ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение принадлежит к области биотехнологии и конкретнее относится к мутантам промоторов гена пируваткарбоксилазы и их применениям.
УРОВЕНЬ ТЕХНИКИ
Аминокислоты, включая лизин, треонин, глутаминовую кислоту и т.д., являются основными веществами, составляющими белки, необходимые для питания животных, и широко используются в таких отраслях, как медицина, здравоохранение, пищевая промышленность, корма для животных и косметика. Аминокислоты в основном продуцируются путем микробиологической ферментации. В настоящее время основные штаммы-продуценты включают такие микроорганизмы, как род Enterobacter и род Corynebacterium. Благодаря своему физиологическому превосходству Corynebacterium стал самым важным штаммом-продуцентом в отрасли. С непрерывным развитием биотехнологии сообщения о метаболической инженерии Corynebacterium для улучшения выхода аминокислот при его применении постепенно растут. Эти инженерные модификации включают повышение экспрессии ферментов, связанных с путями синтеза аминокислот, ослабление экспрессии ферментов, связанных с конкурентными путями, и так далее.
Считается, что пируваткарбоксилаза (кодируемая геном pyc) является ферментом, который играет значительную роль в анаплеротическом пути Corynebacterium, и этот фермент способен катализировать образование щавелевоуксусной кислоты из пировиноградной кислоты и углекислого газа, тем самым входя в цикл ЦТК (цикл трикарбоновых кислот). В многочисленных литературных источниках сообщалось, что сверхэкспрессия пируваткарбоксилазы может увеличить выход аминокислот (таких как лизин, глутаминовая кислота и треонин) при использовании штаммов [1-2]. В настоящее время сверхэкспрессия гена pyc в основном достигается за счет увеличения числа его копий. Однако увеличение числа копий может привести к геномной нестабильности штаммов, тогда как промоторы не имеют вышеуказанного недостатка в регуляции экспрессии генов, и высокоэффективные промоторы также могут применяться для экспрессии различных целевых генов. Следовательно, в данной области техники существует необходимость в разработке промоторных элементов с повсеместной активностью для надлежащего повышения экспрессии генов, таких как pyc, тем самым улучшая продукцию целевого соединения штаммом.
Цитируемые источники:
[1] CN110603321A
[2] CN1275167A
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В первом аспекте настоящего изобретения предложен мутант промотора гена пируваткарбоксилазы, где указанный мутант (i) имеет один или более мутированных нуклеотидов в коровой области, соответствующей положениям с 279 по 317 промотора, имеющего нуклеотидную последовательность, представленную в SEQ ID NO: 21.
Термин «мутант» в контексте настоящего документа относится к полинуклеотиду или полипептиду, который относительно полинуклеотида или полипептида «дикого типа» или «сравнительного» полинуклеотида или полипептида содержит изменение (изменения) (т. е. замену, инсерцию и/или делецию полинуклеотида) в одном или более (например, нескольких) положениях, где замена относится к замене другим нуклеотидом нуклеотида, который занимает одно положение; делеция относится к удалению нуклеотида, который занимает определенное положение; и инсерция относится к добавлению нуклеотида после нуклеотида, прилегающего к и следующего сразу за занятым положением.
В частности, мутант промотора гена пируваткарбоксилазы Corynebacterium glutamicum имеет мутированный нуклеотид(ы) в положениях 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38 или 39 в нуклеотидной последовательности, соответствующей положениям с 279 по 317 нуклеотидной последовательности, представленной в SEQ ID NO: 21.
В некоторых вариантах осуществления мутант содержит замененный нуклеотид. Замена представляет собой мутацию, вызванную заменой одного основания в нуклеотиде другим, отличающимся, основанием, которую также называют заменой основания или точечной мутацией.
В некоторых вариантах осуществления мутант (ii) содержит последовательность, обратно комплементарную нуклеотидной последовательности, представленной в (i).
В некоторых вариантах осуществления мутант (iii) содержит последовательность, обратно комплементарную последовательности, способной к гибридизации с нуклеотидной последовательностью, представленной в (i) или (ii), в высокожестких условиях гибридизации или очень высокожестких условиях гибридизации.
В некоторых вариантах осуществления мутант (iv) содержит нуклеотидную последовательность, обладающую по меньшей мере 90% идентичностью последовательности с нуклеотидной последовательностью, представленной в (i) или (ii). В частности, мутант содержит последовательность, обладающую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с нуклеотидной последовательностью, представленной в (i) или (ii).
Нуклеотидная последовательность мутанта, представленного в любом из (i) - (iv), не является CGATGTTTGATTGGGGGAATCGGGGGTTACGATACTAGG в положениях, соответствующих положениям с 279 по 317 последовательности, представленной в SEQ ID NO: 21; и предпочтительно мутант, представленный в любом из (i) - (iv), обладает повышенной промоторной активностью по сравнению с промотором гена пируваткарбоксилазы, имеющим последовательность, представленную в SEQ ID NO: 21.
В некоторых вариантах осуществления мутант промотора гена пируваткарбоксилазы согласно настоящему изобретению имеет следующую нуклеотидную последовательность, соответствующую положениям с 279 по 317 нуклеотидной последовательности, представленной в SEQ ID NO: 21: NNNNNNTTGATTNNNNNNNNNNNNNNNTANNATNNNNNN, где N выбран из A, T, C или G.
Предпочтительно мутант промотора гена пируваткарбоксилазы обладает промоторной активностью, повышенной в от 1 до 17 раз или более по сравнению с промотором гена пируваткарбоксилазы, имеющим последовательность, представленную в SEQ ID NO: 21. В качестве примера, промоторная активность после мутации повышается по меньшей мере в 1,8 раза, предпочтительно в 3 раза, в 5 раз, в 8 раз, предпочтительно в 10 раз, в 11 раз, в 12 раз, в 13 раз, и более предпочтительно в 14 раз, в 15 раз или в 16 раз, или более.
В некоторых вариантах осуществления нуклеотидная последовательность коровой области промотора мутанта согласно настоящему изобретению представляет собой одну из следующих последовательностей:
CTAATTTTGATTCGTACTGATTTCTGCTACGATGAGTCA;
GGATTGTTGATTTGAGCTTGATGAGCGTACAATCAACTT;
TTCTCCTTGATTGCGCCTTAACCGTGGTATGATTCGATA;
ATTGATTTGATTGGAACCTTACTGTGCTATGATTTGGTA;
TCGAGTTTGATTTCACAACGTGTGTGATAGGATATAATA;
TTGCGTTTGATTAAAGTATGCAAGGGCTAGTATGGTGAT;
ATCATTTTGATTCCGGCGCACATGTGGTAATATGGTATT;
TCGCCATTGATTGCCCGCCATCCATGCTATAATCGGAAG;
TTCCGCTTGATTGTGGCCATAGTATGATATTATTAATTA;
CGGATCTTGATTTTATGATGGGTATTGTATAATCTTGGT;
CGGATATTGATTTGGCCGGTGTTGTGGTAGTATCGTGTT;
AGGGGTTTGATTGGCCGCTCGGTGTGTTATCATGGAGAG;
GAGTTGTTGATTTCGTTGGTGCACGTATACAATGGTTTT;
CTTGGCTTGATTTTTGTTTGAGGGTTGTATAATGTTATT;
GACTAGTTGATTTCCGCCCTTGGTTGATATTATGCTTGA;
ATCCGCTTGATTTAGGCGTACGTTTAATAGTATATTGAA;
CGGGGCTTGATTTCCTTGTCGTGGCGTTATTATAATGGA;
ATGGAGTTGATTATACGATACTACAGATACTATACTGGT;
CCGTAGTTGATTGACTTGGGCAGTATATAGTATAATGAA; или
CGGGCCTTGATTGTAAGATAAGACATTTAGTATAATTAG.
В некоторых вариантах осуществления мутант промотора гена пируваткарбоксилазы согласно настоящему изобретению имеет нуклеотидную последовательность, представленную в любой из SEQ ID NO: 1 - SEQ ID NO: 20.
Мутант промотора гена пируваткарбоксилазы представляет собой молекулу, состоящую из нуклеотидов, обладающую промоторной активностью, и обладает повышенной активностью по сравнению с промотором дикого типа. «Промотор» относится к молекуле нуклеиновой кислоты, которая обычно расположена против хода транскрипции от кодирующей последовательности представляющего интерес гена, обеспечивая сайт распознавания для РНК-полимеразы, и расположена против хода транскрипции от 5’ -направления сайта инициации транскрипции мРНК. Она представляет собой нетранслируемую последовательность нуклеиновой кислоты, с которой связывается РНК-полимераза, чтобы инициировать транскрипцию представляющего интерес гена.
Ген, кодирующий пируваткарбоксилазу, представляет собой ген pyc, который, как полагают, является ферментом, который играет значительную роль в анаплеротическом пути Corynebacterium, и этот фермент способен катализировать образование щавелевоуксусной кислоты из пировиноградной кислоты и углекислого газа. Это один из основных анаплеротических путей цикла трикарбоновых кислот. В многочисленных литературных источниках сообщалось, что сверхэкспрессия пируваткарбоксилазы Pyc может увеличить выход аминокислот (таких как лизин, глутаминовая кислота и треонин) при использовании штаммов.
«Коровая область промотора» относится к последовательности нуклеиновой кислоты, расположенной на промоторе у прокариота, которая представляет собой коровую область последовательности, в которой осуществляется функция промотора, главным образом включающую область -35, область -10, область между областью -35 и областью -10 и сайт инициации транскрипции, область -35 представляет собой сайт распознавания РНК-полимеразы, область -10 представляет собой сайт связывания РНК-полимеразы.
Мутант промотора согласно настоящему изобретению представляет собой улучшенный промотор гена пируваткарбоксилазы Corynebacterium glutamicum, который обладает более высокой промоторной активностью, чем у промотора дикого типа, например, увеличенной по меньшей мере в 1,8 раза, в 3 раза, в 5 раз, в 8 раз, предпочтительно в 10 раз, в 11 раз, в 12 раз, в 13 раз, более предпочтительно в 14 раз или более, в 15 раз или более, в 16 раз или более.
Молекула нуклеиновой кислоты промотора согласно настоящему изобретению может быть выделена или получена с помощью стандартной технологии молекулярной биологии. Например, молекула нуклеиновой кислоты промотора согласно настоящему изобретению может быть выделена с помощью ПЦР с использованием подходящих последовательностей праймеров. Кроме того, молекула нуклеиновой кислоты промотора согласно настоящему изобретению может быть получена стандартным методом синтеза с использованием автоматического синтезатора ДНК.
Молекулы нуклеиновой кислоты с промоторной активностью согласно настоящему изобретению могут применяться для экспрессии другого гена в Corynebacterium или Enterobacterium, включая, не ограничиваясь перечисленным, гены, связанные с синтезом аминокислот, например, пируваткарбоксилазу Pyc, глутаматдегидрогеназу Gdh, аспартаткиназу LysC, треониновый оперон ThrABC, ген аспартат-полуальдегиддегидрогеназы Asd, аспартат-аммоний-лиазу AspB, гомосериндегидрогеназу Hom, гомосерин-O-ацетилтрансферазу MetX, дигидродипиколинатсинтазу DapA, дигидродипиколинатредуктазу DapB, мезо-диаминопимелатдегидрогеназу Ddh, глутаматкиназу ProB, глутамат-5-полуальдегиддегидрогеназу ProA, пирролин-5-карбоксилатдегидрогеназу ProC, пролиндегидрогеназу/пирролин-5-карбоксилатдегидрогеназу PutA, синтазу 5-аминолевулиновой кислоты HemA, фосфоенолпируваткарбоксилазу Ppc, белок-транспортер аминокислот, белок, ассоциированный с системой ptsG, пируватдегидрогеназу AceE, глицеральдегид-3-фосфатдегидрогеназу GapN или лизиндекарбоксилазу CadA или LdcC, и т. д.
Сильные промоторные элементы согласно настоящему изобретению могут применяться для умеренной регуляции экспрессии целевых генов и достижения высокоэффективной продукции целевых продуктов.
Термин «Corynebacterium» в контексте настоящего документа относится к микроорганизмам рода Corynebacterium, включая, не ограничиваясь перечисленным, Corynebacterium glutamicum ATCC 13032, Corynebacterium glutamicum ATCC 13869, Corynebacterium glutamicum B253, Corynebacterium glutamicum ATCC 14067 и производные штаммы, которые продуцируют аминокислоты и получены из штаммов, описанных выше.
Во втором аспекте настоящего изобретения предложены экспрессионная кассета и рекомбинантный вектор, содержащий мутант промотора гена пируваткарбоксилазы.
«Экспрессионная кассета» имеет значение, обычно понимаемое специалистом в данной области техники, т. е. означает элемент, содержащий промотор и представляющий интерес ген, и способна экспрессировать представляющий интерес ген.
Термин «вектор» относится к конструкции ДНК, содержащей последовательности ДНК, функционально лигированные с соответствующими регуляторными последовательностями, для обеспечения экспрессии представляющего интерес гена в подходящем хозяине. Вектор, используемый в настоящем документе, не ограничен каким-либо особым образом и может представлять собой любой вектор, известный в данной области техники, при условии, что он способен реплицироваться в организме-хозяине. То есть вектор включает, не ограничиваясь перечисленным, плазмиду и фаг, например, плазмиду pEC-XK99E, используемую в конкретных примерах настоящего изобретения. После трансформации в подходящего хозяина вектор может реплицироваться и функционировать независимо от генома хозяина или, в некоторых случаях, непосредственно интегрироваться в геном.
В третьем аспекте настоящего изобретения предложена рекомбинантная клетка-хозяин, содержащая мутант промотора гена пируваткарбоксилазы, экспрессионную кассету или рекомбинантный вектор.
Рекомбинантную клетку-хозяина, в частности, получают, например, трансформацией. Термин «трансформация», используемый в настоящем документе, имеет значение, обычно понимаемое специалистом в данной области техники, т. е. означает способ введения экзогенной ДНК в хозяина. Способы трансформации включают любой способ введения нуклеиновой кислоты в клетку. Эти способы включают, не ограничиваясь перечисленным, электропорацию, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), микроинъекцию, метод полиэтиленгликоля (ПЭГ), метод ДЭАЭ-декстрана, метод катионной липосомы и метод ацетата лития-ДМСО.
«Клетка-хозяин» в контексте настоящего документа имеет значение, обычно понимаемое специалистом в данной области техники, т. е. означает клетку, в которую может быть введена нуклеиновая кислота, обладающая промоторной активностью, согласно настоящему изобретению, и которая после введения называется рекомбинантной клеткой-хозяином. Другими словами, в настоящем изобретении может быть использована любая клетка-хозяин, при условии, что она содержит нуклеиновую кислоту, обладающую промоторной активностью, согласно настоящему изобретению, и функционально лигирована с геном для опосредования транскрипции этого гена. Клетка-хозяин согласно настоящему изобретению может представлять собой прокариотическую клетку или эукариотическую клетку. В некоторых вариантах осуществления клетка-хозяин согласно настоящему изобретению может представлять собой штамм любого типа, способный продуцировать целевой продукт, включая штаммы дикого типа и рекомбинантные штаммы. В качестве примера, клетку-хозяина получают из микроорганизмов, подходящих для ферментативного продуцирования целевых продуктов, таких как аминокислоты и органические кислоты, например, Enterobacterium, Corynebacterium, Brevibacterium, Arthrobacterium и Microbacterium, и т. д.
В некоторых предпочтительных вариантах осуществления клетка-хозяин представляет собой Enterobacterium или Corynebacterium, более предпочтительно Corynebacterium glutamicum, включая, не ограничиваясь перечисленным, Corynebacterium glutamicum ATCC 13032, Corynebacterium glutamicum ATCC 13869, Corynebacterium glutamicum B253, Corynebacterium glutamicum ATCC 14067 и производные штаммы, продуцирующие L-аминокислоты, полученные из штаммов, описанных выше.
В одном конкретном варианте осуществления Corynebacterium glutamicum подвергают следующей модификации: последовательность, кодирующую мутацию T311I, вводят в ген, кодирующий аспартаткиназу у Corynebacterium glutamicum, и вводят ген, кодирующий пируваткарбоксилазу, функционально лигированный с мутантом промотора гена пируваткарбоксилазы, описанным выше, для получения лизин-продуцирующего штамма; или экспрессионные кассеты глутаматкиназы, глутамат-5-полуальдегиддегидрогеназы и/или пирролин-5-карбоксилатдегидрогеназы, в которые введена мутация G149K, функционально лигированные с мутантом промотора гена пируваткарбоксилазы согласно настоящему изобретению, интегрируют в ген пролиндегидрогеназы/пирролин-5-карбоксилатдегидрогеназы с получением пролин-продуцирующего штамма; предпочтительно последовательность коровой области мутанта промотора гена пируваткарбоксилазы представляет собой CGGGCCTTGATTGTAAGATAAGACATTTAGTATAATTAG, более предпочтительно последовательность полноразмерного промотора представлена в SEQ ID NO: 20.
В других вариантах осуществления клетка-хозяин может также представлять собой другие типы штаммов, продуцирующих аминокислоты. «Штамм, продуцирующий аминокислоты», описанный в настоящем документе, относится к штамму, который может продуцировать аминокислоты при культивировании бактерий в среде, и может накапливать аминокислоты, или может секретировать аминокислоты в среду, т. е. могут быть получены внеклеточные свободные аминокислоты. Например, штаммы могут представлять собой встречающиеся в природе штаммы, продуцирующие аминокислоты, или сконструированные штаммы, продуцирующие аминокислоты, полученные путем генетических модификаций.
В качестве примера, клетка-хозяин представляет собой лизин-продуцирующую клетку-хозяина. В некоторых вариантах осуществления лизин-продуцирующие клетки-хозяева могут дополнительно включать, не ограничиваясь перечисленным, один или более генов, выбранных из одного или более из следующих, экспрессия которых ослаблена или снижена:
a. ген adhE, кодирующий алкогольдегидрогеназу;
b. ген ackA, кодирующий ацетаткиназу;
c. ген pta, кодирующий фосфатацетилтрансферазу;
d. ген ldhA, кодирующий лактатдегидрогеназу;
e. ген focA, кодирующий транспортер формиата;
f. ген pflB, кодирующий пируватформиатлиазу;
g. ген poxB, кодирующий пируватоксидазу;
h. ген thrA, кодирующий бифункциональный фермент аспартаткиназа I/гомосериндегидрогеназа I;
i. ген thB, кодирующий гомосеринкиназу;
j. ген ldcC, кодирующий лизиндекарбоксилазу; и
h. ген cadA, кодирующий лизиндекарбоксилазу.
В некоторых вариантах осуществления лизин-продуцирующие клетки-хозяева могут дополнительно включать, не ограничиваясь перечисленным, один или более генов, выбранных из одного или более из следующих, которые характеризуются повышенной экспрессией или сверхэкспрессируются:
a. ген dapA, кодирующий дигидродипиридинсинтазу, которая ослабляет ингибирование по типу обратной связи лизином;
b. ген dapB, кодирующий дигидродипиколинатредуктазу;
c. ген ddh, кодирующий диаминопимелатдегидрогеназу;
d. dapD, кодирующий тетрагидродипиколинатсукцинилазу, и dapE, кодирующий сукцинилдиаминопимелатдеацилазу;
e. ген asd, кодирующий аспартат-полуальдегиддегидрогеназу;
f. ген ppc, кодирующий фосфоенолпируваткарбоксилазу;
g. ген pntAB, кодирующий ниацинамидадениндинуклеотид-трансгидрогеназу; и
i. ген lysE, кодирующий белок-транспортер лизина.
В качестве примера, клетки-хозяева представляют собой треонин-продуцирующие клетки-хозяева. В некоторых вариантах осуществления треонин-продуцирующие клетки-хозяева могут представлять собой штаммы, экспрессирующие аспартаткиназу LysC, которая ослабляет ингибирование по типу обратной связи, на основе Corynebacterium glutamicum ATCC 13032. В других вариантах осуществления треонин-продуцирующие клетки-хозяева могут также представлять собой другие типы штаммов, обладающие способностью продуцировать треонин.
В некоторых вариантах осуществления один или более генов, выбранных из группы, состоящей из следующих генов в треонин-продуцирующих клетках-хозяевах, характеризуются повышенной экспрессией или сверхэкспрессируются:
a. ген thrABC, кодирующий треониновый оперон;
b. ген hom, кодирующий гомосериндегидрогеназу, которая ослабляет ингибирование по типу обратной связи;
c. ген GAP, кодирующий глицеральдегид-3-фосфатдегидрогеназу;
d. ген pyc, кодирующий пируваткарбоксилазу;
e. ген mqo, кодирующий малат : хинон-оксидоредуктазу;
f. ген tkt, кодирующий транскетолазу;
g. ген gnd, кодирующий 6-фосфоглюконатдегидрогеназу;
h. ген thE, кодирующий экспортер треонина; и
i. ген eno, кодирующий енолазу.
В качестве примера, клетки-хозяева представляют собой изолейцин-продуцирующие клетки-хозяева. В некоторых вариантах осуществления изолейцин-продуцирующие клетки-хозяева представляют собой штаммы, которые продуцируют L-изолейцин путем замены аминокислоты в положении 323 гена ilvA в L-треониндегидратазе аланином. В других вариантах осуществления изолейцин-продуцирующие клетки-хозяева могут также представлять собой другие типы штаммов, обладающие способностью продуцировать изолейцин.
В качестве примера, клетки-хозяева представляют собой O-ацетилгомосерин-продуцирующие клетки-хозяева. В некоторых вариантах осуществления O-ацетилгомосерин-продуцирующие клетки-хозяева представляют собой штаммы, которые продуцируют O-ацетилгомосерин путем инактивации O-ацетилгомосерин(тиол)-лиазы. В других вариантах осуществления O-ацетилгомосерин-продуцирующие клетки-хозяева также могут представлять собой другие типы штаммов, обладающие способностью продуцировать O-ацетилгомосерин.
В качестве примера, клетки-хозяева представляют собой метионин-продуцирующие клетки-хозяева. В некоторых вариантах осуществления метионин-продуцирующие клетки-хозяева представляют собой штаммы, которые продуцируют метионин путем инактивации регуляторов транскрипции метионина и цистеина. В других вариантах осуществления метионин-продуцирующие клетки-хозяева могут также представлять собой другие типы штаммов, обладающие способностью продуцировать метионин.
В настоящем описании клетки-хозяева могут быть культивированы обычными методами, известными в данной области техники, включая, не ограничиваясь перечисленным, культуру в луночных планшетах, культуру при встряхивании, периодическую культуру, непрерывную культуру, культуру с подпиткой и т. д. Кроме того, различные условия культивирования, такие как температура, время и значение pH среды, могут быть надлежащим образом скорректированы в соответствии с фактическими ситуациями.
В четвертом аспекте настоящего изобретения предложено применение мутанта промотора гена пируваткарбоксилазы согласно первому аспекту, экспрессионной кассеты или экспрессионного вектора согласно второму аспекту или рекомбинантной клетки-хозяина согласно третьему аспекту для по меньшей мере одного из следующих (a) - (c):
(a) повышение уровня транскрипции гена или получение реагента или набора для повышения уровня транскрипции гена;
(b) получение белка или получение реагента или набора для применения при получении белка; и
(c) продуцирование целевого продукта или получение реагента или набора для применения при продуцировании целевого продукта.
В некоторых вариантах осуществления белок представляет собой белок, связанный с синтезом целевого продукта, белок, связанный с мембранным транспортом, или белок, регулирующий экспрессию гена.
В некоторых вариантах осуществления целевой продукт может быть выбран из по меньшей мере одной аминокислоты и ее производных или может быть выбран из других видов соединений, которые могут быть биосинтетически доступны в данной области техники.
В качестве примера, аминокислоты и их производные включают, не ограничиваясь перечисленным, одну или комбинацию двух или более из следующих: пролин, гидроксипролин, лизин, глутаминовая кислота, аргинин, орнитин, глутамин, треонин, глицин, аланин, валин, лейцин, изолейцин, серин, цистеин, метионин, аспарагиновая кислота, аспарагин, гистидин, фенилаланин, тирозин, триптофан, 5-аминолевулиновая кислота, 5-аминовалериановая кислота или производных любой из аминокислот, описанных выше.
В пятом аспекте настоящего изобретения предложен способ повышения экспрессии представляющего интерес гена, включающий функциональное лигирование мутанта промотора гена пируваткарбоксилазы с представляющим интерес геном, предпочтительно повышения экспрессии гена, связанного с синтезом целевого продукта с использованием щавелевоуксусной кислоты в качестве предшественника.
Термин «функционально лигированный» в контексте настоящего документа означает, что мутант промотора гена пируваткарбоксилазы согласно настоящему изобретению функционально лигирован с кодирующим геном для инициирования и опосредования транскрипции гена, что указывает на то, что мутант промотора гена пируваткарбоксилазы согласно настоящему изобретению функционально лигирован с кодирующим геном для контроля транскрипционной активности гена оперона. Способы функционального лигирования могут быть любыми способами, описанными в данной области техники. В случае применения мутанта промотора гена пируваткарбоксилазы согласно настоящему изобретению способ повышения экспрессии целевого гена согласно настоящему изобретению включает способы, обычно применяемые специалистом в данной области техники.
Например, кодирующий ген включает, не ограничиваясь перечисленным, кодирующий ген белка, связанного с синтезом целевого продукта, кодирующий ген белка, связанного с мембранным транспортом, или кодирующий ген белка, регулирующего экспрессию гена.
В некоторых вариантах осуществления мутант промотора гена пируваткарбоксилазы применяют для экспрессии гена, связанного с синтезом аминокислот. Например, мутант промотора гена пируваткарбоксилазы может применяться для экспрессии других генов в Corynebacterium или Enterobacterium, включая, не ограничиваясь перечисленным, гены, связанные с синтезом аминокислот, например, глутаматдегидрогеназу Gdh, ген пируваткарбоксилазы Pyc, аспартаткиназу LysC, треониновый оперон ThrABC, аспартат-полуальдегиддегидрогеназу Asd, аспартат-аммоний-лиазу AspB, гомосериндегидрогеназу Hom, гомосерин-O-ацетилтрансферазу MetX, дигидродипиколинатсинтазу DapA, дигидродипиколинатредуктазу DapB, мезо-диаминопимелатдегидрогеназу Ddh, глутаматкиназу ProB, глутамат-5-полуальдегиддегидрогеназу ProA, пирролин-5-карбоксилатдегидрогеназу ProC, пролиндегидрогеназу/пирролин-5-карбоксилатдегидрогеназу PutA, глутамил-тРНК-редуктазу HemA, фосфоенолпируваткарбоксилазу Ppc, белок-транспортер аминокислот, белок, ассоциированный с системой ptsG, пируватдегидрогеназу AceE, глицеральдегид-3-фосфатдегидрогеназу GapN или лизиндекарбоксилазу CadA или LdcC, и т. д.
В других вариантах осуществления мутант промотора гена пируваткарбоксилазы применяют для экспрессии гена, связанного с синтезом органических кислот, например, белков, связанных с синтезом лимонной кислоты, или генов для кодирования белков, связанных с синтезом янтарной кислоты.
В шестом аспекте настоящего изобретения предложен способ получения белка, включающий стадию экспрессии белка с помощью экспрессионной кассеты или экспрессионного вектора согласно второму аспекту или рекомбинантной клетки-хозяина согласно третьему аспекту; необязательно, белок представляет собой белок, связанный с синтезом целевого продукта, белок, связанный с мембранным транспортом, или белок, регулирующий экспрессию гена; и
необязательно, способ дополнительно включает стадию выделения или очистки белка.
В седьмом аспекте настоящего изобретения предложен способ получения целевого продукта, включающий культивирование клетки-хозяина, содержащей мутант промотора гена пируваткарбоксилазы согласно первому аспекту, который функционально лигирован с геном, связанным с синтезом целевого продукта, и сбор полученного целевого продукта.
Кроме того, в настоящем изобретении предложен способ получения целевого продукта с щавелевоуксусной кислотой в качестве предшественника, включающий культивирование клетки-хозяина, содержащей мутант промотора гена пируваткарбоксилазы, который функционально лигирован с геном, связанным с синтезом целевого продукта с щавелевоуксусной кислотой в качестве предшественника, и сбор полученного целевого продукта. При этом целевой продукт с щавелевоуксусной кислотой в качестве предшественника включает, не ограничиваясь перечисленным, аминокислоты и их производные. Аминокислоты включают аминокислоты семейства аспарагиновой кислоты (лизин, треонин, изолейцин и метионин), аминокислоты семейства глутаминовой кислоты (глутаминовая кислота, пролин, гидроксипролин, аргинин и глутамин) и 5-аминолевулиновую кислоту, и т. д. Производные включают, не ограничиваясь перечисленным, пентандиамин, глутаровую кислоту и т. д. С помощью способов, раскрытых в настоящем документе, выход целевых продуктов с щавелевоуксусной кислотой в качестве предшественника в штаммах может быть увеличен. Более конкретно, ген, связанный с синтезом целевого продукта с щавелевоуксусной кислотой в качестве предшественника, выбран из группы, состоящей из: гена pyc, гена gdh, гена lysC, гена thrABC, гена asd, гена aspB, гена hom, гена metX, гена dapA, гена dapB, гена ddh, гена proB, гена proA, гена proC, гена putA, гена hemA, гена ppc, гена lysE, гена ptsG, гена ptsI, гена aceE, гена gapN, гена cadA, гена ldcC и т. д.
Способ получения целевого продукта с щавелевоуксусной кислотой в качестве предшественника согласно настоящему изобретению представляет собой способ, обычно используемый специалистом в данной области техники, основанный на применении мутанта промотора гена пируваткарбоксилазы, и при этом способ включает стадию извлечения целевого продукта из клетки или культурального раствора. Способ извлечения целевого продукта из клетки или среды хорошо известен в данной области техники и включает, не ограничиваясь перечисленным, фильтрацию, анионообменную хроматографию, кристаллизацию и ВЭЖХ.
Настоящее изобретение обеспечивает следующие преимущества: молекула нуклеиновой кислоты с повышенной промоторной активностью, предложенная в настоящем документе, проявляет более высокую промоторную активность, чем у дикого типа, и пригодна для экспрессии целевого гена, в частности, экспрессии гена, связанного с продукцией аминокислот. Например, при функциональном лигировании с геном, связанным с синтезом целевого продукта, таким как ген pyc, можно повысить интенсивность экспрессии белков, связанных с синтезом целевого продукта, таких как пируваткарбоксилаза, тем самым повышая эффективность продукции аминокислот рекомбинантными штаммами и их производными, и т. д., поэтому он имеет относительно высокое практическое значение.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
ФИГ. 1. Карта плазмидного вектора pEC-XK99E-Ppyc-rfp.
ФИГ. 2. Флуоресцентный скрининг планшета с мутантами промотора pyc.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения терминов
При использовании в сочетании с термином «включая» в формуле изобретения и/или описании, слово в единственном числе может относиться к «одному» или относиться к «одному или более», «по меньшей мере одному» и «одному или более чем одному».
В контексте формулы и описания изобретения подразумевается, что термин «включающий», «имеющий» или «содержащий» является включительным или неограничивающим и не исключает дополнительные или не указанные элементы или способы и стадии.
Во всем документе заявки термин «приблизительно» означает, что значение включает ошибку или стандартное отклонение, вызванное устройством или способом, используемыми для измерения значения.
Применительно к содержанию, раскрытому в настоящем документе, термин «или» определяется только как альтернативы и «и/или», но термин «или», используемый в настоящем документе, относится к «и/или», если явно не указано, что он относится только к альтернативе или взаимному исключению между альтернативами.
Выбранный/необязательный/предпочтительный «числовой диапазон» при использовании в формуле или описании изобретения включает не только числовые конечные точки на обоих концах диапазона, но и все натуральные числа, попадающие между вышеуказанными числовыми конечными точками в отношении этих числовых конечных точек.
В контексте настоящего документа термин «молекула нуклеиновой кислоты» или «полинуклеотид» относится к полимеру, состоящему из нуклеотидов. Молекула нуклеиновой кислоты или полинуклеотид может быть в форме отдельного фрагмента или может быть составной частью более крупной структуры нуклеотидной последовательности, которая получена из нуклеотидной последовательности, которая была выделена по меньшей мере один раз по количеству или концентрации, и может быть распознана, подвергнута манипуляциям и может быть установлена ее последовательность, а также нуклеотидная последовательность, стандартным молекулярно-биологическим методом (например, с использованием вектора клонирования). Когда нуклеотидная последовательность представлена последовательностью ДНК (т. е. A, T, G, C), она также включает последовательность РНК (т. е. A, U, G, C), где «U» заменяет «T». Другими словами, «полинуклеотид» относится к нуклеотидному полимеру, выбитому из дополнительного нуклеотида (отдельному фрагменту или целому фрагменту), или может быть составной частью или компонентом более крупной нуклеотидной структуры, такой как экспрессионный вектор или полицистронная последовательность. Полинуклеотиды включают последовательности ДНК, РНК и кДНК (комплементарная ДНК).
В контексте настоящего документа термины «целевой ген» и «представляющий интерес ген» могут быть использованы взаимозаменяемо.
В контексте настоящего документа термин «дикий тип» относится к объекту, который встречается в природе. Например, полипептид или полинуклеотидная последовательность, присутствующие в организме, которые могут быть выделены из одного источника в природе и не были намеренно модифицированы человеком в лаборатории, являются встречающимися в природе. В контексте настоящего документа термины «встречающийся в природе» и «дикий тип» являются синонимами.
В контексте настоящего документа термины «идентичность последовательностей» и «процентная идентичность» относятся к проценту нуклеотидов или аминокислот, которые являются одинаковыми (т. е. идентичными) в двух или более полинуклеотидах или полипептидах. Идентичность последовательностей между двумя или более полинуклеотидами или полипептидами может быть определена путем выравнивания нуклеотидных последовательностей полинуклеотидов или аминокислотных последовательностей полипептидов и оценки количества положений, в которых нуклеотидные или аминокислотные остатки в выровненных полинуклеотидах или полипептидах идентичны, и сравнения количества этих положений с количеством положений, в которых нуклеотидные или аминокислотные остатки в выровненных полинуклеотидах или полипептидах отличаются. Полинуклеотиды могут отличаться в одном положении, например, содержащем другой нуклеотид (т. е. замену или мутацию), или делецией нуклеотида (т. е. инсерцией или делецией нуклеотида в одном или двух полинуклеотидах). Полипептиды могут отличаться в одном положении, например, содержащем другую аминокислоту (т. е. замену или мутацию), или делецией аминокислоты (т. е. инсерцией или делецией аминокислоты в одном или двух полипептидах). Идентичность последовательностей может быть рассчитана путем деления количества положений, в которых нуклеотидные или аминокислотные остатки идентичны, на общее количество нуклеотидных или аминокислотных остатков в полинуклеотидах или полипептидах. Например, процент идентичности может быть рассчитан путем деления количества положений, в которых нуклеотидные или аминокислотные остатки идентичны, на общее количество нуклеотидных или аминокислотных остатков в полинуклеотидах или полипептидах и умножения результата на 100.
В некоторых вариантах осуществления две или более последовательностей или подпоследовательностей при сравнении и выравнивании при максимальном соответствии алгоритмом выравнивания последовательностей или путем измерения посредством визуального контроля обладают по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% «идентичностью последовательностей» или «процентной идентичностью» нуклеотидов. В некоторых вариантах осуществления последовательность по существу идентична по всей длине одного или обоих сравниваемых биополимеров (например, полинуклеотидов).
В контексте настоящего документа термин «комплементарный» относится к гибридизации или спариванию оснований между нуклеотидами и нуклеотидами, например, между двумя цепями двухцепочечной молекулы ДНК или между олигонуклеотидным праймером и сайтом связывания праймера на одноцепочечном нуклеотиде, подлежащем секвенированию или амплификации.
В контексте настоящего документа термин «высокожесткие условия» означает, что после стандартных процедур блоттинга ДНК зонд длиной по меньшей мере 100 нуклеотидов предварительно гибридизуется или гибридизуется в течение 12-24 часов при 42 °C в 5X SSPE (солевой фосфат натрия-ЭДТА), 0,3% SDS (додецилсульфат натрия), 200 мкг/мл расщепленной и денатурированной ДНК сперматозоида лосося и 50% формамида. Наконец, векторный материал промывают три раза при 65 °С с 2X SSC и 0,2% SDS, каждый раз в течение 15 мин.
В контексте настоящего документа термин «очень высокожесткие условия» означает, что после стандартных процедур блоттинга ДНК зонд длиной по меньшей мере 100 нуклеотидов предварительно гибридизуется или гибридизуется в течение 12-24 часов при 42 °C в 5X SSPE (солевой фосфат натрия-ЭДТА), 0,3% SDS, 200 мкг/мл расщепленной и денатурированной ДНК сперматозоида лосося и 50% формамида. Наконец, векторный материал промывают три раза при 70 °С с 2X SSC и 0,2% SDS, каждый раз в течение 15 мин.
Если не указано иное, все технические и научные термины, используемые в настоящем документе, имеют такие же значения, которые обычно подразумеваются специалистом в области техники, к которой относится настоящее изобретение. Хотя любые методы и материалы, аналогичные или эквивалентные тем, которые описаны в настоящем документе, могут быть использованы для реализации или тестирования настоящего изобретения, методы и материалы, описанные в настоящем документе, являются предпочтительными.
Настоящее изобретение будет дополнительно проиллюстрировано со ссылкой на конкретные примеры. Следует иметь в виду, что эти примеры предназначены только для иллюстрации настоящего изобретения и не предназначены для ограничения объема настоящего изобретения. Экспериментальные методики и экспериментальные методы, используемые в примерах, если не указано иное, представляют собой все обычные методики и методы, например, экспериментальные методы, для которых в следующих примерах не указаны конкретные условия, обычно проводили в соответствии с обычными условиями, такими как условия, описанные Sambrook et al., Molecular Cloning: A Laboratory Manual (New York: Cold Spring Harbor Laboratory Press, 1989), или условиями, которые рекомендованы производителями. Все материалы, реагенты и т. д., использованные в примерах, если не указано иное, являются коммерчески доступными.
Пример 1. Конструирование плазмиды, характеризующей силу промотора гена pyc Corynebacterium glutamicum
Чтобы охарактеризовать силу промотора гена pyc Corynebacterium glutamicum, в настоящем изобретении сначала сконструировали характеризующий вектор, и на основе каркаса плазмиды pEC-XK99E 60 аминокислот на N-конце гена pyc, C-пептид и ген красного флуоресцентного белка были экспрессированы промотором гена pyc. На основе раскрытой геномной последовательности и информации об аннотации гена pyc Corynebacterium glutamicum ATCC13032 были сконструированы праймеры pyc-F/R и был получен фрагмент ДНК размером 180 п. о. промотора гена pyc и N-конца путем ПЦР-амплификации с использованием генома ATCC13032 в качестве матрицы. Плазмиду pEC-XK99E-rfp[3], описанную в литературе, использовали в качестве матрицы, а pEC-F/R - в качестве праймеров для амплификации фрагмента ДНК каркаса плазмиды pEC-XK99E, С-пептида и гена красного флуоресцентного белка. Два вышеуказанных фрагмента лигировали с помощью набора для одностадийного клонирования от Vazyme (Vazyme’s One Step Cloning Kit) с получением характеризующего вектора pEC-XK99E-Ppyc-rfp. Карта плазмидного вектора была такой, как показано на ФИГ. 1. Последовательности праймеров, как использовано выше, были перечислены в таблице 1.
Пример 2. Скрининг и характеристика силы мутантов промотора гена pyc Corynebacterium glutamicum
(1) Конструирование библиотеки мутантов промотора гена pyc Corynebacterium glutamicum
Согласно настоящему изобретению были выполнены мутации коровой области промотора гена pyc Corynebacterium glutamicum:
«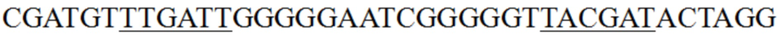 », где подчеркнутые части были основными последовательностями области -35 и области -10 промотора. Согласно настоящему изобретению были выполнены мутации соответствующих положений вышеуказанной коровой области с получением «
», где подчеркнутые части были основными последовательностями области -35 и области -10 промотора. Согласно настоящему изобретению были выполнены мутации соответствующих положений вышеуказанной коровой области с получением «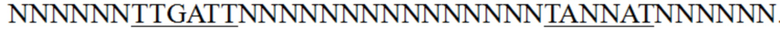 ». Два фрагмента плазмиды амплифицировали с использованием праймеров pyc-M1/ M2 и pyc-M3/M4 соответственно и лигировали с помощью набора для одностадийного клонирования от Vazyme. Все полученные клонированные бактерии собирали и экстрагировали плазмиды с получением библиотеки мутантов промотора гена pyc. Вышеуказанную библиотеку и контроль дикого типа pEC-XK99E-Ppyc-rfp, полученный в примере 1, трансформировали в Corynebacterium glutamicum ATCC13032 соответственно и наносили на планшет TSB. Планшет, на котором были выращены сотни клонов, был сфотографирован для определения флуоресценции с помощью системы флуоресцентной визуализации. Мутанты с улучшенной интенсивностью экспрессии предварительно подвергали скринингу в соответствии с яркостью флуоресценции клонов. Ингредиенты (г/л) среды планшета TSB были следующими: глюкоза, 5 г/л; сухие дрожжи, 5 г/л; соевый пептон, 9 г/л; мочевина, 3 г/л; янтарная кислота, 0,5 г/л; K2HPO4·3H2O, 1 г/л; MgSO4·7H2O, 0,1 г/л; биотин, 0,01 мг/л; витамин B1, 0,1 мг/л; MOPS (3-морфолинопропансульфоновая кислота), 20 г/л; и порошок агара, 15 г/л. Согласно настоящему изобретению был выполнен первоначальный скрининг на более чем 10000 клонов, и, как показано на ФИГ. 2, было получено приблизительно 30 мутантов с повышенной интенсивностью флуоресценции. Последовательности праймеров, использованных выше, были перечислены в таблице 2.
». Два фрагмента плазмиды амплифицировали с использованием праймеров pyc-M1/ M2 и pyc-M3/M4 соответственно и лигировали с помощью набора для одностадийного клонирования от Vazyme. Все полученные клонированные бактерии собирали и экстрагировали плазмиды с получением библиотеки мутантов промотора гена pyc. Вышеуказанную библиотеку и контроль дикого типа pEC-XK99E-Ppyc-rfp, полученный в примере 1, трансформировали в Corynebacterium glutamicum ATCC13032 соответственно и наносили на планшет TSB. Планшет, на котором были выращены сотни клонов, был сфотографирован для определения флуоресценции с помощью системы флуоресцентной визуализации. Мутанты с улучшенной интенсивностью экспрессии предварительно подвергали скринингу в соответствии с яркостью флуоресценции клонов. Ингредиенты (г/л) среды планшета TSB были следующими: глюкоза, 5 г/л; сухие дрожжи, 5 г/л; соевый пептон, 9 г/л; мочевина, 3 г/л; янтарная кислота, 0,5 г/л; K2HPO4·3H2O, 1 г/л; MgSO4·7H2O, 0,1 г/л; биотин, 0,01 мг/л; витамин B1, 0,1 мг/л; MOPS (3-морфолинопропансульфоновая кислота), 20 г/л; и порошок агара, 15 г/л. Согласно настоящему изобретению был выполнен первоначальный скрининг на более чем 10000 клонов, и, как показано на ФИГ. 2, было получено приблизительно 30 мутантов с повышенной интенсивностью флуоресценции. Последовательности праймеров, использованных выше, были перечислены в таблице 2.
(2) Скрининг библиотеки мутантов промотора гена pyc Corynebacterium glutamicum
Все мутанты с повышенной интенсивностью флуоресценции, наблюдаемой в планшете, как описано выше, культивировали в 96-луночном планшете, чтобы охарактеризовать силу промоторов. Ингредиенты (г/л) жидкой среды TSB были следующими: глюкоза, 5 г/л; сухие дрожжи, 5 г/л; соевый пептон, 9 г/л; мочевина, 3 г/л; янтарная кислота, 0,5 г/л; K2HPO4·3H2O, 1 г/л;MgSO4·7H2O, 0,1 г/л; биотин, 0,01 мг/л; витамин B1, 0,1 мг/л; и MOPS, 20 г/л. Штаммы, полученные из планшета, инокулировали зубочистками в 96-луночный планшет, содержащий 200 мкл жидкой среды TSB в каждой лунке. Для каждого штамма отбирали по три образца. Скорость вращения шейкера для планшетов составляла 800 об/мин. После культивирования при 30 °C в течение 24 часов определяли интенсивность флуоресценции штаммов и секвенировали штаммы с улучшенной интенсивностью флуоресценции по сравнению с контролем дикого типа. Результаты приведены в таблице 3. Некоторые из мутантов промотора имеют одинаковую последовательность. Наконец, согласно настоящему изобретению было успешно получило 20 различных мутантов промотора (нуклеотидные последовательности соответствующих мутированных промоторов были пронумерованы как SEQ ID NO: 1-20) с улучшенной интенсивностью экспрессии по сравнению с промотором дикого типа, и диапазон кратности увеличения составлял от 1,8 до 16,1 раза, что могло обеспечить элементы с повсеместной активностью для модификации экспрессии генов, таких как pyc.
Пример 3. Применение мутантов промотора гена pyc Corynebacterium glutamicum для получения целевых продуктов
(1) Конструирование рекомбинантного вектора мутанта промотора гена pyc Corynebacterium glutamicum
Основываясь на известной геномной последовательности Corynebacterium glutamicum ATCC13032, восходящие и нисходящие гомологичные плечи мутантов промотора Ppyc-1, Ppyc-9, Ppyc-16 и Ppyc-20 подвергали ПЦР-амплификации с использованием генома ATCC13032 в качестве матрицы и с использованием pyc-UF/ pyc-UR1 и pyc-DF1/ pyc-DR, pyc-UF/ pyc-UR9 и pyc-DF9/ pyc-DR, pyc-UF/ pyc-UR16 и pyc-DF16/ pyc-DR, и pyc-UF/ pyc-UR20 и pyc-DF20/ pyc-DR в качестве праймеров, соответственно; и в то же время каркас pK18mobsacB амплифицировали с использованием pK18-1/2 в качестве праймеров. Вышеуказанные ПЦР-фрагменты выделяли и затем лигировали с помощью набора для одностадийного клонирования от Vazyme, и получали рекомбинантные векторы pK18- Ppyc-1, pK18- Ppyc-9, pK18- Ppyc-16 и pK18- Ppyc-20 с мутированными промоторами, соответственно. Последовательности праймеров, использованных выше, были перечислены в таблице 4.
(2) Конструирование мутантов промотора гена pyc лизин-продуцирующего штамма Corynebacterium glutamicum
Рекомбинантные векторы pK18- Ppyc-1, pK18- Ppyc-9, pK18- Ppyc-16 и pK18- Ppyc-20, как сконструировано выше, трансформировали в лизин-продуцирующий штамм SCgL30 (в котором Thr в положении 311 аспартаткиназы Corynebacterium glutamicum ATCC13032 был мутирован в Ile [4]) Corynebacterium glutamicum, соответственно. Штамм наносили на твердую среду LBHIS, содержащую 5 г/л глюкозы и 25 мкг/мл канамицина, и культивировали при 30 °С с получением первых рекомбинантных трансформантов. Правильные первые рекомбинантные трансформанты инокулировали в среду LB, содержащую 5 г/л глюкозы соответственно, и культивировали в течение ночи. Затем 1% культуры переносили в среду LB, содержащую 100 г/л сахарозы, и культивировали в течение ночи при 30 °С, а затем наносили соответственно на твердую среду LB, содержащую 100 г/л сахарозы, для скрининга. Проводили секвенирование для подтверждения получения штаммов SCgL33, SCgL34, SCgL35 и SCgL36 с мутированными промоторами pyc, соответственно.
(3) Оценка продуктивности лизина у мутантов промотора гена pyc в лизин-продуцирующем штамме Corynebacterium glutamicum
Чтобы проверить влияние применения мутации промотора pyc в Corynebacterium glutamicum на продуцирование лизина штаммами, были проведены испытания на ферментацию в SCgL30, SCgL33, SCgL34, SCgL35 и SCgL36, соответственно. Ингредиенты ферментационной среды были следующими: глюкоза, 80 г/л; сухие дрожжи, 1 г/л; соевый пептон, 1 г/л; NaCl, 1 г/л; сульфат аммония, 1 г/л; мочевина, 8 г/л; K2HPO4·3H2O, 1 г/л; MgSO4·7H2O, 0,45 г/л; FeSO4·7H2O, 0,05 г/л; биотин, 0,4 мг/л; витамин B1, 0,1 мг/л; MOPS, 40 г/л; и исходный pH 7,2. Штаммы сначала инокулировали в жидкую среду TSB и культивировали в течение 8 ч. Культуры инокулировали в виде инокулянтов в 24-луночный планшет, содержащий 800 мкл ферментационной среды в каждой лунке с начальным значением OD600, контролируемым при приблизительно 0,1, и культивировали при 30 °С в течение 20 ч. Скорость вращения шейкера для планшетов составляла 800 об/мин. Для каждого штамма отбирали по три образца. После ферментации измеряли выходы лизина. Результаты приведены в таблице 5. Все выходы лизина штаммов после мутации промотора pyc были значительно увеличены, и увеличение становилось более значительным по мере увеличения силы промотора.
Пример 4. Применение мутантов промотора гена pyc Corynebacterium glutamicum для продуцирования пролина
(1) Применение Ppyc-20 в конструировании пролин-продуцирующих штаммов
В настоящем изобретении мутацию G149K вводили в штамм Corynebacterium glutamicum ATCC13032, и кодон мутировали из GGT в AAG, тем самым получая штамм SLCgP1. Штамм SLCgP1 дополнительно применяли в настоящем изобретении для интеграции экспрессионной кассеты, которая была сверхэкспрессирована с использованием промотора Ppyc-20, глутаматкиназы proBG149K, которая ослабляет ингибирование по типу обратной связи, глутамат-5-полуальдегиддегидрогеназы proA вместе с пирролин-5-карбоксилатдегидрогеназой proC в ген putA, для получения пролин-продуцирующего штамма, обозначенного как штамм SLCgP2.
Штамм SLCgP2 был специально сконструирован следующим образом: 1) во-первых, конструировали промотор Ppyc-20 на плазмиде pEC-XK99E для экспрессии экспрессионной кассеты proBG149K, proA, наряду с proC. Фрагмент промотора Ppyc-20 амплифицировали с использованием генома штамма SCgL36 в качестве матрицы и pyc-a/b в качестве праймеров; фрагменты proBG149K, proA и proC амплифицировали с использованием генома штамма SLCgP1 в качестве матрицы и proB-1/2, proA-1/2 и proC-1/2 в качестве праймеров, соответственно; и в то же время каркас pEC-XK99E амплифицировали с использованием pEC-1/2 в качестве праймеров. Вышеуказанные 5 фрагментов лигировали с помощью набора для одностадийного клонирования от Vazyme с получением плазмиды pEC- proBG149KproAproC. 2) Конструирование рекомбинантного вектора, содержащего вышеуказанную экспрессионную кассету, вставленную в ген putA на хромосоме. Вышележащие и нижележащие гомологичные плечи, вставленные в ген putA, амплифицировали с использованием генома штамма SCgL36 в качестве матрицы и putA-1/2 и putA-3/4 в качестве праймеров, соответственно; фрагмент экспрессионной кассеты амплифицировали с использованием плазмиды pEC- proBG149KproAproC в качестве матрицы и ABC-F/R в качестве праймеров; и в то же время каркас pK18mobsacB амплифицировали с использованием pK18-1/2 в качестве праймеров. Вышеуказанные ПЦР-фрагменты лигировали с помощью набора для одностадийного клонирования от Vazyme с получением рекомбинантного вектора pK18-proBG149KproAproC. 3) Рекомбинантный вектор pK18-proBG149KproAproC трансформировали в штамм SLCgP1. Штамм наносили на твердую среду LBHIS, содержащую 5 г/л глюкозы и 25 мкг/мл канамицина, и культивировали при 30 °С с получением первых рекомбинантных трансформантов. Правильные первые рекомбинантные трансформанты инокулировали в среду LB, содержащую 5 г/л глюкозы соответственно, и культивировали в течение ночи. Затем 1% культуры переносили в среду LB, содержащую 100 г/л сахарозы, и культивировали в течение ночи при 30 °С, а затем наносили соответственно на твердую среду LB, содержащую 100 г/л сахарозы, для скрининга. Правильные мутанты подтверждали с помощью ПЦР со специфическими праймерами и секвенированием для получения штаммов SLCgP2, соответственно. Последовательности праймеров, использованных выше, были перечислены в таблице 6.
(2) Оценка продуктивности пролина у штаммов, модифицированных мутантом промотора Ppyc-20
Для проверки влияния применения мутанта промотора Ppyc-20 у Corynebacterium glutamicum на продукцию пролина штаммами проводили ферментационные тесты в SLCgP1 и SLCgP2 соответственно. Ингредиенты ферментационной среды были следующими: глюкоза, 72 г/л; сухие дрожжи, 1 г/л; соевый пептон, 1 г/л; NaCl, 1 г/л; сульфат аммония, 1 г/л; мочевина, 10 г/л; K2HPO4·3H2O, 1 г/л; MgSO4·7H2O, 0,45 г/л; FeSO4·7H2O, 0,05 г/л; биотин, 0,4 мг/л; витамин B1, 0,1 мг/л; MOPS, 40 г/л; и исходный pH 7,2. Штаммы сначала инокулировали в жидкую среду TSB и культивировали в течение 8 ч. Культуры инокулировали в виде инокулянтов в 24-луночный планшет, содержащий 800 мкл ферментационной среды в каждой лунке при количестве инокулянта 12 мкл, и культивировали при 30 °С в течение 18 ч. Скорость вращения шейкера для планшетов составляла 800 об/мин. Для каждого штамма отбирали по три образца. После ферментации измеряли выходы пролина. Результаты приведены в таблице 7. Выходы пролина у штаммов были значительно увеличены после того, как промотор Ppyc-20, экспрессирующий экспрессионную кассету proBG149K, proA вместе с proC был вставлен в хромосому, и SLCgP2 продемонстрировал увеличение на 77% по сравнению с SLCgP1.
Приведенные выше результаты свидетельствуют о том, что мутанты с усиленным промотором гена pyc могут применяться для экспрессии различных целевых генов Corynebacterium glutamicum и применены для получения различных продуктов. Во-первых, мутанты с усиленным промотором гена pyc могут применяться для усиления экспрессии непосредственно гена pyc в Corynebacterium glutamicum для повышения активности Pyc, тем самым усиливая синтез из пировиноградной кислоты до щавелевоуксусной кислоты, что может быть использовано для продуцирования целевых продуктов, зависящих от поступления щавелевоуксусной кислоты в качестве предшественника, включая биологическое продуцирование аминокислот с щавелевоуксусной кислотой в качестве основного метаболического предшественника, таких как аминокислоты семейства аспарагиновой кислоты (лизин, треонин, изолейцин и метионин), аминокислоты семейства глутаминовой кислоты (глутаминовая кислота, пролин, гидроксипролин, аргинин и глутамин) и 5-аминолевулиновая кислота. Настоящее изобретение подтвердило в примерах, что мутанты с усиленным промотором гена pyc могут применяться для продуцирования лизина в качестве аминокислоты семейства аспарагиновой кислоты и пролина в качестве аминокислоты семейства глутаминовой кислоты. Было проверено, что повышенная экспрессия и активность Pyc пригодны для продуцирования вышеуказанных продуктов, например: 1) Peters-Wendisch PG et al. сообщили, что сверхэкспрессия Pyc может улучшить выходы глутаминовой кислоты, лизина и треонина [5]; 2) Pyc является основным ферментом в Corynebacterium glutamicum для катализа продуцирования C4 щавелевоуксусной кислоты из C3 (PEP (фосфоенолпировиноградная кислота) или пировиноградная кислота), и сверхэкспрессия и повышенная активность Pyc являются важными целями для продуцирования аминокислот с щавелевоуксусной кислотой в качестве предшественника [6]; 3) в патентном источнике [7] раскрыто, что повышенная активность ферментов (фосфоенолпируваткарбоксилазы или пируваткарбоксилазы), которые облегчают синтез щавелевоуксусной кислоты, может улучшить выход 5-аминолевулиновой кислоты. Все мутанты промотора гена pyc в пируваткарбоксилазе Corynebacterium glutamicum в настоящем изобретении могут применяться для усиления экспрессии и активности Pyc. Следовательно, мутанты промотора гена pyc согласно настоящему изобретению также могут применяться для продуцирования 5-аминолевулиновой кислоты. Далее, примеры настоящего изобретения также демонстрируют, что мутанты с усиленным промотором гена pyc также могут применяться для экспрессии генов, таких как proB, proA и proC, что свидетельствует о том, что мутанты промотора гена pyc согласно настоящему изобретению имеют хорошую универсальность, которую можно использовать для экспрессии большего количества генов и, следовательно, продуцирования большего количества продуктов.
Цитируемые источники
[3] Wang, YC et al. Screening efficient constitutive promoters in Corynebacterium glutamicum based on time-series transcriptome analysis. Chinese Journal of Biotechnology, 2018, 34(11):1760 - 1771.
[4] CN112877269A
[5] Peters-Wendisch P G, Schiel B, Wendisch V F, et al. Pyruvate carboxylase is a major bottleneck for glutamate and lysine production by Corynebacterium glutamicum. [J]. J Mol Microbiol Biotechnol, 2001, 3(2):295 - 300.
[6] Uwe Sauer, Bernhard J. Eikmanns. The PEP-pyruvate-oxaloacetate node as the switch point for carbon flux distribution in bacteria [M]// FEMS Microbiology Reviews.
[7] CN103981203B
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ТЯНЬЦЗИНЬ ИНСТИТЬЮТ ОФ ИНДАСТРИАЛ БАЙОТЕКНОЛОДЖИ, ЧАЙНИЗ
ЭКЭДЕМИ ОФ САЙЕНСИЗ
<120> МУТАНТ ПРОМОТОРА ГЕНА ПИРУВАТКАРБОКСИЛАЗЫ И ЕГО ПРИМЕНЕНИЕ
<130> 6A17-2163278RU
<150> CN202010696983.2
<151> 2020-07-20
<160> 59
<170> SIPOSequenceListing 1.0
<210> 1
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 1
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacct aattttgatt cgtactgatt
300
tctgctacga tgagtcaacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 2
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 2
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacgg attgttgatt tgagcttgat
300
gagcgtacaa tcaacttacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 3
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 3
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaactt ctccttgatt gcgccttaac
300
cgtggtatga ttcgataacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 4
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 4
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacat tgatttgatt ggaaccttac
300
tgtgctatga tttggtaacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 5
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 5
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaactc gagtttgatt tcacaacgtg
300
tgtgatagga tataataacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 6
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 6
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaactt gcgtttgatt aaagtatgca
300
agggctagta tggtgatacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 7
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 7
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacat cattttgatt ccggcgcaca
300
tgtggtaata tggtattacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 8
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 8
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaactc gccattgatt gcccgccatc
300
catgctataa tcggaagacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 9
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 9
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaactt ccgcttgatt gtggccatag
300
tatgatatta ttaattaacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 10
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 10
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaaccg gatcttgatt ttatgatggg
300
tattgtataa tcttggtacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 11
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 11
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaaccg gatattgatt tggccggtgt
300
tgtggtagta tcgtgttacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 12
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 12
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacag gggtttgatt ggccgctcgg
300
tgtgttatca tggagagacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 13
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 13
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacga gttgttgatt tcgttggtgc
300
acgtatacaa tggttttacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 14
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 14
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacct tggcttgatt tttgtttgag
300
ggttgtataa tgttattacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 15
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 15
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacga ctagttgatt tccgcccttg
300
gttgatatta tgcttgaacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 16
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 16
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacat ccgcttgatt taggcgtacg
300
tttaatagta tattgaaacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 17
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 17
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaaccg gggcttgatt tccttgtcgt
300
ggcgttatta taatggaacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 18
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 18
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaacat ggagttgatt atacgatact
300
acagatacta tactggtacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 19
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 19
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaaccc gtagttgatt gacttgggca
300
gtatatagta taatgaaacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 20
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 20
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaaccg ggccttgatt gtaagataag
300
acatttagta taattagacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 21
<211> 373
<212> ДНК
<213> Corynebacterium glutamicum
<400> 21
gaaaacccag gattgctttg tgcactcctg ggttttcact ttgttaagca gttttgggga
60
aaagtgcaaa gtttgcaaag tttagaaata ttttaagagg taagatgtct gcaggtggaa
120
gcgtttaaat gcgttaaact tggccaaatg tggcaacctt tgcaaggtga aaaactgggg
180
cggggttaga tcctgggggg tttatttcat tcactttggc ttgaagtcgt gcaggtcagg
240
ggagtgttgc ccgaaaacat tgagaggaaa acaaaaaccg atgtttgatt gggggaatcg
300
ggggttacga tactaggacg cagtgactgc tatcaccctt ggcggtctct tgttgaaagg
360
aataattact cta
373
<210> 22
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 22
cctgatgcgg tattttctcc gaaaacccag gattgctttg tg
42
<210> 23
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 23
gcggacagct tcagaagcaa aag
23
<210> 24
<211> 84
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 24
ttgcttctga agctgtccgc ggcggtggct ctggaggtgg tgggtccggc ggtggctctg
60
cttcctccga agacgttatc aaag
84
<210> 25
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 25
ggagaaaata ccgcatcagg c
21
<210> 26
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 26
cccgaaaaca ttgagaggaa aacaaaaacn nnnnnttgat tnnnnnnnnn nnnnnntann
60
atnnnnnnac gcagtgactg ctatcaccc
89
<210> 27
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 27
aaccttccat acgaactttg aaacg
25
<210> 28
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 28
caaagttcgt atggaaggtt ccg
23
<210> 29
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 29
ttcctctcaa tgttttcggg c
21
<210> 30
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 30
caggaaacag ctatgacatg gtatcgccat gtatcacgca ctc
43
<210> 31
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 31
tcagtacgaa tcaaaattag gtttttgttt tcctctcaat gttttc
46
<210> 32
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 32
atggccacaa tcaagcggaa gtttttgttt tcctctcaat gttttc
46
<210> 33
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 33
tacgcctaaa tcaagcggat gtttttgttt tcctctcaat gttttc
46
<210> 34
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 34
tatcttacaa tcaaggcccg gtttttgttt tcctctcaat gttttc
46
<210> 35
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 35
ctaattttga ttcgtactga tttctgctac gatgagtcaa cgcagtgact gctatcaccc
60
<210> 36
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 36
ttccgcttga ttgtggccat agtatgatat tattaattaa cgcagtgact gctatcaccc
60
<210> 37
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 37
atccgcttga tttaggcgta cgtttaatag tatattgaaa cgcagtgact gctatcaccc
60
<210> 38
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 38
cgggccttga ttgtaagata agacatttag tataattaga cgcagtgact gctatcaccc
60
<210> 39
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 39
tgtaaaacga cggccagtgc ctaatttgcg aagctcatca ggtg
44
<210> 40
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 40
gcactggccg tcgttttac
19
<210> 41
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 41
catgtcatag ctgtttcctg tgtg
24
<210> 42
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 42
ggagaaaata ccgcatcagg c
21
<210> 43
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 43
ctgttttggc ggatgagaga ag
22
<210> 44
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 44
cctgatgcgg tattttctcc gaaaacccag gattgctttg tg
42
<210> 45
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 45
ttggagatgc gctcacgcat tagagtaatt attcctttca acaagag
47
<210> 46
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 46
atgcgtgagc gcatctccaa c
21
<210> 47
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 47
ttacgcgcgg ctggcgtagt tg
22
<210> 48
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 48
actacgccag ccgcgcgtaa gccttttatg gtgtgatccg ac
42
<210> 49
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 49
ttaaggccta atttgtcctg tgcc
24
<210> 50
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 50
caggacaaat taggccttaa tttgtcgttt tgggccccc
39
<210> 51
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 51
tctctcatcc gccaaaacag ctagcgcttt ccgagttctt cag
43
<210> 52
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 52
caggaaacag ctatgacatg ttctagggca tcgacgaacc ag
42
<210> 53
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 53
gaaattgtta aaagcgcagc gc
22
<210> 54
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 54
gctgcgcttt taacaatttc gaaaacccag gattgctttg tg
42
<210> 55
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 55
tcgaagccgc acgtcatcta g
21
<210> 56
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 56
tagatgacgt gcggcttcga tccgtgaacg cctatctgta cg
42
<210> 57
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 57
tgtaaaacga cggccagtgc gatcgattcc acgcccaaac
40
<210> 58
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 58
gcactggccg tcgttttac
19
<210> 59
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность праймера
<400> 59
catgtcatag ctgtttcctg tgtg
24
<---

Изобретение относится к биотехнологии и представляет собой мутант промотора гена пируваткарбоксилазы Corynebacterium glutamicum. Указанный мутант обладает улучшенной промоторной активностью по сравнению с промотором дикого типа и может применяться для повышения экспрессии целевого гена. Так, функциональное лигирование указанного мутанта с геном пируваткарбоксилазы повышает интенсивность экспрессии пируваткарбоксилазы, тем самым улучшая эффективность продуцирования аминокислот штаммом. 8 н. и 13 з.п. ф-лы, 2 ил., 6 табл., 4 пр.
1. Мутант промотора гена пируваткарбоксилазы Corynebacterium glutamicum, где мутант обладает промоторной активностью и нуклеотидная последовательность мутанта в положениях, соответствующих положениям с 279 по 317 нуклеотидной последовательности, представленной в SEQ ID NO: 21, является следующей:
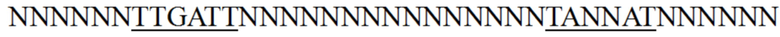 , где N выбран из A, T, C или G;
, где N выбран из A, T, C или G;
где нуклеотидная последовательность мутанта не является CGATGTTTGATTGGGGGAATCGGGGGTTACGATACTAGG в положениях, соответствующих положениям с 279 по 317 последовательности, представленной в SEQ ID NO: 21.
2. Мутант промотора гена пируваткарбоксилазы по п. 1, где мутант обладает промоторной активностью, повышенной в от 1 до 17 раз или более по сравнению с промотором гена пируваткарбоксилазы, имеющим последовательность, представленную в SEQ ID NO: 21.
3. Мутант промотора гена пируваткарбоксилазы по п. 1 или 2, где нуклеотидная последовательность коровой области в положениях с 279 по 317 мутанта представляет собой одну из следующих последовательностей:
1) CTAATTTTGATTCGTACTGATTTCTGCTACGATGAGTCA;
2) GGATTGTTGATTTGAGCTTGATGAGCGTACAATCAACTT;
3) TTCTCCTTGATTGCGCCTTAACCGTGGTATGATTCGATA;
4) ATTGATTTGATTGGAACCTTACTGTGCTATGATTTGGTA;
5) TCGAGTTTGATTTCACAACGTGTGTGATAGGATATAATA;
6) TTGCGTTTGATTAAAGTATGCAAGGGCTAGTATGGTGAT;
7) ATCATTTTGATTCCGGCGCACATGTGGTAATATGGTATT;
8) TCGCCATTGATTGCCCGCCATCCATGCTATAATCGGAAG;
9) TTCCGCTTGATTGTGGCCATAGTATGATATTATTAATTA;
10) CGGATCTTGATTTTATGATGGGTATTGTATAATCTTGGT;
11) CGGATATTGATTTGGCCGGTGTTGTGGTAGTATCGTGTT;
12) AGGGGTTTGATTGGCCGCTCGGTGTGTTATCATGGAGAG;
13) GAGTTGTTGATTTCGTTGGTGCACGTATACAATGGTTTT;
14) CTTGGCTTGATTTTTGTTTGAGGGTTGTATAATGTTATT;
15) GACTAGTTGATTTCCGCCCTTGGTTGATATTATGCTTGA;
16) ATCCGCTTGATTTAGGCGTACGTTTAATAGTATATTGAA;
17) CGGGGCTTGATTTCCTTGTCGTGGCGTTATTATAATGGA;
18) ATGGAGTTGATTATACGATACTACAGATACTATACTGGT;
19) CCGTAGTTGATTGACTTGGGCAGTATATAGTATAATGAA; или
20) CGGGCCTTGATTGTAAGATAAGACATTTAGTATAATTAG.
4. Мутант промотора гена пируваткарбоксилазы по любому из пп. 1-3, где нуклеотидная последовательность мутанта представлена в любой из SEQ ID NO: 1 - SEQ ID NO: 20.
5. Экспрессионная кассета, содержащая мутант промотора гена пируваткарбоксилазы по любому из пп. 1-4.
6. Экспрессионный вектор, содержащий мутант промотора гена пируваткарбоксилазы по любому из пп. 1-4 или экспрессионную кассету по п. 5.
7. Рекомбинантная клетка-хозяин для ферментативного продуцирования целевых продуктов, содержащая мутант промотора гена пируваткарбоксилазы по любому из пп. 1-4, экспрессионную кассету по п. 5 или экспрессионный вектор по п. 6.
8. Рекомбинантная клетка-хозяин по п. 7, где клетка-хозяин принадлежит к роду Enterobacter или роду Corynebacterium, предпочтительно роду Corynebacterium, еще более предпочтительно Corynebacterium glutamicum, более конкретно Corynebacterium glutamicum ATCC 13032, Corynebacterium glutamicum ATCC 13869, Corynebacterium glutamicum B253, Corynebacterium glutamicum ATCC 14067 и их производным штаммам.
9. Применение мутанта промотора гена пируваткарбоксилазы по любому из пп. 1-4, экспрессионной кассеты по п. 5, экспрессионного вектора по п. 6 или рекомбинантной клетки-хозяина по п. 7 или 8 для повышения уровня транскрипции гена или получение реагента или набора для повышения уровня транскрипции гена.
10. Способ повышения экспрессии целевого гена, включающий функциональное лигирование мутанта промотора гена пируваткарбоксилазы по любому из пп. 1-4 с целевым геном или целевой РНК.
11. Способ по п. 10, где целевая РНК включает по меньшей мере одну из тРНК (транспортная РНК) или sРНК (растворимая РНК).
12. Способ по п. 10, где целевой ген включает по меньшей мере один из кодирующего гена белка, связанного с синтезом целевого продукта, кодирующего гена белка, регулирующего экспрессию гена, или кодирующего гена белка, связанного с мембранным транспортом.
13. Способ по п. 10, где целевой ген включает по меньшей мере один из следующих кодирующих генов ферментов: ген pyc пируваткарбоксилазы, ген gdh глутаматдегидрогеназы, ген lysC аспартаткиназы, ген thrABC треонинового оперона, ген asd аспартат-полуальдегиддегидрогеназы, ген aspB аспартат-аммоний-лиазы, ген hom гомосериндегидрогеназы, ген metX гомосерин-O-ацетилтрансферазы, ген dapA дигидродипиколинатсинтазы, ген dapB дигидродипиколинатредуктазы, ген ddh мезо-диаминопимелатдегидрогеназы, ген proB глутаматкиназы, ген proA глутамат-5-полуальдегиддегидрогеназы, ген proC пирролин-5-карбоксилатдегидрогеназы, ген putA пролиндегидрогеназы/пирролин-5-карбоксилатдегидрогеназы, ген hemA глутамил-тРНК-редуктазы, ген ppc фосфоенолпируваткарбоксилазы, ген lysE белка-транспортера аминокислот, кодирующий ген, ассоциированный с системой ptsG, ген aceE пируватдегидрогеназы, ген gapN глицеральдегид-3-фосфатдегидрогеназы и ген cadA/ldcC лизиндекарбоксилазы.
14. Способ получения белка, включающий стадию экспрессии белка с помощью экспрессионной кассеты по п. 5, экспрессионного вектора по п. 6 или рекомбинантной клетки-хозяина по п. 7 или 8.
15 Способ по п. 14, где белок представляет собой белок, связанный с синтезом целевого продукта, белок, связанный с мембранным транспортом, или белок, регулирующий экспрессию гена.
16. Способ по п. 14 или 15, где способ дополнительно включает стадию выделения или очистки белка.
17. Способ получения целевого продукта, включающий культивирование клетки-хозяина, содержащей мутант промотора гена пируваткарбоксилазы по любому из пп. 1-4, который функционально лигирован с геном, связанным с синтезом целевого продукта, и сбор полученного целевого продукта.
18. Способ по п. 17, где целевой продукт представляет собой аминокислоту.
19. Способ по п. 17, где ген, связанный с синтезом целевого продукта, представляет собой ген, связанный с синтезом аминокислоты или ее производного.
20. Способ по п. 19, где аминокислота и ее производное выбраны из одной или комбинации двух или более из: пролина, гидроксипролина, лизина, глутаминовой кислоты, аргинина, орнитина, глутамина, треонина, глицина, аланина, валина, лейцина, изолейцина, серина, цистеина, метионина, аспарагиновой кислоты, аспарагина, гистидина, фенилаланина, тирозина, триптофана, 5-аминолевулиновой кислоты или производных любой из аминокислот, описанных выше.
21. Способ по п. 17, где в качестве предшественника целевого продукта используется щавелевоуксусная кислота; предпочтительно целевой продукт, в качестве предшественника которого используется щавелевоуксусная кислота, включает одно или более из: лизина, треонина, изолейцина, метионина, глутаминовой кислоты, пролина, гидроксипролина, аргинина, глутамина, 5-аминолевулиновой кислоты, пентандиамина, 5-аминовалериановой кислоты или производного любого из вышеперечисленных.
| CN 110951662 A, 03.04.2020 | |||
| Ударно-импульсный механизм | 1986 |
|
SU1425071A2 |
| US 20110129885 A1, 02.06.2011 | |||
| US 20180362991 A1, 20.12.2018 | |||
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2008 |
|
RU2405040C2 |
| Kim et al | |||
| Plant Molecular Biology, 24: 105-117, 1994 | |||
| См | |||
| стр | |||
| Приспособление для останова мюля Dobson аnd Barlow при отработке съема | 1919 |
|
SU108A1 |
| Шкив для канатной передачи | 1920 |
|
SU109A1 |
