Ссылка на родственные заявки
Настоящая заявка родственна предварительной заявке на патент США № 60/435315, поданной 19 декабря 2002 г., предварительной заявке на патент США № 60/375323, поданной 25 апреля 2002 г., и предварительной заявке на патент США № 60/350325, поданной 18 января 2002 г. На основании раздела 35 Кодекса законов США § 119(е)(1) данная заявка претендует на приоритет по указанным предварительным заявкам.
Предпосылки изобретения
Пролиферацию и дифференцировку клеток многоклеточных организмов контролируют гормоны и полипептидные факторы роста. Указанные диффузионные молекулы позволяют клеткам взаимодействовать друг с другом и кооперативно действовать в процессе образования клеток, тканей и органов и восстановления поврежденных тканей. Примеры гормонов и факторов роста включают в себя стероидные гормоны (например, эстроген, тестостерон), паращитовидный гормон, фолликулостимулирующий гормон, интерлейкины, тромбоцитарный фактор роста (PDGF), эпидермальный фактор роста (EGF), колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF), эритропоэтин (ЕРО) и кальцитонин.
Гормоны и факторы роста влияют на клеточный метаболизм в результате связывания с рецепторами. Рецепторы могут быть встроенными в мембрану белками, связанными с путями передачи сигналов в клетке, такими как системы вторичных мессенджеров. Рецепторы других классов являются растворимыми молекулами, такими как факторы транскрипции.
Цитокины обычно стимулируют пролиферацию или дифференцировку клеток гемопоэтической линии или участвуют в механизмах иммунной и воспалительной реакции организма. Примерами цитокинов, влияющих на гемопоэз, являются эритропоэтин (ЕРО), стимулирующий образование эритроцитов; тромбопоэтин (ТРО), стимулирующий образование клеток линии мегакариоцитов; и колониестимулирующий фактор гранулоцитов (G-CSF), стимулирующий образование нейтрофилов. Указанные цитокины способствуют восстановлению нормальных уровней кровяных клеток у пациентов, страдающих анемией, тромбоцитопенией и нейтропенией, или проходящих курс химиотерапии при лечении рака.
Интерлейкины образуют семейство цитокинов, опосредующих иммунологические реакции, включая воспалительные реакции. Интерлейкины опосредуют целый ряд воспалительных патологий. Центральное место в иммунной реакции занимают Т-клетки, которые продуцируют множество цитокинов и вызывают искусственный иммунитет против антигенов. Цитокины, продуцируемые Т-клетками, классифицированы как тип 1 и тип 2 (Kelso, A. Immun. Cell Biol. 76:300-317, 1998). Цитокины типа 1, включая IL-1, IFN-γ, LT-α, участвуют в воспалительных реакциях, вызывают противовирусный иммунитет, иммунитет к внутриклеточным паразитам и отторжение аллотрансплантата. Цитокины типа 2, включая IL-4, IL-5, IL-6, IL-10 и IL-13, участвуют в гуморальных реакциях, вызывают иммунитет к гельминтам и аллергические реакции. Цитокины, занимающие промежуточное положение между типами 1 и 2, включают в себя IL-3, GM-CSF и TNF-α. Имеются данные, позволяющие предположить, что популяции Т-клеток, продуцирующих цитокины типа 1 и типа 2, мигрируют преимущественно в воспаленные ткани разных типов.
Зрелые Т-клетки могут быть активированы, например, антигеном или другим стимулом, в результате чего они начинают продуцировать цитокины, молекулы, осуществляющие биохимическую передачу сигналов, или рецепторы, которые далее влияют на судьбу популяции Т-клеток.
В-клетки могут быть активированы рецепторами на поверхности клеток, включая В-клеточный рецептор и другие вспомогательные молекулы, выполняющие вспомогательные функции клеток, такие как продуцирование цитокинов.
Моноциты/макрофаги и Т-клетки могут быть активированы рецепторами на поверхности клеток, при этом они играют главную роль в иммунной реакции, представляя антиген лимфоцитам, а также действуют в качестве вспомогательных клеток для лимфоцитов, секретируя множество цитокинов.
Естественные клетки-киллеры (NK) имеют общую клетку-предшественник с Т-клетками и В-клетками и играют определенную роль в иммунологическом надзоре. NK-клетки, которые составляют до 15% лимфоцитов крови, не экспрессируют рецепторы антигенов и поэтому не используют функцию распознавания МНС в качестве необходимого условия для связывания с клеткой-мишенью. NK-клетки участвуют в распознавании и уничтожении некоторых опухолевых клеток и вирусинфицированных клеток. Считается, что in vivo NK-клетки требуют активации, однако установлено, что in vitro NK-клетки уничтожают некоторые типы опухолевых клеток без активации.
Рассмотренные действия семейства цитокинов in vivo иллюстрируют большой клинический потенциал и потребность в других цитокинах, агонистах и антагонистах цитокинов. Настоящее изобретение позволяет удовлетворить такие потребности благодаря созданию нового цитокина, стимулирующего клетки гемопоэтической линии, получению композиций, содержащих указанный цитокин, и разработке способов его применения.
Настоящее изобретение относится к таким полипептидам, предназначенным для вышеуказанных и других применений, которые должны быть очевидны специалистам в данной области из приведенного ниже описания изобретения.
Краткое описание чертежей
На фиг.1 представлены результаты сравнительного анализа zcytor17lig человека (SEQ ID NO:2) (zcytor17lig), zcytor17lig мыши (SEQ ID NO:11) (mzcytor17lig), IL-3 мыши (mIL-3) (SEQ ID NO:100) и IL-3 человека (hIL-3) (SEQ ID NO:102).
На фиг.2 представлены результаты сравнительного анализа zcytor17lig человека (SEQ ID NO:2) (zcytor17lig) и zcytor17lig мыши (SEQ ID NO:11) (mzcytor17lig).
На фиг.3 показан график гидрофильности Хоппа-Вудса zcytor17lig человека (SEQ ID NO:2).
Подробное описание изобретения
Подробному описанию изобретения предшествует определение следующих терминов, ознакомление с которыми может способствовать лучшему пониманию изобретения.
Термин «аффинная метка» использован в данном описании для обозначения полипептидного сегмента, который может быть присоединен ко второму полипептиду для очистки или обнаружения второго полипептида или создания сайтов для присоединения второго полипептида к субстрату. В частности, в качестве аффинной метки можно использовать любой пептид или белок, для которого существует антитело или другой специфический связывающий агент. Аффинные метки включают в себя белок А полигистидинового тракта (Nilsson et al., EMBO J. 4:1075, 1985; Nilsson et al., Methods Enzymol. 198:3, 1991), глутатион-S-трансферазу (Smith and Johnson, Gene, 67:31, 1988), аффинную метку Glu-Glu (Grussenmeyer et al., Proc. Natl. Acad. Sci. USA 82:7952-4, 1985), вещество Р, пептид Flag™ (Hopp et al., Biotechnology 6:1204-10, 1988), связывающий стрептавидин пептид или другой эпитоп или связывающий домен. См., например, Ford et al., Protein Expression and Purification 2:95-107, 1991. ДНК-кодирующие аффинные метки можно приобрести у коммерческих поставщиков (например, Pharmacia Biotech, Piscataway, NJ).
Термин «аллельный вариант» использован в данном описании для обозначения любой из двух или большего числа альтернативных форм гена, занимающих один и тот же хромосомный локус. Аллельная вариабельность возникает естественным путем в результате мутации и может вызывать фенотипический полиморфизм в популяциях клеток. Мутации гена могут быть молчащими (не вызывающими изменений в кодированном полипептиде) или могут кодировать полипептиды, имеющие измененную аминокислотную последовательность. Термин «аллельный вариант» использован в данном описании также для обозначения белка, кодированного аллельным вариантом гена.
Термины «аминоконцевой» и «карбоксиконцевой» использованы в данном описании для обозначения положений в полипептидах. В зависимости от контекста, указанные термины использованы применительно к определенной последовательности или части полипептида для обозначения пространственной близости или относительного положения. Например, расположение последовательности у карбоксильного конца контрольной последовательности в полипептиде означает, что данная последовательность находится рядом с карбоксильным концом указанной контрольной последовательности, но необязательно у карбоксильного конца всего полипептида.
Термин «пара комплемент/антикомплемент» означает неидентичные части, образующие нековалентно связанную, устойчивую пару в соответствующих условиях. Например, биотин и авидин (или стрептавидин) являются предполагаемыми членами пары комплемент/антикомплемент. Другие типичные пары комплемент/антикомплемент включают в себя пары рецептор/лиганд, антитело/антиген (или гаптен или эпитоп), смысловой/антисмысловой полинуклеотид и тому подобное. При желании дальнейшего расчленения пары комплемент/антикомплемент следует отметить, что пара комплемент/антикомплемент характеризуется сродством связывания <109 M-1.
Термин «комплементы молекулы полинуклеотида» означает молекулу полинуклеотида, содержащую комплементарную последовательность оснований и имеющую обратную ориентацию по сравнению с контрольной последовательностью. Например, последовательность 5' ATGCACGGG 3' комплементарна последовательности 5' CCCGTGCAT 3'.
Термин «смежная последовательность» означает полинуклеотид, который имеет участок идентичной или комплементарной последовательности, смежный с другим полинуклеотидом. Считается, что смежные последовательности «перекрывают» данный участок полинуклеотидной последовательности на протяжении всей длины или части указанного полинуклеотида. Например, типичными смежными последовательностями для полинуклеотидной последовательности 5'-ATGGCTTAGCTT-3' являются 5'-TAGCTTgagtct-3' и 3'-gtcgacTACCGA-5'.
Термин «вырожденная нуклеотидная последовательность» означает последовательность нуклеотидов, которая включает в себя один или несколько вырожденных кодонов (по сравнению с контрольной молекулой полинуклеотида, кодирующей полипептид). Вырожденные кодоны содержат разные триплеты нуклеотидов, но кодируют один и тот же аминокислотный остаток (то есть каждый триплет GAU и GAC кодирует Asp).
Термин «экспрессирующий вектор» служит для обозначения молекулы ДНК, линейной или кольцевой, которая содержит сегмент, кодирующий представляющий интерес полипептид, функционально связанный с дополнительными сегментами, предназначенными для его транскрипции. Такие дополнительные сегменты содержат промоторную и терминаторную последовательности и могут включать в себя один или несколько ориджинов репликации, один или несколько селектируемых маркеров, энхансер, сигнал полиаденилирования и т.д. Экспрессирующие векторы обычно выделяют из плазмидной или вирусной ДНК, либо они могут содержать элементы ДНК обоих типов.
Термин «выделенный» применительно к полинуклеотиду означает, что данный полинуклеотид был удален из естественной генетической среды и, следовательно, не содержит других периферических или нежелательных кодирующих последовательностей и имеет форму, пригодную для использования в генетически конструируемых системах продуцирования белка. Такие выделенные молекулы представляют собой молекулы, удаленные из естественного окружения, и включают в себя кДНК и геномные клоны. Выделенные молекулы ДНК согласно изобретению не содержат других генов, с которыми они обычно ассоциированы, но могут включать в себя естественные 5'- и 3'-концевые нетранслированные области, такие как промоторы и терминаторы. Идентификация ассоциированных областей должна быть очевидна специалисту в данной области (см., например, Dynan and Tijan, Nature 316:774-78, 1985).
«Выделенный» полипептид или белок является полипептидом или белком, который находится в условиях, отличных от естественного окружения, такого как кровь и ткани животного. В предпочтительной форме выделенный полипептид по существу не содержит других полипептидов, в частности, других полипептидов животного происхождения. Полипептиды предпочтительно должны быть получены в высоко очищенной форме, то есть со степенью чистоты более 95%, более предпочтительно со степенью чистоты более 99%. При использовании в данном контексте термин «выделенный» не исключает наличия такого же полипептида в альтернативных физических формах, таких как димеры либо гликозилированные или производные формы.
Термин «неопластический» применительно к клеткам означает клетки, подвергающиеся новой и аномальной пролиферации, в частности, в ткани, где такая пролиферация является неконтролируемой и прогрессирующей, приводя к возникновению новообразования. Неопластические клетки могут быть злокачественными, то есть инвазивными и метастатическими, или доброкачественными.
Термин «функционально связанный» применительно к сегментам ДНК означает такое расположение сегментов, при котором они кооперативно выполняют присущие им функции, например, транскрипция начинается в промоторе, продолжается в кодирующем сегменте и заканчивается в терминаторе.
Термин «ортолог» означает полипептид или белок, полученный из одного вида, который в функциональном отношении противоположен полипептиду или белку, полученному из другого вида. Различие последовательностей у ортологов является результатом видообразования.
«Паралоги» являются разными, но структурно родственными белками, продуцируемыми организмом. Считается, что паралоги образуются в результате дупликации гена. Например, α-глобин, β-глобин и миоглобин являются паралогами друг друга.
«Полинуклеотид» является одноцепочечным или двухцепочечным полимером дезоксирибонуклеотидных или рибонуклеотидных оснований, считываемых от 5'-конца к 3'-концу. Полинуклеотиды включают в себя РНК и ДНК и могут быть выделены из природных источников, синтезированы in vitro или получены из комбинации природных и синтетических молекул. Величина полинуклеотидов выражается в виде пар нуклеотидов (сокращенно «п.н.»), нуклеотидов («н.») или тысяч пар нуклеотидов («т.п.н.»). В зависимости от контекста два последних термина могут описывать одноцепочечные или двухцепочечные полинуклеотиды. Когда данный термин служит для определения двухцепочечных молекул, он используется для обозначения всей длины и является эквивалентным термину «пары оснований». Специалисту в данной области должно быть понятно, что две цепи в двухцепочечном полинуклеотиде могут иметь немного разную длину, и концы таких цепей могут быть неравномерными в результате ферментативного расщепления, таким образом, не все нуклеотиды в молекуле двухцепочечного полинуклеотида могут быть спарены.
«Полипептид» является полимером аминокислотных остатков, соединенных пептидными связями, которые могут быть продуцированы естественным или синтетическим путем. Полипептиды длиной менее примерно 10 аминокислотных остатков обычно определяются как «пептиды».
Термин «промотор» использован в данном описании в значении, принятом в данной области, и означает часть гена, содержащего последовательности ДНК, которые связывают РНК-полимеразу и инициируют транскрипцию. Промоторные последовательности обычно, но не всегда, находятся в 5'-концевых некодирующих областях генов.
«Белок» представляет собой макромолекулу, содержащую одну или несколько полипептидных цепей. Белок может также содержать непептидные компоненты, такие как углеводородные группы. Углеводороды и другие непептидные заместители могут быть введены в белок клеткой, в которой продуцирован данный белок, и могут изменяться в зависимости от типа клетки. Белки определяются в данном описании с учетом их аминокислотных скелетных структур; заместители, такие как углеводородные группы, обычно не указываются, но, тем не менее, могут присутствовать.
Термин «рецептор» означает ассоциированный с клеткой белок, который связывается с биоактивной молекулой (то есть лигандом) и опосредует действие данного лиганда на клетку. Мембраносвязанные рецепторы характеризуются мультипептидной структурой, включающей в себя внеклеточный лигандсвязывающий домен и внутриклеточный эффекторный домен, который обычно участвует в передаче сигналов. В результате связывания лиганда с рецептором происходит конформационное изменение рецептора, которое вызывает взаимодействие между эффекторным доменом и другой молекулой (молекулами) в клетке. Данное взаимодействие в свою очередь ведет к изменению метаболизма клетки. Метаболические изменения, связанные с взаимодействием рецептор-лиганд, включают в себя транскрипцию гена, фосфорилирование, дефосфорилирование, увеличение продукции циклического АМФ, мобилизацию клеточного кальция, мобилизацию мембранных липидов, клеточную адгезию, гидролиз инозитлипидов и гидролиз фосфолипидов. Как правило, рецепторы могут быть мембраносвязанными, цитозольными или ядерными; мономерными (например, рецептор тироидстимулирующего гормона, бета-адренергический рецептор) или многомерными (например, рецептор PDGF, рецептор гормона роста, рецептор IL-3, рецептор GM-CSF, рецептор G-CSF, рецептор эритропоэтина и рецептор IL-6).
Термин «секреторная сигнальная последовательность» означает последовательность ДНК, кодирующую полипептид («секреторный пептид»), который в качестве компонента более крупного полипептида направляет более крупный полипептид по секреторному пути клетки, в которой он синтезирован. Более крупный полипептид обычно расщепляется для удаления секреторного пептида при прохождении по секреторному пути.
Термин «сплайсированный вариант» использован в данном описании для обозначения альтернативных форм РНК, транскрибированной из гена. Сплайсированный вариант образуется естественным путем в результате использования чередующихся сайтов сплайсинга в молекуле транскрибированной РНК или реже между молекулами отдельно транскрибированной РНК и может вызывать образование нескольких мРНК, транскрибированных из одного гена. Сплайсированные варианты могут кодировать полипептиды, имеющие измененную аминокислотную последовательность. Термин «сплайсированный вариант» использован в данном описании также для обозначения белка, кодируемого сплайсированным вариантом мРНК, транскрибированной из гена.
Вполне понятно, что молекулярные массы и длины полимеров, определяемые неточными аналитическими методами (например, гель-электрофорезом), представляют собой примерные величины. Величина Х, определяемая как «около» Х или «примерно» Х, является точной в пределах погрешности ±10%.
Все приведенные публикации включены в данное описание в качестве ссылки.
В основе настоящего изобретения частично лежит открытие новой последовательности ДНК, которая кодирует белок, имеющий структуру четырехспирального цитокина. В результате выполнения анализов методами клонирования и пролиферации, которые подробно рассмотрены в данном описании, было установлено, что полинуклеотидная последовательность, кодирующая новый полипептидный лиганд, является лигандом с высокой специфичностью в отношении рецептора zcytor17 (SEQ ID NO:5) и имеет по меньшей мере одну дополнительную субъединицу, содержащую бета-рецептор онкостатина М (OSMRbeta) (SEQ ID NO:7) и WSX-1 (SEQ ID NO:9). Данный полипептидный лиганд, получивший название zcytor17lig, был выделен из библиотеки кДНК, созданной из активированных клеток периферической крови человека (hPBC), которые были отобраны при мечении CD3. CD3 является маркером клеточной поверхности, специально предназначенным для клеток лимфоидного происхождения, в частности, Т-клеток.
В нижеследующих примерах для скрининга источника кДНК, кодирующей zcytor17lig, была использована линия клеток, выживание и рост которых при отсутствии других факторов роста зависит от пути, связанного с OSMRbeta и рецептором zcytor17, или от пути, связанного с OSMRbeta, WSX-1 и рецептором zcytor17. Предпочтительной линией клеток, зависящей от фактора роста, которая была использована для трансфекции и экспрессии рецептора zcytor17, была линия клеток BaF3 (Palacios and Steinmetz, Cell 41:727-734, 1985; Mathey-Prevot et al., Mol. Cell. Biol. 6:4133-4135, 1986). Однако для вышеуказанной цели пригодны и другие линии клеток, зависящих от фактора роста, такие как FDC-P1 (Hapel et al., Blood 64:786-790, 1984) и МО7е (Kiss et al., Leukemia 7:235-240, 1993).
Аминокислотная последовательность для рецепторов OSMR, WSX-1 и zcytor17 показала, что кодируемые рецепторы относятся к подсемейству рецепторов цитокина класса I, которое включает в себя, не ограничиваясь ими, рецепторы для IL-2, IL-4, IL-7, Lif, IL-12, IL-15, EPO, TPO, GM-CSF и G-CSF (см. Cosman, "The Hematopoietin Receptor Superfamily" in Cytokine 5(2):95-106, 1993). Рецептор zcytor17 всесторонне описан в принадлежащей на правах общей собственности заявке на патент РСТ № US01/20484 (публикация WIPO № WO 02/00721), и WSX-1 всесторонне описан в патенте США № 5925735. Анализ распределения в тканях мРНК рецептора zcytor17 позволил выявить экспрессию в субпопуляции активированных CD4+ и CD8+ Т-клеток, CD14+-моноцитах и более слабую экспрессию в CD19+ В-клетках. Кроме того, указанная мРНК присутствовала как в покоящихся, так и в активированных линиях клеток моноцитов ТНР-1 (АТСС № TIB-202), U937 (ATCC № CRL-1593.2) и HL60 (ATCC № CCL-240).
Экспрессия WSX-1 является наиболее сильной в тимусе, селезенке, PBL и лимфатическом узле, кроме того, повышенная экспрессия наблюдалась в активированных Т-клетках. Было обнаружено весьма обширное распределение в тканях рецептора OSMRbeta. Распределение в тканях трех вышеуказанных рецепторов позволяет предположить, что мишенью для предполагаемого лиганда zcytor17lig являются клетки гемопоэтической линии, в частности, Т-клетки, моноциты/макрофаги, клетки-предшественники лимфоидных клеток и лимфоидные клетки. Другие известные четырехспиральные цитокины, воздействующие на лимфоидные клетки, включают в себя IL-2, IL-4, IL-7 и IL-15. Для ознакомления с четырехспиральными цитокинами см. Nicola et al., Advances in Protein Chemistry 52:1-65, 1999 and Kelso, A., Immunol. Cell Biol. 76:300-317, 1998.
Кондиционированная среда (СМ), полученная из CD3+-селектированных, РМА/иономицин-стимулированных клеток периферической крови человека, поддерживала рост клеток BaF3, экспрессирующих рецепторы zcytor17, OSMRbeta и WSX-1 и зависящих от IL-3. Кондиционированные клетками среды, которые: 1) не были стимулированы РМА/иономицином или 2) не были CD3-селектированы (при стимуляции РМА/иономицином или без указанной стимуляции), не поддерживали рост клеток BaF3, экспрессирующих рецепторы zcytor17, OSMRbeta и WSX-1 (BaF3/zcytor17/WSX-1/OSMRbeta). Контрольные эксперименты показали, что указанная пролиферативная активность не была присуща другим известным факторам роста и что способность таких кондиционированных сред стимулировать пролиферацию клеток, экспрессирующих рецепторы zcytor17/WSX-1/OSMRbeta, можно было нейтрализовать растворимой формой рецептора zcytor17.
Кондиционированная среда из CD3+-селектированных клеток, активированных РМА/иономицином, также поддерживала рост клеток BaF3, экспрессирующих рецепторы zcytor17 и OSMRbeta (zcytor17/OSMRbeta), в то время как указанная кондиционированная среда не стимулировала клетки BaF3, экспрессирующие только рецепторы zcytor17 и WSХ-1 (zcytor17/WSХ-1) или содержащие только рецептор OSMRbeta.
Пролиферация клеток BaF3, экспрессирующих рецепторы zcytor17/WSX-1/OSMRbeta под действием кондиционированной среды из CD3+-селектированных, РМА/иономицин-стимулированных клеток периферической крови человека, была идентифицирована при визуальном осмотре культур и/или при помощи анализа пролиферации. В данной области известны многие пригодные анализы пролиферации, которые включают в себя восстановление красителя, такого как аламаровый синий (AlamarBlue™) (AccuMed International, Inc. Westlake, Ohio), бромид 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия (Mosman, J. Immunol. Meth. 65:55-63. 1983); 3-(4,5-диметилтиазол-2-ил)-5-3-карбоксиметоксифенил-2Н-тетразолий; гидроксид 2,3-бис(2-метокси-4-нитро-5-сульфофенил)-5-[(фениламино)карбонил]-2H-тетразолия и хлорид цианодитолилтетразолия (которые можно приобрести коммерческим путем в компании Polysciences, Inc., Warrington, PA); анализы митогенеза, такие как измерение включения 3Н-тимидина; вытеснение красителя с использованием, например, нафталинового черного или трипанового синего; поглощение красителя с использованием диацетилфлуоресцеина; и высвобождение хрома. См., например, публикацию Freshney, Culture of Animal Cells: A Manual of Basic Technique, 3rd ed., Wiley-Liss, 1994, которая включена в данное описание в качестве ссылки.
Библиотека кДНК была получена из CD3+-селектированных, РМА- и иономицин-стимулированных первичных клеток периферической крови человека. Библиотеку кДНК CD3+-селектированных, РМА- и иономицин-стимулированных клеток периферической крови человека делили на пулы, содержащие несколько молекул кДНК, и трансфицировали в линию клеток-хозяев, например, клетки ВНК 570 (АТСС, номер доступа 10314). Трансфицированные клетки-хозяева культивировали в среде, которая не содержала экзогенных факторов роста (например, 5% FBS), и собирали кондиционированную среду. Кондиционированные среды анализировали в отношении способности стимулировать пролиферацию клеток BaF3, трансфицированных рецепторами zcytor17, WSX-1 и OSMRbeta. Были идентифицированы пулы кДНК, продуцирующие кондиционированную среду, которая стимулировала клетки BaF3, содержащие рецепторы zcytor17/WSX-1/OSMRbeta. Собранную плазмидную кДНК электропорировали в E. coli. кДНК выделяли из единичных колоний и трансфицировали в клетки ВНК 570. Положительные клоны идентифицировали по положительному результату при выполнении анализа пролиферации клеток BaF3, содержащих рецепторы zcytor17/WSX-1/OSMRbeta, и указанную активность подтверждали нейтрализацией пролиферации с использованием растворимого рецептора zcytor17.
Положительный клон выделяли, и выполняли анализ последовательности, результаты которого показали, что полинуклеотидная последовательность в плазмидной ДНК обладает признаками новизны. Секреторная сигнальная последовательность включает в себя аминокислотные остатки с 1 (Met) по 23 (Ala), и зрелый полипептид содержит аминокислотные остатки с 24 (Ser) по 164 (Thr) (как показано в SEQ ID NO:2). Другой анализ методом секвенирования N-конца очищенного лиганда zcytor17lig из клеток 293Т показал, что N-конец соответствует остатку 27 (Leu), как показано в SEQ ID NO:2, при этом зрелый полипептид содержит аминокислотные остатки с 27 (Leu) по 164 (Thr) (как показано в SEQ ID NO:2).
Как правило, считается, что цитокины имеют структуру, состоящую из четырех альфа-спиралей, в которой спирали А, С и D выполняют особенно важную функцию при взаимодействии лиганд-рецептор и являются более консервативными среди членов данного семейства. На основании аминокислотной последовательности zcytоr17lig человека, показанной в SEQ ID NO:2, был выполнен сравнительный анализ аминокислотных последовательностей zcytоr17lig человека, IL-3 человека и цитокина человека, результаты которого позволили предположить, что спираль А zcytor17lig состоит из аминокислотных остатков 38-52; спираль В состоит из аминокислотных остатков 83-98; спираль С состоит из аминокислотных остатков 104-117, и спираль D состоит из аминокислотных остатков 137-152, как показано в SEQ ID NO:2. Структурный анализ показывает, что петля А/В является длинной, петля В/C является короткой и петля C/D является длинной. Указанная структура петель ведет к организации спиралей по принципу вверх-вверх-вниз-вниз. Благодаря четырехспиральной структуре консервативные остатки цистеина в zcytor17lig соответствуют аминокислотным остаткам 72, 133 и 147 SEQ ID NO:2 и остаткам 74, 137 и 151 описанной ниже SEQ ID NO:11. Сопоставимое расположение цистеина является еще одним подтверждением четырехспиральной структуры. Кроме того, высококонсервативным остатком в zcytor17lig является остаток Glu, показанный в SEQ ID NO:2 в положении остатка 43.
Кроме того, предполагаемая аминокислотная последовательность zcytor17lig мыши на 31% идентична предполагаемому белку человека по всей длине последовательностей (SEQ ID NO:2 и SEQ ID NO:11). В результате сравнения последовательностей человека и мыши консервативные остатки zcytor17lig были обнаружены в областях, предположительно кодирующих альфа-спирали С и D. Соответствующие полинуклеотиды, кодирующие полипептидные области, домены, фрагменты, остатки и последовательности zcytor17lig человека, аналогичны показанным в SEQ ID NO:1.
Хотя спираль D является относительно консервативной в zcytor17lig человека и мыши, наиболее консервативной является спираль С. Хотя у обоих видов в данной области доминируют кислые аминокислоты, расхождения могут быть обусловлены видовой специфичностью взаимодействия между лигандом zcytor17lig и его рецептором, при этом рецептор zcytor17 включает в себя мономерные, гетеродимерные (например, zcytor17/OSMRbeta, WSX-1/OSMRbeta, zcytor17/WSX-1) или многомерные (например, zcytor17/OSMRbeta/WSX-1) рецепторы. Петля А/B и спираль В лиганда zcytor17lig являются маргинально консервативными, и спираль С при переходе петли C/D в спираль D является наиболее консервативной у двух видов; консерватизм данной области позволяет предположить, что она является функционально значимой. Спирали D лиганда zcytor17lig человека и мыши также являются консервативными. Антагонисты рецептора zcytor17 могут быть созданы в результате введения мутаций в спираль D лиганда zcytor17lig. Указанные мутации могут включать в себя усечение белка от остатка Thr156 (SEQ ID NO:2) или сохранение остатков, которые вызывают связывание лиганда с рецептором, но ослабляют активность передачи сигналов.
Четырехспиральные цитокины сгруппированы также по длине спиралей. «Длинноспиральные» цитокины обычно состоят из 24-30 остатков и включают в себя IL-6, ресничный нейтротрофный фактор (CNTF), фактор ингибирования лейкоза (LIF) и гормон роста человека (hGH). «Короткоспиральные» цитокины обычно состоят из 18-21 остатка и включают в себя IL-2, IL-4 и GM-CSF. Считается, что zcytor17lig является новым членом группы короткоспиральных цитокинов. Исследования с использованием CNTF и IL-6 показали, что спираль CNTF может быть заменена эквивалентной спиралью в IL-6, благодаря чему химерам сообщаются CNTF-связывающие свойства. Таким образом, очевидно, что функциональные домены четырехспиральных цитокинов определены на основе структурной гомологии независимо от идентичности последовательностей и могут сохранять функциональную целостность в химере (Kallen et al., J. Biol. Chem. 274:11859-11867, 1999). Следовательно, домены спирали zcytor17lig могут быть пригодны для получения химерных молекул, слитых с другими короткоспиральными цитокинами, для определения и модуляции специфичности связывания с рецептором. Особый интерес представляют слитые белки, созданные с использованием спирали А и/или спирали D, и слитые белки, которые объединяют домены спирали и петли из других короткоспиральных цитокинов, таких как IL-2, IL-4, IL-15, Lif, IL-12, IL-3 и GM-CSF.
Полинуклеотидная последовательность для IL-2 человека показана в SEQ ID NO:161, и соответствующая аминокислотная последовательность показана в SEQ ID NO:162. Секреторная сигнальная последовательность включает в себя аминокислотные остатки с 1 (Met) по 20 (Ser) SEQ ID NO:162; нуклеотиды 48-107 SEQ ID NO:161. Зрелый полипептид содержит аминокислотные остатки с 21 (Ala) по 156 (Thr) SEQ ID NO:162; нуклеотиды 108-515 SEQ ID NO:161. Спираль А IL-2 человека содержит аминокислотные остатки с 27 (Thr) по 48 (Leu) SEQ ID NO:162; нуклеотиды 126-191 SEQ ID NO:161. Спираль В IL-2 человека включает в себя спираль В1 и спираль В2. Спираль В1 IL-2 человека содержит аминокислотные остатки с 73 (Ala) по 80 (Gln) SEQ ID NO:162; нуклеотиды 264-287 SEQ ID NO:161. Спираль В2 IL-2 человека содержит аминокислотные остатки с 83 (Glu) по 92 (Val) SEQ ID NO:162; нуклеотиды 294-323 SEQ ID NO:161. Таким образом, спираль В (включающая в себя спирали В1 и В2) IL-2 представлена аминокислотной последовательностью SEQ ID NO:168 (нуклеотидная последовательность SEQ ID NO:167), в которой аминокислотные остатки 9 и 10 могут быть любой аминокислотой. SEQ ID NO:168 идентична аминокислотам с 73 (Ala) по 92 (Val) SEQ ID NO: 162, в которой аминокислоты 81 и 82 могут быть любой аминокислотой. В предпочтительной форме спираль В IL-2 содержит аминокислоты с 73 (Ala) по 92 (Val) SEQ ID NO:162; нуклеотиды 264-323 SEQ ID NO:161. Спираль С IL-2 человека содержит аминокислотные остатки со 102 (His) по 116 (Val) SEQ ID NO:162; нуклеотиды 351-395 SEQ ID NO:161. Спираль D IL-2 человека содержит аминокислотные остатки со 134 (Thr) по 149 (Gln) SEQ ID NO:162; нуклеотиды 447-494 SEQ ID NO:161.
Полинуклеотидная последовательность для IL-4 человека показана в SEQ ID NO:163, и соответствующая аминокислотная последовательность показана в SEQ ID NO:164. Секреторная сигнальная последовательность содержит аминокислотные остатки с 1 (Met) по 24 (Gly) SEQ ID NO:164; нуклеотиды 64-135 SEQ ID NO:163. Зрелый полипептид содержит аминокислотные остатки с 25 (His) по 153 (Ser) SEQ ID NO:164; нуклеотиды 136-522 SEQ ID NO:163. Спираль А IL-4 человека содержит аминокислотные остатки с 30 (Thr) по 42 (Thr) SEQ ID NO:164; нуклеотиды 151-189 SEQ ID NO:163. Спираль В IL-4 человека содержит аминокислотные остатки с 65 (Glu) по 83 (His) SEQ ID NO:164; нуклеотиды 256-312 SEQ ID NO:163. Спираль С IL-4 человека содержит аминокислотные остатки с 94 (Ala) по 118 (Ala) SEQ ID NO:164; нуклеотиды 343-417 SEQ ID NO:163. Спираль D IL-4 человека содержит аминокислотные остатки со 133 (Leu) по 151 (Cys) SEQ ID NO:164; нуклеотиды 460-516 SEQ ID NO:163.
Полинуклеотидная последовательность для GM-CSF человека представлена в SEQ ID NO:165, и соответствующая аминокислотная последовательность показана в SEQ ID NO:166. Секреторная сигнальная последовательность содержит аминокислотные остатки с 1 (Met) по 17 (Ser) SEQ ID NO:166; нуклеотиды 9-59 SEQ ID NO:165. Зрелый полипептид содержит аминокислотные остатки с 18 (Ala) по 144 (Glu) SEQ ID NO:166; нуклеотиды 60-440 SEQ ID NO:165. Спираль А GM-CSF человека содержит аминокислотные остатки с 30 (Trp) по 44 (Asn) SEQ ID NO:166; нуклеотиды 96-140 SEQ ID NO:165. Спираль В GM-CSF человека содержит аминокислотные остатки с 72 (Leu) по 81 (Gln) SEQ ID NO:166; нуклеотиды 222-251 SEQ ID NO:165. Спираль С GM-CSF человека содержит аминокислотные остатки с 85 (Gly) по 103 (Gln) SEQ ID NO:166; нуклеотиды 261-317 SEQ ID NO:165. Спираль D GM-CSF человека содержит аминокислотные остатки со 120 (Phe) по 131 (Leu) SEQ ID NO:166; нуклеотиды 366-401 SEQ ID NO:165.
Спирали А, В, С и D, содержащие аминокислотные остатки для zcytor17lig, IL-3, IL-2, IL-4 и GM-CSF, показаны в таблице 1.
Настоящее изобретение относится к молекулам полинуклеотидов, включая молекулы ДНК и РНК, кодирующие полипептиды zcytor17lig, рассмотренные в данном описании. Специалистам в данной области должно быть понятно, что, с учетом вырожденности генетического кода, возможны значительные вариации последовательностей в указанных молекулах полинуклеотидов. SEQ ID NO:3 является вырожденной последовательностью ДНК, которая включает в себя все ДНК, кодирующие полипептид zcytor17lig, и его фрагменты SEQ ID NO:2. Специалистам должно быть понятно, что вырожденная последовательность SEQ ID NO:3 также включает в себя все последовательности РНК, кодирующие SEQ ID NO:2, в результате замены остатка Т остатком U. Таким образом, полинуклеотиды, кодирующие полипептид zcytor17lig, которые содержат нуклеотиды с 1 или 70 по 492 SEQ ID NO:3 и их эквивалентные РНК, входят в объем настоящего изобретения. В таблице 2 приведены однобуквенные коды, используемые в SEQ ID NO:3 для обозначения положений вырожденных нуклеотидов. В столбцах «Расшифровка» указаны нуклеотиды, обозначенные кодовым символом. В столбце «Комплемент» указан код для комплементарных нуклеотидов. Например, код Y означает остаток С или Т, и их комплемент R означает остаток А или G, при этом остаток А комплементарен остатку Т, и остаток G комплементарен остатку С.
Вырожденные кодоны, используемые в SEQ ID NO:3, которые включают в себя все возможные кодоны для данной аминокислоты, приведены в таблице 3.
Специалисту должно быть понятно, что существует некоторая неточность в определении вырожденного кодона, типичного для всех возможных кодонов, кодирующих каждую аминокислоту. Например, вырожденный кодон для серина (WSN) в некоторых случаях может кодировать аргинин (AGR), и вырожденный кодон для аргинина (MGN) в некоторых случаях может кодировать серин (AGY). Подобная взаимосвязь существует между кодонами, кодирующими фенилаланин и лейцин. Таким образом, некоторые полинуклеотиды, входящие в определение вырожденной последовательности, могут кодировать вариантные аминокислотные последовательности, но специалист может легко идентифицировать такие вариантные последовательности, обратившись к аминокислотной последовательности SEQ ID NO:2. Функциональную способность вариантных последовательностей можно легко испытать способами, рассмотренными в данном описании.
Специалисту должно быть также понятно, что разные виды могут характеризоваться «предпочтительным использованием кодонов». См., например, Grantham, et al., Nuc. Acids Res. 8:1893-912, 1980; Haas, et al. Curr. Biol. 6:315-24, 1996; Wain-Hobson, et al., Gene 13:355-64, 1981; Grosjean and Fiers, Gene 18:199-209, 1982; Holm, Nuc. Acids Res. 14:3075-87, 1986; Ikemura, J. Mol. Biol. 158:573-97, 1982. В используемом здесь значении термин «предпочтительное использование кодонов» или «предпочтительные кодоны» означает кодоны трансляции белка, которые наиболее часто используются в клетках определенного вида, в результате чего один или несколько возможных кодонов, кодирующих каждую аминокислоту, становятся особенно предпочтительными (см. таблицу 3). Например, аминокислота треонин (Thr) может быть кодирована ACA, ACC, ACG или АСТ, но в клетках млекопитающих АСС является наиболее часто используемым кодоном; в других видах, таких как клетки насекомых, дрожжи, вирусы или бактерии, предпочтение может отдаваться других кодонам Thr. Предпочтительные для определенного вида кодоны могут быть введены в полинуклеотиды согласно изобретению разными методами, известными в данной области. Введение последовательностей с предпочтительными кодонами в рекомбинантную ДНК может, например, усилить продуцирование белка благодаря более эффективной трансляции белка в определенном типе или виде клеток. Поэтому последовательность с вырожденными кодонами, представленная в SEQ ID NO:3, служит в качестве матрицы для оптимизации экспрессии полинуклеотидов в разных типах и видах клеток, обычно используемых в данной области и рассмотренных в данном описании. Последовательности, содержащие предпочтительные кодоны, могут быть испытаны и оптимизированы для экспрессии в определенных видах и исследованы в отношении функциональной способности в соответствии с приведенным описанием.
Как указывалось выше, выделенные полинуклеотиды согласно изобретению включают в себя ДНК и РНК. Способы получения ДНК и РНК хорошо известны в данной области. РНК обычно выделяют из ткани или клетки, которая продуцирует большие количества РНК zcytor17lig. Такие ткани и клетки идентифицируют при помощи нозерн-блоттинга (Thomas, Proc. Natl. Acad. Sci. USA 77:5201, 1980) или исследования активности кондиционированной среды из разных типов клеток в отношении клеток или тканей-мишеней. После идентификации необходимой активности или РНК-продуцирующих клеток или тканей можно получить полную РНК, производя экстракцию изотиоцианатом гуанидиния с последующим выделением путем центрифугирования в градиенте CsCl (Chirgwin et al., Biochemistry 18:52-94, 1979). Поли(А)+ РНК получают из полной РНК способом, описанным Aviv and Leder, Proc. Natl. Acad. Sci. USA 69:1408-12, 1972. Комплементарную ДНК (кДНК) получают из поли(А)+ РНК известными способами. Альтернативно можно выделить геномную ДНК. Затем идентифицируют полинуклеотиды, кодирующие полипептиды zcytor17lig, и выделяют их, например, при помощи гибридизации или полимеразной реакции синтеза цепи (PCR).
Непроцессированный клон, кодирующий zcytor17lig, можно получить обычными методами клонирования. Клоны комплементарной ДНК (кДНК) являются предпочтительными, хотя для некоторых применений (например, для экспрессии в трансгенных животных) может быть желательно использовать геномный клон или модифицировать клон кДНК, так чтобы он включал в себя по меньшей мере один геномный интрон. Способы получения кДНК и геномных клонов хорошо известны специалистам и включают в себя использование описанной здесь последовательности или ее частей для зондирования или инициации библиотеки. Библиотеку экспрессируемых последовательностей можно зондировать антителами к фрагментам zcytor17lig, zcytor17-содержащим растворимым рецепторам или другим специфически связывающимся партнерам.
Полинуклеотидные последовательности zcytor17lig, рассмотренные в данном описании, можно также использовать в качестве зондов или затравок для клонирования 5'-концевых некодирующих областей гена zcytor17lig. Учитывая тканеспецифическую экспрессию, наблюдаемую для zcytor17lig, можно предположить, что область данного гена должна способствовать гемопоэтической и лимфоидспецифической экспрессии. Таким образом, промоторные элементы из гена zcytor17lig можно использовать для того, чтобы направлять тканеспецифическую экспрессию гетерологичных генов в трансгенных животных или пациентах, проходящих курс генотерапии. Клонирование 5'-концевых фланкирующих последовательностей также облегчает продуцирование белков zcytor17lig путем «активации гена», как это описано в патенте США № 5641670. В кратком изложении данный способ включает в себя изменение экспрессии эндогенного гена zcytor17lig в клетке путем введения в локус zcytor17lig конструкции на основе ДНК, содержащей по меньшей мере одну направляющую последовательность, регуляторную последовательность, экзон и неспаренный донорский сайт сплайсирования. Направляющая последовательность является 5'-концевой некодирующей последовательностью zcytor17lig, которая делает возможной гомологичную рекомбинацию конструкции с эндогенным локусом zcytor17lig, в результате чего последовательности в данной конструкции функционально связываются с эндогенной кодирующей последовательностью zcytor17lig. Подобным образом эндогенный промотор zcytor17lig можно заменить или дополнить другими регуляторными последовательностями для достижения повышенной, тканеспецифической или другим образом регулируемой экспрессии.
Настоящее изобретение далее относится к полипептидам и полинуклеотидам из других видов (ортологам). Указанные виды включают в себя, не ограничиваясь ими, млекопитающих, птиц, амфибий, рептилий, рыб, насекомых и других позвоночных и беспозвоночных. Особый интерес представляют полипептиды zcytor17lig из других видов млекопитающих, включая, например, полипептиды, полученные у мышей, свиней, овец, крупного рогатого скота, собак, кошек, лошадей и других приматов. Ортологи zcytor17lig человека можно клонировать, используя информацию и композиции согласно изобретению, в сочетании с известными методами клонирования. Например, кДНК можно клонировать, используя мРНК, полученную из ткани или клетки, экспрессирующей zcytor17lig, описанными здесь способами. Приемлемые источники мРНК можно идентифицировать, зондируя нозерн-блоты зондами, созданными на основе описанных здесь последовательностей. Затем получают библиотеку из мРНК положительной ткани или линии клеток. Zcytor17lig-кодирующую кДНК можно выделить различными способами, такими как зондирование полной или неполной кДНК человека одним или несколькими наборами вырожденных зондов, полученных на основе описанных последовательностей. кДНК можно также клонировать при помощи полимеразной реакции синтеза цепи, или PCR (Mullis, патент США № 4683202), используя затравки, созданные на основе типичной последовательности zcytor17lig человека, рассмотренной в данном описании. В качестве дополнительного способа библиотеку кДНК можно использовать для трансформации или трансфекции клеток-хозяев, при этом экспрессию представляющей интерес кДНК можно обнаружить при помощи антитела к полипептиду zcytor17lig, анализа на связывание или активность. Аналогичные методы можно также использовать для выделения геномных клонов.
Была идентифицирована полинуклеотидная последовательность для ортолога zcytor17lig мыши, которая показана в SEQ ID NO:10 и SEQ ID NO:90, и соответствующая аминокислотная последовательность, показанная в SEQ ID NO:11 и SEQ ID NO:91. Вырожденная полинуклеотидная последовательность, кодирующая полипептид SEQ ID NO:11, показана в SEQ ID NO:12. В аминокислотной последовательности цитокина zcytor17lig мыши спираль А предпочтительно определяется аминокислотными остатками 38-52, спираль В определяется аминокислотными остатками 85-98, спираль С определяется аминокислотными остатками 104-118, и спираль D определяется аминокислотными остатками 141-157, как показано в SEQ ID NO:11 и SEQ ID NO:91. Последовательности мыши и человека идентичны на 31% по всей длине аминокислотных последовательностей (SEQ ID NO:1 и SEQ ID NO:11) zcytor17lig. Зрелая последовательность zcytor17lig мыши предположительно начинается у Met1, как показано в SEQ ID NO:11, что соответствует положению Met1 в последовательности человека, как показано в SEQ ID NO:2. Анализ ткани позволил обнаружить экспрессию zcytor17lig мыши в яичке, головном мозге, CD90+-клетках, клетках предстательной железы, слюнной железе и коже. Дальнейший анализ методом секвенирования N-конца очищенного zcytor17lig из клеток 193Т позволил обнаружить N-конец у остатка 31 (Ala), как это показано в SEQ ID NO:11 и SEQ ID NO:91, при этом зрелый полипептид содержал аминокислотные остатки с 31 (Ala) по 163 (Cys) (как показано в SEQ ID NO:11 и SEQ ID NO:91).
Специалистам должно быть понятно, что последовательность, показанная в SEQ ID NO:1, представляет собой один аллель zcytor17lig человека и что вполне вероятна аллельная вариация и альтернирующий сплайсинг. Аллельные варианты данной последовательности можно клонировать путем зондирования кДНК или геномных библиотек различных субъектов стандартными методами. В объем настоящего изобретения входят аллельные варианты последовательности ДНК, показанные в SEQ ID NO:1, включая варианты, содержащие молчащие мутации, и варианты, в которых мутации вызывают изменение аминокислотной последовательности, а также белки, являющиеся аллельными вариантами SEQ ID NO:2. Кроме того, в объем настоящего изобретения входят кДНК, полученные из альтернативно сплайсированных мРНК, которые сохраняют свойства полипептида zcytor17lig, а также полипептиды, кодированные такими кДНК и мРНК. Аллельные и сплайсированные варианты указанных последовательностей можно клонировать путем зондирования кДНК или геномных библиотек различных субъектов или тканей стандартными методами, известными в данной области.
Настоящее изобретение относится также к реагентам, которые находят применение в диагностике. Например, ген zcytor17lig, зонд, содержащий ДНК или РНК zcytor17lig или их субпоследовательность, можно использовать для определения наличия гена zcytor17lig в хромосоме человека, такой как хромосома 12, или мутации гена. Zсytor17lig расположен в области 12q24.31 хромосомы 12 (пример 13). Обнаруживаемые хромосомные аберрации в локусе гена zcytor17lig включают в себя, не ограничиваясь ими, анеуплоидию, изменение числа копий гена, утрату гетерозиготности (LOH), транслокации, инсерции, делеции, изменения сайта рестрикции и реаранжировки. Такие аберрации можно обнаружить, используя полинуклеотиды согласно изобретению для выполнения молекулярных генетических методов, таких как анализ полиморфизма длины рестрикционного фрагмента (RFLP), анализ короткого тандемного повтора (STR) методами PCR и другие анализы сцепления генов, известные в данной области (Sambrook et al., ibid.; Ausubel et al., ibid.; Marian. Chest 108:255-65, 1995).
Знание точного положения гена может быть полезным для многих целей, которые включают в себя: 1) определение того, является ли некоторая последовательность частью существующей смежной последовательности и получение дополнительных окружающих генетических последовательностей в разных формах, таких как клоны YAC, BAC или кДНК; 2) определение возможного гена, вызывающего наследственное заболевание, который связан с такой же областью хромосомы; и 3) перекрестное сравнение с организмами, представляющими модель заболевания, такими как мыши, которое может оказать помощь при определении функции конкретного гена.
Специалисту должно быть известно, что область 12q24 часто участвует в общих геномных реаранжировках, включая транслокации, делеции, инверсии и дупликации, которые вызывают различные раковые заболевания. В базе данных хромосомных аберраций рака, созданной Мителманом в соответствии с проектом Cancer Genome Anatomy Project, National Institutes of Health, Bethesda, Md, которая размещена в Интернете, перечислено 199 случаев рака с геномными реаранжировками, включающими в себя область 12q24. Большинство указанных случаев являются частью сложных кариотипов с другими реаранжировками; однако в некоторых случаях реаранжировка, включающая в себя область 12q24, является единственным геномным изменением. Принимая во внимание экспрессию рецептора для zcytor17lig в клетках лимфоидной и миелоидной линий, особенно важно отметить, что известны по меньшей мере 4 случая миелоидного лейкоза, описанных в научной литературе, в которых единственным геномным изменением является транслокация (2 случая: Yamagata et al., Cancer Genet. Cytogenet 97:90-93, 1997; Dunphy and Batanian, Cancer Genet Cytogenet 114:51-57, 1999) или дупликация (2 случая: Bonomi et al., Cancer Genet Cytogenet 108:75-78, 1999). Вышеизложенное позволяет предположить, что ген или гены, локализованные в области 12q24, могут принимать непосредственное участие в злокачественной трансформации клеток указанных субъектов. Аномальная сверхэкспрессия zcytor17lig может способствовать злокачественной трансформации, стимулируя аберрантную пролиферацию клеток, несущих рецепторы, под действием аутокринных или паракринных механизмов. Таким образом, ингибирование активности zcytor17lig может тормозить рост таких клеток. Альтернативно, геномная реаранжировка, вызывающая инактивацию гена zcytor17lig, может стимулировать злокачественную трансформацию и/или метастазы в результате устранения иммунорегуляторных функций zcytor17lig. Ген, подавляющий метастазы в случае рака предстательной железы, был картирован в области 12q24-qter (Ichikawa et al., Asian J. Androl 2:167-171, 2000). Если zcytor17lig является геном, расположенным в области, ответственной за подавление метастазов, тогда zcytor17lig сам по себе может иметь терапевтическое значение при лечении рака.
Диагностика может оказать помощь лечащим врачам при определении типа заболевания и выборе соответствующего лечения или выявлении генетической природы заболевания. В таком случае антитела против zcytor17lig, полинуклеотиды и полипептиды согласно изобретению можно использовать в качестве маркеров для обнаружения полипептида zcytor17lig, мРНК или антител против zcytor17lig или непосредственно для выявления генетических или раковых заболеваний при помощи методов, известных в данной области и рассмотренных в настоящем описании. Кроме того, полинуклеотидные зонды к zcytor17lig можно использовать для обнаружения аномалий или генотипов, ассоциированных с делециями и транслокациями в области 12q24.3 хромосомы, вызывающими заболевания человека, или другими транслокациями, способствующими злокачественному развитию опухолей, или других мутаций в области 12q24.3, которые, как предполагается, имеют непосредственное отношение к хромосомным реаранжировкам при злокачественных опухолях или других раковых заболеваниях. Аналогичным образом, полинуклеотидные зонды к zcytor17lig можно использовать для обнаружения аномалий или генотипов, связанных с трисомией 12-й хромосомы и утратой хромосомы, которые вызывают заболевания у человека или самопроизвольный аборт. Так, полинуклеотидные зонды к zcytor17lig можно использовать для обнаружения аномалий или генотипов, ассоциированных с указанными заболеваниями.
Специалисту должно быть понятно, что полинуклеотидные зонды к zcytor17lig особенно пригодны для диагностики общих хромосомных аномалий, обусловленных утратой гетерогенности (LOH), приобретением хромосомы (например, трисомия), транслокацией, амплификацией ДНК и т.д. Известно, что транслокации в хромосомном локусе 12q24.3, где находится ген zcytor17lig, вызывают заболевание у человека. Например, как указывалось выше, делеции и транслокации, дупликации и трисомия в локусе 12q24 вызывают раковые заболевания. Так как ген zcytor17lig картирован в данной критической области, полинуклеотидные зонды к zcytor17lig согласно изобретению можно использовать для обнаружения аномалий или генотипов, обусловленных транслокацией, делецией и трисомией в локусе 12q24, как было описано выше.
Как указывалось выше, дефекты в самом гене zcytor17lig вызывают наследственные заболевания человека. Молекулы согласно изобретению, такие как полипептиды, антагонисты, агонисты, полинуклеотиды и антитела согласно изобретению, могут способствовать обнаружению, диагностике и лечению заболевания, обусловленного генетическим дефектом zcytor17lig. Кроме того, полинуклеотидные зонды к zcytor17lig можно использовать для обнаружения аллельных различий у больных или здоровых субъектов в хромосомном локусе zcytor17lig. Таким образом, последовательности zcytor17lig можно использовать в качестве диагностического средства для выявления чужеродной ДНК.
В данной области известны диагностические методы, используемые для анализа сцепления генов с целью обнаружения генетической аномалии или аберрации у субъекта. Аналитические зонды обычно имеют длину, составляющую по меньшей мере 20 нуклеотидов, хотя иногда можно использовать более короткие зонды (например, 14-17 нуклеотидов). Затравки для PCR имеют длину по меньшей мере 5 нуклеотидов, предпочтительно 15 или более, более предпочтительно 20-30 нуклеотидов. Полинуклеотидный зонд к zcytor17lig, предназначенный для общего анализа генов или хромосомной ДНК, может содержать один или несколько полных экзонов. Экзоны могут быть легко определены специалистом в результате сравнения последовательностей zcytor17lig (SEQ ID NO:1) с геномной ДНК для zcytor17lig мыши (SEQ ID NO:76). Диагностические методы, используемые при выполнении анализа сцепления генов для обнаружения генетической аномалии или аберрации у пациента, хорошо известны в данной области. Большинство диагностических методов включают в себя: (i) получение генетического образца у потенциально больного субъекта или потенциально здорового носителя рецессивного аллеля заболевания; (ii) получение первого продукта реакции путем инкубации генетического образца с полинуклеотидным зондом к zcytor17lig, в процессе которой полинуклеотид гибиридизуется с комплементарной полинуклеотидной последовательностью, как это имеет место при выполнении анализа RFLP, или путем инкубации генетического образца со смысловыми и антисмысловыми затравками при выполнении реакции PCR в приемлемых для данной реакции условиях; (iii) визуализацию первого продукта реакции методом гель-электрофореза и/или другими известными методами, такими как визуализация первого продукта реакции полинуклеотидным зондом к zcytor17lig, в процессе которой полинуклеотид гибридизуется с комплементарной полинуклеотидной последовательностью первого продукта реакции; и (iv) сравнение визуализованного первого продукта реакции со вторым продуктом контрольной реакции, выполненной при использовании генетического образца, полученного у субъекта, несущего ген дикого типа, нормального или контрольного субъекта. Разница между первым продуктом реакции и продуктом контрольной реакции свидетельствует о генетической аномалии у больного или потенциально больного субъекта, или о наличии фенотипа гетерозиготного рецессивного носителя у здорового субъекта, или о наличии генетического дефекта в опухоли у больного субъекта, или о наличии генетической аномалии у плода или эмбриона до имплантации. Например, различия в паттерне рестрикционного фрагмента, длине продуктов PCR, длине повторяющихся последовательностей в локусе гена zcytor17lig и тому подобное являются показателем генетической аномалии, генетической аберрации или аллельного отличия при сравнении с нормальным контрольным субъектом, несущим ген дикого типа. В зависимости от испытания и доступности образцов, контрольные субъекты могут быть не страдающими данным заболеванием членами семьи или посторонними субъектами. Генетические образцы, пригодные для применения в соответствии с настоящим изобретением, включают в себя геномную ДНК, мРНК и кДНК, выделенные из любой ткани или другого биологического образца у субъекта, которые включают в себя, не ограничиваясь ими, кровь, слюну, сперму, эмбриональные клетки, околоплодную жидкость и тому подобное. Полинуклеотидный зонд или затравка могут представлять собой РНК или ДНК и могут содержать часть SEQ ID NO:1, комплемент SEQ ID NO:1 или их эквивалент РНК. Методы выполнения анализа сцепления генов для определения фенотипов заболевания у человека хорошо известны в данной области. Для ознакомления с методами на основе PCR в диагностических целях см., например, Mathew (ed.), Protocols in Human Molecular Genetics (Humana Press, Inc. 1991), White (ed.), PCR Protocols: Current Methods and Applications (Humana Press, Inc. 1993), Cotter (ed.), Molecular Diagnosis of Cancer (Humana Press, Inc. 1996), Hanausek and Walaszek (eds.), Tumor Marker Protocols (Humana Рress, Inc. 1998), Lo (ed.), Clinical Applications of PCR (Humana Press, Inc. 1998), and Meltzer (ed.), PCR in Bioanalysis (Humana Press, Inc. 1998).
Мутации, ассоциированные с локусом zcytor17lig, можно обнаружить при помощи молекул нуклеиновой кислоты согласно изобретению стандартными методами прямого анализа мутаций, такими как анализ полиморфизма длины рестрикционных фрагментов, анализ коротких тандемных повторов методами PCR, системный анализ мутаций, не поддающихся амплификации, обнаружение полиморфизма одноцепочечной структуры, методы расщепления РНКазой, денатурирующий гель-электрофорез в градиенте напряжения, флуоресцентный анализ ошибочного спаривания и другие методы генетических анализов, известные в данной области (см., например, Mathew (ed.), Protocols in Human Molecular Genetics (Humana Press, Inc. 1991), Marian, Chest 108:255 (1995), Coleman and Tsongalis, Molecular Diagnostics (Humana Press, Inc. 1996), Elles (ed.), Molecular Diagnosis of Genetic Diseases (Humana Press, Inc. 1996), Landegren (ed.), Laboratory Protocols for Mutation Detection (Oxford University Press 1996), Birren et al., (eds.), Genome Analysis, Vol. 2: Detecting Genes (Cold Spring Harbor Laboratory Рress 1998), Dracopoli et al. (eds.), Current Protocols in Human Genetics (John Wiley & Sons 1998), and Richards and Ward, "Molecular Diagnostic Testing," in Principles of Molecular Medicine, pages 83-88 (Humana Press, Inc. 1998). Прямой анализ мутации гена zcytor17lig можно выполнить, используя геномную ДНК субъекта. Методы амплификации геномной ДНК, полученной, например, из лимфоцитов периферической крови, хорошо известны специалистам (см., например, Dracopoli et al. (eds.), Current Protocols in Human Genetics, at pages 7.1.6 to 7.1.7 (John Wiley & Sons 1998)).
Положения интронов в гене zcytor17lig мыши были определены путем идентификации геномных клонов с последующим анализом сочленения интрон/экзон. Геномная ДНК мыши показана в SEQ ID NO:76. При обращении к SEQ ID NO:76 видны три кодирующих экзона, разделенных интронами: первый кодирующий экзон находится между нуклеиновыми кислотами 1104-1119 SEQ ID NO:76, второй экзон находится между нуклеиновыми кислотами 1300-1451 SEQ ID NO:76, и третий экзон находится между нуклеиновыми кислотами 2411-2998 SEQ ID NO:76.
В соответствии с вариантами осуществления изобретения, выделенные молекулы нуклеиновой кислоты, кодирующей zcytor17lig, могут гибридизоваться в строгих условиях с молекулами нуклеиновой кислоты, имеющими нуклеотидную последовательность SEQ ID NO:1, молекулами нуклеиновой кислоты, имеющими нуклеотидную последовательность, включающую в себя нуклеотиды 28-519 SEQ ID NO:1, или молекулами нуклеиновой кислоты, имеющими нуклеотидную последовательность, комплементарную SEQ ID NO:1. Как правило, строгие условия выбирают примерно на 5°C ниже температуры плавления (Тm) для конкретной последовательности при определенной ионной силе и рН. Tm означает температуру (при определенной ионной силе и рН), при которой 50% целевой последовательности гибридизуется с образованием полностью спаренного зонда.
Пара молекул нуклеиновых кислот, такая как ДНК-ДНК, РНК-РНК и ДНК-РНК, может гибридизоваться, если нуклеотидные последовательности характеризуются одинаковой степенью комплементарности. Гибриды могут содержать ошибочно спаренные пары оснований в двойной спирали, но степень ошибочного спаривания влияет на устойчивость гибрида. Tm ошибочно спаренного гибрида снижается на 1°C для 1-1,5% ошибочно спаренных пар оснований. Изменяя строгость условий гибридизации, можно контролировать степень ошибочного спаривания, характерного для гибрида. Степень строгости увеличивается по мере повышения температуры гибридизации и уменьшения ионной силы буфера для гибридизации.
Специалист может адаптировать указанные условия применительно к конкретному полинуклеотидному гибриду. Tm для специфической последовательности-мишени является температурой (в установленных условиях), при которой 50% последовательности-мишени гибридизуется с полностью спаренной последовательностью зонда. Указанные условия, влияющие на величину Tm, включают в себя размер и число пар оснований полинуклеотидного зонда, ионную силу раствора для гибридизации и наличие дестабилизирующих агентов в растворе для гибридизации. В данной области известно много уравнений, предназначенных для вычисления Tm, которые являются специфическими для гибридов ДНК, РНК и ДНК-РНК и последовательностей полинуклеотидных зондов разной длины (см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition (Cold Spring Harbor Press 1989); Ausubel et al., (eds.), Current Protocols in Molecular Biology (John Wiley and Sons, Inc. 1987); Berger and Kimmel (eds.), Guide to Molecular Cloning Techniques, (Academic Press, Inc. 1987); and Wetmur, Crit. Rev. Biochem. Mol. Biol. 26:227 (1990)). Программное обеспечение для анализа последовательностей, такое как OLIGO 6.0 (LSR; Long Lake, MN) и Primer Premier 4.0 (Premier Biosoft International; Palo Alto, CA), а также сайты в Интернете весьма пригодны для анализа данной последовательности и вычисления Тm на основании введенных пользователем критериев. Такие программы могут также анализировать данную последовательность в установленных условиях и определять приемлемые последовательности зондов. Более длинные полинуклеотидные последовательности, >50 пар оснований, обычно гибридизуются при температурах, которые примерно на 20-25°C ниже вычисленного значения Tm. Зонды меньшей длины, <50 пар оснований, обычно гибридизуются при Tm или температуре, которая на 5-10°C ниже вычисленного значения Tm. Такие условия обеспечивают максимальную скорость гибридизации для гибридов ДНК-ДНК и ДНК-РНК.
После гибридизации молекулы нуклеиновых кислот можно промыть для удаления негибридизованных молекул нуклеиновых кислот в строгих условиях или очень строгих условиях. Типичные строгие условия промывки включают в себя промывку в 0,5-2-кратном объеме раствора SSC с 0,1% додецилсульфата натрия (SDS) при 55-65°C. То есть молекулы нуклеиновых кислот, кодирующих вариантный полипептид zcytor17lig, гибридизуют с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент) в строгих условиях промывки, в которых строгость промывки эквивалентна 0,5-2-кратному объему раствора SSC с 0,1% SDS при 55-65°C, включая 0,5-кратный объем раствора SSC с 0,1% SDS при 55°C или 2-кратный объем раствора SSC с 0,1% SDS при 65°C. Специалист может легко определить эквивалентные условия, например, заменив раствор SSC раствором SSPE в процессе промывки.
Типичные очень строгие условия промывки включают в себя промывку 0,1-0,2-кратным объемом раствора SSC с 0,1% додецилсульфата натрия (SDS) при 50-65°C. Другими словами, молекулы нуклеиновых кислот, кодирующие вариантный полипептид zcytor17lig, гибридизуют с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент), в очень строгих условиях промывки, в которых строгость промывки эквивалентна 0,1-0,2-кратному объему раствора SSC с 0,1% SDS при 50-65°C, включая 0,1-кратный объем раствора SSC с 0,1% SDS при 50°C или 0,2-кратный объем раствора SSC с 0,1% SDS при 65°С.
Настоящее изобретение относится также к выделенным полипептидам zcytor17lig, которые имеют последовательности, по существу идентичные полипептидам SEQ ID NO:2, или их ортологам. Термин «по существу идентичные последовательности» использован в данном описании для обозначения полипептидов, которые по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или более чем на 99% идентичны последовательностям, показанным в SEQ ID NO:2, или их ортологов. Настоящее изобретение относится также к полипептидам, содержащим аминокислотную последовательность, которая по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или более чем на 99% идентичны последовательности аминокислотных остатков 1-162 или 33-162 SEQ ID NO:2. Настоящее изобретение далее относится к молекулам нуклеиновых кислот, кодирующим такие полипептиды. Способы определения процентного значения идентичности описаны ниже.
Настоящее изобретение относится также к вариантным молекулам нуклеиновых кислот zcytor17lig, которые могут быть идентифицированы по двум критериям: определение сходства между кодированным полипептидом и аминокислотной последовательностью SEQ ID NO:2 и/или выполнение описанного выше анализа методом гибридизации. Такие варианты zcytor17lig включают в себя молекулы нуклеиновых кислот, которые: (1) гибридизуются с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент) в строгих условиях промывки, в которых строгость промывки эквивалентна 0,5-2-кратному объему раствора SSC с 0,1% SDS при 55-65°С; или (2) кодируют полипептид, который по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или более чем на 99% идентичен аминокислотной последовательности SEQ ID NO:2. Альтернативно, варианты zcytor17lig могут быть охарактеризованы как молекулы нуклеиновых кислот, которые (1) гибридизуются с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент) в очень строгих условиях промывки, в которых строгость промывки эквивалентна 0,1-0,2-кратному объему раствора SSC с 0,1% SDS при 50-65°С; и (2) кодируют полипептид, который по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или более чем на 99% идентичен аминокислотной последовательности SEQ ID NO:2.
Процентное значение идентичности последовательности определяют обычными методами. См., например, Altschul et al., Bull. Math. Bio. 48:603 (1986), and Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA 89: 10915 (1992). В кратком изложении данный метод включает в себя сравнительный анализ структуры аминокислотных последовательностей для оптимизации оценок сравнения с использованием 10 штрафных очков за разрыв, 1 штрафного очка за удлинение разрыва и матрицы начисления очков "BLOSUM62" Хеникоффа и Хеникоффа (см. выше), показанной в таблице 4 (аминокислоты указаны стандартными однобуквенными кодами).
Общее число идентичных совмещений
――――――――――――――――――――――――― × 100
[длина более длинной последовательности
плюс число разрывов, введенных
в более длинную последовательность
для согласования двух последовательностей]
Таблица 4

Специалистам должно быть понятно, что существует много алгоритмов, позволяющих сравнить две аминокислотные последовательности. Алгоритм исследования сходства "FASTA" Пиарсона и Липмана является приемлемым методом сравнения белков, предназначенным для исследования уровня идентичности аминокислотной последовательности согласно изобретению и аминокислотной последовательности предполагаемого варианта zcytor17lig. Алгоритм FASTA описан в статьях Pearson and Lipman, Proc. Nat'l Acad. Sci. USA 85:2444 (1988) и Pearson, Meth. Enzymol. 183:63 (1990).
В кратком изложении алгоритм FASTA сначала исследует сходство последовательностей, идентифицируя области, одинаковые для справочной последовательности (например, SEQ ID NO:2) и испытуемой последовательности, которые характеризуются наибольшей плотностью идентичных пар (если переменная ktup равна 1) или числом идентичных пар (если ktup = 2), не учитывая замены, инсерции или делеции консервативных аминокислот. Десять областей с наибольшей плотностью идентичных пар затем вновь оценивают, сравнивая сходство всех спаренных аминокислот с использованием матрицы замены аминокислот, и концы указанных областей «укорачивают» так, чтобы они включали только те остатки, которые соответствуют наивысшей оценке. Если имеется несколько областей с оценками, превышающими значение «отсечения» (вычисленное по заранее определенной формуле на основании длины последовательности и значения ktup), исследуют укороченные исходные области для определения возможности связывания указанных областей с целью приблизительного заполнения разрывов. И наконец, сравнивают области двух аминокислотных последовательностей, получившие наивысшую оценку, используя модифицированный алгоритм Needleman-Wunsch-Sellers (Needleman and Wunsch, J. Mol. Biol. 48:444 (1970); Sellers, SIAM J. Appl. Math. 26:787 (1974)), которые позволяют производить инсерции и делеции аминокислот. Предпочтительными параметрами для анализа FASTA являются: ktup = 1, штрафные очки за разрыв = 10, штрафные очки за удлинение разрыва = 1 и матрица замен = BLOSUM62. Указанные параметры могут быть введены в программу FASTA в результате изменения данных в файле матрицы оценки ("SMATRIX"), как описано в приложении 2 статьи Pearson, Meth. Enzymol. 183:63 (1990).
Программу FASTA можно также использовать для определения идентичности последовательностей молекул нуклеиновых кислот с использованием описанного выше соотношения. При сравнении нуклеотидных последовательностей значение ktup может изменяться от одного до шести, предпочтительно от трех до шести, наиболее предпочтительно указанное значение равно трем, при этом другие параметры вводят в виде значений по умолчанию.
Вариантные полипептиды zcytor17lig или полипептиды, имеющие по существу идентичную последовательность, могут иметь замены, делеции или добавления одной или нескольких аминокислот. Указанные замены предпочтительно являются второстепенными, то есть включают в себя замены консервативных аминокислот (как показано в нижеследующей таблице 5) и другие замены, которые не оказывают существенного влияния на укладку или активность данного полипептида; небольшие делеции, обычно от одной до около 30 аминокислот; амино- или карбоксиконцевые удлиняющие сегменты, такие как аминоконцевой остаток метионина, небольшой линкерный пептид, включающий в себя примерно до 20-25 остатков или аффинную метку. Таким образом, настоящее изобретение относится к полипептидам, содержащим примерно от 108 до 216 аминокислотных остатков, образующих последовательность, которая по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или более чем на 99% идентична соответствующей области SEQ ID NO:2. Полипептиды, включающие в себя аффинные метки, могут далее содержать протеолитический сайт расщепления между полипептидом zcytor17lig и аффинной меткой. Такие сайты предпочтительно включают в себя сайты расщепления тромбина и сайты расщепления Ха-фактора.
Замены консервативных аминокислот
лизин
гистидин
аспарагиновая кислота
аспарагин
изолейцин
валин
триптофан
тирозин
аланин
серин
треонин
метионин
Можно определить аминокислотные остатки, содержащие области или домены, которые имеют важное значение для сохранения структурной целостности. В указанных областях можно определить специфические остатки, которые в большей или меньшей степени допускают замену и сохраняют общую третичную структуру молекулы. Методы анализа структуры последовательности включают в себя, не ограничиваясь ими, сравнительный анализ структуры нескольких последовательностей с высокой степенью идентичности аминокислот или нуклеотидов, анализ допустимости замен во вторичной структуре, анализ двоичных паттернов, анализ комплементарной упаковки и анализ скрытых полярных взаимодействий (Barton, Current Opin. Struct. Biol. 5:372-376, 1995 and Cordes et al., Current Opin. Struct. Biol. 6 :3-10, 1996). Как правило, при внесении изменений в молекулы или идентификации специфических фрагментов определение структуры должно сопровождаться оценкой активности модифицированных молекул.
Изменения в аминокислотные последовательности полипептидов zcytor17lig вносят таким образом, чтобы минимизировать разрушение структуры высшего порядка, имеющей важное значение для биологической активности. Например, когда полипептид zcytor17lig содержит одну или несколько спиралей, аминокислотные остатки следует заменять так, чтобы не нарушить геометрию спиралей и другие компоненты молекулы, где изменение конформации устраняет некоторые важные функции, например, связывание молекулы со связывающими партнерами, в частности, спирали А и D, остатки 43 (Glu), 44 (Glu) и 136 (Phe) SEQ ID NO:2. Влияние замен, вносимых в аминокислотную последовательность, можно прогнозировать, например, при помощи описанного выше компьютерного моделирования или анализа кристаллической структуры (см., например, Laрthorn et al., Nat. Struct. Biol. 2:266-268, 1995). Другие методы, хорошо известные в данной области, включают в себя сравнение укладки вариантного белка со стандартной молекулой (например, нативным белком). Например, можно произвести сравнение паттерна цистеина в вариантной и стандартной молекулах. Масс-спектрометрия и химическая модификация путем восстановления и алкилирования могут быть использованы в качестве методов определения остатков цистеина, которые связаны дисульфидными связями или не имеют таких связей (Bean et al., Anal. Biochem. 201:216-226, 1992; Gray, Protein Sci. 2:1732-1748, 1993; and Patterson et al., Anal. Chem. 66:3727-3732, 1994). Обычно считается, что если в модифицированной молекуле отсутствует такой же паттерн цистеина, что и в стандартной молекуле, нарушается укладка цепи. Другим хорошо известным и общепринятым методом измерения укладки цепи является круговой дихроизм (CD). Обычно производят измерение и сравнение спектров CD модифицированной молекулы и стандартной молекулы (Johnson, Proteins 7:205-214, 1990). Кристаллография является другим хорошо известным методом анализа укладки цепи и структуры. Ядерный магнитный резонанс (ЯМР) и картирование гидролизуемых пептидов и эпитопов также являются хорошо известными методами анализа укладки цепи и структурного сходства между белками и полипептидами (Schaanan et al., Science 257: 961-964, 1992).
Можно создать профиль гидрофильности Хоппа-Вудса белковой последовательности zcytor17lig, показанной в SEQ ID NO:2 (Hopp et al., Proc. Natl. Acad. Sci. 78:3824-3828, 1981; Hopp, J. Immun. Meth. 88:1-18, 1986 and Triquier et al., Protein Engineering 11:153-169, 1998). Указанный профиль получают на основе скользящего окна из шести остатков. Скрытые остатки G, S и Т и доступные остатки Н, Y и W во внимание не принимаются. Например, в zcytor17lig человека гидрофильные области включают в себя аминокислотные остатки 54-59 SEQ ID NO:2, аминокислотные остатки 129-134 SEQ ID NO:2, аминокислотные остатки 53-58 SEQ ID NO:2, аминокислотные остатки 35-40 SEQ ID NO:2 и аминокислотные остатки 33-38 SEQ ID NO:2. Например, в zcytor17lig мыши гидрофильные области включают в себя аминокислотные остатки 34-39 SEQ ID NO:11, аминокислотные остатки 46-51 SEQ ID NO:11, аминокислотные остатки 131-136 SEQ ID NO:11, аминокислотные остатки 158-163 SEQ ID NO:11 и аминокислотные остатки 157-162 SEQ ID NO:11.
Специалистам должно быть понятно, что гидрофильность или гидрофобность должны быть приняты во внимание при внесении изменений в аминокислотную последовательность полипептида zcytor17lig, чтобы не нарушить общий структурный и биологический профиль. Особый интерес для замены представляют гидрофобные остатки, выбираемые из группы, включающей в себя Val, Leu и Ile, или из группы, включающей в себя Met, Gly, Ser, Ala, Tyr и Trp. Например, допускающие замену остатки могут включать в себя Val, Leu и Ile или остатки группы Met, Gly, Ser, Ala, Tyr и Trp, показанные в SEQ ID NO:2. Консервативные остатки цистеина в положениях SEQ ID NO:2 и SEQ ID NO:11 по существу не допускают замены.
Незаменимые аминокислоты можно также идентифицировать при помощи анализа сходства последовательностей IL-3, Lif, IL-12, IL-15, IL-2, IL-4 и GM-CSF c zcytor17lig. При помощи таких методов, как описанный выше анализ "FASTA", можно идентифицировать в семействе белков области, характеризующиеся высоким сходством, и использовать их для анализа аминокислотной последовательности с целью выявления консервативных областей. Альтернативным подходом к идентификации вариантного полинуклеотида zcytor17lig на основе структуры является определение способности молекулы нуклеиновой кислоты, кодирующей возможный вариантный ген zcytor17lig, гибридизоваться с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1, как это было описано выше.
Другими методами идентификации незаменимых аминокислот в полипептидах согласно изобретению являются методы, хорошо известные в данной области, такие как сайт-направленный мутагенез или аланин-сканирующий мутагенез (Cunningham and Wells, Science 244:1081 (1989), Bass et al., Proc. Natl. Acad. Sci. USA 88:4498 (1991), Coombs and Corey, "Site-Directed Mutagenesis and Protein Engineering," in Proteins: Analysis and Design, Angeletti (ed.), pages 259-311 (Academic Press, Inc. 1998)). В соответствии с последним методом, единичные мутации аланина вводят в каждый остаток молекулы, и полученные мутантные молекулы испытывают на биологическую или биохимическую активность, как будет описано ниже, для идентификации аминокислотных остатков, которые оказывают влияние на активность молекулы. См. также Hilton et al., J. Biol. Chem. 271:4699 (1996).
Настоящее изобретение относится также к функциональным фрагментам полипептидов zcytor17lig и молекулам нуклеиновых кислот, кодирующим такие функциональные фрагменты. «Функциональный» zcytor17lig или его фрагмент, в соответствии с определением, принятым в данном описании, характеризуется пролиферативной активностью или дифференцировкой, способностью индуцировать или ингибировать функции специализированных клеток или способностью специфически связываться с антителом против zcytor17lig, антителом к рецептору zcytor17, рецептором zcytor17, WSX-1 или OSMRbeta, гетеродимерами (например, zcytor17/WSX-1 или zcytor17/OSMRbeta) или многомерами (например, zcytor17/WSX-1/OSMRbeta) указанных рецепторов (растворимых или иммобилизованных). Как было описано выше, zcytor17lig характеризуется четырехспиральной структурой, включающей в себя спираль А (аминокислотные остатки 38-52), спираль В (аминокислотные остатки 83-98), спираль С (аминокислотные остатки 104-117) и спираль D (аминокислотные остатки 137-152), как показано в SEQ ID NO:2. Таким образом, настоящее изобретение далее относится к слитым белкам, включающим в себя: (а) молекулы полипептидов, содержащие одну или несколько вышеописанных спиралей, и (b) функциональные фрагменты, содержащие одну или несколько указанных спиралей. Другая полипептидная часть слитого белка может быть образована другим четырехспиральным цитокином, таким как IL-15, IL-2, IL-4 и GM-CSF, или ненативным и/или неродственным секреторным сигнальным пептидом, облегчающим секрецию слитого белка.
Таким образом, настоящее изобретение относится к слитым белкам, содержащим по меньшей мере четыре полипептида, которые характеризуются следующим порядком расположения от N-конца к С-концу: первый полипептид содержит аминокислоты, выбираемые из группы, включающей в себя: (а) аминокислотные остатки 27-48 SEQ ID NO:162 спирали А IL-2; (b) аминокислотные остатки 35-45 SEQ ID NO:102 спирали А IL-3; (c) аминокислотные остатки 30-42 SEQ ID NO:64 спирали А IL-4; (d) аминокислотные остатки 30-44 SEQ ID NO:166 спирали А GM-CSF; и (е) аминокислотные остатки 38-52 SEQ ID NO:2; первый спейсер, состоящий из 6-27 аминокислот; и второй полипептид, содержащий аминокислотные остатки, выбираемые из группы, включающей в себя: (а) аминокислотные остатки SEQ ID NO:168 спирали В IL-2; (b) аминокислотные остатки 65-83 SEQ ID NO:164 спирали В IL-4; (c) аминокислотные остатки 73-86 SEQ ID NO:102 спирали В IL-3; (d) аминокислотные остатки 72-81 SEQ ID NO:166 спирали В GM-CSF; и (е) аминокислотные остатки 83-98 SEQ ID NO:2; второй спейсер, состоящий из 5-11 аминокислотных остатков; третий полипептид, содержащий последовательность аминокислотных остатков, выбираемых из группы, включающей в себя: a) остатки 102-116 SEQ ID NO:162 спирали С IL-2; (b) остатки 94-118 SEQ ID NO:164 спирали С IL-4; (c) остатки 91-103 SEQ ID NO:102 спирали С IL-3; (d) остатки 85-103 SEQ ID NO:166 спирали С GM-CSF; и (е) аминокислотные остатки 104-117 SEQ ID NO:2; третий спейсер, состоящий из 3-29 аминокислотных остатков; и четвертый полипептид, содержащий аминокислотные остатки, выбираемые из группы, включающей в себя (а) аминокислотные остатки 134-149 SEQ ID NO:162 спирали D IL-2; (b) аминокислотные остатки 123-141 SEQ ID NO:102 спирали D IL-3; (c) аминокислотные остатки 133-151 SEQ ID NO:164 спирали D IL-4; (d) аминокислотные остатки 120-131 SEQ ID NO:166 спирали D GM-CSF; и (е) аминокислотные остатки 137-152 SEQ ID NO:2, где по меньшей мере один из четырех полипептидов получен из zcytor17lig. В других вариантах осуществления изобретения спейсерные пептиды выбирают из петель А/B, B/C и C/D zcytor17lig и IL-3, как показано в таблице 1.
Чтобы получить функциональные фрагменты молекулы нуклеиновой кислоты, кодирующей полипептид zcytor17lig, можно выполнить обычные анализы делеций в молекулах нуклеиновых кислот. В качестве примера можно привести метод, в соответствии с которым молекулы ДНК, имеющие нуклеотидную последовательность SEQ ID NO:1 или ее фрагмент, гидролизуют нуклеазой Bal31, получая при этом серию вложенных делеций. Указанные фрагменты ДНК затем встраивают в экспрессирующие векторы в соответствующую рамку считывания, после чего экспрессированные полипептиды выделяют и испытывают на активность zcytor17lig или способность связываться с антителами против zcytor17lig или рецептором zcytor17. Альтернативой гидролиза экзонуклеазой является применение олигонуклеотид-направленного мутагенеза для введения делеций или терминирующих кодонов для продуцирования требуемого фрагмента zcytor17lig. Определенные фрагменты гена zcytor17lig альтернативно можно синтезировать при помощи полимеразной реакции синтеза цепи.
Стандартные методы идентификации функциональных доменов хорошо известны специалистам. Например, результаты исследований по усечению одного или обоих концов интерферонов суммированы в статье Horisberger and Di Marco, Pharmac. Ther. 66:507 (1995). Кроме того, стандартные методы функционального анализа белков описаны, например, в публикациях Treuter et al., Molec. Gen. Genet. 240:113 (1993); Content et al., "Expression and preliminary deletion analysis of the 42 kDa 2-5A synthetase induced by human interferon," in Biological Interferon Systems, Proceedings of ISIR-TNO Meeting on Interferon Systems, Cantell (ed.), pages 65-72 (Nijhoff 1987); Herschman, "Тhe EGF Receptor," in Control of Animal Cell Prolifеration 1, Boynton et al., (eds.) pages 169-199 (Academic Press 1985); Coumailleau et al., J. Biol. Chem. 270:29270 (1995); Fukunaga et al., J. Biol. Chem. 270:25291 (1995); Yamaguchi et al., Biochem. Pharmacol. 50:1295 (1995); and Meisel et al., Plant Molec. Biol. 30:1 (1996).
Многие замены аминокислот могут быть произведены и исследованы при помощи известных методов мутагенеза и скрининга, таких как описанные в публикациях Reidhaar-Olson and Sauer (Science 241:53 (1988)) или Bowie and Sauer (Proc. Nat'l Acad. Sci. USA 86:2152 (1989)). Вышеуказанные авторы описывают методы одновременной рандомизации двух или большего числа положений в полипептиде, выбора функционального полипептида и последующего секвенирования подвергнутых мутагенезу полипептидов для определения совокупности допустимых замен в каждом положении. Другие методы, которые могут быть использованы, включают в себя индикацию фага (например, Lowman et al., Biochem. 30:10832 (1991), Ladner et al., патент США № 5223409, Huse, международная публикация № WO 92/06204) и область-специфический мутагенез (Derbyshire et al., Gene 46:145 (1986), and Ner et al., DNA 7:127 (1988)).
Варианты описанных нуклеотидных и полипептидных последовательностей zcytor17lig могут быть также созданы путем перестановки ДНК методом, описанным в статьях Stemmer, Nature 370:389 (1994), Stemmer, Proc. Natl. Acad. Sci. USA 91:10747 (1994) и в международной публикации № WO 97/20078. Вариантные молекулы ДНК получают методом гомологичной рекомбинации in vitro, произвольной фрагментации родительской ДНК и последующей повторной сборки при помощи PCR, в результате чего получают произвольно введенные точковые мутации. Данный метод может быть модифицирован путем использования семейства молекул родительской ДНК, такого как аллельные варианты или молекулы ДНК из других видов, для введения дополнительной вариабельности в указанный процесс. Выбор или скрининг требуемой активности с последующими дополнительными итерациями мутагенеза и анализа обеспечивают быструю «эволюцию» последовательностей благодаря выбору требуемых мутаций при одновременном исключении разрушительных изменений.
Методы мутагенеза, рассмотренные в данном описании, могут быть объединены с высокопроизводительными, автоматизированными методами скрининга для обнаружения активности клонированных, подвергнутых мутагенезу полипептидов в клетках-хозяевах. Подвергнутые мутагенезу молекулы ДНК, кодирующие биологически активные полипептиды или полипептиды, связывающиеся с антителами против zcytor17lig, растворимым рецептором zcytor17, растворимым рецептором WSX-1, растворимым рецептором OSMR, гетеродимерами или многомерами указанных растворимых рецепторов, могут быть выделены из клеток-хозяев и быстро секвенированы при помощи современной аппаратуры. Указанные методы позволяют быстро определить значение отдельных аминокислотных остатков в представляющем интерес полипептиде и могут быть использованы для исследования полипептидов неизвестной структуры.
Кроме того, белки согласно изобретению (или их полипептидные фрагменты) могут быть связаны с другими биологически активными молекулами, в частности с другими цитокинами, для получения многофункциональных молекул. Например, одна или несколько спиралей zcytor17lig могут быть связаны с другими цитокинами для усиления их биологических свойств или увеличения эффективности продуцирования.
Таким образом, настоящее изобретение относится к серии новых гибридных молекул, в которых сегмент, содержащий одну или несколько спиралей zcytor17lig, слит с другим полипептидом. Слияние предпочтительно осуществляют путем сплайсинга на уровне ДНК, обеспечивающего экспрессию химерных молекул в рекомбинантных продуцирующих системах. У полученных молекул анализируют такие свойства, как улучшенная растворимость, улучшенная устойчивость, более продолжительный период полужизни, более высокие уровни экспрессии и секреции и фармакодинамика. Такие гибридные молекулы могут далее содержать дополнительные аминокислотные остатки (например, полипептидный линкер) между составляющими белками или полипептидами.
Невстречающиеся в природе аминокислоты включают в себя, не ограничиваясь ими, транс-3-метилпролин, 2,4-метанопролин, цис-4-гидроксипролин, транс-4-гидроксипролин, N-метилглицин, аллотреонин, метилтреонин, гидроксиэтилцистеин, гидроксиэтилгомоцистеин, нитроглутамин, гомоглутамин, пипеколевую кислоту, тиазолидинкарбоновую кислоту, дегидропролин, 3- и 4-метилпролин, 3,3-диметилпролин, трет-лейцин, норвалин, 2-азафенилаланин, 3-азафенилаланин, 4-азафенилаланин и 4-фторфенилаланин. В данной области известно несколько методов введения в белки не встречающихся в природе аминокислотных остатков. Например, можно использовать систему in vitro, в которой нонсенс-мутации подавляются химически аминоацилированными супрессорными тРНК. В данной области известны методы синтеза аминокислот и аминоацилирования тРНК. Транскрипцию и трансляцию плазмид, содержащих нонсенс-мутации, обычно осуществляют в бесклеточной системе, содержащей экстракт S30 E. coli, коммерчески производимые ферменты и другие реагенты. Белки очищают при помощи хроматографии. См., например, Robertson et al., J. Am. Chem. Soc. 113:2722 (1991), Ellman et al., Methods Enzymol. 202:301 (1991), Chung et al., Science 259: 806 (1993) and Chung et al., Proc. Nat'l Acad. Sci. USA 90:10145 (1993).
В соответствии со вторым методом, трансляцию осуществляют в ооцитах Xenopus микроинъекцией мутированной мРНК и химически аминоацилированными супрессорными тРНК (Turcatti et al., J. Biol. Chem. 271:19991 (1996)). В соответствии с третьим методом клетки E. coli культивируют без природной аминокислоты, которая должна быть заменена (например, фенилаланин), и в присутствии требуемой невстречающейся в природе аминокислоты (например, 2-азафенилаланин, 3-азафенилаланин, 4-азафенилаланин или 4-фторфенилаланин). Невстречающуюся в природе аминокислоту вводят в белок вместо аналогичной природной аминокислоты. См., Koide et al., Biochem. 33:7470 (1994). Природные аминокислотные остатки можно превратить в их искусственные виды путем химической модификации in vitro. Химическую модификацию можно объединить с сайт-направленным мутагенезом для дальнейшего увеличения возможных замен (Wynn and Richards, Protein Sci. 2:395 (1993). Желательно стабилизировать zcytor17lig для увеличения периода полужизни молекулы, в частности для увеличения метаболической устойчивости в активном состоянии. Для достижения более продолжительного периода полужизни молекулы zcytor17lig могут быть химически модифицированы при помощи описанных методов. Обработка полиэтиленгликолем (PEG) является одним из обычно используемых методов, который, как было показано, увеличивает период полужизни в плазме, повышает растворимость и снижает антигенность и иммуногенность (Nucci et al., Advanced Drug Delivery Reviews 6:133-155, 1991 and Lu et al., Int. J. Peptide Protein Res. 43:127-138, 1994).
Аминокислотные остатки zcytor17lig могут быть заменены ограниченным числом неконсервативных аминокислот, аминокислот, не кодированных генетическим кодом, не встречающихся в природе аминокислот и искусственных аминокислот.
Настоящее изобретение относится также к полипептидным фрагментам или пептидам, содержащим эпитопнесущую часть описанного полипептида zcytor17lig. Такие фрагменты или пептиды могут содержать «иммуногенный эпитоп», который является частью белка, вызывающей реакцию на антитело при использовании всего белка в качестве иммуногена. Иммуногенные эпитопнесущие пептиды можно идентифицировать стандартными методами (см., например, Geysen et al., Proc. Nat'l Acad. Sci. USA 81:3998 (1983)).
В отличие от этого полипептидные фрагменты или пептиды могут содержать «эпитоп», который является областью молекулы белка, с которой может специфически связываться антитело. Некоторые эпитопы имеют линейный или смежный участок аминокислот, и антигенность такого эпитопа не разрушается денатурирующими агентами. В данной области известно, что относительно короткие синтетические пептиды, которые могут имитировать эпитопы белка, можно использовать для стимуляции продуцирования антител против данного белка (см., например, Sutcliffe et al., Science 219:660 (1983). Таким образом, эпитопнесущие пептиды или полипептиды согласно изобретению пригодны для увеличения числа антител (например, нейтрализующих антител), которые связываются с описанными полипептидами. Профили гидрофильности Хоппа-Вудса можно использовать для определения областей, обладающих наиболее сильным антигенным потенциалом (Hopp et al., 1981, ibid. and Hopp, 1986, ibid.). Например, в zcytor17lig человека гидрофильные области включают в себя аминокислотные остатки 54-59 SEQ ID NO:2, аминокислотные остатки 129-134 SEQ ID NO:2, аминокислотные остатки 53-58 SEQ ID NO:2, аминокислотные остатки 35-40 SEQ ID NO:2 и аминокислотные остатки 33-38 SEQ ID NO:2. Например, в zcytor17lig мыши гидрофильные области включают в себя аминокислотные остатки 34-39 SEQ ID NO:11, аминокислотные остатки 46-51 SEQ ID NO:11, аминокислотные остатки 131-136 SEQ ID NO:11, аминокислотные остатки 158-163 SEQ ID NO:11 и аминокислотные остатки 157-162 SEQ ID NO:11.
Антигенные эпитопнесущие пептиды и полипептиды предпочтительно содержат по меньшей мере четыре-десять аминокислот, по меньшей мере десять-четырнадцать аминокислот или около четырнадцати-тридцати аминокислот SEQ ID NO:2 или SEQ ID NO:11. Такие эпитопнесущие пептиды и полипептиды могут быть получены путем фрагментации полипептида zcytor17lig или химического синтеза пептида, представленным в данном описании. Кроме того, эпитопы могут быть выбраны путем индикации фага в библиотеке неспецифических пептидов (см., например, Lane and Stephen, Curr. Opin. Immunol. 5:268 (1993); and Cortese et al., Curr. Opin. Biotechnol. 7:616 (1996)). Стандартные методы идентификации эпитопов и продуцирования антител из коротких пептидов, содержащих эпитоп, описаны, например, в публикации Mole, "Epitope Mapping," in Methods in Molecular Biology, Vol. 10. Manson (еd.), pages 105-116 (The Humana Press, Inc. 1992); Price, "Production and Characterization of Synthetic Peptide-Derived Antibodies," in Monoclonal Antibodies: Production, Engineering, and Clinical Application, Ritter and Ladyman (eds.), pages 60-84 (Cambridge University Press 1995), and Coligan et al. (eds.), Current Protocols in Immunologу, pages 9.3.1-9.3.5 and pages 9.4.1-9.4.11 (John Wiley & Sons 1997).
Независимо от конкретной нуклеотидной последовательности вариантного полинуклеотида zcytor17lig данный полинуклеотид кодирует полипептид, обладающий пролиферативной или дифференцировочной активностью, способностью индуцировать или ингибировать функции специализированных клеток, или способностью специфически связываться с антителом против zcytor17lig или рецептором zcytor17. Более конкретно, вариантные полинуклеотиды zcytor17lig кодируют полипептиды, которые обладают по меньшей мере 50% и предпочтительно по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или более чем 99% активности полипептида, показанного в SEQ ID NO:2.
Для любого полипептида zcytor17lig, включая варианты и слитые белки, специалист может легко создать полностью вырожденную полинуклеотидную последовательность, кодирующую данный вариант, используя информацию, представленную в приведенных выше таблицах 1 и 2.
Настоящее изобретение далее относится к целому ряду других слитых полипептидов (и родственных многомерных белков, содержащих один или несколько слитых полипептидов). Например, полипептид zcytor17lig может быть получен в результате слияния с димеризующим белком, как это описано в патентах США № 5155027 и 5567584. Предпочтительные димеризующие белки включают в себя домены константной области иммуноглобулина. Полипептид zcytor17lig, слитый с иммуноглобулином, может быть экспрессирован в генетически созданных клетках (для продуцирования различных многомерных аналогов zcytor17lig). Вспомогательные домены могут быть слиты с полипептидами zcytor17lig для их направленной доставки в специфические клетки, ткани или макромолекулы. Например, полипептид zcytor17lig или белок может быть направленно доставлен в клетку заранее определенного типа в результате слияния полипептида zcytor17lig с лигандом, который специфически связывается с рецептором на поверхности клетки-мишени. Таким образом, полипептиды и белки могут быть направленно доставлены для терапевтических или диагностических целей. Полипептид zcytor17lig может быть слит с двумя или большим числом частей, таких как аффинная метка для очистки и домен направленной доставки. Слитые полипептиды могут также содержать один или несколько сайтов расщепления, расположенных, в частности, между доменами. См., Tuan et al., Connective Tissue Research 34:1-9, 1996.
Используя описанные методы, специалист может идентифицировать и/или получить разновидность полипептидов, имеющих последовательность, которая по существу идентична остаткам 1-164 или 24-164 SEQ ID NO:2, или их функциональным фрагментам и слитым частям, таким как спирали А-D (остатки 38-152 SEQ ID NO:2), причем такие полипептиды, фрагменты или слитые части сохраняют свойства белка дикого типа, такие как способность стимулировать пролиферацию, дифференцировку, индуцировать функцию специализированной клетки или связываться с рецептором zcytor17 или антителами к zcytor17lig.
Полипептиды zcytor17lig согласно изобретению, включая непроцессированные полипептиды, функциональные фрагменты и слитые полипептиды, могут быть продуцированы в генетически созданных клетках-хозяевах, полученных известными методами. Приемлемыми клетками-хозяевами являются клетки таких типов, которые могут быть трансформированы или трансфицированы экзогенной ДНК и выращены в культуре, и включают в себя бактерии, грибные клетки и культивируемые высшие эукариотические клетки. Предпочтительными являются эукариотические клетки, в частности культивируемые клетки многоклеточных организмов. Методы манипуляции молекулами клонированной ДНК и введения экзогенной ДНК в разнообразные клетки-хозяева описаны в публикациях Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989 and Ausubel et al., eds., Current Protocols in Molecular Biology, John Wiley and Sons, Inc., NY, 1987.
Как правило, последовательность ДНК, кодирующая полипептид zcytor17lig, может быть функционально связана с другими генетическими элементами, необходимыми для ее экспрессии в экспрессирующем векторе, которые обычно включают в себя промотор и терминатор транскрипции. Вектор обычно также содержит один или несколько селектируемых маркеров и один или несколько ориджинов репликации, хотя специалистам должно быть понятно, что в некоторых системах селектируемые маркеры могут быть представлены в отдельных векторах и репликация экзогенной ДНК может быть достигнута встраиванием в геном клетки-хозяина. Выбор промоторов, терминаторов, селектируемых маркеров, векторов и других элементов находится в пределах компетенции специалистов в данной области. Многие такие элементы описаны в научной литературе и могут быть приобретены у коммерческих поставщиков.
Для того чтобы направить полипептид zcytor17lig по секреторному пути клетки-хозяина, в экспрессирующем векторе существует секреторная сигнальная последовательность (известная также как лидерная последовательность, препропоследовательность или препоследовательность). Секреторная сигнальная последовательность может принадлежать zcytor17lig, может быть выделена из другого секретированного белка (например, t-PA) или синтезирована de novo. Секреторная сигнальная последовательность функционально связана с последовательностью ДНК zcytor17lig, то есть две указанные последовательности связаны в правильной рамке считывания и расположены так, чтобы направлять вновь синтезированный полипептид по секреторному пути клетки-хозяина. Секреторные сигнальные последовательности обычно расположены у 5'-конца относительно последовательности ДНК, кодирующей представляющий интерес полипептид, хотя некоторые секреторные сигнальные последовательности могут быть расположены в любом месте представляющей интерес последовательности ДНК (см., например, Welch et al., патент США № 5037743; Holland et al., патент США № 5143830).
Альтернативно, секреторную сигнальную последовательность, находящуюся в полипептидах согласно изобретению, используют для того, чтобы направить другие полипептиды по секреторному пути. В объем настоящего изобретения входят такие слитые полипептиды. Можно получить сигнальный слитый полипептид, в котором секреторная сигнальная последовательность, выделенная из аминокислотных остатков 1-23 SEQ ID NO:2 или остатков 1-23 SEQ ID NO:11, функционально связана с последовательностью ДНК, кодирующей другой полипептид, при помощи методов, известных в данной области и рассмотренных в данном описании. Секреторная сигнальная последовательность, находящаяся в слитых полипептидах согласно изобретению, предпочтительно слита у аминоконца с дополнительным пептидом для того, чтобы направлять данный дополнительный пептид по секреторному пути. Такие конструкции имеют много применений, известных в данной области. Например, указанные новые слитые конструкции секреторной сигнальной последовательности могут направлять секрецию активного компонента обычно несекретируемого белка. Такие слитые конструкции можно использовать in vivo или in vitro для того, чтобы направить пептиды по секреторному пути.
Культивируемые клетки млекопитающих являются приемлемыми хозяевами при осуществлении настоящего изобретения. Методы введения экзогенной ДНК в клетки-хозяева млекопитающих включают в себя трансфекцию, опосредованную фосфатом кальция (Wigler et al., Cell 14:725, 1978; Corsaro and Pearson, Somatic Cell Genetics 7:603, 1981: Graham and Van der Eb, Virology 52:456, 1973), электропорацию (Neumann et al., EMBO J. 1:841-5, 1982), трансфекцию, опосредованную DEAЕ-декстраном (Ausubel et al., см. выше), трансфекцию, опосредованную липосомами (Hawley-Nelson et al., Focus 15:73, 1993; Ciccarone et al., Focus 15:80, 1993), и вирусные векторы (Miller and Rosman, BioTechniques 7:980-90, 1989; Wang and Finer, Nature Med. 2:714-6, 1996). Продуцирование рекомбинантных полипептидов в культивируемых клетках млекопитающих описано, например, в публикациях Levinson et al., патент США № 4713339; Hagen et al., патент США № 4784950; Palmiter et al., патент США № 4579821; и Ringold, патент США № 4656134. Приемлемые культивируемые клетки млекопитающих включают в себя линии клеток COS-1 (АТСС № CRL 1650), COS-7 (ATCC № CRL 1651), ВНК (АТСС № CRL 1632), BHK 570 (ATCC № CRL 10314), 293 (ATCC № CRL 1573; Graham et al., J. Gen. Virol. 36:59-72, 1977) и клетки яичника китайского хомячка (например, СНО-К1; АТСС № CCL 61). В данной области известны дополнительные приемлемые линии клеток, которые могут быть получены из общедоступных депозитариев, таких как Американская коллекция типовых культур, Манассас, шт. Вирджиния. Как правило, предпочтительными являются сильные промоторы транскрипции, такие как промоторы, выделенные из SV-40 или цитомегаловируса. См., например, патент США № 4956288. Другие приемлемые промоторы включают в себя промоторы, полученные из генов металлотионеина (патенты США №№ 4579821 и 4601978), и аденовирусный главный поздний промотор.
Культивируемые клетки млекопитающих, в которые введена чужеродная ДНК, обычно отбирают, используя лекарственное средство. Такие клетки именуются «трансфектантами». Клетки, которые были культивированы в присутствии селективного агента и способны передавать исследуемый ген своему потомству, определяются как «устойчивые трансфектанты». Предпочтительным селектируемым маркером является ген, кодирующий устойчивость к антибиотику неомицину. Отбор производят в присутствии лекарственного средства типа неомицина, такого как G-418 или подобного. Системы отбора можно также использовать для увеличения экспрессии исследуемого гена в ходе процесса, определяемого как «амплификация». Амплификацию осуществляют, культивируя трансфектанты в присутствии селективного агента в низкой концентрации с последующим увеличением количества селективного агента, при этом отбирают клетки, продуцирующие продукты введенных генов на высоком уровне. Предпочтительным амплифицируемым селектируемым маркером является дигидрофолатредуктаза, которая сообщает устойчивость к метотрексату. Можно также использовать другие гены лекарственной устойчивости (например, устойчивости к гигромицину, множественной лекарственной устойчивости, устойчивости к пуромицинацетилтрансферазе). Альтернативные маркеры, которые вводят измененный фенотип, в частности зеленый флуоресцирующий белок или белки клеточной поверхности, такие как CD4, CD8, МНС класса I, щелочная фосфатаза плаценты, можно использовать для отделения трансфицированных клеток от нетрансфицированных клеток такими методами, как сортировка в FACS или отделение магнитными шариками.
Другие высшие эукариотические клетки можно также использовать в качестве хозяев, в том числе растительные клетки, клетки насекомых и клетки птиц. Использование Agrobacterium rhizogenes в качестве вектора для экспрессии генов в растительных клетках было описано в статье Sinkar et al., J. Biosci. (Bangalore) 11:47-58, 1987. Трансформация клеток насекомых и продуцирование в них чужеродных полипептидов описаны в патенте США № 5162222, выданном Гуарино и др., и в публикации WIPO № WO 94/06463. Клетки насекомых могут быть инфицированы рекомбинантным бакуловирусом, обычно выделяемым из вируса ядерного полиэдроза Autographa californicа (AcNPV). См., King, L.A. and Possee, R.D., The Baculovirus Expression System: A Laboratory Guide, London, Chapman & Hall; O'Reilly, D.R. et al., Baculovirus Expression Vectors: A Laboratory Manual, New York, Oxford University Press., 1994; и Richardson, C.D., Ed., Baculovirus Expression Protocols. Methods in Molecular Biology, Totowa, NJ, Humana Press, 1995. Вторым методом получения рекомбинантного бакуловируса является использование системы на основе транспозона, описанной Луковым (Luckow, V.A., et al., J. Virol 67:4566-79, 1993). Данная система входит в набор Bac-to-Bac (Life Technologies, Rockville, MD). В указанной системе использован вектор переноса, pFastBac1™ (Life Technologies), содержащий транспозон Tn7, который предназначен для перемещения ДНК, кодирующей полипептид zcytor17lig, в геном бакуловируса, находящегося в E. coli, в виде большой плазмиды, именуемой «бакмид». В векторе переноса pFastBac1™ использован промотор на основе полигедрина AcNPV для стимуляции экспрессии исследуемого гена, в данном случае zcytor17lig. Однако вектор pFastBac1™ может быть модифицирован в значительной степени. Промотор на основе полигедрина может быть удален и заменен промотором, содержащим основной белок бакуловируса (известный также как промотор Pcor, p6.9 или МР), который ранее был экспрессирован в бакуловирусной инфекции и, как установлено, позволяет эффективно экспрессировать секретированные белки. См., Hill-Perkins, M.S. and Possee, R.D., J. Gen. Virol. 71:971-6, 1990; Bonning, B.C. et al., J. Gen. Virol. 75: 1551-6, 1994; и Chazenbalk, G.D., and Rapoport, B., J. Biol. Chem. 270: 1543-9, 1995. В таких конструкциях вектора переноса можно использовать короткий или длинный вариант промотора в виде основного белка. Кроме того, могут быть созданы векторы переноса, заменяющие нативные секреторные сигнальные последовательности zcytor17lig секреторными сигнальными последовательностями, выделенными из белков насекомых. Например, секреторную сигнальную последовательность, выделенную из экдистероидглюкозилтрансферазы (EGT) мелиттина пчелы (Invitrogen, Carlsbad, CA) или бакуловируса gp67 (PharMingen, San Diego, CA), можно использовать в конструкциях для замены нативной секреторной сигнальной последовательности zcytor17lig. Кроме того, векторы переноса могут быть слиты в рамке считывания с ДНК, кодирующей метку эпитопа у С- или N-конца экспрессированного полипептида zcytor17lig, например метку эпитопа Glu-Glu (Grussenmeyer, T. et al., Proc. Natl. Acad. Sci. 82:7952-4, 1985). При помощи методов, известных в данной области, вектор переноса, содержащий zcytor17lig, вводят в E. coli, и ведут поиск бакмидов, содержащих ген lacZ с разорванной цепью, который является показателем наличия рекомбинантного бакуловируса. Бакмидную ДНК, содержащую геном рекомбинантного бакуловируса, выделяют обычными методами и используют для трансфекции клеток Spodoptera frugiperda, например клеток Sf9. Таким образом продуцируется рекомбинантный вирус, экспрессирующий zcytor17lig. Штаммы рекомбинантных вирусов получают методами, известными в данной области.
Рекомбинантный вирус используют для инфицирования клеток-хозяев, обычно линии клеток, выделенных из «походных червей» Spodoptera frugiperda. См., например, Glick and Pasternak, Molecular Biotechnology: Principles and Applications of Recombinant DNA, ASM Press, Washington, D.C., 1994. Другой приемлемой линией клеток является линия клеток High FiveO™ (Invitrogen), выделенная из Trichoplusia ni (патент США № 5300435).
При осуществлении настоящего изобретения можно также использовать грибные клетки, в том числе дрожжевые клетки. Особенно интересные виды дрожжей включают в себя Saccharomyces cerevisiae, Pichia pastoris и Pichia methanolica. Методы трансформации клеток S. cerevisiae при помощи экзогенной ДНК и продуцирования рекомбинантных полипептидов описаны, например, в следующих патентах: Kawasaki, патент США № 4599311; Kawasaki et al., патент США № 4931373; Brake, патент США № 4870008; Welch et al., патент США № 5037743; и Murray et al., патент США № 4845075. Трансформированные клетки отбирают с учетом фенотипа, определяемого селектируемым маркером, лекарственной устойчивости или способности роста при отсутствии конкретного питательного вещества (например, лейцина). Предпочтительной векторной системой, предназначенной для использования в Saccharomyces cerevisiae, является векторная система РОТ1, описанная Kawasaki et al. (патент США № 4931373), которая позволяет отбирать трансформированные клетки при выращивании в среде, содержащей глюкозу. Приемлемые промоторы и терминаторы, предназначенные для использования в дрожжах, включают в себя аналогичные элементы, выделенные из генов гликолитических ферментов (см., например, Kawasaki, патент США № 4599311; Kingsman et al., патент США № 4615974; и Bitter, патент США № 4977092) и генов спиртовой дегидрогеназы. См. также патенты США №№ 4990446, 5063154, 5139936 и 4661454. В данной области известны системы трансформации других дрожжей, включая Hansenula polymorpha, Schizosaccharomyces pombe, Kluyveromyces lactis, Kluyveromyces fragilis, Ustilago maydis, Pichia partoris, Pichia methanolica, Pichia guillermondii и Candida maltosa. См., например, Gleeson et al., J. Gen. Microbiol. 132:3459-65, 1986 и Cregg, патент США № 4882279. Клетки Aspergillus можно использовать в соответствии с методами, описанными McKnight et al., патент США № 4935349. Методы трансформации Acremonium chrysogenum описаны Sumino et al., патент США № 5162228. Методы трансформации Neurospora описаны Lambowitz, патент США № 4486533.
Применение Pichia methanolica в качестве хозяина для продуцирования рекомбинантных белков описано в публикациях WIPO №№ 97/17450, WO 97/17451, WO 98/02536 и WO 98/02565. Молекулы ДНК, предназначенные для трансформации P. methanolica, обычно получают в виде двухцепочечных кольцевых плазмид, которые предпочтительно переводят в линейную форму до трансформации. Для продуцирования полипептидов в P. methanolica желательно, чтобы промотор и терминатор в плазмиде относились к гену P. methanolica, такому как утилизирующий спирт ген P. methanolica (AUG1 или AUG2). Другие приемлемые промоторы включают в себя промоторы на основе генов дигидроксиацетонсинтазы (DHAS), формиатдегидрогеназы (FMD) и каталазы (САТ). Для облегчения встраивания ДНК в хромосому хозяина желательно иметь полный экспрессирующий сегмент плазмиды, фланкированный с обоих концов последовательностями ДНК хозяина. Предпочтительным селектируемым маркером для использования в Pichia methanolica является ген ADE2 P. methanolica, кодирующий фосфорибозил-5-аминоимидазолкарбоксилазу (AIRC, EC 4.1.1.21), которая позволяет клеткам-хозяевам ade2 расти в отсутствие аденина. В крупномасштабных промышленных процессах, где желательно свести к минимуму применение метанола, предпочтительно используют клетки-хозяева, в которых удалены оба утилизирующих метанол гена (AUG1 и AUG2). Для продуцирования секретированных белков предпочтительными являются клетки-хозяева с отсутствием генов вакуолярной протеазы (РЕР4 и PRB1). Для облегчения введения в клетки P. methanolica плазмиды, содержащей ДНК, кодирующую представляющий интерес полипептид, применяют электропорацию. Желательно трансформировать клетки P. methanolica путем электропорации с использованием экспоненциально затухающего импульсного электрического поля напряженностью от 2,5 до 4,5 кВ/см, предпочтительно около 3,75 кВ/см, и постоянной времени (Ω) от 1 до 40 миллисекунд, наиболее предпочтительно около 20 миллисекунд.
Прокариотические клетки-хозяева, включая штаммы бактерий Escherichia coli, Bacillus и других родов, также являются пригодными клетками-хозяевами при осуществлении настоящего изобретения. Методы трансформации таких хозяев и экспрессии клонированных в них последовательностей чужеродной ДНК хорошо известны в данной области (см., например, Sambrook et al., выше). При экспрессии полипептида zcytor17lig в бактериях, таких как E. coli, данный полипептид может находиться в цитоплазме, обычно в виде нерастворимых гранул, или может быть введен в периплазматическое пространство при помощи секретирующей последовательности бактерий. В первом случае клетки лизируют и гранулы выделяют и денатурируют, используя, например, изотиоцианат гуанидина или мочевину. В денатурированном полипептиде затем можно произвести повторную укладку цепи и димеризацию методом разведения денатуранта, например, путем диализа против раствора мочевины и комбинации восстановленного и окисленного глутатиона с последующим диализом против забуференного солевого раствора. Во втором случае полипептид можно выделить из периплазматического пространства в растворимой и функциональной форме путем разрушения клеток (например, в результате обработки ультразвуком или под действием осмотического шока), в результате чего высвобождается содержимое периплазматического пространства и выделяется белок, что позволяет избежать денатурации и повторной укладки цепи.
Трансформированные или трансфицированные клетки-хозяева культивируют обычными методами в культуральной среде, содержащей питательные вещества и другие компоненты, необходимые для роста выбранных клеток-хозяев. В данной области известны различные приемлемые среды, включая среды определенного состава и комплексные среды, которые обычно содержат источник углерода, источник азота, незаменимые аминокислоты, витамины и минералы. При необходимости среды могут также содержать такие компоненты, как факторы роста или сыворотка. Питательную среду обычно выбирают для клеток, содержащих экзогенно добавленную ДНК, например, на основании лекарственной устойчивости или дефицита незаменимого питательного вещества с использованием селектируемого маркера, находящегося в экспрессирующем векторе или котрансфицированного в клетку-хозяина. Клетки P. methanolica культивируют в среде, содержащей соответствующие источники углерода, азота и питательные микроэлементы при температуре около 25°C-35°C. Жидкие культуры подвергаются достаточной аэрации обычными средствами, такими как встряхивание маленьких колб или барботирование ферментеров. Предпочтительной культуральной средой для P. methanolica является YEPD (2% D-глюкозы, 2% пептона Bacto™ (Difco Laboratories, Detroit, MI), 1% дрожжевого экстракта Bacto™ (Difco Laboratoriеs), 0,004% аденина и 0,006% L-лейцина).
Полипептиды согласно изобретению желательно очистить до степени чистоты ≥80%, более предпочтительно до степени чистоты ≥90%, еще более предпочтительно до степени чистоты ≥95% и особенно предпочтительно до достижения фармацевтически чистого состояния, которое соответствует степени чистоты более 99,9% по отношению к загрязняющим макромолекулам, в частности другим белкам и нуклеиновым кислотам, и предполагает полное отсутствие инфекционных и пирогенных веществ. Очищенный полипептид предпочтительно по существу не содержит других полипептидов, особенно других полипептидов животного происхождения.
Экспрессированные рекомбинантные полипептиды zcytro17lig (или химерные полипептиды zcytor17lig) могут быть очищены при помощи фракционирования и/или обычных методов очистки и сред. Для фракционирования образцов можно использовать осаждение сульфатом аммония и кислотную или хаотропную экстракцию. Типичные стадии очистки могут включать в себя применение гидроксиапатита, вытеснительную хроматографию, жидкостную хроматографию быстрого разрешения (FPLC) и жидкостную высокоэффективную хроматографию с обращенной фазой. Приемлемые хроматографические среды включают в себя производные декстраны, агарозу, целлюлозу, полиакриламид, специальные кремнеземы и тому подобное. Предпочтительными являются производные PEI, DEAE, QAE и Q. Типичными хроматографическими средами являются среды, включающие производные фенильных, бутильных или октильных групп, такие как Phenyl-Sepharose FF (Pharmacia), Toyopearl butyl 650 (Toso Haas, Montgomeryville, PA), Octyl-Sepharose (Pharmacia) и тому подобное; или полиакриловые смолы, такие как Amberchrom CG 71 (Toso Haas) и тому подобное. Приемлемыми твердыми носителями являются стеклянные шарики, смолы на основе кремнезема, целлюлозные смолы, агарозные шарики, сшитые агарозные шарики, полистирольные шарики, сшитые полиакриламидные смолы и пр., которые нерастворимы в условиях применения. Указанные носители могут быть модифицированы реакционно-способными группами, которые делают возможным присоединение белков аминогруппами, карбоксильными группами, сульфгидрильными группами, гидроксильными группами и/или углеводородными частями молекулы. Примеры химических методов связывания включают в себя активацию бромцианом, активацию N-гидроксисукцинимидом, активацию эпоксидом, активацию сульфгидрилом, активацию гидразидом и карбоксильными и аминопроизводными для связывания карбодиимидом. Указанные и другие твердые среды хорошо известны и широко применяются в данной области и могут быть приобретены у коммерческих поставщиков. Методы связывания полипептидов рецепторов с несущими средами хорошо известны в данной области. Выбор определенного метода является обычной практикой и частично определяется свойствами выбранного носителя. См., например, Affinity Chromatography: Principles & Methods, Pharmacia LKB Biotechnology, Uppsala, Sweden, 1988.
Полипептиды согласно изобретению могут быть выделены с учетом их физических или биохимических свойств. Например, для очистки белков с высоким содержанием гистидина, включая белки, содержащие полигистидиновые метки, можно использовать адсорбционную хроматографию с иммобилизованными ионами металла (IMAC). Гель сначала заряжают ионами двухвалентного металла с образованием хелата (Sulkowski, Trends in Biochem. 3:1-7, 1985). Белки с высоким содержанием гистидина адсорбируют на указанной матрице с разной степенью сродства в зависимости от используемых ионов металла, и элюируют путем конкурентного элюирования, уменьшения показателя рН или использования сильных хелатообразующих агентов. Другие методы очистки включают в себя очистку гликозилированных белков путем лектин-аффинной хроматографии и ионообменной хроматографии (Methods in Enzymol., Vol. 182, "Guide to Protein Purification", M. Deutscher, (ed.), Acad Press, San Diego, 1990, pp. 529-39) и применение растворимого рецептора zcytor17. В соответствии с дополнительными вариантами осуществления изобретения, для облегчения очистки может быть произведено слияние представляющего интерес полипептида с аффинной меткой (например, связывающим мальтозу белком, доменом иммуноглобулина).
Кроме того, с помощью методов, описанных в данной области, могут быть получены слитые полипептиды или гибридные белки zcytor17lig при использовании областей или доменов zcytor17lig согласно изобретению и аналогичных элементов других белков семейства цитокинов человека (например, интерлейкинов или GM-CSF) или гетерологичных белков (Sambrook et al., ibid., Altschul et al., ibid., Picard. Cur. Opin. Biology, 5:511-5, 1994, и ссылки, приведенные в указанных публикациях). Указанные методы позволяют определить биологическую значимость более крупных доменов или областей в представляющем интерес полипептиде. Такие гибриды могут изменять кинетику реакции, связывание, ограничивать или расширять специфичность субстрата, или изменять локализацию полипептида в тканях и клетках и могут быть использованы для исследования полипептидов неизвестной структуры.
Слитые белки можно получить методами, известными специалистам в данной области, в соответствии с которыми получают отдельные компоненты слитого белка и конъюгируют их химическими методами. Альтернативно, известными методами можно получить полинуклеотид, кодирующий оба компонента слитого белка в соответствующей рамке считывания, и экспрессировать его способами, рассмотренными в данном описании. Например, часть или вся спираль, определяющая биологическую функцию, может быть заменена в zcytor17lig согласно изобретению функционально эквивалентными спиралями из другого члена семейства, такого как IL-15, IL-2, IL-4 или GM-CSF. Такие компоненты включают в себя, не ограничиваясь ими, секреторную сигнальную последовательность; спирали А, В, С, D; петли A/B, B/C, C/D четырехспиральных цитокинов. Предполагается, что такие слитые белки должны иметь биологический функциональный профиль, аналогичный или подобный полипептидам согласно изобретению или другим известным белкам семейства четырехспиральных цитокинов в зависимости от произведенного слияния. Кроме того, такие слитые белки могут обладать другими свойствами, рассмотренными в данном описании.
Для обмена эквивалентных доменов в полипептиде zcytor17lig и сливаемых полипептидах можно использовать стандартные методы молекулярной биологии и клонирования. Сегмент ДНК, кодирующий представляющий интерес домен, например спирали А-D zcytor17lig, функционально связывают в рамке считывания по меньшей мере с одним другим сегментом ДНК, кодирующим дополнительный полипептид (например, с доменом или областью из другого цитокина, такого как IL-2 или тому подобное), и вводят в соответствующий экспрессирующий вектор, как описано в данном описании. Конструкции ДНК обычно создают таким образом, чтобы несколько сегментов ДНК, кодирующих соответствующие области полипептида, были функционально связаны в рамке считывания с образованием единой конструкции, кодирующей весь слитый белок или его функциональную часть. Например, конструкция ДНК должна кодировать в направлении от N-конца к С-концу слитый белок, содержащий сигнальный полипептид, за которым следует зрелый слитый белок четырехспирального цитокина, содержащий спираль А, затем спираль В, спираль С и спираль D. Такие слитые белки могут быть экспрессированы, выделены и подвергнуты анализу на наличие описанной активности.
Полипептиды zcytor17lig или их фрагменты можно также получить методом химического синтеза. Полипептиды zcytor17lig могут быть мономерами или многомерами, гликозилированными или негликозилированными, обработанными или не обработанными полиэтиленгликолем, и могут содержать или не содержать начальный аминокислотный остаток метионин. Например, указанные полипептиды могут быть получены методом твердофазного синтеза пептидов, например, как описано Merrifield, J. Am. Chem. Soc. 85:2149, 1963.
Активность молекул согласно изобретению можно определить разными анализами, позволяющими измерить пролиферацию и/или связывание с клетками, экспрессирующими рецептор zcytor17. Особый интерес представляют изменения в zcytor17lig-зависимых клетках. Приемлемые линии клеток, которые могут быть сделаны zcytor17lig-зависимыми, включают в себя IL-3-зависимую линию клеток BaF3 (Palacios and Steinmetz, Cell 41:727-734, 1985; Mathey-Prevot et al., Mol. Cell. Biol. 6:4133-4135, 1986), FDC-P1 (Hapel et al., Blood 64:786-790, 1984) и MO7e (Kiss et al., Leukemia 7:235-240, 1993). Линии клеток, зависящие от фактора роста, могут быть созданы в соответствии с опубликованными методами (например, Greenberger et al., Leukemia Res. 8:363-375, 1984; Dexter et al., in Baum et al. Eds., Experimental Hematology Today, 8th Ann. Mtg. Int. Soc. Exp. Hematol. 1979, 145-156, 1980).
Белки согласно изобретению пригодны для стимуляции пролиферации, активации, дифференцировки и/или индукции или ингибирования функций специализированных клеток, участвующих в гомеостазе гемопоэза и иммунной функции. В частности, полипептиды zcytor17lig пригодны для стимуляции пролиферации, активации, дифференцировки, индукции или ингибирования функций специализированных клеток гемопоэтических линий, включая, но не ограничиваясь ими, Т-клетки, В-клетки, моноциты/макрофаги, NK-клетки, нейтрофилы, эндотелиальные клетки, фибробласты, эозинофилы, хондроциты, мастоциты, клетки Лангерганса, моноциты и макрофаги, а также эпителиальные клетки. Эпителиальные клетки включают в себя, например, амелобласты, главные клетки, хроматофоры, аргентофильные клетки, аргентофилподобные клетки, бокаловидные клетки, зернистые клетки, кератиноциты, дендритные клетки, поддерживающие клетки лабиринта, меланоциты, клетки Меркеля, клетки Панета, париетальные клетки, клетки Сертоли и тому подобное. Пролиферацию и/или дифференцировку гемопоэтических клеток можно измерить in vitro, используя культивируемые клетки, или in vivo путем введения молекул согласно изобретению в соответствующую животную модель. Анализы для измерения пролиферации или дифференцировки клеток хорошо известны в данной области. Например, анализы, применяемые для измерения пролиферации, включают в себя анализы химической чувствительности к нейтральному красному красителю (статья Cavanaugh et al., Investigational New Drugs 8:347-354, 1990, включенная в данное описание в качестве ссылки), введение нуклеотидов, меченных радиоактивными изотопами (статья Cook et al., Analytical Biochem. 179:1-7, 1989, включенная в данное описание в качестве ссылки), введение 5-бром-2'-дезоксиуридина (BrdU) в ДНК пролиферирующих клеток (статья Porstmann et al., J. Immunol. Methods 82:169-179, 1985, включенная в данное описание в качестве ссылки) и применение солей тетразолия (статьи Mosmann, J. Immunol. Methods 65:55-63, 1983; Alley et al., Cancer Res. 48:589-601, 1988; Marshall et al., Growth Reg. 5:69-84, 1995 and Scudiero et al., Cancer Res. 48:4827-4833, 1988, включенные в данное описание в качестве ссылки). Анализы, предназначенные для измерения дифференцировки, включают в себя, например, измерение маркеров клеточной поверхности, ассоциированных со стадиеспецифической экспрессией в ткани, ферментативной активности, функциональной активности или морфологических изменений (статьи Watt, FASEB, 5:281-284, 1991; Francis, Differentiation 57:63-75, 1994; Raes, Adv. Anim. Cell Biol. Technol. Bioprocesses, 161-171, 1989, включенные в данное описание в качестве ссылки).
Молекулы согласно изобретению могут быть подвергнуты анализу in vivo с использованием систем вирусной доставки. Типичные вирусы, предназначенные для указанной цели, включают в себя аденовирус, вирус герпеса, ретровирус, вирус коровьей оспы и аденоассоциированный вирус (AAV). Аденовирус, являющийся вирусом с двухцепочечной ДНК, в настоящее время является наиболее исследованным вектором переноса генов для доставки гетерологичной нуклеиновой кислоты (для ознакомления см. T.C. Becker et al., Meth. Cell Biol. 43:161-89, 1984; и J.T. Douglas and D.T. Curiel, Science & Medicine 4:44-53, 1997).
При использовании в качестве лиганда активность полипептида zcytor17lig можно измерить при помощи микрофизиометра с силиконовым биосенсором, который измеряет скорость внеклеточного окисления или экскрецию протонов, обусловленную связыванием с рецептором и последующими физиологическими реакциями клетки. Типичным устройством является микрофизиометр Cytosensor™ компании Molecular Devices, Sunnyvale, CA. При помощи данного метода можно измерить различные реакции клеток, такие как пролиферация клеток, перенос ионов, выделение энергии, воспалительная реакция, активация регуляторной функции и рецепторов и тому подобное. См., например, McConnell, H.M. et al., Science 257:1906-1912, 1992; Pitchford, S. et al., Meth. Enzymol. 228:84-108, 1997; Arimilli, S. et al., J. Immunol. Meth. 212:49-59, 1998; Van Liefde, I. et al., Eur. J. Pharmacol. 346:87-95, 1998.
Кроме того, zcytor17lig можно использовать для идентификации клеток, тканей или линий клеток, реагирующих на zcytor17lig-стимулируемый путь. Микрофизиометр, описанный выше, можно использовать для быстрой идентификации чувствительных к лиганду клеток, таких как клетки, чувствительные к zcytor17lig согласно изобретению. Клетки можно культивировать в присутствии или в отсутствие полипептида zcytor17lig. Клетки, в которых происходит измеряемое изменение внеклеточного окисления в присутствии zcytor17lig, чувствительны к zcytor17lig. Такие клетки или линии клеток можно использовать для идентификации антагонистов и агонистов вышеуказанного полипептида zcytor17lig.
Учитывая широкое распределение рецептора zcytor17 в тканях, агонисты (включая природный zcytor17lig/субстрат/кофактор/и т.д.) и/или антагонисты обладают большими возможностями применения как in vitro, так и in vivo. Соединения, признанные агонистами zcytor17lig, пригодны для стимуляции размножения, пролиферации, активации и дифференцировки клеток и/или для индукции или ингибирования функций специализированных клеток, участвующих в гомеостазе гемопоэза и иммунной функции. Например, zcytor17lig и его агонисты можно использовать в качестве компонентов культуральных сред определенного состава отдельно или в комбинации с другими цитокинами и гормонами для замены сыворотки, которая обычно используется в культуре клеток. Таким образом, агонисты пригодны для специфической стимуляции роста и/или развития Т-клеток, В-клеток, моноцитов/макрофагов, NK-клеток, цитотоксических лимфоцитов и других клеток лимфоидной и миелоидной линий в культуре.
Антагонисты можно также использовать в качестве реагентов для исследования сайтов взаимодействия лиганд-рецептор. Антагонисты пригодны для ингибирования размножения, пролиферации, активации и/или дифференцировки клеток, участвующих в регуляции гемопоэза. Ингибиторы активности zcytor17lig (антагонисты zcytor17lig) включают в себя антитела против zcytor17lig и растворимые рецепторы zcytor17lig, а также другие пептидные и непептидные вещества (включая рибозимы).
Zcytor17lig можно также использовать для идентификации ингибиторов (антагонистов) его активности. Испытуемые соединения добавляют в анализируемую смесь для идентификации соединений, ингибирующих активность zcytor17lig. Помимо анализов, рассмотренных в данном описании, образцы могут быть испытаны на ингибирование активности zcytor17lig в ряде анализов, предназначенных для измерения связывания с рецептором, стимуляции/ингибирования zcytor17lig-зависимых реакций клеток или пролиферации клеток, экспрессирующих рецептор zcytor17.
Полипептид zcytor17lig может быть экспрессирован, будучи слитым с константной областью тяжелой цепи иммуноглобулина, обычно с фрагментом Fc, который содержит два домена константной области и не имеет вариабельной области. Способы осуществления такого слияния описаны в патентах США №№ 5155027 и 5567584. Такие слитые полипептиды обычно секретируются в виде многомерных молекул, в которых части Fc связаны друг с другом дисульфидными связями и два неиммуноглобулиновых полипептида расположены рядом друг с другом. Слитые полипептиды подобного типа можно использовать, например, для димеризации, повышения устойчивости и увеличения периода полужизни in vivo, для аффинной очистки лиганда, в качестве средства выполнения анализа in vitro или антагониста. Применяемые в анализах химеры связывают с носителем при помощи области Fc и используют в формате ELISA.
Zcytor17lig-связывающий полипептид можно также использовать для очистки лиганда. Данный полипептид иммобилизуют на твердом носителе, таком как агарозные шарики, сшитая агароза, стекло, целлюлозные смолы, смолы на основе кремнезема, полистирол, сшитый полиакриламид или подобные материалы, которые являются устойчивыми в условиях применения. Методы связывания полипептидов с твердыми носителями известны в данной области и включают в себя химию аминов, активацию бромцианом, активацию N-гидроксисукцинимидом, активацию эпоксидом, активацию сульфгидрилом и активацию гидразидом. Полученную среду обычно конфигурируют в форме колонки, и текучие среды, содержащие лиганд, пропускают через указанную колонку один или несколько раз для связывания лиганда с полипептидом рецептора. Затем лиганд элюируют, изменяя концентрацию соли, используя хаотропные агенты (HCl гуанидина) или изменяя рН для разрушения связи лиганд-рецептор.
Можно успешно использовать систему анализа на основе лиганд-связывающего рецептора (или антитела, одного члена пары комплемент/антикомплемент) или его связывающего фрагмента при выполнении измерения в коммерчески производимом биосенсорном аппарате (BIAcore, Pharmacia Biosensor, Piscataway, NJ). Такой рецептор, антитело, член пары комплемент/антикомплемент или фрагмент иммобилизуют на поверхности рецепторного чипа. Применение указанного аппарата описано в статьях Karlsson, J. Immunol. Methods 145:229-40, 1991 и Cunningham and Wells, J. Mol. Biol. 234:554-63, 1993. Рецептор, антитело, член пары или фрагмент ковалентно связывают при помощи амина или сульфгидрила с декстрановыми волокнами, которые присоединены к золотой пленке в проточной ячейке. Испытуемый образец пропускают через указанную ячейку. Если в образце присутствует лиганд, эпитоп или противоположный член пары комплемент/антикомплемент, он связывается соответственно с иммобилизованным рецептором, антителом или другим членом пары, вызывая изменение показателя преломления среды, которое определяется как изменение резонанса поверхностного плазмона золотой пленки. Данная система позволяет определить начальные и конечные значения интенсивности, на основании которых можно вычислить сродство связывания и оценить стехиометрию связывания. Альтернативно, связывание лиганд/рецептор можно анализировать методом SELDI(TM) (Ciphergen, Inc., Palo Alto, CA).
Полипептиды лиганд-связывающего рецептора можно также использовать в других системах анализа, известных в данной области. Такие системы включают в себя анализ Скатчарда для определения сродства связывания (см. Scatchard, Ann. NY Acad. Sci. 51:660-72, 1949) и калориметрические анализы (Cunningham et al., Science 253:545-48, 1991; Cunningham et al., Science 245:821-25, 1991).
Полипептиды zcytor17lig можно также использовать для получения антител, связывающихся с эпитопами, пептидами или полипептидами zcytor17lig. Полипептид zcytor17lig или его фрагмент служит в качестве антигена (иммуногена) для инокуляции животного и индукции иммунной реакции. Такие антитела можно использовать для блокирования биологического действия провоспалительного zcytor17lig и в качестве противовоспалительных средств при лечении различных заболеваний, представленных в данном описании. Специалисту должно быть известно, что антигенные, эпитопнесущие полипептиды имеют последовательность, содержащую по меньшей мере 6, предпочтительно по меньшей мере 9 и более предпочтительно по меньшей мере 15-30 смежных аминокислотных остатков полипептида zcytor17lig (например, SEQ ID NO:2). Полипептиды, содержащие большую часть полипептида zcytor17lig, то есть от 30 до 100 остатков и до полной длины аминокислотной последовательности, входят в объем настоящего изобретения. Антигены или иммуногенные эпитопы могут также включать в себя присоединенные метки, адъюванты, наполнители и носители, описанные в данном описании. Приемлемые антигены включают в себя полипептид zcytor17lig, кодированный SEQ ID NO:2 от аминокислоты 24 до аминокислоты 164 или фрагментом, содержащим смежные аминокислоты 9-141. Другие приемлемые антигены включают в себя непроцессированный и зрелый полипептид zcytor17lig, спирали А-D и одну или несколько спиралей А, В, С и D описанной четырехспиральной структуры zcytor17lig. Предпочтительные пептиды, предназначенные для использования в качестве антигенов, являются гидрофильными пептидами, которые могут быть созданы специалистом на основании описанного графика гидрофобности с использованием, например, аминокислотных остатков 114-119, 101-105, 126-131, 113-118 и 158-162 SEQ ID NO:2 и аминокислотных остатков 34-39, 46-51, 131-136, 158-163 и 157-162 SEQ ID NO:11. Кроме того, антигенные эпитопы zcytor17lig, прогнозируемые на основании графика Джеймсона-Вульфа, например, при помощи программы Protean компании DNASTAR (DNASTAR, Inc., Madison, WI), могут служить предпочтительными антигенами и могут быть легко определены специалистом.
Антитела, образующиеся в результате иммунной реакции, возникающей после инокуляции животного указанными антигенами, можно выделить и очистить описанными методами. Методы получения и выделения поликлональных и моноклональных антител хорошо известны в данной области. См., например, Current Protocols in Immunology, Cooligan, et al. (eds.), National Institutes of Health, Jоhn Wiley and Sons, Inc. 1995; Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, NY, 1989; and Haurell, J.G.R., Ed., Monoclоnal Hybridoma Antibodies: Techniques and Applications, CRC Press, Inc. Boca Raton, FL, 1982.
Как должно быть очевидно специалисту, поликлональные антитела могут быть получены в результате инокуляции различных теплокровных животных, таких как лошади, коровы, козы, овцы, собаки, цыплята, кролики, мыши и крысы, полипептидом zcytor17lig или его фрагментом. Иммуногенность полипептида zcytor17lig можно увеличить, используя адъювант, такой как квасцы (гидроксид алюминия), полный или неполный адъювант Фрейнда. Полипептиды, пригодные для иммунизации, включают в себя также слитые полипептиды, например, полученные в результате слияния zcytor17lig или его части с полипептидом иммуноглобулина или связывающим мальтозу белком. Полипептидный иммуноген может быть непроцессированной молекулой или ее частью. Если часть полипептида является «гаптенподобной», такую часть можно успешно присоединить или связать с макромолекулярным носителем (таким, как гемоцианин блюдечка (KLH), альбумин бычьей сыворотки (BSA) или тетанусный токсоид) и использовать для иммунизации.
В используемом здесь значении термин «антитела» включает в себя поликлональные антитела, аффинно очищенные поликлональные антитела, моноклональные антитела и антигенсвязывающие фрагменты, такие как протеолитические F(ab')2 и Fab-фрагменты. В объем данного изобретения входят генетически созданные интактные антитела или их фрагменты, такие как химерные антитела, Fv-фрагменты, одноцепочечные антитела и тому подобное, а также синтетические антигенсвязывающие пептиды и полипептиды. Антитела, отличные от человеческих, могут быть гуманизированы путем трансплантации CDR, отличных от человеческих, в остов и константные области человека или включения всех вариабельных доменов, отличных от человеческих (необязательно «маскируя» их поверхностью, подобной человеческой структуре, путем замены доступных остатков, в результате чего образуется «фанерованное» антитело). В некоторых случаях гуманизированные антитела могут сохранять отличные от человеческих остатки в доменах остова вариабельной области человека для усиления требуемых характеристик связывания. В результате гуманизации антител можно увеличить биологический период полужизни и уменьшить вероятность вредных иммунных реакций при введении человеку. Кроме того, человеческие антитела могут быть продуцированы в трансгенных животных, отличных от человека, которые были созданы для сохранения генов иммуноглобулина человека, как это описано в публикации WIPO № WO 98/24893. Желательно, чтобы эндогенные гены иммуноглобулина в указанных животных были инактивированы или удалены, например, методом гомологичной рекомбинации.
Считается, что антитела специфически связываются, если они 1) обладают пороговым уровнем связывающей активности и 2) по существу перекрестно не реагируют с молекулами родственных полипептидов. Пороговый уровень связывания определяется, если антитела против zcytor17lig связываются с полипептидом, пептидом или эпитопом zcytor17lig со степенью сродства, которая по меньшей мере в 10 раз выше сродства связывания с контрольным полипептидом (не являющимся zcytor17lig). Желательно, чтобы антитела характеризовались сродством связывания (Ka), равным 106 М-1 или выше, предпочтительно 107 М-1 или выше, более предпочтительно 108 М-1 или выше и наиболее предпочтительно 109 М-1 или выше. Специалист может легко определить сродство связывания антитела, например, при помощи анализа Скатчарда (Scatchard, G., Ann. NY Acad. Sci. 51:660-672, 1949).
То что антитела против zcytor17lig по существу перекрестно не реагируют с молекулами родственных полипептидов, можно определить стандартным анализом методом вестерн-блоттинга, при выполнении которого данное антитело обнаруживает полипептид zcytor17lig, а не другие известные родственные полипептиды (Ausubel et al., там же). Примеры известных родственных полипептидов включают в себя полипептиды, ранее описанные в научной литературе, в частности, известные ортологи, паралоги и подобные члены семейства белков. Для скрининга можно также использовать zcytor17lig, отличный от человеческого, и мутантные полипептиды zcytor17lig. Кроме того, антитела могут быть «исследованы в отношении» известных родственных полипептидов с целью отбора популяции, которая специфически связывается с полипептидами zcytor17lig. Например, антитела к zcytor17lig адсорбируют на родственных полипептидах, сцепленных с нерастворимой матрицей; антитела, специфически взаимодействующие с zcytor17lig, будут проходить через матрицу в условиях применения соответствующего буфера. В результате скрининга можно отобрать поликлональные и моноклональные антитела, перекрестно не реагирующие с известными близкородственными полипептидами (Antibodies: A Laboratory Manual, Harlow and Lane (eds.), Cold Spring Harbor Laboratory Press, 1988; Current Protocols in Immunology, Cooligan, et al. (eds.), National Institutes of Health, Jоhn Wiley and Sons, Inc., 1995). Методы скрининга и отбора специфических антител хорошо известны в данной области. См. Fundamental Immunology, Paul (eds.), Raven Press, 1993; Getzoff et al., Adv. in Immunol. 43:1-98, 1988; Monoclonal Antibodies: Principles and Practice, Goding, J.W (eds.), Academic Press Ltd., 1996; Benjamin et al., Ann. Rev. Immunol. 2:67-101, 1984. Специфически связывающиеся антитела против zcytor17lig можно обнаружить рядом методов, известных в данной области и описанных ниже.
Для обнаружения антител, связывающихся с белками или полипептидами zcytor17lig, можно использовать различные анализы, известные специалистам. Типичные анализы подробно описаны в справочнике Antibodies: A Laboratory Manual, Harlow and Lane (Eds.), Cold Spring Harbor Laboratory Press, 1988. Типичные примеры таких анализов включают: одновременный иммуноэлектрофорез, радиоиммуноанализ, радиоиммунопреципитацию, твердофазный иммуноферментный анализ (ELISA), анализ методом дот-блоттинга или вестерн-блоттинга, анализ методом конкурентного ингибирования и анализ методом иммуносэндвича. Кроме того, антитела можно исследовать на связывание с мутантным белком или полипептидом zcytor17lig по сравнению с белком или полипептидом дикого типа.
Антитела к zcytor17lig можно использовать для мечения клеток, экспрессирующих zcytor17lig; для выделения zcytor17lig путем аффинной очистки; для диагностических анализов с целью определения уровней полипептидов zcytor17lig в кровотоке; для обнаружения или количественного определения растворимого zcytor17lig в качестве маркера определенной патологии или заболевания; в аналитических методах с использованием FACS; для скрининга библиотек экспрессируемых последовательностей; для создания антиидиотипических антител и в качестве нейтрализующих антител или антагонистов, блокирующих активность zcytor17lig in vitro и in vivo. Приемлемые прямые маркеры или метки включают в себя радиоизотопы, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные маркеры, хемилюминесцентные маркеры, магнитные частицы и тому подобное; непрямые маркеры или метки могут включать в себя использование пары биотин-авидин или других пар комплемент/антикомплемент в качестве промежуточных продуктов. Антитела могут быть также конъюгированы с лекарственными средствами, токсинами, радиоизотопами и подобными прямыми или непрямыми методами, и указанные конъюгаты могут быть использованы для диагностических или терапевтических применений in vivo. Кроме того, антитела к zcytor17lig или его фрагментам могут быть использованы in vitro для обнаружения денатурированного zcytor17lig или его фрагментов при выполнении таких анализов, как вестерн-блоттинг или другие анализы, известные в данной области.
Приемлемые детектируемые молекулы, которые могут быть присоединены к полипептиду или антителу прямыми или непрямыми методами, включают в себя радиоизотопы, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные маркеры, хемилюминесцентные маркеры, магнитные частицы и тому подобное. Приемлемые цитотоксические молекулы, которые могут быть присоединены к полипептиду или антителу прямыми или непрямыми методами, включают в себя бактериальные или растительные токсины (например, дифтерию, токсин, сапорин, экзотоксин Pseudomonas, рицин, абрин и тому подобное), а также терапевтические радиоизотопы, такие как иод-131, рений-188 или иттрий-90 (прямо присоединенные к полипептиду или антителу или непрямо присоединенные, например, при помощи хелатобразующей части). Полипептиды или антитела могут быть также конъюгированы с цитотоксическими лекарственными средствами, такими как адриамицин. Для непрямого присоединения детектируемой или цитотоксической молекулы указанная детектируемая или цитотоксическая молекула может быть конъюгирована с одним членом пары комплемент/антикомплемент, в которой второй член связан с полипептидом и антителом. Для указанных целей биотин/стрептавидин является типичной парой комплемент/антикомплемент.
Связывающие полипептиды могут также действовать в качестве «антагонистов» zcytor17lig, блокируя связывание zcytor17lig или передачу сигналов in vitro и in vivo. Указанные связывающие полипептиды против zcytor17lig пригодны для ингибирования активности zcytor17lig или связывания с белком.
Белки, полученные в результате слияния полипептида и токсина, или белки, полученные в результате слияния антитела и токсина, можно использовать для ингибирования или уничтожения клеток- или тканей-мишеней (например, для воздействия на раковые клетки или ткани). Альтернативно, если полипептид имеет несколько функциональных доменов (то есть домен активации, домен связывания с рецептором и домен направленной доставки), слитый белок, содержащий только домен направленной доставки, может быть пригоден для направления детектируемой молекулы, цитотоксической молекулы или комплементарной молекулы в представляющую интерес клетку или ткань. В тех случаях, когда домен слитого белка включает в себя комплементарную молекулу, антикомплементарная молекула может быть конъюгирована с детектируемой или цитотоксической молекулой. Такие белки, полученные в результате слияния домена и комплементарной молекулы, содержат генетический носитель для направленной клетко- или тканеспецифической доставки генетических конъюгатов антикомплемент-детектируемой/цитотоксической молекулы.
В другом варианте осуществления изобретения слитые белки zcytor17lig и цитокина или слитые белки антитела и цитокина можно использовать для уничтожения in vivo тканей-мишеней (например, в случае лейкоза, лимфомы, рака легкого, рака ободочной кишки, меланомы, рака поджелудочной железы, рака яичника, рака кожи, крови и костного мозга или других видов рака, в которых экспрессированы рецепторы zcytor17lig) (см., например, Hornick et al., Blood 89:4437-47, 1997). Описанные слитые белки могут направленно доставлять цитокин к требуемому участку воздействия, увеличивая таким образом локальную концентрацию цитокина. Приемлемые полипептиды zcytor17lig или антитела против zcytor17lig направленно воздействуют на нежелательную клетку или ткань (то есть на опухоль или лейкоз), при этом слитый цитокин лучше опосредует лизис клетки-мишени эффекторными клетками. Цитокины, пригодные для указанной цели, включают интерлейкин 2 и колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF).
В другом варианте осуществления изобретения полипептид zcytor17lig или антитело против zcytor17lig, направленно воздействующие на васкулярные клетки или ткани, могут быть конъюгированы с радиоизотопом, в частности с бета-излучающим радиоизотопом, для уменьшения рестеноза. Такой терапевтический подход создает меньшую опасность для врачей, занимающихся лучевой терапией. Например, узкие ленты, пропитанные иридием-192, вводимые в суженные сосуды пациентов до достижения требуемой дозы облучения, вызывали замедление роста тканей в сосуде и увеличение диаметра просвета по сравнению с контрольной группой, которой вводили индифферентные узкие ленты. Кроме того, в испытуемой группе наблюдалась значительно меньшая реваскуляризация и тромбоз, вызванный сужением сосудов. Достижение подобных результатов можно прогнозировать при направленной доставке биологически активного конъюгата, содержащего описанный радиоизотоп.
Описанные биологически активные конъюгаты полипептида или антитела могут быть доставлены к месту действия путем внутривенного, внутриартериального или внутрипротокового введения либо могут быть введены локально непосредственно в предполагаемое место действия.
Кроме того, воспаление является защитной реакцией организма, направленной на уничтожение проникающего агента. Воспаление является каскадно развивающимся событием, в котором участвуют многие клеточные и гуморальные медиаторы. С одной стороны, подавление воспалительных реакций может поставить под угрозу иммунитет хозяина; однако при отсутствии контроля воспаление может привести к серьезным осложнениям, включая хронические воспалительные заболевания (например, ревматоидный артрит, рассеянный склероз, воспалительное заболевание кишечника и тому подобное), септический шок и расстройство многих органов. Важно отметить, что указанные заболевания обусловлены общими воспалительными медиаторами. Все воспалительные процессы являются важной причиной заболеваемости и смертности людей. Поэтому совершенно очевидно, что противовоспалительные антитела и связывающие полипептиды, такие как антитела против zcytor17lig и описанные здесь связывающие полипептиды, могут иметь важное терапевтическое значение для лечения целого ряда заболеваний у человека и животных, начиная с астмы и аллергии и кончая аутоиммунными заболеваниями и септическим шоком. Поэтому противовоспалительные антитела против zcytor17lig и описанные связывающие полипептиды могут найти широкое терапевтическое применение в качестве антагонистов zcytor17lig при лечении таких заболеваний как артрит, эндотоксемия, воспалительное заболевание кишечника, псориаз, родственные заболевания и тому подобное.
1. Артрит
Артрит, в том числе остеоартрит, ревматоидный артрит, артрит суставов вследствие травмы и подобные заболевания, являются обычными воспалительными заболеваниями, при лечении которых могут быть успешно использованы противовоспалительные антитела и связывающие полипептиды, такие как антитела против zcytor17lig и связывающие полипептиды согласно изобретению. Например, ревматоидный артрит (RA) является системным заболеванием, которое поражает все тело, и является одной из наиболее распространенных форм артрита. Данное заболевание характеризуется воспалением выстилки оболочки сустава, которое вызывает боль, тугоподвижность, повышение температуры, покраснение и опухание. Воспалительные клетки высвобождают ферменты, которые могут поражать кость и хрящ. В результате ревматоидного артрита выстилка воспаленного сустава, синовиальная оболочка, может проникать в кость и хрящ и разрушать их, вызывая ухудшение функции сустава и сильную боль наряду с другими физиологическими симптомами. Пораженный сустав теряет свою форму и искривляется, что вызывает боль и утрату подвижности.
Ревматоидный артрит (RA) является иммуноопосредуемым заболеванием, которое прежде всего характеризуется воспалением и последующим разрушением ткани, что ведет к потере трудоспособности и увеличению смертности. В ревматоидных суставах продуцируются различные цитокины. Многочисленные исследования показали, что IL-1 и TNF-альфа, два прототипных провоспалительных цитокина, играют важную роль в механизмах, определяющих синовиальное воспаление и прогрессирующее разрушение сустава. Введение ингибиторов TNF-альфа и IL-1 субъектам, страдающим ревматоидным артритом, вызывало значительное улучшение клинических и биологических симптомов воспаления и уменьшение радиологических признаков эрозии кости и разрушения хряща. Однако, несмотря на обнадеживающие результаты, значительное число субъектов не реагируют на указанные средства, из чего следует, что патологическую физиологию артрита определяют также и другие медиаторы (Gabay, Expert. Opin. Biol. Ther. 2(2):135-149, 2002). Одним из указанных медиаторов может быть zcytor17lig, поэтому молекулы, которые связывает или ингибирует zcytor17lig, в частности антитела против zcytor17lig или связывающие партнеры могут быть ценным лечебным средством, уменьшающим воспаление в случае ревматоидного артрита и других обусловленных артритом заболеваний.
В данной области известно несколько животных моделей ревматоидного артрита. Например, в модели коллаген-индуцированного артрита (CIA) у мышей развивается хронический воспалительный артрит, который близко напоминает ревматоидный артрит человека. Так как CIA характеризуется такими же иммунологическими и патологическими симптомами, что и RA, данная модель является идеальной для поиска потенциальных противовоспалительных соединений для человека. Модель CIA является хорошо известной моделью с использованием мышей, в которой проявляется как иммунная реакция, так и воспалительная реакция. Иммунная реакция включает в себя взаимодействие В-клеток и CD4+ Т-клеток в ответ на воздействие коллагена, вводимого в виде антигена, который вызывает образование антител против коллагена. Воспалительная фаза является результатом реакции тканей, вызываемой медиаторами воспаления, вследствие перекрестной реакции некоторых из указанных антител с нативным коллагеном мыши и активации пути стимуляции комплемента. Преимуществом использования модели CIA является знание основных механизмов патогенеза. Идентифицированы родственные эпитопы Т-клеток и В-клеток в коллагене типа II и определены различные иммунологические (например, гиперчувствительность замедленного типа и антитело против коллагена) и воспалительные (например, цитокины, хемокины и разрушающие матрикс ферменты) параметры, относящиеся к иммуноопосредуемому артриту, благодаря чему указанную модель CIA можно использовать для оценки эффективности испытуемых соединений (Wooley, Curr. Opin. Rheum. 3:407-20, 1999; Williams et al., Immunol. 89:9784-788; Myers et al., Life Sci. 61:1861-78, 1997; and Wang et al., Immunol. 92:8955-959, 1995).
Полипептиды, содержащие растворимый рецептор zcytor17 (включая описанные гетеродимерные и многомерные рецепторы), такие как zcytor17-Fc4 или другие растворимые и слитые белки zcytor17, вводили мышам, используемым в модели CIA, для оценки способности используемого zcytor17 ослаблять симптомы и замедлять развитие болезни. В качестве молекулы, модулирующей иммунную и воспалительную реакцию, zcytor17lig может индуцировать продуцирование SAA, участвующей в патогенезе ревматоидного артрита, при этом антагонисты zcytor17lig могут ослаблять активность SAA in vitro и in vivo; системное или местное введение антагонистов zcytor17lig, таких как антитела против zcytor17lig или связывающие партнеры, полипептиды, содержащие zcytor17 (включая описанные гетеродимерные и многомерные рецепторы), такие как zcytor17-Fc4 или другие растворимые и слитые белки zcytor17, могут подавлять воспалительную реакцию в случае ревматоидного артрита. Другие потенциальные лекарственные средства включают в себя полипептиды zcytor17, полипептиды растворимых гетеродимерных и многомерных рецепторов, антитела против zcytor17lig или связывающие партнеры согласно изобретению и т.п.
2. Эндотоксемия
Эндотоксемия является тяжелым заболеванием, вызываемым инфекционными микроорганизмами, такими как бактерии, и другими инфекционными веществами, вызывающими сепсис, токсический шок, или условно-патогенными инфекциями у субъектов с ослабленной иммунной системой и тому подобными причинами. Терапевтически полезные противовоспалительные антитела и связывающие полипептиды, такие как антитела против zcytor17lig и связывающие полипептиды согласно изобретению, могут предотвращать и лечить эндотоксемию у людей и животных. Другие потенциальные лекарственные средства, включая полипептиды zcytor17, полипептиды растворимых гетеродимерных и многомерных рецепторов, антитела против zcytor17lig или связывающие партнеры согласно изобретению и т.п., могут быть ценными лекарственными средствами для уменьшения воспаления и патологических воздействий в случае эндотоксемии.
Эндотоксемия, вызванная липополисахаридом (LPS), в случае которой задействованы многие провоспалительные медиаторы, оказывающие патологическое действие на инфекционные заболевания и LPS-обусловленную эндотоксемию у грызунов, является широко используемой и приемлемой моделью для исследования фармакологических эффектов потенциальных провоспалительных или иммуномодулирующих средств. LPS, продуцируемый грамотрицательными бактериями, является главной причиной патогенеза септического шока (Glausner et al., Lancet 338:732, 1991). Шокоподобное состояние можно экспериментально вызвать у животных при помощи одной инъекции LPS. Молекулы, продуцируемые клетками под действием LPS, могут осуществлять прямую или непрямую направленную доставку патогенов. Хотя указанные биологические реакции защищают хозяина от проникновения патогенов, они сами могут причинять вред. Так, сильная стимуляция природного иммунитета, происходящая в результате сильной инфекции грамотрицательными бактериями, вызывает чрезмерное продуцирование цитокинов и других молекул и развитие летального синдрома и септического шока, которые сопровождаются лихорадкой, гипотензией, свертыванием крови в сосудах и расстройством многих органов (Dumitru et al., Cell 103:1071-1083, 2000).
Указанные токсические действия LPS связаны главным образом с активацией макрофагов, ведущей к высвобождению множества воспалительных медиаторов. Среди указанных медиаторов TNF, по-видимому, играет решающую роль, о чем свидетельствует предотвращение токсичности LPS в результате введения нейтрализующих антител против TNF (Beutler et al., Science 229:869, 1985). Установлено, что 1 мкг инъекционного раствора LPS E. coli мыши С57В1/6 вызывает значительное увеличение в кровотоке IL-6, TNF-альфа, IL-1 и белков острой фазы (например, SAA) примерно через 2 часа после инъекции. Токсичность LPS, по-видимому, опосредована указанными цитокинами, так как пассивная иммунизация против указанных медиаторов может способствовать снижению смертности (Beutler et al., Science 229:869, 1985). Возможная стратегия иммуноинтервенции для профилактики и/или лечения септического шока включает в себя введение mAb против TNF, антагониста рецептора IL-1, LIF, IL-10 и G-CSF. Так как LPS индуцирует продуцирование провоспалительных факторов, которые, по-видимому, усиливают патологию эндотоксемии, нейтрализация активности zcytor17lig, SAA или других противовоспалительных факторов в результате антагонистического воздействия на полипептид zcytor17lig может уменьшить симптомы эндотоксемии, характерные для эндотоксического шока. Другие потенциальные лекарственные средства включают в себя полипептиды zcytor17, полипептиды растворимых гетеродимерных и многомерных рецепторов, антитела против zcytor17lig или связывающие партнеры согласно изобретению и подобные средства.
3. Воспалительное заболевание кишечника (IBD)
В Соединенных Штатах примерно 500000 человек страдают воспалительным заболеванием кишечника (IВD), которое может поражать ободочную или прямую кишку (неспецифический язвенный колит) или обе вместе, тонкий и толстый кишечник (болезнь Крона). Патогенез указанных заболеваний неясен, но он характеризуется хроническим воспалением пораженных тканей. Потенциальные лекарственные средства, включая полипептиды zcytor17, полипептиды растворимых гетеродимерных и многомерных рецепторов, антитела против zcytor17lig или связывающие партнеры согласно изобретению и т.п., могут уменьшать воспалительные и патологические симптомы в случае IBD и родственных заболеваний.
Неспецифический язвенный колит (UC) является воспалительным заболеванием толстой кишки, обычно именуемой ободочной кишкой, которое характеризуется воспалением и образованием язв в слизистой оболочке или внутренней выстилке ободочной кишки. Подобное воспаление вызывает частое опорожнение ободочной кишки, ведущее к диарее. Симптомы данного заболевания включают в себя частый жидкий стул, сопровождающийся спазмами брюшной полости, лихорадкой и потерей веса. Хотя точная причина возникновения UC неизвестна, недавно выполненные исследования позволяют предположить, что данное заболевание является естественной защитой организма против белков, которые данный организм считает чужеродными («аутоиммунная реакция»). Поскольку данные белки напоминают бактериальные белки в кишечнике, они, вероятно, вызывают или стимулируют воспалительный процесс, который начинает разрушать выстилку ободочной кишки. По мере разрушения выстилки ободочной кишки образуются язвы, выделяющие слизь, гной и кровь. Данное заболевание обычно начинается в ректальной области и постепенно распространяется по всему толстому кишечнику. Повторные вспышки воспаления ведут к утолщению стенки кишечника и прямой кишки рубцовой тканью. Тяжелая форма заболевания может сопровождаться омертвением ткани ободочной кишки или сепсисом. Симптомы неспецифического язвенного колита могут характеризоваться разной степенью тяжести, и их возникновение может быть постепенным или внезапным. Приступы могут быть вызваны многими факторами, включая респираторные инфекции или стресс.
Хотя в настоящее время UC является неизлечимым заболеванием, лечение направлено на подавление аномального воспалительного процесса в выстилке ободочной кишки. Лечение с участием иммуносупрессивных кортикостероидов (например, азатиоприна, меркаптопурина и метотрексата) и аминосалицилатов позволяет ослабить симптомы данного заболевания. Однако длительный прием иммуносупрессоров, таких как кортикостероиды и азатиоприн, может вызвать серьезные побочные эффекты, включая истончение костей, возникновение катаракты, инфекцию и поражение печени и костного мозга. Для пациентов, не восприимчивых к такому лечению, единственным выходом является хирургическая операция. Хирургическая операция связана с удалением всей ободочной и прямой кишки.
Существует несколько животных моделей, которые могут частично имитировать хронический неспецифический язвенный колит. Наиболее широко используемой моделью является колит, вызванный 2,4,6-тринитробензолсульфоновой кислотой/этанолом (TNBS), в результате чего возникает хроническое воспаление и образуются язвы в ободочной кишке. Введение TNBS в ободочную кишку чувствительных мышей при помощи ректальной инстилляции вызывает опосредуемую Т-клетками иммунную реакцию в слизистой оболочке ободочной кишки, что ведет к обширному воспалению слизистой оболочки, характеризующемуся плотной инфильтрацией Т-клеток и макрофагов сквозь стенки толстого кишечника. Кроме того, данная гистопатологическая картина сопровождается клинической картиной снижения веса (истощение), кровавым поносом и утолщением стенок толстого кишечника (Neurath et al., Intern. Rev. Immunol. 19:51-62, 2000).
В другой модели колита использован декстран-сульфат натрия (DSS), который вызывает острый колит, сопровождающийся кровавым поносом, снижением веса, укорочением ободочной кишки и образованием язв в слизистой оболочке при инфильтрации нейтрофилов. DSS-индуцированный колит в гистологическом отношении характеризуется инфильтрацией воспалительных клеток в проприогенный слой, сопровождающейся лимфоидной гиперплазией, поражением кишечных желез и образованием язв в эпителии. Считается, что указанные изменения возникают вследствие токсического воздействия DSS на эпителий и фагоцитоза клеток проприогенного слоя, продуцирования TNF-альфа и IFN-гамма. Несмотря на широкое применение указанной модели, до сих пор не найден ответ на несколько вопросов, касающихся механизмов действия DSS на возникновение данного заболевания у людей. DSS считается моделью, не зависящей от Т-клеток, так как данное заболевание возникает у животных с отсутствием Т-клеток, таких как мыши SCID.
Введение антител против zcytor17lig или связывающих партнеров, в частности полипептидов, содержащих растворимый рецептор zcytor17 (включая гетеродимерные и многомерные рецепторы), такой как zcytor17-Fc4 или другие zcytor17-содержащие растворимые и слитые белки, в модели TNBS или DSS позволяет оценить действие антагонистов zcytor17lig на ослабление симптомов и замедление течения желудочно-кишечного заболевания. Zcytor17lig может играть определенную роль при возникновении воспалительной реакции в случае колита, поэтому нейтрализация активности zcytor17lig благодаря введению антагонистов zcytor17lig является возможным подходом к лечению IBD. Другие потенциальные лекарственные средства включают в себя полипептиды zcytor17, полипептиды растворимых гетеродимерных и многомерных рецепторов, антитела против zcytor17lig или связывающие партнеры согласно изобретению и т.п.
4. Псориаз
Псориаз является хроническим кожным заболеванием, которым страдают более семи миллионов американцев. Псориаз возникает при аномальном росте кожных клеток, в результате чего образуются воспаленные, опухшие и чешуйчатые участки кожи там, где старая кожа не отслаивается достаточно быстро. Бляшкообразующий псориаз, который является наиболее распространенной формой, характеризуется наличием воспаленных участков кожи («поражений»), покрытых серебристо-белыми чешуйками. Псориаз может быть ограничен несколькими бляшками или может поражать средние и обширные участки кожи, возникая главным образом на черепе, коленях, локтях и туловище. Будучи весьма заметным, псориаз не является заразным заболеванием. Патогенез указанного заболевания включает в себя хроническое воспаление пораженных тканей. Полипептиды zcytor17, полипептиды растворимых гетеродимерных и многомерных рецепторов, антитела против zcytor17lig или связывающие партнеры согласно изобретению и тому подобное могут быть ценным лекарственным средством для уменьшения воспалительных и патологических симптомов в случае псориаза, других воспалительных заболеваний кожи, аллергических заболеваний кожи и слизистой оболочки и родственных заболеваний.
Псориаз является опосредуемым Т-клетками воспалительным заболеванием кожи, которое может причинять значительные страдания. Данное заболевание является неизлечимым и поражает людей всех возрастов. Псориазом страдают примерно два процента населения Европы и Северной Америки. Хотя субъекты, страдающие легкой формой псориаза, часто могут контролировать течение болезни при помощи местных средств, более одного миллиона человек во всем мире нуждаются в ультрафиолетовой или системной иммуносупрессорной терапии. К сожалению, неудобство и опасность ультрафиолетового облучения и токсичность многих лекарственных средств ограничивают длительность их применения. Кроме того, у больных псориазом обычно возникают рецидивы заболевания и в некоторых случаях в течение короткого времени после прекращения иммуносупрессорной терапии.
Дифференцировка является прогрессирующим и динамичным процессом, который начинается с плюрипотентных стволовых клеток и заканчивается окончательно дифференцированными клетками. Плюрипотентные стволовые клетки, способные регенерировать без коммитирования с образованием линии клеток, экспрессируют совокупность маркеров дифференцировки, которые утрачиваются после завершения образования линии клеток. Клетки-предшественники экспрессируют совокупность маркеров дифференцировки, которые сохраняются или не сохраняются по мере созревания клеток, образующих линию клеток. Маркеры дифференцировки, экспрессируемые исключительно зрелыми клетками, обычно определяют функциональные свойства, такие как клеточные продукты, ферменты, необходимые для продуцирования клеточных продуктов, и рецепторы. Стадию дифференцировки популяции клеток обычно контролируют, идентифицируя маркеры, присутствующие в популяции клеток.
Вышеизложенное позволяет предположить, что факторы, стимулирующие образование клеток специфических типов вплоть до конечной дифференцировки или дедифференцировки, воздействуют на всю популяцию клеток, образующихся из общей клетки-предшественника или стволовой клетки. Таким образом, настоящее изобретение относится к стимуляции или ингибированию пролиферации лимфоидных клеток, гемопоэтических клеток и эпителиальных клеток.
Zcytor17lig выделяли из ткани, которая, как известно, выполняет важную иммунологическую функцию и содержит клетки, играющие определенную роль в иммунной системе. Zcytor17lig экспрессирован в CD3+-селектированных и активированных клетках периферической крови, при этом установлено, что экспрессия zcytor17lig возрастает после активации Т-клеток. Кроме того, результаты экспериментов, описанных в нижеследующих примерах, позволяют предположить, что полипептиды согласно изобретению могут влиять на рост/увеличение числа моноцитов/макрофагов, Т-клеток, В-клеток, NK-клеток и/или дифференцированное состояние моноцитов/макрофагов, Т-клеток, В-клеток, NK-клеток или клеток-предшественников указанных клеток. Факторы, которые стимулируют пролиферацию клеток-предшественников гемопоэтических клеток и активируют зрелые клетки, как правило, известны, однако пролиферация и активация могут также потребовать дополнительных факторов роста. Например, установлено, что IL-7 и фактор Стила (лиганд с-kit) необходимы для образования колонии клеток-предшественников NK-клеток. IL-15 + IL-2 в комбинации с IL-7 и фактором Стила были более эффективными (Mrozek et al., Blood 87:2632-2640, 1996). Однако для пролиферации специфических субпопуляций NK-клеток и/или клеток-предшественников NK-клеток могут быть необходимы неидентифицированные цитокины (Robertson et al., Blood 76:2451-2438, 1990). Аналогичным образом zcytor17lig может действовать отдельно, кооперативно или синергично с другими цитокинами для усиления роста, пролиферации и модификации дифференцировки моноцитов/макрофагов, Т-клеток, В-клеток или NK-клеток.
Анализы для измерения дифференцировки включают в себя, например, измерение клеточных маркеров, связанных со стадиеспецифической экспрессией ткани, ферментативной активностью, функциональной активностью или морфологическими изменениями (статьи Watt, FASEB, 5:281-284, 1991; Francis, Differentiation 57:63-75, 1994; Raes, Adv. Anim. Cell Biol. Technol. Bioprocesses, 161-171, 1989, которые включены в данное описание в качестве ссылки). Альтернативно, сам полипептид zcytor17lig может служить в качестве дополнительного поверхностного или секретированного маркера, ассоциированного со стадиеспецифической экспрессией ткани. Таким образом, прямое измерение полипептида zcytor17lig или отсутствия его экспрессии в ткани по мере дифференцировки может служить маркером дифференцировки тканей.
Аналогичным образом, прямое измерение полипептида zcytor17lig или отсутствия его экспрессии в ткани можно произвести в ткани или клетках в процессе развития опухоли. Увеличение инвазии и подвижности клеток, возникновение или отсутствие экспрессии zcytor17lig в предраковом или раковом состоянии по сравнению с нормальной тканью может быть использовано для диагностики трансформации, инвазии и образования метастазов в процессе развития опухоли. Представление о стадии развития опухоли или образования метастазов поможет лечащему врачу выбрать наиболее правильное лечение или интенсивность лечения для данного больного раком. Методы измерения возникновения и отсутствия экспрессии (мРНК или белка) хорошо известны в данной области, рассмотрены в данном описании и могут быть применены к измерению экспрессии zcytor17lig. Например, появление или исчезновение полипептидов, регулирующих подвижность клеток, можно использовать для диагностики и прогнозирования рака предстательной железы (Banyard, J. and Zetter, B.R., Cancer and Metast. Rev. 17:449-458, 1999). В качестве эффектора подвижности клеток возникновение или отсутствие экспрессии zcytor17lig может быть использовано для диагностики рака лимфоидных клеток, В-клеток, эпителиальных, гемопоэтических и других типов клеток.
Кроме того, активность zcytor17lig и его влияние на развитие опухоли и образование метастазов можно измерить in vivo. Было создано несколько сингенных моделей с использованием мышей для исследования влияния полипетидов, соединений или других методов лечения на развитие опухоли. В указанных моделях опухолевые клетки, пассированные в культуре, имплантируют мышам такой же линии, что и донор опухоли. Из указанных клеток развиваются опухоли, обладающие такими же характеристиками, что и у мыши-реципиента, и в некоторых моделях образуются метастазы. Соответствующие модели опухолей, используемые в исследованиях авторами данного изобретения, включают в себя рак легкого Льюиса (АТСС № CRL-1642) и меланому В16 (АТСС № CRL-6323) наряду с прочими раковыми заболеваниями. В указанных моделях обычно используются линии опухолей, сингенные мышам С57BL6/J, которые можно легко культивировать и манипулировать in vitro. Опухоли, образующиеся в результате имплантации указанных линий клеток, способны образовывать метастазы в легком мышей С57BL6/J. Модель рака легкого Льюиса с использованием мышей недавно была использована для идентификации ингибитора развития кровеносных сосудов (O'Reilly MS, et al., Cell 79:315-328, 1994). Мышей С57BL6/J подвергали воздействию экспериментального средства в виде ежедневных инъекций рекомбинантного белка, агониста или антагониста или в виде однократной инъекции рекомбинантного аденовируса. Через три дня после инъекции 105-106 клеток имплантировали под кожу спины. Альтернативно, клетки сами могут быть инфицированы рекомбинантным аденовирусом, экспрессирующим zcytor17lig, до имплантации, в результате чего белок синтезируется в месте образования опухоли или внутриклеточно, а не системно. У мышей обычно возникали видимые опухоли в течение 5 дней. Рост опухолей продолжался в течение 3 недель, на протяжении которых их размер достигал 1500-1800 мм3 в лечебной контрольной группе. Размер опухоли и массу тела тщательно контролировали на протяжении всего эксперимента. Затем мышей умерщвляли, опухоль удаляли и взвешивали вместе с легкими и печенью. Было установлено, что масса легких хорошо соответствует массе опухоли с метастазами. В качестве дополнительного измерения подсчитывали метастазы на поверхности легкого. Иссеченную опухоль, легкие и печень подвергали гистопатологическому исследованию, иммуногистохимическому исследованию и гибридизации in situ, используя методы, известные в данной области и рассмотренные в данном описании. Подобным образом можно оценить влияние экспрессированного полипептида, например zcytor17lig, на способность опухоли использовать кровеносные сосуды и образовывать метастазы. Кроме того, помимо использования аденовируса, имплантированные клетки могут быть временно трансфицированы zcytor17lig. Применение устойчивых трансфектантов zcytor17lig и индуцируемых промоторов для активации экспрессии zcytor17lig in vivo известно в данной области и может быть использовано в данной системе для оценки индукции метастазов под действием zcytor17lig. Кроме того, мышам данной модели может быть непосредственно инъецирован очищенный zcytor17lig или zcytor17lig-кондиционированные среды, которые также могут быть использованы в данной системе. В качестве общей ссылки см. O'Reilly MS, et al., Cell 79:315-328, 1994; и Rusciano D, et al. Murine Models of Lever Metastasis. Invasion Metastasis 14:349-361, 1995.
Zcytor17lig или антитела против zcytor17lig пригодны для предотвращения онкогенеза и поэтому могут быть использованы для лечения рака. Zcytor17lig экспрессирован в активированных Т-клетках, моноцитах и макрофагах и связан с областью хромосомы человека, в которой транслокации характерны для лейкозов. Кроме того, установлено, что zcytor17lig действует через рецептор цитокина, zcytor17, который также экспрессирован в активированных Т-клетках, моноцитах и макрофагах. Избыточная стимуляции активированных Т-клеток, моноцитов и макрофагов полипептидом zcytor17lig может привести к заболеванию человека, такому, например, как рак иммунокомпетентных клеток или другие типы рака. Таким образом, идентификация экспрессии zcytor17lig, полипептидов (например, антител против zcytor17lig), растворимых рецепторов zcytor17 (например, рецептора zcytor17), гетеродимеров (например, zcytor17/OSMRbeta, zcytor17/WSX-1), многомеров (например, zcytоr17/OSMbeta/WSX-1)) или других zcytor17lig-связывающих партнеров может служить в качестве диагностического средства и антагонистического действия на пролиферативную активность zcytor17lig. Указанный лиганд можно вводить в комбинации с другими используемыми средствами, включая обычные химиотерапевтические средства и иммунные модуляторы, такие как альфа-интерферон. Установлено, что альфа/бета-интерфероны позволяют эффективно лечить некоторые лейкозы в животных моделях, и ингибирующее действие альфа-интерферона и zcytor17lig может быть аддитивным.
Считается, что NK-клетки играют главную роль в уничтожении метастатических опухолевых клеток, при этом у субъектов, имеющих метастазы и солидные опухоли, наблюдается снижение активности NK-клеток (Whiteside et al., Curr. Top. Microbiol. Immunol. 230:221-244, 1998). Средство, стимулирующее NK-клетки, было бы весьма полезно для уничтожения опухолей.
Настоящее изобретение относится к способу уменьшения пролиферации опухолевых моноцитов/макрофагов, который включает в себя введение млекопитающему с неоплазией моноцитов/макрофагов композиции, содержащей zcytor17lig или антагонист zcytor17lig, в количестве, достаточном для уменьшения пролиферации опухолевых моноцитов/макрофагов. В других вариантах осуществления изобретения данная композиция может содержать по меньшей мере один другой цитокин. Второй цитокин может быть выбран из группы, содержащей лиганд или фактор поддержания стволовых клеток IL-2, IL-3, IL-12, IL-21, IL-22, IL-15, IL-4, GM-CSF, Flt3.
Настоящее изобретение относится к способу ингибирования активации или дифференцировки моноцитов/макрофагов. Моноциты являются неполностью дифференцированными клетками, которые мигрируют в разные ткани, где они созревают и становятся макрофагами. Макрофаги играют главную роль в иммунной реакции, представляя антиген лимфоцитам, и вспомогательную роль в качестве А-клеток для лимфоцитов, секретируя многочисленные цитокины. Активированные макрофаги могут интернализировать внеклеточные молекулы и в результате активации приобретают повышенную способность уничтожать внутриклеточные микроорганизмы и опухолевые клетки. Активированные макрофаги также стимулируют острые или местные воспаления.
Настоящее изобретение относится также к способу уменьшения пролиферации опухолевых В- или Т-клеток, который включает в себя введение млекопитающему с В- или Т-клеточной неоплазией композиции, содержащей антагонист zcytor17lig, в количестве, достаточном для уменьшения пролиферации опухолевых моноцитов/макрофагов. В других вариантах осуществления изобретения данная композиция может содержать по меньшей мере один другой цитокин, который может быть выбран из группы, содержащей лиганд или фактор поддержания стволовых клеток IL-2, IL-3, IL-12, IL-21, IL-22, IL-15, IL-4, GM-CSF, Flt3. Кроме того, антагонист zcytor17lig может быть слитым белком лиганда и токсина.
Слитый токсин zcytor17lig-сапорин можно использовать для лечения лейкозов и лимфом, что позволяет расширить диапазон лейкозов, поддающихся лечению zcytor17lig. Например, такие лейкозы могут быть вызваны сверхэкспрессией рецепторов zcytor17 (таких как рецептор zcytor17, гетеродимеров (например, zcytor17/OSMRbeta, zcytor17/WSX-1), многомеров (например, zcytor17/OSMRbeta/WSX). Опосредуемая слитым токсином активация рецептора zcytor17, гетеродимеров или многомеров zcytor17 (таких, как zcytor19/OSMRbeta, cytor17/WSX-1 или zcytor19/WSX-1/OSMR) предоставляет два независимых способа ингибирования роста клеток-мишеней, первый из которых идентичен действию, присущему одному лиганду, и второй включает в себя доставку токсина вследствие интернализации рецептора. Паттерн ограниченной экспрессии лимфоидных клеток и моноцитов рецептора zcytor17 позволяет предположить, что пациенты толерантны к воздействию конъюгата лиганд-сапорин.
Когда лечение злокачественных опухолей включает в себя аллогенную трансплантацию костного мозга или стволовых клеток, zcytor17lig может усиливать действие трансплантата против опухоли. Zcytor17lig может стимулировать образование литических NK-клеток из клеток-предшественников костного мозга и пролиферацию моноцитов и макрофагов после активации рецепторов антигенов. Поэтому пересадка пациентам аллогенных трансплантатов костного мозга zcytor17lig может усиливать противоопухолевые реакции при вливании донорских лимфоцитов или без такого вливания.
Распределение в тканях рецепторов к данному цитокину является важным индикатором возможных мест действия такого цитокина. Экспрессия zcytor17 была обнаружена в моноцитах и В-клетках при сильном увеличении экспрессии в результате активации CD3+, CD4+ и CD8+ Т-клеток. Кроме того, в двух линиях моноцитов, в частности ТНР-1 (Tsuchiya et al., Int. J. Cancer 26:171-176, 1980) и U937 (Sundstrom et al., Int. J. Cancer 17:565-577, 1976), были получены положительные результаты в отношении экспрессии zcytor17.
Анализ методом нозерон-блоттинга рецептора WSX-1 позволил обнаружить транскрипты во всех исследованных тканях, при этом более высокие уровни экспрессии были обнаружены в селезенке, тимусе, лимфатическом узле, костном мозге и лейкоцитах периферической крови человека. Кроме того, уровни экспрессии WSX-1 увеличивались при активации Т-клеток.
Как показано в научной литературе, экспрессия OSMR является очень обширной (Mosley et al., JBC 271:32635-32643, 1996). Подобное распределение рецепторов zcytor17, WSX-1 и OSM усиливает роль zcytor17lig в иммунных реакциях, в частности, этому способствует увеличение числа Т-клеток в результате активации или роль моноцитов/макрофагов в иммунной системе.
Таким образом, определенные варианты осуществления настоящего изобретения относятся к применению растворимых гетеродимеров zcytor17/WSX-1/OSMR и zcytor17/OSMR в качестве антагонистов для лечения воспалительных и иммунных заболеваний или таких заболеваний, как панкреатит, диабет типа I (IDDM), рак поджелудочной железы, болезнь Грейвса, воспалительное заболевание кишечника (IBD), болезнь Крона, рак ободочной и тонкой кишки, дивертикулез, аутоиммунное заболевание, сепсис, трансплантация органов или костного мозга; воспаление, вызванное травмой, хирургическим вмешательством или инфекцией; амилоидоз, спленомегалия; реакция «трансплантат против хозяина»; для ингибирования воспаления, супрессии иммунной реакции, уменьшения пролиферации гемопоэтических, иммунокомпетентных, воспалительных или лимфоидных клеток, макрофагов, Т-клеток (включая Th1- и Th2-клетки, CD4+ и CD8+-клетки), супрессии иммунной реакции в отношении патогена или антигена. Кроме того, экспрессия zcytor17 в активированных иммунокомпетентных клетках, таких как активированные CD4+ и CD19+-клетки, показывает, что рецептор zcytor17 может участвовать в защитных иммунных реакциях организма против проникновения чужеродных факторов, таких как микроорганизмы и клеточный дебрис, и может играть определенную роль в иммунных реакциях во время воспаления и образования опухоли. Таким образом, антитела и связывающие партнеры согласно изобретению, которые являются агонистами или антагонистами рецептора zcytor17, такими как zcytor17lig, могут быть использованы для модификации иммунной реакции и воспаления.
Структура zcytor17lig и распределение в тканях свидетельствуют о его участии на ранней стадии образования гемопоэтических клеток или тимоцитов и регуляции иммунной реакции или воспаления. Указанные процессы включают в себя стимуляцию пролиферации и дифференцировки клеток в ответ на связывание одного или нескольких цитокинов с родственными рецепторами. Учитывая распределение в тканях zcytor17lig, можно думать об обширных возможностях применения агонистов (включая природные рецепторы) и антагонистов как in vitro, так и in vivo. Соединения, являющиеся агонистами zcytor17lig, пригодны для стимуляции пролиферации и развития клеток-мишеней in vitro и in vivo. Например, агонисты, zcytor17lig или антитела против zcytor17lig могут быть полезными компонентами культуральных сред определенного состава, которые можно использовать отдельно или в комбинации с другими цитокинами и гормонами для замены сыворотки, обычно применяемой в культуре клеток. Таким образом, агонисты пригодны для стимуляции роста и/или развития или активации в культуре моноцитов, Т-клеток, В-клеток и других клеток лимфоидной и миелоидной линий и гемопоэтических клеток.
Zcytor17lig можно использовать для стимуляции опосредуемого клетками иммунитета и пролиферации лимфоцитов, например, при лечении инфекций, подавляющих иммунную реакцию, включая некоторые вирусные инфекции. Дополнительные применения включают в себя подавление развития опухоли, когда злокачественное перерождение вызывает образование опухолевых клеток, являющихся антигенными. Zcytor17lig можно использовать для индукции цитотоксичности, которая может быть опосредована активацией эффекторных клеток, таких как Т-клетки, NK-клетки (природные клетки-киллеры) или LAK (лимфоидные активированные клетки-киллеры), или для прямой индукции по пути апоптоза. Zcytor17lig можно также использовать для лечения лейкопении путем увеличения уровней пораженных клеток и для увеличения регенерации спектра Т-клеток после трансплантации костного мозга; или для усиления пролиферации или активации моноцитов для диагностических и других целей, описанных в данном описании.
Zcytor17lig может найти применение для супрессии иммунной системы, например, при лечении аутоиммунных заболеваний, включая ревматоидный артрит, рассеянный склероз, сахарный диабет, воспалительное заболевание кишечника, болезнь Крона и т.д. Супрессия иммунной системы может быть также полезна для уменьшения отторжения трансплантатов тканей или органов и при лечении лейкозов или лимфом, обусловленных Т-клетками, В-клетками или моноцитами, и других раковых заболеваний, благодаря чему подавляется пролиферация пораженных клеток. Кроме того, zcytor17lig можно использовать для обнаружения моноцитов, макрофагов и активированных Т-клеток, а также для диагностики таких аутоиммунных заболеваний, особенно в тех случаях, когда происходит увеличение числа или активация моноцитов.
Полипептиды, пептиды и антитела против zcytor17lig можно также использовать в диагностических системах для обнаружения уровней zcytor17lig в кровотоке. В родственном варианте осуществления изобретения антитела или другие агенты, которые специфически связываются с полипептидами zcytor17lig, можно использовать для обнаружения полипептидов zcytor17lig в кровотоке. Повышенные или пониженные уровни полипептидов, являющихся лигандами, могут свидетельствовать о патологических состояниях, включая рак. Полипептиды zcytor17lig могут способствовать развитию патологических процессов и могут быть непрямым маркером опосредуемого заболевания.
Кроме того, zcytor17lig можно использовать для обнаружения или направленного воздействия на его рецепторы в случае некоторых заболеваний. Например, повышенные уровни растворимого рецептора IL-2 в сыворотке человека ассоциируются с целым рядом воспалительных и опухолевых заболеваний, таких как инфаркт миокарда, астма, миастения, ревматоидный артрит, острый Т-клеточный лейкоз, В-клеточная лимфома, хронический лимфолейкоз, рак ободочной кишки, рак молочной железы и рак яичника (Heaney et al., Blood 87:847-857, 1996). Аналогичным образом уровень рецептора zcytor17 повышен в активированных моноцитах, поэтому рецептор zcytor17 и/или его растворимые рецепторы могут быть ассоциированы с воспалительными и опухолевыми заболеваниями и могут служить маркером указанных заболеваний. Следовательно, zcytor17lig, включая цитотоксические конъюгаты, можно использовать для обнаружения или направленного воздействия на такие ткани и заболевания.
Молекулы согласно изобретению могут быть использованы для определения активности моноцитов/макрофагов иммунной системы. Известны методы, позволяющие оценить такую активность. Например, гамма-интерферон (IFNγ) является сильным активатором мононуклеарных фагоцитов. Например, увеличение экспрессии zcytor17 при активации клеток ТНР-1 (АТСС № TIB-202) гамма-интерфероном позволяет предположить, что данный рецептор участвует в активации моноцитов. Моноциты являются не полностью дифференцированными клетками, которые мигрируют в разные ткани, где они созревают и становятся макрофагами. Макрофаги играют главную роль в иммунной реакции, представляя антиген лимфоцитам, и вспомогательную роль в качестве А-клеток для лимфоцитов, секретируя многочисленные цитокины. Макрофаги могут интернализировать внеклеточные молекулы и в результате активации приобретают повышенную способность уничтожать внутриклеточные микроорганизмы и опухолевые клетки. Активированные макрофаги также стимулируют острое или местное воспаление. Кроме того, было установлено, что в ряде заболеваний нарушается функционирование моноцитов-макрофагов. См., например, Johnston, R.B., New Eng. J. Med. 318:747-752, 1998.
Специалисту должно быть понятно, что агонисты рецептора zcytor17, такие как zcytor17lig, имеют важное значение. Например, в научной литературе сообщалось, что угнетенная миграция моноцитов характерна для субъектов, предрасположенных к инфекции, в частности для новорожденных детей, больных, проходящих курс лечения кортикостероидами или иммуносупрессивной терапии, больных сахарным диабетом, лиц с ожогами или больных СПИДом. Агонисты рецептора zcytor17, такие как zcytor17lig, могут повышать способность моноцитов мигрировать и, возможно, предотвращать инфекции у таких субъектов. У субъектов, страдающих хроническим грануломатозом, также наблюдается пониженная активность мононуклеарных фагоцитов. Такое явление вызывает образование подкожных абсцессов, а также абсцессов в печени, легких, селезенке и лимфатических узлах. Агонист рецептора zcytor17, такой как zcytor17lig, может корректировать или устранить фагоцитарный дефект. Кроме того, неправильная цитотоксичность моноцитов наблюдается у больных раком и страдающих синдромом Вискотта-Олдрича (экзема, тромбоцитопения и рецидивирующие инфекции). Активация моноцитов агонистами рецептора zcytor17, такими как zcytor17lig, может способствовать лечению указанных заболеваний. Система моноциты-макрофаги участвует в возникновении нескольких заболеваний, связанных с нарушением липидного обмена (сфинголипидозы), таких как болезнь Гоше. Сопротивляемость инфекциям может быть снижена из-за нарушения функции макрофагов, которую можно восстановить при помощи агонистов рецептора zcytor17, таких как zcytor17lig.
Кроме того, специалисту должно быть понятно, что антагонисты zcytor17lig также имеют важное значение. Например, в случае атеросклеротических поражений одним из первых нарушений является локализация моноцитов/макрофагов в эндотелиальных клетках. Указанные поражения можно предотвратить при помощи антагонистов zcytor17lig. В качестве антагонистов zcytor17lig можно также использовать антитела против zcytor17lig (например, zcytor17lig-нейтрализующее антитело), растворимые рецепторы zcytor17, гетеродимеры, многомеры и zcytor17lig-связывающие партнеры. Кроме того, монобластный лейкоз ассоциирован с рядом клинических нарушений, для которых характерно выделение биологических продуктов макрофага, в частности высокие уровни лизоцима в сыворотке и моче и сильная лихорадка. Кроме того, такие лейкозы характеризуются аномальным увеличением моноцитов. Указанные явления, вероятно, можно предотвратить при помощи вышеописанных антагонистов zcytor17lig. Антитела против zcytor17lig могут быть конъюгированы с такими молекулами, как токсины и цитокины, для направленного уничтожения лейкозных моноцитов.
При помощи методов, известных в данной области, специалист может легко оценить активность агонистов и антагонистов zcytor17lig при лечении описанных здесь заболеваний, воспалений, иммунных заболеваний (например, аутоиммунных заболеваний), рака или инфекции, а также других заболеваний, обусловленных моноцитарными клетками. Кроме того, поскольку zcytor17lig специфически экспрессирован в Т-клетках, макрофагах и моноцитах, и указанные заболевания характеризуются нарушениями в моноцитарных клетках, такими как пролиферация, функционирование, локализация и активация клеток, полинуклеотиды, полипептиды и антитела согласно изобретению можно использовать в качестве диагностического средства для обнаружения таких нарушений моноцитарных клеток и выявления наличия заболевания. Такие методы включают в себя взятие у пациента биологического образца, такого как кровь, слюна или ткань, полученная при помощи биопсии, и его сравнение с нормальным контрольным образцом. Для определения относительных уровней или локализации zcytor17lig, или клеток, экспрессирующих zcytor17lig, то есть моноцитов, в образце, полученном у пациента, по сравнению с нормальным контрольным образцом можно использовать гистологические, цитологические, биохимические методы, проточную цитометрию и другие методы. Изменение уровня (увеличение или уменьшение) экспрессии zcytor17lig или изменение числа или локализации моноцитов (например, увеличение или инфильтрация моноцитарных клеток в тканях, где они обычно не встречаются) по сравнению с контрольным образцом является свидетельством заболевания. Такие диагностические методы могут также включать в себя использование радиометрических, флуоресцентных и колориметрических меток, присоединенных к полинуклеотидам, полипептидам или антителам согласно изобретению. Такие методы хорошо известны в данной области и описаны в данном описании.
Аминокислотные последовательности, обладающие активностью zcytor17lig, можно использовать для модуляции иммунной системы путем связывания рецептора zcytor17 и предотвращения, таким образом, связывания zcytor17lig с эндогенным рецептором zcytor17lig. Антагонисты zcytor17lig, такие как антитела против zcytor17lig, можно также использовать для модуляции иммунной системы путем ингибирования связывания zcytor17lig с эндогенным рецептором zcytor17lig. Поэтому настоящее изобретение относится к использованию белков, полипептидов и пептидов, обладающих активностью zcytor17lig (например, полипептидов zcytor17lig), аналогов zcytor17lig (например, антиидиотипических антител против zcytor17lig) и слитых белков zcytor17lig для введения субъекту, у которого наблюдается недостаток указанного полипептида или организм которого продуцирует избыток рецепторов zcytor17. Антагонисты zcytor17 (например, антитела против zcytor17) можно также использовать для лечения субъекта, организм которого продуцирует избыток zcytor17lig или рецепторов zcytor17. Приемлемыми субъектами являются млекопитающие, такие как человек.
Установлено, что zcytor17lig экспрессирован в активированных мононуклеарных клетках и может участвовать в регуляции воспаления. Таким образом, полипептиды согласно изобретению могут быть подвергнуты анализу и использованы для лечения воспаления или в качестве маркера воспаления. Методы определения провоспалительных и противовоспалительных свойств zcytor17lig хорошо известны в данной области и описаны в настоящем описании. Кроме того, zcytor17lig может участвовать в повышении продуцирования активных веществ острой фазы заболевания, таких как сывороточный амилоид А (SAA), α1-антихимотрипсин и гаптоглобин, при этом экспрессию лиганда рецептора zcytor17 можно увеличить при помощи инъекции липополисахарида (LPS) in vivo, который участвует в возникновении воспалительной реакции (Dumoutier, L. et al., Proc. Nat'l. Acad. Sci. 97:10144-10149, 2000). Продуцирование белков острой фазы, таких как SAA, считается кратковременным механизмом выживания, когда воспаление является благоприятным; однако сохранение белков острой фазы в течение более продолжительных периодов времени вызывает хроническое воспаление и может быть вредным для здоровья человека. Для ознакомления с данным вопросом см. Uhlar, CM and Whitehead, AS, Eur. J. Biochem. 265:501-523, 1999, and Baumann H. and Gauldie, J. Immunology Today 15:74-80, 1994. Кроме того, белок острой фазы SAA имеет отношение к патогенезу нескольких хронических воспалительных заболеваний, таких как атеросклероз и ревматоидный артрит, и является предшественником белка амилоида А, характерного для амилоидоза (Uhlar, CM and Whitehead, см. выше). Таким образом, в тех случаях, когда такой лиганд, как zcytor17lig, действует в качестве провоспалительной молекулы и вызывает продуцирование SAA, необходимо использовать антагонисты для лечения воспалительных и других заболеваний, ассоциированных с белками острой фазы, индуцируемыми данным лигандом. Такие антагонисты являются объектом настоящего изобретения. Например, способ уменьшения воспаления включает в себя введение нуждающемуся млекопитающему композиции, содержащей zcytor17lig или антитело против zcytor17lig (например, нейтрализующее антитело), в количестве, достаточном для уменьшения воспаления. Кроме того, способ подавления воспалительной реакции у млекопитающего может включать в себя (1) определение уровня белка сывороточного амилоида А; (2) введение композиции, содержащей полипептид zcytor17lig или антитело против zcytor17lig в приемлемом фармацевтическом носителе; (3) определение уровня белка сывороточного амилоида А после введения композиции; (4) сравнение уровня белка сывороточного амилоида А, измеренного на стадии (1), с уровнем белка сывороточного амилоида А, измеренного на стадии (3), при этом отсутствие увеличения или снижение уровня белка сывороточного амилоида А свидетельствует о подавлении воспалительной реакции.
Рецепторы, связывающие zcytor17lig согласно изобретению, включают в себя по меньшей мере одну субъединицу рецептора zcytor17. Второй полипептид рецептора в структуре гетеродимерного растворимого рецептора относится к подсемейству рецепторов, содержащему субъединицы рецепторов цитокина класса I, в частности OSMRbeta и WSX-1. В соответствии с настоящим изобретением не только полипептид мономерного или гомодимерного рецептора zcytor17, но и гетеродимерный растворимый рецептор zcytor17, содержащий растворимый рецептор zcytor17 + гетеродимерный компонент растворимого рецептора класса I, такой как OSMRbeta или WSX-1, может действовать в качестве антагониста zcytor17lig. Другие варианты осуществления изобретения включают в себя растворимые многомерные рецепторы, содержащие zcytor17, такие как рецептор zcytor17 + многомерный компонент растворимого рецептора класса I, такой как OSMRbeta и WSX-1.
Подобно zcytor17lig, анализ распределения в тканях мРНК, соответствующей кДНК рецептора zcytor17, показал, что уровень мРНК является наиболее высоким в моноцитах и клетках предстательной железы и повышается в активированных моноцитах, активированных CD4+-клетках, активированных CD8+-клетках и активированных CD3+-клетках. Следовательно, рецептор zcytor17 также имеет отношение к индуцированию воспалительной и иммунной реакции. Таким образом, некоторые варианты осуществления настоящего изобретения относятся к применению антител против zcytor17lig, лиганда zcytor17lig и гетеродимеров растворимого рецептора zcytor17 в качестве антагонистов при лечении воспалительных и иммунных заболеваний, таких как панкреатит, диабет типа I (IDDM), рак поджелудочной железы, болезнь Грейвса, воспалительное заболевание кишечника (IBD), болезнь Крона, рак ободочной и тонкой кишки, дивертикулез, аутоиммунное заболевание, сепсис, трансплантация органов или костного мозга; воспаление, вызванное травмой, хирургическим вмешательством или инфекцией; амилоидоз, спленомегалия; реакция «трансплантат против хозяина»; а также для ингибирования воспаления, супрессии иммунной реакции, уменьшения пролиферации гемопоэтических, иммунокомпетентных, воспалительных или лимфоидных клеток, макрофагов, Т-клеток (включая Th1- и Th2-клетки, CD4+ и CD8+-клетки), супрессии иммунной реакции в отношении патогенов или антигенов. Кроме того, наличие рецептора zcytor17 и экспрессии zcytor17lig в активированных иммунокомпетентных клетках, таких как активированные CD3+-клетки, моноциты, CD4+ и CD19+-клетки, показывает, что рецептор zcytor17 может участвовать в защитных иммунных реакциях организма против проникновения чужеродных факторов, таких как микроорганизмы и клеточный дебрис, и может играть определенную роль в иммунных реакциях во время воспаления и образования опухоли. Таким образом, zcytor17lig и антитела против zcytor17lig согласно изобретению, которые являются агонистами или антагонистами рецептора zcytor17, могут быть использованы для модификации иммунной реакции и воспаления.
Кроме того, полипептиды zcytor17lig, связывающиеся с полипептидами рецептора zcytor17, и антитела к указанным полипептидам могут выполнять следующие функции:
1) Оказывать антагонистическое действие или блокировать передачу сигналов zcytor17-содержащими рецепторами в случае острого воспаления, воспаления вследствие травмы, повреждения тканей, хирургического вмешательства, сепсиса или инфекции, хронических воспалительных заболеваний, таких как астма, воспалительное заболевание кишечника (IBD), хронический колит, спленомегалия, ревматоидный артрит, рецидивы острых воспалительных заболеваний (например, туберкулеза), амилоидоз, атеросклероз, болезнь Кастлемана, астма и другие заболевания, обусловленные индукцией реакции в острой фазе.
2) Оказывать антагонистическое действие или блокировать передачу сигналов рецепторами zcytor17 в случае аутоиммунных заболеваний, таких как IDDM, рассеянный склероз (MS), системный эритематоз Люпуса (SLE), миастения, ревматоидный артрит и IBD, для предотвращения или подавления передачи сигналов в иммунокомпетентные клетки (например, лимфоциты, моноциты, лейкоциты) рецептором zcytor17 (Hughes C. et al., J. Immunol. 153:3319-3325, 1994). Альтернативно антитела, такие как моноклональные антитела (MAb) к zcytor17lig, можно также использовать в качестве антагониста для инактивации нежелательных иммунокомпетентных клеток при лечении аутоиммунного заболевания. Астму, аллергию и другие атопические заболевания можно лечить при помощи MAb, например антител против zcytor17lig, растворимых рецепторов zcytor17 или гетеродимеров zcytor17/CRF2-4, которые используют для подавления иммунной реакции или инактивации агрессивных клеток. Блокирование или ингибирование передачи сигналов рецептором zcytor17 с использованием полипептидов и антител согласно изобретению может быть эффективным при лечении заболеваний поджелудочной железы, почек, гипофизарных и нервных клеток. К таким заболеваниям относятся IDDM, NIDDM, панкреатит и рак поджелудочной железы. Рецептор zcytor17 может служить мишенью при лечении рака с помощью MAb, когда антагонистическое действие MAb тормозит рост раковой опухоли и вызывает лизис клеток, опосредуемый иммунокомпетентными клетками. (Holliger P, and Hoogenboom, H: Nature Biotech. 16: 1015-1016, 1998.) Моноклональные антитела (MAb) к мономерам, гомодимерам, гетеродимерам и многомерам растворимого рецептора zcytor17 могут быть также пригодны для лечения нефропатий, таких как гломерулосклероз, мембранозная невропатия, амилоидоз (поражающий почку наряду с другими тканями), почечный артериосклероз, гломерулонефрит разного происхождения, фибропролиферативные заболевания почки, а также нарушение функции почки, обусловленное SLE, IDDM, диабетом типа II (NIDDM), опухолями почки и другими заболеваниями.
3) Оказывать агонистическое действие или инициировать передачу сигналов рецепторами zcytor17 в случае аутоиммунных заболеваний, таких как IDDM, MS, SLE, миастения, ревматоидный артрит и IBD. Zcytor17lig может передавать сигналы в лимфоциты или другие иммунокомпетентные клетки о дифференцировке, замедлении пролиферации или изменении продуцирования цитокинов или белков на поверхности клеток, которые ослабляют аутоиммунную реакцию. В частности, модуляция реакции Т-клеток-хелперов на измененный паттерн секреции цитокинов может направить аутоиммунную реакцию на ослабление симптомов заболевания (Smith J.A. et al., J. Immunol. 160:4841-4849, 1998). Аналогичным образом, zcytor17lig можно использовать для передачи сигналов, вызывающих инактивацию и изменение иммунокомпетентных клеток, опосредующих возникновение астмы, аллергии и атопического заболевания. Передачи сигналов рецептором zcytor17 может оказывать благоприятное влияние на болезни поджелудочной железы, почки, гипофизарных и нервных клеток. К таким заболеваниям относятся IDDM, NIDDM, панкреатит и рак поджелудочной железы. Рецептор zcytor17 может служить мишенью при лечении моноклональными антителами рака поджелудочной железы, когда передача сигналов MAb тормозит рост раковой опухоли и вызывает лизис клеток, опосредуемый иммунокомпетентными клетками (Tutt, A.L. et al., J. Imuunol. 161:3175-3185, 1998). Аналогичным образом, моноклональными антителами (например, нейтрализующим антителом) к zcytor17-содержащим растворимым рецепторам согласно изобретению можно лечить специфические Т-клеточные лейкозы, лимфомы, патологическое изменение плазматических клеток (например, множественную миелому) и карциному.
Антитела против zcytor17lig, мономерные, гомодимерные, гетеродимерные и многомерные полипептиды растворимого рецептора zcytor17 можно использовать для нейтрализации/блокирования активности лиганда рецептора zcytor17 в случае аутоиммунного заболевания, атопического заболевания, NIDDM, панкреатита и нарушения функции почки, как было описано выше. Растворимую форму рецептора zcytor17 можно использовать для стимуляции реакции антитела, опосредуемой Т-клетками, и/или стимуляции продуцирования IL-4 или других цитокинов лимфоцитами или другими иммунными клетками.
Антитела против zcytor17lig и zcytor-17-содержащие растворимые рецепторы можно использовать в качестве антагонистов zcytor17lig. Такое антагонистическое действие может быть достигнуто в результате прямой нейтрализации или связывания природного лиганда. Помимо применения в качестве антагониста растворимые рецепторы могут связывать zcytor17lig и действовать в качестве носителя или белков-носителей для переноса zcytor17lig в другие ткани, органы и клетки организма. Растворимые рецепторы могут быть слиты или связаны с молекулами, полипептидами или химическими частями молекулы, которые направляют комплекс растворимый рецептор-лиганд в определенный участок, такой как ткань, специфические иммунокомпетентные клетки, моноциты или опухоль. Например, в случае острой инфекции или некоторых типов рака благоприятные результаты можно получить в результате индукции воспаления и локальных белков острой фазы. Таким образом, описанные растворимые рецепторы или антитела согласно изобретению можно использовать для того, чтобы направить действие провоспалительного лиганда zcytor17lig. См., Cosman, D. Cytokine 5:95-106, 1993; и Fernandez-Botran, R. Exp. Opin. Invest. Drugs 9:497-513, 2000.
Кроме того, растворимые рецепторы можно использовать для стабилизации zcytor17lig, увеличения биологической доступности, длительности сохранения и/или эффективности лиганда в результате стабилизации лиганда от разрушения или расщепления либо направленной доставки данного лиганда к месту действия в организме пациента. Например, комплекс природный IL-6/растворимый IL-6R стабилизирует IL-6 и может передавать сигналы при помощи рецептора gp130. См., Cosman, D., выше, и Fernandez-Botran, R., выше. Кроме того, zcytor17 можно объединить с родственным лигандом для создания комплекса лиганд/растворимый рецептор. Такие комплексы можно использовать для стимуляции реакций клеток, содержащих субъединицу рецептора. Клеточная специфичность комплексов рецептор zcytor17/zcytor17lig может отличаться от специфичности, характерной для отдельно вводимого лиганда. Кроме того, комплексы могут обладать отличными фармакокинетическими свойствами, такими как период полужизни, соотношение доза/реакция и специфичность органов или тканей. Комплексы zcytor17/лиганд могут оказывать агонистическое действие, направленное на усиление иммунной реакции, или стимуляцию мезангиальных клеток или гепатоцитов. Альтернативно, воздействию могут подвергаться только ткани, экспрессирующие сигнальную субъединицу, которая образует гетеродимер с данным комплексом, аналогично реакциям на комплексы IL6/IL6R (Hirota H. et al., Proc. Na'tl. Acad. Sci. 92:4862-4866, 1995; Hirano, T. in Thomason, A. (Ed.) "The Cytokine Handbook", 3th Ed., p. 208-209). Комплексы растворимый рецептор/цитокин, предназначенные для IL-12 и CNTF, обладают аналогичной активностью.
Zcytor17lig можно также использовать в диагностических системах для определения уровней лиганда в кровотоке и для обнаружения воспалительной реакции в острой фазе. В родственном варианте осуществления изобретения антитела или другие агенты, которые специфически связываются с zcytor17lig, можно использовать для обнаружения полипептидов zcytor17lig в кровотоке, и наоборот, сам zcytor17lig можно использовать для обнаружения полипептидов рецепторов в кровотоке или на отдельных участках. Повышенные или пониженные уровни полипептидов лиганда или рецептора могут указывать на патологическое состояние, включая воспаление или рак. Кроме того, обнаружение белков или молекул острой фазы, таких как zcytor17lig, может указывать на хроническое воспаление в случае некоторых заболеваний (например, ревматоидного артрита). Обнаружение такого состояния помогает лечащему врачу диагностировать заболевание и выбрать правильное лечение.
Полипептиды и белки согласно изобретению можно также использовать ex vivo, например в аутологичной культуре костного мозга. У субъекта берут костный мозг до начала химиотерапии или трансплантации органа и обрабатывают zcytor17lig, необязательно в комбинации с одним или несколькими другими цитокинами. Обработанный костный мозг возвращают пациенту после химиотерапии для ускорения восстановления костного мозга или для подавления реакции «трансплантат против хозяина» после трансплантации. Кроме того, белки согласно изобретению можно также использовать для выращивания ex vivo моноцитов/макрофагов костного мозга или клеток-предшественников периферической крови (PBPC). До обработки костный мозг может быть стимулирован фактором поддержания стволовых клеток (SCF) для высвобождения ранних клеток-предшественников в периферическое кровообращение. Указанные клетки-предшественники можно собрать, концентрировать из периферической крови и обработать zcytor17lig в культуре, необязательно в комбинации с одним или несколькими другими цитокинами, которые включают в себя, не ограничиваясь ими, SCF, IL-2, IL-4, IL-7, Lif, IL-3, IL-12, IL-21 или IL-15, для дифференцировки и пролиферации в культурах лимфоидных клеток высокой плотности, чтобы затем вернуть пациенту после химиотерапии или трансплантации.
Настоящее изобретение относится к способу размножения гемопоэтических клеток и клеток-предшественников гемопоэтических клеток, который включает в себя культивирование клеток костного мозга или периферической крови с композицией, содержащей zcytor17lig в количестве, достаточном для увеличения числа лимфоидных клеток в клетках костного мозга или периферической крови по сравнению с клетками костного мозга или периферической крови, культивируемыми в отсутствие zcytor17lig. В других вариантах осуществления изобретения гемопоэтические клетки и клетки-предшественники гемопоэтических клеток являются лимфоидными клетками. В другом варианте осуществления изобретения лимфоидные клетки являются NK-клетками или цитотоксическими Т-клетками. Кроме того, данная композиция может также содержать по меньшей мере один другой цитокин, выбираемый из группы, содержащей лиганд и фактор поддержания стволовых клеток IL-2, IL-15, IL-4, Lif, IL-3, IL-12, IL-21, GM-CSF, Flt3.
Альтернативно, zcytor17lig может активировать иммунную систему, что имеет важное значение для создания иммунитета к инфекционным заболеваниям, лечения пациентов с ослабленным иммунитетом, например ВИЧ-инфицированных больных, больных раком, или для улучшения вакцин. В частности, стимуляция или размножение моноцитов/макрофагов, Т-клеток, В-клеток, NK-клеток или их клеток-предшественников при помощи zcytor17lig позволяет получить хорошие результаты при лечении вирусной инфекции и в качестве противоопухолевого фактора. Аналогичным образом, стимуляция zcytor17lig иммунной реакции против вирусных и невирусных патогенных микроорганизмов (включая бактерии, простейшие и грибы) является эффективной при лечения таких инфекций благодаря торможению роста указанных инфекций. Присутствие в организме патогена или антигена, такого как опухолевая клетка, можно определить рядом прямых или непрямых методов, известных в данной области и рассмотренных в данном описании.
Настоящее изобретение относится к способу стимуляции иммунной реакции у млекопитающего, подверженного воздействию антигена или патогена, который включает в себя следующие стадии: (1) прямое или непрямое определение уровня антигена или патогена в организме указанного млекопитающего; (2) введение композиции, содержащей полипептид zcytor17lig в приемлемом фармацевтическом носителе; (3) прямое или непрямое определение уровня антигена или патогена в организме указанного млекопитающего; и (4) сравнение уровня антигена или патогена, измеренного на стадии 1, с уровнем антигена или патогена, измеренным на стадии 3, при этом изменение уровня свидетельствует о стимуляции иммунной реакции. В другом варианте осуществления изобретения пациенту повторно вводят композицию, содержащую zcytor17lig. В других вариантах осуществления изобретения антиген является В-клеточной опухолью, вирусом, паразитом или бактерией.
Другим объектом настоящего изобретения является способ стимуляции иммунной реакции у млекопитающего, подверженного воздействию антигена или патогена, который включает в себя (1) определение уровня антиген- или патогенспецифического антитела; (2) введение композиции, содержащей полипептид zcytor17lig в приемлемом фармацевтическом носителе; (3) определение уровня антиген- или патогенспецифического антитела после введения композиции; (4) сравнение уровня антитела, измеренного на стадии (1), с уровнем антитела, измеренным на стадии (3), при этом увеличение уровня антитела свидетельствует о стимуляции иммунной реакции.
Полинуклеотиды, кодирующие полипептиды zcytor17lig, пригодны для применения в генотерапии, когда желательно увеличить или ингибировать активность zcytor17lig. Если у млекопитающего имеется мутированный ген zcytor17lig или такой ген отсутствует, ген zcytor17lig можно ввести в клетки данного млекопитающего. В одном варианте осуществления изобретения ген, кодирующий полипептид zcytor17lig, вводят in vivo в вирусный вектор. Такие векторы содержат ДНК ослабленного или дефектного вируса, включая, но не ограничиваясь ими, вирус простого герпеса (HSV), вирус папилломы, вирус Эпштейна-Барр (EBV), аденовирус, аденоассоциированный вирус (AAV) и пр. Дефектные вирусы, у которых полностью или почти полностью отсутствуют вирусные гены, являются предпочтительными. Дефектный вирус не является инфекционным после введения в клетку. Применение векторов на основе дефектных вирусов позволяет вводить их в клетки на конкретном, локализованном участке, не беспокоясь о том, что вектор может инфицировать другие клетки. Примеры конкретных векторов включают в себя, не ограничиваясь ими, вектор на основе дефектного вируса простого герпеса 1 (HSV1) (Kaplitt et al., Molec. Cell. Neurosci. 2:320-30, 1991); вектор на основе ослабленного аденовируса, такой как вектор, описанный в статье Stratford-Perricaudet et al., J. Clin. Invest. 90:626-30, 1992; и вектор на основе дефектного аденоассоциированного вируса (Samulski et al., J. Virol. 61:3096-101, 1987; Samulski et al., J. Virol. 63:3822-8, 1989).
Ген zcytor17lig можно ввести в ретровирусный вектор способом, описанным в патенте США № 5399346, выданном Anderson et al..; в статье Mann et al. Cell 33:153, 1983; в патенте США № 4650764, выданном Temin et al.; в патенте США № 4980289, выданном Temin et al.; в статье Markowitz et al., J. Virol. 62:1120, 1988; в патенте США № 5124263, выданном Temin et al.; в публикации международного патента № WO 95/07358, Dougherty et al. от 16 марта 1995 г.; в статье Kuo et al., Blood 82:845, 1993. Альтернативно, вектор можно ввести путем липофекции in vivo с использованием липосом. Синтетические катионные липиды можно использовать для получения липосом, предназначенных для трансфекции in vivo гена, кодирующего маркер (Felgner et al., Proc. Natl. Acad. Sci. USA 84:7413-7, 1987; Mackey et al., Proc. Natl. Acad. Sci. USA 85: 8027-31, 1988). Применение липофекции для введения экзогенных генов в конкретные органы in vivo имеет определенные практические преимущества. Направленная молекулярная доставка липосом в специфические клетки является одним из преимуществ. В частности, одним из преимуществ является направленная трансфекция определенных клеток. Например, направленная трансфекция определенных клеток является особенно благоприятной в тканях, состоящих из гетерогенных клеток, таких как иммунная система, поджелудочная железа, печень, почка и головной мозг. Липиды могут быть химически связаны с другими молекулами для достижения направленной доставки. Пептиды-мишени (например, гормоны или нейротрансмиттеры), белки, такие как антитела, или непептидные молекулы, могут быть химически связаны с липосомами.
Клетки-мишени можно удалить из организма, ввести в них вектор в виде плазмиды, содержащей депротеинизированную ДНК, и затем вновь имплантировать трансформированные клетки в организм хозяина. Векторы на основе депротеинизированной ДНК, предназначенные для генотерапии, можно ввести в требуемые клетки-хозяева методами, известными в данной области, такими как трансфекция, электропорация, микроинъекция, трансдукция, слияние клеток, осаждение DEAE-декстраном и фосфатом кальция, применение генного ружья или транспортера векторной ДНК. См., например, Wu et al., J. Biol. Chem. 267:963-7, 1992; Wu et al., J. Biol. Chem. 263:14621-4, 1988.
Метод с применением антисмысловых полинуклеотидов можно использовать для ингибирования транскрипции гена zcytor17lig, например для ингибирования пролиферации клеток in vivo. Полинуклеотиды, комплементарные к сегменту zcytor17lig-кодирующего полинуклеотида (например, полинуклеотид, представленный в SEQ ID NO:1), предназначены для связывания с zcytor17lig-кодирующей мРНК и ингибирования трансляции такой мРНК. Такие антисмысловые полинуклеотиды используют для ингибирования экспрессии генов, кодирующих полипептид zcytor17lig, в культуре клеток или в организме субъекта.
Методами рекомбинантных ДНК можно создать мышей, экспрессирующих ген zcytor17lig, которых именуют «трансгенными мышами», и мышей с полным отсутствием функции гена zcytor17lig, именуемых «дефектными мышами» (Snouwaert et al., Science 257:1983; Lowell et al., Nature 366:740-42, 1993; Capecchi, M.R., Science 244: 1288-1292, 1989; Рalmiter, R.D. et al., Annu Rev. Genet. 20: 465-499, 1986). Например, трансгенных мышей со сверхэкспрессией zcytor17lig, происходящей без внешних стимулов или под воздействием тканеспецифического или тканеограниченного промотора, можно использовать для исследования вопроса о возможности образования фенотипа вследствие указанной сверхэкспрессии. Например, сверхэкспрессия полипептида zcytor17lig дикого типа, фрагмента полипептида или его мутанта может изменять нормальные процессы, происходящие в клетках, с образованием фенотипа, идентифицирующего ткань, в которой экспрессия zcytor17lig является функционально родственной и может представлять собой терапевтическую цель для zcytor17lig, его агонистов или антагонистов. Например, предпочтительной трансгенной мышью является мышь со сверхэкспрессией zcytor17lig (аминокислотные остатки 23-164 SEQ ID NO:2 или 24-163 SEQ ID NO:11). Кроме того, такая сверхэкспрессия может вызывать образование фенотипа, который обладает сходством с болезнями человека. Аналогичным образом, мышей с отсутствием zcytor17lig можно использовать для определения абсолютной необходимости в zcytor17lig in vivo. Фенотип дефектных мышей позволяет прогнозировать результаты воздействия in vivo антагониста zcytor17lig, аналогичные описанным в данном описании. Для создания дефектных мышей можно использовать кДНК zcytor17lig человека или мыши. Указанных мышей можно использовать для исследования гена zcytor17lig и кодируемого им белка в системе in vivo и в моделях in vivo соответствующих болезней человека. Кроме того, у трансгенных мышей могут быть аналогичным образом экспрессированы описанные здесь антисмысловые полинуклеотиды zcytor17lig или рибозимы против zcytor17lig. Исследования могут быть также выполнены при введении очищенного белка zcytor17lig.
Экспериментальные данные подтверждают роль zcytor17lig в развитии заболеваний, поражающих кожу или эпителий внутренних поверхностей, таких, например, как толстый кишечник, тонкий кишечник, поджелудочная железа, легкое, предстательная железа, матка и тому подобное. Как было указано, рецепторы zcytor17, включая бета-рецептор OSM и zcytor17, экспрессированы в клетках нескольких типов, расположенных на поверхности эпителия, включая линии клеток, выделенные из эпителия легкого, фибробласта легкого, эпителия предстательной железы, ободочной кишки, молочной железы, печени, костей и кожи, костных фибробластов и пр. Кроме того, как указано в данном описании, все рассмотренные типы клеток реагируют на активацию лигандом zcytor17lig репортерной конструкции STAT. Кроме того, несколько линий клеток реагируют на стимуляцию zcytor17lig повышением уровней IL-6, IL-8, MCP-1 (хемотаксический фактор). В целом, полученные данные подтверждают роль zcytor17lig в заболеваниях, поражающих эпителий, таких, например, как атопический дерматит, обычный дерматит, псориаз, ревматоидный псориаз, экзема, гингивит, перидонтоз, воспалительное заболевание кишечника (IBD) (например, неспецифический язвенный колит, болезнь Крона), нарушения репродуктивной системы, такие как дисплазия шейки матки, рак шейки матки; другие кожные заболевания, такие как различные типы рака: саркомы, карциномы, меланомы и т.д., то есть не только воспалительные заболевания, так как иммунная система участвует в активации/лечении рака; заболевания, вызываемые нарушением барьерных функций, такие, например, как реакция «трансплантат против хозяина» (GVHD) и воспалительное заболевание кишечника (IBD), и заболевания, поражающие эпителий легкого, такие как астма, эмфизема и тому подобное. Кроме того, высвобождение цитокинов IL-6, IL-8 и МСР-1 клетками, подвергнутыми воздействию zcytor17lig, позволяет предположить, что zcytor17lig участвует в возникновении воспалений. Поэтому регуляция zcytor17lig может быть полезна при лечении аутоиммунных, воспалительных или раковых заболеваний, ассоциированных с тканями, экспрессирующими данный рецептор. Такие заболевания включают в себя, например, простатит, гепатит, остеоартрит и тому подобное. Zcytor17lig может оказывать положительное или отрицательное, прямое или косвенное воздействие на указанные заболевания. Поэтому введение zcytor17lig можно использовать для лечения указанных заболеваний в том виде, как есть, или с молекулами, ингибирующими активность zcytor17lig, которые включают в себя, например, моноклональные антитела к zcytor17lig, моноклональные антитела к zcytor17lig или моноклональные антитела, которые узнают комплекс zcytor17 и бета-рецептора OSM.
Полученные данные также показывают, что zcytor17lig может участвовать в регуляции болезней, опосредуемых Т-клетками ТН2. Zcytor17lig получают, используя субпопуляцию ТН2 активированных Т-клеток. Клетки ТН2 экспрессируют больше zcytor17lig по сравнению с клетками ТН1. Кроме того, по меньшей мере две линии эпителиальных клеток легкого (SK-LU-1, A549) были стимулированы для увеличения мРНК альфа-2-рецептора IL-13 в ответ на стимуляцию лигандом zcytor17. Существует взаимосвязь между цепью альфа-2-рецептора IL-13 и образованием опухоли молочной железы и предстательной железы у человека. Указанные данные позволяют предположить, что zcytor17lig может играть определенную роль в регуляции онкогенности указанных и других типов рака. Поэтому введение антагониста zcytor17lig или прямое использование zcytor17lig может быть эффективным при лечении указанных типов рака, доброкачественных или злокачественных опухолей различной степени тяжести (степени I-IV) и на разных стадиях развития (методы определения стадии заболевания TNM или AJC) у млекопитающих, предпочтительно у человека.
В данной области хорошо известно, что IL-13 участвует в образовании активированных клеток ТН2 и возникновении заболеваний, опосредуемых клетками ТН2, таких как астма, атопический дерматит и тому подобное. Zcytor17lig или антагонисты zcytor17lig можно использовать для лечения заболеваний, опосредуемых Т-клетками ТН2. Такие заболевания включают в себя, например, атопический дерматит, астму, а также другие заболевания, которые обостряются активированными клетками ТН2. Участие zcytor17lig в заболеваниях, таких, например, как атопический дерматит, также подтверждается фенотипом трансгенных мышей со сверхэкспрессией zcytor17lig, у которых появляются симптомы атопического дерматита.
Несмотря на предпочтительную экспрессию zcytor17lig клетками ТН2, наблюдается некоторая экспрессия zcytor17lig в клетках ТН1 и CD8+ Т-клетках. Поэтому zcytor17lig или его антагонисты можно использовать при лечении заболеваний, обусловленных иммунномодуляцией активированных Т-клеток, включая, например, вирусную инфекцию, рак, отторжение трансплантата и тому подобное.
Zcytor17lig может также участвовать в возникновении рака. Обнаружена экспрессия zcytor17 и бета-рецептора OSM в остеосаркоме костных фибробластов человека, меланоме кожных фибробластов человека, эпителиоме ободочной кишки, аденокарциноме, эпителиальной аденокарциноме молочной железы, эпителиальной аденосаркоме предстательной железы, эпителиальной аденокарциноме и карциноме легкого. Поэтому zcytor17lig, его фрагменты или антагонисты zcytor17lig можно использовать для лечения опухолей эпителиального происхождения, которые включают в себя, не ограничиваясь ими, карциному, аденокарциному и меланому. Таким образом, zcytor17lig или антагонист zcytor17lig можно использовать для лечения рака или для ослабления одного или нескольких симптомов рака, включая, но не ограничиваясь ими, плоскоклеточный или эпидермоидный рак, базально-клеточный рак, аденокарциному, папиллярный рак, цистаденокарциному, бронхогенный рак, бронхиальную аденому, меланому, почечно-клеточный рак, злокачественную гепатому, переходно-клеточный рак, хориокарциному, семиному, эмбриональный рак, злокачественную рецидивирующую эпителиому слюнной железы, опухоль Вильмса, незрелую тератому, тератокарциному и другие опухоли, содержащие по меньшей мере некоторые клетки эпителиального происхождения.
Как правило, доза вводимого полипептида zcytor17lig (аналога или слитого белка zcytor17) может изменяться в зависимости от таких факторов, как возраст, масса тела, рост, пол, общее состояние здоровья и анамнез пациента. Реципиенту обычно предпочтительно вводят дозу полипептида zcytor17lig в интервале от около 1 пг/кг до 10 мг/кг (соотношение лекарственное средство/масса тела пациента), хотя, в зависимости от обстоятельств, доза может быть меньше или больше. Специалист легко может определить такие дозы и привести их в соответствие с вышеуказанными факторами, используя методы, известные в данной области.
Полипептид zcytor17lig можно вводить пациенту местно, в виде ингаляции, внутривенно, внутриартериально, внутрибрюшинно, внутримышечно, подкожно, внутриплеврально, подоболочечно, в виде вливания через катетер или в виде прямой инъекции в место поражения. Лекарственные белки можно инъецировать путем непрерывного вливания или введения одного или нескольких болюсов.
Дополнительные способы введения включают в себя пероральное введение, введение через слизистую оболочку, легочное и чрескожное введение. Пероральное введение приемлемо при использовании полиэфирных микросфер, цеиновых микросфер, белковых микросфер, полицианоакрилатных микросфер и систем на основе липидов (см., например, DiBase and Morrel, "Oral Delivery of Microencapsulated Proteins," in Protein Delivery" Physical Systems, Sanders and Hendren (eds.), pages 255-288 (Plenum Press, 1997)). Введение в нос - один из характерных способов введения инсулина (см., например, Hinchcliffe and Illum, Adv. Drug Deliv. Rev. 35:199 (1999)). Сухие или жидкие частицы, содержащие zcytor17lig, могут быть получены и использованы для ингаляции при помощи диспергатора сухого порошка, генераторов жидкого аэрозоля или ингаляторов (например, Pettit and Gombotz, TIBTECH 16:343 (1998); Patton et al., Adv. Drug. Deliv. Rev. 35:235 (1999)). Данный подход можно проиллюстрировать системой лечения диабета AERX, которая представляет собой ручной электронный ингалятор, доставляющий в легкие инсулин в виде аэрозоля. Выполненные исследования показали, что белки в 48000 кДа можно доставлять через кожу в лечебных концентрациях при помощи низкочастотного ультразвука, что иллюстрирует возможность чрескожного введения (Mitragotri et al., Science 269:850 (1995)). Чрескожная доставка при помощи электропорации является другим способом введения молекулы, обладающей zcytor17lig-связывающей активностью (Potts et al., Pharm. Biotechnol. 10:213 (1997)).
Фармацевтическую композицию, содержащую белок, полипептид или пептид, обладающие zcytor17lig-связывающей активностью, можно получить известными методами, предназначенными для приготовления фармацевтических композиций, в которых белки находятся в смеси с фармацевтически приемлемым носителем. Носитель считается «фармацевтически приемлемым», если он не причиняет вреда реципиенту. Одним примером фармацевтически приемлемого носителя является стерильный физиологический раствор с фосфатным буфером. Специалистам хорошо известны другие приемлемые носители. См., например, Gennaro (ed.), Remington's Pharmaceutical Sciences, 19th Edition (Mack Publishing Company, 1995).
В лечебных целях молекулы, обладающие zcytor17lig-связывающей активностью, и фармацевтически приемлемый носитель вводят пациенту в терапевтически эффективном количестве. Считается, что комбинацию белка, полипептида или пептида, обладающего zcytor17lig-связывающей активностью, и фармацевтически приемлемого носителя вводят в «терапевтически эффективном количестве», если вводимое количество является физиологически значимым. Количество лекарственного средства является физиологически значимым, если оно вызывает детектируемое изменение физиологии реципиента. Например, лекарственное средство, используемое для лечения воспаления, является физиологически значимым, если при его введении происходит по меньшей мере частичное ослабление воспалительной реакции.
Фармацевтическую композицию, содержащую zcytor17lig (аналог или слитый белок zcytor17lig), можно вводить в жидкой форме, в виде аэрозоля или в твердой форме. Примерами жидких форм являются инъекционные растворы, аэрозоли, капли, растворы для местного применения и суспензии для перорального введения. Типичные твердые формы включают в себя капсулы, таблетки и формы пролонгированного действия. Примерами форм пролонгированного действия являются миниосмотические насосы и имплантаты (Bremer et al., Pharm. Biotechnol. 10:239 (1997); Ranade, "Implants in Drug Delivery", in Drug Delivery Systems, Ranade and Hollinger (eds.), pages 95-123 (CRC Press 1995); Bremer et al., "Protein Delivery with Infusion Pumps," in Protein Delivery: Physical Systems, Sanders and Hendren (eds.), pages 239-254 (Plenum Press 1997); Yewey et al., "Delivery of Proteins from a Controlled Release Injectable Implant," in Protein Delivery: Physical Systems, Sanders and Hendren (eds.), pages 93-117 (Plenum Press 1997)). Другие твердые формы включают в себя кремы, пасты, другие препараты для местного применения и тому подобное.
Липосомы являются одним из способов доставки лекарственных полипептидов в организм пациента путем внутривенного, внутрибрюшинного, подоболочечного, внутримышечного, подкожного или перорального введения, ингаляции или введения в нос. Липосомы представляют собой микроскопические пузырьки, которые состоят из одного или нескольких липидных слоев, окружающих водные камеры (см., например, Bakker-Woudenberg et al., Eur. J. Clin. Microbiol. Infect. Dis. 12 (Suppl. 1):S61 (1993), Kim, Drugs 46:618 (1993), and Ranade, "Site-Specific Drug Delivery Using Liposomes as Carriers," in Drug Delivery Systems, Ranade and Hollinger (eds.), pages 3-24 (CRC Press 1995)). Липосомы имеют такой же состав, что и клеточные мембраны, благодаря чему липосомы являются безопасными и биологически разлагающимися веществами. В зависимости от способа получения липосомы могут быть однослойными или многослойными и могут иметь диаметр от 0,02 мкм до более чем 10 мкм. В липосомы могут быть инкапсулированы различные лекарственные средства: гидрофобные средства могут находиться между двух слоев, и гидрофильные средства могут находиться во внутреннем водном пространстве (см., например, Machy et al., Liposomes In Cell Biology And Pharmacology (John Libbey 1987) and Ostro et al., American J. Hosp. Pharm. 46:1576 (1989)). Кроме того, можно контролировать терапевтическую доступность инкапсулированного средства, изменяя размер липосомы, число слоев, липидную композицию, а также заряд и поверхностные характеристики липосом.
Липосомы могут адсорбироваться практически всеми типами клеток и медленно высвобождать инкапсулированное лекарственное средство. Альтернативно, адсорбированная липосома может быть поглощена фагоцитарными клетками. Вслед за эндоцитозом происходит внутрилизосомное разложение липосомных липидов и высвобождение инкапсулированных лекарственных средств (Scherphof et al., Ann. N.Y. Acad. Sci. 446:368 (1985)). После внутривенного введения мелкие липосомы (0,1-10 мкм) обычно поглощаются клетками ретикулоэндотелиальной системы, расположенными, главным образом, в печени и селезенке, в то время как липосомы больше 3,0 мкм осаждаются в легких. Такое предпочтительное поглощение более мелких липосом клетками ретикулоэндотелиальной системы используется для доставки химиотерапевтических средств в макрофаги и опухоли печени.
Ретикулоэндотелиальную систему можно обойти несколькими способами, включая насыщение большими дозами липосомных частиц или избирательную инактивацию макрофагов фармакологическими средствами (Claassen et al., Biochim. Biophys. Acta 802:428 (1984)). Кроме того, установлено, что включение в липосомные мембраны фосфолипидов, являющихся производными гликолипида или полиэтиленгликоля, вызывает значительно меньшее поглощение ретикулоэндотелиальной системой (Allen et al., Biochim. Biophys. Acta 1068:133 (1991); Allen et al., Biochim. Biophys. Acta 1150:9 (1993)).
Можно также получить липосомы, направленно воздействующие на определенные клетки или органы, что достигается изменением фосфолипидного состава или введением в липосомы рецепторов или лигандов. Например, липосомы с высоким содержанием неионного поверхностно-активного вещества используют для направленного воздействия на печень (Hayakawa et al., патент Японии 04-244018; Kato et al., Biol. Pharm. Bull. 16:960 (1993)). Такие препараты были получены путем смешивания соевого фосфатидилхолина, α-токоферола и этоксилированного и гидрогенизированного касторового масла (НСО-60) в метаноле, концентрирования полученной смеси в вакууме и восстановления смеси водой. Установлено, что липосомный препарат дипальмитоилфосфатидилхолина (DPPC), содержащий смесь соевого стерилглюкозида (SG) и холестерина (Ch), направленно воздействует на печень (Shimizu et al., Biol. Pharm. Bull. 20:881 (1997)).
Альтернативно, с поверхностью липосомы могут быть связаны различные лиганды направленного действия, такие как антитела, фрагменты антител, углеводороды, витамины и транспортные белки. Например, липосомы могут быть модифицированы производными галактозиллипидов с разветвленной цепью для направленного воздействия на рецепторы азиалогликопротеина (галактозы), которые экспрессированы исключительно на поверхности клеток печени (Kato and Sugiyama, Crit. Rev. Ther. Drug Carrier Syst. 14:287 (1997); Murahashi et al., Biol. Pharm. Bull. 20: 259 (1997)). Аналогичным образом, Wu et al., Hepatology 27:772 (1998) показали, что мечение липосом азиалофетуином сокращает период полужизни липосом в плазме и значительно увеличивает поглощение липосом, меченных азиалофетуином, гепатоцитами. С другой стороны, накопление в печени липосом, содержащих производные галактозиллипидов с разветвленной цепью, можно ингибировать предварительной инъекцией азиалофетуина (Murahashi et al., Biol. Pharm. Bull. 20:259 (1997)). Применение полиаконитилированных липосом сывороточного альбумина человека является еще одним способом доставки липосом в клетки печени (Kamps et al., Proc. Nat'l Acad Sci. USA 94:11681 (1997)). Кроме того, Geho et al. в патенте США № 4603044 описали систему направленной доставки липосом в гепатоциты, которая является специфичной для печеночно-желчных рецепторов, ассоциированных со специализированными метаболическими клетками печени.
Общим подходом к направленному воздействию на ткани является предварительное мечение клеток-мишеней биотинилированными антителами, специфическими для лиганда, экспрессируемого клеткой-мишенью (Harasym et al., Adv. Drug. Deliv. Rev. 32:99 (1998)). После удаления из плазмы свободного антитела вводят стрептавидин-конъюгированные липосомы. В соответствии с другим подходом антитела направленного действия присоединяют непосредственно к липосомам (Harasym et al., Adv. Drug. Deliv. Rev. 32:99 (1998)).
Полипептиды, обладающие zcytor17lig-связывающей активностью, могут быть инкапсулированы в липосомы стандартными методами микроинкапсулирования белка (см., например, Anderson et al., Infect. Immun. 31:1099 (1981), Anderson et al., Cancer Res. 50:1853 (1990), and Cohen et al., Biochim. Biophys. Acta 1063:95 (1991), Alving et al. "Preparation and Use of Liposomes in Immunological Studies," in Liposome Technology, 2nd Edition, Vol. III, Gregoriadis (ed.), page 317 (CRC Press 1993), Wassef et al., Meth. Enzymol. 149:124 (1987)). Как было отмечено выше, терапевтически пригодные липосомы могут содержать различные компоненты. Например, липосомы могут содержать липидные производные полиэтиленгликоля (Allen et al., Biochim. Вiophys. Acta 1150:9 (1993)).
Для поддержания высоких системных уровней лекарственных белков были созданы разрушаемые полимерные микросферы. Указанные микросферы получают из разрушаемых полимеров, таких как сополимер лактида с гликолидом (PLG), полиангидриды, сложные полиортоэфиры, биологически неразрушаемые полиэтилвинилацетаты, в которых белки заключены в полимер (Gombotz and Pettit, Bioconjugate Chem. 6:332 (1995); Ranade, "Role of Polymers in Drug Delivery," in Drug Delivery Systems, Ranade and Hollinger (eds.), pages 51-93 (CRC Press 1995); Roskos and Maskiewicz, "Degragable Controlled Release Systems Useful for Protein Delivery," in Protein Delivery: Physical Systems, Sanders and Hendren (eds.), pages 45-92 (Plenum Press 1997); Bartus et al., Science 281:1161 (1998); Putney and Burke, Nature Biotechnology 16:153 (1998); Putney, Curr. Opin. Chem. Biol. 2:548 (1998)). Наносферы, покрытые полиэтиленгликолем (PEG), также являются приемлемыми носителями для внутривенного введения лекарственных белков (см., например, Gref et al., Pharm. Biotechnol. 10:167 (1997)).
Настоящее изобретение относится также к химически модифицированным полипептидам, обладающим активностью zcytor17lig, таким как полипептид zcytor17lig, агонисты zcytor17lig и антагонисты zcytor17lig, например антитела против zcytor17lig, которые связаны с вышеописанным полимером.
Специалистам должны быть известны другие дозированные лекарственные формы, например, описанные в публикациях Ansel and Popovich, Pharmaceutical Dosage Forms and Drug Delivery Systems, 5th Edition (Lea & Febiger 1990), Gennaro (ed.), Remington's Pharmaceutical Sciences, 19th Edition (Mack Publishing Company 1995), and Ranade and Hollinger, Drug Delivery Systems (CRC Press 1996).
В качестве примера можно отметить, что фармацевтические композиции могут поставляться в виде набора, содержащего емкость, в которой находится полипептид zcytor17lig или антагонист zcytor17lig (например, антитело или фрагмент антитела, который связывается с полипептидом zcytor17lig). Лекарственные полипептиды могут быть получены в виде инъекционного раствора в расчете на одну или несколько доз либо в виде стерильного порошка, который восстанавливают до введения инъекции. Альтернативно, такой набор может включать в себя диспергатор сухого порошка, генератор жидкого аэрозоля или распылитель для введения лекарственного полипептида. Такой набор может дополнительно содержать письменную информацию о назначении и применении лекарственной композиции. Кроме того, такая информация может содержать уведомление о том, что композиция на основе zcytor17lig противопоказана пациентам с повышенной чувствительностью к zcytor17lig.
Первым объектом настоящего изобретения является выделенный полипептид, содержащий последовательность аминокислотных остатков, которая по меньшей мере на 90% идентична последовательности аминокислотных остатков, выбираемых из следующей группы: (а) полипептид от остатка 38 (Val) до остатка 152 (Leu), как представлено в SEQ ID NO:2; (b) полипептид от остатка 27 (Leu) до остатка 164 (Thr), как представлено в SEQ ID NO:2; (с) полипептид от остатка 24 (Thr) до остатка 164 (Thr), как представлено в SEQ ID NO:2; и (d) полипептид от остатка 1 (Met) до остатка 164 (Thr), как представлено в SEQ ID NO: 2. В одном варианте осуществления изобретения выделенный полипептид аналогичен вышеуказанным, при этом аминокислотные остатки 73, 133 и 147 являются цистеином. В другом варианте осуществления изобретения выделенный полипептид аналогичен вышеуказанным, при этом данный полипептид связывается с рецептором zcytor17, представленным в SEQ ID NO:5 или SEQ ID NO:71. В другом варианте осуществления изобретения выделенный полипептид содержит по меньшей мере 14 смежных аминокислотных остатков SEQ ID NO:2 или SEQ ID NO:11. В другом варианте осуществления изобретения выделенный полипептид аналогичен вышеуказанным, при этом аминокислотные остатки выбирают из следующей группы: (а) аминокислотные остатки 38-52 SEQ ID NO:2; (b) аминокислотные остатки 83-98 SEQ ID NO:2; (с) аминокислотные остатки 104-117 SEQ ID NO:2; и (d) аминокислотные остатки 137-152 SEQ ID NO:2.
Вторым объектом настоящего изобретения является слитый белок, содержащий по меньшей мере четыре полипептида, расположенных от N-конца к С-концу в следующем порядке: первый полипептид, содержащий последовательность аминокислотных остатков 38-52 SEQ ID NO:2; первый спейсер, содержащий 6-27 аминокислотных остатков; второй полипептид, содержащий последовательность аминокислотных остатков, выбираемых из следующей группы: (а) аминокислотные остатки спирали В IL-2 SEQ ID NO:168; (b) остатки 65-83 спирали В IL-4 SEQ ID NO:164; (с) остатки 73-86 спирали В IL-3 SEQ ID NO:102; (d) остатки 72-81 спирали В GM-CSF SEQ ID NO:166; и (e) аминокислотные остатки 83-98 SEQ ID NO:2; второй спейсер, содержащий 5-11 аминокислотных остатков; третий полипептид, содержащий последовательность аминокислотных остатков, выбираемых из следующей группы: (а) остатки 102-116 спирали С IL-2 SEQ ID NO:162; (b) остатки 94-118 спирали С IL-4 SEQ ID NO:164; (с) остатки 91-103 спирали С IL-3 SEQ ID NO:102; (d) остатки 85-103 спирали С GM-CSF SEQ ID NO:166; и (е) аминокислотные остатки 104-117 SEQ ID NO:2; третий спейсер, содержащий 3-29 аминокислотных остатков; и четвертый полипептид, содержащий последовательность аминокислотных остатков, выбираемых из следующей группы: (а) остатки 134-149 спирали D IL-2 SEQ ID NO: 162; (b) остатки 123-141 спирали D IL-3 SEQ ID NO:102; (с) остатки 133-151 спирали D IL-4 SEQ ID NO:164; (d) остатки 120-131 спирали D GM-CSF SEQ ID NO:166; и (е) аминокислотные остатки 137-152 SEQ ID NO:2.
Третьим объектом настоящего изобретения является слитый белок, содержащий по меньшей мере четыре полипептида, расположенных от N-конца к С-концу в следующем порядке: первый полипептид, содержащий последовательность аминокислотных остатков, выбираемых из следующей группы: (а) остатки 27-48 спирали А IL-2 SEQ ID NO:162; (b) остатки 30-42 спирали А IL-4 SEQ ID NO:164; (с) остатки 35-45 спирали А IL-3 SEQ ID NO:102; (d) остатки 30-44 спирали А GM-CSF SEQ ID NO:166; и (е) аминокислотные остатки 38-52 SEQ ID NO:2; первый спейсер, содержащий 6-27 аминокислотных остатков; второй полипептид, содержащий последовательность аминокислотных остатков, выбираемых из следующей группы: (а) аминокислотные остатки спирали В IL-2 SEQ ID NO:168; (b) остатки 65-83 спирали В IL-4 SEQ ID NO:164; (с) остатки 73-86 спирали В IL-3 SEQ ID NO:102; (d) остатки 72-81 спирали В GM-CSF SEQ ID NO:166; и (е) аминокислотные остатки 83-98 SEQ ID NO:2; второй спейсер, содержащий 5-11 аминокислотных остатков; третий полипептид, содержащий последовательность аминокислотных остатков, выбираемых из следующей группы: (а) остатки 102-116 спирали С IL-2 SEQ ID NO:162; (b) остатки 94-118 спирали С IL-4 SEQ ID NO:164; (с) остатки 91-103 спирали С IL-3 SEQ ID NO:102; (d) остатки 85-103 спирали С GM-CSF SEQ ID NO:166; и (е) аминокислотные остатки 104-117 SEQ ID NO:2; третий спейсер, содержащий 3-29 аминокислотных остатков, и четвертый полипептид, содержащий последовательность аминокислотных остатков 137-152 SEQ ID NO:2. В другом варианте осуществления изобретения слитый белок аналогичен описанному выше, при этом четвертый полипептид содержит аминокислотные остатки 137-152 SEQ ID NO:2.
Другим объектом настоящего изобретения является молекула выделенного полинуклеотида, содержащего последовательность нуклеотидов, кодирующую вышеописанный полипептид. В одном варианте осуществления изобретения выделенный полинуклеотид аналогичен описанному выше, при этом нуклеотиды выбирают из следующей группы: (а) полинуклеотид, показанный в SEQ ID NO:1 от нуклеотида 139 до нуклеотида 483; (b) полинуклеотид, показанный в SEQ ID NO:1 от нуклеотида 106 до нуклеотида 519; (с) полинуклеотид, показанный в SEQ ID NO:1 от нуклеотида 97 до нуклеотида 519; и (d) полинуклеотид, показанный в SEQ ID NO:1 от нуклеотида 28 до нуклеотида 519.
Другим объектом настоящего изобретения является молекула выделенного полинуклеотида, содержащего последовательность нуклеотидов, кодирующую вышеописанный полипептид.
Другим объектом настоящего изобретения является экспрессирующий вектор, содержащий следующие функционально связанные элементы: (а) промотор транскрипции; (b) сегмент ДНК, кодирующий полипептид, содержащий последовательность аминокислотных остатков, выбираемых из следующей группы: (i) аминокислотные остатки 38-52 SEQ ID NO:2; (ii) аминокислотные остатки 83-98 SEQ ID NO:2; (iii) аминокислотные остатки 104-117 SEQ ID NO:2; (iv) аминокислотные остатки 137-152 SEQ ID NO:2; и (v) комбинацию указанных остатков; и (с) терминатор транскрипции.
Другим объектом настоящего изобретения является экспрессирующий вектор, содержащий следующие функционально связанные элементы: (а) промотор транскрипции; (b) сегмент ДНК, кодирующий полипептид, содержащий последовательность аминокислотных остатков, которые по меньшей мере на 90% идентичны остаткам с 38 (Val) по 152 (Leu), показанным в SEQ ID NO:2; и (с) терминатор транскрипции. В одном варианте осуществления изобретения экспрессирующий вектор аналогичен вышеуказанным и содержит следующие функционально связанные элементы: (а) промотор транскрипции; (b) сегмент ДНК, кодирующий полипептид, содержащий аминокислотные остатки с 38 (Val) по 152 (Leu) SEQ ID NO:2; и (с) терминатор транскрипции.
Другим объектом настоящего изобретения является культивируемая клетка, содержащая вышеописанный экспрессирующий вектор.
Другим объектом настоящего изобретения является способ продуцирования белка, который включает в себя культивирование вышеописанной клетки в условиях, пригодных для экспрессии сегмента ДНК, и выделение белка, кодируемого указанным сегментом ДНК.
Другим объектом настоящего изобретения является способ продуцирования антитела к полипептиду zcytor17lig, который включает в себя введение животному полипептида, выбираемого из следующей группы: (а) полипептид, содержащий 9-141 аминокислоту, который идентичен последовательности смежных аминокислотных остатков SEQ ID NO:2 от аминокислоты 24 (Ser) до аминокислоты 164 (Thr); (b) вышеописанный полипептид; (с) полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, включающую в себя аминокислоты 38-52; (d) полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, включающую в себя аминокислоты 83-98; (е) полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, включающую в себя аминокислоты 104-117, (f) полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, включающую в себя аминокислоты 137-152; (g) полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, включающую в себя аминокислоты 38-152; (h) полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, включающую в себя аминокислоты 24-164; (с) полипептид, содержащий аминокислотную последовательность SEQ ID NO:11, включающую в себя аминокислоты 38-52; (d) полипептид, содержащий аминокислотную последовательность SEQ ID NO:11, включающую в себя аминокислоты 85-98; (е) полипептид, содержащий аминокислотную последовательность SEQ ID NO:11, включающую в себя аминокислоты 104-118; (f) полипептид, содержащий аминокислотную последовательность SEQ ID NO:11, включающую в себя аминокислоты 141-157; (g) полипептид, содержащий аминокислотную последовательность SEQ ID NO:11, включающую в себя аминокислоты 38-157; (h) полипептид, содержащий аминокислотную последовательность SEQ ID NO:11, включающую в себя аминокислоты 24-163; (i) полипептид, содержащий эпитоп, соответствующий профилю гидрофильности Хоппа/Вудса SEQ ID NO:2 или SEQ ID NO:11 на основе скользящего окна из шести остатков. Скрытые остатки G, S и Т и доступные остатки Н, Y и W не учитывались, при этом полипептид вызывает иммунную реакцию у животного, в результате которой образуется антитело; и выделение указанного антитела из организма животного.
Другим объектом настоящего изобретения является антитело (например, нейтрализующее антитело), продуцируемое вышеописанным способом, которое связывается с полипептидом SEQ ID NO:2 или SEQ ID NO:11. В одном варианте осуществления изобретения вышеописанное антитело специфически связывается с полипептидом, показанным в SEQ ID NO:2 или SEQ ID NO:11.
Другим объектом настоящего изобретения является способ стимуляции иммунной реакции у млекопитающего, подверженного воздействию антигена или патогена, который включает в себя следующие стадии: (1) прямое или непрямое определение уровня антигена или патогена в организме указанного млекопитающего; (2) введение композиции, содержащей полипептид zcytor17lig в приемлемом фармацевтическом носителе; (3) прямое или непрямое определение уровня антигена или патогена в организме указанного млекопитающего; и (4) сравнение уровня антигена или патогена, измеренного на стадии 1, с уровнем антигена или патогена, измеренным на стадии 3, при этом изменение уровня свидетельствует о стимуляции иммунной реакции. В одном варианте осуществления изобретения вышеописанный способ стимуляции иммунного ответа у млекопитающего дополнительно включает в себя (5) повторное введение композиции, содержащей полипептид zcytor17lig в приемлемом фармацевтическом носителе; (6) прямое или непрямое определение уровня антигена или патогена в организме указанного млекопитающего; и (7) сравнение уровня антигена или патогена, измеренного на стадии 1, с уровнем антигена, измеренным на стадии 6, при этом изменение уровня свидетельствует о стимуляции иммунной реакции.
Другим объектом настоящего изобретения является способ размножения гемопоэтических клеток и клеток-предшественников гемопоэтических клеток, который включает в себя культивирование клеток костного мозга или периферической крови с композицией, содержащей zcytor17lig в количестве, достаточном для увеличения числа лимфоидных клеток в клетках костного мозга или периферической крови по сравнению с клетками костного мозга или периферической крови, культивируемыми без zcytor17lig. В одном варианте осуществления изобретения способ размножения гемопоэтических клеток и клеток-предшественников гемопоэтических клеток аналогичен вышеописанному, при этом гемопоэтические клетки и клетки-предшественники гемопоэтических клеток являются лимфоидными клетками. В другом варианте осуществления изобретения способ размножения гемопоэтических клеток и клеток-предшественников гемопоэтических клеток аналогичен вышеописанному, при этом лимфоидные клетки являются моноцитами, макрофагами или Т-клетками.
Другим объектом настоящего изобретения является способ стимуляции иммунной реакции у млекопитающего, подверженного воздействию антигена или патогена, который включает в себя (1) определение уровня антиген- или патогенспецифического антитела; (2) введение композиции, содержащей полипептид zcytor17lig в приемлемом фармацевтическом носителе; (3) определение уровня антиген- или патогенспецифического антитела после введения композиции; (4) сравнение уровня антитела, измеренного на стадии (1), с уровнем антитела, измеренным на стадии (3), при этом увеличение уровня антитела свидетельствует о стимуляции иммунной реакции.
Другим объектом настоящего изобретения является способ обнаружения РНК zcytor17lig в биологическом образце, который включает в себя следующие стадии: (а) контактирование зонда нуклеиновой кислоты zcytor17lig в условиях гибридизации с (i) молекулами испытуемой РНК, выделенными из биологического образца, или (ii) молекулами нуклеиновой кислоты, синтезированными из молекул выделенной РНК, при этом зонд имеет нуклеотидную последовательность, содержащую часть нуклеотидной последовательности вышеописанной молекулы нуклеиновой кислоты или ее комплемент, и (b) обнаружение образования гибридов зонда нуклеиновой кислоты с молекулами испытуемой РНК или молекулами синтезированной нуклеиновой кислоты, при этом наличие гибридов свидетельствует о присутствии РНК zcytor17lig в биологическом образце.
Другим объектом настоящего изобретения является способ обнаружения zcytor17lig в биологическом образце, который включает в себя следующие стадии: (а) контактирование биологического образца с вышеописанным антителом или фрагментом антитела, которое осуществляют в условиях, делающих возможным связывание антитела или фрагмента антитела с биологическим образцом, и (b) обнаружение любого связанного антитела или связанного фрагмента антитела.
Другим объектом настоящего изобретения является способ уничтожения раковых клеток, который включает в себя получение у субъекта ex vivo тканевого или биологического образца, содержащего раковые клетки, или идентификацию раковых клеток in vivo; продуцирование полипептида вышеописанным способом; получение препарата, содержащего данный полипептид в фармацевтически приемлемом носителе; и введение указанного препарата пациенту или воздействие данным полипептидом на раковые клетки, в результате которого полипептид уничтожает указанные клетки. В одном варианте осуществления изобретения способ уничтожения раковых клеток аналогичен описанному выше, при этом полипептид далее конъюгирован с токсином. В одном варианте осуществления изобретения антитело аналогично описанному выше, при этом антитело выбирают из следующей группы: (а) поликлональное антитело, (b) моноклональное антитело мыши, (с) гуманизированное антитело, полученное из антитела (b), (d) фрагмент антитела и (е) моноклональное антитело человека.
Другим объектом настоящего изобретения является антитело или фрагмент антитела, которое специфически связывается с полипептидом, содержащим последовательность аминокислотных остатков, выбираемых из следующей группы: (а) полипептид, содержащий остатки с 38 (Val) по 152 (Leu), как представлено в SEQ ID NO:2; (b) полипептид, содержащий остатки с 27 (Leu) по 164 (Thr), как представлено в SEQ ID NO:2; (с) полипептид, содержащий остатки с 24 (Thr) по 164 (Thr), как представлено в SEQ ID NO:2; и (d) полипептид, содержащий остатки с 1 (Met) по 164 (Thr), как представлено в SEQ ID NO:2. В другом варианте осуществления изобретения антитело аналогично описанному выше, при этом указанное антитело дополнительно содержит радиоизотоп, фермент, субстрат, кофактор, флуоресцентный маркер, хемилюминесцентный маркер, пептидную метку, магнитную частицу, лекарственное средство или токсин.
Другим объектом настоящего изобретения является способ ингибирования zcytor17lig-индуцированной пролиферации или дифференцировки гемопоэтических клеток и клеток-предшественников гемопоэтических клеток, который включает в себя культивирование клеток костного мозга или периферической крови с композицией, содержащей вышеописанное антитело в количестве, достаточном для уменьшения пролиферации или дифференцировки гемопоэтических клеток в клетках костного мозга или периферической крови по сравнению с клетками костного мозга или периферической крови, культивируемыми в отсутствие растворимого рецептора цитокина. В одном варианте осуществления изобретения способ ингибирования zcytor17lig-индуцированной пролиферации или дифференцировки гемопоэтических клеток и клеток-предшественников гемопоэтических клеток аналогичен описанному выше, при этом гемопоэтические клетки и клетки-предшественники гемопоэтических клеток являются лимфоидными клетками. В другом варианте осуществления изобретения способ ингибирования zcytor17lig-индуцированной пролиферации или дифференцировки гемопоэтических клеток и клеток-предшественников гемопоэтических клеток аналогичен описанному выше, при этом лимфоидные клетки являются макрофагами или Т-клетками.
Другим объектом настоящего изобретения является способ уменьшения zcytor17lig-индуцированного воспаления, который включает в себя введение млекопитающему, имеющему воспаление, композиции, содержащей вышеописанное антитело в количестве, достаточном для ослабления воспаления.
Другим объектом настоящего изобретения является способ подавления воспалительной реакции у млекопитающего, имеющего воспаление, который включает в себя (1) определение уровня воспалительных молекул; (2) введение композиции, содержащей вышеописанное антитело в приемлемом фармацевтическом носителе; (3) определение уровня воспалительных молекул после введения композиции; (4) сравнение уровня воспалительных молекул, измеренного на стадии (1), с уровнем воспалительных молекул, измеренным на стадии (3), при этом отсутствие увеличения или понижение уровня воспалительных молекул свидетельствует о подавлении воспалительной реакции. В одном варианте осуществления изобретения антитело аналогично описанному выше, при этом указанное антитело дополнительно содержит радиоизотоп, фермент, субстрат, кофактор, флуоресцентный маркер, хемилюминесцентный маркер, пептидную метку, магнитную частицу, лекарственное средство или токсин.
Другим объектом настоящего изобретения является способ ингибирования zcytor17lig-индуцированной пролиферации или дифференцировки гемопоэтических клеток и клеток-предшественников гемопоэтических клеток, который включает в себя культивирование клеток костного мозга или периферической крови с композицией, содержащей вышеописанное антитело в количестве, достаточном для уменьшения пролиферации или дифференцировки гемопоэтических клеток в клетках костного мозга или периферической крови по сравнению с клетками костного мозга или периферической крови, культивируемых в отсутствие растворимого рецептора цитокина. В одном варианте осуществления изобретения способ ингибирования zcytor17lig-индуцированной пролиферации или дифференцировки гемопоэтических клеток и клеток-предшественников гемопоэтических клеток аналогичен описанному выше, при этом гемопоэтические клетки и клетки-предшественники гемопоэтических клеток являются лимфоидными клетками. В другом варианте осуществления изобретения способ ингибирования zcytor17lig-индуцированной пролиферации или дифференцировки гемопоэтических клеток и клеток-предшественников гемопоэтических клеток аналогичен описанному выше, при этом лимфоидные клетки являются макрофагами или Т-клетками.
Другим объектом настоящего изобретения является способ уменьшения zcytor17lig-индуцированного воспаления, который включает в себя введение млекопитающему, имеющему воспаление, композиции, содержащей вышеописанное антитело в количестве, достаточном для уменьшения воспаления.
Другим объектом настоящего изобретения является способ подавления воспалительной реакции у млекопитающего, имеющего воспаление, который включает в себя (1) определение уровня воспалительных молекул; (2) введение композиции, содержащей вышеописанное антитело в приемлемом фармацевтическом носителе; (3) определение уровня воспалительных молекул после введения композиции; (4) сравнение уровня воспалительных молекул, измеренного на стадии (1), с уровнем воспалительных молекул, измеренным на стадии (3), при этом отсутствие повышения или снижение уровня воспалительных молекул свидетельствует о подавлении воспалительной реакции.
Другим объектом настоящего изобретения является способ лечения млекопитающего, страдающего воспалительным заболеванием, опосредованным zcytor17lig, который включает в себя введение млекопитающему антагониста zcytor17lig в количестве, достаточном для уменьшения воспаления, при этом указанный антагонист выбирают из группы, включающей в себя антитело или связывающий полипептид, который специфически связывает полипептид или фрагмент полипептида zcytor17lig (SEQ ID NO:2). В одном варианте осуществления изобретения способ лечения млекопитающего, страдающего воспалительным заболеванием, аналогичен описанному выше, при этом указанное заболевание является хроническим воспалительным заболеванием. В другом варианте осуществления изобретения способ лечения млекопитающего, страдающего воспалительным заболеванием, аналогичен описанному выше, при этом указанное заболевание является хроническим воспалительным заболеванием, выбираемым из группы, включающей в себя воспалительное заболевание кишечника, неспецифический язвенный колит, болезнь Крона, атопический дерматит, экзему и псориаз. В другом варианте осуществления изобретения способ лечения млекопитающего, страдающего воспалительным заболеванием, аналогичен описанному выше, при этом указанное заболевание является острым воспалительным заболеванием. В другом варианте осуществления изобретения способ лечения млекопитающего, страдающего воспалительным заболеванием, аналогичен описанному выше, при этом указанное заболевание является острым воспалительным заболеванием, выбираемым из группы, включающей в себя эндотоксемию, септицемию, синдром токсического шока и инфекционное заболевание. В другом варианте осуществления изобретения способ лечения млекопитающего, страдающего воспалительным заболеванием, аналогичен описанному выше, при этом антитело дополнительно содержит радиоизотоп, фермент, субстрат, кофактор, флуоресцентный маркер, хемилюминесцентный маркер, пептидную метку, магнитную частицу, лекарственное средство или токсин.
Другим объектом настоящего изобретения является способ обнаружения воспаления у пациента, который включает в себя получение у пациента тканевого или биологического образца; инкубацию тканевого или биологического образца с вышеописанным антителом в условиях, при которых антитело связывается с комплементарным полипептидом в тканевом или биологическом образце; визуализацию антитела, связанного с тканевым или биологическим образцом; и сравнение уровней антитела, связанного с тканевым или биологическим образцом, взятым у пациента, с нормальным контрольным тканевым или биологическим образцом, при этом увеличение уровня антитела, связанного с тканевым или биологическим образцом, по сравнению с нормальным контрольным тканевым или биологическим образцом свидетельствует о наличии воспаления у указанного пациента.
Другим объектом настоящего изобретения является способ обнаружения воспаления у пациента, который включает в себя получение тканевого или биологического образца у пациента; мечение полинуклеотида, содержащего по меньшей мере 14 смежных нуклеотидов SEQ ID NO:1 или комплемент SEQ ID NO:1; инкубацию тканевого или биологического образца в условиях, при которых полинуклеотид гибридизуется с комплементарной последовательностью полинуклеотида; визуализацию меченого полинуклеотида в тканевом или биологическом образце; и сравнение уровня гибридизации меченого полинуклеотида с тканевым или биологическим образцом, взятым у пациента, с нормальным контрольным тканевым или биологическим образцом, при этом увеличение уровня гибридизации меченого полинуклеотида с тканевым или биологическим образцом пациента по сравнению с нормальным контрольным тканевым или биологическим образцом свидетельствует о наличии воспаления у указанного пациента.
Данное изобретение далее проиллюстрировано следующими примерами, не ограничивающими объем изобретения.
ПРИМЕРЫ
Пример 1
Создание химеры MPL-полипептид zcytor17: слияние внеклеточного и трансмембранного (ТМ) доменов MPL с внутриклеточным сигнальным доменом zcytor17
5'-Концевой внеклеточный домен рецептора MPL мыши выделяли из плазмиды, содержащей рецептор MPL мыши (плазмида PHZ1/MPL) путем гидролиза EcoRI и BamHI, в результате чего был получен фрагмент длиной 1164 п.н. Гидролиз выполняли в 1% агарозном геле, и указанный фрагмент выделяли при помощи набора для гель-экстракции Qiaquick (Qiagen), в соответствии с инструкциями изготовителя. Остальную часть внеклеточного и трансмембранного домена MPL получали при помощи PCR, используя затравки ZC6673 (SEQ ID NO:13) и ZC29082 (SEQ ID NO:14). Реакцию выполняли в следующих условиях: 15 циклов при 94°C в течение 1 минуты, при 55°C в течение 1 минуты, при 72°C в течение 2 минут; затем при 72°C в течение 7 минут; затем выдерживание при 4°C. Продукт PCR разделяли электрофорезом в 1% агарозном геле, и фрагмент рецептора MРL длиной примерно 400 п.н. выделяли при помощи набора для гель-экстракции Qiaquick™ (Qiagen), в соответствии с инструкциями изготовителя.
Внутриклеточный домен рецептора zcytor17 человека выделяли из плазмиды, содержащей кДНК рецептора zcytor17 (№ 23/pCAP) при помощи PCR, используя затравки ZC29083 (SEQ ID NO:15) и ZC29145 (SEQ ID NO:16). Полинуклеотидная последовательность, соответствующая кодирующей последовательности рецептора zcytor17, показана в SEQ ID NO:5. Условия реакции были аналогичны вышеуказанным. Продукт PCR разделяли электрофорезом в 1% агарозном геле, и фрагмент zcytor17 длиной примерно 320 п.н. выделяли при помощи набора для гель-экстракции Qiaquick, в соответствии с инструкциями изготовителя.
Все вышеописанные выделенные фрагменты PCR смешивали в объемном соотношении 1:1 и использовали при выполнении реакции PCR с применением затравок ZC6673 (SEQ ID NO:13) и ZC29145 (SEQ ID NO:16) для создания химеры MPL-zcytor17 без 5'-концевой части MPL. Реакцию выполняли в следующих условиях: 15 циклов при 94°C в течение 1 минут, при 55°C в течение 1 минуты, при 72°C в течение 2 минут, затем при 72°C в течение 7 минут; затем выдерживание при 4°C. Весь продукт PCR разделяли электрофорезом в 1% агарозном геле и выделяли фрагмент химеры MPL-zcytor17 длиной примерно 700 п.н. при помощи набора для гель-экстракции Qiaquick (Qiagen), в соответствии с инструкциями изготовителя. Фрагмент химеры MPL-zcytor17 гидролизовали BamHI (BRL) и XbaI (Boehringer Mannheim) в соответствии с инструкциями изготовителя. Весь гидролизат разделяли электрофорезом в 1% агарозном геле, и выделяли расщепленную химеру MPL-zcytor17 при помощи набора для гель-экстракции Qiaquick™ (Qiagen), в соответствии с инструкциями изготовителя. Полученную расщепленную химеру MPL-zcytor17 вместе с вышеуказанным 5'-концевым фрагментом MPL EcoRI/BamHI вставляли в экспрессирующий вектор, получая при этом описанный ниже полный химерный рецептор MPL-zcytor17.
Экспрессирующий вектор pZP-7 реципиента гидролизовали EcoRI (BRL) и XbaI (BRL) в соответствии с инструкциями изготовителя и очищали в вышеуказанном геле. Данный фрагмент вектора объединяли при помощи реакции лигирования с EcoRI- и XbaI-расщепленной химерой MPL-zcytor17, выделенной выше при помощи PCR, и выделенным выше 5'-концевым фрагментом MPL EcoRI и BamHI. Лигирование выполняли, используя лигазу Т4 (Epicentre Technologies), при комнатной температуре в течение 1 часа в соответствии с инструкциями изготовителя. Лигированный образец электропорировали в электрокомпетентные клетки DH10B ElectroMAX™ E. coli (25 мкФ, 200 Ом, 1,8 В). Трансформанты культивировали на планшетах, содержащих среду LB с ампициллином, и единичные колонии исследовали при помощи набора Miniprep (Qiagen) и гидролизовали EcoRI для проверки на наличие химеры MPL-zcytor17. В результате гидролиза EcoRI правильных клонов был получен фрагмент длиной около 2 т.п.н. Наличие химерной последовательности MPL-zcytor17 подтверждали анализами последовательности. Вставка имела длину около 3,1 т.п.н. и была непроцессированной.
Пример 2
Пролиферация на основе химеры MPL-zcytor17 при выполнении анализа клеток BAF3 с аламаровым синим
А. Создание клеток BaF3, экспрессирующих химеру MPL-zcytor17
Клетки BaF3, представляющие собой линию интерлейкин-3 (IL-3) зависимых прелимфоидных клеток, выделенных из костного мозга мышей (Placios and Steinmetz, Cell 41:727-734, 1985; Mathey-Prevot et al., Mol. Cell. Biol. 6:4133-4135, 1986), помещали в полную среду (среда RPMI, JRH Bioscience Inc., Lenexa, KS), содержащую 10% термоинактивированной фетальной телячьей сыворотки, 1-2 нг/мл IL-3 мыши (mIL-3) (R&D, Minneapolis, MN), 2 мМ L-glutaMax-1™ (Gibco BRL), 1 мМ пирувата натрия (Gibco, BRL) и антибиотик PSN (Gibco BRL)). До электропорации получали плазмидную ДНК pZP-7/MPL-zcytor17 (пример 1), которую очищали при помощи набора Maxi Prep Qiagen (Qiagen), в соответствии с инструкциями изготовителя. Предназначенные для электропорации клетки BaF3 дважды промывали в среде RPMI и затем вновь суспендировали в среде RPMI при плотности клеток 107 клеток/мл. Один мл повторно суспендированных клеток BaF3 смешивали с 30 мкг плазмидной ДНК pZP-7/MPL-zcytor17 и переносили в отдельные одноразовые камеры для электропорации (GIBCO BRL). При комнатной температуре в аппарате для электропорации (Cyto-Pulse) в клетках вызывали шок, который повторяли 5 раз длительностью 0,1 мсек при напряжении 800 В и затем 5 раз длительностью 2 мсек при напряжении 600 В. Альтернативно, клетки электропорировали двумя последовательными импульсами (800 мкФ/300 В и затем 1180 мкФ/300 В) в аппарате для электропорации Cell-Porator (Gibco BRL). Электропорированные клетки переносили в 50 мл полной среды и помещали в инкубатор на 15-24 часа (37°C, 5% СО2). Затем к клеткам, находящимся в колбе Т-162, добавляли селективную среду (1 мг/мл G418) Geneticin™ (Gibco) для выделения G418-устойчивого пула. Пулы трансфицированных клеток BaF3, далее именуемых клетками BaF3/MPL-zcytor17, анализировали на способность передавать сигналы, как это описано ниже.
В. Испытание на способность передавать сигналы клетками BaF3/MPL-zcytor17 при выполнении анализа пролиферации с аламаровым синим
Клетки BaF3/MPL-zcytor17 центрифугировали и промывали в вышеописанной полной среде, не содержащей mIL-3 (далее именуемой «среда без mIL-3»). Клетки центрифугировали и трижды промывали для полного удаления mIL-3. Затем клетки подсчитывали в гемоцитометре. Клетки помещали на 96-луночный планшет в количестве 5000 клеток/лунку в объеме 100 мкл/лунку, используя среду без mIL-3.
Пролиферацию клеток BaF3/MPL-zcytor17 оценивали, используя тромбопоэтин мышей (mTPO), разведенный средой без mIL-3 до концентраций 200 нг/мл, 100 нг/мл, 50 нг/мл, 25 нг/мл, 12,5 нг/мл, 6,25 нг/мл, 3,1 нг/мл, 1,5 нг/мл. К клеткам BaF3/MPL-zcytor17 добавляли сто микролитров разведенного mTPO. Полный анализируемый объем был равен 200 мкл. Параллельно исследовали отрицательные контрольные образцы, используя только среду без mIL-3 без добавления mTPO. Анализируемые планшеты инкубировали при 37°C, 5% СО2 в течение 3 дней, после чего добавляли аламаровый синий (Accumed, Chicago, IL) в количестве 20 мкл/лунку. Аламаровый синий позволяет получить флуорометрические данные на основе метаболической активности клеток, что является прямым измерением пролиферации клеток по сравнению с отрицательным контрольным образцом. Планшеты снова инкубировали при 37°C, 5% СО2 в течение 24 часов. Планшеты считывали в аппарате для считывания планшетов Fmax™ (Molecular Devices Sunnyvale, CA), используя программу Pro SoftMax™ при длинах волн 544 (возбуждение) и 590 (излучение) или в аппарате для считывания планшетов Wallac Victor 2 (Perkin Elmer Life Sciences, Boston, MA).
Полученные результаты подтвердили способность передавать сигналы внутриклеточной части рецептора zcytor17, так как тромбопоэтин индуцировал пролиферацию, примерно в 9-13 раз превышающую фоновую при концентрациях mTPO, составляющих 50 нг/мл и выше.
Пример 3
Создание вектора, экспрессирующего непроцессированный рецептор zcytor17: pZp7pX/zcytor17
А. Клонирование непроцессированной кДНК zcytor17 для экспрессии в векторе
Для получения непроцессированной кДНК zcytor17 5'- и 3'-концевые продукты PCR выделяли и связывали, используя внутренний сайт PstI. Затравки для PCR были получены с использованием нуклеотидной последовательности SEQ ID NO:4 и включали в себя сайты рестрикции BamHI и XhoI, предназначенные для клонирования.
5'-Концевой продукт PCR был получен с использованием библиотеки кДНК WI-38 в качестве матрицы и олигонуклеотидов ZC29359 (SEQ ID NO:18) и ZC27899 (SEQ ID NO:19) в качестве затравок. WI-38 является библиотекой кДНК, созданной авторами изобретения из линии эмбриональных клеток легкого человека (АТСС CRL-2221). Реакцию PCR 5'-концевого продукта выполняли следующим образом: 30 циклов при 94°C в течение 1 минуты, при 65°C в течение 1 минуты, при 72°C в течение 2 минут, затем при 72°C в течение 7 минут; выдерживание при 10°C. При выполнении реакции PCR использовали примерно 3 мкг плазмиды, полученной из библиотеки кДНК, 20 пмоль каждого олигонуклеотида и пять единиц ДНК-полимеразы PWO (Roche). Примерно 90% 5'-концевого продукта PCR осаждали этанолом, гидролизовали BamHI и PstI и очищали в 1,0% агарозном геле. Полосу длиной около 600 п.н. вырезали и использовали для лигирования с клонирующим вектором pUC18, гидролизованным BamHI и PstI. Полученные трансформанты секвенировали для подтверждения наличия последовательности кДНК zcytor17. Для одного из указанных трансформантов получали плазмидную ДНК и гидролизовали BamHI и PstI. Полученную полосу длиной около 600 п.н. очищали в геле и использовали для лигирования с образованием непроцессированной кДНК.
3'-Концевой продукт PCR был получен с использованием созданной авторами изобретения библиотеки кДНК человека в качестве матрицы и олигонуклеотидов ZC27895 (SEQ ID NO:20) и ZC29122 (SEQ ID NO:21) в качестве затравок. Реакцию PCR 3'-концевого продукта выполняли следующим образом: 30 циклов при 94°C в течение 45 секунд, при 65°C в течение 45 секунд, при 72°C в течение 2 минут, затем при 72°C в течение 7 минут; выдерживание при 10°C. Весь 3'-концевой продукт реакции PCR очищали в 1,0% агарозном геле и вырезали основную полосу длиной 1500 п.н. Данную полосу клонировали в векторе Blunt II TOPO для PCR при помощи набора Zeroblunt TOPO (Invitrogen). Полученные трансформанты секвенировали для подтверждения последовательности кДНК zcytor17. Для одного из указанных трансформантов получали плазмидную ДНК и гидролизовали PstI и XhoI. Полученную полосу длиной около 1500 п.н. очищали в геле. Выполняли трехкомпонентное лигирование, сшивая 5'-концевой BamHI с вышеуказанным фрагментом PstI, 3'-концевой PstI с фрагментом XhoI и экспрессирующий вектор pZp7pX, гидролизованный BamHI и XhoI. Полученная плазмида pZp7pX, содержащая непроцессированную кДНК для zcytor17 (SEQ ID NO:4), получила название pZp7p/zcytor17. Непроцессированная кДНК zcytor17 в плазмиде pZp7p/zcytor17 имеет молчащую мутацию, в соответствии с которой происходит замена остатка Т остатком G в положении 1888 SEQ ID NO:4 (кодирующей остаток Gly в положении остатка 464 SEQ ID NO:5). Так как данная мутация является молчащей, кДНК zcytor17 в pZp7p/zcytor17 кодирует полипептид, как представлено в SEQ ID NO:5. Плазмида pZp7pX является экспрессирующим вектором млекопитающего, содержащим полигенный экспрессирующий кластер, включающий в себя промотор CMV, интрон А, несколько сайтов рестрикции для вставки кодирующих последовательностей и терминатор гормона роста человека. Данная плазмида имеет также ориджин репликации E. coli, экспрессирующую единицу селектируемого маркера млекопитающего, включающую в себя промотор SV40, энхансер и ориджин репликации, ген устойчивости к пуромицину и терминатор SV40.
В. Создание вектора, экспрессирующего непроцессированный рецептор WSX-1
Полный рецептор WSX-1 (SEQ ID NO:9) выделяли из плазмиды, содержащей кДНК рецептора WSX-1 (SEQ ID NO:8) (патент США № 5925735). Плазмидную ДНК hWSX-1/pBluescript SK(+) (Stratagene, La Jolla, CA) гидролизовали EcoRI и XhoI с образованием фрагмента длиной 1075 п.н., который гидролизовали XhoI и XbaI с образованием фрагмента длиной 900 п.н. Оба гидролизата разделяли электрофорезом в 1% агарозном геле и выделяли расщепленные фрагменты WSX-1.
Экспрессирующий вектор pZp7Z реципиента гидролизовали EcoRI и XbaI и очищали в геле, как описано выше. Данный фрагмент вектора объединяли с двумя расщепленными фрагментами zcytor17, выделенными выше, осуществляя реакцию лигирования с использованием лигазы Т4 (BRL). Лигированную смесь инкубировали при комнатной температуре в течение ночи. Образец лигированной смеси электропорировали в электрокомпетентные клетки DH10B electroMAX™ E. coli (25 мкФ, 200 Ом, 2,3 В). Шесть колоний выращивали в культуре и при помощи набора Miniprep получали ДНК, которую гидролизовали для подтверждения наличия правильной непроцессированной вставки WSX-1 длиной 2,0 т.п.н. Полученная плазмида представляет собой pZPZ7Z/WSX-1.
Пример 4
Пролиферация на основе zcytor17 при выполнении анализа клеток BAF3 с аламаровым синим
А. Создание клеток BaF3, экспрессирующих рецептор zcytor17, рецепторы WSХ-1 и OSMR
Клетки BaF3, экспрессирующие непроцессированный рецептор zcytor17, были получены аналогично приведенному выше примеру 2А с использованием 30 мкг вектора экспрессии zcytor17, описанного в примере 3А. Единственным исключением является то, что вместо селективной среды, содержащей генетицин, к трансфицированным клеткам в колбе Т-162 добавляли 2 мкг/мл пуромицина (ClonTech) для выделения пула, устойчивого к пуромицину. Клетки BaF3, экспрессирующие мРНК рецептора zcytor17, получили название BaF3/zcytor17. Для получения клонов клетки BaF3/zcytor17 подсчитывали в гемоцитометре и культивировали на 96-луночных планшетах в количестве 1 клетка/лунку, 0,5 клетки/лунку, 0,1 клетки/лунку и 0,01 клетки/лунку. Пятнадцать клонов переносили в колбы Т75, и пять клонов анализировали в отношении экспрессии zcytor17. Полную РНК выделяли из клеточного дебриса при помощи набора для выделения полной РНК S.N.A.P.™ (In Vitrogen). кДНК первой цепи синтезировали при помощи набора для RT-PCR первой цепи proSTAR™ и выполняли реакцию PCR для скрининга клонов, экспрессирующих zcytor17, используя zcytor17-специфические затравки ZC29180 (SEQ ID NO:22) и ZC29122 (SEQ ID NO:23). Один клон BaF3/zcytor17 №15 был выбран для размножения и трансфекции вектором экспрессии WSX-1.
Клетки BaF3, экспрессирующие zcytor17 и непроцессированный рецептор WSX-1, были получены аналогично приведенному выше примеру 2А с использованием 30 мкг вектора экспрессии WSX-1, именуемого WSX-1/pZp7Z (пример 3В), для электропорации клеток BaF3/zcytor17 №15. Единственным исключением является то, что вместо селективной среды, содержащей генетицин, к трансфицированным клеткам в колбе Т-162 добавляли 200 мкг/мл зеоцина (In Vitrogen) для выделения пула, устойчивого к зеоцину. Клетки BaF3, экспрессирующие zcytor17 и WSX-1, получили название BaF3/zcytor17/hWSX-1. Для получения клонов пулы клеток BaF3/zcytor17/hWSX-1 культивировали при ограниченном разведении на 96-луночных планшетах. Полученные клоны размножали, и выделяли полную РНК при помощи набора для выделения полной РНК S.N.A.P.™ (In Vitrogen). кДНК первой цепи синтезировали при помощи набора для RT-PCR первой цепи proSTAR™, и выполняли реакцию PCR для скрининга клонов, экспрессирующих WSX-1, используя WSX-1-специфические затравки ZC9791 (SEQ ID NO:24) и ZC9793 (SEQ ID NO:25). Один клон, BaF3/zcytor17/hWSX-1 №5, был выбран для дальнейшего размножения и трансфекции вектором экспрессии OSMRbeta.
Клетки BaF3, экспрессирующие zcytor17, WSX-1 и непроцессированный рецептор OSMRbeta, были получены аналогично приведенному выше примеру 2А с использованием 30 мкг вектора экспрессии OSMRbeta, именуемого OSMR/pZp7NX, описанного в примере 29, для электропорации клеток BaF3/zcytor17/hWSX-1 №5. Клетки BaF3, экспрессирующие мРНК zcytor17, WSX-1 и OSMRbeta, получили название BaF3/zcytor17/WSX-1/OSMR. Для получения клонов пулы клеток ВaF3/zcytor17/WSX-1/OSMRbeta культивировали при ограниченном разведении на 96-луночных планшетах. Отдельные клоны размножали, и полную РНК выделяли при помощи набора для выделения полной РНК S.N.A.P.™ (In Vitrogen). кДНК первой цепи синтезировали при помощи набора для RT-PCR первой цепи proSTAR™, и выполняли PCR для скрининга клонов, экспрессирующих zcytor17, WSX-1 и OSMR, используя OSMRbeta-специфические затравки ZC40109 (SEQ ID NO:26) и ZC40112 (SEQ ID NO:27). Был выбран один клон BaF3/zcytor17/WSX-1/OSMR №5, и указанные клетки использовали для скрининга zcytor17lig, описанного в следующих примерах 5 и 6.
В. Создание клеток BaF3, экспрессирующих рецепторы zcytor17 и OSMR
Клетки BaF3, экспрессирующие непроцессированный рецептор zcytor17, были получены аналогично приведенному выше примеру 2А с использованием 30 мкг вектора экспрессии zcytor17, описанного в примере 3А. Единственным исключением является то, что вместо селективной среды, содержащей генетицин, к трансфицированным клеткам в колбе Т-162 добавляли 2 мкг/мл пуромицина (ClonTech) для выделения пула, устойчивого к пуромицину. Клетки BaF3, экспрессирующие мРНК рецептора zcytor17, получили название BaF3/zcytor17. Для получения клонов пулы клеток BaF3/zcytor17 культивировали при ограниченном разведении на 96-луночных планшетах. Указанные клоны размножали в культуре, и полную РНК выделяли при помощи набора для выделения полной РНК S.N.A.P.™ (In Vitrogen). кДНК первой цепи синтезировали при помощи набора для RT-PCR первой цепи proSTAR™ и затем выполняли PCR для скрининга клонов, экспрессирующих zcytor17. Один клон, BaF3/zcytor17 №15, был выбран для выращивания и трансфекции вектором экспрессии OSMRbeta.
Клетки BaF3, экспрессирующие zcytor17 и непроцессированный рецептор OSMRbeta, были получены аналогично приведенному выше примеру 2А с использованием 30 мкг вектора экспрессии OSMRbeta, именуемого OSMR/pZp7NX (пример 29), для электропорации клеток BaF3/zcytor17 №15. Клетки BaF3, экспрессирующие мРНК zcytor17 и OSMRbeta, получили название BaF3/zcytor17/OSMR. Указанные клетки использовали для скрининга zcytor17lig, как это описано в следующем примере 5.
Пример 5
Скрининг zcytor17lig с использованием клеток BaF3/zcytor16/WSX-1/OSMRbeta при выполнении анализа пролиферации с аламаровым синим
А. Активация клеток CCRF-CEM и CCRF-HSB2 для испытания на наличие zcytor17lig
Клетки CCRF-CEM и CCRF-HSB2 были получены из АТСС и стимулированы в культуре для получения кондиционированной среды, используемой для испытания на наличие активности zcytor17lig, как описано ниже. Суспензию клеток высевали в количестве 2 х 105 клеток/мл или 5 х 105 клеток/мл в среду RPMI-1640, содержащую 10% FBS, 2 мМ L-глутамина (GibcoBRL), однократный объем PSN (Gibco BRL), и активировали 10 нг/мл форбол-12-миристат-13-ацетата (РМА) (Calbiochem, San Diego, CA) и 0,5 мкг/мл иономицина™ (Calbiochem) в течение 24 или 48 часов. Супернатант стимулированных клеток использовали для анализа пролиферации клеток BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta, как это описано ниже.
В. Скрининг zcytor17lig с использованием клеток BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta при выполнении анализа пролиферации с аламаровым синим
Клетки BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta центрифугировали и промывали в среде без mIL-3. Клетки трижды центрифугировали и промывали для полного удаления mIL-3. Затем клетки подсчитывали в гемоцитометре. Клетки культивировали на 96-луночном планшете в количестве 5000 клеток/лунку в объеме 100 мкл/лунку, используя среду без mIL-3.
Пролиферацию клеток BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta оценивали, используя кондиционированную среду из активированных клеток CCRF-CEM и CCRF-HSB2 (см. пример 5А). Кондиционированную среду разводили средой без mIL-3 до концентрации, составляющей 50%, 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75% и 0,375%. К клеткам BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta добавляли сто микролитров разведенной кондиционированной среды. Общий анализируемый объем составлял 200 мкл. Анализируемые планшеты инкубировали при 37°C, 5% СО2 в течение 3-5 дней, после чего добавляли аламаровый синий (Accumed. Chicago, IL) в количестве 20 мкл/лунку. Планшеты снова инкубировали при 37°C, 5% СО2 в течение 24 часов. Планшеты считывали в аппарате для считывания планшетов Fmax™ (Molecular Devices), как описано выше (пример 2).
Полученные результаты подтвердили наличие пролиферативной реакции клеток BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta на фактор, присутствующий в среде, кондиционированной активированными клетками CCRF-CEM и CCRF-HSB2. Измеренная реакция была примерно в 10 раз выше фоновой реакции при 25% концентрации. Нетрансфицированные клетки BaF3 не пролиферировали под действием указанного фактора, как и клетки BaF3, трансфицированные рецептором zcytor17 и WSX-1 (клетки BaF3/zcytor17/WSX-1), указывая на то, что данный фактор является специфическим для рецепторов zcytor17/OSMRbeta или zcytor17/OSMRbeta/WSX-1. Кроме того, растворимый рецептор zcytor17 уменьшал указанную пролиферативную активность zcytor17lig в клетках BaF3/zcytor17/WSX-1/OSMRbeta (см. пример 11). Аналогичные результаты предполагается получить в клетках ВaF3/zcytor17/OSMRbeta.
С. Источник первичных клеток человека, используемых для выделения zcytor17lig
У шести доноров были взяты стомиллилитровые пробы крови. Для взятия проб крови использовали десять 10 мл вакуумных пробирок, содержащих гепарин. Кровь, взятую у шести доноров (600 мл), разводили в PBS в отношении 1:1 и разделяли в аппарате Ficoll-Paque® PLUS (Pharmacia Biotech). Выход выделенных первичных клеток человека после разделения в градиенте фиколла составлял 1,2 х 109 клеток.
Клетки суспендировали в 9,6 мл буфера MACS (PBS, 0,5% EDTA, 2 мМ EDTA). Удаляли 1,6 мл суспензии клеток, и добавляли 0,4 мл микрошариков CD3 (Miltenyi Biotec, Auburn, CA). Смесь инкубировали в течение 15 минут при 4°C. Указанные клетки, меченные шариками CD3, промывали 30 мл буфера MACS и затем вновь суспендировали в 2 мл буфера MACS.
Колонку VS+ (Miltenyi) готовили в соответствии с инструкциями изготовителя. Колонку VS+ помещали в магнитное поле VarioMACS™ (Miltenyi). Колонку уравновешивали 5 мл буфера MACS. В колонку вводили выделенные первичные клетки человека. CD3-отрицательные клетки проходили через колонку. Колонку промывали 9 мл (3 х 3 мл) буфера MACS. Затем колонку снимали с магнита и помещали на 15 мл фальконовую трубку. CD3+-клетки элюировали, добавляя в колонку 5 мл буфера MACS, и связанные клетки вымывали при помощи поршня, предоставленного изготовителем. Инкубацию клеток с магнитными шариками CD3, промывку и вышеописанные стадии обработки в колонке VS+ (от инкубации до элюирования) повторяли еще пять раз. Затем собирали фракции CD3+-клеток, полученные в результате шести разделений в колонке. Выход CD3+-селектированных клеток человека составлял в общей сложности 3х108 клеток.
Образец собранных CD3+-селектированных клеток человека окрашивали и сортировали в клеточном сортере с флуоресцентным антителом (FACS) для оценки чистоты клеток. CD3+-селектированные клетки человека на 91% состояли из CD3+-клеток.
CD3+-селектированные клетки человека активировали, инкубируя в среде RPMI, содержащей 5% FBS, 10 нг/мл PMA и 0,5 мкг/мл иономицина (Calbiochem), в течение 13 часов при 37°C. Супернатант указанных активированных CD3+-селектированных клеток человека испытывали в отношении активности zcytor17lig описанным ниже способом. Кроме того, активированные CD3+-селектированные клетки человека использовали для получения библиотеки кДНК, как это описано в следующем примере 6.
D. Испытание супернатанта активированных CD3+-селектированных клеток человека в отношении zcytor17lig с использованием клеток BaF3/zcytor17/WSX-1/OSMRbeta при выполнении анализа пролиферации с аламаровым синим
Клетки BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta центрифугировали и промывали в среде без mIL-3. Клетки трижды центрифугировали и промывали для полного удаления mIL-3. Затем клетки подсчитывали в гемоцитометре. Клетки переносили на 96-луночный планшет в количестве 5000 клеток/лунку в объеме 100 мкл/лунку, используя среду без mIL-3.
Пролиферацию клеток BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta оценивали с использованием кондиционированной среды из активированных CD3+-селектированных клеток человека (см. пример 5С), разведенной средой без mIL-3 до концентрации, составляющей 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75%, 0,375% и 0,187%. К клеткам BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta добавляли сто микролитров разведенной кондиционированной среды. Общий анализируемый объем составлял 200 мкл. Исследуемые планшеты инкубировали и анализировали аналогично примеру 5В.
Полученные результаты подтвердили наличие пролиферативной реакции клеток BaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta на фактор, присутствующий в среде, кондиционированной активированными CD3+-селектированными клетками человека. Измеренная реакция была примерно в 15 раз выше фоновой при 25% концентрации. Нетрансфицированные клетки BaF3 не пролиферировали под действием указанного фактора, как и клетки BaF3, трансфицированные zcytor17 и WSX-1 (клетки ВaF3/zcytor17/WSX-1), указывая на то, что данный фактор является специфическим для рецепторов zcytor17/OSMRbeta или zcytor17/OSMRbeta/WSX-1.
Пример 6
Клонирование zcytor17lig человека из библиотеки CD3+-селектированных клеток человека
Скрининг библиотеки кДНК первичных активированных CD3+-селектированных клеток человека показал, что выделенная кДНК является новым членом семейства четырехспиральных цитокинов. Указанная кДНК кодировала zcytor17lig. кДНК была идентифицирована путем скрининга активности zcytor17lig с использованием рецепторов zcytor17/WSX-1/OSM.
А. Вектор для создания библиотеки CD3+-селектированных клеток
Для создания библиотеки CD3+-селектированных клеток был использован вектор pZP7NX. Вектор pZP7NX был создан следующим образом: кодирующую область для селективного маркера DHFR в векторе pZP7 удаляли путем гидролиза ДНК рестрикционными ферментами NcoI и PstI (Boehringer Mannheim). Гидролизованную ДНК разделяли электрофорезом в 1% агарозном геле, вырезали и очищали в геле при помощи набора для гель-экстракции Qiagen (Qiagen), в соответствии с инструкциями изготовителя. Фрагмент ДНК, представляющий собой кодирующую область селективного маркера зеоцина амплифицировали методом PCR, используя затравки ZC13946 (SEQ ID NO:28), ZC13945 (SEQ ID NO:29) и pZeoSV2(+) в качестве матрицы. В затравке ZC13946 (SEQ ID NO:28) имеются дополнительные сайты рестрикции PstI и BclI и в затравке ZC13945 (SEQ ID NO:29) имеются дополнительные сайты рестрикции NcoI и SfuI. Фрагмент PCR вырезали при помощи рестрикционных ферментов PstI и NcoI и клонировали в векторе pZP7, полученном путем расщепления двумя указанными ферментами с последующей очисткой в геле. Данный вектор получил название pZP7Z. Затем кодирующую область зеоцина удаляли путем гидролиза ДНК вектора pZP7Z рестрикционными ферментами BclI и SfuI. Гидролизованную ДНК разделяли электрофорезом в 1% агарозном геле, вырезали, очищали в геле и лигировали с фрагментом ДНК кодирующей области неомицина, вырезанной из вектора pZem228 (депонирован в Американской коллекции типовых культур (АТСС), Manassas, VA; № депозита АТСС 69446) теми же рестрикционными ферментами (BclI и SfuI).
В новом векторе, получившем название pZP7N, кодирующая область селективного маркера DHFR была заменена кодирующей областью селективного маркера неомицина из вектора pZem228. Заполняющий фрагмент, включающий сайт XhoI, добавляли к pZp7N для создания вектора, пригодного для высокоэффективного направленного клонирования кДНК; новый вектор получил название pZP7NX. Чтобы получить вектор для кДНК, 20 мкг pZР7NX гидролизовали 20 единицами EcoRI (Life Technologies Gaithersberg, MD) и 20 единицами XhoI (Boehringer Mannheim Indianapolis, IN) при 37°C в течение 5 часов, затем при 68°C в течение 15 минут. Гидролизат разделяли электрофорезом в 0,8% агарозном геле 1XTAE с низкой температурой плавления для отделения вставки от вектора. Полосу вектора вырезали и гидролизовали при помощи «бета-агарозы» (New England Biolabs, Beverly, MA), в соответствии с рекомендациями изготовителя. После осаждения этанолом гидролизованный вектор вновь суспендировали в воде до достижения 45 нг/мл для лигирования библиотеки кДНК CD3+-селектированных клеток, как это описано ниже.
В. Получение библиотеки кДНК активированных первичных CD3+-селектированных клеток человека
Примерно 1,5х108 первичных CD3+-селектированных клеток человека, стимулированных в иономицине/PMA, культивировали при 37°C в течение 13 часов и отделяли центрифугированием (пример 5С). Полную РНК выделяли из клеточного дебриса при помощи набора "RNeasy Midi" компании Qiagen, Inc. (Valencia, CA). мРНК выделяли из 225 микрограммов полной РНК при помощи набора для очистки мРНК MPG компании CPG Inc. (Lincoln Park, NJ). 3,4 микрограмма РНК выделяли и превращали в двухцепочечную кДНК следующим способом.
кДНК первой цепи, выделенную из стимулированных CD3+-селектированных клеток человека, синтезировали следующим образом. Девять мкл РНК олиго d(T)-селектированных поли(A) CD3+-клеток при концентрации 0,34 мкг/мл и 1,0 мкг/мл затравки ZC18698 (SEQ ID NO: 30) для получения первой цепи, содержащей сайт рестрикции XhoI, смешивали, нагревали при 65°C в течение 4 минут и охлаждали на льду. Синтез кДНК первой цепи инициировали, добавляя к смеси РНК и затравки 9 мкл буфера для первой цепи (5 х буфер SUPERSCRIPT®; Life Technologies), 4 мкл 100 мМ дитиотреитола и 2 мкл раствора дезоксинуклеотид-трифосфата, содержащего 10 мМ каждого из dATP, dGTP, dTTP и 5-метил-dCTP (Pharmacia Biotech Inc.). Реакционную смесь инкубировали при 45°C в течение 4 минут с последующим добавлением 8 мкл 200 ед/мкл РНКаза Н-обратной транскриптазы SupersctiptII® (Life Technologies). Реакционную смесь инкубировали при 45°C в течение 45 минут, после чего температуру инкубации повышали на 1°C каждые 2 минуты до 50°C и выдерживали реакционную смесь при указанной температуре в течение 10 минут. Для денатурации любой вторичной структуры и дополнительного удлинения кДНК реакционную смесь нагревали до 70°C в течение 2 минут, затем снижали температуру до 55°C в течение 4 минут, после чего добавляли 2 мкл обратной транскриптазы SupersctiptII®, инкубировали еще 15 минут и повышали температуру до 70°C со скоростью 1°C/мин. Невключенные нуклеотиды удаляли из кДНК двукратным осаждением в присутствии 2 мкг гликогенового носителя, 2,0 М ацетата аммония и 2,5 объема этанола с последующей промывкой 100 мкл 70% этанола. кДНК вновь суспендировали в 98 мкл воды для использования в процессе синтеза второй цепи.
Вторую цепь синтезировали на кДНК первой цепи в условиях, стимулирующих праймирование второй цепи первой цепью с образованием петли ДНК. Реакционная смесь, используемая для получения второй цепи, содержала 98 мкл кДНК первой цепи, 30 мкл 5-кратного объема буфера, содержащего полимеразу I (100 мМ трис-буфера с HCl, рН 7,5, 500 мМ KCl, 25 мМ MgCl2, 50 мМ (NH4)2SO4, 2 мкл 100 мМ дитиотреитола, 6 мкл раствора, содержащего 10 мМ дезоксинуклеотид-трифосфата, 5 мкл 5 мМ b-NAD, 1 мкл 3 ед/мкл ДНК-лигазы E. coli (New England Biolabs Inc.) и 4 мкл 10 ед/мкл ДНК-полимеразы I E. coli (New England Biolabs Inc.). Реакционную смесь объединяли при комнатной температуре, инкубировали при комнатной температуре в течение 2 минут и добавляли 4 мкл 3,8 ед/мкл РНКазы Н (Life Technologies). Реакционную смесь инкубировали при 15°C в течение двух часов и затем при комнатной температуре в течение 15 минут. К реакционной смеси добавляли десять микролитров 1 М трис-буфера, рН 7,4, и экстрагировали дважды смесью фенол/хлороформ и один раз хлороформом, органические фазы подвергали обратной экстракции 50 мкл ТЕ (10 мМ трис-буфера, рН 7,4, 1 мМ EDTA), собирали вместе с другой водной фазой и осаждали этанолом в присутствии 0,3 М ацетата натрия. Осадок промывали 100 мкл 70% этанола, подвергали воздушной сушке и вновь суспендировали в 40 мкл воды.
Одноцепочечную ДНК с петлей расщепляли при помощи нуклеазы золотистой фасоли. Реакционную смесь, содержащую 40 мкл кДНК второй цепи, 5 мкл 10-кратного объема буфера с нуклеазой золотистой фасоли (Life technologies), 5 мкл нуклеазы золотистой фасоли (Pharmacia Biotech Corp.), разводили до 1 ед/мкл в однократном объеме буфера с нуклеазой золотистой фасоли. Реакционную смесь инкубировали при 37°C в течение 45 минут. Реакцию прекращали, добавляя 10 мкл 1 М трис-буфера с HCl, pH 7,4, после чего реакционную смесь последовательно экстрагировали смесью фенол/хлороформ и хлороформом, как описано выше. После экстракции кДНК осаждали этанолом в присутствии 0,3 М раствора ацетата натрия. Осадок промывали 100 мкл 70% этанола, подвергали воздушной сушке и вновь суспендировали в 38 мкл воды.
Вновь суспендированную кДНК дефосфорилировали ДНК-полимеразой Т4. кДНК, которую вновь суспендировали в 38 мкл воды, смешивали с 12 мкл пятикратного объема буфера с ДНК-полимеразой Т4 (250 мМ трис-буфера с HCl, рН 8,0, 250 мМ KCl, 25 мМ MgCl2), 2 мкл 0,1 М дитиотреитола, 6 мкл раствора, содержащего 10 мМ каждого дезоксинуклеотид-трифосфата и 2 мкл 1 ед/мкл ДНК-полимеразы Т4 (Boehringer Mannheim Corp.). Реакционную смесь инкубировали при 15°C в течение 45 минут и реакцию прекращали, добавляя 30 мкл ТЕ, после чего реакционную смесь последовательно экстрагировали смесью фенол/хлороформ и хлороформом и подвергали обратной экстракции 20 мкл ТЕ, как описано выше. ДНК осаждали этанолом в присутствии 2 мкл носителя Pellet Paint™ (Novagen) и 0,3 М раствора ацетата натрия и вновь суспендировали в 11 мкл воды.
Адапторы EcoRI лигировали с 5'-концами вышеуказанной кДНК, клонируя ее в экспрессирующем векторе. 11 мкл кДНК и 4 мкл 65 пмоль/мкл гемифосфорилированного адаптора EcoRI (Pharmacia Biotech Corp.) смешивали с 5 мкл пятикратного объема буфера с лигазой (Life Technologies), 2 мкл 10 мМ АТР и 3 мкл 1 ед/мкл ДНК-лигазы Т4 (Life Technologies), 1 мкл 10-кратного объема буфера для лигирования (Promega Corp.) и 9 мкл воды. Дополнительное разведение однократным объемом буфера предотвращало осаждение носителя Pellet Paint. Реакционную смесь инкубировали в течение 9 часов на водяной бане при изменении температуры от 10 до 22°C в течение 9 часов и выдерживали при 25°C в течение 45 минут. Реакцию прекращали путем инкубации при 68°C в течение 15 минут.
Для облегчения направленного клонирования кДНК в экспрессирующем векторе указанную кДНК гидролизовали XhoI с образованием кДНК, имеющей «тупой» 5'-конец EcoRI и «тупой» 3'-конец XhoI. Сайт рестрикции XhoI у 3'-конца кДНК был введен заранее с использованием затравки ZC18698 (SEQ ID NO:30). Гидролиз рестрикционным ферментом осуществляли в реакционной смеси, содержащей 35 мкл вышеописанной лигированной смеси, 6 мкл 10-кратного объема буфера Н (Boehringer Mannheim Corp.), 1 мкл 2 мг/мл BSA (Biolabs Corp.), 17 мкл воды и 1,0 мкл 40 ед/мкл XhoI (Boehringer Mannheim), Гидролиз осуществляли при 37°C в течение 1 часа. Реакцию прекращали, реакционную смесь инкубировали при 68°C в течение 15 минут, осаждали этанолом, промывали, сушили вышеописанным методом и вновь суспендировали в 30 мкл воды.
Повторно суспендированную кДНК нагревали до 65°C в течение 5 минут и охлаждали на льду, добавляли 4 мкл 5-кратного объема красителя для геля (Research Genetics Corp.), кДНК вводили в 0,8% агарозный гель 1Х ТАЕ с низкой температурой плавления (агароза с низкой температурой плавления SEA PLAQUE GTG™; FMC Corp.) и разделяли электрофорезом. Загрязняющие адапторы и кДНК длиной менее 0,6 т.п.н. вырезали из геля. Изменяли полярность электродов, расплавленной агарозой заполняли лунки, буфер заменяли, и кДНК разделяли электрофорезом до достижения концентрации, близкой к началу полосы. Область геля, содержащую концентрированную кДНК, вырезали, помещали в микроцентрифужную пробирку и агарозу расплавляли, нагревая до 65°C в течение 15 минут. Образец уравновешивали до 45°C, добавляли 2 мкл 1 ед/мкл бета-агарозы I (Biolabs, Inc.) и смесь инкубировали при 45°C в течение 90 минут для гидролиза агарозы. После инкубации к образцу добавляли одну десятую объема 3 М раствора ацетата натрия, и смесь инкубировали на льду в течение 15 минут. Образец центрифугировали с ускорением 14000 х g в течение 15 минут при комнатной температуре для удаления негидролизованной агарозы, кДНК осаждали этанолом, промывали в 70% этаноле, подвергали воздушной сушке и повторно суспендировали в 40 мкл воды.
Для определения оптимального отношения кДНК к вектору электропорировали несколько собранных лигированных продуктов. Для этого 2 мкл 5-кратного объема буфера с лигазой Т4 (Life Technologies), 1 мкл 10 мМ АТР, 1 мкл pZP7NX, гидролизованного EcoRI-XhoI, 1 мкл ДНК-лигазы Т4, разведенной до 0,25 ед/мкл (Life Technologies) 10 мкл воды, и 0,5, 1, 2 или 3 мкл кДНК смешивали с получением 4 разных лигированных продуктов, инкубировали при 22°C в течение 4 часов, при 68°C в течение 20 минут, осаждали смесью ацетат натрия-этанол, промывали, сушили и вновь суспендировали в 10 мкл. Один микролитр каждого лигированного продукта электропорировали в 40 мкл электрокомпетентных бактериальных клеток DH10b ElectroMax™ (Life Technologies), используя 0,1 см кювету (Biorad) и импульсный регулятор Genepulser™ (Biorad), настроенный на 2,5 кВ, 251 Ф, 200 Ом. Указанные клетки сразу же повторно суспендировали в 1 мл бульона SOC (Manniatis et al., см. выше) с последующим использованием 500 мкл смеси 50% глицерин/SOC в качестве консерванта. Полученный «глицериновый запас» замораживали в виде нескольких аликвот при -70°C. Аликвоту каждого продукта оттаивали и последовательно культивировали на чашках с агаровой средой LB, содержащей ампициллин в количестве 100 мкг/мл. Число колоний показывало, что оптимальное отношение кДНК CD3+-клеток к вектору pZP7NX составляло 1 мкл к 45 нг; такой лигированный продукт позволил получить 4,5 миллиона первичных клонов.
Для скрининга библиотеки с использованием анализа пролиферации на основе BaF3 (пример 5) полученный выше глицериновый запас разводили с получением жидких культур, содержащих 100 или 250 клонов/пул, на титрационных микропланшетах с глубокими лунками, выращивали при встряхивании в течение 24 часов при 37°C и выделяли плазмиду при помощи набора Qiagen, в соответствии с инструкциями изготовителя. Такую ДНК затем трансфицировали в клетки ВНК, среду кондиционировали в течение 72 часов, клетки собирали и хранили при -80°C, затем их помещали на 5000 клеток ВaF3/zcytor17/WSX-1/OSMRbeta или BaF3/zcytor17/OSMRbeta на 72 часа и оценивали пролиферацию при помощи флуоресцентного анализа с аламаровым синим (пример 5В и пример 2В).
Пример 7
Экспрессирующее клонирование zcytor17lig человека
Глицериновый запас из библиотеки активированных CD3+-селектированных клеток человека (пример 6) добавляли к бульону Super Broth II™ (Becton Dickinson, Cockeysville, MD), содержащему 0,1 мг/мл ампициллина (amp) при концентрации 250 клеток/800 микролитров. Клетки E. coli уравновешивали в течение 24 часов при комнатной температуре. В процессе инокуляции 400 микролитров помещали на планшеты со средой LB, содержащей ампициллин, для определения действительного титра инокуляции. Через 24 часа производили подсчет клеток на планшетах, после чего конечную концентрацию бульона Super Broth II™ и E. coli доводили до 250 клеток/1,2 мл. Два литра трижды инокулировали до достижения 6 литров. Затем среду переносили на 96-луночные планшеты с глубокими лунками (Qiagen). Культивирование производили в 8-канальном диспенсере Q-Fill2™ (Genetix, Christchurch, Dorset, UK). Клетки E. coli выращивали в течение ночи при 37°C, встряхивая со скоростью 250 оборотов/мин на многоярусном шейкере New Brunswick Scientific Innova 4900. Клетки E. coli центрифугировали из раствора со скоростью 3000 оборотов/мин в центрифуге GS-6KR Beckman. Дебрис E. coli замораживали при -20°C или использовали в свежем виде до получения плазмидной ДНК при помощи набора Miniprep. Каждый дебрис содержит примерно 250 клонов кДНК из библиотеки CD3+-селектированных клеток человека.
Пулы, содержащие 250 клонов кДНК, исследовали при помощи набора QIAprep™ 96 Turbo Miniprep (Qiagen). Плазмидную ДНК элюировали, используя 125 мкл ТЕ (10 мМ трис-буфера, рН 8, 12 мМ EDTA). Затем плазмидную ДНК использовали для трансфекции клеток ВНК.
Трансфекция клеток ВНК
Клетки ВНК культивировали на 96-луночных планшетах для тканевой культуры при плотности 12000 клеток/лунку в объеме 100 мкл/лунку. Культуральная среда представляла собой среду DMEM (GibcoBRL), содержащую 5% термоинактивированной фетальной телячьей сыворотки, 2 мМ L-глутамина (GibcoBRL), однократный объем PSN (GibcoBRL), 1 мМ пирувата натрия (GibcoBRL).
На следующий день клетки ВНК промывали один раз 100 мкл SFA. SFA является бессывороточной средой, содержащей DMEM/F12 или DMEM (Gibco/BRL), 2 мМ GlutaMax™ (Gibco/BRL), 1 мМ пирувата натрия, 10 мкг/мл трансферрина, 5 мкг/мл инсулина, 10 мкг/мл фетуина, 2 мкг/мл селена, 25 мМ HEPES (Gibco/BRL), 100 мкМ заменимых аминокислот (Gibco/BRL).
Смесь ДНК/липофектамин™ получали следующим образом: 2,2 мкл реагента липофектамина™ (Gibco/BRL) объединяли со 102,8 мкл SFA при комнатной температуре; затем к смеси липофектамин™/SFA добавляли примерно 5 мкл плазмидной ДНК (200 нг/мкл) с образованием смеси ДНК/липофектамин™, которую инкубировали при комнатной температуре в течение 30 минут. Среду SFA удаляли из клеток ВНК, и клетки инкубировали с 50 мкл смеси ДНК/липофектамин™ в течение 5 часов при 37°C с 5% СО2. Пятьдесят мкл смеси ДНК/липофектамин™ добавляли в каждую из двух лунок, содержащих клетки ВНК, что позволяло дуплицировать трансфекцию.
Клетки ВНК инкубировали со смесью ДНК/липофектамин™ в течение 5 часов, затем смесь ДНК/липофектамин™ удаляли и добавляли 100 мкл культуральной среды. Клетки инкубировали в течение ночи, среду удаляли и заменяли 100 мкл культуральной среды. Клетки культивировали в течение 48-72 часов, кондиционированную среду удаляли, замораживали при -80°C в течение минимум 20 минут и оттаивали, после чего 50 мкл анализировали при помощи анализа пролиферации BaF3, описанного в примере 5, для идентификации пулов из 250 клонов, обладающих активностью лиганда.
Двадцать 96-луночных планшетов исследовали при выполнении одного анализа. Данная величина соответствует примерно 250 кДНК/лунку или в общей сложности 480000 кДНК. Кондиционированная среда примерно из 60 лунок (что соответствует 250 кДНК/лунку) дала положительные результаты при выполнении анализа пролиферации. Один из указанных положительных пулов был расщеплен с выделением единичной кДНК, которая кодировала zcytor17lig. Таким пулом был пул 62А12.
1 мкл ДНК использовали для трансформации клеток DH10B ElectroMax™ (Gibco/BRL) пула 62А12 электропорацией. Трансформанты помещали на планшеты со средой LB, содержащей ампициллин (100 мкг/мл), для получения единичных колоний. Из электропорированного пула 672 отобранных единичных колоний были перенесены при помощи зубочистки на семь 96-луночных планшетов, содержащих 1,2 мл бульона SuperBrothII™ в каждой лунке. Указанные планшеты были пронумерованы с 62.1 по 62.7. Планшеты культивировали в течение ночи, и плазмидную ДНК получали при помощи набора Miniprep, как описано выше. Плазмидную ДНК со всех семи планшетов трансфицировали в клетки ВНК и анализировали на пролиферацию вышеописанным способом, за исключением того, что трансфекции не были дуплицированы.
Из общего числа 672 клонов были идентифицированы два положительных клона 62.6С7 и 62.6Е9. Плазмидную ДНК, полученную при помощи набора Miniprep из клона 62.6Е9, секвенировали и идентифицировали, но из указанных положительных клонов была получена смешанная последовательность. Для дальнейшего выделения кДНК zcytor17lig из единичного клона 1 мкл ДНК из пула 62.6Е9 использовали для электропорации клеток DH10B, и культивировали трансформанты на планшетах со средой LB, содержащей ампициллин (100 мкг/мл), с образованием единичных колоний. Плазмидную ДНК, полученную при помощи набора Miniprep из нескольких колоний, секвенировали, получая точную последовательность ДНК. Полинуклеотидная последовательность zcytor17lig была непроцессированной (SEQ ID NO:1), и соответствующая ей аминокислотная последовательность показана в SEQ ID NO:2.
Пример 8
Создание векторов млекопитающих, экспрессирующих растворимые рецепторы zcytor17: zcytor17CEE, zcytor17CFLG, zcytor17CHIS и zcytor17-Fc4
А. Создание вектора экспрессии zcytor17 млекопитающего, содержащего zcytor17CEE, zcytor17CFLG и zcytor17CHIS
Был получен вектор, экспрессирующий растворимый, внеклеточный домен полипептида zcytor17, pZp9zcytor17CEE, который был предназначен для экспрессии полипептида zcytor17, содержащего прогнозируемый инициирующий метионин, усеченного рядом с прогнозируемым трансмембранным доменом и имеющего С-концевую метку Glu-Glu (SEQ ID NO:32).
Продукт PCR длиной около 1500 п.н. был создан с использованием затравок PCR ZC29451 (SEQ ID NO:33) и ZC29124 (SEQ ID NO:34), предназначенных для введения сайтов рестрикции EcoRI и BamHI. Библиотеку кДНК HPVS человека использовали в качестве матрицы, и амплификацию методом PCR выполняли следующим образом: 30 циклов при 94°C в течение 1 минуты, при 65°C в течение 1 минуты, при 72°C в течение 1,5 минут, затем при 72°C в течение 7 минут; выдерживание при 10°C. Реакционную смесь PCR осаждали этанолом и гидролизовали рестрикционными ферментами EcoRI и BamHI. Гидролизованный продукт PCR очищали в 1,0% агарозном геле и вырезали полосу длиной около 1500 п.н. Данную полосу затем вновь амплифицировали, используя идентичные затравки, в соответствии со следующей цикличностью: 30 циклов при 94°C в течение 1 минуты, при 65°C в течение 1 минуты, при 72°C в течение 3 минут, затем при 72°C в течение 7 минут; выдерживание при 10°C. Реакционную смесь PCR осаждали этанолом и гидролизовали рестрикционными ферментами EcoRI и BamHI. Гидролизованный продукт PCR очищали в 1,0% агарозном геле и вырезали полосу длиной около 1500 п.н. Вырезанную ДНК субклонировали в плазмиде CEEpZp9, которая была разрезана EcoRI и BamHI, с образованием плазмиды, содержащей С-концевой меченый растворимый рецептор GLU-GLU для zcytor17, zcytor17CEEpZp9. Внеклеточный домен в кДНК zcytor17CEE плазмиды zcytor17CEEpZp9 имел молчащую мутацию, которая вызывала замену остатка Т остатком С в положении 1705 SEQ ID NO:4 (кодирующей остаток Pro в положении остатка 403 SEQ ID NO:5). Так как данная мутация была молчащей, кДНК zcytor17 в zcytor17CEEpZp9 кодирует полипептид, представленный в SEQ ID NO:5. Кроме того, в соответствии с используемой конструкцией, пара остатков Gly-Ser была вставлена у С-конца растворимого, внеклеточного домена zcytor17 и до С-конца метки Glu-Glu (SEQ ID NO:3). Таким образом, метка у С-конца внеклеточного домена zcytor17 была меткой Glu-Glu, показанной в SEQ ID NO:17. Плазмида СEEpZp9 является экспрессирующим вектором млекопитающего, содержащим полигенный экспрессирующий кластер, включающий промотор металлотионеин-1 мыши, несколько сайтов рестрикции для вставки кодирующих последовательностей и терминатор гормона роста человека. Данная плазмида также имела ориджин репликации E. coli, экспрессирующую единицу селектируемого маркера млекопитающего, содержащую промотор SV40, энхансер и ориджин репликации, ген DHFR и терминатор SV40. При помощи стандартных методов молекулярной биологии zcytor17CEEpZp9 электропорировали в компетентные клетки DH10B (GIBCO BRL, Gaithersburg, MD), в соответствии с инструкциями изготовителя, культивировали на планшетах со средой LB, содержащей 100 мкг/мл ампициллина, и инкубировали в течение ночи. Колонии исследовали при помощи рестрикционного анализа или PCR, используя ДНК, полученную из единичных колоний. Инсерционную последовательность положительных клонов проверяли при помощи анализа последовательности. Для крупномасштабного получения плазмиды использовали набор Maxi prep QIAGEN® (Qiagen), в соответствии с инструкциями изготовителя.
Аналогичный способ использовали для получения растворимых рецепторов zcytor17 с С-концевой меткой His, состоящей из 6 последовательных остатков His; и с С-концевой меткой FLAG® (SEQ ID NO:36), zcytor17CFLAG. Для создания данных конструкций вышеуказанный вектор имеет метку HIS или FLAG® в месте нахождения метки glu-glu (например, SEQ ID NO:17, SEQ ID NO:32 или SEQ ID NO:35).
В. Создание вектора млекопитающего, экспрессирующего растворимый рецептор zcytor17 человека: zcytor17-Fc4
Экспрессирующий вектор, pЕZE-2 hzcytor17/Fc4, был создан для экспрессии С-концевого Fc4-меченного растворимого варианта hzcytor17 (zcytor17-Fc4 человека) в клетках PF яичника китайского хомячка (СНО). Клетки PF СНО представляют собой линию клеток СНО, адаптированных авторами изобретения для выращивания в безбелковой среде (среда ExCell 325 PF; JRH Biosciences). Линия клеток СНО первоначально была выделена из клеток DG44 СНО (G. Urlaub, J. Mitchell, E. Kas, L.A. Chasin, V.L. Funanage, T.T. Myoda and J.L. Hamlin, "The Effect of Gamma Rays at the Dihydrofolate Reductase Lоcus: Deletions and Inversions," Somatic Cell and Molec. Genet. 12:555-566 (1986). Фрагмент кДНК zcytor17, включающий полинуклеотидную последовательность из внеклеточного домена рецептора zcytor17, был слит в рамке считывания с полинуклеотидной последовательностью Fc4 (SEQ ID NO:37) с образованием слитой последовательности zcytor17-Fc4 (SEQ ID NO:38 и SEQ ID NO:39). Вектор pEZE-2 является экспрессирующим вектором млекопитающего, содержащим полинуклеотидную последовательность Fc4 и сайт клонирования, который позволяет быстро создавать С-концевые слитые последовательности Fc4, используя стандартные методы молекулярной биологии.
При помощи PCR был создан фрагмент из 1566 пар оснований, содержащий внеклеточный домен zcytor17 человека и первые две аминокислоты Fc4 (Glu и Pro) с сайтами FseI и BglII, кодированными соответственно у 5'- и 3'-концов. Данный фрагмент PCR был получен путем амплификации с использованием затравок ZC29157 (SEQ ID NO:40) и ZC29150 (SEQ ID NO:41) из плазмиды, содержащей внеклеточный домен zcytor17 человека (pZр9zcytor17CEE) (пример 8А). Реакцию PCR выполняли в следующих условиях: 25 циклов при 94°C в течение 1 минуты, при 60°C в течение 1 минуты и при 72°C в течение 2 минут; 1 цикл при 72°C в течение 10 минут; затем выдерживание при 4°C. Данный фрагмент гидролизовали рестрикционными эндонуклеазами FseI и BglII, очищали электрофорезом в 1% геле и при помощи набора для гель-экстракции QiaQuick (Qiagen). Полученную очищенную ДНК лигировали в течение 5 часов при комнатной температуре с вектором pEZE-2, предварительно гидролизованным FseI и BglII и содержащим 3'-концевые сайты FseI и BglII последовательности Fc4.
Два мкл лигированной смеси электропорировали в 37 мкл электрокомпетентных клеток DH10B E. coli (Gibco), в соответствии с указаниями изготовителя. Трансформированные клетки разводили в 400 мкл среды LB и культивировали на планшетах со средой LB, содержащей 100 мкг/мл ампициллина. Клоны анализировали на наличие рестрикционных гидролизатов, и положительные клоны отправляли на секвенирование ДНК для подтверждения наличия слитой последовательности. Один микролитр положительного клона вводили в 37 мкл электрокомпетентных клеток DH10B E. coli и высевали на планшет со средой LB, содержащей ампициллин. Единичную колонию удаляли с указанного планшета для получения 250 мл культуры в среде LB, содержащей ампициллин, которую выращивали в течение ночи при 37°C, встряхивая со скоростью 250 оборотов/мин. Данную культуру использовали для получения 750 мкг очищенной ДНК при помощи плазмидного набора Maxi Qiagen (Qiagen).
Пример 9
Трансфекция и экспрессия полипептидов растворимого рецептора zcytor17
Клетки ВНК 570 (АТСС № CRL-10314), клетки DG-44 CHO или другие клетки млекопитающих культивировали в количестве около 1,2 х 106 клеток/лунку (6-луночный планшет) в 800 мкл соответствующей бессывороточной (SF) среды (например, DMEM, Gibco/BRL, с высоким содержанием глюкозы) (Gibco BRL, Gaithersburg, MD). Клетки трансфицировали экспрессирующими плазмидами, содержащими zcytor17CEE, zcytor17СFLG, zcytor17CHIS или zcytor17-Fc4 (пример 8), с использованием липофектина™ (Gibco BRL) в бессывороточной (SF) среде, в соответствии с инструкциями изготовителя. Единичные клоны, экспрессирующие растворимые рецепторы, выделяли, исследовали, выращивали в культуральной среде и очищали стандартными методами.
А. Экспрессия млекопитающим растворимого рецептора zcytor17CEE человека
Клетки ВНК 570 (АТСС № CRL-10314) культивировали в колбах для тканевых культур Т-75 и выращивали примерно до 50-70% слияния при 37°C, 5% СО2 в среде DMEM/FBS (DMEM, Gibco/BRL, с высоким содержанием глюкозы (Gibco BRL, Gaithersburg, MD), содержащей 5% фетальной телячьей сыворотки, 1 мМ L-глутамина (JRH Biosciences, Lenea, KS), 1 мМ пирувата натрия (Gibco BRL)). Затем клетки трансфицировали плазмидой, содержащей zcytor17CEE (пример 8А), с использованием липофектамина™ (Gibco BRL) в бессывороточной (SF) среде (DMEM, 10 мг/мл трансферрина, 5 мг/мл инсулина, 2 мг/мл фетуина, 1% L-глутамина и 1% пирувата натрия). Десять микрограммов плазмидной ДНК pZp9zcytor17CEE (пример 8А) разводили в 15 мл пробирке SF средой до общего конечного объема 500 мкл. Пятьдесят микролитров липофектамина смешивали с 450 мкл SF среды. Смесь с липофектамином добавляли к смеси ДНК и инкубировали в течение примерно 30 минут при комнатной температуре. Четыре мл SF среды добавляли к смеси ДНК:липофектамин. Клетки один раз промывали 5 мл SF среды, среду отсасывали и добавляли смесь ДНК:липофектамин. Клетки инкубировали при 37°C в течение пяти часов и добавляли 5 мл среды DMEM/10%FBS. Колбу инкубировали при 37°C в течение ночи, после чего клетки переносили на 150 мм чашки, содержащие селективную среду (вышеуказанная среда DMEM/FBS с добавлением 1 мкМ метотрексата или 10 мкМ метотрексата (Sigma Chemical Co., St. Lоuis, Mo.) в соотношении 1:2, 1:10 и 1:50. Примерно через 10 дней после трансфекции одну 150 мм чашку, содержащую 1 мкМ колоний, устойчивых к метотрексату, обрабатывали трипсином, клетки собирали и половину клеток снова культивировали в 10 мкМ метотрексата для дальнейшей амплификации экспрессии белка zcytor17CEE. В образце кондиционированной среды из данного пула амплифицированных клеток определяли уровни экспрессии при помощи методов SDS-PAGE и вестерн-блоттинга.
В. Экспрессия млекопитающим растворимого рецептора zcytor17-Fc4 человека
Пять репликатов 200 мкг плазмидной ДНК pEZE-2hzcytor17Fc4 (пример 8) переводили в линейную форму путем рестрикционного гидролиза FspI, являющегося рестрикционным ферментом, который разрезает вектор на одном участке и не разрушает гены, необходимые для экспрессии. К каждому репликату добавляли 200 мкг геномной ДНК клеток СНО в качестве несущей ДНК, после чего ДНК осаждали, добавляя 0,1 объема 3М раствора ацетата натрия, рН 5,2 и 2,2 объема этанола, инкубировали на льду в течение 15 минут и микроцентрифугировали при 4°C. Полученный осадок ДНК промывали в 70% этаноле, подвергали воздушной сушке и вновь суспендировали в 100 мкл безбелковой (PF), неселективной питательной среды для СНО (21 г/л PF среды для клеток 325 СНО/200 мМ L-глутамина (Gibco)/100 мМ пирувата натрия (Gibco)/однократный объем добавки НТ (Gibco). Десять миллионов клеток 61 пассажа клеток СНО в PF среде добавляли к ДНК в 600 мкл PF неселективной питательной среды для клеток СНО, электропорировали в системе для электропорации Gene Pulser II (BioRad) при 950 мкФ и 300 кВ, используя кювету для электропорации с 0,4 см разрывом Gene Pulser (BioRad). Все 5 репликатов электропорированных клеток собирали и отбирали в среде -НТ (21 г/л PF среды для клеток 325 СНО/200 мМ L-глутамина (Gibco)/100 мМ пирувата натрия (Gibco). Клетки отбирали в течение 15 дней в среде -НТ и пассировали в количестве 4 х 105/мл в 50 нм селективной среды МТХ. Через восемь дней клетки высевали в количестве 3,5 х 105 клеток/мл в 200 мМ селективной среды МТХ. Через одну неделю клетки высевали в количестве 4 х 105 клеток/мл в 1 мкМ селективной среды МТХ. Клетки выдерживали в 1 мкМ среды МТХ в течение двух недель и высевали в количестве 1 х 106 клеток/мл в 50 мл для получения кондиционированной среды. Полученную через 72 часа кондиционированную среду анализировали методом вестерн-блоттинга с использованием антитела против Ig человека. Клетки продуцировали белок hzcytor17/Fc4 в количестве около 1 мг/л.
С. Крупномасштабная экспрессия млекопитающим растворимого рецептора zcytor17-Fc4 человека
Двести мкг плазмидной ДНК pEZE-2hzcytor17Fc4 (пример 8В) переводили в линейную форму путем рестрикционного гидролиза FspI, являющегося рестрикционным ферментом, который разрезал вектор pEZE-2 на одном участке и не разрушал гены, необходимые для экспрессии. Двести микрограммов геномной ДНК клеток СНО (собственного изготовления) добавляли в качестве несущей ДНК, после чего ДНК осаждали, добавляя 0,1 объема 3 М раствора ацетата натрия, рН 5,2, и 2,5 объема этанола, и микроцентрифугировали при комнатной температуре. Получали и трансформировали осадки ДНК пяти репликатов. Полученный осадок ДНК промывали в 70% этаноле, подвергали воздушной сушке и вновь суспендировали в 100 мкл безбелковой неселективной питательной среды для клеток СНО (21 г/л PF среды для клеток 325 СНО/200 мМ L-глутамина (Gibco)/100 мМ пирувата натрия (Gibco)/однократный объем добавки НТ (Gibco). Десять миллионов клеток СНО в PF среде добавляли к ДНК в 600 мкл безбелковой неселективной питательной среде для клеток СНО и электропорировали в системе для электропорации Gene Pulser II (BioRad) при 950 мкФ и 300 В, используя кювету для электропорации с 0,4 см разрывом Gene Pulser (BioRad). Электропорированные клетки собирали и переносили в селективную среду -НТ (21 г/л PF среды для клеток 325 СНО/200 мМ L-глутамина (Gibco)/100 мМ пирувата натрия (Gibco). Клетки отбирали в течение 14 дней в среде -НТ и пассировали в количестве 4 х 105/мл в 50 нм селективной среды МТХ. Клетки амплифицировали до 200 нМ в среде МТХ и затем до 1 мкМ в среде МТХ. 50 нМ и 1 мкМ пулы в среде -НТ высевали в количестве 1 х 106 клеток/мл в течение 48 часов и полученную кондиционированную среду анализировали методом вестерн-блоттинга с использованием антитела против Ig человека.
Пример 10
Очистка растворимых рецепторов zcytor17 от клеток ВНК 570 и СНО
А. Временная экспрессия млекопитающим и очистка растворимого рецептора zcytor17-Fc4 человека
Плазмидную ДНК pEZE-2hzcytor17Fc4 (пример 8В) вводили в 40 чашек Maxi с клетками ВНК, используя липофектамин (Gibco BRL), в соответствии с приведенным выше описанием и инструкциями изготовителя. Клетки восстанавливали в течение ночи, затем промывали и подпитывали бессывороточной средой (SL7V4, собственного изготовления). Через 72 часа среду собирали и фильтровали, клетки подпитывали бессывороточной средой. Через 72 часа среду снова собирали и фильтровали.
Бессывороточную кондиционированную среду (2 порции по 1,5 л), полученную из временно трансфицированных клеток BHK, подавали насосом в верхнюю часть 1,5 мл колонки, содержащей белок А-агарозу в 20 мМ трис-буфера, рН 7,5, 0,5 М NaCl. Колонку обильно промывали указанным буфером, после чего связанный белок элюировали 1 мл 0,2 М глицина, рН 2,5, 0,5 М NaCl. Элюированный белок собирали в 0,1 мл 2 М трис-буфера, рН 8,5. Аликвоты собирали для выполнения электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия и основную массу zcytor17-Fc диализовали в течение ночи против PBS. Растворимый рецептор стерилизовали фильтрацией и хранили в виде аликвот при -80°C.
В. Очистка zcytor17-Fc4
Рекомбинантный карбоксиконцевой Fc4-меченный zcytor17 (пример 8 и пример 9) получали из трансфицированных клеток СНО. Клетки СНО трансфицировали методами, известными в данной области. Было собрано примерно пять литров кондиционированной среды, которую стерилизовали фильтрацией, используя 0,2 мкм фильтры Nalgene.
Белок очищали от фильтрованной среды при помощи аффинной хроматографии в колонке с белком А Poros 50 (PerSeptive Biosystems, 1-5559-01, Framingham, MA) и вытеснительной гель-хроматографии в колонке Superdex 200 (Amersham Pharmacia Biotech, Piscataway, NJ). Культуральную среду вводили в колонку с белком А размером 10 х 70 мм (объем слоя 5,5 мл) для аффинной хроматографии со скоростью потока около 3-10 мл/мин. Колонку промывали десятью объемами PBS, и связанный белок элюировали пятью объемами 0,1 М глицина, рН 3,0, со скоростью потока 10 мл/мин). Фракции по 2 мл каждая собирали в пробирки, содержащие 100 мкл 2,0 М трис-буфера, рН 8,0, для нейтрализации элюированных белков. Образцы, взятые из колонки для аффинной хроматографии, анализировали методом SDS-PAGE с окрашиванием кумассии и методом вестерн-блоттинга на наличие zcytor17-Fc4, используя Ig-HRP человека. Фракции, содержащие zcytor17-Fc4, собирали и концентрировали до 1-2 мл в концентраторе Biomax-30 (Millipore) и загружали в колонку для гель-фильтрации Superdex 200 размером 20 х 580 мм. Фракции, содержащие очищенный zcytor17-Fc4, собирали, фильтровали через 0,2 мкм фильтр, делили на аликвоты по 100 мкл каждая и замораживали при -80°C. Концентрацию конечного очищенного белка определяли при помощи анализа ВСА (Pierce, Rockford, IL).
С. Анализ zcytor17/Fc4 методами SDS-PAGE и вестерн-блоттинга
Рекомбинантный zcytor17-Fc4 анализировали методом SDS-PAGE (Nupage 4-12%, Invitrogen, Carlsbad, CA) с окрашиванием кумассии и методом вестерн-блоттинга с использованием Ig-HRP человека. Кондиционированную среду или очищенный белок разделяли электрофорезом, используя миниячейку Xcell II Invitrogen Novex, и переносили на нитроцеллюлозу (0,2 мм; Invitrogen, Carlsbad, CA) при комнатной температуре и перемешивании, используя блот-модуль Xcell II Novex, в соответствии с инструкциями изготовителя. Перенос осуществляли при 500 мА в течение одного часа в буфере, содержащем 25 мМ основного трис-буфера, 200 мМ глицина и 20% метанола. Затем фильтры блокировали 10% обезжиренного сухого молока в PBS в течение 10 минут при комнатной температуре. Нитроцеллюлозу быстро промывали и добавляли антитело против Ig-HRP человека (1:2000) в PBS, содержащем 2,5% обезжиренного сухого молока. Блоты инкубировали, осторожно встряхивая, в течение двух часов при комнатной температуре или в течение ночи при 4°C. После инкубации блоты трижды промывали в PBS в течение 10 минут, затем быстро промывали в Н2О. Блоты проявляли коммерчески производимыми реагентами на хемилюминесцентном субстрате (реагенты 1 и 2 ULTRA SuperSignal®, смешанные в соотношении 1:1, от компании Pierce, Rockford, IL) и улавливали сигнал при помощи программного обеспечения Lumi Analyst 3.0 компании Lumi-Imager (Boehringer Mannheim GmbH, Germany), при этом время воздействия составляло от 10 секунд до 5 минут или соответствовало требуемым условиям.
Очищенный zcytor17-Fc4 был обнаружен в виде одной полосы длиной около 220 кДа при окрашивании кумассии или серебром в невосстановительных условиях и длиной около 120 кДа в восстановительных условиях, из чего следует, что в невосстановительных условиях может быть получена димерная форма zcytor17-Fc4.
Пример 11
Конкурентно-ингибирующий анализ растворимого рецептора zcytor17, включающего растворимый рецептор zcytor17-Fc4
Клетки BaF3/zcytor17/WSX-1/OSMRbeta и BaF3/zcytor17/OSMRbeta центрифугировали и промывали в среде без mIL-3. Клетки трижды центрифугировали и промывали для полного удаления mIL-3. Затем клетки подсчитывали в гемоцитометре. Клетки культивировали на 96-луночном планшете в количестве 5000 клеток/лунку в объеме 100 мкл/лунку, используя среду без mIL-3.
Кондиционированную среду, полученную в результате активации клеток CCRF-CEM, CCRF-HSB2 и CD3+-селектированных клеток человека по примеру 5, добавляли в отдельных экспериментах в концентрациях, составляющих 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75%, 0,375% и 0,187%, с растворимыми рецепторами zcytor17 или без них (zcytor17-Fc4; см. пример 9 и пример 10) в количестве 1-10 мкг/мл. Общий анализируемый объем составлял 200 мкл.
Анализируемые планшеты инкубировали при 37°C, 5% СО2, в течение 3-5 дней, после чего добавляли аламаровый синий (Accumed) в количестве 20 мкл/лунку. Планшеты снова инкубировали при 37°C, 5% СО2, в течение 16-24 часов. Планшеты считывали в аппарате для считывания планшетов Fmax™ (Molecular Devices) аналогично примеру 2. Полученные результаты свидетельствовали о частичном ингибировании роста клеток растворимым рецептором zcytor17-Fc4 при концентрации 10 мкг/мл, из чего следует, что этот фактор в каждом образце является специфическим для рецептора zcytor17.
При помощи вышеуказанного анализа были также получены кривые титрования при разведении растворимого рецептора или гетеродимеров растворимого рецептора, включающих zcytor17/OSMR и zcytor17/WSX-1, на основании которых определяли способность рецепторов zcytor17 полностью ингибировать рост клеток при низких или физиологических концентрациях.
Аналогичные конкурентно-ингибирующие анализы выполняли, используя очищенный zcytor17lig человека (пример 35) и растворимые рецепторы в присутствии люциферазы (пример 20). Полученные результаты показывают, что как гомодимерные zcytor17, так и гетеродимерные zcytor17/OSMR способны ингибировать активность zcytor17lig.
Пример 12
Анализ улавливания секретируемого продукта
Анализ улавливания секретируемого продукта был использован для испытания связывания zcytor17lig с рецепторами, включающими в себя рецептор zcytor17 или гетеродимеры zcytor17/OSMR и zcytor17/WSX-1. Плазмидную ДНК zcytor17lig трансфицировали с клетки COS и использовали для оценки связывания zcytor17lig с рецепторами, включающими в себя рецептор zcytor17, методом улавливания секретируемого продукта нижеописанным способом.
А. Трансфекция клеток COS
Клетки COS трансфицировали следующим образом: 800 нг кДНК zcytor17lig и 4 мкл липофектамина™ смешивали в 80 мкл бессывороточной среды DMEM (55 мг пирувата натрия, 146 мг L-глутамина, 5 мг трансферрина, 2,5 мг инсулина, 1 мкг селена и 5 мг фетуина в 500 мл DMEM) и инкубировали при комнатной температуре в течение 30 минут. Затем добавляли 320 мкл бессывороточной среды DMEM. 500 мкл смеси добавляли к клеткам COS в количестве 2 х 105 клеток/лунку, переносили на 12-луночный планшет для тканевой культуры и инкубировали в течение 5 часов при 37°C. Затем добавляли 500 мкл среды DMEM c 20% FBS (100 мл FBS, 55 мг пирувата натрия и 146 мг L-глутамина в 500 мл DMEM) и инкубировали клетки в течение ночи.
В. Анализ улавливания секретируемого продукта
Секретируемый продукт улавливали следующим образом: клетки вымывали из среды при помощи PBS и фиксировали в течение 15 минут 1,8% формальдегида в PBS. Затем клетки промывали PBS/0,1% BSA и делали проницаемыми при помощи 0,1% тритона-Х в PBS в течение 15 минут и снова промывали PBS/0,1% BSA. Клетки блокировали PBS/0,1% BSA в течение 1 часа. В зависимости от используемого растворимого рецептора, клетки инкубировали в TNB в течение 1 часа с: (А) 1-3 мкг/мл слитого белка zcytor17-Fc4 растворимого рецептора zcytor17 (пример 10); или (В) 1-3 мкг/мл белка растворимого рецептора zcytor17/OSMRbeta. Затем клетки промывали TNT. В зависимости от используемого растворимого рецептора (например, меченного меткой Fc4 (SEQ ID NO:37), С-концевой меткой FLAG (SEQ ID NO:26) или меткой СЕЕ (SEQ ID NO:32; SEQ ID NO:35), клетки инкубировали в течение одного часа с: (А) антителом козы против Ig-HRP человека, разведенным в соотношении 1:200 (Fс-специфический); (В) М2-HRP, разведенным в соотношении 1:1000; (С) антителом против GluGlu-HRP, разведенным в соотношении 1:1000; или (D) смесью стрептавидин-HRP в соотношении 1:300 (набор NEN) в TNB. Клетки снова промывали TNT.
Для обнаружения положительного связывания такой реагент как флуоресцеинтирамид разводили в соотношении 1:50 в буфере для разведения (набор NEN), инкубировали в течение 4-6 минут и промывали TNT. Клетки сохраняли в заливочной среде Vectashield (Vector Labs Burlingame, CA), разведенной TNT в соотношении 1:5. Клетки визуализировали при помощи фильтра FITC в флуоресцентном микроскопе. Результаты данного анализа показали, что zcytor17lig человека не связывается ни с одним из растворимых рецепторов. Указанные данные позволяют предположить, что структура zcytor17lig чувствительна к стадии фиксации данного способа, так как она была способна связываться с рецепторами клеточной поверхности (см., например, данные проточной цитометрии, приведенные в нижеследующем примере 39).
Пример 13
Назначение и расположение генной последовательности zcytor17lig в хромосоме
Генная последовательность zcytor17lig была картирована в хромосоме 12 человека при помощи коммерчески производимого варианта «панели картирования радиационных гибридов Stanford G3» (Research Genetics, Inc., Huntsville, AL). «Панель картирования радиационных гибридов Stanford G3» содержит ДНК каждого из 83 радиационных гибридных клонов всего генома человека и две контрольные ДНК (донор RM и реципиент А3). На общедоступном сервере WWW, расположенном в Интернете по адресу www.stanford.edu, представлена локализация в хромосоме маркеров и генов.
Для картирования генной последовательности zcytor17lig при помощи «панели картирования радиационных гибридов Stanford G3» 20 мкл реакционных смесей помещали на 96-луночный титрационный микропланшет, предназначенный для выполнения PCR (Stratagene, La Jolla, CA), и исследовали в термоячейке для проведения реакций "RoboCycler Gradient 96" (Stratagene). Каждую из 95 реакций PCR выполняли, используя 2 мкл 10-кратного объема буфера для реакции PCR (Qiagen, Inc., Valencia, CA), 1,6 мкл смеси dNTP (по 2,5 мМ каждого, PERKIN-ELMER, Foster City, CA), 1 мкл смысловой затравки ZC41458 (SEQ ID NO:42), 1 мкл антисмысловой затравки ZC41457 (SEQ ID NO:43), 2 мкл реагента "RediLoad" (Research Genetics, Inc., Huntsville, AL), 0,1 мкл ДНК-полимеразы HotStarTaq Qiagen (5 единиц/мкл), 25 нг ДНК из единичного гибридного клона или контрольного образца и дистиллированную воду до достижения общего объема 20 мкл. Реакционные смеси покрывали равным количеством минерального масла и герметично закупоривали. В ячейке для выполнения реакции PCR были созданы следующие условия: 1 начальный цикл денатурации в течение 15 минут при 95°C, 35 циклов денатурации в течение 45 секунд при 95°C, отжиг в течение 1 минуты при 53°C, удлинение цепи в течение 1 минуты 15 секунд при 72°C и 1 конечный цикл удлинения цепи в течение 7 минут при 72°C. Реакционные смеси разделяли электрофорезом в 2% агарозном геле (EM Science, Gibbstown, NJ) и визуализировали, окрашивая бромидом этидия.
Полученные результаты позволили установить связь генной последовательности zcytor17lig с маркером SHGC-83339 хромосомы 12 при оценке LOD >11 и на расстоянии 17 cR_10000 от указанного маркера. При помощи маркера было установлено, что ген zcytor17lig расположен в хромосомной области 12q24.31.
Пример 14
Идентификация и клонирование zcytor17lig мыши
А. Идентификация непроцессированной последовательности zcytor17lig мыши
При использовании пептидной последовательности zcytor17lig человека (SEQ ID NO:2) для поиска в собственной базе данных ДНК, кДНК мыши, № доступа в банке генов АК005939, была идентифицирована как возможная частичная последовательность zcytor17lig мыши. Последовательность кДНК АК005939 использовали для поиска в базе данных, содержащей геномные фрагменты мыши. Была собрана смежная геномная последовательность zcytor17lig мыши (SEQ ID NO:76). Прогнозирование кодирующего потенциала данного геномного фрагмента при помощи программы Genscan позволило выявить подобную последовательность кДНК, имеющую такую же генную структуру, что и zcytor17lig человека. Последовательность кДНК мыши представлена в SEQ ID NO:10, и соответствующая полипептидная последовательность показана в SEQ ID NO:11.
В. Клонирование zcytor17lig мыши из библиотеки кДНК яичка мыши методом PCR
На основании геномной последовательности (SEQ ID NO:76) были созданы две затравки для PCR, которые использовали для идентификации источника кДНК zcytor17lig мыши при помощи PCR. Указанные затравки ZC41498 (SEQ ID NO:86) и ZC41496 (SEQ ID NO:87) были предназначены для предполагаемых 5'- и 3'-концевых нетранслируемых областей последовательностей мыши (SEQ ID NO:76 и SEQ ID NO:10). При помощи PCR было исследовано несколько источников кДНК, включая кДНК Marathon-ready (Clontech) и аликвоты библиотек кДНК, созданных авторами изобретения. Продукты визуализировали в 1% агарозных гелях. Полосы ожидаемого размера наблюдались в реакциях с использованием в качестве матрицы библиотеки кДНК яичка мыши. Реакции PCR были успешно выполнены примерно в 50 мкл объемах с 10% ДМСО или без него при использовании полимеразы pfu turbo (Stratagene), в соответствии с рекомендациями изготовителя, и дополнительном применении горячего воска для инициации реакции с использованием набора Hot Start 50 (Molecular Bioproducts, Inc., San Diego, CA). Термоциклизацию PCR выполняли в следующих условиях: один цикл при 94°C в течение 4 минут, затем 40 циклов при 94°C в течение 30 секунд, при 48°C в течение 30 секунд, при 72°C в течение 50 секунд, и дополнительное конечное удлинение цепи при 72°C в течение 7 минут. Продукты двух реакций PCR собирали и очищали, используя агарозу с низкой температурой плавления и расщепляющий агарозу фермент гелазу (Epicenter, Inc., Madison, WI), в соответствии с рекомендациями изготовителя.
Определение последовательности ДНК указанных продуктов PCR позволило выявить последовательность кДНК zcytor17 мыши (SEQ ID NO:90), которая включала открытую рамку считывания, идентичную SEQ ID NO:10, из чего следует, что SEQ ID NO:10 кодирует полипептид zcytor17lig мыши. Затравки для PCR, ZC41583 (SEQ ID NO:88) и ZC41584 (SEQ ID NO:89), затем использовали для введения сайтов рестрикции FseI и AscI и неполной последовательности Козака в открытую рамку считывания mcytor17lig и терминирующий кодон (SEQ ID NO:92). Термоячейку для проведения реакций Robocycler 40 (Stratagene) использовали для создания градиента температур отжига и циклизации в следующих условиях. Полимеразу pfu turbo (Stratagene) использовали, как было описано выше, но только в 10% ДМСО. Циклизация включала в себя один цикл при 94°C в течение 4 минут, затем 20 циклов при 94°C в течение 30 секунд, градиент от 65°C до 51°C в течение 30 секунд, при 72°C в течение 1 минуты и один цикл удлинения цепи при 72°C в течение 7 минут. В качестве матрицы для второй реакции термоциклизации использовали 1 мкл исходного очищенного в геле продукта PCR mcytor17lig. Продукт PCR, полученный в результате выполнения трех реакций при самых низких температурах, собирали и очищали в геле при помощи вышеописанного метода с использованием гелазы (Epocenter). Очищенный mcytor17lig гидролизовали FseI и AscI и лигировали в вектор pZP7X, модифицированный с включением сайтов FseI и AscI в сайт клонирования. Плазмида pZP7X является экспрессирующим вектором млекопитающего, содержащим полигенный экспрессирующий кластер, включая промотор металлотионеин-1 мыши (МТ-1), несколько сайтов рестрикции для вставки кодирующих последовательностей и терминатор гормона роста человека. Указанная плазмида имеет также ориджин репликации E. coli, экспрессирующую единицу селектируемого маркера млекопитающего, содержащую промотор SV40, энхансер и ориджин репликации, ген DHFR и терминатор SV40. Клонированная последовательность кДНК мыши представлена в SEQ ID NO:90, и соответствующая полипептидная последовательность показана в SEQ ID NO:91 (которая идентична SEQ ID NO:11).
Пример 15
Выделение клона кДНК zcytor17lig мыши из активированной библиотеки селезенки мыши
А. Источник первичных клеток мыши, используемый для выделения zcytor17lig мыши
Селезенки мышей Balb/C собирали и растирали между матированными стеклами для получения суспензии клеток. Выход выделенных первичных клеток мыши составил примерно 6,4 х 108 клеток до описанного ниже отбора.
Клетки селезенки суспендировали в 9,6 мл буфера MACS (PBS, 0,5% EDTA, 2 мМ EDTA). 1,6 мл суспензии клеток удаляли и добавляли 0,4 мл микрошариков CD90 (Thy1.2) (Miltenyi Biotec). Смесь инкубировали в течение 15 минут при 4°C. Клетки, меченные шариками CD90, промывали 30 мл буфера MACS и вновь суспендировали в 2 мл буфера MACS.
Колонку VS+ (Miltenyi) готовили в соответствии с инструкциями изготовителя. Затем колонку VS+ помещали в магнитное поле VarioMACS™ (Miltenyi). Колонку уравновешивали 5 мл буфера MACS. Затем в колонку вводили выделенные первичные клетки мыши. CD90-отрицательные клетки проходили через колонку. Колонку промывали 9 мл (3 х 3 мл) буфера MACS. Затем колонку снимали с магнита и помещали на 15 мл фальконовую трубку. CD90+-клетки элюировали, добавляя в колонку 5 мл буфера MACS, и связанные клетки вымывали при помощи поршня, предоставленного изготовителем. Инкубацию клеток с магнитными шариками CD90, промывку и вышеописанные стадии обработки в колонке VS+ (от инкубации до элюирования) повторяли еще раз. Собирали фракции CD90+, полученные в результате двух разделений в колонке. Выход CD90+-селектированных клеток селезенки мыши составлял в общей сложности примерно 1 х 108 клеток.
Образец собранных CD90+-селектированных клеток мыши удаляли для окрашивания и сортировки в клеточном сортере с флуоресцентным антителом (FACS) для оценки чистоты клеток. Для окрашивания и сортировки CD90+-селектированных клеток использовали РЕ-конъюгированное антитело хомячка против CD3ε мыши (PharMingen). CD90+-селектированные клетки мыши примерно на 93% являются CD3+-клетками, позволяя предположить, что указанные клетки содержат 93% Т-клеток.
CD90+-селектированные клетки мыши активировали путем инкубации 3 х 106 клеток/мл в среде RPMI, содержащей 5% FBS, 10 нг/мл РМА и 0,5 мкг/мл иономицина (Calbiochem), в течение ночи при 37°C. Супернатант активированных CD90+-селектированных клеток мыши испытывали в отношении активности zcytor17lig описанным ниже способом. Кроме того, активированные CD90+-селектированные клетки мыши использовали для получения библиотеки кДНК, как описано в следующем примере 16.
Пример 16
Клонирование zcytor17lig мыши из библиотеки CD90+-селектированных клеток мыши
Скрининг библиотеки кДНК активированных CD90+-селектированных первичных клеток мыши позволил выделить кДНК, которая является новым членом семейства четырехспиральных цитокинов, кодирующих ортолог мыши zcytor17lig человека. Указанная кДНК идентифицирована методом гибридизационного скрининга.
А. Вектор для создания библиотеки CD90+-селектированных клеток
Для создания библиотеки CD3+-селектированных клеток использовали вектор pZP7N (см. пример 6А).
В. Получение библиотеки кДНК активированных CD90+-селектированных первичных клеток мыши
Примерно 1,5 х 108 CD90+-селектированных первичных клеток мыши, стимулированных в иономицине/РМА (пример 15), было выделено центрифугированием. Полную РНК выделяли из клеточного дебриса и превращали в двухцепочечную кДНК в соответствии с примером 6В. Указанную ДНК затем трансфицировали в клетки ВНК по примеру 6В и оценивали пролиферацию при помощи флуоресцентного анализа с «аламаровым синим» (пример 2В).
Для скрининга библиотеки методом клонирования с улавливанием секретируемого продукта необходима комплексная, амплифицированная форма библиотеки для трансфекции клеток COS-7. 4,8 миллиона клонов культивировали на 110 15 см чашках с агаровой средой LB, содержащей 100 мкг/мл ампициллина, 10 мкг/мл метициллина. Бактерии, выращиваемые в чашках в течение ночи при 37°C, собирали соскабливанием и осаждали центрифугированием. Плазмидную ДНК экстрагировали из осажденных бактерий при помощи Nucleobond-giga™ (Clonetech), в соответствии с инструкциями изготовителя. Указанную плазмиду затем использовали для трансфекции клеток COS-7 на предметных стеклах и исследовали нижеописанным методом улавливания секретируемого продукта (пример 17).
С. Скрининг библиотеки кДНК активированных клеток мыши
Примерно 5 х 105 клонов культивировали на 10 чашках Maxi со средой LB, содержащей ампициллин. Колонии собирали, денатурировали, нейтрализовали и сшивали стандартным методом (Sambrook, J. et al., см. выше). Пятьдесят нанограммов 5'-концевого фрагмента длиной 300 п.н., полученного при помощи RACE PCR (пример 14), метили 32Р, используя набор для мечения неспецифических затравок RmT Prime-Itr (Stratagenе). Десять фильтров гибридизовали с указанным меченым зондом при 65°C в течение ночи, используя гибридизационный раствор ExpressHyb™ (Clontech). Затем фильтры трижды последовательно промывали при 60°C в течение 1 часа 0,2-кратным объемом SSC (30 мМ NaCl, 3 мМ цитрата натрия, рН 7,0), 0,1% SDS и затем при 65°C в течение 1 часа. Фильтры экспонировали при -80°C в течение ночи и проявляли рентгеновскую пленку. Вытаскивали агаровые пробки, содержащие положительные колонии, и клоны культивировали на 10 см чашках со средой LB, содержащей ампициллин. Колонии собирали с фильтра и снова гибридизовали вышеописанным способом. Единичные клоны ДНК выделяли и секвенировали стандартными методами для идентификации кДНК мыши.
Пример 17
Отсутствие связывания zcytor17lig мыши с растворимым рецептором zcytor17 человека при выполнении анализа улавливания секретируемого продукта
ДНК zcytor17lig/pZР7 клона мыши трансфицировали в клетки COS и при помощи анализа улавливания секретируемого продукта определяли способность связывания zcytor17, включающего растворимые рецепторы (растворимый рецептор zcytor17 человека zcytor17-Fc4 (пример 10), или гетеродимеров растворимого рецептора (zcytor17/WSX-1 или BaF3/zcytor17/OSMRbeta), с трансфицированными клетками COS (пример 12). Результаты анализа показали, что zcytor17lig мыши не связывается с растворимым рецептором zcytor17 человека.
Клетки COS трансфицировали аналогично примеру 12, используя около 0,7 мкг кДНК zcytor17lig мыши (пример 16) в 3 мкл.
Секретируемый продукт улавливали аналогично примеру 12, используя, например, 1 мкг/мл слитого белка растворимого рецептора zcytor17 и Fc4 (пример 10) (или описанных выше гетеродимеров, содержащих растворимый белок zcytor17) в TNB, и антитело козы против Ig-HRP человека (Fc-специфическое), разведенное в соотношении 1:200 в TNВ, для обнаруживаемого антитела. Положительное связывание растворимого рецептора zcytor17 человека с полученными фиксированными клетками не было обнаружено при выполнении анализа по примеру 12 с использованием такого реагента, как флуоресцеинтирамид. Клетки консервировали и визуализировали аналогично примеру 12.
Полученные результаты показали, что zcytor17lig мыши не связывается с растворимым рецептором zcytor17 человека (или описанными выше гетеродимерами, содержащими растворимый рецептор zcytor17).
Пример 18
Экспрессия zcytor17lig мыши в клетках млекопитающих
Клетки ВНК 570 (АТСС № CRL-10314) культивировали на 10 см чашках для тканевых культур и выращивали примерно до 20% слияния в течение ночи при 37°C, 5% СО2, в среде DMEM/FBS (DMEM, среда с высоким содержанием глюкозы Gibco/BRL; Gibco BRL, Gaithersburg, MD), содержащей 5% фетальной телячьей сыворотки (Hyclone, Logan, UT), 1 мМ L-глутамина (JRH Biosciences, Lenexa, KS), 1 мМ пирувата натрия (Gibco BRL). Клетки трансфицировали плазмидой mzcytor17lig/pZP7X (пример 14) при помощи набора для устойчивой трансфекции клеток млекопитающих с использованием липофектамина (Gibco BRL), в соответствии с инструкциями изготовителя.
Через один день после трансфекции клетки вводили в селективную среду в соотношении 1:10 и 1:20 (среда DMEM/FBS с добавлением 1 мкМ метотрексата (Sigma Chemical Co., St. Louis, MO)) на 150 мм чашках. Среду в клетках заменяли свежей селективной средой на 5-й день после трансфекции. Примерно через 10 дней после трансфекции колонии, устойчивые к метотрексату, обрабатывали трипсином, клетки собирали и вводили в колбы для крупномасштабного культивирования. Клетки, выращенные примерно до 90% слияния, трижды промывали PBS и культивировали в бессывороточной среде ESTEP2 (DMEM (Gibco BRL), 0,11 г/л пирувата натрия, 3,7 г/л NaHCО3, 2,5 мг/л инсулина, 5 мг/л трансферрина, рН 7,0). Кондиционированную среду собирали через три дня и использовали при выполнении анализа пролиферации BaF3 с аламаровым синим в соответствии со следующим примером 19.
Пример 19
Отсутствие активации лигандом zcytor17lig мыши рецептора zcytor17 человека при выполнении анализа BaF3 с аламаровым синим
Пролиферацию клеток BaF3/zcytor17, BaF3/zcytor17/OSMRbeta и BaF3/zcytor17/WSX-1 (примеры 4 и 5В) оценивали, используя бессывороточную кондиционированную среду, полученную из клеток ВНК, экспрессирующих zcytor17lig мыши (пример 18).
Клетки BaF3/zcytor17, BaF3/zcytor17/OSMRbeta и BaF3/zcytor17/WSX-1 центрифугировали, промывали и вводили в среду без mIL-3 в соответствии с примером 5В. Кондиционированную среду, полученную из клеток ВНК, экспрессирующих zcytor17lig мыши (пример 18), разводили средой без mIL-3 до концентраций, составляющих 50%, 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75% и 0,375%. Анализ пролиферации выполняли аналогично примеру 5В. Результаты данного анализа были отрицательными, свидетельствуя о том, что zcytor17lig мыши не активирует zcytor17 человека и комплексы рецепторов zcytor17/OSMRbeta или zcytor17/WSX-1.
Пример 20
Активация лигандом zcytor17lig человека рецептора zcytor17/OSMRbeta человека при выполнении люциферазного анализа
А. Создание линии клеток BaF3/KZ134/zcytor17
Плазмида KZ134 была создана с использованием комплементарных олигонуклеотидов ZC12749 (SEQ ID NO:44) и ZC12748 (SEQ ID NO:45), содержащих связывающие элементы фактора транскрипции STAT из 4 генов, которые включают в себя модифицированный индуцируемый элемент c-fos Sis (m67SIE или hSIE) (Sadowski, H. et al., Science 261:1739-1744, 1993), p21 SIE1 из гена p21 WAF1 (Chin, Y. et al., Science 272:719-722, 1996), реактивный элемент гена β-казеина желез млекопитающего (Schmitt-Ney, M. et al., Mol. Cell. Biol. 11:3745-3755, 1991) и индуцируемый элемент STAT гена Fcg RI (Seidel, H. et al., Proc. Natl. Acad. Sci. 92:3041-3045, 1995). Указанные олигонуклеотиды, имеющие совместимые концы Asp718-XhoI, лигировали стандартными методами с репортерным вектором, содержащим люциферазу светляка, при помощи промотора c-fos (Poulsen, L.K. et al., J. Biol. Chem. 273:6229-6232, 1998), гидролизованного такими же ферментами и содержащего селектируемый маркер неомицина. Плазмиду KZ134 использовали для устойчивой трансфекции клеток BaF3 стандартными методами трансфекции и селекции с получением линии клеток BaF3/KZ134.
Устойчивая индикаторная линия клеток BaF3/KZ134, экспрессирующая непроцессированный рецептор zcytor17 или zcytor17/OSMRbeta, была получена в соответствии с примером 4. Клоны разводили, культивировали и отбирали стандартными методами. Клоны исследовали при помощи люциферазного анализа (см. пример 20В, ниже), используя в качестве индуктора среду, кондиционированную zcytor17lig человека, или очищенный белок zcytor17lig (см. пример 35, ниже). Отбирали клоны с самой высокой реакцией на люциферазу (при использовании люциферазы STAT) и с наименьшей фоновой реакцией. Кроме того, отбирали устойчивые линии клеток трансфектантов. Указанные линии клеток получили название BaF3/KZ134/zcytor17 или BaF3/KZ134/zcytor17/OSMRbeta, в зависимости от рецепторов, трансфицированных в данную линию клеток.
Аналогично, вышеописанным методом были созданы линии клеток ВНК, которые использовали в люциферазных анализах. Указанные линии клеток получили название BHK/KZ134/zcytor17 или BHK/KZ134/zcytor17/OSMRbeta, в зависимости от рецепторов, трансфицированных в линию клеток.
В. Активация лигандом zcytor17lig человека рецептора zcytor17 человека при выполнении люциферазного анализа клеток BaF3/KZ134/zcytor17/OSMRbeta или BHK/KZ134/zcytor17/OSMRbeta
Клетки BaF3/KZ134/zcytor17 и BaF3/KZ134/zcytor17/OSMRbeta осаждали центрифугированием и промывали в среде без mIL-3. Клетки трижды центрифугировали и промывали для полного удаления mIL-3. Затем клетки подсчитывали в гемоцитометре. Клетки культивировали на 96-луночном планшете в количестве около 30000 клеток/лунку в объеме 100 мкл/лунку с использованием среды без mIL-3. Аналогичным способом обрабатывали нетрансфицированные клетки BaF3/KZ134, используемые в качестве контрольного образца в последующем анализе. Клетки BHK/KZ134/zcytor17 или BHK/KZ134/zcytor17/OSMRbeta культивировали на 96-луночном планшете в количестве 15000 клеток/лунку в 100 мкл среды. Родительские клетки BHK/KZ134 использовали в качестве контрольного образца.
STAT-индуцированную активацию клеток ВaF3/KZ134/zcytor17, BaF3/KZ134/zcytor17/OSMRbeta, BHK/KZ134/zcytor17 или BHK/KZ134/zcytor17/OSMRbeta оценивали, используя (1) кондиционированную среду, полученную из клеток ВНК570, трансфицированных zcytor17lig человека (пример 7), (2) кондиционированную среду, полученную из клеток ВНК570, трансфицированных zcytor17lig мыши (пример 18), (3) очищенный zcytor17lig человека (пример 35) или (4) среду без mIL-3, предназначенную для измерения контрольной реакции, вызываемой только средой. Кондиционированную среду разводили средой RPMI без mIL-3 до концентраций, составляющих 50%, 25%, 12,5%, 6,25%, 3,125%, 1,5%, 0,75% и 0,375%. Очищенный zcytor17lig человека разводили до концентраций, составляющих 1200, 600, 300, 150, 75, 37,5, 18,75 или 9,4 пМ. Сто микролитров разведенной кондиционированной среды или белка добавляли к клеткам ВaF3/KZ134/zcytor17, BaF3/KZ134/zcytor17/OSMRbeta, BHK/KZ134/zcytor17 или BHK/KZ134/zcytor17/OSMRbeta. Анализ с использованием кондиционированной среды выполняли параллельно с анализом нетрансфицированных клеток BaF3/KZ134 или BHK/KZ134 в качестве контрольного образца. Общий анализируемый объем составлял 200 мкл. Анализируемые планшеты инкубировали при 37°C, 5% СО2 в течение 24 часов, после чего клетки BaF3 осаждали центрифугированием со скоростью 2000 оборотов/мин в течение 10 минут, среду отсасывали и добавляли 25 мкл буфера для лизиса (Promega). Для линий клеток ВНК стадия центрифугирования была необязательной, так как клетки были сцеплены. Планшеты выдерживали в течение 10 минут при комнатной температуре и измеряли активацию репортерной конструкции STAT в люминометре (Lаbsystems Luminoskan, модель RS), куда добавляли 40 мкл субстрата, используемого в люциферазном анализе (Promega), при интеграции в течение пяти секунд.
Результаты данного анализа показали, что реакция клеток BaF3/KZ134/zcytor17/OSMRbeta и BHK/KZ134/zcytor17/OSMRbeta в репортерной конструкции STAT на zcytor17lig человека при сравнении с клетками BaF3/KZ134/zcytor17, BHK/KZ134/zсytor17, или нетрансфицированными контрольными клетками BaF3/KZ134, или BHK/KZ134 опосредована рецепторами zcytor17/OSMRbeta. Полученные результаты также показали, что zcytor17lig мыши не активирует репортерную конструкцию STAТ при использовании комплекса рецепторов человека.
Пример 21
Активация лиганда zcytor17lig мыши при выполнении анализа с использованием костного мозга мыши
А. Выделение неадгезивных клеток костного мозга с низкой плотностью
Свежий пунктат костного мозга из бедренной кости получали у самцов мышей Balb/C или C57BL/6 в возрасте 6-10 недель. Костный мозг промывали средой RPMI, содержащей 10% FBS (JRH, Lenexa KS; Hyclone, Logan UT), и суспендировали в среде RPMI, содержащей 10% FBS, в виде суспензии цельных клеток костного мозга. Суспензию цельных клеток костного мозга обрабатывали в градиенте плотности (Nycoprep, 1.077, Animal; Gibco BRL) для обогащения преимущественно мононуклеарных клеток с низкой плотностью, выполняемого следующим образом: суспензию цельных клеток костного мозга (около 8 мл) осторожно пипетировали на поверхность примерно 5 мл градиентного раствора Nycoprep в 15 мл конической пробирке и центрифугировали с ускорением 600 х g в течение 20 минут. Межфазный слой, содержащий мононуклеарные клетки с низкой плотностью, удаляли, промывали избытком среды RPMI, содержащей 10% FBS, и осаждали центрифугированием с ускорением 400 х g в течение 5-10 минут. Полученный осадок вновь суспендировали в среде RPMI, содержащей 10% FBS, вводили в колбу Т-75 в количестве около 106 клеток/мл и инкубировали при 37°C, 5% CО2, в течение примерно 2 часов. Клетки, полученные в суспензии, представляли собой неадгезивные клетки костного мозга с низкой плотностью (NA LD).
В. Анализ на 96-луночном планшете
Клетки NA LD костного мозга мыши культивировали в количестве 25000-45000 клеток/лунку на 96-луночных планшетах для тканевых культур в среде RPMI, содержащей 10% FBS, 1 нг/мл фактора поддержания стволовых клеток мыши (mSCF) (R&D Systems, Minneapolis, MN) и 5% кондиционированной среды, полученной из одного из следующих источников, включающих в себя (1) клетки BHK 570, экспрессирующие zcytor17lig мыши (пример 18), (2) клетки ВНК 570, экспрессирующие zcytor17lig человека (пример 7), или (3) контрольные клетки ВНК 570, содержащие вектор и не экспрессирующие никакой лиганд. Указанные клетки затем обрабатывали различными цитокинами для испытания на размножение или дифференцировку гемопоэтических клеток костного мозга. Для выполнения указанного испытания культивируемые клетки NA LD костного мозга мыши подвергали воздействию интерлейкина-15 человека (hIL-15) (R&D Systems) или одного из панели других цитокинов (R&D Sуstems). Серийные разведения hIL-15 или других цитокинов испытывали при двукратном серийном разведении с достижением концентраций от около 50 нг/мл до около 0,5 нг/мл. Через 8-12 дней производили оценку пролиферации клеток при помощи анализа с аламаровым синим в соответствии с примером 5В.
С. Результаты анализа клеток NA LD костного мозга мыши на 96-луночном планшете
Кондиционированные среды, полученные из клеток ВНК, экспрессирующих zcytor17lig мыши и человека, используемый отдельно или в сочетании с другими цитокинами, оказывающими синергичное действие, могут стимулировать увеличение популяции гемопоэтических клеток в клетках NA LD костного мозга мыши по сравнению с контрольной ВНК-кондиционированной средой. Популяция гемопоэтических клеток, выросшая под действием zcytor17lig мыши в присутствии или в отсутствие других цитокинов, и популяция указанных гемопоэтических клеток, выросшая под действием zcytor17lig человека в присутствии или в отсутствие других цитокинов, далее размножалась в культуре клеток. Указанные гемопоэтические клетки окрашивали фикоэритрин-меченным антителом против NK-клеток Pan (PharMingen) и анализировали методом проточной цитометрии, результаты которой показали, что размножившиеся клетки были положительно окрашены в отношении данного маркера естественных клеток-киллеров (NK). Аналогичным образом можно использовать другие специфические маркеры гемопоэтических клеток для определения размножения, например, CD4+ или CD8+ Т-клеток, других популяций Т-клеток, В-клеток и других иммунокомпетентных клеток.
Вышеуказанный анализ выполняли на 96-луночном планшете с использованием свежих клеток костного мозга человека, приобретенных в компании Poietic Technologies, Gaithersburg, MD. Полученные положительные результаты снова показали, что zcytor17lig мыши и человека, используемый отдельно или в комбинации с другими цитокинами, оказывающими синергичное действие, может увеличивать популяцию гемопоэтических клеток, которая положительно окрашивается в отношении специфических клеточных маркеров.
Пример 22
Конструкции для получения трансгенных мышей, экспрессирующих zcytor17lig
Конструкция для экспрессии zcytor17lig человека из промотора МТ-1
Олигонуклеотиды были предназначены для получения фрагмента PCR, содержащего согласованную последовательность Козака и кодирующую область zcytor17lig человека. Указанные олигонуклеотиды имели сайт FseI у 5'-конца и сайт AscI у 3'-конца для облегчения клонирования в (а) рМТ12-8, стандартный трансгенный вектор, или (b) pKFO51, лимфоид-специфический трансгенный вектор (пример 22В).
Реакции PCR выполняли с использованием примерно 200 нг матрицы zcytor17lig человека (SEQ ID NO:1) и олигонуклеотидов, предназначенных для амплификации непроцессированной или активной части zcytor17lig. Условия выполнения реакций PCR определяли методами, известными в данной области. Продукты PCR разделяли электрофорезом в агарозном геле и очищали при помощи набора для гель-экстракции QiaQuick™ (Qiagen). Выделенный фрагмент ДНК правильного размера гидролизовали FseI и AscI (Boehringer-Mannheim), осаждали этанолом и лигировали с вектором pMT12-8, который предварительно гидролизовали FseI и AscI. Плазмида pMT12-8, предназначенная для экспрессии представляющего интерес гена в печени и других тканях трансгенных мышей, содержала полигенный экспрессирующий кластер, фланкированный 5'-концевой ДНК МТ-1 длиной 10 т.п.н. и 3'-концевой ДНК МТ-1 длиной 7 т.п.н. Полигенный экспрессирующий кластер содержал промотор МТ-1, интрон инсулина II крысы, полилинкер для вставки требуемого клона и поли А-последовательность гормона роста человека (hGH).
Примерно один микролитр каждой лигированной реакционной смеси электропорировали в компетентные клетки DH10B ElectroMax™ (GIBCO BRL, Gaithersburg, MD), в соответствии с инструкциями изготовителя, переносили на планшеты со средой LB, содержащей 100 мкг/мл ампициллина, и инкубировали в течение ночи. Колонии собирали и выращивали в среде LB, содержащей 100 мкг/мл ампициллина. ДНК, полученную из собранных клонов при помощи набора Miniprep, подвергали рестрикционному гидролизу только EcoRI или комбинацией FseI и AscI для обнаружения вставки zcytor17lig человека с последующим выполнением электрофореза в агарозном геле. Правильный zcytor17lig человека pMT исследовали при помощи набора Maxiprep. Фрагмент SalI, содержащий 5'- и 3'-концевые фланкирующие последовательности, промотор МТ-1, интрон инсулина II крысы, кДНК zcytor17lig человека и поли А-последовательность hGH использовали для микроинъекции в ооциты оплодотворенной мыши. Для выполнения микроинъекции и создания трансгенных мышей применяли методы, описанные в публикации Hogan, B. et al. Manipulating the Mouse Embryo. 2nd ed., Cold Spring Harbor Laboratory Press, NY, 1994.
В. Конструкция для экспрессии zcytor17lig человека при использовании лимфоид-специфического промотора EµLCK
Олигонуклеотиды были предназначены для получения фрагмента PCR, содержащего согласованную последовательность Козака и кодирующую область zcytor17lig человека. Указанные олигонуклеотиды имели сайт FseI у 5'-конца и сайт AscI у 3'-конца для облегчения клонирования в pKFO51, лимфоид-специфический трансгенный вектор.
Реакции PCR выполняли с использованием примерно 200 нг матрицы zcytor17lig человека (SEQ ID NO:1) и олигонуклеотидов, предназначенных для амплификации непроцессированной или активной части zcytor17lig. Условия выполнения реакций PCR определяли методами, известными в данной области. Выделенный фрагмент ДНК правильного размера гидролизовали FseI и AscI (Boehringer-Mannheim), осаждали этанолом и лигировали с вектором pKFO51, который предварительно гидролизовали FseI и AscI. Трансгенный вектор pKFO51, полученный из p1026X (Iritani, B.M., et al., EMBO J. 16:7019-31, 1997), содержал Т-клеткоспецифический проксимальный промотор lck, В/Т-клеткоспецифический энхансер тяжелой цепи иммуноглобулина µ, полилинкер для вставки требуемого клона и мутированный ген hGH, кодирующий неактивный белок гормона роста (предоставляющий 3'-концевые интроны и сигнал полиаденилирования).
Примерно один микролитр каждой лигированной реакционной смеси электропорировали и культивировали на планшетах, клоны собирали и исследовали описанным выше методом рестрикционного гидролиза для обнаружения вставки zcytor17lig человека. Наличие правильного клона pKFO51-zcytor17lig подтверждали секвенированием и исследованием при помощи набора MaxiPrep. Фрагмент NotI, содержащий проксимальный промотор lck и энхансер иммуноглобулина µ (EµLCK), кДНК zcytor17lig и мутированный ген hGH, использовали для микроинъекции в ооциты оплодотворенной мыши.
С. Конструкция для экспрессии zcytor17lig мыши из промотора EF1альфа
Затравки ZC41498 (SEQ ID NO:86) и ZC41496 (SEQ ID NO:87) использовали для PCR матрицы библиотеки кДНК яичка мыши. Реакции PCR успешно выполняли примерно в 50 мкл объемах с 10% ДМСО или без него, используя полимеразу pfu turbo (Stratagene), в соответствии с рекомендациями изготовителя; при дополнительном применении горячего воска для инициации реакции с использованием набора Hot Start 50 (Molecular Bioproducts, Inc., San Diego, CA). Термоциклизацию PCR выполняли в следующих условиях: один цикл при 94°C в течение 4 минут; затем 40 циклов при 94°C в течение 30 секунд, при 48°C в течение 30 секунд, при 72°C в течение 50 секунд; с дополнительным конечным удлинением цепи при 72°C в течение 7 минут. Продукты двух реакций PCR собирали и очищали, используя агарозу с низкой температурой плавления и расщепляющий агарозу фермент гелазу (Epicenter, Inc., Madison, WI), в соответствии с рекомендациями изготовителя.
В ДНК-секвенированных продуктах PCR была обнаружена последовательность кДНК zcytor17 мыши (SEQ ID NO:90), которая включала в себя открытую рамку считывания, идентичную SEQ ID NO:10, из чего следует, что SEQ ID NO:10 кодирует полипептид zcytor17lig мыши. Затравки для PCR, ZC41583 (SEQ ID NO:88) и ZC41584 (SEQ ID NO:89), затем использовали для введения сайтов рестрикции FseI и AscI и неполной последовательности Козака в открытую рамку считывания mcytor17lig и терминирующий кодон (SEQ ID NO:92). Термоячейку для проведения реакций Robocycler 40 (Stratagene) использовали для создания градиента температур отжига и циклизации в следующих условиях. Полимеразу pfu turbo (Stratagene) использовали, как было описано выше, но только в 10% ДМСО. Циклизация включала в себя один цикл при 94°C в течение 4 минут, затем 20 циклов при 94°C в течение 30 секунд, градиент от 65°C до 51°C в течение 30 секунд, при 72°C в течение 1 минуты и один цикл удлинения цепи при 72°C в течение 7 минут. В качестве матрицы для второй реакции термоциклизации использовали 1 мкл исходного очищенного в геле продукта PCR mcytor17lig. Продукт PCR, полученный в результате выполнения трех реакций при самых низких температурах, собирали и очищали в геле при помощи вышеописанного метода с использованием гелазы (Epicenter). Очищенный фрагмент гидролизовали FseI и AscI и лигировали в вектор pZP7X, модифицированный с включением сайтов FseI и AscI на сайте клонирования. Указанный вектор отправляли на секвенирование для подтверждения правильной структуры последовательности. Клонированная последовательность кДНК мыши представлена в SEQ ID NO:90, и соответствующая полипептидная последовательность показана в SEQ ID NO:91 (которая идентична SEQ ID NO:11).
Выделенный фрагмент ДНК правильного размера гидролизовали FseI и AscI (Boehringer-Mannheim) и субклонировали в плазмиде, содержащей промотор EF1альфа, который предварительно гидролизовали FseI и AscI. Правильный zcytor17lig мыши, полученный при помощи промотора EF1альфа, исследовали с использованием набора MaxiPrep. Полигенный экспрессирующий кластер содержал промотор EF1альфа (с удаленным сайтом FseI), интрон EF1альфа, SUR IRES-подобный сайт для облегчения экспрессии, полилинкер, фланкированный сайтами инсулина II крысы, у 5'-конца которого были добавлены сайты FseI, PmeI, AscI для вставки требуемого клона, и поли А-последовательность гормона роста человека (hGH). Фрагмент NotI длиной 7,5 т.п.н., содержащий промотор EF1альфа, полигенный экспрессирующий кластер и zcytor17lig мыши, был использован для микроинъекции в ооциты оплодотворенной мыши. Плазмида EF1альфа была получена в лаборатории Louis-Marie of the Laboratoire de Differenciation Cellulaire в соответствии с описанием, приведенным в статье Taboit-Dameron et al., 1999, Transgenic Research 8:223-235.
D. Конструкция для экспрессии zcytor17lig мыши из лимфоид-специфического промотора EµLCK
Олигонуклеотиды были предназначены для получения фрагмента PCR, содержащего согласованную последовательность Козака и кодирующую область zcytor17lig мыши. Указанные олигонуклеотиды имели сайт FseI у 5'-конца и сайт AscI у 3'-конца для облегчения клонирования в pKFO51 (см. пример 22В, выше).
Выделенный фрагмент ДНК zcytor17lig правильного размера, использованный в конструкциях с промотором EF1альфа и гидролизованный FseI и AscI (Boehringer-Mannheim), субклонировали в плазмиде, содержащей pKFO51, лимфоид-специфический трансгенный вектор. Трансгенный вектор pKFO51 был получен из p1026X (Iritani, B.M., et al., EMBO J. 16:7019-31, 1997) и содержал Т-клеткоспецифический проксимальный промотор lck, В/Т-клеткоспецифический энхансер тяжелой цепи иммуноглобулина µ, полилинкер для вставки требуемого клона и мутированный ген hGH, кодирующий неактивный белок гормона роста (предоставляющий 3'-концевые интроны и сигнал полиаденилирования). Фрагмент NotI длиной 6,5 т.п.н., содержащий проксимальный промотор lck и энхансер иммуноглобулина µ (EµLCK), кДНК zcytor17lig мыши и мутированный ген hGH использовали для микроинъекции в ооциты оплодотворенной мыши (пример 41).
Пример 23
Создание векторов млекопитающих, экспрессирующих cytor17lig-CEE
А. Создание zcytor17lig-CEE/pZMP21
Экспрессирующая плазмида, содержащая весь или часть полинуклеотида, кодирующего zcytor17lig человека, была создана методом гомологичной рекомбинации. Указанная плазмида получила название zcytor17lig-CEE/pZMP21.
Конструкция zcytor17lig-CEE/pZMP21 была создана путем получения фрагмента zcytor17lig-CEE (SEQ ID NO:95) (ее соответствующая аминокислотная последовательность показана в SEQ ID NO:96) с использованием амплификации методом PCR. В качестве матричной ДНК для получения фрагмента zcytor17lig-CEE был использован zcytor17lig/pZP7nx. Для получения фрагмента zcytor17lig-CEE были использованы следующие затравки: (1) ZC41607 (SEQ ID NO:97) (смысловая последовательность), которая включает от 5'-конца до 3'-конца 28 п.н. фланкирующей последовательности вектора (5'-конец вставки) и 21 п.н., соответствующую 5'-концевой последовательности zcytor17lig; и (2) ZC41605 (SEQ ID NO:98) (антисмысловая последовательность), которая включает от 5'-конца до 3'-конца 37 п.н. фланкирующей последовательности вектора (3'-конец вставки), 3 п.н. терминирующего кодона, 21 п.н., кодирующую С-конец метки ЕЕ, и 21 п.н., соответствующую 3'-концу последовательности zcytor17lig. Фрагмент, полученный в результате амплификации PCR, является копией матричного zcytor17lig с добавлением С-концевой метки ЕЕ, позволяя получить конечный продукт zcytor17lig-CEE.
Реакции PCR выполняли следующим образом: к 100 мкл конечного объема добавляли 10 мкл 10-кратного объема реакционного буфера с Taq-полимеразой, содержащего 15 мМ MgCl2 (Gibco), 1 мкл Taq-ДНК-полимеразы (5 единиц/мкл, Gibco), 3 мкл 10 мМ dNTP, 78 мкл дистиллированной Н2О, 3 мкл 20 пмоль/мкл исходного раствора затравки ZC41607 (SEQ ID NO:97), 3 мкл 20 пмоль/мкл исходного раствора затравки ZC41605 (SEQ ID NO:98) и 2 мкл 0,13 мкг/мкл исходного раствора матричной ДНК zcytor17lig. К смеси добавляли 50 мкл минерального масла. Реакционную смесь нагревали до 94°C в течение 5 минут, затем выполняли 35 циклов при 94°C в течение 1 минут, при 55°C в течение 2 минут; при 72°C в течение 3 минут; удлиняли цепь при 72°C в течение 10 минут и выдерживали продукт реакции при 4°C до сбора реакционной смеси.
Плазмиду pZMP21 подвергали рестрикционному гидролизу ферментом BglII, очищали при помощи набора для очистки продуктов PCR QiaQuick (Qiagen) с использованием микроцентрифуги и использовали для рекомбинации фрагментом PCR. Плазмида pZMP21 была создана из pZMP20, полученной из pZP9 (депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, VA 20110-2209, с присвоением № 98668) с использованием генетических элементов дрожжей, выделенных из pRS316 (депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, VA 20110-2209, с присвоением № 77145), элемента IRES из полиовируса и внеклеточного домена CD8, усеченного у карбоксильного конца трансмембранного домена. PZMP21 является экспрессирующим вектором млекопитающего, содержащим полигенный экспрессирующий кластер, имеющий промотор MРSV, интрон сигнального пептида иммуноглобулина, несколько сайтов рестрикции для вставки кодирующих последовательностей, терминирующий кодон и терминатор гормона роста человека. Указанная плазмида имеет также ориджин репликации E. coli, экспрессирующую единицу селектируемого маркера млекопитающего, содержащую промотор SV40, энхансер и ориджин репликации, ген DHFR, терминатор SV40 и последовательности URA3 и CEN-ARS, необходимые для отбора и репликации в S. cerevisiae.
Пятьдесят микролитров компетентных дрожжевых клеток (S. cerevisiae) независимо объединяли со 100 нг разрезанной плазмиды и 5 мкл ранее описанной смеси PCR и переносили в 0,2 см кювету для электропорации. Смесь дрожжи/ДНК электропорировали при 0,75 кВ (5 кВ/см), бесконечном сопротивлении, 25 мкФ. В каждую кювету добавляли 600 мкл 1,2 М раствора сорбита, дрожжи переносили в виде одной 100 мкл аликвоты и одной 300 мкл аликвоты на две чашки URA-D и инкубировали при 30°C. Примерно через 72 часа трансформанты дрожжей Ura+ из одной чашки вновь суспендировали в 1 мл Н2О и осаждали коротким центрифугированием с образованием осадка дрожжевых клеток. Клеточный дебрис вновь суспендировали в 500 мкл буфера для лизиса (2% тритона Х-100, 1% SDS, 100 мМ NaCl, 10 мМ трис-буфера, рН 8,0, 1 мМ EDTA). 500 мкл лизированной смеси добавляли в пробирку Эппендорфа, содержащую 600 мкм стеклянных шариков, промытых 300 мкл кислоты, и 300 мкл фенол-хлороформа, интенсивно перемешивали два или три раза с интервалами в 1 минуту и центрифугировали в центрифуге Эппендорфа при максимальной скорости в течение 5 минут. Триста микролитров водной фазы переносили в чистую пробирку и ДНК осаждали 600 мкл 100% этанола (EtOH), затем центрифугировали в течение 10 минут при 4°C. Осадок ДНК промывали 500 мкл 70% EtOH и центрифугировали в течение 1 минуты при 4°C. Осадок ДНК вновь суспендировали в 30 мкл Н2О.
Электрокомпетентные клетки E. coli (MC1061) трансформировали 5 мкл ДНК дрожжей и 50 мкл клеток МС1061. Клетки электропорировали при 2,0 кВ, 25 мкФ и 400 Ом. После электропорации добавляли 600 мкл SOC (2% бактотриптона (Difco, Detroit, MI), 0,5% дрожжевого экстракта (Difco), 10 мМ NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкозы). Электропорированные клетки E. coli переносили в виде 200 мкл и 50 мкл аликвот на две чашки со средой LB, содержащей ампициллин (бульон LB (Lennox), 1,8% бактоагара (Difco), 100 мг/л ампициллина). Чашки инкубировали в перевернутом виде в течение примерно 24 часов при 37°C. Произвольно отбирали три колонии, устойчивые к ампициллину, и анализировали последовательность вставки. Плазмидную ДНК выделяли в большом количестве из клона с подтвержденной последовательностью при помощи набора Qiagen Maxi (Qiagen), в соответствии с инструкциями изготовителя.
В. Экспрессия zcytor17lig человека в клетках млекопитающего
Непроцессированный белок zcytor17lig был продуцирован в клетках ВНК, трансфицированных zcytor17lig-CEE/pZMP21 (пример 23А). Клетки ВНК 570 (АТСС CRL-10314) переносили в колбы для тканевой культуры Т75 и выращивали примерно до 50-70% слияния при 37°C, 5% СО2, в питательной среде (SL7V4, 5% FBS, 1% пенициллина/стрептомицина). Затем клетки трансфицировали zcytor17lig-CEE/pZMP21 методом трансфекции, опосредуемой липосомами (с использованием липофектамина™, Life Technologies), в бессывороточной (SF) среде (SL7V4). Плазмиду (16 мкг) разводили бессывороточной средой в 1,5 мл пробирках до общего конечного объема 640 мкл. Тридцать пять микролитров липидной смеси смешивали с 605 мкл бессывороточной среды и полученную смесь инкубировали в течение примерно 15 минут при комнатной температуре. К смеси ДНК:липиды добавляли пять миллилитров бессывороточной среды. Клетки промывали один раз 10 мл PBS, после чего PBS декантировали и добавляли смесь ДНК:липиды. Клетки инкубировали при 37°C в течение пяти часов и в каждую чашку добавляли 15 мл среды (SL7V4, 5% FBS, 1% пенициллина/стрептомицина). Чашки инкубировали при 37°C в течение ночи и на следующий день среду смеси ДНК:липиды заменяли селективной средой (SL7V4, 5% FBS, 1% пенициллина/стрептомицина, 1 мкМ метотрексата). Примерно через 10 дней после трансфекции колонии, устойчивые к метотрексату, из колбы для трансфекции Т75 обрабатывали трипсином, клетки собирали, культивировали в колбе Т-162 и получали крупномасштабную культуру.
Пример 24
Экспрессия растворимого рецептора zcytor17 в клетках E. coli
А. Создание вектора pCMH01, экспрессирующего слитый полипептид huzcytor17/MBP-6H
Экспрессирующая плазмида, содержащая полинуклеотид, кодирующий растворимый рецептор zcytor17, слитый у С-конца со связывающим мальтозу белком (МВР), была получена методом гомологичной рекомбинации. Слитый полипептд содержит N-концевую часть МВР, состоящую примерно из 388 аминокислот, слитую с любыми описанными растворимыми рецепторами zcytor17. Фрагмент кДНК zcytor17 (SEQ ID NO:4) выделяли при помощи PCR. Для получения фрагмента zcytor17 при выполнении стандартной реакции PCR использовали две затравки: (1) одна затравка содержала примерно 40 п.н. фланкирующей последовательности вектора и около 25 п.н., соответствующих аминоконцу zcytor17, и (2) вторая затравка содержала примерно 40 п.н. у 3'-конца, соответствующих франкирующей последовательности вектора, и около 25 п.н., соответствующих карбоксильному концу zcytor17. Два мкл из 100 мкл реакционной смеси PCR разделяли электрофорезом в 1,0% агарозном геле с использованием однократного объема буфера ТВЕ для анализа, при этом был обнаружен ожидаемый фрагмент. Остальную реакционную смесь PCR объединяли с содержимым второй пробирки для PCR и осаждали 400 мкл абсолютного этанола. Осажденную ДНК вводили в SmaI-разрезанный вектор рТАР170 реципиента для получения конструкции, кодирующей слитый фрагмент МВР-zcytor17.
Плазмида рТАР170 была получена из плазмид pRS316 и pMAL-c2. Плазмида pRS316 является шаттл-вектором Saccharomyces cerevisiae (Hieter P. and Sikorski, R., Genetics 122:19-27, 1989). pMAL-C2 (NEB) является экспрессирующей плазмидой E. coli. Указанная плазмида несет промотор tac, стимулирующий MalE (ген, кодирующий МВР), за которым следует метка His, сайт расщепления тромбина, сайт клонирования и терминатор rrnB. Вектор pTAP170 был создан методом гомологичной рекомбинации дрожжей. 100 нг EcoR1-разрезанной плазмиды pMAL-c2 рекомбинировали с 1 мкг PvuI-разрезанного вектора pRS316, 1 мкг линкера и 1 мкг Sca1/EcoR1-разрезнного вектора pRS316. Линкер состоял из олигонуклеотидов zc19372 (SEQ ID NO:157) (100 пмоль), zc19351 (SEQ ID NO:158) (1 пмоль), zc19352 (SEQ ID NO:159) (1 пмоль) и zc19371 (SEQ ID NO:160) (100 пмоль), объединенных в результате выполнения реакции PCR. Реакцию PCR выполняли в следующих условиях: 10 циклов при 94°C в течение 30 секунд, при 50°C в течение 30 секунд и при 72°C в течение 30 секунд; последующее выдерживание при 4°C. Продукты PCR концентрировали, осаждая 100% этанолом.
Сто микролитров компетентных дрожжевых клеток (S. cerevisiae) объединяли с 10 мкл смеси, содержащей примерно 1 мкг вставки zcytor17 человека и 100 нг SmaI-гидролизованного вектора рТАР170, и переносили в 0,2 см кювету для электропорации. Смесь дрожжи/ДНК электропорировали при 0,75 кВ (5 кВ/см), бесконечном сопротивлении и 25 мкФ. В каждую кювету добавляли 600 мкл 1,2 М раствора сорбита. Дрожжи переносили в виде двух 300 мкл аликвот на две чашки-URA D и инкубировали при 30°C.
Примерно через 48 часов трансформанты дрожжей Ura+ собирали с одной чашки, выделяли ДНК и вводили в электрокомпетентнные клетки E. coli (например, МС1061, Casadaban et al., J. Mol. Biol. 138, 179-207) и культивировали на чашках, содержащих ММ/СА+KAN в количестве 25 мкг/л (Pryor and Leiting, Protein Expression and Purification 10:309-319, 1997), в стандартных условиях. Клетки выращивали в ММ/СА с 25 мкг/мл каномиацина, встряхивая, в течение двух часов при 37°C. Один мл культуры индуцировали 1 мМ IPTG. Через два-четыре часа 250 мкл каждой культуры смешивали с 250 мкл промытых кислотой стеклянных шариков, 250 мкл буфера Торнера, содержащего 5% βМЕ и краситель (8М мочевины, 100 мМ трис-буфера, рН 7,0, 10% глицерина, 2 мМ EDTA, 5% SDS). Образцы интенсивно перемешивали в течение одной минуты и нагревали до 65°C в течение 10 минут. 20 мкл образца помещали на одну полосу 4%-12% геля PAGE (NOVEX). Гели разделяли электрофорезом в буфере 1XMES. В положительных клонах, названных рСМН01, анализировали последовательность.
Один микролитр секвенирующей ДНК использовали для трансформации штамма BL21. Клетки электропорировали при 2,0 кВ, 25 мкФ и 400 Ом. После электропорации добавляли 0,6 мл ММ/СА с 25 мкг/л каномиацина. Клетки выращивали в ММ/СА и индуцировали ITPG, как было описано выше. Положительные клоны выращивали для очистки слитого белка huzcytor17/MBP-6H стандартными методами.
В. Очистка растворимого рецептора huzcytor17/MВP-6H от продуктов ферментации E. coli
За исключением особо оговоренных случаев, все операции выполняли при 4°C. Очистку рекомбинантного полипептида растворимого рецептора huzcytor17/MBP-6H производили следующим способом. Клетки E. coli, содержащие конструкцию рСМН01 и экспрессирующие полипептид растворимого рецептора huzcytor17/MBP-6H, получали стандартными методами молекулярной биологии и культивировали в бульоне SuperBroth II (12 г/л казеина, 24 г/л дрожжевого экстракта, 11,4 г/л дикалийфосфата, 1,7 г/л монокалийфосфата (Becton Dickenson, Cockeysville, MD). Полученные клетки собирали и замораживали в 0,5% глицерине. Для очистки белка использовали двадцать граммов замороженных клеток.
Оттаявшие клетки вновь суспендировали в 500 мл буфера для уравновешивания амилозы (20 мМ трис-буфера, 100 мМ NaCl, рН 8,0). Клетки лизировали при помощи системы разрушения клеток French Press (Constant Systems Ltd., Warwick, UK) при температуре от -7°C до -10°C и давлении 43 атм (30000 фунтов/кв.дюйм). Разрушение вновь суспендированных клеток проверяли на основании показаний А600 до и после циклизации в системе French Press. Суспензию лизированных клеток осаждали центрифугированием с ускорением 10000 х g в течение 30 минут. Супернатант собирали с осадка для очистки белка.
Двадцать пять миллилитров амилозной смолы (New England Biolabs, Beverly, MA) выливали в стеклянную колонку Bio-Rad с диаметром 2,5 см и высотой 10 см. Колонку заполняли и уравновешивали под действием силы тяжести 10 объемами колонки (CV) буфера для уравновешивания амилозы. Собранный супернатант клеток порциями загружали в амилозную смолу и выдерживали в течение ночи при покачивании. Загруженную смолу возвращали в стеклянную колонку, промывали 10 объемами колонки буфера для уравновешивания амилозы и элюировали под действием силы тяжести ~2 объемами колонки буфера для элюирования амилозы (буфер для уравновешивания амилозы, 10 мМ мальтозы, Fluka Biochemical, Switzerland). Десять 5 мл фракций собирали в зависимости от профиля элюрования и анализировали оптическую плотность при 280 и 320 нМ. Амилозную смолу регенерировали 1 объемом колонки дистиллированной Н2О, 5 объемами колонки 0,1% (мас./об.) SDS (Sigma), 5 объемами колонки дистиллированной Н2О, 5 объемами колонки буфера для уравновешивания амилозы и, наконец, 1 объемом колонки буфера для хранения амилозы (буфер для уравновешивания амилозы, 0,02% (мас./об.) азида натрия). Регенерированную смолу хранили при 4°C.
Представляющие интерес фракции профиля элюрования собирали и диализовали в камере для диализа 10К (Slide-A-Lyzer, Pierce Immunochemical) против 4 л PBS, рН 7,4 (Sigma), производя 4 замены в течение 8 часов. После диализа собранный материал представлял собой очищенный полипептид huzcytor17/MBP-6H. Очищенный полипептид huzcytor17/ MBP-6H стерилизовали фильтрацией и анализировали методом SDS-PAGE с окрашиванием кумассии для продукта с соответствующей молекулярной массой. Концентрация полипептида huzcytor17/MBP-6H, определенная при помощи анализа ВСА, составляла 0,76 мг/мл.
Очищенный полипептид huzcytor17/MBP-6H использован для получения препарата для иммунизации кроликов и направлен в компанию R&R Research and Development (Stanwood, WA) для получения поликлонального антитела (пример 25, ниже).
Пример 25
Поликлональное антитело против растворимого рецептора zcytor17 человека
А. Получение и очистка
Поликлональные антитела были получены в результате иммунизации двух самок новозеландского белого кролика очищенным рекомбинантным белком huzcytor17/MBP-6H (пример 24). Кроликам сначала внутрибрюшинно (IP) инъецировали 200 мкг очищенного белка в полном адъюванте Фрейнда и затем внутрибрюшинно инъецировали бустер-дозы 100 мкг белка в неполном адъюванте Фрейнда каждые три недели. Через семь-десять дней после введения второй бустер-дозы (всего 3 инъекции) у животных брали кровь и собирали сыворотку. Затем животным вводили бустер-дозу и брали кровь каждые три недели.
Huzcytor17/MBP-6H-специфическую сыворотку кролика предварительно очищали от антител против МВР в колонке для очистки белка CNBr-SEPHAROSE 4B (Pharmacia LKB), которую получали, используя 10 мг неспецифического очищенного рекомбинантного МВР-слитого белка на грамм CNBr-SEPHAROSE. Huzcytor17/MBP-6H-специфические поликлональные антитела очищали аффинной хроматографией от предварительно адсорбированной сыворотки кролика в колонке для очистки белка CNBr-SEPHAROSE 4B (Pharmacia LKB), которую получали, используя 10 мг специфического очищенного от антигена рекомбинантного белка huzcytor17/MBP-6H. После очистки поликлональные антитела диализовали, четырежды заменяя PBS, соответствующий 20-кратному объему антитела, в течение по меньшей мере 8 часов. Huzcytor17-специфические антитела исследовали методом ELISA, используя 500 нг/мл очищенного рекомбинантного белка huzcytor17/MBP-6H в качестве мишени для антитела. Нижний предел обнаружения (LLD) антитела кролика против huzcytor17/MBP-6H, очищенного аффинной хроматографией, составлял 500 пг/мл в отношении специфического очищенного рекомбинантного антигена huzcytor17/MBP-6H.
В. Анализ антитела кролика против zcytor17/MBP-6H человека методами SDS-PAGE и вестерн-блоттинга
Антитело кролика против zcytor17/MBP-6H человека испытывали методом SDS-PAGE (NuPage 4-12%, Invitrogen, Carlsbad, CA) при окрашивании кумассии и методом вестерн-блоттинга с использованием антитела козы против IgG-HRP кролика. Очищенный белок (200-25 нг) или кондиционированную среду, содержащую zcytor17, разделяли электрофорезом с использованием миниячейки Xcell II Invitrogen Novex и переносили на нитроцеллюлозу (0,2 мм; Invitrogen, Carlsbad, CA) при комнатной температуре, используя блот-модуль Xcell Novex при перемешивании в соответствии с инструкциями, содержащимися в руководстве по эксплуатации прибора. Перенос осуществляли при 300 мА в течение одного часа в буфере, содержащем 25 мМ основного трис-буфера, 200 мМ глицина и 20% метанола. Затем фильтр блокировали буфером для вестерн-блоттинга А (собственного изготовления, 50 мМ трис-буфера, рН 7,4, 5 мМ EDTA, рН 8,0, 0,05% Igepal CA-630, 150 мМ NaCl и 0,25% желатина) при осторожном покачивании в течение ночи при 4°C. Нитроцеллюлозу быстро промывали, затем добавляли антитело кролика против zcytor17/MBP-6H человека (1:1000) в буфере для вестерн-блоттинга А. Блот инкубировали, осторожно покачивая, в течение 1,5 часов при комнатной температуре. Блот 3 раза промывали в буфере для вестерн-блоттинга А в течение 5 минут, затем добавляли антитело козы против IgG HRP кролика (1:1000) в буфере для вестерн-блоттинга А. Блот инкубировали, осторожно покачивая, в течение 1,25 часа при комнатной температуре. Блот 3 раза промывали в буфере для вестерн-блоттинга А в течение 5 минут и затем быстро промывали в Н2О. Блот проявляли при помощи коммерчески производимых реагентов для хемилюминесцентного субстрата (детектирующие реагенты для вестерн-блоттинга ECL 1 и 2, смешанные в соотношении 1:1, от компании Amersham Pharmacia Biotech, Вuckinghamshire, England), после чего блот экспонировали на рентгеновской пленке в течение периода времени до 15 минут.
Антитело кролика против zcytor17/MBP-6H человека было способно обнаруживать zcytor17 человека, присутствующий в кондиционированной среде, и очищенный белок zcytor17 в виде полосы, соответствующей 120 кДа, в восстановительных условиях.
Пример 26
Определение распределения zcytor17 мыши в тканевых панелях методом PCR
Панель кДНК из тканей мыши исследовали в отношении экспрессии zcytor17 мыши при помощи PCR. Данная панель была изготовлена авторами изобретения и содержала 94 образца марафонской кДНК и кДНК из различных нормальных и раковых тканей и линий клеток мыши, показанных в нижеследующей таблице 6. Образцы кДНК были получены из созданных авторами изобретения библиотек или марафонских кДНК из собственных препаратов РНК, РНК компании Clontech или РНК компании Invitrogen. Марафонские кДНК мыши получали при помощи набора marathon-Ready™ (Clontech, Palo Alto, CA), производили контроль качества при помощи затравок ZC10651 (SEQ ID NO:46) ZC10565 (SEQ ID NO:47) для рецептора трансферрина и разводили на основании интенсивности полосы трансферрина. Для гарантии качества образцов амплифицированной библиотеки в панели были выполнены три испытания на контроль качества (QC), которые включали в себя (1) оценку качества РНК, используемой для получения библиотек, для чего у полученных авторами изобретения кДНК определяли средний размер вставки при помощи PCR с использованием олигонуклеотидов вектора, специфических к векторным последовательностям библиотеки отдельных кДНК; (2) стандартизацию концентрации кДНК в образцах панели при помощи стандартных методов PCR для амплификации непроцессированного альфа-тубулина или кДНК G3PDH с использованием 5'-концевого олигонуклеотида вектора: ZC14063 (SEQ ID NO:48) и 3'-концевого альфа-тубулин-специфической олгонуклеотидной затравки ZC17574 (SEQ ID NO:49) и 3'-концевой G3PDH-специфической олигонуклеотидной затравки ZC17600 (SEQ ID NO:50); и (3) секвенирование образца для проверки возможного загрязнения рибосомной или митохондриальной ДНК. Панель культивировали на 96-луночном планшете, который включал в себя положительный контрольный образец геномной ДНК мыши (Clontech, Palo Alto, CA). В каждой лунке находилось примерно 0,2-100 пг/мкл кДНК. PCR выполняли с использованием олигонуклеотидов ZC38065 (SEQ ID NO:51) и ZC38068 (SEQ ID NO:52), TaKaRa Ex Taq™ (TAKARA Shuzo Co. LTD, Biomedicals Group, Japan) и красителя Rediload (Research Genetics, Ic., Huntsville, AL). Амплификацию осуществляли в следующих условиях: 1 цикл при 94°C в течение 5 минут; 5 циклов при 94°C в течение 30 секунд, при 68°C в течение 30 секунд; 35 циклов при 94°C в течение 30 секунд, при 56°C в течение 30 секунд и при 72°C в течение 30 секунд, затем 1 цикл при 72°C в течение 5 минут. Примерно 10 мкл продукта реакции PCR разделяли стандартным электрофорезом в 4% агарозном геле. Фрагмент ДНК правильного размера был обнаружен в головном мозге, CD90+-клетках, дендритных клетках, эмбриональных клетках, MEWt№2, линии клеток предстательной железы Tuvak, слюнной железе, коже и яичке.
Фрагмент ДНК для кожи и яичка вырезали и очищали при помощи набора для гель-экстракции (Qiagen, Chatsworth, CA), в соответствии с инструкциями изготовителя. Фрагменты секвенировали для подтверждения того, что они действительно являются zcytor17 мыши.
Пример 27
Экспрессия zcytor17 человека в различных тканях, определяемая количественным анализом методом RT/PCR в реальном времени
А. Затравки и зонды для zcytor17, OSMRbeta, zcytor17lig человека, предназначенные для стандартного и количественного анализа методом RT-PCR
Количественный анализ методом RT-PCR в реальном времени с использованием системы обнаружения последовательностей ABI PRISM 7900 (PE Applied Biosystems, Inc., Foster City, CA) был ранее описан в научной литературе (см. Heid, C.A. et al., Genome Research 6:986-994, 1996; Gibson, U.E.M. et al., Genome Research 6:995-1001, 1996; Sundaresan, S. et al., Endocrinology 139:4756-4764, 1998). Данный метод включает использование геноспецифического зонда, содержащего репортерные и гасящие флуоресцентные красители. Когда зонд находится в интактном состоянии, излучение репортерного красителя подавляется близостью гасящего красителя. При удлинении цепи в результате PCR с использованием дополнительных геноспецифических верхних и нижних затравок зонд расщепляется под действием 5'-концевой нуклеазы Taq-полимеразы, которая высвобождает репортерный краситель из зонда, что вызывает увеличение флуоресцентного излучения.
Затравки и зонды, используемые для количественного анализа экспрессии zcytor17, OSMRbeta и zcytor17lig человека методом RT-PCR в реальном времени, были получены при помощи программного обеспечения создания затравок Primer Express™ (PE Applied Biosystems, Foster City, CA). Затравки для zcytor17 человека были предназначены для заполнения интрон-экзонного сочленения с целью устранения возможной амплификации геномной ДНК. Верхнюю затравку ZC37877 (SEQ ID NO:53) и нижнюю затравку ZC37876 (SEQ ID NO:54) использовали при выполнении реакции PCR в концентрации 200 нМ для синтеза продукта длиной 73 п.н. Соответствующий зонд TaqMan® к zcytor17, обозначенный ZC37776 (SEQ ID NO:55), был синтезирован и помечен в компании PE Applied Biosystems и использован в каждой реакции PCR в концентрации 200 нМ. Зонд ZC37776 (SEQ ID NO:55) был помечен у 5'-конца репортерным флуоресцентным красителем (6-карбоксифлуоресцеин) (FAM) (PE Applied Biosystems) и у 3'-конца гасящим флуоресцентным красителем (6-карбокситетраметилродамин) (TAMRA) (Epoch Biosciences, Вothell, WA).
Затравки для OSMRbeta человека были предназначены для заполнения интрон-экзонного сочленения с целью устранения возможной амплификации геномной ДНК. Верхнюю затравку ZC43891 (SEQ ID NO:122) и нижнюю затравку, ZC43900 (SEQ ID NO:123) использовали при выполнении реакции PCR (описана ниже) в концентрации 200 нМ. Соответствующий зонд TaqMan® к OSMRbeta, обозначенный ZC43896 (SEQ ID NO:124), был синтезирован и помечен в компании PE Applied Biosystems и использован в каждой реакции PCR в концентрации 200 нМ. Зонд ZC43896 (SEQ ID NO:124) был помечен у 5'-конца репортерным флуоресцентным красителем (6-карбоксифлуоресцеин) (FAM) (PE Applied Biosystems) и у 3'-конца нефлуоресцентным гасящим красителем (ECLIPSE) (Epoch Biosciences).
Затравки для zcytor17lig человека были предназначены для заполнения интрон-экзонного сочленения с целью устранения возможной амплификации геномной ДНК. Верхнюю затравку ZC43280 (SEQ ID NO:125) и нижнюю затравку ZC43281 (SEQ ID NO:126) использовали при выполнении реакции PCR (описана ниже) в концентрации около 200 нМ. Соответствующий зонд TaqMan® к zcytor17lig, обозначенный ZC43275 (SEQ ID NO:127), был синтезирован и помечен в компании PE Applied Biosystems и использован в каждой реакции PCR в концентрации 200 нМ. Зонд ZC43275 (SEQ ID NO:127) был помечен у 5'-конца репортерным флуоресцентным красителем (6-карбоксифлуоресцеин) (FAM) (PE Applied Biosystems) и у 3'-конца нефлуоресцентным гасящим красителем (ECLIPSE) (Epoch Вiosciences).
Для контроля целостности и качества испытуемых образцов РНК все образцы РНК исследовали с целью обнаружения рРНК или GUS, используя набор затравок и зондов, заказанных в компании РЕ Applied Biosystems (набор рРНК) или созданных авторами изобретения (GUS). Набор рРНК содержал верхнюю затравку (SEQ ID NO:56), нижнюю затравку для рРНК (SEQ ID NO:57) и зонд TaqMan® к рРНК (SEQ ID NO:58). Зонд к рРНК был помечен у 5'-конца репортерным флуоресцентным красителем VIC (PR Applied Biosystems) и у 3'-конца гасящим флуоресцентным красителем TAMRA (PE Applied Biosystems). Затравки и зонд к GUS были созданы авторами изобретения и использованы в каждой реакции PCR в концентрации, соответственно, 200 нМ и 100 нМ. В качестве верхней затравки использовали затравку ZC40574 (SEQ ID NO:128) и в качестве нижней затравки использовали затравку ZC40575 (SEQ ID NO:129). Зонд ZC43017 к GUS (SEQ ID NO:130) был помечен у 5'-конца репортерным флуоресцентным красителем (Yakima-Yellow) (Epoch Bioscience) и у 3'-конца нефлуоресцентным гасящим красителем (ECLIPSE) (Epoch Biosciences). рРНК и GUS служили также в качестве эндогенного контрольного гена и позволяли нормализовать результаты экспрессии мРНК zcytor17, полученные для испытуемых образцов.
Затравки, предназначенные для выполнения стандартного неколичественного анализа методом RT-PCR, были получены при помощи программного обеспечения создания затравок Primer Express™ (PE Applied Biosystems, Foster City, CA). Затравки для zcytor17 человека образуют продукт из 1000 пар оснований и включают в себя верхнюю затравку ZC28917 (SEQ ID NO:83) и нижнюю затравку ZC28480 (SEQ ID NO:131). Затравки для OSMRbeta человека образуют продукт из 202 пар оснований и включают в себя верхнюю затравку ZC41653 (SEQ ID NO:132) и нижнюю затравку ZC41655 (SEQ ID NO:133). Затравки для zcytor17lig человека образуют продукт из 305 пар оснований и включают в себя верхнюю затравку ZC41703 (SEQ ID NO:134) и нижнюю затравку ZC41704 (SEQ ID NO:135).
В. Затравки и зонды для zcytor17, OSMRbeta и zcytor17lig мыши, предназначенные для выполнения стандартного и количественного анализа методом RT-PCR
Затравки и зонды, используемые для количественного анализа zcytor17, OSMRbeta и zcytor17lig мыши методом RT-PCR в реальном времени, были получены при помощи программного обеспечения создания затравок Primer Express™ (PE Applied Biosystems, Foster City, CA). Затравки для zcytor17 мыши были предназначены для заполнения интрон-экзонного сочленения с целью устранения возможной амплификации геномной ДНК. Верхнюю затравку ZC43272 (SEQ ID NO:136) и нижнюю затравку ZC43273 (SEQ ID NO:137) использовали при выполнении реакций PCR (описаны ниже) в концентрации 300 нМ. Соответствующий зонд TaqMan® к zcytor17, обозначенный ZC43478 (SEQ ID NO:138), был синтезирован и помечен в компании PE Applied Biosystems. Зонд ZC43478 (SEQ ID NO:138) был помечен у 5'-конца репортерным флуоресцентным красителем (6-карбоксифлуоресцеин) (FAM) (PE Applied Biosystems) и у 3'-конца гасящим флуоресцентным красителем (6-карбокситетраметилродамин) (ТАМRА) (PE Applied Biosystems). Зонд ZC43478 (SEQ ID NO:138) использовали в реакциях PCR в концентрации 100 нМ.
Затравки для zcytor17lig мыши были предназначены для заполнения интрон-экзонного сочленения с целью устранения возможной амплификации геномной ДНК. Верхнюю затравку ZC43278 (SEQ ID NO:139) и нижнюю затравку ZC43279 (SEQ ID NO:140) использовали при выполнении реакций PCR в концентрации 500 нМ. Соответствующий зонд TaqMan® к zcytor17lig, обозначенный ZC43276 (SEQ ID NO:141), был синтезирован и помечен в компании PE Applied Biosystems. Зонд ZC43478 (SEQ ID NO:138) был помечен у 5'-конца репортерным флуоресцентным красителем (6-карбоксифлуоресцеин) (FAM) (PE Applied Biosystems) и у 3'-конца нефлуоресцентным гасящим красителем (ECLIPSE) (Epoсh Biosciences). Зонд ZC43276 (SEQ ID NO:141) использовали в реакциях PCR (описаны ниже) в концентрации 200 нМ.
Затравки для OSMRbeta мыши были предназначены для заполнения интрон-экзонного сочленения с целью устранения возможной амплификации геномной ДНК. Верхнюю затравку ZC43045 (SEQ ID NO:142) и нижнюю затравку ZC43046 (SEQ ID NO:143) использовали при выполнении реакций PCR в концентрации 300 нМ. Соответствующий зонд TaqMan® к OSMRbeta, обозначенный ZC43141 (SEQ ID NO:144), был синтезирован и помечен в компании Epoch Biosciences. Зонд ZC43141 (SEQ ID NO:144) был помечен у 5'-конца репортерным флуоресцентным красителем (6-карбоксифлуоресцеин) (FAM) (PE Applied Biosystems) и у 3'-конца нефлуоресцентным гасящим красителем (ECLIPSE) (Epoch Biosciences). Зонд ZC43141 (SEQ ID NO:144) использовали в реакциях PCR (описаны ниже) в концентрации 100 нМ.
Для контроля целостности и качества испытуемых образцов РНК все образцы РНК исследовали для обнаружения GUS или рецептора трансферрина мыши с использованием затравок и зондов, полученных при помощи программы создания затравок Primer Express™ (PE Applied Biosystems Inc., Foster City, CA). Затравки для GUS мыши включают в себя верхнюю затравку ZC43004 (SEQ ID NO:145), нижнюю затравку ZC43005 (SEQ ID NO:146) и зонд TaqMan® ZC43018 (SEQ ID NO:147). Зонд ZC43018 к GUS мыши (SEQ ID NO:147) был помечен у 5'-конца репортерным флуоресцентным красителем Yakima-Yellow (Epoch Biosciences) и у 3'-конца нефлуоресцентным гасящим красителем ECLIPSE (Epoch Biosciences). Затравки для GUS мыши использовали в реакциях PCR в концентрации 300 нМ, и зонд ZC43018 (SEQ ID NO:147) использовали в концентрации 100 нМ. В некоторых случаях вместо GUS использовали рецептор трансферрина мыши в качестве эндогенного контрольного гена. Верхнюю затравку ZC40269 (SEQ ID NO:148) и нижнюю затравку ZC40268 (SEQ ID NO:149) для рецептора трансферрина использовали в концентрации 300 нМ. Зонд к рецептору трансферрина ZC40298 (SEQ ID NO:150) был использован в реакции PCR в концентрации 100 нМ и помечен у 5'-конца репортерным флуоресцентным красителем VIC (PE Applied Biosystems) и у 3'-конца флуоресцентным гасящим красителем (TAMRA) (PE Applied Biosystems). GUS и рецептор трансферрина мыши служили также в качестве эндогенного контрольного гена и позволяли нормализовать результаты экспрессии мРНК zcytor17, OSMRbeta и zcytor17lig, полученные для испытуемых образцов.
Затравки, предназначенные для выполнения стандартного полуколичественного анализа методом RT-PCR, были получены при помощи программного обеспечения создания затравок Primer Express™ (PE Applied Biosystems). Затравки для zcytor17 мыши образуют продукт из 276 пар оснований и включают в себя верхнюю затравку ZC43140 (SEQ ID NO:151) и нижнюю затравку ZC43139 (SEQ ID NO:152). Затравки для OSMRbeta мыши образуют продукт из 575 пар оснований и включают в себя верхнюю затравку ZC41608 (SEQ ID NO:153) и нижнюю затравку ZC41609 (SEQ ID NO:154). Затравки для zcytor17lig мыши образуют продукт из 657 п.н. и включают в себя верхнюю затравку ZC41502 (SEQ ID NO:155) и нижнюю затравку ZC41500 (SEQ ID NO:156).
С. Способы выполнения количественного анализа методом RT-PCR в реальном времени и стандартного полуколичественного анализа методом RT-PCR
Относительные уровни мРНК zcytor17, OSMRbeta и zcytor17lig определяли путем анализа образцов полной РНК методом одностадийной RT-PCR (PE Applied Biosystems). Полную РНК из клеток BAF (человека) или клеток BHK (мыши), трансфицированных zcytor17 и OSMRbeta, выделяли стандартными методами и использовали для построения стандартной кривой, используемой для количественного определения рецепторов zcytor17 и OSMRbeta. Указанная кривая была построена на основании 10-кратных серийных разведений в пределах 100-0,01 нг/мкл, причем каждую точку стандартной кривой анализировали в трех копиях. Аналогичным образом, стандартную кривую для zcytor17lig строили с использованием РНК активированных CD4+ Т-клеток (которая, как было установлено ранее, образует zcytor17lig) в том же интервале 100-0,01 нг/мкл. Полную РНК из клеток человека или мыши анализировали в трех копиях для определения уровней транскриптов zcytor17, OSMRbeta и zcytor17lig человека или мыши и одного из следующих эндогенных контрольных генов: рРНК, GUS или рецептора трансферрина. Каждый образец РНК в объеме 10 мкл подвергали одностадийной реакции RT-PCR, при выполнении которой использовали примерно 50-100 нг полной РНК в предварительно полученном двукратном объеме исходной смеси, содержащей внутренний контрольный краситель (ROX) (карбокси-х-родамин) и ДНК-полимеразу Thermo-Start® (Abgene, Surrey, UK); соответствующие затравки для исследуемого гена (см. части А и В данного примера); соответствующий зонд (см. части А и В для ознакомления с данными концентрации); обратную транскриптазу Superscript® (50 ед/мкл) (PE Applied Biosystems) и соответствующий объем воды без РНКазы. PCR выполняли в следующих условиях термической циклизации: начальная стадия обратной транскрипции (RT) включала один цикл при 48°C в течение 30 минут; стадия активации ферментом Thermo-Start® включала один цикл при 95°C в течение 10 минут; затем 40 циклов амплификации при 95°C в течение 15 секунд и при 60°C в течение 1 минуты. Относительные уровни РНК zcytor17, OSMRbeta и zcytor17lig определяли методом построения стандартной кривой в соответствии с описанием, предоставленным изготовителем, РЕ Biosystems (бюллетень пользователя №2: система обнаружения последовательностей ABI Prism 7700, относительное количественное определение экспрессии генов, 11 декабря 1997 г.). Измерения рРНК, GUS или рецептора трансферрина использовали для нормализации уровней исследуемого гена.
Для полуколичественного анализа методом RT-PCR использовали «одностадийную систему RT-PCR Suprescript с платина-Taq-трансферазой» (Invitrogen, Carlsbad, CA). 25 мкл реакционной смеси включали в себя следующие реагенты: 12,5 мкл двукратного объема реакционного буфера, 0,5 мкл (20 пмоль/мкл) верхней затравки, 0,5 мкл (20 пмоль/мкл) нижней затравки, 0,4 мкл смеси обратная транскриптаза/Taq-полимераза, 5,0 мкл буфера для загрузки геля Redilоad (Invitrogen), 5,1 мкл воды без РНКазы и 1,0 мкл полной РНК (100 нг/мкл). Амплификацию осуществляли следующим образом: один цикл при 45°C в течение 30 минут, затем 35-38 циклов при 94°C в течение 20 секунд; отжиг при разных температурах (см. приведенную ниже таблицу 7) в течение 20 секунд, при 72°C в течение 45 секунд; конечное удлинение цепи при 72°C в течение 5 минут. Восемь-десять микролитров продукта реакции PCR разделяли стандартным электрофорезом в 2% агарозном геле.
D. Выделение РНК из субпопуляций и линий клеток РВМС человека и мыши
У нескольких анонимных доноров брали пробы крови и выделяли мононуклеарные клетки периферической крови (PBMC) стандартным градиентным методом Фиколли. Затем выделяли моноциты при помощи набора для выделения моноцитов и системы магнитного разделения клеток (Miltenyi Biotec, Auburn, CA). Моноциты культивировали на 24-луночных планшетах с ультранизким сцеплением в среде без эндотоксина. Клетки или не стимулировали, или обрабатывали рекомбинантным IFNg человека (R&D Systems Inc.) в концентрации 10 нг/мл. Клетки собирали через 24 и 48 часов. Аналогичным образом, CD4+ и CD8+ Т-клетки выделяли из РВМС, используя магнитные шарики против CD4 или CD8 компании Miltenyi Biotec. Затем клетки активировали в течение 4 или 16 часов на планшетах для тканевых культур, покрытых 0,5 мкг/мл антител против CD3 в среде, содержащей 5 мкг/мл антител против CD-28. NK-клетки также выделяли из PBMC, используя магнитные шарики, покрытые антителом против CD56, компании Miltenyi. Некоторые NK-клетки собирали в нулевой период времени для РНК, и другие клетки культивировали в среде, содержащей ацетат форболмиристата (РМА) (5 нг/мл) и иономицин (0,5 мкг/мл), в течение 24 часов. Кроме того, собирали несколько моноцитоподобных линий клеток человека, U937, THP-1 и HL-60, в состоянии покоя или в активированном состоянии. Клетки U937 активировали РМА (10 нг/мл) в течение ночи. Клетки HL-60 активировали РМА (10 нг/мл) в течение ночи или IFNg (10 нг/мл) в течение 72 и 96 часов для того, чтобы направить их по моноцитарному пути. Клетки ТНР-1 активировали в течение ночи комбинацией LPS (10 нг/мл) и IFNg (10 нг/мл). РНК получали изо всех первичных клеток при помощи набора RNeasy Midiprep™ (Qiagen, Valencia, CA), в соответствии с инструкциями изготовителя. Несущую ДНК удаляли при помощи набора DNA-Free™ (Ambion, Inc., Austin, TX). Концентрацию РНК определяли стандартной спектрофотометрией, и качество РНК определяли при помощи биоанализатора Bioanalyzer 2100 (Agilent Technologies, Palo Alto, CA).
РНК Т-клеток мыши собирали различными способами, хорошо известными в данной области. Первичные селезеночные CD4+ и CD8+ Т-клетки выделяли из селезенки мышей С57BI/6 при помощи магнитных шариков, покрытых антителом, и системы магнитного разделения клеток компании Miltenyi Biotec. CD4+ и CD8+ Т-клетки активировали, культивируя на 24-луночных планшетах, покрытых антителами против CD3 (500 нг/мл), в среде, содержащей антитела против CD28 в концентрации 5 мкг/мл. Клетки собирали для получения РНК через 0, 4 и 16 часов. Аналогичным образом CD4+ Т-клетки выделяли и направляли по пути образования фенотипа Тh1 или Тh2 следующим способом. Так как Т-клетки C57BI/6 уже развиваются в направлении Th1, их нужно было только активировать в течение 6 часов 0,5 мкг/мл РМА и 10 нг/мл иономицина. Фенотип "Th2" был получен путем культивирования наивных CD4+ Т-клеток с 2,5 мкг/мл антител против CD28, 10 нг/мл mIL-2 (R&D Systems Inc.) и 25 нг/мл mIL-4 (R&D Systems) на планшетах, покрытых 0,5 мкг/мл антитела против CD3. После культивирования в течение 2 дней клетки вновь суспендировали в среде, содержащей 10 нг/мл mIL-2 (R&D Systems) и 25 нг/мл mIL-4. Клетки культивировали еще три дня и активировали РМА и иономицином в течение 6 часов.
Одну дополнительную популяцию Т-клеток с фенотипом Th1 и Th2 получали, используя трансгенную линию Т-клеток DO11.10 Т-клеточного рецептора. Все клетки культивировали на планшетах, покрытых антителами против CD3 и CD28. Клетки "Th1" культивировали в среде, содержащей mIL-12 (1 нг/мл) и антитела против IL-4 (10 мкг/мл). Клетки "Th2" культивировали в среде, содержащей mIL-4 (10 нг/мл) и антитела против IFNg (10 мкг/мл). Через 24 часа во все культуры вводили mIL-2 (10 нг/мл). Еще через два дня среду клеток заменяли, добавляли новую среду, содержащую вышеуказанные цитокины, клетки культивировали еще 4 дня и затем собирали.
Вся РНК Т-клеток мыши была получена при помощи набора RNeasy Midiprep™ (Qiagen), при этом загрязняющую ДНК удаляли при помощи набора DNA-free™ компании Ambion.
Е. Выделение РНК из животных моделей панкреатита и воспалительного заболевания кишечника с использованием мышей
Чтобы вызвать заболевание, подобное воспалительному заболеванию кишечника (IBD) человека, использовали гибридную линию мышей С57B16/129S6F1. Мыши были разделены на 4 группы в среднем по шесть мышей в группе. Животных в группе 1, которым не вводили декстран-сульфат натрия (DSS), умерщвляли на 14-ый день. Животным в группе 2 вводили 2% DSS в течение двух дней до того, как их умерщвляли. Животным в группе 3 вводили 2% DSS в течение семи дней до того, как их умерщвляли. Животным в группе 4 вводили 2% DSS в течение семи дней, затем их оставляли выздоравливать в течение семи дней и умерщвляли на 14-ый день. В день умерщвления получали срезы дистальных участков ободочной кишки и помещали в RNAlater™ (Ambion). Срезы ободочной кишки гомогенизировали стандартными методами, и РНК выделяли при помощи набора RNeasy Midiprep™ (Qiagen). Загрязняющую ДНК удаляли при помощи набора DNA-free™ (Ambion), в соответствии с инструкциями изготовителя.
В другом исследовании у самцов мышей CD-1 вызывали острый панкреатит путем инъекции церулейна. Мыши были разделены на три группы (n=8 мышей в группе). Животным в группе 1 делали семь внутрибрюшинных инъекций (1 инъекция в час) носителя (физиологический раствор) и умерщвляли через 12 и 24 часа после первой инъекции. Животным в группах 2 и 3 делали семь внутрибрюшинных инъекций церулейна (Sigma) (№ по каталогу С-9026) в дозе 50 мкг/кг/час в течение шести часов (1 инъекция в час). Животных в группе 2 умерщвляли через 12 часов после первой инъекции и животных в группе 3 умерщвляли через 24 часа после первой инъекции. Поджелудочные железы удаляли во время умерщвления и замораживали для выделения РНК. Ткани гомогенизировали, и РНК выделяли при помощи набора RNeasy Midiprep™ компании Qiagen.
В другом исследовании были получены трансгенные мыши, экспрессирующие zcytor17lig мышей, у которых исследовали фенотипические изменения (см. пример 41). У многих трансгенных мышей наблюдалась пилоэрекция и выпадение шерсти. Четырех трансгенных мышей умерщвляли, брали образцы кожи с нормальных и лишенных шерсти участков и замораживали для последующего выделения РНК. Кроме того, получали срезы кожи у двух нетрансгенных контрольных мышей. Образцы кожи гомогенизировали и гидролизовали протеиназой К (Qiagen) (№ по каталогу 19133) в течение 20 минут при 60°C. РНК выделяли при помощи набора RNeasy Midiprep™ компании Qiagen, в соответствии с инструкциями изготовителя. Несущую ДНК удаляли при помощи набора DNA-free™ компании Ambion.
F. Результаты количественного и полуколичественного анализа zcytor17, OSMRbeta и zcytor17lig человека методом RT-PCR
Экспрессию zcytor17 и OSMRbeta исследовали путем количественного анализа методом RT-PCR в четырех популяциях первичных моноцитов человека, которые находились в состоянии покоя или были активированы IFNg в течение 24 или 48 часов. Экспрессия zcytor17 была ниже порога обнаружения в нестимулированных клетках, но сильно возрастала после 24-часовой активации IFNg и достигала максимальной величины через 48 часов после активации. Экспрессия OSMRbeta не была обнаружена во всех случаях. Экспрессию zcytor17lig не определяли в указанных образцах.
В первичных Т-клетках экспрессия zcytor17 не была обнаружена в покоящихся субпопуляциях CD4+ и CD8+-клеток. Однако после четырехчасовой активации экспрессия zcytor17 повышалась в обеих субпопуляциях и опускалась до более низкого уровня через 16 часов. Экспрессия OSMRbeta не была обнаружена в указанных образцах. Экспрессию zcytor17lig исследовали полуколичественным методом RT-PCR. Экспрессия не была обнаружена в нестимулированных CD4+ и CD8+ Т-клеток. Однако после четырехчасовой активации были обнаружены высокие уровни экспрессии zcytor17lig. Указанный уровень немного снижался через 16 часов.
Экспрессию zcytor17 не исследовали в NK-клетках. Экспрессия OSMRb была ниже порога обнаружения в указанных образцах. Экспрессия zcytor17lig была ниже порога обнаружения в покоящихся NK-клетках, однако активированные NK-клетки генерировали слабый сигнал, из чего следует, что указанные клетки могут продуцировать zcytor17lig в определенных условиях.
В моноцитоподобных линиях клеток человека U937, THP-1 и HL-60 экспрессия OSMRbeta была ниже порога обнаружения во всех покоящихся и активированных образцах, за исключением активированных образцов ТНР-1, в которых был обнаружен слабый сигнал. Экспрессия zcytor17 была высокой в покоящихся линиях клеток U937 и ТНР-1, при этом указанная экспрессия сильно увеличивалась после активации. Экспрессия в клетках U937 была самой высокой среди клеток разных типов. В клетках HL-60 zcytor17 был экспрессирован на среднем уровне в нестимулированных клетках, причем указанная экспрессия уменьшалась при стимуляции РМА. Однако экспрессия zcytor17 сильно увеличивалась в клетках HL-60 при стимуляции IFNg в течение 72 и 96 часов. Все данные об экспрессии человека суммированы в таблице 8, ниже.
G. Результаты количественного и полуколичественного анализа zcytor17, OSMRbeta и zcytor17lig мыши методом RT-PCR
Уровни экспрессии zcytor17, OSMRbeta и zcytor17lig исследовали в нескольких популяциях Т-клеток мыши, и полученные результаты суммированы в приведенной ниже таблице 9. Экспрессия zcytor17 мыши, определяемая полуколичественным методом RT-PCR, была низкой как в покоящихся, так и активированных первичных CD4+ Т-клетках. Экспрессия zcytor17, обнаруженная в покоящихся CD8+ Т-клетках, снижалась после активации антителами против CD3 и CD28 в течение 4 и 16 часов. Экспрессия OSMRbeta, измеренная количественным методом RT-PCR, была обнаружена в покоящихся и активированных CD4+ и CD8+ Т-клетках. Экспрессия OSMRbeta повысилась после четырехчасовой активации и вернулась к нестимулированным уровням в CD4+ и CD8+ Т-клетках через 16 часов. Экспрессия zcytor17lig была обнаружена количественным методом RT-PCR на очень низких уровнях в нестимулированных CD4+ Т-клетках. Однако после четырехчасовой активации экспрессия zcytor17lig сильно увеличилась, а затем несколько уменьшилась по истечении 16 часов. В CD8+ Т-клетках экспрессия zcytor17lig была обнаружена в нестимулированных клетках. Некоторая экспрессия zcytor17lig была обнаружена через 4 часа, но через 16 часов уровни экспрессии опустились ниже порога обнаружения.
В Т-клетках DО11.10 экспрессия zcytor17 была обнаружена в наивных клетках и клетках фенотипа Th2, но не была обнаружена в клетках фенотипа Th1. Экспрессия OSMRbeta была на низком уровне в наивных клетках DO11.10. Наблюдалось значительное увеличение уровней экспрессии OSMRbeta в клетках фенотипа Th1 и среднее увеличение экспрессии в клетках фенотипа Th2. Экспрессия zcytor17lig была обнаружена, главным образом, в субпопуляции клеток фенотипа Th2. Низкие уровни экспрессии были обнаружены в субпопуляции клеток фенотипа Th1, при этом экспрессия полностью отсутствовала в наивных клетках. Полученные результаты суммированы в приведенной ниже таблице 9.
В первичных CD4+ Т-клетках, образующих фенотип Th1 или Th2, экспрессию zcytor17 не определяли. Экспрессия OSMRbeta была обнаружена во всех трех образцах, причем наибольшие уровни были обнаружены в образце Th2. Аналогично результатам, полученным для клеток DO11.10, экспрессия zcytor17lig была обнаружена на высоком уровне в субпопуляции клеток фенотипа Th2 и на низком уровне в субпопуляции клеток фенотипа Th1, причем экспрессия была ниже порога обнаружения в нестимулированных клетках. Результаты суммированы в следующей таблице 9.
В образцах трансгенной кожи, экспрессирующей zcytor17lig, уровни экспрессии zcytor17, OSMRbeta и zcytor17lig определяли количественным методом RT-PCR. Установлено, что zcytor17 присутствует во всех образцах примерно на одинаковых уровнях. Значительно более высокие уровни экспрессии OSMRbeta были обнаружены у нетрансгенных контрольных животных по сравнению с трансгенными животными. Экспрессия zcytor17lig была ниже порога обнаружения у нетрансгенных контрольных животных, и экспрессия от средних уровней до высоких уровней была обнаружена у трансгенных животных. Результаты суммированы в следующей таблице 10.
В другом эксперименте уровни экспрессии zcytor17, OSMRbeta и zcytor17lig измеряли количественным методом RT-PCR в модели острого панкреатита, вызванного у мышей. Экспрессия zcytor17 была ниже порога обнаружения во всех образцах. Была обнаружена низкая экспрессия OSMRbeta в нормальных контрольных образцах (группа 1), при этом экспрессия сильно увеличивалась через 12 часов (группа 2) и несколько уменьшалась через 24 часа (группа 3). Экспрессия zcytor17lig была ниже порога обнаружения у контрольных животных и высокой в группах животных, которым инъецировали церулеин. Полученные данные суммированы в нижеследующей таблице 11.
В другом эксперименте уровни экспрессии zcytor17 и OSMRbeta измеряли в дистальных отделах ободочной кишки мышей, которым вводили DSS. В мышиной модели воспалительного заболевания кишечника уровни экспрессии обоих генов были определены количественным методом RT-PCR и суммированы в следующей таблице 12. Уровни экспрессии zcytor17 увеличивались в зависимости от тяжести заболевания, при этом низкие уровни экспрессии наблюдались у нормальных животных в группе 1 и более высокие уровни экспрессии были обнаружены в группах 2 и 3. У животных в группе 4 уровни экспрессии zcytor17 вернулись к более нормальным уровням. В отличие от экспрессии zcytor17 уровни экспрессии OSMRbeta были самыми высокими у контрольных животных, при этом уровни экспрессии уменьшались во всех трех группах животных, которым вводили DSS.
Пример 28
Распределение в тканях zcytor17lig человека по результатам анализа RT-PCR кДНК первой цепи в нескольких тканях
Экспрессию гена zcytor17lig исследовали при помощи коммерчески производимых нормализованных панелей кДНК первой цепи в нескольких тканях (OriGene Technologies, Inc. Rockville, MD; BD Biosciences Clontеch, Palo Alto, CA). Указанные панели включали «панель человеческих тканей Rapid-Scan™» компании OriGene (№ по каталогу CHSCA-101, содержит 22 различных ткани, костный мозг и лейкоциты плазмы крови) и «панель фракций человеческой крови МТС™» компании BD Biosciences Clontech (№ по каталогу К1428-1, содержит 9 различных фракций крови).
Реакции PCR выполняли с использованием специфических олигонуклеотидных затравок для zcytor17lig, ZC41458 (SEQ ID NO:60) и ZC41457 (SEQ ID NO:61), образующих продукт из 139 п.н., ZC41459 (SEQ ID NO:62) и ZC41460 (SEQ ID NO:63), образующих продукт из 92 п.н., ДНК-полимеразы HotStarTaq компании Qiagen и буфера (Qiagen, Inc., Valencia, CA), дистиллированной Н2О и красителя ReadiLoad™ (Research Genetics, Inc., Huntville. AL). В термоячейке для реакций PCR создавали следующие условия: 1 начальный цикл денатурации при 95°C в течение 15 минут, 35 циклов денатурации при 95°C в течение 45 секунд, отжиг при 53°C в течение 1 минуты или при 56°C в течение 1 минуты и удлинение цепи при 72°C в течение 15 секунд, затем 1 конечный цикл удлинения цепи при 72°C в течение 7 минут. Реакционную смесь разделяли электрофорезом в 2% агарозном геле (EM Science, Gibbstown, NJ) и визуализировали, производя окрашивание бромидом этидия.
Фрагмент ДНК правильного размера был обнаружен при помощи «панели человеческих тканей Rapid-Scan™» компании OriGene в следующих тканях взрослого человека: яичко, лейкоциты плазмы крови (PBL) и костный мозг.
Фрагмент ДНК правильного размера был обнаружен при помощи «панели фракций человеческой крови МТС™» компании BD Biosciences Clontech в следующих фракциях человеческой крови: активированные мононуклеарные клетки (B-и Т-клетки и моноциты), активированные CD8+-клетки ((Т-клетки-супрессоры/цитотоксические), активированные CD4+-клетки (Т-клетки-хелперы/индукторы) и покоящиеся CD8+-клетки.
Пример 29
Клонирование рецептора онкостатина М человека
Бета-рецептор онкостатина М (OSMRbeta) является рецептором цитокина типа 1, который обладает структурным сходством с IL12R-B2. Zcytor17 обладает структурным сходством с IL12R-B1. OSMRbeta и zcytor17 испытывали для выявления возможности их взаимодействия в виде субъединиц сигнального комплекса цитокина и совместного действия в качестве сигнального рецептора или антагониста растворимого рецептора для zcytor17lig.
Для выделения OSMRbeta были созданы олигонуклеотидные затравки для PCR, ZC39982 (SEQ ID NO:64) и ZC39983 (SEQ ID NO:65), предназначенные для амплификации непроцессированной кодирующей области последовательности кДНК бета-рецептора онкостатина М человека (SEQ ID NO:6) (Банк генов, № доступа U60805; Mosley B, JВC Volume 271, Number 50, Issue of December 20, 1996, pp. 32635-32643).
Реакции PCR выполняли на ряде матриц библиотеки кДНК с использованием сильной полимеразы, Advantage II (Clonetech, Palo Alto, CA) для идентификации источника кДНК. Используемая матричная ДНК была получена из библиотек амплифицированной плазмидной кДНК, каждая из которых содержит 5 миллионов независимых клонов кДНК. Реакции выполняли в соответствии с инструкциями изготовителя, используя по 400 фмоль/мкл каждого олигонуклеотида и 2-20 нг/мкл очищенной плазмидной ДНК из библиотеки в качестве матрицы. Библиотеки кДНК были получены из следующих тканей и линий клеток человека: головной мозг плода, гладкие мышцы предстательной железы, костный мозг, RPMI1588, щитовидная железа, WI-38, яички, стимулированные мононуклеарные клетки периферической крови, стимулированные CD3+-клетки, ТНР-1, активированная миндалина, НАСАТ и печень плода. Реакции выполняли в термоячейке в следующих условиях: 30 циклов при 95°C в течение 20 секунд, при 68°C в течение 3 минут. После 30 циклов выполняли один цикл удлинения цепи при 68°C в течение 8 минут. Продукты PCR визуализировали при помощи электрофореза в агарозном геле ТАЕ в присутствии бромида этидия с последующим ультрафиолетовым облучением. Наибольшее количество продукта было обнаружено в библиотеке кДНК гладких мышц предстательной железы. Реакцию PCR с использованием гладких мышц предстательной железы в качестве матрицы и олигонуклеотидов ZC39982 (SEQ ID NO:64) и ZC39983 (SEQ ID NO:65) повторяли, используя менее сильную, но более точную термостойкую ДНК-полимеразу "turboPFu" (Stratagene, La Jolla, CA). Тридцать циклов амплификации выполняли в следующих условиях: денатурация при 94°C в течение 30 секунд, отжиг при 63°C в течение 45 секунд, удлинение цепи при 72°C в течение 3,5 минут. Однополосный продукт очищали в 0,8% агарозном геле ТАЕ.
Данную ДНК затем снова амплифицировали, используя затравки ZC39980 (SEQ ID NO:66) и ZC39981 (SEQ ID NO:67), созданные с возможностью включения последовательностей, узнающих рестрикционные ферменты, для клонирования указанной кДНК в экспрессирующем векторе млекопитающего.
Реакцию PCR выполняли, используя полимеразу "TurboPfu" и очищенный продукт PCR, на протяжении 15 циклов: при 95°C в течение 1 минуты, при 64°C в течение 1 минуты 20 секунд, при 72°C в течение 4,5 минут. Затем реакционную смесь PCR гидролизовали EcoRI и XhoI (Invitrogen, Carlsbad, CA) и очищали в геле вышеописанным способом. Экспрессирующий вектор млекопитающего pZ7NX получали гидролизом EcoRI и XhoI, продукт PCR лигировали с этим вектором и электропорировали в клетки DH10b E. coli. Несколько бактериальных колоний выделяли и секвенировали. Один клон был правильным, за исключением одной неконсервативной мутации. Чтобы заменить указанное основание для достижения соответствия с предполагаемой последовательностью, производили заполняющую мутацию с использованием олигонуклеотида и соседнего сайта рестрикции Pst1 при выполнении реакции PCR с полимеразой "TurboPfu" и pZP7Nx-h. Плазмиду рецептора онкостатина М предварительно секвенировали в качестве матрицы. Амплифицированную ДНК, полученную в результате PCR, гидролизовали Pst1 и Xho1 и клонировали в плазмиде рецептора онкостатина М pZP7Nx-h на месте фрагмента Pst1/Xho1, содержащего неправильную мутацию. Новую плазмиду секвенировали во вновь амплифицированной области от Pst1 до Xho1 для подтверждения правильности последовательности и отсутствия других ошибок, возникающих в процессе амплификации. Указанный анализ подтвердил соответствие данной последовательности ожидаемой последовательности в кодирующей области. Данная последовательность представлена в SEQ ID NO:6, и соответствующая аминокислотная последовательность показана в SEQ ID NO:7.
Пример 30
Конструкции для создания гетеродимера рецептора zcytor17/онкостатин М человека (OSMRbeta)
Система, предназначенная для создания, экспрессии и очистки таких растворимых гетеродимерных рецепторов, известна в данной области и адаптирована для пары рецепторов, включающей онкостатин М человека (OSMRbeta) и zcytor17 человека. Используемый в данной конструкции полинуклеотид для растворимого рецептора OSMRbeta показан в SEQ ID NO:68, и соответствующий полипептид показан в SEQ ID NO:69; а полинуклеотид для растворимого рецептора zcytor17 человека показан в SEQ ID NO:70, и соответствующий полипептид показан в SEQ ID NO:71.
Для получения линии клеток, экспрессирующих секретируемый растворимый гетеродимер hzcytor17/OSMRbeta человека, конструкция была создана таким образом, чтобы гетеродимерный растворимый рецептор включал в себя внеклеточный домен OSMRbeta человека, слитый с тяжелой цепью IgG гамма-1 (Fc4) (SEQ ID NO:37), имеющей метку Glu-Glu (SEQ ID NO:35) у С-конца; при этом внеклеточный домен zcytor17 был слит с Fc4 (SEQ ID NO:37), включающей метку His (SEQ ID NO:72) у С-конца. Для обоих плеч hzcytor17 и OSMRbeta человека гетеродимера был создан спейсер Gly-Ser, состоящий из 12 аминокислот (SEQ ID NO:73), который находился между внеклеточной частью рецептора и N-концом Fc4.
А. Создание части гетеродимера, включающей растворимый рецептор OSMRbeta человека/Fc-4-CEE
Для создания части гетеродимера, включающей в себя растворимый рецептор OSMRbeta человека/Fc4-CEE, выделяли внеклеточную часть OSMRbeta человека при помощи PCR с использованием олигонуклеотидов ZC14063 (SEQ ID NO:48) и ZC41557 (SEQ ID NO:74) в следующих условиях реакции: 30 циклов при 95°C в течение 60 секунд, при 57°C в течение 30 секунд и при 72°C в течение 100 секунд; и при 72°C в течение 7 минут. Продукты PCR очищали при помощи набора для очистки продуктов PCR QIAquick (Qiagen), гидролизовали EcoRI и BglII (Boehringer-Mannheim), разделяли гель-электрофорезом и очищали при помощи набора для гель-экстракции QIAquick (Qiagen).
Полигенный экспрессирующий кластер, остов плазмиды и метка Fc4-GluGlu химеры находились в предварительно созданном авторами изобретения плазмидном векторе. Плазмидный вектор гидролизовали EcoRI и BamHI (Boehringer-Mannheim), разделяли гель-электрофорезом и очищали при помощи набора для гель-экстракции QIAquick (Qiagen). Гидролизованные и очищенные фрагменты плазмиды, содержащей OSMRbeta человека и Fc4-СEE, лигировали друг с другом, используя ДНК-лигазу Т4 (Life Technologies, Bethesda, MD), при помощи стандартных методов лигирования. Лигированные продукты исследовали на наличие вставки EcoRI/SmaI правильного размера (772 п.н.) для растворимого рецептора OSMRbeta, и положительные продукты секвенировали для подтверждения точности выполнения реакции PCR. Новая плазмидная конструкция получила название pZP9-ONCOMR-Fc4CEE.
В. Создание части гетеродимера, включающей растворимый рецептор zcytor17 человека/Fc4-CHIS
Для создания части гетеродимера, включающей в себя hzcytor17/Fc4-CHIS, внеклеточную часть zcytor17 человека выделяли гидролизом плазмиды, ранее содержащей растворимый рецептор zcytor17-Fc4. Указанную плазмиду сначала гидролизовали SalI (New England Biolabs, Beverly, MA), после чего реакционную смесь последовательно экстрагировали фенол-хлороформом и осаждали этанолом. Гидролизованную ДНК затем обрабатывали ДНК-полимеразой Т4 (Boehringer-Mannheim) для заполнения 5'-концевых избыточностей, образовавшихся в результате гидролиза SalI, оставляя тупыми концы ДНК, после чего реакционную смесь последовательно экстрагировали фенол-хлороформом и осаждали этанолом. ДНК с тупыми концами гидролизовали BglII, разрезая у 3'-конца, разделяли гель-электрофорезом и очищали при помощи набора для гель-экстракции QIAquick (Qiagen), в соответствии с инструкциями изготовителя. Полученный фрагмент ДНК, содержащий последовательность, кодирующую внеклеточный домен zcytor17, лигировали с экспрессирующим вектором млекопитающего, содержащим метку Fc4-CHIS, который был получен следующим образом.
Полигенный экспрессирующий кластер, остов плазмиды и метка Fc4-CHIS химеры находились в предварительно созданном авторами изобретения плазмидном векторе. Данный плазмидный вектор гидролизовали EcoR1 (Boehringer-Mannheim), после чего реакционную смесь последовательно экстрагировали фенол-хлороформом и осаждали этанолом. Гидролизованную ДНК затем обрабатывали ДНК-полимеразой Т4 (Boehringer-Mannheim) для заполнения 5'-избыточностей, образовавшихся в результате гидролиза EcoR1, оставляя тупыми концы ДНК, после чего реакционную смесь последовательно экстрагировали фенол-хлороформом и осаждали этанолом. ДНК с тупыми концами гидролизовали BamH1 (Boehringer-Mannheim), разрезая у 3'-конца, разделяли гель-электрофорезом и очищали при помощи набора для гель-экстракции QIAquick (Qiagen). Гидролизованные и очищенные фрагменты плазмиды, содержащей zcytor17 человека и Fc4-CHIS, лигировали друг с другом, используя ДНК-лигазу Т4 (Life Technologies, Bethesda, MD), при помощи стандартных методов лигирования.
Лигированные продукты исследовали методом PCR, используя zcytor17-специфическую смысловую затравку ZC29180 (SEQ ID NO:22) и Fc4-специфическую антисмысловую затравку ZC29232 (SEQ ID NO:75), в следующих условиях реакции: 30 циклов при 94°C в течение 60 секунд, при 68°C в течение 150 секунд и при 72°C в течение 7 минут. Размер продукта, равный 848 п.н., подтверждал правильную сборку плазмиды, получившей название pZEM228 hzcytor17/Fc4HIS.
Была создана вторая конструкция zcytor17-Fc4, предназначенная для получения гомодимерного белка из клеток COS. Кодирующую область полностью слитого белка выделяли, гидролизуя плазмиду, содержащую растворимый рецептор zcytor17-Fc4, при помощи SalI (Boehringer-Mannheim). Реакционную смесь последовательно экстрагировали фенол-хлороформом и осаждали этанолом. Гидролизованную ДНК затем обрабатывали ДНК-полимеразой Т4 (Boehringer-Mannheim) для заполнения 5'-концевых избыточностей, образовавшихся в результате гидролиза EcoRI, оставляя тупыми концы ДНК, после чего реакционную смесь последовательно экстрагировали фенол-хлороформом и осаждали этанолом. ДНК с тупыми концами гидролизовали NotI (Boehringer-Mannheim), разрезая у 3'-конца, разделяли гель-электрофорезом и очищали при помощи набора для гель-экстракции QIAquick (Qiagen). Экспрессирующий вектор млекопитающего, содержащий выделенный из CMV полигенный экспрессирующий кластер, гидролизовали с образованием совместимых концов, и 2 фрагмента лигировали друг с другом. Лигированные продукты исследовали методом PCR, используя вектор-специфическую смысловую затравку ZC14063 (SEQ ID NO:48) и zcytor17-специфическую антисмысловую затравку ZC27899 (SEQ ID NO:19), в следующих условиях реакции PCR: 30 циклов при 94°C в течение 30 секунд, при 64°C в течение 30 секунд, при 70°C в течение 90 секунд и при 72°C в течение 7 минут. Размер ожидаемого продукта примерно в 1000 п.н. подтверждал правильную сборку плазмиды, получившей название pZP7NX-hzcytor17-Fc4. Данную плазмиду затем трансфицировали в клетки COS, используя липофектамин (Gibco/BRL), в соответствии с инструкциями изготовителя. Клетки кондиционировали в течение 60 часов в среде DMEM, содержащей 5% FBS (Gibco/BRL), после чего белок очищали в хроматографической колонке, содержащей белок G-сефароза 4В, в результате чего клетки были пригодны для выполнения биоанализов in vitro, например, аналогичных рассмотренным в данном описании.
С. Создание рецептора zcytor17/онкостатин М человека (OSMRbeta)
Примерно по 16 мкг pZP9-ONCOMR-Fc4CEE и pZEM228 hzcytor17/Fc4HIS котрансфицировали в клетки ВНК-570 (АТСС № CRL-10314), используя липофектамин (Gibco/BRL), в соответствии с инструкциями изготовителя. Трансфицированные клетки отбирали в течение 10 дней в среде DMEM, содержащей 5% FBS (Gibco/BRL), 0,5 мг/мл G418 (Gibco BRL) и 250 нМ метилтрексата (МТХ) (Sigma, St. Louis, MO).
Полученный пул дважды селектированных клеток использовали для создания гетеродимерного белка. На трех клеточных фабриках (Nunc, Denmark) данного пула было получено 10 л бессывороточной кондиционированной среды. Указанную кондиционированную среду пропускали через колонку, заполненную 1 мл белка-А, и элюировали, получая десять 750-микролитровых фракций. Четыре из указанных фракций, которые, как было установлено, характеризовались наибольшей концентрацией, собирали и диализовали (отсекая последовательность с молекулярной массой 10 кДа) против PBS. Требуемый гетеродимерный комплекс растворимого рецептора zcytor17/OSMRbeta и белка отделяли от других компонентов среды, для чего пул пропускали через никелевую колонку и промывали колонку имидазолом с разными концентрациями. Комплекс растворимого рецептора zcytor17/OSMRbeta и белка элюировали при промежуточных концентрациях имидазола, в то время как гомодимер hzcytor17/Fc4HIS элюировали при более высоких концентрациях имидазола.
Пример 31
Распределение в тканях zcytor17 человека, определяемое методами нозерн-блоттинга и PCR с использованием панелей тканей
А. Распределение в тканях zcytor17 человека, определяемое методом нозерн-блоттинга
Нозерн-блоты нескольких тканей человека (12-полосный блот I и II MTN человека и блот II MTN иммунной системы человека; блот II MTN эндокринной системы человека, MTN плода человека, набор нескольких тканей человека) (Clontech), а также полученные авторами изобретения блоты, содержащие различные ткани, зондировали для определения распределения в тканях экспрессии zcytor17 человека. Полученные авторами изобретения блоты содержали мРНК из следующих тканей и линий клеток, включающих в себя клетки SK-Hep-1, клетки THP1, надпочечник (Clontech), почку (Clontech), печень (Clontech и Invitrogen), спинной мозг (Clontech), яичко (Clontech), CD4+ Т-клетки человека, CD8+ Т-клетки человека, CD19+ Т-клетки человека, смесь лимфоцитов человека (MLR), линию клеток ТНР1 (АТСС № TIB-202), линию клеток U937, линию клеток лимфобластов мыши p388D1 (АТСС № CCL-46) при стимуляции или без стимуляции иономицином и линию клеток легкого плода человека WI-38 (ATCC № CRL-2221) при стимуляции или без стимуляции иономицином.
Полученный методом PCR зонд длиной примерно 500 п.н. к zcytor17 (SEQ ID NO:4) амплифицировали, используя в качестве затравок олигонуклеотиды ZC28575 (SEQ ID NO:77) и ZC27899 (SEQ ID NO:19). Амплификацию методом PCR выполняли в следующих условиях: 30 циклов при 94°C в течение 1 минуты, при 65°C в течение 1 минуты и при 72°C в течение 1 минуты, затем 1 цикл при 72°C в течение 7 минут. Продукт PCR визуализировали электрофорезом в агарозном геле, и продукт PCR длиной около 500 п.н. очищали в геле описанным способом. Зонд метили радиоактивными изотопами при помощи набора для мечения неспецифических затравок PRIME IT II™ (Stratagene), в соответствии с инструкциями изготовителя. Зонд очищали в колонке с выталкивающим потоком NUCTRAP™ (Stratagene). Раствор EXPRESSHYB™ (Clontech) использовали для предварительной гибридизации и в качестве гибридизующего раствора для нозерн-блотов. Предварительную гибридизацию осуществляли при 68°C в течение 2 часов. Гибридизацию выполняли в течение ночи при 68°C с использованием меченого зонда, генерирующего примерно 1,5х106 импульсов/мин/мл. Блоты трижды промывали при комнатной температуре 2-кратным объемом SSC, 0,05% SDS, затем 1 раз промывали в течение 10 минут 2-кратным объемом SSC, 0,1% SDS при 50°C. Несколько слабых полос было обнаружено после экспонирования в течение нескольких дней. Транскрипт длиной примерно 9 т.п.н. был обнаружен в трахее, скелетных мышцах и тимусе, транскрипт длиной примерно 2 т.п.н. был обнаружен в клетках PBL, HPV, U937 и ТНР-1, и транскрипт длиной около 1,2 т.п.н. был обнаружен в плаценте, костном мозге, щитовидной железе и клетках HPV и U937. Во всех вышеуказанных тканях интенсивность сигнала была слабой. Данный рецептор, по-видимому, незначительно экспрессирован в большинстве нормальных тканей, из чего следует, что экспрессия zcytor17 может зависеть от активации клеток или тканей, в которых экспрессирован указанный рецептор.
В. Распределение в тканях при анализе методом PCR с использованием панелей тканей
Панель кДНК из тканей человека исследовали на экспрессию zcytor17 методом PCR. Указанная панель была создана авторами изобретения и содержала 94 образца марафонской кДНК и кДНК из разных нормальных и раковых тканей и линий клеток человека, представленных в нижеследующей таблице 13. Образцы кДНК были получены из созданных авторами изобретения библиотек или марафонских кДНК из собственных препаратов РНК, препаратов РНК компании Clontech или РНК компании Invitrogen. Марафонские кДНК, полученные при помощи набора marathon-Ready™ (Clontech, Palo Alto, CA), подвергали контролю качества (QC) с использованием клатриновых затравок ZC21195 (SEQ ID NO:78) и ZC21196 (SEQ ID NO:79) и разводили в соответствии с интенсивностью полосы клатрина. Для гарантии качества образцов панели выполняли три испытания на контроль качества (QC): (1) для оценки качества РНК, используемой для получения библиотек, у полученных авторами изобретения кДНК определяли средний размер вставки методом PCR с использованием олигонуклеотидов вектора, специфических для векторных последовательностей отдельной библиотеки кДНК; (2) стандартизацию концентрации кДНК в образцах панели выполняли стандартными методами PCR для амплификации непроцессированной кДНК альфа-тубулина или G3PDH, используя 5'-концевой олигонуклеотид вектора ZC14063 (SEQ ID NO:48) и 3'-концевую альфа-тубулин-специфическую олигонуклеотидную затравку ZC17574 (SEQ ID NO:49) или 3'-концевую G3PDH-специфическую олигонуклеотидную затравку ZC17600 (SEQ ID NO:50); и (3) образец отправляли на секвенирование для проверки возможного загрязнения рибосомной или митохондриальной ДНК. Панель соответствовала 96-луночному планшету и включала в себя геномную ДНК человека (Clontech, Palo Alto, CA) в качестве положительного контрольного образца. В каждой лунке находилось примерно 0,2-100 пг/мкл кДНК. Реакции PCR выполняли с использованием олигонуклеотидов ZC26358 (SEQ ID NO:80) и ZC26359 (SEQ ID NO:81), TaKaRa Ex Taq™ (TAKARA Shuzo Co. LTD, Biomedicals Group, Japan) и красителя Rediload (Research Genetics, Inc., Huntsville, AL). Амплификацию выполняли в следующих условиях: 1 цикл при 94°C в течение 2 минут, 35 циклов при 94°C в течение 30 секунд, при 66,3°C в течение 30 секунд и при 72°C в течение 30 секунд, затем 1 цикл при 72°C в течение 5 минут. Примерно 10 мкл продукта реакции PCR разделяли стандартным агарозным гель-электрофорезом, используя 4% агарозный гель. Прогнозируемый фрагмент ДНК правильного размера был обнаружен в лимфатическом узле, предстательной железе, щитовидной железе, HPV (эпителий предстательной железы), HPVS (селектированный эпителий предстательной железы), опухоли легкого, опухоли матки и геномной ДНК.
Фрагмент ДНК для ткани предстательной железы (2 образца), HPV (эпителий предстательной железы), HPVS (селектированный эпителий предстательной железы) и геномную ДНК вырезали и очищали при помощи набора для гель-экстракции (Qiagen, Chatsworth, CA) в соответствии с инструкциями изготовителя. Фрагменты секвенировали для подтверждения того, что они действительно представляют собой zcytor17.
иономицин + РМА-стимулированные
С. Анализ экспрессии zcytor17 методом PCR и нозерн-блоттинга
Аннотирование типов клеток и условий роста, влияющих на экспрессию рецептора, является полезным способом определения функции и прогнозирования источника лиганда. С этой целью методом PCR был исследован целый ряд тканей и типов клеток. Термоустойчивую полимеразу Advantage II™ (Clontech, La Jolla, CA) использовали вместе с олигонуклеотидными затравками ZC29180 (SEQ ID NO:22) и ZC29179 (SEQ ID NO:82) и 1-10 нг разных матричных кДНК, указанных ниже для 30 циклов амплификации (94°C, 30 секунд; 66°C, 20 секунд; 68°C, 1 минута 30 секунд). Затем 20% каждой реакционной смеси разделяли электрофорезом в 0,8% агарозных гелях, содержащих ТАЕ/бромид этидия, и визуализировали УФ-светом. Образцы оценивали на основе интенсивности полос. См. приведенную ниже таблицу 14.
Из всех сильных положительных сигналов PCR два принадлежали линиям моноцитов человека U937 и THP1.
Две указанные линии клеток вместе с линией эпителиальных клеток предстательной железы были отобраны для дальнейшего анализа методом нозерн-блоттинга. В предыдущих попытках визуализировать транскрипт методом нозерн-блоттинга с использованием мРНК из разных тканей были получены слабые и размытые сигналы в удивительно большом диапазоне от 7 до 10 т.п.н., что затрудняло интерпретацию указанных данных. При помощи полученного денатурирующего 0,8% агарозного геля, содержащего формальдегид/MOPS (RNA Methodologies, Farrell, RE Academic Press) исследовали 2 мкг полиА+ мРНК для каждого образца вместе с лэддерной последовательностью РНК (Life Technologies, Bethesda, MD). Затем гель переносили в нейлон Hybond (Amersham, Buckinghamshire, UK), сшивали под действием УФ-света, гибридизовали в растворе ExpressHyb (Clontech, La Jolla, CA) при 68°C в течение ночи с использованием зонда к zcytor17 человека, полученного методом PCR с использованием олигонуклеотидов ZC28575 (SEQ ID NO:77) и ZC27899 (SEQ ID NO:19), и метили 32P при помощи набора Megaprime (Amersham). Затем нозерн-блот промывали 0,2-кратным объемом SSC + 0,1% SDS при 65°C в течение 15 минут и экспонировали на пленку при помощи усиливающих экранов в течение 7 дней. Большая полоса длиной 8 т.п.н. была обнаружена в эпителиальных клетках предстательной железы и клетках U937, и более слабая полоса присутствовала в линии клеток ТНР1.
Для оптимизации кДНК, используемой в качестве гибридизационного зонда, четыре разные области непроцессированной последовательности zcytor17 человека амплифицировали методом PCR, метили и гибридизовали вышеописанным способом с саузерн-блотами, содержащими геномную и амплифицированную ДНК из библиотеки кДНК. Четыре зонда, именуемые зондами А-D, амплифицировали с использованием следующих пар затравок: (А) ZC28575 (SEQ ID NO:77), ZC27899 (SEQ ID NO:19); (В) ZC27895 (SEQ ID NO:20), ZC28917 (SEQ ID NO:83); (С) ZC28916 (SEQ ID NO:84), ZC28918 (SEQ ID NO:85) и (D) ZC28916 (SEQ ID NO:84), ZC29122 (SEQ ID NO:21). Геномную ДНК человека вместе с библиотеками амплифицированной кДНК, содержащими zcytor17, как было установлено методом PCR, гидролизовали EcoRI и XhoI с высвобождением вставок и разделяли электрофорезом в дуплицированных 0,8% агарозных гелях с ТАЕ, денатурировали 0,5 М NaOH, 1,5 М NaCl, блоттировали на нейлоне Hybond, сшивали под действием УФ-света и гибридизовали с отдельным зондом. Было установлено, что зонд В характеризуется по меньшей мере неспецифическим связыванием и самым сильным сигналом. Таким образом, зонд В использовали во всех последующих гибридизациях.
С учетом того, что клетки ТНР1 являются великолепной моделью моноцитов, циркулирующих в кровотоке, и экспрессируют zcytor17 на низких уровнях, авторы настоящего изобретения обрабатывали указанные клетки различными соединениями, пытаясь увеличить экспрессию zcytor17. Клетки выращивали до плотности 2е5/мл, промывали и вновь суспендировали в разных стимулирующих средах, затем выращивали в течение четырех или тридцати часов и собирали для получения препаратов РНК. Каждая среда содержала одно из следующих лекарственных средств или два цитокина: 2 мкг/мл LPS (Sigma Chemicals, St. Louis, MO), 2 нг/мл hTNFα (R&D Systems, Minneapolis, MN), 2 нг/мл hGM-CSF (R&D Systems, Minneapolis, MN), 50 нг/мл hIFNγ (R&D Systems, Minneapolis, MN), 1 нг/мл hMCSF (R&D Systems, Minneapolis, MN), 1 нг/мл hIL6 (R&D Systems, Minneapolis, MN), 2 нг/мл hIL1β (R&D Systems, Minneapolis, MN), 50 нг/мл hIFNγ + 0,5 нг/мл hIL4 (R&D Systems, Minneapolis, MN), 50 нг/мл hIFNγ + 1 нг/мл hIL10 (R&D Systems, Minneapolis, MN), 10 нг/мл РМА (Calbiochem, SanDiego, CA) и необработанный контрольный образец. В конце культивирования получали полную РНК при помощи набора RNAeasy Midi (Qiagen, Valencia, CA). ПолиА+РНК отбирали из полной РНК при помощи набора MPG (CPG, Lincoln Park, NJ). 2 мкг полиА+РНК в каждом отдельном случае разделяли электрофорезом в агарозном геле с формальдегидом/MOPS, переносили на нейлон и сшивали под действием УФ-света вышеописанным способом. Указанные нозерн-блоты затем гибридизовали, как указывалось выше, с зондом В при 68°C в течение ночи, промывали в условиях высокой строгости 0,2-кратным объемом SSC, 0,1% SDS при 65°C, экспонировали на пленку в течение ночи и затем экспонировали на фосфорные экраны для количественного определения сигнала. Доминантная мРНК длиной 8 т.п.н. и относительно более слабая полоса длиной 2,8 т.п.н. были видны на всех снимках. 20-кратное увеличение мРНК zcytor17 было обнаружено в РНК, полученной из клеток, обработанных hIFNγ в течение 30 часов, причем указанный эффект был немного меньше при одновременной обработке IL4. Трехкратное увеличение мРНК было обнаружено в РНК, полученной из клеток, обработанных LPS, TNFα и GM-CSF, в то время как MCSF, IL6 и IL1β не влияли на уровни мРНК zcytor17. Полученные данные свидетельствуют об определенной роли рецептора zcytor17 и его лиганда в биологии макрофагов моноцитов и о распространении ряда заболеваний, опосредуемых данными клетками.
Пример 32
Распределение в тканях zcytor17lig человека, определяемое методами нозерн-блоттинга и PCR с использованием панелей тканей
Фрагмент кДНК zcytor17lig человека был получен методом PCR с использованием геноспецифических затравок: смысловой затравки ZC41438 (SEQ ID NO:93) и антисмысловой затравки ZC41437 (SEQ ID NO:94) и матричной кДНК zcytor17lig человека (SEQ ID NO:90). Данный фрагмент очищали стандартными методами, примерно 25 нг метили 32P альфа-dCTP при помощи набора для мечения неспецифических затравок Prime-It RmT (Stratagene), гибридизовали в Ultrahyb (Ambion) и использовали для экспонирования на пленку Biomax/усиливающие экраны в соответствии с рекомендациями изготовителя. Новые, ранее не использовавшиеся блоты, содержащие 12-полосный блот MTN человека Clontech, блот II MTN головного мозга человека, блот IV MTN головного мозга человека, блот II MTN иммунной системы человека и матрицу II МТЕ человека компании Clontech, гибридизовали в течение ночи при 42°C методом Ambion Ultrahyb. Неспецифические радиоактивные импульсы вымывали 0,1 SSC/0,5% SDS при 55°C. Положительные блоты содержали 12-полосный блот MTN человека (Clontech). Из 12 исследованных тканей только плацента дала положительный результат на наличие транскрипта длиной примерно 1,2 т.п.н.
Пример 33
Создание векторов млекопитающих, экспрессирующих zcytor17lig-CEE человека
А. Создание zcytor17lig-CEE/pZMP21
Экспрессирующая плазмида, содержащая весь или часть полинуклеотида, кодирующего zcytor17lig-CEE (SEQ ID NO:95), была создана методом гомологичной рекомбинации. Данная плазмида получила название zcytor17lig-CEE/pZMP21.
В процессе создания zcytor17lig-CEE/pZMP21 путем амплификации методом PCR был получен фрагмент zcytor17lig-CEE. Для получения фрагмента zcytor17lig-CEE была использована матричная ДНК zcytor17lig/pZP7nx. Фрагмент zcytor17lig-CEE был получен при использовании следующих затравок: (1) ZC41607 (SEQ ID NO:97) (смысловая последовательность), содержащая от 5'-конца до 3'-конца: 28 п.н. фланкирующей последовательности вектора (5'-конец вставки) и 21 п.н., соответствующую 5'-концевой последовательности zcytor17lig, и (2) ZC41605 (SEQ ID NO:98) (антисмысловая последовательность), содержащая от 5'-конца до 3'-конца: 37 п.н. фланкирующей последовательности вектора (3'-конец вставки), 3 п.н. терминирующего кодона, 21 п.н., кодирующую С-концевую метку ЕЕ, и 21 п.н., соответствующую 3'-концу последовательности zcytor17lig. Фрагмент, полученный в результате вышеуказанной амплификации методом PCR, представлял собой копию матричного zcytor17lig с добавлением С-концевой метки ЕЕ, что позволило получить конечный продукт zcytor17lig-CEE.
Реакции PCR выполняли следующим образом: для получения реакционной смеси с конечным объемом, равным 100 мкл, добавляли 10 мкл 10-кратного объема Taq-полимеразного реакционного буфера с 15 мМ MgCl2 (Gibco), 1 мкл Taq-ДНК-полимеразы (5 ед/мкл, Gibco), 3 мкл 10 мМ dNTP, 78 мкл дистиллированной Н2О, 3 мкл 20 пмоль/мкл исходного раствора затравки ZC41607 (SEQ ID NO:97), 3 мкл 20 пмоль/мкл исходного раствора затравки ZC41605 (SEQ ID NO:98) и 2 мкл 0,13 мкг/мл исходного раствора матричной ДНК zcytor17lig. К смеси добавляли минеральное масло в объеме 50 мкл. Реакционную смесь нагревали до 94°C в течение 5 минут, затем выполняли 35 циклов при 94°C в течение 1 минуты, при 55°C в течение 2 минут, при 72°C в течение 3 минут, производили удлинение цепи при 72°C в течение 10 минут и выдерживали при 4°C, после чего реакционную смесь собирали.
Плазмиду pZMР21 подвергали рестрикционному гидролизу ферментом BglII, очищали микроцентрифугированием при помощи набора для очистки продуктов PCR QiaQuick (Qiagen) и использовали для рекомбинации с фрагментом PCR. Плазмида pZMP21 была создана из плазмиды pZMP20, которая была получена из pZP9 (депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, VA 20110-2209, под № 98668) с использованием генетических элементов дрожжей из pRS316 (депонирована в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, VA 20110-2209, под № 77145), с использованием элемента IRES из полиовируса и внеклеточного домена CD8, усеченного у карбоксильного конца трансмембранного домена. PZMP21 является экспрессирующим вектором млекопитающего, содержащим полигенный экспрессирующий кластер, включающий в себя промотор MPSV, интрон сигнального пептида иммуноглобулина, несколько сайтов рестрикции для вставки кодирующих последовательностей, терминирующий кодон и терминатор гормона роста человека. Указанная плазмида имеет также ориджин репликации E. coli, экспрессирующую единицу селектируемого маркера млекопитающего, содержащую промотор SV40, энхансер и ориджин репликации, ген DHFR, терминатор SV40, а также последовательности URA3 и CEN-ARS, необходимые для селекции и репликации в S. cerevisiae.
Пятьдесят микролитров компетентных дрожжевых клеток (S. cerevisiae) независимо объединяли со 100 нг разрезанной плазмиды, 5 мкл вышеописанной смеси для PCR и переносили в 0,2 см кювету для электропорации. Смесь дрожжи/ДНК электропорировали при 0,75 кВ (5 кВ/см), ∞ Ом, 25 мкФ. В каждую кювету добавляли 600 мкл 1,2 М раствора сорбита, дрожжи переносили в виде одной 100 мкл аликвоты и одной 300 мкл аликвоты на две чашки URA-D и инкубировали при 30°C Примерно через 72 часа трансформанты дрожжей Ura+ из одной чашки вновь суспендировали в 1 мл Н2О и осаждали коротким центрифугированием с образованием осадка дрожжевых клеток. Клеточный дебрис вновь суспендировали в 500 мкл буфера для лизиса (2% тритона Х-100, 1% SDS, 100 мМ NaCl, 10 мМ трис-буфера, рН 8,0, 1 мМ EDTA). 500 мкл лизированной смеси добавляли в пробирку Эппендорфа, содержащую стеклянные шарики, промытые 300 мкл кислоты, и 300 мкл фенол-хлороформа, интенсивно перемешивали два или три раза с интервалами в 1 минуту и центрифугировали в центрифуге Эппендорфа при максимальной скорости в течение 5 минут. Триста микролитров водной фазы переносили в чистую пробирку, ДНК осаждали 600 мкл 100% этанола (EtOH) и центрифугировали в течение 10 минут при 4°C. Осадок ДНК промывали 500 мкл 70% EtOH и центрифугировали в течение 1 минуты при 4°C. Осадок ДНК вновь суспендировали в 30 мкл Н2О.
Электрокомпетентные клетки E. coli (MC1061) трансформировали 5 мкл препарата дрожжевой ДНК и 50 мкл клеток МС1061. Клетки электропорировали при 2,0 кВ, 25 мкФ и 400 Ом. После электропорации добавляли 600 мкл SOC (2% бактотриптона (Difco, Detroit, MI), 0,5% дрожжевого экстракта (Difco), 10 мМ NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкозы). Электропорированные клетки E. coli переносили в виде 200 мкл и 50 мкл аликвот на две чашки со средой LB, содержащей ампициллин (бульон LB (Lennox), 1,8% бактоагара (Difco), 100 мг/л ампициллина). Чашки инкубировали в перевернутом виде в течение примерно 24 часов при 37°C. Произвольно отбирали три колонии, устойчивые к ампициллину, и анализировали последовательность вставки. Плазмидную ДНК выделяли в крупном масштабе из клона с подтвержденной последовательностью при помощи набора Qiagen Maxi (Qiagen), в соответствии с инструкциями изготовителя.
В. Создание zcytor17lig(m)-CEE/pZMP21 мыши
Экспрессирующая плазмида, содержащая полный полинуклеотид, кодирующий zcytor17lig-CEE мыши (SEQ ID NO:104 и SEQ ID NO:105), также была создана методом гомологичной рекомбинации, описанным в приведенном выше примере 33А. Были использованы следующие затравки: (1) ZC41643 (SEQ ID NO:106) (верхняя смысловая затравка от 5'-конца к 3'-концу), содержащая 28 п.н. вектора, перекрывающего 5'-конец места вставки; 21 п.н. 5'-конца zcytor17lig(m) и (2) ZC41641 (SEQ ID NO:107) (нижняя антисмысловая затравка от 5'-конца к 3'-концу), содержащая 37 п.н. вектора, перекрывающего 3'-конец места вставки, 3 п.н. терминирующего кодона, 21 п.н. С-концевой метки ЕЕ, 24 п.н. 3'-конца zcytor17lig(m)-CEE. Данная плазмида получила название zcytor17lig(m)-CEE/pZMP21. Полинуклеотидная последовательность zcytor17lig(m)-CEE представлена в SEQ ID NO:104, и соответствующая полипептидная последовательность представлена в SEQ ID NO:105.
Пример 34
Трансфекция и экспрессия полипептидов zcytor17lig-CEE
А. Экспрессия zcytor17lig-CEE/pZMP21 человека в клетках 293Т
Zcytor17lig-CEE был временно экспрессирован в клетках 293Т (Stanford University School of Medicine, Stanford, CA, ATCC (SD-3515) для получения исходного очищенного белка. За день до трансфекции клетки 293Т высевали в количестве 6,5 х 104 клеток/см2 в 30 колб для культур Т162, содержащих культуральную среду в общем объеме 30 мл (SL7V4 + 5% FBS + 1% пенициллина/стрептомицина). Клетки инкубировали в течение 24 часов при 37°C.
Смесь ДНК/липосомы была получена следующим образом: две 50 мл конические пробирки заполняли 25 мл среды для трансфекции (SL7V4 + 1% пенициллина/стрептомицина) и добавляли 1,13 мг zcytor17lig-CEE/pZMP21 (пример 33). Отдельный набор из двух 50 мл конических колб заполняли 22 мл среды для трансфекции (указана выше) и добавляли 3 мл липосом (липофектамин, Gibco). В каждом наборе пробирок содержимое одной пробирки с ДНК добавляли к одной пробирке с липосомами, и смесь ДНК/липосомы инкубировали в течение 30 минут. Собирали содержимое двух 50 мл конических пробирок, содержащих смеси ДНК/липосомы (примерно 100 мл), и добавляли 300 мл среды для трансфекции.
30 колб с клетками 293Т декантировали, промывали один раз примерно 15 мл PBS и в каждую колбу добавляли 12,5 мл разведенной смеси ДНК/липосомы. Колбы инкубировали в течение 3 часов при 37°C. После окончания инкубации в каждую колбу Т162 добавляли 25 мл культуральной среды (указана выше). Среду для трансфекции собирали примерно через 96 часов и использовали для очистки белка (пример 35).
В. Экспрессия zcytor17lig-CEE/pZMP21 человека в клетках ВНК
Непроцессированный белок zcytor17lig получали в клетках ВНК, трансфицированных zcytor17lig-CEE/pZMP21 (см. приведенный выше пример 33). Клетки ВНК 570 (АТСС CRL-10314) вводили в колбы для тканевых культур Т75 и выращивали примерно до 50-70% слияния при 37°C, 5% СО2 в питательной среде (SL7V4, 5% FBS, 1% пенициллина/стрептомицина). Затем клетки трансфицировали zcytor17lig-CEE/pZMP21 методом трансфекции, опосредуемой липосомами (с использованием липофектамина™, Life Technologies), в бессывороточной (SF) среде (SL7V4). Плазмиду (16 мкг) разводили бессывороточной средой в 1,5 мл пробирках до общего конечного объема 640 мкл. Тридцать пять микролитров липидной смеси смешивали с 605 мкл бессывороточной среды и полученную смесь инкубировали в течение примерно 15 минут при комнатной температуре. К смеси ДНК/липиды добавляли пять миллилитров бессывороточной среды. Клетки промывали один раз 10 мл PBS, затем PBS декантировали и добавляли смесь ДНК/липиды. Клетки инкубировали при 37°C в течение пяти часов, затем в каждую чашку добавляли 15 мл среды (SL7V4, 5% FBS, 1% пенициллина/стрептомицина. Чашки инкубировали при 37°C в течение ночи, и на следующий день среду смеси ДНК/липиды заменяли селективной средой (SL7V4, 5% FBS, 1% пенициллина/стрептомицина, 1 мкМ метотрексата). Примерно через 10 дней после трансфекции колонии, устойчивые к метотрексату, из колбы для трансфекции Т75 обрабатывали трипсином, клетки собирали, культивировали в колбе Т-162 и переносили в крупномасштабную культуру.
С. Экспрессия zcytor17lig-CEE(m)/pZMP21 мыши в клетках 293Т
Zcytor17lig(m)-CEE мыши временно экспрессировали в клетках 293Т аналогично примеру 34А и использовали культуральную среду для очистки белка (пример 35).
Пример 35
Очистка zcytor17lig-CEE от клеток 293Т
За исключением особо оговоренных случаев, все операции выполняли при 4°C. Zcytor17lig мыши и человека, содержащий С-концевые метки Glu-Glu (EE) (SEQ ID NO:103), очищали следующим способом. Очищали кондиционированную среду, полученную из клеток 293Т, экспрессирующих zcytor17lig-CEE (пример 34). Общие концентрации целевого белка в кондиционированной среде определяли при помощи анализа методом SDS-PAGE и вестерн-блоттинга с антителом против ЕЕ.
5,5 мл смолы с антителом против ЕЕ Poros 50 А (PR BioSystems, Framingham, MA) (полученной следующим способом) выливали в стеклянную колонку Waters AP-1 размером 1 см х 7 см (Waters, Milford, MA). Колонку заполняли проточным методом и уравновешивали в устройстве BioCad Sprint (PE BioSystems, Framingham, MA) физиологическим раствором с фосфатным буфером (PBS), рН 7,4. Кондиционированную среду доводили при помощи NaCl до 0,3 М и рН доводили до 7,2. Затем кондиционированную среду вводили в колонку в течение ночи со скоростью потока около 3 мл/минуту. Колонку промывали 10 объемами (CV) PBS, рН 7,4 и снова 5 раз промывали 3 объемами PBS, pH 7,4, компании Sigma. Элюирование производили 0,5 М ацетата, 0,5 М NaCl, рН 2,5, со скоростью 3 мл/минуту. Пробирки для фракционирования содержали 1 мл основного трис-буфера (без доведения рН) для немедленной нейтрализации элюирования. Колонку снова промывали 5 раз 2 объемами PBS, рН 7,4, компании Sigma для нейтрализации колонки и уравновешивали PBS (рН 7,4). Двухмиллилитровые фракции собирали хроматографией с элюированием, и контролировали оптическую плотность при 280 и 215 нМ; прошедшие через колонку и промытые пулы также сохраняли и анализировали. Максимальные фракции, соответствующие элюированию 5-кратным объемом PBS и кислотой, анализировали на наличие целевого белка при помощи SDS-PAGE с окрашиванием серебром и вестерн-блоттинга с первичным антителом против ЕЕ, конъюгированным с вторичным антителом против HRP мыши. Исследуемые фракции, элюированные кислотой, собирали и концентрировали с 38 мл до 0,8 мл при помощи мембранного центрифужного концентратора с отсечением молекулярной массы 5000 Дальтон (Millipore, Bedford, MA), в соответствии с инструкциями изготовителя.
Для отделения zcytor17lig-CEE от агрегированного материала и любых других загрязняющих совместно очищенных белков, собранные концентрированные фракции подвергали вытеснительной хроматографии в колонке Superdex 75 (Pharmacia, Piscataway, NJ) размером 1,6 х 60 см (120 мл) и вводили в PBS со скоростью потока 1,0 мл/мин при помощи устройства BioCad Sprint. Трехмиллилитровые фракции собирали в течение всей хроматографии, и контролировали оптическую плотность при 280 и 215 нМ. Максимальные фракции исследовали методом SDS-PAGE с окрашиванием серебром, и собирали только самые чистые фракции. Полученный материал представлял собой очищенный белок zcytor16lig-CEE.
При выполнении вестерн-блоттинга и SDS-PAGE с окрашиванием кумассии голубым и серебром zcytor17lig-CEE образовывал одну главную полосу. Концентрацию белка в очищенном материале определяли при помощи анализа ВСА (Pierce, Rockford, IL), после чего отбирали аликвоты белка и хранили при -80°C стандартными методами.
Чтобы получить смолу с антителом против ЕЕ PorosA50, слой Poros A50 объемом 65 мл (PE Biosystems) промывали 100 мл воды и раствором 0,1 М триэтаноламина, рН 8,2 (ТЕА, ICN, Aurora, Ohio), 1 М Na2SO4, рН 8,8, содержащим 0,02% азида натрия, используя фильтровальное устройство с вакуумной колбой. Раствор моноклонального антитела против ЕЕ в концентрации 2 мг/мл в объеме 300 мл смешивали с промытой смолой в объеме 250 мл. Смесь инкубировали в течение ночи при комнатной температуре, и несвязанное антитело удаляли, промывая смолу 5 объемами раствора 200 мМ ТЕА, 1 М Na2SO4, рН 8,8, содержащего 0,02% азида натрия, вышеуказанным способом. Смолу вновь суспендировали в 2 объемах раствора ТЕА, 1 М Na2SO4, рН 8,8, содержащего 0,02% азида натрия, и переносили в подходящую емкость. Добавляли три мл 25 мг/мл (68 мМ) дисукцинимидилсуберата (в ДМСО компании Pierce, Rockford, IL) и раствор инкубировали в течение трех часов при комнатной температуре. Неспецифические участки на смоле блокировали, инкубируя в течение 10 минут при комнатной температуре с 5 объемами 20 мМ этаноламина (Sigma, St. Louis, МО) в 200 мМ ТЕА, рН 8,8, используя фильтровальное устройство с вакуумной колбой. Смолу промывали PBS, рН 7,4, затем 0,1 М глицина, рН 3, и нейтрализовали 10-кратным объемом PBS. После промывки дистиллированной водой смолу, связанную с антителом против ЕЕ, Poros-A50 хранили при 4°C в 20% этаноле.
Пример 36
Секвенирование N-конца zcytor17lig человека и мыши
А. Секвенирование N-конца zcytor17lig человека
Стандартное автоматическое секвенирование N-концевого полипептида (расщепление по Эдману) выполняли с использованием реагентов компании Applied Biosystems. N-концевую последовательность анализировали в системе секвенатора белка модели 494 (Applied Biosystems, Inc., Foster City, CA). Анализ данных производили в системе для секвенирования белка модели 610А, вариант 2.1а (Applied Biosystems).
Был получен очищенный образец zcytor17lig-CEE человека (пример 35). Образец вводили в подготовленный стекловолоконный фильтр для секвенирования N-конца. Стекловолоконный фильтр готовили путем предварительной циклизации биобреном™.
Анализ N-концевой последовательности секретированного полипептида zcytor17lig человека не подтвердил наличия прогнозируемого сайта расщепления сигнальной последовательности, но выявил зрелый инициирующий кодон у остатка 27 (Leu) в SEQ ID NO:2 последовательности-предшественника zcytor17lig человека.
В. Секвенирование N-конца zcytor17lig мыши
Стандартное автоматическое секвенирование N-концевого полипептида (расщепление по Эдману) выполняли с использованием реагентов компании Applied Biosystems. N-концевую последовательность анализировали в системе секвенатора белка модели 494 (Applied Biosystems, Inc., Foster City, CA). Анализ данных производили в системе для секвенирования белка модели 610А, вариант 2.1а (Applied Biosystems).
Очищенный образец zcytor17lig-CEE мыши был получен на шариках, покрытых белком G-сефароза/антителом против ЕЕ (пример 35). Шарики помещали в восстанавливающем буфере для SDS-PAGE на баню с кипящей водой и затем исследовали SDS-PAGE в системе для SDS-PAGE Novex (4-12% бис-трис-буфера MES NuPAGE; Invitrogen) в соответствии с инструкциями изготовителя. Гель электроблоттировали на мембране PVDF Novex (Invitrogen) и окрашивали кумассии голубым (Sigma, St. Louis, MO) стандартными методами. Антитело против ЕЕ анализировали соответствующими методами вестерн-блоттинга для идентификации полосы zcytor17lig с целью секвенирования N-концевого белка. Антитело против ЕЕ, конъюгированное с IgG HRP, было получено авторами настоящего изобретения.
Анализ N-концевой последовательности секретированного полипептида zcytor17lig мыши подтвердил наличие прогнозируемого сайта расщепления сигнальной последовательности с образованием зрелого инициирующего кодона в положении остатка 31 (Ala) при сравнении с SEQ ID NO:11 и SEQ ID NO:91 последовательности-предшественника zcytor17lig мыши.
Пример 37
Анализ связывания с клетками COS
При помощи анализа связывания исследовали связывание zcytor17lig с рецепторами, включая рецептор zcytor17 или гетеродимеры и тримеры рецептора zcytor17 (например, zcytor17/OSMR, zcytor17/WSX-1, zcytor17/OSMR/WSX-1 или другие субъединицы рецепторов цитокина класса 1). Плазмидную ДНК рецептора zcytor17 трансфицировали в клетки COS, и трансфицированные клетки COS использовали для оценки связывания zcytor17lig с рецепторами, включая рецептор zcytor17, следующим способом.
А. Трансфекция клеток COS
Клетки COS трансфицировали следующим образом: 800 нг плазмидной ДНК рецептора смешивали в следующих комбинациях: только pZp7pX/zcytor17; только pZp7Z/WSX-1; только pZp7NX/OSMR; pZp7pX/zcytor17 + pZp7NX/OSMR; pZp7pX/zcytor17 + pZp7Z/WSX-1; рZp7NX/OSMR + pZp7Z/WSX-1; pZp7pX/zcytor17 + pZp7NX/OSMR + pZp7Z/WSX-1) с 4 мкл липофектамина™ в 80 мкл бессывороточной среды DMEM (55 мг пирувата натрия, 146 мг L-глутамина, 5 мг трансферрина, 2,5 мг инсулина, 1 мкг селена и 5 мг фетуина в 500 мл DMEM), инкубировали при комнатной температуре в течение 30 минут и добавляли 320 мкг бессывороточной среды DMEM. Добавляли 400 мкл указанной смеси к 2 х 105 клеток CОS/лунку на 12-луночном планшете для тканевой культуры (покрытом фибронектином) и инкубировали в течение 5 часов при 37°C. Затем добавляли 500 мкл среды DMEM с 20% FBS (100 мл FBS, 55 мг пирувата натрия и 146 мг L-глутамина в 500 мл DMEM) и инкубировали в течение ночи.
В. Анализ связывания
Анализ связывания выполняли следующим образом: среду отмывали от клеток при помощи PBS + 0,1% BSA, после чего клетки блокировали в течение 60 минут тем же раствором. Затем клетки инкубировали в течение 1 часа в PBS + 0,1% BSA с 1,0 мкг/мл очищенного белка zcytor17lig-CEE. Клетки промывали PBS +0,1% BSA и инкубировали еще один час с антителом мыши против GluGlu, разведенным в соотношении 1:1000. Клетки снова промывали PBS + 0,1% BSA и инкубировали в течение 1 часа с конъюгированным антителом козы против HRP мыши, разведенным в соотношении 1:200.
Положительное связывание обнаруживали при помощи реагента флуоресцеинтирамида, разведенного в буфере для разведения (набор NEN) в соотношении 1:50, инкубировали в течение 4-6 минут и промывали PBS + 0,1% BSA. Клетки фиксировали в течение 15 минут 1,8% формальдегида в PBS и промывали PBS + 0,1% BSA. Затем клетки консервировали в заливочной среде Vectashield (Vector Labs Burlingame, CA), разведенной в PBS в соотношении 1:5. Клетки визуализировали при помощи фильтра FITC в флуоресцентном микроскопе.
Положительное связывание было обнаружено в клетках, трансфицированных только zcytor17, zcytor17 + OSMRbeta, zcytor17 + WSX-1 и zсytor17 + OSMRbeta + WSX-1. Связывание не было обнаружено в клетках, трансфицированных WSX-1 + OSMRbeta, только OSMRbeta или только WSX-1.
Пример 38
Активация лигандом zcytor17lig мыши рецептора zcytor17/OSMRbeta мыши при выполнении люциферазного анализа
А. Клонирование непроцессированной последовательности zcytor17 мыши и OSMRbeta мыши с целью экспрессии
Библиотеку кДНК яичка мыши исследовали с целью обнаружения непроцессированного клона zcytor17 мыши. Указанную библиотеку культивировали в количестве 65500 КОЕ/чашку на 24 чашках со средой LB, содержащей ампициллин. Фильтры были получены с использованием нейлона Hybond N (Amersham-Pharmacia Biotech, Inc., Piscataway, NJ) в общей сложности примерно для 1,6 миллиона колоний. Фильтры маркировали горячей иглой для правильной ориентации и денатурировали в течение 6 минут в 0,5 M NaOH и 1,5 М трис-HCl, рН 7,2. Затем фильтры нейтрализовали в 1,5 М NaCl и 0,5 М трис-HCl, рН 7,2, в течение 6 минут. ДНК связывали с фильтрами при помощи ультрафиолетового кросс-линкера (Stratalinker®, Stratagene, La Jolla, CA) при 1200 Дж. Затем фильтры сушили в течение ночи при комнатной температуре.
На следующий день фильтры предварительно промывали при 65°C в буфере для предварительной промывки, содержащем 0,25-кратный объем SSC, 0,25% SDS и 1 мМ EDTA. Клеточный дебрис удаляли вручную при помощи Kimwipes® (Kimberly-Clark), и раствор трижды меняли в течение 1 часа. Фильтры подвергали воздушной сушке и хранили при комнатной температуре. Затем фильтры предварительно гибридизовали в течение примерно 3 часов при 63°C в 20 мл раствора для гибридизации ExpressHyb™ (Clontech, Palo Alto, CA).
Зонд В (пример 31) получали методом PCR из матрицы zcytor17 человека с использованием олигонуклеотидных затравок ZC27895 (SEQ ID NO:20) и ZC28917 (SEQ ID NO:83) и метили 32P, используя коммерчески производимый набор (Система мечения ДНК Megaprime; Amersham Pharmacia Biotech, Piscataway, NJ), в соответствии с инструкциями изготовителя. Зонд очищали в колонке с выталкивающим потоком Stratagene™ (колонка NucTrap®; Stratagene, La Jolla, CA). Зонд денатурировали при 100°C в течение 15 минут и добавляли к раствору для гибридизации ExpressHyb™. Фильтры гибридизовали в 15 мл раствора для гибридизации, содержащего зонд, генерирующий 1,6 х 106 импульсов/мин/мл, при 63°C в течение ночи. Фильтры промывали при 55°C в 2-кратном объеме SSC, 0,1% SDS и 1 мМ EDTA и экспонировали на рентгеновскую пленку при -80°C в течение 4 1/2 дней. Тринадцать положительных образцов удаляли с планшетов в виде пробок и помещали в 1,7 мл пробирки, содержащие 1 мл среды LB с ампициллином. Пробирки выдерживали при 4°C в течение ночи. 13 положительных образцов подвергали еще двум циклам очистки. Планшеты третьего порядка культивировали при 37°C после удаления фильтров, при этом единичные колонии собирали и отправляли на секвенирование. Было установлено, что три из указанных колоний содержат последовательность ортолога zcytor17 мыши.
Кроме того, продукт PCR был получен с использованием кДНК CTLL-2 в качестве матрицы и олигонуклеотидов ZC38239 (SEQ ID NO: 108) и ZC38245 (SEQ ID NO:109) в качестве затравок. CTLL-2 является линией цитотоксических Т-лимфоцитов мыши (АТСС № TIB-214). Реакцию PCR выполняли в следующих условиях: 1 цикл при 95°C в течение 1 минуты, 30 циклов при 95°C в течение 15 секунд, при 68°C в течение 3 минут, затем при 68°C в течение 10 минут; выдерживание при 4°C. При выполнении реакции PCR использовали примерно 0,5 нг кДНК, 20 пмоль каждого олигонуклеотида и 1 мкл полимеразной смеси Asvantage II (ClonTech). Примерно 6% продукта PCR использовали в качестве матрицы при выполнении новой реакции PCR вышеописанным способом с применением олигонуклеотидов ZC38239 (SEQ ID NO:108) и ZC38238 (SEQ ID NO:110). Указанную реакцию PCR выполняли в следующих условиях: 30 циклов при 94°C в течение 45 секунд, при 65°C в течение 45 секунд, при 72°C в течение 1 минуты, затем при 72°C в течение 7 минут; выдерживание при 10°C. Большую часть реакционной смеси PCR вводили в 1,0% агарозный гель, основную полосу длиной около 360 п.н. вырезали, фрагмент ДНК элюировали и ДНК секвенировали.
Последовательность полинуклеотида zcytor17 мыши представлена в SEQ ID NO:111, и соответствующая аминокислотная последовательность показана в SEQ ID NO:112. Кроме того, усеченная растворимая форма полинуклеотида zcytor17 мыши представлена в SEQ ID NO:113, и соответствующая аминокислотная последовательность показана в SEQ ID NO:114.
Для получения непроцессированной кДНК OSMRbeta мыши 5'- и 3'-концевые продукты PCR выделяли и связывали на внутреннем сайте BamHI. Затравки для PCR были созданы с использованием нуклеотидной последовательности SEQ ID NO:119 и включали сайты рестрикции EcoRI и XbaI для клонирования. Геномная последовательность нуклеиновой кислоты OSMRbeta мыши показана в SEQ ID NO:119, в которой кодирующая последовательность включает в себя остатки 780-3692, кодирующие полипептид OSMRbeta, содержащий 970 аминокислот, который показан в SEQ ID NO:120. Вырожденная последовательность нуклеиновой кислоты, которая кодирует полипептид SEQ ID NO:120, показана в SEQ ID NO:121.
5'-Концевой продукт PCR был получен с использованием созданной авторами изобретения библиотеки кДНК 3Т3-L1 (дифференцированные адипоциты мыши) в качестве матрицы и олигонуклеотидов ZC41764 (SEQ ID NO:115) и ZC41598 (SEQ ID NO:116) в качестве затравок. Указанную реакцию PCR для создания 5'-конца выполняли следующим образом: 30 циклов при 95°C в течение 45 секунд, при 55°C в течение 45 секунд, при 72°C в течение 1 минуты 30 секунд, затем при 72°C в течение 7 минут; выдерживание при 4°C. При выполнении реакции PCR использовали примерно 3 мкг плазмиды, полученной из библиотеки кДНК, 20 пмоль каждого олигонуклеотида и пять единиц Pwo-ДНК-полимеразы (Roche). Примерно 90% 5'-концевого продукта PCR гидролизовали EcoRI и BamHI и очищали в 1,0% агарозном геле. Полосу длиной примерно 1446 п.н. вырезали и использовали для лигирования (см. ниже).
3'-Концевой продукт PCR был получен при использовании созданной авторами изобретения библиотеки кДНК из плаценты мыши в качестве матрицы и олигонуклеотидов ZC41948 (SEQ ID NO:117) и ZC41766 (SEQ ID NO:118) в качестве затравок. Указанную реакцию PCR по созданию 3'-конца выполняли следующим образом: 30 циклов при 95°C в течение 45 секунд, при 55°C в течение 45 секунд, при 72°C в течение 1 минуты 30 секунд, затем при 72°C в течение 7 минут; выдерживание при 4°C. При выполнении PCR использовали примерно 3 мкг плазмиды, полученной из библиотеки кДНК, 20 пмоль каждого олигонуклеотида и пять единиц Pwo-ДНК-полимеразы (Roche). Примерно 90% 3'-концевого продукта PCR гидролизовали BamHI и XbaI и очищали в 1,0% агарозном геле. Полосу длиной около 2200 п.н. вырезали и использовали вместе с 5'-концевым продуктом PCR (описан выше) для лигирования с экспрессирующим вектором pZP-5Z, гидролизованным EcoRI и XbaI. Трехкомпонентное лигирование выполняли с использованием вышеуказанного 5'-концевого фрагмента EcoRI-BamHI, 3'-концевого фрагмента BamHI-XbaI и экспрессирующего вектора pZP-5Z, гидролизованного EcoRI и XbaI. В результате выполнения указанной реакции была создана плазмида pZP-5Z, содержащая непроцессированную кДНК для OSMRbeta мыши (нуклеотиды 780-3692 SEQ ID NO:119), получившая название pZP-5Z/OSMRbeta. Непроцессированная кДНК OSMRbeta мыши в плазмиде pZP5Z/OSMRbeta имеет две аминокислотные вставки из SEQ ID NO:120. Обнаружена дупликация аминокислоты глицина в положении 370 и дупликация глутаминовой кислоты в положении 526. Плазмида pZP-5Z является экспрессирующим вектором млекопитающего, содержащим полигенный экспрессирующий кластер, включающий в себя промотор CMV, несколько сайтов рестрикции для вставки кодирующих последовательностей и терминатор гормона роста человека. Указанная плазмида имеет также ориджин репликации E. coli, экспрессирующую единицу селектируемого маркера млекопитающего, включающую в себя промотор SV40, энхансер и ориджин репликации, ген устойчивости к зеоцину и терминатор SV40.
Полученные трансформанты секвенировали для подтверждения наличия последовательности кДНК OSMRbeta мыши.
В. Создание линий клеток BaF3/KZ134/zcytor17m, BaF3/KZ134/cytor17m/OSMRbetam, BHK/KZ134/zcytor17m и BHK/KZ134/zcytor17m/OSMRbetam
Устойчивые линии клеток BaF3/KZ134 и BHK/KZ134 (пример 20) трансфицировали экспрессирующей плазмидой, кодирующей непроцесированный zcytor17 мыши, pZP7P/zcytor17m (пример 38А), для создания, соответственно, клеток BaF3/KZ134/zcytor17m и BHK/KZ134/zcytor17m. Плазмиду экспрессии OSMRbeta мыши pZP-5Z/OSMRbetam (пример 38А) затем трансфицировали в указанные клетки для создания, соответственно, линий клеток BaF3/KZ134/zcytor17m/OSMRbetam и BHK/KZ134/zcytor17m/OSMRbetam. Были использованы способы, описанные в примере 4, за исключением того, что клетки Baf3/KZ134/zcytor17m и BHK/KZ134/zcytor17m отбирали, используя помимо генетицина 2 мкг/мл пуромицина, в то время как клетки Baf3/KZ134/zcytor17m/OSMRbetam и BHK/KZ134/zcytor17m/OSMRbetam отбирали, используя, помимо генетицина, 2 мкг/мл пуромицина и 200 мкг/мл зеоцина.
Клоны разводили, культивировали и отбирали стандартными методами. Клоны исследовали при помощи люциферазного анализа (см. приведенный выше пример 20), используя среду, кондиционированную zcytor17lig мыши, или очищенный белок zcytor17lig мыши (пример 35) в качестве индуктора. Отбирали клоны с самой высокой реакцией на люциферазу (при использовании люциферазы STAT) и самым низким фоновым сигналом. Отбирали устойчивые линии клеток трансфектантов.
С. Активация лигандом zcytor17lig мыши рецептора zcytor17 мыши при выполнении люциферазного анализа с использованием клеток BaF3/KZ134/zcytor17m/OSMRbetam или BHK/KZ134/zcytor17m/OSMRbetam
Линии клеток культивировали для выполнения люциферазных анализов аналогично примеру 20. Активацию люциферазой STAT клеток BaF3/KZ134/zcytor17m, BaF3/KZ134/zcytor17m/OSMRbetam, BHK/KZ134/zcytor17m или BHK/KZ134/zcytor17m/OSMRbeta оценивали при использовании (1) кондиционированной среды, полученной из клеток ВНК570, трансфицированных zcytor17lig человека (пример 7), (2) кондиционированной среды, полученной из клеток ВНК570, трансфицированных zcytor17lig мыши (пример 18), (3) очищенного zcytor17lig мыши и человека (пример 35) и (4) среды без mIL-3 для измерения контрольной реакции, вызываемой только средой. Люциферазные анализы выполняли аналогично примеру 20.
Результаты данного анализа подтверждают наличие реакции, вызываемой репортерной STAT, в клетках BaF3/KZ134/zcytor17m/OSMRbetam и BHK/KZ134/zcytor17m/OSMRbetam на zcytor17lig мыши, по сравнению с клетками BaF3/KZ134/zcytor17m, BHK/KZ134/zcytor17m или нетрансфицированными контрольными клетками BaF3/KZ134 или BHK/KZ134 и показывают, что данная реакция опосредована рецепторами zcytor17/OSMRbeta мыши. Полученные результаты также показывают, что zcytor17lig человека не активирует клетки под действием репортерной STAT при использования комплекса рецепторов мыши.
Пример 39
Связывание лиганда zcytor17lig человека с рецепторами zcytor17 и zcytor17/OSMRbeta, измеренное методом проточной цитометрии
Zcytor17L человека биотинилировали следующим образом: 100 мкл zcytor17 в концентрации 5,26 мг/мл объединяли с 30 мкл 10 мг/мл сульфо-NHS-LC-биотина EZ-link (Pierce, Rockford, IL), растворенного в ddH2O. Реакционную смесь инкубировали в шейкере в течение 30 минут при комнатной температуре. После биотинилирования раствор диализовали в PBS, используя кассету для диализа Slide-A-Lyzer.
Для испытания связывания zcytor17lig человека с другими комбинациями рецепторов клетки BHK и BaF3 трансфицировали экспрессирующими плазмидами при помощи стандартных методов, хорошо известных в данной области. Указанные плазмиды трансфицировали в обе линии клеток в следующих комбинациях: только zcytor17, только OSMRbeta и zcytor17 вместе с OSMRbeta. Трансфекцию выполняли в соответствии с приведенным выше описанием. Нетрансфицированные клетки ВНК и BAF3 использовали в качестве контрольных клеток. Клетки окрашивали FACS следующим образом: клетки 2Е5 окрашивали 2,0 мкг/мл, 100 нг/мл, 10 нг/мл, 1,0 нг/мл, 100 пг/мл, 10 пг/мл, 1,0 пг/мл биотинилированного zcytor17L или оставляли неокрашенными в течение 30 минут на льду в буфере FACS (PBS + 2% BSA + 2% NHS (Gemini) + 2% NGS). Клетки промывали 1,5 раза и окрашивали SA-PE (Jackson Immuno Laboratories) при разведении 1:250 в течение 30 минут на льду. Затем клетки промывали 1,5 раза буфером FACS, вновь суспендировали в буфере FACS и анализировали с использованием буфера FACS в калибраторе BD FACS при помощи программного обеспечения CellQuest (Becton Dickinson, Mountain View, CA).
При исследовании клеток ВНК и BAF3 было установлено, что zcytor17lig связывается как с одним zcytor17, так и в комбинации с OSMRbeta, причем связывание с гетеродимером zcytor17/OSMRbeta является несколько сильнее. Связывание не было обнаружено в линиях клеток, экспрессирующих только OSMRbeta. Связывание zcytor17lig происходило в зависимости от концентрации. Средние значения интенсивности флуоресценции (MFI) для связывания с ВНК приведены в следующей таблице 15.
C17+OSMRbeta
Пример 40
Матричный анализ экспрессии генов в клетках, обработанных zcytor17lig человека
РНК выделяли из клеток А549, обработанных zcytor17lig человека, клеток SK-LU-I, обработанных zcytor17lig, и необработанных контрольных клеток при помощи набора RNeasy Midi (Qiagen, Valencia, CA), в соответствии с инструкциями изготовителя.
Экспрессию генов в клетках, обработанных zcytor17lig, и соответствующих контрольных клетках профилировали при помощи матриц экспрессии кДНК GEArray серии Q (SuperArray Inc., Bethesda, MD). Матрицы экспрессии кДНК серии Q содержат до 96 фрагментов кДНК, ассоциированных со специфическим биологическим путем или генами с подобными функциями или структурными признаками. Сравнение матриц обработанных и контрольных клеток позволяет определить увеличение и уменьшение специфических генов. Мечение зонда, гибридизацию и обнаружение выполняли в соответствии с инструкциями изготовителя. Операции, связанные с обнаружением хемилюминесцентного сигнала и сбором данных, выполняли в рабочей станции Lumi-Imager (Roche, Indianapolis, IN). Полученные графические данные анализировали при помощи программного обеспечения ImageQuant 5.2 (Amersham Biosciences, Inc., Piscataway, NJ) и GEArray Analyser 1.2 (SuperArray Inc., Bethesda, MD).
Анализ результатов, полученных при использовании матриц HS-014N серии Q интерлейкина и рецепторов человека, после нормализации показал примерно 4,7-кратное увеличение сигнала IL13RA2 в клетках SK-LU-1 человека, обработанных zcytor17lig, и примерно 2,2-кратное увеличение сигнала IL13RA2 в клетках А549 человека, обработанных zcytor17lig.
Полученные результаты показывают, что zcytor17lig значительно увеличивает концентрацию IL13RA2 в клетках SK-LU-1 и А549. Вышеуказанные клетки обоих типов являются линиями клеток, выделенных из карциномы легкого человека (Blobel et al., Virchows Arch B Cell Pathol Incl Mol Pathоl., 1984, 45(4):407-29). Более конкретно, клетки А549 можно охарактеризовать как линию эпителиальных клеток легкого человека (Lin, et al., J. Pharm. Pharmacol., 2002 Sep 54(9):1271-8; Martinez et al., Toxicol Sci., 2002 Oct, 69(2):409-23).
Установлено, что интерлейкин-13 (IL13), цитокин, секретируемый активированными Т-лимфоцитами, необходим для экспрессии аллергической астмы и для использования в экспериментальных моделях астмы, которые включают в себя повышенную чувствительность дыхательных путей, рекрутинг эозинофилов и избыточное продуцирование слизи (Wills-Karp et al., Science, 1998, 282:2258-2261). Установлено, что селективная нейтрализация IL13 ослабляет данный фенотип астмы (Grunig et al., Science, 1998, 282:2261-2263). Кроме того, известно, что IL13 увеличивает экспрессию гена муцина MUC8 в эпителии полипа в слизистой оболочке носа человека и вызывает разрастание эпителия в носу (Kimm et al., Acta Otolaryngol, 2002, Sep, 122(6):638-643; Seong et al., Acta Otolaryngol., 2002, Jun, 122(4):401-407). MUC8, главный гликопротеин муцина дыхательных путей, участвует в патогенезе, обусловленном гиперсекрецией слизи в случае хронического синусита, вызываемого полипами (Seong et al., Acta Otolaryngol., 2002, Jun, 122(4):401-407).
В функциональном отношении IL13 передает сигналы через рецепторный комплекс, включающий в себя цепь альфа-1-рецептора интерлейкина-13 (IL13RA1) и альфа-рецептор IL-4 (IL4RA) (Daines and Hershey, J. Biol. Chem., 2002, 22(12):10387-10393). Установлено, что альфа-2-рецептор интерлейкина-13 (IL13RA2) самостоятельно связывается с IL13 с высокой степенью сродства (Daines and Hershey, J. Biol. Chem., 2002, 22(12): 10387-10393). Однако у данного рецептора отсутствует цитоплазматический домен, необходимый для передачи сигналов, поэтому указанный рецептор считается рецептором-западней. Установлено, что IL13RA2 является преимущественно внутриклеточной молекулой, которая может быть быстро мобилизована из внутриклеточного запаса и экспрессирована на поверхности после обработки клетки гамма-интерфероном (IFNγ). Поверхностная экспрессия IL13RA2 после обработки гамма-интерфероном не вызывает синтеза белка и, как следствие этого, уменьшает передачу сигналов IL13 (Daines and Hershey, J. Biol. Chem., 2002, 22(12): 10387-10393).
Результаты анализа матрицы экспрессии генов для zcytor17lig показывают, что действие zcytor17lig является новым по сравнению с действием гамма-интерферона, так как воздействие zcytor7lig на линии клеток, выделенных из эпителия легкого, вызывает значительное увеличение экспрессии гена IL13RA2. Таким образом, воздействие, оказываемое zcytor17lig, может быть благоприятным в тех случаях, когда необходимо длительное увеличение экспрессии IL13RA2 и уменьшение IL13, как это имеет место в случае астмы, гиперактивности дыхательных путей (AHR) и регуляции муцина, включая хронический синусит, вызываемый полипами.
Пример 41
Трансгенные мыши, экспрессирующие zcytor17lig мыши
Для оценки воздействия сверхэкспрессии zcytor17lig in vivo были созданы трансгенные мыши, экспрессирующие мышиную форму гена, полученную при помощи двух разных промоторов: лимфоцитспецифического промотора Eµ/lck и неспецифического промотора EF1α (пример 22). Уровни сывороточного белка составляют примерно 20-300 нг/мл. При помощи промотора Eµ/lck были получены мыши с более высокими уровнями сывороточного белка по сравнению с аналогичными уровнями у трансгенных мышей, экспрессирующих EF1α-zcytor17lig.
У трансгенных мышей, экспрессирующих zcytor17lig, фенотип кожи развивался примерно в возрасте 4-8 недель. Шерсть трансгенных мышей становилась взъерошенной с выраженной пилоэрекцией, при этом наблюдалось выпадение шерсти от среднего до сильного, обычно на спине, боках и вокруг глаз. Данный фенотип был обнаружен у мышей с детектируемыми уровнями белка zcytor17lig в сыворотке. У мышей, экспрессирующих ген, выделенный из Eµ/lck, была отмечена 100% заболеваемость, и у трансгенных мышей, экспрессирующих EF1α-zcytor17lig, была отмечена 50% заболеваемость, что соответствует относительным уровням zcytor17lig, обнаруженным в их сыворотке. Кожа трансгенных мышей, по-видимому, зудела, о чем свидетельствует почесывание мышей, иногда настолько сильное, что оно сопровождалось царапинами и поражениями кожи, которые обычно инфицировались (по меньшей мере Staphуlococcus aureus). Мышей сначала идентифицировали по металлическим ушным меткам, но в большинстве случаев ушные метки были выцарапаны из ушей самими мышами. Такие действия часто сопровождались сильным поражением наружной части уха. Раненые уши часто плохо заживали, на что указывало наличие незаживающих гнойничков и струпьев, при этом у многих животных за ушами и между ушами образовались сочащиеся, постоянно увеличивающиеся раны. У некоторых трансгенных мышей образовались также покрытые струпьями раны на плечах и шее. В данной субпопуляции животных наблюдались поражения кожи, возникающие на участках кожи, где уже происходило выпадение шерсти, которые часто усиливались расчесыванием, производимом мышами.
Количественный анализ методом RT-PCR в реальном времени был использован для обнаружения транскриптов РНК zcytor17lig в образцах трансгенной кожи (но не в образцах нетрансгенной кожи), при этом было установлено что Eµ/lck-трансгенная кожа экспрессирует больше РНК zcytor17lig, чем кожа EF1α-zcytor17lig-трансгенных мышей. Гены, кодирующие субъединицы рецепторов zcytor17, в частности zcytor17 и OSMRbeta, были экспрессированы в коже как нетрансгенных, так и zcytor17lig-трансгенных мышей.
Исследование лимфоидных тканей, полученных из субпопуляции Eµ/lck-трансгенных животных, методом проточной цитометрии показало значительное увеличение концентрации активированных Т-клеток в селезенке и лимфатических узлах указанных мышей. У двух из четырех исследованных мышей были сильно увеличены лимфатические узлы в области шеи, возможно вследствие наличия поражений на шее. Наблюдалось едва заметное увеличение массы селезенки и незначительное увеличение моноцитов и нейтрофилов, циркулирующих в крови трансгенных мышей. Не было отмечено увеличения ряда исследованных цитокинов и изменений в уровнях амилоида А в сыворотке указанных мышей. Указанные воздействия на иммунные клетки у трансгенных мышей могут быть прямым или косвенным результатом действия zcytor17lig или могут представлять собой вторичные явления пораженной кожи.
Гистопатологию исследовали во многих тканях помимо кожи, включая печень, тимус, селезенку, почку и яичко, и в указанных органах не было обнаружено существенных отклонений. Однако анализ трансгенной кожи выявил ряд изменений, которые значительно варьируются в зависимости от источника и локализации кожи (например, нормальная, лишенная шерсти или патологически измененная). Во многих случаях уши трансгенных мышей имели утолщенный эпидермис по сравнению с нетрансгенными контрольными животными (например, примерно 4 слоя по сравнению с 2 слоями), и подстилающие ткани содержали воспалительные клетки от низкого до среднего количества, которые в основном были мононуклеарными при незначительном числе нейтрофилов. Эпидермис на животе во многих местах был немного толще у трансгенных животных, но в подстилающей дерме или подкожном слое не было обнаружено явного увеличения воспалительных клеток. На участках кожи указанных мышей, лишенных шерсти, были обнаружены расширенные волосяные фолликулы, содержащие дебрис при отсутствии волосяного стержня (например, волосы выпали вместе с корнем). На пораженных участках наблюдалось сильное утолщение эпидермиса (акантоз), повышенное содержание кератина на поверхности кожи (гиперкератоз), разбросанные язвы разного размера и значительное число воспалительных клеток в дерме (главным образом нейтрофилов при разном количестве макрофагов и лимфоцитов). Кроме того, дерма содержала большое число мастоцитов, окружающих пораженные участки. Некоторые волосяные стержни на пораженных участках трансгенной кожи находились в активной стадии (анаген), в отличие от многих волосяных стержней на «нормальных» участках, которые находились в инволюционной стадии (катаген) и неактивной стадии (телоген).
Фенотип трансгенных мышей, экспрессирующих zcytor17lig, сильно напоминает фенотип пациентов, страдающих атопическим дерматитом (AD), и животные модели AD с использованием мышей. Атопический дерматит является широко распространенным хроническим воспалительным заболеванием, которое характеризуется наличием гиперактивированных цитокинов субпопуляции 2 (Th2) Т-клеток-хелперов. Zcytor17lig экспрессируют в основном клетки Th2 по сравнению с клетками Th1, что делает данное сравнение еще более достоверным. Хотя точная этиология атопического дерматита неизвестна, данное заболевание обусловлено многими факторами, включая иммунные реакции на гиперактивные клетки Th2, аутоиммунитет, инфекцию, аллергены и генетическую предрасположенность. Главными признаками данного заболевания являются ксероз (сухость кожи), зуд (зудящее состояние кожи), конъюнктивит, воспалительные поражения кожи, инфицирование Staphylococcus aureus, повышенный эозинофильный лейкоцитоз, повышенное содержание IgE и IgG1 в сыворотке и хронический дерматит с инфильтрацией Т-клеток, мастоцитов, макрофагов и эозинофилов. Считается, что инфицирование S. aureus обостряет AD и делает хроническим данное кожное заболевание.
Атопический дерматит часто встречается у пациентов, страдающих астмой и аллергическим ринитом, и является первоначальным признаком аллергического заболевания. Примерно 20% населения в западных странах страдают указанными аллергическими заболеваниями, и заболеваемость AD в развитых странах возрастает по неизвестным причинам. Атопический дерматит обычно начинается в детстве и часто продолжается в подростковом и затем в зрелом возрасте. Существующие методы лечения атопического дерматита включают в себя местное применение кортикостероидов, пероральное введение циклоспорина А, некортикостероидных иммуносупрессоров, таких как такролимус (FK506 в виде мази), и гамма-интерферона. Несмотря на разнообразное лечение AD у многих пациентов симптомы заболевания не пропадают или возникает отрицательная реакция на лекарственные препараты, что требует применения других более эффективных лекарственных средств.
Эпителиальные клетки, экспрессирующие гетеродимерный рецептор для zcytor17lig (zcytor17 и OSMRbeta), расположены на участках (таких как кожа, кишка, легкое и т.д.) проникновения аллергена в организм и тесно взаимодействуют с дендритными клетками (профессиональные антигенпредставляющие клетки) in situ. Дендритные клетки играют важную роль в патогенезе аллергических заболеваний, и zcytor17lig может взаимодействовать со своим рецептором на эпителиальных клетках кожи и легкого и влиять на иммунные реакции в данных органах. Поэтому zcytor17lig и его рецепторы могут способствовать возникновению патогенеза аллергических заболеваний, таких как AD и астма. Кроме того, фенотип трансгенных мышей, экспрессирующих zcytor17lig, позволяет предположить, что данный лиганд может играть определенную роль в заживлении ран, так как у указанных мышей не заживали повреждения ушей и часто возникали незаживающие поражения на спине и боках. Поэтому антагонист zcytor17lig может быть эффективным лекарственным средством для лечения вышеуказанных и других заболеваний.
Пример 42
Люциферазный анализ линий трансформированных эпителиальных клеток человека при временном инфицировании аденовирусным геном-репортером STAT/SRE
Несколько разных линий трансформированных эпителиальных клеток человека (см. приведенную ниже таблицу 16) высевали на 96-луночные планшеты с плоским основанием в количестве 10000 клеток/лунку в обычной питательной среде, определенной для каждого типа клеток. На следующий день клетки инфицировали репортерной конструкцией аденовируса KZ136 с множественностью заражения, равной 5000. Репортер KZ136 содержал элементы STAT помимо элемента, вызывающего реакцию сыворотки. Общий объем составлял 100 мкл/лунку при использовании среды DMEM, содержащей 2 мМ L-глутамина (GibcoBRL), 1 мМ пирувата натрия (GibcoBRL) и однократного объема добавки, включающей в себя инсулин-трансферрин-селен (GibcoBRL) (далее данная среда именуется бессывороточной средой). Клетки культивировали в течение ночи.
На следующий день среду удаляли и заменяли 100 мкл индуцирующей среды. В качестве индуцирующей среды использовали zcytor17lig человека, разведенный в бессывороточной среде в концентрации, составляющей 100 нг/мл, 50 нг/мл, 25 нг/мл, 12,5 нг/мл, 6,25 нг/мл, 3,125 нг/мл и 1,56 нг/мл. Положительную контрольную среду, содержащую 20% FBS, использовали для подтверждения достоверности анализа и гарантии успешного инфицирования аденовирусом. Клетки индуцировали в течение 5 часов, после чего среду отсасывали. Затем клетки промывали в 50 мкл/лунку PBS и лизировали в 30 мкл/лунку однократного объема буфера для лизиса клеток (Promega). После 10-минутной инкубации при комнатной температуре 25 мкл/лунку лизата переносили на матовые 96-луночные планшеты. Затем планшеты считывали в люминометре с 5-секундной интеграцией при инъецировании 40 мкл/лунку люциферазного субстрата (Promega).
Полученные результаты показали способность многих линий эпителиальных клеток реагировать на zcytor17lig, как это показано в следующей таблице 16.
Пример 43
Продуцирование цитокинов линиями эпителиальных клеток человека, культивируемыми с zcytor17lig человека
Пораженные болезнью линии эпителиальных клеток человека (А549, эпителиально-клеточный рак легкого человека; SkLu1, эпителиально-клеточная аденокарцинома легкого человека; DU145, эпителиально-клеточный рак предстательной железы человека; PZ-HPV-7, HPV-трансформированный эпителий предстательной железы человека; U20S, эпителиально-клеточная остеосаркома кости человека) исследовали на продуцирование цитокинов под действием zcytor17lig in vitro. Указанные клетки имеют рецепторы zcytor17 и OSMRbeta, идентифицированные методом RT-PCR, и реагируют на zcytor17lig человека при выполнении анализа с помощью люциферазной репортерной конструкции на основе аденовируса, KZ136 (пример 42). Продуцирование цитокинов указанными линиями клеток под действием zcytor17lig человека определяли в серии из трех экспериментов.
А. Продуцирование цитокинов пораженными болезнью линиями эпителиальных клеток человека, культивируемыми с zcytor17lig человека
Клетки помещали с плотностью 4,5 х 105 клеток/лунку на 6-луноч-ный планшет (Costar) и культивировали в соответствующей питательной среде. Клетки культивировали с испытуемыми реагентами; 100 нг/мл zcytor17lig, 10 нг/мл гамма-интерферона (IFNγ) (R&D Systems, Minneapolis, MN), 10 нг/мл альфа-фактора некроза опухоли (TNF-альфа) (R&D Systems, Minneapolis, MN), 10 нг/мл IL-1beta (R&D Systems, Minneapolis, MN) или 100 мкг/мл липополисахарида (LPS) (Sigma). Супернатанты собирали через 24 и 48 часов и анализировали на наличие цитокинов; GM-CSF (колониестимулирующий фактор гранулоцитов-макрофагов), IL-1b, IL-6, IL-8, MCP-1 (белок-1 хемоаттрактанта макрофагов) и TNFa. Для измерения числа цитокинов в образцах использовали наборы шариков, покрытых антителом, Multiplex компании BioSource International (Camarillo, CA). Анализы выполняли в аппарате Luminex-100 (Luminex, Austin, TX), и полученные данные анализировали при помощи программного обеспечения MasterPlex (MiraiBio, Alameda, CA). Результаты продуцирования цитокина (пг/мл) для каждой линии клеток в образцах, культивируемых в течение 24 часов, представлены в следующей таблице 17.
пг/мл
Все испытуемые линии клеток продуцировали GM-CSF и IL-8 в ответ на стимуляцию контрольными цитокинами IL-1b и TNFa. Большинство линий клеток продуцировали IL-6 и МСР-1 в ответ на стимуляцию IL-1b и TNFa. Zcytor17lig стимулировал продуцирование IL-6 в линии клеток DU145 по сравнению с контрольным образцом (193 пг/мл по сравнению с 71 пг/мл). 3 из 5 линий клеток, стимулированных zcytor17lig, продуцировали IL-8, причем наибольший эффект имел место в клетках А549 (5-кратное увеличение) и двукратное сокращение продуцирования IL-8 наблюдалось в клетках U2OS. Незначительное изменение продуцирования МСР-1 происходило в клетках DU145 и U2OS, культивируемых вместе с zcytor17lig.
В. Продуцирование цитокинов нормальными линиями эпителиальных клеток человека, культивируемыми с zcytor17lig человека
Кроме линий эпителиальных клеток человека испытывали также нормальные эпителиальные клетки бронхов человека (NHBE, Clonetics). Клетки помещали с плотностью 1 х 105 клеток/лунку на 24-луночный планшет и культивировали с испытуемыми реагентами; 1000 нг/мл, 100 нг/мл и 10 нг/мл zcytor17lig (A760F), 10 нг/мл TNFa, 10 нг/мл OSM, 10 нг/мл IFNa, 10 нг/мл TGFb или 10 нг/мл лимфотактина. Супернатанты собирали через 24 и 48 часов и анализировали на наличие цитокинов IL-6, IL-8, MCP-1, MIP-1a, RANTES и эотаксина. Цитокины анализировали описанным выше способом. Результаты продуцирования цитокинов (пг/мл) для всех линий клеток в виде образцов, культивированных в течение 48 часов, приведены в следующей таблице 18.
Клетки DU145 продуцировали IL-6 под воздействием zcytor17lig с таким же результатом, как в примере 43А. Однако только клетки А549 и U2OS продуцировали IL-8 так же, как в примере 43А. Клетки SkLu1 и U2OS продуцировали МСР-1 под воздействием zcytor17lig. Продуцирование цитокинов клетками NHBE было предельным по сравнению с контрольными образцами.
С. Продуцирование цитокинов пораженными болезнью линиями эпителиальных клеток человека, культивируемыми вместе с zcytor17lig человека и гамма-интерфероном
Клетки помещали с плотностью 2 х 105 клеток/лунку на 24-луноч-ный планшет и культивировали вместе с 10 нг/мл гамма-интерферона ± zcytor17lig в количестве 100 нг/мл, 10 нг/мл или 1 нг/мл. Супернатанты собирали через 24 и 48 часов и анализировали на наличие IL-8 и МСР-1 вышеописанным способом. Результаты продуцирования цитокинов (пг/мл) для каждой линии клеток в виде образцов, культивированных в течение 24 часов, показаны в нижеследующей таблице 19.
10 нг/мл IFNg + 10 нг/мл r17L
10 нг/мл IFNg + 1 нг/мл r17L
10 нг/мл IFNg
контрольный образец
75,1
63,6
35,4
36,6
не обнаружен
не обнаружен
не обнаружен
не обнаружен
10 нг/мл IFNg + 10 нг/мл r17L
10 нг/мл IFNg + 1 нг/мл r17L
10 нг/мл IFNg
контрольный образец
92,9
79,9
70,7
79,4
не обнаружен
не обнаружен
не обнаружен
не обнаружен
10 нг/мл IFNg + 10 нг/мл r17L
10 нг/мл IFNg + 1 нг/мл r17L
10 нг/мл IFNg
контрольный образец
194,4
138,7
170,8
203,0
870,7
585,4
652,6
292,3
10 нг/мл IFNg + 10 нг/мл r17L
10 нг/мл IFNg + 1 нг/мл r17L
10 нг/мл IFNg
контрольный образец
108,2
109,9
118,8
146,8
347,7
293,3
159,8
7,0
Клетки А549 продуцировали IL-8 под действием zcytor17lig, однако совместное культивирование клеток с добавлением гамма-интерферона не вызывало никакого эффекта. В клетках U2OS наблюдалось 20-кратное увеличение продуцирования МСР-1 при культивировании с IFNg и 50-кратное увеличение продуцирования МСР-1 при культивировании с гамма-интерфероном + zcytor17lig.
Пример 44
Влияние zcytor17lig на введение 3 Н-TdR в клетки DU145 эпителиально-клеточного рака предстательной железы
Клетки высевали на 96-луночные планшеты для тканевых культур (Falcon) с плотностью 25000 клеток/лунку, в питательной среде МЕМ (Life Technologies), содержащей глутамин, пируват, заменимые аминокислоты (Life Technologies) и 10% фетальной телячьей сыворотки (Hyclone). После слияния клеток (через 24 часа) вместо питательной среды использовали среду для прекращения роста, заменяя сыворотку 0,1% BSA (Life Technologies). Через 48 часов для достижения синхронизации клеток среду для прекращения роста заменяли свежей средой. Затем добавляли рекомбинантный zcytor17lig (испытуемый реагент) человека в разных концентрациях (от 0,24 до 60 нг/мл) (см. приведенную ниже таблицу 16) для исследования влияния белка на репликацию основной ДНК. В некоторые лунки помимо zcytor17lig добавляли 2,5% FBS (Hyclone) для исследования влияния белка на повышенные уровни введения TdR. В качестве положительного контрольного образца использовали 10% FBS и фактор роста ВВ, выделенный из 20 нг/мл тромбоцитов (PDGF-BB) (R&D).
Через восемнадцать часов после добавления zcytor17lig и остальных испытуемых реагентов клетки подвергали импульсному мечению 250 нCi/мл [3H]-тимидина (NEN) в течение 4 часов. После 4-часового импульсного мечения среду декантировали и в каждую лунку добавляли 100 мкл раствора трипсина (Life Technologies) для отсоединения клеток. Для определения радиоактивности клеток DU145 указанные клетки собирали при помощи 196-ячеечного харвестера Packard Filtermate и подсчитывали число включенных меток в сцинтилляционном счетчике для микропланшетов Packard TopCount NXT.
Как показано в нижеследующей таблице 20, zcytor17lig индуцировал введение тимидина в покоящиеся клетки (в 0,1% BSA) в зависимости от концентрации. Достигаемый эффект был в 2,5 раза выше по сравнению с контрольным образцом BSA при самой высокой концентрации, равной 60 нг/мл. Кроме того, воздействие zcytor17lig выражалось в повышении уровня при добавлении 2,5% FBS (которая в данном эксперименте была таким же сильным митогеном, как и 10% FBS). Полученные результаты показывают, что как в исходных, так и стимулированных условиях zcytor17lig может воздействовать на раковые клетки DU145 в качестве митогенного фактора.
В таблице 16 показано воздействие zcytor17lig на включение тимидина в клетки DU145. Результаты выражены в виде числа импульсов/мин/лунку и представляют собой среднее значение ± стандартное отклонение для трех одинаковых лунок.
Пример 45
Экспрессия huzcytor17lig в E. coli
А. Создание вектора pRPS01, экспрессирующего слитый полипептид huzcytor17lig/MBP-6H
Экспрессирующая плазмида, содержащая полинуклеотид, кодирующий huzcytor17lig, слитый у С-конца со связывающим мальтозу белком (МВР), была создана методом гомологичной рекомбинации. N-конец слитого полипептида включает в себя часть МВР, состоящую примерно из 388 аминокислот, которая слита с huzcytor17lig, рассмотренным в данном описании. Фрагмент кДНК huzcytor17lig выделяли вышеописанным методом PCR. Для получения фрагмента zcytor17lig в результате выполнения стандартной реакции PCR использовали две затравки: (1) одна затравка содержала 40 п.н. фланкирующей последовательности вектора и 20 п.н., соответствующих аминоконцу huzcytor17lig, и (2) вторая затравка содержала у 3'-конца 40 п.н., соответствующих фланкирующей последовательности вектора, и 20 п.н., соответствующих карбоксильному концу huzcytor17lig. Два микролитра из 100 мкл реакционной смеси PCR разделяли электрофорезом в 1,0% агарозном геле с использованием однократного объема буфера ТВЕ для анализа, при этом был получен фрагмент с ожидаемой молекулярной массой. Остальную реакционную смесь PCR объединяли с содержимым второй пробирки для РCR и осаждали 400 мкл абсолютного этанола. Осажденную ДНК использовали для рекомбинации в SmaI-разрезанном векторе рТАР98 реципиента с образованием конструкции, кодирующей слитый полипептид MBP-huzcytor17lig, как описано ниже.
Вектор pTAP98 был создан путем гомологичной рекомбинации дрожжей. Сто нанограммов EcoR1-разрезанной pMAL-c2, рекомбинировали, используя 1 мкг Pvu1-разрезанной pRS316, 1 мкг линкера и 1 мкг Sca1/EcoR1-разрезанной pRS316, которые объединяли в реакционной смеси для PCR. Продукты PCR концентрировали, осаждая 100% этанолом. Штамм компетентных дрожжевых клеток (S. cerevisiae) SF838-9Dα объединяли с 10 мкл смеси, содержащей примерно 1 мкг продукта huzcytor17lig, полученного методом PCR (см. выше), и 100 нг SmaI-гидролизованного вектора рТАР98, и электропорировали при 0,75 кВ, 25 мкФ и ∞ Ом. Полученную реакционную смесь помещали на чашки URA-D и инкубировали при 30°C.
Через 48 часов отбирали трансформанты дрожжей Ura+ из одной чашки. ДНК выделяли и вводили в электрокомпетентные клетки E. coli (например, МС1061, Casadaban et al., J. Mol. Biol. 138, 179-207). Полученные клетки E. coli культивировали стандартными методами на чашках со средой ММ/СА, содержащей 100 мг/л ампициллина (Pryor and Leiting, Protein Expression and Purification 10:309-319, 1997). Четыре отдельных клона собирали с чашек и инокулировали в среде ММ/CА со 100 мкг/мл ампициллина в течение двух часов при 37°C. Один миллилитр каждой культуры индуцировали 1 мМ IPTG. Примерно через 2-4 часа 250 мкл каждой индуцированной культуры смешивали с 250 мкл стеклянных шариков, промытых кислотой, и 250 мкл буфера Торнера с 5% βМЕ и красителя (8 М мочевины, 100 мМ трис-буфера, рН 7,0, 10% глицерина, 2 мМ EDTA, 5% SDS). Образцы интенсивно перемешивали в течение одной минуты и нагревали до 65°C в течение 10 минут. Двадцать микролитров каждого образца загружали в одну полосу 4%-12% геля для PAGE (NOVEX). Гели исследовали в однократном объеме буфера MES. В положительных клонах, получивших название pRPS01, выполняли анализ последовательностей.
Один микролитр секвенирующей ДНК использовали для трансформации штамма электрокомпетентных клеток E. coli MC1061. Клетки электропорировали при 2,0 кВ, 25 мкФ и 400 Ом. После электропорации клетки спасали 0,6 мл SOC и выращивали на чашках со средой LB, содержащей 100 мг/л ампициллина, при 37°C в течение ночи. Четыре культуры индуцировали ITPG и исследовали на наличие положительных клонов описанным выше способом. Положительные клоны размножали для очистки слитого белка huzcytor17lig/MBP-6H стандартными методами.
В. Очистка huzcytor17lig/MBP-6H в результате ферментации E. coli
За исключением особо оговоренных случаев, все операции выполняли при 4°C. Очистку рекомбинантного полипептида huzcytor17lig/MBP-6H производили следующим способом. Клетки E. coli, содержащие конструкцию pRPS01 и экспрессирующие huzcytor17lig/MBP-6H, получали стандартными методами молекулярной биологии и культивировали в 50,0 г/л бульона SuperВroth II (12 г/л казеина, 24 г/л дрожжевого экстракта, 11,4 г/л дикалийфосфата, 1,7 г/л монокалийфосфата (Becton Dickenson, Cockeysville, MD), 5 г/л глицерина и 5 мл/л 1 М сульфата магния. Двадцать граммов клеток собирали и замораживали для очистки белка.
Оттаявшие клетки вновь суспендировали в 500 мл буфера для уравновешивания амилозы (20 мМ трис-буфера, 100 мМ NaCl, рН 8,0). Клетки лизировали при помощи системы разрушения клеток French Press (Constant Systems Ltd., Warwick, UK) при температуре от -7°C до -10°C и давлении 43 атм (30000 фунтов/кв.дюйм). Разрушение вновь суспендированных клеток проверяли на основании показаний А600 до и после циклизации в системе French Press. Суспензию лизированных клеток осаждали центрифугированием с ускорением 10000 х g в течение 30 минут для удаления клеточного дебриса, и супернатант собирали для очистки белка.
25 мл амилозной смолы (New England Biolabs, Beverly, MA) (полученной описанным ниже способом) выливали в стеклянную колонку Bio-Rad с диаметром 2,5 см и высотой 10 см. Колонку заполняли и уравновешивали под действием силы тяжести 10 объемами колонки (CV) буфера для уравновешивания амилозы. Супернатант обработанных клеток порциями загружали в амилозную смолу и выдерживали в течение ночи при покачивании. Смолу возвращали в колонку Bio-Rad и промывали 10 объемами буфера для уравновешивания амилозы под действием силы тяжести. Колонку элюировали ~2 объемами буфера для элюирования амилозы (буфер для уравновешивания амилозы + 10 мМ мальтозы, Fluka Biochemical, Switzerland) под действием силы тяжести. Десять 5 мл фракций собирали в соответствии с профилем элюирования и анализировали оптическую плотность при 280 и 320 нМ. Амилозную смолу регенерировали 1 объемом дистиллированной Н2О, 5 объемами 0,1% (мас./об.) SDS (Sigma), 5 объемами дистиллированной Н2О, 5 объемами буфера для уравновешивания амилозы и наконец 1 объемом буфера для хранения амилозы (буфер для уравновешивания амилозы + 0,02% азида натрия). Регенерированную смолу хранили при 4°C.
Представляющие интерес фракции профиля элюирования собирали и диализовали в камере для диализа 10К (Slide-A-Lyzer, Pierce Immunochemical) против 4 х 4 л PBS, рН 7,4 (Sigma) в течение 8 часов для удаления низкомолекулярных загрязняющих примесей, замены буфера и обессоливания. После диализа собранный материал представлял собой очищенный полипептид huzcytor17/MBP-6H. Очищенный полипептид huzcytor17/MBP-6H стерилизовали фильтрацией и анализировали методом SDS-PAGE с окрашиванием кумассии для продукта с соответствующей молекулярной массой. Концентрация полипептида huzcytor17lig/MBP-6H, определенная при помощи анализа ВСА, составляла 1,28 мг/мл.
Пример 46
Поликлональное антитело против zcytor17lig человека
А. Получение и очистка
Поликлональные антитела были получены в результате иммунизации двух самок новозеландского белого кролика очищенным рекомбинантным белком hzcytor17L/MBP-6H (пример 45). Кроликам сначала внутрибрюшинно (IP) инъецировали 200 мкг очищенного белка в полном адъюванте Фрейнда и затем внутрибрюшинно инъецировали бустер-дозы 100 мкг белка в неполном адъюванте Фрейнда каждые три недели. Через семь-десять дней после инъецирования второй бустер-дозы (всего 3 инъекции) у животных брали кровь и собирали сыворотку. Затем животным вводили бустер-дозу и брали кровь каждые три недели.
Hzcytor17L/MBP-6H-специфическую сыворотку кролика предварительно очищали от антител против МВР в колонке для очистки белка CNBr-SЕPНAROSE 4B (Pharmacia LKB), которая была получена с использованием 10 мг неспецифического очищенного рекомбинантного МВР-слитого белка на грамм CNBr-SEPHAROSE. Нzcytor17L/MBP-6H-специфические поликлональные антитела очищали аффинным методом от предварительно адсорбированной сыворотки кролика в колонке для очистки белка CNBr-SEPHAROSE 4B (Pharmacia LKB), которая была получена с использованием 10 мг специфического очищенного от антигена рекомбинантного белка hzcytor17L/MBP-6H. После очистки поликлональные антитела диализовали, четырежды заменяя PBS, соответствующий 20-кратному объему антитела, в течение по меньшей мере 8 часов. Hzcytor17lig-специфические антитела исследовали методом ELISA, используя 500 нг/мл очищенного рекомбинантного белка hzcytor17L/MBP-6H или hzcytor17L-CEE, продуцированного в экспрессирующей системе бакуловируса, в качестве мишени для антитела. Нижний предел обнаружения (LLD) антитела кролика против hzcytor17L/MBP-6H, очищенного аффинным методом, составлял 100 пг/мл в отношении специфического очищенного рекомбинантного антигена hzcytor17L/MBP-6H и 500 пг/г на очищенном рекомбинатном zcytor17L-CEE, продуцированном в экспрессирующей системе бакуловируса.
В. Анализ антитела кролика против zcytor17lig/MBP-6H человека методами SDS-PAGE и вестерн-блоттинга
Антитело кролика против zcytor17lig/MBP-6H человека исследовали методом SDS-PAGE (NuPage 4-12%, Invitrogen, Carlsbad, CA) при окрашивании кумассии и методом вестерн-блоттинга с использованием антитела козы против IgG-HRP кролика. Очищенный белок zcytor17lig человека и мыши (200-25 нг) разделяли электрофорезом в миниячейке Xcell II Invitrogen Novex и переносили на нитроцеллюлозу (0,2 мм; Invitrogen, Carlsbad, CA) при комнатной температуре и перемешивании, используя блот-модуль Xcell Novex, в соответствии с инструкциями, содержащимися в руководстве по эксплуатации прибора. Перенос осуществляли при 300 мА в течение одного часа в буфере, содержащем 25 мМ основного трис-буфера, 200 мМ глицина и 20% метанола. Затем фильтр блокировали буфером для вестерн-блоттинга А (собственного изготовления, 50 мМ трис-буфера, рН 7,4, 5 мМ EDTA, рН 8,0, 0,05% Igepal CA-630, 150 мМ NaCl и 0,25% желатина) при осторожном покачивании в течение ночи при 4°C. Нитроцеллюлозу быстро промывали и добавляли антитело кролика против zcytor17lig/MBP-6H человека (1:1000) в буфере для вестерн-блоттинга А. Блот инкубировали, осторожно покачивая, в течение 1,5 часов при комнатной температуре. Блот 3 раза промывали в буфере для вестерн-блоттинга А в течение 5 минут и добавляли антитело козы против IgG HRP кролика (1:5000) в буфере для вестерн-блоттинга А. Блот инкубировали, осторожно покачивая, в течение 1 часа при комнатной температуре. Блот 3 раза промывали в буфере для вестерн-блоттинга А в течение 5 минут, затем быстро промывали в Н2О. Блот проявляли при помощи коммерчески производимых реагентов для хемилюминесцентного субстрата (детектирующие реагенты для вестерн-блоттинга ECL 1 и 2, смешанные в соотношении 1:1, компании Amersham Pharmacia Biotech, Вuckinghamshire, England), после чего блот экспонировали на рентгеновскую пленку в течение периода времени до 5 минут.
Очищенный zcytor17lig человека был получен в виде большой полосы длиной около 30 кДа и меньшей полосы длиной около 20 кДа в восстановительных условиях. Zсytor17lig мыши не был обнаружен при помощи антитела кролика против zcytor17lig человека.
Пример 47
Влияние zcytor17lig на адгезию моноцитов U937 к монослою трансформированных эндотелиальных клеток костного мозга (TRBMEC)
Трансформированные эндотелиальные клетки костного мозга (TRBMEC) высевали на 96-луночные планшеты для тканевой культуры (Falcon) с плотностью 25000 клеток/лунку в среде М131 (Cascade Biologics), содержащей добавку для роста микрососудов (MVGS) (Cascade Biologics). После слияния клеток (через 24 часа) вышеуказанную среду заменяли средой М199 (Gibco-Life Technologies), содержащей 1% фетальной телячьей сыворотки (Hyclone). Рекомбинантный zcytor17lig человека (испытуемый реагент) добавляли в разных концентрациях (от 0,4 до 10 нг/мл) (cм. приведенную ниже таблицу 21) для определения влияния данного белка на взаимодействие иммунных и эндотелиальных клеток, вызывающее адгезию. В некоторые лунки помимо zcytor17lig добавляли 0,3 нг/мл фактора некроза опухоли (TNFальфа, R&D Systems), известного провоспалительного цитокина, для определения влияния белка на эндотелиальные клетки в воспалительных условиях. TNFальфа в количестве 0,3 нг/мл использовали в качестве положительного контрольного образца, при этом концентрация соответствовала примерно 70% максимальному действию TNFальфа в данной системе, то есть она не вызывала максимального сцепления клеток U937 (линия моноцитоподобных клеток человека) с эндотелием. По этой причине данная установка позволяет обнаружить как увеличение, так и уменьшение воздействия TNFальфа. Исходные уровни адгезии при использовании TNFальфа и без него служили в качестве начала отсчета для оценки влияния испытуемых реагентов.
Эндотелиальные клетки инкубировали в течение ночи с испытуемыми реагентами (zcytor17lig ± TNFальфа), затем клетки U937 окрашивали 5 мкМ флуоресцентного маркера Calcein-AM (Molecular Probes), клетки суспендировали в среде RPMI 1640 (без фенолового красного), содержащей 1% FBS, и культивировали в количестве 100000 клеток/лунку на промытом монослое TRBMEC. Уровни флуоресценции при длинах волн возбуждения/излучения, равных 485/538 нм (аппарат для считывания микропланшетов Molecular Devices, CytoFluor application), измеряли через 30 минут до и после трехкратной промывки лунок теплой средой RPMI 1640 (без фенолового красного) для удаления несцепленных клеток U937. Уровни флуоресценции, соответствующие предварительной промывке (общая) и последующей промывке (адгезионно-специфическая), использовали для определения процентного значения адгезии (истинная адгезия/общая адгезия х 100 = % адгезии).
Как видно из приведенной ниже таблицы 21, добавляемый отдельно zcytor17lig влиял на исходную адгезию клеток U937 с монослоями эндотелиальных клеток в диапазоне используемых концентраций (увеличение меньше чем в 2 раза, р<0,01 при испытании методом ANOVA). Используемый отдельно положительный контрольный образец TNFальфа в количестве 0,3 нг/мл увеличивал адзегию клеток U937 от исходного уровня 5,8%, до 35% (6-кратное увеличение). В присутствии TNFальфа zcytor17lig оказывал синергичное действие и еще больше увеличивал адгезию клеток U937 в зависимости от концентрации в интервале от 0,4 до 10 нг/мл (р<0,01 при испытании методом ANOVA). При концентрации 10 нг/мл zcytor17lig усиливал действие TNFальфа на 62%. Полученные результаты показывают, что zcytor17lig сам по себе является провоспалительным агентом. Zcytor17lig способен оказывать синергичное действие при концентрациях TNFальфа ниже максимальных, усиливающее адгезию моноцитов к эндотелиальным клеткам. Полученные результаты также показывают, что эндотелиальные клетки, особенно под воздействием провоспалительных цитокинов, таких как TNFальфа, по-видимому являются тканью-мишенью для действия zcytor17lig. Следствием воздействия zcytor17lig на эндотелиальные клетки может быть усиление адгезии моноцитов или макрофагов к участку, провоспалительной активности. Активированные моноциты и макрофаги имеют важное значение при многих воспалительных заболеваниях. Поэтому ингибирование адгезии моноцитов/макрофагов может быть терапевтически обоснованным для антагонистов zcytor17lig. Указанные данные подтверждают возможность применения антагонистов лиганда zcytor17 для лечения легочных заболеваний, сосудистых заболеваний, аутоиммунитета, метастазов опухоли, заболеваний, вызванных аллергическими реакциями, для заживления ран и заболеваний кожи, включая контактный, аллергический или неаллергический дерматит или псориаз, и воспалительного заболевания кишечника. В таблице 21 показано воздействие zcytor17lig на адгезию моноцитов U937 к монослоям эндотелиальных клеток TRBMEC. Результаты выражены в процентных значениях адгезии, и приведенные величины представляют собой среднее значение ± стандартное отклонение для трех одинаковых лунок.
Из вышеизложенного должно быть понятно, что несмотря на то, что в данном описании представлены конкретные варианты осуществления изобретения в целях иллюстрации, возможны различные модификации изобретения, не выходящие за пределы объема изобретения. Поэтому данное изобретение ограничено только прилагаемой формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ ЦИТОКИН ZALPHA11-ЛИГАНД | 2000 |
|
RU2258710C2 |
| ИНТЕРФЕРОН-ПОДОБНЫЙ БЕЛОК ZCYTO21 | 2001 |
|
RU2292394C2 |
| ГОМОГЕННЫЕ ПРЕПАРАТЫ IL-28 И IL-29 | 2004 |
|
RU2518324C2 |
| ПОЛИПЕПТИД С АНТИВИРУСНОЙ АКТИВНОСТЬЮ, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2004 |
|
RU2372356C2 |
| ВЫДЕЛЕННЫЙ РАСТВОРИМЫЙ IL-20 РЕЦЕПТОР (ВАРИАНТЫ) | 2000 |
|
RU2279441C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-31 И СПОСОБЫ ПРИМЕНЕНИЯ | 2006 |
|
RU2444528C2 |
| СПОСОБ И НАБОР ДЛЯ ВЫЯВЛЕНИЯ ГЕНА, КОДИРУЮЩЕГО МЕМБРАНОСВЯЗАННЫЙ БЕЛОК, ВЕКТОР (ВАРИАНТЫ) | 1999 |
|
RU2246538C2 |
| ГЕТЕРОДИМЕРНЫЕ ПОЛИПЕПТИДЫ IL-17 A/F И ВОЗМОЖНОСТИ ИХ ЛЕЧЕБНОГО ПРИМЕНЕНИЯ | 2007 |
|
RU2440134C2 |
| ТРОМБОПОЭТИН | 1994 |
|
RU2245365C2 |
| ГЕМОПОЭТИЧЕСКИЙ БЕЛОК (ВАРИАНТЫ), ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, СПОСОБ ПОЛУЧЕНИЯ ГЕМОПОЭТИЧЕСКОГО БЕЛКА, СПОСОБ СЕЛЕКТИВНОЙ ЭКСПАНСИИ КРОВЕТВОРНЫХ КЛЕТОК, СПОСОБ ПРОДУЦИРОВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СПОСОБ СТИМУЛЯЦИИ ПРОДУЦИРОВАНИЯ КРОВЕТВОРНЫХ КЛЕТОК У ПАЦИЕНТА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБ ЛЕЧЕНИЯ | 1997 |
|
RU2245887C2 |


























































































































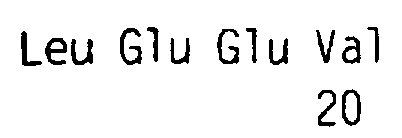




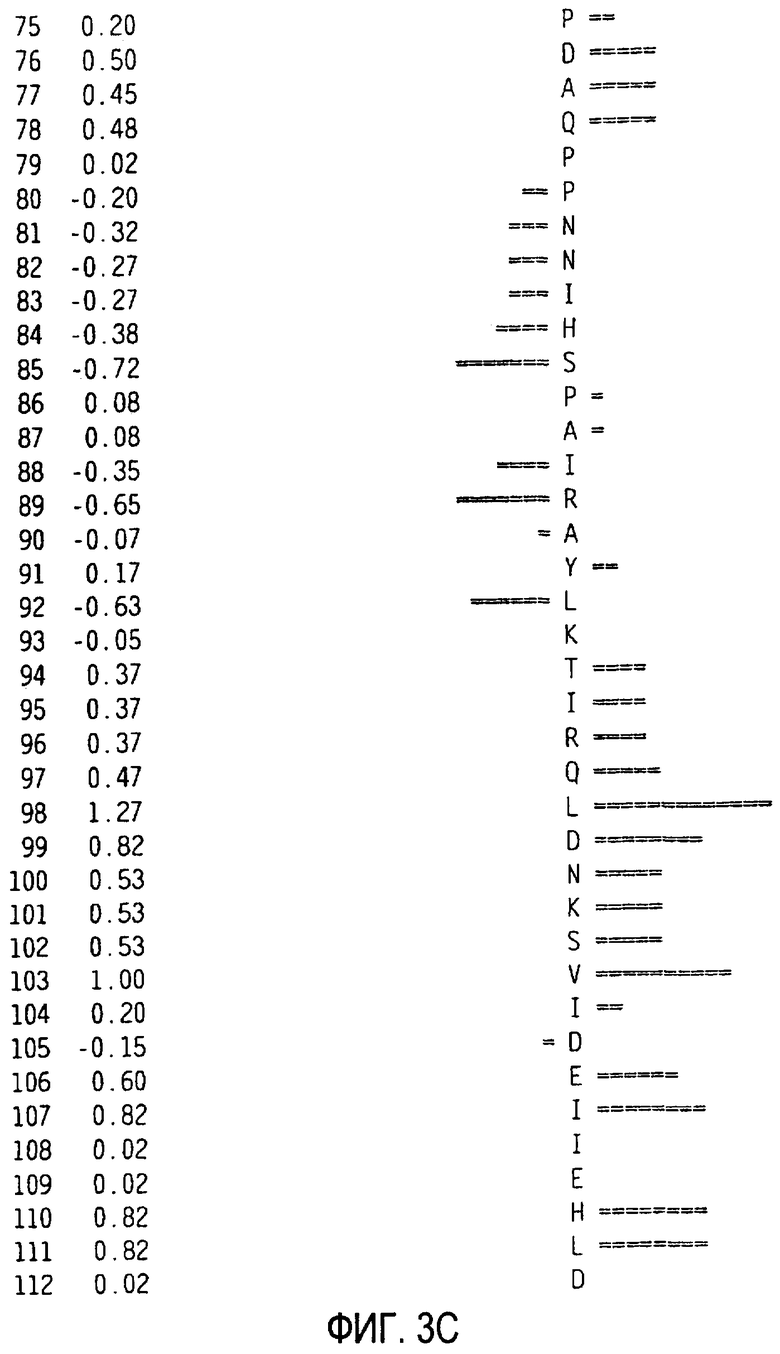

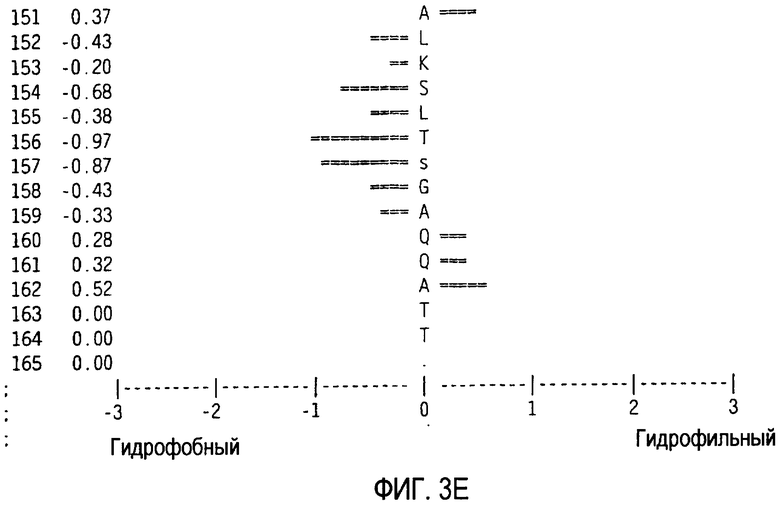
Настоящее изобретение относится к области биотехнологии. Предложен способ лечения млекопитающего, страдающего воспалительным заболеванием, где воспалительное заболевание представляет собой атопический дерматит, экзему или зуд. Способ включает введение млекопитающему выделенного моноклонального антитела или его антиген-связывающего фрагмента, которые получают посредством инокуляции не являющегося человеком животного адъювантом и полипептидом (zcytor17lig), включающим аминокислотные остатки 27-164 SEQ ID NО:2, с последующим выделением указанного антитела из указанного животного. Использование изобретения может найти применение в медицине для лечения атопического дерматита, экземы или зуда. 1 з.п. ф-лы, 7 ил., 21 табл.
1. Способ лечения млекопитающего, страдающего воспалительным заболеванием, который включает введение млекопитающему выделенного моноклонального антитела или его антиген-связывающего фрагмента, которые получают посредством инокуляции не являющегося человеком животного адъювантом и полипептидом, включающим аминокислотные остатки 27-164 SEQ ID NО:2, с последующим выделением указанного антитела из указанного животного, где воспалительное заболевание представляет собой атопический дерматит, экзему или зуд.
2. Способ по п.1, где моноклональное антитело или его антиген-связывающий фрагмент являются гуманизированными.
| WO 2002000721 A2 , 03.01.2002 | |||
| ПРЕПАРАТ С ИММУНОМОДУЛИРУЮЩИМИ СВОЙСТВАМИ | 1995 |
|
RU2102988C1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| VENTER et al | |||
| "The Sequence of the Human Genome Science", 2001, Vol | |||
| СТЕРЕООЧКИ | 1920 |
|
SU291A1 |
| Прибор для записи числа оборотов реверсивных машин | 1926 |
|
SU5507A1 |
| Предохранитель от вывинчивания электрических ламп накаливания | 1924 |
|
SU1304A1 |

