Область техники, к которой относится изобретение
Настоящее изобретение относится к способу продуцирования L-лизина или L-треонина с помощью бактерии Escherichia. L-лизин и L-треонин, как известно, являются незаменимыми аминокислотами и используются в качестве компонентов в фармацевтических композициях и различных питательных смесях, таких как пищевые добавки и животные корма.
Предшествующий уровень техники
L-аминокислоты, такие как L-треонин и L-лизин, производят в промышленных масштабах путем ферментации с использованием продуцирующих L-аминокислоты бактерий, таких как коринебактерии или бактерий Escherichia, которые обладают способностью продуцировать L-аминокислоты. Для повышения продуктивности в качестве бактерии, продуцирующей L-аминокислоты, используют выделенный природный штамм, его мутант, полученный искусственным путем, или рекомбинант, у которых активность фермента биосинтеза L-аминокислот усилена в результате генной рекомбинации. Способ продуцирования L-лизина описан в патентах 1-4. Способ продуцирования L-треонина описан в патентных документах 5-8.
К способам усиления способности продуцирования аминокислот, таких как L-треонин и L-лизин, относится способ усиления энергетической эффективности путем модификации пути дыхательной цепи (патентный документ 13) и способ усиления способности вырабатывать никотинамидадениндинуклеотидфосфат путем амплификации никотинамиднуклеотидтрансдегидрогеназы (патентный документ 9), а также способ повышения уровня экспрессии фермента эндогенного биосинтетического пути.
Кроме того, известны способы модификации общих путей систем биосинтеза аминокислот, которые включают модификацию анаплеротических путей продуцирующих L-аминокислоты бактерий, таких как продуцирующая L-лизин коринебактерия, в которой усилена активность пируваткарбоксилазы (патентный документ 10), продуцирующая L-лизин бактерия Escherichia, дефектная по пируваткиназе (патентный документ 11), и продуцирующая L-лизин коринебактерия, дефектная по малатхининоксидоредуктазе (патентный документ 12).
Малеиновый фермент является одним из ферментов анаплеротического пути. Известно, что у бактерий Escherichia каждый из генов sfcA и b2463 кодируетмалеиновый фермент(непатентный документ 9). Однако независимо от снижения активности малеиновых ферментов, кодируемых генами sfcA и b2463, об эффективности усиления продуцирования L-лизина или L-треонина не сообщалось.
Анализ метаболического потока, также названный как анализ баланса потока, является методом предсказания распределений внутриклеточного метаболического потока путем конструирования стехиометрической модели внутриклеточных биохимических реакций и линейной оптимизации. Этот метод применяется для изучения возможностей систем биохимических реакций микроорганизмов или для предсказания распределений внутриклеточного метаболического потока при различных внешних условиях (непатентные документы 1, 2 и 3). Также имеются сообщения о конструировании стехиометрической модели для Escherichia coli (непатентные документы 4 и 5). Также известен пример использования такой стехиометрической модели в метаболической инженерии продуцирования лизина, разработанной для Corynebacterium glutamicum и используемой в производстве аминокислот (непатентный документ 6). Кроме того, имеются сообщения о большом числе теоретических и экспериментальных методов анализа метаболического потока и их применения (непатентные документы 7 и 8, патентные документы 14, 15 и 16). В патентном документе 14 описывают способ предсказания гена, необходимого для роста на основе стехиометрической модели. В патентном документе 15 описывают метод генетического и эволюционного изменения клеток с целью придания им оптимальных функций в клетках. Далее в патентном документе 16 представляют способ применения ограничений качественной кинетической информации, ограничений качественной контрольной информации и ограничений, основанных на экспериментальных данных, полученных на ДНК-микрочипах, в различных условиях стехиометрической модели. Хотя все они являются способами предсказания более желательных распределений внутриклеточного метаболического потока, не разработали способ теоретического предсказания специфического потока как мишени направленного усиления продуцирования клеточного вещества.
<Патентный документ 1>
Выложенная заявка на патент Японии № 10-165180
<Патентный документ 2>
Выложенная заявка на патент Японии № 11-192088
<Патентный документ 3>
Выложенная заявка на патент Японии № 2000-253879
<Патентный документ 4>
Выложенная заявка на патент Японии № 2001-57896
<Патентный документ 5>
Выложенная заявка на патент Японии № 5-304969
<Патентный документ 6>
Международная публикация № WO98/04715
<Патентный документ 7>
Выложенная заявка на патент Японии № 5-227977
<Патентный документ 8>
Опубликованная заявка на патент США № 2002/0110876
<Патентный документ 9>
Патент Японии № 2817400
<Патентный документ 10>
Выложенная заявка на патент Японии № 2002-508921
<Патентный документ 11>
Международная публикация № WO03/008600
<Патентный документ 12>
Опубликованная заявка на патент США № 2003/0044943
<Патентный документ 13>
Выложенная заявка на патент Японии № 2002-17363
<Патентный документ 14>
Международная публикация № WO00/46405
<Патентный документ 15>
Международная публикация № WO02/061115
<Патентный документ 16>
Международная публикация № WO02/055995
<Непатентный документ 1>
Varma A. and Palsson B.O. Appl. Environ. Microbiol. 60:3724-3731, 1994
<Непатентный документ 2>
Schilling C.H. et al., Biotechnol. Prog., 15:288-295, 1999
<Непатентный документ 3>
Schilling C.H. et al., Biotechnol. Prog., 15:296-303, 1999
<Непатентный документ 4>
Pramanik J. and Keasling J.D., Biotechnol. Bioeng., 56:398-421, 1997
<Непатентный документ 5>
Ibarra R.U. et al., Nature, 420:186-189, 2002
<Непатентный документ 6>
Vallino J.J. and Stephanopoulos G., Biotechnol. Bioeng., 41:633-646, 1993
<Непатентный документ 7>
Wiechert W., Journal of Biotechnology, 94:37-63, 2002
<Непатентный документ 8>
Wiechert W., Metabolic Engineering, 3:195-205, 2001
<Непатентный документ 9>
van der Rest M.E., Frank C., Molenaar D.J., J. Bacteriol., 182(24):6892-6899, 2000
Описание изобретения
В настоящем изобретении предлагается бактерия Escherichia, которая имеет улучшенную способность продуцирования L-лизина или L-треонина, и способ продуцирования L-лизина или L-треонина с помощью бактерии.
Авторы настоящего изобретения для решения проблемы провели тщательное исследование и в результате установили, что производительность метаболического потока, влияющего на продуцирование вещества, может быть определена путем: 1) выбора определенного числа свободных потоков как степени свободы стехиометрической матрицы, рассчитанной по формулам биохимических реакций на субстрате для продуцируемого желательного вещества; 2) расчета распределений метаболического потока на основе выборки случайных комбинаций свободных потоков, достаточной для статистического анализа, основанного на стехиометрической матрице; 3) получения уравнения регрессии, включающего минимальное число свободных потоков, которое коррелирует с продуцированием вещества по рассчитанным распределениям метаболического потока на основе статистического анализа.
Определение метаболических потоков бактерии, продуцирующей L-лизин или L-треонин, с помощью этого способа показало, что модификация, нарушающая нормальную функцию малеинового фермента, вызывает увеличение продуктивности бактерии. Настоящее изобретение выполнено на основе вышеупомянутых данных и предлагает следующее:
1) бактерию Escherichia, которая имеет способность продуцировать L-лизин или L-треонин, и где указанная бактерия модифицирована так, что нарушена нормальная функция малеинового фермента в клетке;
2) бактерию по (1), где ген, кодирующий указанный малеиновый фермент на бактериальной хромосоме, является мутированным и/или мутированной является последовательность, контролирующая экспрессию этого гена, что нарушает нормальную функцию малеинового фермента в клетке;
3) бактерию по (1), где указанный малеиновый фермент не обладает нормальной функцией в результате нарушения гена, кодирующего указанный малеиновый фермент на бактериальной хромосоме;
4) бактерию по (1), где ген, кодирующий указанный малеиновый фермент, содержит sfcA;
5) бактерию по (1), где ген, кодирующий указанный малеиновый фермент, содержит b2463;
6) бактерию по (1), где указанный малеиновый фермент выбран из группы, содержащей:
А) белок, имеющий аминокислотную последовательность, представленную в SEQ ID NO:6;
и
B) белок, который имеет аминокислотную последовательность, содержащую замещение, делецию, инсерцию или добавление одного или нескольких аминокислотных остатков в аминокислотную последовательность, представленную в SEQ ID NO:6, и имеет активность малеинового фермента;
7) бактерию по (1), где указанный малеиновый фермент выбран из группы, содержащей:
С) белок, имеющий аминокислотную последовательность, представленную в SEQ ID NO:8;
и
D) белок, который имеет аминокислотную последовательность, содержащую замещение, делецию, инсерцию или добавление одного или нескольких аминокислотных остатков в аминокислотную последовательность, представленную в SEQ ID NO:8, и имеет активность малеинового фермента;
8) бактерию по (1), где геном, кодирующим указанный малеиновый фермент, является ДНК, выбранная из группы, содержащей:
а) ДНК, имеющую нуклеотидную последовательность, представленную в SEQ ID NO:5;
b) ДНК, которая гибридизируется с нуклеотидной последовательностью, представленной в SEQ ID NO:5, или зонд, который может быть получен из нуклеотидной последовательности, где указанная гибридизация проходит в жестких условиях и где указанная ДНК кодирует белок, имеющий активность малеинового фермента;
9) бактерию по (1), где геном, кодирующим малеиновый фермент, является ДНК, выбранная из группы, содержащей:
c) ДНК, имеющую нуклеотидную последовательность, представленную SEQ ID NO:7;
и
d) ДНК, которая гибридизируется с нуклеотидной последовательностью, представленной в SEQ ID NO:7, или зонд, который может быть получен из нуклеотидной последовательности, где указанная гибридизация проходит в жестких условиях и где указанная ДНК кодирует белок, имеющий активность малеинового фермента;
10) способ продуцирования L-лизина или L-треонина, включающий культивирование бактерии, как определено в любом из (1)-(9), в среде, обеспечивающей продуцирование и секрецию указанных L-лизина или L-треонина, и сбор и выделение L-лизина или L-треонина из среды.
Краткое описание чертежей
Фиг. 1. График продуцирования лизина как функции различных значений свободных потоков посредством использования набора данных 5000 случайных распределений потоков. Выход лизина показан для: а) потока изоцитратлиазы; b) потока малеинового фермента; c) потока PEP-карбоксилазы.
Фиг. 2. График продуцирования лизина как функции величин уравнения 2 для набора данных 5000 случайных распределений потоков. Исходной величиной является поток, выраженный в нм/ч, в расчете на 10 нм/ч потока глюкозы.
Фиг. 3. Представлена структура плазмид pMW118-attL-Tc-attR и pMW118-attL-Cm-attR.
Фиг. 4. Представлена структура плазмиды pMW-intxis-ts.
Лучший способ выполнения изобретения
Далее настоящее изобретение будет объяснено детально.
<I> Бактерия Escherichia настоящего изобретения
Бактерия Escherichia настоящего изобретения является бактерией, принадлежащей к роду Escherichia, которая обладает способностью продуцировать L-лизин или L-треонин и которая модифицирована так, что нарушена нормальная функция малеинового фермента. Бактерия Escherichia настоящего изобретения может иметь способность продуцировать либо L-лизин, либо L-треонин либо может иметь способность продуцировать как L-лизин, так и L-треонин.
Родительский штамм, принадлежащий к роду Escherichia, который использовали для получения бактерии Escherichia настоящего изобретения, включает, но не ограничиваясь ими, штаммы, описанные в книге Neidhardt с сотр. (Neidhardt F.C. с сотр., Escherichia coli and Salmonella Typhimurium, American Society for Microbiology, Washington D. C., 1029, таблица 1). Например, родительским штаммом может быть Escherichia coli. Escherichia coli может быть штаммом W3110 Escherichia coli (ATCC 27325) или штаммом MG1655 Escherichia coli (ATCC 47076), которые оба получены из прототипного штамма K12 дикого типа.
Эти штаммы можно получить, например, из Американской коллекции типовых культур (Адрес: 12301 Parklawn Drive, Rockville, Maryland 20852, United States of America). Штаммам даны соответствующие регистрационные номера. Существует возможность затребовать желательный штамм по его регистрационному номеру. Регистрационные номера, которые соответствуют штаммам, представлены в каталоге Американской коллекции типовых культур.
<1>-1. Придание способности продуцировать L-лизин или L-треонин
Ниже представлен способ придания бактерии Escherichia способности продуцировать L-лизин или L-треонин. Фраза «способность продуцировать L-лизин», как использовано в данном описании, означает способность продуцировать и вызывать накопление или секрецию L-лизина в среде, например, свободного внеклеточного L-лизина, при культивировании бактерии в среде. В частности, фраза «способность продуцировать L-лизин» означает способность вызывать накопление большего количества L-лизина по сравнению с диким типом или родительским штаммом.
Фраза «способность продуцировать L-треонин», как использовано в данном описании, означает способность продуцировать и вызывать накопление или секрецию L-треонина в среде, например, свободного внеклеточного L-треонина, при культивировании бактерии в среде. В частности, эта фраза означает способность вызывать накопление большего количества L-треонина по сравнению с диким типом или родительским штаммом.
Для придания способности продуцировать L-лизин или L-треонин могут быть использованы стандартные способы селекции бактерий Escherichia и коринебактерий, такие как способы получения ауксотрофных мутантных штаммов, штаммов, резистентных к аналогам, или штаммов, мутантных по метаболическому контролю, которые обладают способностью продуцировать L-лизин или L-треонин, и способы продуцирования рекомбинантных штаммов, в которых усилены активности ферментов биосинтеза L-лизина или L-треонина. В селективных бактериях, продуцирующих L-лизин или L-треонин, такие характеристики, как ауксотрофность, резистентность к аналогам и мутации метаболического контроля, могут быть получены по отдельности или в комбинации.
Усиленная активность ферментов биосинтеза L-лизина или L-треонина может быть представлена по отдельности или в комбинации. Далее, получение таких характеристик, как ауксотрофность, резистентность к аналогам и мутации метаболического контроля, может быть комбинировано с усилением активности ферментов биосинтеза L-лизина или L-треонина.
Варианты способов придания или усиления способности продуцировать L-лизин или L-треонин путем усиления активности ферментов биосинтеза L-лизина или L-треонина описаны ниже. Усиление ферментативной активности может быть осуществлено путем, например, введения мутации в ген, кодирующий фермент, или амплификации гена с целью усиления внутриклеточной активности фермента. Это же может быть осуществлено посредством генной рекомбинации.
Гены, кодирующие ферменты биосинтеза L-треонина, включают, но не ограничиваясь ими, ген аспартокиназы III (lysC), ген аспартатполуальдегиддегидрогеназы (asd), аспартокиназу I, кодируемую опероном thr (thrA), ген гомосеринкиназы (thrB), ген треонинсинтазы (thrC). Сокращенное условное обозначение гена показано в круглых скобках. В штамм могут быть введены два или более этих генов. Ген фермента биосинтеза L-треонина может быть введен в бактерию Escherichia, в которой супрессировано расщепление треонина. Примером бактерии Escherichia, в которой супрессировано расщепление треонина, служит штамм TDH6, который является дефектным по активности треониндегидрогеназы (Выложенная заявка на патент Японии № 2001-346578).
Гены, кодирующие ферменты биосинтеза L-лизина, включают, но не ограничиваясь ферментами пути диаминопимелата, такие гены, как ген дигидродипиколинатсинтазы (dapA), ген аспартокиназы (lysC), ген дигидродипиколинатредуктазы (dapB), ген диаминопимелатдекарбоксилазы (lysA), ген диаминопимелатдегидрогеназы (ddh), (все вышеприведенные; Международная публикация № 96/40934), ген фосфоенолпируваткарбоксилазы (ppc) (Выложенная заявка на патент Японии № 60-87788), ген аспартатаминотрансферазы (aspC) (Опубликованная заявка на патент Японии № 6-102028), ген диаминопимелатэпимеразы (dapF) (Выложенная заявка на патент Японии № 2003-135066), ген аспартатполуальдегиддегидрогеназы (asd) (Международная публикация № 00/61723) и ген ферментов пути аминоадипата, такой как ген гомоаконинтатгидратазы (Выложенная заявка на патент Японии № 2000-157276).
Кроме того, бактерия настоящего изобретения может иметь сниженную активность фермента, который катализирует реакцию генерирования соединения, отличающегося от L-лизина, путем ответвления от пути биосинтеза L-лизина или может быть дефектной по такому ферменту. Ферменты, которые катализируют реакцию генерирования соединения, отличающегося от L-лизина, путем ответвления от пути биосинтеза L-лизина, включают гомосериндегидрогеназу и лизиндекарбоксилазу. Штаммы, имеющие сниженные активности ферментов, описаны в патентах WO95/23864 и WO 96/178930.
Усиление активности фермента, кодируемого геном, может быть достигнуто, например, путем амплификации гена биосинтеза L-лизина или L-треонина с плазмидой, автономно реплицирующейся в бактериях Escherichia. Ген биосинтеза может быть интегрирован в бактериальную хромосому. Усиление активности также может быть достигнуто посредством введения гена, который содержит мутацию, вызывающую усиление активности фермента, кодируемого геном. Примерами такой мутации является мутация промоторной последовательности, вызывающей усиление уровня транскрипции гена, и мутация в кодирующей области гена, усиливающая специфическую активность ферментативного белка.
Помимо генной амплификации, как описано выше, экспрессия гена может быть усилена путем замещения последовательности, контролирующей экспрессию, такой как промотор гена на хромосомной ДНК или плазмиде, на функционально более сильную (Международная публикация № WO 00/18935). Сильные промоторы общеизвестны и включают, например, промотор lac, промотор trp, промотор trc, промотор tac и промотор PR бактериофага лямбда.Экспрессия гена может быть усилена путем замещения эндогенного промотора на либо хромосому, либо плазмиду, содержащую функционально более сильный промотор, или путем модификации эндогенного промотора. Модификация последовательности, контролирующей экспрессию, может быть комбинирована с повышением числа копий гена.
Примеры бактерий Escherichia, в которых способность продуцировать L-лизин или L-треонин является приобретенной и которые могут быть использованы в настоящем изобретении, приведены ниже. Однако бактерия настоящего изобретения не ограничена этими примерами, к ней относятся любые бактерии, способные продуцировать L-лизин или L-треонин.
Специфическими примерами штаммов, резистентных к аналогам, или штаммов, мутантных по метаболическому контролю, которые обладают способностью продуцировать L-лизин, являются Escherichia coli AJ11442 (FERM BP-1543, NRRL B-12185; Выложенная заявка на патент Японии № 56-18596 и Патент США № 4346170) и Escherichia coli VL611. Штамм WC196 может быть использован как продуцирующая L-лизин бактерия Escherichia coli (Международная публикация № WO96/17930). Штамм WC196 селектировали путем придания штамму W3110, полученному от штамма K-12 Escherichia coli, резистентности к AEC (S-(2-аминоэтил)цистеину). Этот штамм обозначили как Escherichia coli AJ13069 и депонировали в Национальном институте биологической науки и технологии человека, Агентство по промышленной науке и технике (в настоящее время Национальный институт передовой промышленной науки и технологии, Международная организация по хранению патентов, Tsukuba Central 6, 1-1, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan) от 6 декабря 1994 г. и присвоили инвентарный номер FERM P-14690. Его передали в международный депозитарий по условиям Будапештского соглашения от 29 сентября 1995 г. и присвоили инвентарный номер FERM BP-5252.
Примеры бактерий Escherichia, обладающих способностью продуцировать L-треонин, включают продуцирующий L-треонин мутантный штамм, который является резистентным к 6-диметиламинопурину (Выложенная заявка на патент Японии № 5-304969), рекомбинантные штаммы Escherichia coli, такие как штамм, в котором ген биосинтеза треонина, содержащий введенную мутацию, которая вызывает избыточное продуцирование фермента биосинтеза L-треонина, амплифицирован на плазмиде (Опубликованная заявка на патент Японии № 1-29559 и Выложенная заявка на патент Японии № 5-227977), штамм, в котором оперон треонина амплифицирован на плазмиде (Выложенная заявка на патент Японии № 2-109985), и штамм, в котором гены, кодирующие пируваткарбоксилазу и никотинамиднуклеотидтрансгидрогеназу, амплифицированы (Выложенная заявка на патент Японии № 2002-51787).
Escherichia coli VKPM B-3996 (Патент США № 5175107) также относится к настоящему изобретению. Штамм VKPM B-3996 депонировали в Российской национальной коллекции промышленных микроорганизмов (VKPM), ГНИИ Генетика) от 19 ноября 1987 г. и присвоили инвентарный номер VKPM B-3996. VKPM B-3996 содержит плазмиду pVIC40 (Международная публикация № WO 90/04636), которую получили путем введения генов биосинтеза треонина (треониновый оперон thrABC) в предназначенный для широкого круга хозяев вектор, например плазмиду pAYC32 (Chistoserdov A.Y., Tsygankov Y.D., Plasmid, 1986, 16, 161-167). В pVIC40 десенсибилизировано ингибирование обратной связи посредством L-треонина аспартокиназа I-гомосериндегидрогеназы I, кодируемой thrA в треониновом опероне.
Кроме того, Escherichia coli B-5318 (Патент Европы № 0593792) относится к настоящему изобретению. Штамм B-5318 депонировали в Российской национальной коллекции промышленных микроорганизмов (VKPM), ГНИИ Генетика) от 19 ноября 1987 г. и присвоили инвентарный номер VKPM B-5318. Штамм VKPM является прототрофным по отношению к изолейцину и содержит рекомбинантную плазмидную ДНК. Эту плазмида сконструирована так, что треониновый оперон, включающий гены биосинтеза треонина, содержит дефект в области аттенуации, например в эндогенной области регуляции транскрипции. Оперон расположен ниже температурочувствительного репрессора C1 фага лямбда, промотора PR и N-конца белка Cro и сконструирован таким образом, что экспрессия генов биосинтез треонина находится под контролем репрессора фага лямбда и промотора.
<2> Конструирование бактерии Escherichia настоящего изобретения
Бактерия Escherichia настоящего изобретения является бактерией, принадлежащей к роду Escherichia, которая обладает способностью продуцировать L-лизин или L-треонин и которая модифицирована так, что нарушена нормальная функция малеинового фермента.
В процессе селекции бактерия Escherichia настоящего изобретения либо приобретает способность продуцировать L-лизин или L-треонин, либо несет мутацию, нарушающую нормальную функцию малеинового фермента (EC 1.1.1.38, EC 1.1.1.40). Также бактерия Escherichia, обладающая способностью продуцировать L-лизин или L-треонин, может быть модифицирована так, что нарушена нормальная функция малеинового фермента, и способность продуцировать L-лизин или L-треонин может быть придана бактерии Escherichia, которая пока еще не имеет нормальной функции малеинового фермента.
Фраза «активность малеинового фермента» означает активность, которая катализирует обратимую реакцию продуцирования диоксида углерода и пирувата из малата. Известны малеиновые ферменты, которые используют НАД (EC 1.1.1.38) и НАДФ (EC 1.1.1.40) как коферменты. (EC 1.1.1.38 (S)-малат+НАД+=пируват+CO2+НАДН+H+) (EC 1.1.1.40 (S)-малат+НАДФ+=пируват+CO2+НАДФН+H+). Малеиновый фермент также называют «малатдегидрогеназа» или «малатоксидоредуктаза».
Фраза «модифицированный так, что нарушена нормальная функция малеинового фермента в клетке» означает, что он модифицирован так, что функция малеинового фермента должна быть элиминирована или активность малеинового фермента должна быть снижена или аттенуирована по сравнению с немодифицированным штаммом, таким как штамм дикого типа (родительский). Состоянием, когда нормальная функция малеинового фермента нарушена, может быть, например, состояние ингибирования транскрипции или трансляции гена, кодирующего малеиновый фермент, и, следовательно, его продукта, малеиновый фермент не продуцируется, или его продуцирование снижено, или состояние, когда ген, кодирующий малеиновый фермент на бактериальной хромосоме, мутирован, и/или мутирована последовательность, контролирующая экспрессию гена, и таким образом, активность малеинового фермента снижена или элиминирована. Типичным примером бактерии Escherichia, в которой нарушена функция малеинового фермента, является штамм, несущий разрушенный ген, в котором ген, кодирующий малеиновый фермент на бактериальной хромосоме, разрушен с помощью метода генетической рекомбинации, и мутантный штамм, в котором регулирующая экспрессию последовательность или кодирующая область гена малеинового фермента мутированы, и поэтому функциональный малеиновый фермент не продуцируется длительно.
Фраза «модифицированный так, что активность малеинового фермента аттенуирована» означает, что активность малеинового фермента снижена по сравнению с немодифицированным штаммом, например штаммом дикого типа (родительским) бактерии Escherichia. Предпочтительно активность малеинового фермента снижена не более чем на 50%, более предпочтительно не более чем на 30% и наиболее предпочтительно не более чем на 10% на клетку по сравнению с немодифицированным штаммом.
Примерами бактерии Escherichia, которая может действовать в качестве контроля, являются Escherichia coli W3110 (ATCC 27325) и Escherichia coli MG1655 (ATCC 47076). Эти штаммы дикого типа получены из прототипного штамма K12 дикого типа. Активность малеинового фермента с использованием НАД в качестве кофермента может быть определена по способу Korkes S. с сотр., (Korkes S. с сотр., (1950) J. Biol. Chem. 187, 891-905). Активность малеинового фермента с использованием в качестве кофермента НАДФ может быть определена по способу Ochoa S. (Ochoa S. с сотр., (1947) J. Biol. Chem. 167, 871-872).
Термин «аттенуация» включает, но не ограничиваясь этим, полную элиминацию активности. Каждая активность малеинового фермента с использованием в качестве коферментов НАД или НАДФ может быть аттенуированна индивидуально или совместно. В рамках настоящего изобретения достаточно, что бактерия Escherichia имеет аттенуированную активность малеинового фермента по сравнению со штаммом дикого типа или немодифицированным штаммом. Однако предпочтительно, если бактерия Escherichia настоящего изобретения также имеет усиленную способность вызывать накопление или секрецию L-лизина или L-треонина по сравнению со штаммом дикого типа или немодифицированным штаммом и/или улучшенную продуктивность L-лизина или L-треонина вследствие хорошего роста, главным образом за счет улучшенного выхода продукта за вычетом клеток.
Малеиновый фермент настоящего изобретения содержит белок, имеющий аминокислотную последовательность, представленную в SEQ ID NO:6 или 8. Малеиновый фермент может быть вариантом аминокислотной последовательности, представленной в SEQ ID NO:6 или 8, в которой он может содержать замещение, делецию, инсерцию или добавление одного или нескольких аминокислотных остатков в аминокислотной последовательности, представленной в SEQ ID NO:6 или 8, при условии, что он имеет активность малеинового фермента. Как использовано в данном описании, «нескольких» означает, например, 2-20, предпочтительно 2-10, более предпочтительно 2-5 аминокислотных остатков.
Замещение, делеция, инсерция или добавление одного или нескольких аминокислотных остатков должно быть консервативной мутацией(ями), с тем чтобы сохранялась активность малеинового фермента. Типичной консервативной мутацией является консервативное замещение. Примеры консервативных замещений включают: замещение Ser или Thr на Ala; замещение Gln, His или Lys на Arg; замещение Glu, Gln, Lys, His или Asp на Asn; замещение Asn, Glu или Gln на Asp; замещение Ser или Ala на Cys; замещение Asn, Glu, Lys, His, Asp или Arg на Gln; замещение Asn, Gln, Lys или Asp на Glu; замещение Pro на Gly; замещение Asn, Lys, Gln, Arg или Tyr на His; замещение Leu, Met, Val или Phe на Ile; замещение Ile, Met, Val или Phe на Leu; замещение Asn, Glu, Gln, His или Arg на Lys; замещение Ile, Leu, Val или Phe на Met; замещение Trp, Tyr, Met, Ile или Leu на Phe; замещение Thr или Ala на Ser; замещение Ser или Ala на Thr; замещение Phe или Tyr на Trp; замещение His, Phe или Trp на Tyr и замещение Met, Ile или Leu на Val.
Фраза «модифицированный так, что нарушена нормальная функция малеинового фермента в клетке» может означать снижение числа молекул малеинового фермента на клетку и снижение активности малеинового фермента на молекулу. В частности, модификация может быть проведена путем получения дефектов гена, кодирующего малеиновый фермент на бактериальной хромосоме, или путем модификации последовательности, контролирующей экспрессию, такой как промотор или последовательность Шайна-Дальгарно (SD). Также модификация может быть проведена путем введения замещения аминокислоты (миссенс-мутация) или стоп-кодона (нонсенс-мутация) в кодирующую область, или введения инсерции или делеции 1-2 оснований в кодирующую область (мутация сдвига рамки считывания), или делетирования части гена (Journal of Biological Chemistry 272:8611-8617(1997)).
Примером гена малеинового фермента (ген mez) на хромосоме является ген sfcA, поскольку его ДНК имеет нуклеотидную последовательность, представленную в SEQ ID NO:5. Эта ДНК кодирует фермент, который использует НАД в качестве кофермента. Другим примером является ген b2463, ДНК которого имеет нуклеотидную последовательность, представленную в SEQ ID NO:7. Эта ДНК кодирует фермент, который использует НАДФ в качестве кофермента.
Ген mez может быть ДНК, которая гибридизируется с нуклеотидной последовательностью, представленной в SEQ ID NO:5 или 7, или зондом, полученным из нуклеотидной последовательности в жестких условиях при условии, что он кодирует белок, который имеет активность малеинового фермента. «Жесткие условия» означают условия, при которых формируется специфический гибрид и неспецифический гибрид не формируется. Например, к жестким условиям относится однократное промывание, предпочтительно 2-3-кратное промывание при концентрации соли, соответствующей 1×SSC, 0,1% SDS, предпочтительно 0,1×SSC, 0,1% SDS при 60°C. Подходящая длина зонда может быть выбрана в зависимости от условий гибридизации, как правило, она составляет 100 п.н.-1 т.п.н.
Ген, кодирующий малеиновый фермент, (sfcA, b2643), может быть получен с помощью ПЦР с использованием в качестве матрицы хромосомы Escherichia coli и олигонуклеотидов, синтезированных на основе следующих последовательностей Escherichia coli, зарегистрированных в GenBank в качестве праймеров: sfcA: AAC74552. NAD-связанный малат...[gi:1787754], комплементарный AE000245.1:1208..2932, b2643: AAC75516, предположительно мультимодальный...[gi:1788806], комплементарный AE000333.1:141..2420.
Хромосомная ДНК может быть получена из бактерии для использования в качестве донорной ДНК посредством, например, способа Saito и Miura (ссылка на H. Saito и K. Miura, Biochem. Biophys. Acta, 72, 619 (1963), Text for Bioengineering Experiments, Edited by the Society for Bioscience and Bioengineering, Japan, pp.97-98, Baifukan, 1992) или т.п.
Ген sfcA или b2643, полученный, как описано выше, или его часть может быть использована для разрушения гена. Достаточным условием для гена, используемого для разрушения гена, является степень гомологии, позволяющая проводить гомологичную рекомбинацию генов sfcA или b2463 на хромосоме бактерии Escherichia. Поэтому может быть использован такой гомологичный ген. Степень гомологии, которая должна разрешать гомологичную рекомбинацию, предпочтительно составляет 70% или более, более предпочтительно 80% или более, еще более предпочтительно 90% или более и наиболее предпочтительно 95% или более. Также гомологичная рекомбинация может происходить, если используется ДНК, которая гибридизируется с геном в жестких условиях. «Жесткие условия» являются условиями, в которых формируется специфический гибрид и неспецифический гибрид не формируется. Например, к жестким условиям относится однократное промывание, предпочтительно 2-3-кратное промывание при концентрации соли, соответствующей 1×SSC, 0,1% SDS, предпочтительно 0,1×SSC, 0,1% SDS при 60°C.
Ген sfcA или b2463 может быть разрушен путем, например, получения из гена, как описано выше, гена sfcA или b2463 делеционного типа, в котором частичная последовательность делетирована так, что малеиновый фермент с нормальной функцией не продуцируется. Этим геном делетированного типа или ДНК, содержащей ген, затем может быть трансформирована бактерия Escherichia и вызвана рекомбинация между геном делетированного типа и геном на хромосоме. Разрушение гена посредством подставления гена с помощью гомологичной рекомбинации уже разработано и подтверждено примером с использованием линейной ДНК в способе, разработанном Datsenko К.А. и Wanner B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97, 6640-6645), также названном как "Red-запускаемая интеграция", и способом, основанным на применении плазмиды, несущей температурочувствительную область инициации репликации (Патент США №6303383 и Выложенная заявка на патент Японии №5-7491). Разрушение гена посредством генного вычитания с помощью гомологичной рекомбинации также может быть выполнено посредством использования плазмиды, которая не обладает способностью к репликации в хозяине.
Кроме того, может быть использован способ, основанный на комбинации способа, названного «red-запускаемая интеграция», и эксцизионной системы, полученной из фага лямбда (J.Bacteriol. 2002 Sep; 184(18): 5200-3). Может быть использовано взаимодействие между интегразой и эксцизионазой в фаге лямбда, вырезающее нуклеопротеиновый комплекс (Cho EH, Gumport RI, Gardner JF.), как способ разрушения гена на хромосоме.
В соответствии со способом red-запускаемой интеграции штамм, несущий разрушенный ген, может быть сконструирован в один этап с использованием ПЦР-продукта, который получают с использованием синтетических олигонуклеотидов в качестве праймеров, которые сконструированы так, что содержат часть адресного гена на своем 5'-конце и часть гена резистентности к антибиотику на своем 3'-конце. Кроме того, интегрированный ген резистентности к антибиотику может быть удален путем введения attL и attR, которые являются сайтами прикрепления фага лямбда, и ПЦР-продукта и комбинирования эксцизионной системы, полученной из фага лямбда, со способом red-запускаемой интеграции.
В частности, штамм, в котором адресный ген разрушен и ген резистентности к антибиотику удален, может быть получен с помощью описанного ниже способа.
В первую очередь получили линейную ДНК-кассету, содержащую ген резистентности к антибиотику, сайты прикрепления фага лямбда и адресный ген. Ее получили, как обычно, с помощью ПЦР с использованием специальной полученной матрицы.
Матрицу, в которой attL и attR (SEQ ID NO:9 (GenBank, инвентарный № M12458 и SEQ ID NO:10 (GenBank, инвентарный № M12459)) являются сайтами прикрепления фага лямбда, вводили в соответствующие концы гена резистентности к антибиотику, который использовали как матрицу линейной ДНК-кассеты. Матрица может быть плазмидой, геном, введенным в хромосому, или синтетическим олигонуклеотидом. Хотя геном резистентности к антибиотику предпочтительно является ген резистентности к хлорамфениколу, может быть использован ген резистентности к стрептомицину, или ген резистентности к ампициллину, или ген резистентности к любому другому антибиотику, при условии, что генные функции в качестве гена резистентности к антибиотику в бактериях Escherichia отличаются от маркерного гена, который может содержаться в двух хелперных плазмидах, как описано ниже. Бесспорным подтверждением приобретения резистентности к антибиотику может быть увеличение уровня экспрессии применяемого гена резистентности к антибиотику путем замещения промоторной последовательности и т.п. или усиление активности фермента в результате введения мутации в последовательность структурного гена. Линейную ДНК-кассету получали в следующем порядке, начиная с 5'-конца: (5'-последовательность адресного гена)-(attL)-(ген резистентности к антибиотику)-(attR)-(3'-последовательность адресного гена).
Линейную ДНК-кассету вводили в хромосому. В качестве хелперной плазмиды для интеграции линейной ДНК-кассеты в хромосому может быть использована плазмида pKD46 (Proc. Natl. Acad. Sci. USA, 2000, 97, 6640-6645). pKD46 характеризуется температурочувствительной репликацией и резистентностью к ампициллину и содержит фрагмент 2154 п.н. фрагмента ДНК фага лямбда (GenBank/EMBL, инвентарный № J02459, 31088-33241), который содержит гены (гены γ, β и exo), кодирующие Red-рекомбиназу системы гомологичной рекомбинации λ Red, которая находится под контролем индуцируемого арабинозой промотора ParaB.
pKD46 может быть введена в хозяина с помощью электропорации. pKD46-амплифицированный штамм культивировали с арабинозой. Линейную ДНК-кассету вводили в логарифмической фазе роста и инкубировали при высокой температуре для получения штамма с разрушенным геном, который резистентен к антибиотику благодаря гену резистентности к антибиотику в линейной ДНК-кассете. Подтверждение разрушения гена может быть получено с помощью ПЦР или путем измерения концентрации L-лизина или L-треонина, продуцированного штаммом.
Далее вводили хелперную плазмиду для удаления гена резистентности к антибиотику. Хелперная плазмида содержит ген, кодирующий интегразу (Int) (SEQ ID NO:13, GenBank, инвентарный № J02459. B [gi:215104]), и ген, кодирующий эксцизионазу (Xis) (SEQ ID NO:15, GenBank, инвентарный № J02459 [gi:215104]) фага лямбда, и характеризуется температурочувствительной репликацией. Вследствие введения хелперной плазмиды рекомбинация происходит благодаря узнаванию attL (SEQ ID NO:11) и attR (SEQ ID NO:12) на хромосоме. Ген резистентности к антибиотику, расположенный между attL и attR, вырезается, и в результате структура, которая содержит только последовательности attL или attR, остается на хромосоме. Инкубация при высокой температуре вызывает утрату хелперной плазмиды. Так может быть получен штамм, в котором разрушен адресный ген и элиминирован ген резистентности к антибиотику.
Другой, отличающийся от методов генетической инженерии, способ модификации бактерии, вызывающей нарушение нормальной функции малеинового фермента, может быть примером способа обработки бактерии Escherichia УФ-облучением или мутагенным агентом, обычно используемым для мутагенеза, таким как N-метил-N'-нитро-N-нитрозогуанидин или азотная кислота с последующим отбором бактерий, проявляющих аттенуированную активность малеинового фермента.
Настоящее изобретение было разработано на основании информации о метаболическом потоке. Эта информация была рассчитана с помощью нижеприведенного способа определения влияния метаболического потока на продуцирование вещества с использованием клеток. Однако настоящее изобретение не ограничено способом получения такой информации, то есть способом определения.
Способ определения влияния метаболического потока на продукцию вещества с использованием клеток включает в себя этапы:
1) создание стехиометрической матрицы на основании формул биохимических реакций субстрата с получением желательного вещества;
2) выбор определенного числа независимых метаболических потоков из всей совокупности метаболических потоков в качестве степени свободы стехиометрической матрицы как совокупности свободных потоков;
3) создание достаточного числа случайных комбинаций свободных потоков для статистического анализа и расчета распределения метаболического потока по каждой созданной комбинации на основе стехиометрической матрицы;
4) получение уравнения регрессии, включающего минимальное число свободных потоков, которое показывает корреляцию с продуцированием вещества по рассчитанным распределениям метаболического потока с помощью мультивариантного статистического анализа;
и
5) определение не менее одного метаболического потока, влияющего на продуцирование вещества, на основе коэффициента в полученном уравнении регрессии.
Метаболический поток, использованный в настоящем изобретении, выражается как скорость метаболической реакции (потока), полученная на основе стехиометрической модели внутриклеточных биохимических реакций и закона действия масс между метаболитами; между тем, распределение метаболического потока, как использовано в данном описании, состоит из всех метаболических потоков, где каждый метаболический поток означает отдельную биохимическую реакцию.
На первом этапе способа определения получили стехиометрическую матрицу на основе формул биохимических реакций субстрата с получением желательного вещества в качестве продукта.
Биохимические реакции отражают процесс, в результате которого внутриклеточные метаболиты конвертируют посредством ферментативных реакций в клетке и который был компилирован в различных базах данных в соответствии с типом организма. Например, для ссылки может быть доступна Энциклопедия генов и геномов Kyoto (KEGG, www.genome.ad.jp/kegg/).
Субстрат является веществом, обычно используемым клеткой как источник углерода, и примерами его являются глюкоза, сахароза, фруктоза и т.п.
Вещество в качестве продукта включает не только отдельный вид метаболита, но также агрегаты метаболитов, такие как биомасса (клеточное тело). Продуцирование вещества обычно оценивают по скорости продуцирования вещества. В частности, если желательное вещество является биомассой, ее оценивали по выходу биомассы. Выход биомассы отражает эффективность превращения субстратов, таких как глюкоза, в клеточные компоненты, такие как белок, углевод, нуклеиновая кислота или липид.
Стехиометрическая матрица является матрицей, обычно используемой для анализа метаболического потока, и может быть создана путем составления списков формул биохимических реакций субстрата с образованием желательного вещества в качестве продукта посредством стандартных способов, используемых для анализа метаболического потока. Такие способы, допускающие квазиравновесное состояние внутриклеточного метаболического промежуточного соединения, широко известны (Savinell J.M. and Palsson B.O.J., Theor. Biol., 154:421-454, 1992; Vallino J.J. and Stephanopoulos G., Biotechnol. Bioeng., 41:633-646, 1993). После составления списков формул реакций реакционные пути могут быть упрощены путем допущения серий реакций без разветвления, в виде одной реакции, или допущения метаболитов, конвертируемых с помощью реакции с высокой метаболической скоростью до или после реакции, как одного метаболита и т.д. Когда вещество в качестве продукта является биомассой, стехиометрическая матрица может быть описана путем перечисления биохимических реакций, протекающих в компонентах клетки.
На втором этапе способа определения из общего числа метаболических потоков отобрали определенное число независимых метаболических потоков в качестве свободных потоков, характеризующих степень свободы вышеупомянутой стехиометрической матрицы.
Независимые потоки представляют собой совокупность потоков, каждый из которых должен однозначно соответствовать определенному потоку в системе метаболической сети, как задано стехиометрическим уравнением.
Способ отбора свободных потоков особым образом не ограничен, поскольку может быть отобрано только то число независимых метаболических потоков, которое соответствует степени свободы системы, используемой для анализа. Хотя независимость произвольно выбранных потоков может быть подтверждена, для этого также может быть использована предложенная Reder SIMS-матрица или равновесная внутренняя метаболическая стехиометрическая матрица (Reder C.J., Theor. Biol., 135:175-201, 1988). Согласно этому способу конкретную группу числа метаболических потоков как степень свободы вышеупомянутой стехиометрической матрицы определяли из числа свободных потоков из независимых групп метаболических потоков, определенных на основании вышеупомянутых формул биохимических реакций и метаболический поток выбирали как свободный поток из определенной группы метаболических потоков. Определение специфических групп среди групп потоков гарантирует, что любой поток в группе может быть изменен без влияния на поток других групп. Поэтому становится возможным отбор одного потока из каждой группы как независимого свободного потока. Когда свободный поток отобран из группы потоков, поток замыкает предварительно отобранную точку ветвления.
На третьем этапе способа определения получили случайные комбинации свободных потоков в числе, достаточном для статистического анализа, и распределение метаболических потоков рассчитали для каждой созданной комбинации на основе стехиометрической матрицы.
Случайные комбинации свободных потоков могут быть получены путем подставления случайных значений в свободные потоки, выбранные на предыдущем этапе с целью создания набора данных по комбинациям различных распределений потоков. Способ подставления случайных значений в свободные потоки практически не имеет ограничений, поскольку выбран способ получения комбинаций свободных потоков в специфическом пределе. Указанный специфический предел является набором для придания биологически пригодных значений в последних расчетах. Если число свободных потоков является таким же, как степень свободы специфической стехиометрической матрицы, однозначное распределение метаболического потока может быть разрешено. Для раствора матричная операция с использованием обратной матрицы, в общем, выполнима, и все потоки предпочтительно должны быть нормализованы, например, для определенного количества субстрата. Если субстратом является глюкоза, все значения потока должны быть репрезентативными, например, со значениями для 10 мМ поглощения глюкозы. Растворы распределений метаболического потока, полученные исходя из случайных значений свободного потока, как описано выше, должны быть биологически значимыми. То есть все значения для потоков необратимых реакций должны равняться 0 или более, и значения для биомассы, сформированной потоками, должны равняться 0 или более. Для получения комбинаций более предпочтительных свободных потоков могут быть добавлены условия, основанные на теоретических и/или эмпирических знаниях о продуцировании вещества с помощью клеток. Должно быть получено определенное число комбинаций, то есть рассчитано число биологически значимых распределений потока, не ограниченное практически, пока оно не окажется достаточным для статистического анализа. Для одного свободного потока обычно используют три или пять значений. Поэтому в случае n свободных потоков существует около n степеней ряда значений для одного свободного потока комбинаций. Например, при использовании трех значений для одного свободного потока существует 3 в n-й степени (3n) комбинаций. То есть для семи свободных потоков (n=7) может быть использовано около 2200 комбинаций. В качестве альтернативы, так как число значений для каждого свободного потока в совокупности данных биологически значимых распределений потока может изменяться в зависимости от выбранных свободных потоков или дополнительных условий, число комбинаций, которое может быть использовано, составляет приблизительно от 3 до n-й степени (3n) или от приблизительно 5 до n-й степени (5n), в целом, для n свободных потоков. Для получения растворов биологически значимых распределений потоков в этом ряду обычно начинают с комбинаций случайных свободных потоков с использованием 6-10 значений для одного свободного потока, то есть комбинаций свободных потоков шесть в n-й степени (6n) или 10 в n-й степени (10n).
На четвертом этапе способа определения на основании распределений метаболического потока (совокупность данных по распределениям метаболического потока) и мультивариантного статистического анализа получили уравнение регрессии, включающее минимальное число свободных потоков, которое показывает корреляцию с продуцированием вещества.
Путем проведения мультивариантного статистического анализа совокупности данных по распределениям потока, рассчитанным по случайным комбинациям свободных потоков, полученным на предыдущем этапе, может быть получено уравнение регрессии, включающее минимальное число свободных потоков, которое показывает корреляцию с продуцированием вещества. Мультивариантный статистический анализ (включающий мультивариантный нелинейный регрессионный анализ и мультивариантный линейный регрессионный анализ) может быть выполнен с помощью любого метода, поскольку выбирали метод, с помощью которого можно исследовать корреляции комбинаций свободного потока с продуцированием вещества. Однако более подходящим является мультивариантный линейный регрессионный анализ. Этот метод описан, например, Kachigan S.K., Chapter 4, Regression Analysis in Multivariate Statistical Analysis 2nd Ed., Radius Press, New York, pp.160-193.
Выражение «показывает корреляцию с продуцированием вещества» означает, что коэффициент корреляции является достаточно большим, и «быть значимо большим» обычно означает, что коэффициент корреляции R2 равен 0,8 или более, предпочтительно 0,9 или более.
Уравнение регрессии, содержащее минимальное число свободных потоков (термов), которое показывает корреляцию с продуцированием вещества, может быть получено путем последовательного изменения числа термов, используемых для получения уравнения регрессии. Такое уравнение, которое показывает наиболее высокий коэффициент корреляции, включает в себя каждый терм, и создается возможность отобрать уравнение регрессии, содержащее минимальное число термов со значимо большим коэффициентом корреляции. В качестве альтернативы уравнение регрессии может быть получено способом исключения из общего числа термов одного терма для исследования степени снижения коэффициента корреляции в результате исключения терма; такая же процедура может быть повторена с другими термами за исключением терма, показывающего снижение коэффициента корреляции наименьшее из всех термов; и когда уравнение регрессии, которое показывает корреляцию с продуцированием вещества, может долго не получаться, это позволяет немедленно выбрать уравнение регрессии.
Хотя эти математические операции могут быть запрограммированы по отдельности, они могут быть легко выполнены с помощью серийно выпускаемых математических компьютерных программ, таких как MatLab® (торговое название MathWorks) и Mathematica® (торговое название Wolfram Research).
На пятом этапе способа определения метаболический поток, влияющий на продуцирование вещества, определяли на основании коэффициентов полученного уравнения регрессии.
Вклад свободных потоков в продуцирование вещества с использованием клеток, таких как микроорганизмы, в частности выход биомассы или выход продукта, необходимого для продуцирования вещества, может быть определен путем использования уравнения регрессии, полученного на предыдущем этапе. То есть свободные потоки, представленные в уравнении регрессии, могут быть определены как потоки, влияющие на продуцирование вещества. К тому же, так как коэффициенты уравнения регрессии представляют величину вклада, свободные потоки, имеющие достоверно больший коэффициент (когда потоки нормализованы, свободные потоки имеют большее абсолютное значение относительного коэффициента), могут быть определены как метаболические потоки, значительно влияющие на продуцирование вещества.
Способ определения настоящего изобретения может предоставить информацию, которая имеет важное значение для улучшения бактериальных штаммов, то есть какой свободный поток наиболее влияет на продуцирование адресного вещества и имеет ли свободный поток положительное или отрицательное влияние на продуцирование целевого вещества. Также можно заранее предсказать поток, который требуется изменить для получения благоприятного эффекта на выход адресного продукта и продуктивность.
Как показано в примерах данного описания, можно ожидать, что бактериальные штаммы с улучшенной лизин-продуцирующей способностью могут быть получены путем усиления активности фосфоенолпируваткарбоксилазы при продуцировании лизина с помощью Escherichia coli. В международной публикации № WO01/53459 представлен пример усовершенствования продуцирования лизина путем усиления активности фосфоенолпируваткарбоксилазы. Следовательно, было подтверждено, что бактериальный штамм, обладающий способностью продуцировать вещество, может быть создан на основе способа определения.
<3> Способ получения для продуцирования L-лизина или L-треонина
Способ настоящего изобретения представляет собой способ продуцирования L-лизина или L-треонина и включает в себя этапы: культивирование бактерии, обладающей способностью продуцировать L-лизин или L-треонин в среде; накопление L-лизина или L-треонина в среде или в клетках бактерии; и сбор и выделение L-лизина или L-треонина из среды или из клеток.
Культуральная среда, используемая в настоящем изобретении, может быть средой, обычно применяемой для ферментативного продуцирования L-лизина или L-треонина с помощью микроорганизма. Может быть использована стандартная среда, содержащая источник углерода, источник азота, неорганические ионы и другие органические компоненты, при необходимости. В качестве источника углерода могут быть использованы различные сахариды, такие как глюкоза, сахароза, лактоза, галактоза, фруктоза и гидролизат крахмала, различные спирты, такие как глицерин и сорбит, и различные органические кислоты, такие как фумаровая кислота, лимонная кислота и янтарная кислота. В качестве источника азота могут быть использованы различные неорганические соли аммония, такие как сульфат аммония, хлорид аммония и фосфат аммония, органический азот, такой как соевый гидролизат, газообразный аммиак и водный раствор аммиака и т.п. В качестве следовых количеств органических питательных веществ желательно добавлять необходимые вещества, такие как витамин B1,гомосерин или дрожжевой экстракт и т.п. Кроме того, могут быть добавлены следовые количества фосфата калия, сульфата магния, ион железа, ион марганца. Среда, используемая для культивирования, может быть либо синтетической средой, либо естественной средой, поскольку среда содержит источник углерода и источник азота, неорганические ионы и, если необходимо, следы органических питательных веществ.
Культивирование предпочтительно проводили в аэробных условиях в течение одного-семи дней при температуре 24-37°C и pH 5-9. pH культуры можно регулировать неорганической или органической кислотой или щелочным веществом, например газообразным аммиаком и т.п. Сбор и выделение L-лизина или L-треонина из культуральной среды можно проводить с помощью стандартных способов, таких как способ на основе ионообменной смолы, способ осаждения и других известных способов и их комбинаций. После накопления L-лизина или L-треонина в клетках их можно собирать с помощью способа на основе ионообменной смолы и т.п. из супернатанта, полученного путем разрушения клеток с помощью ультразвука и т.п., и удаления обломков клеток путем центрифугирования.
Примеры
Настоящее изобретение далее детально описано на основании примеров.
Пример 1.
Определение метаболического потока в отношении L-лизина
(1) Создание стехиометрической матрицы
Стехиометрическое уравнение для расчета метаболического потока было получено путем допущения квазиравновесного состояния внутриклеточных метаболических промежуточных соединений (Savinell J.M. and Palsson B.O.J., Theor. Biol., 154:421-454, 1992; Vallino J.J. and Stephanopoulos G., Biotechnol. Bioeng., 41:633-646, 1993). Формулы реакций, включенные в эту модель, показаны в таблице 2. Описание сокращений, применяемых в настоящем изобретении, представлено в таблице 1. Для упрощения формул некоторые реакции без разветвления объединили. Поскольку пентозофосфатный путь является сложным, его представили двумя формулами. Полученные данные использовали для определения соотношения компонентов биомассы (Neidhardt F.C. с сотр., Physiology of the Bacterial Cell., Sinauer Associates, Massachusetts, 1990) и биомассу представили с помощью формулы реакции [68]. Степень свободы стехиометрической матрицы в этой модели равнялась 7.
Список использованных формул реакций. Обратимые реакции обозначены буквой r
(2) Селекция свободных потоков и получение их случайных комбинаций
Специфические группы потоков определили по способу Reder (Reder C.J., Theor. Biol., 135:175-201, 1988). Из каждой группы отобрали поток, замыкающий точку ветвления. В таблице 3 представлены семь отобранных свободных потоков. Путем спецификации этих семи потоков может быть получен уникальный раствор для баланса потоков.
Список свободных потоков для получения случайного распределения потоков
Из почти 300000 комбинаций значений для 7 случайных свободных потоков исключили те, которые нарушали любое ограничение, касающееся обратимой реакционной способности, и те, которые показывали значения как для лизина, так и для биомассы, не превышающие в совокупности пороговые уровни на 20% каждого максимального значения. В результате получили совокупность данных распределений 5000 метаболических потоков в биологически значимой специфической области. Результаты представили с помощью значений, основанных на 10 мМ поглощения глюкозы, и получили матрицу, содержащую 5000 строк, соответствующих распределению случайных потоков, и 68 столбцов, каждый из которых соответствует реакционному потоку.
(3) Корреляционный анализ с помощью мультивариантного анализа и определение метаболических потоков, влияющих на продуцирование вещества
Провели мультивариантную линейную регрессию конденсированной матрицы, включающей Z-меток только столбцов, соответствующих 7 свободным потокам. Для мультивариантной линейной регрессии использовали ступенчатую регрессионную функцию статистической прикладной программы MatLab. С помощью этого метода биомасса или продуцирование лизина может быть выведена из линейной функции 7 свободных потоков. Идентификация этих 7 свободных потоков показывает уникальное определение состояния системы. Поэтому, если все 7 термов использовали в качестве параметров, коэффициент корреляции равен 1, что указывает на полную аппроксимацию. Однако, как правило, можно получить относительно благоприятную аппроксимацию с меньшим числом термов, чем в уравнении. Для проверки различных комбинаций термов уравнение, показывающее наилучшую аппроксимацию для каждого числа содержащихся термов, отбирали с помощью ступенчатой функции программы MatLab. Для выхода биомассы аппроксимацию R2=0,980 получили только с 4 термами - изоцитратлиазой (ICL), малеиновым ферментом (MEZ), PEP-карбоксилазой (PEPC) и АТФазой. Когда число термов в дальнейшем снижали, значение R2 заметноуменьшалось, и любая приемлемая аппроксимация не могла быть получена. Когда реакционные потоки нормализовали по значению на 10 мМ глюкозы и использовали как исходное, точное уравнение представили в следующем виде:
Уравнение 1) Выход биомассы=1,552-0,194 (ICL)+0,184 (MEZ)-0,194 (PEPC)-0,011 (АТФаза)
Выход лизина мог бы быть аппроксимированным согласно модели, содержащий 4 параметра, и в результате получили R2=0,997. Далее, даже когда терм для АТФазы исключили, R2 снизился лишь до 0,856, и аппроксимация была еще положительной. Поэтому в модели лизина использовали следующие 3 параметра.
Уравнение 2) Выход лизина=-1,694+1,176 (ICL) - 1,095 (MEZ)+1,162 (PEPC)
Наконец, общий выход углерода (атомы C) определили по общему числу атомов углерода, направленных в биомассу, и лизин мог бы быть аппроксимирован при R2=0,956 при использовании только терма для АТФазы со следующим уравнением.
Уравнение 3) C-атомы=34,3-0,314 (АТФаза)
Эти результаты показали, что выход биомассы положительно коррелирует с потоком малеинового фермента и что продуцирование лизина положительно коррелирует с потоками PEP-карбоксилазы и изоцитратлиазы (глиоксалатный цикл). Пригодность этого регрессионного анализа может быть показана на фиг. 1 и 2. Когда потоки изоцитратлиазы и малеинового фермента рассматривали отдельно, корреляции с продуцированием лизина не наблюдали, как показано на фиг. 1 (а) и (b). Однако если эти потоки рассматривали как часть уравнения регрессии 2), корреляцию можно было наблюдать, что показано на фиг. 2, и эффект становился очевиден. Таким образом, с помощью данного способа может быть обнаружена невидимая связь между метаболическими потоками. Выход целевого продукта может быть повышен путем усиления регулирующей поток активности, проявляющей положительную корреляцию, и ослаблением регулирующей поток активности, проявляющей отрицательную корреляцию. То есть на основании этих результатов может быть получено руководство для улучшения бактериальных штаммов и усиления активности PEP-карбоксилазы или изоцитратлиазы или аттенуации активности малеинового фермента, проявляющего отрицательную корреляцию с эффективностью продуцирования лизина. В действительности, пример создания бактериального штамма, проявляющего улучшенную способность продуцирования лизина путем усиления активности PEP-карбоксилазы при продуцировании лизина с использованием Escherichia coli, был представлен в Международной публикации № WO01/53459, и, таким образом, была поддержана полезность настоящего изобретения.
Пример 2
Определение метаболического потока в отношении L-треонина
С помощью того же способа, как в примере 1, уравнение, показывающее наибольшую аппроксимацию для каждого числа содержащихся в нем термов, отобрали в отношении L-треонина. Что касается выхода биомассы, аппроксимацию R2=0,986 получили только для 4 термов - изоцитратлиазы (ICL), малеинового фермента (MEZ), PEP-карбоксилазы (PEPC) и АТФазы.
Уравнение 4) Выход биомассы=1,260-0,101 (ICL)+0,093 (MEZ)-0,101 (PEPC)-0,009 (АТФаза)
Выход треонина мог быть аппроксимирован с моделью, содержащей те же 3 параметра, и в результате получили R2=0,937.
Уравнение 5) Выход треонина=-1,432+1,090 (ICL)-1,080 (MEZ)+1,087 (PEPC)
Эти результаты показали, что выход биомассы положительно коррелировал с потоком малеинового фермента, а продуцирование треонина положительно коррелировало с потоками PEP-карбоксилазы и изоцитратлиазы (глиоксилатный цикл). Поэтому в отношении продуцирования треонина также может быть получен руководящий принцип улучшения бактериальных штаммов, и усиление активности PEP-карбоксилазы или изоцитратлиазы или ослабление активности малеинового фермента, проявившего отрицательную корреляцию, является эффективным для продуцирования лизина.
Пример 3
Конструирование продуцирующей L-лизин бактерии, дефектной по малеиновому ферменту
В качестве продуцирующего L-лизин штамма Escherichia coli использовали штамм WC196, который резистентен к AEC (S-(2-аминоэтил)цистеину) (Международная публикация № WO 96/17930).
К малеиновому ферменту из Escherichia coli относится фермент, использующий НАД в качестве кофермента (EC 1.1.1.38) и фермент, использующий НАДФ в качестве кофермента (EC 1.1.1.40). Эти ферменты кодируются генами sfcA и b2463 соответственно.
Гены sfcA и b2463 делетировали путем комбинации способа «red-запускаемой интеграции», который первоначально разработали Datsenko и Wanner (Proc. Natl. Acad. Sci. USA, 2000, 97, 6640-6645), и способа эксцизионной системы, полученной из фага лямбда (J. Bacteriol. 2002 Sep; 184(18): 5200-3). Взаимодействие между интегразой и эксцизионазой в фаге лямбда удаляет нуклеопротеиновый комплекс (Cho EH, Gumport RI, Gardner JF.). В соответствии со способом red-запускаемой интеграции штамм, несущий разрушенный ген, может быть получен в один этап с помощью ПЦР-продукта, полученного путем использования синтетических олигонуклеотидных праймеров, сконструированных так, что они содержат часть адресного гена на своем 5'-конце и часть гена резистентности к антибиотику на своем 3'-конце. Кроме того, интегрированный ген резистентности к антибиотику может быть удален путем дальнейшего комбинирования эксцизионной системы, полученной из фага лямбда, со способом red-запускаемой интеграции.
(1) Разрушение гена sfcA
В качестве ПЦР-матрицы использовали плазмиду pMW118-attL-Cm-attR (ее получение описано ниже). pMW118- attL-Cm-attR является плазмидой, полученной путем инсерции генов attL и attR, которые являются сайтами прикрепления фага лямбда, и гена cat, который является геном резистентности к антибиотику в pMW118 (TaKaRa Bio). Гены ввели в порядке attL-cat-attR. Последовательность attL представлена в SEQ ID NO:11, последовательность attR представлена в SEQ ID NO:12.
ПЦР проводили с использованием праймеров, представленных в SEQ ID NOS:1 и 2 и имеющих последовательности, соответствующие на их 3'-концах attL и attR, и последовательности, соответствующие частям гена sfcA на их 5'-конце соответственно.
Амплифицированный ПЦР-продукт очищали на агарозном геле и вводили в штамм WC196 Escherichia coli, содержащий плазмиду pKD46, характеризующуюся температурочувствительной репликацией, посредством электропорации. pKD46 (Proc. Natl. Acad. Sci. USA, 2000, 97, 6640-6645) содержит 2154 п.н. фрагмент ДНК фага лямбда (GenBank/EMBL инвентарный № J02459, 31088-33241), несущий гены (гены γ, β и exo), кодирующие рекомбиназу Red системы гомологичной рекомбинации λ Red под контролем индуцируемого арабинозой промотора ParaB. pKD46 необходима для интеграции ПЦР-продукта в хромосому штамма WC196.
Компетентные клетки для электропорации получили, как описано ниже. Штамм WC196 Escherichia coli, культивированный в течение ночи при 30°C в среде LB, содержащей 100 мг/л ампициллина, разбавляли в 100 раз 5 мл среды SOB (Sambrook, J. с сотр., "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)), содержащей ампициллин (50 мг/л) и L-арабинозу (1 мМ). Разбавленный продукт культивировали при 30°C в условиях аэрации до тех пор, пока показатель OD600 не равнялся примерно 0,6, затем концентрировали в 100 раз. Для приготовления к электропорации клетки промывали 3 раза 10% глицерином. Электропорацию проводили с использованием 70 мкл компетентных клеток и примерно 100 нг ПЦР-продукта. Перед электропорацией к клеткам добавляли 1 мл среды SOC (Sambrook J. С сотр., "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)). Клетки культивировали при 37°C 2,5 ч, затем помещали на чашки, залитые L-агаром, содержащим 25 мг/л Cm (хлорамфеникол) при 37°C для отбора Cm-резистентного рекомбинанта. Затем для элиминации плазмиды pKD46 клетки субкультивировали дважды при 42°C на Cm-содержащей среде из L-агара. Полученные колонии тестировали на резистентность к ампициллину. Таким образом, получили ампициллин-чувствительный штамм, не содержащий pKD46.
Делецию гена sfcA в мутанте, идентифицированном с помощью гена резистентности к хлорамфениколу, подтвердили с помощью ПЦР. Полученный в результате штамм, дефектный по sfcA, обозначили как WC196ΔsfcA::att-cat.
Для элиминации гена att-cat, который был интегрирован в ген sfcA, использовали хелперную плазмиду pMW-intxis-ts (ее получение описано ниже). pMW-intxis-ts содержит ген, кодирующий интегразу (Int) (SEQ ID NO:13), и ген, кодирующий эксцизионазу (Xis) (SEQ ID NO:15) фага лямбда, и характеризуется температурочувствительной репликацией. В результате введения pMW-intxis-ts, рекомбинация происходит благодаря узнаванию attL (SEQ ID NO:11) и attR (SEQ ID NO:12) на хромосоме, и ген резистентности к антибиотику, расположенный между attL и attR, вырезается, в результате структура, содержащая только последовательности attL или attR, остается на хромосоме.
Компетентные клетки штамма WC196ΔsfcA::att-cat получили в соответствии со стандартным способом путем трансформации их хелперной плазмидой pMW-intxis-ts с последующим культивированием на чашке с L-агаром, содержащим 50 мг/л ампициллина, при 30°C для отбора ампициллин-резистентного штамма.
Для удаления плазмиды pMW-intxis-ts клетки субкультивировали дважды при 42°C на среде, содержащей L-агар. Полученные колонии тестировали на резистентность к ампициллину и резистентность к хлорамфениколу. Получили ампициллин- и хлорамфеникол-чувствительный штамм без att-cat и pMW-intxis-ts. Этот штамм обозначили как WC196ΔsfcA.
(2) Разрушение гена b2463
Делецию гена b2463 в штаммах WC196 и WC196ΔsfcA проводили по способу (1), за исключением того, что праймеры SEQ ID NOS:3 и 4 использовали как праймеры для разрушения b2463. Так получили штаммы WC196Δb2463 и WC196ΔsfcAΔb2463. Полученный штамм WC196ΔsfcAΔb2463 обозначили как WC196Δmez.
(3) Получение ПЦР-матрицы и хелперной плазмиды
ПЦР-матрицу pMW118-attL-Cm-attR и хелперную плазмиду pMW-intxis-ts получили следующим образом:
(3-1) pMW118-attL-Cm-attR
Для конструирования плазмиды pMW118-attL-Cm-attR в качестве исходной использовали плазмиду pMW118-attL-Tc-attR. Лигировали четыре фрагмента ДНК:
BglII-EcoRI - фрагмент ДНК (120 п.н.) (SEQ ID NO:11), несущий attL, который получили с помощью ПЦР-амплификации соответствующей последовательности E. coli W3350 (содержащей профаг λ) хромосомы с использованием олигонуклеотидов P1 и P2 (SEQ ID NOS:17 и 18) в качестве праймеров (эти праймеры содержали вспомогательные сайты узнавания эндонуклеаз BglII и EcoRI);
2) PstI-HindIII - фрагмент ДНК (182 п.н.), несущий attR (SEQ ID NO:12), который получили с помощью ПЦР-амплификации соответствующей последовательности E. coli W3350 (содержащей профаг λ) хромосомы с использованием олигонуклеотидов P3 и P4 (SEQ ID NOS:19 и 20) в качестве праймеров (эти праймеры содержали вспомогательные сайты узнавания эндонуклеаз PstI и HindIII);
3) большой (3916 п.н.) BglII-HindIII фрагмент pMW118-ter_rrnB. pMW118-ter_rrnB получили путем лигирования трех фрагментов ДНК:
большого (2359 п.н.) фрагмента, несущего фрагмент AatII-EcoRIpol pMW118, плазмиду pMW118 переварили рестрикционной эндонуклеазой EcoRI, обработанной фрагментом Кленова ДНК-полимеразы I и затем переварили рестрикционной эндонуклеазой AatII;
маленького (1194 п.н.) фрагмента AatII-BglII pUC19, несущего ген bla резистентности к ампициллину (ApR), полученного путем ПЦР-амплификации соответствующей последовательности плазмиды pUC19 с использованием олигонуклеотидов P5 и P6 (SEQ ID NOS:21 и 22) в качестве праймеров (эти праймеры содержали вспомогательные сайты узнавания эндонуклеаз AatII и BglII);
маленького (363 п.н.) фрагмента BglII-PstIpol терминатора транскрипции ter_rrnB, полученного путем ПЦР-амплификации соответствующей области хромосомы E. coli MG1655 с использованием олигонуклеотидов P7 и P8 (SEQ ID NOS:23 и 24) в качестве праймеров (эти праймеры содержали вспомогательные сайты узнавания эндонуклеаз BglII и PstI);
4) маленький (1388 п.н.) фрагмент EcoRI-PstI (SEQ ID NO:29) pML-Tc-ter_thrL, содержащий ген резистентности к тетрациклину и терминатор транскрипции ter_thrL, pML-Tc-ter_thrL, получили следующим путем:
pML-MSC (2001 #5) переварили рестрикционными эндонуклеазами XbaI и BamHI, затем большой (3342 п.н.) фрагмент лигировали с фрагментом (68 п.н.) XbaI-BamHI, несущим терминатор ter_thrL, который получили путем ПЦР-амплификации соответствующей области хромосомы E. coli MG1655 с использованием олигонуклеотидов P9 и P10 (SEQ ID NOS:25 и 26) в качестве праймеров (эти праймеры содержали вспомогательные сайты узнавания эндонуклеаз XbaI и BamHI), продукт этой реакции представлял собой плазмиду pML-ter_thrL;
затем pML-ter_thrL переварили рестрикционными эндонуклеазами KpnI и XbaI, затем обработали фрагментом Кленова ДНК-полимеразы I и затем лигировали с небольшим (1317 п.н.) EcoRI-Van91I фрагментом pBR322, содержащим ген резистентности к тетрациклину (pBR322 переварили рестрикционными эндонуклеазами EcoRI и Van91I, затем обработали фрагментом Кленова ДНК-полимеразы I), продукт этой реакции представлял собой плазмиду pML-Tc-ter_thrL.
Таким образом, была получена pMW118-attL-Tc-attR.
pMW118-attL-Cm-attR сконструировали путем лигирования большого (4413 п.н.) BamHI-XbaI фрагмента pMW118-attL-Tc-attR и искусственного фрагмента ДНК BglII-XbaI (1162 п.н.), содержащего промотор PA2 (ранний промотор фага T7), ген cat резистентности к хлорамфениколу (CmR), терминатор транскрипции ter_thrL и attR. Искусственный фрагмент ДНК (SEQ ID NO:30) получили следующим путем:
1) pML-MSC (2001 #5) переварили рестрикционными эндонуклеазами KpnI и XbaI и лигировали с маленьким (120 п.н.) фрагментом KpnI-XbaI, который содержал промотор PA2 (ранний промотор фага T7), полученный посредством ПЦР-амплификации соответствующей области ДНК фага T7 с олигонуклеотидами P11 и P12 (SEQ ID NOS:27 и 28) в качестве праймеров (эти праймеры содержали вспомогательные сайты узнавания эндонуклеаз KpnI и XbaI), продукт этой реакции представлял собой плазмиду pML-PA2-MCS;
2) далее сайт XbaI делетировали из pML-PA2-MCS, продукт этой реакции представлял собой плазмиду pML-PA2-MCS(XbaI-);
3) затем маленький (928 п.н.) фрагмент BglII-HindIII плазмиды pML-PA2-MCS(XbaI-), содержащий промотор PA2 (ранний промотор фага T7) и ген cat резистентности к хлорамфениколу (CmR) лигировали с маленьким (234 п.н.) фрагментом HindIII-HindIII плазмиды pMW118-attL-Tc-attR, содержащим терминатор транскрипции ter_thrL и attR;
4) необходимый искусственный фрагмент ДНК (1156 п.н.) получили с помощью ПЦР-амплификации с лигированием с использованием реакционной смеси и олигонуклеотидов P9 и P4 (SEQ ID NOS:25 и 20) в качестве праймеров (эти праймеры содержали вспомогательные сайты узнавания эндонуклеаз HindIII и XbaI).
(3-2) pMW-intxis-ts
Первоначально два фрагмента ДНК амплифицировали с использованием ДНК фага λ ("Fermentas") в качестве матрицы. Первый фрагмент содержал область 37168-38046 п.н. (SEQ ID NO:39) и также содержал ген, кодирующий репрессор cI, промоторы Prm и Pr и лидерную последовательность гена cro. Этот фрагмент получили с использованием олигонуклеотидов P1' и P2' (SEQ ID NOS:31 и 32) в качестве праймеров. Второй фрагмент содержал гены xis-int фага λ и содержал область 27801-29100 п.н. (SEQ ID NO:40). Для этой амплификации в качестве праймеров использовали олигонуклеотиды P3' и P4' (SEQ ID NOS:33 и 34). Все праймеры содержали соответствующие сайты узнавания эндонуклеаз.
Полученный ПЦР-амплифицированный фрагмент, содержащий репрессор cI, переварили рестрикционной эндонуклеазой ClaI, обработали фрагментом Кленова ДНК-полимеразы I и затем переварили рестрикционной эндонуклеазой EcoRI. Второй ПЦР- амплифицированный фрагмент переварили рестрикционными эндонуклеазами EcoRI и PstI. Далее плазмиду pMWPlaclacI-ts переварили эндонуклеазой BglII, обработали фрагментом Кленова ДНК-полимеразы I и затем переварили рестрикционной эндонуклеазой PstI. Векторный фрагмент pMWPlaclacI-ts элюировали из агарозного геля и лигировали с переваренными ПЦР-амплифицированными фрагментами.
Плазмида pMWPlaclacI-ts является производной pMWPlaclacI, которая состоит из следующих частей: 1) BglII-HindIII - искусственный фрагмент ДНК, включающий ген lacI под контролем промотора PlacUV5 и RBS гена 10 бактериофага T7; 2) AatII-BglII - фрагмент ДНК, несущий ген резистентности к ампициллину (ApR), который получили путем ПЦР-амплификации соответствующей последовательности плазмиды pUC19 с использованием олигонуклеотидов P5' и P6' (SEQ ID NOS:35 и 36) в качестве праймеров (эти праймеры содержали вспомогательные сайты узнавания эндонуклеаз AatII и BglII; 3) AatII-HindIII - фрагмент, содержащий фрагмент AatII-PvuI ранее сконструированной рекомбинантной плазмиды pMW118-ter_rrnB. Последнюю плазмиду сконструировали следующим образом: фрагмент ДНК PstI-HindIII, несущий терминатор ter_rrnB, получили посредством ПЦР-амплификации соответствующей области хромосомы E. coli MG1655 с использованием олигонуклеотидов P7' и P8' (SEQ ID NOS:37 и 38), содержащих соответствующие сайты узнавания эндонуклеаз в качестве праймеров. Перед лигированием плазмиду pMW118 и ДНК-фрагмент ter_rrnB (комплементарный SEQ ID NO:41) разрезали эндонуклеазами PvuI и PstI соответственно, обработали фрагментом Кленова ДНК-полимеразы I для получения липких концов и затем разрезали эндонуклеазой AatII или HindIII. Для конструирования варианта pMWPlaclacI-ts фрагмент AatII-EcoRV плазмиды pMWPlaclacI заместили фрагментом AatII-EcoRV плазмиды pMAN997, содержащим локусы par, ori и ген repAts репликона pSC101.
Пример 4
Конструирование продуцирующей L-треонин бактерии, дефектной по малеиновому ферменту
Из штамма VKPM B-5318 получили sfcA- и b2463-дефектные штаммы. Штамм VKPM B-5318 депонировали в Российской национальной коллекции промышленных микроорганизмов (VKPM), (ГНИИ Генетика) от 19 ноября 1987 г. и присвоили инвентарный номер VKPM B-5318.
Штамм, дефектный по одному из генов (sfcA, b2463) малеинового фермента (mez), получили таким же путем, как в примере 3, с помощью способа red-запускаемой интеграции. То есть выполнили таким же образом с помощью способа red-запускаемой интеграции, как в примере 3, за исключением того, что для получения sfcA- или b2463-дефектного штамма как мутанта, идентифицируемого по гену резистентности к хлорамфениколу, вместо штамма WC196 использовали штамм B-5318. Штамм B-5318, в котором разрушили sfcA, обозначили как B-5318ΔsfcA. Штамм B-5318, в котором разрушили b2463, обозначили как B-5318Δb2463. Штамм B-5318 с разрушенными генами sfcA и b2463, B-5318ΔsfcAΔb2463, получили таким же образом, с помощью способа «red-запускаемой интеграции» и эксцизионной системы, как описано в примере 3. Штамм B-5318ΔsfcAΔb2463 обозначили как B-5318mez.
Пример 5
Оценка штамма, дефектного по малеиновому ферменту
<5-1> Оценка продуцирующей L-треонин бактерии, являющейся b2463-дефектным штаммом
Каждый из штаммов B-5318Δb2463 и B-5318 культивировали на среде с LB-агаром (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl и 15 г/л агара), содержащей 20 мг/л сульфата стрептомицина и 25 мг/л сульфата канамицина, при 37°C в течение 24 часов. Бактериальные клетки, взятые с пятой части чашки, вносили в 50 мкл жидкой среды LB (10 г/л триптона, 5 г/л дрожжевого экстракта и 5 г/л NaCl), содержащей 20 мг/л сульфата стрептомицина и 25 мг/л сульфата канамицина, для проведения предварительного культивирования при 40°C и 144 об/мин в течение 3,5 часов.
После завершения предварительного культивирования прекультуральный бульон вносили в 300 мл основной культуральной среды, содержащейся в бродильном ферментере объемом 1 л, в количестве, составляющем 10% объема основной культуральной среды, для проведения основного культивирования при 40°C и pH 7,0. Состав основной культуральной среды приведен ниже.
pH в процессе культивирования поддерживали на уровне 7,0 путем добавления газообразного аммиака.
После расхода добавленного сахара количество L-треонина измеряли с помощью жидкостной хроматографии. Результаты представлены в таблице 5.
При использовании дефектного по b2463 штамма B-5318Δb2463 выход треонина повысился по сравнению с контрольным штаммом B-5318.
<5-2> Оценка продуцирующей L-треонин бактерии, являющейся sfcA-дефектным штаммом
Штаммы B-5318ΔsfcA и B5318 культивировали таким же способом, как описано в <5-1>.
После того как добавленный сахар был использован, количество L-треонина измеряли с помощью жидкостной хроматографии. Результаты представлены в таблице 6.
При использовании дефектного по b2463 штамма B-5318ΔsfcA выход треонина повысился по сравнению с контрольным штаммом B-5318.
<5-3> Оценка продуцирующей L-лизин бактерии, являющейся sfcA- и b2463-дефектным штаммом
Для получения штаммов WC196/pCABD2, WC196ΔsfcA/pCABD2 и WC196Δb2463/pCABD2 штаммы WC196, WC196ΔsfcA и WC196Δb2463 трансформировали в соответствии со стандартным методом с использованием плазмиды pCABD2 для продуцирования лизина, которая содержала гены dapA, dapB и dapC (Международная публикация № WO 01/53459).
Штаммы WC196/pCABD2, WC196ΔsfcA/pCABD2 и WC196Δb2463/pCABD2 культивировали при 37°C в L-среде (как описано ниже), содержащей 20 мг/л стрептомицина до тех пор, пока OD600 на среде не равнялся примерно 0,6. Затем в культуру добавляли эквивалентное количество 40% раствора глицерина. После перемешивания соответствующие аликвоты смеси разливали и хранили при -80°C. Хранящиеся аликвоты назвали глицериновыми штоками.
Глицериновые штоки штаммов размораживали и по 100 мкл каждого штамма равномерно разливали на чашку с L-средой, содержащей 20 мг/л стрептомицина, и культивировали при 37°C в течение 24 часов. С восьмой части полученного высева отбирали бактериальные клетки и вносили в 20 мл ферментативной среды (как описано ниже), содержащей 20 мг/л стрептомицина, и культивировали при 37°C в течение примерно 16 часов на качалке с возвратно-поступательным движением. После культивирования количество накопленного в среде лизина и количество оставшейся глюкозы измеряли с помощью анализатора Biotech AS210 (Sakura Seiki).
Данные по накоплению L-лизина и выхода продукта за вычетом клеток представлены в таблице 7. Выход продукта за вычетом клеток, который представляет собой выход, рассчитанный путем вычитания количества сахара, использованного для формирования бактериальных клеток, рассчитывали на основе допущения, что 50% потребленного сахара использовалось для формирования бактериальных клеток. Результаты показали, что выход продукта за вычетом клеток штаммов WC196(sfcA/pCABD2 и WC196(b2463/pCABD2 увеличился по сравнению с контрольным штаммом WC196/pCABD2.
(%)
Среду, использованную для оценки дефектного по sfcA или b2463 штамма, продуцирующего L-лизин, описали ниже. Использованные реагенты получили от Wako Pure Chemicals или Nakarai Tesque, если не указано иное. pH всей среды регулировали с помощью NaOH или HCl.
Пример 6
Оценка штамма, дефектного по малеиновому ферменту (mez)
<6-1> Оценка продуцирующей L-треонин бактерии, являющейся дефектным по малеиновому ферменту штаммом
Каждый из штаммов B-5318(mez и B-5318 культивировали на среде из LB-агара (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl и 15 г/л агара), содержащей 20 мг/л сульфата стрептомицина и 25 мг/л сульфата канамицина при 37°C в течение 24 часов, и бактериальные клетки брали с одной из чашек и суспендировали в 5 мл жидкой среды LB (10 г/л триптона, 5 г/л дрожжевого экстракта и 5 г/л NaCl). 0.5 мл суспензии вносили в 50 мл жидкой среды LB, содержащей 20 мг/л сульфата стрептомицина и 25 мг/л сульфата канамицина для проведения предварительного культивирования при 39°C и 144 об/мин в течение 4 часов.
После завершения предварительного культивирования прекультуральный бульон вносили в 300 мл основной культуральной среды, содержащейся в бродильном ферментере объемом 1 л, в количестве, составлявшем 10% объема основной культуральной среды, для основного культивирования при 39°C и pH 7.0. Состав основной культуральной среды приведен ниже.
pH в процессе культивирования регулировали до 7,0 путем добавления газообразного аммиака.
После того как добавленный сахар был потреблен и исчерпан, добавляли 600 г/л водного раствора глюкозы.
Через 24 часа в основной культуре измеряли количество L- треонина с помощью жидкостной хроматографии. Результаты представлены в таблице 10.
При использовании дефектного по малеиновому ферменту штамма B-5318Δmez выход треонина повысился по сравнению с контрольным штаммом B-5318.
<6-2> Оценка продуцирующей L-лизин бактерии, являющейся дефектным по малеиновому ферменту штаммом
Для получения штаммов WC196/pCABD2, WC196Δmez/pCABD2 штаммы WC196 и WC196Δmez трансформировали по стандартному методу плазмидой pCABD2 для продуцирования лизина (Международная публикация № WO 01/53459).
Штаммы WC196/pCABD2 и WC196Δmez/pCABD2 культивировали при 37°С в L-среде (аналогичной среде, использованной в примере 5 <5-3>), содержащей 20 мг/л стрептомицина, и культивировали при 37°C в течение примерно 48 часов на качалке с возвратно-поступательным движением. После культивирования количество накопленного в среде лизина и количество оставшейся глюкозы измеряли с помощью анализатора Biotech AS210 (Sakura Seiki).
Результаты по накоплению L-лизина и выхода продукта за вычетом клеток представлены в таблице 11. Выхода продукта за вычетом клеток рассчитывали на основе допущения, согласно которому 50% потребленного сахара использовалось для формирования бактериальных клеток. Результаты показали, что выход продукта за вычетом клеток штамма WC196Δmez/pCABD2 увеличен по сравнению с контрольным штаммом WC196/pCABD2.
Промышленная применимость
В соответствии с настоящим изобретением ферментативный выход L-лизина и/или L-треонина увеличился в способе продуцирования L-лизина или L-треонина путем ферментации с использованием бактерии Escherichia. Кроме того, настоящее изобретение может быть использовано для селекции продуцирующих L-лизин или L-треонин бактерий, относящихся к роду Escherichia.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН pnp, - ПРОДУЦЕНТ L-ТРЕОНИНА, И СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 2005 |
|
RU2330883C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2008 |
|
RU2515044C2 |
| СПОСОБ ПРОИЗВОДСТВА L-АМИНОКИСЛОТ | 2007 |
|
RU2422530C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА И L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН cpxR | 2005 |
|
RU2320723C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА И L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ybiV | 2005 |
|
RU2320719C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН hipA | 2005 |
|
RU2320718C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |
| СПОСОБ ПОЛУЧЕНИЯ ОСНОВНОГО ВЕЩЕСТВА | 2005 |
|
RU2350655C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ ESCHERICHIA | 2005 |
|
RU2304615C2 |
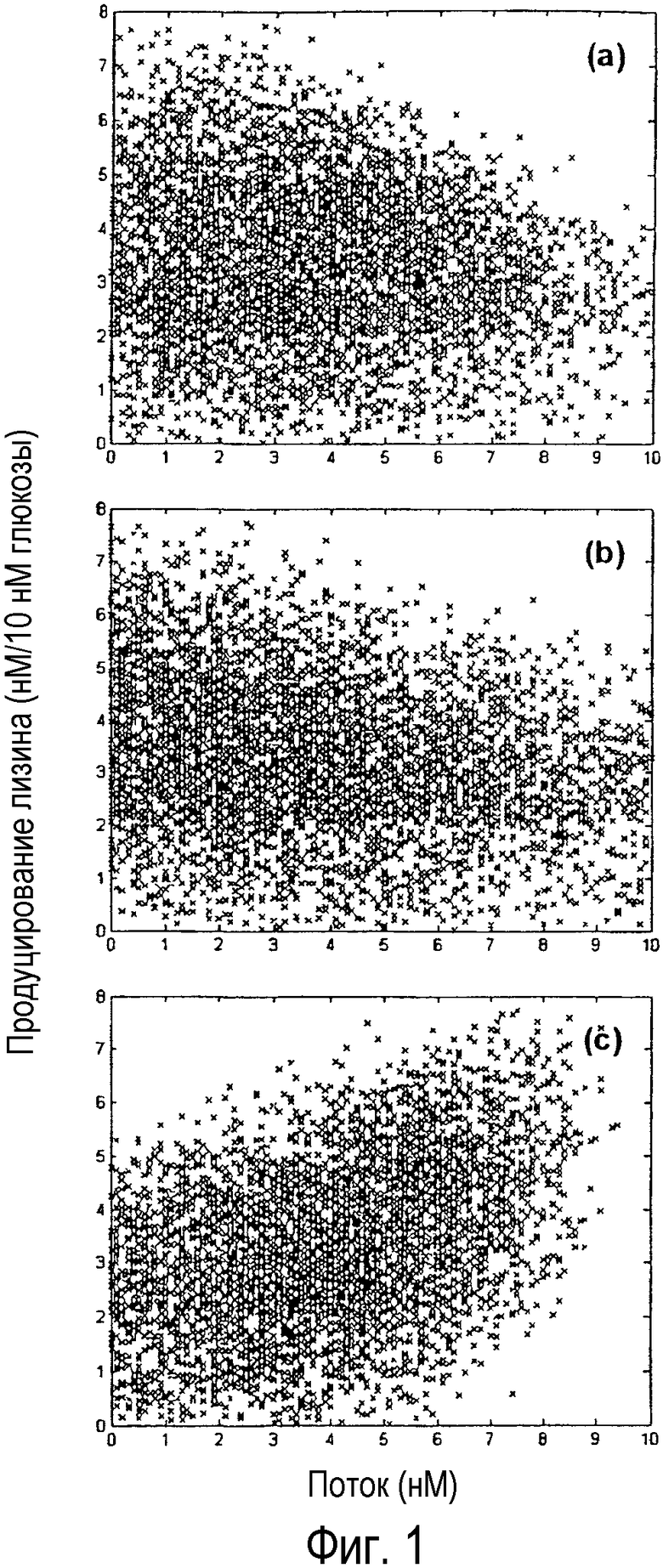
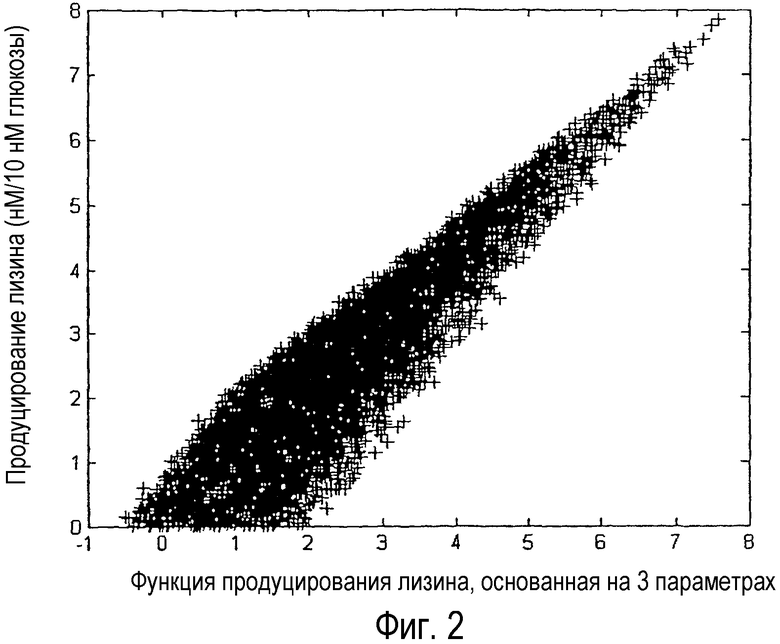
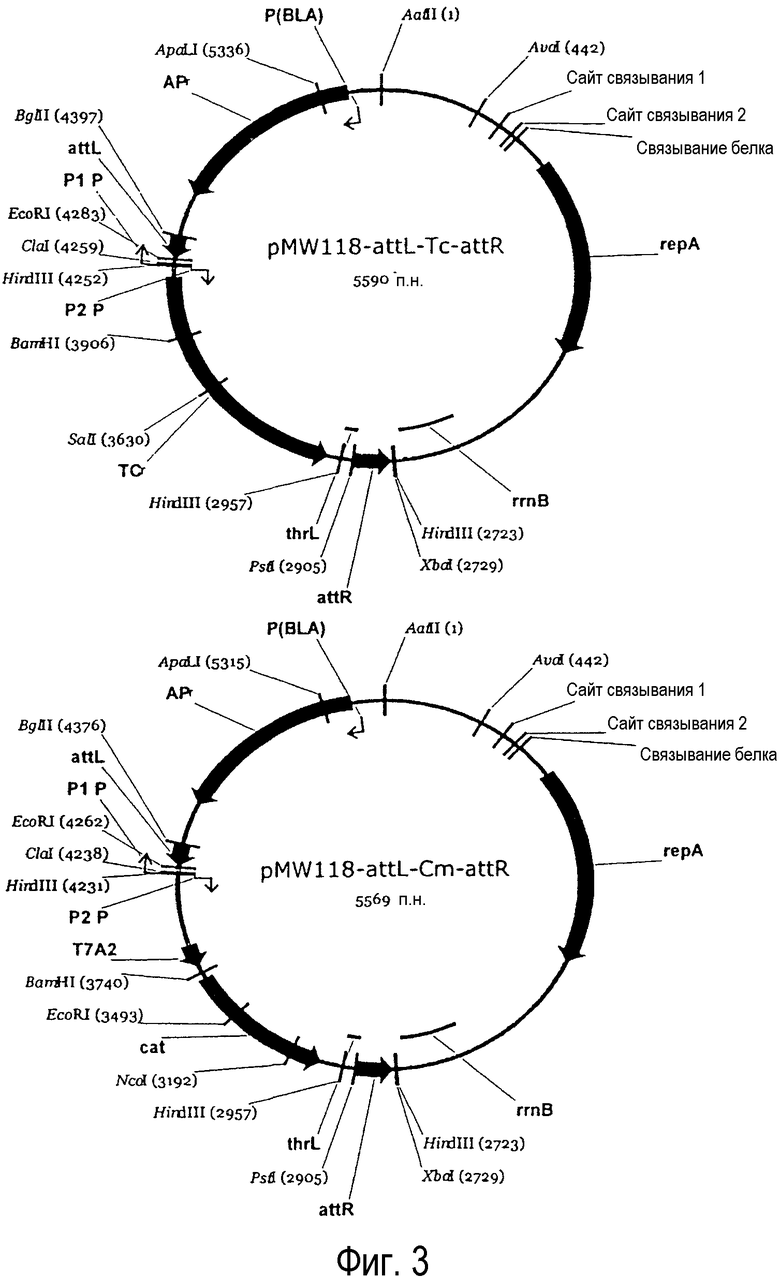
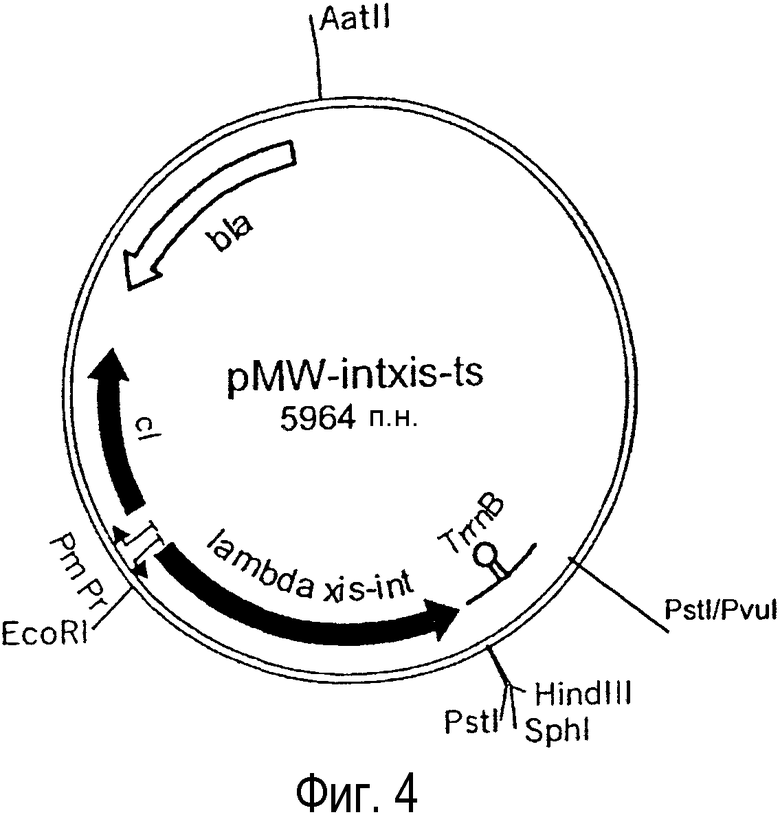






















Изобретение относится к биотехнологии и представляет собой способ продуцирования L-лизина или L-треонина, включающий культивирование бактерии в среде для продуцирования и секреции L-лизина или L-треонина, сбор и выделение L-лизина или L-треонина из среды. При этом указанная бактерия представляет собой бактерию Escherichia, которая обладает способностью продуцировать L-лизин или L-треонин, при этом указанная бактерия модифицирована так, что нарушена нормальная функция малеинового фермента в клетке, при этом малеиновый фермент кодируется геном sfcA и/или геном b2463. Изобретение позволяет получать L-лизин или L-треонин с высокой степенью эффективности. 11 з.п. ф-лы, 4 ил., 11 табл.
(A) белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:6; и
(B) белка, имеющего аминокислотную последовательность, включающего замещение, делецию, инсерцию или добавление одного или нескольких аминокислотных остатков в аминокислотной последовательности, представленной в SEQ ID NO:6, и имеющего активность малеинового фермента.
(C) белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:8;
(D) белка, имеющего аминокислотную последовательность, включающего замещение, делецию, инсерцию или добавление одного или нескольких аминокислотных остатков в аминокислотной последовательности, представленной в SEQ ID NO:8, и имеющего активность малеинового фермента.
(a) ДНК, имеющей нуклеотидную последовательность, представленную в SEQ ID NO:5;
(b) ДНК, которая гибридизируется с нуклеотидной последовательностью, представленной в SEQ ID NO:5, или зонда, который может быть получен из нуклеотидной последовательности, где указанная гибридизация происходит в жестких условиях и где указанная ДНК кодирует белок, имеющий активность малеинового фермента.
(c) ДНК, имеющей нуклеотидную последовательность, представленную в SEQ ID NO:7;
(d) ДНК, которая гибридизируется с нуклеотидной последовательностью, представленной в SEQ ID NO:7, или зонда, который может быть получен из нуклеотидной последовательности, где указанная гибридизация происходит в жестких условиях и указанная ДНК кодирует белок, имеющий активность малеинового фермента.
| VAN DER REST et.al | |||
| Functions of the membrane-associated and cytoplasmic malate dehydrogenases in the citric acid cycle of Escherichia coli | |||
| J Bacteriol | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| Затвор для дверей холодильных камер | 1920 |
|
SU182A1 |
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| RU 2002104983 A, 27.08.2003. | |||

