УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Изучение рака показало значительные достижения, что привело к устойчивому снижению показателей смертности при многих типах злокачественных опухолей. На указанное снижение смертности повлияли усовершенствования раннего обнаружения, прогрессивные хирургические технологии и применение новых видов терапевтических вмешательств. При наличии успеха в снижении показателей смертности, связанных с раком, произошло изменение направления исследований с тем, чтобы сосредоточить внимание на новых видах таргетной терапии рака, среди которых разработка вакцин занимает центральное место. Вакцины высокоэффективны в снижении смертности, вызванной патогенами, благодаря своей способности активировать иммунную систему и обеспечивать иммунитет к чужеродным антигенам. Эффективная вакцинация не только способствует снижению смертности, но вакцины также индуцируют долговременный иммунитет, который защищает против повторной инфекции.
Способность вакцин формировать иммунитет с самого начала привлекала к ним интерес при создании противораковых вакцин. Вне всякого сомнения, противораковые вакцины всех форм продемонстрировали по меньшей мере некоторый успех в стимулировании иммунных ответов против опухолевых антигенов, выбранных в качестве мишеней. Хотя противораковые вакцины не оправдали надежд при использовании в качестве отдельной терапии, особенно на поздних стадиях заболеваний (Terrando et al.; 2007, Vaccine 25,4-16; Burgdorf et al., 2008, Oncol. Rep. 20(6), 1305-131 1), новейшие исследования позволяют предположить, что вакцины на основе дендритных клеток (DC) могут влиять на выживаемость пациентов.
В настоящее время способы, применяемые в иммунотерапии, обычно включают стимулированные ex vivo антиген-представляющие клетки (APC), где предшественники DC культивируют сразу после забора у пациента, и затем нагруженные антигеном DC вводят пациенту непосредственно после получения. Указанные способы создают временные ограничения, что не только ограничивает возможные терапевтические применения DC, но также вызывают необходимость проведения многократного афереза продуктов у пациента во время курса иммунотерапии.
Считают, что помимо презентации антигена, DC при созревании, могут кроме того воздействовать на иммунный ответ путем продуцирования ряда сигнальных молекул, в плане цитокинов и хемокинов. Однако в существовавших ранее методах DC-иммунотерапии не использовали зрелые DC. Кроме того, предыдущая работа по созреванию DC не оптимизировала полностью процесс созревания для использования сигнальной продукции DC. Поскольку нагруженные антигеном зрелые DC уже процессируют антиген и обладают способностью представлять антиген иммунным клеткам, указанные клетки, если они активированы, могут не только быстро вызывать антиген-специфические иммунные ответы, но они также могут продуцировать сигнал для дальнейшего действия иммунного ответа. Нет сомнения, что эффективное криосохранение нагруженных антигеном зрелых DC может обеспечить способ быстрого получения функционально активных клеток для иммунотерапии. Криосохранение таких клеток еще не было продемонстрировано по существу. Таким образом, существует назревшая потребность в данной области в эффективных и управляемых средствах криосохранения активированных DC способом, который сохраняет эффективную антигенную презентацию и профили сигнальной секреции свежеполученных DC. Настоящее изобретение отвечает указанной потребности.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретение включает способ создания нагруженных антигеном, активированных дендритных клеток (DC) для применения в иммунотерапии, способ включает: нагрузку по меньшей мере одним антигеном DC; активациу DC по меньшей мере одним агонистом TLR; криосохранение DC; и размораживание DC; где DC производит эффективное количество по меньшей мере одного цитокина для создания T-клеточного ответа.
В одном варианте осуществления антиген представляет собой опухолевый антиген. В другом варианте осуществления антиген представляет собой микробный антиген. В еще одном варианте осуществления агонист TLR представляет собой LPS. В еще одном варианте осуществления криосохранение включает замораживание DC при температуре приблизительно -70°C или ниже. В еще одном варианте осуществления степень извлечения и жизнеспособность DC после размораживания выше или равны приблизительно 70%. В еще одном варианте осуществления степень извлечения и жизнеспособность DC после размораживания выше или равны приблизительно 80%. В еще одном варианте осуществления DC киосохраняют в течение по меньшей мере приблизительно одной недели. В еще одном варианте осуществления цитокин представляет собой IL12. В еще одном варианте осуществления DC проявляет киллерную функцию, в результате чего DC способны лизировать раковые клетки-мишени.
Изобретение также включает способ активации иммунного ответа у млекопитающего, способ включает введение ранее криосохраненной композиции, содержащей нагруженные антигеном, активированные DC, млекопитающему, которое в этом нуждается, где DC нагружают антигеном и активируют перед тем как их криосохраняют.
В одном варианте осуществления антиген представляет собой опухолевый антиген. В другом варианте осуществления антиген представляет собой микробный антиген. В еще одном варианте осуществления агонист TLR представляет собой LPS. В еще одном варианте осуществления криосохранение включает замораживание DC при температуре приблизительно -70°C или ниже. В еще одном варианте осуществления степень извлечения и жизнеспособность DC после размораживания выше или равны приблизительно 70%. В еще одном варианте осуществления степень извлечения и жизнеспособность DC после размораживания выше или равны приблизительно 80%. В еще одном варианте осуществления DC криосохраняют по меньшей мере в течение приблизительно одной недели. В еще одном варианте осуществления цитокин представляет собой IL12. В еще одном варианте осуществления DC проявляет киллерную функцию, в результате чего DC способны лизировать раковые клетки-мишени.
Изобретение также включает сохраняемую композицию для вызова иммунного ответа у млекопитающего, композиция включает: DC, нагруженную по меньшей мере одним антигеном; где DC активирована путем воздействия по меньшей мере одного агониста TLR; и где DC продуцирует эффективное количество по меньшей мере одного цитокина для развития T-клеточного ответа, независимо от того была ли композиция криосохранена.
В одном варианте осуществления антиген представляет собой опухолевый антиген. В другом варианте осуществления антиген представляет собой микробный антиген. В еще одном варианте осуществления агонист TLR представляет собой LPS. В еще одном варианте осуществления композицию криосохраняют при температуре приблизительно -70°C или ниже. В еще одном варианте осуществления степень извлечения и жизнеспособность DC после размораживания выше или равны приблизительно 70%. В еще одном варианте осуществления степень извлечения и жизнеспособность DC после размораживания выше или равны приблизительно 80%. В еще одном варианте осуществления композицию криосохраняют по меньшей мере в течение по меньшей мере приблизительно одной недели. В еще одном варианте осуществления цитокин является IL12. В еще одном варианте осуществления DC проявляет киллерную функцию, в результате чего DC способны лизировать раковые клетки-мишени.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
С целью иллюстрации изобретения, определенные варианты осуществления изобретения изображаются на чертежах. Однако изобретение не ограничивается определенными схемами и техническими средствами вариантов осуществления, изображенных на чертежах.
На фигуре 1 сравнивают традиционные DC (vac-DC) и ICAIT-DC в отношении продукции панели цитокинов и хемокинов, а также киллерную функцию, при помощи которой указанные клетки способны лизировать клетки линий рака молочной железы.
Фигура 2 иллюстрирует традиционно созревавшие DC и ICAIT-DC, успешно сенсибилизирующие T-клетки против опухолевого антигена, при этом только ICAIT-DC подготавливают T-клетки для фактического распознавания HER-2-экспрессирующих опухолей.
Фигура 3, включающая фигуры 3A и 3B, иллюстрирует аналогичные показатели жизнеспособности и восстановления активности свежих в сравнении с криосохраненными и размороженными ICAIT-DC. Используя тест вытеснения трипанового синего, жизнеспособность и показатель восстановления активности криосохраненных ICAIT DC определяли перед криосохранением и сразу после размораживания и отмывки (с помощью цинтрифугирования) криосохраненных клеток. Фигура 3A иллюстрирует три отдельных случая из набора из двух партий, тогда как фигура 3B иллюстрирует два отдельных случая из набора из двух партий. Сравнивают жизнеспособность свежеполученных и криосохраненных DC1. Степень извлечения криосохраненных DC1 составляет обычно 80-90%, и является значительно выше, чем аналогичный показатель для свежеполученных DC1, вследствие потери клеток при сборе свежеполученных DC1.
Фигура 4, включающая фигуры 4A-4F, иллюстрирует превосходные уровни продукции IL12 DC1 после размораживания при криосохранении. Неприрывную продукцию сигнала 3 (IL12) измеряли методом ELISA. На фигуре 4A показано, что продукция IL12 свежими и криосохраненными цитокин-зависимыми DC (CMDC) является значительно ниже, чем продукция IL12, наблюдаемая у свежих и криосохраненных DC1. Фигура 4B иллюстрирует продукцию IL12 DC1 перед криосохранением, через 2 часа после размораживания и через 12 часов после размораживания. На фигурах 4C-4F показана продукция IL12 криосохраненными DC1 в сравнении с DC1, полученными из криосохраненных моноцитов.
Фигура 5 иллюстрирует уровни INFγ, измеренные для криосохраненных DC1 в сравнении с DC1, полученными из криосохраненных моноцитов. Два образца очищенных аллогенных клеток CD4 (1×106/лунку) культивировали совместно с криосохраненными стимулированными агонистом TLR DC (1×105/лунку) в сравнении с DC1, полученными из криосохраненных моноцитов. Через 9 дней T-клетки собирали и повторно стимулировали на планшетах, покрытых антителом anti-CD3 и anti-CD28. Уровни IFNγ (продуцируемого T-клетками) анализировали в супернатанте через 24 часа.
Фигура 6 иллюстрирует CD4+CD25+T-клетки, ингибирующие пролиферацию иммунореактивных клеток в присутствии незрелых, но не DC1 дендритных клеток. CFSE-меченные нефракционированные реактивные лимфоциты в количестве 2,5×105 культивировали совместно с 1×105 незрелых дендритных клеток (iDC), дендритных клеток, созревавших с применением IFN-γ/LPS (LPS-активированные DC) или дендритных клеток, созревавших с применением подходящего цитокинового коктейля (CMM) в течение 5 дней. Как отмечается, вносили 1,25×105 очищенных сортированных по CD4+CD25+T-клеток (Treg). Пролиферация иммунореактивных клеток показана для CD4-селектированных и CD8-селектированных T-клеток. Представленные данные отражают 10 экспериментов. Также представлена пролиферация CD4-положительных реактивных клеток в присутствии Treg и незрелых дендритных клеток, обработанных в течение короткого периода LPS (15 минут) перед совместным культивированием.
Фигура 7, включающая фигуры 7A-7D, иллюстрирует ингибирование функции Treg дендритными клетками DC1, обусловленное растворимым фактором, но IL-6 и IL-12-независимое. На фигуре 7A показаны 1,25×105 сортированных, очищенных Treg, культивированных совместно с 1×105 незрелых дендритных клеток или LPS-активированных дендритных клеток. Показана экспрессия апоптотических маркеров Аннексин-V и 7-AAD через 24 часа. В столбчатом графике обобщен процент клеток, экспрессирующих оба маркера (+/+), только Аннексин-V (+/-), только 7-AAD (-/+), или ни одного маркера (-/-). На фигуре 7B показаны, как отмечено, 1,25×105 сортированных, очищенных Treg, культивированных совместно с 2,5×105 CFSE-меченных нефракционированных реактивных лимфоцитов в присутствии 1×105 незрелых или LPS-активированных дендритных клеток. Кроме того, 1×105 незрелых или LPS-активированных дендритных клеток добавляли на полупроницаемую мембрану Transwell®, помещенную в лунку с культурой, как отмечено. На фигуре 7C показаны 1,25×105 сортированных, очищенных Treg, культивированных совместно с 2,5×105 CFSE-меченных нефракционированных реактивных лимфоцитов и с 1×105 LPS-активированных дендритных клеток в присутствии 5 мкг/мл нейтрализующего антитела anti-IL-6 или anti-IL-12. Представленные данные отражают по меньшей мере 3 отдельных эксперимента в каждом отдельном случае. На фигуре 7D показаны Treg или CFSE-меченные нефракционированные реактивные лимфоциты, культивированные в 500 мкл культуральной среды и 500 мкл среды, взятой из культур LPS-активированных дендритных клеток приблизительно через 10 часов после добавления LPS (1×106 клеток на мл). Через 24 часа указанные "обработанные" популяции использовали в совместных культурах в стандартном отношении (1:2 Treg:реактивные клетки), как отмечено. Данные отражают 2 отдельных эксперимента.
На фигуре 8, включающей фигуры 8A и 8B, показаны супрессорные CD4+CD25+T-клетки, секретирующие эффекторные цитокины в присутствии дендритных клеток DC1. На фигуре 8 A показаны 2,5×105 сортированных по CD4+CD25+(Treg) или CD4+CD25- (Teff) T-клеток, в сочетании с 2,0×105 незрелых или LPS-активированных дендритных клеток. Супернатанты собирали через 5 дней и использовали метод ELISA для измерения количества IFN-γ, представленного в супернатанте. На 5-й день некоторые культуральные образцы пермеабилизировали и внутриклеточный IFN-γ обнаруживали с помощью проточной цитометрии. 34,4% CD4 T-клеток экспрессировали IFN-γ внутриклеточно (N=3). На фигуре 8B показаны 1,25×105 CD4+CD25+T-клеток, культивированных совместно с 1×105 незрелых или LPS-активированных DC. В некоторые образцы вносили нейтрализующее антитело anti-IL12 (5 мкг/мл). Через 48 часов клетки собирали, пермеабилизировали и внутриклеточную экспрессию T-bet и FoxP3 определяли с помощью внутриклеточного окрашивания. Результаты представлены для CD4-положительных клеток, дающих при сортировке сигнал выше порогового значения (N=3).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к получению и криосохранению зрелых, нагруженных антигеном DC, активированных агонистами Toll-подобного рецептора, для индукции клинически эффективных иммунных ответов, предпочтительно при раннем применении в патологическом процессе. DC настоящего изобретения обладают способностью обуславливать сильные Th1 клеточные ответы, благодаря продукции цитокинов и химокинов, и также обладают способностью вызывать апоптоз опухолевых клеток. Способы развития DC настоящего изобретения также обеспечивают платформу для таргетинга новых молекул и раковых стволовых клеток, что может устранить клетки с высоким метастатическим потенциалом. Настоящее изобретение также относится к криосохранению указанных активированых DC способом, который сохраняет их активность и функциональность при презентации антигена, а также продукцию ими различных цитокинов и хемокинов после размораживания.
Определения
В данном контексте каждый из нижеследующих терминов имеет значение, соответствующее ему в данном разделе.
Неопределенные артикли используются в описании для обозначения одного или более чем одного (т.е. по меньшей мере один) грамматического объекта артикля. В качестве примера, "элемент" означает один элемент или более чем один элемент. Термин "приблизительно" понятен специалистам в данной области, и изменяется до некоторой степени, исходя из контекста, в котором он используется.
Термин "антитело" в данном контексте обозначает молекулу иммуноглобулина, которая способна специфически связываться со специфическим эпитопом антигена. Антитела могут представлять собой интактные иммуноглобулины, полученные из природных источников или из рекомбинантных источников, и могут представлять собой иммуноактивные части интактных иммуноглобулинов. Антитела обычно представляют собой тетрамеры молекул иммуноглобулинов. Антитела в настоящем изобретении могут существовать в различных формах включая, например, поликлональные антитела, моноклональные антитела, Fv, Fab и F(ab)2, а также одноцепочечные антитела и гуманизированные антитела (Harlow et al., 1988; Houston et al., 1988; Bird et al., 1988).
Термин "антиген" или "ag" в данном контексте определяется как молекула, которая вызывает иммунный ответ. Указанный иммунный ответ может включать либо продукцию антител, либо активацию специфических иммунокомпетентных клеток, или оба указанных процесса. Специалисту в данной области понятно, что любая макромолекула, включая практически все белки или пептиды, может служить в качестве антигена. Кроме того, антигены могут быть получены из рекомбинантной или геномной ДНК. Специалисту в данной области понятно, что любая ДНК, которая содержит нуклеотидные последовательности или неполную нуклеотидную последовательность, кодирующую белок, который вызывает иммунный ответ, соответственно, кодирует "антиген", в том виде как этот термин используется в описании. Кроме того, специалисту в данной области понятно, что антиген необязательно кодируется исключительно полноразмерной нуклеотидной последовательностью гена. Совершенно очевидно, что настоящее изобретение включает, но без ограничения, применение неполных нуклеотидных последовательностей более чем одного гена и что указанные нуклеотидные последовательности располагают в различных комбинациях, чтобы вызвать желаемый иммунный ответ. Кроме того, специалисту в данной области понятно, что антиген совсем необязательно должен кодироваться "геном". Совершенно очевидно, что антиген может быть создан или может быть получен из биологического образца. Указанный биологический образец может включать, но без ограничения образец ткани, образец опухоли, клетку или биологичекую жидкость.
"Антигенпрезентирующая клетка" (APC) представляет собой клетку,которая способна активировать T-клетки, и включает, но без ограничения, моноциты/макрофаги, B-клетки и дендритные клетки (DC).
Термин "дендритная клетка" или "DC" относится к любому члену смешанной популяции морфологически сходных клеточных типов, обнаруживаемых в лимфоидной или не лимфоидной тканях. Указанные клетки характеризуются отличительной морфологией и высокими уровнями экспрессии поверхностных молекул MHC класса II. DC можно выделить из ряда тканевых источников. DC обладает высокой способностью к сенсибилизации MHC-рестриктированных T-клеток и является очень эффективной в презентации антигенов T-клеткам in situ. Антигены могут представлять собой аутоантигены, которые экспрессируются во время T-клеточного развития и толерантности, и чужеродные антигены, которые представлены во время обычных иммунных процессов.
В данном контексте "активированная DC" представляет собой DC, которую подвергали воздействию агониста Toll-подобного рецептора. Активированная DC может быть нагружена или не нагружена антигеном.
Термин "зрелая DC" в данном контексте определяется как дендритная клетка, которая экспрессирует молекулы, включая высокие уровни молекул MHC класса II, CD80 (B7. 1) и CD86 (B7.2). Напротив, незрелые дендритные клетки экспрессируют низкие уровни молекул MHC класса II, CD80 (B7.1) и CD86 (B7.2), в то же время еще могут охватывать антиген.
"Нагруженная антигеном APC" или "примированная антигеном APC" включает APC, которую подвергали воздействию антигена и которая активирована антигеном. Например, APC может стать Ag-нагруженной in vitro, например, в процессе культивирования в присутствии антигена. APC также может быть нагружена in vivo в результате воздействия антигена. Обычно "нагруженную антигеном APC" получают одним из двух способов: (1) небольшие пептидные фрагменты, известные как антигенные пептиды, “примируются” непосредственно на наружную поверхность APC; или (2) APC инкубируют с целыми белками или белковыми частицами, которые затем поглощаются APC. Указанные белки перевариваются APC в небольшие пептидные фрагменты и впоследствии транспортируются и презентируются на поверхности APC. Кром того, APC, нагруженная антигеном, также может быть создана путем введения в клетку полинуклеотида, кодирующего антиген.
Термин "аутоиммунное заболевание" в данном контексте определяется как нарушение, которое возникает в результате аутоиммунного ответа. Аутоиммунное заболевание является результатом неадекватного и чрезмерного ответа на аутоантиген. Примеры аутоиммунных заболеваний включают, но без ограничения болезнь Аддисона, очаговая алопеция, анкилозирующий спондилит, аутоиммунный гепатит, аутоиммунный паротит, болезнь Крона, диабет (1-го типа), дистрофический буллезный эпидермолиз, эпидидимит, гломерулонефрит, диффузный тиреотоксический зоб, синдром Гийена-Барре, тиреоидит Хашимото, гемолитическая анемия, системная красная волчанка, рассеянный склероз, злокачественная миастения, pemphigus vulgaris, псориаз, острая ревматическая лихорадка, ревматоидный артрит, саркоидоз, склеродермия, синдром Шегрена, спондилоартропатии, тиреоидит, васкулит, витилиго, myxedema, пернициозная анемия, язвенный колит, наряду с прочим.
В данном контексте термин "аутогенный" предназначен для обозначения любого материала, полученного от того же самого субъекта, которому указанный материал должен быть позднее повторно введен.
Термин "рак" в данном контексте определяется как заболевание, характеризующееся быстрым и неконтролируемым ростом аберрантных клеток. Раковые клетки могут распространяться местно или по кровотоку и лимфатической системе к другим частям организма. Примеры различных раков включают, но без ограничения рак молочной железы, рак предстательной железы, рак яичников, рак шейки матки, рак кожи, рак поджелудочной железы, колоректальный рак, рак почки, рак печени, рак мозга, лимфома, лейкемия, рак легких и т.п.
Термин "криосохраненный" или "криосохранение" в данном контексте относится к клеткам, которые ресуспендируют в криосреде и замораживают при температуре приблизительно -70°C или ниже.
Термин "криосреда" в данном контексте относится к любой среде, смешанной с клеточным образцом при подготовке к замораживанию, такой, что по меньшей мере некоторые из клеток в клеточном образце могут восстанавливаться и остаются жизнеспособными после размораживания.
"Антиген донора" относится к антигену, экспрессированному донорной тканью, которую трансплантируют реципиенту.
"Антиген реципиента" относится к мишени для иммунного ответа для донорного антигена.
В данном контексте "эффекторная клетка" обозначает клетку, которая опосредует иммунный ответ против антигена. Пример эффекторной клетки включает, но без ограничения T-клетку и B-клетку.
В данном контексте "эндогенный" относится к любому материалу, полученному из или произведенному внутри организма, клетки, ткани или системы.
В данном контексте термин "экзогенный" относится к любому материалу, введенному извне или произведенному вне организма, клетки, ткани или системы.
Термин "эпитоп" в данном контексте определяется как небольшая химическая молекула на антигене, которая может вызвать иммунный ответ, индуцируя B- и/или T-клеточные ответы. Антиген может иметь один или более эпитопов. Большинство антигенов имеют много эпитопов; т.е. они являются мультивалентными. В общем, по величине эпитоп представляет собой приблизительно пять аминокислот и/или сахаров. Специалисту в данной области понятно, что, в целом, общая трехмерная структура, скорее чем специфичная линейная последовательность молекулы, является главным критерием антигенной специфичности и следовательно разграничивает один эпитоп от другого.
Термин "хелперная T-клетка" в данном контексте определяется как эффекторная T-клетка, главная функция которой заключается в обеспечении активации и функций других B- и T-лимфоцитов и/или макрофагов. Большинство хелперных T-клеток представляют собой CD4 T-клетки.
В данном контексте "иммуноген" относится к веществу, которое способно стимулировать или вызывать гуморальный антителовисимый и/или клоточно-опосредованный иммунный ответ у млекопитающего. Термин "иммуноглобулин" или "Ig" в данном контексте определяется как класс белков, которые действуют как антитела. Пять членов, входящих в данный класс белков, представляют собой IgA, IgG, IgM, IgD, и IgE. IgA является основным антителом, которое представлено в секретах тела, таких как слюна, слезы, грудное молоко, желудочно-кишечные секреты и слизистые секреты респираторного и мочеполового трактов. IgG является самым распространённым циркулирующим антителом. IgM является главным иммуноглобулином, продуцируемым при первичном иммунном ответе у большинства млекопитающих. IgM является самым эффективным иммуноглобулином в реакциях агглютинации, фиксации комплемента и других гуморальных иммунных ответах, и важен при защите против бактерий и вирусов. IgD представляет собой иммуноглобулин, для которого не известна функция антитела, но может служить в качестве антигенного рецептора. IgE является иммуноглобулином, который опосредует гиперчувствительность немедленного типа, вызывая высвобождение медиаторов из тучных клеток и базофилов после воздействия аллергена.
Термин "главный комплекс гистосовместимости", или "MHC", в данном контексте определяется как специфический кластер генов, многие из которых кодируют эволюционно связанные белки клеточной поверхности, участвующие в презентации антигена, которые принадлежат к наиболее важным детерминантам гистосовместимости. Молекулы класса I MHC, или MHC-I, участвуют, главным образом, в презентации антигена CD8 T-лимфоцитам. Молекулы класса II MHC, или MHC-II, участвуют, главным образом, в презентации антигена CD4 T-лимфоцитам.
В данном контексте термин "модулировать" предназначен для обозначения любого изменения в биологическом состоянии т.е. увеличения, снижения и т.п.
Термин "полипептид" в данном контексте определяется как цепь аминокислотных остатков, обычно характеризующаяся определенной последовательностью. В данном контексте термин полипептид взаимно включает термины "пептид" и "белок".
Термин "аутоантиген" в данном контексте определяется как антиген, который экспрессируется клеткой или тканью хозяина. Аутоантигены могут быть опухолевыми антигенами, но в определенных вариантах осуществления, экспрессируются так в нормальных, так и в опухолевых клетках. Специалистам в данной области понятно, что аутоантиген может экспрессироваться в клетке в повышенных количествах. В данном контексте "по существу очищенная" клетка представляет собой клетку, которая находится в основном без других типов клеток. “По существу очищенная клетка” также относится к клетке, которая отделена от других типов клеток, с которыми она обычно ассоциирована в естественном состоянии. В отдельных случаях популяция по существу очищенных клеток обозначает гомогенную популяцию клеток. В других случаях указанный термин относится просто к клеткам, которые отделили от клеток, с которыми они обычно ассоциированы в естественном состоянии. В некоторых вариантах осуществления клетки являются культурой in vitro. В других вариантах осуществления клетки не культивируют in vitro.
Термин "T-клетка" в данном контексте определяется как клетка тимусного происхождения, которая участвует в ряде клеточно-опосредованных иммунных реакций.
Термин "B-клетка" в данном контексте определяется как клетка, полученная из костного мозга и/или селезенки. B-клетки могут развиваться в плазматические клетки, которые продуцируют антитела.
Термин "Toll-подобный рецептор" или "TLR" в данном контексте определяется как класс белков, который играет роль во врожденной иммунной системе. TLR представляют собой одиночные трансмембранные, некаталитические рецепторы, которые распознают структурно консервативные молекулы микробного происхождения. TLR активируют иммунные клеточные ответы после связывания с лигандом.
Термин "агонисты Toll-подобного рецептора" или "агонисты TLR" в данном контексте определяется как лиганд, который связывается с TLR для активации иммунного клеточного ответа.
В данном контексте "терапевтически эффективное количество" представляет собой количество терапевтической композиции, достаточное, чтобы обеспечить благоприятный эффект у млекопитающего, которому вводят композицию.
Термин "вакцина" в данном контексте определяется как материал, применяемый для возбуждения иммунного ответа после введения материала животному, предпочтительно млекопитающему, и более предпочтительно человеку.
Диапазоны: в настоящем раскрытии, различные аспекты изобретения могут быть представлены в формате диапазонов. Следует понимать, что описание в формате диапазонов используется только для удобства и краткости и не должно истолковываться как жесткое ограничение объема изобретения. Таким образом, описание диапазона следует рассматривать как содержащее конкретно раскрытые все возможные поддиапазоны, а также индивидуальные цифровые значения в пределах указанного диапазона. Например, описание диапазона, такого как от 1 до 6 следует рассматривать как содержащее конкретно раскрытые поддиапазоны, такие как от 1 до 3, от 1 до 4, от 1 до 5, от 2 до 4, от 2 до 6, от 3 до 6, etc., а также отдельные цифровые значения в пределах указанного диапазона, например, 1, 2, 2,7, 3, 4, 5, 5,3 и 6. Указанное выше применимо независимо от ширины диапазона.
Описание
Как предполагается в данном описании, настоящее изобретение предоставляет способ получения и криосохранения DC с улучшенными функциональными свойствами для продукции более сильных сигналов для T-клеток, и что, соответственно, дает в результате более сильную противоопухолевую вакцину на основе DC. Благодаря эффективному криосохранению таких клеток, образцы можно хранить и размораживать для последующего использования, что уменьшает необходимость повторения процессов афереза и элютриации во время приготовления вакцины. Указанные способы также могут быть использованы для непосредственного воздействия на молекулы, вовлеченные в канцерогенные сигнальные пути и на раковые стволовые клетки (CSC).
Настоящее изобретение включает зрелые, нагруженные антигеном DC, активированные агонистами Toll-подобного рецептора, которые вызывают клинически эффективные иммунные ответы, предпочтительно при раннем применении в патологическом процессе. DC настоящего изобретения продуцируют желаемые уровни цитокинов и хемокинов, и, кроме того, обладают способностью вызывать апоптоз опухолевых клеток.
Настоящее изобретение также демонстрирует, что лиганды TLR не только активируют презентирующие клетки, но также ингибируют регуляторные клетки, действие которых ограничивает адаптивные ответы. В определенных вариантах осуществления показано, что передача сигнала с помощью многочисленных Toll-подобных рецепторов, включая TLR-2, TLR-4, TLR-8, и TLR-9, отменяет супрессию под влиянием иммунорегуляторных CD4+CD25+Foxp3+T-клеток (обозначаемых здесь как Treg). В описании показано, что TLR-4-активированные дендритные клетки не только ингибируют эффекты Treg в отношении реактивных клеток, но также, по-видимому, превращают сами регуляторы в IFN-γ продуцирующие эффекторы.
Настоящее изобретение также относится к криосохранению указанных активированных DC способом, который сохраняет их активность и функциональность при представлении антигена, а также продукцию ими различных цитокинов и хемокинов после размораживания, так что криосохраненные и затем размороженные активированные DC являются такими же клинически эффективными как свежеполученные и активированные DC.
Иммунотерапия на основе DC
DC образуются из полипотентных моноцитов, которые выступают в качестве антигенпрезентирующих клеток (APC). DC распространены повсеместно в периферических тканях, где они готовы захватывать антигены. После захвата антигена DC процессируют антиген на небольшие пептиды и двигаются к вторичным лимфоидным органам. В лимфоидных органах DC представляют антигенные пептиды “наивным” T-клеткам, вызывая, таким образом, каскад сигналов которые задают направление T-клеточной дифференцировке. После воздействия DC представляют молекулы антигена, связанные либо со связывающими пептидами MHC класса I либо класса II и активируют CD8+ или CD4+ T-клетки соответственно (Steinman, 1991, Anmi. Rev. Immunol. 9:271-296; Banchereau et al, 1998, Nature392,245-252; Steinman, et al., 2007, Nature 449:419-426; Ginhoux et al., 2007, J. Exp. Med, 204:3133-3146; Banerjee et al., 2006, Blood 108:2655-2661; Sailusto et al., 1999, J. Exp. Med. 189:61 1-614; Reid et al" 2000, Curr. Opin. Immunol.12:1 14-121; Bykovskaia et al., 1999, J. Leukoc. Biol. 66:659-666; Clark et al., 2000, Microbes Infect. 2:257-272).
DC ответственны за индукцию, координацию и регуляцию адаптивного иммунного ответа и также помогают организовать взаимодействие между эффекторами врожденной части и адаптивной части иммунной системы. Указанные особенности делают DC кандидатами, имеющими большие шансы на успех при использовании в иммунотерапии. DC обладают исключительной способностью “брать пробы” из окружающей среды с помощью макропиноцитоза и эндоцитоза, опосредованного рецепторами (Gerner et al., 2008, J. Immunol.1 81:155-164; Stoitzner et al., 2008, Cancer Immunol. Immunother 57:1665-1673; Lanzevecchia A., 1996, Curr. Opin. Immunol.8:348-354; Delamarre et al., 2005, Science, 307(5715):1630-1634).
DC также нуждаются в сигналах созревания для повышения своей антиген-презентирующей способности. DC активируют экспрессию поверхностных молекул, таких как CD80 и CD86 (также известные в качестве молекул второго сигнала) путем предоставления дополнительных сигналов созревания, таких как INF-α, CD40L или агентов кальциевого сигнального пути (Czerniecki et al., 1997. J. Immunol.159:3823-3837; Bedrosian et al. 2000, J. Immunother. 23:31 1-320; Mailiiard et al., 2004, Cancer Res.64,5934-5937; Brossart et al., 1998, Blood 92:4238-4247; Jin et al., 2004, Hum. Immunol. 65:93-103). Установлено, что смесь цитокинов, содержащая TNF-a, IL-1β, IL-6 и простагландин E2 (PGE2), обладает способностью вызывать созревание DC (Jonuleit, et al., 2000, Arch. Derm. Res. 292:325-332). DC также могут созревать с помощью ионофора кальция перед тем как примируются антигеном.
Помимо патоген-распознающих рецепторов, таких как PKR и MDA-5 (Kalali et al., 2008, J. Immunol. 181:2694-2704; Nallagatla et al., 2008, RNA Biol, 5(3):140-144), DC также содержат серии рецепторов, известных как Toll-подобные рецепторы (TLR), которые также способны распознавать опасные патогены. Когда указанные TLR приводятся в действие, индуцируются серии активационных изменений в DC, которые приводят к созреванию и передаче сигналов T-клеток (Boullart et al. 2008, Cancer Immunol. Immunother. 57(11):1589-1597; Kaisho et al., 2003, Curr. Mot. Med. 3(4):373-385; Pulendran et al., 2001, Science 293(5528):253-256; Napolitani et al, 2005, Nat. Immunol. 6(8):769-776). DC могут активировать и усиливать различные ветви клеточно-опосредованного ответа, такие как натуральные киллерные γ-δ T и α-β T-клетки и, после активации DC сохраняют свою иммуногенность (Steinman, 1991, Annu. Rev. Immunol. 9:271-296; Banchereau et al., 1998, Nature 392:245-252; Reid et al., 2000, Curr. Opin. Immunol. 12:114-121 ; Bykovskaia et al., 1999, J. Leukoc. Biol.66:659-666; Clark et al., 2000, Microbes Infect. 2:257-272).
DC-продукция сигнала
Считают, что зрелые DC более эффективны в активации T-клеточно-опосредованных иммунных ответов (Jonuleit et al., 2001, Int. J. Cancer, 93:243-251; Prabakaran et al., 2002, Ann. Surg. Oncol. 9:41 M 18; Xu et al., 2003, J. Immunol. 171:225 1-2261). Зрелые DC способны к формированию более сильного T-клеточного ответа по сравнению с незрелыми DC, отчасти потому что специфические цитокины секретируются зрелыми DC, которые потенцируют более сильный и более действенный T-клеточный ответ. Например, зрелые DC продуцируют IL-12 после взаимодействия с CD4 T-клетками (Koch et al., 1996, J. Exp. Med. 184:741-746; Heifler et al., 1996, Eur. J. Immunol. 26:659-668). DC, которые секретируют Th1-запускающие цитокины, такие как IL-12, IL-18 и IL-23, обозначают как поляризованные DC типа 1, или DC1 (Kalinski, et al., 1999, Immunol. Today 20:561-567; Lanzavecchia et al" 2000, Science 290(5489):92-97).
Цитокины, продуцируемые зрелыми DC, изменяют эффекты на T-клеточный ответ. Например, IL-12, гетородимерный цитокин, продуцируется DC и является основным в образовании IFN-γ-секретирующих CD4+ и CD8+ T-клеток и в усилении антибактериального и противоопухолевого ответов (Gee et al., 2009, Inflamm. Allergy Drug Targets 8:40-52). IL-12 также может ингибировать рост первичной опухоли, а также метастатических опухолевых клеток в мышиных моделях карциномы яичника (OV-HM) (Tatsumi et al., 2001, Cancer Res. 61:7563-7567). IL-12 также может опосредовать образование высоко авидных противоопухолевых T-клеток (Xu et al., 2003, J. Immunol. 171:2251-2261), таким образом, усиливая противоопухолевую T-клеточную функцию. DC также продуцируют хемокины, в качестве четвертого сигнала, который приводит к накоплению T-клеток и затем воздействует на дальнейшие T-клеточные ответы (Xiaoet al., 2003, Cytokine 23:126-132).
DC могут секретировать другие цитокины, которые затем влияют на T-клеточную активацию. Например, DC могут секретировать IL-1, IL-6 и IL-23, которые активируют Th17 клетки. Th17 клетки представляют собой недавно охарактеризованную субпопуляцию провоспалительных T-клеток, которые обеспечивают устранение патогена и тканевое воспаление посредством продукции своего отличительного цитокина, IL-17 (Kikly et al., 2006, Curr, Opin. Immunol. 18:670-675). Продукция IL-12 может приводить к более сильному ответу Th1, тогда как продукция IL-23 может приводить к созреванию Th17 клеток. В сущности, DC, продуцирующие IL-12, могут поляризовать преимущественно Th1 ответ в присутствии IL-23, однако в отличие от этого, DC, которые продуцируют IL-23 в отсутствии IL-12, поляризуют сильный ответ Th17 (Roses et al., 2008, J. Immunol. 181:5120-5127; Acosta-Rodriguez et al., 2007, Nat. Immunol. 8:639-646). Таким образом, поскольку специфические DC-секретированные цитокины оказывают такое сильное влияние на T-клеточную функцию, определение цитокинового профиля зрелых DC является важным показателем, от которого может зависеть образование потенциальных T-клеточных эффекторов. Таким образом, зрелые DC можно охарактеризовать более эффективно по их основной цитокиновой продукции и последующему действию сигнала на T-клетки, чем по более традиционному описанию исключительно в соответствии с экспрессией поверхностных молекул.
Получение нагруженной (примированной) иммунной клетки
Настоящее изобретение включает клетку, которая подвергалась воздействию антигена или иным образом "примирована" антигеном. Например, APC, такая как DC, может стать Ag-нагруженной in vitro, например, при культивировании ex vivo в присутствии антигена, или in vivo при воздействии антигена.
Специалисту в данной области также понятно, что APC может быть "примирована" способом, при котором APC подвергается воздействию антигена в течение времени, достаточного для обеспечения презентации антигена на поверхности APC. Например, APC может подвергаться воздействию антигена в форме небольших пептидных фрагментов, известных как антигенные пептиды, которые "примируются" непосредственно на наружную поверхность APC (Mehta-Damani et al., 1994); или APC можно инкубировать с целыми белками или белковыми частицами, которые затем перевариваются APC. Указанные целые белки перевариваются в небольшие пептидные фрагменты APC и впоследствии переносятся и презентируются на поверхности APC (Cohen et ah, 1994). Антиген в пептидной форме может быть представлен клетке с помощью стандартных технологий “примирования”, описанных здесь.
Вне связи с какой-либо теорией, антиген в форме чужеродного антигена или аутоантигена процессируется APC изобретения, чтобы сохранить иммуногенную форму антигена. Иммуногенная форма антигена подразумевает процессинг антигена с помощью фрагментации для получения формы антигена, которая может распознаваться и стимулировать иммунные клетки, например, T-клетки. Предпочтительно, такой чужеродный антиген или аутоантиген представляет собой белок, который процессируется в пептид APC. Соответствующий пептид, который продуцируется APC, можно выделить и очистить для применения в качестве иммуногенной композиции. Пептиды, процессированные APC, также можно использовать для индукции толерантности к белкам, процессированным APC.
Нагруженные антигеном APC, иначе называемые "примированные APC" изобретения, получают путем воздействия антигена на APC in vitro или in vivo. В случае, когда APC примируется in vitro, APC могут высевать на чашку для культивирования и подвергать воздействию антигена в достаточном количестве и в течение достаточного периода времени, чтобы антиген мог связаться с APC. Количество и время, необходимые для обеспечения связывания антигена с APC, можно определить путем применения методов, известных в данной области или же раскрытых в описании. Другие методы, известные специалистам в данной области техники, например иммунологические анализы или анализы связывания могут использоваться для обнаружения присутствия антигена на APC после воздействия антигена.
В другом варианте воплощения изобретения APC может быть трансфицирована вектором, который предусматривает экспрессию специфического белка APC. Белок, который экспрессируется APC, затем может быть процессирован и представлен на клеточной поверхности. Трансфецированная APC затем может быть использована в качестве иммунногенной композиции для создания иммунного ответа на белок, кодируемый вектором.
Как указано в другом разделе настоящего описания, могут быть изготовлены векторы, которые включают специфический полинуклеотид, который кодирует и экспрессирует белок, на который необходим иммунный ответ. Предпочтительно для инфицирования клетки используют ретровирусные векторы. Более предпочтительно для инфицирования клетки используют аденовирусные векторы.
В другом варианте осуществления вектор можно нацелить на APC путем модификации вирусного вектора для кодирования белка или его частей, которые распознаются рецептором на APC, в результате чего захват рецептора APC вектором будет вызывать эндоцитоз вектора, обеспечивая процессинг и презентацию антигена, кодированного нуклеиновой кислотой вирусного вектора. Нуклеиновая кислота, которая доставляется вирусом, может быть нативной для вируса, если она экспрессируется на APC, кодирует вирусные белки, которые затем процессируются и презентируются на рецепторе MHC APC.
Как предусмотрено в описании, могут быть использованы различные методы для трансфекции полинуклеотида в клетку-хозяин. Методы включают, но без ограничения, осаждение фосфатом кальция, липофекцию, бомбардировку частицами, микроинъекцию, электропорацию, коллоидные дисперсные системы (т.е. макромолекулярные комплексы, нанокапсулы, микросферы, бусины и системы на основе липидов, включающие эмульсии масло-в-воде, мицеллы, смешанные мицеллы и липосомы). Указанные методы подразумеваются в данной области техники и описаны в опубликованной литературе, чтобы предоставить возможность специалисту в данной области осуществить указанные методы.
В другом варианте осуществления полинуклеотид, кодирующий антиген, может быть клонирован в вектор экспрессии, и вектор может быть введен в APC, чтобы получить нагруженную APC другим путем. Различные типы векторов и способы введения нуклеиновых кислот в клетку обсуждаются в доступной опубликованной литературе. Например, вектор экспрессии может быть трансфицирован в клетку-хозяин физическим, химическим или биологическим способами. Смотреть, например, Sambrook et al. (2001, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York), и в Ausubel et al. (1997, Current Protocols in Molecular Biology, John Wiley & Sons, New York). Известно, что введение вектора экспрессии, содержащего полинуклеотид, кодирующий антиген, приводит к образованию примированной клетки. Настоящее изобретение включает различные методы примирования APC, включая, но без ограничения нагрузку APC целым антигеном в форме белка кДНК или мРНК. Однако не следует истолковывать изобретение как ограниченное специфической формой антигена, применяемой для примирования APC. Скорее, изобретение охватывает другие, известные в данной области методы получения антиген-нагруженных APC. Предпочтительно, APC трансфицируют мРНК, кодирующей определенный антиген. мРНК, соответствующая продукту гена, последовательность которого известна, может быть быстро получена in vitro с помощью подходящих праймеров и полимеразной цепной реакции с обратной транскриптазой (RT-PCR) в сочетании с реакциями транскрипции. Трансфекция APC мРНК обеспечивает преимущество по сравнению с другими методами нагрузки антигеном для получения примированной APC. Например, возможность амплифицировать РНК из микроскопического количества ткани, а именно опухолевой ткани, увеличивает применение APC для вакцинации большого числа пациентов.
В отношении антигенной композиции, которую используют в качестве вакцины, антигенная композиция должна вызывать иммунный ответ на антиген у клетки, ткани или млекопитающего (например, человека). В данном контексте "иммунологическа композиция" может содержать антиген (например, пептид или полипептид), нуклеиновую кислоту, кодирующую антиген (например, вектор экспрессии антигена), или клетку, экспрессирующую или презентирующую антиген или клеточный компонент. В конкретных вариантах осуществления антигенная композиция содержит или кодирует весь антиген или часть любого антигена, описанного здесь, или его иммунологически функциональный эквивалент. В других вариантах осуществления антигенная композиция состоит из смеси, которая включает дополнительный иммуностимулирующий агент или нуклеиновые кислоты, кодирующие такой агент. Иммуностимулирующие агенты включают, но без ограничения дополнительный антиген, иммуномодулятор, антигенпрезентирующую клетку или адъювант. В других вариантах осуществления один или более дополнительных агент(ов) ковалентно связывают с антигеном или иммуностимулирующим агентом в любой комбинации. В определенных вариантах осуществления антигенная композиция конъюгирована с или включает аминокислоты якорных мотивов для HLA.
Вакцина, как предусмотрено в описании, может различаться по композиции нуклеиновой кислоты и/или клеточных компонентов. В неограничивающем примере, нуклеиновая кислота, кодирующая антиген, также может быть введена в состав с адъювантом. Разумеется, следует понимать, что различные композиции, описанные в настоящем документе, могут также включать дополнительные компоненты. Например, один или более компонентов вакцины могут быть заключены в липид или липосому. В другом неограничивающем примере вакцина может содержать один или более адъювантов. Вакцина настоящего изобретения и ее различные компоненты могут быть получены и/или введены любым способом, раскрытым здесь, или способом, который должен быть известен специалисту в данной области в свете настоящего раскрытия. Известно, что антигенная композиция настоящего изобретения может быть получена с помощью метода, хорошо известного в данной области техники, включая, но без ограничения химический синтез с помощью твердофазного синтеза и очистку от других продуктов химических реакций с помощью ВЭЖХ, или получение с помощью экспрессии последовательности нуклеиновой кислоты (например, последовательности ДНК), кодирующей пептид или полипептид, содержащий антиген настоящего изобретения в системе трансляции in vitro или в живой клетке. Кроме того, антигенная композиция может содержать клеточный компонент, выделенный из биологического образца. Антигенную композицию выделяют и тщательно диализуют для удаления одной или более нежелательных молекул с низким молекулярным весом и/или лиофилизируют для более легкого включения в желаемую среду для лекарства. Также известно, что дополнительные аминокислоты, мутации, химические модификации и т.п., если имеются, которые производят в компоненте вакцины, предпочтительно не будут существенно препятствовать распознаванию антителами эпитопной последовательности.
Пептид или полипептид, соответствующий одной или более антигенным детерминантам настоящего изобретения, обычно составляет в длину по меньшей мере пять или шесть аминокислотных остатков, и может содержать приблизительно 10, приблизительно 15, приблизительно 20, приблизительно 25, приблизительно 30, приблизительно 35, приблизительно 40, приблизительно 45 или приблизительно 50 остатков или около того. Пептидную последовательность можно синтезировать методами, известными специалистам в данной области, такими как, например, пептидный синтез с применением автоматических пептидных синтезаторов, таких как синтезаторы, поставляемые Applied Biosystems, Inc., Foster City, CA (Foster City, CA).
Более длинные пептиды или полипептиды также могут быть получены, например, рекомбинантными способами. В определенных вариантах осуществления нуклеиновая кислота, кодирующая антигенную композицию и/или компонент, описанный здесь, может быть использована, например, для получения an антигенной композиции in vitro или in vivo для различных композиций и методов настоящего изобретения. Например, в определенных вариантах осуществления нуклеиновая кислота, кодирущая антиген, например, входит в состав, вектора в рекомбинантной клетке. Нуклеиновая кислота может экспрессироваться для продукции пептида или полипептида, содержащего антигенную последовательность. Пептид или полипептид может секретироваться клеткой или может быть включен в виде части клетки или может находиться внутри клетки.
В определенных вариантах осуществления иммунный ответ можно вызвать с помощью трансфекции или инокуляции млекопитающего нуклеиновой кислотой, кодирующей антиген. Одна или более клеток, входящих в млекопитающее-мишень, затем экспрессируют последовательности, кодируемые нуклеиновой кислотой, после введения нуклеиновой кислоты млекопитающему. Вакцина также может быть в форме, например, нуклеиновой кислоты (например, кДНК или РНК), кодирующей целиком всю или часть пептидной или полипептидной последовательности антигена. Экспрессия нуклеиновой кислоты in vivo может происходить, например, с помощью вектора плазмидного типа, вирусного вектора или вектора с вирусно/плазмидной конструкцией. В другом варианте осуществления нуклеиновая кислота включает кодирующую область, которая кодирует все или часть последовательностей, кодирующих соответствующий антиген, или его иммунологически функциональный эквивалент. Разумеется, нуклеиновая кислота может включать в себя и/или кодировать дополнительные последовательности, включая, но без ограничения последовательности, содержащие один или более иммуномодуляторов или адъювантов.
Антигены
Как предусмотрено в описании, настоящее изобретение может включать применение любого антигена, подходящего для нагрузки APC, чтобы вызвать иммунный ответ. В одном варианте осуществления могут использоваться опухолевые антигены. Опухолевые антигены можно разделить на две большие категории: общие опухолевые антигены; и уникальные опухолевые антигены. Общие антигены экспрессируются многими опухолями, тогда как уникальные опухолевые антигены могут возникать вследствие мутаций, вызванных физическими или химическими канцерогенами, и соответственно экспрессируются только индивидуальными опухолями. В определенных вариантах осуществления общие опухолевые антигены нагружают на DC настоящего изобретения. В других вариантах осуществления уникальные опухолевые антигены нагружают на DC настоящего изобретения.
В контексте настоящего изобретения "опухолевый антиген" относится к антигенам, которые являются обычными для специфических гиперпролиферативных нарушений. В определенных аспектах антигены гиперпролиферативного нарушения настоящего изобретения получают из раков, включая, но без ограничения первичную или метастатическую меланому, тимому, лимфому, саркому, рак легких, рак печени, неходжкинскую лимфому, лимфому Ходжкина, лейкемии, рак матки, рак шейки матки, рак мочевого пузыря, рак почки, и аденокарциномы, такие как рак молочной железы, рак предстательной железы, рак яичников, рак поджелудочной железы и т.п.
Злокачественные опухоли экспрессируют ряд белков, которые служат в качестве антигенов-мишеней для иммунной атаки. Указанные молекулы включают, но без ограничения тканеспецифические антигены, такие как MART-1, тирозиназа и GP 100 при меланоме, и простатическая кислая фосфатаза (PAP) и простатоспецифичный антиген (PSA) при раке простаты. Другие молекулы-мишени, принадлежат к группе молекул, связанных с перерождением, такие как онкоген HER-2/Neu/ErbB-2. Еще одна группа антигенов-минешей представляет собой онкофетальные антигены, такие как карциноэмбриональный антиген (CEA). При B-клеточной лимфоме опухоль-специфический идиотип иммуноглобулина составляет истинный опухоль-специфический иммуноглобулиновый антиген, который является уникальным для индивидуальной опухоли. B-клеточные дифференцировочные антигены, такие как CD19, CD20 и CD37, являются другими кандидатами антигенов-мишеней при B-клеточной лимфоме. Некоторые из таких антигенов (CEA, HER-2, CD 19, CD20, идиотип) использовались в качестве мишеней при пассивной иммунотерапии моноклональными антителами с ограниченным успехом. Опухолевый антиген и его антигенные раковые эпитопы могут быть очищены и выделены из естественных источников, например, из первичных клинических изолятов, клеточных линий и т.п. Раковые пептиды и их антигенные эпитопы также могут быть получены с помощью химического синтеза или с помощью технологий рекомбинантной ДНК, известных в данной области. Методики химического синтеза описаны в Steward et al. (1969); Bodansky et al. (1976); Meienhofer (1983); и Schroder et al. (1965). Кроме того, как описано Renkvist et al. (2001), имеются многочисленные антигены, раскрытые в данной области. Хотя аналоги или искусственно модифицированные эпитопы не описываются конкретно, специалисту в данной области понятно, как получить или создать их стандартными способами данной области техники. Другие антигены, идентифицируемые антителами и выявляемые с помощью технологии Serex (смотреть Sahin et al. (1997) и Chen et al, (2000)), указаны в базе данных Ludwig Institute for Cancer Research.
В еще одном варианте осуществления настоящее изобретение может включать микробные антигены для представления с помощью APC. Как предусмотрено в описании, микробные антигены могут быть вирусного, бактериального или грибкового происхождения. Примеры инфекционных вирусов включают: Retroviridae (например, вирусы иммунодефицита человека, такие как HIV-1 (также обозначаемые как HTLV-III, LAV или HTLV-III/LAV, или HIV-III; и другие изоляты, такие как HIV-LP; Picornavhidae (например, полиовирусы, вирус гепатита A; энтеровирусы, вирусы Коксаки человека, риновирусы, ЕСНО-вирусы); Calciviridae (например, штаммы, которые вызывают гастроэнтерит); Togaviridae (например, вирусы энцефалита лошадей, вирусы краснухи); Flaviridae (например, вирусы денге, вирусы энцефалита, вирусы желтой лихорадки); Coronaviridae (например, коронавирусы); Rhabdoviridae (например, вирусы везикулярного стоматита, вирусы бешенства); Filoviridae (например, вирусы Эбола); Paramyxoviridae (например, вирусы парагриппа, вирус эпидемического паротита, вирус кори, респираторно-синтициальный вирус); Orthomyxoviridae (например, вирусы гриппа); Bungaviridae (например, хантавирусы, бунгавирусы, флебовирусы и найровирусы); Arenaviridae (вирусы геморрагической лихорадки); Reoviridae (например, реовирусы, орбивирусы и ротавирусы); Birnaviridae; Hepadnaviridae (вирус гепатита В); Parvovirida (парвовирусы); Papovaviridae (папилломавирусы, полиомавирусы); Adenoviridae (большинство аденовирусов); Herpesviridae (вирус простого герпеса (HSV) 1 и 2, вирус ветряной оспы, цитомегаловирус (CMV), вирус герпеса); Poxviridae (вирусы натуральной оспы, вирусы осповакцины, поксвирусы); и Iridoviridae (например, вирус африканской лихорадки свиней); и неклассифицированные вирусы (например, этиологические агенты Spongiform encephalopathies, агент гепатита дельта (считают дефектным саттелитом вируса гепатита B), агенты гепатита ни-A, ни-B (класс 1 энтерально передающиеся; класс 2 передающиеся парентерально (т.е. гепатит C); Nonvalk и родственные вирусы, и астровирусы). Примеры инфекционных бактерий включают: Helicobacter pyloris, Borelia burgdorferi, Legionella pneumophilia, Mycobacteria sps (например, M. tuberculosis, M. avium, M. intraceliulare, M. kansasii, M. gordonae), Staphylococcus aureus, Neisseria gonorrhoeae, Neisseria meningitidis, Listeria monocytogenes, Streptococcus pyogenes (Group A Streptococcus), Streptococcus agalactiae (Group B Streptococcus), Streptococcus (группа вириданс), Streptococcus faecalis, Streptococcus bovis, Streptococcus (анаэробные виды), Streptococcus pneumoniae, pathogenic Campylobacter sp., Enterococcus sp., Haemophilus influenzae, Bacillus anthracis, corynebacterium diphtheriae, corynebacterium sp., Erysipelothrix rhusiopathiae, Clostridium perfringens, Clostridium tetani, Enterobacter aerogenes, Klebsiella pneumoniae, Pasturella multocida, Bacteroides sp., Fusobacterium nucleatum, Streptobacillus moniliformis, Treponema Treponema pertenue, Leptospira и Actinomyces israelii.
Примеры инфекционных грибов включают: Cryptococcus neoformans, Histoplasma capsufatum, Coccidioides immitis, Blastomyces derniatitidis, Chlamydia trachomatis, и Candida albicans. Другие инфекционные организмы (т.е. протисты) включают: Plasmodium falciparum и Toxoplasma gondii.
Активация DC
Тогда как традиционные вакцины на основе DC (которые раньше преобладали в клинических испытаниях) вызывали созревание DC с помощью коктейльной смеси цитокинов, включающей комбинации TNF, IL-6, PGE2 и IL-1β, которые в конечном итоге стимулировали асептическое воспаление, настоящее изобретение, вместо этого, использует агонист TLR для созревания DC и стимуляции продукции сигнала.
По аспекту настоящего изобретения стимуляция DC комбинацией лигандов TLR приводит к продукции повышенных количеств IL-12. Кроме того, активация DC комбинацией агониста TLR может дать более выраженный CD4 и CD8 T-клеточный ответ (Warger et al., 2006, Blood 108:544-550). Таким образом, DC настоящего изобретения могут секретировать Th1 - запускающие цитокины, такие как IL-12, при воздействии указанных лигандов, которые приводят в действие TLR. Например, добавление poly(I:C), агониста TLR3 к IL-1β, TNF-α, и IFN-γ, может вызвать образование потентной типа 1 поляризованной DC, отличающейся высокими уровнями продукции IL-12 (Heifler et al., 1996, Eur. J. Immunol. 26:659-668). В определенных вариантах осуществления антиген может быть нагружен на DC перед воздействием агониста TLR. В других вариантах осуществления антиген может быть нагружен на DC после воздействия агониста TLR.
По аспекту настоящего изобретения, новый интегральный подход используется для образования высокоэффективных DC, которые производят сильные, противоопухолевые иммунные ответы посредством активации TLR. Данный подход, который также можно назвать иммунной подготовкой, с помощью переноса активированных естественных (аутологичных) клеток (ICAIT), использует DC моноцитарного происхождения, которые специфически активируют биомолекулами, которые имитируют бактериальную инфекцию, тем самым создавая ICAIT-DC. Указанный уникальный способ активации предоставляет DC со свойствами, которые не обнаруживают у DC, которые созревают с помощью цитокинового коктейля из TNF, IL-6, PGE2 и IL-1β ("традиционное созревание"), который также имитирует асептическое воспаление (Lorn bard i et al., 2009, J. Immunol. 182:3372-3379).
В одном варианте осуществления ICAIT-DC настоящего изобретения могут быть активированы комбинацией агониста TLR4, бактериального липополисахарида (LPS), агониста TLR7/8, резиквимод (R848) и/или IFN-γ (Amati et al., 2006, Curr. Pharm. Des 12:4247-4254). При активации DC агонистом TLR4 и бактериальным LPS, получают ICAIT-DC, которые по меньшей мере по существу идентичны (по фенотипу) DC1, полученным с помощью традиционных методов созревания. Указанные ICAIT-DC характеризуются высоким уровнем экспрессии поверхностных молекул, включая CD83, CD80, CD86 и HLA-DR. В других вариантах осуществления могут быть использованы агонисты TLR2, такие как липотейхоевая кислота (LTA), агонисты TLR3, такие как poly(I:C), и/или другие агонисты TLR4, такие как MPL. Как предусмотрено в описании, любой агонист TLR или комбинация агонистов TLR могут быть использованы для активации DC, при условии, что такие лиганды стимулируют продукцию цитокиновых и хемокиновых сигналов активированными DC. Многие другие агонисты TLR известны в данной области техники и их можно найти в списке опубликованной литературы для применения с настоящим изобретением.
Несмотря на сходство фенотипов ICAIT-DC и созревших традиционным путем DC, ICAIT-DC настоящего изобретения обладают многочисленными явными преимуществами. Например, как изображено на фигуре 1, ICAIT-DC продуцируют более высокие уровни TNF, а также высокие уровни IL-12, CCL3 (MIP-1α) и CCL4 (MIP-1β), CCL5 (RANTES), и CXCL10 (IP-10), по сравнению с созревавшими традиционным путем DC.
Каждый из указанных факторов может усиливать аспекты противоопухолевого иммунитета. Например, CXCL 10 (IP-10) является хемоаттрактантом для NK-клеток, которые усиливают отторжение опухоли (Zing et al., 2005, J. Interferon Cytokine Res. 25:103-1 12). TNF и IL-12 являются антиангиогенными агентами и лишают опухоли кровоснабжения (Albini et al, 2009, J. Transl. Med. 7(5)). IL-12 вызывает развитие и рекрутинг IFN-γ-секретирующих Th1-клеток и активирует NK-клетки. В отличие от этого, созревавшие традиционным путем DC характеризуются очень слабой экспрессией всех указанных биомолекул, но вместе с тем, активно экспрессируют CCL17 (TARC), хемокин, ассоциированный с аллергическими реакциями и рекрутингом Th2-клеток (Xiao et al., 2003, Cytokine 23:126-132). Также на фигуре 1 представлена способность ICAIT-DC проявлять уникальную киллерную функцию, которой лишены традиционные DC, в результате чего они способны лизировать клетки линий рака молочной железы. Таким образом, ICAIT-DC настоящего изобретения не только демонстрируют продукцию цитокинов, которые, как известно, являются эффективными противоопухолевыми молекулами, ICAIT-DC также оказывают положительное влияние на качество сенсибилизированных T-клеток. Как изображено на фигуре 2, наряду с тем, что DC, созревшие традиционным путем, и ICAIT-DC могут успешно сенсибилизировать T-клетки против опухолевого антигена, только ICAIT-DC могут подготовить T-клетки для достоверного распознавания RER-2-экспрессирующих опухолей. Указанные факты позволяют предположить, что опухоли обладают механизмами, которые защищают их от распознавания сенсибилизированными T-клетками с помощью ассоциированной с традиционной DC-активацией, но указанные механизмы можно преодолеть с помощью T-клеток, сенсибилизированных и кондиционированных ICAIT-DC. Таким образом, ICAIT-DC воплощают уникальные качества, которыми не обладают обычные DC, и которые предусматривает модель ICAIT-DC для кондиционирования лучшей T-клеточной сенсибилизации, хемотаксической аттракции мультилинейных эффекторных клеток к метастазам опухоли, и участия в прямом разрушении раковых клеток.
Применение ICAIT-DC в иммунотерапии
Несмотря на достигнутый прогресс в создании противораковых вакцин на основе DC, остается значительное количество проблем, которые ограничивают эффективность применения существующих противораковых вакцин на основе DC. Одна из наиболее важных проблем заключалась в преодолении способности опухолей ускользать от иммунной системы. Данную способность можно объяснить тем фактом, что тканеспецифические опухоль-ассоциированные антигены могут быть слабо иммунногенными и, поэтому избегать иммунного ответа хозяина, или иммунный ответ может иммуноредактировать опухоль, устраняя, таким образом, антиген-положительные клетки, оставляя без внимания антиген-отрицательные опухолевые клетки. Указанный процесс иммуноредактирования является способом, в соответствии с которым иммунный ответ может придавать опухоли более агрессивный фентип.
Иммуноредактированием можно объяснить, отчасти, почему таргетинг только отдельных тканеспецифических антигенов приводил к довольно ограниченному клиническому успеху. Наряду с концепцией направленного иммуноредактирования, идея распространения эпитопа также привлекла недавно внимание в качестве другого возможного механизма усиления индукции эффекторных T-клеток.
Большинство клинических испытаний с использованием DC имели своей мишенью только тканеспецифические опухолевые белки/антигены. Однако существует много молекул, которые в случае их повышенной экспрессии ассоциируются с неопластической трансформацией и пока еще не использовались в качестве мишеней. Например, сурвивин, член антиапоптотического семейства, воспринимается как непосредственная расположенная ниже мишень пути сигнального трансдуктора и активатора транскрипции (STAT3) (Gritsko et al., 2006, Clin. Cancer Res. 12:1 1-19). Прямое ингибирование сигнального пути STAT3 блокирует экспрессию белка сурвивина и инициирует апоптоз клеток рака молочной железы. Также полагают, что индукция повышенной экспрессии HER-2/neu активирует сурвивин (Siddiqa et al., 2008, BMC Cancer 8:129). Указанная связь между хорошо определяемым опухолевым антигеном, HER-2/neu, и сигнальным путем гена, сурвивина, предоставляет большой потенциал для развития нового иммунотерапевтического воздействия, которое воздействует на множественные эффекторы в патогенезе рака молочной железы. Установлены другие иммуногенные пептиды гена сурвивина, поэтому создание антисурвивиновых лекарственных препаратов на основе DC с помощью системы и способов настоящего изобретения является весьма вероятным, и в комбинации с общепризнанным anti-HER~2/neu, вакцинация на основе DC может оказать сильное влияние на регрессию, и возможно предотвращение эстроген-независимых раков молочной железы (Reker et al., 2004, Cancer Biol, Ther, 3:180-183).
Белок HER-l/EGFR является еще одним примером новой молекулы, которая может служить мишенью для системы и способа настоящего изобретения. Наряду с сурвивином, HER-l/EGFR также служит в качестве новой нетканеспецифической мишени для противораковых вакцин на основе DC. Повышенная экспрессия или мутация HER-l/EGFR вовлечена в онкогенез различных злокачественных опухолей, которые варьируют от карциномы молочной железы, колоректального рака, мультиформная глиобластома мозга, аденокарцинома поджелудочной железы и до немелкоклеточного рака легких (Hynes et al., 2009, Curr. Opin, Cell Biol, 21:177-1 84). DC могут быть примированы HER-l/EGFR и активированы как описано здесь, и таким образом может развиться anti-HER-1/EGFR T-клеточный ответ.
Муцин 1 (MUC-1) также может служить в качестве мишени для противораковой вакцины. MUC-I представляет собой эпителиально-клеточный гликопротеин, который сверхэкспрессирован в высокой степени и абберантно гликозилирован при многих аденокарциномах, включая аденокарциномы молочной железы, и билиарные и панкреатические аденокарциномы (Vlad et al., 2004, Adv. Immunol. 82:249-293; von Mensdorff-Pouillyet al., 2000, Int. J. Biol. Markers 15,343-356). Сверхпродукция MUC-1 участвует в опухолевой инвазии и метастазировании. Учитывая, что сверхпродукция MUC-1 ассоциирована с рядом гематологических и эпителиальных злокачественных опухолей, комбинирование anti-MUC-1 вакцин на основе DC с вакцинами против тканеспецифических опухолевых мишеней может дать клинически значимые результаты.
Таргетинг молекул может оказать сильный эффект на пролиферацию неопластических клеток, и таким образом остановить прогрессирование заболевания. Поскольку вакцины на основе DC могут содержать белки, такие как сурвивин и HER-l/EGFR, возможно, что прогрессирование заболевания может быть клинически изменено при использовании подхода многоцеловой вакцинации.
Как предусмотрено в описании, CSC также представляют собой иммунотерапевтические мишени для новых вакцин на основе DC настоящего изобретения. Полагают, что субпопуляции стволовых клеток запускают и поддерживают различные новообразования (Wicha et al., 2006, Cancer Res. 66:1883-1890). Считают, что пути, связанные с CSC, испытывают недостаток регуляции и поэтому вызывают неконтролируемое самообновление CSC, которые производят опухоли, которые устойчивы к традиционным видам терапии (Eyler et al., 2008, J, Clin, Oncol. 26:2839-2845). Существующие противораковые воздействия нацелены на дифференцированные опухолевые клетки, но “щадят” популяции CSC (Eyler et al., 2008, J. Clin. Oncol. 26:2839-2845), кроме того, существуют популяции CSC, которые могут инициировать рецидивы заболевания и/или ограничивать благоприятный терапевтический эффект стандартных видов терапии. Таким образом, совершенно необходимо разработать новые стратегии, основанные на лучшем понимании сигнальных путей, которые контролируют аспекты самообновления и выживания в CSC, чтобы определить новые терапевтические мишени в указанных клетках. Маркеры стволовых клеток определяют при ряде злокачественных опухолей человека, включая гематологические злокачественные опухоли и опухоли мозга, предстательной железы, молочной железы, поджелудочной железы, головы и шеи, и толстой кишки. Помимо идентификации маркеров стволовых клеток, пути, которые регулируют самообновление и развитие клеток, какие как Wnt, Notch и Hedgehog также анализируют в мельчайших подробностях (Kakarala et al., 2008, J. Clin, Oncol. 26:2813-2820; Medina et al., 2009, Clin, Transl. Oncol. 11:199-207; Bolos et al., 2009, Clin. Transl. Oncol. 11:11-19; Bisson et al., 2009, Cell Res. 19(6):683-697).
Те же самые механизмы, которые используют при создании направленных на определенную опухоль DC-вакцин, также могут быть использованы для воздействия на молекулы, которые специфичны для CSC. Считают, что не все клетки рака молочной железы одинаковы, и что субпопуляция CSC молочной железы может быть ответственна за развитие инвазии и метастатической болезни. Раки молочной железы человека содержат клеточную популяцию, отличающуюся экспрессией маркеров клеточной поверхности CD44+/CD241ow/lin-, которые демонстрируют свойства стволовых клеток (Al-Hajj et al., 2003, Proc. Natl Acad, Sci. USA 100:3983-3988). Среди молекул, которые регулируют популяции стволовых клеток, есть HER-2/neu. Существует корреляция между экспрессией маркера столовых клеток, альдегиддегидрогеназы 1, и повышенной экспрессией HER-2/neu. Например, в сериях из 477 карцином молочной железы, повышенная экспрессия HER-2/neu нормальными эпителиальными клетками молочной железы человека, а также карциномами молочной железы коррелирует с увеличением доли ALDHl-экспрессирующих стволовых клеток (Ginestier et al., 2007, Cell Stem Cell. 5:555-567). Указанная корреляция между экспрессией HER-2/neu и стволовыми клетками служит ярким примером, как идентификация определенных маркеров CSC может способствовать созданию вакцин на основе DC с применением системы и способа настоящего изобретения, которые не только воздействуют на опухолевые антигены, но также преследуют цель уничтожить саморегулирующиеся клетки, такие как стволовые клетки. Таргетинг молекул, которые заведомо связаны с CSC, может быть эффективен в предотвращении рака отчасти за счет уменьшения клонов плюрипотентных клеток, которые могут служить инициаторами канцерогенеза. Принимая во внимание настоящее изобретение в части разработки вакцин на основе DC, создание противораковой вакцины может таким же образом быть направлено на таргетинг молекул, которые специфичны для CSC. При таргетинге молекул, экспрессированных на CSC, существует возможность уничтожить клоны клеток, которые, возможно, ответственны за большинство системных рецидивов и за отсутствие эффекта существующих противораковых видов терапии. Для достижения этого, идентифицируют молекулы, которые специфичны для CSC, и затем иммунотерапии на основе DC могут быть реализованы, как предусмотрено в описании, чтобы воздействовать на молекулы, которые являются специфичными для стволовых клеток.
Ингибирование регуляторной клеточной функции
Настоящее изобретение также показывает, что лиганды TLR не только активируют презентирующие клетки, но также ингибируют регуляторные клетки, действие которых ограничивает адаптивные ответы. В определенных вариантах осуществления передача сигнала через многочисленные Toll-подобные рецепторы, включая TLR-2, TLR-4, TLR-8 и TLR-9, отменяет супрессию под влиянием Treg. В описании показано, что TLR-активированнные дендритные клетки не только ингибируют эффекты Treg в отношении иммунореактивных клеток, но также, по-видимому, превращает сами регуляторы в эффекторы, продуцирующие IFN-γ.
Дендритные клетки, активированные LPS и IFN-γ, но не DC, созревавшие под действием цитокинов, нейтрализуют ингибиторные эффекты регуляторных T-клеток на ответную клеточную пролиферацию. Указанный эффект не связан с клеточной гибелью, поскольку экспрессия апоптотических маркеров в присутствии TLR-активированных DC не изменяется. Как продемонстрировано в описании, восстановление пролиферации иммунореактивных клеток наблюдали даже когда TLR-активированные дендритные клетки отделяли от регуляторных и иммунореактивных клеток полупроницаемой мембраной. Эффект происходит благодаря растворимому фактору, но не зависит от IL-6 и IL-12. Кроме того, указанный неизвестный растворимый медиатор, по-видимому, действует по меньшей мере отчасти на сами регуляторы, а не на иммунореактивные клетки. Как показано в описании, TLR-активированные дендритные клетки могут индуцировать продукцию цитокинов и эффекторную функцию у регуляторных T-клеток. Регуляторные T-клетки производят значительное количество IFN-γ в присутствии TLR-активированных дендритных клеток, но не незрелых или созревших под действием цитокинов дендритных клеток. Продукция IFN-γ ассоциирована с активацией регулятора транскрипции Th1 T-bet, и значительная фракция IFN-γ-продуцирующих регуляторов коэкспрессирует T-bet и FoxP3. В то время как эффекты LPS-активированной дендритной клетки на ответную клеточную пролиферацию являлись IL-12-независимыми, активация T-bet ингибируется нейтрализующим антителом anti-IL12. Таким образом, DC моноцитарного происхождения, активированные LPS, могут управлять фенотипом иммунного ответа частично с помощью ингибирования супрессорных T-клеток и рекрутинга указанных регуляторов в Th1 эффекторы.
Криосохранение
Как уже разъяснялось, настоящее изобретение предоставляет не только систему и способ для создания более совершенных APC с помощью развития ICAIT-DC, но также предоставляет систему и способ для криосохранения указанных активированных DC путем, который сохраняет их способность производить сигналы, критические для T-клеточной функции после размораживания. Как предусмотрено в описании, настоящее изобретение включает различные методики криосохранения и криосреды, которые известны специалистам в данной области. Например, в определенных вариантах осуществления криосреда для культивированных клеток может включать приблизительно 5-10% DMSO или глицерина и 10-50% сыворотки, например, сыворотки крови человека. В других вариантах осуществления криосреды могут быть бессывороточными. В определенных вариантах осуществления может быть использовано замораживание с контролируемой скоростью, тогда как другие варианты осуществления могут включать применение изотермического контейнера, в котором пузырьки с клетками, смешанными с криосредами помещают в морозильную камеру, например, при температурах в диапазоне приблизительно от -70°C до -80°C. Настоящее изобретение предоставляет способ сохранения активированных ICAIT-DC таким образом, чтобы в дальнейшем облегчить клиническое применение таких клеток и уменьшить потребность в длительных и повторяемых стадиях афереза и элютриации. Как предусмотрено в описании, методики криосохранения могут быть использованы как для небольших, так и для крупных партий.
При рассмотрении практической полезности активированных DC, способность предоставить постоянный запас криосохраненных активированных DC представляет собой важное преимущество, которое может способствовать различным терапевтическим применениям таких клеток, например, крупномасштабная культура активированных DC может быть криосохранена в аликвотах подходящего объема, согласно способам настоящего изобретения, так что отдельные дозы клеток позднее могут быть использованы в любом конкретном иммунотерапевтическом протоколе. В определенных вариантах осуществления активированные DC могут быть криосохранены в течение 2-24 недель при температурах приблизительно -70°C или ниже. При более низких температурах, например, приблизительно - 120°C или ниже, активированные DC могут быть криосохранены по меньшей мере в течение года или больше.
В одном примере осуществления DCs суспендируют в сыворотке крови человека и приблизительно 10% DMSO (об/об). Альтернативно могут быть использованы другие типы сыворотки, такие как фетальная телячья сыворотка. Суспендированные клетки могут быть разделены на аликвоты по небольшим образцам, таким как пузырьки объемом 1,8 мл, и могут храниться приблизительно при -70°C или ниже. В других вариантах осуществления криосреда может включать приблизительно 20% сыворотки и 10% DMSO, и суспендированные клетки могут храниться приблизительно при -180˚C. Другие варианты осуществления могут включать среду, содержащую приблизительно 55% оксиполижелатина, который представляет собой плазмозаменитель, приблизительно 6% оксиэтилированного крахмала, и приблизительно 5% DMSO. Другие показательные криосреды могут включать приблизительно 12% DMSO и приблизительно 25-30% сыворотки.
В то время как настоящее изобретение, как описано здесь, может включать специфические концентрации сыворотки, специалистам в данной области должно быть понятно, что точное количество сыворотки в криосреде может варьировать, и в некоторых вариантах осуществления сыворотка может полностью отсутствовать, но в целом будет находиться в пределах приблизительно от 1% до 30%. Разумеется, любая концентрация сыворотки, которая обеспечивает в результате жизнеспособность клеток приблизительно 50% и/или степень извлечения приблизительно 50%, может быть использована в любой композиции ICAIT-DC настоящего изобретения, а также с любым методом криосохранения, как описано здесь. Предпочтительно, желательны жизнеспособность и степень извлечения клеток по меньшей мере 60%, более предпочтительно по меньшей мере приблизительно 70%, или даже 80% при восстановлении криосохраненных клеток в выбранной криосреде.
Аналогичным образом, несмотря на то, что настоящее изобретение, как описано здесь, может включать конкретные концентрации DMSO, специалисту в данной области понятно, что DMSO может полностью отсутствовать в некоторых вариантах осуществления, тогда как в других вариантах осуществления в криосреде могут использоваться концентрации приблизительно от 5% до 20%, и они включены в методы криосохранения, описанные здесь. В целом, более низкие концентрации DMSO являются предпочтительными, например, приблизительно от 5% до 10%. Однако может быть использована любая концентрация DMSO, которая после размораживания обеспечивает жизнеспособность клеток по меньшей мере 50% и степень извлечения по меньшей мере 50%, и предпочтительно жизнеспособность и степень извлечения клеток по меньшей мере 60%, более предпочтительно приблизительно 70%, более предпочтительно приблизительно 80% и еще более предпочтительно приблизительно 90% и выше.
Несмотря на то, что настоящее изобретение, как описывается здесь, может включать ссылку на замораживание с контролируемой скоростью, специалисту в данной области понятно, что методы замораживания с контролируемой скоростью или неконтролируемой скоростью могут быть использованы обычным путём.
Специалистам в данной области техники также должно быть понятно, что различные среды для криосохранения, которые описаны здесь, могут либо содержать сыворотку, либо быть бессывороточными. Примеры бессывороточных сред могут включать XVIVO 10, XVIVO 15, XVIVO 20, StemPro, а также любые коммерчески доступные бессывороточные среды. Если используют бессывороточную криосреду, методы криосохранения настоящего изобретения обычно не содержат инфекционных агентов, антител и чужеродных белков, которые могут быть антигенными, и любой другой чужеродной молекулы, которую обычно можно обнаружить в криосредах на основе сыворотки.
Криосохранение нагруженных антигеном, активированных DC может происходить в любой момент после активации клеток агонистом TLR. В одном варианте осуществления активированные DC криосохраняют приблизительно через 6-8 часов после воздействия агониста TLR. Предпочтительно, момент времени, выбранный для криосохранения активированных клеток, должен быть обусловлен максимизацией сигнальной продукции клеток, в особенности продукции IL-12.
Терапевтические применения
Настоящее изобретение включает создание нагруженной антигеном, активированной APC, которая продуцирует значительные уровни цитокинов и хемокинов, когда она разморожена после криосохранения, где нагруженная антигеном и активированная APC используется в иммунотерапии млекопитающего, предпочтительно человека. Ответ на антиген, представленный APC, можно измерить с помощью мониторинга индукции цитолитического T-клеточного ответа, хелперного T-клеточного ответа, и/или ответа антител на антиген, с применением способов, хорошо известных в данной области техники.
Настоящее изобретение включает способ усиления иммунного ответа у млекопитающего, включающий стадии: образование незрелых DC из моноцитов, полученных от млекопитающего (например, от пациента); примирование незрелых DCs композицией, содержащей антигенную композицию; активация нагруженных антигеном DC по меньшей мере одним агонистом TLR; криосохранение активированных, нагруженных антигеном DC; размораживание активированных, нагруженных антигеном DC и затем введение активированных, нагруженных антигеном DCs млекопитающему, который в этом нуждается. Композиция включает по меньшей мере антиген, и также может быть вакциной для иммунизации ex vivo и/или терапии in vivo млекопитающего. Предпочтительно, млекопитающее является человеком.
Процедуры ex vivo хорошо известны в данной области и раскрываются более подробно ниже. Кратко, выделяют клетки млекопитающего (предпочтительно, человека). Клетки могут вводиться реципиенту-млекопитающему для обеспечения благоприятного терапевтического эффекта. Реципиент-млекопитающее может быть человеком, и клетки могут быть аутологичными для реципиента.
Альтернативно, клетки могут быть аллогенными, сингенными или ксеногенными для реципиента.
В одном варианте осуществления моноциты периферической крови пациента получают с помощью комбинированного афереза и элютриации. Моноциты можно культивировать в SFM с GM-CSF и IL-4 в течение ночи. На следующий день незрелые DC могут быть примированы антигеном, с последующим взаимодействием DC с IFN-γ и LPS. Активированные DC затем могут быть суспендированы в криосреде и заморожены до готовности для применения в иммунотерапии.
Криосохраненные ICAIT-DC можно культивировать ex vivo в условиях, эффективных для получения процента извлечения и процента жизнеспособности клеток, которые сравнимы с аналогичными показателями свеже-активированных ICAIT-DC. ICAIT-DC, полученные из криосохраненных образцов, могут проявлять аналогичную стабильность, по сравнению со свежеполученными ICAIT-DC. Кроме того, сравнения криосохраненных зрелых DC с теми же свежеполученными DC могут показать по существу идентичные фенотипы, а также профили сигнальной секреции. Как предусмотрено в описании, ICAIT-DC могут быть сохранены как в небольших, так и в крупномасштабных партиях в течение приблизительно от 2 до 24 недель, в различных криосредах, описанных здесь, при температурах приблизительно от -70°C до -80°C. При температурах ниже приблизительно - 120°C, продолжительность хранения может быть увеличена до неопределённо долгого времени, или по меньшей мере до более чем 24 недель без влияния на степень извлечения клеток, жизнеспособность и функциональность DC. Например, в определенных вариантах осуществления активированные клетки могут храниться по меньшей мере в течение одного года и по-прежнему сохранять свою способность продуцировать сигнал после размораживания. Настоящее изобретение предусматривает эффективные профили извлечения и жизнеспособности после размораживания клеток, и, кроме того, условия криосохранения, описанные здесь, не влияют на способность ICAIT-DC сохранять свои сигнальные профили, как объясняется на протяжении всего описания.
В примере осуществления криосохранение может быть выполнено после активации ICAIT-DC с помощью повторного ресуспендирования в предварительно охлажденной сыворотке крови человека и с последующим добавлением приблизительно 10% DMSO к образцу. Затем смесь может быть разделена на аликвоты в 1,8 мл пузырьки и заморожена приблизительно при -80°C в криокамере в течение ночи. На следующий день пузырьки могут быть перенесены в резервуары с жидким азотом. Спустя приблизительно 2-24 недели криосохранения замороженные ICAIT DC можно разморозить и проверить степень извлечения и жизнеспособность. Степень извлечения таких ICAIT DC может быть больше или равной приблизительно 70% с жизнеспособностью больше или равной приблизительно 70%. В других вариантах осуществления DC или даже моноциты могут быть криосохранены перед активацией клеток.
Альтернативно, процедура размножения ex vivo гемопоэтических стволовых и прогениторных клеток описывается в патенте США No. 5199942, который включен в описание посредством ссылки, и может быть применена к клеткам настоящего изобретения. В дополнение к клеточным факторам роста, описанным в патенте США No. 5199942, другие факторы, такие как flt3-L, IL-1, IL-3 и лиганд c-kit, могут использоваться для культивирования и размножения клеток.
Известны различные технологии клеточной селекции для идентификации и выделения клеток из популяции клеток. Например, моноклональные антитела (или другие специфические белки, связывающие клетки) могут быть использованы для связывания с маркерным белком или белком поверхностного антигена, обнаруженного на клетках. Некоторые такие маркеры или антигены клеточной поверхности известны в данной области.
Препараты вакцин
Настоящее изобретение также включает препараты вакцин, подходящие для применения в иммунотерапии. В определенных вариантах осуществления препараты вакцин используют для предотвращения и/или лечения заболевания, такого как рак и инфекционные заболевания. В одном варианте осуществления введение пациенту вакцины согласно настоящему изобретению для предотвращения и/или лечения рака может происходить до или после хирургической процедуры по удалению рака, до или после химиотерапевтической процедуры для лечения рака, и до или после лучевой терапии для лечения рака и любой их комбинации. В других вариантах осуществления препараты вакцины могут быть введены пациенту в сочетании или в комбинации с другой композицией или фармацевтическим продуктом. Следует понимать, что настоящее изобретение также может быть использовано для предотвращения рака у субъектов, не имеющих рака, но которые могут иметь повышенный риск развития рака. Введение противораковой вакцины, приготовленной согласно настоящему изобретению, имеет широкое применение для предотвращения или лечения рака, определяемого отчасти выбором антигенов, образующих часть противораковой вакцины. Раки, которые, соответственно, можно лечить согласно практическим применениям настоящего изобретения включают без ограничения, раки легкого, молочной железы, яичников, шейки матки, толстой кишки, головы и шеи, поджелудочной железы, предстательной железы, желудка, мочевого пузыря, почек, кости, печени, пищевода, мозга, яичка, матки и различные лейкемии и лимфомы.
В одном варианте осуществления вакцины согласно настоящему изобретению могут быть получены из клеток опухоли или рака, который лечат. Например, при лечении рака легкого, клетки рака легкого обрабатывают, как описано выше, для получения вакцины против рака легкого. Аналогичным образом, вакцина против рака молочной железы, вакцина против рака толстой кишки, вакцина против рака поджелудочной железы, вакцина против рака желудка, вакцина против рака мочевого пузыря, вакцина против рака почки и т.п. могут быть произведены и использованы в качестве иммунотерапевтических агентов согласно применениям для предотвращения и/или лечения опухолевой или раковой клетки, из которой была произведена вакцина.
В другом варианте осуществления вакцины согласно настоящему изобретению могут, как указано, также быть приготовлены для лечения различных инфекционных заболеваний, которые поражают млекопитающих, путем отбора подходящих антигенов, распространенных в культуральной среде патогеном. Поскольку существует гетерогенность типов иммуногенных и протективных антигенов, экспрессированных различными видами микроорганизмов, вызывающих одно и то же заболевание, поливалентные вакцины могут быть получены путем создания вакцины из пула организмов, экспрессирующих различные антигены, имеющие значение.
В другом варианте осуществления настоящего изобретения вакцина может быть введена с помощью внутриузловой инъекции в паховые узлы. Альтернативно, и в зависимости от мишени вакцины, вакцина может быть введена внутрикожно или подкожно в конечности, руки и ноги, пациентов, получающих лечение. Хотя обычно данный подход в целом соответствует требованиям при меланоме и других видах рака, включая предотвращение или лечение инфекционных заболеваний, также могут быть использованы другие пути введения, например, внутримышечно или в кровеносное русло.
Кроме того, вакцина может предоставляться вместе с адъювантами и/или иммуномодуляторами для повышения активности вакцины и ответа пациента. Указанные адъюванты и/или иммуномодуляторы известны специалистам в данной области и полностью описаны в имеющейся опубликованной литературе.
Как предусмотрено в описании, и в зависимости от типа вакцины, которую создают, производство вакцины, при необходимости, может быть увеличено путем культивирования клеток в биореакторах или ферментерах или других подобных сосудах или устройствах, подходящих для выращивания клеток в большом объеме. В таком аппарате культуральную среду собирают регулярно, часто или непрерывно, чтобы выделить любые материалы или антигены прежде чем указанные материалы или антигены разрушатся в культуральной среде.
При необходимости устройства или композиции, содержащие вакцину или антигены, произведенные или извлеченные согласно настоящему изобретению, и подходящие для непрерывного или периодического высвобождения, по существу, могут быть имплантированы в организм или нанесены местно для относительно медленного или регулируемого по времени высвобождения таких материалов в организм.
Другие стадии создания вакцин могут быть точно определены для соответствия требованиям конкретных вакцин. Такие дополнительные стадии известны специалистам в данной области, например, некоторые собранные антигенные материалы могут концентрироваться и в некоторых случаях обрабатываются детергентом и подвергаются ультрацентрифугированию для удаления трансплантационных аллогенов.
Указанные методы, описанные здесь, никоим образом не являются всеобъемлющими, и другие методы, которые соответствуют специфическому применению, известны обычному специалисту в данной области. Кроме того, эффективное количество композиций также может быть выражено приближенно по аналогии с соединениями, которые, как известно, оказывают желаемый эффект.
ПРИМЕРЫ
Ниже следует описание изобретения на примерах его осуществления. Указанные Примеры приводятся только с иллюстративной целью, и изобретение не ограничивается указанными Примерами, но скорее охватывает все вариации, которые очевидны благодаря сведениям, предоставленным в описании.
Эксперименты, раскрытые в описании, проводили, чтобы изучить эффекты криосохранения на функциональность нагруженных антигеном, активированных DC, когда их размораживали для применения в иммунотерапии. Размороженные клетки сохраняли способность обуславливать сильные Th1 клеточные ответы, благодаря продукции цитокинов и хемокинов, и также обладают способностью вызывать апоптоз опухолевых клеток.
Далее описываются материалы и методы, используемые в экспериментах и примерах, раскрытых в описании.
Криосохранение ICAIT DC. DC собирали осторожным соскабливанием. Среду и клетки сохраняли на ледяной бане на всех этапах. Клетки осторожно отмывали с помощью центрифугирования приблизительно при 800 RPM в течение 10 мин. 10×106 клеток осторожно суспендировали в сыворотке крови человека и 10% DMSO и переносили в 1,8 мл пузырьки. Клетки замораживали в бане с изопропанолом в контейнере Nalgene® с пеноизоляцией при -80°C. Режим замораживания устанавливался лабораторией в виде снижения температуры на 1°C/мин. Клетки также могут быть заморожены с помощью оборудования для замораживания биологического материала, в котором в качестве источника замораживания используется N2. Через 18-24 часов клетки быстро переносят в парообразную фазу емкости с жидким N2. Парообразная фаза в емкости контролируется извне, так что температура никогда не поднимается выше приблизительно -135°C.
Размораживание DC. Клетки переносили на водяную баню с температурой 37°C. Почти оттаявшие клетки суспендировали в ледяной среде для DC, осторожно опуская пипетку с клетками в среду и по каплям выпуская клетки в среду. Клетки центрифугировали при 800 RPM в течение 5 мин и ресуспендировали. Клетки подсчитывали, и они были готовы к применению.
Подготовка TCAIT DC для криосохранения. Свежевыделенные методом элютриационного центрифугирования миелоидные моноциты культивировали в 6-луночных микропланшетах (12×106 клеток/лунку). Культуральная среда состояла из бессывороточной среды (SFM Invitrogen Carlsbad CA). Конечная концентрация добавленной GMCSF составляла 50 нг/мл, и концентрация IL4 составляла 1000 Ед/мл. Клетки культивировали в течение ночи при 37°C в 5% CO2. В некоторых партиях клетки сенсибилизировали соответствующими пептидами через 16-20 часов и культивировали в течение следующих 6-8 часов, после чего добавляли 1000 Ед/мл IFN-γ. Дендритные клетки созревали в присутствии агониста TLR LPS (TLR 4,10 нг/мл) или R848 (TLR8, 1 пг/мл). Время созревания составляло по меньшей мере приблизительно 6 часов. После этого DC, активированные агонистом TLR, были готовы для криосохранения или немедленного использования.
Далее описываются результаты экспериментов, представленных в описании.
Пример 1. Криосохраненные активированные TCAIT-DC сохраняют сигнальную продукцию на уровнях, сходных с наблюдаемыми у свежеполученных ICAIT-DC
Во-первых, определяли жизнеспособность и степень извлечения, криосохраненных активированных DC. Используя тест вытеснения трипанового синего, определяли жизнеспособность и степень извлечения криосохраненных ICAIT DC перед криосохранением и непосредственно после размораживания и отмывки (с помощью центрифугирования) криосохраненных DC. Как изображено на фигурах 3A и 3B, представлены пять отдельных случаев из двух партий криосохраненных DC. Жизнеспособность свежеполученных и криосохраненных DC1 была сравнимой, и степень извлечения составляла в целом приблизительно 80-90%. Степень извлечения была значительно лучше для криосохраненных образцов, чем для свежеполученных DC 1, вследствие потери клеток при сборе свежеполученных DCI.
Затем сравнивали сигнальную продукцию свежих ICAIT DC и криосохраненных ICAIT DC, а также уровни сигнальной продукции, получаемые при использовании традиционных методик созревания, таких как цитокин-опосредованное созревание DC (CMDC). Как изображено на фигуре 4A, непрерывную продукцию сигнала 3 (IL12) измеряли с помощью теста ELISA. Криосохраненные ICAIT DC, показали сходные значительные уровни продукции IL12. Важно отметить, что продукция IL12 у криосохраненных ICAIT DC была значительно выше, чем у криосохраненных CMDC или у свежих CMDC, что наглядно показывает, что криосохраненные ICAIT DC сохраняют цитокиновые и хемокиновые профили, превосходящие профили традиционно созревавших DC, и поэтому указанные клетки более эффективны в активации T-клеточного ответа. Как изображено на фигуре 4B, уровни продукции IL12 через 2 часа и 12 часов после размораживания были сходными с уровнями продукции IL12, наблюдаемыми перед криосохранением.
Для определения аллосенсибилизации CD4 T-клеток, два образца очищенных аллогенных CD4 клеток (1×106/лунка) культивировали совместно с криосохраненными стимулированными агонистом TLR DC (1×105/лунка), в сравнении с DC1, полученными из криосохраненных моноцитов. Как изображено на фигуре 5, через 9 дней, T-клетки собирали и повторно стимулировали на планшетах, покрытых антителом anti-CD3 и anti-CD28. Уровни IFN-γ (продуцируемые T-клетками) анализировали в супернатанте через 24 часа.
Пример 2: CD4+CD25+T-клетки ингибируют ответную клеточную пролиферацию в присутствии незрелых, но не DC1 дендритных клеток.
Ранее было показано, что очищенные, выделенные методом элютриационного центрифугирования CD 14+моноциты приобретают характеристики активированных DC, если они обработаны сигнальными агентами DC (Czerniecki 1997). Совсем недавно было показано, что указанные дендритные клетки моноцитарного происхождения, несущие опухолевый антиген, полученные с использованием IFN-γ и агониста LPS TLR-4 (LPS-активированные DC), стимулируют направленный иммунный ответ у пациентов с дуктальной карциномой in situ (Czerniecki et al.} 2007, Cancer Res 67:1842-1852). Предыдущие исследования в совокупности показали, что агонисты TLR, включая LPS, способны отменить иммуносупрессию, опосредованную CD4+CD25+Foxp3+T-клетками (ref). Таким образом, предполагается, что TLR-активированная DC может быть способна обратить Treg-опосредованную иммуносупрессию.
Для проверки указанной гипотезы разрабатывали эксперименты по сравнению способности человеческих CD4+CD25+T-клеток ингибировать пролиферацию CD4 и CD8 лимфоцитов при стимуляции TCR (anti-CD3) в присутствии незрелых дендритных клеток (iDC) в сравнении с LPS-активированными DC моноцитарного происхождения. 1,25×105 сортированных CD4+CD25+T-клеток смешивали с 2,5×105 CFSE-меченных нефракционированных лимфоцитов (CD4 и CD8 положительных) и 1×105 незрелых дендритных клеток. Как изображено на фигуре 6, пролиферативный ответ CD4 и CD8 положительных T-клеток на 5-й день ингибировали в присутствие и в отсутствие Treg. Пролиферация сортированных CD4+CD25-T-клеток ингибировалась аналогичных образом, что исключает возможность, что данный результат являлся артефактом в результате использования нефракционированных реактивных клеток (данные не представлены). Напротив, добавление дендритных клеток DC, созревавших при использовании IFN-γ и LPS восстанавливало пролиферацию как CD4 так и CD8 положительных T-клеток, несмотря на присутствие регуляторных клеток. Ответная пролиферация в присутствии Treg и LPS-активированных DC являлась сходной с пролиферацией, наблюдаемой в отсутствие регуляторной популяции. То, что LPS, применяемый для созревания DC моноцитарного происхождения, загрязнял совместную культуру, приводя к указанному результату, исключали путем кратковременной предварительной LPS-обработки iDC (15 минут). Супрессия являлась естественной в данных условиях, что показывает, что популяция DC должна быть формально созревшей, и загрязняющий LPS не являлся причиной получаемого результата. В частности, дендритные клетки, созревавшие с применением общепринятого коктейля для созревания на основе цитокинов (IL-1, IL-6, TNF-a, PGE2) не полностью восстанавливали пролиферативные эффекторы в присутствии регуляторов. Приведенные результаты показывают, что LPS-созревшая DC моноцитарного происхождения ингибирует Treg-опосредуемую супрессию CD4 и CD8 положительных T-клеток, отвечающих на anti-CD3.
Применение CFSE и более конкретно введение метки отдельно в эффекторные клетки представляет собой важнейший аспект данной экспериментальной модели. Пролиферативные тесты, используемые для демонстрации супрессии во многих других исследованиях, страдают от своей зависимости от меченного тритием тимидина. В многочисленных отчетах показано, что регуляторные T-клетки не являются полностью анергичными и в действительности обладают значительным пролиферативным потенциалом, особенно в контексте сигналов воспаления (Brinster et al" 2005, J Immunol 175:7332-7340; Klein et al" 2003, Proc Natl Acad Sci USA 100:8886-8891). Тесты на основе меченного тритием тимидина не могут исключить вероятность того, что отмечаемые различия в пролиферации появляются по меньшей мере отчасти вследствие регуляторного компонента. Тест на основе CFSE специфически устанавливает пролиферацию эффекторных клеток и непосредственно сравнивает пролиферацию указанных эффекторов в присутствии/отсутствии Treg и/или различных популяций дендритных клеток. При этом тест исключает вероятность того, что пролиферация под влиянием регуляторных CD4+CD25+клеток будет способствовать неверной интерпретации результатов.
Пример 3. Ингибирование функции Treg дендритными клетками DC1 обусловлено растворимым фактором, но является IL-6 и IL-12 независимым
Эксперименты разрабатывали, чтобы описать механизм, с помощью которого популяция DC1 ингибирует Treg-опосредованную супрессию. Чтобы определить ингибируют ли дендритные клетки DC1 Treg-опосредованную супрессию путем индукции апоптоза в наборе регуляторных T-клеток, сортированные CD4+CD25+T-клетки культивировали совместно с iDC и с LPS-активированными DC и сравнивали экспрессию апоптотических маркеров Аннексин-V и 7-AAD через 24 часа. В качестве момента времени выбирали день 1 с учетом предположения, что различия в пролиферации, отмечаемые на 5-й день, происходили бы в результате апоптотических явлений значительно раньше. Через 24 часа культивирования, как изображено на фигуре 7A (p>0,2 для групп Аннексин+/7-AAD+ и Аннексин-/7-AAD-), экспрессия Аннексина-V и 7-AAD являлась сходной среди Treg, культивированных совместно с любой из двух популяций дендритных клеток. Указанные данные позволяют предположить, что LPS-активированная популяция DC не изменяет существенно апоптоз Treg, по сравнению с незрелыми дендритными клетками.
После того, как установили, что эффект LPS-активированных DC-клеток моноцитарного происхождения на Treg-опосредованную супрессию осуществлялся не за счет апоптоза, для определения являлся ли данный эффект зависимым от клеточного контакта или опосредовался растворимыми факторами, CFSE-меченные эффекторные клетки культивировали совместно с немечеными CD4+CD25+T-клетками в присутствии незрелых дендритных клеток, затем добавляли другие DC-популяции, отделяя их полупроницаемой мембраной Transwell®. При добавлении дополнительного набора незрелых дендритных клеток к мембране Transwell®, как изображено на фигуре 7B, не наблюдали эффекта на супрессию в присутствии iDC. Однако при добавлении LPS-активированной DC-популяции к мембране Transwell®, наблюдали нарушение Treg-опосредованной супрессии, которое было сходным с нарушением, наблюдаемым при совместном культивировании Tregs непосредственно с указанными DC. Для проверки возможности, что сами DC мигрировали сквозь Transwell® при нарушении супрессии, регуляторные T-клетки культивировали совместно с CFSE-меченными эффекторами в отсутствие дендритных клеток, но добавляли LPS-активированные DC, отделенные мембраной. Отмечали незначительную пролиферацию. Полученные данные в совокупности позволяют предположить, что LPS-активированная популяция DC может ингибировать Treg-опосредованную супрессию способом, независимым от клеточного контакта.
Вне связи с какой-либо конкретной теорией, считают, что двумя наиболее вероятными растворимыми медиаторами отмены супрессии являются IL-12 и IL-6. LPS-активированная популяция DC характеризуется секрецией большого количества IL-12, который главным образом вовлечен в развитие Th1 иммунных ответов (Xu et al., 2003, J Immunol 171:2251-2261). Как было показано, IL-6 занимает центральное место в LPS-опосредуемой отмене Treg-опосредуемой супрессии in vitro (Pasare et al., 2003, Science 299:1033-1036). Проверяли, восстанавливала ли нейтрализация IL-6 или IL-12 ингибиторные эффекты Treg в присутствии LPS-активированных DC, обнаружили, как изображено на фигуре 7C, что пролиферация эффекторов в присутствии Treg и IFN-γ/LPS-активированных DC минимально изменялась при добавлении нейтрализующего антитела anti-IL-12 и по существу не изменялась при добавлении anti-IL-6. Указанные данные в совокупности с результатами экспериментов с применением мембраны Transwell®, позволяют предположить, что растворимый фактор, а не IL-6 и IL-12 опосредует нарушение супрессии, осуществляемое LPS-активированными DC и делает это апоптоз-независимым образом.
В предшествующих экспериментах не представлялось возможным определить, действует ли DC-популяция на сами регуляторы или просто освобождает иммунореактивные клетки от ингибирования Treg. Поскольку эффект DC осуществляется посредством растворимого фактора, чтобы оценить могла ли отменить предварительная обработка регуляторных клеток или реактивных клеток средами, взятыми от LPS-активированных DC Treg-опосредуемую супрессию, сортированные CD4+CD25+T-клетки или CFSE-меченные эффекторные клетки культивировали в равных частях культуральной среды и среды, перенесённой из культур LPS-активированных DC. Указанные клетки собирали и отмывали через 24 часа и затем использовали в совместных культурах. Как изображено на фигуре 7D, предварительная обработка реактивных клеток средами LPS-активированных DC не оказывала эффекта на Treg-опосредуемую супрессию. Предварительная обработка регуляторных клеток перед совместным культивированием, однако, отчасти восстанавливала пролиферацию реактивных клеток в их присутствии. Предварительная обработка Treg только LPS (5 нг/мл) не увеличивала пролиферацию (данные не представлены). Полученные данные в совокупности позволяют предположить, что растворимый фактор может действовать по меньшей мере отчасти на сами регуляторы при отмене супрессии.
Несколько последних исследований показали, что различные Toll-подобные рецепторы, включая TLR-2, TLR-4, TLR-8 и TLR-9 могут отменять Treg-опосредуемую супрессию (Urry et al., 2009, J Clin Invest 1 19:387-398; Pasare et al., 2003, Science 299:1033-1036; Pasare et al., 2003, Science 299:1033-1036; Sutmuller et al., 2006, J Clin Invest 1 16:485-494; Peng et al., 2005, Science 309:1380-1384; Porrett et al., 2008, J Immunol 181:1692-1699; LaRosa et al., 2007, Immunol Lett 108:183-188; Pasare et al., 2003, Science 299:1033-1036). Указанные авторы показали, что дендритные клетки, активированные LPS, секретируют растворимый фактор(ы), который освобождает эффекторные клетки от супрессии с помощью CD4+CD25+T-клеток. Для данного эффекта требовался TL-6, но, по-видимому, он действовал синергично с одним или более цитокинами. Представленные здесь результаты согласуются с приведенными сведениями, но уточняют их в ряде аспектов. Во-первых, эксперименты осуществляли с использованием клеточных популяций человека, включая дендритные клетки, применяемые в настоящее время в иммунотерапии. Наблюдали, что сходный эффект в клеточной популяции человека, применяемой в настоящее время в иммунотерапии опухолей, переводит указанные сведения непосредственно на клиническую площадку. Во-вторых, в то время как указанные авторы добавляли LPS непосредственно к дендритным клеткам, чтобы индуцировать их эффект, эксперименты показали, что дендритные клетки, активированные LPS, затем перенесенные в совместные культуры, содержащие Treg и эффекторные клетки, сохраняют способность к отмене супрессии. Данный факт указывает на продолжительный эффект в отношении APC и позволяет предположить пролонгированную секрецию растворимого медиатора (медиаторов), способного отменить супрессию. В данном исследовании, исключается, что LPS нарушает Treg-опосредованную супрессию непосредственно действуя на супрессорную популяцию или на эффекторы. Скорее, действует LPS-активированная дендритная клетка, которая опосредует эффекты. В-третьих, эксперименты продемонстрировали эффект, который является IL-6 и IL-12-независимым, но, по-видимому, опосредуется растворимым фактором. Возможно, что указанный фактор действует синергично с IL-6 в вышеупомянутом исследовании.
В заключение, эксперименты позволяют предположить, что LPS-активированные дендритные клетки могут секретировать факторы, которые инактивируют сами регуляторы, а не высвобождение эффекторов из контрольных Treg.
Пример 4. Супрессорные CD4+CD25+T-клетки активируют T-bet, подавляют FoxP3, и секретируют эффекторные цитокины в присутствии дендритных клеток DC1
Недавние исследования, проведенные на нескольких экспериментальных моделях, показали, что дендритные клетки с различными фенотипами способны превращать регуляторные T-клетки в антигенспецифические аутоиммунные эффекторы (Baban et al., 2009, J Immunol 1 83:2475-2483, 18; Radhakrishnan et al., 2008, J Immunol 1 81:3 137-3147). Механистически указанный процесс характеризуется подавлением регулятора транскрипции FoxP3 и может вовлекать активацию эффекторных цитокинов. Наиболее последовательно отмечается превращение Treg в IL-17-продуцирующие эффекторные клетки, которые, вероятно, опосредуют Th17 иммунитет (Baban et al., 2009, J Immunol 183:2475-2483, 18; Radhakrishnan et al., 2008, J Immunol 181:3137-3147; Beriou et al., 2009, Blood 1 13:4240-4249). Определяли, отражает ли отмена супрессии простую деактивацию регуляторных T-клеток или их превращение в эффекторные клетки, которые затем усиливали иммунный ответ. Учитывая, что IFN-γ/LPS активированные DC демонстрировали высокий уровень продукции IL-12 и являлись сильными индукторами Th1 иммунитета, предположили, что превращение в эффекторы, скорее, можно было бы охарактеризовать с помощью продукции IFN-γ, нежели чем IL-17. Для проверки данной гипотезы Treg и CD4+CD25-эффекторы культивировали совместно в типичном отношении 1,25:1 и измеряли их цитокиновую продукцию с использованием метода ELISA. Обнаружили, что CD4+CD25+Treg и CD4+CD25-эффекторные клетки, культивированные совместно с незрелыми дендритными клетками, по существу не производят IFN-γ. Однако, как изображено на фигуре 8A, обе популяции производили значительные количества IFN-γ при совместном культивировании с LPS-активированными DC моноцитарного происхождения. Чтобы подтвердить, что измеряемый цитокин продуцировался T-клетками, а не набором дендритных клеток, клетки собирали после совместного культивирования и оценивали внутриклеточную продукцию TFN-γ. Как изображено на фигуре 8A, значительная фракция CD4+T-клеток являлась IFN-γ-положительной. Напротив, значительно меньшее число CD11c-положительных дендритных клеток являлось цитокин-положительными (данные не представлены). Чистота сортированной CD4+CD25+популяции достоверно составляет >99%, исключая вероятность того, что цитокиновая продукция опосредуется контаминирующими клетками (данные не представлены). Однако значительный процент сортированной CD4+CD25+популяции является FoxP3-отрицательным (приблизительно 20%; данные не представлены). Таким образом, возможно, что клетки FoxP3+, которым удачно приписывают супрессию, просто деактивируются, и цитокиновая продукция преимущественно поступает от указанной FoxP3-отрицательной группы клеток.
Исследования, демонстрирующие превращение Treg в эффекторные клетки, устанавливают соответствие указанного фенотипического изменения с супрессией регулятора транскрипции FoxP3 и активацией ряда цитокинов. Известно, что указанная LPS-активированная популяция DC поляризует Th1-иммунный ответ вследствие высокого уровня продукции IL-12, который способствует развитию IFN-γ-продуцирующих T-клеток. Дифференцировка Th1 клеток программируется с помощью действия нескольких факторов транскрипции, включая Tbox фактор T-bet. Предполагали, что переход Treg в IFN-γ-продуцирующие эффекторные клетки в данном исследовании мог быть ассоциирован с подавлением Treg-специфического фактора транскрипции FoxP3 и активацией T-bet. Для проверки данного предположения CD4+CD25+регуляторные клетки культивировали совместно с незрелыми или LPS-активированными DC и растворимым CD3, затем использовали внутриклеточное цитокиновое окрашивание для анализа экспрессии FoxP3 и T-bet. Как изображено на фигуре 8B, по сравнению с незрелыми дендритными клетками, LPS-активированные DC вызывали активацию T-bet у CD4+CD25+T-клеток. Интересно, что активация T-bet была наиболее заметной у FoxP3-положительных клеток. Отмечали, что значительная фракция CD4+CD25+T-клеток на 2-й день экспрессировала как FoxP3 так и T-bet. CD4+CD25+T-клетки, инкубированные в присутствии незрелых дендритных клеток не активировали T-bet, и очень небольшое число клеток оценивали как двойные положительные клетки. Несколько более ранний момент времени (день 2) был выбран в соответствии с предположением, что различия в пролиферации и продукции цитокинов, отмечаемые на 5-й день, отражают ранние изменения в экспрессии фактора транскрипции.
Результаты показывают, что продукция IFN-γ CD4+CD25+T-клетками, культивированными совместно с LPS-активированными DC коррелирует с активацией T-bet, особенно среди FoxP3-положительных клеток. Полученные результаты также делают менее правдоподобной вероятность того, что цитокины, обнаруживаемые с помощью метода ELISA, вырабатываются растущими FoxP3-отрицательными CD4+CD25+T-клетками. Вне связи с какой-либо теорией, считают, что тот факт, что большинство T-bet-положительных клеток также являются FoxP3-положительными, позволяет предположить, что регуляторные T-клетки превращаются в цитокин-продуцирующие эффекторы; менее правдоподобно, что FoxP3-отрицательные клетки развиваются с образованием обнаруживаемого цитокина. О том, что Treg, переходящие в эффекторные клетки, по меньшей мере временно экспрессируют многочисленные регуляторы транскрипции, сообщалось в отношении перехода в Th17 клетки (Beriou et al., 2009, Blood 1 13:4240-4249; Sharma et al., 2009, Blood 1 13:6102-6111).
Поскольку IL-12 является особенно важным для дифференцировки Th1 и последующей продукции IFN-γ, определяли являлось ли превращение регуляторных клеток в IFN-γ-продуцирующие клетки, отмечаемое здесь, IL-12-зависимым. Хотя показали, что указанные DC типа 1 подавляли эффекты Treg на пролиферацию реактивных клеток IL-12-независимым образом, возможно, что другие сигналы обуславливали превращение в эффекторные клетки. Поэтому Treg культивировали совместно с LPS-активированной DC в присутствии нейтрализующего антитела anti-TL-12 и оценивали экспрессию FoxP3 и T-bet. Как изображено на фигуре 8B, обнаружили, что активация T-bet являлась минимальной в присутствии нейтрализующего антитела.
Указанные результаты позволяют предположить новую парадигму интеграции регуляторных T-клеток в иммунную систему. В настоящее время Treg рассматривают в качестве главных медиаторов периферической толерантности, которые помогают смягчить развитие воспалительных иммунных ответов и предотвратить аутоиммунную реакцию. Как инициируется продуктивный иммунный ответ, несмотря на наличие и активность указанных клеток, остается неясным. В настоящее время большое число исследований, демонстрирующих отмену T-клеточной регуляции в контексте воспаления, позволяет предположить одну модель, согласно которой сигналы воспаления, обнаруженные на фоне патогенного повреждения, деактивируют регуляторы при развитии иммунного ответа. Следует отметить, что TLR-активированная DC восстанавливает пролиферацию иммунореактивных клеток, несмотря на присутствие Treg. Недавние исследования, иллюстрирующие превращение Treg в антигенспецифические Th17 клетки, уточняют данную модель, предполагая, что Treg могут становиться частью продуктивного иммунного ответа (Baban et al.(2009, J Immunol 183:2475-2483; Radhakrishnan et al., 2008, J Immunol 181:3 137-3147; Beriou et al., 2009, Blood 113:4240-4249; Sharma et al., 2009, Blood 113:6102-6111) Данные, представленные здесь, указывают, что в присутствии LPS-активированной популяции DC Treg рекрутируются в компартмент, подобный эффекторному компартменту, который преимущественно секретирует IFN-γ и, таким образом, по-видимому, больше соответствуют Th1-клеткам. Интересно, что, данный эффект (в отличие от эффекта, наблюдаемого при пролиферативном ответе) являлся IL-12-независимым. Указанные данные в совокупности позволяют предположить несколько возможностей в отношении интеграции Treg в развитие иммунного ответа. Во-первых, различные сигналы управляют отменой активности Treg и превращением в цитокин-продуцирующие эффекторы. Возможно, что растворимые сигналы, представленные во время инициации иммунного ответа, отчасти влияют на его силу, определяя, будут ли Treg “пассивными наблюдателями” или внесут свой вклад в качестве цитокин-продуцирующих эффекторов. Во-вторых, Treg могут превращаться в различные подтипы (Th1, Th2, Th17) антигенспецифических эффекторов в зависимости от типа иммунного ответа, необходимого в соответствии с природой повреждающего фактора. На молекулярном уровне указанный процесс может осуществляться с помощью активации линиеспецифичных факторов транскрипции (например, T-bet), и клетки могут по меньшей мере временно экспрессировать высокие уровни факторов, которые управляют как регуляторным, так и эффекторным фенотипом. Указанные клетки затем могут действовать синергично с дифференцированными аналогичным образом, “освобожденными” FoxP3-отрицательными клетками для максимального увеличения иммунитета. Указанные результаты также важны для разработки вакцин для иммунотерапии опухолей. Проблематичность применения регуляторных T-клеток доказана попытками ученых стимулировать противоопухолевый иммунитет с 1980-х годов. В некоторых последних исследованиях показано, что различные стратегии вакцинации увеличивают частоту встречаемости и/или активность регуляторных T-клеток (Lacelle et al., 2009, Clin Cancer Res 15:6881-6890; Eggermont, 2009, Clin Cancer Res 15:6745-6747; Francois et al., 2009, Cancer Res 69:4335-4345). Возможно, что особенно важно, обнаружили, что дендритные клетки, созревавшие с применением общепринятого цитокинового коктейля, часто применяемого при получении вакцин, увеличивают количество FoxP3high клеток, которые, по-видимому, обладают супрессорной функцией (Banerjee et al., 2006, Blood 108:2655-2661). Указанный факт может подвергнуть риску развитие и длительность противоопухолевого иммунитета. В свете указанных данных в некоторых современных протоколах совмещают противоопухолевую вакцину с агентами, которые истощают Treg; комбинация агентов часто обладает повышенным эффектом по сравнению с любым агентом по отдельности (Delluc et al., Cancer Immunol Immunother 58:1669-1677; Yamamoto et ai., 2009, Oncol Rep 22:337-343; Morse et al., 2008, Blood 1 12:610-618; Rech et al., 2009, Ann N Y Acad Sci 1 174:99-106; Shimizu et al., 1999, J Immunol 163:521 1-5218; Turk et al" 2004, J Exp Med 200:771-782; Dannull et al" 2005, J Clin Invest 115:3623-3633). Таким образом, оптимизация эффективности вакцин может потребовать замены нормальных процессов регуляции. В связи с этим вакцина на основе LPS-активированных дендритных клеток моноцитарного происхождения является идеальной в том смысле, что она, по-видимому, скорее ингибирует, чем активирует T-клеточно-опосредованную супрессию и может обладать преимуществом превращения указанных клеток в опухоль-реактивные эффекторные клетки.
Хотя не проводили прямого сравнения вакцины на основе DC, активированных LPS, с вакцинами на основе дендритных клеток, созревавших с применением общепринятых протоколов, эффект на функцию Treg, показанный в описании, обладает потенциалом в отношении эффективности вакцины на основе DC. Дополнительным преимуществом является то качество, что вакцина является собственной для организма и может не обладать токсичностями, ассоциированными с рядом экзогенных препаратов, уменьшающих количество Treg, которые используются в настоящее время в комбинированных протоколах.
Раскрытия каждого патента, патентной публикации и публикации, цитируемой здесь, включены в описание посредством ссылки во всей их полноте.
В то время как настоящее изобретение раскрыто со ссылкой на конкретные варианты осуществления, ясно, что другие варианты осуществления и вариации изобретения могут быть разработаны другими специалистами в данной области без отступления от сущности и объема изобретения. Прилагаемые пункты формулы изобретения имеют своей целью включение всех указанных воплощений и эквивалентных вариаций.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ НА ОСНОВЕ ДРОЖЖЕЙ С BRACHYURY | 2012 |
|
RU2619850C2 |
| ИММУНОТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ НА ОСНОВЕ ДРОЖЖЕЙ С BRACHYURY | 2012 |
|
RU2690180C2 |
| ДОСТАВКА БИОМОЛЕКУЛ В КЛЕТКИ ИММУННОЙ СИСТЕМЫ | 2015 |
|
RU2739794C2 |
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| ИММУНОТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ НА ОСНОВЕ ДРОЖЖЕЙ-MUC1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2642300C2 |
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛИРОВАНИЯ СОЗРЕВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СОДЕРЖАЩАЯ ГИБРИДНЫЙ БЕЛОК Rv2299c/ESAT-6 | 2017 |
|
RU2733886C2 |
| ПРИМЕНЕНИЕ МОНОМИКОЛИЛГЛИЦЕРИНА (MMG) В КАЧЕСТВЕ АДЪЮВАНТА | 2008 |
|
RU2479317C2 |
| АГЕНТЫ И СПОСОБЫ, ОСНОВАННЫЕ НА ПРИМЕНЕНИИ ДОМЕНА EDA ФИБРОНЕКТИНА | 2006 |
|
RU2430738C2 |
| ЭКСПРЕСС-АНАЛИЗЫ АКТИВАЦИИ Т-КЛЕТОК, ОСНОВАННЫЕ НА ИЗМЕРЕНИИ РНК С ИСПОЛЬЗОВАНИЕМ ПРОТОЧНОЙ ЦИТОМЕТРИИ | 2013 |
|
RU2671707C2 |
| РАСТВОРИМЫЙ МЕДИАТОР | 2012 |
|
RU2660580C2 |
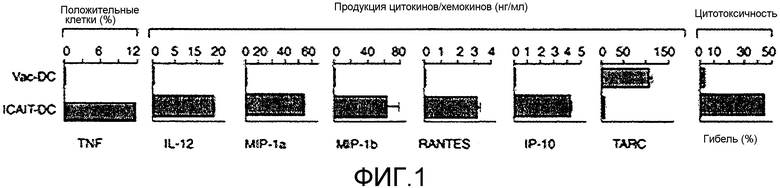
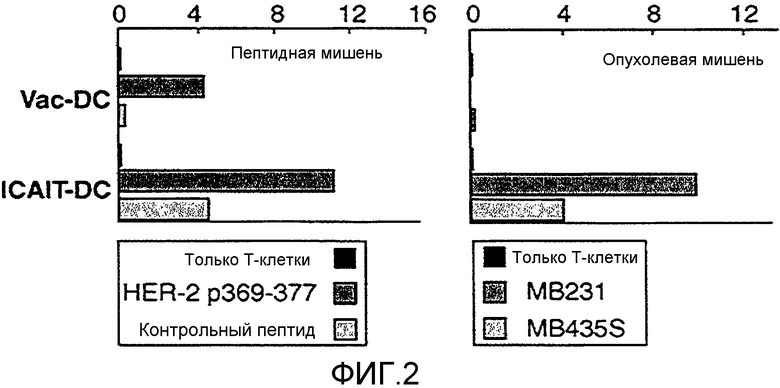
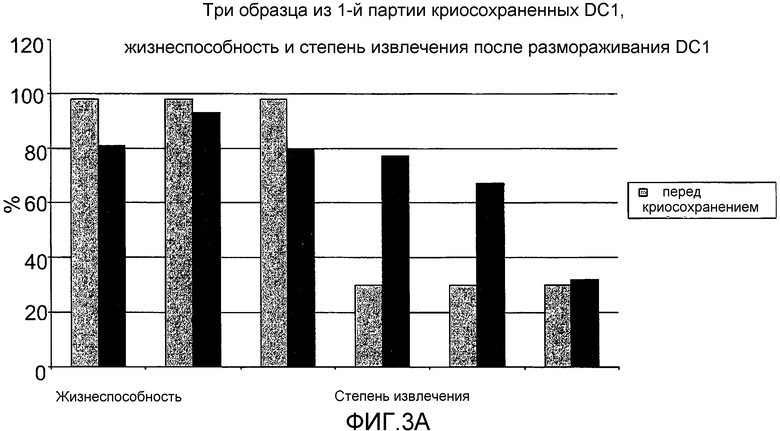
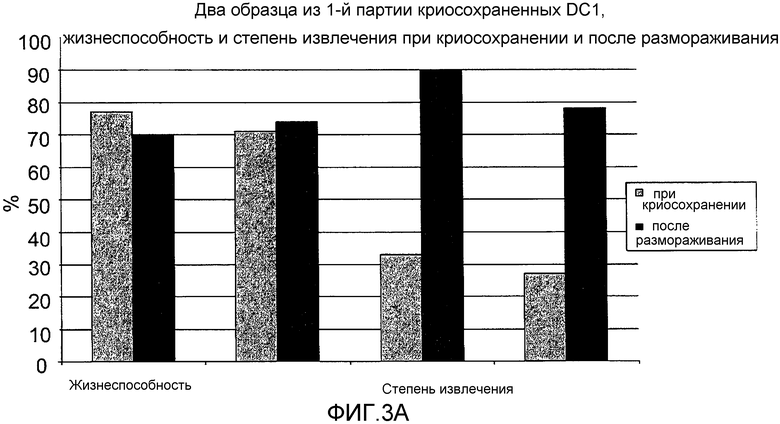
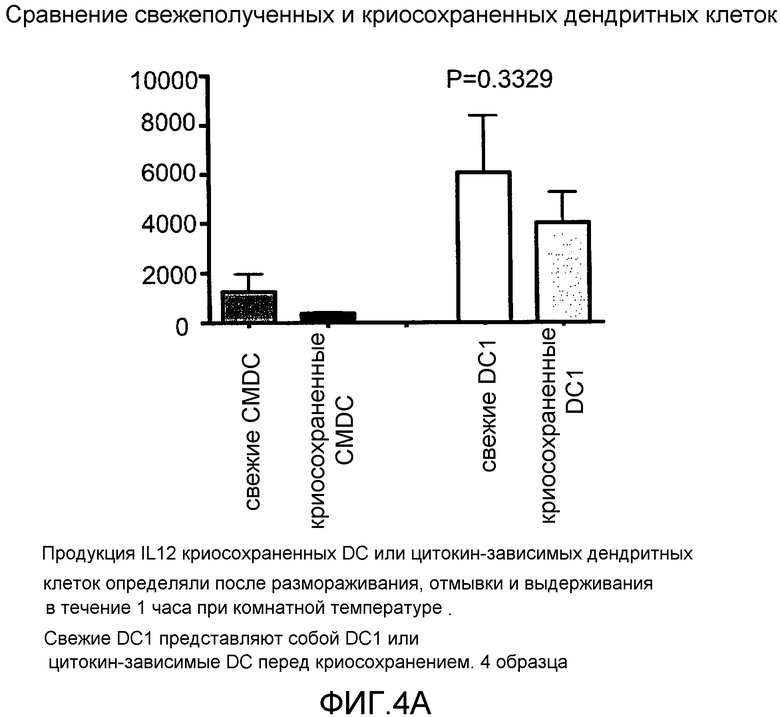
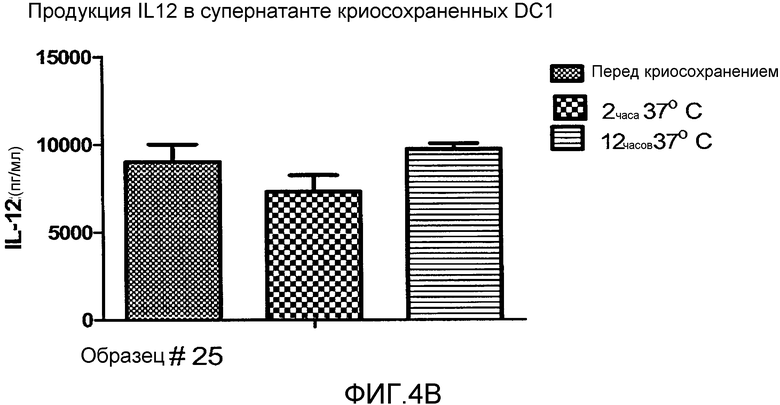
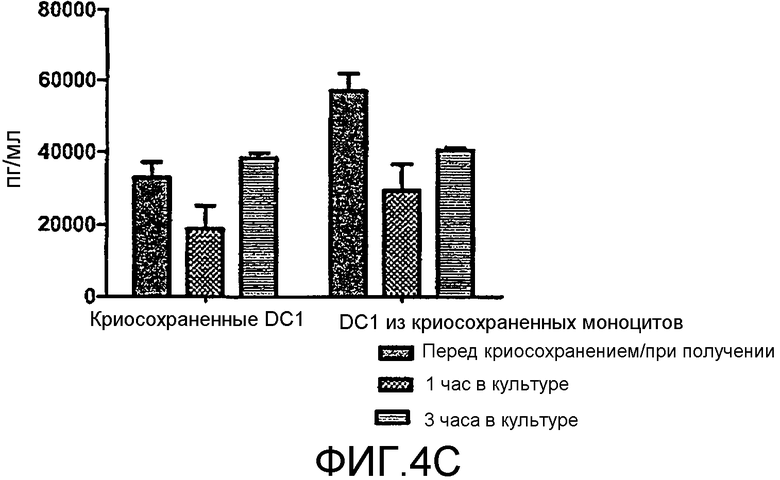
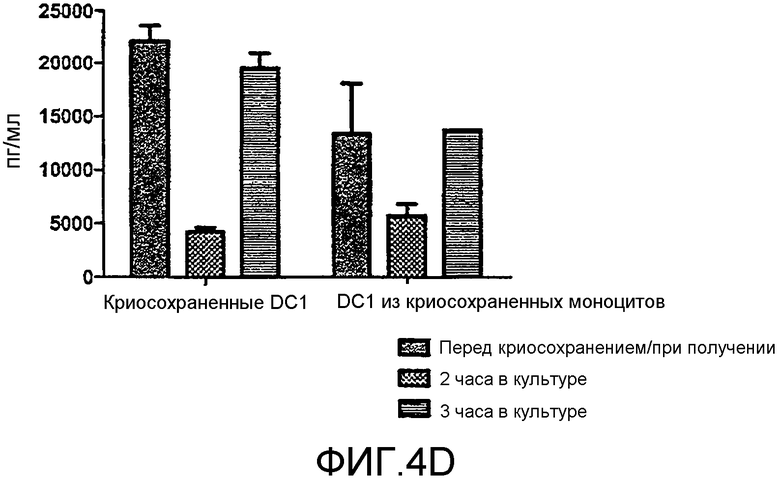
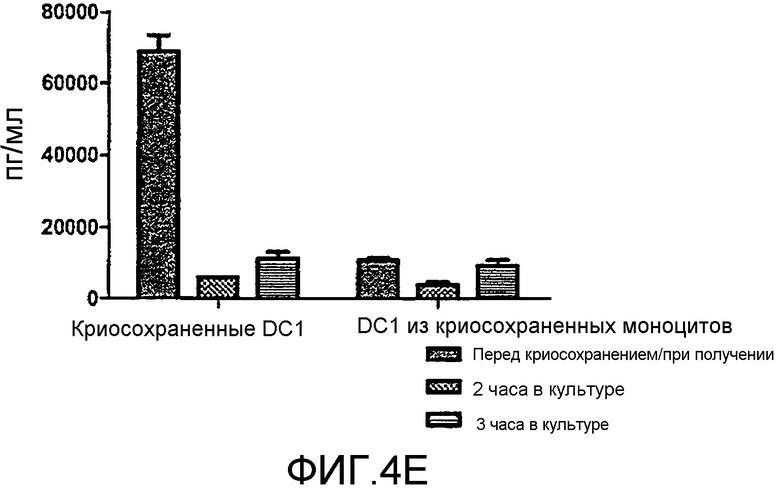
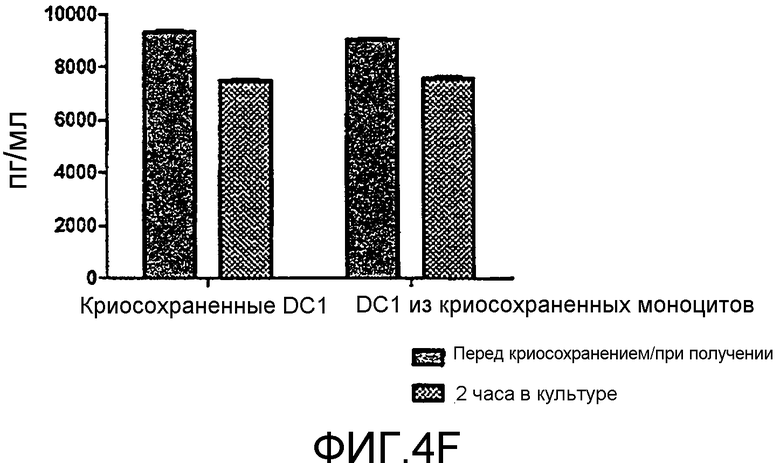
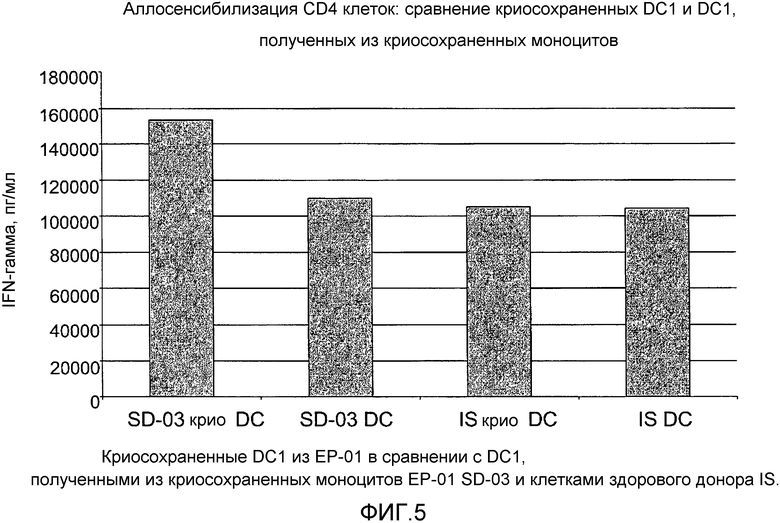
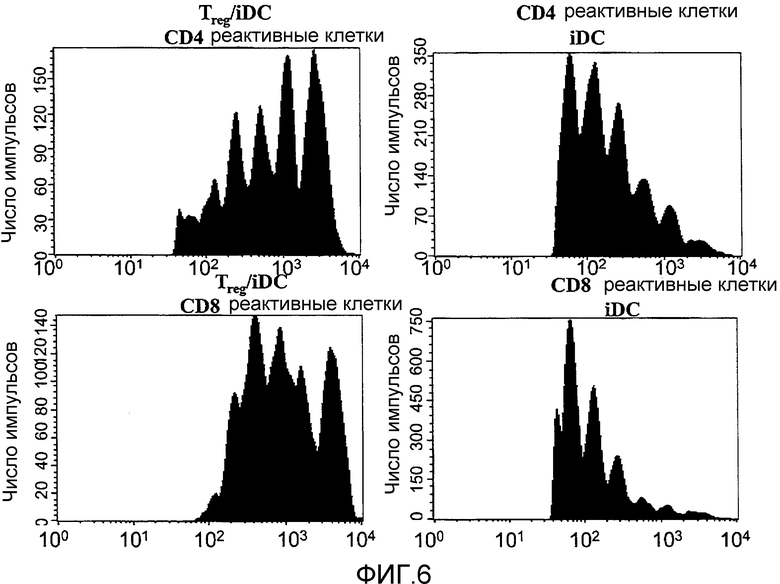
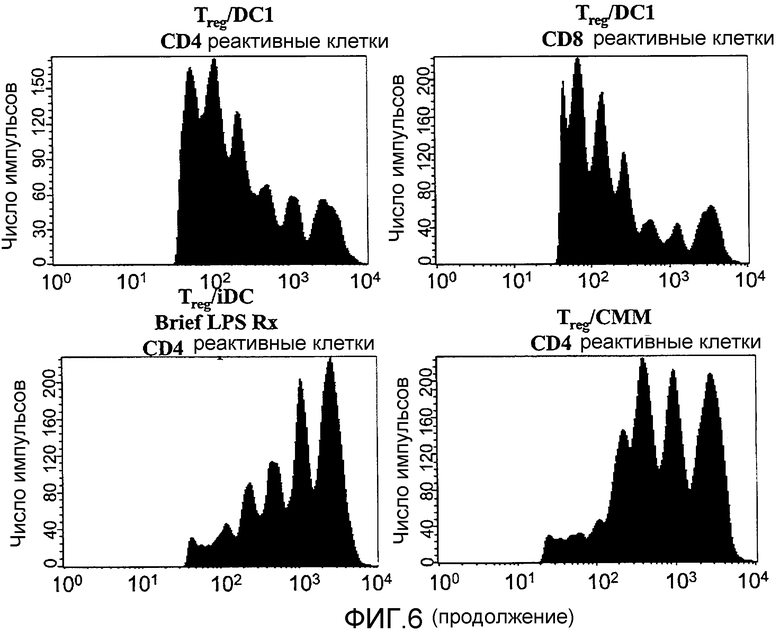
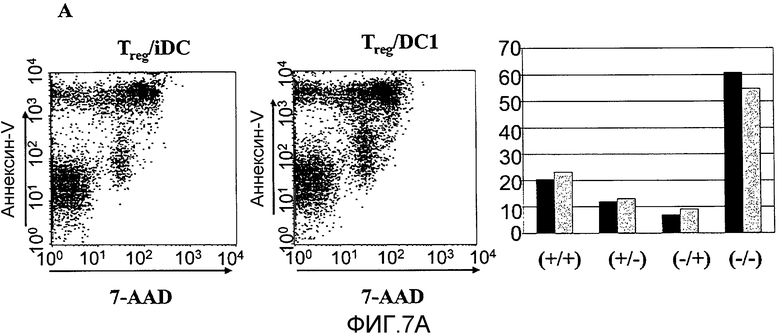
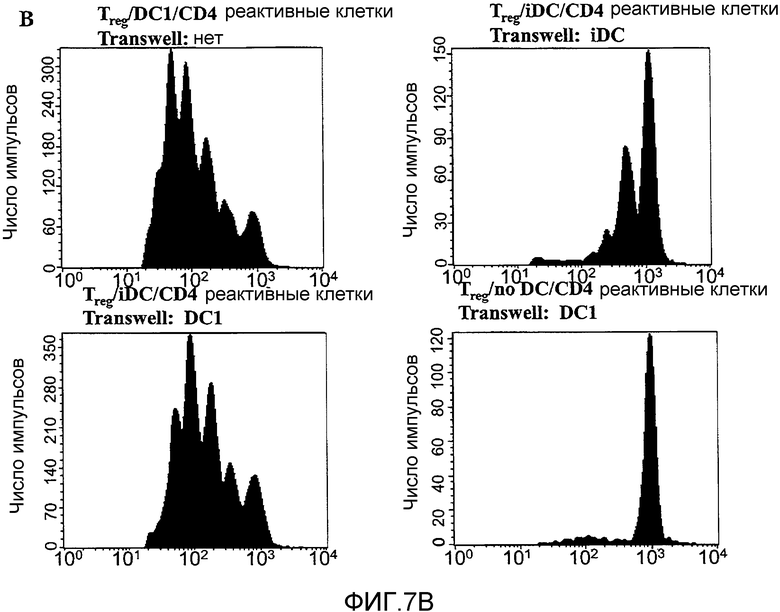
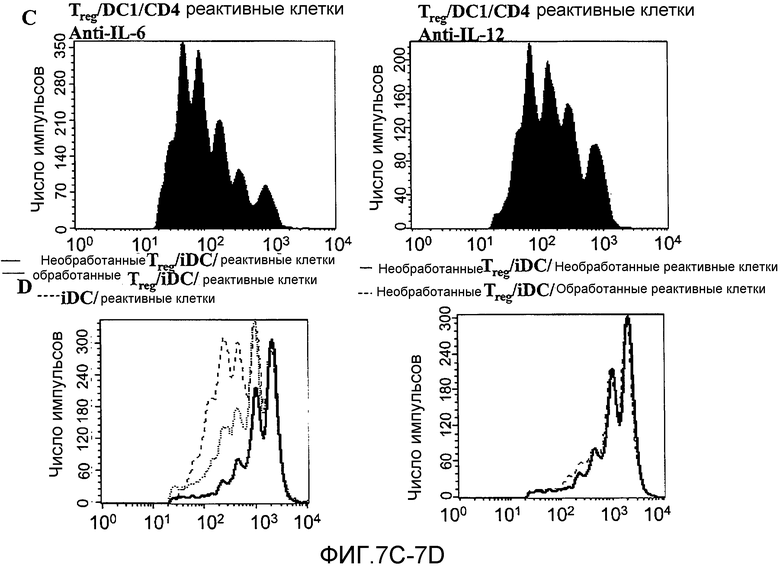
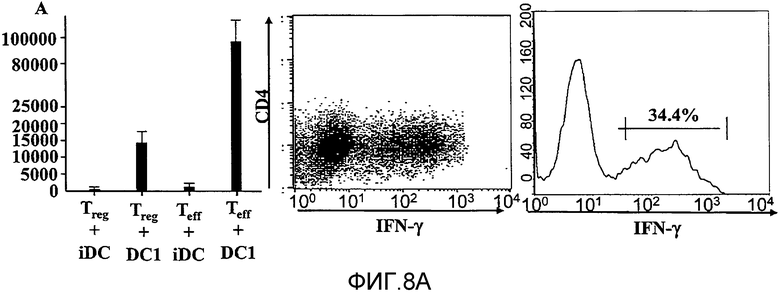
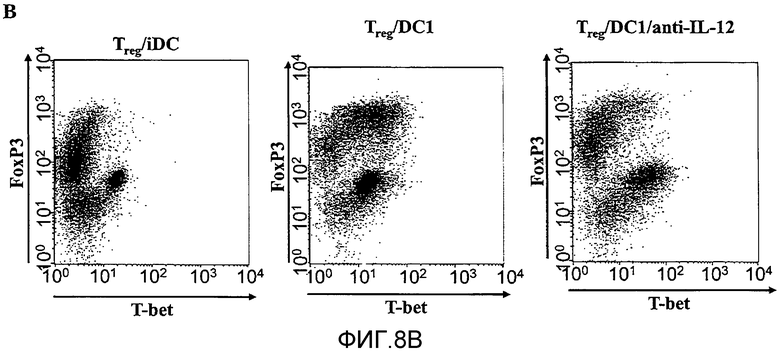
Настоящее изобретение относится к области биотехнологии. Предложен способ получения нагруженных антигеном, активированных дендритных клеток (DC) для применения в иммунотерапии, включающий нагрузку по меньшей мере одним антигеном неактивированных DC; активацию неактивированных DC IFN-γ и липополисахаридом (LPS) для образования активированных DC; криосохранение активированных DC и размораживание активированных DC; где степень извлечения и жизнеспособность активированных DC после размораживания выше или равна приблизительно 70%, и где размороженные активированные DC продуцируют эффективное количество по меньшей мере одного цитокина для создания Т-клеточного ответа. Также рассмотрена криосохраненная композиция, содержащая DC, и способ вызова иммунного ответа у млекопитающего, включающий введение композиции по изобретению. Данное изобретение может найти дальнейшее применение в производстве лекарственных средств на основе DC. 3 н. и 12 з.п. ф-лы, 8 ил., 4 пр.
1. Способ получения нагруженных антигеном, активированных дендритных клеток (DC) для применения в иммунотерапии, включающий:
нагрузку по меньшей мере одним антигеном неактивированных DC;
активацию неактивированных DC IFN-γ и липополисахаридом (LPS) для образования активированных DC;
криосохранение активированных DC; и
размораживание активированных DC;
где степень извлечения и жизнеспособность активированных DC после размораживания выше или равна приблизительно 70%, и где размороженные активированные DC продуцируют эффективное количество по меньшей мере одного цитокина для создания Т-клеточного ответа.
2. Способ по п.1, в котором антиген представляет собой опухолевый антиген или микробный антиген.
3. Способ по п.1, в котором криосохранение включает замораживание DC при температуре -70°C или ниже.
4. Способ по п.1, в котором цитокин представляет собой IL12.
5. Способ по п.1, в котором DC проявляют киллерную функцию, в результате чего DC способны лизировать раковые клетки-мишени.
6. Криосохраненная композиция для вызова иммунного ответа у млекопитающего, где композиция содержит:
DC, нагруженные по крайней мере одним антигеном;
причем DC активированы путем взаимодействия с IFN-γ и LPS; и
причем после размораживания DC продуцируют эффективное количество по меньшей мере одного цитокина для создания Т-клеточного ответа, причем DC были подвергнуты криосохранению в течение по крайней мере одной недели и сохраняли их сигнальные профиля, причем степень извлечения и жизнеспособность зрелых активированных DC после размораживания выше или равна приблизительно 70%.
7. Композиция по п.6, в которой антиген представляет собой опухолевый антиген или микробный антиген.
8. Композиция по п.6, где композицию подвергали криосохранению при температуре -70°C или ниже.
9. Композиция по п.6, в которой цитокин представляет собой IL12.
10. Композиция по п.6, в которой DC проявляют киллерную функцию, в результате чего DC способны лизировать раковые клетки-мишени.
11. Способ вызова иммунного ответа у млекопитающего, включающий ведение композиции по п.6 в организм млекопитающего, нуждающегося в этом.
12. Способ по п.11, в котором антиген представляет собой опухолевый антиген или микробный антиген.
13. Способ по п.11, в котором криосохранение включает замораживание DC при температуре -70°C или ниже.
14. Способ по п.11, в котором цитокин представляет собой IL12.
15. Способ по п.11, в котором DC проявляют киллерную функцию, в результате чего DC способны лизировать раковые клетки-мишени.

