных флуорохромом моноклональных антител (PharMingen), инкубировали на льду в течение 60 минут и затем промывали дважды охлажденным на льду промывным буфером (ЗФР, 1% фетальная телячья сыворотка, 0,1% азид натрия) перед ресуспендированием в 400 мл промывного буфера, содержащего 1 мг/мл 7-AAD (Molecular Probes, Eugene, OR) в качестве маркера жизнеспособности в некоторых пробах. Проточные данные получали на проточном цитометре FACSCaIibur (BD Immunocytometry Systems, San Jose, CA). Как получение, так и анализ выполняли с использованием программного обеспечения CellQuest (BD Immunocytometry Systems).
Трансгенные животные, которые экспрессировали zalpha11-лиганд человека или мыши с высокими уровнями, имели драматически измененные популяции клеток во всех анализированных лимфоидных органах. Наблюдаемые изменения включали в себя полную потерю насыщенности клетками тимуса, полное отсутствие СD45R-позитивных В-клеток и увеличенные размер и насыщенность клетками селезенок. Как селезенка, так и костный мозг имели увеличенные числа клеток миелоидного размера, которые обусловлены увеличениями в количестве моноцитов и нейтрофилов. Маркер pan NK-клеток (DX5) увеличивался во многих популяциях. Умеренно зкспрессирующие основатели имели менее драматические, но все еще значимые изменения, согласующиеся с фенотипом, наблюдаемым в клетках с высокой экспрессией. Мыши с самым низким уровнем экспрессии не имели ни значимого увеличения миелоидных клеток, ни уменьшения чисел В-клеток. Они действительно обнаруживали значимые изменения в популяциях тимоцитов с уменьшениями CD4+CD8+ дважды положительных клеток и увеличениями как в CD4+, так и CD8+ отдельно положительных клеток.
Пример 37
Исследование зависимости ответной реакции от дозы белка очищенного рекомбинантного zalpha11-лиганда человека в нормальных мышах
А. Сущность исследования
Нормальных шестинедельных самок мышей С57В1/6 (Harlan Sprague-Dawley, Indianapolis, IN) обрабатывали внутрибрюшинной инъекцией один раз в день в течение четырех или восьми дней одним из четырех уровней доз очищенного рекомбинантного zalpha11-лиганда человека (пример 24) при 0,1, 0,5, 5 или 50 мкг/мышь/день или носителем в качестве контроля. Веса тела и температуры тела подвергали мониторингу один раз в день. В день 4 или в день 9, четыре из восьми мышей из каждой группы обработки белком и пять из десяти мышей в контрольной группе с носителем умерщвляли. Кровь, костный мозг и ткани собирали и анализировали. Исследовали потенциальные нарушения в лимфоидных тканях, а также обычные физиологические и токсикологические параметры.
Не было доказательства токсичности белка zalpha11-лиганда человека при любой из испытанных доз. Веса тела и температуры тела были неизмененными. Не было видимых изменений в параметрах клинической химии. Однако были стойкие факты, относящиеся к увеличенным процентам клеток миелоидной линии дифференцировки в костном мозгу, селезенке и периферической крови в мышах, обработанных самой высокой дозой zalpha11-лиганда, в сравнении с контролем-носителем. Наблюдалось статичтически значимое увеличение клеток с размером клеток миелоидной линии дифференцировки, идентифицированных проточно-цитометрическим анализом гомогената селезенки в группе с высокой дозой. Селезенки двух групп с наиболее высокой дозой были статистически значимо более крупными, чем селезенки других групп. Однако при гистопатологическом исследовании только несущественное увеличение внемедуллярного гемопоэза наблюдали в группе с наиболее высокой дозой. Статистически значимое увеличение было в отношении миелоидных клеток к эритроидным в костном мозгу в группе с наивысшей дозой в сравнении с другими группами. Наконец, наблюдали увеличения в периферической крови как чисел общих лейкоцитов, так и процента моноцитов в той же самой группе.
В. Приготовление растворов для введения доз
Очищенный рекомбинантный zalpha11-лиганд человека (пример 24) разбавляли в стерильный забуференный фосфатом солевой раствор (Gibco BRL, Grand Island, NY) при концентрациях для доставки 50, 5, 0,5 или 0,1 микрограммов белка в 0,1 мл носителя-ЗФР. Дозы для первых четырех дней готовили в день 0 и замораживали в холодной морозильной камере при -20°С до использования. Дозы для дней 5-8 готовили в день 5 и замораживали, как описано выше. Аликвоты того же самого ЗФР замораживали для обработанной носителем контрольной группы. В день введения соответствующие аликвоты оттаивали и 0,1 мл раствора инъецировали внутрибрюшинно в каждую мышь в течение либо четырех, либо восьми дней.
С. План исследования
Мыши были шестинедельными в начале этого исследования. Каждая группа обработки состояла из восьми мышей за исключением контрольной группы с носителем, которая состояла из десяти мышей. Половину мышей каждой группы умерщвляли после четырех дней обработки, а другую половину после восьми дней.
Перед обработкой каждый день каждую мышь взвешивали и температуру ее тела регистрировали с использованием портативной системы программируемого ноутбука (BMDS, Inc, Maywood, NJ) сканированием мыши на идентификационный номер и температуру тела из имплантированных подкожно ответчиков (IPTT-100, BMDS, Maywood, NJ).
При умерщвлении ткани собирали для оценки популяций лейкоцитов при помощи проточного цитометрического анализа, в том числе костный мозг, тимус и селезенку. FACS-анализ лимфоидных органов и костного мозга выполняли с использованием FACSCalibur (Becton Dickinson, Mansfield, MA). Ткани, собранные для гистологического исследования на признаки токсичности белка, включали в себя: селезенку, тимус, печень, почку, надпочечник, сердце и легкие. Все ткани, фиксированные для гистологии, выдерживали при 4°С в течение ночи в 10% нормальном забуференном солевом растворе (NBF) (Surgipath, Richmond, IL). На следующий день NBF заменяли 70% этанолом и ткани возвращали в условия 4°С до обработки для гистологии.
Ткани обрабатывали и окрашивали гематокислином и эозином в лаборатории и затем отсылали работающему по контракту патологу на гистологический анализ. Кровь собирали для клинического анализа крови (СВС) и показателей химического профиля сыворотки, СВС анализировали у себя в лаборатории при помощи анализатора Cell Dyn 3500 Hematology Analyzer (Abbott Diagnostics Division, Abbott Park, IL) и полученную мануально лейкоцитарную формулу анализировали в лаборатории Phoenix Central Laboratory (Everett, WA). Сыворотку хранили в замороженном виде при -20°С до предоставления в Phoenix Central Laboratory для получения полных панелей химических показателей сыворотки. Для оценки отношений миелоид : эритроид, костный мозг из одной бедренной кости наносили на предметные стекла CytoSpin (CYTOSPIN 3 CYTOCENTRIFUGE и CYTO SLIDES, Shandon, Pittsburgh, PA) и отсылали в Phoenix Central Laboratory для анализа.
D. Результаты исследования
Не было видимых клинических указаний относительно физиологических эффектов или токсичности zalpha11-лиганда человека при дозах 50 мкг/день или более низких дозах. Веса тела и температуры оставались нормальными на протяжении этих обработок. Химические показатели сывороток были в пределах нормы. Числа эритроцитов и тромбоцитов были, по-видимому, нормальными. В мышах, получающих 50 мкг/день в течение 8 дней, мануально полученная лейкоцитарная формула показала, что процент моноцитов был повышенным в периферической крови и было видимое увеличение в общем числе лейкоцитов. В костном мозгу, вымытом из бедренной кости, отношения миелоид-эритроид увеличивались в группе с дозой 50 мкг и в меньшей степени в группе с дозой 5 мкг из 8-дневного предоставления доз. В непараметрическом сравнении множественных столбцов с использованием InStat (InStat MAC; GraphPad Software, Inc., San Diego, CA) это различие было статистически значимым (р=0,0049). Различие между группой с наивысшей дозой и группой носителя было также значимым (р=0,0286). Таким образом, увеличенное число лейкоцитов в периферической крови и значимое увеличение миелоидных предшественников могут быть связанными друг с другом.
Гистологическая оценка следующих тканей не обнаружила видимого доказательства цитологических или структурных изменений, митотических событий или некроза: тимуса, печени, почки, надпочечника, двенадцатиперстной кишки, поджелудочной железы, тощей кишки, слепой кишки, ободочной кишки, мезентериальных лимфатических узлов матки, яичника, слюнных желез, сердца, трахеи, легкого и мозга. Не было видимых различий между группами обработки в весе тимуса, почки, печени или мозга. Из всех испытанных тканей было значимое влияние только на вес селезенки.
Вес селезенки каждой мыши нормализовали относительно веса ее мозга. В группе обработки 50 мкг/день в сравнении с носителем, группами обработки 0,1 мкг и 0,5 мкг, средние веса селезенки были почти на 50% больше после четырех дней обработки и почти на 100% больше после восьми дней, чем средние веса селезенки остальных трех групп. В 4-дневной постановке эксперимента группа 5 мкг/день также имела тенденцию увеличения селезенок в сравнении с контрольной группой и группами с низкими дозами. Различие в отношениях весов селезенки/мозга с данными из 4-дневной и 8-дневной постановок, комбинированных с группой обработки, было статистически значимым (р=0,0072) согласно непараметрическрму ANOVA Kruskall-Wallace, тесту сравнения множественных столбцов с использованием программы InStat (GraphPad Software).
Незначительное увеличение внемедуллярного гемопоэза, в частности, в красной пульпе, было заметно в селезенках мышей из группы с наивысшей дозой, даже в мышах, обработанных в течение четырех дней. Проточный цитометрический анализ селезенок показал значимое увеличение в доле клеток миелоидного размера в группе с наивысшей дозой (р=0,01, t-критерий Стьюдента), представляющей увеличения как моноцитов, так и нейтрофилов. Этот эффект может быть связан с увеличенным процентом мононуклеарных клеток периферической крови, а также видимым увеличением миелоидных предшественников в костном мозгу, описанными выше. Кроме того, трансгенные мыши, полученные инсертированием гена zalpha11 человека, имели увеличенный внемедуллярный гемопоэз в их селезенках, в сравнении с нетрансгенными однопометными животными.
Несколько изменений наблюдали в группе с дозой 50 мкг/день в сравнении с контрольной группой, которые предполагают участие zalpha11-лиганда в образовании и развитии клеток миелоидной линии дифференцировки. Взятые вместе, эти наблюдаемые изменения предполагают, что zalpha11 может быть применим в качестве терапевтического белка в таких медицинских приложениях, как рак и иммунологические нарушения, описанные здесь.
Пример 38
Предварительное исследование элиминации и тканевого распределения очищенного рекомбинантного белка zalpha11-лиганда человека
А. Сущность исследования
Для выяснения характера тканевого распределения и элиминации очищенного rhzalpha11-лиганда было предпринято предварительное фармакокинетическое исследование. Девятинедельные самцы мышей С57В1/6 получали очищенный рекомбинантный белок zalpha11-лиганда человека, меченный 111индием (111ln) (NEN, Boston, MA) одним из трех путей. Каждая мышь получала одну болюсную инъекцию внутривенным (iv), внутрибрюшинным (ip) или подкожным путем (sc). Мышей, инъецированных подкожным или внутрибрюшинным путем, умерщвляли при одном или трех часах после инъекции. Мышей, инъецированных внутривенно, умерщвляли после десяти минут или после одного часа после инъекции. Кровь, плазму и выбранные ткани собирали в различных временных точках и считали гамма-счетчиком для определения полупериода существования в организме и тканевого распределения экзогенно меченного белка. Ткани, которые собирали для счета, а также интервалы умерщвления выбирали на основе сообщений о распределении других цитокинов, меченных радионуклидами.
При умерщвлении собирали ткани для счета радиоактивности, включенной в тимус, селезенку, почку, долю печени, долю легкого и мочевой пузырь. В группе, получающей внутрибрюшинную инъекцию, кишечник также считали для оценки частоты встречаемости введения в кишечник, а в получающей подкожные инъекции мышах считали кожу с подстилающими структурами в зоне инъекции. Число импульсов в минуту для целой печени и целого легкого рассчитывали из участка, который считали, и процента от веса целого органа, представленного этим участком.
После окончания этого исследования собранные ткани, цельную кровь и плазму считали на гамма-счетчике COBRA II AUTO-GAMMA, (Packard Instrument Company, Meriden, CT). Аликвоту исходного меченого раствора для введения доз также считали в конце этого исследования с тканями. Это позволяло рассчитать процент общей инъецированной радиоактивности для каждой мыши и произвести одновременную коррекцию всех чисел имп./мин на радиоактивный распад. Аппроксимации остального объема крови и весов органов показали, что большая часть введенных импульсов была учтена, и, следовательно, проценты имульсов на ткань были разумным представлением распределения импульсов после введения меченого zalpha11-лиганда каждым из путей.
В. Мечение 111индием zalpha11-лиганда
Очищенный рекомбинантный zalpha11-лиганд человека (пример 29) конъюгировали с 10-кратным избытком ДТПА (Pierce, Rockford, II) инкубированием в течение 30 минут при комнатной температуре в ЗФР. Непрореагировавший ДТПА и гидролизаты удаляли сменой буфера на Biomax-5k NMWL (Ultrafree-15, Millipore, Bedford, MA). Пик белка свободного объема концентрировали до 5 мг/мл и брали аликвоту для тестирования в биоанализе (анти-СD40-стимуляция мышиных В-клеток) (пример 44)). При подтверждении, что ДТПА-конъюгат все еще обладает полной биологической активностью, этот конъюгат разбавляли до 0,5 мг/мл 1 М ацетатом натрия рН 6,0. Два мКи 111индия поглощали в 0,5 мл 1 М ацетата натрия рН 6,0 и смешивали с конъюгатом ДТПА-zаlрhа11-лиганд человека в течение 30 минут при комнатной температуре. Невключенный 111индий удаляли во время смены буфера на ЗФР на колонке PD-10 (Pharmacia, Piscataway, NJ). Радиоактивно меченный материал разбавляли немеченым zalpha11-лигандом человека с получением удельной активности 100 мКи/мг, стерильно фильтровали и выдерживали при 4°С в течение ночи. Сто процентов меченого белка удерживались на мембране Biomax-Sk NMWL (Millipore). Меченный 111индием zalpha11-лиганд человека вводили мышам в исследованиях элиминации и фармакокинетических исследованиях. Пятьдесят мкг белка zalpha11-лиганда человека, меченного 5 мкКи меченого zalpha11-лиганда человека, в 0,1 мл носителя-ЗФР вводили в каждое животное.
С. Результаты предварительного исследования распределения
После одного и трех часов после введения всеми тремя путями, наивысшая концентрация 111ln-zаlphа11-лиганда человека была обнаружена в почке, а вторая наивысшая концентрация была в моче и мочевом пузыре, как показано этими тканями, имеющими наивысшие имп/мин. Средние имп/мин, извлеченные из почек, были в 3-8 раз более высокими, чем имп/мин целой печени в зависмости от пути инъекции и временной точки умерщвления. Например, среднее число имп/мин почек при 60 минутах после iv инъекции было в 4,5 раз более высоким, чем средние имп./мин, рассчитанные для целой печени из той же самой группы. В группе, которую умерщвляли через десять минут после внутривенного введения, наивысшее число имп/мин было опять в почке, а второе наивысшее накопление было эквивалентным в печени, мочевом пузыре и моче.
D. Предварительное фармакокинетическое исследование
Взятия крови и плазмы проводили при 10, 30 и 60 минутах после инъекции всеми тремя путями. После инъекции iv путем в отдельных группах мышей пробы крови и плазмы брали при двух, пяти и десяти минутах. В другой группе мышей, которые получали их инъекции ip или sp путем, пробы крови брали при одном, двух и трех часах. В отношении обработки групп см. таблицу 6. Короткие времена сбора охватывают сообщеннный полупериод существования в организме IL-2 после внутривенной инъекции. Сообщенный Т1/2 был в пределах 2,5-5,1 минут. В отношении введения in vivo IL-2, см. Donohue JH and Rosenberg SA J Immunol. 130:2203, 1983. Продолжительные временные точки были выбраны для определения предполагаемой фазы элиминации.
Было показано, что IL-2 элиминируется из сыворотки с полупериодом существования в организме приблизительно три минуты в мышах после iv инъекции. В отношени ссылки см. Donohue, JH and Rosenberg SA supra. После ip и sc инъекции сходных количеств IL-2 продолжительность сохранения активности IL-2 в сыворотке пролонгировалась с 2 единиц/мл в течение менее 30 минут после iv инъекции до более 2 единиц/мл в течение 2 часов после ip и 6 часов после sc инъекций. Основным путем клиренса IL-2, по-видимому, являются почки. Было показано, что zalpha11-лиганд является структурно сходным с IL-2, как обсуждалось здесь. Предварительная оценка элиминации zalpha11-лиганда, по-видимому, согласуется с видимым клиренсом IL-2 почками, на основе накопления имп/мин преимущественно в почках, а затем мочевым пузырем и мочой в данном исследовании.
Были сделаны оценки фармакокинетических параметров на основе некомпартментального анализа данных имп./мин, полученных из плазмы, с использованием программы РК-анализа WinNonLin, Version 1.1. (Scientific Consulting Inc., Gary, NC). Полупериоды существования zalpha-11-лиганда в плазме оценивали с использованием предсказанных констант скоростей терминальной элиминации для внутривенного, подкожного и внутрибрюшинного введения дозы 50 мкг. Фармакокинетические результаты оценивали вследствие ограниченных точек данных в терминальной области элиминации концентрации плазмы против временных профилей. Кроме того, построение терминальной фазы элиминации для введения доз sc и ip требовало применения данных из временных точек, во время которых все еще, по-видимому, имеет место абсорбция 111ln-zalpha11-лиганда человека. Однако оценки полупериодов существования в организме после внутривенного, подкожного и внутрибрюшинного введения доз были 13,6 минут, 18,8 минут и 34,3 минут соответственно. Поскольку диапазон введения доз не оценивался, неясно, имела ли место насыщаемая или активная элиминация (кинетика Михаэлиса-Ментена). Таким образом, эти полупериоды существования в организме являются приблизительными оценками.
Приблизительные расчеты биодоступности меченого белка проводили на основе площадей под кривой (AUC) после подкожного или внутрибрюшинного введения доз в сравнении с площадью под кривой после внутривенного введения доз. Оцененная биодоступность после подкожной и внутрибрюшинной инъекции была 35,8 и 63,9% соответственно. Поскольку исследовали только одну дозу белка, биодоступность не была оценена в зависимости от дозы. Приблизительно расчитанные клиренс и объем распределения (на основе данных из внутривенной инъекции) были 0,48 и 6,1 мл/мин соответственно.
Хотя эти данные являются предварительными, судьба zalpha11-лиганда, вводимого iv, была сходной с судьбой, сообщенной для IL-2, другого цитокина со структурой 4-спирального пучка (Donohue JH and Rosenberg SA, supra). Подобно IL-2 iv вводимый zalpha11-лиганд имеет полупериод существования в плазме только минуты с основным клиренсом в почке. Спустя три часа после инъекции основная часть меченого материала, эестрагированного из почки, все еще удерживалась мембраной Biomax 5K NMLW (Millipore). Поскольку ранее сообщалось, что индий остается связанным с белком даже во время лизосомальной деградации (Staud, F. et al., J. Pharm. Sciences 88:577-585, 1999), zalpha11-лиганд накапливается и может быть деградирован в почке. Настоящее исследование показало также, как наблюдалось и со многими другими белками, в том числе IL-2 (Donahue JH and Rosenberg SA, supra), что введение ip и sc значимо пролонгировало уровни в плазме zalpha11-лиганда.
Пример 39
Выделение и размножение свежей CD34+ фракции MNC костного мозга человека с использованием zalpha11-лиганда для оценки NK-активности
А. Отбор и выделение CD34+ клеток из костного мозга человека
Свежие мононуклеарные клетки (MNC) костного мозга человека получали для обогащения в отношении клеток, имеющих активность МК-клеток (природных клеток-киллеров). Свежие MNC человека получали из Poeitic Technologies (Gaithersburg, MD). 10 мл альфа-МЕМ (JRH, Lenexa, KS), содержащей 10% ФТС HIA (Hyclone, Logan, UT) и антибиотик 1% PSN (Gibco BRL, Grand Island, NY), добавляли к суспензии этих клеток и клетки пропускали через сито 100 мкм. Затем клетки считали, осаждали, промывали 10 мл 3ФР, содержащего 2% ФТС, затем снова осаждали и ресуспендировали в 1 мл ЗФР, содержащего 2% ФТС. Клетки, имеющие маркер клеточной поверхности CD34 (CD34+ клетки), отделяли при помощи магнитных частиц с использованием набора Detachabead с Dynabeads М-450 CD34 (Dynal, Oslo, Norway) в соответствии с инструкциями изготовителя. Как фракции CD34+ клеток, так и фракции CD34- клеток дополнительно анализировали, как описано ниже.
В. Размножение CD34+ клеток с использованием zalpha11-лиганда
Фракцию CD34+ клеток высевали в четыре лунки в 24-луночном планшете. 50000 положительно отобранных клеток, суспендированных в 1 мл среды альфа-МЕМ (JRH), содержащей 10% ФТС HIA (Hyclone) и 1% PSN (Gibco BRL) плюс различные цитокины, описанные ниже, высевали в каждую из 4 лунок (1-4). Различные реагенты использовали для тестирования на индуцированное zalpha11-лигандом размножение CD34+ отобранных MNC костного мозга: реагенты включали в себя: flt3 (R&D, Minneapolis, MN); очищенный zalpha11-лиганд человека (пример 30С и пример 30D); IL-15 человека (R&D). Реагенты комбинировали следующим образом в день 0: В лунку №1 добавляли 2 нг/мл flt3 человека. В лунку №2 добавляли 2 нг/мл flt3 человека и 15 нг/мл очищенного zalpha11-лиганда человека. В лунку №3 добавляли 2 нг/мл flt3 человека и 20 нг/мл IL-15 человека. В лунку №4 добавляли 2 нг/мл flt3 человека, 15 нг/мл очищенного zalpha11-лиганда человека и 20 нг/мл IL-15 человека. После инкубации в течение 18 дней клетки суспензии из каждой лунки осаждали и затем ресуспендировали в 0,5 мл среды альфа MEM (JRH), содержащей 10% ФТС HIA (Hyclone) и 1% PSN (Gibco BRL), и считали для оценки пролиферации фракции CD34+ клеток. Низкий уровень пролиферации наблюдали в присутствии только flt3 (контрольная лунка №1), но присутствие IL-15 или zalpha11 в дополнение к flt3 не оказывало значимого влияния на размножение (лунки №2 и №3). Однако размножение выше контроля flt3 было очевидным в лунке №4, которая содержала IL-15 и zalpha11-лиганд, кроме flt3. Этот результат предполагает, что zalpha11 и IL-15 действуют синергически в размножении популяции CD34+ клеток человека. Кроме того, результаты этого эксперимента подтверждали результаты, наблюдаемые с мышиным zalpha11-лигандом в мышином ВМ-тесте (пример 21).
Затем все популяции клеток тестировали на NK-активность и подвергали проточному цитометрическому анализу, как показано ниже (пример 41).
С. Размножение CD34+ или CD34- клеток с использованием zalpha11-лиганда с задержанным добавлением IL-15
Как CD34+, так и CD34- фракции высевали отдельно в лунки 12-луночного планшета (1-6). Каждая из шести лунок содержала 100000 положительно или отрицательно отобранных клеток в 2 мл альфа MEM, содержащей 10% ФТС HIA и PSN, описанной выше. В лунку №1 добавляли 2 нг/мл flt3 человека в день 0. В лунку №2 добавляли 2 нг/мл flt3 человека в день 0 и после 5 дней инкубации 20 нг/мл IL-15 человека. В лунку №3 добавляли 2 нг/мл flt3 человека и 15 нг/мл zalpha11-лиганда человека в день 0. В лунку №4 добавляли 2 нг/мл flt3 человека и 15 нг/мл zalpha11-лиганда человека в день 0 и после 5 дней инкубации 20 нг/мл IL-15 человека. В лунку №5 добавляли 2 нг/мл flt3 человека и 20 нг/мл IL-15 человека в день 0. В лунку №6 добавляли 2 нг/мл flt3 человека, 15 нг/мл zalpha11-лиганда человека и 20 нг/мл IL-15 в день 0. После инкубации в течение всего 15 дней от начала эксперимента клетки из каждой лунки собирали и считали.
В CD34+ популяции наблюдали низкий уровень пролиферации в присутствии только flt3 (контрольная лунка №1), но присутствие IL-15 или zalpha11, добавляемых в день 0 к flt3, не оказывало значимого действия на размножение (лунки №3 и №5). Добавление IL-15 после 5 дней оказывало некоторое пролиферативное действие в сравнении с flt3-контролем (лунка №2 в сравнении с лункой №1) и пролиферативное действие в присутствии zalpha11 (лунка №4 в сравнении с лункой №3). Однако наибольшее размножение наблюдали в лунке №6, которая содержала IL-15 и zalpha11-лиганд в дополнение к flt3 в день 0.
В CD34- популяции не наблюдали пролиферации в присутствии только flt3 (контрольная лунка №1) и фактически наблюдали уменьшение клеточной популяции. Присутствие zalpha11, добавленного в день 0 в дополнение к flt3 (лунка №3), было сходным с flt3-контролем. Присутствие IL-15, добавленного в день 5, увеличивало пролиферирующее действие на клетки в присутствии (клетка №4) или в отстутствие (клетка №2) zalpha11-лиганда. Опять наивысшее размножение было очевидным в лунке №6, которая содержала IL-15 и zalpha11-лиганд, кроме flt3, в день 0.
Затем все популяции клеток тестировали на МК-активность и подвергали FACS-анализу, как показано ниже (пример 40).
Пример 40
Выделение и размножение свежих мышиных клеток с использованием zalpha11-лиганда человека и мыши для оценки NK-активности и маркеров NK-клеток
А. Выделение и размножение свежих клеток низкой плотности костного мозга с использованием zalpha11-лиганда человека и мыши
Свежие клетки костного мозга мышей выделяли наложением зажимов на оба конца мышиных бедренных костей и промыванием двумя-тремя миллилитрами среды для выращивания (см. ниже) через внутреннее пространство кости в пробирку для сбора. Среда для выращивания была 500 мл среды RPMI 1640 (JRH Biosciences, Lenexa, KS); 5 мл 100х L-глутамина (Gibco BRL, Grand Island, NY); 5 мл 100х Na-пирувата (Gibco BRL); 5 мл ЮОх пенициллина, стрептомицина, неомицина (PSN) (Gibco BRL); и 50 мл инактивированной нагреванием фетальной телячьей сыворотки (ФТС) (Hyclone Laboratories, Logan, UT). Затем клетки костного мозга разрушали пипетированием среды вверх и вниз несколько раз. Затем эти клетки осаждали и промывали один раз средой для выращивания и пропускали через сито 70 микрон. Затем мононуклеарные клетки низкой плотности выделяли подверганием клеток костного мозга градиенту плотности. Клетки костного мозга в пяти-восьми миллилитрах среды для выращивания осторожно пипетировали на верхнюю часть пяти-восьми миллилитров NycoPrep 1,077 Animal (Nycomed, Oslo, Norway) в центрифужной пробирке. Затем этот градиент центрифугировали при 600 × g в течение 20 минут. Мононуклеарные клетки низкой плотности собирали из межфазного слоя между NycoPrep и средой. Затем эти клетки разбавляли до приблизительно 20 миллилитров в среде для выращивания, осаждали и промывали. Затем клетки высевали при приблизительно 0,5-1,5×106 клеток/мл в среде для выращивания в стандартной колбе для культуры ткани и инкубировали при 37°С, 5% CO2 в течение двух часов.
Затем неприкрепленные имеющие низкую плотность (NA LD) клетки костного мозга собирали и высевали при 0,5-2,0×105 клеток на миллилитр в среде для выращивания плюс 2,5 нг/мл мышиного flt3 (R&D Systems, Minneapolis, MN) плюс 25-50 нг/мл интерлейкина 15 (IL-15) человека (R&D Systems) с 50-150 нг/мл zalpha11-лиганда человека или без него; или с 0,12-10 нг/мл мышиного zalpha11-лиганда или без него.
Не было значимого размножения без добавления zalpha11-лиганда человека или мыши. Неприкрепленные клетки размножались в культуре, содержащей мышиный zalpha11-лиганд, всего лишь при 0,12 нг/мл, и в культурах, содержащих zalpha11-лиганд человека, всего лишь 22 нг/мл. В культурах, содержащих как человеческий, так и мышиный zalpha11-лиганд, размножение неприкрепленных клеток увеличивалось с увеличением дозы zalpha11-лиганда, причем насыщающая реакция мышиного лиганда находится при приблизительно 5-10 нг/мл, а zalpha11-лиганд человека не достигает насыщающей реакции даже при наивысшей дозе 200 нг/мл. Zalpha11-лиганд человека является, по-видимому, приблизительно в 20-100 раз более сильным на мышиных клетках, чем мышиный zalpha11-лиганд. Спустя приблизительно пять-десять дней размноженные zalpha11-лигандом мышиные клетки собирали и анализировали проточной цитометрией (FACSCalibur; Becton Dickinson, Mansfield, MA) для определения, какой их процент был положительным в отношении антигенов NK-клеток, причем 46% были положительными в отношении маркера PanNK-клеток DX5 (Pharmingen).
В. Выделение и размножение свежих лишенных линии дифференцировки клеток мышиного костного мозга
Свежие мышиные лишенные линии дифференцировки (lin-) клетки костного мозга выделяли из свежих клеток мышиного костного мозга сначала инкубированием этих клеток со следующими антителами: TER119, Gr-1, B220, MAC-1, CD3e и l-Ab (Pharmingen, San Diego, CA). Затем lin+ клетки удаляли с использованием овечьих антител против крысиного IgG Dynabeads M-450 (Dynal, Lake Success, NY) в соответствии с инструкциями изготовителя.
Затем отрицательно отобранные lin- клетки костного мозга высевали, как описано выше, на среду для выращивания плюс либо 2,5 нг/мл flt3 (R&D Systems) и 25 нг/мл IL-15 (R&D Systems), либо на содержащую flt3, IL-15 и мышиный zalpha11-лиганд, 2-5% ВНК-мышиный zalpha11-лиганд кондиционированную среду. После шести дней выращивания культуру собирали, считали и предоставляли для анализа активности NK-клеток (пример 41). Клетки, выращенные с мышиным zalpha11-лигандом, были приблизительно в два-три раза более эффективными при лизисе клеток-мишеней NK-клеток (YAC-1-клеток), чем клетки, растущие без zalpha11-лиганда.
С. Выделение и размножение CD4- CD8- (дважды негативных или DN) тимоцитов
Свежие мышиные тимоциты выделяли нарезанием и просеиванием тимусов из трех-восьминедельных мышей. Затем CD4- CD8- (DN) клетки негативно отбирали инкубацией тимоцитов с антителами анти-CD4 и анти-CD8 (PharMingen) с удалением затем CD4+ CD8+ клеток с использованием овечьих антител против крысиного IgG Dynabeads M-450 (Dynal) в соответствии с инструкциями изготовителя.
Затем мышиные DN-тимоциты выращивали в среде для выращивания плюс 2,5 нг flt3 (R&D Systems), 25 нг/мл IL-15 (R&D Systems) и 10 нг/мл IL-7 (R&D Systems) с мышиным zalpha11-лигандом или без него, как описано выше. Спустя шесть дней клетки собирали, считали, анализировали проточной цитометрией, как описано выше, и также предоставляли для анализа активности NK-клеток (пример 41).
Культура, росшая с мышиным zalpha11-лигандом, давала приблизительно 480000 клеток, тогда как культура без zalpha11-лиганда давала приблизительно 160000 клеток. Было обнаружено, что культура, росшая с мышиным zalpha11-лигандом, является приблизительно на 16,2% положительной в отношении антигена NK-клеток Pan NK, DX5 (PharMingen). Культура, росшая без zalpha11-лиганда, была на 14,6% положительной в отношении DX5. Клетки, выращенные с zalpha11-лигандом, лизировали клетки-мишени NK-клеток, YAC-1, приблизительно в два раза лучше, чем клетки, выращенные без zalpha11-лиганда. Размноженные клетки не лизировали значимо отрицательную контрольную линию клеток-мишеней, EL4. Эти результаты предполагают, что zalpha11-лиганд избирательно размножает литические NK-клетки.
Пример 41
Активность размноженных zаlрhа11-лигандом человека и мыши клеток и зрелых мышиных NK-клеток в анализах цитотоксичности NK-клеток
А. Анализ NK-клеток
Опосредованный NK-клетками цитолиз испытывали при помощи стандартного теста высвобождения 51Cr. Клетки-мишени (клетки К562 (АТСС No. CRL-243) в анализах человека и клетки YAC-1 (АТСС No. TIB-160) в анализах мыши) лишены молекул главного комплекса гистосовместимости (МНС), что придает им чувствительность к опосредованному NK-клетками лизису. Отрицательной контрольной линией клеток-мишеней в анализе мышей является МНС+ тимома EL4 (АТСС No. TIB-39). Авторы изобретения выращивали клетки К562, EL4 и YAC-1 в среде RP10 (стандартная RPMI 1640 (Gibco BRL, Grand Island, NY), дополненная 10% ФТС (Hyclone, Logan, UT), а также 4 мМ глутамином (Gibco BRL), 100 МЕ/мл пенициллина+100 MCG/мл стрептомицина (Gibco BRL), 50 мкМ β-меркаптоэтанолом (Gibco BRL) и 10 мМ HEPES-буфером (Gibco BRL). В день анализа 1-2×106 клеток-мишеней собирали и ресуспендировали при 2,5-5×106 клеток/мл в среде RP10. Авторы добавляли 50-100 мкл 5 мКи/мл 51Cr-хромата натрия (NEN, Boston, MA) непосредственно к этим клеткам и инкубировали их в течение 1 часа при 37°С, затем промывали их два раза 12 мл ЗФР и ресуспендировали их в 2 мл среды RP10. После счета клеток на гемоцитометре клетки-мишени разбавляли до 0,5-1×105 клеток/мл и 100 мкл (0,5-1×104 клеток) смешивали с эффекторными клетками, как описано ниже.
В анализе человека эффекторные клетки получали из отобранных и размноженных CD34+ ВМ-клеток (пример 39В), которые собирали, промывали, считали, смешивали при различных концентрациях с 51Cr-меченными клетками-мишенями в 96-луночных круглодонных планшетах и инкубировали в течение 4 часов при 37°С. После коинкубирования эффекторных клеток и меченых клеток-мишеней половину супернатанта из каждой лунки собирали и считали в гамма-счетчике в течение 1 минуты на пробу. Процент специфического высвобождения 51Cr рассчитывали из формулы 100 х (X-Y)/(Z-Y), где Х обозначает высвобождение 51Cr в присутствии эффекторных клеток, Y обозначает самопроизвольное высвобождение в отсутствие эффекторов и Z обозначает общее высвобождение 51Cr из клеток-мишеней, инкубированных с 0,5% Тритоном Х-100. Данные строили в виде графика в виде % специфического лизиса в зависимости от отношения эффектор-мишень в каждой лунке.
В. Активность размноженных zalpha11-лигандом человека клеток
Выделенные CD34+ НРС-клетки человека, культивируемые с flt3 +/-zalpha11-лиганд и flt3 + IL-15 +/- zalpha11-лиганд (пример 39), собирали в день 15 для оценки их способности лизировать МНС- К562-клетки в стандартном анализе высвобождения 51Cr, как описано выше, и для анализа фенотипа их поверхности проточной цитометрией. Как ожидалось из предыдущих сообщений, (Mrozek, E et al. Blood 87:2632-2640, 1996; и Yu, H et al. Blood. 92:3647-3657, 1998), одновременное добавление IL-15 и flt3L действительно индуцировало рост небольшой популяции CD56- клеток. Интересно, что, хотя клетки ВМ, культивируемые одновременно с zalpha11-лигандом и flt3L, не размножались значимо, было значимое увеличение в общем числе клеток в культурах, содержащих комбинацию flt3L, zalpha11-лиганда и IL-15 (см. пример 39).
Для оценки фенотипа поверхности этих ВМ-культур человека авторы окрашивали небольшие аликвоты этих клеток для 3-цветного проточного цитометрического анализа mAb анти-СD3-FITC, анти-СD56-РЕ и анти-СD16-CyChrome (все из PharMingen, San Diego, CA) и анализировали их на FACSCalibur с использованием программного обеспечения CellQuest (Becton Dickinson, Mountain View, CA). Этот проточный цитометрический анализ подтвердил, что клетки, выросшие из этих культур, были дифференцированными NK-клетами, так как они были большими и гранулярными и экспрессировали как CD56, так и CD16 и были CD3- (Lanier, LL Annu. Rev. Immunol. 16:359-393, 1998). Кроме того, эти клетки проявляли значимо более высокую эффекторную функцию, чем клетки, росшие с IL-15 и flt3. Более конкретно, клетки, росшие в присутствии всех трех цитокинов, лизировали более 40% мишеней-К562 при отношении эффектор: мишень (Е:Т) 1,5, тогда как клетки, росшие в присутствии IL-15+flt3L, лизировали менее 5% мишеней при Е:Т, равном 2. Эти данные демонстрируют, что, в комбинации с IL-15, zalpha11-лиганд стимулирует дифференцировку NK-клеток из CD34+ ВМ-клеток. С. Активность размноженных мышиным zalpha11-лигандом клеток
Для испытания эффектов zalpha11-лиганда на мышиных гемопоэтических клетках-предшественниках, очищенные отрицательные в отношении линии дифференцировки (Lin-) клетки костного мозга из мышей С57В1/6 размножали в flt3+IL-15+/- zalpha11-лиганд, как описано в примере 40В. В день 6 культуры эти клетки ("эффекторы") собирали и считали, затем суспендировали в 0,4 мл среды RP10 (пример 41А). Две аликвоты (по 0,15 мл каждая) каждой пробы, размноженные с zalpha11-лигандом или без zalpha11-лиганда (пример 41А), разводили серийно 3 раза в двух повторностях в 96-луночных круглодонных планшетах, всего 6 лунок со 100 мкл каждая. Остальные 100 мкл клеток окрашивали на маркеры поверхности NK-клеток с использованием mAb FITC-анти-2В4 и РЕ-анти-ОХ5 (PharMingen) и анализировали проточной цитометрией. Каждая группа клеток, подвергнутая действию flt3+IL-15 в присутствии или в отсутствие zalpha11-лиганда, имела сходные фракции 2B4+DX5+ клеток, в пределах 65-75% положительных в отношении обоих маркеров NK-клеток.
Для анализа NK-лизиса клетки-мишени (YAC-1 и EL4) метили 51Cr, как описано выше. После счета клеток-мишеней на гемоцитометре эти клетки-мишени разбавляли до 0,5-1×105 клеток/мл и 100 мкл YAC-1 или EL4 (0,5-1×104 клеток) смешивали с 100 мкл эффекторных клеток и инкубировали в течение 4 часов при 37°С. Специфический лизис определяли для каждой лунки, как описано выше.
Авторы нашли, что клетки, выращенные в присутствии flt3+IL-15+zalpha11-лиганд, проявляли усиленную литическую активность (приблизительно в 2 раза) против мишеней YAC-1 (но не убивали МНС+-контрольную клеточную линию EL4). При отношении эффектор : мишень (Е:Т) 5, NK-клетки, генерируемые в присутствии всех 3 цитокинов (zаlрhа11-лиганд+flt3+lL-15), лизировали 12% клеток YAC-1, тогда как NK-клетки, размноженные flt3+IL-15, лизировали 6% мишеней YAC-1. Последующие эксперименты подтвердили эту тенденцию.
Во втором подходе к определению биологической активности zalpha11-лиганда на мышиных NK-клетках, авторы выделяли незрелые CD4-CD8- ("дважды отрицательные", DN) мышиные тимоциты, как описано в примере 40С, и культивировали их с IL-15+flt3+IL-7 или IL-15+flt3+IL-2, с zalpha11-лигандом или без zalpha11-лиганда. В день 6 культуры клетки собирали и анализировали на NK-литическую активность на клетках YAC-1 и EL4, как описано выше. Авторы нашли, что клетки, культивируемые в присутствии zalpha11-лиганда, имели наивысшую активность в этом анализе с усиленной литической активностью в сравнении с клетками, культивируемыми в присутствии других цитокинов. Конкретно, DN-тимоциты, выращенные с IL-15+flt3+IL-7, убивали 18% клеток YAC-1 при Е:Т 24, тогда как клетки, выращенные в присутствии IL-15+flt3+IL-7 плюс zalpha11-лиганд, убивали 48% мишеней при том же самом Е:Т. DN-тимоциты, выращенные в IL-15+flt3+IL-2, убивали 15% мишеней YAC-1 при Е:Т 6, тогда как клетки, выращенные с этими 3 цитокинами и zalpha11-лигандом, убивали 35% клеток YAC-1 при Е:Т 9. Проточную цитометрию проводили на культивируемых клетках перед анализом NK-лизиса. Как это было верным для культур костного мозга, несмотря на пролиферативное действие zalpha11-лиганда (числа клеток увеличивались приблизительно в 2 раза при добавлении zalpha11-лиганда) он не увеличивал значимо фракцию DX5+ клеток (17-20% от общего числа клеток в культурах с IL-7 и 35-46% от общего числа клеток в культурах с IL-2). Эти данные предполагают, что zalpha11-лиганд в комбинации с IL-15 и flt3 усиливает литическую активност NK-клеток, полученных из костного мозга или тимуса мышей.
Р. Активность мышиного zalpha11-лиганда на зрелых мышиных NK-клетах
Для испытания действий мышиного zalpha11-лиганда на зрелые NK-клетки авторы изобретения выделяли селезенки из четырех 5-недельных мышей С57В1/6 (Jackson Laboratories, Bar Harbor, ME) и измельчали их между матированными на краях стеклянными предметными стеклами для получения суспензии клеток. Эритроциты удаляли гипотоническим лизисом следующим образом: клетки осаждали и супернатант удаляли аспирацией. Авторы разрушали осадок осторожным перемешиванием на вортексе, затем добавляли 900 мкл стерильной воды при встряхивании, затем быстро (через менее, чем 5 секунд) добавляли 100 мкл 10х HBSS (Gibco BRL). Затем эти клетки ресуспендировали в 10 мл 1х HBSS и дебрис удаляли пропусканием клеток через выстланный найлоновой сеткой деформатор клеток (Falcon). Затем эти лишенные эритроцитов клетки селезенки осаждали и ресуспендировали в MACS-буфере (ЗФР+1% БСА+2 мМ ЭДТА) и считали. Авторы окрашивали 300×106 клеток покрытыми анти-DХ5-магнитными гранулами (Miltenyi Biotec) и положительно отбирали DX5+ МК-клетки пропусканием через MACS VS+ разделительную колонку в соответствии с инструкциями изготовителя, что приводило к извлечению 8,4×106 DX5+ клеток и 251×106 DX5- клеток. Каждую из этих групп клеток культивировали в 24-луночных планшетах (0,67×106 клеток на лунку, 2 лунки на один вариант обработки) в среде RP10 (пример 41А) отдельно или с 1) 30 нг/мл мышиного zalpha11-лиганда, 2) 30 нг/мл рекомбинантного мышиного IL-2 (R&D Systems, Inc., Minneapolis, MN), 3) 30 нг/мл рекомбинантного IL-15 человека (R&D), 4) 30 нг/мл каждого из мышиного zalpha11-лиганда и IL-15 человека, или 5) 30 нг/мл каждого из mlL-2 и hlL-15. Клетки собирали после 21 часа, промывали и ресуспендировали в среде RP10 и считали. Затем клетки анализировали на их способность лизировать 51Cr-меченные клетки- мишени YAC-1 и EL4, как описано в примере 41А.
Обычно была низкая NK-активность из DX5- (не NK-кпетки) групп, но DX5- клетки, культивированные с zalpha11-лигандом и hlL-15, действительно лизировали 25% клеток-мишеней YAC-1 при Е:Т 82. Для сравнения, DX5- клетки, культивированные только с hlL-15, лизировали 14% клеток-мишеней YAC-1 при Е:Т 110. Это предполагает, что zalpha11-лиганд и IL-15 действуют вместе на оставшиеся NK1.1+ NK-клетки в этом препарате клеток. Что касается препаратов DX5- клеток, обработка одним мышиным zalpha11-лигандом не увеличивала значимо их эффекторную функцию (их лизис клеток YAC-1 был сходным с лизисом в необработанной группе). Как ожидалось, как IL-2, так и IL-15 значимо улучшали NK-активность. Однако наивысший уровень лизиса был обнаружен в группе, обработанной zalphall-лигандом и hlL-15 (65% лизис клеток YAC-1 при Е:Т 3,3, против 45% лизиса при Е:Т 4 для группы обработки hlL-15). Взятые вместе, эти результаты предполагают, что, хотя один zalpha11-лиганд не может увеличивать NK-клеточную литическую активность, он действительно усиливает NK-литическую активность зрелых NK-клеток при введении с IL-15.
Пример 42
Пролиферация под действием zalpha11-лиганда человеческих и мышиных Т-клеток в тесте пролиферации Т-клеток
А. Пролиферация мышиным zalpha11-лигандом мышиных Т-клеток
Т-клетки из мышей С57В1/6 (Jackson Laboratories, Bar Harbor, ME) выделяли из объединенных спленоцитов и лимфоцитов из подмышечных, плечевых, паховых, цервикальных и мезентериальных лимфатических узлов (LN). Селезенки разминали матированными на краях стеклянными предметными стеклами для получения суспензии клеток. LN (лимфатические узлы) разрывали на части пинцетом и пропускали через деформатор клеток для удаления дебриса. Объединенные спленоциты и LN-клетки разделяли на субпопуляции CD8+ и CD4+ с использованием двух последовательных MACS магнитных разделительных колонок в соответствии с инструкциями изготовителя (Miltenyi Biotec, Auburn, CA). Целые тимоциты собирали из тех же самых мышей.
Клекти культивировали при 3×105 клеток на лунку (тимоциты) или 105 клеток на лунку (зрелые Т-клетки) с увеличивающимися концентрациями очищенного мышиного zalphall-лиганда (0-30 нг/мл) (пример 24 и пример 29) в 96-луночных плоскодонных планшетах, предварительно покрытых в течение ночи при 4°С различными концентрациями анти-СD3 mAb 2C11 (PharMingen), в течение 3 дней при 37°С. Антитело анти-СD30 служило для активации мышиных Т-клеток через Т-клеточный рецептор. В каждую лунку добавляли 1 мкКи 3H-тимидина в день 2 и планшеты собирали и считали через 16 часов для оценки пролиферации.
При испытании zalpha11-лиганда в тесте пролиферации Т-клеток авторы обнаружили, что он костимулирует анти-СD3-активированные мышиные тимоциты, приводя к ускоренному росту CD8+ CD4+ клеток (большая часть тимоцитов, культивируемых с анти-СD3+zаlрhа11-лиганд, были CD8+ CD4+ в день 3 культуры, тогда как клетки, культивируемые только с анти-СD3, не склонялись значимо к этому фенотипу до дня 5). Авторы не наблюдали значимых уровней пролиферации тимоцитов в ответ на zalpha11-лиганд в отсутствие анти-СD3.
Интересно, что, когда авторы анализировали зрелые мышиные Т-клетки на их способность отвечать на zаlрhа11-лиганд+анти-СD3, они нашли, что только CD8+, но не CD4+ субпопуляция, отвечала зависимым от дозы образом на zalpha11-лиганд. Авторы наблюдали также слабую, но воспроизводимую пролиферацию CD8+ клеток (но не CD4- клеток) в ответ на один zalpha11-лиганд. Интересно, что это не наблюдалось для Т-клеток человека (см. пример 42В ниже).
В. Пролиферация zalpha11-лигандом человека Т-клеток человека
CD4+ и CD8+ Т-клетки человека выделяли из РВМС, как описано в примере 43 (ниже). Клетки культивировали при приблизительно 105 клеток на лунку с увеличивающимися концентрациями очищенного zalpha11-лиганда человека (0-50 нг/мл) (пример 24) в 96-луночных плоскодонных планшетах, предварительно покрытых в течение ночи при 4°С различными концентрациями анти-СD3 mAb UCHT1 (PharMingen), в течение 3 дней при 37°С. В каждую лунку добавляли 1 мкКи 3H-тимидина в день 2 и планшеты собирали и считали через 16 часов для оценки пролиферации. В противоположность результатам авторов с мышиными Т-клетками, предварительные данные предполагают, что zalpha11-лиганд человека костимулирует CD4+, но не CD8+, Т-клеток человека зависимым от дозы образом.
Пример 43
ПНР реального времени показывает экспрессию zalpha11-лиганда в CD4+ клетках
А. Очищенные Т-клетки человека в качестве первичного источника, используемого для оценки экспрессии zalpha11-лиганда человека
Цельную кровь (150 мл) брали из здоровых людей-доноров и смешивали 1:1 с ЗФР в конических пробирках на 50 мл. Затем тридцать мл разбавленной крови наносили на 15 мл фиколла Ficoll Paque Plus (Amersham Pharmada Biotech, Uppsala, Sweden). Эти градиенты центрифугировали 30 минут при 500 g и давали центрифуге остановится без торможения. Лишенные эритроцитов клетки в межфазной границе (РВМС) собирали и промывали 3 раза 3ФР. Выход выделенных мононуклеарных клеток периферической крови (РВМС) был 200×106 перед отбором, описанным ниже.
РВМС суспендировали в 1,5 мл MACS-буфера (3ФР, 0,5% ЭДТА, 2 мМ ЭДТА) и 3×106 клеток отделяли для контрольной РНК и для проточного цитометрического анализа. Затем авторы добавляли 0,25 мл микрогранул с антителом анти-CD8 человека (Miltenyi Biotec) и смесь инкубировали в течение 15 минут при 4°С. Эти клетки, меченные СD8-гранулами, промывали 30 мл MACS-буфера и затем ресуспендировали в 2 мл MACS-буфера. VS+ колонку (Miltenyi) готовили в соответствии с инструкциями изготовителя. Затем VS+ колонку помещали в магнитное поле Vario MACS (Miltenyi). Колонку уравновешивали 5 мл MACS-буфера. Затем выделенные первичные мышиные клетки наносили на колонку. CD8-отрицательным клеткам давали пройти через колонку. Колонку промывали 9 мл (3×3 мл) MACS-буфера. Затем колонку вынимали из магнита и помещали на пробирку falcon на 15 мл. CD8+ клетки элюировали добавлением 5 мл MASC-буфера на колонку и связанные клетки смывали с использованием поршня (плунжера), обеспечиваемого изготовителем. Общий выход CD8+ отобранных клеток периферических Т-клеток был 51×106 клеток. CD8-отрицательные протекающие через колонку клетки собирали, считали, окрашивали покрытыми антителом анти-СD4 гранулами, затем инкубировали и пропускали через новую VS+ колонку при той же самой концентрации, как описано выше. Общий выход CD4+ отобранных периферических Т-клеток был 42×106 клеток.
Пробу каждых из CD8+ и CD4+ отобранных Т-клеток человека удаляли для окрашивания и сортинга на клеточном сортере с активацией флуоресценции (FACS) для оценки их чистоты. РЕ-конъюгированное антитело против CD4 человека, антитело против CD8-FITC Ab и антитело против CD19-CyChrome Ab (все из PharMingen) использовали для окрашивания CD8+ и CD4+ отобранных клеток. CD8+ отобранные клетки в первом эксперименте были на 80% CD8+ клетками, а CD4+ отобранные клетки были на 85% CD4+ клетками. В двух последующих экспериментах (пример 43В) CD8+ очищенные клетки имели чистоту 84% и 81%, а CD4+ клетки имели чистоту 85% и 97% соответственно. В одном эксперименте авторы окрашивали несвязывающиеся (проходящие через колонку) клетки покрытыми aHTH-CD19 человека гранулами (Miltenyi) и пропускали их через третью колонку с магнитными гранулами для выделения CD19+ В-клеток (они имели чистоту 92%).
CD8+, CD4+ и CD19+ отобранные клетки человека активировали инкубированием 0,5×106 клеток/мл в RPMI + 5% ультрасыворотка человека (Gemini Bioproducts, Calabasas, CA) + ФМА 10 нг/мл и Иономицин 0,5 мкг/мл (Calbiochem) в течение приблизительно 4, 16 или 24 часов при 37°С. Т-клетки (2,5×106 на лунку) дополнительно стимулировали в 24-луночных планшетах, предварительно покрытых в течение ночи 0,5 мкг/мл планшет-связанными mAb анти-CD3 UCHT1 (PharMingen) с растворимыми mAb анти-СD28 (PharMingen) при 5 мкг/мл или без них. В каждой временной точке клетки собирали, осаждали, промывали один раз ЗФР и снова осаждали. Супернатант удаляли и осадки сразу же замораживали в бане со смесью сухой лед/этанол, затем хранили при -80°С для получения в последующем РНК.
ПЦР реального времени проводили на этих CD8+, CD4+ и CD19+ отобранных клетках, как описано в примере 43В и 43С ниже, для оценки экспрессии zalpha11-лиганда и zalpha11-рецептора человека.
В. Праймеры и зонды для количественной ОТ-ПЦР для экспрессии zalpha11-лиганда человека
Количественная ОТ-ПЦР реального времени с использованием системы детектирования последовательностей ABI PRISM 7700 (РЕ Applied Biosystems, Inc., Foster City, CA) была описана ранее (См. Heid, C.A. et al., Genome Research 6:986-994, 1996; Gibson, U.E.M. et al., Genome Research 6:995-1001, 1996; и Sundaresan, S. et al., Endocrinology 139:4756-4764, 1998. Этот способ включает в себя применение ген-специфического зонда, содержащего как репортерный, так и гасящий красители. Когда зонд является интактным, излучение репортерного красителя гасится вследствие тесной близости гасящего красителя. Во время ПЦР-удлинения с использованием ген-специфических прямого и обратного праймеров зонд расщепляется 5'-нуклеазной активностью полимеразы Taq, что высвобождает репортерный краситель из зонда, приводя к увеличению флуоресцентного излучения.
Праймеры и зонды, используемые для количественных ОТ-ПЦР-анализов реального времени, сконструированы с использованием программного обеспечения для конструирования праймеров Primer Express™ (PE Applied Biosystems). Праймеры для zalpha11-лиганда человека сконструированы простирающимися с схватыванием интрон-экзонного сочленения для элиминации амплификации геномной ДНК. Прямой праймер, ZC22281 (SEQ ID NO:90), и обратный праймер, ZC22279 (SEQ ID NO:91), использовали в концентрации приблизительно 300 нМ для синтеза продукта 80 п.н. Соответствующий зонд zalpha11-лиганда TaqMan, ZG32 (SEQ ID NO:92) был синтезирован и помечен РЕ Applied Biosystems. Этот зонд метили на 5'-конце репортерным флуресцентным красителем (6-карбоксифлуоресцеином) (FAM) (РЕ Applied Biosystems) и на 3'-конце гасящим флуоресцентным красителем (6-карбокситетраметилродамином) (TAMRA) (РЕ Applied Biosystems). Для тестирования целостности или качества всех проб РНК их подвергали скринингу на рРНК с использованием набора праймеров и зондов, заказанных из РЕ Applied Biosystems (cat. No. 4304483). Репортерным флуоресцентным красителем для этого зонда является VIC (РЕ Applied Biosystems). Результаты по рРНК позволят нормализовать результаты для zalpha11-лигандов.
РНК получали из осадков, полученных в примере 43А, с использованием набора RNeasy Miniprep™ (Qiagen, Valencia, CA) в соответствии с инструкциями изготовителя. Контрольную РНК получали из приблизительно 10 миллионов ВНК-клеток, экспрессирующих zаlрhа11-лиганд человека.
С. Праймеры и зонды для количественной ОТ-ПЦР для экспрессии zalpha11-рецептора человека
ПЦР реального времени выполняли для оценки экспрессии zalpha11-рецептора, как в примере 43В и примере 43D, с использованием клеток, полученных при условиях, подробно описанных в 43А, и зондов, специфических для zalpha11-рецептора. Прямой праймер, ZC22277 (SEQ ID NO:93), и обратный праймер, ZC22276 (SEQ ID NO:94), использовали в ПЦР-реакции (выше) в концентрации приблизительно 300 нМ для синтеза продукта 143 п.н. Соответствующий зонд zalpha11 TaqMan®, названный ZG31 (SEQ ID NO:95) был синтезирован и помечен РЕ Applied Biosystems. РНК из клеток BaF3, экспрессирующую zalpha11-рецептор человека, использовали для генерирования подходящего контроля для стандартных кривых для ПЦР реального времени, описанной в примере 43D ниже.
D. Количественная ОТ-ПЦР реального времени
Относительные уровни РНК zalpha11-лиганда определяли анализом проб тотальной РНК с использованием одностадийного способа ОТ-ПЦР (РЕ Applied Biosystems). РНК из клеток ВНК, экспрессирующих zalpha11-лиганд человека, выделяли стандартными способами и использовали для получения стандартной кривой. Кривая состояла из серийных разведении в диапазоне от 2,5-2,5×10-4 нг для скрининга рРНК и 25-0,0025 нг для скрининга zalpha11-лиганда, причем каждую точку стандартной кривой анализировали в трех повторностях. Пробы тотальной РНК также анализировали в трех повторностях на уровни транскрипта zalpha11-лиганда человека и на уровни рРНК в качестве эндогенного контроля. Каждая одностадийная ОТ-ПЦР-реакция содержала 25 нг тотальной РНК в буфере А (50 мМ KCl, 10 мМ Трис-HCl); внутренний стандартный краситель, (ROX) РЕ Applied Biosystems)); подходящие праймеры (50 нМ для проб рРНК, 300 нМ для проб zalpha11-лиганда) и зонд (50 нМ для рРНК, 100 нМ для zalpha11-лиганда), 5,5 мМ MgCl2; 300 мкМ каждого из d-CTP, d-ATP и d-GTP и 600 мкМ d-UTP; обратную транскриптазу (0,25 Е/мл); ДНК-полимеразу AmpliTaq (0,025 Е/мкл) и ингибитор РНКазы (0,4 Е/мкл) в общем объеме 25 мкл. Условия термоциклирования ПЦР были следующими: начальная стадия обратной транскрипции (ОТ) при 48°С в течение 30 минут; стадия активации AmpliTaq Gold при 95°С в течение 10 минут; с последующими 40 циклами амплификации при 95°С в течение 15 секунд и 60°С в течение 1 минуты. Относительные уровни РНК zalpha11-лиганда определяли с использованием способа стандартных кривых, как описано в User Bulletin No. 2 (РЕ Biosystems; User Bulletin #2: ABI Prism 7700 Sequence Detection System, Relative Quantitation of Gene Expression, December 11, 1997), с использованием измерений рРНК для нормализации уровней zalpha11-лиганда. Пробы сравнивали относительно калибратора в каждом эксперименте. Калибратор произвольно выбирали на основе РНК хорошего качества и уровня экспрессии, с которым другие пробы могли быть подвергнуты значимому сравнению. Результаты этих экспериментов, анализирующих экспрессию zalpha11-лиганда и zalpha11-рецептора в стимулированных и нестимулированных клетках (пример 43А), описаны в примере 43Е ниже.
Е. Экспрессия zalpha11-рецептора и zalpha11-лиганда человека в CD4+, CD8+ и CD19+ клетках
Первый эксперимент использовал ОТ-ПЦР, описанную выше, для оценки экспрессии zalpha11-рецептора в нестимулированных и стимулированных анти-СD3 CD4+ и CD8+ пробах во временных точках 0 ч (нестимулированные ("покоящиеся" клетки) и при 4 ч, 15,5 ч и 24 ч, после стимуляции. Покоящаяся CD4+ проба была произвольно выбрана в качестве калибратора, и ей приписана величина 1,00. Наблюдали приблизительно 4-кратное увеличение экспрессии рецептора в нестимулированных CD4+ клетках от 4 ч до 24 ч культивирования и приблизительно 8-кратное увеличение на протяжении того же самого периода времени в анти-CD3-стимулированных CD4+ клетках. CD8+ клетки обнаружили 7-кратное увеличение экспрессии zalpha11-рецептора, с пиком при 4 ч и снижением на протяжении времени. При анти-CD3-стимуляции CD8+ клетки имели константное 8-кратное увеличение экспрессии рецептора.
Этот первый эксперимент также использовал ОТ-ПЦР для оценки экспрессии zalpha11-лиганда в тех же самых анти-CD3-стимулированных и нестимулированных CD4+ и CD8+ пробах. 4 ч анти-CD3-стимулированная CD8+ проба была произвольно выбрана в качестве калибратора и ей приписана величина 1,00. Результаты показали, что нестимулированные CD4+ и CD8+ клетки не экспрессировали zalpha11-лиганд. Авторы наблюдали значимое повышение экспрессии анти-CD3-стимулированных CD4+ клеток при 4 ч и приблизительно 300-кратное увеличение в сигнале, наблюдаемом при 15,5 ч. CD8+ клетки экспрессировали небольшое количество лиганда при анти-СD3-стимуляции, однако это, возможно, было обусловлено загрязнением CD8+ популяции небольшим числом CD4+ клеток.
Второй эксперимент использовал ОТ-ПЦР для оценки экспрессии zalpha11-рецептора в анти-СD3-стимулированных, ФМА + Иономицин-стимулированных и нестимулированных CD4+ и CD8+ пробах при временных точках 0 ч, и при 3,5 ч, 16 ч и 24 ч после активации. Покоящаяся CD8+ проба была произвольно выбрана в качестве калибратора и ей приписана величина 1,00. Покоящиеся CD4+ и CD8+ клетки не обнаружили значимых количеств экспрессии рецептора. Экспрессия была приблизительно в 3 раза более высокой в ФМА + Иономицин-стимулированных CD4+ пробах при 3,5 ч, 16 ч и 24 ч после стимуляции. Экспрессия в анти-CD3-активированных CD4+ клетках имела максимум в 10 раз выше фоновых уровней при 3,5 ч после стимуляции, затем она снижалась до уровней в 4-раза более высоких, чем фон, при 16 ч после стимуляции. CD8+ клетки обнаружили 4-кратное увеличение экспрессии при 3,5 ч после ФМА + Иономицин-стимуляции, причем экспрессия снижалась в последующих временных точках. Как и в первом эксперименте, анти-CD3-стимулированные CD8+ клетки опять проявляли 8-кратную индукцию выше фона экспрессии рецептора.
Эти пробы из второго эксперимента использовали также для оценки экспрессии zalpha11-лиганда. 24 ч стимулируованную ФМА и Иономицином CD4+ пробу произвольно выбрали в качестве калибратора и ей была приписана величина 1,00. Результаты показали, что нестимулированные клетки не экспрессировали zalpha11-лиганд. Наблюдали приблизительно 30-кратную индукцию экспрессии лиганда в CD4+ клетках, стимулированных антителом анти-CD3, при 3,5 ч, как видно из предыдущего эксперимента (при 4 ч). Однако была только приблизительно 5-кратная индукция при ФМА + Иономицин-стимуляции при 3,5 ч, которая снижалась при последующих временных точках. Опять CD8+ клетки экспрессировали очень малое количество лиганда, что было, возможно, связано с загрязнением CD4+ клетками.
Последний эксперимент использовал ОТ-ПЦР для оценки экспрессии zalpha11-рецептора в анти-CD3- и анти-СD3/анти-СD28-стимулированных и нестимулированных CD4+ и CD8+ пробах во временных точках 0 ч, и 2 ч, 4 ч и 16 ч после стимуляции. CD19+ клетки, активированные ФМА + Иономицин, также подвергали скринингу на экспрессию рецептора при тех же самых временных интервалах. Покоящаяся CD4+ проба была произвольно выбрана в качестве калибратора и ей была приписана величина 1,00. 2 ч анти-СD3-стимутлрованные CD4+ клетки имели только 4-кратную индукцию рецептора в сравнении с 10-кратной индукцией, наблюдаемой при 3,5 ч в предыдущем эксперименте. Комбинация анти-CD3 и анти-CD28 увеличивала экспрессию zalpha11-рецептора до 8-кратной в сравнении с фоном. 16 ч анти-CD3/анти-CD28-стимулированные CD4+ клетки имели очень низкие уровни zalpha11-рецептора, как видно в CD8+ клетках в предыдущих экспериментах (см. выше). CD19+ клетки, стимулированные ФМА + Иономицин, имели наиболее значительную экспрессию zalpha11-рецептора с 19-кратным увеличением при 2 ч, но уровни экспрессии снижались опять до уровней покоящихся клеток при 16 часах.
Эти пробы из последнего эксперимента использовали также для оценки zalpha11-лиганда при помощи ОТ-ПЦР. 16 ч анти-CD3/анти-CD28-стимулированная CD8+ проба была выбрана произвольно в качестве калибратора, и ей была приписана величина 1,00. Эти результаты показали, что при 2 ч CD4+ клетки имели приблизительно 2-кратную индукцию экспрессии zalphall-лиганда с анти-CD3-стимуляцией и 5-кратную индукцию с анти-CD3 плюс анти-CD28 стимуляцией. Эти состояния стимуляции индуцировали экспрессию лиганда во времени с 16 ч стимулированными CD4+ клетками, проявляющими уровни экспрессии в 70 раз более высокие в сравнении с фоном. CD8+ и CD19+ клетки не обнаруживали экспрессию zalpha11-лиганда.
Определенная степень вариации ожидалась между взятиями крови (т.е. множественными пробами в различных временных точках из одного и того же пациента и между множественными пациентами). Таким образом, тенденции данных анализировали в каждом исследовании или из каждой отдельной пробы крови, и три приведенные выше эксперимента сравнивали для общего заключения. Тенденция, полученная из экспериментов ПЦР реального времени, заключается в том, что из всех испытанных типов клеток, CD19+ В-клетки, активированные ФМА + Иономицин, экспрессировали наивысшие уровни РНК zalpha11-рецептора. CD4+ и CD8+ клетки могут быть также стимулированы для экспрессии рецептора, но при более низких уровнях, чем это наблюдается в В-клетках. Zalpha11-лиганд экспрессировался почти исключительно в стимулированных CD4+ Т-клетках (а не в CD8+ Т-клетках или CD19+ В-клетках). Хотя стимуляция ФМА + Иономицин индуцировала хороший сигнал zalpha11-лиганда в этом тесте, значительно более высокий сигнал получали из CD4+ Т-клеток, стимулированных mAb анти-CD3, или комбинацией mAb анти-CD3 и анти-CD28, т.е. в условиях, которые лучше имитируют столкновение антигенов in vivo.
Пример 44
Zalpha11-лиганд-зависимая пролиферация В-клеток, стимулированных антителами анти-СD40 или анти-lgM
А. Очистка В-клеток человека
Флакон, содержащий 1×108 замороженных подвергнутых аферезу мононуклеарных клеток периферической крови человека (РВМС), быстро оттаивали в водяной бане на 37°С и ресуспендировали в 25 мл среды для В-клеток (RPMI-среда 1640 (JRH Biosciences, Lenexa, KS),10% инактивированная нагреванием фетальная телячья сыворотка, 5% L-глутамин, 5% пен/стреп) (Gibco BRL) в пробирке на 50 мл (Falcon VWR, Seattle, WA). Клетки тестировали на жизнеспособность с использованием Трипанового синего (GIBCO BRL). Десять миллилитров смеси Ficoll/Hypaque Plus (Pharmacia LKB Biotechnology Inc., Piscataway, NJ) наслаивали под суспензию клеток и откручивали в течение 30 минут при 1800 об/мин и давали остановиться с выключенным торможением. Затем межфазный слой извлекали и переносили в свежую пробирку Falcon на 50 мл, доводили до конечного объема 40 мл 3ФР и откручивали в течение 10 минут при 1200 об/мин с включенным торможением. Жизнеспособность выделенных В-клеток тестировали с использованием Трипанового синего. Альтернативно, свежую взятую у человека кровь разбавляли 1:1 ЗФР (Gibco BRL) и наслаивали на Ficoll/Hypaque Plus (Pharmacia), откручивали и промывали, как описано выше. Клетки, выделенные из свежего или замороженного источника, давали эквивалентные результаты.
В-клетки очищали из плавающих на фиколле клеток крови здоровых людей-доноров (см. выше) с использованием покрытых анти-СD19 магнитных гранул (Miitenyi Biotec, Auburn, CA) в соответствии с инструкциями изготовителя. Чистоту полученных препаратов подвергали мониторингу при помощи проточного цитометрического анализа с антителами анти-СD22 FITC (Pharmingen, San Diego, CA). Обычно препараты В-клеток имели чистоту >90%.
В. Очистка мышиных В-клеток
Суспензию мышиных спленоцитов получали разрыванием селезенок взрослых мышей С57В1/6 (Charles River Laboratories, Wilmington, MA) согнутыми иглами в среде для В-клеток. Эритроциты удаляли гипотоническим лизисом. CD43 положительные клетки удаляли СD43-магнитными гранулами (Miitenyi Biotec) в соответствии с инструкциями изготовителя. Чистоту полученных препаратов подвергали мониторингу при помощи проточного цитометрического анализа с антителами анти-СD45К FITC Ab (Pharmingen). Обычно препараты В-клеток имели чистоту >90%.
С. Пролиферация анти-СD40-стимулированных В-клеток в присутствии zalpha11-лиганда человека или мыши
В-клетки из человеческого или мышиного источника ресуспендировали при конечной концентрации 1×106 клеток в среде для В-клеток и высевали при 100 мкл на лунку в 96-луночном U-донном планшете (Falcon, VWR), содержащем различные условия стимуляции, с доведением конечного объема до 200 мкл на лунку. Для анти-СD40-стимулированных культур клеток культуры дополняли 1 мкг/мл анти-CD40 человека (Genzyme, Cambridge, MA), а мышиные культуры дополняли 1 мкг/мл анти-мышиный CD40 (Serotec, UK). Zalpha11-лиганд человека или мыши добавляли при разведениях от 1 пг/мл до 100 нг/мл. Специфичность действия zalpha11-лиганда подтверждали ингибированием zalpha11-лиганда 25 мг/мл растворимого рецептора zalpha11CEE человека (пример 10А). Все обработки проводили в трех повторностях. Затем эти клетки инкубировали при 37°С в увлажненном термостате в течение 120 часов (человек) или 72 часов (мышь). За шестнадцать часов перед сбором во все лунки добавляли 1 мкКи 3H-тимидина (Amersham, Piscataway, NJ) для оценки того, произошла ли пролиферация В-клеток. Клетки собирали в 96-луночный фильтровальный планшет (UniFilter GF/C, Packard, Meriden, CT) с использованием харвестера клеток (Packard) и собирали в соответствии с инструкциями изготовителя. Планшеты сушили при 55°С в течение 20-30 минут и дно лунок заделывали непрозрачным герметиком для планшетов. В каждую лунку добавляли 0,25 мл сцинтилляционной жидкости (Microscint-O, Packard) и планшет считывали с использованием сцинтилляционного счетчика для микропланшетов TopCount (Packard).
Инкубирование zalpha11-лиганда при концентрациях 3 нг/мл или более усиливало пролиферацию, индуцированную растворимым анти-СD40, зависимым от дозы образом как в мышиных В-клетках, так и в В-клетках человека в 30 раз. Мышиные и человеческие В-клетки отвечали равным образом также на их соответствующий zalpha11-лиганд. В обоих случаях стимуляция была специфической в отношении zalpha11-лиганда, так как она обращалась (снималась) присутствием растворимого zalpha11-рецептора в культуре.
D Пролиферация анти-lgM-стимулированных В-клеток в присутствии zalpha11-лиганда человека или мыши
В-клетки из человеческого или мышиного источника, как описано выше (пример 44А и пример 44В), высевали, как описано выше (пример 44С). Для анти-lgM-стимуляции клеток человека планшеты предварительно покрывали в течение ночи 10 мг/мл F(ab')2, антителами против lgM человека (Southern Biotech Assiciates, Birmingham, Alabama) и промывали стерильной средой непосредственно перед использованием. Культуры дополняли 0-10 нг/мл hu rlL-4 (R&D Systems, Minneapolis, MN). Для анти-lgM стимуляции мышиных клеток растворимое антитело анти-lgM (Biosource, Camarillo, CA) добавляли к культурам при 10 мг/мл. К каждому из предыдущих условий анти-lgM/L-4 добавляли zalpha11-лиганд человека или мыши при разведениях от 1 пг/мл до 100 нг/мл, как описано выше. Специфичность действия zalpha11-лиганда подтверждали ингибированием растворимым zalpha11-рецептором человека, как описано выше (пример 44С). Все обработки проводили в трех повторностях. Затем эти клетки инкубировали, метили 3H-тимидином, собирали и анализировали, как описано в примере 44С.
Инкубирование с zalpha11-лигандом в концентрациях 0,3 нг/мл или более ингибировало пролиферацию, индуцированную нерастворимым антителом анти-lgM (мышь) или анти-lgM и IL-4 (человека) зависимым от дозы образом. Это ингибирование было специфическим в отношении zalpha11-лиганда, так как оно обращалось присутствием растворимого zalpha11-рецептора в культуре.
Пример 45
Экспрессия растворимого zalpha11-рецептора человека в Е. coli
А. Конструирование экспрессирующего вектора PCZR225, который экспрессирует слитый полипептид huzalphal 1/MBP-6H
Экспрессионную плазмиду, содержащую полинуклеотид, кодирующий растворимый рецептор zalpha11 человека, слитый на С-конце с мальтозусвязывающим белком (МБР), конструировали гомологичной рекомбинацией. Фрагмент кДНК zalpha11 человека (SEQ ID NO:7) выделяли с использованием ПЦР. Полинуклеотидная последовательность для слитого полипептида МВР-растворимый рецептор zalpha11 показана в SEQ ID NO:96. Два праймера использовали в получении фрагмента zalpha11 человека в ПЦР-реакции: (1) Праймер ZC20187 (SEQ ID NO:98), содержащий 40 п.н. векторной фланкирующей последовательности и 25 п.н., соответствующих амино-концу zalpha11 человека, и (2) праймер ZC20185 (SEQ ID NO:99), содержащий 40 п.н. 3'- конца, соответствующих фланкирующей векторной последовательности, и 25 п.н., соответствующих карбоксил-концу zalpha11 человека. Условия ПЦР-реакции были следующими: 25 циклов 94°С в течение 30 секунд, 50°С в течение 30 секунд и 72°С в течение 1 минуты с последующим пропитыванием при 4°С, реакции проводили в двух повторностях. Два мкл из 100 мкл ПЦР-реакции подвергали электрофорезу на 1,0% агарозном геле с 1 х ТВЕ-буфером для анализа и наблюдали ожидаемый фрагмент приблизительно 660 п.н. Оставшиеся 90 мкл ПЦР-реакции объединяла со второй ПЦР-пробиркой, осаждали 400 мкл абсолютного этанола. Осажденную ДНК использовали для рекомбинации в разрезанный Serial реципиентный вектор рТАР98 (пример 31) для получения конструкции, кодирующей слитый МВР-zalpha11. Клоны трансформировали, идентифицировали и выращивали, как описано в примере 31. Положительные клоны были названы pCZR225 и подвергнуты анализу секвенирования. Полинуклеотидная последовательность для МВР-растворимый zalpha11-рецептор показана в SEQ ID NO:96, а соответствующая полипептидная последовательность показана в SEQ ID NO:97. Положительные клоны использовали для выращивания в Е. coli, как описано в примере 34, для очистки слитого белка huzalpha11/MBP-6H (пример 46 ниже).
Пример 46
Очистка слитого белка растворимый рецептор huzalpha11/MBP-6H из ферментации Е. coli
Если нет других указаний, все операции проводили при 4°С. Следующую процедуру использовали для очистки слитого белка растворимый рецепторный полипептид huzalpha11/MBP-6H. Клетки Е. coli, содержащие конструкцию pCZR225 и экспрессирующие растворимый рецептор huzalpha11/MBP-6H (пример 45), выращивали в SuperBroth II (12 г/л казеина, 24 г/л дрожжевого экстракта, 11,4 г/л дикалий-фосфата, 1,7 г/л монокалий-фосфата; Becton Dickinson, Cockeysville, MD) и замораживали в 0,5% глицерине. Двадцать граммов замороженных клеток в SuperBroth II + глицерин использовали для очистки белка. Замороженные клетки оттаивали и разбавляли 1:10 в растворе протеазного ингибитора (буфере для экстракции) перед лизисом этих клеток и высвобождением растворимого рецепторного белка huzalpha11/MBP-6H. Разбавленные клетки содержали конечные концентрации 20 мМ Трис (JT Baker, Philipsburg, NJ), 100 мМ хлорид натрия (NaCl, Mallinkrodt, Paris, KY), 0,5 мМ фенилметилсульфонилфторид (РМСФ, Sigma Chemical Co., St. Louis, МО), 2 мкг/мл лейпептина (Fluka, Switzerland) и 2 мкг/мл апротинина (Sigma). Систему разрушения клеток French Press (Constant Systems Ltd., Warwick, UK) с температурой от -7 до -10°С и 30 кф/кв. дюйм использовали для лизиса клеток. Разбавленные клетки проверяли на разрушение считываниями А600 до и после French Press. Лизированные клетки центрифугировали при 18000 g в течение 45 минут для удаления дебриса разрушенных клеток и супернатант использовали для очистки белка. Концентрации общего белка супернатанта определяли по БСА-способу (Pierce, Rockford, IL) согласно инструкциям изготовителя.
Субстрат колонки (25 мл) Talon Metall Affinity resin (Clontech, Palo Alto, CA) (приготовленный, как описано ниже) выливали в стеклянную колонку Bio-Rad, 2,5 см (диаметр) × 10 см (высота). Колонку упаковывали и уравновешивали под действием силы тяжести 10 колоночными объемами (КО) буфера для уравновешивания Talon (20 мМ Трис, 100 мМ NaCl, рН 8,0). Супернатант периодически наносили на аффинную смолу Talon Metall Affinity resin и качали в течение ночи. Смолу выливали обратно в колонку и промывали 10 колоночными объемами буфера для уравновешивания Talon под действием силы тяжести, затем проводили элюцию под действием силы тяжести 140 мл буфера для злюции (буфер для уравновешивания Talon + 200 мМ имидазол, Fluka Chemical). Talon-колонку очищали 5 колоночными объемами 20 мМ 2-(N-морфолино)этансульфоновой кислоты рН 5,0 (MES, Sigma), 5 колоночными объемами дистиллированной H2O, затем хранили в смеси 20% этанол/0,1% азид натрия. Фракции по четырнадцать мл собирали на протяжении всей элюционной хроматографии и фракции считывали по поглощению при 280 и 320 нм и белок определяли по БСА-способу; проходящий через колонку и промывочный пулы также сохраняли и анализировали. Фракции элюции представляющего интерес белка объединяли и наносили сразу на Амилозную смолу (New England Biolabs, Beverly, MA).
Для получения более чистого полипептида huzalpha11/MBP-6H объединенные фракции элюции с Talon-аффинной колонки подвергали хроматографии на Амилозной смоле (22 мл) при рН 7,4. Колонку Bio-Rad с диаметром 2,5 см и высотой 10 см заливали, упаковывали и уравновешивали в 10 колоночных объемах Амилозного буфера для уравновешивания (20 мМ Трис (JT Baker), 100 мМ NaCl (Mallinkrodt), 1 мМ ПМСФ (Sigma), 10 мМ бета-меркаптоэтанол (ВМЕ, ICN Biomedicals Inc., Aurora, ОН) рН 7,4. Пробу наносили под действием силы тяжести при скорости тока 0,5 мл/мин. Колонку промывали 10 колоночными объемами Амилозного буфера для уравновешивания, затем элюировали примерно 2 колоночными объемами смеси Амилозный буфер для уравновешивания +10 мМ мальтоза (Fluka, Biochemical Switzerland) под действием силы тяжести. Фракции по 5 мл собирали на протяжении всей хроматографии и измеряли поглощение при 280 и 320 нм. Амилозную колонку регенерировали 1 колоночным объемом дистиллированной H2O, 5 колоночными объемами 0,1% (масс/об) ДСН (Sigma), 5 колоночными объемами дистиллированной Н2О и затем 5 колоночными объемами Амилозного буфера для уравновешивания.
Представляющие интерес фракции объединяли и диализовали в Slide-A- Lyzer (Pierce) с 4 × 4л ЗФР рН 7,4 (Sigma) для удаления низкомолекулярных примесей, смены буфера и обессоливания. После смен ЗФР собранный материал представлял собой очищенный полипептид huzalpha11/MBP-6H. Очищенный полипептид huzalpha11/MBP-6H анализировали электрофорезом на ДСН-ПААГ с окрашиванием Кумасси и Вестерн-блот-анализом с антикроличьим антителом, конъюгированным с пероксидазой хрена (HRP) (Rockland, Gilbertsville, PA). Концентрация полипептида huzalpha11/MBP-6H была 1,92 мг/мл, как определено БСА-анализом.
Очищенный полипептид huzalpha11/MBP-6H готовили для инъекции в кроликов и отсылали в R & R Research and Development (Stanwood, WA) для получения антител. Кроликов инъецировали для получения сыворотки против huzalpha11/MBP-6H (пример 47 ниже).
Пример 47
Поликлональные антитела к zalpha11-рецептору
Поликлональные антитела получали иммунизацией двух самок Ново-Зеландских белых кроликов очищенным полипептидом huzalpha11/MBP-6H (пример 46) или очищенным рекомбинантным растворимым рецептором zalpha11CEE (пример 10А). Соответствующие Поликлональные антитела были названы кроличье антитело анти-huzalphа11/МВР-6Н и кроличье антитело анти-huzalpha11CEE-BHK соответственно. Каждый кролик получал исходную внутрибрюшинную инъекцию (IP) 200 мг очищенного белка в полном адъюванте Фрейнда (Pierce, Rockford, IL) с последующими бустерными инъекциями IP 100 мг очищенного белка в неполном адъюванте Фрейнда каждые три недели. Спустя семь-десять недель после введения третьей бустерной инъекции производили кровоизвлечение у животных и собирали сыворотку. Затем кроликам продолжали вводить бустер-инъекции и брали кровь каждые три недели.
Zalpha11-специфические поликлональные антитела очищали аффинно из сывороток кроликов с использованием колонки CNBr-SEPHAROSE 4B protein (Pharmada LKB), которую готовили с использованием 10 мг очищенного полипептида huzalpha11/MBP-6H (пример 32) на грамм CNBr-SEPHAROSE с последующим 20Х диализом в ЗФР в течение ночи. Zalpha11-специфические антитела характеризовали проверкой титра при помощи ELISA с применением 1 мг/мл подходящего белкового антигена в качестве мишени антител. Нижним порогом детектирования (LLD) кроличьего аффинно очищенного антитела анти-huzalpha11/MBP-6H является разведение 500 пг/мл. LLD кроличьего аффинно очищенного антитела анти-huzalpha11-CEE-BHK является разведение 50 пг/мл.
Пример 48
Распределение zalpha11-рецептора
Для оценки распределения zalpha11-рецептора на различных типах клеток авторы изобретения получали кроличьи поликлональные и мышиные моноклональные антитела (mAb), направленные против рецептора человека (пример 35 и пример 47) и конъюгировали эти антитела с биотином для применения в проточной цитометрии. Сначала авторы использовали поликлональные антитела, которые имели относительно низкую аффинность, для окрашивания панели клеточных линий: клетки BaF3 дикого типа IL-3-зависимой мышиной линии В-клеток-предшественников (Palacios and Steinmetz, ibid.4 Mathey-Prevot et al., ibid.); клетки BaF3, трансфицированные zalpha11 человека (пример 4); линии клеток лимфомы Беркитта Raji (ATCC No. CCL-86), Ramos (ATCC No. CRL-1596), RPMI 8226 (ATCC No. CCL-155) и Daudi (ATCC No. CCL-213); линии клеток Jurkat Т-клеточного лейкоза человека (ATCC No. TIB-152); линии клеток Thr-1 миеломоноцитарного лейкоза человека (ATCC No. TIB-202) и UT937 (ATCC No. CRL-1593.2); про-миеломоноцитарные клетки HLT60 человека (ATCC No. CCL-240); линия клеток А20 мышиной В-клеточной лимфомы (ATCC No. TIB-208); и линия клеток EL4 мышиной тимомы (ATCC No. TIB-39).
Эти клетки собирали, промывали один раз промывным буфером FACS с сывороткой (WBS). WBS состояла из сбалансированного солевого раствора Хенкса (Gibco BRL) + 10 mm HEPES (Gibco BRL) + 1% БСА (Sigma) + 10% нормальная козья сыворотка (Gemini Bioproducts, Woodland, CA) + 10% нормальная кроличья сыворотка (Sigma). Промывной буфер (WB) был идентичен WBS за исключением того, что он был бессывороточным. После промывания клетки ресуспендировали в 100 мкл WB, содержащем 10 мкг/мл кроличьих поликлональных антител против zalpha11 (пример 47). Клетки выдерживали на льду с антителом в течение 20 минут, затем промывали WB и ресуспендировали в WB, содержащем козьи антикроличьи-FITC антитела (BioSource, International), инкубировали еще 20 минут на льду, затем промывали и ресуспендировали в 400 мкл WB для анализа на проточном цитометре FACSCalibur (Becton Dickinson). Контрольные пробы окрашивали только вторичным козьим антикроличьим-FITC антителом. Положительное окрашивание определяли в виде смещения выше окрашивания только вторичным антителом. Хотя поликлональные антитела были антителами низкой аффинности, авторы уверенно (значимо) детектировали экспрессию zalpha11 на трансфектанте BaF3/zalpha11, на всех четырех лимфомах Беркитта человека (Raji, Ramos, Daudi и RPMI 8226) и на Т-клетках Jurkat. Покоящиеся (недифференцированные) клетки HL-60 не связывали антитела анти-zalphal 1, но авторы не обнаружили положительного сигнала на клетках HL-60, активированных в течение 20 часов ФМА (Calbiochem, La Jolla, CA), который индуцирует дифференцировку клетки HL-60 в моноцит-подобную клетку. Авторы также наблюдали положительный сигнал на клетках UT937 и Thp-1, хотя этот сигнал мог быть обусловлен неспецифическим связыванием. Поликпональные антитела слабо перкрестно реагировали на мышиной В-клеточной линии А20, но авторы не наблюдали окрашивания мышиной тимомы EL4.
Четыре моноклональных антитела к zalpha11 (пример 35) конъюгировали с биотином и субпопуляцию клеток, описанную выше, подвергали скринингу на экспрессию zalpha11-рецептора (BaF3, BaF3/zalpha11, Raji, Jurkat и покоящиеся HL-60). Клетки собирали, промывали, затем ресуспендировали в 100 мкл WB, содержащего 15 мкг/мл одного из 4 биотинилированных mAb. Клетки инкубировали с mAb в течение 20 минут на льду, затем промывали 1,5 мл WB и осаждали в центрифуге. Супернатант удаляли аспирацией и осадки ресуспендировали в 100 мкл CyCrome-конъюгированного стрептавидина (CYC-SA; PharMingen), затем инкубировали на льду еще в течение 20 минут и промывали и осаждали, как описано ранее. Контрольные пробирки содержали клетки, окрашенные только СуС-SA. Осадки ресуспендировали в 400 мкл WB и проточную цитометрию выполняли, как описано выше. Положительное окрашивание определяли в виде сигнала, превышающего уровень фона окрашивания одним CyC-SA. Применение ВаF3/zаlрhа11-трансфектанта в качестве контроля позволило авторам выстроить эти 4 mAb в связи с их соответствующими средними интенсивностями флуоресценции (MFI), которые могут отражать аффинность антител и/или степень биотинилирования mAb. Эти mAb были выстроены следующим образом, от наивысшей к наинизшей MFI: 249.28.2.1.2.2; 247.10.2.15.4.6; 249.19.2.2.3.5 и 249.15.2.4.2.7. Клетки Raji окрашивались положительно mAb zalpha11. Клетки Jurkat положительно окрашивались mAb zalpha11, но не так сильно, как на В-клетках (Raji). Таким образом, zalpha11-рецептор экспрессировался на этих В- и Т-клеточных линиях. Распределения окрашивания на неактивированных клетках HL60 были идентичными для всех этих mAb, и сигнал был очень слабым. Авторы считают, что этот сигнал не отражает истинной экспрессии zalpha11 клетками HL60, а скорее может быть вызван неспецифическим связыванием мышиных mAb с клетками человека, возможно, через Fc-рецепторы.
Пример 49
Действие zalpha11-лиганда на В-клетки и слитый белок zalpha11-лиганд-токсичный сапорин
Действия zalpha11-лиганда человека испытывали на следующих В-клеточных линиях человека: линиях клеток лимфомы Беркитта Raji (ATCC No. CCL-86) и Ramos (ATCC No. CRL-1596); линии клеток В-клеточной лимфомы EBV человека RPMI 1788 (ATCC No. CRL-156); линии IM-9 миеломы/плазмацитомы человека (ATCC No. CRL159); и EBV-трансформированной В-клеточной линии DAKIKI человека (ATCC No. TIB-206) и линии клеток HS Sultan (ATCC No. CRL-1484). После приблизительно 2-5 дней обработки zalpha11-лигандом изменения экспрессии маркера обнаруживали в клеточных линиях IM-9, Raji, Ramos и RPMI1788, что свидетельствует о том, что эти клетки отвечают на zalpha11-лиганд. В-клеточные линии человека, обработанные zalpha11-лигандом, растут гораздо более медленно, чем необработанные клетки при повторном высеве в чашки для культивирования клеток. Эти клетки имели также увеличенную экспрессию Fas-лиганда, как было определено проточной цитометрией (пример 49D и пример 49Е) и умеренно увеличенной чувствительностью к активирующему Fas-антителу (пример 49А). Этот результат указывает на то, что zalpha11-лиганд мог бы контролировать некторые типы В-клеточных неоплазм индуцированном их к дифференцировке в менее пролиферативное и/или более Fas-лиганд-чувствительное состояние. Кроме того, zalpha11-рецептор экспрессируется на поверхности некоторых из этих клеточных линий (см. пример 48). Таким образом, zalphall-лиганд и конъюгат zalpha11-лиганд человека-иммунотоксин сапорин (пример 49В, ниже) или другой слитый белок zalpha11-лиганд-токсин мог бы использоваться в терапии в В-клеточных лейкозах и лимфомах.
А. Действие zalpha11-лиганда человека на В-клеточные линии
Клетки IM-9 высевали при приблизительно 50000 клеток/мл +/- 50 мкг/мл очищенного zalpha11-лиганда человека (пример 29). После 3 дней роста клетки собирали, промывали и считали и затем повторно высевали при приблизительно 2500 клеток/мл в 96-луночные планшеты в лунки с 0, 0,033, 0,1 или 0,33 мкг/мл антитела против FAS (R&D Systems, Minneapolis). После 2 дней проводили флуоресцентный анализ с Alamar blue (пример 2В) для оценки пролиферации этих клеток.
Обработанные zalpha11-лигандом клетки IM-9 росли только до 27% плотности необработанных клеток в отсутствие антитела анти-FAS. В присутствии 0,33 мкг/мл антитела анти-FAS обработанные zalpha11-лигандом клетки ингибировались дополнительно на 52%, тогда как необработанные клетки ингибировались только на 30%. Общее ингибирование клеточного роста обработкой как zalphall-лигандом, так и 0,33 мкг/мл антитела анти-FAS было 86%.
При предобработке клеток IM-9 в течение трех дней с zalpha11-лигандом или без zalpha11-лиганда и последующем повторном посеве при 100 клеток на лунку и выращивании с антителом анти-FAS или без антитела анти-FAS в течение 6 дней рост необработанных клеток, оцениваемый тестом с Alamar blue (пример 2В), ингибировался только на 25% антителом анти-FAS, тогда как рост обработанных zalpha11-лигандом клеток ингибировался на 95% относительно роста необработанных клеток в отстутствие антитела анти-FAS.
В. Действие слияния zalpha11-лиганд человека-иммунотоксин сапорин на В-клеточные линии
Конструирование и очистка конъюгата zalpha11-лиганд человека-иммунотоксин сапорин (zalpha11L-sap) описаны в примере 50. Zalpha11L человека-sap был гораздо более сильнодействующим, чем один сапорин в ингибировании роста клеток. При повторном посеве обработанных клеток после трех-четырех дней обработки конъюгатом Zalpha11L человека-sap клетки росли очень слабо.
Клетки IM-9, Ramos и К562 (АТСС No. CCL-243) высевали при приблизительно 2500 клеток на лунку в 96-луночные планшеты с 0-250 нг/мл конъюгата Zalpha11L человека-sap или только с 0-250 нг/мл сапорина (Stripe et al., Biotechnology 10:405-412, 1992) в качестве контроля. Планшеты инкубировали 4 дня, затем проводили анализ пролиферации с Alamar blue (пример 5В). При максимальной концентрации конъюгата huzalpha11L-sap рост клеток IM-9 и клеток Ramos ингибировался на 79 и 65%, соответственно. На клетки К562, которые являются низкими/негативными согласно проточной цитометрии в отношении экспрессии zalpha11-рецептора, не было влияния zalpha11L-sap, что свидетельствует о специфичности действия конъюгата.
Клетки IM-9 высевали 50000 клеток/мл в 6-луночные чашки при 0 и 50 нг/мл конъюгата zalpha11L человека-sap. Спустя 3 дня клетки собирали и считали и затем снова высевали от 100 до 0,8 клеток на лунку в 2-кратных серийных разведениях и 12 лунок на одно клеточное разведение без конъюгата zalpha11-лиганд человека-иммунотоксин сапорин. Спустя 6 дней число лунок с ростом при каждом клеточном разведении оценивали в соответствии с результатами анализа пролиферации с Alamar blue (пример 2В).
При оценке числа клеток при помощи анализа с Alamar blue (пример 2В) после 6 дней роста контрольные клетки, посеянные при приблизительно 12,5 и 6,25 клеток на лунку, имели эквивалентный рост с обработанными zalpha11-sap клетками, посеянными при 100 и 50 клеток на лунку, соответственно. Таким образом, рост выживших обработанных клеток IM-9 был заметно ослабленным даже после удаления посредством пересева конъюгата zalpha11-иммунотоксин sap.
Ограниченное тканевое распределение zalpha11-рецептора человека (пример 48) и специфичность действия zalpha11-sap в отношении экспрессирующих рецептор клеточных линий предполагают, что этот конъюгат может быть переносимым in vivo.
С. Действие конъюгата zalpha11-лиганд человека-иммунотоксин сапорин на жизнеспособность В-клеточной линии
Клетки HS Sultan (ATCC No. CRL-1484) высевали при приблизительно 40000 клеток на мл в 12-луночные планшеты и выращивали в течение пяти дней либо без добавления цитокинов, либо с 4 0 нг/мл очищенного zalpha11-лиганда человека (пример 29) или 25 нг/мл конъюгата huzalpha11L-sap (пример 50 ниже) или с 20 нг/мл IFN-альфа (RDI) или zalpha11-лиганда и IFN-альфа. ZalphaH-лиганд ингибировал рост клеток HS Sultan на 63%. IFN-альфа ингибировал рост на 38%. Zalpha11-лиганд плюс IFN-альфа ингибировал рост на 78%, что указывало на то, что ингибирующие эффекты zalpha11-лиганда человека и IFN-альфа могут быть аддитивными. Zalpha11L человека-сапорин(sap) ингибировал рост клеток HS Sultan на 92%.
Результаты, приведенные выше, подтверждают возможное применение zalpha11-лиганда или zalpha11L человека-sap в лечении злокачественностей или других заболеваниий, которые экспрессируют zalpha11-рецептор, в частности заболеваний В-клеточного происхождения. Комбинирование zalpha11-лиганда с IFN-альфа особо предполагается вследствие их аддитивного действия в ингибировании клеток HS Sultan. Некоторые другие типы лимфоидных злокачественностей и заболеваний могут также экспрессировать zalpha11-рецептор, так как активированные Т-клетки также экспрессируют мРНК рецептора (пример 48), и некоторые из этих заболеваний могут быть чувствительными к zalpha11-лиганду при терапии с использованием слияния zalpha11-лиганд-токсин.
D. FAS (СР95)-экспрессия на В-клеточных линиях человека увеличивается стимуляцией zalpha11-лигандом человека
В-клеточные линии человека HS Sultan (ATCC No. CRL-1484), IM-9 (АТСС No. CRL159), RPMI 8226 (АТСС No. CCL-155), RAMOS (ATCC No. CRL-1596), DAKIKI (ATCC No. TIB-206) и RPMI 1788 (ATCC No. CRL-156) обрабатывали 10-50 нг/мл очищенного zalphall-лиганда человека (или без него) (пример 29) в течение 2-8 дней. Затем эти клетки окрашивали РЕ-конъюгированным антителом анти-С095 (PharMingen, San Diego, CA) в соответствии с протоколом изготовителя и анализировали на FACSCalibur (Becton Dickinson, San Jose, CA). Во всех клеточных линиях окрашивание анти-С095 (FAS или АРО-1) увеличивалось в некоторых случаях более чем в 2 раза при обработке zalpha11-лигандом человека.
Е. FAS (СР95)-экспрессия на первичных мышиных В-клетках селезенки увеличивается стимуляцией zalpha11-лигандом человека
Первичные мышиные спленоциты получали измельчением селезенок из 8-12-недельных мышей C57/BL6. Эритроциты лизировали обработкой препарата в течение 5 секунд водой и затем пропускали через сито 70 микрон. Оставшиеся спленоциты промывали и высевали в среду RPMI (JRH Bioscience) плюс 10% HIA-ФТС (Hyclone, Logan, UT). Интерлейкин-2 (IL-2) (R&D Systems) c zalpha11-лигандом человека или без zalphall-лиганда человека, как описано выше. Затем их инкубировали при 37°С в 5% CO2 в течение 5 дней. Спленоциты собирали и окрашивали РЕ-конъюгированным антителом анти-СD95 (PharMingen) и FITC-конъюгированным антителом анти-СD19 (PharMingen) в соответствии с протоколом изготовителя. Клетки анализировали проточной цитометрией на FACScalibur (Becton Dickinson). При установке дискриминирующих окон на мышиные CD19+ В-клетки было обнаружено, что окрашивание антителом анти-СD95 увеличивалось на В-клетках, обработанных IL-2 плюс zalpha11-лиганд человека в сравнении с окрашиванием в одном IL-2. Анти-СD95-окрашивание было равно 37 относительным флуоресцентным единицам (RFU) на В-клетках в одном IL-2 и 55 RFU на В-клетках, культивируемых в IL-2 и zalpha11-лиганде человека.
Пример 50
Конструирование и очистка слияния zalpha11-лиганд-токсин
По контракту о поставке 10 мг zalpha11-лиганда человека (пример 29) послали в Advanced Targeting Systems (ATS, San Diego, CA) для конъюгирования с растительным токсином сапорином (Stripe et al., Biotechnology 10:405-412, 1992). Зимогенетики получали из ATS 1,3 мг белкового конъюгата, содержащего 1,1 молекул сапорина на молекулу zalpha11-лиганда человека, приготовленные в концентрации 1,14 мг/мл в 20 нМ фосфате натрия, 300 нМ хлориде натрия, рН 7,2.
Пример 51
Слияние zalpha11-лиганд-токсин in vivo
А. Испытание конъюгата zalpha11-сапорин в мышах
Конъюгат zalpha11-сапорин (пример 49) вводили мышам C57BL6 (самки, возраст 12 недель, купленные из Taconic) в двух различных дозах: 0,5 и 0,05 мг/кг. Инъекции проводили iv в носителе, состоящем из 0,1% БСА (ICN, Costa Mesa, CA). Три инъекции выполняли на протяжении одной недели (день 0, 2 и 7). Пробы крови брали из мышей в день 0 (перед инъекцией) и в дни 2 и 8 (после инъекции). Кровь собирали в гепаринизированные пробирки (Becton Dickinson, Franklin Lakes, NJ) и числа клеток определяли с использованием автоматического гематологического анализатора (Abbot Cell-Dyn model No. CD-3500CS, Abbot Park, IL). Животных эутанизировали и аутопсировали в день 8 после сбора крови. Селезенку, тимус, печень, почку и костный мозг извлекали для гистопатологии. Селезенку и тимус взвешивали и дополнительную пробу крови собирали в пробирки для отделения сыворотки. Сыворотку отсылали в Pheonix Central Labs, Everett, WA для тестирования в стандартной химической панели. Пробы собирали также для проточного цитометрического анализа, как описано здесь.
Формула циркулирующей крови (гемограмма) и измерения химии сыворотки не отличались значимо между обработанными конъюгатом zalpha11-лиганда мышами и мышами, обработанными эквивалентной дозой неконъюгированного токсина (сапорина). Гистологический анализ тканей в обработанных конъюгатом zalpha11-лиганд-сапорин мышах не показал значимых изменений относительно мышей, обработанных эквивалентной дозой неконъюгированного токсина. Эти результаты показывают, что конъюгат сапорина не был токсичным in vivo.
В. Испытание конъюгата zalpha11-токсин сапорин на происходящих из В-клеток опухолях in vivo
Действия zalpha11-лиганда человека и слияния zalpha11-лиганд человека-токсин сапорин (пример 50) на опухолевые клетки человека испытывали in vivo с использованием мышиной модели опухолевого ксентрансплантата, описанной здесь. Модели ксенотрансплантата сначала испытывали с использованием клеточных линий, выбранных на основе экспериментов in vitro, например линий, описанных в примере 49. Эти клеточные линии включают в себя, но не ограничиваются ими, клеточные линии лимфомы Беркитта человека Raji (ATCC No. CCL-86) и Ramos (ATCC No. CRL-1596); клеточную линию человека RPMI 1788 (ATCC No. CRL-156); линию клеток миеломы/плазмацитомы человека IM-9 (АТСС No. CRL159); клеточную линию человека DAKIKI (ATCC No. TIB-206) и клетки HS Sultan (ATCC No. CRL-1484). Клетки, полученные непосредственно из опухолей человека, могут быть также использованы в этом типе модели. Таким путем, скрининг проб пациента на чувствительность к обработке zalpha11-лигандом или конъюгатом zalpha11-лиганд-токсин сапорин может быть использован для выбора оптимальных показаний для применения zalpha11 в противоопухолевой терапии.
После выбора подходящей модели in vivo ксенотрансплантата, описанной выше, zalpha11-лиганд-индуцированную активность природных клеток-киллеров (NK-клеток) и/или действия zalpha11-лиганда на опухоли В-клеточного происхождения оценивают in vivo. Zalpha11-лиганд человека испытывают на его способность генерировать цитотоксические эффекторные клетки (например, NK-клетки) с активностью против опухолей В-клеточного происхождения с использованием мышиных моделей опухолевых ксенотрансплантатов, описанных здесь. Кроме того, могут оцениваться также прямые действия zalpha11-лиганда человека на опухоли. Модели трансплантата для проведения экспериментов выбирали, как описано выше. Протокол применения стимулированных zalpha11-лигандом клеток человека разрабатывают и испытывают на эффективность в истощении опухолевых клеток и стимуляции выживания у мышей, инокулированных клеточными линиями или первичными опухолями.
Пример 52
Идентификация клонов искусственной хромосомы Р1, содержащей геномную ДНК zalpha11-лиганда человека
кДНК-инсерт zalpha11-лиганда человека амплифицировали при помощи ПЦР с использованием праймеров на основе векторов. ПЦР-продукт 32Р-метили и гибридизовали с фильтрами высокой плотности, представляющими РАС-библиотеку (Искусственной хромосомы Р1). Фильтры и замороженные исходные растворы библиотеки получали из Roswell Park Cancer Institute, Buffalo, New York; сегмент библиотеки представлял собой RPC16 с 4-кратной глубиной покрытия. Фильтры гибридизовали в течение ночи при 65°С в ExpressHyb (Clontech) и промывали в соответствии с рекомендациями изготовителя. Расшифровка позитивных сигналов привела к идентификации четырех РАС-клонов, названных 23Н17, 34А9, 105G9 и 236Н14. ПЦР-анализ с использованием праймеров, специфических для 5'-конца (ZC22452 (SEQ ID NO:100) и ZC22451 (SEQ ID NO:101)) и 3'-конца (ZC22450 (SEQ ID NO:102) и ZC22449 (SEQ ID NO:103)) кодирующего района показала, что РАС 34А9 и РАС 105G9 содержали оба конца, тогда как РАС 23Н17 и РАС 236Н14 содержали только 5'-конец. РАС 23Н17 расщепляли Eco RI и Not I и идентифицировали фрагмент 9 т.п.н., который гибридизуется с кДНК-зондом zalpha11-лиганда. Этот фрагмент выделяли и субклонировали с использованием описанных здесь способов в pBluescript II SK(+) (Stratagene), предварительно расщепленный Eco RI и Not I. Секвенирование выявило, что этот фрагмент содержал приблизительно 380 п.н. промоторного района, экзоны 1, 2 и 3, полные интроны 1 и 2 и заканчивался в интроне 3.
3'-конец гена zalpha11-лиганда человека получали при помощи ПЦР с использованием ДНК из РАС 34А9 в качестве матрицы, с праймерами ZC23771 (SEQ ID NO:104) и ZC22449 (SEQ ID NO:103). Использовали ДНК-полимеразу Taq с поставляемым буфером с добавлением 4% ДМСО. Условия реакции были следующими: 94°С, 5 минут; с последующими 35 циклами 94°С в течение 30 секунд, 52°С в течение 1 минуты, 72°С в течение 3 минут; затем 72°С в течение 7 минут. Это давало фрагмент 2,9 т.п.н., который содержал часть экзона 3, полные интроны 3 и 4, полный экзон 4 и кодирующую часть экзона 5.
Геномная структура гена zalpha11-лиганда человека является следующей от 5' к 3': SEQ ID NO:105, содержащая около 240 п.н. промотора, экзон 1 (номера нуклеотидов 240-455 SEQ ID NO:105), интрон 1 (номера нуклеотидов 456-562 SEQ ID NO:105), зкзон 2 (номера нуклеотидов 563-598 SEQ ID NO:105) и часть интрона 2, содержащая 5' 748 п.н. (номера нуклеотидов 599-1347 SEQ ID NO:105); гэп приблизительно 3 т.п.н.; SEQ ID NO:106, содержащая 3' 718 п.н. интрона 2, экзон 3 (номера нуклеотидов 719-874 SEQ ID NO:106) и часть 5'-конца интрона 3 (номера нуклеотидов 875-1656 SEQ ID NO:106); гэп менее чем приблизительно 800 п.н.; SEQ ID NO: 107, содержащая 644 п.н. интрона 3; гэп менее чем приблизительно 800 п.н.; и SEQ ID NO:108, содержащая 3' 435 п.н. интрона 3, экзон 4 (номера нуклеотидов 436-513 SEQ ID NO:108), интрон 4 (номера нуклеотидов 514-603 SEQ ID NO:108) и часть 5'-конца экзона 5 (номера нуклеотидов 604-645 SEQ ID NO:108).
Пример 53
Исследование связывания 125I-меченного zalpha11-лиганда человека в клеточных линиях
25 микрограммов очищенного zalphall-лиганда человека (пример 29) метили 2 мКи 125I с использованием иодогранул (Pierce, Rockford Illinois) в соответствии с инструкциями изготовителя. Этот меченый белок использовали для оценки связывания zalpha11-лиганда человека с клетками Raji человека (АТСС No. CCL-86) с использованием связывания с мышиными клетками BaF3 дикого типа и клетками BaF3, трансфицированными zalpha11-рецептором (клетками BaF3/hzalpha11), в качестве контролей. Ожидалось связывание zalpha11-лиганда с клетками BaF3/hzalpha11 (положительный контроль), тогда как не ожидалось связывание с клетками BaF3 дикого типа (отрицательный контроль), на основе результатов анализа пролиферации (пример 5). Около 5×105 клеток Raji на лунку, 1×106 BaF3/hzalpha11 и 1×106 клеток BaF3 на лунку высевали отдельно в 96-луночные планшеты. Десять нг/мл меченого zalpha11-лиганда человека добавляли в двух повторностях к этим лункам с серийными разведениями немеченого конкурента-zalpha11-лиганда человека, добавляемыми от 250-кратного молярного избытка в разведениях 1:4 до 0,061-кратного молярного избытка. Каждую точку анализировали в двух повторностях. После добавления меченого zalpha11-лиганда человека к лункам им давали инкубироваться при 4°С в течение 2 часов, для того чтобы позволить лиганду связаться с этими клетками. Затем клетки промывали 3х в буфере для связывания (RPMI-1710 (JRH Bioscience) с 1% БСА (Sigma)) и считали на гамма-счетчике COBRA II AUTO-GAMMA (Packard Instrument Company, Meriden, CT).
Связывание меченого zalpha11-лиганда с клетками наблюдали в клетках Raji и BaF3/hzalpha11. Кроме того, в случае клеток Raji в среднем 250-кратный молярный избыток немеченого zalpha11-лиганда уменьшал связывание в 3 раза в присутствии неспецифического немеченого конкурента (Interferon Gamma из R&D Systems, Minneapolis, MN) и в 3,7 раза относительно варианта без конкурента. Наблюдали зависимую от дозы конкуренцию для специфического немеченого конкурента, zalpha11-лиганда человека. Таким образом, связывание zalpha11-лиганда с клетками Raji было специфическим. Подобным образом, для положительного контроля, клеток BaF3/zalpha11, 250-кратный молярный избыток немеченого zalpha11-лиганда уменьшал связывание в 2 раза относительно неспецифического конкурента и в 3,06 раз относительно варианта без конкурента. Таким образом, связывание zalpha11-лиганда с клетками BaF3/zalpha11 также было специфическим. Не наблюдали конкурентного связывания с клетками BaF3 дикого типа. Таким образом, было показано, что zalpha11-лиганд связывается специфически с клетками Raji и с клетками BaF3/hzalpha11, но не с клетками отрицательного контроля BaF3.
Пример 54
Экспрессия zalpha11-рецептора на клетках крови человека
А. Получение и культивирование клеток периферической крови человека
Свежевзятую кровь человека разбавляли 1:1 с использованием ЗФР (Gibco BRL) и наслаивали на Ficoll/Hypaque Plus (Pharmacia LKB Biotechnology Inc., Piscataway, NJ) и откручивали в течение 30 минут при 1800 об/мин и давали остановиться центрифуге без торможения. Межфазный слой удаляли и переносили в свежую пробирку Falkon на 50 мл (Falcon, VWR, Seattle, WA), доводили до конечного объема 40 мл ЗФР и откручивали в течение 10 минут при 1200 об/мин с включенным тормозом. Жизнеспособность выделенных клеток тестировали с использованием Трипанового синего (Gibco BRL) и клетки ресуспендировали при конечной концентрации 1×106 клеток/мл клеточной среды (среда RPMI 1640, 10% инативированная нагреванием фетальная телячья сыворотка, 5% L-глутамин, 5% пен/стреп) (Gibco BRL).
Клетки культивировали в 6-луночных чашках (Falcon, VWR) в течение 0,4 или 24 часов с различными описанными ниже стимулами. Стимуляцию анти-lgM, анти-С040 и анти-CDS выполняли, как в примере 44 и примере 42. Форболмиристатацетат (ФМА) и иономицин (Sigma, St. Louis, МО) (пример 5С) добавляли к подходящим лункам при 10 нг/мл и 0,5 мг/мл, соответственно. Клетки инкубировали при 37°С в увлажненном термостате в течение различных периодов времени.
В. Окрашивание антителами и анализ
Клетки собирали из чашек, промывали и ресуспендировали в охлажденной на льду окрашивающей среде (HBSS, 1% фетальная телячья сыворотка, 0,1% азид натрия) при концентрации около десять миллионов клеток на миллилитр. Блокирование Fc-рецептора и неспецифического связывания антител с клетками достигалось добавлением 10% нормальной козьей сыворотки (Gemini Bioproducts, Woodland, CA) и 10% нормальной сыворотки человека (Ultraserum, Gemini) к суспензии клеток. Аликвоты клеточных суспензий смешивали с FITC-меченным моноклональным антителом против одного из маркеров линий дифференцировки CD3, CD19 или CD14 (PharMingen, La Jolla, CA) и биотинилированным моноклональным антителом против zalpha11-рецептора человека (huzalpha11) (пример 35). Специфичность окрашивания определяли конкуренцией с использованием расторимого рецептора zalpha11 СЕЕ (пример 10А) при 10-кратном избытке по массе. После инкубирования на льду в течение 60 минут клетки промывали дважды охлажденной на льду окрашивающей средой и ресуспендировали в 50 мл окрашивающей среды, содержащей стрептавидин-РЕ (Caltag, Burlingame, CA). После 30-минутной инкубации на льду клетки промывали дважды охлажденным на льду промывным буфером (ЗФР, 1% телячья сыворотка, 0,1% азид натрия) и ресуспендировали в промывном буфере, содержащем 1 мг/мл 7-AAD (Molecular Probes, Eugene, OR) в качестве маркера жизнеспособности. Данные проточной цитометрии получали на живых клетках при помощи проточного цитометра FACSCalibur (BD Immunocytometry Systems, San Jose, CA). Как получение, так и анализ данных выполняли с использованием программного обеспечения CellQuest (BD Immunocytometry Systems).
Результаты окрашивания антителом анти-zalpha11 показали, что zalpha11-рецептор человека экспрессируется на клетках периферической крови человека, экспрессирующих CDS, CD19 или CD14. Окрашивание на CD3 и CD19 клетках было специфическим, как доказывается абсолютной конкуренцией с растворимым zalpha11-рецептором. Окрашивание на CD14 клетках обнаружило некоторую специфичность в отношении этого лиганда, как доказывается частичной конкуренцией с растворимым рецептором. Активация либо Т-клеток антителом анти-CD3, либо В-клеток антителом анти-CD40 приводила к увеличенному уровню zalpha11 клеточной поверхности при 24 часах. Не наблюдали увеличения уровня экспрессии zalpha11 при 4 часах с любым стимулом на любой популяции клеток. Обработка клеток zalpha11-лигандом приводила к снижению zalpha11-окрашивания на CD3-положительных и CD19-положительных клетках, но не на CD14-положительных клетках как при 4, так и при 24 часах.
Пример 55
Предварительная оценка водной стабильности zalpha11-лиганда человека
Проводили предварительные исследования для оценки характеристик водной стабильности zalpha11-лиганда человека в поддержании биопроцессинга, приготовления композиций и введения in vivo. Преследовались следующие цели: 1) проверка стабильности и извлечения из мининасосов Alzet Minipumps и при обычном хранении и манипулировании, 2) определение детектирующего стабильность характера некоторых аналитических способов, в том числе катионообменной ВЖХ (CX-HPLC), обращеннофазовой ВЖХ (RP-HPLC), гель-фильтрационной ВЖХ (SEC-HPLC) и биоанализа (ВаF3/zаlрhа11R-пролиферации (например, примера 2 и примера 4), и 3) определение ограничивающих стабильность деградационных путей и их кинетических зависимостей.
Аликвоты очищенного zalpha11-лиганда человека (пример 29) готовили разбавлением до 2 мг/мл в ЗФР (рН 7,4) и хранили в криофлаконах из полэтилена низкой плотности (LDPE) (Nalgene, 1,8 мл) при -80°С (контроль), 5°С, 30°С и 37°С. Пробы анализировали периодически на протяжении 29 дней при помощи СХ-, RP-, SEC-HPLC и биоанализа. Аликвоты хранили также при -80°С и подвергали циклическому замораживанию-оттаиванию (з/о) (-80°C/RT; 5x з/о, 10х з/о). Восстановление до нормы zalpha11-лиганда человека определяли в сравнении с контролем -80°С (1 з/о) во всех анализах.
Оставшийся раствор zalpha11-лиганда человека из контрольных -80°С-проб повторно замораживали (-80°С) после анализа. Эту аликвоту (2 з/о) использовали для оценки термической и конформационной стабильности zalpha11-лиганда человека как функции рН с использованием кругового дихроизма (КД). Раствор 2 мг/мл разбавляли до 100 мкг/мл в 3ФР-буферах в диапазоне рН 3,3-8,8. КД-спектры дальнего УФ подвергали мониторингу на протяжении диапазона температур 5-90°С с интервалами 5°С (n=3/рН). Использовали КД-спектрополяриметр Jasco 715 (Jasco, Easton, MD). Термическое развертывание молекулы подвергали мониторингу по изменениям эллиптичности при 222 нм как функции температуры. Оценки Тm получали при предположении модели развертывания с двумя состояниями. Данные строили с использованием программного обеспечения SlideWrite Plus для Windows v4.1 (Advanced Graphics Software; Encinitas, CA).
Извлечение и стабильность из мининасосов Alzet Minipumps (Model No. 1007D; ALZA Corporation, Mountain View, CA) оценивали заполнением насосов 100 мкл раствора 2 мг/мл zalpha11-лиганда человека, помещением насосов в 1,8 мл LDPE, содержащего 1 мл ЗФР (рН 7,4) и хранением их при 37°С. Высвобождение/извлечение zalpha11-лиганда человека из мининасосов оценивали при помощи СХ-, RP-, SEC-HPLC в дни 2, 4 и 7. Активность оценивали при помощи биоанализа в день 7. Данное исследование предназначено для оценки высвобождения из 3 насосов во время взятия проб.
Хроматографические данные позволяли предположить, что СХ- и SEC-HPLC (катионообменная ВЖХ и гель-фильтрационная ВЖХ) были детектирующими стабильность, тогда как обращеннофазовая ВЖХ (RP-HPLC) не обладала таким свойством. По меньшей мере 3 дополнительных пика, указывающих на видимые продукты деградации, наблюдали при помощи CX-HPLC. SEC-HPLC (гель-фильтрационная ВЖХ) разделяла видимые агрегаты zalpha11-лиганда человека, элюирующиеся раньше zalpha11-лиганда человека. Однако не наблюдали значимых дополнительных пиков, элюирующихся после пика zalpha11-лиганда человека. Это предполагает, что продукты деградации, наблюдаемые при помощи катионообменной ВЖХ (CX-HPLC), происходят, наиболее вероятно, из модификаций аминокислот, таких как деамидирование, а не из процессов гидролиза/протеолиза, приводящих к усеченным вариантам. Небольшую степень размытия фронта/образования "хвостов" наблюдали при обращеннофазовой ВЖХ (RP-HPLC) (относительно контроля) в пробах, которые, как было показано, подверглись значимой деградации согласно гель-фильтрационной (SEC-HPLC) и катионообменной ВЖХ (CX-HPLC). Однако видимые продукты деградации не разделялись обращеннофазовой ВЖХ (RP-HPLC). Деградация, наблюдаемая при катионообменной ВЖХ (CX-HPLC), увеличивалась как функция времени-температуры и сопровождалась видимой кинетикой первого порядка. % zalpha11-лиганда человека, извлекаемого при помощи катионообменной ВЖХ (CX-HPLC) после 29 дней при 37°С, 30°С и 5°С, был 39%, 63% и 98% соответственно. Агрегация также увеличивалась зависимым от времени-температуры образом. % агрегата, обнаруженного в препаратах, хранящихся в течение 29 дней при 37°С, 30°С и 5°С, был 7,4, 3,4 и ниже детектируемых пределов (BDL) соответственно. Не наблюдали значимых различий с использованием биоанализа в любой пробе, что позволяет предположить, что продукты деградации обладали активностью, эквивалентной активности интактного zalpha11-лиганда человека. Не наблюдали деградации при любом анализе в пробах, подвергнутых до 10 циклов з/о.
Высвобождение zalpha11-лиганда человека из Alzet Minipumps согласовалось с теоретически ожидаемым объемным высвобождением. Это предполагает, что значительная поверхностная адсорбция не ухудшает доставку zalpha11-лиганда человека с использованием Alzet Minipumps с концентрацией при заполнении 2 мг/мл. Наблюдали деградацию, согласующуюся с ранее отмечаемой деградацией. Чистота в %, определенная с использованием катионообменной ВЖХ zalpha11-лиганда человека, высвобождаемого после 2, 4 и 7 дней, была 96, 90 и 79% соответственно. Следует понимать, что деградация также имеет место, после того как zalpha11-лиганд человека высвобождается в среду высвобождения или разбавляется средой высвобождения. Таким образом, % чистоты в мининасосе может быть несколько отличающимся от %, определенного в среде высвобождения. Биоактивность каждой пробы согласовалась с ожидаемым количеством zalpha11-лиганда человека, высвобождаемого из мининасосов.
КД-спектры в коротковолновой (дальней) УФ-области zalpha11-лиганда человека, как ожидалось, согласовался с интерлейкинами, такими как IL-3 (J. Biochem. 23:352-360, 1991), IL-4 (Biochemistry, 30:1259-1264, 1991) и IL-6-мутантами (Biochemistry, 35:11503-11511, 1996). Не наблюдали больших изменений в дальней УФ-области КД-спектров как функции рН. Результаты показали, что рН максимальной термостабильности/конформационной стабильности был примерно рН 7,4. Анализы кривых развертывания молекулы были основаны на механизме двух состояний развертывания, чтобы сделать возможным сравнение термостабильности/конформационной стабильности в зависимости от рН/состава. Однако могут существовать один или несколько промежуточных продуктов во время процесса развертывания, так как кооперативность была относительно низкой на основе "неглубокости" кривой развертывания. Хотя специальные исследования для определения, происходит ли повторная укладка zalpha11-лиганда человека после термического развертывания до 90°С, не проводились, предварительные данные предполагают, что по меньшей мере частичная повторная укладка происходит после возвращения температуры пробы до 20°С охлаждением.
Эти исследования позволяют идентифицировать аналитический пример для очистки и проверки стабильности zalpha11-лиганда человека. Например, гель-фильтрационную ВЖХ можно использовать для характеристики степени и скорости агрегации в водном растворе. Подобным образом, катионообменную ВЖХ можно использовать для характеристики степени и скорости деградации zalpha11-лиганда человека механизмами, иными, чем агрегация. Биоанализ можно использовать для проверки активности zalpha11-лиганда человека и его продуктов водной деградации. Например, варианты zalpha11-лиганда человека, полученные в водном растворе и разделенные при помощи катионообменной ВЖХ (CX-HPLC), сами могут быть применимы в качестве терапевтических агентов, так как они имеют эквивалентную биологическую активность. Тот факт, что zalpha11-лиганд человека деградируется несколькими различными процессами (агрегацией, модификациями аминокислот), предполагает, что предпочтительная или уникальная композиция, которая минимизирует скорость процесса деградации, может быть необходимой для долгосрочной стабильности продукта в растворе.
Идентификация природы продуктов водной деградации и определение их кинетических зависимостей (рН, концентрации, наполнителей) находятся в процессе исследования. Стабильность zalpha11-лиганда человека в сыворотке/плазме определяется для подтверждения планирования и интерпретации исследований in vivo.
Из вышесказанного должно быть понятно, что, хотя характерные варианты данного изобретения описаны здесь с целью иллюстрации, разнообразные модификации могут быть сделаны без отклонения от идеи и объема данного изобретения. Таким образом, данное изобретение не ограничивается ничем, кроме прилагаемой формулы изобретения.z
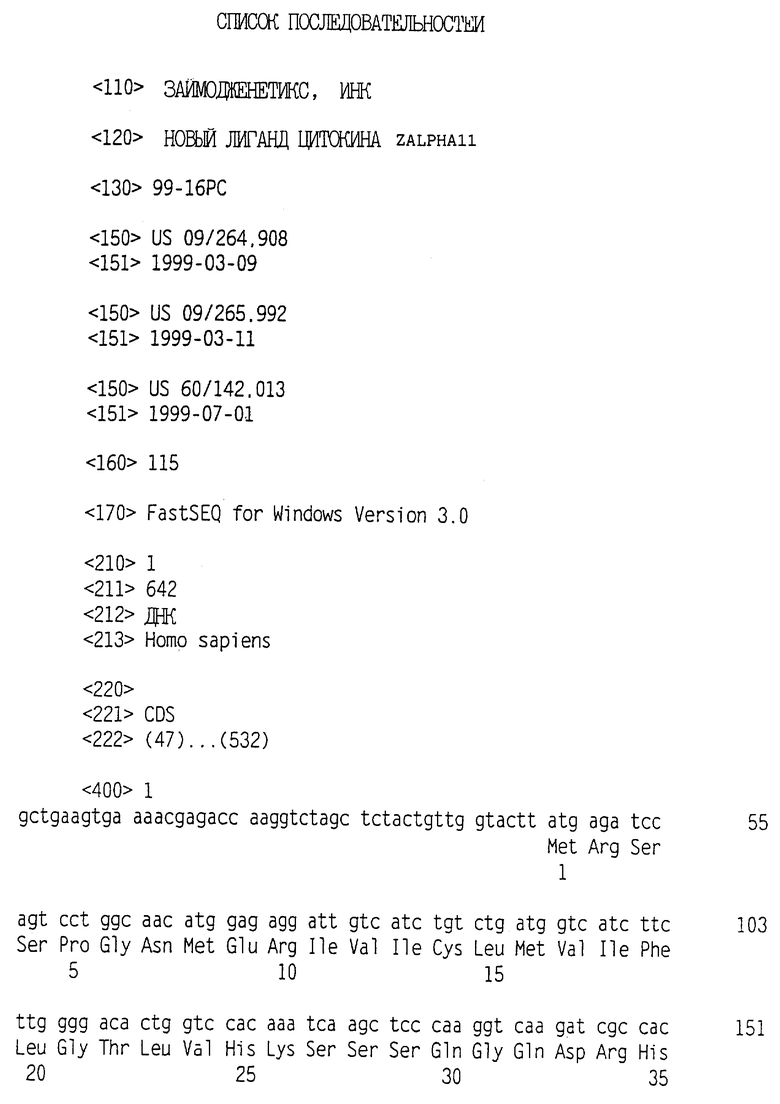

















































| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ ЛИГАНД РЕЦЕПТОРА ЦИТОКИНА ZCYTOR17 | 2003 |
|
RU2490276C2 |
| ВЫДЕЛЕННЫЙ РАСТВОРИМЫЙ IL-20 РЕЦЕПТОР (ВАРИАНТЫ) | 2000 |
|
RU2279441C2 |
| ИНТЕРФЕРОН-ПОДОБНЫЙ БЕЛОК ZCYTO21 | 2001 |
|
RU2292394C2 |
| ГЕМОПОЭТИЧЕСКИЙ БЕЛОК (ВАРИАНТЫ), ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, СПОСОБ ПОЛУЧЕНИЯ ГЕМОПОЭТИЧЕСКОГО БЕЛКА, СПОСОБ СЕЛЕКТИВНОЙ ЭКСПАНСИИ КРОВЕТВОРНЫХ КЛЕТОК, СПОСОБ ПРОДУЦИРОВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СПОСОБ СТИМУЛЯЦИИ ПРОДУЦИРОВАНИЯ КРОВЕТВОРНЫХ КЛЕТОК У ПАЦИЕНТА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБ ЛЕЧЕНИЯ | 1997 |
|
RU2245887C2 |
| МОДИФИЦИРОВАННОЕ АГОНИСТИЧЕСКОЕ АНТИТЕЛО | 2001 |
|
RU2295537C2 |
| ПОЛИПЕПТИД С АНТИВИРУСНОЙ АКТИВНОСТЬЮ, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2004 |
|
RU2372356C2 |
| ГОМОГЕННЫЕ ПРЕПАРАТЫ IL-28 И IL-29 | 2004 |
|
RU2518324C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-21 ЧЕЛОВЕКА | 2008 |
|
RU2504552C2 |
| ДЕГРАДИРОВАННОЕ АНТИТЕЛО, ЯВЛЯЮЩЕЕСЯ АГОНИСТОМ TPO | 2001 |
|
RU2287534C2 |
| ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ЛИГАНД CD86 КОШКИ, ДИАГНОСТИЧЕСКИЙ ОЛИГОНУКЛЕОТИД, КЛОНИРУЮЩИЙ ВЕКТОР, ВАКЦИНА, СПОСОБЫ ИНДУКЦИИ ИЛИ ПОДАВЛЕНИЯ ИММУНИТЕТА У КОШКИ | 1999 |
|
RU2263145C2 |
Изобретение относится к биотехнологии и может быть использовано для получения Zalpha11 лиганда и антител к нему. Zalpha11 лиганд выделяют из библиотеки кДНК, генерированной из активированных клеток периферической крови человека (hPBCs), которые были отобраны на CD3. Zalpha11 лигандом инокулируют животное и получают антитела, которые специфически связываются с эпитопами, пептидами или полипептидами Zalpha11 лиганда. Изобретение позволяет эффективно регулировать пролиферацию и/или развитие гемопоэтических клеток in vitro и in vivo, 11 н. и 7 з.п. ф-лы, 1 ил., 6 табл.
(a) аминокислотных остатков 41-56 SEQ ID NO:2;
(b) аминокислотных остатков 69-84 SEQ ID NO:2;
(c) аминокислотных остатков 92-105 SEQ ID NO:2 и
(d) аминокислотных остатков 135-148 SEQ ID NO:2.
первый полипептид, который содержит последовательность аминокислотных остатков от 41 до 56 SEQ ID NO:2;
первый спейсер из 6-27 аминокислотных остатков;
второй полипептид, который содержит последовательность аминокислотных остатков, выбранных из группы, состоящей из:
(a) остатков 53-75 SEQ ID NO:111 спирали В IL-2;
(b) остатков 65-83 SEQ ID NO:112 спирали В IL-4;
(c) остатков 84-101 SEQ ID NO:113 спирали В IL-15;
(d) остатков 72-81 SEQ ID NO:114 спирали В GM-CSF и
(e) аминокислотных остатков 69-84 SEQ ID NO:2;
второй спейсер из 5-11 аминокислотных остатков;
третий полипептид, который содержит последовательность аминокислотных остатков, выбранную из группы, состоящей из:
(a) остатков 87-99 SEQ ID NO:111 спирали С IL-2;
(b) остатков 95-118 SEQ ID NO:112 спирали С IL-4;
(c) остатков 107-119 SEQ ID NO:113 спирали С IL-15;
(d) остатков 91-102 SEQ ID NO:114 спирали С GM-CSF и
(е) аминокислотных остатков 92-105 SEQ ID NO:2;
третий спейсер из 3-29 аминокислотных остатков и четвертый полипептид, который содержит последовательность аминокислотных остатков, выбранных из группы, состоящей из:
(a) остатков 103-121 SEQ ID NO:111 спирали D IL-2;
(b) остатков 134-157 SEQ ID NO:112 спирали D IL-15;
(c) остатков 134-160 SEQ ID NO:113 спирали D IL-4;
(d) остатков 120-131 SEQ ID NO:114 спирали D GM-CSF и
(e) аминокислотных остатков 135-148 SEQ ID NO:2.
(a) полипептида, состоящего из 9-131 аминокислот, где этот полипептид является идентичным смежной последовательности аминокислотных остатков в SEQ ID NO:2 от номера аминокислоты 32 (Gln) до номера аминокислоты 162 (Ser);
(b) полипептида по п.1;
(c) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 41 (Gln) до номера аминокислоты 148 (Ile);
(d) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 41 (Gln) до номера аминокислоты 56 (Val);
(e) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 69 (Thr) до номера аминокислоты 84 (Leu);
(f) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 92 (Asn) до номера аминокислоты 105 (Arg);
(g) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 135 (Glu) до номера аминокислоты 148 (Ile);
(h) полипептида, содержащего аминокислотную последовательность SEQ ID NO:72;
(i) полипептида, содержащего аминокислотную последовательность SEQ ID NO:73;
(j) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 32 (Gln) до номера аминокислоты 162 (Ser);
(k) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 1 (Met) до номера аминокислоты 162 (Ser);
(1) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 114 до номера аминокислоты 119;
(m) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 101 до номера аминокислоты 105;
(n) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 126 до номера аминокислоты 131;
(о) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 113 до номера аминокислоты 118; и
(р) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2 от номера аминокислоты 158 до номера аминокислоты 162;
причем указанный полипептид индуцирует иммунную реакцию в животном с образованием антитела; и
выделение этого антитела из животного.
| COSMAN D | |||
| The hematopoietin receptor superfamily, Cytokine, 1993, v.5, n.2, p.95-106 | |||
| BRIZZI M.F.et al., Granulocyte-macrophage colony-stimulating factor stimulates JAK2 signaling pathway and rapidly activates p93fes, STAT1 p91, and STAT3 p92 in polymorphonuclear leukocytes, J Biol | |||
| Chem., 1996, v.271, n.7, p.3562-3567 | |||
| RU 95122449 A, 10.10.1998. |

