1. Область изобретения
Настоящее изобретение раскрывает способы и реагенты для диагностики и лечения глаукомы и близких нарушений.
2. Описание области техники
"Глаукомы" представляют группу нарушающих зрение глазных заболеваний, которые являются основной причиной возникновения необратимой слепоты в США и других развитых странах. Первичная открытоугольная глаукома ("POAG") - наиболее распространенная форма глаукомы - характеризуется дегенерацией трабекулярной сети, приводящей к блокаде нормальной способности глазной жидкости к оттоку из глаза без перекрытия пространства (например, "угла") между радужкой и роговицей (Vaughan D. et al. (1992)). Характерным признаком подобной блокады при данном заболевании является повышенное внутриглазное давление ("IOP"), что в конечном итоге при отсутствии соответствующего и своевременного лечения приводит к прогрессирующей потере зрения и слепоте. Установлено, что заболевание поражает 0,4-3,3% взрослых людей в возрасте старше 40 лет (Leske M.C. et al. (1986); Bengtsson B. (1989); Strong N.P. (1992)). Кроме того, распространение заболевания возрастает до 6% у людей в возрасте 75 лет или старше (Strong N.P. (1992)).
Поскольку повышенное IOP является хорошо определяемым признаком глаукомы, то диагностика заболевания в основном проводится по измерению внутриглазного давления (тонометрия) (Strong N.P. (1992); Greve M. et al. (1993)). К сожалению, поскольку пределы давления при глаукоме и нормального давления перекрываются, подобные методы обладают ограниченной ценностью, если только не проводятся многократные определения (Hitchings R.A. (1993); Tuck M.W. et al. (1993); Vaughan D. et al. (1992); Vernon S.A. (1993)). По этой причине часто применяют дополнительные способы, такие как прямое обследование диска зрительного нерва и определение степени потери поля зрения у пациента для повышения точности постановки диагноза (Greve M. et al. (1993)).
При глаукоме поражаются три отдельных ткани в глазе. Повышенное IOP, связанное с POAG, возникает за счет морфологических и биохимических изменений в трабекулярной сети (ТМ), ткани, расположенной в углу между роговицей и радужкой. Большая часть питающей глазной жидкости проходит через передний сегмент глаза по ТМ. Прогрессирующая потеря клеток ТМ и скопление внеклеточного дебриса в ТМ глаза при глаукоме приводит к повышенному сопротивлению оттоку глазной жидкости (Lutjen-Drecoll and Rohen, 1996; Rohen, 1983; Rohen et al., 1993; Grierson and Calthorpe, 1988), в результате чего повышается IOP. Повышенное IOP, а также другие факторы, такие как ишемия, приводят к дегенеративным изменениям в головке глазного нерва (ONH), вызывая прогрессирующее "углубление" ONH (Varma and Minckler, 1996; Hernandez and Gong, 1996; Hernandez et al., 1990; Hernandez and Pena, 1997; Morrison et al., 1990) и потерю ганглиозных клеток сетчатки (Quigley et al., 2000; Quigley, 1999; Quigley et al., 1995; Kerrigan et al., 1997) и аксонов. Детальный молекулярный механизм, ответственный за повреждение ТМ, ONH и ганглиозных клеток сетчатки при глаукоме, остается неизвестным.
Лечение глаукомы в настоящее время направлено на снижение IOP, основного фактора риска для развития и прогрессирования глаукомы. В результате подобного лечения снижается IOP, но оно не направлено непосредственно на механизм патогенеза, и заболевание продолжает прогрессировать. По меньшей мере, у половины пациентов с глаукомой диагноз не поставлен, и со временем, когда у пациентов все-таки глаукома диагностируется, потеря ганглиозных клеток сетчатки уже составляет примерно 40%. Следовательно, имеется потребность в способах более раннего установления и диагностики глаукомы.
В свете значения глаукомы и, по меньшей мере, частичной неадекватности способов диагностики предшествующего уровня техники было бы желательным иметь усовершенствованный, более точный способ диагностики глаукомы на ранних стадиях заболевания. Кроме того, было бы желательным иметь новые терапевтические средства, направленные на механизм патогенеза глаукомы.
Краткое описание изобретения
В настоящем изобретении преодолеваются данные и другие недостатки предшествующего уровня техники обеспечением способов и наборов для ранней диагностики глаукомы, для лечения глаукомы и для идентификации соединений, пригодных для лечения глаукомы.
В некоторых конкретных воплощениях изобретение обеспечивает способ диагностики глаукомы в пробе, полученной из клетки или жидкости организма, детектированием измененной экспрессии гена, члена семейства морфогенных белков кости. В целом способ включает стадии:
а) получения пробы ткани или жидкости от пациента с подозрением на глаукому;
b) экстрагирования ДНК из указанной пробы;
с) получения множества ПЦР-праймеров, при этом каждый из указанных праймеров включает последовательность, состоящую из 18-1547 смежных нуклеотидов последовательностей SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47 или SEQ ID NO:53;
d) амплификации областей экстрагированной ДНК с использованием указанных праймеров с получением продукта ПЦР;
е) выделения продукта ПЦР; и
f) идентификации различий между последовательностью продукта ПЦР и нормальной последовательностью гена;
при этом различие между амплифицированной последовательностью и нормальной последовательностью гена является диагностическим признаком глаукомы.
В основном способы по изобретению могут включать получение пробы от индивидуума и экстрагирование ДНК из указанной пробы. Выбранные ПЦР-праймеры для конкретных членов семейства генов ВМР затем используются для амплификации соответствующих областей экстрагированного гена с получением продукта ПЦР. Продукт ПЦР выделяют методом, с помощью которого можно эффективно установить различия в последовательностях ДНК между нормальной и мутантной формой конкретного гена семейства ВМР, которые подвергаются оценке (экстрагированной ДНК). Установленные различия между последовательностями указывают на наличие глаукомы.
Пробой ткани или жидкости для использования в способах по изобретению может быть кровь или клетки щеки.
Как правило, последовательности праймеров будут иметь длину в пределах примерно от 10, 15 или 18 нуклеотидов до примерно 20 или примерно 30 нуклеотидов. Более длинные последовательности, например, из 40, 50, 80, 90, 95, 100 нуклеотидов, даже до полноразмерных последовательностей, являются даже более предпочтительными для некоторых воплощений. Специалисты в данной области считают допустимыми олигонуклеотиды длиной, по меньшей мере, из 18-20 нуклеотидов, как достаточные для проведения в достаточной мере специфической гибридизации, в качестве молекулярного зонда, описанные Lathe (1985), данный источник специально для этой цели включен здесь для сведения. Предпочтительно нуклеотидная последовательность будет состоять из 20-100 смежных нуклеотидов последовательностей SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47 или SEQ ID NO:53. Также предусматривается, что последовательности праймеров могут состоять из последовательностей, по меньшей мере, из 10, 15 или 18 смежных нуклеотидов последовательностей генов ВМР-рецепторов и связанных с ВМР белков, последовательности которых хорошо известны.
Молекулы нуклеиновой кислоты, имеющие участки из 10, 18, 20, 30, 50, 60, 65 или даже до и включая 100 нуклеотидов, и так далее, комплементарные любой из последовательностей SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47 или SEQ ID NO:53, пригодны в качестве зондов для гибридизации. Специалисты в данной области признают, что праймеры или зонды, имеющие нуклеотидную длину из примерно 18 нуклеотидов, обеспечивают высокую специфичность гибридизации с последовательностью-мишенью. Полный размер фрагмента, а также размер комплементарных участков, в конечном итоге будет зависеть от предназначаемого применения конкретного сегмента нуклеиновой кислоты. В воплощениях гибридизации, как правило, найдут применение меньшие фрагменты, у которых длина комплементарной области может варьировать так, например, в пределах примерно от 10, 18, 20 или 30 и до примерно 50, 60, 70, 80, 90 или 100 нуклеотидов или даже полного размера, соответственно комплементарным последовательностям, которые подвергаются детектированию.
В особо предпочтительных воплощениях праймеры будут состоять из смежных последовательностей из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47 или SEQ ID NO:53. В других предпочтительных воплощениях праймеры будут состоять из смежных последовательностей из генов ВМР-рецепторов (раскрыты у Dijke et al., 1993; Astrom et al., 1999; Nohno et al., 1995, все включены здесь для сведения) или из связанных с ВМР генов, таких как хордин (NCBI NM_029130), гремлин (Murphy et al., 1999; McMahon et al., 2000), фоллистатин (NCBI NM_003892) или бамби (NCBI NM_005791). Наиболее предпочтительно праймеры будут состоять из смежной последовательности из SEQ ID NO:3. В некоторых аспектах, по меньшей мере, некоторые из праймеров могут дополнительно включать детектируемую метку.
В некоторых воплощениях изобретение обеспечивает способ лечения глаукомы введением пациенту, нуждающиеся в этом, композиции, содержащей последовательность, состоящую, по меньшей мере, из одного соединения, выбранного из группы, состоящей из агониста ВМР-2, агониста ВМР-4, агониста ВМР-5, агониста ВМР-7, агониста Smad 1/5, антагониста хордина, антагониста гремлина и антагониста фоллистатина.
В дополнительных аспектах настоящее изобретение обеспечивает способ идентификации терапевтического средства для лечения глаукомы. Терапевтические средства можно идентифицировать, например:
а) получением первой композиции, содержащей популяцию рекомбинантных клеток, экспрессирующих ВМР-2А, ВМР-4, ВМР-5 или ВМР-7;
b) получением соединения-кандидата;
с) инкубацией указанной композиции и указанного соединения-кандидата;
тестированием указанной композиции на ее способность воздействовать на индуцированные ВМР пути передачи сигнала с участием Smad и/или регулируемую ВМР экспрессию гена; и идентификацией соединения-кандидата, которое ингибирует или стимулирует данные эффекты в прямом направлении ВМР.
Другим аспектом изобретения являются диагностические наборы, содержащие последовательности по настоящему изобретению и подходящие реагенты, такие как детектируемая метка, связанная с белком, пептидом или самим антителом. Альтернативно детектируемая метка может быть связана со второй последовательностью, которая избирательно гибридизуется с последовательностью по изобретению.
Близкие воплощения включают терапевтические наборы, которые содержат фармацевтически приемлемые композиции либо последовательностей нуклеиновых кислот, либо последовательностей пептидов или белков, раскрытых здесь. Подобные наборы пригодны для детектирования измененной экспрессии генов ВМР и белков в клинических пробах для диагностики глаукомы.
Краткое описание фигур
Фигуры составляют часть настоящего описания и включены для дополнительной демонстрации некоторых аспектов настоящего изобретения. Изобретение можно лучше понять при обращении к одной или более фигур в сочетании с подробным описанием конкретных воплощений, представленных здесь.
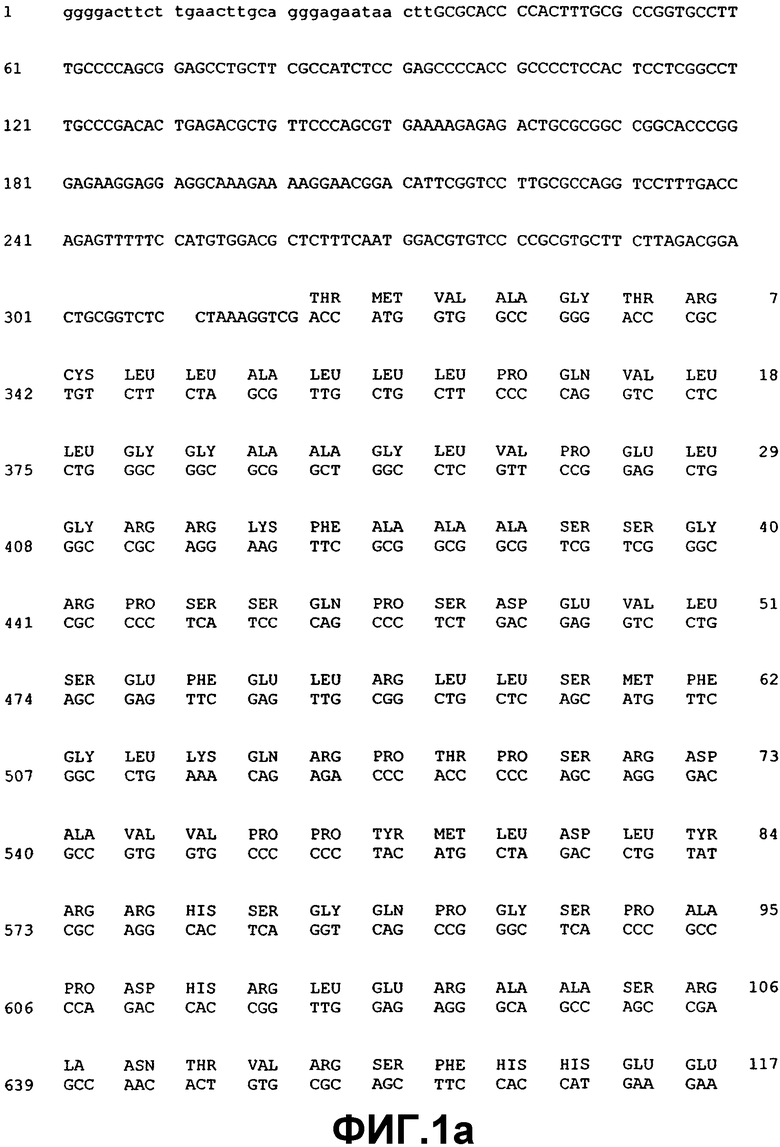
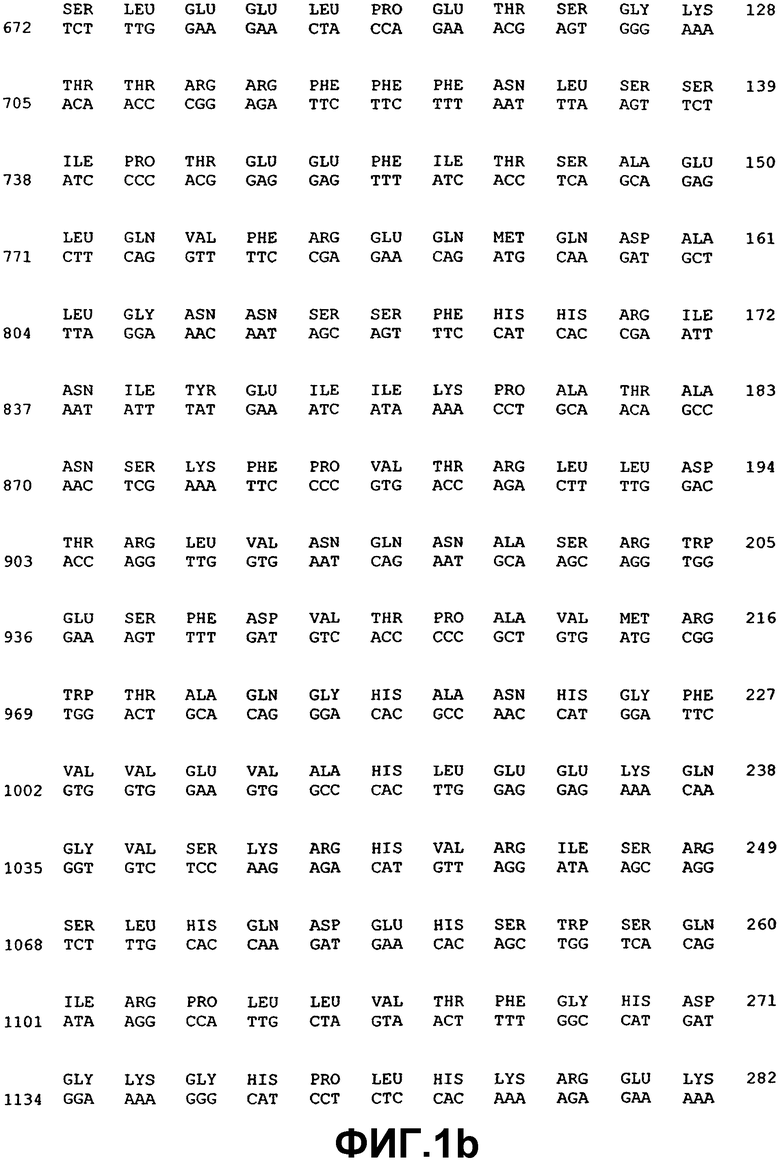
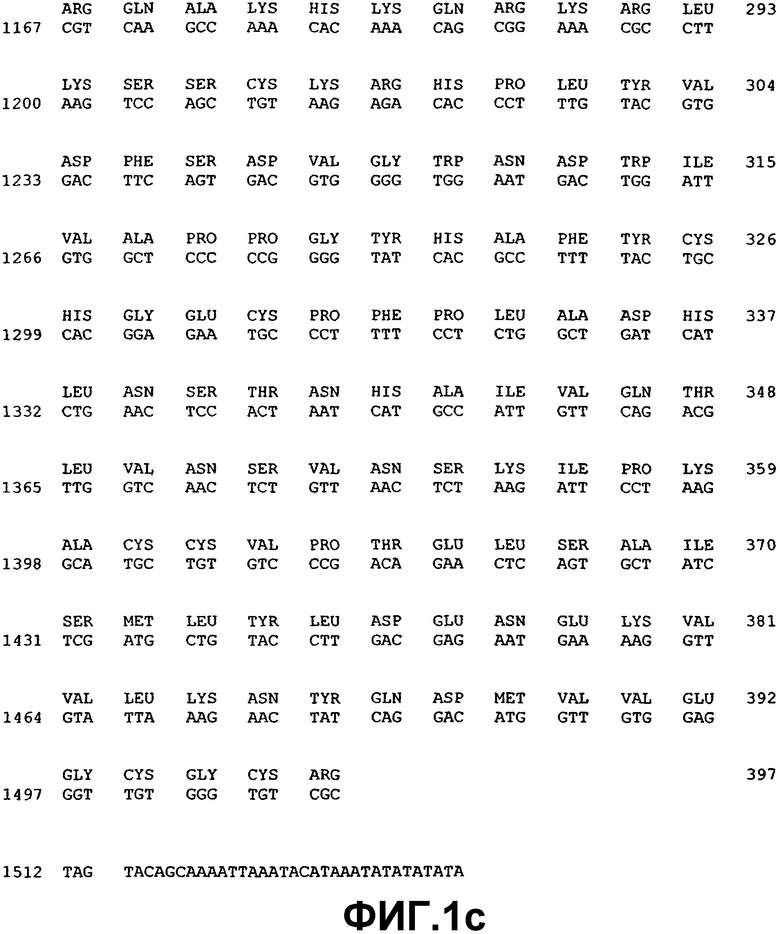
Фиг.1. Нуклеотидная и аминокислотная последовательность ВМР-2А.
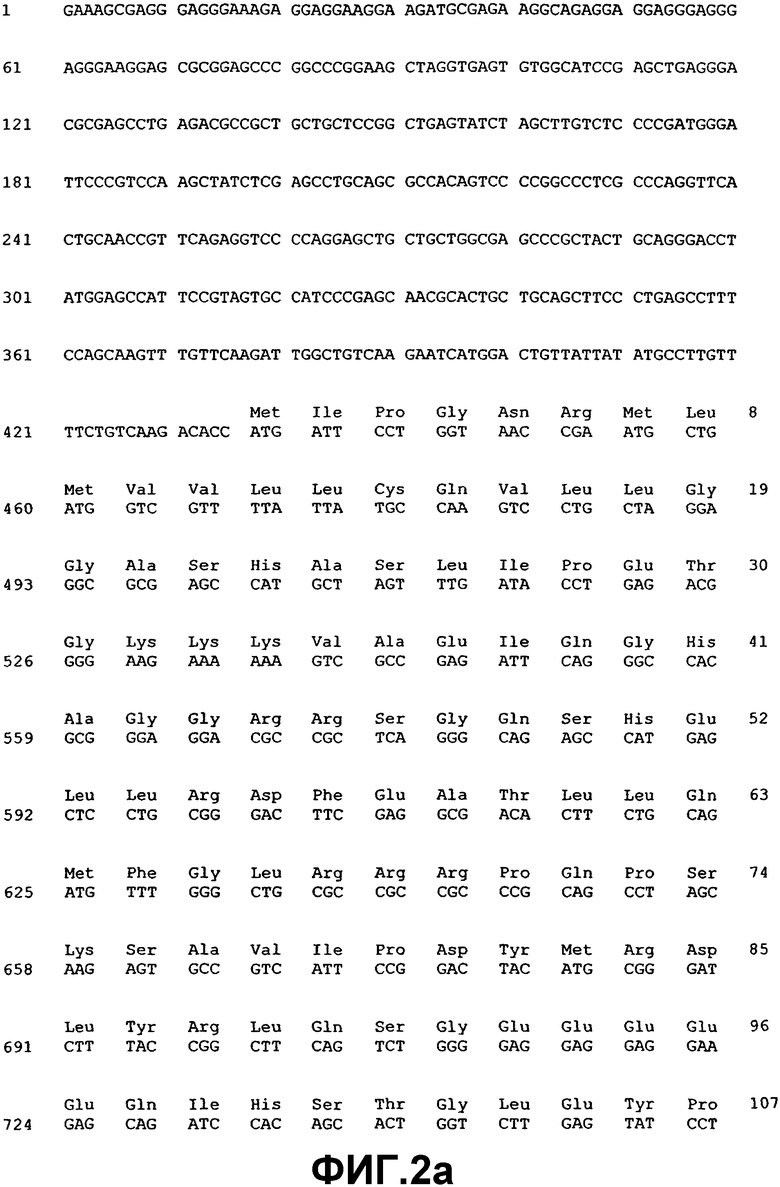
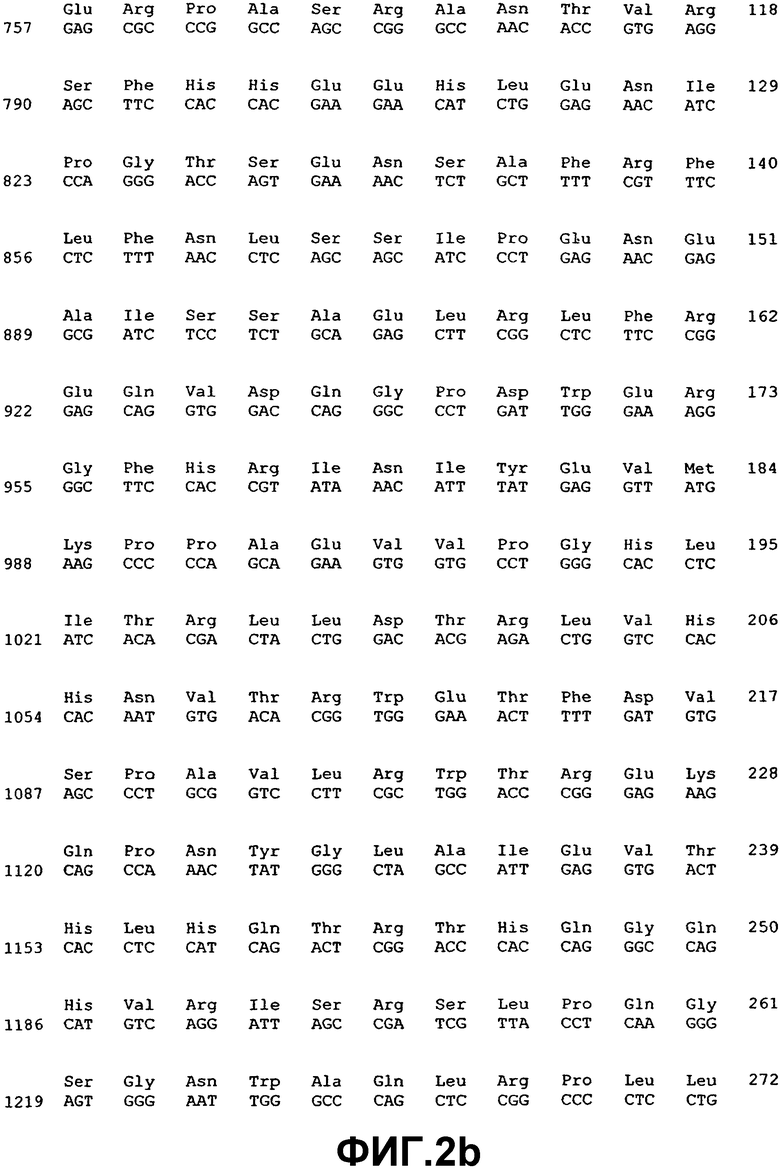
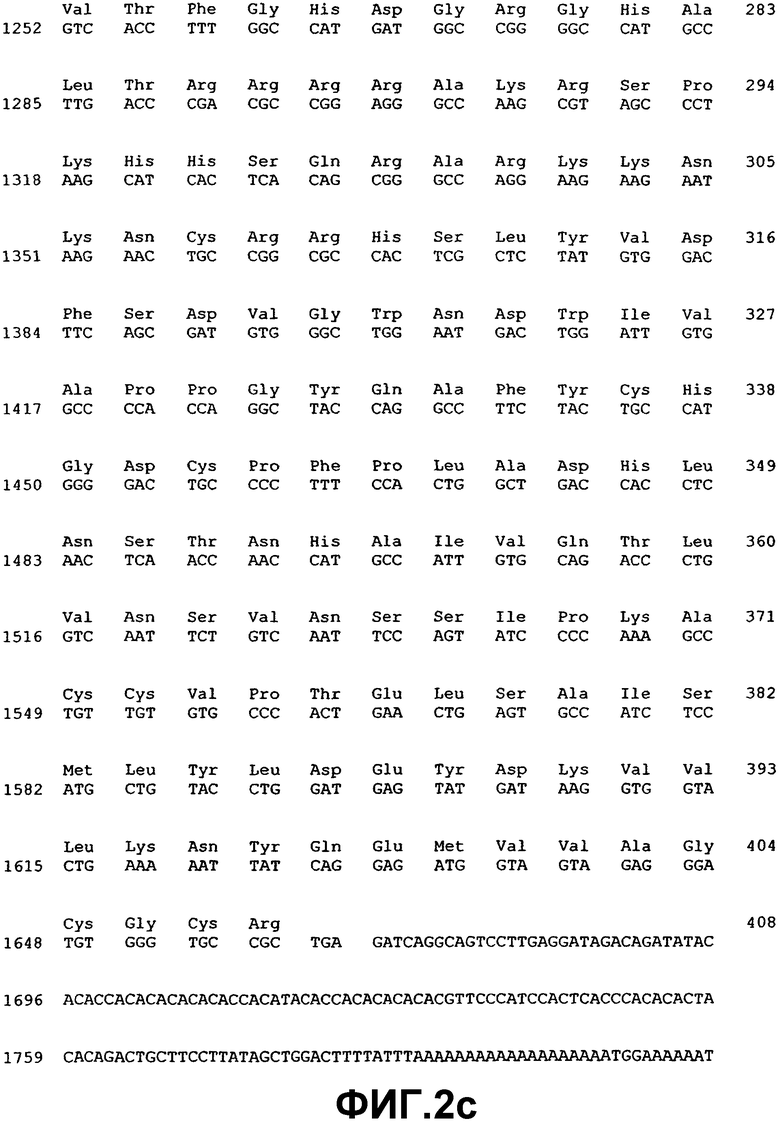
Фиг.2. Нуклеотидная и аминокислотная последовательность ВМР-4.
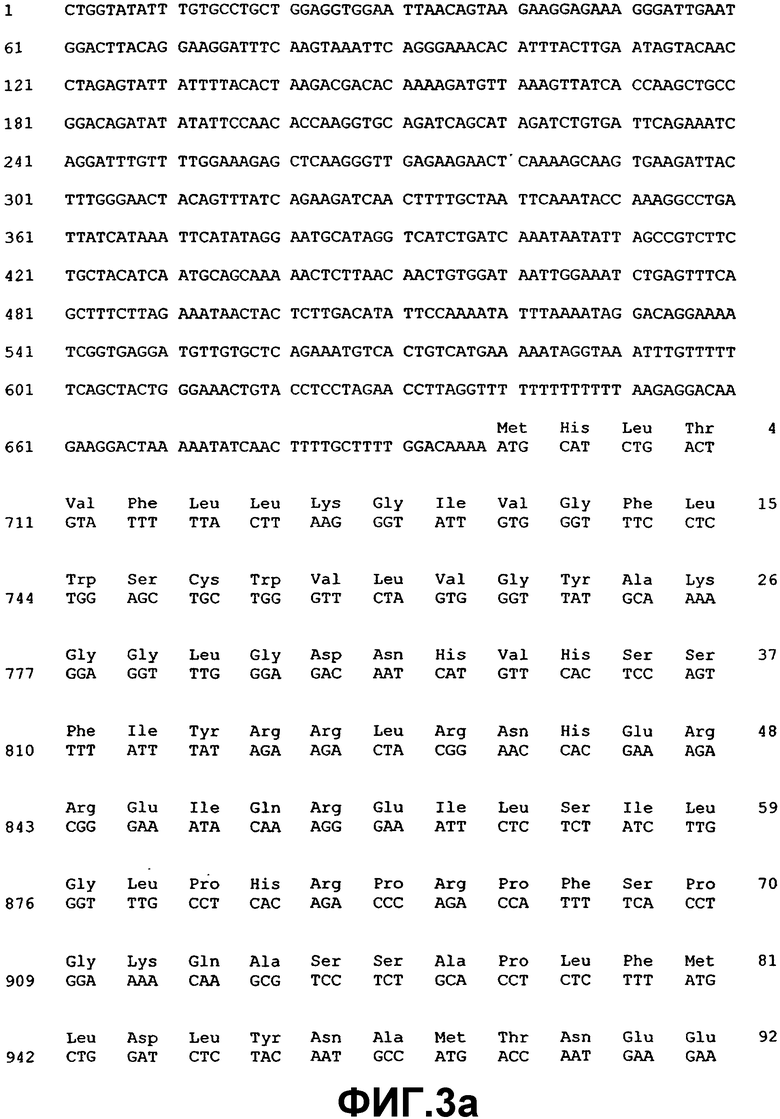
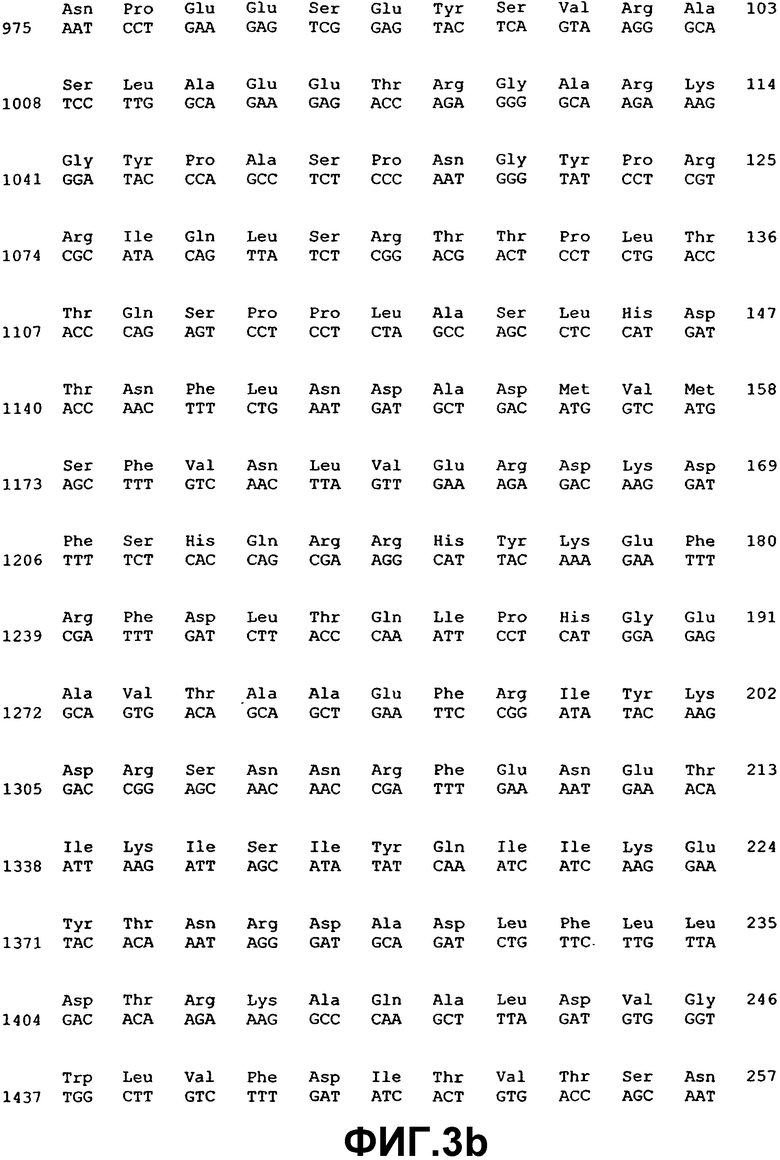
Фиг.3. Нуклеотидная и аминокислотная последовательность ВМР-5.
Фиг.4. Нуклеотидная и аминокислотная последовательность ВМР-7.
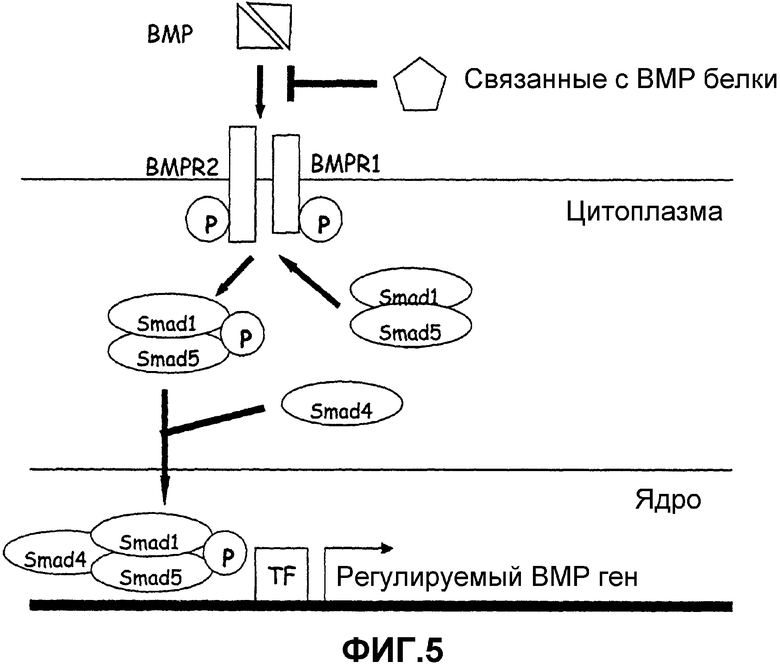
Фиг.5. Путь передачи сигнала с участием морфогенных белков кости. Димеры морфогенных белков кости (ВМР) связываются с мембранным комплексом, состоящим из ВМР-рецепторов 1 и 2, которые представляют серин/треонинкиназы. Регуляторные Smad (Smad 1/Smad 5) фосфорилируются и связываются с со-Smad (Smad 4). Данный образующийся Smad-комплекс проникает в ядро, где он связывается с факторами транскрипции (TF) и регулирует экспрессию гена. Связанные с ВМР белки действуют в качестве антагонистов ВМР, связываясь с ВМР и предупреждая взаимодействие ВМР с ВМР-рецепторами.
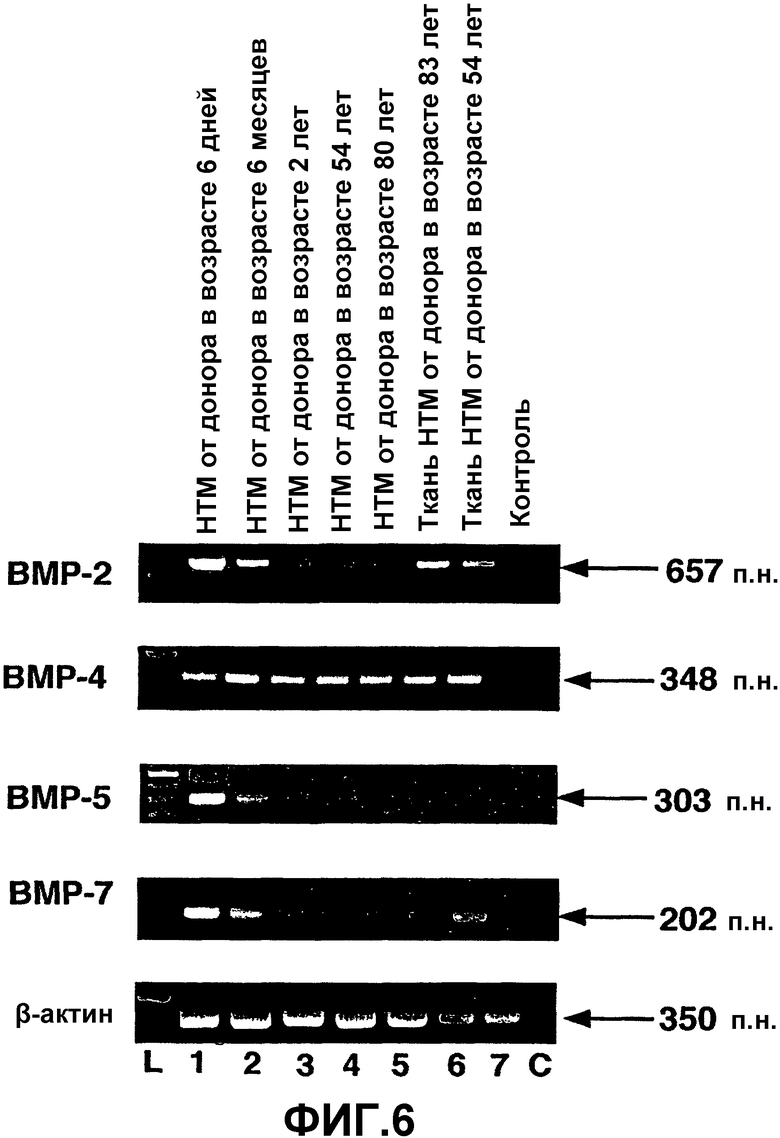
Фиг.6. Экспрессия ВМР в человеческих клетках и тканях ТМ. Окрашенный бромистым этидием агарозный гель продуктов ПЦР для ВМР из проб кДНК, полученных при анализе ОТ-ПЦР экспрессии ВМР в человеческих клетках (полосы 1-5) и тканях (полосы 6-7) ТМ. L = маркеры пар оснований. С = отрицательная контрольная полоса при ПЦР. β-актин использовали в качестве положительного внутреннего контроля при проведении ОТ-ПЦР.
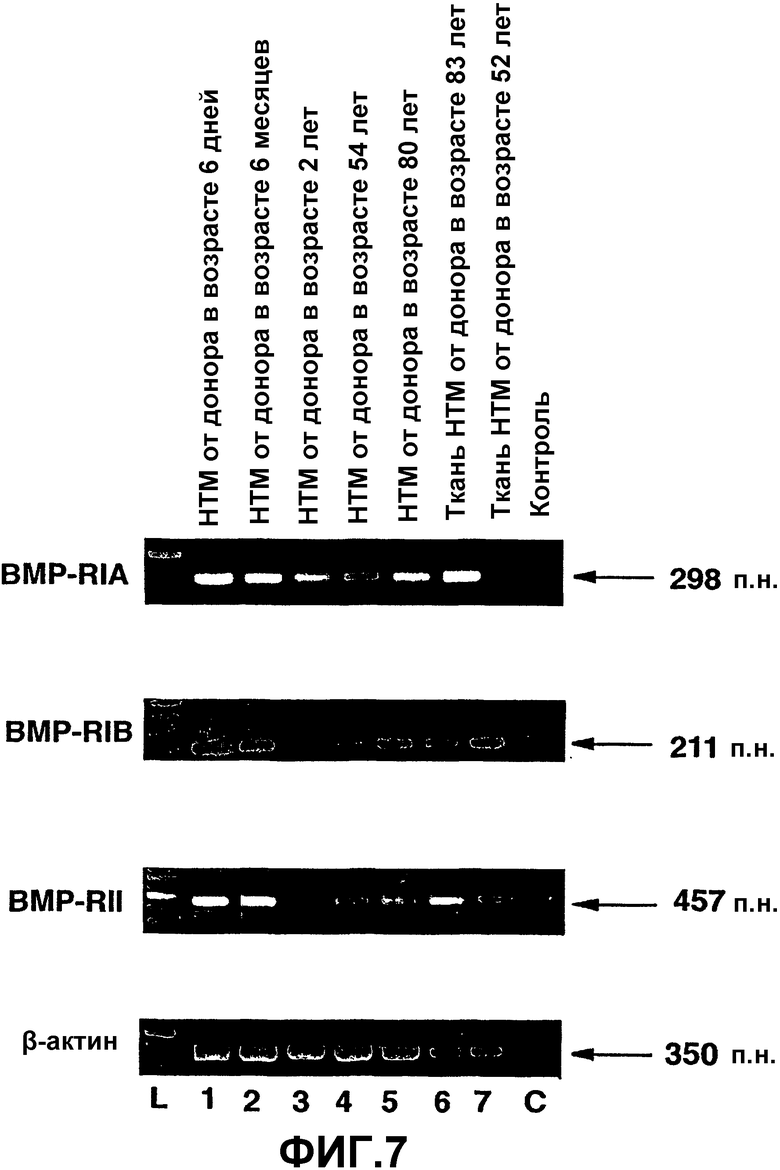
Фиг.7. Экспрессия ВМР-рецептора в человеческих клетках и тканях ТМ. Окрашенный бромистым этидием агарозный гель продуктов ПЦР из проб кДНК, полученных при анализе ОТ-ПЦР экспрессии ВМР-рецепторов в человеческих клетках (полосы 1-5) и тканях (полосы 6-7) ТМ. L = маркеры пар оснований. С = отрицательная контрольная полоса при ПЦР. β-актин использовали в качестве положительного внутреннего контроля при проведении ОТ-ПЦР.
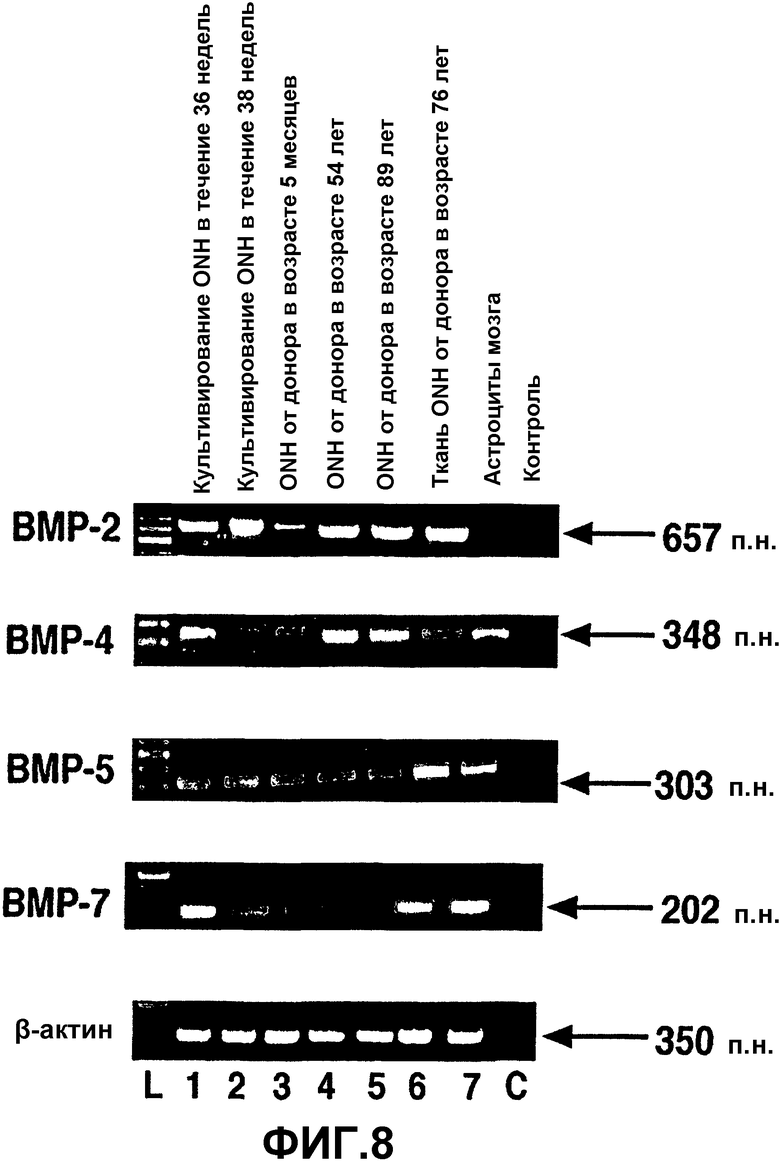
Фиг.8. Экспрессия ВМР в человеческих астроцитах ONH, тканях ONH и астроцитах мозга человека. Окрашенный бромистым этидием агарозный гель продуктов ПЦР из проб кДНК, полученных при анализе ОТ-ПЦР экспрессии ВМР в человеческих астроцитах ONH (полосы 1-5), тканях ONH (полоса 6) и астроцитах мозга человека (полоса 7). L = маркеры пар оснований. С = отрицательная контрольная полоса при ПЦР. β-актин использовали в качестве положительного внутреннего контроля при проведении ОТ-ПЦР.
Фиг.9. Экспрессия ВМР в линиях человеческих клеток решетчатой полости склеры. Окрашенный бромистым этидием агарозный гель продуктов ПЦР из проб кДНК, полученных при анализе ОТ-ПЦР человеческих клеток решетчатой полости склеры (полосы 1-9). L = маркеры пар оснований. С = отрицательная контрольная полоса при ПЦР. β-актин использовали в качестве положительного внутреннего контроля при проведении ОТ-ПЦР.
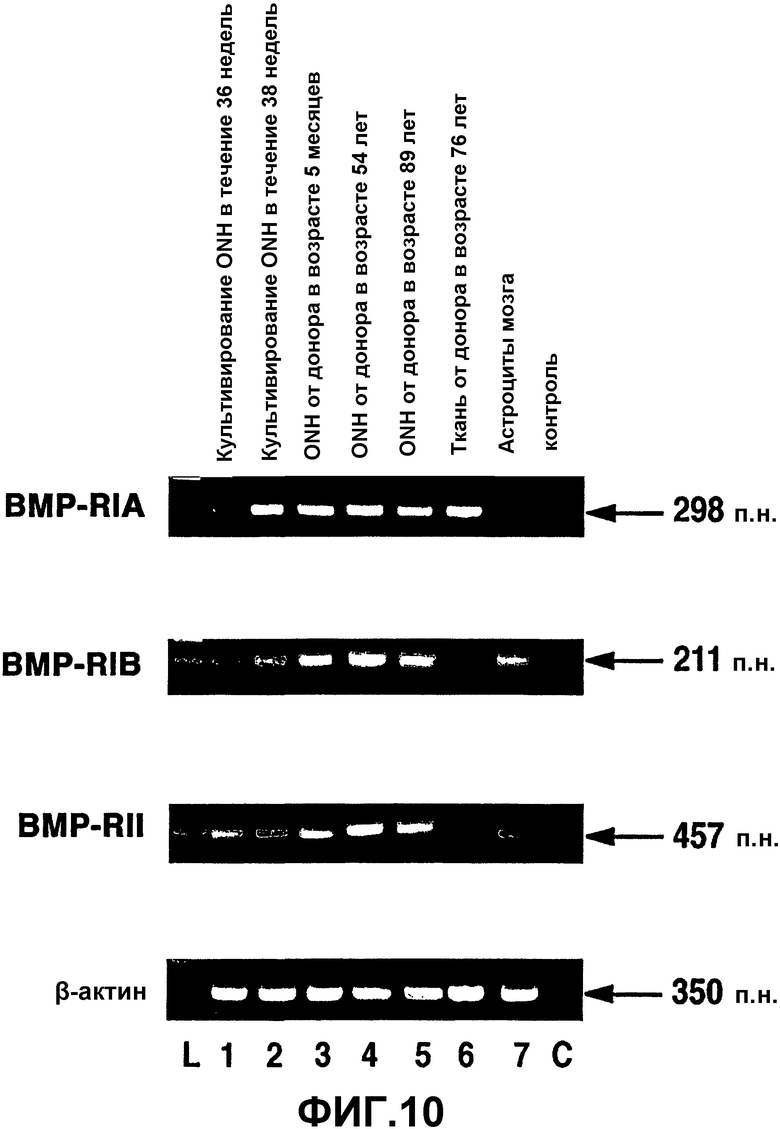
Фиг.10. Экспрессия ВМР-рецептора в человеческих астроцитах ONH, тканях ONH и астроцитах мозга человека. Окрашенный бромистым этидием агарозный гель продуктов ПЦР из проб кДНК, полученных при анализе ОТ-ПЦР экспрессии ВМР-рецепторов в астроцитах головки зрительного нерва человека (ONA) (полосы 1-5), ткани ONH (полоса 6) и астроцитах мозга человека (полоса 7). L = маркеры пар оснований. С = отрицательная контрольная полоса при ПЦР. β-актин использовали в качестве положительного внутреннего контроля при проведении ОТ-ПЦР.
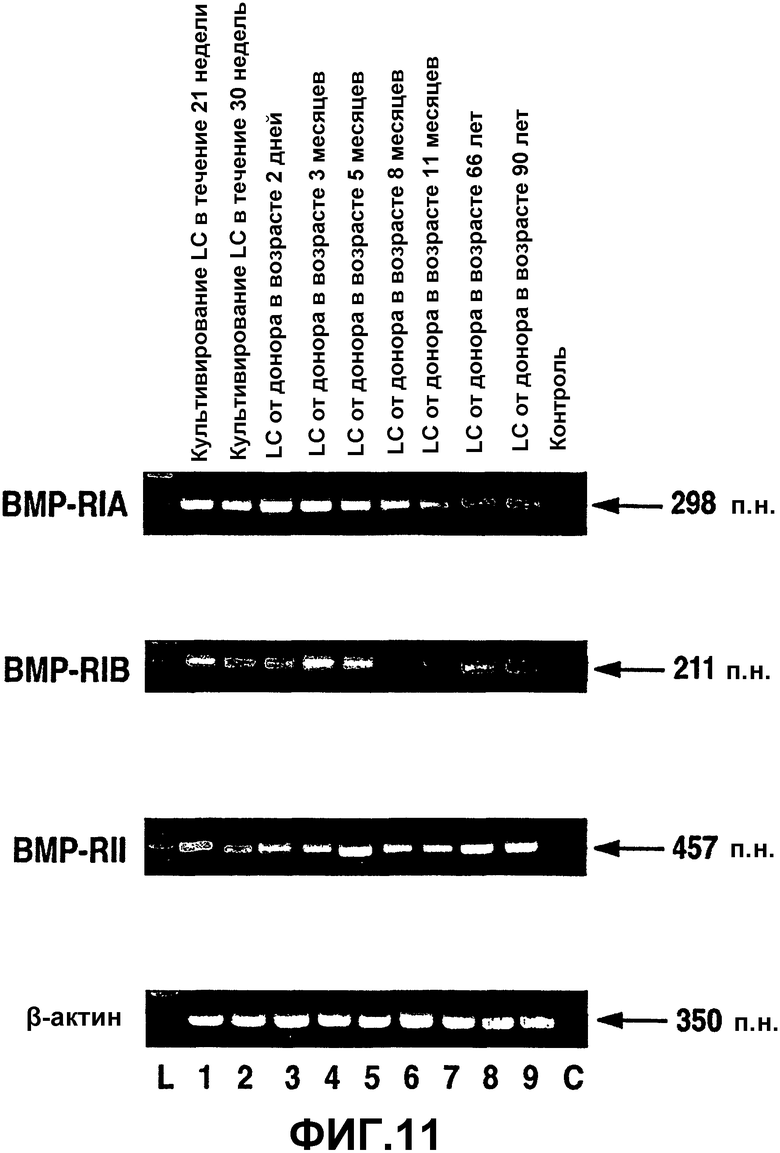
Фиг.11. Экспрессия ВМР-рецепторов в линиях человеческих клеток решетчатой полости склеры. Окрашенный бромистым этидием агарозный гель продуктов ПЦР из проб кДНК, полученных при анализе ОТ-ПЦР человеческих клеток решетчатой полости склеры (полосы 1-9). L = маркеры пар оснований. С = отрицательная контрольная полоса при ПЦР. β-актин использовали в качестве положительного внутреннего контроля при проведении ОТ-ПЦР.
Фиг.12. Вестерн-иммуноблоттинг экспрессии ВМР и ВМР-рецепторов в культивированных человеческих клетках ТМ, астроцитах головки зрительного нерва (ONA) и клетках решетчатой полости склеры. Хемилюминесцентное детектирование ВМР-белков и ВМР-рецепторов в человеческих клетках трабекулярной сети (полосы 1-2), астроцитах ONH (полосы 3-4) и клетках решетчатой полости склеры (полосы 5-6). Масса белков указана в kDa.
Фиг.13. Экспрессия мРНК связанных с ВМР белков в человеческих клетках ТМ. Окрашенный бромистым этидием агарозный гель продуктов ПЦР из проб кДНК, полученных при анализе ОТ-ПЦР человеческих клеток ТМ (полосы 1-5). L = маркеры пар оснований. С = отрицательная контрольная полоса при ПЦР. β-актин использовали в качестве положительного внутреннего контроля при проведении ОТ-ПЦР.
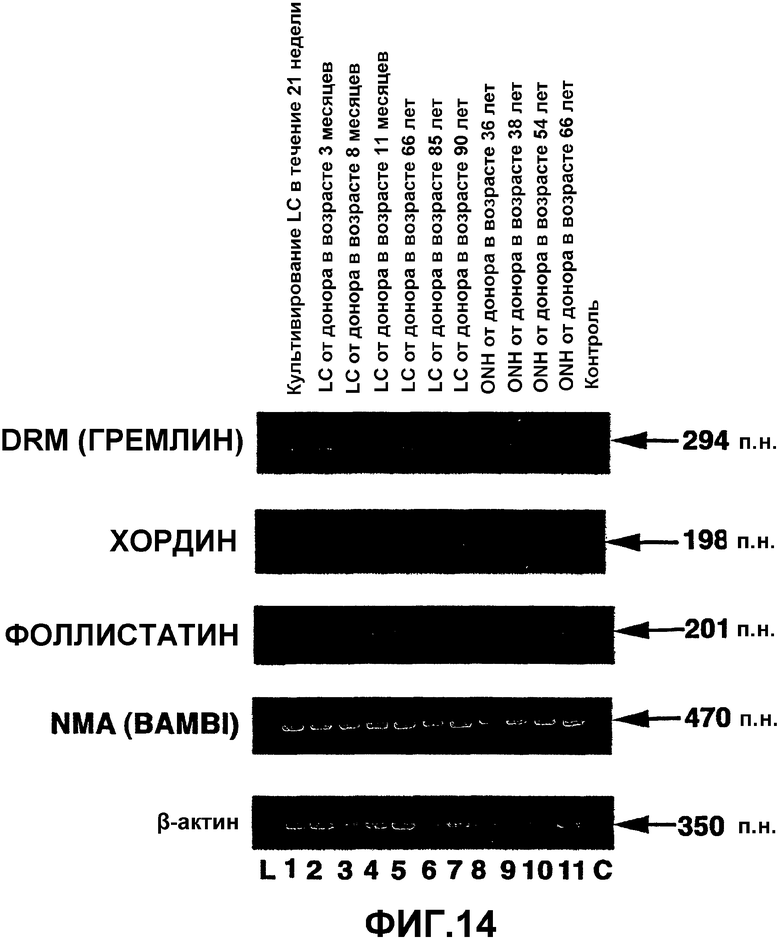
Фиг.14. Экспрессия мРНК связанных с ВМР белков в человеческих клетках решетчатой полости склеры и астроцитах ONH. Окрашенный бромистым этидием агарозный гель продуктов ПЦР из проб кДНК, полученных при анализе ОТ-ПЦР клеток решетчатой полости склеры (LC) (полосы 1-7) и астроцитов ONH (полосы 8-11). L = маркеры пар оснований. С = отрицательная контрольная полоса при ПЦР. β-актин использовали в качестве положительного внутреннего контроля при проведении ОТ-ПЦР.
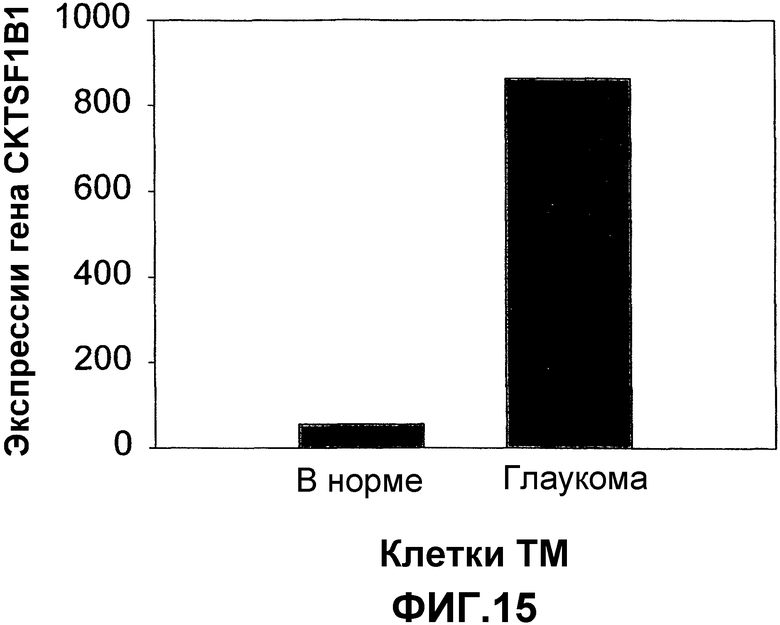
Фиг.15. Иллюстрация повышенной экспрессии антагониста ВМР гремлина (CКTSF1B1) в клетках ТМ при глаукоме. Экспрессию гена оценивали с использованием совокупности генов Affymetrix (генный чип U133А Affymetrix).
Подробное описание предпочтительных воплощений
Было высказано предположение, что трабекулярная сеть играет важную роль в движении глазной жидкости в норме, и полагается, что она представляет основное место сопротивления оттоку в глазе при глаукоме. Клетки трабекулярной сети человека (НТМ) являются специализированными клетками, которые образуют каналы для оттока, по которым глазная жидкость выходит из глаза. Измененная синтетическая функция клеток может участвовать в патогенезе POAG, стероидной глаукомы и других типов глаукомы.
Несмотря на годы интенсивных исследований, точный молекулярный механизм, ответственный за повреждения глаза, возникающие в результате глаукомы, остается неизвестным. В результате недавно проведенных исследований было высказано предположение, что факторы роста могут быть важными в поддержании нормального гомеостаза в тканях глаза, связанных с развитием глаукомы, и изменения факторов роста/рецепторов факторов роста могут играть роль в патогенезе глаукомы. Факторы роста представляют очень большое семейство полипептидов, которые регулируют рост и дифференциацию клеток. Данные молекулы обладают различными специфичными для клеток эффектами в отношении экспрессии генов, состава и отложения внеклеточного матрикса, организации цитоскелета и регуляции функции клеток. ТМ экспрессирует широкий ряд факторов роста, рецепторов факторов роста (Tripathi et al., 1993a; Tripathi et al., 1993b; Tripathi et al., 1994a; Tripathi et al., 1994b; Wordinger et al., 1998; Wordinger et al., 1999), а также нейротрофинов/нейротрофических факторов и их рецепторов (Liu et al., 2001; Wordinger et al., 2000). Астроциты ONH и клетки решетчатой полости склеры, два типа клеток головки зрительного нерва, экспрессируют факторы роста, нейротрофины и их рецепторы (Lambert et al., 2001; Pena et al., 1999). Глазная жидкость также содержит различные факторы роста, включая FGF2, EGF, TGFβ, HGH (Tripathi et al., 1996; Tripathi et al., 1991; Tripathi et al., 1992; Hu and Ritch, 2001), а также нейротрофины (Chundru et al., 2000). Сообщалось о повышенном уровне TGFβ-2 и HGF в глазной жидкости пациентов с POAG (Tripathi et al., 1994c; Inatani et al., 2001; Picht et al., 2001). Факторы роста могут участвовать в развитии глаукомы, изменяя нормальное развитие и/или функцию ТМ и ONH.
Настоящее изобретение частично происходит из представлений о том, что морфогенные белки кости (ВМР) индуцируют образование не только кости и хряща, но являются многофункциональными цитокинами, оказывающими широкий ряд воздействий на многочисленные типы клеток (Hogan, 1996; Reddi, 1997), и экспрессируются как клетками трабекулярной сети (НТМ), так и головки зрительного нерва (ONH) человека (Wordinger et al., 2002). ВМР являются членами суперсемейства TGFβ, и у человека имеется примерно 15-20 генов ВМР, 3 ВМР-рецептора и ряд связанных с ВМР белков, которые функционируют в качестве антагонистов ВМР (Yamashita et al., 1996). ВМР передают сигнал через рецепторный комплекс, состоящий из BMPR-I и BMPR-II. Сообщалось, что члены суперсемейства TGFβ и TGFβR (Agarwal et al., 1997; Lambert et al., 1997) и GDNF и GDNFR (Wordinger et al., 1999; Liu et al., 1999) экспрессируются как клетками НТМ, так и ONH.
ВМР и ВМР-рецепторы экспрессируются в тканях глаза (Obata et al., 1999; You et al., 1999), но предшествующие сообщения в основном касались развития глаза. Функция ВМР является важной для развития глаза, поскольку направленный разрыв генов, кодирующих ВМР у мышей, приводит к тяжелым дефектам развития в сетчатке и хрусталике (Jena et al., 1997; Luo et al., 1995; Dudley et al., 1995). ВМР-2, ВМР-4 и ВМР-7 принимают участие в развитии хрусталика и сетчатки (Jena et al., 1997; Furuta and Hogan, 1998; Reddi, 2000; Trousse et al., 2001). Оказалось, что ВМР-6 и ВМР-7 играют роль в защите нейронов от повреждения в результате гипогликемии или ишемии (Nonner et al., 2001; Liu et al., 2001), и было показано, что ВМР-2 повышает экспрессию нейротрофина в ганглиозных клетках (Zhang et al., 1998). Гетерозиготные "нокаутные" мыши с гаплонедостаточностью по Bmp4 имеют зрительные фенотипы, включающие дисгенез переднего сегмента, повышенное IOP и аномалии зрительного нерва (Chang et al., 2001). Имеется очень ограниченная опубликованная информация, касающаяся роли ВМР для глаза человека в постнатальном периоде.
Mohan и коллеги (1998) сообщали, что ВМР-2-, ВМР-4- и ВМР-рецепторы экспрессировались в клетках роговицы взрослого человека, и предположили, что в функции ВМР может входить воздействие на пролиферацию и апоптоз кератоцитов роговицы. You и коллеги (1999) подтвердили результаты данного исследования и также сообщили об экспрессии ВМР-3, ВМР-5 и ВМР-7 в условиях ex vivo и культивированных клетках эпителия роговицы и стромы. Они сообщили, что уровень транскрипции ВМР был выше в строме, в то время как уровень рецепторов был выше в культивированных эпителиальных клетках роговицы.
С использованием ОТ-ПЦР авторы настоящего изобретения выявили мРНК ВМР, ВМР-рецепторов BMPR-IА, BMPR-IВ и BMPR-II, а также связанных с ВМР белков гремлина, хордина, фоллистатина и бамби в НТМ, решетчатой полости склеры (LC) и клеточных линиях астроцитов и тканях ONH (Wordinger et al., 2002). Кроме того, авторы настоящего изобретения обнаружили, что клетки НТМ и ONH экспрессируют белки ВМР-2, ВМР-4, ВМР-5 и ВМР-7.
Глаукому можно будет диагностировать по обнаружению генетических изменений в генах членов сигнального семейства ВМР. В том смысле, в котором они здесь используются, выражения "ген, член семейства морфогенных белков кости" и "сигнальное семейство ВМР" относится к ВМР, ВМР-рецепторам и связанным белкам. Термин "генетические изменения" хорошо известен специалистам в данной области. Существуют многочисленные примеры заболеваний, связанных с генетическими изменениями в определенных генах (см. примеры у Cummings, 1997; Strachan et al., 1996; Jorde et al., 1999). Генетические изменения в конкретном гене (например, ВМР) можно определить с использованием различных методов, хорошо известных специалистам в данной области, таких как SSCP, DGGE, ASO, RFLP, гетеродуплексный анализ, CCM, PTT и расщепление РНКазой (см. Birren et al., 1998).
Глаукома может быть вызвана измененной экспрессией одного или более генов семейства ВМР в глазе, что приводит к повышенному IOP и/или нейропатии зрительного нерва при глаукоме. "Измененная экспрессия гена ВМР" означает экспрессию продукта данного гена, которая отличается от таковой в норме. Термин может также относится к изменениям в последовательности гена или белка. Ген ВМР в норме был хорошо охарактеризован (см. выше), и сообщалось об экспрессии ВМР в различных тканях, включая ТМ и ONH. Генетические изменения в кодирующей области генов семейства ВМР могут изменять функцию данных белков. Генетические изменения вне кодирующей области также могут привести к глаукоме.
Специалистам в данной области хорошо известно, что "изменения вне" кодирующей области конкретного гена являются важными для регуляции экспрессии гена. Например, известно, что область выше (5') кодирующей области большинства генов служит в качестве промоторной области, которая "способствует" и регулирует экспрессию данного гена. Промоторная область содержит многочисленные нуклеотидные последовательности, распознаваемые различными факторами транскрипции и ДНК-связывающими белками, которые ответственны за активацию или подавление экспрессии гена. Нижние области (3') гена могут также определять полиаденилирование продукта гена, тем самым регулируя процессинг РНК и трансляцию продукта гена.
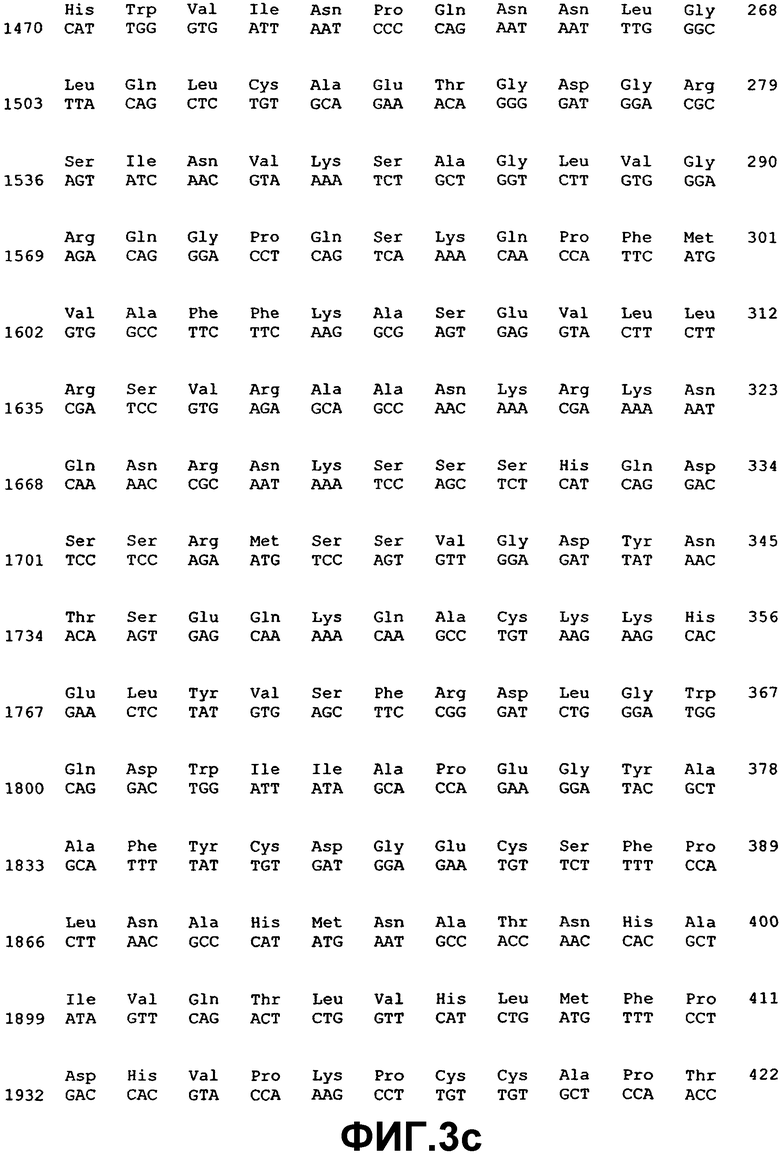
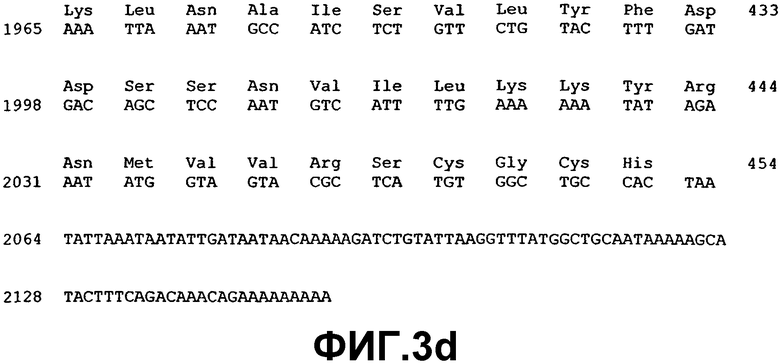
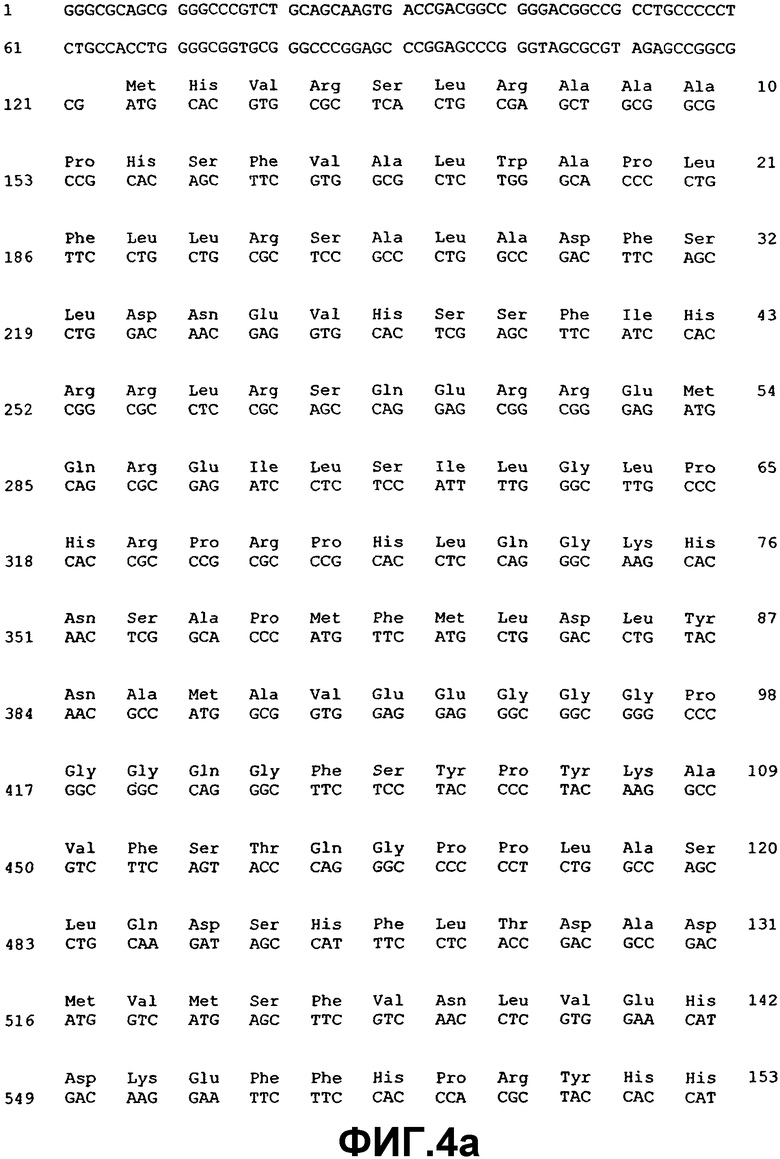
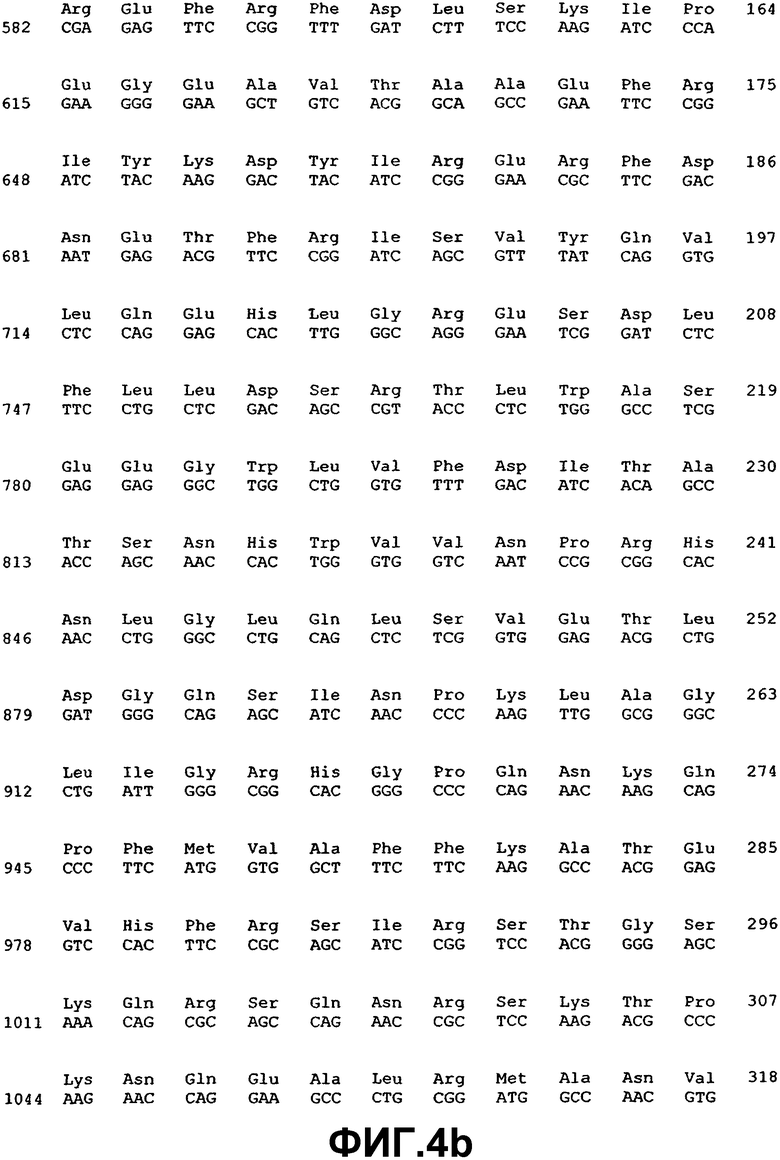
Измененную экспрессию генов ВМР или мутации в последовательности генов, которая указывает на наличие глаукомы, можно детектировать с использованием методов, хорошо известных специалистам в данной области. Например, предусматривается, что можно использовать фрагмент нуклеиновой кислоты практически любой длины, при полной длине предпочтительно ограничивать простотой получения и применения в предназначенном протоколе. Последовательности нуклеиновой кислоты, раскрытые здесь, могут также применяться в качестве зондов или праймеров в воплощениях гибридизации нуклеиновых кислот. Если это так, то предполагается, что сегменты нуклеиновой кислоты, которые включают область последовательности, состоящей из последовательности, по меньшей мере, из 14 смежных нуклеотидов, которая имеет ту же последовательность, или комплементарную последовательность из 14 смежных нуклеотидов ВМР-2А (SEQ ID NO:1), ВМР-4 (SEQ ID NO:3), ВМР-5 (SEQ ID NO:5), ВМР-7 (SEQ ID NO:7), ВМР-RIA (SEQ ID NO:37), ВМР-RIB (SEQ ID NO:39), ВМР-RII (SEQ ID NO:41), хордина (SEQ ID NO:43), гремлина (SEQ ID NO:45), фоллистатина (SEQ ID NO:47) или бамби (SEQ ID NO:53), будет иметь особую применимость. Также в некоторых воплощениях можно использовать смежные идентичные или комплементарные последовательности большей длины, например, примерно из 20, 30, 40, 50, 100, 200, 500, 1000 нуклеотидов (включая все последовательности с промежуточными длинами) и вплоть до полноразмерных последовательностей примерно из 1547 нуклеотидов (для ВМР-2А), 1946 нуклеотидов (для ВМР-4), 2153 нуклеотидов (для ВМР-5) и 1878 нуклеотидов (для ВМР-7), 2932 нуклеотидов (для ВМР-RIA), 2032 нуклеотидов (для ВМР-RIB), 3611 нуклеотидов (для ВМР-RII), 3561 (для хордина), 4049 нуклеотидов (для гремлина), 1386 нуклеотидов (для фоллистатина) и 1523 нуклеотидов (для бамби).
Легко понять, что "последовательности с промежуточными длинами" в данном контексте означают любую длину между указанными пределами, такую как 14, 15, 16, 17, 18, 19, 20 и т.д.; 21, 22, 23 и т.д., 30, 31, 32 и т.д., 50, 51, 52, 53 и т.д., 100, 101, 102, 103 и т.д., 150, 151, 152, 153 и т.д., включая все целые числа в пределах 200-500, 500-1000, 1000-2000, до и включая последовательности из 2001, 2002, 2050, 2051 нуклеотидов и тому подобное.
Способность таких нуклеотидных зондов и праймеров специфически гибридизоваться с кодирующими последовательностями ВМР и праймерами для специфической амплификации последовательностей ВМР позволит использовать их для детектирования наличия комплементарных последовательностей в данной пробе. Однако предусматриваются другие применения, включая применение информации по секвенированию для получения праймеров мутантных видов или праймеров для получения других генетических конструкций.
Молекулы нуклеиновой кислоты, имеющие области последовательности, состоящие из участков смежных нуклеотидов длиной 10, 20, 30, 50 и даже 100-200 нуклеотидов или так далее, идентичные или комплементарные последовательностям ВМР-2А (SEQ ID NO:1), ВМР-4 (SEQ ID NO:3), ВМР-5 (SEQ ID NO:5), ВМР-7 (SEQ ID NO:7), ВМР-RIA (SEQ ID NO:37), ВМР-RIB (SEQ ID NO:39), ВМР-RII (SEQ ID NO:41), хордина (SEQ ID NO:43), гремлина (SEQ ID NO:45), фоллистатина (SEQ ID NO:47) или бамби (SEQ ID NO:53), особенно пригодны для использования в качестве зондов для гибридизации, например, при оценке SNP и в тестах гибридизации на твердой фазе, в дополнение к саузерн- и нозерн-блоттингу. Это позволит проводить анализ структурных и регуляторных генов ВМР в тканях и клетках. Полный размер фрагмента, а также размер комплементарного участка(ов), в конечном итоге будет зависеть от предназначаемого применения конкретного сегмента нуклеиновой кислоты. Меньшие фрагменты, как правило, будут применяться при гибридизации, когда длина смежной комплементарной области может варьировать, например, в пределах примерно от 10 до примерно 100 нуклеотидов, но можно использовать более крупные смежные комплементарные участки вплоть до примерно 1547 нуклеотидов (для ВМР-2А), 1946 нуклеотидов (для ВМР-4), 2153 нуклеотидов (для ВМР-5) и 1878 нуклеотидов (для ВМР-7), 2932 нуклеотидов (для ВМР-RIA), 2032 нуклеотидов (для ВМР-RIB), 3611 нуклеотидов (для ВМР-RII), 3561 (для хордина), 4049 нуклеотидов (для гремлина), 1386 нуклеотидов (для фоллистатина) и 1523 нуклеотидов (для бамби) нуклеотидов, соответственно длине комплементарных последовательностей, которые желательно детектировать.
При применении зонда для гибридизации длиной примерно 10-14 нуклеотидов образуется дуплексная молекула, которая одновременно является стабильной и избирательной. Молекулы, имеющие смежные комплементарные последовательности к участкам длиной более 10 оснований, в основном являются предпочтительными, хотя для повышения стабильности и избирательности гибрида, и тем самым для улучшения качества и степени получаемых конкретных гибридных молекул, как правило, предпочтительно конструировать молекулы нуклеиновых кислот, имеющих комплементарные участки для генов, состоящих из 15-20 смежных нуклеотидов или, при необходимости, длиннее.
Зонды для гибридизации можно выбрать из любого участка любой последовательности, раскрытой здесь. Все, что требуется, это проанализировать представленные последовательности SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47 или SEQ ID NO:53 и выбрать любой смежный участок последовательности длиной примерно от 10 нуклеотидов и вплоть до полноразмерной последовательности, который желательно использовать в качестве зонда или праймера. При выборе последовательностей зонда и праймера можно руководствоваться различными факторами, только в качестве примера, например, желательно ли использовать праймеры к концам целой последовательности или от концов функциональных кодирующих домен последовательностей, для последующей амплификации ДНК.
Способ отбора и получения сегмента нуклеиновой кислоты, который включает смежную последовательность из последовательностей ВМР-2А (SEQ ID NO:1), ВМР-4 (SEQ ID NO:3), ВМР-5 (SEQ ID NO:5), ВМР-7 (SEQ ID NO:7), ВМР-RIA (SEQ ID NO:37), ВМР-RIB (SEQ ID NO:39), ВМР-RII (SEQ ID NO:41), хордина (SEQ ID NO:43), гремлина (SEQ ID NO:45), фоллистатина (SEQ ID NO:47) или бамби (SEQ ID NO:53), альтернативно может быть определен как получение фрагмента нуклеиновой кислоты. Конечно, фрагменты также можно получить другими методами, такими как, например, механическое гидродинамическое фрагментирование ДНК или расщепление рестриктазой. Небольшие сегменты или фрагменты нуклеиновой кислоты можно легко получить, например, прямым синтезом фрагмента химическими методами, как это широко распространено на практике с использованием автоматического синтезатора олигонуклеотидов. Кроме того, фрагменты можно получить применением методов репродукции нуклеиновой кислоты, такой как технология ПЦР™ согласно патенту США № 4683202 и патенту США № 4682195 (каждый включен здесь для сведения), введением отобранных последовательностей в рекомбинантные векторы для рекомбинантной продукции, и другими методами рекомбинантной ДНК, хорошо известными специалистам в области молекулярной биологии.
Следовательно, нуклеотидные последовательности по изобретению можно использовать по их способности к селективному образованию дуплексных молекул с комплементарными участками генов или кДНК ВМР. В зависимости от предполагаемого подхода будет желательно использовать различные степени избирательности гибридизации для достижения различной степени избирательности зонда относительно последовательности-мишени. Для применений, где требуется высокая избирательность, как правило, желательно использовать относительно жесткие условия для образования гибридов, например, следует выбрать условия с относительно низкой концентрацией соли и/или высокой температурой, такие как обеспечиваются при использовании 0,02-0,15 М NaCl при температуре 50-70°С. В подобных условиях имеется небольшая, если вообще таковая существует, вероятность ошибочного спаривания зонда и матрицы или цепи-мишени, и они особенно подходят для исследования генов ВМР.
Конечно, для некоторых применений, например, когда желательно получить или идентифицировать мутанты с использованием цепи мутантного праймера, гибридизованного с лежащей в основе матрицей, или когда необходимо выделить кодирующие последовательности ВМР из близких видов, функциональных эквивалентных вариантов и тому подобное, как правило, требуются менее жесткие условия гибридизации для образования гетеродуплекса. В данных обстоятельствах может быть желательным использовать такие условия, как концентрация соли 0,15-1,0 М при температуре в пределах примерно от 20 до 55°С. Тем самым можно легко идентифицировать перекрестно-гибридизующиеся виды в качестве положительно гибридизующихся сигналов по отношению к контрольной гибридизации. В любом случае в целом предусматривается, что условия можно сделать более жесткими при снижении концентрации NaCl или добавлении возрастающих количеств формамида, который служит для дестабилизации гибридного дуплекса так же, как повышенная температура. Таким образом, можно легко манипулировать условиями гибридизации, и, таким образом, имеется метод выбора в зависимости от желаемых результатов.
В некоторых воплощениях будет преимущественным использовать последовательности нуклеиновых кислот по настоящему изобретению в сочетании с подходящими средствами, такими как метка, для определения результатов гибридизации. В данной области известны самые различные соответствующие индикаторные средства, включая флуоресцентные, радиоизотопные, ферментативные или другие лиганды, такие как авидин/биотин, которые способны воспроизводить детектируемый сигнал. В предпочтительных воплощениях желательно использовать флуоресцентную метку или ферментативную метку, такую как уреаза, щелочная фосфатаза или пероксидаза, вместо радиоактивного или других нежелательных для окружающей среды реагентов. В случае ферментативных меток известны индикаторные субстраты для колориметрии, которые можно использовать для обеспечения средств, видимых человеческому глазу или обнаруживаемых при спектрофотометрии, для идентификации специфической гибридизации с пробами, содержащими комплементарные нуклеиновые кислоты.
В общем предусматривается, что зонды для гибридизации, описанные здесь, будут пригодными в качестве реагентов как для гибридизации в растворе, так и в воплощениях с использованием твердой фазы. В воплощениях с твердой фазой анализируемая ДНК (или РНК) адсорбируется или фиксируется иначе на выбранном матриксе или поверхности. Затем данную фиксированную одноцепочечную нуклеиновую кислоту подвергают специфической гибридизации с отобранными зондами в желаемых условиях. Выбранные условия будут зависеть от конкретных обстоятельств, основанных на конкретных требуемых критериях (в зависимости, например, от содержания G+C, типа нуклеиновой кислоты-мишени, источника нуклеиновой кислоты, размера зонда для гибридизации и т.д.). После промывания гибридизованной поверхности для удаления неспецифически связанных молекул зонда, проводят детектирование специфической гибридизации или даже количественное определение, с помощью метки.
Также следует понимать, что данное изобретение не ограничивается конкретными нуклеотидными и аминокислотными последовательностями ВМР-2А (SEQ ID NO:1), ВМР-4 (SEQ ID NO:3), ВМР-5 (SEQ ID NO:5), ВМР-7 (SEQ ID NO:7), ВМР-RIA (SEQ ID NO:37), ВМР-RIB (SEQ ID NO:39), ВМР-RII (SEQ ID NO:41), хордина (SEQ ID NO:43), гремлина (SEQ ID NO:45), фоллистатина (SEQ ID NO:47) или бамби (SEQ ID NO:53). Так, рекомбинантные векторы и выделенные сегменты ДНК могут включать кодирующие области ВМР сами по себе, "верхние" или "нижние" гены, кодирующие области, несущие выбранные изменения или модификации основной кодирующей области, или они могут кодировать более крупные полипептиды, которые, тем не менее, включают кодирующие области ВМР, или могут кодировать биологически эквивалентные функциональные белки или полипептиды, имеющие вариантные аминокислотные последовательности.
Сегменты ДНК по настоящему изобретению включают биологически эквивалентные функциональные белки и полипептиды ВМР. Данные последовательности можно получить в результате избыточности кодона и функциональной эквивалентности, которые, как известно, имеют место в естественных условиях в последовательностях нуклеиновой кислоты и белках, их кодируемых. Альтернативно, функционально эквивалентные белки или полипептиды можно получить с помощью применения технологии рекомбинантной ДНК, с помощью которой можно воспроизвести изменения в структуре белка, основанные с учетом свойств аминокислот, которые подвергаются замене. Сконструированные человеком изменения можно ввести с помощью методов сайт-направленного мутагенеза, например, для повышения антигенных свойств белка или тестирования мутантов ВМР для исследования связывающей активности на молекулярном уровне.
Терапевтическое средство для лечения глаукомы может быть пептидом или белком, пептидомиметиком, олигонуклеотидом или дериватизированным олигонуклеотидом, или небольшой подобной лекарственному препарату молекулой, которые воздействуют на одну или более сторон зрительных путей с участием ВМР. Предпочтительными терапевтическими средствами являются: (1) агонисты ВМР-2, ВМР-4, ВМР-5 или ВМР-7; (2) антагонисты хордина, гремлина, фоллистатина или бамби; и/или (3) агонисты Smad1, Smad5 и/или Smad4.
Средство можно непосредственно вводить в глаз (например, в виде глазных капель или мазей для местного применения; с помощью устройств с замедленным высвобождением в слепой мешок или имплантированных около склеры или внутри глаза; посредством периокулярной, конъюнктивальной, в теноновую фасцию, в камеру или в стекловидное тело инъекций) или парентерально (например, перорально, с помощью внутривенной, подкожной или внутримышечной инъекций; через кожу и т.д.) с использованием методов, хорошо известных в данной области. Последующее представляет примеры возможных композиций, воплощенных данным изобретением.
Дополнительно предусматривается, что соединения по изобретению можно приготовить в виде устройств для внутриглазной доставки.
А. Анализ терапевтических средств
Данное изобретение также пригодно для обнаружения новых терапевтических средств против глаукомы, которые принимают участие в путях передачи сигналов с ВМР (см. фиг.5). Избирательные лиганды ВМР связываются с ВМР-рецепторами серин/треонинкиназами типа I и типа II (ВМР-RI и ВМР-RII) и передают сигнал через Smad-белки. Сигнал ВМР распространяется Smad через белок-белковые и белок-ДНК взаимодействия (Attisano and Tuen Lee-Hoeflich, 2001). Регуляторные белки Smad1 и Smad5 активируются (посредством фосфорилирования) связанными с лигандом ВМР-рецепторами (Bubnoff and Cho, 2001). Затем данные регуляторные белки Smad взаимодействуют с Smad4 с образованием гетеромерного комплекса, который перемещается в ядро. Данный комплекс способен активировать или подавлять транскрипцию избирательных генов, которые распознают данный транскрипционный комплекс в зависимости от того, какие ядерные кофакторы присутствуют.
Путь передачи сигналов с участием ВМР/Smad отрицательно регулируется несколькими механизмами. Некоторые ВМР-связывающие белки (такие как гремлин, BABMI или фоллистатин) связывают ВМР и подавляют их взаимодействие с ВМР-рецепторами. Кроме того, имеются ингибирующие Smad-белки (например, Smad6 и Smad7), которые связываются и инактивируют ВМР-рецепторы (Kowabata et al., 1998; Itoh et al., 2000; Miyazono, 2000). Авторы настоящего изобретения установили, что человеческие клетки ТМ, астроциты ONH и клетки решетчатой полости склеры экспрессируют мРНК и белок для рецепторного комплекса ВМР. Таким образом, данные клетки могут давать ответную реакцию на эндогенные лиганды ВМР.
Для обнаружения новых терапевтических средств против глаукомы можно использовать различные методы, и данные методы хорошо известны специалистам в данной области. Например, средства на основе пептидов или пептидомиметиков, которые действуют в качестве агонистов или ингибиторов ВМР, можно установить с помощью молекулярного моделирования ВМР/ВМР-рцепторных структур (Nickel et al., 2001). Передача сигнала с участием ВМР включает избирательные группы Smad-белков (Kawabata et al., 1998; Itoh et al., 2000; Attiseno et al., 2000). Избирательные агонисты ВМР и агонисты Smad можно обнаружить с использованием клеточных тестов. Тестируемая клетка должна экспрессировать соответствующие ВМР-рецептор(ы) и располагать соответствующим путем передачи сигнала с участием ВМР. Поскольку основными эффектами передачи сигнала ВМР является изменение экспрессии гена, агонисты ВМР и агонисты Smad можно обнаружить при скрининге ВМР-индуцированных генов. Индукцию регулируемых ВМР генов также можно измерить путем количественного определения уровней мРНК с использованием количественной ОТ-ПЦР (Wang et al., 2001), микросовокупностей ДНК или конструкций репортерного гена. Имеются природные ингибиторы передачи сигнала с участием ВМР, ВМР-связывающие белки (известные как связанные с ВМР белки) такие, как хордин, гремлин и фоллистатин. Антагонисты ингибиторов белков можно обнаружить с использованием тестов связывания лиганда. Например, анализируемые агенты можно добавить к рекомбинантному очищенному гремлину, и идентифицировать те агенты, которые будут связываться с гремлином, с использованием различных методов, известных специалистам в данной области. Для определения того, являются ли данные агенты антагонистами гремлина, используют клеточный тест, аналогичный описанному выше.
Предполагается, что в сочетании с настоящим изобретением можно использовать любую известную модель скрининга in vitro и in vivo для идентификации новых лекарственных средств против глаукомы, обладающих воздействием на семейство генов ВМР. Подобные модели хорошо известны специалистам в данной области и их постановка на практике является общепринятой. Небольшие пептиды или пептидомиметиков можно сконструировать, основываясь на представлениях о структуре/функции ВМР, BMPR и/или продуктов гена, связывающихся с ВМР белков. Можно использовать тесты связывания лиганда для детектирования небольших молекул, которые связываются с ВМР, BMPR или белками, связывающимися с ВМР. При постановке клеточных тестов можно обнаружить действие различных агентов на пути передачи сигнала с участием ВМР. Можно получить клеточные линии "нокин", включающие промоторы генов семейства ВМР, связанные с репортерным геном, для обнаружения средств, которые изменяют экспрессию члена семейства генов ВМР. Данные тесты можно использовать для идентификации как молекул агонистов, так и антагонистов. Тесты ex vivo такие, как культивирование передних сегментов человеческого глаза при перфузии (Clark et al., 1995a; Pang et al., 2000), можно использовать для исследования эффектов средств на IOP и передачу сигнала с участием ВМР в ткани ТМ. Можно создать модели глаукомы на грызунах с использованием хорошо известных методов для создания стабильных трансгенных с членом семейства ВМР, без "нокаутных" или "нокиновых" штаммов мышей и крыс. Данные модели на грызунах можно использовать для скрининга средств, которые изменяют глаукомоподобный фенотип(ы) (например, с помощью тонометрии определяют воздействие на IOP, с помощью гистологии оценивают воздействие на нейрологию зрения при глаукоме).
В. Наборы
Настоящее изобретение обеспечивает способы, композиции и наборы для раннего обнаружения глаукомы. Наборы могут включать сегмент нуклеиновой кислоты, кодирующей полипептид или белок ВМР. Набор может дополнительно включать реагенты для детектирования взаимодействия между пробой и нуклеиновой кислотой или пептидом по настоящему изобретению. Обеспеченный реагент может включать радиоизотопную, флуоресцентную или ферментативную метку. Набор может содержать известный меченый радиоизотопом агент, способный связываться или взаимодействовать с нуклеиновой кислотой или пептидом, или белком по настоящему изобретению.
Реагент набора может быть обеспечен в виде жидкого раствора, связанным с твердой подложкой или в виде сухого порошка. Предпочтительно, когда реагент обеспечивается в виде жидкого раствора, в этом случае жидкий раствор представляет собой водный раствор. Предпочтительно, когда обеспеченный реагент связан с твердой подложкой, в этом случае твердая подложка может быть средой для хроматографии, аналитическим планшетом, имеющим множество лунок, или микроскопическим стеклом. Когда обеспеченный реагент представляет собой сухой порошок, то порошок можно восстановить добавлением подходящего растворителя, который также может находиться в наборе.
В еще дополнительных воплощениях настоящее изобретение касается способов диагностики и связанных наборов для диагностики глаукомы. Предполагается, что связанные с ВМР пептиды и нуклеиновые кислоты по изобретению можно использовать для детектирования полиморфизма или мутаций в нуклеиновых кислотах ВМР из проб пациентов. В основном данные способы будут включать в начале получение пробы с подозрением на присутствие полиморфизма или мутации, контактирование пробы с пептидом или нуклеиновой кислотой по настоящему изобретению в условиях, эффективных для образования комплекса и затем детектирование присутствия комплекса.
В основном детектирование образования комплекса довольно хорошо известно в данной области и его можно проводить при использовании различных подходов. Например, настоящее изобретение предусматривает применение ELISA, RIA, методов непрямой флуоресценции и тому подобное. Обычно образование комплекса будет детектироваться при использовании метки, такой как радиоактивная метка или ферментативная метка (такая как щелочная фосфатаза, пероксидаза хрена и тому подобное). Конечно, можно найти дополнительные преимущества при использовании вторично связывающегося лиганда.
Последующие примеры представляют методы, использованные авторами изобретения для осуществления аспектов настоящего изобретения. Несмотря на то, что данные методы представляют примеры предпочтительных воплощений на практике изобретения, специалисты в данной области в свете настоящего описания должны понимать, что можно сделать многочисленные модификации, не отступая от сущности и предполагаемого объема изобретения.
Пример 1
Культура клеток: человеческие клетки ТМ и клетки ONH получали из глаза доноров, как описано (Steely et al., 1992; Steely et al., 2000; Wilson et al., 1993; Clark et al., 1994; Clark et al., 1995b; Clark et al., 1995c; Clark et al., 1996; Clark et al., 2001a; Clark et al., 2001b; Dickerson et al., 1998; Wordinger et al., 1998; Wordinger et al., 1999; Wordinger et al., 2000; Wordinger et al., 2002; Lambert et al., 2001; Agarwal et al., 1999; Liu et al., 2001). Клетки ТМ культивировали из эксплантатов ТМ от доноров в возрасте от 6 дней до 90 лет. Астроциты головки зрительного нерва и клетки решетчатой полости склеры (LC) человека получали из аккуратно отпрепарированных головок оптического нерва (доноры в возрасте от 2 дней до 90 лет) и охарактеризовали, как описано ранее (Lambert et al., 2001; Clark et al., 1995a). Клетки культивировали до слияния в следующей среде: среда Хэма F10 (JRH Biosciences, Lenexa, KS), содержащая 10% фетальной бычьей сыворотки (FBS) (HyClone, Logan, UT) и антибиотики (Gibco BRL-Life Technologies, Grand Island, NY) для клеток ТМ; модифицированная по Дульбекко среда Игла (DMEM, HyClone), содержащая 10% FBS для клеток LC; и среда для культивирования астроцитов (AGM, Clonetics, San Diego, CA), содержащая 5% FBS для астроцитов ONH.
ОТ-ПЦР: ткани ТМ и ONH также иссекали из глаза доноров (Wordinger et al., 1998; Wang et al., 2001). Общую фракцию РНК выделяли из клеток и тканей ТМ и ONH с использованием экстракции TRIzol (Gibco BRL-Life Technologies) и кДНК получали обратной транскрипцией с использованием обычных методов (Wordinger et al., 1998; Wordinger et al., 1999; Wordinger et al., 2000; Wordinger et al., 2002). Праймеры для ПЦР конструировали с использованием программного обеспечения Oligos 4,0 (см. пары праймеров в таблице 1). Все пары праймеров конструировали таким образом, что в результате амплификации потенциально загрязненных последовательностей геномной ДНК получали продукты ПЦР, мРНК, которые были значительно больше, чем предполагалось, поскольку интронные последовательности, которые вырезали во время процессинга РНК, включались в геномную ДНК. Праймеры ПЦР β-актина AGGCCAACCGCGAGAAGATGACC (верхние) и GAAGTCCAGGGCGACGTAGCAC (нижние) при температуре отжига 55°С давали продукт ПЦР размером 350 п.н.
Реакцию ПЦР проводили как описано (Wordinger et al., 1998; Wordinger et al., 1999; Wordinger et al., 2000; Lambert et al., 2001; Wordinger et al., 2002) с использованием Taq Start Antibody Hot Start при следующих условиях циклов: 2 минуты при 94°С, 2 минуты при 92°С и 40 циклов в течение 30 секунд при оптимальной температуре отжига, удлинения в течение 90 секунд при 72°С и денатурация в течение 45 секунд при 92°С. Амплифицированные продукты ПЦР анализировали горизонтальным электрофорезом в 1,5% агарозном геле. Для обеспечения специфичности продуктов ОТ-ПЦР проводили саузерн-блоттинг с зондами, сконструированными с использованием Oligo 4.0, которые гибридизовали с областью в амплифицированном продукте ПЦР. Продукты ПЦР секвенировали для подтверждения специфичности реакции ПЦР. В таблице 2 представлены члены семейства ВМР, которые экспрессируются в ТМ и ONH человека.

Члены семейства ВМР, экспрессированные в ТМ и ONH человека
Вестерн-иммуноблоттинг: белок выделяли из культивированных клеток с использованием лизирующего буфера, и белки разделяли электрофорезом в полиакриламидном геле в денатурирующих условиях перед электрофоретическим переносом на нитроцеллюлозные мембраны (Lambert et al., 2001). Мембраны блокировали 5% молоком (для ВМР) или 3% желатином (для BMPR) и инкубировали со следующими первичными антителами против: ВМР-2, ВМР-4, ВМР-5, ВМР-7 (все производства Santa Cruz, Santa Cruz, CA) или ВМР-RIA, ВМР-RIB, ВМР-RII (производства Jackson Immuno Research, West Grove, PA). Мембраны промывали, инкубировали со вторичными антителами (козьи анти-мышиный IgG-пероксидаза хрена-антитела для ВМР, Santa Cruz; ослиные анти-козий IgG-пероксидаза хрена-антитела для ВМР-рцепторов, Jackson Immuno Research), и проявляли с использованием хемилюминесцентной системы для иммунодетектирования WesternBreeze (Invitrogen, Carlsbad, CA).
Экспрессия мРНК ВМР, BMPR в человеческих клетках и тканях ТМ: продукты амплификации предлагаемого размера для пар праймеров ВМР-2, ВМР-4, ВМР-5 и ВМР-7 в человеческих клетках и тканях ТМ представлены на фиг.6. При постановке саузерн-блоттинга с использованием специфических зондов было подтверждено, что это были предполагаемые продукты ПЦР. Все человеческие линии клеток и ткани ТМ экспрессировали мРНК для ВМР-2, ВМР-4 и ВМР-7. Однако уровень мРНК для ВМР-5 был низким вплоть до неопределяемого в пробах ткани ТМ человека (фиг.6, полосы 6 и 7). Контрольные реакции без кДНК не давали продуктов амплификации, что указывает на то, что реагенты и праймеры не были загрязнены ДНК или РНК (фиг.6, полоса С).
На фиг.7 показаны продукты амплификации предполагаемого размера для пар праймеров ВМР-RIA, ВМР-RIB и ВМР-RII в клетках и тканях ТМ человека. Все человеческие клетки и ткани ТМ экспрессировали мРНК для рецепторных комплексов ВМР. При постановке саузерн-блоттинга с использованием специфических праймеров было подтверждено, что это были предполагаемые продукты ПЦР. В реакции ПЦР с ВМР-RII детектировали альтернативный продукт амплификации (350 п.н.). Альтернативный продукт амплификации присутствовал во всех клетках и тканях ТМ человека. Данную альтернативную полосу идентифицировали для определения того, соответствует ли она альтернативной сплайсированной форме рецептора. Контрольные реакции без кДНК не давали продуктов амплификации (фиг.7, полоса С), что указывает на отсутствие загрязнения реагентов и праймеров ДНК или РНК.
Экспрессия мРНК ВМР и ВМР-рецепторов в человеческих клетках и тканях ONH: продукты амплификации предлагаемого размера для пар праймеров ВМР-2, ВМР-4, ВМР-5 и ВМР-7 в человеческих астроцитах ONH и тканях ONH представлены на фиг.8. Все астроциты ONH и ткань ONH экспрессировали мРНК для соответствующего ВМР. Астроциты мозга человека использовали в качестве положительной контрольной клеточной линии. При постановке саузерн-блоттинга с использованием специфических зондов было подтверждено, что это были предполагаемые продукты ПЦР. За исключением ВМР-2 все другие ВМР экспрессировались астроцитами мозга человека (фиг.8, полоса 7). Контрольные реакции без кДНК не давали продуктов амплификации (фиг.8, полоса С), что указывает на то, что реагенты и праймеры не были загрязнены ДНК или РНК.
На фиг.9 показаны продукты амплификации предполагаемых размеров для пар праймеров ВМР-2, ВМР-4, ВМР-5 и ВМР-7 в культивированных человеческих клетках LC. Все клеточные линии LC экспрессировали мРНК для каждого ВМР. При постановке саузерн-блоттинга было подтверждено, что это были предполагаемые продукты ПЦР. Контрольные реакции без кДНК не давали продуктов амплификации (фиг.9, полоса С), что указывает на то, что реагенты и праймеры не были загрязнены ДНК или РНК.
Продукты амплификации предполагаемого размера для пар праймеров ВМР-RIA, ВМР-RIB и ВМР-RII в человеческих астроцитах ONH и тканях ONH представлены на фиг.10. Все клеточные линии астроцитов и ткани ONH экспрессировали мРНК для ВМР-RIA и ВМР-RIB. При постановке саузерн-блоттинга с использованием специфических зондов было подтверждено, что это были предполагаемые продукты ПЦР. За исключением ткани ONH (фиг.10, полоса 6), ВМР-RII экспрессировался всеми клеточными линиями астроцитов ONH. мРНК для всех ВМР-рецепторов (фиг.10, полоса 7) экспрессировалась клеточной линией астроцитов мозга человека, которая служила в качестве положительного контроля. Оказалось, что имеются различия в экспрессии ВМР-RII в ткани ONH и клеточных линиях ONH. Низкая экспрессия в ткани ONH может отражать низкий уровень экспрессии. Контрольные реакции без кДНК не давали продуктов амплификации (фиг.5, полоса С), что указывает на то, что реагенты и праймеры не были загрязнены ДНК или РНК.
На фиг.11 показаны продукты амплификации предполагаемых размеров для пар праймер ВМР-RIA, ВМР-RIB и ВМР-RII в культивированных человеческих клетках LC. Все клеточные линии LC экспрессировали мРНК для каждого ВМР-рецептора. При постановке саузерн-блоттинга с использованием специфических зондов было подтверждено, что это были предполагаемые продукты ПЦР. Контрольные реакции без кДНК не давали продуктов амплификации (фиг.11, полоса С), что указывает на то, что реагенты и праймеры не были загрязнены ДНК или РНК.
Экспрессия ВМР-белков и ВМР-рецепторных белков в человеческих клетках и тканях ТМ и ONH: на фиг.12 представлены результаты детектирования белков ВМР-2, ВМР-4, ВМР-5, ВМР-7, ВМР-RIA, ВМР-RIB и ВМР-RII в человеческих клетках и тканях ТМ и ONH с помощью хемилюминесцентного иммуноблоттинга. Все исследованные клеточные линии экспрессировали соответствующие белки ВМР. Белки ВМР детектировали в клеточных линиях со следующими значениями молекулярной массы: 54-56 kDa для ВМР-2, 25-27 kDa для ВМР-4, 55-57 kDa для ВМР-5 и 77 kDa для ВМР-7. Для ВМР-2 и ВМР-4 детектировали множество полос, которые наиболее вероятно соответствуют гликозилированным и частично гликозилированным формам данных ВМР, как показано в других исследованиях. Однако авторы не проводили исследований по гликозилированию, поскольку они находились вне объема данного исследования. ВМР-рецепторные белки детектировали в клеточных линиях со значениями молекулярной массы: 38 kDa для ВМР-RIA, 64 kDa для ВМР-RIB и 57 kDa для ВМР-RII. Для ВМР-RIB и ВМР-RII детектировали множество полос в клетках ТМ, которые наиболее вероятно соответствуют гликозилированным и частично гликозилированным формам, как это следует из результатов других исследований. Оказалось, что уровни экспрессии белков ВМР-рецепторов ниже в клетках ТМ по сравнению с клетками ONH. Например, ВМР-RII не детектировали в клетках ТМ, и ВМР-RIB было значительно снижено.
Экспрессия мРНК связанных с ВМР белков в культивированных человеческих клетках ТМ и человеческих клетках ONH: продукты амплификации предполагаемого размера для пар праймеров белков, связанных с ВМР, в человеческих клеточных линиях ТМ, представлены на фиг.13. Человеческие линии ТМ экспрессировали мРНК для DRM (гремлина), хордина, фоллистатина и NMA (BAMBI). При постановке саузерн-блоттинга с использованием специфических зондов было подтверждено, что это были предполагаемые продукты ПЦР. Явное различие в экспрессии мРНК между клеточными линиями отсутствовало. Все исследованные человеческие клетки ТМ не экспрессировали мРНК для связанных с ВМР белков ноггина и Cer-1. Контрольные реакции без кДНК не давали продуктов амплификации, что указывает на то, что реагенты и праймеры не были загрязнены ДНК или РНК.
Продукты амплификации предполагаемого размера для пар праймеров связанных с ВМР белков в астроцитах ONH и клеточных линиях LC представлены на фиг.14. Все астроциты ONH и клеточные линии LC экспрессировали мРНК для DRM (гремлина), фоллистатина и NMA (BAMBI). При постановке саузерн-блоттинга с использованием специфических зондов было подтверждено, что это были предполагаемые продукты ПЦР. Большая часть клеток LC и астроцитов ONH экспрессировала мРНК для хордина. Все исследованные человеческие астроциты ONH и клетки LC не экспрессировали мРНК связанных с ВМР белков ноггина и Cer-1. Контрольные реакции без кДНК не давали продуктов амплификации, что указывает на то, что реагенты и праймеры не были загрязнены ДНК или РНК.
На фиг.15 показана повышенная экспрессия антагониста ВМР гремлина (CRTSF1B1) в клетках ТМ при глаукоме. Экспрессию гена оценивали с использованием совокупности генов Affymetrix (генные чипы U133A Affymetrix).
Все композиции и/или способы, раскрытые и заявленные в этом документе, можно осуществить и выполнить без излишнего экспериментирования в свете настоящего описания. Несмотря на то, что композиции и способы по данному изобретению описаны здесь в виде предпочтительных воплощений, специалистам в данной области, очевидно, понятно, что можно внести вариации в композиции и/или способы, и стадии или последовательность стадий способа, описанного здесь, не отступая от концепции, сущности и объема изобретения. Конкретнее, очевидно, понятно, что некоторые средства, близкие в химическом и структурном отношении, можно использовать здесь вместо средств, описанных здесь с достижением аналогичных результатов. Предполагается, что все замены и модификации, известные специалистам в данной области, находятся в соответствии с сущностью, объемом и концепцией изобретения, которые определены в прилагаемой формуле изобретения.
Источники литературы
Последующие источники литературы в той степени, в которой они обеспечивают примеры методических и других деталей в дополнение к представленным в этом документе, специально включены здесь для сведения.
Книги


Другие публикации


















































Изобретение относится к медицине, а именно к офтальмологии. Предлагается использовать для лечения глаукомы композиции, содержащие по меньшей мере один агонист ВМР-4. При этом композицию вводят непосредственной доставкой в глаз пациента, например посредством местных глазных капель; глазной мази; устройства замедленного высвобождения, имплантированного в слепой мешок глаза или около склеры глаза или внутри глаза; посредством инъекции. Концентрация ВМР-4 в композиции предпочтительно составляет от 0,01 до 2%. Способ основан на свойстве ВМР-4 снижать внутриглазное давление. 7 з.п. ф-лы, 15 ил., 2 табл.
| СПОСОБ ПРОГНОЗИРОВАНИЯ ПРОГРЕССИРОВАНИЯ ГЛАУКОМЫ | 1998 |
|
RU2139538C1 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ РАЗВИТИЯ КАТАРАКТЫ У БОЛЬНЫХ ПЕРВИЧНОЙ ГЛАУКОМОЙ | 1999 |
|
RU2155346C1 |
| Устройство для транспортирования сыпучих и порошкообразных материалов | 1954 |
|
SU107067A1 |
| ЦЕПНОЙ ЭЛЕВАТОР | 1941 |
|
SU61774A1 |

