Изобретение относится к биотехнологии и касается получения нового штамма, используемого для выделения новой сайт-специфической эндонуклеазы, узнающей и расщепляющей обе цепи нуклеотидной последовательности ДНК, содержащей хотя бы одно С5-метилцитозиновое основание в сайте узнавания 5′-GCNGC-3′, с образованием однонуклеотидного 3′-выступающего конца.
Эндонуклеаза, обладающая данной специфичностью, может быть использована для сайт-специфического гидролиза ДНК, содержащей С5-метилцитозиновые основания, а также для работ по выявлению и расщеплению метилированных участков ДНК, в частности ДНК эукариотических организмов.
Известен штамм Diplococcus pneumoniae, продуцирующий эндонуклеазу рестрикции DpnI, которая узнает и расщепляет метилированную последовательность нуклеотидов 5′-G(m6A)↓TC-3′, где mA - N6-метиладенин [1].
Недостатком известного штамма является то, что продуцируемая им эндонуклеаза рестрикции не способна узнавать сайты, содержащие С5-метилцитозиновые основания.
Известен штамм Glacial ice bacterium I, продуцирующий эндонуклеазу рестрикции GlaI, которая узнает и расщепляет метилированную нуклеотидную последовательность 5′-G(m5C)↓GC-3′, где m5С - С5-метилцитозин [2].
Недостатком известного штамма является то, что продуцируемая им эндонуклеаза рестрикции не способна узнавать метилированную последовательность нуклеотидов 5′-GCNGC-3′, содержащую С5-метилцитозиновые основания.
Наиболее близким к заявляемому штамму - прототипом является штамм Bacillus subtilis 230, продуцирующий эндонуклеазу рестрикции BisI, которая узнает последовательность нуклеотидов 5′-GCNGC-3′ и расщепляет ее перед центральным нуклеотидом (N) с образованием однонуклеотидного 5′ выступающего конца, если хотя бы одно из цитозиновых оснований узнаваемой последовательности метилировано в положении 5 (С5-метилцитозин) [3].
Недостатком известного штамма является то, что продуцируемая им эндонуклеаза рестрикции плохо расщепляет метилированные участки ДНК эукариотических организмов, в которых метилированы цитозиновые основания в динуклеотидной последовательности 5′-CG-3′, в силу низкой удельной активности фермента; а также то, что продуцируемая известным штаммом эндонуклеаза рестрикции расщепляет ДНК с образованием однонуклеотидного 5′-выступающего конца и не способна расщеплять ДНК с образованием однонуклеотидного 3′-выступающего конца.
Технической задачей изобретения является получение бактериального штамма, продуцирующего сайт-специфическую эндонуклеазу, которая узнает и расщепляет обе цепи нуклеотидной последовательности ДНК, содержащей хотя бы одно С5-метилцитозиновое основание в сайте узнавания 5′-GCNGC-3′.
Поставленная техническая задача достигается получением штамма Bacillus simplex 23 - продуцента сайт-специфической эндонуклеазы, узнающей последовательность нуклеотидов на двуцепочечной ДНК и расщепляющей ее с образованием однонуклеотидных 3′-выступающих концов, при условии, что хотя бы одно из цитозиновых оснований узнаваемой последовательности метилировано в положении 5 (С5-метилцитозин):
5′-GCN↓GC-3′
3′-CG↑NCG-5′
(Стрелками указаны позиции расщепления ДНК).
Предлагаемый штамм выделен из природного материала (почвы) в результате целенаправленного систематического поиска.
Полученный штамм Bacillus simplex 23 депонирован в Коллекции культур микроорганизмов НПО "СибЭнзим" под регистрационным номером 7М07, а продуцируемая им сайт-специфическая эндонуклеаза названа Bisl.
Штамм Bacillus simplex 23 характеризуется следующими признаками:
Культурально-морфологические признаки. На агаризованной среде Луриа-Бертрани (ЛБ) образует белые, гладкие, блестящие, непрозрачные, выпуклые, круглые колонии 4 мм в диаметре. В жидкой питательной среде со встряхиванием образует гомогенную муть. Клетки палочковидные, размером 1×(3-5) мкм, одиночные, в парах или в коротких цепочках. Образуют эллиптические центрально-расположенные споры, не раздувающие спорангий. Грамположительные.
Физиолого-биохимические признаки. Облигатно аэробные. Каталазоположительные. Оксидазоотрицательные. Растут при температуре 10-40°С. Оптимальная температура роста равна 30°С, оптимальный рН среды 7,2-7,5. Штамм не растет на среде, содержащей ампицилин (50 мг/л). Содержание гуанина и цитозина в ДНК штамма составляет 35% (определяли при помощи рестриктаз по методу [4]).
Штамм идентифицирован на основе анализа морфологических и биохимических свойств по определителю [5], а также с помощью анализа первичной последовательности фрагмента 16S рибосомной РНК [6] как вид бактерии Bacillus simplex. Продуцируемая заявляемым штаммом сайт-специфическая эндонуклеаза названа по номенклатуре [7].
Хранение штамма осуществляется в лиофильно высушенном состоянии или в растворе 30% глицерина при температуре -60°С.
Для культивирования штамма применяют среду следующего состава (г/л): пептон - 10, дрожжевой экстракт - 5, NaCl - 5, вода - до 1 л. Культивирование проводят при 30°С с аэрацией до достижения стационарной стадии роста.
Выход целевого фермента составляет 200 ед./г сырой биомассы с удельной активностью 1000 ед./мл.
Полученная сайт-специфическая эндонуклеаза BlsI характеризуется следующими свойствами:
1. Узнает и расщепляет последовательность нуклеотидов 5′-GCNGC-3′, в которой хотя бы одно из цитозиновых оснований метилировано в положении 5 (С5-метилцитозин).
2. Расщепляет связи между центральным нуклеотидом (N) и следующим за ним гуаниновым основанием (G) в обеих цепях узнаваемой последовательности.
3. Не расщепляет вышеприведенную последовательность, не содержащую С5-метилцитозиновых оснований (неметилированную).
4. Эффективно расщепляет метилированные участки ДНК эукариотическиских организмов, содержащие С5-метилцитозиновые основания в динуклеотидной последовательности 5′-CG-3′.
5. Проявляет активность в широком температурном диапазоне 20-50°С.
6. Оптимальное значение рН для действия фермента 7,5-8,5.
7. Оптимальная концентрация соли при расщеплении ДНК 100-150 мМ NaCl.
8. Для проявления активности BlsI требуются ионы Mg2+, оптимальная концентрация - 10-20 мМ.
Определяющим отличием предлагаемого штамма от штамма Bacillus subtilis 230 является то, что первый продуцирует сайт-специфическую эндонуклеазу, которая узнает и расщепляет обе цепи нуклеотидной последовательности ДНК, содержащей хотя бы одно С5-метилцитозиновое основание в сайте узнавания 5′-GCNGC-3′ с образованием однонуклеотидного 3′ выступающего конца, а также в отличие от BisI способна эффективно расщеплять метилированные участки эукариотической ДНК. Таким образом, сайт-специфическая эндонуклеаза BlsI представляет собой новый, не имеющий аналогов фермент.
Поскольку предлагаемый штамм получен впервые и для выделения сайт-специфической эндонуклеазы, узнающей и расщепляющей вышеназванную последовательность нуклеотидов в указанной позиции никогда не использовался, можно сделать вывод о соответствии предлагаемого штамма критериям изобретения «новизна» и «изобретательский уровень».
Изобретение иллюстрируется примерами конкретного выполнения.
Пример 1
Выращивание штамма и выделение фермента
Для получения биомассы клетки штамма-продуцента переносят на агаризованную среду ЛБ в чашку Петри и инкубируют в течение 14-16 часов при 30°С. Свежевыращенные колонии переносят стерильной бактериологической петлей в колбы, содержащие жидкую питательную среду ЛБ и культивируют на качалках при 30°С при перемешивании 150 об/мин до достижения стационарной фазы роста. Клетки осаждают центрифугированием при 5000 об/мин при 4°С. Выход биомассы составляет 5 г/л среды. Дезинтеграцию клеток, выделение и очистку фермента проводят по известной методике [8].
Пример 2
Определение специфичности сайт-специфической эндонуклеазы BlsI
Специфичность фермента определяли по картинам специфического расщепления различных ДНК (фигуры 1 и 2). В качестве субстратов для выявления специфичности расщепления использовали различные плазмидные ДНК, а также синтетические олигонуклеотидные ДНК-дуплексы, содержащие или не содержащие метилированные основания (С5-метилцитозин). Расщепление ДНК проводили в реакционной смеси объемом 20 мкл, в которую добавляли 2 мкл препарата фермента, после чего смесь инкубировали в течение 60 мин. Оптимальными условиями для работы фермента являлись: рестрикционный буфер, содержащий 10 мМ TrisHCl, рН 8.0, 10 мМ MgCl2, 100 мМ NaCl, 1 мМ DTT, температура 37°С. В качестве субстратов использовали 1 мкг плазмидной ДНК или 16 нг синтетического олигонуклеотидного дуплекса. Продукты расщепления плазмидной ДНК разделяли путем электрофореза в 1,5% агарозном геле. Фрагменты ДНК, образующиеся в результате расщепления олигонуклеотидных дуплексов, разделяли путем электрофореза в денатурирующем 20% полиакриламидном геле с 7 М мочевиной. Расщепление субстратов происходит только в случае присутствия в них последовательности 5′-GCNGC-3′, в которой хотя бы одно из цитозиновых оснований модифицировано до С5-метилцитозина.
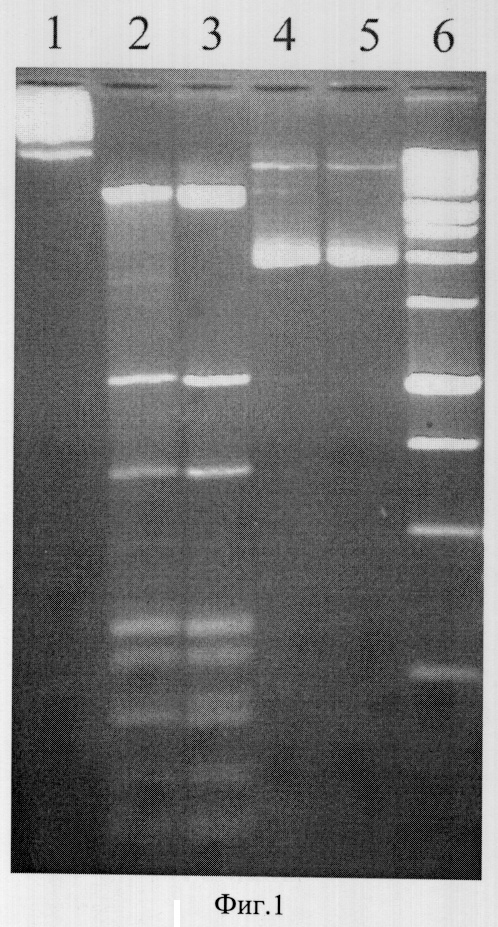
На фигуре 1 представлена электрофореграмма продуктов расщепления ДНК плазмид pUC19 и pFsp4HI1 эндонуклеазами BlsI и BisI.
Описание дорожек на электрофореграмме, изображенной на фиг.1:
1 - ДНК pFsp4HI1;
2 - ДНК pFsp4HI1, обработанная сайт-специфической эндонуклеазой Bisl;
3 - ДНК pFsp4HI1, обработанная эндонуклеазой рестрикции Bisl;
4 - ДНК pUC19;
5 - ДНК pUC19, обработанная сайт-специфической эндонуклеазой Bis1;
6 - маркер молекулярного веса ДНК 1 kb (производство НПО СибЭнзим).
Плазмида pUC19 не содержит метилированных цитозиновых оснований в последовательности 5′-GCNGC-3′. Из фиг.1 видно, что она не расщепляется эндонуклеазой BlsI (дорожка 5). Плазмида pFsp4HI1 включает в себя ген ДНК-метилтрансферазы M.Fsp4HI, которая метилирует первое цитозиновое основание в последовательности 5′-GCNGC-3′. Таким образом, эта плазмида содержит метилированные последовательности 5′-GCNGC-3′. Из фиг.1 также видно, что эндонуклеаза BlsI расщепляет плазмиду pFsp4HI1 (дорожка 2), причем фрагменты ДНК, образовавшиеся в результате гидролиза, идентичны фрагментам, получаемым при расщеплении той же плазмиды рестриктазой BisI (дорожка 3).
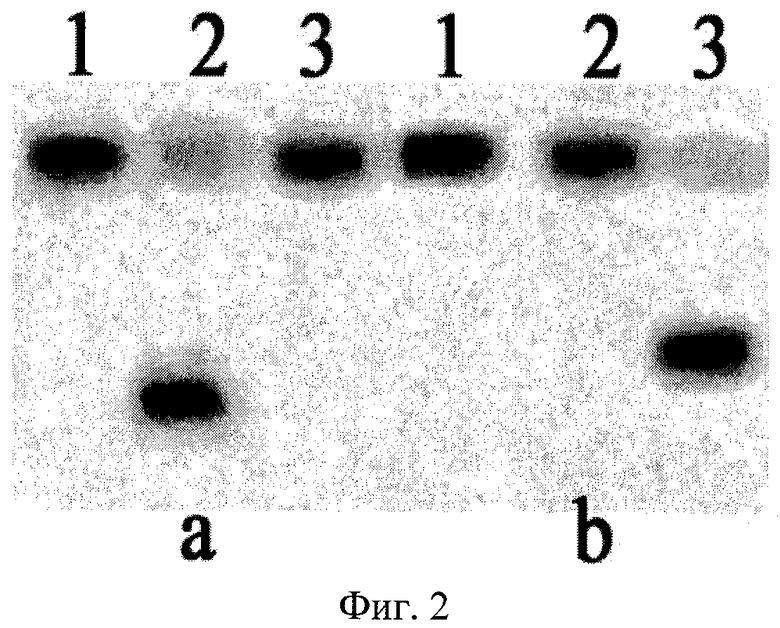
На фиг.2 представлена электрофореграмма продуктов расщепления радиоактивно меченных синтетических олигонуклеотидных дуплексов эндонуклеазами BlsI и BisI. Последовательности олигонуклеотидов приведены в таблице.
Описание дорожек на электрофореграмме, изображенной на фиг.2:
а
1 - исходный дуплекс NN01*/NN02;
2 - дуплекс NN01*/NN02, расщепленный Fsp4HI;
3 - дуплекс NN01*/NN02 после инкубации с BlsI;
b
1 - исходный дуплекс NN1*/NN2;
2 - дуплекс NN1*/NN2 после инкубации с Fsp4HI;
3 - дуплекс NN1*/NN2, расщепленный BlsI.
Олигонуклеотиды, меченные радиоактивной меткой по 5′-концу, помечены знаком *.
Как видно из фиг.2, эндонуклеаза BlsI расщепляет дуплекс NN1*/NN2, содержащий метилированную последовательность 5′-GCNGC-3′, но не расщепляет дуплекс NN01*/NN02, который содержит такую же неметилированную последовательность.
Таким образом, эксперименты, проводимые с олигонуклеотидными дуплексами, содержащими и не содержащими метилированные цитозиновые основания, содержащими сайты узнавания BlsI и Fsp4HI, подтверждают вывод о том, что для расщепления сайт-специфической эндонуклеазой BlsI ДНК требуется присутствие в ней последовательности 5′-GCNGC-3′, содержащей С5-метилцитозиновые основания.
Пример 3
Определение позиций расщепления ДНК сайт-специфической эндонуклеазой BlsI
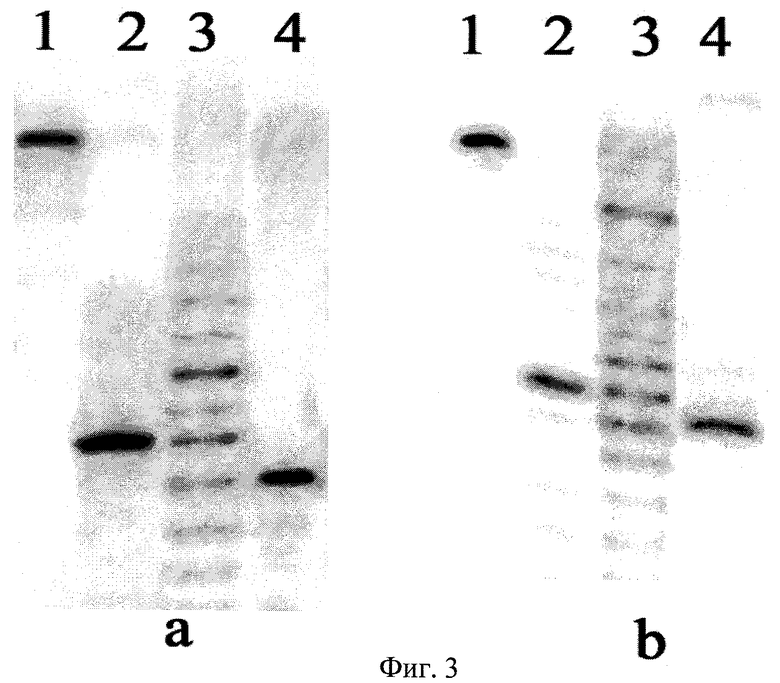
Определение позиций расщепления ДНК сайт-специфической эндонуклеазой BlsI осуществляли путем сравнения длин фрагментов, образующихся при расщеплении олигонуклеотидных дуплексов эндонуклеазами BlsI и BisI (фиг.3). В качестве маркера длин фрагментов использовали продукты частичного расщепления этих же дуплексов экзонуклеазой III из E.coli (ExoIII). Последовательности олигонуклеотидов приведены в таблице.
Описание дорожек на электрофореграмме, изображенной на фиг.3:
а:
1 - исходный дуплекс NN1*/NN2;
2 - дуплекс NN1*/NN2, расщепленный сайт-специфической эндонуклеазой BlsI;
3 - дуплекс NN1*/NN2, обработанный экзонуклеазой III из E.coli;
4 - дуплекс NN1*/NN2, расщепленный эндонуклеазой рестрикции BisI;
b:
1 - исходный дуплекс NN2*/NN1;
2 - дуплекс NN2*/NN1, расщепленный сайт-специфической эндонуклеазой BlsI;
3 - дуплекс NN2*/NN1, обработанный экзонуклеазой III из E.coli;
4 - дуплекс NN2*/NN1, расщепленный эндонуклеазой рестрикции BisI.
Олигонуклеотиды, меченные радиоактивной меткой по 5′-концу, обозначены знаком *.
Из фиг.3 видно, что длины продуктов расщепления ДНК на дорожках 2 и 4 различаются на 1 нуклеотид, причем при расщеплении ДНК эндонуклеазой BlsI образуется более длинный олигонуклеотидный фрагмент, содержащий 5′-меченый конец, что свидетельствует о смещении места расщепления ДНК эндонуклеазой BlsI на 1 нуклеотид в сторону 3′-конца относительно места расщепления ДНК рестриктазой Bisl.
Поскольку известно, что рестриктаза Bisl расщепляет связи на обеих цепях ДНК в сайте узнавания между первым цитозиновым основанием и следующим нуклеотидом (N), то из приведенной электрофореграммы следует, что эндонуклеаза BlsI расщепляет ДНК в сайте узнавания после центрального нуклеотида (N) с образованием однонуклеотидных выступающих 3′-концов.
Пример 4
Расщепление метилированных участков эукариотической ДНК сайт-специфической эндонуклеазой Blsl
В качестве субстрата для расщепления использовали ДНК, выделенные из эукариотических организмов. Для контрольных экспериментов использовали плазмидные ДНК и ДНК фагов, содержащие или не содержащие метилированные основания (С5-метилцитозин) в нуклеотидной последовательности 5′-GCNGC-3′. Расщепление ДНК проводили в оптимальных условиях (37°С, реакционный буфер - 10 мМ TrisHCl, рН 8.0, 10 мМ MgCl2, 100 мМ NaCl, 1 мМ DTT) в течение 3 часов. Продукты расщепления разделяли путем электрофореза в 1% агарозном геле.
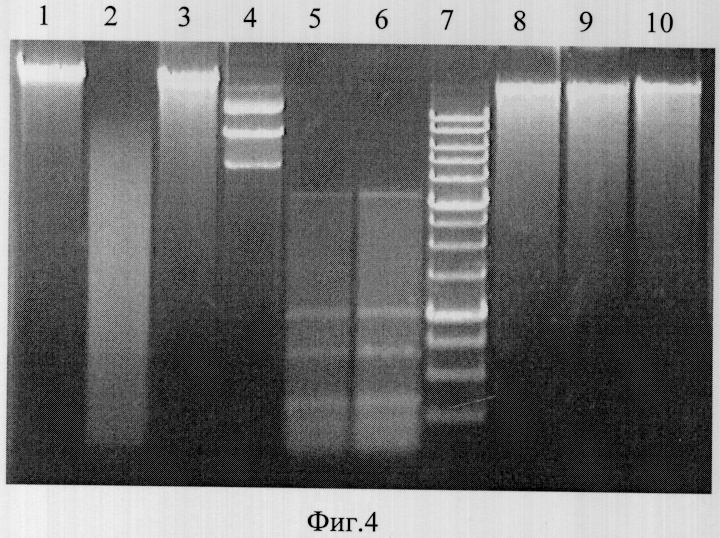
На фиг.4 представлена электрофореграмма продуктов расщепления ДНК из печени крысы линии Wistar, плазмиды pFsp4HI1 и ДНК фага лямбда эндонуклеазами BlsI и BisI.
Описание дорожек на электрофореграмме, изображенной на фиг.4:
1 - ДНК из печени крысы;
2 - ДНК из печени крысы, обработанная сайт-специфической эндонуклеазой BlsI;
3 - ДНК из печени крысы, обработанная эндонуклеазой рестрикции BisI;
4 - ДНК pFsp4HI1;
5 - ДНК pFsp4HI1, обработанная сайт-специфической эндонуклеазой BlsI;
6 - ДНК pFsp4HI1, обработанная эндонуклеазой рестрикции BisI;
7 - маркер молекулярного веса ДНК 1 kb (производство НПО СибЭнзим);
8 - ДНК фага лямбда;
9 - ДНК фага лямбда, обработанная сайт-специфической эндонуклеазой BlsI;
10 - ДНК фага лямбда, обработанная эндонуклеазой рестрикции BisI.
Из фиг.4 видно, что эндонуклеаза BIsI эффективно расщепляет ДНК из печени крысы линии Wistar (дорожка 2), тогда как при обработке этой ДНК эндонуклеазой рестрикции BisI глубокого расщепления субстрата не наблюдается (дорожка 3). При этом оба фермента расщепляют плазмиду pFsp4HI1, содержащую С5-метилцитозиновые основания в сайте 5′-GCNGC-3′, (метилированную) с образованием идентичного набора фрагментов (дорожки 5 и 6) и не расщепляют ДНК фага лямбда, не содержащую С5-метилцитозиновые основания в сайте 5′-GCNGC-3′, (неметилированную) (дорожки 9 и 10).
Выход сайт-специфической эндонуклеазы BlsI определяли по электрофоретической картине расщепления ДНК плазмиды pFsp4HI1. За единицу активности принимали минимальное количество фермента, необходимое для полного расщепления 1 мкг ДНК плазмиды pFsp4HI1 в течение 1 часа при температуре 37°С в 20 мкл реакционной смеси. Выход фермента составляет 200 ед./г сырой биомассы, удельная активность 1000 ед./мл.
Фермент хранится при - 20°С в буфере, содержащем 50% глицерин, 0,2 М NaCl, 10 мМ трисHCl (рН 7,5), 7 мМ β-меркаптоэтанол, 0,1 мМ ЭДТА.
Таким образом получен новый штамм, продуцирующий сайт-специфическую эндонуклеазу BlsI, узнающую и расщепляющую обе цепи нуклеотидной последовательности ДНК, которая содержит хотя бы одно С5-метилцитозиновое основание в сайте узнавания 5′-GCNGC-3′, с образованием однонуклеотидного 3′-выступающего конца, способную эффективно расщеплять метилированные участки эукариотической ДНК, содержащие С5-метилцитозиновые основания в динуклеотидной последовательности 5′-CG-3′.
Данная сайт-специфическая эндонуклеаза может быть использована для выявления метилированной ДНК.
Источники информации
1. Lack, S, Greenberg, В.J. // Biol. Chem. - 1975. - V.250 - P.4060-4066.
2. Чернухин В.А., Наякшина Т.Н., Абдурашитов М.А., Томилова Ю.Э., Мезенцева Н.В., Дедков B.C., Михненкова Н.А., Гончар Д.А., Дегтярев С.Х. // Биотехнология. - 2006 (в печати).
3. Дегтярев С.Х., Чмуж Е.В., Абдурашитов М.А., Каширина Ю.Г., Дедков B.C., Томилова Ю.Э., Мезенцева Н.В., Гончар Д.А. // Патент РФ №2270859, кл. С12N 1/21, опубл. 27.02.2006, Бюл. №6.
4. Дедков B.C. // Биотехнология. - 2004. - №4. - С.77-82.
5. Определитель бактерий Берджи. / Под ред. Дж.Хоулта и др. (9-е издание в 2 томах): Пер. с англ. под ред. акад. РАН Г.А.Заварзина. - М., 1997.
6. Madden, T.L., Tatusov, R.L., Zhang, J. // Meth. Enzymol. - 1996. - V.266. - P.131-141.
7. Smith, H.О., Nathans, D. // J. Mol. Biol. - 1973. - V.81. - P.419-423.
8. Bickle T.A., Pirotta V., Imber R. // Nucl. Acids Res. - 1977. - V.4. - P.2561-2572.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ БАКТЕРИЙ GLACIAL ICE BACTERIUM - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ GLU I | 2006 |
|
RU2322492C1 |
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Bisi | 2004 |
|
RU2270859C1 |
| Штамм бактерий Micrococcus luteus 805 - продуцент сайт-специфической метилзависимой эндонуклеазы MluVI | 2016 |
|
RU2614262C1 |
| ШТАММ БАКТЕРИЙ Microbacterium testaceum 17B - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ MteI | 2011 |
|
RU2475533C1 |
| ШТАММ БАКТЕРИИ Paracoccus carotinifaciens 3K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PcsI. | 2008 |
|
RU2377294C1 |
| ШТАММ БАКТЕРИЙ Plantibacter flavus 3Kz - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PfsI | 2015 |
|
RU2593723C1 |
| ШТАММ БАКТЕРИИ Arthrobacter oxydans - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ AoxI | 2009 |
|
RU2399663C1 |
| ШТАММ БАКТЕРИИ Kocuria rosea - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ KroI | 2009 |
|
RU2394099C1 |
| ШТАММ БАКТЕРИЙ Glacial ice bacterium I - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Gla I | 2005 |
|
RU2287012C1 |
Изобретение относится к биотехнологии и генной инженерии. Предложен штамм Bacillus simplex 23, выделенный из почвы и обеспечивающий получение сайт-специфической эндонуклеазы, узнающей и расщепляющей обе цепи нуклеотидной последовательности ДНК, содержащей хотя бы одно С5-метилцитозиновое основание в сайте узнавания 5′-GCNGC-3′ с образованием однонуклеотидных 3′-выступающих концов. Новый штамм может быть использован для выделения новой сайт-специфической эндонуклеазы, которая может применяться для выявления и расщепления метилированных участков ДНК. 4 ил., 1 табл.
Штамм бактерий Bacillus simplex, депонированный в ККМ НПО «СибЭнзим» под номером 7М07 - продуцент сайт-специфической эндонуклеазы, узнающей и расщепляющей обе цепи нуклеотидной последовательности ДНК, содержащей по меньшей мере одно С5-метилцитозиновое основание в сайте узнавания 5′-GCNGC-3′, с образованием однонуклеотидных 3′-выступающих концов.
| ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Bisi | 2004 |
|
RU2270859C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SCHLEGELII - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ, УЗНАЮЩЕЙ И РАСЩЕПЛЯЮЩЕЙ ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕОТИДОВ 5' - CCNNNNNNN GG-3' | 1994 |
|
RU2073717C1 |
| DNA methylation and epigenetics | |||
| Genetica | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |

