Область техники, к которой относится изобретение
Изобретение относится к молекулярной биологии, биохимии, биоорганической химии, биотехнологии, фармакологии и охране окружающей среды и предоставляет биологический микрочип (биочип), предназначенный для анализа взаимодействия РНК с РНК-связывающимися молекулами, представляющий собой подложку, содержащую иммобилизованные на ней олигорибонуклеотиды, характеризующийся тем, что по одному, нескольким или всем 2'-O- положениям олигорибонуклеотидов присоединены химические защитные группы. Химические защитные группы предохраняют олигорибонуклеотиды от деградации и при необходимости могут быть удалены с иммобилизованных олигорибонуклеотидов прямо на биочипе непосредственно перед его использованием. Изобретение также предлагает способ удаления химических защитных групп с иммобилизованных олигорибонуклеотидов, способ изготовления биочипов, содержащих защищенные от деградации олигорибонуклеотиды, и способ анализа взаимодействия РНК с РНК-связывающимися молекулами.
Уровень техники
Биочипом называется матрица микроячеек, регулярно расположенных на плоской подложке. Размер ячеек и расстояние между ними могут варьироваться от десятков до сотен микрон. В каждой ячейке биочипа иммобилизованы молекулы одного типа (молекулярные зонды), способные с высокой степенью специфичности связывать определенный тип биомолекул исследуемого раствора, наносимого на поверхность биочипа. Взаимодействие биомолекул с зондами регистрируется различными методами, что позволяет детектировать их наличие в анализируемой смеси (Колчинский, A.M., Грядунов, ДА., Лысов, Ю.П., Михайлович, В.М., Наседкина, Т.В., Турыгин, А.Ю., Рубина, А.Ю., Барский, В.Е. и Заседателев, А.С. 2004. Микрочипы на основе трехмерных ячеек геля: история и перспективы. Молекулярная биология, 38: 5-16; Venkatasubbarao, S. 2004. Microarrays-status and prospects. Trends Biotechnol. 22: 630-7).
В настоящее время биочипы с иммобилизованными олигорибонуклеотидами применяются по следующим направлениям:
I. Для исследования кинетики и термодинамики связывания с РНК белков и антибиотиков:
Hendrix, M., Priestley, E.S., Joyce, G.F. and Wong, C.H. 1997. Direct observation of aminoglycoside-RNA interactions by surface plasmon resonance. J. Am. Chem. Soc. 119:
3641-3648.
Wong, C.-H., Hendrix, M., Priestley, E.S. and Greenberg, W. 1998. Specificity of aminoglycoside antibiotics for the A-site of the decoding region of ribosomal RNA. Chem. Biol. 5: 397-406.
Van Ryk, D.I. and Venkatesan, S. 1999. Real-time kinetics of HIV-1 Rev-Rev Response Element Interactions. J. Biol. Chem. 274: 17452-17463.
Xavier, K.A., Eder, P.S. and Giordano T. 2000. RNA as a drug target: methods for biophysical characterization and screening. Trends Biotechnol. 18: 349-356
Kwon, M., Chun, S.-M., Jeong, S. and Yu, J. 2001. In Vitro Selection of RNA against Kanamycin B. Mol. Cells 11: 303-311.
Rajendran, K.S. and Nagy, P.D. 2003. Characterization of the RNA-Binding Domains in the Replicase Proteins of Tomato Bushy Stunt Virus. J. Virol. 77: 9244-9258.
Peters, H., Kusov, Y.Y., Meyer, S., Benie, A.J., Bauml, E., Wolff, M., Rademacher, C., Peters, T. and Gauss-Muller, V. 2005. Hepatitis A virus proteinase 3C binding to viral RNA:
correlation with substrate binding and enzyme dimerization. Biochem. J. 385: 363-370.
Verhelst, S.H., Michiels, P.J., van der Marel, G.A., van Boeckel, C.A. and van Boom, J.H. 2004. Surface plasmon resonance evaluation of various aminoglycoside-RNA hairpin interactions reveals low degree of selectivity. Chembiochem. 5: 937-942.
II. Для исследования пространственной структуры РНК:
Kirn, H.D., Nienhaus, G.U., На, Т., Orr, J.W., Williamson, J.R. and Chu, S. 2002. Mg2+-dependent conformational change of RNA studied by fluorescence correlation and FRET on immobilized single molecules. PNAS 99: 4284-4289.
III. При исследовании каталитических свойств РНК:
Nyholm, Т., Andang, M., Bandholtz, A., Maijgren, С., Persson, В., Hotchkiss, G., Fehniger, Т.Е., Larsson, S. and Ahrlund-Richter, L. 2000. Interaction between hammerhead ribozyme and RNA substrates measured by a surface plasmon resonance biosensor. J. Biochem. Biophys. Methods. 44: 41-57.
IV. Для высокочувствительного определения точечных мутаций с помощью РНКазы H:
Goodrich, T.T., Lee, H.J. and Corn, R.M. 2004. Direct detection of genomic DNA by enzymatically amplified SPR imaging measurements of RNA microarrays. J. Am. Chem. Soc. 126: 4086-4087.
Биочипы с иммобилизованными олигорибонуклеотидами могут быть также применены для исследования термодинамики РНК/РНК дуплексов, РНК/ДНК гибридов и необычных структур нуклеиновых кислот.
Недостатки существующих биочипов, содержащих иммобилизованные олигорибонуклеотиды
На биочипе обычно иммобилизуют олигорибонуклеотиды, которые были энзиматически наработаны или химически синтезированы. В последнем случае защитные группы удаляют с 2'-0- групп сразу после синтеза, до нанесения на биочип. Молекулы РНК в значительно большей степени, чем молекулы ДНК, подвержены химической и энзиматической деградации (Кочетков, Н.К. и Будовский, Э.И. 1970. Органическая химия нуклеиновых кислот. Издательство "Химия", Москва). В частности, условия иммобилизации олигорибонуклеотидов, применяемые при изготовлении биочипов (температура, рН, УФ-облучение), могут приводить к частичной деградации олигорибонуклеотидов. Однако гораздо более общей проблемой, возникающей при производстве и хранении биочипов с иммобилизованными молекулами РНК, является практически повсеместное присутствие в окружающей среде рибонуклеаз (РНКаз), расщепляющих молекулы РНК. РНКазы являются чрезвычайно стабильными ферментами, устойчивыми, в частности, к термоинактивации (Makarov, А.А. and Ilinskaya, O.N. 2003. Cytotoxic ribonucleases: molecular weapons and their targets. FEBS Lett. 540: 15-20). Это требует принятия специальных мер по предотвращению контакта олигорибонуклеотидов с РНКазами в процессе их получения, манипуляций с ними и изготовления биочипов, что не только усложняет процесс, но и приводит к значительному увеличению стоимости. В частности, при работе с олигорибонуклеотидами требуются специально предназначенный для манипуляций с РНК набор дозирующих устройств (например, автоматические пипетки), общелабораторные приборы и устройства (в частности, весы, хроматографическое оборудование, электрофоретическое оборудование, ДНК/РНК синтезатор, и т.д.), наборы реагентов и т.д. Все растворы должны быть свободными от РНКаз, что требует предварительной обработки воды, используемой для приготовления растворов, или, где это допустимо, последующей обработки готовых растворов диэтилпирокарбонатом с последующим автоклавированием с целью стерилизации и разрушения остатков реагента. В идеале все работы с РНК проводятся в специальном отведенном для этих целей боксе, камере или отдельном помещении. (Маниатис, Т., Фрич, Э. и Сэмбрук, Дж. 1984. Методы генетической инженерии. Молекулярное клонирование. Издательство "Мир", глава 6, п.3, 4, стр.188-189).
В основу изобретения положена задача разработать такой биочип с иммобилизованными олигорибонуклеотидами, при изготовлении, хранении и использовании которого деградация иммобилизованных олигорибонуклеотидов была бы сведена к минимуму.
Поставленная задача решается предлагаемым изобретением.
Раскрытие изобретения.
Настоящее изобретение предлагает биологический микрочип, предназначенный для анализа взаимодействия РНК с РНК-связывающимися молекулами, представляющий собой подложку, содержащую иммобилизованные на ней олигорибонуклеотиды, характеризующийся тем, что по меньшей мере по одному из 2'-O- положений олигорибонуклеотидов присоединены химические защитные группы, предотвращающие образование 2',3'-циклофосфата и последующий распад этих олигорибонуклеотидов. При необходимости химические защитные группы могут быть удалены с иммобилизованных олигорибонуклеотидов прямо на биочипе, непосредственно перед его использованием.
При необходимости биочип может дополнительно содержать иммобилизованные олигодезоксирибонуклеотиды.
Химические защитные группы могут быть присоединены ковалентным либо нековалентным способом по 2'-O-положениям иммобилизованных олигорибонуклеотидов.
В качестве химических защитных групп, ковалентно присоединенных по 2'-О-положениям иммобилизованных олигорибонуклеотидов, можно предпочтительно использовать /яре/и-бутилдиметилсилильную или триизопропилсилилоксиметильную защитные группы.
В одном из воплощений биочипа настоящего изобретения олигорибонуклеотиды с защитными группами по 2'-O-положениям иммобилизуют методом сополимеризации в объеме трехмерных гидрогелевых ячеек, расположенных регулярным образом на подложке биочипа.
Настоящее изобретение также предлагает способ удаления химических защитных групп с олигорибонуклеотидов биочипа, проводимый прямо на биочипе непосредственно перед его использованием и предусматривающий инкубацию биочипа с раствором для удаления химических защитных групп в течение времени, достаточного для удаления химических защитных групп. Предпочтительно удаление защитных групп может проводиться в растворе 0,01М-3М тетрабутиламмоний фторида (TBAF) в течение 2-48 часов при температуре 0-60°С.
Настоящее изобретение также обеспечивает способ изготовления биочипа настоящего изобретения, предусматривающий обеспечение олигорибонуклеотидов, содержащих по меньшей мере по одному из 2'-О-положений присоединенные химические защитные группы, и затем изготовление биочипа путем иммобилизации этих олигорибонуклеотидов на подложке биочипа.
При необходимости способ дополнительно предусматривает обеспечение олигодезоксирибонуклеотидов и их иммобилизацию на подложке биочипа.
При изготовлении биочипа можно использовать синтезированные олигорибонуклеотиды, содержащие, по меньшей мере, по одному из 2'-O-положений ковалентно либо нековалентно присоединенные химические защитные группы. Защитные группы предохраняют олигорибонуклеотиды от воздействия рибонуклеаз в процессе постсинтетической обработки, в процессе изготовления и хранения биочипа.
В одном из воплощений способа изготовления биочипа настоящего изобретения иммобилизацию олигорибонуклеотидов проводят методом сополимеризации в объеме трехмерных гидрогелевых ячеек, расположенных регулярным образом на подложке биочипа.
Еще в одном своем воплощении способа изготовления биочипа настоящего изобретения дополнительно проводят процедуру удаления химических защитных групп с иммобилизованных олигорибонуклеотидов на биочипе.
Настоящее изобретение также обеспечивает способ анализа взаимодействия РНК с РНК-связывающимися молекулами, предусматривающий контактирование РНК-связывающихся молекул с биочипом настоящего изобретения.
Далее настоящее изобретение будет подробно раскрыто со ссылкой на чертежи.
Краткое описание чертежей.
Фиг.1. РНК/ДНК биочип. (А) Схема РНК/ДНК биочипа, на которой указаны последовательности и концентрации иммобилизованных олигонуклеотидов для каждой гидрогелевой ячейки в соответствии с расположением ячеек на биочипе. (Б) Картина гибридизации флуоресцентно меченного красителем Техасский красный (TR) олигодезоксирибонуклеотида d(TTCCAT)-TR с олигорибо- и олигодезоксирибонуклеотидами, иммобилизованными в 3D-гидрогелевых ячейках РНК/ДНК биочипа. Изображение получено при -5°С после насыщения флуоресцентных сигналов ячеек в течение 15 минут. Последовательности иммобилизованных олигонуклеотидов указаны в строках. Основания иммобилизованных олигонуклеотидов, приводящие к образованию однократных и двукратных мисматчей при гибридизации олигонуклеотида d(TTCCAT)-TR, подчеркнуты. Последовательности олигорибонуклеотидов указаны слева, а последовательности олигодезоксирибонуклеотидов указаны справа, обозначая расположение соответствующих ячеек относительно среднего столбца гидрогелевых ячеек с пустым гелем. Концентрации иммобилизованных олигонуклеотидов указаны под столбцами ячеек биочипа.
Фиг.2. Изображение 3D-гидрогелевого РНК/ДНК биочипа, инкубированного в течение двух часов с раствором рибонуклеазы биназы, а затем с раствором флуоресцентно меченного олигодезоксирибонуклеотида d(TTCCAT)-TR. Условия гибридизации, последовательности и концентрации иммобилизованных олигонуклеотидов, а также их расположение такое же, как на Фиг.1Б.
Фиг.3. Изучение термодинамических характеристик образования олигонуклеотидных дуплексов на РНК/ДНК биочипе. (А) Нормированные кривые температурной диссоциации (черные символы) и ассоциации (белые символы) совершенных дуплексов, образованных в результате гибридизации флуоресцентно меченного олигонуклеотида d(TTCCAT)-TR с различными олигорибо- и олигодезоксирибонуклеотидами, иммобилизованными в 3D-гидрогелевых ячейках биочипа. Последовательности иммобилизованных олигонуклеотидов указаны на фигуре. (Б) Нормированные равновесные кривые температурной диссоциации совершенных дуплексов, образованных в результате гибридизации флуоресцентно меченного олигонуклеотида d(TTCCAT)-TR с олигорибо- и олигодезоксирибонуклеотидами, иммобилизованными в 3D-гидрогелевых ячейках биочипа. Последовательности иммобилизованных олигонуклеотидов указаны на фигуре.
Осуществление изобретения
Настоящее изобретение предлагает биочип, на подложке которого иммобилизованы олигорибонуклеотиды, содержащие химические защитные группы по одному, нескольким или всем 2'-O-положениям. Удаление этих групп может быть осуществлено непосредственно перед использованием биочипа.
В качестве олигорибонуклеотидов для иммобилизации могут быть использованы олигорибонуклеотиды, содержащие по одному, нескольким или всем 2'-O-положениям любые ковалентно или нековалентно присоединенные химические защитные группы, предотвращающие образование 2',3'-циклофосфата и последующий распад этих олигорибонуклеотидов. К таким группам относятся защитные группы на основе кремния, удаляемые ионами фтора (Wada, Т., Tobe, M-, Nagayama, Т., Furusawa, К. and Sekine, M. 1993. New strategies for oligonucleotide synthesis by use of 2-trimethylsilylethyl and 2-trimethylsilylethoxymethyl as the phosphate and 2'-hydroxyl protecting groups, respectively. Nucleic Acids Symp. Ser. 29: 9-10; Westman, E. and Stromberg, R. 1994. Removal of t-butyldimethylsilyl protection in RNA-synthesis. Triethylamine trihydrofluoride (TEA, 3HF) is a more reliable alternative to tetrabutylammonium fluoride (TBAF). Nucleic Acids Res. 22: 2430-2431), ацетальные группы, удаляемые в кислых условиях (Griffin, В.Е. and Reese, С. В. 1964. Oligoribonucleotide synthesis via 2',5'-protected ribonucleoside derivatives. Tetrahedron Lett. 5: 2925-2931; Rastogi, H. and Usher, D.A. 1995 A new 2'-hydroxyl protecting group for the automated synthesis of oligoribonucleotides. Nucleic Acids Res. 23: 4872-4877) или безводных основных условиях (Umemoto, Т. and Wada, Т. 2004. Oligoribonucleotide synthesis by the use of 1-(2-cyanoethoxy)ethyl (CEE) as a 2'-hydroxy protecting group.Nucleic Acids Symposium Series, 48: 9-10), ортоэфирные защитные группы (Scaringe, S.A., Wincott, F.E. and Caruthers, M.H. 1998. Novel RNA Synthesis Method Using 5'-О-Silyl-2'-О-orthoester Protecting Groups. J. Am. Chem. Soc. 120: 11820-11821; Karwowski, В., Seio, К. and Sekine, M., 2005. 4,5-bis(ethoxycarbonyl)-[1,3]dioxolan-2-yl as a new orthoester-type protecting group for the 2'-hydroxyl function in the chemical synthesis of RNA. Nucleosides Nucleotides Nucleic Acids. 24: 1111-1114). Предпочтительно, в качестве защитных групп по 2'-O-положениям синтезируемых олигорибонуклеотидов могут быть использованы трет-бутилдиметилсилильные (tBDMS) и/или триизопропилсилилоксиметильные (ТОМ, TOM-Protecting-Group™) защитные группы.
Олигорибонуклеотиды могут быть получены различными способами, например химически синтезированы перед нанесением на биочип. В настоящее время разработано несколько различных химических подходов к синтезу олигорибонуклеотидов:
фосфорамидитный метод (Beaucage, S.L. 1993. Oligodeoxyribonucleotides synthesis. Phosphoramidite approach. Methods Mol. Biol. 20: 33-61; Damha, M.J. and Ogilvie, K.K. 1993. Oligoribonucleotide synthesis. The silyl-phosphoramidite method. Methods Mol. Biol. 20: 81-114), Н-фосфонатный (Froehler ВС.1993. Oligodeoxynucleotide synthesis. H-phosphonate approach. Methods Mol. Biol. 20: 63-80), фосфотриэфирный (Christodoulou С.1993. Oligonucleotide synthesis. Phosphotriester approach. Methods Mol. Biol. 20: 19-31), твердофазный (Sinha, N.D. 1993. Large-scale oligonucleotide synthesis using the solid-phase approach. Methods Mol. Biol. 20: 437-463) или проводимый в растворе (Bonora, G.M., Biancotto, G., Maffini, M. and Scremin. C.L. 1993. Large scale, liquid phase synthesis of oligonucleotides by the phosphoramidite approach. Nucleic Acids Res. 21: 1213-1217). Предпочтительно олигорибонуклеотиды с защитными группами синтезируют методом твердофазной фосфорамидитной химии из коммерчески доступных фосфорамидитов, содержащих соответствующие защитные группы (GlenResearch, США).
Олигорибонуклеотиды могут быть наработаны энзиматически, например, с использованием Т7 РНК-полимеразы в ходе in vitro транскрипции короткого фрагмента ДНК, содержащего соответствующий промотор (Milligan, J.F., Groebe, D.R., Witherell, G.W. and Uhlenbeck O.C. 1987. Oligoribonucleotide synthesis using Т7 RNA polymerase and synthetic DNA templates. Nucleic Acids Res. 15: 8783-8798).
Олигорибонуклеотиды также могут быть синтезированы непосредственно в процессе изготовления биочипа, например, методом ступенчатого синтеза олигонуклеотидов заданной последовательности на подложке биочипа, таким как метод фотолитографии (Pease А. С., Solas D., Sullivan E.J., Cronin M.T., Holmes С.Р. and Fodor S.P.A. 1994. Light-generated oligonucleotide arrays for rapid DNA sequence analysis. Proc. Natl. Acad. Sci., 91: 5022-5026).
В качестве биочипов могут быть использованы двумерные (2D) биочипы, олигорибонуклеотиды которых иммобилизованы на модифицированной поверхности подложки, или трехмерные (3D) биочипы, олигорибонуклеотиды которых иммобилизованы в объеме гелевых ячеек, нанесенных на подложку. В качестве подложки может быть использована любая платформа (стекло, пластик, мембрана).
При изготовлении биочипа могут быть использованы различные способы иммобилизации олигонуклеотидов. Олигорибонуклеотиды могут быть иммобилизованы с образованием ковалентных связей между олигорибонуклеотидами, химически модифицированными по 5' или 3' концу, и слоем мономера, полимера или гидрогеля, нанесенным на платформу биочипа (Joos В., Kuster H. and Cone R. (1997). Covalent attachment of hybridizable oligonucleotides to glass supports. Analytical biochemistry, vol. 247, pp.96-101; Goodrich, T.T., Lee, H.J., Corn, R.M. 2004. Direct detection of genomic DNA by enzymatically amplified SPR imaging measurements of RNA microarrays. J. Am. Chem. Soc. 126: 4086-4087; Afanassiev, V., Hanemann, V. and Wolfl, S. 2000; Parfett, C.L., Zhou, G. and Silverman, F. 2005. End-linked amino-modified 50-mer oligonucleotides as RNA profiling probes on nylon arrays: comparison to UV cross-linked DNA probes. BioTechniques 38: 690-694). Олигорибонуклеотиды могут быть иммобилизованы на биочипе нековалентным способом, например, за счет образования связи стрептавидин-биотин, что обычно применяют в производстве сенсорных РНК биочипов для анализа взаимодействия белков и лигандов с РНК методом поверхностного плазменного резонанса (Xavier, K.A., Eder, P.S. and Giordano T. 2000. RNA as a drug target: methods for biophysical characterization and screening. Trends Biotechnol. 18: 349-356; Verhelst, S.H., Michiels, P.J., van der Marel, G.A., van Boeckel, C.A. and van Boom, J.H. 2004. Surface plasmon resonance evaluation of various aminoglycoside-RNA hairpin interactions reveals low degree of selectivity. Chembiochem. 5: 937-942), или, например, посредством физической адсорбции на поверхности нитроцеллюлозной или нейлоновой мембраны (Jones, K.D. September 2001. Membrane immobilization of nucleic acids, Part 2: Probe Attachment Techniques. IVD Technology).
Известно большое число способов изготовления 2D-биочипов, среди которых полезными для осуществления настоящего изобретения могут быть методы ступенчатого синтеза in situ олигонуклеотидов заданной последовательности на поверхности платформы биочипа (микроматрицы), такие как
- метод фотолитографии,
Pease А. С., Solas D., Sullivan E.J., Cronin M.T., Holmes C.P. and Fodor S.P.A. 1994. Light-generated oligonucleotide arrays for rapid DNA sequence analysis. Proc. Natl. Acad. Sci., 91: 5022-5026;
Fodor, Stephen, P. A., Stryer, Lubert, Read, Leighton, J., Pirrung and Michael, C. Methods of making nucleic acid or oligonucleotide arrays, US Patent 6600031, July 29, 2003;
- метод с использованием инжекторной (ink-jet) технологии,
Hughes T.R, Мао М., Jones A.R, Burchard J., Marton M.J., Shannon K.W., Lefkowitz S.M., Ziman М., Schelter J.M., Meyer M.R, Kobayashi S., Davis C., Dai H., He Y.D., Stephaniants S.B., Cavet G., Walker W.L., West A., Coffey E., Shoemaker D.D., Stoughton R., Blanchard A.P., Friend S.H. and Linsley P.S. 2001. Expression profiling using microarrays fabricated by an ink-jet oligonucleotide synthesizer. Nature Biotechnology, 19: 342-347.
В других способах изготовления 2D- и 3D-биочипов, полезных для осуществления настоящего изобретения, синтезированные олигонуклеотиды или их смеси с гелевыми мономерами наносят на модифицированную платформу биочипа с помощью роботов с использованием пинов или пьезоэлектрических распылительных насадок
Afanassiev, V., Hanemann, V. and Wölfl, S. 2000. Preparation of DNA and protein micro arrays on glass slides coated with an agarose film. Nucl. Acids Res. 28: ебб;
Kumar, A. and Liang, Z. 2001. Chemical nanoprinting: a novel method for fabricating DNA microchips. Nucl. Acids Res. 29: e2;
Rubina, A.Y., Pan'kov, S.V., Dementieva, E.I., Pen'kov, D.N., Butygin, A.V., Vasiliskov, V.A., Chudinov, A.V., Mikheikin, A.L., Mikhailovich, V.M. and Mirzabekov, A.D. 2004. Hydrogel drop microchips with immobilized DNA: properties and methods for large-scale production, AnalBiochem, 325: 92-106.
В том случае, когда биочип дополнительно содержит иммобилизованные олигодезоксирибонуклеотиды, они могут быть получены и иммобилизованы на биочипе с использованием принципиально тех же способов, что и олигорибонуклеотиды.
В качестве РНК-связывающихся молекул в рамках настоящего изобретения могут использоваться, без ограничения, молекулы РНК, молекулы ДНК, белки, пептиды, красители, антибиотики, субстраты каталитических молекул РНК (рибозимов), и т.д.
Далее настоящее изобретение будет подробно проиллюстрировано со ссылкой на конкретные примеры, представляющие собой наиболее предпочтительные воплощения данного изобретения. При этом должно быть понятно, что изобретение не ограничивается этими описанными воплощениями. Напротив, предполагается, что оно включает любые альтернативы, модификации или эквиваленты, допустимые с учетом сущности и объема изобретения.
Примеры.
Пример 1. Дизайн РНК/ДНК биочипа, взамодействие флуоресцентно меченного олигонуклеотида d(TTCCAT)-TR с олигорибо- и олигодезоксирибонуклеотидами, иммобилизованными в гелевых ячейках биочипа.
Синтез олигонуклеотидов.
Олигорибо- и олигодезоксирибонуклеотиды с аминолинкером по 3'-концу были синтезированы в количестве 1 мкмоль на синтезаторе Applied Biosystems 394 DNA/RNA synthesizer (Applied Biosystems, США). Для синтеза олигорибонуклеотидов использовали рибофосфорамидиты, содержащие, в частности, 2'-О-трет-бутилдиметилсилильную защитную группу (tBDMS). Рибофософорамидиты (Рас-А-СЕ Phosphoramidite, iPr-Pac-G-CE Phosphoramidite, U-CE Phosphoramidite, Ac-C-CE Phosphoramidite), дезоксирибофосфорамидиты (dA-CE Phosphoramidite, dmf-dG-CE Phosphoramidite, dT-CE Phosphoramidite, Ac-dC-CE Phosphoramidite), 3'-C(7) аминолинкер и 2М триэтиламмоний ацетат (ТЕАА) были закуплены в Glen Research (США).
Защитные группы были удалены с олигорибо- и олигодезоксирибонуклеотидов в соответствии со стандартным протоколом производителя (Glen Research, США). Олигорибонуклеотид r(AUGGAA)* был взят в качестве контроля: все защитные группы с нуклеозидных оснований (аденин, гуанин, цитозин), с фосфатных групп (цианэтильная защитная группа) и с 2'-гидроксильных групп рибозных колец (tBDMS) этого олигорибонуклеотида были удалены непосредственно сразу после синтеза. С остальных олигорибонуклеотидов удалялись все защитные группы, за исключением защитной группы tBDMS, снятие которой не производилось. Все олигонуклеотиды были очищены с помощью обращенной ВЭЖХ на колонке Hypersil ODS (5 мкм, 4,6×250 мм) при 20°С. Олигонуклеотиды были элюированы либо в 0,1 М ТЕАА (рН 7), либо в 0,1 М ТЕАА с градиентом ацетонитрила от 0 до 50% (рН 7,0). Скорость потока составляла 1 мл/мин. Гексадезоксирибонуклеотид d(TTCCAT), используемый для гибридизации с иммобилизованными олигонуклеотидами биочипа, был ковалентно помечен флуоресцентным красителем Texas Red sulfonyl chloride (Molecular Probes, США) в соответствии с протоколом производителя.
Концентрации олигонуклеотидов определяли с помощью спектрофотометра Jasco 550 (Jasco, Япония); при расчете концентраций был взят средний коэффициент экстинкции для гексануклеотидов, равный 60000 M-1см-1.
Изготовление РНК/ДНК биочипов.
Трехмерные гидрогелевые РНК/ДНК биочипы были изготовлены методом сополимеризации по технологии IMAGEChip (Rubina, A.Y., Pan'kov, S.V., Dementieva, E.I., Pen'kov, D.N., Butygin, A.V., Vasiliskov, V.A., Chudinov, A.V., Mikheikin, A.L., Mikhailovich, V.M. and Mirzabekov, A.D. 2004. Hydrogel drop microchips with immobilized DNA: properties and methods for large-scale production. Anal Biochem, 325: 92-106), предоставлены фирмой Биочип-ИМБ (Россия, web-site: www.biochip.ru) и содержали молекулы РНК и ДНК, иммобилизованные в объеме соответствующих гелевых ячеек, расположенных на гидрофобной поверхности стекла. Олигонуклеотиды были иммобилизованы в гелевых ячейках биочипа в двух концентрациях: 1·10-4M и 2·10-4M. Диаметр трехмерных гелевых ячеек полусферической формы составлял 300 мкм с точностью 10%, объем ячеек составлял 2 нл. Расстояние между соседними ячейками составляло 550 мкм.
Удаление защитных групп tBDMS с олигорибонуклеотидов биочипа.
Защитные группы tBDMS удалялись с олигорибонуклеотидов на биочипе, непосредственно перед его использованием, в камере объемом 40 мкл, содержащей раствор 0,01М-3М тетрабутиламмоний фторида (TBAF), в течение 2-48 часов при температуре 0-60°С.
Гибридизация.
Гибридизация олигонуклеотида 5'-TTCCAT-TR в концентрации 4·10-5 М с олигонуклеотидами биочипа и последующая диссоциация дуплексных комплексов проводились в стерильном буфере, содержащем 1 М NaCl, 10 мМ Na-фосфатный буфер (рН 7,0), 1 мМ ЭДТА, 0,1% (об./об.) Tween 20, в камере объемом 40 мкл. Гибридизация проводилась при -5°С в течение 15 мин; в этих условиях флуоресцентные сигналы гелевых ячеек достигали насыщения. Флуоресцентные сигналы гибридизации флуоресцентно меченного олигонуклеотида, кривые температурной диссоциации и ассоциации регистрировались с помощью исследовательского анализатора изображений биочипов (Fotin, A.V., Drobyshev, A.L., Proudnikov, D.Y., Perov, A.N. and Mirzabekov, A.D. 1998. Parallel thermodynamic analysis of duplexes on oligodeoxyribonucleotide microchips. Nucleic Acids Res. 26: 1515-1521).
На Фиг.1А показана схема расположения ячеек РНК/ДНК биочипа. РНК/ДНК биочип представляет собой нанесенную на гидрофобную поверхность стекла матрицу трехмерных гидрогелевых ячеек полусферической формы с иммобилизованными в двух концентрациях олигорибо- и олигодезоксирибонуклеотидами: гексарибонуклеотидом r(AUGGAA), гексарибонуклеотидами с заменами по одному или двум основаниям r(AUGAAA), r(AUAGAA), r(AUAAAA) (положение замен подчеркнуто) и гексадезоксирибонуклеотидами эквиваленной последовательности: d(ATGGAA), d(ATGAAA), d(ATAGAA), d(ATAAAA). Олигодезоксирибонуклеотиды были нанесены на биочип для последующего сравнения термодинамики ДНК дуплексов и РНК/ДНК гибридов на одном биочипе. Иммобилизованные гексарибонуклеотиды содержали защитную группу tBDMS по всем 2'-O-положениям. Защитные группы tBDMS удалялись с олигорибонуклеотидов прямо на биочипе, непосредственно перед его использованием. Гексарибонуклеотид r(AUGGAA)*, отмеченный в тексте звездочкой, был иммобилизован на биочипе в качестве контроля. Этот олигорибонуклеотид был синтезирован идентично олигорибонуклеотиду r(AUGGAA), но все его защитные группы, включая защитную группу tBDMS, были удалены непосредственно сразу после его синтеза, до очистки и иммобилизации в гелевых ячейках биочипа (см. выше).
На Фиг.1Б показана картина флуоресценции ячеек РНК/ДНК биочипа, который вначале был подвергнут процедуре удаления химических защитных групп tBDMS, а затем на его поверхность был нанесен раствор с флуоресцентно меченным гексадезоксирибонуклеотидом d(TTCCAT)-TR. Как видно по флуоресцентным сигналам ячеек, олигонуклеотид d(TTCCAT)-TR образует совершенные и несовершенные дуплексы с олигонуклеотидами, иммобилизованными в объеме гелевых ячеек биочипа. Наибольшие сигналы наблюдаются в случае совершенных дуплексов, менее интенсивные сигналы видны в случае однократных мисматчей. Видно, что связывание практически не происходит в случае двукратных мисматчей, а также то, что гибридизуемый олигонуклеотид не накапливается в ячейках с пустым гелем.
Интенсивность флуоресцентных сигналов совершенных РНК/ДНК гибридных дуплексов и совершенного ДНК дуплекса, образованных в соответствующих гелевых ячейках с одинаковыми концентрациями олигонуклеотидов r(AUGGAA), r(AUGGAA)* и d(ATGGAA), примерно одинаковы, что говорит об одинаковой степени иммобилизации олигорибо- и олигодезоксирибонуклеотидов. На Фиг.1Б наблюдается также зависимость флуоресцентного сигнала от концентрации иммобилизованных олигонуклеотидов.
Пример 2. Взаимодействие рибонуклеазы биназы с олигорибонуклеотидами, иммобилизованными в гелевых ячейках биочипа, и последующая гибридизация флуоресцентно меченного олигонуклеотида d(TTCCAT)-TR.
Олигонуклеотиды, иммобилизованные в гелевых ячейках биочипа, были протестированы на способность быть разрушенными рибонуклеазой биназой, которая, как известно, катализирует гидролиз РНК. Биназа - внеклеточная рибонуклеаза бактерии Bacillus intermedius, штамм 7Р (Яковлев, Г.И., Чепурнова Н.К., Моисеев, Г.П., Бочаров, А.Л. и Лопатнев, С.В. 1987. Специфичность РНКазы Bacillus intermedius 7P в реакциях расщепления полинуклеотидов. Биоорганическая химия, 13: 338-343; Okorokov, A.L., Panov, K.I., Kolbanovskaya, E.Yu., Karpeisky, M.Ya., Polyakov, K.M., Wilkinson, A.J. and Dodson, G.G. 1996. Site-directed mutagenesis of the base recognition loop ofribonuclease from. Bacillus intermedius (binase). FEBS Lett. 384: 143-146).
Вначале РНК/ДНК биочип был подвергнут процедуре удаления химических защитных групп tBDMS (см. Пример 1), затем биочип был отмыт в стерильной воде, после чего биочип был обработан раствором рибонуклеазы биназы при комнатной температуре (20°С) в течение двух часов. Для этого на поверхность биочипа была нанесена капля объемом 50 мкл, представляющая собой раствор фермента биназы (2·10-5 М) в буфере, содержащем 0,1 М NaCl, 50 мМ Tris HCl, 1 мМ ЭДТА, рН 6,5. Концентрация фермента была измерена на спектрофотометре Jasco V-550 (Jasco, Япония); коэффициент экстинкции биназы был взят равным 27400 М-1см-1 при длине волны 280 нм (Schulga, A., Kurbanov, F., Kirpichnikov, М., Protasevich, I., Lobachov, V., Ranjbar, В., Chekhov, V., Polyakov, K., Engelborghs, Y. and Makarov, A. 1998. Comparative study of binase and barnase: experience in chimeric ribonucleases. Protein Eng. 11: 775-782). После инкубирования с биназой биочип был отмыт в стерильной воде, а затем на его поверхность был нанесен раствор с флуоресцентно меченным олигонуклеотидом d(TTCCAT)-TR (см. Пример 1) для проверки результата действия биназы. Результат гибридизации представлен на Фиг.2.
Как видно из Фиг.2, в РНК-содержащих гелевых ячейках РНК/ДНК гибридные дуплексы не образовались, что свидетельствует о том, что биназа разрушила олигорибонуклеотиды, иммобилизованные в ячейках биочипа. Интенсивность флуоресценции этих гелевых ячеек сравнима с интенсивностью флуоресценции гелевых ячеек, содержащих пустой гель. Из Фиг.2 также следует, что фермент не повлиял на иммобилизованные олигодезоксирибонуклеотиды, поскольку ДНК дуплексы образовались в соответствующих ячейках в той же мере, как и в случае, представленном на Фиг.1Б. Для этого достаточно сравнить интенсивность флуоресценции ячеек, содержащих совершенный ДНК дуплекс d(ATGGAA)/d(TTCCAT)-TR, а также однократный мисматч d(ATGAAA)/d(TTCCAT)-TR на обеих фигурах.
Таким образом, РНК биочипы, полученные заявленным способом, сохраняют активность иммобилизованных молекул рибонуклеиновой кислоты и могут применяться для исследования взаимодействия белков и низкомолекулярных соединений с РНК.
Пример 3. Термодинамические характеристики РНК/ДНК биочипа: кривые температурной диссоциации ДНК дуплексов и РНК/ДНК гибридных дуплексов на биочипе.
Измерение кривых температурной диссоциации.
Температурную диссоциацию олигонуклеотидных дуплексов в ячейках биочипа проводили при постепенном увеличении скорости повышения температуры: 1°С/10 мин при низких температурах (от -5 до 15°С), 1°С/7 мин в среднем интервале температур (от 15 до 40°С) и 1°С/2 мин при высоких температурах (от 40 до 60°С). Кривые ассоциации измеряли в тех же интервалах температур, с теми же скоростями (в данном случае при уменьшении температуры) сразу после диссоциации, начиная с 60°С.
Изображения обрабатывались с помощью прецизионной компьютерной программы (OOO Биочип-ИМБ, Россия) по алгоритму, описанному ранее (Fotin, A.V., Drobyshev, A.L., Proudnikov, D.Y., Perov, A.N. and Mirzabekov, A.D. 1998. Parallel thermodynamic analysis of duplexes on oligodeoxyribonucleotide microchips. Nucleic Acids Res. 26; 1515-1521) с одним изменением: квадратная рамка вокруг изображения гелевой ячейки биочипа в данном примере была заменена на круглую.
Температура диссоциации дуплексов определялась по соответствующим зависимостям как температура, при которой в ячейке биочипа происходит диссоциация половины дуплексов.
Теоретический расчет температур диссоциации коротких РНК/ДНК гибридов и ДНК дуплексов на биочипе.
Температуры диссоциации, экспериментально определенные с помощью биочипа, были сопоставлены с соответствующими значениями, рассчитанными для раствора той же ионной силы. В рассмотренных примерах флуоресцентно меченный олигонуклеотид d(TTCCAT)-TR (около 10-9 моль) был гибридизован в избытке по отношению к общему количеству олигонуклеотидов, иммобилизованных в гелевых ячейках биочипа (около 10-12 моль). Поэтому, как описано ранее (Fotin, A.V., Drobyshev, A.L., Proudnikov, D.Y., Perov, A.N. and Mirzabekov, A.D. 1998. Parallel thermodynamic analysis of duplexes on oligodeoxyribonucleotide microchips. Nucleic Acids Res. 26: 1515-1521), равновесная константа образования дуплексов К(Т) на биочипе имеет следующее выражение:
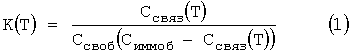
где Т - температура (°К), Ссвяз(Т) - концентрация 5'-TTCCAT-TR, связавшегося с олигонуклеотидами, иммобилизованными в ячейке биочипа, Симмоб - концентрация иммобилизованных олигонуклеотидов и Ссвоб - концентрация гибридизуемого олигонуклеотида 5'-TTCCAT-TR.
Согласно уравнению Вант Гоффа:
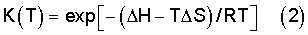
где ΔН и ΔS - изменения стандартной энтальпии и энтропии, соответственно, R - универсальная газовая постоянная.
Поскольку температура диссоциации дуплексов Тдисс определяется как температура, при которой диссоциирует половина двунитевых комплексов в ячейке биочипа, то при этой температуре Ссвяз=Симмоб/2. Принимая во внимание формулы (1) и (2), температура диссоциации дуплексов на биочипе может быть определена по следующей формуле:
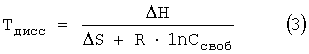
Экспериментальные значения температур диссоциации, полученные с помощью биочипа для дуплексов длиною в 6 пар оснований, были сопоставлены с соответствующими значениями, рассчитанными по формуле (3). Изменения стандартной энтальпии, АН, и энтропии, AS, рассчитывали по алгоритму «ближайших соседей», используя наборы термодинамических параметров Δh и Δs для динуклеотидных дуплексов в растворе 1 М NaCl, указанные в работах Sugimoto, N., Nakano, S., Katoh, М., Matsumura, A., Nakamuta, H., Ohmichi, Т., Yoneyama, М. and Sasaki, М. 1995. Thermodynamic parameters to predict stability of RNA/DNA hybrid duplexes. Biochemistry. 34: 11211-11216; SantaLucia, J., Jr. 1998. A unified view of polymer, dumbbell and oligonucleotide DNA nearest-neighbor thermodynamics. Proc. Natl. Acad. Sci. USA 95: 1460-1465. Отметим, что данный расчет температур диссоциации дуплексов на биочипе не учитывает влияние полиакриламидного геля на процесс образования и диссоциации дуплексных комплексов.
Для исследования термодинамических свойств 3D-гидрогелевых РНК/ДНК биочипов на одном биочипе были сняты кривые температурной диссоциации ДНК дуплексов и РНК/ДНК гибридов и определены соответствующие температуры диссоциации. Как видно из Фиг.3А, кривые диссоциации ДНК дуплексов и РНК/ДНК гибридов совпадают с соответствующими кривыми ассоциации, поэтому процесс диссоциации дуплексов на биочипе проводился в равновесных условиях.
На Фиг.3Б показаны кривые диссоциации совершенных ДНК дуплексов и РНК/ДНК гибридов. Отличие РНК/ДНК гибридных дуплексов r(AUGGAA)*/d(TTCCAT)-TR и r(AUGGAA)/d(TTCCAT)-TR заключается в том, что химические защитные группы tBDMS их олигорибонуклеотидов были удалены, соответственно, до и после изготовления РНК/ДНК биочипа. Защитные группы tBDMS были полностью сняты с уже иммобилизованного гексарибонуклеотида r(AUGGAA), прямо на биочипе, непосредственно перед гибридизацией олигонуклеотида d(TTCCAT)-TR. Кривая диссоциации соответствующего гибридного дуплекса r(AUGGAA)/d(TTCCAT)-TR имеет выраженную S-образную форму. Определенное по этой кривой значение температуры диссоциации такого гибридного дуплекса составляет 27°С. Все защитные группы, включая tBDMS, олигорибонуклеотида r(AUGGAA)* были удалены сразу после его синтеза, до иммобилизации олигорибонуклеотида на биочип. Кривая диссоциации гибридного дуплекса r(AUGGAA)*/d(TTCCAT)-TR, имеет менее пологое верхнее плато и более низкую температуру диссоциации, 25°С. Это означает, что при иммобилизации олигорибонуклеотидов в полиакриламидный гель без защитных групп по 2'-O-положениям происходит частичная деградация олигорибонуклеотидных молекул. Частичный распад олигорибонуклеотидов может быть обусловлен жесткими для РНК условиями полимеризации или результатом действия рибонуклеаз. Отметим, что деградация олигорибонуклеотида r(AUGGAA)* в описанном примере не столь существенна: температура диссоциации дуплекса r(AUGGAA)*/d(TTCCAT) всего на 2 градуса ниже, чем температура диссоциации дуплекса r(AUGGAA)/d(TTCCAT)-TR. Однако в случае олигорибонуклеотидов других последовательностей или длин процент разрушенных молекул РНК в гелевой ячейке может оказаться более значительным. Совершенный ДНК дуплекс d(ATGGAA)/d(TTCCAT)-TR оказался, как и следовало ожидать, менее стабильным, чем совершенный РНК/ДНК гибрид. Его температура диссоциации составила 20°С.
Измеренные на биочипе температуры диссоциации РНК/ДНК гибридного и ДНК дуплексов были сопоставлены с теоретическими значениями, рассчитанными по алгоритму «ближайших соседей» (см. выше). Теоретически рассчитанное и измеренное на биочипе (усредненное по десяти экспериментам) значения температуры диссоциации РНК/ДНК гибридного дуплекса r(AUGGAA)/d(TTCCAT) составили, соответственно, (25,7±0,7)°С и (26,9±0,6)°С. Для ДНК дуплекса d(ATGGAA)/d(TTCCAT) эти величины составили, соответственно, (22,0±1,0)°С и (20,4±0,7)°С. Таким образом, отличие теоретических и экспериментальных значений температуры диссоциации дуплексов на биочипе находится в пределах ошибки измерения. Отметим, что при теоретическом расчете влияние полиакриламидного геля на образование дуплексов не учитывалось.
Это означает, что полиакриламидный гель не оказывает существенного влияния на термодинамические параметры, описывающие взаимодействие этих биомолекул. Сходство диссоциации дуплексов ДНК на полимеризационных биочипах и в растворе отмечалось и ранее (Rubina, A.Y., Pan'kov, S.V., Dementieva, E.I., Pen'kov, D.N., Butygin, A.V., Vasiliskov, V.A., Chudinov, A.V., Mikheikin, A.L., Mikhailovich, V.M. and Mirzabekov, A.D. 2004. Hydrogel drop microchips with immobilized DNA: properties and methods for large-scale production. Anal Biochem, 325: 92-106).
Все патентные документы, публикации, научные статьи и другие документы и материалы, цитируемые или упоминаемые здесь, включены в настоящее описание путем отсылки в такой степени, как если бы каждый из этих документов был включен путем отсылки индивидуально или приведен здесь в его полном виде.
Несмотря на то, что настоящее изобретение было подробно описано со ссылкой на конкретные воплощения, специалисту в данной области будет понятно, что могут быть произведены различные изменения и модификации без отклонения от сущности и объема данного изобретения, которые определяются прилагаемой формулой изобретения.
Дополнительные экспериментальные данные
Дополнение к Примеру 3.
С целью оценить влияние защитных групп tBDMS на формирование РНК/ДНК дуплекса на Фиг.3Б представленной в первоначально поданных материалах заявки также дополнительно приведена кривая температурной диссоциации дуплекса r(AUGGAA)tBDMS/d(TTCCAT)-TR. Перед измерением этой кривой диссоциации процедура удаления защитных групп не была проведена на микрочипе, и иммобилизованный олигорибонуклеотид r(AUGGAA)tBDMS содержал защитные группы tBDMS по всем 2'-O-положениям. Как видно на Фиг.3Б, кривая диссоциации гибридного дуплекса d(TTCCAT)-TR/r(AUGGAA)tBDMS не имеет верхнего плато, а температура плавления такого дуплекса составляет менее -5°С. Таким образом, присутствие защитных групп tBDMS на РНК значительно дестабилизирует РНК/ДНК гибрид, а процедура удаления защитных групп с иммобилизованных олигорибонуклеотидов является необходимой для регенерации микрочипа и последующего изучения на микрочипе взаимодействий РНК с РНК-связывающимися молекулами.
Следует отметить, что эффективность снятия защитных групп с иммобилизованной на микрочипе РНК составляет 100%. После процедуры снятия защитных групп на микрочипе температура диссоциации дуплекса r(AUGGAA)/d(TTCCAT)-TR (с r(AUGGAA) защитные группы снимались во время этой процедуры) оказалась выше, чем температура диссоциации дуплекса r(AUGGAA)*/d(TTCCAT)-TR (иммобилизованный r(AUGGAA)* не содержал защитных групп), и совпала с теоретически рассчитанным значением. Также отметим, что чем короче олигонуклеотиды, тем больше чувствительность кривой диссоциации дуплекса к наличию в олигонуклеотиде модификации/деградации. Например, сложно отличить кривые диссоциации для дуплексов длиной 39 и 40 пар оснований, а кривые диссоциации дуплексов длиной 5 и 6 пар значительно отличаются. И если хотя бы одна защитная группа оставалась на r(AUGGAA) после проведения на микрочипе процедуры снятия защитных групп, то это значительно повлияло бы на кривую температурной диссоциации дуплекса r(AUGGAA)/d(TTCCAT)-TR.
Пример 4. РНК-ДНК биочип, действие рибонуклеазы биназы на не защищенные по 2'-O-положениям РНК последовательности в сравнении с действием этого фермента на РНК последовательности, содержащие tBDMS защитные группы по 2'-О-положениям.
Синтез олигонуклеотидов.
РНК-ДНК олигонуклеотиды с аминолинкером по 3'-концу были синтезированы по той же методике, которая указана в Примере 1, приведенном в первоначально поданном описании заявки.
Все защитные группы с азотистых оснований (аденин, гуанин, цитозин), с фосфатных групп (пианэтильная защитная группа) и с 2'-гидроксильных групп рибозных колец (tBDMS) 40-нуклеотидной РНК-вставки 56-мерной РНК-ДНК d(CAGGGAAA)r(GAGGGUAUAUGUGCGGGUAUAUGUGCGGGUAUAUGUGCAG)*d(A AAGGGAC) были удалены непосредственно сразу после синтеза. С 40-нуклеотидной РНК-вставки 56-мерной РНК-ДНК d(CAGGGAAA)r(GAGGGUAUAUGUGCGGGUAUAUGUGCGGGUAUAUGUGCAG)tBDMS d(AAAGGGAC) удалялись все защитные группы, за исключением защитной группы tBDMS, снятие которой не производилось. Олигонуклеотиды были очищены по методике, указанной в Примере 1. Октадезоксирибонуклеотид d(TTTCCCTG)-TR, используемый для гибридизации с иммобилизованными олигонуклеотидами биочипа, был ковалентно помечен флуоресцентным красителем Texas Red sulfonyl chloride (Molecular Probes, США) в соответствии с протоколом производителя.
Концентрации олигонуклеотидов определяли с помощью спектрофотометра Jasco 550 (Jasco, Япония); при расчете концентраций был взят средний коэффициент экстинкции для одного РНК или ДНК нуклеотида, равный 10000 М-1см-1.
Изготовление РНК-ДНК биочипов.
Трехмерные гидрогелевые РНК-ДНК биочипы были изготовлены. способом, указанным в Примере 1. Олигонуклеотиды были иммобилизованы в гелевых ячейках биочипа в концентрации 0,5·10-5 М.
Гибридизация.
Гибридизацию олигонуклеотида d(TTTCCCTG)-TR в концентрации. 4-10-5 М с олигонуклеотидами биочипа проводили в стерильном буфере А (см. Пример 1), в камере объемом 40 мкл. Гибридизацию проводили при 0°С в течение 30 мин; в этих условиях флуоресцентные сигналы гелевых ячеек достигали насыщения. Флуоресцентные сигналы гибридизации флуоресцентно меченного олигонуклеотида регистрировали по методике, указанной в Примере 1.
РНК-ДНК олигонуклеотиды
d(CAGGGAAA)r(GAGGGUAUAUGUGCGGGUAUAUGUGCGGGUAUAUGUGCAG)*d(AAAGGGAC)-гель (1) и
d(CAGGGAAA)r(GAGGGUAUAUGUGCGGGUAUAUGUGCGGGUAUAUGUGCAG)tBDMS d(AAAGGGAC)-гель (2),
иммобилизованные в гидрогелевых ячейках биочипа, имели одинаковую последовательность. 40-нуклеотидная РНК-вставка иммобилизованного олигонуклеотида (2) содержала защитные группы tBDMS по всем 2'-О-положениям, а 40-нуклеотидная РНК-вставка иммобилизованного олигонуклеотида (1) не содержала защитных групп. 40-нуклеотидные РНК-последовательности образовывали двунитевые структуры - шпильки, а концевые дезокси-участки находились в однонитевой форме, согласно расчетам (Zuker, М. 2003. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids.Ray. 31: 3406-3415).
На Фиг.4А показана картина флуоресценции ячеек РНК-ДНК биочипа, который был инкубирован с раствором флуоресцентно меченного октадезоксирибонуклеотида d(TTTCCCTG)-TR. Олигонуклеотид d(TTTCCCTG)-TR комплементарен 5'-терминирующему октадезоксинуклеотидному участку, d(CAGGGAAA), иммобилизованных олигонуклеотидов. Образование дуплекса d(CAGGGAAA)/d(TTTCCCTG)-TR зависит от целостности 40-нуклеотидной РНК последовательности. Интенсивности флуоресценции ячеек 16 и 26 одинаковы, что говорит об отсутствии видимой деградации незащищенного олигонуклеотида (1) во время производства РНК-ДНК биочипа и его хранения.
Другой РНК-ДНК биочип был вначале обработан раствором рибонуклеазы биназы при комнатной температуре (20°С) в течение 4 часов. Для этого на поверхность биочипа была нанесена капля объемом 50 мкл, представляющая собой раствор фермента биназы (4·10-5 М) в буфере Б. Затем биочип инкубировали с раствором флуоресцентно меченного октадезоксирибонуклеотида d(TTTCCCTG)-TR. На Фиг.4Б показана картина флуоресценции ячеек РНК-ДНК биочипа. Интенсивности флуоресценции ячеек показывают, что фермент значительно разрушил незащищенную 40-нуклеотидную РНК (гелевая ячейка 26, Фиг.4Б) по сравнению с 40-нуклеотидной РНК, содержащей защитные группы tBDMS (гелевая ячейка 16, Фиг.4Б). Таким образом, рибонуклеаза не проявляет биологической активности в отношении РНК последовательности, содержащей защитные группы по 2'-O- положениям, в то время как нативная РНК доступна для действия фермента (см. так же Пример 2, представленный в первоначально поданном описании заявки).
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ И УСТРОЙСТВО ДЛЯ АНАЛИЗА ВЗАИМОДЕЙСТВИЙ БИОЛОГИЧЕСКИХ МОЛЕКУЛ НА БИОЛОГИЧЕСКОМ МИКРОЧИПЕ НА ОСНОВЕ ФЛУОРЕСЦЕНЦИИ АМИНОКИСЛОТНЫХ ОСТАТКОВ ТРИПТОФАНА | 2014 |
|
RU2588816C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ МОДИФИЦИРОВАННЫХ НУКЛЕОТИДОВ РНК | 2012 |
|
RU2522863C2 |
| СПОСОБ МНОЖЕСТВЕННОГО ПАРАЛЛЕЛЬНОГО СКРИНИНГА СПЕЦИФИЧНОСТИ СВЯЗЫВАНИЯ БИОЛОГИЧЕСКИ АКТИВНЫХ СОЕДИНЕНИЙ С НУКЛЕИНОВЫМИ КИСЛОТАМИ С ИСПОЛЬЗОВАНИЕМ БИОЧИПА (ВАРИАНТЫ) | 2000 |
|
RU2182708C2 |
| Способ идентификации генетических полиморфизмов, влияющих на метаболизм противоопухолевых препаратов, с использованием биологических микрочипов | 2018 |
|
RU2697096C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ РНК ВИРУСОВ ГРИППА А И В С ОДНОВРЕМЕННЫМ ОПРЕДЕЛЕНИЕМ ВАРИАНТОВ ГЕМАГГЛЮТИНИНА И НЕЙРАМИНИДАЗЫ ВИРУСА ГРИППА А, ИДЕНТИФИКАЦИЕЙ ГЕНЕТИЧЕСКИХ МАРКЕРОВ ПАТОГЕННОСТИ И УСТОЙЧИВОСТИ К ПРОТИВОГРИППОЗНЫМ ПРЕПАРАТАМ НА БИОЛОГИЧЕСКИХ МИКРОЧИПАХ, БИОЧИП, НАБОР ОЛИГОНУКЛЕОТИДНЫХ ЗОНДОВ, ИСПОЛЬЗУЕМЫЕ В СПОСОБЕ | 2015 |
|
RU2603000C1 |
| НАБОР ДИФФЕРЕНЦИРУЮЩИХ НУКЛЕОТИДОВ И БИОЧИП ДЛЯ ПРИМЕНЕНИЯ В СПОСОБЕ ГЕНОТИПИРОВАНИЯ МАРКЕРОВ ГАПЛОГРУПП Y-ХРОМОСОМЫ ЧЕЛОВЕКА: M130 (C), М145 (DE) | 2012 |
|
RU2539733C2 |
| СПОСОБ ИНТЕГРАЦИИ МНОЖЕСТВЕННЫХ ПОЛИМЕРАЗНЫХ РЕАКЦИЙ АМПЛИФИКАЦИИ С ПОСЛЕДУЮЩИМ АНАЛИЗОМ АМПЛИФИЦИРОВАННЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ НЕПОСРЕДСТВЕННО НА БИОЧИПЕ | 2001 |
|
RU2218414C2 |
| Способ анализа полиморфных маркеров в генах VKORC1, CYP4F2, CYP2C9, CYP2C19, ABCB1, ITGB3 для определения индивидуальной чувствительности к противосвертывающим препаратам | 2018 |
|
RU2689400C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ГЕНОТИПА И ПОДТИПА ВИРУСА ГЕПАТИТА С | 2008 |
|
RU2396355C2 |
| БИОЛОГИЧЕСКИЙ МИКРОЧИП С НАБОРОМ ПРАЙМЕРОВ ДЛЯ АНАЛИЗА ПОЛИМОРФИЗМА В ГЕНАХ AB0, HLA-DQA1, AMEL, DARC, NAT2 | 2014 |
|
RU2582216C2 |
Изобретение относится к молекулярной биологии, биохимии и биоорганической химии и может быть использовано для анализа взаимодействия РНК с РНК-связывающимися молекулами. Биологический микрочип (биочип) представляет собой подложку, содержащую иммобилизованные на ней олигорибонуклеотиды, модифицированные по одному, нескольким или всем 2'-0-положениям химическими защитными группами на основе кремния. Способ изготовления такого биочипа выполняют путем иммобилизации на подложке олигонуклеотидов, содержащих по одному, нескольким или всем 2'-0-положениям химические защитные группы на основе кремния. Для удаления химических защитных групп биочип инкубируют с раствором для удаления химических защитных групп. Данный способ удаления химических защитных групп по 2'-0-положениям: олигорибонуклеотидов, иммобилизованных на биочипе, выполняют непосредственно перед стадией контактирования РНК с РНК-связывающими молекулами для получения биочипа для анализа взаимодействия РНК с РНК-связывающими молекулами и выполнения способа указанного анализа. Применение изобретения позволяет сохранить активность иммобилизованных молекул рибонуклеиновой кислоты. 4 н. и 7 з.п. ф-лы, 4 ил.
1. Биологический микрочип (биочип), предназначенный для получения биочипа для анализа взаимодействия РНК с РНК-связывающимися молекулами, представляющий собой подложку, содержащую иммобилизованные на ней олигорибонуклеотиды, характеризующийся тем, что, по меньшей мере, по одному из 2'-0-положений олигорибонуклеотидов ковалентно присоединены химические защитные группы на основе кремния, предотвращающие образование 2',3'-циклофосфата.
2. Биочип по п.1, в котором подложка дополнительно содержит иммобилизованные на ней олигодезоксирибонуклеотиды.
3. Биочип по п.1, в котором в качестве химической защитной группы по 2'-O-положениям иммобилизованных олигорибонуклеотидов используют трет-бутилдиметилсилильную защитную группу.
4. Биочип по п.1, в котором в качестве химической защитной группы по 2'-O-положениям иммобилизованных олигорибонуклеотидов используют триизопропилсилилоксиметильную защитную группу.
5. Биочип по любому из п.1 или 2, в котором олигорибонуклеотиды с защитными группами по 2'-O-положениям иммобилизованы методом сополимеризации в объеме трехмерных гидрогелевых ячеек, расположенных регулярным образом на подложке биочипа.
6. Способ удаления химических защитных групп по 2'-O-положениям олигорибонуклеотидов, иммобилизованных на биочипе, охарактеризованном в любом из пп.1-5, проводимый прямо на биочипе непосредственно перед его использованием, включающий инкубацию биочипа с раствором для удаления химических защитных групп в течение времени, достаточного для удаления химических защитных групп.
7. Способ по п.6, в котором удаление химических защитных групп на основе кремния с иммобилизованных олигорибонуклеотидов на биочипе проводят в растворе 0,01М-3М тетрабутиламмоний фторида (TBAF) в течение 2-48 ч при температуре 0-60°С.
8. Способ изготовления биочипа, охарактеризованного в п.1, включающий следующие стадии:
а) обеспечивают олигорибонуклеотиды, содержащие, по меньшей мере, по одному из 2'-O-положений ковалентно присоединенные химические защитные группы на основе кремния, предотвращающие образование 2',3'-циклофосфата;
б) изготавливают биочип путем иммобилизации на подложке биочипа олигорибонуклеотидов, обеспеченных на стадии а).
9. Способ по п.8, дополнительно включающий обеспечение олигодезоксирибонуклеотидов и их иммобилизацию на подложке биочипа.
10. Способ по п.8, в котором иммобилизацию проводят методом сополимеризации в объеме трехмерных гидрогелевых ячеек, расположенных регулярным образом на подложке биочипа.
11. Способ анализа взаимодействия РНК с РНК-связывающимися молекулами, предусматривающий:
а) удаление химических защитных групп на основе кремния с олигорибонуклеотидов биочипа, охарактеризованного в любом из пп.1-5, для получения биочипа для анализа взаимодействия РНК с РНК-связывающимися молекулами, причем данную стадию проводят непосредственно перед стадией контактирования РНК-связывающихся молекул с биочипом для анализа взаимодействия РНК с РНК-связывающимися молекулами;
б) контактирование РНК-связывающихся молекул с биочипом для анализа взаимодействия РНК с РНК-связывающимися молекулами.
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Способ регулировки зазора в передачах зацеплением | 1982 |
|
SU1206532A1 |
| КОМПОЗИЦИЯ ДЛЯ ИММОБИЛИЗАЦИИ БИОЛОГИЧЕСКИХ МАКРОМОЛЕКУЛ В ГИДРОГЕЛЯХ, СПОСОБ ПРИГОТОВЛЕНИЯ КОМПОЗИЦИИ, БИОЧИП, СПОСОБ ПРОВЕДЕНИЯ ПЦР НА БИОЧИПЕ | 2001 |
|
RU2206575C2 |

