Изобретение относится к биотехнологии и иммунологии, а именно к препаратам, объединяющим систему доставки антигенов и природных медиаторов, и может быть использовано в области медицины и ветеринарии.
Создание нового поколения вакцин связано с использованием изолированных антигенов, способных обеспечить высокоспецифический иммунный ответ к бактериальным, вирусным патогенам и опухолевым клеткам. Преимущества высокоочищенных субъединичных вакцин по сравнению с традиционными средствами иммунизации характеризуются экономичностью и безопасностью производства, снижением риска возникновения побочных эффектов, возможностью получения препаратов для перорального приема. В то же время иммуногенность большинства индивидуальных антигенов в силу ряда особенностей, таких как низкая молекулярная масса антигена, растворимость, напрямую зависят и опосредуются стимуляцией клеточного звена иммунной системы, а именно Т-хелперных клеток. Поэтому разработка эффективных субъединичных вакцин сопровождается поиском надежных, безопасных и адекватных адъювантов.
Представляют интерес адъюванты липидной природы, используемые в качестве носителей в вакцинных препаратах. Липиды способны связывать одновременно адъюванты и амфифильные антигены и представлять их иммунокомпетентным клеткам в корпускулярной форме. Антигены в составе носителей обладают высокой иммунологической активностью, а сами носители - биодеградируемостью и отсутствием собственной иммуногенности. Комплексы на основе липидной матрицы имеют большие перспективы для решения проблемы адекватного и пролонгированного представления вакцинных антигенов иммунной системе.
Из уровня техники известно, что липидные носители могут существовать в виде эмульсий. Например, описаны водно-масляные растворы обратных мицелл и двухслойных везикул из растительного галактолипида дигалактозидиацилглицерола (ДГДГ) [пат. RU 2127124, опубл. 10.03.1999 г.]. Однако масляные адъюванты имеют ряд недостатков:
- при подкожном и внутримышечном введении вызывают местные реакции воспалительно-некротического характера, а повышенная вязкость затрудняет их введение;
- указанная эмульсия не дает существенного прироста иммуногенности белковых и других антигенов, поскольку сам ДГДГ не обладает иммуноадъювантными свойствами;
- нестабильность при хранении вакцины в готовом виде в результате расслоения фаз и нестандартность качества.
Из уровня техники также известно, что в качестве носителя для представления антигенов используют липосомы, основным структурообразующим компонентом которых является ламеллярный фосфатидилхолин [Allison AG, Gregoriadis G. Liposomes as immunological adjuvants. Nature. 1974. Nov 15; 252(5480):252].
Антигены в составе липосомальных препаратов могут рассматриваться как иммуноадъюванты. В отличие от других адъювантов в месте инъекции липосомальных препаратов образования гранулем не происходит и даже наблюдается предотвращение токсических проявлений инкапсулированных лекарственных средств.
Везикулы липосом могут включать как гидрофобный антигенный материал в липидном бислое, так и гидрофильный во внутренней фазе. Комплексы липосом с антигеном способны стимулировать антителообразование, активизировать фагоциты и поглощаться макрофагами в различных органах и тканях организма, обеспечивать доставку лекарственных средств (антигенов) в клетку. Ограничение их использования является следствием следующих недостатков:
- химическая и физическая нестабильность везикул;
- относительно умеренный иммунный ответ;
- неспособность стимулировать клеточный иммунный ответ, обеспечивающий противовирусную и противоопухолевую защиту.
По технической сущности и достигаемому результату наиболее близким к заявляемому техническому решению является иммуностимулирующий комплекс (ИСКОМ). Это липид-сапониновые частицы, состоящие из смеси растительных тритерпеновых гликозидов Quil А, холестерина и фосфолипидов [пат. RU 2120302, опубл. 20.10.1998 г.] (прототип). Такие комплексы имеют очень специфическую структуру, подтверждаемую данными электронной микроскопии и не являются липидными везикулами или липосомами. Это коллоидные частицы, состоящие из регулярных субъединиц, имеющих открытую сферическую структуру, которые под микроскопом выглядят как кольца. Основная структура ИСКОМ формируется как в присутствии антигена, так и без него. Структурной основой ИСКОМ является сапонин-холестериновый комплекс, способствующий развитию иммунного ответа. ИСКОМ позволяет на порядок снижать дозу вводимых антигенов, индуцировать существенный противовирусный иммунитет и повышать уровень гуморального иммунного ответа в дозах адъюванта намного меньших, чем в обычных вакцинах.
К серьезным недостаткам, препятствующим широкому использованию сапонинов Quil А в качестве адъювантов и системы доставки в составе вакцин против инфекционных заболеваний человека, относятся следующие:
- Quil А представляет собой гетерогенную смесь сапонинов, что является недопустимым для использования в медицине, поскольку смесь сапонинов с разными химическими и биологическими свойствами затрудняет стандартизацию ИСКОМ и может привести к непредсказуемым эффектам in vivo;
- высокая токсичность при внутривенном введении препарата, обусловленная высокой гемолитической активностью;
- воспалительные и болевые реакции у людей при внутримышечном введении, поэтому их использование в качестве неспецифического иммуностимулятора на человеке ограничивается пероральным введением;
- химическая нестабильность в водных растворах по причине гидролиза сложноэфирной связи в одной из боковых углеводородных цепей;
- деацилированные сапонины теряют адъювантные свойства, способность стимулировать Т-клеточный иммунный ответ первого класса и продуцировать антиген-специфические цитотоксические Т-лимфоциты, что негативно сказывается на эффективности вакцин против цитопатических инфекций вирусной или бактериальной природы, противоопухолевых и антипаразитарных вакцин.
Предпринимаемые попытки выделения из Quil А фракции более очищенных сапонинов с низкими цитотоксическими свойствами приводят к тому, что полученные сапонины либо не образуют структуры в виде ИСКОМ, либо не обладают необходимой иммуностимулирующей активностью.
Техническим результатом заявляемого решения является получение носителя антигенов, имеющего высокую иммунологическую эффективность в составе субъединичных вакцин и характеризующегося отсутствием токсичности, воспалительных и болевых реакций при применении.
Технический результат обеспечивается получением носителя антигенов в виде липидного комплекса, состоящего из гликозида, холестерина и липида; в качестве гликозида комплекс содержит голотоксин А1, а в качестве липида моногалактозилдиацилглицериды (МГДГ) из морских макрофитов.
Технический результат оптимальным образом достигается получением носителя антигенов в виде липидного комплекса, в котором весовое содержание голотоксина А1 к холестерину постоянно и равно 3:2, а весовое содержание моногалактозилдиацилглицеридов (МГДГ) из морских макрофитов может изменяться в интервале (2-6).
Полученный липидный комплекс имеет вытянутую нитевидно-тубулярную структуру с длиной частиц от 200 до 300 нм при диаметре тубул около 20 нм, которая отлична от структуры известных иммуностимулирующих комплексов.
Впервые получен носитель антигенов, обладающий высокими иммунологическими и инкорпорирующими характеристиками в отношении амфифильных антигенов, который обеспечивает повышение безопасности вакцин и характеризуется тем, что при его применении достигается снижение токсичности голотоксина A1, входящего в состав комплекса.
Заявляемый носитель антигенов получают посредством смешения голотоксина А1, холестерина и МГДГ, взятых в весовом соотношении 3:2:(2-6) в следующем порядке: смешивают растворы холестерина и МГДГ в хлороформе при весовом соотношении холестерина и МГДГ 2:(2-6), упаривают под вакуумом до удаления хлороформа и добавляют 3 весовые части 0,4 процентного водного раствора голотоксина А1; смесь солюбилизируют, после чего доводят суммарную концентрацию липидов (холестерина и МГДГ) до 2 мг в 1 мл суспензии фосфатно-солевым буфером при pH 7,2. Указанная концентрация липидов является оптимальной для проведения электронно-микроскопических исследований. Полученную суспензию озвучивают с использованием ультразвукового дезинтегратора в течение 5 минут и оставляют на 2 часа до достижения равновесия в системе (пример 1).
Возможность получения носителя антигенов с особой тубулярной суперструктурой при диаметре тубул около 40 нм впервые показана нами в заявке на изобретение «Способ получения антигенов на основе липидов из морских макрофитов и тритерпенового гликозида кукумариозида» при использовании в качестве тритерпенового гликозида кукумариозида А2-2 из кукумарии Cucumaria japonica (заявка на изобретение № 2005131645/13(035476) от 12.10.2005 г.).
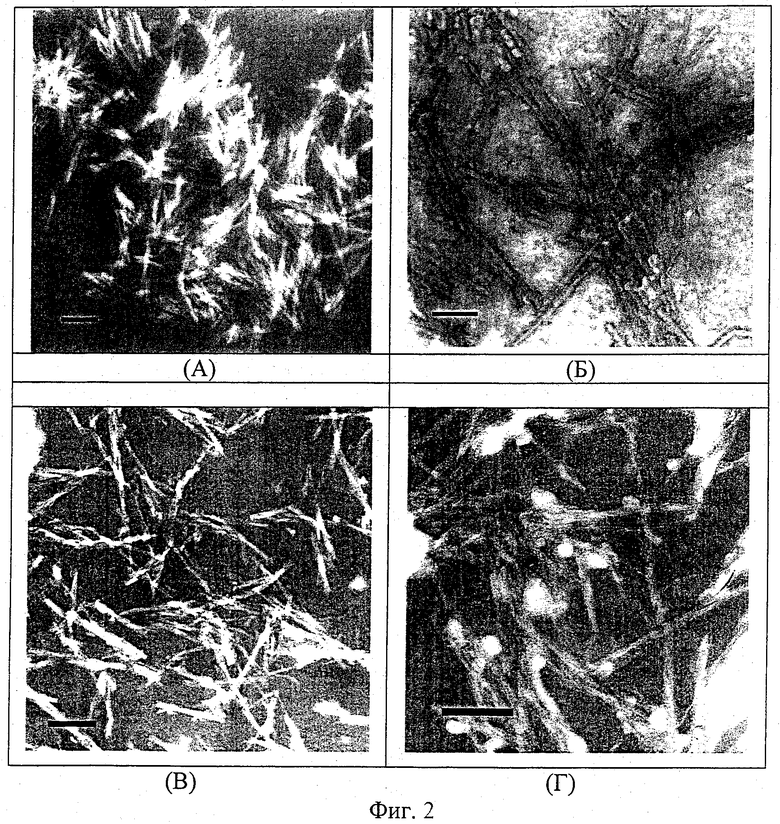
Заявляемый носитель при разных соотношениях компонентов имеет ультрамикроскопические размеры и вытянутую нитевидно-тубулярную структуру с длиной частиц от 200 до 300 нм при диаметре тубул около 20 нм, что подтверждается данными электронной микроскопии (примеры 2,3, фиг.2, снимки А и Б). На фиг.2 приведены электронные микрофотографии липидного носителя антигенов, включающего: голотоксин A1:Хол:МГДГ из Zostera marina в весовых соотношениях 3:2:2 (снимок А) и 3:2:6 (снимок Б). Сравнение снимков А и Б на фиг.2 показывает, что изменение весового содержания МГДГ от 2 до 6 при формировании комплекса носителя антигена приводит к образованию хорошо оформленных тубулярных структур.
Показано также, что количество МГДГ из различных морских макрофитов при получении носителя антигенов заявляемым способом не влияет на его морфологические характеристики, если его весовое соотношение к голотоксину А1 и холестерину изменяется в интервале (2-6), т.е. оптимальным соотношением голотоксина A1, холестерина и МГДГ является 3:2:(2-6) (примеры 3-6, фиг.2, снимки А-Б).
Обнаружено, что комплексы с похожей структурой, но с образованиями меньшего диаметра, могут формироваться и без добавления МГДГ при разных весовых соотношениях голотоксина А1 и холестерина (примеры 7, 8). Это явление обнаружено впервые - при использовании в качестве тритерпенового гликозида кукумариозида А2-2 из кукумарии Cucumaria japonica (заявка на изобретение № 2005131645/13(035476) от 12.09.2005 г.) нами не было зарегистрировано образования комплекса между кукумариозидом А2-2 и холестерином без добавления липидной матрицы.
На снимках В и Г фиг.2 приведены электронные микрофотографии стерин-гликозидных структур, включающих голотоксин А1:Хол в весовых соотношениях 4:1 и 3:2, соответственно. На снимках видны структуры, сходные со структурой комплексных образований на снимках А и В; их диаметр практически в два раза меньше, чем при добавлении МГДГ в структуру комплексов, и составляет 10-12 нм. Увеличение количества холестерина приводит к появлению на снимке Г (фиг.2) белых пятен, свидетельствующих об избытке холестерина.
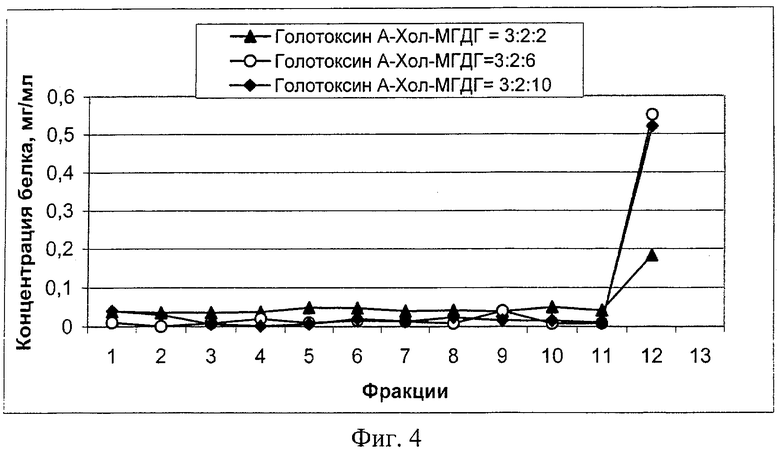
Однако добавление мембранообразующего полярного липида МГДГ усиливает связывание голотоксина А1 со стерином, вызывая существенное снижение гемолитической активности иммуностимулирующего тритерпенового гликозида (пример 9), а также является необходимым условием для самопроизвольного включения амфифильных по своей природе белковых антигенов наружных мембран микроорганизмов. Эффективное весовое содержание МГДГ в системе голотоксин А1:Хол:МГДГ находится в диапазоне (2-6). Исследования с модельным поверхностным белком бактериального происхождения показывают, что оптимальное соотношение голотоксина А1:Хол:МГДГ для эффективного инкорпорирования (более 95%) псевдотуберкулезного порина составляет 3:2:6 (пример 10, фиг.4). На фиг.4 приведено распределение псевдотуберкулезного поринового белка по фракциям после осаждения образцов липид-гликозидных носителей с разным содержанием МГДГ ультрацентрифугированием в фосфатно-солевом буфере.
Снижение содержания МГДГ в комплексе носителя антигенов (менее двух весовых частей) приводит к не полному включению белка в структуру комплекса, а при его увеличении (более 6 весовых частей) не происходит существенного улучшения иммунологических и морфологических характеристик получаемого носителя антигенов.
Примеры 2 и 3 (фиг.2, снимки А, Б) подтверждают, что введение МГДГ в смесь голотоксина A1 и холестерина в указанных выше соотношениях способствует формированию стабильного липидного комплекса, обладающего необходимыми для антигенного носителя свойствами и имеющего стандартную нитевидно-тубулярную структуру.
Оптимальность весового соотношения голотоксина A1 и холестерина (3:2) подтверждается данными электронной микроскопии. При таком соотношении наблюдается образование видимых электронных микроскопических комплексов. Увеличение содержания голотоксина A1 в составе носителя антигенов не рационально по двум причинам:
- это не приводит к изменению структуры комплекса, но приведет к появлению его гемолитической активности;
- голотоксин A1 проявляет свое иммуномодулирующее действие в сверхнизких концентрациях.
Таким образом, экспериментально установлено, что заявляемый липид-холестрин-гликозидный носитель антигенов, полученный при весовом соотношении голотоксин А1:холестерин:МГДГ, равном 3:2:(2-6), имеет оптимальные характеристики и не оказывает токсического действия при внутрибрюшном (в/б) и подкожном (п/к) введении экспериментальным животным (пример 11). По данным электронной микроскопии полученный носитель антигенов имеет вытянутую нитевидно-тубулярную структуру с длиной структурных образований от 200 до 300 нм при диаметре тубул около 20 нм.
Ближайшими структурными аналогами заявляемого носителя являются липид-сапониновые комплексы гликоалкалоидов растений семейства пасленовые [Erik A.J. Keukens, Truus de Vrije, Claudia van den Boom, Pieter de Waard, Henk H. Plasman, Felix Thiel, Vladimir Chupin, Wim M.F.Jongen, Ben de Kruiff. Molekular basis of glicoalkaloid induced membrane disruption // Biochim Biophys Acta. 1995. Vol.1240, P.216-228]. Предложенная модель представляется нам справедливой для заявляемого носителя. Структура заявляемого носителя представляет собой трехслойную тубулу (фиг.1). Внутренний слой (2) структуры состоит из МГДГ, молекулы которого образуют монослой. При этом молекулы МГДГ обращены своими углеводными цепями во внутренний гидратированный канал (1), в то время как их жирнокислотные цепи обращены в противоположную сторону. Над монослоем гликолипидов (2) располагается слой из комплексов холестерин-агликон голотоксина А1 (3). Холестерин-агликоновые комплексы также образуют монослой и ориентированы таким образом, что углеводная цепь молекулы голотоксина A1 экспонирована на внешней стороне носителя (4). Из уровня техники известно, что образование комплекса холестерин-голотоксин А1 обусловлено взаимодействием холестерина и агликона данного гликозида за счет гидрофобных, ван-дер-ваальсовых связей и π-электронов обеих молекул.
Молекулы МГДГ формируют внутренний монослой комплекса за счет гидрофобного взаимодействия жирнокислотных цепей соседних молекул гликолипидов. Сформировавшийся монослой в свою очередь взаимодействует с агликон-холестериновым монослоем, стабилизируя полученную в итоге структуру.
Предположительным механизмом присоединения поринового антигена к заявляемому комплексу является взаимодействие его гидрофобных участков с гидрофобным матриксом носителя.
Голотоксин A1 получают из доступного природного сырья - промысловой голотурии, дальневосточного трепанга Apostichopus japonicus S. [Еляков Г.Б., Мальцев И.И., Калиновский А.И., Стоник В.А. Строение Голотоксина А1 (стихопозида А), основного тритерпенового гликозида тихоокеанской промысловой голотурии Apostichopus japonicus // Биоорг. химия, 1983. Т. 9, № 2. С.280-281].
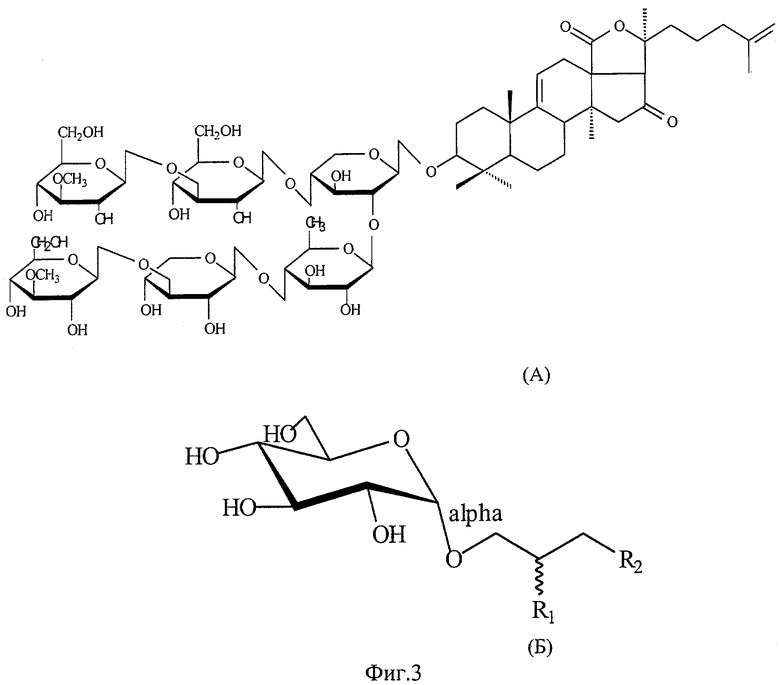
Получаемое сырье включает сумму тритерпеновых гликозидов, основным компонентом которого является голотоксин A1; его структурная формула приведена на фиг.3 А.
Голотоксин A1 в низких дозах обладает адъювантной активностью, оказывая стимулирующее действие на развитие иммунного ответа в разных популяциях иммунокомпетентных клеток при пероральном и парентеральном введении в организм животных. В условиях in vitro голотоксин A1 изменяет функциональную активность иммунокомпетентных клеток, вызывает усиление бластогенеза лимфоцитов селезенки и цитотоксическую активность макрофагов крови. Макрофаги, активированные голотоксином А1, могут играть важную кооперирующую роль в иммуностимулирующей активности этого гликозида [Попов А.М., Атопкина Л.Н., Самошина Н.Ф., Уварова Н.И. Изучение иммуномодулирующей активности тетрациклических тритерпеновых гликозидов даммаранового и голостанового ряда // Антибиотики и химиотерапия. 1994. Т. 39, № 7. С.19-25].
Голотоксин A1 относится к типичным мембранотропным веществам. Мембранная активность голотоксина A1 обусловлена высоким аффинитетом к холестерину. Уже при низких концентрациях он способен с высокой степенью эффективности образовывать нетоксичные гликозид-стериновые или липид-гликозидные структуры [Попов А.М. Сравнительное изучение цитотоксического и гемолитического действия тритерпеноидов женьшеня и голотурий // Изв. РАН. Сер. биол. 2002. № 2. С.155-164].
Таким образом, использование голотоксина A1 в комплексе с холестерином позволяет увеличить эффективность противовирусных вакцин, а именно: повысить иммуногенность антигенной субстанции вследствие корпускулярного представления антигенов в структуре носителя, обладающего иммунными свойствами, а также исключить гемотоксичность голотоксина A1 за счет прочного связывания гликозида с холестерином.
В качестве липидной составляющей заявляемого носителя антигенов был выбран МГДГ, выделенный из морских водорослей, структурная формула которого приведена на фиг.3 (Б). Этот липид сочетает в комплексе структурообразующие, иммунные свойства и способность инкорпорировать поверхностно-мембранные антигены патогенных микроорганизмов. Используемый в качестве липидного компонента МГДГ из Ulva fenestrata (отдел зеленые водоросли - Chlorophyta), Ahnfeltia tobuchiensis (отдел красные водоросли - Rhodophyta), Laminaria japonica (отдел бурые водоросли - Phaeophyta), Zostera marina (зеленая морская трава - отдел Magnoliophyta) был выделен по методу Sanina N., Goncharova S., Kostetsky E. Fatty acid composition of individual polar lipid classes from marine macrophytes // Phytochemistry. 2004. V. 65. №6. Р. 721-730. Препараты липидного комплекса получали по описанной в заявке методике. Для получения конечного продукта использовали МГДГ, выделенный из Zostera marina (пример 3), из Ulva fenestrate (пример 4), Ahnfeltia tobuchiensis (пример 5), Laminaria japonica (пример 6).
Вне зависимости от источника выделения МГДГ из морских макрофитов полученные препараты представляют собой водную суспензию липидного комплекса, имеющего нитевидно-тубулярную структуру длиной 200-300 нм при диаметре тубул около 20 нм.
Этот липид является самым ненасыщенным гликоглицеролипидом и включает широкий спектр полиеновых жирных кислот. Из уровня техники известно влияние ненасыщенных жирных кислот, в частности линоленовой и арахидоновой, на состояние мембран клеток иммунной системы, в особенности макрофагов.
Эффективность использования МГДГ в качестве липида при формировании носителей антигенов, имеющих тубулярную структуру, исследована и обоснована нами в заявке на изобретение 2005131645/13(035476) от 12.10.2005 г. В заявке показано, что МГДГ проявляет самостоятельную иммуноадьювантную активность при в/б и п/к введении, сопоставимую с активностью адъюванта Фрейнда и даже превосходящую его при иммунизации корпускулярным Т-зависимым антигеном - эритроциты барана (ЭБ); обладает способностью подавлять активность эффекторов гиперчувствительности замедленного типа (ГЗТ).
Следует отметить, что новые функции голотоксина А1 и МГДГ, которые они проявляют в заявляемом изобретении, не вытекают с очевидностью из известных для них свойств.
На основе голотоксина А1, холестерина и МГДГ впервые получен носитель антигенов, имеющий нитевидно-тубулярную структуру, позволяющую увеличить эффективность противовирусных вакцин, а именно: повысить иммуногенность антигенной субстанции вследствие корпускулярного представления антигенов в структуре липид-гликозидного носителя, обладающего иммунными свойствами, а также исключить гемотоксичность за счет прочного связывания гликозида с мембранообразующими липидами - холестерином и МГДГ в структуре липид-гликозидного матрикса (пример 9).
Подлинность препарата определяется качественным и количественным составом структурных компонентов (голотоксина A1, холестерина и МГДГ), являющихся исходными при получении носителя антигенов; его структура подтверждается данными электронной микроскопии (фиг.2, снимки А, Б). Препарат стабилен при температуре (-4°С) более трех месяцев.
Предлагаемый липид-гликозидный комплекс при подкожном и внутрибрюшинном введении мышам не вызывает воспалительных, болевых, токсических, а также высоких цитолитических эффектов (пример 11).
Возможность осуществления изобретения подтверждается следующими примерами.
Пример 1. Приготовление препарата липид-гликозидного носителя, состоящего из голотоксина A1, холестерина и МГДГ.
Приготовление растворов исходных компонентов проводят следующим образом: 5 мг МГДГ растворяют в 1 мл хлороформа и 5 мг холестерина растворяют в 1 мл хлороформа, 4 мг голотоксина A1 растворяют в 1 мл дистиллированной воды. Микродозатором отбирают необходимое количество растворенного в хлороформе холестерина и МГДГ, упаривают под вакуумом и солюбилизируют сухой остаток 0,4% раствором голотоксина А1. После тщательного перемешивания в смесь добавляют солевой фосфатный буфер, pH 7,2, до конечной концентрации липидов в смеси 2 мг в 1 мл. Полученную суспензию озвучивают ультразвуковым дезинтегратором SONOPULS Ultraschall - Homogenizatoren HD 2070 (Германия) при частоте 20 кГц, мощность 7 Вт в режиме 7 (0,7 с работа, 0,3 с перерыв) в течение 5 минут, и затем выдерживают в течение 2 часов до полного формирования комплексов.
Пример 2. Приготовление препарата липид-гликозидного носителя, состоящего из трех весовых частей голотоксина A1, двух весовых частей холестерина и двух весовых частей МГДГ из Zostera marina по описанной в примере 1 методике. Микродозатором отбирают 200 мкл раствора холестерина и 200 мкл раствора МГДГ, упаривают под вакуумом досуха и солюбилизируют сухой остаток 0,4% раствором голотоксина А1, объем которого составляет 375 мкл. После тщательного перемешивания в смесь добавляют 625 мкл солевого фосфатного буфера, pH 7,2. Конечная концентрация липидов составляет 2 мг в 1 мл. Суспензию озвучивают 5 минут на ультразвуковом дезинтеграторе. Полученный препарат представляет собой водную суспензию, включающую частицы носителя, имеющего ультрамикроскопическую тубулярную липидную структуру с длиной тубул 100-300 нм. На фиг.2 (снимок А) приведена электронная микрофотография, полученного липид-гликозидного носителя (увеличение Х 50000).
Пример 3. Препарат липидного носителя получали по описанной в примере 1 методике при весовом соотношении исходных компонентов 3:2:6 с использованием МГДГ из Zostera marina. На фиг.2 (снимок Б) приведена электронная микрофотография липидного носителя, полученного с использованием МГДГ из Zostera marina (увеличение Х 50000).
Пример 4. Препарат липидного носителя получали по описанной в примере 3 методике с использованием МГДГ из Ulva fenestrate. Использование МГДГ, выделенного из Ulva fenestrate не изменяло морфологию носителя.
Пример 5. Препарат липидного носителя получали по описанной в примере 3 методике с использованием МГДГ из Ahnfeltia tobuchiensis. Использование МГДГ, выделенного из Ahnfeltia tobuchiensis, не изменяло морфологию носителя.
Пример 6. Препарат липидного носителя получали по описанной в примере 3 методике с использованием МГДГ из Laminaria japonica. Использование МГДГ, выделенного из Laminaria japonica, не изменяло морфологию носителя.
Пример 7. Препарат гликозид-стеринового комплекса получали по описанной в примере 1 методике на основе голотоксина A1 и холестерина при их весовом соотношении 4:1 без добавления МГДГ. По данным электронной микроскопии голотоксин A1 в присутствии холестерина и в отсутствии МГДГ формирует нитевидные структуры длиной 100-200 нм. На фиг.2 (снимок В) приведена электронная микрофотография гликозид-стеринового комплекса, полученного в условиях примера 4 (увеличение Х 50000).
Пример 8. Препарат гликозид-стеринового комплекса получали по описанной в примере 1 методике на основе голотоксина A1 и холестерина при их весовом соотношении 3:2 без добавления МГДГ. По данным электронной микроскопии голотоксин A1 в отсутствии МГДГ и в избытке холестерина формирует нитевидные структуры. На фиг.2 (снимок Г) приведена электронная микрофотография гликозид-стеринового комплекса, полученного в условиях примера 5 (увеличение Х 50000).
Пример 9. Липидные носители, приведенные в примерах 2 и 3, тестировали на гемолитическую активность в условиях in vitro с использованием в качестве тест-системы эритроцитов беспородных мышей. Гепаринизированную кровь, полученную путем декапитации мыши, промывали в изотоническом растворе хлорида натрия 2-кратным центрифугированием при 3000 об/мин в течение 15 мин при комнатной температуре. Из осадка эритроцитов получали 0,5% клеточную суспензию. Исследуемые препараты готовили в фосфатно-солевом буфере, pH 7,4, и последовательно разносили по 10 мкл по лункам планшета методом двукратных серийных разведений с последующим добавлением суспензии эритроцитов по 90 мкл на лунку. В контрольные лунки добавляли по 10 мкл фосфатно-солевого буфера. Инкубирование проводили в течение 1 и 2 часов в термостате при 37°С. Оценку гемолитической активности исследуемых препаратов производили при визуальном осмотре лунок. Минимальную эффективную дозу вещества, вызывающую 100% гемолиз (ЕД100), определяли по появлению прозрачного раствора «лаковой» крови.
Гемолитическое действие голотоксина F1, включенного в комплекс заявляемого носителя антигенов, по отношению к эритроцитам белой мыши в течение двух часов наблюдения в составе липидных носителей снижалось в 9,8 раз по сравнению с действием того же количества индивидуального тритерпенового гликозида.
Пример 10. Полученные по описанной в примере 1 методике препараты липидных носителей смешивали с 0,6 мг псевдотуберкулезного поринового белка. Весовые соотношения структурообразующих компонентов голотоксин А1:Хол:МГДГ составляли 3:2:2, 3:2:6 или 3:2:10. При этом содержание голотоксина во всех исследованных препаратах было постоянно и составляло 0,6 мг. После ультразвукового дезинтегрирования белоксодержащие образцы носителей осаждали на ультрацентрифуге со скоростью 45000×g в течение 30 мин. Из центрифужных пробирок, сверху вниз до осадка, отбирали 12 фракций и в каждой определяли содержание белка по методу Лоури.
В результате эксперимента было показано (фиг.4), что более 95% белка после перемешивания и ультразвуковой обработки включается в состав липидных носителей, содержащих голотоксин A1, холестерин и МГДГ в соотношении 3:2:(6-10). Снижение количества МГДГ в 3-5 раз приводит к частичной утрате инкорпорирующих свойств носителя.
Пример 11. Полученный заявляемым способом (пример 2, 3) липидный носитель антигенов был испытан на группе беспородных мышей (по 10 в контрольной и опытной группе) на его безопасность. В ходе наблюдения у животных контролировали появление воспалительных или болевых реакций, признаков токсичности или гибели.
Однократное внутрибрюшинное введение белым беспородным мышам препарата липидного носителя из расчета 100 и 200 мкг голотоксина A1 на мышь не вызывает гибели животных и проявления токсических эффектов в течение трех дней наблюдения. Испытываемые дозы голотоксина A1 соответствуют 5 и 10 мкг/кг веса. При этом оптимальная иммунотропная доза голотоксина A1 составляет 0,05-50 мкг на 1 кг веса.
Однократное подкожное введение липидных комплексов, приведенных в примерах 2 и 3, из расчета 100 и 200 мкг голотоксина A1 на мышь, не вызывает воспалительных и болевых эффектов у животных в течение трех дней наблюдения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ НОСИТЕЛЯ АНТИГЕНОВ НА ОСНОВЕ ЛИПИДОВ ИЗ МОРСКИХ МАКРОФИТОВ И ТРИТЕРПЕНОВОГО ГЛИКОЗИДА КУКУМАРИОЗИДА | 2005 |
|
RU2319506C2 |
| ИММУНОСТИМУЛИРУЮЩИЙ КОМПЛЕКС, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 2010 |
|
RU2446822C2 |
| СПОСОБ РЕГУЛЯЦИИ ИММУНОГЕННОСТИ АНТИГЕНА | 2010 |
|
RU2440141C1 |
| НОСИТЕЛЬ И АДЪЮВАНТ ДЛЯ АНТИГЕНОВ | 2006 |
|
RU2311926C2 |
| Биологически активная добавка из голотурии и способ её получения | 2021 |
|
RU2755312C1 |
| МИКРОФЛЮИДИЗИРОВАННЫЕ ЭМУЛЬСИИ ТИПА "МАСЛО В ВОДЕ" И ВАКЦИННЫЕ КОМПОЗИЦИИ | 2005 |
|
RU2347586C2 |
| СПОСОБ ПОЛУЧЕНИЯ ФРОНДОЗИДА А И СПОСОБ СТИМУЛИРОВАНИЯ ИММУННОЙ СИСТЕМЫ МЛЕКОПИТАЮЩИХ | 2005 |
|
RU2339644C2 |
| ИММУНОМОДУЛИРУЮЩЕЕ СРЕДСТВО "КУМАЗИД" И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ | 2004 |
|
RU2271820C1 |
| САМОСОБИРАЮЩИЕСЯ СИНТЕТИЧЕСКИЕ БЕЛКИ | 2014 |
|
RU2747857C2 |
| СФИНГОИДНЫЕ ПОЛИАЛКИЛАМИНОВЫЕ КОНЪЮГАТЫ ДЛЯ ВАКЦИНАЦИИ | 2004 |
|
RU2361577C2 |
Изобретение относится к биотехнологии и иммунологии. Предлагается носитель антигенов в виде липидного комплекса, состоящего из гликозида, холестерина и липида. В качестве гликозида он содержит голотоксин A1, а в качестве липида - моногалактозилдиацилглицериды (МГДГ) морских макрофитов, взятые в весовом соотношении 3 (голотоксин А1):2(холестерин):(2-6) (МГДГ). Полученный гликозид-холестерин-липидный носитель имеет вытянутую нитевидно-тубулярную структуру. Применение полученного носителя антигенов позволяет повысить иммуногенную активность вакцинных препаратов, а также понизить или удалить полностью гемолитическую токсичность голотоксина A1 и исключить воспалительные, болевые, токсические и гемолитические эффекты вакцин. 4 ил.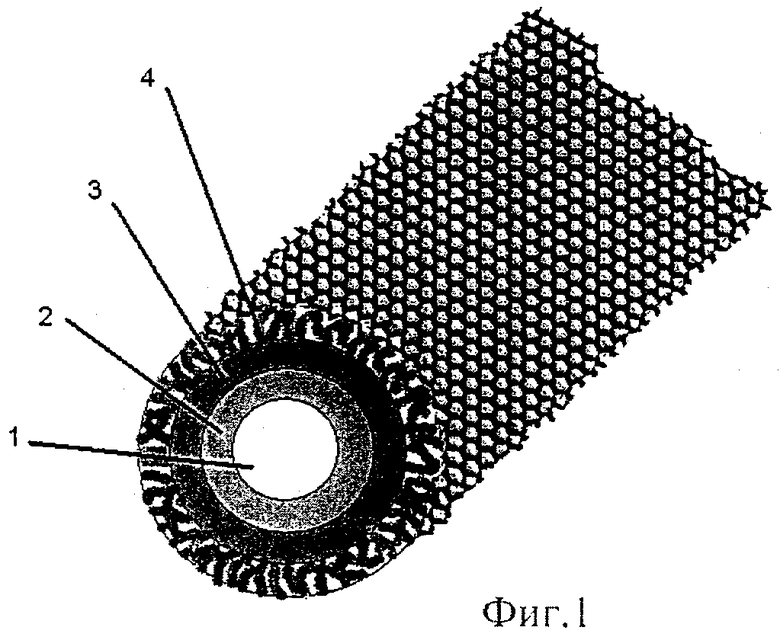
Носитель антигенов в виде липидного комплекса, состоящий из гликозида, холестерина и липида, отличающийся тем, что в качестве гликозида он содержит голотоксин A1, а в качестве липида-моногалактозилдиацилглицериды (МГДГ) из морских макрофитов, взятые в весовом соотношении 3(голотоксин А1):2(холестерин):(2-6) (МГДГ) и представляет собой вытянутую нитевидно-тубулярную структуру.
| ИММУНОМОДУЛИРУЮЩИЙ АГЕНТ | 1989 |
|
RU2120302C1 |
| WO 9205771 A1, 16.04.1992. | |||

