Изобретение относится к области биотехнологии и касается получения нового штамма бактерий Micrococcus luteus 805 - продуцента сайт-специфической метилзависимой эндонуклеазы MluVI, которая может быть использована для крупноблочного сайт-специфического гидролиза ДНК, содержащей С5-метилцитозиновые основания.
На данный момент описано несколько сайт-специфических метилзависимых ДНК-эндонуклеаз (MD-эндонуклеаз), узнающих и расщепляющих ДНК, только при наличии в сайте узнавания 5-метилцитозина.
Например, описан штамм бактерий Glacial ice bacterium I, являющийся продуцентом MD-эндонуклеазы Glal, которая узнает и расщепляет метилированную последовательность нуклеотидов 5'-R(5mC)GY-3', где 5mC - означает 5-метилцитозин, R - пурин, a Y - пиримидин [1, 2].
Известны штаммы бактерий Bacillus simplex 23 [3], Bacillus subtilis 230 [4] и Glacial ice bacterium 24 [5], продуцирующие сайт-специфические эндонуклеазы Blsl, BisI и Glul, соответственно, которые узнают и расщепляют метилированную последовательность 5'-GCNGC-3', при наличии в ней от двух до четырех 5-метилцитозиновых оснований.
Недостатком вышеперечисленных штаммов является то, что продуцируемые ими MD-эндонуклеазы не способны узнавать и расщеплять обе цепи нуклеотидной последовательности ДНК 5'-GCGCNGCGC-373'-CGCGNCGCG-5' имеющей только шесть, семь или восемь 5-метилцитозинов.
Наиболее близким к заявляемому штамму - прототипом, является штамм бактерий Microbacterium testaceum 17В, продуцирующий сайт-специфическую метилзависимую эндонуклеазу MteI, которая узнает метилированную последовательность 5'-GCGCNGCGC-3'/3'-CGCGNCGCG-5' и расщепляет ее перед центральным нуклеотидом (N) с образованием однонуклеотидного 5'-выступающего конца при условии С5-метилирования в ней от пяти до восьми цитозинов [6].
Недостатком штамма-прототипа является то, что продуцируемая им сайт-специфическая эндонуклеаза расщепляет вышеуказанную последовательность, в которой метилированы не менее пяти цитозинов, что в ряде случаев не позволяет использовать ее для получения крупных фрагментов при сайт-специфическом расщеплении ДНК, содержащей С5-метилцитозиновые основания.
В настоящее время не описаны штаммы бактерий, являющиеся продуцентами сайт-специфических метилзависимых ДНК-эндонуклеаз, узнающих и расщепляющих только гиперметилированные последовательности ДНК 5'-GCGCNGCGC-3' при наличии в них не менее шести С5-метилцитозинов.
Задачей изобретения является получение бактериального штамма, продуцирующего сайт-специфическую метилзависимую ДНК-эндонуклеазу, способную узнавать и расщеплять обе цепи нуклеотидной последовательности 5'-GCGCNGCGC-3'/3'-CGCGNCGCG-5' только при наличии в ней не менее шести 5-метилцитозиновых оснований.
Поставленная задача решается путем получения штамма Micrococcus luteus 805 - продуцента сайт-специфической метилзависимой ДНК-эндонуклеазы, узнающей и расщепляющей обе цепи нуклеотидной последовательности 5'-GCGCNGCGC-3'/3'-CGCGNCGCG-5' при условии С5-метилирования в ней не менее шести 5-метилцитозиновых оснований.
Техническим результатом изобретения является получение бактериального штамма, являющегося продуцентом метилзависимой ДНК-эндонуклеазы с новой сайт-специфичностью, обладающей способностью узнавать и расщеплять обе цепи нуклеотидной последовательности 5'-GCGCNGCGC-3'/3'-CGCGNCGCG-5' с образованием однонуклеотидных 5'-выступающих концов, при условии С5-метилирования в ней не менее шести цитозиновых оснований.
Предлагаемый штамм выделен из природного источника (почвы) в результате целенаправленного систематического поиска. Полученный штамм Micrococcus luteus 805 депонирован во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) ФГУП ГосНИИГенетика под регистрационным номером ВКПМ В-12410, а продуцируемая им сайт-специфическая эндонуклеаза названа MluVI.
Штамм Micrococcus luteus 805 характеризуется следующими признаками.
Культурально-морфологические признаки.
Грамположительные неподвижные кокки диметром 0,5-1,5 мкм. Могут образовывать тетрады и нерегулярные скопления. Спор не образуют.
На агаризованной среде Луриа-Бертрани (LB) образует лимонно-желтые, гладкие, блестящие, непрозрачные, выпуклые круглые колонии.
Физиолого-биохимические признаки. Облигатно аэробные. Каталазоположительные. Оксидазоположительные. Массовая доля G/C-нуклеотидов составляет 70-75%. Растут при температуре от 4 до 40°C с температурным оптимумом роста 25-37°C, при рН от 6 до 9.
Штамм идентифицирован на основе анализа морфологических и биохимических свойств по определителю [7], а также с помощью анализа первичной структуры фрагмента гена 16S рРНК по программе BLAST [8] как вид бактерии Micrococcus luteus. Продуцируемая заявляемым штаммом сайт-специфическая метилзависимая эндонуклеаза MluVI названа согласно номенклатуре [9].
Хранение штамма осуществляют в лиофильно высушенном состоянии или в пробирках «Eppendorf» с 22% глицерином, 0,5% желатиной, 20 мМ Трис-HCl (рН 8,1), 60 мМ NaCl, 15 мМ MgCl2, 1 мМ CaCl2 при -50°C.
Для культивирования штамма используют питательную среду следующего состава (г/л): триптон - 10, дрожжевой экстракт - 5, NaCl - 5, рН 7,5. Культивирование осуществляют при 30°C с аэрацией до достижения стационарной стадии роста.
Полученная сайт-специфическая MD-эндонуклеаза MluVI характеризуется следующими свойствами:
1. Узнает и расщепляет в ДНК последовательность нуклеотидов 5'-GCGCNGCGC-3', в которой С5-метилированы не менее шести цитозинов.
2. Расщепляет межнуклеотидные связи перед центральным нуклеотидом (N) в обеих цепях узнаваемой последовательности.
3. Не расщепляет вышеприведенную последовательность, не содержащую С5-метилцитозиновых оснований или содержащую менее шести метилированных цитозинов.
4. Оптимальная температура действия фермента 37°C.
5. Оптимальное значение рН для действия фермента 7,5-9,0.
6. Для проявления активности MluVI требуются ионы Mg2+, оптимальная концентрация - 5-10 мМ.
Определяющим отличием предлагаемого штамма от всех известных в настоящее время штаммов-продуцентов сайт-специфических метилзависимых ДНК-эндонуклеаз (MD-эндонуклеаз) является то, что новый штамм продуцирует сайт-специфическую эндонуклеазу, которая узнает и расщепляет обе цепи метилированной нуклеотидной последовательности ДНК 5'-GCGC^NGCGC-3' только при наличии в ней не менее шести 5-метилцитозинов.
Таким образом, MD-эндонуклеаза MluVI представляет собой новый, не имеющий аналогов фермент, который может быть использован для выявления и анализа участков ДНК, содержащих избыточное количество С5-метилцитозинов.
Поскольку предлагаемый штамм получен впервые и для выделения сайт-специфической эндонуклеазы, узнающей и расщепляющей вышеназванную последовательность нуклеотидов в указанной позиции, никогда не использовался, можно сделать вывод о соответствии предлагаемого штамма критериям изобретения «новизна» и «изобретательский уровень».
Изобретение иллюстрируется следующими примерами конкретного выполнения.
Пример 1. Выращивание штамма-продуцента и выделение фермента MluVI.
Для получения колоний штамм Micrococcus luteus 805 высевают на агаризованную среду LB в чашку Петри и выращивают в течение 48 часов при 22-25°C. Для ферментации штамм выращивают при встряхивании в 0,5-литровых колбах, содержащих по 300 мл среды (1% триптон, 0,5% дрожжевой экстракт, 0,5% NaCl, рН 7,5) при 120-140 об/мин в течение 20 часов при 30°C. Клетки осаждают на центрифуге J2-21 («Весктап», США) в роторе JA-10 при 8000 об/мин и хранят при -20°C. С 1 л среды получают 6-8 г биомассы.
Разрушение клеток ультразвуком, выделение и очистку фермента проводят с использованием модификации известной методики [10]. Фермент хранится при - 20°C в буфере, содержащем 10 мМ Трис-HCl (рН 7,5), 0,2 М NaCl, 0,1 мМ ЭДТА, 0,08% Тритон Х-100, 7 мМ 2-меркаптоэтанол, 50% глицерин.
За единицу активности принимают минимальное количество фермента, необходимое для полного расщепления 1 мкг ДНК плазмиды pHspAI10 [6], дополнительно метилированной метилазой Fsp4HI и линеаризованной эндонуклеазой рестрикции Dril, в течение 1 часа при температуре 37°C в 20 мкл реакционной смеси. Выход фермента составляет 1500 ед/г сырой биомассы, концентрация 2000 ед/мл.
Пример 2. Определение специфичности MD-эндонуклеазы MluVI.
Обработку субстратных ДНК MD-эндонуклеазой MluVI проводят в оптимальных условиях (37°C, реакционный буфер - 33 мМ Трис-ацетат, рН 7,9 (при 25°C), 10 мМ магния ацетат, 66 мМ калия ацетат, 1 мМ дитиотреитол) в течение 1 часа. Продукты расщепления ДНК разделяют путем электрофореза в 1% агарозном геле в буфере ТАЕ, содержащем 0,04 М Трис-HCl (рН 8,4 при 25°C), 0,02 М уксусной кислоты, 1 мМ ЭДТА.
В качестве субстратов для определения специфичности фермента используют различные плазмидные ДНК: метилированные и неметилированные (контрольные ДНК, которые не расщепляются MluVI вследствие отсутствия метилированных сайтов узнавания). Для подтверждения возможности сайт-специфического расщепления последовательности 5'-GCGCNGCGC-3' с различными узорами метилирования используют следующий набор сконструированных плазмид, содержащих ген ДНК-метилтрансферазы HspAI, метилирующей первый цитозин в положении С5 на обеих цепях последовательности 5'-GCGC-3' [6]:
1) Плазмида pHspAI4 содержит единственный гиперметилированный участок:
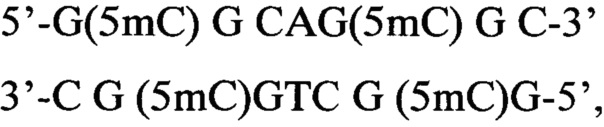
то есть последовательность 5'-GCGCNGCGC-3' с четырьмя 5-метилцитозинами.
2) Плазмида pHspAI10 содержит единственный гиперметилированный участок:
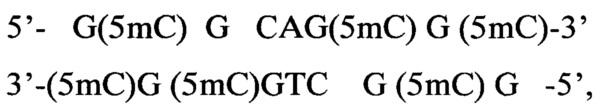
то есть последовательность 5'-GCGCNGCGC-3' с шестью 5-метилцитозинами.
3) Плазмида pHspAI12 содержит единственный гиперметилированный участок:
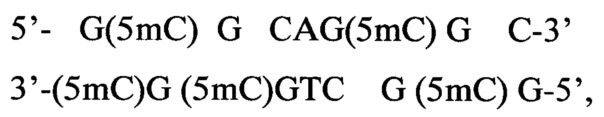
то есть последовательность 5'-GCGCNGCGC-3' с пятью 5-метилцитозинами.
Кроме того, для анализа использовались эти же три плазмиды, pHspAI4, pHspAI10 и pHspAI12, но предварительно метилированые метилазой Fsp4HI по сайту 5'-GCNGC-3', в результате чего в уникальных последовательностях 5'-GCGCNGCGC-3' этих плазмид дополнительно метилировались цитозины перед центральным нуклеотидом на обеих цепях. В результате в последовательностях 5'-GCGCNGCGC-3' этих гиперметилированных плазмид к уже имеющимся 5-метилцитозиновым остаткам добавилось еще по два модифицированных остатка.
Для оценки возможности гидролиза MD-эндонуклеазой MluVI метилированного внутреннего сайта 5'-G(5mC)NGC-3' также использовали плазмиду pFsp4HI3, несущую ген ДНК-метилтрансферазы Fsp4HI [5, 6], метилирующей как раз первый цитозин в последовательности 5'-GCNGC-3'. Поэтому в составе плазмиды pFsp4HI3 встречаются последовательности 5'-GCNGC-3' с двумя, а в случае перекрывания сайтов - с тремя или четырьмя 5-метилцитозинами. Так, в данной плазмиде имеется гиперметилированный участок, содержащий несколько перекрывающихся сайтов узнавания ДНК-метилтрансферазы Fsp4HI:

Следовательно, в плазмиде имеется единственная метилированная последовательность 5'-GCNGC-3' (подчеркнута), в которой все четыре цитозина метилированы, в качестве центрального нуклеотида выступает G/C-пара, и данная последовательность фланкирована с 5'-конца нуклеотидом «С», а с 3'-конца - нуклеотидом «А».
Все используемые плазмиды предварительно были линеаризованы путем расщепления эндонуклеазой рестрикции Dril (сайт узнавания 5'-GACNNN^NNGTC-3').
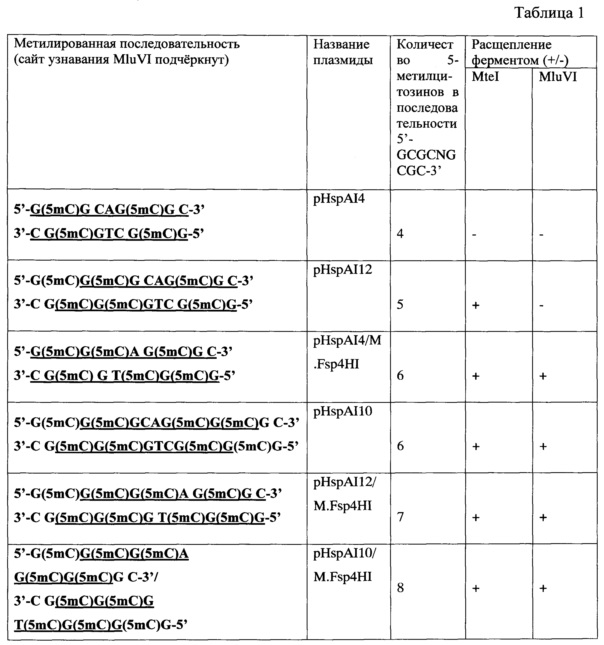
На фиг. 1 представлена электрофореграмма продуктов расщепления MD-эндонуклеазами MluVI и MteI (контроль) ДНК вышеназванных плазмид, в том числе дополнительно метилированных метилазой Fsp4HI.
Описание дорожек на электрофореграмме, изображенной на фиг. 1: 1 - pHspAI4/DriI; 2 - pHspAI4/DriI + MteI; 3 - pHspAI4/DriI + MluVI; 4 - pHspAI10/Dril; 5 - pHspAI1O/Dril + MteI; 6 - pHspAI10/Dril + MluVI; 7 - pHspAI12/DriI; 8 - pHspAI12/DriI + MteI; 9 - pHspAI12/DriI + MluVI; 10 - маркер молекулярного веса ДНК 1 kb; 11 - pHspAI4/DriI + M.Fsp4HI; 12 - pHspAI4/DriI + M.Fsp4HI+MteI; 13 - pHspAI4/DriI + M.Fsp4HI + MluVI; 14 - pHspAI10/Dril + M.Fsp4HI; 15 - pHspAI10/Dril + M.Fsp4HI + MteI; 16 - pHspAI10/Dril + M.Fsp4HI + MluVI; 17 - pHspAI12/DriI + M.Fsp4HI; 18 - pHspAI12/DriI + M.Fsp4HI + MteI; 19 - pHspAI12/DriI + M.Fsp4HI + MluVI; 20 - pFsp4HI3/DriI; 21 - pFsp4HI3/DriI + MluVI; 22 - маркер молекулярного веса ДНК 1 kb.
Как видно из фиг. 1, MD-эндонуклеаза MluVI, подобно MteI, не расщепляет плазмиду, метилированую метилазой HspAI, в которой в последовательности 5'-GCGCNGCGC-3' имеется только четыре 5-метилцитозиновых остатка (pHspAI4, дор. 3). Однако, в отличие от MteI, новый фермент MluVI не расщепляет и плазмиду, в которой в последовательности 5'-GCGCNGCGC-3' присутствует пять 5-метилцитозиновых остатков (pHspAI12, дор. 9).
В то же время MD-эндонуклеаза MluVI расщепляет последовательность 5'-GCGCNGCGC-3' лишь в случаях, если она содержит шесть (pHspAI10, дор. 6 и pHspAI4 + M.Fsp4HI, дор. 13), семь (pHspAI12+M.Fsp4HI, дор. 19) или восемь (pHspAI10 + M.Fsp4HI, дор. 16) 5-метилцитозиновых остатков
Более наглядно эти данные представлены в таблице 1.
Из фиг. 1 также видно, что MD-эндонуклеаза MluVI не расщепляет ДНК плазмиды pFsp4HI3, содержащей последовательности 5'-GCNGC-3' с двумя, тремя и четырьмя С5-метилированными цитозинами. Данный факт свидетельствует о том, что для гидролиза MluVI недостаточно только внутренней метилированной последовательности 5'-GCNGC-3', даже при условии модификации всех цитозинов в ней. Однако данная последовательность является частью сайта узнавания MluVI - 5'-GCGCNGCGC-3'.
Пример 3. Анализ сайт-специфичности и определение позиции гидролиза MluVI на синтетических олигонуклеотидных дуплексах.
Для подтверждения правильности определения узнаваемой новым ферментом последовательности и узора метилирования, а также для определения позиции гидролиза ДНК в сайте узнавания, проводили гидролиз MD-эндонуклеазой MluVI синтетических олигонуклеотидных дуплексов, образованных из олигонуклеотидов следующей структуры:
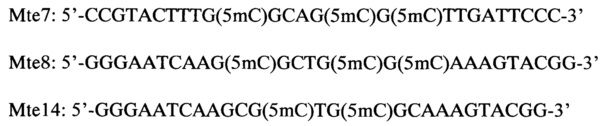
При этом олигонуклеотид Mte7 является комплементарным олигонуклеотидам Mte8 и Mte14, которые, в свою очередь различаются по количеству включенных 5-метилцитозинов (три - в Mte8 и два - в Mte14).
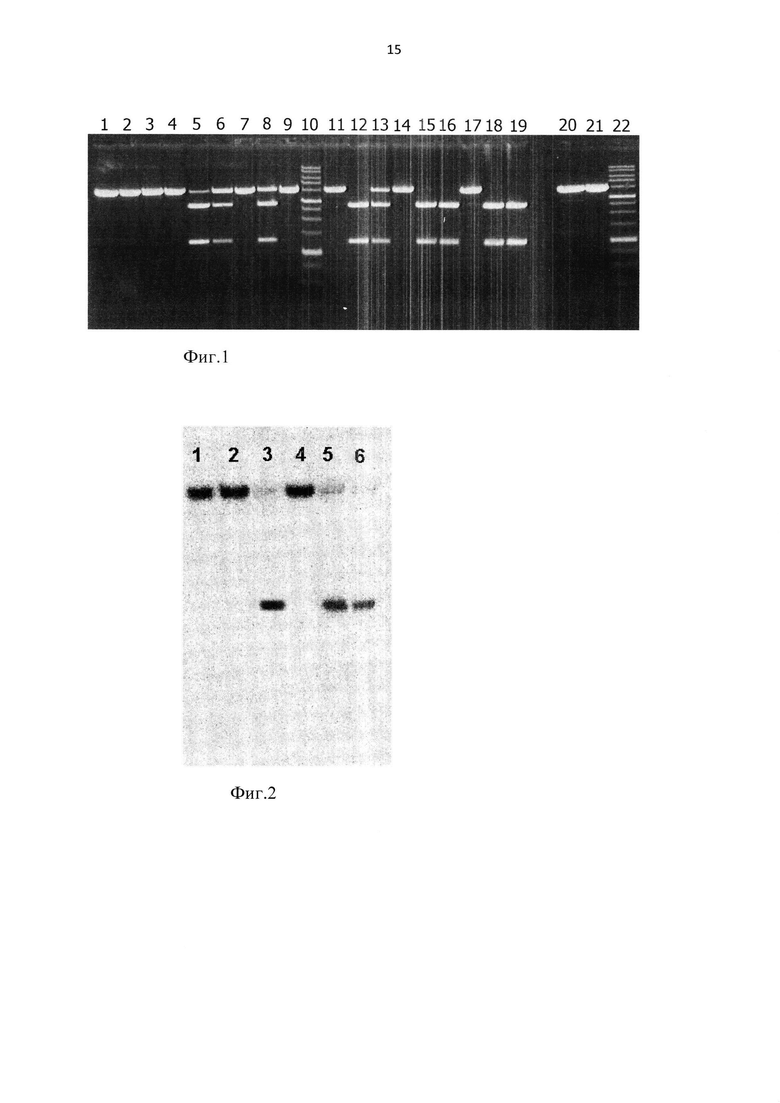
На фиг. 2 приведен радиоавтограф электрофореграммы, полученной в результате расщепления ферментами MluVI или MteI радиоактивно-меченных дуплексов Mte7/Mte14, Mte7/Mte8 и последующего нанесения продуктов на 20% полиакриламидный гель, содержащий 7 М мочевину.
Описание дорожек на электрофореграмме, изображенной на фиг. 2:
1 - исходный дуплекс Mte7*/Mte14; 2 - дуплекс Mte7*/Mte14, обработанный MluVI; 3 - дуплекс Mte7*/Mte14, обработанный MteI; 4 - исходный дуплекс Mte7*/Mte8; 5 - дуплекс Mte7*/Mte8, обработанный MluVI; 6 - дуплекс Mte7*/Mte8, обработанный MteI. Олигонуклеотиды, меченные радиоактивным фосфором Р32 по 5'-концу, обозначены знаком *.
Из фиг. 2 видно, что олигонуклеотидный дуплекс Mte7*/Mte14, содержащий сайт 5'-GCGCNGCGC-3' с пятью 5-метилцитозинами расщепляется MteI (дор. 3), но не расщепляется MluVI (дор. 2).
При этом олигонуклеотидный дуплекс Mte7*/Mte8, содержащий сайт 5'-GCGCNGCGC-3' с шестью 5-метилцитозинами расщепляется как MteI (дор. 6), так и MluVI (дор. 5).
Таким образом, MluVI на олигонуклеотидном дуплексе расщепляет последовательность 5'-GCGCNGCGC-3' с шестью 5-метилцитозинами и не расщепляет эту же последовательность с пятью 5-метилцитозинами.
Из фиг. 2 также видно, что электрофоретические подвижности фрагментов ДНК, образованных в результате гидролиза ферментами MteI и MluVI дуплекса Mte7*/Mte8, совпадают.
Следовательно, можно заключить, что оба фермента имеют одинаковые позиции гидролиза в сайте узнавания, то есть MD-эндонуклеаза MluVI расщепляет обе цепи последовательности узнавания перед центральным нуклеотидом (N), так же, как и MteI, с образованием однонуклеотидных 5'-выступающих концов: 5'-G(5mC)G(5mC)^NG(5mC)G(5mC)-3'/3'-(5mC)G(5mC)GN^(5mC)G(5mC)G-5'.
Использование предлагаемого штамма-продуцента позволит получать новую сайт-специфическую метилзависимую эндонуклеазу MluVI, узнающую и расщепляющую обе цепи нуклеотидной последовательности ДНК 5'-GCGCNGCGC-3', при условии С5-метилирования в ней не менее шести цитозиновых остатков. Данная MD-эндонуклеаза может быть использована в молекулярной биологии, биотехнологии и генной инженерии для крупноблочного сайт-специфического расщепления метилированной ДНК, в частности для выявления и анализа метилированной хромосомной ДНК млекопитающих и человека.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. Чернухин В.А., Наякшина Т.Н., Абдурашитов М.А., Томилова Ю.Э., Мезенцева Н.В., Дедков B.C., Михненкова Н.А., Гончар Д.А., Дегтярев С.Х. // Новая эндонуклеаза рестрикции GlaI узнает метилированную последовательность 5'-GCGC-3'. // Биотехнология. - 2006. - 4. - С. 31-35.
2. G.V. Tarasova, Т.N. Nayakshina, S. Kh. Degtyarev. Substrate specificity of new methyl-directed DNA endonuclease GlaI // BMC Molecular Biology 2008, 9:7.
3. Чернухин В.А., Томилова Ю.Э., Чмуж E.B., Соколова О.О., Дедков B.C., Дегтярев С.Х. Сайт-специфическая эндонуклеаза BIsI узнает последовательности ДНК 5'-G(5mC)NGC-3' и расщепляет ее с образованием 3'-выступающих концов. // Вестник биотехнологии и физико-химической биологии имени Ю.А. Овчинникова. - 2007. - Т. 3. - 1. - С. 28-33.
4. Чмуж Е.В., Каширина Ю.Г., Томилова Ю.Э., Мезенцева Н.В., Дедков B.C., Гончар Д.А., Абдурашитов М.А., Дегтярев С.Х. Новая эндонуклеаза рестрикции BisI из Bacillus subtilis Т30 узнает метилированную последовательность ДНК 5'G(m5C)^NGC-3' // Биотехнология. - 2005. - 3. - C. 22-26.
5. Чернухин В.А., Чмуж Е.В., Томилова Ю.Э., Наякшина Т.Н., Гончар Д.А., Дедков B.C., Дегтярев С.Х. Новая сайт-специфическая эндонклеаза Glul узнает метилированную последовательность ДНК 5'-G(5mC)NG(5mC)-3'/3'-(5mC)GN(5mC)G-5' // Вестник биотехнологии и физико-химической биологии имени Ю.А. Овчинникова. - 2007. - Т. 3. - 2. - С. 13-17.
6. Чернухин В.А., Гончар Д.А., Килева Е.В., Соколова В.А., Голикова Л.Н., Дедков B.C., Михненкова Н.А., Дегтярев С.Х. Новая метилзависимая сайт-специфическая ДНК-эндонуклеаза MteI расщепляет девятинуклеотидную последовательность 5'-G(5mC)G(5mC)NG(5mC)GC-3'/3'-CG(5mC)GN(5mC)G(5mC)G-5'. // Вестник биотехнологии и физико-химической биологии имени Ю.А. Овчинникова. - 2012. - Т. 8. - №1. - С.16-26.
7. Определитель бактерий Берджи. / Под ред. Дж. Хоулта и др.: (9-е издание в 2 томах: Пер. с англ. под ред. акад. РАН Г.А. Заварзина. - М., 1997.
8. Madden, T.L., Tatusov, R.L. & Zhang, J. Applications of network BLAST server. // Meth. Enzymol - 1996. -. V. 266 - P. 131-141.
9. Smith, H.O., Nathans, D. A suggested nomenclature for bacterial host modification and restriction systems and their enzymes. // J. Mol. Biol. - 1973. - Vol. 81. - P. 419-423.
10. Bickle, T.A., Pirrotta, V. and Imber, R. A simple, general procedure for purifying restriction endonucleases. // Nucleic Acids Res. - 1977. - V. 4 - P. 561-2572.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli N42 (pElmI) - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ ElmI | 2015 |
|
RU2597987C1 |
| ШТАММ БАКТЕРИЙ Plantibacter flavus 3Kz - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PfsI | 2015 |
|
RU2593723C1 |
| ШТАММ БАКТЕРИЙ Microbacterium testaceum 17B - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ MteI | 2011 |
|
RU2475533C1 |
| ШТАММ БАКТЕРИИ Paracoccus carotinifaciens 3K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PcsI. | 2008 |
|
RU2377294C1 |
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SIMPLEX - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ BlsI | 2006 |
|
RU2322494C1 |
| ШТАММ БАКТЕРИЙ GLACIAL ICE BACTERIUM - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ GLU I | 2006 |
|
RU2322492C1 |
| ШТАММ БАКТЕРИИ Arthrobacter oxydans - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ AoxI | 2009 |
|
RU2399663C1 |
| ШТАММ БАКТЕРИИ Kocuria rosea - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ KroI | 2009 |
|
RU2394099C1 |
| СПОСОБ САЙТ-СПЕЦИФИЧЕСКОГО ГИДРОЛИЗА ДНК | 2015 |
|
RU2597984C1 |
Изобретение относится к области биотехнологии. Штамм Micrococcus luteus, депонированный под регистрационным номером В-12410 во Всеросиийской Коллекции Промышленных Микроорганизмов ФГУП ГосНИИГенетика, является продуцентом сайт-специфической метилзависимой эндонуклеазы MluVI. Изобретение обеспечивает получение нового фермента MD-эндонуклеазы, который узнает и расщепляет обе цепи последовательности ДНК 5'-GCGC^NGCGC-3'/3'-CGCGN^CGCG-5' перед центральным нуклеотидом (N) с образованием однонуклеотидных 5'-выступающих концов при наличии в данной последовательности не менее шести С5-метилированых цитозинов, и может быть использовано для крупноблочного сайт-специфичного гидролиза ДНК, содержащей С5-метилцитозиновые основания. 2 ил., 1 табл., 3 пр.
Штамм бактерии Micrococcus luteus 805, депонированный под регистрационным номером ВКПМ В-12410, продуцент сайт-специфической метилзависимой ДНК-эндонуклеазы MluVI, узнающей и расщепляющей обе цепи нуклеотидной последовательности 5'-GCGC^NGCGC-3'/3'-CGCGN^CGCG-5' перед центральным нуклеотидом (N), содержащей не менее шести С5-метилированных цитозинов с образованием однонуклеотидных 5'-выступающих концов.
| ШТАММ БАКТЕРИЙ Microbacterium testaceum 17B - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ MteI | 2011 |
|
RU2475533C1 |
| Способ получения биомассы MIcRococcUS LUтеNS ВКПМ В-2836 продуцента рестриктазы MLU 1 | 1989 |
|
SU1738853A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| ЗЕМЛЯНСКАЯ Е.В., ДЕГТЯРЕВ С.Х., Субстратная специфичность и свойства метилзависимых сайт-специфических ДНК-эндонуклеаз.//Молекулярная биология, 2013, т.47, N 6, с.900-913. | |||

