Область и предпосылки изобретения
Настоящее изобретение относится к способам повышения толерантности к абиотическому стрессу и/или биомассы у растений, а более конкретно к растениям, экспрессирующим гены устойчивости к экзогенному абиотическому стрессу.
Условия абиотического стресса (также названного как «экологический стресс»), например засоленность почвы, периоды засухи, наводнения, недостаточно оптимальная температура и токсическое химическое загрязнение являются причиной существенного повреждения сельскохозяйственных растений. У большинства растений существуют развитые стратегии собственной защиты против этих условий. Однако если тяжесть и продолжительность стрессовых условий являются слишком сильными, то влияния на развитие, рост и урожайность растений большинства сельскохозяйственных культур являются глубокими. Кроме того, большинство сельскохозяйственных культур являются очень восприимчивыми к абиотическому стрессу (ABS) и, таким образом, требуют оптимальных условий роста для сбора урожая на коммерческом уровне. Продолжительное воздействие стресса является причиной значительных изменений в метаболизме растений, что приводит, в конечном счете, к гибели клеток, а в результате, к потерям урожая. Таким образом, несмотря на обширные исследования и применение сложных и трудоемких мер по защите урожая, потери вследствие условий абиотического стресса ежегодно составляют миллиарды долларов.
Развитие устойчивых к стрессу растений представляет собой стратегию, обладающую потенциалом для решения или содействия в решении, по крайней мере, некоторых из этих проблем. Однако традиционные стратегии селекции растений, применяемые для развития новых линий растений, проявляющих толерантность к ABS, являются относительно малоэффективными, так как они трудоемки, требуют много времени и не поддаются прогнозированию. Кроме того, значительные проблемы, встречающиеся при традиционном скрещивании, представляют собой ограниченные ресурсы зародышевой плазмы для устойчивости к стрессу и несовместимость при скрещиваниях между отдаленно родственными видами растений. Кроме того, клеточные процессы, приводящие к толерантности к ABS, по своей природе являются комплексными и вовлекают множество механизмов клеточной адаптации и многочисленные метаболические пути.
В уровне техники описаны генно-инженерные исследования, направленные на получение у трансгенных культур толерантности к абиотическому стрессу. В исследованиях Apse and Blumwald (Curr Opin Biotechnol. 13:146-150, 2002), Quesada et al. (Plant Physiol. 130:951-963, 2002), Holmström et al. (Nature 379:683-684, 1996), Xu et al. (Plant Physiol 110:249-257, 1996), Pilon-Smits and Ebskamp (Plant Physiol. 107:125-130, 1995) и Tarczynski et al. (Science 259:508-510, 1993) были предприняты попытки получения устойчивых к стрессу культур.
Кроме того, в нескольких патентах и патентных заявках США также описаны полинуклеотиды, ассоциированные с толерантностью к стрессу, и их применение в создании устойчивых к стрессу культур. В патентах США № 5296462 и 5356816 описаны растения, трансформированные полинуклеотидами, кодирующими белки, вовлеченные в адаптацию к холоду у Arabidopsis thaliana, для обеспечения, таким образом, адаптации к холоду у трансформированных растений.
В патенте США № 6670528 описаны растения, трансформированные полинуклеотидами, кодирующими полипептиды, связанные с элементами, отвечающими на стресс, для повышения, таким образом, толерантности трансформированных растений к абиотическому стрессу.
В патенте США № 6720477 описаны растения, трансформированные полинуклеотидами, кодирующими передающий сигнал, связанный со стрессом белок, способствующий повышению толерантности к абиотическому стрессу у трансформированных растений.
В заявках США №№ 09/938842 и 10/342224 описаны гены, связанные с абиотическим стрессом, и их применение для придания растениям толерантности к абиотическому стрессу.
В заявках США № 10/231035 у растений описана повышенная экспрессия сульфуразы с молибденовым кофактором, для повышения, таким образом, их толерантности к абиотическому стрессу.
Несмотря на то, что указанные выше исследования, по крайней мере отчасти, являлись успешными в создании растений, устойчивых к стрессу, все еще остается необходимость в генах устойчивости к стрессу, которые можно применять для создания растений, устойчивых к широкому спектру условий абиотического стресса.
Во время доведения настоящего изобретения до практического осуществления авторы настоящего изобретения при помощи биоинформатики и лабораторных исследований идентифицировали несколько новых генов толерантности к абиотическому стрессу, которые можно применять для повышения толерантности к абиотическому стрессу и/или биомассы растений.
Сущность изобретения
В одном из аспектов настоящее изобретение относится к способу повышения толерантности растений к абиотическому стрессу. Способ включает в себя экспрессию в растении экзогенного полипептида, включающего в себя аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID №№39-92.
В еще одном аспекте настоящее изобретение относится к растительной клетке, содержащей экзогенный полинуклеотид, по крайней мере, на 90% гомологичный полинуклеотиду, выбранному из группы, состоящей из SEQ ID №№1-18.
В еще одном аспекте настоящее изобретение относится к растительной клетке, содержащей экзогенный полинуклеотид, кодирующий полипептид, включающий в себя аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID №№39-92.
В еще одном аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, включающей в себя полинуклеотид, по крайней мере, на 90% гомологичный нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID №№1-18, и промотор, обеспечивающий управление транскрипцией полинуклеотида в клетке хозяина.
В еще одном аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, включающей в себя полинуклеотид, кодирующий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID №№39-92, и промотор, обеспечивающий управление транскрипцией полинуклеотида в клетке хозяина.
В другом дополнительном аспекте настоящее изобретение относится к выделенному полипептиду, включающему в себя аминокислотную последовательность, по крайней мере, на 90% гомологичный нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID №№ 1-18, и промотор, обеспечивающий управление транскрипцией полинуклеотида в клетке хозяина.
В еще одном аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, включающей в себя полинуклеотид, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID №№ 39-92, и промотор, обеспечивающий управление транскрипцией полинуклеотида в клетке хозяина.
В другом дополнительном аспекте настоящее изобретение относится к выделенному полипептиду, включающему в себя аминокислотную последовательность, по крайней мере, на 90% гомологичную аминокислотной последовательности, кодируемой полинуклеотидом, выбранным из группы, состоящей из SEQ ID №№ 1-18.
В другом дополнительном аспекте настоящее изобретение относится к выделенному полипептиду, включающему в себя аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID №№ 39-92.
Согласно дополнительным признакам в описанных предпочтительных вариантах осуществления экспрессию проводят (i) трансформацией клетки растения экзогенным полинуклеотидом; (ii) генерацией зрелого растения из клетки; и (iii) культивированием зрелого растения в условиях, пригодных для экспрессии экзогенного полинуклеотида в зрелом растении.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления трансформацию проводят введением в растительные клетки конструкции нуклеиновой кислоты, включающей в себя экзогенный полинуклеотид и, по крайней мере, один промотор, обеспечивающий направление транскрипции полинуклеотида в клетке хозяина.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления, по крайней мере, один промотор представляет собой конститутивный промотор.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления конститутивный промотор представляет собой промотор CaMV 35S.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления конститутивный промотор представляет собой At6669 промотор.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления, по крайней мере, один промотор представляет собой индуцибельный промотор.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления индуцибельный промотор представляет собой индуцибельный промотор абиотического стресса.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления, по крайней мере, один промотор представляет собой тканеспецифичный промотор.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления экспрессию проводят инфицированием растения вирусом, включающим в себя экзогенный полинуклеотид.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления вирус представляет собой невирулентный вирус.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления абиотическим стрессом является выбранный из группы, состоящей из засоленности почвы, недостатка воды, низкой температуры, высокой температуры, токсического действия тяжелых металлов, анаэробиоза, недостатка питательных веществ, избытка питательных веществ, загрязнения атмосферы и УФ-излучения.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления растение представляет собой двудольное растение.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления растение представляет собой однодольное растение.
Согласно другим дополнительным признакам в описанных предпочтительных вариантах осуществления растительная клетка формирует часть растения.
Настоящее изобретение успешно обходит недостатки известных в настоящее время конструкций посредством обеспечения способов применения новых генов устойчивости к абиотическому стрессу для повышения толерантности к абиотическому стрессу и/или биомассы растений.
Краткое описание чертежей
Изобретение здесь описано только посредством примера со ссылкой на прилагаемые чертежи. Теперь на конкретном примере подробного описания чертежей подчеркнуто, что сведения представлены посредством примера и только в целях иллюстративного обсуждения предпочтительных вариантов настоящего изобретения и приведены для предоставления того, что, как полагают, является наиболее пригодным и легко понимаемым описанием принципов и концептуальных аспектов изобретения. В этом отношении не делали попыток показать структурные детали изобретения более подробно, чем это необходимо для фундаментального понимания изобретения и для того, чтобы описание, взятое с чертежами, делало очевидным специалистам в данной области, как некоторые части изобретения можно применять на практике.
Фиг.1 представляет собой блок-схему, иллюстрирующую процесс идентификации предполагаемых генов растений устойчивости к стрессу из баз данных последовательностей нуклеиновых кислот.
Фиг.2А-В представляют собой фотографии, иллюстрирующие T2 трансгенное зрелое растение Arabidopsis thaliana на стадии цветения, экспрессирующее трансген экзогенной люциферазы из промотора At6669. Одно и то же растение показано в условиях нормальной освещенности (фиг.2А) и в темноте (фиг.2В). Сильное освещение, указывающее на экспрессию люциферазы, наблюдали на цветках и корневых тканях.
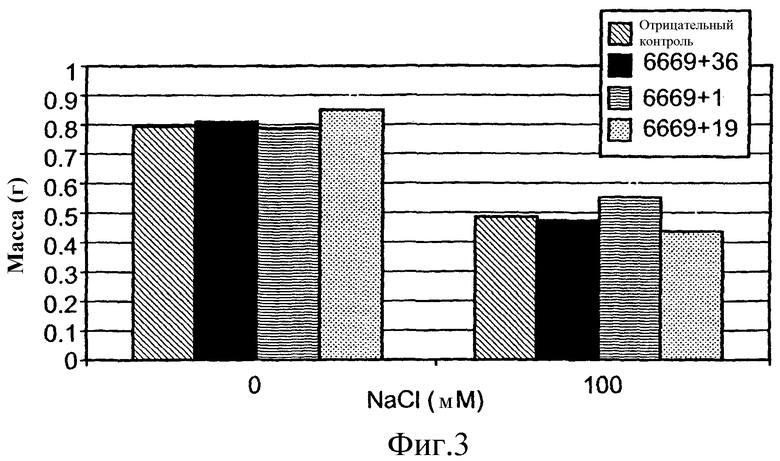
На фиг.3 приведена средняя масса сырой ткани трансгенных растений T1 A. thaliana, выращенных в нормальных или стрессовых условиях (орошение 0 или 100 М раствором NaCl, соответственно). Растения трансформировали предполагаемыми генами толерантности к стрессу, или репортерным геном люциферазы (контроль), помещенными под транскрипционный контроль промотора At6669. Средние значения, обозначенные теми же обозначениями, статистически не отличаются по одностороннему T-тесту ANOVA.
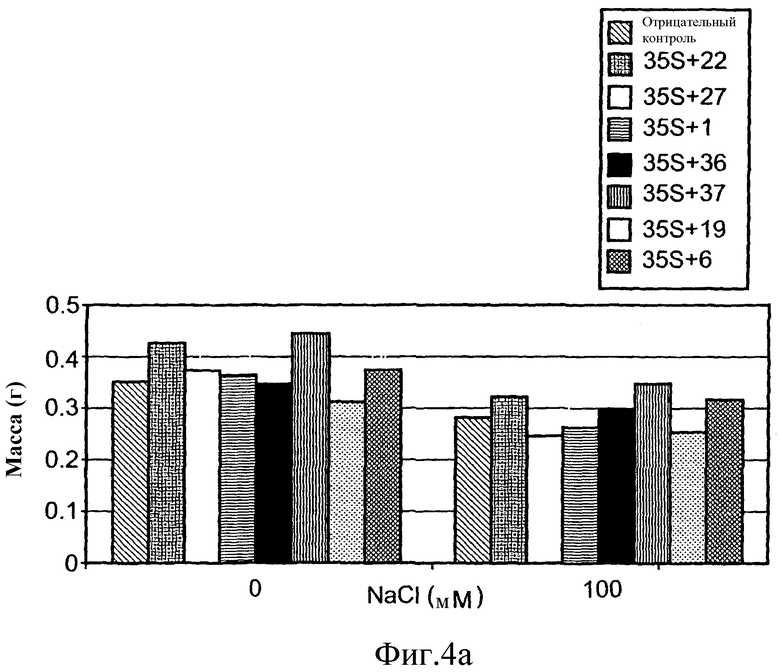
На фиг.4А приведена средняя масса сырой ткани растений T2 A. thaliana выращенных в нормальных или стрессовых условиях (орошение 0 или 100 М раствором NaCl, соответственно). Растения трансформировали предполагаемыми генами толерантности к стрессу по настоящему изобретению, или репортерным геном люциферазы (контроль), помещенными под транскрипционный контроль промотора 35S. Средние значения, обозначенные теми же обозначениями, статистически не отличаются по одностороннему T-тесту ANOVA.
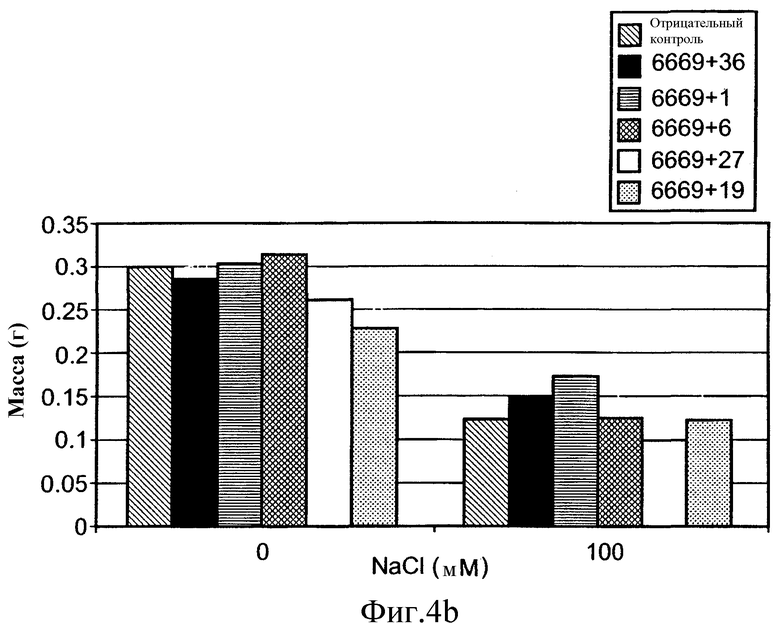
На фиг.4 приведена средняя масса сырой ткани растений T2 A. thaliana выращенных в нормальных или стрессовых условиях (орошение 0 или 100 М раствором NaCl, соответственно). Растения трансформировали предполагаемыми генами толерантности к стрессу по настоящему изобретению, или репортерным геном люциферазы (контроль), помещенными под транскрипционный контроль промотора At6669. Средние значения, обозначенные теми же обозначениями, статистически не отличаются по одностороннему T-тесту ANOVA.
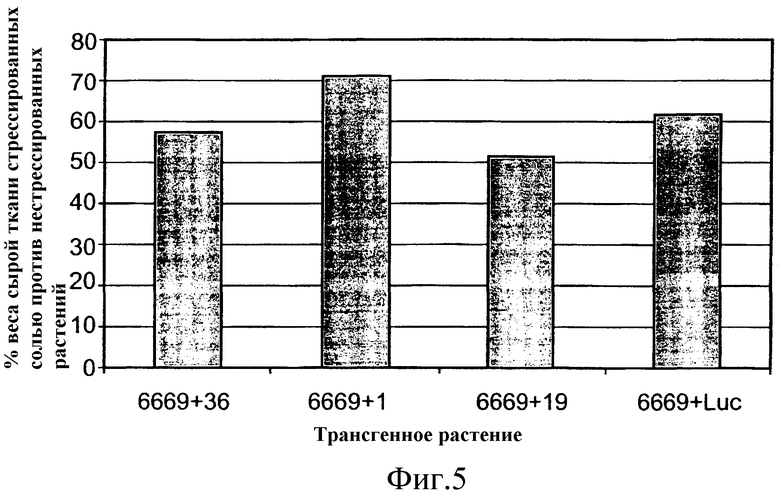
На фиг.5 приведена относительная (процент) масса сырой ткани трансгенных растений T1 A. thaliana, выращенных в стрессовых условиях засоленности почвы (орошение 100 мМ раствором NaCl), по сравнению с подобными растениями, выращенными в нормальных условиях (орошение только водой). Растения трансформировали предполагаемыми генами толерантности к стрессу по настоящему изобретению, или репортерным геном люциферазы (контроль), помещенными под транскрипционный контроль промотора At6669.
Описание предпочтительных вариантов осуществления
Настоящее изобретение относится к способам повышения толерантности растений к абиотическому стрессу и/или биомассы применением новых генов толерантности к абиотическому стрессу и растений, демонстрирующим повышенную толерантность к стрессовым условиям и/или повышенную способность наращивать биомассу.
Принципы и применение настоящего изобретения можно понять лучше со ссылкой на чертежи и сопровождающие описания.
Перед подробным объяснением, по крайней мере, одного варианта осуществления изобретения необходимо понимать, что изобретение не ограничено его применением деталями конструкции и расположением компонентов, указанных в следующем далее описании или проиллюстрированных в чертежах. Изобретение допускает другие варианты осуществления, или применение на практике или выполнение различными способами. Также необходимо понимать, что формулировки и терминологию применяют здесь с целью описания и их не следует рассматривать как ограничивающие.
Во время доведения настоящего изобретения до практического осуществления авторы настоящего изобретения, применяя биоинформатические способы, идентифицировали полинуклеотидные последовательности, кодирующие предполагаемые устойчивые к абиотическому стрессу белки (ABST)(пример 1). Выбранные последовательности выделяли (пример 2), клонировали в экспрессионные векторы (примеры 3-4) и вводили в растения Arabidopsis thaliana (пример 5). Растения, выращенные в стрессовых условиях засоленности почвы или в нормальных условиях, демонстрировали значительно более высокую биомассу по сравнению с подобными растениями, не несущими экзогенные гены ABST (пример 6).
Таким образом, по одному аспекту настоящее изобретение относится к способу повышения толерантности растений к абиотическому стрессу и/или биомассы растений. Способ включает в себя экспрессию в растении экзогенного полинуклеотида, гомологичного, по крайней мере, на 70%, предпочтительно, гомологичного, по крайней мере, на 80%, более предпочтительно, гомологичного, по крайней мере, 85%, наиболее предпочтительно, гомологичного, по крайней мере, на 90% полинуклеотиду, выбранному из группы, состоящей из SEQ ID №№ 1-18. В качестве альтернативы экзогенный полинуклеотид настоящего изобретения кодирует полипептид с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID №№ 39-92.
Используемое здесь идиоматическое выражение «абиотический стресс» употребляется по отношению к любому неблагоприятному действию на метаболизм, рост, репродукцию и/или жизнеспособность растения. Таким образом, абиотический стресс можно индуцировать при помощи субоптимальных условий роста окружающей среды, таких как, например, засоленность почвы, недостаток воды, избыток воды, низкая или высокая температура, токсическое действие тяжелых металлов, анаэробиоз, недостаток питательных веществ, загрязнение атмосферы или УФ-излучение.
Применяемое здесь идиоматическое выражение «толерантность к абиотическому стрессу» относится к способности растения переносить абиотический стресс без существенного изменения в метаболизме, росте, производительности и/или жизнеспособности.
Пригодным для применения в способе настоящего изобретения может быть любое однодольное или двудольное растение, включающее в себя в качестве неограничивающих примеров кукурузу, пшеницу, просо, рожь, овес, рис, сою, арахис, горох, чечевицу и люцерну, хлопок, рапс, канолу, перец, подсолнечник, картофель, табак, томат, баклажан, эвкалипт, растение с древовидным стеблем, декоративное растение, многолетнюю траву и кормовую культуру.
Применяемый здесь термин «экзогенный полинуклеотид» относится к последовательности нуклеиновых кислот, не экспрессируемой растением в природе, но которая при введении в растение в длительных или кратковременных условиях продуцирует, по крайней мере, один полипептидный продукт.
В настоящем изобретении экспрессия экзогенного полинуклеотида растением может быть результатом трансформации одной или более клеток растения экзогенным полинуклеотидом, с последующей генерацией зрелого растения из трансформированных клеток и культивированием зрелого растения в условиях, пригодных для экспрессии экзогенного полинуклеотида в зрелом растении.
Предпочтительно трансформацию проводят введением в растительную клетку конструкции нуклеиновых кислот, включающую в себя экзогенный полинуклеотид настоящего изобретения и, по крайней мере, один промотор, обеспечивающий управление транскрипцией экзогенного полинуклеотида в растительной клетке. Дальнейшие детали применяемых подходов трансформации представлены далее.
Применяемый здесь термин «промотор» относится к области ДНК, расположенной выше области инициации транскрипции гена, с которым связывается РНК-полимераза для инициации транскрипции РНК. Промотор контролирует, где (например, какая часть растения, какой орган в животном и т.д.) и/или когда (например, на какой стадии или условии существования организма) экспрессируется ген.
Любую пригодную промоторную последовательность можно применять при помощи конструкции нуклеиновых кислот настоящего изобретения. Предпочтительно промотор представляет собой конститутивный промотор, тканеспецифичный или индуцибельный промотор абиотического стресса.
Предполагаемые конститутивные промоторы включают в себя, например, промотор CaMV 35S (SEQ ID NO: 19; Odell et al., Nature 313:810-812, 1985); промотор Arabidopsis At6669 (SEQ ID NO: 20); кукурузный Ubi 1 (Christensen et al., Plant Sol. Biol. 18:675-689, 1992); актин риса (McElroy et al., Plant Cell 2:163-171, 1990); pEMU (Last et al., Theor. Appl. Genet. 81:581-588, 1991); и Synthetic Super MAS (Ni et al., The Plant Journal 7:661-76, 1995). Другие конститутивные промоторы включают в себя конститутивные промоторы по патентам США №№ 5659026, 5608149; 5608144; 5604121; 5569597; 5466785; 5399680; 5268463; и 5608142.
Предполагаемые тканеспецифичные промоторы включают в себя в качестве неограничивающих примеров специфичные для листьев промоторами, такими как описано Yamamoto et al., Plant J. 12:255-265, 1997; Kwon et al., Plant Physiol. 105:357-67, 1994; Yamamoto et al., Plant Cell Physiol. 35:773-778, 1994; Gotor et al., Plant J. 3:509-18, 1993; Orozco et al., Plant Mol. Biol. 23:1129-1138, 1993; и Matsuoka et al., Proc. Natl. Acad. Sci. USA 90:9586-9590, 1993.
Предполагаемые индуцибельные промоторы абиотического стресса включают в себя, но не ограничены индуцированными засоленностью промоторами, такими как RD29A (Yamaguchi-Shinozalei et al., Mol. Gen. Genet. 236:331-340, 1993); индуцированными засухой промоторами, такими как промотор кукурузного гена rabl7 (Pla et al., Plant Mol. Biol. 21:259-266, 1993), промотор кукурузного гена rab28 (Busk et al., Plant J. 11:1285-1295, 1997) и промотор кукурузного гена Ivr2 (Pelleschi et al., Plant Mol. Biol. 39:373-380, 1999); и индуцированными повышенной температурой промоторами, такими как тепловой промотор томатов hsp80 из томатов (патент США №5187267).
Конструкция нуклеиновой кислоты настоящего изобретения предпочтительно дополнительно включает в себя соответствующий селективный маркер и/или ориджин репликации. Предпочтительно применяемая конструкция нуклеиновой кислоты представляет собой челночный вектор, способный размножаться в Е.coli (где конструкция содержит соответствующий селективный маркер и ориджин репликации) и пригодный для размножения в клетках. Конструкцией по настоящему изобретению может являться, например, плазмида, бакмида, фагмида, космида, фаг, вирус или искусственная хромосома.
Конструкцию нуклеиновых кислот настоящего изобретения можно применять для устойчивой или кратковременной трансформации растительных клеток. В случае устойчивой трансформации экзогенный полинуклеотид настоящего изобретения встраивали в геном растения, и по существу он представляет устойчивый и наследуемый признак. В случае кратковременной трансформации трансформированные клетки экспрессируют экзогенный полинуклеотид, но он не встроен в геном и по существу представляет кратковременный признак.
Существуют различные способы введения чужеродных генов в однодольные и двудольные растения (Potrykus, I., Annu. Rev. Plant. Physiol., Plant. Mol. Biol. (1991) 42:205-225; Shimamoto et al., Nature (1989) 338:274-276).
Основные способы получения устойчивой интеграции экзогенной ДНК в геномную ДНК растения включают в себя два основных подхода:
(i)Перенос Agrobacterium-опосредованного гена: Klee et al. (1987) Annu. Rev. Plant Physiol. 38:467-486; Klee and Rogers in Cell Culture and Somatic Cell Genetics of Plants, Vol. 6, Molecular Biology of Plant Nuclear Genes, eds. Schell, J., and Vasil, L. K., Academic Publishers, San Diego, Calif. (1989) p.2-25; Gatenby, in Plant Biotechnology, eds. Kung, S. and Arntzen, C. J. , Butterworth Publishers, Boston, Mass. (1989) p.93-112.
(ii) Прямое поглощение ДНК: Paszkowski et al., in Cell Culture and Somatic Cell Genetics of Plants, Vol. 6, Molecular Biology of Plant Nuclear Genes eds. Schell, J., and Vasil, L. K., Academic Publishers, San Diego, Calif. (1989) p.52-68; включающее в себя способы для прямого поглощения ДНК в протопласты, Toriyama, K. et al. (1988) Bio/Technology 6:1072-1074. Поглощение ДНК индуцировали при помощи короткого электрического шока растительных клеток: Zhang et al. Plant Cell Rep. (1988) 7:379-384. Fromm et al. Nature (1986) 319:791-793. Введение ДНК в растительные клетки или ткани при помощи бомбардировки частицами, Klein et al. Bio/Technology (1988) 6:559-563; McCabe et al. Bio/Technology (1988) 6:923-926; Sanford, Physiol. Plant. (1990) 79:206-209; посредством применения систем микропипетирования: Neuhaus and Spangenberg, Physiol. Plant. (1990) 79:213-217; трансформация культур клеток, эмбрионов или костной ткани с применением нитей стекловолокна или карбида кремния, патент США № 5464765, или при помощи прямой инкубации ДНК с зародышевой пыльцой, DeWet et al. in Experimental Manipulation of Ovule Tissue, eds. Chapman, G. P. and Mantell, S. H. and Daniels, W. Longman, London, (1985) p.197-209; и Ohta, Proc. Natl. Acad. Sci. USA (1986) 83:715-719.
Сиситема Agrobacterium включает в себя применение плазмидных векторов, содержащих определенные сегменты ДНК, интегрирующиеся в геномную ДНК растения. Способы инокуляции растительной ткани очень зависят от видов растений и системы доставки Agrobacterium. Способ листовой пластинки представляет собой широко применяемый подход, который можно выполнять для любого эксплантата ткани, который представляет собой хороший источник для инициации дифференцирования целого растения. Horsch et al. in Plant Molecular Biology Manual A5, Kluwer Academic Publishers, Dordrecht (1988) p.1-9. Дополнительный подход использует систему доставки Agrobacterium в комбинации с вакуумной инфильтрацией. Система Agrobacterium является особенно жизнеспособной при создании трансгенных двудольных растений.
Существуют различные способы прямого переноса ДНК в растительные клетки. При электропорации протопласты подвергают кратковременному воздействию сильного электрического поля. При микроинъекции ДНК механически вводят прямо в клетки, применяя очень маленькие микропипетки. При бомбардировке микрочастицами ДНК адсорбируют на бомбардирующие микрочастицы, такие как кристаллы сульфата магния или вольфрамовые частицы, а бомбардирующие микрочастицы с ускорением физически направляют в клетки или растительные ткани.
После устойчивой трансформации проводят размножение растения. Наиболее часто встречающимся способом выращивания растения является посев семян. Однако регенерация посредством размножения семенами имеет тот недостаток, что в результате гетерозиготности существует отсутствие единообразия в культуре, так как растения продуцируют семена в соответствии с генетической изменчивостью, обусловленной законами Менделя. По существу, каждое семя является генетически отличимым, и каждое растет со своими собственными специфическими признаками. Поэтому предпочтительно, что трансформированное растение произведено таким, что восстановленное растение имеет идентичные признаки и свойства родительского трансгенного растения. Таким образом, предпочтительно, что трансформированное растение восстановлено посредством вегетативного размножения, предоставляющего быстрое, устойчивое воспроизведение трансформированных растений.
Вегетативное размножение представляет собой процесс роста нового поколения растений из одного образца ткани, отделенного от отобранного родительского растения или сорта. Этот процесс дает возможность массовому размножению растений с предпочтительной тканевой экспрессией гибридного белка. Новое поколение растений производят генетически идентичными и обладающими всеми свойствами первичного растения. Вегетативное размножение обеспечивает массовую продукцию качественного растительного материала в короткий период времени и предлагает быстрое размножение выбранных сортов при сохранении свойств первично трансгенного или трансформированного растения. Преимуществами размножаемых вегетативным путем растений являются скорость размножения растений, и качество и единообразие продуцируемых растений.
Вегетативное размножение представляет собой многоступенчатый способ, требующий перемены культуральной среды или условий роста между стадиями. Таким образом, процесс вегетативного размножения включает в себя четыре основных стадии: первая стадия - первоначальное культивирование ткани; вторая стадия - размножение тканевой культуры; третья стадия - дифференциация и формирование растения; и четвертая стадия - культивирование в оранжерее и закаливание. В течение первой стадии первоначального культивирования ткани создавали тканевые культуры и подтверждали отсутствие контаминации. В течение второй стадии первоначальную культуру ткани размножили до тех пор, пока не получали достаточного количества образцов ткани для удовлетворения производственных целей. В течение третьей стадии образцы ткани, выращенные во второй стадии, делили и растили в отдельных саженцах. На четвертой стадии трансформированные саженцы переносили в оранжерею для закаливания, где устойчивость растений к свету постепенно повышалась так, что их становилось возможным выращивать в естественной окружающей среде.
Предпочтительно, зрелые трансформированные растения, полученные, как описано выше, далее отбирали по устойчивости к абиотическому стрессу. Соответственно, трансформированные и нетрансформированные (дикого типа) растения подвергали воздействию условиям абиотического стресса, таких как недостаток воды, субоптимальная температура, недостаток питательных веществ, или, предпочтительно, условия солевого стресса. Солевой стресс можно обеспечивать при помощи многих способов, таких как, например, орошение растений гиперосмотическим раствором, культивирование растений способом гидропоники в гиперосмотическом растворе для выращивания (например, раствор Хогланда), или культивирование растений в гиперосмотической питательной среде (например, среда MS). Так как различные растения значительно отличаются по их устойчивости к засоленности, концентрацию соли в орошающей воде, растворе для выращивания или питательной среде предпочтительно уточняют в соответствии со специфическими свойствами специфического сорта или вида растения, для того, чтобы оказать мягкое или среднее действие на физиологию и/или морфологию растений (для руководства в отношении получения подходящих концентраций см. Bernstein and Kafkafi, Root Growth Under Salinity Stress In: Plant Roots, The Hidden Half 3rd ed. Waisel Y, Eshel A and Kafkafi U. (editors) Marcel Dekker Inc., New York, 2002, и ссылки в них). Последующее воздействие стрессовыми условиями на растения часто контролировали до появления значительных физиологических и/или морфологических эффектов в растениях дикого типа. Затем трансформированные растения, не демонстрирующие значительных физиологических и/или морфологических эффектов или демонстрирующие более высокую биомассу, чем растения дикого типа, идентифицировали как растения, устойчивые к абиотическому стрессу.
Хотя в настоящем изобретении предпочтительна стабильная трансформация, в настоящем изобретении также рассматривается транзиторная трансформация клеток листьев, меристематических клеток или целого растения.
Транзиторную трансформацию можно проводить любым из описанных выше способов переноса ДНК или посредством вирусного заражения с применением модифицированных вирусом растений.
Вирусы, для которых показано, что они пригодны для трансформации растений-хозяев, включают в себя CaMV, TMV и BV. Трансформация растений с применением вирусов растений описана в патенте США № 4855237 (BGV), EP-A 67553 (TMV), Japanese Published Application № 63-14693 (TMV), EPA 194809 (BV), EPA 278667 (BV) и Gluzman, Y. et al., Communications in Molecular Biology: Viral Vectors, Cold Spring Harbor Laboratory, New York, pp. 172-189 (1988). Псевдовирусные частицы для применения в экспрессии чужеродной ДНК во многих хозяевах, включая растения, описаны в WO 87/06261.
Предпочтительно, чтобы вирус по настоящему изобретению являлся авирулентным и, таким образом, неспособным вызывать серьезные симптомы, такие как уменьшенная скорость роста, мозаичность, кольцевая пятнистость, скручивание листьев, пожелтение, полосчатость, формирование оспы, формирование опухоли и эрозия. Пригодный авирулентный вирус может представлять собой встречающийся в природе авирулентный вирус или искусственно аттенуированный вирус. Аттенуацию вируса можно проводить с применением хорошо известных в данной области способов, включающих в себя в качестве неограничивающих примеров сублетальное нагревание, химическую обработку или способы прямого мутагенеза, такие как описанные, например, у Kurihara and Watanabe (Molecular Plant Pathology 4:259-269, 2003), Gal-on et al. (1992), Atreya et al. (1992) и Huet et al. (1994).
Пригодные вирусные штаммы можно получить из доступных источников, таких как, например, American Type Culture Collection (ATCC), или выделением из инфицированных растений. Выделение вирусов из инфицированных растительных тканей можно проводить хорошо известными в данной области способами, такими как описанные, например, у Foster and Tatlor, Eds. "Plant Virology Protocols: From Virus Isolation to Transgenic Resistance (Methods in Molecular Biology (Humana Pr), Vol 81)", Humana Press, 1998. В кратком изложении, ткани инфицированного растения, для которых полагают, что они содержат высокую концентрацию пригодного вируса, предпочтительно молодые листья и цветочные лепестки, помещают в буферный раствор (например, раствор фосфатного буфера) для получения инфицированного сока, который можно применять для последующих инокуляций.
Конструирование РНК-вирусов растений для введения в растения и экспрессии в них невирусных экзогенных полинуклеотидных последовательностей продемонстрировано указанными выше ссылками, а также у Dawson, W.O. et al., Virology (1989) 172:285-292; Takamatsu et al. EMBO J. (1987) 6:307-311; French et al. Science (1986) 231:1294-1297 и Takamatsu et al., FEBS Letters (1990) 269:73-76.
Когда вирус представляет, собой ДНК-вирус, пригодные модификации можно проводить с самим вирусом. Альтернативно, для облегчения конструирования желаемого вирусного вектора с чужеродной ДНК вирус сначала можно клонировать в бактериальную плазмиду. Затем вирус можно вырезать из плазмиды. Если вирус представляет собой ДНК-вирус, к вирусной ДНК можно присоединить бактериальный ориджин репликации, а затем реплицировать вирусную ДНК в бактериях. Транскрипция и трансляция этой ДНК продуцирует белок оболочки, инкапсулирующий вирусную РНК. Если вирус представляет собой РНК-вирус, вирус, как правило, клонируют в виде кДНК и вставляют в плазмиду. Затем плазмиду применяют для получения всех конструкций. Затем посредством транскрипции вирусной последовательности плазмиды и трансляции вирусных генов для получения белка(ов) оболочки, инкапсулирующий вирусную РНК, получают РНК-вирус.
Конструирование РНК-вирусов растений для введения в растения и экспрессии в них невирусных экзогенных полинуклеотидных последовательностей, таких как включенные в конструкцию по настоящему изобретению полинуклеотидные последовательности, продемонстрировано указанными выше ссылками, а также в патенте США № 5316931.
В одном из вариантов осуществления представлен полинуклеотид вируса растений, в котором кодирующая нативный белок оболочки последовательность удалена из полинуклеотида вируса, а вставлены кодирующая ненативный белок оболочки вируса растений последовательность и ненативный промотор, предпочтительно субгеномный промотор кодирующей ненативный белок оболочки последовательности, способной к экспрессии в растительном хозяине, пакуя рекомбинантный полинуклеотид вируса растений и обеспечивая системное заражение хозяина рекомбинантным полинуклеотидом вируса растений. Альтернативно, ген белка оболочки можно инактивировать посредством вставки в него ненативной полинуклеотидной последовательности, при которой продуцируется белок. Рекомбинантный полинуклеотид вируса растений может содержать одну или более дополнительных ненативных субгеномных промоторов. Каждый ненативный субгеномный промотор способен направлять транскрипцию или экспрессию прилежащих генов или полинуклеотидных последовательностей в растительном хозяине и не способен к рекомбинации с другими и с нативным субгеномными промоторами. Ненативные (чужеродные) полинуклеотидные последовательности можно вставлять следом за нативным субгеномным промотором вируса растения или нативным и ненативным субгеномными промоторами вирусов растений, если вставлено более одной полинуклеотидной последовательности. Ненативные полинуклеотидные последовательности транскрибируются и экспрессируются в растительном хозяине под контролем субгеномного промотора с получением желательных продуктов.
Во втором варианте осуществления предоставлен рекомбинантный полинуклеотид вируса растения, как и в первом варианте осуществления, за исключением того, что вместо кодирующей ненативный белок оболочки последовательности следом за одним из ненативных субгеномных промоторов белка оболочки помещена последовательность, кодирующая нативный белок оболочки.
В третьем варианте осуществления предоставлен рекомбинантный полинуклеотид вируса растений, в котором нативный ген белка оболочки находится следом за его субгеномным промотором, а в вирусный полинуклеотид вставлены один или несколько ненативных субгеномных промоторов. Вставленные ненативные субгеномные промоторы способны направлять транскрипцию или экспрессию прилежащих генов в растительном хозяине и не способны к рекомбинации с другими и с нативным субгеномными промоторами. Ненативные полинуклеотидные последовательности можно вставлять следом за ненативными субгеномными промоторами вирусов растений так, что последовательности транскрибируются и экспрессируются в растительном хозяине под контролем субгеномных промоторов с получением желательного продукта.
В четвертом варианте осуществления предоставлен рекомбинантный полинуклеотид вируса растения, как и в третьем варианте осуществления, за исключением того, что последовательность, кодирующая нативный белок оболочки, замещена на последовательность, кодирующую ненативный белок оболочки.
Для получения рекомбинантного вируса растений вирусные векторы инкапсулированы посредством белков оболочки, кодируемых рекомбинантным полинуклеотидом вируса растения. Рекомбинантный полинуклеотид вируса растения или рекомбинантный вирус растения применяют для инфицирования соответствующих растений-хозяев. Рекомбинантный полинуклеотид вируса растения способен к репликации в хозяине, системному распространению в хозяине, и транскрипции или экспрессии чужеродного гена(ов) (экзогенный полинуклеотид) в хозяине с продукцией желаемого белка в хозяине.
Способы инокуляции вирусов в растения можно найти в Foster and Taylor, eds. "Plant Virology Protocols: From Virus Isolation to Transgenic Resistance (Methods in Molecular Biology (Humana Pr), Vol 81)", Humana Press, 1998; Maramorosh and Koprowski, eds. "Methods in Virology" 7 vols, Academic Press, New York 1967-1984; Hill, S.A. "Methods in Plant Virology", Blackwell, Oxford, 1984; Walkey, D.G.A. "Applied Plant Virology", Wiley, New York, 1985 и Kado and Agrawa, eds. "Principles and Techniques in Plant Virology", Van Nostrand-Reinhold, New York.
Кроме указанного выше, полинуклеотид по настоящему изобретению также можно вводить в геном хлоропласта, таким образом обеспечивая экспрессию в хлоропласте. Способ введения последовательности экзогенного полинуклеотида в геном хлоропластов известен. Этот способ включает в себя следующие процедуры. Во-первых, клетки растений химически обрабатывают так, чтобы уменьшить количество хлоропластов на клетку приблизительно до одного. Затем в клетки посредством бомбардировки частицами вводят экзогенный полинуклеотид с целью введения в хлоропласты, по меньшей мере, одной экзогенной полинуклеотидной молекулы. Экзогенные полинуклеотиды выбирали так, чтобы они могли интегрироваться в геном хлоропласта посредством гомологичной рекомбинации, которую легко проводить посредством собственных ферментов хлоропласта. С этой целью экзогенный полинуклеотид, кроме представляющего интерес гена, включает в себя, по меньшей мере, один полинуклеотидный фрагмент, полученный из генома хлоропласта. Кроме того, экзогенный полинуклеотид включает в себя селективный маркер, позволяющий последующими процедурами отбора устанавливать, что все или по существу все копии геномов хлоропластов после такого отбора будут включать в себя экзогенный полинуклеотид. Дополнительные детали, относящиеся к этому способу, представлены в патентах США № 4945050 и 5693507, включенные сюда в качестве ссылки. Таким образом, полипептид может продуцироваться посредством системы экспрессии белков хлоропласта и интегрироваться во внутреннюю мембрану хлоропласта.
Так как толерантность к абиотическому стрессу у растений может вовлекать несколько генов, действующих аддитивно или синергично (см., например, в Quesda et al., Plant Physiol. 130:951-063, 2002), в настоящем изобретении рассматривают экспрессию множества экзогенных полинуклеотидов в одном растении-хозяине, для достижения, таким образом, наилучшей толерантности к абиотическому стрессу.
Экспрессию множества экзогенных полинуклеотидов в одном растении-хозяине можно осуществлять совместным введением в одну растительную клетку нескольких конструкций нуклеиновых кислот, где каждая включает в себя отличный экзогенный полинуклеотид. Затем трансформированную клетку можно регенерировать в зрелое растение с применением описанных здесь выше способов.
Альтернативно, экспрессию множества экзогенных полинуклеотидов в одном растении-хозяине можно осуществлять совместным введением в одну растительную клетку единственной конструкции нуклеиновой кислоты, содержащей несколько различных экзогенных полинуклеотидов. Такую конструкцию можно разрабатывать с единственной промоторной последовательностью, способной направлять транскрипцию полицистронной последовательности РНК, включающей в себя различные экзогенные полинуклеотидные последовательности. Для обеспечения совместной трансляции различных полипептидов, кодируемых полицистронными последовательностями РНК, полинуклеотидные последовательности можно связывать между собой посредством последовательности участка внутреннего связывания рибосом (IRES), облегчающей трансляцию полинуклеотидных последовательностей, располагающихся ниже последовательности IRES. В этом случае, транскрибируемая молекула полицистронной РНК, кодирующая различные полипептиды, описанные выше, будет транслироваться с кэпированного 5'-конца и двух внутренних последовательностей IRES молекулы полицистронной РНК, тем самым продуцируя в клетке все различные полипептиды. Альтернативно, конструкция может включать в себя несколько промоторных последовательностей, где каждая связана с отличной экзогенной полинуклеотидной последовательностью.
Растительную клетку, трансформированную конструкцией, включающей в себя несколько различных экзогенных полинуклеотидов, можно регенерировать в зрелое растение с применением способов, описанных здесь выше.
Альтернативно, экспрессию нескольких экзогенных полинуклеотидов в одном растении-хозяине можно осуществлять введением различных конструкций нуклеиновых кислот, включающих в себя различные экзогенные полинуклеотиды, в несколько растений. Затем регенерированные трансформированные растения можно перекрестно скрещивать и получать потомство, отобранное по наилучшей толерантности к абиотическому стрессу и/или характеристикам биомассы с применением обычных способов селекции растений.
Таким образом, настоящая заявка относится к способам применения новых генов толерантности к стрессу для безопасного и экономически эффективного повышения толерантности к абиотическому стрессу и/или биомассы в широком диапазоне хозяйственно-ценных растений.
Дополнительные цели, преимущества и новые особенности настоящего изобретения станут очевидны специалисту в данной области при изучении следующих неограничивающих примеров. Кроме того, каждый из различных вариантов осуществления и аспектов настоящего изобретения, как описано здесь выше и как заявлено в разделе формул изобретения ниже, находит экспериментальное подтверждение в следующих ниже примерах.
Примеры
Теперь приведены ссылки к следующим примерам, которые вместе с приведенными выше описаниями неограниченным образом иллюстрируют изобретение.
Как правило, применяемая здесь номенклатура и применяемые в настоящем изобретении лабораторные процедуры включают в себя молекулярные, биохимические, микробиологические способы и способы рекомбинантных ДНК. Такие способы подробно объяснены в литературе. См., например, "Molecular Cloning: A laboratory Manual" Sambrook et al., (1989); "Current Protocols in Molecular Biology" Volumes I-III Ausubel, R.M., ed. (1994); Ausubel et al., "Current Protocols in Molecular Biology", John Wiley and Sons, Baltimore, Maryland (1989); Perbal, "A Practical Guide to Molecular Cloning", John Wiley & Sons, New York (1988); Watson et al. ,"Recombinant DNA", Scientific American Books, New York; Birren et al. (eds) "Genome Analysis: A Laboratory Manual Series", Vols. 1-4, Cold Spring Harbor Laboratory Press, New York (1998); способы, указанные в патентах США №№ 4666828; 4683202; 4801531; 5192659 и 5272057; "Cell Biology: A Laboratory Handbook", Volumes 1-III Cellis, J.E., ed. (1994); "Current Protocols in Immunology" Volumes 1-III Coligan J.E., ed. (1994); Stites et al. (eds), "Basic and Clinical Immunology" (8th Edition), Appleton & Lange, Norwalk, CT (1994); Mishell and Shiigi (eds), "Selected Methods in Cellular Immunology", W.H. Freeman and Co., New York (1980); доступные иммуноанализы подробно описаны в патентной и научной литературе, см., например, патенты США №№ 3791932; 3839153; 3850752; 3850578; 3853987; 3867517; 3879262; 3901654; 3935074; 3984533; 3996345; 4034074; 4098876; 4879219; 5011771 и 5281521; "Oligonucleotide Synthesis" Gait, M.J., ed. (1984); "Nucleic Acid Hybridization" Hames, B.D., and Higgins S.J., eds. (1985); "Transcription and Translation" Hames, B.D., and Higgins S.J., eds. (1984); "Animal Cell Culture" Freshney, R.I., ed. (1986); "Immobilized Cells and Enzymes" IRL Press, (1986); "A Practical Guide to Molecular Cloning" Perbal, B., (1984) and "Methods in Enzymology" Vol. 1-317, Academic Press;" PCR Protocols: A Guide To Methods And Applications", Academic Press, San Diego, CA (1990); Marshak et al., "Strategies for Protein Purification and Characterization-A Laboratory Course Manual" CSHL Press (1996); все из которых включены сюда в качестве ссылки, как полностью приведенные здесь. Другие основные ссылки приведены на всем протяжении этого документа. Полагают, что приведенные здесь процедуры хорошо известны в данной области и предоставлены для удобства читателя.
Если иначе не определено, все используемые здесь технические и научные термины имеют то же значение, как обычно понимает один из обычных специалистов в данной области, к которой принадлежит это изобретение. Хотя способы и материалы сходны или эквивалентны описанным здесь способам и материалам, можно применять на практике или испытании настоящего изобретения, пригодные способы и материалы описаны ниже.
Пример 1
Идентификация предполагаемых генов толерантности к абиотическому стрессу
Предполагаемые гены толерантности к абиогенному стрессу (ABST) выделяли из баз данных NCBI экспрессируемых маркерных последовательностей (EST) и кДНК томатов. Последовательности баз данных группировали и собирали с применением программного обеспечения LEADS (Compugen). В результате группирования получили более чем 20000 кластеров, где каждый представляет собой отличный ген. Для каждого кластера составили описание профиля экспрессии, объединяя все ключевые слова, включенные в записи последовательностей, содержащих кластер. Затем кластеры скринировали для включения полинуклеотидов, происходящих из библиотек, идентифицированных при помощи ключевых слов, относящихся к ABST. Далее выбранные кластеры фильтровали для исключения любого кластера, включавшего более чем 100 EST на кластер и/или любой кластер, в котором менее чем 50% последовательностей помечают ABST-связанными ключевыми словами.
Гены растений ABST из уровня техники идентифицировали из публикаций Quesada et al. (Plant Physiol. 130:951-963, 2002); Apse and Blumwald (Curr Opin Biotechnol. 13:146-150, 2002); Rontein et al. (Metab Eng 4:49-56, 2002) и ссылок здесь. Известные гены растений ABST выравнивали со сгруппированными последовательностями нуклеиновых кислот томатов с применением программы BLAST. Последовательности томатов, имеющие значение e-шкалы (e-score value) ниже 5, идентифицировали как ортологи ABST. Дополнительные гены томатов ABST предыдущей области идентифицировали при помощи поиска сгруппированных записей последовательностей томатов с применением ключевых слов "root" (корень), "crown gall" (корневой рак), "nutrient" (питательное вещество) "callus" (наплыв), "disease" (заболевание),"pathogen" (патоген), "elicitor" (раздражитель) и "pseudomonas" (синегнойный).
Наконец, все идентифицированные ранее в данной области гены ABST сравнивали (при помощи совмещения последовательности с применением программного обеспечения BLAST) с итоговым набором кластеров генов томатов, выбранных, как описано выше. В результате, примерно 40% генов, выбранных в итоговом наборе кластеров, которые совпадали с генами ABST из предыдущей области, оказались известными генами ABST, указывая на то, что оставшиеся гены выбранных кластеров потенциально способны к повышению толерантности к абиотическому стрессу у растений.
Выбранные полинуклеотидные последовательности (таблица 1a), анализировали на присутствие ORF с применением Vector NTI suite (InforMax, U.K.) версии 6 (Hasting Software, Inc: www.generunner.com/). Идентифицированные в каждой из данных полинуклеотидных последовательностей ORF сравнивали с базой данных последовательностей Genbank с применением Blast(www.ncbi.nlm.nih.gov/BLAST/): для идентификации стартового кодона ATG картировали ORF, показывающие наивысшую степень гомологии с последовательностью или последовательностями GenBank. Затем положение стартового кодона ATG данной ORF сравнивали с положением стартового кодона ATG идентифицированной полинуклеотидной последовательности для подтверждения того, что каждая из описанных здесь восемнадцати последовательностей включает в себя полноразмерную ORF и стартовый кодон ATG (таким образом, определяемые как "предполагаемый ген ABST").
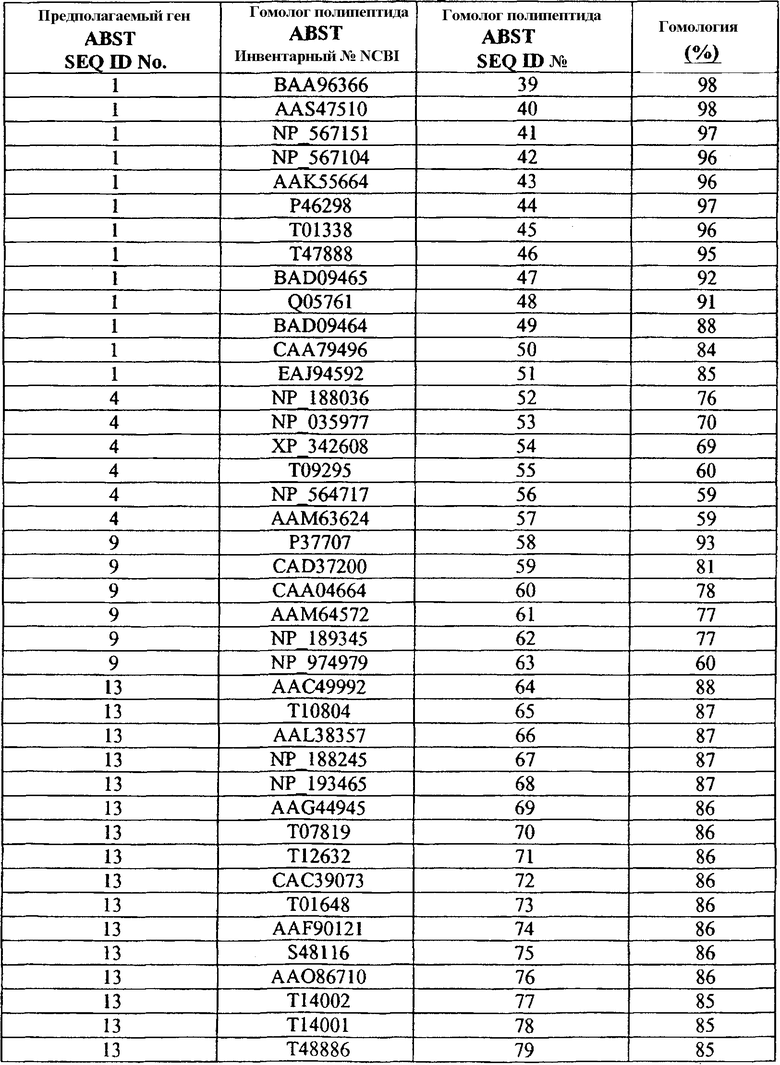
С применением программного обеспечения BLAST из баз данных NCBI идентифицировали гомологи полипептида ABST (Таблица 1b).
Таблица 1b
Гомологи ABST
Пример 2
Выделение предполагаемых генов толерантности к ABS
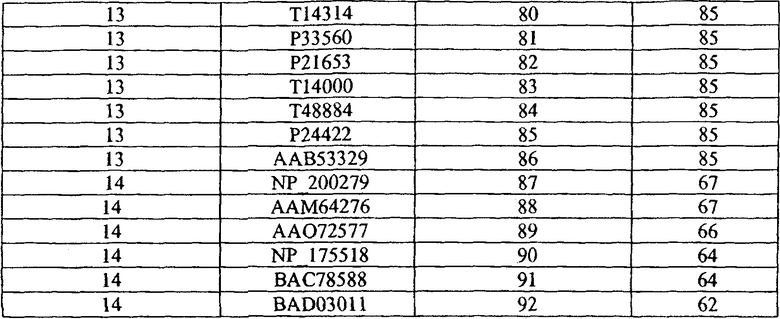
РНК выделяли из корня и листовой ткани 4-недельного томата с применением Tri Reagent (Molecular Research Center, Inc), по протоколу, предоставленному производителем (www.mrcgene.com/tri.htm). Из выделенной мРНК с применением фермента обратной транскриптазы (RT) M-MuLV (Roche) и ДНК-праймера T16NN получали комплементарные молекулы ДНК по инструкциям производителя. Последовательности кДНК, указанные в SEQ ID №№ 1, 4, 8-9 и 12-14, амплифицировали посредством GWH с применением праймеров, описанных в таблице 2, ниже, с применением фермента ДНК-полимеразы с коррекцией считывания PFU (Promega - www.promega.com/pnotes/68/7381_07/7381_07.html), по предоставленному производителем протоколу. Для облегчения клонирования предполагаемых генов толерантности к ABS в бинарные векторы к 5'-концу каждого праймера добавляли дополнительные участки рестрикционных эндонуклеаз.
Таблица 2
ПЦР-праймеры, применяемые для амплификации генов толерантности к ABS tolerance (ABST)
Пример 3
Клонирование предполагаемых генов ABST
Полученные продукты ПЦР с тупыми концами очищали с применением набора PCR Purification Kit (Qiagen, Germany), расщепляли соответствующими рестрикционными ферментами (Roche), а затем вставляли в бинарный плазмидный вектор pPI. Плазмиду pPI конструировали посредством вставки синтетической поли-(A)-сигнальной последовательности, происходящей из плазмидного основного вектора pGL3 (Promega, Acc No U47295; bp 4658-4811), в рестрикционный участок HindIII бинарного вектора pBI1013 (Clontech, Acc. No.U12640).
Полученную плазмиду pPI расщепляли рестрикционными ферментами (BamHI и SacI; MBI Fermentas) и очищали с применением набора PCR Purification Kit (Qiagen, Germany). Затем открытую конструкцию pPI лигировали с каждым из семи описанных выше продуктов ПЦР. Лигирование осуществляли с применением лигирующей смеси, содержащий фермент ДНК-лигазу T4 (Roche) и проводили по инструкциям производителя.
Конструкции pPI, несущие предполагаемые гены ABST, посредством электропорации с применением электропоратора MicroPulser electroporator (Biorad), 0,2 см кювет (Biorad) и программы электропорациии EC-2 (Biorad) вносили в компетентные клетки DH5 E. coli. Обработанные клетки культивировали в жидкой среде LB при 37°C в течение 1 часа, затем наносили на LB-агар, дополненный канамицином (50 мг/л; Sigma) и инкубировали при 37°C в течение 16 часов. Колонии, развивавшиеся на селективной среде, анализировали посредством ПЦР с праймерами, указанными в SEQ ID №№ 35-36, разработанными так, чтобы охватывать вставленную в плазмиду pPI последовательность. Полученные при ПЦР продукты разделяли на 1,5% агарозных гелях, а фрагменты ДНК с предсказанным размером выделяли и проводили секвенировании с применением секвенатора ABI 377 (Amersham Biosciences Inc) для подтверждения того, что в клетки E. coli корректные последовательности ДНК введены правильно.
Пример 4
Получение бинарных векторов, содержащих предсказанные гены ABST и функционально связанные с ними промоторы растений
Получение бинарных векторов, содержащих промотор 35S вируса мозаики цветной капусты: промоторную последовательность вируса мозаики цветной капусты 35S (приведенную в SEQ ID № 19) вставляли выше предполагаемого гена ABST в каждую из описанных выше конструкций pPI. Промотор выделяли из плазмиды pBI121 (Clontech, инвентарный № AF485783) с применением рестрикционных эндонуклеаз HindIII и BamHI (Roche). Выделенный промотор лигировали в конструкции pPI, расщепленные теми же ферментами. Всего получали семь конструкций pPI, где каждая содержала промотор CaMV 35S, расположенный выше предполагаемого гена ABST с последовательностью, приведенной в SEQ ID №№ 1, 4, 8, 9, 12, 13 или 14.
Получение содержащих промотор At6669 бинарных векторов:
в каждую из описанных выше бинарных конструкций pPI выше предполагаемого гена ABST вставляли последовательность промотора At6669 (приведенную в SEQ ID № 20). Промотор выделяли из геномной ДНК Arabidopsis thaliana вар Col0 посредством амплификации ПЦР с применением праймеров, приведенных в SEQ ID №№ 37-38. Продукт ПЦР очищали (Qiagen, Germany) и расщепляли рестрикционными эндонуклеазами HindIII и BamHI (Roche). Полученную промоторную последовательность вводили в открытую бинарную конструкцию, расщепленную теми же ферментами. Всего получали семь конструкций pPI, где каждая содержала промотор At6669, расположенный выше предполагаемого гена ABST с последовательностью, приведенной в SEQ ID № 1, 4, 8, 9, 12, 13 или 14.
Пример 5
Подтверждение активности промотора At6669 в трансгенных Arabidopsis thaliana
Тестировали способность промотора At6669 регулировать транскрипцию генов, перенесенных вектором pPI в растения. Соответственно промотор At6669 вводили в бинарный вектор pPI выше репортерного гена люциферазы. Бинарный вектор вводили в растения Arabidopsis thaliana с применением способа, описанного в примере 6 ниже. Зрелые трансформированные растения T2 Arabidopsis анализировали на биосвечение в темной комнате с применением камеры детекции света сверхнизкой интенсивности (Princeton Instruments Inc., USA) с применением способа, описанного Meissner et al. (Plant J. 22:265, 2000). В цветке и корневых образовательных тканях трансформированных растений наблюдали свечение, указывающее на положительную активность люциферазы (фиг.2).
Пример 6
Трансформация клеток Agrobacterium tumefaciens бинарными векторами, несущими предполагаемые гены ABST
Каждый из описанных выше в Примере 4 бинарных векторов применяли для трансформации клеток Agrobacterium. В качестве отрицательного контроля применяли две дополнительные бинарные конструкции с репортерным геном люциферазы, замещающим ген ABST (расположенный ниже промотора 35S или At6669).
Посредством электропорации вводили бинарные векторы в компетентные клетки GV301 или LB4404 Agrobacterium tumefaciens (примерно 109 клеток/мл). Электропорацию проводили посредством применения электропоратора MicroPulser (Biorad), 0,2 см кюветы (Biorad) и программы электропорации EC-2 (Biorad). Обработанные клетки культивировали в жидкой среде LB при 28°C в течение 3 часов, затем высевали на чашки на агар LB, дополненный гентамицином (50 мг/л для штаммов GV301 Agrobacterium) или стрептомицином (300 мг/л для штамма LB4404 Agrobacterium) и канамицином (50 мг/л) при 28°C в течение 48 часов. Выросшие на селективной среде колонии Agrobacterium анализировали посредством ПЦР с применением приведенных в SEQ ID №№ 35-36 праймеров, разработанных так, чтобы охватывать вставленную последовательность в плазмиду pPI. Полученные продукты ПЦР выделяли и секвенировали так, как описано в примере 4 выше, для подтверждения того, что корректные последовательности ABST введены в клетки Agrobacterium правильно.
Пример 7
Трансформация растений Arabidopsis thaliana предполагаемыми генами ABST
Растения Arabidopsis thaliana Columbia (растения T0) трансформировали с применением процедуры Floral Dip, описанной у Clough and Bent (10) и у Desfeux et al. (11), с небольшими модификациями. В кратком изложении, растения T0 высевали в 250 мл горшки, заполненные основанной на торфе средой для роста. Горшки покрывали алюминиевой фольгой и пластиковым колпаком, выдерживали в течение 2-4 суток при 4°C, затем снимали покрытие и инкубировали в камере для роста при 18-24°C с циклами свет/темнота 16/8 часов. Растения T0 становились готовыми для трансформации за шесть суток до цветения.
Одиночные колонии Agrobacterium, несущие бинарные конструкции, полученные, как описано в примере 6 выше, культивировали в среде LB, дополненной канамицином (50 мг/л) и гентамицином (50 мг/л). Культуры инкубировали в течение 48 часов при 8°C с энергичным перемешиванием, а затем центрифугировали при 4000 об/мин в течение 5 минут. Содержащие клетки Agrobacterium осадки ресуспендировали в среде для трансформации, содержащей Murashig-Skoog (Duchefa) половинной концентрации (2,15 г/л); 0,044 мкМ бензиламинопурина; 112 мкг/л витаминов B5 Gambourg (Sigma); 5% сахарозы и 0,2 мл/л L-77 Silwet (OSI Specialists, CT) в дважды дистиллированной воде при pH 5,7.
Трансформацию растений T0 проводили опрокидыванием каждого растения в суспензию Agrobacterium так, что указанная выше наземная растительная ткань являлась погруженной в течение 3-5 секунд. Каждое инокулированное растение T0 немедленно помещали в пластиковый лоток, покрывали прозрачным пластиковым колпаком для сохранения влажности, а затем в течение 18 часов держали в темноте при комнатной температуре для облегчения заражения и трансформации. Затем с трансформированных (трансгенных) растений снимали покрытие и переносили в теплицу для восстановления и созревания. Трансгенные растения T0 растили в теплице в течение 3-5 недель, пока стручки не становились коричневыми и сухими. Семена собирали из растений и до посева хранили при комнатной температуре.
Для получения несущих гены трансгенных растений T1 и T2 семена собирали из трансгенных растений T0, где поверхность стерилизовали вымачиванием в 70% этаноле в течение 1 минуты, с последующим вымачиванием в 5% гипохлориде натрия и 0,05% тритоне в течение 5 минут. Затем семена со стерилизованной поверхностью тщательно отмывали в стерильной дистиллированной воде, помещали в культуральные планшеты, содержащие Murashig-Skoog (Duchefa) половинной концентрации, 2% сахарозу; 0,8% растительный агар; 50 мМ канамицин и 200 мМ карбенициллин (Duchefa). Культуральные планшеты инкубировали в течение 48 часов при 4°C, затем переносили в теплицу на дополнительную неделю инкубации при 25°C. Зараженные вирусом растения Arabidopsis T1 переносили в свежие культуральные планшеты для дополнительной недели инкубации. После инкубации растения T1 удаляли из культуральных планшетов и растили в среде для роста, содержащейся в 250 мл горшках. Трансгенным растениям позволяли расти в теплице до вызревания. Из растений T1 собирали семена, растения культивировали и растили до созревания в виде растений T2 в тех же условиях, как применяли для культивирования и роста растений T1.
Пример 8
Оценка роста растений, культивируемых при состояниях абиотического стресса
Методы:
Полученные, как описано выше, трансгенные растения T1 или T2 индивидуально пересаживали в горшки, содержащие среду для роста из торфа и вермикулита (соотношение объемов 3:2, соответственно). Горшки на период 24 часа укрывали для закаливания, затем помещали в теплицу в полностью случайном порядке и орошали водопроводной водой (получаемой каждые 3-5 суток со дна горшков) в течение семи суток. Затем половину растений орошали солевым раствором (100 мМ NaCl и 5 мМ CaCl2) для индукции солевого стресса (стрессовые условия). Другую половину растений продолжали орошать водопроводной водой (нормальные условия). Все растения растили в теплице при 100% отн. влажности в течение 28 суток, а затем собирали (указанные выше наземные ткани) и взвешивали (непосредственно после высушивания в печи при 50°C в течение 24 часов).
Результаты:
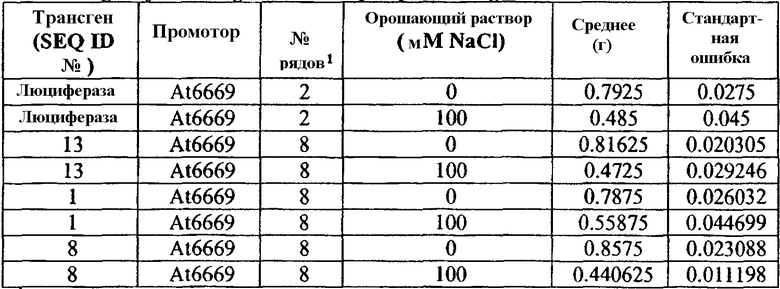
Значимых различий сырой массы растений между растениями T1, трансформированными 3 различными генами ABST, и растениями, трансформированными репортерным геном люциферазы, растущими в нормальных или стрессовых условиях (фиг.3 и таблица 3, ниже), не наблюдали. Однако растения T1, трансформированные SEQ ID № 1, расположенной под контролем регуляторного промотора At6669, сохраняли 71% их сырой массы при воздействии стрессовых условий, тогда как контрольные растения (несущие ген люциферазы, помещенный под регуляторный контроль промотора AT6669) в тех же условиях стресса сохраняли только 61% их сырой массы.
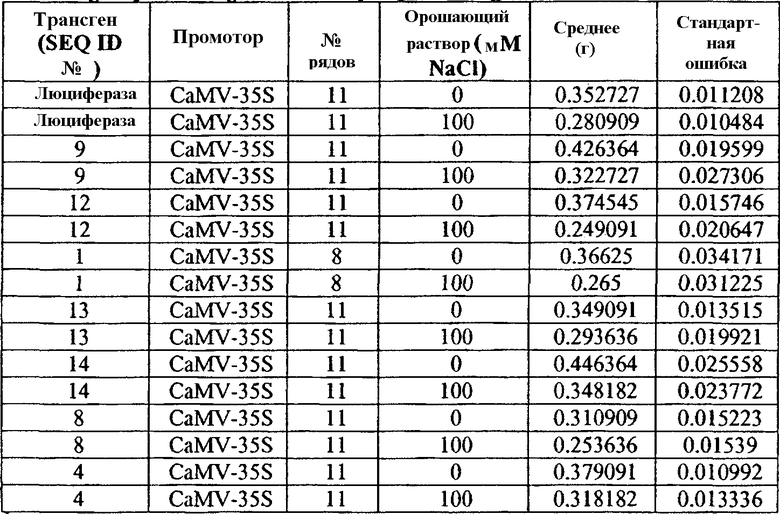
Таблица 3
Сырая масса трансгенных растений Arabidopsis T1 орошаемых водой и солевым раствором
1№ рядов представляет собой количество измеренных растений с независимыми событиями трансформации. Для каждого трансгена применяли 3-5 независимых событий трансформации с 1-3 растениями на одну трансформацию.
Растения T2, трансформированные SEQ ID №№ 7 или 14, расположенными под контролем регуляторного промотора 35S, накапливали значимо большую биомассу, чем контрольные растения, вне зависимости от условий роста. Как показано на фиг.4A и в таблице 4 ниже, средняя сырая масса растений, трансформированных SEQ ID №№ 7 и 14, растущих в стрессовых условиях, составляла 15% и 24%, соответственно, выше, чем средняя сырая масса контрольных растений, растущих при сходных нормальных условиях.
Сходное явление наблюдали для растений T2, трансформированных SEQ ID № 4, расположенной под контролем регуляторного промотора 35S. Таким образом, как показано на фиг.4A и таблице 4 ниже, средняя сырая масса растений, трансформированных SEQ ID № 4 составляла 14% и 7% и была выше, чем средняя сырая масса контрольных растений, растущих в условиях стресса и нормальных условиях, соответственно. Подобным образом растения T2, трансформированные SEQ ID № 4, расположенной под контролем регуляторного промотора At6669, продемонстрировали биомассу на 1,3 и 5% выше, чем контрольные растения, растущие в условиях стресса и в нормальных условиях, соответственно. Однако данные различия не являлись статистически значимыми в экспериментальных условиях.
Таблица 4
Сырая масса трансгенных растений Arabidopsis T2 орошаемых водой и солевым раствором
1№ рядов представляет собой количество измеренных растений с независимыми событиями трансформации. Для каждого трансгена применяли 3-5 независимых событий трансформации с 1-3 растениями на одну трансформацию.
Растения T2, трансформированные SEQ ID №№ 1 и 13, расположенными под контролем регуляторного промотора At6669, и растущие в стрессовых условиях, демонстрировали значимо большую биомассу, чем контрольные растения, растущие в тех же стрессовых условиях. Средняя сырая масса растений T2, трансформированных SEQ ID №№ 1 и 13, расположенных под контролем регуляторного промотора At6669, и растущих в стрессовых условиях, составляла 37% и 21%, соответственно, и являлась выше, чем средняя сырая масса контрольных растений, растущих в сходных стрессовых условиях (фиг.4B и Таблица 5 ниже). Значимого увеличения биомассы по сравнению с контролем, когда данные трансгенные растения (несущие ID №№ 1 и 13, под контролем промотора At6669) росли в нормальных условиях, не наблюдали.
Таблица 5
Сырая масса трансгенных растений Arabidopsis T2 орошаемых водой и солевым раствором
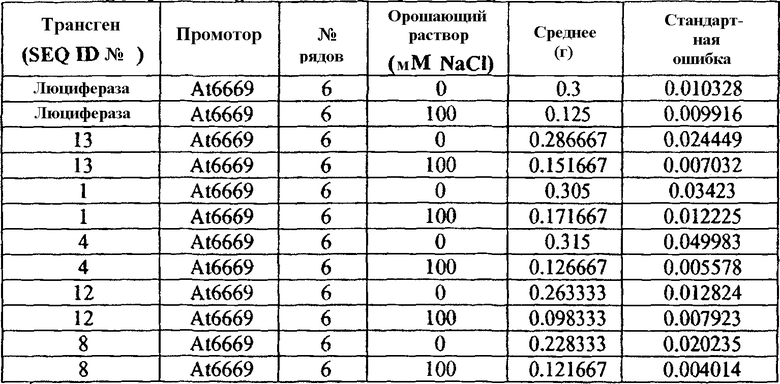
1№ рядов представляет собой количество измеренных растений с независимыми событиями трансформации. Для каждого трансгена применяли 3-5 независимых событий трансформации с 1-3 растениями на одну трансформацию.
Результаты иллюстрируют то, что выделенные предполагаемые гены ABST, приведенные в SEQ ID №№ 1 и 13, способны увеличивать толерантность растений к абиотическому стрессу, такому как стресс засоленности. Кроме того, выделенные предполагаемые гены ABST, приведенные в SEQ ID №№ 7, 14 (а возможно также и в 4), способны существенно увеличивать биомассу растений, растущих в условиях стресса, а также в нормальных условиях.
Следовательно, результаты четко указывают на то, что описанные здесь предполагаемые гены устойчивости к абиотическому стрессу можно легко выделять и применять для значительного увеличения толерантности к абиотическому стрессу и/или биомассы у растений.
Понятно, что определенные отличительные признаки изобретения, которые для ясности описаны в контексте отдельных вариантов осуществления, можно предоставить в сочетании в одном варианте осуществления. Наоборот, различные отличительные признаки изобретения, которые для краткости описаны в контексте одного варианта осуществления, можно также предоставить отдельно или в любой пригодной субкомбинации.
Хотя изобретение описано в связи с его конкретными вариантами осуществления, очевидно, что специалистам в данной области будут видны многие альтернативы, модификации и вариации. Таким образом, оно предназначено охватывать все такие альтернативы, модификации и вариации, которые находятся в пределах сущности и широкого объема прилагаемой формулы изобретения. Все публикации, патенты, патентные заявки и последовательности, идентифицированные их инвентарными номерами, приведенные в данном описании, полностью включены в качестве ссылки в настоящее описание до той же степени, как если бы являлось указанным, что каждая отдельная публикация, каждый отдельный патент или каждая отдельная патентная заявка или последовательность, идентифицированная своим инвентарным номером, конкретно и отдельно включены сюда в качестве ссылки. Кроме того, цитирование или указание любой ссылки в данной заявке не следует рассматривать как допущение того, что такая ссылка доступна как известный по отношению к настоящему изобретению уровень техники.





























































| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ МОДУЛЯЦИИ ПРОВОДИМОСТИ УСТЬИЦА И РАСТИТЕЛЬНЫЕ ЭКСПРЕССИОННЫЕ КОНСТРУКЦИИ ДЛЯ ИХ ОСУЩЕСТВЛЕНИЯ | 2012 |
|
RU2664461C2 |
| УСТОЙЧИВЫЕ К БАКТЕРИЯМ ТРАНСГЕННЫЕ РАСТЕНИЯ | 2012 |
|
RU2640246C2 |
| ТОЛЕРАНТНОЕ К СТРЕССУ ТРАНСГЕННОЕ РАСТЕНИЕ ПШЕНИЦЫ | 2005 |
|
RU2376377C2 |
| УСТОЙЧИВЫЕ К ГЕРБИЦИДАМ РАСТЕНИЯ ПОДСОЛНЕЧНИКА, ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ УСТОЙЧИВЫЕ К ГЕРБИЦИДАМ БЕЛКИ БОЛЬШОЙ СУБЪЕДИНИЦЫ АЦЕТОГИДРОКСИАЦИДСИНТАЗЫ, И СПОСОБЫ ПРИМЕНЕНИЯ | 2005 |
|
RU2415566C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2639275C2 |
| СПОСОБЫ УВЕЛИЧЕНИЯ УРОЖАЯ И СТРЕССОУСТОЙЧИВОСТИ У РАСТЕНИЯ | 2012 |
|
RU2632569C2 |
| УСТОЙЧИВЫЕ К ГЕРБИЦИДАМ РАСТЕНИЯ ПОДСОЛНЕЧНИКА | 2009 |
|
RU2551781C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| СПОСОБЫ ТРАНСФОРМАЦИИ РАСТЕНИЙ ДЛЯ ЭКСПРЕССИИ ДЕЛЬТА-ЭНДОТОКСИНОВ BACILLUS THURINGIENSIS | 1999 |
|
RU2234531C2 |
| БОЛЕЗНЕУСТОЙЧИВЫЕ РАСТЕНИЯ | 2006 |
|
RU2393227C2 |


Клетку растения трансформируют экзогенным полинуклеотидом, экспрессия которого придает клетке повышенную толерантность к абиотическому стрессу, а затем культивируют из нее зрелое растение. Трансформацию осуществляют посредством введения в клетку конструкции, содержащей конститутивный или индуцируемый абиотическим стрессом промотор. Либо растение инфицируют авирулентным вирусом, включающим в себя упомянутый экзогенный полинуклеотид, экспрессия которого в растении придает ему устойчивость к абиотическому стрессу, такому как засоленность почвы, недостаток воды, низкая или высокая температура и др. 5 н. и 32 з.п. ф-лы, 5 ил., 6 табл.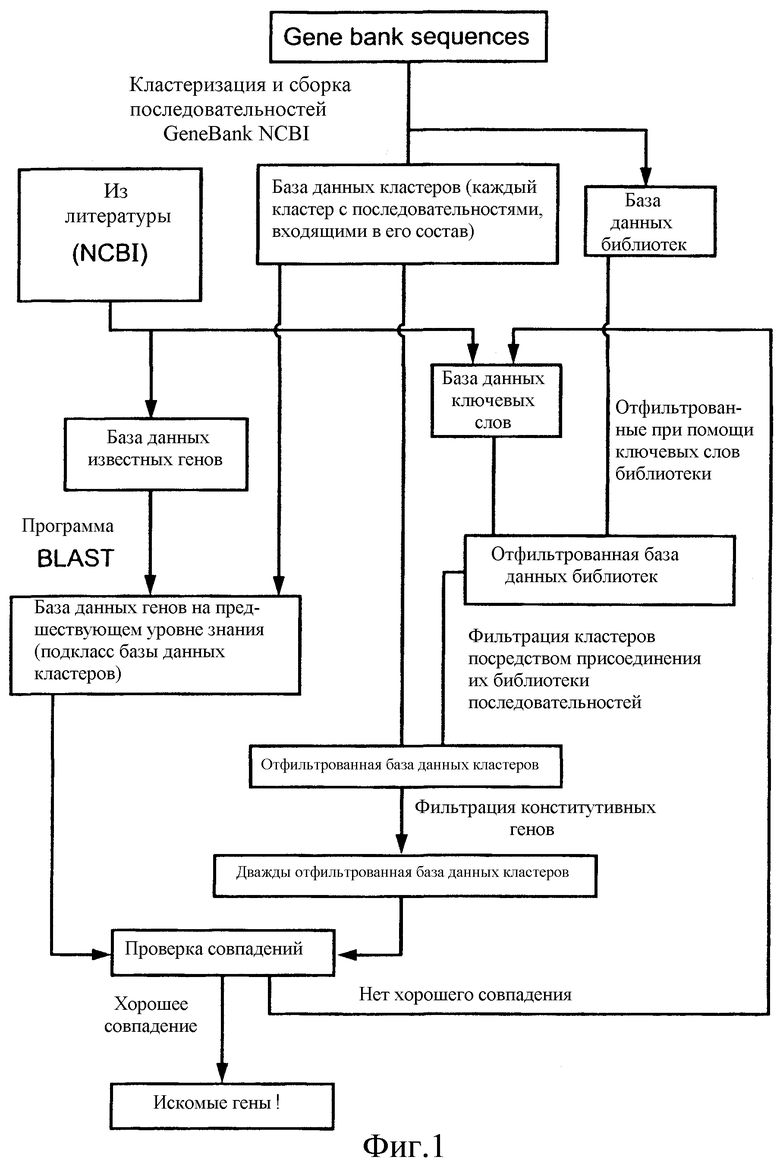
(a) трансформации клетки указанного растения указанным экзогенным полинуклеотидом;
(b) получения из указанной клетки зрелого растения; и
(c) культивирования указанного зрелого растения в условиях, пригодных для экспрессии указанного экзогенного полинуклеотида в указанном зрелом растении.
(a) трансформации клетки указанного растения указанным экзогенным полинуклеотидом;
(b) получения из указанной клетки зрелого растения; и
(с) культивирования указанного зрелого растения в условиях, пригодных для экспрессии указанного экзогенного полинуклеотида в указанном зрелом растении.
| CURRENT OPINION IN BIOTECHNOLOGY, v.12, iss | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| JP 2001186879, 10.07.2001 | |||
| JP 2001054385, 27.02.2001 | |||
| СПОСОБ СОЗДАНИЯ СОЛЕУСТОЙЧИВЫХ ФОРМ МЯГКОЙ ПШЕНИЦЫ | 1998 |
|
RU2138156C1 |

