Изобретение относится к биотехнологии и касается получения нового штамма, используемого для выделения новой эндонуклеазы рестрикции, узнающей последовательность нуклеотидов на двуцепочечной ДНК и расщепляющей ее с образованием тупых концов:
5'-AG↓CT-3'
3'-TC↑GA-5'.
Стрелками указаны позиции расщепления ДНК.
Эндонуклеаза, обладающая данной специфичностью, может быть использована для сайт-специфического гидролиза ДНК, содержащей С5-метилцитозиновые основания, а также при совместном использовании с рестриктазой AluI для работ по выявлению и расщеплению метилированных участков ДНК, как прокариотических, так и эукариотических организмов, а также фагов и вирусов.
Эндонуклеазы рестрикции (рестриктазы, ЭР) являются сайт-специфическими ДНК-эндонуклеазами бактерий и, как правило, входят в состав так называемых систем рестрикции-модификации (РМ систем). В настоящее время описано более 250 различных сайтов узнавания ЭР второго типа и более 2000 различных изотиизомеров рестриктаз, т.е. аналогов ранее обнаруженных ферментов. Хотя изошизомеры имеют одинаковый сайт узнавания с исходным прототипом, они могут отличаться от последнего местом расщепления ДНК или по способности гидролизовать узнаваемую последовательность при наличии в ней метилированных оснований. Использование изошизомеров, отличающихся по способности гидролизовать метилированную ДНК, нашло широкое применение при изучении статуса метилирования природных ДНК [1-4]. Однако известно всего несколько примеров таких пар изошизомеров, например EcoRII - MvaI [5, 6], или HpaII-MspI [7], используемых в практике исследований.
Наиболее близким к заявляемому штамму - прототипом, является штамм Arthrobacter luteus, продуцирующий рестриктазу AluI, которая узнает последовательность нуклеотидов 5'-AGCT-3' и расщепляет ее перед цитозином с образованием тупых концов [8].
Недостатком известного штамма является то, что продуцируемая им сайт-специфическая эндонуклеаза не расщепляет узнаваемую последовательность, если она содержит хотя бы один N4-метилцитозин или 5-метилцитозин [9].
Технической задачей изобретения является получение бактериального штамма, продуцирующего сайт-специфическую эндонуклеазу, которая узнает и расщепляет обе цепи нуклеотидной последовательности ДНК 5'-AGCT-3', как без метилированных оснований, так и в случае, если в сайте узнавания метилирован один или два цитозина в положении С5 или одно основание в положении N4.
Поставленная техническая задача достигается получением штамма Arthrobacter luteus В-продуцента эндонуклеазы рестрикции Alu BI.
Предлагаемый штамм выделен из природного материала (почвы) в результате целенаправленного систематического поиска.
Полученный штамм Arthrobacter luteus В депонирован в Коллекции культур микроорганизмов НПО "СибЭнзим" под регистрационным номером 7М23, а продуцируемая им сайт-специфическая эндонуклеаза названа Alu BI.
Штамм Arthrobacter luteus В характеризуется следующими признаками:
Культурально-морфологические признаки. На среде Лурия-Бертрани (LB) образует белые, гладкие, блестящие, непрозрачные, выпуклые, круглые колонии 4 мм в диаметре. Клетки палочковидные, размером 1х(3-5) мкм, одиночные, в парах или в коротких цепочках. Образуют эллиптические центрально-расположенные споры, не раздувающие спорангий.
Физиолого-биохимические признаки. Грамположительные. Не способны к анаэробному росту, каталазоположительные, оксидазоотрицательные. Растут при температуре 10-40°С.Таксономическая принадлежность данного штамма определена на основе анализа морфологических и биохимических свойств [10].
На основании анализа морфологических и биохимических свойств штамм идентифицирован как вид бактерии Arthrobacter luteus В, а продуцируемая им эндонуклеазу рестрикции названа Alu BI согласно общепринятой номенклатуре [11].
Рестриктаза Alu BI является изошизомером рестриктазы AluI, то есть имеет ту же позицию гидролиза в сайте узнавания, что и AluI. Оба этих фермента не расщепляют узнаваемую последовательность, если она содержит N6-метиладенин [12].
Основным отличием эндонуклеазы, продуцируемой заявляемым штаммом, от известной рестриктазы является то, что Alu BI в отличие от AluI способна эффективно расщеплять двуцепочечную ДНК в случае, когда в сайте узнавания метилирован один или два цитозина в положении С5 или один цитозин в положении N4.
Хранение штамма осуществляют в лиофильно высушенном состоянии или в растворе 30% глицерина при температуре -20°С.
Полученная сайт-специфическая эндонуклеаза Alu BI характеризуется следующими свойствами:
1. Узнает и расщепляет последовательность нуклеотидов 5'-AGCT-3'.
2. Расщепляет связи между гуаниновым (G) и следующим за ним цитозиновым основанием в обеих цепях узнаваемой последовательности.
3. Не расщепляет узнаваемую последовательность, содержащую одно или два N6-метилцитозиновых основания, независимо от того, присутствуют в этой последовательности другие метилированные основания или нет.
4. Не расщепляет узнаваемую последовательность, содержащую два N4-метилцитозиновые основания.
5. Расщепляет вышеприведенную последовательность, содержащую одно N4-метилированное основание и (или) одно или два С5-метилированное основание.
6. Проявляет активность в широком температурном диапазоне 20-50°С.
7. Оптимальное значение рН для действия фермента 7,5-8,5.
8. Для проявления активности Alu BI требуются ионы Mg2+оптимальная концентрация - 10 мМ.
Определяющим отличием предлагаемого штамма от штамма Arthrobacter luteus - продуцента ЭР AluI является то, что предлагаемый штамм продуцирует сайт-специфическую эндонуклеазу Alu BI, которая узнает и расщепляет обе цепи нуклеотидной последовательности ДНК 5'-AGCT-3', содержащей одно или два С3-метилцитозиновых оснований, тогда как AluI не способен гидролизовать метилированные таким образом сайты. Alu BI способен в отличие от AluI, гидролизовать последовательность ДНК 5'-AGCT-3' в случае, если в данной двуцепочечной последовательности метилирован один цитозин в положении N4.
Поскольку предлагаемый штамм получен впервые и для выделения сайт-специфической эндонуклеазы, узнающей и расщепляющей вышеназванную последовательность нуклеотидов в указанной позиции никогда не использовался, можно сделать вывод о соответствии предлагаемого штамма критериям изобретения «новизна» и «изобретательский уровень».
Изобретение иллюстрируется примерами конкретного выполнения.
Пример 1.
Выращивание штамма и выделение фермента.
Для получения биомассы клетки штамма-продуцента переносят на агаризованную среду ЛБ в чашку Петри и инкубируют 3 дня при 22°С. Свежевыращенные колонии переносят стерильной бактериологической петлей в колбы, содержащие 300 мл жидкой питательной среды ЛБ и культивируют на качалках при 30°С при перемешивании - 150 об/мин в течение двух суток до достижения стационарной фазы роста. Клетки осаждают центрифугированием при 5000 об/мин при 4°С. Выход биомассы составляет 5 г/л среды.
Выделение фермента проводили при 4°С путем колоночной хроматографии.
37 г замороженной биомассы суспендировали в 100 мл буфера А (10 мМ Трис-HCl, рН 7,5, 0,1 мМ ЭДТА, 7 мМ β-меркаптоэтанол), содержащего 0,05 М NaCl, 0,3 мг/мл лизоцим, 0,1 мМ фенилметилсульфонилфторид (PSMF), и инкубировали в течение 1 ч при постоянном перемешивании. Далее клетки разрушали ультразвуком и центрифугировали в течение 30 мин при 15000 об/мин.
Супернатант пропускали через колонку с фосфоцеллюлозой Р-11 ("Whatman", Англия) объемом 45 мл, предварительно уравновешенную буфером А, содержащим 0,05 М NaCl, затем линейным градиентом NaCl (0,05 М - 0,6 М) в буфере А объемом 500 мл, собирая фракции по 10 мл. Фракции, содержащие эндонуклеазу, объединяли, диализовали против 20 объемов буфера А и наносили на колонку с 7 мл гепарин-сефарозы ("Bio-Rad", США). Элюцию проводили линейным градиентом NaCl (0,05 М - 0,5 М) в буфере А объемом 120 мл, собирая фракции по 3 мл. Активные фракции объединяли и наносили на колонку с гидроксилапатитом объемом 4 мл, предварительно промытую уравновешивающим буфером (0,01 М K2HPO4, рН 7,2, 7 мМ β-меркаптоэтанол). Колонку промывали 10 мл уравновешивающего буфера и проводили элюцию линейным градиентом, собирая фракции объемом по 2 мл. Активные фракции объединяли и диализовали против 20 объемов концентрирующего буфера (50% глицерин, 10 мМ Трис-HCl, рН 7,55, 0,1 мМ ЭДТА, 7 мМ β-меркаптоэтанол, 0,05 М NaCl). Препарат хранили при -20°С.
Выход фермента составляет 800 ед.акт.фермента с 1 г биомассы с удерльной активностью 5000 ед/мл.
Пример 2.
Определение специфичности эндонуклеазы рестрикции AluI.
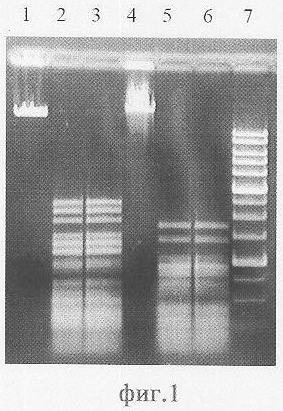
Специфичность фермента определяют по картинам специфического расщепления различных ДНК (фиг.1).
В качестве субстратов для выявления специфичности расщепления используют ДНК фагов λ и Т7. После инкубации в оптимальных условиях (37°С, SE-буфер «Y» - 33 мМ Трис-ацетат, рН 7,9, 10 мМ MgCl2, 66 мМ калия ацетат, 1 мМ DTT) в течение 60 мин при концентрации плазмидной ДНК 0,02 мг/мл продукты реакции разделяют путем электрофореза в 1% агарозном геле.
На фиг.1 представлена электрофореграмма продуктов расщепления ДНК фагов λ и Т7эндонуклеазами рестрикции AluI и Alu BI.
Описание дорожек на электрофореграмме, изображенной на фиг.1:
1 - ДНК фага λ;
2 - ДНК фага λ, обработанная ЭР AluI;
3 - ДНК фага λ, обработанная ЭР Alu BI;
4 - ДНК фага Т7;
5 - ДНК фага Т7, обработанная ЭР AluI;
6 - ДНК фага Т7, обработанная ЭР Alu BI;
7 - маркер молекулярного веса ДНК 1 kb (производство НПО СибЭнзим).
Из фиг.1 видно, что при обработке ДНК фагов λ и Т7 эндонуклеазами рестрикции AluI и Alu BI в результате гидролиза ДНК образуются фрагменты одинаковой длины. Эти результаты говорят о том, что ЭР AluI и Alu BI узнают и расщепляют одни и те же последовательности на ДНК.
Пример 3.
Определение позиций расщепления ДНК сайт-специфической эндонуклеазой Alu BI.
Определение позиций расщепления ДНК сайт-специфической эндонуклеазой Alu BI осуществляли путем сравнения длин фрагментов, образуемых при расщеплении меченых γ[32P олигонуклеотидных дуплексов Alu1*/ Alu2 и Alu2*/ Alu1, имеющих последовательность узнавания AluI эндонуклеазами AluI и Alu BI (подробное описание структуры олигонуклеотидных дуплексов и процедуры внесения метки в олигонуклеотиды приведены в примере 4). В качестве маркера длин фрагментов использовали продукты частичного расщепления этих же дуплексов экзонуклеазой ExoIII. Результаты расщепления данных олигонуклеотидных дуплексов приведены на фиг.2.
Описание дорожек на электрофореграмме, изображенной на фиг.2:
1 - дуплекс Alu1*/Alu2;
2 - дуплекс Alu1*/Alu2, расщепленный экзонуклеазой ExoIII из E.coli;
3 - дуплекс Alu1*/Alu2, расщепленный AluI;
4 - дуплекс Alu1*/Alu2, расщепленный Alu BI;
5 - дуплекс Alu1*/Alu2, расщепленный экзонуклеазой III из E.coli;
6 - дуплекс Alu2*/Alu1;
7 - дуплекс Alu2*/Alu1, расщепленный экзонуклеазой III из E.coli;
8 - дуплекс Alu2*/Alu1, расщепленный AluI;
9 - дуплекс Alu2*/Alu1, расщепленный Alu BI;
10 - дуплекс Alu2*/Alu1, расщепленный экзонуклеазой ExoIII из E.coli.
Как видно из фиг.2 длины фрагментов, образованных при гидролизе олигонуклеотидных дуплексов Alu1*/Alu2 и Alu2*/Alu1 одинаковы. Таким образом, эндонуклеаза рестрикции Alu BI расщепляет сайт узнавания в той же позиции, что и AluI, то есть после гуанина.
Пример 4.
Сравнение чувствительности к метилированию ЭР AluI и Alu BI.
Способность расщеплять метилированные сайты узнавания ЭР AluI и Alu BI изучали на синтетических олигонулеотидных дуплексах, содержащих метилированные основания в сайте узнавания 5'-AGCT-3'.
Для экспериментов брали эндонуклеазы рестрикции AluI (концентация 3000 ед./мкл), AluB I (концентация 3000 ед./мкл), Т4 полинуклеотидкиназу и буферные растворы производства НПО "Сибэнзим" (Россия). Олигодезоксирибонуклеотиды следующего состава, служившие субстратом для эндонуклеаз Alu I и AluB I, были синтезорованы в НПО "Сибэнзим" (Россия):
Alu1: 5'-GGT ATA GGA TGA AGCT TTC GCG GGT ТАА GG-3'
Alu2: 5'-СС ТТА АСС CGC GAA AGCT ТСА ТСС ТАТ ТСС-3'
Alu3: 5'-GGT ATA GGA TGA (M6A)GCT TTC GCG GGT ТАА GG-3'
Alu4: 5'-СС ТТА АСС CGC GAA (M6A)GCT ТСА ТСС TAT ТСС-3'
Alu5: 5'-GGT ATA GGA TGA AG(M5C)T TTC GCG GGT ТАА GG-3'
Alu6: 5'-СС ТТА АСС CGC GAA AG(M5C)T ТСА ТСС TAT ТСС-3'
Alu7: 5'-GGT ATA GGA TGA AG(M4C)T TTC GCG GGT ТАА GG-3'
Alu8: 5'-СС ТТА АСС CGC GAA AG(M4C)T ТСА ТСС ТАТ ТСС-3'
Олигонуклеотиды с четным номером комплементарны олигонуклеотидам с нечетным номером. Все олигонуклеотидные дуплексы имеют одинаковую первичную структуру и отличаются друг от друга наличием или отсутствием метилированного основания в последовательности AGCT (сайт AGCT, узнаваемый AluI и Alu BI, подчеркнут).
Приготовление субстрата для эндонуклеаз AluI и Alu BI.
Одну из цепей олигонуклеотидного дуплекса модифицировали по 5'-концу с помощью Т4-полинуклеотидкиназы и γ-[32Р]АТР. После очистки олигонуклеотида от побочных продуктов реакции к нему добавляли комплементарный немеченый олигонуклеотид и пробирку прогревали 2 минуты при 65°C с последующим охлаждением до комнатной температуры на рабочем столе.
Гидролиз олигонуклеотидных дуплексов эндонуклеазами Alu I и AluB I. Реакцию гидролиза проводили добавлением 1 мкл препарата фермента AluI ил Alu BI, содержащего 3 единицы активности, в 20 мкл реакционной смеси, содержащей SE-буфер «Y» (33 mM Трис ацетат рН 7.9 (при температуре 25°С), 10 mM Mg(СН3СОО)2, 66 mM KCH3СОО, 1 mM DTT) и олигонуклеотидный дуплекс в концентрации 66,7 нМ при температура 37°С в течение 50 мин.
Результаты расщепления метилированных и неметилированных синтетических олигонуклеотидных дуплексов приведены на фиг.3-6.
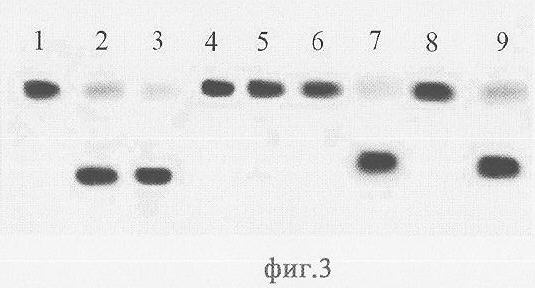
На фиг.3 приведена электрофореграмма продуктов расщепления рестриктазами AluI и Alu BI олигонуклеотидных дуплексов, содержащих сайт узнавания этих ферментов без метилированных оснований или одно метилированное основание (полуметилированный сайт). Как видно из фиг.3, AluI и Alu BI эффективно расщепляют олигонуклеотидный дуплекс, содержащий неметилированный сайт узнавания (дорожки 2, 3). Однако оба фермента не эффективно способны расщеплять ДНК, если в сайте узнавания хотя бы на одной цепи метилирован аденозин (дорожки 4, 5). Если в сайте узнавания, метилирована только одна цепь ДНК в положении С5 или N4, то Alu BI расщепляет неметилированную цепь (дорожки 7, 9), тогда как AluI не расщепляет (дорожки 6, 8).
Описание дорожек на электрофореграмме, изображенной на фиг.3:
1 - дуплекс Alu1*/Alu2;
2 - дуплекс Alu1*/Alu2, расщепленный AluI;
3 - дуплекс Alu1*/Alu2, расщепленный Alu BI;
4 - дуплекс Alu1*/Alu4, обработанный AluI;
5 - дуплекс Alu1*/Alu4, обработанный Alu BI;
6 - дуплекс Alu1*/Alu6, обработанный AluI;.
7 - дуплекс Alu1*/Alu6, расщепленный Alu BI;
8 - дуплекс Alu1*/Alu8, обработанный AluI;
9 - дуплекс Alu1*/Alu8, расщепленный Alu BI.
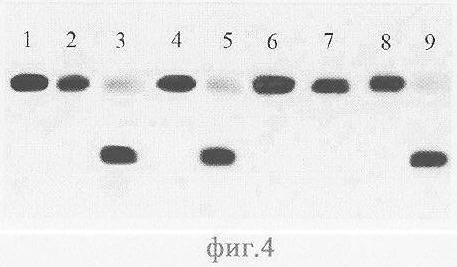
На фиг.4 и 5 приведены электрофореграммы продуктов расщепления ферментами AluI и Alu BI олигонуклеотидных дуплексов, содержащих сайт узнавания этих ферментов с одним или двумя метиилированными основаниями. Как видно из фиг.4, Alu BI расщепляет метилированную цепь в сайте узнавания, если в узнаваемую последовательность входят одно (дорожка 3) или два (дорожка 5) С5-метилцитозиновых оснований; при этом AluI данные метилированные цепи ДНК гидролизовать не может (дорожки 2, 4). Таким образом, Alu BI в отличие от AluI гидролизует обе цепи ДНК в сайте узнавания при наличии в нем одного или двух С5-метилцитозиновых оснований. Однако С5-метилированная цепь не гидролизуется AluI и Alu BI, если вторая цепь ДНК содержит N6-метилированное основание (дорожки 6,7). Alu BI в отличие от AluI гидролизует С5-метилированную цепь при наличии N4-метилированного основания на противоположной цепи (дорожки 8, 9).
Описание дорожек на электрофореграмме, изображенной на фиг.4:
1 - дуплекс Alu5*/Alu2;
2 - дуплекс Alu5*/Alu2, обработанный AluI;
3 - дуплекс Alu5*/Alu2, расщепленный Alu BI;
4 - дуплекс Alu5*/Аlu6, обработанный AluI;
5 - дуплекс Alu5*/Аlu6, расщепленный AluI;
6 - дуплекс Alu5*/Alu4 обработанный Alu BI;
7 - дуплекс Alu5*/Alu4, обработанный AluI;
8 - дуплекс Alu5*/Alu8, обработанный AluI;
9 - дуплекс Alu5*/Alu8, расщепленный Alu BI.
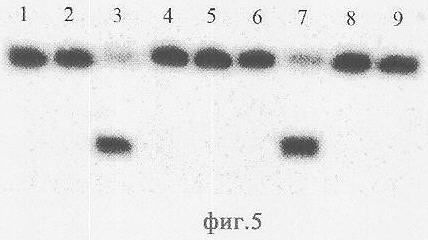
Как видно из фиг.5, Alu BI гидролизует N4-метилированную цепь, если комплементарная цепь не содержит метилированных оснований в узнаваемой последовательности (дорожка 3), тогда как AluI не способен гидролизовать данную цепь (дорожка 2). Однако как AluI, так и Alu BI не гидролизуют сайт узнавания, если он содержит два N4-метилированных основания (дорожки 4, 5). Таким образом, Alu BI в отличие от AluI способен гидролизовать обе цепи ДНК, если в узнаваемую последовательность входит одно N4-метилированное основание. N4-метилированная цепь может гидролизоваться также, если комплементарная цепь узнаваемой последовательности также содержит одно метилированное основание -5m-цитозин (дорожка 7), тогда как AluI не способен гидролизовать данную цепь (дорожка 6). Таким образом, при наличии в сайте узнавания двух метилированных оснований - на одной цепи С5-метилированного основания, а на другой - N4-метилированного основания Alu BI гидролизует обе цепи ДНК, тогда как AluI не гидролизует ни одну из цепей. Цепь ДНК, содержащая в узнаваемой последовательности одно метилированное основание - N4-метилцитозин, не гидролизуется как Alu BI, так и AluI, если в комплементарной цепи в узнаваемой последовательности содержится 6m-аденин (дорожки 8, 9).
Описание дорожек на электрофореграмме, изображенной на фиг.5:
1 - дуплекс Alu7*/Alu2;
2 - дуплекс Alu7*/Alu2, обработанный AluI;
3 - дуплекс Alu7*/Alu2, расщепленный Alu BI;
4 - дуплекс Alu7*/Alu8, обработанный AluI;
5 - дуплекс Alu7*/Alu8, обработанный Alu BI;
6 - дуплекс Alu7*/Alu6, обработанный AluI;
7 - дуплекс Alu7*/Alu6, расщепленный Alu BI;
8 - дуплекс Alu7*/Alu4, обработанный AluI;
9 - дуплекс Alu7*/Alu4, обработанный Alu BI.
Пример 5.
Использование ЭР Alu BI для определения статуса метилирования бактериальных хромосомных ДНК и для избирательного гидролиза С5-метилированных ДНК.
Благодаря различной чувствительности к метилированию цитозина как в положении С5, так и в положении N4, Alu BI вместе с AluI может быть использован для определения наличия или отсутствия С5 и N4-метилированных основания в последовательности 5'-AGCT-3' как в бактериальных, так и эукариотических вирусных и геномных ДНК.
На фиг.6 изображена электрофореграмма хромосомной ДНК Arthrobacter luteus - продуцента ЭР AluI, обработанной рестриктазами AluI и Alu BI.
Известно, что хромосомная ДНК штамма Arthrobacter luteus - продуцента AluI содержит 5-метилцитозин в сайте 5'-AGCT-3' [13]. В соответствии с описанными выше свойствами ферментов Alul и Alu BI, хромосомная ДНК Arthrobacter luteus - продуцента AluI - не гидролизуется рестрктазой AluI, но эффективно расщепляется рестриктазой Alu BI.
Описание дорожек на электрофореграмме, изображенной на фиг.6:
1 - хромосомная ДНК Arthrobacter luteus - продуцента AluI;
2 - хромосомная ДНК Arthrobacter luteus - продуцента AluI, обработанная рестриктазой AluI;
3 - хромосомная ДНК Arthrobacter luteus - продуцента AluI, обработанная рестриктазой Alu BI;
4 - маркер молекулярного веса ДНК 1 kb (производство НПО СибЭнзим).
Таким образом, получен новый штамм, продуцирующий эндонуклеазу рестрикции Alu BI, способную эффективно расщеплять двуцепочечную ДНК в случае, когда в сайте узнавания метилирован один или два цитозина в положении С5 или один цитозин в положении N4.


| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ ESCHERICHIA COLI N16 (PM.ALUBI) - ПРОДУЦЕНТ ДНК-МЕТИЛТРАНСФЕРАЗЫ M.ALUBI | 2015 |
|
RU2603086C1 |
| ШТАММ БАКТЕРИЙ GLACIAL ICE BACTERIUM - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ GLU I | 2006 |
|
RU2322492C1 |
| ШТАММ БАКТЕРИИ Arthrobacter oxydans - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ AoxI | 2009 |
|
RU2399663C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Bisi | 2004 |
|
RU2270859C1 |
| ШТАММ БАКТЕРИЙ BACILLUS SIMPLEX - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ BlsI | 2006 |
|
RU2322494C1 |
| Штамм бактерий Micrococcus luteus 805 - продуцент сайт-специфической метилзависимой эндонуклеазы MluVI | 2016 |
|
RU2614262C1 |
| ШТАММ БАКТЕРИИ Kocuria rosea - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ KroI | 2009 |
|
RU2394099C1 |
| ШТАММ БАКТЕРИЙ Glacial ice bacterium I - ПРОДУЦЕНТ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ Gla I | 2005 |
|
RU2287012C1 |
| ШТАММ БАКТЕРИЙ Microbacterium testaceum 17B - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ MteI | 2011 |
|
RU2475533C1 |
| ШТАММ БАКТЕРИЙ Planomicrobium koreense 78K - ПРОДУЦЕНТ САЙТ-СПЕЦИФИЧЕСКОЙ ЭНДОНУКЛЕАЗЫ PkrI | 2011 |
|
RU2475534C1 |


Изобретение относится к области биотехнологии. Получен штамм бактерий Arthrobacter luteus В, продуцент сайт-специфической эндонуклеазы Alu BI. Изобретение может быть использовано для получения эндонуклеазы рестрикции, которая узнает и расщепляет обе цепи нуклеотидной последовательности ДНК 5'-AGCT-3' как без метилированных оснований, так и в случае, если в сайте узнавания метилирован один или два цитозина в положении С5 или одно основание в положении N4. 6 ил.
Штамм бактерий Arthrobacter luteus В, депонированный в ККМ НПО «СибЭнзим» под номером 7М23 - продуцент сайт-специфической эндонуклеазы Alu BI.
| ROBERTS R.J | |||
| et al | |||
| A specific endonuclease from Haemofilus haemolyticus | |||
| J Mol Biol | |||
| Планшайба для точной расточки лекал и выработок | 1922 |
|
SU1976A1 |
| BUTKUS V et al | |||
| Interaction of Alu I, Cfr6I and Pvu II restriction- modification enzymes with substrates containing either N4- methylcitosine or 5- methylcitosine | |||
| Biochim Biophys Acta | |||
| Кузнечная нефтяная печь с форсункой | 1917 |
|
SU1987A1 |

