Предлагаемая группа изобретений относится к экспериментальной биологии и медицине, а именно - к экспериментальной неврологии, и может использоваться для стимуляции роста мотонейронов коры головного мозга растущего организма млекопитающих.
В перспективе (после проведения соответствующих клинических исследований) возможной областью использования предлагаемой группы изобретений может стать профилактика и лечение детских церебральных параличей, так как (как будет описано ниже) важным патогенетическим фактором этого заболевания является поражение мотонейронов коры головного мозга.
Детский церебральный паралич занимает особое место в патологии детского возраста, является наиболее распространенными заболеванием нервной системы, приводящим к инвалидизации детей, поражая не только двигательную систему, но и вызывая нарушения речи и интеллекта (Семенова К.А.).
Детский церебральный паралич возникает в результате поражения мозга, перенесенного в перинатальный период развития или в период новорожденности. К настоящему времени стало понятно, что родовая травма, кровоизлияние в мозг (которые раньше рассматривались как основная причина заболевания)- явления большей частью вторичные и лишь в относительно в небольшом числе случаев имеют самостоятельное значение в этиологии детского церебрального паралича, возникая в результате различных форм акушерской патологии. К последним относят не только узкий таз матери или неправильное его строение, но и неправильное положение плода. Под действием родовых сил может иметь место деформация головки плода при том или ином предлежании со смещением различных отделов головного мозга и возможность последствий механической травмы в родах. Механическая травма головки плода как правило сопровождается нарушением мозгового кровообращения, кровоизлиянием в мозг. Чаще всего кровоизлияние возникает вследствие разрыва саггитального или поперечного синусов, что в большинстве случаев приводит к смерти плода. При субарахноидальном кровоизлиянии, в результате разрыва мелких сосудов мозга и per diapedesin смертельные исходы наступают реже, чаще развиваются различные формы детских церебральных параличей. Кровоизлияние в мозг может наступить не только в результате механического разрыва сосудов, обусловленного механической травмой в родах, но и асфиксии, начавшейся в родах, хронической гипоксии, связанной с поражением плода до начала акта родов. Оно может быть следствием многих причин, поражающих мозг на разных этапах развития (эмбриогенеза и фетогенеза). Асфиксия в родах может быть лишь конечным результатом поражения и нарушения развития. Однако остается не ясным, почему в одних случаях значительное смещение мозга (и следовательно, достаточная тяжелая травма его) не дают каких-либо нарушений деятельности головного мозга, а в других легкая деформация, казалось бы, не влекущая за собой его повреждения, может рассматриваться как причина церебральной патологии, развивающейся в дальнейшем. В настоящее время не подлежит сомнению, что основной причиной детского церебрального паралича и вторичной микроцефалии в большинстве случаев является тяжелое состояние плода, возникающее под влиянием какого-либо одного или комплекса вредных факторов во время внутриутробного развития. Такие вредные факторы в периоде беременности, как инфекционные агенты, токсины и др., могут проникать через плацентарный барьер, поражать мозговую ткань плода и приводить к ее деструкции, задержке развития мозга.
Ряд авторов придают большое значение в патогенезе детского церебрального паралича аутоиммунному конфликту между матерью и плодом, особенно при тяжелых формах. Начало этого конфликта приходится на разные сроки эмбрио- и фетогенеза. Последствия аутоиммунного конфликта могут сказываться и в постнатальном онтогенезе, проявляясь наличием специфических противомозговых антител (Семенова К.А. и др). Выделяют: 1) раннюю стадию детского церебрального паралича и 2) начальную и позднюю резидуальные стадии детского церебрального паралича. Соответственно, допускаются различные лежащие в их основе процессы, клинические проявления болезни и лечебные подходы.
В первые сутки после рождения в палате соблюдается тишина, нет яркого освещения, придается возвышенное положение в подогреваемой постели, рекомендуется держать холод на расстоянии 2 см от головы ребенка, периодически или постоянно дается увлажненный кислород.
Медикаментозная терапия направлена на дегидратацию мозга (40% глюкоза внутривенно 5-10 мл, сернокислая магнезия 25% по 0,2 мл /кг массы, нативная или сухая плазма 10-15,0 мл внутривенно, викасол 0,002-0,003 г/ сутки, противосудорожные средства, при нарушении глотания кормление через зонд).
В дальнейшем для стимуляции развития мозга назначаются витамин В12, глютаминовая кислота, церебролизин, дибазол, а в начальную и позднюю резидуальную стадию включают в лечение пирогенал, мидокалм, гаммалон, галантамин (или оксазил), АТФ и широко используются физиотерапевтические процедуры, массаж и гимнастика, укладки для конечностей, упражнения для развития речи.
При всяком повреждении (травма, кровоизлияние, инфекция) развивается воспалительный процесс, в том числе и в тканях мозга. Наиболее мощным противовоспалительным действием обладают кортикостероидные гормоны и синтетические препараты, построенные на их основе.
Семенова К.А, Мастюкова Е.М., Смуглин М.Я(1972), Семенова К.А, Махмудова Н.М.(1979. стр 118) применили кортикостероидные препараты для лечения детского церебрального паралича. В комплексе лечебных мероприятий с этой целью использовались в позднем периоде новорожденности (после 10 дней) ингибиторы иммуногенеза - вначале АКТГ от 5 до 10 ед в течение 1-2 месяцев (Семенова К.А. 1972, стр 91), а затем другой группе детей - кортин (представляющий экстракт коры надпочечников убойного скота) по 0,3-0,5 мл внутримышечно через день в сочетании с дексаметазоном по 1/4 таб. (в таблетке 0,5 мг) ежедневно в течение месяца (Семенова К.А и др 1983). Было отмечено, что у детей, не получавших кортин и дексаметазон (1 группа - 10 человек) положительная динамика психомоторного развития наблюдалась в 45,0% случаев, в то время как у лечившихся с применением кортина и дексаметазона (2 группа - 20 человек) - в 54,0% случаев; наблюдались определенные положительные сдвиги в клинической картине, а именно: несколько снизились проявления гипертензионного синдрома, появились установочные рефлексы, снизился мышечный тонус, который до лечения был высоким, появились манипулятивные движения рук. Были отмечены также положительные сдвиги в иммунологическом статусе и улучшилось представительство выделяемых с мочой кортикостероидов.
Кортикостероидные препараты широко используются в преднатальном и постнатальном периоде с целью ослабления дыхательных расстройств при риске преждевременного рождения новорожденных, перенесших внутриутробно и в родах гипоксию, имевших внутричерепные кровоизлияния, при нарушениях гемодинамики (не контролируемой допамином) из-за врожденной слабости надпочечников у преждевременно родившихся детей. С этой целью большей частью использовались кортикостероиды однократно коротким циклом ( гидрокортизон). Повторные курсы рекомендовались только при продленных интубациях и стойкой гипотонии. В качестве альтернативной стратегии рекомендуются препараты стероидов для ингаляций, метилпреднизолон и гидрокортизон (а не дексаметазон).
Вместе с тем, наряду с положительными результатами (снижение частоты преждевременного рождения, повышение выживаемости маловесных детей) обращается внимание на возрастание в таких случаях частоты язвенных энтероколитов, сепсиса у детей, высказываются опасения за последствия, в том числе возможность детского церебрального паралича и задержки нервно-психического развития. Следует заметить, что кортикостероидные препараты, обладая супрессивным действием на иммунокомпетентные клетки, могут существенно повлиять на нейрогенез растущего организма, особенно внутриутробно и в период новорожденности (Rajadurai V.S., Tan К.Н., 2003).
В течение последних десятилетий стало общепринятым, что структурная нейропластичность вовлечена в функциональную адаптацию ЦНС. Клеточная морфология мозга находится в непрерывном переходе всюду по продолжительности жизни как ответ на экологические стимулы. Эффекты до некоторой степени сопряжены с изменяющимся уровнем гонодальных стероидных гормонов. Сегодня ясно, что функции гонадальных стероидов в мозгу простираются вне простого регулирования репродуктивных или нейроэндокринных событий. Важно, что гонадальные стероиды участвуют в формировании развивающегося мозга и в течение всей жизни вовлечены в более высокие мозговые функции типа настроения, познания и памяти (Bovenberg S.A. et al, 2005). Установлены гонадально вынужденные изменения в нейроне: величине, численности синапсов, а также древовидной и синаптической морфологии (Parducz, 2006).
В последние годы возрос интерес к пролиферации новых нейронов из предшественников в поджелудолчковых структурах мозга (зубчатой извилине и гиппокампе), миграции их в другие регионы, которые больше выражены в раннем возрасте, но, уменьшаясь, сохраняются в течение всей жизни.
Dentate gyros взрослого млекопитающего содержат предшественники нервных клеток с мультипотенциалом развития и самовозобновлением. Происхождение и созревание нервных предшественников определены в соответствии с составом и уровнем трофических факторов в их микроокружающей среде (Kosaka N. et al, 2006). Трофические факторы: ресничный нейротрофический фактор (CNTF), глиально-полученный нейротрофический фактор (GDNF), подобные инсулину факторы роста 1 и 2 (IGF-1, IGF-2): наиболее активный из них в продвижении нейрогенеза ресничный фактор (СМТТ) вызывает быстрое увеличение нейрона (Chen H b, 2006).
Экспериментально на культурах тканей мозга была показана роль половых гормонов преимущественно надпочечникового происхождения в нейрогенезе. Обнаружены ферментные системы, обеспечивающие синтез из предшественников (прегненолона, прогестерона, дегигидроэпианростерона) нейрогормонов тестостерона и эстрогенов в тканях мозга. Альфа- и бета-рецепторы эстрогена существенно повышают гиппокампальный нейрогенез; стимуляция сопровождается быстрым увеличением клеток (Massucco, 2006).
Как и в других тканях, нейрогенез модулируется ростовыми факторами, в которых важное место принадлежит трансформирующему фактору роста бета-1 (TGF-beta-1), который синтезируется астропитами, клетками глии. (Galbiati М. Et al, 2005, Chen H. Et al, 2006). Астроциты гиппокампа синтезируют дегидроэпиандростерон (в 3 раза больше, чем астроциты коры), а также тестостерон и эстрогены из прегненолона с участием цитохрома p450c17(Zwain, 1999).
TGF роста бета - запрещает гиппокампальный нейрогенез растущих мышей (Buckwalter, 2006), а дегидроэпиандростерон понижает активность трансформирующего фактора роста бета - 1, который в культурах тормозит образование нейронов из предшественников. Дегидроэпиандростерон стимулирует окислительный механизм энергообразования в мозговых митохондриях развивающихся крыс, значительно увеличивает АТФ-азную активность, уровни дегидрогеназ, ускоряет процесс созревания мозговых митохондрий и тем самым подчеркивается роль дегидроэпиандростерона в мозговом развитии в послеродовой жизни (Patel, 2006).
Отмечены антивоспалительные свойства дегидроэпиандростерона, который может понизить уровни провоспалительных цитокинов - интерлейкина 6 (IL-6) и фактора некроза опухоли -альфа (TNF-alpha) Leowattana W, 2001. Глюкокортикостероиды же запрещают выпуск гонадотропного гормона (GnRH), действуя не только на гипофиз, но непосредственно на гипоталамическом уровне (Calogero A.E. et al.1999).
Подавляющий эффект кортикостерона на выпуск GnRH не моделировался андрогенами (Т uDHT), но устранялся прогестероном, по крайней мере когда кортикостерон использовался в низких концентрациях (10 нитрометаяов), но не высоких (100 нитрометанов); у крыс эффекты кортикостерона соответственны эффектам гидрокортизона у человека.
Вместе с тем отмечены супрессивные эффекты высоких концентраций глюкокортикостероидов на деление предшественников нейронов, их дифференцировку и апаптоз. Изменяя метаболизм нейронов гиппокампа, глюкокортикоиды повышают уязвимость гиппокампа к накапливающимся токсическим продуктам (Robert M Sapolsky, 1985; 2003). Низкие и средние концентрации кортикостероидов в гиппокампальной культуре клеток обладают противовоспалительным действием (снижается выражение IL-1beta и TNF-alfa), в то время как при высоких концентрациях эффекты были противоположными, провоспалительными, повышались концентрации IL-1beta, TNF-alfa (Mac Pherson A et al., 2005).
Дегидроэпиандростерон (DHEA), напротив, защищает гиппокампальные нейроны; антиглюкокортикоидные действия DHEA осуществляются через уменьшение рецепторов к глюкокортикоидным гормонам (Кардоунел А. и др.)
Преднатальное напряжение (стресс) в зависимости от интенсивности увеличивает или подавляет развитие гиппокампальных нейронов: малая продолжительность, умеренное преднатальное напряжение увеличивали нейрогенез и дифференцирование процессов гиппокампальных нейронов, тогда как длительное, серьезное напряжение вредило их морфологии. Рецепторы минералокортикоидов и глюкокортикоидов вносят вклад в вынужденные напряжением морфологические изменения гиппокампальных элементов; минералокортикоидный рецептор повышал нейрогенез и дифференцирование в культуре гиппокампальных нейронов; напротив глюкокортикоидный рецептор был вовлечен в подавление их морфологии ( Fujioka A et al. 2006).
Судороги в первые недели жизни новорожденных приводят к уменьшению нейрогенеза и более поздней дифферинцировке нейронов (Shy X.Y et al., 2006).
В низких концентрациях гидрокортизон и прогестерон усиливают эффекты ростовых факторов (таких как инсулиноподобный фактор роста и др.). В связи с изложенным можно ожидать усиление нейрогенеза при использовании экстрактов коры надпочечников, содержащих комплекс гормоноактивных соединений, оказывающих гормональные и паракринные влияния на метаболические процессы в мозгу. Заслуживает особого рассмотрения возможность использования экстрактов коры свиных и фетальных надпочечников, так как в фетальном возрасте эта железа вместе с плацентой, составляя единый гормональный комплекс, обеспечивают быстрый рост и дифференцировку тканей плода.
Таким образом, использующиеся в настоящее время в клинике способы профилактики и лечения детских церебральных параличей не направлены непосредственно на стимуляцию мотонейронов коры головного мозга. Примененные способы не обеспечивают достаточного эффекта и обладают большим количеством нежелательных побочных проявлений.
В экспериментальной медицине также практически отсутствуют разработки, связанные со стимуляцией роста мотонейронов коры головного мозга. Это направление только начинает развиваться. В известной литературе нами найдена только одна работа, посвященная средству для стимуляции роста мотонейронов коры головного мозга и способу его применения. Эта работа выбрана нами в качестве прототипа - И.Г.Шиленок, И.В.Мухина, И.В.Садовникова, Н.А.Панина. Анаболические эффекты минорных концентраций кортикостероидлов, содержащихся в экстрактах коры надпочечников. // Нижегородский медицинский журнал. - 2004. - №1, с. 92-95. В прототипе для стимуляции роста мотонейронов коры головного мозга (в эксперименте на крысах) предложено использовать экстракт коры фетальных надпочечников в объеме 0,5 мл через день, в течение 5 недель. Несмотря на полученный эффект роста мотонейронов коры головного мозга прототип не лишен недостатков. Прежде всего следует отметить, что фетальные надпочечники брались при вскрытии умерших в родах детей. Очевидно, что использование прототипа в связи с этим очень ограничено, в первую очередь этическими проблемами.
В задачу предлагаемого изобретения положено расширение арсенала средств для стимуляции роста мотонейронов коры головного мозга, обеспечение возможности использования доступного средства.
Поставленная задача достигается применением экстракта коры свиных надпочечников в качестве средства для стимуляции роста мотонейронов коры головного мозга растущего организма млекопитающих.
Поставленная задача в способе стимуляции роста мотонейронов коры головного мозга растущего организма млекопитающих, включающем введение экстракта коры надпочечников, через день, общим курсом 5 недель, достигается тем, что вводят экстракт коры свиных надпочечников в разовой дозе 0,5 мл на 100 г массы тела организма.
Возможность использования экстрактов коры свиных надпочечников в качестве средства для стимуляции роста мотонейронов коры головного мозга растущего организма показана в предлагаемом изобретении впервые. Ранее экстракты коры свиных надпочечников использовались совершенно по другим назначениям, а именно: для стимуляции репаративных процессов в патологически измененной печени (Ru 2185835 С2, 27.07.2002); для коррекции острой почечной недостаточности при токсическом гепаторенальном синдроме (Ru 2290940 С2, 10.01.2007).
В предлагаемом изобретении стимуляция роста мотонейронов коры головного мозга проводится именно на растущем организме, так как предполагалось экстраполировать полученные результаты для использования при лечении пациентов детского возраста.
Существенным признаком являются количественные показатели, введенные в формулу изобретения, а именно - разовая доза и курс введения экстрактов свиных надпочечников. Разовая доза 0,5 мл на 100 г массы определялась целесообразностью испытания минимальных (минорных) концентраций гормоноактивных веществ, обычно имеющихся в крови вне напряжения организма и обладающих биологическим действием (не в миллиграммах и микрограммах, а в нанограммах). Необходимость использования 5-недельного курса объясняется постепенностью развивающегося эффекта и потому требуемого продолжительного времени стимуляции, что обычно применяется в клинических условиях при лечении больных, курсами в течение 4-6 недель.
Необходимость введения экстрактов через день объясняется тем, что при подкожном введении препарат медленно резорбируется и тем самым может достигаться большая равномерность концентраций в течение длительного срока.
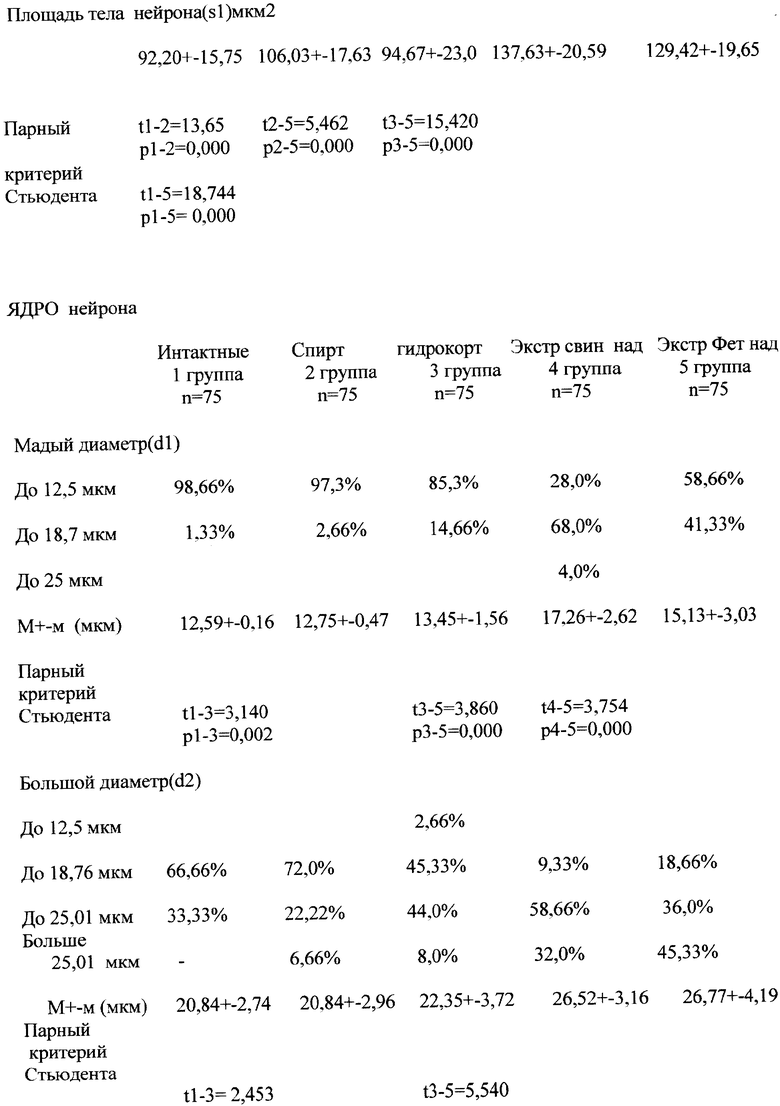
Предлагаемый способ разработан в эксперименте на 25 растущих беспородных крысах-самцах с массой тела в начале опыта 99,8±3,7 г и длиной тела 12,44±0,58 см. В конце опыта животные имели вес 181,7±12,7 г и длину тела 14,0±1,66 см. Животные были разделены на 5 групп, из которых 5 животных были интактными (1 группа контроля), 5 животных получали 10% раствор этилового спирта на физиологическом растворе хлорида натрия (2 группа контроля), 5 животных получали гадрокортизон (10 мг/кг) на основе 10% раствора этилового спирта (3 группа); 5 животных получали экстракт коры свиных надпочечников (4 группа) и 5 животных получали экстракт коры фетальных надпочечников, взятых при вскрытии погибших в родах плодов человека (5 группа).
Экстракты коры надпочечников готовились по принципу органных препаратов, из 3,0 г ткани получали 1 мл экстракта. Препараты в объеме 0,5 мл вводились через день в течение 5 недель. Методом радиоиммунологического анализа в экстрактах коры надпочечников были определены концентрации гидрокортизона, дегидроэпиандростерона и прогестерона. В 0,5 мл экстракта коры свиных надпочечников содержалось гидрокортизона - 65 нмоль, дегидроэпиандростерона - 0,83 нмоль и прогестерона - 1,1 нмоль. В 0,5 мл экстракта коры фетальных надпочечников содержалось гидрокортизона - 5,0 нмоль, дегидроэпиандростерона - 1,15 нмоль и прогестерона - 0,45 нмоль. В 0,5 мл вводимого животным раствора гидрокортизона содержалось 2,6 мкмоль препарата. После 16 инъекций препаратов животные забивались, головной мозг подвергался парафинизации и срезы окрашивались по методу Ниссле. Микроскопические измерения проводились в 4-5 слоях коры при увеличении 7×90 и с помощью шкалы определялись два диаметра (большой и малый) тела и ядра клеток мотонейронов. Ценность деления шкалы была определена с помощью микрометра, одно деление шкалы составляло 16,6 mkm. При расчете площади пользовались формулой
Smkm2=3,14*0,122*(a/2*16,6)*(b/2*16,6), где 0,122 - поправочный коэффициент на оптические среды микроскопа, а - большой диаметр, b - малый диаметр, 16,6 - ценность одного деления шкалы. Диаметры тела и ядра клеток определялись по формуле:
d1,d2mkm=3,14*0,12*a(или b)*16,6. Вычисленные таким образом размеры мотонейронов были сопоставимы с нормативными данными, приводимыми Ю.М.Жаботинским в монографии «Нормальная и патологическая морфология нейрона», 1965.
Исследования представлены в таблице.
Как показали исследования, в группе 2 контроля с введением 10% спирта на изотоническом растворе хлорида натрия полученные данные не расходились с соответствующими измерениями группы 1 контроля у интактных животных или были малозначимыми. Из особенностей воздействия спирта на нейроны головного мозга обращает внимание небольшое увеличение малого диаметра тела нейрона (28,77±4,63 мкм против 24,41±3,08 мкм), что определяло округление клеток и некоторое увеличение площади тела нейрона. Однако при одинаковых параметрах ядра в обеих группах контроля (у интактных и с введением раствора этилового спирта) отношение s2/s1 оказалось сближенным (0,2 против 0,23), что не позволяет думать о существенном возрастании метаболической активности нейрона.
При введении гидрокортизона параметры тела нейрона и его ядра (малый и большой диаметры, площади тела и ядра) были близкими к таковым у интактных животных, хотя статистически можно было подтвердить их различие в размерах ядер (р=0,002; 0,017;0.000). Соотношение площадей ядра и тела соответствовало данным у интактных (1,92).
Значимыми и одной направленности были изменения нейронов головного мозга после введения экстрактов коры свиных и фетальных надпочечников. Все параметры тела и ядра нейронов были достоверно больше, чем у интактных животных и у получивших инъекции гидрокортизона. Из особенностей обращало внимание большее округление тела и ядра клетки после введения экстракта свиных надпочечников (d2/d1 тела =1,54 и ядра =1,54), что может отражать большую активацию нейронов и, возможно, связано с большей концентрацией прогестерона в экстрактах свиных надпочечников (1,1 нмоль против 0,45 нмоль в дозе). При введении экстрактов фетальных надпочечников соотношение большого и малого диаметров тела было ближе к нормативным данным (1,63 против 1,54; у интактных 1,93). Аналогичная направленность изменений большого и малого диаметров была и у ядра (1,77 против 1,54; у интактных 1,65) за счет меньшего возрастания малого диаметра (при равном возрастании большого диаметра, определяющем рост клетки), что позволяет предполагать большее влияние экстрактов фетальных надпочечников на рост нейрона в длину, и возможно, связано с большим содержанием дегидроэпиандростерона (1,15 нмоль в инъекции против 0,83 нмоль в экстрактах свиных надпочечников) и в 13 раз меньшей гидрокортизона (5 нмоль против 65 нмоль).
Следует заметить, что в ранее выполненных нами исследованиях у животных была установлена большая обеспеченность АТФ тканей мозга в условиях гипобарической гипоксии как при введении экстрактов коры фетальных надпочечников, так и экстрактов коры свиных надпочечников. В сравнении с группами контроля и после введения гидрокортизона различия были высоко достоверны. После введения экстрактов фетальных надпочечников концентрация АТФ и соотношение АТФ/АМФ были несколько лучше, чем после введения экстракта свиных надпочечников (2,489±0,109 в сравнении с 2,305±0,134 ; соответственно коэффициент. АТФ/АМФ 10,637±0,463 против 8,801±1,787). Однако в группе гипоксического контроля концентрация АТФ составляла 1,732±0,216, а соотношение АТФ/АМФ всего 4,977±1,265. Все это позволяет считать, что экстракты коры свиных надпочечников мало уступают по стимулирующим эффектам экстрактам коры фетальных надпочечников. При том факте, что сырьевые ресурсы для получения экстрактов свиных надпочечников выше, чем для получения экстрактов коры фетальных надпочечников, возможность использования первых более перспективна.
Следует заметить, обследования экспериментальных животных в динамике опыта показали, что наибольшая прибавка в массе за 5-недельный срок наблюдалась у интактных животных (90 г); животные, которым вводились экстракты свиных и фетальных надпочечников, прибавили в массе 82,4 г и 84,0 г соответственно (в группе с гидрокортизоном - 80,2 г).
Обращает внимание, что прибавка в росте в группе интактного контроля была 1,26 см ; самый высокий прирост отмечен у животных, которым вводились экстракты свиных (2,0 см) и фетальных надпочечников (2,46 см), меньше - при введении гидрокортизона (1,46 см).
Учитывая, что у животных этих групп наблюдались и большие размеры тел и ядер нейронов, мы относим вышеназванные перемены в нейронах растущих животных не столько к проявлениям регуляторной гипертрофии, сколько к истинному приросту за счет всех структур клеток.
Литература
Семенова К.А., Мастюкова Е.М.Смуглин М.Я. Клиника и реабилитационная терапия детских церебральных параличей. Медицина-Москва 1972.
Семенова К.А., Жуковская Е.Д., Яковлева Н.Т., Семенов А.С., Дукарский Ф.Г. Влияние глюкокортикостероидных препаратов на иммунологические показатели и фунгкциональное состояние колры надпочечников у детей раннего возраста с церебральными параличами. Педиатрия. 1983, 12, 25-27.
Bovenberg S.A., van Uum S.H., Hermus A.R. Dehydroepiandrosterone administration in humans:evidence based? Neth J. Med. 2005, 63(8):300-4.
Buckwalter M.S., Yamane M., Coleman B.S.Onnerod B.K., Chin J.T., Wyss-Coray T. Chronically increased transforming growth factor-beta 1 strongly inhibits hippocampal neurogenesis in aged mice. Am J. Pathol, 2006; 169(1): 154-64.
Calogero А.Е., Burello N., Bosboom A.M., Garofalo M.R., Weber R.F., D Agata R.
Glucocorticoids inhibt gonadotropin-releassing hormones by acting directly at the hypothalamic level. Endocrinol. Invest., 1999; 22(9):666-70.
Cardounel A., Regelson W., Kalimi M. Dehydroepiandrosterone protecs hippocampal neurons against neurotoxin-induced cell death: mechanism of action, Proc. Soc/ Exp. Biol., 1999; 222(2): 145-9.
Chen H., Tung Y.C., Li В., Iqbal К., Grundke-Iqbal I. Trophic factors counteract FGF-2-induced inhibition of adult neurogenesis. Neurobiol Aging., 2006,19[Epub ahead of print] Cristi L. Watterberg M.D. Jeffrey S. Gerdes M.D. Xathleen L. Gitibrd R.N. and Hung Mo Lin Ph D Prophylaxis Againts Early Adrenal Insuffiency to Prevent Chronic Lung Disease in Premature Infants/Pediatrics vol 104 No 6, 1999, 1258-1263
Fujioka A., Fujioka T., Ishida Y., Mackawa T., Nakamura S. Differential effects jf prenatal stress on the morphological maturation of hippocampal neurons. Neuroscience, 2006, 25;141(2):907-15.
Galbiati M., Saredi S., Romano N., Martim L., Motta M., Melcangi R.C. Smad proteins are targets of transforming grouth factor betal in immortalized gonadotrophin-releasing hormone releasing neurones. J. Neuroendocrinol. 2005; 17(11):753-60.
Gregory A. Lodigensky M.D., Karin Reademaver M.D., Sewa Zimine Ph.D., Marianne Groenendaal M.D. RhD. Linda S de Vrics M.D. Ph D and Petra S Huppi M.D. Structural and Functional Brain Development Affer Hydrocortisone Treatment for Neonatal chronic Lung. Pediatrics vol 116, No 1, 2005, 1-7.
Rajadurai V.S., Tan K.H. The use abuse of steroids in perinatal medicine. Ann Acad Med Singapore. 2003; 32(3):324-34.
Kosaka N., Kodama M., Sasaki H., Yamamoto Y., Takeshita F., Takahama Y., Sakamoto H., Kato T, Terada M., Ochiya T. FGF-4 regulates neural progenitor cell proliferation and neurosal differentiation. Faseb J., 2006; 20(9): 1484-5.
Leowattana W. DHEA(S): the fountainof youth. J Med. Assoc. Thai. 2001; 84 Suppi 2:S605-12 MacPherson A., Dinkel K., Sapolsky R. Glucocorticoids worsen excitotoxin-induced expression of pro-inflammatory cytokines in hippocampal cultures. Exp Neurol. 2005; 194(2):376-83.
Mazzucco C.A., Lieblich S.E., Bingham B.I., Wi]liamson M.A., Viau V., Galea L.A. Both estrogen receptor alpha and estrogen receptor beta agonists enhance cell proliferation in the dentate gyros of adult female rats. Neuroscience. 2006, 15; 141(4): 1793-800.
Patel M.A., Katyare S.S. Dehydroepiandrosterone (DHEA) treatment stimulates oxidative energy metabolism in the cerebral mitochondria from developing rats. Int. J. Dev Neurosci. 2006; 24(5):327-34.
Robert M.Sapolsky. Glucocorticoid toxicity in the Hippocampus: Temporal aspects of neuronal vulnerability. Bram Research, v 359,1-2,1985: 300-305.
Shi X.Y.,Wang J.W., Sun R.P. Effects and consequence of recurrent seizures of neonatal rat on the hippocampal neurogenesis. Zhonghua Er Ke Za Zhi. 2006; 44(4):289-93.
Zwain I H., Yen S.S. Dehydroepiandrosterone: biosynthesis and metabolism in the brain. Endocrinology, 1999; 140(2): 880-7.


Предлагаемая группа изобретений относится к экспериментальной биологии и медицине, а именно к экспериментальной неврологии, и может быть использовано для стимуляции роста мотонейронов коры головного мозга растущего организма. Предложен способ стимуляции роста мотонейронов коры головного мозга растущего организма млекопитающих в эксперименте, включающий введение экстракта коры надпочечников, через день, общим курсом 5 недель, при этом вводят экстракт коры свиных надпочечников в разовой дозе 0,5 мл на 100 г массы тела. Предложено применение экстракта коры свиных надпочечников в качестве средства для стимуляции роста мотонейронов коры головного мозга растущего организма. Изобретение обеспечивает расширение арсенала средств для стимуляции роста мотонейронов коры головного мозга и возможность использования доступного средства. 2 н.п. ф-лы, 1 табл.
| ШИЛЕНОК И.Г и др | |||
| Анаболические эффекты минорных концентраций кортикостероидов, содержащихся в экстрактах коры надпочесников, Нижегородский медицинский журнал, 2004, №1, с.92-95 | |||
| СПОСОБ ПОЛУЧЕНИЯ КОМПЛЕКСА БИОЛОГИЧЕСКИ АКТИВНЫХ ПОЛИПЕПТИДОВ ДЛЯ НОРМАЛИЗАЦИИ ФУНКЦИЙ ГОЛОВНОГО МОЗГА И ФАРМАЦЕВТИЧЕСКОЕ СРЕДСТВО НА ЕГО ОСНОВЕ | 2004 |
|
RU2275924C2 |
| СПОСОБ СТИМУЛЯЦИИ РЕГЕНЕРАЦИИ ПЕЧЕНОЧНОЙ ТКАНИ | 1999 |
|
RU2185835C2 |
| US 2003224074 A, 04.12 | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| WO 03006038 A1, 23.01 | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |

