 Область изобретения
Область изобретения
Данное изобретение относится к новому подходу в идентификации эпитопов Т-клеток, которые вызывают иммунную реакцию в живом организме-хозяине, который включает расчет значений потенциальных эпитопов Т-клеток для связывающих сайтов молекул MHC класса II в пептиде, посредством автоматизированных методов. Кроме того, изобретение относится к способам получения биологических молекул, прежде всего белков и антител, которые вызывают иммунный ответ при введении в организм-хозяин, предпочтительно человека. С помощью этого способа могут быть получены молекулы, которые неиммунногенны или имеют сниженную иммунногенность, когда подвергаются иммунной системе определенного вида, и в сравнении с соответствующим немодифицированным объектом путем уменьшения или удаления потенциальных эпитопов Т-клеток в пределах последовательности указанных изначально иммуногенных молекул. Таким образом, изобретение относится также к новым биологическим молекулам, которые получены способом согласно изобретению.
Предпосылки изобретения
Терапевтическое применение некоторых пептидов, полипептидов и белков ограничено из-за их иммуногенности у млекопитающих, особенно у людей. Например, при применении мышиных антител к пациентам, которые не имеют иммунодепресантов, большинство таких пациентов проявляют иммунные реакции на введенный инородный объект путем вырабатывания антимышиных антител (НАМА) человека (например, Schroff R.W. и др. (1985), Cancer Res. 45: 879-885; Shawler D.L. и др. (1985) J.Immunol. 135: 1530-1535). Существует два серьезных последствия. Во-первых, антимышиное антитело пациента может связать и уничтожить терапевтическое антитело или иммуноконъюгат до того, как у него появиться возможность связать, например, опухоль и проявить свою терапевтическую функцию. Во-вторых, у пациента может развиться аллергическая чувствительность к мышиному антителу и появиться риск анафилактического шока при любом введении мышиного иммуноглобулина в будущем.
Несколько способов было использовано для решения НАМА проблемы и, таким образом, создана возможность использования на людях терапевтических моноклональных антител (см., например, WO-A-8909622; ЕР-А-0239400; ЕР-А-0438310; WO-A-9106667). Эти подходы на основе рекомбинантной ДНК обычно ослабляют мышиную генетическую информацию в конечной конструкции антитела, усиливая тем самым генетическую информацию человека в полученной конструкции. Тем не менее, полученные "гуманизированные" антитела все же вызывают в ряде случаев иммунный ответ у пациентов (Issacs J.D. (1990) Sem.Jmmunol. 2:449, 456; Rebello, P.R. и др. (1999) Transplantation 68:1417-1420).
Общим аспектом в этих методологиях является введение в терапевтическое антитело, обычно по происхождению от грызунов, остатков аминокислот, даже значительных трактов последовательностей аминокислотных остатков, идентичных тем, которые присутствуют в белках антител человека. Для антител такой способ возможен благодаря относительно высокой степени структурного (и функционального) консерватизма среди молекул антител различных видов. Однако для потенциально терапевтических пептидов, полипептидов и белков, где не может существовать структурной гомологии в хозяйских видах (например, человека) для терапевтического белка, такие способы неприменимы. Кроме того, эти способы предполагают, что основное введение последовательности аминокислотного остатка человека будет делать ремоделированное антитело неиммуногенным. Известно, что определенные короткие пептидные последовательности ("эпитопы Т-клеток") могут высвобождаться во время разрушения пептидов, полипептидов или белков внутри клеток и, следовательно, принимать участие в презентации молекулами основного комплекса гистосовместимости (МНС) для того, чтобы инициировать активацию Т-клеток. Для пептидов, представленных МНС класса II, такая активация Т-клеток может затем вызывать, например, ответ антител путем прямой стимуляции В-клеток для получения таких антител. Соответственно было бы желательно устранить потенциальные эпитопы Т-клеток из пептида, полипептида или белка. Даже белки человеческого происхождения и те, которые имеют существующие у человека аминокислотные последовательности, могут индуцировать иммунный ответ у человека. Показательные примеры включают терапевтическое использование колониестимулирующего фактора гранулоцитов-макрофагов (Wadhwa М. и др. (1999) Clin. Cancer. Res. 5:1353-1361) и интерферон альфа 2 (Russo D. и др. (1996) Bri. J. Haem. 94:300-305; Stein R. и др. (1998) New Engl. J.Med. 318:1409-1413).
Устранение эпитопов Т-клеток из белка было раскрыто ранее (см., например, WO 98/52976, WO 00/34317). Основные способы, раскрытые в предыдущем уровне техники, включают следующие стадии:
(a) Определение аминокислотной последовательности полипептида или его части.
(b) Идентификацию одного или более потенциальных эпитопов Т-клеток в пределах аминокислотной последовательности белка любым способом, включающим определение связывания пептидов с молекулами МНС при использовании способов in vitro или in silico или биологических анализов.
(c) Конструирование новых вариантов последовательности с одной или более аминокислотами в пределах идентифицированных потенциальных эпитопов Т-клеток, модифицированных таким образом для существенного уменьшения или устранения активности эпитопов Т-клеток, как определено путем связывания пептидов с молекулами МНС при использовании способов in vitro или in silico или биологическими анализами. Такие варианты последовательностей разрабатывают таким способом, чтобы избежать образования новых потенциальных эпитопов Т-клеток, путем изменения последовательностей до тех пор, пока такие новые потенциальные эпитопы Т-клеток, в свою очередь, не модифицируют таким способом для существенного снижения или устранения активности эпитопов Т-клеток.
(d) конструирование таких вариантов последовательности с помощью техники рекомбинантных ДНК и тестирование указанных вариантов для того, чтобы идентифицировать один или более вариантов с желаемыми свойствами.
Другие способы, использующие растворимые комплексы рекомбинантных молекул МНС в комбинации с синтетическими пептидами, которые способны к связыванию с клонами Т-клеток образцов периферической крови человека или экспериментальных животных, применяются в области техники (Kern F. и др. (1998) Nature Medicine 4: 975-978; Kwok, W.W. и др. (2001) TRENDS in Immunology 22: 583-588) и могут использоваться в стратегии идентификации эпитопа.
Потенциальные эпитопы Т-клеток обычно определяют как любую последовательность аминокислотных остатков, которая обладает способностью связываться с молекулами МНС класса II. Такие эпитопы Т-клеток можно измерять для установления МНС-связывания. Конечно, "эпитоп Т-клеток" означает эпитоп, который при связывании с молекулами МНС может узнаваться рецептором Т-клеток и который может, по крайней мере, в принципе вызывать активацию этих Т-клеток. Это, однако, обычно понимается как то, что определенные пептиды, которые являются такими, что связываются с молекулами МНС класса II, могут удерживаться в белковой последовательности, поскольку такие пептиды узнаются как "собственные" для организма, в который вводится конечный белок.
Целью настоящего изобретения было побороть практическую реальность, что растворимые белки, введенные в аутологический организм-хозяин с терапевтической целью, могут инициировать иммунный ответ, приводящий к развитию хозяйских антител, которые связывают растворимый белок. Одним среди других примеров является интерферон альфа 2, к которому часть пациентов вырабатывает антитела несмотря на то что этот белок вырабатывается эндогенно [Russo D. и др. (1996) Bri. J. Haem. 94: 300-305; Stein R. и др. (1998) New Engl. J. Med. 318: 1409-1413].
Молекулы МНС класса II представляют собой группу высокополиморфных белков, которые играют центральную роль в отборе хелперных Т-клеток и активации. Человеческая лейкоцитарная антигенная группа DR (HLA-DR) представляет собой преобладающий изотип этой группы белков и является базисной точкой данного изобретения. Однако изотипы HLA-DQ и HLA-DP выполняют сходные функции, таким образом, данное изобретение равно применимо к этим двум изотипам. Молекулы МНС класса II DR понимаются как гомодимеры, в которых каждая "половина" является гетеродимером, состоящим из α и β цепей. Каждый гетеродимер обладает лиганд-связывающим доменом, который связывается с пептидами и имеет длину, варьирующую от 9 до 20 аминокислот, несмотря на то что связывающий желобок может включать максимум 9-11 аминокислот. Лиганд-связывающий домен включает аминокислоты с 1 по 85 α цепи, и аминокислоты с 1 по 94 β цепи. DQ-молекулы, как было показано недавно, имеют гомологичную структуру и DP семейство белков, как ожидается, также является очень подобным. У человека известно около 70 различных аллотипов DR изотипа, для DQ известно 30 различных аллотипов, а для DP - 47 различных аллотипов. Каждая особь несет от двух до четырех аллелей DR, два аллеля DQ и два DP аллеля. Структура ряда DR молекул изучена, и эти структуры указывают на связывающий желобок пептида с открытой концевой структурой с рядом гидрофобных карманов, которые вовлекают гидрофобные остатки (остатки кармана) пептида (Brown и др. Nature (1993) 364: 33, Stern и др. (1994) Nature 368: 215). Полиморфизм, идентифицирующий различные аллотипы класса II молекул, вносит свой вклад в широкое разнообразие различных связывающих поверхностей для пептидов в пределах связывающего желобка пептида и на популяционном уровне обеспечивает максимальную гибкость в отношении способности узнавать чужеродные белки и вызывать иммунный ответ к патогенным организмам.
Существует значительный полиморфизм в пределах лиганд-связывающего домена с четкими "семействами" в пределах различных географических популяций и этнических групп. Этот полиморфизм влияет на характеристики связывания пептид-связывающего домена, и таким образом, различные "семейства" DR молекул будут иметь специфичность для пептидов с различными свойствами последовательности, несмотря на то что они могут в некоторой степени перекрываться. Эта специфичность определяет узнавание Th-клеточных эпитопов (класс II Т-клеточного ответа), которые в конечном счете ответственны за стимулирование выработки антител к В-клеточным эпитопам, которые присутствуют в том же белке, из которого происходит Th-клеточный эпитоп. Таким образом, иммунный ответ на белок в индивидуальном человеческом организме в значительной степени находится под влиянием узнавания Т-клеточного эпитопа, что является функцией пептид-связывающей специфичности такого индивидуального HLA-DR аллотипа. Таким образом, для того чтобы идентифицировать Т-клеточные эпитопы в пределах белка или пептида в контексте глобальной популяции, желательно рассматривать связывающие свойства такого широкого набора HLA-DR аллотипов, насколько это возможно, покрывая таким образом настолько высокий процент мировой популяции, насколько это возможно.
Иммунный ответ на терапевтический белок, такой, как белок в соответствии с изобретением, проходит via путь презентации пептида МНС класса II. Таким образом, для того, чтобы устранить или снизить иммуногенность, желательно идентифицировать и удалять эпитопы Т-клеток из белка.
Немодифицированные биологические молекулы могут быть получены рекомбинантным методом, который per se является хорошо известным в уровне техники, используя некоторое количество различных типов хозяйских клеток.
Однако существует потребность в аналогах указанных биологических молекул с улучшенными свойствами. Желательные усовершенствования включают альтернативные схемы и модальности для экспрессии и очистки указанного терапевтического средства, а также в особенности улучшения биологических свойств белка. Существует особая потребность в улучшении in vivo характеристик при назначении человеку. В этой связи особенно желательно обеспечить выбранные биологические молекулы уменьшенным или отсутствующим потенциалом индукции иммунного ответа в человеческом организме. Ожидается, что такие белки будут показывать повышенное время циркуляции в организме человека и будут особенно полезны в лечении хронических или повторяющихся проявлений болезни, таких как в случае некоторых показаний для указанной биологической молекулы.
Краткое изложение и описание изобретения.
Настоящее изобретение поэтому касается двух основных аспектов, таких как:
(a) удобный и эффективный вычислительный метод для идентификации и расчета эпитопов Т-клеток для универсально различных количеств молекул МНС класса II и, основываясь на этих знаниях, для разработки и конструирования новых вариантов последовательностей биологических молекул с улучшенными свойствами, и
(b) новые биологически активные молекулы для применения особенно к людям и, в частности, для терапевтического применения; указанные биологические молекулы являются согласно данному изобретению иммуногенно модифицированными полипептидами, белками и иммуноглобулинами (антителами), полученными согласно способу данного изобретения, в соответствии с чем модификация приводит к уменьшенной склонности биологической молекулы вызывать иммунный ответ при введении человеку.
В частности, изобретение относится к модификации нескольких в основном известных белков и антител с высоким терапевтическим преимуществом человеческого или нечеловеческого происхождения, полученных методом согласно данному изобретению, что приводит к получению белков, которые являются существенно неиммуногенными или менее иммуногенными, чем любой их немодифицированный эквивалент при использовании in vivo. Ожидается, что молекулы, модифицированные согласно новому методу настоящего изобретения, будут показывать повышенное время циркуляции в организме человека и будут особенно полезны в лечении хронических или повторяющихся проявлений болезни, таких как в случае некоторых показаний. Настоящее изобретение обеспечивает как специфическое воплощение и для демонстрации эффективности метода согласно изобретению, модифицированные формы указанных молекул, которые, как ожидается, покажут улучшенные свойства in vivo. Эти молекулы с модифицированной иммуногенностью, то есть со сниженным иммуногенным потенциалом могут использоваться в фармацевтических композициях. Такие модифицированные молекулы в этом описании названы "иммуногенно" модифицированными.
Способ идентификации эпитопов Т-клеток частично посредством вычислительных средств может использоваться для вычисления теоретических значений эпитопов Т-клеток и, таким образом, идентификации потенциальных связывающих пептидов молекулы МНС класса II в пределах белка; где связывающий сайт включает последовательность аминокислотных сайтов в пределах белка. Идентифицированные пептиды после того могут быть модифицированы без существенного снижения и, возможно, увеличения, терапевтическая ценности белка. Этот вычислительный метод включает выбор области белка, имеющей известную последовательность аминокислотного остатка, последовательно пробуя перекрывающиеся сегменты аминокислотных остатков (окон) предварительно определенной постоянной длины и состоящие из, по крайней мере, трех аминокислотных остатков из выбранной области, вычисляя показатель связывания молекулы МНС класса II для каждого выбранного сегмента и идентифицируя, по крайней мере, один из выбранных сегментов, подходящих для модификации, что основано на рассчитанном показателе связывания молекулы МНС класса II для этого сегмента. Полный показатель связывания молекулы МНС класса II для пептида затем может быть изменен, существенно не уменьшая терапевтическую ценность белка.
Показатель связывания молекулы МНС класса II для выбранного сегмента аминокислотного остатка в одном аспекте данного изобретения рассчитывают, суммируя определенные значения для каждого гидрофобного аминокислотного остатка боковой цепи, присутствующего в выбранном сегменте аминокислотного остатка пептида. Чтобы произвести графический обзор, значение этой суммы затем может быть приписано отдельному аминокислотному остатку в приблизительно средней точке сегмента. Эту процедуру повторяют для каждого из перекрывающихся сегментов (окон) в интересующей области или областях пептида. Определенное значение для каждой присутствующей ароматической боковой цепи равняется приблизительно половине определенного значения для каждого гидрофобного аминокислотного остатка боковой цепи. Гидрофобными аминокислотными остатками боковой цепи являются те, которые присутствуют в валине, лейцине, изолейцине и метионине. Ароматическими боковыми цепями являются те, которые присутствуют в фенилаланине, тирозине и триптофане. Предпочтительное определенное значение для ароматической боковой цепи равно приблизительно 1 и для гидрофобного аминокислотного остатка боковой цепи равно приблизительно 2. Однако могут использоваться другие значения.
Таким образом, в первом аспекте изобретение обеспечивает вычислительный способ, подходящий для идентификации одного или более потенциальных эпитопов Т-клеток пептидов в пределах последовательности аминокислоты биологической молекулы стадиями, включающими определение связывания указанных пептидов с МНС молекулами, используя in vitro или in silico способы или биологические анализы, указанный способ включает следующие стадии:
(а) выбор участка пептида, который имеет известную последовательность аминокислотных остатков;
(b) последовательный отбор перекрывающихся сегментов аминокислотных остатков с предварительно определенным одинаковым размером и состоящих, по крайней мере, из трех аминокислотных остатков выбранного участка;
(c) подсчет показателя связывания молекулы МНС класса II для каждого указанного выбранного сегмента путем суммирования определенных значений для каждого гидрофобного аминокислотного остатка боковой цепи, который присутствует в каждом выбранном сегменте аминокислотного остатка; и
(d) идентификация, по крайней мере, одного из указанных сегментов, подходящих для модификации, основываясь на подсчитанном показателе связывания молекулы МНС класса II этого сегмента, для изменения общего показателя связывания МНС класса II для пептида без существенного уменьшения терапевтической полезности пептида.
В специфическом воплощении изобретение касается способа, в котором стадию (с) выполняют при использовании функции подсчета Вöhm, модифицированной для включения 12-6 составляющей отталкивания энергии комплекса лиганд-белок Ван дер Ваальса и составляющей конформационной энергии лиганда путем:
(1) обеспечения первой базы данных моделей молекулы МНС класса II;
(2) обеспечения второй базы данных возможного пептидного скелета для указанных моделей молекул МНС класса II;
(3) отбора модели из указанной первой базы данных;
(4) отбора возможного пептидного скелета из второй базы данных;
(5) идентификации боковых цепей аминокислотных остатков, которые присутствуют в каждом выбранном сегменте;
(6) определения значения связывающего сродства ко всем боковым цепям, которые присутствуют в каждом выбранном сегменте; и необязательно
(7) повторения стадий от (1) до (5) для каждой указанной модели и каждого указанного скелета.
В следующем воплощении показатель связывания для каждой выбранной последовательности рассчитывают путем (i) обеспечения первой базы данных моделей молекулы МНС класса II; (ii) обеспечения второй базы данных возможного пептидного скелета для моделей молекул указанного МНС класса II; (iii) обеспечения третьей базы данных возможных конформаций аминокислот боковой цепи для каждой из этих двадцати аминокислот в каждом положении каждого скелета; (iv) отбора модели из указанной первой базы данных; (v) отбора возможного пептидного скелета из второй базы данных; (vi) идентификации боковых цепей аминокислотных остатков, которые присутствуют в каждом выбранном сегменте вместе с их возможными конформациями из указанной третьей базы данных; (vii) определения оптимального показателя связывающего сродства ко всем боковым цепям, которые присутствуют в каждом выбранном сегменте в каждой возможной конформаций; (viii) повторения стадий от (v) до (vii) для каждого указанного скелета и определения оптимального показателя связывания; и (ix) повторения стадий от (iv) до (viii) для каждой указанной модели.
Следует понимать, что эти три базы данных, описанные выше, могут быть объединены в одну базу данных, или любые две базы данных могут быть объединены, чтобы обеспечить объединенную базу данных.
Длина сегментов аминокислотного остатка, которые будут выбраны, может изменяться. Предпочтительно выбранные сегменты аминокислотного остатка состоят из приблизительно 10 - приблизительно 15 аминокислотных остатков, более предпочтительно приблизительно 13 аминокислотных остатков.
Выбранные сегменты аминокислотных остатков могут перекрываться до различной степени. Предпочтительно выбранные сегменты аминокислотных остатков перекрываются существенно. Наиболее предпочтительно последовательно выбранные сегменты аминокислотных остатков перекрываются всеми, кроме одного аминокислотного остатка. То есть в сегменте аминокислотного остатка, имеющем n остатков, n-1 остатки перекрываются следующим последовательно выбранным сегментом аминокислотного остатка.
Таким образом, более подробно изобретение, кроме того, касается следующих далее предпочтительных воплощений:
- соответственно указанному способу, в котором указанное значение для каждой ароматической стороны цепи равно приблизительно половине указанного значения для каждой гидрофобной алифатической стороны цепи;
- соответственно указанному способу, в котором выбранный сегмент аминокислотного остатка составлен из 13 аминокислотных остатков;
- соответственно указанному способу, в котором последовательные выбранные сегменты аминокислотного остатка перекрываются одним - пятью аминокислотными остатками;
- соответственно указанному способу, в котором последовательные выбранные сегменты аминокислотного остатка перекрывают друг друга существенно;
- соответственно указанному способу, в котором все, за исключением одного, из аминокислотных остатков в последовательных выбранных сегментах аминокислотного остатка перекрываются.
Во втором основном аспекте настоящее изобретение обеспечивает, модифицированные формы различных биологических молекул с одним или более удаленными эпитопами Т-клеток, где указанная модификация может быть достигнута способами, описанными выше и в формуле изобретения. Молекулы могут также быть получены способами, как описано в вышеуказанном предшествующем уровне техники, однако молекулы, полученные методами данного изобретения, показывают улучшенные свойства. В способах предшествующего уровня техники спрогнозированные эпитопы Т-клеток вычленяются при использовании целесообразных аминокислотных замен в первичной последовательности терапевтического антитела или белка, который не представляет собой антитела, имеющих происхождение как от человека, так и нечеловеческого происхождения.
Настоящее изобретение обеспечивает модифицированные формы белков и иммуноглобулинов, которые, как ожидается, покажут улучшенные свойства in vivo.
Поэтому целью изобретения является обеспечение способа получения иммуногенно модифицированной биологической молекулы, полученной из родительской молекулы, где модифицированная молекула имеет аминокислотную последовательность, отличную от последовательности указанной родительской молекулы, и показывает сниженную иммуногенность относительно родительской молекулы, когда подвергнута иммунной системе определенного вида; указанный способ включает: (i) определение аминокислотной последовательности родительской биологической молекулы или ее части; (ii) идентификацию одного или более потенциальных эпитопов Т-клеток в пределах аминокислотной последовательности белка любым способом, включающим определение связывания пептидов с молекулами МНС при использовании способов in vitro или in silico или биологических анализов; (iii) разработку новых вариантов последовательности путем изменения, по крайней мере, одного аминокислотного остатка в пределах исходно идентифицированных последовательностей эпитопов Т-клеток, указанные варианты модифицируют таким способом, что существенно уменьшают или устраняют активность или некоторое число последовательностей эпитопов Т-клеток и / или некоторое число МНС аллотипов, способных связывать пептиды, производные от указанной биологической молекулы, как определено путем связывания пептида с МНС молекулами при использовании способов in vitro или in silico или биологических анализов или связывания пептид-МНС комплексов с Т-клетками, (iv) конструирование таких вариантов последовательности с помощью техники рекомбинантных ДНК и тестирование указанных вариантов для того, чтобы идентифицировать один или более вариантов с желаемыми свойствами; и (v) необязательное повторение этапов (ii)-(iv), где идентификация последовательностей эпитопов Т-клеток согласно стадии (ii) достигается способом, как указано выше и ниже.
Специфические воплощения стадии (iii) согласно изобретению касаются следующих суммированных стадий:
- соответственно указанному способу, в котором 1-9 аминокислотных остатков, в любом из исходно присутствующих эпитопов Т-клеток изменен;
- соответственно указанному способу, в котором один аминокислотный остаток в любом из исходно присутствующих эпитопов Т-клеток изменен;
- соответственно указанному способу, в котором аминокислотное изменение выполняют в отношении гомологичной последовательности белка и/или в соответствии со способами моделирования in silico.
- соответственно указанному способу, в котором изменение аминокислотных остатков представляет собой замену, делецию или добавление исходно присутствующего(их) аминокислотного(ых) остатка(ов) другим(ми) аминокислотным(ми) остатком(ами) в специфическом(их) положении(ях).
- соответственно указанному способу, в котором дополнительно дальнейшее изменение, предпочтительно заменой, добавлением или делецией определенной(ых) аминокислот(ы), проводится для восстановления биологической активности указанной биологической молекулы.
За исключением стадии (ii) другие стадии раскрытого метода могут быть достигнуты способами и методами, которые являются хорошо известными квалифицированным в данном уровне техники специалистам. Так как модифицированные биологические молекулы получают предпочтительно рекомбинантными способами соответствующих ДНК структур, которые были выведены из последовательности аминокислоты после завершения обмена аминокислотных остатков, идентифицированных способом стадии (i). Рекомбинантные способы, используемые здесь, известны из данного уровня техники (например, Sambrook и другие., 1989, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, NY, USA).
Биологическая молекула, полученная согласно изобретению, является предпочтительно пептидом, белком, антителом, фрагментом антитела или белком слияния. Кроме того, изобретение включает модификации, варианты, мутации, фрагменты, производные, не-, частично или полностью гликозилированные формы указанных молекул, имеющих ту же самую или подобную биологическую и/или фармакологическую активность.
Хотя метод, раскрытый в данном изобретении, не ограничен определенными биологическими молекулами, специфическим воплощением изобретения является обеспечение предпочтительно молекул, которые известны в данном уровне техники, и демонстрация терапевтической полезности и ценности. Таким образом, дополнительной целью изобретения является обеспечение иммуногенно модифицированной биологической молекулы, полученной из родительской молекулы, где модифицированная молекула имеет аминокислотную последовательность, отличную от последовательности указанной родительской молекулы, и показывает сниженную иммуногенность относительно родительской молекулы, когда подвергнута иммунной системе определенного вида, полученная способом согласно изобретению, как подробно раскрыто выше и ниже.
Биологические молекулы, представляющие особый интерес, полученные указанным способом, выбирают из групп:
(а) моноклоналъные антитела:
анти-40kD гликопротеин антиген антитело KS 1/4;
анти-GD2 антитело 14.18;
анти-Неr2 антитело 4D5 (мышиное) и гуманизированная версия (Herceptin®);
анти-Her1 (EGFR) антитело с225 и h425;
анти-IL-2R (анти-Тас) антитело (Zenapax®);
анти-СD52 антитело (САМРАТН®);
анти-СD20 антитела (С2В8, Rituxan®; Bexxar®);
антитело, направленное на С5 комплементный белок человека;
(b) белки человека:
sTNF-R1, sTNF-R2, sTNFR-Fc (Enbrel®);
белок С, асrр30, рицин A, CNTFR лиганды;
субтилизин, GM-CSF, фолликулостимулирующий гормон человека (h-fsh);
β-глюкоцереброзидаза, GLP-1, аполипопротеин А1;
лептин (белок ожирения человека), KGF, GM-CSF;
BDNF, ЕРО, I1-1R антагонист.
Третий основной аспект настоящего изобретения касается последовательностей эпитопов Т-клеток, которые происходят от родительских иммуногенно немодифицированных биологических молекул. Эти эпитопы предпочтительно являются 13-мерными пептидами. В пределах этих пептидов последовательности, имеющие 9 последовательных аминокислотных остатков, являются предпочтительными. Таким образом, другой целью изобретения является обеспечение доступа к таким эпитопам и последовательностям. Более подробно изобретение касается:
- применения потенциального Т-клеточного эпитопа пептида в пределах аминокислотной последовательности родительской иммуногенно немодифицированной биологической молекулы, идентифицированной согласно любому из описанных способов получения биологической молекулы со сниженной иммуногенностью, имеющей ту же самую биологическую активность;
- соответствующего применения потенциального Т-клеточного эпитопа пептида, где указанный эпитоп Т-клеток является 13-мерным пептидом;
- применения последовательности пептида, состоящей, по крайней мере, из 9 последовательных аминокислотных остатков 13-мерного эпитопа Т-клеток, как указано выше, для получения биологической молекулы со сниженной иммуногенностью, имеющей ту же самую биологическую активность по сравнению с родительской немодифицированной молекулой.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
ФИГУРА 1 представляет блок-схему, иллюстрирующую первый аспект настоящего вычислительного метода;
ФИГУРА 2 представляет блок-схему, иллюстрирующую генерирование базы данных для вычислительного метода, воплощаемого настоящим изобретением;
ФИГУРА 3 представляет блок-схему, иллюстрирующую запрос базы данных для создания шаблона пептида для потенциальных эпитопов Т-клеток;
ФИГУРА 4 представляет дополнительную блок-схему, иллюстрирующую вычислительный метод.
ФИГУРА 5 представляет график индекса вероятности эпитопа Т-клеток versus координат (положений) аминокислотного остатка декарбоксилазы глутаминовой кислоты (MW: 65000) изоформ (GAD 65);
ФИГУРА 6 представляет график индекса вероятности эпитопа Т-клеток versus координат (положений) аминокислотного остатка эритропоэтина (ЕРО);
ФИГУРА 7 представляет график индекса вероятности эпитопа Т-клеток versus координат (положений) аминокислотного остатка легкой цепи гуманизированного анти-А33 моноклонального антитела; и
ФИГУРА 8 является графиком индекса вероятности эпитопа Т-клеток versus координат (положений) аминокислотного остатка тяжелой цепи гуманизированного анти-А33 моноклонального антитела.
На вышеупомянутых ФИГУРАХ 5-8, сплошная линия (―) представляет индекс эпитопа Т-клеток, рассчитанный вычислительным способом в соответствии с блок-схемой, показанной в ФИГУРЕ 1, и пунктирная линия (.....) представляет расчетное количество эпитопов Т-клеток, рассчитанное в соответствии с вычислительным методом в соответствии с блок-схемой, показанной на ФИГУРЕ 3, согласно другому аспекту настоящего изобретения.
Детальное описание изобретения
Термин "эпитоп Т-клеток" означает в соответствии с пониманием данного изобретения аминокислотную последовательность, которая способна связываться с приемлемыми эффективными молекулами МСН класса II (или их эквивалентами в нечеловеческих видах), способна стимулировать Т-клетки и/или также связывать (без неизбежной до определенной степени активации) с Т-клетками в комплексе с МСН класса II.
Термин "пептид", как используется в контексте данной заявки и приложенных пунктах формулы, представляет собой соединение, которое включает две или более аминокислот. Аминокислоты связываются вместе с помощью пептидной связи (определено ниже). Существует 20 различных природных аминокислот, вовлеченных в биологическую продукцию пептидов, и любое их количество может связываться в любом порядке для образования пептидной цепи или кольца. Существующие в природе аминокислоты, используемые в биологическом производстве пептидов, все имеют L-конфигурацию. Синтетические пептиды могут быть получены с применением приемлемых способов синтеза при использовании L-аминокислот, D-аминокислот, различных комбинаций аминокислот двух различных конфигураций. Некоторые пептиды содержат только несколько аминокислотных единиц. Короткие пептиды, например, имеющие менее чем десять аминокислотных единиц, иногда называются "олигопептидами". Другие пептиды содержат большое число аминокислотных остатков, например до 100 или более, и называются "полипептидами". По договоренности "полипептидом" можно считать любую пептидную цепь, которая содержит три или более аминокислот, в то время как "олигопептид" обычно считается частным типом "короткого" полипептида. Таким образом, как используется в контексте данной заявки, понятно, что любое упоминание "пептида" также включает и олигопептид. Кроме того, любая ссылка на "пептид" включает полипептиды, олигопептиды и белки. Каждое различное распределение аминокислот образует различные полипептиды или белки. Число полипептидов и отсюда число различных белков, которые могут быть образованы, является практически неограниченным.
Термин "уменьшенная или сниженная иммуногенность", используемый ранее, и здесь далее является относительным термином и касается иммуногенности соответствующей первоначальной исходной молекулы, когда оно подвергается воздействию in vivo тем же видам по сравнению с молекулой, модифицированной согласно изобретению.
Термин "модифицированный белок", как используется в контексте данного изобретения, описывает белок, который имеет уменьшенное количество эпитопов Т-клеток и поэтому выявляет сниженную иммуногенность по сравнению с родительским белком, когда подвергается иммунной системе определенных видов. Термин "немодифицированный белок", как используется в данном изобретении, описывает "родительский" белок, который в сравнении с "модифицированным белком" имеет большее количество эпитопов Т-клеток и поэтому увеличенную иммуногенность относительно модифицированного белка, когда подвергается иммунной системе определенных видов.
"Альфа углерод (Сα)" представляет собой углеродный атом углеводородного (СН) компонента, который находится в пептидной цепи. "Боковая цепь" представляет собой подвешенную группу к Сα, которая может включать простую или сложную группу или остаток и которая имеет физические размеры, которые могут значительно отличаться по сравнению с размерами пептида.
Эпитопы Т-клеток могут быть идентифицированы вычислительным методом данного изобретения путем рассмотрения аминокислотных остатков, важных для связывания специфического эпитопа Т-клеток с молекулами МНС класса II. После идентификации потенциальные эпитопы Т-клеток могут быть вычленены или удалены из последовательности аминокислотного остатка путем изменения, таким как мутирование, ключевых аминокислотных остатков в этой последовательности. Любая модификация, сделанная в последовательности пептида в области, которая, вероятно, будет содержать эпитопы Т-клеток, путем делеции, добавления или замены, приводящая к относительно более низкому полному показателю связывания, будет приводить к менее иммуногенной последовательности аминокислотного остатка. В некоторых случаях может быть желательно увеличить связывание некоторых пептидов с молекулами МНС класса II. Например, было предложено, что переносимость к некоторым аутоантигенам может быть восстановлена у индивидуумов, страдающих от аутоиммунной болезни, если таких индивидуумов лечить пептидными аналогами областей аутоантигена, которые, как известно, содержат эпитопы Т-клеток. Природный эпигон обычно имеет умеренное сродство к молекулам МНС класса II, тогда как пептидный аналог сделан так, что он имеет относительно более высокое сродство к молекулам МНС класса II. Это высокое сродство является важным как для промотирования иммунного наблюдения для очистки таких Т-клеток, представляющих это высокое сродство эпитопа, так и для них, чтобы стать анергическими. Такая модификация в эпитопах Т-клетки может также быть сделана на протеиновом уровне пептида, и весь белок применяется как терапевтическое средство.
Существует целый ряд факторов, которые играют важную роль в определении общей структуры белка или полипептида. Во-первых, пептидная связь, т.е та связь, которая связывает вместе аминокислоты в цепи, является ковалентной связью. Эта связь является плоской по структуре, по сути замещенным амидом. "Амид" представляет собой любую группу органических соединений, содержащих группировку
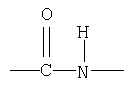
Плоская пептидная связь, соединяющая Сα соседних аминокислот, может быть представлена так, как показано ниже:

Поскольку O=С и C-N атомы лежат в относительно неподвижной плоскости, свободное вращение не происходит вокруг этих осей. Следовательно, плоскость, схематически показанная пунктирной линией, иногда называется как "амидная" или "пептидная плоскость", в которой лежат атомы кислорода (О), углерода (С), азота (N) и водорода (Н) пептидного скелета. В противоположных углах этой амидной плоскости располагаются Сα атомы. Поскольку не происходит существенного вращения вокруг O=С и C-N атомов в пептидной или амидной плоскости, полипептидная цепь, таким образом, включает серию плоскостных пептидных связей, соединяющих Сα атомы.
Вторым фактором, который играет важную роль в определении общей структуры или конформации полипептида или белка, является угол вращения каждой амидной плоскости вокруг общей Сα связи. Термин "угол вращения" и "угол закручивания" употребляются в дальнейшем как эквивалентные термины. Предполагая, что атомы О, С, N и Н остаются в амидной плоскости (что обычно является реальным предположением, несмотря на то что могут существовать некоторые слабые отклонения от плоскости этих атомов для некоторых конформации), эти углы вращения определяют N и R конформацию полипептидного скелета, то есть структуру в том виде, в котором она существует между смежными остатками. Эти два утла известны как φ и ψ. Набор углов φ1 и ψ1, где индекс 1 представляет собой отдельный остаток полипептидной цепи, таким образом, эффективно определяет вторичную структуру полипептида. Договоренности, используемые в определении φ и ψ углов, то есть базисные точки, в которых амидная плоскость образует угол, равный нулю градусов, и определение того, что представляет собой угол φ и что представляет собой угол ψ для данного полипептида, приведено в литературе. Смотри, например, Ramachandran и др. Adv.Prot.Chem. 23:283-437 (1968), а именно, на страницах 285-94, которые введены в данную заявку как ссылки.
Данный способ может быть применен к любому белку и основывается частично на открытии того факта, что у человека анкерное положение первичного кармана 1 связывающих желобков молекулы класса II МНС имеет четко соответствующую специфичность для определенных аминокислотных боковых цепей. Специфичность этого кармана определяется идентичностью аминокислоты в положении 86 бета-цепи молекулы класса II МНС. Этот сайт располагается на дне кармана 1 и определяет размер боковой цепи, которая может подвергаться аккомодации этим карманом (Marshall, K.W., (1994), J.Immunol., 152: 4946-4956). Если этот остаток представляет собой глицин, то все гидрофобные алифатические и ароматические аминокислоты (гидрофобными алифатическими аминокислотами являются: валин, лейцин, изолейцин, метионин, а ароматическими являются: фенилаланин, тирозин и триптофан) могут подвергаться аккомодации в кармане, предпочтение отдается ароматическим боковым цепям. Если этот остаток кармана представляет собой валин, то боковая цепь этой аминокислоты выдается в карман и ограничивает размер боковых цепей пептида, которые могут подвергаться аккомодации таким образом, что только гидрофобные алифатические боковые цепи могут подвергаться аккомодации. Таким образом, в последовательности аминокислотных остатков, где найдена аминокислота с гидрофобной алифатической или ароматической боковой цепью, существует потенциал для присутствующего ограниченного эпитопа Т-клеток класса II МНС. Даже если боковая цепь является гидрофобной алифатической, тем не менее она приблизительно в два раза более ассоциирована с эпитопом Т-клеток, чем ароматическая боковая цепь (даже предполагая приблизительное распределение типов кармана 1 в общей популяции).
Вычислительный способ воплощения параметров данного изобретения касательно вероятности пептидных участков, которые содержат эпитопы Т-клеток, заключается в следующем:
(1) Исследуется первичная последовательность пептидного сегмента предварительно определенной длины и идентифицируются все присутствующие гидрофобные алифатические и ароматические боковые цепи. (2) Гидрофобные алифатические боковые цепи оцениваются значением большим, чем для ароматических боковых цепей; приблизительно в два раза большим значением, чем для ароматический боковых цепей, например, значение 2 для гидрофобной алифатической боковой цепи и значение 1 для ароматической боковой цепи. (3) Определенные значения суммируются для каждого перекрывающегося сегмента аминокислотных остатков (окна) предварительно определенной постоянный длины в пределах пептида, и общее значение для частного сегмента (окна) определяется для одного аминокислотного остатка в промежуточном положении сегмента (окна), предпочтительно для остатка, который находится в центре выборочного сегмента (окна). Эта процедура повторяется для каждого выборочного перекрывающегося сегмента аминокислотных остатков (окна). Таким образом, каждый аминокислотный остаток пептида определяется значением, которое имеет отношение к вероятности эпитопа Т-клеток, который присутствует в этом частном сегменте (окне). (4) Подсчитанные и определенные, как описано в Этапе 3, значения могут быть нанесены против координат аминокислот цельной последовательности аминокислот, которая подвергается анализу. (5) Все части последовательности, которые имеют счет предварительно определенного значения, например, значение 1, считаются вероятно такими, которые содержат эпитоп Т-клеток и могут быть при желании модифицированы. Этот особый аспект данного изобретения обеспечивает общий способ, с помощью которого могут быть описаны участки пептида, вероятностно содержащие эпитопы Т-клеток. Модификации пептида в этих участках имеют потенциал для модификации связывающих характеристик класса II МНС.
В соответствии с другим аспектом данного изобретения эпитопы Т-клеток могут быть предсказаны с большей точностью при использовании более сложного вычислительного способа, который учитывает взаимодействия пептидов с моделями аллелей класса II МНС.
Вычислительное прогнозирование эпитопов Т-клеток, присутствующих в пределах пептида в соответствии с частным аспектом изобретения, рассматривает конструкцию моделей, по крайней мере, 42 аллелей класса II МНС, основываясь на структурах хорошо известных молекул класса II МНС и способе для использования этих моделей в вычислительной идентификации эпитопов Т-клеток, конструкции библиотек пептидных скелетов для каждой модели для того, чтобы получить возможность для известной вариабельности в родственных положениях альфа-углерода (Сα) пептидного скелета, конструкции библиотек структур аминокислотных боковых цепей для каждого скелета с каждой моделью для каждой из 20 аминокислотных альтернатив в критических положениях для каждого взаимодействия между пептидом и молекулой класса II МНС, использовании этих библиотек скелетов и структур боковых цепей в сочетании со счетной функцией для выбора оптимального скелета и структуры боковой цепи для частного пептида, состыкованного с частной молекулой класса II МНС, и получения значения связывания из результатов этого взаимодействия.
Модели молекул класса II МНС могут быть получены посредством гомологичного моделирования из числа подобных структур, находящихся в банке данных белков Брукхавена ("PDB"). Это может быть сделано с использованием полуавтоматического программного обеспечения гомологичного моделирования (Modeller, Sali A. & Bhmdell TL., 1993. J.Mol.Biol. 234: 779-815), которое вводит воспроизводящую отжиговую функцию в сопряжение с CHARMm силовым полем для минимизации энергии (доступно из Molecular Simulations Inc., San Diego, Ca). Альтернативные способы моделирования также могут использоваться.
Настоящий способ значительно отличается от других вычислительных способов, которые используют библиотеки экспериментально полученных данных относительно связывания каждой аминокислотной альтернативы в каждом положении связывающего желобка для небольшого набора молекул класса II МНС (Marshall, K.W и др., Biomed.Pept. Proteins Nucleic Acids, 1 (3): 157-162) (1995)) или даже других вычислительных способов, которые используют простые экспериментальные данные по связыванию для того, чтобы определить характеристики связывания частных типов карманов связывания в пределах бороздки, снова же используя относительно небольшие подмножества молекул класса II МНС и потом "перемешивая и подбирая" типы карманов из этой библиотеки карманов для искусственного создания дальнейших "виртуальных" молекул класса II МНС (Sturniolo Т. И др., Nat. Biotech. 17_(6): 555-561 (1999)). Оба предыдущих способа страдают от основных недостатков, которые благодаря сложности анализов и необходимости синтезировать большое количество вариантов пептидов дают возможность экспериментально исследовать только небольшое количество молекул класса II МНС. Таким образом, первый способ уровня техники может только дать возможность предсказать свойства только для небольшого числа молекул класса II МНС. Второй способ уровня техники также дает возможность сделать предположение, что карман, покрытый подобными аминокислотами в одной молекуле, будет иметь такие же связывающие характеристики в контексте различных аллелей класса II и страдать также теми недостатками, что и те молекулы класса II МНС, которые могут быть виртуально созданы и которые содержат карманы, содержащиеся в пределах библиотеки карманов. Используя моделирующий подход, описанный в данной заявке, структура любого числа и типа исследованных молекул класса II МНС могут быть дедуцированы, и, таким образом, аллели могут быть специфично выбраны как представители общей популяции. В дополнение к этому число исследованных молекул класса II МНС может быть увеличено путем создания дополнительных моделей для получения дополнительных данных посредством комплексных экспериментов.
Использование библиотеки скелетов предусматривает вариации положений Сα атомов различных исследованных пептидов при стыковке с частными молекулами класса II МНС. Это снова-таки отличается от альтернативных вычислительных способов уровня техники, описанных выше, которые полагаются на использование упрощенных пептидных скелетов для исследования аминокислотного связывания в частных карманах. Эти упрощенные скелеты, вероятно, не являются представительными структурами скелета, которые находятся в "реальных" пептидах, что ведет к погрешностям в прогнозировании пептидного связывания. Данная библиотека скелетов создается путем накладывания скелетов всех пептидов, связанных с молекулами класса II МНС, найденными в пределах белкового банка данных и с помощью фиксации исходного значения площади отклонения (RMC) между Сα атомами каждой из одиннадцати аминокислот, размещенных в пределах связывающего желобка. Поскольку эта библиотека может быть получена из небольшого количества приемлемых мышиных или человеческих структур (в настоящее время 13), то для того чтобы предусмотреть возможность даже большей вариабельности, количественные данные RMS для каждого С"-α положения увеличивается на 50%. Потом определяют среднюю величину положения Сα каждой аминокислоты и очерчивают сферу вокруг этой точки, радиус которой равен отклонению RMS в этом положении плюс 50%. Эта сфера представляет все возможные Сα положения. Используя Сα с, по крайней мере, отклонением RMS (те аминокислоты в кармане 1, как упомянуто выше, которые эквивалентны положению 2 одиннадцати остатков в связывающем желобке), сфера имеет пространственную привязку, и каждая вершина в пределах решетки потом используется как возможное расположение для Сα аминокислоты. Полученная амидная плоскость, соответствующая пептиду, связанному с последующей аминокислотой, перемещается на каждый из этих Сα, и углы φ и ψ поворачиваются пошагово на ряд интервалов для того, чтобы переместить последующие Сα. Если последовательные Сα попадают в пределы "сферы разрешенных положений" для этих Сα, то ориентация дипептида принимается, в то время, если она попадает за пределы сферы, то дипептид отвергается. Этот процесс потом повторяют для каждого последующего Сα положения так, что дипептид растет от исходного Сα кармана 1, пока все 9 последующих Сα не будут позиционированы от всех возможных перестановок предыдущих Сα. Процесс потом повторяют еще раз для одного Сα предыдущего кармана 1 для создания библиотеки положений Сα скелета, размещенных в пределах желобка связывания. Количество полученных скелетов зависит от нескольких факторов: размера "сферы возможных положений"; качества привязки "первичной сферы" в положении кармана 1; качества пошагового вращения углов φ и ψ, используемых для позиционирования последующих Сα. При использовании этого процесса может быть создана большая библиотека скелетов. Для большей библиотеки скелетов более вероятно, что она будет такой, что оптимальная подгонка будет найдена для частного пептида в пределах желобка связывания молекулы класса II МНС. Поскольку все скелеты не будут приемлемыми для стыковки со всеми моделями молекул класса II МНС по причине дисгармонии с аминокислотами доменов связывания, то для каждого аллеля создается подмножество библиотеки, включая скелеты, которые могут быть подвергаться аккомодации этими аллелями. Использование библиотеки скелетов в сочетании с моделями молекул класса II МНС создает исчерпывающую базу данных, состоящую из возможных структур боковой цепи для каждой аминокислоты в каждом положении связывающего желобка для каждой молекулы класса II МНС, которая стыкуется с каждым возможным скелетом. Этот набор данных получают при использовании простой функции стерического перекрывания, где молекула класса II МНС стыкуется со скелетом, и аминокислотная боковая цепь перемещается в скелет в желаемом положении. Каждая из способных к вращению связей боковой цепи вращается пошагово на набор интервалов, и полученные положения атомов зависят от записанной связи. Взаимодействие атома с атомами боковых цепей связывающего желобка записывается и положения принимаются либо отвергается в зависимости от следующих критериев: общая сумма перекрывания всех атомов, позиционированных к настоящему моменту времени, не должна превышать предварительно определенного значения. Таким образом, жесткость конформационного поиска является функцией интервала, используемого при пошаговом вращении связи и предварительно определенной границы для общего перекрывания. Таким образом, последнее значение может быть малым, если известно, что определенный карман является жестким, однако жесткость может быть ослаблена, если известно, что положения боковых цепей кармана являются относительно гибкими. Таким образом, может быть сделано допущение для имитации вариаций гибкости в пределах карманов связывающего желобка. Этот конформационный поиск потом повторяют для каждой аминокислоты в каждом положении каждого скелета при стыковке с каждой из молекул класса II МНС для создания исчерпывающей базы данных конформаций боковых цепей.
Приемлемое математическое выражение используется для оценки энергии связывания между моделями молекул класса II МНС в сочетании с конформациями петидных лигандов, которые эмпирически получены путем исследования большой базы данных конформаций скелета/боковых цепей, описанных выше. Таким образом, белок исследуется на потенциальные эпитопы Т-клеток путем исследования каждой возможной длины пептида, которая варьирует между 9 и 20 аминокислотами (несмотря на то что длина сохраняется постоянной для каждого исследования) для следующих вычислений: молекулу класса II МНС выбирают вместе с пептидным скелетом, возможным для этой молекулы и боковых цепей, соответствующих желаемой пептидной последовательности, на которую их переносят. Тождественность атомов и данные относительно межатомного расстояния, относящиеся к определенной боковой цепи в определенном положении в скелете, собирают для каждой возможной конформации этой аминокислоты (полученной из базы данных, описанной выше). Это повторяют для каждой боковой цепи вдоль скелета и получают пептидные значения, используя функцию подсчета. Лучшее значение для этого скелета запоминают и процесс повторяют для каждого возможного скелета для выбранной модели. Значения всех возможных скелетов сравнивают, и самое высокое значение считают как пептидное значение для желаемого пептида в этой модели класса II МНС. Этот процесс потом повторяют для каждой модели с каждым возможным пептидом, который имеет происхождение от исследуемого белка, и показывают значения для пептидов против моделей.
В контексте данного изобретения каждый лиганд, представленный для подсчета сродства связывания, представляет собой аминокислотный сегмент, выбранный из группы пептидов или белков, которые обсуждались выше. Таким образом, лиганд представляет собой выбранную цепь аминокислот, которая содержит от приблизительно 9 до 20 аминокислот в длину и которая имеет происхождение от пептида, полипептида или белка известной последовательности. Термины "аминокислоты" и "остатки" в данной заявке употребляются как эквивалентные термины. Лиганд в форме последовательных аминокислот пептида, который исследуется, перемещают в скелет из библиотеки скелетов, позиционируют в связывающем желобке молекулы класса II МНС из библиотеки моделей молекул класса II МНС через координаты С"-α атомов пептидного скелета и возможные структуры для каждой боковой цепи выбирают из базы данных возможных конформации. Тождественности релевантных атомов и межатомные расстояния также извлекаются из этой базы данных и используются для подсчета пептидного показателя связывания. Лиганды с высокой связывающей аффинностью для связывающего кармана класса II МНС отмечаются как кандидаты для сайт-направленного мутагенеза. Аминокислотные замены делают в отмеченных лигандах (и, таким образом, в белке, который представляет интерес и который потом повторно тестируют, используя счетную функцию для того, чтобы определить изменения, которые уменьшают связывающую аффинность ниже предварительно определенного значения). Эти изменения могут быть потом введены в белок, который представляет интерес, для удаления эпитопов Т-клеток. Связывание между пептидным лигандом и желобком связывания молекул класса II МНС вовлекает нековалентные взаимодействия, включая, но не ограничиваясь, следующие: водородные связи, электростатические взаимодействия, гидрофобные (липофильные) взаимодействия и Ван-дер-ваальсовые взаимодействия. Это включают в пептидную счетную функцию так, как описано более детально ниже. Понятно, что водородная связь представляет собой нековалентную связь, которая может образовываться между полярными или заряженными группами и состоит из водородного атома, поделенного между двумя другими атомами. Водород донора водорода имеет позитивный заряд, в то время как акцептор водорода имеет частичный негативный заряд. Для целей пептидных/белковых взаимодействий доноры водородной связи могут быть или азотами с присоединенным водородом или водородами, присоединенными к кислороду или азоту. Акцептор водородной связи может быть кислородами, не присоединенными к водороду, азотами без присоединенных водородов и одной или двумя связями, или серой только с одной связью. Определенные атомы, такие как кислород, присоединенный к водороду или азоту имина (например, C=NH) могут быть как акцепторами, так и донорами водорода. Энергия водородной связи колеблется от 3 до 7 Ккал/мол и является более сильной, чем Ван-дер-ваальсовые связи, но слабее, чем ковалентные связи. Водородные связи являются также высоко направленными и являются наиболее сильными, когда донорный атом, атом водорода и акцепторный атом колинеарны. Электростатические связи образуются между противоположно заряженными парами ионов, и сила взаимодействия является обратно пропорциональной квадрату расстояния между атомами в соответствии с законом Кулона. Оптимальное расстояние между парами ионов составляет приблизительно 2,8Å. В белок/пептидных взаимодействиях электростатические связи могут образовываться между аргинином, гистидином или лизином и аспартатом или глутаматом. Сила связи будет зависеть от рКа ионизирующих групп и диэлектрической постоянной среды, несмотря на то что они являются приблизительно подобными по силе водородным связям.
Липофильные взаимодействия являются благоприятными гидрофобно-гидрофобными контактами, которые возникают между белком и пептидным дигандом. Обычно они возникают между гидрофобными боковыми аминокислотными цепями пептида, спрятанными внутри карманов желобка связывания, так что они не подвергаются действию растворителя. Подверженность гидрофобных остатков действию растворителя является крайне неблагоприятной, поскольку окружающие молекулы растворителя оказывают влияние на водородную связь с другими клеткоподобными клатрат-формирующими структурами. Полученное в результате уменьшение энтропии является крайне неблагоприятным. Липофильные атомы могут быть серой, которая не является ни полярным, ни водородным акцептором, и атомами углерода, которые не являются полярными.
Ван-дер-ваальсовые связи являются неспецифическими силами, существующими между атомами, которые находятся на расстоянии 3-4Å. Они слабее и менее специфичны, чем водородные и электростатические связи. Распределение электронного заряда вокруг атома меняется со временем, и в любом случае распределение заряда не есть симметричным. Эта кратковременная асимметрия в электронном заряде индуцирует подобную асимметрию в соседних атомах. Проистекающие отсюда силы притяжения между атомами достигают максимума при контактном расстоянии Ван-дер-Ваальса, но очень быстро убывают при расстоянии от приблизительно 1Å до приблизительно 2Å. Наоборот, как только атомы становятся разделенными менее, чем контактным расстоянием, в значительной степени сильные силы отталкивания становятся доминантными тогда, как внешние электронные облака атомов перекрываются. Несмотря на то что силы притяжения являются относительно более слабыми по сравнению с электростатическими и водородными связями (приблизительно 0,6 Ккал/мол), силы отталкивания, в частности, могут быть более важными в определении будет ли пептидный лиганд успешно связываться с белком.
В одном воплощении функция показателя Böhm (SCORE 1 подход) используется для оценки связывающего контакта (Böhm H.J., J.Comput AidedMol. Des 8(3): 243-256 (1994), ссылка введена в данную заявку в своей целостности). В другом воплощении функция показателя (SCORE2 подход) используется для оценки связывающей аффинности как индикатор лиганд-содержащего эпитопа Т-клеток (Böhm H.J., J.Comput Aided Mol. Des. 12(4): 309-323 (1998), ссылка введена в данную заявку в своей целостности). Однако функция оценки Böhm, как описано в указанных выше ссылках, используется для оценки связывающей аффинности лиганда с белком, где заранее известно, что лиганд успешно связывается с белком и комплекс белок/лиганд имеет определенную структуру, определенная структура представлена в банке данных белков ("PDB"). Таким образом, функция подсчета была развита далее с преимуществом известных позитивных данных связывания. Для того чтобы иметь возможность различения между позитивными и негативными связующими, составляющая отталкивания необязательно может прибавляться к равенству. В дополнение более удовлетворительная оценка связывающей энергии достигается путем вычисления липофильных взаимодействий попарным образом, более точно, чем при использовании площади, основанной на составляющей энергии приведенных функций Böhm. Таким образом, в предпочтительном воплощении энергия связывания оценивается при использовании модифицированной счетной функции Böhm. В модифицированной счетной функции Böhm энергия связывания между белком и лигандом (ΔGbind) оценивается при учете следующих параметров: уменьшение энергии связывания благодаря абсолютной потере трансляционной и вращательной энтропии лиганда (ΔG0); вклады идеальных водородных связей (ΔGhb), где, по крайней мере, один партнер является нейтральным; вклады невозмущенных ионных взаимодействий (ΔGionic); липофильные взаимодействия между липофильньши атомами лиганда и липофильными атомами акцептора (ΔGlipo); потеря связывающей энергии благодаря застыванию внутренних степеней свободы в лиганде, то есть свобода вращения вокруг каждой связи С-С уменьшена (ΔGrot); энергия взаимодействия между белком и лигандом (EVdW). Анализ этих составляющих дает равенство 1:
(ΔGbind)=(ΔG0)+(ΔGhb×Nhb)+(ΔGionic×Nionic)+(ΔGlipo×Nlipo)+(ΔGrot+Nrot)+(EVdW).
где N представляет собой количество установленных взаимодействий для специфической составляющей и в одном воплощении, ΔG0, ΔGhb, ΔGionic, ΔGlipo и ΔGrot являются константами, которые имеют значения: 5,4, -4,7, -4,7, -0,17, и 1,4, соответственно.
Составляющую Nhb подсчитывают в соответствии с равенством 2:
Nhb=Σh-bondf(ΔR, Δα)×f(Nneighb)×fpcs
f(ΔR, Δα) представляет собой штрафную функцию, которая подсчитывается для больших отклонений водородных связей от идеальных и рассчитывается в соответствии с равенством 3:
f(ΔR, Δ-α)=f1(ΔR)×f2(Δα)
Где: f1(ΔR)=1, если ΔR<=TOL
или = 1-(ΔR-TOL)/0,4, если ΔR<=0,4+TOL
или = 0, если ΔR>0,4+TOL
И: f2(Δα)=1, если Δα<30°
или = 1-(Δα-30)/50, если Δα<=80°
или = 0, если Δα>80°
TOL представляет собой допустимое отклонение в длине водородной связи = 0,25Å.
ΔR представляет собой отклонение длины H-O/N водородной связи от идеального значения = 1,9Å.
Δα представляет собой отклонение угла водородной связи ∠N/O-H..O/N от идеального значения 180°.
f(Nneighb) проводит отличие между вогнутой и выгнутой частями поверхности белка и, таким образом, устанавливает больший вес для полярных взаимодействий, найденных в карманах, больше, чем таковые для тех, которые найдены на поверхности белка. Эта функция рассчитывается в соответствии с равенством 4, приведенным ниже:
f(Nneighb)=(Nneighb/Nneighb,0)α , где α=0,5
Nneighb представляет собой число неводородных атомов белка, которые находятся ближе, чем 5Å к любому атому белка.
Nneighb,0 представляет собой константу, равную 25.
fpcs является функцией, которая допустима для площади полярной поверхности контакта на одну водородную связь и, таким образом, проводит различия между сильными и слабыми водородными связями, ее значение определяется в соответствии со следующими критериями:
fpcs=β, когда Apolar/NHB<10Å2
или fpcs=1, когда Apolar/NHB>10Å2
Apolar представляет собой размер площади полярного контакта белок-лиганд
NHB - число водородных связей
β является константой, значение которой =1,2
Для осуществления модифицированной функции подсчета Böhm, вклады ионных взаимодействий, ΔGionic подсчитывается тем же способом, что и для водородных связей, описанных выше, поскольку предполагается одинаковая геометрическая зависимость.
Составляющую Nlipo подсчитывают в соответствии с равенством 5, приведенным ниже:
Nlipo =Σ1Lf(r1L)
f(r1L) подсчитывается для всех липофильных лигандных атомов, 1, и всех липофильных белковых атомов, L, в соответствии со следующими критериями:
f(r1L)=1, когда r1l<=R1f(r1L)=(r1l-R1)/(R2-R1), когда R2<r1l>R1
f(r1L)=0, когда f1l>=R2
где: R1=r1VdW+rLVdW+0,5
и R2=R1+3,0
и: r1 VdW представляет собой радиус Ван-дер-Ваальса атома 1
и rL VdW представляет собой радиус Ван-дер-Ваальса атома L
Составляющая Nrot представляет собой число способных к вращению связей аминокислотной боковой цепи и берется для того, чтобы представлять число ациклических sp3-sp3 и sp3-sp2 связей. Вращения терминального конца -СН3 или -NH3 не берутся в расчет.
Конечная составляющая EVdW по дочитывается в соответствии с равенством 6 ниже:
EVdW=ε1ε2((r1VdW+r2VdW)12/(r1VdW+r2 VdW)6/r6, где
где ε1 и ε2 являются константами, зависимыми от идентичности атомов
r1 VdW+r2 VdW являются атомными радиусами Ван-дер-Ваальса
r представляет собой расстояние между парой атомов.
Что касается равенства 6, то в одном воплощении константы ε1 и ε2 имеют следующие значения для атомов: С: 0,245, N: 0,283, О: 0,316, S: 0,316 соответственно (то есть атомы углерода, азота, кислорода и серы соответственно). Что касается равенств 5 и 6, то радиусы Ван-дер-Ваальса имеют следующие значения для атомов: С: 1,85, N: 1,75, О: 1,60, S: 2,00Å.
Понятно, что все предварительно определенные значения и константы, приведенные в равенствах выше, определены в пределах ограниченных условий понимания белок-пептидных взаимодействий в особой связи с типом вычисления, которое проводится в данной заявке. Таким образом, возможно, что эта счетная функция будет усовершенствована в дальнейшем как результат прогресса в области моделирования молекулярных взаимодействий, эти значения и константы могут изменяться, поэтому любое приемлемое числовое значение, которое дает желаемые результаты в составляющих оценки связывающей энергии белка с лигандом, может использоваться и, следовательно, подпадает под объем данного изобретения.
Как описано выше, счетная функция применяется к данным, извлеченным из базы данных конформаций боковых цепей, идентичных атомов и межатомных расстояний. Для целей данного описания число молекул класса II МНС, включенных в эту базу, составляет 42 модели плюс 4 установленные структуры. Из приведенного выше описания понятно, что модульная природа построения вычислительного способа в соответствии с данным изобретением означает, что новые модели могут просто прибавляться и исследоваться совместно с библиотекой пептидных скелетов и функцией поиска конформаций боковых цепей для создания дополнительных наборов данных, которые могут быть обработаны с помощью пептидной счетной функции, как описано выше. Это дает возможность легко увеличить набор исследованных молекул класса II МНС или структур и ассоциированных данных, которые замещаются, если данные являются доступными для создания более точных моделей существующих аллелей.
Понятно, что несмотря на то что описанная выше счетная функция является относительно простой по сравнению с некоторыми сложными методологиями, которые доступны, подсчеты проводятся чрезвычайно быстро. Понятно, что цель не заключается в том, чтобы подсчитать реальную энергию связывания саму по себе для каждого пептида, который стыкуется со связывающим желобком выбранного белка класса II МНС. Основная цель заключается в том, чтобы получить данные относительно энергии связывания для претендентов как цели для прогнозирования размещения эпитопов Т-клеток, основанных на первичной структуре (то есть аминокислотной последовательности) выбранного белка. Относительно высокая энергия связывания или связывающая энергия выбранного значения предполагает присутствие эпитопов Т-клеток в лиганде. Лиганд можно потом подвергать, по крайней мере, одному кругу аминокислотных замен и повторно подсчитывать энергию связывания. Благодаря быстроте подсчета эта обработка пептидной последовательности может быть проведена при использовании компьютерной технологии в пределах интерфейса пользователя программы на эффективной доступной компьютерной технике. Основного вложения в компьютерную технику, таким образом, не требуется.
Любому специалисту в данной области понятно, что другие доступные программные обеспечения могут использоваться для тех же целей. В частности, более сложное программное обеспечение, которое способно к состыковке лигандов в белок-связывающих сайтах, может использоваться в соединение с энергетической минимизацией. Примерами стыковочных программных продуктов являются следующие: DOCK (Kuntz и др., J.Mol.Biol., 161: 269-288 (1982)), LUDI (Böhm H.J, J.Comput Aided Mol. Des., 8: 623-632 (1994) и FLEXX (Rarey M. И др. ISMB, 3: 300-308 (1995)). Примеры программного обеспечения молекулярного моделирования и манипулирования включают: AMBER (Tripos) и CHARMm (Molecular Simulations Inc.). Использование этих вычислительных методов будет строго ограничивать пропускную мощность способа в соответствии с данным изобретением из-за продолжительности времени обработки, которое необходимо для проведения необходимых вычислений. Однако является вероятным, что такие способы могут использоваться как "вторичный скрининг" для получения более точных вычислений энергии связывания для пептидов, которые определены как "позитивные связующие" при использовании способа в соответствии с данным изобретением.
Ограничение времени обработки для сложных молекулярно-механических или молекулярно-динамических вычислений является одной из причин, которая определяется программным обеспечением и возможностями современной технологии компьютерного оборудования. Можно предвидеть, что в будущем с написанием более рационального кода и продолжающегося увеличения скорости процессоров компьютера станет возможным проводить такие вычисления в пределах более поддающихся управлению временных рамок. Дальнейшая информация относительно энергетических функций применимо к макромолекулам и рассмотрение различных взаимодействий, которые имеют место внутри собранной белковой структуры, может быть найдена в: Brooks B.R. и др. J.Comput Chem., 4: 187-217 (1983) и дополнительная информация, касающаяся общих взаимодействий белок-лиганд, может быть найдена в: Dauber-Osguthrpe и др., Proteins 4 (I): 31-47 (1988), эти ссылки введены в данную заявку в своей целостности. Полезная информация также может быть найдена, например, в Fasman G.D. ed.. Prediction of Protein Structure of Protein Conformation, Plenum Press, New York, ISBN: 0-306, 4313-9.
Настоящий способ прогнозирования может быть откалиброван в соответствии с набором данных, включающим большое количество пептидов, сродство которых для различных молекул МНС класса II была ранее экспериментально определена.
Согласно предпочтительному воплощению способа любой из определенных способов прогнозирования, описанных здесь, иди любой другой вычислительный способ прогнозирования взаимодействий пептид-МНС класса II, который приводит к получению числовых показателей для каждой пары пептид/МНС класса II, калибруют в соответствии с набором данных, включающим большое количество пептидов, аффинность которых для различных молекул МНС класса II была ранее экспериментально определена. Путем сравнения подсчитанных данных и экспериментальных данных, как определено выше, можно сократить число значений, если известно, что все экспериментально определенные эпитопы Т-клеток были правильно предсказаны.
В частности, полученный с помощью компьютера числовой показатель рассчитывают для каждой пары пептид/МНС класса II в наборе данных. Показатель рассчитывают так, что более высокий показатель представляет собой увеличенную вероятность связывания. Самый низкий полученный с помощью компьютера показатель для пары пептид/МНС класса II, который найден экспериментально для связывания, взят как граничный. Все полученные с помощью компьютера показатели, которые являются значительно ниже этого граничного показателя, как полагают, представляют несвязывающиеся пары пептид/МНС класса II, в то время как полученные с помощью компьютера показатели выше граничного показателя представляют потенциально связывающуюся пару пептид/МНС класса II. В общем для данного полученного с помощью компьютера оценочного алгоритма будут существовать некоторые комбинации пептид/МНС класса II, которые дают показатели выше граничного показателя, но которые фактически являются несвязывающимися. Таким образом, это предпочтительное воплощение способа может произвести ложные позитивы, но никогда или только редко будет производить ложные негативы.
Это воплощение способа на основе граничного значения особенно полезно, когда цель состоит в том, чтобы устранить путем мутирования наибольшее количество или все эпитопы Т-клеток из белка. В частности, согласно более предпочтительному воплощению способа изобретения наибольшее количество или все эпитопы Т-клеток удаляют из белка следующим образом. Последовательность белка сканируется компьютерным алгоритмом для потенциальных эпитопов Т-клеток. Каждому потенциальному эпитопу Т-клеток присваивают показатель с увеличивающимися показателями, коррелированными по более высокой вероятности связывания с МНС класса II. Каждый сегмент пептида с показателем больше, чем граничный показатель мутируют так, чтобы показатель мутированного сегмента был меньше, чем граничный показатель. Мутации предпочтительно выбирают так, чтобы не уменьшить активность белка ниже активности, необходимой для данной цели. Разрабатывают множественно мутированный белок, не имеющий в основном или вообще прогнозированных компьютером эпитопов Т-клеток. Такой множественно мутированный белок назван "Деиммунизированный белок".
Деиммунизированный белок синтезируют стандартными методами. Например, искусственные последовательности ДНК, кодирующие Деиммунизированный белок, собирают из синтетических олигонуклеотидов, лигированных в вектор экспрессии и функционально связанных с элементами, промотирующими экспрессию Деиммунизированного белка. Деиммунизированный белок затем очищают стандартными методами. Полученный Деиммунизированный белок содержит аминокислоты, мутированные таким образом, что истинные эпитопы Т-клеток устранены. Кроме того, Деиммунизированный белок будет часто содержать в сегментах мутированные аминокислоты, которые, как прогнозируется алгоритмом, являются эпитопами Т-клеток, но которые фактически не являются эпитопами Т-клеток. Однако из мутаций в ложно прогнозированных эпитопах не следуют значительно вредные последствия, потому что мутации выбирают так, чтобы они имели небольшой эффект на активность белка. Кроме того, вредные последствия не следуют из возможного введения новых В-клеточных эпитопов в белок, потому что недостаток Т-клеточных эпитопов предотвращает В-клеточную реакцию на модифицированный белок.
Применение описанной выше методологии к различным пептидам, которые можно рассматривать для деиммунизации, для модификаций для улучшения связывания МНС класса II для терапевтических целей, иллюстрируется ниже.
Изобретение может быть применено к любой биологической молекуле, имеющей определенную биологическую и/или фармакологическую активность с существенно подобными первичными аминокислотными последовательностями, как раскрытые здесь, и поэтому будет включать молекулы, полученные средствами генной инженерии или другими способами. Термин "биологическая молекула" используется здесь для молекул, которые имеют биологическую функцию и вызывают биологический, фармакологический или фармацевтический эффект или активность. Предпочтительно биологическими молекулами согласно изобретениям являются пептиды, полипептиды, белки. Ниже белки, иммуноглобулины являются предпочтительными. Изобретение включает также варианты и другую модификацию определенного полипептида, белка, белка слияния, иммуноглобулина, которые в принципе имеют ту же самую биологическую активность и подобную (сниженную) иммуногенность. Кроме того, включены фрагменты антител, таких как sFv, Fab, Fab', F(ab')2 и Fc, и биологически эффективные фрагменты белков. Антитела человеческого происхождения или гуманизированные антител показывают per se пониженную иммуногенность или отсутствие иммуногенности у людей и не имеют или имеют более низкое количество иммуногенных эпитопов по сравнению с нечеловеческими антителами. Однако есть также потребность в деиммунизации таких молекул, некоторые из которых показали, что они вызывают значительный иммунный ответ у людей. Кроме того, антигены, которые вызывают не желательный и слишком сильный иммунный ответ, могут быть модифицированы согласно методу изобретения и привести к антигенам, которые имеют сниженную иммуногенность, которая, однако, является достаточно сильной, чтобы использовать антиген, например, как вакцину.
Некоторые молекулы, типа лептина, такие как идентифицированные из других млекопитающих источников, имеют в основном многие из пептидных последовательностей настоящего изобретения и имеют в общем много пептидных последовательностей с существенно подобной последовательностью, как те, который раскрыты в списке. Поэтому такие белковые последовательности в равной степени подпадают в область настоящего изобретения.
Изобретение касается аналогов биологических молекул согласно изобретению, в которых замещения, по крайней мере, одного аминокислотного остатка были сделаны в положениях, что приводит к существенному сокращению активности или устранению одного или более потенциальных эпитопов Т-клеток из белка.
Одно или более аминокислотных замещений в специфических точках в пределах любого из потенциальных лигандов МНС класса II, идентифицированные в таблицах примеров, могут приводить к молекуле со сниженным иммуногенным потенциалом при применении человеком в качестве терапевтического средства.
Предпочтительно аминокислотные замещения делают в соответствующих точках в пределах пептидной последовательности, прогнозированной для достижения существенного сокращения или устранения активности эпитопа Т-клетки. Практически соответствующая точка предпочтительно будет приравнена к связыванию аминокислотного остатка в пределах одного из гидрофобных карманов, обеспеченных в пределах связывающей борозды МНС класса II. Аминокислотные остатки в пептиде в положениях, которые приравнены к связыванию в пределах других карманных областей в пределах МНС связывающей щели, также рассматриваются и подпадают в область настоящего изобретения.
Понятно, что отдельные аминокислотные замещения в пределах данного потенциального эпитопа Т-клетки являются наиболее предпочтительным путем, которым эпитоп может быть устранен. Комбинации замещения в пределах отдельного эпитопа могут быть рассмотрены и, например, могут быть особенно приемлемыми, где индивидуально определенные эпитопы перекрываются друг другом. Кроме того, аминокислотные замещения или отдельно в пределах данного эпитопа, или в комбинации в пределах отдельного эпитопа могут быть сделаны в положениях, которые не приравниваются к "карманным остаткам" относительно связывающей бороздки МНС класса II, но в любой точке в пределах последовательности пептида. Все такие замещения подпадают в область настоящего изобретения.
Аминокислотные замещения другие, чем в пределах пептидов, идентифицированных выше, могут быть особенно рассмотрены, когда они сделаны в комбинации с замещением(ями), сделанными в пределах пептида, внесенного в список. Например, изменение может быть предусмотрено для восстановления структуры или биологической активности вариантной молекулы. Такие компенсационные изменения и изменения, включающие делецию или дополнение специфических аминокислотных остатков из молекулы согласно изобретению, приводящие к варианту с желательной активностью, и в комбинации с изменениями в любом из раскрытых пептидов подпадают в область настоящего изобретения.
В другом аспекте настоящее изобретение касается кодирования нуклеиновых кислот указанных биологических молекул, имеющих уменьшенную иммуногенность. Способы для создания генных конструкций и генных продуктов хорошо известны в данном уровне техники. В заключительном аспекте настоящее изобретение касается фармацевтических композиций, включающих указанные биологические молекулы, которые можно получать способами, раскрытыми в настоящем изобретении, и методов терапевтического лечения людей, используя модифицированные молекулы и фармацевтические композиции.
Как может быть видно из следующих примеров, вычислительные методы, описанные здесь выше, обеспечивают очень хороший индикатор того, что эпитопы Т-клеток, вероятно, будут найдены в любом пептиде. Таким образом, это позволяет идентификацию областей последовательностей аминокислотного остатка, которая, если они изменены одним или более изменениями аминокислотных остатков, имеет эффект удаления эпитопов Т-клеток и, таким образом, увеличивает терапевтическую ценность пептида. Посредством этого способа могут быть получены биологические молекулы, такие как пептиды, белки, иммуноглобулины и белки слияния и т.п., имеющие улучшенные свойства и фармакологическую ценность.
Вышеприведенное описание и примеры подразумеваются как иллюстративные и не должны приниматься как ограничение. Тем не менее, другие варианты в пределах сущности и объема данного изобретения возможны и легко представятся специалистами, квалифицированными в данном уровне техники.
ПРИМЕР 1
Этот пример показывает профиль вероятности эпитопа Т-клетки декарбоксилазной изоформы автоантигена глутаминовой кислоты (GAD 65; МВТ: 65.000), который вовлечен в развитие диабета типа 1. Этот специфический белок может быть потенциальной целью для того, чтобы увеличить сродство эпитопов Т-клеток, а также получения хорошего примера для демонстрирования индекса вероятности эпитопа Т-клетки, так как он является относительно длинным белком (585 аминокислотных остатков), и поэтому обеспечивает относительно большой объем выборки для профилирования.
Профиль вероятности эпитопа Т-клетки для GAD 65 показан на ФИГУРЕ 5. Сплошная линия представляет рассчитанный индекс эпитопа Т-клетки с использованием вычислительного метода показанного на ФИГУРЕ 1, и пунктирная линия представляет предсказанный индекс эпитопа Т-клетки с использованием вычислительного метода показанного на ФИГУРЕ 3 и 4.
ПРИМЕР 2
Этот пример показывает профиль вероятности эпитопа Т-клетки эритропоэтина (ЕРО), цитокина, состоящего из 193 аминокислотных остатков, широко используемого как внутривенно (IV) применяемое лекарственное средство для увеличения количества эритроцитов. Он представляет хороший пример биологического лекарственного средства с терапевтической ценностью, но который мог бы вызвать несоответствующие или нежелательные иммунные реакции, особенно при использовании внутривенно и который может поэтому извлечь выгоду из деиммунизации после того, как потенциальные эпитопы Т-клеток идентифицированы.
Профиль вероятности эпитопа Т-клетки для ЕРО показан на ФИГУРЕ 6. Сплошная линия представляет рассчитанный индекс эпитопа Т-клетки с использованием вычислительного метода, показанного на ФИГУРЕ 1, и пунктирная линия представляет предсказанный индекс эпитопа Т-клетки, с использованием вычислительного метода показанного на ФИГУРЕ 3 и 4.
ПРИМЕР 3
ФИГУРЫ 7 и 8 показывают индекс эпитопа Т-клетки для тяжелой и легкой цепи мышиного гуманизированного моноклонального антитела, направленного против А33 антигена. Последний представляет собой трансмембранный гликопротеин, представленный на поверхности >95% раковых образований кишечника и поэтому обладает потенциалом терапевтического антиракововго средства.
На ФИГУРАХ 7 и 8, сплошная линия представляет рассчитанный индекс эпитопа Т-клетки с использованием вычислительного метода показанного на ФИГУРЕ 1 и пунктирная линия представляет предсказанный индекс эпитопа Т-клетки с использованием вычислительного метода показанного на ФИГУРЕ 3 и 4.
ПРИМЕР 4 (лептин):
Одной из этих терапевтически ценных молекул является белок ожирения человека, называемый "лептин". Лептин представляет собой секретированный белок передачи сигналов, состоящий из 146 аминокислотных остатков, вовлеченный в гомеостатические механизмы поддерживающие жировую массу (например, WO 00/40615, WO 98/28427, WO 96/05309). Белок (и его антагонисты) представляет значительный терапевтический потенциал для лечения диабета, высокого кровяного давления и метаболизма холестерина. Белок может быть получен рекомбинантными методами, используя некоторое количество различных типов Т-клеток хозяина. Аминокислотная последовательность лептина (представленная в однобуквенном коде) является следующей:
VPIQKVQDDTKTLIKTIVTRINDISHTQSVSSKQKVTGLDFIPGLHPILTLSKMDQTLAVYQQILTSMPSRNVIQISNDLENLRDLLHVLAFSKSCHLPWASGLETLDSLGGVLEASGYSTEVVALSRLQGSLQDMLWQLDLSPGC
Аминокислотную последовательность, которая является частью последовательности иммуногенно немодифицированного белка ожирения (лептин) человека и которая обладает потенциальной активностью связывания МНС класса II, выбирают из следующей группы идентифицированной согласно способу настоящего изобретения:

Любая из вышеприведенных последовательностей может быть использована для модифицирования путем замены одной или более аминокислот для получения последовательности, которая неиммуногенна или имеет сниженную иммунногенность.
ПРИМЕР 5 (IL-1R антагонист):
IL-1 - важный воспалительный и иммунный цитокин модуляции с плейотропным эффектом на различные ткани, но может привести к патологиям, которые связанны с ревматическим артритом и другими болезнями, которые связанны с местным повреждением ткани. Антагонист IL-1 рецептора, способный ингибировать действие IL-1, был очищен и ген клонирован [Eisenburg S.P. и др. (1990) Nature, 343: 341-346; Carter, D.B. и др. (1990) Nature, 344: 633-637]. Другие обеспечили молекулы IL-IRa [например, US 5075222].
Аминокислотная последовательность IL-IRa (представленная в однобуквенном коде) является следующей:
RPSGRKSSKMQAFRIWDVNQKTFYLRNNQLVAGYLQGPNVNLEEKIDVVPIEPHALFLGIHGGKMCLSCVKSGDETRLQLEAWITDLSENRKQDKRFAFIRSDSGPTTSFESAACPGWFLCTAMEADQPVSLTNMPDEGVMVTKFYFQEDE
Аминокислотную последовательность, которая является частью последовательности иммуногенно немодифицированного IL-IRa и которая обладает потенциальной активностью связывания MHC класса II, выбирают из следующей группы:

Любая из вышеприведенных последовательностей может быть использована для модифицирования, путем замены одной или более аминокислот, для получения последовательности, которая неиммуногенна или имеет сниженную иммунногенность.
ПРИМЕР 6 (BDNF):
Другой терапевтически ценной молекулой является "мозговой нейротрофический фактор (BDNF) человека". BNDF представляет собой гликопротеин семейства белков фактора роста нервов. Зрелый 119 аминокислотный гликопротеин выделяют из большего предшественника, чтобы получить нейтрофический фактор, который промотирует выживание популяций нейронных клеток [Jones KL.R. & Reichardt, L.F. (1990) Proc. Natl. Acad. Sci U.S.A. 87: 8060-8064]. Такие нейронные клетки все находятся или в центральной нервной системе или непосредственно связанны с ней. Рекомбинантные препараты BNDF позволили раскрыть терапевтический потенциал белка для промотирования регенерации нерва и терапии дегенеративного заболевания.
Аминокислотная последовательность мозгового нейротрофического фактора (BDNF) человека (представленная в однобуквенном коде) является следующей:
HSDPARRGELSVCDSISEWVTAADKKTAVDMSGGTVTVLEKVPVSKGQLKQYFYETKCNPMGYTKE
GCRGIDKRHWNSQCRTTQSYVRALTMDSKKRIGWRFIRIDTSCVCTLTIKRGR.
Другие способы обеспечивают модифицированные BDNF молекулы [US 5770577], и направлены на усовершенствование коммерческого производства рекомбинантной BDNF молекулы [US 5986070].
Аминокислотную последовательность, которая является частью последовательности иммуногенно немодифицированного мозгового нейротрофического фактора (BDNF) человека и которая обладает потенциальной активностью связывания МНС класса II, выбирают из следующей группы:

Любая из вышеприведенных последовательностей может быть использована для модифицирования путем замены одной или более аминокислот для получения последовательности, которая неиммуногенна или имеет сниженную иммунногенность.
ПРИМЕР 7 (ЕРО):
Другой терапевтически ценной молекулой является эритропоэтин (ЕРО). ЕРО - гормон гликопротеин, вовлеченный в развитие эритроидных клеток-предшественников в эритроциты. Природный ЕРО вырабатывается печенью в течение эмбриональной жизни и почкой взрослых и циркулирует в крови, чтобы стимулировать продуцирование эритроцитов в костном мозге. Анемия - почти всегда следствие почечной недостаточности вследствие уменьшения продуцирования ЕРО почками. Рекомбинантный ЕРО используется для эффективного лечения анемии вследствие хронической почечной недостаточности. Рекомбинантный ЕРО (выраженный в клетках млекопитающих) имеет аминокислотную последовательность 1-165 эритропоэтина человека [Jacobs, К. и др. (1985) Nature, 313: 806-810; Lin, F. - K. и др. (1985) Proc. Natl. Acad. Sci. U.S.A. 82: 7580-7585] содержащую три N-сцепленных и одну О-сцепленную олигосахаридые цепи имеющие концевые остатки сиаловой кислоты. Последний является важным для обеспечения возможности ЕРО, чтобы избегать быстрого клиренса из кровообращения печеночным асиалогликопротеин связывающим белком.
Аминокислотная последовательность эритропоэтина (ЕРО) (представленная в однобуквенном коде) является следующей:
APPRLICDSRVLERYLLEAKEAENITTGCAEHCSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPWEPLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR
Аминокислотную последовательность, которая является частью последовательности иммуногенно немодифицированного эритропоэтина (ЕРО) человека и которая обладает потенциальной активностью связывания МНС класса II, выбирают из следующей группы:

ПРИМЕР 8 (G-CSF):
Колониестимулирующий гранулоцитарный фактор G-CSF - важный кроветворный цитокин, в настоящее время используемый при лечении показаний, где увеличение нейтрофилов крови будет выгодно. Которые включают терапию рака, различные инфекционные заболевания и связанные с ними состояния, такие как сепсис. G-CSF применяют самостоятельно или в комбинации с другими соединениями и цитокинами при ex vivo экспансии кроветворных клеток для трансплантации костного мозга. Две формы G-CSF человека обычно признаются для этого цитокина. Первая - белок из 177 аминокислот, вторая - белок из 174 аминокислот [Nagata и др. (1986), EMBO J. 5: 575-581], 174 аминокислотная форма, как было установлено, имеет большую специфическую биологическая активность in vivo. Методы рекомбинантной ДНК позволили получать G-CSF в масштабах промышленных количеств, применяя как эукариотические так и прокариотические системы экспрессии клетки-хозяина.
Аминокислотная последовательность колониестимулирующего гранулоцитарного фактора (G-CSF) (представленная в однобуквенном коде) является следующей:
TPLGPASSLPQSFLLKCLEQVRKIQGDGAALQEKLCATYKLCHPEELVLLGHSLGIPWAPLSSCPSQALQLAGCLSQLHSGLFLYQGLLQALEGISPELGPTLDTLQLDVADFATTIWQQMEELGMAPALQPTQGAMPAFASAFQRRAGGVLVASHLQSFLEVSYRVLRHLAQP.
Другие аналоги полипептида и фрагменты пептида G-CSF были ранее раскрыты, включая формы, модифицированные сайт-специфичными заменами аминокислоты и или модификацией химическими аддуктами. Так, US 4810643 раскрывает аналоги со специфическими Cys остатками, замененными другой аминокислотой, и G-CSF с Ala остатком в первом (N-терминалом) положении. ЕР 0335423 раскрывает модификацию по крайней мере одной амино группы в полипептиде, обладающем активностью G-CSF. ЕР 0272703 раскрывает производные G-CSF у которых заменена или удалена аминокислота около N конца. ЕР 0459630 раскрывает производные G-CSF, в которых Cys 17 и Asp 27 заменены остатками Ser. ЕР 0243153 раскрывает G-CSF, модифицированный инактивацией по крайней мере одного сайта обрабатывающего дрожжи КЕХ2 протеаза, для увеличения выработки в рекомбинантной продукции, и US 4904584 раскрывают, лизин измененные белки. WO 90/12874 раскрывает далее Cys измененные варианты и Австралийский патентный документ AU-A-10948/92 раскрывает добавление аминокислот к любому из концов молекулы G-CSF с целью содействия в сворачивании молекулы после прокариотической экспрессии. AU-763 80/91, раскрывает варианты G-CSF в положениях 50-56 для 174 аминокислотной формы G-CSF и в положениях 53-59 для 177 аминокислотной формы. Также были описаны дополнительные изменения в специфическом His остатке.
Аминокислотную последовательность, которая является частью последовательности колониестимулирующего гранулоцитарного фактора (G-CSF) человека и которая обладает потенциальной активностью связывания МНС класса II, выбирают из следующей группы:

Любая из вышеприведенных последовательностей может быть использована для модифицирования путем замены одной или более аминокислот для получения последовательности, которая неиммуногенна или имеет сниженную иммунногенность.

ПРИМЕР 9 (KGF):
Другой терапевтически ценной молекулой является фактор роста кератиноцита (KGF). KGF - член семейства белков фактора роста фибробласта (FGF)/гепарин-связывающего фактора роста. Это - секретированный гликопротеин, выделяемый преимущественно в легком, промотирующий заживание повреждений, стимулируя рост кератиноцитов и других эпителиоцитов [Finch и др. (1989), Science 24:7 52-755; Rubin и др. (1989), Proc. Natl. Acad. Sci. U.S.A. 86: 802-806]. Развитая (преобразованная) форма гликопротеида включает 163 аминокислотных остатка и может быть выделена из условной среды следующей культуры специфических линий клеток [Rubin и др. (1989) там же], или получена, используя рекомбинантные методы [Ron и др. (1993) J.Biol.Chem. 268: 2984-2988]. Белок имеет терапевтическую ценность для стимулирования роста эпителиоцита в ряде серьезных заболеваний и условий регенерации повреждений. Это описание, в частности, касается KGF белка человека, являющегося развитой (преобразованной) формой 163 аминокислотных остатков.
Другие способы обеспечивают молекулы KGF [например, US, 6 008 328; WO 90/08771;] включая модифицированные KGF [Ron и др. (1993) там же; WO 95/01434]. Однако эти методы не признают важности эпитопов Т-клеток для иммуногенных свойств белка, не рассматривают прямого влияния указанных свойств на специфический и контролированный путь в соответствии со схемой данного изобретения.
Аминокислотная последовательность фактора роста кератиноцита (KGF) (представленная в однобуквенном коде) является следующей:
MCNDMTPEQMATNVNCSSPERHTRSYDYMEGGDIRVRRLFCRTQWYLRIDKRGKVKGTQEMKNNYNIMEIRTVAVGIVAIKGVESEFYLAMNKEGKLYAKKECNEDCNFKELILENHYNTYASAKWTHNGGEMFVALNQKGIPVRGKKTKKEQKTAHFLPMAIT
Аминокислотную последовательность, которая является частью последовательности фактора роста кератиноцита (KGF) человека, и которая обладает потенциальной активностью связывания МНС класса II, выбирают из следующей группы:

Любая из вышеприведенных последовательностей может быть использована для модифицирования путем замены одной или более аминокислот для получения последовательности, которая неиммуногенна или имеет сниженную иммунногенность.

ПРИМЕР 10 (растворимый TNF RI):
sTNF-RI (растворимый рецептор фактора некроза опухоли типа I) является производным человеческого рецептора фактора некроза опухоли, описанного ранее [Gray, P.W. и др. (1990) Proc.Nat.Acad.Sci. U.S.A. 87: 7380-7384; Loetschere, Н. и др. (1990) Cell 61: 351-359; Schall, TJ. и др. (1990) Cell 61; 361-370], содержащий внеклеточный домен интактного рецептора и имееющий приблизительную молекулярную массу 30КDа. Дополнительные растворимые TNF ингибиторы и в особенности форма 40 KDa также известны [US 6143866]. Растворимые формы способны связать альфа фактор некроза опухоли с высокой аффинностью и ингибировать цитостатическую деятельность цитокина in vitro. Рекомбинантные препараты sTNF-RI имеют значительную терапевтическую ценность для лечения болезней, где избыточный уровень фактора некроза опухоли причиняет патогенный эффект. Показания, такие как кахексия, сепсис и аутоиммунные нарушения, и, в частности, ревматический артрит, и другие могут объектами для назначения таких терапевтических препаратов sTNF-RI. Другие, включая Brewer и др., US 6143866, обеспечили модифицированные молекулы sTNF-RI.

Любая из вьплеприведенных последовательностей может быть использована для модифицирования путем замены одной или более аминокислот для получения последовательности, которая неиммуногенна или имеет сниженную иммунногенность.
ПРИМЕР 11 (растворимый TNF-R2):
Растворимый рецептор 2 фактора некроза опухоли (sTNF-R2) является производным человеческого рецептора 2 фактора некроза опухоли, описанного ранее [Smith, С.А. и др. (1990) Science 248: 1019-1023; Kohno, Т. и др. (1990) Proc. Nat. Acad. Sci. U.S.A. 87: 8331-8335; Beltmger, C.P. и др. (1996) Genomics 35: 94-100], содержащий внеклеточный домен интактного рецептора. Растворимые формы способны связать фактор некроза опухоли с высокой аффинностью и ингибировать цитостатическую деятельность цитокина in vitro. Рекомбинантные препараты sTNF-R2 имеют значительную терапевтическую ценность для лечения болезней, где избыточный уровень фактора некроза опухоли причиняет патогенный эффект. Специфический рекомбинантный препарат, названный этанерцепт (ethanercept), получил клиническое одобрение для лечения ревматического артрита, и этот и другие подобные агенты могут быть полезными при лечении других показаний, таких как кахексия, сепсис и аутоиммунные нарушения. Этанерцепт - димерный белок слияния содержащий внеклеточный домен молекулы TNFR2 человека в комбинации с Fc доменом молекулы IgGl человека. Димерная молекула включает 934 аминокислоты [US 5395760; US 5605690; US 5945397, US RE36.755].

ПРИМЕР 12 (β-GCR):
Бета-глюкоцереброзидаза (b-D-глюкозил-Н-ацилсфингозин глюкогидролаза, Е.С.3.2.1.45) представляет собой мономерный гликопротеин, состоящий из 497 аминокислотных остатков. Фермент катализирует гидролиз гликолипида глюкоцереброзида до глюкозы и церамида. Недостаток активности GCR приводит к лизосомной болезни накопления, известной как болезнь Гауше (Gaucher disease). Заболевание характеризуется накоплением макрофагов ткани, налитых глюкоцереброзидом, которые накапливаются в печени, селезенке, костном мозге и других органах. Болезнь имеет различные степени серьезности: от типа 1 - болезнь с гематологичными проблемами, но без нейронального поражения, до типа 2 - болезнь проявляющаяся скоро после рождения с обширным нейрональным поражением и общим прогрессированием с фатальным исходом в течение 2 лет. В некоторых классификациях также признан третий тип заболевания, который также характеризуется нейрональным поражением. Ранее единственно полезной терапией болезни Гауше было применение GCR, полученной из человеческой плаценты (известной как аглуцераза), но позже фармацевтические препараты рекомбинантной GCR ("цередаза" и "церезим") показали эффективность при лечении заболевания первого типа [Niederau, С. и др. (1998) Eur. J. Med. Res. 3: 25-30].

ПРИМЕР 13 (Белок C):
Белок С представляет собой витамин К зависимую протеазу серина, вовлеченную в регулирование коагуляции крови. Белок активируется тромбином, чтобы произвести активированный белок С, который в свою очередь подавляет Факторы Va и VIII в каскаде коагуляции. Белок С вырабатывается в печени в виде одиноцепочного предшественника и подвергается ряду обработок, которые заканчиваются молекулой, содержащей легкую и тяжелую цепи, соединенные вместе дисульфидной связью. Белок С активируется отщеплением тетрадекапептида от N-конца тяжелой цепи тромбином. Фармацевтические препараты белка С в нативной или активированной форме являются ценными при лечении пациентов с сосудистыми заболеваниями и/или приобретенным дефицитом в белке С. Такие пациенты включают поэтому индивидуумов, страдающих от тромботического инсульта или дефицита белка С, связанного с сепсисом, процедурами трансплантации, беременностью, серьезными ожогами, радикальной хирургией или другими серьезными травмами. Белок С также используется при лечении индивидуумов с наследственным дефицитом в белке С. Это описание в особенности касается белка С человека, находящегося в зрелой (обработанной) форме, содержащего легкую цепь из 155 аминокислотних остатков и тяжелую цепь из 262 аминокислотних остатков [Foster, D.C. и др. (1985) Proc. Nad. Acad. Set. U.S.A. 82: 4673-4677; Beckman, R.J. и др. (1985) Nucleic Acids Res. 13:5233-5247]. Другими были получены молекулы белка С, включая композиции активированного белка С, и методы применения [US 6159468; US 6156734; US 6037322; US 5618714].

ПРИМЕР 14 (субтилизины)
Субтилизины представляют собой класс фермента протеазы с существенной экономической и промышленной важностью. Они могут использоваться в качестве компонентов моющих или косметических средств или в производстве текстиля и других отраслях промышленности. При воздействии на некоторых людей бактериальные субтилизины могут вызывать нежелательную реакцию повышенной чувствительности. Поэтому существует потребность в аналогах субтилизина с улучшенными свойствами и особенно усовершенствованными биологическими свойствами белка. В связи с этим очень желательно разработать субтилизины с уменьшенным или отсутствующим потенциалом вызывать иммунную реакцию по отношению к человеку. Белки субтилизина идентифицированные из разных источников, включая бактериальные, грибные или позвоночных, а также млекопитающих и человека, имеют в общем многие из последовательностей белка настоящего описания и имеют в общем много последовательностей белка с существенно той же самой последовательностью как те, что раскрыты в списке. Поэтому такие белковые последовательности одинаково подпадают под объем настоящего изобретения. Другими были получены молекулы субтилизина, включая модифицированные субтилизины [US 5700676; US 4914031; US 5397705; US 5972682].




Пример 15 (лиганды CNTF):
Настоящее изобретение обеспечивает модифицированные формы белковых субъединиц, содержащих гетеродимерный лиганд для комплекса рецептора цилиарного нейротрофического фактора (CNTF) в людях. Комплекс рецептора активизирован по крайней мере двумя лигандами, включая CNTF и гетеродимерный комплекс, содержащий кардиотрофин-подобный цитокин (CLC) и растворимый рецептор, подобный цитокину фактор 1 (CLF) [Elson G.С.А. и др. (2000) Nature Neuroscience 3: 867-872]. CLC является белком IL-6 семейства цитокинов, также известный как новый нейротрофин-1/В клетко-стимулирующий фактор-3 [Senaldi, G. и др. (1999) Proc. Nat. Acad. Sci. USA 96: 11458-11463, US 5741772]. CLF гомологичен к белкам семейства рецептора цитокина тип I [Elson, G.С.А. и др. (1998) Journal of Immunol. 161: 1371-1379] и был также идентифицирован как NR6 [Alexander W.S. и др. (1999) Curr. Biol. 9: 605-608]. Гетеродимеры образованные ассоциацией CLC и CLF показали способность непосредственно взаимодействовать с CNTFR, и таким образом сформированный тримерный комплекс способен стимулировать сигнализирование событий в пределах клеток, выражающих другие распознанные компоненты CNTFR комплекса типа gp130 и LIFR [Elson G.С.А. и др. (2000), там же].


ПРИМЕР 16 (фолликулостимулирующий гормон)
Настоящее изобретение обеспечивает модифицированные формы hFSH человека с одним или более удаленными эпитопами Т-клеток. hFSH является гормоном гликопротеином с димерной структурой, содержащей две субъединицы гликопротеина. Белок терапевтически используется при лечении бесплодия человека и рекомбинантные формы белка были предметом множества клинических испытаний [Out, H.J. и др. (1995) Hum. Reprod. 10: 2534-2540; Hedon, В. и др. (1995) Hum. Reprod. 10: 3102-3106; Recombinant Human FSH study Group (1995) Fertil. Steril. 63: 7786; Prevost, R.R. (1998) Pharmacotherapy 18: 1001-1010].

ПРИМЕР 16 (рицин А)
Настоящее изобретение обеспечивает модифицированные формы А-цепи токсина рицина (RTA) с одним или более удаленными эпитопами Т-клеток. Рицин является цитотоксином, изначально выделенным из семян касторового растения, и является примером рибосома-инактивирующего белка типа II (RIP). Нативный зрелый белок является гетеродимером содержащим RTA, который состоит из 267 аминокислотных остатков, на дисульфидной связи с В-цепью рицина, которая состоит из 262 аминокислотных остатков. В-цепь является пектином со связывающим сродством к галактозидам. Нативный белок способен связать клетки посредством В-цепи и войти в клетку эндоцитозом. Внутри клетки RTA выделяется из В-цепи восстановлением дисульфидной связи и выходит из эндосома в цитоплазму посредством неизвестных механизмов. В цитоплазме токсин разрушает рибосомы, действуя как специфичная N-гликозилаза, что быстро приводит к прекращению преобразования белка и гибели клетки. Чрезвычайная цитотоксичность RTA и других RIP-ов, привела к их использованию при экспериментальных терапиях для лечения рака и других заболеваний, где требуется абляция определенной клеточной популяции. Молекулы иммунотоксина, содержащие молекулы антитела в связи с RTA, были получены и использовались в многочисленных клинических испытаниях [Ghetie, M.А. и др. (1991) Cancer Res. 51: 5876-5880; Vitetta, E.S. и др. (1991) CancerRes. 51: 4052-4058; Arnlot, P.L. и др. (1993) Bloody. 2624-2633; Conry, R.M. и др. (1995) J.Immunotlier. Emphasis Tumor Immunol. 18: 231-241; Schnell, R. и др. (2000) Leukaemia 14:129-135]. В иммунотоксине домен антитела обеспечивает связывание с поверхностью желаемой клетки - мишени, и связь с RTA может быть обеспечена посредством химической кросс-связи или как рекомбинанный белок слияния.

ПРИМЕР 17 (адипоцитный комплемент-ассоциированный белок):
Существующее изобретение обеспечивает модифицированные формы Асrр30 человека или мыши с одним или более удаленными эпигонами Т-клеток. Асrр30 представляет собой белок избытка сыворотки с молекулярной массой.приблизительно 30 kDa, выделяемый исключительно адипоцитными (жировыми) клетками [Scherer, Р.Е. и др. (1995) J.Biol. Chez. 270: 26746-26749]. Белковая последовательность человеческого гена Асrр30 описана, например, в US 5869330. Секрецию белка увеличивает инсулин, и уровни белка снижены у страдающих ожирением. Белок вовлечен в регулирование энергетического баланса и в особенности регулирования метаболизма жирных кислот. Четыре последовательности доменов, идентифицированные в белке мыши и человека, содержат расщепленный N-терминальный сигнал, область с нераспознанной гомологией к другим белкам, коллаген подобный домен и глобулярный домен. Глобулярный домен может быть удален из белка мыши обработкой протеазой, чтобы получить gAcrp30. Препараты мышиного gAcrp30 обладают фармацевтическими свойствами и показывают способность уменьшать увеличенные уровни свободных жирных кислот в сыворотке мышей после принятия высоко жирной пищи или внутривенной инъекции липида [Fruebis, J. и др. (2001) Proc. Natl. Acad. Sci. U.S.A. 98: 2005-2010].


ПРИМЕР 18 (анти-С5 антитело):
Настоящее изобретение обеспечивает модифицированные формы моноклональных антител со связывающей специфичностью, направленной на С5 комплементный белок человека. Изобретение обеспечивает модифицированные антитела с одним или более удаленными эпитопами Т-клеток. Антитела со связывающей специфичностью, направленной на С5 комплементный белок, блокируют активацию расщепления С5 конвертазы и таким образом ингибируют продупирование провоспалительных компонентов С5а и С5b-9. Активация комплементной системы является существенным содействующим фактором в патогенезе множества острых и хронических заболеваний, и ингибирование комплементного каскада на уровне С5 предлагает многообещающий терапевтический подход для некоторых из них [Morgan В.Р. (1994) Eur.J.Clin.Invest. 24: 219-228]. Несколько анти-С5 антител и способов их терапевтического использования было описано в уровне техники [Wurzner R. и др. (1991) Complement Inflamm. 8:328-340; Thomas, T.C. и др. (1996) Molecular Immunology 33: 1389-14012; US 5853722; US 607464]. Антитело, обозначенное как 5G1.1 [Thomas, T.C. и др. (1996), там же] и одиноцепочный гуманизированный вариант проходили клинические испытания в отношении нескольких показаний заболеваний, включая экстракорпоральное кровообращение [Fitch, J.C.K. и др. (1999) Circulation 100: 2499-2506] и ревматоидный артрит. Изобретение раскрывает последовательности, идентифицированные в пределах анти-С5 антитела, обозначенного как 5G1.1 [Thomas, T.C. и др.]. Описанные последовательности получены из доменов переменных областей тяжелых и легких цепей последовательности антитела, которые являются потенциальными эпитопами Т-клеток на основании потенциала связывания МНС класса II. В описании далее определяют потенциальные эпитопы в пределах белковых последовательностей одиноцепочного и "гуманизированного" варианта 5G1.1 антитела [Thomas, T.C. и др. (1996), там же].

ПРИМЕР 19 (анти-СD20 антитела):
Настоящее изобретение обеспечивает модифицированные формы моноклонального антитела со связывающей специфичностью, направленной на CD20 антиген человека. CD20 является В-клеточной удельно поверхностной молекулой, вырабатываемой в пре-В и зрелых В-клетках, включая больше чем 90% В-клеток неходжкинской лимфомы (NHL). Моноклональные антитела и радиоиммуноконъюгаты, атакующие CD20, возникли как новые подходы для лечения NHL. Показательные примеры включают моноклональные антитела 2В8 [Reff, M.E. и др. (1994) Blood 83: 435-445] и B1 [US 6090365]. Домены переменных областей 2В8 были клонированы и объединены с доменами постоянных областей человека для получения химерного антитела, обозначенного как С2В8, который продается под названием RituxanTM в США [US 5776456] или MabTheraR (rituximab) в Европе. С2В8 признан как ценный терапевтический агент для лечения NHL и других В-клеточных заболеваний [Maloney, D.G. и др. (1997) J.Clin.Oncol. 15: 3266-3274; Maloney, D.G. и до. (1997) Blood 90: 2188-2195]. В1 антитело подобным образом получило регистрацию для использования как NHL терапевтическое средство, несмотря на то что в этом случае молекула (BexxarTM) является 131I радиоиммуноконъюгатом, хотя нативное (несопряженное) антитело является полезным при ex vivo режимах очистки для терапии аутологической пересадки костного мозга для лимфомы и трудноизлечимой лейкемии [Freedman, A.S. и др. (1990), J.Clin.Oncol. 8: 784]. Несмотря на успех антител типа С2В8 (rituximab) и BexxarTM существует потребность в анти-СD20 аналогах с улучшеными свойствами.

ПРИМЕР 20:
Настоящее изобретение обеспечивает модифицированные формы моноклонального антитела со связывающей специфичностью к рецептору IL-2 человека. Моноклональное антитело обозначенное как анти-Тас и модифицированная форма имеют один или более удаленных эпитопов Т-клеток. Антитело анти-Тас связывается с высокой специфичностью с альфа субъединицей (р55-альфа, CD25 или Тас) человеческого высокоаффинного рецептора IL-2, выраженного на поверхности Т и В лимфоцитов. Связывающее антитело блокирует способность IL-2 связывать рецептор и добиваться активации Т-клетки. Способность анти-Тас антитела выступать в роли антагониста IL-2 обладает существенным клиническим потенциалом для лечения отторжения трансплантата органа. Клинические изучения с использованием антитела мыши показали некоторую начальную полезность для пациентов, которые подверглись почечной пересадке, хотя долговременной выгоды в отношении обычного иммунного подавления не было достигнуто из-за развития ответа НАМА у большинства пациентов [Kirkham, R.L. и др. (1991) Transplantation 51: 107-113]. Было разработано "гуманизированное" анти-Тас антитело, в котором существенные компоненты белка проектировались, чтобы содержать белковую последовательность, идентифицированную из человеческого гена антитела [Queen, С. и др. (1989) Proc. Natl. Acad. Sci. (USA) 86: 10029-10033; US 5530101; US 5585089; US 6013256]. "Гуманизированный" анти-Тас (ZenapaxТМ или daclizumab) был подвергнут клиническим испытаниям как иммуноподавляющее средство для регулирования заболевания острой трансплантации против хозяина и подавления отторжения почечного трансплантата [Anasetti, С. и др. (1994). Blood 84: 1320-1327; Anasetd, С. и др. (1995) Blood 86: Supplement 1: 62a; Eckhoff, D.E. и др. (2000) Transplantation 69: 1867-1872; Ekberg, Н. и др. (1999) Transplant Proc. 31: 267-268].


ПРИМЕР 21 (14.18 антитело):
Если не указано иначе, все аминокислоты в переменной тяжелой и легкой цепях нумеруются, как в Kabat и др., 1991 (Sequences of Proteins of Immunological Interest, US Department of Health and Human Services). Потенциальные эпитопы Т-клеток нумеруются с линейным номером первой аминокислоты эпитопа, считая от первой аминокислоты тяжелой и легкой цепей.
1. Сравнение со Структурами Подгруппы Мыши
Аминокислотные последовательности мышиных 14.18 VH и VK были сравнены с последовательностями общих типичных элементов структуры для Kabat мышиных подгрупп тяжелой и легкой цепи (Kabat и др., 1991). 14.18 VH может быть отнесен к подгруппе IIA тяжелых цепей мыши. Последовательность 14.18 VH показана на SEQ. №1. Сравнение с последовательностью общих типичных элементов структуры для этой подгруппы показывает, что гистидин в положении 81 (обычно глутамин), лизин в положении 82а (обычно серин или аспарагин), валин в положении 93 (обычно аланин) и серин в положении 94 (обычно аргинин) являются несвойственными для этой подгруппы. Остатки в положениях 19, 40 и 66 также нечасто встречаются в этой подгруппе, но, как полагают, они не имеют значительного влияния на структуру и связывание антитела. 14.18 VK может быть отнесен к подгруппе II каппа цепей мыши. Сравнение с последовательностью общих типичных элементов структуры для этой подгруппы показывает, что гистидин в положении 49 является несвойственным для этой подгруппы. Наиболее распространенный остаток для этого положения тирозин.
2. Сравнение со Структурами Человека
Аминокислотные последовательности мышиных 14.18 VH и VK были сравнены с последовательностями справочника зародышевых линий человека (germline) VH (Tomlinson и др., (1992) J.Mol.Biol. 227: 776-798) и VK (Сох и др., (1994) Eur. J. Immunol. 1-4-: 827-83б) последовательностями, а также с областью последовательностей зародышевых линий J человека (Routledge и др., в Protein Engineering of Antibody Molecules for Prophylactic and Therapeutic Applications in Man, Clark, M. ed. Academic Titles, Nottingham, UK, pp 13-44, 1991). Эталонной структурой человека, выбранной для 14.18 VH, была DP25 с JH6 человека. Эта последовательность зародышевой линии была найдена в перестроенном зрелом гене антитела без изменений аминокислот. Для структуры 3 использовалась последовательность зрелого человеческого антитела 29. Эта последовательность идентична мышиной последовательности, непосредственно примыкающей к CDR3. Эталонной структурой человека, выбранной для 14.18VK, была DPK22. Эта последовательность зародышевой линии была найдена в перестроенном зрелом гене антитела без изменений аминокислот. Для структуры 2 использовалась последовательность зрелого человеческого антитела 163.5. Эта последовательность идентична мышиной последовательности, непосредственно примыкающей к CDR2. Последовательностью J области был JK2 человека (Routledge и др., 1993).
3. Разработка "венированных" ("veneered") последовательностей
После идентификации базовых последовательностей эталонных структуры человека некоторые неидентичные аминокислотные остатки в пределах 14.18 VH и VK структур были изменены на соответствующую аминокислоту в базовых последовательностях структуры человека. Остатки, которые, как полагают, являются критическими для структуры антитела и связывания, были исключены из этого процесса и не изменялись. Мышиные остатки, которые были сохранены на этой стадии являлись в значительной степени неповерхностными, внутренними остатками, кроме остатков на N-концах, например, которые являются близкими к CDR-ам в целевом антителе. Этот процесс приводит к последовательности, которая является определенно подобной "венированному" антителу, поскольку поверхностные остатки являются главным образом человеческими, а внутренние остатки, такими, как в первоначальной мышиной последовательности.
4. Пептид проигрывающий анализ
Мышиные и венированные 14.18 VH и VK последовательности были проанализированы с использованием способа согласно изобретению. Последовательности аминокислот были разделены на все возможные тринадцатимеры. Тринадцатимерные белки последовательно представлены моделям связывающей бороздки аллотипов HLA-DR, и значения связывания соотносились каждому пептиду для каждой аллели. Было рассчитано конформационное значение для каждой карман-связанной боковой цепи пептида. Это значение базируется на стерическом наложении, потенциальных водородных связях между белком и остатками в связывающей бороздке, электростатических взаимодействиях и благоприятных контактах между белком и остатками карманна. Конформацию каждой боковой цепи изменили, значение повторно рассчитали. Определив самое высокое конформационное значение, пересчитывали значение связывания основываясь на бороздково-связанных гидрофобных остатках, небороздковых гидрофильных остатках и количестве остатков, которые умещаются в бороздку связывания. Пептиды, которые являются известными связывающими класса II МНС человека, достигают высокого значения связывания почти без неверных результатов. Таким образом, пептиды, которые достигают значительного значения связывания в текущем анализе, как полагают, являются потенциальными эпитопами Т-клеток. Результаты пептид пронизывающего анализа показаны в Таблице 1.
5. Удаление потенциальных эпитопов Т-клеток
Потенциальные эпитопы Т-клеток удаляют заменой аминокислот в специфическом пептиде, который составляет эпитоп. Замены были сделаны введением аминокислот с по возможности подобными физико-химическими свойствами. Однако для того чтобы удалять некоторые потенциальные эпитопы, аминокислоты различного размера, заряда или гидрофобности, возможно, должны быть заменены. Если изменения должны быть сделаны в пределах CDR-ов, которые могли бы иметь эффект связывания, необходимо создать вариант с и без специфического замещения аминокислот. Аминокислотные остатки для замены даны с Kabat номером, указанным в скобках. Потенциальные эпитопы Т-клеток указаны линейным номером первого остатка тринадцатимера.
Необходимые замены аминокислот для удаления эпитопов Т-клеток из венированной 14.18 переменной области тяжелой цепи были следующие:
1. Замена изолейцина на пролин в остатке 41 (Kabat номер 41), объединенная с заменой лейцина на аланин в остатке 50 в CDR2 удаляет потенциальный эпитоп в положении 43.
2. Альтернативно к (1), замена треонина на лейцин в остатке 45 (Kabat номер 45) с пролином в положении 41 (Kabat номер 41) также удаляет потенциальный эпитоп в положении 43.
3. Замена серина на глицин в остатке 66 (Kabat номер 65) в CDR2 и валина на аланин в остатке 68 (Kabat номер 67) удаляет потенциальный эпитоп в положении 58. Серии найден в этом положении в последовательностях антител человека и мыши.
4. Замена изолейцина на лейцина в остатке 70 (Kabat: 69) уменьшает количество МНС аллотипов, которые связываются с потенциальным эпитопом в положении 62 от 11 до 4.
5. Замена аланина на валин в положении 72 (Kabat номер 71) удаляет потенциальный эпитоп в положении 62. Размер аминокислоты в этом положении является критическим и аланин по гидрофобности и размеру подобен валину.
6. Замена треонина на серии в остатке 91 (Kabat номер 87) удаляет потенциальные эпитопы в положениях 81 и 84.
Необходимые замены аминокислот для удаления потенциальных эпитопов Т-клеток из венированной 14.18 переменной области легкой цепи были следующие:
1. Замена серина на аргинин в остатке 32 (Kabat номер 27е) удаляет потенциальный эпитоп в положении 27. Этот остаток находится в пределах CDR2, однако серии часто находят в tWs положении в антителах мыши и человека. Нет никакого изменения с CDR, которое устраняет потенциальный эпитоп Т-клеток.
2. Замена тирозина на гистидин в положении 54 (Kabat номер 49) устраняет потенциальный эпитоп в положении 43. Тирозин - наиболее часто встречающаяся аминокислота, найденная в положении 49 в антителах мыши и человека.
1. Альтернативная замена к (2) для устранения потенциального эпитопа в положении 43, это замена метионина на лейцин в остатке 51 (Kabat номер 46). Метионин подобен лейцину по размеру и гидрофобности.
2. Замена метионина на лейцин в остатке 88 (Kabat номер 83) устраняет потенциальный эпитоп в положении 86.
1. Замена треонина на лейцин в остатке 102 (Kabat номер 96) в CDRH3 в комбинации с заменой глутамина на глицин в положении 105 (Kabat номер 100) уменьшает количество МНС аллотипов, которые связываются с потенциальным эпитопом в положении 97 от 11 до 5.
2. Альтернативная замена к (5), которая устраняет потенциальный эпитоп в положении 97, является заменой пралина на лейцин в остатке 102 (Kabat номер 96).
3. Замена валина на лейцин в остатке 110 (Kabat номер 104) удаляет, устраняет потенциальный эпитоп в положении 100.
6. Разработка деиммунизированных последовательностей
Деиммунизированные последовательности тяжелой и легкой цепи были разработаны с учетом замен, требуемых для удаления потенциальных эпитопов Т-клеток и анализа структуры остатков, которые могли бы быть критичными для структуры антитела и связывания. В дополнение к деиммунизированным последовательностям, которые основаны на венированной последовательности, дополнительная последовательность была разработана для каждого VH и VK, основанного на мышиной последовательности, названные Белок Мыши Прошитая (Мо РТ) версия. Для этой версии замены были сделаны непосредственно в мышиной последовательности, чтобы устранить эпитопы Т-клеток, но изменения сделаны только с CDR-ами, которые не рассматривают как нежелательные для закрепления. Никакой попытки удалять поверхностные (В-клетки) эпитопы не было сделано для этой версии деиммунизированной последовательности.
Первичная деиммунизированная VH включает замены 1, 3, 4, 5 и 6, описанные в разделе 5 выше, и не включает никаких потенциальных эпитопов Т-клеток. Дополнительные 4 деиммунизированных последовательности VH были разработаны, чтобы проверить эффект необходимых различных замен на связывание антитела. Версия 2 является альтернативой к Версии 1, в которой альтернативная замена (2 в Разделе 2.5 выше) использовалась, для удаления того же самого потенциального эпитопа Т-клетки. Совокупные изменения сделаны для первой деиммунизированной последовательности (14.18DIVH1) и оставшиеся потенциальные эпитопы Т-клеток отображены в Таблице 2. Мышиная прошитая версия включена для сравнения.
Первичный деиммунизированный VK включает замены 1, 2, 4, 6 и 7, описанные в разделе 5 выше и не включает никаких потенциальных эпитопов Т-клеток. Дополнительные 5 деиммунизированных последовательности VK были разработаны, чтобы проверить эффект необходимых различных замен, на связывание антитела. Версия 2 является альтернативой Версии 1, в которой использовалась альтернативная замена, чтобы удалить тот же самый потенциальный эпитоп Т-клетки в положении 43. Версия 3 включает альтернативную замену (6 в Разделе 2.5 выше), такая замена уменьшает количество МНС аллотипов, которые связываются с потенциальным эпитопом в положении 97 от 11 до 5. Совокупные изменения сделаны для первичной деиммунизированной последовательности (14.18DIVK1) и оставшиеся потенциальные эпитопы Т-клеток отображены в Таблице 3. Мышиная прошитая версия включена для сравнения.


ПРИМЕР 22 (KS антитело)
1. Сравнение со Структурами Подгруппы Мыши
Аминокислотные последовательности мышиных KS VH и VK были сравнены с последовательностями общих типичных элементов структуры для Kabat мышиных подгрупп тяжелой и легкой цепи (Kabat и др., 1991). KS VH не может быть отнесен к какой либо из подгрупп, но наиболее близок к подгруппе II(А) и V(A). Нетипичные остатки найдены в положении 2, который является обычно валин, 46, который является обычно глутаминовой кислотой, и 68, который является обычно треонином. Остаток 69 является более обычно лейцином или изолейцином. В 82b наиболее часто находят серии. Мышиный KS VK может быть отнесен в Подгруппу VI (Фигура 2). Необычные остатки найдены в 46 и 47, которые оба обычно являются лейцином. Остаток 58 нетипичен с или лейцином, или валином, обычно находящимся в этом положении.
2. Сравнение со Структурами Человека
Аминокислотные последовательности мышиных KS VH и VK были сравнены с последовательностями справочника зародышевых линий человека (germime) VH (Tomlinson и др., (1992)) и VK (Сох и др., (1994)) последовательностями, а также с областью последовательностей зародышевых линий J человека (Routledge и др., (1993)). Эталонной структурой человека, выбранной для KS VH, была DP 10 с JH6 человека. Эта последовательность зародышевой линии была найдена в перестроенном зрелом гене антитела без изменений аминокислот. Эталонной структурой человека, выбранной для KS VK, была В1. Для структуры 2 использовалась последовательность зрелого человеческого антитела IMEV (Kabat и др., 1991). Эта последовательность идентична мышиной последовательности, непосредственно примыкающей к CDR2. Последовательностью J области был JK2 человека (Routledge и др., 1993). Эта последовательность зародышевой линии не была найдена как перестроенная зрелая легкая цепь антитела.
3. Разработка "венированных" ("veneered") последовательностей
После идентификации базовых последовательностей эталонных структур человека некоторые неидентичные аминокислотные остатки в пределах 425 VH и VK структур были изменены на соответствующую аминокислоту в эталонной последовательности человека. Остатки, которые, как полагают, являются критическими для структуры антитела и связывания, были исключены из этого процесса и не изменялись. Мышиные остатки, которые были сохранены на этой стадии, являлись в значительной степени неповерхностными, внутренними остатками, кроме остатков на N-концах, например, которые являются близкими к CDR-ам в целевом антителе. Этот процесс приводит к последовательности, которая является определенно подобной "венированному" антителу, поскольку поверхностные остатки являются главным образом человеческими, а внутренние остатки такими, как в первоначальной мышиной последовательности.
4. Пептид пронизывающий анализ
Мышиные и венированные KS VH и VK последовательности были проанализированы с использованием способа согласно изобретению. Последовательности аминокислот были разделены на все возможные тринадцатимеры. Тринадцатимерные белки последовательно представлены моделям связывающей бороздки аллотипов HLA-DR, и значения связывания соотносились каждому пептиду для каждой аллели. Было рассчитано конформационное значение для каждой кармансвязанной боковой цепи пептида. Это значение базируется на стерическом наложении, потенциальных водородных связях между белком и остатками в связывающей бороздке, электростатических взаимодействиях и благоприятных контактах между белком и остатками карманна. Конформацию каждой боковой цепи изменили, значение повторно рассчитали.
Определив самое высокое конформационное значение, пересчитывали значение связывания основываясь на бороздково-связанных гидрофобных остатках, небороздковых гидрофильных остатках и количестве остатков, которые умещаются в бороздку связывания. Пептиды, которые являются известными связывающими класса II МНС человека, достигают значительного значения связывания почти без неверных результатов. Таким образом, пептиды, которые достигают значительного значения связывания в текущем анализе, как полагают, являются потенциальными эпитопами Т-клеток. Результаты пептид пронизывающего анализа для мышинынных и венированных последовательностей показаны в Таблице 4.
5. Удаление потенциальных эпитопов Т-клеток
Потенциальные эпитопы Т-клеток удаляют заменой аминокислот в специфическом пептиде, который составляет эпитоп. Замены были сделаны введением аминокислот с по возможности подобными физико-химическими свойствами. Однако для того чтобы удалять некоторые потенциальные эпитопы, аминокислоты различного размера, заряда или гидрофобности, возможно, должны быть заменены. Если изменения должны быть сделаны в пределах CDR-ob, которые могли бы иметь эффект связывания, необходимо создать вариант с и без специфического замещения аминокислот. Аминокислотные остатки для замены пронумерованы, как в Kabat. Потенциальные эпитопы Т-клеток указаны линейным номером первого остатка тринадцатимера.
Необходимые замены аминокислот для удаления эпитопов Т-клеток из венированной KS переменной области тяжелой цепи были следующие:
1. замена аргинина на лизин в остатке 38 (Kabat номер 38) удаляет, потенциальный эпитоп в остатке номер 30.
2. замена аланина на лейцин в остатке 72 (Kabat номер 71) и изолейцина на фениаланин в остатке 70 (Kabat номер 69) удаляет потенциальный эпитоп в остатке номер 62. Изолейцин в Kabat номер 69 и аланин в Kabat номер 71 найден в человеческой последовательности VH зародышевой линии, DP10.
3. замена лейцина на аланин в остатке 79 (Kabat номер 78), удаляет потенциальный эпитоп в остатке номер 78.
1. замена треонина на метионин в остатке 91 (Kabat номер 87), удаляет потенциальный эпитоп в остатке номер 89.
2. замена метионина на изолейцин остатке 100 (Kabat номер 96) в CDRH3 удаляет потенциальный эпитоп в остатке 98. Нет никакого изменения с CDRH3, которое удаляет этот потенциальный эпитоп.
Необходимые замены аминокислот для удаления эпитопов Т-клеток из венированной KS переменной области легкой цепи были следующие:
1. Замена изолейцина на метионин в остатке 32 (Kabat номер 33), удаляет потенциальный эпитоп в остатке номер 27. Этот остаток находится в пределах CDR2. Изолейцин обычно находят в этом положении в антителах человека.
2. Потенциальный эпитоп в положении 1 удаляют заменой валина на лейцин в остатке (Kabat номер 3).
3. Замена серина на аланин в остатке 59 (Kabat номер 60), удаляет потенциальный эпитоп в остатке номер 51.
6. Разработка деиммунизированных последовательностей
Деиммунизированные последовательности тяжелой и легкой цепи были разработаны с учетом изменений, требуемых для удаления потенциальных эпитетов Т-клеток и анализа структуры остатков, которые могли бы быть критичными для структуры антитела и связывания. В дополнение к деиммунизированным последовательностям, которые основаны на венированной последовательности, дополнительная последовательность была разработана для каждого VH и VK, основанного на мышиной последовательности, названные Белок Мыши Прошитая (Мо РТ) версия. Для этой версии, замены были сделаны непосредственно в мышиной последовательности, чтобы устранить эпитопы Т-клеток, но изменения сделаны только с CDR-ами, которые не рассматривают как нежелательные для закрепления. Никакой попытки удалять поверхностные (В-клетки) эпитопы не было сделано для этой версии деиммунизированной последовательности.
Первичная деиммунизированная VH включает замены 1-5, описанные в разделе 5 выше, и одно дополнительное изменение в остатке 43 (Kabat номер 43). Лизин, найденный в мышиной последовательности, заменяли глутамином из человеческой структуры. Лизин положительно заряжен и поэтому значительно отличается от глутамина; эта область может быть вовлечена в VH/VL контакты. Первичный деиммунизированный VH не включает никаких потенциальных эпитопов Т-клетки. Дополнительные 4 деиммунизированных последовательности VH были разработаны, чтобы проверить эффект необходимых различных замен на связывание антитела. Совокупные изменения сделаны для первой деиммунизированной последовательности (KSDIVHv1) и оставшиеся потенциальные эпитопы Т-клеток отображены в Таблице 5.
Первичный деиммунизированный VK включает замены 1-3, описанные в разделе 5 выше. Дополнительные 3 деиммунизированных последовательности VK были разработаны, чтобы проверить эффект необходимых различных замен на связывание антитела. Совокупные изменения сделаны для первичной деиммунизированной последовательности (KSDIVKv1) и оставшиеся потенциальные эпитопы Т-клеток отображены в Таблице 6.


| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЕ АНТИ-EGFR АНТИТЕЛА С УМЕНЬШЕННОЙ ИММУНОГЕННОСТЬЮ | 2002 |
|
RU2297245C2 |
| МОДИФИЦИРОВАННЫЕ ЭПИТОПЫ ДЛЯ УСИЛЕНИЯ ОТВЕТОВ CD4+ Т-КЛЕТОК | 2013 |
|
RU2724994C2 |
| ИСКУССТВЕННЫЕ БЕЛКИ С ПОНИЖЕННОЙ ИММУНОГЕННОСТЬЮ | 2002 |
|
RU2363707C2 |
| СПОСОБ КАРТИРОВАНИЯ И УСТРАНЕНИЯ ЭПИТОПОВ Т-КЛЕТОК | 2003 |
|
RU2334235C2 |
| АНТИТЕЛА ПРОТИВ ПРЕДСТАВИТЕЛЯ А5 СЕМЕЙСТВА 19 СО СХОДСТВОМ ПОСЛЕДОВАТЕЛЬНОСТЕЙ И СПОСОБ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2785436C2 |
| ПРЕДСКАЗАНИЕ ИММУНОГЕННОСТИ Т-КЛЕТОЧНЫХ ЭПИТОПОВ | 2014 |
|
RU2724370C2 |
| СПОСОБЫ ПРОГНОЗИРОВАНИЯ ПРИМЕНИМОСТИ СПЕЦИФИЧНЫХ ДЛЯ ЗАБОЛЕВАНИЯ АМИНОКИСЛОТНЫХ МОДИФИКАЦИЙ ДЛЯ ИММУНОТЕРАПИИ | 2018 |
|
RU2799341C2 |
| ФАКТОР КОАГУЛЯЦИИ VIII С УМЕНЬШЕННОЙ ИММУНОГЕННОСТЬЮ | 2013 |
|
RU2631801C2 |
| RA АНТИГЕННЫЕ ПЕПТИДЫ | 2004 |
|
RU2359974C2 |
| НОВЫЕ ИММУНОГЕННЫЕ ПЕПТИДЫ | 2015 |
|
RU2709711C2 |
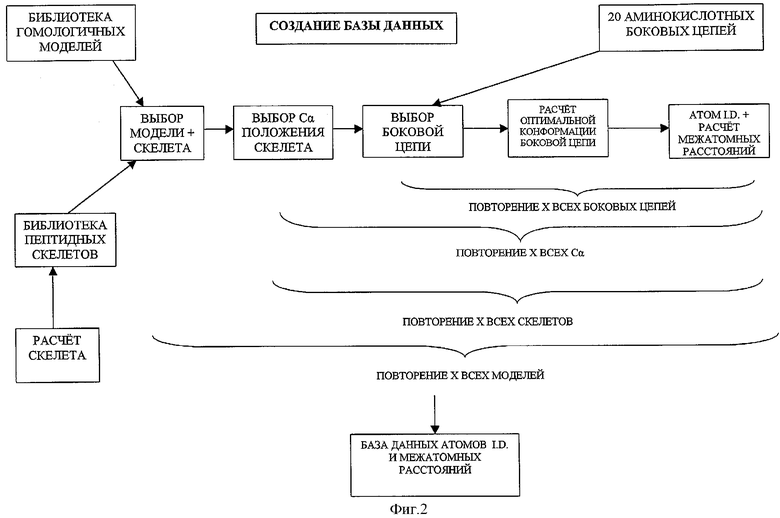



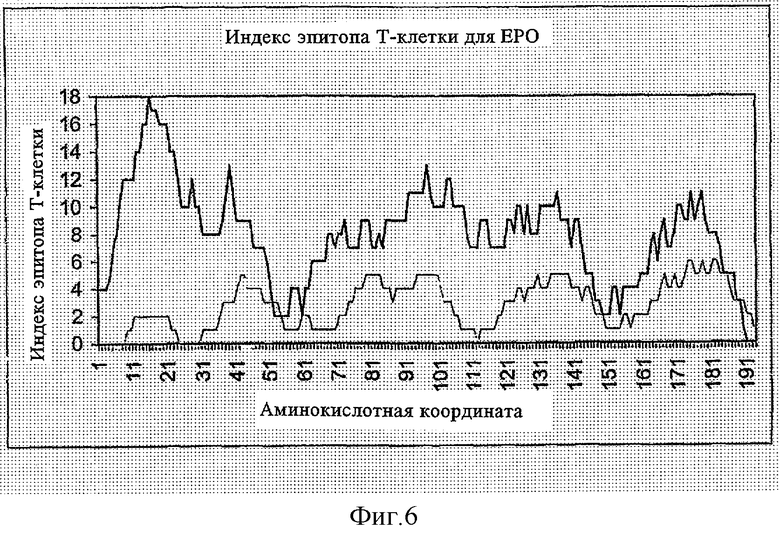
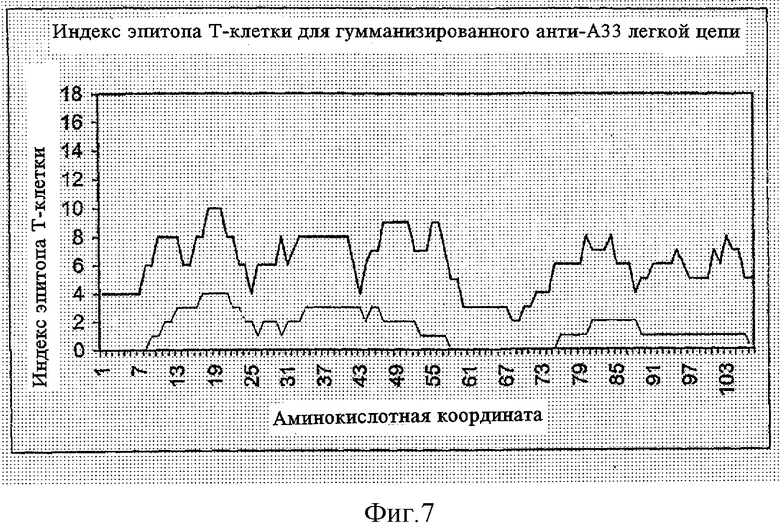

Изобретение относится к биохимии и медицине и касается идентификации эпитопов Т-клеток, которые вызывают иммунную реакцию в живом организме-хозяине. С помощью этого способа могут быть получены биологические соединения, которые неиммунногенны или имеют, по крайней мере, сниженную иммунногенность, и в сравнении с соответствующим немодифицированным объектом путем уменьшения или удаления потенциальных эпитопов Т-клеток в пределах последовательности указанных изначально иммуногенных молекул. Таким образом, изобретение относится также к новым биологическим молекулам, особенно белкам и антителам, которые получены способом согласно изобретению. Изобретение обеспечивает снижение иммуногенности биологических соединений. 5 н. и 12 з.п. ф-лы, 6 табл., 8 ил.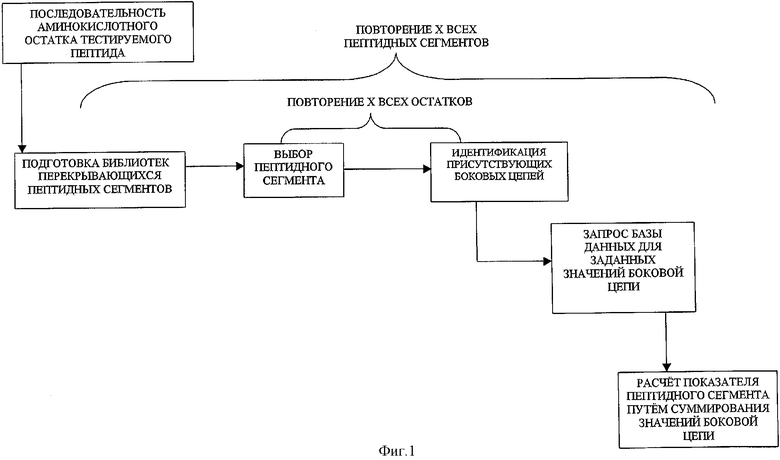
(a) выбор участка пептида, который имеет известную последовательность аминокислотных остатков;
(b) последовательный отбор перекрывающихся сегментов аминокислотных остатков с предварительно определенным одинаковым размером и состоящих, по крайней мере, из трех аминокислотных остатков выбранного участка, причем указанный выбранный сегмент аминокислотных остатков состоит из 13 аминокислотных остатков
(c) подсчет показателя связывания молекулы МНС класса II для каждого указанного выбранного сегмента путем суммирования определенных значений для каждого гидрофобного аминокислотного остатка боковой цепи, который присутствует в указанном выбранном сегменте аминокислотного остатка; и
(d) идентификация, по крайней мере, одного из указанных сегментов, подходящих для модификации, основываясь на подсчитанном показателе связывания молекулы МНС класса II этого сегмента, для изменения общего показателя связывания МНС класса II для пептида без существенного уменьшения терапевтической полезности пептида.
(1) получения компьютерной библиотеки моделей молекулы МНС класса II;
(2) получения компьютерной библиотеки возможной основной цепи пептида для указанных моделей молекул МНС класса II;
(3) отбора модели из указанной первой базы данных;
(4) отбора возможной основной цепи пептида из второй базы данных;
(5) идентификации аминокислотных остатков боковых цепей, которые присутствуют в каждом выбранном сегменте;
(6) определения значения связывающего сродства ко всем боковым цепям, которые присутствуют в каждом выбранном сегменте; и необязательно
(7) повторения стадий от (1) до (5) для каждой указанной модели и каждой указанной боковой цепи.
(i) определение аминокислотной последовательности родительской биологической молекулы или ее части;
(ii) идентификацию одного или более потенциальных эпитопов Т-клеток в пределах аминокислотной последовательности белка любым способом, включающим определение связывания пептидов с молекулами МНС, используя методы in vitro или биологические анализы;
(iii) разработку новых вариантов последовательности путем изменения 1-9 аминокислотных остатков в пределах исходно идентифицированных последовательностей эпитопов Т-клеток, указанные варианты модифицируют таким способом, чтобы существенно уменьшить или устранить активность или количество последовательностей эпитопов Т-клеток и/или количество МНС аллотипов, способных связывать пептиды, полученные от указанной биологической молекулы, как определено путем связывания пептидов с молекулами МНС при использовании способов in vitro или биологических анализов или связывания пептид-МНС комплексов с Т-клетками;
(iv) конструирование таких вариантов последовательности с помощью техники рекомбинантных ДНК и тестирование указанных вариантов для того, чтобы идентифицировать один или более вариантов с желаемыми свойствами; и
(v) необязательное повторение этапов (ii)-(iv),
отличающийся тем, что идентификация последовательностей эпитопов Т-клеток согласно стадии (ii) достигается способом, как указано в любом из пп.1-7.
(a) моноклинальные антитела:
анти-40kD гликопротеин антиген антитело KS 1/4,
анти-GD2 антитело 14.18,
анти-Нer2 антитело 4D5 (мышиное) и гуманизированная версия (Herceptin®),
анти-IL-2R (анти-Тас) антитело (Zenapax®),
анти-CD52 антитело (САМРАТН®),
анти-CD20 антитела (С2В8, Rituxan®; Bexxar®),
антитело, направленное на С5 комплементный белок человека
(b) белки:
sTNF-R1, sTNF-R2, sTNFR-Fc (Enbrel®), белок С, acrp30, рицин A, CNTFR лиганды,
субтилизин, GM-CSF, фолликулостимулирующий гормон человека (h-fsh), β-гликоцереброзидаза, GLP-1, аполипопротеин А1.
| Способ непосредственного перевода с негатива на литографский камень | 1932 |
|
SU34317A1 |
| СИНТЕТИЧЕСКИЙ ПЕПТИД, СИНТЕТИЧЕСКИЙ ЛИПОПЕПТИД, ИММУНОГЕННЫЙ КОНЪЮГАТ (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ОЛИГОМЕРА, ЗАЩИЩЕННЫЙ ТВЕРДЫМ МОНОМЕТИЛОВЫМ ЭФИРОМ ПОЛИЭТИЛЕНГЛИКОЛЯ ПОЛИСАХАРИД | 1993 |
|
RU2141527C1 |
| АНТИГЕННАЯ КОМПОЗИЦИЯ ДЛЯ ИНДУЦИРОВАНИЯ ОТВЕТА ЦИТОТОКСИЧЕСКИХ Т-ЛИМФОЦИТОВ, СПОСОБ ИНДУЦИРОВАНИЯ И СПОСОБ ЛЕЧЕНИЯ БОЛЕЗНИ | 1992 |
|
RU2129439C1 |

