ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к трансгенным растениям и к растительным клеткам, которые включают ген, кодирующий белок NRC1 (NB-LRR, необходимый для HR-ассоциированной гибели клеток 1), интегрированный в геном, и к способам получения таких растений и растительных клеток. В особенности, в настоящем изобретении рассматриваются растения из семейства Solanaceae и части таких растений (семена, плоды, листья и т.п.) с повышенной устойчивостью к заболеваниям. Кроме того, настоящее изобретение относится к выделенным молекулам нуклеиновой кислоты, которые кодируют белки NRC1 согласно настоящему изобретению, к включающим их векторам, а также к выделенным белкам NRC1. Кроме того, в изобретении рассматриваются растительные клетки и растения, включающие одну или несколько мутаций в эндогенной аллели NRC1, где указанные одна или несколько мутаций придают растениям и растительным клеткам повышенную устойчивость к заболеваниям.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Активная система защиты растений, запускаемая при распознавании авирулентного фактора патогена, опосредованная геном устойчивости, соответствует модели ген-на-ген (Dangl and Jones, 2001, Nature 411, 826-833). К настоящему времени были клонированы несколько генов устойчивости из растений (R гены), и на основании структуры белков, которые они кодируют, указанные гены были разделены на несколько групп (Hammond-Kosack and Jones, 1997, Annu. Rev. Plant Physiol. Plant Molec. Biol. 48, 575-607). Большинство R генов кодирует цитоплазматические белки NB-LRR, содержащие сайт связывания нуклеотидов (NB) и обогащенные лейцином повторы (LRR). Данная группа состоит из генов, кодирующих белки CC-NB-LRR, содержащие суперскрученные домены, и генов, которые кодируют белки, содержащие домен, аналогичный рецепторам Toll и интерлейкина (IL) у млекопитающих, так называемые белки TIR-NB-LRR (Hammond-Kosack and Jones, 1997, выше).
Использование таких специфических генов устойчивости в стратегиях разведения с целью достижения длительной устойчивости встречает множество проблем, поскольку патогены легко уходят от распознавания за счет мутаций в своих факторов авирулентности, что препятствует индукции активной системы защиты (Westerink et al., 2004, Mol. Microbiol. 54, 533-545). Аналогично, предполагается существование среди этих белков общих путей, определяющих резистентность (R белки) (Shirasu and Schulze-Lefert, 2000, Plant Mol. Biol. 44, 371-385). В этой связи, идентификация дополнительных генов, необходимых для достижения резистентности, не только дает информацию, как функционируют такие сигнальные пути, но также может позволить идентифицировать гены, которые играют более глобальную роль в резистентности. Так, например, в случае индуцированного вирусом молчания гена (VIGS) в Nicotiana benthamiana было показано, что SGT1 вовлекается в пути реализации множественной устойчивости, такие как N-, Rx- и Pto-опосредованные HR и резистентность, и Cf-4- и Cf-9-опосредованные HR (Peart et al., 2002, Proc. Natl. Acad. Sci. USA 99, 10865-10869; Zhang et al., 2004, Plant J. 40, 213-224). SGT1 представляет собой компонент, функционирующий вместе с SKP1, в виде части SCF E3-лигазного комплекса, который вовлекается в убихитинирование белков, модификацию, запускающую их деградацию (Schwechheimer и Schwager, 2004, Plant Cell Reports 23, 353-364). Была высказана гипотеза, что молчание эссенциального гена в данной системе деградации белков препятствует проведению процесса убихитинирования, что ингибирует деградацию отрицательных регуляторов, которые необходимы для активации защитных механизмов (Azevedo et al., 2002, Science 295, 2073-2076).
Некоторые механизмы резистентности связаны с активацией MAPK (митоген-активированные протеинкиназы) (Zhang and Klessig, 2001, Trends Plant Sci. 6, 520-527; Pedley and Martin, 2005, Curr. Opin. Plant Biol. 8, 541-547). В Cf-9-содержащих растениях табака и клеточных культурах, которые были провоцированы Avr9, активируются NtWIPK (индуцированная повреждением протеинкиназа) и NtSIPK (индуцированная салициловой кислотой протеинкиназа) (Romeis et al., 1999, Plant Cell 11, 273-287). VIGS применительно к NtCDPK (кальций-зависимая протеинкиназа) в N. benthamiana ингибируют Cf-9/Avr9- и Cf-4/Avr4-зависимые HR (Romeis et al., 2001, EMBO J. 20, 5556-5567), а VIGS для LeACIK1 (Avr/Cf-индуцированная каназа 1) в растениях томатов приводит к снижению резистентности к C. fulvum (Rowland et al., 2005, Plant Cell 17, 295-310). Активация киназ при реализации защитных механизмов и сниженная резистентность при «выключении» соответствующих кодирующих генов подтверждают важность их функций в активации системы защиты.
В рамках двунаправленного варианта исследования использовали 21 ген применительно к VIGS в растениях томатов, в отношении которых было известно, что они вовлекаются в сигнальный процесс, связанный с защитой, и было показано, что девять из них вовлекаются в Pto-опосредованную резистентность. Среди них были выявлены два гена, кодирующие MAPKK (LeMEK1 и LeMEK2), и два гена, кодирующие MAPK (LeNTF6 и LeWIPK) (Ekengren et al., 2003, Plant J. 36, 905-917). В рамках другого исследования были клонированы свыше 2400 кДНК из нормализованной библиотеки кДНК N. benthamiana в Х-векторе вируса картофеля и далее они были использованы для VIGS в N. benthamiana. Примерно 3% кДНК оказывали воздействие на Pto-зависимую HR при молчании. В их числе был идентифицирован MAPKKKα в качестве позитивного регулятора резистентности и заболевания (Del Pozo et al., 2004, EMBO J. 23, 3072-3082).
Лу с соавт. (Lu et al. 2003, EMBO J. 22, 5690-5699) получили VIGS с использованием 4992 кДНК из нормализованной библиотеки кДНК N. benthamiana при клонировании в PVX векторе. В числе этих кДНК 79 (1,6%) соответствовали генам, необходимым для Pto-опосредованного HR, при этом молчание только шести из них также нарушало Pto-опосредованную резистентность против Pseudomonas syringae. Состояние VIGS с использованием кДНК, соответствующей HSP90, устраняло не только Pto-опосредованный HR, но также Pto-, Rx- и N-опосредованную резистентность, что указывало на то, что HSP90 необходим для функционирования путей реализации множественной резистентности к заболеваниям. Тот же набор кДНК был использован для достижения VIGS в N-трансгенных N. benthamiana, после чего в растения был инокулирован штамм TMV с GFP-меткой. Резистентность против TMV сильнее всего подавлялась в случае достижения молчания с использованием фрагмента кДНК, полученного из гена, кодирующего CC-NB-LRR, известного как NRG1 (ген 1 требования N) (Peart et al., 2005, Curr. Biol. 15, 968-973). Было показано, что NRG1 специфически необходим для функционирования N гена, то есть белки CC-NB-LRR не только действуют в качестве белков резистентности, вовлекаемых в процесс распознавания факторов авирулентности, но также участвуют в сигнальном пути, инициируемом TIR-NB-LRR белком N, который в итоге приводит к достижению резистентности (Peart et al., 2005, см. выше). Таким образом, несмотря на то, что функции белка NRG1 из растений табака по пути считывания информации в каскаде сигнальной системы защиты растений инициируются белком резистентности, этот путь сопряжен с недостатками, связанными с тем, что он специфически вовлекается в N-опосредованную резистентность против мозаичного вируса табака (TMV) и данный белок не является кофактором резистентности к заболеванию (Rx- и Pto-опосредованная резистентность против PVX и Pseudomonas syringae не затрагиваются при молчании NRG1), что может быть неприемлемо для целей достижения общей системы резистентности к патогенам в культурах, таких как томаты.
Несмотря на возрастающий поток информации о путях достижения болезнеустойчивости растений все еще имеется потребность в идентификации генов и белков, которые могут использоваться для создания растений, обладающих пролонгированной системой резистентности к заболеваниям широкого спектра действия. Целью настоящего изобретения является получение таких нуклеиновых кислот, белков и способов создания растений, в особенности растений, относящихся к семейству Solanaceae, с повышенной резистентностью к заболеваниям.
ОСНОВНЫЕ ОПРЕДЕЛЕНИЯ
Термин "HR" относится к гиперчувствительному ответу, то есть к локальной гибели клеток растений, которая проявляется либо как повреждения, видимые под микроскопом (например, описанные Rivas и Thomas, 2005, Ann Rev Phytopath 43: 395-436), и/или в виде макроскопических повреждений. Гибель гиперчувствительных клеток обычно ассоциирована с другими ответами растений, такими как образование реакционно-активных видов кислорода и активация генов, связанных с системой защиты в клетках, окружающих HR повреждения.
Термин «патогены растений» относится к биотическим агентам, которые способны вызывать заболевание у растений, таким как патогенные грибы, бактерии, вирусы, оомицеты, микоплазменные организмы, нематоды, белокрылки, тли и т.п. В основном, все штаммы, расы или патогенные варианты того вида патогена, который способен вызывать заболевание ткани организма-хозяина, включаются в область настоящего изобретения.
Термин «биотрофные патогены растений» или «биотрофы» относится к патогену, который сохраняет клетки растений организма-хозяина в живом состоянии и существует на живых клетках, нужных для роста и колонизации в ткани.
Термин «хемибиотрофный патоген растений» или «хемибиотроф» относится к растительному патогену, который поддерживают клетки организма-хозяина в живом состоянии в течение по меньшей мере части его жизненного цикла.
Термин «некробиотрофный патоген растения» относится к растительному патогену, который активно уничтожает растительные клетки при колонизации в ткани за счет образования токсичных ферментов, белков или метаболитов, которые уничтожают клетки хозяйского организма.
Термин «элиситор-независимый HR» относится к реакции гиперчувствительности, которая развивается без патогена или патогенного элиситора (например, грибного белка Avr).
В случае растений, экспрессирующих NRC1 белок согласно настоящему изобретению (например, конститутивно активный NRC1 белок), следует также провести разграничение между «конститутивным» HR, в случае которого имеется в виду развитие HR-повреждений в отсутствие патогенов или белков патогенного элиситора, и «индуцированным» HR, где подразумевается развитие HR-повреждений после воздействия индуцирующего стимула (например, после индукции промотора, который запускает экспрессию последовательности нуклеиновой кислоты, кодирующей NRC1 белок или его вариант).
Термин «Solanaceae» относится к роду, виду и сортам растений, принадлежащих к семейству Solanaceae. Они включают виды, относящиеся к роду Solanum (включая Solanum lycopersicum, который известен также как Lycopersicon esculentum), Nicotiana, Capsicum, Petunia, и к другим родам.
Термин «резистентность к заболеванию» в контексте настоящего описания относится к различной степени резистентности или толерантности растения к заболеванию, включая умеренную резистентность и повышенную резистентность или полную резистентность к одному или нескольким патогенам. Указанная резистентность может быть определена и необязательно количественно выражена при сравнении с симптомами, вызванными патогеном (такими как чистота и/или размер HR повреждений, поражений грибным мицелием и т.п.), в сравнении с соответствующими симптомами, выявленными у чувствительных контрольных растений, растущих в идентичных условиях воздействия патогенов. Такие биотесты на заболеваемость могут быть проведены с использованием известных способов. Резистентность к заболеванию может быть измерена опосредованно как повышенный уровень резистентности растений в сравнении с чувствительными растениями, растущими в условиях давления болезнетворного фактора.
Термин «повышенная резистентность к заболеванию» относится к любому статистически значимому повышению болезнеустойчивости растения или растительной ткани в сравнении с соответствующим контролем. В контексте настоящего описания данный термин относится как к качественному (например, переход от чувствительного варианта к резистентному растению), так и к количественному повышению. Также рассматривается снижение частоты заболеваемости (процент инфицированных растений) и/или тяжести заболевания. Предпочтительно, растение, обладающее повышенной болезнеустойчивостью по меньшей мере к одному патогену, представляет собой растение, характеризующееся повышенным по меньшей мере на 1%, 2%, 5%, 10%, 15%, 20%, 30%, 50%, 70%, 80%, 90% или даже на 100% уровнем резистентности к патогену, в сравнении с контрольным растением при сравнении соответствующих результатов, полученных в тестах и/или полевых испытаниях, проводимых для оценки болезнеустойчивости растений.
Термин «широкий спектр» применительно к болезнеустойчивости относится к повышенной резистентности против по меньшей мере двух, трех, четырех или более патогенов разных видов. Так, например, растение-хозяин, обладающее повышенной резистентностью к нескольким биотропным и/или хемибиотропным, и/или некротропным патогенным видам, рассматривается как обладающее широким спектром резистентности.
Термин «симптомы, вызванные патогеном» обозначает любые симптомы заболевания, такие как рост биомассы мицелия на/или внутри хозяйской ткани, рост бактерий/биомассы бактерий, размер и/или частота некротических или хлоротических повреждений растительной ткани, размер и/или частота изъязвлений ткани и т.п.
Термин «последовательность нуклеиновой кислоты» (или молекулы нуклеиновой кислоты) относится к молекуле ДНК или РНК в одноцепочечной или двуцепочечной форме, в частности ДНК, кодирующей белок или белковый фрагмент согласно настоящему изобретению. Термин «выделенная последовательность нуклеиновой кислоты» относится к последовательности нуклеиновой кислоты, которая не будет длиннее, чем в природном среде, из которой она выделена, то есть которая не будет длиннее последовательности нуклеиновой кислоты в бактериальной хозяйской клетке или в ядерном или пластидном геноме растения.
Термины «белок» или «полипептид» используются взаимозаменяемо и относятся к молекулам, состоящим из аминокислотной цепи, независимо от конкретного характера действия, размера, трехмерной структуры или происхождения. Термин «фрагмент» или «часть» применительно к белку может, таким образом, рассматриваться как «белок». Термин «выделенный белок» в контексте настоящего описания относится к белку, который не длиннее, чем в своем природном окружении, например, in vitro или в рекомбинантной бактериальной или растительной хозяйской клетке.
Термин «функциональный» применительно к белкам NRC1 (или вариантам, таким как ортологи или мутанты и фрагменты) относится к способности модифицировать (в количественном и/или качественном отношении) развитие HR-повреждений и/или степень болезнеустойчивости за счет модификации уровня экспрессии гена, кодирующего NRC1 (например, за счет сверхэкспрессии или молчания) в растении. Так, например, функциональность предполагаемого белка NRC1, полученного из растения вида Х, может быть протестирована различными способами. Если белок является функциональным, молчание NRC1 гена, кодирующего данный белок в растении вида Х, при использовании, например, VIGS или векторов с молчащими генами, будет приводить к снижению или супрессии HR-повреждений, индуцированных патогеном или элиситором, и/или к снижению резистентности к патогену, как показано в примерах, относящихся к томату. Кроме того, внесение функционального NRC1 белка способно восстановить связанные с HR повреждения и/или устойчивость к патогенам. Альтернативно, временная или стабильная (супер)экспрессия в растении вида Х гена, кодирующего NRC1 белок (необязательно вместе с посттранскрипционным ингибитором молчания гена), будет приводить к развитию элиситор-независимых HR повреждений и/или к повышению болезнеустойчивости, в особенности против биотрофных и/или хемибиотрофных патогенов. См. также описание, приведенное в разделе «Примеры».
Термин «ген» относится к последовательности ДНК, включающей участок (транскрибированный участок), который транскрибируется c образованием молекулы РНК (например, мРНК) в клетке, оперативно связанной с соответствующими регуляторными участками (например, с промотором). Таким образом, ген может включать несколько оперативно связанных последовательностей, таких как промотор, 5'-лидерная последовательность, включающих, например, последовательности, вовлекаемые в инициацию трансляции, кодирующий участок (для белка) (например, кДНК или геномная ДНК) и 3'-нетранслируемая последовательность, которая включает, например, сайты терминации транскрипции.
Термин «химерный ген» (или рекомбинантный ген) относится к любому гену, который в природном состоянии в норме не обнаруживается в данном виде, в частности ген, в котором присутствует одна или несколько частей последовательности нуклеиновой кислоты, которые не ассоциированы друг с другом в природе. Так, например, промотор не ассоциирован в природе с частью или полностью с транскрибированным участком или с другим регуляторным участком. Термин «химерный ген» относится конструкциям экспрессии, в которых промоторная последовательность или регуляторная последовательность транскрипции оперативно связана с одной или несколькими кодирующими последовательностями, или с антисмысловой последовательностью (последовательность, представляющая собой обратный комплемент смысловой цепи), или с инвертированной повторяющейся последовательностью (смысловая и антисмысловая последовательность, за счет которых РНК транскрипт формирует двуцепочечную РНК при транскрипции).
Термин «3' UTR» или «3'-нетранслируемая последовательность» (также называемая как 3'-нетранслируемый участок или 3'-конец) относится к последовательности нуклеиновой кислоты в направлении считывания информации с кодирующей последовательности гена, которая включает, например, сайт терминации транскрипции и (в большинстве, но не во всех эукариотических мРНК) сигнал полиаденилирования (такой, как, например, AAUAAA или его варианты). После завершения стадии терминации транскрипции транскрипт мРНК может быть расщеплен в направлении считывания информации для сигнала полиаденилирования и может быть добавлен поли(А)фрагмент, который вовлекается в транспортировку мРНК в цитоплазму (где происходит трансляция).
Выражение «экспрессия гена» относится к процессу, в рамках которого участок ДНК, который оперативно связан с соответствующими регуляторными участками, в частности с промотором, транскрибируется в РНК, которая является биологически активной, то есть которая способна транслироваться в биологически активный белок или пептид (или в активный пептидный фрагмент) или которая сама по себе является активной (то есть, например, при посттранскрипционном молчании гена или в случае РНКи). В некоторых вариантах под активным белком понимают белок, который является конститутивно активным. Кодирующая последовательность предпочтительно имеет смысловую ориентацию и кодирует желательный биологически активный белок или пептид или активный пептидный фрагмент. В стратегиях, основанных на молчащих генах, последовательность ДНК предпочтительно присутствует в форме антисмысловой ДНК или в виде инвертированных повторов ДНК, включающих короткую последовательность целевого гена в антисмысловой или в смысловой и антисмысловой ориентации. Термин «эктопическая экспрессия» относится к экспрессии в ткани, в которой данный ген в норме не экспрессируется.
Выражение «регуляторная последовательность транскрипции» в контексте настоящего описания относится к последовательности нуклеиновой кислоты, которая способна регулировать уровень транскрипции (кодирующей) последовательности, оперативно связанной с регуляторной последовательностью транскрипции. Регуляторная последовательность транскрипции в контексте настоящего описания включает все элементы последовательности, необходимые для инициации транскрипции (промоторные элементы), для поддержания и для регуляции транскрипции, включая, например, аттенуаторы или энхансеры. Хотя рассматриваются в основном регуляторные последовательности транскрипции, функционирующие против (в 5'-направлении) направления считывания информации кодирующей последовательности, данное определение относится также к регуляторным последовательностям, локализованным в направлении (в 3'-направлении) считывания информации кодирующей последовательности.
В контексте настоящего описания термин «промотор» относится к фрагментам нуклеиновой кислоты, функция которых связана с контролем транскрипции одного или нескольких генов, расположенных против направления считывания информации относительно направления транскрипции сайта инициации транскрипции гена, и структурно идентифицируется наличием сайта связывания для ДНК-зависимой РНК-полимеразы, сайтов инициации транскрипции и любых других последовательностей ДНК, включающих, без ограничения, сайты связывания фактора транскрипции, сайты связывания белка-репрессора и активатора, а также любые другие нуклеотидные последовательности, известные специалистам в данной области как действующие, непосредственно или опосредованно, в направлении регуляции уровня транскрипции от промотора. Термин «конститутивный» применительно к промотору обозначает промотор, который является активным в большинстве тканей в обычных физиологических условиях и условиях развития. Термин «индуцибельный» применительно к промотору обозначает промотор, который является физиологически регулируемым (например, за счет внешнего внесения некоторых соединений) в ходе развития. «Тканеспецифическим» промотором является промотор, который активен только в специфических типах тканей или в специфических клетках. Выражение «промотор, активный в растениях или растительных клетках» относится к общей способности промотора осуществлять транскрипцию в растении или растительной клетке. При этом не делается никаких ссылок на пространственно-временную активность промотора.
В контексте настоящего описания термин «оперативно связанный» относится к связи полинуклеотидных элементов, находящихся в функциональном родстве. Нуклеиновая кислота рассматривается как «оперативно связанная», когда она находится в функциональной взаимосвязи с последовательностью другой нуклеиновой кислоты. Например, промотор или, скорее, регуляторная последовательность транскрипции оперативно связаны с кодирующей последовательностью, если он/она оказывает эффект на транскрипцию кодирующей последовательности. Выражение «оперативно связанный» означает, что последовательности ДНК, будучи связанными, являются в типичном случае непрерывными и, когда это необходимо для соединения участков, кодирующих два белка, непрерывными и в рамке считывания, с тем чтобы создавался «химерный белок». Термин «химерный белок» или «гибридный белок» представляет собой белок, состоящий из различных белковых «доменов» (или мотивов), которые не встречаются как таковые в природе, но которые объединяются с образованием функционального белка, демонстрирующего функции объединенных доменов (например, могут быть объединены сверхскрученный домен (CC), домен связывания нуклеотидов (NB-ARC) и участок, обогащенный лейциновыми повторами (LRR)). Химерный белок может также представлять собой белок слияния двух или более природных белков. Термин «домен» в контексте настоящего описания относится к одной или нескольким частям или одному или нескольким доменам белка со специфической структурой или функцией, которые могут быть перенесены на другой белок с получением нового гибридного белка, обладающего по меньшей мере функциональными характеристиками данного домена. Специфические домены могут также использоваться для идентификации других NRC1 белков, таких как NRC1 ортологи из растений другого вида.
Термин «целевой пептид» относится к аминокислотным последовательностям, которые направляют белок или белковый фрагмент на внутриклеточные органеллы, такие как пластиды, предпочтительно, хлоропласты, митохондрии, или во внеклеточное пространство или апопласт (сигнальный пептид секреции). Последовательность нуклеиновой кислоты, кодирующая целевой пептид, может быть слита (в рамке считывания) с последовательностью нуклеиновой кислоты, кодирующей амино-конец (N-терминальную часть) белка или белкового фрагмента, или может использоваться с целью замены нативного целевого пептида.
Термин «конструкция нуклеиновой кислоты» или «вектор» в контексте настоящего описания обозначает сконструированную человеком молекулу нуклеиновой кислоты, которую получают по стратегии рекомбинантных ДНК и которая применяется для доставки экзогенной ДНК в хозяйскую клетку. Векторный скелет может представлять собой, например, бинарный или супербинарный вектор (см., например, US 5591616, US 2002138879 и WO95/06722), ко-интегрированный вектор или вектор Т-ДНК, известный в данной области и приведенный в настоящем описании, в который встраивают с последующей интеграцией химерный ген или, если уже имеется подходящая регуляторная последовательность транскрипции, встраивают только нужную последовательность нуклеиновой кислоты (например, кодирующую последовательность, антисмысловую последовательность или инвертированную повторяющуюся последовательность) для интеграции в направлении считывания информации регуляторной последовательности транскрипции. Векторы обычно включают другие генетические элементы, облегчающие их использование при молекулярном клонировании, такие как, например, селектируемые маркеры, сайты множественного клонирования и т.п. (см. приведенное ниже описание).
Термин «хозяйская клетка», или «рекомбинантная хозяйская клетка», или «трансформированная клетка» используется применительно к новой индивидуальной клетке (или организму), которая возникает на основе по меньшей мере одной молекулы нуклеиновой кислоты и которая, в особенности, включает химерный ген, кодирующий желательный белок или последовательность нуклеиновой кислоты, которая при транскрипции дает антисмысловую РНК или инвертированный повтор РНК (или шпилечную РНК) для достижения молчания целевого гена/генного семейства, вводимых в указанную клетку. Хозяйская клетка представляет собой предпочтительно растительную клетку или бактериальную клетку. Указанная хозяйская клетка может содержать конструкцию нуклеиновой кислоты, такую как внехромосомная (эписомальная) реплицирующаяся молекула, или, более предпочтительно, включает химерный ген, интегрированный в ядерный или пластидный геном хозяйской клетки. В тексте настоящего описания термин «хозяйский» относится также к виду растения-хозяина, который патогеноспособен инфицировать или в который он может внедриться, что должно быть ясно из контекста. Растение рассматриваемого вида классифицируют как «хозяйский» или «нехозяйский» вид относительно патогена. «Нехозяйские» виды являются полностью иммунными для инфекции патогеном всех вариантов или штаммов патогена, даже в условиях, оптимальных для развития заболевания. «Хозяйские» виды также описываются «перечнем хозяйских организмов» для патогена и являются иммунным к некоторым (но не ко всем) вариантам патогена.
«Селектируемый маркер» представляет собой термин, известный специалистам в данной области и применяемый для описания любой генетической единицы, которая при экспрессии может использоваться для селекции одной или нескольких клеток, содержащих такой селектируемый маркер. Продукты селектируемого маркерного гена придают, например, резистентность к антибиотикам или, более предпочтительно, устойчивость к гербициду или другому выявляемому и отбираемому признаку, такому как фенотипический признак (например, изменение пигментации) или пищевые потребности. Термин «репортер» в основном используется применительно к визуализируемым маркерам, таким как зеленый флуоресцентный белок (GFP), eGFP, люцифераза, GUS и т.п.
Термин «ортолог» гена или белка в контексте настоящего описания относится к гомологичному гену или белку, обнаруженному в организме другого вида, который имеет ту же функцию, что данный ген или белок, но (обычно) отличается от последовательности за счет дивергенции с того момента времени, когда вид, содержащий данные гены, начал дивергировать (то есть, это гены, возникшие от общего предка путем видообразования).
Ортологи гена томатов NRC1 могут быть, таким образом, идентифицированы в других видах растений на основании сравнения последовательностей (например, при определении процента идентичности последовательностей по всей их длине или по специфическим доменам) и по результатам функционального анализа.
Термины «гомологичный» и «гетерологичный» относятся к родству, имеющемуся между последовательностью нуклеиновой кислоты или аминокислотной последовательностью и соответствующей хозяйской клеткой или хозяйским организмом, в особенности в контексте трансгенных организмов. Гомологичная последовательность представляет собой последовательность, которая в природном состоянии содержится в хозяйском организме того или иного вида (например, в растении томатов, трансформированном геном томатов), тогда как гетерологичная последовательность в природе не встречается в данной хозяйской клетке (например, в случае растения томатов, трансформированного последовательностью, взятой из растений картофеля). В зависимости от контекста термин «гомолог» или «гомологичный» может альтернативно относиться к последовательностям, которые происходят от общей последовательности растения-предка (то есть, они могут быть ортологами).
Термин «жесткие условия гибридизации» может использоваться для идентификации нуклеотидных последовательностей, которые по существу идентичны данной нуклеотидной последовательности. Жесткие условия зависят от используемой последовательности и различаются в разных ситуациях. В основном, жесткие условия выбирают таким образом, чтобы температура поддерживалась примерно на 5°C ниже, чем точка плавления (Тпл) для конкретных последовательностей при заданной ионной силе и заданном значении pH. Показатель Тпл представляет собой температуру (при определенных значениях ионной силы и pH), при которой 50% целевой последовательности гибридизуется с идеально подходящим зондом. В типичном случае жесткие условия выбирают таким образом, чтобы концентрация соли составляла примерно 0,02 моль при pH 7 и температура составляла по меньшей мере 60°C. Снижение концентрации соли и/или повышение температуры усиливают условия жесткости. Жесткие условия для гибридизации РНК-ДНК (нозерн-блоттинг с использованием зонда из примерно 100 нуклеотидов) представляют собой пример таких условий, которые включают по меньшей мере одну промывку в 0,2×SSC при температуре 63°C в течение 20 мин или в эквивалентных условиях. Жесткие условия для гибридизации ДНК-ДНК (саузерн-блоттинг с использованием зонда из примерно 100 нуклеотидов) представляют собой пример таких условий, которые включают по меньшей мере одну промывку (обычно две) в 0,2×SSC при температуре, равной по меньшей мере 50°C, обычно примерно 55°C, в течение 20 минут или в эквивалентных условиях. См. также руководство Самбрука с соавт. (Sambrook et al. (1989) и Sambrook and Russell (2001)).
Параметры «идентичность по последовательности» и «сходство по последовательности» могут быть определены при сопоставлении двух пептидных или двух нуклеотидных последовательностей с использованием алгоритма общего или локального выравнивания. Последовательности могут быть оценены как «по существу идентичные» или «по существу сходные», если они (при проведении оптимального сопоставления, например, с использованием программ GAP или BESTFIT, включающих заданные по умолчанию параметры), характеризуются по меньшей мере некоторым минимальным процентом идентичности по последовательности (см. приведенные ниже определения). В программе GAP используется алгоритм глобального сопоставления Нидлмана и Вунша (Needleman and Wunsch) для сопоставления двух последовательностей по всей длине, максимизации числа спариваний и минимизации числа брешей. В основном, используются в программе GAP заданные по умолчанию параметры, где штраф за образование брешей = 50 (нуклеотидов)/8 (белков) и штраф за расширение брешей = 3 (нуклеотида)/2 (белка). Для нуклеотидов используемая по умолчанию балльная матрица представляет собой nwsgapdna, а для белков заданной по умолчанию балльной матрицей является Blosum 62 (Henikoff & Henikoff, 1992, PNAS 89, 915-919). Выравнивание последовательностей и определение балльных показателей для оценки процента идентичности по последовательности могут быть проведены с использованием компьютерных программ, таких как GCG Wisconsin Package, версия 10.3, доступных от Accelrys Inc., 9685 Scranton Road, San Diego, CA 92121-3752 USA, или EmbossWin, версия 2.10.0 (с использованием программы «needle»). Альтернативно, процент сходства или идентичности может быть определен путем поиска в соответствующих базах данных с использованием алгоритмов, таких как FASTA, BLAST и т.п.
В приведенном документе и в прилагаемой формуле изобретения глагол «включать» и его варианты используются в неограничительном смысле для обозначения того, что охватываются позиции, следующие за данным словом, однако при этом не исключаются позиции, которые конкретно не указаны. Кроме того, ссылка на элемент с использованием единственного числа не исключает возможность того, что может присутствовать более чем один такой элемент, если из контекста явно не следует, что должен быть один и только один такой элемент. Таким образом, единственное число обычно соответствует ситуации «по меньшей мере один». Следует также понимать, что ссылка на «последовательности» в контексте настоящего описания означает в основном ссылку на фактические физические молекулы с определенной последовательностью субъединиц (например, аминокислот).
В контексте настоящего описания термин «растение» включает растительные клетки, растительные ткани или органы, протопласты растений, культуры тканей на основе растительных клеток, из которых данные растения могут быть регенерированы, каллюсы растений, объединения растительных клеток и растительные клетки, которые являются интактными в растениях, или части растений, такие как эмбрионы, пыльца, семяпочки, плоды (например, собранные томаты), цветы, листья, семена, корни, корневые волоски и т.п.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения использовали анализ кДНК-AFLP в сочетании с VIGS (вирус-индуцированное молчание гена) для идентификации генов, которые вовлекаются в Cf-4/Avr4-зависимый HR и резистентность к заболеванию. В числе генов, в случае которых VIGS приводит к подавлению Avr4-индуцированного HR, был идентифицирован один ген из томатов (названный как NRC1), кодирующий аналог белка резистентности типа CC-NB-LRR (обозначаемый в настоящем описании как NRC1 для белка NB-LRR, требуемого для HR-ассоциированной гибели клеток 1). Молчание NRC1 в томатах ухудшает не только развитие Avr4-индуцированного HR, на также резистентность к патогену томатов Cladosporium fulvum. Это указывает на то, что для белка резистентности томатов Cf-4 (внеклеточный белок рецепторного типа) требуется, чтобы цитоплазматический белок NB-LRR был функциональным.
Кроме того, неожиданно было обнаружено, что NRC1 вовлекается в реакцию множественного HR и процесс множественной резистентности к заболеваниям/сигнальные пути клеточной гибели, такие как Cf-9/Avr-9-, LeEix2/Eix-, Pto/AvrPto- и Rx/CP-инициированные HR (см. примеры). Другие тесты проводят для определения, вовлекается ли NRC1 также в другие HR, такие как Mi-опосредованные HR (HR, индуцированные нематодами, белокрылками и тлей; см. US 6613962 и EP0937155B1). Таким образом, NRC1 вовлекается в HR пути, запускаемые как внеклеточными, так и внутриклеточными белками резистентности к заболеваниям, которые принадлежат к различным классам: внеклеточные белки рецепторного типа (RLP, такие как Cf-4, Cf-9 и LeEix2), Ser/Thr протеинкиназы, такие как Pto, и белок CC-NB-LRR (Rx), которые придают резистентность, соответственно, к грибам (Cladosporium fulvum и Trichoderma viride), бактерии (Pseudomonas syringae pv tomato) или вирусу (PVX).
Белок NRC1 (и кодирующий его ген NRC1) может использоваться для придания растениям резистентности или усиления резистентности против множества патогенов, в особенности биотрофных и хемибиотрофных растительных патогенов, а также применительно к некротропным растительным патогенам, таким как виды Botrytis. В особенности, экспрессия NRC1 (или его вариантов или фрагментов, определенных в данном описании) ведет к достижению повышенной резистентности, в особенности против биотрофных и/или хемибиотрофных патогенов, то есть всех патогенов, которые получают питательные вещества от живых клеток. Не ограничивая область настоящего изобретения, следует тем не менее полагать, что ослабление гена (knock-down) (генное молчание) или выключение гена (knick-out) (например, путем TILLING) эндогенных генов NRC1 могут использоваться для придания или повышения резистентности против некротропных патогенов, поскольку затрагивается путь, ведущий к некрозу, а некротропные патогены требуют наличия этого пути. Таким образом, в зависимости от наличия патогена(ов), против которых усиливается резистентность, может использоваться либо повышение, либо понижение экспрессии NRC1 для усиления резистентности. Необязательно, могут быть использованы оба подхода применительно к одному растению, например, под контролем различных промоторов. Так, например, NRC1 может экспрессироваться под контролем промотора, индуцированного (хеми)биотрофным патогеном для придания резистентности к биотрофным и/или хемибиотрофным патогенам листьев, тогда как в то же самое время эндогенный ген NRC1 (или генное семейство) может быть молчащим в некоторых тканях или при индукции некротрофом с использованием промотора, который индуцируется под действием некротофных патогенов или при повреждении.
Кроме того, было показано, что в случае временной продукции конститутивно активного белка NRC1 (NRC1D481V) в растениях томатов, в растительной ткани наблюдается независимая от элиситора гибель клеток (HR), и это указывает на то, что экспрессия функционального белка NRC1 может использоваться для генерирования или усиления резистентности растений к заболеваниям.
Белки и последовательности нуклеиновых кислот согласно настоящему изобретению
Белок NRC1, полученный из растений томатов, характеризуется низкой идентичностью по последовательности (менее 25%) к NRG1 из растений табака. Кроме того, NRC1 также содержит большее число обогащенных лейцином повторов (LRR), чем NRG1. Структура белка NRC1 показана на фиг. 1 и описана в виде SEQ ID NO: 2.
Один вариант настоящего изобретения относится к последовательностям нуклеиновой кислоты и аминокислотным последовательностям белков NRC1 (включая их ортологи), а также к способам выделения или идентификации ортологов NRC1 применительно к другим видам растений, таким как другие представители Solonaceae, предпочтительно для растений картофеля. Дополнительно, в настоящем изобретении описаны способы выделения или идентификации других аллелей NRC1, таких как аллели, выделяемые из других видов томатов, сортов, линий или вариантов.
В одном варианте осуществления настоящего изобретения описываются белки NRC1. Термин «белки NRC1» включает белок, показанный в виде SEQ ID NO: 2 (белок дикого типа) и 4 (конститутивный мутант), а также включает его фрагменты и варианты. Вариант NRC1 включает, например, белки, обладающие по меньшей мере 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 98, 99% или более идентичностью по аминокислотной последовательности (по всей длине) к SEQ ID NO: 2 и/или 4. Идентичность по аминокислотной последовательности определяют при попарном сопоставлении с использованием алгоритма Нидлмана и Вунша (Needleman and Wunsch) и заданных по умолчанию параметров GAP, определенных выше. Варианты NRC1 могут быть получены из различных источников, таких как уже имеющиеся базы данных последовательностей, из других видов растений (в особенности, из других видов Solanaceae, таких как картофель) или из других сортов, или могут быть получены путем синтеза de novo, путем мутагенеза и т.п. Так, например, SEQ ID NO: 4, конститутивно активный мутант NRC1, который представляет собой вариант SEQ ID NO: 2, был получен путем направленного мутагенеза в рамках перекрывающейся ПЦР (см. примеры). Белки NRC1 согласно настоящему изобретению могут быть, таким образом, выделены из природных источников, могут быть синтезированы de novo путем химического синтеза (с использованием, например, пептидного синтезатора, такого как синтезатор, поставляемый компанией Applied Biosystems) или могут быть получены в рекомбинантных хозяйских клетках при экспрессии последовательности нуклеиновой кислоты, содержащей белок NRC1, его фрагмент или вариант.
Варианты NRC1 могут включать консервативные замещения аминокислот, составляющих группу основных аминокислот (например, Arg, His, Lys), кислотных аминокислот (например Asp, Glu), неполярных аминокислот (например, Ala, Val, Trp, Leu, Ile, Pro, Met, Phe, Trp) или полярных аминокислот (например, Gly, Ser, Thr, Tyr, Cys, Asn, Gln). Кроме того, неконсервативные замещения аминокислот также охватываются областью настоящего изобретения.
Функциональная способность любого белка NRC1, его варианта или фрагмента может быть определена с использованием различных способов. Так, например, временная или стабильная суперэкспрессия в растительных клетках может использоваться для выявления, обладает ли данный белок активностью in planta. Функциональную активность предпочтительно тестируют в тех же видах растений, из которых был получен данный белок. Так, например, временная или стабильная экспрессия может использоваться для определения, имеет ли место развитие HR и/или происходит усиление резистентности, что указывает на функциональную активность. Альтернативно, молчание эндогенных генов или генного семейства также будет показывать, является ли данный белок NRC1 функциональным. Так, например, VIGS может использоваться в растениях Solanaceae, таких как растения картофеля, томатов и табака (см. Brigneti et al., 2004, Plant Journal 39: 264; Faivre-Rampant et al. Plant Physiology 134: 1308-1316; Baulcombe 1999, Curr. Opinion. Plant Biol. 2: 109-113; Lu et al. 2003, EMBO J. 22:5690-5699), на модельных организмах, таких как Arabidopsis (Turnage et al. 2002, Plant J. 30: 107-114), а также в однодольных растениях, таких как ячмень (Holzberg et al. 2002, Plant J. 30: 315-327). Альтернативно, молчащие векторы, включающие смысловые и/или антисмысловые фрагменты гена NRC1, могут использоваться для трансформации растительных клеток (см. ниже), в рамках которой далее проводится тест для определения, приводит ли данная способность к развитию связанных с HR повреждений и/или имеет ли место модификация резистентности к заболеванию.
В предпочтительном аспекте осуществления настоящего изобретения рассматриваемые варианты NRC1 включают NRC1 белки, которые являются конститутивно активными в растительных клетках, такие как белок NRC1, описываемый в виде SEQ ID NO: 4, включающий замещение одной аминокислоты в домене MHD (D481V) (см. фиг. 1). Конститутивная активность может быть выявлена при определении, способен ли данный белок демонстрировать HR в растительной ткани в отсутствие элиситора. Так, например, может использоваться конструкция Agroinfiltration 35S:NRC1, описанная в примерах, для инфильтрации тканей листьев. Другие конститутивно активные белки NRC1 могут быть получены либо путем случайного мутагенеза с последующим тестированием активности (как описано в работе Bendahame et al., 2002, р 196), либо путем сайт-направленного мутагенеза единичных аминокислот в домене MHD (любая одна аминокислота в домене VHD или VHDM может быть замещена другой аминокислотой), в домене NB-ARC, например, в домене RNBS-D (аминокислоты FLYFGTFPRGY), либо в одном из 13 LRR доменов (см. фиг. 1). Альтернативно, последовательности нуклеиновых кислот, кодирующие конститутивно активные белки NRC1, могут быть получены из растений, например, путем мутагенеза семян и скрининга их на наличие спонтанного фенотипа с повреждением (например, с микроскопическими повреждениями), см., например, Sharino et al. (2002, The Plant Cell 14: 3149-3162) и приведенное ниже описание.
В одном варианте рассматриваются также химерные белки NRC1. Такие белки включают по меньшей мере домен CC, домен NB-ARC и предпочтительно по меньшей мере 13 LRR. CC-, NB-ARC- и LRR- домены предпочтительно относятся к аминокислотным мотивам, обладающим по меньшей мере 30, 40, 50, 60, 70, 80, 90, 95, 98, 99% или более идентичностью по аминокислотной последовательности к аминокислотам 1-150, к аминокислотам 151-508, или к аминокислотам 509-846 из SEQ ID NO: 2, соответственно. При этом домены могут подвергаться обмену (замена доменов) между белками NRC1 или между белками NRC1 и другими белками CC-NB-LRR или TIR-NB-LRR, при этом главное, чтобы функциональность полученного химерного белка была по существу такой же, как и NRC1, или предпочтительно соответствовала NRC1D481V. Наиболее предпочтительно, химерный белок сохраняет способность придавать или усиливать резистентность к заболеванию при его продукции клетками рекомбинантного растения, как будет описано ниже.
Термин «фрагменты» применительно к белкам NRC1 или к вариантам белков NRC1, приведенным в настоящем описании, включает фрагменты из 100, 150, 200, 300, 400, 500, 600, 700, 800, 850, 855 непрерывных аминокислот или более. Предпочтительно, такие фрагменты являются функциональными в растительной ткани, то есть они способны придавать или усиливать резистентность к патогену, в случае их продукции в растительных клетках. Фрагменты могут также использоваться для создания химерных белков, как было указано выше.
В другом варианте настоящее изобретение относится к выделенным последовательностям нуклеиновой кислоты, кодирующим любой из указанных выше белков, их вариантов или фрагментов, таких как последовательности кДНК, геномной ДНК и РНК. В связи с вырожденностью генетического кода различные последовательности нуклеиновой кислоты могут кодировать одну и ту же аминокислотную последовательность. Любая последовательность нуклеиновой кислоты, кодирующая белки или варианты NRC1, в настоящем описании охватывается термином «NRC1». Рассматриваемые последовательности нуклеиновой кислоты включают природные, искусственные или синтетические последовательности нуклеиновых кислот. Примеры последовательностей нуклеиновых кислот, кодирующих белки NRC1, приведены в виде SEQ ID NO: 1 и 3. Следует понимать, что в том случае, когда последовательности показаны в виде последовательностей ДНК, или в том случае, когда речь идет о РНК, то фактическая последовательность оснований в молекуле РНК идентична указанной последовательности, с той разницей, что в этом случае тимин (T) замещается урацилом (U).
В область настоящего изобретения также включены варианты и фрагменты последовательностей нуклеиновых кислот для NRC1, такие как последовательности нуклеиновых кислот, гибиридизующихся с NRC1 последовательностями нуклеиновой кислоты в жестких условиях гибридизации, определенных в настоящем описании. Варианты NRC1 последовательностей нуклеиновых кислот также включают последовательности нуклеиновой кислоты, которые характеризуются идентичностью к SEQ ID NO: 1 или 3 (по всей длине), составляющей по меньшей мере 50% или более, предпочтительно по меньшей мере 55%, 60%, 70%, 75%, 80%, 90%, 95%, 99%, 99,5%, 99,8% или более. В предпочтительном способе осуществления настоящего изобретения варианты NRC1 кодируют конститутивно активные белки NRC1, приведенные в настоящем описании. Очевидно, что могут использоваться различные методы для идентификации синтеза или выделения вариантов или фрагментов NRC1 последовательностей нуклеиновых кислот, такие как методы гибридизации нуклеиновых кислот, методики на основе ПЦР, анализ in silico и синтез нуклеиновых кислот и т.п.
Последовательность нуклеиновой кислоты, в частности последовательности ДНК, кодирующие белки NRC1 согласно настоящему изобретению, могут быть встроены в векторы экспрессии с целью продукции высоких количеств белков NRC1 (или, например, химерных белков NRC1), приведенных ниже. Для достижения оптимальной экспрессии в организме-хозяине NRC1 последовательности ДНК могут быть оптимизированы по кодонам путем адаптации использования кодонов в растительных генах к наиболее предпочтительному варианту, особенно применительно к генам, которые являются нативными для представляющего интерес рода или вида растения (Bennetzen & Hall, 1982, J. Biol. Chem. 257, 3026-3031; Itakura et al., 1977 Science 198, 1056-1063), с использованием известных таблиц использования кодонов (например, лучше адаптированных для экспрессии в растениях хлопка, соевых бобов или риса). Таблицы использования кодонов для различных видов растений описаны в литературе (Ikemura, 1993, In «Plant Molecular Biology Labfax», Croy, ed., Bios Scientific Publishers Ltd. и Nakamura et al., 2000, Nucl. Acids Res. 28, 292) и приведены в основных базах данных по последовательностям ДНК (например, в базе данных EMBL в Гейдельберге, Германия). Соответственно, синтетические последовательности ДНК могут быть сконструированы таким образом, чтобы создавались одинаковые или по существу одинаковые белки. В научной и патентной литературе описан ряд методик, подходящих для модификации использования кодонов, с тем чтобы сделать их предпочтительными для клеток конкретного организма-хозяина. Однако точный выбор методики модификации использования кодонов не является решающим фактором для осуществления настоящего изобретения.
Небольшие модификации в последовательности ДНК, такие как были описаны выше, могут быть введены стандартными способами, например путем мутагенеза, опосредованного ПЦР (Ho et al., 1989, Gene 77, 51-59., White et al., 1989, Trends in Genet. 5, 185-189). Более выраженные модификации в последовательности ДНК могут быть введены путем синтеза de novo ДНК желательного кодирующего участка с помощью доступных методик.
Кроме того, NRC1 последовательности нуклеиновых кислот могут быть модифицированы таким образом, что N-конец белка NRC1 будет иметь оптимальный характер инициации трансляции за счет добавления или делеции одной или нескольких аминокислот на N-конце белка. Зачастую для достижения оптимальной инициации трансляции предпочтительно, чтобы белки согласно настоящему изобретению экспрессировались в растительных клетках, начиная от Met-Asp или Met-Ala дипептида. Asp или Ala кодон может быть, таким образом, встроен после имеющегося Met, или второй кодон Val может быть замещен кодоном для Asp (GAT или GAC) или Ala (GCT, GCC, GCA или GCG). Последовательности ДНК могут быть также модифицированы с целью удаления неразрешенных сайтов сплайсинга.
Термин «фрагменты» применительно к NRC1 последовательностям нуклеиновых кислот включает фрагменты, содержащие по меньшей мере 10, 12, 15, 16, 18, 20, 30, 40, 50, 100, 200, 500, 1000, 1500, 2000 или более последовательных нуклеотидов из SEQ ID NO: 1 или 3 или из вариантов SEQ ID NO: 1 или 3. Короткие фрагменты могут быть, например, использованы в качестве праймеров для ПЦР или в качестве зондов при гибридизации.
В другом варианте осуществления настоящее изобретение относится к праймерам для ПЦР и/или зондам и наборам для выявления NRC1 последовательностей ДНК или РНК. Вырожденные или специфические праймерные пары для ПЦР с целью амплификации ДНК NRC1 из образцов могут быть синтезированы на основе SEQ ID NO: 1 или 3 (или их вариантов) по известной в данной области методике (см. Dieffenbach и Dveksler (1995) PCR Primer: A Laboratory Manual, Cold Spring Harbor Laboratory Press, and McPherson at al. (2000) PCR-Basics: From Background to Bench, First Edition, Springer Verlag, Germany). Так, например, любой тяж протяженностью в 9, 10, 11, 12, 13, 14, 15, 16, 18 или более непрерывных нуклеотидов из SEQ ID NO: 1 или 3 (или комплементарная цепь) может использоваться в качестве праймера или зонда. Аналогично, фрагменты ДНК из SEQ ID NO: 1 или 3 (или их варианты) могут использоваться в качестве зондов при гибридизации. Набор для выявления NRC1 может включать специфические праймеры NRC1 и/или специфические зонды NRC1 и соответствующий протокол, описывающий использование праймеров или зондов с целью выявления в образце ДНК NRC1. Такого рода набор может, например, использоваться для определения, было ли растение трансформировано геном NRC1 (или его частью) согласно настоящему изобретению. В связи с вырожденностью генетического кода, некоторые аминокислотные кодоны могут быть замещены другими, без изменения аминокислотной последовательности белка.
В еще одном варианте осуществления настоящее изобретение относится к способу идентификации и использования ортологов или аллелей гена NRC1 из томатов (SEQ ID NO: 1 и 3). Данный метод включает следующие стадии:
a) получение или идентификация последовательности нуклеиновой кислоты, обладающей по меньшей мере 70% идентичностью по нуклеиновой кислоте к SEQ ID NO: 1 и/или 3 (или большей процентной идентичностью последовательности, как было указано выше);
b) необязательная модификация последовательности нуклеиновой кислоты, кодирующей конститутивно активный белок NRC1; и
с) использование последовательности нуклеиновой кислоты по пункту a) с целью создания векторов экспрессии и/или молчащих векторов, или использование последовательности нуклеиновой кислоты по пункту b) для создания векторов экспрессии,
d) использование одного или нескольких векторов согласно пункту c) для трансформации растения или одной или нескольких растительных клеток, предпочтительно того вида, растения, из которого была получена нуклеиновая кислота,
e) анализ способности трансформированного растения/растительной ткани способствовать развитию HR-поврежденией (то есть анализ фенотипа HR-повреждения, который может быть необязательно количественно оценен) и/или анализ резистентности к заболеванию полученных трансформантов для того, чтобы определить или подтвердить функцию гена in planta и/или для получения трансгенных растений, обладающих повышенной резистентностью к заболеванию;
f) необязательно отбор для дальнейшего использования тех аллелей или ортологов, которые придают трансгенному растению повышенную резистентность к заболеванию, которые при экспрессии создают слабый фенотип по HR (то есть, не создают фенотип по HR-повреждению или создают сниженный фенотип по HR-повреждению).
Таким образом, с использованием данной методики могут быть идентифицированы NRC1 аллели или ортологи, которые при экспрессии в растениях приводят к снижению частоты и/или уменьшению HR-повреждений, выявляемых при экспрессии SEQ ID NO: 1 или 3 или при экспрессии NRC1 аллелей дикого типа, полученных из вида растения-хозяина, которое было трансформировано. Наиболее предпочтительно, идентифицируют NRC1 аллели или ортологи, которые не вызывают HR-повреждений или по меньшей мере не вызывают макроскопически заметных повреждений при экспрессии, но которые все еще придают повышенную резистентность к заболеванию.
HR-фенотип различных NRC1, аллелей и/или ортологов может быть подвергнут сравнительному анализу при создании векторов экспрессии с использованием тех же промоторов, с последующей трансформацией растительного организма-хозяина указанными векторами и при сравнении фенотипа HR-повреждений в указанных трансгенных растениях. При сравнении различных аллелей в трансгенных растениях Solanaceae (функционирующих, например, под контролем конститутивного или индуцибельного промотора) фенотип HR-повреждения трансформантов, экспрессирующих SEQ ID NO: 1 или 3, предпочтительно используют в качестве эталонного варианта. Альтернативно, в качестве эталонного варианта может использоваться аллель дикого типа, полученная из вида хозяйских организмов, которые были трансформированы. Те аллели, которые обеспечивают достижение сниженной частоты и/или меньших по размеру HR-повреждений в сравнении с SEQ ID NO: 1 или 3, или сниженной частоты и/или меньших HR-повреждений, чем вызванные экспрессией аллели дикого типа, полученной из трансформированных видов организмов, могут быть затем отобраны для дальнейшего использования. Например, экспрессирующие их трансгенные растения могут быть получены по описанной ниже методике.
В частности, аллели из растений томатов и ортологи из растений картофеля могут быть получены или идентифицированы с использованием, например, NRC1-специфических ПЦР праймеров или зондов, или в рамках биоинформационного анализа in silico. Кроме того, может использоваться метод генетического картирования для получения карты локуса NRC1 в геноме растения (например, в растениях томатов или картофеля), и при этом могут быть получены последовательности путем объединения геномной карты с существующими базами данных по геномному секвенированию (например, созданных в рамках проекта секвенирования генома томатов). Такие аллели и/или ортологи могут быть особенно полезны для получения растений с повышенной резистентностью к заболеванию.
При идентификации ортологов NRC1 из растений картофеля в рамках указанного выше способа указанные ортологи (или варианты таких ортологов) предпочтительно используют для создания растений, обладающих повышенной резистентностью к Phytophthora infestans.
Химерные гены, векторы экспрессии и рекомбинантные организмы согласно настоящему изобретению
В одном варианте осуществления настоящего изобретения последовательности нуклеиновых кислот, кодирующие NRC1 белки (включая их варианты или фрагменты), описанные выше, используют для создания химерного гена и векторов, включающих их, для целей переноса химерного гена в хозяйскую клетку и получения одного или нескольких NRC1 белков в хозяйских клетках, таких как клетки, ткани, органы или организмы, полученные из указанных одной или нескольких трансформированных клеток. Векторы, используемые для получения NRC1 белка (или фрагментов или вариантов такого белка) в растительных клетках, обозначаются в рамках настоящего описания как «вектор экспрессии». Хозяйские клетки представляют собой предпочтительно растительные клетки, хотя микробные клетки (бактерии, например, Agrobacterium, дрожжи, грибы и т.п.) также рассматриваются в настоящем изобретении.
В качестве подходящего организма-хозяина может использоваться любое растение, такое как представители однодольных растений или двудольных растений, но наиболее предпочтительно такой растительный организм-хозяин относится к семейству Solanaceae. Так, например, указанное растение принадлежит к роду Solanum (включая Lycopersicon), Nicotiana, Capsicum, Petunia и к другому роду. Приведенные ниже виды организмов-хозяинов могут использоваться согласно настоящему изобретению: табак (виды Nicotiana, например, N. benthamiana, N. plumbaginifolia, N. tabacum, и т.п.), виды овощных растений, такие как томаты (L. esculentum, синоним Solanum lycopersicum), такие как, например, вишневидные томаты, var. cerasiforme или смородиновидные томаты, var. pimpinellifolium) или цифомандра томатное дерево (S. betaceum, синоним Cyphomandra betaceae), картофель (Solanum tuberosum), баклажаны (Solanum melongena), пепино (Solanum muricatum), кокона (Solanum sessiliflorum) и наранхилла (Solanum quitoense), перцы (Capsicum annuum, Capsicum frutescens, Capsicum baccatum), виды декоративных растений (например, Petunia hybrida, Petunia axillaries, P. integrifolia).
Альтернативно, указанное растение может принадлежать к любому другому семейству, такому как Cucurbitaceae или Gramineae. Подходящие растительные организмы-хозяева включают, например, кукурузу (виды Zea), пшеницу (виды Triticum), ячмень (например, Hordeum vulgare), овес (например, Avena sativa), сорго (Sorghum bicolor), рожь (Secale cereale), соевые бобы (Glycine spp, например, G. max), хлопок (виды Gossypium, например, G. hirsutum, G. barbadense), Brassica spp. (например, B. napus, B. juncea, B. oleracea, B. rapa, и т.п.), подсолнечник (Helianthus annus), сафлор красильный, батат, маниока, люцерна (Medicago sativa), рис (виды Oryza, например, O. sativa indica, культивируемая группа или японская культивируемая группа риса), фуражные травы, просо (Pennisetum spp. например, P. glaucum), виды деревьев (Pinus, тополь, пихта, платан и т.п.), растения чая, кофе, масличная пальма, кокосовая пальма, виды овощных растений, такие как горох, цукини, фасоль (например виды Phaseolus), огурцы, артишоки, аспарагус, капуста брокколи, чеснок, лук-порей, латук, лук, редис, турнепс, проростки брюссельской капусты, морковь, цветная капуста, цикорий, сельдерей, шпинат, цикорный салат, фенхель, свекла, растения, несущие мясистые ягоды (виноград, персики, сливы, клубника, манго, яблоко, слива, вишня, абрикос, банан, черная смородина, черника, цитрусовые растения, киви, инжир, лимон, лайм, нектарины, малина, дыня, апельсин, грейпфрут и т.п.), виды декоративных растений (например, розы, петуньи, хризантемы, лилии, виды гербер), травы (мята, петрушка, базилик, тимьян и т.п.), древесные растения (например, виды Populus, Salix, Quercus, Eucalyptus), виды волокнистых растений, например лен (Linum usitatissimum) и конопля (Cannabis sativa) или модельные организмы, используемые в исследованиях, такие как Arabidopsis thaliana.
Предпочтительные хозяйские организмы включают «культурные растения», например виды растений, которые культивируются и выращиваются человеком. Культурные растения могут культивироваться для пищевых целей (например, полевые культуры) или декоративных целей (например, при выращивании цветов, используемых для срезания, трав для лужаек и т.п.). Культурные растения в контексте настоящего описания также включают растения, от которых могут быть получены непищевые продукты, такие как масло для топлива, пластические полимеры, фармацевтические продукты, кора и т.п.
Конструирование химерных генов и векторов для встраивания, предпочтительно стабильного, последовательностей нуклеиновых кислот, кодирующих белок NRC1, в геном хозяйских клеток, в основном известно в данной области. Для получения химерного гена проводят оперативное связывание последовательности нуклеиновой кислоты, кодирующей белок NRC1 (или его вариант или фрагмент), с промоторной последовательностью, подходящей для экспрессии в хозяйских клетках, с использованием стандартных методик молекулярной биологии. Промоторная последовательность может уже присутствовать в векторе, так что нуклеиновая последовательность NRC1 просто вводится в вектор по направлению считывания информации для промоторной последовательности. Затем данный вектор используют для трансформации хозяйских клеток и химерный ген встраивают в ядерный геном или в пластидный, митохондриальный или хлоропластный геном и экспрессируют там с использованием подходящего промотора (см, например, Mc Bride et al., 1995 Bio/Technology 13, 362; US 5693507). В одном варианте химерный ген включает подходящий промотор для экспрессии в растительных клетках или микробных клетках (например, в бактериях), оперативно связанный с последовательностью нуклеиновой кислоты, кодирующей белок NRC1 согласно настоящему изобретению, необязательно с последующей 3'-нетранслируемой последовательностью нуклеиновой кислоты.
Последовательность нуклеиновой кислоты NRC1, предпочтительно химерный ген NRC1, который кодирует функциональный белок NRC1 (или в некоторых вариантах конститутивно активный белок NRC1), может быть стабильно встроена обычным способом в ядерный геном одной растительной клетки, и трансформированная таким образом растительная клетка может использоваться традиционным способом для получения трансформированного растения, которое имеет измененный фенотип, в связи с наличием NRC1 белка в определенных клетках в определенное время. В этой связи может использоваться вектор Т-ДНК, включающий последовательность нуклеиновой кислоты, кодирующей белок NRC1, в Agrobacterium tumefaciens и затем трансформированное растение может быть регенерировано из трансформированной растительной клетки с использованием процедур, описанных, например, в EP 0116718, EP 0270822, в РСТ публикации WO84/02913 и в опубликованной заявке на европейский патент EP 0242246, а также в работе Gould et al. (1991, Plant Physiol. 95, 426-434). Способ конструирования вектора T-ДНК для проведения Agrobacterium-опосредованной трансформации растений известен в данной области. Вектор Т-ДНК может представлять собой бинарный вектор, описанный в EP 0120561 и EP 0120515, или может представлять собой ко-интегрированный вектор, который может интегрироваться в Ti-плазмиду Agrobacterium посредством гомологичной рекомбинации, как описано в EP 0116718.
Предпочтительные Т-ДНК векторы содержат, каждый, промотор, оперативно соединенный с последовательностью нуклеиновой кислоты, кодирующей NRC1 (например, кодирующей SEQ ID NO: 2 или SEQ ID NO: 4), локализованный между пограничными последовательностями Т-ДНК или по меньшей мере расположенный слева от правой пограничной последовательности. Пограничные последовательности описаны в работе Гилен с соавт. (Gielen et al. (1984, EMBO J 3, 835-845). Разумеется, другие типы векторов могут также использоваться для трансформации растительной клетки с использованием процедур, таких как направленный перенос генов (описанный, например, в EP 0223247), трансформация с помощью пыльцы (описанная в EP 0270356 и WO85/01856), протопластная трансформация, описанная, например, в US 4684611, вирус-опосредованная трансформация растительной РНК (описанная, например, в EP 0067553 и US 4407956), трансформация с помощью липосом (описанная, например, в US 4536475), и другие способы. Трансформация растений томатов или табака описана также в работах: An G. et al., 1986, Plant Physiol. 81: 301-305; Horsch R.B. et al., 1988, In: Plant Molecular Biology Manual A5, Dordrecht, Netherlands, Kluwer Academic Publishers, pp 1-9; Koornneef M. et al., 1986, In: Nevins D.J. and R.A. Jones, eds. Tomato Biotechnology, New York, NY, USA, Alan R. Liss, Inc. pp 169-178). Применительно к трансформации картофеля см., например, Sherman and Bevan (1988, Plant Cell Rep. 7: 13-16).
Аналогично, отбор и регенерация трансформированных растений из трансформированных клеток хорошо известны в данной области. Очевидно, что для различных видов и даже для различных сортов или вариантов одного вида протоколы должны быть соответствующим образом адаптированы с целью достижения регенерации трансформантов с высокой частотой.
Кроме трансформации ядерного генома, в настоящем изобретении рассматривается также трансформация пластидного генома, предпочтительно хлоропластного генома. Одним из преимуществ трансформации пластидного генома является снижение риска распространения трансгена(ов). Трансформация пластидного генома может быть проведена по известной в данной области процедуре, см., например, Sidorov VA et al. 1999, Plant J. 19: 209-216 или Lutz KA et al. 2004, Plant J. 37(6):906-13.
Полученное трансформированное растение может использоваться в стандартной схеме выращивания растений для получения трансформированных в большей степени растений, содержащих трансген. Могут быть отобраны трансформанты, содержащие единичную копию, с использованием, например, саузерн-блоттинга или методов на основе ПЦР или теста по процедуре Invader® Technology assay (Third Wave Technologies, Inc.). Трансформированные клетки растения могут быть легко отдифференцированы от нетрансформированных за счет наличия химирного гена. Последовательности растительной ДНК, фланкирующие сайт вставки трансгена, могут быть также секвенированы, при этом может быть разработан соответствующий метод выявления «специфичный для данного события» для целей более удобного применения. См., например, WO0141558, в котором описаны наборы для выявления наиболее подходящих явлений (такие, как наборы для определения по методу ПЦР), которые строятся, например, на основе интегрированной последовательности и фланкирующей (геномной) последовательности.
Последовательность нуклеиновой кислоты NRC1 встраивают в геном растительной клетки, так что встроенная кодирующая последовательность находится по направлению считывания информации (то есть, на 3'-конце) и под контролем промотора, который может направлять экспрессию в растительной клетке. Такой процесс предпочтительно осуществляется путем встраивания химерного гена в геном растительной клетки, в частности в ядерный или пластидный (например, хлоропластный) геном.
Поскольку конститутивная продукция NRC1 белка может приводить к индукции гибели клеток (например, к появлению микроскопических повреждений и/или макроскопических повреждений) и/или может снижать выход белка (см., например, Rizhsky and Mittler, Plant Mol Biol, 2001 46: 313-23), в одном из вариантов осуществления изобретения предпочтительно используют промотор, активность которого индуцируется. Примеры индуцибельных промоторов включают промоторы, индуцируемые повреждением, такие как MPI промотор, описанный Кордером с соавт. (Cordera et al. 1994, The Plant Journal 6, 141), которые индуцируются повреждением растений (такой, который вызывается насекомыми или физическим повреждением), или COMPTII промотор (WO0056897), или PR1 промотор, описанный в US6031151. Альтернативно, указанный промотор может индуцироваться химическим веществом, таким как дексаметазон, как описано Aoyama и Chua (1997, Plant Journal 11: 605-612) и в US6063985 или тетрациклином (TOPFREE или TOP 10 промотор, см., Gatz, 1997, Annu Rev Plant Physiol Plant Mol Biol. 48: 89-108 и Love et al. 2000, Plant J. 21: 579-88). Другие индуцибельные промоторы включают, например, промоторы, индуцируемые изменением температуры, такие как промотор теплового шока, описанный в US 5447858, анаэробными условиями (например, промотор кукурузы ADH1S), световым воздействием (US6455760), патогенами (например, gstl промотор, описанный в EP759085 или vstl промотор, описанным в EP309862), или старением организма (SAG 12 и SAG 13, см. US5689042). Очевидно, что имеется целый перечень других доступных промоторов.
В одном варианте осуществления настоящего изобретения предпочтительно используют индуцируемый патогеном промотор, поскольку в этом случае будет продуцироваться только белок NRC1 (или его вариант или фрагмент) после атаки патогена на растительную ткань. В особенности желательны промоторы генов, которые подвергаются быстрой позитивной регуляции после атаки патогена. Индуцируемые патогеном промоторы включают, например, hsr203J, str246C и sgd24 промоторы из растений табака, промотор EAS4, описанный Еин с соавт. (Yin et al., 1997, Plant Physiology 115(2):437-51), промотор tap1 или tap2 (Mohan et al., 1993, Plant Mol Biol. 1993 22: 475-90), промотор gst1 или их варианты (Martini et al. 1993, Mol. Gen. Gen. 236, 179-186; Hennin C., 1997, Afstudeerwerk, Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen, University of Gent, Belgium), промоторы WRKY (Eulgem et al., EMBO J., 1999, 18(17):4689-99) и химерные промоторы, описанные в WO0029592). Промоторы, индуцируемые конкретным растительным патогеном, могут быть также идентифицированы с использованием известных методов, таких как кДНК-AFLP®.
Предпочтительно, промоторы индуцируются множеством патогенов, например, индуцируются широким перечнем патогенов для хозяйского растительного организма. В случае каждого конкретного вида растительного организма-хозяина может быть наиболее приемлем свой, определенный промотор. Так, например, в случае использования в качестве организма-хозяина растений томата указанный промотор предпочтительно индуцируется по меньшей мере одним, но предпочтительно более чем одним, патогеном томатов. В особенности, предпочтителен промотор, который индуцируется одним или несколькими грибными патогенами растений и/или бактериальными патогенами растений (в особенности, одним или несколькими биотрофными и/или хемибиотрофными патогенами растений).
Подробное описание растительных патогенов растений, симптомов вызываемых ими заболеваний, а также их жизненные циклы описаны для каждого вида растений. Так, например, патогены томатов описаны в «Compendium of Tomato Diseases», Editors Jones, Jones, Stall and Zitter, ISBN 0-89054-120-5, APS Press (http:/www.shopapspress/org). Патогены картофеля описаны в «Compendium of Potato Disease», 2nd edition, Editors Stevenson, Franc and Weingartner, APS Press, ISBN 0-89054-275-9.
Патогены томатов включают, например, следующие виды грибов и бактерий, а также следующие вирусы (приведен неограничивающий перечень): Botrytis cinerea (гриб/некротроф); Colletotrichum coccodes (гриб/некротроф); Alternaria alternata (гриб); Alternaria solani (гриб/некротроф); Stemphylium solani; Phytophthora infestans (оомицет/хемибиотроф); Septoria lycopersici; Cladosporium fulvum, (гриб/хемибиотроф); Phytophthora parasitica; Oidium lycopersicum (биотроф); Fusarium oxysporum; Sclerotium rolfsii; Pythium; Rhizoctonia (гриб/некротроф); Corynebacterium michiganense (бактерия); Pseudomonas syringae pv tomato или pv syringae (бактерия/биотроф); Pseudomonas solanacearum; Pseudomonas corrugate; Clavibacter Xanthomonas campestris (бактерия/биотроф); Verticillium (гриб), вирус пятнистости томата (TSWV); вирус мозаики табака или томата (TobMV, TomMV).
Патогены картофеля включают, например, различные грибы, бактерии, нематоды и вирусы, такие как Phytophthora infestans (оомицет/хемибиотроф), нематоды (биотрофные); Erwinia carotovora (бактерия); Colletotrichum coccodes (грибы); Rhizoctonia solani (грибы/некротроф); Verticillium dahliae (гриб); Streptomyces scabies; Alternaria solani (гриб/некротроф); Pythium; Spongospora subterranean; PVX и PVY; вирус скручивания листьев картофеля (PLRV) и т.п.
На сайте http://www.apsnet.org/online/common/toc.asp описаны заболевания различных видов растений. Таким образом, в одном варианте осуществления настоящего изобретения указанный промотор предпочтительно индуцируется одним или несколькими указанными выше патогенами, более предпочтительно по меньшей мере одним или несколькими указанными биотрофными и/или хемибиотрофными патогенами.
Альтернативно, хозяйское растение может включать различные трансгены NRC1, каждый из которых находится под контролем своего, отличного от других промотора, индуцируемого патогеном, для гарантии того, что белок NRC1 продуцируется после атаки множеством патогенов. Так, например, в случае трансформации растений томатов один промотор может индуцироваться Phytophthora и один может индуцироваться Cladosporium.
Термин «индуцибельный» необязательно требует того, чтобы промотор был полностью неактивным в отсутствие индуцирующего стимула. Может наблюдаться низкий уровень неспецифичной активности, при том, что она не будет приводить к серьезному усилению их образования или к ухудшению качества растения. Таким образом, термин «индуцибельный» предпочтительно относится к повышению активности промотора, что приводит к повышению транскрипции в направлении считывания информации кодирующего участка NRC1 после контакта с индуктором.
Наиболее предпочтительным сочетанием согласно настоящему изобретению является использование индуцируемого патогеном промотора, необязательно соединенного с последовательностью нуклеиновой кислоты NRC1, которая кодирует конститутивно активный белок NRC1, приведенный выше. В этом случае при атаке патогена будет экспрессироваться конститутивно активный NRC1, что скажется на образовании локального HR (ограниченного сайтом атаки патогена) и будет препятствовать дальнейшему росту любого (хеми)биотрофного патогена.
В другом варианте осуществления настоящего изобретения могут использоваться конститутивные промоторы, такие как эффективные конститутивные 35S промоторы или усиленные 35S промоторы («35S промоторы») вируса цветной капусты (CaMV) изолятов CM 1841 (Gardner et al., 1981, Nucleic Acids Research 9, 2871-2887), CabbB-S (Franck et al., 1980, Cell 21, 285-294) и CabbB-JI (Hull and Howell, 1987, Virology 86, 482-493); 35S промотор, описанный Оделлом с соавт. (Odell et al.) (1985, Nature 313, 810-812) или в US5164316, промоторы семейства убихитина (например, промотор убихитина кукурузы, описанный в работах: Christensen et al., 1992, Plant Mol. Biol. 18, 675-689, в EP 0342926, а также Cornejo et al. 1993, Plant Mol. Biol. 23, 567-581), промотор gos2 (de Pater et al., 1992 Plant J. 2, 834-844), промотор emu (Last et al., 1990, Theor. Appl. Genet. 81, 581-588), промоторы актина Arabidopsis, такие как промотор, описанный Ан с соавт. (An et al. 1996, Plant J. 10, 107), промоторы актина риса, такие как промотор, описанный Жангом с соавт. (Zhang et al. 1991, The Plant Cell 3, 1155-1165) и промотор, описанный в US 5641876, или промотор актина 2 риса, описанный в WO070067; промоторы вируса мозаики листовой жилки маниоки (WO 97/48819, Verdaguer et al. 1998, Plant Mol. Biol. 37, 1055-1067), промоторы серии pPLEX из вируса карликовости клевера подземного (WO 96/06932, в частности, S7 промотор), промотор алкогольдегидрогеназы, например, pAdh1S (номера доступа в GenBank Х04049, Х00581), и TR1' промотор и TR2' промотор («TR1' промотор» и «TR2' промотор», соответственно), которые направляют экспрессию 1' и 2' генов, соответственно, из Т-ДНК (Velten et al., 1984, EMBO J 3, 2723-2730), промотор вируса мозаики норичника шишковатого, описанный в US6051753 и в EP426641, промоторы гена гистонов, такие как промотор Ph4a748 из Arabidopsis (PMB 8: 179-191) и другие. В предпочтительном варианте осуществления настоящего изобретения используют AA6 промоторы, описанные в PCT/NL2005/050083 (зарегистрированной 16 декабря 2005 года).
Альтернативно, может использоваться промотор, который не является конститутивным, но, скорее, является специфическим для одного или нескольких видов тканей или органов растений (предпочтительные по ткани/тканеспецифичные, включая промоторы, регулируемые в процессе развития), например, предпочтительные для листа, предпочтительные для эпидермиса, предпочтительные для корня, предпочтительные для ткани цветов, например лепестка или пыльника, предпочтительные для семян, предпочтительны для стручка и т.п.), за счет чего достигается экспрессия гена NRC1 только в клетках, относящихся к одной или нескольким специфическим тканям или к одному или нескольким специфическим органам, и/или только в клетках на определенной стадии развития. Так, например, один или несколько генов NRC1 могут селективно экспрессироваться в листьях растений за счет помещения кодирующей последовательности под контроль светоиндуцируемого промотора, такого как промотор малого субъединичного гена рибулозо-1,5-бифосфаткарбоксилазы из самого растения иди другого растения, такого как горох, согласно US 5254799, или Arabidopsis, как описано в US5034322.
В одном варианте осуществления настоящего изобретения используют промотор эндогенного гена NRC1. Так, например, промотор гена NRC1 из растения томата может быть выделен и оперативно соединен с кодирующим участком, кодирующим белок NRC1, описываемый SEQ ID NO: 2 или 4. Промотор для NRC1 (участок регуляции транскрипции SEQ ID NO: 1 и 3 против направления считывания информации) может быть выделен из растений томата с использованием известных методов, таких как TAIL-PCR (Liu et al. 1995, Genomics 25(3):674-81; Liu et al. 2005, Methods Mol Biol. 286:341-8), линкерная ПЦР или инверсная ПЦР (IPCR).
Кодирующую последовательность для NRC1 предпочтительно встраивают в растительный геном так, чтобы кодирующая последовательность находилась против направления считывания информации (то есть, на 5' конце) соответствующего 3'-концевого нетранслируемого участка («3'-конец» или 3'UTR). Подходящие 3'-концы включают соответствующие участки 35S гена CaMV («3' 35S»), гена нопалинсинтазы («"3' nos») (Depicker et al., 1982 J. Molec. Appl. Genetics 1, 561-573), гена октопинсинтазы («3'ocs») (Gielen et al., 1984, EMBO J 3, 835-845) и гена Т-ДНК 7 («3' ген 7») (Velten and Schell, 1985, Nucleic Acids Research 13, 6981-6998), которые действуют как 3'-нетранслируемые последовательности ДНК в трансформированных растительных клетках и других клетках. В одном варианте осуществления настоящего изобретения используют 3'UTR гена NRC1 из растений томата, как показано в SEQ ID NO: 3, от нуклеотида 2748 до нуклеотида 3168, и как показано в SEQ ID NO: 5. 3'UTR NRC1 также представляет собой один из аспектов настоящего описания, поскольку он также может использоваться в качестве 3'UTR с другими кодирующими участками. Аналогично, также рассматривается любой вариант или фрагмент SEQ ID NO: 5. Вариант SEQ ID NO: 5 включает последовательности нуклеиновой кислоты, обладающие по меньшей мере 40, 50, 60, 70, 80, 90, 95, 98, 99% или большей идентичностью по последовательности нуклеиновой кислоты к SEQ ID NO: 5 (при определении с использованием алгоритма Нидлмана и Вунша (Needleman and Wunsch) и штрафных санкций программы GAP, указанных выше. Рассматриваемые фрагменты включают любые нуклеотидные последовательности, содержащие по меньшей мере 30, 50, 100, 150, 200, 300, 400 или более последовательных нуклеотидов из SEQ ID NO: 5 или варианта SEQ ID NO: 5.
Встраивание Т-ДНК вектора в Agrobacterium может проводиться с использованием известных методов, таких как электропорация или тройное спаривание.
Последовательность нуклеиновой кислоты, кодирующая NRC1, может быть необязательно встроена в растительный геном в качестве последовательности гибридного гена, за счет чего последовательность NRC1 связывается в рамке считывания с геном, кодирующим селектируемый или количественно оцениваемый маркер (US 5254799; Vaeck et al., 1987, Nature 328, 33-37), такой как, например, neo (или nptII) ген (EP 0242236), кодирующий резистентность к канамицину, так что указанное растение экспрессирует белок слияния, который может быть легко выявлен.
Полноразмерная последовательность нуклеиновой кислоты NRC1 или ее часть, кодирующая белок NRC1 (или его вариант или фрагмент), может также использоваться для трансформации микроорганизмов, таких как бактерии (например, Escherichia coli, Pseudomonas, Agrobacterium, Bacillus, и т.п.), грибы или водоросли, или насекомые, или для создания рекомбинантных вирусов. Трансформация бактерий последовательностью нуклеиновой кислоты NRC1 согласно настоящему изобретению, полноразмерной или ее частью, которая встроена в соответствующий носитель для клонирования, может проводиться традиционным способом, предпочтительно с использованием стандартных методик электропорации, описанных, например, в работе Мэйлон с соавт. (Maillon et al. 1989, FEMS Microbiol. Letters 60, 205-210) и в WO 90/06999. Для достижения экспрессия в прокариотической хозяйской клетке, используемый кодон из последовательности нуклеиновой кислоты может быть соответствующим образом оптимизирован (как было описано выше применительно к растениям). Интронные последовательности должны быть удалены и проведены другие необходимые адаптации для достижения оптимальной экспрессии, в соответствии с известными процедурами.
Последовательность ДНК из последовательности нуклеиновой кислоты NRC1 может быть также изменена способом, нейтральным для трансляции, с целью модификации возможных ингибирующих последовательностей ДНК, присутствующих в части гена и/или за счет введения изменений в использование кодонов, например, путем адаптации использования кодона к варианту, наиболее предпочтительному для растений, предпочтительно, специфически для релевантного рода растений, как было описано выше.
В соответствии с одним вариантом осуществления настоящего изобретения, NRC1 белки (или химерные белки) направляются на внутриклеточные органеллы, такие как пластиды, предпочтительно хлоропласты, митохондрии, или секретируются из клетки, и потенциально способны оптимизировать стабильность и/или экспрессию белков. Аналогично, указанный белок может быть нацелен на вакуоли. С этой целью в одном варианте химерные гены согласно настоящему изобретению включают кодирующий участок, кодирующий сигнальный или целевой пептид, соединенный с кодирующим участком белка NRC1 согласно настоящему изобретению. Особенно предпочтительные пептиды, включаемые в белки согласно настоящему изобретению, представляют собой транспортные пептиды, осуществляющие транспортировки в хлоропласты или другие пластиды, в особенности дублицированные участки транспортных пептидов из генов растений, где генный продукт направлен на пластиды, оптимизированный транспортный пептид, описанный Капелладес с соавт. (Capellades et al. US 5635618), транспортный белок ферредоксин-НАДФ+оксидоредуктаза из шпината (Oelmuller et al., 1993, Mol. Gen. Genet. 237, 261-272), транспортный пептид, описанный Вонгом с соавт. (Wong et al. (1992, Plant Molec. Biol. 20, 81-93), и направленные белки, описанные в опубликованной заявке на патент WO 00/26371. Также предпочтительны сигнальные пептиды секреции белка, связывающиеся с таким пептидом за пределами клетки, такие как белок секреции ингибитора II протеиназы картофеля (Keil et al., 1986, Nucl. Acids Res. 14, 5641-5650), сигнал секреции гена альфа-амилазы 3 из риса (Sutliff et al., 1991, Plant Molec. Biol. 16, 579-591) и сигнал секреции белка PR1 из растений табака (Comelissen et al., 1986, EMBO J. 5, 37-40). Особенно часто применяемые сигнальные пептиды согласно настоящему изобретению включают пептид транспорта в хлоропласты (см., например, Van Den Broeck et al., 1985, Nature 313, 358) или оптимизированный пептид транспорта в хлоропласты, описанный в US 5510471 и US 5635618, который запускает транспорт белка в хлоропласты, секреторный сигнальный пептид или пептид, нацеливающий белок на другие пластиды, митохондрии, эндоплазматический ретикулум и другие органеллы. Сигнальные последовательности, нацеливающие на внутриклеточные органеллы или на секрецию за пределы растительной клетки или в клеточную стенку, обнаружены в природных направленных или секретированных белках, предпочтительно белках, описанных Klösgen et al. (1989, Mol. Gen. Genet. 217, 155-161), Klösgen and Weil (1991, Mol. Gen. Genet. 225, 297-304), Neuhaus & Rogers (1998, Plant Mol. Biol. 38, 127-144), Bih et al. (1999, J. Biol. Chem. 274, 22884-22894), Morris et al. (1999, Biochem. Biophys. Res. Commun. 255, 328-333), Hesse et al. (1989, EMBO J. 8, 2453-2461), Tavladoraki et al. (1998, FEBS Lett. 426, 62-66), Terashima et al. (1999, Appl. Microbiol. Biotechnol. 52, 516-523), Park et al. (1997, J. Biol. Chem. 272, 6876-6881), Shcherban et al. (1995, Proc. Natl. Acad. Sci USA 92, 9245-9249).
Для того чтобы секреция белков NRC1 за пределы трансформированной хозяйской клетки была возможна, соответствующий сигнальный пептид секреции может быть слит с аминоконцом (N-терминальный конец) белка NRC1. Предположительные сигнальные пептиды могут быть обнаружены с использованием компьютерного анализа в рамках таких программ, как программа поиска сигнальных пептидов (SignalP V1.1 или 2.0) (Von Heijne, Gunnar, 1986 and Nielsen et al., 1996).
В одном варианте осуществления настоящего изобретения несколько последовательностей нуклеиновой кислоты, кодирующих NRC1, подвергают совместной экспрессии в одном организме-хозяине, необязательно под контролем разных промоторов. Такое растение-хозяин, в котором осуществляется совместная экспрессия, может быть легко получено путем трансформации растения, которое уже экспрессирует белок NRC1 согласно настоящему изобретению, или путем скрещивания растений, трансформированных разными NRC1 согласно настоящему изобретению. Альтернативно, несколько последовательностей нуклеиновой кислоты, кодирующих NRC1 белок, может присутствовать на одном векторе трансформации или может быть трансформировано совместным путем в одно и то же время с использованием отдельных векторов и последующим отбором трансформантов, включающих оба химерных гена. Аналогично, один или несколько генов, кодирующих NRC1, могут быть экспрессированы в одном растении вместе с другими химерными генами, например генами, кодирующими другие белки, которые повышают резистентность к заболеванию или которые вовлекаются в сигнальный путь, определяющий резистентность к заболеванию, и другими.
Следует понимать, что различные белки могут быть экспрессированы в одном и том же растении или каждый из них может быть экспрессирован в одном растении, и затем они могут быть объединены в одном и том же растении путем скрещивания отдельных растений друг с другом. Так, например, при получении гибридных семян каждое родительское растение может экспрессировать один белок. При скрещивании родительские растения продуцируют гибриды и оба белка объединяются в гибридном растении.
Предпочтительно, для целей отбора, а также для контроля размножения сорняков трансгенные растения согласно настоящему изобретению также трансформируют ДНК, кодирующей белок, который придает резистентность к гербициду, такому как гербицид широкого спектра действия, например гербицид на основе глифозината аммония как активного ингредиента (например, Liberty® или BASTA; резистентность создается PAT или bar геном; см. EP 0242236 и EP 0242246) или на основе глифозата (например, RoundUp®; в этом случае резистентность создается генами EPSPS, см., например, EP 0508909 и EP 0507698). Использование генов резистентности к гербициду (или других генов, придающих желательный фенотип) в качестве селектируемого маркера также дает преимущество, заключающееся в том, что можно избежать встраивания генов резистентности к антибиотику.
Альтернативно, могут быть использованы другие селектируемые маркерные гены, такие как гены устойчивости к антибиотику. Поскольку в основном не принято оставлять гены резистентности к антибиотику в трансформированных хозяйских растениях, эти гены могут быть снова удалены после отбора трансформантов. Существуют различные методики удаления трансгенов. Один такой метод удаления включает фланкирование химерного гена сайтами lox и, после селекции, скрещивание трансформированного растения с растением, экспрессирующим CRE-рекомбиназу (см., например, EP506763B1). Сайт-специфическая рекомбинация приводит к исключению маркерного гена. Другая сайт-специфическая система рекомбнации включает систему FLP/FRT, описанную в EP686191 и US5527695. Сайт-специфическая система рекомбинации, такая как CRE/LOX и FLP/FRT, может также использоваться для целей стыковки генов. Кроме того, были описаны однокомпонентные системы исключения, например, в WO9737012 или в WO9500555.
Трансформированные растительные клетки/растения/семена и использованием последовательности нуклеиновой кислоты и белков согласно настоящему изобретению
В приведенном ниже разделе описано использование последовательностей NRC1 согласно настоящему изобретению для получения трансгенных растительных клеток, растений, семян растения и т.п., а также любых их производных/потомства, с фенотипом, характеризующимся повышенной резистентностью к заболеванию.
Трансгенные растения с повышенной резистентностью к заболеванию могут быть получены путем трансформации растительной хозяйской клетки последовательностью нуклеиновой кислоты, кодирующей по меньшей мере один белок NRC1 под контролем соответствующего описанного выше промотора, с последующей регенерацией трансгенного растения из такой клетки.
Предпочтительные промоторы включают промоторы, которые индуцируются внешними биотическими и/или абиотическими стимулами. Особенно предпочтительны промоторы, которые индуцируются патогеном, как было описано выше. Предпочтительный промотор - как сочетание NRC1 - включает
а) промотор, индуцируемый патогеном, - последовательность нуклеиновой кислоты, кодирующая конститутивно активный белок NRC1;
b) промотор, индуцируемый патогеном, - последовательность нуклеиновой кислоты, кодирующая белок NRC1 дикого типа;
с) промотор растительного гена NRC1 (предпочтительно того же вида, который трансформируется) - последовательность нуклеиновой кислоты, кодирующая конститутивно активный белок NRC1;
d) промотор растительного гена NRC1 (предпочтительно того же вида, который трансформируется) - последовательность нуклеиновой кислоты, кодирующая белок NRC1 дикого типа;
e) промотор, индуцируемый биотическим стрессом (например, индуцируемый повреждением насекомыми, индуцируемый патогеном и т.п.) - последовательность нуклеиновой кислоты, кодирующая конститутивно активный белок NRC1;
f) промотор, индуцируемый биотическим стрессом (например, индуцируемый повреждением насекомыми, индуцируемый патогеном и т.п.) - последовательность нуклеиновой кислоты, кодирующая белок NRC1 дикого типа;
g) конститутивный промотор (например, 35S промотор) - последовательность нуклеиновой кислоты, кодирующая белок NRC1 дикого типа;
h) конститутивный промотор (например, 35S промотор) - последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность, обладающую по меньшей мере 70% идентичностью по аминокислотной последовательности к SEQ ID NO:2 по всей длине;
i) промотор, индуцируемый патогеном, - последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность, обладающую по меньшей мере 70% идентичностью по аминокислотной последовательности к SEQ ID NO:2 по всей длине;
j) промотор растительного гена NRC1 - последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность, обладающую по меньшей мере 70% идентичностью по аминокислотной последовательности к SEQ ID NO:2 по всей длине.
В одном варианте осуществления настоящего изобретения указанное трансгенное растение может иметь конститутивные HR-повреждения или индуцируемые HR-повреждения и повышенную резистентность к заболеванию, вызванному одним или несколькими патогенами. Однако в контексте настоящего изобретения рассматривается также вариант, в котором не развиваются HR-повреждения или развиваются «слабые» HR-повреждения (такие, как более мелкие повреждения, например, микроповреждения и/или повреждения с низкой частотой), при том, что растения все еще демонстрируют повышенную резистентность к заболеванию. Аллели или ортологи NRC1, которые при экспрессии в хозяйских растительных организмах под контролем идентичных промоторов приводят к снижению частоты и/или уменьшению размеров HR-повреждений, чем в случае с SEQ ID NO: 1 или 3, или чем при экспрессии NRC1 аллели дикого типа, полученной из того же хозяйского организма, который был трансформирован, являются особенно предпочтительными в рамках настоящего описания, особенно согласно подходам g) и h), описанным выше. Такие алели или ортологи могут рассматриваться как аллели NRC1, придающие «слабый HR фенотип» данному организму-хозяину. Такие аллели или ортологи NRC1 могут быть идентифицированы и/или выделены согласно описанным выше процедурам. HR фенотип разных аллелей и/или ортологов NRC1 можно сравнить путем создания векторов экспрессии и с последующей трансформацией (предпочтительно все нуклеиновые кислоты, которые будут далее сравниваться, должны быть оперативно связаны с идентичными промоторами, например, 35S) растений или растительной ткани указанными векторами и проведения сравнения по фенотипу между указанными растениями. В случае трансформантов Solanaceae фенотип HR-повреждения у трансформантов, экспрессирующих SEQ ID NO: 1 или 3, предпочтительно используют в качестве эталона и любая аллель, приводящая к снижению чистоты и/или уменьшению размеров HR-повреждений при экспрессии под контролем того же промотора, представляет собой аллель, которая придает слабый HR фенотип. Фенотип HR-повреждения можно сравнить и в необязательном варианте количественно охарактеризовать с использованием различных методик, таких как микроскопическое обследование (необязательно с окрашиванием погибших клеток), визуальная оценка по балльной системе, количественный подсчет повреждений, с тем чтобы определить их число на см2, определение диаметра HR-повреждений и т.п.
Предпочтительно, трансгенные растения согласно настоящему изобретению обладают повышенной резистентностью к заболеванию, вызванному одним или несколькими патогенами, особенно биотрофными и/или хемибиотрофными патогенами для данных видов трансгенных растений. Таким образом, например, трансгенное растение томата или картофеля обладает повышенной резистентностью по меньшей мере к одному или нескольким видам грибных, бактериальных видов, к нематодам и/или к вирусным патогенам, перечисленным выше, наиболее предпочтительно, по меньшей мере против одного или нескольких биотрофных и/или хемибиотрофных видов.
Термин «резистентность к заболеванию» или «усиленная/повышенная резистентность к заболеванию» в контексте настоящего описания используется для обозначения повышенной способности трансформантов (в сравнении с растениями дикого типа или контрольными трансформантами) противостоять атаке одного или нескольких растительных патогенов или, иными словами, используется для обозначения выраженного снижения симптомов заболевания у трансформантов в сравнении с нетрансформированными (или трансформированными пустым вектором) контролями. Резистентность к заболеванию или повышенная резистентность к заболеванию может быть определена с использованием множества методов. Часто симптомы заболевания определяют визуально по балльным показателям (либо в рамках биотестов, либо в полевых испытаниях) путем оценки симптомов заболевания в одной или нескольких временных точках после инокуляции патогена или после контакта с патогеном. Альтернативные методы включают такие процедуры, в которых патоген может быть выявлен и необязательно количественно определен. Таким образом, трансгенное растение может демонстрировать повышенную резистентность к заболеванию, если количество выявленного патогена в ткани (или на ткани) существенно снижено в сравнении с контролями, или если патоген распространяется значительно медленнее, чем в контроле. И в итоге значительное повышение среднего показателя выхода трансформантов (например, по меньшей мере на 1%, 2%, 5%, 10% или более), в сравнении с контролями, которые растут в эквивалентных условиях воздействия факторов заболевания (предпочтительно в полевых условиях), представляет возможность непрямого определения повышенной резистентности к заболеванию.
Таким образом, множество трансформантов растений, экспрессирующих NRC1 белок (или конститутивно активный NRC1), демонстрирует повышенную резистентность к заболеванию, если указанные трансформанты характеризуются значительным снижением симптомов заболевания, в сравнении с нетрансформированными контролями или с контролями, трансформированными пустым вектором. Очевидно, что требуется статистический анализ для определения наличия такого значимого снижения. Предпочтительно, в NRC1 трансформантах выраженность одного или нескольких симптомов заболевания в среднем по меньшей мере на 2%, 5%, 10%, 15%, 20%, 30%, 40%, 50% или даже 100% ниже, чем в контрольных растениях. Поскольку для каждого сочетания организм-хозяин - патоген требуются различные тесты на заболевание, нельзя предложить какой-то один конкретный протокол, хотя средний специалист в данной области знает, как определить, демонстрируют ли полученные трансформанты значительно более усиленную резистентность к одному или нескольким патогенам. Биотесты, известные в данной области для каждого сочетания организм-хозяин - патоген, могут использоваться с целью сравнения резистентности трансгенных растений с соответствующими контролями.
Поскольку белок NRC1 может в некоторых вариантах осуществления настоящего изобретения приводить к появлению HR повреждений в отсутствие патогена (например, в том случае, если ген NRC1 находится под контролем конститутивного промотора), в некоторых вариантах осуществления настоящего изобретения может быть важно провести дифференциацию между симптомами, вызванными экспрессией NRC1, и симптомами, определяемыми инфекцией патогеном и его распространением. В этой связи может быть предпочтительно использовать методы, которые позволяют выявить сам патоген (а не вызванный им некроз на растительной ткани) и сравнить количество имеющегося патогена или скорость распространения патогена. Так, например, могут использоваться биотесты, в которых патоген может выявляться путем окрашивания. В приведенных примерах используют трансгенный штамм C.fulvum, который экспрессирует GUS. В этом случае грибной мицелий может быть визуализирован с использованием Х-gluc окрашивания инокулированной ткани растения. Выраженное снижение грибного мицелия в трансгенных растениях в сравнении с контролем указывает на наличие повышенной резистентности к грибам.
Один из вариантов осуществления настоящего изобретения относится также к получению трансгенных растений, которые экспрессируют несколько белков NRC1, предпочтительно под контролем разных промоторов, таких как разные промоторы, индуцируемые патогенами.
Фенотип с резистентностью к заболеваниям может быть достаточно тонко подобран при экспрессии соответствующего количества белка NRC1 в подходящий период времени и в подходящей его локализации. Такой тонкий подбор может быть осуществлен путем определения наиболее подходящего промотора для конкретного сочетания организм-хозяин - патоген, а также при выборе трансгенных «событий», которые демонстрируют желательный уровень экспрессии. Слишком низкий уровень белка NRC1 или слишком медленная индукция продукции белка NRC1 после атаки патогеном могут быть недостаточными для повышения уровня резистентности к заболеванию. С другой стороны, слишком высокий уровень белка или его экспрессии в соответствующие периоды времени и в соответствующей локализации без атаки патогена может приводить к получению агрономически нежелательных фенотипов, таких как повреждение листьев или плодов в отсутствие патогенов, а также к снижению урожая. Однако любой средний специалист в данной области может легко определить, каким образом получать растения, обладающие повышенной резистентностью к заболеванию, но которые в то же время являются агрономически приемлемыми. Оптимальные аллели NRC1 могут быть выделены или идентифицированы в соответствии с приведенным описанием, например, могут быть выделены аллели, обеспечивающие высокий уровень резистентности и лишь слабый фенотип по HR.
Трансформанты, которые экспрессируют желательные уровни белка NRC1, отбирают, например, путем анализа числа копий (саузерн-блот анализ), уровня транскриптов мРНК (например, в рамках ОТ-ПЦР с использованием NRC1 праймерных пар или фланкирующих праймеров) или анализа наличия и уровня белка NRC1 в различных тканях (например, в рамках методов ДСН-ПААГ, ИФТФА и т.п.). Для целей регуляции предпочтительно выбирают однокопийные трансформанты и анализируют последовательности, фланкирующие сайт вставки химерного гена, предпочтительно секвенированного для целей характеристики данного «события». Выбирают трансгенные варианты, характеризующиеся высоким или умеренным уровнем экспрессии NRC1, для дальнейшего скрещивания/обратного скрещивания/самоопыления до получения с высокой частотой элитного варианта, характеризующегося наличием стабильного трансгена NRC1.
Трансформанты, экспрессирующие один или несколько генов NRC1 согласно настоящему изобретению, могут также включают другие трансгены, такие как другие гены, придающие резистентность к заболеванию или определяющие толерантность к другим биотическим и/или абиотическим стрессовым факторам. Для получения таких растений со «сцепленными» трансгенами другие трансгены могут быть либо введены путем интрогрессии в NRC1 трансформанты, либо указанные NRC1 трансформанты могут быть трансформированы впоследствии с использованием одного или нескольких других генов или, альтернативно, могут быть использованы несколько химерных генов для трансформации линии или сорта растений. Так, например, на одном векторе может присутствовать несколько химерных генов или эти несколько химерных генов могут присутствовать на разных векторах, которые используются для совместной трансформации.
В одном варианте осуществления настоящего изобретения указанные ниже гены объединяют с одним или несколькими генами NRC1 согласно настоящему изобретению: известные гены резистентности к заболеванию, в особенности гены, придающие повышенную резистентность к некротрофным патогенам, гены резистентности к вирусу, гены резистентности к насекомым, гены, определяющие резистентность к абиотическому стрессу (например, толерантность к засухе, солевую толерантность, толерантность к высоким температурам или к холоду или т.п.), гены резистентности к гербицидам и т.п. Сцепленные трансформанты могут в этой связи характеризоваться еще более широкой толерантностью к биотическому и/или абиотическому стрессу, резистентностью к патогенам, резистентностью к насекомым, резистентностью к нематодам, к засоленности, к холодовому стрессу, к тепловому стрессу, к водному стрессу и т.п. Кроме того, стратегия на основе молчания NRC1 может быть объединена с экспрессией NRC1 в одном растении. Так, например, суперэкспрессия NRC1 в корнях или клубнях может придавать корню или клубню резистентность к почвенным патогенам или усиливать ее. В то же время отрицательная регуляция NRC1 в надземных частях растений может придавать или усиливать резистентность к некротрофным патогенам (или наоборот).
Возможно также встраивать или вводить путем интрогрессии ген NRC1 при линейном разведении растений, которые уже обладают определенным уровнем резистентности к заболеванию. Для достижения длительной резистентности к заболеваниям в полевых условиях может быть желательно объединить несколько механизмов болезнеустойчивости в растении, при этом предпочтительно, чтобы источники резистентности характеризовались различными молекулярными механизмами действия.
Целые растения, семена, клетки, ткани и потомство (такие, как F1 гибриды, F2 семена/растения и т.п.) любого из указанных выше трансформированных растений охватываются настоящим описанием и могут быть идентифицированы по наличию трангсена в ДНК, например, в рамках ПЦР анализа с использованием суммарной геномной ДНК в качестве матрицы и с использованием NRC1-специфичных праймерных пар для ПЦР. Кроме того, могут быть разработаны методы диагностической ПЦР, «специфичной для события», где используемые для ПЦР NRC1-специфичные праймеры основаны на растительной ДНК, фланкирующей встроенный химерный ген (см. US6563026). Аналогично, могут быть получены специфичные для «события» AFLP карты по типу «отпечатков пальцев» для события или RFLP карты по типу «отпечатков пальцев», которые позволяют идентифицировать трансгенное растения или любое растение, семена, ткани или клетки, полученные из них.
Следует понимать, что трансгенные растения согласно настоящему изобретению предпочтительно не демонстрируют нежелательных фенотипов, таких как снижение урожайности, повышенная чувствительность к заболеванию (в особенности, к некротрофам), нежелательные архитектурные изменения (карликовость, деформации) и т.п., и если такие фенотипы выявляются в первичных трансформантах, они могут быть удалены путем нормального разведения и селекции (скрещивание/обратное скрещивание/самоопыление и т.п.). Любое из указанных в данном описании трансгенных растений может быть гомозиготным или гемизиготным для трансгена.
Подходы, основанные на молчании NRC1 гена, и векторы, включающие молчащий ген
В другом варианте осуществления настоящего изобретения рассматриваются растения с повышенной резистентностью к заболеванию, в особенности против нектротрофных патогенов, за счет трансформации растения вектором, несущим молчащий ген NRC1. Без ограничения области настоящего изобретения следует полагать, что молчание эндогенных NRC1 генов или генных семейств приводит к неспособности трансгенного растения запускать и/или усиливать HR ответ. Поскольку некротрофные патогены требуют для своего роста и развития гибели клеток, такие растения могут демонстрировать усиленную резистентность к одному или нескольким некротрофным патогенам.
Термин «молчание гена» относится к отрицательной регуляции или к полному ингибирования генной экспрессии одного или нескольких целевых генов (например, эндогенных генов NRC1). Использование ингибиторной РНК для снижения или устранения генной экспрессии представляет собой хорошо известный в данной области метод, который описан в ряде обзоров (например, Baulcombe 1996, Stam et al. 1997, Depicker and Van Montagu, 1997). Имеется множество методик, доступных для получения молчащих генов растений, таких как химерные гены, которые продуцируют антисмысловую РНК полноразмерного или части целевого гена (см., например, EP 0140308 B1, EP 0240208 B1 и EP 0223399 B1) или которые продуцируют смысловую РНК (где данный процесс определяется также как ко-супрессия), см. EP 0465572 B1.
Однако самым успешным подходом является получение и смысловой, и антисмысловой РНК целевого гена («инвертированные повторы»), которые формируют двуцепочечную РНК (дцРНК) в клетке и вызывают молчание целевого гена. Методы и векторы, используемые для получения дцРНК и достижения молчания гена, описаны в EP 1068311, EP 983370 A1, EP 1042462 A1, EP 1071762 A1 и EP 1080208 A1. Вектор согласно настоящему изобретению может в этой связи включать участок регуляции транскрипции, который является активным в растительных клетках, оперативно связанный со смысловым и/или антисмысловым фрагментом ДНК NRC1 гена согласно настоящему изобретению. В основном, короткие (смысловые и антисмысловые) тяжи целевой генной последовательности, такие как включающие 17, 18, 19, 20, 21, 22 или 23 нуклеотида из кодирующей или некодирующей последовательности, являются достаточными. Могут также использоваться более длинные последовательности, такие как последовательности из 50, 100, 200 или 250 нуклеотидов или более. Предпочтительно, короткие смысловые и антисмысловые фрагменты разделяются спейсерной последовательностью, такой как интрон, которая формирует петлю (или шпилечную структуру) при формировании дцРНК. Может быть использован любой короткий тяж SEQ ID NO: 1 или 3 или их варианты для создания вектора с молчащим геном NRC1 и трансгенного растения, в котором достигнуто молчание одного или нескольких NRC1 генов во всех или некоторых тканях или органах (в зависимости от используемых промоторов). Удобным способом получения шпилечных конструкций является использование общих векторов, таких как pHANNIBAL и pHELLSGATE, векторов, которые основаны на методике Gateway® (см. Wesley et al. 2004, Methods Mol Biol. 265:117-30; Wesley et al. 2003, Methods Mol Biol. 236:273-86 и Helliwell & Waterhouse 2003, Methods 30(4):289-95), которые все включены в настоящее описание в качестве ссылок.
При выборе консервативных частей нуклеиновой кислоты гена NRC1 может быть достигнуто молчание для NRC1 семейства в хозяйском растении или в частях такого растения. Настоящее изобретение относится также к трансгенным растениям, включающим регуляторный элемент транскрипции, оперативно соединенный со смысловым и/или антисмысловым фрагментом ДНК NRC1 гена и демонстрирующим повышенную резистентность к одному или нескольким патогенам, в особенности, к некротрофным патогенам.
Дополнительно, рассматриваются растения, обладающие повышенной резистентностью к одному или нескольким биотрофным и/или хеми-биотрофным патогенам и к одному или нескольким некротрофным патогенам. Такие растения могут быть получены при выборе соответствующих сочетаний промотор - NRC1 ген. Так, например, функциональный белок NRC1 может быть получен в определенной ткани и в определенное время (например, при его индукции или в надземных частях растений), обеспечивающей резистентность к биотрофным и/или хемибиотрофным патогенам, и при этом достигается молчание одного из нескольких эндогенных генов NRC1 в другой ткани и/или в другое время (например, в проростках, в корнях или клубнях и т.п.), что способствует достижению резистентности к одному или нескольким некротрофным патогенам. В этой связи одно растение может включать и трансген, экспрессирующий химерный NRC1, и молчащий ген по NRC1.
Мутантные аллели и растения согласно настоящему изобретению
В одном из вариантов своего осуществления настоящее изобретение относится также к использованию нетрансгенных методов, например, систем мутагенеза, таких как TILLING (Targeting Induced Local Lesions IN Genomics; McCallum et al., 2000, Nat Biotech 18:455, и McCallum et al. 2000, Plant Physiol. 123, 439-442, где обе работы включены в настоящее описание в качестве ссылок), и соответствующий отбор для получения линий растений, которые продуцируют повышенные уровни одного или нескольких белков NRC1 согласно настоящему изобретению и/или которые продуцируют конститутивно активный белок NRC1 согласно настоящему изобретению. Не ограничивая область настоящего изобретения, следует полагать, что такие растения могут включать точечные/делеционные мутации в промоторе, которые представляют собой сайты связывания для репрессорных белков, что позволяет сделать хозяйский ген NRC1 конститутивным и повысить его экспрессию. Конститутивно активные мутанты NRC1 будут включать мутации в кодирующем участке, таком как MHD. Предпочтительно, уровни NRC1 белка в мутанте или частях мутанта составляют по меньшей мере примерно 2, 5, 10, 15% или более повышения в мутанте, в сравнении с немутантными растениями. В системе TILLING используется традиционный химический мутагенез (например, EMS мутагенез), который сопровождается высокомасштабным скринингом мутаций (например, с использованием Cel1-расщепления гетеродуплексов ДНК мутанта - дикого типа и последующего выявления продуктов путем секвенирования в гелевой системе) (см., например, Henikoff et al. Plant Physiology Preview May 21, 2004). Таким образом, нетрансгенные растения, семена и ткани, характеризующиеся повышенной экспрессией гена NRC1 в одной или нескольких тканях и включающие один или несколько фенотипов NRC1 согласно настоящему изобретению (повышенная резистентность к заболеванию и/или уровень HR повреждений), и способы создания и идентификации таких растений охватываются настоящим изобретением.
Рассматриваемый способ включает в одном из вариантов его осуществления стадии мутагенеза семян растений (например, EMS мутагенез), сбор индивидуальных растений или их ДНК, ПЦР-амплификацию участка, представляющего интерес, образование гетеродуплекса и высокомасштабное выявление, идентификацию мутантного растения и секвенирование ПЦР-продукта мутанта. Следует понимать, что равнозначно могут использоваться другие методы мутагенеза и отбора для получения таких мутантных растений. Так, семена могут быть, например, облучены или химически обработаны и полученные растения могут быть подвергнуты скринингу для определения наличия модифицированного фенотипа, такого как повышенная резистентность к заболеванию и/или наличие HR-повреждений.
В другом варианте осуществления настоящего изобретения используемые материалы растений представляют собой природные популяции видов или родственные виды, которые включают полиморфизмы или вариации в последовательности ДНК в кодирующей и/или регуляторной последовательности ортологов NRC1. Мутации в целевом гене NRC1 могут быть проскринированы, например, с использованием стратегии ECOTILLING (Henikoff et al 2004, выше). В рамках данной методики проводят скрининг природных полиморфизмов при линейном разведении или родственных видов по указанной выше методике TILLING, где используют индивидуальные растения или пулы растений для ПЦР-амплификации NRC1 мишени, получения гетеродуплекса и высокомасштабного анализа. Далее проводится отбор отдельных растений, содержащих нужную мутацию, которые могут быть впоследствии использованы в программах разведения с включением желательной NRC1-ортологичной аллели для целей создания сорта с нужным признаком.
Мутантные растения могут быть выявлены и отделены от немутантных с использованием методов молекулярной биологии, таких как выявление одной или нескольких мутаций, имеющихся в ДНК, определение уровней NRC1 белка, уровней РНК NRC1 и т.п., а также по модифицированным фенотипическим показателям.
Нетрансгенные мутанты могут быть гомозиготными или гетерозиготными по мутации.
Описываемые последовательности
SEQ ID NO 1: кодирующий участок NRC1 гена томата
SEQ ID NO 2: аминокислотная последовательность NRC1 белка томата
SEQ ID NO 3: полноразмерная последовательность кДНК NRC1 гена томата (включающая 5' и 3' UTR)
SEQ ID NO 4: аминокислотная последовательность белка NRC1D481V томата
SEQ ID NO 5: 3'UTR NRC1 гена томата
Описание чертежей
Фиг.1 - Предсказанная последовательность белка NRC1
Первые 150 аминокислотных остатков представляют сверхскрученный (CC) домен и остатки, которые, по прогнозам, формируют СС структуру, подчеркнуты. Остатки 151-508 включают нуклеотид-связывающий домен (NB-ARC), при этом подчеркнуты следующие мотивы (подчеркнуты и выделены): Киназа 1A (P петля), RNBS-A, Киназа 2, RNBS-B, RNBS-C, GLPL, RNBS-D и MHD. Остатки 509-846 включают 13 несовершенных лейцин-обогащенных повторов (LRR); консервативные гидрофобные и пролиновые остатки выделены жирным шрифтом. Под белковой последовательностью показан консенсусный мотив LRR: «l» обозначает консервативный алифатический остаток; «с» обозначает консервативный заряженный остаток и «P» обозначает консервативный пролиновый остаток.
Фиг.2A - NRC1 требуется для полного Cf-4-опосредованного HR томатов к Cladosporium fulvum
Cf-0 растения томата и Cf-4-содержащие растения томата инокулируют указанными TRV-конструкциями и растения анализируют через три недели после проявления VIGS. В листочки TRV-инфицированных Cf-4-содержащих растений томатов инъецируют белок Avr4 и выявляют развитие HR. Количество сайтов, проявляющих HR на TRV:00-инфицированных растениях, было взято за 100%. Каждое приведенное значение ошибки отражает стандартную ошибку, вычисленную по результатам четырех независимых экспериментов.
Фиг.2B - NRC1 требуется для полномасштабной Cf-4-опосредованной резистентности томатов к Cladosporium fulvum
В инфицированные не-TRV и в TRV-инфицированные Cf-4 или Cf-0 растения инокулируют C.fulvum-pGPD::GUS и через две недели после инокуляции оценивают колонизацию листочков в рамках X-gluc теста.
Фиг.3 - Инокуляция Н. benthamiana с использованием TRV:NRC1 воздействует на Cf/Avr-, LeEix2/tvEix-, Pto/AvrPto- и Rx/CP-индуцированный HR
В растения N. benthamiana инокулируют TRV:00 (пустой вектор), TRV:NRC1 и TRV:SGT1. Через три недели листья инфильтруются Agrobacteria, которые экспрессируют HR-индуцирующие белки и оценивают картину через 4 дня после инфильтрации. Первая, вторая и третья колонки: агроинфильтрация листьев N. Benthamiana, экспрессирующих ген резистентности Cf-4, с использованием Avr4 или смесей Cf-9 и Avr9 или смесей LeEix2 и tvEix (в соотношении 1:1), соответственно. В четвертой колонке: показаны листья трансгенных N benthamiana, которые экспрессируют ген резистентности Pto и которые были подвергнуты агроинфильтрации AvrPto. В пятой колонке показано: листья трансгенных растений N. Benthamiana, экспрессирующие ген резистентности Rx, которые были подвергнуты агроинфильтрации геном, экспрессирующим оболочечный белок PVX (CP). Темные кружки обозначают HR, светлые кружки указывают сниженный HR.
Фиг.4 - Конститутивно активный NRC1 индуцирует элиситор-независимый HR и позволяет локализовать NRC1 на сигнальном пути, ведущем к гибели клеток
Растения N. benthamiana, экспрессирующие ген резистентности Cf-4, подвергают агроинфильтрации указанными генами. На панелях А и С показаны растения за три недели до агроинфильтрации, в которые были инокулированы указанные конструкции TRV. Темные кружки указывают HR, светлые кружки обозначают сниженный HR.
(A) Агроинфильтрация генов, кодирующих конститутивно активные MAPKK и MAPK киназы. Первая колонка: агроинфильтрация геном, кодирующим конститутивно активный киназный домен LeMAPKKKα (MAPKKK-KD). Вторая колонка: агроинфильтрация геном, кодирующим конститутивно активную форму LeMEK2 (MEK2DD). Через два дня после инфильтрации индуцируют экспрессию MAPKKK-KD или MEK2DD путем распыления на листья эстрадиола. Картины оценивают через четыре дня после агроинфильтрации.
(B) Агроинфильтрация NRC1 дикого типа (wt) и мутированными формами генов под контролем 35S-промотора, либо в смеси 1:1 c Agrobacterium, направляющим экспрессию гена, кодирующего супрессор молчания pl9 (левая панель), либо одного гена (правая панель). NRC1 K191R (K191R): неактивный P-петлевой мутант NRC1; NRC1 D481V (D481V): конститутивно активный NRC1 (мутированный в MHD мотиве); NRC1K191R/D481v (K191R/D481V): двойной мутант NRC1. Картины оценивают через три дня после агроинфильтрации.
(C) Агроинфильтрация Avr4 и геном, кодирующим конститутивно активный NRC1D481V (D481V). Картины оценивают через три дня после агроинфильтрации.
Фиг.5 - Модель NRC1 опосредованной сигнальной функции, ведущей к гибели клеток
Показана модель эпистазиса в рамках экспериментов, объединяющих тесты по оценке гибели клеток и VIGS в N. benthamiana. Оценивают Cf-4/Avr4-опосредованные сигналы гибели клеток в режиме, независимом от EDS1-, NRC1-, MEK2- и SGT1/RAR1.
Приведенные ниже неограничивающие примеры иллюстрируют различные варианты осуществления настоящего изобретения. Если особо не указанное иное в приведенных примерах, все методики рекомбинантных ДНК проводят в соответствии со стандартными протоколами, описанными в литературе (Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, and Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press, NY; and in Volumes 1 and 2 Ausubel et al. (1994) Current Protocols in Molecular Biology, Current Protocols, USA). Стандартные материалы и методы для проведения молекулярных исследований на растениях описаны в Plant Molecular Biology Labfax (1993), by R.D.D. Croy, где указанная работа представляет собой совместную публикацию BIOS Scientific Publications Ltd (UK) и Blackwell Scientific Publications, UK.
ПРИМЕРЫ
1. МАТЕРИАЛЫ И МЕТОДЫ
1.1 VIGS в N. benthamiana. Агроинфильтрация, HR и тестирование заболеваний
Четырехнедельные растения N. benthamiana подвергают агроинфильтрации смесью, в соотношении 1:1, полученных на основе pTV00 конструкций (бинарный вектор TRV RNA2) и pBintra6 (бинарный вектор TRV RNA1) (Ratcliff et al., 2001 Plant J. 25, 237-245) или смесью, в соотношении 1:1, полученных на основе pTRV-RNA2 конструкций и pTRV-RNA1 (Liu et al., 2002, Plant J. 31, 777-786; Liu et al., 2002, Plant J. 30, 415-429). Используют следующие конструкции TRV: TRV:NRC1, TRV:Cf-4 и TRV:SGT1 (Peart et al., 2002, Proc. Natl. Acad. Sci. USA 99, 10865-10869), которые, все, входят в вектор TRV, как описано Ратклиффом с соавт. (Ratcliff et al. (2001, см. выше) и TRV:EDS1, TRV:MEK2, TRV:RAR1 и TRV:NDR1 (Ekengren et al., 2003, Plant J. 36, 905-917), все в векторе TRV, как описано Лью с соавт. (Liu et al. (2002, Plant J. 30, 415-429). Для каждой конструкции TRV в каждом эксперименте используют четыре растения. AvrPto и CP подвергают агроинфильтрации в TRV-инфицированные растения N. benthamiana, экспрессирующие ген резистентности Pto (N. benthamiana:Pto (линия 38-12) (Rommens et al., 1995, Plant Cell 7, 1537-1544)) (Pedley and Martin, 2003, Annu. Rev. Phytopathol. 41, 215-243) и Rx (N. benthamiana:Rx (линия Rx-18) (Bendahmane et al., 1999, Plant Cell 11, 781-791), соответственно. Во всех других случаях агроинфильтрацию проводят с использованием растений N. benthamiana, экспрессирующих гены резистентности Cf-4 (N. benthamiana:Cf-4). Через три недели после инокуляции TRV проводят провокацию третьего, четвертого и пятого листов выше инокулированных листьев с использованием Agrobacterium tumefaciens, которые направляют экспрессию AvrPto (OD600=0,06) (Tang et al., 1996, Science 274, 2060-2063), CP (OD600=0,12) (Bendahmane et al., 1999, см. выше), Avr4 (OD600=0,03), Cf-9 и Avr9 (смесь в соотношении 1:1, OD600=0,2) (Van der Hoorn et al., 2000 Mol. Plant-Microbe Interact. 13, 439-446), LeEix2 и tvEix (смесь в соотношении 1:1, OD600=1) (Ron and Avni, 2004, Plant Cell 16, 1604-1615), ген β-глюкуронидазы (GUS) (OD600=2) (Van der Hoorn et al., 2000, Mol. Plant-Microbe Interact. 13, 439-446), NRC1 и p19 (смесь в соотношении 1:1, OD600=l) (Voinnet et al., 2003, Plant J. 33, 949-956), конститутивно активного NRC1 D481V или неактивного двойного мутанта NRC1K191R/D481V (OD600=2), LeMAPKKKα KD, LeMAPKKKα KD- (Del Pozo et al., 2004, EMBO J. 23, 3072-3082) (оба имеют значение при OD600=0,12) или LeMEK2DD и LeMEK2 (Del Pozo et al., 2004, см. выше) (оба имеют значение при OD600=0,25). Через два дня после инфильтрации LeMAPKKKa KD , LeMAPKKKa KD -, LeMEK2DD или LeMEK2 на листья распыляют 7,5 мкМ раствор β-эстрадиола в воде, содержащий силвет (4 мкл/100 мл) (Del Pozo et al., 2004, см. выше). Для инъекций белка белок с фрагментом Avr4-HIS-FLAG обрабатывают энтерокиназой EK-max, в соответствии с инструкциями производителя (Invitrogen, Breda, NL), и используют для инъекций 5 мкМ Avr4 белка в воде с добавкой 0,2% (об./об.) твина. Через три-пять дней после агроинфильтрации или инъекции белка листья осматривают для оценки развития HR или определения активности β-глюкуронидазы (GUS).
1.2 VIGS в растениях томатов, тесты на HR и заболевания
Для получения VIGS в растениях томатов используют векторы pTRV-RNA1 и pTRV-RNA2, описанные Лью с соавт. (Liu et al. (2002, Plant J. 30, 415-429). Фрагменты Cf-4 и NRC1 вырезают путем расщепления pTV00 с использованием BamHI/Asp718 и встраивают в BamHI/Asp718-расщепленный pTRV-RNA2 (pYL156) (Liu et al., 2002, Plant J. 31, 777-786). Для конструирования TRV:222-UTR часть 3'-UTR из NRC1 амплифицируют с использованием праймеров 222-3'UTR-F (5'-GTGGATCCGCAGGTTCAACCAGCCTGGT-3'; сайт BamHI подчеркнут) и 222-3'UTR-R (5'-GTGGTACCCAAGTGACTTGTTCTGCTGT-3'; сайт Asp718 подчеркнут) и для конструирования TRV:222-LRR проводят амплификацию части участка NRC1, кодирующего LRR с использованием праймеров
222-LRR-F (5'-GTGGATCCGTTAAGAGGCTGCAATTTCT-3'; сайт BamHI подчеркнут) и 222-LRR-R (5'-GTGGTACCGATCTTCTCAAGTTTATCAC-3'; сайт Asp718 подчеркнут). ПЦР фрагменты расщепляют BamH1/Asp718 и встраивают в BamHI/Asp718-расщепленный pTRV-RNA2. Конструирование TRV:Prf было описано в литературе (Ekengren et al., 2003, Plant J. 36, 905-917). Все плазмиды трансформируют A. tumefaciens, штамм GV3101 путем электропорации (Takken et al., 2000, Plant J. 24, 275-283). Для установления VIGS в томатах семяпочки десяти-двенадцатидневных проростков томатов подвергают агроинфильтрации смесью pTRV-RNA1- и pTRV-RNA2-расщепленных конструкций (объединенные в соотношении 1:1) (Liu et al., 2002, см. выше). Для каждой конструкции TRV используют либо четыре Cf-4-содержащих растения томатов (Cf-0 растения, трансформированные Hcr9-4D (Cf-4)) (Thomas et al., 1997, Plant Cell 9, 2209-2224), резистентные к C. fulvum, экспрессирующему Avr4, либо четыре Cf-9-содержащих растения томатов (Cf-0 растения, трансформированные Hcr9-9C (Cf-9)) (Jones et al., 1994, Science 266, 789-793), резистентные к C. fulvum, экспрессирующему Avr9. В качестве контроля используют Cf-0 растения томатов (MM-Cf-0), полностью чувствительные к C. fulvum, при инокуляции или TRV:00, или TRV:NRC1. Для тестирования заболеваний через три недели после TRV-инокуляции в Cf-0- и Cf-4-содержащие растения инокулируют C. fulvum (De Wit, 1977, Neth. J. Plant Path. 83, 109-122). Используют C. fulvum штамм 5-pGPD::GUS (который экспрессирует Avr4 и ген β-глюкуронидазы под контролем конститутивного промотора GPD). Колонизацию листочков оценивают через две недели после окрашивания в тесте на X-gluc. Параллельно анализируют листочки второго, третьего или четвертого сложного листа в рамках ОТ-ПЦР анализа для тестирования «выключения» интересующего гена (см. приведенные ниже данные). Для тестов на HR в листочки третьего сложного листа TRV-инфицированных Cf-4- или Cf-9-содержащих растений инъецируют Avr4 или Avr9 соответственно. Оба элиситора инъецируют в листочки с помощью микрошприца (Ito Corporation, Fuji, Japan). Avr4 инъецируют в концентрации 10 мкМ, в десяти сайтах на листочек и в четыре листочка на растение. В случае Avr9, апопластическую жидкость в восьмикратном разбавлении, содержащую примерно 10 мкМ Avr9, выделенную после совместимого взаимодействия между расой 5 C. fulvum и Cf-0-растениями, инъецируют в восьми сайтах на листочек и в четыре листочка на растение. Резистентность против Pseudomonas syringae pv. tomato оценивают в растениях томатов RG-PtoR (Pto/Pto, Prf/Prf), при инокуляции TRV:00, TRV:Prf или TRV:NRC1. Процедуру инокуляции и определения колонизации бактерий на листьях проводят по описанной ранее методике (Ekengren et al., 2003, см. выше).
1.3 Конструирование бинарного вектора 35S:NRC1 и мутагенез
Полноразмерную кДНК NRC1 подвергают ПЦР-амплификации с использованием праймеров
222-Старт-F (5'-GGGATCCATGGTTGATGTAGGGGTTGA-3') и
222-Стоп-R (5'-GTCACTGCAGACCTTTCTAAGAAGCTGTCTG-3'), в ходе которой достигается встраивание сайта рестрикции NcoI и PstI, соответственно (сайты рестрикции подчеркнуты). ПЦР фрагмент представляет собой NcoI/PstI-расщепленный продукт, который встраивают в NcoI/PstI-расщепленный pRH80 (Van der Hoorn et al., 2000, Mol. Plant-Microbe Interact. 13, 439-446). Далее конструкцию расщепляют с использованием XbaI/KpnI и полученный фрагмент, содержащий 35S промотор, открытую рамку считывания для NRC1 и NOS терминатор (tNOS) клонируют в XbaI/KpnI-расщепленном бинарном векторе pMOG800 (Honée et al., 1998, Plant Physiol. 117, 809-820) с образованием плазмиды NRC1(wt). Для создания конститутивно активного бинарного NRC1 D481V встраивают мутацию D481V в рамках перекрывающейся реакции ПЦР-расширения (Higuchi et al., 1988, Nucleic Acids Res. 16, 7351-7367) с использованием плазмиды NRC1wt в качестве матрицы и фланкирующих праймеров 222-Старт-F и 222-Стоп-R и ошибочных праймеров
222MHD-F (5'-CAAAACTTGTCGTGTTCATGTCATGTTGTATGAG-3') и
222MHD-R (5'-CCAGCAAAACTCATACAACATGACATGAACACGAC-3')
(мутация подчеркнута).
Фрагмент, который представляет собой продукт расщепления NcoI/PstI, встраивают в pRH80, вырезают фрагмент 35S-NRC1 D481V -tNOS и встраивают в pMOG800, описанный выше. Аналогичным образом получают P-петлевой мутант NRC1 K191R и неактивный двойной мутант NRC1 K191R/D481V. При этом мутацию K191R встраивают с использованием неправильных праймеров
222петля-F (5'-GGAATGCCTGGTCTTGGCAGAACTACACTAGC-3') и
222петля-R (5'-GCTAGTGTAGTTCTGCCAAGACCAGGCATTCC-3') (мутация подчеркнута) с использованием в качестве матрицы плазмид NRC1(wt) и NRC1 D481V, соответственно. Все конструкции были подтверждены путем секвенирования и трансформированы в A. tumefaciens штамм GV3101.
1.4 Гель-блот анализ ДНК
Геномную ДНК из N. benthamiana выделяют с использованием протокола экстракции ДНК QIA-Gen (Qiagen, Venlo, NL), а для растений томатов используют стандартный протокол, описанный в руководстве Самбрука и Расселла (Sambrook and Russell, 2001, Molecular cloning: A Laboratory Manual, 3rd ed. (Cold Spring Harbor, NY, U.S.A.: Cold Spring Harbor Laboratory Press). ДНК расщепляют с использованием BamHI, HindIII, EcoRI, EcoRV или XbaI. Гель-блотты из N. benthamiana гибридизуют с 32P-меченым фрагментом размером 252 п.н. (Prime-a-gene Labeling System, Promega, Madison, WI), присутствующим в векторе TRV:NRC1, и гель-блот ДНК из томатов гибридизуют с 32P-меченым зондом из 1293 оснований, соответствующим нуклеотидам 1876-3168 в полноразмерной кДНК для NRC1. Используемые для рестрикционных ферментов сайты не присутствуют в зондах. Условия низкой жесткости относятся к промывке при температуре 55°C в 2x SSC и 0,5% ДСН. Условия высокой жесткости включают промывку при температуре 65°C в 0,5x SSC и 0,5% ДСН.
1.5 ОТ-ПЦР, проводимая для демонстрации молчания гена NRC1 в томатах
Четыре листовых диска (примерно по 100 мг ткани в целом) собирают со второго, третьего или четвертого сложного листа из TRV-инфицированных растений. Суммарную ДНК экстрагируют с использованием протокола экстракции QIA-Gen RNAeasy (Qiagen, Venlo, NL) и обрабатывают не содержащей РНКазы ДНКазой (Bio-Rad, Veenendaal, NL). Первую цепь кДНК синтезируют на основе 1 мкг суммарной РНК с использованием набора для синтеза кДНК Био-Рад (Bio-Rad, Veenendaal, NL) и проводят ОТ-ПЦР с использованием следующих циклов: 95°C в течение 15 секунд, 60°C в течение 45 секунд и 72°C в течение 60 секунд. Используют следующие праймеры:
(222F: 5'-TGAGGTATATTGCTTTCTCATCTGAC-3' и
222R: 5,-AGCTATTTTCCCACGGATGCCCAG-3')
которые не охватывают фрагмент, встроенный в TRV:NRC1. Используют актиновые праймеры
(АктинFnrl82: 5'-TATGGAAACATTGTGCTCAGTGG-3' и
АктинRnrl83: 5'-CCAGATTCGTCATACTCTGCC-3')
для подтверждения наличия равных количеств кДНК в реакциях ПЦР.
Пример 2 - Результаты
2.1 NRC1 томатов: белок CC-NB-LRR
Проводят анализ по методу кДНК-AFLP и затем VIGS идентифицированных фрагментов из томатов для N. benthamiana:Cf-4. Идентифицируют 20 фрагментов кДНК, в которых VIGS затрагивает Cf-4/Avr4-индуцированный HR. В случае одного из них, NRC1, выделяют полноразмерную кДНК, описываемую в виде SEQ ID NO: 3. Открытая рамка считывания, которая показана в виде SEQ ID NO: 1, кодирует белок NRC1, представленный в виде SEQ ID NO: 2.
Предсказанная первичная структура белка NRC1 (SEQ ID NO: 2) в типичном случае отражает структуру белков резистентности CC-NB-LRR (фиг. 1). NRC1 содержит амино-концевой суперскрученный домен (CC), NB-ARC домен (нуклеотид-связывающий адаптер, общий для Apaf-1, R белков и для CED4) (Van der Biezen and Jones, 1998, Curr. Biol. 8, R226-R227; Aravind et al., 1999, Trends Biochem. Sci. 24, 47-53) и 13 несовершенных лейцин-обогащенных повторов (LRR). Как показано на фиг. 1, сравнение с гомологичными доменами NB-ARC выявляет наличие мотива киназы 1A или P-петлевого мотива, четыре RNBS мотива (нуклеотид-связывающий сайт, определяющий резистентность) и GLPL и MHD мотив (Meyers et. al., 1999, Plant J. 20, 317-332; Meyers et al., 2003, Plant Cell 15, 809-834).
Фрагмент кДНК-AFLP размером 252 п.н., присутствующий в TRV:NRC1 векторе и используемый для VIGS, кодирует аминокислоты на участке 599-681, которые локализованы на четвертом-седьмом LRR.
Гель-блот анализ ДНК в условиях низкой жесткости геномной ДНК томатов, расщепленных BamHI-, HindIII-, EcoRI-, EcoRV- и XbaI проводят при гибридизации с фрагментом кДНК для NRC1 размером 1293 п.н. (участок нуклеотидов с 1876 по 3168 из SEQ ID NO: 3), который охватывает последовательность NRC1, присутствующую в конструкциях TRV:NRC1-, TRV:NRC1-LRR- и TRV:NRC1-UTR (см. ниже), в качестве зонда. Указанный саузерн-блоттинг выявляет только одну выраженную полосу после промывки в условиях высокой жесткости, указывая на то, что NRC1 представляет однокопийный ген в растениях томатов.
Гель-блоты BamHI-, HindIII-, EcoRI-, EcoRV- и XbaI-расщепленной геномной ДНК N. benthamiana зондируют фрагментом кДНК-AFLP для NRC1, присутствующим в векторе TRV, и выявляют две-три гибридизующиеся полосы (результаты не показаны) (0,5 х SSC, 0,5% ДСН, 65°C). Полученный результат позволяет полагать, что имеется по меньшей мере два-три ортолога NRC1 в геноме N. benthamiana, которые могут стать молчащими при инокуляции TRV:NRC1.
2.2 NRC1-молчащие растения томата поражаются при Cf-4-опосредованном HR и разистентности к заболеванию
Для исследования функции NRC1 в HR-сигнальной активности и резистентности к C. fulvum, авторы получили VIGS в растениях томатов, поскольку указанные растения являются единственным хозяином для данного гриба. Десятидневные сеянцы томатов подвергают агроинфильтрации с использованием TRV:NRC1 и через три недели после инфильтрации выделяют РНК из листочков с потенциально молчащим геном и проводят анализ по методу ОТ-ПЦР. Уровни транскриптов NRC1 варьировали в различных TRV:NRC1 -инфицированных растениях, но в большинстве случаев они были ниже, чем в TRV:00-инфицированных растениях, указывая на то, что имеет место «выключение» экспрессии NRC1 (данные не показаны).
Для исключения возможности того, что наблюдаемый в растениях томатов фенотип вызван молчанием дополнительных белков NB-LRR, авторы изобретения провели VIGS в томатах с использованием фрагмента NRC1 размером 360 п.н., направленного на восьмой-двенадцатый LRR (TRV:NRC1-LRR), и фрагмент, состоящий из 297 п.н. из 3'-нетранслируемого участка (UTR) NRC1 (TRV:NRC1-UTR). С использованием указанных конструкций авторы проверили, необходим ли NRC1 для Cf-4-опосредованного HR в растениях томатов, путем инъекции Avr4 белка в TRV:222-LRR и TRV:222-UTR инфицированные Cf-4-содержащие растения томатов. Молчание NRC1 (с использованием каждой из трех конструкций) приводит к получению смягченного фенотипа, поскольку растения томата, по всей видимости, демонстрируют меньший эффект, чем TRV:00- или TRV:Cf-4-инфицированные растения (данные не приведены). В качестве контролей использовали варианты, в которых Avr4 белок был инъецирован в TRV:00- и TRV:Cf-4-инфицированные растения. В случае TRV:Cf-4-инфицированных растений, процент отзывающихся на инъекцию Avr4 сайтов составлял 52% (фиг. 2), указывая на то, что снижение HR связано с молчанием Cf-4. В случае TRV:222-LRR- и TRV:222-UTR-инфицированных растений указанный процент был ниже (56% и 48%, соответственно) (фиг. 2), подтверждая функцию NRC1 в Cf-4/Avr4-индуцированном HR также и в томатах. Аналогичные результаты были получены при оценке VIGS для NRC1 в Cf-9-содержащих растениях томатов и последующих инъекциях апопластической жидкости, содержащей Avr9 (результаты не показаны). Для проведения VIGS с использованием Cf-9 в Cf-9-содержащих растениях томатов авторы использовали конструкцию TRV:Cf-4, поскольку фрагмент Cf-4 размером 404 п.н. кодирует высококонсервативные LRR с 15 по 21, позволяя тем самым достичь молчания Cf-4 и гомологичного гена резистентности Cf-9 (Van der Hoorn et al., 2001, см. выше.).
Кроме того, исследовали, требуется ли NRC1 также для достижения полной резистентности растений томатов к C. fulvum. В Cf-0 и Cf-4-растения инокулируют TRV:00, TRV:Cf-4 и TRV:NRC1, и через три недели в растения с молчанием гена инокулируют штамм C. fulvum, экспрессирующий Avr4 и ген β-глюкуронидазы (GUS), позволяющий визуализировать рост гриба. Через две недели после инокуляции C. fulvum листья проводят тест с окрашиванием по X-gluc. В листочках Cf-4 растений, инфицированных TRV:00, не наблюдается рост C. fulvum, тогда как очажки синего окрашивания в TRV:Cf-4-инфицированных Cf-4 растениях указывают на сниженную Cf-4-опосредованную резистентность (результаты не показаны). Кроме того, в TRV:NRC1-инфицированных растениях маленькие очажки синего окрашивания указывают на полную потерю резистентности против данного гриба. Микроскопирование выявляет межклеточный рост грибных гифов в TRV:Cf-4- и TRV:NRC1-инфицированных растениях, но не в TRV:00-инфицированных контрольных растениях. Все Cf-0 растения демонстрируют обширную колонизацию C. fulvum, указывая на то, что ни TRV инфекция сама по себе, ни VIGS с использованием TRV:NRC1 не затрагивают чувствительность данных растений к грибу.
2.3 VIGS для NRC1 затрагивает HR, индуцированную разными сочетаниями R гена спаривания /Avr гена
Кроме снижения Cf-4/Avr4-индуцированного HR при VIGS с использованием NRC1, было также показано, что HR, индуцированный Infl элиситором патогенного оомицета Phytophthora infestans, снижается в случае VIGS с использованием NRC1 в N. benthamiana.
Для дальнейшего исследования специфичности NRC1 в сигнальной защитной функции авторы изобретения исследовали ее значимость для HR, индуцированного другими сочетаниями R/Avr. В качестве контролей были использованы TRV:00 (пустой вектор) и TRV:SGT1, поскольку известно, что SGT1 необходим для HR, индуцированного несколькими сочетаниями R/Avr (Peart et al., 2002, Proc. Natl. Acad. Sci. USA 99, 10865-10869).
Агроинфильтрация смесью Cf-9 и Avr9 (Van der Hoorn et al., 2000, см. выше) или смесью LeEix2 и tvEix (Ron and Avni, 2004, Plant Cell 16, 1604-1615) TRV:NRC1-инфицированных растений N. benthamiana приводит к снижению HR, тогда как TRV:00-инфицированные растения демонстрируют развитие нормального HR. В TRV:SGT1-инфицированных растениях HR полностью устраняется, подтверждая результаты Пирта с соавт. (Peart et al. 2002, см. выше) (фиг. 3). Кроме того, AvrPto из бактериального патогена Pseudomonas syringae pv tomato и ген, кодирующий белок оболочки (CP) вируса картофеля X (PVX), подвергают агроинфильтрации в TRV-инфицированные N. benthamiana, экспрессирующие ген резистентности Pto (Pedley and Martin, 2003, Annu. Rev. Phytopathol. 41, 215-243) и Rx (Bendahmane et al., 1999, Plant Cell 11, 781-791), соответственно. В обоих случаях растения, инфицированные TRV:00, демонстрируют HR, тогда как HR устраняется в TRV:SGT1-инфицированных растениях. TRV:NRC1-инфекция приводит к серьезному снижению Pto/AvrPto-, а также Rx/CP-индуцированного HR, и это указывает на то, что в N. benthamiana белок NRC1 необходим для HR сигнальной функции, активируемой несколькими генными сочетаниями R/Avr (фиг. 3).
Для исключения возможности того, что недостаточно выраженный HR в TRV:NRC1-инфицированных растениях N. benthamiana определяется сниженной эффективностью трансформации Agrobacterium, авторы провели инфильтрацию TRV:00- и TRV:NRC1-инфицированных N. benthamiana:Cf-4 с использованием Agrobacterium, экспрессирующим ген β-глюкуронидазы (GUS) (Van der Hoorn et al., 2000, см. выше). Через три дня после инфильтрации в TRV:00- и TRV:NRC1-инфицированных растениях была выявлена аналогичная интенсивность синего окрашивания, подтверждающая, что трансформация растений Agrobacterium не затрагивается (данные не показаны). Кроме того, TRV:NRC1-инфицированные растения также демонстрируют HR при инъекции белка Avr4, тогда как TRV:00-инфицированные растения демонстрируют четкое развитие HR в течение 2 дней.
2.4 NRC1 действует в направлении считывания информации EDSl и против направления считывания информации в каскадном пути MAPK в сигнальном пути гибели клеток
Поскольку NRC1 требуется не только для Cf-4/Avr4-индуцированного HR, но также для HR, индуцируемого некоторыми дополнительными R/Avr сочетаниями, следует полагать, что NRC1, по-видимому, вовлекается в общий путь HR сигнальной активности. Типичным ответом организма-хозяина, который предшествует инициации HR, является MAPK каскад (Romeis et al., 2001, EMBO J. 20, 5556-5567; Del Pozo et al., 2004, EMBO J. 23, 3072-3082; Pedley and Martin, 2005, Plant Biol. 8, 541-547).
Для исследования потребности в NRC1 для HR инициированных MAPK проводят эксперименты по эпистазису в растениях N. benthamiana. В растения инокулируют TRV:00, TRV:SGT1 и TRV:NRC1 и далее проводят агроинфильтрацию генами, кодирующими киназный домен LeMAPKKKα (LeMAPKKKαKD ) или конститутивно активный LeMEK2 (LeMEK2DD) (Yang et al., 2001, Proc. Natl. Acad. Sci. USA 98, 741-746; Del Pozo et al., 2004, EMBO J. 23, 3072-3082). Через два дня после агроинфильтрации индуцируют экспрессию генов путем опрыскивания инфильтрованных листьев эстардиолом. Временная экспрессия каждого из генов приводит к появлению HR в TRV:00-инфицированных растениях, тогда как в TRV:SGT1-инфицированных растениях HR снижается (фиг 4А). В TRV:NRC1-инфицированных растениях HR, вызванная обеими конститутивно активными киназами, не затрагивается (фиг. 4А). Агроинфильтрация соответствующих негативных контролей LeMAPKKKa KD- и LeMEK2 дикого типа не приводит к HR в любом из TRV-инфицированных растений (данные не показаны). Полученные результаты означают, что SGT1 функционирует в направлении считывания информации для данных MAPK, при этом MAPK действуют либо по пути считывания информации NRC1, либо независимо от NRC1.
2.5 Временная суперэкспрессия NRC1 и конструирование конститутивно активного белка NRC1
Для дальнейшего выяснения, какие гены необходимы для HR сигнальной функции применительно к CC-NB-LRR белку, исследуют эффект суперэкспрессии NRC1. В этой связи кодирующую последовательность (SEQ ID NO: 1) из кДНК конденсируют с конститутивным 35S промотером и встраивают в бинарный вектор. Агроинфильтрация данной конструкции в вектор N. benthamiana не приводит к развитию HR, тогда как экспрессия смеси NRC1 и ингибитора молчания p19 (Voinnet et al., 2003, Plant J. 33, 949-956) провоцирует образование элиситор-независимого HR (фиг. 4B). Агрофильтрация конструкции, кодирующей Р-петлевой мутант NRC1 (K191R), разрушает Р-петлевой мотив, затрагивая гидролиз АТФ (Tameling et al., 2002, Plant Cell 14, 2929-2939) либо с использованием pl9, либо без него, и это не приводит к развитию HR (фиг. 4B).
Приведенные выше данные указывают, что посттранскрипционное молчание гена (PTGS) применительно к гену NRC1 может препятствовать развитию HR в ткани, демонстрирующей суперэкспрессию NRC1. Кроме того, разрушение Р-петлевого мотива приводит к образованию неактивного белка NRC1.
Поскольку мутации в MHD мотиве в NB-LRR белков резистентности Rx (D460V) (Bendahmane et al., 2002; Tameling et al., 2002) и I-2 (D495V) (Bendahmane et al., 2002, Plant J. 32, 195-204; Tameling et al., 2002, Plant Cell 14, 2929-2939; Van Bentem et al., 2005, Plant J. 43, 284-298) приводят к достижению конститутивной активности, авторы изобретения получили аналогичный мутант для NRC1 (NRC1D481V). Фактически, агроинфильтрация NRC1D481V приводит к появлению элиситор-независимых HR в листьях N. benthamiana в течение трех дней после инфильтрации, и снова при агроинфильтрации двойного мутанта NRC1K191R/D481V не наблюдается HR (фиг 4B). Кроме того, не индуцируется HR при экспрессии NRC1D481V в растениях с молчащим SGT1 (см. ниже). Приведенные результаты показывают, что ответ, индуцированный при агроинфильтрации NRC1D481V, является специфичным в связи с конститутивной активностью белка NRC1 и что функция NRC1 в каскаде передачи сигнала приводит к развитию HR.
2.6 Эксперименты по эпистазису с использованием NCR1 белка
Эксперименты по эпистазису с использованием NRC1D481V проводят для дальнейшего выяснения, какие гены необходимы для HR-сигнальной функции указанного белка и в этой связи для определения его предположительной роли в пути HR. Кроме VIGS генов, в отношении которых известно, что они вовлекаются в HR сигнальную функцию, таких как SGT1 и RAR1 (требуемые для резистентности к Mlal2) (Shirasu and Schulze-Lefert, 2003, Trends Plant Sci. 8, 252-258), в N. benthamiana:Cf-4 было достигнуто молчание для NDR1 (резистентность к заболеванию, неспецифичная по штамму) (Century et al., 1995, Proc. Natl. Acad. Sci. USA 92, 6597-6601), EDSl (повышенная чувствительность к заболеванию (Aarts et al., 1998, Proc. Natl. Acad. Sci. USA 95,10306-10311) и MEK2 (MAPKK) (Ekengren et al., 2003, Plant J. 36, 905-917), и впоследствии проведена агроинфильтрация NRC1D481V или Avr4. Кроме того, VIGS с использованием TRV:00, TRV:Cf-4 и TRV:NRC1 были включены в качестве контролей. Снижение NRC1D481V- или Avr4-индуцированного HR указывает на выключение гена, нужного для NRC1- или Cf-4/Avr4-индуцированного сигнального HR, соответственно.
Как и ожидалось, HR, индуцированный при агроинфильтрации Avr4, снижается в TRV:Cf-4- и TRV:NRC1-инфицированных растениях. Cf-4-опосредованная сигнальная функция также требует присутствия EDS1, так как растения, молчащие по данному гену, проявляют менее выраженный Avr4-индуцированный HR. Дополнительно, авторы обнаружили сниженный HR при агроинфильтрации Avr4 в растения с молчанием по MEK2, RAR1 и SGT1 (фиг. 4C; светлые кружки). Avr4-индуцированный HR не снижается в TRV:00- и TRV:NDR1-инфицированных растениях (фиг. 4C; темные кружки), указывая на то, что NDR1 не требуется для осуществления Cf-4-опосредованной сигнальной функции. Аналогично, NRC1D481v-индуцированный HR не ухудшается в TRV:00- и TRV:NDR1-инфицированных растениях, а также не снижается в TRV:Cf-4-инфицированных растениях. Интересно отметить, что, в отличие от Avr4, NRC1D481V продолжает индуцировать HR в TRV:EDS1-инфицированных растениях, указывая на то, что NRC1 функционирует в направлении считывания информации в EDS1 (фиг. 4C; темные кружки). NRC1D481v-индуцированный HR снижается в растениях с молчанием по MEK2, и это указывает на то, что NRC1 требует наличия MAP-киназного каскада для его сигнальной функции и может быть расположен против направления считывания информации для данных киназ. VIGS для RAR1 и SGT1 также ухудшает D481V-индуцированный HR, аналогично HR, который индуцируется Avr4 (фиг 4C; светлые кружки). Таким образом, NRC1 необходим для HR сигнальной функции, инициируемой Cf-4, и может быть расположен против направления считывания информации по MAPK каскаду и по направлению считывания информации для EDS1.
См. фиг. 5, на которой показана модель NRC1, опосредованной сигнальной функции клетки.
Пример 3 - Потребность в NRC1 для Mi-опосредованной резистентности
Для того чтобы определить, необходим ли NRC1 для Mi-опосредованной резистентности против нематод, белокрылки и тли, конститутивно активную формую Mi подвергают агроинфильтрации (см. US 6613962 и EP0937155B1) в NRC1-молчащие растения. Сниженный HR в NRC1-молчащих растениях указывает на то, что NRC1 также необходим для Mi-опосредованного HR и что (супер)экспрессия NRC1 может использоваться для получения трансгенных растений, обладающих повышенной резистентностью против нематод, белокрылок и тли.











| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОВЫШЕНИЯ РЕЗИСТЕНТНОСТИ ТРАНСГЕННЫХ РАСТЕНИЙ К ПАТОГЕННЫМ ВОЗДЕЙСТВИЯМ | 2007 |
|
RU2428480C9 |
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ПРИДАНИЯ РАСТЕНИЯМ УСТОЙЧИВОСТИ К ЗАБОЛЕВАНИЯМ | 1996 |
|
RU2203320C2 |
| АУТОАКТИВИРУЮЩИЙСЯ БЕЛОК УСТОЙЧИВОСТИ | 2006 |
|
RU2375453C2 |
| КЛОНИРОВАНИЕ И ИСПОЛЬЗОВАНИЕ ФУНКЦИОНАЛЬНОГО R-ГЕНА ИЗ SOLANUM CHACOENSE | 2010 |
|
RU2586509C2 |
| УСТОЙЧИВЫЕ К БАКТЕРИЯМ ТРАНСГЕННЫЕ РАСТЕНИЯ | 2012 |
|
RU2640246C2 |
| ГЕН, ПРИДАЮЩИЙ УСТОЙЧИВОСТЬ К PHYTOPHTHORA INFESTANS (ФИТОФТОРОЗУ) В СЕМЕЙСТВЕ SOLANACEAE | 2003 |
|
RU2361920C2 |
| НОВЫЙ ИММУНОГЕННЫЙ ЭКСТРАКТ ГРИБОВ И ПАТТЕРН-РАСПОЗНАЮЩИЙ РЕЦЕПТОР РАСТЕНИЙ | 2013 |
|
RU2700621C2 |
| ПРИМЕНЕНИЕ ДЕНДРИМЕРНОЙ НАНОТЕХНОЛОГИИ ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ | 2010 |
|
RU2571928C2 |
| ТРАНСГЕННЫЕ РАСТЕНИЯ С УЛУЧШЕННЫМИ ХАРАКТЕРИСТИКАМИ РОСТА | 2009 |
|
RU2582260C2 |
| ПОЛУЧЕНИЕ ПИКОРНАВИРУСОПОДОБНЫХ ЧАСТИЦ В РАСТЕНИЯХ | 2013 |
|
RU2693431C2 |

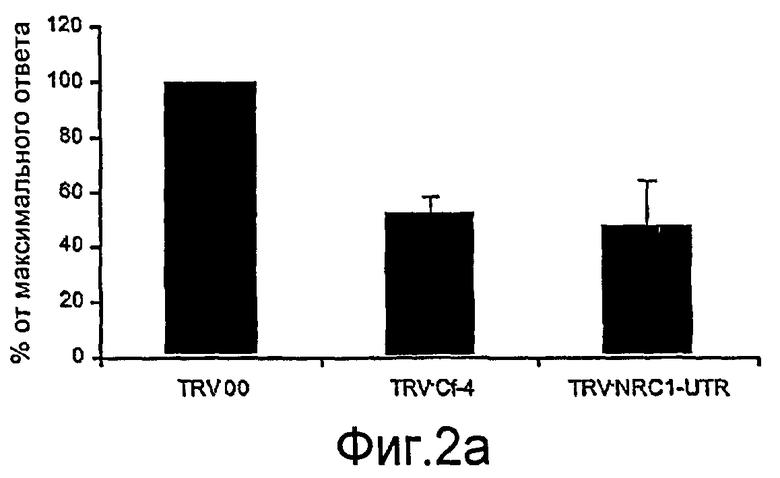
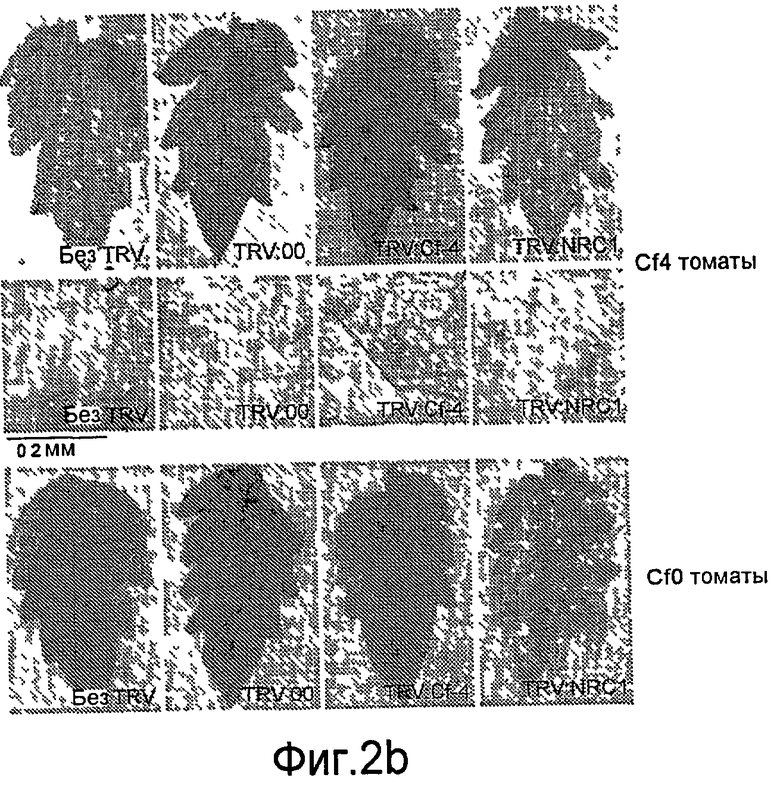
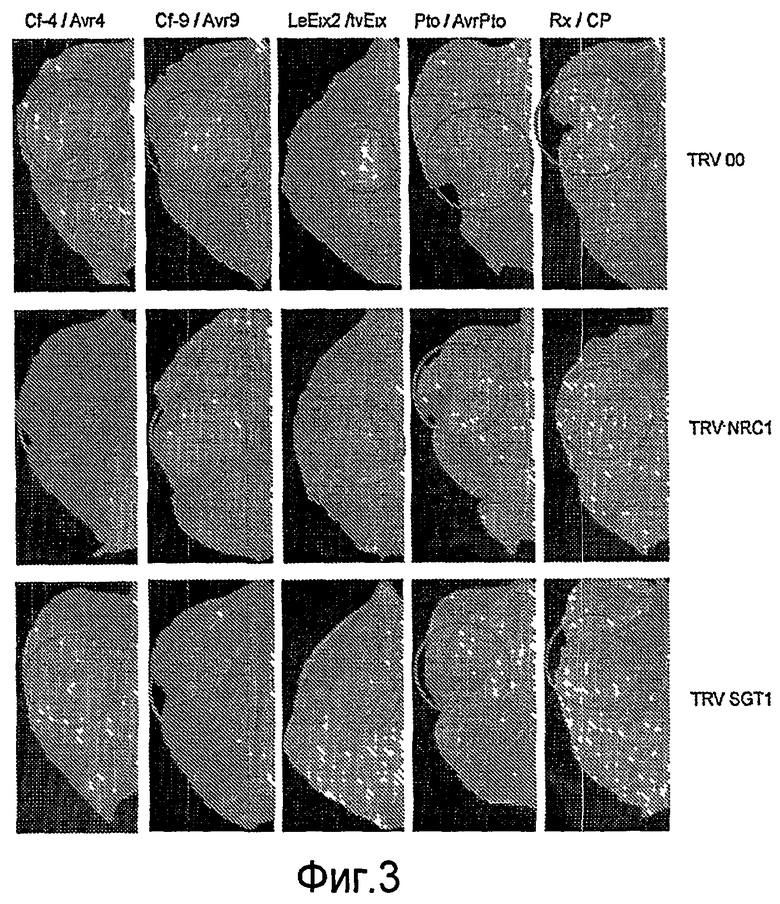
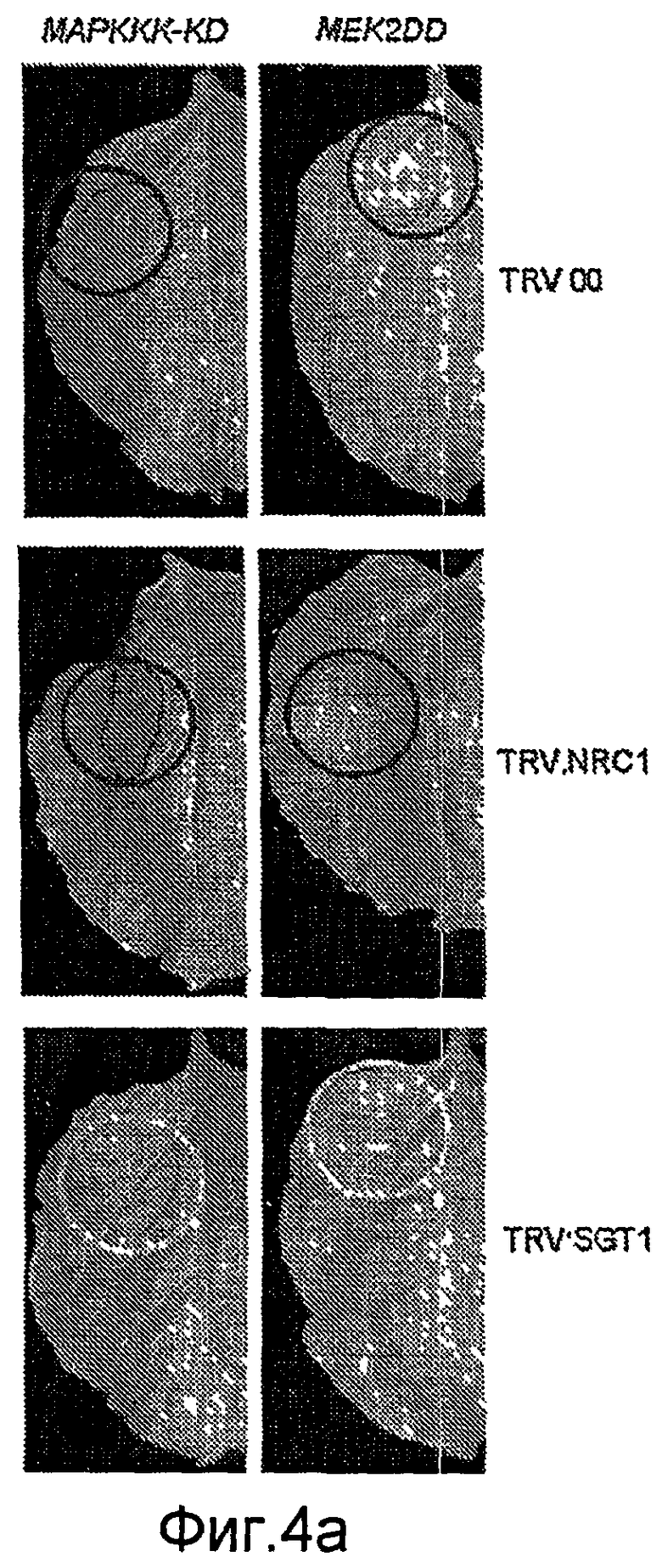
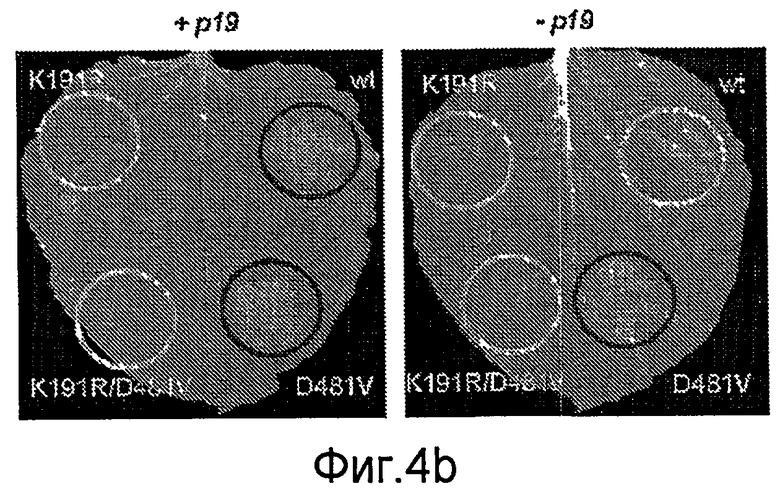
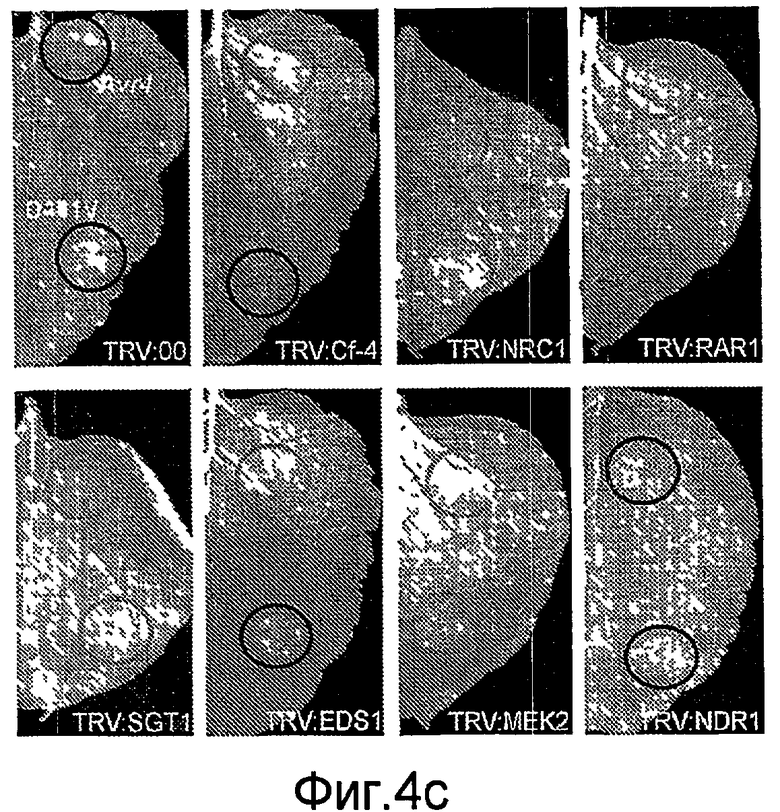
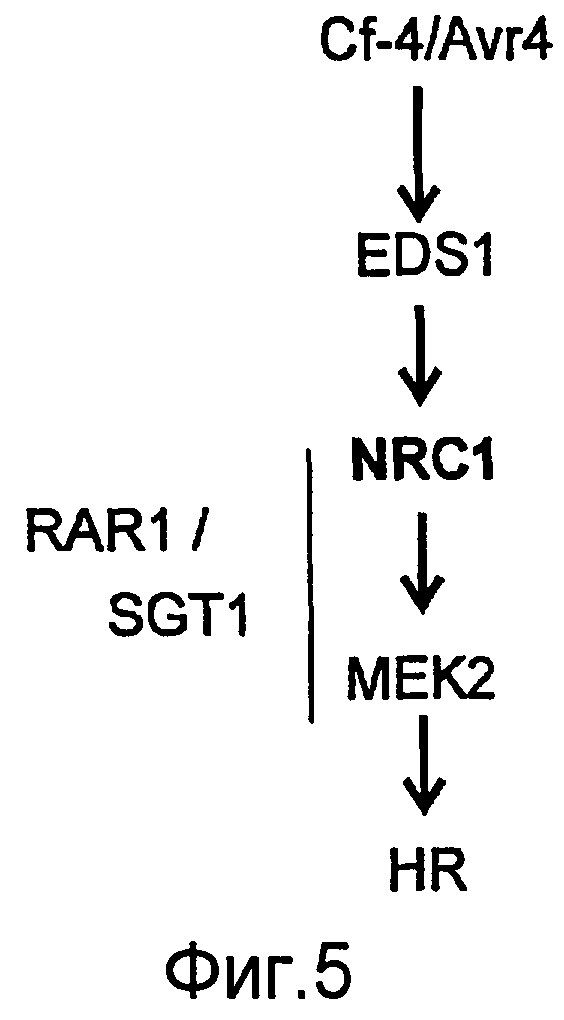
Растения или растительные клетки трансформируют нуклеотидной последовательностью, кодирующей белок, обозначенный как NRC1. Экспрессия указанного белка в растениях придает им повышенную устойчивость к патогенам. 7 н. и 5 з.п. ф-лы, 8 ил.
1. Способ получения трансгенного растения, обладающего повышенной резистентностью к патогену в сравнении с нетрансгенным контрольным растением, причем указанный способ включает стадии
(a) трансформации растения или клетки растения нуклеотидной последовательностью, кодирующей белок NRC1, оперативно связанной с промотором, активным в растительных клетках, причем указанный белок NRC1 включает аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:4 или аминокислотную последовательность, обладающую, по меньшей мере, 70%-ной идентичностью по аминокислотной последовательности с SEQ ID NO:2 по всей ее длине
(b) регенерации растения.
2. Способ по п.1, где указанная нуклеотидная последовательность интегрирована в геном указанного растения.
3. Способ по п.1 или 2, дополнительно включающий стадию
(c) скрининга регенерированного растения или растения, полученного из него путем самоопыления или скрещивания, на резистентность к одному или нескольким растительным патогенам и идентификации растения, обладающего повышенной резистентностью к одному или нескольким указанным патогенам растений.
4. Способ по п.1, где указанный промотор представляет собой промотор, индуцируемый патогеном.
5. Способ по п.1, где растение принадлежит к семейству Solanaceae.
6. Способ по п.5, где растение относится к роду Solanum.
7. Трансгенное растение, клетка растения, семя или плод, включающие нуклеотидную последовательность, кодирующую белок NRC1, причем указанный белок NRC1 включает аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:4 или аминокислотную последовательность, обладающую, по меньшей мере, 70%-ной идентичностью по аминокислотной последовательности с SEQ ID NO:2 по всей ее длине.
8. Выделенный белок, включающий аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:4 или аминокислотную последовательность, обладающую, по меньшей мере, 70%-ной идентичностью по аминокислотной последовательности с SEQ ID NO:2 по всей ее длине, где указанный белок сообщает растению повышенную резистентность к патогену.
9. Выделенная молекула нуклеиновой кислоты, кодирующая белок по п.8.
10. Химерный ген, включающий промотор, активный в растительных клетках, оперативно связанный с молекулой нуклеиновой кислоты по п.9 и, необязательно, дополнительно оперативно связанный с молекулой 3'-нетранслируемой нуклеиновой кислоты.
11. Вектор экспрессии, включающий химерный ген по п.10.
12. Применение молекулы нуклеиновой кислоты, кодирующей белок NRC1, для получения растений, резистентных к заболеванию, где указанный белок NRC1 включает аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:4 или аминокислотную последовательность, обладающую, по меньшей мере, 70%-ной идентичностью по аминокислотной последовательности с SEQ ID NO:2 по всей ее длине.
| CURRENT BIOLOGY, vol.15, no.10 | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |

