Изобретение относится к технологиям стабилизации изгибов и петлеобразных структур синтетических пептидов и может использоваться в фармакологии для получения препаратов нового поколения, предназначенных для диагностики, терапии и вакцинопрофилактики инфекционных заболеваний и патологических состояний человека и животных.
Синтетические пептидные антигены все шире используются как ключевые компоненты качественно новых препаратов для диагностики, терапии и вакцинопрофилактики, так как они лишены недостатков, характерных для традиционно используемых рекомбинантных и природных антигенов, таких как патогенность, остаточная вирулентность, неполная инактивация, пирогенность, сопутствующие неспецифические антигены и т.д. Тем не менее синтетические пептиды имеют ряд специфических особенностей, затрудняющих их успешное использование в диагностике, терапии и вакцинопрофилактике, а именно:
1. Низкомолекулярные синтетические пептиды, неконъюгированные с носителем, как правило, не способны вызывать иммунный ответ [Hermanson GT, Mallia AK, Smith PK. «Immobilized Affinity Ligand Techniques» San Diego: Academic Press; 1992].
2. Низкомолекулярные синтетические пептиды требуют особых условий сорбции на поверхности носителя, используемого в иммунодиагностических системах [Gomara M.J., Riedemann S., Vega I., Ibarra H., Ercilla G., Haro I. «Use of linear and multiple antigenic peptides in the immunodiagnosis of acute hepatitis A virus infection» // J. Immunological Methods 234, 23-34; 2000].
3. Низкомолекулярные синтетические пептиды, как правило, не способны формировать стабильную конформацию, аналогичную той, которую имеют имитируемые пептидами фрагменты нативного антигена [Rizo J, Gierasch LM. Constrained Peptides: Models of Bioactive Peptides and Protein Substructures. // Annual Review of Biochemistry Vol.61: 387-416; 1992].
Для улучшения иммунологических характеристик пептидных антигенов их конъюгируют с каким-либо высокомолекулярным носителем. В качестве коньюгатов используются как природные и рекомбинантные белки [Wong SS: Chemistry of Protein Conjugation and Crosslinking. Boca Raton: CRC Press; 1991], так и носители небелковой природы: микрочастицы, липосомы, вирусоподобные частицы и др. [Tam JP, Spetzler JC: Multiple Antigen Peptide System. Methods Enzymol. 289:612-636; 1997]. Пептидные антигены коньюгируются с носителем посредством формирования как ковалентных связей, так и нековалентных взаимодействий. С целью увеличения эффективности коньюгирования, основанного на нековалентных взаимодействиях с современньми носителями (микрочастицы, липосомы), синтетические пептиды модифицируют гидрофобными углеводородами, имеющими высокое сродство к гидрофобным частям микрочастиц и липосом [Haro I, Perez S, Garcia M, Chan WC, Ercilla G. «Liposome entrapment and immunogenic studies of a synthetic lipophilic multiple antigenic peptide bearing VP1 and VP3 domains of the hepatitis A virus: a robust method for vaccine design» //FEBS Lett. Apr. 10; 540(1-3):133-40; 2003].
С целью увеличения эффективности сорбции пептидных антигенов на поверхностях носителей, используемых в иммунодиагостических системах, поверхности гидрофобных полимеров модифицируют различными способами. Либо изменяют сам пептидный антиген, связывая его ковалентно с поверхностью носителя, либо с высокомолекулярной структурой, способной эффективно сорбироваться на поверхностях носителей, используемых в иммунодиагостических системах [Gomara MJ, Riedemann S, Vega I, Ibarra H, Ercilla G, Haro I. «Use of linear and multiple antigenic peptides in the immunodiagnosis of acute hepatitis A virus infection» // J. Immunological Methods 234, 23-34; 2000].
Синтетические пептиды, используемые в фармакологии, как правило, имитируют поверхностные фрагменты нативных антигенов. Поверхность нативных белковых антигенов содержит большое количество изгибов и петлеобразных участков [Kuntz ID. «Protein-Folding» // J.Amer. Chem. Soc.94 4009 (1972)]. Синтетические пептиды, как правило, не способны формировать какую - либо стабильную конформацию в растворе, что приводит к потере аффинности связывания противопептидных антител с нативным антигеном и антител от нативных антигенов с синтетическим пептидами и, в свою очередь, сильно снижает активность субстанций, основанных на пептидных антигенах.
Известен способ стабилизации вторичной петлеобразной структуры с помощью дисульфидной связи [Broun JR. «Struktural origins of mammalian albumin» // Fed. Proc 35 2141 (1976)]. В этом случае с помощью включения N- и С-концевых цистеинов в состав пептида с последующим замыканием дисульфидной связи можно получить стабильные петлеобразные структуры. Для реализации этого способа в последовательности целевого пептида с N- и С-концов включают аминокислоту цистеин. Аминокислота цистеин может быть включена в последовательность как непосредственно конечная, так и локализована в непосредственной близости от концевых аминокислот. На заключительной стадии формирования циклического пептида тиоловые группы цистеинов окисляют, что приводит к формированию внутримолекулярной дисульфидной связи.
Этот способ стабилизации имеет следующие недостатки: существует высокая вероятность формирования нецелевых межмолекулярных дисульфидных связей; накладываются ограничения на аминокислотный состав целевого пептида, поскольку структура пептида не должна содержать внутренних цистеинов, иначе возможно формирование нецелевых внутримолекулярных дисульфидных связей; полученный таким образом пептид не является готовым прекурсором для связывания его с носителем.
Известен также способ стабилизации бета-изгибов коротких пептидов, который основан на склонности аминокислотных последовательностей к формированию тех или иных вторичных структур пептидной цепи, стабилизированных внутримолекулярными нековалентными взаимодействиями (водородными связями, гидрофобными взаимодействиями, солевыми мостиками). Гидрофобные взаимодействия при нормальных условиях в водной среде самые сильные из всех нековалентных взаимодействий. Одним из примеров стабилизации бета-изгибов в коротких пептидах является триптофановый зиппер [A.G.Cochran, N.J.Skelton, M.A.Starovasnik "Triptophan zippers: stable, monomeric β-hairpins". PNAS, 98, (10), p.5578-5583; 2001]. Стабилизация бета-изгиба обуславливается гидрофобными взаимодействиями боковых групп пар молекул триптофана, расположенных симметрично относительно центра изгиба. Этот способ принят за прототип изобретения.
Прототип обладает следующими недостатками:
- стабилизация изгиба пептидной цепи зачастую требует вмешательства в аминокислотную последовательность пептида, что может привести к потере аффиности комплексов антиген-антитело;
- полученный пептид не является готовым прекурсором для связывания его с носителем;
- способ пригоден только для стабилизации бета-изгибов.
Изобретение решает задачу создания способа стабилизации изгибов и петлеобразных структур пептидов, позволяющего не затрагивать аминокислотную последовательность пептида, а также создавать пептиды, являющиеся готовым прекурсором для связывания их с носителями, при этом универсального, пригодного и для стабилизации изгибов и петлеобразных структур пептидов, имеющих произвольную аминокислотную последовательность.
Поставленная задача решается тем, что предлагается способ стабилизации вторичной петлеобразной структуры синтетических пептидов путем нековалентного взаимодействия гидрофобных углеводородов или их радикалов между собой, в соответствии с которым гидрофобные углеводороды или их радикалы располагают на противоположных N и С концах пептида, либо в непосредственной близости от них, причем названные гидрофобные углеводороды или их радикалы также нековалентно связывают с гидрофобным носителем.
При ковалентном связывании не менее чем двух сильно гидрофобных углеводородных радикалов, находящихся с противоположных N и С концов пептидной цепи, происходит изгибание пептидной цепи за счет сближения ее N и С концов и формирование нековалентного взаимодействия между N и С концевыми гидрофобными углеводородными радикалами, а также формирование нековалентного взаимодействия названых гидрофобных углеводородных радикалов с гидрофобньм носителем.
Гидрофобные углеводородные радикалы могут быть ковалентно связаны с реакционноспособными группами пептида, содержащимися на его противоположных N и С концах или в непосредственной близости от них, причем реакционноспособными группами пептида могут являться карбоксильные группы, или амидные группы, или тиоловые группы, или иные реакционноспособные группы, способные обеспечить селективное ковалентное связывание гидрофобных углеводородных радикалов с молекулой пептида.
Носитель может быть выполнен из полимера, им может быть высокомолекулярная структура из амфипатических молекул, а также возможны иные решения.
Присоединяемые к концам пептидов гидрофобные углеводородные радикалы могут быть алифатическимим углеводородами или ароматическими углеводородами.
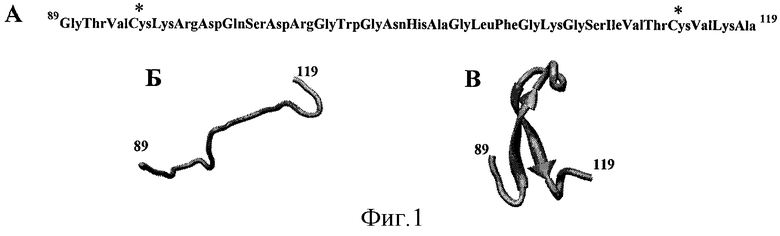
На фиг.1А приведена аминокислотная последовательность пептида, обозначенного здесь как пептид №1, соответствующая фрагменту 89-119 белка Е вируса клещевого энцефалита (штамм 205). Звездочкой помечены аминокислоты Cys в положениях 92 и 116, по тиольным группам которых проводилось модифицирование пептида.
На фиг.1Б приведено схематическое изображение пространственной структуры пептида №1.
На фиг.1В приведена пространственная структура фрагмента 89-119 белка Е вируса клещевого энцефалита, соответствующая нативной, полученной с помощью рентгеноструктурного анализа монокристалла белка [Rey FA, Heinz FX, Mandl C, Kunz С, Harrison SC. «The envelope glycoprotein from tick-bome encephalitis virus at 2 A resolution.» // Nature. May 25; 375(6529):291-8; 1995].
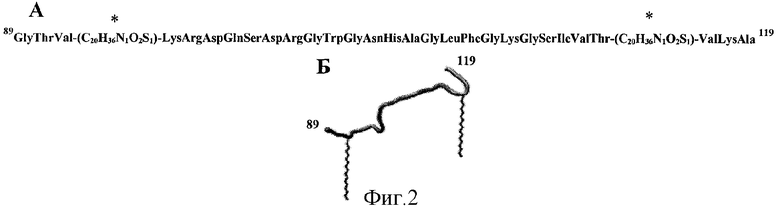
На фиг.2А приведена первичная структура пептида, обозначенного здесь №1-М - модифицированного пальмитиновыми кислотами пептида №1. Звездочкой помечены модифицированные пальмитиновой кислотой аминокислоты Cys в положениях 92 и 116.
На фиг.2Б схематично изображена пространственная структура пептида №1-М.
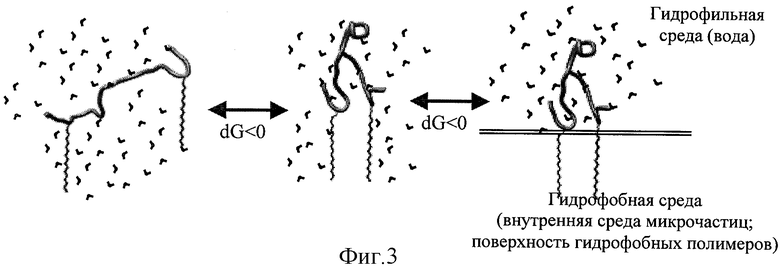
На фиг.3 схематично изображен самопроизвольный процесс стабилизации петлеобразной структуры пептида №1-М, аналогичной нативной структуре фрагмента 89-119 белка Е вируса клещевого энцефалита. Самопроизвольность процесса обеспечивается уменьшением площади контакта гидрофобных радикалов с водой [Chothia С. "Hydrophobic bonding and accessible surface area in proteins." // Nature. Mar 22;248(446):338-9; 1974].
Синтетический пептид модифицируют путем ковалентного связывания производных гидрофобных углеводородов с карбоксильными, амидными либо тиоловыми группами, содержащимися на N и С концах пептида (пример 1). Обязательным условием такой модификации является формирование пары сильно гидрофобных углеводородных радикалов на противоположных концах пептидной цепи, как показано на фиг.2. Такая модификация, с одной стороны, приводит к формированию нековалентной внутримолекулярной связи между N- и С- концами пептида, а с другой стороны, позволяет осуществить нековалентное связывание полученного модифицированного пептида с гидрофобным носителем, как показано на фиг.3. В предлагаемом способе происходит стабилизация изгибов или петлеобразных структур пептида за счет внедрения дополнительных концевых молекул, способных осуществить внутримолекулярную связь, что подтверждается высокой аффинностью антипептид №1-М антител к нативному антигену (Пример 6), а также происходит связывание синтетического пептида с гидрофобными носителями за счет нековалентных взаимодействий (пример 3).
Таким образом, предлагаемый способ имеет следующие преимущества:
- возможность стабилизации как изгибов пептидной цепи, так и петлеобразных структур за счет нековалентных взаимодействий;
- возможность связывания пептида с гидрофобными носителями и поверхностями;
- способность образовывать высокомолекулярные структуры (мицелы) без участия дополнительных высокомолекулярных носителей.
Ниже следуют примеры, раскрывающие сущность изобретения.
Пример 1. Твердофазный синтез пептида №1.
Аминокислотная последовательность пептида №1 изображена фиг.1А - она соответствует фрагменту 89-119 белка Е вируса клещевого энцефалита. Этот фрагмент 89-119 в нативном белке Е вируса клещевого энцефалита представляет собой петлеобразный изгиб, как показано на фиг.1В. Пространственная структура нативного белка Е вируса клещевого энцефалита была получена с помощью рентгеноструктурного анализа монокристалла белка [Rey FA, Heinz FX, Mandl C, Kunz С, Harrison SC. «The envelope glycoprotein from tick-borne encephalitis virus at 2 A resolution» // Nature. May 25;375 (6529):291-8; 1995]. Схематичное изображение пространственной структуры пептида №1 представлено на фиг. 1Б.
Синтез пептида, аминокислотная последовательность которого имитирует петлеобразный фрагмент 89-119 поверхности белка Е вируса клещевого энцефалита, проводили ручным твердофазным методом на 4-гидроксиметилфеноксиметильном стирол-дивинилбензольном (1%) сополимере (смола Ванга) [Albericio F, Barany G. «Application of N,N-dimethylformamide dineopentyl acetal for efficient anchoring of N alpha-9-fluorenylmethyloxycarbonylamino acids as p-alkoxybenzyl esters in solid-phase peptide synthesis» // Int J Pept Protein Res. Apr; 23(4):342-9; 1984]. В качестве реактора использовали стандартный 10 мл полипропиленовый шприц, снабженный пористой полипропиленовой пластинкой. Пептид синтезировали Nsc/tBu-методом [Sabirov AN, Kirn YD, Kim HJ, Samukov VV. «FMOC- and NSC-groups as a base labile N(alpha)-amino protection: A comparative study in the automated SPPS.» // Protein Peptide Lett. 4, 307-312; 1997]. В качестве постоянных защитных групп применялись Nγ-Trt для Asn и Gln; Nim-Trt для His; S-Trt для Cys; tBu для Thr и Ser; tBu-эфир для Asp; NG-Pbf для Arg; Nε-Boc для Lys. Для присоединения первой аминокислоты 200 мг полимера, 6 мл дихлорэтана, 2 мл N-метилпирролидона, 0,4 ммоль Nsc-Ala-OH, 1,0 ммоль дициклогексилкарбодиимида и 0,1 ммоль 4-диметиламинопиридина перемешивали 24 ч. Полученный аминоацил-полимер промывали DMF, метанолом, эфиром и хлороформом, сушили в вакууме. Выход 220 мг, содержание аминогрупп 500 мкмоль/г по тесту с пикриновой кислотой [Gisin BF. «The monitoring of reactions in solid-phase peptide synthesis with picric acid.» // Anal Chim Acta. 1972 Jan; 58(1):248-9]. Наращивание пептидной цепи проводили однократными конденсациями. По окончании сборки с пептида удаляли N-концевую Nsc-защиту, пептидил-полимер (1,4 г) промывали хлороформом, метанолом, эфиром и сушили в вакууме. Полученный пептидил-полимер 60 мин обрабатывали смесью 4 мл TFA, 0,4 мл тиоанизола, 0,1 мл этандитиола и 0,1 мл мета-крезола, полимер отделяли фильтрованием. Пептид осаждали из фильтрата большим объемом холодного эфира. Осадок сырого пептида отфильтровывали, промывали эфиром и сушили в вакууме. Выход неочищенного пептида 150 мг. Очистку пептида осуществляли полупрепаративной обращенно-фазовой ВЭЖХ с использованием хроматографа Pharmacia-LKB 2249 (Швеция): аналитическую - на колонке 4×150 мм с сорбентом Lichrosorb RP18, 5 мкм (Merck) при скорости потока 1 мл/мин в линейном градиенте концентрации ацетонитрила (0-80%) в воде с 0,5% Н3PO4; препаративную - на колонке 10×250 мм с фазой SynChropak RPP 100, 10 мкм (Eichrom Ind.), при скорости потока 4 мл/мин в линейном градиенте концентрации ацетонитрила (0-66%) в воде с 0,1% трифторуксусной кислоты. Детекцию осуществляли спектрофотометрически по поглощению при 226 нм с обработкой результатов с использованием системы MultiChrom (Ampersand). Фракции, содержащие основное вещество, упаривали в вакууме с изопропанолом и лиофилизовали из воды, содержащей 10% уксусной кислоты. Выход 42 мг (13%, для TFA-соли с м.в. 3250). Содержание SH-групп по реакции с 2,2'-дипиридилдисульфидом 0,58 мкмоль/мг (94,5%). При помощи препаративной ВЭЖХ был выделен мажорный продукт с чистотой около 85%, идентичность которого была доказана при помощи аминокислотного анализа. Элементный анализ проводили на CHN-анализаторе Perkin-Elmer, аминокислотный - на приборе Biotronik LC5001 после гидролиза пробы в 6 н. HCl, 110°, 24 ч. Количественное определение сульфгидрильных групп осуществляли при помощи теста с 2,2'-дипиридилдисульфидом (Carlsson et al., 1978), качественное - по реакции с нитропруссидом натрия.
Таким образом был изготовлен пептид, аминокислотная последовательность которого соответствует фрагменту 89-119 белка Е вируса клещевого энцефалита. Этот пептид является прекурсором для получения модифицированного пальмитиновыми аминокислотами пептида, способного формировать стабильную петлеобразную структуру.
Пример 2. Синтез модифицированного пептида №1-М.
Пептид №1-М, как уже упоминалось, это модифицированный пальмитиновыми кислотами пептид №1. Первичная структура пептида №1-М изображена на фиг.2А. Модифицирование пептида №1 проводилось по тиольным группам аминокислот Cys в положениях 92 и 116, эти положения на фиг.1А помечены звездочками.
Для изготовления модифицированного пептида №1-М смесь 1,0 г (10 ммоль) малеинового ангидрида и 2,4 г (10 ммоль) цетиламина в 20 мл диоксана кипятили над 2 г молекулярных сит (4Å) 5 ч, охлаждали до комнатной температуры и отфильтровывали выпавший осадок N-гексадецилмалеамида. Выход 2,7 г (80%), т.пл. 58-59°С. Часть полученного продукта (1 г) нагревали в открытой пробирке в токе аргона 1 ч при 120°С, охлаждали, растворяли в 10 мл гексана и отфильтровывали от нерастворившегося остатка. Фильтрат упаривали и сушили в вакууме. Выход 0,8 г (85%). Найдено: С 74,02; Н 11,02; N 4,18. С20Н35NO2. Вычислено: С 74,12; Н 10,97; N 4,36. Растворяли 20 мг пептида №1 в смеси 4 мл этанола и 0,5 мл воды, добавляли раствор 10 мг N-гексадецилмалеимида в 0,2 мл этанола и выдерживали при 35°С до негативного теста на SH-группы с нитропруссидом натрия (5 ч). Реакционную массу упаривали в вакууме, промывали эфиром, сухим изопропанолом, лиофилизовали из воды с добавлением 10% уксусной кислоты. Выход модифицированного пептида №1-М 18 мг.
Таким образом был изготовлен пептид №1-М, содержащий в непосредственной близости от N и С концов гидрофобные радикалы, представляющие собой остатки пальмитиновой кислоты. Схематическое изображение пространственной структуры пептида №1-М представлено на фиг.2Б. Гидрофобные молекулы (радикалы) в гидрофильной среде (вода) стремятся формировать плотные контакты, исключающие наличие молекул гидрофильной среды (вода) в области контакта. Формирование плотных контактов между гидрофобными радикалами приводит к уменьшению площади контакта с гидрофильной средой (вода), этот процесс именуется «гидрофобные взаимодействия», является энергетически выгодным и протекает самопроизвольно. Гидрофобное взаимодействие является достаточно сильным, что позволяет стабилизировать структуры даже крупных белковых молекул [Chothia С. «Hydrophobic bonding and accessible surface area in proteins» // Nature. Mar 22; 248 (446):338-9; 1974]. В изготовленном пептиде №1-М гидрофобные радикалы локализованы в непосредственной близости от N и С концов пептидной цепочки фиг.2Б. Гидрофобное взаимодействие между N и С концами пептида стремится сблизить их, тем самым изогнуть пептидную цепочку и стабилизировать изгиб синтетического пептида, как показано на фиг.3. Стоит отметить, что аминокислотная последовательность пептида вносит несущественный вклад в процесс формирования изгиба пептидной цепочки, иными словами, этот способ стабилизации изгибов приемлем для любых синтетических пептидов.
Пример 3. Коньюгирование модифицированного пептида №1-М.
Помимо того, что гидрофобные радикалы, входящие в состав пептида №1-М, обеспечивают стабилизацию изгиба пептидной цепочки, они же обеспечивают нековалентное связывание молекул модифицированного пептида с другими гидрофобными средами. Так при физиологических значениях рН через 1-2 суток в растворе пептида №1-М формируются высокомолекулярные структуры (эмульсия). Эти высокомолекулярные структуры представляют собой агрегировавшие молекулы пептида №1-М за счет нековалентного взаимодействия гидрофобных радикалов, входящих в состав соседних молекул пептида. Пептид №1-М обладал более высокой по сравнению с немодифицированным пальмитиновыми кислотами пептидом №1 способностью к адсорбции на поверхности гидрофобного полимера - полистирола.
Кроме того, гидрофобные радикалы, входящие в состав пептида №1-М, способны фиксировать пептид на поверхности гидрофобных микрочастиц, как показано на фиг.3.
Микрочастицы, представляющие собой мелкие катионные мицеллы, были изготовлены на основе коммерческого катионного амфифила DDAB - /Dimethyl-dioctadecyi - ammonium bromide (ICN)/ и неионного амфифила Tween 80. По данным спектротурбидиметрического анализа микрочастицы имели размер 200 нм. Коньюгирование модифицированного пептида №1-М с микрочастицами проводили инжектированном смеси пептида и микрочастиц через пористую мембрану с диаметром пор 0,5 мкм. Полноту коньюгирования пептида №1-М с микрочастицами подтверждали данными электрофореза на содержание свободного пептида. Схематическое изображение самопроизвольного процесса стабилизации петлеобразной структуры пептида №1-М, аналогичной нативной структуре фрагмента 89-119 белка Е вируса клещевого энцефалита, представлено на фиг.3.
Таким образом было проведено коньюгирование пептида №1-М как с катионньми микрочастицами и гидрофобной поверхностью полистирола, так и молекул пептида №1-М между собой с образованием эмульсий. Способность пептида №1-М формировать высокомолекулярные структуры (эмульсии) и самопроизвольно связываться с гидрофобньми микрочастицами позволяет с легкостью изготавливать субстанции, содержащие пептидный антиген и способные вызывать иммунный ответ.
Пример 4. Приготовление поликлональных антипептидных антител.
Аффинность взаимодействия антиген-антитело очень чувствительна к структуре антигена. Это свойство антиген-антительного взаимодействия использовалось для подтверждения того, что предлагаемый способ стабилизации действительно позволяет стабилизировать изгиб или петлеобразную структур пептида, аналогичную нативной. Для этого были получены антипептидные антитела мышей.
Мыши линии BALB/c в возрасте 5-6 недель иммунизировались внутрибрюшинно трехкратно изготовленными пептидами в составе катионных микрочастиц, разведенными в фосфатном буферном растворе. Мышам вводилось по 0,2 мл раствора пептидов в дозе 50 мкг/гол, вторая и третья иммунизации проводились через 2 и 4 недели, соответственно, после первой иммунизации. Контрольной группе животных вводили раствор пептида, мимикрирующего антигенный участок белка gp120 вируса иммунодифицита человека, по той же схеме и в тех же дозировках. Через 10 дней после последней иммунизации у животных из ретроорбитальной вены брали пробу крови для определения титра специфических антител в сыворотке. Для определения титра специфических антител в лунках микропланшетов для иммуноферментного анализа («Медполимер», г.Москва) иммобилизировали следующие антигены: 1) пептид №1 или 2) пептид №1-М. Пептидные антигены растворяли в 0,05 М карбонат-бикарбонатном буферном растворе, рН 9,6, до конечной концентрации 10 мкг/мл, вносили в каждую лунку микропланшета по 0,1 мл раствора и инкубировали в течение ночи при температуре 37°С. Лунки промывали 3 раза фосфатно-буферным раствором с твин 20 и блокировали неспецифическое связывание 0,5%-ным раствором бычьего сывороточного альбумина. Двукратные разведения проб сывороток крови мышей в трис-HCl буферном растворе наносили по 0,1 мл в лунки с иммобилизованным антигеном и инкубировали при 37°С в течение 30 мин, после чего лунки промывали 5 раз фосфатным буферным раствором. Образовавшиеся комплексы антиген-антитело выявляли путем нанесения в лунки микропланшета по 0,1 мл фосфатного буферного раствора, содержащего конъюгированные пероксидазой хрена антитела козы против иммуноглобулинов класса G мыши (Sigma), инкубирования при 37°С в течение 30 мин и добавления субстратной смеси, содержащей орто-фенилендиамин. Ферментную реакцию останавливали добавлением раствора серной кислоты, результат реакции учитывали по оптической плотности, измеренной с помощью планшетного ридера «Мультискан ЕХ» на длине волны 492 нм. Титром сыворотки считали максимальное разведение, при котором оптическая плотность в 2 раза превышала соответствующий сигнал в лунке с отрицательным контролем. В качестве контроля использовали сыворотки животных из контрольной группы.
Результаты определения титра антипептидных антител в сыворотке крови мышей представлены в таблице 1.
Таким образом, трехкратная внутрибрюшинная иммунизация мышей пептидами, мимикрирующими фрагмент белка Е ВКЭ вызывает образование сывороточных антител, специфичных к пептидам (иммунизирующий антиген).
Пример 5. Приготовление нативного антигена вируса клещевого энцефалита.
Для получения нативного антигена вируса КЭ, штамм 205, осветленную вирусную суспензию очищали ультрацентрифугированием в комбинированном градиенте глицерина и тартрата калия, в котором верхнюю часть образует 30%-ный (вес/вес) раствор глицерина в STE-буфере (50 мМ трис-HCl, 1 мМ ЭДТА (рН 7,5)), а нижнюю (более плотную) 40%-ный (вес/вес) раствор тартрата калия в STE-буфере (рН 8,5). На приготовленный градиент наслаивали вирусную суспензию и центрифугировали при 100000 g в течение 3 часов при температуре +4°С на центрифуге L8-70 (Beckman). Фракцию, содержащую вирус («бэнд»), разводили в равном объеме STE буфера (рН 7,5) и центрифугировали в течение 2 часов при 100000 g при температуре +4°С. Осадок вируса растворяли в боратном буфере (0,15М NaCl, 50 mM Н3ВО3, рН 9,0) и хранили при -70°С. Концентрацию общего белка оценивали при помощи набора "Bio-Rad Protein Assay Kit". Чистоту выделения вируса проверяли методом электрофореза в 12% полиакриламидном геле с 0,1% додецилсульфата натрия (Саминский, 1966). Окраску гелей проводили при помощи красителя Кумасси G-250. Две полосы на электрофореграмме соответствовали белкам Е (54 кД) и NS1 (41 kD) вируса КЭ. Для получения антигенов вируса КЭ очищенную вирусную суспензию обрабатывали 0,2% раствором додецилсульфата натрия.
Пример 6. Определение аффинности антипептидных антител к нативному антигену.
Аффинность взаимодействия антиген-антитело очень чувствительна к структуре антигена. Высокие значения аффинности антител однозначно указывают на то, что антиген представлен в той пространственной структуре, на которую были получены антитела. Иными словами, та структура пептидного антигена, которая вызовет формирование антисыворотки, обладающей наибольшей аффинностью к нативному антигену, имеет пространственную структуру, наиболее сходную с нативной.
Для анализа взаимодействия специфических антипептидных антисывороток на полистероловые планшеты наносили 1 мкг нативного антигена вируса клещевого энцефалита в 100 мкл 0,05 М раствора двухзамещенного фосфатного буферного раствора, рН 8, и инкубировали при 37°С в течение 1 часа. Места неспецифического связывания насыщали 0,5% раствором казеина при 20°С в течение 60 минут. Затем инкубировали со специфическими антипептидными антисыворотками 1 час при 37°С. Образовавшиеся комплексы антиген-антитело выявляли путем нанесения в лунки микропланшета по 0,1 мл фосфатного буферного раствора, содержащего конъюгированные пероксидазой хрена антитела козы против иммуноглобулинов класса G мыши (Sigma), инкубирования при 37°С в течение 30 мин. Ферментную реакцию останавливали добавлением раствора серной кислоты, результат реакции учитывали по оптической плотности, измеренной с помощью планшетного ридера «Мультискан ЕХ» на длине волны 492 нм.
Аффинность взаимодействие антиген-антитело характеризуется значением константы диссоциации комплекса антиген-антитело (Kd) - чем меньше Kd тем выше аффинность. Для определения Kd результаты измерения оптических плотностей при различных разведениях сыворотки представляли в координатах Скэтчарда (А, А/С), где А - оптическая плотность, а С - концентрация антител (сыворотки). Предполагалось, что имеет место взаимодействие антител с двумя типами связывающих центров: специфическое связывание антител с антигеном и неспецифическое взаимодействие антитела с пластиком. Тангенсы углов наклона касательных к кривой Скэтчарда соответствуют значениям Kd1 -1 специфического взаимодействия антиген-антитело и (Kd1 -1+Kd1 -2) специфического и неспецифического взаимодействия. Для двух типов связывающих центров, при условии, что один из них является высокоспецифичным, а второй неспецифическим, уравнение взаимодействия в координатах Скэтчарда имеет вид [Ворфоломеев С.Д., Зайцев С.В. «Кинетические методы в биохимических исследованиях». - Издательство Московского университета, 1982]:
у=(((R1-x)/K1+(R2-x)/K2+(((R1-x)/K1+(R2-x)/K2)^2+4*x*(R1+R2-x)/(K1*K2)^0,5)/2),
где у - количество комплексов антиген-антитело/концентрация антител;
х - количество высокоспецифических комплексов и количество низкоспецифических комплексов;
K1, K2 - константы диссоциации высокоспецифических и низкоспецифических комплексов;
R1, R2 - количество высокоспецифических и низкоспецифических центров связывания.
Рассчитанные с помощью уравнения Скэтчарда значения констант диссоциации высокоспецифического взаимодействия антипептидных антисывороток с нативным антигеном представлены в таблице 2.
Таким образом, антисыворотки, полученные на пептид №1-М, обладают меньшей Kd, или большей аффинностью к нативному антигену. Следовательно, N и С концевые модификации приводят к формированию пространственной структуры, сходной с нативной петлеобразной структурой района 89-119 белка Е вируса клещевого энцефалита. Иными словами, модифицирование N и С концов, приводящие к формированию гидрофобных радикалов по концам пептида, обеспечивает стабилизацию петлеобразной пространственной структуры пептида, аналогичной нативной петлеобразной структуре мимикрируемого района.
Из приведенных примеров видно, что предлагаемый способ стабилизации изгибов и петлеобразных структур пептидов позволяет не затрагивать аминокислотную последовательность пептида, а также создавать пептиды, являющиеся готовым прекурсором для связывания их с носителями. При этом способ универсален, пригоден и для стабилизации изгибов и петлеобразных структур пептидов, имеющих произвольную аминокислотную последовательность.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОГЕННАЯ БИБЛИОТЕКА ХИМЕРНЫХ ПЕПТИДОВ, МИМИКРИРУЮЩАЯ ГЕНЕТИЧЕСКОЕ МНОГООБРАЗИЕ ГИПЕРВАРИАБЕЛЬНОГО РАЙОНА V3 БЕЛКА ОБОЛОЧКИ GP120 ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА | 2002 |
|
RU2237065C2 |
| СИНТЕТИЧЕСКИЙ ПЕПТИД, СИНТЕТИЧЕСКИЙ ЛИПОПЕПТИД, ИММУНОГЕННЫЙ КОНЪЮГАТ (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ОЛИГОМЕРА, ЗАЩИЩЕННЫЙ ТВЕРДЫМ МОНОМЕТИЛОВЫМ ЭФИРОМ ПОЛИЭТИЛЕНГЛИКОЛЯ ПОЛИСАХАРИД | 1993 |
|
RU2141527C1 |
| Пептиды-иммуногены и вакцина "ЭпиВакЭбола" против лихорадки Эбола с использованием указанных пептидов | 2017 |
|
RU2635998C1 |
| Рекомбинантная плазмида pHis6-flagG-protE, обеспечивающая синтез рекомбинантного химерного белка, включающего эпитопы гликопротеина Е вируса клещевого энцефалита и флагеллин G S.typhii и используемого в качестве основы для вакцины против вируса клещевого энцефалита | 2018 |
|
RU2702716C2 |
| КОНЪЮГАТ АНГИОТЕНЗИНОВОЙ ПЕПТИДНОЙ СОСТАВЛЯЮЩЕЙ С НОСИТЕЛЕМ, КОМПОЗИЦИЯ ВАКЦИНЫ, СПОСОБ ИММУНИЗАЦИИ ЖИВОТНОГО И СПОСОБ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ ФИЗИЧЕСКОГО НАРУШЕНИЯ, АССОЦИИРОВАННОГО С АКТИВИРУЕМОЙ РЕНИНОМ АНГИОТЕНЗИНОВОЙ СИСТЕМОЙ | 2002 |
|
RU2350622C2 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРЕЗЕНТАЦИИ МНОЖЕСТВЕННЫХ АНТИГЕНОВ, ОТНОСЯЩИЕСЯ К НЕЙ СПОСОБЫ И ПРИМЕНЕНИЯ | 2012 |
|
RU2619176C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ АДЪЮВАНТНЫХ ВИРОСОМ И АДЪЮВАНТНЫЕ ВИРОСОМЫ, ПОЛУЧАЕМЫЕ УКАЗАННЫМИ СПОСОБАМИ | 2014 |
|
RU2694367C2 |
| Пептидные иммуногены и вакцинная композиция против коронавирусной инфекции COVID-19 с использованием пептидных иммуногенов | 2020 |
|
RU2738081C1 |
| МОДИФИЦИРОВАННЫЙ БИОТИН-СВЯЗЫВАЮЩИЙ БЕЛОК, СЛИТЫЕ БЕЛКИ НА ЕГО ОСНОВЕ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2632651C2 |
| Вакцинная композиция против коронавирусной инфекции COVID-19 | 2020 |
|
RU2743595C1 |
Изобретение относится к технологиям стабилизации петлеобразной структуры синтетических пептидов и может использоваться в фармакологии для получения препаратов нового поколения, предназначенных для диагностики, терапии и вакцинопрофилактики инфекционных заболеваний и патологических состояний человека и животных. Предлагается способ стабилизации петлеобразной структуры синтетических пептидов путем нековалентного взаимодействия гидрофобных углеводородов или их радикалов между собой, причем гидрофобные углеводороды или их радикалы располагают на противоположных N и С концах пептида, либо в непосредственной близости от них и названные гидрофобные углеводороды или их радикалы также нековалентно связывают с гидрофобным носителем. Изобретение решает задачу осуществления стабилизации петлеобразных структур пептидов, не затрагивая аминокислотную последовательность пептида, а также обеспечивает возможность нековалентного связывания стабилизированного синтетического пептидного антигена с носителем, при этом способ является универсальным и может использоваться для стабилизации различных структур. 8 з.п. ф-лы, 3 ил., 2 табл.
| ЭПИТОПЫ, ОБРАЗОВАННЫЕ НЕКОВАЛЕНТНЫМ СВЯЗЫВАНИЕМ КОНЪЮГАТОВ | 2000 |
|
RU2259194C2 |
| ЦИКЛИЧЕСКИЕ ПЕПТИДЫ ИЛИ ИХ ФАРМАЦЕВТИЧЕСКИ ПРИЕМЛЕМЫЕ СОЛИ, СПОСОБ ИХ ПОЛУЧЕНИЯ | 1992 |
|
RU2095368C1 |
| A.Cochran et al, "Tryptophan zippers:Stable, monomeric-hairpins" PNAS, 08.05.2001, v.98, № 10, p.5578-5583. | |||

