ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
«Перечень последовательностей», поданный одновременно с данным документом в электронном виде в соответствии с § 1821 раздела 37 Свода законов США (C.F.R.) в машиночитаемой форме (CRF) с помощью системы EFS-Web в виде файла под именем 200606_415WO_SEQUENCE_LISTING.txt, включен в данный документ в качестве ссылки. Электронная копия Перечня последовательностей была создана 16 января 2015 года, размер файла 851 килобайт.
ОБЛАСТЬ ТЕХНИКИ
Данное описание относится к новым микроорганизмам для повышенного продуцирования аминокислот и кормовым продуктам, полученным из рекомбинантных метаболизирующих C1 микроорганизмов, содержащих генетически модифицированный метаболический путь для повышенного продуцирования аминокислот, и к связанным с ними композициям и способам.
УРОВЕНЬ ТЕХНИКИ
Прогресс в эффективности использования животного корма за последние несколько десятилетий был достигнут благодаря использованию кормовых добавок. Такие добавляемые вещества повышают содержание питательных веществ, энергетическую ценность композиций животного корма и/или их способность бороться с болезнями. Растущим вызовом для коммерческих производителей животноводческой продукции является увеличивающаяся стоимость зерна. Растущая стоимость частично вызвана конкурентным спросом на зерно для производства биотоплива и продуктов питания. С увеличением стоимости зерна и белковых компонентов, в сочетании с ограниченной доступностью земель для производства продуктов питания, альтернативные дешевые кормовые продукты для животных с полезными питательными свойствами и способностью повышать сопротивляемость болезням становятся весьма желательными.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В первом варианте реализации настоящее изобретение обеспечивает рекомбинантный метаболизирующий С1 микроорганизм, содержащий первую экзогенную нуклеиновую кислоту, выбранную из группы, состоящей из экзогенной нуклеиновой кислоты, кодирующей фермент биосинтеза L-аминокислоты, и экзогенной нуклеиновой кислоты, кодирующей контрольную последовательность экспрессии, функционально связанную с нуклеиновой кислотой, кодирующей нативный фермент биосинтеза L-аминокислоты, где рекомбинантный метаболизирующий С1 микроорганизм способен конвертировать углеродное сырье, получаемое из природного газа, в требуемую L-аминокислоту, и где δ13С рекомбинантного метаболизирующего C1 микроорганизма имеет значение менее -30%o.
В другом варианте реализации рекомбинантный метаболизирующий C1 микроорганизм по настоящему изобретению дополнительно содержит вторую экзогенную нуклеиновую кислоту, кодирующую cache-полипептид. В дополнительных вариантах реализации настоящее изобретение обеспечивает биомассу, получаемую из культуры рекомбинантного метаболизирующего С1 микроорганизма по настоящему изобретению, и композицию, содержащую биомассу по настоящему изобретению. В другом варианте реализации настоящее изобретение обеспечивает композицию аминокислот, содержащую одну или несколько аминокислот, экстрагированных из биомассы по настоящему изобретению.
В дополнительном варианте реализации настоящее изобретение обеспечивает животный корм, содержащий биомассу или композицию аминокислот по настоящему изобретению. В некоторых вариантах реализации животный корм дополнительно содержит добавку, выбранную из группы, состоящей из материала растительного происхождения, материала животного происхождения, и материала, полученного из микроорганизмов.
В дополнительных вариантах реализации настоящее изобретение обеспечивает способ продуцирования требуемой L-аминокислоты, включающий культивацию рекомбинантного метаболизирующего C1 микроорганизма по настоящему изобретению в присутствии углеродного сырья, получаемого из природного газа, в условиях, достаточных для продуцирования требуемой L-аминокислоты. В некоторых вариантах реализации L-аминокислота демонстрирует значение δ13С менее -30%o.
В дополнительном варианте реализации настоящее изобретение обеспечивает способ продуцирования биомассы, обогащенной L-аминокислотой, включающий культивацию рекомбинантного метаболизирующего С1 микроорганизма по настоящему изобретению в питательной среде в присутствии углеродного сырья, получаемого из природного газа, в условиях, достаточных для промотирования роста рекомбинантного метаболизирующего С1 микроорганизма с получением биомассы.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 представляет собой графическое изображение уровня продуцирования внеклеточного L-лизина в Metylococcus capsulatus штамм Bath, который сверхэкспрессирует индивидуальные гены из оперона биосинтеза лизина, как описано в Примере 4. Для каждого штамма представлена концентрация внеклеточного L-лизина, детектируемая в клеточном супернатанте штаммов, гетерологически экспрессирующих указанный ген, деленная на оптическую плотность (OD600нм) культуры. Генотипы, обозначенные номерами 34 и 1067, представляют собой контрольную пустую плазмиду и контроль, экспрессирующий посторонний ген (флуоресцентный белок) в контексте того же промотора, соответственно. График показывает, что сверхэкспрессия указанных генов приводит к повышению внеклеточного L-лизина по сравнению с контрольной пустой плазмидой (генотип номер 34) и контроль, экспрессирующий посторонний ген (зеленый флуоресцентный белок).
Фигура 2 представляет собой графическое изображение концентрации клеточно-ассоциированного L-лизина, детектируемого в образцах клеточной биомассы штаммов, гетерологически экспрессирующих разные кодон-оптимизированные последовательности, каждая из которых кодирует один и тот же cache-белок (полипептид с высоким содержанием лизина), как описано в Примере 5. Генотип номер 34 представляет собой контрольную пустую плазмиду. Столбики отображают усредненные значения для восьми клонов каждого экспериментального генотипа.
Фигура 3 представляет собой графическое изображение уровня клеточно-ассоциированного L-лизина, определяемого в пересчете на абсолютно сухую (dry weight) биомассу культур штаммов, гетерологически экспрессирующих cache-полипептид с повышенным содержанием лизина (HighK) и dapA, как описано в Примере 6. Столбики отображают усредненные значения для восьми клонов каждого экспериментального генотипа. Все гены HighK в экспериментальных штаммах 1186-1193 кодируют одну и ту же аминокислотную последовательность, но имеют разное использование кодонов. График показывает, что коэкспрессия этих генов приводит к диапазону значений повышенного продуцирования клеточно-ассоциированного L-лизина.
Фигура 4 представляет собой графическое изображение уровня внеклеточного L-лизина, детектируемого в супернатанте культуры образцы штаммов, гетерологически экспрессирующие cache-полипептид с повышенным содержанием лизина (HighK) и dapA, как описано в Примере 6. Столбики отображают усредненные значения для восьми клонов каждого экспериментального генотипа. График показывает, что экспрессия комбинации генов приводит к (росту) внеклеточного L-лизина от 8- до 23-кратного по сравнению с контролем (генотип номер 1067), при расчете по средним значениям для каждого набора клонов.
ПОДРОБНОЕ ОПИСАНИЕ
Данное описание обеспечивает новые рекомбинантные метаболизирующие C1 микроорганизмы, обладающие способностью утилизировать относительно дешевое углеродное сырье в качестве источника энергии и, дополнительно, обеспечивает требуемые связанные с ними биомассу, питательные продукты, композиции. Рекомбинантные микроорганизмы по настоящему изобретению генетически модифицированы для повышенного продуцирования L-аминокислот, являющихся коммерчески требуемыми. Такие рекомбинантные микроорганизмы, а также получаемые из них биомасса и композиции L-аминокислот, пригодны для использования в качестве источника корма для животных (таких как, например, домашний скот, рыба, домашняя птица и т.п.), а также культивируемые или ферментируемые микроорганизмы.
В одном варианте реализации настоящее изобретение обеспечивает рекомбинантный метаболизирующий C1 микроорганизм, причем рекомбинантный метаболизирующий C1 микроорганизм содержит экзогенную нуклеиновую кислоту, выбранную из группы, состоящей из экзогенной нуклеиновой кислоты, кодирующей фермент биосинтеза L-аминокислоты, и экзогенной нуклеиновой кислоты, кодирующей контрольную последовательность экспрессии, функционально связанную с нуклеиновой кислотой, кодирующей нативный фермент биосинтеза L-аминокислоты, при том, что рекомбинантный метаболизирующий С1 микроорганизм способен конвертировать углеродное сырье природного газа в требуемую L-аминокислоту, и где рекомбинантный метаболизирующий C1 микроорганизм демонстрирует δ13С менее -30%o, и часто менее -35%o. Типично, рекомбинантный метаболизирующий С1 микроорганизм представляет собой нефотосинтезирующий метаболизирующий C1 микроорганизм, и более типично, представляет собой метанотрофный микроорганизм.
В таких вариантах реализации рекомбинантные микроорганизмы по настоящему изобретению генетически модифицированы для превращения получаемого из природного газа сырья, представляющего собой относительно дешевый и обильный ресурс (например, природный газ, или субстрат C1, выделенный из природного газа, такой как, например, метан) по сравнению с более дорогими углеводами, в более ценные питательные вещества. В используемом в данном документе значении, термин «получаемое из природного газа сырье» относится к природному газу или любым компонентам, выделенным из природного газа (включая субстраты С1) или полученным путем конверсии природного газа (например, сингаз).
Термин «природный газ» относится в данном документе к встречающимся в природе газовым смесям, которые могут быть получены обычными способами (например, бурение и заводнение поровых коллекторов) или нетрадиционными способами (например, гидроразрыв, горизонтальное бурение или направленное бурение формаций, имеющих низкую газопроницаемость). Газовые смеси состоят из метана и других соединений, включая другие соединения С1, а также другие газообразные легкие алканы (такие как, например, этан, пропан, бутан, пентан и т.п.), двуокись углерода, азот, сероводород и т.п., и их комбинации. Нетрадиционный природный газ может быть получен из таких источников, как, например, плотные газовые пески, образующиеся в песчанике или карбонате, метан угольных пластов, образующийся в угольных месторождениях и адсорбированный частицами угля, сланцевый газ, образующийся в тонкозернистых сланцевых породах и адсорбированный частицами глины или удерживаемый в мелких порах или микротрещинах, метангидраты, представляющие собой кристаллические комбинации природного газа и воды, образующиеся при низкой температуре и высоком давление в таких местах, как глубины океанов и вечная мерзлота.
В используемом в данном документе значении, термин «субстрат C1» или «соединение C1» относится к любой углерод содержащей молекуле или композиции, не имеющей углерод-углеродной связи. Типичные примеры субстратов C1 включают сингаз, метан, метанол, формальдегид, муравьиную кислоту или ее соль, окись углерода, двуокись углерода, метилированные амины (например, метиламин, диметиламин, триметиламин и т.д.), метилированные тиолы, метилгалогениды (например, бромметан, хлорметан, йодметан, дихлорметан и т.д.), цианид или любую их комбинацию.
В определенных вариантах реализации настоящего изобретения, получаемое из природного газа сырья часто является природным газом, субстратом С1 из природного газа, или представляет собой сингаз. Типично субстрат C1 представляет собой метан. Рекомбинантные метаболизирующие C1 микроорганизмы по изобретению, использующие в качестве сырья полученный из природного газа углеродный субстрат, демонстрируют характерную изотопную подпись углерода, которая описана более детально ниже. Эта характерная изотопная подпись углерода также наблюдается у продуктов таких рекомбинантных микроорганизмов (например, биомасса, L-аминокислота, композиции L-аминокислот и т.п.).
В используемом в данном документе значении, термины «метаболизирующий C1 микроорганизм» или «метаболизирующий C1 нефотосинтезирующий микроорганизм» относятся к любому микроорганизму, обладающему способностью использовать субстрат C1 в качестве источника энергии или в качестве своего основного источника энергии и биомассы, и может использовать или не использовать другие углеродные субстраты (такие как сахара и сложные углеводы) для получения энергии и биомассы. Например, метаболизирующий C1 микроорганизм может окислять субстрат С1, такой как метан или метанол. Метаболизирующие С1 микроорганизмы включают бактерии (такие как метанотрофы и метилотрофы) и дрожжи. В определенных вариантах реализации метаболизирующий С1 микроорганизм не включает фотосинтезирующие микроорганизмы, такие как водоросли. В некоторых вариантах реализации метаболизирующий С1 микроорганизм будет «облигатным метаболизирующим C1 микроорганизмом», что означает, что его единственным источником энергии являются субстраты С1. В дополнительных вариантах реализации метаболизирующий C1 микроорганизм (например, метанотроф) будет культивироваться в присутствии сырьевого субстрата С1 (т.е. с использованием субстрата C1 в качестве источника энергии).
Рекомбинантные метаболизирующие C1 микроорганизмы по настоящему изобретению генетически модифицированы для повышенного продуцирования требуемой L-аминокислоты путем модификации нуклеиновой кислоты, кодирующей фермент биосинтеза L-аминокислоты (АВ), или ее экспрессии. Термины «фермент биосинтеза L-аминокислоты» и «АВ-фермент» используются в данном документе взаимозаменяемо по отношению к ферменту, принимающему участие в продуцировании L-аминокислоты рекомбинантным метаболизирующим C1 микроорганизмом-хозяином.
В одном варианте реализации настоящее изобретение обеспечивает рекомбинантный метаболизирующий C1 микроорганизм, содержащий первую экзогенную нуклеиновую кислоту, выбранную из группы, состоящей из экзогенной нуклеиновой кислоты, кодирующей фермент биосинтеза L-аминокислоты, и экзогенной нуклеиновой кислоты, кодирующей контрольную последовательность экспрессии, функционально связанную с нуклеиновой кислотой, кодирующей нативный фермент биосинтеза L-аминокислоты. Типично, δ13С рекомбинантного метаболизирующего C1 микроорганизма имеет значение менее -30%o, как описано более детально ниже. Обычно рекомбинантный метаболизирующий С1 микроорганизм представляет собой метанотрофный микроорганизм.
В некоторых вариантах реализации рекомбинантный метаболизирующий C1 микроорганизм по настоящему изобретению дополнительно содержит вторую экзогенную нуклеиновую кислоту, кодирующую cache-полипептид. В используемом в данном документе значении, термин «cache-полипептид» относится к полипептиду, имеющему аминокислотную последовательность, которая: (1) имеет длину по меньшей мере около 10 аминокислотных остатков; и (2) содержит L-аминокислоту, содержание которой должно быть увеличено на уровне, составляющем по меньшей мере 10% (по видам) от общего количества аминокислотных остатков в аминокислотной последовательности. Типично, cache-полипептид не является ферментом АВ (биосинтеза аминокислоты).
Cache-полипептиды пригодны для обеспечения повышенного продуцирования требуемых L-аминокислот в рекомбинантных метаболизирующих С1 микроорганизмах по настоящему изобретению, как продемонстрировано в примерах. В некоторых вариантах реализации аминокислотная последовательность cache-полипептида может (1) иметь длину по меньшей мере около 10 амино(кислотных) остатков, по меньшей мере около 15, по меньшей мере около 20, по меньшей мере около 30, по меньшей мере около 40, по меньшей мере около 50, по меньшей мере около 60, по меньшей мере около 70, по меньшей мере около 80, по меньшей мере около 90, по меньшей мере около 100, по меньшей мере около 150, по меньшей мере около 200, по меньшей мере около 250, по меньшей мере около 300, по меньшей мере около 350, по меньшей мере около 400, по меньшей мере около 450 или по меньшей мере около 500 аминокислотных остатков; и (2) включать L-аминокислоту, содержание которой должно быть увеличено на уровне, составляющем по меньшей мере около 10% (по видам) от общего количества аминокислотных остатков в аминокислотной последовательности, или уровне, составляющем по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 13%, по меньшей мере около 14%, по меньшей мере около 15%, по меньшей мере около 16%, по меньшей мере около 17%, по меньшей мере около 18%, по меньшей мере около 19%, по меньшей мере около 20%, по меньшей мере около 21%, по меньшей мере около 22%, по меньшей мере около 23%, по меньшей мере около 24%, по меньшей мере около 25%, по меньшей мере около 26%, по меньшей мере около 27%, по меньшей мере около 28%, по меньшей мере около 29%, по меньшей мере около 30%, по меньшей мере около 31%, по меньшей мере около 32%, по меньшей мере около 33%, по меньшей мере около 34%, по меньшей мере около 35%, по меньшей мере около 36%, по меньшей мере около 37%, по меньшей мере около 38%, по меньшей мере около 39%, по меньшей мере около 40%, по меньшей мере около 41%, по меньшей мере около 42%, по меньшей мере около 43%, по меньшей мере около 44%, по меньшей мере около 45%, по меньшей мере около 46%, по меньшей мере около 47%, по меньшей мере около 48%, по меньшей мере около 49%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, по меньшей мере около 75%, по меньшей мере около 80%, по меньшей мере около 85%, по меньшей мере около 90% или по меньшей мере около 95%, от общего количества аминокислотных остатков в аминокислотной последовательности. Cache-полипептид, используемый в практике настоящего изобретения, может быть природным полипептидом или неприродным полипептидом. Вторая экзогенная нуклеиновая кислота, кодирующая cache-полипептид, типично является кодон-оптимизированной для оптимальной экспрессии рекомбинантным метаболизирующим C1 микроорганизмом-хозяином. Типичным примером cache-полипептида является пептидаза семейства S9A (SEQ ID NO: 152), как показано в Примерах 5 и 6. Последовательности нуклеиновой кислоты, кодирующие пептидазу семейства S9A, которая была оптимизирована по использованию кодонов для оптимальной экспрессии в Methylococcus capsulatus Bath, представлены как SEQ ID NO: 152, 154, 156, 158, 160, 162, 164 и 166. В используемом в данном документе значении, термины «полипептид» и «белок» используются взаимозаменяемо по отношению к полимеру аминокислотных остатков.
Рядовой специалист в данной области техники может легко определить cache-полипептиды, пригодные для использования в настоящем изобретении, путем проведения поиска белков, в которых по меньшей мере 10% от общего содержания аминокислот приходится на требуемую L-аминокислоту. Это может быть сделано, например, путем направления запроса в базу данных последовательностей, такую как, например, GENBANK (интернет-адрес ncbi.nlm.nih.gov/genbank/) по всем белкам организма, представляющего интерес, и затем расчета отношения [число вхождений требуемой аминокислоты] / [длина белковой последовательности] для каждого белка. Результат этого анализа может быть подвергнут фильтрации и сортировке по величине отношения для определения пригодных белков.
Экзогенные нуклеиновые кислоты, кодирующие АВ-ферменты и cache-полипептиды, используемые в практике настоящего изобретения, типично являются кодон-оптимизированными для оптимальной экспрессии рекомбинантным метаболизирующим С1 микроорганизмом-хозяином. Они могут кодировать фермент или белок, которые или являются нативными для вида, гетерологичного по отношению к C1 микроорганизму-хозяину, или кодировать мутант (т.е. вариант) полипептида, существующего в природе. В тех случаях, когда первая экзогенная нуклеиновая кислота кодирует АВ-фермент, экзогенная нуклеиновая кислота может быть нуклеиновой кислотой, кодирующей АВ-ферменты, принимающие участие в биосинтезе L-аминокислоты, выбранной из группы, состоящей из L-лизина, L-триптофана, L-метионина, L-цистеина, L-треонина, L-гистидина, L-изолейцина, L-лейцина, L-фенилаланина, L-валина, L-аланина, L-аргинина, L-аспарагина, L-аргининовой кислоты, L-глутаминовой кислоты, L-глутамина, L-глицина, L-орнитина, L-пролина, L-серина и L-тирозина. В некоторых вариантах реализации L-аминокислоту выбирают из группы, состоящей из L-лизина, L-триптофана, L-метионина, L-цистеина, L-треонина и т.п. В некоторых случаях L-аминокислоту выбирают из группы, состоящей из L-лизина, L-триптофана, L-метионина, L-цистеина и L-треонина. Пригодные экзогенные нуклеиновые кислоты включают кодирующие фермент биосинтеза L-аминокислоты, выбранный из группы, состоящей из:
(i) фермента биосинтеза L-лизин, такого как, например: лизин-чувствительная аспартокиназа III (lysC), аспартаткиназа, аспартат-семиальдегид-дегидрогеназа (asd), дигидродипиколинатсинтаза (dapA), дигидродипиколинатредуктаза (dapB), 2,3,4,5-тетрагидропиридин-2,6-карбоксилат-N-сукцинилтрансфераза (dapD), ацетилорнитин/сукцинилдиаминопимелатаминотрансфераза (argD), сукцинил-диаминопимелат-десукцинилаза (dapE), сукцинилдиаминопимелаттрансаминаза, диаминопимелатэпимераза (dapF), диаминопимелатдикарбоксилаза (lysA), и т.п.;
(ii) фермента биосинтеза L-триптофана, такого как, например, хоризмат-пируват-лиаза (ubiC), антранилатсинтазы компонент I (trpE), антранилатсинтазы компонент II (frpG), антранилат-фосфорибозилтрансфераза (trpD), фосфорибозилантранилат-изомераза (trpC), белок биосинтеза триптофана (trpC), N-(5'-фосфорибозил)антранилат-изомераза (trpF), индол-3-глицеролфосфатсинтаза, триптофансинтазы альфа-цепь (trpA), триптофансинтазы бета-цепь (trpB), и т.п.;
(iii) фермента биосинтеза L-метионина, такого как, например, гомосерин-О-сукцинилтрансфераза (metA), цистатионин-гамма-синтаза (metB), белок MalY, цистатионин бета-лиаза (metC), В12-зависимая метионинсинтаза (metH), 5-метилтетрагидроптероилтриглутамат-гомоцистеин-S-метилтрансфераза (metE), и т.п.;
(iv) фермента биосинтеза L-цистеина, где фермент биосинтеза цистеина выбирают из группы, состоящей из серин-ацетилтрансферазы (CysE), цистеинсинтазы А, цистеинсинтазы В, и т.п.; и
(v) фермента биосинтеза L-треонина, такого как, например, аспартаттрансаминаза, PLP-зависимая аминотрансфераза, аспартатаминотрансфераза, аспартаткиназа, аспартат-семиальдегид-дегидрогеназа, гомосериндегидрогеназа, гомосеринкиназа, треонинсинтаза, и т.п.
Вышеперечисленные ферменты могут присутствовать в ряде гетерологичных видов, включая микроорганизмы, такие как, например, Е. coli и Corynebacterium glutamicum. В определенных конкретных вариантах реализации экзогенная нуклеиновая кислота кодирует фермент биосинтеза L-аминокислоты, имеющий последовательность по любому из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 88, 90, 92, 94, 96, 98, 100, 102, 104, 106, 108, 110, 112, 114, 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148 или 150, приведенных в Таблице А ниже. Как было описано выше, экзогенная нуклеиновая кислота типично является кодон-оптимизированной для оптимальной экспрессии в рекомбинантном метаболизирующем С1 микроорганизме. Типичные примеры последовательностей нуклеиновой кислоты, кодирующих такие АВ-ферменты, также представлены в Таблице А. Эти последовательности нуклеиновой кислоты были оптимизированы по использованию кодонов для экспрессии в Methylococcus capsulatus Bath.
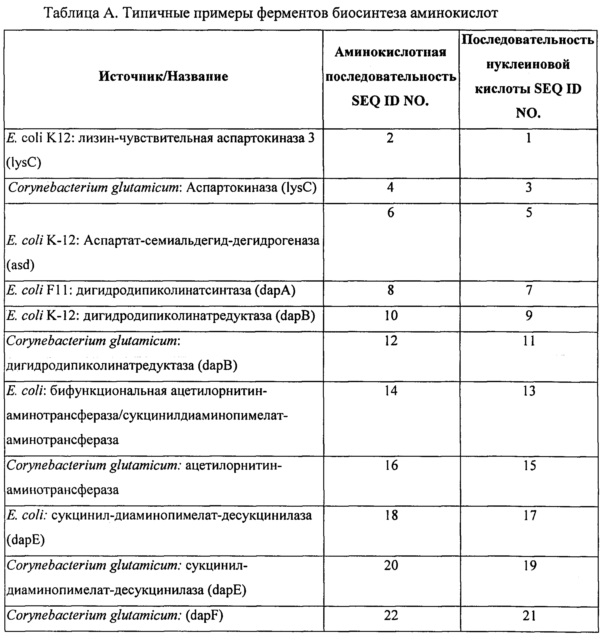
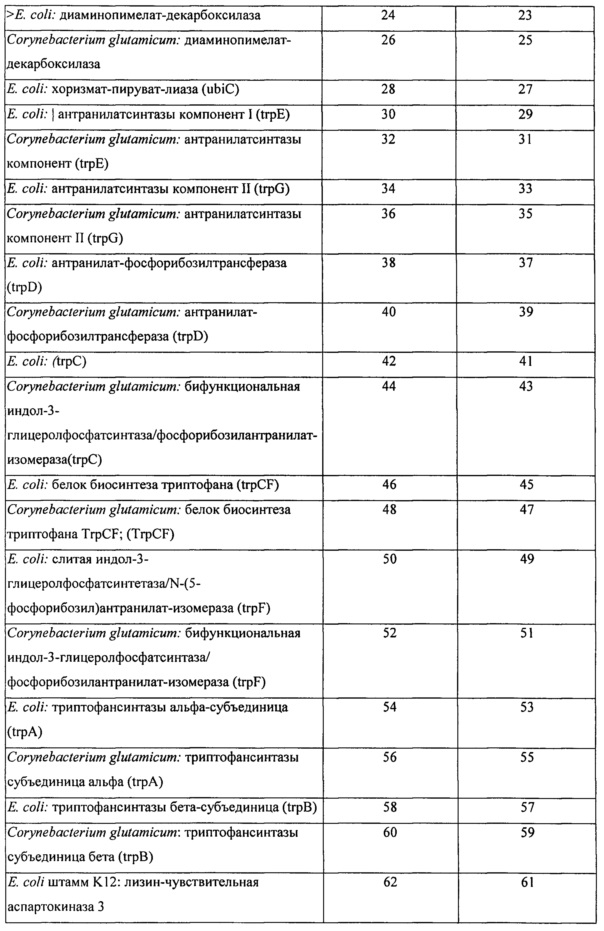
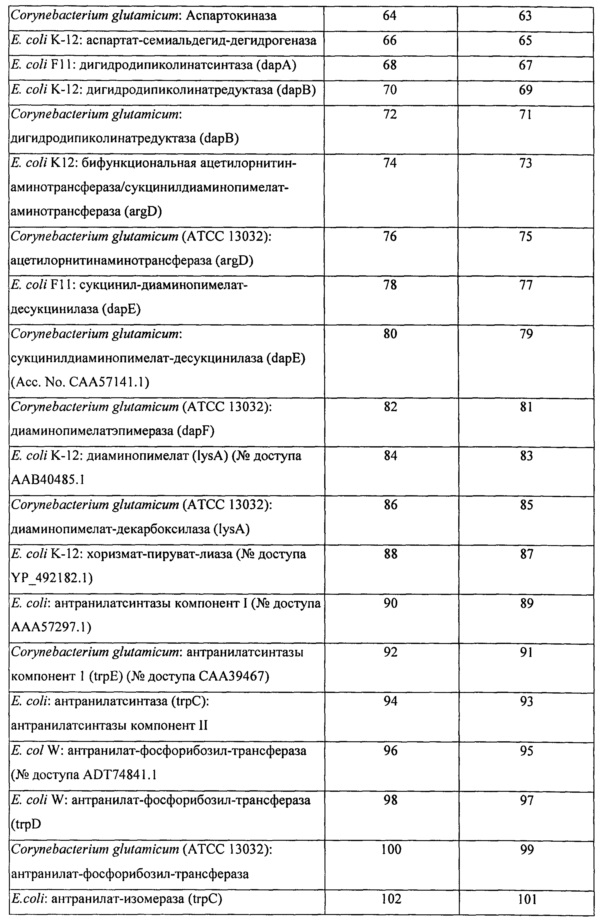
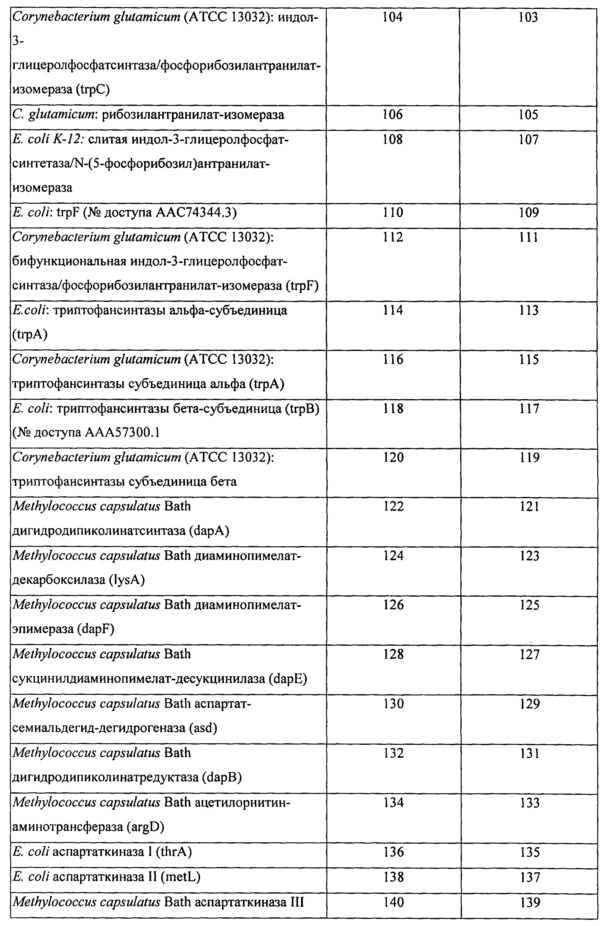

Пригодные экзогенные нуклеиновые кислоты, используемые в практике настоящего изобретения, включают кодирующие вариант последовательности АВ-фермента, который по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичен референсной полипептидной последовательности или исходной полипептидной последовательности дикого типа, такой как, например, референсная последовательность, соответствующая любому из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 88, 90, 92, 94, 96, 98, 100, 102, 104, 106, 108, 110, 112, 114, 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148 или 150, при условии, что вариант сохраняет активность фермента биосинтеза L-аминокислоты, представляющую интерес. В определенных вариантах реализации варианты полипептидов АВ-фермента будут включать по меньшей мере одно замещение аминокислоты (например, 1, 2, 3, 5, 6, 7, 8, 9 или 10 или больше или до 20, 25 или 30 замещений) в предварительно определенном положении по сравнению с референсным АВ-ферментом или АВ-ферментом исходного дикого типа, при условии, что вариант сохраняет активность АВ-фермента, представляющую интерес. Вариант полипептида АВ-фермента может дополнительно содержать одно или несколько консервативных замещений. «Консервативное замещение» понимается в данной области техники как замещение одной аминокислоты на другую аминокислоту, обладающую схожими свойствами. Типичные примеры консервативных замещений хорошо известны специалистам (см., например, WO 97/09433, р. 10; Lehninger, Biochemistry, 2nd Edition; Worth Publishers, Inc. NY:NY (1975), pp,71-77; Lewin, Genes IV, Oxford University Press, NY and Cell Press, Cambridge, MA (1990), p. 8, которые включены в данный документ в качестве ссылок).
Способы получения пригодных экзогенных нуклеиновых кислот, кодирующих такие варианты ферментов, описаны более детально ниже.
«Процент идентичности» между двумя или больше последовательностями нуклеиновых кислот или аминокислотными последовательностями является функцией числа идентичных позиций, общих для последовательностей (т.е. % идентичности = число идентичных позиций/общее число позиций × 100), с учетом числа пробелов, и длины каждого пробела, необходимого для оптимизации выравнивания двух или больше последовательностей. Сравнение последовательностей и определение процента идентичности между двумя или больше последовательностями может быть осуществлено с использованием математического алгоритма, такого как программы BLAST и Gapped BLAST с параметрами по умолчанию (например, Altschul et al., J. Mol. Biol. 275:403, 1990; см. также BLASTN по интернет-адресу ncbi.nlm.nih.gov/BLAST, которые включены в данный документ в качестве ссылки).
Как указано выше, экзогенные нуклеиновые кислоты, используемые в практике настоящего изобретения, могут быть оптимизированы по использованию кодонов для экспрессии в метаболизирующем C1 микроорганизме. Экспрессия рекомбинантных белков может быть затруднена вне их исходного хозяина. Например, наблюдались вариации в смещении использования кодонов для разных видов бактерий (Sharp et al., Nucl. Acids. Res. 33:1141, 2005, который включен в данный документ в качестве ссылки). Сверхэкспрессия рекомбинантных белков даже в их нативном хозяине может также быть затруднена. В определенных вариантах реализации нуклеиновая кислота, предназначенная для введения в хозяина, как описано в данном документе, может быть подвергнута оптимизации кодонов перед введением в хозяина для обеспечения эффективной или повышенной экспрессии белка. Оптимизация кодонов относится к изменению кодонов в генах или кодирующих участках нуклеиновых кислот перед трансформацией для соответствия типичному использованию кодонов в хозяине без изменения полипептида, кодируемого неприродной молекулой ДНК. Способы оптимизации кодонов для оптимальной генной экспрессии в гетерологичных хозяевах были описаны ранее (см., например, Welch et al., PLoS One 4:e7002, 2009; Gustafsson et al., Trends Biotechnol. 22:346, 2004; Wu et al., Nucl. Acids Res. 35:D76, 2007; Villalobos et al., BMC Bioinformatics 7:285, 2006; патентные публикации США №№2011/0111413 и 2008/0292918; раскрытие указанных способов включено в данный документ в качестве ссылки, в полном объеме). Экзогенные нуклеиновые кислоты, кодирующие АВ-ферменты, пригодные для использования в практике настоящего изобретения, включают имеющие последовательность нуклеиновой кислоты, по меньшей мере на около 85% идентичную референсной последовательности нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87, 89, 91, 93, 95, 97, 99, 101, 103, 105, 107, 109, 111, 113, 115, 117, 119, 121, 123, 125, 127, 129, 131, 133, 135, 137, 139, 141, 143, 145, 147 и 149. Иллюстративные экзогенные нуклеиновые кислоты, кодирующие фермент СВ, пригодный для использования в практике изобретения, включают последовательности, которые были оптимизированы по использованию кодонов для оптимальной экспрессии в Methylococcus capsulatus Bath, такие как, например, любая из SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87, 89, 91, 93, 95, 97, 99, 101, 103, 105, 107, 109, 111, 113, 115, 117, 119, 121, 123, 125, 127, 129, 131, 133, 135, 137, 139, 141, 143, 145, 147 и 149.
Аналогично, молекулы экзогенной нуклеиновой кислоты в соответствии с настоящим описанием, кодирующие варианты полипептидов, могут быть сконструированы с использованием филогенетических способов, описанных в указанных выше ссылках (патент США №8005620; Gustafsson et al.; Welch et al.; Villalobos et al.; Minshull et al., все из которых включены в данный документ в качестве ссылок).
Экзогенная нуклеиновая кислота, кодирующая фермент биосинтеза L-аминокислоты, включает полинуклеотиды, которые кодируют полипептид, полипептидный фрагмент, пептид или слитый полипептид, имеющий или сохраняющий соответствующую активность фермента биосинтеза L-аминокислоты. Способы определения наличия у полипептида конкретной активности путем измерения способности полипептида к превращению субстрата в продукт известны специалистам.
В некоторых вариантах реализации экзогенная нуклеиновая кислота кодирует контрольную последовательность экспрессии, функционально связанную с нуклеиновой кислотой, кодирующей нативный углевод фермент биосинтеза. Типично, контрольная последовательность экспрессии приводит к сверхэкспрессии нативного фермента биосинтеза L-аминокислоты.
В тех случаях, когда первая экзогенная нуклеиновая кислота кодирует АВ-фермент, она типично функционально связана с экзогенной контрольной последовательностью экспрессии. Экзогенная контрольная последовательность экспрессии может быть функционально связана с экзогенной нуклеиновой кислотой, кодирующей фермент биосинтеза L-аминокислоты для усиления продуцирования требуемой L-аминокислоты. Экзогенная контрольная последовательность экспрессии типично является кодон-оптимизированной для оптимальной экспрессии клеткой-хозяином метаболизирующего C1 микроорганизма.
В некоторых вариантах реализации первая экзогенная нуклеиновая кислота кодирует контрольную последовательность экспрессии, функционально связанную с нуклеиновой кислотой, кодирующей нативный фермент биосинтеза L-аминокислоты. Типичные примеры контрольных последовательностей экспрессии включают промоторы, описанные ниже. В таких вариантах реализации экспрессия нативного фермента биосинтеза L-аминокислоты типично повышена (т.е. имеет место сверхэкспрессия). Повышенная экспрессия или активность включает повышение экспрессии или активности гена или белка выше уровня дикого типа (нативного или генетически немодифицированного) контрольного или референсного микроорганизма. Ген или белок сверхэкспрессируется, если экспрессия или активность наблюдаются в микроорганизме, в котором он нормально не экспрессируется или не проявляет активность. Ген или белок сверхэкспрессируется, если экспрессия или активность продлевается или присутствует дольше в рекомбинантном микроорганизме, чем в контроле дикого типа или референсном микроорганизме. В используемом в данном документе значении, термин «сверхэкспрессируемый» и «сверхэкспрессия» по отношению к гену или белку означает увеличение экспрессии или активности гена или белка. Термины «нуклеиновая кислота», «полинуклеотид» и «ген» используются в данном документе взаимозаменяемо по отношению к полимеру остатков нуклеиновой кислоты (например, дезоксирибонуклеотидов или рибонуклеотидов) в одно- или двухцепочечной форме.
В дополнение к экзогенным нуклеиновым кислотам, описанным выше, рекомбинантные метаболизирующие C1 микроорганизмы по настоящему изобретению могут содержать дополнительные генетические модификации, которые усиливают продуцирование требуемой L-аминокислоты. Например, рекомбинантный метаболизирующий С1 микроорганизм по настоящему изобретению может дополнительно содержать экзогенную контрольную последовательность экспрессии, функционально связанную с эндогенной нуклеиновой кислотой, кодирующей эндогенный фермент, утилизирующий один или несколько из тех же субстратов, которые утилизируются ферментами биосинтеза L-аминокислот, или утилизирующий требуемую L-аминокислоту в качестве субстрата (т.е., «конкурирующий» эндогенный фермент). Это может быть сделано для понижающей регуляции конкурирующего эндогенного фермента.
В некоторых вариантах реализации может быть желательным снижение или ингибирование активности конкурирующего эндогенного фермента путем мутирования конкурирующего эндогенного фермента для удаления или ослабления его активности. Термин «ингибировать» или «ингибированный», в используемом в данном документе значении, относится к изменению, уменьшению, понижающей регуляции, нейтрализации или удалению, прямо или косвенно, экспрессии гена-мишени или активности молекулы-мишени по сравнению с контрольной, эндогенной или референсной молекулой, причем изменение, уменьшение, понижающая регуляция или нейтрализация являются статистически, биологически, промышленно или клинически значимыми.
Различные способы понижающей регуляции, инактивации, нокаута или делеции функции эндогенного гена в метаболизирующих C1 микроорганизмах известны специалистам. Например, разрыв гена-мишени является эффективным способом понижающей регуляции гена, когда экзогенный полинуклеотид вставлен в структурный ген для нарушения транскрипции. Генетические кассеты, содержащие экзогенную инсерционную ДНК (например, генетический маркер), фланкированную последовательностью, имеющей высокую степень гомологии с частью гена-мишени хозяина, предназначенные для разрыва, вводят в метаболизирующий С1 микроорганизм-хозяин. Экзогенная ДНК разрывает ген-мишень хозяина по механизмам репликации нативной ДНК. Аллельный обмен для конструирования делеционных/инсерционных мутантов в метаболизирующих C1 микроорганизмах, включая метанотрофные бактерии, был описан, например, в Toyama and Lidstrom, Microbiol. 144:183, 1998; Stoylar et al., Microbiol. 145:1235, 1999; Ali et al., Microbiol. 152:2931, 2006; Van Dien et al., Microbiol. 149:601, 2003; Martin and Murrell, FEMS Microbiol. Lett. 127:243, 2006, все из которых включены в данный документ в качестве ссылок. Например, ферменты, принимающие участие в других путях, таких как путь синтеза углеводов, могут также быть мишенями понижающей регуляции для того, чтобы сосредоточить метаболическую активность микроорганизма-хозяина на биосинтезе L-аминокислоты.
Рекомбинантный метаболизирующий С1 микроорганизм может быть, таким образом, генетически модифицирован для создания способности продуцировать требуемую L-аминокислоту с повышенными уровнями. В некоторых из этих вариантов реализации рекомбинантный метаболизирующий C1 микроорганизм продуцирует требуемую L-аминокислоту на уровне по меньшей мере на около 10% выше, чем при продуцировании нативным метаболизирующим С1 микроорганизмом, и до около 2 раз больше, до около 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, и до около 500 или около 1000 раз больше уровня, продуцируемого нативным метаболизирующим C1 микроорганизмом, при культивации в присутствии получаемого из природного газа сырья (например, природного газа, метана и т.п.) при по меньшей мере одном наборе условий культивации. В других вариантах реализации рекомбинантный метаболизирующий C1 микроорганизм продуцирует требуемую L-аминокислоту на уровне, который на по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, по меньшей мере около 75%, по меньшей мере около 80%, по меньшей мере около 85%, по меньшей мере около 90%, или по меньшей мере около 95%, превышает продуцирование нативным метаболизирующим C1 микроорганизмом, и до около 2 раз больше, до около 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, до около 500 или около 1000 раз больше уровня, продуцируемого нативным метаболизирующим C1 микроорганизмом, при культивации в присутствии получаемого из природного газа сырья при по меньшей мере одном наборе условий культивации. Типично, повышенный уровень продуцирования требуемой L-аминокислоты рекомбинантным метаболизирующим C1 микроорганизмом по настоящему изобретению по меньшей мере около в 2 раза, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 раз больше, чем у нативного метаболизирующего C1 микроорганизма, при культивации в присутствии получаемого из природного газа сырья при по меньшей мере одном наборе условий культивации.
Рекомбинантные способы экспрессии экзогенных нуклеиновых кислот в бактериальных организмах хорошо известны специалистам. Описание таких способов можно найти, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, Third Ed., Cold Spring Harbor Laboratory, New York (2001); и Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, MD (1999), все из которых включены в данный документ в качестве ссылок. Генетические модификации молекул нуклеиновой кислоты, кодирующих ферменты, или их функциональных фрагментов могут придавать рекомбинантной клетке биохимическую или метаболическую способность, измененную по сравнению с ее природным состоянием.
В используемом в данном документе значении, термины «эндогенный» и «нативный», при использовании по отношению к нуклеиновой кислоте, полипептиду, такому как фермент, соединению или активности, относятся к нуклеиновой кислоте, полипептиду, соединению или активности, которые нормально присутствуют в клетке-хозяине. Термин «гомологичный» или «гомолог» относится к молекуле или активности из экзогенного (ненативного) источника, которые являются такими же или схожими с молекулой или активностью, присутствующими в или выделенными из клетки-, вида- или штамма-хозяина.
В используемом в данном документе значении, термины «гетерологичный» и «экзогенный», при использовании по отношению к молекуле нуклеиновой кислоты, конструкту или последовательности, относятся к молекуле нуклеиновой кислоты или части последовательности молекулы нуклеиновой кислоты, которая на является нативной для клетки, в которой она экспрессируется, молекуле нуклеиновой кислоты или части молекулы нуклеиновой кислоты, нативной для клетки-хозяина, которая была изменена или мутирована, или молекуле нуклеиновой кислоты с измененной экспрессией по сравнению с нативными уровнями экспрессии в схожих условиях. Например, гетерологичная контрольная последовательность (например, промотор, энхансер) может быть использована для регуляции экспрессии гена или молекулы нуклеиновой кислоты способом, отличным от гена или молекулы нуклеиновой кислоты, нормально экспрессируемых в природе или в культуре. В определенных вариантах реализации, гетерологичная молекула нуклеиновой кислоты может быть гомологичной по отношению к нативному гену клетки-хозяина, но может иметь измененный уровень экспрессии или иметь отличную от него последовательность, или оба эти признака. В других вариантах реализации гетерологичные или экзогенные молекулы нуклеиновой кислоты могут не быть эндогенными по отношению к клетке-хозяину или геному хозяина, и вместо этого могут быть добавлены в клетку-хозяина путем конъюгации, трансформации, трансфекции, электропорации и т.п., причем добавленная молекула может интегрироваться в геном хозяина или может существовать как внехромосомный генетический материал (например, плазмида или другой самореплицирующийся вектор). Термин «гетерологичный» при использовании по отношению к организму касается вида, отличного от клетки-хозяина.
В определенных вариантах реализации, несколько гетерологичных или экзогенных молекул нуклеиновой кислоты может быть введено в клетку-хозяина в виде отдельных молекул нуклеиновой кислоты, в виде полицистронной молекулы нуклеиновой кислоты, в виде отдельной молекулы нуклеиновой кислоты, кодирующей гибридный белок, или любой их комбинации, и будет считаться несколькими гетерологичныыми или экзогенными нуклеиновыми кислотами. Например, метаболизирующий С1 микроорганизм может быть модифицирован для экспрессии двух или больше гетерологичных или экзогенных молекул нуклеиновой кислоты, которые могут быть одинаковыми или разными, которые кодируют один или несколько ферментов биосинтеза L-аминокислоты, как раскрыто в данном документе. В определенных вариантах реализации, в клетку-хозяина вводят множество копий полинуклеотидной молекулы, кодирующей фермент биосинтеза L-аминокислоты, которые могут быть двумя, тремя, четырьмя, пятью, шестью, семью, восемью, девятью, десятью или больше копиями полинуклеотидов, кодирующих один и тот же фермент биосинтеза L-аминокислоты или разные ферменты биосинтеза L-аминокислоты.
Клетки-хозяева и способы трансформации
При осуществлении настоящего изобретения на практике экзогенные нуклеиновые кислоты, описанные выше, трансформируются в клетку-хозяина, которая представляет собой метаболизирующий C1 микроорганизм. Используемый метаболизирующий C1 микроорганизм может быть природным, адаптированным штаммом (например, путем проведения ферментации для селекции штаммов с повышенной скоростью роста и повышенным общим выходом биомассы по сравнению с исходным штаммом), или рекомбинантно модифицированным для продуцирования или сверхэкспрессирования фермента биосинтеза L-аминокислоты, представляющего интерес, и/или обеспечения повышенной скорости роста. Типично, метаболизирующий C1 микроорганизм представляет собой нефотосинтезирующий C1 микроорганизм (например, не является водорослями или растением).
В определенных вариантах реализации настоящее изобретение использует метаболизирующие C1 микроорганизмы, являющиеся прокариотами или бактериями, такие как Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium, Methanomonas, Methylophilus, Methylobacillus, Methylobacterium, Hyphomicrobium, Xanthobacter, Bacillus, Paracoccus, Nocardia, Arthrobacter, Rhodopseudomonas или Pseudomonas.
В дополнительных вариантах реализации используемые метаболизирующие C1 бактерии представляют собой метанотрофа или метилотрофа. Типичные примеры метанотрофов включают Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium, Methanomonas, Methylocella или их комбинацию. Типичные примеры метилотрофов включают Methylobacterium extorquens, Methylobacterium radiotolerans, Methylobacterium populi, Methylobacterium chloromethanicum, Methylobacterium nodulans или их комбинацию. В используемом в данном документе значении, термин «метилотрофные бактерии» относится к любым бактериям, способным окислять любое соединение в любой форме (например, твердой, жидкой, газообразной), содержащее по меньшей мере один (атом) углерода и не содержащее углерод-углеродных связей. В определенных вариантах реализации метилотрофная бактерия может быть метанотрофом. Например, «метанотрофные бактерии» относятся к любым метил отрофным бактериям, которые обладают способностью окислять метан в качестве источника углерода и энергии, который может быть основным источником углерода и энергии. Типичные примеры метанотрофных бактерий включают Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium или Methanomonas.
Метанотрофные бактерии делятся на три группы на основе их путей ассимиляции углерода и строения внутренней мембраны: тип I (гамма-протеобактерии), тип II (альфа-протеобактерии и тип X (гамма-протеобактерии). Метанотрофы типа I используют путь рибулозамонофосфата (RuMP) для ассимиляции углерода, в то время как метанотрофы типа II используют сериновый путь. Метанотрофы типа X используют путь RuMP, но также экспрессируют низкие уровни ферментов серинового пути. Метанотрофные бактерии, используемые в практике настоящего изобретения, включают облигатные метанотрофы, которые могут утилизировать только субстраты С1 для получения углерода и источников энергии, и факультативные метанотрофы, которые от природы обладают способностью утилизировать некоторые многоуглеродные субстраты в качестве источника углерода и энергии.
Типичные примеры факультативных метанотрофов, используемых в практике настоящего изобретения, включают некоторые виды Methylocella, Methylocystis и Methylocapsa (например, Methylocella silvestris, Methylocella palustris, Methylocella tundrae, Methylocystis daltona штамм SB2, Methylocystis bryophila и Methylocapsa aurea KYG), Methylobacterium organophilum (ATCC 27,886), Methylibium petroleiphilum, или их быстрорастущие варианты. Типичные примеры облигатных метанотрофных бактерий, пригодных для использования в практике настоящего изобретения, включают Methylococcus capsulatus Bath (NCIMB 11132), Methylomonas sp. 16a (ATCC РТА 2402), Methylosinus trichosporium OB3b (NRRL B-11,196), Methylosinus sporium (NRRL B-11,197), Methylocystis parvus (NRRL B-11,198), Methylomonas methanica (NRRL B-11,199), Methylomonas albus (NRRL B-11,200), Methylobacter capsulatus Y (NRRL B-11,201), Methylomonas flagellata sp. AJ-3670 (FERM P-2400), Methylacidiphilum infernorum, Methylacidiphilum fumariolicum, Methylomicrobium alcaliphilum, Methyloacida kamchatkensis, или их быстрорастущие варианты.
Пригодные метаболизирующие C1 микроорганизмы, пригодные для использования в практике настоящего изобретения, включают бактерии, метаболизирующие сингаз, такие как, например, Clostridium, Moorella, Pyrococcus, Eubacterium, Desulfobacterium, Carboxydothermus, Acetogenium, Acetobacterium, Acetoanaerobium, Butyribaceterium, Peptostreptococcus, и т.п. Типичные примеры бактерий, метаболизирующих сингаз, включают Clostridium autoethanogenum, Clostridium ljungdahli, Clostridium ragsdalei, Clostridium carboxydivorans, Butyribacterium methylotrophicum, Clostridium woodii, Clostridium neopropanologen, и т.п.
Другие пригодные метаболизирующие C1 микроорганизмы, пригодные для использования в практике настоящего изобретения, включают эукариоты, такие как, например, дрожжи, включая Candida, Yarrowia, Hansenula, Pichia, Torulopsis, Rhodotorula, и т.п.
Каждый из микроорганизмов в соответствии с настоящим описанием может быть выращен в виде изолированной культуры, с гетерологичным организмом, который может способствовать росту, или одна или несколько таких бактерий могут быть скомбинированы для получения смешанной культуры. В дополнительных вариантах реализации, метаболизирующие C1 нефотосинтезирующие микроорганизмы в соответствии с настоящим описанием являются облигатными метаболизирующими C1 нефотосинтезирующими микроорганизмами, такими как облигатный метанотроф или метилотроф.
Любой из вышеупомянутых метаболизирующих С1 микроорганизмов может быть использован в качестве исходной или референсной клетки-хозяина для получения рекомбинантных метаболизирующих С1 микроорганизмов в соответствии с настоящим описанием. В используемом в данном документе значении, термин «рекомбинантный» относится к не встречающемуся в природе организму, микроорганизму, клетке, молекуле нуклеиновой кислоты, или вектору, которые имеют по меньшей мере одно генетическое изменение или были модифицированы путем введения гетерологичной молекулы нуклеиновой кислоты, или относится к клетке, которая была изменена таким образом, чтобы экспрессия эндогенной молекулы нуклеиновой кислоты или гена могла контролироваться. Рекомбинантный также относится к клетке, которая была получена от клетки или потомства клетки, имеющих одну или несколько таких модификаций. Генетические изменения включают, например, модификации, вводящие экспрессируемые молекулы нуклеиновой кислоты, кодирующие белки или ферменты, или другие добавления, делеции, замещения молекулы нуклеиновой кислоты, или другие функциональные изменения генетического материала клетки. Например, рекомбинантные клетки могут экспрессировать гены или другие молекулы нуклеиновой кислоты, не встречающиеся в идентичной форме в нативной клетке (т.е. немодифицированной клетке или клетке дикого типа), или могут обеспечивать измененный характер экспрессии эндогенных генов, таких генов, которые могут иначе сверхэкспрессироваться, иметь пониженную экспрессию, минимальную экспрессию, или вообще не экспрессироваться.
Любые рекомбинантные метаболизирующие С1 микроорганизмы или метанотрофные бактерии, описанные в данном документе, могут быть трансформированы для включения по меньшей мере одной экзогенной нуклеиновой кислоты для обеспечения хозяина с новой или повышенной активностью (например, ферментативной активностью), или могут быть генетически модифицированы для удаления или значительного снижения функции эндогенного гена с использованием различных способов, известных специалистам.
Трансформация относится к введению молекулы нуклеиновой кислоты (например, экзогенной или гетерологичной молекулы нуклеиновой кислоты) в клетку-хозяина. Трансформированная клетка-хозяин может нести экзогенную или гетерологичную молекулу нуклеиновой кислоты внехромосомно или интегрированной в хромосому. Интеграция в геном клетки-хозяина и самореплицирующиеся векторы обычно приводят к генетически стабильному наследованию трансформированной молекулы нуклеиновой кислоты. Клетки-хозяева, содержащие трансформированные молекулы нуклеиновой кислоты, называются «неприродными» или «генетически модифицированными», или «рекомбинантными», или «трансформированными», или «трансгенными» клетками (например, бактериями).
Системы экспрессии и экспрессионные векторы, пригодные для экспрессии гетерологичных нуклеиновых кислот в метаболизирующих C1 микроорганизмах (например, метанотрофные бактерии), являются известными.
Электропорация метаболизирующих C1 бактерий описана в данном документе и была ранее описана, например, в Toyama et al., FEMS Microbiol. Lett. 166:1, 1998; Kim and Wood, Appl. Microbiol. Biotechnol. 48:105, 1997; Yoshida et al., Biotechnol. Lett. 23:787, 2001, и публикации патентной заявки США № 2008/0026005.
Бактериальная конъюгация, которая относится к определенному типу трансформации, предусматривающему непосредственный контакт клеток донора и реципиента, чаще используется для переноса молекул нуклеиновой кислоты в метаболизирующие С1 бактерии. Бактериальная конъюгация предусматривает смешивание клеток «донора» и «реципиента» вместе в тесном контакте друг с другом. Конъюгация происходит путем образования цитоплазматических соединений между бактериями-донорами и реципиентами, с однонаправленным переносом заново синтезированных донорных молекул нуклеиновой кислоты в клетки-реципиенты. Реципиент в реакции конъюгации представляет собой любую клетку, которая может принимать нуклеиновые кислоты путем горизонтального переноса от бактерии-донора. Донор в реакции конъюгации представляет собой бактерию, которая содержит конъюгативную плазмиду, конъюгативный транспозон или мобилизованную плазмиду. Физический перенос донорной плазмиды может происходить с помощью самопередающейся плазмиды или с помощью «хелперной» плазмиды. Конъюгация с участием метаболизирующих С1 бактерий описана в данном документе и была описана ранее в Stolyar et al., Mikrobiologiya 64:686, 1995; Motoyama et al., Appl. Micro. Biotech. 42:61, 1994; Lloyd et al., Arch. Microbiol. 171:364, 1999; публикации PCT № WO 02/18617; и Ali et al., Microbiol. 152:2931, 2006.
Контрольные последовательности экспрессии, пригодные для использования в практике настоящего изобретения, включают, например, промоторы, терминаторы, энхансеры, репрессоры, индукторы и т.п. Промоторы, пригодные для использования в практике настоящего изобретения, могут быть конститутивными, просачивающимися или индуцируемыми, и нативными или ненативными для используемой клетки-хозяина. Типичные примеры промоторов включают промотор пируват-декарбоксилазы (PDC), промотор дезоксиксилулозы-фосфатсинтазы, промотор метанолдегидрогеназы (MDH) (такой как, например, промотор в вышележащей межгенной области гена mxaF Methylococcus capsulatus Bath (№ доступа MCA0779) или промотор MDH М. extorquens (см. Springer et al., FEMS Microbiol. Lett. 160:119 (1998)), промотор гексулоза-6-фосфатсинтазы, промотор рибосомального белка S16, промотор серингидроксиметилтрансферазы, промотор серин-глиоксилатаминотрансферазы, промотор фосфоенолпируваткарбоксилазы, промотор Т5, промотор Trc, промотор синтеза PHA (Foellner et al., Appl. Microbiol. Biotechnol. 40:284, 1993), промотор пируват-декарбоксилазы (Tokuhiro et al., Appl. Biochem. Biotechnol. 131:195, 2006), промотор lac-оперона Plac (Toyama et al., Microbiol. 143:595, 1997), гибридный промотор, такой как Ptrc (Brosius et al., Gene 27:161, 1984), промоторы, идентифицированные в нативных плазмидах метилотрофов (ЕР 296484), метанотрофов, и т.п.
Дополнительно, могут быть использованы пригодные гомологичные или гетерологичные промоторы для высокой экспрессии молекул экзогенной нуклеиновой кислоты. Например, патент США № 7098005 описывает использование промоторов для высокой экспрессии в присутствии метана или метанола гетерологичной кодирующей нуклеиновой кислоты в метаболизирующих С1 бактериях.
В определенных вариантах реализации регулируемая экспрессия экзогенных нуклеиновых кислот, кодирующих фермент биосинтеза L-аминокислоты, может быть требуемой для оптимизации скорости роста неприродного метаболизирующего C1 микроорганизма, и может ускорять бактериальный рост в условиях различных источников углерода. Это может быть достигнуто путем использования системы индуцируемого промотора.
В определенных вариантах реализации нуклеиновая кислота, кодирующая АВ-фермент, функционально связана с индуцируемым промотором. Системы индуцируемого промотора, используемые в практике настоящего изобретения, известны специалистам и включают тетрациклиновую систему индуцируемого промотора; систему индуцируемого промотора оперона IPTG/lac, систему индуцируемого промотора теплового шока; системы чувствительных к металлам промоторов; нитратную систему индуцируемого промотора; систему индуцируемого светом промотора; систему экдизон-индуцируемого промотора, индуцируемую/регулируемую систему, описанную для использования в метилотрофных и метанотрофных бактериях (см., например, патентную заявку США № US 2010/0221813, которая включена в данный документ в качестве ссылки), и т.п. Например, в одном варианте реализации неприродный метаболизирующий C1 микроорганизм (например, метанотроф, метилотроф) содержит: (1) экзогенную нуклеиновую кислоту, кодирующую АВ-фермент, функционально связанную с промотором, фланкированным последовательностями оператора lacO, и (2) экзогенную нуклеиновую кислоту, кодирующую репрессорный белок lacI, функционально связанный с конститутивным промотором (например, промотором гексулоза-6-фосфатсинтазы). Индукция инициируется, когда репрессорный белок LacI связывается с последовательностями оператора lacO, фланкирующими LDH или другой промотор, препятствуя транскрипции. IPTG связывает репрессор lacI и освобождает его от последовательностей lacO, обеспечивая возможность транскрипции. Благодаря использованию системы индуцируемого промотора, синтез лактата можно контролировать путем добавления индуктора.
Системы экспрессии и экспрессионные векторы, используемые в практике настоящего изобретения, необязательно содержат генетические элементы, такие как, например, один или несколько сайтов связывания рибосом для инициации трансляции и сайт терминации транскрипции, сигналы полиаденилирования, сайты рестрикции ферментами, сайты множественного клонирования, другие кодирующие сегменты и т.п. В определенных вариантах реализации промоторы и/или оптимизация кодонов (описана более детально выше) используются для высокой конститутивной экспрессии экзогенных полинуклеотидов, кодирующих один или несколько ферментов биосинтеза углеводов в метанотрофных бактериях-хозяевах. Может быть использована также регулируемая экспрессия экзогенной нуклеиновой кислоты в метанотрофной бактерии-хозяине. Например, может быть использована индуцируемая/регулируемая система рекомбинантной экспрессии белка в метилотрофных и метанотрофных бактериях, как описано, например, в патентной заявке США № US 2010/0221813.
В определенных вариантах реализации промоторы или оптимизация кодонов (описана более детально выше) используются для высокой конститутивной экспрессии экзогенных полинуклеотидов, кодирующих один или несколько ферментов биосинтеза L-аминокислоты в метанотрофных бактериях-хозяевах. Также может быть использована регулируемая экспрессия экзогенной нуклеиновой кислоты в метанотрофной бактерии-хозяине. Например, может быть использована индуцируемая/регулируемая система рекомбинантной экспрессии белка в метилотрофных и метанотрофных бактериях, как описано, например, в патентной заявке США № US 2010/0221813.
Способы продуцирования требуемой L-аминокислоты
Настоящее изобретение обеспечивает способ продуцирования L-аминокислоты, который включает культивацию рекомбинантного метаболизирующего С1 микроорганизма в присутствии углеродного сырья, получаемого из природного газа, в условиях, достаточных для продуцирования L-аминокислоты, причем метаболизирующий C1 микроорганизм содержит экзогенную нуклеиновую кислоту, кодирующую фермент биосинтеза L-аминокислоты. Типично, полученное из природного газа углеродное сырье представляет собой природный газ, метан или сингаз. Иллюстративные условия, пригодные для культивации микроорганизмов по настоящему изобретению, описаны в примерах.
Различные методики культивации могут быть использованы для микроорганизмов, описанных в данном документе. Например, метаболизирующие C1 микроорганизмы (такие как метанотрофные или метилотрофные бактерии) могут выращиваться по методикам периодической культуры или непрерывной культуры. В определенных вариантах реализации культуры выращивают в устройстве для контролируемой культивации, таком как ферментер, биореактор, ячейка с пустотелыми волокнами и т.п. Обычно клетки в лог-фазе часто отвечают за массовое продуцирование продукта или промежуточного соединения, представляющего интерес, в некоторых системах, в то время как в других системах может быть получена стационарная или пост-экспоненциальная фаза продуцирования.
Классический способ периодической культивации представляет собой замкнутую систему, в которую помещают композицию среды при запуске культуры и которая не изменяется в процессе культивации. Это означает, что среду инокулируют в начале процесса культивации одним или несколькими выбранными микроорганизмами и затем позволяют расти без добавления чего-либо в систему. В используемом в данном документе значении, «периодическая» (порционная) культура является указанием на то, что количество конкретного источника углерода, добавленного в начальный момент времени, остается неизменным, в то время как контроль факторов, таких как рН и концентрация кислорода, может проводиться с внесением изменений в процессе культивации. В периодических системах, состав метаболита и биомассы в системе постоянно изменяются до времени завершения культивации. В периодических культурах, клетки (например, бактерии, такие как метилотрофы) будут обычно переходить от статической латентной фазы к логарифмической фазе быстрого роста и к стационарной фазе, где скорость роста снижается или останавливается (и со временем приходят к гибели клеток, если условия не изменятся).
Периодическая система с подпиткой является вариантом стандартной периодической системы, в которой углеродный субстрат, представляющий интерес, добавляют порциями по мере развития культуры. Периодические системы с подпиткой полезны в случаях, когда клеточный метаболизм вероятно будет ингибироваться в результате катаболитной репрессии, и когда желательно ограничивать количество субстрата в среде. Поскольку фактическую концентрацию субстрата в периодических системах с подпиткой измерить трудно, проводят оценку на основе изменений измеримых факторов, таких как рН, растворенный кислород и парциальное давление отходящих газов. Способы периодической культивации и периодической культивации с подпиткой являются обычными и известны специалистам (см., например, Thomas D. Brock, Biotechnology: A Textbook of Industrial Microbiology, 2nd Ed. (1989) Sinauer Associates, Inc., Sunderland, MA; Deshpande, Appl. Biochem. Biotechnol. 36:221, 1992).
Непрерывные культуры являются «открытыми» системами в том смысле, что питательные среды определенного состава непрерывно добавляют в биореактор, причем равное количество использованной («кондиционной») среды одновременно удаляется для переработки. Непрерывные культуры обычно поддерживают клетки при постоянной высокой плотности жидкой фазы, в которой клетки преимущественно находятся на логарифмической фазе роста. Альтернативно, может быть реализована непрерывная культура с иммобилизованными клетками (например, биопленкой), в которой углерод и питательные вещества непрерывно добавляют, и ценные продукты, побочные продукты и отходы непрерывно удаляют из клеточной массы. Иммобилизация клеток может быть достигнута с помощью широкого спектра твердых носителей, состоящих из природных материалов, синтетических материалов или их комбинации.
Непрерывная или полунепрерывная культура позволяет регулировать один или несколько факторов, влияющих на рост клеток или концентрацию конечного продукта. Например, один способ может поддерживать фиксированное значение лимитирующего питательного вещества (например, источника углерода, азота) и позволять всем другим параметрам изменяться со временем. В других вариантах реализации несколько факторов, влияющих на рост, могут непрерывно изменяться, в то время как концентрация клеток, определяемая по мутности среды, поддерживается постоянной. Целью системы непрерывной культуры является поддержание условий стабильного роста, причем потеря клеток вследствие отбора среды уравновешивается скоростью роста клеток. Способы регулирования питательных веществ и факторов роста для процессов непрерывной культивации и методы максимизации скорости образования продукта хорошо известны специалистам (см. Brock, 1992).
Биореакторы с жидкой фазой (например, с перемешиваемой емкостью, насадкой, одной жидкой фазой, двумя жидкими фазами, мембраной из полых волокон) хорошо известны специалистам и могут быть использованы для выращивания неприродных микроорганизмов и биокатализа.
При использовании биореакторов с газовой фазой субстраты для биопродуцирования абсорбируются неприродными микроорганизмами, клеточными лизатами или их бесклеточными фракциями из газа, а не из жидкости. Использование биореакторов с газовой фазой с микроорганизмами известно специалистам (например, патенты США №№ 2793096; 4999302; 5585266; 5079168; и 6143556; правовая публикация США (U.S. Statutory Invention Registration) H1430; публикация патентной заявки США №2003/0032170; Emerging Technologies in Hazardous Waste Management III, 1993, eds. Tedder and Pohland, стр. 411-428). Типичные примеры биореакторов с газовой фазой включают прямоточную систему, систему замкнутого контура с принудительной подачей и реактор с псевдоожиженным слоем. Благодаря использованию биореакторов с газовой фазой, метан или другие газообразные субстраты будут легкодоступными для биокатализа полипептидами, например, с активностью монооксигеназы. В определенных вариантах реализации, способы конверсии газа в L-аминокислоту осуществляются в биореакторах с газовой фазой. В дополнительных вариантах реализации способы конверсии газа в L-аминокислоту осуществляются в реакторах с псевдоожиженным слоем. В реакторе с псевдоожиженным слоем флюид (т.е. газ или жидкость) проходит вверх через слой дисперсного носителя, обычно, песка, гранулированного активированного угля или диатомовой земли, на котором микроорганизмы могут закрепляться и расти. Скорость флюида является такой, чтобы слой дисперсного носителя с прикрепившимися микроорганизмами находился в суспендированном состоянии (т.е. чтобы происходило ожижение слоя). Микроорганизмы, прикрепившиеся к дисперсным частицам носителя, свободно циркулируют во флюиде, что обеспечивает возможность эффективного массопереноса субстратов из флюида к микроорганизмам и усиление бактериального роста. Типичные примеры реакторов с псевдоожиженным слоем включают реакторы идеального вытеснения и реакторы полного смешения. Использование реакторов с псевдоожиженным слоем с бактериальными биопленками известно специалистам (например, Pfluger et al., Bioresource Technol. 102:9919, 2011; Fennell et al., Biotechnol, Bioengin. 40.1218, 1992; Ruggeri et al., Water Sci. Technol. 29:347, 1994; патенты США №№4032407; 4009098; 4009105; и 3846289).
Рекомбинантные метаболизирующие C1 микроорганизмы, описанные в данном документе, могут выращиваться в виде изолированной чистой культуры, с гетерологичным неметаболизирующим C1 микроорганизмом (микроорганизмами), которые могут способствовать росту, или один или несколько разных штаммов или видов метаболизирующих C1 микроорганизмов могут быть объединены для получения смешанной культуры.
В определенных вариантах реализации L-аминокислоты по настоящему изобретению продуцируются на определенной фазе роста клеток (например, латентной фазе, лог-фазе, стационарной фазе или фазе гибели). Может быть желательной конверсия углерода из сырья в L-аминокислоту, а не рост и поддержание метаболизирующего C1 микроорганизма. В некоторых вариантах реализации неприродный метаболизирующий C1 микроорганизм (например, метанотрофы, метилотрофы), предусматриваемый в данном документе, культивируют при плотности клеток от низкой до средней (OD600) и затем инициируют продуцирование L-аминокислоты. В некоторых вариантах реализации L-аминокислота продуцируется, когда метанотрофные бактерии прекращают делиться или делятся очень медленно. В некоторых вариантах реализации L-аминокислота продуцируется только на стационарной фазе. В некоторых вариантах реализации L-аминокислота продуцируется на лог-фазе и стационарной фазе.
Композиция ферментера, содержащая L-аминокислоту, продуцируемую рекомбинантным метаболизирующим С1 микроорганизмом (например, метанотрофами, метилотрофами), предусматриваемая в данном документе, может дополнительно содержать другие органические соединения, ассоциированные с процессами биологической ферментации. Например, биологические побочные продукты ферментации могут включать один или несколько спиртов, эпоксидов, альдегидов, кетонов, сложных эфиров или их комбинацию. В определенных вариантах реализации, композиция ферментера может содержать один или несколько из следующих спиртов: метанол, этанол, бутанол или пропанол. Другие соединения, такие как Н2О, СО, СО2, СО N2, Н2, О2, и неутилизированное углеродное сырье, такое как метан, этан, пропан и бутан, также могут присутствовать в отходящем газе ферментера.
В определенных вариантах реализации рекомбинантные метаболизирующие С1 микроорганизмы (например, метанотрофы, метилотрофы), предусматриваемые в данном документе, продуцируют L-аминокислоту по настоящему изобретению в количестве от около 0,001 г/л культуры до около 500 г/л культуры. В некоторых вариантах реализации количество продуцируемой L-аминокислоты составляет от около 1 г/л культуры до около 100 г/л культуры. В некоторых вариантах реализации количество продуцируемой L-аминокислоты составляет около 0,001 г/л, 0,01 г/л, 0,025 г/л, 0,05 г/л, 0,1 г/л, 0,15 г/л, 0,2 г/л, 0,25 г/л, 0,3 г/л, 0,4 г/л, 0,5 г/л, 0,6 г/л, 0,7 г/л, 0,8 г/л, 0,9 г/л, 1 г/л, 2,5 г/л, 5 г/л, 7,5 г/л, 10 г/л, 12,5 г/л, 15 г/л, 20 г/л, 25 г/л, 30 г/л, 35 г/л, 40 г/л, 45 г/л, 50 г/л, 60 г/л, 70 г/л, 80 г/л, 90 г/л, 100 г/л, 125 г/л, 150 г/л, 175 г/л, 200 г/л, 225 г/л, 250 г/л, 275 г/л, 300 г/л, 325 г/л, 350 г/л, 375 г/л, 400 г/л, 425 г/л, 450 г/л, 475 г/л или 500 г/л.
Продукты
Настоящее изобретение обеспечивает другие полезные продукты в дополнение к рекомбинантным метаболизирующим C1 клеткам по настоящему изобретению. В одном варианте реализации, настоящее изобретение обеспечивает биомассу, получаемую из культуры рекомбинантного метаболизирующего C1 микроорганизма по настоящему изобретению. В конкретном варианте реализации настоящее изобретение обеспечивает биомассу, содержащую рекомбинантный метаболизирующий С1 микроорганизм, который содержит экзогенную нуклеиновую кислоту, кодирующую фермент биосинтеза L-аминокислоты, и при этом рекомбинантный метаболизирующий C1 микроорганизм способен конвертировать получаемое из природного газа сырье в требуемую L-аминокислоту. В дополнительном варианте реализации настоящее изобретение обеспечивает биомассу, содержащую рекомбинантный метаболизирующий C1 микроорганизм, причем, рекомбинантный метаболизирующий С1 микроорганизм содержит экзогенную нуклеиновую кислоту, кодирующую фермент биосинтеза L-аминокислоты, и при этом, рекомбинантный метаболизирующий C1 микроорганизм способен конвертировать метан в требуемую L-аминокислоту.
В используемом в данном документе значении, «биомасса» относится к органическому материалу, имеющему биологическое происхождение, который может включать что-то одно или несколько из цельных клеток, лизированных клеток, внеклеточного материала и т.п. Например, материал, собранный из культивируемых микроорганизмов (например, бактериальной или дрожжевой культуры) считается биомассой, которая может включать клетки, клеточные мембраны, клеточную цитоплазму, тельца включения, продукты, секретируемые или экскретируемые в питательную среду, или любую их комбинацию. В определенных вариантах реализации биомасса включает метаболизирующие C1 микроорганизмы в соответствии с настоящим описанием вместе с питательными культурами, в которых метаболизирующие С1 микроорганизмы в соответствии с настоящим описанием были выращены. В других вариантах реализации биомасса включает метаболизирующие C1 микроорганизмы (цельные или лизированные или вместе) в соответствии с настоящим описанием, выделенные из культуры, выращенной на субстрате С1 (например, природный газ, метан и т.п.). В других вариантах реализации биомасса включает супернатант истощенной среды из культуры метаболизирующего С1 микроорганизма, выращиваемой на субстрате С1. Такая культура может считаться возобновляемым ресурсом. Биомасса по настоящему изобретению является обогащенной по отношению к уровням требуемой L-аминокислоты.
Рекомбинантные метаболизирующие С1 микроорганизмы по настоящему изобретению, которые обеспечиваются полученным из природного газа субстратом для роста клеток, отличаются по своему углеродному отпечатку пальцев, представленному значениями δ13С (так же, как и продукты, полученные от таких рекомбинантных метаболизирующих C1 микроорганизмов). В качестве пояснения укажем, что измерения стабильных изотопов и подходы, основанные на массовом балансе, широко используются для оценки глобальных источников и стоков метана (см. Whiticar and Faber, Org. Geochem. 70:759, 1986; Whiticar, Org. Geochem. 16: 531, 1990). Для использования значений δ13C остаточного метана для определения количества окисленного, необходимо знать степень изотопного фракционирования, вызванную бактериальным окислением метана. Например, аэробные метанотрофы могут метаболизировать метан с помощью специфического фермента, метанмонооксигеназы (ММО). Метанотрофы превращают метан в метанол и затем в формальдегид. Формальдегид может быть дополнительно окислен до СО2 для обеспечения клетки энергией в форме восстановительных эквивалентов (NADH), или включен в биомассу посредством RuMP или серинового циклов (Hanson and Hanson, Microbiol. Rev. 60:439, 1996), которые прямо аналогичны путям ассимиляции углерода в фотосинтезирующих организмах. Более конкретно, метанотроф типа I использует путь RuMP для синтеза биомассы и генерирует биомассу целиком из СН4, в то время как метанотроф типа II использует сериновый путь, который ассимилирует 50-70% углерода клетки из СН4 и 30-50% из СО2 (Hanson and Hanson, 1996). Способы измерения изотопного состава углерода приведены, например, Templeton et al. (Geochim. Cosmochim. Acta 70:1739, 2006), и настоящим включены в качестве ссылок в полном объеме. Пример 2 описывает определение характеристик распределения стабильных изотопов углерода в клетках разных метаболизирующих C1 микроорганизмов. Высокие отрицательные значения δ13С для клеток аналогично отражаются в значениях δ13С соединений, экстрагированных из этих клеток, как описано в примерах. Значения δ13С продуктов по изобретению, описанных в данном документе (т.е., рекомбинантного метаболизирующего C1 микроорганизма по настоящему изобретению (как описано выше), связанной с ним биомассы и получаемых из него L-аминокислот и композиций) может меняться в зависимости от источника и чистоты используемого субстрата C1, как показано в Примере 3.
В определенных вариантах реализации рекомбинантный метаболизирующий C1 микроорганизм по настоящему изобретению и связанная с ним биомасса и L-аминокислоты и композиция, получаемые из него, демонстрируют значение δ13С менее -30%o, менее -31%o, менее -32%o, менее -33%o, менее -34%o, менее -35%o, менее -36%o, менее -37%o, менее -38%o, менее -39%o, менее -40%o, менее -41%o, менее -42%o, менее -43%o, менее -44%o, менее -45%o, менее -46%o, менее -47%o, менее -48%o, менее -49%o, менее -50%o, менее -51%o, менее -52%o, менее -53%o, менее -54%o, менее -55%o, менее -56%o, менее -57%o, менее -58%o, менее -59%o, менее -60%o, менее -61%o, менее -62%o, менее -63%o, менее -64%o, менее -65%o, менее -66%o, менее -67%o, менее -68%o, менее -69%o, или менее -70%o.
В определенных вариантах реализации рекомбинантный метаболизирующий C1 микроорганизм по настоящему изобретению, и связанная с ним биомасса и получаемые из него L-аминокислоты и композиция, ним биомасса и L-аминокислоты и композиция, получаемые из него, демонстрируют значение δ13С, равное от около -35%o до около -50%o, от -45%o до около -35%o, или от около -50%o до около -40%o, или от около -45%o до около -65%o, или от около -60%o до около -70%o, или от около -30%o до около -70%o.
В дополнительных вариантах реализации биомасса метаболизирующего С1 нефотосинтезирующего микроорганизма имеет δ13С менее чем около -30%o, или в диапазоне значений от около -40%o до около -60%o. В определенных вариантах реализации биомасса содержит рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм вместе с истощенной средой, или биомасса содержит композицию супернатанта истощенной среды из культуры рекомбинантного метаболизирующего С1 нефотосинтезирующего микроорганизма, причем величина δ13С биомассы имеет значение менее чем около -30%o. В определенных других вариантах реализации композицию L-аминокислот экстрагируют или концентрируют из биомассы, которая может содержать рекомбинантные метаболизирующие С1 нефотосинтезирующие микроорганизмы вместе с истощенной средой из культуры, или композицию супернатанта истощенной среды из культуры рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма.
В определенных вариантах реализации композиция L-аминокислот, получаемая из метаболизирующего C1 микроорганизма (которая может необязательно быть экстрактом или изолятом из биомассы метаболизирующего C1 микроорганизма) содержит атомы водорода, кислорода и углерода, составляющие по меньшей мере от около 50% до около 80% от массы композиции, и величина δ13С композиции имеет значение менее чем около -35%o, или менее чем около -36%o, или менее чем около -37%o, или менее чем около -38%o, или менее чем около -39%o, или менее чем около -40%o. В определенных вариантах реализации получаемая из него композиция L-аминокислот содержит молекулы, имеющие атомы водорода, кислорода и углерода, где атомы водорода, кислорода и углерода составляют по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, или по меньшей мере 80%, или по меньшей мере 90%, или по меньшей мере 95%, от массы композиции, и где величина δ13С композиции находится в диапазоне значений от около -30%o до около -70%o, или где значение δ13С в биомассе снижается с увеличением плотности клеток на величину от около -5%o до около -20%o, или где значение δ13С биомассы выше, чем у СО2, продуцируемого в то же время в среднем на величину от 5%o до 15%o при культивации в присутствии меди или без нее.
Определение значений δ13С для некоторых метаболизирующих C1 микроорганизмов, культивируемых в присутствии получаемого из природного газа сырья, проиллюстрировано в примерах ниже.
Настоящее изобретение дополнительно обеспечивает животный корм, содержащий рекомбинантный метаболизирующий C1 микроорганизм, связанную с ним биомассу, и/или композицию L-аминокислот по настоящему изобретению. Как предусматривается в практике настоящего изобретения, животный корм может быть кормом для домашнего скота (таким как, например, корм для свиней, корм для крупного рогатого скота, корм для овец и т.п.), кормом для домашней птицы (таким как, например, корм для курей, корм для индеек и т.п.), или рыбным кормом (таким как, например, корм для лососевых, корм для моллюсков и ракообразных и т.п.). Животный корм может дополнительно содержать добавку, такую как, например, растительный материал (включая, например, полученный из зерновых культур, таких как, например, кукуруза, ячмень, овес, рис, рожь, пшеница, сорго, пивная дробина и т.п.; и материалы, полученные из бобовых, таких как, например, люцерна, клевер, горох, бобы, чечевица, соя и т.п.), материал животного происхождения (такой как, например, рыбная мука), и/или материал, полученный из микроорганизмов (включая, например, биомассу гетерологичных микроорганизмов которые могут быть, например, бактериями, дрожжами или водорослями). В некоторых вариантах реализации добавка материала растительного происхождения представляет собой соевую муку или белок гороха.
В дополнительном варианте реализации настоящее изобретение обеспечивает среду для культивации или ферментации, содержащую рекомбинантный метаболизирующий С1 микроорганизм, связанную с ним биомассу и/или композицию L-аминокислот по настоящему изобретению. Типично, среда для культивации или ферментации дополнительно содержит углевод (например, сахар и т.п.) и/или воду. В дополнительном варианте реализации настоящее изобретение обеспечивает композицию клеточной культуры, содержащую среду для культивации или ферментации в соответствии с изобретением, и второй микроорганизм. Типично, второй микроорганизм представляет собой бактериальную, дрожжевую или водорослевую клетку.
Варианты реализации изобретения включают следующие:
1. Биомасса, получаемая из культуры рекомбинантного метаболизирующего C1 микроорганизма, причем, рекомбинантный метаболизирующий C1 микроорганизм содержит экзогенную нуклеиновую кислоту, кодирующую фермент биосинтеза L-аминокислоты, при этом метаболизирующий С1 микроорганизм способен конвертировать углеродное сырье, получаемое из природного газа, в L-аминокислоту, и величина δ13С биомассы имеет значение менее -40%o.
2. Биомасса по варианту реализации 1, отличающаяся тем, что рекомбинантный метаболизирующий С1 микроорганизм представляет собой нефотосинтезирующий метаболизирующий C1 микроорганизм.
3. Биомасса по любому из вариантов реализации 1-2, отличающаяся тем, что экзогенная нуклеиновая кислота кодирует фермент, выбранный из группы, состоящей из фермента биосинтеза лизина, фермента биосинтеза триптофана, фермента биосинтеза метионина, фермента биосинтеза цистеина и фермента биосинтеза треонина.
4. Биомасса по любому из вариантов реализации 1-3, отличающаяся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза лизина, выбранный из группы, состоящей из лизин-чувствительной аспартокиназы III (lysC), аспартаткиназы, аспартат-семиальдегид-дегидрогеназы (asd), дигидродипиколинатсинтазы (dapA), дигидродипиколинатредуктазы (dapB), 2,3,4,5-тетрагидропиридин-2,6-карбоксилат-N-сукцинилтрансферазы (dapD), ацетилорнитин/сукцинилдиаминопимелат-аминотрансферазы (argD), сукцинил-диаминопимелат-десукцинилазы (dapE), сукцинилдиаминопимелат-трансаминазы, диаминопимелат-эпимеразы (dapF) и диаминопиметат-дикарбоксилазы (lysA).
5. Биомасса по варианту реализации 4, отличающаяся тем, что L-аминокислота представляет собой L-лизин.
6. Биомасса по любому из вариантов реализации 1-3, отличающаяся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза триптофана, выбранный из группы, состоящей из хоризмат-пируват-лиазы (ubiC), антранилатсинтазы компонента I (trpE), антранилатсинтазы компонента II (trpG), антранилат-фосфорибозилтрансферазы (trpD), фосфорибозилантранилат-изомеразы (trpC), белка биосинтеза триптофана (trpC), N-(5'-фосфорибозил)антранилат-изомеразы (trpF), индол-3-глицеролфосфатсинтазы, триптофансинтазы альфа-цепи (trpA) и триптофансинтазы бета-цепи (trpB).
7. Биомасса по варианту реализации 6, отличающаяся тем, что L-аминокислота представляет собой L-триптофан.
8. Биомасса по любому из вариантов реализации 1-3, отличающаяся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза метионина, выбранный из группы, состоящей из гомосерин-О-сукцинилтрансферазы (metA), цистатионин-гамма-синтазы (metB), белка MalY, цистатионин бета-лиазы (metC), В12-зависимой метионинсинтазы (metH) и 5-метилтетрагидроптероилтриглутамат-гомоцистеин-S-метилтрансферазы (metE).
9. Биомасса по варианту реализации 8, отличающаяся тем, что L-аминокислота представляет собой L-метионин.
10. Биомасса по любому из вариантов реализации 1-3, отличающаяся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза цистеина, выбранный из группы, состоящей из серин-ацетилтрансферазы (CysE) цистеинсинтазы А и цистеинсинтазы В.
11. Биомасса по варианту реализации 10, отличающаяся тем, что L-аминокислота представляет собой L-цистеин.
12. Биомасса по любому из вариантов реализации 1-3, отличающаяся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза треонина, выбранный из группы, состоящей из аспартат-трансаминазы, PLP-зависимой аминотрансферазы, аспартат-аминотрансферазы, аспартаткиназы, аспартат-семиальдегид-дегидрогеназы, гомосерин-дегидрогеназы, гомосеринкиназы и треонинсинтазы.
13. Биомасса по варианту реализации 12, отличающаяся тем, что L-аминокислота представляет собой L-треонин.
14. Биомасса по любому из вариантов реализации 1-13, отличающаяся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза L-аминокислоты, являющийся эндогенным по отношению к микроорганизму, выбранному из группы, состоящей из Е. coli и С. glutamicum.
15. Биомасса по любому из вариантов реализации 1-2, отличающаяся тем, что экзогенная нуклеиновая кислота кодирует последовательность фермента биосинтеза L-аминокислоты, выбранную из группы, состоящей из любой из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40,42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 88, 90, 92, 94, 96, 98, 100, 102, 104, 106, 108, 110, 112, 114, 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148, или 150.
16. Биомасса по любому из вариантов реализации 1-15, отличающаяся тем, что последовательность экзогенной нуклеиновой кислоты оптимизирована по использованию кодонов для оптимальной экспрессии рекомбинантным метаболизирующим C1 микроорганизмом.
17. Биомасса по любому из вариантов реализации 1-16, отличающаяся тем, что экзогенная нуклеиновая кислота, кодирующая фермент биосинтеза L-аминокислоты, функционально связана с контрольной последовательностью экспрессии.
18. Биомасса по варианту реализации 17, отличающаяся тем, что контрольная последовательность экспрессии является экзогенной контрольной последовательностью экспрессии.
19. Биомасса по любому из вариантов реализации 1-18, отличающаяся тем, что метаболизирующий C1 микроорганизм дополнительно содержит делецию активности эндогенного фермента.
20. Биомасса по любому из вариантов реализации 1-19, отличающаяся тем, что метаболизирующий С1 микроорганизм является метанотрофом.
21. Биомасса по варианту реализации 20, отличающаяся тем, что метанотроф представляет собой Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium, Methanomonas, Methylocella или Methylocapsa.
22. Биомасса по варианту реализации 20, отличающаяся тем, что метанотроф выбирают из группы, состоящей из Methylococcus capsulatus штамм Bath, Methylomonas methanica 16a (АТСС РТА 2402), Methylosinus trichosporium OB3b (NRRL B-11,196), Methylosinus sporium (NRRL B-11,197), Methylocystis parvus (NRRL B-11,198), Methylomonas methanica (NRRL B-11,199), Methylomonas albus (NRRL B-11,200), Methylobacter capsulatus (NRRL B-11,201), Methylobacterium organophilum (ATCC 27,886), Methylomonas sp. AJ-3670 (FERM P-2400), Methylocella silvestris, Methylocella palustris (ATCC 700799), Methylocella tundrae, Methylocystis daltona штамм SB2, Methylocystis bryophila, Methylocapsa aurea KYG, Methylacidiphilum infernorum, Methylibium petroleiphilum и Methylomicrobium alcaliphilum.
23. Биомасса по любому из вариантов реализации 1-22, отличающаяся тем, что полученное из природного газа углеродное сырье выбирают из группы, состоящей из природного газа, сингаза, метана, метанола, формальдегида, муравьиной кислоты, окиси углерода, двуокиси углерода, цианида, метиламина, метилтиола, метилгалогена и любой комбинации или двух или больше из них.
24. Биомасса по варианту реализации 23, отличающаяся тем, что полученное из природного газа углеродное сырье является природным газом.
25. Биомасса по варианту реализации 23, отличающаяся тем, что полученное из природного газа углеродное сырье является метаном.
26. Биомасса по варианту реализации 23, отличающаяся тем, что полученное из природного газа углеродное сырье является сингазом.
27. Биомасса по варианту реализации 23, отличающаяся тем, что метаболизирующий C1 микроорганизм является бактериями, метаболизирующими сингаз.
28. Биомасса по варианту реализации 27, отличающаяся тем, что бактерии, метаболизирующие сингаз, выбирают из группы, состоящей из Clostridium autoethanogenum, Clostridium ljungdahli, Clostridium ragsdalei, Clostridium carboxydivorans, Butyridbacterium methylotrophicum, Clostridium woodii и Clostridium neopropanologen.
29. Биомасса по любому из вариантов реализации 1-28, отличающаяся тем, что δ13С биомассы имеет значение менее -50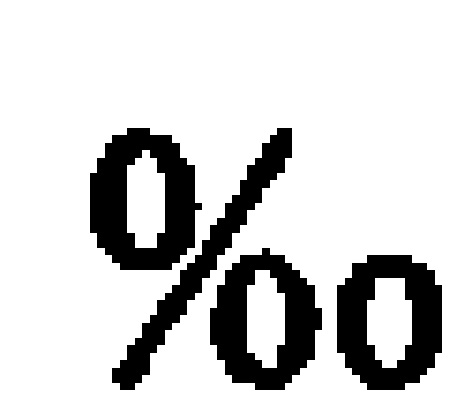 .
.
30. Композиция, содержащая L-аминокислоту, которая демонстрирует δ13С менее -40.
31. Композиция по варианту реализации 30, в которой L-аминокислоту выбирают из группы, состоящей из L-лизина, L-триптофана, L-метионина, L-цистеина и L-треонина.
32. Животный корм, содержащий биомассу по любому из вариантов реализации 1-29, или композиция по любому из вариантов реализации 30-31.
33. Животный корм по варианту реализации 32, дополнительно содержащий материал растительного происхождения.
34. Животный корм по варианту реализации 33, отличающийся тем, что материал растительного происхождения выбирают из группы, состоящей из соевой муки и белка гороха.
35. Среда для культивации или ферментации, содержащая биомассу по любому из вариантов реализации 1-27, или композиция по любому из вариантов реализации 40-42.
36. Рекомбинантный метаболизирующий C1 микроорганизм, содержащий экзогенную нуклеиновую кислоту, кодирующую фермент биосинтеза L-аминокислоты, где метаболизирующий С1 микроорганизм способен конвертировать углеродное сырье, получаемое из природного газа, в L-аминокислоту, и где δ13С биомассы имеет значение менее -40.
37. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 36, отличающийся тем, что рекомбинантный метаболизирующий C1 микроорганизм представляет собой нефотосинтезирующий метаболизирующий C1 микроорганизм.
38. Рекомбинантный метаболизирующий С1 микроорганизм по любому из вариантов реализации 36-37, отличающийся тем, что экзогенная нуклеиновая кислота кодирует фермент, выбранный из группы, состоящей из фермента биосинтеза лизина, фермента биосинтеза триптофана, фермента биосинтеза метионина, фермента биосинтеза цистеина и фермента биосинтеза треонина.
39. Рекомбинантный метаболизирующий С1 микроорганизм по любому из вариантов реализации 36-38, отличающийся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза лизина, выбранный из группы, состоящей из лизин-чувствительной аспартокиназы III (lysC), аспартаткиназы, аспартат-семиальдегид-дегидрогеназы (asd), дигидродипиколинатсинтазы (dapA), дигидродипиколинатредуктазы (dapB), 2,3,4,5-тетрагидропиридин-2,6-карбоксилат-N-сукцинилтрансферазы (dapD), ацетилорнитин/сукцинилдиаминопимелат-аминотрансферазы (argD), сукцинил-диаминопимелат-десукцинилазы (dapE), сукцинилдиаминопимелат-трансаминазы, диаминопимелат-эпимеразы (dapF) и диаминопиметат-дикарбоксилазы (lysA).
40. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 39, отличающийся тем, что L-аминокислота представляет собой L-лизин.
41. Рекомбинантный метаболизирующий С1 микроорганизм по любому из вариантов реализации 36-38, отличающийся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза триптофана, выбранный из группы, состоящей из хоризмат-пируват-лиазы (ubiC), антранилатсинтазы компонента I (trpE), антранилатсинтазы компонента II (trpG), антранилат-фосфорибозилтрансферазы (trpD), фосфорибозилантранилат-изомеразы (trpC), белка биосинтеза триптофана (trpC), N-(5'-фосфорибозил)антранилат-изомеразы (trpF), индол-3-глицеролфосфатсинтазы, триптофансинтазы альфа-цепи (trpA) и триптофансинтазы бета-цепи (trpB).
42. Рекомбинантный метаболизирующий С1 микроорганизм по варианту реализации 41, отличающийся тем, что L-аминокислота представляет собой L-триптофан.
43. Рекомбинантный метаболизирующий С1 микроорганизм по любому из вариантов реализации 36-38, отличающийся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза метионина, выбранный из группы, состоящей из гомосерин-О-сукцинилтрансферазы (metA), цистатионин-гамма-синтазы (metB), белка MalY, цистатионин-бета-лиазы (metC), В12-зависимой метионинсинтазы (metH), и 5-метилтетрагидроптероилтриглутамат-гомоцистеин-S-метилтрансферазы (metE).
44. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 43, отличающийся тем, что L-аминокислота представляет собой L-метионин.
45. Рекомбинантный метаболизирующий C1 микроорганизм по любому из вариантов реализации 36-38, отличающийся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза цистеина, выбранный из группы, состоящей из серин-ацетилтрансферазы (CysE), цистеинсинтазы А и цистеинсинтазы В.
46. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 45, отличающийся тем, что L-аминокислота представляет собой L-цистеин.
47. Рекомбинантный метаболизирующий C1 микроорганизм по любому из вариантов реализации 36-38, отличающийся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза треонина, выбранный из группы, состоящей из аспартат-трансаминазы, PLP-зависимой аминотрансферазы, аспартат-аминотрансферазы, аспартаткиназы, аспартат-семиальдегид-дегидрогеназы, гомосерин-дегидрогеназы, гомосеринкиназы и треонинсинтазы.
48. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 47, отличающийся тем, что L-аминокислота представляет собой L-треонин.
49. Рекомбинантный метаболизирующий C1 микроорганизм по любому из вариантов реализации 36-48, отличающийся тем, что экзогенная нуклеиновая кислота кодирует фермент биосинтеза L-аминокислоты, являющийся эндогенным по отношению к микроорганизму, выбранному из группы, состоящей из Е. coli и С. glutamicum.
50. Рекомбинантный метаболизирующий C1 микроорганизм по любому из вариантов реализации 36-37, отличающийся тем, что экзогенная нуклеиновая кислота кодирует последовательность фермента биосинтеза L-аминокислоты, выбранную из группы, состоящей из любой из SEQ ID NO: 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 88, 90, 92, 94, 96, 98, 100, 102, 104, 106, 108, 110, 112, 114, 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148 и 150.
51. Рекомбинантный метаболизирующий C1 микроорганизм по любому из вариантов реализации 36-50, отличающийся тем, что последовательность экзогенной нуклеиновой кислоты оптимизирована по использованию кодонов для оптимальной экспрессии рекомбинантным метаболизирующим С1 микроорганизмом.
52. Рекомбинантный метаболизирующий C1 микроорганизм по любому из вариантов реализации 36-51, отличающийся тем, что экзогенная нуклеиновая кислота, кодирующая фермент биосинтеза L-аминокислоты, функционально связана с контрольной последовательностью экспрессии.
53. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 52, отличающийся тем, что контрольная последовательность экспрессии является экзогенной контрольной последовательностью экспрессии.
54. Рекомбинантный метаболизирующий С1 микроорганизм по любому из вариантов реализации 36-53, отличающийся тем, что метаболизирующий С1 микроорганизм дополнительно содержит делецию активности эндогенного фермента.
55. Рекомбинантный метаболизирующий С1 микроорганизм по любому из вариантов реализации 36-54, в котором метаболизирующий C1 микроорганизм является метанотрофом.
56. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 55, отличающийся тем, что метанотроф представляет собой Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium, Methanomonas, Methylocella или Methylocapsa.
57. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 55, отличающийся тем, что метанотроф выбирают из группы, состоящей из Methylococcus capsulatus штамм Bath, Methylomonas methanica 16a (АТСС РТА 2402), Methylosinus trichosporium OB3b (NRRL B-11,196), Methylosinus sporium (NRRL B-11,197), Methylocystis parvus (NRRL B-11,198), Methylomonas methanica (NRRL B-11,199), Methylomonas albus (NRRL B-11,200), Methylobacter capsulatus (NRRL B-11,201), Methylobacterium organophilum (ATCC 27,886), Methylomonas sp. AJ-3670 (FERM P-2400), Methylocella silvestris, Methylocella palustris (ATCC 700799), Methylocella tundrae, Methylocystis daltona штамм SB2, Methylocystis bryophila, Methylocapsa aurea KYG, MethylaC1diphilum infernorum, Methylibium petroleiphilum, и Methylomicrobium alcaliphilum.
58. Рекомбинантный метаболизирующий C1 микроорганизм по любому из вариантов реализации 36-57, отличающийся тем, что полученное из природного газа углеродное сырье выбирают из группы, состоящей из природного газа, сингаза, метана, метанола, формальдегида, муравьиной кислоты, окиси углерода, двуокиси углерода, цианида, метиламина, метилтиола, метилгалогена и любой комбинации или двух или больше из них.
59. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 58, отличающийся тем, что полученное из природного газа углеродное сырье является природным газом.
60. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 58, отличающийся тем, что полученное из природного газа углеродное сырье является метаном.
61. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 58, отличающийся тем, что полученное из природного газа углеродное сырье является сингазом.
62. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 61, отличающийся тем, что метаболизирующий C1 микроорганизм представляет собой бактерии, метаболизирующие сингаз.
63. Рекомбинантный метаболизирующий C1 микроорганизм по варианту реализации 62, отличающийся тем, что бактерии, метаболизирующие сингаз, выбирают из группы, состоящей из Clostridium autoethanogenum, Clostridium ljungdahli, Clostridium ragsdalei, Clostridium carboxydivorans, Butyridbacterium methylotrophicum, Clostridium woodii, и Clostridium neopropanologen.
64. Рекомбинантный метаболизирующий C1 микроорганизм по любому из вариантов реализации 36-63, в котором δ13С биомассы имеет значение менее -50%o.
65. Способ продуцирования L-аминокислота, включающий культивацию рекомбинантного метаболизирующего С1 микроорганизма по любому из вариантов реализации 36-64 в присутствии углеродного сырья, получаемого из природного газа, в условиях, достаточных для продуцирования L-аминокислоты.
66. L-аминокислота, продуцируемая способом по варианту реализации 65, отличающийся тем, что L-аминокислота демонстрирует δ13С, имеющий значение менее -40%o.
Вышеперечисленные и другие аспекты изобретения могут быть более понятны в сочетании со следующими неограничивающими Примерами.
ПРИМЕРЫ
ПРИМЕР 1
УСЛОВИЯ КУЛЬТИВАЦИИ И ВЫРАЩИВАНИЯ В БИОРЕАКТОРЕ ДЛЯ МЕТАБОЛИЗИРУЮЩИХ С1 МИКРООРГАНИЗМОВ
Типичные примеры метаболизирующих С1 микроорганизмов в соответствии с данным описанием (метанотрофы, метилотрофы, клостридии) культивировали в пробирках, во флаконах, в бутылях, на планшетах, или в биореакторе (ферментация). Условия роста, среды, и источники углерода для различных микроорганизмов описаны в данном Примере.
Methylosinus trichosporium штамм OB3b (NCIMB 11131); Methylomonas sp. штамм 16а (ATCC РТА-2402); или Methylomonas methanica
Для сывороточных бутылок, бактерии культивировали при 30°С в минимальной среде Хиггинса с нитратными солями (NSM; Cornish et al., J. Gen. Microbiol. 130:2565, 1984; Park et al., Biotechnol. Bioeng. 38:423, 1991) или среде MM-W1. Состав в свободном пространстве доводили до соотношения метан:воздух, равного 1:1 по объему. Бутылки встряхивали со скоростью 200-250 об/мин. Альтернативно, культуру поддерживали на планшетах со средой NSM, содержащей 1,5% мас./об. агара, выращивали в герметично закрытой камере, содержащей газовую смесь 1:1 (об./об.) метан:воздух, или в присутствии паров метанола (с помощью 0,5 мл метанола в крышке герметизованных парафилмом планшетов) или на планшетах со средой NSM с добавкой 0,5% метанола. Планшеты инкубировали в перевернутом положении в увлажненной камере при 30°С.
Используемая среда NSM имела следующий состав: 1,0 г MgSO4*7H2O, 0,20 г CaCl2*6H2O, 2,0 мл раствор хелата железа (0,1 г железа(III)-аммония цитрата или 0,5 г железа(III) хлорида; 0,2 г ЭДТА, натриевой соли; 0,3 мл HCl, концентрированной; 100,0 мл дистиллированной деионизированной H2O), 1,0 г KNO3, 0,5 мл раствора следовых элементов (500,0 мг ЭДТА, 200,0 мг FeSO4. 7H2O, 10,0 мг ZnSO4*7H2O, 3,0 мг MnCl2*4H2O, 30,0 мг Н3ВО3, 20,0 мг CoCl2*6H2O, 1,0 мг CaCl2*2H2O, 2,0 мг NiCl2*6H2O, 3,0 мг Na2MoO4*2H2O, 1,0 л дистиллированной воды), 0,272 г KH2PO4, 0,717 г Na2HPO4*12H2O, необязательно 12,5 г очищенного агара (например, агара Oxoid L28 или Bacto™; используется при приготовлении планшетов), 1,0 л дистиллированной деионизированной воды, рН доводят до 6,8 и автоклавируют при 121°С в течение 15 минут.
Для ферментации, 2-литровый биореактор, содержащий 1 л стерилизованной среды MM-W1 определенного состава инокулировали клетками из сывороточной бутылки периодической культуры (10-20% об./об.), выращенной в MM-W1 при подаче смеси 1:1 (об./об.) метана и воздуха. Использованная среда MM-W1 имела следующий состав: 0,8 мМ MgSO4*7H2O, 10 мМ NaNO3, 0,14 мМ CaCl2, 1,2 мМ NaHCO3, 2,35 мМ KH2PO4, 3,4 мМ K2HPO4, 20,7 мкМ Na2MoO4*2H2O, 1 мкМ CuSO4*5H2O, 10 мкМ FeIII-Na-ЭДТА, и 1 мл на литр раствора следовых металлов (содержащего, на литр, 500 мг FeSO4*7H2O, 400 мг ZnSO4*7H2O, 20 мг MnCl2*7H2O, 50 мг CoCl2*6H2O, 10 мг NiCl2*6H2O, 15 мг H3BO3, 250 мг ЭДТА). Фосфат, бикарбонат и FeIII-Na-ЭДТА прибавляли после автоклавирования и охлаждения среды. Бикарбонат прибавляли до 0,1% (мас./об.) для некоторых ферментации. Содержимое реактора перемешивали с помощью погружной лопастной мешалки при постоянной скорости 750 об/мин. Культуру подпитывали постоянным продуванием метана при расходе от около 60 мл/мин до около 120 мл/мин, в то время как концентрированный кислород (по меньшей мере 85%) подавали при переменном расходе около 10-100 мл/мин для поддержания уровня растворенного кислорода от около 40% до около 80% (по отношению к насыщению среды воздухом).
Температуру в биореакторе поддерживали при 30°С и рН поддерживали равным 7,1±0,1 путем автоматического добавления 0,5М NaOH и 0,5М HCl, вместе с другими добавками, в культуру через каждые от около 4 часов до около 24 часов (соответствует увеличению OD600 приблизительно на 5 единиц оптической плотности (OD)). Другими добавками были поочередно добавка металлов (конечные концентрации 10 мкМ CuSO4, 5 мкМ FeSO4, 5 мкМ FeIII-Na-ЭДТА) и добавка питательных веществ (5,75 мМ KxHyPO4, 10 мМ NaNO3). В таких условиях наблюдался по существу линейный рост, с эффективной скоростью образования биомассы от около 2,7 до около 3,3 грамм сухой массый клеток на литр в день до OD600 более 20. Культуру биомассы собирали центрифугированием, промывали один раз средой MM-W1, и выделенную биомассу или замораживали при -80°С, или немедленно использовали для фракционирования клеточных компонентов (например, экстракция липидов).
Метод полунепрерывной ферментации также может быть использован для поддержания продуктивности биомассы и уменьшения времени, связанного с остановкой и запуском ферментации (т.е., оборачиваемость или время производственного цикла).
Сбор бактериальной биомассы осуществляют с интервалами приблизительно 12-24 часа, когда плотность культуры приближается к (но не доходит до) стационарной фазе. Отбирали приблизительно половину объема биореактора путем перемещения в отдельный контейнер с помощью центробежного насоса. Равный объем стерилизованной или рециркулированной среды затем возвращали в биореактор таким образом, чтобы оптическая плотность реактора составляла приблизительно половину от его начального значения. Ферментацию в биореакторе продолжали в соответствии с вышеописанным протоколом, так чтобы множество циклов роста и выделения биомассы могло осуществляться на протяжении одной ферментации.
Methylococcus capsulatus Bath (NCIMB 11132)
Бактерии культивировали при 42°С в сывороточных бутылках, содержащих минимальную среду Хиггинса с нитратными солями (NSM) или среду MM-W1. Состав газов в свободном пространстве над жидкостью доводили до 1:1 смеси метан:воздух по объему. Бутылки встряхивали со скоростью 200-250 об/мин. Альтернативно, культуру поддерживали на планшетах со средой NSM, отвержденной с помощью 1,5% мас./об. агара, выращивали в герметичной камере, содержащей газовую смесь 1:1 (об./об.) метан:воздух. Планшеты инкубировали в перевернутом положении в камере при 42°С.
Для ферментации, 3-литровый биореактор, содержащий 1,25 л стерилизованной среды MMF1.1, инокулировали клетками из сывороточной бутылки с периодической культурой (10-20% об./об.), выращенной в той же среде с добавкой 1:1 (об./об.) смеси метана и воздуха. Среда MMF1.1 имела следующий состав: 0,8 мМ MgSO4*7H2O, 40 мМ NaNO3, 0,14 мМ CaCl2, 6 мМ NaHCO3, 4,7 мМ KH2PO4, 6,8 мМ K2HPO4, 20,7 мкМ Na2MoO4*2H2O, 6 мкМ CuSO4*5H2O, 10 мкМ FeIII-Na-ЭДTA, и 1 мл на литр раствора следовых металлов (содержащего, на литр 500 мг FeSO4*7H2O, 400 мг ZnSO4*7H2O, 20 мг MnCl2*7H2O, 50 мг CoCl2*6H2O, 10 мг NiCl2*6H2O, 15 мг H3BO3, 250 мг ЭДТА). Фосфат, бикарбонат и FeIII-Na-ЭДТА прибавляют после автоклавирования и охлаждения среды. Содержимое реактора перемешивали с помощью погружной лопастной мешалки при постоянной скорости 750 об/мин. Культуру подпитывали постоянным продуванием метана со скоростью от около 60 до около 200 мл/мин, в то время как концентрированный кислород (>85%) подавали с переменным расходом в диапазоне значений 15-90 мл/мин и уровень содержания растворенного кислорода поддерживали ниже 10% (по отношению к насыщению среды воздухом).
Температуру в биореакторе поддерживали равной 44°С и рН поддерживали на уровне 7,0±0,1 путем автоматического добавления 0,5М NaOH и 0,5М HCl, вместе с добавками меди и железа (конечная концентрация 5 мкМ CuSO4, 5 мкМ FeSO4, 10 мкМ FeIII-Na-ЭДTA) в культуру через каждые 3-6 часов (соответствует увеличению значения OD600 приблизительно на 3-5 единиц OD после достижения OD 5). В таких условиях наблюдался по существу линейный рост, с эффективной скоростью образования биомассы более 5 грамм сухой массы клеток на литр в день до значения CD600 больше 10. Культуру биомассы собирали центрифугированием, клетки промывали один раз средой MM-W1 и клеточный осадок или замораживали при -80°С, или использовали немедленно для фракционирования клеточных компонентов.
Истощение питательных веществ было определено как фактор, который может ограничивать урожай биомассы в процессе ферментации. Во избежание ограничения питательных веществ, преимущественно азота и фосфата, инициировали подачу питательных веществ, состоящих из 2-кратной концентрированной MMF1.1, после того как значение OD600 культуры превышало 5. Подачу питательных веществ инициировали при степени разбавления, соответствующей приблизительно половине скорости роста культуры во избежание вымывания и для поддержания увеличения OD при расширении объема культура. Ферментацию в биореакторе продолжали в соответствии с вышеописанным протоколом таким образом, чтобы множество циклов роста и выделения биомассы можно было провести в ходе одной ферментации.
Methylobacterium extorquens или Methylosinus trichosporium штамм OB3b (NCIMB 11131)
Бактерии культивировали при 30°С в пробирках, содержащих минимальную среду Хиггинса с нитратными солями (NSM) с добавкой 0,5% метанола. Пробирки встряхивали со скоростью 200-250 об/мин. Альтернативно, культуры поддерживаемые на планшетах со средой NSM, содержащей 1,5% мас./об. агара, выращивали в присутствии паров метанола (с помощью 0,5 мл метанола в крышке планшетов, герметизованных с помощью пленки парафилм (parafilm)), или с добавкой 0,5% метанола. Планшеты инкубировали в перевернутом положении в увлажненной камере с нормальной атмосферой при 30°С.
Для ферментации, 2-литровый биореактор, содержащий 1 л среды MM-W1 определенного состава, инокулировали клетками из пробирки с периодической культурой (10-20% об./об.). Среда MM-W1 имела состав, описанный выше. Содержимое реактора перемешивали с помощью погружной лопастной мешалки при постоянной скорости 800 об/мин. Культуру подпитывали начальной увеличенной дозой метанола до конечной концентрации 0,5% и метанолом с переменной скоростью подачи, в то время как чистый кислород подавали с переменным расходом в диапазоне 30-100 мл/мин для поддержания уровня растворенного кислорода 60-90% (по отношению к насыщению среды воздухом).
Температуру в биореакторе поддерживали равной 30°С, и рН поддерживали на уровне 7,1±0,1 путем автоматического добавления 0,5М NaOH и 1М HCl, вместе с добавками металлов и питательных веществ, как описано выше. В таких условиях, наблюдается по существу линейный рост, с эффективной скоростью образования биомассы от 2,7 до 3,3 грамм сухой массы клеток на литр в день до величины OD600 более 20. Культуру биомассы собирали центрифугированием, клетки промывали один раз в среде MM-W1 и клеточный осадок или замораживали при -80°С, или использовали немедленно для фракционирования клеточных компонентов.
Методика полунепрерывной ферментации также может быть применена для поддержания продуктивности биомассы и уменьшения времени на остановку и запуск ферментации (т.е. оборачиваемость или время производственного цикла).
Сбор накопленной бактериальной биомассы проводят с интервалами приблизительно 12-24 часов, когда плотность культуры приближается к (но не доходит до) стационарной фазе. Отбирают приблизительно половину объема биореактора путем перемещения в отдельный контейнер с помощью центробежного насоса. Равный объем свежей или рециркулированной среды затем возвращали в биореактор таким образом, чтобы оптическая плотность в реакторе составляла приблизительно половину от ее начального значения. Ферментацию в биореакторе продолжают в соответствии с вышеописанным протоколом таким образом, чтобы множество циклов роста и выделения биомассы осуществлялись за время одной ферментации.
Clostridium autoethanogenum и Clostridium ljungdahlii
Бактерии Clostridium культивировали анаэробно в 100 мл модифицированной среде РЕТС (среда АТСС 1754) при 37°С в бутылках Schott Duran® GL45 на 500 мл с пластиковым покрытием с пробками из бутилкаучука и отходящим газом сталеплавильного завода под давлением 200 кПа. Рост контролировали путем измерения оптической плотности при 600 нм (OD600).
Модифицированная среда РЕТС содержит (на литр) 1 г NH4Cl, 0,4 г KCl, 0,2 г MgSO4*7H2O, 0,8 г NaCl, 0,1 г KH2PO4, 20 мг CaCl2*2H2O, 10 мл раствора следовых элементов (см. ниже), 10 мл витаминного раствора Вульфа (Wolfe's) (см. ниже), 2 г NaHCO3 и 1 мг ресазурина. После доведения рН до 5,6 среду кипятят, анаэробно дозируют и автоклавируют при 121°С в течение 15 мин. Отходящий газ сталеплавильного завода (состав: 44% СО, 32% N2, 22% CO2, 2% Н2) или эквивалентные синтетические смеси используются в качестве источника углерода. Среда имеет конечное значение рН 5,9 и восстанавливается цистеином-HCl и Na2S в концентрации 0,008% (мас./об.).
Раствор следовых элементов содержит 2 г нитрилотриуксусной кислоты (доведена до рН 6 с помощью КОН перед добавлением остальных ингредиентов), 1 г MnSO4, 0,8 г Fe(SO4)2(NH4)2*6H2O, 0,2 г CoCl2*6H2O, 0,2 мг ZnSO4*7H2O, 20 мг CuCl2*2H2O, 20 мг NiCl2*6H2O, 20 мг Na2MoO4*2H2O, 20 мг Na2SeO4, и 20 мг Na2WO4 на литр.
Витаминный раствор Вульфа (Wolin et al., J. Biol. Chem. 238:2882, 1963) содержит (на литр) 2 мг биотина, 2 мг фолиевой кислоты, 10 мг пиридоксина гидрохлорида, 5 мг тиамина-HCl, 5 мг рибофлавина, 5 мг никотиновой кислоты, 5 мг D-(+)-пантотената кальция, 0,1 мг витамина В12, 5 мг п-аминобензойной кислоты и 5 мг тиоктовой кислоты.
а. Ферментация Clostridium autoethanogenum
Ферментация Clostridium autoethanogenum проводится с использованием способов, аналогичных описанным, например, в патентной заявке США № 2011/0300593. Вкратце, 2-литровый биореактор, содержащий 1,3 л Раствора А (3,083 г NH4Ac; 0,61 г MgCl2*6H2O; 0,294 г CaCl2*2H2O; 0,15 г KCl; 0,12 г NaCl (необязательно); доводят до 1 л дистиллированной водой) продувают газообразным N2. Прибавляют 85% раствор Н3РО4 (2,025 мл, 30 мМ) и рН доводят до 5,3 с помощью концентрированного водного NH4OH. Затем прибавляют 13,5 мл Раствора В (20,0 мг биотина; 20,0 мг фолиевой кислоты; 10,0 мг пиридоксина HCl; 50,0 мг тиамина*HCl; 50,0 мг рибофлавина; 50,0 мг никотиновой кислоты; 50,0 мг D-(*)-пантотената кальция; 50,0 мг витамина В12; 50,0 мг п-аминобензойной кислоты; 50,0 мг тиоктовой кислоты; доводят до 1 л дистиллированной водой) и раствор продувают газообразным N2. Прибавляют хлорид хрома(II) для того, чтобы снизить окислительно-восстановительный потенциал (ОВП) раствора до приблизительно -200 мВ, после чего прибавляют ресазурин (1,35 мл 2 г/л раствора). Прибавляют полисульфид натрия (5,4 мл 3М раствора, см. ниже) и раствор продувают N2 и затем СО-содержащим газом (1% Н2; 13% N2; 71% СО; 15% CO2). Прибавляют раствор сульфида металла (150 мл, см. ниже) и раствор продувают еще 30 минут перед инокуляцией культурой активно растущего С. autoethanogenum в количестве приблизительно 5% (об./об.).
Раствор полисульфида натрия готовят в колбе на 500 мл, в которую загружают Na2S (93,7 г, 0,39 моль) и 200 мл H2O. Раствор перемешивают до растворения соли и серу (25 г, 0,1 моль) прибавляют под постоянным током N2. После перемешивания при комнатной температуре в течение 2 часов раствор полисульфида натрия (около 4М по Na и около 5М по сере), представляющий собой прозрачную красновато-коричневую жидкость, переносят в продутые N2 сывороточные бутылки и заворачивают в алюминиевую фольгу.
Раствор хрома(II) готовят в трехгорлой колбе на 1 л, оснащенной газонепроницаемыми входным и выходным отверстиями для обеспечения возможности работы под инертным газом и последующего переноса требуемого продукта в пригодную колбу для хранения. В колбу загружают CrCl3*6H2O (40 г, 0,15 моль), гранулы цинка [20 меш (0,841 мм)] (18,3 г, 0,28 моль), ртуть (13,55 г, 1 мл, 0,0676 моль) и 500 мл дистиллированной воды. После продувки N2 в течение одного часа смесь нагревают до около 80°С для инициации реакции. После двух часов перемешивания под постоянным током N2, смесь охлаждают до комнатной температуры и непрерывно перемешивают еще 48 часов, за это время реакционная смесь превращается в темно-синий раствор. Раствор переносят в продутые N2 сывороточные бутылки и хранят при 4°С для последующего использования.
Раствор сульфида металла готовят путем прибавления около 950 мл Раствора А в ферментер на 1 л и продувки газообразным N2. Прибавляют 85% раствор H3PO4(1,5 мл, 30 мМ) и рН доводят до 5,3 с помощью концентрированного водного NH4OH. Прибавляют раствор В (10 мл) и раствор продувают N2. Прибавляют хлорид хрома(II), чтобы снизить окислительно-восстановительный потенциал (ОВП) раствора до приблизительно -200 мВ, после чего прибавляют ресазурин (1 мл 2 г/л раствора). Прибавляют раствор С (1/10; 10 мл FeCl3; 5 мл CoCl2; 5 мл NiCl2; 1 мл Н3ВО3; 1 мл Na2MoO4; 1 мл MnCl2; 1 мл Na2WO4; 1 мл ZnCl2; 1 мл Na2SeO3; в 1 л среды), затем прибавляют полисульфид натрия (2 мл 3М раствора), и затем раствор продувают газообразным N2.
Ферментация субстрата, содержащего СО, с использованием С. autoethanogenum в условиях периодического процесса в присутствии полисульфида приводит к значительно повышенной скорости накопления и накоплению конечного количества биомассы приблизительно 4 г/л за период 2-3 дней. Например, после короткой латентной фазы продолжительностью приблизительно 1 день, биомасса может нарасти от около 0,5 г/л до по меньшей мере 3,5 г/л за приблизительно 36 часов ферментации. Кроме того, ацетат не продуцируется в фазе роста в присутствии полисульфида (как это типично происходит при периодической ферментации) и в определенных обстоятельствах некоторая часть ацетата расходуется, так что происходит суммарное снижение количества ацетата в ферментере. Культуру биомассы собирали центрифугированием, клетки промывали один раз в среде и клеточный осадок или замораживали при -80°С, или использовали немедленно для фракционирования клеточных компонентов.
Метод полунепрерывной ферментации также может быть использован для поддержания продуктивности биомассы и уменьшения времени, связанного с остановкой и запуском ферментации (т.е. оборачиваемости или времени производственного цикла).
Сбор накопленной бактериальной биомассы осуществляют с интервалами приблизительно 12-24 часов, когда плотность культуры приближается к (но не достигает) стационарной фазе. Отбирают приблизительно половину объема биореактора путем перемещения в отдельный контейнер с помощью центробежного насоса. Равный объем свежей или рециркулированной среды затем возвращают в биореактор таким образом, чтобы оптическая плотность в реакторе составляла приблизительно половину от исходного значения. Ферментацию в биореакторе продолжают в соответствии с вышеописанным протоколом таким образом, чтобы множество циклов роста и выделения биомассы осуществлялось за одну ферментацию.
b. Ферментация Clostridium ljungdahlii
Ферментация Clostridium ljungdahlii проводится с использованием способов, аналогичных описанным, например, в патентах США №№5173429 и 5593886. Вкратце, периодическую ферментацию проводят с использованием биологически чистой культуры С. ljungdahlii. Приготовление среды ((1) 80,0 мл соли, содержащей КН2РО4 3,00 г/л, K2HPO4 3,00 г/л, (NH4)2SO4 6,00 г/л, NaCl 6,00 г/л, MgSO4*2H2O 1,25 г/л; (2) 1,0 г дрожжевого экстракта; (3) 1,0 г триптиказы; (4) 3,0 мл раствора следовых металлов PFN (Pfenning), содержащего FeCl2*4H2O 1500 мг, ZnSO4*7H2O 100 мг, MnCl2*4H2O 30 мг, Н3ВО3 300 мг, CoCl2*6H2O 200 мг, CuCl2*H2O 10 мг, NiCl2*6H2O 20 мг, NaMoO4*2H2O 30 мг, Na2SeO3 10 мг, и дистиллированную воду до 1 л; (5) 10,0 мл витаминов В, включая пиридоксаль-HCl 10 мг, рибофлавин 50 мг, тиамин-HCl 50 мг, никотиновую кислоту 50 мг, Ca-D-пантотеинат 50 мг, липоевую кислоту 60 мг, п-аминобензойную кислоту 50 мг, фолиевую кислоту 20 мг, биотин 20 мг, цианокобаламин 50 мг и дистиллированную воду до 1 л; (6) 0,5 г цистеина-HCl; (7) 0,06 г CaCl2*2H2O; (8) 2,0 г NaHCO3; (9) 1,0 мл ресазурина (0,01%); и (10) 920,0 мл дистиллированной воды) осуществляют анаэробно в атмосфере 80% азота и 20% CO2. рН среды контролируют в процессе ферментации и поддерживают равным 5,0 с помощью HCl. При необходимости регулировку рН проводят с помощью стерильных растворов 10% NaOH или 1,0% уксусной кислоты. Среду переносят в сывороточные бутылки на 157,5 мл и закрывают пробками из бутилкаучука и алюминиевыми колпачками. Бутылки затем автоклавируют при 121°С в течение 20 минут.
Приблизительно за 48 часов до начала эксперимента готовят посевную культуру из маточной культуры С. ljungdahlii в бутылке аналогично тому, как описано выше. Посевную культуру выращивают в инкубаторе-шейкере при 37°С и встряхивают при 100 об/мин. Восстановительные растворы (2,0 мл Na2S, 2,5% раствор, и 2,0 мл цистеина-HCl, 3,5% раствор) прибавляют к культуре, которую помещают в инкубатор-шейкер приблизительно на 15 минут для обеспечения полного удаления кислорода и температурной адаптации. В отличие от процедуры, используемой для выделения биологически чистой культуры организма, добавление ингибиторов метана не требуется при периодической ферментации.
Ферментация С. ljungdahlii проводится в 2,5-литровом ферментере New Brunswick Scientific Bioflow IIc, содержащем питательную среду при 37°С, и поддерживают постоянный уровень жидкости, равный 1,5 литра, причем жидкость перемешивают с переменной скоростью до 1000 оборотов в минуту при подаче газа со скоростью приблизительно 500 кубических сантиметров в минуту. Оптимальное время удержания газа составляет около трех минут. Газовое сырье меняется в процессе его поглощения бактериями, которое в свою очередь является функцией плотности клеток.
Сбор накопленной бактериальной биомассы осуществляют с интервалами приблизительно 12-24 часов, когда плотность культуры приближается к (но не достигает) стационарной фазе. Отбирают приблизительно половину объема биореактора путем перемещения в отдельный контейнер с помощью центробежного насоса. Равный объем свежей или рециркулированной среды затем возвращают в биореактор таким образом, чтобы оптическая плотность в реакторе составляла приблизительно половину от начального значения. Ферментацию в биореакторе продолжают в соответствии с вышеописанным протоколом таким образом, чтобы множество циклов роста и выделения биомассы осуществлялось за время одной ферментации.
ПРИМЕР 2
РАСПРЕДЕЛЕНИЕ СТАБИЛЬНЫХ ИЗОТОПОВ УГЛЕРОДА В ЛИПИДАХ МЕТАБОЛИЗИРУЮЩИХ C1 МИКРООРГАНИЗМОВ
Сухие образцы биомассы М. trichosporium и липидных фракций анализировали на содержание углерода и азота (% сухой массы), и соотношения стабильных изотопов углерода (13С) и азота (15N) с помощью элементного анализа (analyzer)/масс-спектрометрии изотопного соотношения в непрерывном потоке с использованием элементного анализатора CHNOS (vario ISOTOPE cube, Elementar, Hanau, Germany)/ сопряженного с IsoPrime100 IRMS (Isoprime, Чидл, Великобретания). Образцы метанотрофной биомассы, культивируемой в ферментерах или сывороточных бутылках, центрифугировали, ресуспендировали в деионизированной воде, и объемы, соответствующие 0,2-2 мг углерода (около 0,5-5 мг сухой массы клеток) переносили в оловянные капсулы 5×9 мм (Costech Analytical Technologies, Inc., Valencia, CA) и высушивали при 80°C в течение 24 часов. Аналогично, предварительно экстрагированные липидные фракции суспендировали в хлороформе и объемы, содержащие 0,1-1,5 мг углерода, переносили в оловянные капсулы и выпаривали досуха при 80°С в течение 24 часов. Стандарты, содержащие 0,1 мг углерода, давали достоверные значения δ13С.
Изотопное соотношение выражали в виде показателя «дельта» (%o), где изотопный состав материала определяется по сравнению со стандартом как отклонение на миллион (per million deviation basis), выраженное как δ13С (или δ15N) = (Rобразец/RСтандарт-1)×1000, где R обозначает молярное отношение тяжелой формы изотопа к легкой. Стандартом для углерода является Vienna Pee Dee Belemnite (V-PDB), и для азота - воздух. В качестве калибровочного стандарта использовали предложенный NIST (Национальный институт стандартов и технологий) SRM (стандартный эталонный материал) №1547, листья персика. Все изотопные анализы проводили в Центре биогеохимии стабильных изотопов (Center for Stable Isotope Biogeochemistry) Университета Калифорнии (University of California, Беркли). Долгосрочная воспроизводимость результатов для изотопных анализов С и N составляла 0,10%o и 0,15%o, соответственно.
М. trichosporium штамм OB3b выращивали на метане в трех разных экспериментах по ферментации, М. capsulatus Bath выращивали на метане в двух разных экспериментах по ферментации, и Methylomonas sp. 16а выращивали на метане в одном эксперименте по ферментации. Биомассу каждой из этих культур анализировали на распределение стабильных изотопов углерода (значения δ13С; см. Таблицу 1).
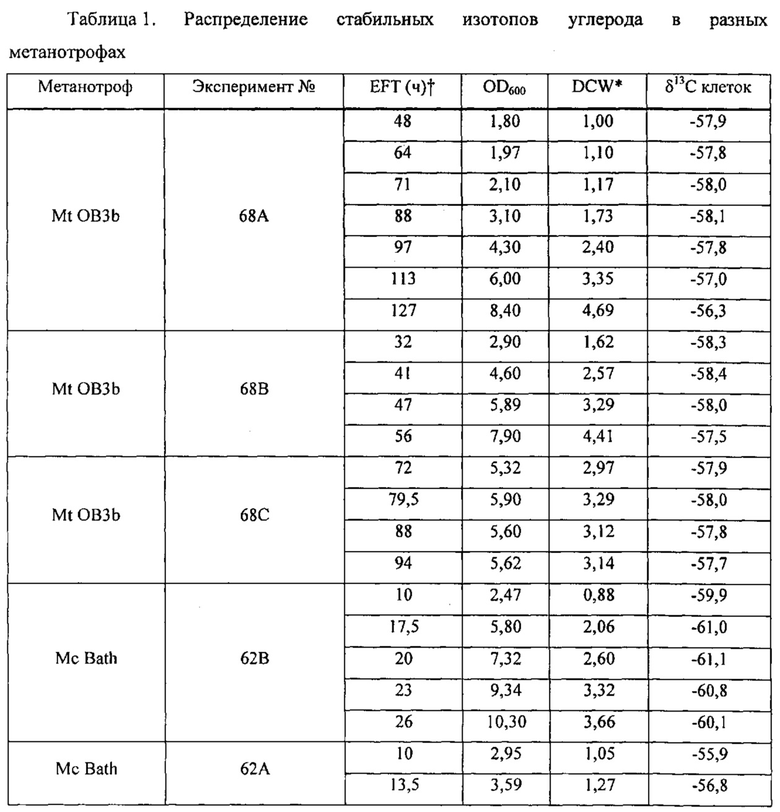
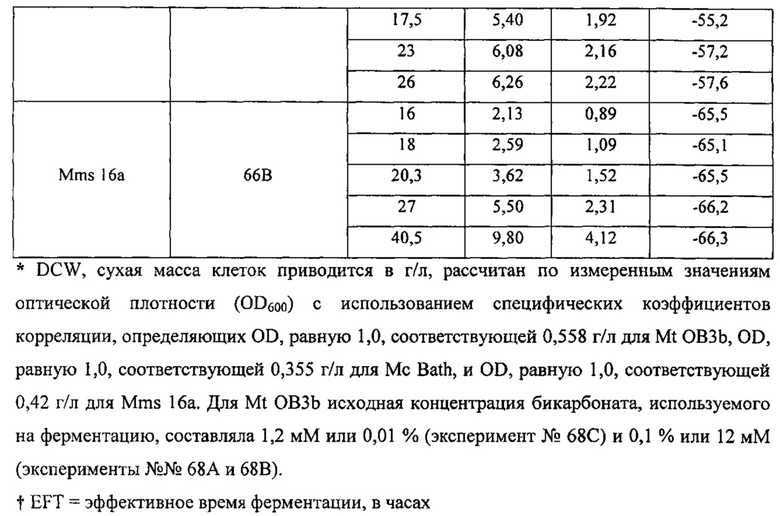
Кроме этого, был проведен анализ стабильных изотопов углерода для биомассы и соответствующих липидных фракций (см. Таблица 2) штаммов Methylosinus trichosporium OB3b (Mt OB3b), Methylococcus capsulatus Bath (Mc Bath), и Methylomonas sp. 16a (Mms 16a), выращенных на метане в биореакторах, как описано в Примере 1.

Биомассу штаммов Mt OB3b, Mc Bath и Mms 16а собирали в моменты времени 94 ч (3,14 г DCW/л), 26 ч (2,2 г DCW/л) и 39 ч (1,14 г DCW/л), соответственно. Значения δ13С для липидов в Таблице 4 представляют собой среднее для двух параллельных измерений.
ПРИМЕР 3
ЭФФЕКТ ИСТОЧНИКА МЕТАНА И ЧИСТОТЫ НА РАСПРЕДЕЛЕНИЕ СТАБИЛЬНЫХ ИЗОТОПОВ УГЛЕРОДА В ЛИПИДАХ
Для исследования роста метанотрофов на компонентах природного газа, содержащих метан, ряд 0,5-литровых сывороточных бутылок, содержащих 100 мл среды MMS1.0 определенного состава, инокулировали Methylosinus trichosporium OB3b или Methylococcus capsulatus Bath из периодической культуры в сывороточных бутылках (5% об./об.), выращенных в той же среде с добавкой 1:1 (об./об.) смеси метана и воздуха. Среда MMS1.0 имела следующий состав: 0,8 мМ MgSO4*7H2O, 30 мМ NaNO3, 0,14 мМ CaCl2, 1,2 мМ NaHCO3, 2,35 мМ KH2PO4, 3,4 мМ K2HPO4, 20,7 мкМ Na2MoO4*2H2O, 6 мкМ CuSO4*5H2O, 10 мкМ FeIII-Na-ЭДTA, и 1 мл на литр раствора следовых металлов (содержащего, на л: 500 мг FeSO4*7H2O, 400 мг ZnSO4*7H2O, 20 мг MnCl2*7H2O, 50 мг CoCl2*6H2O, 10 мг NiCl2*6H2O, 15 мг H3BO3, 250 мг ЭДТА). Фосфат, бикарбонат и FeIII-Na-ЭДТА прибавляют после автоклавирования и охлаждения среды. Конечный рН среды был равен 7,0±0,1.
Инокулированные бутылки закрывали герметизующими резиновыми пробками и вводили путем инъекции с помощью шприца 60 мл газообразного метана через стерильный 0,45 мкм фильтр и стерильные иглы 27G. В два параллельных образца культуры вводили инъекцией 60 мл объема (А) метана 99% чистоты (сорт 2.0, Praxair через Alliance Gas, San Carlos, CA), (В) метана 70% чистоты, соответствующего стандарту природного газа (Sigma-Aldrich; также содержит 9% этана, 6% пропана, 3% метилпропана, 3% бутана и другие незначительные углеводородные компоненты), (С) метана 85% чистоты, поставляемого в виде 1:1 смеси источников метана А и В; и (D) метана >93% (сорт 1.3, Specialty Chemical Products, South Houston, TX; анализ собственными силами показал содержание метана >99%). Культуры инкубировали при 30°С (М trichosporium штамм OB3b) или 42°С (М capsulatus Bath) с вращательным встряхиванием со скоростью 250 об/мин, и рост измеряли с интервалами приблизительно 12 часов путем отбора 1 мл образцов для определения OD600. В эти моменты времени бутылки вентилировали и свободное пространство над содержимым заполняли 60 мл метана из соответствующего источника (А, В, С, или D) и 60 мл концентрированного кислорода (по меньшей мере 85% чистоты). С интервалами около 24 часа отбирали по 5 мл образцов, клетки выделяли центрифугированием (8000 об/мин, 10 минут), и затем хранили при -80°С до проведения анализа.
Проводили анализы содержания углерода и азота (% сухой массый), и соотношений стабильных изотопов углерода (13С) и азота (15N) для метанотрофной биомассы, полученной от М. trichosporium штамма OB3b и М. capsulatus Bath. В Таблице 3 приведены результаты анализа стабильных изотопов углерода для биомассы образцов М. capsulatus Bath, выращенных на метане с разной степенью чистоты и в разных партиях бутылочных культур.
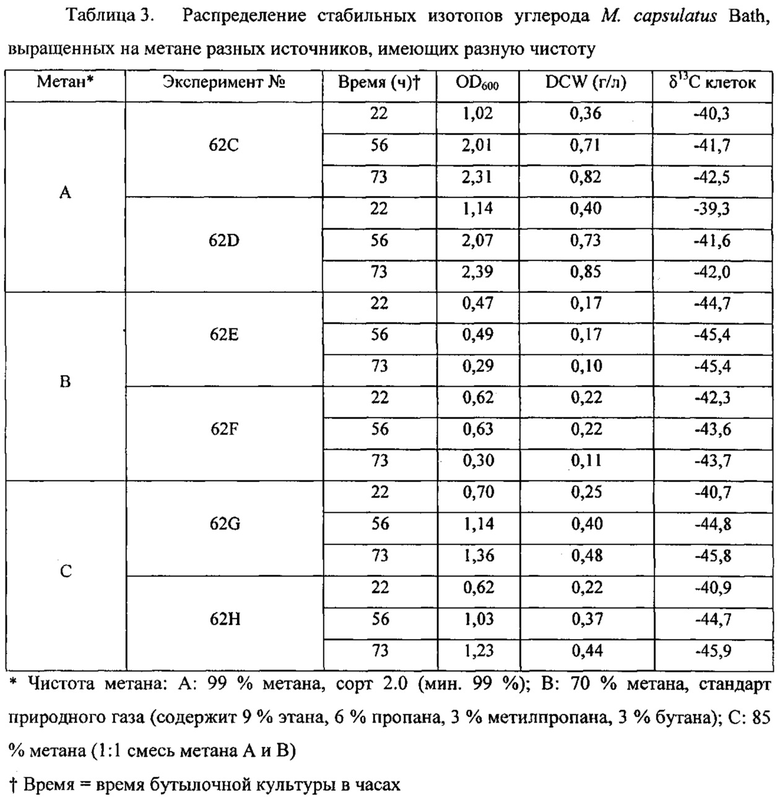
Среднее δ13С для М. capsulatus Bath, выращенных на одном источнике метана (А, 99%), было равно -41,2±1,2, в то время как среднее δ13С для М. capsulatus Bath, выращенных на разных источниках метана (В, 70%), составляло -44,2±1,2. В тех случаях, когда источники метана А и В смешивали, было получено промежуточное среднее значение δ13С, равное -43,8± 2,4. Эти данные показывают, что δ13С клеточного материала, выращенного на метане источников А и В, значительно отличаются друг от друга из-за различий в δ13С исходного метана. Однако клетки, выращенные на смеси двух газов, предпочтительно утилизировали 12С и, таким образом, демонстрировали тенденцию к более отрицательным значениям δ13С.
Аналогичный эксперимент проводили для исследования того, будут ли два разных метанотрофа, Methylococcus capsulatus Bath и Methylosinus trichosporium OB3b, выращенные на метане разных источников и в разных партиях бутылочных культур, демонстрировать различия в распределении δ13С (см. Таблицу 4).
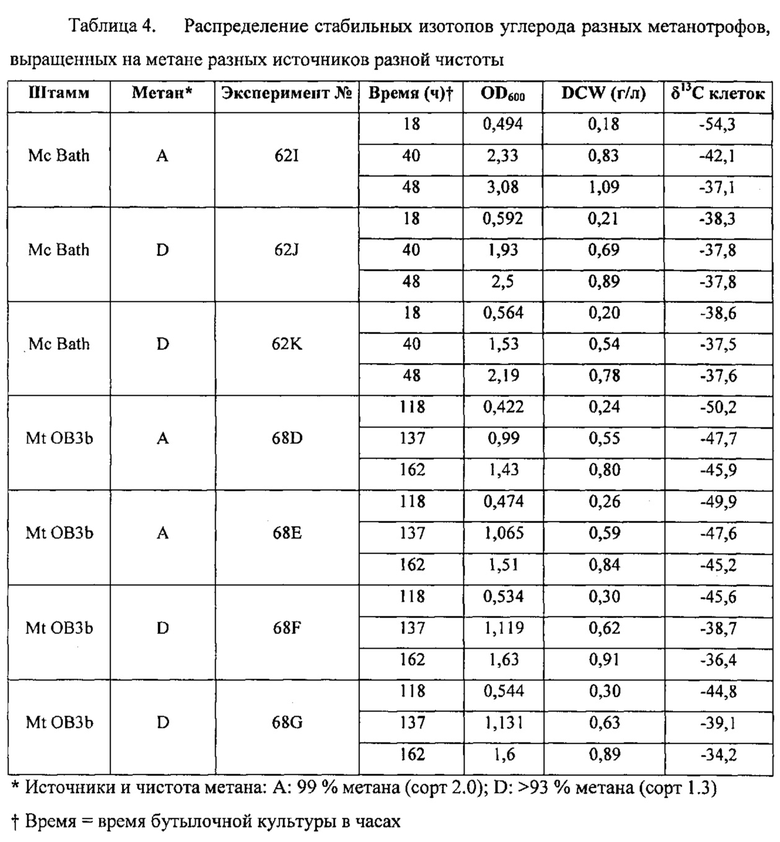
Среднее δ13С для М. capsulatus, выращенных на первом источнике метана (А), составляло -44,5±8,8, тогда как среднее δ13С для М. trichosporium было равно -47,8±2,0 при выращивании на том же источнике метана. Среднее δ13С для М. capsulatus, выращенного на втором источнике метана (В), было равно -37,9±0,4, тогда как среднее δ13С для М. trichosporium составляло -39,8±4,5. Эти данные показывают, что δ13С клеточного материала, выращенного на источнике метана, очень близко к значению δ13С клеточного материала другого штамма, выращенного на том же источнике метана. Таким образом, наблюдаемый δ13С клеточного материала, по-видимому, преимущественно зависит от состава подаваемого газа, а не от свойств конкретного исследуемого бактериального штамма.
ПРИМЕР 4
КЛОНИРОВАНИЕ И ЭКСПРЕССИЯ ОДНОГО ИЛИ ДВУХ ГЕНОВ L-ОПЕРОНА БИОСИНТЕЗА ЛИЗИНА В METHANOCOCCUS CAPSULATUS ШТАММ BATH
Для создания метанотрофного бактериального штамма, который сверхпродуцирует аминокислоту L-лизин, метанотрофный экспрессионный вектор, содержащий ген, кодирующий аминокислотную последовательность фермента дигидродипиколинатсинтазы (dapA) (SEQ ID NO 121), или диаминопимелат-декарбоксилазы (lysA) (SEQ ID NO: 123) и диаминопимелат-эпимеразы (dapF) (SEQ ID NO: 125) или сукцинилдиаминопимелат-десукцинилазы (dapE) (SEQ ID NO: 127), или аспартат-семиальдегид-дегидрогеназы (asd) (SEQ ID NO: 129), или дигидропиколинат-редуктазы (dapB) (SEQ ID NO: 131), или ацетилорнитин-аминотрансферазы (argD) (SEQ ID NO: 133), вводили в Methylococcus capsulatus Bath путем конъюгативного скрещивания. Ферменты являются нативными по отношению к Methylococcus capsulatus Bath. Эписомальная экспрессионная плазмида (содержащая последовательности, кодирующие начало репликации, начало переноса, маркер резистентности к лекарственному средству (канамицину), и сайты множественного клонирования), была использована для клонирования каждой из этих полинуклеотидных последовательностей ниже генетически модифицированного IPTG-индуцируемого (LacIq) промотора метанолдегидрогеназы (MDH). Колонии конъюгация-компетентного штамма Е. coli (S-17), содержащие или плазмиду с геном пути, несвязанный ген в той же самой системе экспрессии, или плазмиду без описанного промоторного конструкта (донорные штаммы), инокулировали в жидкую (среду) LB (Луриа-Бертани), содержащую канамицин (30 мкг/мл), и выращивали при 37°С в течение ночи. Одну часть жидкой донорной культуры инокулировали в 100 частей свежей LB, содержащей канамицин (30 мкг/мл), на 3-5 ч до использования для скрещивания с реципиентными метанотрофными штаммами. Метанотрофный (реципиентный) штамм инокулировали в жидкую среду MM-W1 (Pieja et al., 2011, Microbial Ecology 62:564-573) с около 40 мл метана на 1-2 дня перед скрещиванием до достижения ими логарифмической фазы роста (OD600 около 0,3).
Бипарентальное скрещивание проводили путем приготовления штаммов реципиента и донора в таком объеме, чтобы отношение CD600 было равно 1:1 (например, 1 мл метанаотрофа с величиной OD600 1,5 и 1 мл донора с OD600, равной 1,5). Эти клетки затем собирали центрифугированием в течение 60 с при 13200 об/мин. Супернатант удаляли и клеточный осадок осторожно ресуспендировали в 500 мкл MM-W1. Для донорного штамма Е. coli, центрифугирование и ресуспендирование повторяли еще 2 раза для обеспечения удаления антибиотиков. Равные объемы ресуспендированных клеток штаммов реципиента и донора затем объединяли и перемешивали осторожным пипетированием. Композицию для скрещивания центрифугировали в течение 60 с при 13,2k об/мин, и удаляли максимально возможную часть супернатанта. Осадок клеток затем осторожно перемешивали и наносили одной каплей на агар для скрещивания (полная среда MM-W1, содержащая стерильный 0,5% дрожжевой экстракт). Планшеты для скрещивания инкубировали в течение 48 ч в герметично закрытой камере, содержащей метан и воздух (в соотношении 1:1) при 37°С. После 48 ч периода инкубации, клетки с планшетов для скрещивания собирали путем прибавления на планшеты 1 мл среды MM-W1 и переноса суспендированных клеток в пробирку Эппендорфа на 2 мл. Клетки осаждали центрифугированием и ресуспендировали с использованием 100 мкл свежей MM-W1 перед высеиванием на планшеты для селекции (полная MM-W1 агаровая среда, содержащая 7,5 мкг/мл канамицина) для селекции клеток, стабильно поддерживающих конструкты. Несущие плазмиду метанотрофы появлялись на этих планшетах спустя около 1 неделю инкубации при 42°С. Колонии затем выращивали в инкубаторе-шейкере при 42°С в 2,5 мл жидкой среды (MMS1) в герметично закрытых сосудах, содержащих метан с воздухом в соотношении 1:1. Спустя 24 часа, прибавляли IPTG до конечной концентрации 5 мМ для индуцирования экспрессии целевого гена. Спустя еще 72 часа культуры анализировали на продуцирование аминокислоты с использованием способа, описанного в Примере 9. В Таблице 5 приведено краткое описание штаммов.

Результаты представлены на Фигуре 1, которая представляет собой график зависимости концентрации внеклеточного L-лизина (детектируемого в клеточном супернатанте штаммов, гетерологически экспрессирующих указанный ген), деленной на оптическую плотность (OD600нм) культуры в зависимости от штамма. Генотипы, обозначенные номерами 34 и 1067, представляют контрольную пустую плазмиду и контроль, экспрессирующий посторонний ген (кодирующий зеленый флуоресцентный белок) в контексте того же промотора. Столбики отображают среднее значение для от одного до восьми образцов (некоторые клоны не росли). График показывает, что сверхэкспрессия указанных генов приводит к увеличению внеклеточного L-лизина от 10 до 16 раз по сравнению с контрольной пустой плазмидой (генотип номер 34) и от 1,2 до 2,1 раза по сравнению с контрольным посторонним геном (генотип номер 1067). Результаты демонстрируют повышенное продуцирование внеклеточного L-лизина в результате экспрессии фермента биосинтеза указанной нуклеиновой кислоты, кодирующей L-аминокислоту, находящейся под контролем экзогенного промотора.
ПРИМЕР 5
КЛОНИРОВАНИЕ И ЭКСПРЕССИЯ ГЕНА, КОДИРУЮЩЕГО БЕЛОК С ВЫСОКИМ СОДЕРЖАНИЕМ L-ЛИЗИНА (CACHE-БЕЛОК) В METHANOCOCCUS CAPSULATUS BATH
Для создания метанотрофного бактериального штамма, который сверхпродуцирует аминокислоту L-лизин в ассоциированной с клетками биомассе, метанотрофный экспрессионный вектор, содержащий один из нескольких вариантов гена, кодирующего аминокислотную последовательность белка с повышенным содержанием L-лизина (пептидаза семейства S9A, обозначенная в данном документе как HighK (SEQ ID NO: 152)) вводили в Methylococcus capsulatus Bath путем конъюгативного скрещивания. Эписомальную экспрессионную плазмиду (содержащую последовательности, кодирующие начало репликации, начало переноса, маркер резистентности к лекарственному средству (канамицину), и сайты множественного клонирования), использовали для клонирования полинуклеотидных последовательностей HighK (SEQ ID NO: 151, 153, 155, 157, 159, 161, 163 и 165, все из которых являются разными кодон-оптимизированными последовательностями Methylococcus capsulatus Bath), расположенными ниже генетически модифицированного IPTG-индуцируемого промотора (LacIq) метанолдегидрогеназы (MDH). Колонии конъюгация-компетентного штамма Е. coli (S-17), содержащего плазмиду с экспериментальным геном или плазмиду «отрицательного контроля» без описанного конструкта промотор-ген (донорные штаммы) инокулировали в жидкую среду LB, содержащую канамицин (30 мкг/мл), и выращивали при 37°С в течение ночи. Одну часть жидкой донорной культуры инокулировали в 100 частей свежей LB, содержащей канамицин (30 мкг/мл), на 3-5 ч перед их использованием для скрещивания с реципиентными метанотрофными штаммами. Метанотрофный (реципиентный) штамм инокулировали в жидкую среду MM-W1 (Pieja et al., 2011, Microbial Ecology 62:564-573) с около 40 мл метана на 1-2 дня перед скрещиванием до достижения ими логарифмической фазы роста (OD600 около 0,3).
Бипарентальное скрещивание проводят путем приготовления штаммов реципиента и донора в таком объеме, чтобы соотношение OD600 составляло 1:1 (например, 5 мл метанотрофа с OD600, равной 0,3, и 5 мл донора с OD600, равной 0,3). Эти клетки затем собирали центрифугированием в течение 60 с при 13,2k об/мин. Супернатант удаляли, и клеточный осадок осторожно ресуспендировали в 500 мкл MM-W1. Для донорного штамма Е. coli, центрифугирование и ресуспендирование повторяли еще 2 раза для обеспечения удаления антибиотиков. Равные объемы ресуспендированных клеток штаммов реципиента и донора затем объединяли и перемешивали осторожным пипетированием. Композицию для скрещивания центрифугировали в течение 60 с при 13,2k об/мин, и удаляли максимально возможную часть супернатанта. Осадок клеток затем осторожно перемешивали и наносили одной каплей на агар для скрещивания (полная среда MM-W1, содержащая стерильный 0,5% дрожжевой экстракт). Планшеты для скрещивания инкубировали в течение 48 ч в герметично закрытой камере, содержащей метан и воздух (в соотношении 1:1) при 37°С. После 48 ч периода инкубации, клетки с планшетов для скрещивания собирали путем прибавления на планшеты 1 мл среды MM-W1 и переноса суспендированных клеток в пробирку Эппендорфа на 2 мл. Клетки осаждали центрифугированием и ресуспендировали с использованием 100 мкл свежей MM-W1 перед высеиванием на планшеты для селекции (полная MM-W1 агаровая среда, содержащая 7,5 мкг/мл канамицина) для селекции клеток, стабильно поддерживающих конструкты. Несущие плазмиду метанотрофы появлялись на этих планшетах спустя около 1 неделю инкубации при 42°С. Колонии затем выращивали в инкубаторе-шейкере при 42°С в 2,5 мл жидкой среды (MMS1) в герметично закрытых сосудах, содержащих метан с воздухом в соотношении 1:1. Спустя 24 часа, прибавляли IPTG до конечной концентрации 5 мМ для индуцирования экспрессии целевого гена. Через заданный период времени (дополнительно 48 или 72 часа, в зависимости от условий культивации), культуры анализировали на продуцирование аминокислоты с использованием способа, описанного в Примере 9. В Таблице 6 приведено краткое описание штаммов.

Концентрация клеточно-ассоциированного L-лизина, детектируемого в образцах клеточной биомассы штаммов, гетерологически экспрессирующих) к указанные гены, изображены на Фигуре 2. Генотип номер 34 представляет собой контрольную пустую плазмиду. Столбики отображают усредненные значения для восьми клонов каждого экспериментального генотипа. Все гены кодируют один и тот же белок пептидазы семейства S9A (HighK), но имеют все разные последовательности, соответствующие различному использованию кодонов. График показывает, что экспрессия указанных генов приводит к диапазону значений содержания клеточно ассоциированного L-лизина. Генотип номер 1122 не демонстрирует увеличения этого значения по сравнению с контролем (34), в то время как генотип номер 1115 демонстрирует рост на 53% по сравнению с контролем.
ПРИМЕР 6
КЛОНИРОВАНИЕ И КОЭКСПРЕССИЯ ДИГИДРОПИКОЛИНАТСИНТАЗЫ И CACHE-БЕЛКА В METHANOCOCCUS CAPSULATUS BATH
Для создания метанотрофного бактериального штамма, который сверхпродуцирует аминокислоту L-лизин, метанотрофный экспрессионный вектор, содержащий ген, кодирующий аминокислотную последовательность фермента дигидродипиколинатсинтазы (dapA) (SEQ ID NO: 121) и один из нескольких вариантов гена, кодирующего аминокислотную последовательность белка с повышенным содержанием лизина (пептидаза семейства S9A, обозначенная в данном документе как HighK (SEQ ID NO: 152)), вводили в Methylococcus capsulatus Bath путем конъюгативного скрещивания. Эписомальную экспрессионную плазмиду (содержащую последовательности, кодирующие начало репликации, начало переноса, маркер резистентности к лекарственному средству (канамицину), и сайты множественного клонирования), использовали для клонирования dapA (SEQ ID NO: 121) и полинуклеотидных последовательностей HighK (SEQ ID NO: 151, 153, 155, 157, 159, 161, 163 и 165, все из которых являются разными кодон-оптимизированными последовательностями Methylococcus capsulatus Bath) для Methylococcus capsulatus Bath) ниже генетически модифицированного IPTG-индуцируемого промотора (LacIq) метанолдегидрогеназы (MDH). Колонии конъюгация-компетентных штаммов Е. coli (S-17), содержащих или плазмиду с экспериментальными генами, или плазмиду «отрицательного контроля» без описанного конструкта промотор-гены (донорные штаммы), инокулировали в жидкую среду LB, содержащую канамицин (30 мкг/мл), и выращивали при 37°С в течение ночи. Одну часть жидкой донорной культуры инокулировали в 100 частей свежей LB, содержащей канамицин (30 мкг/мл), на 3-5 ч перед их использованием для скрещивания с реципиентными метанотрофными штаммами. Метанотрофный (реципиентный) штамм инокулировали в жидкую среду ММ-W1 (Pieja et al., 2011, Microbial Ecology 62:564-573) с около 40 мл метана на 1-2 дня перед скрещиванием до достижения ими логарифмической фазы роста (OD600 около 0,3).
Бипарентальное скрещивание проводят путем приготовления штаммов реципиента и донора в таком объеме, чтобы соотношение OD600 составляло 1:1 (например, 5 мл метанотрофа с OD600, равным 0,3, и 5 мл донора с OD600, равным 0,3). Эти клетки затем собирали центрифугированием в течение 60 с при 13,2k об/мин. Супернатант удаляли, и клеточный осадок осторожно ресуспендировали в 500 мкл MM-W1. Для донорного штамма Е. coli центрифугирование и ресуспендирование повторяли еще 2 раза для обеспечения удаления антибиотиков. Равные объемы ресуспендированных клеток штаммов реципиента и донора затем объединяли и перемешивали осторожным пипетированием. Композицию для скрещивания центрифугировали в течение 60 с при 13,2k об/мин и удаляли максимально возможную часть супернатанта. Осадок клеток затем осторожно перемешивали и наносили одной каплей на агар для скрещивания (полная среда MM-W1, содержащая стерильный 0,5% дрожжевой экстракт). Планшеты для скрещивания инкубировали в течение 48 ч в герметично закрытой камере, содержащей метан и воздух (в соотношении 1:1) при 37°С. После 48 ч периода инкубации клетки с планшетов для скрещивания собирали путем прибавления на планшеты 1 мл среды ММ-W1 и переноса суспендированных клеток в пробирку Эппендорфа на 2 мл. Клетки осаждали центрифугированием и ресуспендировали с использованием 100 мкл свежей MM-W1 перед высеиванием на планшеты для селекции (полная MM-W1 агаровая среда, содержащая 7,5 мкг/мл канамицина) для селекции клеток, стабильно поддерживающих конструкты. Несущие плазмиду метанотрофы появлялись на этих планшетах спустя около 1 неделю инкубации при 42°С. Колонии затем выращивали в инкубаторе-шейкере при 42°С в 2,5 мл жидкой среды (MMS1) в герметично закрытых сосудах, содержащих метан с воздухом в соотношении 1:1. Спустя 24 часа, прибавляли IPTG до конечной концентрации 5 мМ для индуцирования экспрессии целевого гена. Через заданный период времени (дополнительно 48 или 72 часа, в зависимости от условий культивации), культуры анализировали на продуцирование аминокислоты. В Таблице 7 приведено краткое описание штаммов.

Результаты приведены в Таблице 8 ниже и на Фигурах 3 и 4, как описано ниже.
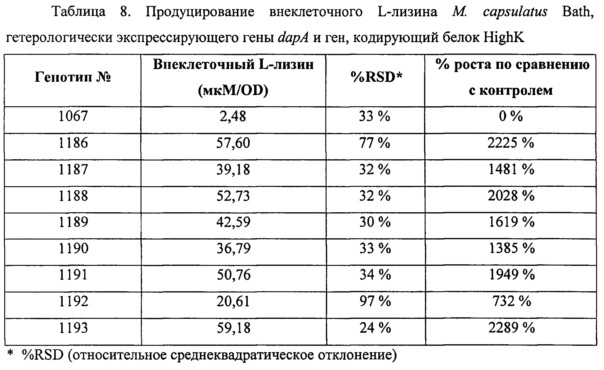
Концентрации клеточно-ассоциированного L-лизина, определяемые в культурах штаммов, гетерологически экспрессирующих указанные гены, представлены на Фигуре 3. На графике, генотип номер 1067 обозначает контроль, экспрессирующий посторонний ген (флуоресцентный репортерный ген) в контексте того же промотора. Столбики отображают усредненные значения для восьми клонов каждого экспериментального генотипа. Экспериментальные штаммы (1186-1193) содержат dapA, а также ген, кодирующий белок с повышенным содержанием L-лизина, обозначенный HighK. Все гены HighK кодируют одну и ту же аминокислотную последовательность, но имеют разное использование кодонов. Ожидалось, что различное использование кодонов будет влиять на количество продуцируемого полипептида HighK. График показывает, что экспрессия указанных генов приводит к диапазону значений клеточно-ассоциированного L-лизина; генотип номер 1186 демонстрирует увеличение этого значения на 18% по сравнению с контролем (1067), в то время как генотип номер 1188 демонстрирует 36% роста по сравнению с контролем (1067).
Концентрации внеклеточного L-лизина, детектируемого в супернатантах культур образцов штаммов, гетерологически экспрессирующих указанные гены, представлены на Фигуре 4. Как и на Фигуре 3, генотип номер 1067 обозначает контроль, экспрессирующий посторонний ген (флуоресцентный репортерный ген), в контексте того же промотора. Столбики отображают усредненные значения для восьми клонов каждого экспериментального генотипа. График показывает, что экспрессия указанных генов приводит к увеличению внеклеточного L-лизина от 8 до 23 раз по сравнению с контролем (генотип номер 1067), при расчете по средним значениям для каждого набора клонов.
Результаты, представленные на Фигурах 3 и 4, демонстрируют повышенное продуцирование внеклеточного и клеточно-ассоциированного L-лизина в результате экспрессии указанных экзогенных последовательностей нуклеиновой кислоты.
ПРИМЕР 7
КЛОНИРОВАНИЕ И КОЭКСПРЕССИЯ ГЕНОВ ИЗ ОПЕРОНОВ БИОСИНТЕЗА АМИНОКИСЛОТЫ В METHANOCOCCUS CAPSULATUS BATH ДЛЯ ПРОДУЦИРОВАНИЯ L-ЛИЗИНА
Для создания метанотрофного бактериального штамма, который сверхпродуцирует аминокислоту L-лизин, различные метанотрофные экспрессионные векторы, содержащие определенные комбинации генов («опероны»), кодирующие аминокислотные последовательности ферментов путей биосинтеза аминокислоты Methylococcus capsulatus Bath и/или Е. coli, были сконструированы и введены в Methylococcus capsulatus Bath путем конъюгативного скрещивания. Эти опероны содержали следующие гены:
Оперон А: аспартаткиназа I (thrA; SEQ ID NO: 135); дигидродипиколинатсинтаза (dapA; SEQ ID 121); диаминопимелат-декарбоксилаза (lysA; SEQ ID NO: 123)
Оперон В: аспартаткиназа II (metL; SEQ ID NO: 137); дигидродипиколинатсинтаза (dapA; SEQ ID NO: 121); диаминопимелат-декарбоксилаза (lysA; SEQ ID NO: 123)
Оперон С: аспартаткиназа III (lysC; SEQ ID NO: 139); гомосерин-дегидрогеназа (hom; SEQ ID NO: 141); дигидродипиколинатсинтаза (dapA; SEQ ID NO: 121); диаминопимелат-декарбоксилаза (lysA; SEQ ID NO: 123)
Оперон D: гомосерин-О-сукцинилтрансфераза (metA; SEQ ID NO: 143); O-сукцинил-L-гомосерин-сульфгидрилаза (metZ; SEQ ID NO: 145); гомоцистеин-трансметилаза (metE; SEQ ID NO: 147); метионинсинтаза (metH; SEQ ID NO: 149)
Оперон F: аспартаткиназа II (metL; SEQ ID NO: 137); гомосерин-О-сукцинилтрансфераза (metA; (SEQ ID NO: 143); гомоцистеин-трансметилаза (metE; SEQ ID NO: 147)
Оперон G: аспартаткиназа III (lysC; SEQ ID NO: 139); гомосерин-дегидрогеназа (hom; SEQ ID NO: 141); гомосерин-О-сукцинилтрансфераза (metA; SEQ ID NO: 143); гомоцистеин-трансметилаза (metE; SEQ ID NO: 147)
Эписомальную экспрессионную плазмиду (содержащую последовательности, кодирующие начало репликации, начало переноса, маркер резистентности к лекарственному средству (канамицин), и сайты множественного клонирования), использовали для клонирования полинуклеотидных последовательностей вышеуказанных оперонов ниже генетически модифицированного IPTG-индуцируемого промотора (LacIq) метанолдегидрогеназы (MDH). Колонии конъюгация-компетентных штаммов Е. coli (S-17), содержащих плазмиду с опероном или плазмиду без описанного конструкта промотор-оперон (донорные штаммы) инокулировали в жидкую среду LB, содержащую канамицин (30 мкг/мл), и выращивали при 37°С в течение ночи. Одну часть жидкой донорной культуры инокулировали в 100 частей свежей LB, содержащей канамицин (30 мкг/мл), на 3-5 ч перед их использованием для скрещивания с реципиентными метанотрофными штаммами. Метанотрофный (реципиентный) штамм инокулировали в жидкую среду MM-W1 (Pieja et al., 2011, Microbial Ecology 62:564-573) с около 40 мл метана на 1-2 дня перед скрещиванием до достижения ими логарифмической фазы роста (OD600 около 0,3).
Бипарентальное скрещивание проводят путем приготовления штаммов реципиента и донора в таком объеме, чтобы соотношение OD600 составляло 1:1 (например, 5 мл метанотрофа с OD600, равной 0,3 и 5 мл донора с OD600, равной 0,3). Эти клетки затем собирали центрифугированием в течение 60 с при 13,2k об/мин. Супернатант удаляли и клеточный осадок осторожно ресуспендировали в 500 мкл MM-W1. Для донорного штамма Е. coli центрифугирование и ресуспендирование повторяли еще 2 раза для обеспечения удаления антибиотиков. Равные объемы ресуспендированных клеток штаммов реципиента и донора затем объединяли и перемешивали осторожным пипетированием. Композицию для скрещивания центрифугировали в течение 60 с при 13,2k об/мин, и удаляли максимально возможную часть супернатанта. Осадок клеток затем осторожно перемешивали и наносили одной каплей на агар для скрещивания (полная среда MM-W1, содержащая стерильный 0,5% дрожжевой экстракт). Планшеты для скрещивания инкубировали в течение 48 ч в герметично закрытой камере, содержащей метан и воздух (в соотношении 1:1) при 37°С. После 48 ч периода инкубации клетки с планшетов для скрещивания собирали путем прибавления на планшеты 1 мл среды MM-W1 и переноса суспендированных клеток в пробирку Эппендорфа на 2 мл. Клетки осаждали центрифугированием и ресуспендировали с использованием 100 мкл свежей MM-W1 перед высеиванием на планшеты для селекции (полная MM-W1 агаровая среда, содержащая 7,5 мкг/мл канамицина) для селекции клеток, стабильно поддерживающих конструкты. Несущие плазмиду метанотрофы появлялись на этих планшетах спустя около 1 неделю инкубации при 42°С. Колонии затем выращивали в инкубаторе-шейкере при 42°С в 2,5 мл жидкой среды (MMS1) в герметично закрытых сосудах, содержащих метан с воздухом в соотношении 1:1. Спустя 24 часа прибавляли IPTG до конечной концентрации 5 мМ для индуцирования экспрессии целевых генов. Через заданный период времени (дополнительно 48 или 72 часа, в зависимости от условий культивации), культуры анализировали на продуцирование аминокислоты.
Результаты, приведенные в Таблице 9, демонстрируют повышенное продуцирование внеклеточного L-лизина в результате экспрессии указанных экзогенных последовательностей нуклеиновых кислот.
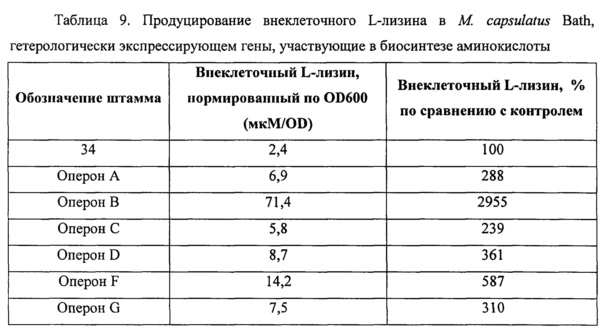
Для каждой последовательности в Таблице 9 приведено наибольшее детектируемое определенное по результатам анализа значение для набора клонов, предположительно содержащих указанные последовательности. В качестве нижнего предела при определении рекомбинантных клеток как обладающих повышенным продуцированием лизина была установлена концентрация внеклеточного лизина, в 2 раза превышающая значение, определенное по результатам анализа для неизмененного контроля (т.е. штамм отрицательного контроля). В нескольких случаях, когда набор тестируемых клонов предположительно содержал одну и ту же комбинацию гетерологичных генов, не наблюдалось увеличения концентрации внеклеточного лизина по сравнению с контролем. Было установлено, что на рост М. capsulatus Bath отрицательно влияют даже низкие концентрации внеклеточных аминокислот (Eroshin, Harwood and Pirt, 1968, J. appl. Bact., 31, 560-567), что позволяет предположить, что изменение биосинтеза аминокислоты вероятно будет являться нежелательным для нормального роста клеток. Не желая ограничиваться какой-либо теорией, укажем, что, как считается, изменение активности нативного биосинтеза аминокислоты индуцирует высокое селекционное давление на экспрессирующие клетки, направленное на мутирование или иную инактивацию введенных генов. Таким образом, отсутствие активности выше пороговой может быть вызвано рядом факторов (как наблюдались в нескольких клонах, идентифицированных как имеющие показатели ниже порога обнаружения), включая мутации в последовательностях нуклеиновой кислоты или потерю плазмиды, и не является убедительным доказательством присутствия или отсутствия функции фермента в данном случае.
ПРИМЕР 8
КЛОНИРОВАНИЕ И КОЭКСПРЕССИЯ ГЕНОВ ИЗ ОПЕРОНОВ БИОСИНТЕЗА АМИНОКИСЛОТЫ В METHANOCOCCUS CAPSULATUS BATH ДЛЯ ПРОДУЦИРОВАНИЯ L-МЕТИОНИНА
Для создания метанотрофного бактериального штамма, который сверхпродуцирует аминокислоту L-метионин, различные метанотрофные экспрессионные векторы, содержащие четыре разные комбинации генов («опероны»), кодирующие аминокислотные последовательности ферментов Methylococcus capsulatus Bath и/или Е. coli пути биосинтеза аминокислоты, были сконструированы и введены в Methylococcus capsulatus Bath путем конъюгативного скрещивания. Эти опероны содержали следующие гены:
Оперон В: аспартаткиназа II (metL; SEQ ID NO: 137); дигидродипиколинатсинтаза (dapA; SEQ ID NO: 121); диаминопимелат-декарбоксилаза (lysA; SEQ ID NO: 123)
Оперон D: гомосерин-О-сукцинилтрансфераза (metA; SEQ ID NO: 143); O-сукцинил-L-гомосерин-сульфгидрилаза (metZ; SEQ ID NO: 145); гомоцистеин-трансметилаза (metE; SEQ ID NO: 147); метионинсинтаза (metH; SEQ ID NO: 149)
Оперон F: аспартаткиназа II (metL; SEQ ID NO: 137); гомосерин-О-сукцинилтрансфераза (metA; SEQ ID NO: 143); гомоцистеин-трансметилаза (metE; SEQ ID NO: 147)
Оперон G: аспартаткиназа III (lysC; SEQ ID NO: 139); гомосерин-дегидрогеназа (hom; SEQ ID NO: 141); гомосерин-О-сукцинилтрансфераза (metA; SEQ ID NO: 143); гомоцистеин-трансметилаза (metE; SEQ ID NO: 147)
Эписомальную экспрессионную плазмиду (содержащую последовательности, кодирующие начало репликации, начало переноса, маркер резистентности к лекарственному средству (канамицину), и сайты множественного клонирования), использовали для клонирования полинуклеотидных последовательностей этих оперонов (см. выше) ниже генетически модифицированного IPTG-индуцируемого промотора (LacIq) метанолдегидрогеназы (MDH). Колонии конъюгация-компетентного штамма Е. coli (S-17), содержащего плазмиду с опероном или плазмиду без описанного конструкта промотор-оперон (донорные штаммы), инокулировали в жидкую среду LB, содержащую канамицин (30 мкг/мл), и выращивали при 37°С в течение ночи. Одну часть жидкой донорной культуры инокулировали в 100 частей свежей LB, содержащей канамицин (30 мкг/мл), на 3-5 ч перед их использованием для скрещивания с реципиентными метанотрофными штаммами. Метанотрофный (реципиентный) штамм инокулировали в жидкую среду MM-W1 (Pieja et al., 2011, Microbial Ecology 62:564-573) с около 40 мл метана на 1-2 дня перед скрещиванием до достижения ими логарифмической фазы роста (OD600 около 0,3).
Бипарентальное скрещивание проводят путем приготовления штаммов реципиента и донора в таком объеме, чтобы соотношение OD600 составляло 1:1 (например, 5 мл метанотрофа с OD600, равной 0,3 и 5 мл донора с OD600, равной 0,3). Эти клетки затем собирали центрифугированием в течение 60 с при 13,2k об/мин. Супернатант удаляли и клеточный осадок осторожно ресуспендировали в 500 мкл MM-W1. Для донорного штамма Е. coli, центрифугирование и ресуспендирование повторяли еще 2 раза для обеспечения удаления антибиотиков. Равные объемы ресуспендированных клеток штаммов реципиента и донора затем объединяли и перемешивали осторожным пипетированием. Композицию для скрещивания центрифугировали в течение 60 с при 13,2k об/мин, и удаляли максимально возможную часть супернатанта. Осадок клеток затем осторожно перемешивали и наносили одной каплей на агар для скрещивания (полная среда MM-W1, содержащая стерильный 0,5% дрожжевой экстракт). Планшеты для скрещивания инкубировали в течение 48 ч в герметично закрытой камере, содержащей метан и воздух (в соотношении 1:1) при 37°С. После 48 ч периода инкубации клетки с планшетов для скрещивания собирали путем прибавления на планшеты 1 мл среды MM-W1 и переноса суспендированных клеток в пробирку Эппендорфа на 2 мл. Клетки осаждали центрифугированием и ресуспендировали с использованием 100 мкл свежей MM-W1 перед высеиванием на планшеты для селекции (полная MM-W1 агаровая среда, содержащая 7,5 мкг/мл канамицина) для селекции клеток, стабильно поддерживающих конструкты. Несущие плазмиду метанотрофы появлялись на этих планшетах спустя около 1 неделю инкубации при 42°С. Колонии затем выращивали в инкубаторе-шейкере при 42°С в 2,5 мл жидкой среды (MMS1) в герметично закрытых сосудах, содержащих метан с воздухом в соотношении 1:1. Спустя 24 часа прибавляли IPTG до конечной концентрации 5 мМ для индуцирования экспрессии целевых генов. Через заданный период времени (дополнительно 48 или 72 часа, в зависимости от условий культивации), культуры анализировали на продуцирование аминокислоты.
Результаты, представленные в Таблице 10, демонстрируют повышенное продуцирование внеклеточного L-метионина в результате экспрессии указанных экзогенных последовательностей нуклеиновой кислоты.
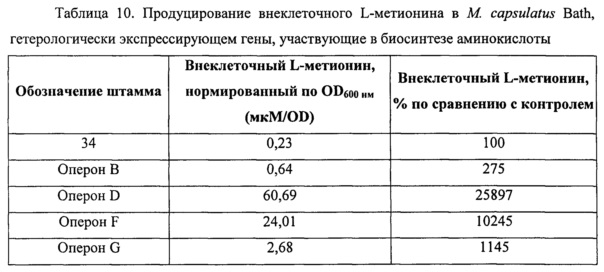
Для каждой последовательности в Таблице 10 приведено наибольшее детектируемое определенное по результатам анализа значение для набора клонов, предположительно содержащих указанные последовательности. В качестве нижнего предела при определении рекомбинантных клеток как обладающих повышенным продуцированием L-метионина была установлена концентрация внеклеточного лизина, в 2 раза превышающая значение, определенное по результатам анализа для неизмененного контроля (т.е. штамм отрицательного контроля). В нескольких случаях, когда набор тестируемых клонов предположительно содержал одну и ту же комбинацию гетерологичных генов, не наблюдалось увеличения концентрации внеклеточного лизина по сравнению с контролем. Было установлено, что на рост М. capsulatus Bath отрицательно влюяют даже низкие концентрации внеклеточных аминокислот (Eroshin, Harwood and Pirt, 1968, J. appl. Bact., 31, 560-567), что позволяет предположить, что изменения биосинтеза аминокислоты вероятно будет являться нежелательным для нормального роста клеток. Не желая ограничиваться какой-либо теорией, укажем, что, как считается, изменение активности нативного биосинтеза аминокислот индуцирует высокое селекционное давление на экспрессирующие клетки, направленное на мутирование или иную инактивацию введенных генов. Таким образом, отсутствие активности выше пороговой может быть вызвано рядом факторов (как наблюдались в нескольких клонах, идентифицированных как имеющие показатели ниже порога обнаружения), включая мутации в последовательностях нуклеиновой кислоты или потерю плазмиды, и не является убедительным доказательством присутствия или отсутствия функции фермента в данном случае.
ПРИМЕР 9
АНАЛИЗЫ ПРОДУЦИРОВАНИЯ L-ЛИЗИНА И L-МЕТИОНИНА METHANOCOCCUS CAPSULATUS BATH
Были получены образцы культуры штамма М. capsulatus Bath, содержащего вектор с IPTG-индуцируемым (LacIq) конструктом промотор-dapA или вектор без такого конструкта (оба выращенные в присутствии 5 мМ IPTG). Эти образцы центрифугировали в течение 1,5 минут при 13,2k об/мин и как супернатант, так и клеточный осадок анализировали на продуцирование аминокислоты. Бесклеточные супернатанты анализировали методом ГХ-МС (GC-MS) после дериватизации метилхлорформиатом (эталон, Sue et. al). Для анализа аминокислотного состава биомассы клеточный осадок сначала гидролизовали в 6N НСl в течение 24 ч при 100°С при непрерывном перемешивании с последующей нейтрализацией, дериватизацией метилхлороформиатом, и последующим ГХ-МС анализом. Количественный анализ проводят с использованием ГХ-МС системы Agilent 6890/5972. ГХ оснащен капиллярной колонкой HP-5MS с размерами 0,25 мм × 30 м × 0,25 мкМ и использует в качестве газа-носителя гелий с объемным расходом 1 мл/мин. Программа температуры печи начинается с 55°С в течение 3 минут, постепенно повышается до 325°С со скоростью 20°С/мин и выдерживается в течение 2 минут. Впрыскивают 1 мкл образцов с помощью автоматического шприца-пробоотборника Hamilton 10мкл. Входное отверстие образца поддерживается при температуре 250°С, имеет коэффициент деления 15:1 и облицовано вкладышем для инжектора Restek Sky с прецизионным низким перепадом давления, набитым стекловатой.
Производное L-лизина элюируется из колонки в момент времени 11,51 минут и количественно охарактеризовано с использованием характеристического иона 142 m/z. Идентификация соединения подтверждается путем контроля квалифицирующего иона при 88 m/z и 26% относительного содержания по сравнению с ионом-мишенью. Калибровочные стандарты были приготовлены из L-лизина дигидрохлорида в деионизированной воде. Калибровочную кривую для L-лизина аппроксимировали с использованием 1/х2 взвешенной линейной регрессии.
Производное L-метионина элюируется из колонки в момент времени 9,954 минут и количественно охарактеризовано с использованием характеристического иона 61 m/z. Идентификация соединения подтверждается путем контроля квалифицирующих ионов при 115 m/z (относительное содержание 52%) и 147 m/z (относительное содержание 34%). Калибровочные стандарты были приготовлены из чистого метионина в деионизированной воде. Калибровочную кривую для L-метионина аппроксимировали с использованием 1/х2 взвешенной линейной регрессии.
ПРИМЕР 10
РАСПРЕДЕЛЕНИЕ СТАБИЛЬНЫХ ИЗОТОПОВ УГЛЕРОДА В БИОМАССЕ И ПРОДУКТЫ, ПОЛУЧЕННЫЕ ИЗ МЕТАБОЛИЗИРУЮЩИХ С1 МИКРООРГАНИЗМОВ
Полученная из метана биомасса генетически модифицированных штаммов М. capsultus Bath, продемонстрировавших продуцирование повышенных уровней аминокислот, в частности. L-лизина (Пример 1), была проанализирована на соотношение стабильных изотопов углерода (13С) путем элементного анализа/изотопной масс-спектрометрии (IRMS) в непрерывном потоке с использованием элементного анализатора Costech, сопряженного с прибором Thermo Delta IRMS. Образцы метанотрофной биомассы, культивируемой в сывороточных бутылках, центрифугировали, промывали один раз в деионизированной воде, ресуспендировали в деионизированной воде и объемы, соответствующие 0,2-2 мг углерода (около 0,5-5 мг сухой массы клеток) переносили в оловянные капсулы 5×9 мм (Costech Analytical Technologies, Inc., Valencia, CA) и высушивали при 80°C в течение по меньшей мере 60 часов. В качестве контролей, образцы М. capsulatus Bath дикого типа и генетически модифицированный М. capsulatus штамм Bath, экспрессирующий последовательности нуклеиновой кислоты, кодирующие флуоресцентный белок, параллельно культивировали, промывали и ресуспендировали в деионизированной воде, переносили в оловянные капсулы 5×9 мм и высушивали при 80°С в течение по меньшей мере 60 часов. Достоверные значения δ13С получали для образцов, обеспечивающих отклик от ~100 до 10000 мВ в окне детектирования IRMS. Диапазон отклика IRMS для проанализированных образцов биомассы составлял от 3195 до 8827 мВ. Каждый образец анализировали с двумя параллельными измерениями и средние соотношения стабильных изотопов углерода (13С) приведены в Таблице 11.
Изотопное соотношение выражали как показатель «дельта» (), где изотопный состав материала определяется по сравнению со стандартом как отклонение на миллион, записанное как δ13С = (Rобразец/RСтандарт-1)×1000, где R обозначает молярное отношение тяжелой формы изотопа (13С) к легкой (12С). Стандартом для углерода является Vienna Рее Dee Belemnite (V-PDB). Изотопные анализы проводила фирма Beta Analytic Inc. (Miami, FL) под строгим наблюдением и контролем качества (аккредитация на проведение испытаний в соответствии с ISO/IEC 17025:2005; сертификат PJLA №59423). IRMS калибровали по NIST-8541 (графит: -15,90 +/- 0,25) и NIST-8542 (сахароза: -10,47 +/-0,13). Перед и после измерений неизвестных образцов проводили измерения рабочего стандарта, откалиброванного по указанным стандартам NIST (ацетанилид: -33,4 +/- 0,3). После коррекции по результатам холостого измерения, значения, измеренные для указанного вторичного стандарта, составляли -33,22%o и -33,22%o. Затем проводили нормирование, внося в результаты поправку на 0,2%o для систематизации результатов в соответствии с первичными эталонами NIST. Завышенная ошибка, равная +/- 0,3%o (абсолютная), приписывается к результатам для учета неопределимых ошибок. Хотя производитель указывает, что детекторы имеют воспроизводимость в пределах 0,02%o, отклонение между индивидуальными измерениями могут колебаться от 0,3 до 0,5%o, и поэтому Beta Analytics Inc. указывает результаты с точностью до десятых долей, а не до сотых, и указывает величину абсолютной ошибки +/- 0,3%o.
Отдельные штаммы М. capsulatus Bath, сконструированные для индуцируемой экспрессии нуклеиновых кислот, кодирующих dapA или зеленый флуоресцентный белок (см. Таблицу 11), выращивали на метане в 0,5 л сывороточных бутылках, содержащих 110 мл среды MMS1.0 определенного состава с добавкой 50 мкг/мл канамицина. Канамицин не добавляли в среду, используемую для культивации М. capsulatus Bath дикого типа. Штаммы инокулировали из периодических культур в 25 мл сывороточных бутылках, выращенных в той же среде, с добавкой приблизительно 1:1 (об./об.) смеси метана и воздуха (например, 60 мл метана в свободном пространстве, содержащем 75 мл воздуха). Среда MMS1.0 имела следующий состав: 0,8 мМ MgSO4*7H2O, 30 мМ NaNO3, 0,14 мМ CaCl2, 1,2 мМ NaHCO3, 2,35 мМ KH2PO4, 3,4 мМ K2HPO4, 20,7 мкМ Na2MoO4*2H2O, 6 мкМ CuSO4*5H2O, 10 мкМ FeIII-Na-ЭДTA, и 1 мл на литр раствора следовых металлов (содержащего, на л: 500 мг FeSO4*7H2O, 400 мг ZnSO4*7H2O, 20 мг MnCl2*7H2O, 50 мг CoCl2*6H2O, 10 мг NiCl2*6H2O, 15 мг H3BO3, 250 мг ЭДТА). Фосфат, бикарбонат и FeIII-Na-ЭДТА прибавляют после автоклавирования и охлаждения среды. Конечный рН среды был равен 7,0±0,1. Инокулировали объем 110 мл культуры из сывороточных бутылок до начальной OD600, равной 0,1, герметизировали резиновыми пробками и вводили инъекцией 120 мл газообразного метана (чистота 99%; сорт 2.0, Praxair, поставляется Alliance Gas, San Carlos, CA), который набирали в шприц через стерильный фильтр 0,45 мкм и стерильные иглы 27G. Культуры инкубировали вертикально при 42°С с вращательным встряхиванием со скоростью 250 об/мин и рост измеряли путем отбора образцов 1 мл для определения OD600. В тех случаях, когда культуры достигали OD600, равной по меньшей мере 0,6, бутылки вентилировали и газы свободного пространства заменяли на 60 мл метана и 120 мл концентрированного кислорода (чистота по меньшей мере 85%). Газы свободного пространства заменяли таким образом через каждые 8-12 часов. В тех случаях, когда OD600 достигала значения 1,6 (приблизительно через 48 часов роста), все культуры индуцировали для инициации сверхэкспрессии целевых генных продуктов путем прибавления стерилизованного на фильтре IPTG до конечной концентрации 5 мМ. Культуры выращивали еще 48 часов с заменой газов свободного пространства, как описано выше. Конечные образцы биомассы собирали центрифугированием (8000 об/мин, 10 минут), и готовили для анализа стабильного изотопа 13С, как описано выше.
В Таблице 11 приведены краткие сведения о распределении стабильных изотопов углерода (значения δ13С) для генетически модифицированных штаммов М. capsulatus Bath, сверхэкспрессирующих dapA, для обеспечения повышенного L-продуцирования лизина (Генотип №1057), а также контрольных штаммов, экспрессирующих гены, кодирующие флуоресцентный белок Dasher (Генотип №1067), и исходного штамма дикого типа. Результаты показывают, что δ13С имеет сильно отрицательные значения, и указывают на высокую степень обогащения 12С полученной из метана биомассы и продуктов штаммов, генетически модифицированных для сверхэкспрессии аминокислот, флуоресцентного белка, а также штамма М. capsulatus Bath дикого типа.
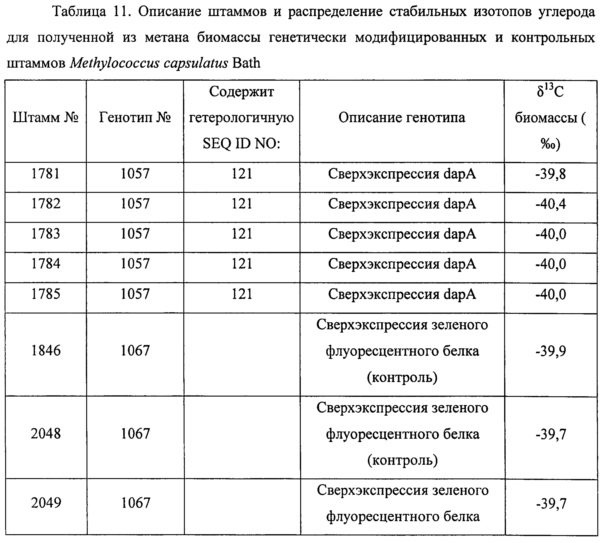

Различные варианты реализации, описанные выше, могут быть объединены для обеспечения дополнительных вариантов реализации. Все патенты США, публикации патентных заявок США, патентные заявки США, иностранные патенты, иностранные патентные заявки и непатентные публикации, упоминаемые в данном описании и/или перечисленные в библиографических данных заявки, включая, без ограничений, временную заявку США № 61/928401, поданную 16 января 2014 года, включены в данный документ в качестве ссылок в полном объеме.
В общем, в приведенной ниже формуле изобретения, используемые термины не должны истолковываться как ограничивающие притязания конкретными вариантами реализации, раскрытыми в описании заявки и формуле изобретения, но должны рассматриваться как включающие все возможные варианты реализации вместе с полным объемом эквивалентов, на которые распространяются такие притязания. Соответственно, формула изобретения не ограничена описанием.
| название | год | авторы | номер документа |
|---|---|---|---|
| СИСТЕМА, СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ БИОПЕРЕРАБОТКИ | 2013 |
|
RU2723620C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БИОЛОГИЧЕСКОГО ПОЛУЧЕНИЯ ЛАКТАТА ИЗ С1-СОЕДИНЕНИЙ С ПРИМЕНЕНИЕМ ТРАНСФОРМАНТОВ ЛАКТАТ ДЕГИДРОГЕНАЗЫ | 2014 |
|
RU2710714C2 |
| СИСТЕМЫ ФЕРМЕНТАЦИИ С ПОДАЧЕЙ ГАЗА | 2013 |
|
RU2639542C2 |
| РЕАКТОРЫ, СИСТЕМЫ И ПРОЦЕССЫ ФЕРМЕНТАЦИИ С ПОДАЧЕЙ ГАЗА, ИСПОЛЬЗУЮЩИЕ ЕМКОСТИ РАЗДЕЛЕНИЯ ГАЗА/ЖИДКОСТИ | 2018 |
|
RU2769130C2 |
| Способ культивирования аэробных метанассимилирующих микроорганизмов | 2021 |
|
RU2768401C1 |
| Штамм Methylococcus capsulatus - продуцент высокобелковой биомассы | 2022 |
|
RU2787202C1 |
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ОСНОВАННОГО НА РЕКОМБИНАЦИИ ПОЛУЧЕНИЯ СОЕДИНЕНИЙ ШАФРАНА | 2012 |
|
RU2676730C2 |
| БИОЛОГИЧЕСКИЙ СИНТЕЗ ДИФУНКЦИОНАЛЬНЫХ АЛКАНОВ ИЗ АЛЬФА-КЕТОКИСЛОТ | 2009 |
|
RU2496880C2 |
| МИКРОБИОЛОГИЧЕСКАЯ КОНВЕРСИЯ МЕТАНА | 2014 |
|
RU2658440C2 |
| РЕКОМБИНАНТНЫЕ МИКРООРГАНИЗМЫ, ПРОДУЦИРУЮЩИЕ МЕТИОНИН | 2006 |
|
RU2447146C2 |














































































































































































































































































































































































































































































































































































































































































































































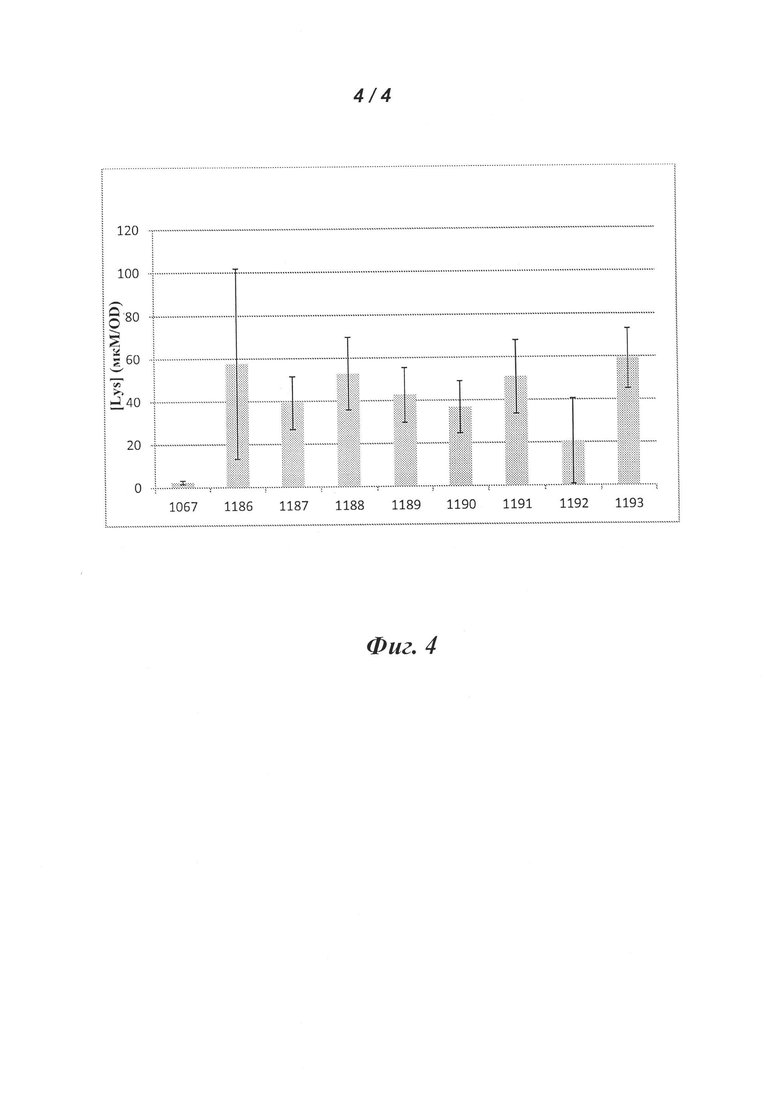
Настоящее описание относится к биотехнологии, в частности к рекомбинантным микроорганизмам, генетически модифицированным для усиления продуцирования требуемой аминокислоты. Изобретение представляет собой рекомбинантный метаболизирующий C1 микроорганизм, содержащий первую экзогенную нуклеиновую кислоту и вторую экзогенную нуклеиновую кислоту, где указанная первая нуклеиновая кислота выбрана из группы, состоящей из экзогенной нуклеиновой кислоты, кодирующей фермент биосинтеза L-аминокислоты, способный продуцировать требуемую L-аминокислоту, и экзогенной нуклеиновой кислоты, кодирующей контрольную последовательность экспрессии, функционально связанную с нуклеиновой кислотой, кодирующей нативный фермент биосинтеза L-аминокислоты, способный продуцировать указанную требуемую L-аминокислоту, где указанная вторая нуклеиновая кислота кодирует полипептид, содержащий аминокислотную последовательность длиной по меньшей мере около 10 аминокислотных остатков, в котором по меньшей мере 10% от общего количества аминокислотных остатков приходится на указанную требуемую L-аминокислоту, и отличающийся тем, что рекомбинантный метаболизирующий C1 микроорганизм способен конвертировать углеродное сырье, получаемое из природного газа, в указанную требуемую L-аминокислоту, имеющую значение δ13С менее - 30 %o. 2 н. и 22 з.п. ф-лы, 4 ил., 11 табл., 10 пр.
1. Рекомбинантный метаболизирующий C1 микроорганизм, содержащий первую экзогенную нуклеиновую кислоту и вторую экзогенную нуклеиновую кислоту,
где указанная первая нуклеиновая кислота выбрана из группы, состоящей из экзогенной нуклеиновой кислоты, кодирующей фермент биосинтеза L-аминокислоты, способный продуцировать требуемую L-аминокислоту, и экзогенной нуклеиновой кислоты, кодирующей контрольную последовательность экспрессии, функционально связанную с нуклеиновой кислотой, кодирующей нативный фермент биосинтеза L-аминокислоты, способный продуцировать указанную требуемую L-аминокислоту,
где указанная вторая нуклеиновая кислота кодирует полипептид, содержащий аминокислотную последовательность длиной по меньшей мере около 10 аминокислотных остатков, в котором по меньшей мере 10% от общего количества аминокислотных остатков приходится на указанную требуемую L-аминокислоту, и
отличающийся тем, что рекомбинантный метаболизирующий C1 микроорганизм способен конвертировать углеродное сырье, получаемое из природного газа, в указанную требуемую L-аминокислоту, имеющую значение δ13С менее - 30 %o.
2. Рекомбинантный метаболизирующий C1 микроорганизм по п. 1, отличающийся тем, что первая экзогенная нуклеиновая кислота кодирует фермент биосинтеза L-аминокислоты.
3. Рекомбинантный метаболизирующий C1 микроорганизм по любому из пп. 1, 2, отличающийся тем, что получаемое из природного газа сырье представляет собой природный газ.
4. Рекомбинантный метаболизирующий C1 микроорганизм по п. 3, отличающийся тем, что получаемое из природного газа сырье представляет собой метан.
5. Рекомбинантный метаболизирующий C1 микроорганизм по любому из пп. 1, 2, отличающийся тем, что рекомбинантный метанотрофный микроорганизм содержит способность продуцировать указанную требуемую L-аминокислоту в количестве по меньшей мере на примерно 10% больше, чем продуцируется нативным метаболизирующим С1 микроорганизмом.
6. Рекомбинантный метаболизирующий C1 микроорганизм по любому из пп. 1, 2, отличающийся тем, что первая экзогенная нуклеиновая кислота кодирует фермент биосинтеза L-аминокислоты, выбранный из группы, состоящей из фермента биосинтеза лизина, фермента биосинтеза триптофана, фермента биосинтеза метионина, фермента биосинтеза цистеина, и фермента биосинтеза треонина.
7. Рекомбинантный метаболизирующий C1 микроорганизм по п. 6, отличающийся тем, что указанная требуемая L-аминокислота представляет собой лизин, и тем, что первая экзогенная нуклеиновая кислота кодирует фермент биосинтеза лизина, выбранный из группы, состоящей из лизин-чувствительной аспартокиназы III (lysC), аспартаткиназы, аспартат-семиальдегид-дегидрогеназы (asd), дигидродипиколинатсинтазы (dapA), дигидродипиколинатредуктазы (dapB), 2,3,4,5-тетрагидропиридин-2,6-карбоксилат-N-сукцинилтрансферазы (dapD), ацетилорнитин/сукцинилдиаминопимелат-аминотрансферазы (argD), сукцинил-диаминопимелат-десукцинилазы (dapE), сукцинилдиаминопимелат-трансаминазы, диаминопимелат-эпимеразы (dapF), и диаминопиметатдикарбоксилазы (lysA).
8. Рекомбинантный метаболизирующий C1 микроорганизм по п. 7, отличающийся тем, что первая экзогенная нуклеиновая кислота кодирует дигидродипиколинатсинтазу (dapA).
9. Рекомбинантный метаболизирующий C1 микроорганизм по п. 6, отличающийся тем, что указанная требуемая L-аминокислота представляет собой L-метионин, и тем, что первая экзогенная нуклеиновая кислота кодирует фермент биосинтеза метионина, выбранный из группы, состоящей из гомосерин-О-сукцинилтрансферазы (metA), цистатионин-гамма-синтазы (metB), белка MalY, цистатионин-бета-лиазы (metC), В12-зависимой метионинсинтазы (metH), и 5-метилтетрагидроптероилтриглутамат-гомоцистеин-S-метилтрансферазы (metE).
10. Рекомбинантный метаболизирующий C1 микроорганизм по п. 6, отличающийся тем, что указанная L-аминокислота представляет собой L-цистеин, и тем, что первая экзогенная нуклеиновая кислота кодирует фермент биосинтеза цистеина, выбранный из группы, состоящей из серин-ацетилтрансферазы (CysE), цистеинсинтазы А, и цистеинсинтазы В.
11. Рекомбинантный метаболизирующий C1 микроорганизм по п. 6, отличающийся тем, что указанная требуемая L-аминокислота представляет собой L-треонин, и тем, что первая экзогенная нуклеиновая кислота кодирует фермент биосинтеза треонина, выбранный из группы, состоящей из аспартат-трансаминазы, PLP-зависимой аминотрансферазы, аспартат-аминотрансферазы, аспартаткиназы, аспартат-семиальдегид-дегидрогеназы, гомосерин-дегидрогеназы, гомосеринкиназы, и треонинсинтазы.
12. Рекомбинантный метаболизирующий C1 микроорганизм по п. 6, отличающийся тем, что указанная требуемая L-аминокислота представляет собой L-триптофан, и тем, что первая экзогенная нуклеиновая кислота кодирует фермент биосинтеза триптофана, выбранный из группы, состоящей из хоризмат-пируват-лиазы (ubiC), антранилатсинтазы компонента I (trpE), антранилатсинтазы компонента II (trpG), антранилат-фосфорибозилтрансферазы (trpD), фосфорибозилантранилат-изомеразы (trpC), белка биосинтеза триптофана (trpC), N-(5'-фосфорибозил)антранилат-изомеразы (trpF), индол-3-глицеролфосфатсинтазы, триптофансинтазы альфа-цепи (trpA), и триптофансинтазы бета-цепи (trpB).
13. Рекомбинантный метаболизирующий C1 микроорганизм по любому из пп. 1, 2, отличающийся тем, что первая экзогенная нуклеиновая кислота кодирует фермент биосинтеза L-аминокислоты, имеющий аминокислотную последовательность, которая по меньшей мере на 90% идентична референсной последовательности, выбранной из группы, состоящей из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 88, 90, 92, 94, 96, 98, 100, 102, 104, 106, 108, 110, 112, 114, 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148, и 150.
14. Рекомбинантный метаболизирующий C1 микроорганизм по п. 1, отличающийся тем, что первая экзогенная нуклеиновая кислота кодирует контрольную последовательность экспрессии, функционально связанную с нуклеиновой кислотой, кодирующей нативный фермент биосинтеза L-аминокислоты.
15. Рекомбинантный метаболизирующий C1 микроорганизм по любому из пп. 1, 2, отличающийся тем, что последовательность первой экзогенной нуклеиновой кислоты оптимизирована по использованию кодонов для оптимальной экспрессии рекомбинантным метаболизирующим C1 микроорганизмом.
16. Рекомбинантный метаболизирующий С1 микроорганизм по любому из пп. 1, 2, отличающийся тем, что последовательность второй экзогенной нуклеиновой кислоты оптимизирована по использованию кодонов для оптимальной экспрессии рекомбинантным метаболизирующим C1 микроорганизмом.
17. Рекомбинантный метаболизирующий C1 микроорганизм по любому из пп. 1, 2, отличающийся тем, что первая экзогенная нуклеиновая кислота кодирует фермент биосинтеза L-аминокислоты, и тем, что первая экзогенная нуклеиновая кислота содержит последовательность нуклеиновой кислоты, которая по меньшей мере на 85% идентична референсной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87, 89, 91, 93, 95, 97, 99, 101, 103, 105, 107, 109, 111, 113, 115, 117, 119, 121, 123, 125, 127, 129, 131, 133, 135, 137, 139, 141, 143, 145-147 и 149.
18. Рекомбинантный метаболизирующий C1 микроорганизм по любому из пп. 1, 2, отличающийся тем, что указанная требуемая L-аминокислота демонстрирует значение δ13С от -30%o до -70%о.
19. Рекомбинантный метаболизирующий С1 микроорганизм по любому из пп. 1, 2, отличающийся тем, что метаболизирующий C1 микроорганизм является метанотрофом или метилотрофом.
20. Рекомбинантный метаболизирующий C1 микроорганизм по п. 19, отличающийся тем, что метаболизирующий C1 микроорганизм является метанотрофом.
21. Рекомбинантный метаболизирующий C1 микроорганизм по п. 19, отличающийся тем, что метаболизирующий C1 микроорганизм выбран из группы, состоящей из Methylococcus capsulatus Bath, Methylomonas methanica 16a, Methylosinus trichosporium OB3b, Methylosinus sporium, Methylocystis parvus, Methylomonas methanica, Methylomonas albus, Methylobacter capsulatus, Methylobacterium organophilum, Methylomonas sp AJ-3670, Methylocella silvestris, Methylocella palustris, Methylocella tundrae, Methylocystis daltona strain SB2, Methylocystis bryophila, Methylocapsa aurea KYG, Methylacidiphilum infernorum, Methylibium petroleiphilum и Methylomicrobium alcaliphilum.
22. Способ продуцирования биомассы, включающий культивацию рекомбинантного метаболизирующего C1 микроорганизма по любому из пп. 1-21 в устройстве для контролируемой культивации в присутствии субстрата C1 в условиях и в течение времени, достаточных для продуцирования биомассы.
23. Способ по п. 22, отличающийся тем, что указанное устройство для культивации представляет собой ферментер, биореактор или ячейку с пустотелыми волокнами.
24. Способ по любому из пп. 22, 23, отличающийся тем, что субстрат C1 содержит природный газ или метан.
| US 20030166174 A1, 04.09.2003 | |||
| US 7217543 B2, 15.05.2007 | |||
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2368659C2 |

