Область техники, к которой относится изобретение
Настоящее изобретение относится к способу лечения неврологических состояний у млекопитающего путем введения гематопоэтического фактора роста, такого как гранулоцитарный колониестимулирующий фактор (GCSF), гранулоцитарно-макрофагальный колониестимулирующий фактор (GMCSF), и/или других гематопоэтических факторов, например MCSF, за исключением эритропоэтина (EPO). Настоящее изобретение также относится к способам скрининга соединений, связывающихся с рецептором GCSF или MCSF, обнаруженным на поверхности нервной клетки; и которые обеспечивают нейропротективную, нейропролиферативную активность и/или активность по активации STAT гена.
Уровень техники
Факторы роста представляют собой белки, которые по существу участвуют в регуляции жизнеспособности, пролиферации, созревания и роста развивающихся нейрональных клеток. Например, экспрессия большого числа факторов роста повышается в ответ на различные типы мозговых инсультов. Многие факторы обладают эндогенными нейропротективными и нейротропными эффектами (см. Arvidsson A et al., Neurosciense 2001; 106:27-41; Larsson E. et al., J .Cereb. Blood Flow Metab 1999;19:1220-8; Mattison MP. et al., J Neurotrauma 1994; 11:3-33; Semkova I, et al., Brain Res Rev 1999; 30:176-88). О таких эффектах также сообщали после экзогенного введения in vitro и in vivo после травмы головного мозга и инсульта (см. Semkova I., et al., Brain Res Rev 1999; 30:176-88; Fisher M., et al., J.Cereb. Blood Flow Metab 1995;15:953-9; Schabitz WR, et al., Stroke 2001; 32:1226-33; Schabitz WR, et al., Stroke 2000; 31:2212-7). После связывания с высокоаффинными мембранными рецепторами воздействие факторов роста опосредуется каскадом событий внутриклеточной передачи сигналов (Kernie SG, et. al., Arch Neurol. 2000; 57:654-7), стимулирующим рост и дифференцировку клеток или обеспечивающим питание для жизнеобеспечения клеток.
Гранулоцитарный колониестимулирующий фактор (GCSF) представляет собой белок размером 20 кДа, который вместе с фактором некроза опухоли-α (TNF-α) и интерлейкинами является представителем цитокинового семейства факторов роста. GCSF представляет собой основной фактор роста, участвующий в продукции нейтрофильных гранулоцитов.
GCSF проявляет свою функцию посредством активации мембранного рецептора (рецептора GCSF), принадлежащего к суперсемейству рецепторов гематопоэтинов, обозначаемых также как рецепторы цитокинов I класса (de Koning and Touw, Curr. Opin. Hematol., 1996, 3, 180-4).
Обнаружено, что ряд рецепторов лимфокинов, гематопоэтических факторов роста и молекул, родственных гормону роста, обладают общим связывающим доменом. Данные рецепторы называют рецепторами гематопоэтинов, а соответствующие лиганды - гематопоэтинами. Кроме того, гематопоэтины подразделяются на две основных структурных группы: большие/длинные и малые/короткие гематопоэтины. Одна подгруппа отдельных рецепторных цепей, которые являются частью рецепторных комплексов крупных гематопоэтинов, содержит в своих внеклеточных частях общие структурные элементы: иммуноглобулин-подобный домен, домен рецептора гематопоэтина и три домена фибронектина типа III (2 в лептиновом рецепторе). Данную подгруппу обозначили как "семейство рецепторов gp130" (Mosley, et.al., J.Biol. Chem. 1996, 271, 32635-43), и она включает в себя лептиновый рецептор (LPTR), рецептор гранулоцитарного колониестимулирующего фактора (GCSFR), общую бета-цепь интерлейкина-6/-11/LIF/OSM/CNTF (GP130), рецептор ингибирующего лейкоз фактора (LIFR), бета-цепь рецептора онкостатина-М (OSMR), бета-1-цепь рецептора интерлейкина-12 (IL12RB1), бета-2-цепь рецептора интерлейкина-12 (IL12RB2). Данные рецепторные цепи при связывании с узнаваемым цитокином образуют гомодимеры (GCSFR, GP130, LPTR) или гетеродимеры (GP130 с LIFR или OSMR, IL12RB1 с IL12RB2). Кроме того, характеристикой данного семейства рецепторов является консенсусная структура prosite:
N-x(4)-S-x(28,35)-[LVIM]-x-W-x(0,3)-P-x(5,9)-[YF]-x(1,2)-[VILM]-x-W (SEQ ID № 1)
GCSF стимулирует пролиферацию, жизнеспособность и созревание клеток, коммитированных в направлении линии нейтрофильных гранулоцитов посредством связывания со специфическим рецептором GCSF (GCSFR) (см. Hartung T., et al., Curr. Opin. Hematol. 1998; 5:221-5). Опосредованный GCSFR сигнальный путь активирует семейство белков переносчиков сигнала и активаторов транскрипции (STAT), перемещающихся в ядро и регулирующих транскрипцию (Darnell JE Jr., Science 1997; 277:1630-5). GCSF, как правило, применяют для лечения различных видов нейтропении у людей. Он представляет собой один из немногих факторов роста, одобренных для клинического применения. В частности, его применяют для снижения вызванной химиотерапией (ХТ) цитопении (Viens et al., J. of Clin. Oncology, Vol. 20, No. 1, 2002:24-36). GCSF также применяют для лечения инфекционных заболеваний как потенциальное дополнительное средство (Hubel et al., J. of Infectious Diseases, Vol. 185:1490-501, 2002). Сообщалось, что GCSF кристаллизован до некоторой степени (EP 344 796) и что предположена общая структура GCSF, но только и в грубом приближении (Bazan, Immunology Today 11:350-354 (1990); Parry et al., J. Molecular recognition 8: 107-110 (1988)).
В последние годы ряд факторов роста, таких как bFGF, и перспективные в фармацевтическом плане вещества, такие как блокаторы адгезии тромбоцитов типа анти-GP IIb/IIa и Abcizimab, протестированы в клинических испытаниях на наличие у них нейропротективного эффекта. К сожалению, ни одно из указанных средств не обладало достаточной нейропротективной эффективностью. В частности, антагонисты NMDA, уловители свободных радикалов и антагонисты глутамата либо являлись неэффективными, либо оказывали серьезные побочные эффекты. Перечень веществ, таких как анти-ICAM или ингибиторы опосредованной глутаматом NO-синтетазы, не способствующие росту, представлен в (De Keyser, et al., (1999), Trends Neurosci, 22, 535-40).
Большинство исследований ишемии головного мозга и тестирования фармакологических веществ in vivo сосредоточены только на непосредственных эффектах лекарственного средства на исследуемую модель (например, размер инфаркта через 24 часа после индукции инсульта). Однако более надежным параметром истинной эффективности конкретного вещества является длительное действие на восстановление функциональной активности, что также отражено в исследованиях инсульта у человека, где шкалы клинической оценки (например, скандинавская шкала оценки, шкала оценки национального института здравоохранения США (NIH), индекс Бартеля) также отражают способность осуществлять ежедневную жизнедеятельность. Восстановление в первые несколько суток после очаговых поражений может происходить вследствие разрешения отека или реперфузии области "ишемической полутени". Большая часть восстановления функции, достигаемая после острой фазы, вероятно, происходит вследствие пластичности головного мозга, где смежные зоны коры головного мозга берут на себя выполнение тех функций, которые ранее выполняли поврежденные участки (Chen R, Cohen LG, Hallett M, Neuroscience 2002; 111(4): 761-73). Два основных механизма, предложенных для объяснения такой перестройки, представляют собой демаскирование ранее существовавших, но функционально неактивных связей и рост новых связей, таких как коллатеральные пучки (Chen R, Cohen LG, Hallett M, Neuroscience 2002; 111(4): 761-73). Краткосрочные пластические изменения опосредованы снятием ингибирования возбуждающих синапсов, что, вероятно, происходит вследствие сниженного ГАМК-эргического ингибирования (Kaas JH. Annu Rev Neurosci. 1991; 14:137-67; Jones EG. Cereb Cortex. 1993 Sep-Oct; 3(5):361-72). Происходящие в течение более длительного периода времени пластические изменения в дополнение к демаскировке латентных синапсов включают в себя такие механизмы, как долговременная потенциация (LTP), требующая активации рецептора NMDA и повышенной внутриклеточной концентрации кальция (Hess and Donoghue, J. Neurophysiol. 1994, 71(6): 2543-7). Долговременные изменения также включают в себя регенерацию аксонов и ростки с изменениями в форме, количестве, размере и типе синапсов (Kaas JH, Annu Rev Neurosci. 1991; 14: 137-67, 3:).
Инсульт представляет собой третью ведущую причину смертности и основную причину инвалидизации на Западе. Он представляет собой большую социально-экономическую проблему. Этиология может являться или ишемической (в большинстве случаев), или геморрагической. Причиной ишемического инсульта, как правило, является эмболия или тромб. До настоящего времени эффективного лечения для большинства пациентов с инсультом не существует. Единственным клинически подтвержденным лекарственным средством до настоящего времени является тканевой активатор плазминогена (ТАП) и аспирин. После массовой гибели клеток непосредственно в очаге инфаркта в связи с недостатком глюкозы и кислорода зона инфаркта со временем расширяется под действием вторичных механизмов, таких как опосредованная глутаматом эксайтотоксичность, механизмы апоптоза и образование свободных радикалов.
Боковой амиотрофический склероз (ALS; болезнь Лу-Герига; болезнь Шарко) представляет собой нейродегенеративное заболевание с частотой заболеваемости от 0,4 до 1,76 на 100000 человек в год (Adams et al., Principles of Neurology, 6th ed., New York, pp. 1090-1095). Он представляет собой наиболее общую форму заболевания моторных нейронов с типичными проявлениями генерализованных фасцикуляций, прогрессирующей атрофии и слабости скелетных мышц, спастичности и пирамидных симптомов, дизартрии, дисфагии и диспноэ. По существу патология заключается в потере нервных клеток в переднем роге спинного мозга и в двигательных ядрах нижней части ствола головного мозга, а также может включать в себя моторные нейроны первого порядка в коре головного мозга. Патогенез данного изнуряющего заболевания до сих пор почти совершенно неизвестен, хотя хорошо изучена роль мутантов по супероксиддисмутазе (SOD1) в семейных случаях, что соответствует гипотезе окислительного стресса. До настоящего времени в белке SOD1 описаны более 90 мутаций, способных служить причиной ALS (Cleveland and Rothstein (2001), Nat Rev Neurosci, 2, 806-19). Кроме того, при данном заболевании показана роль нервных волокон. Эксайтотоксичность, механизм, индуцируемый избыточной стимуляцией глутаматом, также является важным фактором, иллюстрируемым эффективным действием рилузола у человеческих пациентов. Как наиболее убедительно показано у мутантов SOD1, активация каспаз и апоптоза, по-видимому, представляет собой общий заключительный этап патогенеза ALS (Ishigaki et al., (2002), J. Neurochem, 82, 576-84., Li, et al., (2002), Science, 288, 335-9). Следовательно, по-видимому, ALS также удовлетворяет той же общей патогенетической модели, которая также работает при других нейродегенеративных заболеваниях и при инсульте, например участие глутамата, окислительный стресс и программируемая гибель клеток.
Болезнь Паркинсона представляет собой наиболее частое двигательное расстройство, которым страдают приблизительно 1 миллион пациентов в Северной Америке; приблизительно 1 процент населения в возрасте старше 65 лет. Основными симптомами заболевания являются мышечная ригидность, тремор и акинезия (Adams et al., Principles of Neurology, 6th ed., New York, pp. 1090-1095). Этиология болезни Паркинсона неизвестна. Тем не менее, значительная часть биохимических данных, полученных на основании исследований образцов аутопсии головного мозга человека и на животных моделях, указывают на происходящий процесс окислительного стресса в черном веществе, который может инициировать дофаминэргическую нейродегенерацию. Окислительный стресс, как индуцируемый нейротоксинами 6-гидроксидофамином и MPTP (N-метил-4-фенил-1,2,3,6-тетрагидропиридином), применяли на животных моделях для изучения процесса нейродегенерации. Несмотря на существование симптоматической терапии (например, L-ДОФА совместно с ингибитором декарбоксилазы; агонисты дофамина, такие как бромокриптин, перголид; и антихолинергические средства, такие как тригексилфенидил (артан)), существует очевидная потребность в каузативном лечении, например в нейропротективной терапии, которая действительно останавливает прогрессирование заболевания. Данные животные модели применяли для исследования эффективности акцепторов радикалов, хелаторов железа, агонистов дофамина, ингибиторов синтазы оксида азота и определенных антагонистов кальциевых каналов. Отчетливо видно, что в животных моделях, а также у пациентов действуют механизмы апоптоза (Mochizuki et al. (2001), Proc. Natl. Acad. Sci. USA, 98, 10918-23, Xu et al. (2002), Nat. Med. 8, 600-6, Vismanath, et al. (2001), J. Neurosci. 21, 9519-28, Hartmann, et al. (2002), Neurology, 58, 308-10). Данная патофизиология с участием окислительного стресса и апоптоза также включает болезнь Паркинсона в число других нейродегенеративных расстройств и инсульта.
Ишемия головного мозга может возникать в результате множества причин, нарушающих мозговой кровоток (CBF) и ведущих к недостаточности кислорода и глюкозы. С другой стороны, травматическое повреждение головного мозга (TBI) включает в себя участие первичного механического воздействия, которое обычно вызывает перелом черепа и грубо разрушает паренхиму головного мозга с деформацией и разрывом кровеносных сосудов и ткани головного мозга. Это, в свою очередь, запускает каскад событий, характеризующихся активацией молекулярных и клеточных реакций, что ведет ко вторичному повреждению. Развитие такого вторичного повреждения представляет собой активный процесс, в который вовлечено множество биохимических путей (Leker and Shohami (2002), Brain Res. Rev., 39, 55-73). Выявлены многочисленные совпадения между патологическими механизмами, ведущими ко вторичной гибели клеток в зоне частичной ишемии, и патологическими механизмами в области, подвергавшейся вторичному посттравматическому повреждению (например, эксайтотоксичность, при избыточном высвобождении глутамата, оксида азота, реакционно-способных форм кислорода, воспаление и апоптоз (Leker and Shohami (2002), Brain Res. Rev., 39, 55-73)). Кроме того, сообщалось, что после травматического повреждения мозга возникают ранние эпизоды ишемии, добавляя компонент ишемии в первичное механическое повреждение.
Сердечно-сосудистые заболевания представляют собой основную причину смерти в западных развитых странах. В Соединенных Штатах Америки случается примерно один миллион смертельных случаев каждый год, из них примерно 50% связано с внезапной смертью и происходит вне больницы (Zheng, et al. (2001), Circulation, 104, 2158-63). Ежегодно проводят 40-90 сердечно-легочных реанимаций (CPR) на 100000 человек, а восстановление самостоятельного кровообращения (ROSC) достигают у 25-50% указанных пациентов. Однако количество выписавшихся из больницы после удачной ROSC пациентов составляет лишь 2-10% (Bottiger, et al. (1999), Heart, 82, 674-9). Таким образом, в подавляющем большинстве случаев остановок сердца, наблюдаемых ежегодно в Соединенных Штатах, не проводится успешного лечения. Основной причиной низкого уровня выживания после удачной CPR, например уровня внутрибольничной смертности после остановки сердца, является стойкое повреждение головного мозга. Повреждение мозга после остановки кровообращения связано с небольшим периодом толерантности к гипоксическому стрессу и со специфическими расстройствами после реперфузии (Safar (1986), Circulation, 74, IV138-53, Hossmann (1993), Resuscitation, 26, 225-35). На начальной стадии после остановки сердца большее число пациентов можно стабилизировать гемодинамически, однако многие из них умирают в результате поражения центральной нервной системы. Личностные, социальные и экономические последствия повреждения головного мозга, наступающие после остановки сердца, разрушительны. Следовательно, одной из наиболее важных проблем в исследовании остановки сердца и реанимации ("полная ишемия организма и реперфузия") является реанимация головного мозга и повреждение головного мозга после остановки сердца (Safar (1986), Circulation, 74, IV138-53, Safar, et al. (2002), Crit Care Med, 30, p. 140-4). В настоящее время никакими терапевтическими мерами после остановки сердца невозможно снизить первичное повреждение нейронов, вызываемое гипоксией, возникающей при остановке сердца. Основные патофизиологические исходы включают в себя гипоксию и последующий некроз, реперфузионное повреждение с образованием свободных радикалов, приток кальция в клетки, высвобождение аминокислот, вызывающих возбуждение, реперфузионные расстройства церебральной микроциркуляции и программированную гибель нейронов или апоптоз (Safar (1986), Circulation, 74, IV138-53, Safar, et al. (2002), Crit Care Med, 30, p. 140-4).
Предприняты несколько безуспешных клинических испытаний с целью улучшения неврологического исхода после остановки сердца. Тестировали терапевтическое применение барбитуратов (для усиления нейропротекции) или применение блокаторов кальциевых каналов (для снижения реперфузионного повреждения при ишемии) (Group (1986), Am. J. Emerg. Med., 4, 72-86, Group (1986), N. Engl. J. Med. 314, 397-403, Group (1991), Control.Clin. Trials 12, 525-45, Group (1991), N. Engl. J. Med. 324, 1225-31). В настоящее время не доступно каких-либо терапевтических средств после остановки сердца для улучшения неврологических исходов после остановки деятельности сердца в клинических условиях (с возможным исключением слабой гипотермии и тромболиза, для исследования которых в настоящее время проводятся крупномасштабные рандомизированные и контролируемые клинические испытания, результаты которых ожидают с большим вниманием) (Safar, et al. (2002), Crit Care Med., 30, p. 140-4). Следовательно, новые подходы к лечению для улучшения неврологического исхода после остановки сердца являются очень важными.
Рассеянный склероз представляет собой модельное воспалительное аутоиммунное заболевание центральной нервной системы, риск развития которого составляет один к 400, и потенциально он является наиболее частой причиной неврологической инвалидности у совершеннолетних молодых людей. Во всем мире насчитывается примерно 2-5 миллионов пациентов, страдающих от данного заболевания (Compston and Coles (2002), Lancet, 359, 1221-31). Как и при всех комплексных синдромах, данное заболевание является результатом взаимодействия еще не выявленных факторов внешней среды и генов предрасположенности. Вместе данные факторы запускают каскад событий, включающих в себя вовлечение иммунной системы, острое воспалительное повреждение аксонов и глии, восстановление функции и структурную репарацию, глиоз после воспаления и нейродегенерацию. Последовательное вовлечение указанных процессов лежит в основе клинического течения, характеризующегося случаями с восстановлением, случаями со стойкой недостаточностью и с вторичной прогрессией. Целью лечения является снижение частоты и уменьшение длительного воздействия рецидивов, ослабление симптомов, профилактика инвалидности, возникающей при развитии заболевания, и содействие восстановлению ткани.
Депрессия является распространенным психическим расстройством, характеризующимся унынием, потерей интереса к активной жизни и уменьшением активной деятельности. Депрессия отличается от нормальных изменений настроения степенью тяжести, симптомами и длительностью расстройства. Суицид остается одним из самых распространенных и часто неизбежных последствий депрессии. Если заболевание включает чередование депрессивных эпизодов с эйфорией или раздражительностью, его относят к биполярному расстройству. Депрессивные расстройства и шизофрения являются причиной 60% всех суицидов. Депрессия может быть вызвана разными причинами. На начало и течение депрессивных эпизодов могут оказывать влияние психосоциальные факторы, такие как неблагоприятные жизненные условия. Генетические и биологические факторы также могут вносить определенный вклад.
По расчетам в настоящее время от депрессии страдает 121 миллион человек. Депрессия является ведущей причиной инвалидности по данным оценки параметра YLD (годы, прожитые с инвалидностью) и является 4-м по важности фактором, определяющим суммарный поражающий эффект (DALY = длительность жизни, скорректированная с учетом инвалидности; сумма лет потенциальной жизни, потерянных в связи с преждевременной смертью, и годы продуктивной жизни, потерянные из-за инвалидности) в 2000 году. По оценкам, ежегодно от приступов депрессии страдают 5,8% мужчин и 9,5% женщин. Прогнозируют, что к 2020 году депрессия достигнет второго места в шкале DALY, рассчитываемой для всех возрастов и обоих полов. В развитых странах депрессия займет одно из ведущих мест в ряду всех причин заболеваний.
В настоящее время первая линия лечения большинства людей с депрессией включает применение антидепрессантов, психотерапии или их сочетания. Антидепрессанты эффективны на всем диапазоне тяжести большинства приступов депрессии. В настоящее время эффективная антидепрессивная терапия тесно связана с модуляцией или регулировкой серотонинергической нейротрансмиссии. Лекарственные средства, повышающие уровень серотонина в головном мозге, являются самыми сильными из известных антидепрессантов (такие как флуоксетин, прозак (Prozac®) или флуктин (Fluctin®)). Способы лечения с антидепрессивными эффектами у пациентов также включают в себя, например, фармакологические антидепрессанты, такие как литий, электросудорожную терапию и физические упражнения. Другие способы воздействия включают в себя создание систем поддержки для уязвимых индивидуумов, семей и групп. Менее убедительны доказательства профилактического лечения депрессии, лишь в нескольких отдельных исследованиях показали эффективность предлагаемых для профилактики депрессии способов воздействия. Важно иметь в виду, что существующие лекарственные средства направлены на облегчение симптомов заболевания, но не воздействуют на основные патофизиологические механизмы, вызывающие данное заболевание. В этой связи имеется потребность в новом лечении, которое было бы специфически направлено на новые выявленные причины депрессии.
Шизофрения является самым распространенным психическим заболеванием. Шизофрения поражает приблизительно 1 из 100 человек (1% населения). Это расстройство распространено во всем мире, охватывая все расы и культуры. Шизофрения поражает и мужчин, и женщин в равной мере, хотя в среднем у мужчин шизофрения развивается в более раннем возрасте, чем у женщин. Как правило, первые признаки шизофрении у мужчин появляются в середине 20-летнего возраста, а у женщин первые признаки - в конце 20-летнего возраста. Шизофрения сопряжена с огромными экономическими потерями, так, например, в США связанные с нею убытки составляют 32,5 миллиарда долларов в год. Шизофрения характеризуется наличием ряда симптомов: бред, галлюцинации, спутанные мышление и речь, негативные симптомы (отсутствие социальной адаптации, отсутствие эмоций и их выражения, снижение энергии, мотивации и активности), кататония. Главным способом лечения шизофрении является применение нейролептиков, таких как хлорпромазин, галоперидол, оланзапин, клозапин, тиоридазин и другие. Однако при лечении нейролептиками часто не удается ослабить все симптомы шизофрении. Кроме того, антипсихотическое лечение оказывает серьезные побочные эффекты, такие как поздняя дискинезия. Этиология шизофрении не ясна, хотя предполагают, что на нее сильное воздействие оказывают генетические факторы. В последнее время стало ясно, что у шизофрении существуют, по меньшей мере, несколько аспектов нейродегенеративного заболевания. В частности, в исследованиях с применением MR у больных шизофренией выявлена быстрая потеря серого вещества в коре головного мозга (Thompson, et al. (2001), Proc Natl Acad Sci USA 98, 11650-5; Cannon, et al. (2002), Proc Natl Acad Sci USA, 99, 3228-33). Следовательно, обосновано лечение больных шизофренией нейропротективным лекарственным средством, таким как GCSF, или GMCSF, или другие гематопоэтические факторы.
У людей существует потребность в способах повышения познавательной способности и усиления интеллекта. "Интеллект" в современном понимании не ограничивается просто логическими или семантическими способностями. Так, например, теория множественного интеллекта Говарда Гарднера оценивает интеллект с точки зрения эволюционной и антропологической перспективы и выдвигает более широкую концепцию, которая включает в себя атлетические, музыкальные, артистические и эмпатические способности, а также лингвистические/логические способности, которые чаше всего связывают с интеллектом и определяют с помощью тестов IQ. Такое расширенное понимание интеллекта распространяется и на область творчества. Кроме того, существует непатологическое состояние, известное как возрастное снижение памяти у людей (ARML), или легкое когнитивное нарушение (MCI), или возрастное снижение познавательной способности (ARCD), которое, как правило, начинается приблизительно в возрасте 40 лет и отличается от ранних симптомов болезни Альцгеймера.
В течение всего периода жизни взрослого человека происходит физиологическое снижение количества нервных клеток, составляющее, по подсчетам, 100000 нейронов в сутки. В течение указанного периода отмечается постепенное снижение веса и объема мозга. Оно составляет примерно 2% в 10 лет. В отличие от более ранних представлений указанное снижение не усиливается после 50 лет, но продолжается примерно с одной и той же скоростью, начиная с раннего периода совершеннолетия. Аккумулятивные эффекты указанного процесса, как правило, не заметны до старости.
Хотя головной мозг уменьшается в размерах, этого не происходит равномерно. Определенные структуры больше подвержены такому уменьшению. Например, часто становятся меньше гиппокамп и лобные доли, две структуры, вовлеченные в механизмы памяти. Частично это происходит вследствие потери нейронов и частично - вследствие атрофии некоторых нейронов. Размер многих других структур головного мозга не уменьшается. Снижение интеллектуальной деятельности может происходить вследствие повреждения нейронов, когда они разрушаются, уменьшаются или утрачивают связи. Такое снижение числа полностью функционирующих нейронов делает необходимым привлечение дополнительных сетей нейронов для управления видами умственной деятельности, которые в ином случае выполнялись бы проще или автоматически. Таким образом, процесс замедляется. Часть лобной доли, называемая префронтальная кора, участвует в управлении и контроле мыслей и действий. Атрофия, происходящая в этом участке головного мозга, может являться причиной испытываемой пожилыми людьми трудности в подборе слова. Она также может являться причиной забывания места нахождения ключей от машины или общей рассеянности. Полагают, что за затруднения с памятью ответственно уменьшение лобной доли и гиппокампа. Таким образом, также сохраняется необходимость в улучшении или усилении познавательной способности индивидуума.
В связи с указанным выше существует необходимость лечения неврологических и/или психиатрических состояний, таких как неврологические заболевания, связанные с усилением пластичности и восстановлением функций или клеточной гибелью в нервной системе. В частности, имеется потребность в лечении неврологических заболеваний путем создания нейропротекции для вовлеченных нервных клеток.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Таким образом, одной из целей настоящего изобретения является предоставление способа лечения неврологического или психиатрического состояния у млекопитающего посредством введения данному млекопитающему гематопоэтического фактора, такого как GMCSF, производное GMCSF, GCSF, производное GCSF и их сочетания, или клеток, секретирующих GCSF, или GMCSF, или их производные, для лечения указанного состояния.
Другой целью настоящего изобретения является предоставление способа лечения неврологического состояния у млекопитающих посредством кондиционирования композиции нейрональных стволовых клеток с гематопоэтическим фактором, таким как GMCSF, производное GMCSF, GCSF, производное GCSF и их сочетаний, с последующим введением нейрональных стволовых клеток млекопитающему для лечения указанного состояния.
Другой целью настоящего изобретения является предоставление способа идентификации соединения, которое связывается с рецептором гранулоцитарного колониестимулирующего фактора (GCSFR) на нервных клетках и которое активирует экспрессию гена STAT в нервной клетке, посредством взаимодействия нервной клетки с данным соединением и измерения степени повышения активации STAT относительно активации STAT в нервной клетке, которая не контактировала с данным соединением, и определения степени активации относительно, например, опосредованной GCSF активации STAT. Кроме того, соединения, получаемые этим способом, а также способы применения данных соединений для лечения неврологических состояний представляют собой дополнительные объекты настоящего изобретения.
Другой целью настоящего изобретения является предоставление способа идентификации соединения, которое связывается с рецептором гранулоцитарно-макрофагального колониестимулирующего фактора (GMCSFR) на нервных клетках и/или которое активирует экспрессию гена STAT в нервной клетке, путем взаимодействия нервной клетки с данным соединением и измерения степени повышения уровня активации STAT относительно активации гена STAT в нервной клетке, которая не контактировала с данным соединением. Кроме того, соединение, получаемое этим способом, а также способы применения данных соединений для лечения неврологических состояний представляют собой дополнительные объекты настоящего изобретения.
Другой целью настоящего изобретения является предоставление способа идентификации агониста рецептора GCSF с повышенной нейропротективной активностью посредством взаимодействия соединения с нервной клеткой, несущей рецептор GCSF, измерения нейропротективного эффекта соединения на нервную клетку и сравнения эффекта соединения с эффектом GCSF. Кроме того, соединения, получаемые этим способом, а также способы применения данных соединений для лечения неврологических состояний представляют собой дополнительные объекты настоящего изобретения.
Другой целью настоящего изобретения является предоставление способа идентификации агониста рецептора GMCSF с улучшенной нейропротективной активностью путем осуществления взаимодействия соединения с нервной клеткой, несущей рецептор GMCSF, измерения нейропротективного эффекта соединения на нервную клетку и сравнения эффекта соединения с эффектом GMCSF. Кроме того, соединения, получаемые этим способом, а также способы применения данных соединений для лечения неврологических состояний представляют собой дополнительные объекты настоящего изобретения.
Другой целью настоящего изобретения является предоставление способа идентификации соединения с повышенной активностью в качестве агониста в отношении рецептора GCSF посредством взаимодействия соединения с нервной клеткой, несущей рецептор GCSF, сравнения уровня экспрессии гена STAT в нервной клетке со второй нервной клеткой, контактировавшей с GCSF. Кроме того, соединения, получаемые этим способом, а также способы применения данных соединений для лечения неврологических состояний представляют собой дополнительные объекты настоящего изобретения.
Другой целью настоящего изобретения является предоставление способа идентификации соединения с улучшенной активностью в качестве агониста в отношении рецептора GMCSF посредством взаимодействия соединения с нервной клеткой, несущей рецептор GMCSF, сравнения уровня экспрессии гена STAT в нервной клетке со второй нервной клеткой, контактировавшей с GMCSF. Кроме того, соединения, получаемые этим способом, а также способы применения данных соединений для лечения неврологических состояний представляют собой дополнительные объекты настоящего изобретения.
Другой целью настоящего изобретения является предоставление способа стимуляции экспрессии GCSF или GMCSF и/или их высвобождения в эндогенных нервных клетках посредством применения агонистов рецептора GCSF или GMCSF, расположенного на клетке. В другом варианте осуществления клетки приводят в контакт с веществом, которое повышает экспрессию и/или высвобождение GCSF или GMCSF. Такие способы можно применять в тестах, выявляющих высвобождение или повышение экспрессии GCSF или GMCSF в нервных клетках, например в культурах нервных клеток (например, анализы ПЦР и/или ELISA).
Другой целью настоящего изобретения является предоставление способа лечения неврологического состояния у млекопитающего путем применения агонистов рецептора GMCSF, рецептора GCSF или обоих рецепторов для лечения неврологического состояния.
Другой целью настоящего изобретения является предоставление способа повышения жизнеспособности клеток, трансплантированных в организм млекопитающего посредством введения в клетку одного или нескольких полинуклеотидов, которые кодируют GMCSF, производное GMCSF, GCSF, производное GCSF и/или их сочетания, перед трансплантацией клетки в организм млекопитающего, посредством чего достигается экспрессия в клетке гематопоэтического фактора в количестве, достаточном для повышения жизнеспособности клетки в сравнении с показателем жизнеспособности клетки до введения полинуклеотидов.
Другой целью настоящего изобретения является предоставление способа повышения жизнеспособности культуры нервных клеток посредством введения GMCSF, производного GMCSF, GCSF, производного GCSF и/или их сочетаний для повышения жизнеспособности культуры нервных клеток в сравнении с жизнеспособностью культуры до введения гематопоэтического фактора. В таком способе гематопоэтические факторы можно применять для взаимодействия с клетками культуры или их можно предоставлять с применением полинуклеотидов, которые кодируют и экспрессируют гематопоэтические факторы.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показана эффективная нейропротекция in vivo и in vitro при применении GCSF. А - уровень нейропротективного воздействия GCSF после очаговой ишемии головного мозга (с применением модели нитевидной структуры; окклюзии средней мозговой артерии, MCAO), как измерено посредством окрашивания ТТС. Значения по оси Y означают процент инфаркта от всего полушария (данные приведены как среднее значение ± SD; T-тест; p<0,05). В - тест жизнеспособности клеток, проведенный на клетках РС12, обработанных NGF в условиях повышающегося окислительного стресса под действием H2O2 (0 мкМ, 400 мкМ, 750 мкМ). Добавление GCSF приводит к резкому повышению жизнеспособности клеток. Для сравнения представлена жизнеспособность клеток после обработки клеток эритропоэтином (EPO), известным нейропротективным веществом. Ось Y: относительная (отн.) жизнеспособность клеток (световые единицы люциферазной активности).
На фиг.2А показана ОТ-ПЦР, специфичная для мышиного GCSFR. РНК GCSF-R обнаружена в ткани головного мозга и имела ожидаемый размер 567 п.н. Идентичность подтверждена определением последовательности продукта ПЦР.
На фиг.4 приведены результаты иммуногистохимического анализа, показывающие распределение рецептора GCSF-R по различным участкам головного мозга мыши (парафиновые срезы, 2 мкм). a-d: локализация GCSF-R в гиппокампе. Следует отметить, что антитело преимущественно окрашивает нейроны в зоне СА3 (a,b), с наличием четкой границы между участками СА3 и СА2 (с, стрелка). GCSF-R распределен по всем клеткам, а также отросткам нейронов (b, стрелка). Следует отметить наличие рецептора в районе ворот и слоях базальных клеток зубчатой извилины (d, стрелка). Рецептор GCSF также выявляется в областях коры головного мозга: в пириформной коре головного мозга (е) и в периринальной коре головного мозга (f) в качестве примеров. В мозжечке метятся клетки Пуркинье (g, стрелка). Кроме того, положительными по GCSF-R являются некоторые из крупных митральных клеток обонятельной луковицы (h, стрелка). Сильное окрашивание проявляют передние рога спинного мозга (i, j), а при большом увеличении как позитивные по GCSF-R выявляются крупные двигательные нейроны (k, l). Следует отметить, что отростки нейронов также сильно метятся. В среднем мозге нейроны черного вещества демонстрируют положительное окрашивание по GCSF-R (m). Особенно сильно метятся все нейроны в компактной части (SNC) (стрелки в m и n). Кроме того, несколько нейронов в сетчатой части экспрессируют GCSF-R (o). Кроме нейронов, также окрашены олигодендроциты в пучках белого вещества, например, в передней спайке (р, стрелка). Неожиданным является окрашивание лиганда GCSF (антитело sc13102, Santa Cruz) локализуется совместно с экспрессией его рецептора (антитело sc694, Santa Cruz) (фиг.4 q-u). Полученные данные свидетельствуют об аутокринном механизме как защитном механизме нейронов против повреждающих факторов. В гиппокампе для лиганда GCSF наблюдают такую же специфичность к подструктурам (q: GCSFR, r: GCSF, стрелки указывают на границу между подструктурами СА2-3, са3 и са2 метки указывают подструктуру). Указанная специфичность совпадает с известными различиями в чувствительности данных участков к ишемическому повреждению и является дополнительным аргументом в пользу нейропротективной функции системы GCSF. Кроме того, в зубчатой извилине наблюдают такой же интересный профиль экспрессии в воротах и в зоне под зернистым слоем (фиг.4s, рецептор GCSF, фиг.4t, GCSF; стрелки указывают на один нейрон в зоне под зернистым слоем, метки: s: зона под зернистым слоем, h: ворота зубчатой извилины), которая подчеркивает важность системы GCSF для нейрогенеза и очень хорошо соответствует экспрессии рецептора и лиганда на нейросферах (см. фиг.13). Интересно отметить, что GCSF также экспрессируется в больших мотонейронах спинного мозга (фиг.4u), где также экспрессируется его рецептор (фиг.4i-l).
На фиг.3 показаны характерные иммуноблоты GCSF-R. С применением антисыворотки к GCSF-R в ткани коры головного мозга крысы выявлена полоса размером примерно 130 кДа (трек 1). Дополнительные полосы с меньшим молекулярным весом, вероятно, относятся к продуктам распада. После предварительной абсорбции антисыворотки соответствующим пептидом указанные полосы уже не выявляются (трек 2).
На фиг.5 показано окрашивание GCSFR на культивируемых нейронах коры головного мозга (a, b) и нейронах гиппокампа (c, d). Рецептор присутствует и на телах клеток, и на отростках нейронов. Не все нейроны в препарате являются мечеными. Предварительная инкубация антитела с соответствующим пептидом, как и отсутствие первичного антитела, не приводит к специфическому окрашиванию (результаты не приведены).
На фиг.6 и 7 показаны результаты иммунохимического анализа с использованием антитела против STAT3. Фиг.6: иммуногистохимия STAT3. Следует отметить, что в области "полутени" коры головного мозга у крыс в группе, обработанной GCSF (b, стрелки), в сравнении с периинфарктной зоной в коре головного мозга животных после введения плацебо положительно окрашены множество ядер нейронов (а, стрелки; слева: зона инфаркта; справа: область "полутени"; исходное увеличение х200). Фиг.7: Подсчитано количество нейронов коры головного мозга на непораженной контралатеральной стороне (CL) и на ипсилатеральной стороне рядом с зоной инфаркта (IL) как у животных, обработанных GCSF, так и у контрольных животных. Происходила значимая активация STAT3 в нейронах вблизи зоны инфаркта у животных в группе, обработанной GCSF, по результатам подсчета нейронов с транслокацией STAT3 в ядро (*p<0,05, t-тест).
На фиг.8 показаны эффект обработки GCSF в фототромботической модели ишемии головного мозга с применением бенгальского розового. А - поведение на вращающемся барабане (ротароде); В - неврологическая шкала, включающая в себя способность поддержания баланса на барабане; С, D: - тест с удалением лейкопластыря, измеряемый как на ипси- (С), так и на контралатеральной стороне (D). Подписи: ишемия: группа крыс с ишемией, не подвергнутых обработке; ишемия + GCSF: группа крыс с ишемией, обработанных GCSF; симуляция + GCSF: животные с ложной операцией, обработанные GCSF; симуляция: не обработанные животные после ложной операции; L: тест с удалением лейкопластыря на левой лапе; R - тест с удалением лейкопластыря на правой лапе. Следует отметить, что в тесте с удалением липкой ленты эффект у животных присутствует на обеих сторонах (C, D), что, вероятно, вызвано доминирующим двигательным дефицитом в случае наличия ленты на ипсилатеральной стороне и доминирующим сенсорным дефицитом, когда лента находится на контралатеральной стороне.
На фиг.9 показана повышающая регуляция рецептора GCSF на модели с применением бенгальского розового через 48 часов после индукции фототромбоза на контралатеральной стороне относительно зоны ишемии. a - схема коронарный срез головного мозга крысы, образцы ткани 3 и 4 применяли для количественного определения мРНК рецептора GCSF в сравнении с образцами той же ткани у крыс после ложной операции. b - количественное определение мРНК рецептора GCSF в образцах из области "полутени" коры головного мозга (образцы 3 и 4 на фиг.9а). За начальной повышающей регуляцией через 6 часов после индукции ишемии на ипсилатеральной стороне следует повышающая регуляция на контралатеральной стороне через 48 часов после инфаркта. Данную повышающую регуляцию на контралатеральной стороне также наблюдали и для рецептора GMCSF (см. ниже).
На фиг.10 показаны результаты выравнивания GCSF разных видов с применением алгоритма ClustalW (SEQ ID №№ 28-33) (MEGALIGNTM, Lasergene, Wisconsin).
На фиг.11 показано выравнивание рецепторов GCSF мыши и человека и фрагмента рецептора крысы с применением алгоритма ClustalW (SEQ ID №№ 34-36) (MEGALIGNTM, Lasergene, Wisconsin).
На фиг.12 показано присутствие рецептора GCSF на нейрональных стволовых клетках взрослых животных (nsc) способом ОТ-ПЦР (ПЦР с обратной транскрипцией). Показан агарозный гель. Трек 1: маркер размера; трек 2: продукты ПЦР из нейрональных стволовых клеток (nsc), видна специфичная для GCSFR крысы полоса (279 п.н.); трек 3: отрицательный контроль.
На фиг.13 показано присутствие рецептора GCSF и GCSF на нейрональных стволовых клетках. а - DAPI-окрашивание нейросферы для визуализации всех клеток, b - окрашивание той же нейросферы антителом против рецептора GCSF. c, d - нейросфера, окрашенная с использованием DAPI (с) и антитела против GCSF (d).
На фиг.14 показано быстрое всасывание биотинилированного GCSF после внутрибрюшинной инъекции мышам. А - вестерн-блоттинг сыворотки от мышей, умерщвленных через 1, 2, 4, 6, 20, 28 часов после инъекции 7,5 мкг GCSF/мышь. В - ELISA сывороточного GCSF после в/б инъекции. Происходит быстрое всасывание GCSF из брюшины с достижением пиковых уровней в сыворотке через 2 часа, что указывает на применимость этого способа введения.
На фиг.15 показано выявление рецептора GMCSF в виде повышенной экспрессии мРНК после индукции фототромбоза (модель с использованием бенгальского розового) на ипсилатеральной и контралатеральной стороне зоны ишемии. На части а схематически показан коронарный срез головного мозга мыши, а интересующие зоны помечены серым цветом; на части в показан срез геля RMDD, на котором транскрипт рецептора GMCSF идентифицируется как регулируемый. На треках представлены продукты ОТ-ПЦР, проводимой с использованием образцов РНК из головного мозга мыши. Образцы отбираются из области "полутени" коры головного мозга в различные временные точки после инсульта (3 и 4 на части а соответственно).
На фиг.16 приведен контроль повышения экспрессии рецептора GMCSF в модели с применением бенгальского розового при количественной ОТ-ПЦР через 48 часов с применением LightCyclerSystem. Образцы отбирали через 6 часов, 48 часов и 21 сутки после индукции фототромбоза, а уровни индукции сравнивали с животными после ложной операции. На ипсилатеральном повышенная экспрессия рецептора GMCSF максимальна сразу после ишемии коры головного мозга, а затем монотонно снижается до 21 суток. В образце коры головного мозга из соответствующей контралатеральной стороны повышенная экспрессия лучше всего видна через 2 суток после инфаркта и остается все еще умеренно повышенной к 21 суткам. Такой профиль регуляции на контралатеральной стороне является реминисцентной для рецептора GCSF (см. выше).
На фиг.17 показано выравнивание рецепторов GMCSF человека, мыши и последовательности крысы, идентифицированной как транскрипт с повышающейся регуляцией (SEQ ID №№ 22, 23 и 24) (алгоритм ClustalW MEGALIGNTM Lasergene, Wisconsin). На основании данной гомологии сделан вывод о том, что идентифицированная последовательность представляет собой последовательность рецептора GMCSF крысы.
На фиг.18 показаны результаты сравнения образцов GMCSF из образцов человека, мыши и крысы (SEQ ID №№ 25, 26 и 27) (алгоритм ClustalW MEGALIGNTM Lasergene, Wisconsin).
На фиг.19 приведены результаты иммуногистохимического анализа распределения рецептора GMCSF-альфа (a-d) и GMCSF (e-g) в разных участках головного мозга мыши (парафиновые срезы, 2 мкм). а: - в мозжечке, метятся клетки Пуркинье (стрелка), b-d: локализация GMCSF-R-альфа в гиппокампе. Необходимо отметить присутствие рецептора в воротах зубчатой извилины (b, стрелка). Антитело преимущественно окрашивает нейроны в зоне СА3 (с) с наличием четкой границы между участками СА3 и СА2 (с, стрелка). GMCSF-R-альфа распределен по всем клеткам (CA3), а также отросткам нейронов (СА2). Рецептор GMCSF также выявляли в энторинальной области коры головного мозга (d). Распределение GMCSF аналогично картине распределения рецептора GMCSF-альфа. Следует отметить, что антитело к GMCSF окрашивает также клетки Пуркинье (е, стрелка), нейроны в зоне СА3 (f) с наличием резкой границы между участками СА3 и СА2 (f, стрелка) и нейроны в энторинальной области коры головного мозга (g). Фиг.19h-m: показана неожиданная совместная локализация рецептора GMCSF и его лиганда в нейронах; h - локализация рецептора GMCSF (антитело sc690, Santa Cruz) в нейронах подполе гиппокампа СА3, стрелка указывает на отчетливую экспрессию на границе с прилегающим участком СА2. i - для лиганда GMCSF (антитело sc13101) показан тот же специфичный для подполя характер экспрессии, стрелка указывает на один нейрон в области СА3. j - экспрессия рецептора GMCSF в воротах и зоне под зернистым слоем зубчатой извилины. Стрелка указывает на нейрон в зоне под зернистым слоем. k - экспрессия лиганда GMCSF в данной области. Здесь у лиганда выявили немного иной характер экспрессии, чем у рецептора. Видна четкая экспрессия в области СА3 (стрелка), которая, однако, ниже, чем в области зубчатой извилины. l, m: экспрессия рецептора GMCSF (l) и лиганда (m) в больших двигательных нейронах спинного мозга. Неожиданная экспрессия является явным указанием на терапевтическую применимость системы GMCSF для заболеваний моторных нейронов, в частности бокового амиотрофического склероза (ALS).
На фиг.20 показано окрашивание GMCSFR-альфа в культурах нервных клеток коры головного мозга. Рецептор присутствует как на клетках, так и на отростках нейронов (подтверждено двойным мечением антителом к ядерному эпитопу NeuN и антителом к синаптофизину, что не показано на чертеже). Как предварительная инкубация антитела с соответствующим пептидом, так и отсутствие первичного антитела не приводило к специфичному окрашиванию (результаты не показаны).
На фиг.21 показано наличие рецептора GMCSF-альфа на нейрональных стволовых клетках (nsc) взрослого индивидуума и на клетках PC12 (PC12) посредством ОТ-ПЦР. Показан агарозный гель. Трек 1: маркера размера (М); трек 2: ПЦР с нейрональными стволовыми клетками (nsc); трек 3: ПЦР с клетками РС12 (РС12); трек 4: отрицательный контроль (neg). На треках 2 и 3 видна полоса, специфичная для GMCSFR-альфа крысы (176 п.н.).
На фиг.22 показано присутствие рецептора GMCSF-альфа в нейрональных стволовых клетках (nsc) взрослого посредством иммуногистохимии. Показана одна нейросфера, окрашенная DAPI (A - окрашиваются все клеточные ядра) и антителом, специфичным к рецептору GMCSF (В) (увеличение 10х).
На фиг.23 показано эффективное нейропротективное действие GMCSF in vitro. Тест на жизнеспособность клеток проведен с использованием обработанных NGF клеток РС12 в условиях повышающегося окислительного стресса, вызванного H2O2 (0 мкМ, 400 мкМ, 750 мкМ). Обработка GMCSF приводит к резкому повышению жизнеспособности клеток. Для сравнения приведены показатели жизнеспособности клеток после их обработки эритропоэтином (EPO; 0,5 Ед/мл) - известным нейропротективным средством. Ось Y - относительный показатель жизнеспособности клеток (световые единицы люциферазной активности).
На фиг.24 показан эффект внутривенного введения GCSF на размер области инфаркта в крысиной модели MCAO при применении через 120 минут после появления ишемии в сравнении с введением C-CSF (контроль).
На фиг.25 показано окрашивание гиппокампа и коры головного мозга ТТС для выявления рецептора GCSG и GCSF. Приведенные данные показывают, что рецептор GCSF (a, d, g) локализован вместе со своим лигандом (b, e, h) как в гиппокампе (a-c: зубчатая извилина, d-f: ворота), так и в коре головного мозга (g-i).
На фиг.26 показана совместная экспрессия рецепторов GCSF и GMCSF на нейронах в зубчатой извилине (a-c) и в воротах (d-f) гиппокампа, а также в коре головного мозга (g-i). Рецептор GCSF (a, d, g) и рецептор GMCSF (b, e, h) экспрессируются на одних и тех же нейронах как в гиппокампе, так и в коре головного мозга (c, f, i).
На фиг.27 показано, что GCSF оказывает антиапоптотическое действие путем активации в нейронах stat3.
На фиг.28 (части I и II) в части А показана очень сильная повышающая регуляция GCSF через 2 часа и 6 часов после ишемии на ипси- и контралатеральной сторонах, тогда как для рецептора (В) через 6 часов отмечается умеренная регуляция. Индуцированная экспрессия GCSF не специфична для модели MCAO, но может отмечаться, хотя и в меньшей степени, при общей ишемии (С).
На фиг.29 показаны GCSF и его рецептор в области "полутени" инфаркта (бенгальский розовый).
На фиг.30 показана экспрессия рецепторов GCSF (a, d, g) и doublecortin (b, e, h) на нейронах в зубчатой извилине (а-f) и в воротах (g-i) гиппокампа.
На фиг.31 показана сильная и зависимая от концентрации повышающая регуляция нейрональных маркеров NSE и бета-III-тубулина через 4 суток после введения GCSF. PLP и GFAP подвергаются умеренной регуляции в зависимости от концентрации GCSF. Отрезки, означающие ошибки, означают стандартные отклонения, рассчитанные на основе 3-кратных последовательно разведенных образцов кДНК, и отражают достоверность проведенных измерений.
На фиг.32 показана совместная локализация в головном мозге GMCSF и его рецептора. Рецептор GMCSF (a, d, g) и сам GMCSF (b, e, h) локализованы вместе на нейронах в зубчатой извилине (a-с), в воротах (d-f) и в коре головного мозга (g-i).
На фиг.33 показано, что GMCSF и его рецептор подвергаются повышающей регуляции под действием ишемии головного мозга.
На фиг.34 показана экспрессия рецептора GMCSF (a, d, g) и doublecortin (b, e, h) на нейронах в зубчатой извилине (a-с) и в воротах (d-i) гиппокампа.
На фиг.35 показаны потенциал нейрональных стволовых клеток к дифференцировке и экспрессия специфических маркеров дифференцированных клеток и показано, что обработка нейрональных стволовых клеток 10 нг/мл GMCSF приводит к значимо индуцированной экспрессии бета-III-тубулина (n=3; p<0,05; двусторонний t-тест) и в меньшей степени - NSE, маркера зрелых нейронов (В).
На фиг.36 показано, что GCSF проходит через гематоэнцефалический барьер (ГЭБ).
На фиг.37 показано, что GMCSF проходит через гематоэнцефалический барьер (ГЭБ).
На фиг.38 проиллюстрирована эффективность применения GCSF при лечении бокового амиотрофического склероза (ALS).
На фиг.39 показана эффективность применения GCSF при лечении болезни Паркинсона (PD) на моделях грызунов.
На фиг.40 показано, что GMCSF оказывает в нейронах антиапоптотическое действие путем активации путей передачи сигнала с участием stat3, на части I чертежа показано, что GMCSF ингибирует расщепление PARP, являющегося специфическим признаком апоптоза. Гибель клеток индуцировали в первичных нейронах применением NOR-3 как донора NO. На части II чертежа показано, что применение GMCSF не приводит к повышению уровня фосфорилирования STAT1 ("pSTAT1") или STAT5 ("pSTAT5"), хотя сами белки экспрессируются в нейронах. На части IIIA чертежа показано, что применение GMCSF приводит к зависимой от времени активации STAT3, которая достигает максимума через 5 минут после введения GMCSF. В - количественный результат трех независимых экспериментов. С - активацию STAT3 посредством фосфорилирования можно ингибировать введением ингибитора JAK2-AG490. На части IV чертежа показано, что GMCSF сильно индуцирует экспрессию генов-мишеней stat3 - Bc12 и BclXI. Известно, что эти гены являются антиапоптотическими.
На фиг.41 показано значительное снижение размера зоны инфаркта у животных после введения GMCSF как в начале (часть I), так и в конце временного окна длительностью 3 часа (часть II).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
GCSF
Гранулоцитарный колониестимулирующий фактор (GCSF) представляет собой хорошо известный фактор роста. Варианты GCSF, которые можно применять в способах по настоящему изобретению, представляют собой полноразмерные кодирующие последовательности, белковые последовательности и различные функциональные варианты, мутеины или миметики, которые уже известны и доступны. В приведенном ниже обсуждении их обозначают как производные GCSF.
Структура кодирующей ДНК и последовательности белка известна, а также известны способы получения полипотентного гранулоцитарного колониестимулирующего фактора млекопитающих рекомбинантными способами (WO 87/01132; патент США 4810643). Например, несколько аминокислотных последовательностей, соответствующих GCSF, показаны на фиг.10 и в перечне последовательностей, например SEQ ID №№ 28, 29, 30, 31, 32 и 33.
В одном варианте осуществления в настоящем изобретении можно применять описываемые здесь белки, которые, по меньшей мере, на 70%, предпочтительно, по меньшей мере, на 80%, более предпочтительно, по меньшей мере, на 90%, идентичны полноразмерным аминокислотным последовательностям человеческого GCSF, например SEQ ID № 28. В другом варианте осуществления настоящего изобретения варианты GCSF, подходящие для применения, включают в себя варианты, кодируемые полинуклеотидной последовательностью, которая, по меньшей мере, на 70%, предпочтительно, по меньшей мере, на 80%, более предпочтительно, по меньшей мере, на 90%, 95% и 97%, идентична полноразмерной кодирующей последовательности человеческого GCSF дикого типа, например полинуклеотид, кодирующий SEQ ID № 28, данные полинуклеотиды гибридизуются в строгих условиях с полноразмерной кодирующей полинуклеотидной последовательностью человеческого GCSF дикого типа. Термины "строгие условия" или "условия строгой гибридизации" включают в себя указание на условия, при которых полинуклеотид гибридизуется со своей последовательностью-мишенью в заметно большей степени, чем другие последовательности (например, по меньшей мере, в 2 раза превышая фоновый уровень). Строгие условия представляют собой применение концентрации солей менее чем приблизительно 1,5 М иона Na, как правило, приблизительно от 0,01 до 1,0 М концентрации иона Na (или других солей) при pH 7,0 - 8,3 и температуре, по меньшей мере, равной приблизительно 30°C для коротких зондов (например, длиной примерно от 10 до 50 нуклеотидов) и, по меньшей мере, примерно 60°C для длинных зондов (например, более чем 50 нуклеотидов), например, очень строгие условия включают в себя гибридизацию в присутствии 50% формамида, 1 M NaCl, 1% SDS при 37°C и отмывку в 0,1Х SSC при температуре от 60 до 65°C (см. Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, Part I, Chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays"; Elsevier, New York (1993); и Current Protocols in Molecular Biology, Chapter 2, Ausubel, et al., Eds., Greene Publishing and Wiley-Interscience, New York (1995)). Идентичность, гомологию и/или сходство аминокислот и полинуклеотидов можно определять с применением алгоритма ClustalW (MEGALIGNTM, Lasergene, Wisconsin).
Примеры различных функциональных вариантов, мутеинов и миметиков GCSF включают в себя функциональные фрагменты и варианты (например, структурно и биологически близкие к белку дикого типа и содержащие по меньшей мере один биологически эквивалентный домен), химические производные GCSF (например, содержащие дополнительные химические группы, такие как полиэтиленгликоль и производные полиэтиленгликоля и/или его гликозилированные формы, такие как ЛеногастримTM (LenogastrimTM)) и пептидомиметики GCSF (например, низкомолекулярное соединение, которое имитирует пептид по структуре и/или по функции (см., например, Abell, Advances in Amino Acid Mimetics and Peptidomimetics, London: JAI Press (1997); Gante, Peptidmimetica - massgeschneiderte Enzyminhibitoren Angew. Chem. 106: 1780-1802 (1994); и Olson et al., J. Med. Chem. 36: 3039-3049 (1993)).
Дополнительные примеры производных GCSF включают гибридный белок альбумина и GCSF (AlbugraninTM) или другие модификации, полученные по типу слияния, такие как описанные в патенте США № 6261250); конъюгаты PEG-GCSF и другие PEGилированные формы; формы, описанные в WO 00/44785 и в работе Viens et al., J. of Clin. Oncology, VI., Nr. 1, 2002: 24-36; норлейциновые аналоги GCSF, например, описанные в патенте США № 5599690; миметики GCSF, например, описанные в WO 99/61445, WO 99/61446 и в работе Tian et al., Science, Vol. 281, 1998: 257-259; описанные в патентах США №№ 5214132 и 5218092 мутеины GCSF, в которых модифицированы, удалены или вставлены одна или несколько аминокислот; производные GCSF, описанные в патенте США № 6261550 и в патенте США № 4810643; а также химерные молекулы, которые содержат полную последовательность GCSF или ее часть в сочетании с другими фрагментами последовательности, например Леридистим (Leridistim) (см., например, Streeter et al. (2001), Exp. Hematol., 29, 41-50, Monahan, et al. (2001) Exp. Hematol., 29, 416-24; Hood et al. (2001), Biochemistry, 40, 13598-606, Farese et al. (2001) Stem Cells, 19, 514-21; Farese et al. (2001) Stem Cells, 19, 522-33; MacVittie, et al. (2000) Blood, 95, 837-45). Кроме того, производные GCSF включают в себя такие варианты, где цистеиновые остатки в положениях 17, 36, 42, 64 и 74 (из числа вариантов, содержащих 174 аминокислоты (SEQ ID № 37) или из числа вариантов, содержащих 175 аминокислот, где дополнительная аминокислота, представляющая собой N-концевой метионин (SEQ ID № 38), заменены другой аминокислотой (такой как серин), как описано в патенте США № 6004548, GCSF, содержащий аланин в первом (N-концевом) положении; вариант, содержащий модификацию, по меньшей мере, одной аминогруппы в полипептиде, обладающем активностью GCSF, как описано в EP 0 335 423; производные GCSF, содержащие замещенную или удаленную на N-концевом участке белка аминокислоту, как описано в EP 0 272 703; производные природных GCSF, обладающие, по меньшей мере, одним из биологических свойств природного GCSF и стабильностью в растворах в концентрации, по меньшей мере, 35% при 5 мг/мл, где в указанном производном, по меньшей мере, один Cys17 в природной последовательности замещен остатком Ser17, и Asp27 в природной последовательности замещен остатком Ser27, как описано в EP 0459630; модифицированную последовательность ДНК, кодирующую GCSF, в которой N-конец модифицирован с целью усиления экспрессии белка в рекомбинантных клетках хозяина, без изменения аминокислотной последовательности белка, как описано в EP 0459630; GCSF, модифицированный путем инактивации, по меньшей мере, одного участка расщепления дрожжевой протеазы KEX2, с целью повышения выхода рекомбинантной продукции при использовании дрожжей, как описано в ЕР 0 243 153; белки, измененные лизином, как описано в патенте США № 4904584; варианты белков, измененные цистеиновым остатком, как описано в патенте США WO/9012874 (US 5166322); варианты с добавлением аминокислот к любому из концов молекулы GCSF для облегчения укладки молекулы после экспрессии в прокариотах, как описано в AU-A-10948/92; варианты с замещением последовательности Leu-Gly-His-Ser-Leu-Gly-Ile (SEQ ID № 11) в положении 50-56 в молекуле GCSF, содержащей 174 аминокислоты (SEQ ID № 37), и в положении 53-59 в молекуле GCSF, содержащей 177 аминокислот (SEQ ID № 39), и/или, по меньшей мере, одного из четырех гистидиновых остатков в положениях 43, 79, 156 и 170 в зрелом GCSF, содержащем 174 аминокислоты (SEQ ID № 37), или в положениях 46, 82, 159 или 173 в зрелом GCSF, содержащем 177 аминокислот (SEQ ID № 39), как описано в AU-A-76380/91; и синтетическую последовательность нуклеиновой кислоты, кодирующую GCSF, в которую встроены сайты рестрикции для облегчения кассетного мутагенеза выбранных областей и фланкирующие участки рестрикции для облегчения вставки гена в желательную систему экспрессии, как описано в GB 2 213 821. Дополнительные примеры аналогов GCSF включают в себя SEQ ID №№ 64 и 65 и другие, описанные в патенте США № 6632426. Содержание указанных выше документов включено в настоящее описание в качестве ссылки.
Различные функциональные производные, варианты, мутеины и/или миметики GCSF предпочтительно сохраняют, по меньшей мере, 20%, предпочтительно 50%, более предпочтительно, по меньшей мере, 75% и/или наиболее предпочтительно - по меньшей мере, 90% биологической активности GCSF млекопитающего дикого типа, уровень биологической активности включает в себя 25%, 30%, 35%, 40%, 45%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 95%; а также все значения и поддиапазоны между ними. Кроме того, функциональные производные, варианты, мутеины и/или миметики GCSF могут также обладать 100% или большей биологической активностью в сравнении с активностью GCSF млекопитающего дикого типа, уровень биологической активности включает в себя, по меньшей мере, 105%, по меньшей мере, 110%, по меньшей мере, 125%, по меньшей мере, 150% и, по меньшей мере, 200%.
Для определения биологической активности GCSF можно применять несколько известных анализов как по отдельности, так и в сочетании. Один из примеров определения функции GCSF проиллюстрирован в примере 1. Известны также другие способы определения функции GCSF, которые включают в себя анализ колониеобразующей активности с применением клеток костного мозга мышей; стимуляцию пролиферации клеток костного мозга индуцированную GCSF; специфические биологические анализы с применением клеточных линий, рост которых зависит от GCSF или которые отвечают на GCSF (например, AML-193; 32D; BaF3; GNFS-60; HL-60, M1; NFS-60; OCI/AML1a; и WEHI-3B). Эти и другие анализы описаны в Braman et al., Am. J. Hematology 39: 194-201 (1992); Clogston CL et al., Anal. Biochem. 202: 375-83 (1992); Hattori K et al., Blood 75: 1228-33 (1990); Kuwabara Т et al., Journal of Pharmacobiodyn 15: 121-9 (1992); Motojima H et al., Journal of Immunological Methods 118; 187-92 (1989); Sallerfors B and Olofsson, European Journal of Haematology 49: 199-207 (1992); Shorter SC et al., Immunology 75: 468-74 (1992); Tanaka H and Kaneko, Journal of Pharmacobiodyn 15: 359-66 (1992); Tie F et al., Journal of Immunological Methods 149; 115-20 (1992); Watanabe M et al., Anal. Biochem. 195: 38-44 (1991).
В одном варианте осуществления настоящего изобретения GCSF модифицирован, введен в состав соответствующей композиции или существует в виде миметика GCSF, за счет чего достигается повышение его способности проходить через гематоэнцефалический барьер или смещается его коэффициент распределения в сторону мозговой ткани. Примером такой модификации является добавление последовательностей PTD или TAT (Cao et al. (2002) J. Neurosci. 22: 5423-5431; Mi et al. (2000) Mol. Ther. 2: 339-347; Morris et al. (2001) Nat Biotechnol 19: 1173-1176; Park et al. (2002) J. Gen Virol 83: 1173-1181). Указанные последовательности также можно применять в мутированных формах и добавлять с дополнительными аминокислотами к амино- или карбоксиконцу белков. Кроме того, добавление брадикинина или аналогичных веществ при внутривенном введении любого препарата GCSF способствует его доставке в головной мозг или спинной мозг (Emerich et al. (2002) Clin Pharmacokinet 40: 105-123; Siegal et al. (2002) Clin Pharmacokinet 41: 171-186).
GMCSF
Гранулоцитарно-макрофагальный колониестимулирующий фактор (GMCSF) представляет собой хорошо известный фактор роста (см., например, фиг.18, SEQ ID №№ 25, 26 и 27). GMCSF, пригодные для применения по способам настоящего изобретения, представляют собой полноразмерные кодирующие последовательности, белковые последовательности и различные функциональные варианты, химерные белки, мутеины и миметики, которые известны и доступны, например PEGилированные формы или формы, связанные с альбумином. Известна структура кодирующей ДНК и белка, а также способы рекомбинантного получения полипотентного гранулоцитарно-макрофагального колониестимулирующего фактора (патент США 5641663). Также известен рецептор GMCSF и описан, например, в патенте США 5629283.
В одном варианте осуществления в настоящем изобретении можно применять белки, которые, по меньшей мере, на 70%, предпочтительно, по меньшей мере, на 80%, более предпочтительно, по меньшей мере, на 90%, идентичны аминокислотным полноразмерным последовательностям человеческого GMCSF. В другом варианте осуществления настоящего изобретения GMCSF, которые можно применять в способах по настоящему изобретению, включают в себя формы, кодируемые полинуклеотидной последовательностью, которая, по меньшей мере, на 70%, предпочтительно, по меньшей мере, на 80%, более предпочтительно, по меньшей мере, на 90%, 95%, 97% и 98% идентична полноразмерной кодирующей последовательности человеческого GMCSF дикого типа, например полинуклеотидом, который кодирует SEQ ID № 25, указанные полинуклеотиды гибридизуются в строгих условиях с полноразмерной кодирующей полинуклеотидной последовательностью человеческого GMCSF дикого типа. Термины "строгие условия" или "условия строгой гибридизации" включают в себя указание на условия, при которых полинуклеотид гибридизуется со своей последовательностью-мишенью в заметно большей степени, чем другие последовательности (например, по меньшей мере, в 2 раза превышая фоновый уровень). Строгие условия представляют собой применение концентрации солей менее чем приблизительно 1,5 М иона Na, как правило, приблизительно от 0,01 до 1,0 М концентрации иона Na (или других солей) при pH 7,0-8,3 и температуре, по меньшей мере, равной приблизительно 30°C для коротких зондов (например, длиной примерно от 10 до 50 нуклеотидов) и, по меньшей мере, примерно 60°C для длинных зондов (например, более чем 50 нуклеотидов), например, очень строгие условия включают в себя гибридизацию в присутствии 50% формамида, 1 M NaCl, 1% SDS при 37°C и отмывку в 0,1Х SSC при температуре от 60 до 65°C (см. Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, Part I, Chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays"; Elsevier, New York (1993); и Current Protocols in Molecular Biology, Chapter 2, Ausubel, et al., Eds., Greene Publishing and Wiley-Interscience, New York (1995)). Идентичность, гомологию и/или сходство аминокислот и полинуклеотидов можно определять с применением алгоритма ClustalW (MEGALIGNTM, Lasergene, Wisconsin).
Различные функциональные производные, варианты, мутеины и/или миметики GMCSF предпочтительно сохраняют, по меньшей мере, 20%, предпочтительно 50%, более предпочтительно, по меньшей мере, 75% и/или наиболее предпочтительно, по меньшей мере, 90%, биологической активности GCSF млекопитающего дикого типа, уровень биологической активности включает в себя 25%, 30%, 35%, 40%, 45%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 95%; а также все значения и поддиапазоны между ними. Кроме того, функциональные производные, варианты, мутеины и/или миметики GCSF могут также обладать 100% или большей биологической активностью в сравнении с активностью GCSF млекопитающего дикого типа, уровень биологической активности включает в себя, по меньшей мере, 105%, по меньшей мере, 110%, по меньшей мере, 125%, по меньшей мере, 150% и, по меньшей мере, 200%.
Для практического осуществления настоящего изобретения предпочтительны производные GMCSF, более предпочтительны миметики GMCSF, которые сохраняют свой потенциал в отношении защиты нейронов и которые обладают сниженным уровнем воздействия на лейкоциты, за счет чего минимизируются возможные неблагоприятные эффекты. Производные GMCSF, предпочтительно миметики GMCSF, можно тестировать в нейропротективном анализе in vitro, таком как анализ, описанный в примере 17. Вещества, демонстрирующие положительный нейропротективный эффект в данном анализе, далее можно анализировать на наличие у них иммуномодулирующей активности.
Для измерения уровня биологической активности GMCSF можно применять несколько известных анализов как по отдельности, так и в сочетании. Подлежащие анализу функции GMCSF включают в себя его известные иммуномодулирующие функции и одну или несколько функций, связанных с его ролью в нейропротекции. Другие способы определения функции GMCSF включают в себя, например, анализ колониеобразования посредством формирования колоний, содержащих макрофаги, нейтрофилы, эозинофилы и мегакариоциты; а также специфические биологические анализы с применением клеточных линий, рост которых зависит от наличия GMCSF или которые отвечают на данный фактор (например, клеточные линии AML-193; B6SUt-A; BAC1.2F5; BCL1; Da; FDCP1; CF-D8; GM/SO; IC-2; MO7E; NFS-60; PT-18; TALL-103; TF-1; UT-7). Эти и другие анализы описаны в Cebon J. et al., Blood 72: 1340-7 (1988); Katzen NA et al., European Cytokine Network 3: 365-72 (1992); Lewis CE et al., Journal of Immunological Methods 127; 51-9 (1990); Mortensen BT et al., Experimental Hematology 21: 1366-70 (1993); Oez S et al., Experimental Hematology 18: 1108-11 (1990); Roncaroli F et al., Journal of Immunological Methods 158; 191-6 (1993); Sallerfors B and Olofsson, European Journal of Haematology 49: 199-207 (1992); Zenke G et al., Journal of Immunoassay 12; 185-206 (1991).
Также предпочтительны модификации или композиции GMCSF или вещества-миметики, которые повышают его способность проходить через гематоэнцефалический барьер или сдвигают его коэффициент распределения в сторону мозговой ткани. Примером такой модификации является добавление домена трансдукции белка (PTD) или последовательностей TAT (Cao G et al. (2002) J. Neurosci. 22: 5423-5431; Mi Z. et al. (2000) Mol. Ther. 2: 339-347; Morris et al. (2001) Nat Biotechnol 19: 1173-1176; Park et al. (2002) J. Gen Virol 83: 1173-1181). Указанные последовательности можно также применять в мутантных формах и добавлять с дополнительными аминокислотами на амино- или карбоксиконец белков. Кроме того, добавление брадикинина или аналогичных веществ к любому препарату GMCSF при внутривенном введении способствует его доставке в головной мозг или спинной мозг (Emerich et al. (2002) Clin Pharmacokinet 40: 105-123; Siegal et al. (2002) Clin Pharmacokinet 41: 171-186). Примерами различных пригодных форм и производных являются сарграмостим (лейкин (Leukine®), прокин (Prokine®), лейкотропин (Leucotropin®)) или молграмостим (лейкомакс (Leucomax®)).
мРНК GMCSFR выявлена в выделенной микроглии, астроцитах, олигодендроцитах и нейронах (Sawada et al. (1993), Neurosci Lett, 160, 131-40; Baldwin et al. (1993), Blood, 82, 3279-82). Показано, что GMCSF стимулирует пролиферацию астроцитов (Guillemin et al. (1996), Glia, 16, 71-80). GMCSF также индуцирует высвобождение интерлейкина-6 из микроглии (Suzumura et al. (1996), Brain Res, 713, 192-8). Недавно появилось сообщение о том, что интерлейкин-1 повышает продукцию белка GMCSF в нейрональной клеточной линии NT2 (Dame et al. (2002), Eur. Cytokine Netw, 13, 128-33). Присутствие GMCSFR в головном мозге было показано при исследовании материала человеческого эмбриона в рамках направленного поиска экспрессии рецептора GMCSF в эмбриональных тканях (Dame et al. (1999), Pediatr Res, 46, 358-66). Dame et al на основе своих наблюдений сделали вывод о том, что GMCSF может играть определенную роль в развитии нервной системы эмбриона и в иммунном контроле в головном мозге взрослого. Важно, что они не упомянули о его возможной связи с развитием заболеваний, особенно в контексте его участия в нейропротекции. GMCSF in vitro повышает активность холинацетилтрансферазы (клеточная линия SN6.10.2.2 и культивируемые мышиные нейроны перегородки), а in vivo повышает жизнеспособность поврежденных холинергических нейронов перегородки крысы после перерезания по лини бахромка-свод (Kamegai et al. (1990), Brain Res, 532, 323-5; Konishi et al. (1990), Brain Res, 609, 29-35). Важно, что данные наблюдения относятся только к определенному подтипу нейронов и не обобщены авторами на другие типы нейронов или нервных клеток. Из полученных данных они также не выявили основного нейропротективного свойства GMCSF, а также не упомянули о какой-либо возможности его терапевтического применения. GMCSF активируется в астроцитах при добавлении конечных продуктов с более высоким уровнем гликозилирования (AGE) (Li et al. (1998), Mol. Med., 4, 46-60). Было показано, что GMCSF ускоряет пролиферацию микроглии in vitro (Lee et al. (1994), Glia, 12, 309-18). Недавно обнаружено, что уровень GMCSF повышается в цереброспинальной жидкости (csf) у пациентов с болезнью Альцгеймера (Tarkowski et al. (2001), Acta Neurol Scand, 103, 166-74). GMCSF также исследовали в CSF у пациентов с инсультом (Tarkowski et al. (1999), Stroke, 30, 321-7). В последнем исследовании было показано, что уровень FAS-лиганда, проапоптозного белка, прямо коррелирует с уровнем GMCSF в цереброспинальной жидкости у пациентов с инсультом на 21-26 сутки и спустя 3 месяца. Однако при этом не был сделан вывод относительно терапевтической применимости данных наблюдений для пациентов с инсультом, а также не упоминалось о какой-либо возможной нейропротивной роли GMCSF. Высвобождение GMCSF после инсульта по существу может иметь несколько причин и не позволяет сделать какие-либо прогнозы относительно его функций. GMCSF проходит через гематоэнцефалический барьер (McLay et al. (1997), Brain, 120, 2083-91). Однако это исследование не проводили с целью продемонстрировать возможность терапевтического применения GMCSF при неврологических заболеваниях и в его контексте не содержится каких-либо упоминаний о возможной применимости GMCSF в случае нейродегенеративных заболеваний или инсульта.
Обобщая, следует отметить, что указанные литературные ссылки предоставляют доказательства наличия рецептора и лиганда GMCSF в нервной системе, но не показывают какого-либо общего нейропротективного свойства GMCSF. Данные исследования, безусловно, не предполагают применения GMCSF для лечения нейродегенеративных и ишемических расстройств, таких как инсульт.
ДРУГИЕ ГЕМАТОПОЭТИЧЕСКИЕ ФАКТОРЫ РОСТА
В других вариантах осуществления настоящего изобретения можно применять сочетания препаратов GCSF и/или GMCSF, которые поддерживают терапевтическое действие каждого из них, предпочтительно нейропротивное действие. Эффект, достигаемый при применении данных сочетаний, может быть кумулятивным или супераддитивным/синергическим. В одном варианте различные производные GCSF и/или GMCSF можно использовать в сочетании друг с другом. Аналогично GCSF и/или GMCSF можно использовать в сочетании с одним или несколькими дополнительными гематопоэтическими факторами роста, такими как эритропоэтин и его производные, которые, как недавно было показано, оказывают сильное нейропротективное воздействие (например, Brines et al. (2000), Proc Natl Acad Sci USA 97: 10526-10531; Cerami et al. (2002), Nephrol Dial Transplant, 17: 8-12; Siren AL, Ehrenhreich H (2001) Eur Arch Psychiatry Clin Neurosci, 251: 179-184). Кроме того, GMCSF и/или GCSF можно применять в сочетании, например, с различными колониестимулирующими факторами (такими как M-CSF), SCF (фактор роста стволовых клеток), SCPF (фактор пролиферации стволовых клеток), различными интерлейкинами (IL1, IL3, IL4, IL5, IL6, IL11, IL12), LIF, TGF-β, MIP-1-α, TNF-α, а также многими другими низкомолекулярными факторами.
В другом варианте осуществления настоящего изобретения различные гематопоэтические факторы роста, за исключением эритропоэтина, можно применять самостоятельно без GMCSF и/или GCSF. В другом варианте, например, в случае инсульта GCSF и/или GMCSF можно сочетать с веществами, обладающими тромболитической активностью, например тканевым активатором плазминогена (ТАП), стрептокиназой, урокиназой и/или анкродом (Ancrod). GCSF и/или GMCSF также можно сочетать с ацетилсалициловой кислотой (аспирином) или гепарином, с мелатонином и/или веществами, которые препятствуют сигналам апоптоза (например, ингибиторами каспаз). Также, а особенно для лечения бокового амиотрофического склероза (ALS), хотя и не только его, применимы сочетания GCSF и/или GMCSF с рилузолом (Rilutek®), с витаминами, такими как витамин E, Q10 или с антиоксидантами. Для лечения болезни Паркинсона можно применять сочетания GCSF и/или GMCSF с известными лекарственными средствами, применяемыми при лечении болезни Паркинсона, такими как тригексилфенидил, селегилин, L-ДОФА, перголид и другие. Другие вещества, которые могут сочетать с гематопоэтическими факторами роста, например с GCSF и/или GMCSF, включают в себя bFGF, антитела к GPIIb/IIa, антитела к ICAM, ингибиторы оксида азота, тромболитики, ингибиторы ротамазы, ингибиторы кальциневрина и циклоспорин. Дополнительно при лечении нейродегенеративных расстройств может практиковаться введение одного или более средств, оказывающих воздействие на уровень нейротрансмиттеров. Кроме того, в случае лечения когнитивных состояний можно вводить одно или несколько холинергических средств, ингибиторов повторного поглощения катехоламинов и т.п.
Также можно применять сочетания с известными антидепрессантами, где пригодно, например для лечения депрессии у индивидуума. Неограничивающие примеры антидепрессантов включают в себя: SSRI (селективные ингибиторы повторного всасывания серотонина): паксил (Paxil) (пароксетин); прозак (Prozac) (флуоксетин); золофт (Zoloft) (сертралин); целекса (Celexa) (циталопрам); лексапро (Lexapro) (оксалат эсциталопрама); лувокс (Luvox) (флувоксамин); ИМАО (MOAI) (ингибиторы моноаминоксидазы): нардил (Nardil) (фенелзин); парнат (Parnate) (транилципромин); ТЦА (трициклические антидепрессанты): адапин (Adapin) (доксепин); анафранил (Anafranil) (кломипрамин); элавил (Elavil) (амитриптилин); эндеп (Endep) (амитриптилин); людиомил (Ludiomil) (мапротилин); норпрамин (Norpramin) (дезипрамин); памелор (Pamelor) (нортриптилин); пертофран (Pertofrane) (дезипрамин); синекван (Sinequan) (доксепин); сурмонтил (Surmontil) (тримипрамин); тофранил (Tofranil) (имипрамин); вивактил (Vivactil) (протриптилин) или другие типы лекарственных средств: эффексор (Effexor) (венлафаксин); цимбалта (Cymbalta) (дулоксетин); дезирел (Desyrel) (тразодон); буспар (Buspar) (буспирон); эдронакс (Edronax), вестра (Vestra) (ребоксетин); ремерон (Remeron) (миртазапин); серзон (Serzone) (нефазодон); веллбутрин (Wellbutrin) (дупропион).
НЕВРОЛОГИЧЕСКИЕ СОСТОЯНИЯ
Неврологические состояния, лечение которых можно проводить в соответствии с настоящим изобретением, как правило, относятся к трем классам: заболевания с ишемическими или гипоксическими механизмами; нейродегенеративные заболевания (см. Adams et al., Principles of Neurology, 1997, 6th Ed., New York, p. 1048 ff) и неврологические и психиатрические заболевания, сопровождающиеся гибелью нервных клеток. Другие неврологические состояния, которые можно лечить по настоящему изобретению, включают в себя также усиление познавательной способности, лечение опухолей головного мозга, таких как глиобластомы, астроцитомы, менингиомы и невриномы.
Заболевания с ишемическими или гипоксическими механизмами далее можно подразделить на заболевания общего характера и ишемию головного мозга. Примеры таких заболеваний общего характера, в которые вовлечены ишемические или гипоксические механизмы, включают в себя инфаркт миокарда, сердечную недостаточность, порок сердца, застойную сердечную недостаточность, миокардит, перикардит, перимиокардит, коронарную болезнь сердца (стеноз коронарных артерий), стенокардию, врожденный порок сердца, шок, ишемию конечностей, стеноз почечных артерий, диабетическую ретинопатию, тромбоз, ассоциированный с малярией, наличие искусственных клапанов сердца, анемии, гиперспленический синдром, эмфизему, фиброз легкого и отек легкого. Примеры ишемической болезни мозга включают в себя инсульт (а также геморрагический инсульт), церебральную микроангиопатию (болезнь мелких сосудов), интрапартальную ишемию головного мозга, ишемию головного мозга, возникающую во время/после остановки сердца или реанимации, ишемию головного мозга, связанную с проблемами в ходе операции, ишемию головного мозга при операции на сонной артерии, хроническую ишемию головного мозга, вызванную стенозом артерий, поддерживающих кровоток головного мозга, синусовый тромбоз или тромбоз вен головного мозга, врожденные пороки сосудов головного мозга и диабетическую ретинопатию.
Примеры нейродегенеративных заболеваний включают в себя боковой амиотрофический склероз (ALS), болезнь Паркинсона, болезнь Хантингтона, болезнь Уилсона, множественную атрофию, болезнь Альцгеймера, болезнь Пика, болезнь Леви, болезнь Халлервордена-Шпатца, торсионную дистонию, наследственные сенсомоторные невропатии (HMSN), болезнь Герстманна-Штраусслера-Шанкера, болезнь Крейтцфельд-Якоба, болезнь Мачадо-Джозефа, атаксию Фридрайха, атаксию не-Фридрайха, синдром Жилль-де-ля-Туретта, семейный тремор, оливопонтоцеребеллярную дегенерацию, паранеопластические церебральные синдромы, наследственные спастические параплегии, наследственную невропатию зрительного нерва (болезнь Лебера), пигментный ретинит, болезнь Штаргардта и синдром Киернса-Сайре.
Примеры неврологических и психических заболеваний, связанных с гибелью нервных клеток, включают в себя септический шок, внутримозговое кровотечение, субарахноидальное кровотечение, деменцию, возникшую в результате множественных инфарктов, воспалительные заболевания (такие как васкулит, рассеянный склероз и синдром Гийена-Барре), травмы нервной системы (такие как травма спинного мозга и травма головного мозга), периферические невропатии, полиневропатии, эпилепсии, шизофрению, депрессию, метаболические энцефалопатии и инфекции центральной нервной системы (вирусные, бактериальные, грибковые).
Под термином "лечение" понимают замедление, прерывание, остановку или приостановку прогрессирования заболевания или состояния, что необязательно требует полного устранения всех симптомов и признаков заболевания. Термин "профилактика" применяется в контексте предупреждения неврологического заболевания, включая любую степень отдаления времени начала развития или подавления тяжести, признаков или симптомов заболевания, состояния, в том числе, без ограничения, полное предупреждение развития заболевания или состояния.
Поскольку была выявлена сильная экспрессия рецептора GCSF и рецептора GMCSF на больших моторных нейронах спинного мозга (см. фиг.4i-l) и показано, что GCSF эффективен при очаговой ишемии головного мозга (см. фиг.1), в которую вовлечены те же основные патогенетические механизмы, что и в случае ALS и других нейродегенеративных заболеваний, таких как, например, вовлечение глутамата, окислительный стресс и программируемая гибель клеток, то GCSF особенно подходит для длительного лечения хронического состояния, такого как ALS, поскольку он хорошо переносится пациентами при длительном применении (Ozer et al. (2000), J. Clin. Oncol, 18, 3558-85). Таким образом, в одном варианте осуществления настоящего изобретения для лечения ALS можно применять гематопоэтические факторы роста, такие как GCSF и GMCSF, отдельно, в сочетании друг с другом и/или в сочетании с одним или несколькими дополнительными факторами.
Ассоциированная с болезнью Паркинсона патофизиология, такая как вовлечение окислительного стресса и апоптоза, размещает болезнь Паркинсона среди других нейродегенеративных расстройств и инсульта. GCSF оказывает сильный нейропротективный эффект на вызванную H2O2 гибель клеток в клеточной линии РС12 (фиг.1b). Клетки РС12 проявляют признаки дофаминэргических клеток, например, присутствием функционального переносчика дофамина (Maruyama et al. (2001), Arch Toxicol, 75, 209-13) и используются в качестве моделей in vitro для изучения важных аспектов болезни Паркинсона, например токсичности для MPTP метаболита MPP+ (Bai et al. (2002), Neurosci. Lett, 321, 81-4). H2O2 оказывает вредное воздействие на клетки PC12, четко моделируя на них некоторые аспекты болезни Паркинсона. Например, H2O2 является одним из промежуточных продуктов в цепи клеточных событий, вызванных МРТР (Fabre et al., 1999, J Physiol Biochem 55(4): 325-31). Существует выраженное перекрывание каскада клеточных событий и механизмов передачи сигнала, вовлеченных в опосредованную MPTP и H2O2 клеточную гибель у дофаминэргических нейронов (Chun HS et al., J Neurochem, 2001, Feb; 76(4): 1010-21; Lai et al., Biochem Pharmacol, 1993, Feb 24; 45(4): 927-33). H2O2 действует посредством продукции реакционно-способных видов кислорода (ROS), что ведет к окислительному стрессу и апоптозу. Повреждение кислородными радикалами представляет собой одно из важнейших патофизиологических событий при болезни Паркинсона (Bonnet and Houeto (1999), Biomed Pharmacother, 53, 117-21, Beal (2001), Nat Rev Neurosci, 2, 325-34), и для таких пациентов эффективно лечение антиоксидантами (Beal (2002), Free Radic Res, 36, 455-60, Shults et al. (2002), Arch Neurol, 59, 1541-50). Таким образом, эффективность GCSF, при вызванной Н2О2 гибели клеток в клеточной линии РС12, модели для важных патофизиологических механизмов окислительного стресса и апоптоза, позволяет прогнозировать эффективность его применения при лечении болезни Паркинсона (PD). Неожиданно выявили экспрессию рецептора GCSF в области, поражаемой при болезни Паркинсона, в черном веществе (SN), в частности в компактной части черного вещества (SNC) (см. фиг.4m-o). Эффективность на клеточной модели, локализация рецепторов in vivo и перекрывание патофизиологических механизмов с ишемией головного мозга (кислородные радикалы/апоптоз) позволяют обоснованно полагать, что при лечении болезни Паркинсона можно применять GCSF и/или другие факторы роста, например GMCSF. Тестирование эффективности на моделях грызунов можно проводить, как проиллюстрировано в примере 5. Таким образом, в другом варианте осуществления настоящего изобретения гематопоэтические факторы роста, такие как GCSF и GMCSF, отдельно, в сочетании друг с другом и/или в сочетании с одним или несколькими дополнительными факторами, можно применять для лечения болезни Паркинсона.
Авторы настоящего изобретения на основании содержащегося здесь описания показали, что GCSF и GMCSF обладают способностью усиливать нейрогенез и улучшают поведенческие последствия ишемического повреждения. Нейрогенез представляет собой один из механизмов, который может приводить к повышению пластичности нейронных сетей и может замещать постепенную потерю нейронов. Таким образом, один вариант настоящего изобретения относится к обеспечению усиления, улучшения или повышения познавательной способности у индивидуума, страдающего от некоторого снижения познавательной способности, индивидуума демонстрирующего его и/или у которого предполагают его наличие, за счет введения такому индивидууму одной или нескольких описанных здесь композиций в соответствии с описанным здесь режимом введения. В альтернативном варианте осуществления усиление познавательной способности может также являться полезной для тех индивидуумов, у которых нет патологических состояний, например у тех индивидуумов, у которых отсутствуют признаки ухудшения познавательной способности. Способы определения уровня познавательной способности и в этой связи ее улучшения известны специалистам в данной области. Когда измеряют повышение или усиление познавательной способности, их сравнивают до введения композиции по изобретению и после введения (в некоторых вариантах осуществления можно определять во время введения) по тому же тесту, например, с теми же критериями, параметрами и т.п.
Кроме того, применение композиций по настоящему изобретению для усиления познавательной способности не ограничено случаями непатологического снижения интеллекта, но его также можно применять для усиления нормальной физиологической совокупности умственных способностей, например для улучшения памяти, улучшения двигательной координации и/или усиления способностей к логическому мышлению.
Депрессия характеризуется унынием, потерей интереса к активной жизни и уменьшением активной деятельности. Другие симптомы включают в себя утрату уверенности и самоуважения, чувство неоправданной вины, мысли о смерти и суициде, сниженную концентрацию и нарушение сна и аппетита. Также могут присутствовать множество соматических симптомов. Хотя депрессивные ощущения являются обычной реакцией, особенно после столкновения с жизненными препятствиями, такие состояния диагностируются как депрессивные расстройства только в тех случаях, когда симптомы достигают порогового значения и длятся, по меньшей мере, две недели. Депрессия может варьировать по тяжести от умеренной до очень тяжелой.
Депрессия может поражать людей на любой стадии жизни, хотя наибольшая частота отмечается в среднем возрасте. Однако все чаще депрессия выявляется у подростков и у совершеннолетних молодых людей (Lewinsohn et al. (1993), J Abnorm Psychol, 102, 110-20). Депрессия по существу представляет собой эпизодически повторяющееся расстройство, где каждый эпизод длится, как правило, от нескольких месяцев до нескольких лет, с периодами нормального самочувствия между ними. Однако примерно в 20% случаев депрессия протекает в хронической форме, без ремиссии (Thornicroft and Sartorius (1993), Psychol Med, 23, 1023-32), особенно в тех случаях, когда отсутствует адекватное лечение. Частота рецидивов у тех, кто восстановился после первого периода, составляет около 35% в течение 2 лет и примерно 60% в течение 12 лет. Частота рецидивов выше у лиц старше 45 лет. Одним из самых трагических исходов депрессивного расстройства является суицид. Примерно 15-20% пациентов с депрессией заканчивают жизнь совершением самоубийства. Суицид остается одним из наиболее частых и неизбежных последствий депрессии. Обобщая, можно сказать, что депрессия представляет собой часто встречающееся психическое расстройство, приводящее к очень высокому уровню тяжести заболевания и прогнозируемой тенденцией к росту в предстоящие 20 лет.
Депрессия может поражать пациента постоянно (монополярное расстройство) или в режиме биполярного характера (маниакальная или биполярная депрессия). Биполярная депрессия характеризуется экстремальными колебаниями настроения от "подъемов" в виде избыточной энергии и эйфории до "спадов" в виде крайнего отчаяния и апатии. Маниакальную депрессию часто лечат литием, который выравнивает колебания настроения.
Депрессия может представлять собой сезонное аффективное расстройство (SAD). Это такой тип депрессии, который в основном совпадает с приближением зимы, начиная с сентября, и длящийся до прихода весны, приносящей более длинные сутки и больше солнечного света.
Депрессия может представлять собой постнатальную депрессию, которая поражает детей, начиная примерно с 2-х недельного возраста до 2 летнего возраста.
Для определения характера депрессии у детей и подростков предприняты попытки классификации депрессии по степени тяжести (см., например, Abdel-Khalek (2003), Psychol Rep, 93, 544-60), в соответствии с которой она была распределена по восьми основным группам, таким как пессимизм, слабая концентрация внимания, проблемы со сном, ангедония, усталость, чувство одиночества, низкий уровень самоуважения и соматические жалобы.
Депрессия может протекать как идиопатическое заболевание (без связанного с нею соматического заболевания) или может развиваться как психический симптом соматического заболевания, в особенности ряда нейродегенеративных расстройств. Депрессивные расстройства (DD) представляют собой частые психические сочетанные заболевания при неврологических расстройствах, таких как рассеянный склероз, инсульт, деменция, мигрень, болезнь Паркинсона и эпилепсия. Клинические проявления DD при таких расстройствах идентичны их проявлениям при идиопатических расстройствах настроения. При нейродегенеративных расстройствах на начальной стадии заболевания часто выявляют "классические" психические симптомы. Симптоматика депрессии в контексте нейродегенеративного расстройства может отличаться от депрессии в чистом виде. Ниже перечислены некоторые нейродегенеративные расстройства, которые включают в качестве симптома депрессию:
1. Рассеянный склероз
2. Травматическое повреждение головного мозга: депрессия может поражать пациентов с травматическим повреждением головного мозга (TBI): от 37% до 38% пациентов с TBI можно классифицировать как пребывающих в депрессии (Seel and Kreuzer 03)
3. Инсульт: Примерно 30% пациентов с инсультом пребывают в клинически выраженной депрессии (Williams 86)
4. Деменция и болезнь Альцгеймера
5. Мигрень
6. Болезнь Паркинсона: Депрессивные симптомы встречаются примерно у половины пациентов с PD и являются существенной причиной функциональных нарушений у пациентов с PD. Накапливается все больше доказательств того, что депрессия при PD является вторичной относительно лежащей в ее основе нейроанатомической дегенерации, а не просто представляет собой реакцию на психологический стресс и инвалидность. Частота депрессии коррелирует с изменениями серотонинергической функции центральной нервной системы и нейродегенерацией специфических кортикальных и субкортикальных путей. Таким образом, понимание депрессии как сопутствующего патологического процесса при PD может помочь в понимании нейроанатомической основы меланхолии.
7. Эпилепсия: Эпилепсия представляет собой хроническое состояние, которое оказывает комплексное воздействие на социальные, профессиональные и психологические функции. Показано, что у пациентов с эпилепсией ряд психических расстройств встречается чаще по сравнению с населением в целом. Депрессия, по всей видимости, является наиболее частым сопутствующим психическим заболеванием, хотя тревожность и другие диагностические признаки не исследованы в достаточной мере. Однако при эпилепсии, депрессивные расстройства часто могут также встречаться в сочетании с клиническими характеристиками, отличаясь при этом от характеристик идиопатической депрессии, и не соответствуют критериям, описанным в руководстве по диагностике и статистическому анализу психических заболеваний (Diagnostics and Statistical Manual of Psychiatric Disorders-Fourth Edition). В результате ряда исследований показано, что депрессия или физиологическое расстройство могут служить сильнейшими параметрами прогноза относящегося к здоровью качества жизни, даже включая сюда частоту и тяжесть приступов, способность работать или управлять механизмами (Gilliam, et al., (2003), Epilepsy Behav, 4, Suppl 4, 26-30). Сообщается, что люди с эпилепсией, у которых наблюдается клинически выраженная картина депрессии, более тяжело и беспокойно воспринимают приступы, а также имеют значительно больше проблем при общем восстановлении от приступов, чем люди с аналогичными типами приступов, но без депрессии. Всестороннее влияние депрессивных симптомов на сообщения об активности приступов дает основание полагать, что людей с эпилепсией следует подвергать скринингу на депрессию. Приведенные данные демонстрируют важность выявления и лечения депрессии у людей с эпилепсией (Gilliam, Hecimovic and Sheline (2003), Epilepsy Behav, 4 Suppl 4, 26-30).
8. Болезнь Хантингтона: Болезнь Хантингтона (HD) представляет собой нейродегенеративный процесс, который проявляется как снижение контроля двигательной активности индивидуума, снижение познавательной способности и эмоциональной стабильности. Эмоциональная нестабильность отражается во множестве симптомов, таких как изменение личности, тревожное состояние и раздражительность. Ослабление познавательной функции может предшествовать двигательным симптомам, наблюдаемым при болезни Хантингтона (HD). Депрессия широко распространена при HD, а также связана с ослаблением познавательной способности. Депрессивное настроение и предполагаемое время наступления заболевания, определяемое с применением длины мутации ДНК, являются важными факторами прогноза функционирования оперативной памяти. Полученные результаты соответствуют имеющимся данным, внося дополнительный вклад в существующие исследования индивидуумов с симптомами, предшествующими HD, что позволяет идентифицировать потенциально поправимые параметры, способствующие ухудшению познавательной способности (например, депрессивное настроение (Nehl, et.al. (2001) J Neuropsychiatry Clin Neurosci, 13, 342-6). Депрессия также может наступать в связи с другими соматическими заболеваниями, которые в данном перечне не приведены.
В последнее время достигнут прогресс в изучении патогенеза депрессии, в соответствии с которым депрессия связана с нейродегенеративными явлениями в структурах головного мозга и с возможной неспособностью к коррекции у взрослых нейрогенеза в структурах гиппокампа. Таким образом, лечение гематопоэтическими факторами роста, например GCSF и/или GMCSF, которые оказывают антиапоптотические и способствующие регенерации воздействия (включая стимуляцию эндогенных нейрональных стволовых клеток у взрослых), как описано здесь, можно применять для лечения депрессии.
Ниже описано исследование, относящееся к нейродегенеративной концепции депрессии (обзор Kempermann and Kronenberg (2003), Biol Psychiatry, 54, 499-503).
Изучение морфологических изменений у пациентов с депрессией выявило наличие структурных изменений в гиппокампе, включая изменения в сером веществе (Sheline (2000), Biol, Psychiatry, 48, 791-800). Исследования рецидивирующей депрессии с ранним началом, депрессии в позднем возрасте, связанной с неврологическими расстройствами, и биполярного заболевания выявили наличие структурных изменений головного мозга в пределах нейроанатомического контура. Указанный контур обозначен как лимбикокортикостриопаллидарноталамический тракт и состоит из структур, которые в высшей степени взаимосвязаны. В исследованиях трехмерной магнитно-резонансной томографией при аффективном заболевании выявлено, что у многих структур, составляющих данный тракт, снижен объем или присутствуют структурные аномалии. Механизмы, предложенные для объяснения потери объема при депрессии, включают в себя вызывающую потерю нейронов нейротоксичность, снижение нейрогенеза и потерю пластичности. Данные аспекты можно лечить воздействием GCSF или GMCSF. Одна из преимущественных гипотез является нарушение нейрогенеза в гиппокампе взрослого (Kempermann and Kronenberg (2003), Biol Psychiatry, 54, 499-503, Jacobs, et al., (2000), Mol. Psychiatry, 5, 262-9, Jacobs (2002) Brain Behar Immun, 16, 602-9). Следовательно, депрессия и другие расстройства настроения представляют собой "расстройство стволовых клеток". Последние исследования выявили новые аспекты процесса регуляции нейрогенеза в гиппокампе взрослого. Одним из факторов, который сильно подавляет нейрогенез у взрослого, является стресс, возможно, из-за повышенного высвобождения глюкокортикоидов (Jacobs, Praag and Gage (2000), Mol. Psychiatry, 5, 262-9).
Нейрогенез гиппокампа у взрослого впервые описан в 1965 году Altman and Das и претерпел несколько повторных открытий. Широкий интерес к данному явлению впервые проявлен в ранних 80-х годах в связи с сообщениями о корреляции активности с нейрогенезом в головном мозге взрослых канареек. Мечение пролиферирующих клеток головного мозга бромдезоксиуридином (BrdU) в сочетании с конфокальной лазерной сканирующей микроскопией, вместо более обременительных способов применения меченого тритием тимидина с последующей ауторадиографической оценкой, позволили провести более эффективный количественный анализ нейрогенеза у взрослого. Подмножество новых клеток выживает, мигрирует в зернистый клеточный слой и дифференцируется в нейроны. Они образуют аксональные проекции до зоны СА3 вдоль волокнистых трактов, так же как и другие зернистые клетки в слое зернистых клеток, становясь визуально неотличимыми от окружающих более старых клеток.
Выявление того факта, что длительное лечение антидепрессантами позволяет компенсировать снижение нейрогенеза у взрослого, поддерживает описанную выше гипотезу (Benninghoff, et al., (2002), J Neural Transm, 109, 947-62, Dremencov, et al., (2003), Prog Neuropsychopharmacol Biol Psychiatry, 27, 729-39, D'Sa and Duman (2002), Bipolar Disord, 4, 183-94, Duman, et al., (2001), J Pharmacol Exp Ther, 299, 401-7, Duman, et al., (1999), Biol Psychiatry, 46, 1181-91).
Таким образом, в другом варианте осуществления настоящего изобретения гематопоэтические факторы роста, например GCSF и GMCSF, отдельно, в сочетании друг с другом, в сочетании с дополнительными факторами, как описано здесь, можно применять для лечения депрессии и/или для проведения профилактической терапии депрессии, например, путем их введения пациенту, предрасположенному к депрессии или у которого ожидают развитие симптомов депрессии.
В одном варианте осуществления настоящего изобретения депрессию можно лечить композициями GCSF и/или GMCSF, обладающими увеличенными периодами полувыведения из плазмы, описанными здесь выше, например, с применением PEGилированных форм (PEG-филграстим), форм, связанных с альбумином (альбугранин). Лечение не ограничено идиопатической депрессией, а его можно также применять для лечения симптоматических депрессий и депрессии, представляющей собой сопутствующую патологию. В другом варианте осуществления настоящего изобретения лечение можно проводить еженедельно путем подкожных инъекций, которые можно сочетать с перорально доступными агонистами GCSF или рецепторами GMCSF в головном мозге. Применяемые для лечения дозы могут быть такими, как указано выше, а в одном предпочтительном варианте осуществления они могут составлять от 0,1 до 1000 мкг/кг массы тела GCSF или GMCSF ежедневно. Сходное нейрохимическое окружение вокруг очага ишемии и участка травмы в сочетании со сходно измененной генной транскрипцией позволяют полагать, что сходные нейропротективные подходы, направленные на создание препятствий действию повреждающих механизмов, могут являться эффективными при ишемии головного мозга и травматическом повреждении головного мозга. Целью такой терапии в случае обоих типов повреждений является минимизация активации токсических путей при усилении активности эндогенных механизмов нейропротекции, поскольку баланс между указанными путями и будет в итоге определять судьбу такой ткани с повышенным риском. Действительно, показано, что большинство нейропротективов, эффективные в моделях экспериментального инсульта, эффективны также в моделях травматического поражения головного мозга (TBI).
В свете общности патологических и защитных процессов, активных при ишемии головного мозга и травматическом поражении головного мозга, а также общего ответа на применение нейропротективных подходов в лечении можно считать, что терапия с применением GCSF будет эффективна и при травматическом повреждении головного мозга. Существует одно исследование, в котором изучали GCSF в условиях травматического повреждения головного мозга ( Heard , et al , (1998), Crit. Care Med., 26748-54). Однако в данном исследовании стремились не к какому-либо нейропротективному эффекту GCSF (филграстим), а просто к снижению параметров инфекции (основные задачи исследования: повышение абсолютного числа нейтрофилов, безопасность филграстима и частота развития внутрибольничных инфекций (пневмония, бактериемия и инфекция мочевых путей). В данном исследовании не выявлено улучшения показателей смертности. В контексте клинической безопасности данное исследование продемонстрировало, что введение GCSF безопасно для TBI, подтвердив безопасность практического применения GCSF с целью нейропротекции по настоящему изобретению. Таким образом, в другом варианте настоящего изобретения гематопоэтические факторы, например GCSF или GMCSF, отдельно, в сочетании друг с другом, в сочетании с одним или более дополнительными факторами, можно применять для лечения ишемии головного мозга и травматического повреждения головного мозга, например, посредством предоставления профилактического способа защиты нервных клеток у таких пациентов с травмой.
Так как основные патофизиологические механизмы, действующие при ишемии головного мозга, вызванной сердечной недостаточностью и реанимацией, сравнимы с механизмами, действующими при ишемии головного мозга, вызванной окклюзией кровеносных сосудов (см. пример 1), терапия с применением GCSF также может являться эффективной для нейропротекции в условиях кардиологических осложнений. Таким образом, в другом варианте настоящего изобретения гематопоэтические факторы роста, например GCSF или GMCSF, отдельно, в сочетании друг с другом, в сочетании с одним или более дополнительными факторами, можно применять для лечения ишемии, возникшей в результате кардиологических осложнений/заболеваний, и/или для проведения профилактической нейропротективной терапии. Лечение можно начинать сразу же, как только приступают к неотложной реанимации. Альтернативно, у пациентов, входящих в известные группы риска в связи с осложнениями сердечной функции (первичный инфаркт миокарда, высокий уровень холестерина в крови, высокое кровяное давление, диабет, табакокурение), можно проводить длительное профилактическое лечение с применением гематопоэтических факторов роста, например GCSF, например, с использованием форм фактора(ов) с замедленным высвобождением.
Также указанные выше подходы применимы к большой группе пациентов, подвергающихся хирургическому вмешательству, у которых впоследствии развивается ишемия головного мозга. В частности, операция на сердце (Hogue et al. (1999), Circulation, 100, 642-7) и операция на крупных кровеносных сосудах, питающих головной мозг (например, эндартерэктомии сонной артерии), сопряжены с высоким риском развития неврологических осложнений. Объективный ретроспективный обзор 358 случаев эндартериэктомий, проведенных на сонной артерии в нейрохирургических блоках управления обучением в образовательном центре университета в Торонто в 1982 году, выявил уровень периоперационных инсультов в размере 3,9% и уровень смертности в размере 1,5%. Большая часть (82%) неврологических осложнений во время операции происходит непосредственно после послеоперационного периода (24 часа). Такая высокая частота инсульта с задержкой позволяет полагать, что большая часть периоперационных инсультов имеет эмболическую, а не гемодинамическую природу. При проведении эндартериэктомии на сонной артерии следует ожидать сочетанного уровня заболеваемости и смертности в размере 5-6% (Group (1986), Stroke, 17, 848-52). Эти и другие данные демонстрируют явную потребность в профилактической нейропротективной терапии при проведении данных процедур. Таким образом, в другом варианте осуществления настоящего изобретения гематопоэтические факторы роста, например GCSF или GMCSF, отдельно, в сочетании друг с другом, в сочетании с одним или более дополнительными факторами, можно применять для лечения ишемии, возникающей как результат индуцированной операцией ишемии головного мозга, и/или для проведения профилактической нейропротективной терапии. В одном варианте осуществления настоящего изобретения лечение у пациентов в группе риска начинают перед проведением основного хирургического вмешательства.
В другом варианте осуществления настоящего изобретения гематопоэтические факторы роста, например, такие как GCSF или GMCSF, отдельно, в сочетании друг с другом и/или в сочетании с одним или более дополнительными факторами, можно применять для лечения рассеянного склероза (РС) и/или для проведения профилактической нейропротективной терапии у пациентов с рассеянным склерозом. Данный способ основан на наличии рецептора GCSF на олигодендроцитах, что подтверждает эффективность непосредственного воздействия GCSF на первичные клетки-мишени при РС. Кроме того, рецептор GCSF присутствует на нервных клетках и их отростках, которые нарушены на поздних стадиях заболевания и могут коррелировать с развитием длительной нетрудоспособности (Cid, et al. (2002), J Neurol Sci, 193, 103-9). Действительно, в последнее время наблюдается сдвиг парадигмы в терапевтических концепциях для рассеянного склероза от иммуномодуляции к нейропротекции, поскольку считается, что нейродегенеративные механизмы наиболее важны с точки зрения развития инвалидизирующих аспектов рассеянного склероза (Bo, et al. (2003), Mult Scler, 9, 323-31, Bjartmar, et al., (2003), J. Neurol Sci, 206, 165-71, Wujek, et al. (2002), J. Neuropathol Exp Neurol, 61, 23-32, Bjartmar, et al. (2001), Neurology, 57, 1248-52, Peterson, et al., (2001), Ann. Neurol, 50, 389-400, Bjartmar and Trapp (2001), Curr Opin Neurol, 14, 271-8, Bjartmar, et al. (2000), Ann. Neurol, 48, 893-901, Bjartmar, et al. (1999), J. Neurocytol, 28, 383-95, Trapp, et al. (1999), Curr Opin Neurol, 12, 295-302, Trapp, et al. (1998), N. Engl J Med, 338, 278-85, Neuhaus, et al. (2003), Trends Pharmacol Sci, 24, 131-8, Pryce, et al. (2003), Brain, 126, 2191-202, Waubant (2003), Expert Opin Emerg Drugs, 8, 145-61, Graumann, et al. (2003), Brain Pathol, 13, 554-73, Golde, et al. (2003), Eur J Neurosci, 18, 2527-37). Даже в тех зонах головного мозга, которые кажутся нормальными с точки зрения изменений в белом веществе, выявляют признаки нейродегенерации. Таким образом, патология аксонов и нейродегенерация являются важнейшими мишенями для лечения при рассеянном склерозе. Как показано в настоящем изобретения, GCSF или GMCSF с их антиапоптотической активностью в отношении нейронов и их способствующий регенерации потенциал (за счет повышения нейрогенеза и пластичности) подтверждают, что описанные здесь композиции можно применять в виде нового терапевтического подхода для лечения рассеянного склероза.
Кроме того, патофизиологические механизмы рассеянного склероза перекрываются с важными механизмами развития ишемии головного мозга, например с вовлечением оксида азота (Smith, et al. (2001), Ann Neurol, 49, 470-6) и с вовлечением токсического возбуждения глутаматом (Pitt, et al. Nat Med, 6, 67-70). В свете данной информации гематопоэтические факторы роста, такие как GCSF или GMCSF, представляют собой новую альтернативу для лечения рассеянного склероза и воспалительных расстройств головного мозга.
Другой вариант осуществления настоящего изобретения относится к нейропротективному лечению шизофрении. В последние годы получены неожиданные данные, указывающие на прогрессирующее снижение серого вещества у шизофреников. Эти результаты вначале были получены с использованием новых способов магнитно-резонансной томографии. Хотя нейродегенеративные процессы при шизофрении на молекулярном уровне неизвестны, нейропротективная защита при шизофрении с использованием GCSF или GMCSF представляет собой новый подход к лечению данного заболевания.
Для тестирования эффективности гематопоэтических факторов роста, таких как GCSF, для защиты первичных нейронов нейроны можно получать следующим образом. Материал коры головного мозга можно получить из эмбрионов 10-12 крыс на стадии Е18 (18 сутки развития эмбриона). Ткань можно подвергнуть диссоциации обработкой трипсином [10 мг/мл]/ЭДТА/ДНКазой [5 мг/мл] (Roche Diagnostics, Mannheim, Германия) в HBSS (сбалансированный раствор Хэнкса, BioWithakker). Расщепление может быть остановлено с использованием 4 частей среды (нейробазальная среда + 1 мл 50Х добавки В-27 (Invitrogen) + 0,5 мМ L-глютамина + 25 мкМ глутамата) с последующим центрифугированием при комнатной температуре в течение 5 минут при 800 g. Осадок можно растворять в 5 мл среды, а количество клеток определять путем подсчета (предметное стекло Neubauer). Клетки при плотности 250 000 клеток на лунку 24-луночных планшетов для микротитрования можно размещать на покровных стеклах, которые могут быть покрыты поли-L-лизином. Затем данные нейроны можно обрабатывать сочетаниями защитного стимула (GCSF) и вредного стимула (глутамат, 100 мкМ). GCSF применяют за 30 минут до обработки клеток глутаматом. Контрольные группы не обрабатывают или GCSF (только солевой раствор), или глутаматом. Через 24 часа можно определять гибель нервных клеток с использованием анализа на LDH (Roche Diagnostics, Mannheim, Германия) в соответствии с рекомендациями производителя. Альтернативно, можно применять другие вредные стимулы, известные своей способностью индуцировать гибель клеток, например NMDA и глицин, 3-нитропропионовую кислоту (3-NPA), H2O2, стауроспорин, гипоксию/недостаточность глюкозы, потерю калия, MPP+, интерлейкин-1-бета, TNF-альфа, FAS-лиганд или другие вредные для клеток и нейронов стимулы. Для оценки уровня гибели клеток или их относительной жизнеспособности можно применять другие анализы, например анализ гибели клеток ELISA (Roche Diagnostics), окрашивание аннексином/иодидом пропидия с последующим анализом лазерной сканирующей цитометрией (Kamenstky (2001), Methods Cell Biol., 63, 51-87, Compucyte, Cambridge, Massachusetts), подсчет клеточных ядер, содержащих апоптозные структуры при окрашивании DAPI или HOECHST33342 (конденсация, фрагментация), подсчет клеток, положительных на активированную каспазу 3, после иммунного окрашивания с использованием антитела, специфичного к расщеплению (например, антитела к каспазе 3 Promega), или анализ активности каспазы 3 в клеточных лизатах (анализ ApoOne, Promega; вестерн-блот-анализ, Elisas), или любой другой анализ, пригодный для определения жизнеспособности клеток или наличия в них характерных апоптозных свойств. Альтернативно, можно применять другие клетки, например дифференцированные РС12 клетки, NH33 клетки, SHSY5 клетки, первичные нейроны гиппокампа, первичные двигательные нейроны, первичные сенсорные ганглиозные клетки, первичные культуры клеток среднего мозга, нейрональные стволовые клетки, дифференцированные клетки ES или другие известные нейроноподобные клетки или клетки, обладающие одним или несколькими известными нейрональными фенотипами. Временные и концентрационные характеристики, приведенные в примерах, могут также варьировать, например, GCSF может применяться одновременно со стимулирующим фактором, до или после его воздействия. Также можно применять изменяющиеся концентрации GCSF, например, от 0,1 до 100 мкг/мл. В принципе, данный анализ можно также адаптировать для использования в культурах срезов головного мозга. Для тестирования эффективности гематопоэтических факторов роста, таких как GCSF, на модели травмы головного мозга (контролируемый удар по коре головного мозга) можно проводить следующую процедуру. Экспериментальные протоколы может принимать местный этический комитет. Двадцать самцов крыс Wistar (Charles River, Германия) весом от 280 до 320 грамм можно случайным образом распределить по следующим группам: А - (контрольная группа, n=10, травматическое повреждение головного мозга (TBI), обработка 2 мл 0,9% физиологического раствора в течение 90 минут, начиная с 30 минуты TBI); В - (группа с введением GCSF, n = 10, ишемия в течение 90 минут, лечение 60 мкг/кг массы тела рекомбинантным GCSF (Neupogen®, Amgen, Europe B.V., Нидерланды), растворенным в 2 мл 0,9% физиологического раствора в течение 90 минут, которое начинают за 30 минут до TBI); С - (контрольная группа GCSF с ложной операцией, n = 10, ложная операция, лечение 60 мкг/кг веса тела рекомбинантным GCSF (Neupogen®, Amgen , Europe B.V., Нидерланды), растворенным в 2 мл 0,9% физиологического раствора в течение 90 минут, которое начинают через 30 минут после TBI).
Животных анестезируют путем внутрибрюшинной инъекции 100 мг/кг массы кетамингидрохлорида (WDT, Garbsen, Германия). При необходимости анестезию можно поддерживать на уровне 50 мг/кг веса тела. В правую бедренную артерию для непрерывного мониторинга среднего артериального давления, наличия газов в крови, гематокрита, подсчета лейкоцитов и уровня глюкозы в крови можно вставлять полиэтиленовую трубку PE-50. В правую бедренную вену можно ввести канюлю с применением трубки PE-50 для инфузии. В ходе эксперимента можно контролировать ректальную температуру и поддерживать ее на уровне 37°C с помощью термостатически контролируемой нагреваемой прокладки (Fohr Medical Instruments, Германия).
У животных в группе с TBI можно разрезать кожу вокруг зонда и открыть и очистить череп. TBI можно осуществлять с использованием устройства типа падающего груза непрямого воздействия, модифицированного для достижения совместимости с микродиализом (устройство весом 150 г падает с высоты 40 см на цилиндр из ПВХ с тефлоновым покрытием диаметром 2,0 мм). Контрольных животных с ложной операцией можно готовить идентично крысам, получающим TBI, но без травмы.
У всех животных результат исследователь может оценить слепым способом в виде смертности, а также шкалой тяжести неврологических повреждений (Neurological Severity Scores (NSS)), проводя оценку ежедневно в течение 1 недели после травматического повреждения головного мозга. Функционирование нервной системы можно ранжировать по шкале от 0 до 16 (нормальный балл - 0; балл максимального дефицита - 16). NSS представляет собой сочетание двигательного, сенсорного и рефлексного тестов и включает в себя тест на поддержание баланса на барабане. При ранжировании тяжелых повреждений 1 балл по указанной шкале присваивается в случае неспособности выполнять тест или в случае отсутствия исследуемого рефлекса; таким образом, чем выше балльный показатель, тем более тяжелым является повреждение.
Через неделю после TBI крыс анестезируют с использованием 150 мг/кг массы тела кетамина и декапитируют. Мозг крыс удаляют и фиксируют 4% параформальдегидом в 0,1 моль/л фосфатном буфере в течение 24 часов. После погружения в парафин можно получить срезы толщиной 1 мкМ и применять для окрашивания Г/Э, окрашивания по Нисслю и для иммуногистохимического анализа.
Иммуногистохимическое исследование можно проводить с использованием антисыворотки к миелопероксидазе (DAKO, США) и GCSFR (Santa Cruz Biothechnology Inc., США). Антисыворотки можно получать от кроликов, иммунизированных выделенным человеческим белком (антимиелопероксидаза), или с применением синтетического пептида, соответствующего карбоксиконцу GCSFR мышиного происхождения соответственно. Для поиска антигена срезы, приготовленные для иммуногистохимического анализа GCSFR, можно нагревать в течение 20 минут в 10 мМ цитратном буфере при 99°C. Далее срезы инкубируют в нормальной сыворотке свиньи (10% в фосфатно-солевом буфере) в течение 30 минут и затем в первичной антисыворотке в течение ночи при 4°С. Первичные антитела можно разбавлять 1:150 (миелопероксидаза) или 1:400 (GCSFR). Иммунореактивность можно выявлять способом образования комплекса авидин-биотин (Vectastain, Vector Laboratories, США). В срезах можно получать окраску в 0,02% диаминобензидине (DAB) с добавкой 0,02% перекиси водорода. Интенсивность продукта реакции усиливают добавлением 0,02% хлорида кобальта и сульфата аммония-никеля. Жизнеспособность нейронов после TBI может быть определена путем количественного подсчета нейронов под микроскопом (увеличение Х40) в гиппокампе животных после введения GCSF и у контрольных животных. Инвазию нейтрофильных гранулоцитов (НГ) можно измерять полуколичественно по четырехбалльной шкале (0 - отрицательный результат MPO, 1 - низкая экспрессия МРО, 2 - средняя экспрессия МРО, 3 - сильная экспрессия МРО).
СТАТИСТИЧЕСКИЙ АНАЛИЗ
Результаты выражают в виде среднего значения ±SD. После получения всех данных коды можно нарушить. Тест способом ANOVA, а затем защищенный тест Фишера post hoc по оценке наименее значимого различия можно применять для определения статистической значимости различий непрерывных переменных, таких как физиологические параметры. Для сравнения повреждения нейронов и иммуногистохимических данных можно применять t-тест. Также для анализа непараметрических данных, таких как уровень смертности и результаты иммуногистохимического анализа для оценки МРО можно проводить U-тест Манна-Уитни (Mann-Whitney U test). Величину р<0,05 рассматривают как статистически значимую.
На основании действия гематопоэтических факторов, таких как GMCSF или GCSF, и эффектов агонистов соответствующих рецепторов на нервных клетках, другой вариант осуществления настоящего изобретения относится к лечению опухолей головного мозга или других видов неврологического рака путем антагонистов рецепторов GMCSF и/или GCSF на раковых клетках.
НЕЙРОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ
В последнее время признана важность формирования новых нервных клеток (нейрогенеза) для лечения неврологических заболеваний. В отличие от многих других тканей зрелый мозг имеет ограниченную способность к регенерации, а его необычная степень специализации клеток ограничивает величину, до которой оставшаяся здоровая ткань может приобретать функции поврежденного участка головного мозга. Однако нейроны головного мозга происходят из клеток-предшественников, которые постоянно присутствуют в головном мозге взрослого, так что стимуляция эндогенных предшественников нервных клеток в головном мозге у взрослого может иметь терапевтический потенциал.
Нейрогенез происходит в дискретных участках головного мозга взрослого, включая ростральную субвентрикулярную зону (SVZ) латеральных желудочков и зону под зернистым слоем (SGZ) зубчатой извилины (DG). У взрослых животных происходит нейрогенез, в особенности после некоторых неврологических событий (например, ишемии головного мозга (Jin, et al. (2001), Proc, Natl. Acad. Sci. USA, 98, 4710-5, Jiang, et al. (2001), Stroke, 32, 1201-7, Kee, et al. (2001), Exp. Brain Res., 136, 313-20, Perfilieva, et al. (2001), J Cereb. Blood Flow Metab., 21, 211-7). Нейрогенез также показан у людей (Eriksson, et al. (1998) Nat. Med, 4, 1313-7) и показано, что он фактически ведет к появлению функционирующих нейронов (van Praag, et al. (2002), Nature, 415, 1030-7). В частности, зона под зернистым слоем зубчатой извилины и ворота обладают способностью образовывать у взрослого новые нейроны (Gage, et al. (1998) J Neurobiol. 36, 249-66). Удивительно, что рецептор GCSF экспрессируется в данной области (фиг.4 a, d). Вместе с неожиданными данными, демонстрирующими улучшение функционального исхода после лечения GCSF (фиг.8), и тем фактом, что GCSF является фактором для стволовых клеток в другой системе (гематопоэз), можно ожидать, что GCSF проявит часть своей активности, особенно долговременные воздействия (фиг.8) посредством своей стимулирующей функции на стволовые клетки у взрослого, по меньшей мере, в зубчатой извилине.
Настоящая заявка подтверждает присутствие рецептора GCSF на нейрональных стволовых клетках взрослого, выделенных из области гиппокампа крысы, окружающего зубчатую извилину (фиг.12 и 13). Важная роль GCSF в нейрогенезе дополнительно обосновывает его применимость и пользу при лечении всех вариантов нейродегенеративного заболевания и всех состояний, связанных с гибелью нейронов. Кроме случаев применения, которые определяются воздействием на эндогенные стволовые клетки в мозге при лечении неврологических состояний, GCSF может быть также использован in vitro для манипуляций со стволовыми клетками, например, с целью их дифференцировки и пролиферации. Лечение людей стволовыми клетками в настоящее время проводится при многих заболеваниях, в частности при болезни Паркинсона и инсульте. Желательно, в случае лечения болезни Паркинсона, дифференцировать стволовые клетки в культуре на конкретные типы нервных клеток, например дофаминэргические клетки. Далее дифференцированные или иным способом адаптированные к новому окружению клетки вводят различными способами в организм. При болезни Паркинсона, например, стволовые клетки инъецируют непосредственно в головной мозг для восполнения потери дофаминэргических нейронов в черном веществе ("заместительная терапия") (Arenas (2002), Brain Res. Bull, 57, 795-8-8, Barker (2002), Mov. Disord., 17, 233-41).
В этой связи один вариант настоящего изобретения относится к стимуляции роста и дифференцировки нейрональных стволовых клеток или подготовительной обработке нейрональных стволовых клеток перед имплантацией в организм млекопитающих с использованием гематопоэтических факторов роста и их производных. Другой вариант настоящего способа относится к применению таких нейрональных стволовых клеток в способах лечения неврологического заболевания, описанных выше, предпочтительно в способах, которые обеспечивают нейропротективный эффект в тех случаях, когда нейрональные стволовые клетки вводят индивидууму.
В одном варианте осуществления настоящего изобретения стволовые клетки могут быть введены внутривенно или внутриартериально. Было показано, например, в случае ишемии головного мозга или травматического повреждения головного мозга, что стромальные клети костного мозга, инъецированные в/в, находят свой путь к зонам мишени в головном мозге (Mahmood, et al. (2001), Neurosurgery, 49, 1196-203; обсуждение 1203-4, Lu, et al. (2001), J Neurotrauma, 18, 813-9, Lu, et al. (2002), Cell Transplant, 11, 275-81, Li, et al. (2002), Neurology, 59, 514-23). Таким образом, стволовые клетки могут быть обработаны GCSF, GMCSF или другими гематопоэтическими факторами и затем инъецированы различными способами пациентам, имеющим любое из описанных выше заболеваний.
В одном варианте осуществления настоящего изобретения трансплантируемые стволовые клетки подвергают генетическим манипуляциям для экспрессии факторов, которые они будут секретировать, повышая при этом жизнеспособность окружающих клеток, или для усиления пролиферации и/или дифференцировки эндогенных стволовых клеток организма в состоянии зрелости. Например, стволовые клетки могут быть подвергнуты генетическим манипуляциям для достижения стабильной экспрессии GCSF, GMCSF и/или одного или более дополнительных гематопоэтических факторов, после чего могут быть введены в центральную нервную систему для постоянной секреции GCSF, или GMCSF, или других гематопоэтических факторов роста в локальное окружение.
Стволовые клетки могут быть обработаны GCSF, или GMCSF, или другими агонистами рецептора гематопоэтического фактора. Стволовые клетки, которые могут при этом использоваться, включают иммортализованные стволовые клетки (например, иммортализованный онкоген), нейросферы и нервные клетки, полученные из эмбриональных стволовых клеток (ES) (Gottlieb (2002), Annu Rev Neurosci, 25, 381-407), а также включать клетки типа стромальных клеток костного мозга или клетки почечного канатика (Lu, et al. (2002), Cell Transplant, 11, 275-81, Li, et al. (2002), Neurology, 59, 514-23). Трансплантация стволовых клеток различных типов является одним из подходов в лечении множества неврологических заболеваний, включая болезнь Паркинсона (Isacson (2002), Brain Res Bull, 57, 839-46) и инсульт (Kondziolka, et al. (2002), J. Clin Neurosci, 9, 225-30).
Стволовые клетки, например нейрональные стволовые клетки человека, могут быть обработаны факторами, позволяющими улучшить их состояние перед трансплантацией. Один из примеров таких кондиционирующих агентов включает факторы роста. Примером такого процесса кондиционирования является дифференцировка клеток в желательном направлении, например, до нейронов (Svendsen, et al. (1999), Brain Pathol, 9, 499-513). Присутствие рецептора GCSF на стволовых клетках указывает на важность агонистов этого рецептора в процессе кондиционирования таких клеток. Нейрональные стволовые клетки из организма в состоянии зрелости могут быть обработаны различными концентрациями GCSF или других агонистов рецептора GCSF и далее могут быть проанализированы на наличие повышенного потенциала дифференциации по методике количественной ПЦР, например путем количественного определения соотношения нейрональных маркеров (Map2, NSE (нейрон-специфичная энолаза), нейрофиламент, NeuN) и маркеров нейрональных стволовых клеток (нестин). Повышенный коэффициент соотношения после указанной обработки сигнализирует об усиленной дифференцировке стволовых клеток в направлении нейрональной линии. Для обработки стволовых клеток перед трансплантацией в случае неврологического заболевания могут быть использованы соответствующие показатели концентрации и временного диапазона.
В другом варианте осуществления настоящего изобретения могут использоваться GCSF, GMCSF, их производные, а также агонисты рецептора GCSF и GMCSF для облегчения культивирования нервных клеток, таких как, например, нейрональные стволовые клетки. В рамках данного способа GCSF, GMCSF, их производные, а также агонисты рецептора GCSF и GMCSF могут быть внесены в среду и перемешаны перед добавлением к клеткам или могут быть внесены в среду, в которой уже культивируются клетки. В другом варианте осуществления данного способа нервные клетки трансфицируют полинуклеотидом, который кодирует GCSF, GMCSF и их производные, которые далее, после трансфекции, экспрессируют соответствующие факторы в клетке.
СПОСОБ ВВЕДЕНИЯ/КОМПОЗИЦИЯ/ДОЗИРОВКА
GCSF, GMCSF и другие факторы, производные, гены и их сочетания могут быть введены в виде множества дозированных форм, которые включают, без ограничения, жидкие растворы или суспензии, таблетки, пилюли, порошки, суппозитории, полимерные микрокапсулы или микровезикулы, липосомы и инъецирумые или инфузируемые растворы. Предпочтительная форма определяется способом введения и видом терапевтического применения.
Композиция может иметь вид жидкости, взвеси или стерильного твердого вещества, которое может быть растворено в стерильной инъецируемой среде перед использованием. Парентеральное введение включает предпочтительно внутривенное введение. Данная инъекция может быть осуществлена шприцем или с помощью аналогичного устройства. Используемая в таком случае форма может содержать фармацевтически приемлемый носитель. Альтернативно, композиции, например, содержащие GCSF, могут быть введены через слизистую в соответствующей дозе и в жидкой форме. В случае перорального введения композиции, например, содержащие GCSF, могут быть введены в жидкой или твердой форме с использованием соответствующего носителя.
Млекопитающее, подлежащее лечению, может представлять собой, например, морскую свинку, собаку, кошку, крысу, мышь, лошадь, корову, овцу, обезьяну или шимпанзе. В одном варианте осуществления настоящего изобретения данное млекопитающее представляет собой человека. Аналогично, в одном варианте осуществления настоящего изобретения гематопоэтические факторы, такие как GCSF или GMCSF, используемые для лечения или профилактики, включают человеческий фактор или фактор, полученный из организма человека как источника.
Терапевтически эффективное количество гематопоэтических факторов, используемых в способах лечения неврологического заболевания, при использовании указанных факторов по отдельности или в сочетании, представляет собой то количество, которое приводит к достижению нейропротективного эффекта. Такое количество может варьировать от примерно 100 нг до примерно 10 мг/кг веса тела указанного фактора или его сочетания и может быть определено с учетом возраста, расы, пола и других показателей конкретного пациента. Например, количество GCSF, используемого в настоящем способе, будет составлять от примерно 5 до примерно 60 мкг/кг, а в случае GMCSF данное количество может варьировать от примерно 5 до примерно 750 мкг/кг веса тела. В случае введения факторов в сочетании они могут быть предварительного смешаны перед введением, введены одновременно или введены порознь в серии.
В некоторых вариантах лечения неврологических состояний, приведенных в настоящем описании, может быть особенно полезно использование повышенных доз GCSF и GMCSF, например, может использоваться по меньшей мере 1 мг, по меньшей мере 2 мг или по меньшей мере 3 мг, при этом предпочтителен диапазон от 1 до 3 мг. Указанные повышенные дозы трудно вводить при использовании доступных в настоящее время фасованных дозированных форм. В этой связи один вариант осуществления настоящего изобретения относится к фасованным дозированным формам, в особенности к дозам в диапазоне от 20 до 250 мкг/кг веса тела, которые могут вводиться, например, внутривенно или подкожно с целью достижения нейропротективного эффекта согласно настоящему описанию. Примеры фасованных дозированных форм, используемых для доставки высоких доз композиций, включают, например, флаконы с однодозовой единицей и шприцы-тюбики, содержащие 0,75-25 мг GCSF и/или GMCSF в полном объеме 1,0 мл или 2,0 мл. В предпочтительном варианте фасованные дозированные формы могут также содержать инструкции, или этикетки, или отдельно напечатанный материал, описывающие применение композиции(й), содержащейся(ихся) внутри.
Предпочтительны также шприцы-тюбики для введения GCSF и/или GMCSF, откалиброванные по шкале кг/вес тела, что позволяет осуществлять точную и быструю дозировку для каждого конкретного пациента в соответствии с его весом. Указанные фасованные дозированные формы особенно полезны в отделениях неотложной помощи, например, при инсульте, где пациентов необходимо лечить очень быстро после поражения. Соответственно, другой вариант настоящего изобретения относится к нейрозащите, в частности, применительно к пациенту с инсультом или имеющему другое нейротравматическое состояние, путем введения одной или нескольких композиций из числа приведенных в описании в течение короткого периода времени после повреждения. В альтернативном варианте композиции, описанные в изобретении, такие как шприцы-тюбики, могут вводиться/использоваться в любое время после повреждения, без конкретного ограничения необходимостью использовать лишь в течение короткого периода времени после повреждения.
В контексте данного варианта осуществления изобретения фраза "в течение короткого периода времени после повреждения" означает, что воздействие осуществляется примерно в течение 10 минут, 15 минут, 20 минут, 25 минут, 30 минут или даже примерно до 1 часа после повреждения или после события, вызвавшего повреждение. Композиции могут вводиться медицинским работником или в ряде случаев не медицинским работником, который имеет доступ к фасованным дозированным формам, содержащим соответствующие дозировки фактора(ов). В случае медицинского работника, такого как технический сотрудник автомобиля скорой помощи, медсестра, врач и т.п., введение может быть осуществлено в том месте, где пациент получил нейротравму, такую как, например, инсульт, в автомобиле скорой помощи или в другом транспортном средстве во время транспортировки в больницу (или другое медицинское учреждение) или, например, в палату неотложной помощи. Без ограничения какой-либо конкретной теорией следует отметить, что введение одного или нескольких гематопоэтических факторов, таких как композиции, содержащие GCSF и/или GMCSF, в течение короткого времени после повреждения, позволяет достичь большего нейропротективного эффекта.
Другой вариант осуществления настоящего изобретения относится также к устройству, которое особенно хорошо подходит для медленного высвобождения и длительного постоянного применения и которое может быть имплантировано в виде мини-насоса, предпочтительно имплантированного подкожно (см., например, описание в Edith Mathiowitz; (Ed.), Encyclopedia of Controlled Drug Delivery, John Wiley & Sons vol. 2, pp. 896-920, 1999). Такие насосы известны по применению в инсулиновой терапии. Примеры таких насосов включают устройства, производимые/распространяемые различными компаниями (Animas, Dana Diabecare, Deltec Cozm, Disetronic Switzerland, Medtonic и Nipro Amigo), а также устройства, описанные, например, в патентах США №№: 5474552; 6558345; 6122536; 5492534 и 6551276, соответствующее содержание которых включено в настоящее описание в качестве ссылок.
Настоящее изобретение также относится к устройству, которое позволяет измерять эндогенные сывороточные уровни GCSF или GMCSF и на основе полученных результатов с учетом возраста и веса пациентов вычислить и затем осуществить инъекцию соответствующего количества лекарственного средства (см., например, описание в работе Renard E., Curr Opin Phermacol. 2002 Dec; 2(6): 708-16). Такой процесс может быть осуществлен на постоянной основе, например, путем использования описанных выше мини-насосов или в случае острой необходимости. Примером такого устройства является устройство, производимое компанией Медтроник Минимед (Medtronic MiniMed, 18000 Devonshire Street, Northridge, CA 91325-1219), используемое при инсулиновой терапии. Для измерения сывороточного уровня GCSF или GMCSF могут использоваться белковые чипы или матрицы, описанные, например, в патентах США №№ 6630358; 6537749 и 6475809, соответствующее содержание которых включено в настоящее описание в качестве ссылок.
Указанные выше варианты доставки лекарственного средства с использованием насосов и/или в сочетании с определением сывороточных уровней GCSF и/или GMCSF могут быть особенно полезны при лечении хронических нейродегенеративных заболеваний, например болезни Паркинсона, бокового амиотрофического склероза (ALS), деменции и других состояний.
Способ введения включает типичные способы, такие как пероральный, подкожный, чрескожный, внутрикожный, ректальный, вагинальный, внутримышечный, внутривенный способы, а также введение путем непосредственной инъекции в головной мозг и парентеральный способ. Дополнительно, в некоторых случаях может использоваться внутрилегочное введение, например, с использованием легочных аэрозолей и других вдыхаемых форм.
В дополнение к легочным аэрозолям интраназальный (ИН) способ доставки (например, с использованием назальных аэрозолей) представляет собой предпочтительный способ доставки композиций по настоящему изобретению в случае неврологических/психических состояний. Интраназальная доставка хорошо подходит как способ введения белков и пептидов и очень удобна для использования, особенно в случае длительных курсов лечения. Примеры использования интраназальной доставки (назальные аэрозоли) для введения пептидов или белков в головной мозг описаны в литературе (Lyritis and Trovas (2002), Bone, 30, 71S-74S, Dhillo and Bloom (2001), Curr Opin Pharmacol, 1, 651-5, Thorne and Frey (2001), Clim Pharmacokinet, 40, 907-46, Tirucherai, et al. (2001), Expert Opin Bio[ Ther, 1, 49-66, Jin et al., (2003), Ann Neurol, 53, 405-9, Lemere et al. (2002), Neurobiol Aging, 23, 991-1000, Lawrence (2002), Lancet, 359, 1674, Liu et al. (2001), Neurosci Lett, 308, 91-4). Для целей внутриназального введения GCSF/GMCSF и другие композиции, приведенные в настоящем описании, могут быть объединены с растворителями, детергентами и веществами, которые повышают проникновение в назальный эпителий или доставку в кровеносные сосуды, такими как лекарственные средства, которые расширяют кровеносные сосуды носа, усиливают перфузию и т.п.
На основе выбранного способа введения и с учетом соответствующей фармакокинетики препарата, используемые дозы могут быть далее модифицированы, например, доза, выбранная для непосредственной инъекции в головной мозг, может быть снижена, а требуемое количество может быть определено в абсолютных единицах, на основании данных по локальной доступности GCSF, GMCSF или их производных (например, в суммарной дозе 5 мкг). Предпочтительно GCSF, GMCSF и другие гематопоэтические факторы и их производные вводят внутривенно, подкожно или путем внутримозговой инъекции, которая может быть осуществлена с помощью осмотического насоса.
В другом варианте осуществления настоящего изобретения GCSF, GMCSF и другие гематопоэтические факторы и их производные могут быть доставлены в организм пациента путем введения одной или более нуклеиновых кислот, которые кодируют указанные факторы. Кодирующую последовательность нуклеиновой кислоты предпочтительно вводят в виде рекомбинантного вектора, такого как вирусный вектор. Методики выбора соответствующего вектора и последовательностей, контролирующих экспрессию, а также конструирования векторов известны. Примеры вирусных векторов включают аденовирусный вектор (Acsadi et al. Hum. Gene. Ther. 7(2): 129-140 (1996); Quantin et al. PNAS USA 89(7): 2581-2584 (1992); и Ragot et al. Nature 361 (6413): 647-650 (1993), аденоассоциированный вирусный вектор (Rabinowitz et al. Curr Opin Biotechnol. 9(5): 470-475, (1998)), ретровирусный вектор (Federico, Curr Opin Biotechnol. 10(5): 448-453 (1999)), вектор на основе Herpes simplex (Latchman, Gene 264(1): 1-9 (2001)), лентивирусный вектор, вектор на основе вируса Sindbis или вирусный вектор на основе Semliki forest. Подходящими векторами являются также липосомы, содержащие белки, которые присоединяют к нервным клеткам, например вирусные эпитопы, и которые содержат любую нуклеиновую кислоту, кодирующую GCSF или GMCSF, или белок, или олигонуклеотиды. Примером такой транспортной системы является HVJ-липосома (Kaneda, et al. (2002), Mol. Ther, 6, 219-26; Kaneda (1999), Mol. Membr. Biol, 16, 119-22). Предпочтительно выделенная и очищенная нуклеиновая кислота, кодирующая и экспрессирующая белок или полипептид, оперативно связана с промотором, который подходит для экспрессии в нервных клетках. Указанные и другие подходящие векторы приведены в литературе (обзор Kay et al. Nature Medicine 7: 33-40 (2001); Somia et al. Nature Reviews, 1: 91-99 (2000) и van Deutekom et al. Neuromiscular Disorders 8: 135-148 (1998)).
Подходящие промоторы для оперативного соединения с выделенной и очищенной нуклеиновой кислотой известны в данной области техники. Предпочтительно выделенная и очищенная нуклеиновая кислота, кодирующая полипептид, оперативно связана с промотором, выбранным из группы, состоящей из промотора мышечной креатинкиназы (МСК) (Jaynes, et al. Mol Cell Biol, 6: 2855-2864 (1986), промотора цитомегаловируса (CMV), промотора, регулируемого тетрациклином/доксициклином (Gossen, et al. PNAS USA 89: 5547-5551 (1992)).
В основном для обеспечения эффективной транспортировки векторов согласно настоящему изобретению берут от примерно 1 до примерно 5000 копий вектора на клетку, используемую для взаимодействия на основе их приблизительного количества, определяемого данным способом введения, и еще более предпочтительно в каждую клетку вводят от примерно 3 до примерно 3000 БОЕ. Указанное количество вирусных частиц может варьировать в зависимости от потребностей и от того, в каких условиях, in vitro или in vivo, осуществляется процедура. Фактические показатели дозы и режимы могут также варьировать в зависимости от того, вводят ли данную композицию в сочетании с другими композициями, например с фармацевтическими композициями, или в зависимости от индивидуальных различий в фармакокинетике, распределении лекарственного средства и метаболизме. Аналогично, эти количества могут варьировать в случаях применения in vitro, в зависимости от конкретного типа клеток или от способа, посредством которого осуществляется перенос вектора.
Описанные выше белки или их производные, другие вещества или нуклеиновые кислоты могут быть введены в состав композиции для достижения определенных медицинских целей в соответствии со стандартными процедурами, доступными в данной области, например, может быть добавлен фармацевтически приемлемый носитель (или эксципиент). Носитель, или эксципиент может представлять собой твердый, полутвердый или жидкий материал, который может служить в качестве носителя или жидкой среды для активного ингредиента. Соответствующие формы и способ введения могут быть выбраны в зависимости от конкретных характеристик выбранного продукта, заболевания или состояния, подлежащего лечению, стадии заболевания или состояния и других релевантных условий (Remington's Pharmaceutical Sciences, Mack Publishing Co. (1990)). Вводимая пропорция фармацевтически приемлемого носителя или эксципиента и его природа определяются по растворимости и химическим свойствам вещества, выбранного для данного способа введения, и в соответствии со стандартными нормами фармацевтической практики. Фармацевтический препарат может быть адаптирован для перорального, парентерального или местного применения и может вводиться пациенту в виде таблеток, капсул, суппозиториев, растворов, суспензий или т.п. Факторы роста, их производные, соответствующая кодирующая последовательность нуклеиновой кислоты по настоящему изобретению, будучи эффективными сами по себе, могут быть изготовлены и введены в виде фармацевтически приемлемых солей, таких как аддитивные соли кислоты или аддитивные соли основания, для достижения целей стабильности, легкости кристаллизации, повышенной растворимости и т.п.
Для эффективной терапии некоторых неврологических заболеваний, особенно ишемических заболеваний, чрезвычайно важно не задерживать начало лечения. В предпочтительном варианте настоящее изобретение относится к способу, в рамках которого GCSF, GMCSF, или их производные, или вещество, активирующее STAT белки, или агонист рецепторов GCSF или GMCSF вводят в течение 24, предпочтительно в течение 10 и наиболее предпочтительно в течение 3-6 часов после окклюзии кровеносного сосуда, или начала развития неврологических симптомов, или проявляемого иным образом начала развития ишемии. Поскольку GCSF и GMCSF эффективны по достижению длительного функционального улучшения, возможно также начать лечение через некоторое время после ишемического инсульта (например, в период с 1 по 7 сутки после инсульта). Лечение может быть продолжено в течение нескольких суток и недель после ишемического события в качестве способа улучшения функционального восстановления пациента по аналогии с функциональным улучшением, наблюдаемым на модели длительного инфаркта коры головного мозга (см. фиг.8). Лечение может включать введение нескольких доз в сутки (например, коротких в/в инфузий) для достижения стабильных уровней GCSF, GMCSF или родственных факторов. Лечение может также включать непрерывную медленную инфузию описанных веществ (например, с помощью перфузора) для гарантии достижения стойких сывороточных уровней.
В случае хронических нейродегенеративных процессов, таких как болезнь Паркинсона или боковой амиотрофический склероз, лечение, скорее всего, будет включать однократное введение суточной дозы GCSF или родственных факторов или предпочтительно использование композиции с медленным высвобождением или более стабильных производных GCSF или родственных факторов.
СКРИНИНГ НЕЙРОПРОТЕКТИВНЫХ ВЕЩЕСТВ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С РЕЦЕПТОРОМ GCSF ИЛИ GMCSF НА НЕРВНЫХ КЛЕТКАХ
В практике осуществления настоящего изобретения предпочтительны производные GCSF, GMCSF (например, миметики GCSF или GMCSF) и/или другие гематопоэтические факторы, которые сохраняют свой нейропротективный потенциал и характеризуются сниженным действием на лейкоциты, минимизируя тем самым возможные неблагоприятные эффекты. В этой связи один вариант настоящего изобретения относится к скринингу соединений, которые связываются с рецептором GCSF или GMCSF на нервнных клетках и которые стимулируют нейропротективный эффект, наблюдаемый при использовании GCSF или GMCSF, описанных в данной заявке.
Производные GCSF, например миметики GCSF, могут быть проанализированы в тесте на нейропротективный эффект in vitro, как описано, в частности, в примере 10. Данный тест на нейропротективный эффект может быть модифицирован с сохранением основного принципа данного теста путем адаптации его, например, (i) к другому вредному фактору, такому как интерлейкин-1, кислородная недостаточность, пептиды А4, FAS лиганд, или (ii) к другим клеткам, например клеткам, которые подобны нейрональным, типа SH-SY5Y клеток или РС12 клеток, или (iii) для оценки других показателей жизнеспособности клеток или гибели клеток, таких как фрагментация ДНК, ядерная конденсация, активность совместно трансфицированной бета-галактозидазы, обнаружение флуоригенного субстрата каспаз и т.п. Все указанные многочисленные формы адаптации известны и могут также использоваться в широкомасштабных системах скрининга. Методика широкомасштабного скрининга часто используется для определения быстрого процессинга клеток в крупных масштабах. Вещества, демонстрирующие позитивный нейропротекторный эффект в данном тесте, могут быть далее проанализированы на наличие у них гранулопоэтической активности, как описано, например, в работе Тиана с соавт. (Tian et al., Science, 281, 257-259). Выбор производных GCSF предпочтительно основан на сравнении указанных двух специфических эффектов, проявляемых исследуемыми веществами, с эффектами, проявляемыми известными формами GCSF. Аналогично, производные GMCSF, например миметики GMCSF, могут быть проанализированы в тесте на нейропротективный эффект in vitro, по методике, описанной для GCSF.
Другие нейропротективные вещества могут быть получены при определении наличия активированных факторов транскрипции, таких как STAT белки, включая STAT-3 и STAT-5, в нейрональных или нейроноподобных клетках (например, PC12, SH-SY5Y, hNT, NT2, hn33) после воздействия на них исследуемого вещества. В этой связи в другом варианте осуществления настоящего изобретения способность конкретного вещества или соединения действовать в качестве агониста к рецептору GCSF или GMCSF может быть оценена по активации STAT белков, например STAT3 и/или STAT5, в нейроноподобных клетках, как показано в настоящем описании, и с использованием соответствующих методов анализа экспрессии гена, таких как количественная ПЦР в системах ЛайтЦиклер (LightCyclerTM) или Такман (TaqmanTM).
Активируемые STAT белки фосфорилируются (pSTAT) и могут быть далее выявлены с использованием коммерчески доступного pSTAT-специфического антитела (Santa Cruz Biotechnology) методами вестерн-блоттинга или иммуногистохимического анализа или ИФТФА.
Альтернативно, STAT активность может быть определена с использованием репортерной конструкции, которая включает STAT-отзывчивый промоторный элемент (например, мультимеризованный STAT-связывающий элемент, такой как мультимеризованный STAT-3- или STAT-5-связывающий элемент), присоединенный к репортерному гену, такому как ген люциферазы, SEAP (секретируемой щелочной фосфатазы), хлорамфениколтрансферазы (CAT), зеленого флуоресцентного белка (GFP), или к другим часто применяемым репортерам генной экспрессии. Клетки после их трансфекции репортерной конструкцией клетки подвергают взаимодействию с исследуемым веществом и идентифицируют уровень экспрессии репортера. Данный метод измерения уровня STAT может быть адаптирован для широкомасштабного применения.
Типичный репортерный тест может быть проведен с использованием, например, коммерчески доступной системы анализа (Mercury Pathway Profiling System от Clontech; Path Detect-System от Stratagene). Ниже описан пример проведения такой процедуры.
Клетки культивируют в многоячеечном планшете, например по 20000 клеток на ячейку в 96-ячеечном планшете. Через двое суток после высева клеточной культуры среду заменяют на бессывороточную среду (40 мкл на ячейку) и клетки трансфицируют репортерной плазмидой, кодирующей STAT-связывающий элемент и репортерный белок (STAT-3-плазмида, кодирующая люциферазу светлячков; 50 нг/ячейку), и плазмидой контроля трансфекции (плазмида, кодирующая экспрессию люциферазы рениллы; 10-20 нг/ячейку) в условиях, оптимально подобранных для каждого типа клеток (например, РС12 клетки могут быть трансфицированы липофектином с использованием Липофектамина2000 (LIPOFECTAMINE2000TM Invitrogen, по инструкции производителя). Через 48-72 часов после трансфекции может быть проведен оценочный тест. Клетки стимулируют исследуемым веществом, взятым в широком диапазоне разных концентраций. В тесте используют множественные временные точки, начиная от 5 минут после стимуляции. При использовании люциферазного теста результат анализа оценивают в люминометре плоского формата (Berthold, Германия), измеряя постадийно активность люциферазы рениллы и светлячков. Выявление двух люциферазных активностей проводят при использовании коммерчески доступного набора на люциферазу (Dual luciferase reporter assay kit, Promega; Luciferase reporter assay kit, Clonetech). Далее показатели активности люциферазы светлячка нормализуют относительно значений активности люциферазы рениллы и вычисляют относительную индукцию репортерного гена.
В альтернативном варианте метода скрининга может быть использована и измерена агонистская активность в отношении рецептора GCSF или GMCSF на нейрональных клетках. Например, гомодимеризация при связывании лиганда может быть оценена по методу определения флуоресцентного резонансного переноса энергии (FRET или BRET (метод биолюминесцентного резонансного переноса энергии)) (Siegel RM et al. Sci STKE (2000) 2000 (38); L1; Xu Y et al. Proc Natl Acad Sci USA, (199), 96(1); 151-6) или с использованием репортерных систем для явлений димеризации, например двухходовой системы (DE 10211063.8), бета-gal репортерной системы (Rossi F et al., Proc Natl Acad Sci USU, (1997) 94(16): 8405-10) или системы расщепления убихитина (WO 954/29195, US 5 585 245 или Johnsson N and Varshavsky, Proc Natl Acad Sci USA, (1994) 91(22): 10340-4).
В других альтернативных вариантах скрининга нейропротективных веществ, которые присоединяются к GCSF или GMCSF на нейрональных клетках, настоящее изобретение относится к индукции эндогенных уровней GCSF или GMCSF в головном мозге. Как описано и продемонстрировано в соответствующих примерах, неожиданно было обнаружено, что GCSF или GMCSF являются эндогенными лигандами головного мозга. Указанные лиганды индуцируются несколькими ишемическими состояниями мозга. В этой связи они действуют как часть эндогенной нейропротективной системы мозга. Они действуют в том же направлении, как и экзогенно добавленные GCSF или GMCSF, которые пересекают гематоэнцефалический барьер и оказывают эффективное воздействие на широкий перечень состояний, при которых нейрозащита является важным терапевтическим подходом, и поэтому лекарственные средства, которые повышают в головном мозге эндогенные уровни GCSF/GMCSF, могут быть полезны в таких состояниях.
Система скрининга, позволяющая идентифицировать такие вещества, может быть установлена в основном путем добавления веществ (например, из большой библиотеки малых молекул) к клеткам в культуре, например нейрональным клеткам, т.е. первичным клеткам или нейроноподобным иммортализованным клеткам, таким как РС12 клетки, SHSY5-клетки и другие, с последующим измерением уровней GCSF и GMCSF.
Измерение уровня GCSF/GMCSF может быть осуществлено посредством анализа самих белков (методом вестерн-блоттинга, ИФТФА и других методик, известных специалистам в данной области), посредством измерения уровня мРНК, кодирующих данные белки (таких как метод количественной ПЦР, нозерн-блоттинг, тест на РНК-азную защиту, РНК-дот-блоттинг и другие методики, известные специалистам в данной области) или путем анализа активности регуляторных элементов генов GCSF/GMCSF. Предпочтительно активность регуляторных элементов может быть оценена с использованием репортерных конструкций, включающих сегменты ДНК из промотора, энхансера и/или интронных элементов, связанных с кДНК, которые кодируют репортеры (такие как люцифераза, бета-галактозидаза, зеленый флуоресцентный белок или другие репортерные гены, которые могут быть легко выявлены). Указанные репортерные конструкции могут быть трансфицированы в клетку либо в стабильном или временном режиме.
Клетки могут быть разрушены в физиологических условиях или дополнительно подвергнуты воздействию вредных факторов (таких как гипоксия, недостаточность глюкозы, доноры NO, избыток глутамата и другие) для выявления эффектов добавления соответствующих веществ к системе тестирования.
Идентифицированные вещества могут быть далее оценены путем введения их животным и измерения содержания GCSF/GMCSF в мозге (например, путем количественной ПЦР).
В другом варианте осуществления настоящего изобретения повышенные уровни экспрессии рецепторов GCSF или GMCSF могут быть полезны для целей повышения отзывчивости клеток к лигандам. В этой связи описанные выше методы скрининга могут быть также адаптированы для выявления повышенного уровня экспрессии рецептора.
На основе указанного выше описания настоящего изобретения далее приводится его пояснение с помощью некоторых конкретных примеров, которые даны лишь для целей иллюстрации и никоим образом не ограничивают область настоящего изобретения.
ПРИМЕРЫ
Эксперименты, проведенные в рамках настоящего изобретения, показывают, что GCSF является нейропротективным веществом in vitro и что GCSF демонстрирует значительные эффекты по уменьшению инфаркта после временной очаговой ишемии головного мозга. Нейроны на периферии инфарктной зоны, а также на контралатеральной стороне проявляют способность к специфическому связыванию антитела против GCSFR, что показательно для рецептора GCSF. Присутствие GCSF на нейронах показано впервые и было подтверждено методом вестерн-блоттинга, ОТ-ПЦР и детальным иммуногистохимическим анализом образцов мозга, а также методами ПЦР и иммуногистохимии в культивируемых первичных нейронах. Кроме того, активация STST-3, как следует из результатов исследования ядерной транслокации STAT-3, значительно повышается в нейронах в затемненной области у GCSF-обработанных животных в сравнении с контрольными, позволяя полагать, что имеется механизм действия, опосредованный GCSFR/STAT. В эксперименте с ишемией in vivo (окклюзия средней церебральной артерии, MCAO) не было выявлено воздействия на мозговой кровоток, по данным лазерной допплер-флуометрии (ldf), при сравнении обеих групп. Между обеими группами при проведении MCAO эксперимента не отмечалось также существенных различий по физиологическим параметрам и по снижению веса. Уровень смертности был значительно снижен у животных, которым вводили GCSF, в сравнении с контролями в рамках MCAO эксперимента. Количество нейтрофилов в крови (NGC) значительно возросло через 24 часа у животных, которым вводили GCSF, в сравнении с контролями. Миелопероксидазное (МРО) окрашивание как мера оценки внедряющихся нейрофильных гранулоцитов (НГ) в ишемизированное полушарие существенно не различалось между животными, подвергнутыми лечению GCSF, и контролями.
Доза, в/в доставленного GCSF (60 мкг/кг/веса тела), используемая в экспериментах, была сравнима с дозами, применяемыми в других экспериментальных условиях. В более раннем пилотном проекте была исследована его безопасность и впоследствии также не было выявлено существенных побочных эффектов. Указанная доза (60 мкг/кг/веса тела) в шесть раз превышает разрешенную дозу для лечения миелобластных и других заболеваний человека и, как было показано на модели крыс, не оказывает заметных побочных эффектов.
Эффект по снижению инфаркта на 50%, достигаемый при использовании GCSF, сравним с результатом использования других факторов роста, таких как bFGF или IGF в режиме системного введения (Fisher M, et al. J Cereb. Blood. Flow Metab. 1995; 15:963-9; Schabitz WR, et al. Stroke, 2001; 32: 1226-33). Можно полагать, что недостаточность глюкозы и процесс токсического возбуждения с последующей перегрузкой клеток Ca2+, а также апоптоз, наличие реакционно-способных видов кислорода и снижение энергетических резервов в ситуации повышенных потребностей (например, в связи с развитием депрессии) являются основными причинами гибели нейрональных клеток после ишемии (Lee JM, et al. Nature, 1999; 399 (Suppl): A7-14). Как было показано в приведенных ниже примерах, GCSF защищает in vitro клетки нейронального типа (NGF-дифференцированные РС12 клетки) от H2O2-индуцированной гибели. H2O2 вызывает окислительный стресс путем образования реакционно-способных видов кислорода (ROS), которые приводят к гибели клеток. H2O2-опосредованный клеточный стресс в клетках млекопитающих хорошо описан с точки зрения клеточного фенотипа, доз, временных характеристик и участвующих в нем сигнальных путей. Широко распространенное использование H2O2 в большом числе исследований подтверждает апоптозные механизмы эффекта H2O2 на клетки (см., например, FASEB J. 2002 Jan; 16(1): 113-3; J Cell Biochem 2001; 82(3): 437-44). Например, были убедительно продемонстрированы функци проапоптозной киназы ASK1 и FAS лиганда в H2O2-опосредованной гибели клеток (Tobiume K, EMBO Rep 2001 Mar; 2(3): 222-8; Kwon, D., J Neuroimmunol, 2001 Feb 1; 113(1):1-9; Facchinetti, F., J Neorosci Res, 2002 Jul 15; 69(2): 178-88). В этой связи вполне вероятно, что GCSF препятствует действию апоптозных механизмов, вовлеченных в ишемию головного мозга. Воздействие GCSF, по всей видимости, происходит через связывание с высокоафинным рецептором GCSFR. Взаимодействие с данным рецептором активирует семейство киназ Janus (JAK) и белки STAT (Darnell JE Jr. Science, 1997, 277: 1630-5). JAK включает тирозинпротеинкиназы нерецепторного типа, которые активируются при димеризации рецептора, индуцированного лигандом. GCSF-индуцированная активация JAK приводит к фосфорилированию STAT по консервативному тирозиновому остатку, которое далее индуцирует димеризацию STAT (Quelle FW, et al. Mol. Cell. Biol. 1994; 14; 4335-41). Кроме того, STAT перемещается в ядро и впоследствии регулирует генную экспрессию (Shuai K, et al. Nature, 1993; 366:580-3; Shuai K, Oncogene, 2000; 19: 2638-44). STAT3 представляет собой главный STAT-белок, активируемый под действием GCSFR (Shuai K, Oncogene, 2000; 19: 2638-44) STAT3 вовлечен в антиапоптозный механизм через активацию bcl-2, индуцируя пролиферацию и дифференцировку гранулоцитов посредством позитивной регуляции c-myc гена (Fukada T, Immunity, 1996; 5: 449-60; Shimozaki K, Biol Chem, 1997; 272: 25184-9). Как было показано авторами, при использовании методов иммуногистохимии, ОТ-ПЦР и вестерн-блоттинга GCSFR существует не только на гематопоэтических клетках, но также на нейронах, клетках глии, клетках нейронального типа и нерональных стволовых клетках. Кроме того, GCSF-индуцированная позитивная регуляция STAT3 в нейронах из затемненной области может опосредовать антиапоптозные эффекты, такие как позитивная регуляция bcl-2, показанная для BDNF и bFGF, (Schabitz WR, et al., Stroke, 2001; 32: 1226-33) и обеспечивать трофную поддержку нейронов, повышая их жизнеспособность. Мечение плотной ядерной массы на STAT3 в затемненной области инфаркта может отражать опосредованное рецептором мембранное перемещение STAT3 из цитоплазмы в ядро, продемонстрированное уже в активированной микроглии после ишемии головного мозга (Planas AM, et al., Eur, J, Neurosci. 1996; 8: 2612-8). Известно, что GCSF стимулирует высвобождение, усиление эффекторной функции и увеличение времени жизни путем задержки апоптозной гибели нейтрофильных гранулоцитов (NGC) как первой линии защиты организма от разных видов инфекции (Hartung T., Curr Opin Hematol, 1998; 5: 221-5). Нейтрофилы могут вызвать закупорку микрососудов с последующей инвазией лейкоцитов, которая запускает высвобождение протеолитических ферментов, свободных кислородных радикалов, интерлейкинов и THF-α, то есть развитие процессов, которые, как известно, увеличивают размер инфаркта и ухудшают последствия ишемии головного мозга (Del Zoppo GJ, Stroke 1991; 22:1276-83; Jean WC., et al., Neurosurgery 1998; 43: 1382-96; Matsuo Y., et al., Brain Res. 1994; 656: 344-52). GCSF, напротив, оказывает выраженное противовоспалительное действие: на моделях инфекции периферических органов GCSF снижает уровень TNF-α, IL-1β, IL-2, IL-6 и IL-8 и повышает уровень антагонистов рецептора IL-1β (Gorgen I., et al., J Immunol 1992; 149: 918-24; Heard SO, et al., Crit. Care Med. 1999; 27: 1019-21; Heard SO, et al., Crit Care Med. 1998; 26: 748-54; Hebert JC, et al., Arch Surg 1990; 125: 1075-8; Lundblad R., et al., Crit Care Med. 1996; 24: 820-6; Squadrito F., et al., Br J Pharmacol 1990; 120:333-9). GCSF снижает высвобождение TNF-α in vitro и in vivo у здоровых волонтеров и повышает уровень антагонистов TNF, IL-6, IL-8 и IL-1β (Hartung T., Curr Opin Hematol, 1998; 5: 221-5; Gorgen I., et al., J Immunol 1992; 149: 918-24; Heard SO, et al., Crit Care Med. 1999; 27: 1019-21). Кроме того, наблюдается сниженная инфильтрация нейтрофилов в легких и в повздошной кишке на модели висцеральной ишемии и реперфузии через 15 минут после введения GCSF и реперфузии тонкого кишечника (Squadrito F., et al., Br J Pharmacol 1990; 120"333-9). Однако при этом не было выявлено повышения уровня инфильтрации нейтрофилов в ишемизированное полушарие, хотя общее количество нейтрофильных гранулоцитов (NGC) повышается после лечения GCSF.
Другой возможный механизм действия факторов роста, в частности bFGF, включает воздействие на мозговой кровоток. Лечение с использованием bFGF расширяет коллатерали вокруг ишемизированной зоны, даже в дозах, которые не стимулируют системную гипотензию, повышая таким образом кровоток в затемненной области (Tanaka R., et al., Stroke, 1995; 26: 2154-8; обсуждение: 2158-9). Однако, как показано в настоящем изобретении, лечение с использованием GCSF не снижает системного кровяного давления или не изменяет мозговой кровоток в сравнении с контрольной группой, по данным оценки LDF.
На фототромбической модели с использованием красителя бенгальского розового внутривенное введение GCSF после ишемического события явно улучшает сенсоромоторную функцию через шесть недель после фототромботического инсульта, дополнительно подтверждая предположение о том, что факторы роста индуцируют процесс восстановления и регенерации после травматических повреждений головного мозга in vivo. Два других соединения, основной фактор роста фибробластов (bFGF) и остеогенный белок-1 (OP-1), ранее были описаны как средства, способные улучшить сенсомоторную функцию и индуцировать рост нейрональных клеток после очаговой ишемии головного мозга (Kawamata et al., Proc, Natl. Acad. Sci USA, 1997; 94: 8179-84); Kawamata et al., Neuroreport 1998; 9: 1441-5; Ren et al., Neuropharmacology, 2000; 39: 860-5). Чаще всего в указанных исследованиях факторы роста вводят в желудочки мозга, что клинически нерелевантно. Только Рамирес с соавт. (Ramirez et al., (1999), Neuroreport., 10, 1201-4) показали, что внутривенное введение bFGF поддерживает в гиппокампе рост, индуцированный повреждением, но авторы не изучили функциональные показатели таких последствий. Представленные в данном описании результаты указывают на то, что процедура введения клинически релевантной дозы GCSF индуцирует функциональное восстановление после ишемии головного мозга. Способность к усилению пластичности не ограничивается зоной ишемического повреждения головного мозга, но также может рассматриваться применительно к нейродегенеративным заболеваниям, таким как болезнь Паркинсона и боковой амиотрофический склероз (ALS), тринуклеотидные повторные заболевания, ишемии головного мозга, связанные с реанимацией или интранатальными проблемами, и, вероятно, также применительно к деменциям, таким как болезнь Альцгеймера, а также к нейродегенеративным аспектам шизофрении.
Обзор способов, применяемых в экспериментах с ишемией головного мозга
Ишемию индуцируют с использованием модели окклюзии путем наложения шва на среднюю мозговую артерию (90 минут) у крыс. Через 30 минут после индукции ишемии животным (n=12 на группу) вводят внутривенно в течение указанных 90 минут 60 мкг/кг веса тела GCSF или носитель. Объем инфарктной зоны вычисляют по результатам теста с красителем ТТС (2,3,5-трифенитатразолийхлорид) в течение 24 часов после ишемии. Экспрессию GCSFR исследуют иммуногистохимически, подтверждая результатами ОТ-ПЦР и иммуноблоттинга. Экспрессию STAT3 оценивают методами иммуногистохимии. Эффективность GCSF по функциональному восстановлению исследуют на фототромботической модели с красителем бенгальского розового.
Статистический анализ
Представленные в данном исследовании показатели выражают в виде среднего значения ±СКО. После получения всех данных были расшифрованы коды, использованные для рандомизации. Для определения статистической значимости различий полученных in vitro данных и физиологических параметров, а также функциональных последствий исследуемых схем был использован метод ANOVA и затем post hoc тест по методу Фишера или методика коррекции по Бонферрони. Для сравнения объемов инфарктной зоны после посмертного осмотра, результатов анализа МРО и иммуногистохимического анализа STAT3 используют тест на соответствие по критерию Стьюдента (t-тест). Тест на соответствие по критерию хи-квадрат был проведен для обработки данных по уровню смертности. Значение p<0,05 рассматривалось как статистически значимое.
Результаты
GCSF снижает объем инфарктной зоны до 131,96 мм3±112,7 мм3 в сравнении с 278,9 мм3±91,56 мм3 (p<0,05) в контрольной группе. Результаты иммуногистохимического анализа, вестерн-блоттинга и ОТ-ПЦР показали наличие рецепторов GCSF в нейронах и глиальных клетках. GCSF существенно активирует STAT3 по периферии инфарктной зоны в сравнении с контролем (p<0,05). GCSF эффективен в плане улучшения функционального восстановления после ишемии на модели фототромбоза с использованием красителя бенгальского розового.
В этой связи было продемонстрировано, что GCSF оказывает выраженный нейропротективный эффект в клеточной культуре и после очаговой мозговой ишемии. Данный эффект, по всей видимости, опосредован взаимодействием с GCSF и активацией STAT3.
Пример 1. Очаговая мозговая ишемия
Процедура индукции очаговой мозговой ишемии (МСАО, окклюзия средней мозговой артерии)
Процедуры эксперимента были утверждены местным этическим комитетом. Двадцать четыре самца крыс Вистар (Wistar) (Charles River, Германия) весом 280-320 были случайным образом распределены по следующим группам: А (контрольная группа, n=12, ишемия в течение 90 минут, лечение с использованием 2 мл 0,9% раствора в течение 90 минут, начиная с 30 минут после окклюзии сосуда); В - (группа введения GCSF, n=12, ишемия в течение 90 минут, лечение с использованием 60 мкг/кг веса тела рекомбинантного человеческого GCSF (Neupogen®, Amgen, Europe B.V., Нидерланды), растворенного в 2 мл 0,9% солевого раствора в течение 90 минут, которое начинают через 30 минут после окклюзии сосуда. Альтернативно, могут быть использованы любой GCSF, или его производное, или композиция от другого источника (другого производителя, например LenogastrimTM от Roche, или GranocyteTM от Chugai, или AlbugraninTM от HGS, или NeulastaTM от Roche/Amgen).
Затем животных подвергают анестезии путем внутрибрюшинной инъекции 100 мг/кг веса тела гидрохлорида кетамина (WDT, Garbsen, Германия). Анестезию поддерживают на уровне 50 мг/кг веса тела, при необходимости. Полиэтиленовую трубку (ПЭ) вставляют в правую бедренную артерию для непрерывного мониторинга среднего артериального давления крови, уровня газов в крови, гематокрита, подсчета лейкоцитов и определения уровня глюкозы в крови. Правая бедренная вена может быть канюлирована с использованием трубки ПЭ-50 для лечения методом инфузии. В ходе эксперимента отслеживают ректальную температуру и поддерживают ее на уровне 37°С за счет термостатически контролируемой нагретой прокладки (Fohr Medical Instruments, Германия). Временную очаговую ишемию головного мозга индуцируют при моделировании окклюзии путем наложения шва, как детально описано в литературе (Zea Longa et al. (Stroke 1989; 20: 84-91)). Вкратце, процедура состоит в том, что правую общую сонную артерию и правую внешнюю сонную артерию обнажают через срединный разрез горла. Затем нейлоновую моноволокнистую нитку для шва, покрытую силиконом (Bayer, Германию), вставляют посредством артериэктомии в общую сонную артерию, осторожно подтягивают внутреннюю сонную артерию и располагают примерно в 17 мм от разветвления сонной артерии. В рамках данной методики верхняя часть шва закупоривает в одностороннем режиме проксимальную переднюю часть мозговой артерии, исходный участок МСА и заднюю соединяющую часть артерии. При этом в типичном случае создается крупный инфаркт в пределах МСА. Реперфузию проводят путем снятия закупоривающей нити через 90 минут после окклюзии сосуда. Животных с ложной операцией подвергают тем же процедурам, что и описаны выше, но нейлоновую нить не затягивают над общей сонной артерией, так что инфаркт не происходит. После хирургии катетер удаляют и животным позволяют восстановиться после анестезии, дают пищу и воду ad libitum.
Измерение регионального кровотока мозга
Лазерную Допплер-флоуметрию (LDF) (Periflux 4001 Master, Perimed AB, Швеция) используют для оценки мозгового кровотока (CBF) до, во время и после окклюзии MCA. После помещения животных в стереотаксическую рамку обнажают череп животного и просверливают отверстие диаметром 5 мм под микроскопом с правой стороны, в 4 мм латерально и в 2 мм каудально от брегмы. Твердую мозговую оболочку оставляют интактной и в просверленное отверстие помещают LDF зонд (диаметром 1,4 мм). Площадь, выбранная для CBF мониторинга, соответствует ишемизированной полутеневой зоне на модели крыс с MCA окклюзией.
Вычисление объема инфаркта
Через 24 часа после окклюзии МСА крыс анестезируют кетамином в дозе 150 мг/кг веса тела и подвергают декапитации. Мозг животного рассекают на пять коронарных срезов толщиной 2 мм, инкубируют в 2% растворе 2,3,5-трифенилтетразолия хлорида (ТТС) при 37°С в течение 30 минут и фиксируют погружением в 10% забуференный раствор формалина. Срезы, окрашенные ТТС, фотографируют с использованием камеры с присоединенным считывающим устройством (EDH-1000HR Computer Camera, Electrim Corporation, Princetown, NJ, США). Исследователь, работающий в режиме слепой пробы, измеряет размер инфарктных зон с помощью присоединенного к компьютеру анализатора (Bio Scan Optimas, Edmonds, WA). Для компенсации эффекта, вызванного отеком мозга, вычисляют откорректированный объем инфарктной зоны, как ранее было описано более подробно: корректированную площадь инфаркта получают при вычитании площади оставленного нетронутым левого полушария (площадь инфаркта получают при вычитании площади оставленного нетронутым правого полушария) (Schabitz WR, et al., Stroke 2001; 32: 1226-33). Объем инфаркта выражают в виде процента от контралатерального полушария.
Эксперимент по ишемии
GCSF оказывает мощный нейропротективный эффект после очаговой мозговой ишемии. Средний объем инфаркта у животных с внутрибрюшинным введением GCSF составляет 131,96 мм3±112,7 мм3 в сравнении с 278,9 мм3±91,56 мм3 в контрольной группе или 9,96±8,31% (n=12) в сравнении с 22,7 ± 6,69% относительно всего полушария (p<0,05; фиг.1).
Применение GCSF существенно снижает уровень смертности: четыре животных в контрольной группе и одно животное в группе введения GCSF умерли в течение 24-часового периода реперфузии (p<0,05). Не отмечалось статистических различий между контрольной группой и группой введения GCSF по ректальной температуре, pH, pCO2, pO2, гематокриту (hct), уровню глюкозы в крови, частоте сердечных сокращений, среднему значению артериального давления и весу тела для всех животных (таблица 1). Количество лейкоцитов в периферической крови существенно повышалось через 24 часа после ишемии у животных в группе введения GCSF в сравнении с контрольной группой (p<0,05, таблица).
LDF-мониторинг не выявляет статистически значимых различий между двумя исследуемыми группами (данные не приведены).
Пример 2. Иммуногистохимическое исследование очаговой мозговой ишемии
Использованные методы иммуногистохимии
Для проведения морфологического анализа активации STAT3 (фиг.6) и распределения GCSFR в инфарктном мозге и подсчета числа нейтрофильных гранулоцитов срезы мозга толщиной 2 мм из GCSF-обработанных животных и из контрольных животных фиксируют погружением в 4% параформальдегид в 0,1 моль/л фосфатного буфера на 24 часа (n=5 на группу). После погружения в парафин нарезают срезы толщиной 1 мкм и далее используют их для окрашивания красителями H&E, Nissl и для иммуногистохимического анализа.
Иммуногистохимические исследования проводят с использованием антисыворотки против миелопероксидазы (DAKO, Carpinteria, CA, США), фибриллярного кислого белка глии (GFAP) (DAKO, Carpinteria, CA, США), GCSFR (Santa Cruz Biotechnology Inc., Santa Cruz, CA, США) и STAT3 (Santa Cruz Biotechnology Inc., Santa Cruz, CA, США). Для выявления антигена срезы, подготовленные для гистохимического анализа GCSF и STAT3, нагревают в течение 10 минут в 10 мМ цитратном буфере при 99°С. Затем срезы инкубируют в нормальной сыворотке свиньи (в 10% фосфатно-буферном растворе) в течение 30 минут и затем в первичной антисыворотке в течение ночи при 4°С. Первичные антитела разбавляют 1:150 (для миелопероксидазы), 1:400 (для GFAP), 1:400 (для GCSFR) и 1:100 (для STAT3) соответственно. Иммунореактивность визуализируют по методу образования комплекса авидин-биотин (Vectastain, Vector Laboratories, США). Срезы обрабатывают 0,02% диаминобензидином (ДАБ (DAB)) с добавлением 0,02% перекиси водорода. Интенсивность окрашивания реакционного продукта усиливают за счет добавления 0,02% хлорида кобальта и сульфата никеля-аммония. В подмножестве контрольных препаратов предварительная сорбция антисыворотки к GCSFR на соответствующем пептиде не приводит к развитию иммунологического окрашивания (данные не показаны). При отсутствии первичной сыворотки иммунологическое окрашивание вообще не развивается (данные не приведены).
Инвазию нейтрофильных гранулоцитов (НГ) оценивают при подсчете количества НГ в инфарктном полушарии. Экспрессию STAT3 белка количественно оценивают при осмотре в двух перекрывающихся полях: в ростро-каудальном направлении в районе инфаркта теменной части коры головного мозга и на соответствующей контралатеральной стороне (увеличение х400). В этом случае подсчитывают количество нейронов с ядерной транслокацией и выражают результат в виде процента STAT3-позитивных нейронов относительно всех нейронов.
Результаты иммуногистохимических экпериментов с очаговой церебральной ишемией
Окрашивание с использованием миелопероксидазы (МРО) не выявляет нейтрофильных гранулоцитов (НГ) в неишимизированных полушариях в обеих группах. МРО окрашивание незначительно отличалось между животными из группы введения GCSF и контрольными животными по данным количественного определения МРО-позитивных клеток в ишемизированном полушарии (14±17,6 в сравнении с 14,3±12,5, н/о).
Иммунореактивность по GFAP (ИР) выявляется в процессах, связанных с астроцитами, рассеянными по коре головного мозга, полосатому телу и белому веществу в неинфарктном полушарии. Не было выявлено каких-либо различий в характере и интенсивности GFAP-окрашивания в кортикальной околоинфарктной зоне как у контрольных крыс, так и у крыс в группе введения GCSF. В частности, ИР по GCSF не увеличивается в кортикальной части полутеневой зоны ни в группе плацебо, ни в группе введения GCSF (данные не приведены). Внутри инфарктного ядра не выявляется GFAP-иммунореактивность рассеянных астроцитов (данные не приведены).
Иммуногистохимически выявляется окрашивание по GCSFR в рассеянных кортикальных нейронах и нейритах (данные приведены) как в группе необработанных животных, так и в группе введения GCSF. Глиальные клетки также окрашиваются антителом к GCSF-R (данные не приведены). В инфарктном ядре не выявляются клетки, иммунореактивные по GCSF-R. Не наблюдается различий между двумя экспериментальными группами в характере и интенсивности ИР по GCSF-R.
ИР по STAT3 отмечается в рассеянных ядрах нейронов и в глиальных клетках в не пораженном инфарктом полушарии как в группе плацебо, так и в группе введения GCSF. Некоторое цитоплазматическое окрашивание отмечается также в ряде рассеянных нейронов. Экспрессия STAT3 белка после обработки GCSF значительно повышается в полутени инфарктной зоны в сравнении с необработанными контрольными животными (34,4±7,05 в сравнении с 13,7±4,4; p<0,0003) (фиг.6,7). Не отмечалось различий на контралатеральной стороне (16,2±6,9 в сравнении с 13,3±6,9, н/о).
Пример 3. Вестерн-блоттинг и ПЦР при очаговой мозговой ишемии
Вестерн-блотинг (фиг.3)
Для проведения иммуноблоттинга ткани мозга (временная ишемия в течение 2 часов) подвергают лизису в 20 объемах (вес/объем) гомогенизирующего буфера (320 мМ сахарозы, 0,1 мМ фенилметилсульфонилфторида и 2 мкг/мл пепстатина) при 4°С. Гомогенаты центрифугируют при 9200 G в течение 15 минут при 4°С. После ресуспендирования осадка в 1/10 объема гомогенизирующего буфера отбирают аликвоты для определения белка (анализ белка по методу Bio-Rad, Munich, Германия) и образцы быстро замораживают и хранят в окиси азота при -70°С. Для каждой полосы вносят по 15 мкг белка в 8% ДСН-полиакриламидный гель, содержащий 4 М мочевины, и проводят электрофорез в стандартных условиях. Белки электрофоретически переносят на мембраны Immobilon-PTM (Millipore Corp., Eschborn, Германия) путем полусухого блоттинга. После блокирования в 3% нежирном сухом молоке в TBST (20 мМ Трис-основания, pH 7,6, 137 мМ NaCl и 0,05% Твин-20) в течение 1 часа при комнатной температуре (КТ) мембраны инкубируют с первичным антителом к GCSFR (1:500) в течение ночи при 4°С. После промывки в TBST мембраны инкубируют в течение 1 часа при КТ и в разведении 1:2000 с соответствующим вторичным антителом, конъюгированным с пероксидазой хрена. Иммунореактивные полосы визуализируют в линейном диапазоне по усиленной хемилюминесценции (Amersham Intl., Braunschweig, Германия).
В экспериментах по иммуноблоттингу с использованием кортикальных экстрактов (фиг.3) антисыворотка к GCSF-R выявляет белковую полосу размером примерно 130 кД, соответствующую расчетной молекулярной массе (Fukunaga R., et al., J Biol Chem, 1990; 265: 14008-15). Дополнительно отмечается несколько полос более низкого молекулярного веса, что, вероятно, отражает наличие продуктов распада. После предварительной абсорбции антисыворотки к GCSFR на соответствующем пептиде полосы исчезают (фиг.3).
ПЦР для рецептора GCSF в мозге (фиг.2А)
После глубокой анестезии крыс и транскардиальной перфузии мозг быстро рассекают. РНК экстрагируют из мозга с использованием набора для выделения РНК (AGS, Heidelberg, Германия) в соответствии с инструкциями производителя. В общей сложности 10 мкг РНК подвергают обратной транскрипции с использованием обратной транскриптазы MMLV и случайных гексамеров. Для проведения ПЦР используют следующие праймеры от экзонов 5 и 7 мышиного GCSFR: смысловой: 5'-CCC CTC AAA CCT ATC CTG CCT C-3' (SEQ ID NO: 5) и антисмысловой: 5'-TCC AGG CAG AGA TCA GCG AAT G-3' (SEQ ID NO: 6) (Ashihara E., et al., J Cell Physiol, 1997; 171: 343-56). ПЦР проводят по следующей схеме: 3 минуты при 94°С; 1 минута при 94°С; 1 минута при 58°С и 1 минута при 72°С (40 циклов). Продукт анализируют в 2% агарозном геле.
С использованием ОТ-ПЦР, специфичной для мышиного GCSF, мРНК для GCSF-R выявляется в ткани мозга (фиг.2А). ПЦР продукт имеет ожидаемый размер 567 п.н. Идентичность подтверждается секвенированием ПЦР продукта (фиг.2).
Пример 4. Эффективность GCSF на моделях с ALS
Тест на выживание в рамках моделирования ALS на мышах
Предшествующие эксперименты показали, что модели ALS с использованием мышей SOD1 являются прогностическими для оценки результатов лечения у людей (Cleveland and Rothstein (2001), Nat Rev Neurosci, 2, 806-19). Первичные временные точки в таких анализах включают начало появления моторных симптомов и уровень смертности. Так, например, начало моторных симптомов может быть определено как первые сутки, когда мышь не может в течение 7 минут оставаться на движущемся со скоростью 20 об/мин барабане (ротароде) (Li, et al. (2000), Science, 288, 335-9). Уровень смертности подсчитывается в сутки смерти или в сутки, когда отмечаемая недостаточность становится настолько серьезной, что мышь должна быть умерщвлена (например, апатия, неспособность самой взбираться на стержень). Дополнительные параметры определяются путем измерения двигательных усилий по тестам на прочность захвата, путем подсчета двигательных нейронов в спинном мозге, определения толщины нервов (например, седалищного нерва, диафрагмального нерва) и по наличию апоптозного окрашивания в двигательных нейронах спинного мозга. GCSF может быть подвергнут инфузии с помощью осмотического насоса в желудочки мозга в заданной дозе, например 60 мкг/кг веса тела/сутки. Альтернативно, GCSF вводят в/в или в/б инъекцией в дозе 60 мкг/кг веса тела в сутки или в более высоких дозах. Альтернативно, вводят форму GCSF с медленным высвобождением, такую как PEG-композиция (см. выше), или альбуминовая композиция (см. выше), или другие композиции с медленным высвобождением. Альтернативно, используют любой GCSF, или его производное, или композицию, полученную из другого источника (другого производителя, например LENOGASTRIMTM от Roche, GRANOCYTETM от Chugai Pharma, Co., Ltd., ALBUGRANINTM от Human Genome Science или NEULASTATM от Roche/Amgen). Введение начинают на 60 сутки поздней предсимптоматической стадии, характерной для мутанта SOD1 G93A. У не подвергнутых обработке мышей с наследственным ALS моторные нарушения появляются в возрасте 12-14 недель, тогда как паралич не отмечается ранее чем в 20 недель. Ожидаемая продолжительность жизни составляет 140-170 суток. Эффективное лечение продолжают в течение всей жизни, длительность которой более чем на 15% превышает соответствующий показатель для контрольной группы (Cleveland and Rothstein (2001), Nat Rev Neurosci, 2, 806-19). В качестве контрольной группы для оценки результатов лечения используют животных, которым вводят и носитель, и zVADfmk (мощный ингибитор каспазы, который продемонстрировал эффективность в данной модели). Каждая группа включает по 10 животных.
Пример 5. Эффективность GCSF на моделях болезни Паркинсона
Имеются различные модели болезни Паркинсона с использованием грызунов, которые подходят для исследования эффективности GCSF (Grunblatt et al. (2000), J Neurol, 247, Suppl 2, 1195-102). Одна хорошо описанная модель представляет собой использование 1-метил-4-фенил-1,2,3,6-тетрагидропиридина (MPTP). 4 дозы MPTP-HCl (15 мг/кг на дозу) могут вводиться 8-недельным самцам мышей (n=20) в/б инъекцией с интервалами 2 часа. Животным с ложной обработкой вводят солевой раствор. Для 20 животных проводят введение и MPTP, и ежедневной дозы GCSF (в/б, 60 мкг/кг веса тела), через семь суток после введения последней дозы MPTP мышей умерщвляют. До указанного момента времени мышей подвергают анализу для оценки двигательных параметров (поведение на движущемся стержне (ротароде), свободная локомоция). Мозг от 10 мышей подвергают обработке для иммуногистохимического анализа всей совокупности нейронов в черном веществе, которые являются позитивными на тирозингидроксилазу или на переносчик допамина (с использованием коммерчески доступных антител), для определения числа позитивных по апоптозу нейронов (окрашивание по методу TUNEL, окрашивание по каспазе-3). Оставшиеся допиминергические нейроны после MPTP-обработки сравнивают с их количеством в случае введения MPTP плюс GCSF и в группе с ложной обработкой.
Оставшихся 10 животных в каждой группе подвергают транскардиальной перфузии солевым раствором, умерщвляют и вырезают полосатое тело. Затем полосатое тело гомогенизируют на льду и измеряют содержание допамина методом ВЭЖХ при электрохимическом обнаружении. Сравнение трех групп обеспечивает хороший критерий для оценки истощения по допамину из-за потери клеток в черном веществе.
Данный эксперимент может быть также проведен с использованием различных схем дозирования MPTP (например, 40 мг/кг веса тела один раз; 30 мг/кг веса тела два раза и т.п.).
Пример 6. GCSF улучшает сенсоромоторную функцию после фототромботической мозговой ишемии (фиг.8)
Экспериментальные группы
Экспериментальные процедуры были одобрены местным комитетом по этике. Самцов крыс Вистар (Wistar) (Charles River, Германия) весом от 280 до 320 грамм были случайным образом распределены по следующим группам: А (контрольная группа, n=6), ишемия, лечение с использованием 0,5 мл 0,9% солевого раствора в виде в/в инфузии болюсом, начиная с 1 часа после ишемии; В - (группа введения GCSF, n=6), ишемия, лечение с использованием 5 мкг GCSF (Amgen), растворенного в 0,5 мл 0,9% солевого раствора в виде в/в болюса, начиная с 1 часа после ишемии. Альтернативно, в данном способе может использоваться любой GCSF, или его производное, или композиция, полученная из другого источника (другой производитель, например LenogastrimTM от Roche, или GranocyteTM от Chugai, или AlbugraninTM от HGS, или NeulastaTM от Roche/Amgen). Затем проводят повторные в/в инфузии болюсом через хвостовую вену в период с 1 по 5 сутки. С - (группа животных с ложной операцией, n=6), ложная операция, без ишемии, лечение с использованием 0,5 мл 0,9% солевого раствора в виде в/в инфузии болюсом, начиная с 12 часа после ишемии.
Очаговая мозговая ишемия, индуцируемая фототромбозом
Животных анестезируют путем внутримышечной инъекции 100 мг/кг веса тела гидрохлорида кетамина (WDT, Garbsen, Германия). Анестезию поддерживают на уровне 50 мг/кг веса тела, при необходимости. Полиэтиленовую трубку ПЭ-50 вводят в правую бедренную артерию для непрерывного мониторинга среднего артериального давления крови и уровня газов в крови. Правую бедренную вену канюлируют с использованием трубки ПЭ-50 для инфузии. В ходе эксперимента отслеживают ректальную температуру и поддерживают ее на уровне 37°С с использованием термостатически контролируемой нагретой прокладки (Fohr Medical Instruments, Германия).
Фототромботическую ишемию индуцируют в правой теменной части коры головного мозга крысы по методу Уотсона с соавт. (Watson BD, Dietrich WD, Busto R, Wachtel MS, Ginsberg MD. Induction of reproducible brain infarction by photochemically initiated thrombosis. Ann Neurol., 1985; 17: 497-504). Животных анестезируют с использованием гидрохлорида кетамина и помещают в стереотаксическую камеру, рассекают скальп для вскрытия поверхности черепа. Для освещения оптоволоконный тяж с отверстием 1,5 мм помещают стереотаксически в череп в направлении 4 мм назад от брегмы и на расстоянии 4 мм латерально от средней линии. Затем череп освещают лучом холодного белого света (150 Вт) в течение 20 минут. В течение первых 2 минут освещения инъецируют внутривенно краситель бенгальский розовый (0,133 мл/кг веса тела, 10 мг/мл солевого раствора). Животных с ложной операцией подвергают тем же экспериментальным процедурам, что и описаны выше, но без инфузии красителем бенгальского розового и без освещения. После хирургической процедуры удаляют катетеры и животных оставляют восстанавливаться от анестезии и дают пищу и воду ad libitum.
Бихевиоральное тестирование
На всех животных перед индукцией ишемии, на базовом уровне, а также через 2, 3, 4, 5 и 6 недель после ишемии исследователь, который работает в режиме слепой пробы в отношении распределения экспериментальных групп, осуществляет целую серию бихевиоральных тестов. Для проведения теста с движущимся стержнем (ротародом) крыс помещают на ускоряющийся цилиндр ротарода и измеряют время, в течение которого животные остаются на данном ротароде (Hamm et al., J Neurotrauma, 1994 Apr; 11(2):187-96, Chen J, Li Y. Wang L, Zhang Z, Lu D, Lu M, Chopp M. Therapeutic Benefit of Intravenous Administration of Bone Marrow Stromal Cells After Cerebral Ischemia in Rats. Stroke. 2001; 32: 1005). Скорость медленно повышают от 4 до 40 об/мин в течение 5 минут. Испытание заканчивают, когда все животные падают со ступеньки или хватаются за устройство и крутятся два последовательных вращения без попыток взобраться на ступеньки. Для крыс устанавливают произвольный лимит времени, равный 500 секунд, на движущемся цилиндре, в процессе обучения, а также в процедурах тестирования. Животных обучают 3 суток перед индукцией ишемии. Средняя длительность (в секундах) пребывания на устройстве подсчитывается при 3 измерениях на вращающемся устройстве за 1 сутки до проведения хирургической процедуры. Данные по моторному тесту представляют в виде процента от средней длительности (3 испытания) нахождения на вращающемся устройстве в сравнении с базовым уровнем, характерным для внутреннего контроля на базовом уровне (до хирургической процедуры).
В тесте с удалением липкой ленты определяют соматосенсорный дефицит как до, так и после ишемии (Schallert T, Kozlowski DA, Humm JL, Cocke RR. Use-dependent structural events in recovery of function. Adv. Neurol. 1997;73: 229-238; Chen J, Li Y., Wang L, Zhang Z, Lu D, Lu M, Chopp M. Therapeutic Benefit of Intravenous Administration of Bone Marrow Stromal Cells After Cerebral Ischemia in Rats. Stroke. 2001; 32: 1005). Всем крысам дают освоиться с окружающей средой в условиях тестирования. В первоначальном тесте два маленьких бумажных кружочка с липкой основой (равного размера, по 113,1 мм2) используют в качестве двустороннего тактильного раздражителя, занимающего дистально-радиальный участок на запястье каждой передней лапы. Затем каждую из крыс возвращают в свою клетку. Время удаления каждого из раздражителей с передних лап определяют в ходе пяти испытаний в сутки для каждой из передних лап. Отдельные испытания разделяют по меньшей мере пятью минутами. Перед проведением хирургического вмешательства животных обучают в течение 3 суток. Как только крысы оказываются способными снимать кружочки в течение 10 секунд, их подвергают ишемии.
Используют модифицированную шкалу для оценки тяжести неврологических симптомов (NSS) (Chen J, Li Y, Wang L, Zhang Z, Lu D, Lu M, Chopp M. Therapeutic Benefit of Intravenous Administration of Bone Marrow Stromal Cells After Cerebral Ischemia in Rats. Stroke. 2001; 32: 1005; и Schallert T, Kozlowski DA, Humm JL, Cocke RR. Use-dependent structural events in recovery of function. Adv. Neurol. 1997;73: 229-238). Выраженность неврологической функции ранжируют по шкале от 0 до 16 (нормальный балл - 0; балл максимального дефицита -16). NSS представляет собой сложный показатель, определяемый на основе теста по оценке двигательной, сенсорной активности, рефлексов и баланса (Chen J, Li Y, Wang L, Zhang Z, Lu D, Lu M, Chopp M. Therapeutic Benefit of Intravenous Administration of Bone Marrow Stromal Cells After Cerebral Ischemia in Rats. Stroke. 2001; 32: 1005; Germano AF, Dixon CE, d'Avella D, Hayes RL, Tomasello F. Behavioral deficits following experimental subarachnoid hemorrhage in the rat. J Neurotrauma. 1994; 11: 345-353). В соответствии с данной шкалой оценки тяжести повреждения показатель в 1 балл присваивается неспособности выполнять тест или в случае потери исследуемого рефлекса. Таким образом, чем выше балл, тем более тяжелым является повреждение.
Результаты
Не наблюдается статистических различий между группами по ректальной температуре, pH, pCO2, pO2, гематокриту (hct), уровню глюкозы в крови, частоте сердечных сокращений, среднему артериальному давлению, весу тела и уровню смертности для всех животных (данные не приведены).
Функциональное восстановление животных в группе введения GCSF с течением времени происходит значительно лучше, чем во всех других группах. Животные, которым вводят GCSF, характеризуются значительно меньшими баллами по шкале NSS, включая результат лучевого баланса, в сравнении с контрольной группой (p<0,001, фиг.8B), что сказывается на значительно лучшем поведении животных на вращающемся стержне (ротароде) в сравнении с контролем (p<0,05, фиг.8А). Сенсоромоторная функция, по результатам теста на удаление липкой ленты, также лучше у животных в группе введения GCSF в сравнении с контролем как на контралатеральной передней лапе с течением времени, так и на ипсилатеральной передней лапе к 6 неделе (см. Фиг.8 C, D).
Пример 7. Рецептор GCSF подвергается позитивной регуляции на фототромботической модели мозговой ишемии
РНК выделяют в соответствии со стандартными процедурами (Chomczynski and Sacchi (1987), Anal Biochem, 162, 156-159) с последующей очисткой с использованием мини-набора Qiagen RNeasyTM из образцов кортикальной полутеневой зоны, взятых из участков, расположенных на ипси- и контралатеральной стороне относительно участка повреждения (см. фиг.9а с изображением локализации образцов ткани; в данной иллюстрации: 3 против 4). кДНК синтезируют на основе 1 мкг суммарной РНК с помощью олигоdT праймеров, обратной транскриптазы Superscript II (Gibco) в стандартных условиях. Количественную ПЦР проводят с использованием системы ЛайтЦиклер (LightCycler®) (Roche Diagnostics, Mannheim, Германия) при окрашивании двуцепочечной ДНК красителем SYBR-зеленый. Циклическую обработку проводят по следующей схеме: 5 минут - 95°С, 5 секунд - 95°С, 7 секунд - 66°С, 30 секунд - 72°С, 9 секунд - 84°С, в течение 55 циклов. Кривые плавления строят по следующим параметрам: 95°С, охлаждение до 50°С; подъем до 99°С со скоростью 0,2°С в секунду. Используют следующие пары праймеров: "крысиный GCSF-frag-32s" CCATTGTCCATCTTGGGGATC (SEQ ID NO: 7) и "крысиный GCSF-frag-265as" CCTGGAAGCTGTTGTTCCATG (SEQ ID NO: 8). ПЦР в системе ЛайтЦиклер (LightCycler®) проводят с использованием основной смеси, включающей краситель SYBR-зеленый и далее в соответствии с рекомендациями производителя (Roche Diagnostics). Специфичность продукта гарантируется анализом точки плавления и электрофоретическим разделением в агарозном геле. Содержание кДНК в образцах нормализуют до уровня экспрессии циклофилина (праймеры: "cyc5" ACCCCACCGTGTTCTTCGAC (SEQ ID NO: 9); "acyc300" CATTTGCCATGGACAAGATG (SEQ ID NO: 10). Относительные уровни регуляции получают после нормализации до уровня циклофилина и сравнения с животными из группы с ложной операцией. На фиг.9b показана позитивная регуляция GCSF чрез 48 часов на контралатеральной стороне, стрелки ошибок указывают на стандартные отклонения, которые вычисляют на основе трехкратного анализа серийно разбавленных образцов кДНК, и отражают достоверность измерений.
Пример 8. Рецептор GCSF присутствует на нейрональных клетках в первичных культурах: данные иммуноцитохимии
Получение нейронов
10-12 образцов коркового вещества получают из эмбрионов на стадии Е18 (18 эмбриональные сутки). Ткань подвергают расщеплению с использованием трипсина [10мг/мл]/ЭДТА/ДНК-азы [5 мг/мл] (Roche Diagnostics, Mannheim, Германия) в HBSS (сбалансированный раствор Хэнкса, BioWithakker). Расщепление может быть остановлено с использованием 4 частей среды (нейробазальная среда + 1 мл 50Х В-27 добавки (Invitrogen) + 0,5 мМ L-глютамина + 25 мкМ глютамата) с последующим центрифугированием при комнатной температуре в течение 5 минут при 800 g. Осадок растворяют в 5 мл среды и далее определяют количество клеток путем подсчета (в препарате Neubauer). Клетки помещают в 24-ячеечные микротитрационные планшеты по 250 000 клеток в ячейки на покровные стекла, которые далее могут быть покрыты слоем поли-L-лизина.
Иммуногистохимия
Через 14 суток после получения нейроны промывают ФБР (Gibco) (37°C) и фиксируют в 2% параформальдегиде в течение 10 минут на льду. Затем клетки промывают ФБР (4°C) и хранят при 4°C. Клетки инкубируют в течение 10 минут в 50 мМ глицине и затем промывают ФБР. Клетки подвергают обработке на льду для усиления проницаемости с использованием 0,2% Тритона Х-100 (Sigma) в ФБР и инкубируют с блокирующим раствором (0,2% Тритон Х-100, 4% нормальная козья сыворотка (NGS) (Jackson Immunoresearch Laboratories) в ФБР) при комнатной температуре. Первичное антитело (кроличье антитело к рецептору GCSF, направленное против С-конца мышиного GCSFR, М-20; sc-694; Santa Cruz Biothechnology, Inc.) используют в разбавлении 1:800 (0,1% ТритонХ-100/2% NGS) и инкубируют в течение ночи при 4°С. Затем клетки обрабатывают 1% NGS/ФБР и инкубируют в течение 30 минут со вторичным антителом (антикроличий FITC, 1:400; dianova) при комнатной температуре. Клетки быстро промывают 1% NGS/ФБР и окрашивают Hoechst 33342 (Molecular Probes) (1:10000 в ФБР) для контрастного окрашивания ядер. В конце процедуры покровные стекла быстро промывают два раза в 1% NGS/ФБР, два раза в ФБР и один раз в течение 10 минут в 10 мМ Трис-HCl, pH 7,6. Покровные стекла погружают с использованием Aquamount (Polysciense).
Фотографии получают на цифровом аппарате с использованием микроскопа OlympusIX81 и проводят анализ с пакетом прикладных программ "Analysis" (Soft Imaging Systems, Stuttgart, Германия).
Пример 9. Рецептор GCSF присутствует на нейрональных стволовых клетках: ПЦР и иммуноцитохимия (фиг.12,13); рецептор GCSF присутствует на РС12 клетках: ПЦР (фиг.2В)
Получение нейрональных стволовых клеток
Нейрональные стволовые клетки выделяют из участков головного мозга, образуемых в результате известного процесса спонтанного нейрогенеза, например, из гиппокампа, обонятельной луковицы и субвентрикулярной зоны, у 4-6-недельных самцов крыс Вистар (Wistar), как описано в литературе (Ray J et al. (1993), Proc Natl Acad Sci USA 90, 3602-6). Используемые процедуры соответствуют принципам работы с животными, зафиксированным в правилах Национального Исследовательского Совета США, и удовлетворяют требованиям Германского законодательства. Вкратце, процедура состоит в том, что животных анестезируют 1 об.% изофлурана, 70% N2О, 29% кислорода и умерщвляют декапитацией. Головные мозги удаляют и промывают 50 мл охлажденного льдом фосфатно-буферного раствора Дульбекко (DPBS) с добавкой 4,5 г/л глюкозы (DPBS/Glc). Гиппокамп, обонятельную луковицу и субвентрикулярную зону от шести животных разрезают, промывают в 10 мл DPBS/Glc и центрифугируют в течение 5 минут при 1600 х g при 4°С. После удаления супернатанта ткань гомогенизируют с помощью ножниц и скальпелей. Кусочки такни промывают средой DPBS/Glc в течение 5 минут при 800 х g при 4°С и 3 осадка ресуспендируют в 0,01% (вес/объем) папаина, 0,1% (вес/объем) диспазы II (нейтральная протеаза), 0,01% (вес/объем) ДНК-азы I и 12,4 мМ сульфата марганца в сбалансированном солевом растворе Хэнкса (HBSS). Ткань растирают наконечниками пластиковых пипеток и инкубируют в течение 40 минут при комнатной температуре, но при этом каждые 10 минут раствор хорошо перемешивают. Суспензию центрифугируют при 800 х g в течение 5 минут при 4°С и осадки промывают три раза в 10 мл среды DMEM-Ham F-12 с добавкой 2 мМ L-глютамина, 100 единиц/мл пенициллина и 100 единиц/мл стрептомицина. Затем клеточные осадки ресуспендируют в 1 мл нейробазальной среды с добавкой В27 (Invitrogen, Carlsbad, CA, США), 2 мл L-глютамина, 100 единиц/мл пенициллина и 100 единиц/мл стрептомицина, 20 нг/мл EGF, 20 нг/мл FGF-2 и 2 мкг/мл гепарина. Клетки помещают в стерильных условиях в 6-гнездные чашки в концентрации 25000-100000 клеток/мл. Чашки инкубируют при 37°С в среде с содержанием 5% СО2. Культуральные среды для клеток меняют один раз в неделю, при этом около одной трети сред замещается (Ray J et al. (1993), Proc Natl Acad Sci USA 90, 3602-6).
Процедура проведения ОТ-ПЦР
РНК выделяют в соответствии со стандартными процедурами из стволовых клеток гиппокампа (фиг.12), которые подвергали размножению в течение 3 недель в культуре после оттаивания из замороженных образцов для хранения, в соответствии с рекомендациями производителя (набор RNeasy, Qiagen). кДНК синтезируют с использованием праймеров олигоdT, обратной транскриптазы Superscript II (Gibco) в стандартных условиях. Полимеразно-цепьевую реакцию (ПЦР) проводят по следующей схеме циклической обработки: 3 минуты - 94°С, 30 секунд - 94°С, 30 секунд - 60°С, 1 минута - 72°С, в течение 32 циклов; с использованием праймерных пар "крысиный GCSFR-frag-8s" GCGGGCAAATCAGGATCTCAC (SEQ ID NO: 2) и "крысиный GCSFR-frag-287as" CGAAGCTCAGCTTGATCCAGG (SEQ ID NO: 3). Праймеры получают из фрагмента GCSFR, выделенного и идентифицированного из геномных баз данных при поисках в режиме TBLASTX (см. фиг.11). Условия реакции: 2 мМ MgCl2, 200 мкМ дНТФ, 200 нМ каждого праймера, 1 единица Taq полимеразы (Invitrogen)/25 мкл. ПЦР продукт разрешают в 1,5% геле агарозы. Длина специфического ПЦР продукта составляет 279 п.н., с последовательностью (SEQ ID NO: 4), которая приведена ниже (праймерные последовательности подчеркнуты).
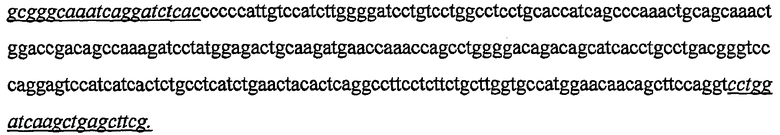
Для проведения ПЦР на клетках РС12 (фиг.2В) используют указанную выше процедуру.
Иммуноцитохимия нейросфер:
Нейросферы, состоящие из нейрональных стволовых клеток, наносят пипеткой на стеклянную пластину, накрывают покровным стеклом и препарат оставляют при -80°C в течение по меньшей мере 30 минут. Покровное стекло удаляют, а препарат сразу же помещают в 4% ПФА (параформальдегид) в 0,1 М фосфатном буфере, pH 7,4. Клетки фиксируют в течение 20 минут. Затем клетки промывают 3х5 мин в ФБР (pH 7,4) с добавкой 1% ФСТ и 0,02% NaN3. Клетки подвергают обработке для усиления проницаемости в течение 10 минут в ФБР, содержащем 0,5% ТХ-100. Антигены блокируют выдерживанием в течение 60 минут в ФБР (pH 7,4), содержащем ФСТ и 0,02% NaN3. Клетки окрашивают в течение 20 минут ядерным красителем DAPI в концентрации 0,001 мг/мл в ФБР. Далее клетки промывают 2х5 минут ФБР (pH 7,4), содержащим 1% ФСТ и 0,02% NaN3. Первое антитело наносят на 2 часа при разбавлении ФБР (pH 7,4), содержащим 1% ФСТ и 0,02% NaN3, при соблюдении следующих концентраций: анти-GCSF - 1:1000, анти-GCSF-R - 1:1000, анти-GMCSF-R - 1:1000 (Santa Cruz) соответственно. Клетки промывают 3х5 мин ФБР (pH 7,4), содержащем 1% ФСТ и 0,02% NaN3. Второе антитело (козий IgG-FITC против антигена кролика (DAKO)) выдерживают в течение 60 минут в ФБР (pH 7,4), содержащем 1% ФСТ и 0,02% NaN3, в концентрации 1:30. Клетки промывают 3х5 минут в ФБР (pH 7,4), содержащем 1% ФСТ и 0,02% NaN3. В конце процедуры клетки вносят в Aquamount и закрывают покровным стеклом.
Пример 10. GMCSFR-альфа подвергается позитивной регуляции в рамках фототромботической модели мозговой ишемии (выявлено по RMDD)
Экспериментальные процедуры одобрены местным комитетом по этическим вопросам. Самцов крыс Вистар (Wistar) (Charles River, Германия) весом 280-320 грамм случайным образом распределяют по следующим группам: а) ишемия в разные временные точки (6 часов, 48 часов и 21 сутки); b) животные с ложной операцией без ишемии, с оценкой в соответствующие временные точки (6 часов, 48 часов и 21 сутки), n=2 для каждой временной точки и для каждого вида обработки.
Очаговая мозговая ишемия, индуцируемая фототромбозом
Животных анестезируют внутримышечной инъекцией 100 мг/кг веса тела гидрохлоридом кетамина (WDT, Garbsen, Германия). Анестезию поддерживают на уровне 50 мг/кг веса тела, при необходимости. Полиэтиленовую трубку ПЭ-50 вставляют в правую бедренную артерию для непрерывного мониторинга среднего артериального давления крови и уровня газов в крови. Правую бедренную вену канюлируют с использованием трубки ПЭ-50 для проведения инфузии. В ходе эксперимента отслеживают ректальную температуру и поддерживают ее на уровне 37°С за счет термостатически контролируемой нагретой прокладки (Fohr Medical Instruments, Германия).
Фототромботическую ишемию индуцируют в правой теменной части коры головного мозга крысы по методу Уотсона с соавт. (Watson BD, Dietrich WD, Busto R, Wachtel MS, Ginsberg MD. Induction of reproducible brain infarction by photochemically initiated thrombosis. Ann Neurol, 1985; 17: 497-504). Животных анестезируют с использованием гидрохлорида кетамина и помещают в стереотаксическую камеру и надрезают скальп для вскрытия поверхности черепа. Для освещения помещают оптоволоконный тяж с отверстием 1,5 мм стереотаксически в череп в направлении 4 мм назад от брегмы и на расстоянии 4 мм латерально от средней линии. Далее череп освещают лучом холодного белого света (150 Вт) в течение 20 минут. В течение первых 2 минут освещения инъецируют внутривенно краситель бенгальский розовый (0,133 мл/кг веса, 10 мг/мл солевого раствора). Животных с ложной операцией подвергают тем же экспериментальным процедурам, описанным выше, но без инфузии красителя бенгальского розового и без освещения. После окончания хирургической процедуры удаляют катетеры и животным позволяют восстановиться от анестезии и дают пищу и воду ad libitum. Животных умерщвляют в различные временные точки (6 часов, 48 часов и 21 сутки после ишемии и ложной операции соответственно) и отбирают препараты из полутеневой зоны коры головного мозга с ипси- и контралатеральной стороны, как известно специалистам в данной области техники.
Выделение РНК и RMDD
РНК выделяют в соответствии со стандартными процедурами (Chomczynski and Sacchi (Anal Biochem, (1987), 162, 156-9) с последующей очисткой при использовании мини-набора Qiagen RNeasyTM) из образцов полутеневой зоны коры головного мозга, взятых из участков, расположенных на ипси- и контралатеральной стороне относительно локализации повреждения (см. фиг.13а с изображением локализации образцов ткани; в данном случае: 3 против 4). Синтез кДНК проводят на основе 1 мкг суммарной РНК в соответствии с процедурой RMDD (дифференциальная процедура, включающая рестрикцию), описанной в ЕР 0743367 А2 и в US 5876932. После синтеза первой нити и второй нити проводят MboI-расщепление на участке адапторного лигирования. Осуществляют две реакции ПЦР с подмножеством праймерных сочетаний. Далее ПЦР продукты вносят на денатурированный гель и переносят блоттингом на нейлоновую мембрану (GATC Biotech AG, Konstanz, Германия). Меченные биотином полосы визуализируют при проведении обычной реакции с образованием комплекса стрептавидин-пероксидаза. ПЦР-образцы из полутеневой зоны коры головного мозга наносят на гель в следующем порядке: ипсилатеральные образцы: нативные (необработанные), ложная обработка - 6 часов, ложная обработка - 48 часов, ложная обработка - 21 сутки и через 6 часов, 48 часов и 21 сутки после фототромбоза; контралатеральные образцы: ложная обработка - 6 часов, ложная обработка - 48 часов, ложная обработка - 21 сутки и через 6 часов, 48 часов и 21 сутки после фототромбоза. Полосы, имеющие разную интенсивность на ипсилатеральном и контралатеральном участках, вырезают из нейлоновой мембраны и проводят повторную амплификацию соответствующего ПЦР продукта. Амплифицированные продукты клонируют в векторе pCR-BluntII-TOPO (Invitrogen GmbH, Karlsruhe, Германия) и секвенируют с использованием праймеров T7 и M13rev (ABI 3700). Полученные последовательности сравнивают с компонентами, имеющимися в базе данных EMBL. Идентифицируют последовательность, которая подверглась позитивной регуляции через 48 часов как на ипси-, так и на контралатеральном участке полутеневой зоны коры головного мозга (фиг.14). Идентифицированную EST последовательность расширяют при проведении поисков в базах данных EST в режиме BLASTN и идентифицируют гомологичную последовательность мышей, кодирующую мышиный GMCSFR-альфа, в базах данных EST и геномных базах данных (ensembl; www.ensembl. org) с использованием программ скрининга (BLAST, TBLASTN (Altschul, et al. (1997), Nucleic Acids Res, 25, 3389-402)).
Скрининги проводят в библиотеках кДНК крысы, полученных по методике клонирования продуктов ПРЦ (Shepard (1997) Nucleic Acids Res 25: 3183-3185), для подтверждения корректности полученной последовательности крысы. Указанный метод основан на гибридизации молекул кДНК, полученных из плазмидной библиотеки, с олигонуклеотидной последовательностью, связанной с биотином. После экстракции плазмиды при использовании магнитных шариков, связанных со стрептавидином, получают результат, который подтверждают диагностической ПЦР и двухкратной репликацией на стадиях, следующих после повторной трансформации полученных плазмид до восстановления единственного клона. Используют следующие праймерные сочетания:
5'-bloc-2.clb4-4-4: CGGGATCCGGGACCGCGTATCTGATGACGAGCGTGTCAA (SEQ ID NO: 12)
25bio-2.clb4-4-4: CTCGGAGACGCTGAGGAAGGACCTG (SEQ ID NO: 13)
3'bloc-2.clb4-4-4: CTGCGGCCCTAGACCACGCCCACCGCTCCCCGTGACGTCG (SEQ ID NO: 14)
(ОРС определяют для единственного клона, полученная при этом последовательность показана как SEQ ID NO: 40, а соответствующая аминокислотная последовательность показана в виде SEQ ID NO: 41).
Пример 11. Рецептор GMCSF-альфа подвергается позитивной регуляции в рамках фототромботической модели мозговой ишемии (подтверждение методом количественной ПЦР)
РНК выделяют в соответствии со стандартными процедурами (Chomczynski and Sacchi (Anal Biochem (1987), 162, 156-9) с последующей очисткой при использовании мини-набора Qiagen RntasyTM) из образцов полутеневой зоны коры головного мозга крысы на ипси- и контралатеральной стороне относительно локализации повреждения (см. фиг.13а с изображением локализации образцов ткани; в данном случае: 3 против 4). Синтез кДНК проводят на основе 1 мкг суммарной РНК с использованием олигоdT праймеров, обратной транскриптазы Superscript II (Gibco) в стандартных условиях. Количественную ПЦР проводят в системе ЛайтЦиклер (LightCycler®) (Roche Diagnostics, Mannheim, Германия) при окрашивании двуцепочечных ДНК красителем SYBR-зеленый. Циклическую обработку проводят по следующей схеме: 5 минут - 95°С, 5 секунд - 95°С, 7 секунд - 62°С, 30 секунд - 72°С, 9 секунд 80°С, в течение 50 циклов. Кривые плавления строят по следующим параметрам: 95°С, охлаждение до 50°С; подъем до 99°С со скоростью 0,2°С в секунду. Используют следующие праймеры: "крысиный BR4-4s96" ACGTCGTTGGCTCAGTTATGTC (SEQ ID NO: 15) и "крысиный BR4-4as272" ATTTATGTCAGAGATGGAGGATGG (SEQ ID NO: 16). ПЦР в системе ЛайтЦиклер (LightCycler®) проводят с использованием основной смеси, включающей SYBR зеленый, и далее по инструкции производителя (Roche Diagnostics). Специфичность продукта подтверждают анализом точки плавления и электрофоретическим разделением в агарозном геле. Содержание кДНК в образцах нормализуют до уровня экспрессии циклофилина (праймеры: "cyc5" ACCCCACCGTGTTCTTCGAC (SEQ ID NO: 17); "acyc300" CATTTGCCATGGACAAGATG (SEQ ID NO: 18)). Относительные уровни регуляции получают после нормализации до уровня циклофилина и при сравнении с животными из группы с ложной операцией. На фиг.14А показана позитивная регуляция GMCSFR-альфа через 48 часов на ипси- и на контралатеральной стороне. Не обнаруживается существенной регуляции через 21 сутки после индукции фототромбоза (фиг.14 B). Стрелки ошибок указывают на стандартные отклонения, которые вычисляют на основе трехкратного анализа серийно разбавленных образцов кДНК, и отражают достоверность измерений.
Пример 12. Рецептор GMCSF присутствует на нейрональных клетках в первичных кортикальных культурах: получение нейронов
10-12 образцов коры головного мозга получают из эмбрионов на стадии Е18 (18 сутки развития эмбриона). Ткань подвергают разволокнению с использованием трипсина [10 мг/мл]/ЭДТА/ДНК-азы [5 мг/мл] (Roche Diagnostics, Mannheim, Германия) в HBSS (сбалансированный раствор Хэнкса, BioWithakker). Расщепление останавливают с использованием 4 частей среды (нейробазальная среда + 1 мл 50Х В-27 добавки (Invitrogen) + 0,5 мМ L-глютамина + 25 мкМ глютамата) с последующим центрифугированием при комнатной температуре в течение 5 минут при 800 х g. Осадок растворяют в 5 мл среды и определяют количество клеток путем подсчета (в препарате Neubauer). Клетки вносят с плотностью 250000 клеток на ячейку в 24-ячеечные планшеты на покровные стекла, которые далее закрывают слоем поли-L-лизина.
Иммуногистохимия
Через 1 неделю после получения нейроны промывают ФБР (Gibco) (37°C) и фиксируют 2% параформальдегидом в течение 10 минут на льду. Затем клетки промывают ФБР (4°C) и инкубируют в течение 10 минут в 50 мМ глицине в ФБР, после чего снова промывают ФБР. Клетки подвергают обработке для усиления проницаемости с использованием 0,2% ТритонХ-100 (Sigma) в ФБР на льду и инкубируют с блокирующим раствором (0,2% ТритонХ-100, 4% нормальная козья сыворотка (NGS) (Jackson Immunoresearch Laboratories) в ФБР) при комнатной температуре. Первичное антитело (кроличье антитело против рецептора GMCSF, направленное против С-конца мышиного GMCSFR, М-20; sc-694; (Santa Cruz Biothechnology, Inc.) используют в разбавлении 1:300 (0,1% ТритонХ-100/2% NGS) и инкубируют в течение ночи при 4°С. Затем клетки обрабатывают 1% NGS/ФБР и инкубируют 30 минут со вторичным антителом (анти-кроличий FITC, 1:400; dianova) при комнатной температуре. Клетки быстро промывают в 1% NGS/ФБР и окрашивают Hoechst 33342 (Molecular Probes) (1:10000 в ФБР) для контрастного окрашивания ядер. В конце процедуры покровные стекла быстро промывают два раза в 1% NGS/ФБР, два раза в ФБР и один раз в течение 10 минут в 10 мМ Трис-HCl, pH 7,6. Покровные стекла погружают с использованием Aquamount (Polysciense).
Фотографии получают на цифровом аппарате с использованием микроскопа Olympus IX81 и проводят анализ с использованием пакета прикладных программ "Analysis" (Soft Imaging Systems, Stuttgart, Германия).
Пример 13. Рецептор GMCSF присутствует на нейрональных стволовых клетках: получение нейрональных стволовых клеток (фиг.13)
Нейрональные стволовые клетки выделяют из участков головного мозга, образуемых в результате известного процесса спонтанного нейрогенеза, например, из гиппокампа, обонятельной луковицы и субвентрикулярной зоны, у 4-6-недельных самцов крыс Вистар (Wistar), как описано в литературе (Ray J et al. (1993), Proc Natl Acad Sci USA 90, 3602-6). Используемые процедуры соответствуют принципам работы с животными, зафиксированным в правилах Национального Исследовательского Совета США, и удовлетворяют требованиям Германского законодательства. Вкратце, процедура состоит в том, что животных анестезируют 1 об.% изофлурана, 70% N2О, 29% кислорода и умерщвляют декапитацией. Головные мозги удаляют и промывают 50 мл охлажденного льдом фосфатно-буферного раствора Дульбекко (DPBS) с добавкой 4,5 г/л глюкозы (DPBS/Glc). Гиппокамп, обонятельную луковицу и субвентрикулярную зону от 6 животных разрезают, промывают в 10 мл DPBS/Glc и центрифугируют в течение 5 минут при 1600 х g при 4°С. После удаления супернатанта ткань гомогенизируют с помощью ножниц и скальпелей. Кусочки такни промывают средой DPBS/Glc в течение 5 минут при 800 х g при 4°С и три осадка ресуспендируют в 0,01% (вес/объем) папаина, 0,1% (вес/объем) диспазы II (нейтральная протеаза), 0,01% (вес/объем) ДНК-азы I и 12,4 мМ сульфата марганца в сбалансированном солевом растворе Хэнкса (HBSS). Ткань растирают наконечниками пластиковых пипеток и инкубируют в течение 40 минут при комнатной температуре, но при этом каждые 10 минут раствор хорошо перемешивают. Суспензию центрифугируют при 800 х g в течение 5 минут при 4°С и осадки промывают три раза в 10 мл среды DMEM-Ham F-12 с добавкой 2 мМ L-глютамина, 100 единиц/мл пенициллина и 100 единиц/мл стрептомицина. Затем клеточные осадки ресуспендируют в 1 мл нейробазальной среды с добавкой В27 (Invitrogen, Carlsbad, CA, США), 2 мл L-глютамина, 100 единиц/мл пенициллина и 100 единиц/мл стрептомицина, 20 нг/мл EGF, 20 нг/мл FGF-2 и 2 мкг/мл гепарина. Клетки помещают в стерильных условиях в 6-гнездные чашки в концентрации 25000-100000 клеток/мл. Чашки инкубируют при 37°С в среде с содержанием 5% СО2. Культуральные среды для клеток меняют один раз в неделю, при этом около одной трети сред замещается (Ray J et al. (1993), Proc Natl Acad Sci USA 90, 3602-6).
Процедура проведения ОТ-ПЦР
РНК выделяют в соответствии со стандартными процедурами из стволовых клеток гиппокампа (фиг.12), которые подвергали размножению в течение 3 недель в культуре после оттаивания из замороженных образцов для хранения, в соответствии с рекомендациями производителя (набор RNeasy, Qiagen). кДНК синтезируют с использованием праймеров олигоdT, обратной транскриптазы Superscript II (Gibco) в стандартных условиях. Полимеразно-цепьевую реакцию (ПЦР) проводят по следующей схеме циклической обработки: 3 минуты - 94°С, 30 секунд - 94°С, 30 секунд - 60°С, 1 минута - 72°С, в течение 32 циклов; с использованием праймерных пар "крысиный BR4-4s96" ACGTCGTTGGCTCAGTTATGTC (SEQ ID NO: 19) и "крысиный BR4-4as272" ATTTATGTCAGAGATGGAGGATGG (SEQ ID NO: 20). Условия реакции: 2 мМ MgCl2, 200 мкМ дНТФ, 200 нМ каждого праймера, 1 единица Taq полимеразы (Invitrogen)/25 мкл. ПЦР продукт разрешают в 1,5% геле агарозы. Длина специфического продукта ПЦР составляет 176 п.н. и имеет следующую последовательность (праймерные последовательности подчеркнуты)

Пример 14: Тест для определения периода полувыведения GCSF/GMCSF из сыворотки и прохождения через гематоэнцефалический барьер
Желательно знать, проходят ли GCSF и GMCSF через гематоэнцефалический барьер. GCSF/GMCSF подвергают биотинилированию с целью использования высокочувствительного авидин-биотинового взаимодействия для обнаружения хемокинов в ткани мозга. GCSF (Neupogen, Amgen) биотинилируют с использованием биотина-XX-SE (Molecular Probes B 1606). GCSF разбавляют в 20 мМ натрий-карбонатном буфере, pH 8, содержащем 250 мМ сорбита и 0,004% Твин-80, и добавляют биотин-XX-SE. После выдерживания в течение 1 часа при комнатной температуре добавляют Трис-буфер, pH 8 до 50 мМ концентрации для гашения непрореагировавшего меченого реагента. Образец откручивают в течение 30 минут со скоростью 45000 об/мин в роторе TLA 100 (Beckman Instruments) для удаления агрегатов.
7,5 мкг биотинилированного GCSF инъецируют внутрибрюшинно мышам в нулевой временной точке (200 мкл 20 мМ натрий-карбонатного буфера, pH 8, содержащего 250 мМ сорбита и 0,004% Твин-80). Мышей анестезируют хлоралгидратом в указанные временные точки и отбирают образцы крови (примерно 200 мкл) из правой камеры сердца. Добавляют ЭДТА до 5 мМ и образец центрифугируют в течение 30 минут при 1000 g с получением сыворотки. К сыворотке добавляют 4 х буферных образца, белки подвергают денатурации при нагревании до 95°С в течение 5 минут и 20 мкл наносят на минигель. Белки переносят на нитроцеллюлозу, блокируют и инкубируют со стрептавидином-HRP (Amersham) в TBST. После промывки выявляют сигналы хемилюминесцирующего реагента.
Для проведения анализа методом ИФТФА сывороточные образцы разбавляют в пропорции 1:20 буфером для тестирования и проводят анализ по инструкции производителя (IBL, Hamburg, Германия).
Данный тест может быть адаптирован к условиям исследования цереброспинальной жидкости (scf) или гомогената тканей мозга для выявления прохождения GCSF через гематоэнцефалический барьер.
Пример 15. Тест на нейропротективное действие GCSF, GMCSF (фиг.1B)
Нейропротективное действие GCSF/GMCSF определяют in vitro на NGF-обработанных РС12 клетках. РС12 клетки высевают в 96-ячеечные планшеты, покрытые поли-L-лизином (0,01% конечная концентрация) с плотностью 40000 клеток/ячейку. Клетки выдерживают в среде DMEM, содержащей 1000 мг глюкозы/л, 10% ЛС (лошадиная сыворотка), 5% ФСТ (фетальная сыворотка теленка), 1% пенициллина/стрептомицина. Клетки подвергают трансфекции с использованием pSV40-RL (ген рениллы, кодирующий люциферазу ) с использованием трансфицирующего агента Липофектамин2000® (Lipofectamine2000®) (Gibco, BRL) (0,2 мкг ДНК/ячейку) в соответствии с рекомендациями производителя. Сразу же после трансфекции добавляют NGF (ростовой фактор нервных клеток) в концентрации 40 нг/мл для индукции дифференциации РС12 клеток. Через 24 часа после обработки РС12 клетки развивают нейроно-подобную морфологию с вытянутыми структурами. Далее клетки обрабатывают H2O2 в варьирующих концентрациях (фиг.1B) и GCSF в варьирующих концентрациях (1-100 нг/мл). Добавляют ЕРО в качестве позитивного контроля, как вещество с известной нейропротективной эффективностью in vitro (Cerami, et al. (2002), Nephrol Dial Transplant, 17: 8-12; Kawakami, et al. (2001), J Biol Chem, 276, 39469-75; Sinor and Greenberg (2000), Neurosci Lett, 290, 213-5; Chong, et al. (2002), J Cereb Blood Flow Metab. 22, 503-14), в концентрациях от 0,01 Ед/мл до 1 Ед/мл. Через 24 часа супернатант среды отбрасывают и клетки лизируют с использованием буфера для пассивного лизиса (Promega). Измеряют люциферазную активность рениллы с помощью люминометра (Mithras, Berthold) и результаты выражают в виде относительных световых единиц. Данный тест позволяет измерить уровень выживания клеток в соответствии с выявляемым количеством люциферазы. В этой связи чем выше показатель относительных световых единиц, тем больше клеток выжило. В данном тесте GCSF продемонстрировал зависимое от дозы нейропротективное действие на РС12 клетки, что характеризуется более мощным эффектом, чем в случае эритропоэтина.
Пример 16. Рецептор GCSF экспрессируется в различных участках мозга, важных с точки зрения неврологического заболевания (фиг.4); GMCSF экспрессируется в различных важных участках мозга (фиг.19)
Для систематической оценки распределения рецептора GCSF в нормальном мозге мышей С57/bl6 (возраст 2-3 месяца ) подвергают анестезии в/в инъекцией Ромпуна® (Rompun®) и Кетанеста® (Ketanest®). Затем мышей подвергают транскардиальной перфузии с использованием 20 мл сбалансированного солевого раствора Хэнкса (HBSS) и далее 20 мл 4% параформальдегида (ПФА) в ФБР (pH 7,4). Мозг вырезают и хранят в течение ночи в 2% растворе ПФА. Погруженные в парафин участки ткани нарезают с получением срезов (2 мкм), которые наносят на предварительно обработанные предметные стекла (DAKO, Glostrup, Дания), сушат на воздухе в течение ночи и далее подвергают депарафинизации. После микроволновой обработки (цитратный буфер; 500 Вт, 10 минут) в ткани вносят антитело против GCSFR (1:400) и далее инкубируют в течение 1 часа при комнатной температуре в увлажненной камере. Антительную метку визуализируют с использованием обычной методики ABC и ДАБ (DAB) в качестве хромогена в соответствии с рекомендациями производителя (DAKO, Glostrup, Дания). Отрицательные контроли включают аналогичным образом обработанные срезы, в которых первичное антитело полностью отсутствует, а также срезы, для которых используют соответствующую нормальную сыворотку (Dianova, Hamburg, Германия). Локализация GCSF-R отмечается в гиппокампе (фиг.4а-d) с преобладающим окрашиванием нейронов на участке СА3 (фиг.4а, b), при наличии резкой границы между участками СА3 и CA2 (фиг.4с, стрелка). GCSF-R распределяется по всей соме, как и процессы, определяемые нейронами (фиг.4b, стрелка). Рецептор присутствует в области ворот и в слоях базальных клеток зубчатой извилины (фиг.4d, стрелка). Рецептор GCSF-R также выявляется в кортикальных зонах: в пириформной коре (фиг.4е, стрелка) и в периринальной коре (f) в качестве примеров. В мозжечке метятся клетки Пуркинье (фиг.4g, стрелка). Кроме того, некоторые из крупных митральных клеток в обонятельной луковице также являются позитивными по GCSF-R (фиг.4h, стрелка). Сильное окрашивание выявляется в передних столбах спинного мозга (фиг.4i, j), а при большем увеличении выявляются крупные двигательные нейроны в виде положительных по GCSF-R структур (фиг.4k, l). Следует отметить, что нейрональные процессы сильно метятся. В среднем мозге нейроны в черном веществе демонстрируют позитивность относительно GCSF-R (фиг.4m, n, o). Кроме нейронов, окрашиваются также олигодендроциты в проходах белого вещества, например в передней спайке (фиг.4р, стрелка).
Процедура указанного примера была использована для определения локализации GMCSFR (фиг.19). В этом случае окрашивание отмечается в гиппокампе, в коре, в мозжечке и в хороидном сплетении. При этом средний мозг и спинной мозг не исследовались.
Пример 17. Тест на нейропротективное действие GCSF, GMCSF (фиг.1 B, фиг 23)
Нейропротективное действие GCSF/GMCSF определяют in vitro на NGF-обработанных РС12 клетках. РС12 клетки высевают в 96-ячеечные планшеты, покрытые поли-L-лизином (0,01% конечная концентрация) с плотностью 40000 клеток/ячейку. Клетки выдерживают в среде DMEM, содержащей 1000 мг глюкозы/л, 10% ЛС (лошадиная сыворотка), 5% ФСТ (фетальная сыворотка теленка), 1% пенициллина/стрептомицина. Клетки подвергают трансфекции с использованием pSV40-RL (ген рениллы, кодирующий люциферазу) с использованием трансфицирующего агента Липофектамин2000® (Lipofectamine2000®) (Gibco, BRL) (0,2 мкг ДНК/ячейку) в соответствии с рекомендациями производителя. Сразу же после трансфекции добавляют NGF (ростовой фактор нервных клеток) в концентрации 40 нг/мл для индукции дифференциации РС12 клеток. Через 24 часа после обработки РС12 клетки развивают нейроно-подобную морфологию с вытянутыми структурами. Далее клетки обрабатывают H2O2 в варьирующих концентрациях (фиг.1B) и GCSF в варьирующих концентрациях (1-100 нг/мл). Добавляют ЕРО в качестве позитивного контроля как вещество с известной нейропротективной эффективностью in vitro (Cerami et al. (2002), Nephrol Dial Transplant, 17: 8-12; Kawakami, et al. (2001), J Biol Chem, 276, 39469-75; Sinor and Greenberg (2000), Neurosci Lett, 290, 213-5; Chong, et al. (2002), J Cereb Blood Flow Metab. 22, 503-14), в концентрациях от 0,01 Ед/мл до 1 Ед/мл. Через 24 часа супернатант среды отбрасывают и клетки лизируют с использованием буфера для пассивного лизиса (Promega). Измеряют люциферазную активность рениллы с помощью люминометра (Mithras, Berthold) и результаты выражают в виде относительных световых единиц. Данный тест позволяет измерить уровень выживания клеток в соответствии с выявляемым количеством люциферазы. В этой связи чем выше показатель относительных световых единиц, тем больше клеток выжило. В данном тесте GCSF продемонстрировал зависимое от дозы нейропротективное действие на РС12 клетки, что характеризуется более мощным эффектом, чем в случае эритропоэтина.
Пример 18. Тромэмболическая мозговая ишемия
Процедуры эксперимента были утверждены местным комитетом по этическим вопросам. Сорок самцов крыс Вистар (Wistar) (Charles River, Германия) весом 280-320 г случайным образом распределяют по следующим группам: А) ранний тромболизис под действием 10 мг rt-PA/кг веса тела в течение 1 часа, через 1 час после тромбоэмболической окклюзии сосуда; B) поздний тромболизис под действием 10 мг rt-PA/кг веса тела в течение 1 часа, через 3 часа после тромбоэмболической окклюзии сосуда; С) без тромболизиса, но при обработке 60 мкг/кг веса тела рекомбинантным GCSF (Neupogen®, Amgen, Europe B.V., Нидерланды) в 2 мл 0,9% солевого раствора в течение 90 минут, которую начинают через 30 минут после тромбоэмболической ишемии; D) обработка 60 мкг/кг веса тела рекомбинантным GCSF (Neupogen®, Amgen, Europe B.V., Нидерланды) в 2 мл 0,9% солевого раствора в течение 90 минут, которую начинают через 30 минут после тромбоэмболической ишемии в сочетании с поздним тромбозом под действием 10 мг rt-PA/кг веса тела в течение 1 часа, через 3 часа после тромбоэмболической окклюзии сосуда.
Животных анестезируют, например, внутрибрюшинной инъекцией 100 мкг/кг веса гидрохлорида кетамина (WDT, Garbsen, Германия). Анестезию поддерживают на уровне 50 мг/кг веса тела, при необходимости. Полиэтиленовую трубку ПЭ-50 вставляют в правую бедренную артерию для непрерывного мониторинга среднего артериального давления крови, уровня газов в крови, гематокрита, подсчета лейкоцитов и определения уровня глюкозы в крови. Правую бедренную вену канюлируют с использованием трубки ПЭ-50 для проведения инфузии. В ходе эксперимента отслеживают ректальную температуру и поддерживают ее на уровне 37°С за счет термостатически контролируемой нагретой прокладки (Fohr Medical Instruments, Германия).
Тромбоэмболический инсульт индуцируют по модифицированной методике Буша с соавт. (Busch et al., Brain Res 1997 Dec 5: 778(1): 16-24). В общих чертах процедура состоит в том, что правую общую сонную артерию (CCA), внутреннюю сонную артерию (ICA) и внешнюю сонную артерию (ECA) обнажают через срединный разрез шеи. Дальнейшее рассечение позволяет идентифицировать начало крылонебной артерии (PPA). ECA и PPA лигируют в постоянном режиме шелковой нитью 6-0. ССА подвергают лишь временному лигированию на период эмболизации. Катетер вставляют в ECA проксимально к месту ее лигирования и инъецируют 12 сгустков крови (каждый имеет 0,35 мм в диаметре и 3 мм в длину), что приводит к эмболизации в правой средней мозговой артерии (MCA).
Развитие инфаркта отслеживают методом МР-фотографии через 1, 2, 4 и 24 часа с использованием диффузной, перфузионной и Т2-опосредованной визуализации. У всех животных результат выражают через уровень смертности, а также при описании неврологических последствий по пятибалльной шкале через 24 часа после ишемии. Через 24 часа после ишемии животных анестезируют кетамином в дозе 150 мг/кг веса тела и подвергают декапитации. Мозг животных удаляют и фиксируют 4% параформальдегидом в 0,1 моль/л фосфатного буфера в течение 24 часов. После погружения в парафин нарезают срезы толщиной 1 мкм и используют их для окрашивания красителями ТТС, H&E и Nissl, а также для иммуногистохимического анализа.
Статистический анализ
Результаты выражают в виде среднего значения ±СКО. После получения всех данных расшифровывают коды рандомизации. Тест по методу ANOVA и затем post hoc тест Фишера по оценке наименее значимого различия могут быть использованы для определения статистической значимости различий непрерывных переменных, таких как физиологические параметры и объем инфарктной зоны. Проводят U-тест Манна-Уитни (Mann-Whitney U test) для обработки непараметрических данных, таких как уровень смертности. Полученное значение р<0,05 рассматривается как статистически значимое.
Пример 19. GCSF эффективен при введении в широком временном диапазоне в случае окклюзии средней мозговой артерии (MCAO) как модели инсульта на грызунах
На фиг.24 показан эффект внутривенного введения GCSF на объем инфаркта у крыс в рамках модели MCAO в случае введения его через 120 минут после начала ишемии. Эксперимент проводят по методике примера 1, с тем исключением, что анестезию индуцируют ингаляцией (1% галотана, 30% О2 и 70% N2O). В данном случае используют дозу 60 мкг GCSF (Neupogen®, AMGEN)/кг веса тела крысы. Отмечается существенная защита при сравнении объемов инфаркта, как следует по результатам ТТС-окрашивания (см. фиг.25). Операции проводят в другом составе исследователей, как показано в примере 1, демонстрируя эффективность в других лабораторных условиях. Указанный пример дает дополнительное подтверждение полезности GCSF в качестве способа лечения инсульта и других ишемических расстройств нервной системы.
Пример 20. Совместная локализация GCSF и его рецептора в головном мозге
Использованные иммуногистохимические методы
Срезы погруженных в парафин тканей (2 мкм) подвергают депарафинизации путем обработки их 2 раза по 5 минут ксилолом, 2 раза по 2 минуты 100% этанолом и затем снижающимися концентрациями этанола от 96% до 70%. В конце процедуры срезы промывают дистиллированной водой и подвергают микроволновой обработке (цитратный буфер при 600 Вт в течение 15 минут). После этого срезы промывают дистиллированной водой и антигены блокируют при обработке 3 раза по 5 минут в 1 х TBS (pH 7,4), содержащем 0,2% БСА. Антисыворотку к рецептору GCSF (SC694; Santa Cruz Biotechnology, Santa Cruz, CA, США; 1:100), разбавленную в 1 x TBS (pH 7,4), содержащем 0,2% БСА, инкубируют при комнатной температуре в течение 1 часа в увлажненной камере. Затем срезы промывают 3 раза по 2 минуты в 1 х TBS (pH 7,4), содержащем 0,2% БСА. Затем добавляют второе антитело (козий Fab-FITC против антигена кролика, dianova, 1:50) и выдерживают в течение ночи при 4°С в 1 х TBS (pH 7,4), содержащем 0,2% БСА. После промывки срезов 3 раза по 2 минуты 0,2% БСА в 1 х TBS (pH 7,4) вносят антисыворотку к GCSF (SC13102; Santa Cruz Biotechnology, Santa Cruz, CA, США), разбавленную в пропорции 1:100 0,2% БСА в 1 х TBS (pH 7,4), на 1 час при комнатной температуре. Срезы промывают 3 раза, как было описано ранее, и инкубируют в течение 30 минут с биотинилированным козьим IgG против антигена кролика (Vector, Laboratories, США), разбавленным в пропорции 1:200 с использованием 1 x TBS (pH 7,4), содержащего 0,2% БСА. И вновь после промывания срезов вносят TRITC-конъюгированный стрептавидин (dianova; 1:200) на 1,5 часа при комнатной температуре. Далее срезы промывают 3 раза по 2 минуты в 1 х TBS и окрашивают в течение 10 минут ядерным красителем DAPI, разбавленным в пропорции 1:10000 в 1 x TBS. В конце процедуры срезы промывают 3 раза по 2 минуты 1 х TBS и вносят в среду для измерения флуоресценции (Vectashield, Vector Laboratories, США). Фотографии получают на цифровом аппарате с использованием микроскопа Olympus IX81 и проводят анализ с пакетом прикладных программ "Analysis" (Soft Imaging Systems, Stuttgart, Германия).
Все эксперименты с двойной оценкой флуоресценции контролируют параллельным однократным окрашиванием, которое используют для подтверждения отсутствия флуоресцирующего проводника во втором канале. В качестве второго контроля все процедуры окрашивания при двукратной флуоресценции проводят при переключении хромофоров на второе антитело.
Результаты
Для рецептора GCSF (фиг.25a, d, g) показана совместная локализация с его лигандом (фиг.25b, e, h) как в гиппокампе (фиг.25a-c в зубчатой извилине, так и в d-f, ворота), так и в коре головного мозга (фиг.25g-i). Неожиданным является тот факт, что некоторые нейроны экспрессируют и рецептор, и лиганд, что позволяет полагать наличие аутокринного сигнального механизма в отношении GCSF, который поддерживает новую эндогенную нейропротективную нервную систему.
Пример 21. Совместная локализация рецептора GCSF и рецептора GMCSF в мозге
Иммуногистохимические методы
Иммуногистохимическое исследование проводят по методике примера 20. После инкубации срезов с антисывороткой к GCSFR и козьим Fab-FITC против антигена кроликов срезы промывают 3 раза по 2 минуты 0,2% БСА в 1 х TBS (pH 7,4). После этого вносят антисыворотку к GMCSFR (SC690; Santa Cruz Biotechnology, Santa Cruz, CA, США, 1:100) на 1 час при комнатной температуре. Последующую процедуру, включая инкубирование с биотинилированным козьим IgG против кролика (Vector, Laboratories, США) и TRITC-конъюгированным стрептавидином (dianova; 1:200), проводят по методике, описанной в примере 2 применительно к выявлению GCSF.
Все эксперименты с двойной оценкой флуоресценции контролируют параллельным однократным окрашиванием, которое используют для подтверждения отсутствия флуоресцирующего проводника во втором канале. В качестве второго контроля все процедуры окрашивания при двукратной флуоресценции проводят при переключении хромофоров на второе антитело.
Результаты
Рецептор GCSF (фиг 26a, d, g) и рецептор GMCSF (фиг.26b, e, h) экспрессируются на одних и тех же нейронах как в гиппокампе, так и в коре головного мозга (фиг.26c, f, i). На фиг.26 показана совместная экспрессия обоих рецепторов на нейронах в зубчатой извилине (a-c) и в воротах (d-f) гиппокампа, а также в коре головного мозга (g-i). Данный результат дополнительно иллюстрирует заявление о том, что GCSF и GMCSF могут использоваться в сочетании для лечения неврологических состояний и что нейропротективные свойства гематопоэтических факторов могут быть объединены при совместном применении.
Пример 22. GCSF действует антиапоптозно путем активации stat в нейронах
Результаты
Первичные культивируемые нейроны являются инструментом для вскрытия механизма действия. В этой связи авторы проверили, может ли рецептор GCSF экспрессироваться на нейронах гиппокампа или на кортикальных нейронах в культуре после 21 суток. Фактически, авторы подтвердили наличие экспрессии путем ПЦР (предыдущие примеры) и методами иммунохимии как в кортикальных, так и в большинстве нейронов гиппокампа. Одним из наиболее важных механизмов в патофизиологии инсульта является задержанная гибель нейрональных клеток (Choi (1996, Curr Opin Neurobiol, 6, 667-72, Schneider, et al., (1999), Nat. Med., 5, 554-9, Mattson (2000), Nat Rev Mol Cell Biol, 1, 120-9). В этой связи авторы предположили, что GCSF может препятствовать каскадам апоптозных явлений в нейронах. В первичных культурах нейрональных клеток, которые, как было показано, экспрессируют рецептор GCSF, воздействие окиси азота, представляющего собой релевантную модель событий, происходящих во время мозговой ишемии, ведет к зависимому от дозы повышению программированной гибели клеток, что следует по расщеплению PARP (данные не приведены). Лечение с использованием 50 нг/мл GCSF резко снижает апоптозную гибель клеток после обработки NOR3, как видно на фиг.27. В клетках гематопоэтической линии рецептор GMCSF переносит свой антиапоптозный сигнал через киназу 2 Януса (JAK2) и белки сигнальной трансдукции и сигнальной трансактивации (stat) (Jak-stat путь) (см., например, Epling-Burnette, et al (2001), J. Immunol, 166, 7486-95, Sakamoto, et al., (2003), Int J Hematol, 77, 60-70)). Хотя авторы не смогли пока выявить активацию фосфорилирования stat1 или stat5 (фиг.27, часть III), показано, что stat3 сильно фосфорилируется при добавлении GCSF, зависимым от времени образом (фиг.27, часть IV), что типично для кинетики типа JAK-stat клетках других видов (фиг.27, часть V; взято и адаптировано из Kuroki, M.& O'Flaherty, J. T. Extracellular signal-regulated protein kinase (ERK)-dependent and ERK-independent pathways target STAT3 on serine-727 in human neutrophils stimulated by chemotactic factors and cytokines. Biochem J 341 (Pt3), 691-6 (1999). Фосфорилирование stat3 tyr705, осуществляемое при добавлении GCSF к культуральной среде, специфически опосредовано через рецептор GCSF/JAK2 путь, поскольку блокирование связи JAK2/stat3 ингибитором AG490 ведет к резкому снижению сигналов в присутствии GCSF. Активация stat3 ведет к индукции антиапоптозных белков из семейства bcl (Yoshida, et al. (2002), J Exp Med, 196, 641-53, Sakai and Kraft (1997), J Biol Chem, 272, 12350-8, Nielsen, et al. (1999), Leukemia, 13, 735-8). Добавление GCSF к культуре нейрональных клеток ведет к зависимому от времени повышению уровня Bcl-XI и Bcl-2, мощных антиапоптозных белков в нейронах, и во время ишемии (фиг.27, часть VI). Интересно, что в недавно опубликованном отчете предполагается роль stat3 в качестве обязательного белка для выживания двигательных нейронов после повреждения нервов (Schweizer, et al. (2002), J Cell Biol, 156, 287-97). В этой связи можно полагать, что GCSF действует по пути, отличному от предложенного для EPO антиапоптозного механизма, включающего активацию stat5 и NF-kB (Digicaylioglu and Lipton (2001), Nature, 412, 641-7).
Методы
Первичные кортикальные нейроны крыс получают следующим образом: десять - двенадцать участков коры головного мозга вырезают из крысиных эмбрионов Е18. Ткань подвергают разволокнению с использованием 10 мг/мл трипсина, 5 мг/мл ЭДТА/ДНКазы (Roche Diagnostics, Mannheim, Германия) в HBSS (BioWhitakker, Taufkirchen, Германия). Процесс разложения останавливают при использовании 4 частей нейробазальной среды, содержащей добавку 1 Х В-27 (Invitrogen, Karlsruhe, Германиия). После центрифугирования осадок растворяют в 5 мл среды и клетки вносят с плотностью 250000 клеток на ячейку в 24-ячеечный планшет на покровные стекла и наносят слой поли-L-лизина. В типичном случае среда содержит: 50 мл нейробазальной среды (Life Technologies, Karlsruhe, Германия), 1 мл добавки В27 (Life Technologies), 5 мкл bFGF (Sigma, 19 мкг растворяют в 100 мкл 10 мМ Трис/HCl, pH 7) и 50 мкл пенициллина/стрептомицина (Life Technologies).
Выявление STAT1, pSTAT1, STAT5, pSTAT5, GCSFR, PARP, расщепленного PARP, каспазы3, расщепленной каспазы3, Bcl2 и Bcl xl проводят по методу вестерн-блоттинга. В общих чертах, процедура состоит в том, что нейроны обрабатывают в течение 24 часов 150 мМ NOR-3 (Sigma-Aldrich, Seelze, Германия) и 50 нг/мл GCSF (Neupogen, Amgen, Thousand Oaks, CA, США), как указано. Клетки соскребают с планшета, откручивают при 800 об/мин в течение 5 минут и промывают охлажденным на льду ФБР, содержащим 2,5 мг/мл пепстатина (Sigma-Aldrich, Seelze, Германия) и апротинина (1:1000, Sigma-Aldrich). Осадки ресуспендируют в 100 мкл раствора бензоназы (содержащем 10 мкл 10 х ФБР, 79,5 мкл H2O, 10 мкл 10% ДСН, 0,5 мкл 100 мМ MgCl2, 0,1 мкл апротинина, 0,1 мкл лейпептина, 0,1 мкл пепстатина, 0,1 мкл PMSF, 0,1 мкл бензоназы (Roche Diagnostics, Mannheim, Германия)). После солюбилизации добавляют 1 объем ФБР и определяют концентрацию белка (тест на БСА, Pierce, Rockford, IL, США). После денатурации при 90°С в течение 5 минут 100 мкг анализируют в 8%-12% ДСН-полиакриламидных гелях. Белки переносят на нитроцеллюлозные мембраны (Protan BA79, Schleicher & Schuell, Dassel, Германия) с использованием полусухой блоттинг-камеры (Whatman Biometra, Gottingen, Германия). Блоты блокируют в 5% порошковом молоке в ФБР/0,02 Твине-20, промывают три раза ФБР/0,02% Твин-20 и инкубируют в течение ночи при 4°С с соответствующим первичным антителом (антитело против расщепленного PARP, Cell Signaling, 1:1000; анти-PARP, Cell Signaling, 1:1000; против расщепленной каспазы3, Cell Signaling, 1:200; против каспазы3, Cell Signaling, 1:200; против Bcl2, BD Transduction Laboratories, 1:500; против Bcl XI, BD Transduction Laboratories, 1:500; против pSTAT3tyr, Cell Signaling, 1:500; против STAT3, Cell Signaling, 1:500; против GCSF, Santa Cruz, 1:200; против pSTAT5, Cell Signaling, 1:500; против stat5, Cell Signaling, 1:200; против pSTAT1, Cell Signaling, 1:500). После промывки блоты инкубируют с соответствующим вторичным антителом (антикроличья или антимышиная антисыворотка, связанная с пероксидазой хрена, Dianova, Hamburg, Германия, 1:4000) в течение 1 часа при комнатной температуре. Сигналы оценивают с использованием суперчувствительной системы выявления хемилюминесценции (Pierce, Rockford, США) и переносят на пленки Hyperfilm-ECL (Amersham Pharmacia Biotech, Piscataway, NJ, США). Уровень расщепления PARP определяют количественно по методу ауторадиографического сканирования с использованием программы Windows ImageJ v1.29 (http://rsb.info.nih.gov/ij/index.html).
Пример 23. GCSF и его рецептор индуцируются мозговой ишемией
MCAO как модель мозговой ишемии
Процедуру индукции очаговой мозговой ишемии (MCAO, окклюзия средней мозговой артерии) проводят по методике примера 1.
Модель общей ишемии
Экспериментальные процедуры на животных, рандомизированных по группам ишемия/реперфузия, проводят по ранее подробно описанной методике (Brambrink, et al. (2000), J. Cereb Blood Flow Metab, 20, 1425-36). В общих чертах, процедура состоит в том, что крыс подвергают анестезии (хлоралгидрат), после чего проводят пероральную интубацию и механическую вентиляцию (Paco2: 35-40 мм Hg, Paco2: 95-110 мм Hg, в течение всего эксперимента). Вскрывают обе общих сонных артерии (ССА) и в левую сторону вставляют катетер в виде полиэтиленовой трубки (ПЭ-50) для отбора образцов крови и непрерывного мониторинга среднего артериального давления (MABP) (Sirecust 310, Siemens, Danvers, MA, США). На правой стороне нейлоновой ниткой делают несильную перетяжку вокруг ССА для последующего затягивания ее с целью индукции временной мозговой ишемии. Голову животного фиксируют в стереотаксической рамке для измерения ЭЭГ и мозгового кровотока (CBF) (подробное описание см. в литературе) (Brambrink, Schneider, Noga, Astheimer, Gotz, Korner, Heimann, Welschof and Kepski (2000), J Cereb Blood Flow Metab, 20, 1425-36)). Свод черепа вскрывают путем срединного разреза и делают две выемки для введения хлорсеребряных штифтовых ЭЭГ электродов (высокоскоростная стоматологическая бормашина, расположенная над левым соматосенсорным участком головного мозга и передним синусом, по атласу головного мозга (в соответствии с Praxinos and Watson (1986) brain atlas)). Над правым полушарием из кости черепа размером примерно 24 мм2 удаляют тонкую костную пластину (1,3-5,3 мм латерально вправо и 1,5-7,5 мм в сторону затылка от брегмы). Используют допплер-флуометрический зонд (длина волны 780 мкм, BPM 403A, TSI Inc., St. Paul, MN, США) для определения в компьютерном варианте контроля с микроманипулятором регионального мозгового кровотока (rCBF) в 30 положениях, по методике сканирования (Soehle, et al. (2000), Acta Neurochir Suppl, 76, 181-4). На контралатеральной стороне (3,3-5,3 мм латерально влево и 5,0-7,0 мм в сторону затылка от брегмы) устанавливают меньшее (4 мм2) окно для измерения местного мозгового кровотока (lCBF) с использованием стационарного зонда для лазерной допплер-фуометрии (длина волны: 780 мкм, BPM2, Vasamedices Inc., St. Paul, MN, США). Температурные зонды помещают, как описывалось ранее (Brambrink, et al. (1999), J Neurosci Methods, 92, 111-22), в правую височную мышцу через левую аурикулярную трубку и на 6 см вглубь прямой кишки. Температуру ядерной зоны (термостатически контролирумое нагреваемое одеяло) и температуру в височной мышце (нагревающий радиатор возле инфарктной зоны) контролируют на уровне 37,5±0,1°C в ходе всех экспериментальных процедур. Нижнюю часть тела животного помещают в воздухонепроницаемую камеру. Указанный подход позволяет установить гипобарическое давление с помощью регулируемого электронного вакуумного насоса, за счет чего достигается снижение артериального давления у животного (венозное депо) контролируемым образом («гипобарическая гипотензия»). По прошествии 30-минутного периода стабилизации после хирургической процедуры инициируют временную глобальную ишемию головного мозга путем затягивания нейлоновой нитки, присоединенной к грузу определенного веса, вокруг правой сонной артерии для окклюзии сосуда (артериальное давление измеряют катетером, помещенным в левую сонную артерию); среднее артериальное давление (MABP) сразу снижается до 35 мм Hg при использовании методики индукции гипобарической гипотензии. Ишемию мозга подтверждают при проведении непрерывных измерений местного мозгового кровотока (ICBF), регионального мозгового кровотока (rCBF) и ЭЭГ. После 15 минут общей мозговой ишемии нитку разрезают, прекращают вакуумный отсос, позволяя реперфузию. После 90 минут восстановительного периода удаляют катетер из ССА, зашивают разрезы и животных отключают от системы вентиляции. При экстубации (приблизительно после 110 минут реперфузии) при наличии стабильных признаков жизни крыс возвращают в клетки и используют нагретое одеяло для предупреждения потери тепла.
Множественная ПЦР
РНК выделяют в соответствии со стандартными процедурами (Chomczynski and Sacchi (1987), Anal Biochem, 162, 156-159), с последующей очисткой при использовании мини-набора QiagenRNeasyTM. Образцы тканей отбирают из объединенного мозга крыс после общей мозговой ишемии как на ипси-, так и на контралатеральных сторонах относительно локализации повреждения в различные временные точки после очаговой мозговой ишемии соответственно. Синтез кДНК проводят на основе 10 мкг суммарной РНК с использованием олигоdT праймеров, обратной транскриптазы Superscript II (Gibco) в стандартных условиях. Количественную ПЦР проводят в системе ЛайтЦиклер (LightCycler®) (Roche Diagnostics, Mannheim, Германия) окрашивании двуцепочечной ДНК красителем SYBR-зеленый. Циклическую обработку проводят по следующей схеме: GCSFR: 5 минут - 95°С, 5 секунд - 95°С, 10 секунд - 66°С, 30 секунд - 72°С, 10 секунд - 84°С, в течение 50 циклов; GCSF: 5 минут - 95°С, 5 секунд - 95°С, 10 секунд - 64°С, 30 секунд - 72°С, 10 секунд - 88°С, в течение 50 циклов. Кривые плавления строят по следующим параметрам: 95°С, охлаждение до 50°С; подъем до 99°С со скоростью 0,2°С в секунду. Используют следующие праймерные пары: "крысиный GCSFR-frag-32a" CCA TTG TCC ATC TTG GGG ATC (SEQ ID NO: 42), "крысиный GCSFR-frag-265as" CCT GGA AGC TGT TGT TCC ATG (SEQ ID NO: 43), "крысиный GCSF-345s" CAC AGC GGG CTC TTC CTC TAC CAA (SEQ ID NO: 44) и "крысиный GCSF-862as" AGC AGC GGC AGG AAT CAA TAC TCG (SEQ ID NO: 45).
ПЦР в системе ЛайтЦиклер (LightCycler®) проводят с использованием основной смеси, включающей краситель SYBR-зеленый, и далее в соответствии с инструкциями производителя (Roche Diagnostics). Специфичность продукта гарантируют анализом точки плавления и электрофоретическим разделением в агарозном геле. Содержание кДНК в образцах нормализуют до уровня экспрессии циклофилина (праймеры: "cyc5" ACC CCA CCG TGT TCT TCG AC (SEQ ID NO: 46); "acyc300" CATTTGCCATGGACAAGATG (SEQ ID NO: 47). Определяют относительные уровни регуляции после нормализации до циклофилина и при сравнении с животными из группы с ложной операцией. Стрелки ошибок указывают на стандартные отклонения, которые вычисляют на основе трехкратного анализа серийно разбавленных образцов кДНК, и отражают достоверность измерений.
На фиг.28А (часть I и II) показана очень сильная позитивная регуляция GCSF через 2 и 6 часов после очаговой ишемии на ипси- и контралатеральной стороне, тогда как рецептор подвергается умеренной регуляции (фиг.28 B) через 6 часов.
Индуцированная экспрессия GCSF не специфична для модели МСАО, но может также отмечаться, хотя и в меньшей степени, при общей ишемии (фиг.28С). Данное наблюдение поддерживает гипотезу, согласно которой GCSF является эндогенным для мозга нейропротективным лигандом, так что в лечении GCSF или GMCSF фактически используется эндогенный принцип нейропротекции.
Пример 24. GCSF и его рецептор подвергаются позитивной регуляции в полутеневой зоне кортикального инфаркта
Методы
Ишемическую модель фототромбической кортикальной ишемии осуществляют по методике, описанной в приведенных выше примерах.
Иммуногистохимия: Срезы погруженных в парафин тканей (2 мкм) подвергают депарафинизации и микроволновой обработке (цитратный буфер при 500 Вт, в течение 10 минут). Далее срезы инкубируют при комнатной температуре с антисывороткой к GCSF или рецептору GCSF (SC13102 или SC694; Santa Cruz Biotechnology, Santa Cruz, СА, США, 1:500) в течение 1 часа в увлажненной камере. Окрашивание визуализируют с использованием методики ALS и красителя ДАБ в качестве хромогена (DAKO, Glostrup, Дания). Отрицательные контроли готовят без добавления первичной антисыворотки.
Результаты
Чрез 6 часов после индукции фототромботической ишемии отмечаются явные признаки усиления окрашивания нейронов в окружающей зоне вокруг первичного ишемического поражения ("полутеневая зона"), как показано стрелками на фиг.29, и для рецептора, и для лиганда. Данное наблюдение подчеркивает обнаруженный авторами принцип, связанный с наличием новой эндогенной нейропротективной системы защиты нервной системы. Оно также подчеркивает тот факт, что первичный механизм действия GCSF в полутеневой зоне действительно больше всего напоминает антиапоптозный механизм.
Пример 25. Совместная локализация GCSF и doublecortin в гиппокампе
Использованные иммуногистохимические методы
Данный эксперимент проводят по методике примера 21. Вместо антисыворотки к GCSF используют поликлональное антитело против doublecortin из морской свинки (DCX) (Chemicon International, Германия), разбавленное в пропорции 1:200 0,2% БСА в 1 х TBS (pH 7,4). Срезы после промывки 3 раза по 2 минуты 0,2% БСА в 1 х TBS (pH 7,4) инкубируют в течение 30 минут с биотинилированным козьим IgG против антигенов морской свинки (Vector, Laboratories, США) в разбавлении 1:200 в 1 х TBS (pH 7,4), содержащем 0,2% БСА. И вновь после промывки срезов добавляют TRITC-конъюгированный стрептавидин (dianova; 1:200) на 1,5 часа при комнатной температуре с последующим проведением процедуры, описанной в примере 21.
Результаты
На фиг.30 показана экспрессия рецептора GCSF (a, d, g) и doublecortin (b, e, h) на нейронах в зубчатой извилине (a-f) и в воротах (g-I) гиппокампа. Перекрытие экспрессии рецептора GCSF и раннего нейронального маркера doublecortin (Jin, et al. (2001), Proc Natl Acad Sci USA, 98, 4710-5) (фиг.30 c, f, i) указывает на функциональную роль GCSF на ранней стадии нейрональной дифференциации.
Пример 26. GCSF индуцирует нейрональную дифференциацию зрелых нейрональных стволовых клеток
Получение нейрональных стволовых клеток (NSC)
Получение зрелых нейрональных стволовых клеток крысы осуществляют по методике примера 9.
39 культивируемых в DIV нейросфер, полученных из субвентрикулярной зоны (SVZ), однократно стимулировали следующими концентрациями GCSF: 10 нг/мл, 100 нг/мл и 500 нг/мл соответственно. Через 4 суток после добавления рекомбинантного человеческого GCSF (Neupogen®, Amgen, Europe B.V., Нидерланды) клетки собирали и из них выделяли РНК. Необработанные клетки служили в качестве контроля.
Количественная ПЦР
РНК из обработанных GCSF и необработанных нейросфер из SVZ выделяли с использованием мини-набора Qiagen RNeasyTM по процедуре производителя. Синтез кДНК проводят на основе 2 мкг суммарной РНК с помощью олигоdT праймеров, обратной транскриптазы Superscript II (Gibco) в стандартных условиях. Количественную ПЦР проводят в системе ЛайтЦиклер (LightCycler®) (Roche Diagnostics, Mannheim, Германия) при окрашивании двуцепочечной ДНК красителем SYBR-зеленый. Циклическую обработку проводят в следующих условиях:
Нестин и НСЭ (NSE) (нейрон-специфичная энолаза): 3 минуты - 95°С, 5 секунд - 95°С, 10 секунд - 58°С, 30 секунд - 72°С, 10 секунд - 81°С, в течение 50 циклов; бета III-тубулин: 3 минуты - 95°С, 5 секунд - 95°С, 10 секунд - 65°С, 30 секунд - 72°С, 10 секунд - 87°С, в течение 50 циклов; PLP: 3 минуты - 95°С, 5 секунд - 95°С, 10 секунд - 62°С, 30 секунд - 72°С, 10 секунд - 84°С, в течение 50 циклов; GFAP: 3 минуты - 95°С, 5 секунд - 95°С, 10 секунд - 60°С, 30 секунд -72°С, 10 секунд - 81°С, в течение 50 циклов. Кривые плавления строят по следующим параметрам: 95°С, охлаждение до 50°С; подъем до 99°С со скоростью 0,2°С в секунду. Используют следующие праймерные пары: "крысиный нестин-плюс" AGG AAG AAG CTG CAG GAG AG (SEQ ID NO: 48), "крысиный нестин-минус" TTC ACC TGC TTG GGC TCT AT (SEQ ID NO: 49), "крысиная NSE-плюс " GGC AAG GAT GCC ACT AAT GT (SEQ ID NO: 50), "крысиная NSE-минус" AGG GTC AGC AGG AGA CTT GA (SEQ ID NO: 51), "крысиный бета III-tub-716s" CCA CCT ACG GGG ACC TCA ACC AC (SEQ ID NO: 52), "крысиный бета III-tub-1022as" GAC ATG CGC CCA CGG AAG ACG (SEQ ID NO: 53), "крысиный PLP-518s" TCA TTC TTT GGA GCG GGT GTG (SEQ ID NO: 54), "крысиный PLP-927as" TAA GGA CGG CAA AGT TGT AAG TGG (SEQ ID NO: 55), "крысиный GFAP3'-1123s" ССТ ТТС ТТА TGC ATG TAC GGA G (SEQ ID NO: 56), "крысиный GFAP3'-1245as" GTA CAC TAA TAC GAA GGC ACT C (SEQ ID NO: 57). ПЦР в системе ЛайтЦиклер (LightCycler®) проводят с использованием основной смеси, включающей краситель (SYBR-зеленый) и далее в соответствии с рекомендациями производителя (Roche Diagnostics). Специфичность продукта гарантируют анализом точки плавления и электрофоретическим разделением в агарозном геле. Содержание кДНК в образцах нормализуют до уровня экспрессии циклофилина (праймеры: "cyc5" ACC CCA CCG TGT TCT TCG AC (SEQ ID NO: 58); "acyc300" CAT TTG CCA TGG ACA AGA TG (SEQ ID NO: 59)). Относительные уровни регуляции получают после нормализации до уровня циклофилина и при сравнении с необработанными клетками. На фиг.31 показана сильная, зависимая от концентрации позитивная регуляция нейрональных маркеров НСЭ (NSE) и бета III-тубулина через 4 суток после обработки GCSF. PLP и GFAP подвергаются умеренной регуляции, зависимой от концентрации GCSF. Стрелки ошибок указывают на стандартные отклонения, которые вычисляют на основе результатов трехкратного анализа серийно разбавленных образцов кДНК, и отражают достоверность измерений.
Пример 27. Совместная локализация GMCSF и его рецептора в мозге
Иммуногистохимические методы
Срезы погруженных в парафин тканей (2 мкм) подвергают депарафинизации путем обработки их 2 раза по 5 минут ксилолом, 2 раза по 2 минуты 100% этанолом и затем снижающимися концентрациями этанола от 96% до 70%. В конце процедуры срезы промывают дистиллированной водой и подвергают микроволновой обработке (цитратный буфер при 600 Вт в течение 15 минут). После этого срезы промывают дистиллированной водой и антигены блокируют при обработке 3 раза по 5 минут в 1 х TBS (pH 7,4), содержащем 0,2% БСА. Антисыворотку к рецептору GMCSF (SC690; Santa Cruz Biotechnology, Santa Cruz, CA, США; 1:100), разбавленную в 1 x TBS (pH 7,4), содержащем 0,2% БСА, инкубируют при комнатной температуре в течение 1 часа в увлажненной камере. Затем срезы промывают 3 раза по 2 минуты в 1 х TBS (pH 7,4), содержащем 0,2% БСА. Затем добавляют второе антитело (козий Fab-FITC против антигена кролика, dianova, 1:50) и выдерживают в течение ночи при 4°С в 1 х TBS (pH 7,4), содержащем 0,2% БСА. После промывки срезов 3 раза по 2 минуты 0,2% БСА в 1 х TBS (pH 7,4) вносят антисыворотку к GMCSF (SC13101; Santa Cruz Biotechnology, Santa Cruz, CA, США), разбавленную в пропорции 1:100 0,2% БСА в 1 х TBS (pH 7,4), на 1 час при комнатной температуре. Срезы промывают 3 раза, как было описано ранее, и инкубируют в течение 30 минут с биотинилированным козьим IgG против антигена кролика (Vector, Laboratories, США), разбавленным в пропорции 1: 200 с использованием 1 x TBS (pH 7,4), содержащего 0,2% БСА. И вновь после промывания срезов вносят TRITC-конъюгированный стрептавидин (dianova; 1:200) на 1,5 часа при комнатной температуре. Далее срезы промывают 3 раза по 2 минуты в 1 х TBS и окрашивают в течение 10 минут ядерным красителем DAPI, разбавленным в пропорции 1:10000 в 1 x TBS. В конце процедуры срезы промывают 3 раза по 2 минуты 1 х TBS и вносят в среду для измерения флуоресценции (Vectashield, Vector Laboratories, США). Фотографии получают на цифровом аппарате с использованием микроскопа Olympus IX81 и проводят анализ с пакетом прикладных программ "Analysis" (Soft Imaging Systems, Stuttgart, Германия).
Все эксперименты с двойной оценкой флуоресценции контролируют параллельным однократным окрашиванием, которое используют для подтверждения отсутствия флуоресцирующего проводника во втором канале. В качестве второго контроля все процедуры окрашивания при двукратной флуоресценции проводят при переключении хромофоров на второе антитело.
Результаты
Рецептор GMCSF (фиг.32a, d, g) и GMCSF (фиг.32b, e, h) локализованы вместе на нейронах в зубчатой извилине (фиг.32a-c), в воротах (фиг.32d-f) и в коре головного мозга (фиг.32g-i). Приведенные данные подтверждают представление о том, что GMCSF представляет собой аутокринный лиганд.
Пример 28. GMCSF и его рецептор подвергаются позитивной регуляции при церебральной ишемии
МСАО как модель церебральной ишемии
Процедуру индукции очаговой мозговой ишемии (МСАО, окклюзия средней мозговой артерии) проводят по методике, описанной в примере 1.
Модель общей ишемии
Подготовку к хирургической операции и индукцию временной общей мозговой ишемии проводят по описанной ранее методике.
Количественная ПРЦ
Выделение РНК и синтез кДНК проводят по ранее описанной методике. Циклическую обработку проводят в следующих условиях: GMCSFR: 5 минут - 95°С, 5 секунд - 95°С, 10 секунд - 62°С, 30 секунд - 72°С, 10 секунд - 80°С, в течение 50 циклов; GMCSF: 5 минут - 95°С, 5 секунд - 95°С, 10 секунд - 60°С, 30 секунд - 72°С, 10 секунд - 81°С, в течение 50 циклов. Кривые плавления строят по следующим параметрам: 95°С, охлаждение до 50°С; подъем до 99°С со скоростью 0,2°С в секунду. Используют следующие праймерные пары: крысиный GMCS "BR4-4s96" ACG TCG TTG GCT CAG TTA TGT C (SEQ ID NO: 60), "BR4-4as272" ATT TAT GTC AGA GAT GGA GGA TGG (SEQ ID NO: 61), "крысиный GMCSF-723s" GGA GCT CTA AGC TTC TAG ATC (SEQ ID NO: 62) и "крысиный GMCSF-908as" GGC TCA ATG TGA TTT CTT GGG (SEQ ID NO: 63).
ПЦР в системе ЛайтЦиклер (LightCycler®) проводят с использованием основной смеси, включающей краситель SYBR-зеленый, и далее в соответствии с рекомендациями производителя (Roche Diagnostics). Специфичность продукта гарантируют анализом точки плавления и электрофоретическим разделением в агарозном геле. Содержание кДНК в образцах нормализуют до уровня экспрессии циклофилина (праймеры: "cyc5" ACC CCA CCG TGT TCT TCG AC (SEQ ID NO: 58); "acyc300" CAT TTG CCA TGG ACA AGA TG (SEQ ID NO: 59)). Относительные уровни регуляции получают после нормализации до уровня циклофилина и при сравнении с животными из группы с ложной операцией. Стрелки ошибок указывают на стандартные отклонения, которые вычисляют на основе результатов трехкратного анализа серийно разбавленных образцов кДНК, и отражают достоверность результатов измерения.
Очень сильная позитивная регуляция GMCSF через 2 часа и 6 часов после очаговой ишемии на ипси- и контралатеральной стороне показана на фиг.33А. Индукция GMCSF может быть продемонстрирована также при общей ишемии, хотя и в меньшей степени (фиг.33 В). Даже рецептор GMCSF подвергается незначительной позитивной регуляции через 6 часов после общей ишемии (фиг.33 С). Указанные данные поддерживают гипотезу о том, что GMCSF представляет собой часть эндогенного нейропротективного механизма.
Пример 29. Совместная локализация экспрессии GCSFR doublecortin в гиппокампе
Использованные иммуногистохимческие методы
Эксперимент проводят по описанной ранее методике. Вместо антисыворотки к GMCSF вносят поликлональное антитело из морской свинки против doublecortin (DCX) (Chemicon International, Германия), разбавленное 1:200 в 0,2% БСА в 1 х TBS (pH 7,4). Срезы после промывки 3 раза по 2 минуты 0,2% БСА в 1 х TBS (pH 7,4) инкубируют в течение 30 минут с биотинилированным козьим IgG против антигенов морской свинки (Vector, Laboratories, США) в разбавлении 1:200 в 1 х TBS (pH 7,4), содержащем 0,2% БСА. И вновь после промывки среза вносят TRITC-конъюгированный стрептавидин (dianova; 1:200) на 1,5 часа при комнатной температуре по описанной выше процедуре.
Результаты
На фиг.34 показана экспрессия рецептора GMCSF (a, d, g) и doublecortin (b, e, h) на нейронах в зубчатой извилине (a-с) и в воротах (d-i) в гиппокампе. Перекрывание экспрессии рецептора GMCSF и раннего нейронального маркера doublecortin (Jin, Minami, Lan, Mao, Batteur, Simon and Greenberg (2001), Proc Natl Acad Sci USA, 98, 4710-5) (фиг.34 c, f, i) указывает на функциональную роль GMCSF на ранней стадии нейрональной дифференциации и подчеркивают применимость GMCSF в случае всех нейродегенеративных заболеваний, при которых воздействие на нейрогенез может быть важной терапевтической мишенью, таких как инсульт, болезнь Паркинсона, ALS и многие другие.
Пример 30. GMCSF индуцирует дифференциацию зрелых нейрональных стволовых клеток
Получение нейрональных стволовых клеток
Зрелые нейрональные стволовые клетки крыс получают по методике, описанной в примере 9. Клетки подвергают пассажу каждые две-три недели, после чего клетки центрифугируют в течение 5 минут при 800 х g, удаляют супернатант и промывают один раз 1 х ФБР. После ресуспендирования в 1 мл аккутазы (Sigma) и последующей инкубации при 37°С в течение 15 минут клетки подсчитывают и помещают с плотностью 1,5 - 2,0 х 105/ячейку в 6-ячеечный планшет. Нейрональные стволовые клетки, полученные из гиппокампа, стимулируют после 4-го пассажа путем добавления 10 нг/мл GMCSF (Leukine®, Berlex, Schering AG Германия) и собирают через 3 суток для выделения ДНК. Необработанные клетки используются в качестве контроля.
Пример 30. Количественная ПЦР
РНК из GMCSF-обработанных и необработанных нейрональных стволовых клеток SVZ выделяют с использованием мини-набора Qiagen RNeasyTM по процедуре производителя. Синтез кДНК проводят на основе 2 мкг суммарной РНК с помощью олигоdT праймеров, обратной транскриптазы Superscript II (Gibco) в стандартных условиях. Количественную ПЦР проводят в системе ЛайтЦиклер (LightCycler®) (Roche Diagnostics, Mannheim, Германия) при окрашивании двуцепочечной ДНК красителем SYBR-зеленый. Проведение ПЦР в системе ЛайтЦиклер (LightCycler®), условия циклической обработки и праймерные пары, использованные для бета III-тубулина, НСЭ (NSE), PLP и GFAP, описаны ранее. Относительный уровень регуляции получают при нормализации до уровня циклофилина и при сравнении с необработанными клетками. На фиг.35А схематически показан потенциал дифференциации нейрональных стволовых клеток и специфических маркеров экспрессии дифференцированных клеток. Обработка нейрональных стволовых клеток 10 нг/мл GMCSF приводит к значительной индукции экспрессии бета III-тубулина (n=3; p<0,05, двуфакторный t-тест), в меньшей степени НСЭ (NSE), представляющей собой маркер зрелых нейронов (фиг.35 В). Не отмечается изменений в уровне экспрессии PLP и GFAP.
Данный пример подчеркивает применимость GMCSF для модуляции нейрогенеза. Указанная обработка полезна как для генерирования дифференцированных нейронов in vitro, так и в плане воздействия на эндогенные стволовые клетки и применима при широком перечне заболеваний человека, в особенности нейродегенеративных заболеваний.
Пример 31. GCSF проходит через гематоэнцефалический барьер (ГЭБ)
Большая часть доступных в литературе данных содержит информацию о сывороточных уровнях GCSF и родственных цитокинов. Одним из важнейших параметров для эффективного использования GCSF в лечении неврологических заболеваний является прохождение белка через гематоэнцефалический барьер (ГЭБ). Данный вопрос представляет особый интерес, поскольку известно, что многие лекарственные компоненты белковой природы не проходят через природный барьер между кровью и нейронами. С другой стороны, в отношении ряда белков, включая гематопоэтические факторы типа эритропоэтина и фактора, стимулирующего образование колоний гранулоцитов и макрофагов (GMCSF), показано, что они могут проходить через ГЭБ и эффективно достигать нейронов в ЦНС (McLay, et al. (1997), Brain, 120, 2083-91); Brines, et al. (2000), Proc. Natl. Acad. Sci. USA, 97, 10526-31).
Для доказательства того, что GCSF может проходить через гематоэнцефалический барьер, авторы исследовали появление инъецированного GCSF в мозге крысы. С этой целью был проведен чрезвычайно чувствительный тест по методу иодирования. GCSF и БСА в качестве контроля подвергают радиоактивному мечению 131I (Amersham Biosciences, Freiburg, Германия) иодогеном (Iodogen) (Sigma, Taufkirchen, Германия) и очищают на колонках с сефадексом G-25 (Amersham Biosciences, Freiburg, Германия). Очищенные белки элюируют немечеными стандартными соединениями в виде одного пика с нужным молекулярным весом, по данным анализа методом вытеснительной хроматографии. Радиоактивно меченные белки инъецируют через хвостовую вену самкам крыс Спрэг-Доли (Sprague-Dawley) (250-300 г). Для исследования конкурентного поглощения охлажденные белки смешивают с немеченым соединением и совместно инъецируют. Непосредственно перед вскрытием крыс подвергают анестезии ромпуном/кетанестом (Rompun/Ketanest) и проводят перфузию 100 мл солевого раствора через нижнюю брюшную аорту (через 1, 4 и 24 часов после инъекции) для удаления крови. Полный мозг и кровь извлекают, промакивают досуха и взвешивают. Кровь центрифугируют с получением сыворотки. После инъекции измеряют радиоактивность на γ-счетчике (LB 951G, Berthold, Германия) по всему образцу для вычисления показателя %ID/г ткани. Результаты показывают, что GCSF присутствует в мозге и в сравнении с альбумином прохождение через гематоэнцефалический барьер повышается в 4-5 раз с течением времени. Образцы, которые были взяты через 24 часа после инъекции радиоактивно меченного GCSF, демонстрируют повышенный уровень в сравнении с результатами при отборе через 1 и 4 часа (фиг.36). Данные наблюдения показывают, что GCSF, инъецированный внутривенно, в крови пересекает ГЭБ и может, в принципе, активировать специфические рецепторы на нейронах ЦНС.
Количество радиоактивно меченного GCSF в сыворотке и мозге измеряют, строят график зависимости коэффициента соотношения уровня в мозге/сыворотке в зависимости от времени. В качестве контроля используют бычий сывороточный альбумин. Для предупреждения загрязнения мозговой ткани кровью перед извлечением крыс подвергают перфузии 100 мл солевого раствора.
Пример 32. GMCSF проходит через гематоэнцефалический барьер (ГЭБ)
Для демонстрации того, что GMCSF может проходить через гематоэнцефалический барьер, авторы инъецируют крысам иодированный GMCSF по методике примера 31, описанной для GCSF. Проведенный эксперимент дал аналогичные результаты, а сравнение уровня GMCSF в головном мозге крысы после внутривенного введения показывает, что уровни GMCSF (соотношение уровня в мозге и в сыворотке) в 3-4 раза выше, чем в случае альбумина. Приведенные данные показывают, что GMCSF может проходить через гематоэнцефалический барьер (фиг.37).
На фиг.37 измеряют количество радиоактивно меченного GMCSF в сыворотке и строят график зависимости соотношения уровня в мозге/сыворотке от времени. В качестве контроля используют бычий сывороточный альбумин. Для предупреждения загрязнения мозговой ткани кровью крыс перед вскрытием подвергают перфузии 100 мл солевого раствора.
Пример 33. Применение GCSF для лечения бокового амиотрофического склероза (ALS)
Идея о том, что GCSF и GMCSF могут оказывать благоприятный эффект при лечении ALS имеет два обоснования. Во-первых, оба: и рецептор, и лиганд GCSF и GMCSF экспрессируются в крупных двигательных нейронах в переднем отростке спинного мозга. Во-вторых, доказательство того, что активность GCSF частично реализуется через противодействие апоптозным каскадам, является весьма привлекательным в свете данных о том, что апоптозные механизмы работают при ALS (Li, et al. (2000), Science, 288, 335-9). Самой распространенной моделью ALS на мышах являются трансгенные мыши, несущие мутацию в гене SOD1, который, как было показано, ответственен за наследственный ALS у человека. Наиболее часто используемой при этом является трансгенная линия SOD1 (G93A). В типичном случае данные мыши характеризуются сниженной продолжительностью жизни и демонстрируют прогрессирующие признаки слабости моторной функции, которые могут быть легко оценены бихевиоральными тестами (например, в рамках теста на силу хватательного рефлекса, теста на вращающемся стержне (ротароде)). На данных мышах было проведено множество тестов, важных для оценки терапевтической эффективности, и продемонстрирована активность в плане продления жизни таких веществ, как рилузол, миноциклин, карнитин (Riluzole, Minocycline, Carnitine) и другие. Считается, что данные мыши обладают клинически предсказуемой релевантностью, поскольку рилузол как единственное разрешенное для применения людьми лекарственное средство указанного типа оказывает защитное действие на данных мышей, тогда как BDNF, в отношении которого не было получено нужных результатов в испытаниях на людях, был также неэффективным. В этой связи авторы решили выяснить, увеличивает ли GCSF ожидаемую продолжительность жизни или функциональные последствия у SOD1-трансгенных мышей, как модели ALS. Мышей SOD1, как и мышей дикого типа, подвергают подкожной инъекции с использованием 10 мкг/кг веса тела GCSF или носителя, начиная с 60 суток постнатального периода и осматривают в течение всего времени. Результаты указывают на явную тенденцию к увеличению ожидаемой продолжительности жизни животных в группе введения GMCSF (фиг.38А), а также улучшению двигательной функции (тест на силу хватательного рефлекса (фиг.38В)), которые достигают статистической значимости, по данным измерений в нескольких временных точках. Кроме того, отмечается тенденция к увеличению веса животных в испытуемой группе.
Фиг.38А: Мышам SOD1-tg инъецируют 10 мкг/кг веса тела, начиная с 60 суток после рождения. Отмечается четкая тенденция к увеличению продолжительности жизни у мышей SOD1-tg в группе введения GCSF (сплошные линии против прерывистой линии). В нескольких временных точках данное различие достигает статистической значимости.
Фиг.38В: Мышам SOD1-tg инъецируют 10 мкг/кг веса тела, начиная с 60 суток после рождения. Тест на определение силы хватательного рефлекса демонстрирует улучшение двигательной силы у мышей SOD1-tg после введения GCSF (открытые квадраты против открытых треугольников). В нескольких временных точках данные различия достигают статистической значимости.
Пример 34: Эффективность GCSF на моделях грызунов при болезни Паркинсона (PD)
Модель MPTP
Хорошо описанная модель болезни Паркинсона (PD) была разработана с использованием нейротоксина: 1-метил-4-фенил-1,2,3,6-тетрагидропиридина (MPTP). Для исследования эффективности GCSF на модели болезни Паркинсона авторы вводят MPTP 8-недельным самцам мышей. Мышам в каждой группе (n=15) делают повторную в/б инъекцию MPTP-HCl или солевого раствора (один раз в сутки в течение 5 последовательных суток в концентрации 30 мг/кг, 5 мл/кг) и повторно п/к вводят (один раз в сутки в течение 22 последовательных суток) буфер, GCSF (0,03 мг/кг, 5 мл/кг) или миноциклин (45 мг/кг, 5 мл/кг). При первом применении GCSF вводят сразу после MPTP (или солевого раствора в группе 0), тогда как моноциклин вводят через 30 минут в связи с возможными взаимодействиями обоих соединений. Всех животных в каждой из групп умерщвляют на 22 сутки. До указанного момента мышей анализируют на локомоторную активность (ускоренный тест на ротароде) и проводят ежедневное определение веса тела. Далее мозг каждого животного подвергают анализу методом ВЭЖХ с электрохимической оценкой определяемой концентрации допамина, 3,4-дигидроксифенилуксусной кислоты (DOPAC) и гомованилиновой кислоты (HVA) и в полосатом теле и в nucleus accumbens.
Однако наиболее важным и прямым параметром для оценки нейропротективного эффекта лекарственного средства в данной модели является количество выживших нейронов после воздействия токсина. В этой связи проводят иммуногистохимический анализ с тирозингидроксилазой (ТГ) и стереологическое количественное определение ТГ-позитивных нейронов в срезах среднего мозга каждого животного (Triarhou, et al., (1988), J Neurocytol, 17, 221-32).
Полученные результаты указывают на тенденцию к увеличению веса животных в группе введения GCSF (данные не приведены). Кроме того, наблюдается усиление локомоторной активности при тестировании по методу оценки поведения на ротароде (данные не приведены). Недавнее исследование, проведенное с MPTP-индуцированной болезнью Паркинсона на мышах, показало что миноциклин предотвращает допаминергическую нейродегенерацию полосатого тела в черном веществе у таких животных (Du, et al. (2001), Proc Natl Acad Sci USA, 98, 14669-74). В настоящем исследовании авторы отобрали миноциклин как стандартное нейропротективное соединение для оценки полученных результатов. Применение МРТР в подостром режиме в течение 5 последовательных суток существенно снижает уровень нейронов, позитивных по тирозингидролазе (ТГ) в pars compacta черного вещества (фиг.39 А). Обработка GCSF и миноциклином приводит почти к одинаковой эффективности в противодействии указанному снижению уровня ТГ-позитивных нейронов в черном веществе (фиг.39В). Кроме того, GCSF и миноциклин демонстрируют практически одинаковую терапевтическую эффективность, если учитывать уровни допамина в полосатом теле (фиг.39 В) и его метаболитов.
Пример 35. Эффективность GMCSF на модели болезни Паркинсона
Аналогично описанию для случая GCSF, эффективность GMCSF на модели болезни Паркинсона также оценивают в рамках соответствующего, приведенного ниже исследования (см. предыдущий пример).
Мышам в каждой из групп (n=15) проводят повторную в/б инъекцию MPTP-HCl или солевого раствора (один раз в сутки в течение 5 последовательных суток в концентрации 30 мг/кг, 5 мл/кг) и повторно вводят п/к (один раз в сутки в течение 22 последовательных суток) буфер, GMCSF (0,03 мг/кг, 5 мл/кг) или миноциклин (45 мг/кг, 5 мл/кг). При первом применении GCSF вводят сразу после MPTP (или солевого раствора в группе 0), тогда как миноциклин вводят через 30 минут в связи с возможными взаимодействиями обоих соединений. Всех животных в каждой из групп умерщвляют на 22 сутки.
Наблюдается улучшение в плане набора веса тела и локомоторной активности в тесте с ускоряющимся стержнем (ротародом) после применения GMCSF, аналогично по балльной оценке тому, что наблюдалось при использовании GCSF (данные не приведены). В этой связи терапевтическая эффективность GMCSF будет более сравнима с GCSF, если принимать во внимание уровни ТГ-позитивных нейронов в черном веществе (фиг.39 В). Снижение уровня допамина и его метаболитов в полосатом теле может рассматриваться как противодействие ситуации, развивающейся после введения GMCSF, что сравнимо с тем, что происходит при введении GMCSF (данные не приведены).
Пример 36. Эффективность GCSF и GMCSF по 6-гидроксидопамину (6OHDA) на модели болезни Паркинсона у крыс
Как приведено в указанном выше примере, GCSF, как и GMCSF обладают сильными нейропротективными свойствами на МРТР-модели мышей. Указанный факт является очень сильным аргументом в пользу применимости в случае болезни Паркинсона у человека, поскольку МРТР является токсином, который был обнаружен по его способности вызывать PD у людей.
Одной из дополнительных моделей для изучения эффективности гематопоэтических факторов применительно к болезни Паркинсона является 6-OHDA модель. Данная модель основана на инъекции 6-OHDA непосредственно в черное вещество или полосатое тело. Лекарственное средство селективно накапливается в допаминергических нейронах и ведет к апоптозу данных клеток. У крыс 6-OHDA является эффективным нейротоксином, который преимущественно используют для получения односторонних повреждений. Уровень истощения по допамину может быть оценен при оценке поведения животных в тесте на вращение в ответ на введение амфетамина или апоморфина (Ungerstedt 1971). Легкий и количественно хорошо выявляемый моторный дефицит составляет основное достоинство данной модели. Дополнительно к поведенческому параметру могут быть также определены иммуногистохимическим методом уровень в полосатом теле нейронов, позитивных по тирозингидроксилазе (ТГ), и уровень допамина и его метаболитов методом ВЭЖХ. 6-OHDA может использоваться для оценки эффективности GCSF и GMCSF на модели PD у крыс. Взрослые крысы Спрэг-Доли (Spraque-Dawley) (вес тела 250 г) имеют одностороннее повреждение после стереотаксической инъекции 8 мкг в объеме 2 мкл 6-OHDA в черное вещество или в полосатое тело. Различные дозы GMCSF (0,03 мг/кг, 0,1 мг/кг или другие) могут вводиться подкожно ежедневно сразу после нанесения повреждения в течение 2 недель. Другим группам экспериментальных животных вводят в полосатое тело или в черное вещество однократную дозу GCSF или GMCSF (300 мкг/кг) сразу после инъекции 6-OHDA. Как и в случае МРТР-модели, миноциклин может использоваться в качестве стандартного нейропротективного соединения (45 мг/кг один раз в сутки, п/к). Животные с ложной операцией и животные с повреждениями, которым вводят буфер, используются в качестве контрольной группы. Через две недели после нанесения повреждения животных подвергают бихевиоральному тесту при вращении. Крысам инъецируют п/к апоморфин, помещают в сферическую клетку и фиксируют число контралатеральных вращений в течение 1 часа. Количество вращений для каждого животного сравнивают методами стандартной статистической обработки результатов. После бихевиорального тестирования животных умерщвляют и головной мозг обрабатывают для определения общего числа ТГ-позитивных нейронов иммугистонохимическим методом и уровня допамина методом ВЭЖХ.
Пример 37. GMCSF действует антиапоптозно на нейроны за счет активации пути через stat3
Как уже было показано выше (пример 22), GCSF обладает мощной активностью по ингибированию апоптоза в первичных нейронах. Авторы полагают, что GMCSF может также обладать сильным антиапоптозным механизмом действия. В этой связи был проведен эксперимент, аналогичный описанному выше в примере 22, но с использованием GMCSF. В данной ситуации концентрация применительно к нейронам составляет 50 нг GMCSF (Leukine, Immunex)/мл среды.
На фиг.40, часть I, показано, что GMCSF ингибирует расщепление PARP, что является специфическим признаком апоптоза. Гибель клеток индуцируется в первичных нейронах при использовании NOR-3 как донора NO.
На фиг.40, часть II, показано, что применение GMCSF не приводит к повышению уровня фосфорилирования STAT1 ("pSTAT1") или STAT5 ("pSTAT5"), хотя сами белки экспрессируются в нейронах.
На фиг.40, часть IIIA, показано, что применение GMCSF приводит к зависимой от времени активации STAT3, которая является максимальной к 5 минуте после стимулирования GMCSF. К 60 минуте уровень STAT3 падает ниже исходного уровня, что представляет известную для клеток гематопоэтической системы кинетику ответной реакции. B: количественное определение в трех независимых экспериментах. С: активация STAT3 за счет фосфорилирования может быть подавлена ингибитором JAK2 - AG490. Через 5 минут после внесения GMCSF уровень STAT3 будет совершенно другим в присутствии ингибитора.
На фиг.40, часть IV, показано, что GMCSF сильно индуцирует экспрессию целевых генов для stat3: Bcl2 и BclXI. Указанные гены известны как антиапоптозные.
Данный эксперимент показывает, что: 1) GMCSF действует антиапоптозно на нейроны; 2) Указанное действие опосредовано механизмом, реализуемым через stat3-путь. В этой связи показана также возможность использования stat3 в нейронах как инструмент скрининга новых нейропротективных лекарственных соединений, включая новые миметики GCSF или GMCSF.
Пример 38. GMCSF снижает объем инфаркта на моделях инсульта у грызунов
Авторы провели эксперименты с GMCSF на основе самой известной и распространенной модели для изучения инсульта у крыс: на модели окклюзии средней мозговой артерии (МСАО).
В принципе, эксперимент проводят по методике, описанной в примерах 1 и 19. В общих чертах процедура состоит в том, что самцов крыс Вистар (Wistar) подвергают анестезии методом ингаляции (галотан/N2O/O2). Обнажают сонную артерию, в общую сонную артерию вставляют покрытую нейлоном нить и затягивают до блокирования кровотока через MCA. Правильное положение нити подтверждают лазерной допплер-флуометрией. Через 90 минут после окклюзии нить убирают и проводят реперфузию. GMCSF (Leukine, Immunex) вводят крысам в дозе 250 мкг/кг веса тела через внутривенный катетер (бедренная вена) в течение примерно 20 минут с помощью инфузионного насоса через 30 минут или 3 часа после начала ишемии. Объемы инфарктной зоны определяют через 24 часа по стандартной методике ТТС-окрашивания и с помощью компьютерной волюметрии.
На фиг.41 показано, что наблюдается значительно снижение инфарктного объема у животных после введения GMCSF как при ранней оценке (фиг.41, часть I), так и в случае более позднего временного интервала в 3 часа (фиг.41, часть II).
Полученные результаты иллюстрируют применимость GMCSF в случае нейродегенеративных заболеваний в целом, и в частности, для лечения инсульта.
Все цитированные в описании материалы, в том числе патенты, патентные заявки и публикации, включены в настоящее описание полностью в качестве ссылок.
Вполне очевидно, что возможны различные модификации и вариации настоящего изобретения в свете приведенного описания. В этой связи следует понимать, что в рамках тематики, определяемой прилагаемой формулой изобретения, настоящее изобретение может осуществляться иначе, нежели это конкретно описано в данной заявке.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ ГАМКергических НЕЙРОНОВ in vitro ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ НЕВРОЛОГИЧЕСКИХ РАССТРОЙСТВ | 2004 |
|
RU2380410C2 |
| НОВОЕ ПРИМЕНЕНИЕ | 2004 |
|
RU2416426C2 |
| ИНГИБИТОРЫ ПЕРЕДАЧИ СИГНАЛА ТРАНСФОРМИРУЮЩИХ ФАКТОРОВ РОСТА (TGF-R) ДЛЯ ЛЕЧЕНИЯ РАССТРОЙСТВ ЦНС | 2005 |
|
RU2385933C2 |
| ПОЛУЧЕНИЕ ТЕРМИНАЛЬНО ДИФФЕРЕНЦИРОВАННЫХ ДОФАМИНЕРГИЧЕСКИХ НЕЙРОНОВ ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2004 |
|
RU2345133C2 |
| ПРИМЕНЕНИЕ СТВОЛОВЫХ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ МЕЗЕНХИМАЛЬНЫЕ И НЕЙРОНАЛЬНЫЕ МАРКЕРЫ, И ИХ КОМПОЗИЦИЙ ДЛЯ ЛЕЧЕНИЯ НЕВРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ (ВАРИАНТЫ) | 2017 |
|
RU2742828C2 |
| ЛЕЧЕНИЕ НЕВРОЛОГИЧЕСКОЙ ДИСФУНКЦИИ С ПРИМЕНЕНИЕМ СУЛЬФАМАТОВ ФРУКТОПИРАНОЗЫ И ЭРИТРОПОЭТИНА | 2002 |
|
RU2317086C2 |
| СТВОЛОВЫЕ КЛЕТКИ, ЭКСПРЕССИРУЮЩИЕ МЕЗЕНХИМАЛЬНЫЕ И НЕЙРОНАЛЬНЫЕ МАРКЕРЫ, ИХ КОМПОЗИЦИИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2017 |
|
RU2741839C2 |
| КОМПОЗИЦИЯ ДЛЯ НЕЙРОПРОТЕКЦИИ И СТИМУЛЯЦИИ НЕЙРОРЕГЕНЕРАЦИИ ГОЛОВНОГО МОЗГА ПОСЛЕ ПОВРЕЖДЕНИЯ, СРЕДСТВО НА ЕЕ ОСНОВЕ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2022 |
|
RU2803286C1 |
| ТРАНСПЛАНТАЦИЯ НЕРВНЫХ КЛЕТОК ДЛЯ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ СОСТОЯНИЙ | 2005 |
|
RU2434636C2 |
| Способ получения глиальных производных индуцированных плюрипотентных стволовых клеток с повышенной экспрессией BDNF для терапии ишемического инсульта | 2023 |
|
RU2829361C1 |
Изобретение относится к области медицины и касается способов лечения неврологических состояний с применением введения гематопоэтических факторов роста. Сущность изобретения включает G-CSF или его функционально активный вариант, который на 90% идентичен последовательности G-CSF человека, представленной в SEQ ID NO: 28 для применения в способе лечения амиотрофического бокового склероза. Преимущество изобретения заключается в создании нейропротекции вовлеченных в заболевание нейронных клеток. 1 з.п. ф-лы, 1 табл. 41 ил.
1. G-CSF или его функционально активный вариант, который на 90% идентичен последовательности G-CSF человека, представленной как SEQ ID NO: 28, для применения в способе лечения амиотрофического бокового склероза.
2. G-CSF по п.1, где G-CSF представляет собой G-CSF человека.
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| US 20020151488 A1, 17.10.2002 | |||
| DE 10033219 A, 24.01.2002 | |||
| KONISHI Y | |||
| et al | |||
| Trophic effect of erythropoietin and other hematopoietic factors on central cholinergic neurons in vitro and in vivo, Brain Res, 1993 Apr 23; 609 (1-2), 29-35. | |||

