ЗАЯВЛЕНИЕ В ОТНОШЕНИИ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Перечень последовательностей, относящийся к настоящей заявке, приведен в формате текстового файла вместо бумажной копии, и тем самым включен в описание изобретения посредством ссылки. Название текстового файла, содержащего перечень последовательностей - 200206_404_SEQUENCE_LISTING.txt. Указанный текстовый файл имеет размер 146 Кбайт, создан 12 июля 2013 года и подан в электронном виде через EFS-Web.
УРОВЕНЬ ТЕХНИКИ
Область техники
Настоящее изобретение относится к биотехнологическим способам получения биотоплива, и в частности к применению реакторной системы на основе метаболизирующего C1 микроорганизма для превращения C1 субстратов, таких как метан или метанол, в биомассу и затем в биотопливо, биопластики или т.п.
Уровень техники
Учитывая постоянно растущее истощение месторождений ископаемого топлива, увеличение выбросов парниковых газов и недавнюю озабоченность в отношении изменения климата, усилия в промышленности направлены на замену ископаемых топлив на биотоплива (например, этанол, биодизельное топливо). Однако получаемые в настоящее время виды биотоплива характеризуются своими трудностями и проблемами. Биотопливо первого поколения получают из растений (например, крахмала, тростникового сахара и кукурузного, рапсового, соевого, пальмового и других растительных масел), но такие топливные зерновые культуры конкурируют с зерновыми культурами, выращиваемыми для потребления человеком и животными. Количество имеющихся сельскохозяйственных земель недостаточно для удовлетворения глобальных потребностей одновременно как в продовольствии, так и в топливе. Поэтому биотопливо второго поколения получают, например, из целлюлозы или водорослей. Тем не менее, в связи с наличием технических трудностей при производстве, а также с высокой стоимостью производства, биотопливо второго поколения не стало более экономически эффективным или доступным.
Таким образом, существует потребность в биотопливе третьего или следующего поколения, которое можно было бы получать из альтернативного сырья (т.е., без применения сахара, кукурузы, водорослей). В этом отношении, метан представляет собой один из наиболее распространенных отечественных видов углеродного сырья, источником которого является главным образом природный газ. Недавний рост внутреннего производства метана (от 48 млрд. фут3/день (1359 млрд. л) в 2006 году до 65 млрд. фут3/день (1841 млрд. л) в 2012 году) довел стоимость природного газа до рекордно низкого уровня (от примерно $14,00/млн. БТЕ в 2006 году до примерно $2,50/млн. БТЕ в 2012 году). Природный газ внутри страны в основном получают путем гидроразрыва пласта («фрэкинга»), но метан можно также получить из других источников, таких как мусорные свалки и сточные воды. Кроме того, освоение метановых источников принесет значительную пользу для окружающей среды, поскольку парниковый эффект метана в 23 раза больше по сравнению с CO2.
Однако летучесть метана делает его транспортировку и непосредственное применение в качестве топлива проблематичными. По этой причине имеется сильный стимул для превращения газа в жидкую форму, что позволит облегчить транспортировку к месту применения. В настоящее время используют два подхода: сжижение, приводящее к получению сжиженного природного газа (СПГ), и химическое превращение с конвертированием газа в жидкость (ГВЖ) (Patel, 7th World Congress of Chemical Engineering, Глазго, Шотландия, Соединенное Королевство, 2005). Метод Фишера-Тропша (Ф-Т) является в настоящее время наиболее распространенным ГВЖ-способом для превращения метана из природного газа в углеводороды более высокого порядка (Patel, 2005). Следует отметить, что в методе Ф-Т в качестве сырья используют синтез-газ, который получают из природного газа путем парового риформинга (источником синтез-газа также может быть газификация угля, при высокотемпературной реакции с водой и кислородом). В процессе Ф-Т получают нефтепродукты, которые соответствуют сегодняшним требованиям к поставке топлива, однако указанный способ имеет ряд недостатков, в том числе низкие выходы, плохую селективность (что делает сложной утилизацию при переработке), и требует значительных капитальных затрат и больших масштабов для обеспечения экономически выгодного производства (Spath и Dayton, December 2003 NREL/TP-510-34929). Большой масштаб, необходимый для функционирования установки Ф-Т (для типичной установки капитальные затраты составляют более $2 миллиардов $ [Patel, 2005]) также приводит к значительному ограничению вследствие большого количества исходного метана, требующегося для непрерывной работы такой установки. Так как во многих случаях стоимость транспортировки метана чрезвычайно высокая, указанную установку следует размещать вблизи или большого газового источника, или трубопровода. Дополнительные затраты и коэффициент масштабирования определяют собой экономику технологий мокрой очистки газа (Spath и Dayton, 2003), так как Ф-Т катализаторы проявляют высокую чувствительность к обычным загрязняющим веществам в природном газе, которые успешно выдерживают процесс превращения синтез-газа.
Установки Ф-Т работают в полунепрерывном режиме с 1938. В настоящее время несколько компаний изучают возможность введения новых установок с учетом текущей доступности и цены метана, приведенной выше. Однако, несмотря на значительное количество научных исследований и разработок, сделанных за последние 70 с лишним лет, недостатки технологии Ф-Т препятствуют широкому внедрению промышленных способов ГВЖ. Требование свободного доступа к большим объемам чистого газа в сочетании с внушительными капитальными вложениями ограничивают в настоящее время применение установок Ф-Т на основе природного газа их успешным функционированием только в нескольких местах по всему миру (Spath и Dayton, 2003). Строгое требование к минимальной обработке для установки ГВЖ или СПГ в сочетании с высокой стоимостью транспортировки приводят к применению более маленьких источников метана, именуемых «труднодоступным» газом (например, природного газа, добываемого на нефтяных скважинах в открытом море, или метана, выделяющегося из мусорных свалок). При текущем отсутствии эффективных маломасштабных технологий превращения такие источники труднодоступного газа, как правило, выбрасываются в атмосферу или сжигаются на факелах, так как накопление метана несет значительную угрозу безопасности.
В связи с ограничениями, связанными с производством биотоплива первого, второго и следующего поколений, в данной области техники имеется явная потребность в новых способах эффективного и рентабельного получения альтернативных видов топлива без воздействия на окружающую среду или без конкуренции с производством продуктов питания. Настоящее изобретение решает эту проблему путем обеспечения эффективных и рентабельных способов получения биотоплива и других продуктов с применением биоинженерии.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Согласно одному из аспектов, в настоящем изобретении предложен способ получения топлива путем переработки масляной композиции, полученной из метаболизирующего C1 нефотосинтезирующего микроорганизма (например, в устройстве переработки), с получением топлива. Кроме того, в настоящем изобретении предложен способ получения топлива путем превращения биомассы из культуры, в основном содержащей метаболизирующий C1 нефотосинтезирующий микроорганизм, в масляную композицию и переработки масляной композиции с получением топлива. Согласно еще одному аспекту, в настоящем изобретении предложена установка для биопереработки, содержащая блок обработки, в котором масляную композицию получают из метаболизирующего C1 нефотосинтезирующего микроорганизма; и устройство переработки для переработки масляной композиции с получением топлива. Согласно другому аспекту, в настоящем изобретении предложена масляная композиция или биотопливная композиция, содержащая молекулы, содержащие атомы водорода и углерода, составляющие по меньшей мере от примерно 50% до примерно 99% относительно массы композиции, при этом δ13C композиции составляет от примерно - 35‰ до примерно - 50‰, от -45‰ до примерно - 35‰, или от примерно - 50‰ до примерно - 40‰, или от примерно - 45‰ до примерно - 65‰, или примерно - 60‰ до примерно - 70‰, или от примерно - 30‰ до примерно - 70‰.
Согласно некоторым вариантам реализации, в настоящем изобретении предложены метаболизирующие C1 микроорганизмы, которые представляют собой прокариоты или бактерии, такие как Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium, Methanomonas, Methylophilus, Methylobacillus, Methylobacterium, Hyphomicrobium, Xanthobacter, Bacillus, Paracoccus, Nocardia, Arthrobacter, Rhodopseudomonas или Pseudomonas. Согласно другим вариантам реализации изобретения, метаболизирующие C1 бактерии представляют собой метанотроф или метилотроф. Типичные метанотрофы включают Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium, Methanomonas или их комбинацию.
Типичные виды метанотрофов включают Methylomonas sp. 16a (ATCC РТА 2402), Methylosinus trichosporium OB3b (NRRL В-11, 196), Methylosinus sporium (NRRL В-11, 197), Methylocystis parvus (NRRL В-11, 198), Methylomonas methanica (NRRL В-11, 199), Methylomonas albus (NRRL В-11, 200), Methylococcus capsulatus Bath (NCIMB 11132), Methylobacter capsulatus Y (NRRL B-11, 201), Methylobacterium organophilum (ATCC 27, 886), Methylomonas sp. AJ-3670 (FERM P-2400), Methylomicrobium alcaliphilum, Methylocella silvestris, Methylacidiphilum infernorum, Methylibium petroleiphilum, Methylococcus capsulatus (Bath) или их быстрорастущие варианты.
Типичные виды метилотрофов включают Methylobacterium extorquens, Methylobacterium radiotolerans, Methylobacterium populi, Methylobacterium chloromethanicum, Methylobacterium nodulans или их комбинацию.
Согласно другим вариантам реализации в настоящем изобретении предложены метаболизирующие C1 микроорганизмы, которые представляют собой бактерии, метаболизирующие синтез-газ, такие как Clostridium, Moorella, Pyrococcus, Eubacterium, Desulfobacterium, Carboxydothermus, Acetogenium, Acetobacterium, Acetoanaerobium, Butyribaceterium, Peptostreptococcus или любую их комбинацию. Типичные бактерии, метаболизирующие синтез-газ, включают Clostridium autoethanogenum, Clostridium ljungdahli, Clostridium ragsdalei, Clostridium carboxydivorans, Butyribacterium methylotrophicum, Clostridium woodii, Clostridium neopropanologen или любую их комбинацию.
Согласно некоторым другим вариантам реализации изобретения, метаболизирующие C1 микроорганизмы представляют собой эукариоты, такие как дрожжи, в том числе Candida, Yarrowia, Hansenula, Pichia, Torulopsis или Rhodotorula.
Согласно другим вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой рекомбинантный микроорганизм, содержащий гетерологичный полинуклеотид, кодирующий фермент синтеза жирных кислот, фермент ассимиляции формальдегида или их комбинацию. Согласно некоторым вариантам реализации изобретения, гетерологичный полинуклеотид кодирует тиоэстеразу, малонил-КоА: ацилпереносящий белок-трансацилазу, ацетил-КоА-карбоксилазу или любую их комбинацию. Например, тиоэстераза может представлять собой кодон-оптимизированный tesA E. coli, в котором отсутствует последовательность нацеливания на периплазму; малонил-КоА: ацилпереносящий белок-трансацилаза может представлять собой кодон-оптимизированный fabD E. coli; и ацетил-КоА-карбоксилаза может представлять собой кодон-оптимизированный ассА, ассВ, ассС, accD E. Coli, или любую их комбинацию. Согласно некоторым другим вариантам реализации изобретения, метаболизирующий C1 микроорганизм дополнительно содержит мутацию, которая минимизирует или устраняет активность жирная кислота: КоА-лигазы.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 показана типичная концептуальная модель реакторной системы на основе метаболизирующего C1 микроорганизма, предназначенная для улавливания метана и превращения его в алкановое топливо согласно некоторым вариантам реализации настоящего изобретения.
На фиг. 2 показана типичная концептуальная модель реакторной системы на основе метаболизирующего C1 микроорганизма, предназначенная для улавливания метана и превращения его в биодизельное топливо согласно некоторым вариантам реализации настоящего изобретения.
На фиг. 3A и 3B показано, что рекомбинантный Methylobacter capsulatus, экспрессирующий TesA' (ген TesA из Е. coli с удаленной последовательностью нацеливания на периплазму) приводит (A) к увеличению продуцирования свободных жирных кислот, и (B) указанное увеличение основном обусловлено повышенными уровнями C16:0 и C18:0 липидов.
На фиг. 4A и 4B показаны ГХ/МС-хроматограммы масляной композиции, экстрагированной из М. Trichosporium, до (A) и после (B) гидролиза и переэтерификации с применением KOH в смеси толуол : метанол.
На фиг. 5A и 5B показаны ГХ/МС-хроматограммы масляной композиции, экстрагированной из M. Capsulatus, до (A) и после (B) гидролиза и переэтерификации с применением KOH в смеси толуол : метанол.
На фиг. 6A и 6B показаны ГХ/МС-хроматограммы масляной композиции, экстрагированной из Methylomonas sp. 16a, до (A) и после (B) гидролиза и переэтерификации с применением KOH в смеси толуол : метанол.
На фиг. 7 показана схема распределения δ13C в разных источниках углерода.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложены композиции, способы и системы для получения биотоплив и биопластика, в которых культивируют метаболизирующие C1 микроорганизмы с получением биомассы, характеризующейся максимальным накоплением биомасла. Например, предложен способ ферментации, включающий превращение метана в биотопливо и представляющий собой масштабируемый промышленный процесс. Такой новый подход позволяет использовать, например, метилотрофные или метанотрофные бактерии в качестве новой системы-хозяина для продуцирования биомассы с получением биотоплива в форме, например, эстерифицированного биодизельного топлива или алканового топлива для гидрообработки или для биопластика в форме полигидроалканоатов (ПГА). Кроме того, представляющую интерес масляную композицию можно получить из метилотрофных или метанотрофных бактерий, поскольку указанные организмы могут аккумулировать значительные количества мембранных липидов в условиях, описанных в настоящей заявке и, более того, указанные микроорганизмы продуцируют высокое содержание мембран.
В качестве справочной информации, метан из различных источников, в том числе природного газа, представляет собой имеющийся в изобилии отечественный ресурс. Химические подходы, направленные на разработку технологии превращения газа в жидкости (ГВЖ) для улучшения использования метана в качестве топлива имеют на сегодняшний день только ограниченный успех, несмотря на значительные капиталовложения. Напротив, на развитие современных биотехнологических подходов в отношении разработки способов ГВЖ были потрачены небольшие усилия. Некоторые ограничения, в первую очередь, стоимость сахарного сырья, препятствовали экономически выгодному производству биотоплива с применением микробных систем. Эксплуатация дешевого, имеющегося в изобилии внутри страны углеродного сырья, такого как метан, является экономически рациональной альтернативой производства биотоплива. С помощью новых биотехнологических инструментов и методов были разработаны новые промышленные микроорганизмы, которые обеспечивают биопроцесс ГВЖ в промышленном масштабе, как описано в настоящей заявке. Кроме того, характеристики топлива после переработки и повышения качества экстрагированных липидов демонстрируют падающий потенциал для таких применений, как дизельное топливо, газолин, топливо для реактивных двигателей или олефины.
Согласно одному из аспектов, в настоящем изобретении предложен способ получения топлива в устройстве переработки путем переработки масляной композиции, полученной из метаболизирующего C1 нефотосинтезирующего микроорганизма, с получением топлива. Кроме того, в настоящем изобретении предложен способ получения топлива путем превращения биомассы из культуры, в основном содержащей метаболизирующий C1 нефотосинтезирующий микроорганизм, в масляную композицию и обработку масляной композиции с получением топлива. В другом аспекте, в настоящем изобретении предложена установка для биопереработки, содержащая блок обработки, в котором масляную композицию получают из метаболизирующего C1 нефотосинтезирующего микроорганизма; и устройство переработки для переработки масляной композиции с получением топлива.
Согласно другому аспекту, в настоящем изобретении предложена масляная композиция или полученная из нее биотопливная композиция, содержащая молекулы, содержащие атомы водорода и углерода, составляющие по меньшей мере от примерно 50% до примерно 99% относительно массы композиции, при этом δ13C композиции составляет менее - 30‰ или составляет от примерно - 70‰ до примерно - 35‰, или, при смешивании с компонентом топлива с получением топливного продукта, составляет от примерно - 37‰ до примерно - 10‰. В соответствующем аспекте, в настоящем изобретении предложена биомасса, δ13C которой составляет менее - 30‰ или находится в диапазоне от примерно - 35‰ до примерно - 50‰, от - 45‰ до примерно - 35‰, или от примерно - 50‰ до примерно - 40‰, или от примерно - 45‰ до примерно - 65‰, или от примерно - 60‰ до примерно - 70‰, или от примерно - 30‰ до примерно - 70‰.
Перед более подробным изложением настоящего изобретения, может быть полезным для его понимания дать определения некоторым терминам, применяемым настоящей заявке. Дополнительные определения приведены по всему тексту настоящего описания.
В настоящем описании подразумевают, что любой диапазон концентраций, диапазон процентных отношений, диапазон соотношений или диапазон целочисленных значений включает величину любого целочисленного значения в пределах приведенного диапазона и, когда это целесообразно, его доли (такие как одна десятая и одна сотая целого числа), если не указано иное. Кроме того, подразумевают, что любой диапазон чисел, приведенный в настоящей заявке в отношении любого физического свойства, такого как элементарные звенья полимера, размер или толщина, включает любой целочисленное значение в пределах приведенного диапазона, если не указано иное. В настоящей заявке, термин «примерно» означает ±20% от указанного диапазона, значения или структуры, если не указано иное. Термин «состоящий в основном из» ограничивает объем формулы изобретения конкретными материалами или стадиями или теми материалами или стадиями, которые по существу не влияют на основные и новые характеристики заявленного изобретения. Следует иметь в виду, что в настоящей заявке термины в единственном числе относятся к «одному или более» перечисленных компонентов. Следует понимать, что применение альтернативного варианта (например, «или») означает любой один, оба или любая комбинация альтернативных вариантов. В настоящей заявке, термины «включают», «имеют» и «содержат» применяют синонимично, что предполагает толкование указанных терминов и их вариантов как неограничивающих.
В настоящей заявке, «C1 субстрат» или «C1 соединение» относится к любой углеродсодержащей молекуле или композиции, у которой отсутствует связь углерод-углерод. Типичные C1 субстраты включают природный газ, нетрадиционный природный газ, синтез-газ, метан, метанол, формальдегид, муравьиную кислоту или ее соль, моноксид углерода, диоксид углерода, метилированные амины (например, метиламин, диметиламин, триметиламин, и т.п.), метилированные тиолы, метилгалогены (например, бромметан, хлорметан, иодметан, дихлорметан, и т.п.), цианид или любую их комбинацию.
В настоящей заявке, «метаболизирующий C1 микроорганизм» или «метаболизирующий C1 нефотосинтезирующий микроорганизм» относится к любому микроорганизму, обладающему способностью использовать C1 субстрат в качестве источника энергии или в качестве своего основного источника энергии и биомассы, и который может использовать или не использовать другие углеродные субстраты (такие как сахара и сложные углеводы) для энергии и биомассы. Например, метаболизирующий C1 микроорганизм может окислять C1 субстрат, такой как метан или метанол. Метаболизирующие C1 микроорганизмы включают бактерии (такие как метанотрофы и метилотрофы) и дрожжи. Согласно некоторым вариантам реализации изобретения, метаболизирующий C1 микроорганизм не включает фотосинтезирующий микроорганизм, такой как водоросль. Согласно некоторым вариантам реализации изобретения, метаболизирующий C1 микроорганизм будет представлять собой «облигатный метаболизирующий C1 микроорганизм», что означает, что его единственным источником энергии являются C1 субстраты. Согласно другим вариантам реализации изобретения, метаболизирующий C1 микроорганизм (например, метанотроф) будет расти в культурной среде в присутствии сырья, представляющего собой C1 субстрат (т.е., при применении C1 субстрата в качестве источника энергии).
В настоящей заявке, термин «метилотрофные бактерии» относится к любым бактериям, способным окислять любое соединение в любой форме (например, твердое вещество, жидкость, газ), которое содержит по меньшей мере один углерод и которое не содержит связи углерод-углерод. Согласно некоторым вариантам реализации изобретения, метилотрофная бактерия может представлять собой метанотроф. Например, «метанотрофные бактерии» относятся к любым метилотрофным бактериям, которые обладают способностью окислять метан в качестве источника углерода и энергии, который может представлять собой основной источник углерода и энергии. Типичные метанотрофные бактерии включают Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium или Methanomonas. Согласно некоторым вариантам реализации изобретения, метилотрофная бактерия представляет собой «облигатную метилотрофную бактерию», относящуюся к бактериям, которые ограничены применением C1 субстратов для выработки энергии. Согласно некоторым вариантам реализации изобретения, метилотрофные бактерии представляют собой «факультативные метанотрофные бактерии», которые наряду с C1 субстратами по природе способны использовать субстраты, содержащие несколько атомов углерода, такие как ацетат, пируват, сукцинат, малат или этанол, в качестве их источника углерода и энергии. Факультативные метанотрофы включают некоторые виды Methylocella, Methylocystis, Methylocapsa (например, Methylocella silvestris, Methylocella palustris, Methylocella tundrae, Methylocystis daltona SB2, Methylocystis bryophila, и Methylocapsa aurea KYG), и Methylobacterium organophilum (ATCC 27, 886).
В настоящей заявке, термин «утилизирующая CO бактерия» относится к бактерии, которая по природе обладает способностью окислять моноксид углерода (CO) в качестве источника углерода и энергии. Можно использовать моноксид углерода из «синтез-газа» или «синтез-газа», смеси моноксида углерода и водорода, образующейся при газификации любого органического сырья, такого как уголь, угольная нефть, природный газ, биомасса или органические отходы. Утилизирующие CO бактерии не включают бактерии, которые должны быть генетически модифицированы для выращивания на CO в качестве источника углерода.
В настоящей заявке, «синтез-газ» относится к смеси, содержащей моноксид углерода (CO) и водород (H2). Синтез-газ может также включать CO2, метан, и другие газы в меньших количествах относительно CO и H2.
«Рост» определяют как увеличение клеточной массы. Такое увеличение может происходить путем деления клеток (репликации) и образования новых клеток при «сбалансированном росте» или при «несбалансированном росте», когда клеточная масса увеличивается вследствие аккумулирования конкретного соединения или полимера, такого как некоторые липиды. В последнем случае рост может проявляться в виде увеличения размера клеток вследствие аккумулирования биополимера внутри клетки.
Во время «сбалансированного клеточного роста» все сырье (электронные доноры и акцепторы электронов) и все питательные вещества присутствуют в соотношениях, необходимых для создания всех макромолекулярных компонентов клетки. То есть, сырье или питательное вещество не ограничивает синтез белков, сложных углеводных полимеров, жиров или нуклеиновых кислот. Напротив, во время «несбалансированного клеточного роста» сырье или питательное вещество, необходимое для построения одной или более макромолекул клетки, не присутствует в количестве или соотношении, требуемом для сбалансированного роста. Соответственно, такое сырье или питательное вещество становится ограничивающим фактором и называется «ограничивающим питательным веществом».
Некоторые клетки могут все еще обеспечивать чистый рост при несбалансированных условиях, но такой рост является несбалансированным, и полимеры, которые могут синтезироваться в отсутствии ограничивающего сырья или питательного вещества, будут аккумулироваться. Указанные полимеры включают липиды или продукты, хранящиеся внутри клеток, такие как полигидроксиалканоаты (ПГА), в том числе полигидроксибутират (ПГБ), полигидроксивалерат (ПНВ), и полигидроксигексаноат (ПГГ)-гликоген, или секретируемые материалы, такие как внутриклеточный полисахарид. Указанные масляные композиции можно применять при производстве биопластика.
Типичные сбалансированные и несбалансированные условия роста могут различаться по содержанию азота в средах. Например, азот составляет примерно 12% относительно сухой массы клеток, что означает, что для выращивания 100 мг/л сухой массы клеток необходимо обеспечить 12 мг/л азота (например, в форме нитрата или соли аммония вместе с сырьем и другими питательными веществами в требуемых стехиометрических соотношениях). Не желая быть связанными теорией, предполагают, что фиксация атмосферного азота с образованием аммиака (т.е. посредством фиксации азота) не является важным источником азота для биосинтетических промежуточных соединений или клеточных компонентов. Если другое сырье и питательные вещества доступны в количествах, необходимых для получения 100 мг/л сухой массы клеток, но поступает меньше 12 мг/л азота, то может происходить несбалансированный клеточный рост с аккумулированием полимеров, не содержащих азот. Если впоследствии подают азот, накопленный полимер может служить в качестве сырья для клетки, обеспечивая сбалансированный рост с репликацией и образованием новых клеток.
В настоящей заявке термин «ростовой цикл», применимый к клетке или микроорганизму, относится к метаболическому циклу, посредством которого клетка или микроорганизм развивается в условиях культивирования. Например, цикл может включать различные стадии, такие как лаг-фаза, экспоненциальная фаза, конец экспоненциальной фазы и стационарная фаза.
Термин «экспоненциальный рост», «экспоненциальная фаза роста», «логарифмическая фаза» или «логарифмическая фаза роста» относятся к скорости, с которой микроорганизмы растут и делятся. Например, во время логарифмической фазы, микроорганизмы растут с их максимальной скоростью с учетом их генетического потенциала, природы среды и условий, при которых их выращивают. Скорость роста микроорганизма является постоянной во время экспоненциальной фазы, и микроорганизм делится и его количество удваивается через равные промежутки времени. Клетки, которые являются «активно растущими», представляют собой клетки, которые растут в логарифмической фазе. Напротив, «стационарная фаза» относится к точке в цикле роста, во время которой рост культуры клеток замедляется или даже прекращается. Термин «окружающая среда, изменяющая рост» относится к энергии, химическим веществам или живым существам, обладающим способностью подавлять клеточный рост или уничтожать клетки. Ингибиторы могут включать мутагены, лекарственные препараты, антибиотики, УФ свет, экстремальную температуру, pH, метаболические побочные продукты, органические химические вещества, неорганические химические вещества, бактерии, вирусы или т.п.
В настоящей заявке, «быстрорастущий вариант» относится к организму, микроорганизму, бактерии, дрожжам или клетке, способной расти с помощью C1 субстрата, такого как метан или метанол, в качестве единственного или основного источника углерода и энергии и которая характеризуется скоростью роста в экспоненциальной фазе, более быстрой, чем в случае родительского, эталонного или дикого типа организма, микроорганизма, бактерии, дрожжей или клетки - то есть, быстрорастущий вариант имеет более быстрое время удвоения и, следовательно, высокую скорость роста и высокий выход клеточной массы на грамм метаболизируемого C1 субстрата по сравнению с родительской клеткой (см., например, патент США №6689601).
В настоящей заявке, «биотопливо» относится к топливу, по меньшей мере частично полученному из «биомассы».
В настоящей заявке «биомасса» или «биологический материал» относится к органическому материалу биологического происхождения, который может включать одну или более цельных клеток, лизированных клеток, внутриклеточный материал или т.п. Например, материал, собранный из культивируемого микроорганизма (например, бактериальной или дрожжевой культуры) считается биомассой, которая может включать клетки, клеточные мембраны, клеточную цитоплазму, тельца включений, продукты, секретируемые или экскретируемые в культуральную среду, или любую их комбинацию. Согласно некоторым вариантам реализации изобретения, биомасса содержит метаболизирующие C1 микроорганизмы согласно настоящему изобретению вместе с культивирующими средами, в которых выращивали метаболизирующие C1 микроорганизмы согласно настоящему изобретению. Согласно другим вариантам реализации изобретения, биомасса содержит метаболизирующие C1 микроорганизмы (цельные или лизированные или и то, и другое) согласно настоящему изобретению, извлеченные из культуры, растущей на C1 субстрате (например, природном газе, метане). Согласно другим вариантам реализации изобретения, биомасса содержит супернатант отработавшей среды из культуральной жидкости метаболизирующего C1 микроорганизма, культивируемого на C1 субстрате. Такую культуру можно рассматривать как возобновляемый ресурс.
В настоящей заявке, «масляная композиция» относится к липидному содержанию биомассы (например, бактериальной культуры), в том числе к содержанию жирных кислот, сложных эфиров жирных кислот, триглицеридов, фосфолипидов, полигидроксиалканоатов, изопренов, терпенов или т.п. Масляную композицию биомассы можно экстрагировать из остатка материала биомассы с помощью способов, описанных в настоящей заявке, таких экстракция гексаном или хлороформом. Кроме того «масляную композицию» можно обнаружить в любой одной или более участках культуры, включая клеточную мембрану, клеточную цитоплазму, тельца включений, секретируемые или экскретируемые в культуральную среду, или любую их комбинацию. Масляная композиция не является ни природным газом, ни сырой нефтью.
В настоящей заявке, термин «хозяин» относится к клетке или микроорганизму (например, метанотрофу), который можно генетически модифицировать с помощью молекулы экзогенной нуклеиновой кислоты с образованием представляющего интерес полипептида (например, тиоэстеразы [tesA], ацетил-КоА-карбоксилазы [accABCD], малонил-КоА: АСР-трансацилазы [fabD]). Согласно некоторым вариантам реализации изобретения, клетка-хозяин может уже обладать другими генетическими модификациями или быть модифицирована таким образом, чтобы включать другие генетические модификации, которые придают требуемые свойства, связанные или несвязанные с биосинтезом липидов (например, клетка может быть модифицирована путем исключения, изменения или процессирования длинноцепочечной жирной кислоты КоА-лигазы [fadD]). Например, клетка-хозяин может обладать генетическими модификациями, которые минимизируют или уменьшают расщепление жирных кислот, минимизируют или уменьшают продуцирование ингибиторов роста клетки-хозяина, обеспечивают быстрый рост, устойчивость к загрязняющим веществам или конкретным условиям культивирования (например, кислотоустойчивость, устойчивость к биоцидам), способность метаболизировать дополнительные углеродные субстраты или способность синтезировать дополнительные необходимые продукты или промежуточные вещества.
В настоящей заявке, «рекомбинантный» или «искусственный» относится к организму, микроорганизму, клетке, молекуле нуклеиновой кислоты или вектору, который имеет по меньшей мере одно генетическое изменение или был модифицирован путем введения молекулы гетерологичной нуклеиновой кислоты, или относится к клетке, которая была изменена таким образом, чтобы можно было регулировать экспрессию молекулы эндогенной нуклеиновой кислоты или гена. Рекомбинантный также относится к клетке, которую получают из клетки или которая является потомком клетки, содержащей одну или более таких модификаций. Генетические изменения включают, например, модификации, вносящие способные к экспрессии молекулы нуклеиновой кислоты, кодирующие белки или ферменты, или другие присоединения, делеции, замещения молекул нуклеиновой кислоты или другое функциональное изменение генетического материала клетки. Например, рекомбинантные клетки могут экспрессировать гены или другие молекулы нуклеиновой кислоты, которые не встречаются в идентичной форме в нативной клетке (т.е., немодифицированной клетке или клетке дикого типа), или могут обеспечить измененный профиль экспрессии эндогенных генов, таких генов, которые можно иным образом сверхэкспрессировать, недостаточно экспрессировать, минимально экспрессировать или не экспрессировать совсем.
Рекомбинантные способы экспрессии экзогенных или гетерологичных нуклеиновых кислот в микробных организмах хорошо известны в данной области техники. Описание таких способов можно найти, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, Third Ed., Cold Spring Harbor Laboratory, New York (2001); и Ausubel et al., Current Protocols in Molecular Biology, John Wiley и Sons, Baltimore, MD (1999). Типичные экзогенные белки или ферменты, подлежащие экспрессии, включают тиоэстеразу, одну или более ацетил-КоА-карбоксилаз, малонил-КоА: ACP-трансацилазу или любую их комбинацию. Генетические модификации молекул нуклеиновых кислот, кодирующих ферменты или их функциональные фрагменты, могут придавать биохимическую или метаболическую способность рекомбинантной клетке, которая изменена по сравнению с ее естественным состоянием.
В настоящей заявке, термин «эндогенный» или «нативный» относится к гену, белку, соединению или активности, которая обычно присутствует в клетке-хозяине. Термин «гомологичный» или «гомолог» относится к молекуле или активности, относящейся к экзогенному (ненативному) источнику, который представляет собой такую же или похожую молекулу или активность, обнаруживаемую в клетке, виде или штамме хозяина или получаемую из такой клетки, вида или штамма.
В настоящей заявке, молекула, конструкт или последовательность «гетерологичной» нуклеиновой кислоты относится к молекуле нуклеиновой кислоты или части последовательности молекулы нуклеиновой кислоты, которая не является нативной по отношению к клетке, в которой ее экспрессируют, к молекуле нуклеиновой кислоты или части молекулы нуклеиновой кислоты, нативной по отношению к клетке-хозяину, которая была изменена или подверглась мутации, или к молекуле нуклеиновой кислоты с измененной экспрессией по сравнению с уровнями нативной экспрессии в похожих условиях. Например, гетерологичную управляющую последовательность (например, промотор, энхансер) можно использовать для регулирования экспрессии гена или молекулы нуклеиновой кислоты, которая в какой-то мере отличается от гена или молекулы нуклеиновой кислоты, обычно экспрессируемой в природе или культуре. Согласно некоторым вариантам реализации изобретения, молекула гетерологичной нуклеиновой кислоты может быть гомологичной относительно гена нативной клетки-хозяина, но может иметь измененный уровень экспрессии или содержать другую последовательность или и то, и другое. Согласно другим вариантам реализации изобретения, молекулы гетерологичной или экзогенной нуклеиновой кислоты могут быть не эндогенными относительно клетки-хозяина или генома хозяина, но в качестве альтернативы их можно было внести в клетку-хозяина путем конъюгации, трансформации, трансфекции, электропорации или т.п., при этом внесенная молекула может встраиваться в геном хозяина или может существовать в виде экстрахромосомного генетического материала (например, плазмиды или другого самореплицирующегося вектора).
Согласно некоторым вариантам реализации изобретения, более чем одна молекула гетерологичной или экзогенной нуклеиновой кислоты может быть введена в клетку-хозяина в виде отдельных молекул нуклеиновой кислоты, в виде полицистронной молекулы нуклеиновой кислоты, в виде одной молекулы нуклеиновой кислоты, кодирующей гибридный белок, или любой их комбинации, и все еще может рассматриваться как более чем одна гетерологичная или экзогенная нуклеиновая кислота. Например, метаболизирующий C1 микроорганизм можно модифицировать таким образом, чтобы он экспрессировал две или более молекулы гетерологичной или экзогенной нуклеиновой кислоты, которые могут быть одинаковыми или разными, которые кодируют одну или более тиоэстераз, как описано в настоящей заявке. Согласно некоторым вариантам реализации изобретения, многократные копии полинуклеотидной молекулы кодирующей тиоэстеразу (ТЕ), вводят в клетку-хозяина, которая может представлять собой две, три, четыре, пять, шесть, семь, восемь, девять, десять или более копий полинуклеотидов, кодирующих одну и ту же ТЕ или другую ТЕ.
Когда две или более молекул экзогенной нуклеиновой кислоты вводят в метаболизирующий C1 микроорганизм хозяина, понятно, что еще две молекулы экзогенной нуклеиновой кислоты можно ввести в виде одной молекулы нуклеиновой кислоты (например, на один вектор), на отдельные вектора, встроенные в хромосому хозяина на одном сайте или нескольких сайтах, и каждый из указанных вариантов реализации изобретения все еще следует рассматривать как две или более молекулы экзогенной нуклеиновой кислоты. Таким образом, количество упомянутых молекул гетерологичной нуклеиновой кислоты или активность белка относится к количеству кодирующих молекул нуклеиновой кислоты или показателю активности белка, не к количеству отдельных молекул нуклеиновой кислоты, введенных в клетку-хозяина.
«Идентичность, выраженная в процентах» двух или более последовательностей нуклеиновых кислот представляет собой функцию количества идентичных позиций, совместно используемых последовательностями (т.е., идентичность, % = количество идентичных позиций / общее количество позиций × 100), с учетом количества разрывов и длины каждого разрыва, который необходимо внести, чтобы оптимизировать выравнивание двух или более последовательностей. Сравнение последовательностей и установление идентичности в процентах двух или более последовательностей можно выполнить, используя математический алгоритм, такой как программы BLAST и Gapped BLAST при их параметрах, используемых по умолчанию (например, Altschul et al., J. Mol. Biol. 275:403, 1990; см. также BLASTN на сайте www.ncbi.nlm.nih.gov/BLAST).
В данной области техники «консервативным замещением» считается замещение одной аминокислоты на другую аминокислоту, имеющую похожие свойства. Типичные консервативные замещения хорошо известны в данной области техники (см., например, WO 97/09433, р. 10; Lehninger, Biochemistry, 2nd Edition; Worth Publishers, Inc. NY:NY (1975), pp. 71-77; Lewin, Genes IV, Oxford University Press, NY и Cell Press, Cambridge, MA (1990), p. 8).
В настоящей заявке «сверхэкспрессируемый» в отношении гена или белка означает увеличение экспрессии или активности гена или белка. Повышенная экспрессия или активность включает экспрессию или активность гена или белка, повышенную относительно уровня контрольного микроорганизма дикого типа (не созданного методами генетической инженерии) или эталонного микроорганизма. Ген или белок сверхэкспрессируется, если экспрессия или активность имеет место в микроорганизме, где он обычно не экспрессируется или не активен. Ген или белок сверхэкспрессируется, если экспрессия или активность расширена или протекает дольше в рекомбинантном микроорганизме, чем в контрольном микроорганизме дикого типа или эталонном микроорганизме.
В настоящей заявке «ингибирование» или «ингибированный» относится, непосредственно или косвенно, к изменению, уменьшению, угнетающей регуляции, устранению или делеции при экспрессии гена-мишени или активности молекулы-мишени (например, длинноцепочечная жирная кислота: КоА-лигазы) по сравнению с контрольной, эндогенной или эталонной молекулой, при этом изменение, уменьшение, угнетающая регуляция или устранение является статистически, биологически, промышленно или клинически значимым.
В настоящей заявке, «установка для биопереработки» относится к средствам производства, которые объединяют оборудование и способы превращения биомассы для получения топлива из биомассы.
В настоящей заявке «установка по переработке» относится к нефтеперерабатывающему заводу или его аспектам, в котором можно обрабатывать масляные композиции (например, биомассу, биотопливо или ископаемые топлива, такие как сырая нефть, уголь или природный газ). Типичные процессы, выполняемые на нефтеперерабатывающих заводах, включают крекинг, переэтерификацию, риформинг, дистилляцию, гидроочистку, изомеризацию или любую их комбинацию.
Системы для производства биотоплива
Системы для получения биотоплива согласно настоящему изобретению могут включать отдельные блоки (например, близко расположенные или прилегающие друг к другу или не прилегающие), комплексные блоки или сама система может быть объединенной и частично или полностью комплексной. В системах согласно настоящему изобретению можно использовать биомассу микроорганизма, выращенного в комплексной установке для биопереработки с получением топливных композиций и топливных продуктов, в частности биотоплива. Согласно некоторым вариантам реализации изобретения, в установке для биопереработки в качестве биомассы используют одну биомассу или смешанную биомассу с получением топлива (например, дизельного топлива, топлива для реактивных двигателей, газолина), такую как метаболизирующий C1 микроорганизм (например, метанотроф, такой как Methylosinus trichosporium OB3b, Methylococcus capsulatus Bath, Methylomonas sp. 16a, Methylomonas methanica, Methylomicrobium alcaliphilum или их быстрорастущие варианты).
Типичная система биопереработки проиллюстрирована на фиг. 1. В такой системе можно выполнить одну или более из следующих стадий: культивирование представляющего интерес штамма микроорганизма (например, метанотрофа, метилотрофа или дрожжей), которые могут иметь одно или более улучшенных свойств (например, быть рекомбинантными, иметь более быструю скорость роста, способность расти при высоком pH, проявлять улучшенную усвояемость питательных веществ, термоустойчивость, повышенный выход биомассы), выделение из микроорганизма продукта, такого как масляная композиция (например, жирные кислоты, триглицериды, фосфолипиды, изопрены, терпены, РНА или любая их комбинация), и обработку масляной композиции с получением предшественников пластмассы или одного или более видов топлива, таких как топливо для реактивных двигателей, дизельное топливо, газолин или их комбинация. С помощью указанной системы можно одновременно или последовательно получить различные биотопливные композиции и продукты. Например, система может включать установку или блок гидрообработки, в которой можно превратить масляную композицию в топливо для реактивных двигателей и дизельное топливо. Система может также включать установку по переработке нефти, в которой можно превратить сырую нефть и продукты, получаемые на установке гидрообработки, в газолин. Например, при производстве топлива для реактивных двигателей и дизельного топлива можно получить дополнительные продукты, такие как нафта и легкие углеводороды, в том числе, пропан, которые затем используют для получения газолина. Типичные легкие углеводороды включают метан, этан, пропан, бутан, пентан, бутанол и изобутанол. В другом примере при производстве газолина можно получить дополнительные продукты, такие как дизельное топливо, которые можно использовать для получения топлива для реактивных двигателей.
Альтернативная типичная система биопереработки показана на фиг. 2. В такой системе можно выполнить одну или более из следующих стадий: культивирование представляющего интерес штамма микроорганизма (например, метанотрофа, метилотрофа или дрожжей), которые могут иметь одно или более улучшенных свойств (например, быть рекомбинантными, иметь более быструю скорость роста, способность расти при высоком pH, проявлять улучшенную усвояемость питательных веществ, термоустойчивость, повышенный выход биомассы), выделение из микроорганизма продукта, такого как масляная композиция (например, жирных кислот, сложных эфиров жирных кислот, триглицеридов, фосфолипидов, изопренов, терпенов, РНА или любой их комбинации), и модифицирование масляной композиции с получением биодизельной топливной композиции. Например, система может включать установку или блок эстерификации, в котором можно превратить масляную композицию в биодизельной топливо посредством реакции со спиртом. Типичные спирты включают метанол, этанол, пропанол, бутанол или жирные спирты с более длиной цепью.
Согласно некоторым вариантам реализации изобретения, в системах, описанных в настоящей заявке, в качестве микроорганизма используют бактерии, такие как метилотрофы или метанотрофы или дрожжи. Бактерии или дрожжи можно собрать и отделить от культуральных сред (если их не выращивают, например, в виде биопленки), получив в результате бактериальную или дрожжевую пасту. Перед получением масляной композиции из биомассы бактериальную или дрожжевую биомассу можно высушить. Согласно некоторым вариантам реализации изобретения, бактериальная или дрожжевая биомасса остается до некоторой степени влажной и ее не нужно полностью высушивать перед получением, отделением или экстрагированием масляной композиции. Бактериальные или дрожжевые масляные композиции можно экстрагировать из биомассы и отделить от бактериальной или дрожжевой твердой фазы или шлама.
Экстракцию масляной композиции можно выполнить с применением других различных способов или растворителей (например, полярного растворителя, неполярного растворителя, нейтрального растворителя, кислого растворителя, щелочного растворителя, гексана или их комбинации), таких как гексан или кислый метанол или смесь хлороформ/метанол, в способах, таких как способы, более подробно описанные в настоящей заявке, или с применением других способов экстракции, известных в данной области техники.
Согласно некоторым вариантам реализации изобретения, масляную композицию метаболизирующего C1 микроорганизма (например, метанотрофа), содержащуюся в собранной биомассе, отделяют от биомассы путем приведения в контакт с большими сдвиговыми усилиями с органическим растворителем (например, гексаном) и облагораживающим реагентом. В качестве справочной информации, масло растворяется в гексане или других похожих растворителях, образуя раствор мисцеллы, тогда как вода и твердое клеточное вещество не растворяются и могут быть отделены от мисцеллы. Несмешиваемость воды и гексана используют для достижения требуемого отделения. Согласно некоторым вариантам реализации изобретения, после смешивания с большими сдвиговыми усилиями, смесь масляной композиции/гексана/воды направляют в декантатор, где ее разделяют на две различные жидкости: фазу (мисцеллу), содержащую более легкие гексан и масляную композицию, и фазу, содержащую более тяжелые воду и отработавшее твердое вещество. Согласно другим вариантам реализации изобретения, мисцеллу из декантатора направляют в процесс дистилляции, в котором масляную композицию отделяют от растворителя, что позволяет извлекать и повторно использовать растворитель и очищает масло до той степени, при которой оно готово для последующей обработки. При дистилляции, например, используют преимущество, состоящее в разнице между точками кипения растворителя и масла, для разделения указанных двух компонентов.
Согласно некоторым вариантам реализации изобретения, масляную композицию, предложенную в настоящем изобретении, подвергают переработке. Переработка может включать крекинг, переэтерификацию, риформинг, дистилляцию, гидроочистку, изомеризацию или их комбинацию. При необходимости, переработка может включать удаление загрязняющих веществ. Например, гетероатомы и металлы можно удалить путем гидрообработки (например, гидродеазотирования (HDN), гидродеоксигенирования (HDO), гидродесульфирования (HDS), гидродеметаллизации (HDM)). Гидрообработка также может включать насыщение олефинов, гидрообработку дистиллятов, гидрообработку вакуумного газойля, гидрообработку остатка в неподвижном слое или их комбинацию. При гидрообработке масляной композиции можно получить топливо для реактивных двигателей или дизельное топливо. Масляную композицию можно также очистить с помощью крекинга, такого как каталитический крекинг, с получением газолина. Типичные способы крекинга могут включать каталитический крекинг, флюид-каталитический крекинг, паровой крекинг, гидрокрекинг, термический крекинг, термический каталитический крекинг или их комбинацию. Переработку путем гидрообработки и крекинга можно осуществлять одновременно (протекают оба процесса) или альтернативно (протекает один или другой процесс). Процессы переработки также можно осуществлять последовательно один за другим, например, продукты, полученные при гидрообработке, затем можно обработать посредством крекинга. Продукты, полученные с применением одного способа переработки (например, H2), можно также в дальнейшем использовать в другом способе переработки. Процессы переработки можно выполнять в отдельных блоках системы или в одном и том же блоке. Более того, бактериальное или дрожжевое твердое вещество или шлам можно использовать для получения топлива, корма для животных или энергии, такого как метан, выделяющийся при разложении твердого вещества или шлама.
Согласно некоторым вариантам реализации в настоящем изобретении предложена установка для биопереработки, содержащая: (а) блок обработки, в котором масляную композицию получают из метаболизирующего C1 нефотосинтезирующего микроорганизма; и (b) устройство переработки для переработки масляной композиции с получением топлива. Согласно другим вариантам реализации изобретения, установка для биопереработки может дополнительно содержать блок регулируемого культивирования для выращивания метаболизирующего C1 нефотосинтезирующего микроорганизма в присутствии сырья, содержащего C1 субстрат, при этом культивируемые бактерии продуцируют масляную композицию.
Типичные блоки регулируемого культивирования включают ферментатор, биореактор, половолоконную ячейку, биореактор с фильтрующим наполнителем или т.п. Согласно другим вариантам реализации изобретения, культуру можно выращивать в форме жидкофазной ферментации или твердофазной ферментации. Например, бактерии, такие как метилотрофы или метанотрофы, можно культивировать в биореакторе, содержащим сбалансированные среды или несбалансированные среды, которые содержат ограничивающие количества фосфора, азота, микроэлементов, кислорода или любой их комбинации, так что некоторые липиды или другие представляющие интерес полимеры (например, ПГА) аккумулируются в клетках.
Согласно некоторым вариантам реализации изобретения, культуры включают бактериальное сообщество, в том числе, различные метилотрофы или метанотрофы, которые продуцируют самые высокие уровни представляющей интерес масляной композиции (т.е., с высоким соотношением, масс./масс., липидов к биомассе). Можно использовать целый ряд конфигураций биореактора, в том числе мембранные биореакторы для секвенирования и конфигурацию для непрерывного многостадийного роста в суспензии. Согласно некоторым вариантам реализации изобретения, биореактор функционирует таким образом, что в нем происходит отбор бактерий, эффективно продуцирующих представляющую интерес масляную композицию из метана, например, условия в биореакторе позволяют отсеять те бактерии, которые или не продуцируют представляющую интерес масляную композицию из метана или продуцируют такую композицию неэффективно.
Согласно другим вариантам реализации в настоящем изобретении предложен блок регулируемого культивирования, в котором при ферментации в твердой фазе C1 субстрат (например, метан или синтез-газ) доставляют в газовой фазе к микробным биопленкам. Согласно другим вариантам реализации изобретения, при ферментации в твердой фазе устанавливают сбалансированные или несбалансированные условия роста. Согласно другим вариантам реализации изобретения, метилотрофы или метанотрофы выращивают при сбалансированных условиях роста, собирают и отделяют от жидкой фазы и переносят в камеру для проведения ферментации в твердой фазе, куда C1 субстрат поставляют при несбалансированных условиях (например, не включают азот), при этом бактерии потребляют субстрат и вырабатывают представляющую интерес масляную композицию.
Согласно некоторым вариантам реализации в настоящем изобретении предложена установка для биопереработки, содержащая (а) блок регулируемого культивирования для выращивания метаболизирующего C1 нефотосинтезирующего микроорганизма в присутствии сырья, содержащего C1 субстрат, при этом культивируемые бактерии продуцируют масляную композицию; (b) блок обработки, в котором масляную композицию получают или экстрагируют из метаболизирующего C1 нефотосинтезирующего микроорганизма; и (с) устройство переработки для переработки масляной композиции с получением топлива. Согласно другим вариантам реализации изобретения, сырьевой C1 субстрат, применяемый в установке для биопереработки, представляет собой метан, метанол, формальдегид, муравьиную кислоту или ее соль, моноксид углерода, диоксид углерода, синтез-газ, метиламин, метилтиол или метилгалоген.
Согласно другим вариантам реализации установки для биопереработки метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой метанотроф или метилотроф, сырьевой C1 субстрат представляет собой природный газ или метан, и бактерии культивируют в аэробных условиях. Согласно другим вариантам реализации изобретения, метанотроф представляет собой Methylosinus trichosporium OB3b, Methylococcus capsulatus Bath, Methylomonas sp. 16a, Methylomonas methanica, Methylomicrobium alcaliphilum, любую их комбинацию или их быстрорастущий вариант, и метилотроф представляет собой Methylobacterium extorquens, Methylobacterium radiotolerans, Methylobacterium populi, Methylobacterium chloromethanicum, Methylobacterium nodulans, любую их комбинацию или их быстрорастущий вариант. Согласно некоторым другим вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой облигатный метаболизирующий C1 нефотосинтезирующий микроорганизм, такой как облигатный метанотроф или метилотроф.
Согласно другим вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой рекомбинантный микроорганизм, содержащий гетерологичный полинуклеотид, кодирующий фермент синтеза жирных кислот, фермент ассимиляции формальдегида или их комбинацию. Например, биосинтез свободных жирных кислот (СЖК), которые можно использовать в качестве предшественников для получения топлива или других ценных химических веществ, можно усилить путем введения гена тиоэстеразы (ТЕ) в метаболизирующий C1 нефотосинтезирующий микроорганизм согласно настоящему изобретению (например, Methylosinus trichosporium OB3b, Methylococcus capsulatus Bath, Methylomonas sp. 16a, Methylomonas methanica). Биосинтез СЖК также можно усилить путем введения более чем одного гена ТЕ, гена малонил-КоА: ацилпереносящий белок-трансацилазы (FabD, также называемого МСТ), одного или более генов оперона ацетил-КоА-карбоксилазы (AccABCD) или любой их комбинации. Согласно некоторым вариантам реализации изобретения, продуцирование СЖК можно улучшить путем сверхэкспрессии малонил-КоА: ацилпереносящий белок-трансацилазы (FabD, также называемого МСТ), поскольку первой обязательной стадией биосинтеза жирных кислот является превращение ацетил-КоА в малонил-КоА под действием аденозинтрифосфат (АТФ)-зависимой ацетил-КоА-карбоксилазы с последующим превращением малонил-КоА в малонил-АСР посредством FabD фермента.
Согласно другим вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой рекомбинантный микроорганизм, содержащий генетическую модификацию, которая минимизирует или уменьшает расщепление жирных кислот. Например, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой рекомбинантный микроорганизм, содержащий одну или более мутаций, которые процессируют или нокаутируют активность длинноцепочечная жирная кислота: КоА-лигазы, кодированную одним или более эндогенными генами fadD.
Последовательности нуклеиновых кислот, кодирующие белки FadD дикого типа, представляют собой эталонную исходную точку для конструирования мутантных генов fadD. Например, последовательность белка FadD дикого типа, кодированная М. trichosporium OB3b, М. capsulatus Bath, M. methanica, M. extorquens и С. ljungdahlii, находится в банке данных GenBank Accession №№ EFH00931.1, YP_114021.1, YP_004512148.1, YP_002964871.1 и YP_003782065.1, соответственно. Согласно некоторым вариантам реализации изобретения, молекулу нуклеиновой кислоты гена fadD, кодирующего любой из перечисленных выше белков, отдельно модифицируют, чтобы подвергнуть мутации fadD. В примере 2, описанном в настоящей заявке, ген fadD из разного метаболизирующего C1 микроорганизма синтезировали для внедрения нескольких мутаций остановки и сдвигов рамки в 5'-область гена M. trichosporium OB3b (SEQ ID №: 1), M. methanica (SEQ ID №: 35), M. extorquens (SEQ ID №: 52) и С. ljungdahlii (SEQ ID №: 85). Для гена fadD M. capsulatus, синтезировали молекулу нуклеиновой кислоты, содержащую внутриклональную делецию, чтобы оставшиеся 5' и 3' концы гена могли соединиться с сохранением первоначальной рамки считывания (SEQ ID №: 18).
Для некоторых метаболизирующих C1 микроорганизмов, в которых последовательность гена fadD не известна (например, Clostridium autoethanogenum), геном можно секвенировать и fadD, гомологичный Е. coli, идентифицируют путем поиска с применением TBLASTN (поиск транслированных нуклеотидных последовательностей гена с последовательностью белка FadD Е. coli). Например, синтезируют молекулу нуклеиновой кислоты гена fadD С. autoethanogenum для введения нескольких мутаций остановки и сдвигов рамки в 5'-область гена.
Согласно некоторым вариантам реализации изобретения, молекулу нуклеиновой кислоты мутантного fadD клонируют в плазмидном экспрессионном векторе (и, возможно, не имеющем точки начала репликации метаболизирующего C1 микроорганизма и кодирующем устойчивость к антибиотику) для конъюгации, электропорации или трансформации в метаболизирующем C1 микроорганизме с применение способов, описанных в настоящей заявке. Согласно некоторым вариантам реализации изобретения, мутант fadD встраивается в геном клетки-хозяина путем гомологической рекомбинации и образует рекомбинантные клетки, в которых отсутствует или которые проявляют минимальную активность длинноцепочечная жирная кислота: КоА-лигазы.
Согласно некоторым вариантам реализации изобретения, любой или все гены ТЕ, МСТ и Acc, введенные в метаболизирующие C1 микроорганизмы согласно настоящему изобретению, могут быть сверхэкспрессированы, при этом метаболизирующие C1 микроорганизмы могут содержать мутацию, которая минимизирует или устраняет активность жирная кислота: КоА-лигазы (например, нокаут fadD).
Согласно некоторым вариантам реализации изобретения, в блоке обработки для биопереработки можно получить масляную композицию с помощью влажной экстракции, сверхкритической флюидной экстракции, сухой экстракции, термической экстракции (например, посредством отгонки водяным паром, гидротермического сжижения, варки под давлением), ферментативного гидролиза (например, клеточной стенки), экстракции в импульсном электрическом поле, микропузырьков, экстракции в полых волокнах или т.п. Согласно другим вариантам реализации изобретения, влажная экстракция включает применение полярного растворителя, неполярного растворителя, нейтрального растворителя, кислого растворителя, основного растворителя, гексана или их комбинации. Согласно некоторым вариантам реализации изобретения, масляную композицию получают или экстрагируют из клеточной мембраны метаболизирующего C1 нефотосинтезирующего микроорганизма или указанную композицию можно извлечь из супернатанта культуры при секретировании или экскретировании, или с применением их комбинации. Согласно другим вариантам реализации изобретения, установка для биопереработки дополнительно содержит второй блок обработки, представляющий собой блок обработки отходов, в котором перерабатывают остаток от очищенной масляной композиции и которое включает установку для анаэробного разложения, установку для аэробного разложения или и то, и другое. Согласно другим вариантам реализации изобретения, установка для биопереработки дополнительно содержит трубопровод для подачи по меньшей мере одного продукта из блока обработки отходов для применения при культивировании или поддержании метаболизирующего C1 нефотосинтезирующего микроорганизма.
Согласно другим вариантам реализации изобретения, блок обработки для биопереработки дополнительно содержит блок регулируемого культивирования, представляющий собой блок для ферментации в твердой фазе, в котором может происходить культивирование и обработка (например, экстракция) в одном и том же блоке или даже в одной и той же камере. Согласно некоторым вариантам реализации изобретения, установка для биопереработки, объединяющая устройство для культивирования/обработки, включает сверхкритическую флюидную экстракцию, например, посредством сверхкритической жидкости, содержащей CO2, метанол или H2O.
В некоторых аспектах, любые из описанных выше установок для биопереработки являются комплексными.
Метаболизирующие C1 микроорганизмы
Метаболизирующие C1 микроорганизмы согласно настоящему изобретению могут быть природными, представлять собой адаптированный штамм (например, проводит ферментацию для выбора штаммов с улучшенными скоростями роста и повышенным общим выходом биомассы по сравнению с родительским штаммом) или могут быть рекомбинантно модифицированными таким образом, чтобы продуцировать представляющие интерес липиды (например, генетически измененными таким образом, чтобы экспрессировать фермент синтеза жирных кислот, фермент ассимиляции формальдегида или их комбинацию) или иметь повышенные скорости роста или и то, и другое. Согласно некоторым вариантам реализации изобретения, метаболизирующие C1 микроорганизмы не представляют собой такие метаболизирующие C1 нефотосинтезирующие микроорганизмы, как водоросли или растения.
Согласно некоторым вариантам реализации, в настоящем изобретении предложены метаболизирующие C1 микроорганизмы, которые представляют собой прокариоты или бактерии, такие как Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium, Methanomonas, Methylophilus, Methylobacillus, Methylobacterium, Hyphomicrobium, Xanthobacter, Bacillus, Paracoccus, Nocardia, Arthrobacter, Rhodopseudomonas или Pseudomonas.
Согласно другим вариантам реализации изобретения, метаболизирующие C1 бактерии представляют собой метанотроф или метилотроф. Типичные метанотрофы включают Methylomonas, Methylobacter, Methylococcus, Methylosinus, Methylocystis, Methylomicrobium, Methanomonas, Methylocella или их комбинацию. Типичные метилотрофы включают Methylobacterium extorquens, Methylobacterium radiotolerans, Methylobacterium populi, Methylobacterium chloromethanicum, Methylobacterium nodulans или их комбинацию.
Согласно некоторым вариантам реализации изобретения, метанотрофные бактерии созданы методами генетической инженерии со способностью превращать сырье, представляющее собой C1 субстрат, в масляную композицию. Метанотрофные бактерии обладают способностью окислять метан как источник углерода и энергии. Метанотрофные бактерии классифицируют на три группы на основе их путей ассимиляции углерода и структуры внутренней мембраны: I тип (гамма протеобактерии), II тип (альфа протеобактерии) и Х тип (гамма протеобактерии). Для ассимиляции углерода метанотрофы I типа используют рибулозомонофосфатный (РМФ) путь, тогда как метанотрофы II типа используют сериновый путь. Метанотрофы Х типа используют РМФ путь, но также экспрессируют низкие уровни ферментов серинового пути. Метанотрофные бактерии включают облигатные метанотрофы, которые могут только утилизировать C1 субстраты как источники углерода и энергии, и факультативные метанотрофы, которые по природе обладают способностью утилизировать некоторые субстраты, содержащие более одного атома углерода, в качестве источника углерода и энергии.
Типичные факультативные метанотрофы включают некоторые виды Methylocella, Methylocystis и Methylocapsa (например, Methylocella silvestris, Methylocella palustris, Methylocella tundrae, Methylocystis daltona штамм SB2, Methylocystis bryophila u Methylocapsa aurea KYG), Methylobacterium organophilum (ATCC 27, 886), Methylibium petroleiphilum или их быстрорастущие варианты. Типичные облигатные метанотрофные бактерии включают Methylococcus capsulatus Bath (NCIMB 11132), Methylomonas sp. 16a (ATCC РТА 2402), Methylosinus trichosporium OB3b (NRRL B-11, 196), Methylosinus sporium (NRRL B-11, 197), Methylocystis parvus (NRRL B-11, 198), Methylomonas methanica (NRRL B-11, 199), Methylomonas albus (NRRL B-11, 200), Methylobacter capsulatus Y (NRRL B-11, 201), Methylomonas flagellata sp. AJ-3670 (FERM P-2400), Methylacidiphilum infernorum и Methylomicrobium alcaliphilum или их быстрорастущие варианты.
Согласно другим вариантам реализации в настоящем изобретении предложены метаболизирующие C1 микроорганизмы, которые представляют собой бактерии, метаболизирующие синтез-газ, такие как Clostridium, Moorella, Pyrococcus, Eubacterium, Desulfobacterium, Carboxydothermus, Acetogenium, Acetobacterium, Acetoanaerobium, Butyribaceterium, Peptostreptococcus или любая их комбинация. Типичные бактерии, метаболизирующие синтез-газ, включают Clostridium autoethanogenum, Clostridium ljungdahli, Clostridium ragsdalei, Clostridium carboxydivorans, Butyribacterium methylotrophicum, Clostridium woodii, Clostridium neopropanologen или любую их комбинацию.
Согласно некоторым другим вариантам реализации изобретения, метаболизирующие C1 микроорганизмы представляют собой эукариоты, такие как дрожжи, в том числе Candida, Yarrowia, Hansenula, Pichia, Torulopsis или Rhodotorula.
Согласно некоторым другим вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой облигатный метаболизирующий C1 нефотосинтезирующий микроорганизм, такой как облигатный метанотроф или метилотроф. Согласно другим вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой рекомбинантный микроорганизм, содержащий гетерологичный полинуклеотид, кодирующий фермент синтеза жирных кислот, фермент ассимиляции формальдегида или их комбинацию. Согласно некоторым вариантам реализации изобретения, любой или все гены ТЕ, МСТ и Acc, введенные в метаболизирующий C1 микроорганизм согласно настоящему изобретению, могут быть сверхэкспрессированы и метаболизирующие C1 микроорганизмы могут иметь мутацию, которая минимизирует или устраняет активность жирная кислота: КоА-лигазы (например, нокаут fadD).
Каждый из микроорганизмов согласно настоящему изобретению можно выращивать как изолированную культуру, вместе с гетерологичным организмом, который может содействовать росту, или одну или более из указанных бактерий можно объединять с получением смешанной культуры. Согласно другим вариантам реализации изобретения, метаболизирующие C1 нефотосинтезирующие микроорганизмы согласно настоящему изобретению представляют собой облигатные метаболизирующие C1 нефотосинтезирующие микроорганизмы.
Любой из перечисленных выше метаболизирующих C1 микроорганизмов можно использовать в качестве родительской или эталонной клетки-хозяина для создания рекомбинантных метаболизирующих C1 микроорганизмов согласно настоящему изобретению.
Оптимизация кодонов
Экспрессия рекомбинантных белков может быть трудной за пределами их первоначального хозяина. Например, изменчивость в отклонении частоты использования кодона наблюдалась во всех различных видах бактерий (Sharp et al., Nucl. Acids. Res. 33:1141, 2005). Сверхэкспрессия рекомбинантных белков даже внутри их нативного хозяина также может быть трудной. Согласно некоторым вариантам реализации изобретения, молекулы нуклеиновых кислот (например, нуклеиновых кислот, кодирующих тиоэстеразу, fabD, accABCD), которые должны быть введены в хозяина, как описано в настоящей заявке, можно подвергнуть оптимизации кодонов перед введением в хозяина для обеспечения эффективной или усиленной экспрессии белка. Оптимизация кодонов относится к изменению кодонов в генах или кодирующих областях нуклеиновых кислот перед трансформацией для отражения типичной частоты использования кодона хозяина без изменения полипептида, кодированного искусственной молекулой ДНК. Способы оптимизации кодонов для достижения оптимальной экспрессии генов в гетерологичных хозяевах были описаны ранее (см., например, Welch et al., PLoS One 4:е7002, 2009; Gustafsson et al., Trends Biotechnol. 22:346, 2004; Wu et al., Nucl. Acids Res. 35:D76, 2007; Villalobos et al., BMC Bioinformatics 7:285, 2006; публикации патентов США №№2011/0111413 и 2008/0292918; описание предложенных в указанных ссылках способов в полном объеме включено в настоящую заявку посредством ссылки).
Подобным образом, можно сконструировать молекулы экзогенных нуклеиновых кислот согласно настоящему изобретению, кодирующие варианты полипептида, используя филогенетические способы, описанные в ссылках, приведенных выше (патент США №8005620; Gustafsson et al.; Welch et al.; Villalobos et al.; Minshull et al.). Каждый вариантный полипептид, получаемый с помощью указанных способов, будет сохранять По меньшей мере 50% активность (предпочтительно 100% или более активность) и содержать последовательность полипептидов, которая по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% идентична или на 100% идентична эталонной или родительской полипептидной последовательности дикого типа. Согласно некоторым вариантам реализации изобретения, вариантные полипептиды будут включать по меньшей мере одну аминокислотную замену (например, 1, 2, 3, 5, 6, 7, 8, 9 или 10 или более или до 20, 25 или 30 замен) в заранее определенном положении относительно эталонного или родительского фермента дикого типа, при условии, что вариант сохраняет представляющую интерес активность (например, активность тиоэстеразы или продуцирование жирных кислот).
Согласно некоторым вариантам реализации изобретения, тиоэстераза Е. coli, Cinnamomum camphorum, Umbellularia californica, Streptoccus pyogenes, Ricinius communis или Jatropha curcus является кодон-оптимизированной для экспрессии в метаболизирующем C1 микроорганизме согласно настоящему изобретению (например, метанотрофе, метилотрофе, клостридии). Согласно другим вариантам реализации изобретения, любую одну или более из последовательностей кодон-оптимизированной тиоэстеразы вводят (например, путем трансформации, конъюгации, электропорации) в метаболизирующий C1 микроорганизм согласно настоящему изобретению. Типичные последовательности кодон-оптимизированной тиоэстеразы приведены в (1) SEQ ID №№: 3-13 для М. trichosporium OB3b; (2) SEQ ID №№: 20-30 для М. capsulatus Bath; (3) SEQ ID №№: 37-47 для М. methanica; (4) SEQ ID №№: 54-64 for М. extorquens; (5) SEQ ID №№: 70-80 для С. autoethanogenum; и (6) SEQ ID №№: 87-97 для С. ljungdahlii.
Согласно некоторым вариантам реализации изобретения, последовательность малонил-КоА: ацилпереносящий белок-трансацилазы (fabD) в Е. coli является кодон-оптимизированной для экспрессии в метаболизирующем C1 микроорганизме согласно настоящему изобретению (например, метанотрофе, метилотрофе, клостридии). Согласно другим вариантам реализации изобретения, любую одну или более из последовательностей кодон-оптимизированного fabD вводят (например, путем трансформации, конъюгации, электропорации) в метаболизирующий C1 микроорганизм согласно настоящему изобретению. Типичные последовательности кодон-оптимизированного fabD приведены в (1) SEQ ID №: 2 для М. trichosporium OB3b; (2) SEQ ID №: 19 для М. capsulatus Bath; (3) SEQ ID №: 36 для М. methanica; (4) SEQ ID №: 53 для М. extorquens; (5) SEQ ID №: 69 для С. autoethanogenum; и (6) SEQ ID №: 86 для С. ljungdahlii.
Согласно некоторым вариантам реализации изобретения, одна или более последовательностей ацетил-КоА-карбоксилазы (например, ассА, ассВ, ассС и accD из Е. coli) являются кодон-оптимизированными для экспрессии в метаболизирующем C1 микроорганизме согласно настоящему изобретению (например, метанотрофе, метилотрофе, клостридии). Согласно другим вариантам реализации изобретения, любую одну или более из последовательностей кодон-оптимизированного Acc вводят (например, путем трансформации, конъюгации, электропорации) в метаболизирующий C1 микроорганизм согласно настоящему изобретению. Согласно другим вариантам реализации изобретения, вводят кодон-оптимизированный ассА или вводят кодон-оптимизированный accABCD. Типичные последовательности кодон-оптимизированных ассА, ассВ, ассС и accD приведены, соответственно, в (1) SEQ ID №№: 14-17 для М. trichosporium OB3b; (2) SEQ ID №№: 31-34 для М. capsulatus Bath; (3) SEQ ID №№: 48-51 для М methanica; (4) SEQ ID №№: 65-68 для М. extorquens; (5) SEQ ID №№: 81-84 для С. autoethanogenum; и (6) SEQ ID №№: 98-101 для С. ljungdahlii.
Способы трансформации
Любые из рекомбинантных метаболизирующих C1 микроорганизмов или метанотрофных бактерий, описанных в настоящей заявке, можно трансформировать таким образом, чтобы они содержали по меньшей мере одну экзогенную нуклеиновую кислоту для придания хозяину новой или усиленной активности (например, ферментативной активности), или можно генетически модифицировать для удаления или по существу понижения функции эндогенного гена с применением любых различных способов, известных в данной области техники.
Трансформация относится к введению молекулы нуклеиновой кислоты (например, молекулы экзогенной или гетерологичной нуклеиновой кислоты) в клетку-хозяина. Трансформированная клетка-хозяин может нести молекулу экзогенной или гетерологичной нуклеиновой кислоты, экстрахромосомную или встроенную в хромосому. Интеграция в геном клетки-хозяина и самореплицирующиеся векторы в целом приводит к генетически стабильному наследованию трансформированной молекулы нуклеиновой кислоты. Клетки-хозяина, содержащие трансформированные молекулы нуклеиновых кислот, называют «искусственными» или «созданными методами генетической инженерии» или «рекомбинантными» или «трансформированными» или «трансгенными» клетками (например, бактериями).
Известны экспрессионные системы и экспрессионные векторы, применимые для экспрессии гетерологичных нуклеиновых кислот в метаболизирующих C1 микроорганизмах (например, метанотрофных бактериях).
Электропорация метаболизирующих C1 бактерий описана в настоящей заявке и было ранее описана, например, в Toyama et al., FEMS Microbiol. Lett. 166:1, 1998; Kim и Wood, Appl. Microbiol. Biotechnol. 48:105, 1997; Yoshida et. al., Biotechnol. Lett. 23:787, 2001 и в патенте США Appl. Pub. №2008/0026005.
Бактериальную конъюгацию, которая относится к конкретному типу трансформации, включающему непосредственный контакт клеток донора и реципиента, более часто используют для переноса молекул нуклеиновых кислот в метаболизирующие C1 бактерии. Бактериальная конъюгация включает смешивание клеток «донора» и «реципиента» в тесном контакте друг с другом. Конъюгация происходит при образовании цитоплазматических связей между бактериями донора и реципиента, при однонаправленном переносе вновь синтезированных молекул-доноров нуклеиновых кислот в клетки реципиента. Реципиент в реакции конъюгации представляет собой любую клетку, которая может принять нуклеиновые кислоты посредством горизонтального переноса из бактерии-донора. Донор в реакции конъюгации представляет собой бактерию, которая содержит конъюгативную плазмиду, конъюгативный транспозон или мобилизованную плазмиду. Физический перенос плазмиды-донора может произойти посредством самотрансмиссивной плазмиды или с помощью плазмиды «помощника». Конъюгации, включающие метаболизирующие C1 бактерии, описаны в настоящей заявке и были ранее описаны в Stolyar et al., Mikrobiologiya 64-686, 1995; Motoyama et al., Appl. Micro. Biotech. 42:67, 1994; Lloyd et al., Arch. Microbiol. 171:364, 1999; публикации РСТ № WO 02/18617; и Ali et al., Microbiol. 152:2931, 2006.
Экспрессия гетерологичных нуклеиновых кислот в метаболизирующих C1 бактериях известна в данной области техники (см., например, патент США №6818424; публикацию заявки на патент США №2003/0003528). Была описана основанная на мю-транспозоне трансформация метилотрофных бактерий (Akhverdyan et al., Appl. Microbiol. Biotechnol. 91:857, 2011). Была описана система мини-Tn7 транспозона для одной и мультикопийной экспрессии гетерологичных генов без инсерционной инактивации генов хозяина в Methylobacterium (публикация заявки на патент США №2008/0026005).
Можно использовать различные способы инактивации, нокаута или делеции функции эндогенного гена в метаболизирующих C1 бактериях. Аллельный обмен с помощью суицидных векторов с созданием мутантов с делецией/вставкой при медленной росте метаболизирующих C1 бактерий был также описан в настоящей заявке и, например, в Toyama и Lidstrom, Microbiol. 144:183, 1998; Stolyar et al., Microbiol. 145-1235, 1999; Ali et al., Microbiol. 152:2931, 2006; Van Dien et al., Microbiol. 149:601, 2003.
Для сильной экспрессии молекул экзогенных нуклеиновых кислот можно использовать подходящие гомологичные или гетерологичные промоторы. Например, в патенте США №7098005 описано применение промоторов, которые сильно экспрессируются в присутствии метана или метанола, для экспрессии гетерологичных генов в метаболизирующих C1 бактериях. Дополнительные промоторы, которые можно использовать, включают промотор оперона деокси-ксилулоза-фосфат-синтаза метанол-дегидрогеназы (Springer et al., FEMS Microbiol. Lett. 160:119, 1998); промотор для синтеза PHA (Foellner et al., Appl. Microbiol. Biotechnol. 40-284, 1993); промотор пируватдекарбоксилаза (Tokuhiro et al., Appl. Biochem. Biotechnol. 131:795, 2006); или промоторы, идентифицированные по нативной плазмиде в метилотрофах (ЕР 296484). Ненативные промоторы включают промотор lac-оперон-Plac (Toyama et al., Microbiol. 143:595, 1997) или гибридный промотор, такой как Ptrc (Brosius et al., Gene 27:161, 1984).
Согласно некоторым вариантам реализации изобретения, промоторы или оптимизацию кодонов применяют для сильной конститутивной экспрессии экзогенных полинуклеотидов, кодирующих один или более ферментов, продуцирующих лактат, в метанотрофных бактериях-хозяевах. Можно также использовать регулируемую экспрессию экзогенной нуклеиновой кислоты в метанотрофной бактерии-хозяине. Согласно некоторым вариантам реализации изобретения, для оптимизации продуцирования липидов искусственными метанотрофными бактериями может быть необходима регулируемая экспрессия экзогенных нуклеиновых кислот, кодирующих один или более ферментов тиоэстеразы, ацетил-КоА-карбоксилазы или малонил-КоА: АСР-трансацилазы. Например, можно использовать индуцируемую/регулируемую систему экспрессии рекомбинантных белков в метилотрофных и метанотрофных бактериях, как описано, например, в заявке на патент США №2010/0221813.
Рекомбинантные метаболизирующие C1 микроорганизмы
Как указано в настоящей заявке, для создания рекомбинантных метаболизирующих C1 микроорганизмов в качестве родительской или эталонной клетки-хозяина можно использовать любой из рекомбинантных метаболизирующих C1 микроорганизмов (например, метанотрофные бактерии), описанных в настоящей заявке. Согласно некоторым вариантам реализации в настоящем изобретении предложен рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм, содержащий последовательность гетерологичных нуклеиновых кислот, связанную с биосинтезом жирных кислот, причем экспрессия последовательности гетерологичных нуклеиновых кислот приводит к аккумулированию повышенного уровня жирных кислот или сверхэкспрессии жирных кислот в рекомбинантном метаболизирующем C1 микроорганизме по сравнению с родительским или эталонным метаболизирующим C1 нефотосинтезирующим микроорганизмом.
Согласно некоторым вариантам реализации изобретения, рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм содержит гетерологичный полинуклеотид, кодирующий фермент синтеза жирных кислот, фермент ассимиляции формальдегида или любую их комбинацию. Согласно другим вариантам реализации изобретения, гетерологичный полинуклеотид кодирует тиоэстеразу, малонил-КоА: ацилпереносящий белок-трансацилазу, ацетил-КоА-карбоксилазу или любую их комбинацию. Например, тиоэстераза может представлять собой тиоэстеразу Е. coli, Cinnamomum camphorum, Umbellularia californica, Streptoccus pyogenes, Ricinius communis или Jatropha curcus. Типичные гены fabD, accA, accB, accC и accD могут быть из Е. coli или любого другого выбранного организма.
Согласно другим вариантам реализации изобретения, рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм содержит последовательность гетерологичных нуклеиновых кислот, кодон-оптимизированную для эффективной экспрессии в метаболизирующем C1 нефотосинтезирующем микроорганизме. Согласно некоторым вариантам реализации изобретения, любую одну или более из тиоэстеразы, fabD, accA, accB, accC и accD подвергают оптимизации кодонов для метаболизирующего C1 нефотосинтезирующего микроорганизма. Согласно одному из вариантов реализации изобретения кодон-оптимизированная тиоэстераза представляет собой tesA Е. coli, в которой отсутствует последовательность нацеливания на периплазму.
Согласно другим вариантам реализации изобретения, любой из описанных выше рекомбинантных метаболизирующих C1 нефотосинтезирующих микроорганизмов дополнительно содержит мутацию, которая минимизирует или устраняет активность жирная кислота: КоА-лигазы.
Типичные организмы, применяемые при конструировании рекомбинантных метаболизирующих C1 нефотосинтезирующих микроорганизмов согласно настоящему изобретению, включают бактерии или дрожжи. Согласно некоторым вариантам реализации изобретения, родительская или эталонная метаболизирующая C1 бактерия, применяемая для создания рекомбинантной метаболизирующей C1 бактерии согласно настоящему изобретению, представляет собой метанотроф или метилотроф, такой как Methylomonas sp. 16а (АТСС РТА 2402), Methylosinus trichosporium OB3b (NRRL В-11, 196), Methylosinus sporium (NRRL В-11, 197), Methylocystis parvus (NRRL В-11, 198), Methylomonas methanica (NRRL В-11, 199), Methylomonas albus (NRRL В-11, 200), Methylobacter capsulatus Y (NRRL B-11, 201), Methylococcus capsulatus Bath (NCIMB 11132), Methylobacterium organophilum (АТСС 27, 886), Methylomonas sp. AJ-3670 (FERM P-2400), Methylomicrobium alcaliphilum, Methylocella silvestris, Methylacidiphilum infernorum, Methylibium petroleiphilum, Methylobacterium extorquens, Methylobacterium radiotolerans, Methylobacterium populi, Methylobacterium chloromethanicum, Methylobacterium nodulans или любую их комбинацию.
Согласно другим вариантам реализации изобретения, родительская или эталонная метаболизирующая C1 бактерия, применяемая для создания рекомбинантной метаболизирующей C1 бактерии согласно настоящему изобретению, представляет собой бактерию, метаболизирующую синтез-газ, такую как Clostridium autoethanogenum, Clostridium ljungdahli, Clostridium ragsdalei, Clostridium carboxydivorans, Butyribacterium methylotrophicum, Clostridium woodii, Clostridium neopropanologen или их комбинацию.
Способы культивирования и способы получения масляных композиций
Для выращивания микроорганизмов, бактерий и дрожжей, описанных в настоящей заявке, можно использовать различные методологии культивирования. Например, метаболизирующие C1 микроорганизмы (такие как метанотрофные или метилотрофные бактерии) можно выращивать с применением методик периодического культивирования или непрерывного культивирования. Согласно некоторым вариантам реализации изобретения, указанные культуры выращивают в блоке регулируемого культивирования, таком как ферментатор, биореактор, половолоконная ячейка или т.п. В целом в некоторых системах клетки в логарифмической фазе часто обеспечивают массовое продуцирование продукта или представляющего интерес промежуточного вещества, тогда как в других системах можно осуществить продуцирование в стационарной или пост-экспоненциальной фазе.
Классический способ периодического культивирования представляет собой закрытую систему, в которой задают состав среды в начале культивирования и не меняют его в процессе культивирования. То есть, в начале процесса культивирования среду инокулируют одним или более выбранными микроорганизмами и затем оставляют расти без добавления в систему каких-либо компонентов. В настоящей заявке, термин «периодическое» культивирование используют со ссылкой на неизменность количества конкретного источника углерода, добавленного в начале, при этом в процессе культивирования можно контролировать и изменять такие факторы, как pH и концентрация кислорода. В системах периодического действия, составы метаболитов и биомассы в системе изменяются постоянно до времени завершения культивирования. В периодических культурах, клетки (например, бактерии, такие как метилотрофы) будут в целом перемещаться со статической лаг-фазы на логарифмическую фазу быстрого роста и на стационарную фазу, на которой скорость роста уменьшается или рост прекращается (что, в конце концов, приведет к некрозу клеток, если условия не изменятся).
Система периодического действия с подпиткой представляет собой вариацию стандартной системы периодического действия, в которую по мере развития культуры добавляют в возрастающих количествах представляющий интерес углеродный субстрат. Систему периодического действия с подпиткой можно использовать, когда метаболизм клеток, вероятно, может быть ингибирован посредством катаболитной репрессии и когда желательно иметь ограниченные количества субстрата в среде. Поскольку в системе периодического действия с подпиткой трудно измерить фактическую концентрацию субстрата, оценку делают на основе изменений измеримых факторов, таких как pH, растворенный кислород и парциальное давление отходящих газов. Способы периодического культивирования и периодического культивирования с подпиткой являются общепринятыми и известны в данной области техники (см., например, Thomas D. Brock, Biotechnology: A Textbook of Industrial Microbiology, 2nd Ed. (1989) Sinauer Associates, Inc., Sunderland, MA; Deshpande, Appl. Biochem. Biotechnol. 36:227, 1992).
Непрерывные культуры представляют собой «открытые» системы в том смысле, что определенную культуральную среду непрерывно добавляют в биореактор, при этом равное количество используемой («кондиционированной») среды одновременно удаляют для обработки. В целом в непрерывных культурах клетки поддерживаются при постоянной высокой плотности жидкой фазы, при этом клетки в основном находятся в фазе логарифмического роста. Согласно альтернативному варианту реализации изобретения непрерывное культивирование можно осуществить на практике с помощью иммобилизованных клеток (например, биопленки), при этом непрерывно добавляют углерод и питательные вещества, а ценные продукты, побочные продукты и отходы непрерывно удаляют из клеточной массы. Иммобилизацию клеток можно осуществить с помощью широкого ряда твердых подложек, состоящих из природных материалов, синтетических материалов или их комбинации.
Непрерывное или полунепрерывное культивирование позволяет модулировать один или более факторов, которые влияют на клеточный рост или концентрацию конечного продукта. Например, один из способов позволяет поддерживать лимитирующее питательное вещество при фиксированной скорости (например, источник углерода, азот) и позволяет изменять со временем все другие параметры. Согласно другим вариантам реализации изобретения, несколько факторов, влияющих на рост, можно непрерывно изменять, тогда как концентрацию клеток, измеряемую посредством измерения мутности среды, поддерживают постоянной. Цель системы непрерывного культивирования состоит в поддержании условий роста, обеспечивающих стационарную фазу, при уравновешивании потери клеток вследствие удаления среды относительно скорости клеточного роста. Способы модулирования питательных веществ и факторов роста для процессов и технологий непрерывного культивирования для максимизирования скорости получения продукта хорошо известны в данной области техники (см. Brock, 1992).
Согласно некоторым вариантам реализации изобретения, культуральная среда включает углеродный субстрат в качестве источника энергии для метаболизирующего C1 микроорганизма. Подходящие субстраты включают C1 субстраты, такие как метан, метанол, формальдегид, муравьиная кислота (формиат), моноксид углерода, диоксид углерода, метилированные амины (метиламин, диметиламин, триметиламин и т.п.), метилированные тиолы или метилгалогены (бромметан, хлорметан иодметан, дихлорметан и т.п.). Согласно некоторым вариантам реализации изобретения, культуральные среды могут содержать один C1 субстрат в качестве единственного источника углерода для метаболизирующего C1 микроорганизма или могут содержать смеси из двух или более C1 субстратов (композицию смешанных C1 субстратов) в качестве нескольких источников углерода для метаболизирующего C1 микроорганизма.
Кроме того, как известно, некоторые C1 метаболизирующие организмы для обеспечения метаболической активности утилизируют вещества, не относящиеся к C1 субстратам, такие как сахар, глюкозамин или различные аминокислоты. Например, некоторые виды Candida могут метаболизировать аланин или олеиновую кислоту (Sulter et al., Arch. Microbiol. 753:485, 1990). Methylobacterium extorquens AM1 способна расти на ограниченном количестве C2, C3 и C4 субстратов (Van Dien et al., Microbiol. 149:601, 2003). Согласно альтернативному варианту реализации изобретения метаболизирующий C1 микроорганизм может представлять собой рекомбинантный вариант, обладающий способностью утилизировать альтернативные углеродные субстраты. Поэтому, предполагают, что источник углерода в культуральной среде может содержать смесь углеродных субстратов, с соединениями, содержащими один и несколько атомов углерода, в зависимости от выбранного метаболизирующего C1 микроорганизма.
Согласно некоторым вариантам реализации в настоящем изобретении предложен способ получения топлива, включающий превращение биомассы из культуры, в основном содержащей метаболизирующий C1 нефотосинтезирующий микроорганизм, в масляную композицию и обработку масляной композиции с получением топлива. Согласно некоторым вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой облигатный метаболизирующий C1 нефотосинтезирующий микроорганизм, такой как облигатный метанотроф или метилотроф. Согласно другим вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой рекомбинантный микроорганизм, содержащий гетерологичный полинуклеотид, кодирующий фермент синтеза жирных кислот, фермент ассимиляции формальдегида или их комбинацию. Согласно некоторым вариантам реализации изобретения, любой или все гены ТЕ, МСТ и Acc, введенные в метаболизирующие C1 микроорганизмы согласно настоящему изобретению, можно сверхэкспрессировать, при этом метаболизирующие C1 микроорганизмы могут содержать мутацию, которая минимизирует или устраняет активность жирная кислота: КоА-лигазы (например, нокаут fadD). Согласно другим вариантам реализации изобретения, масляную композицию получают или экстрагируют из клеточной мембраны метаболизирующего C1 нефотосинтезирующего микроорганизма (например, метилотрофа, метанотрофа, дрожжей) или ее можно извлечь из супернатанта культуры при секретировании или экскретировании, или с применением их комбинации.
Согласно другим вариантам реализации изобретения, стадия превращения биомассы в масляную композицию включает экстрагирование масляной композиции, например, с применением влажной экстракции, сверхкритической флюидной экстракции, сухой экстракции, термической экстракции (например, отгонки водяным паром, гидротермического сжижения, варки под давлением), ферментативного гидролиза (например, клеточной стенки), экстракции в импульсном электрическом поле, микропузырьков, экстракции в полых волокнах или т.п. Типичные способы экстракции известны в данной области техники, такие как способ Фолча с применением смеси хлороформ : метанол (2:1 об./об.) (раствор ХМ) (см. Folch et al., J. Biol. Chem. 226:497, 1957) или его модифицированный способ (см. пример 3); способ экстракции Hara и Radin с применением смеси гексан : изопропанол (ГИП) (см. Hara and Radin, Anal. Biochem. 90:420, 1978); способ Bligh и Dyer с применением смеси хлороформ : метанол : вода (см. Bligh and Dyer, Canadian J. Biochem. Physiol. 37:911, 1959); или т.п. Другие типичные способы экстракции включают способы с применением колонки для твердофазной экстракции (Pinkart et al., J. Microbiol. Meth. 34:9, 1998), одностадийную реактивную экстракцию (Nelson, All Graduate Theses and Dissertations. Paper 642, digitalcommons.usu.edu/etd/642), экстракцию α-гидроксисульфоновой кислотой (публикация патента США №2013/0144078), экстракцию при высоких температуре и давлении (публикация патента США №2012/0110898) или ускоренную экстракцию растворителем (ASE), экстракцию в аппарате Сокслета, ультразвуковую экстракцию и осциллирующую экстракцию (см. Liu et al., J. Earth Sci. 27:300, 2010). Каждый из указанных способов экстракции в полном объеме включен в настоящую заявку посредством ссылки и может быть использован в любом из упомянутых выше способов или систем биопереработки, описанных в настоящей заявке.
Согласно некоторым вариантам реализации в настоящем изобретении предложен способ получения топлива путем переработки масляной композиции (например, в устройстве переработки) с получением топлива, при этом масляную композицию получают из метаболизирующего C1 нефотосинтезирующего микроорганизма, такого как метилотроф или метанотроф. Согласно другим вариантам реализации изобретения, указанный способ дополнительно включает экстрагирование масляной композиции или применение блока обработки для экстрагирования масляной композиции из метаболизирующего C1 нефотосинтезирующего микроорганизма. Согласно другим вариантам реализации изобретения, указанный способ включает: (а) культивирование метаболизирующих C1 бактерий в присутствии сырья, содержащего C1 субстрат, в блоке регулируемого культивирования, при этом культивируемые бактерии продуцируют масляную композицию; (b) экстрагирование масляной композиции из культивируемых бактерий или экстрагирование масляной композиции в блоке обработки; и (с) обработку экстрагированной масляной композиции или обработку масляной композиции в устройстве переработки с получением топлива. Согласно некоторым вариантам реализации изобретения, исходный C1 субстрат представляет собой метан, метанол, формальдегид, муравьиную кислоту, моноксид углерода, диоксид углерода, метиламин, метилтиол или метилгалоген.
В любом из описанных выше способов получения топлива или биотоплива метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой метанотроф, метилотроф или клостридий, исходный C1 субстрат представляет собой природный газ, синтез-газ или метан и бактерии культивируют в аэробных или анаэробных условиях. Согласно другим вариантам реализации изобретения, метанотроф представляет собой Methylosinus trichosporium OB3b, Methylococcus capsulatus Bath, Methylomonas sp. 16a, Methylomonas methanica, Methylomicrobium alcaliphilum, любую их комбинацию или их быстрорастущий вариант; метилотроф представляет собой Methylobacterium extorquens, Methylobacterium radiotolerans, Methylobacterium populi, Methylobacterium chloromethanicum, Methylobacterium nodulans, любую их комбинацию или их быстрорастущий вариант; и клостридий представляет собой Clostridium autoethanogenum, Clostridium ljungdahli, Clostridium ragsdalei, Clostridium carboxydivorans, Clostridium woodii, Clostridium neopropanologen или любую их комбинацию или их быстрорастущий вариант. Согласно некоторым другим вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой облигатный метаболизирующий C1 нефотосинтезирующий микроорганизм, такой как облигатный метанотроф, метилотроф или клостридий.
В любом из описанных выше способов получения топлива или биотоплива метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой метанотроф, исходный C1 субстрат представляет собой природный газ или метан и бактерии культивируют в аэробных условиях. Согласно другим вариантам реализации изобретения, метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой метанотроф, C1 субстрат представляет собой природный газ или метан и бактерии культивируют с применением ограничивающих количеств фосфора, азота, микроэлементов, кислорода или любой их комбинации.
Топливные композиции и топливные продукты
В качестве справочной информации, для оценки глобальных источников и стоков метана широко используют измерения устойчивых изотопов и подходы, основанные на балансе масс (см. Whiticar и Faber, Org. Geochem. 10:759, 1986; Whiticar, Org. Geochem. 16:531, 1990). Для использования значений δ13C остаточного метана для определения количества окисленного метана необходимо знать степень фракционирования изотопов, вызванную микробным окислением метана. Например, аэробные метанотрофы могут метаболизировать метан с помощью конкретного фермента, метанмонооксигеназы (ММО). Метанотрофы превращают метан в метанол и затем в формальдегид. Далее формальдегид можно окислить до CO2, что обеспечивает клетке энергию в форме восстанавливающих эквивалентов (NADH), или ввести в биомассу посредством или РМФ, или серинового циклов (Hanson и Hanson, Microbiol. Rev. 60:439, 1996), которые прямо аналогичны путям ассимиляции углерода в фотосинтезирующих организмах. Более конкретно, метанотроф I типа использует РМФ путь для синтеза биомассы и генерирует биомассу полностью из CH4, тогда как метанотроф II типа использует сериновый путь, который ассимилирует 50-70% клеточного углерода из CH4 и 30-50% из CO2 (Hanson и Hanson, 1996). Способы измерения изотопных составов углерода приведены, например, Templeton et al. (Geochim. Cosmochim. Acta 70:1739, 2006), при этом указанные способы тем самым в полном объеме включены посредством ссылки. Устойчивое отношение углерода 13C/12C в масляной композиции, полученной из биомассы (обозначаемое как значение «дельта» ‰, δ13C), может меняться в зависимости от источника и чистоты применяемого C1 субстрата (см., например, фиг. 7).
Масляные композиции, полученные при использовании метаболизирующих C1 нефотосинтезирующих микроорганизмов и способов, описанных в настоящей заявке, а также полученные из них биотопливные композиции, могут отличаться с точки зрения «отпечатков пальцев» углерода от масла и топлив, полученных из нефтепродуктов или из фотосинтезирующих микроорганизмов или растений. Согласно некоторым вариантам реализации изобретения, δ13C биомассы, масляной композиции или биотоплива, полученного из биомассы или масляной композиции, составляет менее - 30‰, менее - 31‰, менее - 32‰, менее - 33‰, менее - 34‰, менее - 35‰, менее - 36‰, менее - 37‰, менее - 38‰, менее - 39‰, менее - 40‰, менее - 41‰, менее - 42‰, менее - 43‰, менее - 44‰, менее - 45‰, менее - 46‰, менее - 47‰, менее - 48‰, менее - 49‰, менее - 50‰, менее - 51‰, менее - 52‰, менее - 53‰, менее - 54‰, менее - 55‰, менее - 56‰, менее - 57‰, менее - 58‰, менее - 59‰, менее - 60‰, менее - 61‰, менее - 62‰, менее - 63‰, менее - 64‰, менее - 65‰, менее - 66‰, менее - 67‰, менее - 68‰, менее - 69‰ или менее - 70‰.
Согласно некоторым вариантам реализации изобретения, биомасса метаболизирующего C1 микроорганизма содержит масляную композицию, при этом δ13C биомассы составляет от примерно - 35‰ до примерно - 50‰, от - 45‰ до примерно - 35‰, или от примерно - 50‰ до примерно - 40‰, или от примерно - 45‰ до примерно - 65‰, или от примерно - 60‰ до примерно - 70‰, или от примерно - 30‰ до примерно - 70‰. Согласно другим вариантам реализации изобретения, масляная композиция биомассы содержит по меньшей мере 50% жирных кислот или содержит по меньшей мере 50% свободных жирных кислот. Согласно другим вариантам реализации изобретения, масляная композиция биомассы содержит смесь диацилглицеридов и триацилглицеридов. Согласно другим вариантам реализации изобретения, большая часть (более 50% масс./масс.) жирных кислот в масляной композиции биомассы содержит углеродную цепь длиной от C14 до C18 или от C16 до C18, или большая часть жирных кислот содержит углеродную цепь длиной менее C16. Согласно другим вариантам реализации изобретения, масляная композиция биомассы содержит более 50% масс./масс. терпеноидных или изопреноидных соединений, при этом терпеноид может представлять собой фарнезен или лимонен.
Согласно другим вариантам реализации изобретения, биомасса метаболизирующего C1 нефотосинтезирующего микроорганизма имеет δ13C, составляющую менее примерно - 30‰ или от примерно - 40‰ до примерно - 60‰. Согласно некоторым вариантам реализации изобретения, биомасса содержит рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм вместе с отработавшей средой или биомасса содержит композицию супернатанта отработавшей среды из культуральной жидкости рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма, при этом δ13C биомассы составляет менее примерно - 30‰. Согласно некоторым другим вариантам реализации изобретения, масляную композицию экстрагируют или концентрируют из биомассы, которая может содержать рекомбинантные метаболизирующие C1 нефотосинтезирующие микроорганизмы вместе с отработавшей средой из культуральной жидкости или композицией супернатанта отработавшей среды из культуральной жидкости рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма.
Согласно некоторым вариантам реализации изобретения, биомасса рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма содержит гетерологичный полинуклеотид, кодирующий фермент синтеза жирных кислот, фермент ассимиляции формальдегида или любую их комбинацию. Согласно другим вариантам реализации изобретения, гетерологичный полинуклеотид кодирует тиоэстеразу, малонил-КоА: ацилпереносящий белок-трансацилазу, ацетил-КоА-карбоксилазу или любую их комбинацию. Например, тиоэстераза может представлять собой тиоэстеразу Е. coli, Cinnamomum camphorum, Umbellularia californica, Streptoccus pyogenes, Ricinius communis или Jatropha curcus. Типичные гены fabD, accA, accB, accC и accD могут быть из Е. coli или любого другого выбранного организма.
Согласно другим вариантам реализации изобретения, биомасса состоит из рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма, содержащего последовательность гетерологичных нуклеиновых кислот, кодон-оптимизированную для эффективной экспрессии в метаболизирующем C1 нефотосинтезирующем микроорганизме. Согласно некоторым вариантам реализации изобретения, любой один или более из генов, выбранных из тиоэстеразы, fabD, accA, accB, ассС и accD, подвергают оптимизации кодонов для метаболизирующего C1 нефотосинтезирующего микроорганизма. Согласно одному из вариантов реализации изобретения кодон-оптимизированная тиоэстераза представляет собой tesA Е. coli, в которой отсутствует последовательность нацеливания на периплазму.
Согласно другим вариантам реализации изобретения, любая из описанной выше биомассы рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма дополнительно содержит мутацию, которая минимизирует или устраняет активность жирная кислота: КоА-лигазы.
Типичные организмы, применяемые для получения биомассы рекомбинантных метаболизирующих C1 нефотосинтезирующих микроорганизмов согласно настоящему изобретению включают бактерии или дрожжи. Согласно некоторым вариантам реализации изобретения, биомасса представляет собой биомассу метаболизирующих C1 бактерий, выбранных из метанотрофа или метилотрофа, таких как Methylomonas sp. 16a (АТСС РТА 2402), Methylosinus trichosporium OB3b (NRRL В-11, 196), Methylosinus sporium (NRRL В-11, 197), Methylocystis parvus (NRRL В-11, 198), Methylomonas methanica (NRRL B-11, 199), Methylomonas albus (NRRL B-11, 200), Methylobacter capsulatus Y (NRRL B-11, 201), Methylococcus capsulatus Bath (NCIMB 11132), Methylobacterium organophilum (ATCC 27, 886), Methylomonas sp. AJ-3670 (FERM P-2400), Methylomicrobium alcaliphilum, Methylocella silvestris, Methylacidiphilum infernorum, Methylibium petroleiphilum, Methylobacterium extorquens, Methylobacterium radiotolerans, Methylobacterium populi, Methylobacterium chloromethanicum, Methylobacterium nodulans или любая их комбинация.
Согласно другим вариантам реализации изобретения, биомасса метаболизирующих C1 бактерий из рекомбинантных метаболизирующих C1 бактерий согласно настоящему изобретению представляет собой бактерию, метаболизирующую синтез-газ, такую как Clostridium autoethanogenum, Clostridium ljungdahli, Clostridium ragsdalei, Clostridium carboxydivorans, Butyribacterium methylotrophicum, Clostridium woodii, Clostridium neopropanologen или их комбинацию.
Согласно некоторым вариантам реализации изобретения, δ13C масляной композиции составляет от примерно - 35‰ до примерно - 50‰, от - 45‰ до примерно - 35‰, или от примерно - 50‰ до примерно - 40‰, или от примерно - 45‰ до примерно - 65‰, или от примерно - 60‰ до примерно - 70‰, или от примерно - 30‰ до примерно - 70‰. Согласно другим вариантам реализации изобретения, масляная композиция содержит по меньшей мере 50% масс./масс. жирных кислот или содержит по меньшей мере 50% масс./масс. свободных жирных кислот. Согласно другим вариантам реализации изобретения, масляная композиция содержит смесь диацилглицеридов и триацилглицеридов. Согласно другим вариантам реализации изобретения, масляная композиция содержит жирные кислоты, большая часть которых имеет углеродную цепь длиной от C14 до C18 или от C16 до C18, или жирные кислоты, большая часть которых имеет углеродную цепь длиной менее C16. Согласно другим вариантам реализации изобретения, масляная композиция содержит более 50% масс./масс. терпеноидных или изопреноидных соединений, при этом терпеноидные соединения могут представлять собой фарнезен, лимонен или и то, и другое.
Согласно некоторым вариантам реализации изобретения, биотопливо, полученное из биомассы или масляной композиции, имеет δ13C, составляющее от примерно - 35‰ до примерно - 50‰, от - 45‰ до примерно - 35‰, или от примерно - 50‰ до примерно - 40‰, или от примерно - 45‰ до примерно - 65‰, или от примерно - 60‰ до примерно - 70‰, или от примерно - 30‰ до примерно - 70‰. Согласно некоторым другим вариантам реализации изобретения, δ13C биотоплива, полученного из масляной композиции, составляет от примерно - 35‰ до примерно - 50‰, от - 45‰ до примерно - 35‰, или от примерно - 50‰ до примерно - 40‰, или от примерно - 45‰ до примерно - 65‰, или от примерно - 60‰ до примерно - 70‰, или от примерно - 30‰ до примерно - 70‰.
Согласно другим вариантам реализации изобретения, биотопливо содержит по меньшей мере 50% масс./масс. сложных метиловых эфиров жирных кислот (СМЭЖК). Согласно связанным вариантам реализации изобретения, биотопливо содержит по меньшей мере 50% СМЭЖК, при этом большая часть СМЭЖК имеет углеродные цепи длиной C14-C18, C16-C18 или менее C16. Согласно другим вариантам реализации изобретения, биотопливо содержит по меньшей мере 50% масс./масс. сложных этиловых эфиров жирных кислот (СЭЭЖК). Согласно связанным вариантам реализации изобретения, биотопливо содержит по меньшей мере 50% СЭЭЖК, при этом большая часть СЭЭЖК имеет углеродные цепи длиной C14-C18, C16-C18 или менее C16. Согласно другим вариантам реализации изобретения, биотопливо содержит по меньшей мере 50% масс./масс. гидрогенизованных терпеноидов, таких как фарнезан или лимонан. Согласно некоторым вариантам реализации изобретения, большинство гидрогенизованных терпеноидов состоят из фарнезана, лимонана или и того и другого. Согласно некоторым вариантам реализации изобретения, биотопливо содержит гидрогенизованную биомассу. Согласно некоторым вариантам реализации изобретения, большая часть гидрогенизованной биомассы содержит смесь линейных и разветвленных алканов. Согласно некоторым вариантам реализации изобретения, биотопливо содержит жирные кислоты, большая часть которых имеет углеродную цепь длиной от C14 до C18 или от C16 до C18, или жирные кислоты, большая часть которых имеет углеродную цепь длиной менее C16. Согласно другим вариантам реализации изобретения, биотопливо содержит более 50% масс./масс. терпеноидных или изопреноидных соединений, при этом терпеноид может представлять собой фарнезен или лимонен.
Согласно некоторым вариантам реализации изобретения, масляная композиция метаболизирующего C1 микроорганизма (которая может представлять собой экстракт или изолят из биомассы метаболизирующего C1 микроорганизма) содержит атомы водорода и углерода, составляющие по меньшей мере от примерно 50% до примерно 80% относительно массы композиции, при этом δ13C композиции составляет менее примерно - 35‰, или менее примерно - 36‰, или менее примерно - 37‰, или менее примерно - 38‰, или менее примерно - 39‰, или менее примерно - 40‰. Согласно некоторым вариантам реализации изобретения, масло или полученная из него биотопливная композиция содержит молекулы, содержащие атомы водорода и углерода, составляющие по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75% или по меньшей мере 80% или по меньшей мере 90% или по меньшей мере 95% относительно массы композиции, при этом δ13C композиции составляет от примерно - 30‰ до примерно - 70‰, или δ13C в биомассе уменьшается при увеличении плотности клеток на от примерно - 5‰ до примерно - 20‰, или δ13C биомассы было выше, чем в случае одновременного полученного CO2 в среднем на от 5‰ до 15‰ при культивировании в присутствии или при отсутствии меди.
Согласно другим вариантам реализации изобретения, масляная композиция метаболизирующего C1 микроорганизма согласно настоящему изобретению (которую можно экстрагировать или выделить из биомассы метаболизирующего C1 микроорганизма) содержит атомы водорода и углерода, составляющие примерно по меньшей мере 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% относительно массы композиции. Согласно некоторым вариантам реализации изобретения, масляная композиция или полученная из нее биотопливная композиция содержит молекулы, содержащие атомы водорода и углерода, составляющие по меньшей мере примерно 90% относительно массы композиции, и при этом δ13C композиции составляет от примерно - 40‰ до примерно - 55‰.
Компонент топлива, описанный в настоящей заявке и известный в данной области техники, может представлять собой ископаемое топливо или смешивающуюся смесь для получения топливного продукта. Например, смесь для получения топливной или биотопливной смеси может представлять собой углеводородную смесь, подходящую для смешивания с другой углеводородной смесью с получением топливного или биотопливного продукта. Например, смесь легких алканов может не иметь определенное октановое число, соответствующее данному типу топлива; однако, указанную смесь можно смешать со смесью с высоким содержанием октана с получением топливного продукта. Согласно некоторым вариантам реализации изобретения, биомасса, масляная композиция или полученное из нее биотопливо согласно настоящему изобретению представляет собой после переработки топливный или биотопливный компонент.
Согласно некоторым вариантам реализации изобретения, биотопливная композиция содержит молекулы, содержащие атомы водорода и углерода, составляющие по меньшей мере 80% относительно массы композиции, причем распределение δ13C композиции составляет от примерно - 37% до примерно - 10%, или распределение δ13C в биомассе увеличивается при увеличении плотности клеток от - 22% до - 9%, или δ13C композиции биомассы было выше, чем в случае одновременного полученного CO2 в среднем на от 5‰ до 15‰ при культивировании в присутствии или при отсутствии меди.
Биотопливный продукт, описанный в настоящей заявке, представляет собой продукт, полученный путем смешивания масляной композиции или полученной из нее биотопливной композиции согласно настоящему изобретению с топливным или биотопливным компонентом. В некоторых случаях, распределение δ13C биотопливного продукта составляет более - 60‰, или более - 50‰, или более - 40‰, или более - 30‰, при условии, что часть смеси, представляющая собой масляную композицию или полученную из нее биотопливную композицию, не получают из фотосинтезирующего микроорганизма или растения. Согласно некоторым вариантам реализации изобретения, компонент топлива, применяемый для смешивания, представляет собой композицию на основе нефти или присадку к топливу (например, оксигенаты, такие как метанол, этанол, изопропанол; простые эфиры, такие как метил-трет-бутиловый эфир, трет-амилметиловый эфир; противоокислители, такие как бутилированный гидрокситолуол, этилендиамин; антидетонаторы, такие как тетраэтилсвинец, ферроцен толуол; уловители свинца, такие как трикрезилфосфат; красители; или т.п.). Например, масляную композицию метаболизирующего C1 микроорганизма можно смешать с компонентом топлива перед переработкой (например, переэтерификацией; крекингом) с получением топливного продукта, описанного в настоящей заявке. Согласно другим вариантам реализации изобретения, масляная композиция представляет собой жидкость или твердое вещество и ее можно очистить с получением присадки к топливу, применяемой для получения биотопливного продукта. Согласно некоторым вариантам реализации изобретения, масляная композиция содержит терпен, терпеноид, изопрен или изопреноид. Согласно другим вариантам реализации изобретения, биотопливный продукт имеет октановое число, составляющее от 85 до 120, или октановое число больше 90.
ПРИМЕРЫ
ПРИМЕР 1
УСЛОВИЯ КУЛЬТИВИРОВАНИЯ И В БИОРЕАКТОРЕ ДЛЯ ПОЛУЧЕНИЯ ЛИПИДОВ С ПОМОЩЬЮ МЕТАБОЛИЗИРУЮЩИХ C1 МИКРООРГАНИЗМОВ
Типичные метаболизирующие C1 микроорганизмы согласно настоящему изобретению (метанотрофы, метилотрофы, клостридии) культивировали в пробирках, во флаконах, в склянках, на планшетах или в биореакторе (ферментация). В настоящем примере описаны условия роста, среды и источник углерода для различных микроорганизмов.
Methylosinus trichosporium штамм OB3b (NCIMB 11131); Methylomonas sp. штамм 16a (ATCC PTA-2402); или Methylomonas methanica
Для приготовления флаконов с сывороткой указанные бактерии культивировали при 30°C в среде Хиггинза с минимальным содержанием нитратных солей (NSM; Cornish et al., J. Gen. Microbiol. 130:2565, 1984; Park et al., Biotechnol. Bioeng. 38:423, 1991) или в среде MM-W1. Состав газа в свободном пространстве регулировали до объемного соотношения метан : воздух 1:1. Флаконы встряхивали при скорости 200-250 об/мин. Согласно альтернативному варианту реализации изобретения культуры находились на планшетах со средой NSM, содержащих 1,5% масс./об агара, выращенного в газонепроницаемой камере, содержащей газовую смесь 1:1 (об./об.) метан : воздух, или в присутствии пара метанола (путем применения 0,5 мл метанола в крышечке запечатанных парафильмом планшетов) или на планшетах со средой NSM с добавкой 0,5% метанола. Планшеты инкубировали перевернутыми в увлажнительной камере при 30°C.
Состав применяемой среды NSM был следующим: 1,0 г MgSO4×7H2O, 0,20 г CaCl2×6H2O, 2,0 мл раствора хелатного железа (0,1 г цитрата аммония железа (III) или 0,5 г хлорида железа (III); 0,2 г ЭДТК, натриевой соли; 0,3 мл HCl, концентрированной; 100,0 мл дистиллированной деионизированной H2O), 1,0 г KNO3, 0,5 мл раствора микроэлементов (500,0 мг ЭДТК, 200,0 мг FeSO4. 7H2O, 10,0 мг ZnSO4×7H2O, 3,0 мг MnCl2×4H2O, 30,0 мг H3BO3, 20,0 мг CoCl2×6H2O, 1,0 мг CaCl2×2H2O, 2,0 мг NiCl2×6H2O, 3,0 мг Na2MoO4×2H2O, 1,0 л дистиллированной воды), 0,272 г KH2PO4, 0,717 г Na2HPO4×12H2O, возможно 12,5 г очищенного агара (например, агара Oxoid L28 или Bacto™; применяемого при изготовлении планшетов), 1,0 л дистиллированной деионизированной воды, устанавливали pH 6,8 и стерилизовали в автоклаве при 121°C в течение 15 минут.
Для проведения ферментации, 2-литровый биореактор, содержащий 1 л стерилизованной заданной среды MM-W1, инокулировали клетками из периодических культур, содержащихся во флаконах с сывороткой (10-20% об./об.), выращенных в среде MM-W1, в которую подавали смесь 1:1 (об./об.) метана и воздуха. Состав применяемой среды MM-W1 был следующим: 0,8 мМ MgSO4×7H2O, 10 мМ NaNO3, 0,14 мМ CaCl2, 1,2 мМ NaHCO3, 2,35 мМ KH2PO4, 3,4 мМ K2HPO4, 20,7 мкМ Na2MoO4×2H2O, 1 мкМ CuSO4×5H2O, 10 мкМ FeIII-Na-ЭДТК, и 1 мл на литр раствора металлических микроэлементов (содержащего, на литр 500 мг FeSO4×7H2O, 400 мг ZnSO4×7H2O, 20 мг MnCl2×7H2O, 50 мг CoCl2×6H2O, 10 мг NiCl2×6H2O, 15 мг H3BO3, 250 мг ЭДТК). После автоклавирования и охлаждения среды добавляли фосфат, бикарбонат и FeIII-Na-ЭДТК. При некоторых ферментациях добавляли бикарбонат до концентрации 0,1% (масс./об). Содержимое реактора перемешивали с помощью подвесной мешалки при постоянной скорости 750 об/мин. Культуры подкармливали при постоянном барботировании метана со скоростью от примерно 60 мл/мин до примерно 120 мл/мин, при этом подавали концентрированный кислород (по меньшей мере 85%) с переменной скоростью от примерно 10 до 100 мл/мин для поддержания уровня растворенного кислорода от примерно 40% до примерно 80% (относительно насыщения среды воздухом).
Температуру в биореакторе поддерживали при 30°C, а pH поддерживали при 7,1±0,1 путем автоматизированного добавления в культуру 0,5 М NaOH и 0,5 М HCl вместе с другими добавками примерно каждые от 4 часов до примерно 24 часов (что соответствовало увеличению OD600 на приблизительно 5 единиц оптической плотности (OD)). При добавлении других компонентов чередовали добавление металлов (конечные концентрации 10 мкМ CuSO4, 5 мкМ FeSO4, 5 мкМ FeIII-Na-ЭДТК) и добавление питательных веществ (5,75 мМ KxHyPO4, 10 мМ NaNO3). При указанных условиях наблюдался по существу линейный рост, при этом эффективная скорость образования биомассы составляла от примерно 2,7 до примерно 3,3 граммов сухой массы клеток на литр в день при достижении OD600 более 20. Культивируемую биомассу собирали путем центрифугирования, промывали один раз в среде MM-W1, и извлеченную биомассу либо замораживали при -80°C, либо сразу же применяли для фракционирования клеточных компонентов (например, экстракции липидов).
Подход на основе полунепрерывной ферментации можно также применять для поддержания продуктивности биомассы и уменьшения времени, связанного с остановкой и запуском ферментации (т.е., времени оборота или времени производственного цикла).
Сбор бактериальной биомассы выполняли с промежутками приблизительно 12-24 часа, когда плотность культуры достигала стационарной фазы (но до ее наступления). Приблизительно половину объема биореактора удаляли путем переноса в отдельный контейнер с помощью центробежного насоса. Затем равный объем стерилизованной или повторно используемой среды возвращали в биореактор, так что оптическая плотность реактора составляла приблизительно половину от ее исходного значения. Ферментацию в биореакторе продолжали согласно приведенному выше протоколу таким образом, чтобы можно было выполнить несколько циклов выращивания и извлечения биомассы в течение одного цикла ферментации.
Methylococcus capsulatus Bath (NCIMB 11132)
Бактерии культивировали при 42°C во флаконах с сывороткой, содержащих среду Хиггинза с минимальным содержанием нитратных солей (NSM) или среду MM-W1. Состав газа в свободном пространстве регулировали до объемного соотношения метан : воздух 1:1. Флаконы встряхивали при скорости от 200 до 250 об/мин. Согласно альтернативному варианту реализации изобретения культуру содержали на планшетах со средой NSM, затвердевших под действием 1,5% масс./об агара, выращенного в газонепроницаемой камере, содержащей газовую смесь 1:1 (об./об.) метан : воздух. Планшеты инкубировали перевернутыми в камере при 42°C.
Для проведения ферментации 3-литровый биореактор, содержащий 1,25 л стерилизованной среды MMF1.1 инокулировали клетками из периодических культур, содержащихся во флаконах с сывороткой (10-20% об./об.), выращенных в одинаковой среде, в которую подавали смесь 1:1 (об./об.) метана и воздуха. Состав среды MMF1.1 был следующим: 0,8 мМ MgSO4×7H2O, 40 мМ NaNO3, 0,14 мМ CaCl2, 6 мМ NaHCO3, 4,7 мМ KH2PO4, 6,8 мМ K2HPO4, 20,7 мкМ Na2MoO4×2H2O, 6 мкМ CuSO4×5H2O, 10 мкМ FeIII-Na-ЭДТК и 1 мл на литр раствора металлических микроэлементов (содержащего на литр 500 мг FeSO4×7H2O, 400 мг ZnSO4×7H2O, 20 мг MnCl2×7H2O, 50 мг CoCl2×6H2O, 10 мг NiCl2×6H2O, 15 мг H3BO3, 250 мг ЭДТК). После автоклавирования и охлаждения среды добавляли фосфат, бикарбонат и FeIII-Na-ЭДТК. Содержимое реактора перемешивали с помощью подвесной мешалки при постоянной скорости 750 об/мин. Культуры подкармливали при постоянном барботировании метана со скоростью от примерно 60 до примерно 200 мл/мин, при этом концентрированный кислород (>85%) подавали с переменной скоростью от 15 до 90 мл/мин, и уровень растворенного кислорода поддерживали ниже 10% (относительно насыщения среды воздухом).
Температуру в биореакторе поддерживали при 44°C, а pH поддерживали при 7,0±0,1 путем автоматизированного добавления в культуру 0,5 М NaOH и 0,5 М HCl вместе с добавлениями меди и железа (конечная концентрация 5 мкМ CuSO4, 5 мкМ FeSO4, 10 мкМ FeIII-Na-ЭДТК) каждые 3-6 часа (что соответствует увеличению OD600 на приблизительно 3-5 единиц оптической плотности после достижения значения 5 единиц оптической плотности). При указанных условиях по существу наблюдался линейный рост, при этом эффективная скорость образования биомассы составляла более 5 граммов сухой массы клеток на литр в день до достижения OD600 больше 10. Культивируемую биомассу собирали путем центрифугирования, клетки промывали один раз в среде MM-W1 и конгломераты клеток либо замораживали при -80°C, либо сразу же применяли для фракционирования клеточных компонентов.
Исчерпание питательных веществ было признано проблемой, которая могла ограничить выход биомассы в процессе ферментации. Чтобы избежать недостатка питательных веществ, главным образом, азота и фосфата, подачу питательных веществ, содержащих 2-кратно концентрированную MMF1.1, начинали после того, как OD600 культуры превышала 5. Подачу питательных веществ начинали при скоростях разбавления, соответствующих приблизительно половине скорости роста культуры, чтобы избежать вымывания и поддержать увеличение OD при расширении объема культуры. Ферментацию в биореакторе продолжали согласно приведенному выше протоколу таким образом, чтобы можно было выполнить несколько циклов выращивания и извлечения биомассы в течение одного цикла ферментации.
Methylobacterium extorquens или Methylosinus trichosporium штамм OB3b (NCIMB 11131)
Бактерии культивировали при 30°C в пробирках, содержащих среду Хиггинза с минимальным содержанием нитратных солей (NSM) с добавкой 0,5% метанола. Пробирки встряхивали при скорости от 200 до 250 об/мин. Согласно альтернативному варианту реализации изобретения культуры содержали на планшетах со средой NSM, содержащих 1,5% масс./об агара, выращенного в присутствии пара метанола (путем применения 0,5 мл метанола в крышечке запечатанных парафильмом планшетов) или с добавкой 0,5% метанола. Планшеты инкубировали перевернутыми в увлажнительной камере при нормальной атмосфере при 30°C.
Для проведения ферментации, 2-литровый биореактор, содержащий 1 л заданной среды MM-W1, инокулировали клетками из содержащейся в культуральной пробирке периодической культуры (10-20% об./об.). Использовали состав среды MM-W1, описанный выше. Содержимое реактора перемешивали с помощью подвесной мешалки при постоянной скорости 800 об/мин. Культуру подпитывали посредством начального болюса метанола до конечной концентрации 0,5% и переменной подачи метанола, при этом чистый кислород подавали с переменной скоростью от 30 до 100 мл/мин для поддержания уровня растворенного кислорода от 60 до 90% (относительно насыщения среды воздухом).
Температуру в биореакторе поддерживали при 30°C, а pH поддерживали при 7,1±0,1 путем автоматизированного добавления 0,5 М NaOH и 1 М HCl вместе с добавлениями металлов и питательных веществ, как описано выше. При указанных условиях наблюдался по существу линейный рост, при этом эффективная скорость образования биомассы составляла от 2,7 до 3,3 граммов сухой массы клеток на литр в день до достижения величины OD600 более 20. Культивируемую биомассу собирали путем центрифугирования, клетки промывали один раз в среде MM-W1 и конгломераты клеток либо замораживали при -80°C, либо сразу же применяли для фракционирования клеточных компонентов.
Подход на основе полунепрерывной ферментации можно также применять для поддержания продуктивности биомассы и уменьшения времени, связанного с остановкой и запуском ферментации (т.е., времени оборота или времени производственного цикла).
Сбор аккумулированной бактериальной биомассы выполняли с промежутками приблизительно 12-24 часа, когда плотность культуры достигала стационарной фазы (но до ее наступления). Приблизительно половину объема биореактора удаляли путем переноса в отдельный контейнер с помощью центробежного насоса. Затем равный объем свежей или повторно используемой среды возвращали в биореактор, так что оптическая плотность реактора составляла приблизительно половину от ее исходного значения. Ферментацию в биореакторе продолжали согласно приведенному выше протоколу таким образом, чтобы было выполнено несколько циклов роста и извлечения биомассы в течение одного цикла ферментации.
Clostridium autoethanogenum и Clostridium ljungdahlii
Бактерии рода клостридий культивировали в анаэробных условиях в 100 мл модифицированной среды РЕТС (среда АТСС 1754) при 37°C в покрытых пластмассой 500 мл флаконах Schott Duran® GL45 с бутилрезиновыми пробками, содержащих отходящие газы сталелитейного завода при давлении 200 кПа. Рост контролировали путем измерения оптической плотности при 600 нм (OD600).
Модифицированная среда РЕТС содержала (на литр) 1 г NH4Cl, 0,4 г KCl, 0,2 г MgSO4×7H2O, 0,8 г NaCl, 0,1 г KH2PO4, 20 мг CaCl2×2H2O, 10 мл раствора микроэлементов (см. ниже), 10 мл витаминного раствора Вольфа (см. ниже), 2 г NaHCO3 и 1 мг резазурина. После установления pH 5,6, среду кипятили, распределяли в анаэробных условиях и стерилизовали в автоклаве при 121°C в течение 15 мин. В качестве источника углерода применяли отходящие газы сталелитейного завода (состав: 44% CO, 32% N2, 22% CO2, 2% H2) или эквивалентные синтетические смеси. Указанная среда имела конечное значение pH 5,9 и восстанавливалась посредством цистеина-HCl и Na2S при концентрации 0,008% (масс./об).
Раствор микроэлементов содержал 2 г нитрилотриацетной кислоты (устанавливали pH 6 с помощью KOH перед добавлением оставшихся ингредиентов), 1 г MnSO4, 0,8 г Fe(SO4)2(NH4)2×6H2O, 0,2 г CoCl2×6H2O, 0,2 мг ZnSO4×7H2O, 20 мг CuCl2×2H2O, 20 мг NiCl2×6H2O, 20 мг Na2MoO4×2H2O, 20 мг Na2SeO4 и 20 мг Na2WO4 на литр.
Витаминный раствор Вольфа (Wolin et al., J. Biol. Chem. 235:2882, 1963) содержал (на литр) 2 мг биотина, 2 мг фолиевой кислоты, 10 мг пиридоксина гидрохлорида, 5 мг тиамина-HCl, 5 мг рибофлавина, 5 мг никотиновой кислоты, 5 мг кальция D-(+)-пантотената, 0,1 мг витамина B12, 5 мг n-аминобензойной кислоты и 5 мг липоевой кислоты.
а. Ферментация Clostridium autoethanogenum
Ферментацию Clostridium autoethanogenum проводили с применением способов, аналогичных способам, описанным, например, в заявке на патент США №2011/0300593. Короче, 2-литровый биореактор, содержащий 1,3 л раствора А (3,083 г NH4Ac; 0,61 г MgCl2×6H2O; 0,294 г CaCl2×2H2O; 0,15 г KCl; 0,12 г NaCl (необязательно); доводили до 1 л дистиллированной водой) барботировали газообразным N2. Добавляли 85% раствор H3PO4 (2,025 мл, 30 мМ) и устанавливали pH 5,3, используя концентрированный водный NH4OH. Затем добавляли 13,5 мл раствора В (20,0 мг биотина; 20,0 мг фолиевой кислоты; 10,0 мг пиридоксина HCl; 50,0 мг тиамина × HCl; 50,0 мг рибофлавина; 50,0 мг никотиновой кислоты; 50,0 мг кальция D-(*)-пантотената; 50,0 мг витамина B12; 50,0 мг n-аминобензойной кислоты; 50,0 мг липоевой кислоты; доводили до 1 л дистиллированной водой), и полученный раствор барботировали газообразным N2. Добавляли хлорид хрома (II) до тех пор, пока окислительно-восстановительный потенциал (ORP) раствора не уменьшился до приблизительно - 200 мВ, затем добавляли резазурин (1,35 мл 2 г/л раствора). Добавляли полисульфид натрия (5,4 мл 3 М раствора, см. ниже), и полученный раствор барботировали N2 и затем газом, содержащим CO (1% H2; 13% N2; 71% CO; 15% CO2). Добавляли раствор сульфида металла (150 мл, см. ниже), и полученный раствор барботировали в течение дополнительных 30 минут, перед инокуляцией посредством активно растущей культуры С. autoethanogenum при уровне приблизительно 5% (об./об.).
Готовили раствор полисульфида натрия в 500 мл колбе, в которую загружали Na2S (93,7 г, 0,39 моль) и 200 мл H2O. Раствор перемешивали до растворения соли и добавляли серу (25 г, 0,1 моль) при постоянном потоке N2. После перемешивания при комнатной температуре в течение 2 часов раствор полисульфида натрия (примерно 4 М относительно Na и примерно 5 М относительно серы), теперь прозрачная красновато-коричневая жидкость, переносили в прошедшие продукву N2 флаконы с сывороткой и заворачивали в алюминиевую фольгу.
Готовили раствор хрома (II) в 1 л трехгорлой колбе, снабженной газонепроницаемыми входным и выходным отверстиями, что позволяло работать в атмосфере инертного газа и осуществить последующий перенос требуемого продукта в подходящую колбу для хранения. Колбу загружали CrCl3×6H2O (40 г, 0,15 моль), гранулами цинка [20 меш] (18,3 г, 0,28 моль), ртутью (13,55 г, 1 мл, 0,0676 моль) и 500 мл дистиллированной воды. После продукви N2 в течение одного часа, смесь нагревали до примерно 80°C, чтобы инициировать реакцию. После двух часов перемешивания при постоянном потоке N2, смесь охлаждали до комнатной температуры и непрерывно перемешивали в течение еще 48 часов, за время которых реакционная смесь превращалась в темно-голубой раствор. Раствор переносили в прошедшие продукву N2 флаконы с сывороткой и хранили при 4°C для будущего применения.
Раствор сульфида металла готовили путем добавления примерно 950 мл раствора А в 1 л ферментатор и барботирования газообразным N2. Добавляли 85% раствор H3PO4 (1,5 мл, 30 мМ) и устанавливали pH 5,3, используя концентрированный водный NH4OH. Добавляли раствор В (10 мл), и полученный раствор барботировали с помощью N2. Добавляли хлорид хрома (II) до тех пор, пока окислительно-восстановительный потенциал (ORP) раствора не уменьшился до приблизительно - 200 мВ, затем добавляли резазурин (1 мл 2 г/л раствора). Добавляли раствор C (1/10; 10 мл FeCl3; 5 мл CoCl2; 5 мл NiCl2; 1 мл H3BO3; 1 мл Na2MoO4; 1 мл MnCl2; 1 мл Na2WO4; 1 мл ZnCl2; 1 мл Na2SeO3; в 1 л среды), затем добавляли полисульфид натрия (2 мл 3 М раствора) и далее полученный раствор барботировали газообразным N2.
Ферментация субстрата, содержащего CO, посредством С. autoethanogenum в статических условиях в присутствии полисульфида приводила к значительному увеличению скорости аккумулирования, при этом конечное аккумулирование биомассы составляло приблизительно 4 г/л на протяжении периода от 2 до 3 дней. Например, после короткой лаг-фазы, составляющей приблизительно 1 день, биомасса могла увеличиться от примерно 0,5 г/л до по меньшей мере 3,5 г/л в течение приблизительно 36 часов ферментации. Кроме того, ацетат не образовывался во время фазы роста в присутствии полисульфида (который обычно находят при периодической ферментации) и в некоторых случаях определенное количество ацетата потреблялось, так что имело место чистое уменьшение количества ацетата в ферментаторе. Культивируемую биомассу собирали путем центрифугирования, клетки промывали один раз в среде и конгломерат клеток либо замораживали при -80°C, либо сразу же применяли для фракционирования клеточных компонентов.
Подход на основе полунепрерывной ферментации можно также применять для поддержания продуктивности биомассы и уменьшения времени, связанного с остановкой и запуском ферментации (т.е., времени оборота или времени производственного цикла).
Сбор аккумулированной бактериальной биомассы выполняли с промежутками приблизительно 12-24 часа, когда плотность культуры достигала стационарной фазы (но до ее наступления). Приблизительно половину объема биореактора удаляли путем переноса в отдельный контейнер с помощью центробежного насоса. Затем равный объем свежей или повторно используемой среды возвращали в биореактор, так что оптическая плотность реактора составляла приблизительно половину от ее исходного значений Ферментацию в биореакторе продолжали согласно приведенному выше протоколу таким образом, чтобы было выполнено несколько циклов роста и извлечения биомассы, в течение одного цикла ферментации.
b. Ферментация Clostridium ljungdahlii
Ферментацию Clostridium ljungdahlii выполняли с применением способов, аналогичных способам, описанным, например, в патентах США №№5173429 и 5593886. Короче, периодические ферментации проводили с применением биологически чистой культуры С. ljungdahlii. Приготовление среды ((1) 80,0 мл соли, содержащей KH2PO4 3,00 г/л, K2HPO4 3,00 г/л, (NH4)2SO4 6,00 г/л, NaCl 6,00 г/л, MgSO4×2H2O 1,25 г/л; (2) 1,0 г дрожжевого экстракта; (3) 1,0 г триптиказы; (4) 3,0 мл раствора металлических микроэлементов PFN (Pfenning), содержащего FeCl2×4H2O 1500 мг, ZnSO4×7H2O 100 мг, MnCl2×4H2O 30 мг, H3BO3 300 мг, CoCl2×6H2O 200 мг, CuCl2×H2O 10 мг, NiCl2×6H2O 20 мг, NaMoO4×2H2O 30 мг, Na2SeO3 10 мг и дистиллированную воду до 1 л; (5) 10,0 мл витаминов В, содержащих пиридоксаль HCl 10 мг, рибофлавин 50 мг, тиамин HCl 50 мг, никотиновую кислоту 50 мг, Ca-D-пантотенат 50 мг, липоевую кислоту 60 мг, n-аминобензойную кислоту 50 мг, фолиевую кислоту 20 мг, биотин 20 мг, цианокобаламин 50 мг и дистиллированную воду до 1 л; (6) 0,5 г цистеина HCl; (7) 0,06 г CaCl2×2H2O; (8) 2,0 г NaHCO3; (9) 1,0 мл резазурина (0,01%); и (10) 920,0 мл дистиллированной воды) выполняли в анаэробных условиях в атмосфере 80% азота и 20% CO2. В процессе ферментации регулировали показатель pH среды, который поддерживали равным 5,0 посредством HCl. При необходимости, регулировку pH осуществляли с помощью стерильного 10% NaOH или 1,0% раствора уксусной кислоты. Среду переносили в 157,5 мл флаконы с сывороткой и герметично закрывали с помощью бутилрезиновых пробок и алюминиевых прокладок. Затем флаконы стерилизовали в автоклаве при 121°C в течение 20 минут.
За приблизительно 48 часов до начала эксперимента готовили затравочную культуру из исходной культуры С. ljungdahlii во флаконе, аналогичном флаконам, описанным выше. Затравочную культуру выращивали в вибрационном инкубаторе при 37°C и встряхивали при 100 об/мин. Добавляли восстанавливающие растворы (2,0 мл Na2S, 2,5% раствор и 2,0 мл цистеин-HCl, 3,5% раствор) к культуре, которую помещали в вибрационный инкубатор на приблизительно 15 минут для полного удаления кислорода и температурной акклиматизации. В отличие от процедуры, применяемой для выделения биологически чистой культуры организма, при периодической ферментации не требуется добавление ингибиторов метана.
Ферментацию с применением С. ljungdahlii выполняли в 2,5-литровом ферментаторе New Brunswick Scientific Bioflow IIc, содержащем питательную среду при 37°C, и поддерживали постоянный уровень жидкости 1,5 литров при перемешивании жидкости при переменных скоростях, составляющих до 1000 оборотов в минуту, с помощью газа, введенного со скоростью приблизительно 500 кубических сантиметров в минуту. Оптимальные времена удерживания газа находились в диапазоне трех минут. Подачу газа изменяли при поглощении его бактериями, которое в свою очередь являлось функцией плотности клеток.
Сбор аккумулированной бактериальной биомассы выполняли с промежутками приблизительно 12-24 часа, когда плотность культуры достигала стационарной фазы (но до ее наступления). Приблизительно половину объема биореактора удаляли путем переноса в отдельный контейнер с помощью центробежного насоса. Затем равный объем свежей или повторно используемой среды возвращали в биореактор, так что оптическая плотность реактора составляла приблизительно половину от ее исходного значения. Ферментацию в биореакторе продолжали согласно приведенному выше протоколу таким образом, чтобы было выполнено несколько циклов роста и извлечения биомассы в течение одного цикла ферментации.
ПРИМЕР 2
МЕТАБОЛИЗИРУЮЩИЕ C1 МИКРООРГАНИЗМЫ, СКОНСТРУИРОВАННЫЕ ДЛЯ УСИЛЕНИЯ ПРОДУЦИРОВАНИЯ ЛИПИДОВ
Клетки-хозяина были сконструированы таким образом, что они обладают генетическими модификациями, минимизирующими или уменьшающими расщепление жирных кислот - путем нокаута активности длинноцепочечная жирная кислота: КоА-лигазы, кодированной эндогенным геном fadD. Кроме того, биосинтез свободных жирных кислот (СЖК) был усилен путем введения гена тиоэстеразы (ТЕ) в метанотроф согласно настоящему изобретению (Methylococcus capsulatus). Такие рекомбинантные изменения дополнительно описаны в настоящем примере.
Молекулы рекомбинантных нуклеиновых кислот
Последовательности нуклеиновых кислот, кодирующие белки FadD дикого типа, представляли собой эталонную исходную точку для конструирования мутантных генов fadD. Например, последовательность белка FadD дикого типа, кодированная М. trichosporium OB3b, М. capsulatus Bath, М. methanica, М. extorquens и С. ljungdahlii, находится в банке данных GenBank Accession №№ EFH00931.1, YP_114021.1, YP_004512148.1, YP_002964871.1 и YP_003782065.1, соответственно. В результате, молекула нуклеиновой кислоты генов fadD, кодирующих указанные выше белки, была отдельно синтезирована таким образом, что содержит несколько мутаций остановки и сдвиги рамки в 5'-области гена из М. trichosporium OB3b (SEQ ID №: 1), М. methanica (SEQ ID №: 35), М. extorquens (SEQ ID №: 52) и С. ljungdahlii (SEQ ID №: 85). Для гена fadD М. capsulatus, была синтезирована молекула нуклеиновой кислоты, содержащая внутриклональную делецию, так что оставшиеся 5' и 3' концы гена могли соединиться с сохранением первоначальной рамки считывания (SEQ ID №: 18).
В случае С. autoethanogenum геном являлся секвенированным и fadD, гомологичный Е. coli, идентифицировали путем поиска с помощью TBLASTN (поиск транслированных нуклеотидных последовательностей генов с последовательностью белка FadD Е. coli). Молекулу нуклеиновой кислоты гена fadD С. autoethanogenum синтезировали для внедрения нескольких мутаций остановки и сдвигов рамки в 5'-область указанного гена.
Молекулы нуклеиновых кислот мутантного fadD по отдельности клонировали в плазмидном векторе (не имеющем точки начала репликации метанотрофа или клостридии и кодирующем устойчивость к канамицину) для конъюгации, электропорации или трансформации в метаболизирующий C1 микроорганизм с применением способов, описанных в настоящей заявке. Такой вектор (который не реплицирует в метаболизирующем C1 микроорганизме) гарантирует, что любой канамициноустойчивый метаболизирующий C1 микроорганизм будет иметь ген устойчивости, встроенный в клеточный геном хозяина в результате гомологической рекомбинации и замещения эндогенного гена fadD указанными выше мутантами fadD (так что рекомбинантные клетки будут отсутствовать или проявлять минимальную активность длинноцепочечная жирная кислота: КоА-лигазы).
Кроме того одна или более выбранных последовательностей тиоэстеразы, последовательность малонил-КоА: ацилпереносящий белок-трансацилазы (fabD) и последовательность ацетил-КоА-карбоксилазы (например, ассА, accB, ассС и accD из Е. coli) подвергали оптимизации кодонов и синтезировали с помощью подходящих промоторов. Затем один или более генов тиоэстеразы и ген ацетил-КоА-карбоксилазы (например, ассА или accABCD) клонировали в подходящем экспрессирующем векторе и конъюгировали, электропорировали или трансформировали в метаболизирующие C1 микроорганизмы дикого типа или метаболизирующие C1 микроорганизмы с нокаутированным fadD, описанные в настоящей заявке.
Кодон-оптимизированные последовательности тиоэстеразы приведены в (1) SEQ ID NOS.: 3-13 для М. trichosporium OB3b; (2) SEQ ID NOS.: 20-30 для М. capsulatus Bath; (3) SEQ ID NOS.: 37-47 для М. methanica; (4) SEQ ID NOS.: 54-64 для М. extorquens; (5) SEQ ID NOS.: 70-80 для С. autoethanogenum; и (6) SEQ ID NOS.: 87-97 для С. ljungdahlii. Кодон-оптимизированные последовательности fabD приведены в (1) SEQ ID №: 2 для М. trichosporium OB3b; (2) SEQ ID №: 19 для М capsulatus Bath; (3) SEQ ID №: 36 для М. methanica; (4) SEQ ID №: 53 для М. extorquens; (5) SEQ ID №: 69 для С. autoethanogenum; и (6) SEQ ID №: 86 для С. ljungdahlii. Кодон-оптимизированные последовательности ассА, ассВ, ассС и accD описаны, соответственно, в (1) SEQ ID NOS.: 14-17 для М. trichosporium OB3b; (2) SEQ ID NOS.: 31-34 для М. capsulatus Bath; (3) SEQ ID NOS.: 48-51 для М. methanica; (4) SEQ ID NOS.: 65-68 для М. extorquens; (5) SEQ ID NOS.: 81-84 for С autoethanogenum; и (6) SEQ ID NOS.: 98-101 для С. ljungdahlii.
Конъюгация
Процедура конъюгирования плазмид из Escherichia coli в М. trichosporium OB3b или М. methanica была основана на способе, разработанном Martin и Murrell (FEMS Microbiol. Lett. 727:243, 1995), тогда как процедура конъюгирования плазмид из Е. coli в M. capsulatus была основана на способе, описанном Ali и Murrell (Microbiology 755:761, 2009).
Короче, мобилизуемая плазмида, содержащая один или более представляющих интерес генов (например, мутантный fadD, МСТ, один или более ТЕ, один или более Acc) и кодирующая устойчивость к канамицину, была сначала трансформирована в S17-1 Е. coli с применением стандартных способов электропорации. Трансформация была подтверждена выбором устойчивых к канамицину колоний на LB-агаре, содержащем 30 мкг/мл канамицина. Трансформированные колонии инокулировали в среду LB, содержащую 30 мкг/мл канамицина, и встряхивали всю ночь при 37°C. Затем 10 мл аликвоту полученной за ночь культуры собирали на стерильный 47 мм нитроцеллюлозный фильтр (размер пор 0,2 мм). Донорные клетки Е. coli промывали на фильтре с помощью 50 мл стерильной среды NSM для удаления остаточной среды и антибиотика.
Параллельно, пробу штаммов-реципиентов M. trichosporium OB3b, M. methanica или M. capsulatus Bath по отдельности инокулировали в 100 мл флаконы с сывороткой, содержащие от 20 до 50 мл среды NSM. Затем свободное пространство во флаконах заполняли смесью 1:1 кислорода и метана и флаконы герметично закрывали бутилрезиновыми крышками и обжимали. Флаконы непрерывно встряхивали в инкубаторе при 30°C (M. trichosporium OB3b, M. methanica) или 45°C (M. capsulatus Bath) до достижения OD600 приблизительно 0,3. Далее клетки собирали на такой же фильтр, что и штамм-донор Е. coli. Фильтр снова промывали с помощью 50 мл стерильной среды NSM. Указанный фильтр помещали (клетками вверх) на планшет с агаровой средой NSM, содержащий 0,2% дрожжевого экстракта, и инкубировали в течение 24 ч при 30°C (M. trichosporium OB3b, M. methanica) или 37°C (M. capsulatus Bath) в присутствии смеси 1:1 метана и воздуха. Через 24 ч перед концентрированием путем центрифугирования клетки повторно суспендировали в 10 мл стерильной среды (NSM). Собранные клетки повторно суспендировали в 1 мл стерильной среды NSM и аликвоты (100 мкл) распределяли по планшету с агаровой средой NSM, содержащему 10 мкг/мл канамицина.
Планшеты инкубировали в герметичных камерах, содержащих смесь 1:1 метана и воздуха, и выдерживали при 30°C (M. trichosporium OB3b, M. methanica) или 45°C (M. capsulatus Bath). Газовую смесь пополняли каждые 2 дня до формирования колоний, как правило, через 7-14 дней. Колонии засеивали штрихом на планшетах с NSM, содержащих канамицин, для подтверждения устойчивости к канамицину, а также для дополнительного выделения трансформированных метанотрофных клеток из остаточных клеток-доноров Е. coli.
Электропорация - Methanobacterium
Процедура введения плазмид в М. extorquens была основана на процедуре, описанной Ueda et al., Appl. Environ. Microbiol. 57:924, 1991. Короче, М. extorquens дикого типа (wt) культивировали при 30°C в среде NSM с добавкой 0,5% метанола. Клетки NR-2 М. extorquens, выращенные до фазы середины логарифмического роста (1,4×109/мл), собирали путем центрифугирования при 6000 × г в течение 10 мин и промывали электропорационным буферным раствором (10 мМ Tris-HCl, 2 мМ MgCl2, 6H2O, 10% [масс./об.] сахарозы [pH 7,5]). Клетки повторно суспендировали в том же буферном растворе при концентрации клеток 7,0×1010/мл. Клеточную суспензию и вектор (70 мкг/мл) смешивали при соотношении 9:1 (об./об.) в пробирке и затем 10 мкл переносили пространство между электродами камеры, где указанную суспензию приводили в равновесие в течение 3 минут. После воздействия 10 импульсами 10 кВ/см электрического поля в течение 300 мкс/импульс, 5 мкл аликвоту смеси переносили в чистую пробирку и добавляли 0,2 мл среды NSM. Затем клеточную суспензию инкубировали в течение 2 ч при 30°C для протекания экспрессии генов, устойчивых к антибиотикам, перед культивированием на планшетах со средой NSM, содержащих 0,5 метанола и 20 мкг/мл канамицина.
Указанные планшеты инкубировали при 30°C до образования колоний. Колонии засеивали штрихом на одинаковых планшетах для подтверждения устойчивости к канамицину, а также для дополнительного выделения трансформированных метанотрофных клеток из остаточных донорных клеток Е. coli.
Электропорация - Clostridium
Способы трансформации для С. autoethanogenum или С. ljungdahlii выполняли, как описано в публикации патента США №2011/0236941, или с применением модифицированного протокола для С. tyrobutyricum (Zhu et al., Biotechnol. Bioeng. 90:154, 2005). Короче, для получения компетентных клеток 50 мл культуры С. autoethanogenum пересеивали в свежую среду в течение 3 последовательных дней согласно условиям культивирования, описанным в настоящей заявке. Указанные клетки применяли для инокулирования 50 мл среды РЕТС, содержащей 40 мМ DL-треонина, при OD600 0,05. Когда OD600 культуры достигла 0,4, клетки переносили в анаэробную камеру и собирали при 4700 × г и 4°C. Культуру дважды промывали ледяным электропорационным буферным раствором (270 мМ сахарозы, 1 мМ MgCl2, 7 мМ фосфата натрия, pH 7,4) и в заключении суспендировали в свежем электропорационном буферном растворе объемом 600 мкл. Указанную смесь переносили в предварительно охлажденную кювету для электропорации с 0,4 см зазором между электродами, содержащую 1 мкг вектора (не имеющего точки начала репликации Clostridium и содержащего представляющую интерес молекулу нуклеиновой кислоты и кодирующего устойчивость к кларитромицин) и сразу же воздействовали импульсами, используя систему электропорации Gene pulser Xcell (Bio-Rad) при следующих установочных параметрах: 2,5 кВ, 600 мкл и 25 мкФ. Обеспечивали постоянные времени от 3,7 до 4,0 мс. Культуру переносили в 5 мл свежей среды. Регенерацию клеток контролировали при длине волны 600 нм, используя спектрофотометр Spectronic Helios Epsilon (Thermo), оборудованный держателем для пробирок. После начального уменьшения биомассы, клетки снова возобновили рост. После того, как количество биомассы удвоилось относительно указанной точки, клетки собирали, суспендировали в 200 мкл свежей среды и высеивали на планшетах с селективной средой РЕТС (содержащих 1,2% Bacto™ Agar (BD)) с 4 мкг/мкл кларитромицина. После 4-5 дней инкубирования с применением газа от сталелитейного завода при давлении 30 psi (207 кПа) при 37°C, колонии были ясно видны.
Согласно альтернативному варианту реализации изобретения после электропорационного импульса клетки переносили в 5 мл предварительно нагретой среды в пробирке Хангейта и инкубировал при 37°C до тех пор, пока рост не становился видимым (измерения проводили в пробирках Хангейта в фотометре). Аликвоты трансформантов инокулировали в 5 мл жидкой среды и распределяли по содержащим кларитромицин планшетам для выращивания мутантных колоний.
Выбранные рекомбинантные колонии применяли для инокулирования 2 мл среды РЕТС, содержащей 4 мкг/мкл кларитромицина. Когда происходил рост, объем культуры увеличивали в 5 мл и затем в 50 мл среды РЕТС, содержащей 4 мкг/мкл кларитромицина и газ от сталелитейного завода при давлении 30 psi (207 кПа) в качестве источника углерода.
Рекомбинантные метаболизирующие C1 бактерии
Трансформация была подтверждена устойчивостью клеток к выбранному антибиотику и экспрессия генов была подтверждена методами ПЦР (полимеразной цепной реакцией), нозерн-блоттинга, вестерн-блоттинга или ELISA. Например, для подтверждения переноса можно выделить плазмидную ДНК и подвергнуть ее ПЦР, используя гранулы illustra PuReTaq Ready-To-Go™ ПЦР Beads (GE Healthcare) и стандартные условия (95°C в течение 5 мин; 32 цикла при 95°C в течение 30 с, 50°C в течение 30 с и 72°C в течение 1 мин; 72°C в течение 10 мин). В качестве дополнительного контроля, 1 мкл каждой из изолированных плазмид повторно трансформировали в XL1-Blue MRF' Kan E. coli (Stratagene, Ла-Холья, Калифорния), откуда плазмиды можно аккуратно выделить и верифицировать с помощью рестриктазных фрагментов.
Способы идентификации актов гомологичной рекомбинации надежно отработаны в данной области техники, например, ПЦР и секвенирование с применением уникальных праймеров в геноме и векторе для подтверждения соответствующей вставки. Далее рекомбинантные бактерии, имеющие согласно идентификации соответствующую вставку, выращивали в отсутствии селективного давления (например, без канамицина или кларитромицина) в течение нескольких поколений и чувствительные к канамицину клоны идентифицировали методом отпечатков (или эквивалентным методом). Приблизительно 50% чувствительных к канамицину ревертантов должно обладать мутировавшей формой гена-мишени вместо дикого типа, что подтверждалось посредством ПЦР и секвенирования. Потерю экспрессии или функции fadD можно подтвердить с помощью одного или более методов, выбранных из (1) ПЦР и секвенирования, (2) Нозерн-блоттинга и (3) анализа на активность ацил-КоА синтетазы.
Для анализа активности ацил-КоА синтетазы можно использовать, например, способ Kameda et al. (J. Biol. Chem. 256:5702, 1981) путем выращивания клеток до фазы середины логарифмического роста в среде NSM с антибиотиками, при необходимости, сбора клеток путем центрифугирования, промывания дважды с помощью NSM, суспендирования клеток до плотности 1,2×109 клеток/мл в 10 мМ Tris-HCl, pH 7,5 и затем лизирования при трех циклах обработки ультразвуком на льду. Готовили реакционные смеси в общем объеме 0,5 мл, содержащие 200 мМ Tris-HCl, pH 7,5, 2,5 мМ АТФ, 8 мМ MgCl, 2 мМ ЭДТК, 20 мМ NaF, 0,1% Triton® X-100, 10 пМ [3H]олеата, 0,5 мМ кофермента А и клеточный экстракт. Ферментативные реакции инициировали путем добавления кофермента А, инкубировали при 35°C в течение 10 минут и заканчивали путем добавления 2,5 мл изопропилового спирта : н-гептана : 1М H2SO4 (40:10:1). Радиоактивную олеиновую кислоту удаляли путем органической экстракции с применением н-гептана, тогда как олеоил-КоА, образовавшийся во время реакции, оставался в водной фракции для количественного определения посредством сцинтилляционного счета. Концентрации белка в ферментных экстрактах определяли методом Бредфорда, используя бычий сывороточный альбумин в качестве стандарта.
Получение жирных кислот из C1 субстратов (CH4 и CO)
Для метанотрофов применяли М. trichosporium OB3b, М. methanica, M. extorquens или М. capsulatus Bath, дикого типа или с нокаутированным fadD, трансформированные вектором, содержащим гены, кодирующие один или более генов тиоэстеразы или сверхэкспрессирующие гены ацетил-КоА-карбоксилазы, для инокулирования 100 мл флаконов с сывороткой или культуральных пробирок, содержащих от 20 до 50 мл среды NSM и 10 мкг/мл канамицина. Для М. extorquens в среду добавляли 0,5% метанола в качестве источника углерода, тогда как для М. trichosporium OB3b, М. methanica и М. capsulatus Bath свободное пространство во флаконе заполняли смесью 1:1 кислорода и метана в качестве источника углерода. Флаконы герметично закрывали бутилрезиновыми крышками и обжимали. Затем флаконы или пробирки непрерывно встряхивали при скорости 200-250 об/мин во время инкубирования при 30°C (М. trichosporium OB3b, М. methanica, М. extorquens) или 42-45°C (М. capsulatus Bath).
В случае клостридии применяли С. autoethanogenum или С. ljungdahlii дикого типа или с нокаутированным fadD, трансформированные вектором, содержащим гены, кодирующие один или более ферментов тиоэстеразы, и с содержанием или без генов ацетил-КоА-карбоксилазы, для инокулирования 2 мл среды РЕТС, содержащей 4 мкг/мкл кларитромицина. Когда происходил рост, объем культуры увеличивали в 5 мл и затем в 50 мл среды РЕТС, содержащей 4 мкг/мкл кларитромицина и газ от сталелитейного завода при давлении 30 psi (207 кПа) в качестве источника углерода. Затем флаконы непрерывно встряхивали при скорости от 200 до 250 об/мин во время инкубирования при 37°C.
Количественное определение жирных кислот, продуцируемых рекомбинантными метаболизирующими C1 бактериями, выполняли с применением газового хроматографа/масс-спектрометра (ГХ/МС). Жирные кислоты в клеточной культуре экстрагировали путем интенсивного перемешивания с бутилацетатом, содержащим ундекановую кислоту в качестве внутреннего стандарта для ГХ/МС анализа экстракта. После недолгого центрифугирования смеси небольшую часть органической фазы переносили в отдельный флакон, с последующим добавлением равного объема N,O-бис(триметилсилил)трифторацетамида. Пробу анализировали посредством ГХ с масс-спектрометрическим детектором (HP 5792), используя ГХ/МС колонку Agilent HP-5MS (толщина пленки 30,0 м × 250 мкм × 0,25 мкм). Для инжектора применяли отношение деления потока 20:1 при 250°C, и в качестве газа-носителя использовали гелий при скорости потока 1,2 мл/мин. Температуру термостата поддерживали при 60°C в течение 1 минуты, с последующим увеличением градиента температуры на 19°C/мин до достижения температуры 250°C. Концентрацию жирных кислот в клеточной культуре рассчитывали в режиме селективного детектирования ионов на основе калибровочных кривых стандартов жирных кислот. Поскольку метан был единственным источникам углерода, поставляемым в клетки, все продуцируемые жирные кислоты должны были быть получены из метана.
Результаты
Профиль жирных кислот М. capsulatus Bath изменяли посредством нокаута fadD и путем введения и экспрессии гена тиоэстеразы Е. coli. Сначала синтезировали ген тиоэстеразы Е. coli с удаленной последовательностью нацеливания на периплазму (TesA'), используя три различные композиции кодонов (TesA'-3, SEQ ID NO: 102; TesA'-37, SEQ ID NO: 103; и TesA'-20, SEQ ID NO: 104), сконструированные для получения вариантов с различными уровнями экспрессии. Варианты TesA' клонировали в плазмиду на основе IncP (содержащую Inc-P oriV и oriT) и функционально связывали с промотором, который функционировал в метанотрофах. Рекомбинантный экспрессирующий вектор, содержащий TesA', трансформировали в М. capsulatus, как описано в настоящей заявке. Культуры М. capsulatus выращивали в течение 5 дней в 5 мл объеме в 150 мл герметично закрытых флаконах с сывороткой с использованием 40 мл метана и 80 мл кислорода. После стадии роста 1 мл каждой культуры анализировали на концентрацию и состав жирных кислот, используя ГХ/МС, как описано в настоящей заявке. Измеренные концентрации свободных жирных кислот нормализировали до OD600 с помощью культуры. Отметим, что фракция C16:1 состояла из по меньшей мере трех различных изомеров, при этом наиболее распространенной была Δ9-цис-пальмитолеиновая кислота (данные не показаны).
Параллельно, гомолог ацилкофермент А (КоА)-синтетазы (fadD) Е. coli рекомбинантно нокаутировали SEQ ID NO: 18 в геноме М. capsulatus, как описано в настоящей заявке, и подтверждали с помощью анализа ПЦР. Было показано, что нокаутированный FadD в некоторых других микробных штаммах увеличивал уровни свободных жирных кислот (см., например, Lennen et al., Trends Biotechnol. 72:659, 2012). Нокаутированный мутант fadD М. capsulatus не проявлял значительного увеличения уровней свободных жирных кислот, что указывает на то, что один или более дополнительных гомологов FadD может присутствовать в геноме М. capsulatus, но липидный профиль был сдвинут, поскольку имело место увеличение содержания C18:0 липидов.
Пул свободных жирных кислот в трансформированных клетках резко увеличился (см. фиг. 3A), причем указанное увеличение в основном связано с повышенными уровнями С16:0 и С18:0 липидов (см. фиг. 3B).
ПРИМЕР 3
ЭКСТРАКЦИЯ ЛИПИДОВ ИЗ МЕТАБОЛИЗИРУЮЩИХ C1 МИКРООРГАНИЗМОВ
Масляную композицию, содержащуюся в собранной бактериальной биомассе, экстрагировали согласно модифицированной версии протокола экстракции Фолча (Folch et al., J. Biol. Chem. 226:497, 1957), выполняемого при 20°C (т.е., комнатной температуре) в экстрактном растворе, приготовленном из одного объема метанола в двух объемах хлороформа (раствор ХМ). Для экстракции использовали примерно 5 г влажной клеточной массы (ВКМ) из каждой свежей бактериальной биомассы (или бактериальной биомассы, которую хранили при -80°C и затем оттаивали). К клеточному материалу добавляли 100 мл раствора ХМ и смесь энергично экстрагировали в делительной воронке. Через по меньшей мере 10 минут три фазы растворялись. Органическая фаза, содержащая экстрагированные липиды, оседала в нижней части делительной воронки, которую сливали в чистый стеклянный флакон. Средний слой содержал в основном лизированный клеточный материал, который можно было отделить от легкой водной фазы, содержащей соли и другие растворимые клеточные компоненты.
При необходимости, твердые вещества в водной фазе можно сконцентрировать, используя центрифугу или другое механическое оборудование для концентрирования. Воду, удаленную из твердой фазы, можно повторно использовать, тогда как твердые вещества с некоторым количеством остаточной воды можно загрузить в устройство для обработки твердой фазы.
Для повышения эффективности экстракции липидов выполняли вторую стадию экстракции путем добавления дополнительных 100 мл свежего раствора ХМ непосредственно в делительную воронку, содержащую оставшуюся лизированную клеточную массу и остаточную воду. Смесь опять тщательно перемешивали, оставляли фазы разделяться и объединяли нижние органические фазы, полученные в результате двух экстракций. Затем объединенные органические фазы промывали в делительной воронке 100 мл деионизированной воды для удаления любого остаточного водорастворимого материала. Снова сливали отделенную органическую фракцию из нижней части делительной воронки и удаляли растворитель путем вращательного испарения при нагревании, предпочтительно, в отсутствии кислорода или путем испарения при 55°C в потоке азота.

Затвердевшие масляные композиции, экстрагированные из собранных культур М. trichosporium OB3b, Methylococcus capsulatus Bath и Methylomonas sp. 16a, взвешивали, и результаты взвешивания приведены в таблице 1 в виде массовой доли относительно первоначальной сухой массы клеток (СМК). Полученные данные показывают, что значительная часть СМК из указанных метаболизирующих C1 микроорганизмов состоит из липидов.
Масляную композицию из биомассы Methylomonas sp. 16a также экстрагировали способом экстракции Hara и Radin с применением смеси гексан : изопропанол (ГИП) (Anal. Biochem. 90:420, 1978). Анализ масляной композиции, экстрагированной способом с применением ГИП, показал, что указанная масляная композиция была по существу идентична масляной композиции, экстрагированной с применением модифицированного способа Фолча (данные не показаны).
ПРИМЕР 4
ПРЕВРАЩЕНИЕ ЛИПИДОВ ИЗ МЕТАБОЛИЗИРУЮЩИХ C1 МИКРООРГАНИЗМОВ В СЛОЖНЫЕ МЕТИЛОВЫЕ ЭФИРЫ ЖИРНЫХ КИСЛОТ
Липидные фракции, экстрагированные из культивируемой биомассы М. capsulatus Bath, М. trichosporium OB3b и Methylomonas sp. 16a в форме сухого твердого вещества, подвергали по отдельности гидролизу с применением гидроксида калия (KOH) и превращали в сложные метиловые эфиры жирных кислот (СМЭЖК) посредством реакции с метанолом в одну стадию. Примерно 5 г экстрагированных твердых липидов в 10 мл стеклянном флаконе растворяли в 5 мл 0,2 М KOH раствора толуола : метанола (1:1 об./об.). Флакон энергично встряхивали и затем перемешивали при 250 об/мин при 42°C в течение 60 минут, после чего раствор оставляли охлаждаться до температуры окружающей среды и переносили в делительную воронку. В делительную воронку добавляли приблизительно 5 мл дистиллированной воды и 5 мл раствора ХМ, перемешивали и затем оставляли фазы разделяться под действием силы тяжести или путем центрифугирования (3000 об/мин, 25°C) в течение 5 минут. Удаляли верхний водный слой, содержащий растворенные сложные эфиры глицеринфосфата, тогда как тяжелую масляную фазу (нижнюю) собирали и концентрировали до сухости путем вращательного испарения или с применением постоянного потока азота.
Анализ СЖК и СМЭЖК, обнаруженных в липидах из каждой метанотрофной культуры, выполняли с применением газового хроматографа/масс-спектрометра (ГХ/МС). Твердую фазу, собранную перед и после стадии гидролиза/переэтерификации растворяли в 300 мкл бутилацетата, содержащего ундекановую кислоту в качестве внутреннего стандарта для ГХ/МС анализов. Полученный раствор центрифугировали в течение 5 минут при 14000 об/мин для удаления нерастворимых остатков. К супернатанту, полученному на стадии центрифугирования, добавляли одинаковый объемный эквивалент N,O-бис(триметилсилил)трифторацетамида и быстро встряхивали. Пробы помещали в ГХ, оборудованный масс-спектрометрическим детектором (HP 5792), и применяли ГХ/МС колонку Agilent HP-5MS (толщина пленки 30,0 м × 250 мкм × 0,25 мкм) для разделения СЖК и СМЭЖК. Идентичность СЖК и СМЭЖК подтверждали на основе времени удерживания и масс-спектров электронной ионизации их стандартов. В ГХ/МС способе в качестве газа-носителя использовали гелий при скорости потока 1,2 мл/мин. Температуру в отверстии для ввода пробы поддерживали при 250°C при отношении деления потока 20:1. Температуру термостата поддерживали при 60°C в течение 1 минуты при последующем градиенте температуры, включающем увеличение на 8°C/мин до 300°C. Площадь в % каждой СЖК и СМЭЖК рассчитывали на основе общего количество ионов из показания масс-детектора.
Твердый остаток, собранный перед и после гидролиза/переэтерификации, анализировали на содержание СЖК и СМЭЖК с применением ГХ/МС (см. таблицу 2). Кроме того, хроматограммы, полученные при анализе ГХ/МС, приведены на фиг. 4-6.
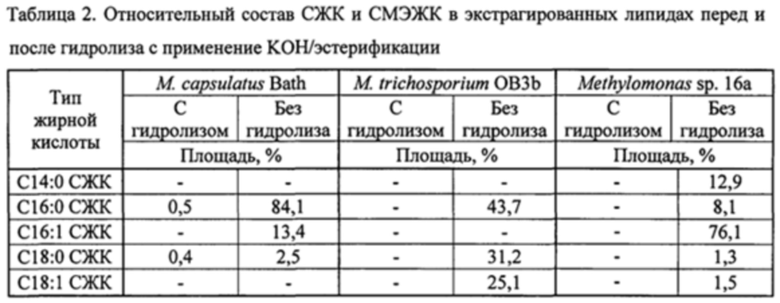
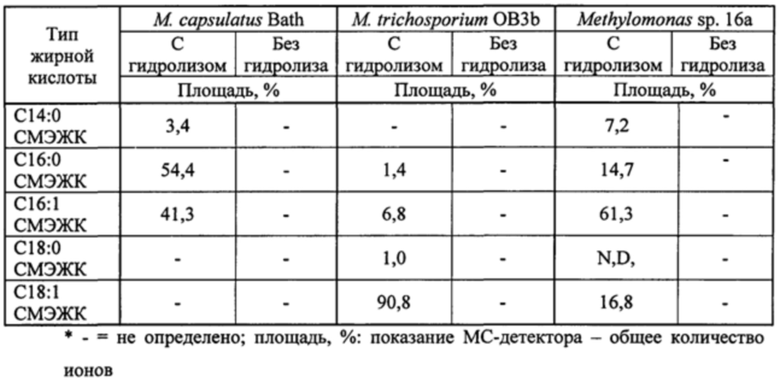
Как видно из таблицы 2 и фиг. 4-6, экстрагированные масляные композиции перед гидролизом/переэтерификацией содержали в изобилии свободные жирные кислоты и дополнительные жирные кислоты, присутствующие в виде (наиболее вероятно) ди- и триацилглицеридов (которые не были обнаружены при следовом ГХ/МС анализе), но после гидролиза/переэтерификации СЖК превращались в сложные метиловые эфиры жирных кислот различной длины. Полученные данные указывают, что масляные композиции из метаболизирующих C1 микроорганизмов согласно настоящему изобретению можно очистить и применять для получения высокоценных молекул.
ПРИМЕР 5
ПОЛУЧЕНИЕ БИОТОПЛИВА С ПОМОЩЬЮ МАСЛЯНЫХ КОМПОЗИЦИЙ ИЗ МЕТАБОЛИЗИРУЮЩИХ C1 МИКРООРГАНИЗМОВ
Экстрагированные масляные композиции из метаболизирующих C1 микроорганизмов можно обработать на расположенной вблизи установке для переработки или транспортировать на удаленную установку для переработки. Установку для переработки применяли для превращения триглицеридов из биовозобновляемого сырья (такого как жиры, животные жиры и метанотрофные масла) в смесь жидких углеводородных топлив, в основном биодизельного топлива и биотоплива для реактивных двигателей, высококачественного синтетического парафинированного керосина (СПК). Указанный способ требует водорода, который можно получить на месте посредством риформинга метана или который обеспечивают с применением аппарата для ферментации, расположенного вблизи существующей установки для переработки.
Установка для переработки может работать в смешенном режиме, при этом выходной продукт представлял собой смесь биодизельного топлива и биотоплива для реактивных двигателей, или в режиме дизельного топлива, при этом выходной продукт представлял собой в основном биодизельное топливо.
Во время переработки жирные кислоты и глицериды превращали в СПК за три стадии. Сначала сырье обрабатывали для удаления загрязняющих веществ-катализаторов и воды, при необходимости. На второй стадии цепи жирных кислот трансформировали в н-парафины в гидроочистителе. Примером является превращение олеиновой кислоты в н-октадекан посредством реакций гидрирования и дезоксигенирования в гидроочистителе. Для большинства биомасел, жиров и животных жиров полученный в гидроочистителе жидкий продукт представлял собой, главным образом, композицию C15-C18 н-парафинов. На третьей стадии способа указанные парафины с длинными прямыми цепями подвергали гидрокрекингу с получением более коротких разветвленных парафинов. Продукты гидрокрекинга главным образом соответствовали интервалу температур кипения керосина.
Полученный СПК предпочтительно соответствовал или превосходил все соответствующие целевому назначению спецификации топлива для реактивных двигателей, за исключением плотности. Высокое отношение H к C в СПК, придающее ему отличную термическую устойчивость и низкий показатель выбросов твердых частиц, означает углеводородную композицию с пониженной плотностью: 760-770 кг/м3 по сравнению с минимальным значением 775 кг/м3, приведенным в спецификации ASTM (Американского общества по испытанию материалов)' Но такая пониженная плотность не является проблемой в случае смесей нефтяного топлива для реактивных двигателей: СПК (например, 50/50).
ПРИМЕР 6
РАСПРЕДЕЛЕНИЕ СТАБИЛЬНОГО ИЗОТОПА УГЛЕРОДА В ЛИПИДАХ ИЗ МЕТАБОЛИЗИРУЮЩИХ C1 МИКРООРГАНИЗМОВ
Сухие пробы биомассы М. trichosporium и липидных фракций анализировали на содержание углерода и азота (% сухой массы) и отношения стабильных изотопов углерода (13C) и азота (15N) с помощью элементного анализатора/масс-спектрометрии изотопных отношений при непрерывном потоке, используя элементный анализатор CHNOS (vario ISOTOPE cube, Elementar, Ханау, Германия), соединенный с IsoPrime100 IRMS (Isoprime, Чидл, Соединенное Королевство). Пробы метанотрофной биомассы, культивируемой в ферментаторах или флаконах с сывороткой, центрифугировали, ресуспендировали в деионизированной воде и объемы, соответствующие от 0,2 до 2 мг углерода (примерно от 0,5 до 5 мг сухой массы клеток), переносили в 5×9 мм оловянные капсулы (Costech Analytical Technologies, Inc., Валенсия, Калифорния) и высушивали при 80°C в течение 24 часов. Подобным образом, предварительно экстрагированные липидные фракции суспендировали в хлороформе и объемы, содержащие от 0,1 до 1,5 мг углерода, переносили в оловянные капсулы и испаряли до сухости при 80°C в течение 24 часов. Стандарты, содержащие 0,1 мг углерода, обеспечивали надежные значения δ13C.
Изотопное отношение выражали с помощью условного обозначения «дельта» (‰), при этом изотопный состав материала относительно состава стандартного образцам расчете на миллион отклонений обозначали как δ13C (или δ15N)=(RПроба/RСтандарт-1)×1000, где R представлял собой молекулярное отношение тяжелой формы изотопа к легкой форме. Стандартным образцом для углерода являлся венский белемнит Pee Dee (V-PDB), а для азота - воздух. В качестве калибровочного стандартного образца применяли предложенный NIST (Национальным институтом стандартов и технологии) SRM (стандартный эталонный материал) №1547, листья персика. Все изотопные анализы проводили в Центре биогеохимии стабильных изотопов при Калифорнийском университете, Беркли. Долгосрочная внешняя точность для анализов С и N изотопов составляла 0,10‰ и 0,15‰, соответственно.
Штамм М. trichosporium OB3b выращивали на метане в трех различных ферментируемых партиях, М. capsulatus Bath выращивали на метане в двух различных ферментируемых партиях и Methylomonas sp. 16a выращивали на метане в одной ферментируемой партии. Биомассу из каждой из указанных культур анализировали на распределение стабильного изотопа углерода (значения δ13C; см. таблицу 3).
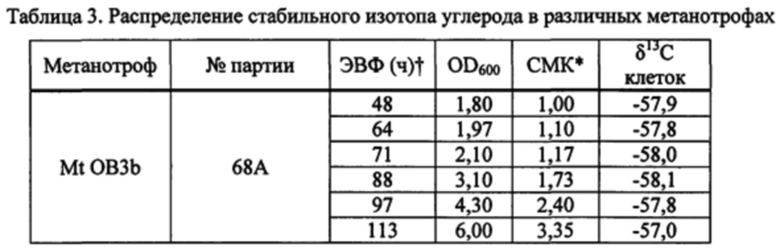
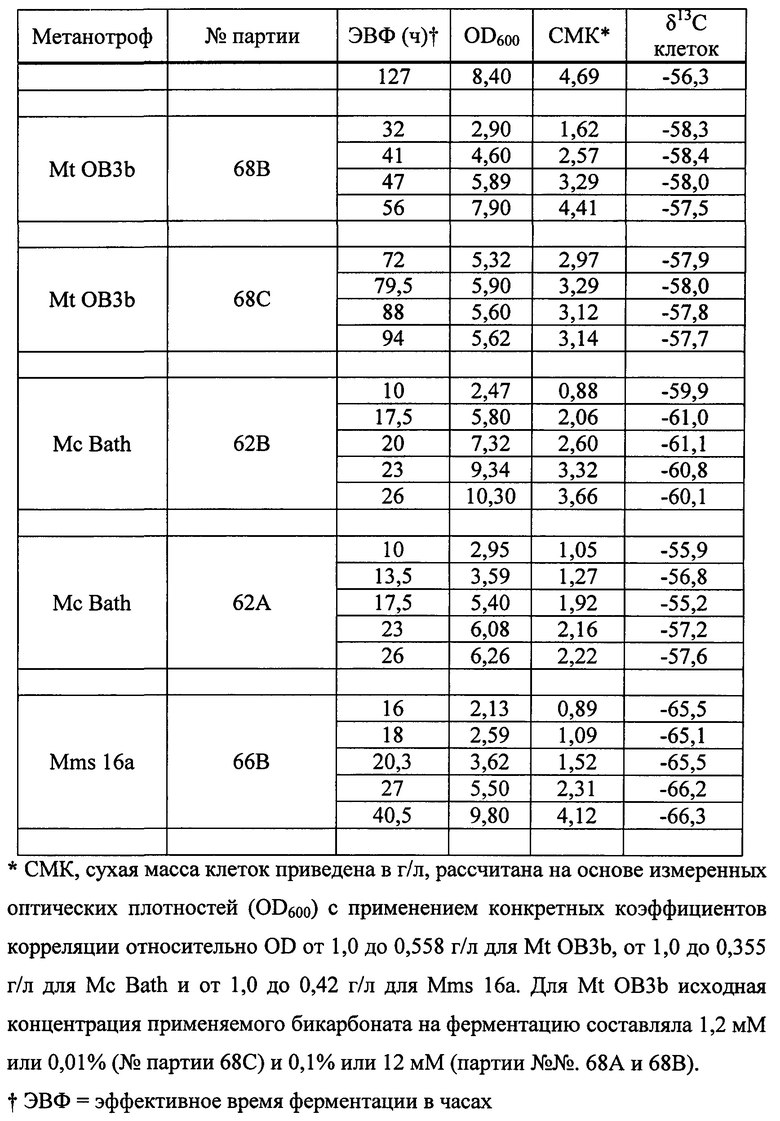
Кроме того, выполняли анализ на стабильный изотоп углерода для биомассы и соответствующих липидных фракций (см. таблицу 4) из штаммов Methylosinus trichosporium OB3b (Mt OB3b), Methylococcus capsulatus Bath (Mc Bath) и Methylomonas sp. 16a (Mms 16a), выращенных на метане в биореакторах, как описано в примере 1.

Биомассу из штаммов Mt OB3b, Me Bath и Mms 16a собирали через 94 ч (3,14 г СМК/л), 26 ч (2,2 г СМК/л) и 39 ч (1,14 г СМК/л), соответственно. В таблице 4 значения δ13C для липидов представляют собой среднее значение повторных определений.
ПРИМЕР 7
ВЛИЯНИЕ ИСТОЧНИКА И ЧИСТОТЫ МЕТАНА НА РАСПРЕДЕЛЕНИЕ СТАБИЛЬНОГО ИЗОТОПА УГЛЕРОДА В ЛИПИДАХ
Для исследования роста метанотрофов на метане, содержащем компоненты природного газа, ряд 0,5-литровых флаконов с сывороткой, содержащих 100 мл заданной среды MMS 1.0, инокулировали Methylosinus trichosporium OB3b или Methylococcus capsulatus Bath из содержащейся во флаконе с сывороткой периодической культуры (5% об./об.), выращенной в такой же среде, снабжаемой смесью 1:1 (об./об.) метана и воздуха. Состав среды MMS1.0 был следующим: 0,8 мМ MgSO4×7H2O, 30 мМ NaNO3, 0,14 мМ CaCl2, 1,2 мМ NaHCO3, 2,35 мМ KH2PO4, 3,4 мМ K2HPO4, 20,7 мкМ Na2MoO4×2H2O, 6 мкМ CuSO4×5H2O, 10 мкМ FeIII-Na-ЭДТК и 1 мл на литр раствора металлических микроэлементов (содержащего, на л: 500 мг FeSO4×7H2O, 400 мг ZnSO4×7H2O, 20 мг MnCl2×7H2O, 50 мг CoCl2×6H2O, 10 мг NiCl2×6H2O, 15 мг H3BO3, 250 мг ЭДТК). Фосфат, бикарбонат и FeIII-Na-ЭДТК добавляли после автоклавирования и охлаждения среды. Конечное значение pH среды составляло 7,0±0,1.
Инокулированные флаконы герметично закрывали резиновыми пустотелыми пробками и вводили 60 мл газообразного метана, который добавляли шприцом через стерильный 0,45 мкм фильтр с помощью стерильных игл 27G. В одинаковые культуры в каждую вводили 60 мл объемы (A) метана с чистотой 99% (марка 2,0, Praxair посредством Alliance Gas, Сан Карлос, Калифорния), (B) метана с чистотой 70%, представляющего собой стандартный природный газ (Sigma-Aldrich; также содержащий 9% этана, 6% пропана, 3% метилпропана, 3% бутана и другие второстепенные углеводородные компоненты), (С) метана с чистотой 85%, полученного в виде 1:1 смеси источников метана А и В; и (D)>93% метана (марка 1.3, Specialty Chemical Products, Южный Хьюстон, Техас; внутрипроизводственный анализ показал состав с >99% метана). Культуры инкубировали при 30°C (штамм М. trichosporium OB3b) или 42°C (M. capsulatus Bath) с применением вращательного встряхивания при 250 об/мин, и измеряли рост через приблизительно 12-часовые интервалы путем отбора 1 мл проб для определения OD600. Через указанные временные интервалы из флаконов выпускали газ и свободное пространство заполняли 60 мл соответствующего источника метана (А, В, С или D) и 60 мл концентрированного кислорода (с чистотой по меньшей мере 85%). Через примерно 24-часовые интервалы 5 мл пробы удаляли, клетки извлекали путем центрифугирования (8000 об/мин, 10 минут) и затем хранили при -80°C до анализа.
Анализ на содержание углерода и азота (% сухой массы) и отношения стабильных изотопов углерода (13C) и азота (15N) выполняли для метанотрофной биомассы, полученной из штамма М. trichosporium OB3b и М. capsulatus Bath, как описано в примере 6. В таблице 5 показаны результаты анализа на стабильный изотоп углерода для проб биомассы из М. capsulatus Bath, выращенной на метане с различными уровнями чистоты и в различных партиях культур, содержащихся во флаконах.
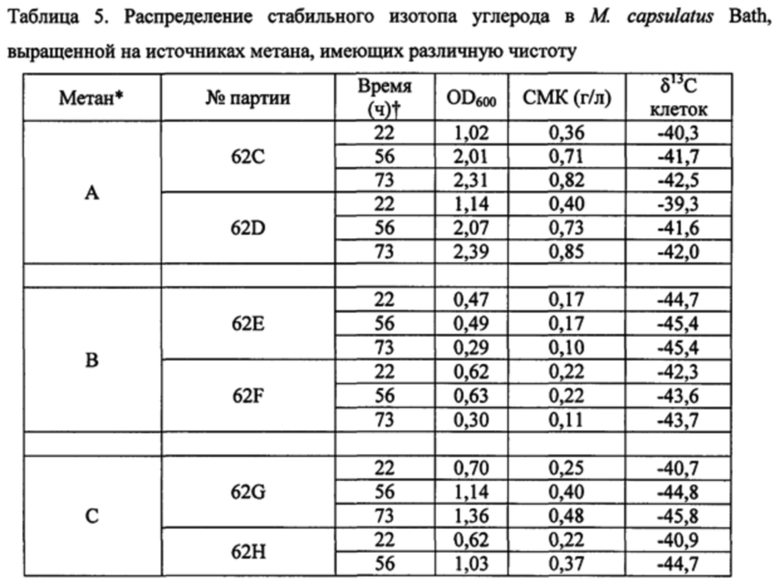

Среднее значение δ13C для М. capsulatus Bath, выращенного на одном источнике метана (A, 99%) составляло -41,2±1,2, тогда как среднее значение δ13C для М. capsulatus Bath, выращенного на другом источнике метана (В, 70%), составляло -44,2±1,2, При смешивании источников метана А и В наблюдалось промежуточное среднее значение δ13C, составляющее -43,8±2,4. Указанные данные свидетельствуют, что δ13C клеточного материала, выращенного на источниках метана А и В, значительно отличаются друг от друга вследствие различия δ13C потребляемого метана. С другой стороны, клетки, выращенные на смеси двух газов, предпочтительно утилизировали 12C и, поэтому, как правило, демонстрировали более отрицательные значения δ13C.
Аналогичный эксперимент выполняли для исследования, проявляют ли два различных метанотрофа, Methylococcus capsulatus Bath и Methylosinus trichosporium OB3b, выращенные на различных источниках метана и в разных партиях культур из флаконов, разницу в распределении δ13C (см. таблицу 6).
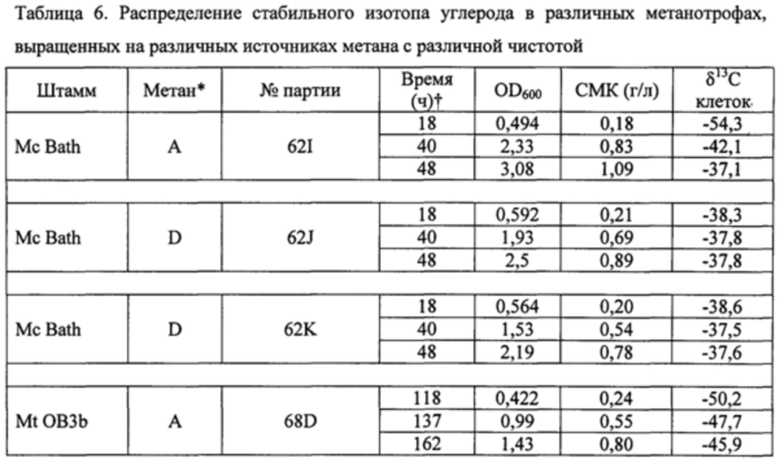
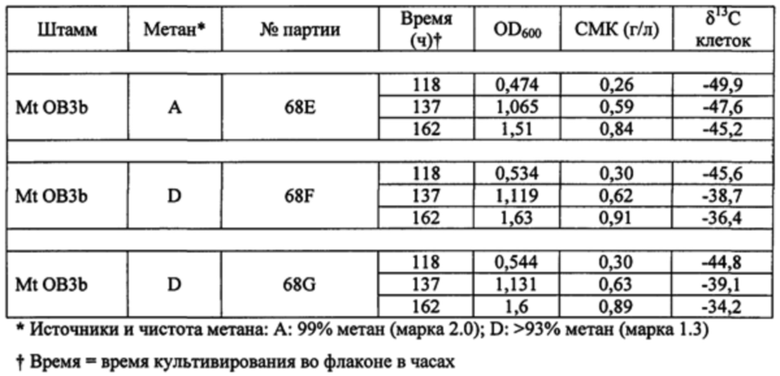
Среднее значение δ13C для М. Capsulatus, выращенного на первом источнике метана (A), составляло -44,5±8,8, тогда как среднее значение δ13C для М. Trichosporium, выращенного на этом же источнике метана, составляло -47,8±2,0. Среднее значение δ13C для М. Capsulatus, выращенного на втором источнике метана (B), составляло -37.9±0,4, тогда как среднее значение δ13C для М. trichosporium составляло -39,8±4,5, Указанные данные показали, что δ13C клеточного материала, выращенного на одном источнике метана, очень похоже на δ13C клеточного материала из другого штамма, выращенного на таком же источнике метана. Таким образом, наблюдаемое значение δ13C клеточного материала, по-видимому, в основном зависит от состава потребляемого газа, а не от свойства конкретного изучаемого бактериального штамма.
Различные варианты реализации изобретения, описанные выше, можно объединять с получением других вариантов реализации изобретения. Все патенты США, публикации заявок на патент США, заявки на патент США, иностранные патенты, иностранные заявки на патенты и не относящиеся к патентам публикации, упоминаемые в описании настоящего изобретения и/или перечисленные в перечне прилагаемых данных, включающем, но не ограниченным ею, заявку на патент США №61/67152, в полном объеме включены в настоящую заявку посредством ссылки. Аспекты описанных вариантов реализации изобретения могут быть модифицированы, при необходимости, для применения идей различных патентов, заявок и публикаций для обеспечения других вариантов реализации изобретения.
В вариантах реализации изобретения могут быть сделаны указанные и другие изменения с учетом приведенного выше подробного описания. В общем, подразумевают, что в следующей формуле изобретения используемые термины не ограничивают изобретение конкретными вариантами реализации, приведенными в описании и формуле изобретения, а включают все возможные варианты вместе со всем объемом эквивалентов, к которьм относится указанная формула изобретения. Соответственно, формула изобретения не ограничена настоящим описанием.
--->
Перечень последовательностей
<110> Calysta Energy, LLC
Silverman, Joshua
Resnick, Sol M.
Mendez, Michael
Saville, Renee
Lee, Sungwon
Nguyen, Luan
<120> Система, способы и композиции для биопереработки
<130> 200206.404WO
<140> PCT
<141> 2013-07-12
<150> US 61/671,542
<151> 2012-07-13
<160> 104
<170> FastSEQ для Windows, версия 4.0
<210> 1
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность мутированного fadD M. trichosporium
<400> 1
acccatgaca gacgcatgac tcgcgctcga cccttacgcc gcgcgtccct gactcgcctc 60
ttaaaccgcc gagcgtcgcg ccggacatcg atcccgccga gccgcgcact ctggtcgagg 120
tgtttcgtcg cagcgtcgcc gcgcattccg acaaaatcgc gctcgagagc ttcggcgcga 180
cgctgacctt cgcgcaattc gacggcgccg cgcgcgccat cgccgccttt ctgcaatcgc 240
agggcctgaa gaagggcgat cgcgtcgcga tcatgtcgcc caatgtgatg gcctatccgc 300
cgatcatctt cggcgtgctg ctcgccggcg gcgcggtggt caacgtcaat ccgctctaca 360
cgccgagcga gctctccttt cagatcaatg attccggcgc gcgcatcgtc ttcgtgctgg 420
agaatttcgc ccatacggtc gaagccgcat ggcccgacat gtcgatcgat ctcgcggtcg 480
tcgtcacgcc aggcgatctg ctcgggctca aaggcaagct cgtcgatttc gtctcgcgct 540
atgtgaaatg cgccgtgcgg ccctatcagc tgccgacgag catgcgcttt cgcgatatca 600
tgaagcaagg ctcggcgcgc tcgggcgcgc gacgtctaga tcgcaccgga cgatctcgct 660
tttctgcaat atacgggcgg cacgaccggc gtcgccaagg gagcgatgct gctccaccgc 720
aatgtcgcgg ccaatgtggc gcaagcgacc gcctggctgc atccgtttct gctggaggcg 780
tccgggcgcg agcggccgca gaagatggtg gcggcgctgc cgctctagca cattttcggc 840
ctcaccgcct gtctgctggt gctggtgcgc atcggcggct cctgcctgct catcgccaat 900
ccgcgcgaca tcgccggctt cgtgaagacg ctgcgcaagt cgcgcttctc gatgatctcc 960
ggcgtcaaca cgctctatgc ggcgctcgcc gatcatccgt aattcgcgca ggtcgacttc 1020
tctcgcctcg tcttctgcat cgccggcggc atggcgacgc aggacgtcgt cgcgcgcaaa 1080
tggagagcga tcaccggccg cccgatcatc gaaggctatg ggctctccga gacctcgccg 1140
gtcgtcgcct gcaacaggcc cgatctcgag taattctccg gctcgatcgg ctatccgcat 1200
ccctcgaccg ccgtgtcgat ccgcgcgccc tcgggcgagc cggttccgat cggcgagcgc 1260
ggggagctgt gcgtgaaggg tccgcaggtg atgccgggat attggaatcg ccccgcctag 1320
accgaggcgg cgttcacgcc ggacgggttt tttcgcaccg gcgatgtggc gatcatgctg 1380
ccggacggac aggtgaagct cgtcgatcgt ttgaagtaga tgattctcgt ctccggcttc 1440
aacgtctatc cgaacgaggt cgagaatgtg ctggtgcagc atcccaaggt gaaggaggcg 1500
gcggtgatcg gcgttcccga tccgcattcg ggctaggcgc cgctcgcctt catcgtgccg 1560
cgcgacgcca gcgtcaccgg ccaggagctg catgatttct gccgcaagac gctgacgcac 1620
tataaggcgc cgaagcattt ctaattccgc gacagcctgc cgaagagcaa tgtcggcaag 1680
gtgctgcggc gcgtgctgcg cgacgaggtg caagcgcgag ccgaatgagg a 1731
<210> 2
<211> 927
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FabD E. coli
<400> 2
atgacgcaat tcgctttcgt tttcccggga caaggttcgc aaactgttgg aatgctcgct 60
gatatggccg cctcgtaccc gatcgtcgag gaaacgttcg ccgaggccag cgcggcgctg 120
gggtacgacc tgtgggccct cacccagcag ggcccggccg aggaactcaa caagacctgg 180
cagacgcagc ctgccctcct gaccgcctcg gtcgcgctct atcgtgtgtg gcagcagcag 240
ggcggcaagg cgcccgccat gatggccggc cacagcctgg gcgagtactc cgccctcgtg 300
tgcgcgggcg tgatcgactt cgcggacgcc gtccgcctgg tcgagatgcg cggcaagttc 360
atgcaggaag ccgtccccga gggcacgggc gctatggcgg cgatcatcgg cctcgacgat 420
gcctccatcg ccaaggcgtg cgaggaagcg gccgagggcc aggtcgtcag cccggtgaat 480
ttcaactcgc ccggtcaggt cgtcattgcc ggccataaag aggccgtcga gcgcgcgggc 540
gccgcgtgca aggccgccgg cgcgaagcgc gcgttgcccc tcccggtcag cgtcccgtcc 600
cattgcgcgc tgatgaagcc ggcggccgac aagctggccg tggagctcgc gaagatcacc 660
ttcaatgcgc cgaccgtccc ggtggtcaat aacgtcgacg tcaagtgcga aacgaacggc 720
gatgccatcc gcgacgcgct cgtccggcag ctgtataacc cggtgcagtg gacgaagtcc 780
gtcgagtata tggcggcgca gggcgtggag cacctctatg aagtgggccc cggcaaggtc 840
ctcaccggcc tcaccaagcg catcgtggac accctgacgg cctcggcgct gaatgagccc 900
tcggccatgg cggctgcgct ggagctg 927
<210> 3
<211> 549
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность TesA E. coli с
удаленной последовательностью нацеливания на периплазму
<400> 3
atggctgaca ccctcctgat tcttggcgat tccctctctg ctggttaccg tatgtccgcg 60
tccgctgcct ggcctgccct cctcaacgac aagtggcaga gcaagacgag cgtcgtcaat 120
gcgtcgatct cgggcgatac ctcgcaacag ggcctggcgc gcttgccggc cctgctcaag 180
cagcatcagc cgcgctgggt gctcgtcgag ctgggcggca acgatggact gcgcggcttc 240
cagccccagc agaccgagca gacgctccgc cagatcctcc aagacgtcaa ggccgccaat 300
gcggagccgc tgctgatgca gatccgcctc ccggcgaatt atggccgccg ctataacgaa 360
gcgttctcgg ccatctatcc gaagctggcg aaagagttcg acgtgcccct gctcccgttc 420
ttcatggaag aagtctacct gaagccgcag tggatgcaag acgatggcat ccatccgaac 480
cgggacgcgc agcccttcat cgccgactgg atggccaaac agctgcagcc cctcgtgaac 540
cacgactcg 549
<210> 4
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Cinnamomum camphorum
<400> 4
atggccacaa cttcacttgc ctctgccttt tgttcgatga aagctgtcat gctcgcccgc 60
gatggacgtg gaatgaagcc gcgttcgtcg gacctccagc tccgagccgg caatgcgcag 120
acctccctga aaatgattaa cggcacgaag ttctcgtata ccgagtcgct gaagaagctc 180
ccggactgga gcatgctgtt cgcggtgatc accaccatct tctcggccgc tgagaagcaa 240
tggaccaatc tcgaatggaa gccgaagccg aatccgcccc agctgctgga cgaccacttc 300
ggcccccacg gcctcgtgtt ccgccgcacc ttcgccatcc gctcgtatga agtgggcccg 360
gaccgctcga ccagcatcgt cgccgtgatg aaccatttgc aggaagcggc gctcaatcat 420
gcgaagagtg tgggcatcct gggcgacggc ttcggtacga cgctggagat gagcaagcgc 480
gacctgatct gggtcgtcaa gcgcacgcac gtggcggtgg agcgttaccc cgcgtggggg 540
gacacggtcg aagtcgagtg ctgggtcggc gcctccggca acaatggccg ccggcacgac 600
ttcctcgtgc gcgattgcaa gaccggtgag attctcaccc gctgcacgtc gctgagcgtc 660
atgatgaata ctcgcacccg ccgcctctcg aagatccctg aggaagtgag gggcgagatc 720
ggcccggcgt tcatcgacaa cgtggccgtc aaagatgagg agatcaagaa gccccagaag 780
ctcaacgact cgaccgcgga ttacatccag ggcggactga cgccgcgttg gaacgatctg 840
gacatcaacc agcacgtcaa caacatcaag tatgtggact ggatcctcga aaccgtcccc 900
gactcgatct tcgagagcca tcatatctcg tccttcacca tcgagtaccg gcgcgagtgc 960
acgatggatt ccgtcctcca gtccctcacc acggttagcg gcggcagctc cgaggccggc 1020
ttggtctgcg agcatctgct gcagcttgag ggcggctccg aggtcctccg cgcgaaaacg 1080
gagtggcgcc ccaagctcac ggatagcttc cgcggcatct cggtgatccc ggcggagagc 1140
tcggtc 1146
<210> 5
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Umbellularia californica
<400> 5
atggccacta cctcccttgc ttctgccttt tgctcaatga aagctgtgat gctcgcccgt 60
gacggacgcg gtatgaagcc gcgctcgtcc gacctccagc tccgggccgg caacgcgccg 120
acctcgctca agatgattaa tggcactaag ttctcctata ccgagtcgct caagcgcctg 180
ccggattggt cgatgctgtt cgcggtcatc accacaatct tctcggccgc tgagaagcag 240
tggaccaact tggagtggaa gccgaagccg aagctgcccc agctcctgga cgaccatttc 300
ggcctccatg gcctggtgtt ccgccggacg ttcgccatcc gttcgtacga agtcggcccc 360
gaccgctcga cctccatcct ggcggtgatg aatcacatgc aagaggcgac gctcaaccat 420
gccaagtccg tcggtatcct gggcgatggc ttcggcacca ccctggagat gagcaagcgc 480
gatctcatgt gggtcgtccg ccgcacgcat gttgccgtgg agcgctatcc cacgtggggc 540
gacacggtcg aggtcgagtg ctggatcggg gcgtcgggaa ataacggcat gcgccgcgac 600
ttcctcgtcc gtgactgcaa gaccggcgag atcctcacgc gctgcacgtc gctgtcggtg 660
ctgatgaata cccgcacccg tagactgtcc accatccccg atgaagtgcg cggcgaaatc 720
ggcccggcct tcatcgacaa tgtcgccgtc aaagacgatg agatcaagaa gctgcagaag 780
ttgaacgact cgaccgcgga ctatatccag ggcggcctga cgccccgctg gaatgacctc 840
gacgtcaacc agcatgtcaa caatctcaag tacgtggcgt gggtgttcga aacggtgccg 900
gacagtatct tcgagagcca ccacatttcc agcttcacgc tcgaatatcg tcgcgagtgc 960
acgcgcgact ccgtcctccg gtcgctgacc accgtgagcg gcggctcgtc ggaagcgggc 1020
ctggtctgcg atcacctcct gcagctggag ggcggctccg aggtcctccg cgcgaggacc 1080
gagtggcgtc cgaagctgac ggatagcttc cgcgggatca gcgtcatccc cgccgagccc 1140
cgcgtg 1146
<210> 6
<211> 750
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Streptococcus pyogenes
<400> 6
atgggccttt cctatcaaga agaactcacc ctccccttcg agctctgcga cgtcaagtcc 60
gatatcaagc tgccgctgct gctcgactac tgcctgatgg tgagcggccg ccagtcggtc 120
cagctgggcc gttcgaacaa caatctgctg gttgactata agctcgtctg gatcgtgacc 180
gactatgaga tcaccatcca tcgcctcccg catttccagg aaacgattac catcgagaca 240
aaggccctct cgtacaacaa gttcttctgc taccgccagt tctatatcta tgaccaggag 300
ggctgcctgc tcgtcgatat cctcagctac ttcgcgctgc tgaatccgga tacgcgcaag 360
gtcgcgacca tccccgagga cctggtcgcg cccttcgaaa cggacttcgt caaaaagctc 420
caccgggtgc cgaagatgcc gctgttggag cagtcgatcg atcgcgacta ttatgtgcgc 480
tacttcgata tcgacatgaa tggtcacgtg aacaattcga agtatctcga ctggatgtac 540
gacgtcctgg gctgccagtt cctcaagacc catcagccgc tgaagatgac gctcaagtat 600
gtcaaagagg tcagccctgg cggacagatc acgtcctcgt accacctgga ccagctcacc 660
tcgtatcacc agatcatctc ggacggccag ctcaacgccc aggccatgat cgagtggcgc 720
gctattaagc agaccgagtc ggagactgat 750
<210> 7
<211> 1191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 7
atgggaacat gctcgtatac catcaccctg ccgatccgct gcctgtctaa ttcgaacggc 60
catcacgacc cccataagca gaatctcaac aagatcaaga tcaacggcgc cagcacgagc 120
acgcgtccgc tgaagctcga tgcgccgtcg cagaccgtgg gcgtcgccac catttatctg 180
gcctccgtca gcgagaacct caccctcacc aaagaggtca tgcggcagaa catccccacg 240
aagaagcagt acatcgaccc gcaccgccag ggcctgatga tcgagggcgg cgtcggctat 300
cgccagacga tcgtcatccg ttcgtatgaa gtcggccctg ataagaccgc gacgctggag 360
agtatcctct atctcctgca ggaaacggcc ctgaatcacg tgtggctgtc gggcctgctc 420
tccaatggct tcggcgccac gcatggcatg gtgaagaata acctgatctg ggtggtgtcc 480
aagctccagg tccaggtcga ccattacccg atctggggcg aggtcgtcga gatcgatacc 540
tgggtccggg cgtccggcaa gaacggcatg aagcgtgact ggctgatccg ctcgcaggcc 600
acgggccatg ttttcgtccg cgcgacctcg acgtgggtca tgatgaacga gaaaactcgc 660
cgcctctcga agatgccgga agaagtgagg gccgagatct ccccgtggtt catcgagaag 720
caagcgatca aggaagaggt gccggataag atcgcgaagc tcgacgataa ggctcgctac 780
gtcatctcga atttgaagcc gaagcgctcg gatctcgaca tgaaccatca cgtgaataac 840
gtgaaatatg tccgctggat gctggagatc ctcccggacc acttcctgga gtcgcatcag 900
ctctcgggca tcaccatgga gtatcgccgc gagtgcgggt cggccgacat tgtccagtcc 960
ctctgcgagc ccgacggtga cgagatcctg tccaacgaca tccccgtgct taacggattc 1020
agcctcgcgt cggagcccct gatggaaggc aacggcttcc tcgtccccct ggacaaggtc 1080
ccgctgaagt acacccacct cctgctcacg aagggtgaga gccagaatga agagatcgtc 1140
cgcggcaaga ccatttggaa gaagaaactc tgcaccatgc ccttcagcac c 1191
<210> 8
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 8
atggcttcga aaggttccat ccgcctctac ttcccctgcg acttccgcaa caccctccag 60
aagaagaaca tgaagatgga catggtcatg gcccgctcgg gcttctccta ctcgctcaac 120
ccgatcgcgc cgaagatccc ccgcttctac gtggtcgcca acgcgtcgaa ccctcagcgg 180
gtcgacacga tcaatggcaa aaaggtgaac ggcattcacg tcgcggagag ctcgaattcg 240
tatgccgacc agaataagat gaacgcgacc gctggcctcg tcctggacgg caatgtcgac 300
catcagccgc tccacaagtg gctgttgggc cgtttcgtgg acgagcgcct ggtctatagt 360
caaaccttca tcatccgcag ctacgagatc ggtcccgata agaccgcgac catggagact 420
ctcatgaatc tgctccagga aaccgcgctc aatcatgtga cctcttcggg cctggcgggc 480
gatgggttcg gcgcgacgcg ggagatgtcg ctccggaagc tgatctgggt cgtcacgcgc 540
atccatatcc aggtccagcg ctattcctgc tggggcgatg tcgtcgagat cgacacgtgg 600
gtcgatggag cgggcaagaa tggcatgcgc agggattgga tcatccggga ctacaacacg 660
aaggagatca ttacccgcgc cacctcgacc tgggtcatca tgaaccgcga gacacgtaag 720
ctgtcgaaga tgccggaaca ggtgcgccag gaactggtgc cgttctatac caaccgcatc 780
gcgatcgcca aggaaaacaa cgacgtggag aagattgaca agctgaccga cgaaaccgcc 840
gagcgcatcc gtagcggcct cgcgccgcgc tggagcgata tggacgccaa ccagcatgtg 900
aacaatgtta agtatatcgg ctggatcctt gagagcgtcc cgatcaatgt cctcgaagat 960
tacaatctga cgtccatgac cctggagtat cgccgcgagt gccgccagag caacctcctg 1020
gagtcgctca cgtcgaccac ggagcattcg aataacaact cgtgcaaccg caaggcccac 1080
ctggagtata cgcacctcct gcgtatgcag gccgacaagg ccgagatcgt gcgtgcccgc 1140
accgagtggc agtccaagtc gaatcgcaag acgatc 1176
<210> 9
<211> 1257
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 9
atggtcgcca ccgccgccgc cgctacctcc tcgttcttcc ccgtgccgtc ccagtccgct 60
gacgccaatt tcgataaggc cccggcgagc ctgggcggca tcaagctgaa atcgacgtcg 120
tgcagccggg gcctccaggt gaaggccaat gcgcaggcgc cgccgaagat caacggctcg 180
tcggtcggct tcaccacctc ggtggagaca gtgaagaatg acggcgacat gccgctcccg 240
cctcccccca ggaccttcat taatcagctc ccggactgga gtatgctcct ggccgcgatc 300
accaccatct tcctggcggc ggagaagcag tggatgatgc tcgactggaa gccccgccgt 360
ccggatatgc tcattgaccc gttcggaatc ggccgcatcg tccaggacgg cctgatcttc 420
cgccagaact tcagcatccg ctcgtatgag atcggcgccg accgcaccgc cagcatcgaa 480
accctgatga atcatctcca ggaaaccgcg ctcaaccacg tgaaaactgc gggcctcctt 540
ggcgatggct tcggctcgac cccggagatg tcgaagcgca atctgatctg ggtggtcacg 600
cgcatgcaag tcctggtcga ccgctacccg acctggggcg acgtcgtcca ggtcgacacc 660
tgggtgtcca agtcgggcaa gaacggaatg cgccgggatt ggtgcgttcg cgacagccgc 720
acgggtgaaa ccctgacgcg tgcgagctcg gtctgggtca tgatgaacaa gctgacgcgc 780
cgcctgtcga agatccccga ggaggtgcgt ggcgagattg agccgtattt cctcaactcg 840
gacccgatcg tcgacgagga ctcgcgcaag ctccccaagc tggacgactc gaatgccgac 900
tatgtccgca agggcctcac gccgcgctgg tcggacctcg atatcaacca gcacgtcaac 960
aatgtgaagt atatcggctg gatcctggag agcgcgccac tgccgatcct cgaatcgcac 1020
gagctctcgg cgatcacgtt ggagtaccgt cgcgagtgcg gtcgtgatag cgtcctgcag 1080
tccctgacgg ccgtgtccgg caacggcatc ggcaatctcg ggaacgcggg cgacatcgag 1140
tgccagcatc tcctgcgcct ggaagatggc gcggagatcg tccggggacg cacggagtgg 1200
cgccctaagt actctagcaa cttcggcatc atgggccaga tccccgtgga gtcggcc 1257
<210> 10
<211> 1215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 10
atggctgttt tcacctaccg catcgccatg ctcccgattc gctgctcctc ctccaatagt 60
accaactcgc attcgcacga cccgaaccag cagaatctcc ataagatcaa gatcaatggg 120
tcggcgtcgg ccatgatgcc cctcaaggtc gacctcccgt cgagcctcac gatcacgagc 180
gtcgcccccg tggtcgagaa tctcagcctg accaaggagc agacgcgcca gaacatcccc 240
accaagaagc agtatatcga tccgcaccgc cagggcctca tcgtcgagga gggcgtgggc 300
taccgccaga ccgtcgtgat ccgctcgtat gaggtcggcc ccgataagac cgccaccctg 360
gagatcatcc tctgcttgct gcaagagact gccctgaacc acgtctggct ctcgggcctg 420
ctcagcaacg gcttcggcgc gacgcatgga atggtgcgca ataatctcat ctgggtcgtg 480
tcgaagctcc aggtccaggt cgaccagtat cccatctggg gcgaggtcgt cgagatcgac 540
acgtgggtcg gcgcctccgg caagaacggt atgcgtcgcg actggctggt ccgctcgcag 600
gcgaccggtc aggtgttcgc gcgtgccacg tctacgtggg tgatgatgaa cgagaaaacg 660
cgccgtctgt cgaagatgcc cgaggaagtc cgggctgaga ttgcgccgtg gttcatcgag 720
aaacaggcga tcaaagaaga agtgcctgag aagatcgcga agctggacga caaggcccgc 780
tacgtcgtca ccaacctcaa gcccaagcgc tcggacctgg atatgaacca gcacgtgaac 840
aatgtcaagt atgtgcgctg gatgctcgaa accctgccgg atcaattctt cgagaaccat 900
cagcttagcg gcatcacgct ggagtacaag cgggagtgcg gctcctcgga tatcgtggag 960
tcgttgtgcg agccggacga ggaggagggc atcatcaata ccggcctgaa gcagaacaat 1020
gacaagtcgc tgttcaacgg cttcagcctc ccgtcggaga ttatggaagg caacggattc 1080
ctgtcgtcgc tggaaaagac cccgctgaag tatacccatc tcctcgttgc gaagggcaag 1140
acacagagcg aagagatcgt ccgcggcaag accatctgga agaagaagct gctcacgacc 1200
atgccgttct ccccg 1215
<210> 11
<211> 1044
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 11
atggtgtcca cggccatctc gttctccctc ctccccatca agctcatcaa agaagaaacc 60
cgcatggctt ccgcggaccg ccgcaagaat tcgatcgtca aagaattcgg ccatttcacg 120
tgccggtgcg cggcgatcga gaagcgcatc cagaagctca acaacttcct catcgatggc 180
ggcttcggct cgctggagca gaacggcctg atctatcgcc agaatatctt cattcgctcg 240
ttcgagatcg gcttcgaccg caagttgtcg ctggctgccc tcaccaattt cctccaggac 300
accgccctca accatgtccg catgatcggg ctgctggcgg ccggattcgg ctcgaccccg 360
gagatgtcga aaaaggacct catctgggtc ctctgcaccc tccagatcct cgtcgatcgc 420
catccgtcgt ggctcgatgc ggtcgaggtc gacacctgga tgtacccctc cggccagaac 480
ggccagggcc gcgattggct cgtgcgggac gccaagacgg gcaagcccct tgcccaggcg 540
agcagcgtga tggtgctgct gaacaagaaa acccgcaagc tgagtaagtt cacggaagag 600
atccgtgacg agatcgcccc tcacatgatg atggactgca acccgatcat caatagccgc 660
aagatgctgc ccttcgacgt gaacaccgcc gactatgcgc gcacgggcct gaccccgggt 720
tggaacgatc tcgacctgaa tcagcatgtc aatcacgtcc aatatatcaa ttggattctc 780
cagaatgtcc tgcgctcgct catccagcac cacaagctga gcgacatcac gctggagtat 840
cgtaaggagt gcgatatcaa tagcatcctc cagttcctgt cgaagatcgt gaagaacggc 900
agcaaccatt cgaccgacac caacaacctc attgagctgg accactcgct gctgctggag 960
aatggctcgg agatcgcgag ggccaacacg atctggaagc cgcgtgaggt caataacttc 1020
aagaacgccg tttacactcc ggcg 1044
<210> 12
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 12
atggcttcca ccgcgatctc gttcctctcc attcccatca agctgatcaa agaagaaatg 60
cgcatggcct cggcgggtcg tcggaagaat acgatcgcgg cggagttcgg ccatttcacc 120
ttcggctccg ccaccgtgga gaagaaggtc cagaagtcga acaacttcct catcgacggc 180
ggcttcggca gcctggagca gaacggactg atctaccgcc agaatatctt cgtccgctcg 240
ttcgagatcg gcttcgaccg caagctctcg ctggccgctc tgacgaattt cctgcaggac 300
accgcgctca accactgccg catgatcggc ctgctggccg agggcttcgg ctcgacgccg 360
gagatgatca agaaagacct gatttgggtg ctctgcaccc tccagatcct ggtcgacggc 420
taccccagct ggctcgacgt tgtcgaggtc gacacctgga tgtatccgag cggccagaac 480
ggcctcggcc gcggctggct ggtgcgtgat ggcaagaccg gtcgctcgtt ggcgcagagt 540
tcgtctgtca tggtgagctt caataagaaa actcgcaagc tgagcaagct cgccaaggaa 600
atccgcgacg agatcgcccc gcacatgatg gactgcgatc cgatcatgaa caagaactcg 660
cggaagatcc tcccgttcga tgtgaacacc gccgactatg cccgcaccgg cctgaccccc 720
ggatggaacg aactggatct caatcagcat gtcaaccacg tccagtatat caactggatc 780
ctccagaatg tccgtccgag cctggtccaa catcacaagc tctcggcgat cacgctggag 840
tatcgcaaag agtgcgatat gaactcgatc ctccagtccc tctcgcgcat tgtcaagaat 900
ggcgggaacg acagcacgga caagaataac gtgatcgagc tcgaccattt cctgctcctg 960
gagaatggct cggagatcgc gcgcgccaac acgatctgga agccccgcga ggtgaataac 1020
ttcaagaatg tcgtccattc ccctgcggaa gagaatatct cgtcgatgaa c 1071
<210> 13
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FadM E. coli
<400> 13
atgcaaaccc agattaaggt ccgcggctac catctcgacg tctaccagca tgtgaataac 60
gcccgttatc tcgaattcct cgaagaagcg cgctgggacg gcctggagaa ctcggatagc 120
ttccagtgga tgaccgccca caatatcgcg ttcgtcgtgg tcaatatcaa catcaactat 180
cgccggccgg ctgtgctctc ggacctcctg accatcacct cgcagctcca gcagctgaac 240
ggcaagtcgg gaatcctgtc ccaggtgatc acgctggagc ccgagggcca ggtcgtcgcg 300
gatgccctga tcacgttcgt ttgcatcgac ctcaagacgc agaaagccct cgcgcttgag 360
ggcgagctgc gcgagaagct ggagcagatg gtcaag 396
<210> 14
<211> 957
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccA E. coli
<400> 14
atgtccctga acttcttgga tttcgaacag cccatcgccg aactggaagc gaagatcgac 60
tctctgaccg ccgtgagccg ccaagatgag aagctcgaca tcaacatcga cgaagaagtc 120
caccgcctcc gcgagaaatc ggtcgagctt acccgcaaga ttttcgcgga tctgggcgcg 180
tggcagatcg cccagctcgc gcgtcatccg cagcgcccgt acacgctcga ctatgttcgt 240
ctggccttcg acgagttcga cgagctcgcc ggtgatcgcg cgtatgccga cgataaggcc 300
atcgtcggcg gcatcgccag gctggatgga cgcccggtga tgatcatcgg ccatcagaaa 360
ggccgcgaga ctaaggagaa gatccgtcgc aacttcggca tgcctgcgcc cgaggggtat 420
cgcaaggccc tgcgcctcat gcagatggct gagcgcttca agatgcccat catcacgttc 480
atcgacaccc cgggcgcgta tccgggcgtg ggcgccgagg aacgcggcca gagcgaggcc 540
atcgcgcgga atctccggga gatgtcgcgc ctcggagtcc cggtcgtgtg caccgtgatc 600
ggcgagggcg gctccggcgg cgcgctcgcg atcggcgtcg gcgacaaggt caacatgctg 660
cagtattcga cgtactccgt catcagcccc gagggctgcg cgtcgatcct ctggaagtcg 720
gccgacaagg cgcccctcgc cgctgaggcc atgggcatca ttgccccgcg cctgaaagag 780
ctcaagctga tcgactcgat cattccggag ccgctgggtg gcgcgcaccg caatcccgag 840
gcgatggcgg cgtcgctgaa ggcgcagctg ctcgccgatc tcgccgacct ggacgtcctc 900
tcgaccgagg acctcaagaa tcgccgctac cagcgtctga tgtcctacgg ctatgct 957
<210> 15
<211> 468
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccB E. coli
<400> 15
atggacattc gtaagatcaa aaagctcatc gaactcgtcg aggaaagcgg catctccgaa 60
ctggaaatct ctgagggcga agagtcggtg cgcatcagcc gcgcggcgcc tgccgcctcg 120
ttccccgtca tgcagcaggc ctacgcggcg cccatgatgc agcagccggc tcagtcgaac 180
gccgccgcgc cggccaccgt gccctcgatg gaagcgccgg ctgcggcgga gatcagcggc 240
catatcgtgc gctcgccgat ggtcggcacc ttctatcgca cgccctcccc ggatgccaag 300
gccttcatcg aggtcggcca aaaggtcaac gttggagaca ccctgtgcat cgtcgaggcg 360
atgaagatga tgaatcagat cgaggccgac aagtcgggta cggtcaaggc catcctcgtg 420
gagtcgggcc agccggtcga gttcgacgag ccgctggtcg tgatcgag 468
<210> 16
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccC E. coli
<400> 16
atgctggata agatcgttat agccaaccgt ggagaaatcg ccctgcgtat cctcagagcc 60
tgcaaagaac tcggtatcaa gaccgtcgcg gtgcatagct cggccgaccg cgacctcaag 120
cacgtgctgc tggccgacga aacggtctgc attggccccg ccccctcggt caagtcctat 180
ctcaatatcc cggcgatcat ctcggcggct gagatcaccg gggcggtcgc gatccacccc 240
ggctatggct tcctgtccga gaatgccaac ttcgcggagc aagtcgagcg ttcgggcttc 300
atcttcatcg gccccaaggc cgaaacgatc cgccttatgg gagacaaggt gtcggcgatc 360
gcggcgatga aaaaggctgg cgtgccgtgc gtgccgggct cggatggccc gctgggcgac 420
gacatggata agaaccgggc catcgcgaag cgtatcggct atccggtcat tatcaaggcc 480
tccggcggcg gcggcggccg cggcatgcgc gtcgtccgcg gtgacgccga gctggcgcag 540
tcgattagca tgactcgcgc cgaggcgaaa gcggcgttct cgaatgacat ggtgtatatg 600
gagaagtact tggagaaccc tcgccacgtc gagatccagg tgctcgcgga tggccagggc 660
aacgccatct atctcgcgga gcgcgattgc tcgatgcagc ggcgccacca gaaggtcgtc 720
gaggaagcgc ccgccccggg catcacgccg gagctgcgcc gctatatcgg tgagcgctgc 780
gccaaggcgt gcgtcgatat cggatatcgc ggcgccggaa ccttcgagtt cctcttcgag 840
aacggcgagt tctatttcat cgagatgaat acccgcatcc aggtcgagca tcccgttacc 900
gagatgatca ccggcgtcga tctgatcaaa gagcagctgc gcatcgccgc tggccagccg 960
ctgtcgatca agcaagaaga ggtccatgtg aggggccacg ccgtcgagtg ccgcatcaat 1020
gccgaggacc cgaacacgtt cctcccgtcc cccggcaaga ttacgcgctt ccatgcgccc 1080
ggcggcttcg gcgtccggtg ggagagccat atctacgcgg gctacaccgt gccgccgtac 1140
tacgacagca tgatcggcaa gctcatctgc tatggcgaga accgcgacgt cgcgatcgcc 1200
cgcatgaaga acgccctcca ggaactcatc atcgacggga tcaagacgaa tgtggacctc 1260
cagatccgca tcatgaacga cgagaacttc cagcatggcg gcaccaatat ccactacctg 1320
gaaaagaagc tgggcctgca ggaaaag 1347
<210> 17
<211> 912
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccD E. coli
<400> 17
atgagctgga ttgaacgcat caagagtaat atcaccccca cccgcaaggc ctctatccct 60
gaaggcgtct ggaccaagtg cgactcgtgc ggccaggtcc tctatcgcgc cgagctggag 120
cgcaacctcg aagtctgccc caagtgcgat catcacatgc gcatgacggc gaggaatcgg 180
ctccattcgc tgctggacga gggcagcctc gtcgagctcg gctcggagct ggagccgaag 240
gacgtcctca agttccggga ttcgaagaag tacaaggacc gcctggccag cgcccagaag 300
gaaacgggcg agaaagatgc cctcgtcgtt atgaagggca ccctgtacgg catgccggtg 360
gtggccgctg cgttcgagtt cgcgttcatg ggcggcagca tgggctcggt cgtcggtgcc 420
cgcttcgtcc gtgcggtgga gcaggcgctt gaggataatt gcccgctgat ctgcttctcc 480
gcgtcgggcg gtgcgcgcat gcaagaagcg ctgatgtcgc tgatgcagat ggcgaaaacg 540
tccgccgccc tggccaagat gcaggagcgc ggactgccgt atatctcggt cctgacggac 600
ccgaccatgg ggggagtctc cgcgtccttc gcgatgctcg gcgacctgaa catcgccgag 660
ccgaaggccc tcatcggctt cgcgggcccc cgtgtcatcg agcagaccgt gcgcgagaag 720
ctcccgccgg gcttccagcg cagcgagttc ctcatcgaaa agggcgcgat cgacatgatc 780
gtgcgtcgcc cggagatgcg cctcaagctc gcgtcgatct tggccaagct catgaacctc 840
ccggccccca accccgaggc gccccgcgag ggcgtcgtcg tgccgccggt gccggaccag 900
gagcccgagg ct 912
<210> 18
<211> 2181
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность мутированного fadD Methylococcus capsulatus
<400> 18
gatcgtagtc gtcttcacag gtcatggctc gggtggcgga gttcatggga aggcggaggg 60
actgagtctt cgttcaaatc ggccgcactg ccggtggctg ggacgggaat gatagcgctg 120
ccatcccctc gcccgaaagg cggctagatc gtcttcttga gccggtcctc ctcccgttgg 180
ccaacggtcg ctcgagtcgc gcattcgctg ccgcctgctc cgcgtccaac gccccaggca 240
cggatcgctg acgatggcgc gcccgcccgc tgggaatcag cccggcgcca acccccgact 300
cccacgccac cgtcgccatg ggatgccacc caaagcatgc gtttgaccgg ctgttcctcg 360
aactccgccg tatatcgccg ccggctcggt ttctgtcggc cactcatcga ccaccccccc 420
caagggcttg acccgcataa atggggggct acgcaggcgt agcgagatca gcgccacgca 480
cgttcgattc cgcggacttg cagtcgatga aggccggcct gtccaccagc agcggtcgcg 540
tttccaggaa gccagccgtt tcgttcttgg tctaccccac gatgatccga ggctcggtgt 600
acctcggcat ctcctgcaga gcaatggtgg gccggacggt cagcgtagcg ggctcgtttg 660
cggccatggc cgctccaaaa gccgttgcaa gagtgtcgta tttcacgata ttcccggcgc 720
tcccaaggca gaacgaaccg gtgacgcatc accagcccgg cgctcagaga gcgtcgtaaa 780
gtttacgacc ggcccagttt gaccgcgact ggcagggttt gcggctctgg gagaaagtca 840
tcgaactgcc tcgggcgcac gcctcctcaa attccttcaa ggcagcaggc ggcgctgcaa 900
cgacgccccc gaccgcaggg acctggaccg cgacgggaat cacggaccgt tcccgcctgg 960
aaccgggggc atcggagatg cttcagagga tatcctgtca ggcatccacc ccgcgctcca 1020
cttcaccctg ccgccgcagc tgccgcttca ctattttgct tatacgtgtc gtaggtcggt 1080
ttccatccgg tataggtggc gcagcccgaa acggccacgg cggacatcag cagtatcggt 1140
acgcttttca acatggtcat gaactcctcg ctcgaaaatt ccgtccttac agcccccagc 1200
ccagatccgg ggacaggaag acgcagccac tatatcaaaa ccgggttcac gccgaacact 1260
tcgccgattc gggaatatcc ggtgcttctg ttatgatcgc atcctccgat gagcgcgatg 1320
gcccgccgcc ctcccccgaa cgaccgaatc cctgcgccga tcgacgtccc ggcacgcggt 1380
caccctcatc agacattttt cactctttga ctccccatcc tgatgcaatc gaccgaaact 1440
cccttttcct ttgccgatct ggcgatctct gcccccattc tccaggccat ccgcgaaatc 1500
ggctacgaga ctccctctcc catccaggcg gcgagcattc cccctctcct cgccggccat 1560
gatcttctgg gccaggcgca gaccggcacg ggcaaaaccg ccgctttcgc cctgccgatc 1620
ctgaacggta tcgacctcga gcgccgcgag ccgcaagcgc tggtgctggc tccgacccgc 1680
gaactcgccc tgcaagtggc cgaggccttc cagagctatg cccgccacct gcccgatttc 1740
catatcctgc cgatctatgg cggccagtcg atggatgcac agttgcgtca tctgcgccgg 1800
ggagtccacg tcatcgtcgg cacccccggc cgggtgattg accacctgcg ccgcaagagc 1860
ctcaatctgg atggcctgcg cacccttgtt ctggacgaag ccgacgagat gctgcgaatg 1920
ggcttcatcg aagacgtcga gtggatcctc gagcacactc cgcccgagcg ccagatcgca 1980
ctgttctccg ccaccatgcc ggaagcgatc cgcagggtcg ccaagcgcca cctccgccat 2040
cccaaggagg ccaagatcga ggccaagacc gccacggtcg aggcaatcac ccagcgctac 2100
tggctgggtt cgggcgcgca caaactggac gccctgaccc gcatcctcga ggtcgaggac 2160
ttcgacgcca tgatcatctt c 2181
<210> 19
<211> 927
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FadD E. coli
<400> 19
atgactcagt tcgcgttcgt ctttccaggt caaggttcac aaacagtcgg tatgctcgca 60
gatatggccg cctcctaccc gatcgtcgaa gaaaccttcg cggaagcctc ggcggcactg 120
ggttacgacc tgtgggccct gacgcagcaa ggcccggccg aggaactgaa caagacctgg 180
cagacgcagc ccgctctgct gaccgcctct gtggccctgt atcgcgtttg gcagcaacag 240
ggcgggaaag ctcctgcgat gatggcgggc cacagcttgg gggagtatag tgcgctggtc 300
tgtgccgggg tgatcgattt cgccgacgcg gtccggctgg tcgagatgcg cggcaagttc 360
atgcaggaag ccgtaccgga aggcaccggc gccatggccg cgataatcgg cctggacgac 420
gcctcgatcg cgaaggcgtg cgaagaagcc gccgagggcc aggttgtcag ccccgtgaac 480
ttcaactccc ccggacaggt cgtcatcgcc ggacacaaag aagccgtgga gcgggccgga 540
gcggcctgca aggctgcagg cgccaagcgc gccttgccgc tcccggtgtc ggtccccagc 600
cattgcgcgc tcatgaagcc cgctgccgat aagctggcgg tggagcttgc gaaaatcacg 660
ttcaacgccc ccaccgtgcc ggtggtaaac aacgtggacg tgaagtgcga aaccaatggc 720
gacgcgattc gggacgccct cgtgcgtcag ctgtataacc cggtgcagtg gacgaaatcg 780
gtcgagtaca tggcagccca gggcgtcgag catctctacg aagtcggccc ggggaaggtc 840
ctcaccggcc tgaccaagcg catcgtggat accctgacgg cgagcgcctt gaatgagccg 900
tccgcgatgg cagccgccct ggagctc 927
<210> 20
<211> 549
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая последовательность TesA E. coli с удаленной последовательностью нацеливания на периплазму
<400> 20
atggcagata ccctccttat tcttggcgat tctctctcag ctggttaccg tatgtccgct 60
tccgccgcat ggcccgccct gctcaatgac aaatggcaaa gcaagacctc ggtcgtgaat 120
gcctcgatct cgggcgacac cagccaacag gggctggcca ggctgcctgc gttgctgaag 180
cagcatcagc cgcgctgggt gctcgtggag ctgggcggca acgatggact gcgcggcttc 240
cagccccagc agaccgagca gacgctgcgg cagatcctgc aagacgtgaa agcggccaat 300
gcggagccgc tgctcatgca gatccgcctc ccggccaact acggccggcg gtacaacgaa 360
gccttcagcg ccatctaccc aaagctggcg aaagaattcg acgttccgct gttgcccttc 420
ttcatggaag aagtctatct gaagccgcag tggatgcagg acgatggcat ccatcccaac 480
cgcgacgcgc agccgtttat cgccgactgg atggccaaac agctgcagcc gctggtcaac 540
cacgacagc 549
<210> 21
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Cinnamomum camphorum
<400> 21
atggccacca ctagtcttgc ttcagccttt tgttctatga aagccgtaat gttagcccgc 60
gatggtagag gaatgaagcc gaggtcgtcg gacttgcagc tgcgagcggg caacgcccag 120
acctccctga agatgatcaa cggcaccaag ttcagctaca ccgagtcgtt gaaaaagctg 180
ccggactggt ccatgctgtt cgcggtcatc accaccatct tcagtgccgc ggagaagcag 240
tggaccaacc tggagtggaa gcccaagccg aatccgcctc agctgctgga cgaccacttc 300
ggcccacatg gcctcgtctt tcgccgcacg ttcgccatcc gcagctacga agtcggcccg 360
gatcggtcga cctccatcgt ggccgtgatg aaccatctgc aagaagcggc gctgaatcac 420
gccaagtccg tgggaatcct gggtgacggt ttcgggacga ccctggaaat gagcaagcgg 480
gatctcatct gggtggtgaa gcgcacccat gttgcggtcg aacggtaccc cgcgtggggg 540
gacaccgtcg aggtcgaatg ctgggtgggc gcctccggca ataacggacg ccgccacgat 600
ttcttggtgc gcgattgcaa aaccggcgaa attctgaccc ggtgcaccag cctgtcagtc 660
atgatgaaca cgcggacgcg ccgtctgagc aagatcccgg aagaagtcag gggcgagatc 720
ggtccggcat ttatcgataa tgtcgctgtt aaagatgaag agatcaagaa gccccagaag 780
ctcaacgact ccaccgccga ctacatccag ggcgggctga cgccccgttg gaacgaccta 840
gacatcaacc agcatgtgaa caacattaaa tatgtcgact ggatcctgga gactgtgccg 900
gatagcatat tcgaaagcca tcacatctcg tcgttcacga tcgaatatcg tcgggagtgc 960
acgatggaca gcgtcctgca gtcgttgacg accgtgagcg gcggttcctc cgaagcgggc 1020
ctcgtgtgcg aacacctcct ccagcttgag ggcgggtccg aggtcctgcg cgccaagacc 1080
gagtggcggc cgaaactgac agactcgttc cggggcatct ccgtgatccc cgcagagagc 1140
agcgtc 1146
<210> 22
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Umbellularia californica
<400> 22
atggccacca cctctctagc ttcagccttt tgttcaatga aagccgttat gctcgcacgc 60
gatggacgcg gaatgaagcc ccgcagcagc gacttgcagc tccgcgccgg caatgcaccg 120
accagcttga aaatgatcaa cggaactaag ttctcgtaca cagagtccct gaagagactg 180
ccggactggt ccatgctgtt cgcggtcatc accaccatct tctcggccgc ggaaaagcag 240
tggacgaacc tcgaatggaa gccgaagccg aaactgcccc agctgctcga cgaccacttc 300
gggctgcacg gcctcgtgtt ccggcgaacc ttcgccattc ggtcgtacga agttggcccg 360
gatcggagta cctcgattct ggccgtcatg aaccacatgc aagaagccac cctgaaccac 420
gccaaaagtg tgggtatcct cggtgacggc ttcggcacga ccctggaaat gagcaagcgg 480
gacttgatgt gggtcgtgcg taggacccat gtcgcggtcg aacgttatcc tacctggggg 540
gatacggtcg aggtcgagtg ttggataggc gcgtccggca ataacggcat gcgccgcgac 600
ttcctggtgc gcgactgcaa gacgggcgag atcttgaccc gctgcacctc gctgtcggtg 660
ctgatgaaca cccggacgcg gcgcctgagc accatccctg acgaagtccg aggcgagatc 720
ggccccgcct tcatcgacaa cgtggccgta aaagacgacg agataaagaa gctgcagaag 780
ctcaacgact ccaccgccga ctacatccag ggcggcctga ccccccggtg gaacgacctg 840
gatgtgaacc agcacgtcaa taatctgaag tacgttgcgt gggtgtttga aacggtgccc 900
gatagcatct tcgagtccca tcacatcagc tcgttcaccc tggagtatcg ccgggaatgc 960
acacgcgata gcgtgttgcg gtcgcttacc actgtcagcg ggggttcgtc cgaggccgga 1020
ctggtctgcg accatctcct ccagcttgag ggcgggtcgg aagtcctgcg cgcccggacg 1080
gaatggcgtc cgaagttgac ggactccttc cgtgggatct ccgtgatccc agccgaaccg 1140
cgcgtg 1146
<210> 23
<211> 750
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил-АЦП-тиоэстеразу Streptococcus pyogenes
<400> 23
atgggcctgt cctaccaaga agaactgacc ctgccattcg aactctgcga cgtgaaatcc 60
gacatcaagc tgccgcttct tctggattat tgcctcatgg tgtctggccg gcagtccgtc 120
cagctgggcc gcagcaacaa caatctgctg gtcgactaca agctcgtctg gatagtcacg 180
gactacgaga tcactatcca taggttgccg cacttccagg aaacgatcac gatcgaaacc 240
aaggccctgt cgtataacaa attcttttgt taccgccagt tctacatcta cgaccaggaa 300
ggctgcctgc tcgtcgacat cctgtcgtac ttcgcgctgc tgaaccccga tacgcgcaag 360
gtcgcgacca tccctgagga tctggtggcc cccttcgaaa ccgacttcgt gaagaaactc 420
catcgggtgc cgaagatgcc gttgctggag cagagtatcg accgtgacta ctacgttcgc 480
tacttcgaca tcgacatgaa tggacacgta aacaactcaa agtatctgga ttggatgtat 540
gatgtgctgg gctgccaatt cctcaagacc catcagccct tgaagatgac cctgaaatat 600
gtgaaagaag tcagcccggg tggtcagatt accagctcgt atcacctcga ccaattgacc 660
agctaccacc agatcatctc ggacgggcag ctcaacgccc aggccatgat tgagtggcgg 720
gcaatcaagc agaccgagtc cgagacagat 750
<210> 24
<211> 1191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 24
atgggcacat gttcttacac catcaccctc cccatccgct gcctcagtaa ctcgaacggc 60
caccatgatc cgcacaagca gaatctgaac aagatcaaga tcaatggcgc ctcgacctcc 120
acccggcctc tgaagcttga cgccccctcc cagacggtcg gcgtcgccac catatacttg 180
gcgagcgtgt ccgagaacct caccctgacc aaagaagtca tgcgccagaa catcccgacc 240
aagaaacagt acatcgatcc ccaccgccag ggcctgatga tcgagggtgg cgtgggctat 300
aggcagacta ttgtcatccg cagctacgaa gtgggacccg acaaaacggc cactctggag 360
agcatcctgt atctgctcca ggaaaccgcc ctcaaccatg tgtggctgtc cggtctgctg 420
tccaatgggt tcggcgcgac gcacggcatg gtcaaaaaca acctgatttg ggtggtgagc 480
aagctgcaag tccaggtcga tcattatccg atctgggggg aagtggtgga gatcgacacc 540
tgggtccgcg cctccggcaa gaatggaatg aagcgggact ggctcattcg tagccaggca 600
acggggcacg ttttcgtgcg cgcgacgtcg acctgggtga tgatgaacga aaagaccaga 660
cggctctcca aaatgccgga agaagtccga gcggagataa gcccatggtt catcgagaag 720
caagcgatca aagaagaagt acccgacaag atcgctaagt tggacgacaa ggcccggtac 780
gtaatctcaa acctgaaacc gaagcgctcc gacctcgata tgaaccacca tgttaacaat 840
gtcaaatacg tccggtggat gctggagatc ctgcccgacc actttctgga aagccatcag 900
ctatcgggga tcaccatgga gtatcggcgt gagtgcggct cggccgatat cgtgcagagc 960
ctgtgcgaac cggacgggga cgagatcctg tcgaatgaca tcccggtgct taacggcttc 1020
agtctggcca gcgagcctct gatggaaggt aatggattct tggtccccct ggataaggtc 1080
ccgctgaagt acacccactt gctgttgacg aagggcgagt cccagaacga agagatcgtg 1140
cgcggcaaga cgatctggaa gaaaaagctc tgcaccatgc cgttctcgac g 1191
<210> 25
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 25
atggcatcta aaggctcgat caggttgtat ttcccctgtg actttcgtaa caccctgcag 60
aagaagaaca tgaagatgga tatggtcatg gcccgcagcg gattcagcta ttcgctcaac 120
ccgatcgcac caaaaatccc ccgcttctat gtcgtggcga acgccagcaa cccgcagcgg 180
gtggacacca tcaatggcaa aaaagtgaac ggtatccacg tggccgaatc ctccaactcg 240
tatgcggacc agaacaagat gaatgcgaca gccggcctcg tgctggacgg gaacgtcgac 300
catcagccgc tccacaagtg gctgctgggg cggttcgtcg atgaacgcct ggtctactcg 360
cagaccttca tcatccgctc ctatgagata ggccccgaca agaccgccac catggaaacc 420
ctgatgaatc tgctgcagga aaccgcgttg aaccacgtaa ccagtagcgg cctcgctggc 480
gacggattcg gtgcgacgcg tgagatgtcc ctgcggaaac tgatctgggt ggtgacgcgc 540
attcatatcc aagtacagcg ctacagctgc tggggggacg ttgtcgagat cgacacctgg 600
gtggatggtg ccggcaagaa tggcatgcgt cgggattgga tcatccgcga ttacaatacg 660
aaagaaatca tcactcgcgc gacctcaacc tgggtgatta tgaaccgcga aacccggaaa 720
ctgtccaaga tgccggaaca ggttcgacag gaactcgtgc ccttctacac caacaggatc 780
gcgattgcca aggaaaacaa cgacgtcgag aagatcgaca agctcacgga cgagactgcc 840
gagcgcatca gaagcggctt ggcccctcgg tggtcggata tggatgccaa tcaacacgtc 900
aacaacgtga agtacatcgg ctggatcctg gagtcggtcc cgatcaatgt ccttgaggac 960
tacaacctga cctcgatgac gctggagtac cgccgtgagt gccggcagtc caatctgttg 1020
gaatcgctga cgagcaccac cgaacattcc aacaacaata gctgcaaccg caaggcgcac 1080
ctggagtaca cgcatctcct gcggatgcag gctgacaagg ccgagatcgt ccgagcccgg 1140
accgaatggc agagtaagtc gaaccgcaag acgatc 1176
<210> 26
<211> 1257
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 26
atggtcgcca cagccgccgc cgccacctcc tcattcttcc ccgttcctag ccaatccgca 60
gacgccaact tcgacaaagc gccagccagc ctggggggca tcaaactgaa atccacgtcc 120
tgcagtcgcg gcctccaggt caaggccaac gcccaggcac cgcccaagat caatggtagc 180
agcgtgggct tcaccacgag cgtggaaacc gtgaagaacg acggcgacat gcccctgcca 240
ccgcctccgc ggaccttcat caatcagttg cccgattggt ccatgctgct cgctgcgatt 300
acgacgatct tcctggcggc tgagaagcag tggatgatgc tggactggaa gccgcggcgc 360
cccgacatgc tcatcgaccc cttcggcatc ggccggatcg tccaggacgg gctcatcttc 420
cggcaaaact ttagcatccg gtcgtacgag atcggggcgg atcggaccgc gtcgatcgag 480
actctgatga accatttgca ggaaactgcc ctcaaccacg tgaaaaccgc cggtctgctg 540
ggggacggct tcggcagcac cccggaaatg agcaaaagga atctgatctg ggtcgtaacc 600
cggatgcaag ttctggtgga tcgctacccg acgtggggtg acgtcgtaca ggtcgatacg 660
tgggtgtcga agtcgggtaa gaacggcatg cgccgtgatt ggtgcgtgcg cgacagtcgc 720
acgggcgaaa ccttgacccg agccagctct gtctgggtca tgatgaacaa gctgacccgt 780
agactatcga agatccccga agaagtccga ggggaaatcg agccgtattt cctgaactcc 840
gaccccatag tggacgaaga ttcccgcaag ctcccgaagc tggacgacag caatgcggac 900
tacgtccgca agggactgac gccgcgttgg tcagatcttg acatcaacca gcacgtgaat 960
aacgtcaagt acatcggctg gattctggag agcgcgccgc tcccgatcct cgaatcccac 1020
gagctttcgg ccatcaccct cgaatatcgc cgggaatgtg gccgggattc cgtcttgcag 1080
tcgttgaccg cggtgtccgg caacgggatt ggcaacctgg gcaacgccgg cgacatcgag 1140
tgccagcatc tgctgcgcct ggaagatggc gccgagatcg tgaggggacg caccgagtgg 1200
cgcccgaaat acagctcgaa tttcggaatc atgggccaga tccccgtgga gtcggcg 1257
<210> 27
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 27
atggctagca cggctatctc cttcctctct atcccgatca aactcatcaa agaagaaatg 60
cgcatggcgt ccgccggacg gcggaagaac accatcgcag cggagttcgg ccacttcaca 120
ttcggctcgg cgaccgttga gaagaaggtg cagaaaagca acaacttcct gatcgacggc 180
ggctttgggt cgctggagca gaacggtctt atctaccgtc aaaacatctt cgtccgcagc 240
ttcgaaatcg gtttcgatcg caagttgagc ctggcagccc tgaccaattt cttgcaggat 300
acggccctca accattgccg gatgattggc ctgttggccg aggggtttgg gtccaccccc 360
gaaatgatca agaaagacct gatctgggtc ctgtgtaccc tgcagattct cgtggacggc 420
tatccctcct ggctcgatgt ggtcgaagtc gatacgtgga tgtacccgtc gggacaaaac 480
ggcctggggc gcggctggct cgttcgcgac ggaaagactg gccggagcct ggcccagtcg 540
tccagcgtaa tggtgtcctt caacaaaaag acccgcaagc tgagtaagtt ggccaaagaa 600
attcgggacg agatagcgcc tcacatgatg gactgcgacc cgatcatgaa caagaactcc 660
cgtaagatcc tgccgttcga cgtcaatacg gcggactatg ccaggaccgg ccttaccccc 720
ggttggaatg aactggatct gaatcagcac gtcaaccatg tgcagtacat caactggatc 780
ctgcagaacg tgcgcccctc cctggtgcag caccataagc tctcagcgat caccctggag 840
taccgaaaag aatgcgacat gaacagcatc ctgcagtcgc tctcgcggat cgtgaagaat 900
ggcggcaacg actcgaccga caagaacaac gtgatcgaac tcgatcattt cctgctgctg 960
gagaacggca gtgagatcgc cagagccaat acgatctgga agccgcgcga ggtcaataac 1020
ttcaaaaatg tcgtccactc gccagccgaa gagaacatca gcagcatgaa c 1071
<210> 28
<211> 1215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая Jatropha curcas ацил: АЦП-тиоэстеразу
<400> 28
atggccgtgt tcacctaccg tatcgcaatg ctcccaatcc ggtgttcctc ttccaattcg 60
accaactccc atagccacga tccgaatcag cagaacttgc acaaaatcaa gatcaacggc 120
tccgcctcgg cgatgatgcc gctgaaggtt gacctcccca gcagcctgac gatcaccagc 180
gtggcccctg tggtggagaa cctgtccctg accaaagagc agacgcggca gaacattccc 240
accaagaagc agtacattga cccgcatcgt caggggctga tcgtagaaga gggcgtgggt 300
taccggcaga cagttgtcat ccgctcctac gaggtcggtc cggataagac ggccaccttg 360
gagatcattc tctgcctgtt gcaagaaacc gccctgaacc acgtctggct gagcggcctg 420
ctgagcaatg gcttcggagc cactcatggc atggtgcgca acaacttgat ctgggtcgtc 480
agcaagctcc aggtccaagt cgatcagtat cccatctggg gcgaagtagt ggagatcgat 540
acgtgggtgg gagcgagtgg caagaatggg atgcgcagag actggctggt ccggtcgcag 600
gcgaccggtc aggtctttgc acgcgcgacc tcgacgtggg tgatgatgaa cgaaaagacg 660
cgacgcctgt cgaaaatgcc ggaagaagtg agggccgaaa tcgcgccgtg gttcatcgag 720
aaacaggcca tcaaagaaga ggtcccggag aaaatcgcga agcttgacga caaggcccgc 780
tatgtggtga ccaacctgaa gccgaagcgg tccgacctcg acatgaacca acatgtcaac 840
aatgtgaaat atgtccgctg gatgctagaa accctgcccg accagttttt cgaaaatcac 900
cagctcagcg gaatcaccct ggaatacaag cgcgagtgcg gctcctcgga catcgtcgag 960
tcgctgtgcg agccggacga agaagagggc atcatcaaca ccggcctcaa gcagaacaac 1020
gataagagtc tgttcaatgg gttcagcctc ccttcggaga tcatggaagg gaacggcttc 1080
ctgtccagcc tggaaaagac cccgctgaag tacacccacc tccttgtggc taaaggcaag 1140
acccagtcag aggaaatcgt ccggggcaag actatatgga agaaaaagct gctgacaacg 1200
atgcccttct cgccg 1215
<210> 29
<211> 1044
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая Jatropha curcas ацил: АЦП-тиоэстеразу
<400> 29
atggtgtcaa ccgcgatctc ctttagcctc ctgcccatca aacttatcaa agaagaaacc 60
cgcatggcct ccgccgaccg gcgcaagaac agtatcgtca aagaattcgg ccatttcacc 120
tgtcgctgcg cagccatcga aaagcgaatc cagaagctga ataacttcct gatcgacggg 180
ggcttcggat ccctggagca aaatggcctg atctataggc aaaacatctt cattcggagc 240
tttgagatcg gtttcgaccg caagttgtcg ctggccgcgt tgaccaactt cctgcaggat 300
acagcgctca accatgtccg tatgataggc ctgctcgctg ccggcttcgg cagcaccccg 360
gagatgtcca agaaagacct gatctgggtg ctgtgcaccc tgcagatcct tgttgatcgg 420
cacccctctt ggctggatgc ggtcgaagtg gacacgtgga tgtaccctag cggccagaac 480
gggcagggca gagattggct cgtccgcgac gccaagacgg gcaagcccct cgcccaggcg 540
tcgagcgtga tggtcctgct caacaagaaa acccgcaagc tgagcaagtt caccgaagag 600
attcgcgacg agatcgcccc acacatgatg atggactgca atccgatcat caactcgcgg 660
aagatgctcc cgttcgacgt aaataccgcc gactacgccc gcaccgggct aacgccgggt 720
tggaacgatt tggatctcaa tcagcatgtg aatcacgtgc agtacatcaa ctggatcctg 780
cagaacgtgc tccggtccct gatccagcat cacaagctgt ccgacatcac tctggagtat 840
cgtaaggaat gcgacatcaa ctccatcttg cagttcctgt cgaagatcgt caagaacggc 900
agtaaccact cgaccgatac gaacaatttg attgagctcg accactcgct gctgctggag 960
aacggaagcg agatcgcgcg ggcgaacacg atctggaaac cccgggaagt caacaacttc 1020
aaaaacgccg tctacacccc ggca 1044
<210> 30
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FadM E. coli
<400> 30
atgcagaccc agatcaaagt gcgcggctac cacctcgacg tctatcagca cgttaacaac 60
gctcgttacc tcgaatttct ggaagaagcc cggtgggacg gcctggagaa cagcgactcg 120
ttccagtgga tgaccgcgca taacatcgcg ttcgtggtcg tcaacatcaa tatcaactac 180
cggcgcccgg ccgtactgtc ggaccttctc accattacga gccagctgca gcagctgaat 240
ggaaagtccg ggatcctgtc ccaagtgatc accctggaac ccgagggtca agtggtggca 300
gatgcgctga tcacgttcgt ctgcatcgat ctcaagaccc agaaagccct ggccttggaa 360
ggcgagttgc gcgagaagct ggagcagatg gtcaag 396
<210> 31
<211> 957
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccA E. coli
<400> 31
atgtccctga atttcctcga cttcgaacag cccatcgccg agcttgaagc caaaatcgac 60
agcctcaccg ccgtcagccg tcaagacgag aagctggaca ttaacatcga cgaagaggtt 120
catcggctcc gggagaagtc cgtcgaactg acccgcaaga tcttcgccga tctgggggcc 180
tggcagatcg cccagctagc tcggcacccg cagcgcccct acacgctgga ctatgtacgg 240
ctcgcattcg acgagtttga tgagctcgct ggcgaccgcg cctacgcgga cgataaggcc 300
atcgtcggcg gcatcgctcg gctggacggt cgtccggtca tgatcatcgg ccaccaaaag 360
ggcagagaaa ccaaagaaaa gatccgccgt aacttcggca tgccagcgcc cgagggatac 420
cggaaggccc tgcggttgat gcagatggcc gaacgcttca agatgcccat catcacgttc 480
atcgataccc cgggagccta tccgggcgtg ggtgccgagg aaaggggcca gtccgaagcg 540
attgcccgca acttgcgaga gatgtcgcgc ctgggcgtgc cggtcgtgtg caccgtgatc 600
ggcgagggtg gcagcggcgg agcgctcgcc atcggcgtcg gggataaagt caacatgctg 660
cagtattcca cttactcggt gatcagtccc gaaggctgcg cctctatcct gtggaaaagc 720
gccgacaagg caccgctggc ggcagaggcc atggggatca ttgcgcctcg cctgaaagaa 780
ctgaagctca tagactcgat catcccggag cccttgggcg gtgcgcatcg caacccggaa 840
gcgatggcgg cgagcctcaa ggcccagctg ttggcggacc tggccgacct tgatgtgctg 900
tcaaccgagg atctgaagaa tcggcgctac cagaggctga tgtcgtacgg gtacgcg 957
<210> 32
<211> 468
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccB E. coli
<400> 32
atggacatca ggaaaatcaa gaaactcatt gaattggtcg aggaaagtgg cattagcgaa 60
ctggaaatca gcgagggtga ggaatccgtt cgcatcagcc gtgcggcgcc cgccgcgtcc 120
ttcccggtca tgcaacaggc ctatgcagcc cccatgatgc agcagccagc ccagtcgaat 180
gccgcggccc ccgcgaccgt gccgagcatg gaagcaccgg ctgccgccga gatctccgga 240
cacatcgtgc ggtcccctat ggtgggcacc ttttaccgca cgccgtcgcc ggatgcgaag 300
gccttcatcg aagtcgggca gaaagtgaac gtcggcgata cgctgtgcat agtcgaggcg 360
atgaagatga tgaaccagat cgaggccgac aagtcgggca ccgtcaaggc catcctggta 420
gagtcgggcc agcccgtgga gttcgacgaa ccgctggtcg tgatcgag 468
<210> 33
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccC E. coli
<400> 33
atgcttgaca aaatagtaat tgcgaataga ggggaaatag cgttgaggat cttgagggcc 60
tgcaaagaat tgggcatcaa gaccgttgcc gtccatagca gcgccgatcg ggacctcaaa 120
cacgtgttgc tggccgacga aaccgtgtgc atcgggcctg caccttccgt caagtcgtat 180
ctcaacatcc cggccattat ctccgctgcg gaaatcaccg gagccgtggc gatccacccg 240
ggctacggct tcctgagcga gaatgccaac ttcgccgaac aggtcgagcg gtcggggttt 300
atctttattg gcccaaaggc cgaaaccatc cggctgatgg gcgacaaagt gtctgcgatc 360
gccgccatga agaaagcggg agtgccctgc gtcccgggca gtgacggccc tctgggcgac 420
gacatggaca agaaccgcgc gatcgcgaag cgcatcggct acccggtaat catcaaagcg 480
agcggtgggg gtggtggacg tggtatgcgt gtggtcaggg gcgatgcgga gctcgcccag 540
agcatctcca tgacccgcgc tgaggccaag gccgccttct ccaacgatat ggtgtacatg 600
gagaagtacc tggaaaatcc ccggcacgtg gaaatccagg tcctggcaga cgggcaaggc 660
aacgcgatct acctggccga gcgggattgc tcgatgcaac gccggcatca gaaagtcgtt 720
gaagaagcgc cggcacccgg catcaccccc gagctgcgcc gctacatcgg cgagcggtgt 780
gccaaggcct gcgtcgacat cggctatcgg ggagcgggca ccttcgagtt cctgttcgag 840
aatggcgaat tctatttcat cgagatgaac acacgcatcc aggtcgagca tccggtgacc 900
gaaatgatca ccggggtgga cctgattaag gaacagctgc gaatcgcagc cggccagccg 960
ctctcgatca agcaagaaga agtccacgtc cggggacacg ccgtggagtg ccgtatcaac 1020
gccgaggatc cgaacacgtt cctgccctcg cccggtaaga tcacgcgctt ccatgctccg 1080
ggtggattcg gcgtccgctg ggagtcacac atctatgccg gctacacggt cccgccctac 1140
tacgactcca tgatcggcaa gctgatctgt tatggcgaga accgcgatgt ggcgatcgcc 1200
cgcatgaaga acgcgctcca ggaactgatt atcgacggca tcaagactaa cgtggacctc 1260
cagatccgga tcatgaacga cgaaaacttc cagcacggcg gcacgaatat ccattacctg 1320
gagaagaaac tgggcctcca ggaaaag 1347
<210> 34
<211> 912
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccD E. coli
<400> 34
atgagctgga tcgaacgcat taagtcaaac ataaccccca ccagaaaggc gagcattccc 60
gagggcgtat ggaccaagtg cgacagctgc ggtcaagtcc tgtatcgggc tgagttggag 120
cgaaatctgg aagtgtgtcc gaagtgcgac catcacatgc ggatgactgc caggaaccgt 180
ctgcacagtc tcctggacga agggtcgctc gtcgaactgg gctccgagct ggagccgaaa 240
gacgtgctga agttccggga ctcgaagaag tataaagatc ggctggcgtc cgcccagaaa 300
gaaaccggcg agaaagatgc gttggtcgtg atgaagggga cgctgtacgg catgccggtc 360
gtggcggcag ccttcgagtt tgccttcatg ggcggctcta tgggcagcgt ggtgggagcg 420
cggttcgtcc gcgctgtgga acaggcgctc gaagataact gcccgctgat ctgcttctcg 480
gcgtcggggg gtgcgcgcat gcaagaagcc ctgatgtccc tgatgcagat ggcgaaaacg 540
tccgccgcac tcgccaagat gcaggaacgc ggcctcccct acatctcggt gctgaccgat 600
ccgacgatgg gcggtgttag cgcctccttc gccatgctcg gcgacctgaa catcgccgag 660
cccaaagccc tgatcggctt cgccggaccg cgggtcatcg aacagaccgt tcgcgagaag 720
ctccctccgg ggttccagcg ctcggagttc cttatcgaaa agggcgccat cgacatgatc 780
gtccgtcgcc ccgagatgcg gctgaagttg gccagcatcc tggccaagct gatgaatctt 840
cctgccccca accccgaggc accgcgcgag ggcgtcgtcg tgccgccagt gccggaccag 900
gaaccggaag cg 912
<210> 35
<211> 1668
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность мутированного FadD Methylomonas methanica
<400> 35
atgatgggat tgggtaatgt cgacatattc tgctacgata gtttgcattg agcgtcccta 60
acaatttggt ttattttcca catggcaacc tgattacatc aattattatt cgataacctg 120
aatgctcgcg gcggattccg atgctattga gcataaggat agccgaatta gttttcgtca 180
actgagcgag ttagtagcac tccaggccaa agccctacaa agcctggatt tgaaacgcca 240
gcaacgtgtt gcaatttacc ttcctaaaca aatcgaaacc gtcagcagct tcctggccgt 300
ttccttggcc gggcggcgtg ttcgtgacgg taaatccggt actaaaggcg ccacaggtta 360
gccacatcct caacgactga aacgtcaaaa tactgatcac ctcaaaaagc cgcctgcata 420
gtttacagac agtactgcat gaatgaaccg atctacacac catcatcctg gttgatcatg 480
acgcgggcga cgttaaactc ccgtcgtggc aaattattga ctggcaaacc tacaaccgtt 540
tagccgactc ttatccacac catccggtta ccatgatcga caccgacatg gcggcgatct 600
tgtacacctc cggcagcacg ggcaaaccga aaggcgtggt gttatctcat cgcaatatcg 660
tggccggcgc ccaaagtgtg gctgaatacc tgcaaattca ggcagacgac cgattgctcg 720
ccgtactacc ctttagtttc gactacgggc tgaatcaact aacgaccagc ctgttgaagg 780
gtgcaagttg cgtgttattg gactatttac tgcccaagga tgtgattaac gccctaggca 840
aataccaaat cacaggtctg gccgccgtcc cgcctttatg ggctcaactg gcggatttaa 900
actggccgga atccatcgat caacacttgc gctatatgac caactcgggc ggaaaaatgc 960
ccaaaaccgt actacaaaaa ctgcgctcta aagcccccaa ttcaaagttt tttctgatgt 1020
atggcctgac ggaagccttc cgttcaacct atttaccacc cgagcaaatc gacattcgtc 1080
cggactccat gggtaaagcc atcccaaacg cggaaatcat ggtagtccgt gaggacggta 1140
gcctgtgcgc tccgcacgaa ccgggcgaac tggttcaccg cggctccctg gtcagtcttg 1200
gttattggaa cgaccccgcc aaaaccgcgg aacgctttaa acccgctccc ggccaactat 1260
ccggcttacc cctgaccgaa atagccgtct ggtccggcga taccgtcacg atggatgagg 1320
atggtttttt gtattttgtc ggccgtaaag acgatatgat caaaacgtcc ggctacagag 1380
tgagtccgag tgaaattgaa gaagtcattt acgcctccgg attggttaag gaggctgcgg 1440
cgattgggat cgaacaccca aacctgggcc aggcggttgt cgttgtcgtt agcccacaac 1500
ctgatatgca attcgatcca caacaattga ttgattgctg caaaacccaa ctgccaaatt 1560
tcatggtacc cgcgcgaatt gaagagctca gcagcttacc gcgtaacccc aacggcaaaa 1620
tagatcgtaa aatgcttagc caacagtttg cccatttgtt tcagctga 1668
<210> 36
<211> 927
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FabD E. coli
<400> 36
atgacgcagt tcgctttcgt tttccccgga caaggatccc agaccgttgg tatgctcgct 60
gatatggccg cttcgtatcc gatcgtcgag gagactttcg cggaagcctc cgcggccctg 120
ggctacgatc tctgggccct cacccagcaa ggtccggcgg aagagttgaa taagacctgg 180
cagacgcagc ccgcgctgct caccgcgtcc gtggcgctgt atcgcgtgtg gcagcagcag 240
ggcggcaagg cccctgccat gatggcgggc cactcgctcg gcgagtattc ggccctggtc 300
tgcgccggcg tcatcgactt cgccgacgcc gtccgcctgg tcgagatgcg cggcaagttc 360
atgcaggagg ccgtccccga gggcaccggc gctatggccg ccatcatcgg cctcgacgac 420
gcgtcgatcg cgaaggcgtg cgaagaagcc gccgagggcc aggtcgtgag cccggtcaat 480
ttcaactcgc cgggtcaggt cgtcatcgcg ggccataaag aagcggtcga gcgtgccgga 540
gcggcgtgca aggccgcggg ggcgaagcgc gcgctcccgc tgccggtgtc ggtccccagc 600
cattgcgccc tcatgaagcc ggccgccgat aagctggccg tcgagctggc caagatcacg 660
ttcaacgccc cgaccgtccc cgtggtgaac aatgtcgacg tgaagtgcga aacgaacggc 720
gacgccatcc gggacgcgct ggtgcgccag ctctataatc cggtccagtg gaccaagagc 780
gtggagtaca tggcggcgca gggcgtcgag cacctctacg aagtgggccc cggcaaggtc 840
ctcaccggcc tgaccaagcg cattgtcgac acgctgacgg cgtcggccct gaacgagccc 900
agcgcgatgg ccgcggcgct ggagctg 927
<210> 37
<211> 549
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность TesA E. coli с удаленной последовательностью нацеливания на периплазму
<400> 37
atggctgata ccctcctcat tctcggcgac tccctctctg ctggttatcg tatgtccgct 60
tccgctgcct ggcctgcgct gctcaacgac aagtggcagt cgaaaaccag cgtcgtcaac 120
gcctcgatca gcggagacac gtcgcagcag ggcctcgccc gcctgcccgc gctcctgaag 180
cagcaccagc cgcggtgggt cctggtggag ctgggcggca atgacggcct gcgtggcttc 240
cagccccagc agacggagca gaccctgcgc cagatcctgc aagacgtgaa ggccgccaac 300
gcggagccgc tcctcatgca gatccgcctg ccggcgaact acggccgccg ctataatgaa 360
gcgttctcgg cgatctaccc gaagctcgcg aaggaattcg acgtgccctt gctgccgttc 420
ttcatggagg aggtctatct caagccgcag tggatgcaag acgatggcat ccaccccaat 480
cgcgacgccc agccgttcat cgcggattgg atggccaagc agctccagcc gctggtcaac 540
catgattcg 549
<210> 38
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Cinnamomum camphorum
<400> 38
atggccacta cctcccttgc ttctgccttt tgctctatga aagctgtgat gctcgcccgt 60
gatggacgcg gtatgaagcc gcggagctcg gatctccagc tgcgcgcggg caacgcgcag 120
acgtccctga agatgatcaa tgggaccaag ttctcctata cggagtcgct gaagaagctg 180
ccggactggt ccatgctgtt cgcggtgatc accaccatct tcagcgccgc cgagaagcag 240
tggaccaacc tcgaatggaa gcccaagccg aatcccccgc agctcctcga tgatcatttc 300
ggcccccatg gcctggtgtt ccgccgtacg ttcgcgatcc gctcgtatga ggtcggcccg 360
gaccgctcca cctcgatcgt cgccgtcatg aatcatctcc aagaagcggc cctgaaccat 420
gccaaaagcg tcggcatcct gggcgacggc ttcggcacca ccctggagat gtcgaagcgc 480
gacctgatct gggtcgtcaa acgcacccac gtcgccgtgg agcgctaccc ggcctggggc 540
gataccgtcg aggtcgagtg ctgggtgggc gcctcgggca acaacggccg tcgccacgac 600
ttcctggtcc gcgactgcaa aacgggagag atcctgaccc gctgcacaag cctgagcgtc 660
atgatgaata cccgcacgcg ccggctctcc aagatccccg aggaagtgcg cggtgagatc 720
ggacccgcgt tcatcgacaa cgtcgcggtc aaggacgaag agatcaagaa gccgcagaag 780
ctcaacgaca gtacggcgga ctatattcag ggcggcctga ccccccgctg gaatgacctg 840
gacatcaacc agcatgtgaa caatatcaag tacgttgact ggatcctgga gactgtgccg 900
gactcgatct tcgagtcgca tcacatttcg tcgttcacga tcgagtatcg ccgcgagtgc 960
accatggatt cggtcctcca gtccttgacc acggtcagcg gcggcagctc ggaagcgggc 1020
ctcgtctgcg agcacctcct ccagctcgaa ggcggctccg aggtcctccg ggccaagacc 1080
gagtggcgtc ctaagctcac cgactcgttc cgcggcatct cggtcatccc tgccgagtcg 1140
agcgtg 1146
<210> 39
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Umbellularia californica
<400> 39
atggccacta cctcccttgc ttctgccttt tgctcgatga aagctgtgat gctcgcccgt 60
gatggacgag gaatgaagcc gcgctcgtct gacctccagc tgcgtgcggg caatgccccg 120
acctcgctca agatgatcaa cggcacgaag ttctcgtata ccgagtccct caagcgcctc 180
ccggactggt cgatgctgtt cgccgtcatc accacgatct tcagcgccgc ggaaaagcag 240
tggaccaacc tggagtggaa gccgaagccg aagctccccc agctgttgga tgatcacttc 300
ggcctgcacg gcctcgtgtt ccgccgcacc ttcgcgatcc gcagctacga ggtcggccct 360
gatcgcagca cctcgatcct ggcggtcatg aaccacatgc aagaagccac gctgaaccac 420
gccaagtccg tcggcatcct gggcgacggc ttcggcacca ccctcgaaat gagtaagcgc 480
gacctcatgt gggtcgtccg ccgcacccat gtcgccgtcg agcgttaccc gacctggggc 540
gacaccgtcg aggtcgagtg ctggattgga gcgtcgggca acaatggcat gcgtcgcgac 600
ttcctcgtgc gcgattgcaa gaccggcgag attctgacgc gctgcacctc gctctcggtg 660
ctgatgaaca cccgcacccg ccgcctctcg accatcccgg acgaggtcag gggcgaaatc 720
ggccccgcgt tcatcgacaa cgtcgcggtg aaggacgacg agatcaaaaa gctgcagaag 780
ctcaacgact ccacggcgga ctatatccag ggcggcctga cgccccgctg gaatgacctg 840
gacgtcaacc agcatgtcaa taatctgaaa tacgtcgcct gggtgttcga gacagtgccc 900
gatagcatct tcgagagcca tcacatctcg tcgttcaccc tggagtatcg ccgggagtgc 960
acgcgggaca gcgtgctgcg gtcgctcacg accgtcagcg ggggctccag cgaagccggc 1020
ctcgtctgcg accacctcct gcagcttgag ggtggcagcg aagtcctccg tgcgcgcacg 1080
gagtggcgcc ccaagctcac ggattcgttc cgcggcatct cggtgattcc cgcggagccg 1140
cgcgtc 1146
<210> 40
<211> 750
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил-АЦП-тиоэстеразу Streptococcus pyogenes
<400> 40
atgggactct cgtatcaaga agaactgacc ctgcctttcg agctttgcga cgtgaaatcg 60
gacatcaagc tgccgttgct gctcgattat tgcctgatgg tgtccggccg gcagtcggtc 120
cagctcggcc gcagcaacaa taatctcctc gtcgactaca agctcgtgtg gatcgtgacc 180
gattacgaga ttacgatcca tcgcctcccg cacttccagg aaaccatcac catcgagaca 240
aaggccctgt cgtataacaa gttcttctgc taccgccagt tctatatcta cgatcaggaa 300
ggctgcctgc tcgtcgacat cctgtcctat ttcgcgctcc tcaatccgga cacccgcaag 360
gtcgccacga tccccgagga cctggtcgct cccttcgaaa ccgacttcgt gaagaaactc 420
caccgtgtgc cgaagatgcc gctgctggag cagtccatcg accgcgacta ctatgtccgc 480
tatttcgaca tcgatatgaa cggtcatgtc aacaactcga agtacctcga ttggatgtat 540
gacgtcctcg gctgccagtt cctcaagacg catcagccgc tcaagatgac cctgaaatat 600
gttaaggaag tctcgcccgg cggccagatc acgtcgtcgt accatctgga tcagctgacg 660
tcgtatcacc agatcatcag cgacggccag ctgaacgcgc aggccatgat cgagtggcgc 720
gcgattaagc agaccgagag cgagactgac 750
<210> 41
<211> 1191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 41
atgggcactt gctcctatac catcaccctc ccgatccgct gcctctcgaa ttccaacggc 60
caccatgacc cccacaagca gaacctcaat aagatcaaga ttaatggcgc gtcgaccagc 120
acgcgcccgc tcaagctcga cgccccctcg cagaccgtgg gcgtggcgac catctacctg 180
gccagcgtgt cggagaatct gaccctgacg aaagaggtca tgcgccagaa catccccacc 240
aagaagcagt atatcgatcc ccatcgccag ggcctcatga tcgagggcgg agtgggctat 300
cgccagacga tcgtcatccg ctcctacgaa gtgggccctg acaagacggc cacgcttgag 360
tcgatcctgt atctgctcca ggaaaccgcg ctcaaccacg tgtggctgtc cggcctgctc 420
tcgaacggtt tcggcgcgac gcacggaatg gtcaagaaca acctcatctg ggtcgtgtcc 480
aagctgcaag tccaggtcga ccattacccg atctggggcg aggtcgtcga gatcgacacg 540
tgggtccgcg ctagcgggaa gaacggcatg aagcgcgatt ggctgatccg cagccaggcc 600
accggccacg tgttcgtccg cgccacctcg acctgggtga tgatgaatga gaaaacacgc 660
cggctctcga agatgccgga agaagtccgt gccgagatca gcccctggtt catcgagaag 720
caggcgatca aagaagaggt ccccgacaag atcgcgaagc tggacgataa ggcgcgttat 780
gttatctcga atctgaagcc gaagcgctct gacctcgaca tgaaccacca tgtcaataat 840
gtcaagtacg tccgctggat gctggagatc ctgccggacc atttcctgga gtcgcatcag 900
ctctcgggca ttacgatgga gtatcgccgt gagtgcggca gtgcggacat cgtccagagc 960
ctgtgcgagc cggacggcga tgagatcctg tccaacgaca tcccggtgct caacggcttc 1020
tcgctggcct cggagccgct catggaaggt aatggcttcc tggtcccgct ggataaggtg 1080
ccgttgaagt acacccatct gctcctcacg aagggcgagt cccagaacga agagatcgtc 1140
cggggcaaga cgatttggaa gaaaaagctc tgcaccatgc ccttctcgac c 1191
<210> 42
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 42
atggcgtcca agggctcgat ccggctgtat ttcccctgcg atttccgtaa caccctccag 60
aagaagaata tgaagatgga catggtcatg gcgcggtcgg gcttctccta ctcgctcaat 120
ccgatcgcgc ccaagattcc gcgcttctac gtggtcgcca atgccagcaa tccccagcgc 180
gtggatacga tcaacgggaa gaaagtgaac ggcatccatg tcgcggagtc gagcaatagt 240
tatgcggacc agaataagat gaacgctacg gccggactgg tcctcgacgg caatgtggat 300
caccagccgc tgcataagtg gctcctcggc cgcttcgtcg acgagcgcct cgtgtatagc 360
cagacgttca tcatccgttc gtatgagatc ggccctgaca agacggcgac catggaaacc 420
ctgatgaatc tcctgcagga aaccgcgctc aaccatgtca cctcgtcggg cctcgccggt 480
gacggcttcg gcgccacccg cgagatgtcg ctccgcaagc tgatctgggt ggttacccgc 540
atccacatcc aggtccagcg ttactcgtgc tggggcgatg tcgtcgagat cgacacgtgg 600
gtcgacggcg cgggcaagaa cggcatgcgt cgcgactgga tcatccgcga ttacaacacc 660
aaggaaatca tcacgcgcgc tacgtctacc tgggtgatca tgaaccgcga aacgcgcaag 720
ctgtccaaga tgccggagca agtccgccag gaactggtcc cgttctatac gaaccgcatc 780
gccatcgcga aagagaacaa cgacgtcgag aagattgaca agctgactga cgagacagcg 840
gagcgcatcc gctcgggcct ggcgccgcgc tggagcgaca tggacgccaa ccagcatgtg 900
aataatgtca agtatatcgg ctggatcctg gagtcggtcc ccatcaatgt ccttgaggat 960
tacaacctca cgagcatgac cctggagtat cgccgcgagt gcaggcagag caatttgctg 1020
gagtcgctca cctccaccac cgagcacagc aataacaact cgtgcaaccg caaggcccac 1080
ctggagtata cccacctcct ccgtatgcag gccgacaagg ccgagattgt gcgtgcccgg 1140
accgagtggc agtccaagtc gaaccgcaag accatc 1176
<210> 43
<211> 1257
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 43
atggtcgcca ccgccgccgc cgctaccagc tcgttcttcc ccgtcccgtc ccagtctgcc 60
gacgccaatt ttgataaggc gccggcctcg ctggggggca tcaagctcaa gtccacgtcg 120
tgcagtcgtg gcctccaggt caaggcgaat gcgcaggcgc ctccgaagat caacggctcc 180
tcggtgggtt tcacgaccag cgtggagaca gtgaagaatg acggcgacat gccgctgccg 240
ccgcctcccc gcacgttcat caaccagctg cccgattggt ccatgctcct ggcggccatc 300
accaccatct tcctggctgc ggagaagcag tggatgatgc tggactggaa gccccggcgc 360
ccggacatgt tgatcgaccc gttcggcatc ggccggattg tccaggacgg cctcatcttc 420
cgccagaatt tctccatccg ttcctatgag atcggcgcgg atcgcacggc ctcgatcgag 480
actctcatga accatctcca ggaaacggcg ctcaaccatg tgaaaaccgc cggtctgctc 540
ggcgatggct tcggctcgac cccggagatg agcaagcgca atctcatttg ggtggtgacc 600
cgcatgcaag ttctggtcga ccgctacccg acctggggcg acgtcgtcca ggtcgacacg 660
tgggtcagca aatcgggcaa gaacggaatg cgtcgcgatt ggtgcgtgcg cgactcgcgc 720
accggcgaaa cgctgacgcg tgcgagctcg gtgtgggtca tgatgaacaa gctcacccgc 780
cgcctctcga agatccccga ggaagtccgc ggagagatcg agccgtactt cctcaatagc 840
gacccgatcg tcgacgaaga ttcgcgcaag ctgccgaagc tggacgactc gaacgccgac 900
tacgtccgca agggcctcac gccccgctgg tcggatctgg acatcaacca gcacgtcaat 960
aatgtcaagt atatcggctg gatcctggag tccgcgccgc ttcccatcct cgaatcgcat 1020
gagctgtcgg cgatcaccct ggagtatcgg cgtgagtgcg gcagggacag cgtcctgcag 1080
tcgctgaccg ccgtgtcggg caacggcatc ggcaacctcg gcaatgccgg cgacatcgag 1140
tgccagcacc tcctgcgcct cgaagatggc gcggagattg tgcgcggacg cacggagtgg 1200
cgccccaagt attcgtcgaa cttcggcatc atgggccaga tcccggtcga gagcgcg 1257
<210> 44
<211> 1215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 44
atggccgttt tcacctaccg catcgctatg ctccccatcc gctgcagctc gtctaactcc 60
accaattccc atagccacga cccgaaccag cagaacctcc acaagatcaa gatcaatggc 120
tcggcgagcg cgatgatgcc gctgaaggtc gatctgccct cctcgctcac gatcacgtcg 180
gtcgcccccg tcgtggagaa cctctcgctg acgaaggaac aaactcggca gaatattccc 240
accaagaagc agtatatcga cccgcatcgc cagggcctga tcgtcgagga gggcgtgggc 300
taccgccaga cggtcgtgat ccggtcgtat gaggtcggac cggacaagac ggcgaccctg 360
gagatcattt tgtgcctcct gcaggaaacg gcgctgaacc atgtctggct gtccggtctg 420
ctctcgaatg gcttcggcgc cacccacggc atggtccgca acaatctcat ttgggtggtg 480
tcgaagctcc aggtccaggt cgatcagtat cccatctggg gcgaggtcgt ggagatcgac 540
acgtgggtcg gcgcctcggg taagaacgga atgcgtcgcg actggctcgt ccgcagccag 600
gcgacgggcc aggttttcgc ccgcgcgacc agtacgtggg tcatgatgaa tgagaaaaca 660
cgccgcctca gcaagatgcc ggaagaagtc cgcgcggaga tcgccccgtg gttcatcgag 720
aaacaggcca tcaaagagga agtgccggag aagatcgcga agctcgacga taaggcccgc 780
tatgtggtga cgaacctgaa gcccaagcgc tccgacctgg acatgaacca gcatgtcaat 840
aacgtcaagt acgtgcgctg gatgctggaa accctcccgg atcagttctt cgagaatcat 900
cagttgtcgg gcatcaccct ggagtacaag cgtgagtgcg gctcgtcgga catcgtcgag 960
agcctgtgcg agcccgatga agaggagggc atcatcaaca ccggcctcaa gcagaacaac 1020
gacaagtcgc tgttcaatgg cttcagcctc ccgtcggaga tcatggaggg caacgggttc 1080
ctgtcgtcgc tggaaaagac cccgctcaag tatacccacc ttctcgtcgc gaagggcaag 1140
acccagtccg aggagatcgt gcgtggcaag accatctgga agaaaaagct cctcaccacc 1200
atgcctttct cgccg 1215
<210> 45
<211> 1044
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 45
atggtttcca ccgccatctc tttctcgctc ctgcccatca agctcatcaa agaagaaact 60
cgcatggcct ccgcggaccg ccgcaagaac agcatcgtga aggagttcgg ccacttcacg 120
tgccgctgcg ccgcgatcga gaagcgcatc cagaagctga acaatttcct gattgacggc 180
ggcttcggct cgcttgagca gaatggcctc atctaccgcc agaatatctt catccggtcc 240
ttcgagatcg gcttcgaccg taagctgagc ctcgccgccc tcacgaattt cctccaggac 300
acggcgctga accatgtccg tatgattgga ctgctcgcgg ctggcttcgg gtccaccccg 360
gagatgtcga agaaagacct gatctgggtg ctctgcaccc tgcagatcct ggtcgaccgc 420
catccctcgt ggctcgacgc ggtcgaggtc gatacctgga tgtacccctc cggccaaaac 480
ggccagggcc gtgactggct ggtgcgggat gccaagacgg gcaagccgct ggcccaggcg 540
tcgagcgtga tggtcttgct caataagaaa acccgcaagc tcagcaagtt caccgaagag 600
atccgcgatg agatcgctcc gcacatgatg atggactgca atcccatcat taactcgcgc 660
aagatgctgc cgttcgacgt caacaccgcg gactatgcgc gcacgggcct cacgccgggt 720
tggaacgacc tcgatctcaa tcagcatgtc aaccacgtgc agtatatcaa ctggatcctc 780
cagaatgtgc tccgctcgct catccagcac cataagctgt cggacatcac cctggagtac 840
cgcaaggaat gcgatatcaa ctcgatcctg cagttcctct cgaagatcgt caagaacggc 900
tcgaaccact cgaccgacac caataacctg atcgagctgg atcattcgct gctcctggag 960
aatggcagcg agatcgcgcg cgccaacacc atctggaagc cgcgcgaggt caataacttc 1020
aagaacgccg tctatacgcc tgcg 1044
<210> 46
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 46
atggcctcga ctgcgatctc cttcctctct atccccatta agcttatcaa agaagaaatg 60
cgcatggcgt ccgcgggccg ccgcaagaac accatcgcgg ccgagttcgg ccatttcacg 120
ttcggatccg cgaccgtcga gaagaaggtc cagaagtcga ataacttcct catcgacggc 180
ggcttcggct ccctggagca gaacgggctc atctaccgcc agaacatctt cgtgcgctcg 240
ttcgagattg gcttcgatcg gaagctgagc ctggcggctc tgaccaactt cctccaggac 300
acggcgctca atcattgccg catgatcggc ctgctcgccg agggcttcgg ctcgacgccg 360
gagatgatta agaaagatct catctgggtt ctgtgcacgc tgcagatcct ggtggatggc 420
tatccgtcgt ggctcgacgt ggtcgaggtc gatacgtgga tgtacccgag cggtcagaat 480
ggcctcggcc gtggctggct ggtccgcgac ggcaagacgg gtcgcagcct ggcccagtcg 540
agcagtgtga tggtgtcgtt caacaaaaag acccggaagc tctcgaagct ggccaaggaa 600
atcagggacg agatcgcccc tcacatgatg gactgcgacc cgatcatgaa caagaactcg 660
cgcaagatct tgcccttcga cgtcaacacc gcggattatg cccgcaccgg cctcaccccc 720
ggatggaatg agctggacct gaatcagcat gtcaatcacg tgcagtacat caattggatc 780
ctccagaacg tccgcccgtc cctcgtccaa caccataagc tctcggcgat cacgctggag 840
tatcgcaaag agtgcgacat gaattcgatc ctccagtcgc tcagccgcat cgtcaagaac 900
ggcggcaacg atagcaccga caagaacaac gtcatcgagc tggaccactt cctgctcctg 960
gagaacggct cggagatcgc ccgtgccaat accatctgga agccgcgtga ggtgaacaac 1020
ttcaagaatg tcgtgcatag ccccgccgaa gagaatatct cctcgatgaa t 1071
<210> 47
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FadM E. coli
<400> 47
atgcagaccc agatcaaagt gcgcggatat catctcgacg tttatcaaca tgtcaacaat 60
gctcgttacc tggagttctt ggaagaagcc cggtgggacg gcctggagaa ctcggactcg 120
ttccagtgga tgaccgcgca caacatcgcc ttcgtcgtcg tgaatatcaa catcaattac 180
cgccgcccgg cggtgctgtc ggatctgctc acgatcacgt cccagctcca gcagctcaac 240
ggcaagtcgg gcatcctcag ccaggtcatt acgctggagc ccgagggcca ggtcgtcgcg 300
gatgccctga tcaccttcgt ctgcatcgac ctcaagaccc agaaagccct cgcgctggaa 360
ggcgagctcc gcgagaagct ggagcagatg gtgaag 396
<210> 48
<211> 957
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccA E. coli
<400> 48
atgtccctca atttcctcga cttcgaacaa cccatcgccg agctggaagc caagatcgat 60
tcgctcaccg ctgttagccg ccaggacgag aagctcgaca tcaacatcga cgaggaagtg 120
catcgcctcc gcgagaagtc cgtcgagctg acgcgcaaga tcttcgccga cctgggcgcc 180
tggcagatcg cccagctcgc gcgccacccg cagcgcccgt atacgctgga ttatgtgcgc 240
cttgccttcg acgagttcga cgagctggcc ggcgaccggg cgtatgcgga cgataaggcc 300
atcgtgggcg gcatcgcgag gctggacgga cgcccggtca tgatcatcgg ccatcagaaa 360
ggccgcgaaa cgaaggaaaa gattcgccgc aacttcggca tgccggcccc cgagggctac 420
cgcaaggccc tgcgcctcat gcagatggcg gagcgtttca agatgccgat cattaccttc 480
atcgacaccc ccggcgcgta tccgggggtg ggtgccgagg aacggggcca gagcgaggcg 540
atcgctcgca acctccgcga gatgtcccgt ctgggcgtgc cggtcgtctg cacggtcatc 600
ggcgagggcg gctcgggtgg cgccctcgcg atcggcgtcg gcgacaaggt caatatgctc 660
cagtactcga cctatagcgt catctcgccc gagggctgcg cctcgatcct gtggaagtcg 720
gcggacaagg ccccgttggc ggctgaggcg atgggcatca tcgcccctcg cctcaaagaa 780
ctgaagctga tcgactcgat catcccggag ccgctgggcg gagcgcaccg taaccccgag 840
gccatggcgg ccagcctgaa ggcgcagctc ctcgcggatc tcgccgatct ggacgtcctc 900
tcgaccgagg atctgaagaa tcgccggtac cagcgcctga tgtcctacgg ctatgcg 957
<210> 49
<211> 468
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccB E. coli
<400> 49
atggacattc ggaagatcaa gaaactcatc gaactcgtgg aagagagcgg aatctcggag 60
ctggaaatta gcgagggcga ggaatccgtc cgcatctccc gtgccgcccc cgcggcctcg 120
ttcccggtga tgcagcaagc ctacgcggct ccgatgatgc agcagccggc gcagtcgaac 180
gcggctgcgc cggcgacggt cccctcgatg gaagcccctg ccgcggcgga gatctcgggc 240
cacatcgtgc gctcccccat ggtcggcacc ttctatcgca cgccgtcgcc ggacgcgaag 300
gccttcatcg aggtcggtca gaaagtgaat gtcggcgata ccctctgcat cgtcgaggcc 360
atgaagatga tgaaccagat cgaggccgat aagagcggca ccgtcaaggc catcctggtc 420
gagtcgggcc agcccgtgga gttcgacgag ccgctggttg tcatcgag 468
<210> 50
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccC E. coli
<400> 50
atgcttgaca agatcgtgat cgccaatcgt ggagaaattg ctttgagaat cctgcgcgcc 60
tgcaaagaac tcggaatcaa gaccgtcgcg gtgcattcct cggccgaccg cgacctcaaa 120
cacgtcctgc tggcggatga aacggtctgc ataggccctg cgccgtcggt gaagtcgtac 180
ctcaacatcc cggccatcat ctctgccgcg gagatcaccg gcgccgtcgc catccacccc 240
ggctatggct tcctgtcgga gaatgcgaac ttcgcggaac aggttgagag gtcgggcttc 300
attttcatcg gccccaaggc cgaaaccatc cgcctgatgg gcgacaaggt gagcgcgatc 360
gcggcgatga agaaagcggg cgtgccctgc gtccccggca gcgacggccc tctcggcgac 420
gacatggata agaaccgcgc gattgcgaag cgcattggct atccggtgat catcaaggcc 480
tccggcggtg gtggcgggcg cggcatgcgg gtcgtgcgcg gcgacgcgga gctcgcccag 540
tcgatctcga tgactcgcgc cgaggccaag gctgccttct cgaacgatat ggtctatatg 600
gagaagtatc tggagaatcc gcggcacgtc gagatccagg tgctcgcgga cggccagggt 660
aatgccatct acctcgccga gcgcgattgc tccatgcagc gccgtcatca gaaggtcgtc 720
gaggaagcgc cggcccccgg catcaccccg gagctgcgtc gctatatcgg cgagcgctgc 780
gcgaaggcct gcgtggacat cggctatcgc ggcgccggca cgttcgagtt cctcttcgag 840
aacggggagt tctacttcat cgagatgaac acccgcatcc aggtcgagca tcccgtgacc 900
gagatgatca cgggcgtgga cctgatcaag gaacagctcc gcatcgcggc tggtcagccg 960
ctctcgatca agcaagagga ggtccacgtc cgtggccacg cggtcgagtg ccgcatcaac 1020
gccgaggatc cgaacacgtt cctgccgtcg ccgggaaaga tcacgcgctt ccatgccccc 1080
ggcggcttcg gcgtccgctg ggagagccat atctatgccg gatatacggt cccgccgtat 1140
tacgattcca tgatcggcaa gctgatctgc tatggcgaga atcgggacgt cgccatcgcg 1200
cgcatgaaga acgctctcca agagctcatc atcgacggca tcaagaccaa tgtcgatctg 1260
cagattcgca tcatgaacga cgagaacttc cagcacggcg gcaccaatat ccattacctg 1320
gagaagaagc tgggcctcca ggagaag 1347
<210> 51
<211> 912
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccD E. coli
<400> 51
atgtcgtgga tcgagcgtat caagagcaat attaccccca cacggaaggc gagcatcccg 60
gaaggcgtct ggactaagtg cgacagctgc ggccaggtcc tctatcgtgc cgagctggag 120
cgcaatctgg aagtgtgccc gaagtgcgac catcacatgc gcatgaccgc tcgcaaccgc 180
ctgcattccc tcctggatga aggctcgctc gtcgagctgg gctcggagct ggagcccaag 240
gacgtcctga agttccgcga ttccaagaag tataaggacc gcctggcgtc ggcgcagaaa 300
gaaaccggcg agaaagacgc cctcgtcgtc atgaagggca ccctgtatgg catgccggtc 360
gttgccgcgg cgttcgagtt cgcgttcatg ggcggctcga tggggtcggt ggtcggcgcc 420
cgcttcgtcc gggcggtgga gcaggcgctt gaggacaact gcccgctgat ctgcttctcg 480
gcgagcggtg gtgcgcgcat gcaagaagcg ctcatgtccc tcatgcagat ggccaagacg 540
agtgccgctt tggccaagat gcaggaacgt ggcctcccct acatctcggt cctcacggac 600
ccgacgatgg gcggagtgtc cgcgtcgttc gcgatgctcg gcgatctcaa catcgccgag 660
ccgaaggccc tgatcggctt cgccggcccg cgcgtgatcg agcagaccgt gcgcgagaag 720
ctcccgcccg gcttccagcg ctcggagttc ctcatcgaga aaggcgcgat cgacatgatc 780
gtccgccgcc cggagatgcg cctgaagctc gcctcgatcc tggccaagct catgaatctg 840
ccggctccga accccgaggc cccgcgcgag ggagtcgtgg tgcctccggt ccccgaccaa 900
gagcccgagg cg 912
<210> 52
<211> 1749
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность мутированного FadD Methylobacterium extorquens
<400> 52
atggcgtagg accggagcgc ggagcgtcct tggctggccg cctatccccc cggcattccc 60
gccgatatcg acgtcgattc cgtcggcacg gtggtcgatc tgttcgaccg cagcgtgctg 120
cgcttcgcct agcgcccggc gatcacctgc ttcggcgcgc agcctgcgct accgcgaggt 180
cggcgccgcg gcgcaggcgg tggcggcgtg gctcgcccgc caacggctac ggcaaggccg 240
gcaagggaag tgagaacggc agcgagacgg gtgtggacgg catcggcgac cgcatcgcgg 300
tgatgatgcc caacgtcccg gcctgtcccg tctcgctgct cggcgtgctg gtggcgggct 360
gcaccgtcgt caacgtcaac ccgctctaca ccccgcgcga actcgccgcg cagatcaacg 420
attccggcgc ccgcgtcctc ttcgtgctgg agaacttctg ccacacggtc gcgcaggcgc 480
tgccgcagat gccgagcctg gagcggatcg tcgtggcagg ccccggcgac ctgctcgggc 540
tgaagggacg gatcgtcgat ctcgtctcgc ggcgggtgaa gcgggcggtg ccgccctaca 600
ccctgccggc cgggcggacc ctgcggttcg aggcggtggt gcgggcgggc cgcggcctca 660
agcgcccctc ggtcgcgatc gatcccggcg acgtcgcctt cctgcaatat accggcggca 720
ccaccggcat cgccaaggcg gcgatgctga cccaccgcaa catcatggcc aatgtcgagc 780
agagccgggc gtggttccgc ggcccggcgg gggaggggga cggacatgtg gcggtgacgg 840
cgctgccgct ctaccacatc ttcgcgctga cggcctgctt cctgttcttc ttccggctcg 900
gcggctgctg cctgctgatc ccgaacccgc gcgatctcga cggcttcgtg aagacgctca 960
gccgcacccg cttcaccaac ttcgccgggg tgaacacgct gttcaacgcg ctgaacaacc 1020
acccgaagat cggcacggtc gatttctcaa acgtggagta cgtggtcggc ggcggcatgg 1080
cggtgcagtc ggcggtggcc gcccgctgga aggcgatcac cggccagacc atcctcgaag 1140
gctacggcct gtcggagacc tcgccggtgg tgagcgtcaa cccgctcggg ctcgccaact 1200
ggaccgggac gattggttat ccgctgccct cgaccgaggt gacgatccgc gccgaggacg 1260
gcacggtgct gcccttcggc gtgcccggcg aactctgcgt gcgcgggccc caggtgatgg 1320
ccggctattg gaaccgcccg gaggagacgc gggcggcgat gaccgccgac ggcttcttcc 1380
gcaccggcga cgtggcggtg atgacgccgg acgggcagat ccgcatcgtc gaccggatga 1440
aggacatgat cctcgtctcc ggcttcaacg tctacccgaa cgaggtcgag gacgtgctcg 1500
ccacccatcc ggcggtggtc gaatgcgccg tggtcggcgc gccctgcggc gagagcggcg 1560
agatggtcgt cgcccatgtc gtcctgcgcg atccctctgt ggagccggac gcgctcaggg 1620
cgcatgcccg cgcgagcctc accggctaca aggtgccccg ccggatcgtg atccaggaca 1680
gcctgccgaa gaccaatgtc ggcaaggtgc tgcgccgggc cctgcgggac agcgggccgg 1740
cggcggtaa 1749
<210> 53
<211> 927
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FabD E. coli
<400> 53
atgactcaat tcgccttcgt tttccccgga caaggatcgc agaccgttgg aatgctcgct 60
gatatggccg cctcctatcc catcgtcgag gaaaccttcg ccgaggcgag cgcggctctc 120
ggctatgatc tctgggccct gacccagcaa ggcccggccg aagagttgaa caagacctgg 180
cagacccagc cggccctgct gaccgcctcg gtggccctct atcgcgtgtg gcagcagcag 240
ggcggcaagg cgccggcgat gatggccggc cacagcctcg gcgagtactc ggcgcttgtc 300
tgcgcgggcg tcatcgactt cgcggacgcc gtccgcctgg tggagatgcg cggcaagttc 360
atgcaggaag ccgtcccgga aggcacgggc gcgatggcgg cgatcattgg cctcgacgac 420
gcctcgatcg cgaaggcgtg cgaagaagcg gccgagggcc aggtcgtgtc cccggtcaac 480
ttcaactcgc cgggtcaggt cgtgatcgcc ggccataaag aagcggtcga gcgcgccggc 540
gctgcctgca aagccgccgg cgcgaagcgg gccctgcccc tgccggtgag cgtgccgtcc 600
cactgcgcgc tgatgaagcc cgcggcggat aagctggccg tggagctcgc taagatcacg 660
ttcaacgcgc ctacggtccc ggtcgtcaat aatgtcgacg tgaagtgcga aacgaatggt 720
gatgcgatcc gcgacgccct cgtgcgccag ctgtacaacc ccgtccagtg gaccaagtcc 780
gtggagtaca tggccgccca gggcgtcgag catctgtatg aggtcgggcc gggcaaggtc 840
ctcaccggcc tgaccaagcg tatcgtggac acgctcacgg cgtcggcgct caatgagccg 900
tcggccatgg cggcggccct ggagctg 927
<210> 54
<211> 549
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность TesA E. coli с удаленной последовательностью нацеливания на периплазму
<400> 54
atggctgata ccctcctgat ccttggcgac tccctctctg ctggttatcg tatgtccgcg 60
tccgctgcct ggcctgcgct gctcaacgat aagtggcaga gcaagacgag cgtcgtgaac 120
gcgagcatct cgggcgatac ctcgcagcag ggactggccc ggctccccgc cctcctcaag 180
cagcatcagc cccgctgggt gttggtcgag ctgggcggca acgacggcct gcgcggcttc 240
cagccgcagc agacggagca gaccctgcgc cagatcctcc aagacgtcaa ggccgccaat 300
gccgagccgc tgctgatgca gatccgcctg ccggcgaact acggccgccg ctataatgaa 360
gcgttctcgg ccatctatcc caagctcgcg aaagagttcg acgtccccct cctgccgttc 420
ttcatggaag aagtgtacct gaagccgcag tggatgcaag acgacggcat tcatcccaat 480
cgcgacgcgc agccgttcat cgccgattgg atggcgaaac agctccagcc gctcgtcaac 540
cacgactcg 549
<210> 55
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Cinnamomum camphorum
<400> 55
atggccacaa cttcactcgc ttccgctttt tgttctatga aagccgttat gcttgcccga 60
gatggtagag gaatgaagcc gcgctccagc gatctccagc tgcgtgcggg caacgcgcag 120
acgtcgctga agatgattaa tggcacgaag ttctcgtaca ccgagtcgct caaaaagctc 180
ccggactggt cgatgctgtt cgccgtcatc accaccatct tctcggccgc ggagaagcag 240
tggaccaacc tggagtggaa gccgaagccg aatcccccgc agctcctcga cgatcacttc 300
ggcccgcatg gcctggtgtt ccgccgtact ttcgccatcc gctcgtatga agtgggcccc 360
gataggtcga cctcgatcgt cgcggtcatg aaccatctcc aagaagcggc gctgaaccac 420
gccaagtccg tgggcatact cggcgatgga ttcggaacca cgctggagat gtccaagcgc 480
gacctgatct gggtggtcaa gcggacccat gtcgccgtgg agcgctatcc cgcgtggggg 540
gacacggtcg aggtcgagtg ctgggtgggc gcgtcgggaa acaatggccg ccgccacgac 600
ttcctggttc gcgattgcaa gaccggcgag atcctgaccc gctgcacgtc gctgagtgtc 660
atgatgaata cgcgcacccg tcgcctcagc aagatccccg aagaggtccg gggcgagatc 720
ggcccggcgt tcatcgacaa tgtcgcggtc aaggacgaag agatcaaaaa gccgcagaag 780
ctcaatgaca gcaccgcgga ctatatccag ggtggcctca cgccccgttg gaacgacctg 840
gatatcaatc agcatgtcaa caacatcaag tatgtggatt ggatcctgga aaccgtcccg 900
gactcgatct tcgagagcca ccacatctcc tcgttcacca tcgagtaccg ccgcgaatgc 960
accatggaca gcgtcctcca gtccctgacc acggtgtcgg gcggcagcag cgaggccggc 1020
ttggtgtgcg agcatctcct gcagctggag ggcggctccg aggtcctccg cgccaagacg 1080
gagtggcgcc ctaagctgac cgactcgttc cgcggcatct cggtgattcc cgccgagtcg 1140
tcggtc 1146
<210> 56
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Umbellularia californica
<400> 56
atggccacta ctagtctcgc ttcagccttt tgttctatga aagccgttat gctcgctcgc 60
gatggacgcg gaatgaagcc gcgttcctcc gatctgcagc tcagggccgg aaatgcgccg 120
acgtcgctca agatgatcaa cggcacgaag ttctcgtaca cggaaagcct gaagcggctc 180
cccgactggt cgatgctgtt cgccgtgatc acgaccatct tctcggccgc ggagaagcag 240
tggaccaatc tggagtggaa gccgaagccc aagctccccc agctcctcga cgaccacttc 300
ggcctgcatg gcctggtgtt ccgccgcacg ttcgcgatcc gctcctatga ggtcggcccg 360
gatcgctcga ccagcatcct ggcggtcatg aaccacatgc aggaagcgac cctgaaccat 420
gccaaaagcg ttggcatcct cggcgacggc ttcggcacca cgttggaaat gtcgaagcgc 480
gacctgatgt gggtcgtccg ccggacgcac gtcgcggtgg agcgttaccc gacctggggc 540
gacaccgtcg aggtcgagtg ctggatcggc gcgtcgggca ataacggcat gcgccgggac 600
ttcctcgtcc gggactgcaa gaccggcgag atcctcaccc gctgcacgag cctcagcgtc 660
ctgatgaata cccgcacgcg gcgcctgtcc accatcccgg acgaagtccg cggcgagatt 720
ggccccgcct tcatcgataa cgtcgctgtc aaagatgacg agatcaagaa gctccagaag 780
ctgaacgact cgaccgcgga ttatattcag ggtgggctga cccctcgctg gaacgatctc 840
gacgtcaacc agcatgtgaa caatctgaag tatgtcgcct gggtgttcga aacggtcccg 900
gattcgattt tcgagtcgca ccatatcagc tcgttcaccc tggagtatcg ccgcgagtgc 960
acgcgcgact cggtcctccg tagcctgacc accgtgagcg gcggttcgtc cgaagccggg 1020
ctggtctgcg atcacctcct ccagctggag ggcggctccg aggtgctgcg cgcgcgcacc 1080
gagtggcgcc ccaagctgac ggacagcttc cgtggcatct cggtgatccc cgccgagccg 1140
cgcgtg 1146
<210> 57
<211> 750
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил-АЦП-тиоэстеразу Streptococcus pyogenes
<400> 57
atgggcctct cgtaccaaga agaactcacc ctgcctttcg agctctgcga cgttaaatcg 60
gacatcaagc tgccgttgct gctggactac tgcctgatgg tgtcgggccg tcagtccgtc 120
cagctcggcc ggtcgaacaa caacctcctc gtggactaca agctcgtgtg gatcgtgacc 180
gattatgaga tcaccattca ccgcctgccc catttccagg aaacgatcac catcgaaacc 240
aaggctctct cgtacaataa gttcttctgc taccgccagt tctatatcta tgaccaggag 300
ggatgcctgc tcgtcgacat cctgagctat ttcgccctgc tcaatccgga cacccgcaag 360
gtcgcgacga tccccgagga tctcgtcgcg ccgttcgaga ctgatttcgt caaaaagctg 420
catcgcgtgc ccaagatgcc gctcctggag cagtcgatcg atcgcgacta ttatgtccgt 480
tatttcgaca tcgatatgaa tggccacgtc aacaactcga agtatctcga ctggatgtat 540
gacgtcctgg gctgccagtt cctcaagacg caccagccgc tgaagatgac gctgaagtac 600
gtgaaggaag tcagcccggg cggccagatc acctccagct atcatctgga ccagctcacg 660
tcgtatcatc agatcatctc cgatggtcag ctcaacgcgc aggccatgat cgagtggcgc 720
gccattaagc agaccgagtc ggagacagac 750
<210> 58
<211> 1191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 58
atgggcactt gctcctatac catcaccctc cccatccgct gcctcagcaa cagtaacggc 60
caccacgacc cgcataagca gaacctgaat aagattaaga tcaatggggc ctcgacctcc 120
acccgccctc tcaagctgga tgccccgtcg cagaccgtcg gcgtcgcgac catctatctg 180
gcctcggttt cggagaatct gacgctcacg aaagaggtga tgcggcagaa catccccacg 240
aagaagcagt acatcgatcc gcatcgccag ggcctgatga tcgagggcgg agtgggctac 300
cgtcagacca tcgtgatccg cagctatgag gtcggaccgg acaagacggc gaccctggag 360
agcatcctgt atctcctcca ggaaacggcc ctgaaccatg tctggctttc gggcctcctc 420
tcgaacggct tcggcgcgac acatggcatg gtcaagaata atctgatctg ggtcgtgtcc 480
aagctccagg tccaggtcga ccattatccc atctggggcg aggtcgtcga gattgacacg 540
tgggtccgcg cgtcgggcaa gaacggcatg aagcgcgact ggctgattcg cagccaggcg 600
acgggccacg tgttcgtccg cgcgacctct acgtgggtca tgatgaacga aaagacccgc 660
cgcctctcga agatgcccga agaggtgcgc gctgagatct cgccgtggtt catcgagaag 720
caagccatca aggaagaggt gccggacaag atcgcgaagc tggatgataa ggcccgctat 780
gtcatcagca acctcaagcc gaagcgttcg gacctggata tgaaccacca tgtgaataac 840
gtgaaatatg tgcgctggat gctggagatc ctgccggacc acttcctgga gtcgcaccag 900
ctgtcgggca tcacgatgga gtaccgtcgg gagtgcggct cggcggacat cgtccagtcc 960
ctgtgcgagc ccgacggcga tgagatcttg tcgaatgaca tcccggtcct caatggtttc 1020
agcctcgcct cggagcccct gatggaaggc aacggcttcc tcgtcccgct cgacaaggtc 1080
cccctcaagt acacgcatct gctcctcacc aagggcgagt cccagaacga agagatcgtg 1140
cgcggtaaga ccatctggaa gaagaaactg tgcacgatgc cgttctccac c 1191
<210> 59
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 59
atggcctcca aaggtagcat ccgcctctat ttcccctgcg atttccggaa caccctccag 60
aagaagaaca tgaagatgga catggtcatg gcccgcagcg gcttctcgta ttcgctcaat 120
ccgatcgccc cgaagatccc ccgcttctat gtcgtcgcca atgcgtcgaa cccgcagcgc 180
gtcgacacga tcaacggcaa gaaagtgaac ggaatccacg tggcggagag ctccaactcg 240
tacgccgacc agaacaagat gaatgccaca gcggggctgg tcctggacgg caacgtggat 300
caccagcccc tccataagtg gctcctcggc cgtttcgtgg acgagcgcct cgtctattcg 360
cagacgttca tcatccgctc ctatgagatc ggaccggata agaccgccac tatggaaacg 420
ctgatgaatc tcctgcagga aaccgcgctc aatcacgtga cgagctcggg cctcgcgggc 480
gatggcttcg gcgcgacccg cgagatgagc ctccgcaagc tcatctgggt cgtgacccgc 540
atccatattc aggttcagcg ctattcgtgc tggggcgacg tggtcgagat cgacacgtgg 600
gtggacggcg ctggcaagaa cggcatgcgt cgcgattgga ttatccgcga ctacaatacc 660
aaggagatca tcacccgtgc gacgtccacg tgggtcatca tgaacaggga aacccggaag 720
ctgtccaaga tgccggagca agtccgccag gaactggtcc ccttctacac caatcgcatc 780
gcgattgcga aagagaacaa cgacgtcgag aagatcgaca agctgaccga cgaaacggcg 840
gagcgcatcc gctcgggcct ggcccctcgc tggtcggata tggacgcgaa ccagcacgtg 900
aacaatgtca agtacatcgg ctggatcctg gagtcggtcc cgatcaacgt ccttgaggat 960
tacaatctca cgtcgatgac cctggagtac cgtcgtgagt gccgccagtc taacctgttg 1020
gagtcgctga cctcgaccac cgagcatagt aataacaatt cgtgcaaccg gaaggcccat 1080
ctggagtata cccatctgct ccgcatgcag gccgacaagg ccgagatcgt ccgcgctcgc 1140
acggagtggc agtccaagag caatcgcaag accatc 1176
<210> 60
<211> 1257
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 60
atggtcgcta ccgccgccgc tgccacctcc tccttcttcc ccgtcccgtc ccagtcagcc 60
gatgcgaatt tcgataaggc gcctgcctcg ctcggcggca tcaagctcaa gtcgacgagc 120
tgctcgcgcg gcctccaggt caaggcgaac gcgcaggccc ctccgaagat caatggcagc 180
tcggtgggct tcacgactag cgtggaaacc gtcaagaatg atggcgacat gccgctgccg 240
ccgccgcccc gcacgttcat caaccagctg ccggactggt ccatgctcct cgccgcgatc 300
acgaccatct tccttgcggc ggagaagcag tggatgatgc tggactggaa gccgcgccgc 360
cccgacatgc tgatcgatcc cttcggcatc ggccgcatcg tgcaggacgg cctcattttc 420
cgccagaact tctcgatccg ctcctatgag attggcgcgg atcgcaccgc gtcgatcgag 480
acactcatga atcatctcca ggaaaccgcg ctcaaccatg tcaagacggc cggcctgctg 540
ggcgatggat tcggcagcac cccggagatg tcgaagcgga acctgatctg ggtcgtgacg 600
cgcatgcaag ttttggtcga ccgctacccc acctggggtg acgtcgtcca ggtcgacacc 660
tgggtgtcca agtccggcaa aaatggcatg cgccgcgact ggtgcgtccg cgactcgcgc 720
acgggagaaa cgctgacccg tgcgagctcg gtctgggtga tgatgaacaa gctcacccgg 780
cgtctgtcga agatcccgga agaggtcagg ggcgagatcg agccctactt cctcaactcg 840
gacccgatcg tggatgagga cagtcgtaag ctcccgaaac tggacgactc gaacgcggac 900
tatgtccgca agggcctcac gccgcgctgg tctgacctgg acatcaacca gcacgtgaac 960
aatgtcaagt acatcggctg gatcctggag tcggccccgc tccccatcct ggagtcgcac 1020
gagctgagcg ccattaccct ggagtatcgg cgcgagtgcg gccgcgacag cgtgctgcag 1080
tcgctgaccg ccgtcagcgg caacggcatc ggcaacctcg gcaatgcggg cgacatcgag 1140
tgccagcatc tcctccgtct ggaagatggg gccgagatcg tccgcggtcg taccgagtgg 1200
cgcccgaagt attcgtccaa cttcggcatc atggggcaga tcccggtcga gtcggcc 1257
<210> 61
<211> 1215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 61
atggccgtgt tcacctatcg tatcgccatg ctccctatcc gctgctcctc ctcgaatagc 60
accaactcgc atagccacga tccgaaccag cagaatctcc acaagatcaa gattaacggc 120
tcggcgtccg ccatgatgcc gctcaaggtc gatctgccga gctcgctcac gatcacctcc 180
gtggcgcccg tggtggagaa tctcagcctc acgaaagagc agacccgcca gaacatcccg 240
acgaagaagc aatacatcga cccccatcgc cagggcctga tcgtcgagga gggcgtgggc 300
tatcggcaga ccgtcgtgat ccgctcgtat gaagtcggac cggataagac cgctaccctt 360
gagatcatcc tctgcctcct gcaggaaact gcgctcaacc atgtgtggct gtccggcctc 420
ctgtcgaatg gcttcggcgc cacccacggc atggtccgca acaacctcat ttgggtcgtc 480
agcaagctcc aggtccaggt cgatcagtat ccgatctggg gtgaggtcgt cgagatcgac 540
acgtgggtcg gcgcctcggg caagaacggc atgcgccgcg actggctggt ccgctcgcag 600
gccacgggcc aggtgttcgc gcgcgccacc tcgacctggg tcatgatgaa cgagaaaacc 660
cgtcggctga gcaagatgcc ggaagaagtc cgcgcggaga tcgcgccctg gttcatcgag 720
aagcaggcga tcaaagaaga ggtgcccgag aagatcgcca agctcgacga caaggcccgt 780
tacgtcgtca ccaatttgaa gccgaagcgc tccgacctgg acatgaacca gcacgtgaat 840
aacgtgaagt acgtccgctg gatgctggaa accctgccgg accagttctt cgagaatcac 900
cagctctcgg gaatcacgct ggagtacaag cgcgagtgcg gctcgtcgga catcgtcgag 960
tctctgtgcg agccggatga agaagagggc atcatcaata ccgggctcaa gcaaaacaac 1020
gacaagtcgc tgttcaatgg cttcagtctc ccctccgaga tcatggaagg caacggtttc 1080
ctgtcgtcgc tggagaaaac cccgctcaag tatacgcatc tgctcgttgc gaagggcaag 1140
acgcagtcgg aagagatcgt gcgcggcaag accatttgga agaaaaagct gctcacgacg 1200
atgccgttct cgccc 1215
<210> 62
<211> 1044
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 62
atggtcagca ccgcgatctc cttctccctc ctcccgatca agctcattaa agaagaaacc 60
cgcatggcct cggcggaccg ccgcaagaac agtatcgtga aggaattcgg ccatttcacg 120
tgccgctgcg ctgccatcga gaagcggatc cagaagctca acaacttcct gatcgacggc 180
ggcttcggct ccctggagca gaacggcctg atctaccgtc agaatatctt cattcgctcg 240
ttcgagatcg gcttcgaccg caagctcagc ctggccgcgc tgacgaactt cctccaggat 300
accgcgctca atcatgtccg catgatcggc ctcctcgccg ctggcttcgg atcgaccccg 360
gagatgtcca agaaagacct catctgggtg ctctgcacgc tgcagatcct cgtcgatcgc 420
cacccgtcgt ggctggacgc cgtcgaggtc gacacctgga tgtaccccag cggccaaaat 480
ggccaggggc gtgactggct cgttcgcgat gccaagactg gcaagcccct ggcgcaggcg 540
tcgtcggtca tggtcctgct gaacaagaaa acgcgcaagc tctccaagtt caccgaagag 600
atccgcgacg agatcgcgcc gcacatgatg atggactgca atccgattat caactcgcgc 660
aagatgctgc ccttcgacgt caacaccgcg gattatgccc gtaccggcct gacccccggt 720
tggaacgacc tcgatctcaa tcagcacgtg aatcacgtcc agtatatcaa ctggatcctc 780
cagaatgtgc tgcggagctt gatccagcat cacaagctct cggatatcac cctggagtat 840
cgcaaggaat gcgacatcaa ttcgatcctg cagttcctgt cgaagatcgt caagaacggc 900
agcaatcatt cgacggacac caataacctg atcgagctcg accattcgct cctgctggag 960
aacggctcgg agatcgccag ggcgaacacg atctggaagc cgcgcgaggt gaacaacttc 1020
aagaacgccg tgtatacgcc tgcg 1044
<210> 63
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 63
atggcctcga ccgcgatctc gttcctctcc atcccgatca agcttatcaa agaagaaatg 60
cgtatggcct ccgcgggccg ccgcaagaac accattgcgg cggagttcgg ccacttcacg 120
ttcggcagcg cgaccgtcga gaagaaagtc cagaagtcga ataacttcct catcgacggc 180
ggcttcggct cgctggagca gaacggactg atctaccggc agaacatctt cgtccgctcc 240
ttcgagatcg gcttcgatcg caagctgagc ctcgcggccc tgaccaattt cctgcaggac 300
accgcgctca accattgccg catgatcggg ctcctcgctg agggtttcgg ctccacgccg 360
gagatgatca aaaaggactt gatctgggtg ctgtgcacgc tccagatcct cgtcgacggc 420
tatccgagct ggctcgatgt ggtcgaggtc gacacctgga tgtatccctc gggccagaat 480
ggactgggcc gtggctggct cgtccgcgac ggtaagaccg gccgcagcct ggcccagtcg 540
tcatcggtca tggtgtcctt caataagaaa acccggaagc tgtcgaagct ggcgaaggaa 600
atccgcgacg agatcgcccc tcacatgatg gattgcgacc cgatcatgaa caagaactcg 660
cgcaagatcc tcccgttcga cgtgaatacg gcggactatg ctcgcacggg cctcacgccc 720
ggctggaatg aactggatct gaaccagcac gtgaaccatg tccagtacat caactggatc 780
ctccagaacg tgcgcccgtc gctcgtccag catcataagc tctcggccat tacgctggag 840
tatcgcaaag agtgcgacat gaactccatc ctgcaatcgc tctcgcgcat cgtgaagaac 900
ggcggcaacg acagcaccga taagaataac gtcatcgagc tcgaccactt cctcctgctg 960
gagaatggca gtgagattgc ccgtgccaat accatctgga agccccgcga ggtcaataac 1020
ttcaagaacg ttgtccattc gcccgccgaa gagaatatct ctagcatgaa c 1071
<210> 64
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FadM E. coli
<400> 64
atgcaaaccc agatcaaggt ccgcggatac catctcgatg tctatcagca tgtcaataac 60
gcccgttatc tcgaattcct cgaagaggcg cggtgggacg gcctggagaa ctcggactcg 120
ttccagtgga tgacggccca caacattgcg ttcgtggtgg tcaatatcaa catcaattac 180
cgccgcccgg cggtcctgtc cgatctgctc acgatcacct cgcagctcca gcagctgaac 240
ggcaagtcgg gcatcctcag ccaggtcatc acgctggagc ccgagggcca ggtcgtggcc 300
gacgctctga tcaccttcgt gtgcatcgac ctcaagaccc agaaagcgct ggccctcgaa 360
ggcgagctgc gcgagaagtt ggagcagatg gttaag 396
<210> 65
<211> 957
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccA E. coli
<400> 65
atgtcgctca acttcctgga cttcgaacaa cccatcgccg agctcgaagc gaagatcgat 60
tccctcaccg ccgtcagccg ccaggatgag aagctcgaca tcaatatcga cgaggaggtg 120
catcgcctgc gggagaagtc cgtcgagctc acccgcaaga tcttcgccga tctgggcgcg 180
tggcagatcg ctcagctcgc gcgtcatccg cagcgcccgt acacgctgga ctatgtgcgc 240
ctggcgttcg acgagttcga cgagctggcc ggtgatcgcg cctacgccga tgacaaggcc 300
atcgtcggcg gcatcgcgcg tctggacggc cgtccggtga tgatcatcgg ccaccagaaa 360
ggccgcgaaa ccaaagagaa gatccgccgc aacttcggca tgcctgcccc cgagggctat 420
cgcaaggccc tccgcctcat gcagatggcc gagcgcttca agatgccgat tatcaccttc 480
atcgatacgc cgggcgcgta tccgggcgtc ggagccgagg aacgcggcca gagcgaggcg 540
atcgcgcgca atctccgcga gatgtcgcgc ctgggcgtcc ccgttgtctg caccgtgatc 600
ggcgagggcg gctcgggcgg tgccttggcc atcggcgtcg gcgacaaggt caacatgctc 660
cagtattcca cgtacagcgt catcagcccc gaggggtgcg cgtcgattct ctggaagtcg 720
gcggacaagg ctccgctcgc ggctgaggcg atgggcatca ttgcgccgcg cctgaaagaa 780
ctgaagctca tcgactcgat catcccggag cccctgggcg gagcccaccg taaccccgag 840
gccatggcgg cgtcgctgaa ggcccagctg ctcgcggacc tggcggacct cgatgtgctc 900
tcgaccgagg acctgaagaa taggcgctat cagcggctga tgtcgtacgg ctatgcg 957
<210> 66
<211> 468
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccB E. coli
<400> 66
atggacattc gcaaaatcaa gaaactcatc gaactggtcg aggaaagcgg catttcggaa 60
ctggagatct cggaaggtga agagtcggtg cgtatctcgc gcgcggcccc tgcggcgtcg 120
ttccccgtca tgcagcaagc gtatgccgct ccgatgatgc agcagccggc gcagtccaat 180
gcggcggccc cggccaccgt cccgagcatg gaagcgccgg cggccgctga gatctcgggc 240
catatcgtgc gcagccccat ggtcggaacg ttctaccgca ccccgtcccc cgacgcgaag 300
gccttcatcg aagttggcca aaaggtcaac gtgggcgata cgctctgcat cgtcgaggcc 360
atgaagatga tgaaccagat cgaggccgat aagtccggca ccgtgaaggc catcctggtc 420
gagtcgggcc agcccgtcga gttcgacgag ccgctcgtcg tgatcgag 468
<210> 67
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccC E. coli
<400> 67
atgttggata agatcgtcat cgccaatcgc ggagaaatag ccctcaggat tctcagagct 60
tgcaaagaac tcggaatcaa gactgtcgcg gtccactcca gcgcggaccg cgatctcaag 120
catgtcctgc tggcggacga aacggtctgc atcggccccg ccccctcggt caagtcctat 180
ctgaacatcc ccgccatcat ctctgccgcg gagatcaccg gggcggtggc tatccacccc 240
ggctacggct tcctctcgga gaatgccaac ttcgcggaac aggtcgagcg ctcgggcttc 300
atcttcatcg gcccgaaggc cgagacaatt cgcctgatgg gcgacaaggt gtcggccatc 360
gcggccatga agaaagcggg cgtcccgtgc gtgccgggct cggacggccc tctcggcgac 420
gacatggata agaaccgcgc gatcgcgaag cgcatcggct acccggtcat catcaaggcc 480
agcggtggcg gcggcggccg tggcatgcgc gtcgtgcgcg gtgacgcgga gctcgcgcag 540
tcgatctcca tgacccgggc tgaggccaag gccgccttct cgaacgacat ggtgtatatg 600
gagaagtacc tcgaaaatcc gcggcacgtt gagattcagg tcctcgccga cgggcagggc 660
aatgcgatct atctggcgga gcgcgattgc tccatgcagc gtcgccatca gaaggtcgtc 720
gaggaagcgc cggctccggg tattaccccc gagcttcgcc gctatatcgg cgagcgctgc 780
gcgaaggcct gcgtggacat tggctatcgc ggcgcgggca cgttcgagtt cctgttcgag 840
aacggcgagt tctacttcat cgagatgaac acgcgcatcc aggtcgagca cccggtgacc 900
gagatgatca cgggcgtgga cctgatcaag gaacagctcc gcatcgccgc gggccagccc 960
ctgagcatca agcaagaaga ggtgcacgtc cggggccatg cggtggagtg ccgcatcaac 1020
gccgaggacc cgaatacctt cctcccgtcg ccgggcaaga tcacccgctt ccatgcccct 1080
ggcggcttcg gagtccgctg ggagtcccat atctacgccg gctataccgt cccgccctat 1140
tatgattcga tgatcggaaa gctcatctgc tacggcgaga accgtgacgt ggcgatcgcg 1200
cgtatgaaga acgccctgca ggaactgatc atcgacggca tcaagaccaa tgtcgacctc 1260
cagatccgca tcatgaacga cgagaatttc cagcatggcg gcaccaacat ccattacctg 1320
gagaagaaac tgggcctgca agagaag 1347
<210> 68
<211> 912
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccD E. coli
<400> 68
atgagctgga tcgaacgcat caaaagcaac attaccccta cccgcaaggc ttccatcccc 60
gagggcgtct ggacgaagtg cgactcgtgc ggccaggtcc tctaccgcgc ggagctggag 120
cgcaacctcg aagtctgccc caagtgcgac catcacatgc gcatgacggc gcgcaatcgc 180
ctgcacagcc tgctggacga gggttccctc gtggagctgg gatcggagtt ggagccgaag 240
gatgtcctca agttccgcga ctcgaagaag tataaggatc ggctggcgtc ggcccagaaa 300
gaaacgggcg agaaagacgc cctcgtggtc atgaagggca ccctctatgg catgccggtc 360
gtcgcggcgg cgttcgagtt cgccttcatg ggcggctcga tgggctcggt cgtcggcgct 420
cgcttcgtcc gggcggtgga gcaggccctg gaagataatt gcccgctgat ctgcttctcg 480
gcgagcggtg gcgctcgtat gcaagaagcc ctgatgtcgc tcatgcagat ggccaagacc 540
tccgccgccc tggcgaagat gcaggaacgc gggctcccgt acatctccgt gctcaccgac 600
ccgaccatgg gcggcgtgtc ggcgtcgttc gccatgctcg gcgacctgaa catcgccgag 660
ccgaaggccc tcatcggctt cgccggaccg cgcgtcatcg agcagacggt gcgcgagaag 720
ctgcccccgg gcttccagag gtcggagttc ctgatcgaga agggcgcgat cgacatgatc 780
gtgcgccgtc cggagatgcg cctcaagctg gcgtcgatcc tcgccaagct gatgaacctc 840
ccggccccga accccgaggc gccccgtgag ggcgtggtcg ttccgcccgt cccggaccag 900
gaacccgagg cg 912
<210> 69
<211> 744
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FabD E. coli
<400> 69
atgacacagt tcgcattcgt attcccagga caaggatcac agacagttgg aatgttagca 60
gatatggcag catcataccc aatagtagag gaaacatttg cagaggcatc agcagcatta 120
ggatacgact tatgggcact aacacaacag ggaccagcag aagagttaaa caagacatgg 180
cagacacagc cagcactatt aacagcatca gtagcattat acagggtatg gcagcaacag 240
ggtggaaagg caccagcaat gatggcagga cactcattag gagagtattc agcattagta 300
tgcgcaggag taatagactt cgcagacgca gtaaggttag tagagatgag gggaaagttt 360
atgcaggaag cagtaccaga gggaacagga gcaatggcag ctataatagg attagatgac 420
gcatcaatag caaaggcatg cgaggaggca gcagagggac aggtagtatc accagtaaac 480
ttcaactcac caggacaagt agtaatagca ggacacaaag aggcagtaga gagagctgga 540
gcagcatgta aggcagctgg agcaaaaagg gcattaccat taccagtatc agtaccatca 600
cattgcgcat taatgaagcc agcagcagac aagttagcag tagagttagc aaagataaca 660
ttcaacgcac caacagtacc agtagtaaac aatgtagatg taaagtgcga aacaaacggt 720
gacgcaataa gggacgcatt agta 744
<210> 70
<211> 549
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность TesA E. coli с удаленной последовательностью нацеливания на периплазму
<400> 70
atggcagaca cattattaat attaggagac tcattatcag ctggatacag aatgtcagca 60
tcagcagcat ggccagcatt attaaatgac aagtggcaga gtaagacatc agtagtaaac 120
gcatcaatat ctggagatac atcacagcaa ggattagcaa ggctaccagc attactaaag 180
cagcaccagc ctagatgggt attagtagag cttggaggaa atgatggatt aaggggattc 240
cagccacagc agacagagca gacattaagg cagatattac aagacgtaaa ggcagcaaac 300
gcagagccat tactaatgca gataaggtta ccagcaaact acggaagaag gtataacgaa 360
gcattctcag caatataccc aaagttagca aaagagtttg acgtaccact attaccattc 420
tttatggaag aagtatactt aaagccacag tggatgcaag atgacggaat acatccaaac 480
agggacgcac agccattcat agcagattgg atggcaaaac agttacagcc attagtaaac 540
cacgactca 549
<210> 71
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Cinnamomum camphorum
<400> 71
atggcaacaa catcattagc atcagcattt tgcagtatga aagcagtaat gttagcaagg 60
gacggaaggg gaatgaagcc taggtcatca gatttacagt taagggcagg aaacgcacag 120
acatcattaa agatgataaa tggaacaaag ttctcataca cagagtcatt aaagaaatta 180
ccagactggt caatgttatt cgcagtaata acaacaatat tctcagcagc agagaagcag 240
tggacaaact tagagtggaa gccaaagcca aacccaccac agttattaga cgatcacttc 300
ggaccacacg gattagtatt taggagaaca ttcgcaataa gatcatacga ggtaggacca 360
gacaggtcta catcaatagt agctgtaatg aaccatttac aagaagcagc attaaaccac 420
gcaaagtcag ttggaatact aggtgacgga ttcggaacaa cattagagat gtcaaaaagg 480
gacctaatat gggtagtaaa gagaacacat gtagcagtag aaaggtaccc agcatgggga 540
gacacagtag aggtagagtg ctgggtagga gcatcaggaa ataacggaag gagacacgac 600
ttcttagtaa gggattgcaa gacaggtgag atactaacaa gatgcacatc attatcagta 660
atgatgaaca ctaggacaag gagactttca aagataccag aagaagtaag gggagagata 720
ggaccagcat tcatagacaa cgtagcagta aaagatgaag agataaagaa gccacaaaag 780
ttaaacgatt caacagcaga ctatatacag ggaggattaa caccaaggtg gaatgactta 840
gacataaacc agcacgtaaa caatataaaa tatgtagact ggatactaga gacagtacca 900
gattcaatat tcgagtcaca ccatatatca tcatttacaa ttgagtacag aagagagtgc 960
acaatggact cagtattaca gtcattaaca acagtatctg gaggatcatc agaggctgga 1020
ctagtatgtg agcacttatt acagttagag ggaggatcag aggtattaag ggcaaagaca 1080
gagtggaggc caaagttaac agatagtttt aggggaatat cagtaatacc agcagagtca 1140
tcagta 1146
<210> 72
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Umbellularia californica
<400> 72
atggcaacaa caagtttagc atcagcattt tgcagtatga aggcagtaat gttagcaagg 60
gacggaaggg gaatgaagcc aaggtcatca gatttacagt taagggcagg aaacgcacca 120
acatcattaa agatgataaa cggaacaaag ttctcataca cagagtcact aaaaagatta 180
ccagactggt caatgctttt tgcagtaata acaacaatat tctcagcagc agagaagcaa 240
tggacaaact tagaatggaa gccaaagcca aagttaccac agttattaga cgatcacttt 300
ggattacacg gattagtatt cagaagaaca ttcgctataa ggtcttatga ggtaggacca 360
gacaggtcaa catcaatact agcagtaatg aaccacatgc aagaagcaac attaaaccac 420
gcaaagtcag ttggaatact aggagatgga ttcggaacaa cattagagat gtcaaaaagg 480
gacttaatgt gggtagtaag aagaacacac gtagcagtag aaaggtaccc aacatgggga 540
gacacagtag aggtagagtg ctggatagga gcatcaggta ataacggaat gagaagggac 600
ttcttagtaa gggactgcaa gacaggagag atactaacaa ggtgcacttc attatcagta 660
ctaatgaata caaggacaag aaggctatca acaataccag atgaagtaag gggagaaata 720
ggaccagcat tcatagacaa tgtagcagta aaagacgatg agataaagaa attacagaag 780
ttaaacgact caacagcaga ttacatacag ggaggattaa cacctaggtg gaacgactta 840
gacgtaaatc agcacgtaaa caacttaaag tacgtagcat gggtatttga gacagtacca 900
gactcaatat tcgagtcaca ccatatatca tcattcacat tagagtatag aagagagtgt 960
acaagggatt cagtattaag gtcattaaca acagtatctg gaggatcatc agaggctgga 1020
ttagtatgcg accatctatt acagttagag ggaggatcag aggtattaag ggcaaggaca 1080
gagtggaggc caaagttaac agactcattc aggggaatat cagtaatacc agcagagcca 1140
agggta 1146
<210> 73
<211> 750
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил-АЦП-тиоэстеразу Streptococcus pyogenes
<400> 73
atgggattat catatcaaga agagttaaca ttaccatttg agttatgcga cgtaaagtca 60
gacataaagt taccattact attagattat tgcttaatgg tatctggaag gcaatcagta 120
cagttaggaa gatcaaacaa caacttatta gtagactata agttagtatg gatagtaaca 180
gactacgaga taacaataca caggttacca cacttccaag agacaataac aatagagaca 240
aaggcattat catacaacaa gtttttctgt tacaggcagt tctacatata cgaccaagag 300
ggatgcttac tagtagacat actatcatac ttcgcattac taaacccaga tacaagaaag 360
gtagcaacaa taccagagga tttagtagca ccattcgaaa cagactttgt aaagaaatta 420
cacagggtac caaagatgcc attactagag cagtcaatag atagggacta ctatgtaagg 480
tacttcgaca tagacatgaa tggacatgta aacaactcaa agtacttaga ctggatgtac 540
gacgtattag gttgccagtt cttaaagaca caccagccat taaagatgac attaaaatac 600
gtaaaagaag tatcacctgg aggacagata acttcatcat atcatttaga tcagttaaca 660
tcataccacc agataatatc agatggacag ttaaatgcac aggcaatgat agagtggaga 720
gcaataaagc agacagagag tgagacagac 750
<210> 74
<211> 1191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 74
atgggaacat gctcatacac aataacatta ccaataagat gcttatcaaa ctcaaacgga 60
caccacgacc cacataagca gaacttaaac aagataaaga taaatggagc atctacatca 120
acaaggccat taaagttaga cgcaccatca cagacagttg gagtagcaac aatatactta 180
gcatcagtat cagagaactt aactttaaca aaagaagtaa tgagacagaa tataccaaca 240
aagaagcagt acatagatcc acacaggcag ggattaatga tagagggtgg agtaggatat 300
aggcagacaa tagtaataag atcttacgag gtaggaccag acaagacagc aacattagag 360
tcaatactat acttattaca agagacagca ttaaaccacg tatggttatc aggactatta 420
tcaaacggat tcggagcaac acatggaatg gtaaagaata acctaatatg ggtagtatca 480
aagttacagg tacaggtaga ccactaccca atatggggag aggtagtaga gatagacaca 540
tgggtaaggg caagtggaaa gaacggaatg aaaagggact ggttaataag atcacaagca 600
acaggacatg tatttgtaag ggcaacatca acatgggtaa tgatgaatga gaaaactaga 660
aggctatcaa agatgccaga ggaagtaagg gcagagatat caccatggtt catagagaag 720
caagcaataa aagaagaagt acctgataag attgctaagt tagacgacaa ggcaaggtat 780
gtaatatcaa acttaaagcc aaaaagatca gacttagaca tgaaccatca cgtaaacaat 840
gtaaaatatg taaggtggat gttagagata ttaccagatc acttcttaga gtcacaccag 900
ttaagtggaa taacaatgga gtacaggaga gagtgtggat cagcagacat agtacagtca 960
ttatgcgagc cagacggaga tgagatacta tcaaacgata taccagtact aaatggattt 1020
tcattagcat cagagccatt aatggaagga aacggattct tagtaccatt agataaggta 1080
ccattaaagt acacacactt actattaaca aagggagagt cacagaacga agagatagta 1140
aggggaaaga caatatggaa gaaaaagtta tgcacaatgc cattctcaac a 1191
<210> 75
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 75
atggcatcaa agggatcaat aaggctatac tttccatgcg acttcaggaa cacactacag 60
aagaagaata tgaagatgga catggtaatg gcaaggtcag gattttcata ctcattaaac 120
ccaatagcac caaagatacc aaggttctat gtagtagcaa atgcatcaaa tccacagagg 180
gtagatacaa taaacggaaa gaaagtaaac ggaatacacg tagcagagtc atcaaactca 240
tacgcagacc agaataagat gaacgcaaca gcaggattag tattagacgg aaacgtagat 300
catcagccat tacacaagtg gttattaggt aggttcgtag acgagaggtt agtatattca 360
cagacattca taataagatc atacgagata ggaccagaca agacagcaac aatggagaca 420
ttaatgaact tattacaaga gacagcatta aaccacgtaa catcaagtgg attagcagga 480
gatggattcg gagcaacaag agagatgtca ttaagaaagc taatatgggt agtaacaagg 540
atacacatac aggtacaaag gtactcatgc tggggagatg tagtagagat agatacatgg 600
gtagacggag ctggaaagaa cggaatgaga agggactgga taataaggga ttacaacaca 660
aaagaaataa taacaagggc aacatcaaca tgggtaataa tgaataggga aactagaaag 720
ttatcaaaga tgccagaaca ggtaaggcag gaattagtac cattctacac aaataggata 780
gcaatagcaa aagagaataa cgacgttgag aagatagata agttaacaga cgagacagca 840
gaaaggataa ggtctggatt agcaccaaga tggagtgaca tggacgcaaa ccagcacgta 900
aacaacgtaa agtacatagg atggatacta gagtcagtac caataaacgt attagaggac 960
tataacttaa cttcaatgac attagagtac agaagagagt gcaggcagtc aaacttatta 1020
gagtcattaa catcaacaac agagcattca aacaacaatt cttgtaacag gaaagcacac 1080
ttagagtata cacacttact aagaatgcaa gcagacaagg cagagatagt aagggcaagg 1140
acagagtggc agtcaaagtc aaacagaaag acaata 1176
<210> 76
<211> 1257
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 76
atggtagcaa cagcagcagc agcaacatca tcattctttc cagtaccatc acagtcagca 60
gatgcaaact tcgataaggc accagcatca ttaggaggaa taaagttaaa gtcaacttca 120
tgctcaaggg gactacaggt taaggcaaac gcacaggcac caccaaagat aaacggatca 180
tctgtaggat ttacaacatc agtagagaca gtaaagaacg acggagacat gccattacca 240
ccaccaccaa gaacattcat aaaccagtta ccagattggt caatgttatt agcagcaata 300
acaacaatat tcttagcagc agagaagcaa tggatgatgt tagattggaa gccaagaagg 360
ccagacatgt taatagaccc atttggaata ggaaggatag tacaagacgg attaatattc 420
aggcagaatt tctcaataag atcatacgag ataggagcag acaggacagc atcaatagag 480
acattaatga atcacttaca ggaaacagca ttaaaccatg taaagacagc aggactatta 540
ggtgacggat tcggatcaac accagagatg tcaaaaagaa acctaatatg ggtagtaaca 600
agaatgcaag tattagtaga taggtatcca acatggggag atgtagtaca ggtagacaca 660
tgggtatcaa agagtggaaa gaacggaatg agaagggact ggtgcgtaag ggactcaagg 720
acaggagaga cactaacaag ggcatcatca gtatgggtaa tgatgaataa gttaacaaga 780
aggctatcaa agataccaga agaagtaagg ggagagatag agccatactt cttaaactca 840
gacccaatag tagacgaaga ttctaggaaa ttaccaaagt tagacgattc aaacgcagac 900
tacgtaagga aaggattaac acctaggtgg agtgatttag acataaatca gcacgtaaac 960
aatgtaaagt acataggatg gattttagag tcagcaccat taccaatact agagtcacac 1020
gagttatcag caataacatt agagtataga agagagtgcg gaagggactc agtattacag 1080
tcattaacag cagtatcagg aaacggaata ggaaacttag gaaacgctgg agacatagaa 1140
tgtcagcact tactaaggtt agaggacgga gcagagatag taaggggaag gacagagtgg 1200
aggccaaagt actcatcaaa ctttggaata atgggacaga taccagtaga gtctgca 1257
<210> 77
<211> 1215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 77
atggcagtat ttacatacag aatagcaatg ttaccaataa gatgttcatc atcaaactca 60
acaaattcac actcacacga cccaaaccaa cagaatttac acaagataaa gataaacgga 120
tcagcttcag caatgatgcc attaaaggta gacttaccat catcattaac aataacatca 180
gtagcaccag tagtagagaa cttatcttta acaaaagagc agacaaggca gaacatacca 240
acaaagaagc agtatataga cccacatagg cagggactaa tagtagagga gggtgtagga 300
tacaggcaga cagtagtaat aagatcatac gaggtaggac cagacaagac agcaacatta 360
gagataatac tatgcttatt acaagagaca gcattaaacc acgtatggtt atcaggatta 420
ctatcaaacg gatttggagc aacacacgga atggtaagaa ataacctaat atgggtagta 480
tcaaagttac aggtacaggt agaccagtac ccaatatggg gagaggtagt agagatagac 540
acatgggtag gagcatctgg aaagaacgga atgagaaggg attggttagt aaggtcacag 600
gcaactggac aggttttcgc aagggcaaca agtacatggg taatgatgaa tgagaaaaca 660
agaaggctat caaagatgcc agaggaagta agggcagaga tagcaccatg gttcatagag 720
aagcaagcaa taaaagaaga agtaccagag aagatagcaa agttagacga taaggcaagg 780
tatgtagtaa caaacttaaa gcctaagagg tcagatttag acatgaatca acatgtaaac 840
aacgtaaaat acgtaaggtg gatgttagag acattaccag accagttctt cgagaaccac 900
cagttaagtg gaataacttt agagtataag agagagtgcg gatcatcaga tatagtagag 960
tcattatgcg agccagatga agaagaggga ataataaaca caggattaaa gcagaacaac 1020
gacaagtcat tattcaatgg attctcatta ccatcagaga taatggaagg aaacggattc 1080
ttatcatcat tagaaaagac accattaaag tacacacact tattagtagc aaagggaaag 1140
acacagtcag aggaaatagt aaggggaaag acaatatgga agaaaaagtt actaacaaca 1200
atgccatttt cacca 1215
<210> 78
<211> 1044
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил: АЦП-тиоэстеразу Jatropha curcas
<400> 78
atggtatcaa cagcaatatc attttcacta ttaccaataa agttaataaa agaagagaca 60
agaatggcat cagcagatag gagaaagaac tcaatagtaa aagaattcgg acacttcaca 120
tgcagatgtg cagcaataga gaagagaata caaaagttaa acaacttctt aatagacgga 180
ggattcggat cattagagca aaatggatta atatataggc agaacatatt cataaggtca 240
ttcgagatag gattcgacag aaagttatca ttagcagcat taactaattt cttacaggat 300
acagcattaa accacgtaag aatgatagga ttactagctg caggatttgg atcaacacca 360
gagatgtcaa agaaagactt aatatgggta ttatgcacat tacagatatt agtagacagg 420
cacccatctt ggttagacgc agtagaggta gacacatgga tgtacccatc aggacagaac 480
ggacagggaa gggactggtt agtaagggac gcaaagacag gaaagccatt agcacaggca 540
tcaagtgtaa tggtattatt aaacaaaaag acaaggaagc tatcaaagtt cacagaagag 600
ataagggacg agatagcacc acacatgatg atggactgca acccaataat aaattctagg 660
aagatgttac cattcgatgt aaatacagca gattacgcaa ggacaggatt aacaccaggt 720
tggaacgatt tagacttaaa tcagcatgta aaccatgtac agtacataaa ctggatatta 780
cagaacgtat taaggtcact aatacagcat cacaagttat cagatataac attagagtac 840
agaaaggaat gcgacataaa ctcaatacta cagtttctat caaagatagt taagaatgga 900
tcaaaccact caacagacac aaataactta atagagttag accactcatt actattagag 960
aacggatcag agatagcaag ggcaaacaca atatggaagc caagagaagt aaacaacttt 1020
aagaacgcag tatatacacc agca 1044
<210> 79
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил:АЦП-тиоэстеразу Jatropha curcas
<400> 79
atggcatcaa ctgcaatatc attcttaagt ataccaataa agttaataaa agaagagatg 60
agaatggcat ctgcaggaag gagaaagaac acaatagcag cagagttcgg acattttaca 120
ttcggatcag caacagtaga gaaaaaagta cagaagtcaa ataacttcct aatagacgga 180
ggatttggat cattagagca gaacggatta atatacaggc agaacatatt tgtaaggtca 240
tttgagatag gattcgatag aaagttatca ttagcagcat taacaaactt cttacaggac 300
acagcattaa accactgcag aatgatagga ttactagcag agggattcgg atcaacacca 360
gagatgataa agaaagattt aatatgggta ttatgtacat tacagatatt agtagacgga 420
tatccaagtt ggctagacgt agtagaggta gatacatgga tgtacccatc aggacaaaac 480
ggattaggta ggggatggtt agtaagggac ggaaagacag gaaggtcatt agcacagtca 540
tcatcagtaa tggtatcatt caataagaaa acaaggaaac tatcaaagtt agcaaaggag 600
ataagggacg agatagcacc acacatgatg gactgcgacc caataatgaa caagaactca 660
agaaagatat taccattcga cgtaaataca gctgattatg caaggacagg tttaacacca 720
ggatggaacg aattagactt aaaccaacac gtaaatcacg tacagtacat aaactggata 780
ttacagaatg taaggccatc attagttcag catcacaagt tatcagcaat tacattagag 840
tacagaaaag agtgcgacat gaattctata ctacagtcat tatcaaggat agtaaagaac 900
ggaggaaacg attcaacaga caagaacaac gtaatagagt tagaccactt cttactatta 960
gagaacggat cagagatagc aagggcaaac acaatatgga agccaagaga agtaaacaac 1020
tttaagaatg tagtacactc accagcagaa gagaacatat catcaatgaa t 1071
<210> 80
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FadM E. coli
<400> 80
atgcagacac agataaaggt aaggggatat cacttagacg tataccaaca cgtaaacaac 60
gcaaggtact tagagttctt agaagaagca agatgggatg gattagagaa ctcagattca 120
ttccagtgga tgacagcaca taacatagca tttgttgtag taaacataaa tataaactac 180
agaaggccag cagtattatc agacctatta acaataacat cacagttaca gcagttaaat 240
ggaaagtcag gaatactatc acaggtaata acattagagc cagagggaca ggtagtagca 300
gacgcactaa taacattcgt atgcatagac ttaaagacac aaaaggcatt agcattagag 360
ggagagttaa gggagaaatt agagcagatg gtaaag 396
<210> 81
<211> 957
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccA E. coli
<400> 81
atgtcattaa acttcttaga tttcgagcag ccaatagcag agttagaggc aaagatagac 60
tcattaacag cagtatcaag gcaagatgag aagttagaca taaacataga cgaagaagta 120
cataggttaa gggagaagtc agtagagtta acaagaaaga tattcgcaga cttaggagca 180
tggcagatag cacagctagc aaggcaccca cagaggccat acacattaga ctatgtaagg 240
ttagcattcg acgagttcga tgagcttgca ggagacaggg catacgcaga cgataaggca 300
atagtaggag gaatagcaag gttagacgga aggccagtaa tgataatagg acaccaaaag 360
ggaagggaga caaaagagaa gataagaaga aactttggaa tgcctgctcc agagggatac 420
agaaaggcat taaggttaat gcagatggca gaaaggttta agatgccaat aataacattc 480
atagacacac caggagcata cccaggagta ggagcagagg aaaggggaca gagtgaggca 540
atagcaagaa acttaaggga aatgtcaagg ttaggtgtac cagtagtatg cacagtaata 600
ggagagggag gatctggagg agcattagca ataggagtag gtgacaaggt aaacatgtta 660
cagtactcaa catattcagt aatatcacca gagggatgcg catcaatact atggaaatca 720
gcagataagg caccattagc tgcagaggca atgggaataa tagcaccaag gctaaaagaa 780
ctaaagttaa tagactcaat aataccagag ccattaggag gagcacacag aaatccagag 840
gcaatggcag catcattaaa ggcacagcta ttagcagatt tagcagactt agatgtatta 900
tcaacagagg acttaaagaa tagaagatac cagaggttaa tgtcatacgg atatgca 957
<210> 82
<211> 468
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccB E. coli
<400> 82
atggacataa gaaagataaa gaaattaata gagttagtag aagagtcagg aatatcagag 60
ttagaaatat cagagggaga ggaatcagta agaatatcaa gggcagctcc agcagcatca 120
ttcccagtaa tgcaacaggc atacgcagca ccaatgatgc agcagcctgc acagagtaac 180
gcagcagcac cagcaacagt accatcaatg gaagcaccag cagcagcaga gatatctgga 240
cacatagtaa ggtcaccaat ggtaggaaca ttctatagga caccatcacc agacgcaaag 300
gcattcatag aggtaggaca aaaggtaaac gttggagata cattatgcat agtagaggca 360
atgaagatga tgaatcagat agaggcagat aagtcaggaa cagtaaaggc aatactagta 420
gagtcaggac agccagtaga gtttgacgag ccattagtag taatagag 468
<210> 83
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccC E. coli
<400> 83
atgttagaca agatagtaat agcaaatagg ggagagatag cattaagaat actaagggca 60
tgcaaagagt taggaataaa gacagtagct gtacactcat cagcagacag ggacttaaaa 120
catgtattac tagcagatga gacagtttgt ataggaccag caccatcagt aaagtcatac 180
ttaaacatac cagcaataat atcagcagca gagataacag gagcagtagc aatacaccca 240
ggatacggat tcttatcaga aaacgcaaat ttcgcagagc aagtagagag atcaggattc 300
atattcatag gaccaaaggc agagacaata aggctaatgg gagacaaggt atcagcaata 360
gcagcaatga agaaagctgg agtaccatgc gtaccaggat cagacggacc attaggtgat 420
gacatggata agaatagggc aatagcaaaa aggataggat acccagtaat aataaaggca 480
agtggaggag gtggaggaag gggaatgaga gtagtaaggg gagatgcaga gttagcacag 540
tcaatatcaa tgacaagggc agaggcaaag gcagcattct caaatgacat ggtatacatg 600
gagaagtact tagagaaccc aaggcacgta gagatacagg tattagcaga cggacaggga 660
aacgcaatat acttagcaga gagggactgc tcaatgcaga gaaggcacca gaaagtagta 720
gaagaagcac cagctccagg aataacacca gagttaagaa gatacatagg agagaggtgc 780
gcaaaggcat gcgtagacat aggatatagg ggagcaggaa catttgagtt cttattcgag 840
aacggagagt tttacttcat agagatgaac acaaggatac aggtagagca cccagtaaca 900
gagatgataa caggagtaga cttaataaag gaacagttaa ggattgcagc aggacagcca 960
ttatctataa agcaagaaga agtacacgta aggggacatg cagtagagtg caggataaac 1020
gcagaggatc ctaacacatt tctaccatca ccaggaaaga taacaagatt ccatgcacca 1080
ggaggattcg gagtaaggtg ggagtcacac atatatgcag gatacacagt accaccatat 1140
tacgattcaa tgataggtaa gctaatatgc tacggagaga acagggacgt agcaatagca 1200
agaatgaaga acgcattaca ggaattaata atagacggaa taaagacaaa tgtagattta 1260
cagataagga taatgaacga cgagaacttt cagcacggag gaacaaacat acactattta 1320
gagaaaaagt taggattaca agagaag 1347
<210> 84
<211> 912
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccD E. coli
<400> 84
atgtcatgga tagagaggat aaagtcaaac ataacaccaa caagaaaggc atcaatacct 60
gagggagtat ggacaaagtg cgactcatgc ggacaggtac tatatagggc agaattagag 120
agaaacttag aggtatgccc aaagtgcgat catcacatga gaatgacagc aaggaatagg 180
ttacactcat tattagatga aggatcatta gtagagttag gatcagagtt agagccaaag 240
gacgtactaa agtttaggga cagtaagaag tacaaggaca ggttagcatc agcacagaaa 300
gagacaggag agaaagacgc attagtagta atgaagggaa cattatacgg aatgccagta 360
gtagcagcag catttgagtt cgcattcatg ggaggatcaa tgggatcagt agtaggagca 420
aggttcgtaa gggcagtaga gcaagcttta gaggacaact gcccattaat atgtttctca 480
gcatctggag gtgcaagaat gcaagaagca ttaatgtcat taatgcagat ggcaaagaca 540
tcagcagcat tagcaaaaat gcaggaaagg ggactaccat acatatcagt attaacagat 600
ccaacaatgg gaggagtatc agcatcattc gcaatgttag gagacttaaa catagctgag 660
ccaaaggcac taataggatt cgcaggacca agggtaatag agcagacagt aagggagaag 720
ttaccaccag gatttcagag gtcagagttc ttaatagaaa agggagcaat agacatgata 780
gttagaaggc cagagatgag attaaagtta gcatcaatac tagcaaagtt aatgaattta 840
ccagcaccaa acccagaggc accaagagag ggagtagtag taccaccagt accagatcag 900
gaaccagagg ca 912
<210> 85
<211> 1716
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность мутированного FadD Clostridium ljungdahlii
<400> 85
atggatgaaa ttaaaataaa tgatagccaa aaaagatagt atgaaaaaat gggctattgg 60
ggtaaaaaaa cactctaaga ttactggcat acgtcggtaa aaaagtatag agataaggaa 120
tttgtagtag atgatagagg atatcgttac acctagggac aattggatga aaaagcaggt 180
attgttgctt cctattttct tagtattgga gtaaagcctc tagatgttat ttcttttcaa 240
ataccaatat ggagtgaatt tgtaattgtt agtattgcat gcatgaaggt ggggagctgt 300
tgttaaccca attggcatgt gctatagtgg tccgggaagt ttcatatctt ttgaatttat 360
gtaaaagtaa agtttttctc tgccctacat ggtacaataa gacaaattat gaaaaattga 420
ttctatcagt caaaaaggat gtaaaaagtt taaaacacat tgtattactt gacaatctta 480
aagaaaaaga aagtaacagt attactttaa aacatatatt atcatcatac ttttccttga 540
acataaaaga tgaggtaatg gttgacagta atgatgtagc agctatcctt tgcacatccg 600
gtactactgg atgtgcaaag ggagccatgc tgactcataa taacataata tttagtgaaa 660
aatactttaa taaggaactt ggcattacaa aagatgatat tatgtttatg ccggcacctt 720
taaatcatgc aactggcttt catcatggaa ttattgcacc tatgcttatt ggatcaaaag 780
tggttttaca gcagaagttt aaaagtaaaa aggcaataga acttatgaat agagaaaaat 840
gcacctggtc aatgggagct acaccattta tatatgatat tttgaaaaac attagagagg 900
atgaagttta ccttagttcg ttaaaatttt atctttgtgg tggagctgta gtaccagaag 960
aaatggtaag gcaagcctat gaatatggaa taaaattatg tgaagtttat ggatctacag 1020
aaagtgttcc acatgtattt gtaagaccag atgaaaatat tgagttaaca tttggcactg 1080
caggaagggc tatggaaggc gtagaagtta aaattgtaga tgaaaataga aaagagatac 1140
tacctggaaa tctaggggaa gaagtatcga gaggtccaaa tgtatttgta ggatatatag 1200
gtgataaatc agctacaaat aaggtactgg atgatgaagg atggttttat agtggcgatt 1260
tatgtgtaag tgatataagt ggcaatatta atatcattgg gagaaaaaaa gacataattg 1320
ttagaggcgg tgaaaatctc aactcgaatc atataagtca gtatatttca aaatttccac 1380
taattaaaga tgaggcagtt attggaatgc ctgataagcg tcttggtgaa cgaatttgtg 1440
cttatgtcgt attgaaaaaa gaagttaatt ctctgaaatt agaagaactt ttagagtaca 1500
tggaaaagga aaaaatccct aaaaggtatt ggccggaaca cctggagatt atagataaaa 1560
ttcccagaac tgacagtgga aaggtgaaaa aaaatttatt ggcaaaggat ttaaaagttc 1620
gaatgagtag acaggaggag tcaagttgga agggcgagaa ttcggtccaa gaagatcaag 1680
agaaacaaca aaaccaaggg ttgtgtgccc caataa 1716
<210> 86
<211> 927
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FabD E. coli
<400> 86
atgactcaat tcgcgttcgt ttttccaggg caaggttctc agaccgtagg tatgttagca 60
gatatggcag catcatatcc aattgtagag gaaacctttg cagaagcatc agcggcgtta 120
ggctatgact tatgggcgct tacccaacag ggcccagcag aggaacttaa taagacttgg 180
cagacccagc cagcgttact aaccgcatca gttgcgcttt atagagtttg gcagcaacag 240
ggcggcaagg cgccagcgat gatggcaggg cattcattag gtgagtattc agcgcttgtg 300
tgtgcgggcg tcattgactt cgcggacgcg gtgagactag ttgaaatgag aggtaagttc 360
atgcaagaag cagtaccaga gggtaccgga gcaatggcag ccattattgg gcttgacgat 420
gcgagtatag cgaaagcgtg tgaagaagcg gccgagggtc aggttgtgtc accagtcaat 480
ttcaattcac caggacaagt tgttattgcc ggtcataaag aagcggttga gagagctggc 540
gcagcatgta aagcagcggg agcaaagaga gcacttccat taccagtttc agttccttca 600
cattgtgcct taatgaagcc tgcggcggac aagttagcgg ttgagttagc gaagattacc 660
ttcaatgcgc caactgtacc agttgtcaat aatgttgacg ttaaatgtga aaccaatggc 720
gacgcgatta gagatgcact tgtgagacaa ctttataatc cagttcaatg gaccaaatca 780
gttgagtata tggcggcaca gggtgttgaa catctttatg aagtcggccc aggtaaagta 840
cttaccggcc ttaccaagag aattgtagac acccttactg cgtcagcgtt aaatgagcct 900
tcagcgatgg cggcagcgct tgagtta 927
<210> 87
<211> 549
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность TesA E. coli с удаленной последовательностью нацеливания на периплазму
<400> 87
atggcagata ccttacttat tcttggcgac agtttatcag cgggttatag aatgtcagca 60
tcagcagcgt ggcctgcgtt actaaatgac aaatggcaat caaagacctc agttgttaat 120
gcatcaatat ctggtgacac ttcacaacag ggattagcga gacttccagc gttacttaag 180
cagcatcagc caagatgggt attagttgag cttggtggga atgacggcct tagaggtttc 240
caaccacagc aaaccgagca aaccttaaga cagattcttc aagatgtgaa agcggcgaat 300
gcggagcctc ttcttatgca gattagactt cctgcgaatt atggcagaag atataatgaa 360
gcgttttcag ccatttatcc aaagttagca aaagaattcg acgttccatt acttccattt 420
ttcatggaag aagtctatct aaagccacag tggatgcaag atgacggcat tcatccaaat 480
agagatgcac aaccattcat tgcggactgg atggcaaaac agcttcagcc attagtaaat 540
catgactca 549
<210> 88
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Cinnamomum camphorum
<400> 88
atggcaacca ccagtttagc gagtgcgttt tgttcaatga aagccgttat gttagcaaga 60
gatgggagag gaatgaaacc aagatcatca gaccttcaac ttagagcggg caatgcgcag 120
acctcattaa agatgattaa tggtaccaaa ttctcatata ccgaatcatt aaagaaactt 180
ccagactggt caatgctttt cgcagttatt accactattt tctcagccgc ggaaaagcaa 240
tggaccaatt tagagtggaa acctaagcca aatcctccac agttattaga tgaccatttc 300
ggtccacatg gtttagtatt cagaagaact tttgcgatta gatcttatga agttggccca 360
gacagatcaa cctcaatagt tgcggtcatg aatcatttac aagaagcagc gcttaatcat 420
gcgaaatcag ttggcattct aggcgacggt ttcggcacta cccttgaaat gtcaaagaga 480
gatcttattt gggttgtgaa gagaacccat gtagcagttg agagatatcc agcgtgggga 540
gataccgttg aagttgaatg ttgggttgga gcatcaggga ataatggtag acgacatgac 600
ttcttagtaa gagactgtaa aactggcgag attcttacca gatgtacttc actttcagta 660
atgatgaata ccagaaccag aagattatca aagattccag aagaagtgag aggtgagatt 720
ggtccagcgt ttattgataa tgttgcggtg aaagatgaag agattaagaa gccacaaaag 780
cttaatgatt caaccgcaga ctatattcag ggtggcctta cccctagatg gaatgactta 840
gacattaatc agcatgtcaa taatattaaa tatgtcgact ggattcttga aactgttcca 900
gactcaattt tcgagtcaca tcatatttca tcatttacca ttgaatatag aagagagtgt 960
accatggact cagtcctaca atcattaacc accgtttctg gcgggtcatc agaggcaggc 1020
cttgtgtgtg agcatcttct acagcttgag ggcggatcag aagttttaag agcgaaaacc 1080
gagtggaggc caaagcttac tgactcattc agaggtattt cagtaatacc agcggagtca 1140
tcagtt 1146
<210> 89
<211> 1146
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая тиоэстеразу Umbellularia californica
<400> 89
atggcaacta cttcattagc gtcagcattt tgttcaatga aagccgtaat gttagcaaga 60
gatgggagag gaatgaagcc aagatcaagt gacttacagt taagagcggg taatgcgcca 120
acttcactta agatgataaa tggtaccaaa ttctcatata ccgaatcatt aaagagactt 180
ccagactggt caatgttatt cgcggtgatt accaccattt tctcagcggc cgagaaacaa 240
tggaccaatc tagagtggaa acctaagcca aaacttccac aactacttga cgaccatttt 300
ggcttacatg gtcttgtttt cagaagaacc tttgcgatta gatcttatga agttggccct 360
gacagatcaa cttctattct tgcggttatg aatcacatgc aagaagcaac tcttaatcat 420
gcaaagtcag ttggtattct tggtgacggt ttcgggacca ctcttgaaat gtcaaagaga 480
gacttaatgt gggtagtaag aagaactcat gtcgcggttg aaagatatcc aacctgggga 540
gacaccgttg aggttgaatg ttggattggt gcgtcaggca ataatggcat gagaagagac 600
tttcttgtga gagattgtaa gaccggagag attcttacca gatgtacctc actttcagtt 660
cttatgaata cccgtaccag aagattatca accatacctg acgaagtgag aggcgaaatt 720
gggccagcgt tcattgacaa tgttgcggtt aaagatgatg agattaaaaa gcttcaaaag 780
cttaatgact caaccgcaga ctatattcag ggcggcttaa ccccaagatg gaatgacctt 840
gacgttaatc agcatgtcaa taatctaaaa tatgtagcgt gggttttcga aaccgttcca 900
gactcaattt tcgaatcaca tcatatttca tcattcacct tagagtatag aagagaatgt 960
accagagatt cagttttaag gtcattaacc accgtaagtg gtggttcatc agaggcaggc 1020
cttgtctgtg atcatctttt acagttagag ggcggatcag aggtgcttag agcgagaacc 1080
gagtggcgac caaagttaac tgattcattc agaggcattt cagttattcc agcggagcca 1140
agagtt 1146
<210> 90
<211> 750
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая ацил-АЦП-тиоэстеразу Streptococcus pyogenes
<400> 90
atgggcttat catatcaaga agagttaacc ttaccatttg aattatgtga tgttaagtca 60
gacattaaac ttccattact tttagactat tgtcttatgg tttcaggcag acagtcagtg 120
cagcttggta gatcaaataa taatctttta gtcgactata aacttgtatg gatagttact 180
gactatgaga ttaccattca tagattacca catttccaag aaaccattac cattgaaacc 240
aaagcgctaa gttataataa gttcttttgt tatagacaat tctatattta tgaccaagag 300
ggctgtcttt tagttgacat tctatcatat ttcgcgcttc ttaatccaga taccagaaag 360
gttgcaacta ttccagagga cttagtcgcc ccattcgaga ctgactttgt taagaaatta 420
catagagttc caaaaatgcc tttacttgag cagtcaattg acagagacta ttatgtaaga 480
tatttcgata ttgacatgaa tggacatgtt aataattcaa agtatcttga ctggatgtat 540
gatgttttag gctgtcaatt ccttaagacc catcagccac ttaagatgac ccttaaatat 600
gtgaaagaag tatcacctgg tggtcaaatt acctcatcat atcatcttga ccagttgacc 660
tcatatcatc agattatatc agatgggcag cttaatgcac aggcgatgat tgaatggaga 720
gcgattaagc aaaccgagtc tgaaaccgat 750
<210> 91
<211> 1191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 91
atggggacct gttcatatac cattaccctt ccaattcgat gtttatcaaa ttcaaatggc 60
catcatgatc cacataagca gaatcttaat aagataaaga taaatggcgc gtcaactagt 120
accagaccac ttaagttaga cgcaccttca caaaccgttg gtgttgcgac catttatcta 180
gcatcagttt cagaaaatct tacccttacc aaagaagtca tgagacagaa tattccaacc 240
aagaaacagt atattgaccc acatagacaa gggttaatga ttgagggtgg agttggttat 300
agacaaacta ttgtcattag aagttatgaa gttggcccag acaaaaccgc gactttagag 360
tcaattcttt atcttttaca ggaaaccgcc ttaaatcatg tatggctttc aggtctactt 420
tcaaatggct tcggggcgac ccatggcatg gtaaagaata atttgatttg ggtagtgtct 480
aagcttcagg ttcaagtgga tcattatcca atttggggag aggttgtaga gattgacacc 540
tgggttagag catcaggtaa gaatggtatg aaaagagact ggcttattag atcacaagca 600
accggccatg tatttgttag agcgacctca acttgggtta tgatgaatga gaaaaccaga 660
agattatcaa agatgccaga agaagtcaga gcggagattt caccatggtt cattgaaaag 720
caagcgatta aagaagaagt gccagacaaa attgcaaagt tagacgacaa ggcgagatat 780
gttatttcaa atctaaaacc taaaagatca gaccttgata tgaatcatca tgtgaataat 840
gtcaaatatg ttagatggat gcttgagatt ttaccagatc atttcttaga gtctcatcag 900
ttatcaggca taactatgga gtatagaaga gaatgtggat cagcagatat tgttcaatca 960
ttatgtgagc ctgacggtga cgagattctt tcaaatgaca ttcctgttct taatggtttc 1020
tcacttgcgt cagaaccatt aatggaaggc aatggtttcc ttgtaccatt agataaggtt 1080
ccattaaagt atacccatct tttacttacc aaaggcgagt cacagaatga agagattgtt 1140
agaggaaaaa ccatttggaa gaagaaactt tgtactatgc cattttcaac c 1191
<210> 92
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 92
atggcatcaa agggctcaat tagattatat tttccatgtg acttcagaaa tactttacag 60
aaaaagaata tgaaaatgga catggtaatg gcgagatcag gattctcata ttcacttaat 120
ccaattgcgc caaagattcc aagattctat gttgtcgcta atgcgtcaaa tccacagaga 180
gttgatacca ttaatggcaa aaaagttaat ggtatacatg ttgcagagtc atcaaattct 240
tatgcggatc agaataagat gaatgcaacc gcgggtttag tcttagacgg caatgtagat 300
catcagccac ttcataagtg gcttttaggt agattcgttg acgaaagact tgtttattca 360
caaaccttca ttattagaag ttatgagatt ggtccagata agaccgcgac tatggaaacc 420
ttaatgaatt tacttcaaga aaccgcactt aatcatgtta cctcatcagg ccttgcggga 480
gatggttttg gtgcgactag agagatgtca ctaagaaaac taatttgggt ggtaactaga 540
attcatattc aagtccagag gtattcatgt tggggggatg tggttgaaat agacacctgg 600
gttgacggcg caggcaagaa tggaatgaga agagactgga ttattagaga ctataatacc 660
aaagaaataa ttactagagc aacctcaacc tgggtcatta tgaatagaga aactagaaag 720
ctttcaaaga tgccagagca agttagacaa gagttagttc cattctatac caatagaatt 780
gcaattgcga aagagaataa tgacgttgag aaaattgaca agttaaccga cgaaaccgcg 840
gagagaattc gatcagggct tgcgcctaga tggtcagaca tggacgccaa tcaacatgtg 900
aataatgtta agtatattgg ctggattctt gaatcagtac ctattaatgt acttgaggac 960
tataatctta cctcaatgac ccttgagtat agaagagaat gtagacagtc aaatctttta 1020
gagtcattaa ccagtaccac cgaacattca aataataatt cttgtaatag aaaggcccat 1080
ttagaatata cccatttact tagaatgcaa gcggataaag cggaaattgt tagagcaaga 1140
actgagtggc agtcaaagtc aaatagaaaa accatt 1176
<210> 93
<211> 1257
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая пальмитоил: ацилпереносящий белок-тиоэстеразу Ricinus communis
<400> 93
atggtggcga ccgcagcagc agcaacctca tcatttttcc cagtcccatc acaatcagcc 60
gacgcgaatt tcgacaaagc accagcgtca cttggcggca ttaaactaaa atcaacttca 120
tgttctagag gattacaggt taaagcaaat gcacaagcgc ctccaaagat aaatggatca 180
tcagttggtt tcaccacttc agtagaaacc gttaagaatg acggtgatat gcctttacct 240
ccaccaccaa gaaccttcat taatcagtta ccagactggt ctatgttact tgcggccatt 300
accaccattt tcttagcggc tgaaaagcag tggatgatgc ttgattggaa gccacgtaga 360
ccagacatgc ttattgatcc atttggcatt ggaagaattg tccaagacgg gttaatattt 420
agacagaatt tctcaattag aagttatgag attggtgcag acagaaccgc gtcaattgag 480
actttaatga atcatcttca agaaaccgcg ttaaatcatg ttaaaaccgc gggtctttta 540
ggtgacggtt tcggttcaac tccagaaatg agtaaaagaa atctaatttg ggttgtcacc 600
agaatgcaag ttttagtaga cagatatcca acctggggtg atgttgttca ggttgatact 660
tgggtttcaa aatctggtaa gaatggcatg agaagagact ggtgtgtgag agactcaaga 720
accggcgaaa cccttacccg agcgtcatca gtatgggtaa tgatgaataa gcttaccaga 780
agattatcaa agattccaga agaagtgaga ggcgaaattg agccatattt ccttaattca 840
gacccaattg tagacgaaga ttcaagaaag cttcctaaac ttgacgactc aaatgcggac 900
tatgttagaa aaggtttaac ccctagatgg tcagatcttg acataaatca acatgtgaat 960
aatgttaagt atattggttg gattcttgag tcagcgccat taccaattct agagtcacat 1020
gaactttcag cgattaccct tgagtataga agagaatgtg ggagagactc agttcttcag 1080
tcacttaccg cggtttcagg caatggcatt ggcaatcttg gcaatgcggg ggatattgag 1140
tgtcagcatt tacttagatt agaagatgga gcagagattg ttagaggtag aactgagtgg 1200
aggccaaagt attcatcaaa tttcggcatt atgggccaaa ttccagtcga gtcagcg 1257
<210> 94
<211> 1215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая Jatropha curcas ацил: АЦП-тиоэстеразу
<400> 94
atggcagtat tcacctatag aattgcgatg ttaccaatta gatgttcatc atcaaattca 60
accaattcac attcacatga cccaaatcaa cagaatttac ataagattaa gattaatggc 120
tcagcgtcag cgatgatgcc tcttaaggtt gatttaccaa gttcattgac gattacctca 180
gttgcaccag ttgtagagaa tctttcatta actaaagaac aaaccagaca gaatattcca 240
accaagaagc agtatattga cccacataga caaggtctta ttgtcgaaga gggcgttggt 300
tatagacaga ccgtcgttat tagatcttat gaagttggcc ctgacaaaac cgcaaccctt 360
gagattatac tttgtctact tcaagagact gcgttaaatc atgtttggtt atcaggctta 420
ctttcaaatg gttttggggc gacccatgga atggtgagaa ataatcttat ttgggttgtt 480
tcaaaacttc aggtacaagt tgatcaatat ccaatttggg gcgaagttgt cgagattgac 540
acttgggttg gcgcgagtgg caagaatggt atgcgaagag actggcttgt tagatcacag 600
gcgaccggcc aggtatttgc aagggccacc tcaacctggg ttatgatgaa tgagaaaact 660
agaagattat caaaaatgcc agaagaagtg agagcggaaa ttgcgccatg gttcattgag 720
aagcaagcga ttaaagaaga agtgccagag aagattgcaa aactagacga taaggcgaga 780
tatgtagtta ccaatcttaa gcctaaaaga tcagaccttg acatgaatca gcatgtgaat 840
aatgtcaaat atgttagatg gatgcttgaa accttaccag atcaattctt cgagaatcat 900
cagttatcag gtattacttt agagtataag agagaatgtg gatcatcaga tatagtagag 960
tcattatgtg agccagacga agaggaaggg attataaata ccggacttaa gcagaataat 1020
gacaaatcac tattcaatgg tttctcactt ccatcagaga ttatggaagg taatggtttc 1080
ctttcatcat tagaaaagac ccctcttaag tatactcatc ttttagttgc aaaagggaaa 1140
acccaatctg aggaaattgt tagaggcaag accatttgga aaaagaaatt acttaccacc 1200
atgccatttt cacca 1215
<210> 95
<211> 1044
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая Jatropha curcas ацил: АЦП-тиоэстеразу
<400> 95
atggtttcaa ccgcgatttc attttcactt ttaccaatta agttaattaa agaagaaact 60
agaatggcct cagcggacag aagaaagaat tcaattgtaa aagaattcgg tcatttcacc 120
tgtagatgtg cagcgattga aaagagaatt caaaaactta ataatttctt aatagacggc 180
ggcttcggct cattagagca aaatggtctt atatatagac agaatatttt cattagatca 240
ttcgagattg gattcgacag aaagctttca ttagcagcgt tgaccaattt ccttcaggac 300
accgcgctta atcatgttag aatgattggc ttacttgcag cgggtttcgg cagtactcca 360
gaaatgtcaa agaaagacct tatttgggtt ttatgtacct tacagattct tgttgacaga 420
catccttcat ggttagatgc agttgaggtc gacacctgga tgtatccatc aggccagaat 480
gggcaaggta gagactggct tgttagagat gcgaaaaccg gaaagccact tgcgcaggca 540
tcttcagtta tggttctatt aaataaaaag accagaaaac tttctaagtt taccgaagag 600
attagagatg aaattgcgcc acatatgatg atggactgta atccaattat taattcaaga 660
aaaatgctac catttgacgt gaatactgcg gattatgcaa gaaccggtct taccccaggt 720
tggaatgacc ttgaccttaa tcagcatgta aatcatgtcc aatatattaa ttggatttta 780
caaaatgttt taagatcact tattcaacat cataagttat cagatattac cttagagtat 840
agaaaagagt gtgacataaa ttcaattctt cagttccttt caaagattgt gaaaaatggg 900
tcaaatcatt caaccgatac caataatcta attgagttag accattcact tcttttagaa 960
aatggctcag agattgcgcg agcgaatact atttggaagc caagagaagt aaataatttt 1020
aagaatgcag tttatacccc tgcc 1044
<210> 96
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность, кодирующая Jatropha curcas ацил: АЦП-тиоэстеразу
<400> 96
atggcatcaa ccgcgattag tttcctttca attccaatta agcttattaa agaagagatg 60
agaatggcgt cagcgggaag aagaaagaat actattgcag cagaattcgg ccatttcacc 120
ttcggctcag cgactgttga gaaaaaagtc caaaagtcaa ataatttctt aattgatggc 180
gggtttggct cattagaaca gaatgggtta atatatagac agaatatttt cgtgaggtca 240
ttcgagattg gttttgaccg aaagttatca ttagcagcgc ttaccaattt tctacaggac 300
actgcgctta atcattgtag aatgattggc ttattagccg agggtttcgg atcaacccca 360
gaaatgatta agaaagacct tatttgggtt ctttgtaccc ttcaaattct tgttgacgga 420
tatccatcat ggcttgacgt tgttgaggtc gacacctgga tgtatccttc aggtcaaaat 480
ggtcttggca gaggttggtt agtaagagat ggcaagactg gcagatcatt agcgcagtca 540
tcatcagtta tggtgtcatt caataagaaa accagaaaac tttctaagtt agcgaaagaa 600
attagagatg agattgcacc acacatgatg gattgtgacc caattatgaa taagaattca 660
agaaagatac ttccattcga tgtaaatacc gcagactatg cgagaaccgg ccttacccca 720
ggttggaatg aattagactt aaatcagcat gttaatcatg tccagtatat taattggatt 780
ttacaaaatg ttagaccttc acttgttcaa catcataagc tttcagcgat taccttagaa 840
tatagaaaag agtgtgacat gaattcaatt ctacaatcac tttcaagaat tgttaagaat 900
ggtgggaatg atagtaccga caagaataat gttattgagt tagaccattt tcttcttcta 960
gagaatggtt cagaaattgc gagagcgaat accatttgga aaccaagaga agtaaataat 1020
ttcaaaaatg tagtgcattc accagcagaa gagaatatat cttcaatgaa t 1071
<210> 97
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность FadM E. coli
<400> 97
atgcaaactc agattaaggt tagaggctat catctagacg tttatcaaca tgtgaataat 60
gcgagatatc ttgagttcct tgaagaagcc agatgggacg gtttagagaa ttcagattca 120
tttcagtgga tgaccgcaca taatattgca ttcgtagtgg tcaatattaa tataaattat 180
agacgaccag cggtcttatc agacctttta accattacct cacaacttca acagttaaat 240
ggaaagtcag ggattctttc acaggttatt acccttgaac cagagggtca ggttgtagca 300
gatgcgctta ttaccttcgt ttgtattgac cttaaaactc aaaaagcgtt agcgcttgaa 360
ggcgagttaa gagaaaagtt agagcagatg gttaaa 396
<210> 98
<211> 957
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccA E. coli
<400> 98
atgtcattaa atttcttaga ctttgaacaa ccaatagcag aacttgaagc gaaaattgat 60
tcattaaccg cagtttcaag acaagatgaa aagttagaca ttaatataga cgaagaagtc 120
catagactta gagaaaaatc agttgagtta actagaaaga ttttcgcaga ccttggtgca 180
tggcagattg cgcaacttgc aagacatcca caaagaccat ataccttaga ctatgtaaga 240
ttagcatttg acgagttcga cgagcttgcg ggtgaccgag cgtatgcgga cgataaagca 300
attgttggtg gcattgccag attagacgga aggccagtta tgattattgg acatcagaaa 360
gggagagaga ctaaggaaaa gattagaaga aatttcggca tgccagcacc agagggctat 420
agaaaggcac ttagacttat gcagatggcg gaaagattca agatgccaat tataaccttc 480
attgataccc ctggagcgta tccaggcgtt ggcgccgaag agagaggcca atcagaggcg 540
attgcgagaa atcttagaga gatgtcaaga ctaggtgtgc ctgtagtatg taccgttatt 600
ggcgagggtg gaagtggcgg tgcccttgcg attggggttg gtgacaaagt gaatatgtta 660
cagtattcaa cctattcagt catttcacct gagggctgtg cgtcaattct ttggaaatca 720
gcggacaaag cgccattagc ggcagaggcg atggggatta ttgcaccaag acttaaagaa 780
ctaaagctta ttgattcaat tattccagag ccacttggtg gtgcgcatag aaatccagaa 840
gcgatggcag cgtcattaaa ggcgcagtta ttagcggatc tagcggactt agacgttctt 900
tcaaccgaag atcttaagaa tagaagatat cagagactta tgtcttatgg ttatgca 957
<210> 99
<211> 468
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccB E. coli
<400> 99
atggatatta gaaagataaa gaaactaatt gagttagtcg aagagagtgg tatatcagag 60
ttagaaattt cagagggtga agaatcagtt agaatttcaa gagcggcacc agcggcctct 120
ttcccagtca tgcagcaagc gtatgcggca ccaatgatgc agcaaccagc ccagtcaaat 180
gcggcagcgc cagcgaccgt accatcaatg gaagcgccag cagcagcgga gatttcaggc 240
catattgtta gatcacctat ggttggtacc ttctatagaa ccccttcacc agacgcgaaa 300
gcgtttattg aagtgggaca aaaggttaat gtaggcgata ccctttgtat tgttgaggca 360
atgaaaatga tgaatcagat tgaagcggac aaatcaggca ctgttaaggc aattcttgta 420
gagtcagggc aaccagttga gttcgacgaa ccacttgtgg ttattgaa 468
<210> 100
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccC E. coli
<400> 100
atgttagaca agattgttat tgcaaataga ggggaaattg cgcttagaat tcttagagcg 60
tgtaaagaat taggcattaa gactgttgca gttcattcat cagcagacag agatcttaaa 120
catgttttac ttgcggacga aaccgtttgt attggtccag caccatcagt aaagtcatat 180
cttaatattc cagccattat ttcagccgcg gagattaccg gagcggtagc gattcatcca 240
ggttatggct tcttatcaga gaatgcgaat ttcgcagaac aagttgagag atcaggattc 300
attttcattg gtccaaaggc agaaaccatt agattaatgg gcgacaaggt ttcagcgatt 360
gcggcgatga aaaaggcagg cgtcccatgt gttccaggct cagatggccc attaggtgac 420
gatatggaca agaatagagc catagcgaaa agaatagggt atccagtaat tataaaggcg 480
tctgggggtg gcggtggtag aggtatgaga gtagtcagag gcgacgcaga gctagcacaa 540
tcaatttcaa tgaccagagc ggaagcaaaa gcggcattct caaatgacat ggtgtatatg 600
gagaagtatt tagagaatcc aagacatgtg gagattcaag ttcttgcgga cggccagggt 660
aatgcgattt atcttgcgga aagagactgt agtatgcaga gaaggcatca gaaagttgtt 720
gaagaagcac cagcgccagg tattacccct gaacttagaa gatatatagg cgagagatgt 780
gcaaaggcgt gtgttgacat tggttataga ggggcaggga cctttgagtt tctttttgag 840
aatggagagt tctatttcat tgaaatgaat accagaattc aagttgaaca tcctgttacc 900
gagatgatta ccggcgtgga cttaattaaa gagcaattaa gaattgcagc gggccagcca 960
ctttcaatta agcaagaaga agtccatgtt agaggacatg cggtcgagtg tagaattaat 1020
gcggaagatc ctaatacttt cttaccatca ccaggcaaga ttaccagatt ccatgcacct 1080
ggtggtttcg gtgtaagatg ggaatcacat atttatgcgg gctatactgt tccaccatat 1140
tatgactcaa tgattggtaa acttatttgt tatggagaga atagagatgt agcgattgcg 1200
agaatgaaaa atgctttaca agaactaatt attgacggta ttaagaccaa tgtggatctt 1260
cagattcgaa ttatgaatga tgagaatttt cagcatggcg gcaccaatat acattatctt 1320
gagaagaaac ttggattaca ggaaaaa 1347
<210> 101
<211> 912
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность AccD E. coli
<400> 101
atgtcatgga tagaaagaat taagtcaaat attaccccaa ccagaaaggc gtcaattcca 60
gagggcgtat ggaccaagtg tgactcatgt ggtcaggttc tttatagagc cgaattagaa 120
agaaatcttg aggtttgtcc aaaatgtgac catcacatga gaatgaccgc aagaaataga 180
ttacattcac ttttagacga aggctcactt gtggaattag gctcagagct tgaaccaaaa 240
gacgttctta agttcagaga cagtaagaag tataaagata gattagcatc agcacagaaa 300
gagactgggg agaaagatgc gttagtggtc atgaaaggta ccttatatgg aatgcctgtc 360
gttgcagctg cgttcgaatt cgcgtttatg ggcggttcaa tgggttcagt tgtaggcgcg 420
agattcgtta gagcagttga acaggcgtta gaagataatt gtccactaat ttgtttttca 480
gcgtctggtg gtgcaagaat gcaagaagca ctaatgtcac ttatgcaaat ggcgaaaact 540
tcagcggcgc ttgcaaagat gcaagagaga gggttaccat atatttcagt tcttaccgac 600
ccaaccatgg ggggtgtctc agcgtcattc gcgatgttag gcgacttaaa tattgccgag 660
ccaaaagcac ttattggctt cgcgggacca agagtgattg aacagaccgt tagggaaaag 720
cttccacctg gatttcagag atcagagttc cttattgaaa aaggcgcgat tgatatgata 780
gttagacgac cagagatgag attaaagctt gcgtcaattc ttgcaaagtt aatgaatctt 840
ccagcgccta atccagaagc accaagagag ggtgttgtag tacctccagt tccagaccaa 900
gagccagagg cg 912
<210> 102
<211> 568
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность TesA E. coli с удаленной последовательностью нацеливания на периплазму
<400> 102
ggtctctaat ggcagacaca cttttgatct tgggcgactc actttccgcg ggctatcgta 60
tgagtgcgtc cgcggcgtgg cccgcgctgc tgaacgacaa gtggcagagc aagacctccg 120
tcgtcaacgc gtccatctcg ggcgacacga gccagcaggg actggcccgc ctcccggcgc 180
tgctcaagca gcatcagcca cgctgggtgc tcgtcgagct gggtggcaac gacgggctcc 240
gcggcttcca accgcagcag accgaacaga ccctccggca aattctccaa gacgtcaaag 300
cggccaacgc ggagcccctg ctgatgcaga tccgcctgcc ggcgaactac ggtcggcgtt 360
ataatgaggc gttcagcgcc atctatccga agctggccaa ggaattcgac gtgcccctcc 420
tcccgttttt catggaagaa gtgtacctca agccccagtg gatgcaagac gacggcatcc 480
acccgaatcg cgacgcgcag ccgttcatcg cggactggat ggcgaagcag ctgcagccgc 540
tcgtaaacca cgactcgtaa aagagacc 568
<210> 103
<211> 568
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность TesA E. coli с удаленной последовательностью нацеливания на периплазму
<400> 103
ggtctctaat ggctgatacc ctcttgattc ttggtgattc ccttagcgcc ggctaccgca 60
tgtccgcgag cgccgcatgg ccggcgctgc tgaatgacaa gtggcagtcc aagacctcgg 120
tcgtcaatgc gagcatcagc ggcgacacct cgcagcaggg cctcgcgcgg ctgccggccc 180
tcctgaagca acatcagccg cgctgggtac tcgtggagct cggcggcaat gatgggctgc 240
gtggcttcca gccccaacag accgagcaga cgttgaggca gatcctccaa gacgtgaagg 300
cagcgaatgc ggagcctctc ctcatgcaga tccggctgcc cgcgaattat ggccgccgtt 360
acaacgaggc cttctcggcg atctatccga agctcgcgaa agagttcgac gtcccgctgc 420
tgcccttctt tatggaggag gtgtacctga agccgcaatg gatgcaggat gacggcatcc 480
acccgaaccg ggacgcgcag cccttcatcg ccgattggat ggcgaagcag ctgcagccgc 540
tggttaatca tgacagttaa aagagacc 568
<210> 104
<211> 568
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная последовательность TesA E. coli с удаленной последовательностью нацеливания на периплазму
<400> 104
ggtctctaat ggccgatacc ctgctgatcc tcggcgattc acttagcgct ggataccgca 60
tgagcgcctc ggctgcgtgg cctgcactcc tcaacgacaa gtggcagagc aagacctcgg 120
tggtcaacgc gtccatttcg ggcgacacca gtcagcaggg cctcgccagg ctgcctgccc 180
tgctgaagca acatcagccg cgctgggtcc tggtcgagct tggcggcaac gacggcctcc 240
gggggttcca gccgcaacag acggagcaga cgttgcggca gattctccag gatgtaaagg 300
cggcgaatgc cgagccgctc ctgatgcaga tcaggctccc ggccaactat ggtcgccgct 360
ataacgaggc gttctccgca atctacccca agctggcaaa agaattcgac gtgccgctgc 420
tgccattttt catggaagaa gtgtacctca agccgcagtg gatgcaagat gatggtatcc 480
accccaaccg ggacgcccaa cccttcatcg ccgactggat ggcgaagcag ctgcagcccc 540
ttgtcaatca cgactcctaa aagagacc 568
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МИКРООРГАНИЗМЫ ДЛЯ ПОВЫШЕННОГО ПРОДУЦИРОВАНИЯ АМИНОКИСЛОТ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2015 |
|
RU2688486C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БИОЛОГИЧЕСКОГО ПОЛУЧЕНИЯ ЛАКТАТА ИЗ С1-СОЕДИНЕНИЙ С ПРИМЕНЕНИЕМ ТРАНСФОРМАНТОВ ЛАКТАТ ДЕГИДРОГЕНАЗЫ | 2014 |
|
RU2710714C2 |
| СИСТЕМЫ ФЕРМЕНТАЦИИ С ПОДАЧЕЙ ГАЗА | 2013 |
|
RU2639542C2 |
| РЕАКТОРЫ, СИСТЕМЫ И ПРОЦЕССЫ ФЕРМЕНТАЦИИ С ПОДАЧЕЙ ГАЗА, ИСПОЛЬЗУЮЩИЕ ЕМКОСТИ РАЗДЕЛЕНИЯ ГАЗА/ЖИДКОСТИ | 2018 |
|
RU2769130C2 |
| СИСТЕМА И СПОСОБ (ВАРИАНТЫ) ИНТЕНСИФИКАЦИИ ПРОИЗВОДСТВА БИОМАССЫ | 2017 |
|
RU2747305C2 |
| РЕАКТОРЫ, СИСТЕМЫ И СПОСОБЫ ФЕРМЕНТАЦИИ С ПОДАЧЕЙ ГАЗА С ПРИМЕНЕНИЕМ ЗОНЫ ВЕРТИКАЛЬНОГО ПОТОКА | 2018 |
|
RU2761409C2 |
| КОМПОЗИЦИЯ ПИТАТЕЛЬНОЙ СРЕДЫ И УЛУЧШЕННЫЕ СПОСОБЫ ПОЛУЧЕНИЯ БИОМАССЫ И ПРОДУКТА С ДОБАВЛЕННОЙ СТОИМОСТЬЮ | 2019 |
|
RU2811519C2 |
| ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИНТЕРЛЕЙКИН-12 (IL12), И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2769316C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ УЛУЧШЕНИЯ ПРИЗНАКОВ РАСТЕНИЙ | 2018 |
|
RU2797537C2 |
| Способ продуцирования рекомбинантного белка | 2017 |
|
RU2742006C2 |
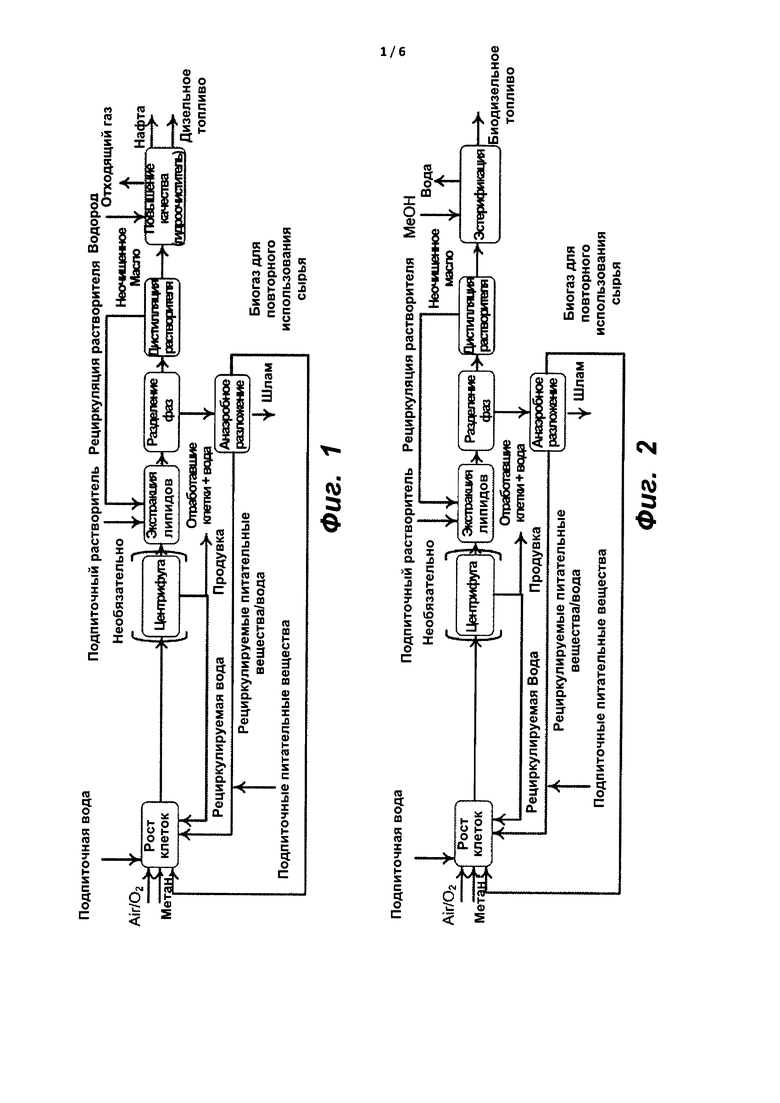
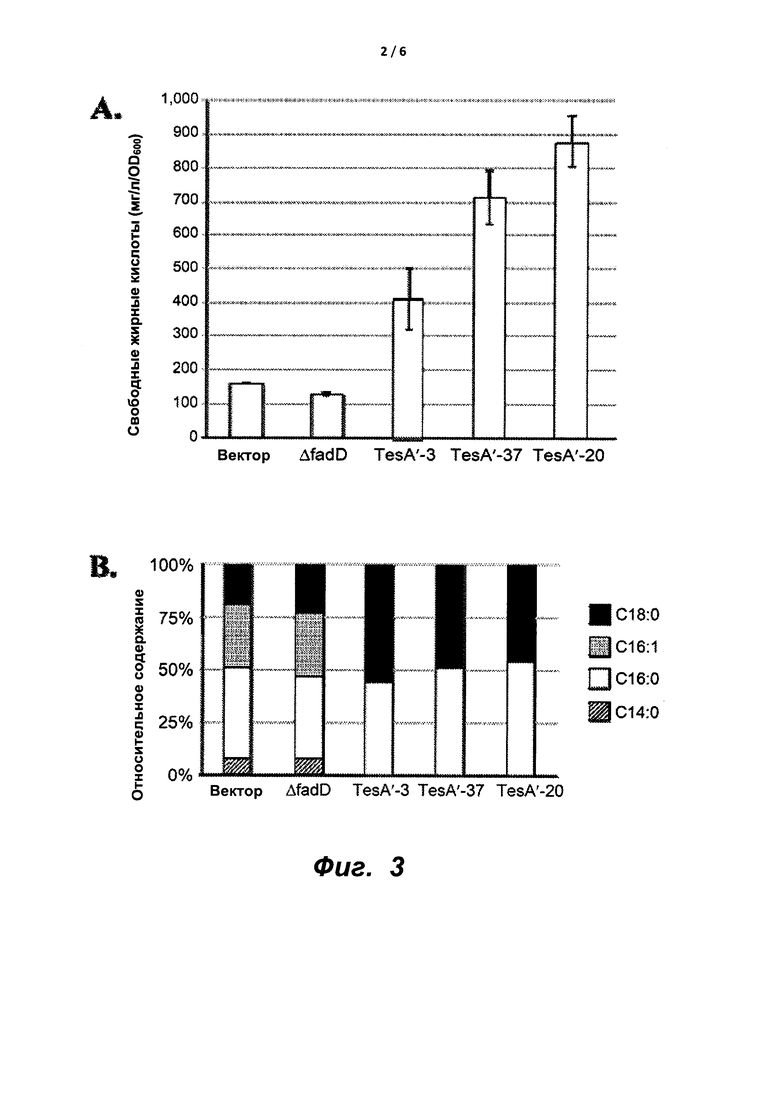
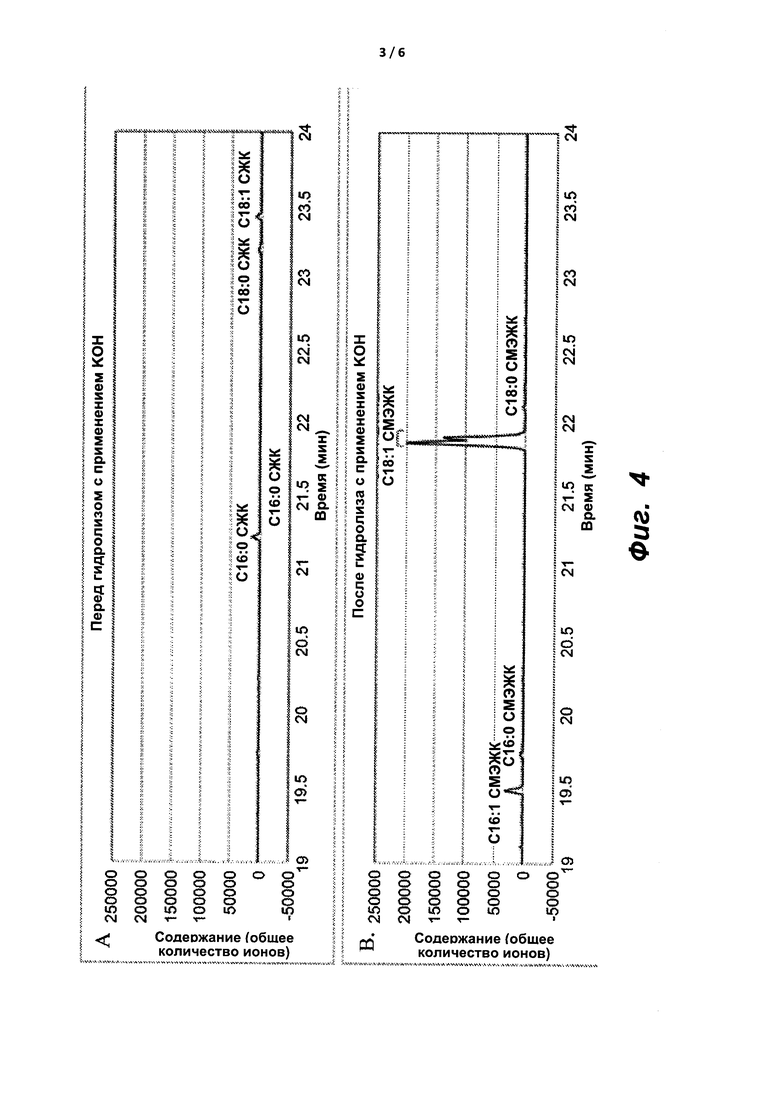
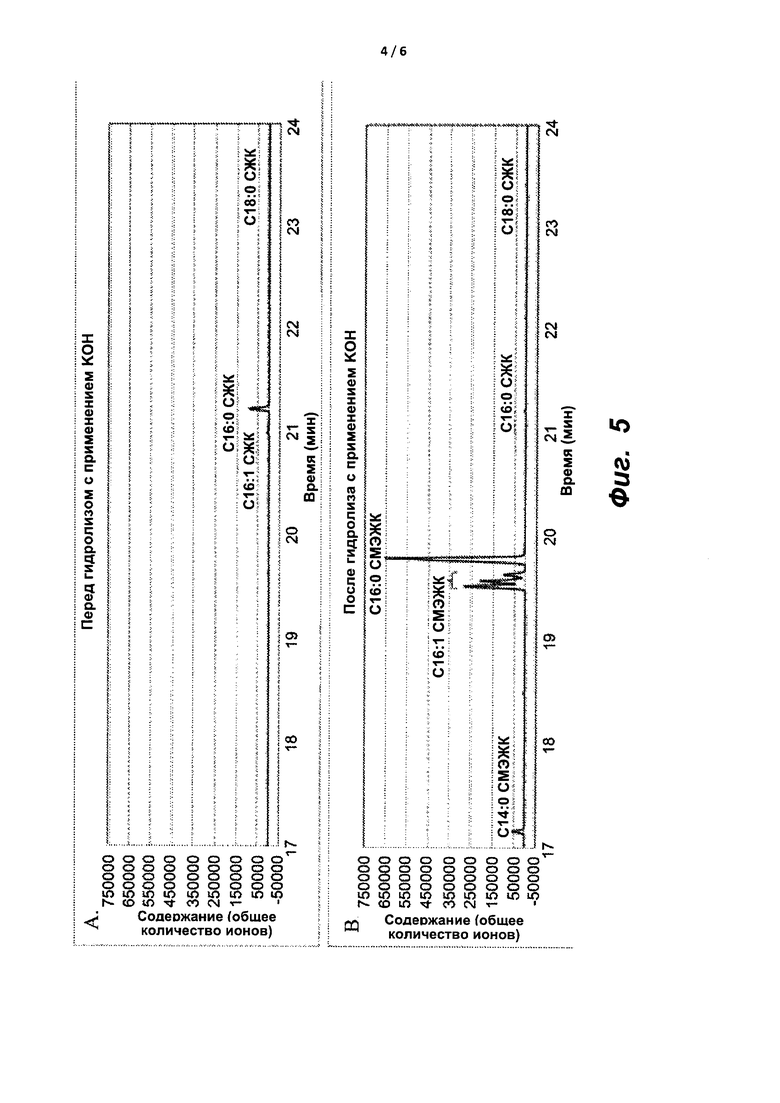
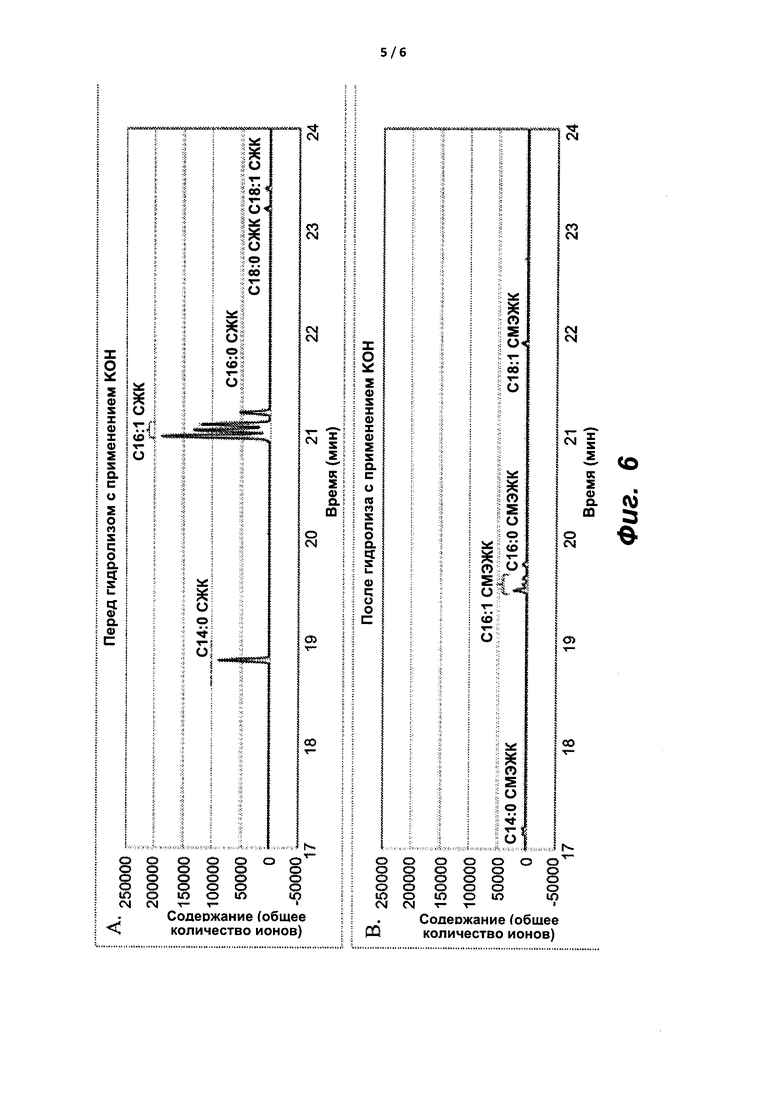
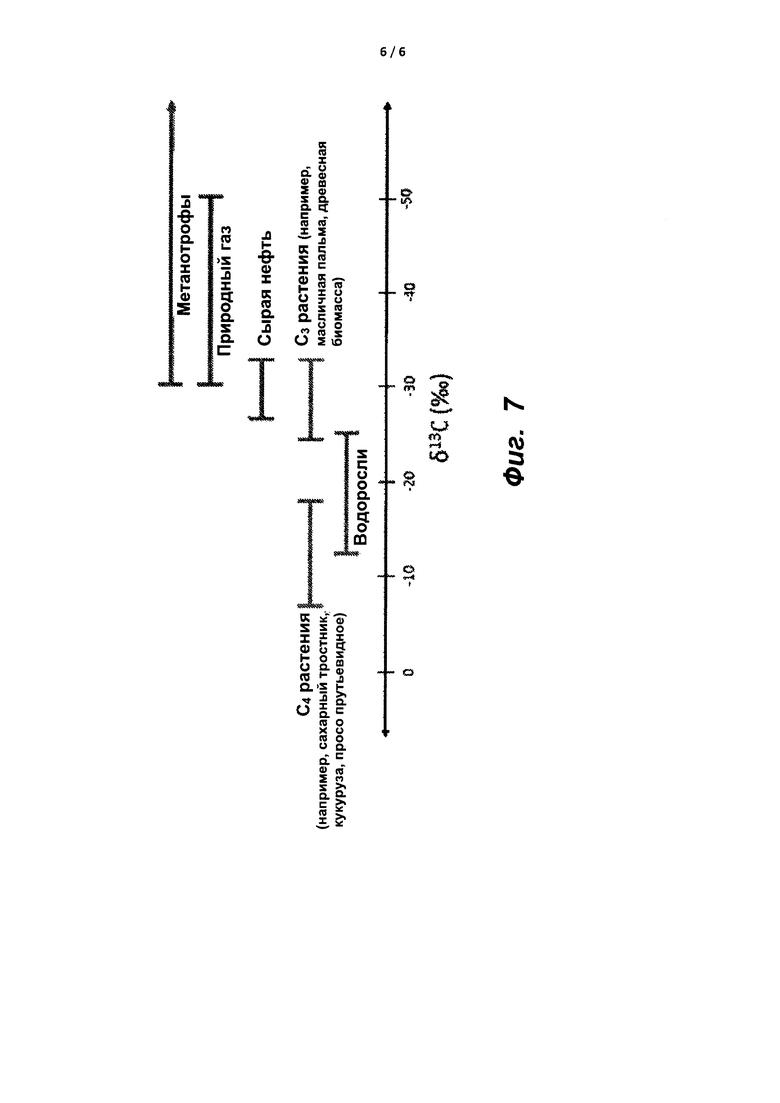
Группа изобретений относится к биотехнологии. Предложен рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм, содержащий гетерологичный полинуклеотид, кодирующий тиоэстеразу, малонил-КоА: ацилпереносящий белок-трансацилазу, ацетил-КоА-карбоксилазу или любую их комбинацию. При этом экспрессия указанного гетерологичного полинуклеотида приводит к аккумуляции в вышеуказанном микроорганизме жирных кислот на повышенном уровне по сравнению с родительским метаболизирующим C1 нефотосинтезирующим микроорганизмом. Вышеуказанный микроорганизм представляет собой метилотрофную и/или метанотрофную бактерию. Предложена также композиция биомассы вышеуказанного микроорганизма, содержащая жирные кислоты на повышенном уровне по сравнению с биомассой родительского метаболизирующего C1 нефотосинтезирующего микроорганизма, предназначенная для применения для получения топлива, топливной добавки, корма для животных, энергии или высокоэнергетической молекулы, которая выбрана из жирных кислот, сложных эфиров жирных кислот, триглицеридов, фосфолипидов, изопренов, терпенов, полигидроксиалканоатов и жирных спиртов. Предложены также способ получения биомассы, включающий культивирование вышеуказанного микроорганизма, масляная композиция вышеуказанного микроорганизма, предназначенная для применения для получения топлива, топливной добавки, корма для животных, энергии или высокоэнергетической молекулы, которая выбрана из жирных кислот, сложных эфиров жирных кислот, триглицеридов, фосфолипидов, изопренов, терпенов, полигидроксиалканоатов и жирных спиртов, способ получения указанной масляной композиции, а также применение вышеуказанного микроорганизма. Группа изобретений может быть использована в биотехнологических способах получения биотоплива. 7 н. и 10 з.п. ф-лы, 7 ил., 6 табл., 7 пр.
1. Рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм, при этом указанный рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм содержит гетерологичный полинуклеотид, кодирующий тиоэстеразу, малонил-КоА: ацилпереносящий белок-трансацилазу, ацетил-КоА-карбоксилазу или любую их комбинацию, и при этом экспрессия указанного гетерологичного полинуклеотида приводит к аккумуляции в указанном рекомбинантном метаболизирующем C1 нефотосинтезирующем микроорганизме жирных кислот на повышенном уровне по сравнению с родительским метаболизирующим C1 нефотосинтезирующим микроорганизмом, при этом указанный рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой метилотрофную и/или метанотрофную бактерию.
2. Рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм по п. 1, отличающийся тем, что:
(i) тиоэстераза является кодон-оптимизированной для указанного метаболизирующего C1 нефотосинтезирующего микроорганизма;
(ii) малонил-КоА: ацилпереносящий белок-трансацилаза представляет собой кодон-оптимизированный fabD E. coli;
(iii) ацетил-КоА-карбоксилаза представляет собой кодон-оптимизированный accA, accB, accC, accD E. coli или любую их комбинацию; или
(iv) в указанном рекомбинантном метаболизирующем C1 микроорганизме дополнительно минимизирована или устранена активность жирная кислота: КоА-лигазы.
3. Рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм по любому из пп. 1 или 2, отличающийся тем, что указанный рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм представляет собой метилотрофную бактерию.
4. Рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм по п. 3, отличающийся тем, что указанная родительская бактерия представляет собой Methylobacterium extorquens, Methylobacterium radiotolerans, Methylobacterium populi, Methylobacterium chloromethanicum или Methylobacterium nodulans.
5. Рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм по п. 1 или 2, отличающийся тем, что указанная рекомбинантная метаболизирующая C1 бактерия представляет собой метанотрофную бактерию.
6. Рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм по п. 5, отличающийся тем, что указанная родительская бактерия представляет собой Methylomonas sp. 16a, Methylosinus trichosporium OB3b, Methylosinus sporium, Methylocystis parvus, Methylomonas methanica, Methylomonas albus, Methylobacter capsulatus Y, Methylococcus capsulatus Bath, Methylobacterium organophilum, Methylomonas sp. AJ-3670, Methylomicrobium alcaliphilum, Methylocella silvestris или Methylacidiphilum infernorum.
7. Композиция биомассы метаболизирующего C1 нефотосинтезирующего микроорганизма, содержащая жирные кислоты на повышенном уровне по сравнению с биомассой родительского метаболизирующего C1 нефотосинтезирующего микроорганизма, для применения для получения топлива, топливной добавки, корма для животных, энергии или высокоэнергетической молекулы, которая выбрана из жирных кислот, сложных эфиров жирных кислот, триглицеридов, фосфолипидов, изопренов, терпенов, полигидроксиалканоатов и жирных спиртов, при этом указанная композиция содержит:
(а) культуру рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма по любому из пп. 1-6 совместно с культуральной средой, в которой выращен указанный микроорганизм;
(b) рекомбинантный метаболизирующий C1 нефотосинтезирующий микроорганизм по любому из пп. 1-6, выращенный в культуральной среде и извлеченный из указанной культуральной среды; или
(с) супернатант отработавшей среды, полученный из культуры указанного рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма по подпункту (а),
и при этом δ13C указанной биомассы составляет менее приблизительно -30‰.
8. Биомасса по п. 7, отличающаяся тем, что δ13C биомассы составляет от приблизительно -40‰ до приблизительно -60‰.
9. Способ получения биомассы, включающий культивирование рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма по любому из пп. 1-6 в блоке регулируемого культивирования в присутствии сырья, содержащего субстрат С1 при условиях и времени, которые являются достаточными для получения биомассы, при этом δ13C указанной биомассы составляет от приблизительно -70‰ до приблизительно -30‰, причём указанная биомасса содержит жирные кислоты на повышенном уровне по сравнению с биомассой родительского метаболизирующего C1 нефотосинтезирующего микроорганизма.
10. Масляная композиция метаболизирующего C1 нефотосинтезирующего микроорганизма по любому из пп. 1-6 для применения для получения топлива, топливной добавки, корма для животных, энергии или высокоэнергетической молекулы, которая выбрана из жирных кислот, сложных эфиров жирных кислот, триглицеридов, фосфолипидов, изопренов, терпенов, полигидроксиалканоатов и жирных спиртов, при этом указанная композиция содержит
(а) по меньшей мере 50% (масс./масс.) жирных кислот; или
(b) по меньшей мере 50% (масс./масс.) свободных жирных кислот.
11. Масляная композиция по п. 10, отличающаяся тем, что указанный терпеноид представляет собой фарнезин, лимонен или и то, и другое.
12. Способ получения масляной композиции, при этом указанный способ включает:
(i) выращивание микроорганизма по любому из пп. 1-6 на субстрате С1 путем периодического или непрерывного культивирования, и
(ii) экстракцию указанной масляной композиции из клеточной мембраны указанного микроорганизма или путем получения указанной масляной композиции из супернатанта культуры, или их комбинацию.
13. Способ по п. 12, дополнительно включающий очистку указанной масляной композиции путем гидрообработки для получения реактивного топлива, дизельного топлива или их комбинации, причем указанная гидрообработка приводит к образованию легких углеводородов, выбранных из метана, этана, пропана, бутана, пентана, бутанола и изобутанола.
14. Способ по п. 9 или 12, отличающийся тем, что указанный субстрат С1 представляет собой природный газ, метан или метанол.
15. Применение блока регулируемого культивирования, в котором C1 субстрат, необязательно представляющий собой природный газ или синтез-газ, при твердофазной ферментации доставляют в газовой фазе к микробным биопленкам, при этом микробные биопленки содержат рекомбинантный метаболизирующий C1 микроорганизм по любому из пп. 1-6 для получения масляной композиции по п. 10.
16. Применение рекомбинантного метаболизирующего C1 нефотосинтезирующего микроорганизма по любому из пп. 1-6 для получения масляной композиции, которая содержит:
(а) по меньшей мере 50% (масс./масс.) жирных кислот; или
(b) по меньшей мере 50% (масс./масс.) свободных жирных кислот.
17. Применение по п. 16, отличающееся тем, что указанную масляную композицию применяют для получения топлива, топливной добавки, корма для животных, энергии или высокоэнергетической молекулы, которая выбрана из жирных кислот, сложных эфиров жирных кислот, триглицеридов, фосфолипидов, изопренов, терпенов, полигидроксиалканоатов и жирных спиртов.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| SUMMONS R.E | |||
| ET AL | |||
| Carbon isotopic fractionation in lipids from methanotrophic bacteria: Relevance for interpretation of the geochemical record of biomarkers | |||
| Geochimica et Cosmochimica Acta, 1994, vol | |||
| Способ окисления боковых цепей ароматических углеводородов и их производных в кислоты и альдегиды | 1921 |
|
SU58A1 |
| Насос | 1917 |
|
SU13A1 |
| МНОГОХОДОВОЙ ПРОБОЧНЫЙ КРАН | 1925 |
|
SU2853A1 |
| US 20080305540 А1, 11.12.2008 | |||
| ПРИСПОСОБЛЕНИЕ ДЛЯ УКРЕПЛЕНИЯ НА РАБОЧЕМ КОНТАКТНОМ ПРОВОДЕ ВСПОМОГАТЕЛЬНОГО ПРОВОДА, СЛУЖАЩЕГО ДЛЯ УПРАВЛЕНИЯ ПУТЕВЫМИ СТРЕЛКАМИ И Т. П. | 1928 |
|
SU16258A1 |
| JAHNKE L.L | |||
| The effects of | |||

