Область техники, к которой относится изобретение.
Изобретение относится к области биомедицинских измерительных технологий, а именно к способам оценки фагоцитарного звена иммунитета при ряде физиологических и патологических состояний. Необходимость проведения подобных исследований закреплена в «Номенклатуре клинических лабораторных исследований» (п.6.15.5. - Признаки реактивности нейтрофилов), утвержденной Приказом Министерства здравоохранения РФ №64 от 21.02.2000 г.
В частности, изобретение предназначено для одновременного определения двух наиболее значимых и взаимосвязанных параметров фагоцитарной реакции: 1) интенсивности киллинга фагоцитируемых клеток-мишеней, являющегося наиболее интегральным показателем завершенности фагоцитоза; 2) активации кислородзависимого метаболизма фагоцитов, характеризующего состоятельность их бактерицидных систем и лежащего в основе развития киллингового эффекта.
Таким образом, заявляемый способ предназначен для использования в лабораториях биологического и медицинского профиля, решающих вопросы оценки иммунного статуса организма и уровня его противоинфекционной резистентности с целью выявления их возможных нарушений и контроля эффективности соответствующих коррекционных мероприятий.
Уровень техники.
Реализация бактерицидного потенциала фагоцитов и обусловленный этим киллинг (умерщвление) бактериальных клеток после поглощения специализированными клетками крови являются одними из завершающих и наиболее важных этапов фагоцитарного процесса [1]. В связи с этим результаты их исследования служат интегральными показателями фагоцитоза, нарушения которого ведут к развитию пиогенных инфекций различной степени тяжести.
Для оценки бактерицидности/киллинга предложен достаточно большой спектр различных способов и вариантов их выполнения.
К наиболее простым из них следует отнести микроскопический метод, с использованием которого проводится подсчет обычных (жизнеспособных) и дегенеративных (разрушенных) микроорганизмов после совместной инкубации с фагоцитами в присутствии опсонизирующей сыворотки при 37°С в течение 1 часа и последующей окраски по Романовскому-Гимзе или иными красителями [2]. Однако, как и любая другая биомедицинская технология с визуальной оценкой, данный способ является весьма субъективным, а потому неточным.
Более надежен микробиологический метод, заключающийся в определении числа микроорганизмов, выживших после контакта с фагоцитами и за счет этого сохранивших способность к формированию изолированных колоний при выращивании на плотных питательных средах [3]. Для его реализации готовят смесь фагоцитов и опсонизированных микроорганизмов в соотношениях от 1:1 до 1:100, инкубируют при 37°С и через различные промежутки времени отбирают пробы, из которых, предварительно разрушив фагоциты, делают высевы на питательный агар. Еще через 18-24 часа инкубации подсчитывают количество выросших колоний, сравнивая их с таковым в контроле (посева из взвеси микроорганизмов без фагоцитов), после чего выражают киллинговую способность фагоцитов как % погибших микробных клеток. Данный способ чрезвычайно надежен, в связи с чем часто используется в качестве «золотого стандарта» при оценке киллинговой способности фагоцитов, однако достаточно трудоемок и позволяет получать результат не ранее суток от момента начала исследования.
Наиболее современные методы оценки фагоцитоза основаны на введении в фагоцитарную систему дополнительных «меток», инструментальное исследование которых позволяет получить объективное и экспрессное представление об изучаемых явлениях.
В частности, известен радиоизотопный метод оценки фагоцитоза, основанный на предварительном маркировании микроорганизмов 3Н-тимидином [4], 3Н-глюкозой или 14С-аминокислотами. При этом о степени разрушения бактериальных клеток при фагоцитозе судят на основе учета распределения радиоактивности в супернатанте и клеточном осадке на различных сроках совместной инкубации микроорганизмов с фагоцитами. К преимуществам данного метода следует отнести его высокую чувствительность и объективность, к недостаткам же - необходимость использования радиоактивных веществ, а также относительно высокую стоимость используемого оборудования и расходных материалов.
В свою очередь, цитофлюориметрический метод позволяет получить представление об интенсивности киллинга бактериальных клеток при ведении в фагоцитарную систему различных флюорохромов (карбокифлюоресцеина, пропидиума иодида и др.) - флюоресцирующих красителей, связывающимся с телами микробных клеток и при внешнем облучении сообщающих различный характер свечения их живым и убитым вариантам [5]. Применение подобного подхода с использованием специального прибора - проточного цитофлюориметра - позволяет в одном измерении анализировать несколько тысяч клеток и, тем самым, получать максимально объективные результаты. К причинам же, ограничивающим ее широкое использование, вновь следует отнести высокую стоимость используемого оборудования и расходных материалов.
По совокупности признаков наиболее близким к заявляемому и в этой связи характеризуемым как способ-прототип является биолюминесцентный метод оценки фагоцитоза [6], основанный на использовании в качестве фагоцитируемых объектов рекомбинантных микроорганизмов вида Escherichia coli с клонированными в них генами биолюминесценции (lux+). Соответственно, снижение уровня биолюминесценции E.coli lux+ в процессе их взаимодействия с нейтрофилами периферической крови человека на 0, 10, 30 и 60 минутах инкубации по сравнению с контролем позволяет осуществить расчет фагоцитарной активности (ФА) по формуле ФА=100·(X1-X2)/X1, где X1 - интенсивность биолюминесценции контрольной пробы, Х2 - интенсивность биолюминесценции опытной пробы.
Однако попытки использования данного способа заставляют констатировать существование ряда причин, препятствующих получению требуемого результата, а именно:
1) в формуле и описании способа-прототипа отсутствует процедура опсонизации бактериальных клеток, являющаяся одним из важнейших условий для воспроизведения фагоцитарной реакции [7];
2) в формуле способа-прототипа и примере его использования описывается проведение взаимодействия бактерий с суспензией нейтрофилов, содержащей 25% аутоплазмы, которая потенциально может вести к развитию самостоятельных киллинговых эффектов, не связанных с фагоцитозом, но определяемых присутствием в ней гуморальных бактерицидных факторов [8];
3) в формуле и описании способа-прототипа в качестве объекта для фагоцитоза упоминается генно-инженерный штамм E.coli lux+ без указания источника (донора) генетического материала, клонированного в данном микроорганизме и кодирующего ферменты, необходимые для формирования биолюминесценции. Вместе с тем рекомбинантные микроорганизмы на основе одного и того же штамма E.coli с клонированными lux-оперонами разных люминесцирующих бактерий при развитии бактерицидного эффекта демонстрируют различный, в том числе противоположный характер изменения уровня биолюминесценции [9].
Таким образом, отсутствие процедуры опсонизации, наличие в фагоцитарной системе дополнительных бактерицидных агентов, а также отсутствие четких требований к генно-инженерным люминесцирующим микроорганизмам, используемым в качестве объектов фагоцитоза, существенно снижает точность определения фагоцитарной активности нейтрофилов по способу-прототипу и делает его результаты сложноинтерпретируемыми.
Еще одним недостатком способа-прототипа является невозможность получения информации об активности фагоцитарных систем, лежащих в основе киллингового эффекта и по современным представлениям определяемых существованием кислородзависимых и кислороднезависимых механизмов [1], из которых первые являются ведущими. В их основе лежит активация гексозомонофосфатного шунта с окислением кислорода НАДФН-оксигеназной системой и образованием активных форм кислорода (АФК) - так называемый «респираторный взрыв» фагоцитов. При этом интенсивность респираторного взрыва может быть оценена по выраженности другого типа свечения - хемилюминесценции, возникающего при введении в фагоцитарную систему специальных люминофоров (например, люминола), при взаимодействии с АФК переходящих в возбужденное состояние с испусканием кванта света [10].
Вместе с тем способы раздельной одновременной или последовательной оценки хеми- и биолюминесценции в фагоцитарной системе, характеризующих как активность кислородзависимых систем фагоцитов, так и выраженность обусловленного их действием киллингового эффекта в отношении бактериальных клеток-мишеней, неизвестны. Альтернативные подходы, ведущие к устранению данных недостатков, в доступной нам литературе также не описаны.
Таким образом, заявляемый способ не известен из уровня техники, т.е. является новым.
Сущность изобретения.
Основной задачей, на решение которой направлен заявляемый способ, является одновременная раздельная оценка хеми- и биолюминесценции в фагоцитарной системе, позволяющая получить комплексное представление как об уровне активации кислородзависимых бактерицидных систем фагоцитов (по показателям люминолзависимой хемилюминесценции), так и выраженность обусловленного их действием киллингового эффекта в отношении бактериальных клеток-мишеней (по показателям биолюминесценции). Дополнительной задачей, решаемой при осуществлении заявляемого способа оценки фагоцитоза, является повышение его точности за счет адаптированной процедуры опсонизации бактериальных клеток-мишеней, исключения из фагоцитарной системы иных бактерицидных факторов, а также использования биолюминесцирующего штамма с определенными биологическими (генетическими) характеристиками.
Сущностью заявляемого способа, сформулированной на уровне функционального обобщения и лежащей в его основе, является следующее:
1) снижение уровня биолюминесценции при фагоцитозе, пропорциональное развитию киллингового эффекта, регистрируется при использовании в качестве фагоцитируемых объектов рекомбинантных люминесцирующих бактерий Escherichia coli с клонированными luxCDABE генами Photobacterium leiognathi; при этом для исключения не связанных с фагоцитозом киллинговых эффектов их опсонизация проводится нормальным иммуноглобулином человека в конечной концентрации 6-10 мг/мл в течение 10 мин при 37°С.
2) раздельный учет интенсивности люминолзависимой хемилюминесценции фагоцитов и биолюминесценции бактериальных клеток-мишеней в фагоцитарной системе возможен в одной пробе путем оценки интенсивности свечения в двух различных диапазонах спектра (<430 нм при оценке хемилюминесценции и >540 нм при оценке биолюминесценции) или в двух отдельных пробах, в одну из которых люминесцирующие бактерии вносят после дополнительной инкубации при 42°С в течение 10 минут (для подавления биолюминесценции), а в другую не вносят люминол (для исключения хемилюминесценции).
Соответственно, при реализации заявляемого способа оценки фагоцитоза характеристика действий, порядок их выполнения и условия осуществления в сравнении со способом-прототипом представляются следующим образом (таблица 1).
На первом (подготовительном) этапе готовят компоненты реакции: фагоцитируемые объекты (рекомбинантные люминесцирующие бактерии) и фагоцитирующие клетки (нейтрофилы периферической крови человека). При этом в соответствии с формулой заявляемого способа в качестве фагоцитируемых объектов используют микроорганизмы вида Escherichia coli с клонированными luxCDABE генами Photobacterium leiognathi. В частности, в качестве подобной бактериальной культуры могут использоваться генно-инженерный штамм E.coli, производимый МГУ им. М.В.Ломоносова под названием «Эколюм-9» [11], или E.coli Z905, депонированный в «Коллекции культур светящихся бактерий» Института биофизики Сибирского отделения Российской академии наук [12] и производимый ИБСО РАН под названием «Микробиосенсор E.coli Z905». Оба названных микроорганизма выпускаются в лиофилизированном состоянии, из которого перед выполнением исследований их восстанавливают добавлением холодной дистиллированной воды и выдерживают в течение 30 минут при 4°С для стабилизации уровня биолюминесценции. После этого микроорганизмы смешивают в равных объемах с коммерческим препаратом нормального иммуноглобулина человека, так чтобы его конечная концентрация составляла 6-10 мг/мл, а конечная концентрация бактерий равнялась 5×108/мл. Полученную смесь выдерживают при 37°С в течение 10 мин. Для получения фагоцитов цельную кровь в количестве 2-4 мл забирают из локтевой вены в силиконированную или пластиковую пробирку, содержащую гепарин (50 Ед/мл), после чего выдерживают в течение 2 часов при 4°С для разделения эритроцитов (нижняя фаза) и обогащенной лейкоцитами плазмы (верхняя фаза). Содержимое верхней фазы наслаивают на двойной градиент фиколл-верографина с плотностями 1,077 г/мл и 1,112 г/мл, после чего центрифугируют в течение 45 мин при 1500 об/мин. После разделения нейтрофилы собирают с нижней интерфазы, отмывают холодным физиологическим раствором и ресуспендируют в среде 199 до концентрации 5×106/мл.
Особенностями этапа, принципиально отличающими его от такового по способу-прототипу, являются: 1) использование в качестве объектов фагоцитоза рекомбинантных штаммов Escherichia coli с определенной характеристикой клонированных в них luxCDABE генов из природного морского люминесцирующего микроорганизма вида Photobacterium leiognathi, для которых показана количественная зависимость снижения уровня биолюминесценции от выраженности киллингового эффекта (см. ниже); 2) проведение опсонизации данного микроорганизма в режиме, обеспечивающем его последующий фагоцитоз, но исключающем развитие не связанного с ним бактерицидного эффекта (см. ниже). При выполнении способа по п.3 формулы заявляемого изобретения, кроме того, делят пробу опсонизированных люминесцирующих бактерий на две равные части, одну из которых дополнительно инкубируют при 42°С в течение 10 минут для подавления активности ферментной системы генерации свечения.
На втором (основном) этапе выполнения способа формируют фагоцитарную систему путем смешивания 1 объема опсонизированных люминесцирующих бактерий с концентрацией 5×108/мл и 9 объемов фагоцитов с концентрацией 5×106/мл, так что достигаемое соотношение «бактерии:фагоциты» составляет 10:1. Полученную смесь инкубируют в течение 60 минут при 37°С в термостате или измерительной ячейке люминометра (если последний имеет функцию термостатирования).
Содержание этапа - фагоцитоз с активацией кислородзависимых бактерицидных систем фагоцитов и формированием киллингового эффекта в отношении бактериальных клеток-мишеней - также имеет отличия от такового в способе-прототипе, т.к.: 1) в смесь не добавляют аутоплазму, способную инициировать не связанные с фагоцитозом киллинговые эффекты (см. ниже) и, тем самым, исказить результаты исследования; 2) в смесь добавляют люминол в конечной концентрации 10-3М, при окислении генерируемыми фагоцитами активными формами кислорода формирующий собственное, отличное от биолюминесценции свечение - хемилюминесценцию. При выполнении способа по п.3 формулы заявляемого изобретения параллельно готовят две фагоцитарные системы, в одну из которых вносят опсонизированные люминесцирующие бактерии, но не вносят люминол, а в другую вносят опсонизированные люминесцирующие бактерии, дополнительно инкубированные при при 42°С в течение 10 минут, и люминол.
На завершающем этапе осуществления заявляемого способа проводят измерение интенсивности свечения в фагоцитарной системе с использованием прибора - люминометра, на основании чего рассчитывают показатели, характеризующие активность фагоцитоза. При этом свечение измеряется на 0, 10, 30 и 60 мин или в непрерывном режиме (в случае инкубации пробы в измерительной термостатируемой ячейке). В частности, в качестве измерительного прибора при реализации заявляемого способа может использоваться двухканальный биохемилюминометр 8802М2К (СКТБ «Наука», Красноярск), обеспечивающий термостатирование изучаемой пробы и непрерывное измерение интенсивности ее свечения, в том числе в двух диапазонах длин волн или одновременно в двух анализируемых пробах. При выполнении способа по п.3 формулы заявляемого изобретения также могут использоваться биохемилюминометр БХЛ-6 (НПО «Биофармавтоматика», Нижний Новгород), БЛМ 8802 М (СКТБ «Наука», Красноярск), а также прочие отечественные и зарубежные люминометры, обеспечивающие возможность количественного измерения интенсивности свечения в области спектра 380-600 нм. В свою очередь, итоговый расчет показателей активности фагоцитоза проводят по формуле: 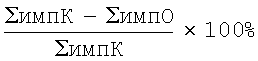 , где ΣимпК - сумма импульсов, испускаемых фагоцитарной системой в контроле (в том числе на 0 секунде исследования), ΣимпО - сумма импульсов, испускаемых фагоцитарной системой в опыте.
, где ΣимпК - сумма импульсов, испускаемых фагоцитарной системой в контроле (в том числе на 0 секунде исследования), ΣимпО - сумма импульсов, испускаемых фагоцитарной системой в опыте.
Основным отличием данного этапа от такового в способе-прототипе, в значительной степени определяющим новизну всего изобретения, является то, что интенсивность свечения фагоцитарной системы оценивается раздельно в двух диапазонах спектра: <430 нм и >540 нм.

Проведение измерений в первом диапазоне позволяет оценить интенсивность люминолзависимой хемилюминесценции и на ее основе сформировать представление о степени активации кислородзависимых бактерицидных систем фагоцитов, а во втором - на основе изменения уровня биолюминесценции бактериальных клеток-мишеней количественно оценить выраженность развивающегося в отношении них киллингового эффекта. При выполнении же способа по п.3 формулы заявляемого изобретения аналогичные заключения делают на основании измерения интенсивности свечения, проводится во всем спектре длин волн (380-600 нм), но в двух пробах: 1) с добавлением люминола и инкубированных при 42°С бактерий, в которой формируется только хемилюминесценция; 2) без добавления люминола с опсонизированными люминесцирующими бактериями, в которой формируется только биолюминесценция.
Возможность получения технического результата при выполнении перечисленных действий в указанных интервалах значений определяется следующим комплексом причинно-следственных связей:
1) в качестве объекта для фагоцитоза при осуществлении биохемилюминесцентного способа его оценки может использоваться не любой рекомбинантный (генно-инженерный) штамм Escherichia coli с клонированными в нем генами люминесцентной системы, в наиболее общем виде обозначаемый E.coli lux+ (как в способе-прототипе), а только рекомбинантные штаммы с определенной генетической характеристикой, а именно - с клонированными luxCDABE генами морских люминесцирующих Photobacterium leiognathi. Обоснованием для подобного утверждения являются полученные экспериментальные данные о том, что именно данные микроорганизмы демонстрируют в фагоцитарной системе пропорциональность между снижением уровня биолюминесценции и выраженностью развивающегося в отношении них киллингового эффекта, отсутствующую у других генно-инженерных вариантов E.coli lux+.
Так рекомбинантные штаммы E.coli с клонированными luxCDABE генами Р.leiognathi, представленные двумя независимыми вариантами, выпускаемыми под коммерческими названиями «Микробиосенсор E.coli Z905» (ИБСО РАН) и «Эколюм-9» (МГУ им. М.В.Ломоносова), при введении в фагоцитарную систему характеризовались прогрессирующим угнетением уровня биолюминесценции, что сопровождалось пропорциональным снижением количества жизнеспособных микроорганизмов при высеве на плотные питательные среды (коэффициент корреляции r=0,96; t=7,l 1; Р<0,001). В то же время другой генно-инженерный штамм E.coli с клонированными luxCDABE генами почвенного люминесцирующего микроорганизма Photorhabdus luminescence, выпускаемый под коммерческими названиями «Эколюм-8» (МГУ им. М.В.Ломоносова), в тех же условиях отвечал стимуляцией уровня свечения, противоположной динамике развития киллингового эффекта.
2) опсонизация фагоцитируемых частиц является одним из основных условий для эффективного фагоцитоза, однако проведение этой процедуры по способу-прототипу не предусмотрено. Причиной этого является невозможность рутинной процедуры опсонизации с использованием сыворотки/плазмы крови, так как в отличие от ее использования для опсонизации грамположительных микроорганизмов данная гуморальная система способна оказать на жизнеспособность грамотрицательных бактерий (в т.ч. E.coli) самостоятельное, предшествующее фагоцитозу, бактерицидное воздействие. При обосновании подходов к решению данного вопроса был изучен ряд факторов, потенциально обладающих опсонизирующим эффектом, а именно: плазма крови, сыворотка крови, термоинактивированная (+56°С, 10 мин) сыворотка крови, комплемент (комплемент морской свинки), иммуноглобулины (нормальный иммуноглобулин человека). В качестве критериев использовались: а) отсутствие выраженного ингибирующего влияния на биолюминесценцию бактериальных клеток; б) отсутствие киллингового эффекта в отношении бактериальных клеток; в) способность активировать фагоцитоз бактериальных клеток (собственно опсонизирующий эффект).
Полученные результаты, обобщенные в таблице 2, свидетельствовали о том, что большинство изученных факторов оказывали на уровень биолюминесценции и жизнеспособность бактериальных клеток E.coli выраженный ингибирующий эффект, негативно влияющий на их последующий опсонофагоцитоз. На этом фоне единственным фактором, применимым для заявленной цели, оказался нормальный иммуноглобулин человека, инкубация в контакте с которым в конечной концентрации 6-10 мг/мл в течение 10 мин при 37°С вела к умеренно выраженной (<20% от исходного уровня) стимуляции биолюминесценции, не вызывала развития бактерицидного эффекта, но позволяла достичь последующей активации фагоцитов, более чем в 5 раз превышающей фоновый уровень их хемилюминесценции и в 4 раза превышающей таковую при использовании интактных (неопсонизированных) бактерий.
При этом возможность опсонизации рекомбинантных люминесцирующих бактерий E.coli с использованием нормального иммуноглобулина человека определяется отсутствием в его составе белков системы комплемента и других гуморальных факторов, способных оказать бактерицидный эффект и, как следствие, вызвать ингибицию уровня их свечения. Одновременно формирование опсонизирующего эффекта определяется присутствием в составе нормального иммуноглобулина человека антител, специфичных к липополисахаридам энтеробактерий и после связывания с ними способных взаимодействовать с Fc-рецепторами на мембране нейтрофилов периферической крови человека.
3) приведенные выше результаты экспериментальных исследований также свидетельствуют о невозможности получения результата по способу-прототипу, так как введение в фагоцитарную систему 25% аутоплазмы обуславливает развитие самостоятельного (не связанного с фагоцитозом) киллингового эффекта и, как следствие, не связанного с фагоцитозом снижение уровня биолюминесценции бактериальных клеток-мишеней.
4) возможность раздельной оценки люминолзависимой хемилюминесценции, формируемой в результате активации (окислительного взрыва) фагоцитов, и биолюминесценции бактериальных клеток-мишеней основана на экспериментальных данных о различиях в спектрах их световой эмиссии (таблица 3).
Полученные результаты свидетельствуют о существовании достаточно существенной зоны «перекрывания» двух сравниваемых спектров. Однако спектр хемилюминесценции люминола сдвинут в сторону коротких волн, так что его отдельный учет возможен в диапазоне <430 нм. В свою очередь, спектр бактериальной биолюминесценции характеризуется сдвигом в сторону более длинных волн, так что ее отдельный учет возможен в диапазоне >540 нм.
5) возможность другого подхода к раздельной оценке люминолзависимой хемилюминесценции фагоцитов и биолюминесценции бактериальных клеток-мишеней основана на выявленной возможности термоинактивации (42°С, 10 мин) клонированной в E.coli ферментной системы генерации свечения P.leiognathi без изменения жизнеспособности реципиентных бактериальных клеток (таблица 4). Кроме того, установлено, что в исследованном интервале температур это невозможно для другого генно-инженерного штамма E.coli, содержащего ферментную систему генерации свечения почвенного люминесцирующего микроорганизма P.luminescence, что еще раз указывает на возможность осуществления способа только с использованием рекомбинантных штаммов с определенной генетической характеристикой (см. выше).
Полученные результаты явились обоснованием возможности проведения исследований во всем спектре длин волн (380-600 нм), но в двух пробах: 1) фагоциты + люминол + инкубированные при 42°С опсонизированные бактерии (учитывается только хемилюминесценция); 2) фагоциты + опсонизированные люминесцирующие бактерии (учитывается только биолюминесценция).
Таким образом, резюмируя приведенные выше материалы о сущности заявляемого способа, характеристике действий, порядке их выполнения и условия осуществления, можно констатировать, что заявляемый способ не следует из уровня техники и по этому показателю должен быть оценен как соответствующий критерию «изобретательский уровень».
Сведения, подтверждающие возможность осуществления изобретения.
Последовательность выполнения действий при осуществлении заявляемого способа, их условия и режимы являются следующими:
1) Для получения фагоцитов цельную кровь в количестве 2-4 мл забирают из локтевой вены в силиконированную или пластиковую пробирку, содержащую гепарин (50 Ед/мл), после чего выдерживают в течение 2 часов при 4°С для разделения эритроцитов (нижняя фаза) и обогащенной лейкоцитами плазмы (верхняя фаза). Содержимое верхней фазы наслаивают на двойной градиент фиколл-верографина с плотностями 1,077 г/мл и 1,112 г/мл, после чего центрифугируют в течение 45 мин при 1500 об/мин. После разделения нейтрофилы собирают с нижней интерфазы, отмывают холодным физиологическим раствором и ресуспендируют в среде 199 до концентрации 5×106/мл.
2) Для получения фагоцитируемых объектов готовят суспензию живых люминесцирующих бактерий - рекомбинантнтных E.coli с клонированными luxCDABE генами морских люминесцирующих Photobacterium leiognathi, используя для этого лиофилизированый препарат под коммерческим названием «Микробиосенсор E.coli Z 905» или «Эколюм-9». Непосредственно перед проведением исследований микроорганизмы в составе этих препаратов восстанавливают из лиофилизированного состояния добавлением холодной дистиллированной воды, после чего выдерживают в течение 30 минут при 4°С для стабилизации уровня биолюминесценции.
3) Для проведения опсонизации фагоцитируемых объектов микроорганизмы смешивают в равных объемах с коммерческим препаратом нормального иммуноглобулина человека, так чтобы его конечная концентрация составляла 6-10 мг/мл, а конечная концентрация бактерий равнялась 5×108/мл. Полученную смесь выдерживают при 37°С в течение 10 мин.
При выполнении исследований по п.3 формулы изобретения полученную взвесь опсонизированных микроорганизмов разделяют на две равные части, одну из которых дополнительно инкубируют при 42°С в течение 10 мин для подавления активности ферментной системы генерации свечения.
4) В измерительную кювету люминометра последовательно вносят 0,88 мл нейтрофилов периферической крови человека, суспендированных в среде 199 до концентрации 5×106/мл, раствор люминола до конечной концентрации 10-3М, а также 0,1 мл взвеси опсонизированных люминесцирующих бактерий с концентрацией 5×108/мл, так что достигаемое соотношение «бактерии:фагоциты» составляет 10:1.
При выполнении исследований по п.3 формулы изобретения готовят две кюветы, в одну из которых в указанных количествах вносят нейтрофилы и опсонизированные люминесцирующие бактерии, но не вносят люминол, а в другую вносят нейтрофилы, люминол и опсонизированные бактерии с термоинактивированной ферментной системой генерации свечения.
5) Кювету помещают в термостатируемое кюветное отделение двухканального биохемилюминометра, имеющее выходы на два оптических регистратора (фотоэлектронных умножителя), перед первым из которых установлен светофильтр с областью пропускания <430 нм, а перед вторым светофильтр с областью пропускания >540 нм.
При выполнении исследований по п.3 формулы изобретения кюветы помещают в кюветное отделение двухканального биохемилюминометра, имеющее два отдельных гнезда - каждое с изолированным выходом на отдельный оптический регистратор (фотоэлектронный умножитель). Светофильтры в этом случае не устанавливают.
6) Пробы инкубируют при 37°С в течение 60 мин, осуществляя измерение интенсивности свечения на каждом канале биохемилюминометра в непрерывном режиме.
7) По результатам измерения производят расчет активации кислородзависимых бактерицидных систем фагоцитов и определяемой их действием киллинговой активности в отношении бактериальных клеток по формуле:
где ΣимпК - сумма импульсов? испускаемых фагоцитарной системой в контроле (на 0 секунде исследования), ΣимпO - сумма импульсов, испускаемых фагоцитарной системой в опыте.
При выполнении исследований по п.2 формулы изобретения для проведения расчетов используют величины, зафиксированные в двух диапазонах спектра: <430 нм (при оценке люминолзависимой хемилюминесценции фагоцитов) и >540 нм (при оценке киллингового эффекта в отношении люминесцирующих клеток-мишеней).
При выполнении исследований по п.3 формулы изобретения для расчета активации кислородзависимых бактерицидных систем фагоцитов используют величины, зафиксированные при исследовании пробы, содержащей нейтрофилы, люминол и опсонизированные бактерии с термоинактивированной ферментной системой генерации свечения, а для расчета киллингового эффекта используют величины, зафиксированные при исследовании пробы, содержащей нейтрофилы, опсонизированные люминесцирующие бактерии, но не содержащей люминол.
Пример.
В хирургическое отделение Отделенческой клинической железнодорожной больницы ст. Оренбург 12.08.07 г. поступила больная З., 58 лет, с диагнозом «Некротическая флегмона левой стопы». После предоперационной подготовки больная была оперирована, произведено вскрытие клетчаточных пространств, некрэктомия с наложением глухого шва на рану на дренажно-промывной системе, а также поставлен катетер в артерию конечности для проведения внутриартериальных инфузий. Однако несмотря на проведенное лечение и начатую антибиотикотерапию, послеоперационный период протекал неблагоприятно, что явилось основой для постановки вопроса о состоятельности иммунной системы пациента, в том числе его фагоцитарного звена.
Для проведения исследования цельную кровь пациентки в количестве 2-4 мл забрали из локтевой вены в силиконированную или пластиковую пробирку, содержащую гепарин (50 Ед/мл), выдержали в течение 2 часов при 4°С, после чего отделившуюся верхнюю фазу разделили центрифугированием при 1500 об/мин в течение 45 мин на двойном градиенте фиколл-верографина с плотностями 1,077 г/мл и 1,112 г/мл. Нейтрофилы собрали с нижней интерфазы, отмыли холодным физиологическим раствором и ресуспендировали в среде 199 до концентрации 5×106/мл.
Параллельно провели регидратацию лиофилизированных рекомбинантных люминесцирующих бактерий E.coli с клонированными luxCDABE генами P.leiognathi («Эколюм-9») добавлением во флакон 10 мл холодной дистиллированной воды с последующим выдерживанием в течение 30 минут при 4°С для стабилизации уровня биолюминесценции. После этого микроорганизмы в равных объемах смешали с коммерческим препаратом нормального иммуноглобулина человека, так чтобы его конечная концентрация составляла 8 мг/мл, а конечная концентрация бактерий равнялась 5×108/мл. Полученную смесь инкубировали при 37°С в течение 10 мин.
На следующем этапе 0,88 мл взвеси нейтрофилов, 0,02 мл раствора люминола (10-3М), а также 0,1 мл взвеси опсонизированных люминесцирующих бактерий последовательно внесли в измерительную кювету, которую сразу после этого поместили в термостатируемое (37°С) кюветное отделение двухканального биохемилюминометра 8802М2К (СКТБ «Наука», Красноярск), имеющее выходы на два отдельных фотоэлектронных умножителя, перед первым из которых был установлен светофильтр с областью пропускания <430 нм, а перед вторым светофильтр с областью пропускания >540 нм. Измерение интенсивности свечения на каждом канале провели в течение 60 мин в непрерывном режиме.
После расчета полученных результатов по формуле:
где ΣимпК - сумма импульсов, испускаемых фагоцитарной системой в контроле (на 0 минуте исследования), ΣимпО - сумма импульсов испускаемых фагоцитарной системой в опыте (в интервале до 60 мин от начала исследования), были констатированы недостаточная выраженность «респираторного взрыва» фагоцитов (стимуляция на 258% при нормативных значениях >500%)), а также низкая эффективность киллинга бактериальных клеток (подавление на 37% при нормативных значениях>75%), что позволило сделать заключение об имеющемся дефекте фагоцитарного звена иммунитета.
При анализе причин подобной ситуации пациентке было проведено дополнительное исследование уровня глюкозы и гликозилированного гемоглобина в крови, показавшее величины 8,1 ммоль/л и 7% НbA1с соответственно, что явилось основанием для постановки дополнительного диагноза «Сахарный диабет II типа». Для коррекции данного состояния были назначены инъекции инсулина пролонгированного действия в дозе 32 ЕД/сут.
Проведенный через 5 суток после начала инсулинотерапии биохемилюминесцентный анализ показал рост активности кислородзависимых бактерицидных систем фагоцитов до 580%, а киллинга бактериальных клеток-мишеней до 88%.
Рана зажила первично, больная выписана на двадцатые сутки.
Таким образом, положительный эффект от использования способа проявился в комплексном и точном анализе состояния фагоцитарного звена иммунитета, что позволило провести оперативную коррекцию комплекса лечебных мероприятий.
Источники информации
1. Хаитов P.M., Игнатьева Г.А., Сидорович И.Г. Иммунология. - М.:Медицина, 2000. - 432 с.
2. Хаитов P.M., Пинегин Б.В. Современные подходы к оценке основных этапов фагоцитарного процесса // Иммунология. - 1995. - № 3. - С.3-8.
3. Рудик Д.В., Тихомирова Е.И. Методы изучения процесса фагоцитоза и функционально-метаболического состояния фагоцитирующих клеток. - Саратов: СГУ, 2006. - 112 с.
4. Verhoef J., Peterson P.K., Quie P.G. Kinetics of staphylococcal opsonization, attachment, ingestion and killing by human polymorphonuclear leukocytes: a quantitative assay using [3H]thymidine labeled bacteria // J. Immunol. Methods. - 1977. V.14. - № 3-4. - Р.303-311.
5. Martin E., Bhakdi S. Flow cytometric assay for quantifying opsonophagocytosis and killing of Staphylococcus aureus by peripheral blood leukocytes // J. Clin. Microbiol. - 1992. - V.30. - № 9. - P.2246-2255.
6. Патент RU2292553. Способ определения фагоцитарной активности нейтрофилов периферической крови человека по степени гашения биолюминесценции. Опубл. 27.01.2007, Бюл. № 3.
7. Verhoev J., Peterson P.K., Kim Y., Sabath L.D., Quie P.G. Opsonic requirements for staphylococcal phagocytosis. Heterogeneity among strains // Immunology. - 1977. - V.33. - № 2. - P.191-197.
8. Дерябин Д.Г., Поляков Е.Г. Причины, определяющие характер изменения бактериальной люминесценции при воздействии сыворотки крови // Микробиология. - 2005. - № 2. - С.191-197.
9. Дерябин Д.Г., Поляков Е.Г. Влияние сыворотки крови человека на уровень свечения природных и рекомбинантных люминесцирующих бактерий // Бюлл. экспер. биол. мед. - 2004. - № 9. - С.311-315.
10. Albrecht D., Jungi T.W. Luminol-enhanced chemiluminescence induced in peripheral blood-derived human phagocytes: obligatory requirement of myeloperoxidase exocytosis by monocytes // J.Leucocyte Biol. - 1993. - V.54. - № 4. - P.300-306.
11. Данилов B.C., Зарубина А.П., Ерошников Г.Е. и др. Сенсорные биолюминесцентные системы на основе lux-оперонов разных видов люминесцентных бактерий // Вестник МГУ. Сер. 16. Биология. - 2002. - № 3. - С.20-24.
12. Каталог светящихся бактерий / под ред. Э.К.Родичевой. - Новосибирск: Наука, 1997. - 125 с.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ ФАГОЦИТАРНОЙ АКТИВНОСТИ НЕЙТРОФИЛОВ ПРИ ПОМОЩИ МИКРОБИОЛОГИЧЕСКОГО ПОДХОДА | 2019 |
|
RU2727880C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ФАГОЦИТАРНОЙ АКТИВНОСТИ НЕЙТРОФИЛОВ ПЕРИФЕРИЧЕСКОЙ КРОВИ ЧЕЛОВЕКА ПО СТЕПЕНИ ГАШЕНИЯ БИОЛЮМИНЕСЦЕНЦИИ | 2005 |
|
RU2292553C1 |
| СПОСОБ ОЦЕНКИ ФАГОЦИТАРНОЙ АКТИВНОСТИ НЕЙТРОФИЛОВ | 2023 |
|
RU2824345C1 |
| СПОСОБ БИОХЕМИЛЮМИНЕСЦЕНТНОЙ ОЦЕНКИ ТОКСИЧНОСТИ РУБЦОВОЙ ЖИДКОСТИ IN VITRO | 2013 |
|
RU2603104C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ НЕСПЕЦИФИЧЕСКОЙ АНТИБАКТЕРИАЛЬНОЙ РЕЗИСТЕНТНОСТИ ОРГАНИЗМА | 1992 |
|
RU2068997C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ БИОЛОГИЧЕСКОЙ (АНТИМИКРОБНОЙ) АКТИВНОСТИ ДЕФЕНСИНОВ И ИХ ПРОИЗВОДНЫХ | 2009 |
|
RU2405835C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ БИОТОКСИЧНОСТИ НАНОУГЛЕРОДА | 2010 |
|
RU2437938C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ БАКТЕРИЦИДНЫХ СВОЙСТВ СЫВОРОТКИ КРОВИ | 2011 |
|
RU2489489C1 |
| Способ определения тактики лечения хронического рецидивирующего бактериального простатита | 2020 |
|
RU2723997C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ В ЭКСПЕРИМЕТНЕ СПОСОБНОСТИ НЕЙТРОФИЛОВ ПРОДУЦИРОВАТЬ АКТИВИРОВАННЫЕ МЕТАБОЛИТЫ КИСЛОРОДА | 1993 |
|
RU2089907C1 |
Изобретение относится к области лабораторной диагностики и может быть использовано для определения фагоцитарной активности нейтрофилов периферической крови человека. Сущность способа: в качестве объектов фагоцитоза используют рекомбинантные люминесцирующие бактерии, а именно микроорганизмы Escherichia coli с клонированными luxCDABE генами Photobacterium leiognathi, предварительно опсонизированные в течение 10 мин нормальным иммуноглобулином человека в конечной концентрации 6-10 мг/мл, а также тем, что в пробу дополнительно вносят люминол до конечной концентрации 10-3М. Проводят раздельный учет интенсивности люминолзависимой хемилюминесценции фагоцитов и биолюминесценции бактериальных клеток-мишеней путем измерения свечения в двух различных диапазонах спектра: <430 нм при оценке хемилюминесценции и >540 нм при оценке биолюминесценции. Использование способа позволяет провести комплексную оценку уровня активации кислородзависимых бактерицидных систем фагоцитов, выраженность обусловленного их действием киллингового эффекта в отношении бактериальных клеток-мишеней, а также позволяет повысить точность проводимого исследования. 4 табл.
Способ определения фагоцитарной активности нейтрофилов периферической крови человека путем измерения интенсивности свечения в смесях рекомбинантных люминесцирующих бактерий с фагоцитами в соотношении 10:1 в течение 60 мин при 37°С, отличающийся тем, что в качестве рекомбинантных люминесцирующих бактерий используют микроорганизмы Escherichia coli с клонированными luxCDABE генами: Photobacterium leiognathi, предварительно опсонизированные в течение 10 мин нормальным иммуноглобулином человека в конечной концентрации 6-10 мг/мл, а также тем, что в пробу дополнительно вносят люминол до конечной концентрации 10-3М, после чего проводят раздельный учет интенсивности люминолзависимой хемилюминесценции фагоцитов и биолюминесценции бактериальных клеток-мишеней путем измерения свечения в двух различных диапазонах спектра: <430 нм при оценке хемилюминесценции и >540 нм при оценке биолюминесценции.
| СПОСОБ ОПРЕДЕЛЕНИЯ ФАГОЦИТАРНОЙ АКТИВНОСТИ НЕЙТРОФИЛОВ ПЕРИФЕРИЧЕСКОЙ КРОВИ ЧЕЛОВЕКА ПО СТЕПЕНИ ГАШЕНИЯ БИОЛЮМИНЕСЦЕНЦИИ | 2005 |
|
RU2292553C1 |
| СПОСОБ АЛЛЕРГОДИАГНОСТИКИ ПО ПОКАЗАТЕЛЯМ ХЕМИЛЮМИНЕСЦЕНТНОГО СВЕЧЕНИЯ НЕЙТРОФИЛОВ | 2004 |
|
RU2289138C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ВОЗДЕЙСТВИЯ ЭЛЕКТРОМАГНИТНОГО ИЗЛУЧЕНИЯ С ПОМОЩЬЮ БИОЛЮМИНЕСЦЕНЦИИ БАКТЕРИЙ | 2005 |
|
RU2291196C1 |
| US 2003134348 A1, 17.07.2003 | |||
| US 5900362 A1, 10.04.1996 | |||
| ARISAWA F | |||
| et al | |||
| MCLA-dependent chemiluminescence suggests that singlet oxygen plays a pivotal role in myeloperoxidase-catalysed bactericidal action in neutrophil phagosomes | |||
| Luminescence, 2003, Jul-Aug; | |||

