Область техники
Настоящее изобретение относится к области биотехнологии, а именно к антителам или их антигенсвязывающим фрагментам, а также их применению. Более конкретно, настоящее изобретение относится к моноклональным антителам, которые специфически связываются с GITR (Glucocorticoid-induced TNFR-related protein/индуцируемый глюкокортикоидами рецептор фактора некроза опухолей/TNFRSF18/tumor necrosis factor receptor superfamily, member 18/мембранный белок, рецептор из надсемейства рецепторов фактора некроза опухоли). Изобретение также относится к нуклеиновой кислоте, кодирующей данное антитело или его антигенсвязывающему фрагменту, вектору экспрессии, способу получения антитела и применению антитела для лечения заболеваний или нарушений, связанных с GITR.
Уровень техники
TNFRSF18, GITR (англ. Glucocorticoid-induced TNFR-related protein/индуцируемый глюкокортикоидами рецептор фактора некроза опухолей/TNFRSF18/tumor necrosis factor receptor superfamily) - мембранный белок, рецептор из надсемейства рецепторов фактора некроза опухоли.
GITR представляет собой трансмембранный белок типа I, который состоит из 216 аминокислот, молекулярная масса его составляет 26 кДа. N-концевой внеклеточный домен содержит 3 повтора TNFR-Cys и участок N-гликозилирования. 3 цистеин-богатых домена и цитоплазматический хвост GITR имеет значительную гомологию с 4-1BB, OX40 и CD27 (Nocentini, et al. (1997) Proc. Natl. Acad. Sci. 94:6216-6221).
Человеческий GITR экспрессируется на низких уровнях иммунореактивными Т-клетками, при этом клетки CD4+ демонстрируют более высокую экспрессию по сравнению с клетками CD8+. Экспрессия GITR значительно повышается на несколько дней после активации Т-клеток. GITR постоянно экспрессируется на высоких уровнях в регуляторных Т-клетках (Treg), таких как клетки CD4+CD25+ или CD8+CD25+, и дополнительно повышается при активации этих клеток (Nocentini and Riccardi (2005) Е. J. Immunol. 35:1016)
При этом экспрессия GITR не ограничивается исключительно Т-клетками. В некоторых работах также указывалось, что GITR экспрессируется NK-клетками, макрофагами, В-клетками, дендритными клетками, тучными клетками и моноцитами (Nocentini and Riccardi (2005) Е. J. Immunol. 35:1016-1022).
GITR экспрессируется в лимфатических узлах, лейкоцитах периферической крови и в меньшей степени - в селезенке, конституитивно экспрессируется в высоких количествах на Treg, в низких количествах - на наивных Т-клетках и клетках памяти.
Однако GITR экспрессируется не только на иммунных, но и на опухолевых клетках. Был проведен RNA-Seq анализ данных в отношении экспрессии GITR в 33 типах опухолей, высокий уровень экспрессии выявлен в ПРГШ (плоскоклеточный рак головы и шеи), НМРЛ (немелкоклеточный рак легкого), РМЖ (рак молочной железы), раке пищевода и РМП (рак мочевого пузыря).
Подобные результаты получены в ходе RNA-Seq анализа образцов 24 типов опухолей. Таким образом, GITR экспрессируется не только на иммунных клетках, но и на мембране опухолевых клеток. В образцах опухолей GITRL-Fc усиливал экспрессию генов, ассоциированных с T-клетками, CD8 T клетками, цитотоксичностью, Th1 клетками, ИФН-гамма, NK, Teff, и T-клеточными маркерами активации.
Экспрессия GITR и его лиганда не ограничивается гематопоэтическими клетками. GITR также экспрессируется на кератиноцитах, предшественниках остеокластов, GITRL - на эндотелиальных клетках.
GITRL представляет собой трансмембранный белок типа II, что типично для большинства представителей семейства лигандов TNF. Проводимые на данный момент исследования показывают, что человеческий GITRL, как правило, существует в виде тримера, хотя он также может находиться в виде мономера или образовывать другие мультимерные формы (Chattopadhyay, et al. (2007) Proc. Natl. Acad. Sci. 104:19452-19457; Zhou, et al. (2008) Proc. Natl. Acad. Sci. 105:635-640). Существуют данные, позволяющие предположить, что вырабатывается также и растворимая форма GITRL (Baltz, et al. (2008) Blood 112:3735-3743; Mahesh, et al. (2006) Eur. J. Immunol. 36: 2128-2138). GITRL экспрессируется главным образом на антигенпрезентирующих клетках (АПК), включая макрофаги, В-клетки, дендритные клетки и эндотелиальные клетки, которые могут функционировать как АПК (Nocentini and Riccardi (2005) Е. J. Immunol. 35:1016-1022; Agostini, et al. (2005) Infect. Immun. 73:7502-7508; и Nocentini, et al. (2007) E. J. Immunol. 37:11651169).
Связывание GITRL на АПК с GITR на иммунореактивных Т-клетках запускает сигнализацию GITR, что костимулирует иммунореактивные Т-клетки и ингибирует супрессивную активность клеток регуляторных Т-клеток. Сигнализация GITR служит сигналом коактивации как для CD4+, так и CD8+ наивных Т-клеток, тем самым индуцируя или усиливая пролиферацию и эффекторную функцию, в частности, когда стимуляция рецепторов Т-клеток (РТК) близка к оптимальной (Schaer, et al. (2012) Curr. Opin. Immunol. 24:217224). Более конкретно, GITR может оказывать несколько видов действия на эффекторные Т-клетки и регуляторные Т-клетки, включая: костимуляцию и активацию эффекторных Т-клеток так, что они становятся более устойчивыми к ингибированию, ингибирование регуляторных Т-клеток, снижение чувствительности эффекторных Т-клеток к супрессии регуляторными Т-клетками и частичное удаление регуляторных Т-клеток из циркуляции (Nocentini, et al. (2007) Eur. J. Immunol. 37:1165-1169).
Основными функциями GITR, а значит и основными эффектами анти-GITR антитела, являются усиление пролиферации и функционирования эффекторных Т-клеток, а также ингибирующее влияние на супрессивное действие Tregs. Тeff клетки генерируются изначально с неспособностью противостоять ингибиторному опухолевому микроокружению и супрессии Treg - клетками. Стимуляция GITR на повторных стадиях прайминга и экспансии, через анти-GITR агонист, растворимый GITR лиганд или DC вакцину, модулирует Teff и Treg опухолевый ответ в пользу первых, что приводит к регрессии опухоли. Таким образом, анти-GITR обеспечивает устойчивость Teff к супресссии Treg.
Вместе вышеуказанные функции, в частности, костимуляция иммунореактивных Т-клеток и нейтрализация супрессорной активности регуляторных Т-клеток, означают, что активация GITR приводит к повышению иммунного ответа. Такая активация потенциально может приводить к восстановлению иммунных ответов на инфекции и опухоли. Соответственно, молекулы, способные активировать GITR, имели бы ценность как иммуностимулирующие агенты в условиях, когда необходимо вызвать повышенный иммунный ответ.
Антитело должно обладать свойствами агониста рецептора GITR, с эффекторными, цитотоксическими свойствами в отношении Treg лимфоцитов.
Из уровня техники известны различные антитела к GITR (например, WO2015187835, WO2015031667, WO2017068186, WO2017096189, WO2017214548).
На данный момент на стадии доклинических и клинических исследований находится 23 агониста к GITR (антител/рекомбинантных GITRL). Только 2 антитела находятся на 2 фазе КИ (TRX518, INCAGN1876), на 1 фазе КИ - AMG228, MEDI1873, MK-4166. Представлено очень мало клинических данных.
Однако, на данный момент, в мире нет ни одного антитела, которое специфически связывается с GITR, одобренного к терапевтическому использованию.
В связи с вышесказанным, актуальным является создание новых агонистических антител, которые взаимодействует с GITR, активирует рецептор, подавляет супрессорное действие Т-регуляторных лимфоцитов, ингибирует/деплетирует Т-супрессорное (регуляторное) звено иммунитета за счет ADCC эффекторных свойств.
BCD-166 представляет собой агонистическое моноклональное антитело, которое взаимодействует с GITR, активирует рецептор, подавляет супрессорное действие Т-регуляторных лимфоцитов, ингибирует/деплетирует Т-супрессорное (регуляторное) звено иммунитета за счет ADCC эффекторных свойств, тем самым увеличивая количество CD8+ и CD4+ эффекторных клеток и активируя Т-эффекторное звено иммунитета в микроокружении опухоли.
Краткое описание изобретения
В одном из аспектов настоящее изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему фрагменту, которое специфично связывается с GITR, включающему:
(a) вариабельный домен тяжелой цепи, содержащему:
(i) CDR1 с аминокислотной последовательностью, выбранной из группы:
NYGMH (SEQ ID NO: 1) или YYWMY (SEQ ID NO: 12);
(ii) CDR2 с аминокислотной последовательностью, выбранной из группы:
VIWFDGSNKFYTDSVKG (SEQ ID NO: 2) или AISWNGGRTYYAESMKG (SEQ ID NO: 13);
(iii) CDR3 с аминокислотной последовательностью, выбранной из группы:
ELGGYYYDSSGFRPYYYGMDV (SEQ ID NO: 3) или NRYYSDPNYGMNL (SEQ ID NO: 14), и
(b) вариабельный домен легкой цепи, содержащему:
(i) CDR1 с аминокислотной последовательностью, выбранной из группы:
RASQSIGSWLA (SEQ ID NO: 7) или TGTSTDIGTYKYIS (SEQ ID NO: 17);
(ii) CDR2 с аминокислотной последовательностью, выбранной из группы:
AASTLQR (SEQ ID NO: 8) или GVSHRPS (SEQ ID NO: 18);
(iii) CDR3 с аминокислотной последовательностью, выбранной из группы:
QQSHSHPLT (SEQ ID NO: 9) или SSYTSSGTVV (SEQ ID NO: 19).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3, соответственно;
- вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14, соответственно;
- вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 4.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 4.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 15.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 15.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 10.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 10.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 20.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 20.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 4;
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 10.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 4;
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 10.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, которой содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 15;
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 20.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 15;
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 20.
В некоторых вариантах моноклональное антитело, которое специфично связывается с GITR, представляет собой полноразмерное антитело IgG.
В некоторых вариантах моноклональное антитело IgG относится к изотипу IgG1, IgG2, IgG3 или IgG4 человека.
В некоторых вариантах моноклональное антитело IgG относится к изотипу IgG1 человека.
В некоторых вариантах моноклональное антитело, которое специфично связывается с GITR, включает мутацию E345R в Fc фрагменте для увеличения агонистических свойств, антителозависимой клеточной цитотоксичности (ADCC), но не комплементзависимой цитотоксичности (CDC).
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 5.
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 5.
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 6.
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 6.
В некоторых вариантах моноклональное антитело включает легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 11.
В некоторых вариантах моноклональное антитело включает легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11.
В некоторых вариантах моноклональное антитело включает:
- тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 5;
- легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 11.
В некоторых вариантах моноклональное антитело включает:
- тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 5;
- легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11.
В некоторых вариантах моноклональное антитело включает:
- тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 6;
- легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 11.
В некоторых вариантах моноклональное антитело включает:
- тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 6;
- легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11.
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 16.
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 16.
В некоторых вариантах моноклональное антитело включает легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 21.
В некоторых вариантах моноклональное антитело включает легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 21.
В некоторых вариантах моноклональное антитело включает:
- тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 16;
- легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 21.
В некоторых вариантах моноклональное антитело включает:
- тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 16;
- легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 21.
В одном из аспектов настоящее изобретение относится к выделенной нуклеиновой кислоте, которая кодирует любое вышеуказанное антитело или его антигенсвязывающий фрагмент.
В некоторых вариантах нуклеиновая кислота представляет собой ДНК.
В одном из аспектов настоящее изобретение относится к экспрессионному вектору, содержащему вышеуказанную нуклеиновую кислоту.
В одном из аспектов настоящее изобретение относится к способу получения клетки-хозяина для получения вышеуказанного антитела или его антигенсвязывающего фрагмента, который включает трансформирование клетки вышеуказанным вектором.
В одном из аспектов настоящее изобретение относится к клетке-хозяину для получения вышеуказанного антитела или его антигенсвязывающего фрагмента, содержащему вышеуказанную нуклеиновую кислоту.
В одном из аспектов настоящее изобретение относится к способу получения вышеуказанного антитела или его антигенсвязывающего фрагмента, заключающемуся в культивировании вышеуказанной клетки-хозяина в культуральной среде в условиях, достаточных для получения указанного антитела, при необходимости, с последующим выделением и очисткой полученного антитела.
В одном из аспектов настоящее изобретение относится к фармацевтической композиции для лечения заболевания или нарушения, опосредуемого GITR, содержащей вышеуказанное антитело или его антигенсвязывающий фрагмент в терапевтически эффективном количестве, в сочетании с одним или несколькими фармацевтически приемлемыми эксципиентами.
В некоторых вариантах фармацевтическая композиция предназначена для лечения заболевания или нарушения, опосредуемого GITR, выбранного из группы: РШМ (рак шейки матки), рак головы и шеи, рак желудка, РМЖ (рак молочной железы), почечно-клеточный рак, КРР (колоректальный рак), РЯ (рак яичника), НМРЛ (немелкоклеточный рак легкого).
В одном из аспектов настоящее изобретение относится к фармацевтической композиции для лечения заболевания или нарушения, опосредуемого GITR, содержащей вышеуказанное антитело или его антигенсвязывающий фрагмент в терапевтически эффективном количестве и по меньшей мере одно терапевтически активное противоопухолевое соединение в терапевтически эффективном количестве.
В некоторых вариантах фармацевтическая композиция предназначена для лечения заболевания или нарушения, опосредуемого GITR, выбранного из группы РШМ (рак шейки матки), рак головы и шеи, рак желудка, РМЖ (рак молочной железы), почечно-клеточный рак, КРР (колоректальный рак), РЯ (рак яичника), НМРЛ (немелкоклеточный рак легкого).
В некоторых вариантах фармацевтическая композиция включает терапевтически активное противоопухолевое соединение, которое выбирают из химиотерапевтического средства, антитела или противогормонального средства.
В некоторых вариантах фармацевтическая композиция включает терапевтически активное противоопухолевое соединение представляет антитело, которое выбирают из группы: антитела к PD1, антитела к PD-L1, антитела к CTLA4, антитела к анти 4-1BB, антитела к OX40 или их комбинации.
В некоторых вариантах фармацевтическая композиция включает терапевтически активное противоопухолевое соединение, которое представляет собой малую молекулу.
В некоторых вариантах фармацевтическая композиция включает терапевтически активное противоопухолевое соединение, которое выбирают из группы активаторов врожденного или приобретенного иммунитета.
В некоторых вариантах фармацевтической композиции вышеуказанное антитело и по меньшей мере одно терапевтически активное противоопухолевое соединение вводятся последовательно.
В некоторых вариантах фармацевтической композиции вышеуказанное антитело и по меньшей мере одно терапевтически активное противоопухолевое соединение вводятся одновременно.
В одном из аспектов настоящее изобретение относится к способу ингибирования биологической активности GITR у субъекта, нуждающемуся в таком ингибировании, включающему введение субъекту эффективного количества вышеуказанного антитела или его антигенсвязывающего фрагмента.
В одном из аспектов настоящее изобретение относится к способу лечения заболевания или нарушения, опосредованного GITR, включающему введение субъекту вышеуказанного антитела или его антигенсвязывающего фрагмента или вышеуказанной фармацевтической композиции, нуждающемуся в таком лечении, в терапевтически эффективном количестве.
В некоторых вариантах способ лечения включает заболевание или нарушение, которое выбрано из группы: РШМ (рак шейки матки), рак головы и шеи, рак желудка, РМЖ (рак молочной железы), почечно-клеточный рак, КРР (колоректальный рак), РЯ (рак яичника), НМРЛ (немелкоклеточный рак легкого).
В одном из аспектов настоящее изобретение относится к применению вышеуказанного антитела или его антигенсвязывающего фрагмента или вышеуказанной фармацевтической композиции для лечения у субъекта, нуждающегося в таком лечении, заболевания или нарушения, опосредуемого GITR.
В некоторых вариантах применение включает заболевание или нарушение, которое выбрано из группы: РШМ (рак шейки матки), рак головы и шеи, рак желудка, РМЖ (рак молочной железы), почечно-клеточный рак, КРР (колоректальный рак), РЯ (рак яичника), НМРЛ (немелкоклеточный рак легкого).
Краткое описание чертежей
Фиг. 1. Плазмида для наработки белка pEE-GITR-TEV-Fc lama.
Фиг. 2. Плазмида для наработки белка pEE-GITR -Fc.
Фиг. 3. Карта плазмиды pEE-humGITR-ligand-foldon-EPEA.
Фиг. 4. SDS-гель-электрофорез в 4-20% PAGE
1. Актемра 2,5 мг
2. маркер Fermentas unstained PW
3. -
4. GITR-TEV-Fc-lama +β-ME 10 мл
5. Ангиопоетин 2-H6F + β-ME 10 мл
6. -
7. GITR-TEV-Fc-lama -β -ME 10 мл
8. Ангиопоетин 2-H6F - β-ME 10 мл
Фиг. 5. SDS-гель-электрофорез
1. маркер Fermentas unstained PW
2. -
3. hGITR лиганд -EPEA среда до очистки 5 мл
4. hGITR лиганд -EPEA среда после очистки 5 мл
5. hGITR лиганд -EPEA
Фиг. 6. Схема синтеза комбинаторной наивной библиотеки человека.
Фиг. 7. Карта фагмиды pH5 для клонирования Fab фаговых дисплейных библиотек.
Фиг. 8. Карта экспрессионной плазмиды pLL для наработки секретируемых Fab.
Фиг. 9. Карта экспрессионного вектора pEE-BCD166-01-001-VH-HC кодирующего тяжелую цепь антител для транзиентной продукции антител в клетках млекопитающих
Фиг. 10. Карта экспрессионного вектора pEE-BCD166-01-001-VK-CK кодирующего легкую цепь антител для транзиентной продукции антител в клетках млекопитающих
Фиг. 11. SDS-гель-электрофорез в 12% PAGE + β-ME
1. Актемра 5 мл
2. маркер Fermentas unstained PW
3. -
4. анти-GITR антитело lot#1161 среда до очистки 5 мл
5. анти-GITR антитело lot#1161 среда после очистки 5 мл
6. анти-GITR антитело lot#1161 5 мл
7. анти-GITR антитело lot#1162 5 мл
8. анти-GITR антитело lot#1163 5 мл
9. анти-GITR антитело lot#1164 5 мл
10. анти-GITR антитело lot#1165 5 мл
11. анти-GITR антитело lot#1166 среда до очистки 5 мл
12. анти-GITR антитело lot#1166 среда после очистки 5 мл
13. анти-GITR антитело lot#1166 5 мл
14. анти-GITR антитело lot#1167 5 мл
Фиг. 12. SDS-гель-электрофорез в 12% PAGE + β-ME
1. маркер Fermentas unstained PW
2. анти-GITR антитело lot#1208 среда до очистки 10 мл
3. анти-GITR антитело lot#1208 среда после очистки 10 мл
4. анти-GITR антитело lot#1208 5 мл
5. анти-GITR антитело lot#1209 5 мл
6. анти-GITR антитело lot#1210 5 мл
7. анти-GITR антитело lot#1211 5 мл
8. анти-GITR антитело lot#1212 5 мл
9. анти-GITR антитело lot#1213 5 мл
10. анти-GITR антитело lot#1214 среда до очистки 10 мл
11. анти-GITR антитело lot#1214 среда после очистки 10 мл
12. анти-GITR антитело lot#1214 5 мл
Фиг. 13. График с результатами проверки специфической агонистической активности анти-GITR моноклональных антител лотов 1161 (BCD166-01-01), 1210 BCD166-01-011) и 1213 BCD166-01-014) в тесте на репортерной клеточной линии.
Фиг. 14. График с результатами проверки специфического связывания антитела BCD166-01-01 с GITR человека, GITR мыши и GITR обезьяны циномолгус.
Фиг. 15. График с результатами проверки специфического связывания антитела BCD166-01-011 с GITR человека, GITR мыши и GITR обезьяны циномолгус.
Фиг. 16. График с результатами проверки специфического связывания антитела BCD166-01-014 с GITR человека, GITR мыши и GITR обезьяны циномолгус.
Фиг. 17. График с результатами уровня активации в клеточном агонистическом тесте исследуемых BCD166-01-01, BCD166-02-01, BCD166-01-014.
Фиг. 18. График с результатами величины EC50 в клеточном ADCC тесте исследуемого BCD166-02-01, BCD166-01-01, BCD166-01-014, BCD166-02-014.
Фиг. 19. График с результатами уровня CDC исследуемого BCD166-01-01, BCD166-02-01, BCD166-01-014.
Фиг. 20. График с результатами влиянии исследуемых anti-GITR кандидатов на иммунореактивные клетки (NK) в сравнении с негативным контролем.
Фиг. 21. График с результатами влиянии исследуемых anti-GITR кандидатов на иммунореактивные клетки (B) в сравнении с негативным контролем.
Фиг. 22. График с результатами влиянии исследуемых anti-GITR кандидатов на иммунореактивные клетки (CD3) в сравнении с негативным контролем.
Фиг. 23. График с результатами влиянии исследуемых anti-GITR кандидатов на иммунореактивные клетки (CD8+ T клетки) в сравнении с негативным контролем.
Фиг. 24. График с результатами влиянии исследуемых anti-GITR кандидатов на иммунореактивные клетки (CD4+ T клетки) в сравнении с негативным контролем.
Фиг. 25. Анализ антитело-зависимой деплеции nTreg под действием изучаемых anti-GITR кандидатов для клеточного материала донора 1.
Фиг. 26. Анализ антитело-зависимой деплеции nTreg под действием изучаемых anti-GITR кандидатов для клеточного материала донора 2.
Фиг. 27. Анализ антитело-зависимой деплеции iTreg под действием изучаемых anti-GITR кандидатов для клеточного материала донора 1 и донора 2
Фиг. 28. Результаты анализа влияния анти-GITR антитела на уровень секреции провоспалительных цитокинов IL-2 и IFN- γ, обуславливающих антираковых эффект.
Фиг. 29. Таблица данных констант аффинности связывания BCD166-02-01 кандидата с гамма рецепторами.
Фиг. 30. График по стабильности BCD166-02-01 при 72-часовой инкубации при 50°С.
Фиг. 31. График по стабильности BCD166-01-014 при 72-часовой инкубации при 50°С.
Фиг. 32. Средние значения объемов опухолей в группах в течение эксперимента.
Фиг. 33. Показатель торможения роста опухоли на 33 сутки.
Описание изобретения
Определения и общие методы
Если иное не определено в настоящем документе, научные и технические термины, используемые в связи с настоящим изобретением, будут иметь значения, которые обычно понятны специалистам в данной области.
Кроме того, если по контексту не требуется иное, термины в единственном числе включают в себя термины во множественном числе, и термины во множественном числе включают в себя термины в единственном числе. Как правило, используемая классификация и методы культивирования клеток, молекулярной биологии, иммунологии, микробиологии, генетики, аналитической химии, химии органического синтеза, медицинской и фармацевтической химии, а также гибридизации и химии белка и нуклеиновых кислот, описанные в настоящем документе, хорошо известны специалистам и широко применяются в данной области. Ферментативные реакции и способы очистки осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как описано в настоящем документе.
Определения, связанные с антителом
TNFRSF18, GITR (англ. Glucocorticoid-induced TNFR-related protein/индуцируемый глюкокортикоидами рецептор фактора некроза опухолей/TNFRSF18/tumor necrosis factor receptor superfamily) - мембранный белок, рецептор из надсемейства рецепторов фактора некроза опухоли. GITR представляет собой трансмембранный белок типа I, который состоит из 216 аминокислот, молекулярная масса его составляет 26 кДа. N-концевой внеклеточный домен содержит 3 повтора TNFR-Cys и участок N-гликозилирования. 3 цистеин-богатых домена и цитоплазматический хвост GITR имеет значительную гомологию с 4-1BB, OX40 и CD27 (Nocentini, et al. (1997) Proc. Natl. Acad. Sci. 94:6216-6221).
Амплификация гена GITR и/или сверхэкспрессия его белка были обнаружены при многих раковых заболеваниях, в том числе при РШМ (раке шейки матки), раке головы и шеи, раке желудка, РМЖ (раке молочной железы), почечно-клеточном раке, КРР (колоректальном раке), РЯ (раке яичника), НМРЛ (немелкоклеточном раке легкого).
Термин «связывающая молекула» включает в себя антитела и иммуноглобулины.
Термин «антитело» или «иммуноглобулин» (Ig), как использовано в данном описании, включает целые антитела и любой антигенсвязывающий фрагмент (т.е. «антигенсвязывающую часть») или его отдельные цепи. Термин “антитело» относится к гликопротеину, содержащему по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, взаимосвязанные дисульфидными связями, или его антигенсвязывающей части. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно называемую в данном описании как VH) и константную область тяжелой цепи. Известно пять типов тяжелых цепей антител млекопитающих, которые обозначают греческими буквами: α, δ, ε, γ и µ. Присутствующий тип тяжелой цепи определяет класс антитела; указанные цепи обнаружены в антителах типа IgA, IgD, IgE, IgG и IgM соответственно. Различные тяжелые цепи отличаются по размеру и составу; α и γ содержат примерно 450 аминокислот, а µ и ε состоят примерно из 550 аминокислот. Каждая тяжелая цепь содержит две области, т.е. константную область и вариабельную область. Константная область является идентичной во всех антителах одного и того же изотипа, но отличается в антителах различного изотипа. Тяжелые цепи γ, α и δ содержат константную область, которая состоит из трех константных доменов CH1, СН2 и CH3 (выстроены в ряд) и шарнирной области, которая придает гибкость (Woof J., Burton D., Nat Rev Immunol 4, 2004, cc.89-99); тяжелые цепи µ и ε содержат константную область, которая состоит из четырех константных доменов CH1, СН2, CH3 и CH4. У млекопитающих известно только два типа легких цепей, которые обозначают как лямбда (λ) и каппа (κ). Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно называемой в данном описании как VL) и константной области легкой цепи. Примерная длина легкой цепи составляет 211-217 аминокислот. Предпочтительно легкая цепь представляет собой легкую каппа (κ)-цепь, а константный домен CL предпочтительно представляет собой С-каппа (κ).
«Антитела» согласно изобретению могут представлять собой антитела любого класса (например, IgA, IgD, IgE, IgG и IgM, предпочтительно IgG) или подкласса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2, предпочтительно IgG1).
Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), разбросанные между областями, которые являются более консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат домен связывания, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями хозяина или факторами, включая различные клетки иммунной системы (например, эффекторными клетками), и первый компонент (Clq) классической системы комплемента.
Термин «антигенсвязывающая часть» антитела или «антигенсвязывающий фрагмент» (или просто «часть антитела» или «фрагмент антитела»), как использовано в данном описании, относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфически связываться с антигеном. Было показано, что антигенсвязывающая функция антитела может выполняться фрагментами полноразмерного антитела. Примеры связывающих фрагментов, включенных в термин “антигенсвязывающая часть» антитела включают (i) Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH 1; (ii) F(ab’)2-фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; (iii) Fd- фрагмент, состоящий из доменов VH и CH 1; (iv) Fv-фрагмент, состоящий из доменов VL и VH в едином плече антитела, (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH/VHH; и (vi) выделенная определяющая комплементарность область (CDR). Кроме того, две области Fv-фрагмента, VL и VH, кодируются разными генами, они могут быть соединены при помощи рекомбинантных способов с использованием синтетического линкера, который дает возможность получать их в виде единой белковой цепи, в которой области VL и VH спариваются с образованием одновалентных молекул (известных как одноцепочечный Fv (scFv); см., например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Предполагается, что такие одноцепочечные молекулы также включены в термин “антигенсвязывающая часть» антитела. Такие фрагменты антител получают с использованием общепринятых способов, известных специалистам в данной области, и эти фрагменты подвергают скринингу таким же образом, как и интактные антитела.
Предпочтительно CDR антигенсвязывающего участка или весь антигенсвязывающий участок антител по изобретению имеет происхождение из мыши, ламы или донорской человеческой библиотеки или по существу человеческое происхождение с определенными аминокислотными остатками, измененными, например, замещенными разными аминокислотными остатками с тем, чтобы оптимизировать конкретные свойства антитела, например KD, koff, IC50, EC50, ED50. Предпочтительно каркасные участки антитела по изобретению имеют человеческое происхождение или по существу человеческое происхождение (по крайней мере на 80, 85, 90, 95, 96, 97, 98 или 99% человеческое происхождение).
В других вариантах осуществления антигенсвязывающий участок антитела по изобретению может происходить из других нечеловеческих видов, включая мыши, ламы, кролика, крысу или хомяка, но не ограничиваясь ими. Альтернативно, антигенсвязывающий участок может происходить из человеческих видов.
Термин «вариабельный» относится к тому факту, что определенные сегменты вариабельных доменов широко отличаются в последовательности среди антител. Домен V опосредует связывание антигена и определяет специфичность конкретного антитела к его конкретному антигену. Однако вариабельность неравномерно распределяется на участке вариабельных доменов из 110 аминокислот. Напротив, V области состоят из инвариантных фрагментов, называемых каркасными областями (FR) из 15-30 аминокислот, разделенных более короткими участками чрезвычайной вариабельности, называемых «гипервариабельными областями» или CDR. Каждый вариабельный домен нативных тяжелых и легких цепей содержит четыре FR, в основном принимающих конфигурацию бета-листов, связанных тремя гипервариабельными областями, которые образуют петли, связывающие, и в некоторых случаях являющиеся частью бета-складчатой структуры. Гипервариабельные области в каждой цепи удерживаются вместе в тесной близости с помощью FR и с гипервариабельными областями другой цепи вносят вклад в образование антигенсвязывающего сайта антител. Константные домены не принимают непосредственного участия в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности (АЗКЦ, ADCC).
Термин «гипервариабельная область» по данному описанию относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Обычно гипервариабельная область содержит аминокислотные остатки из «области, определяющей комплементарность» или «CDR», и/или такие остатки из «гипервариабельной петли».
В некоторых случаях может также быть предпочтительным изменение одного или более остатков аминокислот CDR-участков с целью повышения аффинности связывания с целевым эпитопом. Это известно, как «созревание аффинности» и в некоторых случаях может выполняться в связи с гуманизацией, например, в ситуациях, когда гуманизация антитела приводит к снижению специфичности или аффинности связывания, и не представляется возможным в достаточной степени улучшить специфичность или аффинность связывания с помощью только обратных мутаций. Различные методы созревания аффинности известны в данной области техники, например, способ in vitro сканирующего насыщающего мутагенеза, описанный Burks et al., Proc Natl Acad Sci USA, 94:412-417 (1997), и способ пошагового in vitro созревания аффинности, предложенный Wu et al., Proc Natl Acad Sci USA 95:6037 6042 (1998).
«Каркасные области» (FR) представляют собой остатки вариабельного домена, отличные от CDR остатков. Обычно каждый вариабельный домен имеет четыре FR, определяемые как FR1, FR2, FR3 и FR4. Если CDR определяются согласно Kabat, FR остатки легкой цепи локализуются, приблизительно, в области остатков 1-23 (LCFR1), 35-49 (LCFR2), 57-88 (LCFR3) и 98-107 (LCFR4), а остатки FR тяжелой цепи локализуются, приблизительно, в области остатков 1-30 (HCFR1), 36-49 (HCFR2), 66-94 (HCFR3) и 103- 113 (HCFR4) в тяжелой цепи. Если участки CDR содержат аминокислотные остатки из гипервариабельных петель, FR остатки легкой цепи локализуются, приблизительно, в остатках 1-25 (LCFR1), 33-49 (LCFR2), 53-90 (LCFR3) и 97-107 (LCFR4) в легкой цепи, а FR остатки тяжелой цепи локализуются, примерно, в остатках 1-25 (HCFRI), 33-52 (HCFR2), 56-95 (HCFR3) и 102-113 (HCFR4) в остатках тяжелой цепи. В некоторых примерах, когда CDR содержит аминокислоты как из CDR по Kabat, так и аминокислоты из гипервариабельной петли, FR соответствующим образом корректируются. Например, когда CDRH1 включает аминокислоты Н26-Н35, остатки FR1 тяжелой цепи находятся в положениях 1-25, а остатки FR2 находятся в положениях 36-49.
Кристаллизующийся фрагмент иммуноглобулина (англ. fragment crystallizable region, Fc region, Fc) - это концевая часть молекулы иммуноглобулина, которая взаимодействует с Fc-рецептором на поверхности клетки и с некоторыми белками системы комплемента. Данное свойство позволяет антителам активировать иммунную систему. Fc-участок IgG, IgA и IgD изотипов состоит из двух одинаковых белковых фрагментов, соответственно, второго и третьего константных доменов двух тяжелых цепей; в случае изотипов IgM и IgE Fc содержит три константных домена тяжелых цепей (домены CH 2-4) в каждой полипептидной цепочке.
Антитело по данному изобретению, «которое связывает» целевой антиген, представляет собой антитело, которое связывает антиген с достаточной аффинностью так, что антитело можно применять в качестве диагностического и/или терапевтического агента при нацеливании на белок или клетку, или ткань, экспрессирующую антиген, и в незначительной степени перекрестно реагирует с другими белками. По данным аналитических методов: сортинга флуоресцентно-активированных клеток (FACS), радиоиммунопреципитации (RIA) или ИФА (ELISA), в таких вариантах изобретения степень связывания антитела с белком, не являющимся «мишенью» (с «нецелевым белком»), составляет менее 10% от связывания антитела с конкретным белком-мишенью. По отношению к связыванию антитела с молекулой-мишенью термин «специфическое связывание» или выражения «специфически связывается с» или «специфический к» конкретному полипептиду или эпитопу на конкретном полипептиде-мишени означает связывание, которое заметно (измеримо) отличается от неспецифического взаимодействия (например, в случае bH1-44 или bH1-81 неспецифическое взаимодействие представляет собой связывание с бычьим сывороточным альбумином, казеином, фетальной бычьей сывороткой или нейтравидином).
Специфическое связывание можно определять количественно, например, определяя связывание молекулы по сравнению со связыванием контрольной молекулы. Например, специфическое связывание можно определять конкурентной реакцией с другой молекулой, аналогичной мишени, например, с избытком немеченой мишени. В этом случае специфическое связывание указывается, если связывание меченой мишени с зондом конкурентно ингибируется избытком немеченой мишени. В данном описании термин «специфическое связывание» или выражения «специфически связывается с» или «специфический к» конкретному полипептиду или эпитопу на конкретном полипептиде-мишени можно характеризовать на примере молекулы, имеющей Kd к мишени по меньшей мере около 200 нМ, или же по меньшей мере около 150 нМ, или же по меньшей мере около 100 нМ, или же по меньшей мере около 60 нМ, или же по меньшей мере около 50 нМ, или же по меньшей мере около 40 нМ, или же по меньшей мере около 30 нМ, или же по меньшей мере около 20 нМ, или же по меньшей мере около 10 нМ, или же по меньшей мере около 8 нМ, или же по меньшей мере около 6 нМ, или же по меньшей мере около 4 нМ, или же по меньшей мере около 2 нМ, или же по меньшей мере около 1 нМ или выше. В одном варианте изобретения термин «специфическое связывание» относится к связыванию, при котором молекула связывается с конкретным полипептидом или эпитопом на конкретном полипептиде, практически не связываясь с каким-либо другим полипептидом или эпитопом на полипептиде.
Термин «Ka», как использовано в данном описании, относится к скорости ассоциации конкретного взаимодействия антитело-антиген.
Термин «Kd», как использовано в данном описании, относится к скорости диссоциации конкретного взаимодействия антитело-антиген.
«Аффинность связывания» обычно относится к силе совокупных нековалентных взаимодействий между единичным сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указано иначе, «аффинность связывания» относится к внутренней (характерной, истинной) аффинности связывания, которая отражает 1:1 взаимодействие между членами пары связывания (например, антителом и антигеном). Аффинность молекулы Х к своему партнеру Y обычно можно представить константной диссоциации (Kd). Желательно, чтобы величина Kd составляла, примерно, 200 нМ, 150 нМ, 100 нМ, 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 8 нМ, 6 нМ, 4 нМ, 2 нМ, 1 нМ или менее. Аффинность можно измерять обычными методами, известными в уровне техники, включая методы по данному описанию. Низкоаффинные антитела обычно медленно связываются с антигеном и имеют тенденцию легко диссоциировать, тогда как высокоаффинные антитела обычно быстрее связывают антиген и имеют тенденцию дольше оставаться в связанном состоянии. В уровне техники известны различные методы измерения аффинности связывания, любой из этих методов можно использовать для целей настоящего изобретения.
В одном варианте изобретения «Kd» или «величину Kd» по данному изобретению измеряют методами поверхностного плазмонного резонанса на приборе BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ) при 25°С, используя чипы с иммобилизованным антигеном СМ5 при ~10 относительных единицах (единицах отклика, RU). Коротко говоря, биосенсорные чипы с карбоксиметилдекстраном (СМ5, BIAcore Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламинопропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями производителя. Антиген разводят 10 мМ раствором ацетата натрия, рН 4.8, до концентрации 5 мкг/мл (~0.2 мкМ), а затем вводят (инжекция) при скорости потока 5 мкл/минута до достижения, примерно, 10 относительных единиц (RU) связанного белка. После введения антигена вводят 1 М раствор этаноламина, чтобы блокировать непрореагировавшие группы. Для кинетических измерений двукратные серийные разведения Fab (например, от 0.78 нМ до 500 нМ) вводят в PBS с 0.05% Tween 20 (PBST) при 25°С при скорости потока, примерно, 25 мкл/мин. Величины скорости ассоциации (kon) и скорости диссоциации (koff) рассчитывают, применяя простую модель Ленгмюра для связывания один-плюс-один (BIAcore Evaluation Software версия 3.2), с помощью одновременного получения сенсограммы ассоциации и диссоциации. Константу равновесной диссоциации (Kd) рассчитывают как отношение koff/kon. См., например, Chen, Y., et al., (1999) J. Mol. Biol. 293: 865-881. Если по данным вышеуказанного метода поверхностного плазмонного резонанса скорость ассоциации превышает 106 М-1 сек-1, тогда ее можно определять методом тушения флуоресценции, который измеряет увеличение или уменьшение интенсивности флуоресцентной эмиссии (возбуждение=295 нм; эмиссия (излучение)=340 нм, полоса 16 нм) при 25°С раствора антитела против антигена (Fab форма) с концентрацией 20 нМ в PBS, рН 7.2, в присутствии увеличивающихся концентраций антигена, измеряемых с помощью спектрометра, такого как спектрофотометр остановленного потока (Aviv Instruments) или спектрофотометр SLM-Aminco (ThermoSpectronie) серии 8000 с кюветой с перемешиванием.
Термин «koff» относится к константе скорости диссоциации конкретного взаимодействия связывающей молекулы и антигена. Константу скорости диссоциации koff можно измерить посредством биослойной интерферометрии, например, с помощью системы Octet™.
«Скорость ассоциации» («on-rate») или «kon» по данному изобретению можно также определять тем же самым описанным выше методом поверхностного плазмонного резонанса на приборе BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ) при 25°С, используя чипы с иммобилизованным антигеном СМ5 при ~10 относительных единицах (единицах отклика, RU). Коротко говоря, биосенсорные чипы с карбоксиметилдекстраном (СМ5, BIAcore Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламино пропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями производителя. Антиген разводят 10 мМ раствором ацетата натрия, рН 4.8, до концентрации 5 мкг/мл (~0.2 мкМ), а затем вводят (инжекция) при скорости потока 5 мкл/минута до достижения, примерно, 10 относительных единиц (RU) связанного белка. После введения антигена вводят 1 М раствор этаноламина, чтобы блокировать непрореагировавшие группы.
Если специально не указано иначе, выражения «биологически активный», и «биологическая активность», и «биологические характеристики», по отношению к полипептиду по данному изобретению, означают обладание способностью связываться с биологической молекулой.
Выражение «биологическая молекула» относится к нуклеиновой кислоте, белку, углеводу, липиду и их комбинации. В одном варианте изобретения биологическая молекула существует в природе.
Участки антител, такие как Fab- и F (аb') 2-фрагменты, могут быть получены из целых антител с использованием традиционных методов, таких как папаиновый или пепсиновый гидролиз целых антител. Более того, антитела, части антител и молекулы иммуноадгезии могут быть получены с использованием стандартных методов рекомбинантной ДНК, например, как описано в настоящем документе.
Термин «рекомбинантное антитело» означает антитело, которое экспрессируется в клетке или клеточной линии, содержащей нуклеотидную последовательность (нуклеотидные последовательности), которая кодирует антитела, при этом указанная нуклеотидная последовательность (нуклеотидные последовательности) не ассоциирована с клеткой в природе.
Термин «вариантное» антитело, используемый в данном документе, относится к антителу, имеющему аминокислотную последовательность, которая отличается от аминокислотной последовательности его «родительского» антитела путем добавления, удаления и/или замены одного или более аминокислотных остатков относительно последовательности родительского антитела. В предпочтительном варианте осуществления изобретения вариантное антитело содержит по меньшей мере одно или более (например, от одного до двенадцати, например, два, три, четыре, пять, шесть, семь, восемь или девять, десять, одиннадцать или двенадцать; и в некоторых вариантах осуществления изобретения от одного до примерно десяти) добавлений, делеций и/или замен аминокислот относительно родительского антитела. В некоторых вариантах осуществления изобретение добавления, делеции и/или замены осуществляются на CDR-участках вариантного антитела. Идентичность или гомология по отношению к последовательности вариантного антитела определяется в настоящем документе как процент аминокислотных остатков в последовательности вариантного антитела, которые идентичны остаткам родительского антитела, после выравнивания последовательностей и введения гэпов, если это необходимо, для достижения максимального процента идентичности последовательности. Вариантное антитело сохраняет способность связываться с тем же антигеном, и предпочтительно эпитопом, с которым связывается родительское антитело, и в некоторых вариантах осуществления изобретения по меньшей мере одно свойство или биологическая активность превосходит аналогичные свойства родительского антитела. Например, вариантное антитело может иметь, например, более выраженную аффинность связывания, более длительный период полувыведения, более низкое значение ИК50 или повышенную способность подавлять биологическую активность антигена по сравнению с родительским антителом. Особый интерес в настоящем документе представляет вариантное антитело, показывающее биологическую активность, превышающую по меньшей мере в 2 раза (предпочтительно, по меньшей мере в 5 раз, 10 раз или 20 раз) биологическую активность родительского антитела.
Термин «биспецифическое антитело» означает антитело, содержащее антигенсвязывающий домен или антигенсвязывающие домены, которые способны к специфическому связыванию с двумя различными эпитопами на одной биологической молекуле или способны к специфическому связыванию с эпитопами на двух различных биологических молекулах. Биспецифичное антитело также упоминается в настоящем документе, как обладающее «двойной специфичностью» или как являющееся антителом с «двойной специфичностью».
Термин «химерное антитело» относится в широком смысле к антителу, которое содержит одну или более областей из одного антитела, и одну или более областей из одного или нескольких других антител, как правило, антитело, частично человеческого происхождения и частично нечеловеческого происхождения, то есть полученное частично из не относящегося к человеку животного, например, мыши, крысы или другого грызуна или верблюдовых, таких как лама или альпака. Химерные антитела являются предпочтительными по сравнению с нечеловеческими антителами для того, чтобы снизить риск иммунного ответа, направленного против антител у человека, например, ответа, направленного против мышиных антител у человека в случае мышиного антитела. Примером типичного химерного антитела является то, в котором последовательности вариабельного участка являются мышиными, в то время как последовательности константного участка являются человеческими. В случае химерного антитела нечеловеческие части могут быть подвергнуты дальнейшему изменению с целью гуманизации антитела.
Термин «гуманизация» относится к факту, что когда антитело имеет полностью или частично нечеловеческое происхождение, например, антитело мыши или ламы, полученное при иммунизации мышей или лам, соответственно, с представляющим интерес антигеном, или является химерным антителом на основе такого антитела мыши или ламы, можно заменить некоторые аминокислоты, например, в каркасных областях и константных доменах тяжелой и легкой цепей, с тем чтобы избежать или свести к минимуму иммунный ответ у человека. Специфичность взаимодействия антитела с антигеном-мишенью присуща главным образом аминокислотным остаткам, расположенных в шести CDR-участках тяжелой и легкой цепи. Поэтому аминокислотные последовательности внутри CDR-участков, являются гораздо более вариабельными между отдельными антителами, по сравнению с последовательностями вне CDR-участков. Поскольку последовательности CDR участков отвечают за большинство антитело-антиген взаимодействий, можно экспрессировать рекомбинантные антитела, которые имитируют свойства специфического природного антитела, или в более общем плане какого-либо специфического антитела с данной аминокислотной последовательностью, например, путем конструирования экспрессионных векторов, которые экспрессируют последовательности CDR-участков из специфического антитела и каркасные последовательности другого антитела. В результате, можно «гуманизировать» нечеловеческое антитело и в значительной степени сохранить специфичность связывания и аффинность исходного антитела. Несмотря на то, что невозможно точно предсказать иммуногенность и тем самым иммунный ответ, направленный против антитела у человека на конкретное антитело, нечеловеческие антитела, как правило, более иммуногенны, чем человеческие антитела. Химерные антитела, у которых инородные (например, грызуна или верблюда) константные участки были заменены последовательностями человеческого происхождения, показали в целом более низкую иммуногенность, чем антитела полностью инородного происхождения, и существует тенденция использовать в терапевтических антителах гуманизированные или полностью человеческие антитела. Химерные антитела или другие антитела нечеловеческого происхождения, таким образом, могут быть гуманизированы, чтобы снизить риск иммунного ответа, направленного против антитела, у человека.
Для химерных антител, гуманизация обычно включает в себя модификацию каркасных участков последовательностей вариабельного участка. Аминокислотные остатки, которые являются частью участков, определяющих комплементарность (CDR участков), чаще всего не будут изменяться в связи с гуманизацией, хотя в некоторых случаях это может быть желательным, чтобы изменить отдельные аминокислотные остатки CDR-участка, например, чтобы удалить участок гликозилирования, участок дезамидирования, участок изомеризации аспартата или нежелательный остаток цистеина или метионина. N-связанное гликозилирование происходит путем присоединения олигосахаридной цепи к остатку аспарагина в трипептидной последовательности Asn-X-Ser или Asn-X-Thr, где Х может быть любой аминокислотой, кроме Pro. Удаление участка N-гликозилирования может быть достигнуто путем мутирования Asn или Ser/Thr остатка другим остатком, предпочтительно путем консервативной замены. Дезамидирование остатков аспарагина и глутамина может происходить в зависимости от таких факторов, как рН и обнажение поверхности. Остатки аспарагина особенно восприимчивы к дезамидированию, прежде всего, если они присутствуют в последовательности Asn-Gly, и в меньшей степени в других дипептидных последовательностях, таких как Asn-Ala. При наличии такого дезамидированного участка, например, Asn-Gly в последовательности CDR-участка, может быть предпочтительным удалить этот участок, как правило, путем консервативной замены для удаления одного из вовлеченных остатков.
В данной области техники известны многочисленные способы гуманизации последовательности антитела. Одним из наиболее часто используемых методов является трансплантация CDR-участков. Трансплантация CDR участка может быть основана на определениях CDR-участков по Kabat, хотя в более поздней публикации (Magdelaine-Beuzelin et al., Crit Rev.Oncol Hematol. 64:210 225 (2007)) предполагается, что определение по IMGT® (the international ImMunoGeneTics information system®, www.imgt.org) может улучшить результат гуманизации (см Lefranc et al., Dev. Comp Immunol. 27:55-77 (2003)). В некоторых случаях, трансплантация CDR-участка может уменьшить специфичность и аффинность связывания, и, следовательно, биологическую активность, в CDR трансплантированном нечеловеческом антителе, по сравнению с родительским антителом, из которого получены CDR-участки. Обратные мутации (иногда именуемые «ремонт каркасного участка»), могут применяться в выбранных положениях CDR трансплантированного антитела, как правило, в каркасных участках, для того, чтобы восстановить специфичность и аффинность связывания родительского антитела. Определение позиций для возможных обратных мутаций может быть выполнено с использованием информации, имеющейся в литературе и в базах данных антител. Аминокислотные остатки, которые являются кандидатами для обратных мутаций, как правило, расположены на поверхности молекулы антитела, в то время как остатки, которые углублены или имеют низкую степень обнажения поверхности обычно не будут подвержены изменениям. Метод гуманизации, альтернативный трансплантации CDR-участка и обратной мутации, представляет собой изменение поверхности, при котором неэкспонированные на поверхности остатки нечеловеческого происхождения, сохраняются, в то время как экспонированные на поверхности остатки изменяются в человеческие остатки.
Существует две технологии получения полностью человеческих антител: с использованием in vitro собранных фаговых библиотек или in vivo иммунизацией гуманизированных животных (мышей, крыс и т.д.).
Конструирование комбинаторных фаговых библиотек антител начинается с выбора источника генного репертуара, в зависимости от которого можно выделить несколько видов библиотек антител: наивные, иммунные или синтетические. Наивные и иммунные библиотеки конструируют, используя естественным образом реорганизованные гены, кодирующие вариабельные домены иммуноглобулинов здоровых или иммунных к какому-либо антигену доноров, соответственно. Для этого выделяют мРНК клеток лимфоидного ряда, продуцирующих антитела. Чаще всего это лимфоциты периферической крови, но в некоторых случаях используют спленоциты [Sheets MD, Amersdorfer P, Finnern R, Sargent P, Lindquist E, Schier R, et al. Efficient construction of a large nonimmune phage antibody library: the production of high-affinity human single-chain antibodies to protein antigens. Proc Natl Acad Sci U S A 1998,95:6157-6162 и de Haard HJ, van Neer N, Reurs A, Hufton SE, Roovers RC, Henderikx P, et al. A large non-immunized human Fab fragment phage library that permits rapid isolation and kinetic analysis of high affinity antibodies. J Biol Chem 1999,274:18218-18230.], клетки миндалин или лимфоциты костного мозга [Vaughan TJ, Williams AJ, Pritchard K, Osbourn JK, Pope AR, Earnshaw JC, et al. Human antibodies with sub-nanomolar affinities isolated from a large non-immunized phage display library. Nat Biotechnol 1996,14:309-314.]. На основе мРНК синтезируют кДНК, при этом для праймирования реакции могут быть взяты олиго-dT праймеры и статистические гексаолигонуклеотиды, что позволяет получать кДНК копии всех возможных вариантов генов, кодирующих вариабельные домены антител [Улитин АБ, Капралова МВ, Ламан АГ, Шепеляковсткая АО, Булгакова ЕВ, Фурсова КК, et al. Библиотека миниантител человека в формате фагового дисплея. Создание и апробация. ДАН: Изд-во «Наука»; 2005.].
Сразу могут использоваться один или несколько праймеров, ограничивающих набор амплифицируемых генов до одного или нескольких семейств генов вариабельных доменов или изотипов антител уже на уровне кДНК [Marks JD, Hoogenboom HR, Bonnert TP, McCafferty J, Griffiths AD, Winter G. Bypassing immunization. Human antibodies from V-gene libraries displayed on phage. J Mol Biol 1991,222:581-597]. Праймеры, используемые для амплификации генов, кодирующих иммуноглобулины, комплементарны их наиболее консервативным участкам. Их последовательности выбирают из коллекций генов, которые организованы в базы данных, такие как база данных Kabat или V BASE. Дизайн праймеров также предусматривает наличие в них внутренних сайтов рестрикции, позволяющих клонировать ПЦР продукты в состав соответствующих векторов.
Конструирование синтетических библиотек основано на замене природных CDR на набор случайных последовательностей, что позволяет создавать огромное разнообразие антигенсвязывающих сайтов.
Фаговый дисплей является первой и самой широко распространенной in vitro технологией для поиска антител. В 1985 году Смит обнаружил, что последовательности чужеродной ДНК могут быть клонированы в нитевидный бактериофаг M13 таким образом, что клонированные последовательности генов экспрессируются на поверхности фаговых частиц как слитые белки (Smith GP: Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228:1315-1317.). Таким образом, можно проводить селекцю интересующих нас слитых белков на основе их способности связывать другие белки. Это открытие было скомбинировано с методами ПЦР-амплификации, что позволило клонировать кДНК репертуар генов иммуноглобулинов для создания разнообразных фаговых библиотек, содержащих вариабельные домены, которые могут быть использованы для быстрого поиска мишень-специфичных моноклональных антител. Репертуар фаговых библиотек отражает репертуар антител B-лимфоцитов каждого человека или животного, кровь которого была использована при создании библиотеки. В 1995 году две статьи сообщили о создании генетически сконструированных мышей, которые экспрессировали полностью человеческие антитела, репертуар которых может быть соспоставим с полученных гибридомной технологией (Lonberg N, Taylor LD, Harding FA, Trounstine M, Higgins KM, Schramm SR, Kuo CC, Mashayekh R, Wymore K, McCabe JG et al.: Antigen-specific human antibodies from mice comprising four distinct genetic modifications. Nature 1994, 368:856-859) У этих животных были целенаправленно разрушены гены своих собственных эндогенных тяжелых и k легких цепей иммунноглобулинов и введены трансгены, представляющие собой сегменты генов тяжелых и k легких цепей человека .Оказалось, что репертуар генов человека может быть использован мышиной иммунной системой для создания высокоспецифичных и высокоаффинных антител ко большему разнообразию антигенов. Несмотря на то, что трансгенные мыши экспрессируют B-клеточные рецепторы, которые по существу являются гибридными мышиных и человеческих (человеческий иммуноглобулин, мышиные Igα, Igβ и другие сигнальные молекулы), их B-клетки нормально развиваются и созревают.
В некоторых случаях может также быть предпочтительным изменение одного или более остатков аминокислот CDR-участков с целью повышения аффинности связывания с целевым эпитопом. Это известно, как «созревание аффинности» и в некоторых случаях может выполняться в связи с гуманизацией, например, в ситуациях, когда гуманизация антитела приводит к снижению специфичности или аффинности связывания, и не представляется возможным в достаточной степени улучшить специфичность или аффинность связывания с помощью только обратных мутаций. Различные методы созревания аффинности известны в данной области техники, например, способ in vitro сканирующего насыщающего мутагенеза, описанный Burks et al., Proc Natl Acad Sci USA, 94:412-417 (1997), и способ пошагового in vitro созревания аффинности, предложенный Wu et al., Proc Natl Acad Sci USA 95:6037 6042 (1998).
Термин «моноклональное антитело» или «mAb» относится к антителу, которое синтезировано и выделено отдельной клональной популяцией клеток. Клональная популяция может быть клональной популяцией иммортализованных клеток. В некоторых вариантах осуществления изобретения иммортализованные клетки в клональной популяции являются гибридными клетками, гибридомами, которые обычно получают путем слияния отдельных В-лимфоцитов от иммунизированных животных с отдельными клетками лимфоцитарной опухоли. Гибридомы представляют собой тип сконструированных клеток и не встречаются в природе.
«Нативные антитела» обычно являются гетеротетрамерными гликопротеидами с молекулярной массой примерно 150000 дальтон, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, тогда как количество дисульфидных связей между тяжелыми цепями варьирует в разных изотипах иммуноглобулинов. Каждая тяжелая и легкая цепь также имеет равномерно расположенные внутрицепочечные дисульфидные мостики. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), за которым следует несколько константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен на другом конце. Константный домен легкой цепи выровнен с первым константным доменом тяжелой цепи, и вариабельный домен легкой цепи выровнен с вариабельным доменом тяжелой цепи. Полагают, что конкретные аминокислотные остатки образуют поверхность раздела между вариабельными доменами легкой цепи и тяжелой цепи.
Определение «выделенный» («изолированный»), применяемое для описания различных антител по данному описанию, означает антитело, идентифицированное и выделенное и/или регенерированное из клетки или клеточной культуры, в которой оно экспрессируется. Примеси (загрязняющие компоненты) из природной среды представляют собой материалы, которые, как правило, мешают диагностическому или терапевтическому применению полипептида, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах изобретения антитело очищают (1) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности, при использовании секвенатора с вращающейся стеклянной чашечкой (секвенатора Эдмана), или (2) до гомогенности методом SDS-PAGE в невосстанавливающих или восстанавливающих условиях с применением окрашивания Кумасси бриллиантовым голубым или, предпочтительно, серебром. Выделенное антитело включает антитела in situ внутри рекомбинантных клеток, так как по меньшей мере один компонент природной среды полипептида отсутствует. Обычно выделенный полипептид получают в результате по меньшей мере одной стадии очистки.
«Выделенная» молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая идентифицирована и отделена от по меньшей мере одной молекулы нуклеиновой кислоты-примеси, с которой она обычно связана в естественном источнике нуклеиновой кислоты антитела. Выделенная молекула нуклеиновой кислоты отличается от той формы или набора, в которых она находится в естественных условиях. Таким образом, выделенная молекула нуклеиновой кислоты отличается от молекулы нуклеиновой кислоты, существующей в клетках в естественных условиях. Однако выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, находящуюся в клетках, в которых в норме происходит экспрессия антитела, например, в случае, если молекула нуклеиновой кислоты имеет локализацию в хромосоме, отличную от ее локализации в клетках в естественных условиях.
Термин «эпитоп» при использовании в данном документе относится к части (детерминанте) антигена, который специфически связывается со связывающей молекулой (например, и антитело или родственная молекула, такие как биспецифичная связывающая молекула). Эпитопные детерминанты обычно состоят из химически активных поверхностных групп молекул, таких как аминокислоты или углеводы, или боковые цепи сахаров, и, как правило, имеют специфические трехмерные структурные характеристики, а также специфические характеристики зарядов. Эпитоп может быть «линейным» или «конформационным.» В линейном эпитопе, все точки взаимодействия между белком (например, антиген) и взаимодействующей молекулой (такой как антитело) происходит линейно вдоль первичной аминокислотной последовательности белка. В конформационном эпитопе, точки взаимодействия происходят через аминокислотные остатки на белке, отделенные друг от друга в первичной аминокислотной последовательности. Когда желаемый эпитоп антигена определен, можно генерировать антитела к этому эпитопу с использованием методик, хорошо известных в данной области техники. Кроме того, генерация и характеристика антител или других связывающих молекул могут пролить свет на информацию о желательных эпитопах. Основываясь на этой информации, можно затем конкурентно скринировать связывающие молекулы для связывания с теми же или аналогичными эпитопами, например, путем проведения исследований конкуренции, чтобы найти связывающие молекулы, которые конкурируют за связывание с антигеном.
Термин «пептидный линкер» в настоящем документе означает любой пептид с возможностью соединения доменов с длиной в зависимости от доменов, которые он связывает между собой, содержащий любую аминокислотную последовательность. Предпочтительно пептидный линкер имеет длину более 5 аминокислот и состоит из любого набора аминокислот, выбранного из G, A, S, P, E, T, D, K.
Термин «in vitro» относится к биологическому объекту, биологическому процессу или биологической реакции вне организма, смоделированному в искусственных условиях. Например, рост клеток in vitro должен пониматься как рост клеток в среде вне организма, например, в пробирке, культуральном флаконе или микропланшете.
Термин «IС50» (50% ингибирующая концентрация) относится к концентрациям препарата, при которых измеряемая активность или отклик, например, рост или пролиферация клеток, таких как опухолевые клетки, ингибируется на 50%. Значение IС50 может оцениваться c помощью соответствующих кривых зависимости ответа от логарифма дозы, с использованием специальных статистических программ для обработки кривых.
Термин GI50 (50% ингибирование роста) относится к концентрациям препарата, при которых пролиферация клеток, таких как опухолевые клетки, ингибируется на 50%.
Термин ED50 (EC50) (50% эффективная доза/концентрация) относится к концентрациям препарата, при которых измеряемый биологический эффект достигается на 50% (может включать цитотоксичность).
Термин «антипролиферативное действие» подразумевает остановку или ингибирование роста пролиферирующих клеток, таких как раковые клетки.
Понятие «эффекторная функция» антитела относится к видам биологической активности, связанным с Fс-областью (нативной последовательностью Fc-области или с вариантами аминокислотной последовательности Fc-области) антитела, и варьируют в зависимости от изотипа антитела. Примерами эффекторных функций антитела являются: Clq- связывание; комплементзависимая цитотоксичность; связывание Fc-рецептора; антитело-зависимая клеточно-опосредованная цитотоксичность (ADCC); фагоцитоз; понижающая регуляция рецепторов клеточной поверхности (например, В-клеточного рецептора, BCR) и В-клеточная активация.
«Антитело-зависимая клеточная цитотоксичность» или «ADCC» относится к опосредованному клетками ответу, при котором неспецифичные цитотоксические клетки, которые экспрессируют рецепторы Fc (FcR) (например, природные клетки-киллеры (NK), нейтрофилы и макрофаги), узнают связанное антитело на клетке-мишени и затем вызывают лизис или фагоцитоз клетки-мишени. Первичные клетки для опосредования ADCC, NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках суммирована в таблице 3 на странице 464 в публикации Ravetch and Kinet, Annu. Rev. Immunol 9: 457-92 (1991). Чтобы оценить активность в ADCC представляющей интерес молекулы можно осуществить анализы ADCC in vitro, такие как анализы, описанные в патентах США № 5500362 или 5821337. Применимые эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (PBMC) и природные клетки-киллеры (NK). Альтернативно или дополнительно ADCC-активность представляющей интерес молекулы можно оценить in vivo, например, в животной модели, такой как модель, описанная в Clynes et al. PNAS (USA) 95: 652-656 (1998).
«Эффекторными клетками человека» являются лейкоциты, которые экспрессируют один или несколько FcR и осуществляют эффекторные функции. Предпочтительно клетки экспрессируют, по меньшей мере, FcγRIII и осуществляют ADCC-эффекторную функцию. Примеры лейкоцитов человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (PBMC), природные клетки-киллеры (NK), моноциты, цитотоксические T-клетки и нейтрофилы; при этом предпочтительны PBMC и NK-клетки. Эффекторные клетки могут быть выделены из их природного источника, например из крови или PBMC, как описано в настоящей публикации.
Термины «Fc-рецептор» и «FcR» используют для описания рецептора, который связывается с Fc-областью антитела. Предпочтительным FcR является FcR человека с нативной последовательностью. Кроме того, предпочтительным FcR является FcR, который связывает IgG-антитело (гамма-рецептор), и к предпочтительным рецепторам относятся рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и альтернативно сплайсируемые формы указанных рецепторов. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), которые имеют сходные аминокислотные последовательности, которые отличаются главным образом своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит в своем цитоплазматическом домене основанный на тирозине мотив активации иммунорецептора (ITAM). Ингибирующий рецептор FcγRIIB содержит в своем цитоплазматическом домене основанный на тирозине мотив ингибирования иммунорецептора (ITIM) (см. обзор в Daeron, Annu. Rev. Immunol. 15: 203-234 (1997)). Обзор, посвященный FcR, представлен в Ravetch and Kinet, Annu. Rev. Immunol 9: 457-92 (1991). Другие FcR, включая FcR, которые будут идентифицированы в будущем, включены в настоящем описании в термин «FcR». Термин также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG в плод.
«Комплемент-зависимая цитотоксичность» и «CDC» относятся к способности молекулы лизировать мишень в присутствии комплемента. Путь активации комплемента инициируется связыванием первого компонента системы комплемента (C1q) с молекулой (например, антителом) в комплексе со своим антигеном. Чтобы оценить активацию комплемента можно осуществить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996).
Термин «идентичность» или «гомологичность» следует толковать как означающее процентное содержание остатков аминокислот в кандидатной последовательности, которые идентичны остаткам соответствующей последовательности, с которой ее сравнивают, после сравнения последовательностей и введения «брешей», если необходимо достичь максимального процента идентичности для полной последовательности и не учитывая любые консервативные замещения как часть идентичности последовательности. Ни N- или С-концевой удлиняющей, ни инсерционные сегменты не следует толковать как уменьшающие идентичность или гомологичность. Методы и компьютерные программы для сравнения хорошо известны. Идентичность последовательности можно определить, используя программное обеспечение для анализа последовательности (например, Sequence Analysis Software Package, Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Ave., Madison, WI 53705). Данное программное обеспечение подходит для подобных последовательностей путем определения степени гомологичности для разнообразных замещений, делеций (элиминирований) и других модификаций.
Фразу «гомологичный», что касается полипептидной последовательности антитела, следует толковать как антитело, проявляющее по крайней мере, 70%-ную, предпочтительно 80%-ную, более предпочтительно 90%-ную и наиболее предпочтительно 95%-ную идентичность последовательности относительно полипептидной последовательности. Термин в отношении последовательности нуклеиновой кислоты следует толковать как последовательность нуклеотидов, проявляющих, по крайней мере, 85%-ную, предпочтительно 90%-ную, более предпочтительно 95%-ную и наиболее предпочтительно 97%-ную идентичность последовательности относительно последовательности нуклеиновой кислоты.
Предлагается модификация(и) аминокислотных последовательностей антител, описанных в настоящей публикации. Например, может быть желательным улучшение аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотной последовательности антитела получают введением соответствующих изменений нуклеотидов в нуклеиновую кислоту антитела или пептидным синтезом. Такие модификации включают, например, делеции, и/или инсерции, и/или замены остатков в аминокислотных последовательностях антитела. Осуществляют любое сочетание делеции, инсерции и замены, чтобы получить конечную конструкцию, при условии, что конечная конструкция обладает требуемыми характеристиками. Изменения аминокислот также могут изменять посттрансляционные процессы в антителе, такие как изменение количества или положения сайтов гликозилирования.
Вариант модификации аминокислотных последовательностей антител с помощью аминокислотных замен. Такой вариант представляет собой замену, по меньшей мере, одного аминокислотного остатка в молекуле антитела на другой остаток. Места, представляющие наибольший интерес для мутагенеза путем замен, включают гипервариабельные области или CDR, но также предполагаются изменения и в области FR или Fc. Консервативные замены показаны в таблице А под заголовком «предпочтительные замены». Если такие замены приводят к изменению биологической активности, то могут быть введены дополнительные существенные изменения, названные «примерами заменам» в таблице А, или изменения, дополнительно описанные ниже при описании классов аминокислот, и может быть проведен скрининг продуктов.
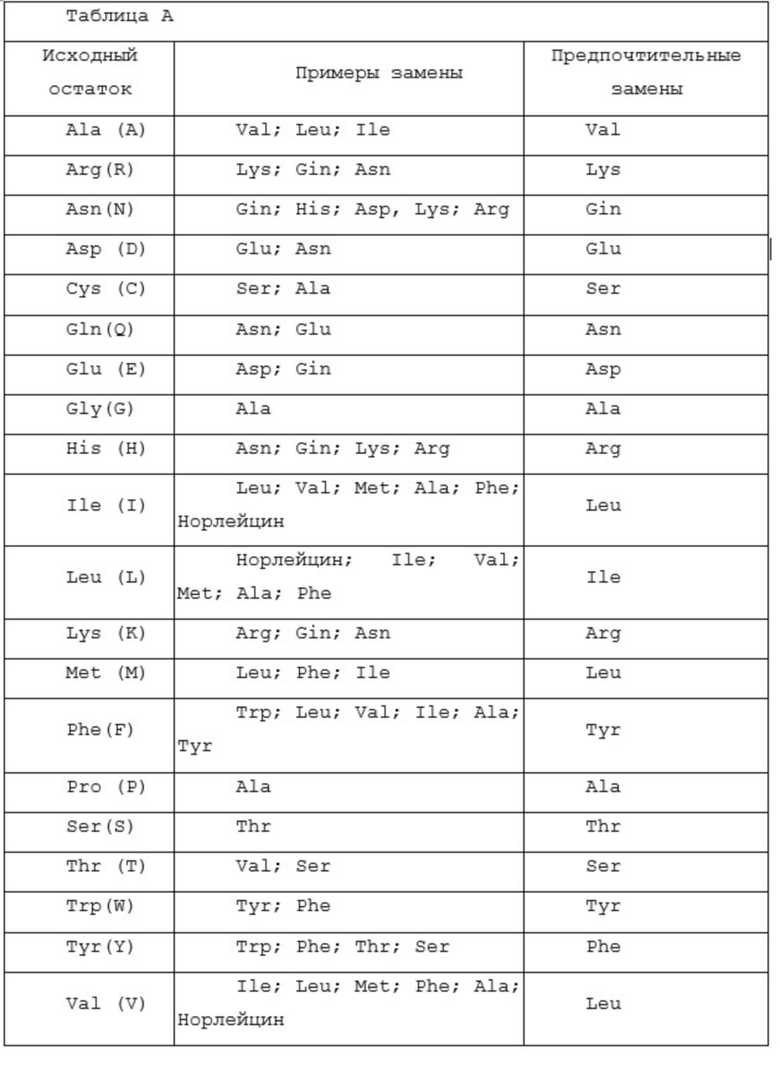
Термины «нуклеиновая кислота», «нуклеиновая последовательность» или «нуклеиновокислотная последовательность», «полинуклеотид», «олигонуклеотид», «полинуклеотидная последовательность» и «нуклеотидная последовательность», которые используются равнозначно в данном описании, обозначают четкую последовательность нуклеотидов, модифицированных или не модифицированных, определяющую фрагмент или участок нуклеиновой кислоты, содержащую или не содержащую неприродные нуклеотиды и являющуюся либо двухцепочечной ДНК или РНК, либо одноцепочечной ДНК или РНК, либо продуктами транскрипции указанных ДНК.
Здесь также следует упомянуть, что данное изобретение не относится к нуклеотидным последовательностям в их природной хромосомной среде, т.е. в природном состоянии. Последовательности данного изобретения были выделены и/или очищены, т.е. были взяты прямо или косвенно, например, путем копирования, при этом их среда была по меньшей мере частично модифицирована. Таким образом, также здесь следует подразумевать изолированные нуклеиновые кислоты, полученные путем генетической рекомбинации, например, с помощью принимающих клеток (клеток-хозяев), или полученные путем химического синтеза.
Ссылка на нуклеотидную последовательность охватывает его комплимент, если не указано иное. Таким образом, ссылка на нуклеиновую кислоту, имеющую определенную последовательность следует понимать как охватывающие ее комплементарную цепь с ее комплементарной последовательностью.
Выражение «контролирующие последовательности» относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в определенном организме-хозяине. Пригодные для прокариот контролирующие последовательности представляют собой, например, промотор, необязательно оператор и сайт связывания рибосомы. Как известно, в эукариотических клетках присутствуют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота «функционально связана», если она находится в функциональной связи с другой нуклеотидной последовательностью. Например, ДНК предпоследовательности или секреторной лидерной последовательности функционально связывают с ДНК полипептида, если он экспрессируется в виде предпротеина, который принимает участие в секреции полипептида; промотор или энхансер функционально связывают с кодирующей последовательностью, если он оказывает воздействие на транскрипцию последовательности; сайт связывания рибосомы функционально связывают с кодирующей последовательностью, если он расположен так, что может облегчать трансляцию. Как правило, «функционально связан» обозначает, что связанные последовательности ДНК являются смежными, а в случае секреторной лидерной последовательности являются смежными и находятся в фазе считывания. Однако энхансеры не обязательно должны быть смежными.
Термин «вектор» при использовании в настоящем документе означает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой она соединена. В некоторых вариантах осуществления изобретения вектор представляет собой плазмиду, т.е. кольцевую двухцепочечную часть ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. В некоторых вариантах осуществления изобретения вектор представляет собой вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. В некоторых вариантах осуществления изобретения векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальный сайт инициации репликации и эписомные векторы млекопитающих). В других вариантах осуществления изобретения векторы (например, неэписомальные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина, и таким образом реплицируются вместе с геном хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально соединены. Такие векторы упоминаются в данном документе как «рекомбинантные экспрессирующие векторы» (или просто «экспрессирующие векторы»).
Термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») при использовании в данном документе означает клетку, в которую введен рекомбинантный экспрессионный вектор. Настоящее изобретение относится к клеткам-хозяевам, которые могут включать, например, вектор в соответствии с настоящим изобретением, описанным выше. Настоящее изобретение относится также к клеткам-хозяевам, которые включают, например, нуклеотидную последовательность, кодирующую тяжелую цепь или ее антигенсвязывающие части, нуклеотидную последовательность, кодирующую легкую цепь или ее антигенсвязывающие части, или обе из них, первого связывающего домена и/или второго связывающего домена связывающей молекулы по данному изобретению. Следует понимать, что «рекомбинантная клетка-хозяин» и «клетка-хозяин» означают не только конкретную заявленную клетку, но также и потомство такой клетки. Поскольку модификации могут проходить в последующих поколениях вследствие мутации или воздействий окружающей среды, такое потомство не может, на самом деле, быть идентичным родительской клетке, но такие клетки по-прежнему включены в объем термина «клетка-хозяин» при использовании в настоящем документе.
Термин «эксципиент» используется в данном документе для описания любого ингредиента, отличающегося от соединения(ий) по данному изобретению.
Термин «заболевание или нарушение, опосредованное GITR» подразумевает все заболевания или нарушения, которые либо прямо, либо косвенно связаны с GITR, включая этиологию, развитие, прогресс, персистентность или патологию заболевания или нарушения.
«Лечить», «лечение» и «терапия» относятся к методу смягчения или устранения биологического расстройства и/или по меньшей мере одного из сопутствующих ему симптомов. Используемый в данном документе, термин «облегчить» болезнь, заболевание или состояние, означает уменьшение тяжести и/или частоты возникновения симптомов заболевания, расстройства или состояния. Кроме того, содержащиеся в данном документе ссылки на «лечение» включает ссылки на лечебную, паллиативную и профилактическую терапию.
В одном аспекте субъект лечения или пациент является млекопитающим, предпочтительно человеческим субъектом. Вышеупомянутый субъект может быть мужского или женского пола любого возраста.
Термин «нарушение» означает любое состояние, которое можно улучшить в результате лечения по настоящему изобретению. В определение данного термина входят хронические и острые нарушения или заболевания, включающие в себя патологические состояния, которые вызывают предрасположенность млекопитающего к возникновению данного нарушения.
Термины «рак» или «раковый» относятся к физиологическому состоянию или описывают физиологическое состояние у млекопитающих, которое, как правило, характеризуется нерегулируемым(/ой) ростом/пролиферацией клеток. Это определение охватывает доброкачественные и злокачественные раковые заболевания. Примеры раковых заболеваний включают, но без ограничения, карциному, лимфому, бластому, саркому и лейкоз. Более конкретные примеры таких раковых заболеваний включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого, плоскоклеточную карциному легкого, рак брюшины, печеночноклеточный рак, рак желудка, включая рак ЖКТ, рак поджелудочной железы, глиобластому, глиому, цервикальный рак, рак яичника, рак печени, рак мочевого пузыря, рак молочной железы, рак толстой кишки, колоректальный рак, карциному эндометрия и матки, карциному слюнных желез, рак почки (почечноклеточную карциному), рак предстательной железы (рак простаты), рак наружных женских половых органов, рак щитовидной железы, печеночную карциному, карциному анального канала, карциному пениса, меланому и различные типы рака головы и шеи.
Термины «иммунный ответ», «аутоиммунная реакция», «аутоиммунное воспаление» относятся, например, к действию лимфоцитов, антиген-представляющих клеток, фагоцитирующих клеток, гранулоцитов и растворимых макромолекул, вырабатываемых указанными клетками или клетками печени (включая антитела, цитокины и комплемент, образующиеся в результате селективного повреждения, разрушения или элиминации из человеческого организма инвазивных патогенов, клеток или тканей, инфицированных патогенами, раковых клеток или, в случаях аутоиммунитета или патологического воспаления, нормальных человеческих клеток или тканей).
«Терапевтически эффективным количеством» считается количество вводимого в процессе лечения терапевтического агента, которое избавит в определенной степени от одного или нескольких симптомов заболевания, по поводу которого проводится лечение.
Понятие «хроническое» применение относится к непрерывному (продолжительному) применению агента(ов) в противоположность острому (кратковременному) пути введения, так чтобы поддерживать первоначальное терапевтическое действие (активность) в течение длительного периода времени.
«Прерывистое» применение обозначает лечение, которое не осуществляют последовательно без перерывов, но которое скорее по своей природе является периодическим.
В настоящем описании и в последующей формуле изобретения, если контекстом не предусмотрено иное, слова «иметь», «включать» и «содержать» или их вариации, такие как «имеет», «имеющий», «включает», «включающий», «содержит» или «содержащий», следует понимать как включение указанного целого или группы целых, но не исключение любого другого целого или группы целых.
Подробное описание изобретения
Антитело
Настоящее изобретение относится к моноклональному антителу, которое специфично связывается с GITR.
В одном из аспектов настоящее изобретение относится к моноклональному антителу или его антигенсвязывающему фрагменту, которое специфично связывается с GITR, включающему:
(a) вариабельный домен тяжелой цепи, содержащему:
(i) CDR1 с аминокислотной последовательностью, выбранной из группы:
NYGMH (SEQ ID NO: 1) или YYWMY (SEQ ID NO: 12);
(ii) CDR2 с аминокислотной последовательностью, выбранной из группы:
VIWFDGSNKFYTDSVKG (SEQ ID NO: 2) или AISWNGGRTYYAESMKG (SEQ ID NO: 13);
(iii) CDR3 с аминокислотной последовательностью, выбранной из группы:
ELGGYYYDSSGFRPYYYGMDV (SEQ ID NO: 3) или NRYYSDPNYGMNL (SEQ ID NO: 14), и
(b) вариабельный домен легкой цепи, содержащему:
(i) CDR1 с аминокислотной последовательностью, выбранной из группы:
RASQSIGSWLA (SEQ ID NO: 7) или TGTSTDIGTYKYIS (SEQ ID NO: 17);
(ii) CDR2 с аминокислотной последовательностью, выбранной из группы:
AASTLQR (SEQ ID NO: 8) или GVSHRPS (SEQ ID NO: 18);
(iii) CDR3 с аминокислотной последовательностью, выбранной из группы:
QQSHSHPLT (SEQ ID NO: 9) или SSYTSSGTVV (SEQ ID NO: 19).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3, соответственно;
- вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14, соответственно;
- вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19, соответственно.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSS (SEQ ID NO: 4).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSS (SEQ ID NO: 4).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности
QVQLVQSGGGLVQPGGSLRLSCAASGFTFSYYWMYWVRQAPGKGLEWVSAISWNGGRTYYAESMKGRFTISRDNAQNTLYLQMNSLKSEDTAVYYCAKNRYYSDPNYGMNLWGKGTTVTVSS (SEQ ID NO: 15).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность QVQLVQSGGGLVQPGGSLRLSCAASGFTFSYYWMYWVRQAPGKGLEWVSAISWNGGRTYYAESMKGRFTISRDNAQNTLYLQMNSLKSEDTAVYYCAKNRYYSDPNYGMNLWGKGTTVTVSS (SEQ ID NO: 15).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности
DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIK (SEQ ID NO: 10).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIK (SEQ ID NO: 10).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности QSALTQPASVSGSPGQSITISCTGTSTDIGTYKYISWYQQHPGKAPKLIIYGVSHRPSGVSDRFSGSKSDNTASLTISGLQAEDEADYYCSSYTSSGTVVFGGGTKVTVL (SEQ ID NO: 20).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность QSALTQPASVSGSPGQSITISCTGTSTDIGTYKYISWYQQHPGKAPKLIIYGVSHRPSGVSDRFSGSKSDNTASLTISGLQAEDEADYYCSSYTSSGTVVFGGGTKVTVL (SEQ ID NO: 20).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSS (SEQ ID NO: 4);
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIK (SEQ ID NO: 10).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSS (SEQ ID NO: 4);
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIK (SEQ ID NO: 10).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, которой содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности QVQLVQSGGGLVQPGGSLRLSCAASGFTFSYYWMYWVRQAPGKGLEWVSAISWNGGRTYYAESMKGRFTISRDNAQNTLYLQMNSLKSEDTAVYYCAKNRYYSDPNYGMNLWGKGTTVTVSS (SEQ ID NO: 15);
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности QSALTQPASVSGSPGQSITISCTGTSTDIGTYKYISWYQQHPGKAPKLIIYGVSHRPSGVSDRFSGSKSDNTASLTISGLQAEDEADYYCSSYTSSGTVVFGGGTKVTVL (SEQ ID NO: 20).
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент включает:
- вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность QVQLVQSGGGLVQPGGSLRLSCAASGFTFSYYWMYWVRQAPGKGLEWVSAISWNGGRTYYAESMKGRFTISRDNAQNTLYLQMNSLKSEDTAVYYCAKNRYYSDPNYGMNLWGKGTTVTVSS (SEQ ID NO: 15);
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность QSALTQPASVSGSPGQSITISCTGTSTDIGTYKYISWYQQHPGKAPKLIIYGVSHRPSGVSDRFSGSKSDNTASLTISGLQAEDEADYYCSSYTSSGTVVFGGGTKVTVL (SEQ ID NO: 20).
В некоторых вариантах моноклональное антитело, специфичное к GITR, представляет собой полноразмерное антитело IgG.
В некоторых вариантах моноклональное антитело IgG относится к изотипу IgG1, IgG2, IgG3 или IgG4 человека.
В некоторых вариантах моноклональное антитело IgG относится к изотипу IgG1 человека.
В некоторых вариантах моноклональное антитело, специфичное к GITR, включает мутацию E345R в Fc фрагменте для увеличения агонистических свойств, антителозависимой клеточной цитотоксичности (ADCC), но не комплементзависимой цитотоксичности (CDC).
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 5).
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 5).
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPRRPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 6).
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPRRPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 6).
В некоторых вариантах моноклональное антитело включает легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 11).
В некоторых вариантах моноклональное антитело включает легкую цепь, содержащую аминокислотную последовательность DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 11).
В некоторых вариантах моноклональное антитело включает:
- тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 5);
- легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 11).
В некоторых вариантах моноклональное антитело, которое специфично связывается с GITR, представляет собой BCD166-01-001.
Моноклональное антитело BCD166-01-001 включает:
- тяжелую цепь, содержащую аминокислотную последовательность EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 5);
- легкую цепь, содержащую аминокислотную последовательность DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 11).
В некоторых вариантах моноклональное антитело включает:
- тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPRRPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 6);
- легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 11).
В некоторых вариантах моноклональное антитело, которое специфично связывается с GITR, представляет собой BCD166-02-001.
BCD166-02-001 отличается от BCD166-01-001 одной мутацией E345R в Fc фрагменте для увеличения агонистических свойств, антителозависимой клеточной цитотоксичности (ADCC), но не комплементзависимой цитотоксичности (CDC).
Моноклональное антитело BCD166-02-001 включает:
- тяжелую цепь, содержащую аминокислотную последовательность EVQLVQSGGGVVQPGKSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVAVIWFDGSNKFYTDSVKGRFTISRDNSKDTLSLQMNSLRAEDTAVYYCARELGGYYYDSSGFRPYYYGMDVWGQGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPRRPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 6);
- легкую цепь, содержащую аминокислотную последовательность DVVMTQSPSSVSASVGDRVTITCRASQSIGSWLAWYQQKPGEAPKLLIYAASTLQRGVPSRFSGGGYGTEFTLTISSLQPEDFATYFCQQSHSHPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 11).
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности QVQLVQSGGGLVQPGGSLRLSCAASGFTFSYYWMYWVRQAPGKGLEWVSAISWNGGRTYYAESMKGRFTISRDNAQNTLYLQMNSLKSEDTAVYYCAKNRYYSDPNYGMNLWGKGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 16).
В некоторых вариантах моноклональное антитело включает тяжелую цепь, содержащую аминокислотную последовательность QVQLVQSGGGLVQPGGSLRLSCAASGFTFSYYWMYWVRQAPGKGLEWVSAISWNGGRTYYAESMKGRFTISRDNAQNTLYLQMNSLKSEDTAVYYCAKNRYYSDPNYGMNLWGKGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 16).
В некоторых вариантах моноклональное антитело включает легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности QSALTQPASVSGSPGQSITISCTGTSTDIGTYKYISWYQQHPGKAPKLIIYGVSHRPSGVSDRFSGSKSDNTASLTISGLQAEDEADYYCSSYTSSGTVVFGGGTKVTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS (SEQ ID NO: 21).
В некоторых вариантах моноклональное антитело включает легкую цепь, содержащую аминокислотную последовательность QSALTQPASVSGSPGQSITISCTGTSTDIGTYKYISWYQQHPGKAPKLIIYGVSHRPSGVSDRFSGSKSDNTASLTISGLQAEDEADYYCSSYTSSGTVVFGGGTKVTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS (SEQ ID NO: 21).
В некоторых вариантах моноклональное антитело включает:
- тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности QVQLVQSGGGLVQPGGSLRLSCAASGFTFSYYWMYWVRQAPGKGLEWVSAISWNGGRTYYAESMKGRFTISRDNAQNTLYLQMNSLKSEDTAVYYCAKNRYYSDPNYGMNLWGKGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 16).
- легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности QSALTQPASVSGSPGQSITISCTGTSTDIGTYKYISWYQQHPGKAPKLIIYGVSHRPSGVSDRFSGSKSDNTASLTISGLQAEDEADYYCSSYTSSGTVVFGGGTKVTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS (SEQ ID NO: 21).
В некоторых вариантах моноклональное антитело, которое специфично связывается с GITR, представляет собой BCD166-01-014.
Моноклональное антитело BCD166-01-014 включает:
- тяжелую цепь, содержащую аминокислотную последовательность QVQLVQSGGGLVQPGGSLRLSCAASGFTFSYYWMYWVRQAPGKGLEWVSAISWNGGRTYYAESMKGRFTISRDNAQNTLYLQMNSLKSEDTAVYYCAKNRYYSDPNYGMNLWGKGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 16).
- легкую цепь, содержащую аминокислотную последовательность QSALTQPASVSGSPGQSITISCTGTSTDIGTYKYISWYQQHPGKAPKLIIYGVSHRPSGVSDRFSGSKSDNTASLTISGLQAEDEADYYCSSYTSSGTVVFGGGTKVTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS (SEQ ID NO: 21).
Молекулы нуклеиновых кислот
Настоящее изобретение также относится к молекулам нуклеиновых кислот, а именно к последовательностям, кодирующим моноклональное антитело, которое специфически связывается с GITR, по данному изобретению, которые описаны в данном документе, необязательно включающим любую последовательность соединяющего их пептидного линкера.
Ссылка на нуклеотидную последовательность, охватывает его комплимент, если не указано иное. Таким образом, ссылка на нуклеиновую кислоту, имеющую определенную последовательность следует понимать, как охватывающие ее комплементарную цепь с ее комплементарной последовательностью. Термин «полинуклеотид», упоминаемый в данном документе, означает полимерную форму нуклеотидов, по меньшей мере 10 оснований в длину, либо рибонуклеотидов, либо дезоксинуклеотидов или модифицированную форму любого типа нуклеотида. Термин включает в себя одно- и двухцепочечные формы.
В одном из аспектов настоящее изобретение относится к молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует аминокислотную последовательность, выбранную из SEQ ID NO: 1-21. Молекула нуклеиновой кислоты может также содержать любую комбинацию указанных нуклеотидных последовательностей.
В одном из аспектов настоящее изобретение относится к нуклеиновой кислоте, которая содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с GITR, и включает:
(a) вариабельный домен тяжелой цепи, содержащему:
(i) CDR1 с аминокислотной последовательностью, выбранной из группы:
NYGMH (SEQ ID NO: 1) или YYWMY (SEQ ID NO: 12);
(ii) CDR2 с аминокислотной последовательностью, выбранной из группы:
VIWFDGSNKFYTDSVKG (SEQ ID NO: 2) или AISWNGGRTYYAESMKG (SEQ ID NO: 13);
(iii) CDR3 с аминокислотной последовательностью, выбранной из группы:
ELGGYYYDSSGFRPYYYGMDV (SEQ ID NO: 3) или NRYYSDPNYGMNL (SEQ ID NO: 14), и
(b) вариабельный домен легкой цепи, содержащему:
(i) CDR1 с аминокислотной последовательностью, выбранной из группы:
RASQSIGSWLA (SEQ ID NO: 7) или TGTSTDIGTYKYIS (SEQ ID NO: 17);
(ii) CDR2 с аминокислотной последовательностью, выбранной из группы:
AASTLQR (SEQ ID NO: 8) или GVSHRPS (SEQ ID NO: 18);
(iii) CDR3 с аминокислотной последовательностью, выбранной из группы:
QQSHSHPLT (SEQ ID NO: 9) или SSYTSSGTVV (SEQ ID NO: 19).
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3, соответственно.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14, соответственно.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9, соответственно.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует вариантах моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19, соответственно.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает:
- вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3, соответственно;
- вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9, соответственно.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает:
- вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14, соответственно;
- вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 c аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19, соответственно.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 4.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 4.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 15.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 15.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 10.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 10.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 20.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 20.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает:
- вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 4;
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 10.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает:
- вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 4;
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 10.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает:
- вариабельный домен тяжелой цепи, которой содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 15;
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 20.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело или его антигенсвязывающий фрагмент, которое включает:
- вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 15;
- вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 20.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое специфично связывается с GITR, которое представляет собой полноразмерное антитело IgG.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело IgG, которое относится к изотипу IgG1, IgG2, IgG3 или IgG4 человека.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело IgG, которое относится к изотипу IgG1 человека.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое специфично связывается с GITR, которое включает мутацию E345R в Fc фрагменте для увеличения агонистических свойств, антителозависимой клеточной цитотоксичности (ADCC), но не комплементзависимой цитотоксичности (CDC).
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 5.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 5.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 6.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 6.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 11.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает:
- тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 5;
- легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 11.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает:
- тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 5;
- легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает:
- тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 6;
- легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 11.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает:
- тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 6;
- легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 16.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 16.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 21.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 21.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает:
- тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 16;
- легкую цепь, содержащую аминокислотную последовательность по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 21.
В некоторых вариантах молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует моноклональное антитело, которое включает:
- тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 16;
- легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 21.
В некоторых вариантах нуклеиновая кислота представляет собой ДНК.
В любом из указанных выше вариантах осуществления изобретения молекулы нуклеиновых кислот могут быть выделенными.
Молекула нуклеиновой кислоты по данному изобретению может быть выделена из любого источника, который продуцирует моноклональное антитело, которое специфически связывается с GITR. В определенных вариантах осуществления изобретения молекула нуклеиновой кислоты данному изобретению может быть синтезирована, а не выделена.
В одном варианте осуществления изобретения молекулы нуклеиновых кислот, кодирующие домены VH (SEQ ID NO: 4 или SEQ ID NO: 15) или VL (SEQ ID NO: 10 или SEQ ID NO: 20) преобразуются в гены антитела по всей длине путем вставки в экспрессионный вектор, уже кодирующей константные домены тяжелой цепи (СН) или легкой цепи (CL), соответственно, так что VH сегмент функционально соединен с CH сегментом(-ами) в векторе и/или VL сегмент оперативно соединен с CL сегментом в векторе. В другом варианте осуществления изобретения молекулы нуклеиновых кислот, кодирующие VH и/или VL домены преобразуются в гены по всей длине антитела путем соединения, например, лигирования, молекулы нуклеиновой кислоты, кодирующей VH и/или VL домены, к молекуле нуклеиновой кислоты, кодирующей CH и/или CL домены с использованием стандартных молекулярно-биологических методов. Молекулы нуклеиновых кислот, кодирующие по всей длине тяжелую и/или легкую цепи, могут затем экспрессироваться из клетки, в которую они были введены.
Молекулы нуклеиновой кислоты могут использоваться для экспрессии большого количества рекомбинантного моноклонального антитела, которое специфически связывается с GITR.
Вектор
В еще одном аспекте настоящее изобретение относится к вектору, подходящему для экспрессии любой из нуклеотидных последовательностей, описанных в настоящем документе.
Настоящее изобретение относится к векторам, содержащим молекулы нуклеиновых кислот, которые кодируют любую из аминокислотных последовательностей моноклонального антитела, которое специфически связывается с GITR, или их частей (например, последовательностей тяжелой цепи первого и/или тяжелой и/или легкой цепи второго связывающих доменов) как описано в настоящем документе. Настоящее изобретение далее относится к векторам, содержащим молекулы нуклеиновых кислот, кодирующих слитые белки, модифицированные антитела, фрагменты антител.
В некоторых вариантах осуществления изобретения, моноклональное антитело, которое специфически связывается с GITR, по данному изобретению экспрессируются путем вставки ДНК, кодирующей частично или полностью последовательность первого и второго связывающего домена (например, последовательности тяжелой и легкой цепи, где связывающий домен содержит последовательности тяжелой и легкой цепи), полученный, как описано выше, в экспрессионных векторах таким образом, что гены функционально соединены с необходимыми последовательностями, контролирующими экспрессию, такими как транскрипционные и трансляционные контрольные последовательности. Экспрессионные векторы включают плазмиды, ретровирусы, аденовирусы, аденоассоциированные вирусы (AAV), вирусы растений, такие как вирус мозаики цветной капусты, вирусы табачной мозаики, космиды, YAC, EBV полученные эписомы и тому подобное. Молекулы ДНК могут быть лигированы в вектор таким образом, что последовательности, контролирующие транскрипцию и трансляцию в векторе, выполняют предусмотренную функцию регуляции транскрипции и трансляции ДНК. Экспрессионный вектор и последовательности контроля экспрессии могут быть выбраны таким образом, чтобы быть совместимыми с используемой экспрессирующей клеткой-хозяином. Молекулы ДНК, кодирующие частично или по всей длине последовательности первого и второго связывающих доменов (например, последовательности тяжелой и легкой цепи, где связывающий домен содержит последовательность тяжелой и легкой цепи) могут быть введены в отдельные векторы. В одном варианте осуществления изобретения любая комбинация указанных выше молекул ДНК вводится в тот же экспрессионный вектор. Молекулы ДНК могут быть введены в экспрессионный вектор стандартными способами (например, лигированием комплементарных сайтов рестрикции на фрагменте гена антитела и вектора или лигированием тупых концов, если сайты рестрикции отсутствуют).
Подходящим вектором является тот, который кодирует функционально законченные последовательности CH или CL человеческого иммуноглобулина с конструированием соответствующего места рестрикции так, что любая последовательность VH или VL может быть легко включена и экспрессирована, как описано выше. HC - и LC-кодирование генов в таких векторах может содержать интронные последовательности, что приводит к общему увеличению белковых продуктов антитела путем стабилизации соответствующей мРНК. Интронные последовательности находятся в окружении сплайс-донора и сплайс-акцептора сайтов, которые определяют, где будет происходить сплайсинг РНК. Расположение интронных последовательностей может быть либо в вариабельных или константных участках цепей антитела, или как в вариабельных, так и константных участках, когда используются несколько интронов. Прекращение полиаденилирования и транскрипции может произойти вниз по ходу сайта нативной хромосомы кодируемых участков. Рекомбинантный экспрессионный вектор также может кодировать сигнальный пептид, который облегчает выработку цепочки антитела клеткой-хозяином. Ген цепочки антитела может быть клонирован в вектор таким образом, что сигнальный пептид соединен с рамкой считывания аминоконца цепи иммуноглобулина. Сигнальным пептидом может быть сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (то есть, сигнальный пептид белка не иммуноглобулиновой природы).
Помимо цепочки генов антител, рекомбинантная экспрессия векторов по данному изобретению может нести регулирующие последовательности, которые контролируют экспрессию генов цепи антител в клетке-хозяине. Специалистам в этой области будет понятно, что дизайн экспрессионного вектора, включая выбор регулирующих последовательностей, может зависеть от таких факторов, как селекция клетки-хозяина для трансформации, уровень экспрессии желаемого белка, и т.д. Предпочтительные регулирующие последовательности для экспрессирующей клетки-хозяина млекопитающих включают вирусные элементы обеспечивающие высокий уровень экспрессии белков в клетках млекопитающих, таких как промоторы и/или энхансеры, полученные из ретровирусной LTR, цитомегаловируса (CMV) (например, CMV промотора/энхансера), обезьяньего вируса 40 (SV40) (например, SV40 промотора/энхансера), аденовируса, (например, большого позднего промотора аденовируса (AdMLP)), вирус полиомы, а также сильных промоторов млекопитающих, таких как промотор нативных иммуноглобулинов или промотор актина. Для дальнейшего описания вирусных регулирующих элементов и их последовательностей см., например, патенты США 5,168,062, 4,510,245 and 4,968,615. Способы экспрессии связывающих молекул, таких как антитела растений, в том числе описание промоторов и векторов, а также трансформация растений, известны в данной области техники. См., например, патент США 6,517,529. Методы экспрессии полипептидов в бактериальных клетках или клетках грибов, например, дрожжевых клетках, также хорошо известны в данной области техники.
В дополнение к генам цепи антитела и регулирующим последовательностям, рекомбинантные векторы экспрессии изобретения могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, точки начала репликации) и гены селектируемого маркера. Ген селектируемого маркера облегчает селекцию клеток-хозяев, в которые был введен вектор (см., например, патенты США 4,399,216, 4,634,665 и 5,179,017). Например, обычно ген селектируемого маркера придает устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат, клетке-хозяину, в которую вектор введен. Например, гены селектируемого маркера включают ген дигидрофолат редуктазы (DHFR) (для использования в dhfr-клетках-хозяевах при селекции/амплификации метотрексата), ген нео (для селекции G418) и ген синтетазы глутамата.
Термин «последовательность контроля экспрессии», используемый в данном описании, означает полинуклеотидные последовательности, которые необходимы для воздействия на экспрессию и процессинг кодирующих последовательностей, к которым они лигированы. Контролирующие экспрессию последовательности включают соответствующие последовательности инициации транскрипции, терминации, промотора и энхансера; эффективные сигналы процессинга РНК, такие как сплайсинг и сигналы полиаденилирования; последовательности, которые стабилизируют цитоплазматическую мРНК; последовательности, которые повышают эффективность трансляции (т.е. консенсусная последовательность Козака); последовательности, которые повышают стабильность белка; и, при желании, последовательности, которые усиливают секрецию белка. Характер таких контролирующих последовательностей различается в зависимости от организма-хозяина; в прокариотах такие контролирующие последовательности, как правило, включают промотор, сайт связывания рибосомы, а также последовательности терминации транскрипции; в эукариотах, как правило, такие контролирующие последовательности включают промоторы и последовательности терминации транскрипции. Термин «контролирующие последовательности» включает, как минимум, все компоненты, наличие которых имеет важное значение для экспрессии и процессинга, и может также включать дополнительные компоненты, чье присутствие является полезным, например, лидирующие последовательности и последовательности слившихся клеток.
Клетки-хозяева
Дополнительный аспект настоящего изобретения относится к способам получения моноклонального антитела, которое специфически связывается с GITR, по данному изобретению. Один вариант осуществления изобретения относится к способу получения моноклонального антитела, которое специфически связывается с GITR, как определено в настоящем документе, содержащему получение рекомбинантной клетки-хозяина, способной экспрессировать моноклональное антитело, которое специфически связывается с GITR, культивированию указанной клетки-хозяина в условиях, подходящих для экспрессии продукции моноклонального антитела, которое специфически связывается с GITR, и выделение полученного моноклонального антитела, которое специфически связывается с GITR. Моноклональное антитело, которое специфически связывается с GITR, полученное такой экспрессией в таких рекомбинантных клетках-хозяевах упоминается в данном документе как «рекомбинантные моноклональное антитело, которое специфически связывается с GITR». Изобретение также относится к потомству клеток таких клеток-хозяев и моноклональному антителу, которое специфически связывается с GITR, полученному аналогично.
Молекулы нуклеиновой кислоты, кодирующие моноклональное антитело, которое специфически связывается с GITR, по изобретению и векторы, содержащие эти молекулы нуклеиновой кислоты, могут быть использованы для трансфекции подходящего млекопитающего или его клетки, растения или его клетки, бактериальной или дрожжевой клетки-хозяина. Преобразование может происходить любым известным способом для введения полинуклеотидов в клетку-хозяина. Способы введения гетерологичных полинуклеотидов в клетки млекопитающих хорошо известны в данной области и включают декстран-опосредованную трансфекцию, трансфекцию комплексом нуклеиновой кислоты и позитивно заряженного полимера, трансфекцию преципитатом нуклеиновой кислоты и фосфата кальция, полибрен-опосредованную трансфекцию, слияние протопластов, трансфекцию инкапсулированными в липосомы полинуклеотидами и прямую микроинъекцию ДНК в ядра. В дополнение, молекулы нуклеиновых кислот могут быть введены в клетки млекопитающих вирусными векторами. Способы трансфекции клеток хорошо известны в данной области техники. См., например, патенты США, 4,399,216, 4,912,040, 4,740,461 и 4,959,455. Способы трансформации клеток растений хорошо известны в данной области, включая, например, Agrobacterium-опосредованную трансформацию, биолистическую трансформацию, прямую инъекцию, электропорацию и вирусную трансформацию. Методы трансформации клеток бактерий и дрожжей также хорошо известны в данной области.
Клеточные линии млекопитающих, используемые в качестве хозяев для трансформации, хорошо известны в данной области и включают множество иммортализованных доступных клеточных линий. К ним относятся, например, клетки яичников китайского хомячка (CHO), NS0 клетки, клетки SP2, HEK-293T клетки, 293 Фристайл клетки (Invitrogen), NIH-3T3 клетки, клетки HeLa, клетки почек хомячка (BHK), клетки почек африканских зеленых мартышек (COS), клетки гепатоцеллюлярной карциномы человека (например, Hep G2), A549 клетки и ряд других клеточных линий. Клеточные линии выбираются путем определения, какие клеточные линии имеют высокие уровни экспрессии и обеспечивают необходимые характеристики продуцируемого белка. Другими клеточными линиями, которые могут быть использованы, являются клеточные линии насекомых, такие как Sf9 или Sf21 клетки. Когда векторы рекомбинантной экспрессии, кодирующие моноклональное антитело, которое специфически связывается с GITR вводятся в клетки-хозяева млекопитающих, антитела продуцируются путем культивирования клеток-хозяев в течение времени, достаточного для экспрессии антител в клетках-хозяевах или, предпочтительнее, выделения антител в питательную среду, в которой выращиваются клетки-хозяева. Моноклональное антитело, которое специфически связывается с GITR, может быть выделено из питательной среды с использованием стандартных методов очистки белка. Клетки-хозяева растений, например, включают Nicotiana, Arabidopsis, ряску, кукурузу, пшеницу, картофель и т.д. Клетки бактерий хозяина включают виды Escherichia и Streptomyces. Дрожжевые клетки-хозяева включают Schizosaccharomyces pombe, Saccharomyces cerevisiae и Pichia pastoris.
Кроме того, уровень продукции моноклонального антитела, которое специфически связывается с GITR, по данному изобретению из продуцирующей клеточной линии можно усилить с помощью ряда известных методов. Например, система экспрессии гена глутамин синтетазы (система GS) является достаточно распространенной для усиления экспрессии при определенных условиях. Система GS обсуждается в целом или частично в связи с патентами ЕР 0216846, 0256055, 0323997 и 0338841.
Вполне вероятно, что моноклональное антитело, которое специфически связывается c GITR, различных клеточных линий или трансгенные животные будут отличаться друг от друга профилем гликозилирования. Однако моноклональное антитело, которое специфически связывается с GITR, кодируемое молекулами нуклеиновой кислоты, описанными в данном документе, или содержащее аминокислотные последовательности, приведенные в настоящем документе, являются частью данного изобретения, независимо от состояния гликозилирования связывающих молекул и в целом, независимо от наличия или отсутствия пост-трансляционных модификаций.
Получение антител
Изобретение относится также к способам и процессам получения моноклонального антитела, которое специфически связывается с GITR, и их антигенсвязывающих фрагментов.
Моноклональные антитела
Моноклональные антитела можно создавать, например, с помощью метода на основе гибридом, который впервые описан Kohler и др., Nature, 256, 1975, с. 495, или с помощью методов рекомбинантной ДНК (US 4816567).
При использовании метода на основе гибридом мышь или другое пригодное животное-хозяин, такое как хомячок, иммунизируют согласно описанной выше методике, для того, чтобы вызывать образование лимфоцитов, которые продуцируют или могут продуцировать антитела, обладающие способностью специфически связываться с белком, применяемым для иммунизации. Согласно другому варианту лимфоциты можно получать в результате иммунизации in vitro. После иммунизации лимфоциты выделяют и затем сливают с клеточной линией миеломы с помощью приемлемого связывающего агента, такого как полиэтиленгликоль, с получением клетки гибридомы.
Полученные таким образом клетки гибридомы высевают и выращивают в пригодной культуральной среде, которая предпочтительно содержит одно или несколько веществ, ингибирующих рост или выживание неслитых родительских клеток миеломы. Например, если родительские клетки миеломы не содержат фермента гипоксантингуанинфосфорибозилтрансферазы (HGPRT или HPRT), то культуральная среда для гибридом, как правило, должна включать гипоксантин, аминоптерин и тимидин (НАТ-среда), т.е. вещества, которые препятствуют росту клеток с дефицитом HGPRT.
Предпочтительными применяемые в качестве компонента для слияния клетками миеломы являются клетки, которые легко поддаются слиянию, поддерживают стабильный высокий уровень производства антител выбранными продуцирующими антитела клетками и чувствительны к избирательной среде, на которой происходит отбор несвязанных родительских клеток. Предпочтительными линиями клеток миелом являются линии мышиной миеломы, такие как созданные на основе мышиных линий опухолевых клеток линии МОРС-21 и МРС-11, которые можно получать из Salk Institite Cell Disrtibution Center, Сан-Диего, шт. Калифорния, США, и линии SP-2 или Х63- Ag8-653, которые можно получать из Американской коллекции типовых культур, Роквилл, шт. Мэриленд, США. Описано также применение линий клеток человеческой миеломы и гетеромиеломы типа «мышь-человек» для производства моноклональных антител (Kozbor, J. Immunol., 133, 1984, с. 3001).
Предпочтительно специфичность связывания моноклональных антител, полученных с использованием клеток гибридомы, определяют методом иммунопреципитации или с помощью анализа связывания in vitro, такого как радиоиммунный анализ (РИА) или твердофазный имммуноферментный анализ (ELISA).
Аффинность связывания моноклонального антитела можно, например, определять с помощью Скэтчард-анализа, описанного у Munson и др., Anal. Biochem., 107, 1980, с. 220.
После выявления клеток гибридомы, которые продуцируют антитела требуемой специфичности, аффинности и/или активности, клоны можно субклонировать с помощью метода лимитирующих разведений и выращивать стандартными методами. Пригодные для этой цели среды включают, например, среду DMEM или RPMI-1640. Кроме того, клетки гидрибомы можно выращивать in vivo в виде асцитных опухолей в организме животных, например, путем внутрибрюшинной (i.p.) инъекции клеток мышам.
Моноклональные антитела, секретируемые субклонами, можно отделять от культуральной среды, асцитной жидкости или сыворотки с помощью обычных методов очистки антител, например, аффинной хроматографией (например, с использованием протеин А- или протеин Q-сефарозы), или ионообменной хроматографии, хроматографии на гидроксилапатитах, гель-электрофореза, диализа и т.д.
ДНК, кодирующую моноклональные антитела, можно легко выделять и секвенировать с помощью общепринятых процедур (например, с использованием олигонуклеотидных зондов, которые обладают способностью специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител). В качестве предпочтительного источника такой ДНК служат клетки гибридомы. После выделения ДНК можно включать в экспрессионные векторы, которыми затем трансфектируют клетки-хозяева, такие как клетки Е. coli, обезьяньи COS- клетки, клетки яичника китайского хомячка (СНО) или клетки миеломы, которые без трансфекции не продуцируют белок антитела, что приводит к синтезу моноклональных антител в рекомбинантных клетках-хозяевах.
Согласно еще одному варианту осуществления изобретения моноклональные антитела или фрагменты антител можно выделять из фаговых библиотек антител, созданных с помощью методов, описанных у McCafferty и др., Nature, 348, 1990, cc. 552-554 . У Clackson и др., Nature, 352, 1991, cc. 624- 628 и Marks и др., J. Mol. Biol., 222, 1991, cc. 581-597 описано выделение мышиных и человеческих антител соответственно с помощью фаговых библиотек. В последующих публикациях было описано производство высокоаффинных (нМ-диапазон) человеческих антител с помощью перестановки цепи (Marks и др., Bio/Technology, 10, 1992, cc. 779-783), а также комбинаторная инфекция и рекомбинация in vivo в качестве стратегии конструирования очень больших фаговых библиотек (Waterhouse и др., Nucl. Acids. Res., 21, 1991, cc. 2265-2266). Таким образом, эти методы представляют собой реальную альтернативу традиционным методам выделения моноклональных антител на основе гибридом моноклональных антител.
ДНК, кодирующую антитело, можно также модифицировать, например таким образом, чтобы получать химерные или слитые полипептиды антител, например, путем замены последовательностей константных областей тяжелой и легкой цепи (Сн и CL) на гомологичные мышиные последовательности (US 4816567 и Morrison и др., Proc. Natl. Acad. Sci. USA: 81, 1984, с. 6851) или с помощью ковалентного связывания кодирующей последовательности иммуноглобулина со всей или с частью кодирующей последовательности полипептида, не относящегося к иммуноглобулинам (гетерологичного полипептида). Не относящиеся к иммуноглобулинам полипептидные последовательности можно заменять на константные области антитела или заменять на вариабельные области антигенсвязывающего центра антитела, создавая химерное бивалентное антитело, которое содержит один антигенсвязывающий центр, обладающий специфичностью в отношении антигена, и другой антигенсвязывающий центр, обладающий специфичностью в отношении другого антигена.
Человеческие антитела и методика, основанная на применении фаговой дисплейной библиотеки
В настоящее время можно получать трансгенных животных (например, мышей), которые после иммунизации могут продуцировать полный спектр человеческих антител без производства эндогенного иммуноглобулина. Например, было описано, что гомозиготная делеция областей стыка гена тяжелой цепи антитела (Jн-сегмента) в организме химерных мышей и мышей с мутацией в зародышевой линии приводит к полному ингибированию производства эндогенного антитела. Перенос набора зародышевой линии гена человеческого иммуноглобулина в такую мутантную зародышевую линию мышей приводит к производству человеческих антител после контрольного заражения антигеном (US 5545806, 5569825, 5591669 (все на имя фирмы GenPharm); 5545807; и WO 97/17852).
В альтернативном варианте для получения человеческих антител и фрагментов антител in vitro из спектра генов вариабельной области (V) иммуноглобулина из организма иммунизированных доноров можно использовать фаговую дисплейную технологию (McCafferty и др., Nature, 348, 1990, сс. 552-553). Согласно этой методике гены V-области антитела клонируют в рамке считывания либо с основным, либо с минорным геном оболочечного белка нитчатого бактериофага, такого как М13 или fd, и презентируют в виде функциональных фрагментов антитела на поверхности фаговой частицы. Поскольку нитчатая частица содержит копию одноцепочечной ДНК фагового генома, селекции по признаку функциональных свойств антитела, также приводят к отбору гена, кодирующего антитело, которое обладает указанными свойствами. Таким образом, фаг имитирует некоторые свойства В-клетки. Фаговую презентацию можно осуществлять в различных форматах. Для фаговой презентации можно использовать различные источники сегментов V-генов. Clackson и др., Nature, 352, 1991, сс. 624-628 выделили различные наборы антител к оксазолону из небольшой произвольной комбинаторной библиотеки V-генов, полученной из селезенки иммунизированных мышей. Можно конструировать спектр V-генов, полученных из организма иммунизированных людей-доноров, и антитела к различному набору антигенов (включая аутоантигены) можно выделять в целом согласно методам, описанным у Marks и др., J. Mol. Biol., 222, 1991, cc. 581-597.
Как описано выше, человеческие антитела могут продуцироваться также in vitro активированными В-клетками (см. US 5567610 и 5229275).
Фрагменты антител
В определенных обстоятельствах целесообразно применять фрагменты антител, а не полные антитела. Меньший размер фрагментов способствует их быстрому клиренсу и может способствовать лучшему проникновению в плотные опухоли.
Для получения фрагментов антител разработаны различные методы. Традиционно эти фрагменты получали путем протеолитического расщепления интактных антител. Однако в настоящее время эти фрагменты можно получать непосредственно с помощью рекомбинантных клеток-хозяев. Fab-, Fv- и scFv-фрагменты антител можно экспрессировать и секретировать из Е. coli, что позволяет облегчать производство больших количеств указанных фрагментов. Фрагменты антител можно выделять из описанных выше фаговых библиотек антител. Согласно другому варианту Fab'-SH-фрагменты можно непосредственно выделять из Е. coli и химически сшивать с получением F(аb')2-фрагментов (Carter и др., Bio/Technology, 10, 1992, сс. 163-167). Согласно другому подходу F(аb')2 - фрагменты можно выделять непосредственно из культуры рекомбинантных клеток-хозяев. Fab- и F(аb')2-фрагмент с повышенным временем полужизни in vivo, в которых сохранены остатки эпитопсвязывающего рецептора, описаны в US 5869046.Специалистам в данной области должны быть очевидны другие методики получения фрагментов антител. В других вариантах осуществления изобретения выбранное антитело представляет собой одноцепочечный Fv- фрагмент (scFv) (см. WO 93/16185; US 5571894 и US США 5587458). Fv и sсFv представляют собой единственные виды с интактными связывающими сайтами, лишенные константных областей; в результате их можно применять для пониженного неспецифического связывания при применении in vivo. Слитые белки, несущие sсFv, можно конструировать для получения слияния эффекторного белка либо на N-, либо на С-конце sсFv. Фрагмент антитела может представлять собой также «линейное антитело», например, описанное в US 5641870. Такие фрагменты линейного антитела могут быть моноспецифическими или биспецифическими.
Фармацевтические композиции
Другим аспектом изобретения является фармацевтическая композиция, содержащая в качестве активного ингредиента (или в качестве единственного активного ингредиента) моноклональное антитело, которое специфически связывается с GITR.
Фармацевтическая композиция может включать по меньшей мере одно моноклональное антитело, которое специфически связывается с GITR и, по крайней мере, один из компонентов, выбранных из группы, состоящей из фармацевтически приемлемых и фармакологически совместимых эксципиентов.
Фармацевтическая композиция может включать по меньшей мере одно моноклональное антитело, которое специфически связывается с GITR, и одну или более дополнительных связывающих молекул (например, антител), которые нацелены на один или более соответствующих поверхностных рецепторов. В некоторых вариантах осуществления изобретения композиции предназначены для улучшения, профилактики или лечения нарушений, которые могут быть связаны с GITR.
«Фармацевтическая композиция» обозначает композицию, включающую в себя моноклональное антитело, которое специфически связывается с GITR, согласно изобретению и, по крайней мере, один из компонентов, выбранных из группы, состоящей из фармацевтически приемлемых и фармакологически совместимых эксипиентов, таких как наполнители, растворители, разбавители, носители, вспомогательные, распределяющие, средства доставки, консерванты, стабилизаторы, эмульгаторы, суспендирующие агенты, загустители, регуляторы пролонгированной доставки, выбор и соотношение которых зависит от природы и способа назначения и дозировки. Фармацевтические композиции по настоящему изобретению и способы их изготовления будут бесспорно очевидными для специалистов в этой области. Производство фармацевтических композиций предпочтительно должно соответствовать требованиям GMP (надлежащей производственной практики). Композиция может включать буферную композицию, тонические агенты, стабилизаторы и солюбилизаторы. Пролонгированное действие композиции может быть обеспечено с помощью агентов, замедляющих абсорбцию активного фармацевтического ингредиента, например, моностеарат алюминия и желатин. Примерами подходящих носителей, растворителей, разбавителей и средств доставки являются вода, этанол, полиспирты, а также их смеси, масла и инъекционные органические сложные эфиры.
«Лекарственное средство (препарат)» - вещество или смесь веществ в виде фармацевтической композиции в виде таблеток, капсул, порошков, лиофилизатов, инъекций, инфузий, мазей и др. готовых форм, предназначенное для восстановления, исправления или изменения физиологических функций у человека и животных, а также для лечения и профилактики болезней, диагностики, анестезии, контрацепции, косметологии и прочего. Любой способ введения пептидов, белков или антител, принятый в данной области, может соответствующим образом использоваться для моноклонального антитела, которое специфически связывается с GITR, по данному изобретению.
Термин «фармацевтически приемлемый» означает один или несколько совместимых жидких или твердых компонентов, которые подходят для введения млекопитающему, предпочтительно человеку.
Термин «эксципиент» или «вспомогательное вещество» используется в данном документе для описания любого компонента, отличающегося от ранее описанных по данному изобретению. Это вещества неорганического или органического происхождения, используемые в процессе производства, изготовления лекарственных препаратов для придания им необходимых физико-химических свойств.
Под термином «буфер», «буферная композиция», «буферный агент» понимается раствор, способный сохранять значение pH, благодаря взаимодействию кислотных и щелочных компонентов, входящих в его состав, который дает возможность препарату моноклонального антитела, которое специфически связывается с GITR, проявлять устойчивость к изменениям рН. В общем случае, преимущественными являются значения рН фармацевтической композиции от 4,0 до 8,0. В качестве буферных агентов могут быть использованы, например, ацетатный, фосфатный, цитратный, гистидиновый, сукцинатный и т.п. буферные растворы, но, не ограничиваясь ими.
Термины «тонический агент», «осмолитик» или «осмотический агент» в том виде, как они здесь использованы, относятся к эксципиенту, который может подводить осмотическое давление жидкого препарата антитела. «Изотоничный» препарат представляет собой препарат, который имеет осмотическое давление, эквивалентное давлению человеческой крови. Изотоничные препараты обычно имеют осмотическое давление от примерно 250 до 350 мОсм/кг. В качестве изотонических агентов могут быть использованы полиолы, моно- и дисахара, аминокислоты, соли металлов, например, хлорид натрия, и т.п., но, не ограничиваясь ими.
Под «стабилизатором» понимается вспомогательное вещество или смесь двух и более вспомогательных веществ, которые обеспечивают физическую и/или химическую стабильность активного агента. В качестве стабилизаторов могут быть использованы аминокислоты, например, аргинин, гистидин, глицин, лизин, глутамин, пролин, но, не ограничиваясь ими; поверхностно-активные вещества, например, полисорбат 20 (торговое наименование Tween 20), полисорбат 80 (торговое наименование Tween 80), полиэтилен-полипропилен гликоль и его кополимеры (торговые наименования Полоксамер (Poloxaner), Плуроник (Pluronic)), натрия додецилсульфат (SDS), но, не ограничиваясь ими; антиоксиданты, например, метионин, ацетилцистеин, аскорбиновая кислота, монотиоглицерол, соли серистых кислот, и т.п., но не ограничиваясь ими; хелатирующие агенты, например, этилендиаминтетрауксусная кислота (ЭДТА), диэтилентриаминпентауксусная кислота (ДТПА), цитрат натрия и т.п., но не ограничиваясь ими.
Фармацевтическая композиция является «стабильной», если активный агент сохраняет свою физическую стабильность и/или химическую стабильность и/или биологическую активность в течение заявленного срока годности при температуре хранения, например, при 2-8 °С. Предпочтительно, чтобы активный агент сохранял и физическую, и химическую стабильность, а также биологическую активность. Период хранения выбирается на основании результатов исследования стабильности при ускоренном и естественном хранении.
Фармацевтическая композиция по данному изобретению может изготавливаться, упаковываться или широко продаваться в виде единичной стандартной дозы или множества единичных стандартных доз в виде готовой лекарственной формы. Используемый в данном документе термин «единичная стандартная доза» означает дискретное количество фармацевтической композиции, содержащей заранее определенное количество активного ингредиента. Количество активного ингредиента обычно равно дозировке активного ингредиента, который будет вводиться субъекту, или удобной части такой дозировки, например, половине или трети такой дозировки.
Фармацевтические композиции по настоящему изобретению, как правило, пригодны для парентерального введения в виде стерильных лекарственных средств, предназначенных для введения в организм человека с нарушением целостности кожных покровов или слизистых оболочек, минуя желудочно-кишечный тракт путем инъекций, инфузий или имплантации. Например, предполагается, что парентеральное введение включает, помимо прочего, подкожную, внутрибрюшинную, внутримышечную, внутривенную, внутриартериальную, интратекальную, внутрижелудочковую, интрауретральную, внутричерепную, внутрисуставную, трансдермальную инъекцию или инфузию; и почечные диализные инфузионные методики. Внутриопухолевая доставка, например, внутриопухолевая инъекция, также может быть применима. Также предусмотрена региональная перфузия. Предпочтительные варианты осуществления изобретения включают внутривенный и подкожный пути. Любой способ введения пептидов или белков, принятый в данной области, может соответствующим образом использоваться для моноклонального антитела, которое специфически связывается с GITR, по данному изобретению.
Инъекционные лекарственные средства могут быть изготовлены, упакованы или проданы в стандартной лекарственной форме, например, в ампулах, флаконах, полимерных контейнерах, преднаполненных шприцах, устройствах для автоинжекции, но, не ограничиваясь ими. Лекарственные средства для парентерального введения включают, помимо прочего, суспензии, растворы, эмульсии в масляных или водных основах, пасты и тому подобное.
Еще одним вариантом осуществления изобретения является лекарственное средство для парентерального введения, где фармацевтическая композиция предоставлена в сухой форме, то есть, порошка или гранул для растворения в подходящем растворителе (например, стерильной апирогенной воде) перед введением. Такое лекарственное средство может быть получено, например, с помощью лиофилизации, т.е. процесса, известного в данной области техники как сушка из замороженного состояния, включающая в себя замораживание препарата и последующее удаление растворителя из замороженного содержимого.
Моноклональное антитело, которое специфически связывается с GITR, по данному изобретению может также вводиться интраназально или ингаляционно, самостоятельно, в виде смеси с подходящим фармацевтически приемлемым наполнителем из ингалятора, такого как аэрозольный контейнер под давлением, помпа, спрей, распылитель) или небулайзер, в котором используется или не используется подходящий пропеллент, или в виде назальных капель или спрея.
Лекарственное средство для парентерального введения может быть немедленного или модифицированного высвобождения. Лекарственные средства с модифицированным высвобождением включают отсроченное, замедленное, пульсирующее, контролируемое, нацеленное и программируемое высвобождение.
Терапевтическое применение моноклонального антитела, которое специфически связывается с GITR, по данному изобретению
В одном аспекте моноклональное антитело, которое специфически связывается с GITR, по данному изобретению применяется в лечении нарушений, которые связаны (опосредованы) с активностью GITR.
В одном аспекте субъект лечения или пациент является млекопитающим, предпочтительно человеческим субъектом. Вышеупомянутый субъект может быть мужского или женского пола и любого возраста.
В некоторых вариантах моноклональное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с GITR, применяют для лечения заболевания или нарушения, опосредуемого GITR, где заболевание или нарушение выбрано из группы: РШМ (рак шейки матки), рак головы и шеи, рак желудка, РМЖ (рак молочной железы), почечно-клеточный рак, КРР (колоректальный рак), РЯ (рак яичника), НМРЛ (немелкоклеточный рак легкого).
В случае опухоли (например, раковой опухоли) терапевтически эффективное количество антитела или фрагмента антитела (например, антитела или фрагмента антитела, которое специфически связывает GITR, может уменьшать число раковых клеток; уменьшать начальный размер опухоли; ингибировать (т.е. до некоторой степени замедлять и, предпочтительно, прекращать) инфильтрацию раковыми клетками периферических органов; ингибировать (т.е. до некоторой степени замедлять и, предпочтительно, прекращать) метастазирование опухоли; ингибировать, до некоторой степени, рост опухоли; и/или облегчать, до некоторой степени, один или более симптомов, обусловленных расстройством. Антитело или фрагмент антитела может, до некоторой степени, предупреждать рост и/или убивать имеющиеся раковые клетки, оно может вызывать цитостатический и/или цитотоксический эффект. При терапии рака эффективность in vivo можно определять, например, оценивая продолжительность жизни, время до прогрессирования заболевания (ТТР), частоту ответа опухоли на лечение (RR), продолжительность ответа и/или качество жизни.
Используемые в данном документе термины «совместное назначение», «совместно назначенный» и «в сочетании с» относятся к моноклональному антителу, которое специфически связывается с GITR, с одним или более другими терапевтическими агентами, как предполагается, означают, ссылаются или включают:
1) одновременное введение такой комбинации моноклонального антитела, которое специфически связывается с GITR, по данному изобретению и терапевтического агента пациенту, который нуждается в лечении, когда такие компоненты сформулированы вместе в одной лекарственной форме, из которой указанные компоненты высвобождаются практически одновременно указанному пациенту,
2) одновременное введение такой комбинации моноклонального антитела, которое специфически связывается с GITR, по данному изобретению и терапевтического агента пациенту, который нуждается в лечении, когда такие компоненты сформулированы отдельно в разных лекарственных формах, введение которых происходит практически в одно и то же время указанному пациенту, после чего указанные компоненты высвобождаются практически одновременно указанному пациенту,
3) последовательное введение такой комбинации моноклонального антитела, которое специфически связывается с GITR, по данному изобретению и терапевтического агента пациенту, который нуждается в лечении, когда такие компоненты сформулированы отдельно друг от друга в отдельных лекарственных формах, которые принимаются в последовательно по времени указанным пациентом со значимым временным интервалом между каждым введением, после чего указанные компоненты высвобождаются в практически разное время указанному пациенту; а также
4) последовательное введение такой комбинации моноклонального антитела, которое специфически связывается с GITR, по данному изобретению и терапевтического агента пациенту, который нуждается в лечении, когда такие компоненты сформулированы вместе в единый лекарственной форме, из которой высвобождение указанных компонентов происходит контролируемым образом, после чего они одновременно, последовательно или совместно высвобождаются в одно и то же время и/или разное время указанному пациенту, где каждая часть может быть введена одним или разными путями.
Моноклональное антитело, которое специфически связывается с GITR, по данному изобретению могут назначаться без дополнительного терапевтического лечения, т.е. в качестве самостоятельной терапии. Кроме того, лечение моноклональным антителом, которое специфически связывается с GITR, по данному изобретению может включать по меньшей мере одно дополнительное терапевтическое лечение (комбинированная терапия). В некоторых вариантах осуществления изобретения, моноклональное антитело, которое специфически связывается с GITR, может вводиться совместно или быть сформулирована с другим медикаментом/препаратом для лечения рака.
Термин «цитотоксическое средство» в используемом в настоящем описании смысле относится к веществу, которое ингибирует или предотвращает функционирование клеток и/или вызывает разрушение клеток. Подразумевается, что термин включает радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивные изотопы Lu), химиотерапевтические средства и токсины, такие как низкомолекулярные токсины или ферментативно активные токсины, имеющие происхождение из бактерий, грибов, растений или животных, включая их фрагменты и/или варианты.
«Химиотерапевтическим средством» является химическое соединение, применимое для лечения злокачественной опухоли. Примеры химиотерапевтических средств включают алкилирующие агенты, такие как тиотепа и циклофосфамид (CYTOXAN®); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбохинон, метуредопа и уредопа; этиленимины и метилмеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилмеламин; ацетогенины (например, буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, MARINOL®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; камптотецин (включая синтетический аналог топотекан (HYCAMTIN®), CPT-11 (иринотекан, CAMPTOSAR®), ацетилкамптотецин, скополектин и 9-аминокамптотецин); бриостатин; каллистатин; CC-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); подофиллотоксин; подофиллиновую кислоту; тенипозид; криптофицины (например, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги KW-2189 и CB1-TM1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид оксида мехлорэтамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урамустин; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, например, калихеамицин гамма II и калихеамицин омега II (см., например, Agnew, Chem. Intl. Ed. Engl., 33: 183-186 (1994)); динемицин, включая динемицин A; эсперамицин; а также хромофор неокарциностатина и родственные хромофоры хромопротеинов - энедииновые антибиотики, аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (включая ADRIAMICIN®, морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин, доксорубициноНСl в инъецируемых липосомах (DOXOL®), липосомный доксорубицин TLC D-99 (MYOCET®), пегилированный липосомный доксорубицин (CAELYX®) и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин C, микофеноловую кислоту, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат, гемцитабин (GEMZAR®), тегафур (UFTORAL®), капецитабин (XELODA®), эпотилон и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; средства, подавляющие функции надпочечников, такие как аминоглютетимид, митотан, трилостан; компенсатор фолиевой кислоты, такой как фолиновая кислота; ацеглатон; гликозид альдофосфамида; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазихон; элфорнитин; ацетат эллиптиния; этоглуцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2»-трихлортриэтиламин; трихотецены (например, токсин T-2, верракурин A, роридин A и ангуидин); уретан; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («ara-C»); тиотепа; таксоид, например паклитаксел (TAXOL®), препарат паклитаксела на основе сконструированных связанных с альбумином наночастиц (ABRAXANETM) и доцетаксел (TAXOTERE®); хлорамбуцил; 6-тиогуанин; меркаптопурин; метотрексат; средства на основе платины, такие как цисплатин, оксалиплатин и карбоплатин; алкалоиды барвинка, которые предотвращают полимеризацию тубулина из образующихся микротрубочек, включая винбластин (VELBAN®), винкристин (ONCOVIN®), виндезин (ELDISINE®, FILDESIN®) и винорелбин (NAVELBINE®); этопозид (VP-16); ифосфамид; митоксантрон; лейковорин; новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота, включая бексаротен (TARGRETIN®); бифосфонаты, такие как клодронат (например, BONEFOS® или OSTAC®), этидронат (DIDROCAL®), NE-58095, золедроновую кислоту/золедронат (ZOMETA®), алендронат (FOSAMAX®), памидронат (AREDIA®), тилудронат (SKELID®) или ризендронат (ACTONEL®); троксацитабин (1,3-диоксолановый нуклеозидный аналог цитозина); антисмысловые олигонуклеотиды, например олигонуклеотиды, которые ингибируют экспрессию генов в путях передачи сигналов, вовлеченных в пролиферацию аберрантных клеток, таких как, например, PKC-альфа, Raf, H-Ras и рецептора эпидермального фактора роста (EGF-R); вакцины, такие как вакцина THERATOPE® и вакцины для генной терапии, например вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; ингибитор топоизомеразы 1 (например, LURTOTECAN®); rmRH (например, ABARELIX®); BAY439006 (сорафениб; Bayer); SU-11248 (Pfizer); перифосин, ингибитор ЦОГ-2 (например, целекоксиб или эторикоксиб), ингибитор протеосом (например, PS341); бортезомиб (VELCADE®); CCI-779; типифарниб (811577); орафениб, ABT510; ингибитор Bcl-2, такой как облимерсен натрия (GENASENSE®); пиксантрон; ингибиторы EGFR (см. определение ниже); ингибиторы тирозинкиназ (см. определение ниже); и фармацевтически приемлемые соли, кислоты или производные любого из указанных выше средств; а также сочетания двух или более указанных выше средств, такие как CHOP, сокращенное название комбинированной терапии циклофосфамидом, доксорубицином, винкристином и преднизолоном, и FOLFOX, сокращенное название схемы лечения оксалиплатином (ELOXATINTM) в сочетании с 5-FU и лейковорином.
Также в указанное определение включены противогормональные средства, которые действуют, регулируя или ингибируя действие гормонов на опухоли, такие как антиэстрогены со смешанным профилем агонист/антагонист, включая тамоксифен (NOLVADEX®), 4-гидрокситамоксифен, триоксифен, торемифен (FARESTON®); идоксифен, дролоксифен, ралоксифен (EVISTA®), триоксифен, кеоксифен и избирательные модуляторы рецепторов эстрогена (SERM), такие как SERM3; чистые антиэстрогены без агонистических свойств, такие как фулвестрант (FASLODEX®) и EM800 (такие средства могут блокировать димеризацию рецепторов эстрогена (ER), ингибировать связывание ДНК, усиливать метаболизм ER и/или снижать уровни ER); ингибиторы ароматазы, включая стероидные ингибиторы ароматазы, такие как форместан и эксеместан (AROMASIN®), и нестероидные ингибиторы ароматазы, такие как анастразол (ARIMIDEX®), летрозол (FEMARA®) и аминоглутетимид и другие ингибиторы ароматазы, включая ворозол (RIVISOR®), ацетат мегестрола (MEGASE®), фадрозол, имидазол; агонисты рилизинг-гормона лютеинизирующего гормона, включая лейпролид (LUPRON® и ELIGARD®), гозерелин, бузерелин и триптерелин; половые стероиды, включая прогестины, такие как ацетат мегестрола и ацетат медроксипрогестерона, эстрогены, такие как диэтилстилбестрол и премарин и андрогены/ретиноиды, такие как флуоксиместерон, полностью трансретиноевая кислота и фенретинид; онапристон; антипрогестероны; понижающие регуляторы рецепторов эстрогенов (ERD); антиандрогены, такие как флутамид, нилутамид и бикалутамид; тестолактон; и фармацевтически приемлемые соли, кислоты или производные любого из указанных выше средств; а также сочетания двух или более указанных выше средств.
Другими терапевтическими средствами, которые могут применяться в комбинации с антителом, которое специфически связывается с GITR, по данному изобретению могут быть ингибиторы функции фактора роста, например, такие ингибиторы включают антитела фактора роста и антитела рецептора фактора роста (например, анти-erbB2 антитело трастузумаб [Herceptin], анти-EGFR антитело панитумумаб, анти-erbB1 антитело цетуксимаб [Erbitux, C225] и любые антитела факторов роста или рецепторов фактора роста, раскрытые Stern et al. Critical reviews in oncology/haematology, 2005, Vol. 54, pp11-29); антиангиогенные агенты, такие как ингибирующие эффекты сосудистого эндотелиального фактора роста, [например, антитело против фактора роста сосудистых эндотелиальных клеток бевацизумаб (Avastin)], антитела против рецепторов сосудистого эндотелиального фактора роста, такие как анти-KDR антитела и анти-flt1 антитела; антисмысловые терапии, например, направленные на вышеперечисленные мишени, такие как ISIS 2503, анти-ras антисмысловые средства или G3139 (Genasense), анти-bcl2 антисмысловые средства; подходами генной терапии, включая, например, подходы с заменой аберрантных генов, таких как аберрантный p53 или аберрантный BRCA1 или BRCA2, GDEPT (ген-направленная ферментная пролекарственная терапия) подходы с использованием цитозиндеаминазы, тимидинкиназы или фермента бактериальной нитроредуктазы, и подходы, направленные на повышение толерантности пациента к химиотерапии или лучевой терапии, такие как генная терапия мультилекарственной резистентности; иммунотерапевтические подходы, включая, например, лечение Алемтузумабом (campath-1H), моноклональным антителом, направленным на CD52, или лечение антителами, направленными на CD22, ex vivo и in vivo подходы по повышению иммуногенности опухолевых клеток пациента, трансфекцию цитокинами, такими как интерлейкин 2, интерлейкин 4 или гранулоцит-макрофаг колониестимулирующий фактор, подходы, направленные на снижение анергии T-клеток, такие как лечение моноклональными антителами, ингибирующими функцию CTLA-4, подходы с использованием трансфицированных иммунных клеток, таких как цитокин-трансфицированные дендритные клетки, подходы с использованием цитокин-трансфицированных опухолевых клеточных линий и подходы, использующие анти-идиотипические антитела, адаптивный перенос T-клеток с использованием T-клеток, подвергнутых неспецифической активации или нацеленных на конкретный антиген, представляющий интерес, ex vivo; ингибиторы деградации белка, такие как ингибитор протеасом, такой как Велкаде (бортезомид); биотерапевтические терапевтические подходы, например, использующие пептиды или белки (такие как антитела или растворимые конструкты внешних рецепторных доменов), которые секвестируют рецепторы лигандов, блокируют связывание лиганда с рецептором или ослабляют сигнализацию рецептора (например, вследствие повышенной деградации рецептора или сниженных уровней экспрессии).
Другим терапевтическим средством, которое может применяться в комбинации с антителом, которое специфически связывается с GITR, по данному изобретению может быть антитело, которое выбирают из группы: антитела к PD1, антитела к PD-L1, антитела к CTLA4, антитела к анти 4-1BB, антитела к OX40 или их комбинации.
Другим терапевтическим средством, которое может применяться в комбинации с антителом, которое специфически связывается с GITR, по данному изобретению может быть терапевтически активное противоопухолевое соединение, которое выбирают из группы активаторов врожденного или приобретенного иммунитета.
Подразумевается, что моноклональное антитело, которое специфически связывается с GITR, по данному изобретению могут использоваться в способах лечения, как описано выше, могут использоваться в лечении, как описано выше, и/или могут использоваться в производстве медикаментов для лечения, как описано выше.
Дозы и пути введения
Моноклональное антитело, которое специфически связывается с GITR, по данному изобретению будет вводиться в количестве, эффективном для лечения состояния, о котором идет речь, т.е. в дозах и в течение периодов времени, необходимых для достижения желаемого результата. Терапевтически эффективное количество может изменяться в зависимости от таких факторов, как конкретное состояние, по поводу которого проводится лечение, возраста, пола и веса пациента, а также является ли введение моноклонального антитела, которое специфически связывается с GITR, самостоятельным лечением или оно проводится в комбинации с одним или более дополнительных препаратов или методов лечения.
Схемы приема лекарственных средств можно регулировать, чтобы обеспечить оптимальный желаемый ответ. Например, может быть введен один болюс, несколько разделенных доз могут быть введены в течение некоторого времени, или доза могут быть пропорционально уменьшена или увеличена в зависимости от остроты терапевтической ситуации. Особенно полезным является изготовление парентеральных композиций в стандартной лекарственной форме для простоты введения и однородности дозирования. Стандартная лекарственная форма при использовании в данном документе, относится к физически дискретным единицам, пригодным в качестве единичных доз для пациентов/субъектов, подлежащих лечению; каждая единица содержит заданное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта в сочетании с требуемым фармацевтическим носителем. Спецификация для стандартных лекарственных форм по настоящему изобретению, как правило, диктуется и непосредственно зависит от (a) уникальных характеристик химиотерапевтического агента и конкретного терапевтического или профилактического эффекта, которые должны быть достигнуты, и (b) ограничений, присущих в технике компаундирования такого активного соединения для лечения чувствительности у субъектов.
Таким образом, квалифицированным специалистам понятно, исходя из раскрытия, представленного в данном документе, что дозы и режим дозирования корректируются в соответствии со способами, хорошо известными в терапевтической области. Это означает, что может быть легко установлена максимально переносимая доза и может быть также определено эффективное количество, обеспечивающее обнаруживаемый терапевтический эффект для пациента, так же как и требования к времени введения каждого агента для достижения видимого терапевтического эффекта для пациента. Таким образом, хотя некоторые дозы и схемы режима введения приведены в качестве примеров в данном документе, эти примеры никоим образом не ограничивают дозы и режимы введения, которые могут понадобиться для пациента в практике применения настоящего изобретения.
Следует отметить, что значения дозировки могут изменяться, в зависимости от типа и тяжести состояния, которое следует облегчить, и может включать одну или более доз. Кроме того, необходимо понимать, что для любого конкретного пациента, конкретные схемы введения должны быть скорректированы через некоторое время согласно индивидуальной потребности и на усмотрение медицинского работника, который осуществляет введение или контролирует введение композиций, и что диапазоны концентрации, приведенные в данном описании, приведены только в качестве примера и не предназначены для ограничения объема или практики заявленных композиций. Кроме того, режим дозирования с композициями по данному изобретению может быть основан на различных факторах, включая тип заболевания, возраст, вес, пол, состояния здоровья пациента, тяжесть состояния, путь введения и конкретное используемое моноклональное антитело, которое специфически связывается с GITR. Таким образом, режим дозирования может широко варьироваться, но может определяться регулярно с помощью стандартных методов. Например, дозы могут быть скорректированы на основе фармакокинетических и фармакодинамических параметров, которые могут включать клинические эффекты, такие как токсические эффекты или лабораторные значения. Таким образом, настоящее изобретение охватывает индивидуальное повышение дозы, которое определяется квалифицированным специалистом. Определение необходимой дозы и режимы хорошо известны в соответствующей области техники и будут понятны специалисту в данной области после ознакомления с идеями, раскрытыми в данном документе.
Примеры подходящих способов введения предусмотрены выше.
Предполагается, что подходящая доза моноклонального антитела, которое специфически связывается с GITR, по данному изобретению будет в диапазоне от 0,1-200 мг/кг, предпочтительно 0,1-100 мг/кг, в том числе около 0,5-50 мг/кг, например, около 1-20 мг/кг. Моноклональное антитело, которое специфически связывается с GITR, может быть введено, например, в дозе по меньшей мере 0,25 мг/кг, например, по меньшей мере 0,5 мг/кг, в том числе не менее 1 мг/кг, например, по меньшей мере 1,5 мг/кг, например, также как не менее 2 мг/кг, например, по меньшей мере 3 мг/кг, в том числе по меньшей мере 4 мг/кг, например, по меньшей мере 5 мг/кг; и например вплоть до максимально 50 мг/кг, в том числе вплоть до максимально 30 мг/кг, например, вплоть до максимально 20 мг/кг, в том числе вплоть до максимально 15 мг/кг. Введение будет повторяться обычно в подходящие промежутки времени, например, раз в неделю, раз в две недели, один раз каждые три недели или один раз каждые четыре недели, и так долго, как будет сочтено целесообразным ответственным врачом, который может в некоторых случаях увеличить или уменьшить дозу при необходимости.
Диагностическое использование и композиции
Моноклональное антитело, которое специфически связывается с GITR, по настоящему изобретению также используются в диагностических целях (например, in vitro, ex vivo). Например, данное моноклональное антитело, которое специфически связывается с GITR, по данному изобретению может использоваться для обнаружения или измерения уровня GITR в образцах, полученных от пациента, например, образец ткани или образец жидкости тела, такой как воспалительный экссудат, кровь, сыворотка крови, жидкость кишечника, слюна или моча. Подходящие методы обнаружения и измерения включают иммунологические методы, такие как проточная цитометрия, твердофазный иммуноферментный анализ (ИФА), хемилюминесцентный анализ, радиоиммуноанализ и иммуногистология. Изобретение далее включает наборы, например, диагностические наборы, содержащие моноклональное антитело, которое специфически связывается с GITR, по настоящему изобретению, описанное в данном документе.
Примеры
Для наилучшего понимания изобретения приводятся следующие примеры. Эти примеры приведены только в иллюстративных целях и не должны толковаться как ограничивающие сферу применения изобретения в любой форме.
Все публикации, патенты и патентные заявки, указанные в этой спецификации включены в данный документ путем отсылки. Хотя вышеупомянутое изобретение было довольно подробно описано путем иллюстрации и примера в целях исключения двусмысленного толкования, специалистам в данной области на основе идей, раскрытых в данном изобретении, будет вполне понятно, что могут быть внесены определенные изменения и модификации без отклонения от сущности и объема прилагаемых вариантов осуществления изобретения.
Материалы и общие методы
Общая информация, касающаяся нуклеотидных последовательностей легких и тяжелых цепей человеческого иммуноглобулина, представлена у: Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991. Аминокислоты цепей антител пронумерованы согласно нумерации EU (Edelman G.M. и др., Proc. Natl. Acad. Sci. USA 63, 1969, ее. 78- 85; Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991).
Методы рекомбинантной ДНК
Для манипуляций с ДНК использовали стандартные методы, описанные у Sambrook J. и др., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Реагенты для молекулярной биологии использовали согласно инструкциям производителей.
Синтез генов
Требуемые сегменты генов получали из олигонуклеотидов, созданных путем химического синтеза. Генные сегменты длиной от 300 до 4000 т.п.н., которые фланкированы уникальными сайтами рестрикции, собирали путем отжига и лигирования олигонуклеотидов, включая ПЦР-амплификацию и последующее клонирование через указанные сайты рестрикции. Последовательности ДНК субклонированных генных фрагментов подтверждали путем секвенирования ДНК.
Определение последовательностей ДНК
Последовательности ДНК определяли путем секвенирования по Сенгеру.
Анализ последовательностей ДНК и белков и обработка данных о последовательностях
Применяли пакет программ фирмы Infomax's Vector NT1 Advance suite, версия 8.0 для создания, картирования, анализа, аннотирования и иллюстрации последовательностей.
Экспрессионные векторы
Для экспрессии описанных антител и антигенов применяли варианты экспрессионных плазмид, предназначенных для экспрессии в клетках прокариот (E.coli) кратковременной экспрессии в клетках эукариот (например, в клетках CHO). Помимо кассеты экспрессии антитела векторы содержали: сайт инициации репликации, обеспечивающий репликацию указанной плазмиды в Е.coli, гены, придающие устойчивость в Е.coli к различным антибиотикам (например, к ампициллину и канамицину).
Слияние генов, содержащих описанные цепи антитела, как указано ниже, осуществляли с помощью ПЦР и/или синтеза и сборки генов с использованием известных методов и процедур рекомбинации путем соединения соответствующих сегментов нуклеиновых кислот, например, с использованием уникальных сайтов рестрикции в соответствующих векторах. Субклонированные нуклеотидные последовательности подтверждали секвенированием ДНК. Для кратковременных трансфекций создавали большие количества плазмид посредством получения плазмид из трансформированных культур E. coli.
Пример 1
Продукция рекомбинантных антигенов и антител в суспензионной культуре клеток млекопитающих
Для приготовления рекомбинантных антигенов на основе последовательности экстраклеточной части GITR человека и ортологов был создан ряд конструкций, содержащих экстраклеточный домен антигена human GITR (https://www.uniprot.org/uniprot/Q9Y5U5), mcyGITR Macaca mulatta (https://www.uniprot.org/uniprot/Q1PBC4) и musGITR Mus musculus (https://www.uniprot.org/uniprot/O35714). Последовательности генов были синтезированы с помощью ПЦР с матрицы, кодирующий полноразмерный антигена GITR человека из плазмидного вектора RDC0359 (https://www.rndsystems.com/products/human-gitr-tnfrsf18-np_004186-versaclone-cdna_rdc0359) и собраны из синтетических олигонуклеотидов для генов ортологов GITR. Последовательность гена GITR клонировали в плазмиду для наработки белка в клетках млекопитающих с тагом Fc IgG1 Lama glama (фиг. 1) по сайтам рестрикции SalI/NotI. Кроме того, последовательность антигена отделена от Fc протеаз-лабильным сайтом TEV протеазы, который также использовался для отщепления Fc фрагмента обработкой с TEV протеазой и получении безтаговой версии антигена. Последовательность также клонировали в плазмиду, содержащую FLAG-EPEA-tag (C-tag GE Healthcare) на С-конце белка (фиг. 2) по сайтам рестрикции SalI/NotI. Кроме того, провели аналогичное клонирование рекомбинантного GITR-лиганда человека (GITRL) https://www.uniprot.org/uniprot/Q9UNG2) в плазмиду, содержащую гомотримеризующий домен foldon [1] и EPEA-tag на C-terminus GITRL (фиг. 3). Плазмиды нарабатывали в необходимых количествах в клетках E.Coli и очищали при помощи набора Maxiprep Qiagen.
Антитела и антигены продуцировали в клетках постоянной клеточной линии, полученной из клеток яичника китайского хомячка (линия CHO-K1), согласно опубликованным протоколам [Biotechnol Bioeng. 2005 Sep 20; 91(6):670-677, Liao Metal., 2004; Biotechnol Lett. 2006 Jun;28(11):843-848; Biotechnol Bioeng. 2003 Nov 5;84(3):332-342]. Использовали клетки, конститутивно экспрессирующие ген белка EBNA1 (Epstein-Barrvirus nuclear antigen 1). Суспензионное культивирование осуществляли в колбах на орбитальном шейкере, с использованием бессывороточных сред производства компании Life Technologies Corporation и согласно инструкциям производителя. Для транзиентной экспрессии клетки в концентрации 2*106/мл трансфецировали с помощью линейного полиэтиленимина (ПЭИ «MAX», компания «Polysciences»). Соотношение ДНК/ПЭИ составляло 1:3/1:10. Через 5-7 дней после трансфекции культуральную среду центрифугировали при 2000 g в течение 20 минут и фильтровали через фильтр с размером пор 0,22 мкм. Целевые белки выделяли из культуральной жидкости с помощью аффинной хроматографии.
Рекомбинантные белки GITR и GITRL, содержащий EPEA-tag (glutamic acid-proline-glutamic acid-alanine) на С-конце белка, выделяли и очищали из культуральной жидкости, используя сорбент CaptureSelect C-tag Affinity Matrix. Культуральную жидкость пропускали через хроматографическую колонку, предварительно заполненную 5 мл C-tag сорбента, затем промывали колонку 25 мл ФСБ, чтобы вымыть неспецифически связывающиеся компоненты. Связанный антиген элюировали в мягких условиях, используя 20 mM Tris, 2M MgCl2 pH7,0-7,4. Далее белок переводили в ФСБ (pH 7,4) с помощью диализа, используя полупроницаемую диализную мембрану, фильтровали (0,22 мкм), переносили в пробирки и хранили при -70°С.
Рекомбинантный антиген GITR-TEV-Fc из культуральной жидкости выделяли и очищали, используя колонку для аффинной хроматографии протеин А. Осветленную культуральную жидкость пропускали через колонку HiTrap rProtein A Sepharose FF объемом 5 мл (GE Healthcare), уравновешенную фосфатно-солевым буфером (ФСБ, pH 7,4). Затем колонку промывали 5 колоночными объемами ФСБ, чтобы вымыть неспецифически связывающие компоненты. Связанный антиген элюировали, используя 0,1 М глициновый буфер рН 3. Главный протеиновый пик элюции собирали и доводили его рН до нейтральности с помощью 1 М буфера Tris (рН 8). Все стадии проводили при скорости потока 110 см/ч. Далее белок переводили в ФСБ (pH 7,4) с помощью диализа, используя технологию SnakeSkin Dialysis Tubing, фильтровали (0,22 мкм), переносили в пробирки и хранили при -70°С.
Все рекомбинантные антитела IgG очищали на колонке Hi Trap rProteinA FF объемом 1 мл (GE Healthcare) согласно методике, описанной выше для антигенов. Чистоту полученного раствора белка оценивали с помощью SDS-гель-электрофореза (пример фиг. 4 и 5).
Пример 2
Создание наивной Fab-библиотеки человеческих антител MeganLibTM
Суммарную РНК В-лимфоцитов из индивидуальных образцов крови более тысячи человеческих доноров выделяли с помощью набора RNeasy Mini Kit согласно предлагаемому протоколу (от QIAGEN). Концентрацию РНК определяли с помощью набора Nanovue (от GE Healthcare) и проверяли качество выделенной РНК с помощью электрофореза в 1,5%-ном агарозном геле.
Реакцию обратной транскрипции проводили с использованием набора MMLV RT kit (от Evrogen) согласно рекомендуемому протоколу с использованием обратной транскриптазы MMuLV и рандом-гексамерных олигонуклеотидов в качестве затравки.
Продукты обратной транскрипции использовали в качестве матрицы в двухступенчатой полимеразной цепной реакции для получения генов вариабельных доменов, фланкированных сайтами рестрикции, с использованием набора олигонуклеотидов по протоколам авторов [J Biol Chem. 1999 Jun 25; 274(26): 18218-30].
Полученный ДНК препарат VL-CK-VH (фиг. 6) обрабатывали эндонуклеазами рестрикции NheI/Eco91I и лигировали в оригинальную фагмиду pH5 (фиг. 7). Продукты лигирования трансформировали в электрокомпетентные клетки штамма SS320, приготовленные согласно протоколам [Methods Enzymol. 2000;328: 333-63.]. Репертуар комбинаторной фаговой Fab-дисплейной библиотеки MeganLibTM составлял 1011 трансформантов. Препараты фага Fab-библиотек готовили согласно процедуре, описанной ранее [J Mol Biol. 1991 Dec 5;222(3): 581-97].
Пример 3
Создание иммуных Fab-библиотек гибридных лама-человеческих антител (CHARM).
Суммарную РНК В-лимфоцитов из индивидуальных образцов крови иммунизированных рекомбинантными GITR антигенами Lama Glama, которые продемонстрировали максимальный специфический титр сыворотки, выделяли с помощью набора RNeasy Mini Kit согласно предлагаемому протоколу (от QIAGEN). Концентрацию РНК определяли с помощью набора Nanovue (от GE Healthcare) и проверяли качество выделенной РНК с помощью электрофореза в 1,5%-ном агарозном геле.
Реакцию обратной транскрипции проводили с использованием набора MMLV RT kit (от Evrogen) согласно рекомендуемому протоколу с использованием обратной транскриптазы MMuLV и рандом-гексамерных олигонуклеотидов в качестве затравки.
Продукты обратной транскрипции использовали в качестве матрицы в двухступенчатой полимеразной цепной реакции для получения генов вариабельных доменов, фланкированных сайтами рестрикции, с использованием набора олигонуклеотидов по протоколам авторов [J Biol Chem. 1999 Jun 25; 274(26): 18218-30].
Полученный ДНК препарат VL-CK-VH (фиг. 6) обрабатывали эндонуклеазами рестрикции NheI/Eco91I и лигировали в оригинальную фагмиду pH5 (фиг. 7). Продукты лигирования трансформировали в электрокомпетентные клетки штамма SS320, приготовленные согласно протоколам [Methods Enzymol. 2000;328: 333-63.]. Репертуар комбинаторной фаговой Fab-дисплейной библиотеки MeganLibTM составлял 1011 трансформантов. Препараты фага Fab-библиотек готовили согласно процедуре, описанной ранее [J Mol Biol. 1991 Dec 5;222(3): 581-97]. Суммарная таблица препаратов иммунных библиотек приведена в таблице 1.
Таблица 1.
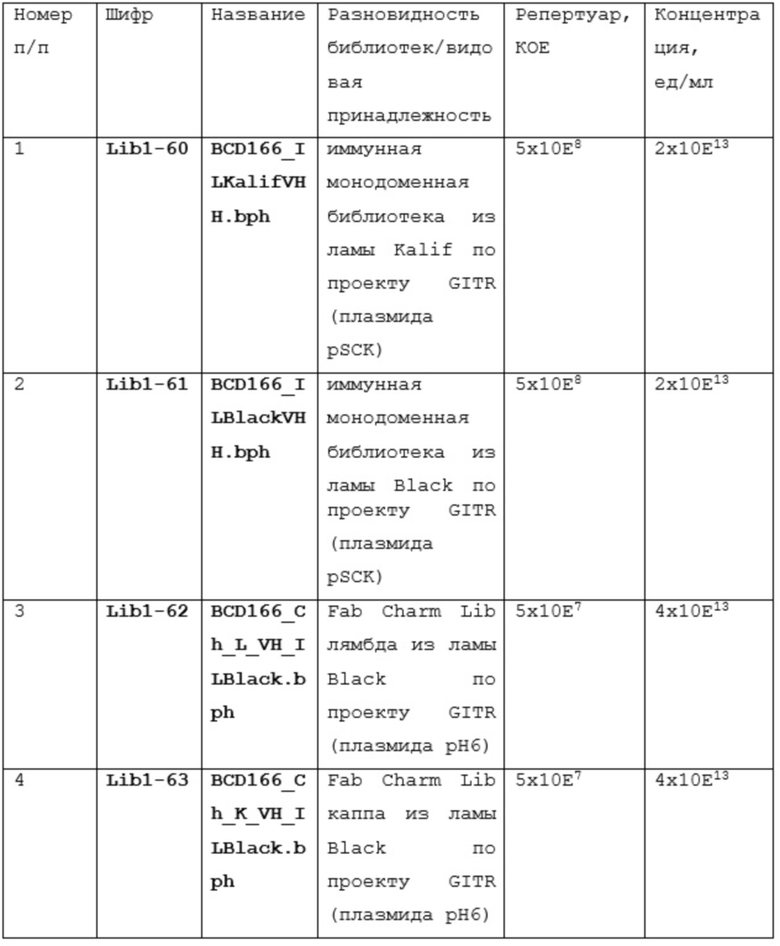
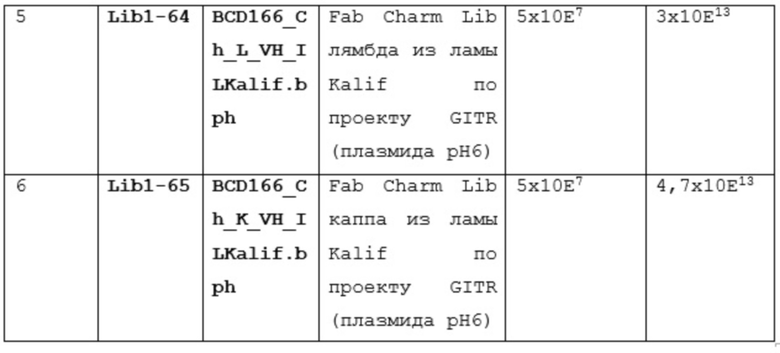
Пример 4
Селекция Fab-библиотек фаговых антител
Специфичные фаговые Fab-антитела человека против GITR получали из комбинаторных фаговой Fab-дисплейной библиотеки MeganLibTM и иммунных CHARM библиотек, описанных в примерах 2 и 3. Селекцию проводили на рекомбинантных антигенах GITR человека методом фагового дисплея [Nat Biotechnol. 1996 Mar;14(3):309-14; J Mol Biol.1991 Dec 5;222(3): 581-97].
Селекция иммунных фаговых библиотек проводили на EIA/RIA Tube High Binding 5 ml immunotube. Антиген (0.5 мл 2 мкг\мл) сорбировали в карбонатном буфере(0.1 M NaHCO3, pH 9.5) в течение ночи при+ 4С. Затем отмыли пробирки PBST (10 mM фосфатно-солевой буфер pH7,3-7,5, 137 mM NaCl и 2,7 mM KCl, 0,1% Polysorbate 20) и добавили полный объем (5 мл) 0,5% сухого молока в PBST для забивки. Забивка чередуется от раунда к раунду, например, если на 1 раунде это было 0,5% молоко на PBST, то на 2-м - 1% BSA на PBST. Инкубировали в течение часа при комнатной температуре на качалке. Отмыли пробирки PBST и добавили в них по 2 мл забивочного буфера и фаговые библиотеки до концентрации примерно 2*1012 фаговых частиц на мл. Инкубировали с фагами в течение 1-2 часов на комнатной температуре. На последующих раундах использовали 4 мл фагового супернатанта с предыдущего раунда, предварительно осветленного на центрифуге при 17000g 10-15 минут. Отмыли пробирки 20 раз PBST. Элюировали фаги с поверхности пробирок 0,5 мл 0,1М глицинового буфера рН 2,2: добавили в пробирки глициновый буфер и несильно помешивали 15 минут при комнатной температуре. Затем перенесли раствор фагов в чистые не сорбирующие пробирки со 100 мкл 1М ТрисHCl рН8,0 для нейтрализации. Держали пробирки во льду до заражения клеток.
Бактериофаг М13 после селекции культивируют, используя штамм Е.соli TG1 в качестве хозяина. Амплификацию проводят посредством заражения фагами культуры штамма-хозяина и последующим ростом в течение 12-15 часов.
0,5-1 мл фагов после селекции смешивали с культурой клеток (OП6oo=0,3-0,4) и инкубировали 1.5 часа на 37°С. Затем клетки осаждали на 3000-4000 об\мин в течение 10-15 минут, ресуспендировали в 1 мл среды и растирали на чашки Петри с антибиотиком для селекции (ампициллин). Наращивали колонии в термостате на 30°С. Спустя 12-15 часов рассчитали количество колоний и смыли клетки с чашек 5-10 мл LB среды. 100 мкл клеточной суспензии добавили к 20 мл среды c антибиотиком (ампициллин). Нарастили до OП6oo=0,35-0,5 на 37. Добавили фаг-хэлпер К07 из расчета 1 мкл на 10 мл культуры (1010 частиц на мл) и инкубировали на 37°С 1,5 часа без качания. Затем к клеточной культуре добавили равный объем среды с однократным ампициллином (100 мкг\мл) и двукратным канамицином (40 мкг/мл) и двукратным IPTG (0,2 мМ). Переставили на качалку и нарабатывали фаг 4-5 часов при 30°С. Культуру клеток центрифугировали 25-30 минут 17000g и отобрали супернатант в пробирки. Затем полученный супернатант использовали в следующем раунде селекции или выделяли фаг осаждением для последующего хранения.
Выделяли фаги следующим способом: добавляли к супернатанту 1/6 объема раствора, содержащего 20% полиэтиленгликоля и 2,5M хлорида натрия, интенсивно перемешали. Инкубировали раствор во льду не менее 3 часов. Затем раствор центрифугировали 10 минут на 8000g, образовавшийся осадок, содержащий фаги растворили в 1 мл буфера TBS (Трис - Боратный Буфер).
Бактериофаги после двух-трех раундов выделяли и проводили анализ специфического связывания поликлонального фага с GITR человека и неспецифических антигенов.
Пример 5
Анализ поликлонального фага, полученных после 2 и 3 раундов селекции.
После проведения 3 раундов селекции 2 наивных библиотек MeganLib и 6 иммунных библиотек на целевом антигене проверили специфическое и неспецифическое связывания поликлональных фагов полученных после 2 и 3 раундов с помощью ИФА.
На пластик планшета для ИФА сорбировали в течение ночи в карбонатном буфере (0.1 M NaHCO3, pH 9.5) целевой антиген GITR для анализа наличия специфически связывающихся фагов (2 мкг/мл) или ГКСФ, hIL6R-Fc, интерферон альфа 2b, Rituximab (2 мкг/мл) для анализа неспецифически связывающихся фагов. Затем отмыли планшеты PBST (10 mM фосфатно-солевой буфер pH7,3-7,5, 137 mM NaCl и 2,7 mM KCl, 0,1% Polysorbate 20) и добавили по 300 мкл/лунку 0,5% сухого молока в PBST для забивки. Инкубировали в течение часа при комнатной температуре на качалке. Отмыли планшеты PBST и добавили в них по 50 мкл растворов фагов, полученных после 2 и 3 раундов селекции, разведенные забивочным буфером от 2 до 256 раз. Инкубировали в течение часа при комнатной температуре на качалке. Отмыли планшеты PBST и нанесли анти-M13 антитело, конъюгированное пероксидазой хрена, в забивочном буфере. После инкубации в течение часа отмыли планшеты и добавили по 50 мкл субстрата реакции (Н2О2-0,02% и ТМБ) в ацетатном буфере рН-5.0. Инкубировали при комнатной температуре в темноте до появления легкого фона в отрицательных контролях (но не более 20-ти мин). Остановили реакцию добавлением 25 мкл на лунку 10% Н2SO4, и измерили ОД в лунках на 450 нм.
По результатам анализа было выявлено специфическое (5Х превышение над фоновым сигналом) связывание 1 наивной Fab человека, и 6 иммунных Fab CHARM библиотек после 3-его раунда селекции на целевом антигене и отсутствие значимого (менее 2Х превышение) неспецифического связывания.
Гены вариабельных доменов антител из данных библиотек были реклонированы в экспрессионный вектор pLL-Fab (Фиг. 8) для наработки секретирумой растворимой формы Fab в клетках Е.coli.
Пример 6
Скрининг Fab, специфически связывающих GITR человека
ИФА (ELISA) использовали для поиска Fab, секретирующихся в среду из моноклонов продуцентов E.coli полученных из из примера 5, которые специфически связывают GITR человека. В качестве позитивного контроля использовали Fab с опубликованной последовательностью Fab-gitr3215 (заявка на патент). Для анализа специфического связывания лунки планшетов ELISA (medium binding от Greiner bio one) покрывали 50 мкл GITR (0,2 мкг/мл в 1× карбонатном буфере), герметично закрывали и инкубировали в течение ночи при 4°С. Все последующие этапы проводили по стандартному протоколу ИФА с использованием высокопроизводительной автоматизированной платформы на базе роботизированных систем Genetix Qpix2xt (от Molecular Device) и Tecan Freedom EVO 200 (от Tecan). Для блокирования неспецифического связывания добавляли блокирующий буфер BB (200 мкл 0,5% нежирного молока в ФСБ). Планшеты инкубировали на шейкере в течение часа при комнатной температуре. После отмывок ФСБ-Твином добавляли по 50 мкл на лунку тестируемого клеточного супернатанта, содержащего исследуемый Fab, смешанного с равным объемом BB. Планшеты снова инкубировали, встряхивая, один час при комнатной температуре, после чего каждую лунку планшетов три раза промывали буфером ФСБ-Твин. После промывания добавляли (50 мкл/лунку) анти-человеческого Fab HRP-конъюгированного вторичного антитела (от Pierce-ThermoScientific) в соотношении 1:5000 в ФСБ-Твин. Планшеты встряхивали на ротационном шейкере (50 мин, комнатная температура) и промывали три раза буфером ФСБ-Твин, как описано выше. Колориметрический сигнал развивали добавлением ТМВ (50 мкл/лунку) до насыщения (в среднем 3-5 мин), затем дальнейшее развитие останавливали добавлением стопового раствора (30 мкл/лунку, 10% серная кислота). Цветовой сигнал измеряли при длине волны 450 нм, используя подходящий планшет-ридер Tecan-Sunrise (от Tecan). Степень связывания антитела была пропорциональна продуцированию цветового сигнала. Клоны, у которых цветовой сигнал 5Х превышал фоновый сигнал и был соизмерим с сигналом контрольного антитела Fab-gitr3215, проверяли в ИФА на неспецифическое связывание.
Пример 7
Анализ неспецифического связывания отобранных Fab с другими антигенами
ИФА (ELISA) использовали также анализа неспецифического связывания исследуемых Fab-фрагментов с другими антигенами. Исследование проводили, как описано выше, но в качестве антигенов для иммобилизации использовали IL6R-Fc, INFα2b, PCSK9-VG-FE, PD-1-Fc (2,5 мкг/мл в 1× карбонатном буфере). В качестве контроля специфического связывания использовали GITR-TEV-Fc (0,2 мкг/мл в 1× карбонатном буфере). Все последующие этапы проводили по стандартному протоколу ИФА с использованием высокопроизводительной автоматизированной платформы на базе роботизированных систем Genetix Qpix2xt (от Molecular Device) и Tecan Freedom EVO 200 (от Tecan). Клоны, у которых цветовой сигнал неспецифического связывания не превышал фонового сигнала и составлял 5Х меньшие значения от специфического связывания считались позитивными и их гены были секвенированы для определения последовательности генов вариабельных доменов антител и определения уникальности. В результате секвенирования было отобрано 30 уникальных последовательностей для конверсии в полноразмерный формат IgG1 антител.
Пример 8
Продукция рекомбинантных антител в суспензионной культуре клеток млекопитающих
Клонирование генов вариабельных доменов антител из примера 7 проводили по стандартной методике. Для этого, наработали ПЦР-продукты, содержащие гены вариабельных доменов тяжелой и легкой цепей антител. Вариабельный домен тяжелой цепи клонировали в вектор pEE-Hc IgG1 по сайтам рестрикции SalI/NheI, схематичная карта которого приведена на фиг.9). Вариабельный домен легкой цепи лигировали в вектор pEE-CK по сайтам рестрикции SalI/BsiWI схематичная карта которого приведена на (фиг.10).
Антитела продуцировали в клетках постоянной клеточной линии, полученной из клеток яичника китайского хомячка (линия CHO-K1), согласно опубликованным протоколам [Biotechnol Bioeng. 2005 Sep 20; 91(6):670-677, Liao Metal., 2004; Biotechnol Lett. 2006 Jun;28(11):843-848; Biotechnol Bioeng. 2003 Nov 5;84(3):332-342]. Использовали клетки, конститутивно экспрессирующие ген белка EBNA1 (Epstein-Barrvirus nuclear antigen 1).
Для наработки антител в системе транзиентной экспрессии использовали клетки линии CHO-K1-S. Клетки культивировали в колбах с отбойниками (125, 250, 500, 1000 и 3000 мл) в смеси сред CHO-S-SFM II и FreeStyle CHO (1:1) с 4 мМ глутамином, 0,05 мг/мл гентамицином и 10 мкг/мл ципрофлоксацином в орбитальных шейкерах-инкубаторах при +37°С в присутствии 5%СО2, при оборотах 110 или 150 rpm в зависимости от размера колбы. Клетки пересевали 3 раза в неделю, плотность засева 0,2*106 клеток /мл.
Для трансфекции клетки засевали накануне с плотностью 0,8*106 клеток /мл, трансфекцию проводили через сутки, на клеточной культуре с плотностью 2*106 клеток /мл. Для приготовления трансфекционной смеси использовали среду RPMI-1640 с 2 мМ глутамина и 0,05 мг/мл гентамицина. Отдельно в среде разводили вектора из расчета 0,75 мкг/мл, отдельно - трансфецирующий агент полиэтиленимин (ПЭИ), из расчета ПЭИ:ДНК 7:1 по массе. Разведения векторов и ПЭИ смешивали и инкубировали в течение 10 минут при комнатной температуре, после чего вносили трансфекционную смесь к клеткам и культивировали клетки при стандартных условиях.
На следующие сутки после трансфекции к клеткам вносили смесь сред CHO-S-SFM II и FreeStyle CHO (1:1) с 0,05 мг/мл гентамицином и 10 мкг/мл ципрофлоксацином, 1 мМ вальпроата натрия и 10% фидинга F12.7 и культивировали клетки в орбитальных шейкерах-инкубаторах при +34°С в присутствии 5%СО2, при оборотах 110 или 150 rpm в зависимости от размера колбы. На 3-4 день после трансфекции к клеткам вносили 10% фидинга F12.7.
На 7 сутки после трансфекции отбирали образцы клеточной жидкости для измерения концентрации нарабатываемого белка на чипах Protein-A Biosense на приборе OctetRed 96 согласно инструкции компании ForteBio (таблица 2). Рекомбинантные антитела из культуральной жидкости выделили и очистили, используя колонку для аффинной хроматографии протеин А. Осветленную культуральную жидкость пропускали через колонку Hi Trap rProteinA FF объемом 1 мл (GE Healthcare), которая была уравновешена фосфатно-солевым буфером (ФСБ, pH 7,4). Затем колонку промыли 5 колоночными объемами ФСБ, чтобы вымыть неспецифически связывающие компоненты. Связанный антитело элюировали, используя 0,1 М глициновый буфер рН 3. Главный протеиновый пик элюции собрали и довели его рН до нейтральности с помощью 1 М буфера Tris (рН 8). Все стадии проводили при скорости потока 110 см/ч. Далее белок перевели в ФСБ (pH 7,4) с помощью диализа, используя технологию SnakeSkin Dialysis Tubing, профильтровали (0,22 мкм), перенесли в пробирки и хранили при -70°С.
Чистоту полученного раствора белка оценивали с помощью SDS-гель-электрофореза фиг.11 и фиг. 12. Все антитела удовлетворяют чистоте необходимой для исследования физико-химических свойств и активности для клеточных тестов.
Таблица 2.
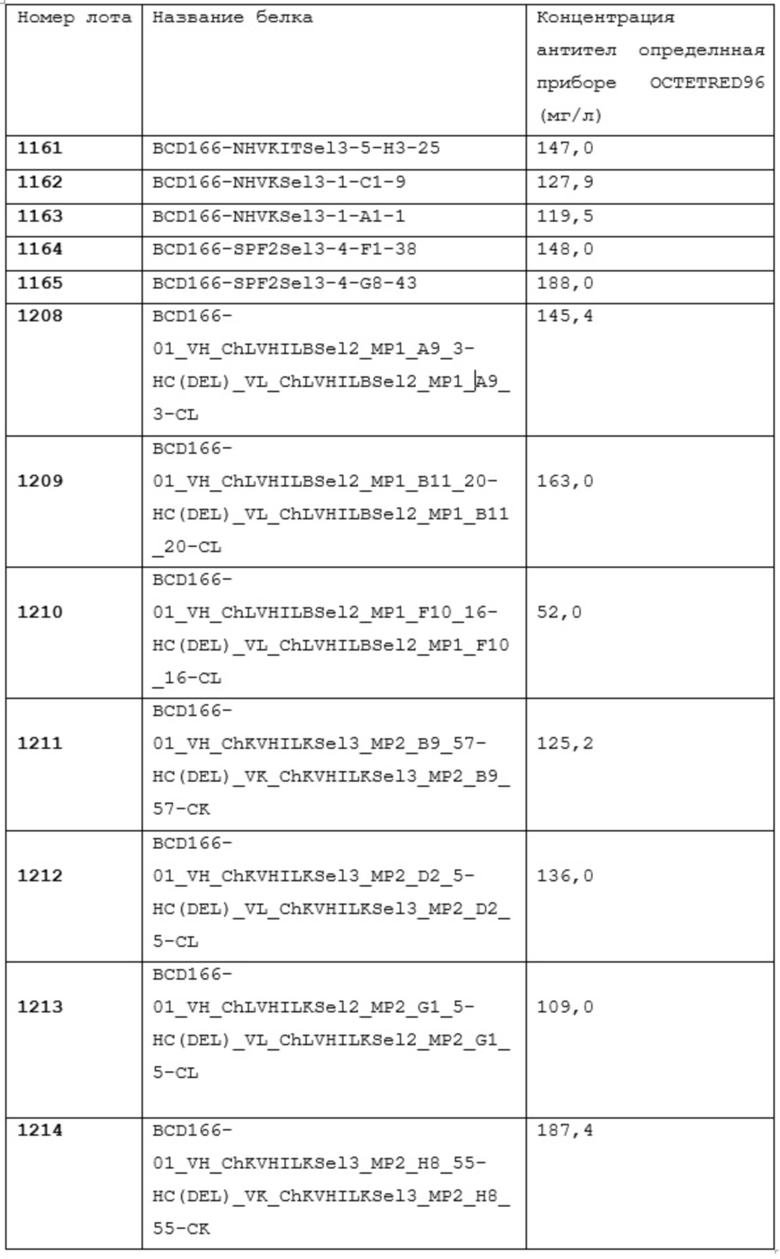
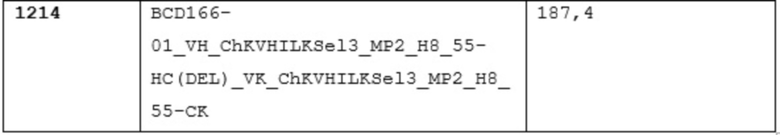
Пример 9
Кинетический анализ взаимодействия anti-GITR IgG1 антител с GITR человека
Константы аффинности связывания анти-GITR антител с GITR человека, макаки резус и яванской макаки получили с помощью прибора OctetRed 96 согласно инструкциям компании ForteBio. Антиген 25 мкг/мл был неспецифически иммобилизован на поверхность аминореактивных сенсоров второго поколения (от AR2G) (ForteBio по стандартному протоколу согласно инструкции производителя для подготовки и иммобилизации AR2G сенсоров). анти-GITR антител Антитела добавлялись с концентрацией Анализ был произведен при 30°С c использованием PBS, содержащим 0,1% Твин-20 и 0,1% БСА в качестве рабочего буфера.
Полученные сенсограммы с вычетом базового сигнала анализировали с помощью программы Octet Data Analysis (версия 8.0) согласно стандартной процедуре с использованием модели взаимодействия 1:1. Полученные константы аффинности приведены в таблице 3. Все исследованные антитела является высокоаффинными и специфичными к GITR человека.
Таблица 3. Константы аффинности анти-GITR антител при взаимодействии с GITR human.
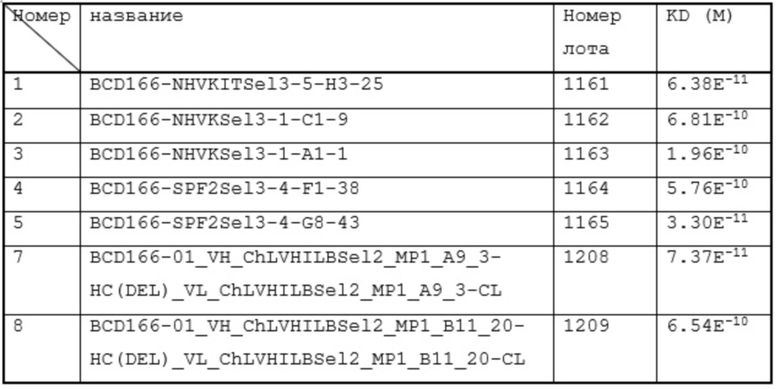
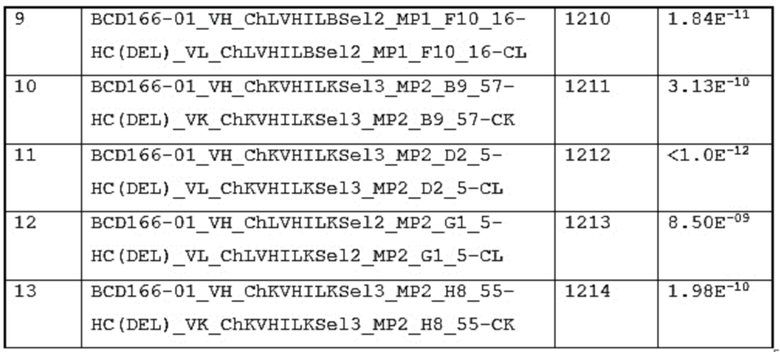
Пример 10
Тест на определение анти-GITR специфической агонистической активности.
Для теста использовали клеточную линию HEK293-GITR-NFkB-Luc, созданную на основе клеточной линии Hek-293, стабильно экспрессирующую на поверхности GITR и содержащую ген, кодирующий люциферазы светляка, под контролем NFkB-промотора.
Тест проводили в культуральном 96-луночном планшете. Суспензия в каждой лунке содержала клетки HEK293-GITR-NFkB-Luc и исследуемое антитело в указанной на графике концентрации. Все компоненты суспензии готовили в питательной среде для культивирования клеток. После добавления всех компонентов планшет инкубировали при 37°С, 5% CO2 и далее, с помощью набора для оценки люминесценции и планшетного ридера, измеряли интенсивность люциферазы в лунках.
Результаты проверки (фиг.13) специфической активности анти-GITR моноклональных антител обнаружил что антитела лотов 1161, 1210 и 1213 являются функционально активными и проявляют значительную агонистическую активность с уровнем верхнего плато соизмеримым как с контрольным антителом анти-GITR3215 антителом так и с GITRL. Эти антитела были переименованы согласно таблице 4.
Таблица 4.

Пример 11
Иммуноферментный анализ взаимодействий анти-GITR антител с GITR из разных организмов
ИФА (ELISA) использовали для измерения сравнительной аффинности антител к GITR-Fc разных организмов. Для анализа связывания лунки планшетов ELISA (medium binding от Greiner bio one) покрывали 50 мкл GITR-Fc человека, циномолгуса, мыши, (0,5 мкг/мл в 1× карбонатном буфере), герметично закрывали и инкубировали в течение ночи при 4°С. Все последующие этапы проводили по стандартному ИФА протоколу, описанному в примере 6. Анти-GITR антитело BCD166-01-01 специфически связывалось с GITR человека и обезьяны циномолгус и не связывалось с мышиным GITR (фиг. 14). Антитело BCD166-01-011 специфически связывалось с GITR человека, но не показало связывания с GITR обезьяны циномолгус и мыши (фиг. 15). Антитело BCD166-01-014 специфически связывалось с GITR человека, гораздо слабее было связывание с GITR обезьяны циномолгус и полностью отсутствовало для GITR мыши (фиг. 16). Таким образом, кандидат BCD166-01-011 был исключен из дальнейшей работы из-за отсутствия видимой специфичности к GITR макаки. Кандидаты BCD166-01-01 и BCD166-01-014 были финализированы для последующей разработки.
Пример 12
Кинетический анализ взаимодействия anti-GITR IgG1 антител с macaca mulatta GITR и macaca Cynomolgus GITR
Константы аффинности связывания анти-GITR антител с GITR макаки резус и яванской макаки получили с помощью прибора OctetRed 96 согласно инструкциям компании ForteBio. Антиген 25 мкг/мл был неспецифически иммобилизован на поверхность аминореактивных сенсоров второго поколения (от AR2G) (ForteBio по стандартному протоколу согласно инструкции производителя для подготовки и иммобилизации AR2G сенсоров). анти-GITR антител Антитела добавлялись с концентрацией Анализ был произведен при 30°С c использованием PBS, содержащим 0,1% Твин-20 и 0,1% БСА в качестве рабочего буфера.
Полученные сенсограммы с вычетом базового сигнала анализировали с помощью программы Octet Data Analysis (версия 8.0) согласно стандартной процедуре с использованием модели взаимодействия 1:1. Полученные константы аффинности приведены в таблице 5 и 6. Все исследованные антитела является высокоаффинными и специфичными к GITR макаки, что является основанием для исследования в дальнейшем на этой релевантной in vivo животной модели.
Таблица 5.
Константы аффинности анти-GITR антител при взаимодействии с macaca mulatta GITR.
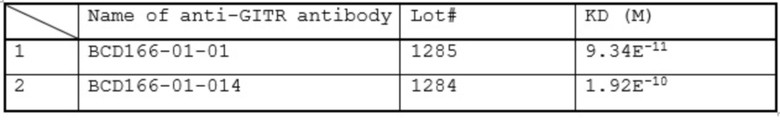
Таблица 6.
Константы аффинности анти-GITR антител при взаимодействии с macaca Cynomolgus GITR.
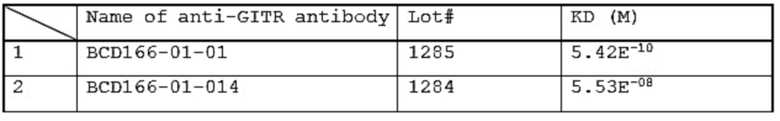
Пример 13
Клеточный in vitro тест на определение анти-GITR специфической агонистической активности диких и мутантного кандидата с усиленной агонистической активностью.
Для возможного усиления агонистической активности anti-GITR антитела BCD166-01-01, производного человека, на основании работ по изучению таких замен приводящих в олигомеризации антитела в работах WO2005047327 , WO2006104989 (A2), WO2007005612 (A2) и Science. 2014 Mar 14;343(6176):1260-3, была выбрана и введена мутация E345R в IgG1 Fc регион антитела. Было показано что такая мутация может приводить к олигомеризации антитела на поверхности клеток после связывания с антигеном, и усилению различных эффекторных способностей тк ADCC, ADCP, CDC а также фармакокинетики (PK). Целью было получить антитело с усиленной агонистической способностью, ADCC, но не CDC. Мутагенез производили согласно стандартным генно-инженерным протоколам и рекомбинантное антитело синтезировали согласно примеру 8. Антитело было названо и используется далее как BCD166-02-01. В тест также были взяты и два родительских антитела BCD166-01-01, BCD166-01-014.
Тест проводили аналогично тесту в Примере 10.
Результаты приведены на фиг.17 свидетельствуют об 5-кратном увеличении уровня активации в клеточном агонистическом тесте исследуемого BCD166-02-01 в сравнении с предшественником BCD166-01-01, в который была введена мутация E345R, что обуславливается антиген-зависимой олигомеризацией anti-GITR BCD166-02-01 антитела на клетке мишени. Причем закономерно увеличился не только уровень верхнего плато активации но и величина EC50 изменилась более чем в 20 раз в сторону усиления.
Пример 14
Клеточный in vitro тест на определение антителозависимой клеточной цитотоксичности (ADCC) для анти-GITR антител.
В тесте исследовали ADCC активность для кандидатов BCD166-02-01. В тест также были взяты и два антитела BCD166-01-01, BCD166-01-014. Кроме того, по аналогии с кандидатом BCD166-01-01, мутацию E345R ввели в кандидата BCD166-01-014 (который назвали BCD166-02-014), чтобы повысить ADCC активность в сравнении с дикой формой.
В тесте использовали клеточную линию Jurkat-NFAT-Luc-CD16, созданную на основе клеточной линии Jurkat, стабильно экспрессирующую на поверхности СD16 и содержащую ген, кодирующий люциферазу светляка, под контролем NFAT-промотора; а также клеточную линию Hek-293-GITR, созданную на основе клеточной линии Hek-293, стабильно экспрессирующую на поверхности GITR.
Тест проводили в культуральном 96-луночном планшете. Суспензия в каждой лунке содержала смесь клеток-эффекторов Jurkat-NFAT-Luc-CD16 и клеток мишеней Hek-293-GITR, а также анализируемые антитела в указанной на графике концентрации. Все компоненты суспензии готовили в питательной среде для культивирования клеток. После добавления всех компонентов планшеты инкубировали при 37°С, 5% CO2 и далее, с помощью набора для оценки люциферы и планшетного ридера, измеряли интенсивность люминесценции в лунках.
Результаты приведены на фиг.18 свидетельствуют об 40-кратном усилении величины EC50 в клеточном ADCC тесте исследуемого BCD166-02-01 в сравнении с предшественником BCD166-01-01, в который была введена мутация E345R, что обуславливается антиген-зависимой олигомеризацией anti-GITR BCD166-02-01 антитела на клетке мишени. Также было достоверно определено 5-кратное усиление величины EC50 в клеточном ADCC тесте исследуемого BCD166-02-14 в сравнении с предшественником BCD166-01-14, в который была введена мутация E345R. Вместе совокупные данные явно свидетельствуют о значительном позитивном эффекте замены E345R в Fc части IgG1 антител на разные эффекторные характеристики необходимые для создания высокоактивного агониста GITR.
Пример 15
Клеточный in vitro тест на определение комплемент зависимой цитотоксичности (CDC) для анти-GITR антител.
В данном тесте были исследованы антитела BCD166-01-01, BCD166-01-014, BCD166-02-01 и контрольное антитело anti-GITR-3215.
Тест проводили в культуральном 96-луночном планшете. Суспензия в каждой лунке содержала по клетки HEK-293-GITR, а также анализируемые антитела в указанной концентрации и комплемент сыворотки крови человека.. Планшет с описанной суспензией инкубировали в течение 4 ч при 37°С, 5% СО2. Далее в каждую лунку вносили раствор аламара синего, и далее инкубировали при 37°С, 5% СО2. Далее, с помощью планшетного ридера, измеряли в лунках величину флуоресценции (длина волны возбуждения 544 нм, длина волны испускания - 590 нм),
Результаты приведены на фиг.19 свидетельствуют об незначимой активности CDC , те не более 10% до концентрации 1 мкг/мл, которая является граничной для оценочной терапевтической дозы предполагаемой для человека, для антител BCD166-01-01, BCD166-02-01 и контрольного антитела anti-GITR-3215. Для кандидата BCD166-01-14 активности при концентрации 1 мкг/мл CDC составила около 30% лизиса, что свидетельствует о видимом эффекте, но возможно слабо проявляющемся in vivo. Вместе совокупные данные явно свидетельствуют незначительном эффекте замены E345R в Fc части IgG1 кандидатов anti-GITR.
Пример 16
Клеточный in vitro тест на определение цитотоксичности anti-GITR кандидатов.
Для оценки негативного или позитивного влияния изучаемых anti-GITR кандидатов на изменение количества различных популяций иммунореактивных клеток был проведен данный тест. В данном тесте были исследованы антитела BCD166-01-01, BCD166-01-014, BCD166-02-01 и контрольное антитело anti-CD20 (rituximab).
Клетки PBMCs были изолированы из цельной крови здоровых доноров путем центрифугирования в градиенте плотности фиколла. Тест проводили в культуральном 96-луночном планшете. Суспензия в каждой лунке содержала клетки PBMCs и указанное на графике антитело в концентрации 25, 1 и 0 мкг/мл,.. После смешивание PBMCs и антител планшет инкубировали в течение 16 ч при 37°С, 5% CO2. Далее оценивали в суспензиях долю CD56+, CD19+, CD3+, CD4+ и CD8+ субпопуляций PBMCs c помощью прямого окрашивания суспензий флуоресцентно-меченными антителами против соответствующих CD и последующего анализа клеток на проточном цитофлуориметре. Для CD56+, CD19+, CD3+ клеток на графиках приведена доля от всех клеток анализируемой суспензии, для CD4+, CD8+ - доля от CD3+ клеток.
Результаты представленные на фиг. 20-24 свидетельствуют об незначимом негативном влиянии изученных anti-GITR кандидатов, те не более 10% уменьшения количества иммунореактивных клеток в сравнении с негативным контролем. В случае популяции NK клеток мы наблюдали приблизительно 30-70% увеличение количества клеток в зависимости от дозы. При этом контрольное антитело anti-CD20 показало также увеличение процента NK клеток, но практически закономерное полную деплецию популяции CD20+ B клеток. Таким образом, все рассматриваемые кандидаты не обладают неспецифической цитотоксичностью in vitro на клетках крови человека.
Пример 17
Клеточный in vitro тест на определение антитело-зависимой деплеции nTreg клеточной популяции человека.
Одним из основных предполагаемых механизмов терапевтического анти-онкогенного действия anti-GITR антител предполагается ADCC зависимая деплеция GITR+ клеток nTreg популяции. В данном тесте были исследованы антитела BCD166-01-01, BCD166-01-014, BCD166-02-01 и контрольное антитело anti-CTLA4 (ipilimumab).
Мононуклеарные клетки перифеческой крови (PBMC) были изолированы из цельной крови здоровых доноров центрифугированием в градиенте плотности фиколла. PBMC были обогащены nTreg с помощью «CD4+ CD25+ Regulatory T Cell Isolation Kit, human» (Miltenyi Biotec, Германия). Полученные клетки активировали с помощью магнитных частиц с иммобилизованными анти-CD3 и анти-CD28 антителами.
NK клетки были изолированы из PBMC с помощью «NK Cell Isolation Kit, human» (Miltenyi Biotec, Германия).
В культуральный планшет внесли растворы антител, суспензию NK клеток и суспензию nTreg. Планшет инкубировали при 37°С 5% CO2 в течение 16 часов.
Окрашивали образцы клеточных суспензий с помощью флуоресцентно меченных антител против CD3, CD4, CD25, FoxP3 (Biolegend, США). Образцы окрашенных клеточных суспензий анализировали с помощью проточной цитофлюориметрии.
Результаты анализа антитело-зависимой деплеции nTreg под действием изучаемых антител представленные на фиг. 25 для клеточного материала донора 1 и на фиг. 26 донора 2 свидетельствуют о значимом 70-100% уменьшении популяции клеток, т.е. деплетирующей активности изученных anti-GITR кандидатов особенно для материала донора 2. Кроме того, сравнительный анализ с контрольным anti-CTLA4 антителом ipilimumab демонстрирует большую активность в пользу anti-GITR антител особенно для финального кандидата BCD166-02-01.
Пример 18
Клеточный in vitro тест по анализу антитело-зависимой деплеции iTreg клеточной популяции человека.
Одним из основных предполагаемых механизмов терапевтического анти-онкогенного действия anti-GITR антител предполагается ADCC зависимая деплеция GITR+ клеток iTreg популяции. В данном тесте были исследованы антитела BCD166-01-01, BCD166-01-014, BCD166-02-01.
PBMC были изолированы из цельной крови здоровых доноров центрифугированием в градиенте плотности фиколла. Выделяли из PBMC моноциты, как популяцию клеток, связывающихся с культуральным пластиком. Для получения дендритных клеток моноциты инкубировали в присутствии 1000 Ед/мл GM-CSF (Peprotech, США) и 500 Ед/мл IL-4 (Thermo Scientific, США) при 37°С 5% CO2 в течение 120 часов. Добавили в культуральную среду раствор LPS (Sigma, США) до концентрации 0,5 мкг/мл, инкубировали клетки при 37°С 5% CO2 в течение 48 часов.
В культуральный планшет внесли растворы разведений антител, сусензию дендритных клеток и PBMC. Планшет инкубировали при 37°С 5% CO2 в течение 120 часов.
Окрашивали образцы клеточных суспензий с помощью флуоресцентно меченных антител против CD3, CD4, CD25, FoxP3 (Biolegend, США). Образцы окрашенных клеточных суспензий анализировали с помощью проточной цитофлюориметрии. Антитела против GITR BCD166-01-01, BCD166-01-14 и BCD166-02-01 вызывают деплецию iTreg in vitro.
Результаты анализа антитело-зависимой деплеции iTreg под действием изучаемых антител представленные на фиг.27 для клеточного материала донора 1 и донора 2 свидетельствуют о значимой 2-5Х кратной деплетирующей активности изученных anti-GITR кандидатов особенно для материала донора 1. Финальный кандидат BCD166-02-01 демонстрирует достаточную iTreg деплетирующую активность.
Пример 19
Клеточный in vitro тест на определение секреции провоспалительных цитокинов IL-2 и IFN- γ под действием анти-GITR антитела в клеточной популяции человека.
Клетки PBMCs были изолированы из цельной крови здоровых доноров путем центрифугирования в градиенте плотности фиколла.
Иммобилизовали в лунках 96-луночного планшета aнтиCD3 и антиGITR антитела. Для этого внесли в соответствующие лунки планшета по 100 мкл раствора DPBS, содержащего антиGITR антитело BCD166-02-01 в указанной на графике концентрации и aнтиCD3 антитело в субоптимальной концентрации, инкубировали планшет 16 часов при комнатной температуре.
Тест проводили 96-луночном планшете с предварительно иммобилизованными в лунках aнти-CD3 и анти-GITR антителами. Суспензия в каждой лунке содержала 40 000 клеток PBMCs, а также aнти-CD28 антитело в субоптимальной концентрации. Все компоненты суспензии готовили в среде RPMI-1640, содержащей 10% FBS. После добавления клеточной суспензии, планшет инкубировали в течение 6 дней при 37°С, 5% CO2. На 4-й день инкубации из лунок отбирали аликвоты культуральной жидкости. Далее определяли в культуральной жидкости 4-го и 6-го дня инкубации концентрацию IL-2 и IFN- γ с помощью ИФА.
Результаты анализа влияния анти-GITR антитела на уровень секреции провоспалительных цитокинов IL-2 и IFN-γ, обуславливающих антираковых эффект, представлены на фиг.28 демонстрируют значительное повышение концентрации этих веществ в культуральной жидкости. Таким образом, кандидат BCD166-02-01 демонстрирует высокую активность в активации цитокиновой секреции, что может свидетельствовать о значимом анти-онкогенном действии.
Пример 20
Анализ связывания антитела BCD166-02-01 с FcRn, FcgRIIIa158V, FcgRIIIa158F, FcgRIIa131H, FcgRIIa131R, FcgRIIb и FcgRIa рецепторами человека.
Константы аффинности связывания антитела BCD166-02-01 к FcRn, FcgRIIIa158V, FcgRIIIa158F, FcgRIIa131H, FcgRIIa131R, FcgRIIb и FcgRIa человека получили с помощью прибора OctetRed 96 (от ForteBio). Биотинилированные рецепторы иммобилизировали на поверхности стрептавидиновых сенсоров (SA). Провели и проанализировали ассоциацию и диссоциацию рецепторов и антитела в рабочем буфере (PBS, содержащим 0,1% Твин-20 и 0,1% БСА: с рецепором FcRn с рН6, с остальными рецепторами - в буфере с рН7,4) при 30°С. Полученные сенсограммы проанализировали в соответствии с моделями 1:1 или 2:1 и рассчитали константы аффинности, используя программное обеспечение Octet Data Analysis Software 8.0 User Guide Copyright 2011©.
Результаты анализа аффинности BCD166-02-01 к FcRn, FcgRIIIa158V, FcgRIIIa158F, FcgRIIa131H, FcgRIIa131R, FcgRIIb и FcgRIa человека представлены на Фиг. 29. Они демонстрируют значения кратно усиленные по сравнению с литературными данными и данными многократно измеренными в нашей компании для антител IgG1 изотипа, что свидетельствует о заметном но не качественном влиянии мутации E345R в составе Fc фрагмента на кинетику мономерных антител адсорбирующихся на этих рецепторах из раствора.
Пример 21
Определение коллоидной и термической стабильности по точке агрегации белка методом динамического светорассеяния
Для определения температуры агрегации исследуемых образцов методом динамического светорассеяния получали зависимость размера частиц в среде от температуры с помощью прибора DynaPro® Plate Reader II (Wyatt Technology Corp.) при постепенном нагревании от 40 до 85°С. Результаты представлены в таблице 7.
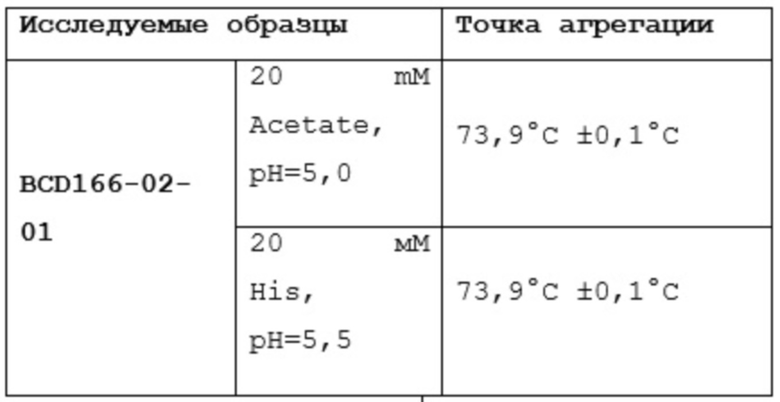
Таблица 7. Точка агрегации BCD166-02-01
Можно заключить, что молекула BCD166-02-01 имеет высокую термоколлоидную стабильность (точка агрегации в 20mM Acetate, pH=5,0 и 20мМ His, pH=5,5 буферных растворах составляет > 65).
Аналогичные данные были получены для BCD166-01-014.
Пример 22
Определение термической стабильности при термострессе 50°С
Исследуемые образцы устанавливали в воздушный термостат и термостатировали при 50°С в течение 72 часов. После окончания прогрева интактные и стрессированные образцы передавали на анализ методом эксклюзионной ВЭЖХ (SEC HPLC) с УФ-детектором и методом капиллярного изоэлектрического фокусирования. Хроматографию проводили на системе ВЭЖХ Agilent 1100 на колонке Tosoh TSK-Gel G3000SWXL, детектирование проводили при длине волны 220 нм. Определение гетерогенности по заряду методом капиллярного изоэлектрического фокусирования проводился на приборе Labchip GX II, Caliper. Приготовление рабочих растворов и подготовку чипа проводили по стандартной методике с использованием наборов HT Protein Charge Variant Labeling Kit и Protein Charge Variant Buffer Kit, PerkinElmer.
Результирующие данные стабильности BCD-166 (для BCD166-02-01 и BCD166-01-014) при инкубировании при 50°С представлены в табл. 8 и на фиг. 30 (для BCD166-02-01) и 31 (для BCD166-01-014) представлены совмещенные хроматограммы при 72-часовой инкубации при 50°С.
Общий вывод: образец обладает высокой коллоидной и термической стабильностью.
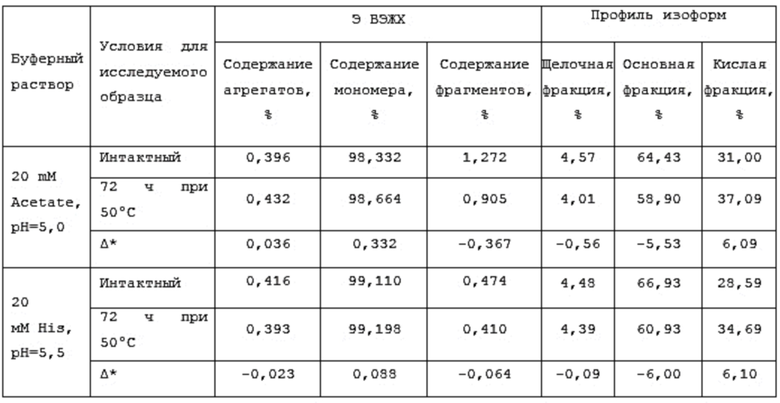
* Δ - разница чистоты стрессированного образца и чистоты интактного образца (входного контроля), %.
Таблица 8. Зависимость содержания мономера по эксклюзивной ВЭЖХ и ЭФ для BCD-166.
Пример 23
Оценка противоопухолевой активности препаратов BCD-166 на модели подкожных ксенографтов
Для оценки противоопухолевой активности препаратов BCD-166 использовали модель подкожных опухолевых ксенографтов. Для этого гуманизированным мышам huNOG-EXL подкожно трансплантировали 3×106 клеток линии A2058 в смеси с Matrigel®. На 7 день после инокуляции клеток животных распределяли по группам, как представлено в таблице 9.
Таблица 9. Распределение животных по группам в эксперименте по определению противоопухолевой активности препаратов BCD-166
Введение препаратов проводили внутрибрюшинно в дозе 20 мг/кг на 1, 5, 8, 12 сутки после распределения животных по группам. Группе отрицательного контроля вводили гистидиновый буфер. В течение эксперимента измеряли массу тела мышей и линейные размеры опухоли. Для вычисления объема опухоли использовали формулу V=L×W×H×π/6. В качестве критерия эффективности исследуемого препарата использовали показатель торможения роста опухоли (ТРО), который рассчитывается с учетом среднего объема опухоли в группе отрицательного контроля (Vc) и группе препарата (Vt) по формуле
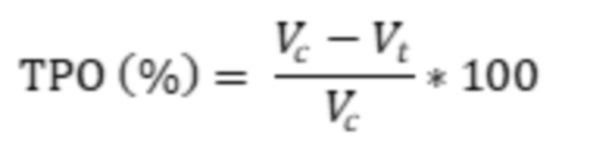
По результатам эксперимента препараты BCD-166-01-014 и BCD-166-02-01 показали значимую противоопухолевую активность, препарат BCD-166-01-01 не показал значимой противоопухолевой активности. Результаты показаны на фигурах 32 и 33.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЗАО «БИОКАД»
<120> Моноклональное антитело, которое специфически связывается с GITR
<160> 21
<170> BiSSAP 1.3.6
<210> 1
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность HCDR1 BCD166-01/02-001
<400> 1
Asn Tyr Gly Met His
1 5
<210> 2
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность HCDR2 BCD166-01/02-001
<400> 2
Val Ile Trp Phe Asp Gly Ser Asn Lys Phe Tyr Thr Asp Ser Val Lys
1 5 10 15
Gly
<210> 3
<211> 21
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность HCDR3 BCD166-01/02-001
<400> 3
Glu Leu Gly Gly Tyr Tyr Tyr Asp Ser Ser Gly Phe Arg Pro Tyr Tyr
1 5 10 15
Tyr Gly Met Asp Val
20
<210> 4
<211> 130
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность BCD166-01/02-001_VH
<400> 4
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Lys
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Phe Asp Gly Ser Asn Lys Phe Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asp Thr Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Leu Gly Gly Tyr Tyr Tyr Asp Ser Ser Gly Phe Arg Pro
100 105 110
Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Met Val Thr Val
115 120 125
Ser Ser
130
<210> 5
<211> 460
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность BCD166-01-001_HC
<400> 5
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Lys
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Phe Asp Gly Ser Asn Lys Phe Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asp Thr Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Leu Gly Gly Tyr Tyr Tyr Asp Ser Ser Gly Phe Arg Pro
100 105 110
Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Met Val Thr Val
115 120 125
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
130 135 140
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
145 150 155 160
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
165 170 175
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
180 185 190
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
195 200 205
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
210 215 220
Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
225 230 235 240
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
245 250 255
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
260 265 270
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
275 280 285
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
290 295 300
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
305 310 315 320
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
325 330 335
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
340 345 350
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
355 360 365
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
370 375 380
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
385 390 395 400
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
405 410 415
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
420 425 430
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
435 440 445
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455 460
<210> 6
<211> 460
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность BCD166-02-001_HC
<400> 6
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Lys
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Phe Asp Gly Ser Asn Lys Phe Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asp Thr Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Leu Gly Gly Tyr Tyr Tyr Asp Ser Ser Gly Phe Arg Pro
100 105 110
Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Met Val Thr Val
115 120 125
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
130 135 140
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
145 150 155 160
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
165 170 175
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
180 185 190
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
195 200 205
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
210 215 220
Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
225 230 235 240
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
245 250 255
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
260 265 270
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
275 280 285
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
290 295 300
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
305 310 315 320
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
325 330 335
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
340 345 350
Lys Gly Gln Pro Arg Arg Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
355 360 365
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
370 375 380
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
385 390 395 400
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
405 410 415
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
420 425 430
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
435 440 445
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455 460
<210> 7
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность LCDR1 BCD166-01/02-001
<400> 7
Arg Ala Ser Gln Ser Ile Gly Ser Trp Leu Ala
1 5 10
<210> 8
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность LCDR2 BCD166-01/02-001
<400> 8
Ala Ala Ser Thr Leu Gln Arg
1 5
<210> 9
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность LCDR3 BCD166-01/02-001
<400> 9
Gln Gln Ser His Ser His Pro Leu Thr
1 5
<210> 10
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность BCD166-01/02-001_VL
<400> 10
Asp Val Val Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Glu Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Gly Gly Tyr Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser His Ser His Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 11
<211> 214
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность BCD166-01/02-001_LC
<400> 11
Asp Val Val Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Glu Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Gly Gly Tyr Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser His Ser His Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 12
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность HCDR1 BCD166-01-014
<400> 12
Tyr Tyr Trp Met Tyr
1 5
<210> 13
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность HCDR2 BCD166-01-014
<400> 13
Ala Ile Ser Trp Asn Gly Gly Arg Thr Tyr Tyr Ala Glu Ser Met Lys
1 5 10 15
Gly
<210> 14
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность HCDR3 BCD166-01-014
<400> 14
Asn Arg Tyr Tyr Ser Asp Pro Asn Tyr Gly Met Asn Leu
1 5 10
<210> 15
<211> 122
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность BCD166-01-014_VH
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Trp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Trp Asn Gly Gly Arg Thr Tyr Tyr Ala Glu Ser Met
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Gln Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asn Arg Tyr Tyr Ser Asp Pro Asn Tyr Gly Met Asn Leu Trp
100 105 110
Gly Lys Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 16
<211> 452
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность BCD166-01-014_HC
<400> 16
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Trp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Trp Asn Gly Gly Arg Thr Tyr Tyr Ala Glu Ser Met
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Gln Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asn Arg Tyr Tyr Ser Asp Pro Asn Tyr Gly Met Asn Leu Trp
100 105 110
Gly Lys Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 17
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность LCDR1 BCD166-01-014
<400> 17
Thr Gly Thr Ser Thr Asp Ile Gly Thr Tyr Lys Tyr Ile Ser
1 5 10
<210> 18
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность LCDR2 BCD166-01-014
<400> 18
Gly Val Ser His Arg Pro Ser
1 5
<210> 19
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность LCDR3 BCD166-01-014
<400> 19
Ser Ser Tyr Thr Ser Ser Gly Thr Val Val
1 5 10
<210> 20
<211> 110
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность BCD166-01-014_VL
<400> 20
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Ile Gly Thr Tyr
20 25 30
Lys Tyr Ile Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Ile Ile Tyr Gly Val Ser His Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Asp Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Gly Thr Val Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 21
<211> 216
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность BCD166-01-014_LC
<400> 21
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Ile Gly Thr Tyr
20 25 30
Lys Tyr Ile Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Ile Ile Tyr Gly Val Ser His Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Asp Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Gly Thr Val Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО К PD-L1, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2778085C2 |
| ИММУНОЦИТОКИН, ВКЛЮЧАЮЩИЙ ГЕТЕРОДИМЕРНЫЙ БЕЛКОВЫЙ КОМПЛЕКС НА ОСНОВЕ IL-15/IL-15Rα, И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2753282C2 |
| Моноклональное антитело или его антигенсвязывающий фрагмент, которое специфически связывается с BCMA, и его применение | 2022 |
|
RU2820350C2 |
| Выделенное биспецифическое антитело, которое специфически связывается с CD47 и PD-L1 | 2021 |
|
RU2815823C2 |
| АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2815280C1 |
| Двухвалентное биспецифическое химерное антитело, включающее гетеродимер на основе MHC или MHC-подобных белков | 2021 |
|
RU2820683C2 |
| НАЦЕЛЕННЫЕ НА uPARAP КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2017 |
|
RU2740311C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ CD3E/BCMA И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800164C2 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С СОРТИЛИНОМ И ПОДАВЛЯЮТ СВЯЗЫВАНИЕ ПРОГРАНУЛИНА | 2016 |
|
RU2735639C2 |
| Иммунотерапия с применением антител, связывающих лиганд 1 белка программируемой смерти клеток (PD-L1) | 2017 |
|
RU2766582C2 |
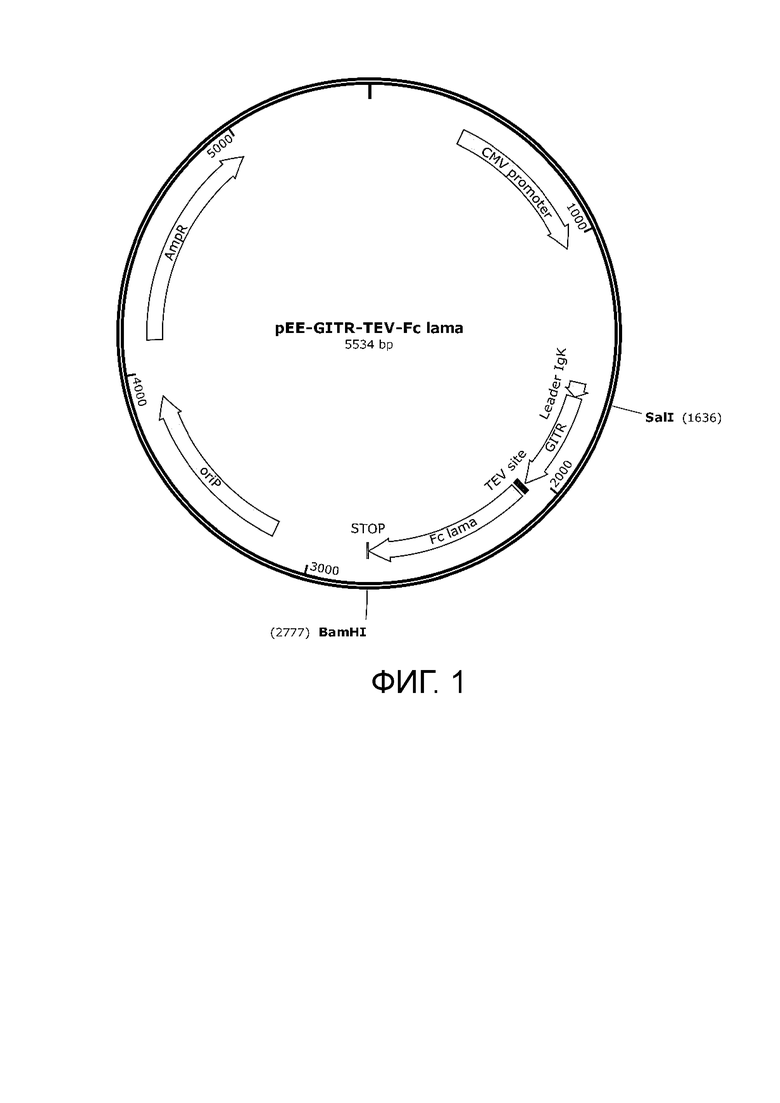
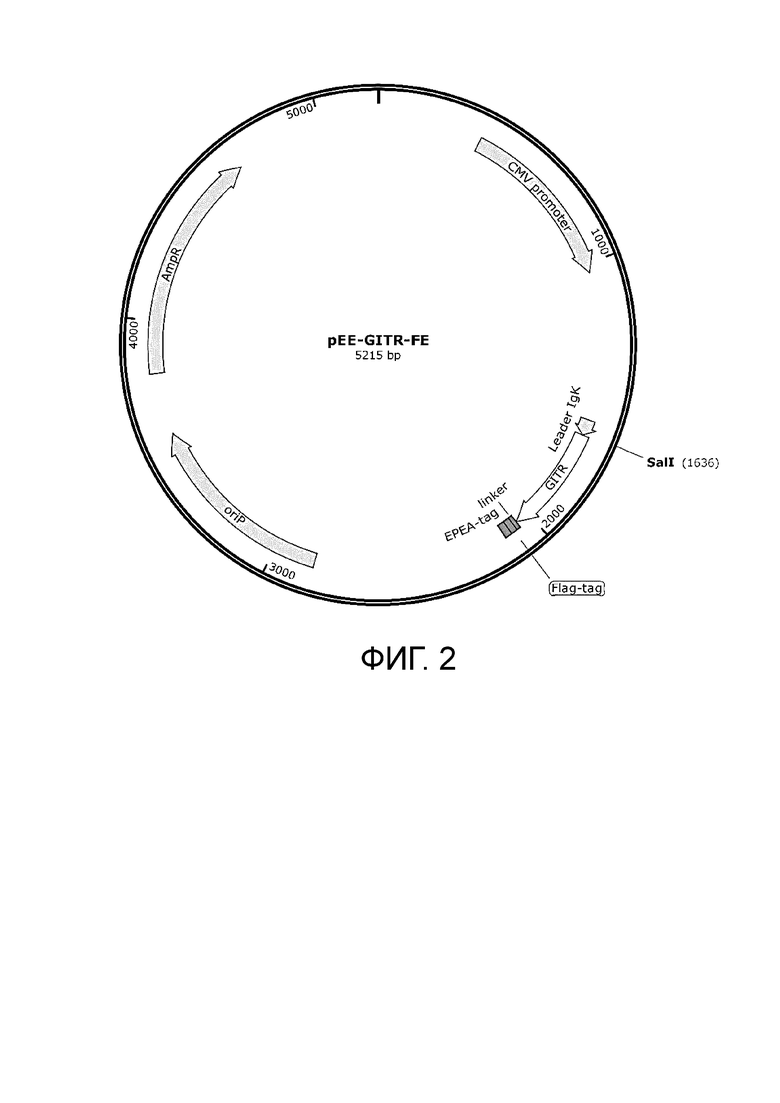
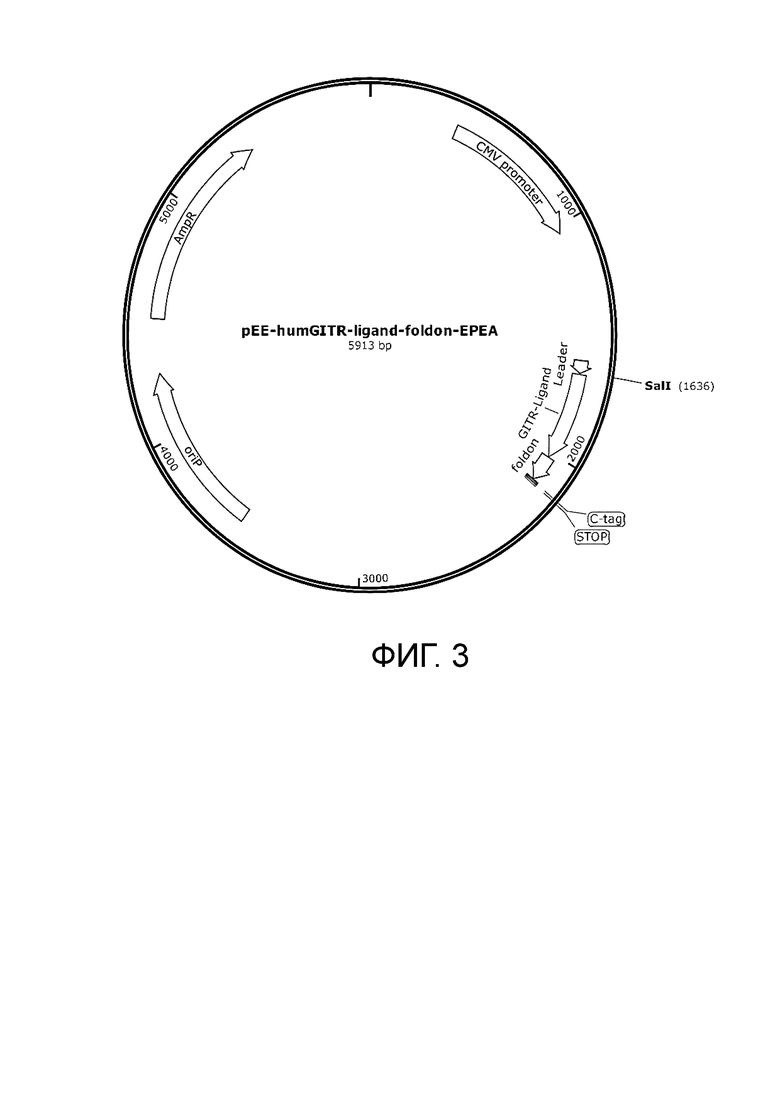
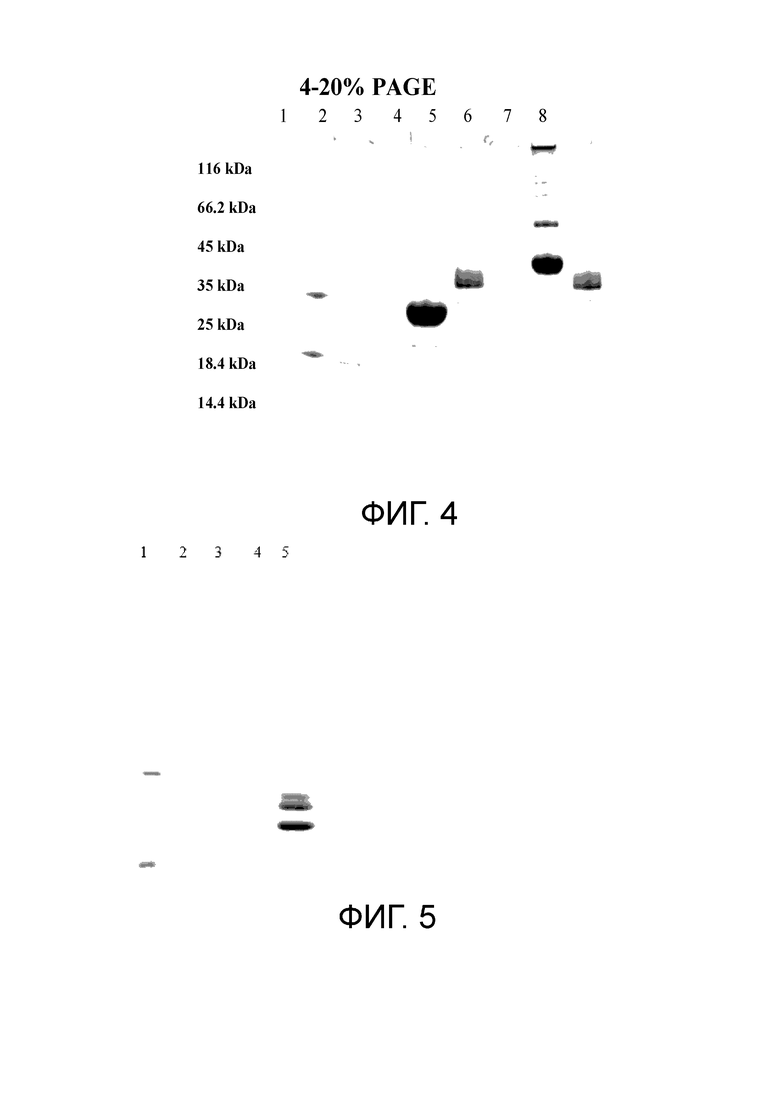
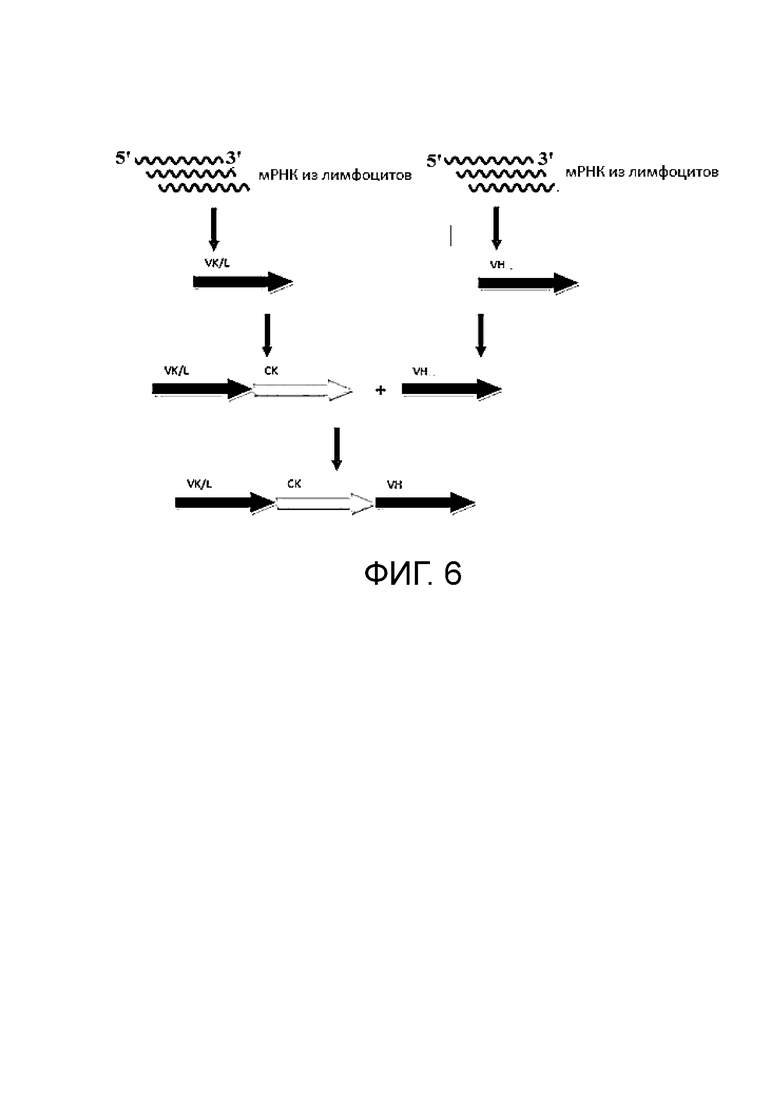
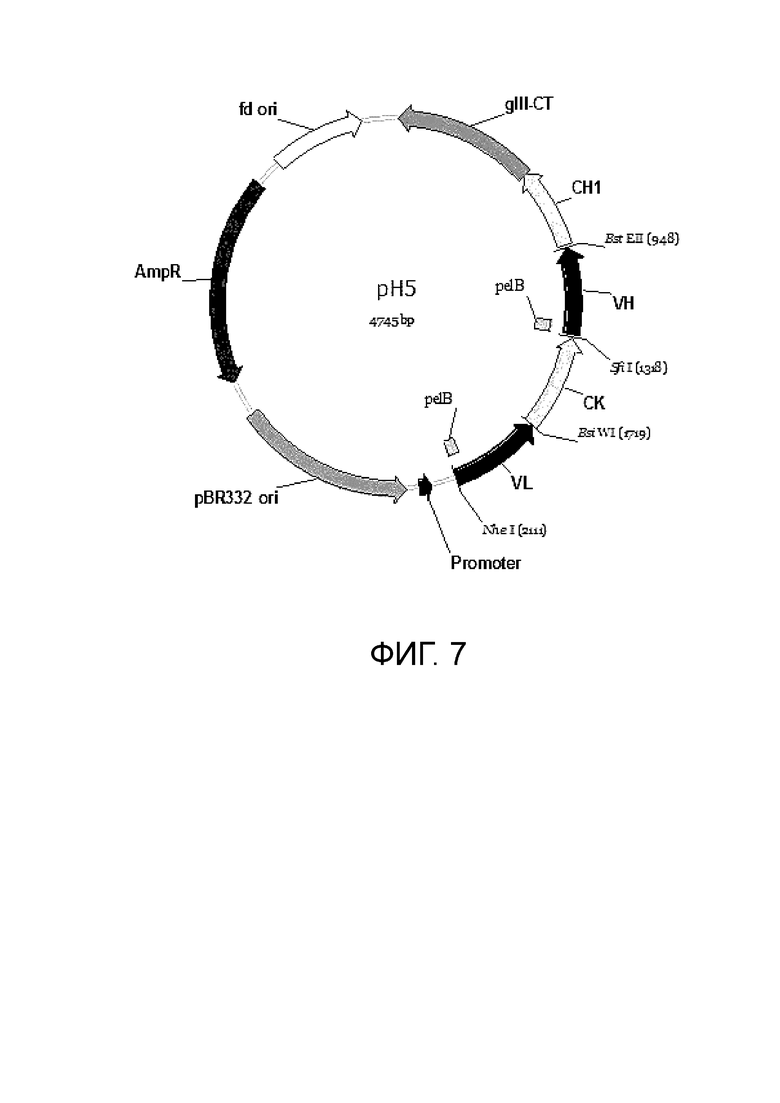
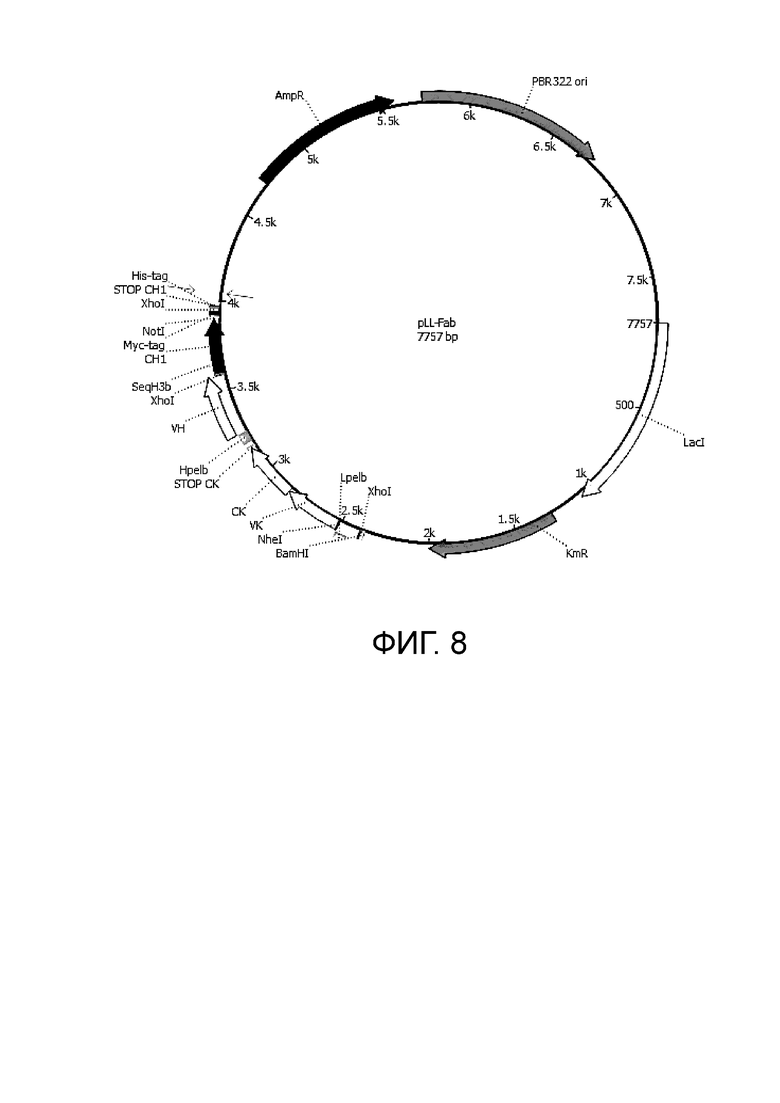
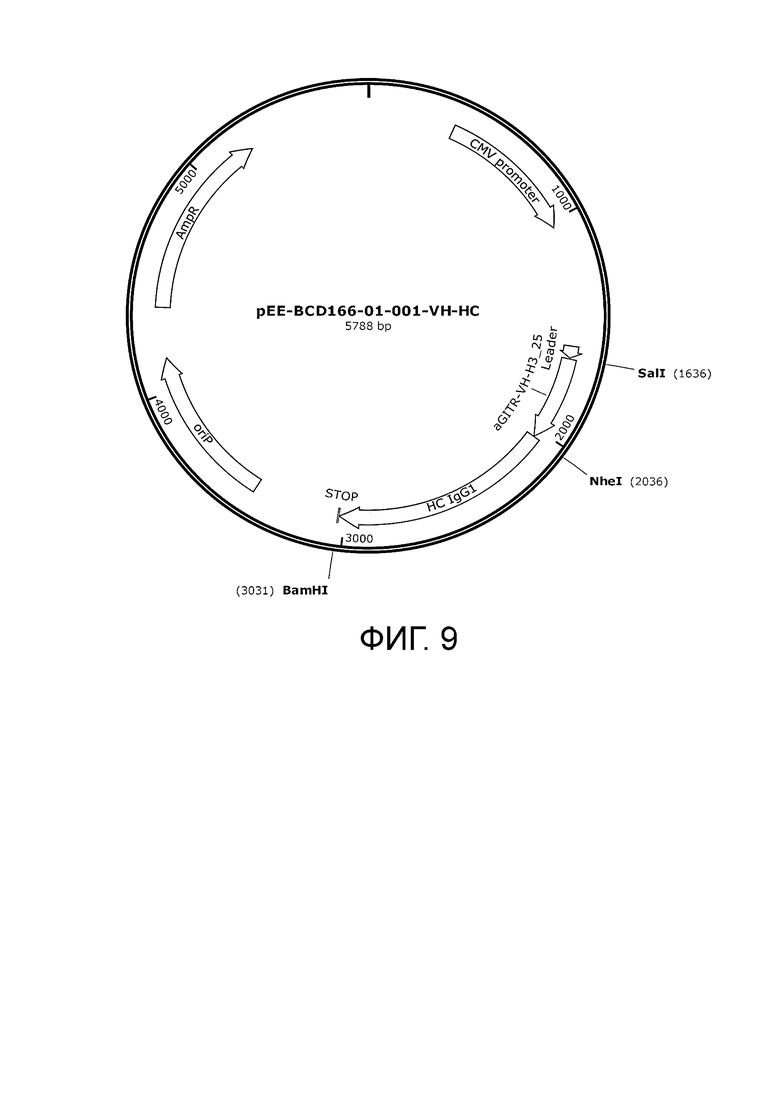
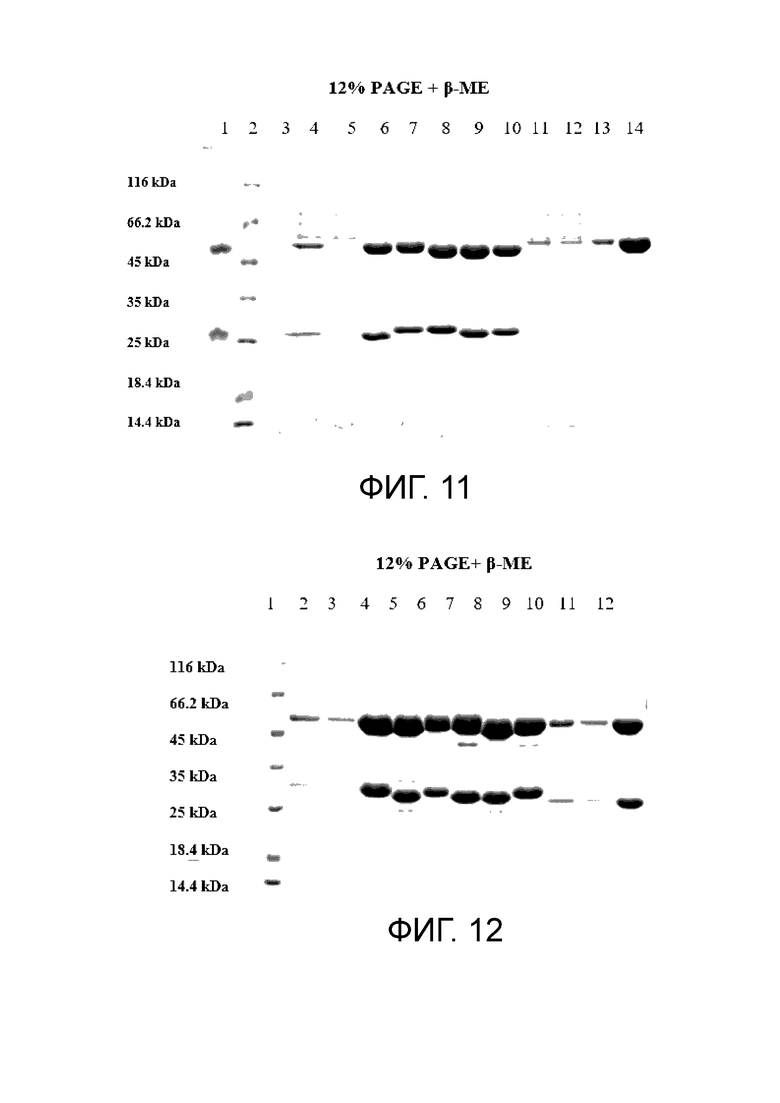
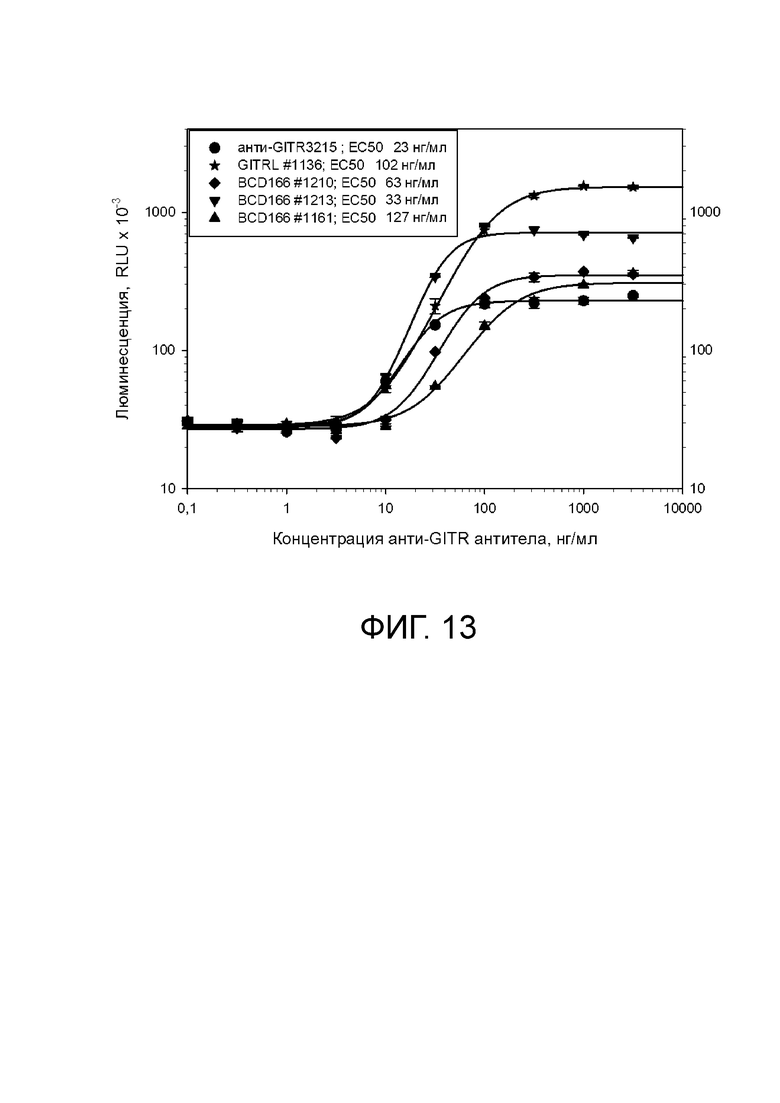
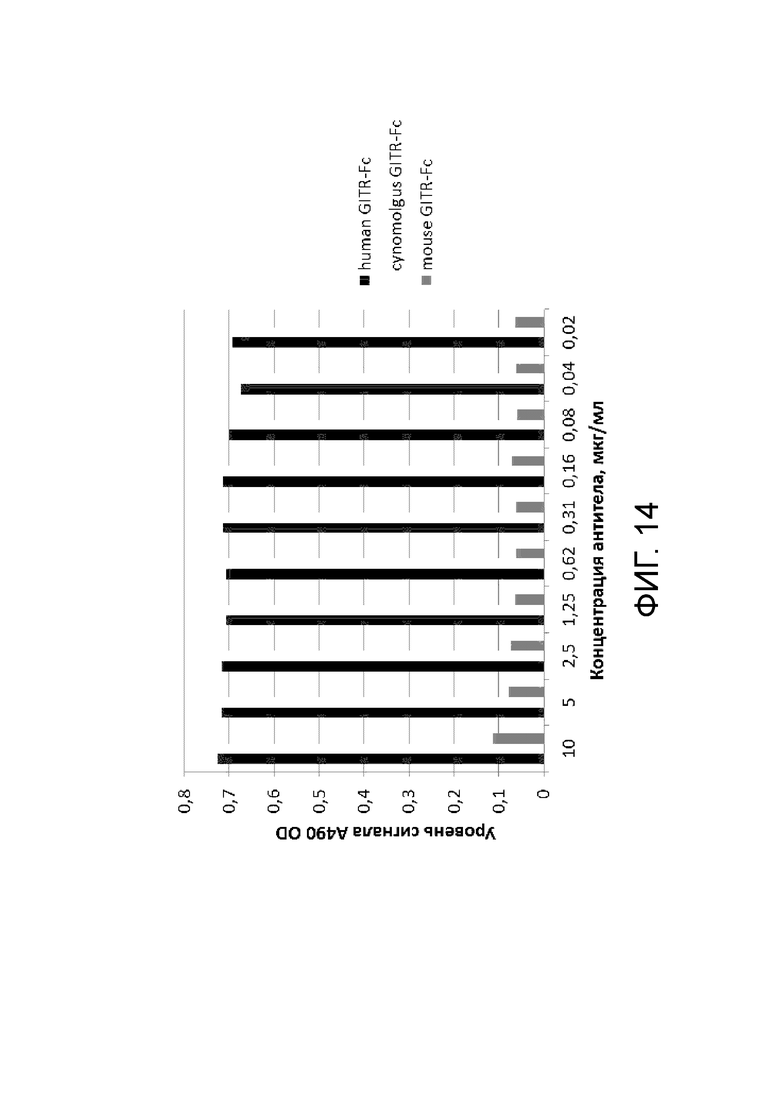
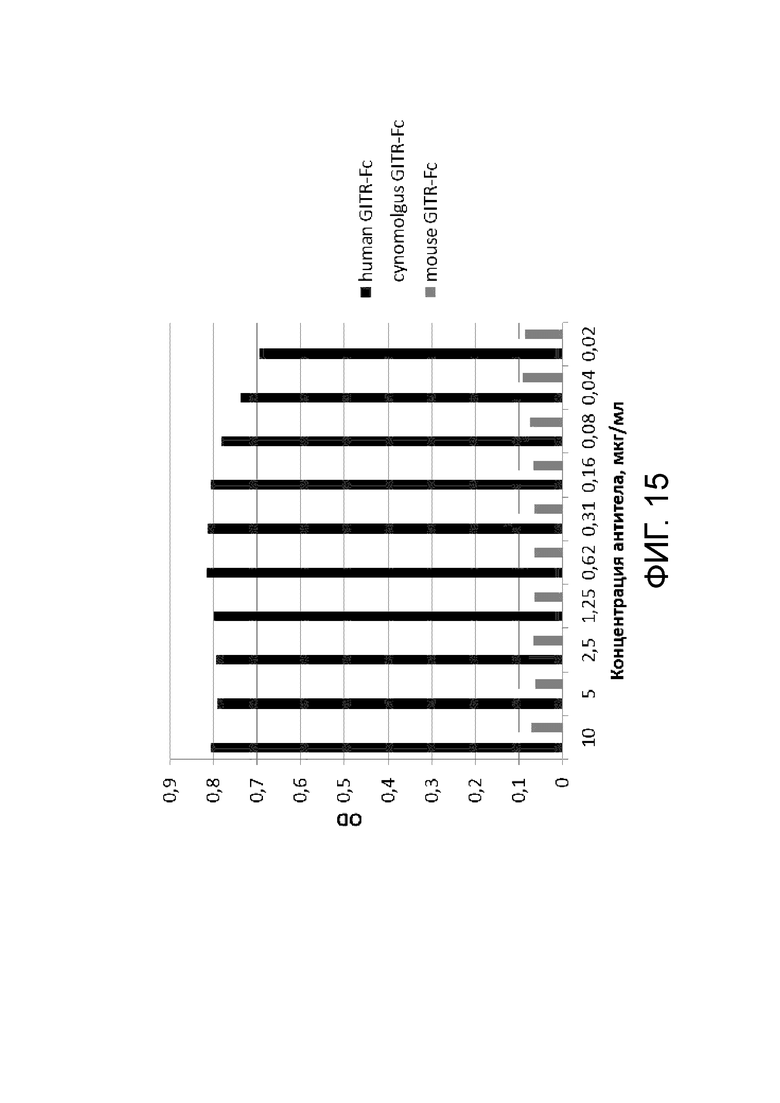
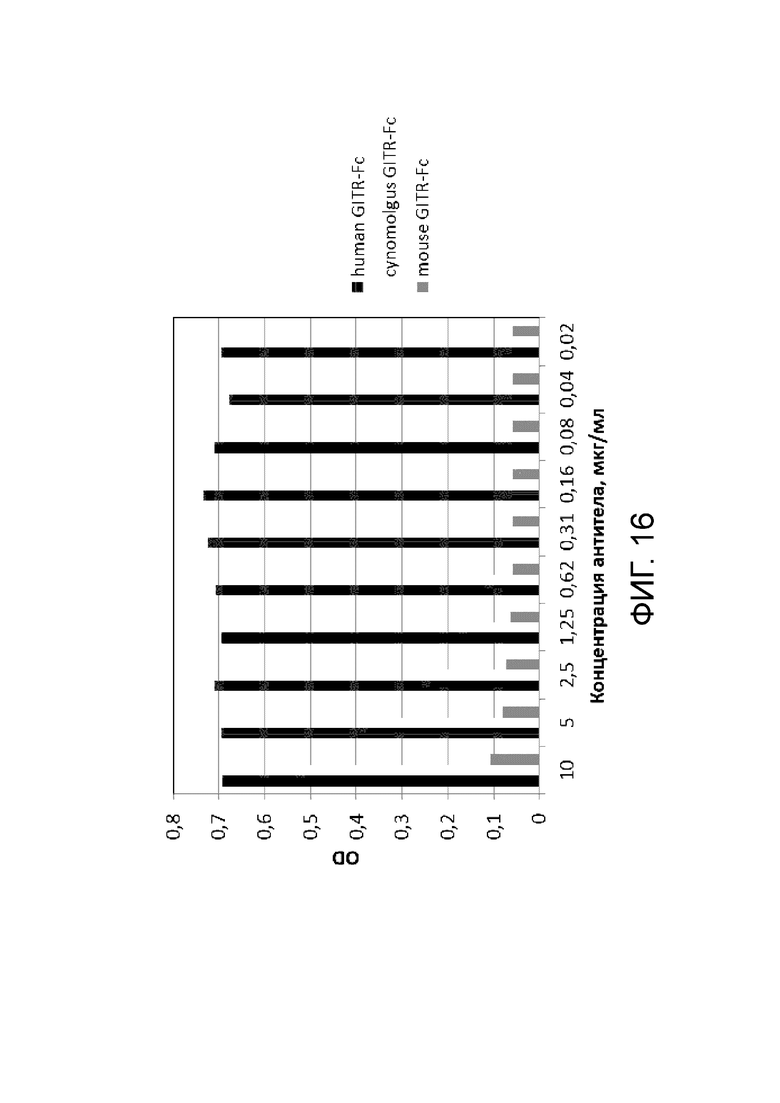
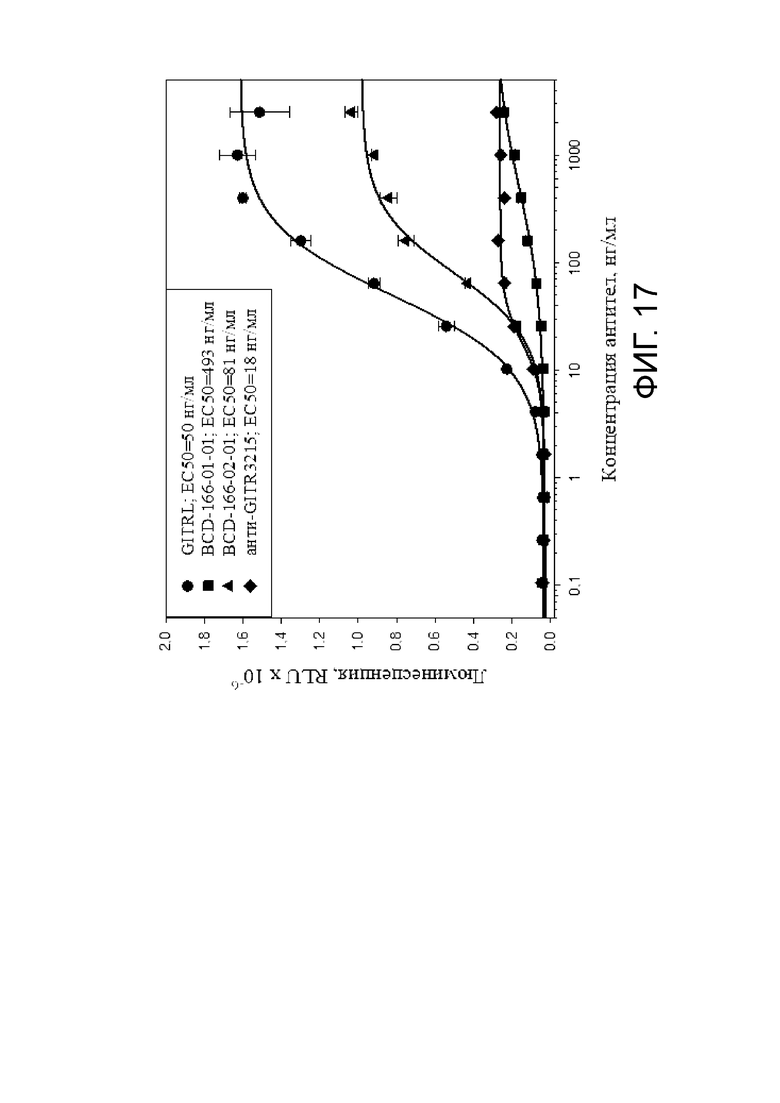
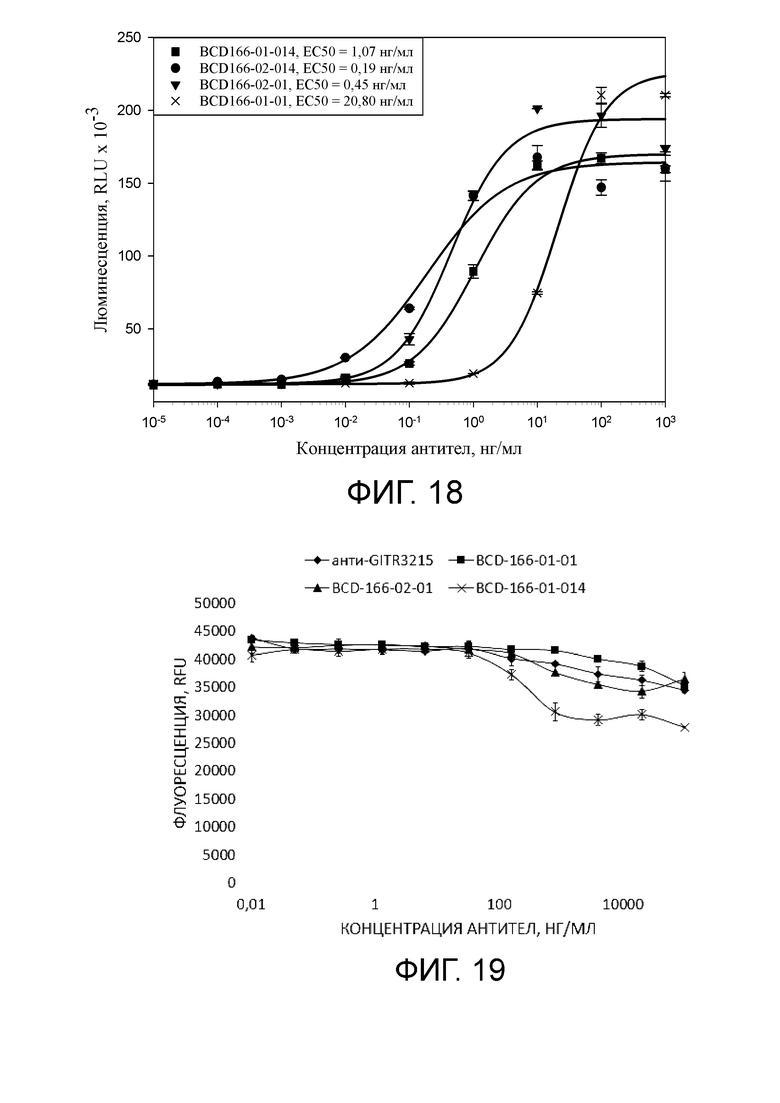
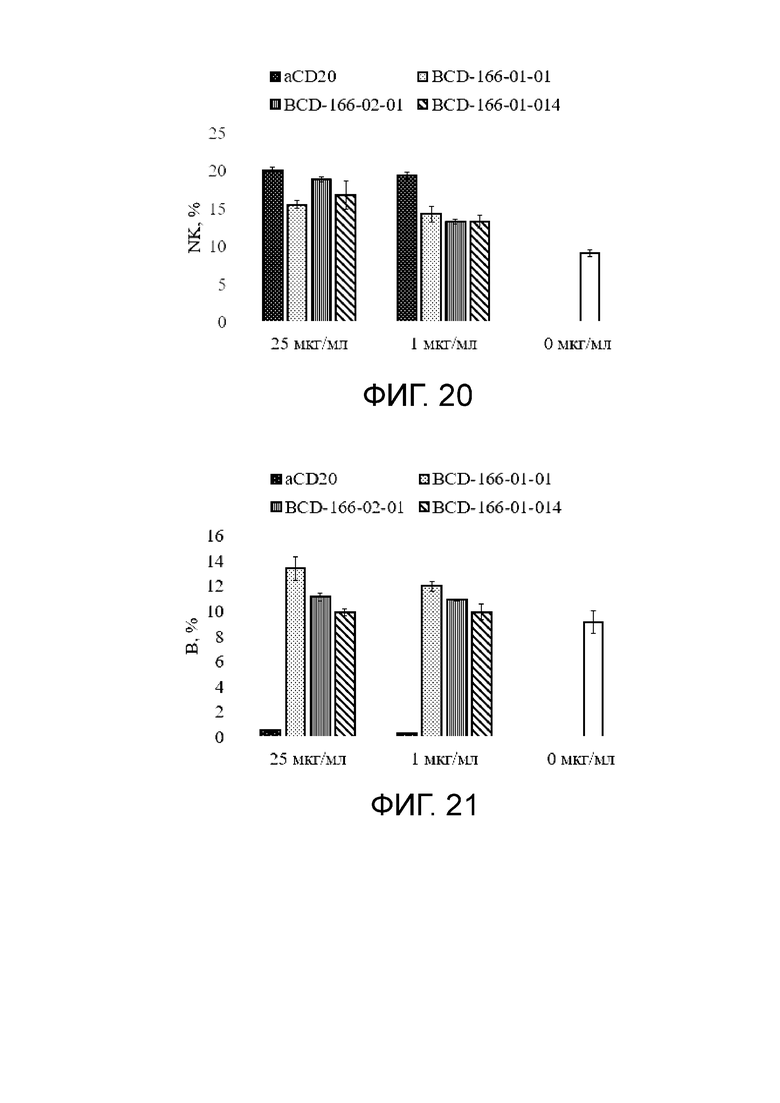
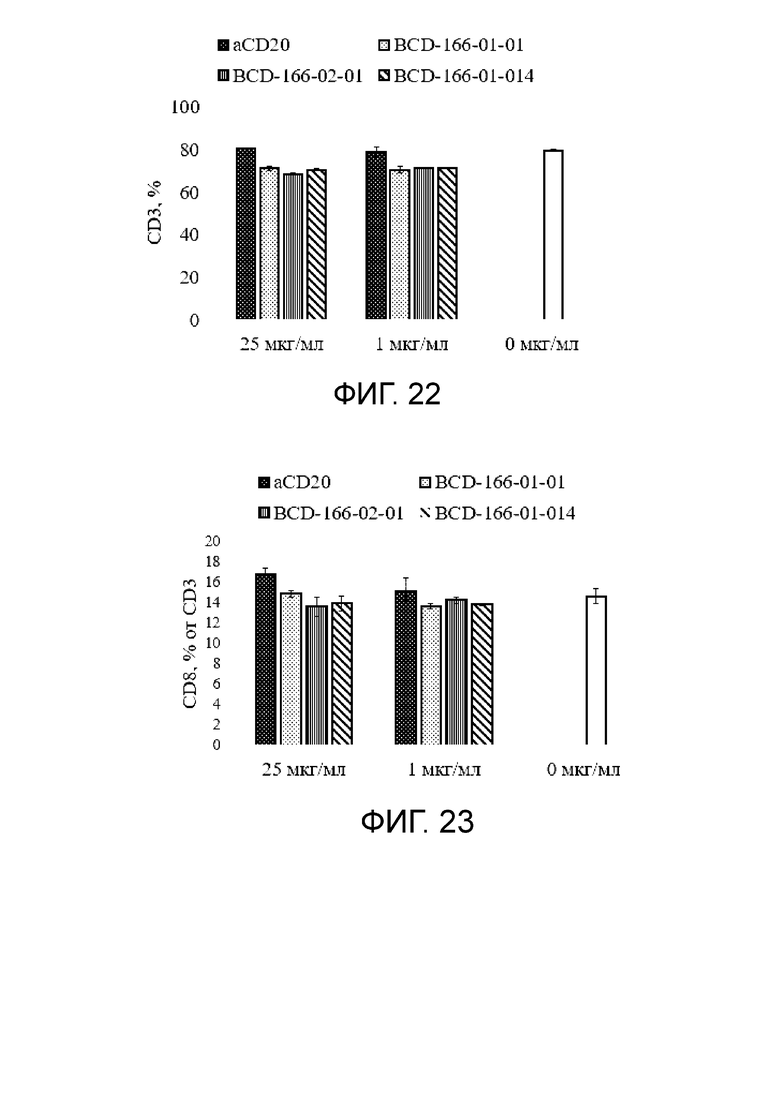
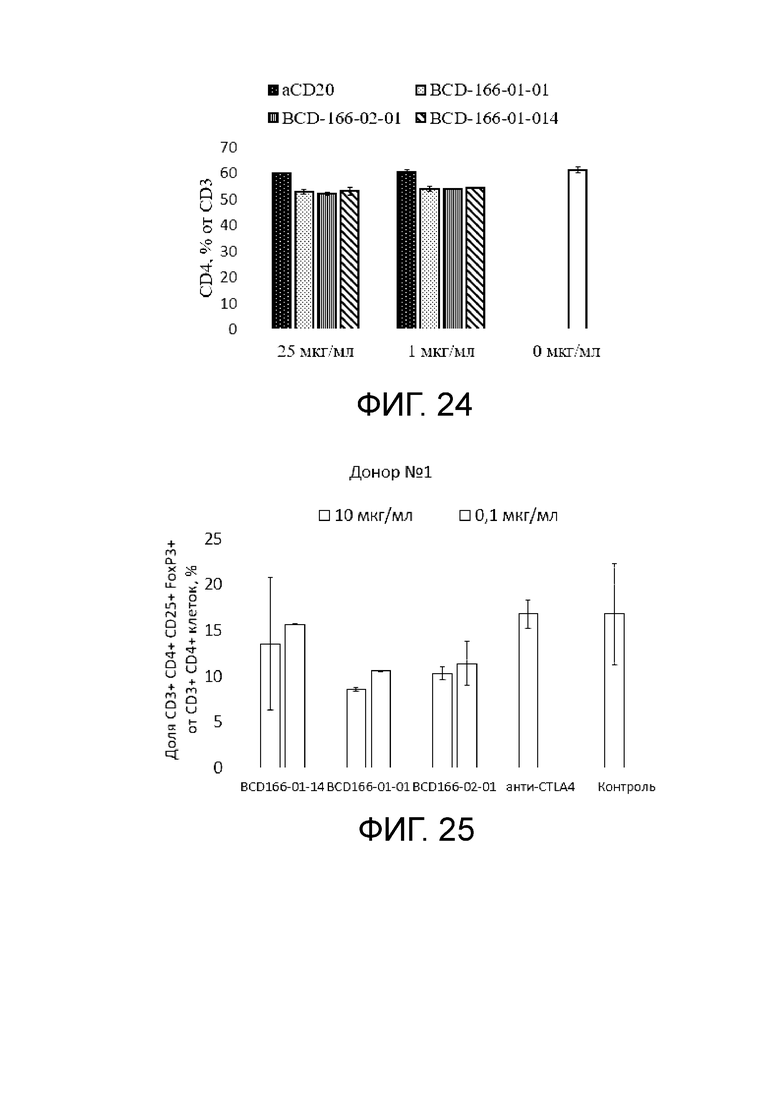
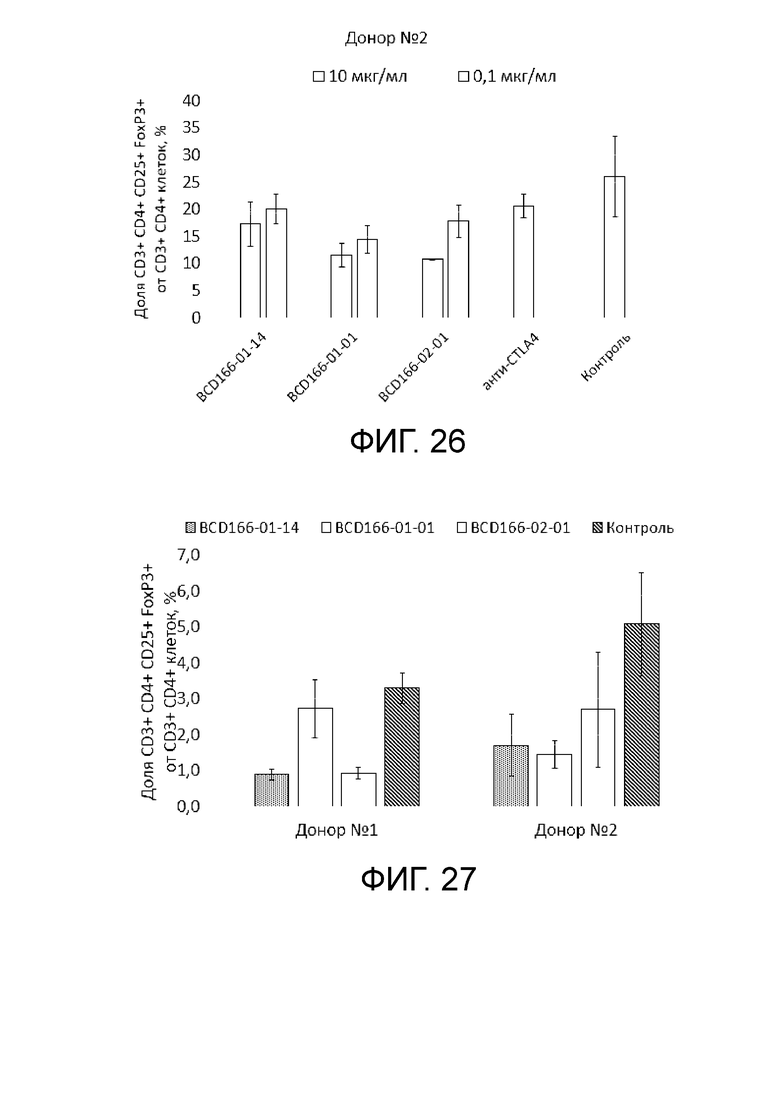
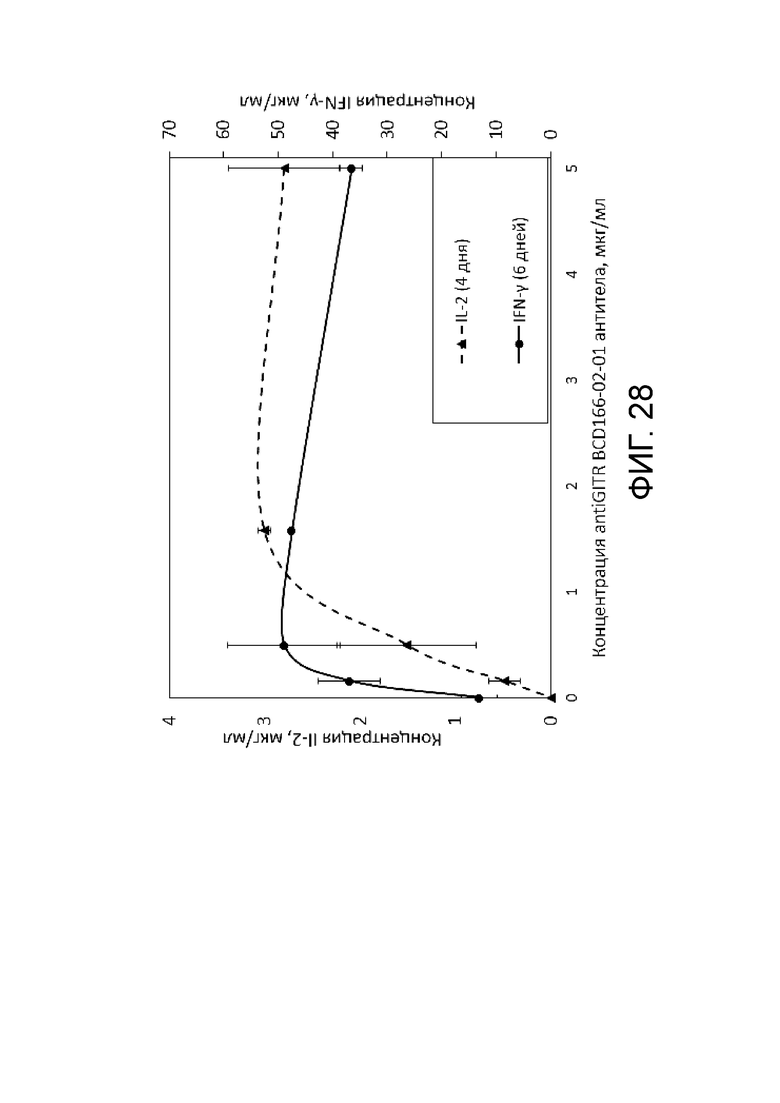

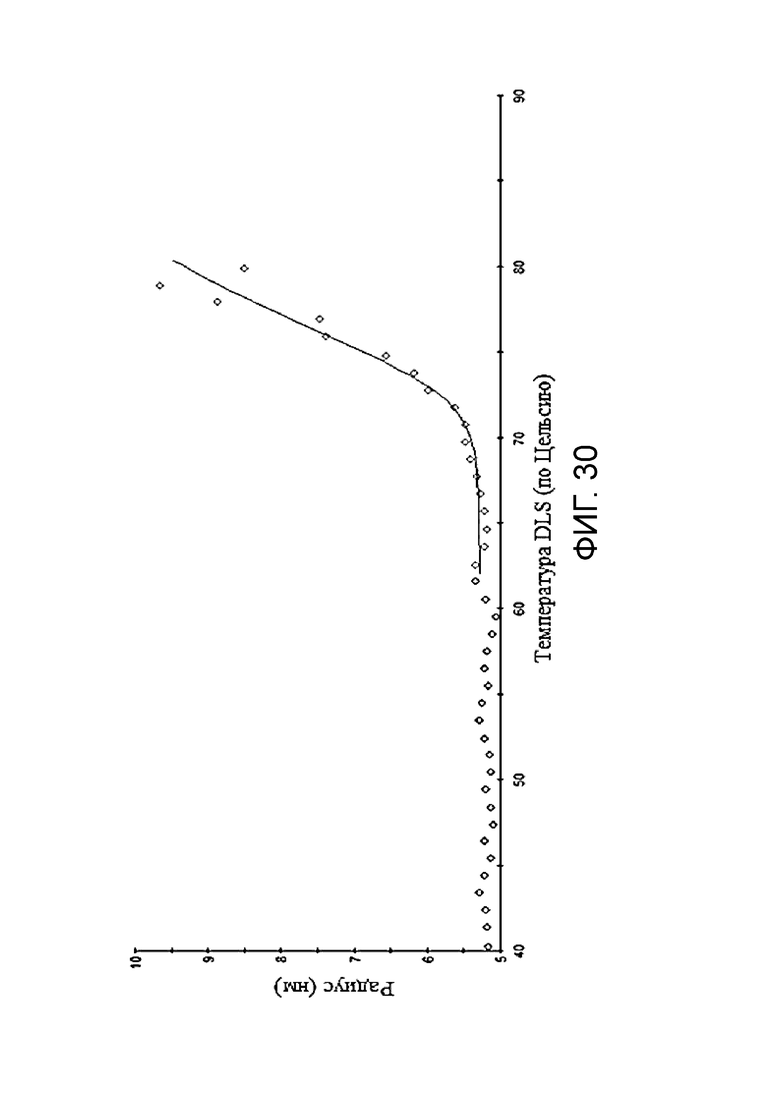
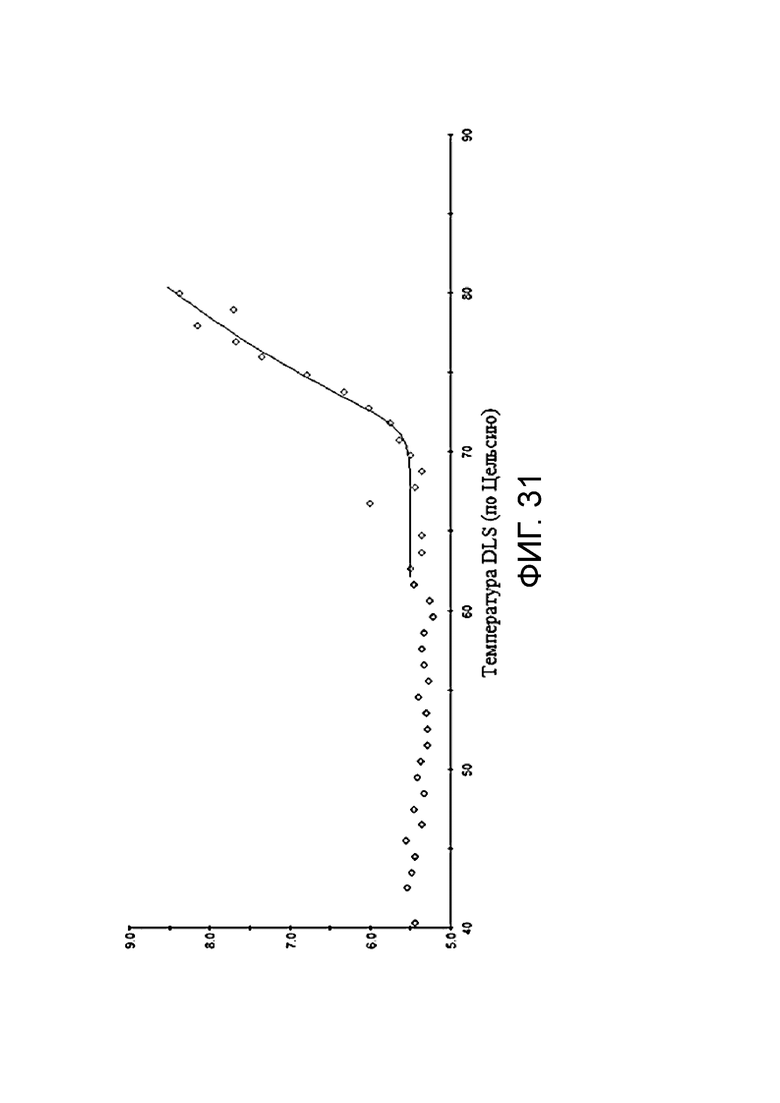
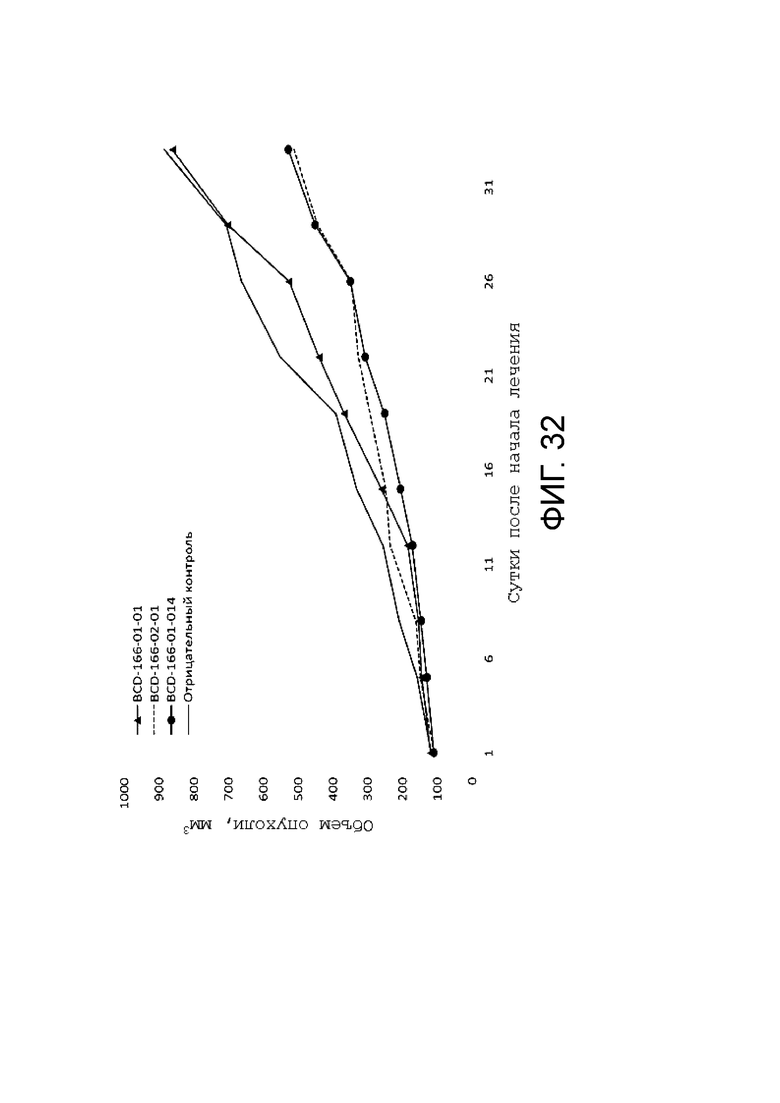
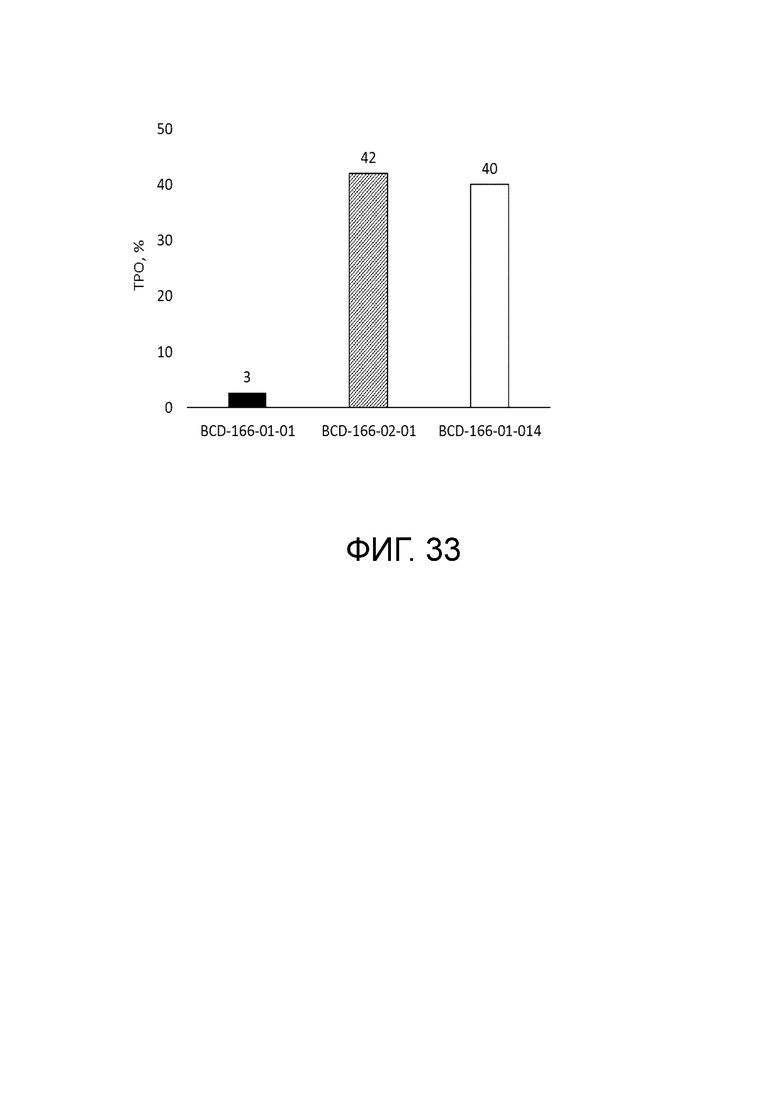
Настоящее изобретение относится к области иммунологии. Предложено антитело или его антигенсвязывающий фрагмент против GITR, нуклеиновая кислота, экспрессионный вектор, способ получения клетки-хозяина, клетка-хозяин. Также рассмотрен способ получения антитела или его антигенсвязывающего фрагмента, фармацевтические композиции. Кроме того, предложен способ ингибирования биологической активности GITR, способ лечения, применение антитела или его антигенсвязывающего фрагмента. Данное изобретение может найти дальнейшее применение в терапии рака, опосредуемого GITR. 11 н. и 49 з.п. ф-лы, 33 ил., 10 табл., 23 пр.
1. Выделенное моноклональное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с GITR, включающее:
а) вариабельный домен тяжелой цепи, который содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3 соответственно, или с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14 соответственно, и
(b) вариабельный домен легкой цепи, который содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9 соответственно, или с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19 соответственно,
при этом, если вариабельный домен тяжелой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3 соответственно, то вариабельный домен легкой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9 соответственно,
если вариабельный домен тяжелой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14 соответственно, то вариабельный домен легкой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19 соответственно.
2. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельный домен тяжелой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3 соответственно.
3. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельный домен тяжелой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14 соответственно.
4. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельный домен легкой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9 соответственно.
5. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельный домен легкой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19 соответственно.
6. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где
вариабельный домен тяжелой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 1, 2 и 3 соответственно;
вариабельный домен легкой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 7, 8 и 9 соответственно.
7. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где
вариабельный домен тяжелой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 12, 13 и 14 соответственно;
вариабельный домен легкой цепи содержит CDR 1, 2 и 3 с аминокислотными последовательностями, представленными последовательностями SEQ ID NO: 17, 18 и 19 соответственно.
8. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 4.
9. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 5, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 4.
10. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 15.
11. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 5, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 15.
12. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельный домен легкой цепи содержит аминокислотную последовательность, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 10.
13. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 12, где вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 10.
14. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельный домен легкой цепи содержит аминокислотную последовательность, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 20.
15. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 12, где вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 20.
16. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где
вариабельный домен тяжелой цепи содержит аминокислотную последовательность, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 4;
вариабельный домен легкой цепи содержит аминокислотную последовательность, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 10.
17. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 16, где
вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 4;
вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 10.
18. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, где
вариабельный домен тяжелой цепи содержит аминокислотную последовательность, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 15;
вариабельный домен легкой цепи содержит аминокислотную последовательность, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 20.
19. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 18, где
вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 15;
вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 20.
20. Моноклональное антитело по п. 1, где антитело, которое специфично связывается с GITR, представляет собой полноразмерное антитело IgG.
21. Моноклональное антитело по п. 20, где полноразмерное антитело IgG относится к изотипу IgG1, IgG2, IgG3, IgG4 человека.
22. Моноклональное антитело по п. 21, где полноразмерное антитело IgG относится к изотипу IgG1 человека.
23. Моноклональное антитело по п. 1, где антитело, которое специфично связывается с GITR, включает мутацию E345R в Fc фрагменте для увеличения агонистических свойств, антителозависимой клеточной цитотоксичности (ADCC), но не комплементзависимой цитотоксичности (CDC).
24. Моноклональное антитело по п. 1, включающее тяжелую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 5.
25. Моноклональное антитело по п. 1, включающее тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 5.
26. Моноклональное антитело по п. 1, включающее тяжелую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 6.
27. Моноклональное антитело по п. 1, включающее тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 6.
28. Моноклональное антитело по п. 1, включающее легкую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 11.
29. Моноклональное антитело по п. 1, включающее легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11.
30. Моноклональное антитело по п. 1, включающее:
тяжелую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 5;
легкую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 11.
31. Моноклональное антитело по п. 1, включающее:
тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 5;
легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11.
32. Моноклональное антитело по п. 1, включающее:
тяжелую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 6;
легкую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 11.
33. Моноклональное антитело по п. 1, включающее:
тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 6;
легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11.
34. Моноклональное антитело по п. 1, включающее тяжелую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 16.
35. Моноклональное антитело по п. 1, включающее тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 16.
36. Моноклональное антитело по п. 1, включающее легкую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 21.
37. Моноклональное антитело по п. 1, включающее легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 21.
38. Моноклональное антитело по п. 1, включающее:
тяжелую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 16;
легкую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% гомологичную последовательности SEQ ID NO: 21.
39. Моноклональное антитело по п. 1, включающее:
тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 16;
легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 21.
40. Выделенная нуклеиновая кислота, которая кодирует антитело или его антигенсвязывающий фрагмент по любому из пп. 1-39.
41. Нуклеиновая кислота по п. 40, где нуклеиновая кислота представляет собой ДНК.
42. Экспрессионный вектор, содержащий нуклеиновую кислоту по любому из пп. 40, 41.
43. Способ получения клетки-хозяина для получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-39, включающий трансформирование клетки вектором по п. 42.
44. Клетка-хозяин для получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-39, содержащая нуклеиновую кислоту по любому из пп. 40, 41.
45. Способ получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-39, заключающийся в культивировании клетки-хозяина по п. 44 в культуральной среде в условиях, достаточных для получения указанного антитела, при необходимости, с последующим выделением и очисткой полученного антитела.
46. Фармацевтическая композиция для лечения заболевания или нарушения, опосредуемого GITR, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-39 в терапевтически эффективном количестве в сочетании с одним или несколькими фармацевтически приемлемыми эксципиентами.
47. Фармацевтическая композиция по п. 46, предназначенная для лечения заболевания или нарушения, опосредуемого GITR, выбранного из группы: РШМ (рак шейки матки), рак головы и шеи, рак желудка, РМЖ (рак молочной железы), почечно-клеточный рак, КРР (колоректальный рак), РЯ (рак яичника), НМРЛ (немелкоклеточный рак легкого).
48. Фармацевтическая композиция для лечения заболевания или нарушения, опосредуемого GITR, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-39 в терапевтически эффективном количестве и по меньшей мере одно терапевтически активное противоопухолевое соединение в терапевтически эффективном количестве.
49. Фармацевтическая композиция по п. 48, предназначенная для лечения заболевания или нарушения, опосредуемого GITR, выбранного из группы РШМ (рак шейки матки), рак головы и шеи, рак желудка, РМЖ (рак молочной железы), почечно-клеточный рак, КРР (колоректальный рак), РЯ (рак яичника), НМРЛ (немелкоклеточный рак легкого).
50. Фармацевтическая композиция по п. 48, где терапевтически активное противоопухолевое соединение выбирают из химиотерапевтического средства, антитела или противогормонального средства.
51. Фармацевтическая композиция по п. 50, где терапевтически активное противоопухолевое соединение представляет собой антитело, которое выбирают из группы: антитела к PD1, антитела к PD-L1, антитела к CTLA4, антитела к анти 4-tlBB. антитела к ОХ40 или их комбинации.
52. Фармацевтическая композиция по п. 50, где терапевтически активное противоопухолевое соединение представляет собой малую молекулу.
53. Фармацевтическая композиция по п. 48, где терапевтически активное противоопухолевое соединение выбирают из группы активаторов врожденного или приобретенного иммунитета.
54. Фармацевтическая композиция по пп. 50-53, где антитело по пп. 1-39 и по меньшей мере одно терапевтически активное противоопухолевое соединение вводятся последовательно.
55. Фармацевтическая композиция по пп. 50-53, где антитело по пп. 1-39 и по меньшей мере одно терапевтически активное противоопухолевое соединение вводятся одновременно.
56. Способ ингибирования биологической активности GITR у субъекта, нуждающегося в таком ингибировании, включающий введение субъекту эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп. 1-39.
57. Способ лечения заболевания или нарушения, опосредованного GITR, включающий введение субъекту антитела или его антигенсвязывающего фрагмента по любому из пп. 1-39 или фармацевтической композиции по п. 46 или 48, нуждающемуся в таком лечении, в терапевтически эффективном количестве.
58. Способ лечения заболевания или нарушения по п. 57, где заболевание или нарушение выбрано из группы: РШМ (рак шейки матки), рак головы и шеи, рак желудка, РМЖ (рак молочной железы), почечно-клеточный рак, КРР (колоректальный рак), РЯ (рак яичника), НМРЛ (немелкоклеточный рак легкого).
59. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-39 или фармацевтической композиции по п. 46 или 48 для лечения у субъекта, нуждающегося в таком лечении, заболевания или нарушения, опосредуемого GITR.
60. Применение по п. 59, где заболевание или нарушение выбрано из группы: РШМ (рак шейки матки), рак головы и шеи, рак желудка, РМЖ (рак молочной железы), почечно-клеточный рак, КРР (колоректальный рак), РЯ (рак яичника), НМРЛ (немелкоклеточный рак легкого).
| АНТИ-GITR-АНТИТЕЛА | 2010 |
|
RU2646139C1 |
| АНТИ-GITR-АНТИТЕЛА | 2010 |
|
RU2595409C2 |
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| КАДАГИДЗЕ З.Г | |||
| "Новые подходы к регуляции противоопухолевого иммунитета", Маммология, 2007, No | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| BIEDERBICK KD, SCHMIDT-WOLF IGH "Efficacy of cytokine induced killer cells targeting CD40 and GITR", ONCOLOGY LETTERS, 2019, Vol | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| Слуховой прибор | 1925 |
|
SU2425A1 |

