Изобретение относится к биотехнологии и касается получения нового штамма, используемого для выделения новой эндонуклеазы рестрикции Gla I, узнающей и расщепляющей (гидролизующей) последовательность нуклеотидов на двуцепочечной ДИК, при условии, что первое цитозиновое основание узнаваемой последовательности метилировано в положении 5 (С5-метилцитозин, 5mC) на обеих цепях ДНК в последовательности
Вследствие гидролиза образуются тупые концы ДНК.
Эндонуклеаза рестрикции (рестриктаза), обладающая данной специфичностью, может быть использована для работ по выявлению и гидролизу метилированных участков ДНК, в частности ДНК эукариотических организмов, содержащих С5-метилцитозин.
В настоящее время известно небольшое число эндонуклеаз рестрикции, узнающих и гидролизующих только те специфические последовательности ДНК, которые имеют метилированные основания.
Известен штамм Diplococcus pneumoniae, продуцирующий рестриктазу DpnI, которая узнает и гидролизует метилированную нуклеотидную последовательность 5' - G mA Т С - 3', где mA - N6-метиладенин (Lacks S., Greenberg В. J. Biol. Chem., 1975, v.250, p.4060-4066).
Рестриктаза DpnI не способна узнавать сайты, содержащие С5-метилцитозиновые основания.
Наиболее близким к заявляемому штамму - прототипом, является штамм Haemophilus haemolyticus, продуцирующий рестриктазу HhaI, узнающую и расщепляющую последовательность нуклеотидов 5'-GCGC-3', не содержащую метилированных оснований (Roberts R.J., Myers P.A., Morrison A., Murray К. J. Mol. Biol., 1976, v.103, p.199-208).
Недостатком известного штамма является то, что продуцируемая им рестриктаза не способна расщеплять сайты узнавания, содержащие С5-метилцитозиновые основания.
Технической задачей изобретения является выявление бактериального штамма, продуцирующего эндонуклеазу рестрикции, которая узнает и расщепляет обе цепи нуклеотидной последовательности 5'-GCGC-3', обязательно содержащей С5-метилцитозиновые основания.
Поставленная техническая задача достигается получением штамма Glacial ice bacterium I - продуцента эндонуклеазы рестрикции Gla I. Таксономическая принадлежность данного штамма была определена на основе анализа последовательности 16 S рибосомной РНК. Данный вид бактерий относится к классу Actinobacteria, порядку Actinomycetales, семейству Microbacteriaceae, к группе неклассифицированных Microbacteriaceae.
Предлагаемый штамм выделен из природного материала (пресноводный водоем) в результате целенаправленного систематического поиска.
Полученный штамм Glacial ice bacterium I депонирован в Коллекции культур микроорганизмов НПО "СибЭнзим" под регистрационным номером 6М57, а продуцируемая им рестриктаза названа Gla I.
- Штамм Glacial ice bacterium I характеризуется следующими признаками:
Клетки представляют собой грамм-положительные аэробные нерегулярные, мелкие палочки размером 0,5-(1-1,5) мкм, одиночные и образующие палисадные скопления, неподвижные, не образующие спор. На среде Лурия-Бертрани (ЛБ) образует светло-коричневые, полупрозрачные, небольшие (1-2 мм), округлые, с ровными краями колонии. Температура для роста - от 10 до 30°С. Клетки продуцируют каталазу. Оксидаза не обнаружена.
Оптимальная температура выращивания штамма 30°С, рН 7,0-7,2.
Хранение штамма осуществляют в лиофильно высушенном состоянии или в растворе 30% глицерина при температуре - 60°С.
Для культивирования штамма применяют среду следующего состава, г/л: пептон - 10, дрожжевой экстракт - 5, NaCl - 5. Культивирование проводят при 30°С с аэрацией до достижения стационарной стадии роста.
За единицу активности принимали минимальное количество фермента, необходимое для полного расщепления 1 мкг ДНК плазмиды pHspAIM в течение 1 часа при температуре 37°С в 20 мкл реакционной смеси. Выход фермента составляет 10 ед/г сырой биомассы, удельная активность 200 ед/мл.
Фермент хранят при - 20°С в буфере, содержащем 50% глицерин, 0,2 М NaCl, 10 мМ трис-HCl (рН 7,5), 7 мМ β-меркаптоэтанол, 0,1 мМ ЭДТА.
Полученная эндонуклеаза рестрикции Gla I характеризуется следующими свойствами:
1. Узнает и гидролизует связи между первым цитозиновым остатком (С) и следующим за ним гуанином в обеих комплементарных цепях последовательности нуклеотидов 5' - G С G С - 3', в которой первое цитозиновое основание на каждой цепи ДНК узнаваемой последовательности метилировано в положении 5:
2. Не расщепляет эту же последовательность, не содержащую С5-метилцитозиновых оснований (неметилированную);
3. Фермент проявляет активность в широком температурном диапазоне: 27-50°С;
4. Оптимальное значение рН активности фермента 7,5-8,5;
5. Для проявления активности Gla I требуются ионы Mg2+, оптимальная концентрация - 10 мМ.
Определяющим отличием предлагаемого штамма от штамма Haemophilus haemolyticus является то, что первый продуцирует одну рестриктазу, которая узнает и расщепляет обе цепи нуклеотидной последовательности 5'-GCGC-3', обязательно содержащей С5-метилцитозиновые основания.
Поскольку предлагаемый штамм получен впервые и для выделения рестриктазы, узнающей и гидролизующей вышеназванную последовательность нуклеотидов, никогда не использовался, можно сделать вывод о соответствии предлагаемого штамма критериям изобретения "новизна" и "изобретательский уровень".
Изобретение иллюстрируется примерами конкретного выполнения.
Пример 1
Выращивание штамма и выделение фермента.
Культуру хранят в 30%-м растворе глицерина при -60°С. Для получения биомассы клетки штамма-продуцента переносят на агаризованную среду ЛБ в чашку Петри и инкубируют в течение ночи при 30°С. Свежевыращенные колонии переносят стерильной бактериологической петлей в колбы, содержащие жидкую питательную среду ЛБ, и культивируют на качалках при 30°С при перемешивании - 150 об/мин, до достижения стационарной фазы роста. Клетки осаждают центрифугированием при 5000 об/мин при 4°С. Выход биомассы составляет 5 г/л среды. Дезинтеграцию, выделение и очистку фермента проводят по известной методике (Bickle T.A., Pirotta V., Imber R. Nucl.Fcids Res., 1977, v.4, p.2561-2572).
Пример 2
Определение специфичности эндонуклеазы рестрикции Gla I
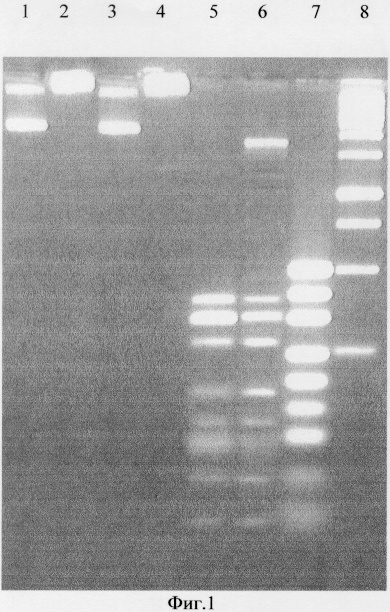
Специфичность фермента определяют по картинам специфического расщепления различных ДНК (фиг.1). В качестве субстратов для выявления специфичности расщепления использовали различные плазмидные ДНК, а также синтетические олигонуклеотидные ДНК-дуплексы, содержащие или не содержащие метилированные основания (С5-метилцитозин или А6-метиладенин). После инкубации в оптимальных условиях в течение 30 мин продукты реакции разделяли путем электрофореза в 2% агарозном геле. Расщепление субстратов происходит только в случае присутствия в них последовательности 5' - G С G С - 3', в которой первое цитозиновое основание модифицировано в С5-метилцитозин в обоих комплиментарных цепях ДНК сайта узнавания.
На фиг.1 представлена электрофореграмма продуктов расщепления ДНК плазмид pUC19 и pHspAIM эндонуклеазами рестрикции Gla I и HspAI Плазмида pHspAIM получена в результате клонирования фрагмента геномной ДНК штамма бактерии Haemophilus species AI длиной 1432 п.о. в линеаризованную по сайту рестрикции EcoRI плазмиду pUC19. Клонированный фрагмент ДНК содержит ген ДНК-метилтрансферазы HspAI, которая метилирует первый цитозин в обеих цепях палиндромной последовательности 5'-GCGC-3' в положении С5.
Описание дорожек на электрофореграмме, изображенной на фиг.1:
1, 2 - ДНК pUC19 и pHspAI соответственно;
3 - ДНК pUC19, обработанные эндонуклеазой рестрикции Gla I;
4 - ДНК pHspAIM, обработанная эндонуклеазой рестрикции HspAI;
5 - ДНК pUC19, обработанная эндонуклеазой рестрикции HspAI;
6 - ДНК pHspAI, обработанная эндонуклеазой рестрикции Gla I;
7 - маркер молекулярного веса ДНК pUC19/Msp I (производство НПО "СибЭнзим"),
8 - маркер молекулярного веса ДНК 1 kb (производство НПО "СибЭнзим").
Поскольку плазмида pUC19 не содержит метилированных оснований в последовательности 5'-GCGC-3', она гидролизуется рестриктазой HspAI и не гидролизуется рестриктазой Gla I. Плазмида pHspAIM, несущая ген ДНК-метилтрансферазы HspAI, содержит метилированную по первому цитозину последовательность 5' - G (mC) G С - 3'. Это приводит к тому, что ДНК pHspAIM гидролизуется рестриктазой Gla I, но не гидролизуется эндонуклеазой рестрикции HspAI. Длины фрагментов ДНК, образуемые при гидролизе Gla I плазмиды pHspAIM соответствуют гидролизу по сайтам 5'-G(mC)GC-3'. Как видно из фиг.1, идентичные части плазмид pUC19 и pHspAIM образуют фрагменты одинаковой длины при гидролизе эндонуклеазами рестрикции HspAI и Gla I соответственно.
Пример 3
Определение позиций гидролиза ДНК эндонуклеазой рестрикции Gla I
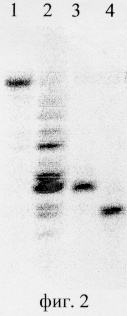
Определение места гидролиза ДНК рестриктазой Gla I осуществляли путем сравнения длины фрагментов, образуемых при расщеплении олигонуклеотидных дуплексов эндонуклеазами рестрикции Gla I и HspAI (фиг.2). Последовательности олигонуклеотидов приведены в таблице.
Первичная структура олигонуклеотидов, используемых для определения места гидролиза рестриктазой Gla I.
На фиг.2 изображен радиоавтограф электрофореграммы продуктов расщепления дезоксирибоолигонуклеотидных радиоактивно меченых дуплексов, в 20% полиакриламидном геле, содержащем 8-молярную мочевину.
Описание дорожек на электрофореграмме, изображенной на фиг.2:
1 - исходный дуплекс 1*/2;
2 - дуплекс 1*/2, обработанный экзонуклеазой ExoIII
3 - дуплекс 1*/2, расщеленный рестриктазой Gla I;
4 - дуплекс 3*/4, расщепленный рестриктазой HspAI
В качестве маркера длин фрагментов использовали продукты частичного расщепления этих же дуплексов экзонуклеазой ExoIII (дорожка 2).
Из фиг.2 видно, что продукты гидролиза ДНК на дорожках 3 и 4 имеют разные длины, отличающиеся на один нуклеотид. Поскольку известно, что рестриктаза HspAI расщепляет связи между первым гуаниновым и первым цитозиновым остатком в сайте узнавания на обеих цепях, то из приведенной электрофореграммы следует, что эндонуклеаза рестрикции Gla I расщепляет ДНК в сайте узнавания между первым цитозиновым и вторым гуаниновым остатком, образуя у гидролизованных продуктов тупые концы.
Пример 4
Идентификация штамма с помощью анализа первичной последовательности 16S рибосомной РНК.
Проведено секвенирование большей части фрагмента 16S рибосомной РНК, полученного с помощью полимеразной цепной реакции. Из примерно 1540 нуклеотидов 16S рибосомной РНК была определена первичная последовательность фрагмента длиной 1417 нуклеотидов (фиг.3). Используя программу BLAST, было установлено, что наибольшую гомологию данная последовательность имеет с фрагментом 16S рибосомной РНК штамма Glacial ice bacterium G50-TB8 (Christner, B.C. Thesis, 2002, Ohio State University, 484 W. 12th Avenue, Columbus, USA). Из 1417 нуклеотидов обнаружено несовпадение только по одному нуклеотиду. Данный штамм относится к порядку Actinomycetales, семейству Microbacteriaceae. Из известных родов семейства Microbacteriaceae наибольшую гомологию отсеквенированная нами последовательность имеет к последовательностям 16S рибосомной РНК представителей рода Mycetocola (Tsukamoto Т., Takeuchi M., Shida О., Murata H., Shirata A. Int. J. Syst. Evol. Microbiol., 2001, v.51, p.937-944).
Таким образом, получен новый штамм, продуцирующий эндонуклеазу рестрикции Gla I, узнающую и гидролизующую обе цепи нуклеотидной последовательности, обязательно содержащей С5-метилцитозиновые основания в двуцепочечной последовательности ДНК 5'-GCGC-3'. Данная рестриктаза может быть использована для выявления метилированной ДНК.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |
| ШТАММ БАКТЕРИЙ GLACIAL ICE BACTERIUM - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ GLU I | 2006 |
|
RU2322492C1 |
| Штамм бактерий Micrococcus luteus 805 - продуцент сайт-специфической метилзависимой эндонуклеазы MluVI | 2016 |
|
RU2614262C1 |
| ШТАММ БАКТЕРИЙ Microbacterium testaceum 17B - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ MteI | 2011 |
|
RU2475533C1 |
| ШТАММ БАКТЕРИИ Arthrobacter oxydans - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ AoxI | 2009 |
|
RU2399663C1 |
| ШТАММ БАКТЕРИИ Paracoccus carotinifaciens 3K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PcsI. | 2008 |
|
RU2377294C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SIMPLEX - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ BlsI | 2006 |
|
RU2322494C1 |
| ШТАММ БАКТЕРИЙ Plantibacter flavus 3Kz - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PfsI | 2015 |
|
RU2593723C1 |
| ШТАММ БАКТЕРИИ Kocuria rosea - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ KroI | 2009 |
|
RU2394099C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Bisi | 2004 |
|
RU2270859C1 |

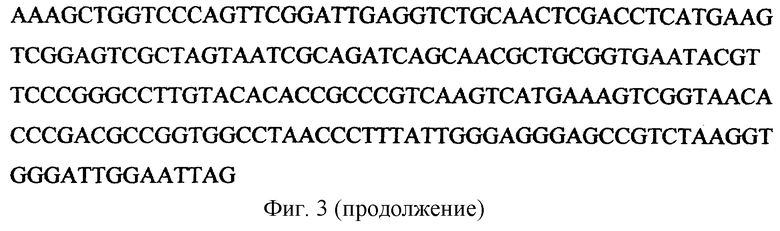
Изобретение относится к биотехнологии и генной инженерии и касается получения нового штамма, используемого для выделения новой эндонуклеазы рестрикции Gla I. Такая эндонуклеаза может быть использована для выявления метилированной ДНК. Штамм Glacial ice bacterium I выделен из водоема в результате поиска продуцентов рестриктаз и обеспечивает получение эндонуклеазы рестрикции Gla I, узнающей и расщепляющей обе цепи нуклеотидной последовательности ДНК, содержащей С5-метилцитозиновые основания в сайте узнавания 5'-GCGC-3'. Заявленное изобретение позволяет получить выход фермента 10 ед/г сырой биомассы, при удельной активности целевого фермента 200 ед/мл. 3 ил., 1 табл.
Штамм бактерий Glacial ice bacterium I KKM НПО "СибЭнзим" 6M57 - продуцент эндонуклеазы рестрикции, узнающей и расщепляющей обе цепи нуклеотидной последовательности 5'-GCGC-3', содержащей С5-метилцитозиновые основания.
| ROBERTS RJ et.al | |||
| A specific endonuclease from Haemophilus haemolyticus | |||
| J Mol Biol., 1976 May 5; 103(1), p.199-208 | |||
| LACKS S et.al | |||
| A deoxyribonuclease of Diplococcus pneumoniae specific for methylated DNA., J Biol Chem | |||
| Сплав для отливки колец для сальниковых набивок | 1922 |
|
SU1975A1 |
| ШТАММ БАКТЕРИИ BACILLUS STEAROTHERMOPHILUS - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ, УЗНАЮЩЕЙ И РАСЩЕПЛЯЮЩЕЙ ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕОТИДОВ 5'-GGTNACC-3' | 1996 |
|
RU2115728C1 |
| Штамм бактерий BacILLUS SрнаеRIсUS-продуцент рестриктазы BSI 1 | 1991 |
|
SU1784642A1 |
| Штамм бактерий BacILLUS SUвFILIS - продуцент эндонуклеазы рестрикции BSU 15 I | 1987 |
|
SU1449583A1 |
| ШТАММ БАКТЕРИЙ BACILLUS SCHLEGELII - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ, УЗНАЮЩЕЙ И РАСЩЕПЛЯЮЩЕЙ ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕОТИДОВ 5' - CCNNNNNNN GG-3' | 1994 |
|
RU2073717C1 |

