Изобретение относится к биотехнологии и касается получения нового штамма, используемого для выделения новой сайт-специфической эндонуклеазы, узнающей и расщепляющей обе цепи метилированной нуклеотидной последовательности ДНК 5'-^GG (m5C)C-3', где m5C - 5-метилцитозин.
Эндонуклеаза, обладающая данной специфичностью, может быть использована для сайт-специфического гидролиза ДНК, содержащей С5-метилцитозиновые основания.
К настоящему времени известны несколько сайт-специфических эндонуклеаз, узнающих и расщепляющих ДНК, только при наличии в сайте узнавания 5-метилцитозина.
Известен штамм Glacial ice bacterium I, продуцирующий эндонуклеазу GlaI, которая узнает и расщепляет метилированную нуклеотидную последовательность 5'-G(m5C)↓GC-3', где m5C - 5-метилцитозин [1].
Известны штаммы Bacillus simplex 23 [2], Bacillus subtilis 230 [3] и Glacial ice bacterium 24 [4], продуцирующие сайт-специфические эндонуклеазы BlsI, BisI и Glul, которые узнают и расщепляют метилированную нуклеотидную последовательность 5'-GCNGC-3', при наличии в сайте узнавания 5-метилцитозиновых оснований.
Недостатком известных штаммов является то, что продуцируемые ими эндонуклеазы рестрикции не способны узнавать и расщеплять обе цепи нуклеотидной последовательности ДНК 5'-^GG (m5C)C-3'/3'-C(m5C)GG^-5'.
Наиболее близким к заявляемому штамму - прототипом является штамм Haemophilus aegyptius, продуцирующий рестриктазу HaeIII, которая узнает последовательность нуклеотидов 5'-GGCC-3' и расщепляет ее перед цитозином с образованием тупых концов [5]. Недостатком известного штамма является то, что продуцируемая им сайт-специфическая эндонуклеаза не расщепляет узнаваемую последовательность, если центральные цитозины метилированы в положении С5.
Однако в настоящее время не описаны штаммы бактерий, являющихся продуцентами сайт-специфических эндонуклеаз и узнающих ДНК 5'-GGCC-3' только при наличии в ней С5-метилцитозинов.
Технической задачей изобретения является получение бактериального штамма, продуцирующего сайт-специфическую эндонуклеазу, которая узнает и расщепляет обе цепи нуклеотидной последовательности ДНК 5'-^GG (m5C)C-3'/3'-C(m5C)GG^-5'.
Поставленная техническая задача решается путем получения штамма Arthrobacter oxydans 25K - продуцента сайт-специфической эндонуклеазы, узнающей и расщепляющей метилированную последовательность нуклеотидов:
5'-↓G G(m5C)C-3'
3'-С(m5C) GG↑-5',
где m5C - 5-метилцитозин (стрелками указаны позиции расщепления ДНК).
Предлагаемый штамм выделен из природного материала (почвы) в результате целенаправленного систематического поиска.
Полученный штамм Arthrobacter oxydans 25K депонирован в Коллекции культур микроорганизмов НПО "СибЭнзим" под регистрационным номером 7М89, а продуцируемая им сайт-специфическая эндонуклеаза названа Aoxl.
Штамм Arthrobacter oxydans 25K характеризуется следующими признаками:
Культурально-морфологические признаки. На агаризованной среде Луриа-Бертрани (ЛБ) образует оранжево-красные, гладкие, блестящие, полупрозрачные, выпуклые, круглые колонии 1-2 мм в диаметре. В жидкой питательной среде со встряхиванием образует гомогенную муть. Клетки палочковидные, размером 0,3-0,7 мкм в диаметре и 1-3 мкм в длину. Спор не образуют. Грамположительные.
Физиолого-биохимические признаки. Облигатно аэробные. Каталазоположительные. Растут при температуре от 4 до 33°С, при pH от 6 до 9.
Штамм идентифицирован на основе анализа морфологических и биохимических свойств по определителю [6], а также с помощью анализа первичной структуры фрагмента 16S РНК по программе BLAST [7] как вид бактерии Arthrobacter oxydans 25K. Продуцируемая заявляемым штаммом сайт-специфическая эндонуклеаза Aoxl названа согласно номенклатуре [8].
Хранение штамма осуществляется в лиофильно высушенном состоянии или в растворе 30% глицерина при температуре - 60°С.
Для культивирования штамма применяют среду следующего состава (г/л): пептон - 10, дрожжевой экстракт - 5, NaCl - 5. Культивирование проводят при 28°С с аэрацией до достижения стационарной стадии роста.
Выход целевого фермента составляет 100 ед/г сырой биомассы.
Полученная сайт-специфическая эндонуклеаза Aoxl (рестриктаза) характеризуется следующими свойствами:
1. Узнает и расщепляет метилированную последовательность нуклеотидов 5'-^GG (m5C)C-3'/3'-C(m5C)GG^-5', где m5C - 5-метилцитозин.
2. Расщепляет связи перед первым гуанином в обеих цепях узнаваемой последовательности.
3. Не расщепляет вышеприведенную последовательность, не содержащую двух центральных С5-метилцитозиновых оснований.
4. Оптимальная температура действия 37°С.
5. Оптимальное значение pH для действия фермента 7,5-8,5.
6. Оптимальная концентрация соли при расщеплении ДНК 50-100 мМ NaCl.
7. Для проявления активности AoxI требуются ионы Mg2+, оптимальная концентрация - 5-10 мМ.
Определяющим отличием предлагаемого штамма от всех известных в настоящее время штаммов-продуцентов сайт-специфических эндонуклеаз является то, что данный штамм продуцирует сайт-специфическую эндонуклеазу, которая узнает и расщепляет обе цепи метилированной нуклеотидной последовательности ДНК 5'-^GG(m5C)C-3'/3'-C(m5C)GG^-5', где m5C - 5-метилцитозин. Таким образом, сайт-специфическая эндонуклеаза AoxI представляет собой новый, не имеющий аналогов фермент.
Поскольку предлагаемый штамм получен впервые и для выделения сайт-специфической эндонуклеазы, узнающей и расщепляющей вышеназванную последовательность нуклеотидов в указанной позиции, никогда не использовался, можно сделать вывод о соответствии предлагаемого штамма критериям изобретения «новизна» и «изобретательский уровень».
Изобретение иллюстрируется следующими примерами конкретного выполнения.
Пример 1
Выращивание штамма и выделение фермента
Для получения биомассы клетки штамма-продуцента переносят на агаризованную среду ЛБ в чашку Петри и инкубируют в течение ночи при 28°С. Свежевыращенные колонии переносят стерильной бактериологической петлей в колбы, содержащие жидкую питательную среду ЛБ, и культивируют на качалках при 28°С при перемешивании - 150 об/мин, до достижения стационарной фазы роста. Клетки осаждают центрифугированием при 5000 об/мин при 4°С на центрифуге "Beckman" (США) в роторе JA-10. Выход биомассы составляет 5 г/л среды. Дезинтеграцию клеток, выделение и очистку фермента проводят по известной методике [9].
Пример 2
Сайт-специфический гидролиз С5-метилированной плазмидной ДНК эндонуклеазой AoxI
Расщепление ДНК проводят в оптимальных условиях (37°С, реакционный буфер - 10 мМ TrisHCl, pH 8.0, 10 мМ MgCl2, 100 мМ NaCl, 1 мМ DTT) в течение 60 мин. Продукты расщепления плазмидной ДНК разделяют путем электрофореза в 1% агарозном геле.
В качестве субстратов для выявления специфичности расщепления используют различные метилированные и неметилировнные плазмидные и фаговые ДНК: фагов лямбда и Т7, плазмид pHspAI1 (содержит метилированные последовательности 5'-G(m5C)GC-3′/3′-CG(m5C)G-5' [1]), pFsp4HI3 (содержит метилированные последовательности 5'-G(5mC)NGC-3'/3'-CGN(5mC)G-5' [4]) и полученной нами плазмиды pMHaeIII (содержит метилированные последовательности 5'-GG(5mC)C-3'/3'-C(5mC)GG-5').
Плазмида pMHaeIII была получена путем встраивания в плазмиду pUC19 ПЦР-фрагмента (992 п.н.) геномной ДНК Haemophilus aegyptius no сайтам гидролиза эндонуклеаз рестрикции PstI и BamHI. Данный ПЦР-фрагмент был получен с использованием следующих праймеров (Р1 и Р2):
Р1: 5'-GACCGCACTTATAGAAATCAATTTGTTTAG-3'
Р2: 5'-TGCTCTGCCAGGATCCTTACTTTTATTTGAC-3'
Данный ПЦР-фрагмент включает ген ДНК-метилтрансферазы М.HaeIII, метилирующей первый цитозин в обеих цепях последовательности 5'-GGCC-3' в положении С5.
Благодаря встроенному гену, кодирующему М.HaeIII плазмида pMHaeIII, содержит метилированные последовательности 5'-GG(5mC)C-3'/3'-C(5mC)GG-5'.
На фиг.1 представлена электрофореграмма продуктов расщепления ДНК фагов λ и Т7, а также плазмид pFsp4HI3, pHspAI1, pMHaeIII и pUC19 эндонуклеазой AoxI.
Описание дорожек на электрофореграмме, изображенной на фиг.1:
1 - ДНК фага лямбда;
2 - ДНК фага лямбда, обработанная эндонуклеазой AoxI;
3 - ДНК фага Т7;
4 - ДНК фага Т7, обработанная эндонуклеазой AoxI;
5 - ДНК pFsp4HI3;
6 - ДНК pFsp4HI3, обработанная эндонуклеазой AoxI.
7 - ДНК pHspAI1;
8 - ДНК pHspAI1, обработанная эндонуклеазой AoxI;
9 - ДНК pMHaeIII;
10 - ДНК pMHaeIII, обработанная рестриктазой HaeIII;
11 - ДНК pMHaeIII, обработанная эндонуклеазой AoxI;
12 - ДНК pUC19;
13 - ДНК pUC19, обработанная рестриктазой AoxI;
14 - ДНК pUC19, обработанная эндонуклеазой HaeIII;
М - маркер молекулярного веса ДНК 1 kb (производство НПО СибЭнзим).
Как видно из фиг.1, AoxI не расщепляет ДНК фагов лямбда и Т7, а также ДНК плазмид pHspAI, pFsp4HI3 и pUC19, в которых отсутствует метилированная последовательность 5'-GG(5mC)C-3'/5'-GG(5mC)C-3'.
Рестриктаза HaeIII расщепляет неметилированные последовательности 5'-GGCC-3' в плазмиде pUC19. Теоретически рассчитанные длины фрагментов ДНК при этом расщеплении: 587, 458, 434, 298, 267, 257, 174, 102, 80, 18, 11 п.н.
HaeIII не расщепляет плазмиду pMHaeIII вследствие метилирования центральных цитозинов в последовательности 5'-GGCC-3'. В противоположность этому эндонуклеаза AoxI, наоборот, не расщепляет ДНК pUC19, но расщепляет pMHaeIII. Теоретически рассчитанные длины фрагментов ДНК, образующиеся при полном расщеплении плазмиды pMHaeIII по последовательности 5'-GGCC-3': 1283, 587, 458, 434, 298, 267, 257, 174, 102, 80, 18, 11 п.н., что соответствует полученной картине гидролиза ДНК.
Пример 3
Определение позиции гидролиза ДНК эндонуклеазой AoxI
Для подтверждения правильности определения узнаваемой последовательности и определения позиции гидролиза ДНК в сайте узнавания проводили гидролиз синтетических олигонуклеотидных дуплексов, образованных из олигонуклеотидов следующей структуры:
3М: 5'-ACGGATCGAGGAGCGGGCCCTCGTGAATTCACG-3'
4М: 5'-CGTGAATTCACGAGGGCCCGCTCCTCGATCCGT-3'
3M(m5C): 5'-ACGGATCGAGGAGCGGG(m5C)CCTCGTGAATTCACG-3'
4M(m5C): 5'-CGTGAATTCACGAGGG(m5C)CCGCTCCTCGATCCGT-3'
Олигонуклеотиды 3М и 3M(5mC) комплементарны олигонуклеотидам 4М и 4(5mC).
Олигонуклеотидные дуплексы 3М*/4М и 3M(5mC)*/4M(5mC) имеют сходную последовательность нуклеотидов и отличаются друг от друга только наличием или отсутствием метальных групп в первом цитозине последовательности 5'-GGCC-3'.
Определение места гидролиза ДНК рестриктазой AoxI осуществляли путем сравнения длин фрагментов, образуемых при расщеплении рестриктазой HaeIII неметилированного дуплекса 3М*/4М и AoxI метилированного дуплекса 3M(5mC)*/4M(5mC). В качестве маркера длин фрагментов использовали продукты частичного расщепления этих же дуплексов экзонуклеазой ExoIII из E.coli.
На фиг.2 изображен радиоавтограф электрофореграммы продуктов расщепления радиоактивно меченных дуплексов 3М*/4М и 3M(5mC)*/4M(5mC) в 20% полиакриламидном геле, содержащем 7-молярную мочевину.
Олигонуклеотиды, меченные радиоактивным фосфором 32P по 5'-концу, обозначены знаком *.
Описание дорожек на электрофореграмме, изображенной на фиг.2:
1 - дуплекс 3М*/4М;
2 - дуплекс 3M(5mC)*/4M(5mC);
3 - дуплекс 3М*/4М, обработанный эндонуклеазой AoxI;
4 - дуплекс 3M(5mC)*/4M(5mC), обработанный рестриктазой HaeIII;
5 - дуплекс 3М*/4М, обработанный рестриктазой HaeIII;
6 -дуплекс 3M(5mC)*/4M(5mC), обработанный экзонуклеазой ExoIII;
7 - дуплекс 3M(5mC)*/4M(5mC), обработанный эндонуклеазой AoxI;
Из фиг.2 видно, что фрагмент ДНК, образующийся при гидролизе дуплекса 3М*/4М рестриктазой HaeIII на дорожке 5, длиннее на два нуклеотида фрагмента, образующегося при гидролизе дуплекса 3M(5mC)*/4M(5mC) эндонуклеазой AoxI.
На основе полученных результатов можно сделать вывод, что в палиндромной последовательности 5'-GG(5mC)C-3'/3'-GG(5mC)C-5' эндонуклеаза AoxI расщепляет ДНК перед первым гуанином с образованием с образованием четырехнуклеотидных 5'-выступающих концов.
Выход сайт-специфической эндонуклеазы AoxI определяют по электрофоретической картине расщепления ДНК плазмиды pMHaeIII.
За единицу активности принимают минимальное количество фермента, необходимое для полного расщепления 1 мкг ДНК данной плазмиды в течение 1 часа при температуре 37°С в 20 мкл реакционной смеси. Выход фермента составляет 100 ед/г сырой биомассы, концентрация 200 ед/мл.
Фермент хранится при - 20°С в буфере, содержащем 50% глицерин, 0,08% тритон
XI 00, 0,2 М NaCl, 10 мМ трисHCl (pH 7,5), 7 мМ β-меркаптоэтанол, 0,1 мМ ЭДТА.
Таким образом, получен новый штамм, продуцирующий сайт-специфическую эндонуклеазу AoxI, узнающую и расщепляющую обе цепи нуклеотидной последовательности ДНК, которая содержит четыре С5-метилцитозиновых оснований в сайте узнавания 5'-GG(5mC)C-3'/3'-C(5mC)GG-5', с образованием четырехнуклеотидных 5'-выступающих концов. Данная сайт-специфическая эндонуклеаза может быть использована для выявления и анализа метилированной ДНК.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. Чернухин В.А., Наякшина Т.Н., Абдурашитов М.А., Томилова Ю.Э., Мезенцева Н.В., Дедков B.C., Михненкова Н.А., Гончар Д.А., Дегтярев С.Х. // Новая эндонуклеаза рестрикции GlaI узнает метилированную последовательность 5'-GCGC-3'. // Биотехнология. - 2006. - №4. - С.31-35.
2. Чернухин В.А., Томилова Ю.Э., Чмуж Е.В., Соколова О.О., Дедков B.C., Дегтярев С.Х. Сайт-специфическая эндонуклеаза BlsI узнает последовательности ДНК 5'-G(5mC)NGC-3' и расщепляет ее с образованием 3'-выступающих концов. // Вестник биотехнологии и физико-химической биологии имени Ю.А.Овчинникова. - 2007. - Т.3. - №1. - С.28-33.
3. Чмуж Е.В., Каширина Ю.Г., Томилова Ю.Э., Мезенцева Н.В., Дедков B.C., Гончар Д.А., Абдурашитов М.А., Дегтярев С.Х. Новая эндонуклеаза рестрикции BisI из Bacillus subtilis T30 узнает метилированную последовательность ДНК 5'G(m5C)^NGC-3' // Биотехнология. - 2005. - №3. - С.22-26.
4. Чернухин В.А., Чмуж Е.В., Томилова Ю.Э., Наякшина Т.Н., Гончар Д.А., Дедков B.C., Дегтярев С.Х. Новая сайт-специфическая эндонклеаза Glul узнает метилированную последовательность ДНК 5'-G(5mC)NG(5mC)-3'/3'-(5mC)GN(5mC)G-5' // Вестник биотехнологии и физико-химической биологии имени Ю.А.Овчинникова. - 2007. - Т.3. - №2. - С.13-17.
5. Roberts R.J., Vineze Т., Posfai J., Macelis D. 2003. REBASE - restriction enzymes and methylases. Nucl. Acids Res. 31, 418-420.
6. Определитель бактерий Берджи. / Под ред. Дж.Хоулта и др.: (9-е издание в 2 томах: Пер. с англ. под ред. акад. РАН Г.А.Заварзина. - М., 1997.
7. Madden, T.L., Tatusov, R.L. & Zhang, J. Applications of network BLAST server. // Meth. Enzymol - 1996. -. V.266 - P.131-141.
8. Smith, H.O., Nathans, D. A suggested nomenclature for bacterial host modification and restriction systems and their enzymes. // J. Mol. Biol. - 1973. - Vol.81. - P.419-423.
9. Bickle, T.A., Pirrotta, V. and Imber, R. A simple, general procedure for purifying restriction endonucleases. // Nucleic Acids Res. - 1977. - V.4 - P.561-2572.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |
| ШТАММ БАКТЕРИЙ Microbacterium testaceum 17B - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ MteI | 2011 |
|
RU2475533C1 |
| ШТАММ БАКТЕРИИ Paracoccus carotinifaciens 3K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PcsI. | 2008 |
|
RU2377294C1 |
| ШТАММ БАКТЕРИИ Kocuria rosea - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ KroI | 2009 |
|
RU2394099C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli N42 (pElmI) - ПРОДУЦЕНТ МЕТИЛЗАВИСИМОЙ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ ElmI | 2015 |
|
RU2597987C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Bisi | 2004 |
|
RU2270859C1 |
| ШТАММ БАКТЕРИЙ Glacial ice bacterium I - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Gla I | 2005 |
|
RU2287012C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SIMPLEX - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ BlsI | 2006 |
|
RU2322494C1 |
| ШТАММ БАКТЕРИЙ GLACIAL ICE BACTERIUM - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ GLU I | 2006 |
|
RU2322492C1 |
| СПОСОБ САЙТ-СПЕЦИФИЧЕСКОГО ГИДРОЛИЗА ДНК | 2015 |
|
RU2597984C1 |
Изобретение относится к области биотехнологии, а именно к штамму бактерий Arthrobacter oxydans 25K. Штамм бактерий Arthrobacter oxydans 25К выделяют из почвы. Данный штамм обеспечивает получение сайт-специфической эндонуклеазы Aoxl, узнающей и расщепляющей обе цепи нуклеотидной последовательности ДНК 5'-GGCC-3/3'-CCGG-5', в которых два центральных цитозина метилированы в положении С5, с образованием четырехнуклеотидных 5'-выступающих концов. Представленное изобретение позволяет получить бактериальный штамм, продуцирующий сайт-специфическую эндонуклеазу, которая узнает и расщепляет обе цепи нуклеотидной последовательности ДНК 5'-^GG (m5C)C-3/3'-C(m5C)GG^-5'. 2 ил.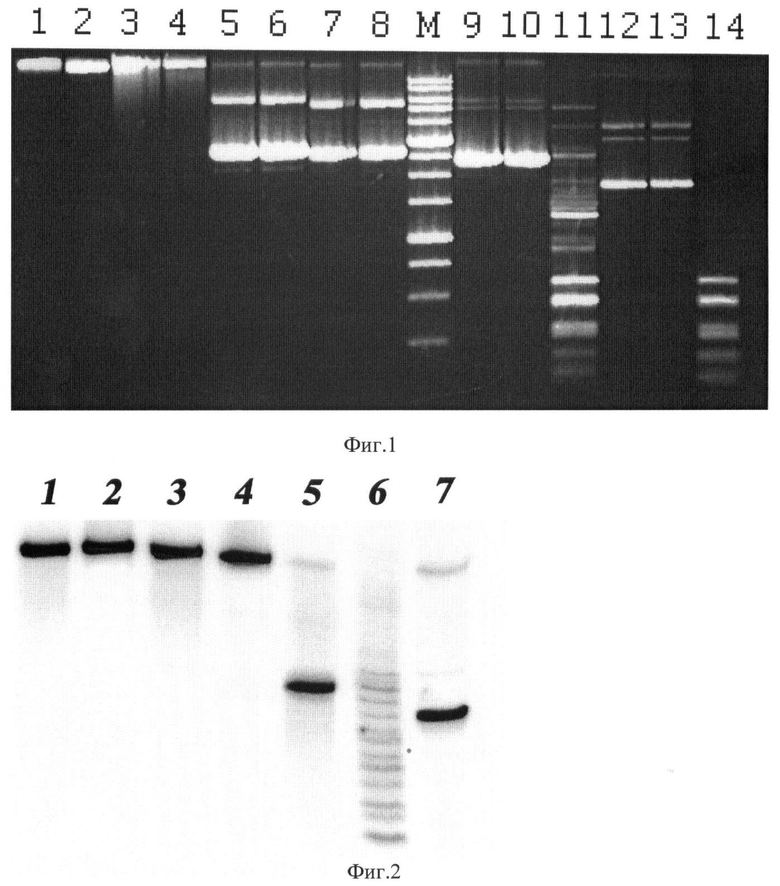
Штамм бактерий Arthrobacter oxydans 25K KKM НПО «СибЭнзим» 7М89 - продуцент сайт-специфической эндонуклеазы, узнающей и расщепляющей обе цепи нуклеотидной последовательности ДНК 5'-GGCC-3'/3'-CCGG-5', в которых два центральных цитозина метилированы в положении С5, с образованием четырехнуклеотидных 5'-выступающих концов.
| ROBERTS R.J | |||
| et al | |||
| REBASE: restriction enzymes and methyltransferases | |||
| Nucleic Acids Res | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| ЧЕРНУХИН В.А | |||
| и др | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Биотехнология, 2006, №4, с.31-35 | |||
| ЧЕРНУХИН В.А | |||
| и др | |||
| Сайт-специфическая эндонуклеаза BIsI узнает | |||

