Область техники, к которой относится изобретение
Настоящее изобретение относится к способам предсказания Т-клеточных эпитопов. В частности, настоящее изобретение относится к способам предсказания, являются иммуногенными ли или нет модификации в пептидах или полипептидах, таких как опухоль-ассоциированные неоантигены. Способы согласно изобретению полезны, в частности, при получении вакцин, которые являются специфическими в отношении опухоли пациента и, таким образом, в контексте персонализированных противораковых вакцин.
Сведения о предшествующем уровне техники
Персонализированные противораковые вакцины представляют собой специально изготовленные терапевтические вакцины для направленного воздействия на опухоль-специфические мутации, которые являются уникальными для данного пациента. Такое лечение дает большую надежду онкопациентам, так как не наносит вред здоровым клеткам и имеет потенциал для обеспечения ремиссии в течение всего периода жизни. Тем не менее, не каждую мутацию, которая экспрессируется опухолью, можно использовать в качестве мишени для вакцины. В действительности, большинство вызывающих рак соматических мутаций не будут приводить к иммунному ответу при вакцинировании (J.С. Castle et al., Exploiting the mutanome for tumor vaccination. Cancer Research 72, 1081 (2012)). Поскольку опухоли могут кодировать не менее 100000 соматических мутаций (М.R. Stratton, Science Signalling 331, 1553 (2011)), а вакцины направленно действуют только на незначительное число эпитопов, очевидно, что главной целью иммунотерапии рака является идентификация мутаций, которые могут быть иммуногенными.
С биологической точки зрения, для того, чтобы соматическая мутация генерировала иммунный ответ, должно выполняться несколько критериев: аллель, содержащий мутацию, должен экспрессироваться клеткой; мутация должна быть в кодирующей белок области и несинонимичной; транслированный белок должен расщепляться протеасомой; и эпитоп, содержащий мутацию, должен быть презентирован комплексом МНС, при этом презентированный эпитоп должен распознаваться Т-клеточным рецептором (TCR); и, в заключение, комплекс TCR-pMHC должен запускать сигнальный каскад, который активирует Т-клетки (S. Whelan, N. Goldman, Molecular biology and evolution 18, 691 (2001)). Таким образом, до сих пор не существует алгоритма, с помощью которого можно предсказывать с высокой степенью достоверности, какие мутации могут удовлетворять этим критериям. В настоящем документе рассматривается несколько факторов, которые могут способствовать иммуногенности, проводится сравнение этих факторов с экспериментальными данными и предлагается простая модель для идентификации иммуногенных мутаций.
Предсказание связывания с МНС: Существующий уровень техники
Более 20 лет назад было установлено, что в пептиде, связывающемся с МНС, имеются положения, которые в большей степени содействуют способности к связыванию, чем другие положения (например, A. Sette et al., Proceedings of the National Academy of Sciences 86, 3296 (1989)). Идентификация и описание этих якорных положений позволило обнаружить паттерны пептидов, связывающихся с МНС, и, таким образом, послужило основой для разработки методов предсказания. В последние годы были достигнуты значительные успехи в области моделей in silico механизма процессинга антигена (Antigen Processing Machinery). Два новаторских подхода, которые были разработаны в конце 1990-х годов, BIMAS (K.С. Parker, М.A. Bednarek, J.Е. Coligan, The Journal of Immunology 152, 163 (1994)) и SYFPEITHI (H.-G. Rammensee, J. Bachmann, N. P. N. Emmerich, O.A. Bachor, S.  Immunogenetics 50, 213 (1999)), основаны на знании якорных положений и на выявленных аллель-специфических мотивах. По мере того, как становилось доступным все большее количество экспериментальных данных, касающихся связывания пептида с МНС, все большее количество инструментов разрабатывалось с использованием разнообразных статистических и компьютерных технологий (для обзора смотри фигуру 1). Так называемые методы на основе матриц используют позиционные весовые матрицы (position-specific scoring matrices) для определения совпадения последовательности пептида со связывающим мотивом конкретного аллеля МНС. Другой класс методов предсказания связывания с МНС использует методы компьютерного осмысления, такие как искусственные нейронные сети (Artificial Neural Networks) или метод опорных векторов (Support Vector Machines) (смотри фигуру 1). Выполнение этих алгоритмов в значительной степени зависит от количества и качества доступных обучающих баз данных для каждой модели аллеля (например, «HLA-A*02:01», «H2-Db» и т.п.) для «изучения» основных паттернов/отличительных признаков, которые обладают способностью предсказывать связывание. Недавно появились методы, основанные на структуре, которые обходят проблемы, связанные с большим обучающим набором, так как опираются исключительно на кристаллических структурах пептид-МНС и оценочных функциях (scoring functions) (например, различных энергетических функциях) для предсказания взаимодействий пептид-МНС путем, например, минимизации энергии (смотри фигуру 1). Однако точность этих подходов все еще сильно отстает от способов, основанных на последовательностях. Сравнительные исследования показывают, что инструмент NetMHC на основе искусственной нейронной сети (С. Lundegaard et al., Nucleic Acids Research 36, W509 (2008)) и алгоритм SMM на основе матриц (В. Peters, А. Sette, ВМС bioinformatics 6, 132 (2005)) исполняются наилучшим образом на тестированных оценочных данных (В. Peters, A. Sette, ВМС bioinformatics 6, 132 (2005); Н.Н. Lin, S. Ray, S. Tongchusak, E.L. Reinherz, V. Brusic, BMC immunology 9, 8 (2008)). Оба подхода интегрированы в так называемые консенсусные методы IEDB, доступные в базе данных иммуноэпитопов (Immune Epitope Database) (Y. Kim et al., Nucleic Acids Research 40, W525 (2012)). Моделирование взаимодействий связывания пептид-МНС II является более сложным, чем для МНС I, так как молекулы МНС II обладают связывающей бороздкой с открытыми концами с каждой стороны, обеспечивая связывание пептидов различной длины. Поскольку связывание пептидов с МНС I ограничивается главным образом 8-12 аминокислотами, эта длина может сильно отличаться для пептидов МНС II (9-30 аминокислот). Недавнее сравнительное исследование показало, что существующие в настоящее время способы предсказания для МНС II характеризуются ограниченной точностью по сравнению с предсказанием для МНС I (Н.Н. Lin, S. Ray, S. Tongchusak, E.L. Reinherz, V. Brusic, BMC immunology 9, 8 (2008)).
Immunogenetics 50, 213 (1999)), основаны на знании якорных положений и на выявленных аллель-специфических мотивах. По мере того, как становилось доступным все большее количество экспериментальных данных, касающихся связывания пептида с МНС, все большее количество инструментов разрабатывалось с использованием разнообразных статистических и компьютерных технологий (для обзора смотри фигуру 1). Так называемые методы на основе матриц используют позиционные весовые матрицы (position-specific scoring matrices) для определения совпадения последовательности пептида со связывающим мотивом конкретного аллеля МНС. Другой класс методов предсказания связывания с МНС использует методы компьютерного осмысления, такие как искусственные нейронные сети (Artificial Neural Networks) или метод опорных векторов (Support Vector Machines) (смотри фигуру 1). Выполнение этих алгоритмов в значительной степени зависит от количества и качества доступных обучающих баз данных для каждой модели аллеля (например, «HLA-A*02:01», «H2-Db» и т.п.) для «изучения» основных паттернов/отличительных признаков, которые обладают способностью предсказывать связывание. Недавно появились методы, основанные на структуре, которые обходят проблемы, связанные с большим обучающим набором, так как опираются исключительно на кристаллических структурах пептид-МНС и оценочных функциях (scoring functions) (например, различных энергетических функциях) для предсказания взаимодействий пептид-МНС путем, например, минимизации энергии (смотри фигуру 1). Однако точность этих подходов все еще сильно отстает от способов, основанных на последовательностях. Сравнительные исследования показывают, что инструмент NetMHC на основе искусственной нейронной сети (С. Lundegaard et al., Nucleic Acids Research 36, W509 (2008)) и алгоритм SMM на основе матриц (В. Peters, А. Sette, ВМС bioinformatics 6, 132 (2005)) исполняются наилучшим образом на тестированных оценочных данных (В. Peters, A. Sette, ВМС bioinformatics 6, 132 (2005); Н.Н. Lin, S. Ray, S. Tongchusak, E.L. Reinherz, V. Brusic, BMC immunology 9, 8 (2008)). Оба подхода интегрированы в так называемые консенсусные методы IEDB, доступные в базе данных иммуноэпитопов (Immune Epitope Database) (Y. Kim et al., Nucleic Acids Research 40, W525 (2012)). Моделирование взаимодействий связывания пептид-МНС II является более сложным, чем для МНС I, так как молекулы МНС II обладают связывающей бороздкой с открытыми концами с каждой стороны, обеспечивая связывание пептидов различной длины. Поскольку связывание пептидов с МНС I ограничивается главным образом 8-12 аминокислотами, эта длина может сильно отличаться для пептидов МНС II (9-30 аминокислот). Недавнее сравнительное исследование показало, что существующие в настоящее время способы предсказания для МНС II характеризуются ограниченной точностью по сравнению с предсказанием для МНС I (Н.Н. Lin, S. Ray, S. Tongchusak, E.L. Reinherz, V. Brusic, BMC immunology 9, 8 (2008)).
Первое крупномасштабное и систематическое использование этих алгоритмов для обнаружения Т-клеточных эпитопов было предпринято Moutaftsi et al. (М. Moutaftsi et al., Nature Biotechnology 24, 817 (2006)), в котором были объединены различные инструменты для предсказания возможных вакцин-кандидатов для инфицированных вирусом оспорвакцины мышей C57BL76, выделения спленоцитов и измерения CD8+ Т-клеточных ответов относительно топового 1% предсказанных пептидов. Было идентифицировано 49 (из 2256) пептидов, которые индуцировали Т-клеточный ответ. После этого было опубликовано много исследований, использующих различные инструменты предсказания связывания с МНС для поиска Т-клеточных эпитопов в качестве кандидатов для вакцины, главным образом, для патогенов, например, Leishmania major (С. Herrera-Najera, R.  F. Xacur-Garcia, M.J. Ramirez-Sierra, E. Dumonteil, Proteomics 9, 1293 (2009)). Однако использование только инструментов предсказания связывания с МНС I для предсказания иммуногенности является ошибочным, так как эти инструменты нацелены на предсказание, обладает ли заданный пептид потенциалом к связыванию с заданным аллелем МНС. Рациональным объяснением использования предсказаний связывания с МНС для предсказания иммуногенности является предположение о том, что пептиды, связывающиеся с высокой аффинностью с соответствующим аллелем МНС, с большой долей вероятности являются иммуногенными (A. Sette et al., The Journal of Immunology 153, 5586 (1994)). Однако существует ряд исследований, которые указывают на то, что низкая аффинность связывания с МНС также может привести к высокой иммуногенности (М.С. Feltkamp, М.P. Vierboom, W.М. Kast, С.J. Melief, Molecular Immunology 31, 1391 (1994)), и что стабильность пептид-МНС может быть более эффективным предиктором иммуногенности, чем аффинность пептидов (М. Harndahl et al., European Journal of Immunology 42, 1405 (2012)). По этой причине предсказание иммуногенности до настоящего времени было не очень точным, что отражалось в низких показателях успешности (success rates) предсказания иммуногенности. Однако пептидное связывание является необходимым, но недостаточным условием распознавания Т-клеточного эпитопа, и эффективное предсказание может значительно снизить количество пептидов, подлежащих экспериментальному тестированию.
F. Xacur-Garcia, M.J. Ramirez-Sierra, E. Dumonteil, Proteomics 9, 1293 (2009)). Однако использование только инструментов предсказания связывания с МНС I для предсказания иммуногенности является ошибочным, так как эти инструменты нацелены на предсказание, обладает ли заданный пептид потенциалом к связыванию с заданным аллелем МНС. Рациональным объяснением использования предсказаний связывания с МНС для предсказания иммуногенности является предположение о том, что пептиды, связывающиеся с высокой аффинностью с соответствующим аллелем МНС, с большой долей вероятности являются иммуногенными (A. Sette et al., The Journal of Immunology 153, 5586 (1994)). Однако существует ряд исследований, которые указывают на то, что низкая аффинность связывания с МНС также может привести к высокой иммуногенности (М.С. Feltkamp, М.P. Vierboom, W.М. Kast, С.J. Melief, Molecular Immunology 31, 1391 (1994)), и что стабильность пептид-МНС может быть более эффективным предиктором иммуногенности, чем аффинность пептидов (М. Harndahl et al., European Journal of Immunology 42, 1405 (2012)). По этой причине предсказание иммуногенности до настоящего времени было не очень точным, что отражалось в низких показателях успешности (success rates) предсказания иммуногенности. Однако пептидное связывание является необходимым, но недостаточным условием распознавания Т-клеточного эпитопа, и эффективное предсказание может значительно снизить количество пептидов, подлежащих экспериментальному тестированию.
Очевидно, что при разработке модели, которая предсказывает иммуногенность, следует также учитывать распознавание Т-клеточного рецептора (TCR), а также центральную толерантность, то есть негативную и положительную селекцию Т-клеток в процессе развития в тимусе.
Существует потребность в модели предсказания, которая способна моделировать все аспекты, упомянутые выше, для точного предсказания иммуногенности эпитопа, а не только связывания.
Краткое описание изобретения
В одном аспекте настоящее изобретение относится к способу предсказания иммуногенных аминокислотных модификаций, при этом способ включает следующие стадии:
a) определение показателя связывания модифицированного пептида с одной или несколькими молекулами МНС, и
b) определение показателя связывания немодифицированного пептида с одной или несколькими молекулами МНС, и/или
c) определение показателя связывания модифицированного пептида в случае присутствия в комплексе МНС-пептид с одним или несколькими Т-клеточными рецепторами.
В одном варианте осуществления модифицированный пептид содержит фрагмент модифицированного белка, при этом указанный фрагмент, содержащий модификацию(и), присутствует в белке. В одном варианте осуществления немодифицированный пептид или белок содержит аминокислоту зародышевой линии в положении(ях), соответствующем положению(ям) модификации(й) в модифицированном пептиде или белке.
В одном варианте осуществления немодифицированный пептид или белок и модифицированный пептид или белок являются идентичными, за исключением модификации(й). Предпочтительно, немодифицированный пептид или белок и модифицированный пептид или белок имеют одинаковую длину или/и последовательность (за исключением модификации(й)).
В одном варианте осуществления немодифицированный пептид и модифицированный пептид имеют от 8 до 15, предпочтительно от 8 до 12 аминокислот в длину.
В одном варианте осуществления одна или несколько молекул МНС включают различные типы молекул МНС, в частности, различные аллели МНС. В одном варианте осуществления одна или несколько молекул МНС представляют собой молекулы МНС класса I или молекулы МНС класса II. В одном варианте осуществления одна или несколько молекул МНС содержат набор аллелей МНС, например, набор аллелей МНС индивидуума или его поднабор.
В одном варианте осуществления показатель связывания с одной или несколькими молекулами МНС определяют с помощью процесса, включающего сравнение последовательности с базой данных МНС-связывающих мотивов.
В одном варианте осуществления стадия а) включает определение, удовлетворяет ли указанный показатель предварительно определенной пороговой величине связывания с одной или несколькими молекулами МНС, и/или стадия b) включает определение, удовлетворяет ли указанный показатель предварительно определенной пороговой величине связывания с одной или несколькими молекулами МНС. В одном варианте осуществления пороговая величина, применяющаяся на стадии а), отличается от пороговой величины, применяющейся на стадии b). В одном варианте осуществления предварительно определенная пороговая величина связывания с одной или несколькими молекулами МНС отражает вероятность связывания с одной или несколькими молекулами МНС.
В одном варианте осуществления один или несколько Т-клеточных рецепторов включают набор Т-клеточных рецепторов, такой как набор Т-клеточных рецепторов индивидуума или его поднабор. В одном варианте осуществления стадия с) включает допущение того, что указанный набор Т-клеточных рецепторов не включает Т-клеточные рецепторы, которые связываются с немодифицированным пептидом в случае присутствия в комплексе МНС-пептид, и/или не включают Т-клеточные рецепторы, которые связываются с высокой аффинностью с немодифицированным пептидом в случае присутствия с комплексе МНС-пептид.
В одном варианте осуществления стадия с) включает определение показателя химического и физического сходства между немодифицированными и модифицированными аминокислотами. В одном варианте осуществления стадия с) включает определение, удовлетворяет ли указанный показатель предварительно определенной пороговой величине химического и физического сходства между аминокислотами. В одном варианте осуществления указанная предварительно определенная пороговая величина химического и физического сходства между аминокислотами отражает вероятность того, что аминокислоты являются химически и физически сходными. В одном варианте осуществления показатель химического и физического сходства определен на основании вероятности того, что аминокислоты являются взаимозаменяемыми в природе. В одном варианте осуществления, чем чаще аминокислоты взаимозаменяются в природе, тем более сходными аминокислоты считаются, и наоборот. В одном варианте осуществления химическое и физическое сходство определяют с использованием эволюционных матриц логарифмических различий (log-odds).
В одном варианте осуществления, если немодифицированный пептид имеет показатель связывания с одной или несколькими молекулами МНС, который удовлетворяет пороговой величине, свидетельствуя о связывании с одной или несколькими молекулами МНС, и модифицированный пептид имеет показатель связывания с одной или несколькими молекулами МНС, который удовлетворяет пороговой величине, свидетельствуя о связывании с одной или несколькими молекулами МНС, то модификация или модифицированный пептид предсказан как иммуногенный, если немодифицированные или модифицированные аминокислоты имеют показатель химического и физического сходства, который удовлетворяет пороговой величине, свидетельствующей о химическом и физическом несходстве.
В одном варианте осуществления, если немодифицированный пептид связывается с одной или несколькими молекулами МНС, или имеет вероятность связывания с одной или несколькими молекулами МНС, и модифицированный пептид связывается с одной или несколькими молекулами МНС, или имеет вероятность связывания с одной или несколькими молекулами МНС, то модификация или модифицированный пептид предсказан как иммуногенный, если немодифицированные и модифицированные аминокислоты являются химически и физически несходными, или имеют вероятность являться химически или физически несходными.
В одном варианте осуществления модификация не находится в якорном положении для связывания с одной или несколькими молекулами МНС.
В одном варианте осуществления, если немодифицированный пептид имеет показатель связывания с одной или несколькими молекулами МНС, удовлетворяющий пороговой величине, свидетельствующей об отсутствии связывания с одной или несколькими молекулами МНС, и модифицированный пептид имеет показатель связывания с одной или несколькими молекулами МНС, удовлетворяющий пороговой величине, свидетельствующей о связывании с одной или несколькими молекулами МНС, то модификация или модифицированный пептид является предсказанным как иммуногенный.
В одном варианте осуществления, если немодифицированный пептид не связывается с одной или несколькими молекулами МНС, или имеет вероятность не связываться с одной или несколькими молекулами МНС, и модифицированный пептид связывается с одной или несколькими молекулами МНС или имеет вероятность связывания с одной или несколькими молекулами МНС, то модификация или модифицированный пептид является предсказанным как иммуногенный.
В одном варианте осуществления модификация находится в якорном положении для связывания с одной или несколькими молекулами МНС.
В одном варианте осуществления способ согласно изобретению включает выполнение стадии а) на двух или более различных модифицированных пептидах, при этом указанные два или более различных модифицированных пептидов содержат одинаковую модификацию(и). В одном варианте осуществления два или более различных модифицированных пептидов, содержащих одинаковую модификацию(и), содержат различные фрагменты модифицированного белка, при этом указанные различные фрагменты содержат одинаковую модификацию(и), присутствующую в белке. В одном варианте осуществления два или более различных модифицированных пептидов, содержащих одинаковую модификацию(и), содержат все потенциальные связывающиеся с МНС фрагменты модифицированного белка, при этом указанные фрагменты содержат одинаковую модификацию(и), присутствующую в белке. В одном варианте осуществления способ согласно изобретению дополнительно включает отбор модифицированного пептида(ов) из двух или более различных модифицированных пептидов, содержащих одинаковую модификацию(и), имеющих вероятность или имеющих наиболее высокую вероятность связывания с одной или несколькими молекулами МНС. В одном варианте осуществления два или более различных модифицированных пептидов, содержащих одинаковую модификацию(и), отличаются по длине и/или положению модификации(й).
В одном варианте осуществления способ согласно изобретению включает исполнение стадии а) и необязательно одной или обеих стадий b) и с) на двух или более различных модифицированных пептидах. В одном варианте осуществления указанные два или более различных модифицированных пептидов содержат одинаковую модификацию(и) и/или содержат различные модификации. В одном варианте осуществления различные модификации присутствуют в одинаковых и/или различных белках. Набор из двух или более различных модифицированных пептидов, используемых на стадии а), и необязательно на одной или обеих стадиях b) и с), может быть одинаковым или различным. В одном варианте осуществления набор из двух или более различных модифицированных пептидов, используемых на стадии b) и/или на стадии c), представляет собой поднабор набора из двух или более различных модифицированных пептидов, используемых на стадии а). Предпочтительно, указанный поднабор включает пептид(ы), оцененные лучше на стадии а).
В одном варианте осуществления способ согласно изобретению включает сравнение показателей двух или более указанных различных модифицированных пептидов. В одном варианте осуществления способ согласно изобретению включает ранжирование двух или более указанных различных модифицированных пептидов. В одном варианте осуществления показатель связывания модифицированного пептида с одной или несколькими молекулами МНС оценен выше, чем показатель связывания модифицированного пептида с одним или несколькими Т-клеточными рецепторами в случае присутствия в комплексе МНС-пептид; предпочтительно показатель химического и физического сходства между немодифицированными и модифицированными аминокислотами, и показатель связывания модифицированного пептида с одним или несколькими Т-клеточными рецепторами в случае присутствия с комплексе МНС-пептид, предпочтительно показатель химического или физического сходства между немодифицированными и модифицированными аминокислотами оценен выше, чем показатель связывания немодифицированного пептида с одной или несколькими молекулами МНС.
В одном варианте осуществления способ согласно изобретению дополнительно включает идентификацию несинонимичных мутаций в одной или нескольких кодирующих белок областях.
В одном варианте осуществления модификации идентифицированы в соответствии с изобретением путем частичного или полного секвенирования генома или транскриптома одной или нескольких клеток, таких как одна или несколько раковых клеток, и необязательно одной или нескольких нераковых клеток, и идентификации мутаций в одной или нескольких кодирующих белок областях.
В одном варианте осуществления указанные мутации представляют собой соматические мутации. В одном варианте осуществления указанные мутации представляют собой раковые мутации.
В одном варианте осуществления способ согласно изобретению используется для изготовления вакцины. В одном варианте осуществления вакцина получена из (а) модификации(й) или (а) модифицированного пептида(ов), предсказанного как иммуногенный с помощью способов согласно изобретению.
В следующем аспекте в настоящем изобретении предлагается способ обеспечения вакцины, включающий стадию:
идентификации (а) модификации(й) или (а) модифицированного пептида(ов), предсказанного как иммуногенный с помощью способов согласно изобретению.
В одном варианте осуществления способ дополнительно включает стадию:
обеспечения вакцины, содержащей пептид или полипептид, содержащий модификацию(и) или модифицированный пептид(ы), предсказанный как иммуногенный, или нуклеиновую кислоту, кодирующую пептид или полипептид.
В следующем аспекте настоящее изобретение обеспечивает вакцину, которая может быть получена с использованием способов в соответствии с изобретением. Предпочтительные варианты осуществления таких вакцин описаны в настоящем документе.
Вакцина, обеспеченная в соответствии с изобретением, может содержать фармацевтически приемлемый носитель, и необязательно может содержать один или несколько адъювантов, стабилизаторов и т.д. Вакцина может быть представлена в форме терапевтической или профилактической вакцины.
Другой аспект относится к способу индукции иммунного ответа у пациента, включающему введение пациенту вакцины, обеспеченной в соответствии с изобретением.
Другой аспект относится к способу лечения онкопациента, включающему следующие стадии:
(a) обеспечение вакцины с помощью способов в соответствии с изобретением; и
(b) введение указанной вакцины пациенту.
Другой аспект относится к способу лечения онкопациента, включающему введение пациенту вакцины в соответствии с изобретением.
В следующих аспектах изобретение обеспечивает вакцины, описанные здесь, для применения в способах лечения, описанных здесь, в частности, для применения в лечении или предупреждении рака.
Терапии рака, описанные здесь, могут быть комбинированы с хирургической резекцией и/или лучевой терапией, и/или стандартной химиотерапией.
Другие отличительные признаки и преимущества настоящего изобретения станут очевидными из последующего подробного описания и формулы изобретения.
Подробное описание изобретения
Несмотря на то, что настоящее изобретение подробно описано далее, следует понимать, что данное изобретение не ограничивается конкретными методологиями, протоколами и реагентами, описанными здесь, поскольку они могут варьироваться. Также, следует понимать, что используемая здесь терминология предназначена только для описания конкретных вариантов осуществления изобретения и не предназначена для ограничения объема настоящего изобретения, который будет ограничен только прилагаемой формулой изобретения. Если не определено иначе, все технические и научные термины, используемые здесь, имеют те же значения, которые обычно понимаются специалистами в данной области.
Далее будут описаны элементы настоящего изобретения. Эти элементы перечислены с помощью конкретных вариантов осуществления, однако следует понимать, что они могут быть комбинированы любым способом и в любом количестве для создания дополнительных вариантов осуществления изобретения. По-разному описанные примеры и предпочтительные варианты осуществления не должны рассматриваться как ограничивающие настоящее изобретение только однозначно описанными вариантам осуществления. Данное описание следует понимать как поддерживающее и охватывающее варианты осуществления, которые объединяют конкретно описанные варианты осуществления с любым числом раскрытых и/или предпочтительных элементов. Кроме того, любые перестановки и комбинации всех описанных элементов в этой заявке должны считаться раскрытыми с помощью описания настоящей заявки, если контекст не указывает иначе.
Предпочтительно, используемым здесь терминам дают определения, как описано в руководстве "A multilingual glossary of biotechnological terms: (IUPAC Recommendations)", H.G.W. Leuenberger, B. Nagel, and H.  Eds., (1995) Helvetica Chimica Acta, CH-4010 Basel, Switzerland.
Eds., (1995) Helvetica Chimica Acta, CH-4010 Basel, Switzerland.
При практическом осуществлении настоящего изобретения будут задействованы, если не указано иначе, общепринятые методы биохимии, иммунологии и методики рекомбинантной ДНК, которые описаны в литературе в данной области (смотри, например, Molecular Cloning: A Laboratory Manual, 2nd Edition, J. Sambrook et al. eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor 1989).
Во всем данном описании и пунктах формулы изобретения, которые следуют далее, если контекст не требует иного, под словом «содержать» и вариациями, такими как «содержит» и «содержащий», будет подразумеваться включение указанного члена, целого числа или стадии, или группы членов, целых чисел или стадий, но не исключение любого другого члена, целого числа или стадии, или группы членов, целых чисел или стадий, хотя в некоторых вариантах осуществления такой другой член, целое число или стадия, или группа членов, целых чисел или стадий может быть исключена, то есть объект состоит во включении указанного члена, целого числа или стадии, или группы членов, целых чисел или стадий. В данном документе слова, употребляемые в единственном числе, также включают и множественное число, если иное не указано или явно не следует из контекста. Перечисление диапазонов значений в данном документе служит просто для того, чтобы выступать в качестве укороченного метода задания ссылок отдельно на каждое значение, попадающее в диапазон. Если не указано иное, каждое отдельное значение включается в описание так, как если бы оно было отдельно перечислено в данном документе.
Все способы, описанные здесь, могут быть выполнены в любом подходящем порядке, если не указано иное здесь, или иным образом ясно не продиктовано контекстом. Использование любого или всех примеров, или образного языка (например, «такой как»), в настоящем документе предназначено просто для того, чтобы лучше разъяснять изобретение, и не налагает никаких ограничений на область применения изобретения, если не заявлено иное. Ничто в данном описании не должно истолковываться как указание какого-либо незаявленного элемента в качестве важнейшего для применения изобретения на практике.
Несколько документов процитировано в тексте данного описания. Каждый из документов, процитированных здесь (включая все патенты, заявки на патент, научные публикации, технические требования изготовителя, инструкции и т.п.), или выше или ниже, тем самым включен путем отсылки в полном объеме. Ничто здесь не должно толковаться как допущение того, что изобретение не обладает правом предвосхищать такое раскрытие в связи с тем, что изобретение имеет более раннюю дату создания.
В соответствии с настоящим изобретением термин «пептид» относится к веществам, содержащим две или более, предпочтительно 3 или более, предпочтительно 4 или более, предпочтительно 6 или более, предпочтительно 8 или более, предпочтительно 10 или более, предпочтительно 13 или более, предпочтительно 16 или более, предпочтительно 21 или более, и предпочтительно вплоть до 8, 10, 20, 30, 40 или 50, в частности, 100 аминокислот, ковалентно связанных пептидными связями. Термин «полипептид» или «белок» относится к крупным пептидам, предпочтительно к пептидам, содержащим более 100 аминокислотных остатков, но обычно термины «пептид», «полипептид» и «белок» являются синонимами и используются здесь взаимозаменяемо.
В соответствии с изобретением термин «модификация» в отношении пептидов, полипептидов или белков относится к изменению последовательности в пептиде, полипептиде или белке по сравнению с родительской последовательностью, такой как последовательность пептида, полипептида или белка дикого типа. Термин включает варианты аминокислотных инсерций, варианты аминокислотных добавлений, аминокислотных делений и варианты аминокислотных замен, предпочтительно варианты аминокислотных замен. Все эти изменения последовательностей в соответствии с изобретением могут потенциально приводить к образованию новых эпитопов.
Варианты аминокислотных инсерций включают вставки одной или более аминокислот в конкретную аминокислотную последовательность.
Варианты аминокислотных добавлений включают амино- и/или карбокси-концевые слияния с одной или несколькими аминокислотами, например, 1, 2, 3, 4 или 5, или более аминокислот.
Варианты аминокислотных делеций характеризуются удалением одной или нескольких аминокислот из последовательности, например, удалением 1, 2, 3, 4 или 5, или более аминокислот.
Варианты аминокислотных замен характеризуются, по меньшей мере, одним остатком в последовательности, который удален, и другим остатком, который вставлен не его место.
В соответствии с изобретением модификация или модифицированный пептид, используемый для тестирования в способах согласно изобретению, может быть получен из белка, содержащего модификацию.
Термин «полученный» в соответствии с изобретением означает, что конкретная молекулярная единица, в частности, конкретная пептидная последовательность, присутствует в объекте, из которого она получена. В случае аминокислотных последовательностей, в особенности конкретных участков последовательностей, «полученный», в частности, означает, что релевантная аминокислотная последовательность получена из аминокислотной последовательности, в которой она присутствует.
Белок, содержащий модификацию, из которого может быть получена модификация или модифицированный пептид, используемый для тестирования в способах согласно изобретению, может представлять собой неоантиген.
В соответствии с изобретением термин «неоантиген» относится к пептиду или белку, включающему одну или несколько аминокислотных модификаций по сравнению с родительским пептидом или белком. Например, неоантиген может представлять собой опухоль-ассоциированный неоантиген, при этом термин «опухоль-ассоциированный неоантиген» включает пептид или белок, включающий аминокислотные модификации вследствие опухоль-ассоциированных мутаций.
В соответствии с изобретением термин «опухоль-специфическая мутация» или «рак-специфическая мутация» относится к соматической мутации, которая присутствует в нуклеиновой кислоте опухолевой или раковой клетки, но отсутствует в нуклеиновой кислоте, соответствующей нормальной, то есть неопухолевой или нераковой клетки. Термины «опухоль-специфическая мутация» и «опухолевая мутация», и термины «рак-специфическая мутация» и «раковая мутация» используются здесь взаимозаменяемо.
Термин «иммунный ответ» относится к общему ответу организма на мишень, такую как антиген, и предпочтительно относится к клеточному иммунному ответу или клеточному, а также гуморальному иммунному ответу. Иммунный ответ может быть защитным/превентивным/профилактическим и/или терапевтическим.
«Индукция иммунного ответа» может означать, что иммунный ответ отсутствовал до индукции, но также может означать, что некоторый уровень иммунного ответа присутствовал до индукции, а после индукции указанный иммунный ответ усилился. Таким образом, «индукция иммунного ответа» также включает «усиление иммунного ответа». Предпочтительно, чтобы после индукции иммунного ответа у субъекта указанный субъект был защищен от развития заболевания, такого как раковое заболевание, или заболевание было ослабленным в виду индукцированного иммунного ответа. Например, иммунный ответ на экспрессированный опухолью антиген может быть индуцирован у пациента, имеющего раковое заболевания, или у субъекта, имеющего риск развития ракового заболевания. Индукция иммунного ответа в этом случае может означать, что заболевание у субъекта является ослабленным, что у субъекта не развиваются метастазы, или что у субъекта, имеющего риск развития ракового заболевания, не развивается раковое заболевание.
Термины «клеточный иммунный ответ» и «клеточный ответ» или аналогичные термины относятся к иммунному ответу, направленному на клетки, характеризующиеся презентацией антигена посредством молекул МНС класса I или класса II, с участием Т-клеток или Т-лимфоцитов, которые действуют как «хелперы» или «киллеры». Хелперные Т-клетки (также называемые CD4+ Т-клетками) играют центральную роль путем регулирования иммунного ответа, и клетки-киллеры (также называемые цитотоксическими Т-клетками, цитолитическими Т-клетками, CD8+ Т-клетками или CTL) уничтожают больные клетки, такие как раковые клетки, предотвращая выработку большего количества больных клеток. В предпочтительных вариантах осуществления настоящее изобретение включает стимуляцию противоопухолевого ответа CTL против опухолевых клеток, экспрессирующих один или несколько опухоль-ассоциированных антигенов, и предпочтительно презентирующих такие экспрессированные опухолью антигены посредством МНС класса I.
«Антиген» в соответствии с изобретением включает любое вещество, предпочтительно пептид или белок, которое является мишенью и/или вызывает иммунный ответ, такой как специфическая реакция с антителами или Т-лимфоцитами (Т-клетками). Предпочтительно, антиген содержит, по меньшей мере, один эпитоп, такой как Т-клеточный эпитоп. Предпочтительно, антиген в контексте настоящего изобретения представляет собой молекулу, которая необязательно после процессинга индуцирует иммунную реакцию, которая предпочтительно является специфической в отношении антигена (включая клетки, экспрессирующие антиген). Антиген или его Т-клеточный эпитоп предпочтительно презентированы клеткой, предпочтительно антигенпрезентирующей клеткой, которая включает больную клетку, в частности, раковую клетку в контексте молекул МНС, что приводит к иммунному ответу против антигена (включая клетки, экспрессирующие антиген).
В одном варианте осуществления антиген представляет собой опухолевый антиген (также называемый здесь как экспрессируемый опухолью антиген), то есть часть опухолевой клетки, такая как белок или пептид, экспрессируемый в опухолевой клетке, которая может происходить из цитоплазмы, клеточной поверхности или клеточного ядра, в частности, такие, которые в первую очередь образуются внутриклеточно, или в виде поверхностных антигенов опухолевых клеток. Например, опухолевые антигены включают карциноэмбриональный антиген, α1-фетопротеин, изоферритин и фетальный сульфогликопротеин, α2-Н-ферропротеин и γ-фетопротеин. В соответствии с настоящим изобретением опухолевый антиген предпочтительно включает любой антиген, который экспрессируется опухолевыми клетками и необязательно является характерным в отношении типа и/или уровня экспрессии для опухолей или типов рака, а также для опухолевых или раковых клеток, то есть опухоль-ассоциированный антиген. В одном варианте осуществления термин «опухоль-ассоциированный антиген» относится к белкам, которые в нормальных условиях специфически экспрессируются в ограниченном числе тканей и/или органов, или на определенных стадиях развития, например, опухоль-ассоциированные антигены в нормальных условиях могут специфически экспрессироваться в ткани желудка, предпочтительно в слизистой оболочке желудка, в репродуктивных органах, например, в семеннике, трофобластической ткани, например, в плаценте или клетках зародышевой линии, и экспрессируются или аберрантно экспрессируются в одной или нескольких опухолевых или раковых тканях. В этом контексте «ограниченное число» предпочтительно означает не более, чем 3, более предпочтительно не более, чем 2. В контексте настоящего изобретения опухолевые антигены включают, например, дифференцировочные антигены, предпочтительно дифференцировочные антигены, специфичные для типов клеток, то есть белки, которые в нормальных условиях специфически экспрессируются в определенных типах клеток на определенной стадии дифференцировки; раково-тестикулярные антигены, то есть белки, которые в нормальных условиях специфически экспрессируются в семеннике и иногда в плаценте; и антигены, специфические к зародышевой линии. Предпочтительно, опухолевый антиген или аберрантная экспрессия опухолевого антигена идентифицирует раковые клетки. В контексте настоящего изобретения опухолевый антиген, который экспрессируется раковой клеткой у субъекта, например, пациента, страдающего раковым заболеванием, предпочтительно представляет собой собственный белок указанного субъекта. В предпочтительных вариантах осуществления опухолевый антиген в контексте настоящего изобретения экспрессируется в нормальных условиях специфически в ткани или органе, который является неосновным, то есть тканях или органах, которые при повреждении иммунной системой не приводят к смерти субъекта, или в органах или структурах организма, которые недоступны или труднодоступны для иммунной системы.
В соответствии с изобретением термины «опухолевый антиген», «опухоль-ассоциированный антиген», «раковый антиген» и «рак-экспрессирующий антиген» являются эквивалентами и используются здесь взаимозаменяемо.
Термин «иммуногенность» относится к относительной эффективности в отношении индукции иммунного ответа, который предпочтительно связан с терапевтическими видами лечения, такими как терапии против рака. Используемый здесь термин «иммуногенный» относится к свойству обладания иммуногенностью. Например, термин «иммуногенная модификация» при использовании в отношении пептида, полипептида или белка относится к эффективности указанного пептида, полипептида или белка в отношении индукции иммунного ответа, который вызван и/или направлен против указанной модификации. Предпочтительно, немодифицированный пептид, полипептид или белок не вызывает иммунный ответ, вызывает другой иммунный ответ, или вызывает другой уровень, предпочтительно низкий уровень иммунного ответа.
Термины «главный комплекс гистосовместимости» и сокращение «МНС» включают молекулы МНС класса I и МНС класса II, и относятся к комплексу генов, который есть у всех позвоночных. Белки или молекулы МНС являются важными для передачи сигнала между лимфоцитами и антигенпрезентирующими клетками или больными клетками в иммунных реакциях, при этом белки или молекулы МНС связываются с пептидами и презентируют их для распознавания Т-клеточными рецепторами. Белки, кодируемые МНС, экспрессируются на поверхности клеток и представляют собственные антигены (пептидные фрагменты из самой клетки) и несобственные антигены (например, фрагменты инвазирующих микроорганизмов) Т-клеткам.
Область МНС подразделяется на три подгруппы, класс I, класс II, и класс III. Белки МНС класса I содержат α-цепь и β2-микроглобулин (не часть МНС, кодированную хромосомой 15). Они презентируют фрагменты антигена цитотоксическим Т-клеткам. На поверхности большинства клеток иммунной системы, в особенности на поверхности антигенпрезентирующих клеток, белки МНС класса II содержат α- и β-цепи, и они презентируют фрагменты антигена Т-хелперным клеткам. Область МНС класса III кодирует другие компоненты иммунной системы, такие как компоненты комплемента и некоторые, которые кодируют цитокины.
МНС является как полигенным (имеется несколько генов МНС класса I и МНС класса II), так и полиморфным (имеется множество аллелей каждого гена).
Используемый здесь термин «гаплотип» относится к аллелям HLA, расположенным на одной хромосоме, и кодируемым ими белкам. Гаплотип может также относиться к аллелю, который находится в любом локусе в пределах МНС. Каждый класс МНС представлен несколькими локусами: например, антигены HLA-A (Human Leukocyte Antigen-A - человеческий антиген лейкоцитов), HLA-B, HLA-C, HLA-E, HLA-F, HLA-G, HLA-H, HLA-J, HLA-K, HLA-L, HLA-P и HLA-V для класса I, и HLA-DRA, HLA-DRB1-9, HLA-, HLA-DQA1, HLA-DQB1, HLA-DPA1, HLA-DPB1, HLA-DMA, HLA-DMB, HLA-DOA и HLA-DOB для класса II. Термины «аллель HLA» и «аллель МНС» используются здесь взаимозаменяемо.
Гены МНС отличаются высокой степенью полиморфизма: в пределах человеческой популяции в каждом генетическом локусе имеется большое число гаплотипов, содержащих различные аллели. Различные полиморфные аллели МНС обоих классов, класса I и класса II, обладают различными специфичностями в отношении пептидов: каждая аллель кодирует белки, которые связываются с пептидами, проявляющими паттерны конкретных последовательностей.
В одном предпочтительном варианте осуществления всех аспектов изобретения молекула МНС представляет собой молекулу HLA.
В контексте настоящего изобретения термин «пептид, связывающийся с МНС» включает пептиды, связывающиеся с МНС класса I и/или класса II, или пептиды, которые могут быть процессированы с получением пептидов, связывающихся с МНС класса I и/или класса II. В случае комплексов МНС класса I/пептид связывающиеся пептиды имеют длину, которая обычно составляет 8-12, предпочтительно 8-10 аминокислот, хотя более длинные или более короткие пептиды могут быть эффективными. В случае комплексов МНС класса II/пептид связывающиеся пептиды имеют длину, которая обычно составляет 9-30, предпочтительно 10-25 аминокислот и, в частности, 13-18 аминокислот, при этом более длинные и более короткие пептиды могут быть эффективными.
В случае, если пептид должен быть презентирован напрямую, то есть без процессинга, в частности, без расщепления, то он имеет длину, которая является подходящей для связывания с молекулой МНС, в частности, молекулой МНС класса I, и предпочтительно составляет 7-30 аминокислот, такую как 7-20 аминокислот, более предпочтительно 7-12 аминокислот, более предпочтительно 8-11 аминокислот, в частности, 9 или 10 аминокислот.
В случае, если пептид является частью более крупной молекулярной единицы, содержащей дополнительные последовательности, например, последовательность вакцины или полипептида, и должен быть презентирован после процессинга, в частности, после расщепления, то пептид, полученный путем процессинга, имеет длину, которая является подходящей для связывания с молекулой МНС, в частности, молекулой МНС класса I, и предпочтительно составляет 7-30 аминокислот, такую как 7-20 аминокислот, более предпочтительно 7-12 аминокислот, более предпочтительно 8-11 аминокислот, в частности, 9 или 10 аминокислот. Предпочтительно, последовательность пептида, которая должна быть презентирована после процессинга, получена из аминокислотной последовательности антигена или полипептида, используемого для вакцинации, то есть его последовательность по существу соответствует и предпочтительно является полностью идентичной фрагменту антигена или полипептида.
Таким образом, пептид, связывающийся с молекулами МНС, в одном варианте осуществления содержит последовательность, которая по существу соответствует и предпочтительно является полностью идентичной фрагменту антигена.
Термин «эпитоп» относится к антигенной детерминанте в молекуле, такой как антиген, то есть к части в молекуле или фрагменту молекулы, который распознается иммунной системой, например, который распознается Т-клеткой, в частности, при презентации в контексте молекул МНС. Эпитоп белка, такой как опухолевый антиген, предпочтительно содержит непрерывную или прерывающуюся часть указанного белка и имеет длину, которая составляет от 5 до 100, предпочтительно от 5 до 50, более предпочтительно от 8 до 30, наиболее предпочтительно от 10 до 25 аминокислот, например, эпитоп может предпочтительно иметь длину, которая составляет 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 аминокислот. Особенно предпочтительно, чтобы эпитоп в контексте настоящего изобретения представлял собой Т-клеточный эпитоп.
В соответствии с изобретением эпитоп может связываться с молекулами МНС, такими как молекулы МНС на поверхности клетки и, таким образом, может представлять собой «пептид, связывающийся с МНС».
Используемый здесь термин «неоэпитоп» относится к эпитопу, который не присутствует в референсной клетке, такой как нормальная нераковая клетка или клетка зародышевой линии, но обнаруживается в раковых клетках. Это включает, в частности, ситуации, когда в нормальной нераковой клетке или клетке зародышевой линии обнаруживается соответствующий эпитоп, однако вследствие одной или нескольких мутаций в раковой клетке последовательность эпитопа изменяется таким образом, что приводит к образованию неоэпитопа.
Используемый здесь термин «Т-клеточный эпитоп» относится к пептиду, который связывается с молекулой МНС в конфигурации, распознаваемой Т-клеточным рецептором. Обычно Т-клеточные эпитопы презентированы на поверхности антигенпрезентирующих клеток.
Используемый здесь термин «предсказание Т-клеточных эпитопов» относится к предсказанию, будет ли пептид связываться с молекулой МНС и будет ли распознаваться Т-клеточным рецептором. Термин «предсказание Т-клеточных эпитопов» является по существу синонимом выражению «предсказание, является ли пептид иммуногенным».
В соответствии с изобретением Т-клеточный эпитоп может присутствовать в вакцине как часть более крупного объекта, такого как вакцинная последовательность и/или полипептид, содержащей более одного Т-клеточного эпитопа. Представленный пептид или Т-клеточный эпитоп получен после подходящего процессинга.
Т-клеточные эпитопы могут быть модифицированы по одному или нескольким остаткам, которые не являются важными для распознавания Т-клеточными рецепторами (TCR) или для связывания с МНС. Такие модифицированные Т-клеточные эпитопы могут считаться иммунологически эквивалентными.
Предпочтительно, Т-клеточный эпитоп, в случае презентации МНС и распознавании Т-клеточным рецептором, способен индуцировать в присутствии подходящих костимулирующих сигналов клональное размножение Т-клеток, несущих Т-клеточный рецептор, специфически распознающий комплекс пептид/МНС.
Предпочтительно, Т-клеточный эпитоп содержит аминокислотную последовательность, по существу соответствующую аминокислотной последовательности фрагмента антигена. Предпочтительно, указанный фрагмент антигена представляет собой пептид, презентированный МНС класса I и/или класса II.
Т-клеточный эпитоп в соответствии с изобретением предпочтительно относится к части или фрагменту антигена, который способен стимулировать иммунный ответ, предпочтительно клеточный ответ против антигена или клеток, характеризующихся экспрессией антигена и предпочтительно презентацией антигена, таких как больные клетки, в частности, раковые клетки. Предпочтительно, Т-клеточный эпитоп способен стимулировать клеточный ответ против клетки, характеризующейся презентацией антигена с участием МНС класса I, и предпочтительно способен стимулировать отвечающие на антиген цитотоксические Т-лимфоциты (CTL).
«Процессинг антигена» или «процессинг» относится к деградации пептида, полипептида или белка на продукты процессинга, которые представляют собой фрагменты указанного пептида, полипептида или белка (например, деградации полипептида на пептиды), и ассоциации со специфическими Т-клетками одного или нескольких из этих фрагментов (например, путем связывания) с молекулами МНС для презентации клетками, предпочтительно антигенпрезентирующими клетками.
«Антигенпрезентирующие клетки» (АРС) представляют собой клетки, которые презентируют пептидные фрагменты белковых антигенов в ассоциации с молекулами МНС на их клеточной поверхности. Некоторые АРС могут активировать антиген-специфические Т-клетки.
«Профессиональные» антигенпрезентирующие клетки являются очень эффективными в отношении интернализации антигена либо путем фагоцитоза или рецептор-опосредованного эндоцитоза, и затем представляют фрагмент этого антигена на их мембране в комплексе с молекулами МНС класса II. Т-клетка распознает и взаимодействует с комплексом антиген-молекула МНС класса II на мембране антигенпрезентирующей клетки. После этого дополнительный костимулирующий сигнал продуцируется антигенпрезентирующей клеткой, приводя к активации Т-клетки. Экспрессия этих костимулирующих молекул является характерной чертой «профессиональных» антигенпрезентирующих клеток.
Основными типами «профессиональных» антигенпрезентирующих клеток являются дендритные клетки, которые презентируют наиболее широкий ряд антигенов и, вероятно, являются наиболее важными антигенпрезентирующими клетками; макрофаги; В-клетки; и некоторые активированные эпителиальные клетки. Дендритные клетки (DC) представляют собой популяции лейкоцитов, которые презентируют антигены, захваченные в периферических тканях, Т-клеткам посредством обоих путей презентации антигена, МНС класса II и I. Хорошо известно, что дендритные клетки являются мощными индукторами иммунных ответов, и активация этих клеток является важной стадией для индукции противоопухолевого иммунитета. Дендритные клетки обычно подразделяются на «незрелые» и «зрелые» клетки, что можно использовать в качестве простого способа проведения различия между двумя хорошо охарактеризованными фенотипами. Однако эта номенклатура не должна предназначаться для исключения всех возможных промежуточных стадий дифференцировки. Незрелые дендритные клетки характеризуются как антигенпрезентирующие клетки, обладающие высокой способностью поглощать и процессировать антиген, которая коррелирует с высокой экспрессией Fcγ-рецептора и рецептора маннозы. Зрелый фенотип обычно характеризуется более низкой экспрессией этих маркеров, но отличается высокой экспрессией молекул клеточной поверхности, ответственных за активацию Т-клеток, таких как молекулы МНС класса I и класса II, молекулы адгезии (например, CD54 и CD11) и костимулирующие молекулы (например, CD40, CD80, CD86 и 4-1 ВВ). Созревание дендритных клеток называется статусом активации дендритных клеток, при котором такие антигенпрезентирующие дендритные клетки приводят к примированию Т-клеток, при этом презентация незрелыми дендритными клетками приводит к толерантности. Созревание дендритных клеток вызвано, главным образом, биомолекулами с микробиологическими характеристиками, детектированными рецепторами врожденного иммунитета (бактериальная ДНК, вирусная РНК, эндотоксины и т.п.), провоспалительными цитокинами (TNF, IL-1, IFN), взаимодействием CD40 с CD40L на поверхности дендритной клетки, и веществами, высвобождающимися из клеток, подвергающихся стрессовой клеточной гибели. Дендритные клетки могут быть получены путем культивирования клеток костного мозга in vitro с цитокинами, такими как гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF) и фактор некроза опухоли-альфа.
«Непрофессиональные» антигенпрезентирующие клетки не экспрессируют конститутивно белки МНС класса II, необходимые для взаимодействия с нативными Т-клетками; они экспрессируются только при стимуляции «непрофессиональных» антигенпрезентирующих клеток определенными цитокинами, такими как IFNγ.
Антигенпрезентирующие клетки могут быть загружены пептидами, презентированными МНС класса I, путем трансдукции клеток нуклеиновой кислотой, предпочтительно РНК, кодирующей пептид или полипептид, содержащий пептид, подлежащий презентации, например, нуклеиновой кислотой, кодирующей антиген или полипептид, используемый для вакцинации.
В некоторых вариантах осуществления пациенту можно вводить фармацевтическую композицию или вакцину, содержащую доставляющий нуклеиновую кислоту носитель, которая направленно воздействует на дендритную или другую антигенпрезентирующую клетку, приводя к трансфекции, которая происходит in vivo. Как правило, трансфекция in vivo дендритных клеток может быть выполнена, например, с использованием любых способов, известных в данной области, например таких способов, которые описаны в WO 97/24447, или подхода «генной пушки», описанного в Mahvi et al., Immunology and cell Biology 75: 456-460, 1997.
В соответствии с изобретением термин «антигенпрезентирующая клетка» также включает клетки-мишени.
«Клетка-мишень» означает клетку, которая является мишенью для иммунного ответа, такого как клеточный иммунный ответ. Клетки-мишени включают клетки, которые презентируют антиген, то есть пептидный фрагмент, происходящий из антигена, и включают любую нежелательную клетку, такую как раковая клетка. В предпочтительных вариантах осуществления клетка-мишень представляет собой клетку, экспрессирующую антиген, как описано здесь, и предпочтительно презентирующую указанный антиген с участием МНС класса I.
Термин «доля» относится к фракции. В отношении конкретной структуры, такой как аминокислотная последовательность или белок, термин его «доля» может обозначать непрерывную или прерывающуюся фракцию указанной структуры. Предпочтительно, доля аминокислотной последовательности содержит по меньшей мере 1%, по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, предпочтительно по меньшей мере 40%, предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 70%, даже более предпочтительно по меньшей мере 80%, и наиболее предпочтительно по меньшей мере 90% аминокислот указанной аминокислотной последовательности. Предпочтительно, если доля представляет собой прерывающуюся фракцию, то указанная прерывающаяся фракция состоит из 2, 3, 4, 5, 6, 7, 8 или более частей структуры, при этом каждая часть является непрерывным элементом структуры. Например, прерывающаяся фракция аминокислотной последовательности может состоять из 2, 3, 4, 5, 6, 7, 8 или более, предпочтительно из не более 4 частей указанной аминокислотной последовательности, при этом каждая часть предпочтительно содержит по меньшей мере 5 непрерывных аминокислот, по меньшей мере 10 непрерывных аминокислот, по меньшей мере 20 непрерывных аминокислот, по меньшей мере 30 непрерывных аминокислот аминокислотной последовательности.
Термины «часть» и «фрагмент» используются здесь взаимозаменяемо и относятся к непрерывному элементу. Например, часть структуры, такой как аминокислотная последовательность или белок, относится к непрерывному элементу указанной структуры. Доля, часть или фрагмент структуры предпочтительно обладает одним или несколькими функциональными свойствами указанной структуры. Например, доля, часть или фрагмент эпитопа, пептида или белка является предпочтительно иммунологически эквивалентным эпитопу, пептиду или белку, из которого он происходит. В контексте настоящего изобретения «часть» структуры, такой как аминокислотная последовательность, предпочтительно содержит, предпочтительно составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 94%, по меньшей мере 96%, по меньшей мере 98%, по меньшей мере 99% всей структуры или аминокислотной последовательности.
Термин «иммунореактивная клетка» в контексте настоящего изобретения относится к клетке, которая проявляет эффекторные функции во время иммунной реакции. «Иммунореактивная клетка» предпочтительно способна к связыванию с антигеном или клеткой, характеризующейся презентацией антигена или его пептидного фрагмента (например, Т-клеточного эпитопа) и опосредованием иммунного ответа. Например, такие клетки секретируют цитокины и/или хемокины, секретируют антитела, распознают раковые клетки и необязательно уничтожают такие клетки. Например, иммунореактивные клетки включают Т-клетки (цитотоксические Т-клетки, хелперные Т-клетки, инфильтрующие опухоль Т-клетки), В-клетки, естественные клетки киллеры, нейтрофилы, макрофаги и дендритные клетки. Предпочтительно, в контексте настоящего изобретения «иммунореактивные клетки» представляют собой Т-клетки, предпочтительно CD4+ и/или CD8+ Т-клетки.
Предпочтительно «иммунореактивная клетка» распознают антиген или его пептидный фрагмент с некоторой степенью специфичности, в частности, в случае презентации в контексте молекул МНС, например, на поверхности антигенпрезентирующих клеток или больных клеток, таких как раковые клетки. Предпочтительно, указанное распознавание позволяет клетке, которая распознает антиген или его пептидный фрагмент, быть отвечающей или реактивной. Если клетка представляет собой хелперную Т-клетку (CD4+ Т-клетку), несущую рецепторы, которые распознают антиген или его пептидный фрагмент в контексте молекул МНС класса II, такая отвечаемость или реактивность может включать высвобождение цитокинов и/или активацию CD8+ лимфоцитов (CTL) и/или В-клеток. Если клетка представляет собой CTL, то такая отвечаемость или реактивность может включать уничтожение клеток, презентированных в контексте молекул МНС класса I, то есть клеток, характеризующихся презентацией антигена с участием МНС класса I, например, посредством апоптоза или перфорин-опосредованного лизиса клеток. В соответствии с изобретением, отвечаемость CTL может включать устойчивые потоки ионов кальция, клеточное деление, выработку цитокинов, таких как IFN-γ и TNF-α, повышение экспрессии маркеров активации, таких как CD44 и CD69, и специфическое цитолитическое уничтожение экспрессирующих антиген клеток-мишеней. Отвечаемость CTL можно также определить с использованием искусственного репортера, который достоверно показывает отвечаемость CTL. Такие CTL, которые распознают антиген или фрагмент антигена, и являются отвечающими или реактивными, также называются здесь «антиген-отвечающими CTL». Если клетка представляет собой В-клетку, то такая отвечаемость может включать высвобождение иммуноглобулинов.
Термины «Т-клетка» и «Т-лимфоцит» используются здесь взаимозаменяемо и включают Т-хелперные клетки (CD4+ Т-клетки) и цитотоксические Т-клетки (CTL, CD8+ Т-клетки), которые содержат цитолитические Т-клетки.
Т-клетки принадлежат группе белых клеток крови, известных как лимфоциты, и играют центральную роль в опосредованном клетками иммунитете. Они отличаются от других типов лимфоцитов, таких как В-клетки и естественные клетки-киллеры, присутствием специального рецептора на своей клеточной поверхности, называемого Т-клеточным рецептором (TCR). Тимус является важным органом, который отвечает за созревание Т-клеток. Было открыто несколько различных поднаборов Т-клеток, при этом каждый поднабор выполняет отдельную функцию.
Т-хелперные клетки содействуют другим белым клеткам крови в иммунологических процессах, включая, помимо других функций, созревание В-клеток в плазматические клетки и активацию цитотоксических Т-клеток и макрофагов. Эти клетки также известны как CD4+ Т-клетки, так как они экспрессируют белок CD4 на своей поверхности. Хелперные Т-клетки становятся активированными, когда они презентированы с пептидными антигенами молекулами МНС класса II, которые экспрессируются на поверхности антигенпрезентирующих клеток (АРС). После активации они быстро делятся и секретируют небольшие белки, называемые цитокинами, которые регулируют или способствуют активному иммунному ответу.
Цитотоксические Т-клетки разрушают инфицированные вирусом клетки и опухолевые клетки, а также вовлечены в отторжение трансплантата. Эти клетки также известны как CD8+ Т-клетки, поскольку они экспрессируют гликопротеин CD8 на своей поверхности. Эти клетки распознают свои мишени путем связывания с антигеном, ассоциированным с МНС класса I, который презентирован на поверхности почти каждой клетки организма.
Большинство Т-клеток несут Т-клеточный рецептор (TCR), существующий в виде комплекса из нескольких белков. Фактический Т-клеточный рецептор состоит из двух отдельных пептидных цепей, которые получены из независимых генов Т-клеточного рецептора альфа и бета (TCRα и TCRβ) и называются α- и β-TCR-цепи. γδ-Т-клетки (гамма-дельта-Т-клетки) представляют небольшой поднабор Т-клеток, которые содержат особый Т-клеточный рецептор (TCR) на своей поверхности. Однако, в γδ-Т-клетках, TCR состоит из одной γ-цепи и одной δ-цепи. Эта группа Т-клеток является гораздо менее распространенной (2% от общего числа Т-клеток), чем αβ-Т-клетки.
Первый сигнал к активации Т-клеток обеспечивается путем связывания Т-клеточного рецептора с коротким пептидом, презентированным МНС на другой клетке. Это гарантирует активацию только Т-клетки с TCR, специфическим к этому пептиду. Клетка-партнер обычно представляет собой антигенпрезентирующую клетку, такую как «профессиональная» антигенпрезентирующая клетка, обычно дендритная клетка в случае нативных ответов, хотя В-клетки и макрофаги могут являться важными АРС.
В соответствии с настоящим изобретением молекула способна связываться с мишенью, если она обладает значительной аффинностью в отношении указанной предварительно определенной мишени и связывается с указанной предварительно определенной мишенью в стандартных анализах. «Аффинность» или «аффинность связывания» часто измеряется равновесной константой диссоциации (KD). Молекула (по существу) неспособна связываться с мишенью, если она не обладает достаточной аффинностью в отношении указанной мишени и не связывается в значительной степени с указанной мишенью в стандартных анализах.
Цитотоксические Т-лимфоциты могут быть получены in vivo путем встраивания антигена или его пептидного фрагмента в антигенпрезентирующие клетки in vivo. Антиген или его пептидный фрагмент может быть представлен в виде белка, ДНК (например, в пределах вектора), или РНК. Антиген может быть процессирован для получения пептидного партнера для молекулы МНС, при этом его фрагмент может быть презентирован без необходимости дополнительного процессинга. Последнее, в частности, относится к случаю, когда они указанный фрагмент может связываться с молекулами МНС. Как правило, введение пациенту путем интрадермальной инъекции является возможным. Однако введение можно также осуществлять интранодально в лимфатический узел (Maloy et al. (2001), Proc Natl Acad Sci USA 98:3299-303). Полученные клетки презентируют представляющий интерес комплекс и распознаются аутологичными цитотоксическими Т-лимфоцитами, которые затем размножаются.
Специфическую активацию CD4+ или CD8+ Т-клеток можно детектировать различными способами. Способы детекции специфической активации Т-клеток включает детекцию пролиферации Т-клеток, выработки цитокинов (например, лимфокинез) или генерирования цитолитической активности. Для CD4+ Т-клеток предпочтительным способом детекции специфической активации Т-клеток является детекция пролиферации Т-клеток. Для CD8+ Т-клеток предпочтительным способом детекции специфической активации Т-клеток является детекция генерирования цитолитической активности.
Выражение «клетка, характеризующаяся презентацией антигена» или «клетка, презентирующая антиген», или аналогичные выражения означают клетку, такую как больная клетка, например раковая клетка, или антигенпрезентирующую клетку, презентирующую антиген, который она экспрессирует, или фрагмент, полученный из указанного антигена, например путем процессинга антигена, в контексте молекул МНС, в частности, молекул МНС класса I. Аналогично, выражение «заболевание, характеризующееся презентацией антигена» означает заболевание, в которое вовлечены клетки, характеризующиеся презентацией антигена, в частности, с участием МНС класса I. Презентация антигена клеткой может быть достигнута путем трансфекции клетки нуклеиновой кислотой, такой как РНК, кодирующей антиген.
Выражение «фрагмент антигена, который презентирован» или сходные выражения означают, что фрагмент может быть презентирован МНС класса I или класса II, предпочтительно МНС класса I, например, при непосредственном добавлении в антигенпрезентирующие клетки. В одном варианте осуществления фрагмент представляет собой фрагмент, который естественным образом презентирован клетками, экспрессирующими антиген.
Термин «иммунологически эквивалентный» означает, что иммунологически эквивалентная молекула, такая как иммунологически эквивалентная аминокислотная последовательность, проявляет такие же или по существу такие же иммунологические свойства, и/или проявляет такие же или по существу такие же иммунологические эффекты, например, в отношении типа иммунологического эффекта, такого как индукция гуморального и/или клеточного иммунного ответа, силы и/или продолжительности индуцированной иммунной реакции, или специфичности индуцированной иммунной реакции. В контексте настоящего изобретения термин «иммунологически эквивалентный» предпочтительно используется в отношении иммунологических эффектов или свойств пептида, используемого для иммунизации. Например, аминокислотная последовательность является иммунологически эквивалентной референсной аминокислотной последовательности, если указанная аминокислотная последовательность при воздействии иммунной системы субъекта индуцирует иммунную реакцию, обладающую специфичностью реакции, индуцируемой референсной аминокислотной последовательностью.
Термин «иммунные эффекторные функции» в контексте настоящего изобретения включает любые функции, опосредованные компонентами иммунной системы, которые приводят, например, к уничтожению опухолевых клеток или ингибированию роста опухоли и/или ингибированию развития опухоли, включая ингибирование диссеминации опухоли и метастазов. Предпочтительно, иммунные эффекторные функции в контексте настоящего изобретения представляют собой эффекторные функции, опосредованные Т-клетками. Такие функции в случае хелперной Т-клетки (CD4+ Т-клетки) включают распознавание антигена или фрагмента антигена в контексте молекул МНС класса II Т-клеточными рецепторами, высвобождение цитокинов и/или активацию CD8+ лимфоцитов (CTL), и/или В-клеток, и в случае CTL распознавание антигена или фрагмента антигена в контексте молекул МНС класса I Т-клеточными рецепторами, уничтожение клеток, презентированных в контексте молекул МНС класса I, то есть клеток, характеризующихся презентацией антигена с участием МНС класса I, например, посредством апоптоза или перфорин-опосредованного лизиса клеток, выработку цитокинов, таких как IFN-γ и TNF-α, и специфическое цитолитическое уничтожение экспрессирующих антиген клеток-мишеней.
В соответствии с изобретением термин «показатель» относится к результату теста или обследования, обычно выраженному в виде цифры. Термины, такие как «показатель лучше» или «самый лучший показатель» относится к результату, который лучше, или самому лучшему результату теста или обследования.
Термины, такие как «предсказывать», «предсказывание» или «предсказание», относятся к определению вероятности.
В соответствии с изобретением определение показателя связывания пептида с одной или несколькими молекулами МНС включает определение вероятности связывания пептида с одной или несколькими молекулами МНС.
Показатель связывания пептида с одной или несколькими молекулами МНС может быть определен путем использования любых прогностических инструментов для предсказания связывания пептид:МНС. Например, можно использовать анализ базы данных иммунных эпитопов (IEDB-AR: http://tools.iedb.org)
Предсказания обычно проводят в отношении набора молекул МНС, такого как набор различных аллелей МНС, таких как все возможные аллели МНС, или набор или поднабор аллелей МНС, обнаруженных у пациента, предпочтительно имеющих модификацию(и), иммуногенность которых подлежит определению в соответствии с изобретением.
В соответствии с изобретением определение показателя связывания модифицированного пептида, в случае присутствия в комплексе МНС-пептид, с одним или несколькими Т-клеточными рецепторами включает определение вероятности связывания пептида, в случае присутствия в комплексе с молекулой МНС, с Т-клеточными рецепторами.
Предсказания могут быть сделаны в отношении одного Т-клеточного рецептора, такого как Т-клеточный рецептор, обнаруженный у пациента, или предпочтительно в отношении набора Т-клеточных рецепторов, такого как неизвестный набор различных Т-клеточных рецепторов, или набор или поднабор Т-клеточных рецепторов, обнаруженных у пациента, предпочтительно имеющих модификацию(и), иммуногенность которых подлежит определению в соответствии с изобретением.
Кроме того, предсказания обычно проводятся в отношении набора молекул МНС, такого как набор различных аллелей МНС, таких как все возможные аллели МНС, или набора или поднабора аллелей, обнаруженного у пациента, предпочтительно имеющих модификацию(и), иммуногенность которых подлежит определению в соответствии с изобретением.
Показатель связывания модифицированного пептида, в случае присутствия в комплексе МНС-пептид, с одним или несколькими Т-клеточными рецепторами может быть определен путем оценки эффекта модификации на связывание комплекса Т-клеточный рецептор-пептид-МНС при условии (неизвестного) репертуара Т-клеточного рецептора. Показатель связывания модифицированного пептида, в случае присутствия в комплексе МНС-пептид, с одним или несколькими Т-клеточными рецепторами может быть, в целом, определен как показатель распознавания данного комплекса пептид-молекула МНС соответствующим Т-клеточным рецептором.
Показатель связывания модифицированного пептида, в случае присутствия в комплексе МНС-пептид, с одним или несколькими Т-клеточными рецепторами может быть определен путем установления физико-химических различий между модифицированной и немодифицированной аминокислотой. Например, могут быть использованы матрицы замен. Такие матрицы описывают скорость, при которой одна аминокислота в последовательности изменяется на другие аминокислотные состояния во времени.
Например, могут быть использованы матрицы логарифмических различий (лог-различий, log-odds), такие как эволюционные матрицы log-odds: замена с низким показателем log-odds имеет больше шансов найти совпадающий Т-клеточный рецептор из пула (неизвестных) молекул Т-клеточного рецептора, чем замена с высоким показателем log-odds (вследствие негативной селекции Т-клеточного рецептора, совпадающего с немодифицированными пептидами). Однако существуют другие способы определения этого показателя. Например, рассмотрение положения мутации в пептиде (некоторые положения могут иметь более низкое воздействие на связывание, чем другие), принятие во внимание ближайшего окружения замещенной аминокислоты (которое может влиять на вторичную структуру замещенной аминокислоты), принятие во внимание всей пептидной последовательности, принятие во внимание полной информации о структуре пептида в молекуле МНС, и т.д. Определение показателя может также включать определение репертуара Т-клеточного рецептора (такого как репертуар Т-клеточного рецептора пациента или его поднабор), например, с использованием секвенирования следующего поколения (NGS) и выполнения симулирования докинга комплексов Т-клеточный рецептор-пептид-МНС.
Настоящее изобретение может также включать выполнение способа согласно изобретению на различных пептидах, содержащих одинаковую модификацию(и) и/или различные модификации.
Термин «различные пептиды, содержащие одинаковую модификацию(и)» в одном варианте осуществления относится к пептидам, содержащим или состоящим из различных фрагментов модифицированного белка, при этом указанные различные фрагменты содержат одинаковую модификацию(и), присутствующую в белке, но различаются по длине и/или положению модификации(й). Если белок имеет модификацию в положении х, то два или более фрагментов указанного белка, каждый содержащий различное окно последовательности указанного белка, покрывающее указанное положение х, считаются различными пептидами, содержащими одинаковую модификацию(и).
Термин «различные пептиды, содержащие различные модификации» в одном варианте осуществления относится к пептидам одинаковой и/или различной длины, содержащим различные модификации одинаковых и/или различных белков. Если белок имеет модификации в положениях х и у, два фрагмента указанного белка, каждый содержащий окно последовательности указанного белка, покрывающее положение х или положение у, считаются различными пептидами, содержащими различные модификации.
Настоящее изобретение может также включать разрывание белковых последовательностей, имеющих модификации, иммуногенность которых подлежит определению в соответствии с изобретением, на пептиды соответствующий длины для связывания с МНС, и определение показателей связывания с одной или несколькими молекулами МНС различных модифицированных пептидов, содержащих одинаковые и/или различные модификации одинаковых и/или различных белков. Результаты могут быть ранжированы и могут состоять из списка пептидов и их предсказанных показателей, указывающих на вероятность их связывания.
Стадия определения показателя связывания немодифицированного пептида с одной или несколькими молекулами МНС, и/или стадия определения показателя связывания модифицированного пептида, в случае присутствия в комплексе МНС-пептид, с одним или несколькими Т-клеточными рецепторами может быть последовательно выполнена со всеми различными модифицированными пептидами, содержащими одинаковые и/или различные модификации, их поднабором, например, модифицированными пептидами, содержащими одинаковые и/или различные модификации, имеющими более высокий показатель связывания с одной или несколькими молекулами МНС, или только с одним модифицированным пептидом, имеющим лучший показатель связывания с одной или несколькими молекулами МНС.
После указанных дополнительных стадий результаты могут быть ранжированы и могут состоять из списка пептидов и их предсказанных показателей, указывающих на вероятность того, что они являются иммуногенными.
Предпочтительно, чтобы в таком ранжировании показатель связывания модифицированного пептида с одной или несколькими молекулами МНС был оценен выше, чем показатель связывания модифицированного пептида, в случае присутствия в комплексе МНС-пептид, с одним или несколькими Т-клеточными рецепторами, предпочтительно показатель химического и физического сходства между немодифицированными и модифицированными аминокислотами, и показатель связывания модифицированного пептида, в случае присутствия в комплексе МНС-пептид, с одним или несколькими Т-клеточными рецепторами, предпочтительно показатель химического и физического сходства между немодифицированными и модифицированными аминокислотами был оценен выше, чем показатель связывания немодифицированного пептида с одной или несколькими молекулами МНС.
Аминокислотные модификации, иммуногенность которых подлежит определению в соответствии с настоящим изобретением, могут являться результатом мутаций в нуклеиновой кислоте клетки. Такие мутации могут быть идентифицированы известными методами секвенирования.
В одном варианте осуществления мутации представляют собой рак-специфические соматические мутации в образцах опухоли ракового пациента, которые могут быть определены путем идентификации различий последовательностей между геномом, экзомом и/или транскриптомом образца опухоли и геномом, экзомом и/или транскриптомом неонкогенного образца.
В соответствии с изобретением образец опухоли относится к любому образцу, такому как физиологический образец, полученный от пациента, содержащий или предположительно содержащий опухоль или раковые клетки. Физиологический образец может представлять собой образец любой ткани, такой как кровь, образец ткани, полученный из первичной опухоли или опухолевых метастазов, или любой другой образец, содержащий опухолевые или раковые клетки. Предпочтительно, физиологический образец представляет собой кровь, и рак-специфические соматические мутации или различия последовательностей определяют в одной или нескольких циркулирующих опухолевых клетках (СТС), содержащихся в крови. В другом варианте осуществления образец опухоли относится к одной или нескольким изолированным опухолевым или раковым клеткам, таким как циркулирующие опухолевые клетки (СТС), или образцу, содержащему одну или несколько изолированных опухолевых или раковых клеток, таких как циркулирующие опухолевые клетки (СТС).
Неонкогенный образец относится к любому образцу, такому как физиологический образец, полученный от пациента или другого индивидуума, который предпочтительно относится к такому же виду, что и пациент, предпочтительно здорового индивидуума, не содержащего или предположительно не содержащего опухолевые или раковые клетки. Физиологический образец может представлять собой образец любой ткани, такой как кровь, или образец, полученный из неонкогенной ткани.
Изобретение может включать определение сигнатуры раковой мутации пациента. Термин «сигнатура раковой мутации» может относиться ко всем раковым мутациям, присутствующим в одной или нескольких раковых клетках пациента, или может относиться только к части раковых мутаций, присутствующих в одной или нескольких раковых клетках пациента. Таким образом, настоящее изобретение может включать идентификацию всех рак-специфических мутаций, присутствующих в одной или нескольких раковых клетках пациента, или может включать идентификацию только части рак-специфических мутаций, присутствующих в одной или нескольких раковых клетках пациента. В целом, способы согласно изобретению предназначены для идентификации количества мутаций, которые обеспечивают достаточное количество модификаций или модифицированных пептидов, подлежащих включению в способы согласно изобретению.
Предпочтительно, мутации, идентифицированные в соответствии с настоящим изобретением, представляют собой несинонимичные мутации, предпочтительно несинонимичные мутации белков, экспрессирующихся в опухолевой или раковой клетке.
В одном варианте осуществления рак-специфические соматические мутации или различия последовательностей определяют в геноме, предпочтительно во всем геноме образца опухоли. Таким образом, изобретение может включать идентификацию мутационной сигнатуры ракового генома, предпочтительно всего генома одной или нескольких раковых клеток. В одном варианте осуществления стадия идентификации рак-специфических соматических мутаций в образце опухоли ракового пациента включает идентификацию профиля раковых мутаций по всему геному.
В одном варианте осуществления рак-специфические соматические мутации или различия последовательностей определяют в экзоме, предпочтительно во всем экзоме образца опухоли. Таким образом, изобретение может включать идентификацию мутационной сигнатуры ракового экзома, предпочтительно всего экзома одной или нескольких раковых клеток. В одном варианте осуществления стадия идентификации рак-специфических соматических мутаций в образце опухоли ракового пациента включает идентификацию профиля раковой мутации по всему экзому.
В одном варианте осуществления рак-специфические соматические мутации или различия последовательностей определяют в транскриптоме, предпочтительно во всем транскриптоме образца опухоли. Таким образом, изобретение может включать идентификацию мутационной сигнатуры ракового транскриптома, предпочтительно всего транскриптома одной или нескольких раковых клеток. В одном варианте осуществления стадия идентификации рак-специфических соматических мутаций в образце опухоли ракового пациента включает идентификацию профиля раковых мутаций по всему транскриптому.
В одном варианте осуществления стадия идентификации рак-специфических соматических мутаций или идентификации различия последовательностей включает секвенирование единичных клеток одной или нескольких, предпочтительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 раковых клеток или даже более. Таким образом, изобретение может включать идентификацию сигнатуры раковой мутации указанной одной или нескольких раковых клеток. В одном варианте осуществления раковые клетки представляют собой циркулирующие опухолевые клетки. Раковые клетки, такие как циркулирующие опухолевые клетки, могут быть изолированы перед секвенированием единичных клеток.
В одном варианте осуществления стадия идентификации рак-специфических соматических мутаций или идентификации различий последовательностей включает использование секвенирования следующего поколения (NGS).
В одном варианте осуществления стадия идентификации рак-специфических соматических мутаций или идентификации различий последовательностей включает секвенирование геномной ДНК и/или РНК образца опухоли.
Для выявления рак-специфических соматических мутаций или различий последовательностей, информацию о последовательности, полученную из образца опухоли, предпочтительно сравнивают с эталоном, таким как информация о последовательности, полученная в результате секвенирования нуклеиновой кислоты, такой как ДНК или РНК нормальных неонкогенных клеток, таких как клетки зародышевой линии, которые могут быть получены от пациента или другого индивидуума. В одном варианте осуществления нормальная геномная ДНК зародышевой линии получена из мононуклеарных клеток периферической крови (РВМС).
Термин «геном» относится к общему количеству генетической информации в хромосомах организма или клетки.
Термин «экзом» относится к части генома организма, образованной экзонами, которые являются кодирующими участками экспрессирующихся генов. Экзом обеспечивает генетический код, используемый в синтезе белков и других функциональных генных продуктов. Он представляет наиболее функционально значимую часть генома и, следовательно, с наибольшей вероятностью влияет на фенотип организма. Экзом генома человека предположительно составляет 1,5% от всего генома (Ng, PC et al., PLoS Gen., 4(8): 1-15, 2008).
Термин «транскриптом» относится к набору молекул РНК, включая мРНК, рРНК, тРНК, и других некодирующих РНК, синтезируемых одной клеткой или популяцией клеток. В контексте настоящего изобретения транскриптом означает набор всех молекул РНК, синтезируемых одной клеткой, популяцией клеток, предпочтительно популяцией раковых клеток или всех клеток данного индивидуума в определенный момент времени.
«Нуклеиновая кислота» в соответствии с изобретением предпочтительно представляет собой дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК), более предпочтительно РНК, наиболее предпочтительно транскрибированную in vitro РНК (IVT RNA) или синтетическую РНК. В соответствии с изобретением нуклеиновые кислоты включают геномную ДНК, кДНК, мРНК, рекомбинантно полученные и химически синтезированные молекулы. В соответствии с изобретением нуклеиновая кислота может быть представлена в виде одноцепочечной или двухцепочечной, и линейной или ковалентно циклически замкнутой молекулы. Нуклеиновая кислота в соответствии с изобретением может быть изолированной. Термин «изолированная нуклеиновая кислота» означает, в соответствии с изобретением, что нуклеиновая кислота (i) была амплифицирована in vitro, например посредством полимеразной цепной реакции (PCR), (ii) была получена рекомбинантно путем клонирования, (iii) была очищена, например, путем расщепления и разделения с помощью гель-электрофореза, или (iv) была синтезирована, например, путем химического синтеза. Нуклеиновую кислоту можно использовать для встраивания внутрь, то есть трансфекции клеток, в частности, в форме РНК, которая может быть получена путем транскрипции in vitro на матрице ДНК. Кроме того, РНК может быть модифицирована до использования путем стабилизации последовательностей, кэпирования и полиаденилирования.
Термин «генетический материал» относится к изолированной нуклеиновой кислоте, ДНК или РНК, участку двойной спирали, участку хромосомы или полному геному организма или клетки, в частности, его экзому или транскриптому.
Термин «мутация» относится к изменению последовательности нуклеиновой кислоты или различию в последовательности нуклеиновой кислоты (нуклеотидному замещению, добавлению или делеции) по сравнению с эталоном. «Соматическая мутация» может возникать в любой клетке организма, за исключением зародышевых клеток (спермиев и яйцеклеток) и, следовательно, не передается детям. Эти изменения могут (но не всегда) вызывать рак или другие заболевания. Предпочтительно мутация представляет собой несинонимичную мутацию. Термин «несинонимичная мутация» относится к мутации, предпочтительно нуклеотидному замещению, которое приводит к аминокислотному изменению, такому как аминокислотная замена в продукте трансляции.
В соответствии с изобретением термин «мутация» включает точечные мутации, инсерции-делеции (Indels), слияния, хромотрипсис и редактирование РНК.
В соответствии с изобретением термин «инсерции-делеции (Indels)» описывает специальный класс мутации, определенный как мутация, приводящая к одновременной инсерции и делеции, и чистому наращиванию или утрате нуклеотидов. В кодирующих областях генома, если длина Indel не кратна трем, они вызывают мутацию сдвига рамки считывания (frameshift mutation). Indels отличаются от точечной мутации; там, где Indels вставляют и удаляют нуклеотиды из последовательности, точечная мутация представляет собой форму замещения, при которой происходит замена одного из нуклеотидов.
Слияния могут генерировать гибридные гены, сформированные из двух ранее отдельных генов. Это может происходить в результате транслокации, интерстициальной делеции или хромосомной инверсии. Часто гены слияния представляют собой онкогены. Онкогенные гены слияния могут приводить к образованию генного продукта, обладающего новой функцией или функцией, отличающейся от двух партнеров слияния. Альтернативно, протоонкоген является слитым с сильным промотором, и тем самым онкогенная функция настраивается на функционирование путем повышенного регулирования, вызванного сильным промотором, расположенным выше по ходу транскрипции (в 3'-5' направлении) от партнера слияния. Онкогенные транскрипты слияния также могут быть вызваны событиями транс-сплайсинга или сквозного прочитывания (read-through).
В соответствии с изобретением термин «хромотрипсис» относится к генетическому явлению, посредством которого специфические области генома разрываются на части, а затем объединяются между собой посредством одного губительного события.
В соответствии с изобретением термин «редакция РНК» или «редактирование РНК» относится к молекулярным процессам, в ходе которых информация, содержащаяся в молекуле РНК, изменяется путем химической модификации оснований. Редактирование РНК включает в себя модификации нуклеозидов, например, дезаминирование цитидина (С) в уридин (U) и аденозина (А) в инозин (I), а также добавления и инсерций нуклеотидов без матрицы. Редактирование РНК в случае мРНК значительно изменяет аминокислотную последовательность кодируемого белка, таким образом, он может сильно отличаться от закодированного в геномной ДНК.
Термин «раковая мутационная сигнатура» относится к набору мутаций, которые присутствуют в раковых клетках по сравнению с нераковыми эталонными клетками.
В соответствии с изобретением «эталон» может быть использован для корреляции и сравнения с результатами, полученными в способах согласно изобретению от образцов опухоли. Обычно «эталон» может быть получен на основе одного или нескольких нормальных образцов, в частности, образцов, которые не поражены раковым заболеванием или получены от пациента, или одного или нескольких разных индивидуумов, предпочтительно здоровых индивидуумов, в частности, индивидуумов одного вида. «Эталон» может быть определен эмпирически путем тестирования достаточно большого количества нормальных образцов.
Любой подходящий метод секвенирования может быть использован в соответствии с изобретением для определения мутаций, при этом предпочтительными являются технологии секвенирования следующего поколения (NGS). Методы секвенирования третьего поколения могут быть заменой для технологии NGS в будущем для ускорения в методе стадии секвенирования. Для прояснения: термины «секвенирование следующего поколения, Next Generation Sequencing» или «NGS» в контексте настоящего изобретения означают все новые высокопроизводительные технологии секвенирования, которые в отличие от «традиционных» методов секвенирования, известных как химия Сэнгера, читают матрицы нуклеиновых кислот в произвольном порядке одновременно по всему геному путем разрушения всего генома на малые части. Такие технологии NGS (также известные как технологии массивного параллельного секвенирования) способны доставлять информацию о нуклеотидной последовательности всего генома, экзома, транскриптома (все транскрибированные последовательности генома) или метилома (все метилированные последовательности генома) за очень короткие промежутки времени, например, в течение 1-2 недель, предпочтительно в течение 1-7 дней, и наиболее предпочтительно в течение менее 24 часов, и обеспечивают, в целом, подходы к секвенированию единичных клеток. Множество платформ NGS, которые являются коммерчески доступными или описаны в литературе, могут быть использованы в контексте настоящего изобретения, например такие, которые описаны подробно в Zhang et al. 2011: The impact of next-generation sequencing on genomics. J. Genet Genomics 38 (3), 95-109; or in Voelkerding et al. 2009: Next generation sequencing: From basic research to diagnostics. Clinical chemistry 55, 641-658. Неограничивающими примерами таких технологий/платформ NGS являются:
1) Технология секвенирования путем синтеза (sequencing-by-synthesis), известная как пиросеквенирование, реализованная, например, на платформе GS-FLX 454 Genome Sequencer™ производства 454 Life Sciences фирмы Roche (Branford, Connecticut), впервые описанная в Ronaghi et al. 1998: A sequencing method based on real-time pyrophosphate". Science 281 (5375), 363-365. В данной технологии используется эмульсионная PCR, в которой шарики, на которые иммобилизуются одноцепочечные фрагменты ДНК, путем энергичного перемешивания заключаются в водные капельки, окруженные маслом и содержащие реактивы, необходимые для проведения PCR-амплификации. В ходе процесса пиросеквенирования квант света, испускаемый фосфатными молекулами во время встраивания нуклеотида, регистрируется, как только полимераза синтезирует цепь ДНК.
2) Подходы к секвенированию путем синтеза, разработанные компанией Solexa (в настоящее время часть компании Illumina Inc., San Diego, California), которые основаны на обратимой терминации с использованием красителей и реализованы, например, на секвенаторе Genome Analyzer™ компании Illumina/Solexa и секвенаторе HiSeq 2000 Genome Analyzer™ компании Illumina. В данной технологии все четыре нуклеотида добавляют одновременно в олиго-примированные фрагменты кластера в каналы проточных кювет вместе с ДНК-полимеразой. Мостиковая амплификация удлиняет кластерные цепи со всеми четырьмя флуоресцентно мечеными нуклеотидами для секвенирования.
3) Подходы секвенирования путем лигирования (sequencing-by-ligation), например, реализованные на платформе SOLid™ компании Applied Biosystems (в настоящее время Life Technologies Corporation, Carlsbad, California). В этой технологии пул всех возможных олигонуклеотидов фиксированной длины мечены в соответствии с секвенированным положением. Олигонуклеотиды ренатурируют и лигируют; предпочтительное лигирование с участием ДНК-лигазы для совпадающих последовательностей приводит к сигналу, содержащему информацию о нуклеотиде в этом положении. Перед секвенированием ДНК амплифицируют путем эмульсионной PCR. Полученный шарик, каждый, содержащий только копии одной и той же молекулы ДНК, осаждают на покровное стекло. В качестве второго примера, в платформе Polonator™ G.007 компании Dover Systems (Salem, New Hampshire) также используется подход к секвенированию путем лигирования путем использования эмульсионной PCR на основе случайным образом расположенных шариков для амплификации фрагментов ДНК для параллельного секвенирования.
4) Технологии одномолекулярного секвенирования (single-molecule sequencing), такие как, например, реализованные на системе PacBio RS компании Pacific Biosciences (Menlo Park, California) или на платформе HeliScope™ компании Helicos Biosciences (Cambridge, Massachusetts). Отличительной характеристикой этой технологии является ее способность секвенировать единичные молекулы ДНК или РНК без амплификации, названной одномолекулярным секверированием ДНК в режиме реального времени (Single-Molecule Real Time, SMRT). Например, HeliScope использует высокочувствительную систему детекции флуоресценции для прямой детекции каждого нуклеотида по мере его синтезирования. Сходный подход, основанный на флуоресцентном резонансном переносе энергии (FRET), был разработан фирмой Visigen Biotechnology (Houston, Texas). Другие одномолекулярные технологии на основе флуоресценции доступны от компании U.S. Genomics (GeneEngine™) и Genovoxx (AnyGene™).
5) Нанотехнологии для одномолекулярного секвенирования, в которых используются различные наноструктуры, которые, например, расположены на чипе для контроля движения молекулы полимеразы на единичной цепи во время репликации. Неограничивающие примеры подходов на основе нанотехнологий представляют собой платформу GridON™ компании Oxford Nanopore Technologies (Oxford, UK), платформы для секвенирования через нанопоры (hybridization-assisted nano-pore sequencing) (HANS™), разработанные компанией Nabsys (Providence, Rhode Island), и патентованная платформа Ligase-based DNA Sequencing с технологией DNA nanoball (DNB), называемая Combinatorial probe-anchor ligation (cPAL™).
6) Технологии на основе электронной микроскопии для одномолекулярного секвенирования, например такие, которые разработаны компанией LightSpeed Genomics (Sunnyvale, California) и Halcyon Molecular (Redwood City, California).
7) Ионное полупроводниковое секвенирование, которое основано на обнаружении ионов водорода, выделяющихся при полимеризации ДНК. Например, в системе Ion Torrent Systems (San Francisco, California) используются микрочипы, содержащие высокоплотную матрицу микроячеек для выполнения этого биохимического процесса массивно-параллельным путем. Каждая ячейка содержит различную ДНК-матрицу. Под ячейками находится ионно-чувствительный слой и под ним патентованный ионный сенсор.
Предпочтительно, препараты ДНК и РНК служат в качестве исходного материала для NGS. Такие нуклеиновые кислоты могут быть легко получены из образцов, таких как биологический материал, например, свежие, свежезамороженные или фиксированные формалином заключенные в парафин (FFPE) ткани опухоли, или из свежеизолированных клеток, или из СТС, которые присутствуют в периферической крови пациентов. Нормальная немутированная геномная ДНК или РНК может быть экстрагирована из нормальной, соматической ткани, однако клетки зародышевой линии являются предпочтительными в контексте настоящего изобретения. ДНК или РНК зародышевой линии может быть экстрагирована из мононуклеарных клеток периферической крови (РВМС) у пациентов с негематологическими злокачественными новообразованиями. Хотя нуклеиновые кислоты, экстрагированные из FFPE тканей или свежеизолированных единичных клеток, являются высокофрагментированными, они являются подходящими для применения в NGS.
В литературе описано несколько методов таргетного NGS для секвенирования экзома (для обзора смотри, например, Teer and Mullikin 2010: Human Mol Genet 19 (2), R145-51), все из которых можно использовать в сочетании с настоящим изобретением. Многие из этих методов (описанных, например, как захват генома, разделение генома, обогащение генома, и т.п.) используют методы гибридизации и включают подходы к гибридизации на основе микрочипов (например, Hodges et al. 2007: Nat. Genet. 39, 1522-1527) и жидкости (например, Choi et al. 2009: Proc. Natl. Acad. Sci USA 106, 19096-19101). Коммерческие наборы для приготовления образцов ДНК и последующего захвата экзома также являются доступными: например, Illumina Inc. (San Diego, California) предлагает набор TruSeq™ DNA Sample Preparation Kit и набор Exome Enrichment Kit TruSeq™ Exome Enrichment Kit.
Для сокращения количества ложноположительных результатов при детекции рак-специфических соматических мутаций или различий последовательностей при сравнении, например, последовательности образца опухоли с последовательностью эталонного образца, такой как последовательность образца зародышевой линии, предпочтительно определить последовательность одного или обоих образцов этих типов в нескольких повторах. Таким образом, определение последовательности эталонного образца, такой как последовательность образца зародышевой линии, предпочтительно проводить два раза, три раза или более. Альтернативно или дополнительно, последовательность образца опухоли определяют два раза, три раза или более. Также, возможно определение последовательности эталонного образца, такой как последовательность образца зародышевой линии, и/или последовательности образца опухоли более одного раза путем определения не менее одного раза последовательности в геномной ДНК и определения не менее одного раза последовательности в РНК указанного эталонного образца и/или указанного образца опухоли. Например, путем определения вариаций между повторами эталонного образца, такого как образец зародышевой линии, можно определить предполагаемую частоту ложноположительных результатов (FDR) для соматических мутаций в качестве статистического количества. Технические повторы образца будут давать идентичные результаты, и любая обнаруженная мутация в этом «сравнении сам против себя» является ложноположительным результатом. В частности, для определения частоты ложноположительных результатов для детекции соматической мутации в образце опухоли по сравнению с эталонным образцом, может быть использован технический повтор эталонного образца в качестве эталона для определения числа ложноположительных результатов. Кроме того, различные связанные с качеством показатели (например, охват или качество SNP) могут быть объединены в один показатель качества с использованием подхода компьютерного обучения. Для заданной соматической вариации могут быть подсчитаны все другие вариации с превышающим показателем качества, которые позволяют ранжировать все вариации в наборе данных.
В контексте настоящего изобретения термин «РНК» относится к молекуле, которая содержит, по меньшей мере, один рибонуклеотидный остаток и предпочтительно полностью или в основном состоит из рибонуклеотидных остатков. «Рибонуклеотид» относится к нуклеотиду, содержащему гидроксильную группу в 2'-положении β-D-рибофуранозильной группы. Термин «РНК» включает двухцепочечную РНК, одноцепочечную РНК, выделенную РНК, такую как частично или полностью очищенная РНК, по существу чистая РНК, синтетическую РНК, и полученную рекомбинантными методами РНК, такую как модифицированная РНК, которая отличается от природной РНК за счет внесения в нее добавки, делеции, замены и/или изменения одного или нескольких нуклеотидов. Такие изменения могут включать добавление материала ненуклеотидной природы, такие как добавки, содержащиеся на одном или обоих концах молекулы РНК или введенные внутрь молекулы, например, локализованные в одном или нескольких нуклеотидах в составе РНК. Нуклеотиды в составе молекул РНК могут также включать нестандартные нуклеотиды, такие как неприродные нуклеотиды или химически синтезированные нуклеотиды, или дезоксинуклеотиды. Указанные измененные РНК могут рассматриваться как аналоги или аналоги природных РНК.
В соответствии с настоящим изобретением термин «РНК» включает и предпочтительно относится к «мРНК». Термин «мРНК» означает «информационная РНК» и относится к «транскрипту», который создан путем использования матрицы ДНК и кодирует пептид или полипептид. Обычно мРНК содержит 5'-UTR, кодирующую белок область, и 3'-UTR. мРНК обладает лишь ограниченным периодом полужизни в клетках и in vitro. В контексте настоящего изобретения мРНК может быть создана путем транскрипции in vitro матрицы ДНК. Методика транскрипции in vitro является известной для специалистов в данной области. Например, существует множество различных коммерчески доступных наборов для транскрипции in vitro.
В соответствии с изобретением стабильность и эффективность трансляции РНК можно необходимым образом модифицировать. Например, РНК можно стабилизировать и ее трансляцию увеличить с помощью одной или нескольких модификаций, обладающих стабилизирующими эффектами и/или повышающих эффективность трансляции РНК. Такие модификации описаны, например, в РСТ/ЕР2006/009448, включенном здесь путем отсылки. Для того, чтобы увеличить экспрессию РНК, используемую в соответствии с настоящим изобретением, ее можно модифицировать в пределах кодирующей области, то есть последовательности, кодирующей экспрессирующийся пептид или белок, предпочтительно без изменения последовательности экспрессирующегося пептида или белка, чтобы тем самым увеличить содержание GC-пар для повышения стабильности мРНК и выполнить оптимизацию кодонов и, таким образом, усилить трансляцию в клетках.
Используемый в настоящем изобретении термин «модификация» в контексте РНК включает любую модификацию РНК, которая естественным образом не присутствует в указанной РНК.
В одном варианте осуществления РНК, используемая в соответствии с изобретением, не содержит некэпированных 5'-трифосфатов. Удаление таких некэпированных 5'-трифосфатов может быть достигнуто путем обработки РНК фосфатазой.
РНК в соответствии с изобретением может содержать модифицированные рибонуклеотиды для повышения ее стабильности и/или уменьшения цитотоксичности. Например, в одном варианте осуществления в РНК, используемой в соответствии с изобретением, 5-метилцитидин является замещенным частично или полностью, предпочтительно полностью, на цитидин. Альтернативно или дополнительно, в одном варианте осуществления в РНК, используемой в соответствии с изобретением, псевдоуридин является замещенным частично или полностью, предпочтительно полностью, на уридин.
В одном варианте осуществления термин «модификация» относится к обеспечению РНК 5'-кэпом или аналогом 5'-кэпа. Термин «5'-кэп» относится к кэп-структуре, находящейся на 5'-конце молекулы мРНК и, как правило, состоит из гуанозинового нуклеотида, соединенного с мРНК посредством нестандартной 5'-5' трифосфатной связи. В одном варианте осуществления этот гуанозин является метилированным в 7-положении. Термин «общепринятый 5'-кэп» относится к 5'-кэпу РНК природного происхождения, предпочтительно к 7-метилгуанозиновому кэпу (m7G). В контексте настоящего изобретения термин «5'-кэп» включает аналог 5'-кэпа, который образует кэп-структуру РНК и является модифицированным для проявления способности стабилизировать РНК и/или усиливать трансляцию РНК в случае присоединения к ней, предпочтительно in vivo и/или в клетке.
Обеспечение РНК 5'-кэпом или аналогом 5'-кэпа может быть достигнуто путем транскрипции in vitro матрицы ДНК в присутствии указанного 5'-кэпа или аналога 5'-кэпа, при этом указанный 5'-кэп является котранскрипционно встроенным в генерированную цепь РНК, или РНК может быть генерирована, например, путем транскрипции in vitro, и 5'-кэп может быть присоединен к РНК посттранскрипционно с использованием кэпирующих ферментов, например, кэпирующих ферментов вируса осповакцины.
РНК может содержать дополнительные модификации. Например, дополнительная модификация РНК, используемая в настоящем изобретении, может представлять собой удлинение или усечение природного поли(А) хвоста, или изменение 5'- или 3'-нетранслируемых областей (UTR), такое как введение UTR, которая не относится к кодирующей области указанной РНК, например, замену существующей 3'-UTR или инсерцию одной или нескольких, предпочтительно двух копий 3'-UTR, полученных их гена глобулина, такого как альфа2-глобулина, альфа1-глобулина, бета-глобулина, предпочтительно бета-глобулина, более предпочтительно бета-глобулина человека.
РНК, несущая активную (немаскированную) поли-А-последовательность, транслируется более эффективно, чем РНК, несущая неактивную (маскированную) поли-А-последовательность. Термин «поли(А)-хвост» или «поли-А-последовательность» относится к последовательности остатков аденила (А), которые обычно расположены на 3'-конце молекулы РНК, и «активная (немаскированная) поли-А-последовательность» означает, что поли-А-последовательность на 3'-конце молекулы РНК оканчивается остатками аденила (А) поли-А-последовательности, за которыми не следуют нуклеотиды, отличные от остатков аденила (А), расположенных на 3'-конце, то есть в направлении ниже по ходу транскрипции, поли-А-последовательности. Кроме того, длинная поли-А-последовательность, состоящая из около 120 пар оснований, приводит к оптимальной стабильности транскрипта и эффективности трансляции РНК.
Таким образом, для увеличения стабильности и/или экспрессии РНК, используемой в соответствии с настоящим изобретением, ее можно модифицировать таким образом, чтобы она присутствовала в сочетании с поли-А-последовательностью, предпочтительно имеющей длину от 10 до 500, более предпочтительно от 30 до 300, даже более предпочтительно от 65 до 200 и в особенности от 100 до 150 остатков аденозина. В особенно предпочтительном варианте осуществления поли-А-последовательность имеет длину приблизительно 120 аденозиновых остатков. Для дополнительного увеличения стабильности и/или экспрессии РНК, используемой в соответствии с изобретением, последовательность поли-А может быть немаскированной.
Кроме того, встраивание 3'-нетранслируемой области (UTR) в 3'-нетранслируемую область молекулы РНК может привести к усилению эффективности трансляции. Синергический эффект может быть достигнут путем встраивания двух или более таких 3'-нетранслируемых областей. 3'-Нетранслируемые области могут быть аутологичными или гетерологичными в отношении РНК, в которую они вводятся. В одном конкретном варианте осуществления 3'-нетранслируемая область получена из человеческого гена β-глобулина.
Комбинация описанных выше модификаций, то есть встраивание поли-А-последовательности, демаскирование поли-А-последовательности и встраивание одной или нескольких 3'-нетранслируемых областей, имеет синергическое влияние на стабильность РНК и повышение эффективности трансляции.
Термин «стабильность» РНК относится к «полужизни» РНК. «Полужизнь» относится к периоду времени, которое необходимо для устранения половины активности, объема или количества молекул. В контексте настоящего изобретения полужизнь РНК является показателем стабильности указанной РНК. Полужизнь РНК может влиять на «продолжительность экспрессии» РНК. Можно предположить, что РНК, имеющая длинный период полужизни, будет экспрессироваться в течение продолжительного периода времени.
Разумеется, если согласно настоящему изобретения желательно уменьшить стабильность РНК и/или эффективность трансляции РНК, то можно модифицировать РНК таким образом, чтобы препятствовать функции элементов, описанных выше, повышающих стабильность и/или эффективность трансляции РНК.
Термин «экспрессия» используется в соответствии с изобретением в его самом общем значении и включает продуцирование РНК и/или пептидов, полипептидов или белков, например, путем транскрипции и/или трансляции. В отношении РНК термин «экспрессия» или «трансляция» относится, в частности, к продуцированию пептидов, полипептидов или белков. Он также включает частичную экспрессию нуклеиновых кислот. Кроме того, экспрессия может быть временной или стабильной.
В соответствии с изобретением термин экспрессия также включает «аберрантную экспрессию» или «ненормальную экспрессию». «Аберрантная экспрессия» или «ненормальная экспрессия» означает в соответствии с изобретением, что экспрессия является измененной, предпочтительно увеличенной, по сравнению с эталоном, например, состоянием у субъекта, не имеющего заболевания, связанного с аберрантной или ненормальной экспрессией определенного белка, например, опухолевого антигена. Увеличение экспрессии относится к увеличению по меньшей мере на 10%, в частности, по меньшей мере на 20%, по меньшей мере на 50% или по меньшей мере на 100% или более. В одном варианте осуществления экспрессия обнаруживается только в больной ткани, тогда как экспрессия в здоровой ткани является репрессированной.
Термин «специфически экспрессирующийся» означает, что белок экспрессируется преимущественно только в определенной ткани или органе. Например, опухолевый антиген, специфически экспрессирующийся в слизистой оболочке желудка, означает, что указанный белок в первую очередь экспрессируется в слизистой оболочке желудка и не экспрессируется в других тканях или не экспрессируется на значительном уровне в тканях или органах других типов. Таким образом, белок, который экспрессируется исключительно в клетках слизистой оболочки желудка и на значительно меньшем уровне в любой другой ткани, такой как семенник, специфически экспрессируется в клетках слизистой оболочки желудка. В некоторых вариантах осуществления опухолевый антиген может также специфически экспрессироваться в нормальных условиях в более чем одном типе ткани или органе, например, в 2 или 3 типах ткани или органах, но предпочтительно не более чем в 3 различных типах ткани или органах. В этом случае опухолевый антиген затем специфически экспрессируется в этих органах. Например, если опухолевый антиген экспрессируется в нормальных условиях предпочтительно на приблизительно одинаковом уровне в легких и желудке, то указанный опухолевый антиген специфически экспрессируется в легких и желудке.
В контексте настоящего изобретения термин «транскрипция» относится к процессу, в котором генетический код в последовательности ДНК транскрибирован в РНК. Затем РНК может быть транслирована в белок. В соответствии с настоящим изобретением термин «транскрипция» включает «транскрипцию in vitro», при этом термин «транскрипция in vitro» относится к процессу, в котором РНК, в частности мРНК, синтезируется in vitro в бесклеточной системе, предпочтительно с использованием соответствующих клеточных экстрактов. Предпочтительно, для создания транскриптов используют вектора клонирования. Эти вектора клонирования, как правило, разработаны в качестве векторов транскрипции и в соответствии с настоящим изобретением охвачены термином «вектор». В соответствии с настоящим изобретением РНК, используемая в настоящем изобретении, предпочтительно является in vitro транскрибированной РНК (IVT-RNA) и может быть получена путем in vitro транскрипции соответствующей матрицы ДНК. Промотором для контроля транскрипции может быть любой промотор для любой РНК-полимеразы. Конкретными примерами РНК-полимеразы являются РНК-полимеразы Т7, Т3 и SP6. Предпочтительно, транскрипция in vitro в соответствии с изобретением контролируется промотором Т7 или SP6. ДНК-матрица для транскрипции in vitro может быть получена путем клонирования нуклеиновой кислоты, в частности кДНК, и встраивания ее в соответствующий вектор для транскрипции in vitro. кДНК может быть получена путем обратной транскрипции РНК.
Термин «трансляция» в соответствии с изобретением относится к процессу, происходящему в рибосомах клетки, в ходе которого цепь информационной РНК направляет сборку последовательности аминокислот на получение пептида, полипептида или белка.
Последовательности, участвующие в контроле экспрессии или регуляторные последовательности, которые в соответствии с изобретением могут быть связаны функционально с нуклеиновой кислотой, могут быть гомологичными или гетерологичными по отношению к нуклеиновой кислоте. Кодирующая последовательность и регуляторная последовательность связаны вместе «функционально», если они соединяются ковалентной связью, таким образом, что транскрипция или трансляция кодирующей последовательности находится под контролем или под влиянием регуляторной последовательности. Если кодирующая последовательность подлежит трансляции в функциональный белок, с функциональной связью регуляторной последовательности с кодирующей последовательностью, то индукция регуляторной последовательности приводит к транскрипции кодирующей последовательности без вызывания сдвига рамки считывания в кодирующей последовательности или невозможности трансляции кодирующей последовательности в желательный белок или пептид.
Термин «последовательность, участвующая в контроле экспрессии» или «регуляторная последовательность» в соответствии с изобретением включает промоторы, связывающие рибосомы последовательности, и другие контрольные элементы, которые контролируют транскрипцию нуклеиновой кислоты или трансляцию полученной РНК. В определенных вариантах осуществления изобретения регуляторные последовательности могут быть контролируемыми. Конкретная структура регуляторных последовательностей может варьироваться в зависимости от вида или типа клетки, но, как правило, содержит 5'-нетранскрибируемые и 5'- и 3'-нетранслируемые последовательности, которые участвуют в инициации транскрипции или трансляции, такие как ТАТА-бокс, кэпирующая последовательность, СААТ-последовательность и т.п. В частности, такие 5'-нетранскрибируемые регуляторные последовательности содержат промоторную область, которая содержит последовательность промотора для транскрипционного контроля функционально связанного гена. Такие регуляторные последовательности могут также содержать энхансерные последовательности или активаторные (3''-5'') последовательности.
В соответствии с изобретением предпочтительно введение РНК, подлежащей экспрессии в клетке, в указанную клетку. В одном варианте осуществления способов согласно изобретению РНК, которая подлежит введению в клетку, получена путем in vitro транскрипции соответствующей матрицы ДНК.
В соответствии с изобретением термины, такие как «РНК, способная к экспрессии» и «РНК, кодирующая» используются взаимозаменяемо в настоящем документе и в отношении конкретного пептида или полипептида означают, что РНК, в случае присутствия в соответствующей среде, предпочтительно в пределах клетки, может экспрессироваться с получением указанного пептида или полипептида. Предпочтительно, РНК в соответствии с изобретением может также взаимодействовать с клеточным механизмом трансляции для обеспечения экспрессии пептида или полипептида.
Термины, такие как «перенос», «введение» или «трансфекция», используются здесь взаимозаменяемо и относятся к введению нуклеиновых кислот, в частности, экзогенных или гетерологичных нуклеиновых кислот, в частности РНК, в клетку. В соответствии с настоящим изобретением клетка может формировать часть органа, ткани и/или организма. В соответствии с настоящим изобретением введение нуклеиновой кислоты осуществляется либо в виде голой нуклеиновой кислоты или в сочетании с введением реагента. Предпочтительно, введение нуклеиновых кислот осуществляют в форме голых нуклеиновых кислот. Предпочтительно, РНК вводят в комбинации со стабилизирующими веществами, такими как ингибиторы РНКазы. В настоящем изобретении также рассматривается повторное введение нуклеиновых кислот в клетки для обеспечения непрерывной экспрессии в течение продолжительных периодов времени.
Клетки могут быть трансфицированы с использованием любых носителей, с которыми может быть ассоциирована РНК, например, путем формирования комплексов с РНК или формирования везикул, в которые заключена или инкапсулирована РНК, что приводит к повышенной стабильности РНК по сравнению с голой РНК. Носители, подходящие в соответствии с изобретением, включают, например, липид-содержащие носители, такие как катионные липиды, липосомы, в частности, катионные липосомы, и мицеллы, и наночастицы. Катионные липиды могут формировать комплексы с отрицательно заряженными нуклеиновыми кислотами. Любой катионный липид может быть использован в соответствии с изобретением.
Предпочтительно, введение РНК, которая кодирует пептид или полипептид, в клетку, в частности, в клетку, присутствующую in vivo, приводит к экспрессии указанного пептида или полипептида в клетке. В конкретных вариантах осуществления нацеливание нуклеиновых кислот на конкретные клетки является предпочтительным. В таких вариантах осуществления носитель, который применяется для введения нуклеиновой кислоты в клетку (например, ретровирус или липосома), представляет собой нацеливающую молекулу. Например, молекула, такая как антитело, которая является специфической к поверхностному мембранному белку на клетке-мишени или лиганду для рецептора на клетке-мишени, может быть введена в носитель нуклеиновой кислоты или может быть присоединена к ней. В случае, когда нуклеиновую кислоту вводят с помощью липосом, белки, которые связываются с поверхностным мембранным белком путем эндоцитоза, могут быть введены в состав липосом для обеспечения направленного воздействия и/или облегчения поглощения. Такие белки включают капсидные белки или их фрагменты, которые являются специфическими для определенного типа клеток, антитела против белков, которые проходят интернализацию в цикле, белки, которые воздействуют на внутриклеточную локализацию и т.д.
Термин «клетка» или «клетка-хозяин» предпочтительно представляет собой интактную клетку то есть клетку с интактной мембраной, которая не высвобождает свои нормальные внутриклеточные компоненты, такие как ферменты, органеллы или генетический материал. Интактная клетка предпочтительно представляет собой жизнеспособную клетку, то есть живую клетку, способную выполнять свои нормальные метаболические функции. Предпочтительно, указанный термин в соответствии с изобретением относится к любой клетке, которая может быть трансформирована или трансфицирована экзогенной нуклеиновой кислотой. Термин «клетка» в соответствии с изобретением включает прокариотические клетки (например, Е. Coli) или эукариотические клетки (например, дендритные клетки, В-клетки, клетки СНО, клетки COS, клетки K562, клетки НЕK293, клетки HELA, клетки дрожжей и клетки насекомых). Экзогенная нуклеиновая кислота может быть обнаружена внутри клетки (i) в свободном диспергированном состоянии, (ii) введена в рекомбинантный вектор, или (iii) интегрирована в геном клетки-хозяина или митохондриальную ДНК. Клетки млекопитающего являются особенно предпочтительными, такие как клетки человека, мышей, хомяков, свиней, коз и приматов. Клетки могут быть получены из большого числа типов тканей и включают первичные клетки и клеточные линии. Конкретные примеры включают кератиноциты, лейкоциты периферической крови, стволовые клетки костного мозга и эмбриональные стволовые клетки. В других вариантах осуществления клетка представляет собой антигенпрезентирующую клетку, в частности, дендритную клетку, моноцит или макрофаг.
Клетка, которая содержит молекулу нуклеиновой кислоты, предпочтительно экспрессирует пептид или полипептид, кодированный нуклеиновой кислотой.
Термин «клональное размножение» относится к процессу, в ходе которого специфическая единица размножается. В контексте настоящего изобретения указанный термин предпочтительно используется в контексте иммунологического ответа, в котором лимфоциты, стимулированные антигеном, пролиферируют, и специфический лимфоцит, распознающий указанный антиген, является амплифицированным. Предпочтительно, клональное размножение приводит к дифференциации лимфоцитов.
Термины, такие как «уменьшение» или «ингибирование», относятся к способности вызывать общее уменьшение на уровне, предпочтительно 5% или выше, 10% или выше, 20% или выше, более предпочтительно 50% или выше, и наиболее предпочтительно 75% или выше. Термин «ингибировать» или сходные выражения включают полное или по существу полное ингибирование, то есть уменьшение до нуля или по существу до нуля.
Термины, такие как «увеличение», «усиление», «содействие» или «пролонгирование» предпочтительно относятся к увеличению, усилению, содействию или пролонгированию по меньшей мере на 10%, предпочтительно, по меньшей мере на 20%, предпочтительно, по меньшей мере на 30%, предпочтительно, по меньшей мере на 40%, предпочтительно, по меньшей мере на 50%, предпочтительно, по меньшей мере на 80%, предпочтительно, по меньшей мере на 100%, предпочтительно, по меньшей мере на 200%, и в особенности по меньшей мере на 300%. Эти термины могут также относиться к увеличению, усилению, содействию или пролонгированию от нуля или неизмеряемого или недетектируемого уровня до уровня больше чем нулевой, или уровня, который является измеряемым или детектируемым.
Настоящее изобретение обеспечивает вакцины, такие как раковые вакцины, разработанные на основе аминокислотных модификаций или модифицированных пептидов, предсказанных как иммуногенные с помощью способов настоящего изобретения.
В соответствии с изобретением термин «вакцина» относится к фармацевтическому препарату (фармацевтической композиции) или продукту, который при введении вызывает иммунный ответ, в частности, клеточный иммунный ответ, который распознает и атакует патоген или больную клетку, такую как раковая клетка. Вакцину можно применять для предупреждения или лечения заболевания. Термин «персонализированная вакцина против рака» или «индивидуализированная вакцина против рака» относится к конкретному онкопациенту и означает, что вакцина против рака адаптирована к потребностям или конкретным обстоятельствам индивидуального онкопациента.
В одном варианте осуществления вакцина, обеспеченная в соответствии с изобретением, может содержать пептид или полипептид, содержащий одну или несколько аминокислотных модификаций, или один или несколько модифицированных пептидов, предсказанных как иммуногенные с помощью способов согласно изобретению, или нуклеиновую кислоту, предпочтительно РНК, кодирующую указанный пептид или полипептид.
Вакцины против рака, обеспеченные в соответствии с изобретением, при введении пациенту обеспечивают один или несколько Т-клеточных эпитопов, подходящих для стимуляции, примирования и/или распространения Т-клеток, специфичных для опухоли пациента. Т-клетки предпочтительно направлены против клеток, экспрессирующих антигены, из которых получены Т-клеточные эпитопы. Таким образом, описанные здесь вакцины предпочтительно способны индуцировать или стимулировать клеточный ответ, предпочтительно цитотоксическую Т-клеточную активность, против ракового заболевания, характеризующегося презентацией одного или нескольких опухоль-ассоциированных неоантигенов с участием МНС класса I. Поскольку вакцина, обеспеченная в соответствии с настоящим изобретением, будет направленно воздействовать на раковые специфические мутации, она будет специфической по отношению к опухоли пациента.
Вакцина, обеспеченная в соответствии с изобретением, относится к вакцине, которая при введении пациенту предпочтительно обеспечивает один или несколько Т-клеточных эпитопов, таких как 2 или более, 5 или более, 10 или более, 15 или более, 20 или более, 25 или более, 30 или более, и предпочтительно до 60, до 55, до 50, до 45, до 40, до 35 или до 30 Т-клеточных эпитопов, введение аминокислотных модификаций или модифицированных пептидов, предсказанных как иммуногенные с помощью способов согласно изобретению. Такие Т-клеточные эпитопы также называются здесь как «неоэпитопы». Презентация этих эпитопов клетками пациента, в частности, антигенпрезентирующими клетками, предпочтительно приводит к Т-клеткам, направленно воздействующим на эпитопы при связывании с МНС и, таким образом, на опухоль пациента, предпочтительно первичную опухоль, а также метастазы опухоли, экспрессирующие антигены, из которых происходят Т-клеточные эпитопы, и презентирующие эти эпитопы на поверхности опухолевых клеток.
Способы согласно изобретению могут включать дополнительную стадию определения применимости идентифицированных аминокислотных модификаций или модифицированных пептидов для вакцинирования против рака. Таким образом, дополнительные стадии могут включать одну или несколько из следующих: (i) определение, локализованы ли модификации в известных или предсказанных презентированных МНС эпитопах; (ii) тестирование in vitro и/или in silico, локализованы ли модификации в презентированных МНС эпитопах, например, тестирование, являются ли модификации частью пептидных последовательностей, которые процессированы и/или представлены в виде презентированных МНС эпитопов; и (iii) тестирование in vitro, способны ли предполагаемые модифицированные эпитопы, особенно в случае присутствия в контексте своей природной последовательности, например, в случае фланкирования аминокислотными последовательностями, также фланкирующими указанные эпитопы в природном белке, и в случае экспрессии в антигенпрезентирующих клетках, стимулировать Т-клетки, такие как Т-клетки пациента, обладающие желательной специфичностью. Такие фланкирующие последовательности, каждая, могут содержать 3 или более, 5 или более, 10 или более, 15 или более, 20 или более и предпочтительно до 50, до 45, до 40, до 35 или до 30 аминокислот, и могут фланкировать последовательность эпитопа на N-конце и/или С-конце.
Модифицированные пептиды, определенные в соответствии с изобретением, могут быть ранжированы на их применимость в качестве эпитопов для вакцинации против рака. Таким образом, в одном аспекте способ согласно изобретению включает неавтоматизированный или основанный на применении компьютера аналитический метод, в котором проводится анализ и отбор идентифицированных модифицированных пептидов на их применимость в соответствующей подлежащей получению вакцине. В предпочтительном варианте осуществления указанный аналитический метод представляет собой метод на основе вычислительного алгоритма. Предпочтительно, указанный аналитический метод включает определение и/или ранжирование эпитопов в соответствии с предсказанием их способности являться иммуногенными.
Неоэпитопы, идентифицированные в соответствии с изобретением и обеспеченные вакциной согласно изобретению, предпочтительно представлены в форме полипептида, содержащего указанные неоэпитопы, такие как полиэпитопный полипептид или нуклеиновая кислота, в особенности РНК, кодирующая указанный полипептид. Кроме того, неоэпитопы могут присутствовать в полипептиде в форме вакцинной последовательности, то есть присутствовать в контексте своей природной последовательности, например, фланкированные аминокислотными последовательностями, также фланкируют указанные эпитопы в природном белке. Такие фланкирующие последовательности, каждая, могут содержать 5 или более, 10 или более, 15 или более, 20 или более и предпочтительно до 50, до 45, до 40, до 35, до 30 аминокислот, и могут фланкировать последовательность эпитопа на N-конце и/или С-конце. Таким образом, последовательность вакцины может содержать 20 или более, 25 или более, 30 или более, 35 или более, 40 или более и предпочтительно до 50, до 45, до 40, до 35 или до 30 аминокислот. В одном варианте осуществления неоэпитопы и/или вакцинные последовательности расположены в полипептиде в направлении «голова-к-хвосту».
В одном варианте осуществления неоэпитопы и/или вакцинные последовательности разделены линкерами, в частности, нейтральными линкерами. Термин «линкер» в соответствии с изобретением относится к пептиду, добавленному между двумя пептидными доменами, такими как эпитопы или вакцинные последовательности, для соединения указанных пептидных доменов. Не существует конкретного ограничения в отношении последовательности линкера. Однако предпочтительно, чтобы последовательность линкера уменьшала стерическое затруднение между двумя пептидными доменами, хорошо транслировалась и способствовала или обеспечивала возможность процессинга эпитопов. Кроме того, линкер не должен содержать или должен содержать незначительное количество элементов иммуногенной последовательности. Предпочтительно, линкеры не должны вызывать образование неэндогенных неоэпитопов, таких как неоэпитопы, генерированные из линии соединения соседних неоэпитопов, которые могут генерировать нежелательные иммунные реакции. Таким образом, полиэпитопная вакцина предпочтительно должна содержать последовательности линкера, которые способны уменьшать число нежелательных эпитопов линии соединения МНС-связывания. В работе Hoyt et al. (EMBO J. 25(8), 1720-9, 2006) и Zhang et al. (J. Biol. Chem., 279(10), 8635-41, 2004) показано, что богатые глицином последовательности ухудшают протеасомальный процессинг и, таким образом, использование богатых глицином последовательностей линкера уменьшает до минимума количество линкер-содержащих пептидов, которые могут быть процессированы протеасомой. Кроме того, наблюдалось, что глицин ингибирует сильное связывание в положениях бороздки МНС-связывания (Abastado et al., J. Immunol. 151(7), 3569-75, 1993). В работе Schlessinger et al. (Proteins, 61(1), 115-26, 2005) было показано, что аминокислоты глицин и серин, включенные в аминокислотную последовательность, приводят к более гибкому белку, который более эффективно транслируется и процессируется протеасомой, обеспечивая лучший доступ к кодированным неоэпитопам. Каждый линкер может содержать 3 или более, 6 или более, 9 или более, 10 или более. 15 или более. 20 или более и предпочтительно до 50, до 45, до 40, до 35 или до 30 аминокислот. Предпочтительно линкер является обогащенным аминокислотами глицином и/или серином. Предпочтительно, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% аминокислот линкера представляют собой глицин и/или серин. В одном предпочтительном варианте осуществления линкер по существу состоит из аминокислот глицина и серина. В одном варианте осуществления линкер содержит аминокислотную последовательность (GGS)a(GSS)b(GGG)c(SSG)d(GSG)e в которой а, b, с, d и e независимо представляют собой число, выбранное из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20, и где a+b+c+d+e не равно 0 и предпочтительно равно 2 или более, 3 или более, 4 или более, или 5 или более. В одном варианте осуществления линкер содержит последовательность, описанную здесь, включающую последовательности линкера, описанные в примерах, такие как последовательность GGSGGGGSG.
В одном особенно предпочтительном варианте осуществления полипептид, включающий один или несколько неоэпитопов, такой как полиэпитопный полипептид, в соответствии с настоящим изобретением вводят пациенту в форме нуклеиновой кислоты, предпочтительно РНК, такой как транскрибированная in vitro или синтетическая РНК, которая может экспрессироваться в клетках пациента, таких как антигенпрезентирующие клетки, с получением полипептида. В настоящем изобретении также рассматривается введение одного или нескольких мультиэпитопных полипептидов, которые для целей настоящего изобретения описаны термином «полиэпитопный полипептид», предпочтительно в форме нуклеиновой кислоты, предпочтительно РНК, такой как транскрибированная in vitro или синтетическая РНК, которая может экспрессироваться в клетках пациента, таких как антигенпрезентирующие клетки, с получением одного или нескольких полипептидов. В случае введения более чем одного мультиэпитопного полипептида, неоэпитопы, обеспеченные различными мультиэпитопными полипептидами, могут быть различными или частично перекрывающимися. В случае присутствия в клетках пациента, таких как антигенпрезентирующие клетки, полипептид в соответствии с изобретением процессирован с получением неоэпитопов, идентифицированных согласно изобретению. Введение вакцины, обеспеченной в соответствии с изобретением, может обеспечить презентированные МНС класса II эпитопы, которые способны вызывать ответ CD4+ хелперных Т-клеток против клеток, экспрессирующих антигены, из которых получены эпитопы, презентированные МНС. Альтернативно или дополнительно, введение вакцины, обеспеченной в соответствии с изобретением, может обеспечить эпитопы, презентированные МНС класса I, которые способны вызывать ответ CD8+ Т-клеток против клеток, экспрессирующих антигены, из которых получены эпитопы, презентированные МНС. Кроме того, введение вакцины, обеспеченной в соответствии с изобретением, может обеспечить один или несколько неоэпитопов (включая известные неоэпитопы и неоэпитопы, идентифицированные в соответствии с изобретением), а также один или несколько эпитопов, не содержащих рак-специфических соматических мутаций, но экспрессирующихся раковыми клетками и предпочтительно индуцирующих иммунный ответ против раковых клеток, предпочтительно рак-специфический иммунный ответ. В одном варианте осуществления введение вакцины, обеспеченной в соответствии с изобретением, обеспечивает неоэпитопы, которые представляют собой эпитопы, презентированные МНС класса II и/или способные вызывать ответ CD4+ хелперных Т-клеток против клеток, экспрессирующих антигены, из которых получены эпитопы, презентированные МНС, а также эпитопы, не содержащие рак-специфических соматических мутаций, которые представляют собой эпитопы, презентированные МНС класса I и/или способные вызывать ответ CD8+ Т-клеток против клеток, экспрессирующих антигены, из которых получены презентированные МНС эпитопы. В одном варианте осуществления эпитопы, не содержащие рак-специфических соматических мутаций, получены из опухолевого антигена. В одном варианте осуществления неоэпитопы и эпитопы, не содержащие рак-специфических соматических мутаций, обладают синергическим эффектом в лечении рака. Предпочтительно, вакцина, обеспеченная в соответствии с изобретением, является полезной для полиэпитопной стимуляции цитотоксических и/или хелперных Т-клеточных ответов.
Вакцина, обеспеченная в соответствии с изобретением, может представлять собой рекомбинантную вакцину.
Термин «рекомбинантная» в контексте настоящего изобретения означает «созданная посредством генной инженерии». Предпочтительно, «рекомбинантная единица», такая как рекомбинантный полипептид, в контексте настоящего изобретения не имеет природного происхождения и предпочтительно является результатом комбинирования единиц, таких как аминокислотные или нуклеотидные последовательности, которые не являются комбинированными в природе. Например, рекомбинантный полипептид в контексте настоящего изобретения может содержать несколько аминокислотных последовательностей, таких как неоэпитопные или вакцинные последовательности, полученные из различных белков или различных частей одного и того же белка, слитых вместе, например, при помощи пептидных связей или соответствующих линкеров.
Используемый здесь термин «природный» относится к тому факту, что объект может существовать в природе. Например, пептид или нуклеиновая кислота, которая присутствует в организме (включая вирусы) и может быть выделена из источника, существующего в природе, и которая не была специально модифицирована человеком в лаборатории, является природной.
Агенты, композиции и способы, описанные здесь, могут быть использованы для лечения субъекта, имеющего заболевание, например, заболевание, характеризующееся присутствием больных клеток, экспрессирующих антиген и презентирующих его фрагмент. Особенно предпочтительными заболеваниями являются раковые заболевания. Агенты, композиции и способы, описанные здесь, могут быть использованы для иммунизации или вакцинации для предупреждения описанного здесь заболевания.
В соответствии с изобретением термин «заболевание» относится к любому патологическому состоянию, включая раковые заболевания, в частности, такие формы раковых заболеваний, которые описаны в настоящем документе.
Термин «нормальный» относится к удовлетворительному состоянию здоровья или состояниям у здорового субъекта или ткани, то есть непатологическим состояниям, при этом «здоровый» предпочтительно означает нераковый.
«Заболевание, в которое вовлечены клетки, экспрессирующие антиген» означает в соответствии с изобретением, что детектирована экспрессия антигена в клетках больной ткани или органа. Экспрессия в клетках больной ткани или органа может быть повышенной по сравнению с состоянием в здоровой ткани или органе. Повышение относится к повышению по меньшей мере на 10%, в частности, по меньшей мере на 20%, по меньшей мере на 50%, по меньшей мере на 100%, по меньшей мере на 200%, по меньшей мере на 500%, по меньшей мере на 1000%, по меньшей мере на 10000%, или даже больше. В одном варианте осуществления экспрессия обнаруживается только в больной ткани, тогда как экспрессия в здоровой ткани является репрессированной. В соответствии с изобретением заболевания, в которые вовлечены клетки или которые ассоциированы с клетками, экспрессирующими антиген, включают раковые заболевания.
В соответствии с изобретением термин «опухоль» или «опухолевое заболевание» относится к ненормальному росту клеток (называемых неопластическими клетками, онкогенными клетками или опухолевыми клетками), предпочтительно формирующих набухание или поражение. Под «опухолевой клеткой» понимается ненормальная клетка, которая растет путем быстрой неконтролируемой клеточной пролиферации и продолжает расти после прекращения действия стимула, вызвавшего новый рост. Опухоли показывают частичное или полное отсутствие структурной организации и функциональной координации с нормальной тканью и обычно формируют отдельную массу ткани, которая может быть либо доброкачественной, предраковой или злокачественной.
Рак (медицинский термин: злокачественная неоплазма) представляет собой класс заболеваний, в которых группа клеток проявляет неконтролируемый рост (деление, выходящее за рамки нормальных пределов), инвазию (проникновение и разрушение соседних тканей) и иногда метастазирование (распространение в другие локализации в организме через лимфатические узлы или кровь). Эти три свойства злокачественности рака отличают их от доброкачественных опухолей, которые являются самоограничивающимися и не инвазируют или метастазируют. Большинство видов рака формируют опухоли, но некоторые, такие как лейкоз, нет. Злокачественность, злокачественная неоплазма и злокачественная опухоль являются по существу синонимами рака.
Неоплазма представляет собой ненормальную массу ткани, которая возникает в результате неоплазии. Неоплазия (в переводе с греческого языка означает новый рост) представляет собой аномальную пролиферацию клеток. Рост клеток превышает нормальный уровень и не согласуется с уровнем в нормальных окружающих тканях. Рост продолжается таким же чрезмерным образом даже после прекращения действия стимула. Это обычно вызывает образование уплотнения или опухоли. Неоплазмы могут быть доброкачественными, предраковыми или злокачественными.
«Рост опухоли» или «опухолевый рост» в соответствии с изобретением относится к тенденции опухоли увеличивать свой размер и/или тенденции опухолевых клеток к пролиферации.
Для целей настоящего изобретения термины «рак» и «раковое заболевание» используются взаимозаменяемо с терминами «опухоль» и «опухолевое заболевание».
Рак классифицируется по типу клеток, из которых образуется опухоль и, следовательно, тканей, из которых предположительно развивается опухоль. Это гистология и локализация, соответственно.
Термин «рак» в соответствии с изобретением включает карциномы, аденокарциномы, бластомы, лейкозы, семиномы, меланомы, тератомы, лимфомы, нейробластомы, глиомы, рак прямой кишки, рак эндометрия, рак почек, рак надпочечников, рак щитовидной железы, рак крови, рак кожи, рак головного мозга, рак шейки матки, интестинальный рак, рак печени, рак толстой кишки, рак желудка, рак кишечника, рак головы и шеи, рак желудочно-кишечного тракта, рак лимфатического узла, рак пищевода, колоректальный рак, рак поджелудочной железы, рак уха, носа и гортани (ENT), рак молочной железы, рак предстательной железы, рак матки, рак яичников и рак легкого, и их метастазы. Их примерами являются карцинома легкого, карцинома молочной железы, карцинома предстательной железы, карцинома толстой кишки, почечно-клеточный рак, карцинома матки, или метастазы рака или опухолей, описанных выше. Термин рак в соответствии с изобретением также включает метастазы рака и рецидив рака.
Под «метастазами» понимается распространение раковых клеток из их первоначального месторасположения в другую часть организма. Формирование метастазов является очень сложным процессом и включает отделение опухолевых клеток от первичной опухоли, инвазию во внеклеточный матрикс, проникновение через эндотелиальные базальные мембраны с поступлением в полость организма и сосуды, и затем, после переноса кровью, инфильтрацию органов-мишеней. В заключение, рост новой опухоли, то есть вторичной опухоли или метастатической опухоли в целевом участке зависит от ангиогенеза. Метастазы опухоли часто возникают даже после удаления первичной опухоли, так как опухолевые клетки или компоненты могут остаться и развивать метастатический потенциал. В одном варианте осуществления термин «метастазы» в соответствии с изобретением относится к «отдаленным метастазам», которые относятся к метастазам, происходящим из первичной опухоли и системы региональных лимфатических узлов.
Клетки вторичной или метастатической опухоли сходны с клетками исходной опухоли. Это означает, например, что если рак яичников метастазирует в печень, вторичная опухоль состоит из ненормальных клеток яичника, а не ненормальных клеток печени. Поэтому опухоль в печени называется метастатическим раком яичников, а не раком печени.
Термин «циркулирующие опухолевые клетки» или «СТС» относится к клеткам, которые отрываются от первичной опухоли или опухолевых метастазов и циркулируют в кровотоке. СТС могут оставлять «семена» для последующего роста дополнительных опухолей (метастазов) в различных тканях. У пациентов с метастатическим заболеванием циркулирующие опухолевые клетки обнаруживаются с концентрацией порядка 1-10 СТС на мл общей крови. Разработаны методы исследования для изолирования СТС. Некоторые методы исследования изолирования СТС описаны в данной области, например, методики, в которых используется тот факт, что эпителиальные клетки обычно экспрессируют белок клеточной адгезии ЕрСАМ, который отсутствует в нормальных клетках крови. Улавливание на основе иммуномагнитных частиц включает обработку проб крови антителом к ЕрСАМ, которое конъюгировано с магнитными частицами, с последующим разделением меченых клеток в магнитном поле. Изолированные клетки затем окрашивают антителом к другому эпителиальному маркеру, цитокератину, а также общему лейкоцитарному антигену CD45 для того, чтобы отличить редкие СТС от контаминирующих белых клеток крови. Этот простой и полуавтоматизированный подход позволяет идентифицировать СТС со средним выходом, составляющим приблизительно 1 СТС/мл, и чистотой 0,1% (Allard et al., 2004: Clin Cancer Res 10, 6897-6904). Во втором способе изоляции СТС используется микрожидкостное устройство для улавливания СТС, которое обеспечивает прокачивание цельной крови через камеру, загруженную 80000 микростолбиков, которые были наделены функциональностью путем покрытия антителом к ЕрСАМ. СТС затем окрашивают вторичными антителами против цитокератина или тканеспецифических маркеров, таких как PSA в случае рака предстательной железы, или HER2 в случае рака молочной железы, и визуализируют путем автоматизированного сканирования микростолбиков в разных проекциях по трем пространственным координатам. СТС-чипы способны идентифицировать цитокератин-положительные цикрулирующие опухолевые клетки у пациентов со средним выходом 50 клеток/мл и чистотой в диапазоне 1-80% (Nagrath et al., 2007: Nature 450, 1235-1239). Другой возможностью для изолирования СТС является использование теста CellSearch™ Circulating Tumor Cell (CTC) Test от компании Veridex, LLC (Raritan, NJ), который улавливает, идентифицирует и подсчитывает СТС в пробирке с кровью. Система CellSearch™ представляет собой методику количественного определения СТС в цельной крови, одобренную Управлением США по санитарному надзору за качеством пищевых продуктов и медикаментов (U.S. Food and Drug Administration, FDA), которая основана на комбинировании иммуномагнитного мечения и автоматизированной цифровой микроскопии. Существуют другие методы изолирования СТС, которые описаны в литературе, и могут быть использованы в сочетании с настоящим изобретением.
Рецидив или повторное возникновение происходит, когда у пациента вновь возникает заболевание, которым он страдал в прошлом. Например, если пациент страдал опухолевым заболеванием, получал успешное лечение указанного заболевания, и у него вновь развилось указанное заболевание, то указанное вновь развившееся заболевание может считаться рецидивом или повторным возникновением заболевания. Однако, в соответствии с изобретением, рецидив или повторное возникновение опухолевого заболевания необязательно возникает в участке первоначального опухолевого заболевания. Таким образом, например, если пациент страдал опухолью яичника и получал успешное лечение, рецидив или повторное возникновение может представлять собой возникновение опухоли яичника или возникновение опухоли в участке, отличном от яичника. Рецидив или повторное возникновение опухоли также включает ситуации, при которых опухоль возникает в участке, отличном от участка первоначальной опухоли, а также в участке первоначальной опухоли. Предпочтительно, первоначальная опухоль, по поводу которой пациент получал лечение, представляет собой первичную опухоль, и опухоль в участке, отличном от участка первоначальной опухоли, является вторичной или метастатической опухолью.
Под «лечением» понимается введение соединения или композиции, описанной здесь, субъекту для предупреждения или устранения заболевания, включая уменьшение размера опухоли или количества опухолей у субъекта; купирование или замедление развития заболевания у субъекта; ингибирование или замедление развития нового заболевания у субъекта; снижение частоты или тяжести симптомов и/или повторного возникновения у субъекта, который на текущий момент имеет или ранее имел заболевание; и/или продление, то есть увеличение продолжительности жизни субъекта. В частности, термин «лечение заболевания» включает излечение, сокращение продолжительности, ослабление, предупреждение, замедление или ингибирование прогрессирования или ухудшения, или предупреждение или задержку начала развития заболевания или его симптомов.
Под «имеющий риск» понимается субъект, то есть пациент, который идентифицирован как имеющий более высокий, чем нормальный шанс развития заболевания, в частности рака, по сравнению с общей популяцией. Кроме того, субъект, который имел или на текущий момент имеет заболевание, в частности рак, представляет собой субъекта, который имеет повышенный риск развития заболевания, поскольку у такого субъекта заболевание может продолжать развиваться. Субъекты, которые на текущий момент имеют или которые имели рак, также имеют повышенный риск метастазирования рака.
Термин «иммунотерапия» относится к лечению, включающему активацию специфической иммунной реакции. В контексте настоящего изобретения термины, такие как «защищать», «предупреждать», «профилактический», «превентивный» или «протективный», относятся к предупреждению или лечению, или предупреждению и лечению возникновения и/или распространения заболевания у субъекта и, в частности, сведению к минимуму вероятности того, что у субъекта будет развиваться заболевание, или задержке развития заболевания. Например, лицо, имеющее риск развития опухоли, как описано выше, будет считаться кандидатом на терапию для предупреждения возникновения опухоли.
Профилактическое проведение иммунотерапии, например, профилактическое введение вакцины согласно изобретению, предпочтительно защищает реципиента от развития заболевания. Терапевтическое проведение иммунотерапии, например, терапевтическое введение вакцины согласно изобретению, может привести к ингибированию прогрессирования/роста заболевания. Это включает замедление прогрессирования/роста заболевания, в частности, остановку прогрессирования заболевания, что предпочтительно приводит к устранению заболевания.
Иммунотерапия может проводиться с применением различных способов, при которых предлагаемые здесь агенты действуют в направлении удаления больных клеток у пациента. Такое удаление может иметь место в результате усиления или индукции иммунного ответа у пациента, специфичного по отношению к антигену или клетке, экспрессирующей антиген.
В определенных вариантах осуществления иммунотерапия может представлять собой активную иммунотерапию, при которой лечение основано на стимуляции in vivo эндогенной иммунной системы хозяина для вызывания реакции против больных клеток путем введения средств, модифицирующих иммунные реакции (таких как полипептиды и нуклеиновые кислоты, описанные здесь).
Агенты и композиции, представленные здесь, можно применять отдельно или в сочетании со стандартными терапевтическими режимами, такими как хирургия, лучевая терапия, химиотерапия и/или трансплантация костного мозга (аутологичная, сингенная, аллогенная или неродственная).
Термин «иммунизация» или «вакцинация» описывает процесс лечения субъекта с целью индукции иммунного ответа по терапевтическим или профилактическим причинам.
Термин «in vivo» относится к ситуации у субъекта.
Термины «субъект», «индивидуум», «организм» или «пациент» используются взаимозаменяемо и относятся к позвоночным, предпочтительно млекопитающим. Например, млекопитающие в контексте настоящего изобретения представляют собой человека, приматов нечеловеческого происхождения, домашних животных, таких как собаки, кошки, овцы, крупный рогатый скот, козы, свиньи, лошади и т.д., лабораторных животных, таких как мыши, крысы, кролики, морские свинки и т.д., а также животных в неволе, такие как животные в зоопарках. Термин «животное», используемый здесь, также включает человека. Термин «субъект» может также включать пациента, то есть животное, предпочтительно человека, имеющего заболевание, предпочтительно заболевание, описанное здесь.
Термин «аутологичный» используется для описания чего-либо, что происходит от одного и того же субъекта. Например, «аутологичный трансплантат» относится к трансплантату ткани или органов, полученных от одного и того же субъекта. Такие процедуры являются предпочтительными, так как они преодолевают иммунологический барьер, который в ином случае приводит к отторжению.
Термин «гетерологичный» используется для описания чего-либо, состоящего из множества различных элементов. В качестве примера, перенос костного мозга одного индивидуума другому индивидууму представляет гетерологичный трансплантат. Гетерологичный ген представляет собой ген, происходящий из источника, отличного от субъекта.
В качестве части композиции для иммунизации или вакцинации, предпочтительно один или несколько агентов, описанных здесь, вводить вместе с одним или несколькими адъювантами для индукции иммунного ответа или для повышения иммунного ответа. Термин «адъювант» относится к соединениям, которые продлевают или усиливают, или ускоряют иммунный ответ. Композиция согласно настоящему изобретению предпочтительно проявляет свой эффект без добавления адъювантов. Кроме того, композиция согласно настоящей заявке может содержать любой известный адъювант. Адъюванты содержат гетерогенную группу соединений, таких как масляные эмульсии (например, адъюванты Фрейнда), минеральные соединения (такие как квасцы), бактериальные продукты (такие как токсин Bordetella pertussis), липосомы и иммуностимулирующие комплексы. Примерами адъювантов являются монофосфорил-липид-А (MPL SmithKline Beecham). Сапонины, такие как QS21 (SmithKline Beecham), DQS21 (SmithKline Beecham; WO 96/33739), QS7, QS17, QS18 и QS-L1 (So et al., 1997, Mol. Cells 7: 178-186), неполные адъюванты Фрейнда, полные адъюванты Фрейнда, витамин Е, монтанид, квасцы, олигонуклеотиды CpG (Krieg et al., 1995, Nature 374: 546-549), и различные эмульсии вода-в-масле, которые получены из биологически разлагаемых масел, таких как сквален и/или токоферол.
Также, можно вводить другие вещества, которые стимулируют иммунный ответ пациента. Например, для вакцинации возможно применение цитокинов благодаря их регуляторному влиянию на лимфоциты. Такие цитокины включают, например, интерлейкин-12 (IL-12), который, как было показано, увеличивает защитные действия вакцин (смотри Science 268:1432-1434, 1995), GM-CSF и IL-18.
Существует ряд соединений, которые усиливают иммунный ответ и которые, следовательно, можно применять для вакцинации. Указанные соединения содержат костимулирующие молекулы, представленные в форме белков или нуклеиновых кислот, такие как В7-1 и В7-2 (CD80 и CD86, соответственно).
В соответствии с изобретением физиологический образец может представлять собой образец ткани, включая физиологические жидкости, и/или клеточный образец. Такие физиологические образцы могут быть получены обычным способом, таким как биопсия ткани, включая прицельную биопсию с помощью биопсийных щипцов, и путем взятия крови, бронхиального аспирата, мокроты, мочи, кала или других физиологических жидкостей. В соответствии с изобретением термин «образец» также включает обработанные образцы, такие как фракции или изоляты биологических образцов, например, нуклеиновую кислоту или клеточные изоляты.
Агенты, такие как вакцины и композиции, описанные здесь, можно вводить любым стандартным путем, включая инъекцию или инфузию. Введение может быть выполнено, например, перорально, внутривенно, интраперитонеально, внутримышечно, подкожно или трансдермально. В одном варианте осуществления введение проводится интранодально, например, путем инъекции в лимфатический узел. Другие формы введения предусматривают трансфекцию in vitro антигенпрезентирующих клеток, таких как дендритные клетки, с нуклеиновыми кислотами, описанными здесь, с последующим введением антигенпрезентирующих клеток.
Агенты, описанные здесь, вводят в эффективных количествах. «Эффективное количество» относится к количеству, которое обеспечивает достижение желаемой реакции или желаемого эффекта отдельно или вместе с дополнительными дозами. В случае лечения конкретного заболевания или конкретного состояния, желательная реакция предпочтительно относится к ингибированию хода течения заболевания. Это включает замедление прогрессирования заболевания и, в частности, прерывание или изменение прогрессирования заболевания. Желательной реакцией в лечении заболевания или состояния может также являться задержка начала или предупреждение начала развития указанного заболевания или указанного состояния.
Эффективное количество агента, описанного здесь, будет зависеть от состояния, подлежащего лечению, тяжести заболевания, индивидуальных параметров пациента, включая возраст, физиологическое состояние, размер и массу, продолжительности лечения, типа сопутствующей терапии (в случае присутствия), специфического пути введения и аналогичных факторов. Соответственно, дозы вводимых агентов, описанных здесь, могут зависеть от таких различных параметров. В случае, если при введении начальной дозы реакция у пациента является недостаточной, могут применяться более высокие дозы (или эффективные более высокие дозы, которые достигаются с помощью различных, более локализованных путей введения).
Фармацевтические композиции, описанные здесь, являются предпочтительно стерильными и содержат эффективное количество терапевтически активного вещества для генерирования желательной реакции или желательного эффекта.
Фармацевтические композиции, описанные здесь, как правило, вводят в фармацевтически совместимых количествах и в фармацевтически совместимом препарате. Термин «фармацевтически совместимый» относится к нетоксичному материалу, который не нарушает действие активного компонента фармацевтической композиции. Препараты данного вида обычно содержат соли, буферные вещества, консерванты, носители, дополнительные повышающие иммунитет вещества, такие как адъюванты, например, олигонуклеотиды CpG, цитокины, хемокины, сапонин, GM-CSF и/или РНК, и, при необходимости, другие терапевтически активные соединения. При использовании в медицине соли должны быть фармацевтически совместимыми. Однако соли, которые не являются фармацевтически совместимыми, могут быть использованы для приготовления фармацевтически совместимых солей, и включены в изобретение. Фармакологически и фармацевтически совместимые соли этого типа включают, но без ограничения, такие соли, которые получены из следующих кислот: соляной, бромистоводородной, серной, азотной, фосфорной, малеиновой, уксусной, салициловой, лимонной, муравьиной, малоновой, янтарной и т.п. Фармацевтически совместимые соли могут быть также получены в виде солей щелочных металлов или солей щелочно-земельных металлов, таких как соли натрия, соли калия или соли кальция.
Фармацевтическая композиция, описанная здесь, может содержать фармацевтически совместимый носитель. Термин «носитель» относится к органическому или неорганическому компоненту природного или синтетического происхождения, с которым комбинируют активный компонент для содействия применению. В соответствии с изобретением термин «фармацевтически совместимый носитель» включает один или несколько совместимых твердых или жидких наполнителей, разбавителей или инкапсулирующих веществ, которые являются подходящими для введения пациенту. Компоненты фармацевтической композиции согласно изобретению обычно представляют собой такие компоненты, которые не вызывают возникновения реакций, которые значительно не влияют на желательную фармацевтическую эффективность.
Фармацевтические композиции, описанные здесь, могут содержать подходящие буферные вещества, такие как уксусная кислота в форме соли, лимонная кислота в форме соли, борная кислота в форме соли, и фосфорная кислота в форме соли.
Фармацевтические композиции могут при необходимости также содержать подходящие консерванты, такие как бензалкония хлорид, хлорбутанол, парабен и тимеросал.
Фармацевтические композиции обычно представлены в виде стандартной лекарственной формы и могут быть приготовлены известным способом per se. Фармацевтические композиции согласно изобретению могут быть представлены в форме капсул, таблеток, пластинок, растворов, суспензий, сиропов, эликсиров или, например, в форме эмульсий.
Композиции, подходящие для парентерального введения, обычно содержат стерильные водные или неводные препараты активного соединения, которые предпочтительно являются изотоничными крови реципиента. Примерами совместимых носителей и растворителей являются раствор Рингера и изотоничный раствор хлорида натрия. Кроме того, в качестве раствора или суспензионной среды обычно используют стерильные нелетучие масла.
Настоящее изобретение описано далее подробно с помощью фигур и примеров, которые используются только в иллюстративных целях и не предназначены для ограничения изобретения. На основе описания и примеров для специалиста в данной области будет ясно, что возможны дополнительные варианты осуществления, которые могут быть включены в изобретение.
ФИГУРЫ
Фигура 1. Общая схема предсказания связывания с МНС
Фигура 2. Анализ иммуногенности как функции показателя Mmut для 50 приоритетных мутаций B16F10 и 82 приоритетных мутаций CT26.WT (всего 132 мутации), из которых 30 мутаций были иммуногенными. Все вакцинирования выполняли с использованием РНК. Для B16F10 иммуногенность оценивали путем введения РНК в BMDC и измерения иммунного ответа спленоцитов с помощью ELISPOT и FACS. Для CT26.WT иммуногенность оценивали путем введения по отдельности РНК и пептидов в BMDC, и измерения иммунного ответа спленоцитов с помощью ELISPOT; мутация считалась иммуногенной, если пептид или РНК регистрировали иммунный ответ. А Кумулятивное распределение иммуногенных мутаций как функция показателя Mmut. На графике показано общее количество мутаций ниже заданного показателя Mmut (красный), количество мутаций, которые были иммуногенными (синий), и процент иммуногенных мутаций от общего числа (черный). В Гистограмма процента иммуногенных мутаций на интервал Mmut для следующих диапазонов: ≤0,3, (0,3, 1), >1. Показанные ошибки представляют собой стандартные ошибки.
Фигура 3. Анализ иммуногенности B16F10 и CT26.WT как функция Mmut. Кумулятивное распределение иммуногенных мутаций как функция показателя Mmut, для В16 (А) и СТ26 (С). Гистограмма процента иммуногенных мутаций на интервал Mmut для следующих диапазонов: [0,1, 0,3], (0,3, 1], (1, ∞) для В16 (В) и СТ26 (D). Фигуры А и В основаны на анализе 50 приоритетных мутаций B16F10, из которых 12 были иммуногенными. Фигуры С и D основаны на анализе 82 приоритетных мутаций B16F10, из которых 30 были иммуногенными. Для более подробной информации смотри фигуру 2. Ошибки представляют собой стандартные ошибки.
Фигура 4. Модели иммуногенности и контрольные гипотезы. Иммуногенность класса I, обозначенная НA, предполагает, что оба эпитопа WT и MUT презентированы клетками, и что мутация в достаточной степени изменила физико-химические свойства аминокислоты, таким образом, что иммунная система зарегистрировала это изменение и генерировала иммунный ответ (обозначены в виде молнии). Гипотеза Hn, которая служит в качестве контроля для НA, представляет собой всего лишь обратную гипотезу HA, состоящую в том, что мутация значительно не изменила физико-химические свойства аминокислоты и поэтому имеет более низкую вероятность быть «детектированной» иммунной системой и генерировать иммунный ответ. Что касается иммуногенности класса II (HB в сравнении с HC), эпитоп WT не презентирован, а эпитоп MUT является презентированным. НB и НC различаются по показателю Т, то есть высокий (T>τ) против низкого (Т≤τ), соответственно. Следует учесть, что для α*=α, модель иммуногенности HBC1 (Mmut<β) представляет собой композицию всех четырех групп: HBCl=U[HA,HB,HC,Hn].
Фигура 5. Гипотетическое связь показателя Т с иммуногенностью. В соответствии с моделью иммуногенности класса I, во время развития Т-клеток были удалены Т-клеточные рецепторы (TCR), которые сильно связаны с эпитопом дикого типа. Оставшиеся TCR должны проявлять только слабую аффинность связывания с эпитопом дикого типа (А) или ее отсутствие. Эпитопы, которые содержат аминокислотную замену, которая имеет высокий показатель Т, имеют сходные с аминокислотой дикого типа физико-химические свойства и поэтому вероятно будут иметь незначительное воздействие на аффинность связывания с оставшимися TCR (В). Эпитопы, которые содержат аминокислотную замену с низким показателем Т, имеют более высокий шанс увеличить аффинность связывания с конкретными TCR и поэтому большую вероятность быть иммуногенными (С). В этой схематической иллюстрации кодирование цветом используют для спаривания Т-клеток с совпадающим пептидом. Оранжевые/желтые мутации представляют собой мутации с высокими показателями Т (аналогично WT), тогда как синие/бордовые мутации представляют собой мутации с низкими показателями Т (значительное физико-химическое различие по сравнению с WT).
Фигура 6. Кумулятивное распределение иммуногенных мутаций как функция Mmut. А Сравнение процента иммуногенных мутаций, которые удовлетворяют исходной контрольной гипотезе HBC1: {Mmut≤β}, с процентом иммуногенных мутаций, которые удовлетворяют частичной гипотезе HA': 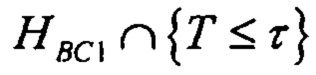 , частичной гипотезе НВС2:
, частичной гипотезе НВС2: 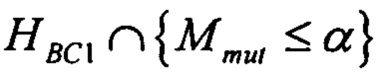 и полной гипотезе HA:
и полной гипотезе HA: 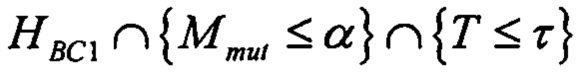 , для α=1, τ=1. В Сравнение процента иммуногенных мутаций, которые удовлетворяют исходным контрольным гипотезам HBC1: {Mmut≤β}, с процентом иммуногенных мутации, которые удовлетворяют обратным частичным гипотезам:
, для α=1, τ=1. В Сравнение процента иммуногенных мутаций, которые удовлетворяют исходным контрольным гипотезам HBC1: {Mmut≤β}, с процентом иммуногенных мутации, которые удовлетворяют обратным частичным гипотезам: 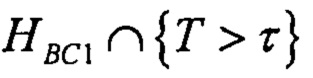 и
и 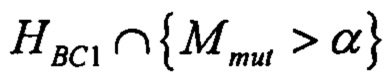 . Анализы в А и В основаны на наборах данных для B16F10 и CT26.WT, содержащих 132 мутации, из которых 30 являются иммуногенными. Каждая экспериментальная точка на графике основана на ≥4 мутациях.
. Анализы в А и В основаны на наборах данных для B16F10 и CT26.WT, содержащих 132 мутации, из которых 30 являются иммуногенными. Каждая экспериментальная точка на графике основана на ≥4 мутациях.
Фигура 7. Кумулятивное распределение иммуногенных мутаций как функция показателя Mmut. Сравнение процента иммуногенных мутаций, которые удовлетворяют исходной контрольной гипотезе HBC1: {Mmut≤β}, с процентом иммуногенных мутаций, которые удовлетворяют частичной гипотезе НА': 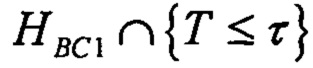 , частичной гипотезе НВС2:
, частичной гипотезе НВС2: 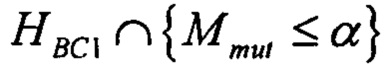 и полной гипотезе HA:
и полной гипотезе HA: 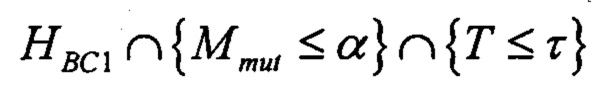 , при условии, что α=1, τ=1 для В16 (А) и СТ26 (С). Сравнение процента иммуногенных мутаций, которые удовлетворяют исходной контрольной гипотезе HBC1: {Mmut≤β}, с процентом иммуногенных мутаций, которые удовлетворяют обратным частичным гипотезам:
, при условии, что α=1, τ=1 для В16 (А) и СТ26 (С). Сравнение процента иммуногенных мутаций, которые удовлетворяют исходной контрольной гипотезе HBC1: {Mmut≤β}, с процентом иммуногенных мутаций, которые удовлетворяют обратным частичным гипотезам: 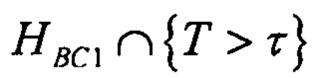 и
и 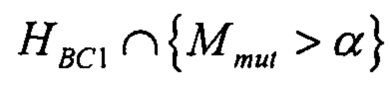 для В16 (В) и СТ26 (D). Фигуры А и В основаны на анализах 50 приоритетных мутаций B16F10, из которых 12 являются иммуногенными. Фигуры С и D основаны на 82 приоритетных мутациях B16F10, из которых 30 являются иммуногенными. Каждая экспериментальная точка на графике основана на ≥4 мутациях.
для В16 (В) и СТ26 (D). Фигуры А и В основаны на анализах 50 приоритетных мутаций B16F10, из которых 12 являются иммуногенными. Фигуры С и D основаны на 82 приоритетных мутациях B16F10, из которых 30 являются иммуногенными. Каждая экспериментальная точка на графике основана на ≥4 мутациях.
Фигура 8. Контроль иммуногенности WT. Для проверки, имеет ли влияние пропуск решений MUT+/WT+ на эти обнаружения, из набора данных были исключены 9 MUT+/WT+ мутаций и 2 мутации, в отношении которых WT не был измерен, оставляя всего 121 мутацию (43 В16 и 78 СТ26), из которых 19 были MUT+/WT- (5 для В16 и 14 для СТ26). Снова были обнаружены такие же тенденции, как в полном наборе данных, а именно, высоко нелинейный ответ как функция показателя Mmut, превосходство гипотезы На над частичной гипотезой, и нахождение на более низкой ступени обратных гипотез по сравнению с исходной контрольной гипотезой HBC1. А Кумулятивное распределение иммуногенности как функция показателя Mmut. В Гистограмма процента иммуногенных мутаций на интервал Mmut. С Сравнение процента иммуногенных мутаций, которые удовлетворяют исходным контрольным гипотезам HBC1, с HA', HBC2 и HA. D Сравнение процента иммуногенных мутаций, которые удовлетворяют исходным контрольным гипотезам НBC1, с обратными гипотезами. Смотри фигуру 5 для дополнительной информации.
Фигура 9. Фракция иммуногенных мутаций как функция RPKM. Красный: все 50 мутаций В16 и 82 мутации СТ26 без фильтрации (всего 132 мутации). Синий: мутации, удовлетворяющие гипотезе НА с α=1, β=0.5, τ=1. B. Процент иммуногенных мутаций для различных диапазонов RPKM без фильтраций. Интервалы RPKM: 1=(0,1],2=(1,5],3=(5,50],4=(50,∞). С Процент иммуногенных мутаций для различных диапазонов RPKM согласно гипотезе НА с α=1, β=0.5, τ=1 Интервалы RPKM: 1=(0,1], 2=(1,∞). Ошибки представляют собой стандартные ошибки.
Фигура 10. Мутированные в якорных и неякорных положениях иммуногенные эпитопы класса II. Мотивы якорных положений анализировали с использованием SYFPEITHI.
Фигура 11. Предложенные модели для иммуногенных опухоль-ассоциированных эпитопов.
Фигура 12. Пример способа взвешенной оценки рангового положения мутаций. Для каждой мутации ранговое положение в списке ранжированных мутаций можно дополнительно подвергнуть Взвешенной оценке на число решений, для которых комбинация типов HLA для пациента, возможные длины окон для типа HLA и положение мутации в пределах эпитопа привели к решению с низким показателем Mmut, или к классификации HA и/или HBUHC. Так как все решения на мутацию потенциально могут быть представлены параллельно, этот взвешенный коэффициент может являться важным фактором для ранжирования положения мутации.
Фигура 13. Пример диаграммы разброса для всех решений эпитопов для мутации chr14_52837882 из СТ26 против Mmut и ΔМ=Mmut-Mwt.
ПРИМЕРЫ
Методики и способы, используемые здесь, описаны в настоящем документе и проводятся известным способом per se и как описано, например, в работе Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Edition (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. Все способы, включая использование наборов и реагентов, осуществляют в соответствии с инструкциями производителя, если специально не указано иное.
Пример 1: Построение модели для предсказания иммуногенности Т-клеточных эпитопов
Ранее авторы использовали иммуногенность 50 соматических мутаций, идентифицированных в клеточной линии мышиной меланомы B16F10 (J.С. Castle et al., Exploiting the mutanome for tumor vaccination. Cancer Research 72, 1081 (2012)). Эти 50 мутаций были отобраны из пула 563 экспрессирующихся несинонимичных соматических мутаций в первую очередь для максимального увеличения экспрессии МНС класса I (J.С. Castle et al., Exploiting the mutanome for tumor vaccination. Cancer Research 72, 1081 (2012)) (смотри также пример 2). Для каждой мутации был предсказан минимальный эпитоп, то есть эпитоп, имеющий самый низкий консенсусный показатель (consensus score) для МНС класса I (Y. Kim et al., Nucleic Acids Research 40, W525 (2012)) (определенный здесь как Mmut) при поиске множества всех возможных аллелей МНС класса I, потенциальных длин эпитопов и окон последовательностей (где положение мутации) (J.С. Castle et al., Exploiting the mutanome for tumor vaccination. Cancer Research 72, 1081 (2012)). Измерение иммуногенности этих мутаций с использованием вакцинации РНК с последующим прочтением пептидов (смотри пример 2) подтвердило полученные ранее данные исследования с использованием вакцинации пептидов (J.С. Castle et al., Exploiting the mutanome for tumor vaccination. Cancer Research 72, 1081 (2012)) и показало, что только 12 из 50 мутаций (24%) были иммуногенными (таблица 1) с последовательностями MUT+/WT-, содержащими только 10% для всех тестированных мутаций.

* Две мутации MUT+ СТ26 были исключены из этой таблицы, так как реактивность их WT еще не была измерена. Всего было измерено 18 MUT+ мутаций из 82 мутаций СТ26 с получением, таким образом, показателя успешности 22%
Результаты тестирования мышиной линии B16F10 показывают, что наивный отбор экспрессирующихся несинонимичных мутаций с низкими показателями Mmut (≤3,9) дает достаточно низкие показатели успешности для предсказания иммуногенности. Таким образом, для того, чтобы персонализированные вакцины, направленно воздействующие на опухоль-специфические неоантигены, стали эффективными терапиями, требуется лучшее понимание механизмов, управляющих иммуногенностью. Для обнаружения дополнительных переменных, которые способствуют иммуногенности, авторы использовали иммуногенность экспрессирующихся несинонимичных соматических мутаций, идентифицированных в мышиной клеточной линии колоректального рака CT26.WT. Всего было отобрано 96 мутаций на основе их показателей Mmut (низкий в сравнении с высоким), среднего значения RPKM (низкий в сравнении с высоким) и клеточной локализации (внутриклеточной в сравнении с внеклеточной), и проведено тестирование на иммуногенность с использованием вакцинации РНК с прочтением как пептида, так и РНК (смотри пример 2 для дополнительной информации). Вместе с клеточной линией B16F10 набор данных состоял из 132 эпитопов, иммуногенность которых была измерена ex vivo на мышиных спленоцитах.
Консенсусный показатель (consensus score) МНС. Для исследования зависимости иммуногенности от Mmut на график наносили кумулятивный процент иммуногенных мутаций как функцию Mmut, то есть процент мутаций, имеющих показатель Mmut меньше, чем заданное пороговое значение (обозначенное как β), которые были иммуногенными. Анализ объединенных наборов данных для В16 и СТ26, охватывающих всего 132 мутации, выявил высоко нелинейную зависимость показателя успешности иммуногенности от Mmut (фигура 2А). На фигуре 2А показано, что иммуногенные мутации богаты крайне низкими показателями Mmut (≤~0,2). Для Mmut≤0,1 процент иммуногенных мутаций достигает максимального значения при ~60%, и быстро уменьшается по мере возрастания Mmut, опускаясь ниже ~25% для Mmut≥2. Процент иммуногенных мутаций с Mmut≤0,3 в сравнении с>0,3 составил 44,4% по сравнению с 17,1%, статистически значимым различием (значение Р=0,004, односторонний точный критерий Фишера). Гистограмма процента иммуногенных мутаций для трех интервалов Mmut: ≤0,3, (0,3, 1] и >1 показывает, что процент иммуногенных мутаций падает по мере увеличения Mmut (фигура 2В). На фигуре 2В различия между показателями успешности самого низкого интервала (Mmut≤0,3), 44,4%, и обоими, центрального, 20,7%, и самого высокого интервала (Mmut>1), 15,8%, были статистически значимыми (величина Р=0,05 и 0,004, соответственно, односторонний точный критерий Фишера), что указывает на то, что для Mmut>~0,3 показатель успешности падает статистически значимым образом. Сходная тенденция показателя успешности также наблюдается при раздельном анализе конкретных раковых мутаций (мутаном, mutanomes) В16 и СТ26 (фигура 3).
Таким образом, критерий для отбора мутаций направлен на презентацию и наблюдалось, что ограничение показателя связывания с МНС мутированного эпитопа обеспечивает возможность предсказания иммуногенных эпитопов с точностью до 60%. Однако презентация является необходимым, но недостаточным условием для индукции иммуногенности. Дополнительно улучшить точность предсказания возможно путем идентификации дополнительного критерия для узнавания TCR. Предложено два взаимоисключающих механизма для управления иммуногенностью, которые названы моделями иммуногенности класса I и класса II.
Иммуногенность класса I. Для того, чтобы репертуар TCR узнавал мутированный эпитоп и генерировал иммунный ответ, была предложена гипотеза, состоящая в том, что должно выполняться три условия (HA, фигура 4): (i) эпитоп дикого типа в некоторой точке в процессе развития организма презентирован иммунной системе, приводя к делеции совпадающих TCR посредством сильного связывания TCR/pMHC; (ii) мутированный эпитоп является презентированным, и (iii) мутированная аминокислота по физико-химическим свойствам в достаточной степени «отличается» от аминокислоты дикого типа (по некоторым показателям, которые будут определены ниже), таким образом, что репертуар TCR способен «детектировать» или «регистрировать» это изменение. Условия (i) и (ii) гарантируют то, что иммунная система фактически подвергается изменению, то есть мутации. Условие (iii) требует, чтобы мутация значительно изменяла физико-химическую характеристику аминокислоты дикого типа таким образом, что аффинность связывания мутированного эпитопа с сохранившимся (неудаленным) TCR потенциально возрастает, запуская тем самым сигнальный каскад, который приводит к иммунному ответу (фигура 5).
Показатель распознавания TCR. Для моделей иммуногенности класса I требуются критерий для установления физико-химического различия между двумя аминокислотами. В теории молекулярной эволюции хорошо известно, что аминокислоты, которые часто взаимозаменяются, вероятно обладают химическим и физическим сходством, тогда как аминокислоты, которые редко взаимозаменяются, вероятно обладают различными физико-химическими свойствами. Вероятность возникновения данной замены в природе по сравнению с вероятностью случайного возникновения этой замены измеряется с помощью матриц логарифмических различий (log-odds). Паттерны, наблюдаемые в матрицах log-odds в условиях естественного отбора, «отражают сходство функций аминокислотных остатков, состоящее в их слабых взаимодействиях друг с другом в трехмерной конформации белков» (М.О. Dayhoff, R.М. Schwartz, В. С. Orcutt, А model for evolutionary change. MO Dayhoff, ed. Atlas of protein sequence and structure Vol. 5, 345 (1978)). Таким образом, были использованы эволюционные матрицы логарифмических различий (log-odds), которые называются здесь как «показатели Т», для отражения распознавания TCR, в качестве эффективных оценивающих матриц для аминокислотных замен, ассоциированных с раком. Замены с положительными показателями Т (то есть log-odds) вероятно возникают в природе и, следовательно, соответствуют двум аминокислотам, которые обладают сходными физико-химическими свойствами. Модель класса I предсказывает, что замены с положительными показателями Т будут иметь более низкую вероятность быть иммуногенными. Напротив, замены с отрицательными показателями Т отражают замены, которые маловероятно возникают в природе и поэтому соответствуют двум аминокислотам, которые имеют значительно отличающиеся физико-химические свойства. В соответствии с предложенной моделью, такие замены будут иметь больший шанс являться иммуногенными. Проведено сравнение различных способов определения матриц log-odds и обнаружено, что результаты являются в значительной степени робастными для конкретного выбранного метода. Оказалось, что подход на основе оценки максимальной вероятности (ML), известный как WAG (S. Whelan, N. Goldman, Molecular biology and evolution 18, 691 (2001)), в котором используется эволюционное расстояние РАМ (число фиксированных точковых мутаций на 100 звеньев цепи (point accepted mutation)), равное 250 единицам, лучше отделяет предсказанные иммуногенные мутации от неиммуногенных и поэтому представлены результаты, полученные с использованием этой матрицы (смотри пример 2 для дополнительной информации).
Имуногенность класса II. В модели иммуногенности класса II авторы выдвинули гипотезу о том, что мутация, по-видимому, является иммуногенной, если иммунная система никогда ранее не видела эпитоп дикого типа и поэтому заразилась мутированным эпитопом. Таким образом, для того, чтобы мутация была иммуногенной, в этой модели авторы выдвинули гипотезу о том, что должно выполняться два условия: (i) эпитоп дикого типа никогда не был презентирован иммунной системе; (ii) мутированный пептид является презентированным. Эти условия могут выполняться одновременно, если, например, мутация поражает якорное положение, изменяя тем самым эпитоп «несвязующее» на «связующее». Формально, иммуногенность класса II можно разделить на две подгипотезы: высокие показатели Т (HB на фигуре 4) и низкие показатели Т (HC на фигуре 4). Однако, так как условием является то, что эпитоп дикого типа не является презентированным, то можно предположить, что природа аминокислотной замены не будет влиять на распознавание TCR, и поэтому иммуногенность класса II можно приравнять объединенной гипотезе: Нв U HC.
Тестирование иммуногенности класса I. Предположения иммуногенности класса I (HA на фигуре 4) могут быть представлены математически следующим образом: требуется, чтобы эпитоп дикого типа был презентирован (Mwt≤α), мутированный эпитоп был презентирован (Mmut≤β), и аминокислотная замена являлась нетривиальной (Т≤τ), где Mwt определен как консенсусный показатель МНС мутированного эпитопа (такая же аллель HLA и длина окна), заменяя мутированную аминокислоту на аминокислоту дикого типа, и Т обозначает показатель Т. Так как все три условия являются необходимыми, можно предположить, что точность классификатора HA будет выше по сравнению с классификатором, основанным только на Mmut (HBC1 на фигуре 4), или по сравнению с частичными гипотезами: 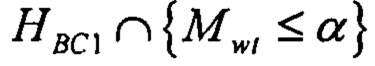 и
и 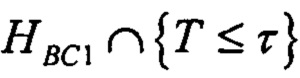 . Таким образом, был рассчитан процент иммуногенных мутаций (количество истинных положительных результатов, деленное на сумму истинных положительных результатов и ложных положительных результатов) как функция β для HBC1, для частичной гипотезы HA':
. Таким образом, был рассчитан процент иммуногенных мутаций (количество истинных положительных результатов, деленное на сумму истинных положительных результатов и ложных положительных результатов) как функция β для HBC1, для частичной гипотезы HA': 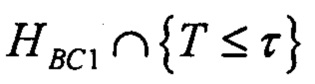 и для частичной гипотезы НВС2:
и для частичной гипотезы НВС2: 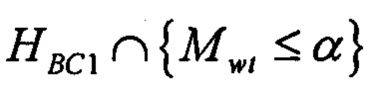 . Было обнаружено, что стабильное пороговое значение τ в интервале примерно от 0,5 до 1 выполнялось лучше (диапазон матрицы WAG250 составляет от -5,1 (замена F↔G) до +5,4 (замена F↔Y). Также, было обнаружено, что значение α может быть консервативно ограничено по сравнению с β, составляя α≈1. Более того, на фигуре 6А показано, что при рассмотрении объединенных мутаном В16 и СТ26, классификаторы на основе НВС2, и HA' достигали большей точности, чем исходная контрольная гипотеза HBC1. Кроме того, классификатор на основе полной гипотезы HA достигал большей точности, чем частичные гипотезы HBC1, и HBC2, демонстрируя тем самым аддитивный эффект. Такие же заключения могут быть сделаны при раздельном анализе наборов данных для В16 и СТ26 (фигура 7).
. Было обнаружено, что стабильное пороговое значение τ в интервале примерно от 0,5 до 1 выполнялось лучше (диапазон матрицы WAG250 составляет от -5,1 (замена F↔G) до +5,4 (замена F↔Y). Также, было обнаружено, что значение α может быть консервативно ограничено по сравнению с β, составляя α≈1. Более того, на фигуре 6А показано, что при рассмотрении объединенных мутаном В16 и СТ26, классификаторы на основе НВС2, и HA' достигали большей точности, чем исходная контрольная гипотеза HBC1. Кроме того, классификатор на основе полной гипотезы HA достигал большей точности, чем частичные гипотезы HBC1, и HBC2, демонстрируя тем самым аддитивный эффект. Такие же заключения могут быть сделаны при раздельном анализе наборов данных для В16 и СТ26 (фигура 7).
Поскольку условия Mwt≤α и T≤τ постулированы как необходимые условия иммуногенности, будет исключено, что классификатор на основе условия 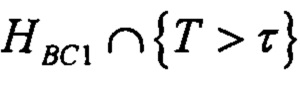 или условия
или условия 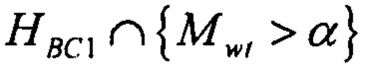 (то есть отрицание второго условия), будет исполняться хуже, чем НBC1. На самом деле, было обнаружено, что это действительно так для В16 и СТ26 при их совместном (фигура 6В) или раздельном (фигура 7) анализе. Таким образом, можно сделать вывод о том, что оба набора данных для В16 и СТ26 поддерживают вместе или по отдельности гипотезу НА. Пропуск мутаций, где WT РНК также показал реактивность, не повлиял на эти заключения (фигура 8).
(то есть отрицание второго условия), будет исполняться хуже, чем НBC1. На самом деле, было обнаружено, что это действительно так для В16 и СТ26 при их совместном (фигура 6В) или раздельном (фигура 7) анализе. Таким образом, можно сделать вывод о том, что оба набора данных для В16 и СТ26 поддерживают вместе или по отдельности гипотезу НА. Пропуск мутаций, где WT РНК также показал реактивность, не повлиял на эти заключения (фигура 8).
Контроль гипотезы HA. Несмотря на то, что мутации с высокими показателями Т все еще могут оставаться иммуногенными, гипотеза, которая богата такими мутациями, должна быть статистически богатой неиммуногенными мутациями. Таким образом, если сравнивать гипотезу НА (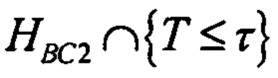 ) с ее обратным вариантом,
) с ее обратным вариантом, 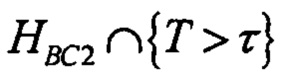 (Hn на фигуре. 4), то будет наблюдаться статистически значимая деплеция иммуногенных мутаций. Действительно, в таблице 2 показано, что для Mmut≤β=0,5, Mwt≤α=1 и Т≤τ=1, HA превосходит Hn с показателем успешности 52,5% (n=21) по сравнению с 21,4% (n=14; Р=0,068, односторонний точный критерий Фишера).
(Hn на фигуре. 4), то будет наблюдаться статистически значимая деплеция иммуногенных мутаций. Действительно, в таблице 2 показано, что для Mmut≤β=0,5, Mwt≤α=1 и Т≤τ=1, HA превосходит Hn с показателем успешности 52,5% (n=21) по сравнению с 21,4% (n=14; Р=0,068, односторонний точный критерий Фишера).
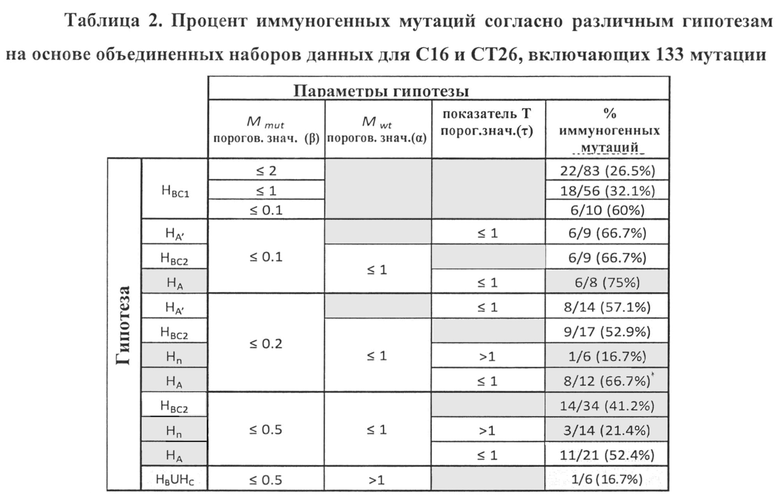
Гипотеза HA также выполняется лучше, чем исходная контрольная гипотеза HBC2, которая достигает 41,2% (n=35). Так как было снижено β, различие между показателями успешности HA и Hn становится больше, так как чем жестче условие по β, тем больше ложноположительных результатов удаляется из группы HA. Например, для β=0,25 показатель успешности группы HA составил 67% (n=14) по сравнению с показателем успешности 17% (n=6) для группы Hn (Р=0,066, односторонний точный критерий Фишера) - смотри таблицу 3.
Таблица 3. Ранжированный список из 133 измеренных мутаций B16F10/CT26.WT, которые удовлетворяют основной контрольной гипотезе НBC1 (Мmut≤0,25), разбитый на три непересекающихся класса гипотез: гипотеза НA для иммуногенных мутаций (Mwt≤0,8, Т≤0,5), Hn/обратная гипотеза НА, богатая неиммуногенными мутациями (Mwt≤0,8, Т>0,5), и гипотеза HbUHC для иммуногенных мутаций (Mwt>0,8). Было предложено ранжировать кандидаты НA и HbUHC на основе относительной важности различительных переменных. Для НА предложенный порядок представляет собой: Мmut (убывание) → показатель Т (убывание) → Mwt (убывание). Для HBUHC предложенный порядок представляет собой: Мmut (убывание) → MWT (возрастание). Ошибки представляют собой стандартные ошибки
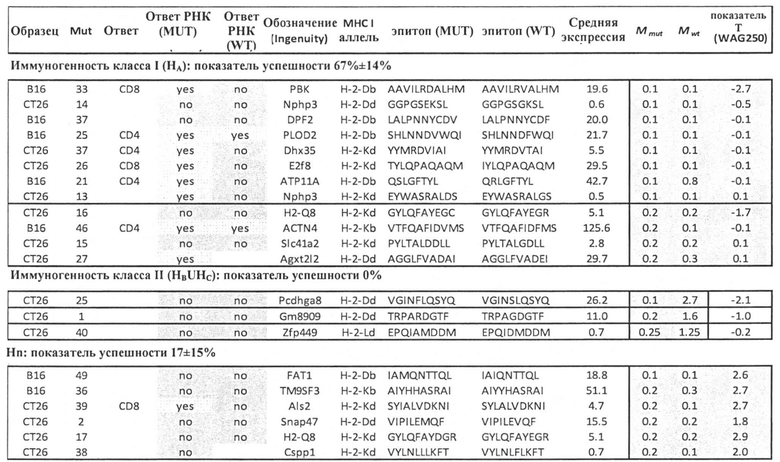
yes - да
no - нет
Пример дополнительных весовых коэффициентов, которые могут дополнительно улучшить ранжирование иммуногенности, представлен в примере 3.
В более общем смысле, список мутаций, которые удовлетворяют HBC1 (Mmut≤β), может быть классифицирован на три категории: НА, Hn и HBUHC (таблица 3), где HA богата иммуногенными мутациями, Hn богата неиммуногенными мутациями. В случае В16 и СТ26, все три кандидата в группе HBUHC были неиммуногенными вопреки ожиданиям. Однако, в случае выбора более правдоподобного порогового значения α* для Mwt, такого как α*>>α, предсказаний, которые могли бы быть тестированы для HBUHC, не будет.
Максимальная точность классификаторов иммуногенности. В соответствии с таблицей 1 средний показатель успешности предсказания иммуногенности в объединенных наборах данных В16 и СТ26 составил 22,7% (=30/132). Путем применения самого жесткого порогового значения к показателю Mmut (β=0.1), точность классификатора иммуногенности увеличилась до 60% (=6/10; HBC1 в таблице 2). Путем объединения HBC1 с условием Mwt≤α или условием T≤τ (α=1, τ=1) точность увеличилась до 66,7% (=6/9). Классификатор на основе НA, который объединяет оба критерия, приводит к аддитивному ответу, который увеличивает точность до 75% (=6/8) (таблица 2).
Эпитоп MUT33 В16. Эпитоп HA-класса, который был ранжирован как самый высокий всеми эволюционными моделями (за исключением матрицы РАМ) в объединенном наборе данных В16/СТ26, представлял собой MUT33 В16 (смотри таблицу 3). Дополнительный анализ выявил, что MUT33 фактически вызвал рестриктированный по МНС класса I ответ CD8+-клеток и проявил ex vivo иммуногенность по сравнению с минимально предсказанным эпитопом (данные не показаны).
Роль генной экспрессии. Нанесение на график фракции иммуногенных мутаций (количество иммуногенных мутаций со значениями RPKM ниже заданного порогового значения относительно общего количества иммуногенных мутаций) как функции значений RPKM для В16 и СТ26 показывает, что это соотношение в некоторой степени находится в стагнации при очень низких значениях RPKM (фигура 9А). Этот эффект наблюдается независимо от того, применяется ли критерий НA или нет. Нанесение на график процента иммуногенных мутаций для различных интервалов значений RPKM (фигура 9В и С) дает основание предположить, что значения RPKM≤ ~1 имеют более низкий показатель успешности (как с применением, так и без применения фильтрующей гипотезы HA), хотя предполагается, что при этом эти результаты находятся в пределах диапазона ошибки.
Анализ опубликованных эпитопов CD8+. Далее авторов заинтересовало, удовлетворяют ли опубликованные опухолевые антигены, распознаваемые Т-клетками, с единичными аминокислотными заменами, вызывающие рестриктированный ответ CD8+, предложенным в настоящем документе моделям иммуногенности. Из опубликованных 17 эпитопов (P. Van der Bruggen, V. Stroobant, N. Vigneron, В. Van den Eynde. (Cancer Immun, http://www.cancerimmunity.org/peptide/, 2013)) (таблица 4) пять удовлетворяло критерию для НА (α=0,7, β=0,2, τ=0,5), четыре удовлетворяло критерию для HCUHB (α=2,2, β=0,4) и два удовлетворяло критерию для Hn (α=0,6, β=0,3, τ=1,7).
Таблица 4. Опубликованные эпитопы с единичной аминокислотной заменой, генерирующие CD8+-ответы. Список ссылочных материалов представлен в примере 2. Мутации якорных положений в группе HBUHC выделены красным цветом.
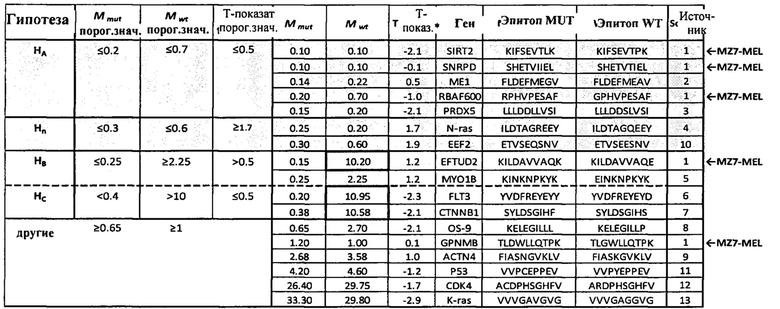
* Based on WAG250 log-odds matrix, color legend: T≤0.5, 0.5<T≤1, T>1
* Ha основе матрицы log-odds WAG250, цветовое обозначение: T≤0,5, 0,5<Т≤1, Т>1
Таким образом, гипотезы НА и HCUHB вместе насчитывают примерно 50% опубликованных эпитопов. Примечательно, что 3 из 4 опубликованных эпитопов, которые удовлетворяют условию HCUHB (красные рамки в таблице 4), имели показатель Mwt, который был больше 10 вследствие мутаций якорных положений (фигура 10). Поскольку требование для гипотезы HCUHB заключается в том, что вероятность презентации эпитопа дикого типа какой-либо клеткой в ходе развития организма сохраняется ничтожно низкой, предполагается, что пороговое значение для Mwt будет сохраняться высоким, то есть  . Действительно, при увеличении α от 0,8 до >3 ложноположительные результаты для В16/СТ26 в таблице 3 исчезают. Таким образом, более правдоподобное пороговое значение для Mwt в условиях гипотезы HcUHb может находиться в интервале от 3 до 10. Клеточная линия MZ7-MEL. Для тестирования способности предложенных в настоящем документе моделей иммуногенности предсказывать иммуногенные эпитопы на модели опухоли человека, использовали клеточную линию MZ7-MEL, полученную в 1988 году из метастазов в селезенке пациента со злокачественной меланомой (V. Lennerz et al., Proceedings of the National Academy of Sciences of the United States of America 102, 16013 (2005)). Скрининг библиотеки кДНК из клеток MZ7-MEL с аутологичными опухоль-реактивными Т-клетками выявил, по меньшей мере, пять неоантигенов, способных генерировать CD8+-ответы (V. Lennerz et al., Proceedings of the National Academy of Sciences of the United States of America 102, 16013 (2005)). На сегодняшний день это представляет самый крупный набор неоантигенов CD8+, полученных от пациента. При применении предложенных в настоящем документе моделей иммуногенности к этим эпитопам было обнаружено, что три неоантигена классифицировались как HA эпитопы, и один неоантиген, мутация якорного положения, классифицировался как HbUHc эпитоп (стрелки в таблице 4, и фигура 10). Таким образом, четыре из пяти эпитопов могут быть объяснены предложенными в настоящем документе моделями иммуногенности.
. Действительно, при увеличении α от 0,8 до >3 ложноположительные результаты для В16/СТ26 в таблице 3 исчезают. Таким образом, более правдоподобное пороговое значение для Mwt в условиях гипотезы HcUHb может находиться в интервале от 3 до 10. Клеточная линия MZ7-MEL. Для тестирования способности предложенных в настоящем документе моделей иммуногенности предсказывать иммуногенные эпитопы на модели опухоли человека, использовали клеточную линию MZ7-MEL, полученную в 1988 году из метастазов в селезенке пациента со злокачественной меланомой (V. Lennerz et al., Proceedings of the National Academy of Sciences of the United States of America 102, 16013 (2005)). Скрининг библиотеки кДНК из клеток MZ7-MEL с аутологичными опухоль-реактивными Т-клетками выявил, по меньшей мере, пять неоантигенов, способных генерировать CD8+-ответы (V. Lennerz et al., Proceedings of the National Academy of Sciences of the United States of America 102, 16013 (2005)). На сегодняшний день это представляет самый крупный набор неоантигенов CD8+, полученных от пациента. При применении предложенных в настоящем документе моделей иммуногенности к этим эпитопам было обнаружено, что три неоантигена классифицировались как HA эпитопы, и один неоантиген, мутация якорного положения, классифицировался как HbUHc эпитоп (стрелки в таблице 4, и фигура 10). Таким образом, четыре из пяти эпитопов могут быть объяснены предложенными в настоящем документе моделями иммуногенности.
Для тестирования способности предсказывать эти эпитопы de novo в клеточной линии MZ7-MEL проводили секвенирование экзома клеточной линии MZ7-MEL (смотри раздел «Способы»). Всего было идентифицировано 743 экспрессирующихся несинонимичных мутаций. Были обнаружены все пять мутаций, ранее идентифицированных Lennerz et al. (V. Lennerz et al., Proceedings of the National Academy of Sciences of the United States of America 102, 16013 (2005)). Затем для каждой мутации рассчитывали показатель Т, Mmut, и Mwt, регистрируя также аллель HLA и эпитоп, которые привели к минимальному консенсусному показателю МНС для заданной мутации. Мутации классифицировали в одну из трех групп: HA, HBUHC и Hn с использованием пороговых значений α=0,8, β=0,2, τ=0,5 (и условии RPKM>0,2), и затем ранжировали на основе их потенциала быть иммуногенными, как поясняется в таблице 3. Было обнаружено, что из 743 мутаций 32 мутации удовлетворяли критерию НA (таблица 5), 12 удовлетворяли критерию HBUHC (таблица 6) и 15 удовлетворяли критерию Hn.
Таблица 5. HA-классифицированные мутации клеток MZ7-MEL. 32 из 743 экспрессирующихся несинонимичных мутаций в MZ7-MEL классифицировали как HA-иммуногенные с использованием пороговых значений: α=0,8, β=0,2 и τ=0,5. Ранжирование основано на схеме сортировки Mmut (убывание) → показатель Т (убывание). Иммуногенные неоантигены, идентифицированные Lennerz et al., выделены желтым цветом. Кроме того, требовалось, чтобы значение RPKM превышало 0,2.

* Rank is based on Mmut and the T score
** T score is based on the WAG250 log-odds matrix
* Ранг основан на Mmut и показателе Т
** Показатель Т основан на матрице log-odds WAG250
Таблица 6. HBUHC-классифицированные мутации клеток MZ7-MEL. 12 из 743 экспрессирующихся несинонимичных мутаций в MZ7-MEL классифицировали как HBUHC-иммуногенные с использованием пороговых значений: α*=0,8, β=0,2 и RPKM>2. Ранжирование основано на схеме сортировки Mmut (убывание)→Mwt (возрастание). Иммуногенные неоантигены, идентифицированные Lennerz et al., выделены желтым цветом.
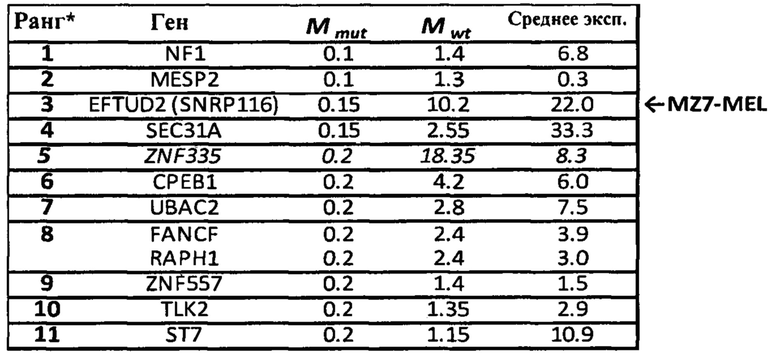
* Rank is based on Mmut and Mwt
* Ранг основан на Mmut и Mwt
Из 32 мутаций, классифицированных как На, три мутации HA-класса, идентифицированные Lennerz et al. (SIRT2, SNRPD1 и RBAF600), ранжировали во 2-ое, 4-ое и 13-ое положения из 18 классов ранжирования с использованием схемы ранжирования Mmut → показатель Т (смотри таблицу 3). Из 12 мутаций, классифицированных как HBUHC, четвертую мутацию согласно Lennerz et al. (SNRP116) ранжировали в 3-е положение. Кроме того, если использовали более высокое (более правдоподобное) пороговое значение для Mwt (например, α*~5), то четвертую мутацию согласно Lennerz et al. ранжировали в 1-е положение (вместе с только одной дополнительной мутацией якорного положения - таблица 7). В заключение, четыре мутации согласно Lennerz et al. были предсказаны как имеющие соответствующий аллель HLA, длину эпитопа и положение мутации, как описано авторами.
Таблица 7. HBUHC-классифицированные мутации клеток MZ7-MEL. 2 из 743 экспрессирующихся несинонимичных мутаций в MZ7-MEL классифицировали как HBUHC-иммуногенные с использованием пороговых значений: α*=5, β=0,2 и RPKM>2. Ранжирование основано на схеме сортировки Mmut (убывание) → Mwt (возрастание). Иммуногенные неоантигены, идентифицированные Lennerz et al., выделены желтым цветом.

* Rank is based on Mmut and Mwt
* Ранг основан на Mmut и Mwt
Выводы
Анализ наборов данных для В16 и СТ26 поддерживает модель, в которой иммуногенность сообщается при выполнении трех условий: пептид дикого типа является презентированным, мутированный пептид является презентированным и аминокислотная замена имеет достаточно низкий показатель log-odds (фигура 11А). Эта модель иммуногенности, которая называется как иммуногенность класса I, дополнительно поддерживается в модели клеточной линии меланомы человека, MZ7-MEL. Модель MZ7-MEL и опубликованные CD8+-рестриктированные неоантигены поддерживают вторую модель, которая называется иммуногенностью класса II, в которой эпитоп дикого типа не презентирован, но замена (например, в якорном положении) приводит к значительному увеличению консенсусного показателя МНС (от >5 до 10), что приводит к новому, никогда ранее не наблюдавшемуся эпитопу (фигура 11В). Эта концепция определения иммуногенности включает схему классификации по трем переменным (Mmut, Mwt, показатель Т). С помощью этой схемы классификации авторы смогли сократить 743 мутации MZ7-MEL до списка из 34 мутаций, при этом 3 из 5 эпитопов согласно Lennerz et al. были ранжированы в первых 5 классах.
В таблице 7 показано, что иммуногенные мутации класса II являются редкими. Из 743 мутаций только 2 были классифицированы как иммуногенные класса II (с использованием правдоподобного порогового значения для Mwt) по сравнению с примерно 30 иммуногенными мутациями класса I. Недостаточность мутаций HBUHC-класса также наблюдалось в мышиных моделях меланомы (таблица 8). Это наблюдение подчеркивает важность иммуногенных мутаций класса I для персонализированных вакцин, которые, как предполагается, являются доминирующим типом обнаруженных в образцах от пациентов мутаций, которые могут применяться для вакцинации. В то же время, тот факт, что один из пяти эпитопов, обнаруженных Lennerz et al., относился к классу II иммуногенности, может указывать на то, что иммуногенные мутации класса II являются более сильными или каким-либо образом отобранными иммунной системой.
Таблица 8. Количество кандидатных мутаций HA и HBUHC в различных моделях опухолей.
(α=0,8, α*=5, β=0,2, τ=0,5)
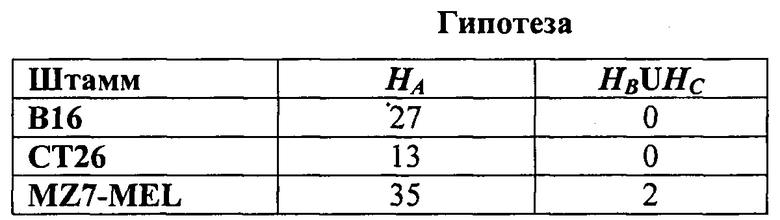
Пример 2: Материалы и методы
Материалы и методы, используемые в примере 1, описаны ниже:
Животные
Мышей C57BL/6J и Balb/cJ (CRL) содержали в соответствии с федеральной и государственной политикой по проведению исследований на животных в университете Майнца (University of Mainz).
Клетки для мышиной модели меланомы и колоректальной опухоли
Клеточную линию меланомы B16F10 (Продукт: АТСС CRL-6475, Номер партии: 58078645) и клеточную линию карциномы толстой кишки CT26.WT (Продукт: АТСС CRL-2638, Номер партии: 58494154) получали в 2010 году из Американской коллекции типовых культур (American Type Culture Collection). Ранние пассажи (3-й, 4-й) клеток использовали для экспериментов по секвенированию. Клетки тестировали обычным способом на микоплазму. Повторную аутотентификацию клеток не выполняли после получения. Клеточную линию MZ7-MEL (полученную в январе 1988 года) и аутологичную В-клеточную линию, трансформированную вирусом Эпштейна-Барр, получали от Dr. Thomas  (Department of Medicine, Hematology Oncology, Johannes Gutenberg University).
(Department of Medicine, Hematology Oncology, Johannes Gutenberg University).
Синтетические пептиды
Пептиды получали от компании Jerini Peptide Technologies (Berlin, Germany) или синтезировали из пептидов TRON. Синтетические пептиды представляли собой 27 аминокислот вместе с мутировавшей (MUT) аминокислотой или аминокислотой дикого типа (WT) в положении 14.
Иммунизация мышей
Мышиным самкам C57BL/6 или Balb/c сопоставимого возраста инъецировали внутривенно 20 мкг транскрибированной in vitro мРНК, формулированной с 20 мкл реагента Lipofectamine™ RNAiMAX (Invitrogen) в PBS в общем инъекционном объеме 200 мкл (3 мыши на группу). Мышей иммунизировали на день 0, 3, 7, 14 и 18. Через двадцать три дня после первой инъекции мышей умерщвляли, и спленоциты выделяли для иммунологического тестирования (смотри анализ ELISPOT). ДНК-последовательности, представляющие одну (Моноэпитоп) или две мутации (Биэпитоп) конструировали с использованием последовательности из 27 аминокислот (аа) с мутацией в положении 14, и клонировали в основную цепь pST1-2BgUTR-A120 (S. Holtkamp et al., Blood 108, 4009 (2006)). Транскрипция in vitro с этой матрицы и очистка были описаны ранее (S. Kreiter et al., Cancer Immunology, Immunotherapy 56, 1577 (2007)).
Иммуноферментный спот-анализ
Иммуноферментный спот-анализ (ELISPOT) (S. Kreiter et al., Cancer Research 70, 9031 (2010)) и генерация сингенных дендритных клеток, происходящих из костного мозга (BMDC), в качестве стимуляторов описаны ранее (L. MB et al., J. Immunol. Methods 223, 77 (1999)). Для модели B16F10 клетки BMDC сенсибилизировали пептидом (6 мкг/мл) с указанной мутацией, соответствующим пептидом дикого типа или контрольным пептидом (VSV-NP). Для модели СТ26 дополнительно к рестимуляции пептидами BMDC трансфицировали соответствующей транскрибированной in vitro мРНК и использовали также для рестимуляции. Для анализа 5×104 BMDC коинкубировали с 5×105 свежевыделенных спленоцитов в микротитровальном планшете, покрытом анти-IFN-γ-антителом (10 мкг/мл, клон AN18; Mabtech). Через 18 часов при 37°C секрецию цитокина детектировали с помощью анти-IFN-γ антитела (клон R4-6A2; Mabtech). Число пятен подсчитывали и анализировали с использованием анализатора ImmunoSpot® S5 Versa ELISPOT, программного обеспечения ImmunoCaptureTM Image Acquisition и программного обеспечения ImmunoSpot® Analysis версии 5. Статистический анализ выполняли с использованием t-критерия Стьюдента и теста Манна-Уитни (непараметический тест). Ответы считались значимыми при p-величине <0,05.
Анализ внутриклеточных цитокинов
Аликвоты спленоцитов, приготовленные для анализа ELISPOT, подвергали анализу на внутриклеточную выработку цитокинов с помощью проточной цитометрии. Для этого 2×106 спленоцитов на образец вносили в культуральную среду (RPMI+10% FCS), дополненную ингибитором транспорта белков в аппарат Гольджи - брефелдином А (10 мкг/мл), в 96-луночном планшете. Клетки от каждого животного рестимулировали в течение 5 ч при 37°C с 2×105 BMDC, сенсибилизированных пептидом или трансфицированных РНК. После инкубации клетки промывали PBS, ресуспендировали в 50 мкл PBS и внеклеточно окрашивали следующими антимышиными антителами в течение 20 мин при 4°C: анти-CD4 FITC, анти-CD8 АРС-Су7 (BD Pharmingen). После инкубации клетки промывали PBS и затем ресуспендировали в 100 мкл раствора Cytofix/Cytoperm (BD Bioscience) в течение 20 мин при 4°C для пермеабилизации внешней мембраны. После пермеабилизации клетки промывали буфером Perm/Wash (BD Bioscience), ресуспендировали в 50 мкл/образец в буфере Perm/Wash и внутриклеточно окрашивали следующими анти-мышиными антителами в течение 30 мин при 4°С: анти-IFN-γ РЕ, анти-TNF-α РЕ-Су7, анти-IL2 АРС (BD Pharmingen). После промывания буфером Perm/Wash клетки ресуспендировали в PBS, содержащем 1% параформальдегида, для анализа методом проточной цитометрии. Образцы анализировали с использованием проточного цитометра BD FACSCanto™ II и программного обеспечения FlowJo (Версия 7.6.3).
Секвенирование следующего поколения
Экстрагирование нуклеиновой кислоты: ДНК и РНК из клеточной массы и ДНК из мышиных тканей экстрагировали с использованием набора Qiagen DNeasy Blood and Tissue kit (для ДНК) и набора Qiagen RNeasy Micro kit (для РНК)
Секвенирование экзома ДНК: Захват экзома для B16F10, C57BL/6J и CT26.WT, и ресеквенирование ДНК для Balb/cJ выполняли в трех повторах, как описано ранее (J.С. Castle et al., Exploiting the mutanome for tumor vaccination. Cancer Research 72, 1081 (2012)). Захват экзома для МZ7-МЕL/ресеквенирование ДНК для EBV-B выполняли в двух повторах с использованием набора для захвата на основе решения Agilent XT Human all Exon 50 Mb, разработанного для захвата всех белок-кодирующих областей. 3 мкг очищенной геномной ДНК (гДНК) фрагментировали на 150-200 пар оснований с использованием ультразвукового прибора Covaris S2. Концы фрагментов «затупляли» (end repaired), 5'-фосфорилировали и 3'- аденилировали в соответствии с инструкциями изготовителя. К фрагментам геномной ДНК лидировали адаптеры «Agilent indexing specific paired-end adapters» с использованием молярного соотношения 10:1 адаптера к геномной ДНК. Перед захватом выполняли 4 цикла амплификации с использованием праймеров Agilent's InPE 1.0 и SureSelect indexing pre-capture PCR primers, и HerculaseII polymerase. 500 нг лигированного адаптера, PCR-обогащенные фрагменты геномной ДНК гибридизировали с зондами (baits) для захвата экзома фирмы Agilent в течение 24 ч при 65°С. Гибридизированные комплексы геномная ДНК/РНК-зонд удаляли с использованием покрытых стрептавидином магнитных шариков, промывали, и РНК-зонды (baits) отщепляли во время элюирования элюирующим буфером SureSelect. Элюированные фрагменты геномной ДНК амплифицировали при помощи PCR после захвата в течение 10 циклов с использованием праймеров SureSelect Indexing Post-Capture PCR и индексных праймеров PCR, и HerculaseII polymerase. Все операции по очистке выполняли с помощью 1,8-кратного объема магнитных шариков Agencourt AMPure ХР. Все оценки качества проводили при помощи набора Qubit HS (Invitrogen), и размер фрагментов определяли при помощи HS DNA assay на анализаторе 2100 Bioanalyzer (Agilen). Обогащенные экзомом библиотеки геномной ДНК кластеризовали на системе cBot с помощью набора Truseq SR cluster kit v2.5 с использованием библиотеки 7 рМ, и 1×100 пар оснований секвенировали на Illumina HiSeq2000 с использованием наборов Truseq SBS kits.
Определение профиля экспрессии генов РНК (RNA-Seq): Библиотеки штрихкодированной кДНК mRNA-seq готовили в двух повторах из 5 мкг общей РНК (модифицировали протокол Illumina mRNA-seq с использованием реагентов NEB). мРНК изолировали с использованием магнитных частиц Seramag Oligo(dT) (Thermo Scientific) и фрагментировали с использованием двухвалентных катионов и нагрева. Полученные фрагменты (160-220 пар оснований) конвертировали в кДНК с использованием случайных праймеров и обратной транскриптазы SuperScriptII (Invitrogen) с последующим синтезом второй цепи с использованием ДНК-полимеразы I и RNaseH. Проводили репарацию концов кДНК, 5'-фосфорилировали и 3'-аденилировали в соответствии с инструкциями к набору библиотеки NEB RNA library kit. Специфические мультиплексные адаптеры Illumina с 3'-выступающими концами, содержащими остатки Т, лигировали с помощью ДНК-лигазы Т4 при молярном соотношении 10:1 адаптера к вставке кДНК. Библиотеки кДНК очищали, и отбирали по размеру, составляющему 300 пар оснований (E-Gel 2% SizeSelect gel, Invitrogen). Обогащение, добавление последовательностей «Illumina six base index» и последовательностей, специфических к проточной кювете проводили с помощью PCR с использованием ДНК-полимеразы Phusion и специфических PCR-праймеров для Illumina. Все операции по очистке до этой стадии выполняли с помощью 1,8-кратного объема магнитных частиц Agencourt AMPure ХР. Все контроли качества выполняли с использованием набора Qubit HS (Invitrogen), и размер фрагментов определяли с использованием набора HS DNA assay на биоанализаторе Agilent 2100 Bioanalyzer. Штрихкодированные RNA-Seq библиотеки кластеризовали, и 50 пар оснований секвенировали, как описано выше.
Анализ данных NGS, экспрессия генов: Полученные в результате секвенирования риды из образцов РНК предварительно обрабатывали в соответствии со стандартным протоколом Illumina, включая фильтрацию ридов низкого качества. Риды, полученные в результате секвенирования, выравнивали на последовательность референсного генома mm9 (А.Т. Chinwalla et al., Nature 420, 520 (2002)) или hg18 (F. Collins, E. Lander, J. Rogers, R. Waterston, I. Conso, Nature 431, 931 (2004)) с использованием программы Bowtie (версия 0.12.5) (В. Langmead, С. Trapnell, М. Pop, S.L. Salzberg, Genome Biol 10, R25 (2009)). Для геномных выравниваний допускалось два несовпадения, и регистрировали только лучшее выравнивание («-v2 -best»), для выравниваний транскриптов использовали параметры по умолчанию. Риды, которые не могут быть выровнены на геномную последовательность, выравнивали с базой данных всех возможных последовательностей экзон-экзонных соединений известных генов UCSC (F. Hsu et al., Bioinformatics 22, 1036 (2006)). Величины экспрессии определяли по пересечению координат рида с координатами транскриптов RefSeq, путем подсчета перекрывающихся ридов экзона и соединения, и нормализации к единицам экспрессии RPKM (Reads which map per Kilobase of exon model per million mapped reads) (A. Mortazavi, B.A. Williams, K. McCue, L. Schaeffer, B. Wold, Nature methods 5, 621 (2008)).
Данные анализа NGS, обнаружение соматических мутаций: Соматические мутации идентифицировали, как описано ранее (J.С. Castle et al., Exploiting the mutanome for tumor vaccination. Cancer Research 72, 1081 (2012)). Риды последовательностей выравнивали на референсный геном mm9 или hg18 с использованием программы для выравнивания bwa (параметры по умолчанию, версия 0.5.8с) (Н. Li, R. Durbin, Bioinformatics 25, 1754 (2009)). «Двусмысленные» риды, которые картируются на множество мест в геноме, были удалены. Мутации идентифицировали с использованием консенсуса двух компьютерных программ: samtools (version 0.1.8) (Н. Li, Bioinformatics 27, 1157 (2011)) и SomaticSniper (A. McKenna et al., Genome Research 20, 1297 (2010)). GATK также включали для B16F10 и C57BL/6J (A. McKenna et al., Genome Research 20, 1297 (2010)). Потенциальным соматическим вариациям, идентифицированным во всех соответствующих репликатах присваивали величину достоверности, «долю ложноположительных результатов» (FDR) (М.  et al., PLoS computational biology 8, e1002714 (2012)) (только CT26 и MZ7-MEL).
et al., PLoS computational biology 8, e1002714 (2012)) (только CT26 и MZ7-MEL).
Отбор и валидация мутаций
Критерии для отбора 50 мутаций B16F10 для тестирования иммуногенности были описаны ранее (J.С. Castle et al., Exploiting the mutanome for tumor vaccination. Cancer Research 72, 1081 (2012)). Эти критерии для мутаций включали: (i) присутствие во всех трех репликатах B16F10 и отсутствие во всех трипликатах C57BL/6; (ii) появление в транскрипте RefSeq; (iii) вызывание несинонимичного изменения; (iv) появление в генах, экспрессирующихся в B16F10 (медиана RPKM по репликатам >10, экспрессия экзона >0); и (v) для каждой мутации показатель Mmut (смотри ниже) должен быть <5. Из 59 оставшихся мутаций был сформирован продукт квантильных рангов показателя МНС класса I, показателя МНС класса II и экспрессии транскрипта, и первые 50 мутаций (0,1≤Mmut≤3,9) были отобраны для подтверждения с помощью PCR (смотри (J.С. Castle et al., Exploiting the mutanome for tumor vaccination. Cancer Research 72, 1081 (2012)) для более подробной информации). Критерии для 96 мутаций CT26.WT, отобранных для тестирования иммуногенности, были дополнительно оптимизированы и включали следующие: (i) присутствие во всех трех репликатах CT26.WT и отсутствие во всех трех репликатах Balb/cJ; (ii) FDR≤0,05; (iii) появление в транскрипте UCSC Known Gene; (iv) вызывание несинонимичного изменения; (v) отсутствие в базе данных dbSNP; и (vi) отсутствие в геномной области повтора. Из оставшихся 493 мутаций восемь 12-членных групп оптимизировали согласно трем признакам: показатель Mmut (самый низкий - [0.1,1.9] против самого высокого - [3.9-20.3]), компартмент белка (внеклеточный, внутриклеточный) и экспрессия генов (выше и ниже медианы RPKM 7,1), отбирая мутации в соответствии с жадным алгоритмом и устанавливая соответствующие пороговые значения. Из 96 полученных мутаций 94 подтверждали при помощи PCR с последующим секвенированием Sanger.
Критерии для отбора мутаций MZ7-ML для анализа включали: (i) присутствие в двух репликатах MZ7-MEL и отсутствие в двух аутологичных репликатах EBV-B, с последующими критериями с (ii) по (vi), описанными выше для CT26.WT. Применение критериев (i)-(vi) сократило первоначальный список из ~8000 мутаций до 743.
Предсказание связывания с МНС и расчет показателя Mmut
Предсказания связывания с МНС выполняли с использованием IEDB Analysis Resource Consensus Tool (http://tools.immuneepitope.org/analyze/html/mhc_binding.html) (Y. Kim et al., Nucleic Acids Research 40, W525 (2012)), который объединяет методы наилучшего выполнения предсказаний, основанные на сравнительных исследованиях (Н. Н. Lin, S. Ray, S. Tongchusak, E. L. Reinherz, V. Brusic, BMC immunology 9, 8 (2008); B. Peters et al., PLoS computational biology 2, e65 (2006)) из ANN (С. Lundegaard et al., Nucleic Acids Research 36, W509 (2008); M. Nielsen et al., Protein Science 12, 1007 (2009)), SMM (B. Peters, A. Sette, BMC bioinformatics 6, 132 (2005)), и для некоторых аллельных моделей также comblib (J. Sidney et al., Immunome Research 4, 2 (2008)). Консенсусный подход объединяет показатели предсказаний всех инструментов путем генерации ранга процентиля, который отражает показатели предсказания связывания заданного пептида относительно пептидных показателей пяти миллионов случайных пептидов из SWISSPROT.
Для каждой мутации рассчитывали предсказанный консенсусный показатель МНС для всех возможных (i) окон последовательности (там, где находится мутация), (ii) длин эпитопов и (iii) мышиных аллелей МНС класса I. Минимальный из всех консенсусных показателей МНС был определен как показатель Mmut.
Расчет матриц log-odds и показателя Т
Марицы log-odds могут быть определены из сравнений выравнивания последовательностей баз данных крупных белков. Ранние матрицы log-odds были основаны на парном сравнении последовательностей (BLOSUM62 (S. Kreiter et al., Cancer Immunology, Immunotherapy 56, 1577 (2007))) и методе оценки максимального приближения (MP) (например, РАМ250 (М. О. Dayhoff, R. М. Schwartz, В.С. Orcutt, А model for evolutionary change. MO Dayhoff, ed. Atlas of protein sequence and structure Vol.5, 345 (1978)), JTT250 (S. Q. Le, O. Gascuel, Molecular biology and evolution 25, 1307 (2008)), и матрица Gonnet (С.С. Dang, V. Lefort, V. S. Le, Q. S. Le, O. Gascuel, Bioinformatics 27, 2758 (2011))). Позднее были разработаны методы на основе максимального правдоподобия (ML) (например, VT160 (P. G. Higgs, Т. K. Attwood, Bioinformatics and molecular evolution. (Wiley-Blackwell, 2009)), WAG (S. Whelan, N. Goldman, Molecular biology and evolution 18, 691 (2001)) и LG (V. Lennerz et al., Proceedings of the National Academy of Sciences of the United States of America 102, 16013 (2005))). Так как ML не ограничивается сравнением только близкородственных последовательностей, как в случае с подходами на основе MP, предполагается, что этот подход к оценке является наиболее точным.
Расчет матрицы log-odds описан подробно в различных источниках (С.С. Dang, V. Lefort, V. S. Le, Q. S. Le, O. Gascuel, Bioinformatics 27, 2758 (2011)). Вкратце, стандартная модель замен аминокислот предполагает Марковскую, с непрерывным временем, реверсивную модель, представленную 20×20 матрицей вероятности (rate matrix) Qij, где qij(i≠j) представляет собой количество замен аминокислот i на j за единицу времени, и где выбранные диагональные элементы удовлетворяют 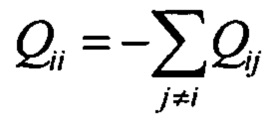 . Q может быть разложено таким образом, что Qij=Sij⋅πj для i≠j, где Si,j представляет собой симметричную матрицу взаимозаменяемости, и πi представляет собой вероятность наблюдать аминокислоту i (С.С. Dang, V. Lefort, V. S. Le, Q. S. Le, O. Gascuel, Bioinformatics 27, 2758 (2011)). В заключение, Q нормализуют таким образом, что
. Q может быть разложено таким образом, что Qij=Sij⋅πj для i≠j, где Si,j представляет собой симметричную матрицу взаимозаменяемости, и πi представляет собой вероятность наблюдать аминокислоту i (С.С. Dang, V. Lefort, V. S. Le, Q. S. Le, O. Gascuel, Bioinformatics 27, 2758 (2011)). В заключение, Q нормализуют таким образом, что  , так что единица времени t=1,0 соответствует 1,0 предполагаемой замене на сайт, или одной «принятой или фиксированной мутации (accepted point mutation)» на сайт, обозначенной как эволюционное расстояние РАМ, равное 100 (М.О. Dayhoff, R.М. Schwartz, В.С. Orcutt, A model for evolutionary change. MO Dayhoff, ed. Atlas of protein sequence and structure Vol. 5, 345 (1978); S. Q. Le, O. Gascuel, Molecular biology and evolution 25, 1307 (2008); С.C. Dang, V. Lefort, V. S. Le, Q. S. Le, O. Gascuel, Bioinformatics 27, 2758 (2011)). Вероятность того, что аминокислота i будет заменена на кислоту j через время t, Pr(i→j|t)=Pij(t), описывается 20×20 матрицей вероятности P(t)=etQ (с символами, означающими экспонирование матрицы). Матрица log-odds, рассчитанная для времени t, описывается 20×20 матрицей log-odds
, так что единица времени t=1,0 соответствует 1,0 предполагаемой замене на сайт, или одной «принятой или фиксированной мутации (accepted point mutation)» на сайт, обозначенной как эволюционное расстояние РАМ, равное 100 (М.О. Dayhoff, R.М. Schwartz, В.С. Orcutt, A model for evolutionary change. MO Dayhoff, ed. Atlas of protein sequence and structure Vol. 5, 345 (1978); S. Q. Le, O. Gascuel, Molecular biology and evolution 25, 1307 (2008); С.C. Dang, V. Lefort, V. S. Le, Q. S. Le, O. Gascuel, Bioinformatics 27, 2758 (2011)). Вероятность того, что аминокислота i будет заменена на кислоту j через время t, Pr(i→j|t)=Pij(t), описывается 20×20 матрицей вероятности P(t)=etQ (с символами, означающими экспонирование матрицы). Матрица log-odds, рассчитанная для времени t, описывается 20×20 матрицей log-odds 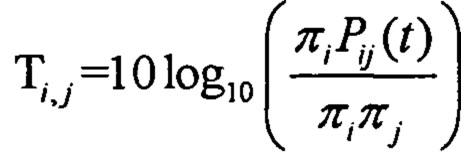 (М.О. Dayhoff, R.М. Schwartz, В.С. Orcutt, A model for evolutionary change. MO Dayhoff, ed. Atlas of protein sequence and structure Vol.5, 345 (1978, 1978)). Реверсивная (time-reversible) означает, что πiPij(t)=πjPji(t) и следовательно, Ti,j является симметричным (P.G. Higgs, Т.K. Attwood, Bioinformatics and molecular evolution. (Wiley-Blackwell, 2009)).
(М.О. Dayhoff, R.М. Schwartz, В.С. Orcutt, A model for evolutionary change. MO Dayhoff, ed. Atlas of protein sequence and structure Vol.5, 345 (1978, 1978)). Реверсивная (time-reversible) означает, что πiPij(t)=πjPji(t) и следовательно, Ti,j является симметричным (P.G. Higgs, Т.K. Attwood, Bioinformatics and molecular evolution. (Wiley-Blackwell, 2009)).
Показатель T для замены i↔j определен здесь как Ti,j и зависит от эволюционной модели и времени t. Были использованы различные модели и эволюционные расстояния РАМ для показателя Т, включая РАМ, BLOSUM62, JTT, VT160, Gonnet, WAG, WAG* и LG (смотри ссылки выше). Фигуры в этом документе были созданы с использованием показателя Т на основе модели WAG и эволюционного расстояния РАМ 250. Такое большое эволюционное расстояние РАМ означает, что существует значительная вероятность замены аминокислоты (P.G. Higgs, Т.K. Attwood, Bioinformatics and molecular evolution. (Wiley-Blackwell, 2009)), и является полезным для детекции взаимосвязей между последовательностями на расстоянии, в которых остатки могут не быть идентичными, но сохраняются физико-химические свойства аминокислот (М.О. Dayhoff, R.М. Schwartz, В.С. Orcutt, A model for evolutionary change. MO Dayhoff, ed. Atlas of protein sequence and structure Vol. 5, 345 (1978); P.G. Higgs, Т.K. Attwood, Bioinformatics and molecular evolution. (Wiley-Blackwell, 2009)).
С использованием статистических данных теста t-распределения сравнивали средние показатели Т иммуногенных эпитопов относительно неиммуногенных эпитопов из таблицы 3 для матрицы WAG с использованием различных единиц РАМ (1, 10, 25, 50, 100, 150, 200 и 250). Анализ статистических данных тестирования показал, что величина Р постепенно уменьшалась с эволюционным расстоянием РАМ, означая, что расстояние РАМ 250 было оптимальным решением, как предполагалось (данные не показаны). Классификация на На и Hn была одинаковой для всех матриц, за исключением матрицы РАМ, которая является наименее достоверной из всех эволюционных моделей. Из всех эволюционных моделей модель WAG250 приводит к максимальному разделению между эпитопами HA и Hn в таблице 3, оценивая разделение с помощью статистики критерия: [максимальный показатель Т (HA)-минимальный показатель Т(Hn)]/σ(показатель Т (НА), показатель Т (Hn)) (данные не показаны). Такая же статистика критерия была также максимальной для расстояния РАМ 250 по сравнению с меньшим расстоянием.
Опубликованные эпитопы CD8+
Эпитопы CD8+с аминокислотами, имеющими одиночные мутации, собирали из списка опухолевых антигенов, возникших в результате мутаций, опубликованного в журнале Cancer Immunity Journal (P. Van der Bruggen, V. Stroobant, N. Vigneron, B. Van den Eynde. (Cancer Immun, http://www.cancerimmunity.org/peptide/, 2013) (http://cancerimmunity.org/peptide/mutations/). Аллели HLA были взяты из опубликованной таблицы или из оригинального документа, если последний был более точным. Ссылки, перечисленные в таблице 4, представляют собой следующие: (1) Lennerz et al. PNAS 102 (44), pp. 16013-16018 (2005); (2) Karanikas et al. Cancer Res 61 (9), pp. 3718-3724 (2001); (3) Sensi et al. Cancer Res 65 (2), pp.632-640 (2005); (4) Linard et al. J. Immunol 168 (9), pp. 4802-4808 (2002); (5) Zorn et al. Eur. J. Immunol 29 (2), pp. 592-601 (1999); (6) Graf et al. Blood 109 (7), pp.2985-2988 (2007); (7) Robbins et al. J. Exp. Med 183 (3), pp. 1185-1192 (1996); (8) Vigneron et al. Cancer Immun 2, pp. 9 (2002); (9) Echchakir et al. Cancer Res 61 (10), pp. 4078-4083 (2001); (10) Hogan et al. Cancer Res 58 (22), pp. 5144-5150 (1998); (11) Ito et al. Int. J. Cancer 120 (12), pp. 2618-2624 (2007); (12) et al. Science 269 (5228), pp. 1281-1284 (1995); (13) Gjertsen et al. Int. J. Cancer 72 (5), pp. 784-790 (1997).
Пример 3. Пример схемы взвешенной оценки показателей мутации для улучшения приоритизации иммуногенных мутаций
РНК, которую вводят в клетку, после трансляции и расщепления на короткие пептиды может быть презентирована на различных типах HLA в пределах клетки. Таким образом, есть все основания полагать, что чем больше типов HLA, которые согласно предсказанию должны иметь низкий консенсусный (или сходный) показатель МНС, тем больше вероятности того, что заданная мутация будет иммуногенной, поскольку она потенциально может размещаться на более чем одном типе HLA одновременно. Таким образом, взвешенная оценка мутаций по количеству типов HLA, для которых мутация классифицирована как HA и/или HBUHC, или даже взвешенная оценка каждой мутации только по количеству типов HLA, которые имеют низкий показатель Mmut, может улучшить ранжирование иммуногенности. В наиболее общем решении, при введении в клетку 27-мерной РНК или пептида существует не только возможность отбора типа HLA, но также длины пептида и положения мутации в пределах этого пептида. Таким образом, можно сканировать все возможные типы HLA, все возможные длины окон и все возможные положения для мутации в пределах окна, и рассчитать число решений (на заданную мутацию), которые классифицируются как HA и/или HBUHC (фигура 12). Это может являться важным взвешивающим коэффициентом для приоритизации мутаций для отбора наиболее эффективных эпитопов для вакцинации. Пример графика разброса всех этих решений как функции Mmut и ΔМ=Mmut-Mwt показан на фигуре 13.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> BioNTech AG et al.
<120> DIAGNOSIS AND THERAPY OF CANCER INVOLVING CANCER STEM CELLS
<130> 342-79 PCT
<150> PCT/EP2013/002272
<151> 2013-07-31
<160> 12
<170> PatentIn version 3.5
<210> 1
<211> 220
<212> PRT
<213> Homo sapiens
<400> 1
Met Ala Ser Ala Gly Met Gln Ile Leu Gly Val Val Leu Thr Leu Leu
1 5 10 15
Gly Trp Val Asn Gly Leu Val Ser Cys Ala Leu Pro Met Trp Lys Val
20 25 30
Thr Ala Phe Ile Gly Asn Ser Ile Val Val Ala Gln Val Val Trp Glu
35 40 45
Gly Leu Trp Met Ser Cys Val Val Gln Ser Thr Gly Gln Met Gln Cys
50 55 60
Lys Val Tyr Asp Ser Leu Leu Ala Leu Pro Gln Asp Leu Gln Ala Ala
65 70 75 80
Arg Ala Leu Cys Val Ile Ala Leu Leu Val Ala Leu Phe Gly Leu Leu
85 90 95
Val Tyr Leu Ala Gly Ala Lys Cys Thr Thr Cys Val Glu Glu Lys Asp
100 105 110
Ser Lys Ala Arg Leu Val Leu Thr Ser Gly Ile Val Phe Val Ile Ser
115 120 125
Gly Val Leu Thr Leu Ile Pro Val Cys Trp Thr Ala His Ala Ile Ile
130 135 140
Arg Asp Phe Tyr Asn Pro Leu Val Ala Glu Ala Gln Lys Arg Glu Leu
145 150 155 160
Gly Ala Ser Leu Tyr Leu Gly Trp Ala Ala Ser Gly Leu Leu Leu Leu
165 170 175
Gly Gly Gly Leu Leu Cys Cys Thr Cys Pro Ser Gly Gly Ser Gln Gly
180 185 190
Pro Ser His Tyr Met Ala Arg Tyr Ser Thr Ser Ala Pro Ala Ile Ser
195 200 205
Arg Gly Pro Ser Glu Tyr Pro Thr Lys Asn Tyr Val
210 215 220
<210> 2
<211> 220
<212> PRT
<213> Homo sapiens
<400> 2
Met Ala Ser Ala Gly Met Gln Ile Leu Gly Val Val Leu Thr Leu Leu
1 5 10 15
Gly Trp Val Asn Gly Leu Val Ser Cys Ala Leu Pro Met Trp Lys Val
20 25 30
Thr Ala Phe Ile Gly Asn Ser Ile Val Val Ala Gln Val Val Trp Glu
35 40 45
Gly Leu Trp Met Ser Cys Val Val Gln Ser Thr Gly Gln Met Gln Cys
50 55 60
Lys Val Tyr Asp Ser Leu Leu Ala Leu Pro Gln Asp Leu Gln Ala Ala
65 70 75 80
Arg Ala Leu Cys Val Ile Ala Leu Leu Val Ala Leu Phe Gly Leu Leu
85 90 95
Val Tyr Leu Ala Gly Ala Lys Cys Thr Thr Cys Val Glu Glu Lys Asp
100 105 110
Ser Lys Ala Arg Leu Val Leu Thr Ser Gly Ile Val Phe Val Ile Ser
115 120 125
Gly Val Leu Thr Leu Ile Pro Val Cys Trp Thr Ala His Ala Val Ile
130 135 140
Arg Asp Phe Tyr Asn Pro Leu Val Ala Glu Ala Gln Lys Arg Glu Leu
145 150 155 160
Gly Ala Ser Leu Tyr Leu Gly Trp Ala Ala Ser Gly Leu Leu Leu Leu
165 170 175
Gly Gly Gly Leu Leu Cys Cys Thr Cys Pro Ser Gly Gly Ser Gln Gly
180 185 190
Pro Ser His Tyr Met Ala Arg Tyr Ser Thr Ser Ala Pro Ala Ile Ser
195 200 205
Arg Gly Pro Ser Glu Tyr Pro Thr Lys Asn Tyr Val
210 215 220
<210> 3
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 3
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ile Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 4
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 4
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Leu His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Val Tyr Ser
35 40 45
Thr Ser Asn Leu Pro Ser Gly Val Pro Ala Arg Phe Gly Gly Ser Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ile Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 5
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 5
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Phe Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 6
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 6
Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Cys Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 7
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 7
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Ile Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Phe Tyr Cys
85 90 95
Ala Arg Asp Phe Gly Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 8
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 8
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Thr Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 9
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 9
Glu Val Gln Leu Gln Gln Ser Arg Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Leu Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Ser Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Tyr Val Phe Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 10
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 10
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Met His
20 25 30
Trp Phe Gln Leu Lys Pro Gly Thr Ser Pro Lys Leu Leu Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Asn Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 11
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 11
Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Gly Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 12
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 12
Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Ser Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, ПРИГОДНЫЕ В ДИАГНОСТИКЕ РАКА | 2018 |
|
RU2815883C1 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2782261C2 |
| ИНДИВИДУАЛИЗИРОВАННЫЕ ПРОТИВООПУХОЛЕВЫЕ ВАКЦИНЫ | 2012 |
|
RU2779946C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ ПРОТИВ КЛАУДИНА 18.2 ДЛЯ ЛЕЧЕНИЯ РАКА | 2014 |
|
RU2792932C2 |
| ТЕРАПЕВТИЧЕСКАЯ РНК ДЛЯ РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ | 2020 |
|
RU2838161C2 |
| ТЕРАПЕВТИЧЕСКАЯ РНК ДЛЯ РАКА ЯИЧНИКА | 2020 |
|
RU2832172C2 |
| ДИАГНОСТИКА И ЛЕЧЕНИЕ РАКА, СВЯЗАННОГО С РАКОВЫМИ СТВОЛОВЫМИ КЛЕТКАМИ | 2014 |
|
RU2749867C2 |
| ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА, ВКЛЮЧАЮЩАЯ АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 | 2014 |
|
RU2832015C2 |
| СОСТАВЫ ВАКЦИН ПРОТИВ НЕОПЛАЗИИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2016 |
|
RU2753246C2 |
| ПЕРСОНАЛИЗИРОВАННАЯ ВАКЦИНА | 2018 |
|
RU2779987C2 |

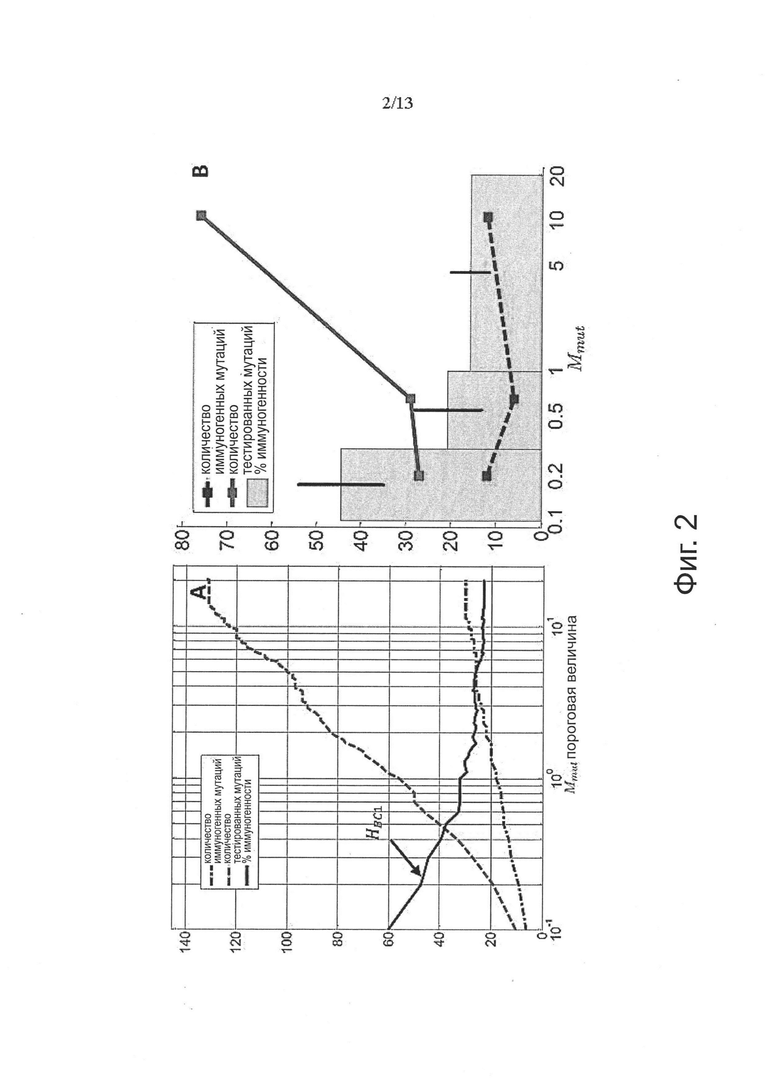
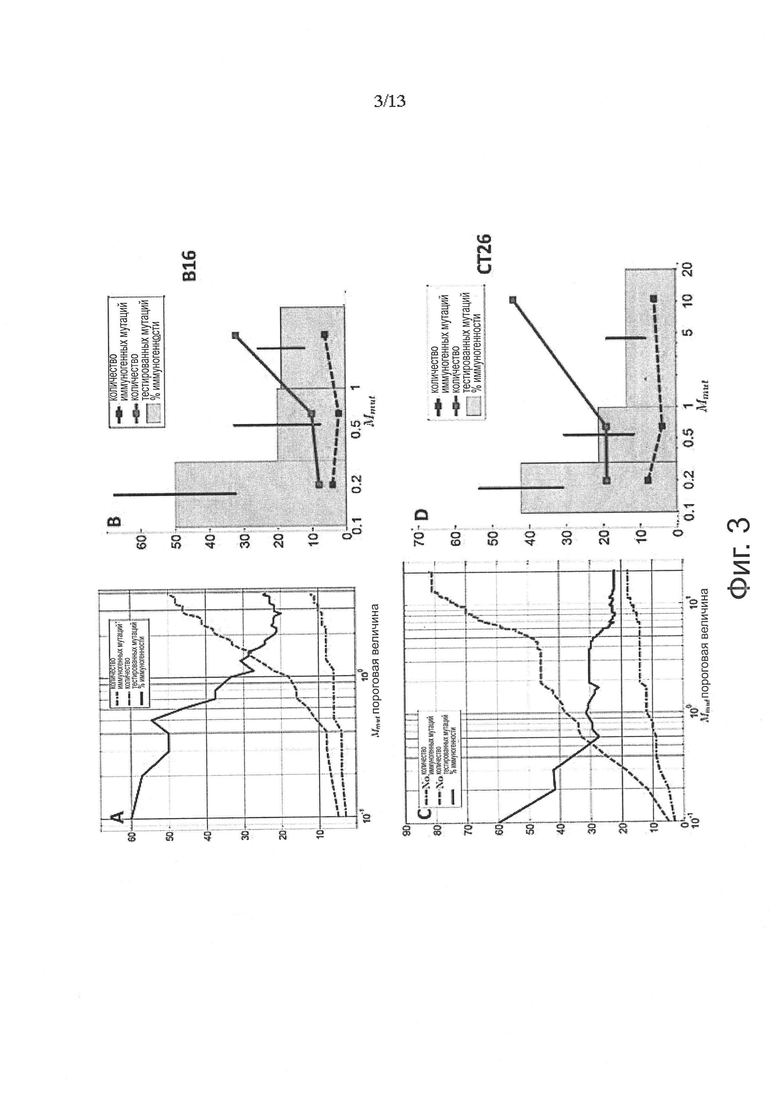
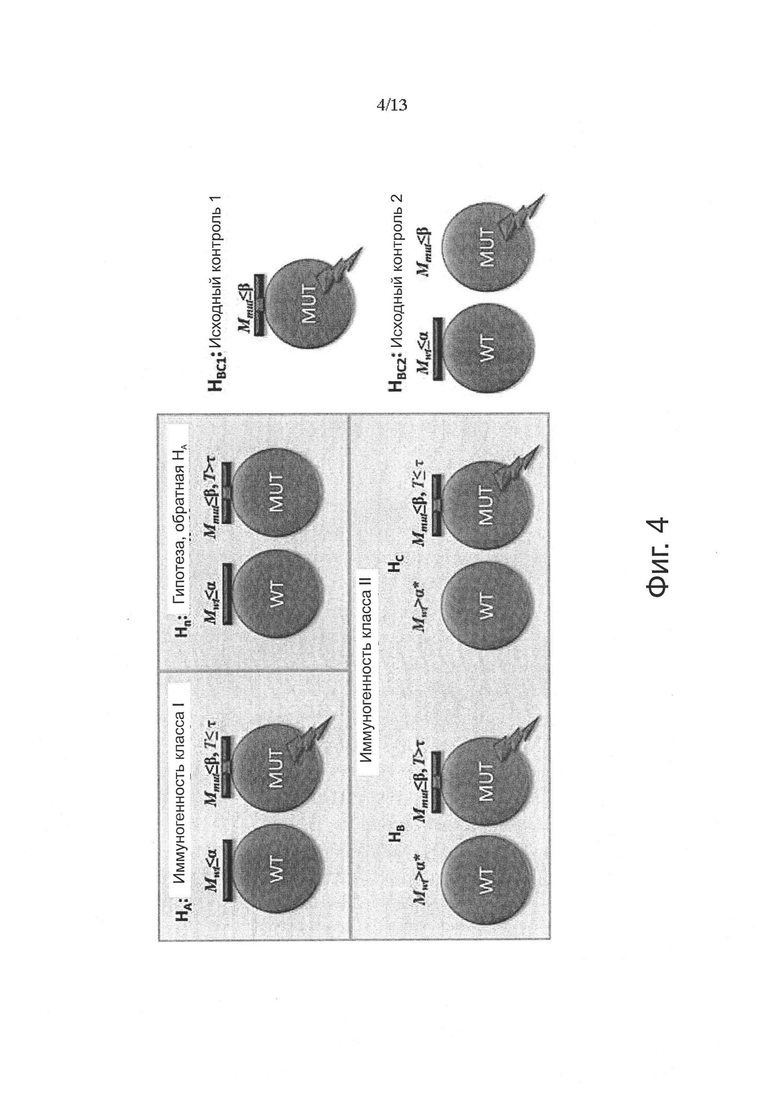


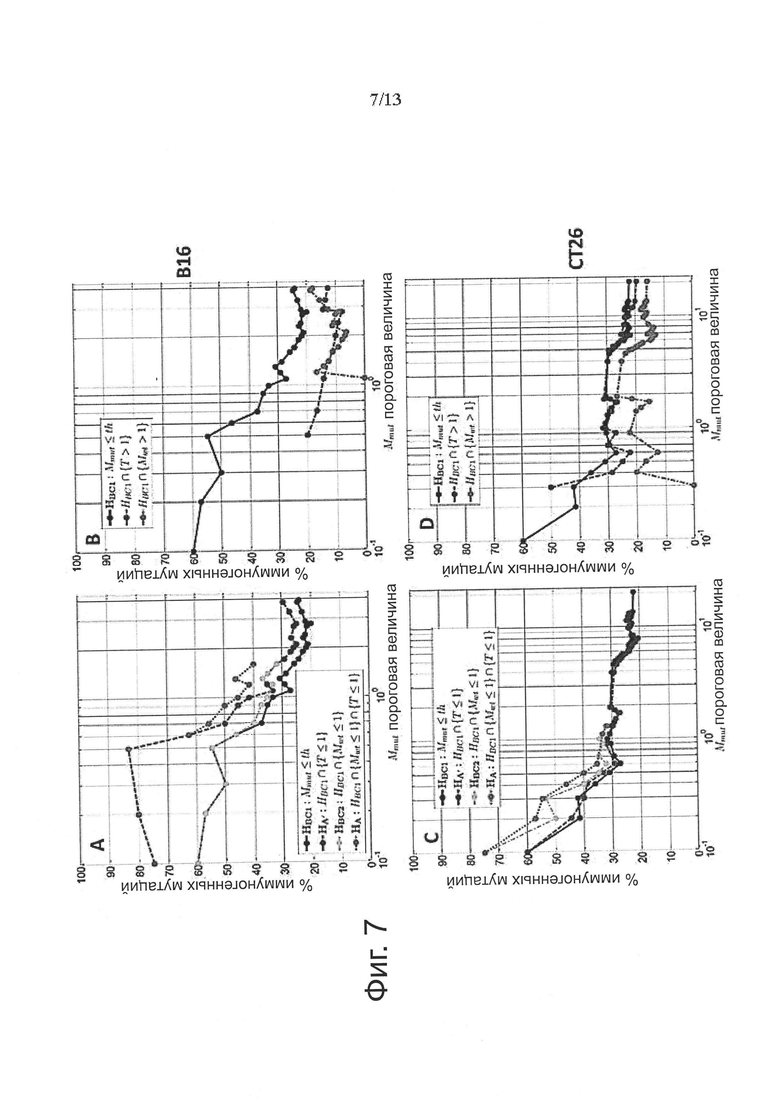
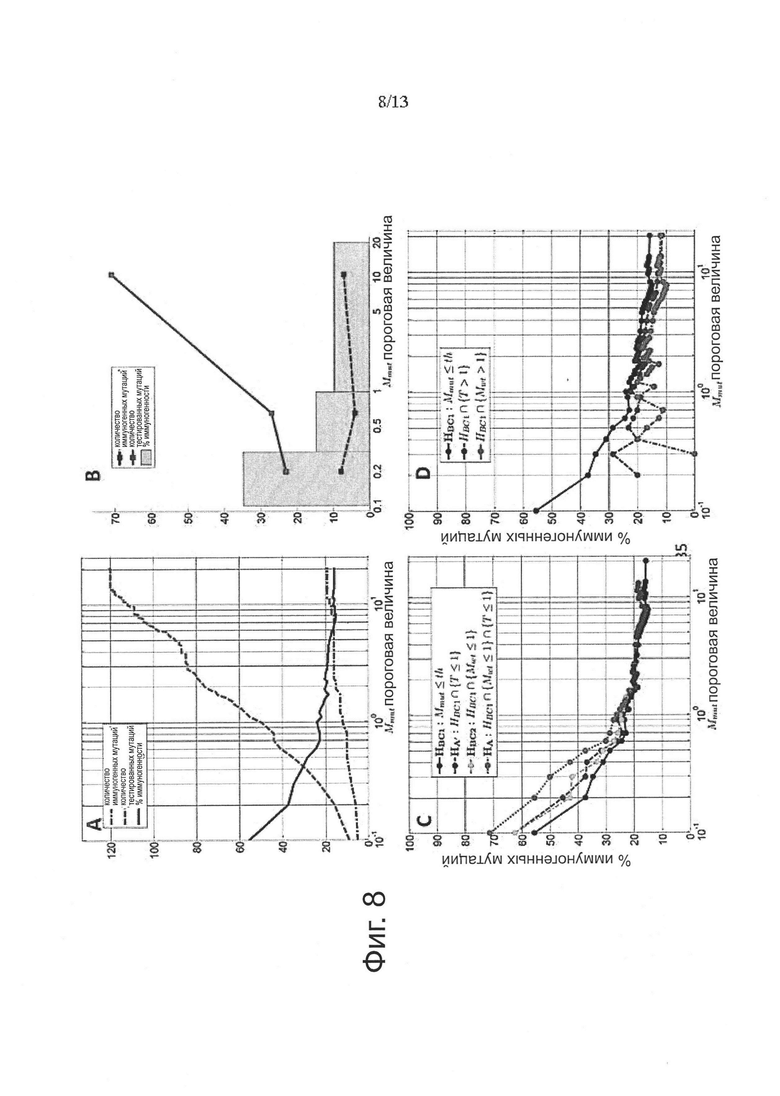

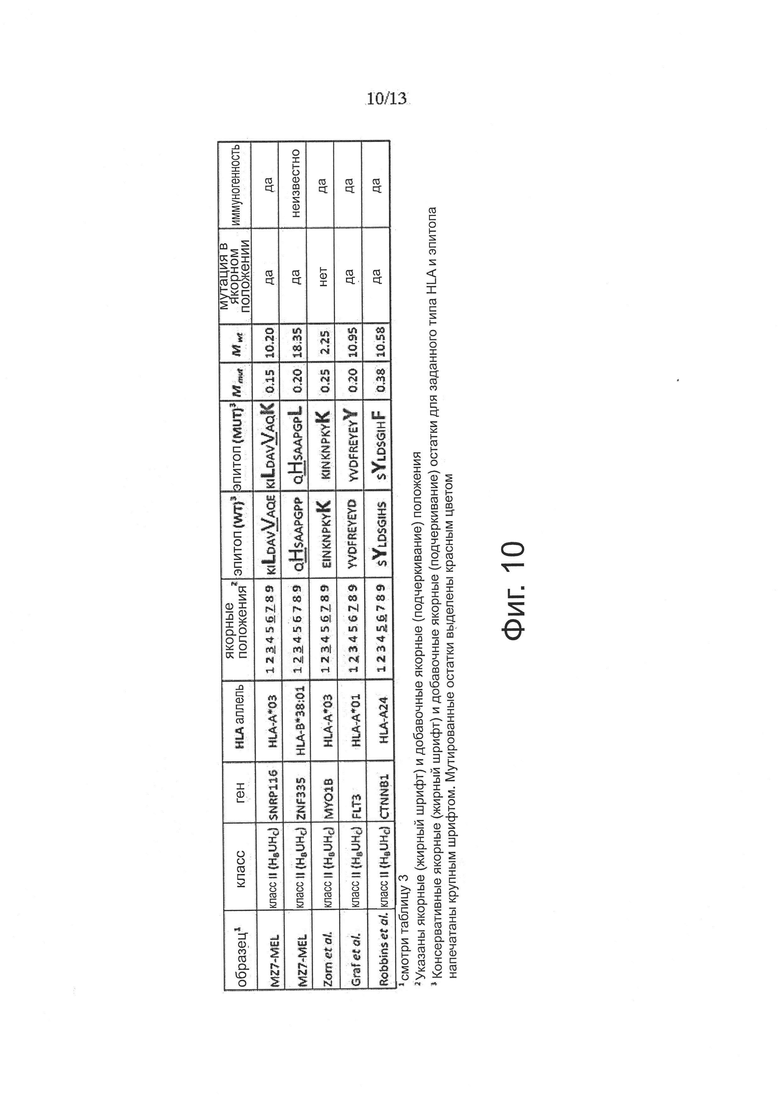
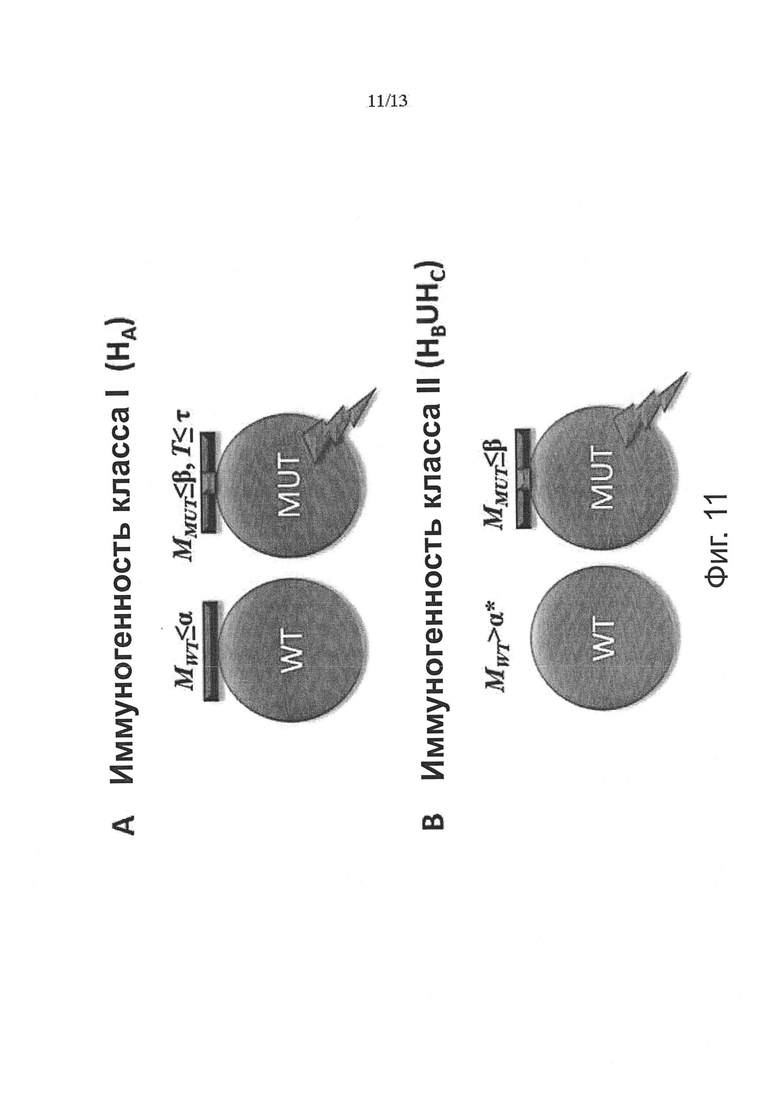

Изобретение относится к области биотехнологии, конкретно к способу предсказания иммуногенных модифицированных пептидов, содержащих аминокислотные модификации, и может быть использовано в медицине. Способ предсказания иммуногенных модифицированных пептидов может быть использован для создания вакцин. 28 з.п. ф-лы, 13 ил., 8 табл., 2 пр.
1. Способ предсказания иммуногенных модифицированных пептидов, содержащих аминокислотные модификации, причем указанные модификации представляют собой аминокислотные замены, где способ включает следующие стадии:
а) идентификация модификаций в одной или более областях, кодирующих белки, путем частичного или полного секвенирования генома, экзома или транскриптома одной или более раковых клеток и необязательно одной или более нераковых клеток;
b) определение показателя связывания с одной или несколькими молекулами МНС модифицированного пептида, содержащего модификацию, идентифицированную на стадии а),
c) определение показателя связывания с одной или более молекулами МНС немодифицированного пептида,
где модифицированный пептид и немодифицированный пептид идентичны во всем за исключением указанной идентифицированной модификации,
где указанные одна или более молекул МНС представляют собой молекулы МНС класса I и/или молекулы МНС класса II,
d) определение показателя химического и физического сходства между немодифицированными и модифицированными аминокислотами, где показатель химического и физического сходства определен на основании вероятности того, что аминокислоты являются взаимозаменяемыми в природе, где чем чаще аминокислоты являются взаимозаменяемыми в природе, тем более похожими эти аминокислоты считаются,
где делают предсказание, что указанный модифицированный пептид является иммуногенным если (i) показатель связывания указанного немодифицированного пептида с одной или несколькими молекулами МНС соответствует порогу, указывающему на связывание с одной или несколькими молекулами МНС, и (ii) показатель связывания указанного модифицированного пептида с одной или несколькими молекулами МНС соответствует порогу, указывающему на связывание с одной или несколькими молекулами МНС, и (iii) немодифицированные и модифицированные аминокислоты имеют показатель химического и физического сходства, соответствующий порогу, свидетельствующему о химическом и физическом несходстве.
2. Способ по п. 1, отличающийся тем, что модифицированный пептид содержит фрагмент модифицированного белка, при этом указанный фрагмент содержит модификацию(и), присутствующую(ие) в белке.
3. Способ по п. 1, отличающийся тем, что немодифицированный пептид содержит аминокислоту зародышевой линии в положении(ях), соответствующем(их) положению(ям) модификации(й) в модифицированном пептиде.
4. Способ по п. 1, отличающийся тем, что немодифицированный пептид и модифицированный пептид имеют длину от 8 до 15, предпочтительно от 8 до 12 аминокислот.
5. Способ по п. 1, отличающийся тем, что одна или несколько молекул МНС включают различные типы молекул МНС, в частности различные аллели МНС.
6. Способ по п. 1, отличающийся тем, что показатель связывания с одной или более молекулами МНС определен с помощью процесса, включающего сравнение последовательности с базой данных МНС-связывающихся мотивов.
7. Способ по п. 1, отличающийся тем, что стадия b) включает определение, удовлетворяет ли указанный показатель предварительно определенной пороговой величине связывания с одной или более молекулами МНС.
8. Способ по п. 1, отличающийся тем, что стадия c) включает определение, удовлетворяет ли указанный показатель предварительно определенной пороговой величине связывания с одной или более молекулами МНС.
9. Способ по п. 1, отличающийся тем, что стадия b) и стадия с) включают определение, удовлетворяет ли указанный показатель предварительно определенной пороговой величине связывания с одной или более молекулами МНС, и пороговая величина, применяющаяся на стадии b), отличается от пороговой величины, применяющейся на стадии c).
10. Способ по п. 1, отличающийся тем, что стадия b) и/или стадия c) включают определение, удовлетворяет ли указанный показатель предварительно определенной пороговой величине связывания с одной или несколькими молекулами МНС, и предварительно определенная пороговая величина связывания с одной или более молекулами МНС отражает вероятность связывания с одной или более молекулами МНС.
11. Способ по п. 1, отличающийся тем, что стадия d) включает определение, удовлетворяет ли указанный показатель предварительно определенной пороговой величине химического и физического сходства между аминокислотами.
12. Способ по п. 1, отличающийся тем, что химическое и физическое сходство определяют с использованием эволюционных матриц логарифмических различий log-odds.
13. Способ по п. 1, отличающийся тем, что модификация не находится в якорном положении для связывания с одной или более молекулами МНС.
14. Способ по п. 1, отличающийся тем, что модификация находится в якорном положении для связывания с одной или более молекулами МНС.
15. Способ по п. 1, который включает выполнение стадии а) на двух или более различных модифицированных пептидах, при этом указанные два или более различных модифицированных пептида содержат одинаковую(ые) модификацию(и).
16. Способ по п. 15, отличающийся тем, что два или более различных модифицированных пептида, содержащие одинаковую(ые) модификацию(и), содержат различные фрагменты модифицированного белка, при этом различные фрагменты содержат одинаковую(ые) модификацию(и), присутствующую(ие) в белке.
17. Способ по п. 15, отличающийся тем, что два или более различных модифицированных пептида, содержащие одинаковую(ые) модификацию(и), содержат все потенциальные фрагменты модифицированного белка, связывающиеся с МНС, при этом, указанные фрагменты содержат одинаковую(ые) модификацию(и), присутствующую(ие) в белке.
18. Способ по п. 15, дополнительно включающий отбор модифицированного(ых) пептида(ов) из двух или более различных модифицированных пептидов, содержащих одинаковую(ые) модификацию(и), обладающих вероятностью или обладающих самой высокой вероятностью связывания с одной или несколькими молекулами МНС.
19. Способ по п. 15, отличающийся тем, что два или более различных модифицированных пептидов, содержащих одинаковую(ые) модификацию(и), отличаются по длине и/или положению модификации(й).
20. Способ по п. 1, который включает выполнение стадии а) и необязательно одну или обе из стадий b) и с) на двух или более различных модифицированных пептидах.
21. Способ по п. 20, отличающийся тем, что указанные два или более различных модифицированных пептида содержат одинаковую(ые) модификацию(и) и/или содержат различные модификации.
22. Способ по п. 21, отличающийся тем, что различные модификации присутствуют в одинаковых и/или различных белках.
23. Способ по п. 15, который включает сравнение показателей двух или более указанных различных модифицированных пептидов.
24. Способ по п. 22, отличающийся тем, что показатель связывания модифицированного пептида с одной или более молекулами МНС является более высоким по весу, чем показатель химического и физического сходства между немодифицированными и модифицированными аминокислотами, и показатель химического или физического сходства между немодифицированными и модифицированными аминокислотами является более высоким по весу, чем показатель связывания немодифицированного пептида с одной или несколькими молекулами МНС.
25. Способ по п. 1, дополнительно включающий идентификацию несинонимичных мутаций в одной или нескольких кодирующих белок областях.
26. Способ по п. 1, отличающийся тем, что указанные мутации представляют собой соматические мутации.
27. Способ по п. 1, отличающийся тем, что указанные мутации представляют собой раковые мутации.
28. Способ по любому из пп. 1-27, который применяют для изготовления вакцины.
29. Способ по п. 28, отличающийся тем, что вакцина получена на основании модификации(й) или модифицированного(ых) пептида(ов), предсказанного(ых) как иммуногенный(ые) с помощью указанного способа.
| CASTLE J | |||
| C | |||
| et al., Exploiting the mutanome for tumor vaccination, Cancer research, 2012, V | |||
| Термосно-паровая кухня | 1921 |
|
SU72A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| WO 2012159754, 29.11.2012 | |||
| MICHIELIN O | |||
| at al., Binding free energy differences in a TCR-peptide-MHC complex induced by a peptide mutation: a simulation analysis, Journal of molecular biology, 2002, V | |||
| Телефонный аппарат, отзывающийся только на входящие токи | 1921 |
|
SU324A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| WAN S | |||

