Изобретение относится к биотехнологии и медицине, а именно иммунологии. Предлагается носитель и адъювант для антигенов, имеющий эффективную адъювантную форму и объединяющий систему доставки антигенов и природных медиаторов.
В современных технологиях изготовления вакцинных препаратов большое внимание уделяют проблеме поиска и конструирования различных носителей для антигенов, назначение которых оптимальная форма доставки антигена иммунокомпетентным клеткам, направленный транспорт, защита антигена от биодеградирующих механизмов (протеаз, нуклеаз и др.), адъювантный эффект за счет собственной иммуномодулирующей активности носителя и процессинг антигена соответствующими клетками.
Наиболее известный аналог таких носителей - полный адъювант Фрейнда (ПАФ) [Freund J., Am. J. Clin. Pathol., 21, 645 (1951)]. Это суспензия микобактерий Mycobacterium tuberculosis на основе эмульсии минерального масла и детергента - эмульгатора. Подобные носители часто оказываются токсичными, вызывают образование гранулем в месте инъекции и в связи с этим непригодны для приготовления препаратов, в том числе вакцин, клинического или ветеринарного назначения.
Делаются многочисленные попытки разработки адъювантов с активностью, сравнимой с ПАФ, но без нежелательных побочных эффектов. Большое число исследований посвящено различным липидным носителям, объединяющим свойства носителя антигена и источника для иммунологически высокореактогенных производных продуктов метаболизма липидов.
Известны липидные носители антигенов - липосомы, которые являются везикулами с гидрофобной и гидрофильной фазами, способными инкорпорировать или включать в мембрану антиген для транспортирования и представления его иммунокомпетентным клеткам [Allison A.G., Gregoriadis G. Liposomes as immunological adjuvants. Nature. 1974. Nov 15; 252(5480):252].
Основным структурообразующим компонентом липосом является ламеллярный фосфатидилхолин. Такие носители со встроенным антигеном способны стимулировать антителообразование, активизировать фагоциты и поглощаться макрофагами в различных органах и тканях организма, обеспечивать доставку антигена в клетку. Образующийся комплекс антигена с липосомами может рассматриваться как иммуноадъювантный комплекс. Однако широкое использование вакцинных препаратов на основе липосом ограничивается рядом недостатков, к которым относятся химическая и физическая нестабильность везикул, относительно умеренный иммунный ответ, а также неспособность стимулировать клеточный иммунный ответ, обеспечивающий противовирусную и противоопухолевую защиту.
Наиболее близким к заявляемому техническому решению по технической сущности и достигаемому при использовании техническому результату является иммуностимулирующий комплекс (ИСКОМ), представляющий собой липид-сапониновые частицы, состоящие из смеси растительных тритерпеновых гликозидов QuilA из растения Quillajia saponaria, холестерина и фосфолипидов в соотношении 3:2:6 [пат. RU 2120302, опубл. 20.10.1998 г.]. Такие комплексы не являются липидными везикулам или липосомами и имеют специфическую морфологическую структуру, подтверждаемую данными электронной микроскопии. ИСКОМы - это коллоидные частицы, состоящие из регулярных субъединиц, имеющих открытую сферическую структуру, которые под микроскопом выглядят как кольца. QuilA сапонины в составе носителя обладают иммуностимулирующими свойствами и необходимы для его структурирования. Адъювантная активность ИСКОМ позволяет индуцировать существенный специфический иммунный ответ гуморального и клеточного типа при использовании доз антигена, намного меньших, чем в обычных вакцинах.
Основным недостатком ИСКОМ как системы доставки антигенов является токсичность при парентеральном введении препарата, главной причиной которой является относительно высокая гемолитическая активность сапонинов QuilA. Парентеральное введение ИСКОМ сопровождается воспалительными и болевыми реакциями; в связи с этим в настоящее время использование в клинике QuilA сапонинов в качестве адъюванта ограничивается пероральным введением.
Группа изобретений, лежащих в основе данной заявки, относится к иммунологии. Предлагаются: носитель и адъювант для антигенов, имеющий эффективную адъювантную форму и объединяющий систему доставки антигенов и природных медиаторов; вакцинный препарат, полученный на основе данного носителя, способ его получения, а также применение, обеспечивающее повышение безопасности (отсутствие токсичности, воспалительных и болевых реакций) и эффективности вакцинации.
Задачей заявляемого технического решения является получение липид-сапонинового носителя и адъюванта индивидуальных белковых антигенов, состоящего из смеси тритерпеновых гликозидов кукумариозида А2-2 (КД), холестерина и полярного липида моногалактозилдиацилглицерида (МГДГ) их морских макрофитов, взятых в весовом соотношении 3:2:6.
Задачей заявляемого технического решения являются также получение антиген-содержащих липид-сапониновых комплексов, включающих указанный выше липид-сапониновый носитель и мономерный порин.
Задачами заявляемого технического решения являются также способ получения антигенсодержащих липид-сапониновых комплексов и их применение в качестве вакцинных препаратов.
Поставленные задачи оптимальным образом решаются следующим образом.
На основе природных липидов получают гидрофобный носитель антигенов, объединяющий адъювантный эффект и систему доставки антигенов и природных медиаторов. А именно получают носитель в виде липид-сапониновой системы, в которой структурообразующей основой является нетоксичный комплекс из тритерпенового гликозида кукумариозида А2-2 дальневосточной голотурии Cucumaria japonica (препарат КД) с холестерином в постоянном весовом соотношении КД к холестерину 3:2, и моногалактозилдиацилглицеридов (МГДГ) из морских макрофитов, содержание которых в комплексе КД: холестерин: МГДГ оптимально при весовом соотношении 3:2:6 (тубулярно-искомный комплекс или ТИ-комплекс).
Заявляемое изобретение обеспечивает повышение эффективности вакцин на основе индивидуальных или субъединичных антигенов, которые по тем или иным причинам не способны индуцировать эффективный иммунный ответ в отношении инфекционных агентов разной этиологии (патогенных вирусов, бактерий) и опухолевых клеток.
Липид-сапониновые комплексы получают следующим образом. Растворы холестерина и МГДГ в хлороформе при весовом соотношении холестерина и МГДГ 2:6 упаривают под вакуумом до удаления хлороформа и добавляют 3 вес.ч. 0,4 процентного водного раствора кукумариозида, смесь солюбилизируют, после чего доводят суммарную концентрацию липидов (холестерина и МГДГ) до 2 мг в 1 мл суспензии фосфатно-солевым буфером при рН 7,2, после чего озвучивают полученную суспензию ультразвуковым дезинтегратором в течение 5 мин. Полученные препараты представляют собой водную суспензию липид-сапониновых комплексов, стабильную при температуре (-4°С) более трех месяцев. Подлинность препарата определяется качественным и количественным составом исходных структурных компонентов (КД, холестерина и МГДГ).
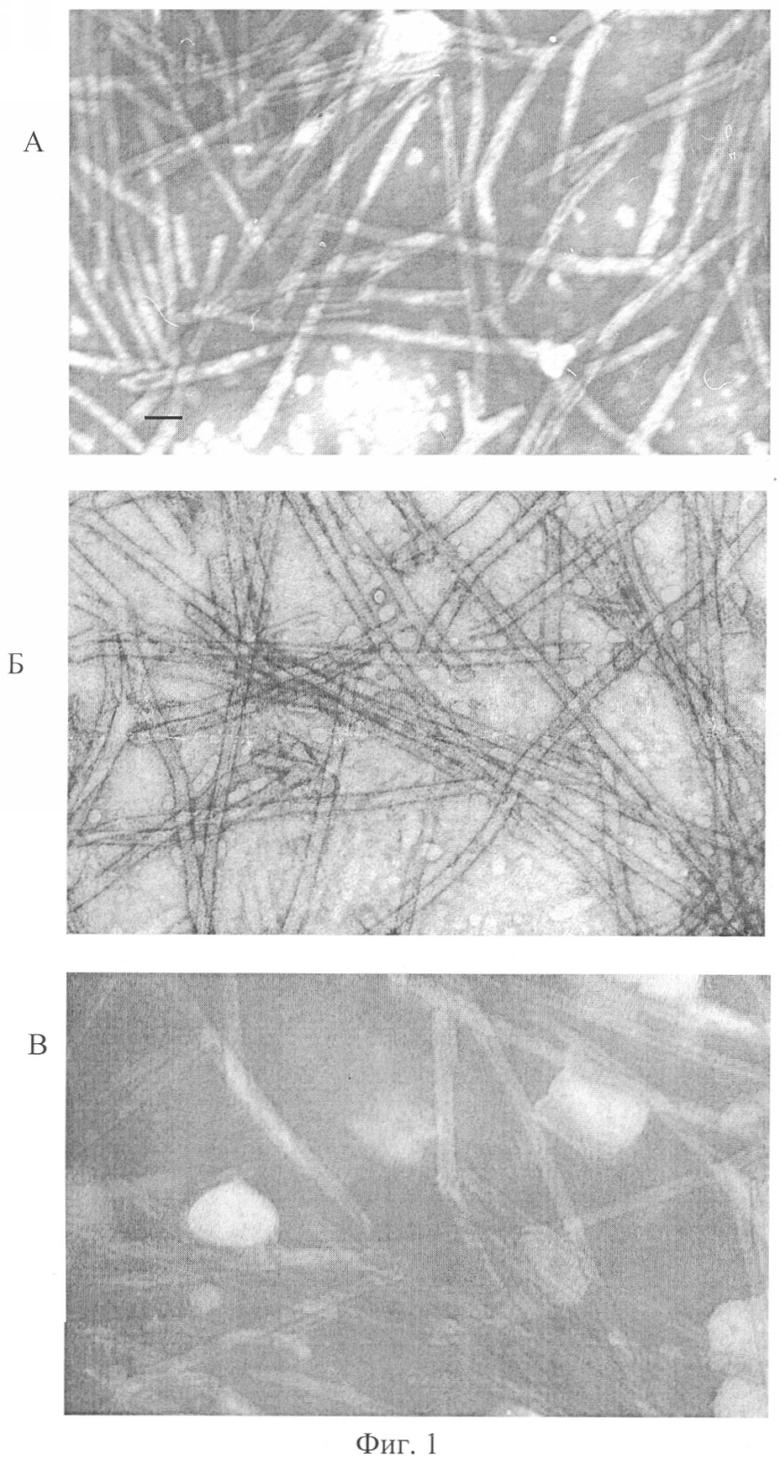
Образующийся ТИ-комплекс (пример 1) оценивают электронно-микроскопически (фиг.1, снимок А). На снимке А представлена электронно-микроскопическая фотография матрицы ТИ-комплекса при соотношении компонентов в системе КД: холестерин: МГДГ (3:2:6). Длина масштабной линии 100 нм, увеличение ×50000.
Полученные комплексы - это суперструктуры, состоящие из комплексов с ультромикроскопической тубулярной структурой при диаметре тубул около 40 нм, сформированных искомоподобными структурами сходного диаметра при наличии последних между тубулами. Получен липид-сапониновый носитель антигенов, имеющий структуру, отличную от структуры ранее описанных иммуностимулирующих комплексов.
Эти суперструктуры, названные нами тубулярно-искомными комплексами (ТИ-комплексами), являются морфологически устойчивыми; их назначение - представление антигена иммунокомпетентным клеткам в составе ТИ-комплекса и устранение ряда негативных эффектов, характерных для ИСКОМ, таких как гемолитический эффект и соответственно токсичность при парентеральном введении.
Ранее авторами показано, что замена в ИСКОМах фосфатидилхолина (ФХ) на МГДГ не вызывает изменения структуры ИСКОМ. [Acta Biochimica Polonica. Vol.51 №1 (2004) 263-272]. Однако замена в структуре ИСКОМ ФХ на МГДГ, a QuilA на КД приводит к появлению новой тубулярно-искосомальной структуры.
Присутствие в структуре ТИ-комплексов тубулярных структур наряду с искомоподобными структурами может приводить к пролонгированному высвобождению искомоподобных структур, способных фагоцитироваться в иммунизированном организме, и, следовательно, усилению адъювантных и иммуномодулирующих свойств ТИ-комплекса.
Для оценки иммуннологических свойств ТИ-комплекса в качестве объектов сравнения, способных переносить антиген и обеспечивать адъювантный эффект, использовали широко известный из уровня техники полный адъювант Фрейнда, иммуностимулирующий комплекс (ИСКОМ) на основе сапонина QuilA, холестерина и фосфатидилхолина в соотношении QuilA: Хол: ФХ = 3:2:6 и ТИ-комплекс, в котором МГДГ заменен на ФХ (ТИ-ФХ-ком) в соотношении КД:Хол:ФХ=3:2:6. Подобная замена проведена для оценки вклада, прежде всего состава жирно-кислотных остатков на иммуннологические свойства.
На первом этапе была проведена серия экспериментов для анализа собственной адъювантной активности структурообразующих единиц ТИ-комплекса: КД и МГДГ.
Из уровня техники известно, что препарат КД не проявляет эмбриотоксических и мутагенных свойств, повышает неспецифическую устойчивость к бактериальным и вирусным инфекциям в чрезвычайно низких нетоксичных для организма дозах. Эффект кукумариозида связывают со стимуляцией фагоцитарной активностью тканевых макрофагов, влиянием на уровень пролиферативной активности стволовых клеток, усилением кооперативного взаимодействия иммунокомпетентных клеток (Т- и В-лимфоцитов) в процессе развития специфического иммунного ответа и, вероятно, прямым действием на стадии взаимодействия вирус - клетка.
Биологическую активность МГДГ морских макрофитов обоснованно связывают как со структурой углеводной составляющей, так и с присутствием большого количества полиненасыщенных жирных кислот (ПНЖК). Используемый нами в качестве липидного компонента МГДГ содержит до 93% омега-3 ПНЖК, которые согласно известным данным обладают иммуномодулирующим действием, оказывая влияние на активацию Т-хелперов 1 и 2 класса, стимулируют Т-клеточную пролиферацию и образование цитокинов, влияют на состояние мембран клеток иммунной системы, в частности макрофагов, и обладают избирательной цитотоксичностью в отношении опухолевых клеток, усиливая действие цитотоксических агентов.
Следует отметить, что новые функции КД и МГДГ, которые они проявляют в заявляемом изобретении, не вытекают с очевидностью из известных для них свойств.
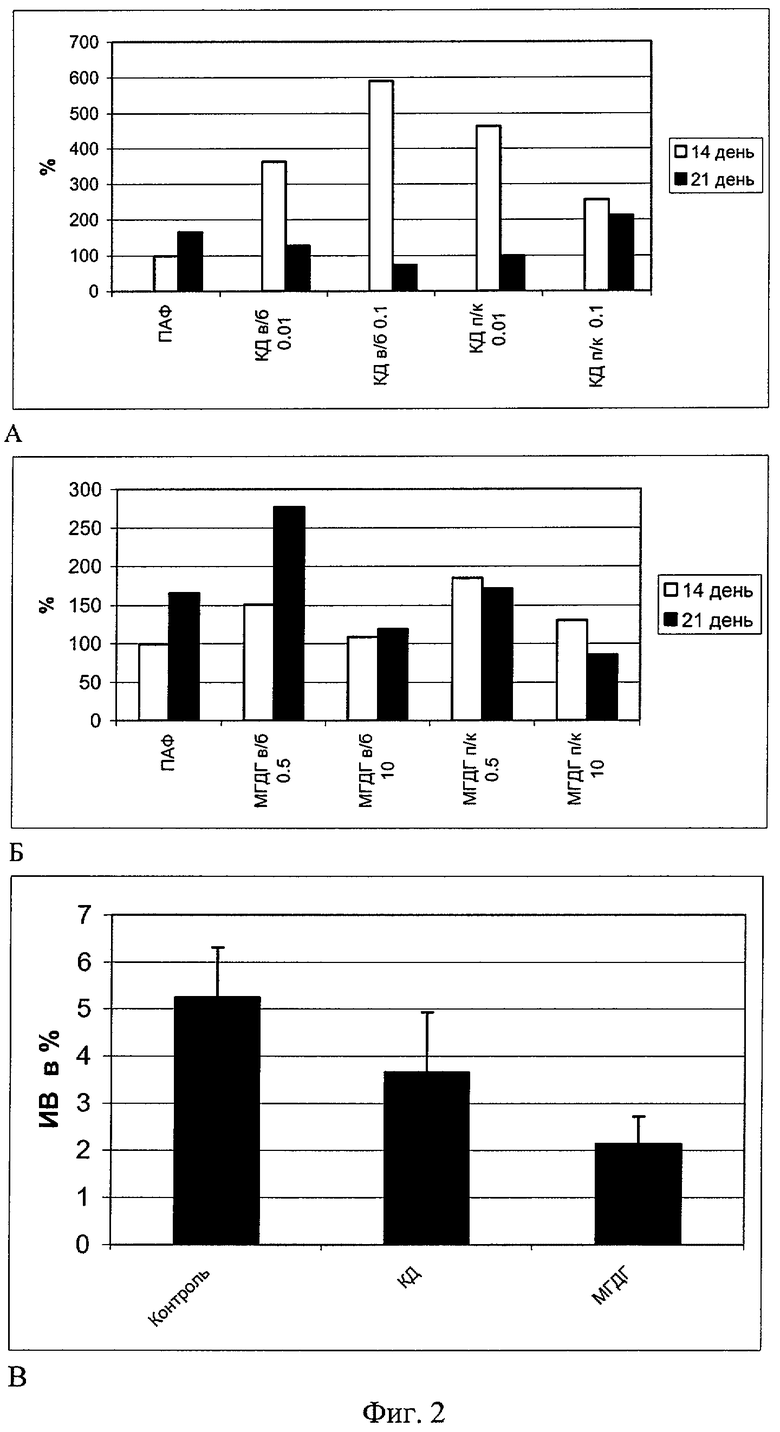
Экспериментально показано, что препараты КД и МГДГ проявляют самостоятельную иммуноадъювантную активность (пример 3, фиг.2, снимки А и Б). На фиг.2 представлено адъювантное действие структурообразующих компонентов ТИ-комплекса на 14 и 21 день после внутрибрюшинной (в/б) или подкожной (п/к) иммунизации эритроцитами барана (ЭБ) беспородных мышей: А - КД в дозах 0,01 и 0,1 мкг/мышь, Б - МГДГ в дозах 0,5 и 10 мкг/мышь. По оси ординат - процент прироста титра гемагглютинирующих антител по отношению к контролю. По оси абсцисс - экспериментальные группы животных. Представленные данные демонстрируют наличие иммуноадъювантных свойств у КД (в дозах 0.01 и 0.1 мкг/мышь) и МГДГ (в дозах 0.5 и 10 мкг/мышь) при их в/б и п/к введении. Причем эти свойства более выражены в сравнении с ПАФ. Особенно заметен процент прироста (до 600%) гемаглютинирующих антител к ЭБ в первые 14 дней после иммунизации животных, получивших КД одновременно с ЭБ. МГДГ также показал адъювантный эффект, превосходящий ПАФ, при в/б и п/к введении в дозах 0.5 мкг/мышь. Таким образом, оба компонента ТИ-комплекса характеризуются способностью проявлять иммуноадъювантные свойства, сопоставимые с активностью адъюванта Фрейнда и даже превосходящие его при иммунизации корпускулярным Т-зависимым антигеном - ЭБ.
Оба препарата обладают также способностью подавлять активность эффекторов гиперчувствительности замедленного типа (ГЗТ), что свидетельствует об их влиянии на клеточно-опосредованные иммунные реакции. При постановке классической реакции ГЗТ против эритроцитов барана, введенных в/б, КД и МГДГ при в/б введении снижали индекс воспаления (ИВ) ГЗТ (пример 3, фиг.2, снимок В). На фиг.2, снимок В приведена оценка реакции ГЗТ у беспородных белых мышей, иммунизированных ЭБ при совместном в/б введением КД в дозе 0,1 мкг/мышь или МГДГ в дозе 2 мкг/мышь. По оси ординат приведен индекс воспаления ГЗТ в процентах, а по оси абсцисс - экспериментальные группы животных. Особенно заметно активность эффекторов ГЗТ снижалась в группе животных, получивших МГДГ, что, видимо, является результатом влияния полиненасыщенных жирно-кислотных метаболитов МГДГ, выступающих в роли медиаторов, снижающих интенсивность местных воспалительных процессов. Эти результаты подтверждают иммуномодулирующий потенциал МГДГ и КД и позволяют предположить их положительное влияние в составе ТИ-комплекса на безопасность иммунизации (за счет снижения местных клеточно-опосредованных воспалительных реакций).
Для оценки свойств ТИ-комплекса как носителя и адъюванта для антигена получен антигенсодержащий липид-сапониновый комплекс, в котором в качестве амфифильного мембранного антигена использован порин. В частности, в примере 2 используют термически обработанную, гидрофобную, мономерную форму порообразующего белка наружной мембраны возбудителя псевдотуберкулеза Yersinia pseudotuberculosis с молекулярной массой около 36 КД. Этот белок обладает видовой специфичностью, иммуногенностью, отсутствием гемолитической активности и других видов токсичности.
Способ получения таких поринсодержащих ТИ-комплексов включает получение липидного комплекса из КД, холестерина и МГДГ, взятых в весовом соотношении 3:2:6, с последующим добавлением поринового белка в необходимой концентрации. Сначала смешивают растворы холестерина и МГДГ в хлороформе при весовом соотношении холестерина и МГДГ 2:6, упаривают под вакуумом до удаления хлороформа и добавляют 3 вес.ч. 0,4 процентного водного раствора кукумариозида, смесь солюбилизируют, после чего доводят суммарную концентрацию липидов (холестерина и МГДГ) до 2 мг в 1 мл суспензии фосфатно-солевым буфером при рН 7,2. К полученному ТИ-комплексу добавляют порин, который вводят в фосфатно-солевом буфером растворе (рН 7,2) в дозах, соответствующих 0,1-1-10 мкг белка на 1 мкг содержания КД, а именно 75 мкг, 750 мкг или 7500 мкг на 1 мл полученной смеси. Полученную смесь озвучивают ультразвуковым дезинтегратором в течение 5 мин и оставляют на 3 ч. Амфифильные мембранные антигены, подобные мономерному пориновому белку из Y.Pseudotuberculosis, включаются самопроизвольно в ТИ-комплекс.
Получают продукт, поринсодержащий ТИ-комплекс, готовый для иммунизации, интегрирующий адъювантный эффект, систему доставки антигенов и природных медиаторов и позволяющий при применении обеспечить безопасность иммунизации, уменьшить иммуносупрессорные свойства порина и обеспечить более высокий уровень гуморального и клеточного иммунного ответа при использовании незначительных доз антигена.
Полученный препарат представляет собой водную суспензию порин-липид-сапониновых комплексов, стабильную при температуре (-4°С) более трех месяцев. Подлинность препарата определяется качественным и количественным составом исходных структурных компонентов (КД, холестерина и МГДГ). Для оценки связывания порина с ТИ-комплексом проводят ультрацентрифугирование при 54000×g в течение 45 мин.
Содержание несвязанного белка определяют в надосадочной жидкости методом Лоури. Количество белка, включенного в ТИ-комплекс, составляет не менее 95%. На фиг.1, снимках Б и В представлены электронно-микроскопические фотографии матрицы ТИ-комплекса с порином при соотношении компонентов в системе КД: холестерин: МГДГ: порин (3:2:6:0,3) и (3:2:6:30) соответственно. Длина масштабной линии 100 нм, увеличение ×50000.
При электронно-микроскопическом исследовании установлено, что процесс взаимодействия порина с ТИ-комплексом не нарушает морфологическую структуру ТИ-комплекса. Таким образом, получался комплексный препарат порина, инкорпорированного в структуру матрицы ТИ-комплекса.
Полученные ТИ-комплексы с антигеном - это суперструктуры, отличные от известных иммуностимулирующих комплексов; они имеют ультрамикроскопическую тубулярную структуру, представленную тубулами с диаметром поперечного сечения около 40 нм, сформированными искомоподобными структурами сходного диаметра при наличии последних между тубулами.
На втором этапе на мышах разных линий проведена серия экспериментов по оценке эффективности иммунизации субъединичным белком порином в индивидуальном виде и в составе ТИ-комплексов в сравнении с известными носителями и адъювантами: ПАФ, ИСКОМ, ТИ-ФХ-ком. Проведена оценка специфического клеточного иммунного ответа (ГЗТ к порину), гуморального (уровень антипориновых антител в сыворотке крови животных), уровня общих иммуноглобулинов классов М, G, А в сыворотке крови и цитокинов - интерферон-гамма (IFNg) и интерлейкин-1-бета (IL-1b), имеющих важный иммунорегуляторный и эффекторный потенциал. В экспериментах использованы в/б и п/к способы введения препаратов; количество порина - в следующем диапазоне доз: 0,1-1-10-30-50-100 мкг/мышь.
Как оптимальные были отобраны дозы 0,1, 1 и 10 мкг, так как более высокие дозы не приводят к усилению гуморального иммунного ответа.
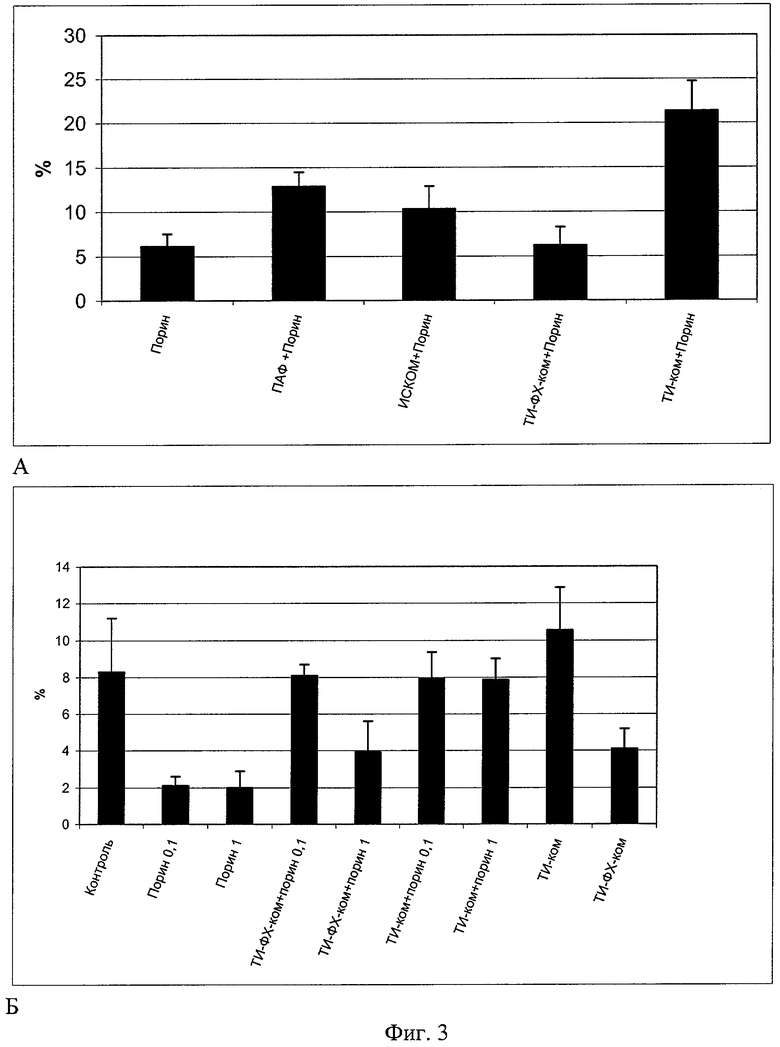
Серия экспериментов по оценке эффективности специфического клеточного иммунного ответа (ГЗТ к порину), выполненная на беспородных белых мышах с использованием субъединичного белка порина и белка в составе ТИ-комплексов в сравнении с известными носителями и адъювантами, показывает, что индивидуальный порин и порин в составе липидных комплексов по-разному влияют на показатели специфического иммунного ответа клеточного типа. При в/б иммунизации порином ИВ эффекторов ГЗТ, специфичных к порину, оказывается значительно выше при использовании антигена в составе ТИ-комплекса (пример 4, фиг.3 снимок А, Б). На фиг.3 снимок А приведены показатели ГЗТ беспородных белых мышей после в/б иммунизации индивидуальным порином и порином в составе адъювантных носителей в дозе 10 мкг/мышь. По оси ординат приведен ИВ ГЗТ в процентах, а оси абсцисс - экспериментальные группы животных.
ТИ-комплекс с порином способен переносить, представлять антиген и генерировать развитие специфического иммунного ответа клеточного типа более эффективно, чем индивидуальный порин и комплексы порина с ПАФ, ИСКОМами и ТИ-ФХ-комплексами. Это доказывает перспективность использования ТИ-комплекса и его конкурентноспособность в сравнении с другими известными липидными носителями.
Как известно из уровня техники, порин является белком с умеренно выраженной иммуногенностью, вызывающим модуляцию механизмов клеточного и гуморального иммунитета. В наших экспериментах порин существенно подавляет активность эффекторов ГЗТ при иммунизации мышей в диапазоне доз 0,1 -1 -10 мкг/мышь, т.е. оказывается супрессором в отношении этого механизма клеточного иммунного ответа. Применение порина в тех же дозах, но в составе ТИ-комплекса, характеризуется устранением супрессорного эффекта порина в отношении эффекторов ГЗТ (фиг.3, снимки А, Б). При этом следует отметить, что сам по себе ТИ-комплекс обладает способностью повышать ИВ ГЗТ, демонстрируя тем самым стимулирующее действие в отношении клеточных механизмов иммунного ответа. Данный эффект ТИ-комплекса как носителя порина превосходит по своей эффективности действие комплекса ТИ-ФХ-ком с порином (фиг.3Б). На фиг.3, снимок Б приведены показатели ГЗТ беспородных белых мышей, иммунизированных в/б индивидуальным порином и порином в составе адъювантных носителей в дозах 0,1 и 1 мкг/мышь. По оси ординат приведен ИВ ГЗТ в процентах, а оси абсцисс - экспериментальные группы животных.
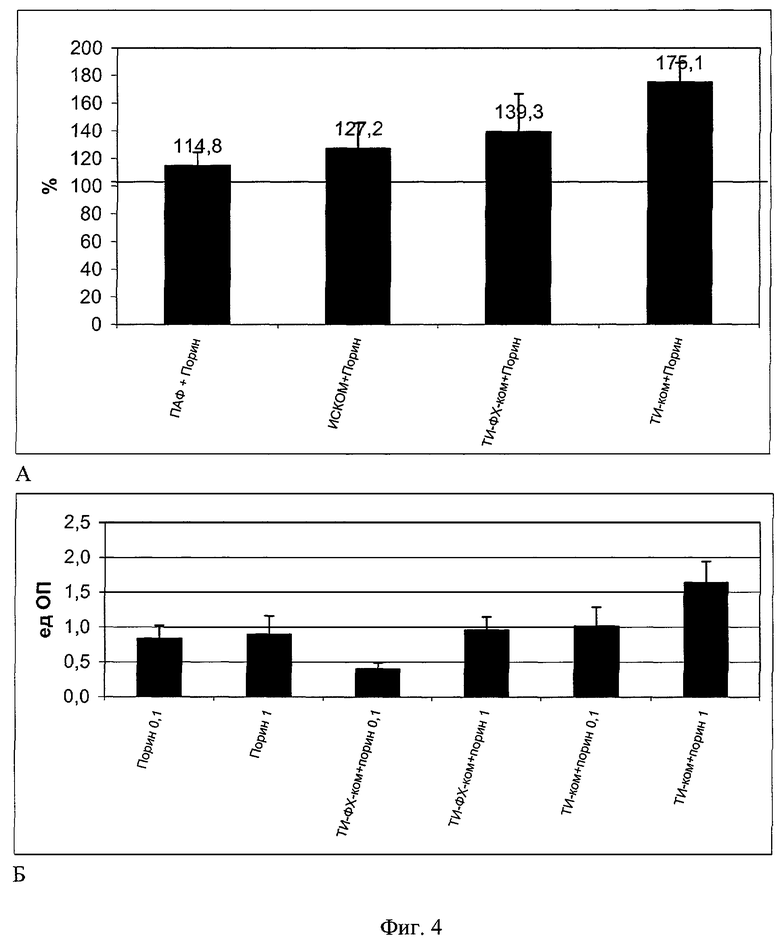
Для исследования специфического иммунного ответа гуморального типа (уровень антипориновых антител в сыворотке крови животных) проведена серия экспериментов по оценке эффективности иммунизации индивидуальным порином и белком в составе ТИ-комплексов в сравнении с известными носителями и адъювантами (пример 4, фиг.4, снимки А, Б). Уровень гуморального иммунного ответа при иммунизации порином оценивается методом иммуноферментного анализа по концентрации специфических антипориновых антител в сыворотке крови беспородных белых мышей и выражается величиной сдвига (в процентах) относительно показателей животных, иммунизированных индивидуальным порином, и в единицах оптической плотности при длине волны 492 нм (фиг.4, снимки А, Б). На фиг.4А приведены данные, характеризующие различия концентраций антипориновых антител в сыворотке крови беспородных белых мышей после иммунизации порином в составе различных адъювантных носителей. По оси ординат приведены относительные показатели содержания антипориновых антител в сыворотке крови мышей, иммунизированных порином в составе различных адъювантных носителей, в процентах к показателям сыворотки крови мышей, иммунизированных только порином (принятым за 100% и показанным на фигуре горизонтальной линией). По оси абсцисс приведены экспериментальные группы животных. При в/б введении порина в дозе 10 мкг/мышь содержание специфических антипориновых антител оказалось значительно выше при использовании ТИ-комплекса, чем при использовании индивидуального антигена. Следовательно, ТИ-комплекс с порином способен переносить, представлять антиген и генерировать развитие специфического иммунного ответа гуморального типа более эффективно, чем комплексы порина с ПАФ, ИСКОМами и ТИ-ФХ-комплексами. ТИ-комплекс с порином обеспечивает прирост специфических антител на 75% в сравнении с животными, иммунизированными индивидуальным порином, тогда как комплексы порина с ПАФ, ИСКОМ и ИСКОМ-КД обеспечивают прирост специфических антител в диапазоне 15-39% (фиг.4А). Это доказывает перспективность использования ТИ-комплекса и его преимущества в сравнении с другими известными липидными носителями.
Из уровня техники известно, что порин является иммуногенным белком. Однако это слабый антиген, характеризующийся рядом негативных эффектов, в том числе иммуносупрессорного свойства. Это особенно подчеркивает необходимость правильного сочетания при иммунизации: повышение иммуногенности и снижение дозы антигена. Данные, представленные на фиг.4, снимок Б, демонстрируют, что дозы порина в 0,1 и 1 мкг/мышь обеспечивают гуморальный иммунный ответ практически на одном уровне, тогда как иммунизация порином в этих же дозах, но в составе ТИ-комплексов, обеспечивает более высокие показатели. Этот прирост особенно значителен в отношении дозы порина в 1 мкг/мышь. Очевидно именно в таком соотношении порина и ТИ-комплекса достигается оптимальный баланс сочетания адъювантных свойств носителя и его способности к экранированию (нейтрализации) негативных свойств индивидуального антигена. При этом доза порина в 0,1 мкг/мышь в составе ТИ-комплекса обеспечивает более высокий гуморальный иммунный ответ, чем при использовании индивидуального порина в дозе 1 мкг/мышь. Это позволяет говорить о способности ТИ-комплекса на порядок повышать эффективность иммунизации в сравнении с индивидуальным порином. Это с большой вероятностью доказывает преимущества иммунизации с использованием ТИ-комплексного носителя. Сравнение действия порина в составе ТИ-комплекса и порина в составе комплекса ТИ-ФХ-ком также демонстрируют явное преимущество ТИ-комплекса, который обеспечивает адъювантный эффект, тогда как комплекс ТИ-ФХ-ком обеспечивает практически только экранирование порина, ингибировав специфический гуморальный иммунный ответ (фиг.4, снимок Б). На этом снимке приведена концентрация антител к порину в сыворотке крови беспородных белых мышей при двукратной иммунизации индивидуальным порином и порином в составе различных адъювантных носителей в дозах 0,1 и 1 мкг/мышь. По оси ординат приведены показатели содержания антипориновых антител, определенных методом иммуноферментного анализа (ИФА) и выраженных в единицах оптической плотности при длине волны 492 нм (разведение сыворотки крови 1:160). По оси абсцисс приведены экспериментальные группы животных.
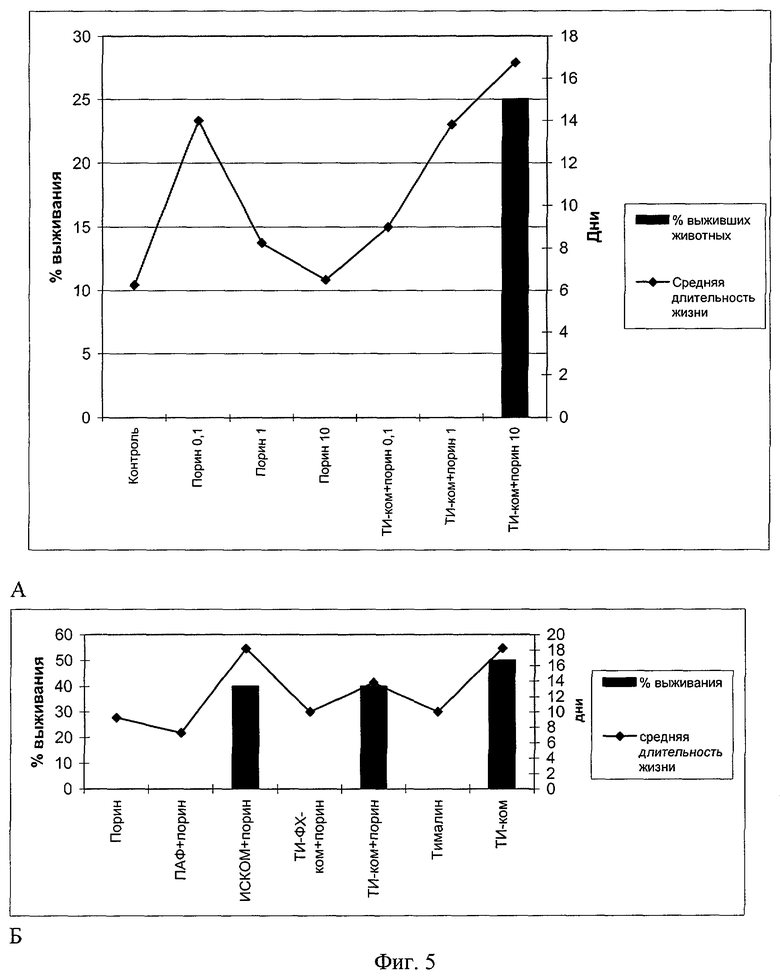
Эксперименты по оценке выживаемости мышей, инфицированных Y.pseudotuberculosis, после иммунизации порином в составе ТИ-комплексов дали следующие результаты.
Супрессорное действие порина в отношении клеточных механизмов иммунного ответа нарушает адекватное развитие иммунологических реакций и дает в итоге негативную динамику выживаемости животных, иммунизированных порином в диапазоне доз 0,1-1-10 мкг/мышь при модельной инфекции Y.pseudotuberculosis (пример 5, фиг.5, снимки А, Б). При этом имеет место обратная дозозависимость - повышение средней длительности жизни зараженных животных при уменьшении иммунизирующей дозы. Доза порина в 10 мкг/мышь обеспечивает наименьший защитный эффект как у мышей линии СВА, так и у беспородных белых мышей (фиг.5, снимки А, Б). Это доказывает негативные побочные эффекты порина, проявляющиеся после иммунизации этим белком в чистом виде. Применение порина в составе ТИ-комплексов дает обратную картину: повышение показателей выживаемости животных при увеличении дозы. Порин в составе ТИ-комплекса при иммунизации мышей линии СВА приводит к повышению средней длительности жизни и 25%-ной выживаемости мышей (фиг.5, снимок А). На этом снимке приведен процент выживания и средняя продолжительность жизни мышей линии СВА, зараженных в/б 10 LD50 Y.pseudotuberculosis, после иммунизации индивидуальным порином и порином в составе ТИ-комплексов в дозах 0,1,1 и 10 мкг/мышь. По оси ординат, слева, приведен процент выживших животных по истечении 21 суток наблюдения; справа - средняя продолжительность жизни мышей. По оси абсцисс - экспериментальные группы животных.
В экспериментах на беспородных белых мышах иммунизация порином в составе ТИ-комплекса в дозе порина 10 мкг/мышь тоже приводит к повышению средней длительности жизни и 40%-ной выживаемости (фиг.5, снимок Б). Эти показатели сопоставимы с результатами иммунизации порином в составе ИСКОМ и значительно превосходят соответствующие показатели с использованием порина в составе ПАФ и ТИ-ФХ-ком, которые не обеспечивают никакого протективного эффекта. Тималин - известный иммуномодулятор, обладающий протективной активностью в отношении ряда инфекционных заболеваний, в условиях нашего эксперимента также не дает заметного защитного эффекта при самостоятельном применении, тогда как самостоятельное применение ТИ-комплекса (без порина) обеспечивает значительный защитный эффект (увеличение средней длительности жизни и 50%-ный показатель выживаемости мышей). На фиг.5, снимок Б приведен процент выживаемости и средняя продолжительность жизни беспородных мышей, зараженных в/б 1 LD 100 Y.pseudotuberculosis после иммунизации индивидуальным порином (в/б в дозе 10 мкг/мышь) и порином в составе) адъювантных носителей. По оси ординат, слева, процент выживших животных по истечении 21 суток наблюдения, справа - средняя продолжительность жизни мышей в днях. По оси абсцисс - экспериментальные группы животных.
Применение для иммунизации ТИ-комплексов как носителей порина приводит к устранению негативных свойств индивидуального антигена и улучшению прогноза модельного заболевания, т.е. к улучшению результативности вакцинации. Эти результаты коррелируют с данными экспериментов, демонстрирующих способность ТИ-комплекса увеличивать содержание сывороточных иммуноглобулинов класса А (пример 6, фиг.6) и активировать систему Интерферона (пример 7, фиг.7).
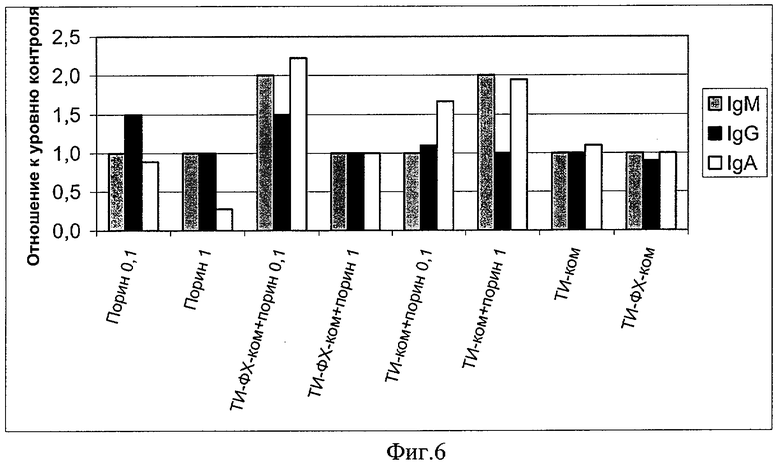
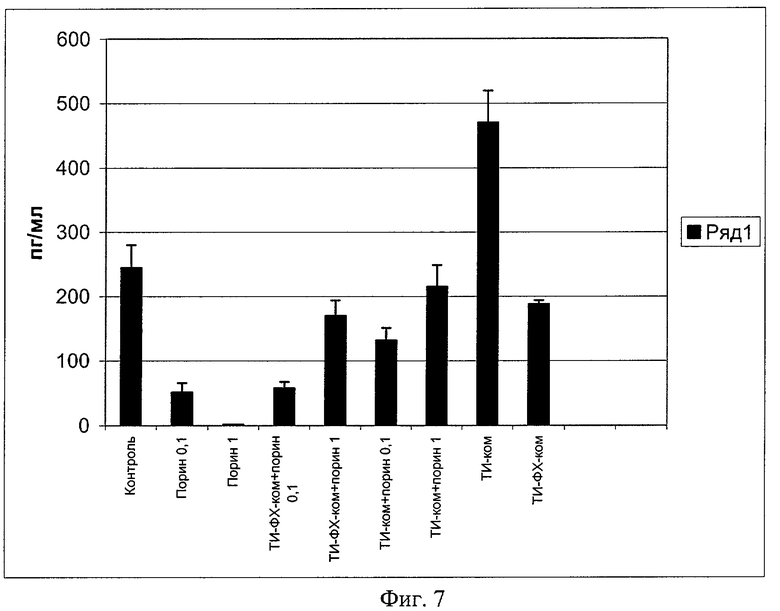
Под влиянием ТИ-комплекса повышается уровень иммуноглобулинов классов M, G, A в сыворотке крови иммунизированных мышей, т.е. срабатывает механизм гуморального неспецифического иммунитета. Порин в дозах 0,1 и 1 мкг/мышь практически не изменяет уровень иммуноглобулинов классов М, увеличивает уровень IgG (в дозе 0,1 мкг/мышь) и приводит к глубокому угнетению IgA при в/б способе введения (фиг.6). На этой фигуре приведен относительный уровень в сыворотке крови беспородных белых мышей, иммунизированных в/б в дозах 0,1 и 1 мкг/мышь. Уровень контроля принят за 1. По оси ординат приведена величина изменения уровня иммуноглобулинов сыворотки крови относительно показателей контрольных животных. По оси абсцисс - экспериментальные группы животных. Данный способ иммунизации приводит еще к одной заметной сопутствующей негативной особенности - значительному угнетению интерферонового статуса. Уровень IFNg в сыворотке крови мышей падает до близких к нулевым значениям (пример 6, фиг.7), что свидетельствует о негативных результатах иммунизации порином, приводящих к нарушению эффекторных и регуляторных механизмов. Применение порина в составе ТИ-комплекса устраняет эти негативные эффекты: поднимает уровень иммуноглобулинов класса А до уровней, вдвое превышающих соответствующие показатели интактных животных (фиг.6), и нейтрализует супрессорное действие порина в отношении системы интерферона (фиг.7). ТИ-комплекс сам по себе оказался индуктором интерферона, превосходящим по активности комплекс ТИ-ФХ-ком (фиг.7). На фиг.7 приведена концентрация интерферона - гамма (IFNg) в сыворотке крови беспородных белых мышей, иммунизированных в/б в дозах 0,1 и 1 мкг/мышь индивидуальным порином и порином в составе различных адъювантных носителей. По оси ординат приведены титры IFNg в пг/мл, а по оси абсцисс - экспериментальные группы животных.
Позитивное действие ТИ-комплекса в отношении системы интерферона и иммуноглобулинов класса А может считаться основой для дальнейшего испытания в качестве носителя для вирусных антигенов.
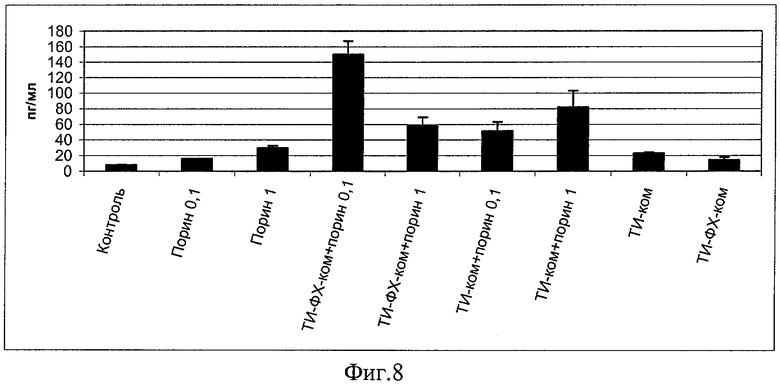
Порин в составе ТИ-комплекса дал более эффективную активацию интерлейкин-1-бета (IL-1b), важного для запуска первичных иммунных реакций, реакций воспаления и межклеточной кооперации цитокина (пример 7, фиг.8). На фиг.8 приведена концентрация интерлейкина-1-бета (IL-1b) в сыворотке крови беспородных белых мышей, иммунизированных индивидуальным порином (в/б в дозах 0,1 и 1 мкг/мышь) и порином в составе различных адъювантных носителей. По оси ординат приведены титры IL-1b в пг/мл, по оси ординат - экспериментальные группы животных.
При иммунизации порином в составе ТИ-комплекса активация IL-1b существенно выше, чем при иммунизации порином без носителя. Сам по себе ТИ-комплекс вызывает достаточно умеренную стимуляцию уровней IL-1b, демонстрируя способность обеспечивать более высокую функциональную способность (преддиспозицию) иммунной системы к ответу на антигенное раздражение.
Разработка и применение предлагаемого ТИ-комплекса в качестве носителя и адъюванта для антигенов позволяет:
- получать иммунные ТИ-комплексы, взаимодействие которых с мембранным белковым антигеном не нарушает их суперструктуры;
- доказать наличие адъювантной активности компонентов ТИ-комплекса - МГДГ и КД в отношении гуморального звена иммунного ответа и их супрессорные свойства в отношении антипориновых эффекторов ГЗТ;
- выявить положительное влияние КД и МГДГ на безопасность иммунизации (за счет снижения местных клеточно-опосредованных воспалительных реакций);
- доказать действие ТИ-комплекса как системы, обладающей свойствами носителя антигена и адъюванта, обеспечивающих усиление специфического иммунного ответа при иммунизации белком порином и способность активировать антипориновые эффекторы ГЗТ в отличие от супрессорных свойств индивидуального порина;
- установить, что применение порина в составе ТИ-комплексов при иммунизации животных более эффективно, чем имммунизация животных индивидуальным порином, и позволяет устранить или минимизировать негативные эффекты индивидуального порина (супрессорное действие в отношении эффекторов ГЗТ, иммуноглобулинов класса А и системы интерферона) и обеспечить более интенсивный синтез специфических антипориновых антител;
- установить, что системное действие ТИ-поринового комплекса способно нейтрализовать негативные эффекты порина в отношении важнейших эффекторных и регуляторных иммунологических реакций;
- установить, что порин в составе ТИ-комплексов слабо повышает содержание в сыворотке крови IL-1b, как свидетельство отсутствия у ТИ-комплекса негативных провоспалительных свойств;
- установить противоположно дозозависимый позитивный эффект от иммунизации порином в составе ТИ-комплексов в отношении модельной инфекции, вызванной заражением животных Y.Pseudotuberculosis: эффект усиливается при увеличении иммунизирующей дозы порина;
- установить преимущество использования ТИ-комплекса при сравнительной оценке иммунологического действия различных липидных носителей порового белка с чистым порином; комплекс стабильно превосходит по своим адъювантным свойствам полный адъювант Фрейнда (ПАФ), ИСКОМы и ТИ-ФХ-ком;
- установить, что ТИ-комплекс без антигена обладает иммуномодулирующей активностью, активирует эффекторы ГЗТ, является индуктором IFNg и IL-1b, обеспечивает протективную активность (значительно выше, чем у известного иммуномодулятора тималина) при модельной инфекции на экспериментальных животных.
Пример 1. Приготовление препарата ТИ-комплекса, состоящего из 3 весовых частей КД, двух весовых частей холестерина и шести весовых частей МГДГ.
5 мг МГДГ растворяют в 1 мл хлороформа; 5 мг холестерина растворяют в 1 мл хлороформа, а 4 мг КД растворяют в 1 мл дистиллированной воды. Микродозатором отбирают 27 мкл раствора холестерина и 81 мкл раствора МГДГ, упаривают под вакуумом досуха, добавляют 3 вес.ч.4% раствора кукумариозида в объеме 50 мкл и солюбилизируют остаток раствором КД, объем которого 500 мкл. В смесь добавляют 500 мкл фосфатного буфера, рН 7,2. Конечная концентрация липидов составляет 2 мг в 1 мл. Суспензию озвучивают 5 мин ультразвуковым дезинтегратором. Полученный препарат представляет собой водную суспензию, включающую частицы комплекса.
Пример 2. ТИ-комплекс с порином получают следующим образом: к ТИ-комплексу, полученному в примере 1, добавляют субъединичный, гидрофобный, термически обработанный пориновый белок с молекулярной массой около 36 КД из порообразующего белка наружной мембраны возбудителя псевдотуберкулеза Y.pseudotuberculosis. Порин добавляют в ТИ-комплекс (КД: холестерин: МГДГ (3:2:6)) в фосфатно-солевом буферном растворе (рН 7,2) в дозах, соответствующих 0,1 мкг белка (фиг.1, снимок Б) или 10 мкг белка (фиг.1 снимок В) на 1 мкг содержания КД. Полученную смесь ТИ-комплекса с порином озвучивают ультразвуковым дезинтегратором в течение 5 мин и оставляют на 3 ч. Амфифильные мембранные антигены, подобные мономерному пориновому белку из Y.Pseudotuberculosis, включаются самопроизвольно в ТИ-комплекс, образуя порин-содержащий Ти-комплекс.
Пример 3. Перед иммунизацией животных комплекс разводят фосфатным буфером до рабочих концентраций из расчета 1 мкг КД/мышь в объеме 0,2 мл. При этом используют ТИ-комплекс (КД: Хол: МГДГ = 3:2:6) и систему антигена с ТИ-комплексом (КД: Хол: МГДГ: порин = 3:2:6:0,3-3-30). Эмульсию мышам вводят в/б и п/к в дозе 1 мкг КД/мышь в объеме 0,2 мл.
Для оценки собственной иммуномодулирующей активности компонентов ТИ-комплекса производят оценку влияния МГДГ и КД на интенсивность реакций клеточного и гуморального иммунного ответа при иммунизации беспородных белых мышей корпускулярным Т-зависимым антигеном - эритроцитами барана (ЭБ). Мышей иммунизируют в/б в дозе 5×106 клеток в объеме 0,5 мл физиологического раствора. Одновременно вводят в/б или п/к КД в дозах 0,01 и и 0,1 мкг/мышь или МГДГ в дозах 0,5 и 10 мкг/мышь. Препараты вводят в объеме 0,2 мл фосфатно-солевого буфера (ФСБ). В качестве положительного контроля используют полный адъювант Фрейнда (ПАФ), который вводят п/к в объеме 0,2 мл. Разрешающую дозу ЭБ для ГЗТ вводят на 6-й день после иммунизации мышам в подушечку одной из лапок в половинной дозе от иммунизирующей в объеме 0,02 мл ФСБ. В контрлатеральную лапку вводят такой же объем ФСБ. Эвтаназию животных производят на 7-й день после иммунизации, с последующим определением эффекторов гиперчувствительности замедленного типа (ГЗТ), специфичных к ЭБ, методом расчета индекса воспаления (ИВ) ГЗТ. ИВ оценивают как разницу веса опытной и контрольной лапок, в %. На 14-21 день после иммунизации у мышей соответствующих опытных групп получают сыворотку крови и определяют содержание в сыворотке крови специфических антител против ЭБ в реакции прямой гемагглютинации. За титр антител принимают обратную величину, соответствующую максимальному разведению сыворотки, при которой наблюдалась реакция агглютинации ЭБ.
Установлено иммуноадъювантное действие КД и МГДГ, которое оказалось более выраженным, чем эффект от адъюванта Фрейнда при иммунизации корпускулярным Т-зависимым антигеном - ЭБ. (фиг.2, снимок А, Б). Особенно заметен процент прироста (250-600%) гемаглютинирующих антител к ЭБ при введении КД в первые 14 дней после иммунизации. МГДГ показал наилучший эффект, превосходящий адьювант Фрейнда при в/б и п/к введении в дозах 0.5 мкг/мышь. Оба препарата обладают также способностью подавлять активность эффекторов ГЗТ. При постановке классической реакции ГЗТ против эритроцитов барана КД и МГДГ при в/б введении снижает индекс воспаления ГЗТ (фиг.2, снимок В). Особенно значимо активность эффекторов ГЗТ снижалась в группе животных, получивших МГДГ.
Пример 4. Беспородных белых мышей иммунизируют в/б порином в дозах 0,1-1-10 мкг/мышь в объеме 0,2 мл ФСБ и порином в тех же дозах в составе ТИ-комплексов. В качестве групп контроля используют порин в смеси с адъювантом Фрейнда в составе ИСКОМ и ТИ-ФХ-комплексов. Мышей иммунизируют дважды: повторную иммунизацию производят через 14 дней после первой. Данный режим иммунизации был выбран как оптимальный на основании данных, известных из уровня техники. На 6-й день после второй иммунизации мышам в подушечку одной из лапок вводят разрешающую дозу порина в половинной дозе от иммунизирующей в объеме 0,02 мл ФСБ. В контрлатеральную лапку вводят такой же объем ФСБ. Забой животных производят на 7 день после второй иммунизации. Оценивают ИВ эффекторов гиперчувствительности замедленного типа как разницу веса опытной и контрольной лапок в процентах. Получают сыворотку крови и определяют содержание специфических антипориновых антител методом иммуноферментного анализа. Измерение оптической плотности производят при длине волны 492 нм. Определяют относительную величину содержания антипориновых антител по отношению к показателям интактных животных, которую выражают в процентах. Данный эксперимент доказывает, что ТИ-комплексы наиболее эффективно способны переносить, представлять антиген и генерировать развитие специфического иммунного ответа. На фиг.3 снимок А, Б и фиг.4, снимки А, Б показано, что ТИ-комплекс с порином обеспечивает прирост показателей специфического иммунного ответа как клеточного, так и гуморального типа. Данный эффект ТИ-комплекса с порином оказался более выраженным в сравнении с адъювантом Фрейнда, ИСКОМами и ТИ-ФХ-комплексом.
Пример 5. Модельную инфекцию вызывают у мышей линии СВА и беспородных белых мышей путем в/б введения взвеси иерсиний в количестве, составляющем соответственно 10LD50 и 1LD100. В течение последующих 21 дня (срок наблюдения) оценивают процент выживших животных и среднюю продолжительность жизни (срок жизни выживших животных оценивали как 21 день). Данные показатели оценивают у интактных животных и животных, иммунизированных порином в дозах 0,1-1-10 мкг/мышь. Иммунизацию производят 2-кратно по стандартной схеме (14+7 дней). Заражение животных производят через 21 день от даты первой иммунизации порином, который вводят в свободном виде, в составе ТИ-комплексов и других липидных носителей. В данных экспериментах оценивают также самостоятельную протективную активность ТИ-комплекса без порина в сравнении с действием известного неспецифического иммуностимулятора тималина, примененного также по 2-кратной схеме в/б в дозах 1 мкг/мышь.
Пример 6. Изучают влияние иммунизации индивидуальным порином и порином в составе ТИ-комплексов и других липидных носителей на состояние системы гуморального иммунитета у беспородных белых мышей. Применяют в/б способ введения чистого порина и порина в комплексе с липидными носителями в дозах 0,1-1 мкг/мышь. Содержание иммуноглобулинов классов М, G, А в сыворотке крови мышей, иммунизированных порином самостоятельно и порином в составе ТИ-комплексов, определяют методом радиальной иммунодиффузии с использованием готовых диагностических наборов производства МР Biomedicals, США в соответствии с приложенными методическими рекомендациями. Величину радиальной иммунодиффузии оценивают в мм и определяют концентрацию иммуноглобулинов по калибровочной кривой. Рассчитывают относительные показатели содержания иммуноглобулинов в сыворотке крови опытных животных по сравнению с показателями интактных животных. Показатели интактных животных принимают за единицу. Оценивают положительные или отрицательные отклонения содержания иммуноглобулинов классов М, G, А в опытных группах по сравнению с контролем.
Пример 7. Цитокины определяют в сыворотке крови беспородных белых мышей, иммунизированных порином в/б в дозах 0,1, 1 и 10 мкг/мышь самостоятельно и порином в составе ТИ-коплексов. Проводят сравнительную оценку действия порина в составе ТИ-комплекса, а также цитокин-индуцирующую активность самого ТИ-комплекса. Титры цитокинов выражают в пг/мл.
Содержание интерферона-гамма (IFNg) и интерлейкина-1-бета (IL-1b) определяют методом твердофазного иммуноферментного анализа (ИФА) с использованием стандартных наборов производства BD Bioscience, США в соответствии с методическими рекомендациями.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОСТИМУЛИРУЮЩИЙ КОМПЛЕКС, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 2010 |
|
RU2446822C2 |
| СПОСОБ ПОЛУЧЕНИЯ НОСИТЕЛЯ АНТИГЕНОВ НА ОСНОВЕ ЛИПИДОВ ИЗ МОРСКИХ МАКРОФИТОВ И ТРИТЕРПЕНОВОГО ГЛИКОЗИДА КУКУМАРИОЗИДА | 2005 |
|
RU2319506C2 |
| НОСИТЕЛЬ АНТИГЕНОВ | 2006 |
|
RU2322259C1 |
| СПОСОБ РЕГУЛЯЦИИ ИММУНОГЕННОСТИ АНТИГЕНА | 2010 |
|
RU2440141C1 |
| СПОСОБ УСИЛЕНИЯ ИММУННОГО ОТВЕТА | 2010 |
|
RU2442604C1 |
| Вакцина и способ повышения иммуногенности и специфической активности средств профилактики инфекций | 2020 |
|
RU2741834C1 |
| СПОСОБЫ ПОЛУЧЕНИЯ АДЪЮВАНТНЫХ ВИРОСОМ И АДЪЮВАНТНЫЕ ВИРОСОМЫ, ПОЛУЧАЕМЫЕ УКАЗАННЫМИ СПОСОБАМИ | 2014 |
|
RU2694367C2 |
| СПОСОБ ИММУНОПРОФИЛАКТИКИ ЭКСПЕРИМЕНТАЛЬНОГО МЕЛИОИДОЗА ИНКАПСУЛИРОВАННЫМИ АНТИГЕНАМИ Burkholderia pseudomallei | 2008 |
|
RU2373955C1 |
| АДЪЮВАНТНАЯ КОМПОЗИЦИЯ ДЛЯ ИНЪЕКЦИОННЫХ ВАКЦИН ПРОТИВ ТКАНЕВЫХ ГЕЛЬМИНТОЗОВ | 2007 |
|
RU2348427C2 |
| СПОСОБ ПОВЫШЕНИЯ ПРОТЕКТИВНОСТИ МЕЛИОИДОЗНЫХ АНТИГЕНОВ ЦИТОКИНАМИ | 2008 |
|
RU2376031C1 |
Изобретение относится к биотехнологии и иммунологии и медицине. Предложен липид-сапониновый иммуностимулирующий комплекс, являющийся носителем для белковых антигенов, который состоит из смеси тритерпенового гликозида кукумариозида А2-2 (КД), холестерина и полярного липида моногалактозилдиацилглицерида (МГДГ) из морских макрофитов, взятых в весовом соотношении 3:2:6. Указанный комплекс представляет собой ультрамикроскопические тубулы с диаметром тубул около 40 нм. Описан также антигенсодержащий липид-сапониновый комплекс, структура которого не отличается от структуры исходного комплекса, состоящий из носителя для белковых антигенов и мономерного порина. Раскрыт способ его получения. Способ включает смешивание раствора холестерина и МГДГ в хлороформе при весовом соотношении холестерина и МГДГ 2:6. Упаривание досуха под вакуумом. Затем добавляют 3 весовые части 0,4 процентного раствора кукумариозида. Солюбилизируют смесь и доводят суммарную концентрацию холестерина и МГДГ до 2 мг на 1 мл фосфатно-солевого буфера. Добавляют раствор мономерного порина в дозах 0,1-10 мкг белка на 1 мкг содержания КД. Полученную суспензию озвучивают ультразвуковым дезинтегратором в течение 5 мин и выдерживают в течение трех часов для полного формирования поринового комплекса. Описано применение указанного комплекса в качестве вакцинного препарата на мыши. Использование изобретения обеспечивает усиление иммуногенности бактериального антигена как в отношении специфического, так и неспецифического иммунного ответа при отсутствии токсических свойств, что может найти применение в медицине в качестве адъюванта. 4 н. и 1 з.п. ф-лы, 19 ил.
| ИММУНОМОДУЛИРУЮЩИЙ АГЕНТ | 1989 |
|
RU2120302C1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| BATRAKOV et al., Triterpene glycosides with antifungal activity isolated from the sea cucumber, Cucumaria japonica, Antibiotiki, 1980, 25(6), реф. | |||

