Родственные заявки
Данная заявка заявляет приоритет предварительной заявки на патент США №60/141031, поданной 25 июня 1999 года. Данная заявка заявляет также приоритет заявки на патент Германии №19931454.3, поданной 8 июля 1999 года, заявки на патент Германии №19931478.0, поданной 8 июля 1999 года, заявки на патент Германии №19931563.9, поданной 8 июля 1999 года, заявки на патент Германии №19932122.1, поданной 9 июля 1999 года, заявки на патент Германии №19932124.8, поданной 9 июля 1999 года, заявки Германии на патент №19932125.6, поданной 9 июля 1999 года, заявки Германии на патент №19932128.0, поданной 9 июля 1999 года, заявки на патент Германии №19932180.9, поданной 9 июля 1999 года, заявки на патент Германии №19932182.5, поданной 9 июля 1999 года, заявки на патент Германии №19932190.6, поданной 9 июля 1999 года, заявки на патент Германии №19932191.4, поданной 9 июля 1999 года, заявки на патент Германии №19932209.0, поданной 9 июля 1999 года, заявки на патент Германии №19932212.0, поданной 9 июля 1999 года, заявки на патент Германии №19932227.9, поданной 9 июля 1999 года, заявки на патент Германии №19932228.7, поданной 9 июля 1999 года, заявки на патент Германии №19932229.5, поданной 9 июля 1999 года, заявки на патент Германии №19932230.9, поданной 9 июля 1999 года, заявки на патент Германии №19932927.3, поданной 14 июля 1999 года, заявки на патент Германии №19933005.0, поданной 14 июля 1999 года, заявки на патент Германии №19933006.9, поданной 14 июля 1999 года, заявки на патент Германии №19940764.9, поданной 27 августа 1999 года, заявки на патент Германии №19940765.7, поданной 27 августа 1999 года, заявки на патент Германии №19940766.5, поданной 27 августа 1999 года, заявки на патент Германии №19940830.0, поданной 27 августа 1999 года, заявки на патент Германии №19940831.9, поданной 27 августа 1999 года, заявки на патент Германии №19940832.7, поданной 27 августа 1999 года, заявки на патент Германии №19940833.5, поданной 27 августа 1999 года, заявки на патент Германии №19941378.9, поданной 31 августа 1999 года, заявки на патент Германии №19941379.7, поданной 31 августа 1999 года, заявки на патент Германии №19941395.9, поданной 31 августа 1999 года, заявки на патент Германии №19942077.7, поданной 3 сентября 1999 года, заявки на патент Германии №19942078.5, поданной 3 сентября 1999 года, заявки на патент Германии №19942079.3, поданной 3 сентября 1999 года, и заявки на патент Германии №19942088.2, поданной 3 сентября 1999 года. Полные содержания всех вышеуказанных заявок включены здесь в виде ссылки.
Предпосылки изобретения
Некоторые продукты и побочные продукты природно встречающихся метаболических процессов в клетках имеют применение в широком списке отраслей промышленности, в том числе в кормовой, пищевой, косметической и фармацевтической отраслях промышленности. Эти молекулы, совокупно называемые "химическими продуктами тонкого органического синтеза", включают органические кислоты, как входящие в состав белков (протеиногенные), так и не входящие в состав белков (непротеиногенные) аминокислоты, нуклеотиды и нуклеозиды, липиды и жирные кислоты, диолы, углеводы, ароматические соединения, витамины и кофакторы и ферменты. Их получение наиболее удобно выполнять посредством крупномасштабной культуры бактерий, разработанной для продуцирования и секреции больших количеств одной или более желаемых молекул. Одним особенно применимым организмом для этой цели является Corynebacterium glutamicum, грамположительная, не патогенная бактерия. Посредством отбора штаммов был получен ряд мутантных штаммов, которые продуцируют ряд желаемых соединений. Однако, отбор штаммов, улучшенных в отношении продуцирования конкретной молекулы, является трудоемким процессом, отнимающим много времени.
Сущность изобретения
Данное изобретение представляет новые бактериальные молекулы нуклеиновых кислот, которые имеют множество применений. Эти применения включают идентификацию микроорганизмов, которые могут быть использованы для получения химических продуктов тонкого органического синтеза, модуляции образования химических продуктов тонкого органического синтеза в С. glutamicum или родственных бактериях, типирования или идентификации С. glutamicum или родственных бактерий, в качестве ссылочных точек для картирования генома С. glutamicum и в качестве маркеров для трансформации. Эти новые молекулы нуклеиновых кислот кодируют белки, называемые здесь белками конструирования мембран и мембранного транспорта (МСТ).
С. glutamicum является грамположительной, аэробной бактерией, которую обычно используют в промышленности для крупномасштабного получения множества химических продуктов тонкого органического синтеза, а также для расщепления углеводородов (например, в разливах нефти) и для окисления терпеноидов. Таким образом, молекулы нуклеиновых кислот МСТ данного изобретения могут быть использованы для идентификации микроорганизмов, которые могут быть использованы для получения химических продуктов тонкого органического синтеза, например, посредством ферментационных процессов. Модуляция экспрессии нуклеиновых кислот МСТ данного изобретения или модификация последовательности молекул нуклеиновых кислот МСТ данного изобретения может использоваться для модуляции образования одного или нескольких химических продуктов тонкого органического синтеза из микроорганизма (например, для улучшения выхода или получения одного или нескольких химических продуктов тонкого органического синтеза из видов Corynebacterium или Brevibacterium).
Нуклеиновые кислоты МСТ данного изобретения могут быть также использованы для идентификации организма как являющегося Corynebacterium glutamicum или близкородственной бактерией или для идентификации присутствия С. glutamicum или родственной бактерии в смешанной популяции микроорганизмов. Данное изобретение представляет последовательности нуклеиновых кислот ряда генов С. glutamicum; зондированием экстрагированной геномной ДНК культуры уникальной или смешанной популяции микроорганизмов при строгих условиях зондом, охватывающим участок гена С. glutamicum, который является уникальным в отношении этого организма, можно определить, присутствует ли данный организм. Хотя сама бактерия Corynebacterium glutamicum является непатогенной, она является родственной видам, патогенным в человеке, таким как Corynebacterium diphtheriae (возбудитель дифтерии); выявление таких организмов имеет важное клиническое значение.
Молекулы нуклеиновых кислот МСТ данного изобретения могут также служить в качестве ссылочных точек для картирования генома С. glutamicum или геномов родственных организмов. Подобным образом, эти молекулы, или их варианты или части, могут служить в качестве маркеров для генетически сконструированных видов Corynebacterium или Brevibacterium.
Белки МСТ, кодируемые новыми молекулами нуклеиновых кислот данного изобретения, способны, например, выполнять функцию, участвующую в метаболизме (например, биосинтезе или расщеплении) соединений, необходимых для биосинтеза мембран, или способствовать трансмембранному транспорту одного или большего количества соединений в клетку или из клетки. При условии доступности клонирующих векторов для применения в Corynebacterium glutamicum, таких как векторы, описанные в Sinskey et al., U.S. Patent №4649119, и способов для генетической манипуляции С. glutamicum и родственных видов Brevibacterium (например, lactofermentum) (Yoshihama et al., J. Bacteriol. 162: 591-597 (1985); Katsumata et al., J. Bacteriol. 159: 306-311 (1984) и Santamaria et al., J. Gen. Microbiol. 130: 2237-2246 (1984)), молекулы нуклеиновых кислот данного изобретения могут быть использованы в генетической инженерии этого организма, чтобы сделать его лучшим или более эффективным продуцентом одного или нескольких химических продуктов тонкого органического синтеза. Это улучшенное продуцирование или улучшенная эффективность продуцирования химического продукта тонкого органического синтеза могут быть обусловлены прямым действием манипулирования геном данного изобретения или могут быть обусловлены непрямым действием такого манипулирования.
Существует ряд механизмов, при помощи которых изменение белка МСТ данного изобретения может непосредственно влиять на выход, продуцирование и/или эффективность продуцирования химического продукта тонкого органического синтеза из штамма С. glutamicum, включающего в себя такой измененный белок. Белки МСТ, участвующие в экспорте молекул химических продуктов тонкого органического синтеза из клетки, могут быть увеличены в числе или усилены в активности, так что большие количества этих соединений секретируются во внеклеточную среду, из которой они более легко извлекаются. Подобным образом, белки МСТ, участвующие в импорте питательных веществ, необходимых для биосинтеза одного или нескольких химических продуктов тонкого органического синтеза (например, фосфатных, сульфатных, азотсодержащих соединений и т.д.), могут быть увеличенными в концентрации или активности, так что эти предшественники, кофакторы или промежуточные соединения увеличиваются в концентрации в клетке. Далее, жирные кислоты и липиды сами являются желательными химическими продуктами тонкого органического синтеза; посредством оптимизации активности или увеличения числа одного или нескольких белков МСТ данного изобретения, участвующих в биосинтезе этих соединений, или посредством нарушения активности одного или нескольких белков МСТ, участвующих в расщеплении этих соединений, можно увеличить выход, продуцирование и/или эффективность продуцирования молекул жирных кислот и липидов из С. glutamicum.
Мутагенез одного или нескольких генов МСТ данного изобретения может также приводить к белкам МСТ, имеющим измененные активности, которые опосредованно влияют на продуцирование одного или нескольких желательных химических продуктов тонкого органического синтеза из С. glutamicum. Например, белки МСТ данного изобретения, участвующие в экспорте отработанных продуктов клетки (возможно, увеличенных в количестве вследствие сверхпродуцирования желательного химического продукта тонкого органического синтеза), эффективно экспортируются, прежде чем они способны повредить нуклеотиды и белки в клетке (что уменьшало бы жизнеспособность данной клетки) или помешать биосинтетическим путям химических продуктов тонкого органического синтеза (что уменьшало бы выход, продуцирование или эффективность продуцирования желательного химического продукта тонкого органического синтеза). Далее, относительно большие внутриклеточные количества желательного химического продукта тонкого органического синтеза может само быть токсичным для клетки, так что посредством увеличения активности или числа переносчиков (транспортеров), способных экспортировать это соединение из клетки, можно увеличить жизнеспособность данной клетки в культуре, что, в свою очередь, приводит к большему числу клеток в культуре, продуцирующих желательный химический продукт тонкого органического синтеза. Белки МСТ данного изобретения могут быть подвергнуты манипулированию таким образом, что продуцируются относительно измененные количества различных молекул липидов и жирных кислот. Это может оказывать сильное влияние на состав липидов мембраны клетки. Поскольку каждый тип липида имеет различные физические свойства, изменение в липидном составе мембраны может существенно изменять текучесть мембраны. Изменения в текучести мембраны могут влиять на транспорт молекул через мембрану, а также на целостность клетки, причем оба эти свойства имеют сильное влияние на продуцирование химических продуктов тонкого органического синтеза из С. glutamicum в крупномасштабной ферментационной культуре.
Данное изобретение представляет новые молекулы нуклеиновых кислот, которые кодируют белки, называемые здесь белками МСТ, которые способны, например, участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. Молекулы нуклеиновых кислот, кодирующие белок МСТ, называют здесь молекулами нуклеиновых кислот МСТ. В предпочтительном варианте белок МСТ участвует в метаболизме соединений, необходимых для построения (конструирования) клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. Примеры таких белков включают белки, кодируемые генами, представленными в таблице 1.
Таким образом, один аспект данного изобретения относится к выделенным молекулам нуклеиновых кислот (например, кДНК, ДНК или РНК), содержащим нуклеотидную последовательность, кодирующую белок МСТ или его биологически активные части, а также фрагментам нуклеиновых кислот, пригодным в качестве праймеров или гибридизационных зондов для обнаружения или амплификации МСТ-кодирующей нуклеиновой кислоты (например, ДНК или мРНК). В особенно предпочтительных вариантах выделенная молекула нуклеиновой кислоты содержит одну из нуклеотидных последовательностей, представленных в виде имеющих нечетные номера SEQ ID NO в Списке последовательностей (например, SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7...), или кодирующий район или его комплемент одной из этих нуклеотидных последовательностей. В других особенно предпочтительных вариантах выделенная молекула нуклеиновой кислоты данного изобретения содержит нуклеотидную последовательность, которая гибридизуется с нуклеотидной последовательностью или является по меньшей мере на приблизительно 50%, предпочтительно по меньшей мере на приблизительно 60%, более предпочтительно по меньшей мере на приблизительно 70%, 80% или 90% и даже более предпочтительно по меньшей мере на приблизительно 95%, 96%, 97%, 98%, 99% или более гомологичной нуклеотидной последовательности, представленной в виде имеющих нечетные номера SEQ ID NO в Списке последовательностей (например, SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7...), или ее части. В других предпочтительных вариантах выделенная молекула нуклеиновой кислоты кодирует одну из аминокислотных последовательностей, представленных в виде имеющих четные номера SEQ ID NO в Списке последовательностей (например, SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8...). Предпочтительные белки МСТ данного изобретения также предпочтительно имеют по меньшей мере одну из описанных здесь активностей МСТ.
В другом варианте выделенная молекула нуклеиновой кислоты кодирует белок или его часть, причем этот белок или его часть включает аминокислотную последовательность, которая является достаточно гомологичной аминокислотной последовательности данного изобретения (например, последовательности, имеющей четный номер SEQ ID NO: в Списке последовательностей), например, достаточно гомологичной аминокислотной последовательности данного изобретения, так что этот белок или его часть сохраняет активность МСТ. Предпочтительно, белок или его часть, кодируемые этой молекулой нуклеиновой кислоты, сохраняют способность участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. В одном варианте белок, кодируемый указанной молекулой нуклеиновой кислоты, является по меньшей мере на приблизительно 50%, предпочтительно по меньшей мере на приблизительно 60% и более предпочтительно по меньшей мере на приблизительно 70%, 80% или 90% и наиболее предпочтительно по меньшей мере на приблизительно 95%, 96%, 97%, 98%, 99% или более гомологичным аминокислотной последовательности данного изобретения (например, полной аминокислотной последовательности, выбранной из аминокислотных последовательностей, имеющих четные номера SEQ ID NO: в Списке последовательностей). В другом предпочтительном варианте этот белок является полноразмерным белком С. glutamicum, который является по существу гомологичным полной аминокислотной последовательности данного изобретения (кодируемой открытой рамкой считывания, показанной в соответствующих имеющих нечетные номера SEQ ID NO в Списке последовательностей (например, SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7...).
В другом предпочтительном варианте выделенная молекула нуклеиновой кислоты происходит из С. glutamicum и кодирует белок (например, слитый белок МСТ), который включает биологически активный домен, который по меньшей мере на приблизительно 50% или более гомологичен одной из аминокислотных последовательностей данного изобретения (например, последовательности одной из имеющих четные номера SEQ ID NO: в Списке последовательностей) и способен участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны, или имеет одну или несколько активностей, представленных в таблице 1, и также включает гетерологичные последовательности нуклеиновых кислот, кодирующие гетерологичные полипептид или регуляторные участки.
В другом варианте выделенная молекула нуклеиновой кислоты имеет длину по меньшей мере 15 нуклеотидов и гибридизуется при строгих условиях с молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность данного изобретения (например, последовательность, имеющая нечетный номер SEQ ID NO: в Списке последовательностей). Предпочтительно, выделенная молекула нуклеиновой кислоты соответствует природно встречающейся молекуле нуклеиновой кислоты. Более предпочтительно, выделенная нуклеиновая кислота кодирует природно встречающийся белок МСТ С. glutamicum или его биологически активную часть.
Другой аспект данного изобретения относится к векторам, например, рекомбинантным экспрессирующим векторам, содержащим молекулы нуклеиновых кислот данного изобретения, и клеткам-хозяевам, в которые были введены такие векторы. В одном варианте такую клетку-хозяина используют для получения белка МСТ культивированием клетки-хозяина в подходящей среде. Затем белок МСТ может быть выделен из этой среды или из клетки-хозяина.
Еще один аспект данного изобретения относится к генетически измененному микроорганизму, в котором ген МСТ был введен или изменен. В одном варианте геном микроорганизма был изменен введением молекулы нуклеиновой кислоты данного изобретения, кодирующей последовательность МСТ дикого типа или мутированную последовательность МСТ, в качестве трансгена. В другом варианте эндогенный ген МСТ в геноме микроорганизма был изменен, например, функционально нарушен, гомологичной рекомбинацией с измененным геном МСТ. В другом варианте эндогенный или введенный ген МСТ в микроорганизме был изменен одной или несколькими точковыми мутациями, делециями или инверсиями, но все еще кодирует функциональный ген белка МСТ. Еще в одном варианте один или несколько регуляторных участков (например, промотор, репрессор или индуктор) гена МСТ в микроорганизме был изменен (например, делецией, укорочением, инверсией или точковой мутацией), так что экспрессия гена МСТ является модулированной. В предпочтительном варианте этот микроорганизм принадлежит к роду Corynebacterium или Brevibacterium, причем особенно предпочтительным является Corynebacterium glutamicum. В предпочтительном варианте этот микроорганизм используют также для получения желаемого соединения, такого как аминокислота, причем особенно предпочтительным является лизин.
В другом аспекте данное изобретение представляет способ идентификации присутствия или активности Corynebacterium diphtheriae в субъекте. Этот способ включает обнаружение одной или нескольких последовательностей нуклеиновых кислот или аминокислотных последовательностей данного изобретения (например, последовательностей, представленных в Списке последовательностей в виде SEQ ID NO:1-676), в субъекте, детектированием тем самым присутствия или активности Corynebacterium diphtheriae в субъекте.
Еще один аспект данного изобретения относится к выделенным белку МСТ или его части, например, его биологически активной части. В предпочтительном варианте выделенные белок МСТ или его часть может участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. В другом предпочтительном варианте выделенные белок МСТ или его часть являются достаточно гомологичными аминокислотной последовательности данного изобретения (например, последовательности, имеющей четный номер SEQ ID NO: в Списке последовательностей), так что этот белок или его часть сохраняет способность участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны.
Данное изобретение представляет также выделенный препарат белка МСТ. В предпочтительных вариантах белок МСТ содержит аминокислотную последовательность данного изобретения (например, последовательность, имеющая четный номер SEQ ID NO: Списка последовательностей). В другом предпочтительном варианте данное изобретение относится к выделенному полноразмерному белку, который является по существу гомологичным полной аминокислотной последовательности данного изобретения (например, последовательности, имеющей четный номер SEQ ID NO: Списка последовательностей) (кодируемому открытой рамкой считывания, приведенной в соответствующих имеющих нечетные номера SEQ ID NO: Списка последовательностей). Еще в одном варианте этот белок является по меньшей мере на приблизительно 50%, предпочтительно по меньшей мере на приблизительно 60% и более предпочтительно по меньшей мере на приблизительно 70%, 80% или 90% и наиболее предпочтительно по меньшей мере на приблизительно 95%, 96%, 97%, 98%, 99% или более гомологичным полной аминокислотной последовательности данного изобретения (например, последовательности, имеющей четный номер SEQ ID NO: Списка последовательностей). В других вариантах выделенный белок МСТ содержит аминокислотную последовательность, которая является по меньшей мере на приблизительно 50% или более гомологичной одной из аминокислотных последовательностей данного изобретения (например, последовательности, имеющей четный номер SEQ ID NO: Списка последовательностей), и способен участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum в транспорте молекул через клеточную мембрану, или имеет одну или несколько активностей, приведенных в таблице 1.
Альтернативно, выделенный белок МСТ может содержать аминокислотную последовательность, которая кодируется нуклеотидной последовательностью, которая гибридизуется, например, гибридизуется при строгих условиях, с нуклеотидной последовательностью, или является по меньшей мере на приблизительно 50%, предпочтительно по меньшей мере на приблизительно 60%, более предпочтительно по меньшей мере на приблизительно 70%, 80% или 90% и даже более предпочтительно по меньшей мере на приблизительно 95%, 96%, 97%, 98%, 99% или более гомологичной нуклеотидной последовательности одной из имеющих четные номера SEQ ID NO:, представленных в Списке последовательностей. Также предпочтительно, чтобы предпочтительные формы белков МСТ имели также одну или несколько из биологических активностей МСТ, описанных здесь.
Полипептид МСТ или его биологически активная часть могут быть функционально связаны с полипептидом не-МСТ с образованием слитого белка. В предпочтительных вариантах этот слитый белок имеет активность, которая отличается от активности одного белка МСТ. В других предпочтительных вариантах этот слитый белок участвует в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. В особенно предпочтительных вариантах интеграция этого слитого белка в клетку-хозяина модулирует продуцирование желаемого соединения из данной клетки.
В другом аспекте данное изобретение представляет способы скрининга молекул, которые модулируют активность белка МСТ, либо посредством взаимодействия с самим белком или субстратом или партнером связывания белка МСТ, либо посредством модуляции транскрипции или трансляции молекулы нуклеиновой кислоты МСТ данного изобретения.
Другой аспект данного изобретения относится к способу получения химического продукта тонкого органического синтеза. Этот способ предусматривает культивирование клетки, содержащей вектор, направляющий экспрессию молекулы нуклеиновой кислоты МСТ данного изобретения таким образом, что образуется химический продукт тонкого органического синтеза. В предпочтительном варианте этот способ дополнительно включает стадию получения клетки, содержащей такой вектор, в которой клетку трансфицируют вектором, направляющим экспрессию нуклеиновой кислоты МСТ. В другом предпочтительном варианте этот способ дополнительно предусматривает стадию извлечения химических продуктов тонкого органического синтеза из культуры. В особенно предпочтительном варианте эта клетка является клеткой из рода Corynebacterium или Brevibacterium или выбрана из штаммов, приведенных в таблице 3.
Другой аспект данного изобретения относится к способам модуляции продуцирования молекулы из микроорганизма. Такие способы включают контактирование клетки с агентом, который модулирует активность белка МСТ или экспрессию нуклеиновой кислоты МСТ, так что связанная с клеткой активность является измененной относительно той же самой активности в отсутствие этого агента. В предпочтительном варианте клетку модулируют в отношении одного или нескольких метаболических путей С. glutamicum для компонентов клеточной мембраны или модулируют в отношении транспорта соединений через такие мембраны, так что выходы или скорость продуцирования желательного химического продукта тонкого органического синтеза этим микроорганизмом улучшается. Агент, который модулирует активность белка МСТ, может быть агентом, стимулирующим активность белка МСТ или экспрессию нуклеиновой кислоты МСТ. Примеры агентов, стимулирующих активность белка МСТ или экспрессию нуклеиновой кислоты МСТ, включают небольшие молекулы, активные белки МСТ и нуклеиновые кислоты, кодирующие белки МСТ, которые были введены в клетку. Примеры агентов, ингибирующих активность МСТ или экспрессию, включают небольшие молекулы и антисмысловые молекулы нуклеиновых кислот МСТ.
Другой аспект данного изобретения относится к способам модуляции выходов желаемого соединения из клетки, включающим введение гена МСТ дикого типа или мутантного МСТ в клетку, либо сохраняемого на отдельной плазмиде, либо интегрируемого в геном клетки-хозяина. При интегрировании в геном такая интеграция может быть случайной или она может происходить посредством гомологичной рекомбинации, так что нативный ген заменяется вводимой копией, обусловливая модуляцию продуцирования желаемого соединения из этой клетки. В предпочтительном варианте указанные выходы увеличиваются. В другом предпочтительном варианте указанный химический продукт является химическим продуктом тонкого органического синтеза. В конкретном предпочтительном варианте указанный химический продукт тонкого органического синтеза является аминокислотой. В особенно предпочтительном варианте указанная аминокислота является L-лизином.
Подробное описание изобретения
Данное изобретение представляет молекулы нуклеиновой кислоты и белка МСТ, которые участвует в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте соединений через эти мембраны. Молекулы данного изобретения могут быть использованы в модуляции получения химических продуктов тонкого органического синтеза из микроорганизмов, таких как С. glutamicum, непосредственно (например, когда сверхэкспрессия или оптимизация белка, участвующего в биосинтезе жирных кислот, оказывает прямое действие на выход, продуктивность и/или эффективность продуцирования жирных кислот из модифицированного С. glutamicum) или могут иметь опосредованное действие, которое тем не менее приводит к увеличению выхода, продуцирования и/или эффективности продуцирования желаемого соединения (например, когда модуляция метаболизма компонентов клеточной мембраны приводит к изменениям в выходе, продуцировании и/или эффективности продуцирования или составе клеточной мембраны, что, в свою очередь, влияет на продуцирование одного или большего количества химических продуктов тонкого органического синтеза). Аспекты данного изобретения дополнительно объясняются ниже.
I. Химические продукты тонкого органического синтеза
Термин «химический продукт тонкого органического синтеза» является признанным в данной области и включает молекулы, продуцируемые организмом, которые имеют применения в различных отраслях промышленности, таких как, но не только, фармацевтическая, сельскохозяйственная и косметическая отрасли промышленности. Такие соединения включают органические кислоты, такие как винная кислота, итаконовая кислота и диаминопимелиновая кислота, как протеиногенные, так и непротеиногенные аминокислоты, пуриновые и пиримидиновые основания, нуклеозиды и нуклеотиды (описанные, например, в Kuninaka, А. (1996) Nucleotides and related compounds, p.561-612, in Biotechnology vol.6, Rehm et al., eds. VCH: Weinheim, и содержащихся в них ссылках), липиды, насыщенные и ненасыщенные жирные кислоты (например, арахидоновая кислота), диолы (например, пропандиол и бутандиол), углеводы (например, гиалуроновая кислота и трегалоза), ароматические соединения (например, ароматические амины, ванилин и индиго), витамины и кофакторы (описанные в Ullmann's Encyclopedia of Industrial Chemistry, vol.A27, "Vitamins", p.443-613 (1996) VCH: Weinheim и ссылках в них; и Ong, A.S., Niki, E. and Packer, L. (1995) "Nutrition, Lipids, Health, and Disease" Proceedings of the UNESCO/Confederation of Scientific and Technological Associations in Malaysia, and Society for Free Radical Research-Asia, held Sept.1-3, 1994 at Penang, Malaysia, AOCS Press, (1995)), ферменты, поликетиды (Cane et al., (1998) Science 282: 63-68) и все другие химические продукты, описанные в Gutcho (1983) Chemicals by Fermentation, Noyes Data Corporation, ISBN: 0818805086 и имеющихся в этой работе ссылках. Метаболизм и применения некоторых из этих химических продуктов тонкого органического синтеза дополнительно раскрываются ниже.
А. Метаболизм и применения аминокислот
Аминокислоты составляют основные структурные единицы всех белков и как таковые являются важными для нормального клеточного функционирования во всех организмах. Термин «аминокислота» является признанным в данной области. Протеиногенные аминокислоты, которыми являются 20 видов, служат в качестве структурных единиц для белков, в которых они связаны пептидными связями, тогда как непротеиногенные аминокислоты (сотни которых известны) обычно не обнаруживаются в белках (см. Ullmann's Encyclopedia of Industrial Chemistry, vol.A2, p.57-97 VCH: Weinheim (1985)). Аминокислоты могут быть в D- или L-конфигурации, хотя L-аминокислоты являются обычно единственным типом, обнаруживаемым в природно встречающихся белках. Биосинтетические и пути разложения каждой из 20 протеиногенных аминокислот были хорошо охарактеризованы как в прокариотических, так и в эукариотических клетках (см., например, Stryer, L. Biochemistry, 3rd edition, pages 578-590 (1988)). «Незаменимые» аминокислоты (гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан и валин), названные так, поскольку они обычно являются необходимыми в питании вследствие сложности их биосинтеза, легко превращаются посредством простых биосинтетических путей в остальные 11 «не-незаменимых» аминокислот (аланин, аргинин, аспарагин, аспартат, цистеин, глутамат, глутамин, глицин, пролин, серин и тирозин). Высшие животные действительно сохраняют способность синтезировать некоторые из этих аминокислот, но незаменимые аминокислоты должны предоставляться из пищевого рациона, для того, чтобы имел место нормальный синтез белков.
Помимо их функции в биосинтезе белков, эти аминокислоты сами по себе представляют интерес, и было обнаружено, что многие из них имеют различные применения в кормовой, пищевой, химической, косметической, сельскохозяйственной и фармацевтической отраслях промышленности. Лизин является важной аминокислотой в питании не только человека, но также моногастрических животных (животных с однокамерным желудком), таких как домашняя птица и свинья. Глутамат наиболее часто используется в качестве вкусовой добавки (мононатрий-глутамат, MSG) и широко применяется в пищевой промышленности, так же как и аспартат, фенилаланин, глицин и цистеин. Глицин, L-метионин и триптофан используются в фармацевтической промышленности. Глутамин, валин, лейцин, изолейцин, гистидин, аргинин, пролин, серин и аланин применяют как в фармацевтической, так и в косметической промышленности. Треонин, триптофан и D/L-метионин являются общепринятыми пищевыми добавками (Leuchtenberger, W. (1996) Amino acids - technical production and use, p.466-502 in Rehm et al., (eds.) Biotechnology vol.6, chapter 14a, VCH: Weinheim). Кроме того, было обнаружено, что эти аминокислоты применимы в качестве предшественников для синтеза синтетических аминокислот и белков, таких как N-ацетилцистеин, S-карбоксиметил-L-цистеин, (S)-5-гидрокситриптофан и другие, описанные в Ullmann's Encyclopedia of Industrial Chemistry, vol.A2, p.57-97 VCH: Weinheim, 1985.
Биосинтез этих природных аминокислот в организмах, способных их продуцировать, таких как бактерии, был хорошо охарактеризован (в отношении обзора бактериального биосинтеза аминокислот и его регуляции см., например, Umbarger, H.E. (1978) Ann. Rev. Biochem. 47: 533-606). Глутамат синтезируется восстановительным аминированием α-кетоглутарата, промежуточного продукта цикла лимонной кислоты. Глутамин, пролин и аргинин, каждый, образуются затем из глутамата. Биосинтез серина является трехстадийным процессом, начинающимся с 3-фосфоглицерата (промежуточного продукта в гликолизе) и приводящим к этой аминокислоте после стадий окисления, переаминирования и гидролиза. Как цистеин, так и глицин образуются из серина; первый посредством конденсации гомоцистеина с серином, а последний переносом β-углеродного атома боковой цепи к тетрагидрофолату, в реакции, катализируемой серин-трансгидроксиметилазой. Фенилаланин и тирозин синтезируются из предшественников гликолитического и пентозофосфатного пути эритрозо-4-фосфата и фосфоенолпирувата в 9-стадийном биосинтетическом пути, который отличается только на конечных двух стадиях после синтеза префената. Триптофан также образуется из этих двух исходных молекул, но его синтез является 11-стадийным путем. Тирозин может быть также синтезирован из фенилаланина, в реакции, катализируемой фенилаланингидроксилазой. Аланин, валин и лейцин - все являются биосинтетическими продуктами пирувата, конечного продукта гликолиза. Аспартат образуется из оксалоацетата, промежуточного продукта цикла лимонной кислоты. Аспарагин, метионин, треонин и лизин, каждый, образуются преобразованием аспартата. Изолейцин образуется из треонина. Сложный 9-стадийный путь приводит к образованию гистидина из 5-фосфорибозил-1-пирофосфата, активированного сахара.
Аминокислоты в превышающем потребности белкового синтеза клетки количестве не могут запасаться и вместо этого разрушаются с образованием промежуточных продуктов для основных метаболических путей клетки (в отношении обзора см. Stryer, L. Biochemistry 3rd ed. Ch.21 "Amino Acid Degradation and. the Urea Cycle" p.495-516 (1998)). Хотя клетка способна превращать нежелательные аминокислоты в полезные метаболические промежуточные продукты, получение аминокислот является дорогостоящим в отношении энергии, молекул предшественников и ферментов, необходимых для их синтеза. Таким образом, неудивительно, что биосинтез аминокислот регулируется ингибированием по типу обратной связи, в котором присутствие конкретной аминокислоты служит для замедления или полной остановки ее собственного образования (в отношении обзора механизмов по типу обратной связи в путях биосинтеза аминокислот см. Stryer, L. Biochemistry 3rd ed. Ch.24: "Biosynthesis of Amino Acids and Heme" p.575-600 (1988)). Таким образом, выход любой конкретной аминокислоты лимитирован количеством этой аминокислоты, присутствующим в клетке.
В. Метаболизм и применения витаминов, кофакторов и нутрацевтических веществ (пищевых добавок)
Витамины, кофакторы и нутрацевтические вещества (пищевые добавки) составляют другую группу молекул, которые высшие животные не синтезируют вследствие утраты способности их синтеза и которые, следовательно, должны приниматься животными с пищей, хотя они легко синтезируются другими организмами, такими как бактерии. Эти молекулы либо сами являются биологически активными веществами, либо они являются предшественниками биологически активных веществ, которые могут служить в качестве носителей электронов или промежуточных продуктов в многочисленных метаболических путях. Помимо их питательной ценности, эти соединения имеют также значительную промышленную ценность в качестве красящих агентов, антиоксидантов и катализаторов или других технологических вспомогательных средств. (В отношении обзора структуры, активности и промышленных применений этих соединений см., например, Ullmann's Encyclopedia of Industrial Chemistry, "Vitamins" vol.A27, p.443-613 VCH: Weinheim, 1996). Термин "витамин" является признанным в данной области и включает в себя питательные вещества, которые необходимы организму для нормального функционирования, но которые этот организм не может сам синтезировать. Группа витаминов может охватывать кофакторы и добавочные питательные (нутрацевтические) соединения (пищевые добавки). Термин "кофактор" включает в себя небелковые соединения, требуемые для того, чтобы имела место нормальная ферментативная активность. Такие соединения могут быть органическими или неорганическими; молекулы кофакторов данного изобретения являются предпочтительно органическими. Термин "нутрацевтическое вещество" включает в себя пищевые добавки, полезные для здоровья растений и животных, в частности, людей. Примерами таких молекул являются витамины, антиоксиданты и также некоторые липиды (например, полиненасыщенные жирные кислоты).
Биосинтез этих молекул в организмах, способных их продуцировать, таких как бактерии, был в значительной степени охарактеризован (Ullmann's Encyclopedia of Industrial Chemistry, "Vitamins" vol.A27, p.443-613, VCH: Weinheim, 1996; Michal, G. (1999) Biochemical Pathways: An Atlas of Biochemistry and Molecular Biology, John Wiley and Sons; Ong, A.S., Niki, E and Packer, L. (1995) "Nutrition, Lipids, Health, and Disease" Proceedings of the UNESCO/Confederation of Scientific and Technological Associations in Malaysia, and Society for Free Radical Research-Asia, held Sept.1-3, 1994 at Penang, Malaysia, AOCS Press: Champaign, IL X, 374 S).
Тиамин (витамин B1) образуется химическим связыванием пиримидиновой и тиазоловой частей молекулы. Рибофлавин (витамин В2) синтезируется из гуанозин-5'-трифосфата (ГТФ) и рибозо-5'-фосфата. Рибофлавин, в свою очередь, используется для синтеза флавинмононуклеотида (FMN) и флавинадениндинуклеотида (FAD). Семейство соединений, совокупно называемых «витамином Be» (например, пиридоксин, пиридоксамин, пиридоксаль-5'-фосфат и коммерчески используемый пиридоксингидрохлорид), включает соединения, являющиеся производными общей структурной единицы, 5-гидрокси-6-метилпиридина. Пантотенат (пантотеновая кислота, (R)-(+)-N-(2,4-дигидрокси-3,3-диметил-1-оксобутил)-β-аланин) может быть получен химическим синтезом или ферментацией. Конечные стадии в биосинтезе пантотената состоят из АТФ-управляемой конденсации β-аланина и пантоевой кислоты. Ферменты, ответственные за стадии биосинтеза для превращения в пантоевую кислоту, в β-аланин и для конденсации в пантотеновую кислоту, являются известными. Метаболически активной формой пантотената является Кофермент А, биосинтез которого протекает в виде 5 ферментативных стадий. Пантотенат, пиридоксаль-5'-фосфат, цистеин и АТФ являются предшественниками Кофермента А. Эти ферменты катализируют не только образование пантотената, но также образование (R)-пантоевой кислоты, (R)-пантолактона, (R)-пантенола (провитамина В5) (и его производных) и кофермента А.
Биосинтез биотина из молекулы-предшественника пимелоил-КоА в микроорганизмах был подробно исследован, и были идентифицированы несколько участвующих в нем генов. Было обнаружено, что многие из соответствующих белков участвуют также в синтезе Fe-кластера и являются членами белков nifS-класса. Липоевую кислоту получают из октановой кислоты и она служит в качестве кофермента в энергетическом обмене, где становится частью пируватдегидрогеназного комплекса и α-кетоглутарат-дегидрогеназного комплекса. Фолаты являются группой веществ, которые все являются производными фолиевой кислоты, которая, в свою очередь, образуется из L-глутаминовой кислоты, п-аминобензойной кислоты и 6-метилптерина. Биосинтез фолиевой кислоты и ее производных, начинающийся с промежуточных продуктов метаболизма гуанозин-5'-трифосфата (ГТФ), L-глутаминовой кислоты и п-аминобензойной кислоты, был подробно исследован в некоторых микроорганизмах.
Корриноиды (такие как кобаламины и, в частности, витамин В12) и порфирины принадлежат к группе химических продуктов, характеризующихся системой тетрапиролльного кольца. Биосинтез витамина B12 является достаточно сложным, так что он еще не был полностью охарактеризован, но многие участвующие в нем ферменты и субстраты в настоящее время известны. Никотиновая кислота (никотинат) и никотинамид являются производными пиридина, которые называют также "ниацином". Ниацин является предшественником важных коферментов НАД (никотинамидадениндинуклеотида) и НАДФ (никотинамидадениндинуклеотидфосфата) и их восстановленных форм.
Крупномасштабное получение этих соединений в значительной степени основано на бесклеточных химических синтезах, хотя некоторые из этих химических продуктов были также получены крупномасштабным культивированием микроорганизмов, такие как рибофлавин, витамин В6, пантотенат и биотин. Только витамин B12 получают исключительно ферментацией вследствие сложности его синтеза. Методологии in vitro требуют значительных затрат материалов и времени, часто при высокой стоимости.
С. Метаболизм и применения пуринов, пиримидинов, нуклеозидов и нуклеотидов
Гены метаболизма пуринов и пиримидинов и их соответствующие белки являются важными мишенями для терапии опухолевых заболеваний и вирусных инфекций. Термины "пурин" или "пиримидин" включают в себя азотистые основания, которые являются составляющими нуклеиновых кислот, коферментов и нуклеотидов. Термин "нуклеотид" включает в себя основные структурные единицы молекул нуклеиновых кислот, которые состоят из азотистого основания, пентозного сахара (в случае РНК этот сахар является рибозой; в случае ДНК этот сахар является D-дезоксирибозой) и фосфорной кислоты. Термин "нуклеозид" включает в себя молекулы, которые служат в качестве предшественников для нуклеотидов, но которые не содержат части, являющейся фосфорной кислотой, которую имеют нуклеотиды. Ингибированием биосинтеза этих молекул или их мобилизации для образования молекул нуклеиновых кислот можно ингибировать синтез РНК и ДНК; ингибированием этой активности нацеленным на раковые клетки образом можно ингибировать способность опухолевых клеток делиться и размножаться. Кроме того, имеются нуклеотиды, которые не образуют молекул нуклеиновых кислот, а служат в качестве запасов энергии (т.е. АМФ) или в качестве коферментов (т.е. FAD и NAD).
В нескольких публикациях было описано применение этих химических продуктов для медицинских показаний, посредством влияния на метаболизм пуринов и/или пиримидинов (например, Christopherson, R.I. and Lyons, S.D. (1990) "Potent inhibitors of de novo pyrimidine and purine biosynthesis as chemotherapeutic agents." Med. Res. Reviews 10: 505-548). Исследования ферментов, участвующих в метаболизме пуринов и пиримидинов, было сосредоточено на развитии новых лекарственных средств, которые могут быть использованы, например, в качестве иммунодепрессантов или антипролиферантов (Smith, J.L., (1995) "Enzymes in nucleotide synthesis." Curr. Opin. Struct. Biol. 5: 752-757; (1995) Biochem. Soc. Transact. 23: 877-902). Однако, пуриновые и пиримидиновые основания, нуклеозиды и нуклеотиды имеют другие применения: в качестве промежуточных продуктов в биосинтезе некоторых химических продуктов тонкого органического синтеза (например, тиамина, S-аденозилметионина, фолатов или рибофлавина), в качестве энергоносителей для клетки (например, АТФ или ГТФ) и для самих химических продуктов, обычно используемых в качестве усилителей аромата (например, ИМФ или ГМФ) или для некоторых медицинских применений (см., например, Kuninaka, A. (1996) Nucleotides and Related Compounds in Biotechnology vol.6, Rehm et al., eds. VCH: Weinheim, p.561-612). Кроме того, ферменты, участвующие в метаболизме пуринов, пиримидинов, нуклеозидов или нуклеотидов, все больше используются в качестве мишеней, против которых разрабатываются химические продукты для защиты урожая сельскохозяйственных культур, в том числе фунгициды, гербициды и инсектициды.
Метаболизм этих соединений в бактериях был охарактеризован (в отношении обзора см., например, Zaikin, H. and Dixon, J.E. (1992) "de novo purine nucleotide biosynthesis", in: Progress in Nucleic Acid Research and Molecular Biology, vol. 42, Academic Press: p. 259-287 и Michal, G. (1999) "Nucleotides and Nucleosides", Chapter 8 in: Biochemical Pathways: An Atlas of Biochemistry and Molecular Biology, Wiley: New York). Метаболизм пуринов был объектом интенсивного исследования и он является существенным для нормального функционирования клетки. Нарушенный метаболизм пуринов в высших животных может вызывать серьезное заболевание, такое как подагра. Пуриновые нуклеотиды синтезируются из рибозо-5-фосфата, в ряде стадий через промежуточное соединение инозин-5'-фосфат (ИМФ), приводя к образованию гуанозин-5'-монофосфата (ГМФ) или аденозин-5'-монофосфата (АМФ), из которых легко образуются трифосфатные формы, используемые в качестве нуклеотидов. Эти соединения используются также в качестве энергозапасов, и таким образом их разложением обеспечивается энергия для многих различных биохимических процессов в клетке. Биосинтез пиримидинов протекает посредством образования уридин-5'-монофосфата (УМФ) из рибозо-5-фосфата. УМФ, в свою очередь, превращается в цитидин-5'-трифосфат (ЦТФ). Дезоксиформы всех этих нуклеотидов образуются в одностадийной реакции восстановления из дифосфатрибозной формы нуклеотида в дифосфатную дезоксирибозную форму этого нуклеотида. При фосфорилировании эти молекулы способны участвовать в синтезе ДНК.
D. Метаболизм и применения трегалозы
Трегалоза состоит из двух молекул глюкозы, связанных в α,α-1,1-связи. Ее обычно используют в пищевой промышленности в качестве подслащивающего вещества, добавки для высушенных или замороженных пищевых продуктов и в напитках. Однако, она применяется также в фармацевтической, косметической и биотехнологической отраслях промышленности (см., например, Nishimoto et al., (1998) U.S. Patent №5759610; Singer, M.A. and Lindquist, S. (1998) Trends Biotech. 16: 460-467; Paiva, C.L.A. and Panek, A.D. (1996) Biotech. Ann. Rev.2: 293-314; и Shiosaka, M. (1997) J. Japan. 172: 97-102). Трегалоза образуется ферментами из многих микроорганизмов и природно высвобождается в окружающую среду, из которой она может быть собрана с использованием способов, известных в данной области.
II. Биосинтез мембран и трансмембранный транспорт
Клеточные мембраны обслуживают различные функции в клетке. Во-первых и прежде всего, мембрана дифференцирует содержимое клетки от окружающей среды, создавая таким образом целостность клетки. Мембраны могут также служить в качестве барьеров для вхождения вредных или нежелательных соединений, а также выхода желательных соединений. Клеточные мембраны являются по их природе непроницаемыми для необлегченной диффузии гидрофильных соединений, таких как белки, молекулы воды и ионы, вследствие их структуры: бислоя липидных молекул, в котором группы полярных головок обращены наружу (в направлении внешнего и внутреннего пространства клетки, соответственно), а неполярные хвосты обращены внутрь в центре бислоя, образуя гидрофобное основание (в отношении общего обзора структуры и функции мембран см. Gennis, R.B. (1989) Biomembranes, Molecular Structure and Function, Springer: Heidelberg). Этот барьер позволяет клеткам поддерживать относительно более высокую концентрацию желательных соединений и относительно более низкую концентрацию нежелательных соединений, в сравнении с окружающей средой, так как диффузия этих соединений эффективно блокируется мембраной. Однако, мембрана представляет также эффективный барьер для импорта желательных соединений и экспорта отработанных молекул. Для преодоления этой трудности клеточные мембраны включают многочисленные типы белков-переносчиков (транспортеров), которые способны облегчать трансмембранный транспорт различных типов соединений. Существуют два основных класса этих транспортных белков: поры или каналы и переносчики (транспортеры). Первые являются интегральными мембранными белками, иногда комплексами белков, которые образуют регулируемое отверстие через мембрану. Эта регуляция, или «воротный механизм» является обычно специфическим в отношении транспортируемых порой или каналом молекул, что делает эти трансмембранные конструкции селективно проницаемыми для специфического класса субстратов; например, калиевый канал сконструирован таким образом, что только ионы, имеющие заряд и размер, аналогичные заряду и размеру калия, могут проходить через него. Белки каналов и пор склонны иметь дискретные гидрофобные и гидрофильные домены, так что гидрофобная поверхность белка может связываться с внутренней частью мембраны, тогда как гидрофильная поверхность выстилает внутреннее пространство канала, обеспечивая таким образом защищенную гидрофильную среду, через которую может проходить выбранная гидрофильная молекула. Многие такие поры/каналы известны в данной области, в том числе каналы для ионов калия, кальция, натрия и хлорида.
Эта опосредуемая порами и каналами система облегченной диффузии ограничена очень небольшими молекулами, такими как ионы, так как поры или каналы, достаточно крупные для прохождения целых белков посредством облегченной диффузии, были бы неспособны предотвращать прохождение также и более мелких гидрофильных молекул. Транспорт молекул посредством этого процесса называют иногда "облегченной диффузией", так как для этого транспорта необходима движущая сила градиента концентрации. Пермеазы также делают возможной облегченную диффузию более крупных молекул, таких как глюкоза или другие сахара, в клетку, когда концентрация этих молекул на одной стороне мембраны является более высокой, чем концентрация на другой стороне (также называемой "унипортом", т.е. транспортом одного растворенного вещества через мембрану). В противоположность порам или каналам, эти интегральные мембранные белки (часто имеющие 6-14 простирающихся через мембрану α-спиралей) не образуют открытых каналов через мембрану, а связываются с молекулой-мишенью на поверхности мембраны и затем подвергаются конформационному сдвигу, так что молекула-мишень высвобождается на противоположной стороне мембраны.
Однако, клеткам необходим импорт или экспорт молекул против существующего градиента концентрации ("активный транспорт"), ситуации, в которой не может происходить облегченная диффузия. Существует два основных механизма, используемых клетками для такого мембранного транспорта: симпорт или антипорт, и сопряженный с энергией транспорт, например, опосредованный АВС-транспортерами. Системы симпорта и антипорта сопрягают движение двух различных молекул через мембрану (посредством пермеаз, имеющих два отдельных сайта связывания для двух различных молекул); в симпорте, обе молекулы транспортируются в одном и том же направлении, тогда как в антипорте одна молекула импортируется, тогда как другая экспортируется. Это является энергетически возможным, так как одна из двух молекул движется в соответствии с градиентом концентрации, и это энергетически благоприятное событие допускается только при сопутствующем движении желательного соединения против преобладающего градиента концентрации. Отдельные молекулы могут транспортироваться через мембрану против градиента концентрации в использующем энергию процессе, например, процессе, используемом АВС-транспортерами. В этой системе транспортный белок, расположенный в мембране, имеет АТФ-связывающую кассету; при связывании молекулы-мишени АТФ превращается в АДФ+Pi, и полученное высвобождение энергии используется для проведения перемещения молекулы-мишени на противоположную сторону мембраны, облегченного этим транспортером. В отношении более подробных описаний всех этих транспортных систем см.: Bamberg, E. et al., (1993) "Charge transport of ion pumps on lipid bilayer membranes", Q. Rev. Biophys. 26: 1-25; Findlay, J.B.C. (1991) "Structure and function in membrane transport systems", Curr. Opin. Struct. Biol. 1:804-810; Higgins, C.F. (1992) "ABC transporters from microorganisms to man", Ann. Rev. Cell Biol. 8: 67-113; Gennis, R.B. (1989) "Pores, Channels and Transporters", in: Biomembranes, Molecular Structure and Function, Springer: Heidelberg, p.270-322; и Nikaido, H. and Saier, H. (1992) "Transport proteins in bacteria: common themes in their design", Science 258: 936-942, и содержащиеся в каждой из этих работ ссылки.
Синтез мембран является хорошо охарактеризованным процессом, в котором участвует ряд компонентов, наиболее важными из которых являются липидные молекулы. Синтез липидов может быть разделен на две части: синтез жирных кислот и их присоединение к sn-глицерол-3-фосфату и добавление или модификация полярной группы головки. Типичные липиды, используемые в бактериальных мембранах, включают фосфолипиды, гликолипиды, сфинголипиды и фосфоглицериды. Синтез жирных кислот начинается с превращения ацетил-КоА либо в малонил-КоА ацетил-КоА-карбоксилазой, либо в ацетил-АСР ацетилтрансацилазой. После реакции конденсации эти две полученные молекулы образуют вместе ацетоацетил-АСР, который превращается посредством ряда реакций конденсации, восстановления и дегидратации с образованием молекулы жирной кислоты, имеющей желательную длину цепи. Образование ненасыщенных жирных кислот из таких молекул катализируется специфическими десатуразами либо аэробно, с использованием молекулярного кислорода, либо анаэробно (в отношении ссылки по синтезу жирных кислот см. F.C. Neidhardt et al. (1996) E. coli and Salmonella. ASM Press: Washington, D.C., p.612-636 и содержащиеся в этой работе ссылки; Lengeler et al. (eds) (1999) Biology of Procaryotes. Tieme: Stuttgart, New York, и содержащиеся в этой работе ссылки; и Magnuson, К. et al. (1993) Microbiological Reviews 57: 522-542, и ссылки в этих работах). Циклопропан-жирные кислоты (CFA) синтезируются специфической CFA-синтазой с использованием SAM в качестве ко-субстрата. Жирные кислоты с разветвленной цепью синтезируют из аминокислот с разветвленной цепью, которые деаминируют с получением 2-оксокислот с разветвленной цепью (см. Lengeler et al. (eds) (1999) Biology of Procaryotes. Thieme: Stuttgart, New York, и содержащиеся в этой работе ссылки). Другой существенной стадией в синтезе липидов является перенос жирных кислот на группы полярной головки при помощи, например, глицерофосфат-ацилтрансфераз. Комбинирование различных молекул-предшественников и биосинтетических ферментов приводит к получению различных молекул жирных кислот, которые оказывают сильное действие на состав мембраны.
III. Элементы и способы изобретения
Данное изобретение основывается, по меньшей мере частично, на обнаружении новых молекул, называемых здесь молекулами нуклеиновых кислот и белковыми молекулами МСТ, которые регулируют продуцирование клеточных мембран в С. glutamicum и управляют движением молекул через такие мембраны. В одном варианте молекулы МСТ участвуют в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. В предпочтительном варианте активность молекул МСТ данного изобретения в отношении регуляции продуцирования компонентов мембран и мембранного транспорта оказывает воздействие на продуцирование желательного химического продукта тонкого органического синтеза этим организмом. В особенно предпочтительном варианте молекулы МСТ данного изобретения модулируются в активности таким образом, что метаболические пути С. glutamicum, которые регулируют белки МСТ данного изобретения, модулируются в отношении выхода, продуцирования и/или эффективности продуцирования, и транспорт соединений через мембраны изменяется в эффективности, что либо прямо, либо опосредованно модулирует выход, продуцирование и/или эффективность продуцирования желаемого химического продукта тонкого органического синтеза С. glutamicum.
Выражение «белок МСТ» или «полипептид МСТ» включает белки, которые участвуют в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. Примеры белков МСТ включают белки, кодируемые генами МСТ, представленными в таблице 1 и имеющими нечетные номера SEQ ID NO:. Термины «ген МСТ» или «последовательность нуклеиновой кислоты МСТ» включают последовательности нуклеиновых кислот, кодирующих белок МСТ, которые состоят из кодирующего района, а также из соответствующих нетранслируемых 5'- и 3'-участков последовательности. Примеры генов МСТ включают гены, приведенные в таблице 1. Термины «продуцирование» или «продуктивность» являются признанными в данной области и включают в себя концентрацию продукта ферментации (например, желаемого химического продукта тонкого органического синтеза), образованного за конкретное время и в конкретном объеме ферментации (например, кг продукта в час на литр). Термин «эффективность продуцирования» включает в себя время, необходимое для конкретного уровня продуцирования, который должен быть достигнут (например, время, которое необходимо для клетки для достижения конкретной скорости выхода химического продукта тонкого органического синтеза). Термин «выход» или «выход продукт/углерод» является признанным в данной области и включает в себя эффективность превращения источника углерода в продукт (т.е. в химический продукт тонкого органического синтеза). Это обычно выражается как, например, кг продукта на кг источника углерода. Посредством увеличения выхода или продуцирования соединения количество извлеченных молекул этого соединения в конкретном количестве культуры на протяжении конкретного периода времени увеличивается. Термины «биосинтез» или «биосинтетический путь» являются признанными в данной области и включают в себя синтез соединения, предпочтительно органического соединения, клеткой из промежуточных соединений, который может включать в себя многостадийный и высокорегулируемый процесс. Термины «разложение» или «путь разложения» являются признанными в данной области и включают в себя распад соединения, предпочтительно органического соединения, клеткой до продуктов деградации (говоря в общем, меньших по размеру или менее сложных молекул), который может включать в себя многостадийный и высокорегулируемый процесс. Термин «метаболизм» является признанным в данной области и включает в себя всю сумму биохимических реакций, которые имеют место в организме. Тогда метаболизм конкретного соединения (например, метаболизм аминокислоты, например, глицина) включает в себя общие биосинтетические пути, пути модификации и разложения в клетке, связанные с этим соединением.
В другом варианте, молекулы МСТ данного изобретения способны модулировать продуцирование желаемой молекулы, такой как химический продукт тонкого органического синтеза, в микроорганизме, таком как С. glutamicum. Существуют ряд механизмов, при помощи которых изменение белка МСТ данного изобретения может непосредственно влиять на выход, продуцирование и/или эффективность продуцирования химического продукта тонкого органического синтеза из С. glutamicum, включающего такой измененный белок. Белки МСТ, участвующие в экспорте молекул химических продуктов тонкого органического синтеза из клетки, могут быть увеличенными в числе или активности, так что большие количества этих соединений секретируются во внеклеточную среду, из которой они могут быть более легко извлечены. Подобным образом, белки МСТ, участвующие в импорте питательных веществ, необходимых для биосинтеза одного или нескольких химических продуктов тонкого органического синтеза (например, фосфатных, сульфатных или азотсодержащих соединений и т.д.), могут быть увеличены в числе или активности, так что эти соединения-предшественники, соединения-кофакторы или промежуточные соединения увеличиваются в концентрации в клетке. Далее, сами жирные кислоты и липиды являются желательными химическими продуктами тонкого органического синтеза; оптимизацией активности или увеличением числа одного или нескольких белков МСТ данного изобретения, которые участвуют в биосинтезе этих соединений, или нарушением активности одного или нескольких белков МСТ, которые участвуют в разложении этих соединений, можно увеличивать выход, продуцирование и/или эффективность продуцирования молекул жирных кислот и липидов из С. glutamicum.
Мутагенез одного или более генов МСТ данного изобретения может также обуславливать наличие белков МСТ, имеющих измененные активности, которые опосредованно влияют на продуцирование одного или более желательных химических продуктов тонкого органического синтеза из С. glutamicum. Например, белки МСТ данного изобретения, участвующие в экспорте отработанных продуктов клетки, могут быть увеличены в количестве или активизированы таким образом, что отработанные продукты метаболизма клетки (возможно, увеличенных в количестве вследствие сверхпродуцирования желательного химического продукта тонкого органического синтеза), эффективно экспортируются, прежде чем они способны повредить нуклеотиды и белки в клетке (что уменьшало бы жизнеспособность данной клетки) или помешать биосинтетическим путям химических продуктов тонкого органического синтеза (что уменьшало бы выход, продуцирование или эффективность продуцирования желательного химического продукта тонкого органического синтеза). Далее, относительно большие внутриклеточные количества желательного химического продукта тонкого органического синтеза может само быть токсичным для клетки, так что посредством увеличения активности или числа транспортеров, способных экспортировать это соединение из клетки, можно увеличить жизнеспособность данной клетки в культуре, что, в свою очередь, приводит к большему числу клеток в культуре, продуцирующих желательный химический продукт тонкого органического синтеза. Белки МСТ данного изобретения могут быть подвергнуты манипулированию таким образом, что продуцируются относительные количества различных молекул липидов и жирных кислот. Это может оказывать сильное влияние на состав липидов мембраны клетки. Поскольку каждый тип липида имеет отличающиеся физические свойства, изменение в липидном составе мембраны может существенно изменять текучесть мембраны. Изменения в текучести мембраны могут влиять на транспорт молекул через мембрану, а также на целостность клетки, причем оба эти свойства имеют сильное влияние на продуцирование химических продуктов тонкого органического синтеза из С. glutamicum в крупномасштабной ферментационной культуре.
Выделенные последовательности нуклеиновых кислот данного изобретения содержатся в геноме штамма Corynebacterium glutamicum, доступного в Американской Коллекции Типовых культур, с номером депонирования АТСС 13032. Нуклеотидная последовательность выделенных ДНК МСТ С. glutamicum и предсказанные аминокислотные последовательности белков МСТ С. glutamicum показаны в Списке последовательностей в виде имеющих нечетные номера SEQ ID NO: и четные номера SEQ ID NO:, соответственно. Были проведены компьютерные анализы, которые классифицировали и/или идентифицировали эти нуклеотидные последовательности как последовательности, кодирующие белки, участвующие в метаболизме компонентов клеточных мембран, или белки, участвующие в транспорте соединений через такие мембраны.
Данное изобретение относится также к белкам, имеющим аминокислотную последовательность, которая является по существу гомологичной аминокислотной последовательности данного изобретения (например, последовательности, имеющей четный номер SEQ ID NO: Списка последовательностей). В применении здесь, белок, имеющий аминокислотную последовательность, которая является по существу гомологичной выбранной аминокислотной последовательности, является по меньшей мере на приблизительно 50% гомологичным выбранной аминокислотной последовательности, например, полной выбранной аминокислотной последовательности. Белок, имеющий аминокислотную последовательность, которая является по существу гомологичной выбранной аминокислотной последовательности, может также быть по меньшей мере на приблизительно 50-60%, предпочтительно по меньшей мере на приблизительно 60-70% и более предпочтительно по меньшей мере на приблизительно 70-80%, 80-90% или 90-95% и наиболее предпочтительно по меньшей мере на приблизительно 96%, 97%, 98%, 99% или более гомологичным выбранной аминокислотной последовательности.
Белок МСТ или его биологически активная часть или биологически активный фрагмент данного изобретения может участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum, или в транспорте молекул через эти мембраны, или может иметь одну или большее количество активностей, приведенных в таблице 1.
Различные аспекты данного изобретения описаны с дополнительными подробностями в следующих подразделах:
А. Выделенные молекулы нуклеиновых кислот
Один аспект данного изобретения относится к выделенным молекулам нуклеиновых кислот, которые кодируют полипептиды МСТ или их биологически активные части, а также фрагментам нуклеиновых кислот, достаточным для применения в качестве гибридизационных зондов или праймеров для идендификации или амплификации кодирующей МСТ нуклеиновой кислоты (например, ДНК МСТ). В применении здесь, термин "молекула нуклеиновой кислоты" включает в себя молекулы ДНК (например, кДНК или геномную ДНК) и молекулы РНК (например, мРНК) и аналоги ДНК или РНК, полученные с использованием нуклеотидных аналогов. Этот термин включает в себя также нетранслируемую последовательность, локализованную по обоим 3'- и 5'-концам кодирующего участка этого гена: по меньшей мере приблизительно 100 нуклеотидов последовательности выше (против хода транскрипции) от 5'-конца кодирующего участка и по меньшей мере приблизительно 20 нуклеотидов ниже (по холу транскрипции) от 3'-конца кодирующего участка этого гена. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но предпочтительно является двухцепочечной ДНК. "Выделенная" молекула нуклеиновой кислоты является молекулой нуклеиновой кислоты, которая отделена от других молекул нуклеиновых кислот, которые присутствуют в природном источнике этой нуклеиновой кислоты. Предпочтительно, "выделенная" нуклеиновая кислота не содержит последовательностей, которые природно фланкируют эту нуклеиновую кислоту (т.е. последовательностей, локализованных по 5'- и 3'-концам этой нуклеиновой кислоты) в геномной ДНК организма, из которого получена эта нуклеиновая кислота. Например, в различных вариантах выделенная молекула нуклеиновой кислоты МСТ может содержать менее, чем приблизительно 5 т.п.н., 4 т.п.н., 3 т.п.н., 2 т.п.н., 1 т.п.н., 0,5 т.п.н. или 0,1 т.п.н. нуклеотидных последовательностей, которые природно фланкируют эту молекулу нуклеиновой кислоты в геномной ДНК клетки, из которой получена эта нуклеиновая кислота (например, клетки С. glutamicum). Кроме того, "выделенная" молекула нуклеиновой кислоты, например, молекула ДНК, может по существу не содержать другого клеточного материала или культуральной среды при получении рекомбинантными способами или при химическом синтезе химических предшественников или других химических продуктов.
Молекула нуклеиновой кислоты данного изобретения, например, молекула нуклеиновой кислоты, имеющая нуклеотидную последовательность имеющих нечетные номера SEQ ID NO: Списка последовательностей, или ее часть, может быть выделена с использованием стандартных способов молекулярной биологии и информации последовательности, представленной здесь. Например, ДНК МСТ С. glutamicum может быть выделена из библиотеки С. glutamicum с использованием всей или части одной из имеющих нечетные номера SEQ ID NO: последовательностей Списка последовательностей в качестве гибридизационного зонда и стандартных способов гибридизации (например, описанных в Sambrook J., Fritsh, E.F., and Maniatis, Т. Molecular Cloning. A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989). Кроме того, молекула нуклеиновой кислоты, охватывающая полную последовательность или часть одной из последовательностей нуклеиновых кислот данного изобретения (например, имеющих нечетные номера SEQ ID NO: Списка последовательностей), может быть выделена полимеразной цепной реакцией с использованием олигонуклеотидных праймеров, сконструированных на основе этой последовательности (например, молекула нуклеиновой кислоты, охватывающая всю последовательность или часть одной из последовательностей нуклеиновых кислот данного изобретения (например, имеющих нечетные номера SEQ ID NO: Списка последовательностей), может быть выделена полимеразной цепной реакцией с использованием олигонуклеотидных праймеров, сконструированных на основе той же самой последовательности). Например, мРНК может быть выделена из нормальных эндотелиальных клеток (например, процедурой экстракции гуанидинийтиоцианатом Chirgwin et al. (1979) Biochemistry 18: 5294-5299), а ДНК может быть получена с использованием обратной транскриптазы (например, обратной транскриптазы Moloney MLV, доступной из Gibco/BRL, Bethesda, MD; или обратной транскриптазы AMV, доступной из Seikagaku America, Inc., St. Petersburg, FL). Синтетические олигонуклеотидные праймеры для амплификации при помощи полимеразной цепной реакции могут быть сконструированы на основе одной из нуклеотидных последовательностей, показанных в Списке последовательностей. Нуклеиновая кислота данного изобретения может быть амплифицирована с использованием кДНК или, альтернативно, геномной ДНК, в качестве матрицы и подходящих олигонуклеотидных праймеров в соответствии со стандартными способами ПЦР-амплификации. Амплифицированная таким образом нуклеиновая кислота может быть клонирована в подходящий вектор и охарактеризована анализом последовательности ДНК. Кроме того, олигонуклеотиды, соответствующие нуклеотидной последовательности МСТ, могут быть получены стандартными синтетическими способами, например, с использованием автоматического ДНК-синтезатора.
В предпочтительном варианте выделенная молекула нуклеиновой кислоты данного изобретения содержит одну из нуклеотидных последовательностей, показанных в Списке последовательностей. Последовательности нуклеиновых кислот данного изобретения, приведенные в Списке последовательностей, соответствуют ДНК МСТ Corynebacterium glutamicum данного изобретения. Эта ДНК содержит последовательности, кодирующие белки МСТ (т.е. "кодирующий участок", показанный в каждой имеющей нечетный номер SEQ ID NO: последовательности в Списке последовательностей), а также 5'-нетранслируемые последовательности и 3'-нетранслируемые последовательности, также показанные в каждой имеющей нечетный номер SEQ ID NO: последовательности в Списке последовательностей. Альтернативно, молекула нуклеиновой кислоты может содержать только кодирующий участок любой из последовательностей нуклеиновых кислот Списка последовательностей.
Для целей данной заявки, должно быть понятно, что каждая из последовательностей нуклеиновых кислот или аминокислотных последовательностей, приведенных в Списке последовательностей, имеет идентифицирующий номер RXA, RXN, RXS или RXC, имеющий обозначение "RXA", "RXN", "RXS" или "RXC", с последующими 5 цифрами (т.е. RXA02099, RXN03097, RXS00148 или RXC01748). Каждая из этих последовательностей нуклеиновых кислот содержит до трех частей: верхний 5'-участок (против хода транскрипции), кодирующий участок и нижний участок (по ходу транскрипции). Каждый из этих трех участков идентифицирован тем же самым обозначением RXA, RXN, RXS или RXC для избежания путаницы. Тогда ссылка на "одну из имеющих нечетные номера последовательностей Списка последовательностей" относится к любой из последовательностей нуклеиновых кислот в Списке последовательностей, которые могут быть также различены отличающимися обозначениями RXA, RXN, RXS или RXC. Кодирующий участок каждой из этих последовательностей транслируется в соответствующую аминокислотную последовательность, которая также представлена в Списке последовательностей, в виде имеющих четные номера SEQ ID NO: непосредственно после соответствующей последовательности нуклеиновой кислоты. Например, кодирующий участок для RXA03097 представлен в SEQ ID NO:1, тогда как аминокислотная последовательность, которую он кодирует, представлена в виде SEQ ID NO:2. Последовательности молекул нуклеиновых кислот данного изобретения идентифицируются теми же самыми обозначениями RXA, RXN, RXS или RXC, что и молекулы аминокислот, которые они кодируют, так что они могут быть легко соотнесены друг с другом. Например, аминокислотные последовательности, обозначенные RXA02099, RXN03097, RXS00148 или RXC01748, являются трансляциями кодирующего участка нуклеотидных последовательностей молекул нуклеиновых кислот RXA02099, RXN03097, RXS00148 или RXC01748, соответственно. Соответствие между нуклеотидными и аминокислотными последовательностями RXA, RXN, RXS и RXC данного изобретения и соответствующими SEQ ID NO: показано в таблице 1. Например, как показано в таблице 1, нуклеотидная последовательность RXA00104 является SEQ ID NO:5, а аминокислотная последовательность RXA00104 является SEQ ID NO:6.
Несколько генов данного изобретения являются "F-обозначенными генами». F-обозначенный ген включает гены, представленные в таблице 1, которые имеют "F" перед обозначением RXA, RXN, RXS или RXC. Например, последовательность SEQ ID NO:11, обозначенная, как указано в таблице 1, как "F RXA02581", является F-обозначенным геном, как и SEQ ID NO:31, 33 и 43 (обозначенные в таблице 1 как "F RXA02487", "F RXA02490" и "F RXA02809", соответственно).
В одном варианте молекулы нуклеиновых кислот данного изобретения не включают молекулы нуклеиновых кислот, представленные в таблице 2. В случае гена dapD, последовательность для этого гена была опубликована Wehrmann, A., et al., (1998) J. Bacteriol. 180(12): 3159-3165. Однако, последовательность, полученная авторами данной заявки, является значительно более длинной, чем опубликованная версия. Авторы считают, что опубликованная версия основывается на неправильном стартовом (инициирующем) кодоне и, следовательно, представляет только фрагмент истинного кодирующего участка.
В другом предпочтительном варианте выделенная молекула нуклеиновой кислоты данного изобретения содержит молекулу нуклеиновой кислоты, которая является комплементом одной из нуклеотидных последовательностей данного изобретения (например, последовательности, имеющей нечетный номер SEQ ID NO: Списка последовательностей), или ее часть. Молекула нуклеиновой кислоты, которая комплементарна одной из нуклеотидных последовательностей данного изобретения, является последовательностью, которая достаточно комплементарна одной из нуклеотидных последовательностей в Списке последовательностей (например, последовательности, имеющей нечетный номер SEQ ID NO:), так что она может гибридизоваться с одной из нуклеотидных последовательностей данного изобретения с образованием в результате стабильного дуплекса.
Еще в одном предпочтительном варианте выделенная молекула нуклеиновой кислоты данного изобретения содержит нуклеотидную последовательность, которая является по меньшей мере на приблизительно 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59% или 60%, предпочтительно по меньшей мере на приблизительно 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69% или 70%, более предпочтительно по меньшей мере на приблизительно 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79% или 80%, 81%, 82%, 84%, 85%, 86%, 87%, 88%, 89% или 90%, или 91%, 92%, 94%, или даже более предпочтительно на по меньшей мере приблизительно 95%, 96%, 97%, 98%, 99% или более гомологичной нуклеотидной последовательности данного изобретения (например, последовательности, имеющей нечетный номер SEQ ID NO: Списка последовательностей), или ее часть. Подразумевается, что диапазоны и величины идентичности, промежуточные относительно указанных выше диапазонов (например, идентичные на 70-90% или идентичные на 80-95%), также охватываются данным изобретением. Например, подразумевается, что включены также диапазоны величин идентичности, использующие комбинацию любых из цитированных выше величин в качестве верхнего и/или нижнего пределов. В дополнительном предпочтительном варианте выделенная молекула нуклеиновой кислоты данного изобретения содержит нуклеотидную последовательность, которая гибридизуется, например, гибридизуется при строгих условиях, с одной из нуклеотидных последовательностей данного изобретения или с ее частью.
Кроме того, молекула нуклеиновой кислоты данного изобретения может содержать только часть кодирующего участка последовательности одной из имеющих нечетные номера SEQ ID NO: Списка последовательностей, например, фрагмент, который может быть использован в качестве зонда или праймера, или фрагмента, кодирующего биологически активную часть белка МСТ. Эти нуклеотидные последовательности, определенные из клонирования генов МСТ из С. glutamicum, позволяют получать зонды и праймеры, сконструированные для применения в идентификации и/или клонировании МСТ-гомологов в других клеточных типах и организмах, а также МСТ-гомологов из других Corynebacteria или родственных видов. Зонд/праймер обычно содержит по существу очищенный олигонуклеотид. Олигонуклеотид обычно содержит участок нуклеотидной последовательности, который гибридизуется при строгих условиях по меньшей мере с приблизительно 12, предпочтительно приблизительно 25, более предпочтительно приблизительно 40, 50 или 75 последовательными нуклеотидами смысловой цепи одной из нуклеотидных последовательностей данного изобретения (например, последовательностей, имеющих нечетные номера SEQ ID NO: Списка последовательностей), антисмысловой последовательностью одной из этих последовательностей или их природно встречающимися мутантами. Праймеры на основе нуклеотидной последовательности данного изобретения могут быть использованы в ПЦР-реакциях для клонирования МСТ-гомологов. Зонды на основе нуклеотидных последовательностей МСТ могут быть использованы для обнаружения транскриптов или геномных последовательностей, кодирующих те же самые или гомологичные белки. В предпочтительных вариантах такой зонд дополнительно содержит группу метки, присоединенную к нему, например, эта группа метки может быть радиоактивным изотопом, флуоресцентным соединением, ферментом или кофактором фермента. Такие зонды могут быть использованы как часть диагностического тест-набора для идентификации клеток, которые имеют нарушенную экспрессию белка МСТ, например, измерением уровня кодирующей МСТ нуклеиновой кислоты в пробе клеток, например, детектированием уровней мРНК МСТ, или определением, был ли геномный ген МСТ мутирован или делетирован.
В одном варианте молекула нуклеиновой кислоты данного изобретения кодирует белок или его часть, которые включают аминокислотную последовательность, которая является достаточно гомологичной аминокислотной последовательности данного изобретения (например, последовательностей, имеющих четные номера SEQ ID NO: Списка последовательностей), так что этот белок или его часть сохраняет способность участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. В применении здесь, термин "достаточно гомологичные" относится к белкам или их частям, которые имеют аминокислотные последовательности, которые включают минимальное число идентичных или эквивалентных (например, аминокислотный остаток, который имеет такую же боковую цепь, что и аминокислотный остаток в последовательности одной из имеющих четные номера SEQ ID NO: Списка последовательностей) аминокислотных остатков относительно аминокислотной последовательности данного изобретения, так что этот белок или его часть способны участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. Белковые члены таких метаболических путей компонентов мембран или систем мембранного транспорта, как описано здесь, могут играть роль в продуцировании и секреции одного или более химических продуктов тонкого органического синтеза. Примеры таких активностей также описаны здесь. Таким образом, "функция белка МСТ" вносит вклад, прямо или опосредованно, в выход, продуктивность и/или эффективность продуцирования одного или более химических продуктов тонкого органического синтеза. Примеры активностей белков МСТ приведены в таблице 1.
В другом варианте этот белок является по меньшей мере на приблизительно 50-60%, предпочтительно по меньшей мере на приблизительно 60-70% и более предпочтительно по меньшей мере на приблизительно 70-80%, 80-90%, 90-95% и наиболее предпочтительно по меньшей мере на приблизительно 96%, 97%, 98%, 99% или более гомологичным полной аминокислотной последовательности данного изобретения (например, последовательности из имеющих четные номера SEQ ID NO: Списка последовательностей).
Части белков, кодируемые молекулами нуклеиновых кислот МСТ данного изобретения, являются предпочтительно биологически активными частями одного из белков МСТ. В применении здесь, термин "биологически активная часть белка МСТ" включает часть, например, домен/мотив, белка МСТ, которая участвует в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны, или имеет активность, приведенную в таблице 1. Для определения, может ли белок МСТ или его биологически активная часть участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны, может быть выполнен анализ ферментативной активности. Такие способы анализов хорошо известны в данной области специалистам с обычной квалификацией в данной области, как подробно описано в примере 8 раздела иллюстрации примерами.
Дополнительные фрагменты нуклеиновых кислот, кодирующие биологически активные части белка МСТ, могут быть получены выделением части одной из аминокислотных последовательностей данного изобретения (например, последовательности, имеющей четный номер SEQ ID NO: Списка последовательностей), экспрессией кодируемой части белка или пептида МСТ (например, рекомбинантной экспрессией in vitro) и оценкой активности кодируемой части этого белка или пептида МСТ.
Далее, данное изобретение охватывает молекулы нуклеиновых кислот, которые отличаются от нуклеотидных последовательностей данного изобретения (например, последовательности, имеющей нечетный номер SEQ ID NO: Списка последовательностей) (или их части) вследствие вырожденности генетического кода и, следовательно, кодируют тот же самый белок МСТ, что и кодируемый нуклеотидными последовательностями данного изобретения. В другом варианте выделенная молекула нуклеиновой кислоты данного изобретения имеет нуклеотидную последовательность, кодирующую белок, имеющий аминокислотную последовательность, показанную в Списке последовательностей (например, имеющую четный номер SEQ ID NO:). Еще в одном варианте молекула нуклеиновой кислоты данного изобретения кодирует полноразмерный белок С. glutamicum, который является по существу гомологичным аминокислотной последовательности данного изобретения (кодируемой открытой рамкой считывания, показанной в имеющих нечетные номера SEQ ID NO: Списка последовательностей).
Специалисту в данной области должно быть понятно, что в одном варианте последовательности данного изобретения не должны включать последовательности предыдущего уровня техники, такие как последовательности Genbank, приведенные в таблицах 2 или 4, которые были известны до данного изобретения. В одном варианте данное изобретение включает нуклеотидные и аминокислотные последовательности, имеющие процентную идентичность относительно нуклеотидной или аминокислотной последовательности данного изобретения, более высокую, чем у последовательностей предыдущего уровня техники (например, последовательности Genbank (или белка, кодируемого такой последовательностью), приведенных в таблицах 2 или 4). Например, данное изобретение включает нуклеотидную последовательность, которая является более, чем на 38%, и/или по меньшей мере на 38% идентичной нуклеотидной последовательности, обозначенной RXA01420 (SEQ ID NO:7), нуклеотидную последовательность, которая является более, чем на 43%, и/или по меньшей мере на 43% идентичной нуклеотидной последовательности, обозначенной RXA00104 (SEQ ID NO:5), и нуклеотидную последовательность, которая является более, чем на 45%, и/или по меньшей мере на 45% идентичной нуклеотидной последовательности, обозначенной RXA02173 (SEQ ID NO:25). Специалист с обычной квалификацией в данной области сможет рассчитать низший предел процентной идентичности для любой конкретной последовательности данного изобретения исследованием оценок рассчитанной с использованием программы GAP процентной идентичности, приведенных в таблице 4 для каждого из трех наивысших совпадений для конкретной последовательности, и вычитанием наивысшей рассчитанной с использованием программы GAP процентной идентичности из 100 процентов. Специалисту в данной области будет также понятно, что последовательности нуклеиновых кислот и аминокислотные последовательности, имеющие процентные идентичности, большие, чем нижний предел, рассчитанный таким образом (например, по меньшей мере на 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59% или 60%, предпочтительно по меньшей мере на приблизительно 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69% или 70%, более предпочтительно по меньшей мере на приблизительно 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79% или 80%, 81%, 82%, 84%, 85%, 86%, 87%, 88%, 89% или 90%, или 91%, 92%, 94%, или даже более предпочтительно по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или более идентичные), также охватываются данным изобретением.
Специалистам в данной области должно быть понятно, что, кроме нуклеотидных последовательностей МСТ С. glutamicum, приведенных в Списке последовательностей в виде имеющих нечетные номера SEQ ID NO:, в популяции (например, в популяции С. glutamicum), могут существовать полиморфизмы последовательности ДНК, которые приводят к заменам в аминокислотных последовательностях белков МСТ. Такой генетический полиморфизм в гене МСТ может существовать среди индивидуумов в популяции вследствие природной изменчивости. В применении здесь, термины "ген" и "рекомбинантный ген" относятся к молекулам нуклеиновых кислот, содержащих открытую рамку считывания, кодирующую белок МСТ, предпочтительно белок МСТ С. glutamicum. Такие природные вариации могут обычно приводить к 1-5% дисперсии в нуклеотидной последовательности гена МСТ. Подразумевается, что любые и все такие вариации нуклеотидов и полученные полиморфизмы аминокислот в МСТ, которые являются результатом природной изменчивости и которые не изменяют функциональную активность белков МСТ, входят в объем данного изобретения.
Молекулы нуклеиновых кислот, соответствующие природным вариантам и не относящимся к С. glutamicum гомологам ДНК МСТ С. glutamicum данного изобретения, могут быть выделены на основе их гомологии описанной здесь нуклеиновой кислоте МСТ С. glutamicum с использованием ДНК С. glutamicum или ее части в качестве гибридизационного зонда в соответствии со стандартными способами гибридизации при строгих условиях гибридизации. Таким образом, в другом варианте выделенная молекула нуклеиновой кислоты данного изобретения имеет длину по меньшей мере 15 нуклеотидов и гибридизуется при строгих условиях с молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность, имеющую нечетный номер SEQ ID NO: Списка последовательностей. В других вариантах эта нуклеиновая кислота имеет длину по меньшей мере 30, 50, 100, 250 или более нуклеотидов. В применении здесь, термин "гибридизуется при строгих условиях" относится к условиям гибридизации и промывки, при которых нуклеотидные последовательности, по меньшей мере на 60% гомологичные друг другу, обычно остаются гибридизованными друг с другом. Предпочтительно, эти условия являются такими, что последовательности, по меньшей мере приблизительно на 65%, более предпочтительно по меньшей мере приблизительно на 70% и даже более предпочтительно по меньшей мере на приблизительно 75% или более гомологичные друг другу, обычно остаются гибридизованными друг с другом. Такие строгие условия известны специалистам в данной области и могут быть найдены в Current Protocols in Molecular Biology, John Wiley and Sons, N.Y. (1989), 6.3.1-6.3.6. Предпочтительным, неограничительным примером строгих условий гибридизации являются условия гибридизации в 6Х смеси хлорид натрия/цитрат натрия (SSC) при приблизительно 45°С с последующими одной или несколькими промывками в 0,2×SSC, 0,1 ДСН при 50-65°С. Предпочтительно, выделенная молекула нуклеиновой кислоты данного изобретения, которая гибридизуется при строгих условиях с нуклеотидной последовательностью данного изобретения, соответствует природно встречающейся молекуле нуклеиновой кислоты. В применении здесь, «природно встречающейся» молекулой нуклеиновой кислоты называют молекулу РНК или ДНК, имеющую нуклеотидную последовательность, которая встречается в природе (например, кодирует природный белок). В одном варианте эта нуклеиновая кислота кодирует природный белок МСТ С. glutamicum.
Специалисту в данной области будет также понятно, что кроме природно встречающихся вариантов последовательности МСТ, которые могут существовать в популяции, в нуклеотидную последовательность данного изобретения могут быть введены изменения посредством мутации, что приведет к изменениям в аминокислотной последовательности кодируемого белка МСТ, без изменения функциональной способности белка МСТ. Например, в нуклеотидной последовательности данного изобретения могут быть произведены замены нуклеотидов, приводящие к заменам аминокислот в «не-незаменимых» аминокислотных остатках. «Ненезаменимым» аминокислотным остатком является остаток, который может быть изменен в сравнении с последовательностью дикого типа одного из белков МСТ (например, имеющих четные номера SEQ ID NO: Списка последовательностей) без изменения активности указанного белка МСТ, тогда как "незаменимый" аминокислотный остаток является необходимым для активности белка МСТ. Однако, другие аминокислотные остатки (например, остатки, которые являются неконсервативными или только полуконсервативными в домене, имеющем активность МСТ) могут не быть незаменимыми для активности и, следовательно, будут, вероятно, доступными для изменения без изменения активности МСТ.
Таким образом, другой аспект данного изобретения относится к молекулам нуклеиновых кислот, кодирующим белки МСТ, которые содержат изменения в аминокислотных остатках, которые являются не-незаменимыми для активности МСТ. Такие белки МСТ отличаются по аминокислотной последовательности от последовательностей, имеющих четные номера SEQ ID NO: Списка последовательностей, но все еще сохраняют по меньшей мере одну из описанных здесь активностей МСТ. В одном варианте выделенная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую белок, который содержит аминокислотную последовательность, по меньшей мере на приблизительно 50% гомологичную аминокислотной последовательности данного изобретения, и способен участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны, или имеет одну или несколько активностей, приведенных в таблице 1. Предпочтительно, белок, кодируемый этой молекулой нуклеиновой кислоты, является по меньшей мере на приблизительно 50-60% гомологичным аминокислотной последовательности одной из имеющих нечетные номера SEQ ID NO: Списка последовательностей, более предпочтительно по меньшей мере на приблизительно 60-70% гомологичным одной из этих последовательностей, даже более предпочтительно по меньшей мере на приблизительно 70-80%, 80-90%, 90-95% гомологичным одной из этих последовательностей, и наиболее предпочтительно по меньшей мере на приблизительно 96%, 97%, 98% или 99% гомологичным одной из аминокислотных последовательностей данного изобретения.
Для определения процентной гомологии двух аминокислотных последовательностей (например, одной из аминокислотных последовательностей данного изобретения и ее мутантной формы) или двух нуклеиновых кислот, эти последовательности сопоставляют для целей оптимального сравнения (например, в последовательность одного белка или нуклеиновой кислоты могут быть введены гэпы (бреши) для оптимального сопоставления с другим белком или другой нуклеиновой кислотой). Затем сравнивают аминокислотные остатки или нуклеотиды в соответствующих положениях аминокислот или нуклеотидов. Когда положение в одной последовательности (например, в одной из аминокислотных последовательностей данного изобретения) занято тем же самым аминокислотным остатком или нуклеотидом, что и соответствующее положение в другой последовательности (например, в мутантной форме этой аминокислотной последовательности), тогда эти молекулы являются гомологичными в данном положении (т.е., в применении здесь, "гомология" аминокислоты или нуклеиновой кислоты эквивалентна "идентичности" аминокислоты или нуклеиновой кислоты). Процентная гомология между двумя последовательностями является функцией числа идентичных положений, общих у двух последовательностей (т.е. %-ная гомология=число идентичных положений/общее число положений×100).
Выделенная молекула нуклеиновой кислоты, кодирующая белок МСТ, гомологичный последовательности белка данного изобретения (например, последовательностей, имеющих четные номера SEQ ID NO: Списка последовательностей), может быть создана введением одной или более нуклеотидных замен, добавлений или делеций в нуклеотидную последовательность данного изобретения, так что одна или более замен, добавлений или делеций аминокислот вводятся в кодируемый белок. Мутации могут быть введены в одну из нуклеотидных последовательностей данного изобретения стандартными способами, такими как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Предпочтительно, производят консервативные замены аминокислот в одном или более предсказанных не-незаменимых аминокислотных остатках. "Консервативной заменой аминокислоты" является замена, в которой аминокислотный остаток заменяют аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, известны из уровня техники. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, предсказанный не-незаменимый аминокислотный остаток в белке МСТ предпочтительно заменяют другим аминокислотным остатком из того же самого семейства боковых цепей. Альтернативно, в другом варианте мутации могут быть введены случайным образом вдоль всей или части кодирующей последовательности МСТ, например, насыщающим мутагенезом, и полученные мутанты могут быть подвергнуты скринингу на активность МСТ, описанную здесь, для идентификации мутантов, которые сохраняют активность МСТ. После мутагенеза одной из нуклеотидных последовательностей, имеющей нечетный номер SEQ ID NO: Списка последовательностей, кодируемый белок может быть экспрессирован рекомбинантно, и активность этого белка может быть определена с использованием, например, анализов, описанных здесь (см. пример 8 раздела "Примеры").
Кроме молекул нуклеиновых кислот, кодирующих описанные выше белки МСТ, другой аспект данного изобретения относится к выделенным молекулам нуклеиновых кислот, которые являются антисмысловыми по отношению к ним. "Антисмысловая" нуклеиновая кислота содержит нуклеотидную последовательность, которая комплементарна "смысловой" нуклеиновой кислоте, кодирующей белок, например, комплементарна кодирующей цепи двухцепочечной молекулы кДНК или комплементарна последовательности мРНК. Таким образом, антисмысловая нуклеиновая кислота может связываться водородной связью со смысловой нуклеиновой кислотой. Антисмысловая нуклеиновая кислота может быть комплементарной полной кодирующей цепи МСТ или только ее части. В одном варианте антисмысловая молекула нуклеиновой кислоты является антисмысловой относительно "кодирующего участка" кодирующей цепи нуклеотидной последовательности, кодирующей белок МСТ.
Термин "кодирующий участок" относится к участку нуклеотидной последовательности, содержащей кодоны, которые транслируются в аминокислотные остатки (например, полный кодирующий участок SEQ ID NO:5 (RXA00104) содержит нуклеотиды 1-756). В другом варианте антисмысловая молекула нуклеиновой кислоты является антисмысловой относительно «некодирующего участка» кодирующей цепи нуклеотидной последовательности, кодирующей МСТ. Термин «некодирующий участок» относится к 5'- и 3'-последовательностям, которые фланкируют кодирующий участок, которые не транслируются в аминокислоты (т.е. их называют также 5'- и 3'-нетранслируемыми участками).
При наличии последовательностей кодирующей цепи, кодирующих МСТ, описанных здесь (например, последовательности, представленные в имеющих нечетные номера SEQ ID NO: в Списке последовательностей), антисмысловые нуклеиновые кислоты данного изобретения могут быть сконструированы согласно правилам спаривания оснований Уотсона и Крика. Антисмысловая молекула нуклеиновой кислоты может быть комплементарна полному кодирующему участку мРНК МСТ, но более предпочтительно она является олигонуклеотидом, который является антисмысловым относительно только части кодирующего или некодирующего участка мРНК МСТ. Например, антисмысловой олигонуклеотид может быть комплементарным участку, окружающему стартовый сайт трансляции мРНК МСТ. Антисмысловой олигонуклеотид может иметь длину, например, приблизительно 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 нуклеотидов. Антисмысловая нуклеиновая кислота данного изобретения может быть сконструирована при помощи химического синтеза и реакциями ферментативного лигирования с использованием процедур, известных в данной области. Например, антисмысловая нуклеиновая кислота (например, антисмысловой олигонуклеотид) может быть химически синтезирована с использованием природно встречающихся нуклеотидов или разнообразно модифицированных нуклеотидов, сконструированных для увеличения биологической стабильности этих молекул или для увеличения физической стабильности дуплекса, образуемого между антисмысловой и смысловой нуклеиновыми кислотами, например, могут быть использованы фосфоротиоатные производные и замещенные акридином нуклеотиды. Примеры модифицированных нуклеотидов, которые могут быть использованы для получения антисмысловой нуклеиновой кислоты, включают 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-иодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидроксиметил)урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D-галактозилквеозин, инозин, N6-изопентениладенин, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеозин, 5'-метоксикарбоксиметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентениладенин, урацил-5-оксиуксусную кислоту (v), вибутоксозин, псевдоурацил, квеозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, метиловый эфир урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусную кислоту (v), 5-метил-2-тиоурацил, 3-(3-амино-3-N-2-карбоксипропил)урацил, (acp3)w и 2,6-диаминопурин. Альтернативно, антисмысловая нуклеиновая кислота может быть получена биологически с использованием экспрессирующего вектора, в который была субклонирована нуклеиновая кислота в антисмысловой ориентации (т.е. РНК, транскрибированная со встроенной нуклеиновой кислоты, будет иметь антисмысловую ориентацию относительно представляющей интерес нуклеиновой кислоты-мишени, описанной далее в следующем подразделе).
Молекулы антисмысловых нуклеиновых кислот данного изобретения обычно вводят в клетку или генерируют in situ таким образом, что они гибридизуются или связываются с клеточной мРНК и/или геномной ДНК, кодирующей белок МСТ, для ингибирования посредством этого экспрессии белка, например, посредством ингибирования транскрипции и/или трансляции. Гибридизация может происходить посредством общепринятой комплементарности нуклеотидов с образованием стабильного дуплекса или, например, в случае антисмысловой молекулы нуклеиновой кислоты, которая связывается с ДНК-дуплексами, через специфические взаимодействия в большой бороздке двойной спирали. Антисмысловая молекула может быть модифицирована таким образом, что она специфически связывается с рецептором или антигеном, экспрессируемым на поверхности выбранной клетки, например, посредством связывания молекулы антисмысловой нуклеиновой кислоты с пептидом или антителом, которые связываются с рецептором поверхности клетки или антигеном. Молекула антисмысловой нуклеиновой кислоты может также доставляться в клетки с использованием описанных здесь векторов. Для получения достаточных внутриклеточных концентраций антисмысловых молекул предпочтительными являются векторные конструкции, в которые молекулу антисмысловой нуклеиновой кислоты помещают под контроль сильного, предпочтительно, прокариотического, вирусного или эукариотического промотора.
Еще в одном варианте молекула антисмысловой нуклеиновой кислоты данного изобретения является α-аномерной молекулой нуклеиновой кислоты. α-Аномерная молекула нуклеиновой кислоты образует специфические двухцепочечные гибриды с комплементарной РНК, в которых, в противоположность обычным β-единицам, цепи идут параллельно друг другу (Gaultier et al. (1987) Nucleic Acids Res. 15:6625-6641). Молекула антисмысловой нуклеиновой кислоты может также содержать 2'-о-метилрибонуклеотид (Inoue et al. (1987) Nucleic Acids Res.15: 6131-6148) или химерный аналог РНК-ДНК (Inoue et al. (1987) FEBS Lett. 215: 327-330).
Еще в одном варианте антисмысловая нуклеиновая кислота данного изобретения является рибозимом. Рибозимы являются каталитическими молекулами РНК с рибонуклеазной активностью, которые способны расщеплять одноцепочечную нуклеиновую кислоту, такую как мРНК, к которой они имеют комплементарный участок. Таким образом, рибозимы (например, молотоголовые рибозимы (описанные в Haselhoff and Gerlach (1988) Nature 334:585-591)) могут быть использованы для каталитического расщепления мРНК-транскриптов МСТ с ингибированием тем самым трансляциии мРНК МСТ. Рибозим, имеющий специфичность в отношении кодирующей МСТ нуклеиновой кислоты, может быть сконструирован на основе нуклеотидной последовательности описанной здесь ДНК МСТ (т.е. SEQ ID NO:5 (RXA00104)). Например, может быть сконструировано производное РНК Tetrahymena L-19 IVS, в котором нуклеотидная последовательность активного сайта является комплементарной нуклеотидной последовательности, которая должна быть расщепелена в кодирующей МСТ мРНК. См., например, Cech et al. U.S. Patent №4987071 и Cech et al. U.S. Patent №5116742. Альтернативно, мРНК МСТ может быть использована для отбора каталитической РНК, имеющей специфическую рибонуклеазную активность, из пула молекул РНК. См., например, Bartel, D. and Szostak, J.W. (1993) Science 261: 1411-1418.
Альтернативно, экспрессия гена МСТ может ингибироваться нацеливающими нуклеотидными последовательностями, комплементарными регуляторному району нуклеотидной последовательности МСТ (например, промотору и/или энхансерам МСТ) с образованием тройных спиральных структур, которые препятствуют транскрипции гена МСТ в клетках-мишенях. См., в общем, Helene, С. (1991) Anticancer Drug Des. 6(6): 569-84; Helene, С. et al. (1992) Ann. N.Y. Acad. Sci. 660:27-36; и Maher, L.J. (1992) Bioassays 14(12):807-15.
В. Рекомбинантные экспрессирующие векторы и клетки-хозяева
Другой аспект данного изобретения относится к векторам, предпочтительно экспрессирующим векторам, содержащим нуклеиновую кислоту, кодирующую белок МСТ (или его часть). В применении здесь, термин "вектор" относится к молекуле нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой она была связана. Одним типом вектора является "плазмида", которая является кольцевой двухцепочечной ДНК-петлей, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, причем дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в который их вносят (например, бактериальные векторы, имеющие бактериальный сайт инициации репликации, и эписомные векторы млекопитающих). Другие векторы (например, не-эписомные векторы млекопитающих) интегрируются в геном клетки-хозяина при введении в клетку-хозяина и поэтому реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы называют здесь "экспрессирующими векторами". Обычно, экспрессирующие векторы, используемые в способах рекомбинантных ДНК, часто находятся в форме плазмид. В данном описании термины "плазмида" и "вектор" используются взаимозаменяемо, так как плазмида является наиболее обычно используемой формой вектора. Однако, данное изобретение предполагает включение таких других форм экспрессирующих векторов, как вирусные векторы (например, ретровирусы с дефектной репликацией, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
Рекомбинантные экспрессирующие векторы данного изобретения содержат нуклеиновую кислоту данного изобретения в форме, пригодной для экспрессии этой нуклеиновой кислоты в клетке-хозяине, что означает, что эти рекомбинантные экспрессирующие векторы включают одну или более регуляторных последовательностей, выбранных на основе используемых для экспрессии клеток-хозяев, которые функционально связаны с экспрессируемой последовательностью нуклеиновой кислоты. В рекомбинантном экспрессирующем векторе, "функционально связанные" означает, что представляющая интерес нуклеотидная последовательность связана с регуляторной последовательностью (регуляторными последовательностями) таким образом, что возможна экспрессия этой нуклеотидной последовательности (например, в системе транскрипции/трансляции in vitro или в клетке-хозяине, при введении этого вектора в клетку-хозяина). Термин "регуляторная последовательность" включает промоторы, энхансеры и другие элементы регуляции экспрессии (например, сигналы полиаденилирования). Такие регуляторные последовательности описаны, например, в Goeddel: Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Регуляторные последовательности включают последовательности, которые управляют конститутивной экспрессией нуклеотидной последовательности во многих типах клетки-хозяина и которые управляют экспрессией нуклеотидной последовательности только в определенных клетках-хозяевах. Предпочтительными регуляторными последовательностями являются, например, промоторы, такие как cos-, tac-, trp-, tet-, trp-tet, lpp-, lac-, lpp-lac-, lacIq-, T7-, T5-, T3-, gal-, trc-, ara-, SP6-, arny, SPO2, λ-РR- или λ PL, которые используют предпочтительно в бактериях. Дополнительными регуляторными последовательностями являются, например, промоторы из дрожжей и грибов, такие как ADC1, MFα, AC, Р-60, CYC1, GAPDH, TEF, rp28, ADH, промоторы из растений, такие как CaMV/35S, SSU, OCS, lib4, usp, STLS1, B33, nos или убиквитин- или фазеолин-промоторы. Можно также использовать синтетические промоторы. Специалисту в данной области будет понятно, что конструкция экспрессирующего вектора может зависеть от таких факторов, как выбор трасформируемой клетки-хозяина, уровень экспрессии желаемого белка и т.д. Экспрессирующие векторы данного изобретения могут быть введены в клетки-хозяева для получения при помощи них белков или пептидов, в том числе слитых белков или пептидов, кодируемых нуклеиновыми кислотами, описанными здесь (например, белков МСТ, мутантных форм белков МСТ, слитых белков и т.д.).
Рекомбинантные экспрессирующие векторы данного изобретения могут быть сконструированы для экспрессии белков МСТ в прокариотических или эукариотических клетках. Например, гены МСТ могут быть экспрессированы в бактериальных клетках, таких как С. glutamicum, клетках насекомых (с использованием бакуловирусных экспрессирующих векторов), дрожжах и других грибных клетках (см. Romanes, M.A. et al. (1992) "Foreign gene expression in yeast: a review". Yeast 8: 423-488; van den Hondel, C.A.M.J.J. et al. (1991) "Heterologous gene expression in filamentous fungi" in: More Gene Manipulations in Fungi, J.W.Bennet and L.L.Lasure, eds., p.396-428: Academic Press: San Diego; и van den Hondel, C.A.M.J.J. and Punt, P.J. (1991) "Gene transfer systems and vektor development for filamentous fungi, in: Applied Molecular Genetics of Fungi, Peberdy, J.F. et al., eds., p.1-28, Cambridge University Press: Cambridge), водорослях и клетках многоклеточных растений (см. Schmidt, R. and Willmitzer, L. (1988) High efficiency Agrobacterium tumefaciens-mediated transformation of Arabidopsis thaliana leaf and cotyledon explants" Plant Cell Rep.: 583-586) или клетках млекопитающих. Подходящие клетки-хозяева обсуждаются дополнительно в Goeddel: Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Альтернативно, рекомбинантный экспрессирующий вектор может быть транскрибирован и транслирован in vitro, например, с использованием промоторных регуляторных последовательностей Т7 и полимеразы Т7.
Экспрессию белков в прокариотах наиболее часто проводят с векторами, содержащими конститутивные или индуцибельные промоторы, направляющие экспрессию слитых или неслитых белков. Слитые векторы добавляют ряд аминокислот к кодируемому в них белку, обычно к амино-концу рекомбинантного белка, но также и к С-концу, или сливают с подходящими участками в белке. Такие слитые векторы обычно служат для достижения трех целей: 1) для увеличения экспрессии рекомбинантного белка; 2) для увеличения растворимости рекомбинантного белка и 3) для того, чтобы способствовать очистке рекомбинантного белка посредством действия в качестве лиганда в аффинной очистке. Часто, в слитых экспрессирующих векторах, вводится сайт протеолитического расщепления в месте соединения слитой части молекулы и рекомбинантного белка для обеспечения возможности отделения рекомбинантного белка от слитой части молекулы после очистки слитого белка. Такие ферменты и их родственные последовательности узнавания включают фактор Ха, тромбин и энтерокиназу.
Типичные слитые экспрессирующие векторы включают pGEX (Pharmacia Biotech Inc; Smith, D.B. and Johnson, K.S. (1988) Gene 67:31-40), pMAL (New England Biolabs, Beverly, MA) и pRIT5 (Pharmacia, Piscataway, NJ), которые сливают глутатион S-трансферазу (GST), мальтозу Е-связывающий белок, или белок А, соответственно, с рекомбинантным белком-мишенью. В одном варианте кодирующую последовательность белка МСТ клонируют в экспрессирующий вектор pGEX для создания вектора, кодирующего слитый белок, содержащий, от N-конца к С-концу, GST-сайт расщепления тромбином-Х белок. Этот слитый белок может быть очищен аффинной хроматографией с использованием глутатион-агарозной смолы. Рекомбинантный белок МСТ, не слитый с GST, может быть извлечен расщеплением слитого белка тромбином.
Примеры подходящих индуцибельных неслитых экспрессирующих векторов Е. coli включают pTrc (Amann et al., (1988) Gene 69:301-315), pLG338, pACYC184, pBR322, pUC18, pUC19, рКС30, pRep4, pHS1, pHS2, pPLc236, pMBL24, pLG200, pUR290, pIN-III113-B1, λgt11, pBdC1 и pET 11d (Studier et al., Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, California (1990) 60-89; и Pouwels et al., (1985) Cloning Vectors, Elsevier: New York IBSN 0 444 904018). Экспрессия гена-мишени из вектора pTrc основана на транскрипции РНК-полимеразой хозяина от гибридного trp-lac слитого промотора. Экспрессия гена-мишени из вектора pET 11d основана на транскрипции от слитого промотора gn10-lac T7, опосредованной коэкспрессируемой вирусной РНК-полимеразой (gnl Т7). Эта вирусная полимераза поставляется штаммами-хозяевами BL21(DE3) или HMS174(DE3) из резидентного профага λ, несущего ген gnl T7 под транскрипционным контролем промотора lacUV 5. Для трансформации других разновидностей бактерий, могут быть выбраны соответствующие векторы. Например, известно, что плазмиды pIJ101, pIJ364, pIJ702 и pIJ361 применимы в трансформации Streptomyces, тогда как плазмиды pUB110, рС194 или pBD214 пригодны для трансформации видов Bacillus. Несколько плазмид, применимых в переносе генетической информации в Corynebacterium, включают рНМ1519, pBL1, p3A77 или pAJ667 (Pouwels et al., (1985) Cloning Vectors, Elsevier: New York IBSN 0 444 904018).
Одна стратегия максимизации экспрессии рекомбинантного белка состоит в экспрессии этого белка в бактерии-хозяине с нарушенной способностью протеолитически расщеплять рекомбинантный белок (Gottesman, S., Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, California (1990) 119-128). Другая стратегия состоит в изменении последовательности нуклеиновой кислоты, которая должна быть встроена в экспрессирующий вектор, таким образом, что индивидуальные кодоны для каждой аминокислоты являются кодонами, преимущественно используемыми в бактерии, выбранной для экспрессии, такой как С. glutamicum (Wada et al. (1992) Nucleic Acids Res. 20:2111-2118). Такое изменение последовательностей нуклеиновых кислот данного изобретения может проводиться стандартными способами синтеза ДНК.
В другом варианте экспрессирующий белок МСТ вектор является дрожжевым экспрессирующим вектором. Примеры векторов для экспрессии в дрожжах S. cerevisiae включают pYepSec1 (Baldari, et al., (1987) EMBO J. 6:229-234), 2 μ, pAG-1, Yep6, Yep13, pEMBLYe23, pMFa (Kurjan and Herskowitz, (1982) Cell 30:933-943), pJRY88 (Schuiz et al., (1987) Gene 54:113-123) и pYES2 (Invitrogen Corporation, San Diego, CA). Векторы и способы конструирования векторов, пригодных для применения в других грибах, таких как нитчатые грибы, включают способы и векторы, подробно описанные в van den Hondel, C.A.M.J.J. and Punt, P.J. (1991) "Gene transfer systems and vector development for filamentous fungi, in: Applied Molecular Genetics of Fungi, J.F. Peberdy, et al., eds., p.1-28, Cambridge University Press: Cambridge, и Pouwels et al., (1985) Cloning Vectors, Elsevier: New York IBSN 0 444 904018).
Альтернативно, белки МСТ могут экспрессироваться в клетках насекомых с использованием бакуловирусных экспрессирующих векторов. Бакуловирусные векторы, пригодные для экспрессии белков в культивируемых клетках насекомых (например, в клетках Sf9), включают серию рАС (Smith et al., (1983) Mol. Cell Biol. 3:2156-2165) и ряд pVL (Lucklow and Summers (1989) Virology 170:31-39).
В другом варианте белки МСТ данного изобретения могут экспрессироваться в клетках одноклеточных растений (таких как водоросли) и в растительных клетках из высших растений (например, семенных растений, таких как сельскохозяйственные культуры). Примеры экспрессирующих векторов растений включают векторы, подробно описанные в Becker, D., Kemper, E., Schell, J. and Masterson, R. (1992) "New plant binary vectors with selectable markers located proximal to the left border", Plant Mol. Biol. 20: 1195-1197; и Bevan, M.W. (1984) "Binary Agrobacterium vectors for plant transformation", Nucl. Acid. Res. 12: 8711-8721, и включают pLGV23, pGHlac+, pBIN19, pAK2004 и pDH51 (Pouwels et al., (1985) Cloning Vectors, Elsevier: New York IBSN 0 444 904018).
Еще в одном варианте нуклеиновая кислота данного изобретения экспрессируется в клетках млекопитающих с использованием экспрессирующего вектора млекопитающего. Примеры экспрессирующих векторов млекопитающих включают pCDM8 (Seed, В. (1987) Nature 329:840) и pMT2PC (Kaufman et al., (1987) EMBO J. 6:187-195). При использовании в клетках млекопитающего регуляторные функции этого экспрессирующего вектора часто обеспечиваются вирусными регуляторными элементами. Например, обычно используемые промоторы происходят из полиомы, аденовируса 2, цитомегаловируса и вируса обезьян 40. В отношении подходящих экспрессирующих систем для прокариотических и эукариотических клеток см. главы 16 и 17 Sambrook J., Fritsh, E.F., and Maniatis, T. Molecular Cloning. A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
В другом варианте рекомбинантный экспрессирующий вектор млекопитающих способен направлять экспрессию нуклеиновой кислоты преимущественно в конкретном типе клеток (например, используют тканеспецифические регуляторные элементы для экспрессии нуклеиновой кислоты). Тканеспецифические регуляторные элементы известны в данной области. Неограничивающие примеры подходящих тканеспецифических промоторов включают промотор альбумина (печеночно-специфический; Pinkert et al., (1987) Genes Dev. 1:268-277), лимфоидно-специфические промоторы (Calame and Eaton (1988) Adv. Immunol. 43:235-275), в частности, промоторы Т-клеточных рецепторов (Winoto and Baltimore (1989) EMBO J. 8:729-733) и иммуноглобулинов (Banerji et al., (1983) Cell 33:729-740; Queen and Baltimore (1983) Cell 33:741-748), нейрон-специфические промоторы (например, промотор нейрофиламента; Byrne and Ruddle (1989) PNAS 86:5473-5477), специфические для поджелудочной железы промоторы (Ed-lund et al. (1985) Science 230:912-916) и специфические для молочной железы промоторы (например, промотор молочной сыворотки; Патент США №4 873 316 и Публикация Европейской заявки №264 166). Включены также регулируемые стадией развития промоторы, например, мышиные промоторы hox (Kessel and Gruss (1990) Science 249:374-379) и промотор α-фетопротеина (Campes and Tilghman (1989) Genes Dev. 3:537-546).
Далее, данное изобретение представляет рекомбинантный экспрессирующий вектор, содержащий молекулу ДНК данного изобретения, клонированную в этот экспрессирующий вектор в антисмысловой ориентации. Т.е., эта молекула ДНК функционально связана с регуляторной последовательностью таким образом, что возможна экспрессия (транскрипцией этой молекулы ДНК) молекулы РНК, которая является антисмысловой относительно мРНК МСТ. Могут быть выбраны регуляторные последовательности, функционально связанные с нуклеиновой кислотой, клонированной в антисмысловой ориентации, которые направляют непрерывную экспрессию этой антисмысловой молекулы РНК в многочисленных клеточных типах, например, вирусные промоторы и/или энхансеры, или могут быть выбраны регуляторные последовательности, которые направляют конститутивную, тканеспецифическую или специфическую для определенного типа клеток экспрессию антисмысловой РНК. Антисмысловой экспрессирующий вектор может быть в форме рекомбинантной плазмиды, фагмиды или аттенуированного вируса, в котором антисмысловые нуклеиновые кислоты продуцируются под контролем высокоэффективного регуляторного участка, активность которого может определяться типом клеток, в которые вводят этот вектор. В отношении обсуждения регуляции экспрессии генов с использованием антисмысловых генов см. Weintraub, H. et al., Antisense RNA as a molecular tool for genetic analysis. Reviews - Trends in Genetics, Vol.1(1) 1986.
Другой аспект данного изобретения относится к клеткам-хозяевам, в которые вводили рекомбинантный экспрессирующий вектор данного изобретения. Термины «клетка-хозяин» и «рекомбинантная клетка-хозяин» используются здесь взаимозаменяемо. Понятно, что такие термины относятся не только к конкретной рассматриваемой клетке, но и к потомству или потенциальному потомству такой клетки. Поскольку некоторые модификации могут встречаться в последующих поколениях вследствие мутации или влияний окружающей среды, такое потомство на самом деле может не быть идентичным родительской клетке, но оно все-таки включено в рамки этого термина, в применении здесь.
Клетка-хозяин может быть любой прокариотической или эукариотической клеткой. Например, белок МСТ может быть экспрессирован в бактериальных клетках, таких как С. glutamicum, клетках насекомых, клетках дрожжей или млекопитающих (таких как клетки яичника Китайского хомячка (СНО) или клетки COS). Другие подходящие клетки-хозяева известны специалисту в данной области. Микроорганизмы, родственные Corynebacterium glutamicum, которые могут быть легко использованы в качестве клеток-хозяев для молекул нуклеиновых кислот и белков данного изобретения, приведены в таблице 3.
Вектор ДНК может быть введен в прокариотические или эукариотические клетки посредством общепринятых способов трансформации или трансфекции. В применении здесь, термины "трансформация" и "трансфекция", "конъюгация" и "трансдукция" относятся к различным признанным в данной области способам введения чужеродной нуклеиновой кислоты (например, линейной ДНК или РНК (например, линеаризованного вектора или генной конструкции отдельно без вектора) или нуклеиновой кислоты в форме вектора (например, плазмиды, фага, фазмиды, фагмиды, транспозона или другой ДНК) в клетку-хозяина, в том числе совместным осаждением фосфатом кальция или хлоридом кальция, опосредованной ДЭАЭ-декстраном трансфекцией, липофекцией, естественной конкуренцией химически опосредованным переносом или электропорацией. Подходящие способы для трансформации или трансфекции клеток-хозяев могут быть найдены в Sambrook, et al., (Molecular Cloning. A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989) и других лабораторных руководствах-справочниках.
В отношении стабильной трансфекции клеток млекопитающих известно, что, в зависимости от используемого экспрессирующего вектора и способа трансфекции, только небольшая фракция клеток может интегрировать чужеродную ДНК в их геном. Для идентификации и отбора этих интегрантов ген, который кодирует селектируемый маркер (например, ген устойчивости к антибиотикам), вводят обычно в клетки-хозяева вместе с представляющим интерес геном. Предпочтительные селектируемые маркеры включают маркеры, придающие устойчивость к лекарственным средствам, таким как G418, гигромицин и метотрексат. Нуклеиновая кислота, кодирующая селектируемый маркер, может быть введена в клетку-хозяина на том же самом векторе, что и вектор, кодирующий белок МСТ, или может быть введена на отдельном векторе. Клетки, стабильно трансфицированные введенной нуклеиновой кислотой, могут быть идентифицированы отбором с лекарственным средством (например, клетки, которые включили ген селектируемого маркера, выживают, тогда как другие клетки погибают).
Для создания гомологичного рекомбинантного микроорганизма получают вектор, который содержит по меньшей мере часть гена МСТ, в который была введена делеция, добавление или замена, для изменения посредством этого, например, функционального нарушения, гена МСТ. Предпочтительно, этот ген является геном МСТ Corynebacterium glutamicum, но он может быть гомологом из родственной бактерии или даже из млекопитающего, дрожжей или насекомого. В предпочтительном варианте этот вектор конструируют таким образом, что, при гомологичной рекомбинации, эндогенный ген МСТ функционально разрушается (т.е. уже больше не кодирует функциональный белок; этот вектор называют также «нокаут»-вектором). Альтернативно, вектор может быть сконструирован таким образом, что, при гомологичной рекомбинации, эндогенный ген МСТ мутируется или иным образом изменяется, но все еще кодирует функциональный белок (например, регуляторный участок против хода транскрипции может быть изменен для изменения посредством этого экспрессии эндогенного белка МСТ). В векторе гомологичной рекомбинации, изменяемая часть гена МСТ фланкирована на ее 5'- и 3'-концах дополнительной нуклеиновой кислотой гена МСТ, чтобы сделать возможной гомологичную рекомбинацию между экзогенным геном МСТ, переносимым этим вектором, и эндогенным геном МСТ в микроорганизме. Дополнительная фланкирующая нуклеиновая кислота МСТ имеет достаточную длину для успешной гомологичной рекомбинации с эндогенным геном. Обычно, несколько т.п.н. фланкирующей ДНК (как на 5'-, так и на 3'-концах) включают в этот вектор (см., например, Thomas, K.R. and Capecchi, M.R. (1987) Cell 51: 503 в отношении описания векторов гомологичной рекомбинации). Вектор вводят в микроорганизм (например, электропорацией), и клетки, в которых введенный ген МСТ был гомологично рекомбинирован с эндогенным геном МСТ, отбирают с использованием известных в данной области способов.
В другом варианте могут быть получены рекомбинантные микроорганизмы, которые содержат выбранные системы, которые делают возможной регулируемую экспрессию введенного гена. Например, включение гена МСТ на векторе, с помещением его под контроль lac-оперона делает возможной экспрессию гена МСТ только в присутствии IPTG. Такая регуляторная система хорошо известна в данной области.
В другом варианте эндогенный ген МСТ в клетке-хозяине разрушают (например, гомологичной рекомбинацией или другими генетическими способами, известными в данной области), так что экспрессия его белкового продукта не происходит. В другом варианте эндогенный или введенный ген МСТ в клетке-хозяине был изменен одной или более точковыми мутациями, делециями или инверсиями, но все еще кодирует функциональный белок МСТ. Еще в одном варианте один или более регуляторных участков (например, промотор, репрессор или индуктор) гена МСТ в микроорганизме был изменен (например, делецией, укорочением, инверсией или точковой мутацией), так что экспрессия гена МСТ является модулированной. Специалисту в данной области будет понятно, что клетки-хозяева, содержащие более чем одну из описанных модификаций гена и белка МСТ, могут быть легко получены с использованием способов данного изобретения, и подразумевается, что они включены в данное изобретение.
Клетка-хозяин данного изобретения, например, прокариотическая или эукариотическая клетка-хозяин в культуре, может быть использована для продуцирования (т.е. экспрессии) белка МСТ. Таким образом, данное изобретение представляет, далее, способы получения белков МСТ с использованием клеток-хозяев данного изобретения. В одном варианте этот способ включает культивирование клетки-хозяина данного изобретения (в которую был введен рекомбинантный экспрессирующий вектор, кодирующий белок МСТ, или в геном которой был введен ген, кодирующий белок МСТ дикого типа или измененный белок МСТ) в подходящей среде до тех пор, пока не продуцируется белок МСТ. В другом варианте этот способ дополнительно включает выделение белков МСТ из среды или из клетки-хозяина.
С. Выделенные белки МСТ
Другой аспект данного изобретения относится к выделенным белкам МСТ и их биологически активным частям. "Выделенный" или "очищенный" белок или его биологически активная часть по существу не содержит клеточного материала при получении способами рекомбинантных ДНК или при химическом синтезе химических предшественников или других химических продуктов. Выражение "по существу не содержащие клеточного материала" включает препараты белка МСТ, в которых белок отделен от клеточных компонентов клеток, в которых он природно или рекомбинантно продуцировался. В одном варианте выражение "по существу не содержащие клеточного материала" включает препараты белка МСТ, имеющие менее, чем приблизительно 30% (по сухому весу) белка не-МСТ (называемого здесь также "загрязняющим (примесным) белком"), более предпочтительно менее, чем приблизительно 20% белка не-МСТ, еще более предпочтительно менее, чем приблизительно 10% белка не-МСТ, и наиболее предпочтительно менее, чем приблизительно 5% белка не-МСТ. При рекомбинантном получении белка МСТ или его биологически активной части он предпочтительно является по существу не содержащим культуральной среды, т.е. культуральная среда представляет менее, чем приблизительно 20%, более предпочтительно менее, чем приблизительно 10%, и наиболее предпочтительно менее, чем приблизительно 5% объема препарата белка. Выражение "по существу не содержащие химических предшественников или других химических продуктов" включает препараты белка МСТ, в которых этот белок отделен от химических предшественников или других химических продуктов, которые участвуют в синтезе этого белка. В одном варианте выражение "по существу не содержащие химических предшественников или других химических продуктов" включает препараты белка МСТ, имеющие менее, чем приблизительно 30% (по сухому весу) химических предшественников или химических продуктов не-МСТ, более предпочтительно менее, чем приблизительно 20% химических предшественников или химических продуктов не-МСТ, еще более предпочтительно менее, чем приблизительно 10% химических предшественников или химических продуктов не-МСТ, и наиболее предпочтительно менее, чем приблизительно 5% химических предшественников или химических продуктов не-МСТ. В предпочтительных вариантах выделенные белки или их биологически активные части не содержат загрязняющих (примесных) белков из того же самого организма, из которого получен белок МСТ. Обычно такие белки получают рекомбинантной экспрессией, например, белка МСТ С. glutamicum, в микроорганизме, например, С. glutamicum.
Выделенный белок МСТ или его часть данного изобретения может участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны, или имеет одну или более активностей, приведенных в таблице 1. В предпочтительных вариантах этот белок или его часть содержит аминокислотную последовательность, которая является достаточно гомологичной аминокислотной последовательности данного изобретения (например, последовательностей, имеющих четные номера SEQ ID NO: Списка последовательностей), так что этот белок или его часть сохраняет способность участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны. Часть этого белка является предпочтительно биологически активной частью, как описано здесь. В другом предпочтительном варианте белок МСТ данного изобретения имеет аминокислотную последовательность, представленную в виде имеющих четные номера SEQ ID NO: Списка последовательностей. Еще в одном предпочтительном варианте белок МСТ имеет аминокислотную последовательность, которая кодируется нуклеотидной последовательностью, которая гибридизуется, например, гибридизуется при строгих условиях, с нуклеотидной последовательностью данного изобретения (например, последовательностей, имеющих нечетные номера SEQ ID NO: Списка последовательностей). Еще в одном предпочтительном варианте белок МСТ имеет аминокислотную последовательность, которая кодируется нуклеотидной последовательностью, которая является по меньшей мере на приблизительно 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59% или 60%, предпочтительно по меньшей мере на приблизительно 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69% или 70%, более предпочтительно по меньшей мере на приблизительно 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79% или 80%, 81%, 82%, 84%, 85%, 86%, 87%, 88%, 89% или 90%, или 91%, 92%, 94%, или даже более предпочтительно на по меньшей мере приблизительно 95%, 96%, 97%, 98%, 99% или более гомологичной нуклеотидной последовательности данного изобретения, или ее часть. Подразумевается, что диапазоны и величины идентичности, промежуточные относительно указанных выше диапазонов (например, идентичные на 70-90% или идентичные на 80-95%), также охватываются данным изобретением. Например, подразумевается, что включены также диапазоны величин идентичности, использующие комбинацию любых из цитированных выше величин в качестве верхнего и/или нижнего пределов. Предпочтительные белки МСТ данного изобретения также предпочтительно обладают по меньшей мере одной из описанных здесь активностей МСТ. Например, предпочтительный белок МСТ данного изобретения включает аминокислотную последовательность, кодируемую нуклеотидной последовательностью, которая гибридизуется, например, гибридизуется при строгих условиях, с нуклеотидной последовательностью данного изобретения, и может участвовать в метаболизме соединений, необходимых для конструирования клеточных мембран в С. glutamicum или в транспорте молекул через эти мембраны, или имеет одну или несколько активностей, приведенных в таблице 1.
В других вариантах белок МСТ является по существу гомологичным аминокислотной последовательности данного изобретения (например, последовательностей, имеющих четные номера SEQ ID NO: Списка последовательностей) и сохраняет функциональную активность белка одной из аминокислотных последовательностей данного изобретения, несмотря на то, что он отличается по аминокислотной последовательности вследствие природной изменчивости или мутагенеза, как описано подробно в подразделе I выше. Таким образом, в другом варианте белок МСТ является белком, который содержит аминокислотную последовательность, которая является по меньшей мере на приблизительно 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59% или 60%, предпочтительно по меньшей мере на приблизительно 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69% или 70%, более предпочтительно по меньшей мере на приблизительно 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79% или 80%, 81%, 82%, 84%, 85%, 86%, 87%, 88%, 89% или 90%, или 91%, 92%, 94%, или даже более предпочтительно по меньшей мере на приблизительно 95%, 96%, 97%, 98%, 99% или более гомологичной полной аминокислотной последовательности данного изобретения, и который имеет по меньшей мере одну из описанных здесь активностей МСТ. Подразумевается, что диапазоны и величины идентичности, промежуточные относительно указанных выше диапазонов (например, идентичные на 70-90% или идентичные на 80-95%), также охватываются данным изобретением. Например, подразумевается, что включены также диапазоны величин идентичности, использующие комбинацию любых из цитированных выше величин в качестве верхнего и/или нижнего пределов. В другом варианте данное изобретение относится к полноразмерному белку С. glutamicum, который является по существу гомологичным полной аминокислотной последовательности данного изобретения.
Биологически активные части белка МСТ включают пептиды, содержащие аминокислотные последовательности, полученные из аминокислотной последовательности белка МСТ, например, аминокислотной последовательности, имеющей четный номер SEQ ID NO: Списка последовательностей, или аминокислотную последовательность белка, гомологичного белку МСТ, которая содержит меньше аминокислот, чем полноразмерный белок МСТ или полноразмерный белок, который гомологичен белку МСТ, и проявляют по меньшей мере одну активность белка МСТ. Обычно, биологически активные части (пептиды, например, пептиды, которые имеют длину, например, 5, 10, 15, 20, 30, 35, 36, 37, 38, 39, 40, 50, 100 или более аминокислот) содержат домен или мотив с по меньшей мере одной активностью белка МСТ. Кроме того, другие биологически активные части, в которых делетированы другие участки этого белка, могут быть получены рекомбинантными способами и оценены на одну или несколько описанных здесь активностей. Предпочтительно, биологически активные части белка МСТ включают один или несколько доменов/мотивов или их частей, имеющих биологическую активность.
Белки МСТ предпочтительно получают способами рекомбинантных ДНК. Например, молекулу нуклеиновой кислоты, кодирующую этот белок, клонируют в экспрессирующий вектор (как описано выше), экспрессирующий вектор вводят в клетку-хозяин (как описано выше) и белок МСТ экспрессируется в клетке-хозяине. Затем белок МСТ может быть выделен из клеток с использованием подходящей схемы очистки при помощи стандартных способов очистки белков. Альтернативно рекомбинантной экспрессии, белок, полипептид или пептид МСТ может быть синтезирован химически при помощи стандартных способов синтеза пептидов. Кроме того, нативный белок МСТ может быть выделен из клеток (например, эндотелиальных клеток), например, с использованием анти-МСТ-антитела, которое может быть получено стандартными способами, использующими белок МСТ или его фрагмент данного изобретения.
Данное изобретение представляет также химерные или слитые белки МСТ. В применении здесь, "химерный белок" или "слитый белок" МСТ содержит полипептид МСТ, функционально связанный с полипептидом не-МСТ. "Полипептид МСТ" обозначает полипептид, имеющий аминокислотную последовательность, соответствующую белку МСТ, тогда как "полипептид не-МСТ" обозначает полипептид, имеющий аминокислотную последовательность, соответствующую белку, который не является по существу гомологичным белку МСТ, например, белку, который отличается от белка МСТ и который получен из того же самого или другого организма. В слитом белке, термин "функционально связанный" означает, что полипептид МСТ и полипептид не-МСТ слиты в рамке считывания друг с другом. Полипептид не-МСТ может быть слит с N-концом или С-концом полипептида МСТ. Например, в одном варианте слитый белок является слитым белком GST-MCT, в котором последовательности МСТ слиты с С-концом последовательностей GST. Такие слитые белки могут облегчать очистку рекомбинантных белков МСТ. В другом варианте слитый белок является белком МСТ, содержащим гетерологичную сигнальную последовательность на его N-конце. В некоторых клетках-хозяевах (например, в клетках-хозяевах млекопитающих) экспрессия и/или секреция белка МСТ может быть увеличена посредством применения гетерологичной сигнальной последовательности.
Предпочтительно, химерный или слитый белок МСТ данного изобретения получают стандартными способами рекомбинантных ДНК. Например, фрагменты ДНК, кодирующие различные полипептидные последовательности, лигируют вместе в рамке считывания в соответствии с общепринятыми способами, например, посредством использования тупых концов или ступенчатых концов для лигирования, расщепления рестриктазами для обеспечения подходящих концов, достраивания липких концов, если нужно, обработки щелочной фосфатазой для избежания нежелательного связывания и ферментативного лигирования. В другом варианте слитый ген может быть синтезирован общепринятыми способами, в том числе с использованием автоматических синтезаторов ДНК. Альтернативно, может быть проведена ПЦР-амплификация фрагментов гена с использованием якорных праймеров, которые вызывают появление выступов между двумя последовательными генными фрагментами, которые могут быть затем подвергнуты отжигу и повторно амплифицированы с образованием химерной последовательности гена (см., например, Current Protocols in Molecular Biology, eds. Ausubel et al., John Wiley and Sons: 1992). Кроме того, коммерчески доступными являются многие экспрессирующие векторы, которые уже кодируют необходимую для слияния часть (например, полипептид GST). Кодирующая МСТ нуклеиновая кислота может быть клонирована в такой экспрессирующий вектор таким образом, что сливаемая часть связывается в рамке считывания в белком МСТ.
Гомологи белка МСТ могут быть получены мутагенезом, например, дискретной точковой мутацией или укорочением белка МСТ. В применении здесь, термин «гомолог» относится к вариантной форме белка МСТ, которая действует как агонист или антагонист активности белка МСТ. Агонист белка МСТ может сохранять по существу те же самые биологические активности, или субпопуляцию биологических активностей белка МСТ. Антагонист белка МСТ может ингибировать одну или более активностей природно встречающейся формы белка МСТ, например, конкурентным связыванием со стоящим ниже или выше по ходу процесса членом метаболического каскада компонентов клеточных мембран, который включает белок МСТ, или связыванием с белком МСТ, который медиирует транспорт соединений через такие мембраны, предотвращая тем самым осуществление транслокации.
В альтернативном варианте гомологи белка МСТ могут быть идентифицированы скринингом комбинаторных библиотек мутантов, например, укороченных мутантов, белка МСТ на агонистическую или антагонистическую активность относительно белка МСТ. В одном варианте смешанная библиотека вариантов МСТ генерируется комбинаторным мутагенезом на уровне нуклеиновых кислот и кодируется библиотекой смешанных (разнообразных) генов. Смешанная библиотека вариантов МСТ может быть получена, например, ферментативным лигированием смеси синтетических олигонуклеотидов в последовательности генов таким образом, что вырожденный набор потенциальных последовательностей МСТ может экспрессироваться в виде индивидуальных полипептидов, или альтернативно в виде набора более крупных слитых белков (например, для фаг-дисплея (фагового представления)), содержащих в них ряд последовательностей МСТ. Существуют различные способы, которые могут быть использованы для получения библиотек потенциальных гомологов МСТ из вырожденной олигонуклеотидной последовательности. Химический синтез вырожденной последовательности гена может проводиться в автоматическом синтезаторе ДНК, и синтетический ген затем лигируют в подходящий экспрессирующий вектор. Применение вырожденного набора генов позволяет обеспечить, в одной смеси, все последовательности, кодирующие желаемый набор потенциальных последовательностей МСТ. Способы синтеза вырожденных олигонуклеотидов известны в данной области (см., например, Narang, S.A. (1983) Tetrahedron 39:3; Itakura et al., (1984) Annu. Rev. Biochem. 53:323; Itakura et al., (1984) Science 198:1056; Ike et al., (1983) Nucleic Acid Res.11:477.
Кроме того, библиотеки фрагментов кодирующей белок МСТ последовательности могут быть использованы для генерирования смешанной популяции фрагментов МСТ для скрининга и последующего отбора гомологов белка МСТ. В одном варианте библиотека фрагментов кодирующей последовательности может быть получена обработкой двухцепочечного ПЦР-фрагмента кодирующей МСТ последовательности нуклеазой в условиях, в которых происходит образование одноцепочечных разрывов только приблизительно один раз на молекулу, денатурацией двухцепочечной ДНК, ренатурацией этой ДНК с образованием двухцепочечной ДНК, которая может включать в себя смысловые/антисмысловые пары из различных разорванных продуктов, удалением одноцепочечных частей из повторно образованных дуплексов обработкой нуклеазой S1 и лигированием полученной библиотеки фрагментов в экспрессирующий вектор. При помощи этого способа может быть получена экспрессионная библиотека, которая кодирует N-концевые, С-концевые и внутренние фрагменты различного размера белка МСТ.
Несколько способов известны в данной области для скрининга генных продуктов комбинаторных библиотек, приготовленных с использованием точковых мутаций или укорочения, и для скрининга библиотек кДНК на генные продукты, имеющие выбранное свойство. Такие способы можно приспособить для быстрого скрининга библиотек генов, генерируемых комбинаторным мутагенезом гомологов МСТ. Наиболее широко распространенные способы, которые поддаются высокопроизводительному анализу, для скрининга больших библиотек генов, обычно включают клонирование библиотеки генов в реплицируемые экспрессирующие векторы, трансформацию подходящих клеток полученной библиотекой векторов и экспрессию комбинаторных генов в условиях, в которых обнаружение желаемой активности облегчает выделение вектора, кодирующего ген, продукт которого был детектирован. Рекурсивный комплексный мутагенез (REM), новый способ, который усиливает частоту встречаемости функциональных мутантов в библиотеках, может быть использован в сочетании с анализами скрининга для идентификации гомологов МСТ (Arkin and Yourvan (1992) PNAS 89:7811-7815; Delgrave et al., (1993) Protein Engineeribg 6(3):327-331).
В другом варианте анализы на основе клеток могут быть использованы для анализа смешанной библиотеки МСТ с применением способов, хорошо известных в данной области.
D. Применения и способы изобретения
Молекулы нуклеиновых кислот, белки, белковые гомологи, слитые белки, праймеры, векторы и клетки-хозяева, описанные здесь, могут быть использованы в одном или большем количестве из следующих способов: идентификации С. glutamicum и родственных организмов; картировании геномов организмов, родственных С. glutamicum; идентификации и локализации представляющих интерес последовательностей С. glutamicum; эволюционных исследований; определении участков белка МСТ, необходимых для функции; модуляции активности белка МСТ, модуляции метаболизма одного или более компонентов клеточных мембран; модуляции трансмембранного транспорта одного или нескольких соединений и модуляции клеточного продуцирования желаемого соединения, такого как химический продукт тонкого органического синтеза.
Молекулы нуклеиновых кислот МСТ данного изобретения имеют множество применений. Во-первых, они могут быть использованы для идентификации организма как являющегося Corynebacterium glutamicum или близкородственным ему организмом. Они могут быть также использованы для идентификации присутствия С. glutamicum или родственного ему организма в смешанной популяции микроорганизмов. Данное изобретение представляет последовательности нуклеиновых кислот ряда генов С. glutamicum; зондированием экстрагированной геномной ДНК культуры уникальной или смешанной популяции микроорганизмов при строгих условиях зондом, охватывающим участок гена С. glutamicum, который является уникальным в отношении этого организма, можно определить, присутствует ли данный организм.
Хотя сама бактерия Corynebacterium glutamicum является непатогенной, она является родственной патогенным видам, таким как Corynebacterium diphtheriae. Corynebacterium diphtheriae является возбудителем дифтерии, быстро развивающейся, острой, протекающей с повышением температуры инфекции, которая включает как местную, так и системную патологию. В этом заболевании местное повреждение развивается в верхних дыхательных путях и включает некротическое повреждение эпителиальных клеток; бациллы секретируют токсин, который диссеминируется через это повреждение к дистальным чувствительным тканям тела. Дегенеративные изменения, вызываемые ингибированием белкового синтеза в этих тканях, которые включают сердце, мышцы, периферические нервы, надпочечники, почки, печень и селезенку, приводят к системной патологии этого заболевания. Дифтерия продолжает встречаться с высокой частотой во многих частях света, в том числе в Африке, Азии, Восточной Европе и независимых государствах бывшего Советского Союза. Продолжающиеся эпидемии дифтерии в двух последних районах привели по меньшей мере к 5000 случаям со смертельным исходом с 1990 года.
В одном варианте данное изобретение представляет способ идентификации присутствия или активности Corynebacterium glutamicum в субъекте. Этот способ включает обнаружение одной или большего количества последовательностей нуклеиновых кислот или аминокислотных последовательностей данного изобретения (например, последовательностей, приведенных в имеющих нечетные или четные номера SEQ ID NO:, соответственно, в Списке последовательностей) в субъекте, с обнаружением посредством этого присутствия или активности Corynebacterium diphtheriae в этом субъекте. С. glutamicum и С. diphtheriae являются родственными бактериями, и многие из молекул нуклеиновых кислот и белковых молекул в С. glutamicum являются гомологичными молекулам нуклеиновых кислот и белковым молекулам Corynebacterium diphtheriae и, следовательно, могут быть использованы для обнаружения Corynebacterium diphtheriae в субъекте.
Молекулы нуклеиновых кислот и белковые молекулы данного изобретения могут также служить в качестве маркеров для специфических участков генома. Это находит применение не только в картировании генома, но также для функциональных исследований белков С. glutamicum. Например, для идентификации участка генома, с которым связывается конкретный связывающий ДНК С. glutamicum белок, геном С. glutamicum можно было бы подвергнуть расщеплению и фрагменты можно было бы инкубировать с ДНК-связывающим белком. Фрагменты, которые связывают этот белок, могут быть дополнительно зондированы молекулами нуклеиновых кислот данного изобретения, предпочтительно с легко обнаруживаемыми метками; связывание такой молекулы нуклеиновой кислоты с фрагментом генома позволяет определить местоположение этого фрагмента на карте генома С. glutamicum и, при проведении этого много раз с различными ферментами, облегчает быстрое определение последовательности нуклеиновой кислоты, с которой связывается данный белок. Далее, молекулы нуклеиновых кислот данного изобретения могут быть достаточно гомологичными последовательностям родственных видов, так что эти молекулы нуклеиновых кислот могут служить в качестве маркеров для конструирования геномной карты в родственных бактериях, таких как Brevibacterium lactofermentum.
Молекулы нуклеиновых кислот МСТ данного изобретения применимы также для эволюционных исследований и исследований структуры белков. Метаболические и транспортные процессы, в которых участвуют молекулы данного изобретения, используется в большом разнообразии прокариотических и эукариотических клеток; посредством сравнения последовательностей молекул нуклеиновых кислот данного изобретения с молекулами нуклеиновых кислот, кодирующими сходные ферменты из других организмов, может оцениваться эволюционное родство этих организмов. Подобным образом, такое сравнение позволяет оценить, какие участки этой последовательности являются консервативными, а какие не являются консервативными, что может способствовать определению тех участков этого белка, которые являются незаменимыми для функционирования данного фермента. Этот тип определения является ценным для исследований по конструированию белков и может дать указание на то, какой белок может выносить условия мутагенеза без утраты функции.
Манипулирование молекулами нуклеиновых кислот МСТ данного изобретения может приводить к получению белков МСТ, имеющих функциональные различия в сравнении с белками МСТ дикого типа. Эти белки могут быть улучшены по эффективности или активности, могут присутствовать в более высоких количествах в клетке, чем обычно, или могут иметь уменьшенную эффективность или активность.
Данное изобретение представляет способы для скрининга молекул, которые модулируют активность белка МСТ, либо посредством взаимодействия с самим этим белком или субстратом или партнером связывания белка МСТ, либо модуляцией транскрипции или трансляции молекулы нуклеиновой кислоты МСТ данного изобретения. В таких способах микроорганизм, экспрессирующий один или более белков МСТ данного изобретения, контактируют с одним или более испытуемыми соединениями и оценивают действие каждого тест-соединения на активность или на уровень экспрессии данного белка МСТ.
Существует ряд механизмов, при помощи которых белок МСТ данного изобретения может непосредственно влиять на выход, продуцирование и/или эффективность продуцирования химического продукта тонкого органического синтеза из штамма С. glutamicum, несущего такой измененный белок. Извлечение химических продуктов тонкого органического синтеза из крупномасштабных культур С. glutamicum значительно улучшается, если С. glutamicum секретирует эти желательные соединения, так как такие соединения могут быть легко очищены из культуральной среды (в противоположность экстракции из массы клеток С. glutamicum). Посредством увеличения либо числа, либо активности молекул-транспортеров, которые экспортируют химические продукты тонкого органического синтеза из клетки, можно увеличить количество продуцируемого химического продукта тонкого органического синтеза, который присутствует во внеклеточной среде, обеспечивая более легкие сбор и очистку. Напротив, для эффективного сверхпродуцирования одного или нескольких химических продуктов тонкого органического синтеза могут требоваться увеличенные количества кофакторов, молекул-предшественников и промежуточных соединений для подходящих биосинтетических путей. Таким образом, увеличением числа и/или активности белков-транспортеров, участвующих в импорте питательных веществ, таких как источники углерода (т.е. сахаров), источники азота (т.е. аминокислоты, соли аммония), фосфат и сера, можно улучшать продуцирование химического продукта тонкого органического синтеза вследствие устранения каких-либо лимитирующих факторов в питательных веществах в биосинтетическом процессе. Далее, жирные кислоты и липиды сами являются желательными химическими продуктами тонкого органического синтеза, так что оптимизацией активности или увеличением числа одного или нескольких белков МСТ данного изобретения, которые участвуют в биосинтезе этих соединений, или нарушением активности одного или нескольких белков МСТ, которые участвуют в разложении этих соединений, можно увеличивать выход, продуцирование и/или эффективность продуцирования молекул жирных кислот и липидов из С. glutamicum.
Конструирование одного или более генов МСТ данного изобретения может также приводить к белкам МСТ, имеющим измененные активности, которые косвенно влияют на продуцирование одного или более химических продуктов тонкого органического синтеза из С. qlutamicum. Например, нормальные биохимические процессы метаболизма приводят к продуцированию разнообразных отработанных продуктов (например, пероксида водорода и других реакционноспособных разновидностей кислорода), которые могут активно взаимодействовать с теми же самыми метаболическими процессами (например, известно, что пероксинитрит осуществляет нитрование боковых цепей тирозина, инактивируя некоторые ферменты, имеющие в активном сайте тирозин (Groves, J.T. (1999) Curr. Opin. Chem. Biol. 3(2): 226-235). Хотя эти отработанные продукты обычно экскретируются, штаммы С. glutamicum, используемые для крупномасштабного ферментативного производства, являются оптимизированными в отношении сверхпродуцирования одного или более химических продуктов тонкого органического синтеза и, следовательно, могут продуцировать больше продуктов-отходов, чем это является типичным для С. glutamicum дикого типа. Оптимизацией активности одного или более белков МСТ данного изобретения, участвующих в экспорте молекул-отходов, можно улучшать жизнеспособность клетки и поддерживать эффективную метаболическую активность. Присутствие высоких внутриклеточных уровней желаемого химического продукта тонкого органического синтеза может быть также фактически токсичным для клетки, так что увеличением способности этой клетки секретировать эти соединения можно улучшить жизнеспособность клетки.
Далее, белки МСТ данного изобретения могут быть подвергнуты манипулированию таким образом, что относительные количества различных молекул липидов и жирных кислот изменяются. Это может оказывать сильное влияние на состав липидов мембраны клетки. Поскольку каждый тип липида имеет различные физические свойства, изменение в липидном составе мембраны может существенно изменять текучесть мембраны. Изменения в текучести мембраны могут влиять на транспорт молекул через мембрану, что, как объяснялось ранее, может модифицировать экспорт продуктов-отходов или продуцируемого химического продукта тонкого органического синтеза или импорт необходимых питательных веществ. Такие изменения текучести мембраны могут также сильно влиять на целостность клетки; клетки с относительно более слабыми мембранами являются более чувствительными в среде крупномасштабного ферментера к механическим стрессам, которые могут повреждать или убивать клетку. При манипулировании белками МСТ, участвующими в продуцировании жирных кислот и липидов для построения мембран, таким образом, что полученная мембрана имеет состав мембраны, более адаптируемый к условиям окружающей среды, существующим в настоящее время в культурах, используемых для производства химических продуктов тонкого органического синтеза, большая доля клеток С. glutamicum будет выживать и размножаться. Более высокие количества клеток С. glutamicum в культуре будут давать более высокие выходы, продуцирование или эффективность продуцирования химического продукта тонкого органического синтеза из данной культуры.
Вышеупомянутые стратегии мутагенеза для белков МСТ с целью получения увеличенных выходов химического продукта тонкого органического синтеза из С. glutamicum не должны рассматриваться как органичительные; вариации в таких стратегиях будут вполне очевидными специалисту с обычной квалификацией в данной области. С использованием таких стратегий и посредством включения описанных здесь механизмов молекулы нуклеиновых кислот и белков данного изобретения могут быть использованы для генерирования С. glutamicum или родственных штаммов бактерий, экспрессирующих мутированные молекулы нуклеиновых кислот и белков МСТ, так что выход, продуцирование и/или эффективность продуцирования желаемого соединения улучшается. Это желаемое соединение может быть любым природным продуктом, продуцируемым С. glutamicum, который включает конечные продукты путей биосинтеза и промежуточные продукты природно встречающихся метаболических путей, а также молекулы, которые не встречаются природно в метаболизме С. glutamicum, но которые продуцируются штаммом С. glutamicum данного изобретения.
Данное изобретение иллюстрируется далее следующими примерами, которые не должны рассматриваться как ограничивающие. Содержания всех ссылок, патентных заявок, патентов, опубликованных патентных заявок, таблиц и Списка последовательностей, цитируемые в данной заявке, включены здесь в качестве ссылки.
ПРИМЕРЫ
Пример 1: Получение общей геномной ДНК Corynebacterium glutamicum ATCC 13032
Культуру Corynebacterium glutamicum (ATCC 13032) выращивали в течение ночи при 30°С с сильным встряхиванием в среде BHI (Difco). Клетки собирали центрифугированием, супернатант отбрасывали и клетки ресуспендировали в 5 мл буфера-I (5% исходного объема культуры - все указанные объемы рассчитывали для 100 мл объема культуры). Состав буфера-I: 140,34 г/л сахарозы, 2,46 г/л MgSO4×7Н2O, 10 мл/л раствор К2HPO4 (100 г/л, доведенный до рН 6,7 с помощью КОН), 50 мл/л концентрата М12 (10 г/л (NH4)2SO4, 1 г/л NaCl, 2 г/л MgSO4×7Н2О, 0,2 г/л CaCl2, 0,5 г/л дрожжевого экстракта (Difco), 10 мл/л смеси микроэлементов (200 мг/л FeSO4×Н2О, 10 мг/л ZnSO4×7Н2О, 3 мг/л MnCl2×4Н2О, 30 мг/л Н3ВО3, 20 мг/л CoCl2×6Н2О, 1 мг/л NiCl2×6H2O, 3 мг/л Na2MoO4×2Н2О, 500 мг/л комплексообразующего агента (ЭДТА или лимонная кислота), 100 мл/л смеси витаминов (0,2 мг/л биотина, 0,2 мг/л фолиевой кислоты, 20 мг/л п-аминобензойной кислоты, 20 мг/л рибофлавина, 40 мг/л Са-пантотената, 140 мг/л никотиновой кислоты, 40 мг/л пиридоксолгидрохлорида, 200 мг/л миоинозита). К этой суспензии добавляли лизоцим до конечной концентрации 2,5 мг/мл. После приблизительно 4 часов инкубации при 37°С клеточную стенку разрушали и полученные протопласты собирали центрифугированием. Осадок промывали один раз 5 мл буфера-I и один раз 5 мл ТЭ-буфера (10 мМ Трис-HCl, 1 мМ ЭДТА, рН 8). Осадок ресуспендировали в 4 мл ТЭ-буфера и добавляли 0,5 мл раствора ДСН (10%) и 0,5 мл раствора NaCl (5 М). После добавления протеиназы К до конечной концентрации 200 мкг/мл суспензию инкубировали в течение приблизительно 18 часов при 37°С. ДНК очищали экстракцией фенолом, смесью фенол-хлороформ-изоамиловый спирт и смесью хлороформ-изоамиловый спирт с использованием обычных процедур. Затем ДНК осаждали добавлением 1/50 объема 3 М ацетата натрия и 2 объемов этанола с последующими 30-минутной инкубацией при -20°С и 30-минутным центрифугированием при 12000 об/мин в высокоскоростной центрифуге с использованием ротора SS34 (Sorvall). ДНК растворяли в 1 мл ТЭ-буфера, содержащего 20 мкг/мл РНКазы А, и диализовали при 4°С против 1000 мл ТЭ-буфера в течение по меньшей мере 3 часов. Это время буфер заменяли 3 раза. К аликвотам 0,4 мл диализованного раствора ДНК добавляли 0,4 мл 2 М LiCl и 0,8 мл этанола. После 30 мин инкубации при -20°С ДНК собирали центрифугированием (13000 об/мин, Biofuge Fresco, Heraeus, Hanau, Germany). Осадок ДНК растворяли в ТЭ-буфере. ДНК, полученную этой процедурой, можно было использовать для всех целей, в том числе для Саузерн-блоттинга или конструирования геномных библиотек.
Пример 2: Конструирование геномных библиотек Corynebacterium glutamicum ATCC 13032 в Escherichia coli
С использованием ДНК, полученной, как описано в примере 1, космидные и плазмидные библиотеки конструировали в соответствии с известными и хорошо установленными способами (см., например, Sambrook et al. (1989) "Molecular Cloning. A Laboratory Manual", Cold Spring Harbor Laboratory Press или Ausubel, F.M. et al. (1994) "Current Protocols in Molecular Biology", John Wiley and Sons).
Можно использовать любую плазмиду или космиду. Особенно применимыми были плазмиды pBR322 (Sutcliffe, J.G. (1979) Proc. Natl. Acad. Sci. USA, 75:3737-3741); pACYC177 (Change and Cohen (1978) J. Bacteriol. 134:1141-1156), плазмиды серии pBS (pBSSK+, pBSSK- и другие; Stratagene, La Jolla, USA) или космиды в виде SuperCos1 (Stratagene, La Jolla, USA) или Lorist6 (Gibson, T.J., Rosenthal A. and Waterson, R.H. (1987) Gene 53:283-286. Библиотеки генов специально для применения в С. glutamicum могут быть сконструированы с использованием плазмиды pSL109 (Lee, H.-S. and A.J.Sinskey (1994) J. Microbiol. Biotechnol. 4: 256-263).
Пример 3: Секвенирование и вычислительный функциональный анализ ДНК
Геномные библиотеки, описанные в примере 2, использовали для секвенирования ДНК в соответствии со стандартными способами, в частности, при помощи способа терминации цепи с использованием установки для секвенирования ABI377 (см., например, Fleischman, R.D. et al., (1995) "Whole-genom Random Sequencing and Assembly of Haemophilus Influenzae Rd., Science, 269:496-512). Использовали секвенирующие праймеры со следующими последовательностями: 5'-GGAAACAGTATGACCATG-3' (SEQ ID NO:677) или 5'-GTAAAACGACGGCCAGT-3' (SEQ ID NO:678).
Пример 4: Мутагенез in vivo
Мутагенез in vivo Corynebacterium glutamicum может проводиться пассажем ДНК плазмиды (или другого вектора) через Е. coli или другие микроорганизмы (например, виды Bacillus или дрожжи, такие как Saccharomyces cerevisiae), у которых была нарушена их способность поддерживать целостность их генетической информации. Типичные мутированные штаммы имеют мутации в генах для системы репарации ДНК (например, mutHLS, mutD, mutt, и т.д.; в отношении ссылки см. Rupp, W.D. (1996) DNA repair mechanisms, in: Escherichia coli and Salmonella, p.2277-2294, ASM, Washington). Такие штаммы хорошо известны специалисту в данной области. Применение таких штаммов иллюстрируется, например, в Greener, A. and Callahan, M. (1994) Strategies 7: 32-34.
Пример 5: Перенос ДНК между Escherichia coli и Corynebacterium glutamicum
Некоторые виды Corynebacterium и Brevibacterium содержат эндогенные плазмиды (например, рНМ1519 или pBL1), которые реплицируются автономно (в отношении обзора см, например, Martin, J.F. et al. (1987) Biotechnology, 5:137-146). Челночные векторы для Escherichia coli и Corynebacterium glutamicum могут быть легко сконструированы с использованием стандартных векторов для Е. coli (Sambrook et al. (1989) "Molecular Cloning. A Laboratory Manual", Cold Spring Harbor Laboratory Press или Ausubel, F.M. et al. (1994) "Current Protocols in Molecular Biology", John Wiley and Sons), к которым добавляют сайт инициации репликации (ориджин) и подходящий маркер из Corynebacterium glutamicum. Такие сайты инициации репликации предпочтительно берутся из эндогенных плазмид, выделенных из видов Corynebacterium и Brevibacterium. Особенно применимы в качестве маркеров трансформации для этих видов гены устойчивости к канамицину (такие, как полученные из транспозонов Tn5 или Tn903) или к хлорамфениколу (Winnacker, E.L. (1987) "From Genes to Clones - Introduction to Gene Technology, VCH, Weinheim). В литературе имеются многочисленные примеры конструирования большого разнообразия челночных векторов, которые реплицируются как в Е. coli, так и в С. glutamicum и которые могут быть использованы для нескольких целей, в том числе для сверхэкспрессии генов (в отношении ссылки см, например, Yoshihama, M. et al. (1985) J. Bacteriol. 162:591-597, Martin, J.F. et al. (1987) Biotechnology, 5:137-146 и Eikmanns, B.J. et al. (1991) Gene, 102:93-98).
С использованием стандартных способов можно клонировать представляющий интерес ген в один из описанных выше челночных векторов и вводить такие гибридные векторы в штаммы Corynebacterium glutamicum. Трансформация С. glutamicum может быть достигнута трансформацией протопластов (Kastsumata, R. et al. (1984) J. Bacteriol. 159306-311), электропорацией (Liebl, E. et al. (1989) FEMS Microbiol. Letters, 53:399-403) и в случаях, когда используют специальные векторы, также конъюгацией (как описано, например, в Schafer, A. et al. (1990) J. Bacteriol. 172:1662-1666). Можно также переносить челночные векторы для С. glutamicum в Е. coli получением плазмидной ДНК из С. glutamicum (с использованием стандартных способов, хорошо известных в данной области) и трансформацией ее в Е. coli. Эту стадию трансформации можно проводить с использованием стандартных способов, но предпочтительно использовать Мсг-недостаточный штамм Е. coli, такой как NM522 (Gough and Murray (1983) J. Mol. Biol. 166:1-19).
Гены могут быть сверхэкспрессированы в штаммах С. glutamicum с использованием плазмид, содержащих pCG1 (патент США №4617267) или ее фрагменты, и необязательно ген устойчивости к канамицину из TN903 (Grindley, N.D. and Joyce, C.M. (1980) Proc. Natl. Acad. Sci. USA 77(12): 7176-7180). Кроме того, гены могут быть сверхэкспрессированы в штаммах С. glutamicum с использованием плазмиды pSL109 (Lee, H.-S. and A.J.Sinskey (1994) J. Microbiol. Biotechnol. 4: 256-263).
Помимо применения реплицирующихся плазмид, сверхэкспрессия генов может быть также достигнута интеграцией их в геном. Геномная интеграция в С. glutamicum или других видах Corynebacterium или Brevibacterium может выполняться хорошо известными способами, такими как гомологичная рекомбинация с участком (участками) генома, опосредованная рестрикционными эндонуклеазами интеграция (REMI) (см., например, патент DE Patent 19823834), или посредством использования транспозонов. Можно также модулировать активность представляющего интерес гена модификацией регуляторных участков (например, промотора, репрессора и/или энхансера) посредством модификации последовательности, инсерцией или делецией с использованием сайт-направленных способов (таких как гомологичная рекомбинация) или способов на основе случайных событий (таких как транспозонный мутагенез или REMI). Последовательности нуклеиновых кислот, которые функционируют как терминаторы транскрипции, могут быть также встроены по положению 3' от кодирующего участка одного или более генов данного изобретения; такие терминаторы хорошо известны в данной области и описаны, например, в Winnacker, E.L. (1987) "From Genes to Clones - Introduction to Gene Technology, VCH, Weinheim.
Пример 6: Оценка экспрессии мутантного белка
Наблюдения активности мутированного белка в трансформированной клетке-хозяине основываются на том факте, что мутантный белок экспрессируется сходным образом и в сходном количестве в сравнении с белком дикого типа. Применимым способом для определения уровня транскрипции мутантного гена (индикатора количества мРНК, доступного для трансляции генного продукта) является проведение Нозерн-блоттинга (в отношении ссылки см., например, Ausubel, F.M. et al. (1994) "Current Protocols in Molecular Biology", Wiley; New York), в котором праймер, сконструированный для связывания с представляющим интерес геном, метят детектируемой меткой (обычно радиоактивной или хемилюминесцентной), так что при экстракции общей РНК культуры организма, подвергании ее электрофорезу на геле, переносе на стабильный матрикс и инкубировании с этим зондом связывание и количество связывания этого зонда указывает на присутствие, а также показывает количество мРНК для этого гена. Эта информация показывает степень транскрипции мутантного гена. Общая клеточная РНК может быть получена из Corynebacterium glutamicum при помощи нескольких способов, которые хорошо известны в данной области, например, описанных в Bormann, E.R. et al. (1992) Mol. Microbiol. 6: 317-326.
Для оценки присутствия или относительного количества белка, транслируемого из этой мРНК, могут быть использованы стандартные способы, такие как Вестерн-блоттинг (см., например, Ausubel, F.M. et al. (1994) "Current Protocols in Molecular Biology", Wiley: New York). В этом способе экстрагируют общие клеточные белки, разделяют их гель-электрофорезом, переносят на матрикс, такой как нитроцеллюлоза, и инкубируют с зондом, таким как антитело, которое специфически связывается с желаемым белком. Этот зонд обычно является меченым хемилюминесцентной или колорметрической меткой, которая может легко детектироваться. Присутствие и количество наблюдаемой метки показывает присутствие и количество желаемого мутантного белка, присутствующего в данной клетке.
Пример 7: Культивирование генетически модифицированного Corynebacterium glutamicum - среды и условия культуры
Генетически модифицированные Corynebacteria культивируют в синтетических или природных средах для выращивания. Ряд различных сред для выращивания для Corynebacteria являются как хорошо известными, так и легко доступными (Lieb et al. (1989) Appl. Microbiol. Biotechnol., 32:205-210; von der Osten et al. (1998) Biotechnology Letters, 11:11-16; Patent DE 4120867; Liebl (1992) "The Genus Corynebacterium, in: The Procaryotes, Volume II, Balows, A. et al., eds. Springer-Verlag). Эти среды состоят из одного или более источников углерода, источников азота, неорганических солей, витаминов и микроэлементов. Предпочтительными источниками углерода являются сахара, такие как моно-, ди- или полисахариды. Например, очень хорошими источниками углерода являются глюкоза, фруктоза, манноза, галактоза, рибоза, сорбоза, рибулоза, лактоза, мальтоза, сахароза, рафиноза, крахмал или целлюлоза. Сахар можно также предоставлять в среду через комплексные соединения, такие как мелассы или другие побочные продукты очистки сахаров. Предпочтительно, можно также предоставлять смеси различных источников углерода. Другими возможными источниками углерода являются спирты и органические кислоты, такие как метанол, этанол, уксусная кислота или молочная кислота. Источниками азота обычно являются органические или неорганические соединения азота или материалы, содержащие эти соединения. Примеры источников азота включают газообразный аммиак или аммониевые соли, такие как NH4Cl, (NH4)2SO4, NH4OH, нитраты, мочевина, аминокислоты или комплексные источники азота, такие как жидкий кукурузный экстракт, соевая мука, соевый белок, дрожжевой экстракт, мясной экстракт и другие.
Соединения, являющиеся неорганическими солями, которые могут быть включены в эти среды, включают хлоридные, фосфористые или сульфатные соли кальция, магния, натрия, кобальта, молибдена, калия, марганца, цинка, меди и железа. Хелатообразующие соединения могут быть добавлены к среде для поддержания ионов металлов в растворе. Особенно применимые хелатообразующие соединения включают дигидроксифенолы, такие как катехин или прокатехуат, или органические кислоты, такие как лимонная кислота. Для этих сред является типичным, что они также содержат другие факторы роста, такие как витамины или стимуляторы роста, примеры которых включают биотин, рибофлавин, тиамин, фолиевую кислоту, никотиновую кислоту, пантотенат и пиридоксин. Факторы роста и соли часто происходят из комплексных компонентов для сред, таких как дрожжевой экстракт, мелассы, жидкий кукурузный экстракт и другие. Точный состав соединений сред сильно зависит от конкретного эксперимента и решение о составе принимается для каждого особого случая. Информация об оптимизации сред доступна в руководстве "Applied Microbiol. Physiology. A Practical Approach (eds. P.M.Rhodes, P.F.Stanbury, IRL Press (1997) pp.53-73, ISBN 0 19 963577 3). Возможен также выбор сред для выращивания от коммерческих поставщиков, таких как стандарт 1 (Merck) или BHI (инфузионный раствор для мозга и сердца, Difco) или другие.
Все компоненты среды стерилизуют либо нагреванием (20 мин при 1/5 бар и 121°С), либо стерильным фильтрованием. Эти компоненты могут стерилизоваться в начале культивирования или они могут необязательно добавляться непрерывно или периодически.
Условия культивирования определяют отдельно для каждого эксперимента. Температура должна быть в диапазоне между 15°С и 45°С. Температура может поддерживаться на постоянном уровне или может меняться во время эксперимента. рН Среды должен быть в диапазоне 5-8,5, предпочтительно около 7,0, и может поддерживаться добавлением буферов к средам. Примером буфера для этой цели является калий-фосфатный буфер. Альтернативно или одновременно могут быть использованы синтетические буферы, такие как MOPS, HEPES, ACES и другие. Можно также поддерживать постоянный рН культуры добавлением NaOH или NH4OH во время культивирования. При использовании комплексных компонентов среды, таких как дрожжевой экстракт, необходимость в дополнительных буферах может уменьшаться вследствие того факта, что многие соединения комплекса имеют высокие буферные способности. При использовании ферментера для культивирования микроорганизмов рН может регулироваться при помощи газообразного аммиака.
Время инкубации обычно находится в диапазоне от нескольких часов до нескольких дней. Это время выбрано для получения возможности накопления максимального количества продукта в бульоне. Описанные эксперименты с культивированием могут проводиться в различных сосудах, таких как микротитрационные планшеты, стеклянные пробирки, стеклянные колбы или металлические ферментеры разных размеров. Для скрининга большого количества клонов микроорганизмы должны культивироваться в микротитрационных планшетах, стеклянных пробирках или встряхиваемых колбах с перегородками или без перегородок. Предпочтительно, используют встряхиваемые колбы на 100 мл, заполненные 10% (по объему) требующейся среды для выращивания. Колбы должны встряхиваться на ротационном шейкере (амплитуда 25 мм) с использованием диапазона скоростей 100-300 об/мин. Потери, связанные с испарением, могут быть уменьшены поддержанием влажной атмосферы; альтернативно должна проводиться математическая коррекция на потери от испарения.
При испытании генетически модифицированных клонов должен также испытываться немодифицированный контрольный клон или контрольный клон, содержащий основную плазмиду без какой-либо вставки. Среду инокулируют до OD600 0,5-1,5 с использованием клеток, выращенных на чашках с агаром, таких как СМ-чашки (10 г/л глюкозы, 2,5 г/л NaCl, 2 г/л мочевины, 10 г/л полипептона, 5 г/л дрожевого экстракта, 5 г/л мясного экстракта, 22 г/л агара, рН 6,8 с 2 М NaOH), которые инкубировали при 30°С. Инокуляцию сред выполняют либо введением суспензии клеток С. glutamicum в солевом растворе из СМ-чашек, либо добавлением жидкой предварительной культуры этой бактерии.
Пример 8 - Анализ in vitro функции мутантных белков
Определение активностей и кинетических параметров ферментов является хорошо разработанным в данной области. Эксперименты для определения активности любого конкретного измененного фермента должны выполняться относительно удельной активности фермента дикого типа, что находится вполне в рамках способностей специалиста в данной области. Обзоры по ферментам вообще, а также конкретные детали, касающиеся структуры, кинетики, принципов, способов, применений и примеров для определения активностей многих ферментов, могут быть найдены в следующих ссылках: Dixon, M. and Webb, B.C., (1979) Enzymes, Longmans: London; Fersht, (1985) Enzyme Structure and Mechanism, Freeman: New York; Walsh, (1979) Enzymatic Reaction Mechanisms. Freeman: San Francisco; Price, N.C., Stevens, L. (1982) Fundamentals of Enzymology, Oxford Univ, Press: Oxford; Boyer, P.O., ed. (1983) The Enzymes, 3rd ed. Academic Press: New York; Bisswanger, H., (1994) Enzymkinetik, 2nd ed. VCH: Weinheim (IBSN 3527300325); Bergmeyer, H.U., Bergmeyer, J., Grassi, M., eds. (1983-1986) Methods of Enzymatic Analysis, 3rd ed., vol.I-XII, Verlag Chemie: Weinheim; и Ullmann's Encyclopedia of Industrial Chemistry (1987) vol.A9, "Enzymes", VCH: Weinheim, p.352-363.
Активность белков, которые связываются с ДНК, может быть измерена несколькими установленными способами, такими как анализы смещения полос ДНК (также называемые гель-ретардационными анализами (анализами замедления миграции в геле)). Действие таких белков на экспрессию других молекул может быть измерено с использованием анализов с репортерными генами (например, описанными в Kolmar, H. et al. (1995) EMBO J. 14: 3895-3904 и цитируемых в этой работе ссылках). Тест-системы с репортерными генами хорошо известны и разработаны для применений как в прокариотических, так и эукариотических клетках, с использованием ферментов, таких как бета-галактозидаза, зеленый флуоресцентный белок и некоторые другие.
Определение активности белков мембранного транспорта может проводиться в соответствии со способами, такими как способы, описанные в Gennis, R.B. (1989) "Pores, Channels and Transporters", in Biomemranes, Molecular Structure and Function, Springer: Heidelberg, p.85-137; 199-234 и 270-322.
Пример 9: Анализ влияния мутантных белков на продуцирование желаемого продукта
Влияние генетической модификации в С. glutamicum на продуцирование желаемого соединения (такого как аминокислота) может быть оценено культивированием модифицированного микроорганизма при подходящих условиях (таких как условия, описанные выше) и анализом среды и/или клеточного компонента на увеличенное продуцирование желаемого продукта (т.е. аминокислоты). Такие способы анализа являются хорошо известными специалисту в данной области и включают спектроскопию, тонкослойную хроматографию, способы окрашивания разного рода, ферментативные и микробиологические способы и аналитическую хроматографию, например, высокоэффективную жидкостную хроматографию (см., например, Ullmann, Encyclopedia of Industrial Chemistry, vol.A2, p.89-90 и p.443-613, VCH: Weinheim (1985); Fallen, A. et al., (1987) "Applications of HPLC in Biochemistry" in: Laboratory Techniques in Biochemistry and Molecular Biology, vol.17; Rehm et al. (1993) Biotechnology, vol.3, Chapter III: "Product recovery and purification", page 469-714, VCH: Weinheim; Belter, P.A. et al. (1988) Bioseparations: downstream processing for biotechnology, John Wiley and Sons; Kennedy, J.F. and Cabral, J.M.S. (1992) Recovery processes for biological materials, John Wiley and Sons; Shaeiwitz, J.A. and Henry, J.D. (1988) Biochemical separations, in: Ullmann's Encyclopedia of Industrial Chemistry, vol.B3, Chapter 11, page 1-27, VCH: Weinheim; и Dechow, F.J. (1989) Separation and purification techniques in biotechnology, Noyes Publications).
Кроме измерения конечного продукта ферментации, можно также анализировать другие компоненты метаболических путей, используемых для продуцирования желаемого соединения, такие как промежуточные продукты и побочные продукты, для определения общей эффективности продуцирования соединения. Способы анализа включают измерения уровней питательных веществ в среде (например, сахаров, углеводородов, источников азота, фосфата и других ионов), измерения состава биомассы и роста, анализ продуцирования обычных метаболитов биосинтетических путей и измерение газов, продуцируемых во время ферментации. Стандартные способы для этих измерений описаны в "Applied Microbiol. Physiology. A Practical Approach (eds. P.M.Rhodes and P.F.Stanbury, eds., IRL Press p.103-129; и 165-192 (ISBN 0 199635773) и цитируемые ссылки.
Пример 10: Очистка желаемого продукта из культуры С. glutamicum
Извлечение желаемого продукта из клеток С. glutamicum или супернатанта описанной выше культуры может быть выполнено различными способами, хорошо известными в данной области. Если желаемый продукт не секретируется из клеток, клетки могут быть собраны из культуры низкоскоростным центрифугированием, эти клетки могут быть лизированы стандартными способами, такими как механические способы или обработка ультразвуком. Клеточный дебрис удаляют центрифугированием, а фракцию супернатанта, содержащую растворимые белки, сохраняют для последующей очистки желаемого соединения. Если продукт секретируется из клеток С. glutamicum, то эти клетки удаляют из культуры низкоскоростным центрифугированием, а фракцию супернатанта сохраняют для последующей очистки.
Фракцию супернатанта из любого способа очистки подвергают хроматографии с подходящей смолой, в которой либо желаемая молекула удерживается на хроматографической смоле, тогда как примеси в пробе не удерживаются, либо примеси удерживаются этой смолой, тогда как проба не удерживается. Такие стадии хроматографии могут быть повторены, если требуется, с использованием той же самой смолы или отличающихся смол. Специалист в данной области должен быть вполне сведущим в выборе подходящих хроматографических смол и в их наиболее эффективном применении для конкретной подлежащей очистке молекулы. Очищенный продукт может быть сконцентрирован фильтрованием или ультрафильтрацией и может храниться при температуре, при которой стабильность этого продукта является максимизированной.
Имеется большое число способов очистки, известных в данной области, и предыдущий способ очистки не должен рассматриваться как ограничивающий изобретение. Такие способы очистки описаны, например, в Bailey, J.E. and Oilis, D.F. Biochemical Engineering Fundamentals, McGraw-Hill: New York (1986).
Идентичность и чистота выделенных соединений могут оцениваться способами, стандартными в данной области. Они включают высокоэффективную жидкостную хроматографию (ВЭЖХ), спектроскопические способы, способы окрашивания, тонкослойную хроматографию, NIRS, ферментативный анализ или микробиологические способы. Такие способы анализа рассматриваются в Patek et al. (1994) App. Environ. Microbiol. 60: 133-140; Malakhova et al. (1996) Biotekhnologiya 11: 27-32 и Schmidt et al. (1998) Bioprocess Engineer, 19: 67-70, Ullmann's Encyclopedia of Industrial Chemistry, (1996) vol. A27, VCH: Weinheim, p.89-90, p.521-540, p.540-547, p.559-566, 575-581 и р.581-587; Michal, G. (1999) Biochemical Pathways: An Atlas of Biochemistry and Molecular Biology, John Wiley and Sons; Fallen, A. et al.(1987) Applications of HPLC in Biochemistry in: Laboratory Techniques in Biochemistry and Molecular Biology, vol.17.
Пример 11: Анализ последовательностей генов изобретения
Сравнение последовательностей и определение процента гомологии между двумя последовательностями являются известными в данной области способами и могут быть выполнены с использованием математического алгоритма, такого как алгоритм Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87:2264-68, модифицированный, как описано в Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-77. Такой алгоритм включен в программы NBLAST и XBLAST (версия 2.0) Altschul, et al. (1990) J. Mol. Biol. 215:403-10. blast-поиски нуклеотидов могут выполняться с программой NBLAST, оценка=100, длина слова=12, для получения нуклеотидных последовательностей, гомологичных молекулам нуклеиновых кислот МСТ данного изобретения, blast-поиски белков могут выполняться с программой XBLAST, оценка=50, длина слова=3, для получения аминокислотных последовательностей, гомологичных молекулам белка МСТ данного изобретения. Для получения имеющих гэпы сопоставлений для целей сравнения может быть использована программа Gapped BLAST, описанная в Altschul, et al. (1997) Nucleic Acids Res. 25(17): 3389-3402. При использовании программ BLAST и Gapped BLAST специалист в данной области должен знать, как оптимизировать параметры этой программы (например, XBLAST и NBLAST) для конкретной анализируемой последовательности.
Другим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Meyers and Miller (1998) Comput. Appl. Biosci. 4: 11-17). Такой алгоритм включен в программу ALIGN (версия 2.0), которая является частью GCG-пакета программ сопоставления последовательностей. При использовании программы ALIGN для сравнения аминокислотных последовательностей могут быть использованы таблица РАМ120 остатков весовых коэффициентов, штраф длины гэпа 12 и штраф гэпа 4. Дополнительные алгоритмы для анализа последовательностей известны в данной области и включают ADVANCE и ADAM, описанные в Torelli and Robotti (1994) Comput. Appl. Biosci. 10:3-5; и FASTA, описанную в Pearson and Lipman (1988) P.N.A.S. 85:2444-8.
Определение процента гомологии между двумя аминокислотными последовательностями может также выполняться с использованием программы GAP GCG-пакета программ (доступной в http://www.gcg.com), использующей либо матрицу Blosum 62, либо матрицу РАМ250 и весовой коэффициент гэпа 12, 10, 8, 6 или 4 и весовой коэффициент длины 1, 3 или 4. Процент гомологии между двумя последовательностями нуклеиновых кислот может быть определен с использованием программы GAP в GCG-пакете программ, с применением стандартных параметров, таких как весовой коэффициент гэпа 50 и весовой коэффициент длины 3.
Сравнительный анализ последовательностей генов данного изобретения с последовательностями, присутствующими в Genbank, выполняли с использованием способов, известных в данной области (см., например, Bexevanis and Oullette, eds. (1998) Bioinformatics: A Practical Guide to the Analysis of Genes and Proteins. John Wiley and Sons: New York). Последовательности генов данного изобретения сравнивали с генами, присутствующими в Genbank, в трехстадийном процессе. В первой стадии выполняли анализ BLASTN (например, локальный анализ сопоставления) для каждой из последовательностей данного изобретения в отношении нуклеотидных последовательностей, присутствующих в Genbank, и высшие 500 совпадений использовали для дальнейшего анализа. Последующий поиск FASTA (например, объединенный локальный и общий анализ сопоставления, в котором сопоставляли лимитированные участки этих последовательностей) проводили на этих 500 совпадениях. Каждую последовательность гена данного изобретения затем сопоставляли в целом с каждым из наивысших трех совпадений FASTA с использованием программы GAP в GCG-пакете программ (с использованием стандартных параметров). Для получения точных результатов длину последовательностей, извлеченных из Genbank, корректировали относительно длины исследуемых последовательностей при помощи способов, хорошо известных в данной области. Результаты этого анализа приведены в таблице 4. Полученная информация идентична информации, которая была бы получена, если бы проводили только анализ GAP (общий) на каждом из генов данного изобретения в сравнении с каждой из ссылочных последовательностей в Genbank, но требовалось значительно уменьшенное время вычисления в сравнении со временем вычисления анализа GAP (общего) с широкой базой данных. Последовательности данного изобретения, для которых не получали сопоставлений выше величин отсечения, показаны в таблице 4 отсутствием информации сопоставления. Кроме того, специалисту в данной области будет понятно, что проценты гомологии сопоставления GAP, приведенные в таблице 4 под заголовком «%-ная гомология (GAP)», перечислены в Европейском цифровом формате, где "," (запятая) обозначает десятичную точку. Например, величина "40,345" в этом столбце обозначает именно "40,345%".
Пример 12: Конструирование и эксплуатация микроматриц ДНК
Последовательности данного изобретения могут быть дополнительно использованы в конструировании и применении микроматриц ДНК (конструирование, методология и применения матриц ДНК хорошо известны в данной области и описаны, например, в Schena, M. et al. (1995) Science 270: 467-470, Wodicka, L. et al. (1997) Nature Biotechnology 15: 1359-1367; DeSaizieu, A. et al. (1998) Nature Biotechnology 16: 45-48 и DaRisi, J.L. et al. (1997) Science 278: 680-686).
Микроматрицы ДНК являются твердыми или гибкими подложками, состоящими из нитроцеллюлозы, нейлона, стекла, силикона или других материалов. Молекулы нуклеиновых кислот могут быть присоединены к поверхности упорядоченным образом. После подходящего мечения другие нуклеиновые кислоты или смеси нуклеиновых кислот могут быть гибридизованы с иммобилизованными молекулами нуклеиновых кислот, и метка может быть использована для мониторинга и измерения индивидуальных интенсивностей сигнала гибридизованных молекул в определенных участках. Эта методология позволяет проводить одновременное определение относительного или абсолютного количества всех или выбранных нуклеиновых кислот в нанесенной пробе или смеси нуклеиновых кислот. Таким образом, микроматрицы ДНК позволяют проводить анализ экспрессии множественных (до 6800 или более) нуклеиновых кислот параллельно (см., например, Schena, M. (1996) BioEssays 18(5): 427-431).
Последовательности данного изобретения могут быть использованы для конструирования олигонуклеотидных праймеров, которые способны амплифицировать определенные участки одного или более генов С. glutamicum посредством реакции амплификации нуклеиновых кислот, такой как полимеразная цепная реакция. Выбор и конструирование 5'- или 3'-олигонуклеотидных ПЦР-праймеров или подходящих линкеров делают возможным ковалентное присоединение полученных продуктов к поверхности среды-носителя, описанной выше (и описанной также, например, Schena, M. (1995) Science 270: 467-470).
Микроматрицы нуклеиновых кислот могут быть также сконструированы синтезом олигонуклеотидов in situ, как описано Wodicka, L. et al. (1997) Nature Biotechnology 15: 1359-1367. При помощи фотолитографических способов точно определенные участки матрицы экспонируются свету. Защитные группы, которые являются фотолабильными, активируются посредством этого и подвергаются добавлению нуклеотидов, тогда как участки, которые маскированы от света, не подвергаются никакой модификации. Последующие циклы защиты и активации светом делают возможным синтез различных олигонуклеотидов в определенных положениях. Небольшие, определенные участки генов данного изобретения могут быть синтезированы на микроматрицах посредством твердофазного синтеза олигонуклеотидов.
Молекулы нуклеиновых кислот данного изобретения, присутствующие в пробе или смеси нуклеотидов, могут быть гибридизованы с микроматрицами. Эти молекулы нуклеиновых кислот могут быть помечены в соответствии со стандартными способами. Вкратце, молекулы нуклеиновых кислот (например, молекулы мРНК или молекулы ДНК) метят включением изотопно или флуоресцентно меченых нуклеотидов, например, во время обратной транскрипции или синтеза ДНК. Гибридизация меченых нуклеиновых кислот с микроматрицами описана (например, в Schena, M. et al. (1995) supra; Wodicka, L. et al. (1997) supra и DeSaizieu, A. et al. (1998) supra). Детектирование и количественное определение гибридизованной молекулы выполняются в соответствии со специфической включенной меткой. Радиоактивные метки могут быть детектированы, например, как описано в Schena, M. et al. (1995 supra), а флуоресцентные метки могут быть детектированы, например, по способу Shalon et al. (1996) Genome Research 6: 639-645).
Применение последовательностей данного изобретения к технологии микроматриц ДНК, как описано выше, делает возможным сравнительный анализ различных штаммов С. glutamicum или других Corynebacteria. Например, исследования вариаций между штаммами на основе профилей индивидуальных транскриптов и идентификация генов, которые являются важными для специфических и/или желательных свойств штаммов, таких как патогенность, продуктивность и устойчивость к стрессам, облегчаются методологиями микроматриц нуклеиновых кислот. Сравнения профиля экспрессии генов данного изобретения в ходе реакции ферментации также являются возможными с использованием технологии матриц нуклеиновых кислот.
Пример 13: Анализ динамики клеточных популяций белков (протеомики)
Гены, композиции и способы данного изобретения могут применяться для исследования взаимодействий и динамики популяций белков, называемой "протеомикой". Представляющие интерес популяции белков включают, но не ограничиваются, общую популяцию белков С. glutamicum (например, в сравнении с популяциями белков других организмов), белки, которые являются активными при специфических условиях окружающей среды или метаболических условиях (например, во время ферментации, при высокой или низкой температуре или при высоком или низком рН), или другие белки, которые являются активными во время конкретных фаз роста и развития.
Популяции белков можно анализировать разнообразными хорошо известными способами, такими как гель-электрофорез. Клеточные белки могут быть получены, например, лизисом или экстракцией, и могут быть отделены друг от друга с использованием различных электрофоретических способов. Электрофорез в полиакриламидном геле с додецилсульфатом натрия (электрофорез в ДСН-ПААГ) разделяет белки в значительной степени на основе их молекулярной массы. Электрофорез в полиакриламидном геле с изоэлектрическим фокусированием (ИЭФ-электрофорез в ПААГ) разделяет белки по их изоэлектрической точке (которая отражает не только аминокислотную последовательность, но также посттрансляционные модификации данного белка). Другим, более предпочтительным способом анализа белков является последовательная комбинация ИЭФ-электрофореза в ПААГ и электрофореза в ДСН-ПААГ, известная как 2-D-гель-электрофорез (описанная, например, в Hermann et al. (1998) Electrophoresis 19: 3217-3221; Fountoulakis et al. (1998) Electrophoresis 18: 1184-1192; Antelmann et al. (1997) Electrophoresis 19:1193-1202; Langen et al. (1997) Electrophoresis 18: 1451-1463). Другие способы разделения могут быть также использованы для разделения белков, такие как капиллярный гель-электрофорез; подобные способы хорошо известны в данной области.
Белки, разделенные с использованием этих методологий, могут быть визуализированы стандартными способами, такими как окрашивание или мечение. Подходящие красители известны в данной области и включают Кумасси бриллиантовый синий, содержащий серебро краситель (серебряный краситель) или флуоресцентные красители, такие как Sypro Ruby (Molecular Probes). Включение радиоактивно меченых аминокислот или других белковых предшественников (например, 35S-метионина, 35S-цистеина, 14С-меченых аминокислот, 15N-аминокислот, 15NO3 или 15NH4 + или 13С-меченых аминокислот) в среду С. glutamicum позволяет проводить мечение белков из этих клеток перед их разделением. Подобным образом могут быть использованы флуоресцентные метки. Меченые белки могут быть экстрагированы, выделены и разделены в соответствии с описанными ранее способами.
Белки, визуализированные этими способами, можно дополнительно анализировать посредством измерения количества используемых красителя или метки. Количество конкретного белка может быть определено количественно с использованием, например, оптических методов и может сравниваться с количеством других белков в том же самом геле или в других гелях. Сравнения белков на гелях могут производиться, например, оптическим сравнением, спектроскопией, сканированием изображений и анализом гелей или посредством применения фотографических пленок и экранов. Такие способы хорошо известны в данной области.
Для определения идентичности любого конкретного белка могут быть использованы прямое секвенирование или другие стандартные способы. Например, может быть использовано секвенирование N- и/или С-концевых аминокислот (например, разложение по Эдману), а также масс-спектрометрия (а именно, способах MALDI или ESI (см., например, Langen et al. (1997) Electrophoresis 18: 1184-1192)). Белковые последовательности, представленные здесь, могут быть использованы для идентификации белков С. glutamicum при помощи этих способов.
Информация, полученная этими способами, может быть использована для сравнения параметров присутствия белков, активности или модификации между различными пробами из различных биологических условий (например, различных организмов, временных точек ферментации, условий сред или различных биотопов, среди прочих). Данные, полученные только из таких экспериментов, или в комбинации с другими способами, могут быть использованы для различных применений, например, для сравнения поведения различных организмов в конкретной (например, метаболической) ситуации, для увеличения продуктивности штаммов, которые продуцируют химические продукты тонкого органического синтеза, или для увеличения эффективности продуцирования химических продуктов тонкого органического синтеза.
Эквиваленты
Специалистам в данной области будут очевидны многие эквиваленты или они будут способны получить с использованием не большего труда, чем этого требует рутинное экспериментирование, многочисленные эквиваленты относительно конкретных вариантов описанного здесь изобретения. Предполагается, что такие эквиваленты охватываются следующей формулой изобретения.




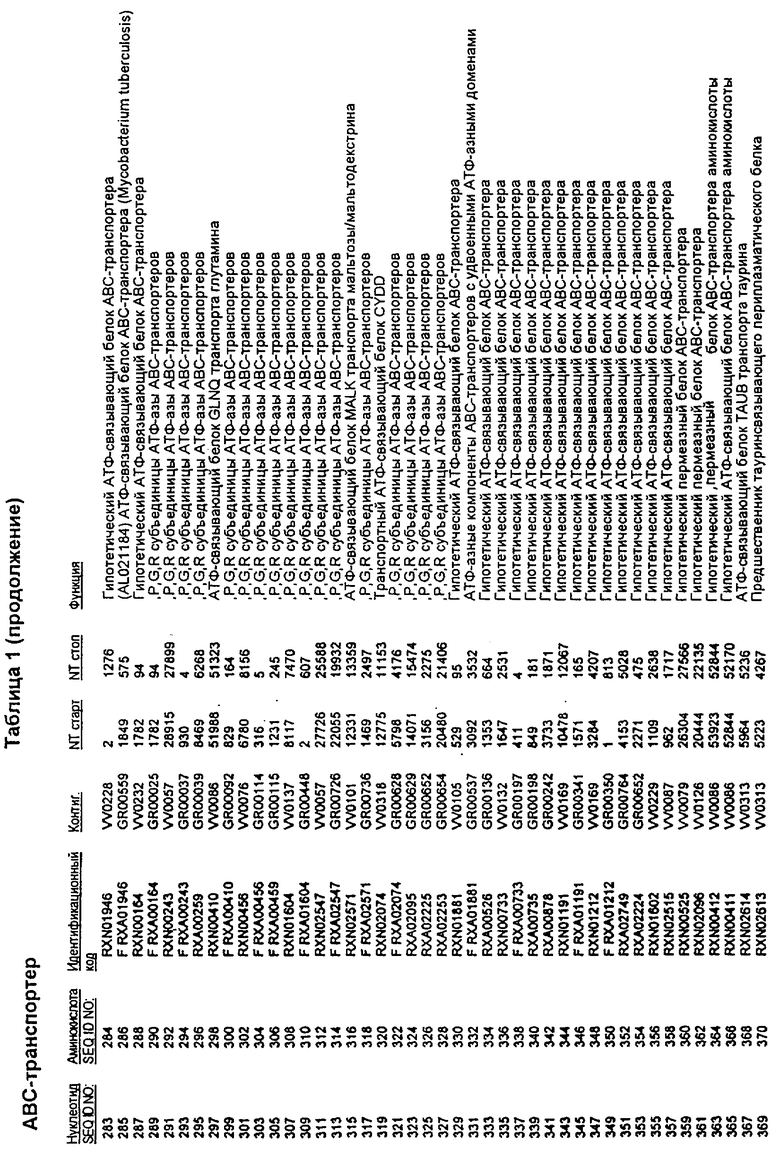




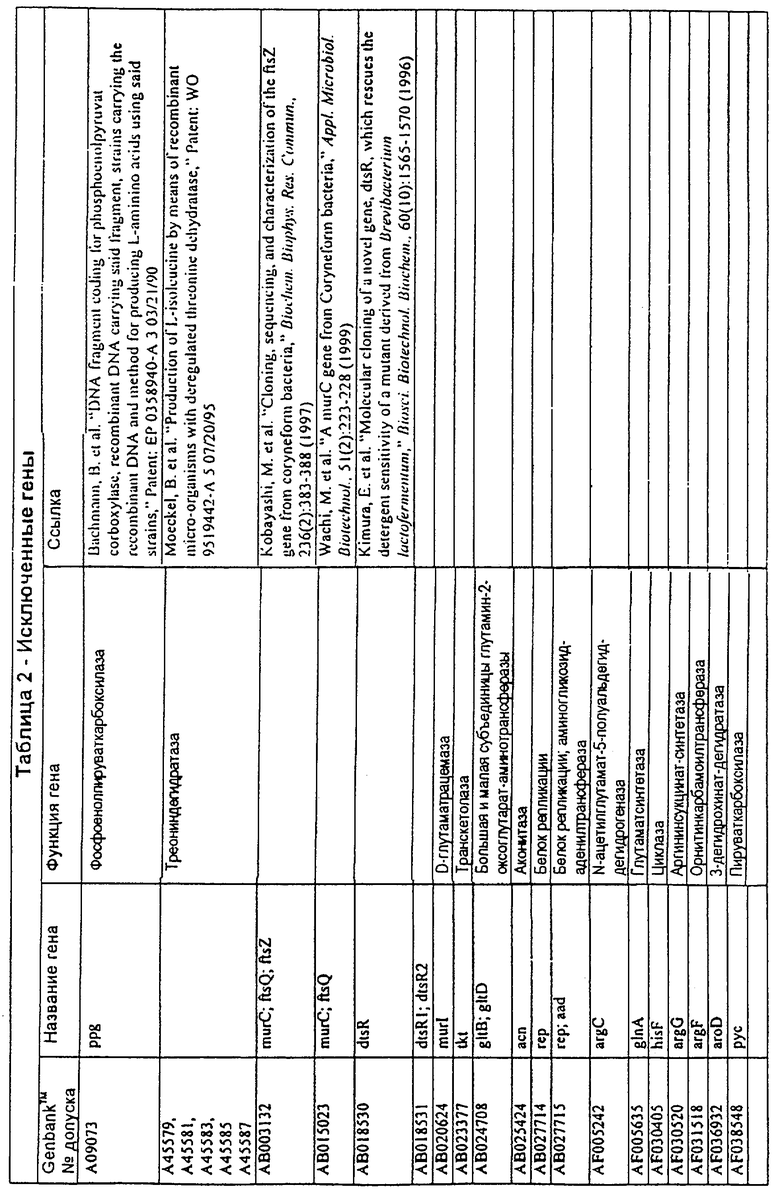
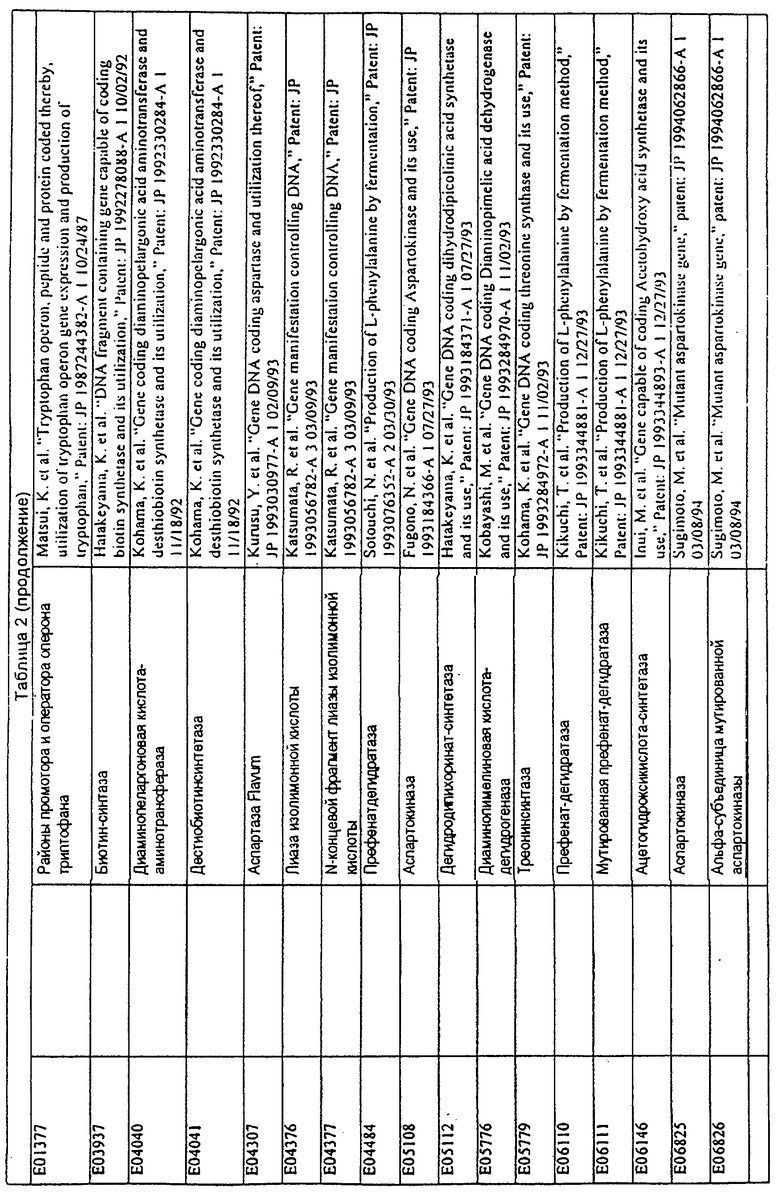
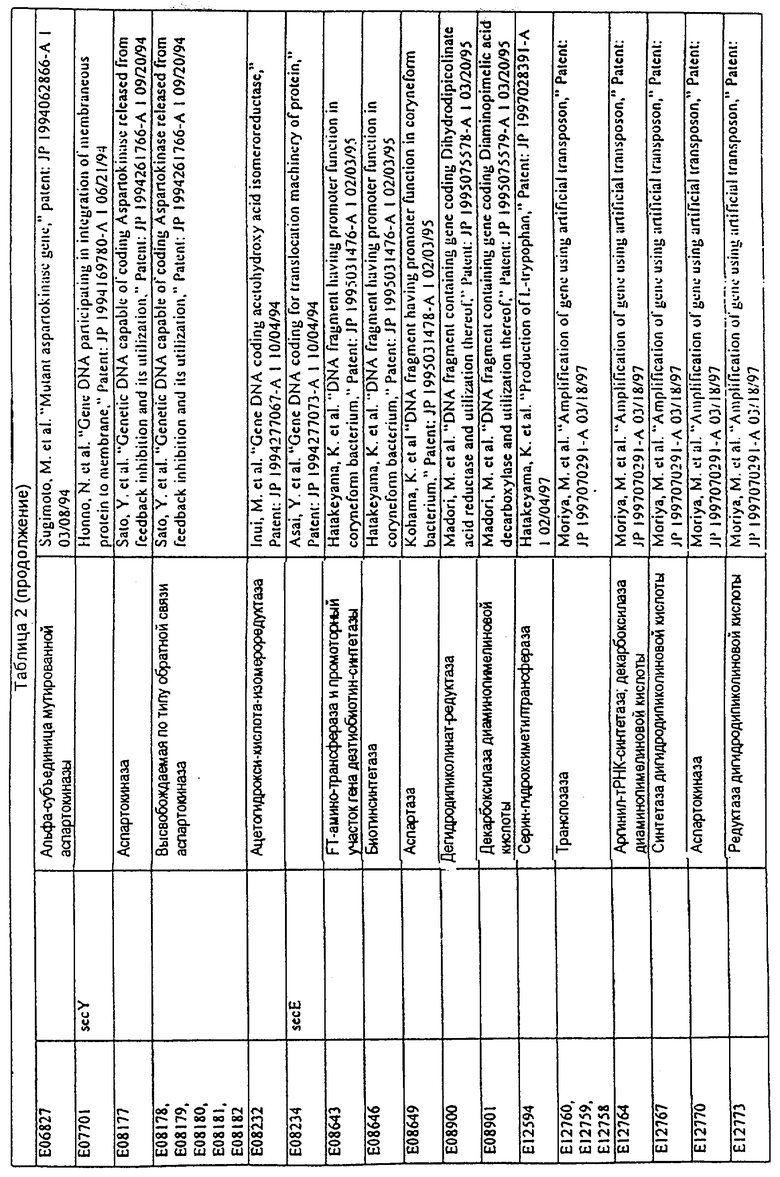
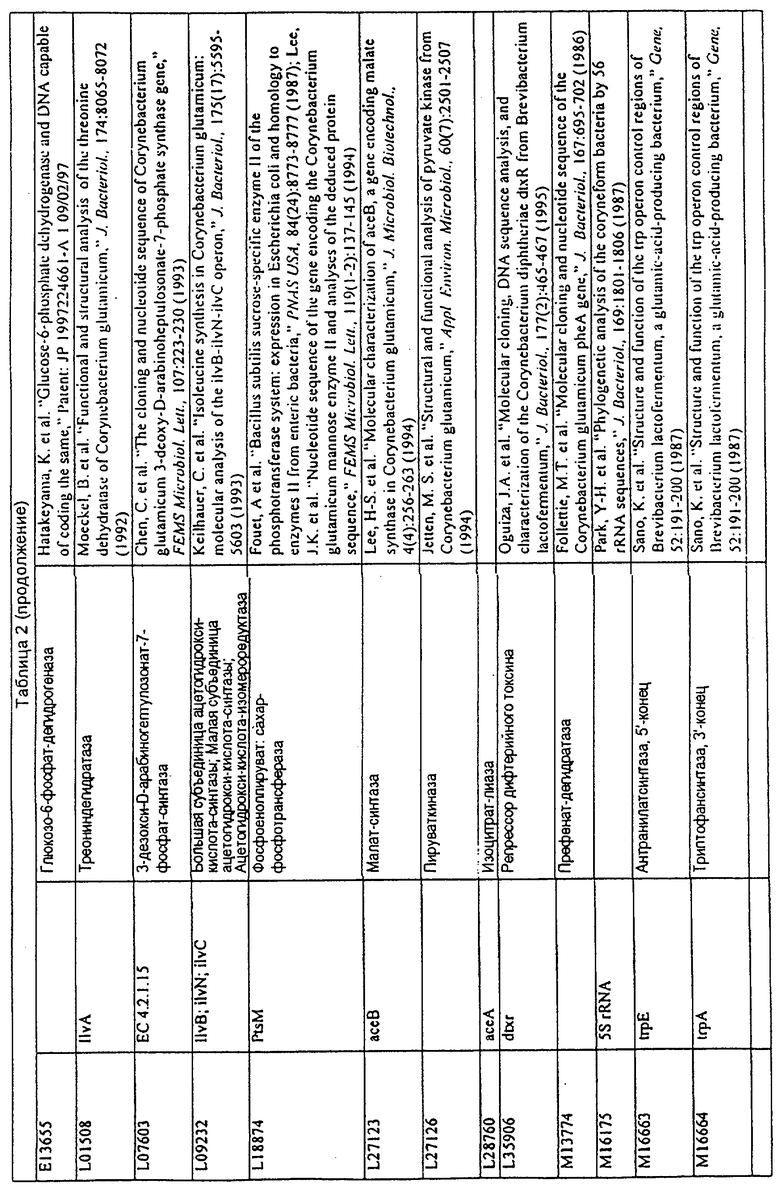
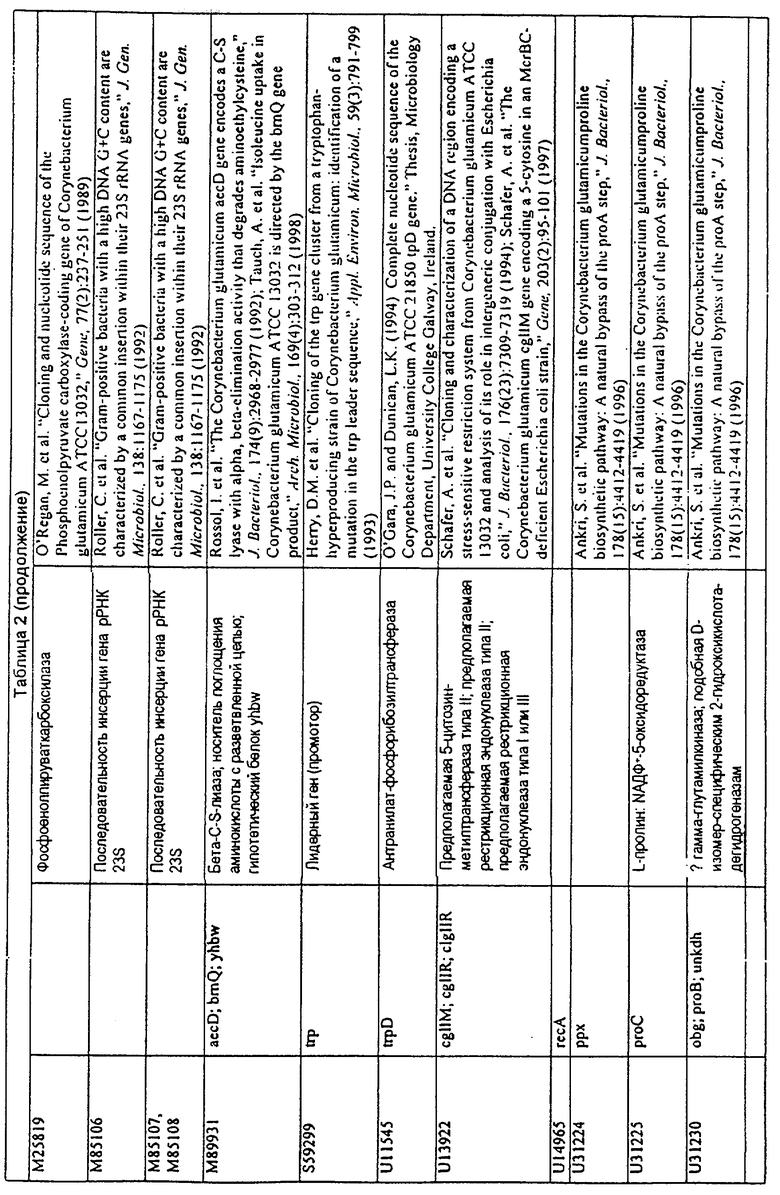
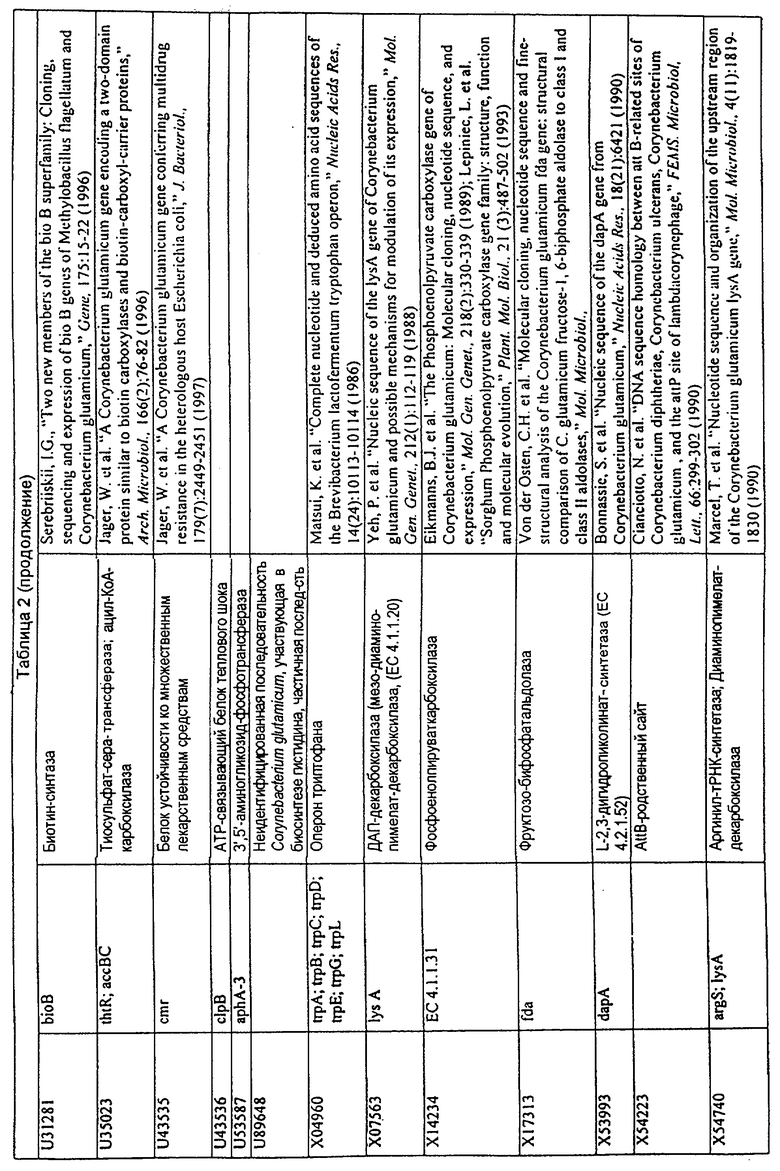
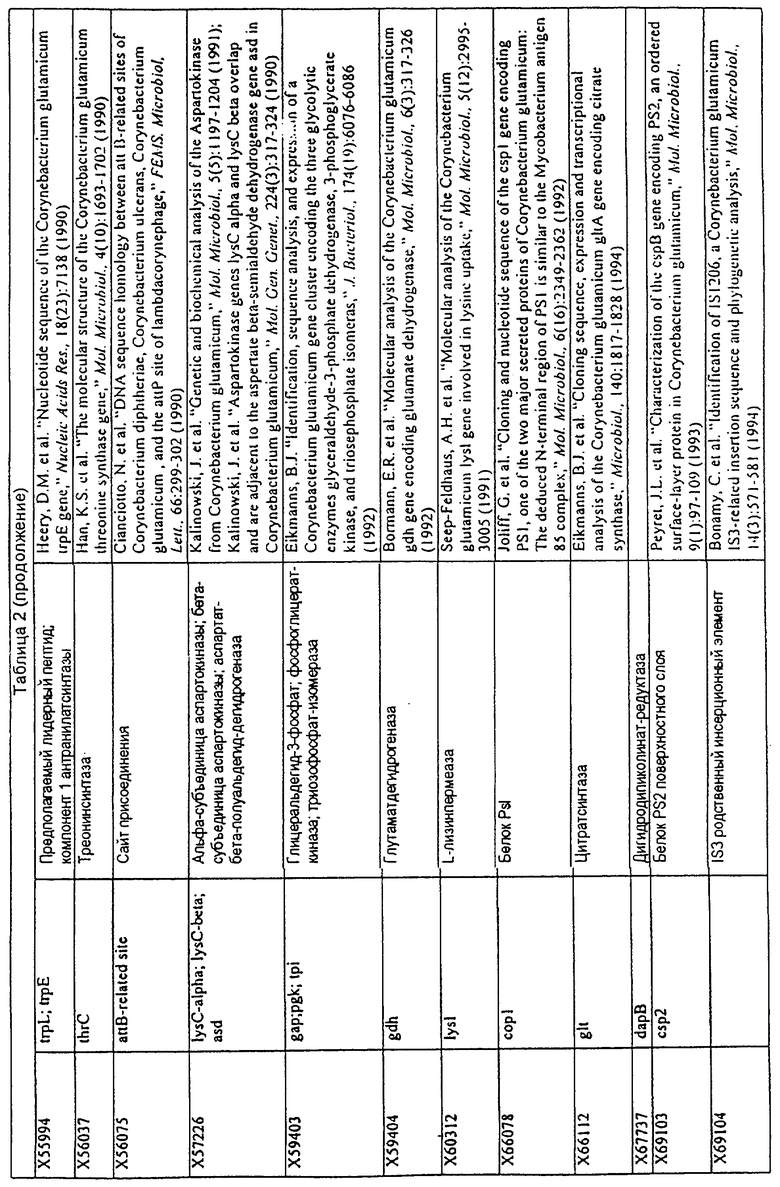
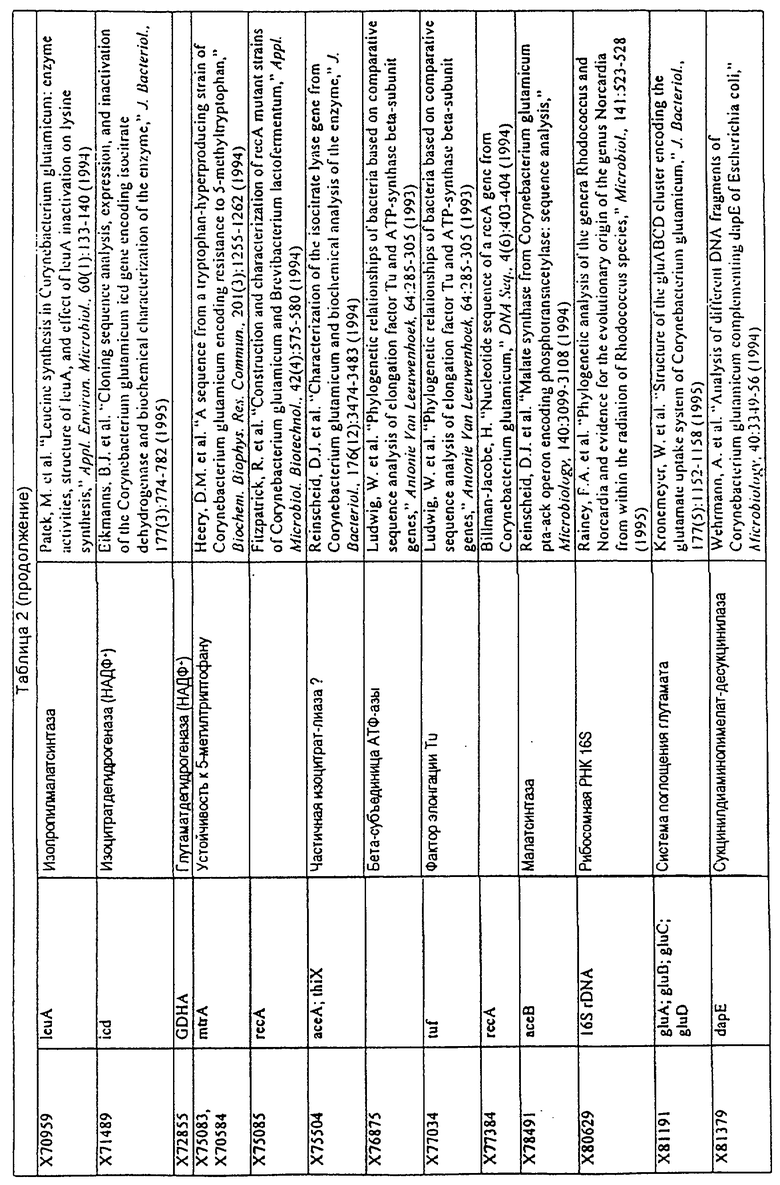
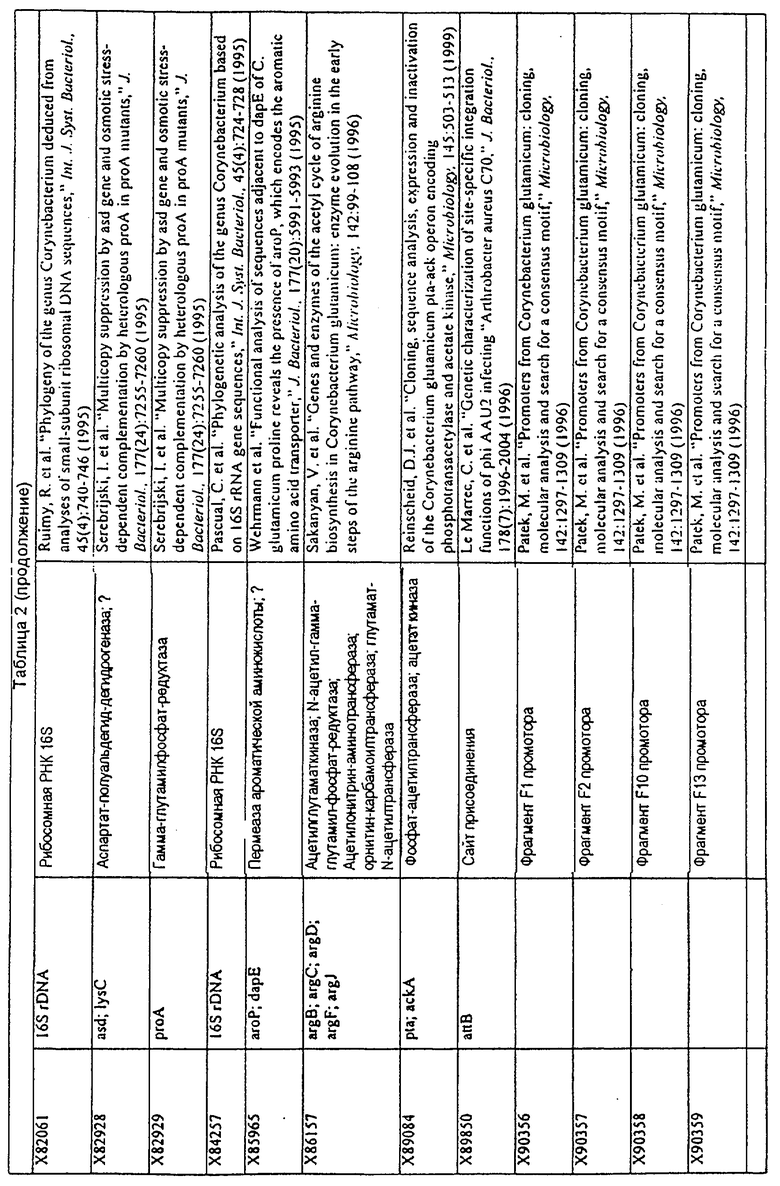
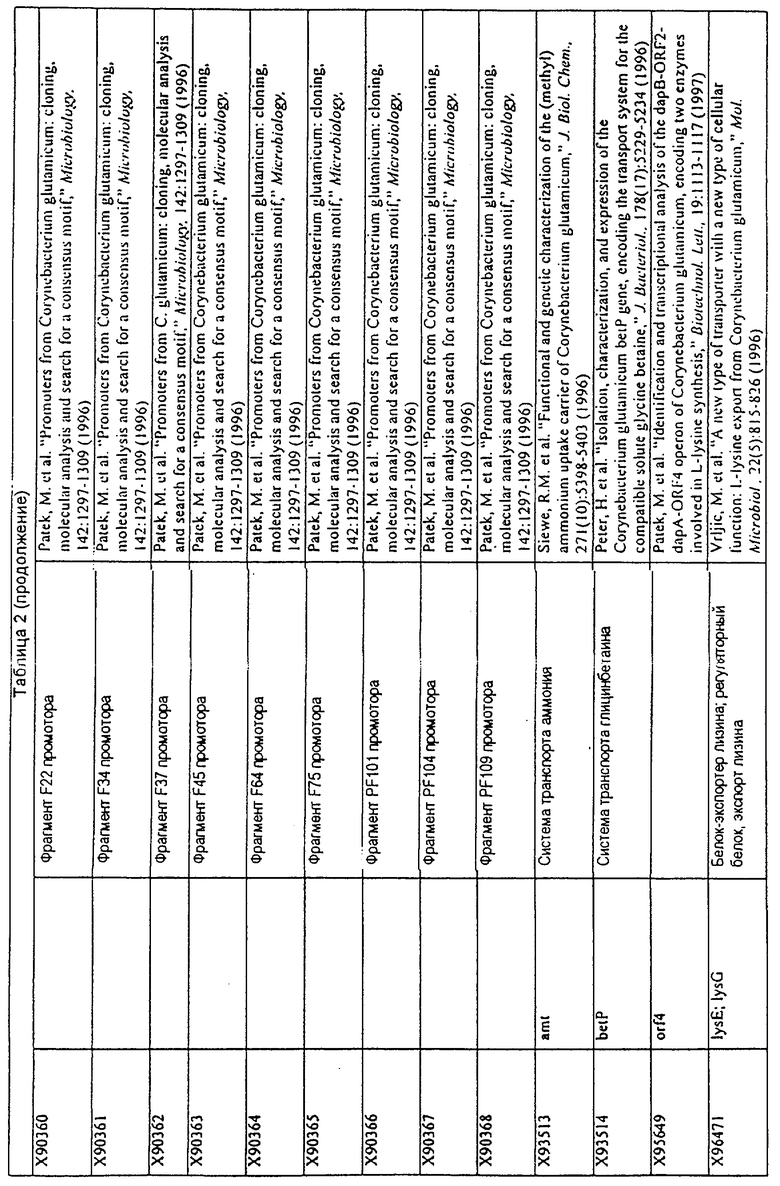
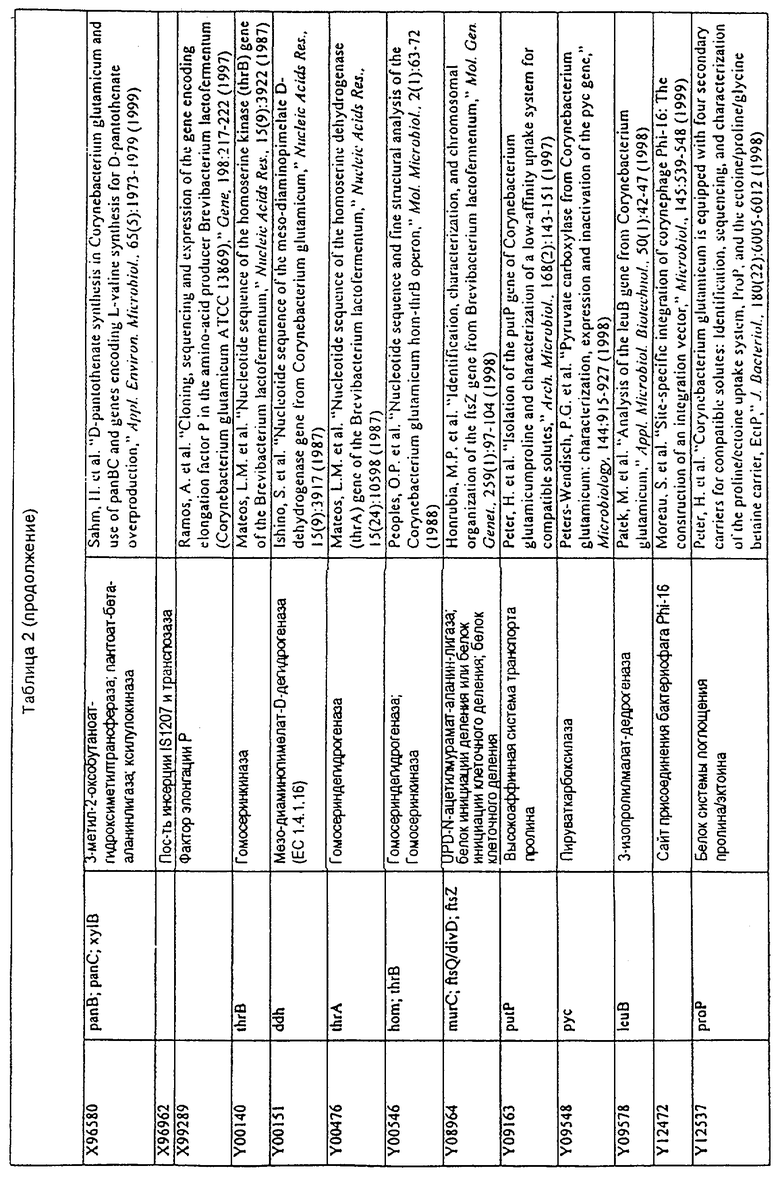
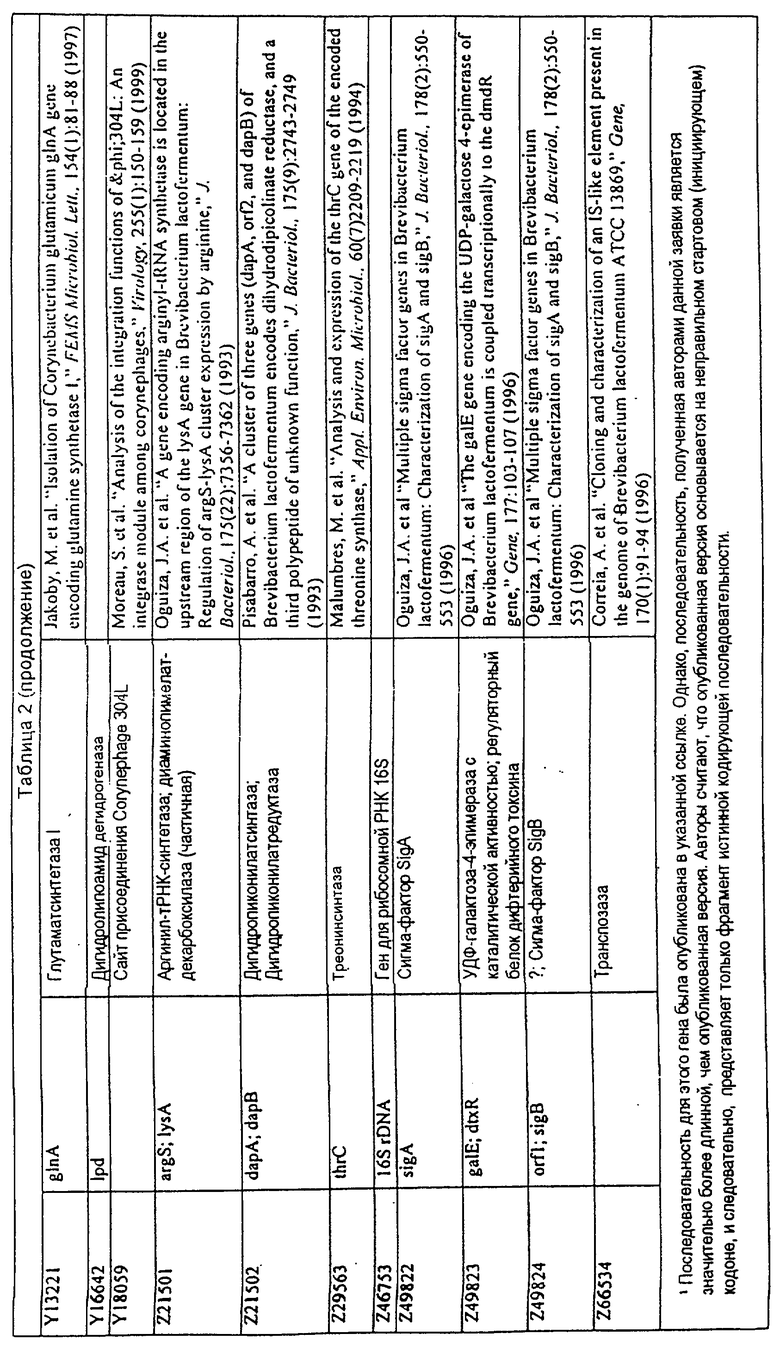
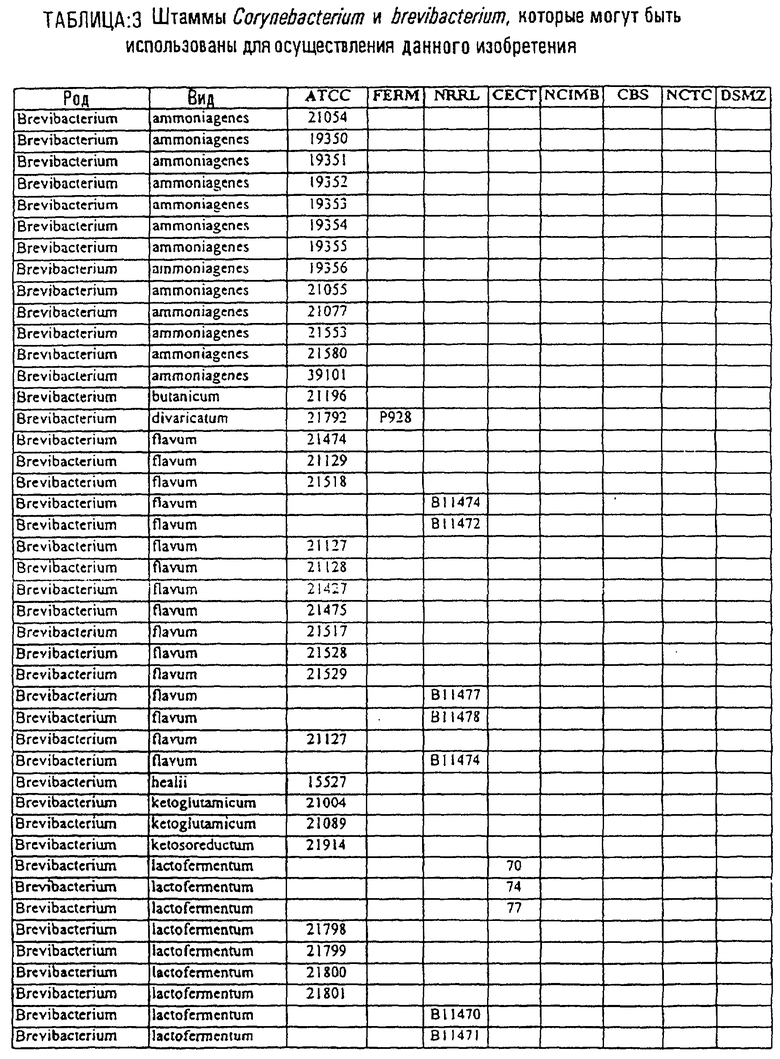
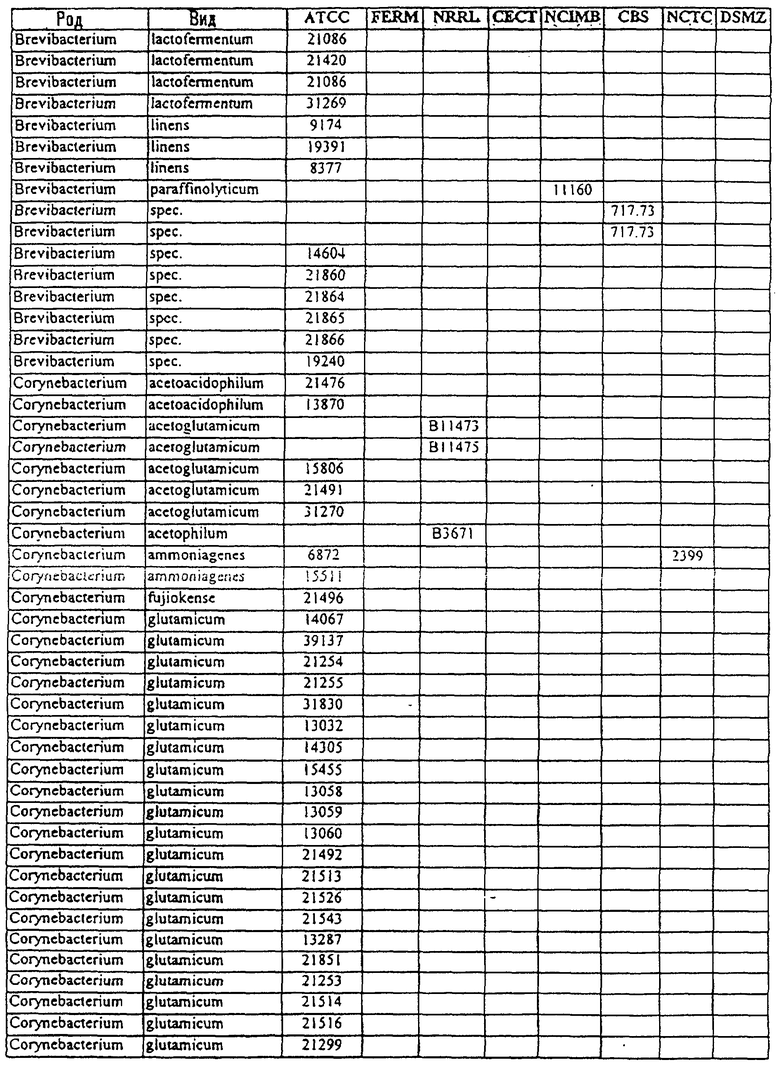
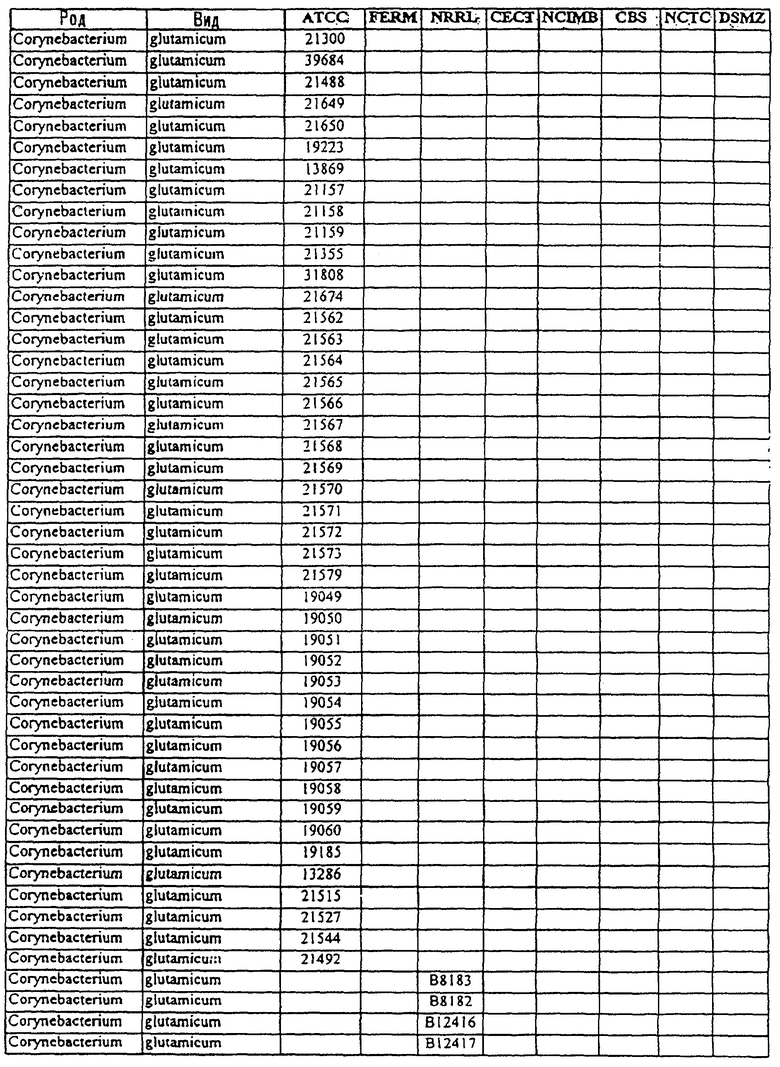
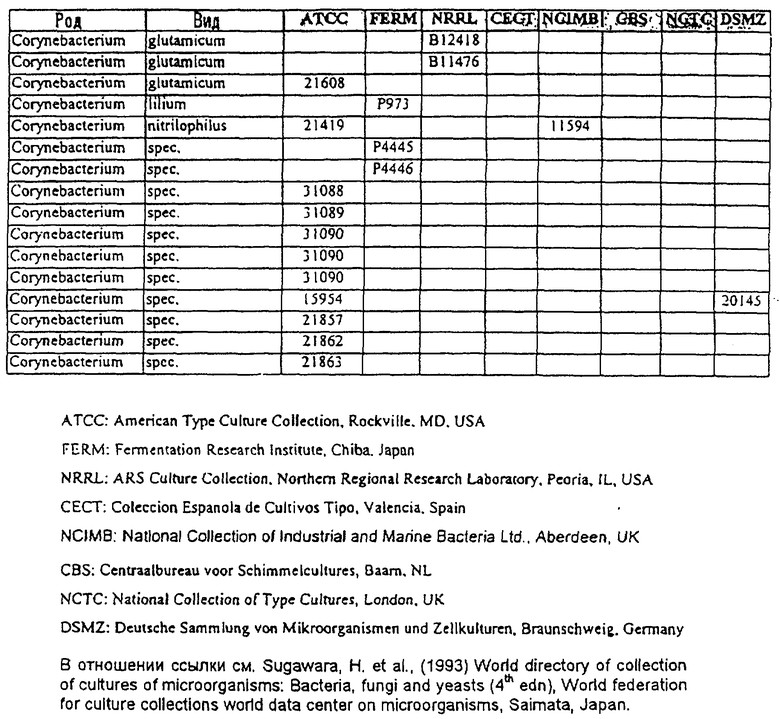





















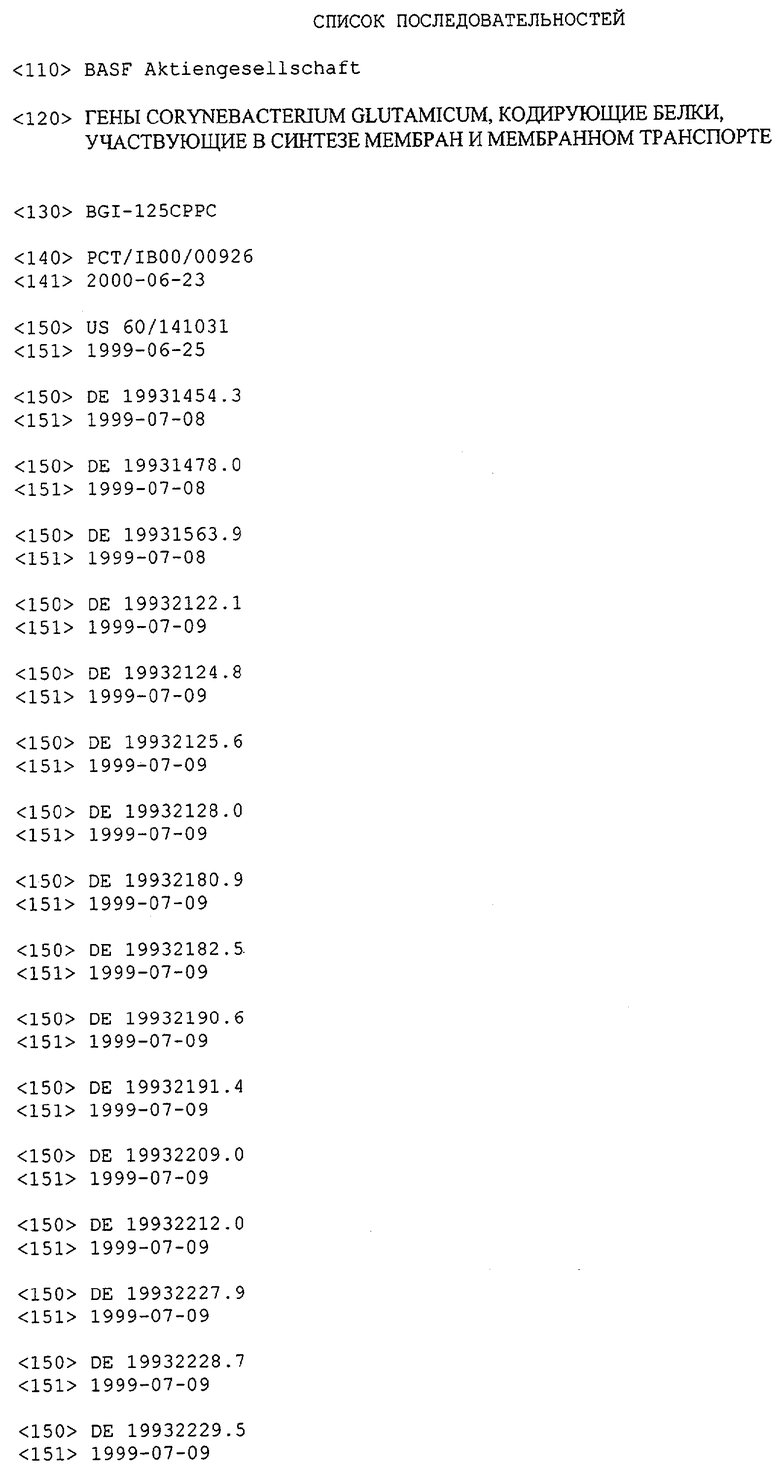
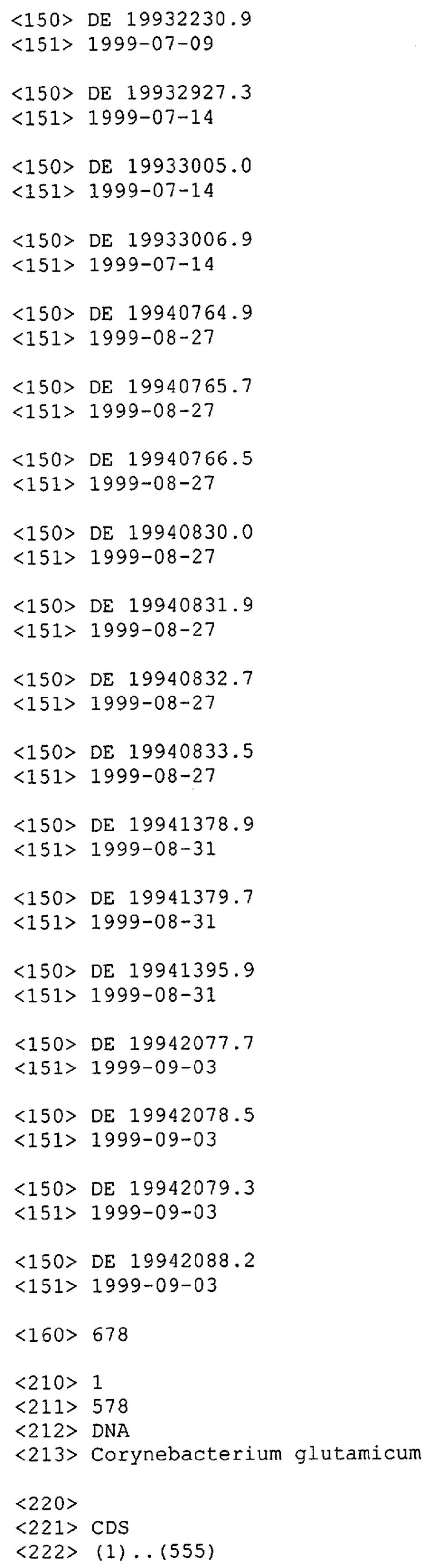






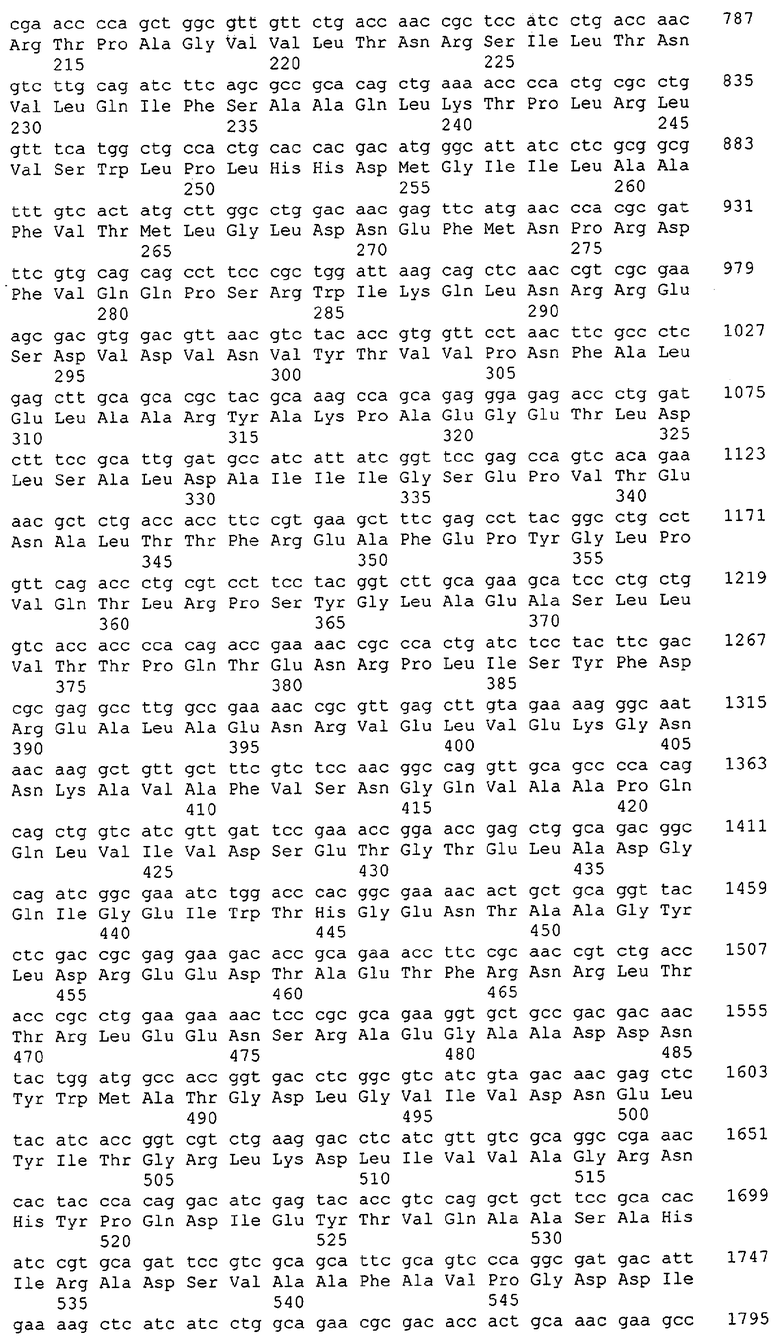

















































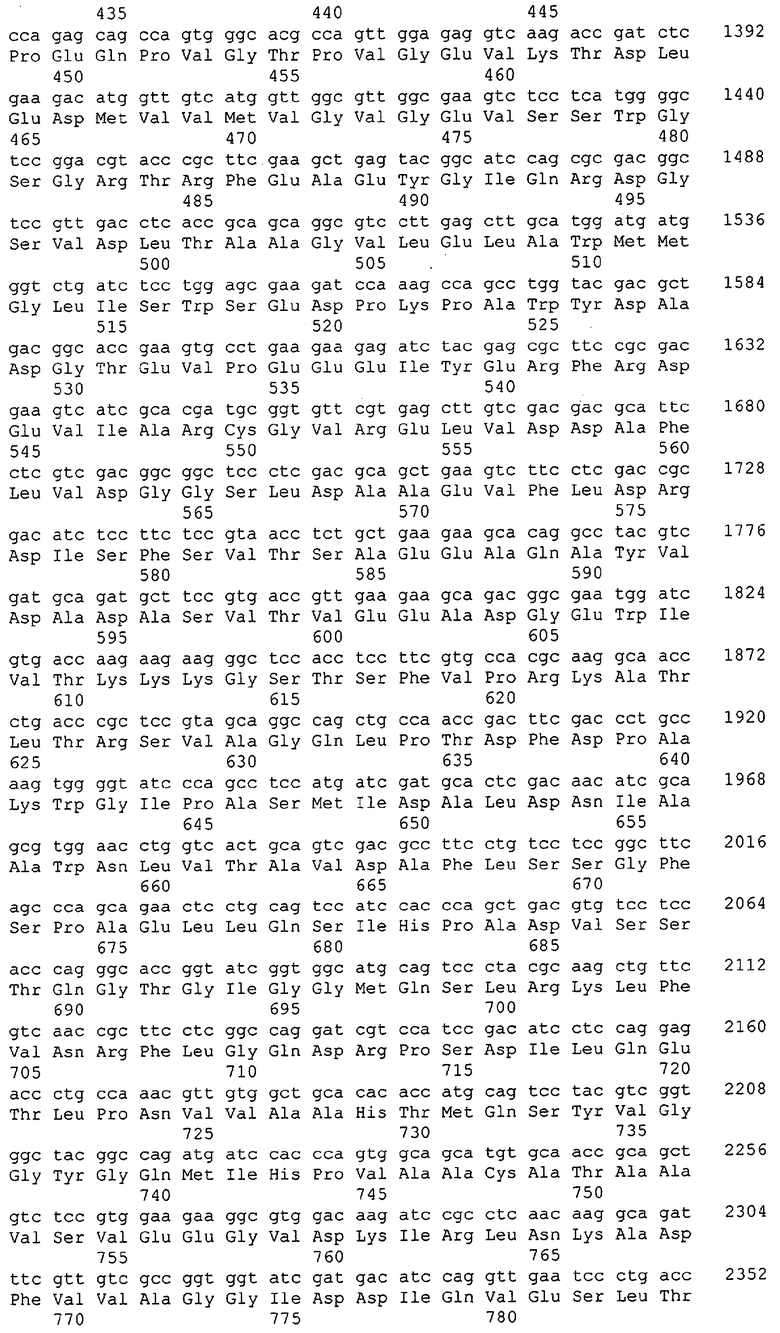




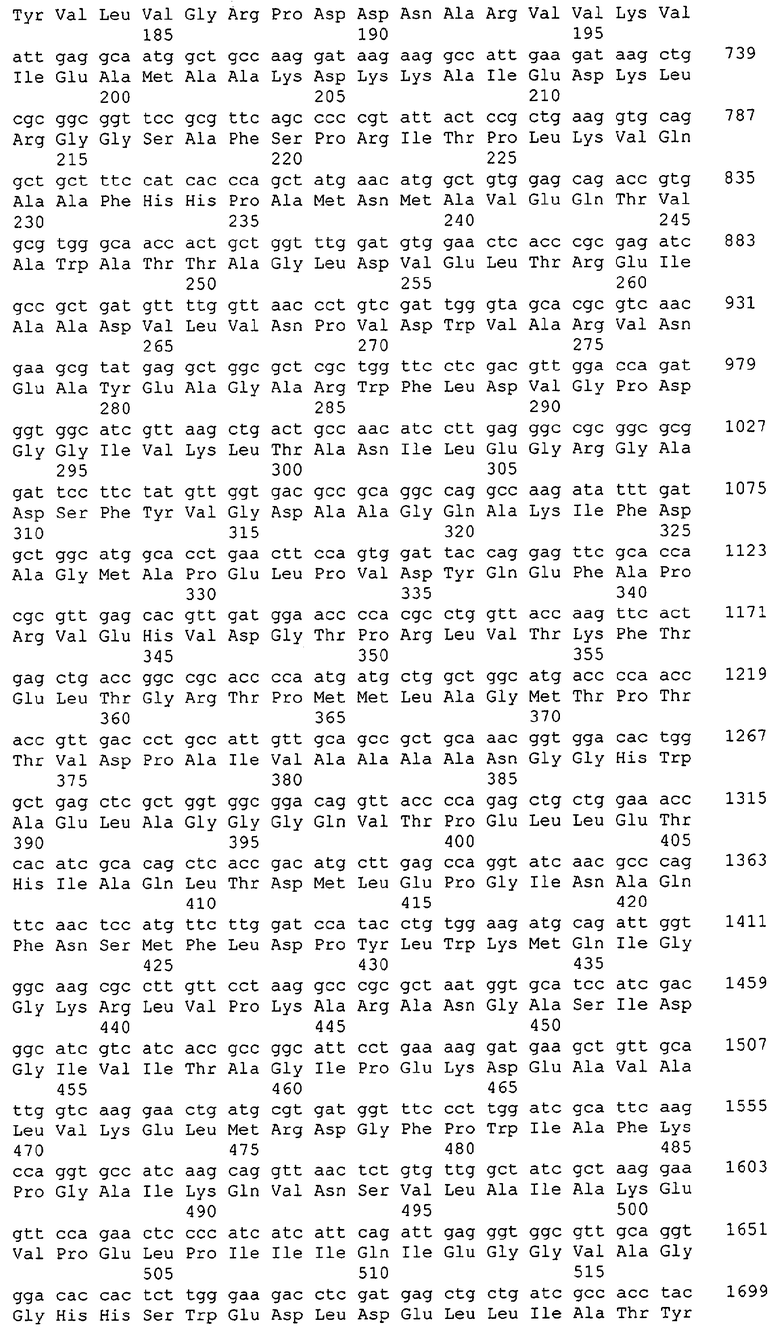
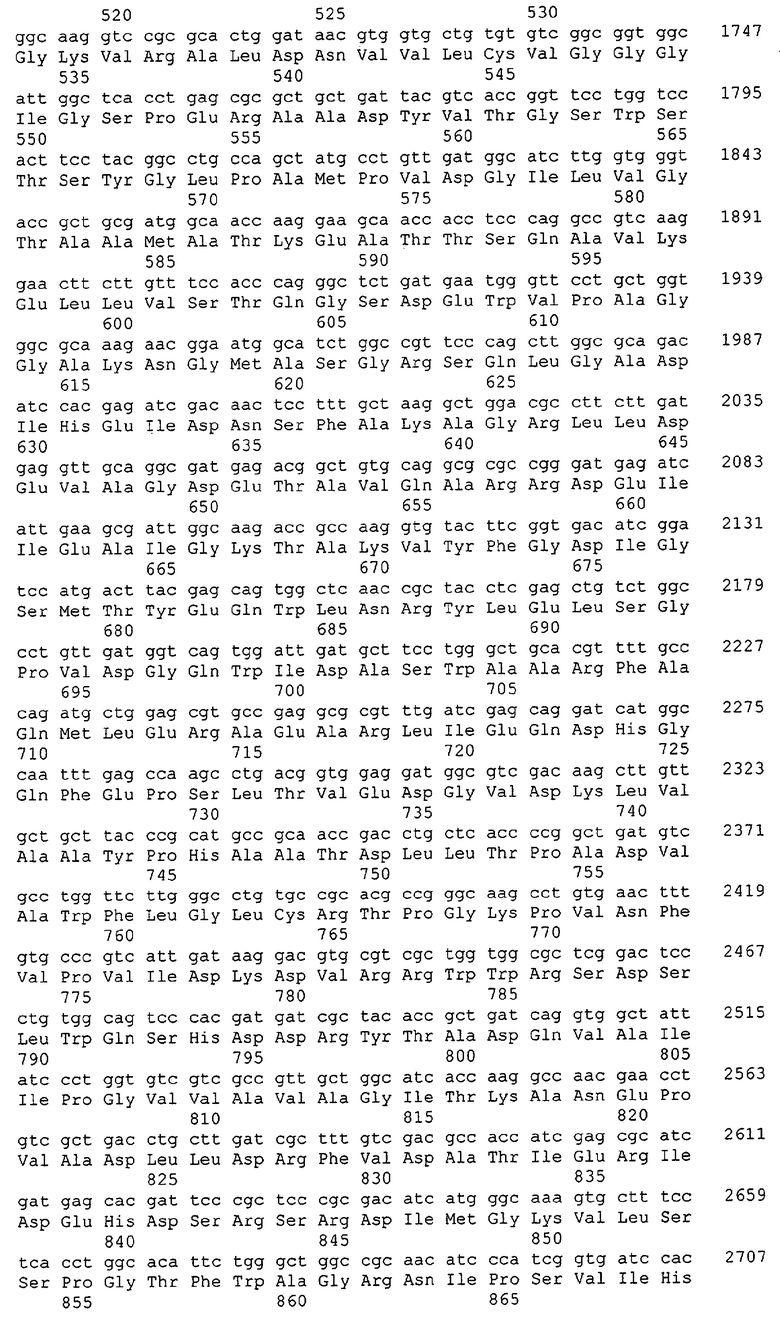


































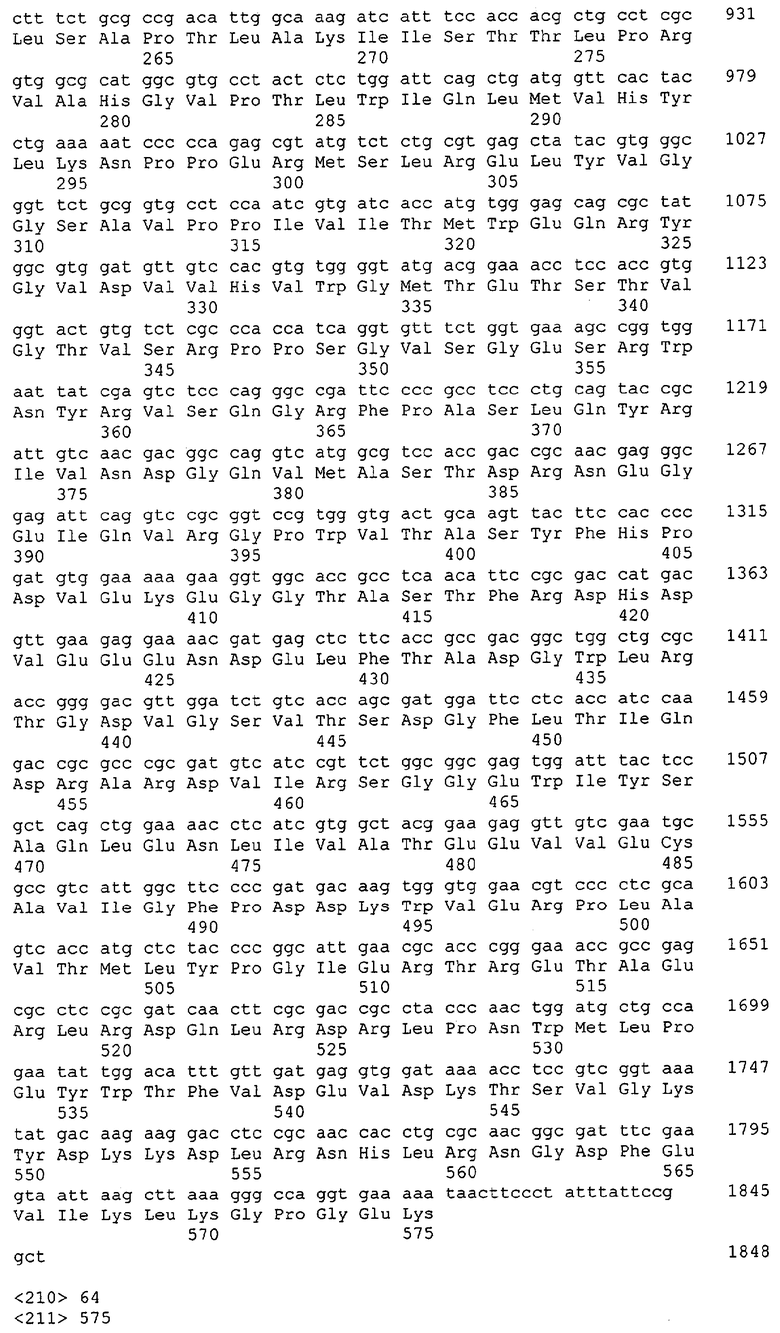







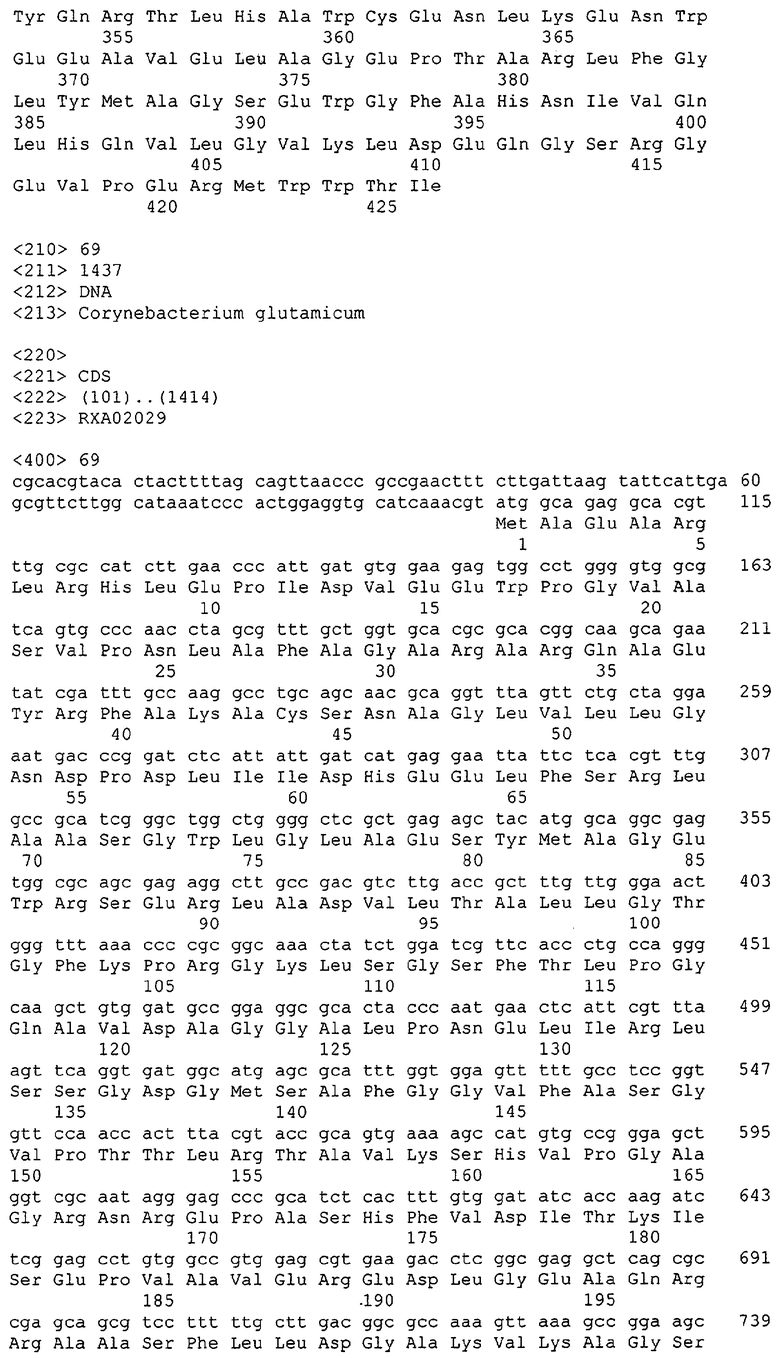










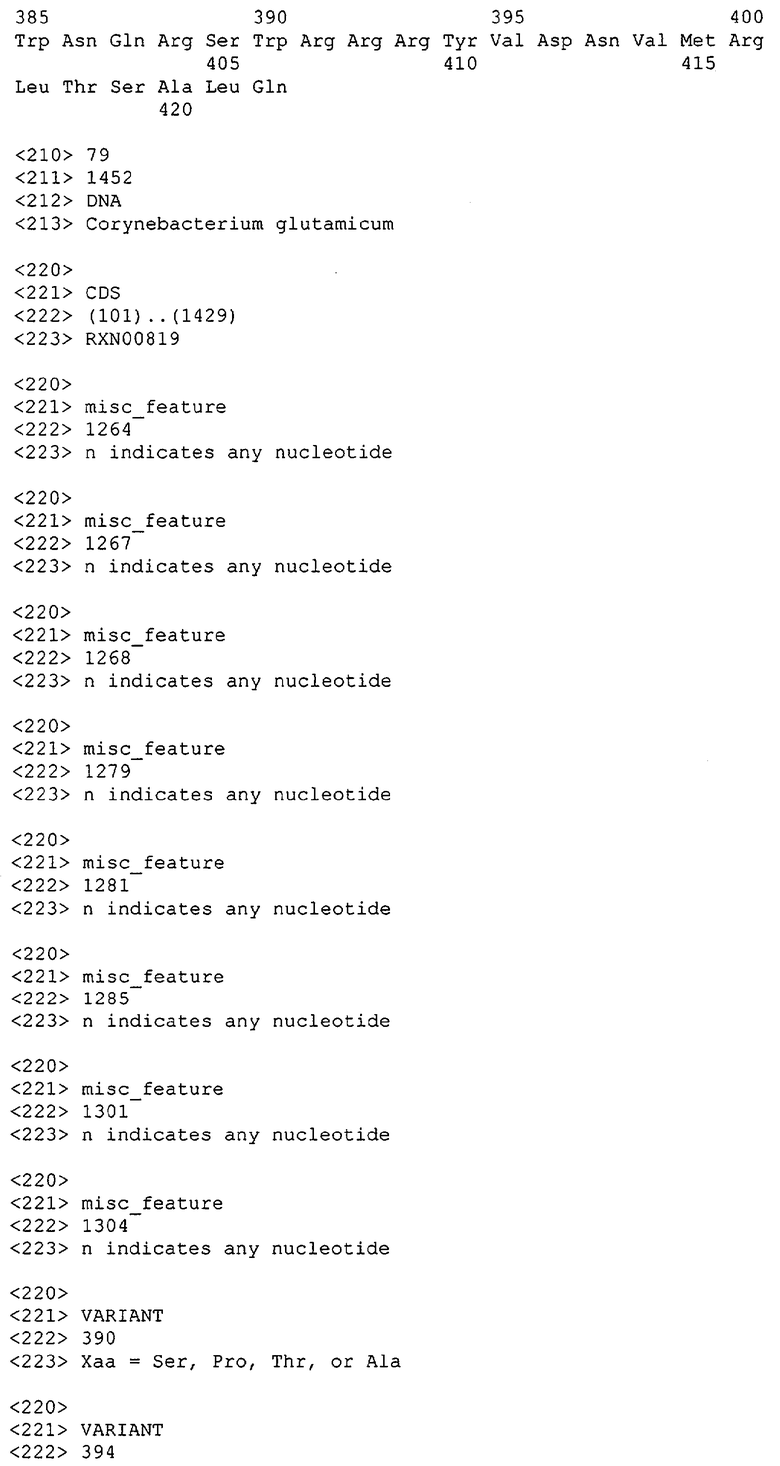
















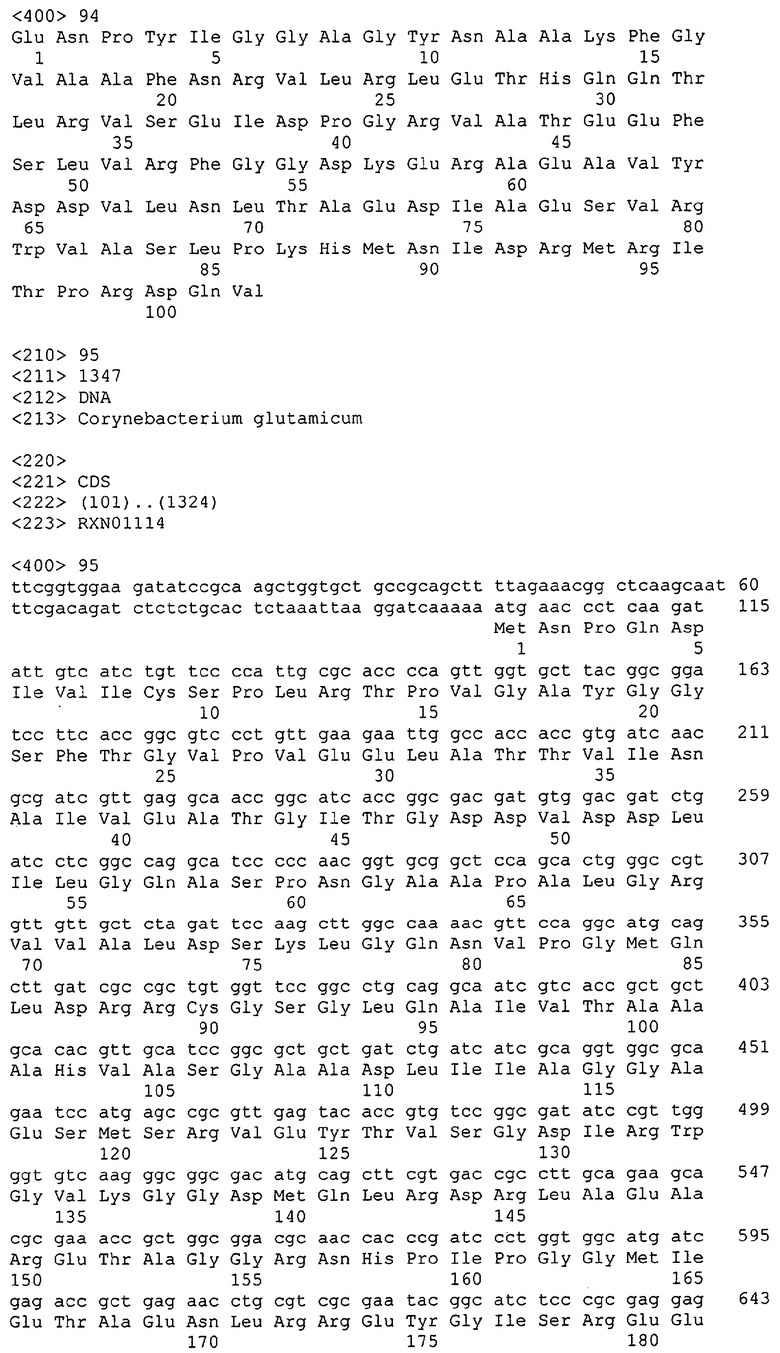




























































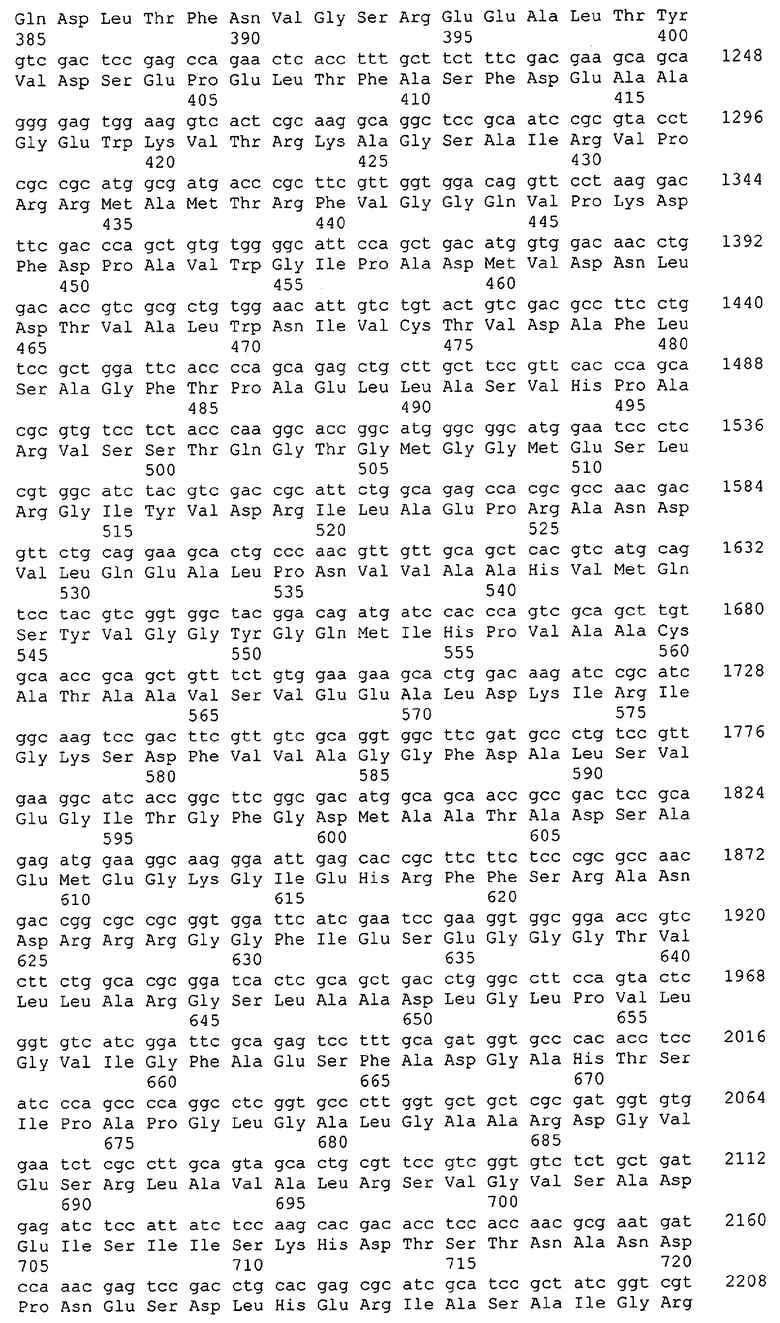

















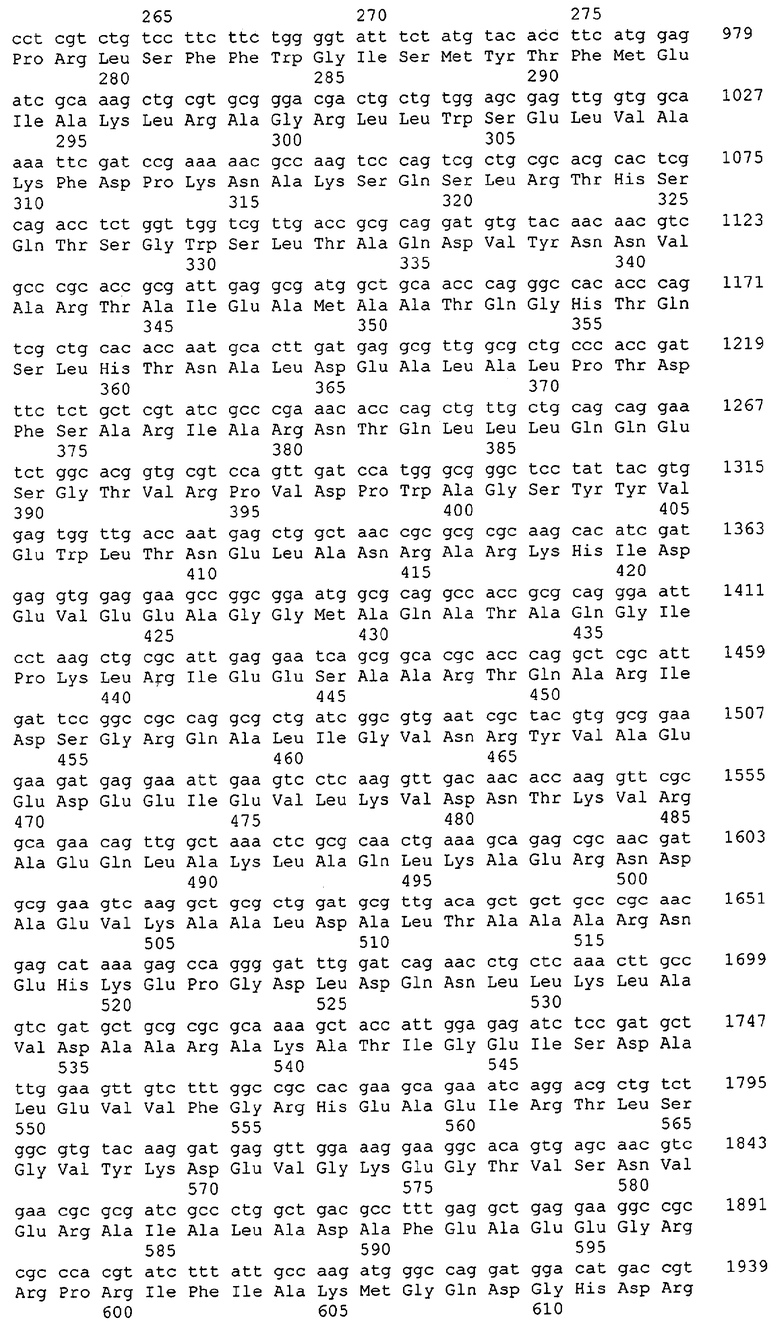



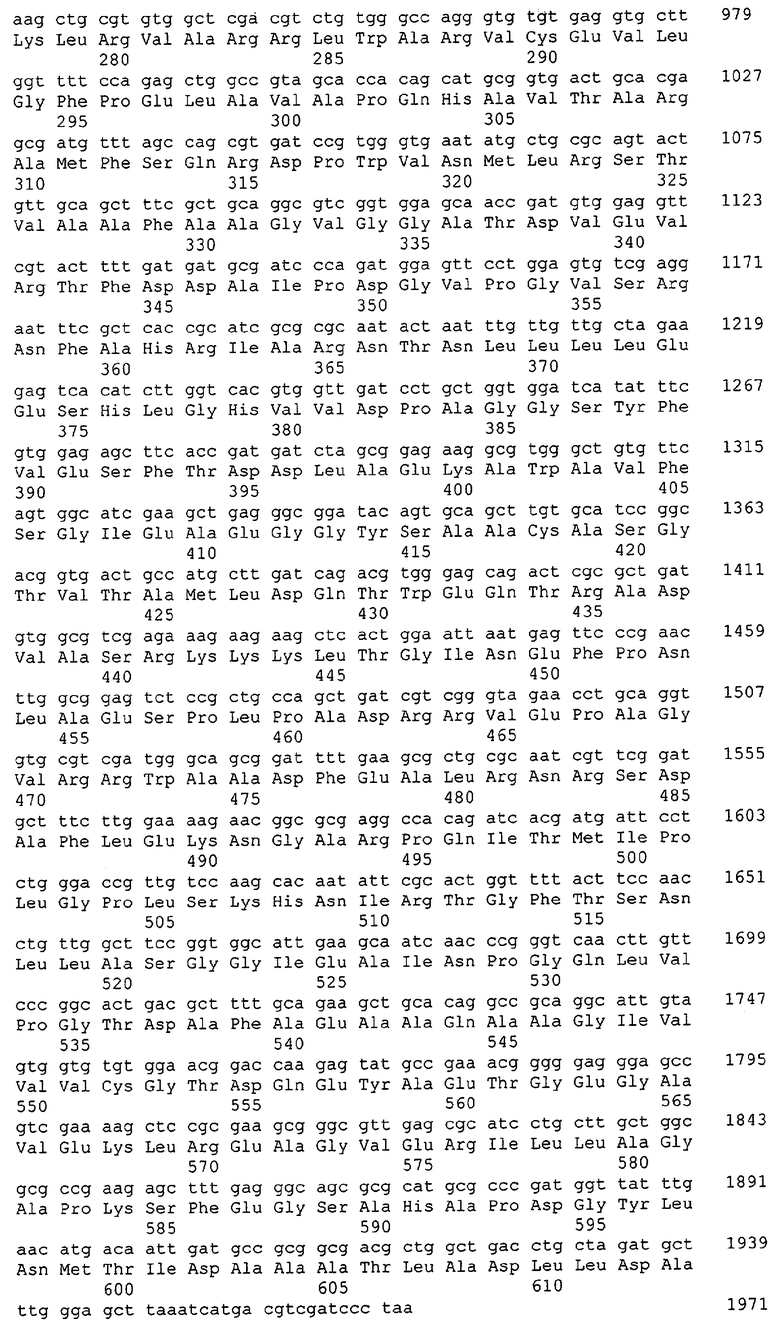





































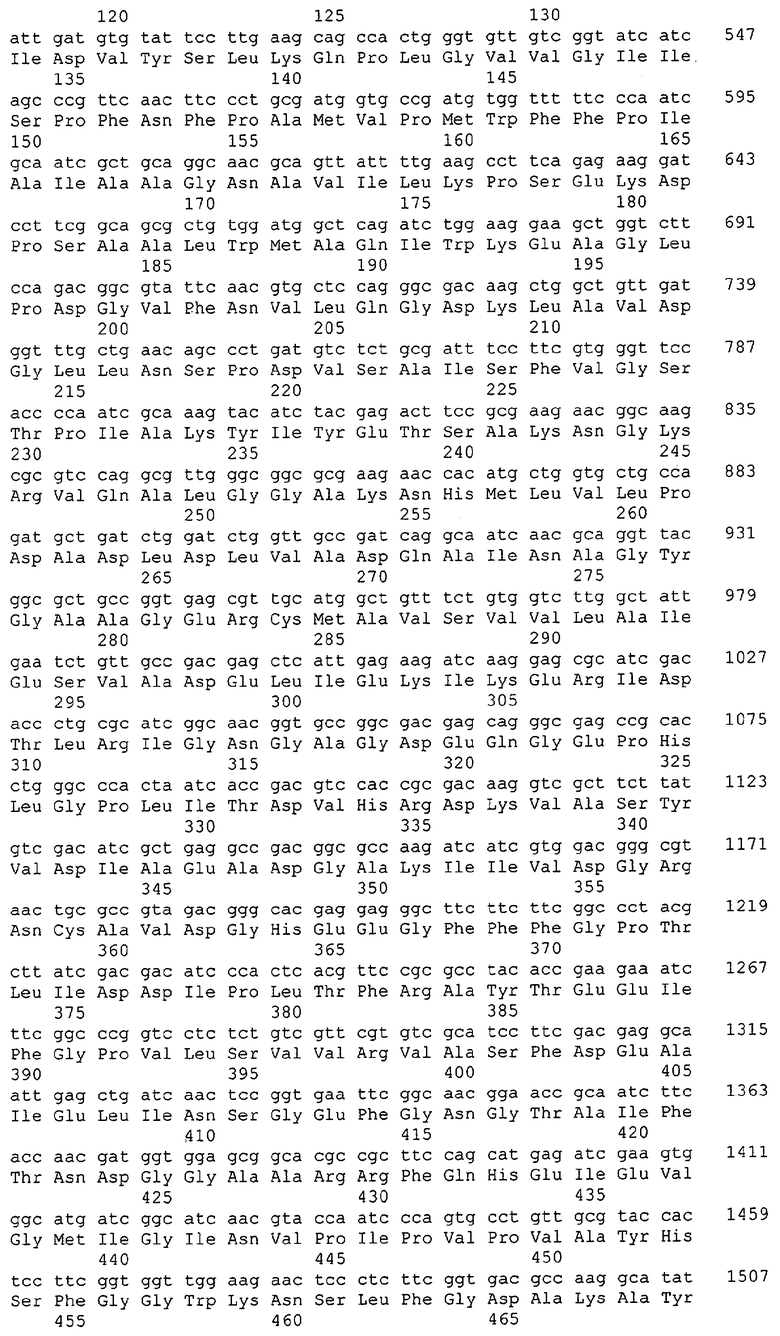






























































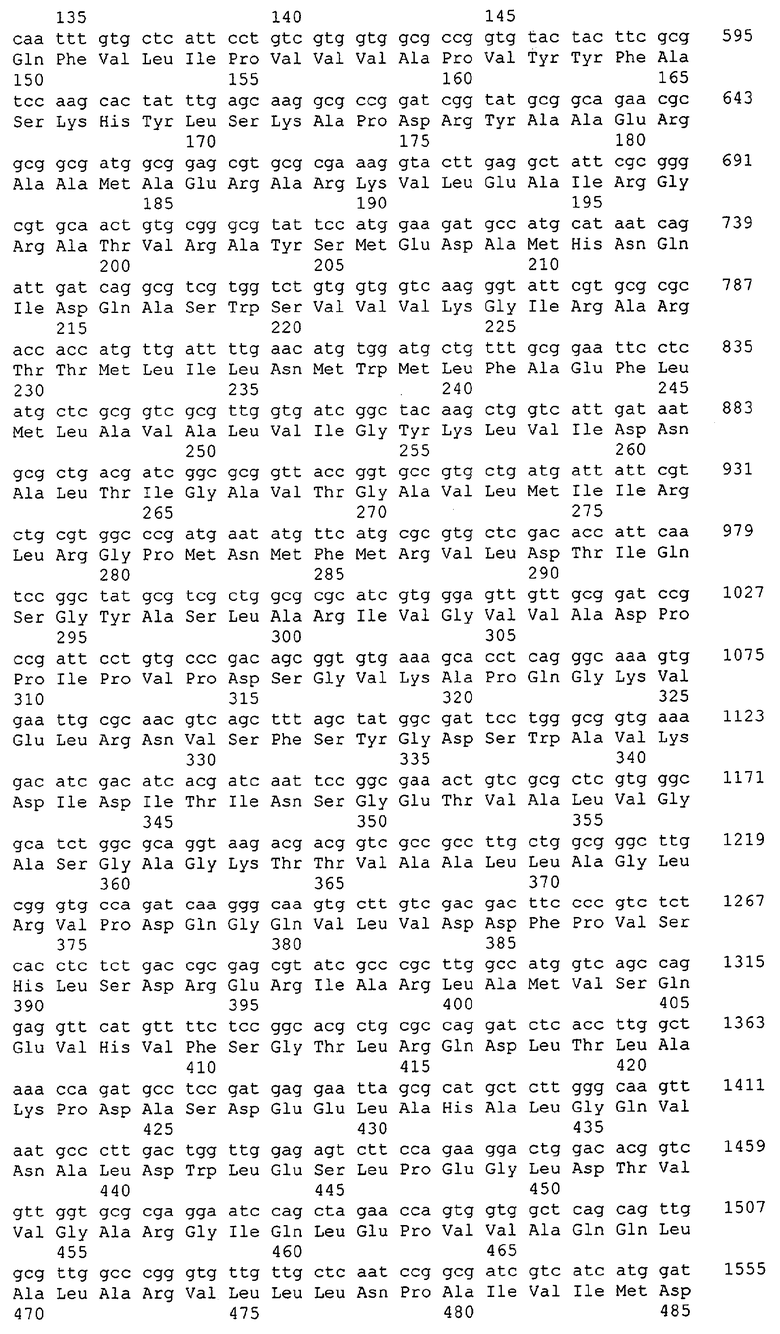





























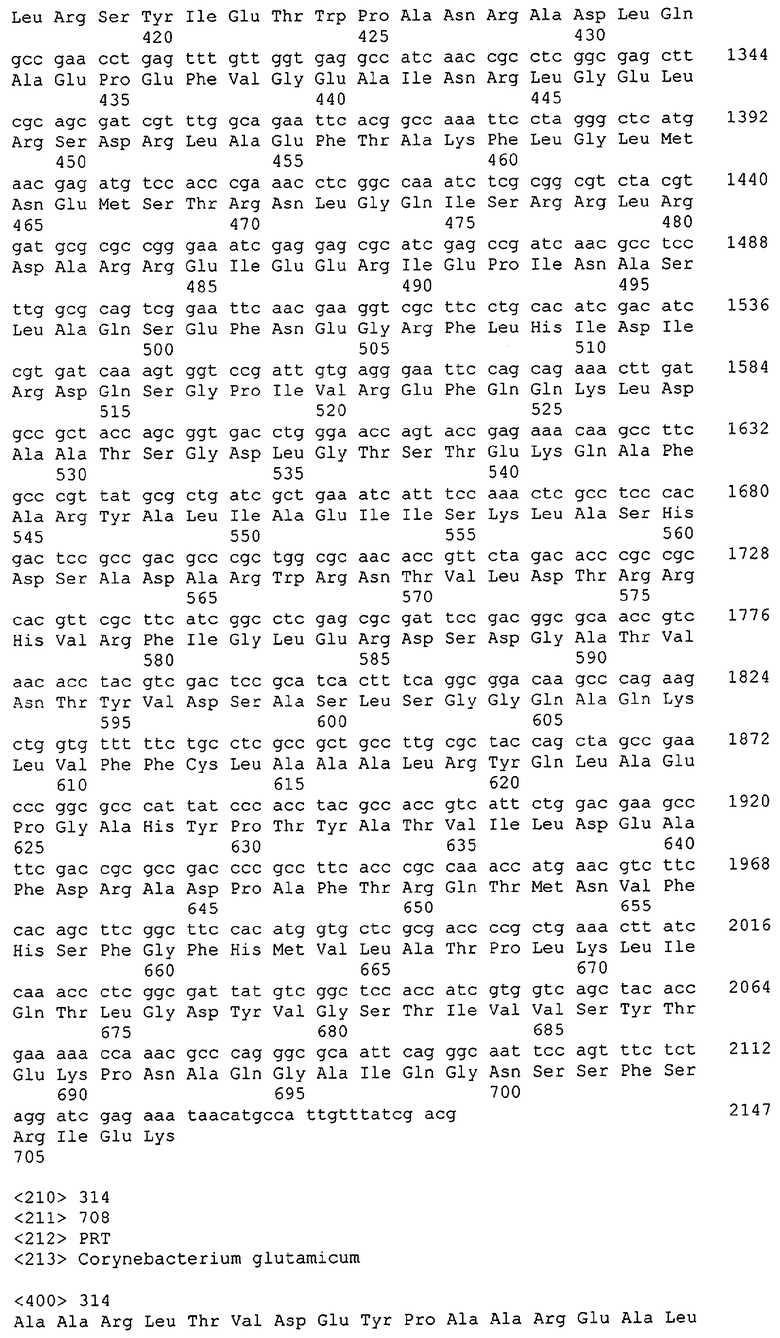



























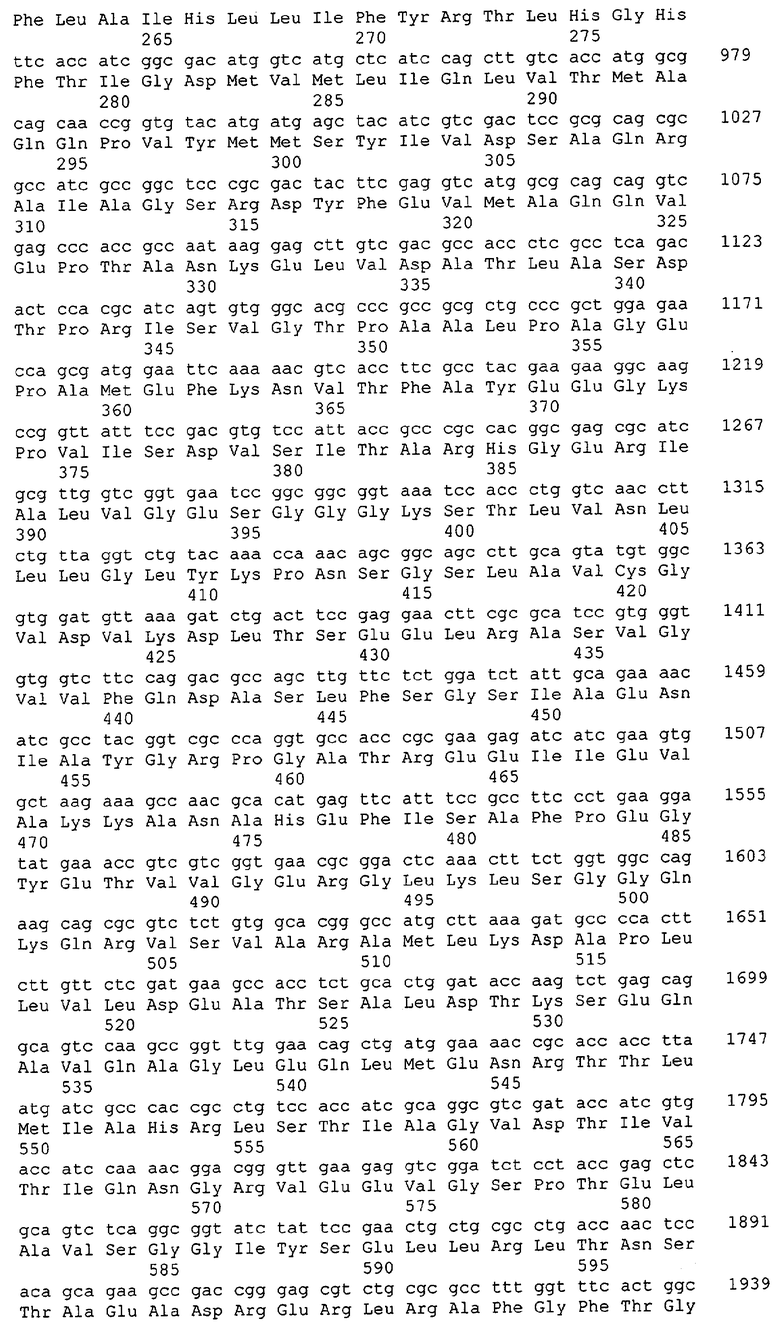













































































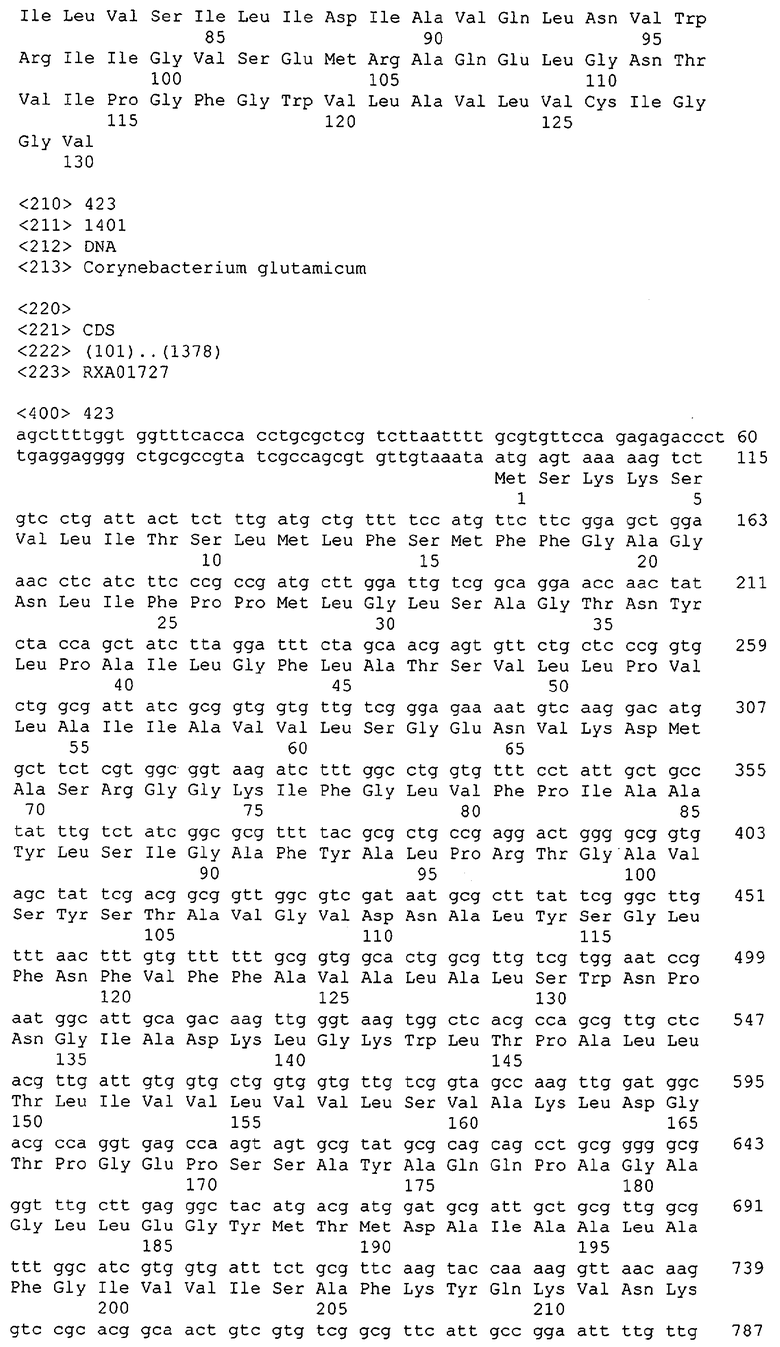






















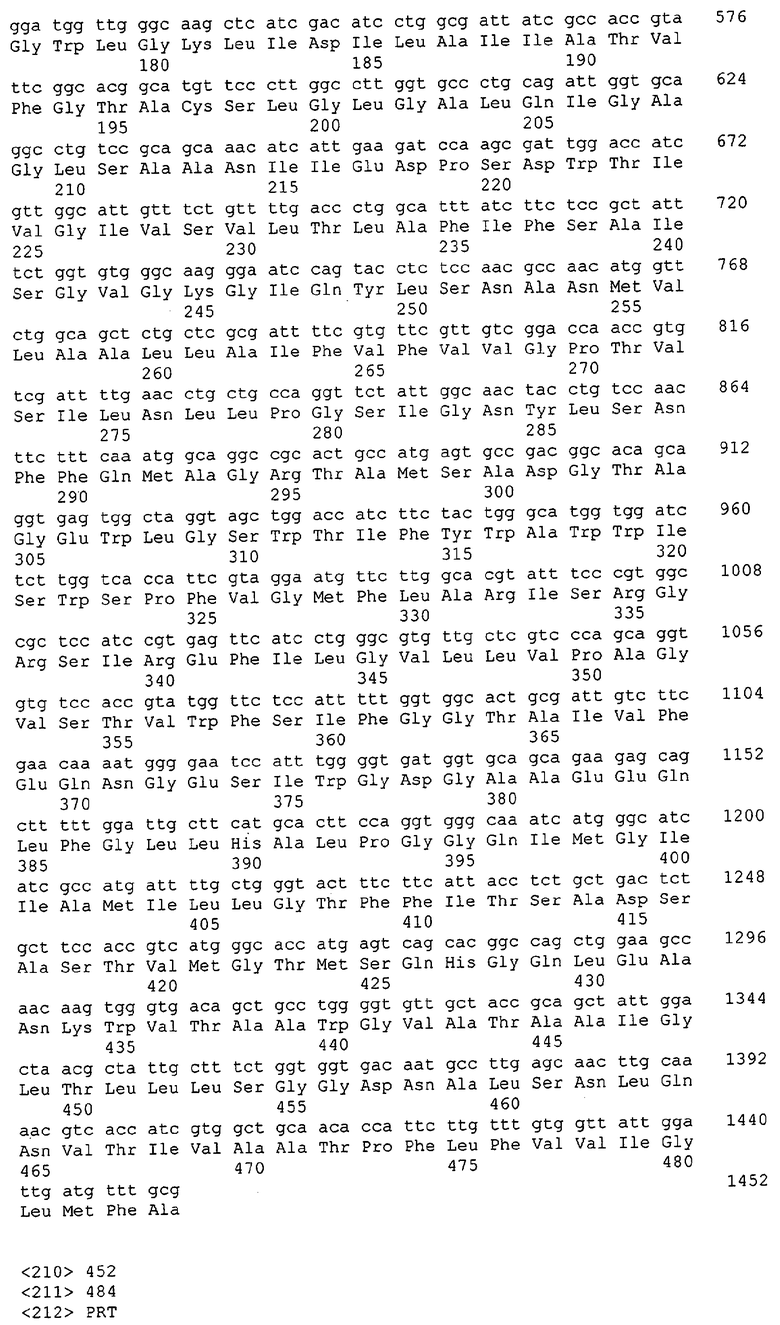



































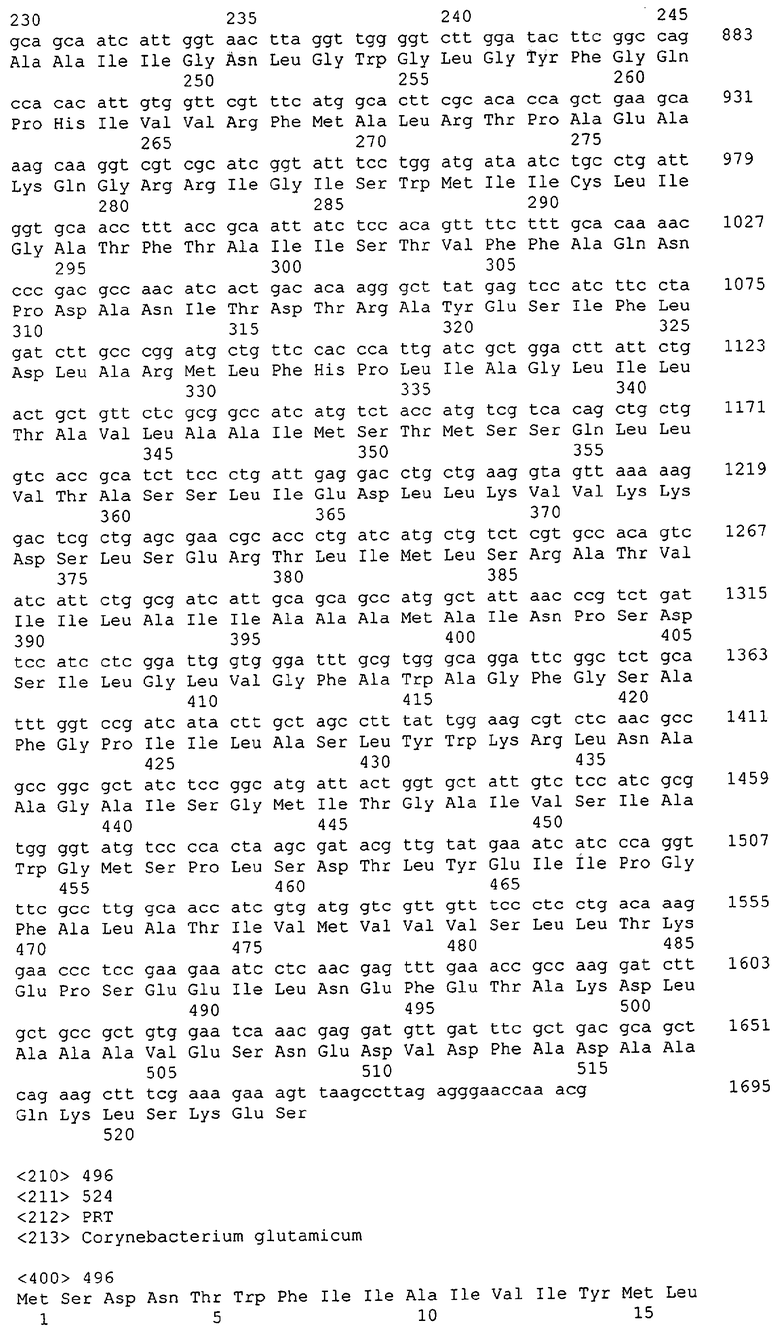










































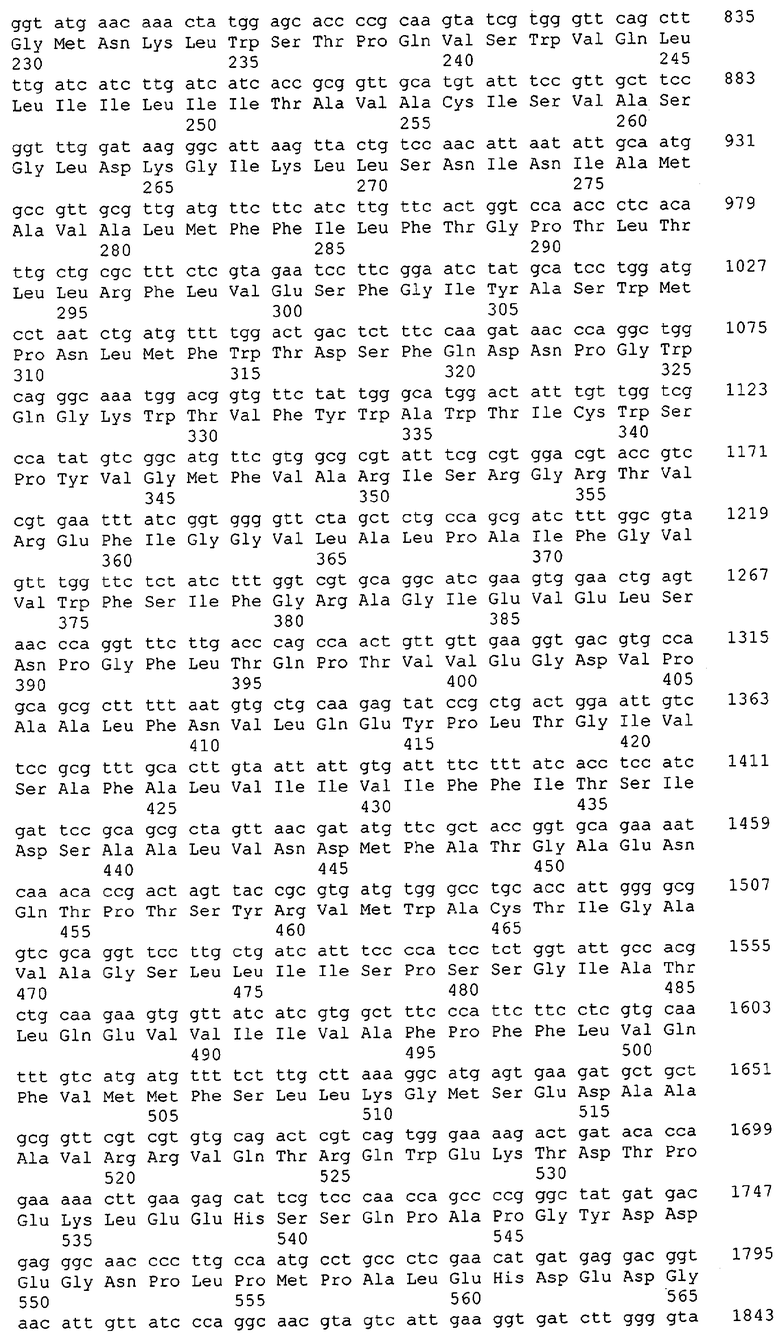




























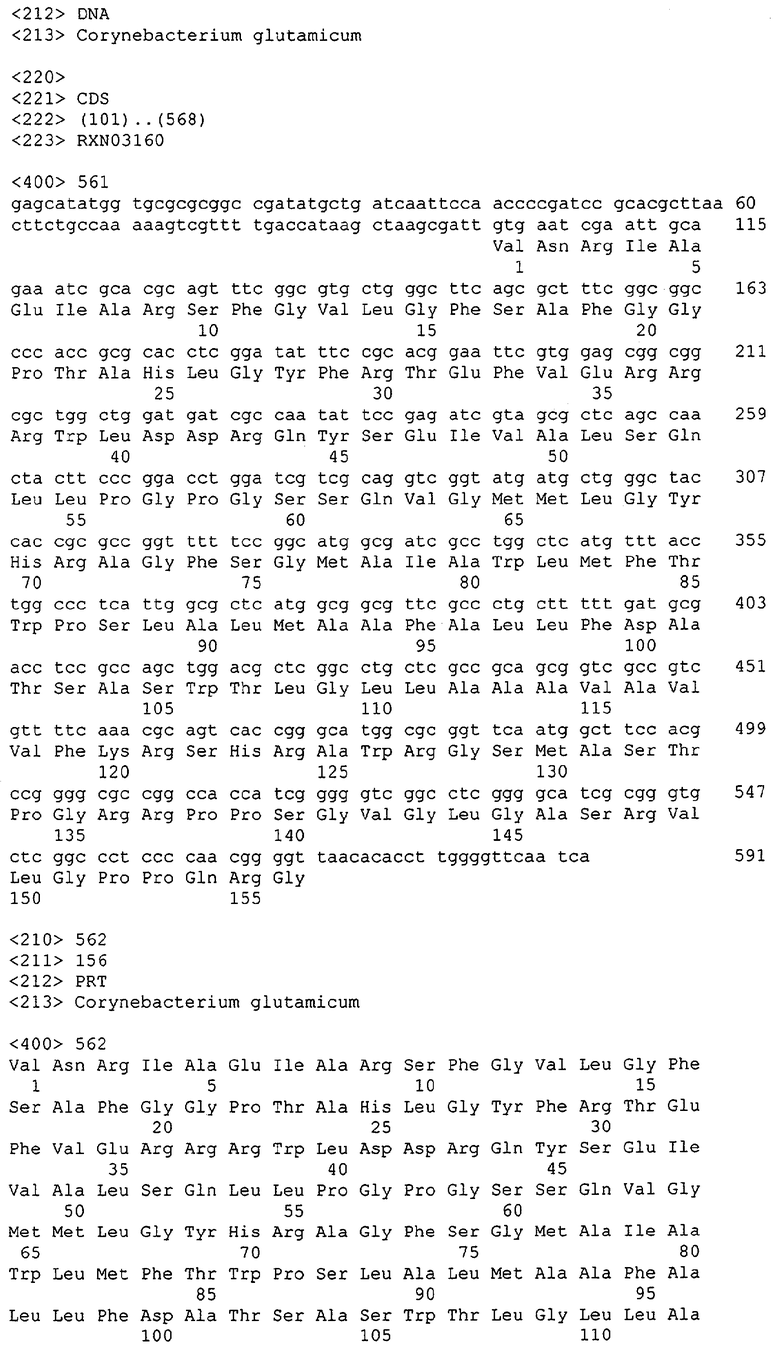










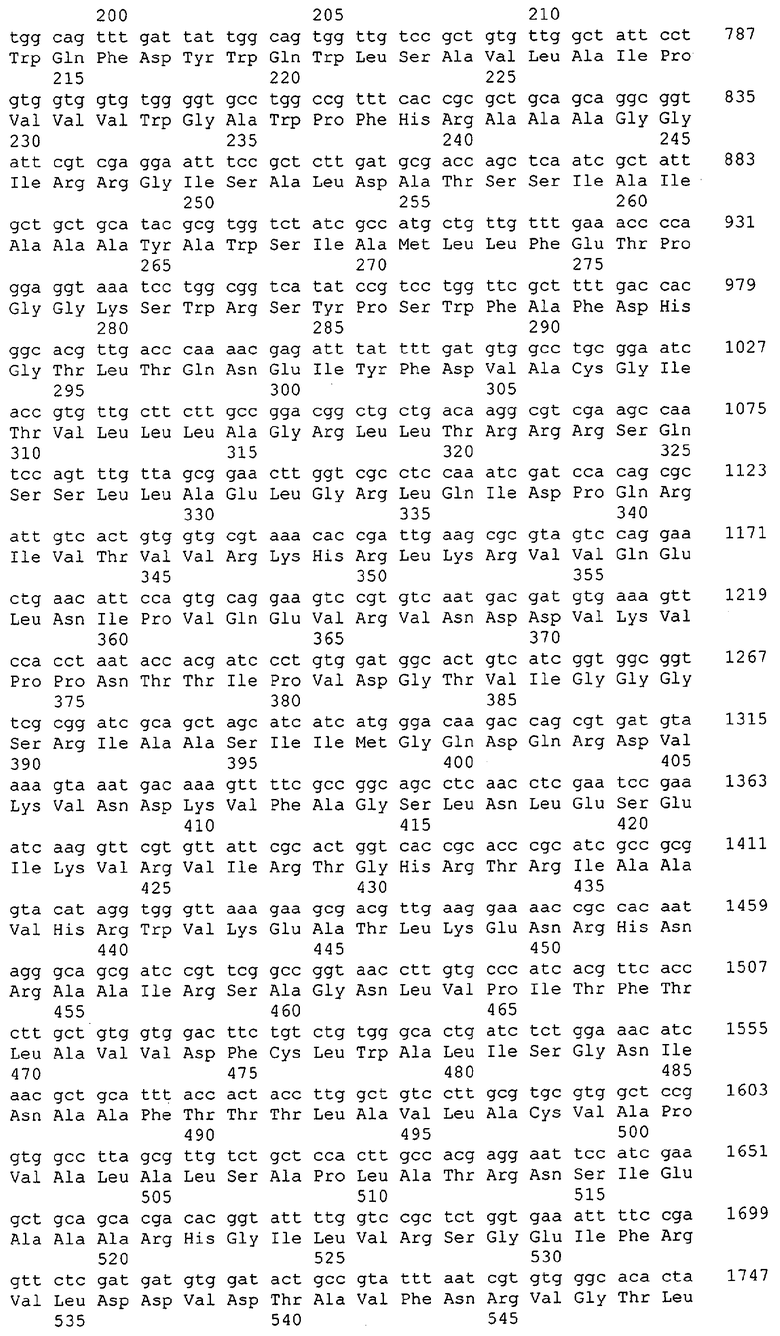






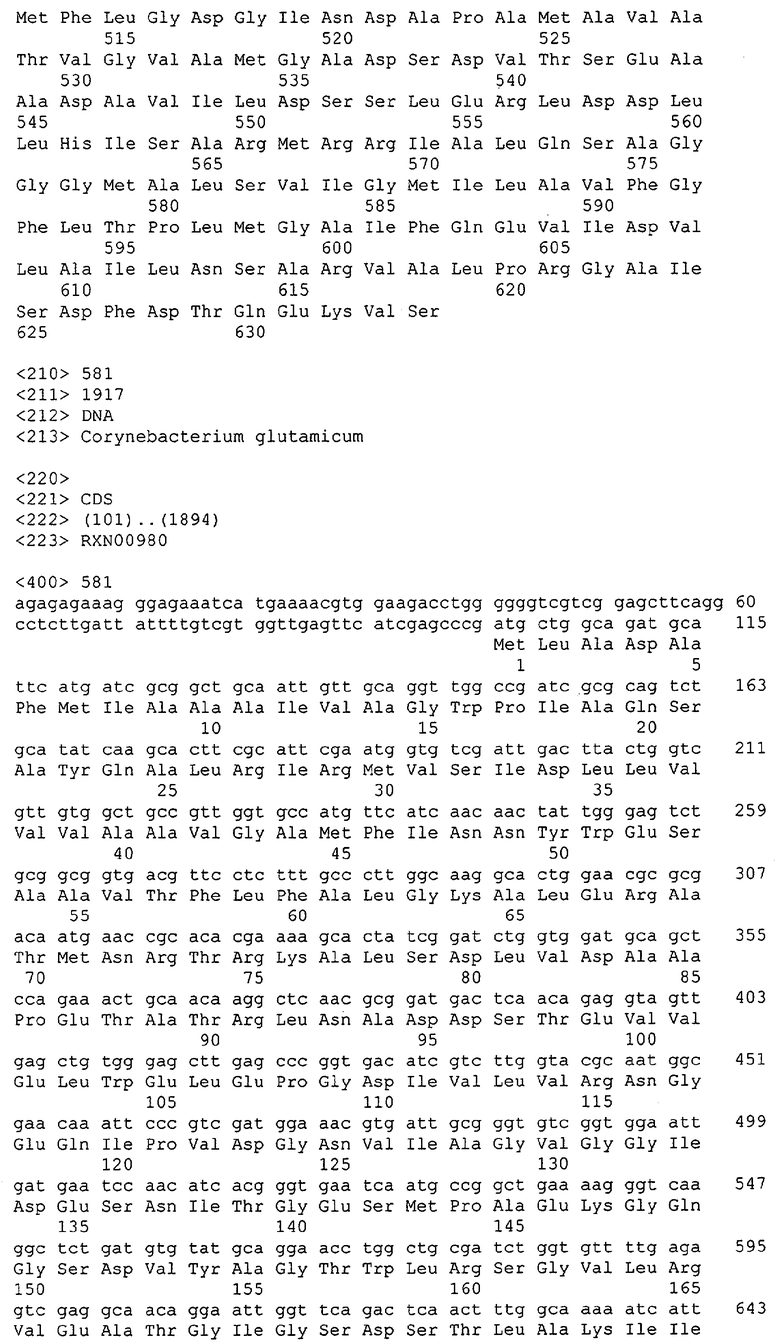
































































































Изобретение относится к биотехнологии и представляет собой выделенные молекулы нуклеиновой кислоты Corynebacterium glutamicum, которые кодируют полипептид с активностью CysQ. Изобретение относится к также рекомбинантным экспрессирующим векторам, содержащим такие молекулы нуклеиновых кислот, и клеткам-хозяевам, в которые были введены эти экспрессирующие векторы. Данное изобретение касается также способа получения аминокислот при помощи культивирования указанных клеток. Изобретение позволяет расширить ассортимент ферментов и получать аминокислоты с высокой степенью эффективности. 23 н. и 13 з.п. ф-лы, 4 табл.
Приоритеты по пунктам:
| Способ изготовления носителей информации на полосковых магнитных доменах | 1978 |
|
SU752472A1 |
| ANDREW F.NEUWALD cysQ, a gene needed for cysteine synthesis in Escherichia coli K-12 only during aerobic growth | |||
| J Bacteriol | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ПОЛИПЕПТИД С АКТИВНОСТЬЮ НИТРИЛГИДРАТАЗЫ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ НИТРИЛГИДРАТАЗЫ, ШТАММ БАКТЕРИЙ ESHCERICHIA COLI - ПРОДУЦЕНТ ПОЛИПЕПТИДА СО СВОЙСТВАМИ НИТРИЛГИДРАТАЗЫ, СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДА СО СВОЙСТВАМИ НИТРИЛГИДРАТАЗЫ | 1991 |
|
RU2081173C1 |

