Изобретение относится к биотехнологии и касается получения нового штамма, используемого для выделения новой сайт-специфической эндонуклеазы, узнающей и расщепляющей обе цепи нуклеотидной последовательности ДНК 5'-G(m5C)↓NG(m5C)-3'.
Эндонуклеаза, обладающая данной специфичностью, может быть использована для сайт-специфического гидролиза ДНК, содержащей С 5-метилцитозиновые основания.
Известен штамм Diplococcus pneumoniae, продуцирующий эндонуклеазу рестрикции DpnI, которая узнает и расщепляет метилированную последовательность нуклеотидов 5'-G(m6A)↓TC-3', где m6А - N6-метиладенин [1].
Недостатком известного штамма является то, что продуцируемая им эндонуклеаза рестрикции не способна узнавать сайты, содержащие С5-метилцитозиновые основания.
Известен штамм Glacial ice bacterium I, продуцирующий эндонуклеазу рестрикции GlaI, которая узнает и расщепляет метилированную нуклеотидную последовательность 5'-G(m5C)↓GC-3', где m5С - С5-метилцитозин [2].
Недостатком известного штамма является то, что продуцируемая им эндонуклеаза рестрикции не способна узнавать метилированную последовательность нуклеотидов 5'-GCNGC-3', содержащую С5-метилцитозиновые основания.
Известен штамм Bacillus simplex 23, продуцирующий сайт-специфическую эндонуклеазу BlsI, которая узнает и расщепляет метилированную нуклеотидную последовательность 5'-GCN↓GC-3', если она содержит хотя бы одно С5-метилцитозиновое основание [3].
Недостатком известного штамма является то, что продуцируемая им сайт-специфическая эндонуклеаза расщепляет метилированную нуклеотидную последовательность 5'-GC↓NGC-3' с любым количеством и расположением С5-метилцитозинов, что в ряде случаев не позволяет использовать ее для получения крупных фрагментов при сайт-специфическом расщеплении ДНК, содержащей С5-метилцитозиновые основания; а также то, что продуцируемая известным штаммом сайт-специфическая эндонуклеаза расщепляет ДНК с образованием однонуклеотидных 3'-выступающих концов и не способна расщеплять ДНК с образованием однонуклеотидных 5'-выступающих концов.
Наиболее близким к заявляемому штамму - прототипом, является штамм Bacillus subtilis 230, продуцирующий эндонуклеазу рестрикции BisI, которая узнает последовательность нуклеотидов 5'-GC↓NGC-3' и расщепляет ее перед центральным нуклеотидом (N) с образованием однонуклеотидных 5'-выступающих концов, если хотя бы одно из цитозиновых оснований узнаваемой последовательности метилировано в положении 5 (С5-метилцитозин) [4].
Недостатком известного штамма является то, что продуцируемая им сайт-специфическая эндонуклеаза расщепляет метилированную нуклеотидную последовательность 5'-GC↓NGC-3' с любым количеством и расположением С5-метилцитозинов, что в ряде случаев не позволяет использовать ее для получения крупных фрагментов при сайт-специфическом расщеплении ДНК, содержащей С5-метилцитозиновые основания.
Технической задачей изобретения является получение бактериального штамма, продуцирующего сайт-специфическую эндонуклеазу, которая узнает и расщепляет обе цепи нуклеотидной последовательности ДНК 5'-G(m5C)↓NG(m5C)-3'.
Поставленная техническая задача достигается получением штамма Glacial ice bacterium 24 - продуцента сайт-специфической эндонуклеазы, узнающей и расщепляющей метилированную последовательность нуклеотидов:
5'-G(m5C)↓N G(m5C)-3'
3'-(m5C)GN↓(m5C)G-5',
где m5C - С5-метилцитозин. (Стрелками указаны позиции расщепления ДНК).
Таксономическая принадлежность заявляемого штамма была определена на основе анализа первичной последовательности 16S рибосомной РНК [5]. Данный вид бактерий относится к классу Actinobacteria, порядку Actinomycetales, семейству Microbacteriaceae, к группе неклассифицированных Microbacteriaceae.
Предлагаемый штамм выделен из природного материала (почвы) в результате целенаправленного систематического поиска.
Полученный штамм Glacial ice bacterium 24 депонирован в Коллекции культур микроорганизмов НПО "СибЭнзим" под регистрационным номером 6М74, а продуцируемая им сайт-специфическая эндонуклеаза названа GluI.
Штамм Glacial ice bacterium 24 характеризуется следующими признаками:
Культурально-морфологические признаки. На агаризованной среде Луриа-Бертрани (ЛБ) образует бледно-желтые, гладкие, блестящие, полупрозрачные, выпуклые, круглые колонии 1-2 мм в диаметре. В жидкой питательной среде со встряхиванием образует гомогенную муть. Клетки палочковидные, размером 0,4×1 мкм, одиночные и в небольших группах располагаются палисадно. Неподвижные. Спор не образуют. Грамположительные.
Физиолого-биохимические признаки. Облигатно-аэробные. Каталазоположительные. Оксидазоотрицательные. Растут при температуре 4-30°С, при 0,5-1,5% NaCl. Содержание гуанина и цитозина в ДНК штамма составляет 65% (определяли при помощи рестриктаз по методу [6]).
Хранение штамма осуществляется в лиофильно-высушенном состоянии или в растворе 30% глицерина при температуре -60°С.
Для культивирования штамма применяют среду следующего состава (г/л): пептон - 10, дрожжевой экстракт - 5, NaCl - 5. Культивирование проводят при 25-28°С с аэрацией до достижения стационарной стадии роста.
Выход целевого фермента составляет 200 ед/г сырой биомассы с удельной активностью 1000 ед/мл.
Полученная сайт-специфическая эндонуклеаза GluI характеризуется следующими свойствами:
1. Узнает и расщепляет метилированную последовательность нуклеотидов 5'-G(m5C)↓NG(m5C)-3', где m5С - С5-метилцитозин.
2. Расщепляет связи между первым цитозиновым основанием (С) и следующим за ним нуклеотидом (N) в обеих цепях узнаваемой последовательности.
3. Не расщепляет вышеприведенную последовательность, не содержащую С5-метилцитозиновых оснований (неметилированную).
4. Оптимальная температура действия 33-40°С.
5. Оптимальное значение рН для действия фермента 7,5-8,5.
6. Оптимальная концентрация соли при расщеплении ДНК 100-150 мМ NaCl.
7. Для проявления активности GluI требуются ионы Mg2+, оптимальная концентрация - 5-10 мМ.
Определяющим отличием предлагаемого штамма от штамма Bacillus subtilis 230 является то, что первый продуцирует сайт-специфическую эндонуклеазу, которая узнает и расщепляет обе цепи метилированной нуклеотидной последовательности ДНК 5'-G(m5C)↓NG(m5C)-3', то есть метилирование хотя бы одного цитозинового основания в сайте узнавания не является достаточным условием для расщепления ДНК. Таким образом, сайт-специфическая эндонуклеаза GluI представляет собой новый, не имеющий аналогов фермент.
Поскольку предлагаемый штамм получен впервые и для выделения сайт-специфической эндонуклеазы, узнающей и расщепляющей вышеназванную последовательность нуклеотидов в указанной позиции никогда не использовался, можно сделать вывод о соответствии предлагаемого штамма критериям изобретения «новизна» и «изобретательский уровень».
Изобретение иллюстрируется примерами конкретного выполнения.
Пример 1.
Выращивание штамма и выделение фермента.
Для получения биомассы клетки штамма-продуцента переносят на агаризованную среду ЛБ в чашку Петри и инкубируют в течение ночи при 30°С. Свежевыращенные колонии переносят стерильной бактериологической петлей в колбы, содержащие жидкую питательную среду ЛБ и культивируют на качалках при 25-28°С при перемешивании - 135-140 об/мин, до достижения стационарной фазы роста. Клетки осаждают центрифугированием при 8000 об/мин при 4°С. Выход биомассы составляет 5 г/л среды. Дезинтеграцию клеток, выделение и очистку фермента проводят по известной методике [7].
Пример 2.
Определение специфичности сайт-специфической эндонуклеазы GluI.
Специфичность фермента определяют по картинам специфического расщепления различных ДНК (фигуры 1 и 2). В качестве субстратов для выявления специфичности расщепления используют различные плазмидные ДНК, а также синтетические олигонуклеотидные ДНК-дуплексы, содержащие или не содержащие метилированные основания (С5-метилцитозин). Расщепление ДНК проводят в оптимальных условиях (37°С, реакционный буфер - 10 мМ TrisHCl, pH 8.0, 10 мМ MgCl2, 100 мМ NaCl, 1 мМ DTT) в течение 60 мин. Продукты расщепления плазмидной ДНК разделяют путем электрофореза в 1,2% агарозном геле. Фрагменты ДНК, образующиеся в результате расщепления олигонуклеотидных дуплексов, разделяют путем электрофореза в денатурирующем 20% полиакриламидном геле с 7 М мочевиной. Расщепление субстратов происходит только в случае присутствия в них метилированной последовательности 5'-G(m5C)↓NG(m5C)-3'.
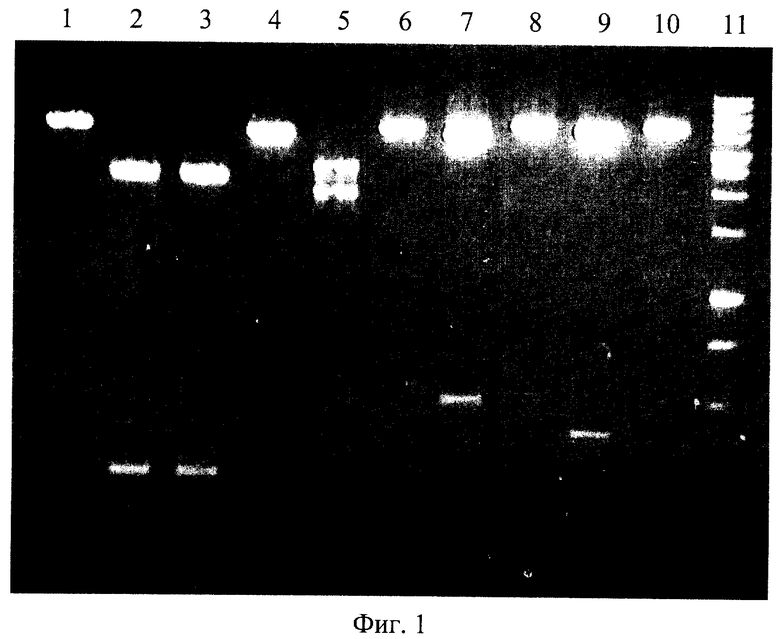
На фигуре 1 представлена электрофореграмма продуктов расщепления ДНК плазмид pFsp4HI2 и pFsp4HI3 эндонуклеазами GluI, BlsI и BisI.
Описание дорожек на электрофореграмме, изображенной на фиг.1:
1 - ДНК pFsp4HI2, линеаризованная ЭР BglII;
2 - ДНК pFsp4HI2, линеаризованная ЭР BglII, обработанная эндонуклеазой рестрикции BlsI;
3 - ДНК pFsp4HI2, линеаризованная ЭР BglII, обработанная сайт-специфической эндонуклеазой BlsI;
4 - ДНК pFsp4HI2, линеаризованная ЭР BglII, обработанная сайт-специфической эндонуклеазой GluI;
5 - ДНК pFsp4HI3, линеаризованная ЭР BglII, обработанная сайт-специфической эндонуклеазой GluI;
6 - ДНК pFsp4HI3, линеаризованная ЭР BglII;
7 - ДНК pFsp4HI3, линеаризованная ЭР DseDI, обработанная сайт-специфической эндонуклеазой GluI;
8 - ДНК pFsp4HI3, линеаризованная ЭР DseDI;
9 - ДНК pFsp4HI3, линеаризованная ЭР Pcil, обработанная сайт-специфической эндонуклеазой GluI;
10 - ДНК pFsp4HI3, линеаризованная ЭР Pcil;
11 - маркер молекулярного веса ДНК 1 kb (производство НПО СибЭнзим).
Плазмида pFsp4HI2 включает в себя ген ДНК-метилтрансферазы M.Fsp4HI, которая метилирует первое цитозиновое основание в последовательности 5'-GCNGC-3'. Таким образом, эта плазмида содержит метилированные последовательности 5'-GCNGC-3'. Из фиг.1 видно, что эндонуклеазы BisI и BlsI расщепляют плазмиду pFsp4HI2 с образованием идентичных фрагментов ДНК (дорожки 2, 3), тогда как эндонуклеаза GluI не расщепляет ДНК этой плазмиды (дорожка 4). Плазмида pFsp4HI3 отличается от pFsp4HI2 четырехнуклеотидной заменой в районе полилинкера, в результате которой образуется последовательность нуклеотидов 5'-GCCGCGGCAGC-3', представляющая собой 3 перекрывающихся сайта узнавания 5'-GCNGC-3'. Плазмида также включает в себя ген ДНК-метилтрансферазы M.Fsp4HI. В результате метилирования приведенной выше последовательности данной метилазой формируется полностью метилированная последовательность нуклеотидов 5'-G(m5C)NG(m5C)-3'/3'-(m5C)GN(m5C)G-5'. Из фиг.1 видно, что эта плазмида расщепляется эндонуклеазой Glul в одном месте (дорожка 5). Локализацию места расщепления ДНК проводили посредством гидролиза ДНК плазмиды сайт-специфической эндонуклеазой Glul совместно с рестриктазами DseDI и PciI (дорожки 7 и 9 соответственно). Длины выщепляемых фрагментов ДНК соответствуют расчетным в случае расщепелния ДНК эндонуклеазой GluI в сайте 5'-G(m5C)↓NG(m5C)-3'.
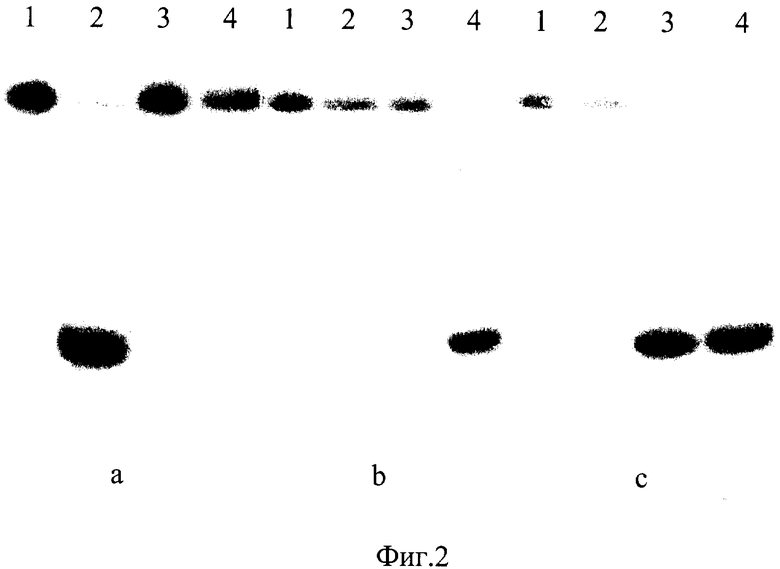
На фиг.2 представлена электрофореграмма продуктов расщепления радиоактивно меченных синтетических олигонуклеотидных дуплексов эндонуклеазами GluI, BlsI и BisI. Последовательности олигонуклеотидов приведены в таблице.
3'
Описание дорожек на электрофореграмме, изображенной на фиг.2:
а
1 - исходный дуплекс NN01*/NN02;
2 - дуплекс NN01*/NN02, расщепленный Fsp4HI;
3 - дуплекс NN01*/NN02 после инкубации с GluI;
4 - дуплекс NN01*/NN02 после инкубации с BisI;
b
1 - исходный дуплекс NN1*/NN2;
2 - дуплекс NN1*/NN2 после инкубации с Fsp4HI;
3 - дуплекс NN1*/NN2 после инкубации с GluI;
4 - дуплекс NN1*/NN2, расщепленный BisI;
с
1 - исходный дуплекс DD1*/DD2;
2 - дуплекс DD1*/DD2 после инкубации с Fsp4HI;
3 - дуплекс DD1*/DD2, расщепленный GluI;
4 - дуплекс DD1*/DD2, расщепленный BisI.
Олигонуклеотиды, меченные радиоактивной меткой по 5'-концу, помечены знаком *.
Как видно из фиг.2, эндонуклеаза GluI расщепляет дуплекс DD1*/DD2, содержащий полностью метилированную последовательность 5'-GCNGC-3', но не расщепляет дуплекс NN1*/NN2, содержащий ту же последовательность с 2 метилированными цитозиновыми основаниями, и дуплекс NN01*/NN02, который содержит такую же неметилированную последовательность.
Таким образом, эксперименты, проводимые с олигонуклеотидными дуплексами, содержащими и не содержащими метилированные цитозиновые основания, содержащими сайты узнавания GluI, BisI и Fsp4HI, подтверждают вывод о том, что для расщепления ДНК сайт-специфической эндонуклеазой GluI требуется присутствие в ней полностью метилированной нуклеотидной последовательности 5'-G(m5C)NG(m5C)-3'.
Пример 3.
Определение позиций расщепления ДНК сайт-специфической эндонуклеазой GluI
Определение позиций расщепления ДНК сайт-специфической эндонуклеазой GluI осуществляли путем сравнения длин фрагментов, образующихся при расщеплении олигонуклеотидных дуплексов эндонуклеазами GluI и BisI (фиг.3). В качестве маркера длин фрагментов использовали продукты частичного расщепления этих же дуплексов экзонуклеазой III из Е.coli (ExoIII). Последовательности олигонуклеотидов приведены в таблице.
Описание дорожек на электрофореграмме, изображенной на фиг.3:
а:
1 - исходный дуплекс DD1*/DD2;
2 - дуплекс DD1*/DD2, расщепленный эндонуклеазой рестрикции BisI;
3 - дуплекс DD1*/DD2, обработанный экзонуклеазой III из E.coli;
4 - дуплекс DD1*/DD2, расщепленный сайт-специфической эндонуклеазой GluI;
b:
1 - исходный дуплекс DD2*/DD1;
2 - дуплекс DD2*/DD1, расщепленный эндонуклеазой рестрикции BisI;
3 - дуплекс DD2*/DD1, обработанный экзонуклеазой III из E.coli;
4 - дуплекс DD2*/DD1, расщепленный сайт-специфической эндонуклеазой GluI.
Олигонуклеотиды, меченные радиоактивной меткой по 5'-концу, обозначены знаком *.
Из фиг.3 видно, что длины продуктов расщепления ДНК на дорожках 2 и 4 совпадают, что свидетельствует о совпадении места расщепления ДНК эндонуклеазам BisI и Glul. Поскольку известно, что рестриктаза BisI расщепляет связи на обеих цепях ДНК в сайте узнавания между первым цитозиновым основанием и следующим нуклеотидом (N), то из приведенной электрофореграммы следует, что эндонуклеаза GluI также расщепляет ДНК в сайте узнавания после первого цитозинового основания (С) перед центральным нуклеотидом (N), с образованием однонуклеотидных 5'-выступающих концов.
Выход сайт-специфической эндонуклеазы GluI определяют по электрофоретической картине расщепления ДНК плазмиды pFsp4HI3. За единицу активности принимают минимальное количество фермента, необходимое для полного расщепления 1 мкг ДНК линеаризованной плазмиды pFsp4HI3 в течение 1 часа при температуре 37°С в 50 мкл реакционной смеси. Выход фермента составляет 200 ед/г сырой биомассы, удельная активность 1000 ед/мл.
Фермент хранится при - 20°С в буфере, содержащем 50% глицерин, 0,2 М NaCl, 10 мМ трисHCl (рН 7,5), 7 мМ 3-меркаптоэтанол, 0,1 мМ ЭДТА.
Таким образом, получен новый штамм, продуцирующий сайт-специфическую эндонуклеазу Glul, узнающую и расщепляющую обе цепи нуклеотидной последовательности ДНК 5'-G(m5C)↓NG(m5C)-3', с образованием однонуклеотидных 5'-выступающих концов.
Данная сайт-специфическая эндонуклеаза может быть использована для выявления метилированной ДНК.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. Lack, S., Greenberg, B.J. // Biol. Chem. - 1975. - V.250 - P.4060-4066.
2. Чернухин В.А., Наякшина Т.Н., Абдурашитов М.А., Томилова Ю.Э., Мезенцева Н.В., Дедков B.C., Михненкова Н.А., Гончар Д.А., Дегтярев С.Х. // Биотехнология. - 2006 (в печати).
3. Чернухин В.А., Томилова Ю.Э., Чмуж Е.В., Соколова О.О., Дедков B.C., Дегтярев С.Х. // Заявка на выдачу патента РФ №2006130604, кл. С12N 1/21, дата подачи 24.08.2006 г.
4. Дегтярев С.Х., Чмуж Е.В., Абдурашитов М.А., Каширина Ю.Г., Дедков B.C., Томилова Ю.Э., Мезенцева Н.В., Гончар Д.А. // Патент РФ №2270859, кл. С12N 1/21, опубл. 27.02.2006, Бюл.№6.
5. Madden, T.L., Tatusov, R.L., Zhang, J. // Meth. Enzymol. - 1996. - V.266. - P.131-141.
6. Дедков B.C. // Биотехнология. - 2004. - №4. - С.77-82.
7. Bickle T.A., Pirotta V., Imber R. // Nucl. Acids Res. - 1977. - V.4. - P.2561-2572.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ БАКТЕРИЙ Microbacterium testaceum 17B - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ MteI | 2011 |
|
RU2475533C1 |
| ШТАММ БАКТЕРИИ Paracoccus carotinifaciens 3K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PcsI. | 2008 |
|
RU2377294C1 |
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |
| ШТАММ БАКТЕРИИ Kocuria rosea - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ KroI | 2009 |
|
RU2394099C1 |
| ШТАММ БАКТЕРИИ Arthrobacter oxydans - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ AoxI | 2009 |
|
RU2399663C1 |
| Штамм бактерий Micrococcus luteus 805 - продуцент сайт-специфической метилзависимой эндонуклеазы MluVI | 2016 |
|
RU2614262C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SIMPLEX - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ BlsI | 2006 |
|
RU2322494C1 |
| ШТАММ БАКТЕРИЙ Plantibacter flavus 3Kz - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PfsI | 2015 |
|
RU2593723C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Bisi | 2004 |
|
RU2270859C1 |
| ШТАММ БАКТЕРИЙ Glacial ice bacterium I - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Gla I | 2005 |
|
RU2287012C1 |

Изобретение относится к области биотехнологии и генной инженерии и может быть использовано для анализа ДНК. Из почвы выделен штамм Glacial ice bacterium, обеспечивающий получение сайт-специфической эндонуклеазы, узнающей и расщепляющей обе цепи метилированной нуклеотидной последовательности ДНК 5′-G(m5C)↓NG(m5C)-3′. Применение изобретения обеспечивает получение новой специфической эндонуклеазы, специфичной к метилированной последовательности ДНК. 3 ил., 1 табл.
Штамм бактерий Glacial ice bacterium KKM НПО «СибЭнзим» 6М74 - продуцент сайт-специфической эндонуклеазы, узнающей и расщепляющей обе цепи метилированной нуклеотидной последовательности ДНК 5′-G(m5C)↓NG(m5C)-3′ с образованием однонуклеотидных 5′-выступающих концов.
| ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Bisi | 2004 |
|
RU2270859C1 |
| LEUNG D.W., LUI A.C., MERILEES H., MCBRIDE В.С., SMITH M.A restriction enzyme from Fusobacterium nucleatum 4H which recognizes GCNGC | |||
| Nucleic Acids Res | |||
| Дверной замок, автоматически запирающийся на ригель, удерживаемый в крайних своих положениях помощью серии парных, симметрично расположенных цугальт | 1914 |
|
SU1979A1 |
| LUBYS A., JANULAITIS A | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Gene | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |

