Область техники настоящего изобретения
Изобретение, главным образом, относится к области молекулярной биологии. В частности, настоящее изобретение относится к новому пептиду, применимому для идентификации, очистки или выделения содержащих его рекомбинантных белков. Кроме того, настоящее изобретение относится к нуклеотидной последовательности, кодирующей указанную пептидную метку. Настоящее изобретение также относится к вектору для клонирования, содержащему нуклеотидную последовательность, кодирующую интересующий рекомбинантный белок, слитую с сохранением одной рамки считывания с нуклеотидной последовательностью, кодирующей указанный новый пептид.
Предшествующий уровень техники настоящего изобретения
В ходе лабораторных, в частности молекулярно-биологических, исследований зачастую возникает необходимость детекции рекомбинантных белков. Для этой цели обычно используют специфичные антитела. Однако ввиду высокой вариабельности структуры белков такой подход к детекции требует получения специфичных антител для каждого конкретного белка.
Альтернативой является использование пептидной эпитопной метки, которую вводят в рекомбинантный белок, и антител, специфично узнающих эту метку, для детекции рекомбинантных белков, содержащих эту метку. Метка в комплексе с узнающими ее антителами также позволяет проводить и аффинную очистку содержащих ее рекомбинантных белков.
В качестве метки часто используют короткие пептиды. Использование коротких аминокислотных последовательностей позволяет минимизировать эффект присутствия посторонних аминокислот на свойства рекомбинантного белка. Размер таких эпитопных меток может варьироваться в пределах от нескольких аминокислотных остатков до десятков аминокислотных остатков. Метка может располагаться как на N-конце рекомбинантного белка (например, метка FLAGTM, 8 аминокислотных остатков, Sigma-Aldrich, Inc., каталожный номер F3290; Норр ТР et al. (1988) Bio/Technology 6:1204-10; Chubet R and Brizzard В (1996) BioTechniques 20:136-141; Hernan R et al. (2000) BioTechniques 28:789-93), так и на С-конце (например, метка recА, 144 аминокислотных остатка; Krivi GG et al. (1985) J. Biol. Chem. 260:10263-7), однако возможно и расположение метки на 3'-конце. Кроме того, метка может быть встроена в неконцевых положениях аминокислотной последовательности рекомбинантного белка.
Для получения рекомбинантных белков с меткой используют плазмидные или другие векторы, которые содержат кодирующую метку нуклеотидную последовательность. Клонирование в такие векторы кДНК интересующего белка с сохранением одной рамки считывания белка и метки позволяет экспрессировать рекомбинантный белок с меткой. Как правило, для этих целей в различных экспрессирующих системах (например, в клетках Е.coli или млекопитающих с использованием различных контролирующих экспрессию вспомогательных последовательностей) используют различные векторы. Альтернативой является введение нуклеотидной последовательности, кодирующей метку, в кДНК с использованием олигонуклеотидов с помощью полимеразной цепной реакции с последующим клонированием продукта полимеразной цепной реакции в желаемый вектор для продукции рекомбинантных белков.
В качестве ближайшего аналога технического решения, составляющего основу настоящего изобретения, можно привести метку FLAGTM.
Краткое описание чертежей
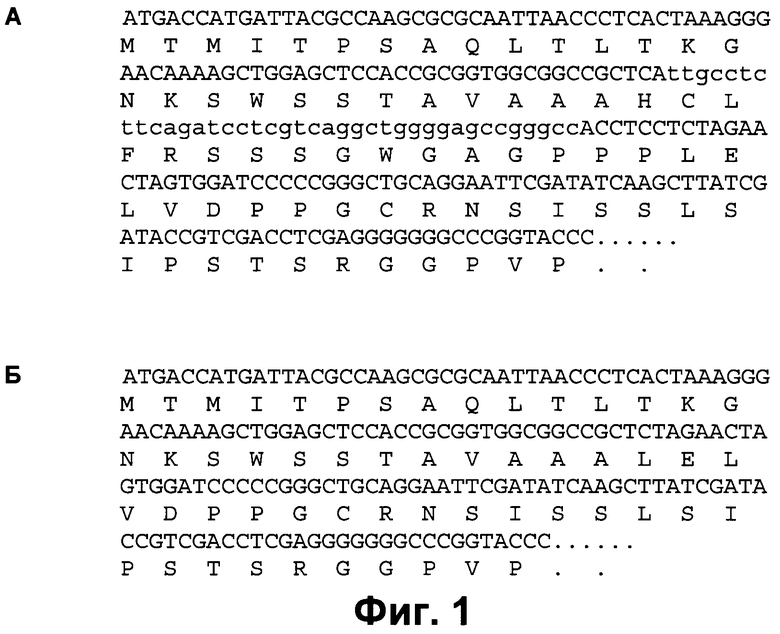
Фиг.1. Приведены нуклеотидная последовательность района полилинкера вектора pDED (фиг.1А) и для сравнения - аналогичного района вектора pBluescript SKII(+) (фиг.1Б). Показана рамка считывания кДНК lacZ, которая не нарушается в результате вставки нуклеотидной последовательности, кодирующей пептид DED. Для вектора pDED нуклеотидная последовательность, комплементарная последовательности, кодирующей идентификационный пептид DED, выделена строчными буквами.
Фиг.2. Фрагмент последовательности белка Рабаптин-5-гамма мыши, включающий последовательность, идентичную последовательности пептида DED, и фрагмент последовательности белка Рабаптин-5, в котором последовательность, идентичная последовательности пептида DED, разделена на две части 43 аминокислотными остатками. Последовательность, идентичная последовательности пептида DED в белке Рабаптин-5-гамма, и фрагменты последовательности, идентичной последовательности пептида DED в белке Рабаптин-5, показаны жирным шрифтом.
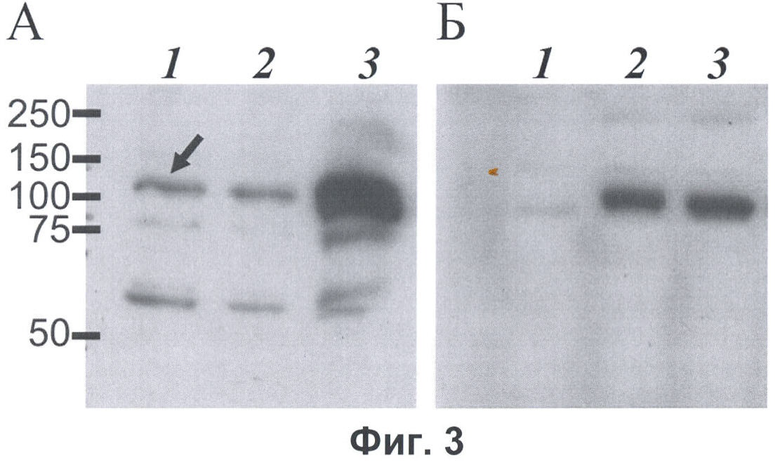
Фиг.3. Вестерн-блот анализ с моноклональными антителами мыши, специфически узнающими пептид DED (А) и поликлональными анти-FLAG антителами кролика (Sigma) (Б) лизатов клеток линии COS-7 (1), лизатов клеток линии COS-7, продуцирующих белок Рабаптин-5 с С-концевым FLAG-эпитопом (2), и лизатов клеток линии COS-7, продуцирующих белок Рабаптин-5-гамма с С-концевым FLAG-эпитопом (3). Белки разделяли в 7,5% ДСН-полиакриламидном геле. Детекцию первичных антител мыши проводили с помощью конъюгированных с пероксидазой хрена антител против иммуноглобулинов мыши (GE Healthcare, Великобритания) и системы ECL+ (GE Healthcare), детекцию первичных антител кролика проводили с помощью конъюгированных с пероксидазой хрена антител против иммуноглобулинов кролика (GE Healthcare,) и системы ECL+(GE Healthcare). Слева показаны положения маркеров молекулярных весов белков в кДа. Эндогенно продуцируемый клетками линии COS-7 белок Рабаптин-5-гамма отмечен стрелкой.
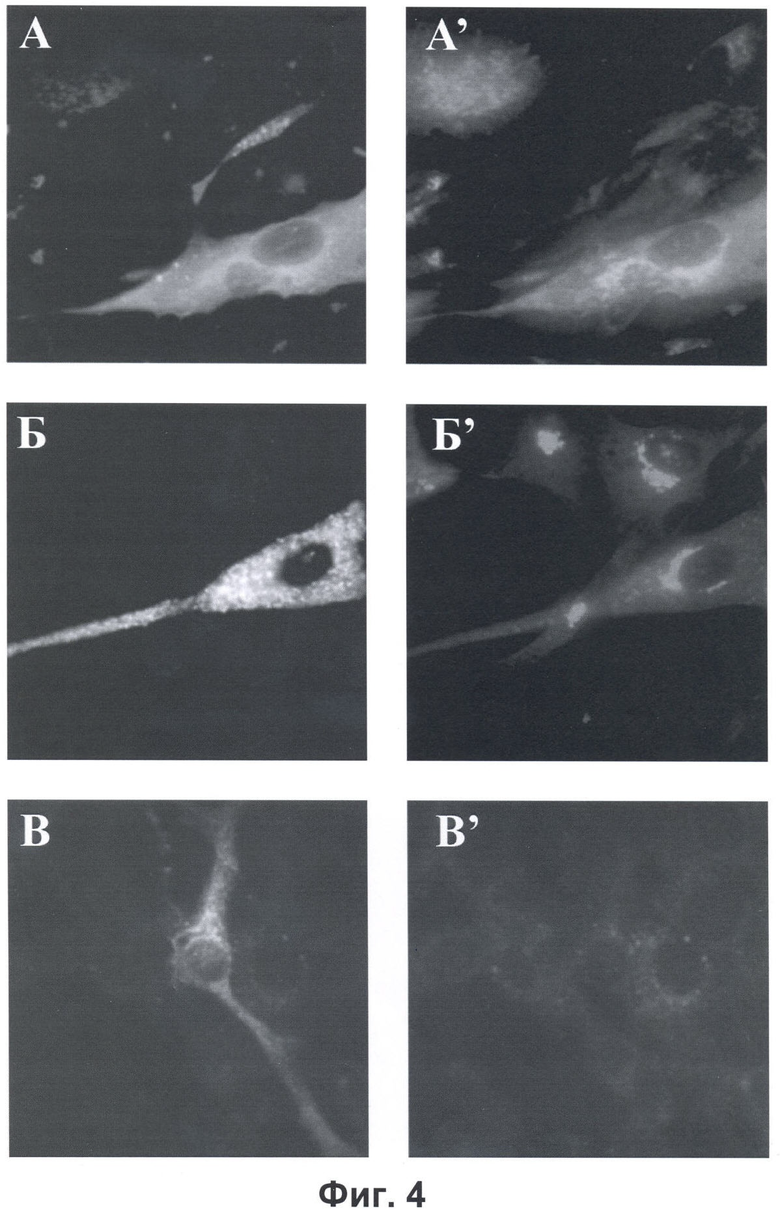
Фиг.4. Результаты иммуноцитохимической детекции химер зеленого флуоресцентного белка с Рабаптином-5-гамма (А-А') и Рабаптина-5 (Б-Б'), продуцируемых клетками линии ВНК-21, с помощью моноклональных антител, специфически узнающих пептид DED, и химеры зеленого флуоресцентного белка с Рабаптином-5-гамма с помощью моноклональных антител, специфически узнающих пептид DED, которые были предварительно проинкубированы с 10-кратным молярным избытком пептида DED (В-В'). Показана флуоресценция зеленого флуоресцентного белка (А, Б, В) и флуоресценция вторичных антител, связанных с иммобилизованными антителами к пептиду DED (А', Б', В').
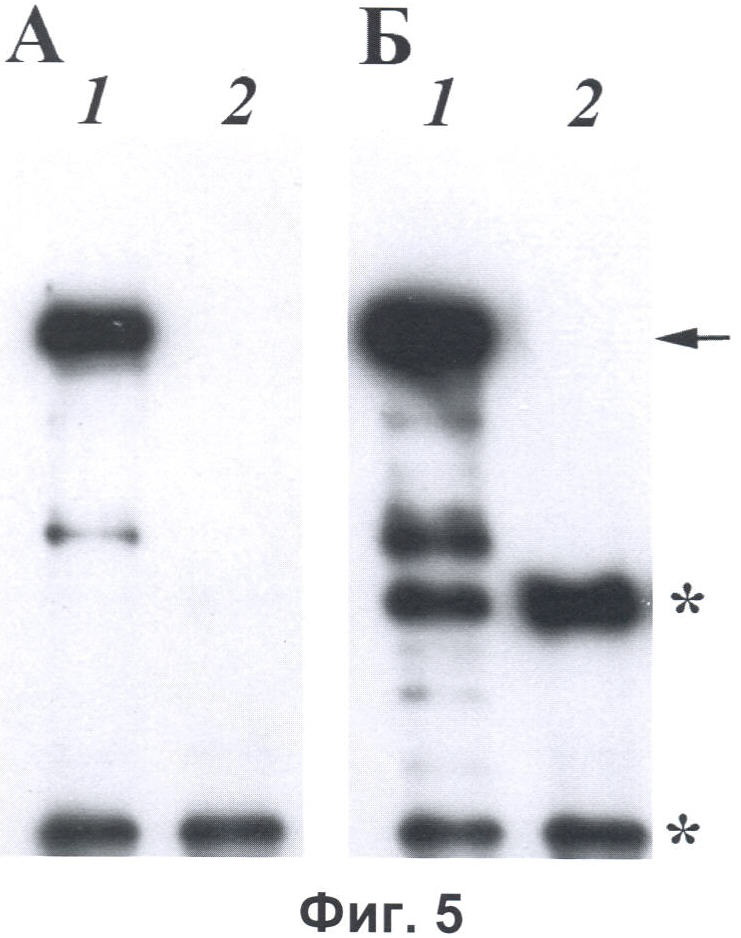
Фиг.5. Вестерн-блот анализ с анти-FLAG антителами кролика (А) и моноклональными антителами мыши к пептиду DED (Б) белков, очищенных с помощью антител к пептиду DED, иммобилизованных на Протеин G-Сефарозе из лизатов клеток линии COS-7, продуцирующих белок Рабаптин-5-гамма мыши (1) или белок Рабаптин-5 мыши (2) с С-концевыми FLAG-эпитопами. Белки разделяли в 7,5% ДСН-полиакриламидном геле. Детекцию первичных антител мыши проводили с помощью конъюгированных с пероксидазой хрена антител против иммуноглобулинов мыши и системы ECL+, детекцию первичных антител кролика проводили с помощью конъюгированных с пероксидазой хрена антител против иммуноглобулинов кролика и системы ECL+. Положения тяжелых и легких цепей иммуноглобулинов отмечены звездочками, положение белка Рабаптин-5-гамма с С-концевым FLAG-эпитопом отмечено стрелкой.
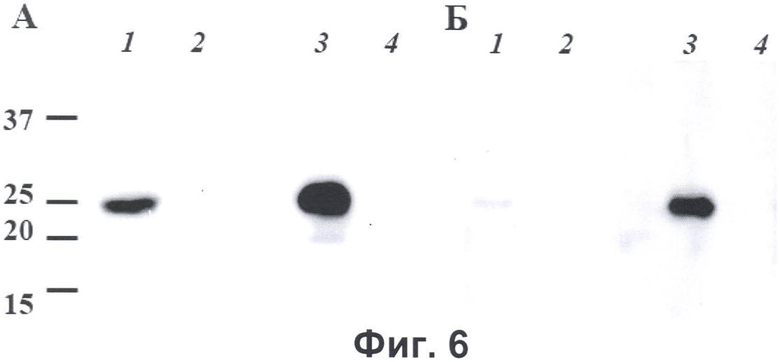
Фиг.6. Детекция белка hVEGF-165 с С-концевым идентификационным пептидом DED в кондиционной среде клеток линии СНО, трансфицированных плазмидой pVEGF-DED. Аликвоты кондиционных сред клеток линии СНО, трансфицированных плазмидой pVEGF-DED (1) и нетрансфицированных клеток линии СНО (2), а также осажденные из них белки (3 и 4, соответственно), разделяли в 12,5% ДСН-полиакриламидном геле, белки переносили на поливинилхлоридную мембрану и проводили Вестерн-блот анализ с использованием моноклональных антител мыши, специфически узнающих идентификационный пептид (А) или поликлональных антител козы к белку hVEGF-165 (Sigma) (Б). Детекцию первичных моноклональных антител мыши проводили с помощью конъюгированных с пероксидазой хрена антител против иммуноглобулинов мыши и системы ECL+, детекцию первичных антител козы проводили с помощью конъюгированных с пероксидазой хрена антител против иммуноглобулинов козы и системы ECL+. Слева показаны положения маркеров молекулярных весов белков в кДа.
Раскрытие настоящего изобретения
Существует несколько необходимых условий для создания эффективной системы детекции и очистки на основе гибридных полипептидов. Первым основным требованием к добавляемой маркерной последовательности является отсутствие влияния метки на нативную укладку белка, к которому эта метка присоединяется. Во-вторых, маркерная пептидная последовательность должна быть гидрофильной и экспрессироваться по существу на поверхности белка с тем, чтобы она могла надежно взаимодействовать со своим лигандом (например, антителом). Кроме того, желательно, чтобы использование метки позволяло применять для аффинной очистки относительно простую и недорогую методику. И наконец, также предпочтительно, чтобы пептидная метка при необходимости могла быть легко удалена с получением нативного белка (Einhauer A. and Jungbauer A. (2001) J. Biochem. Biophys. Methods 49:455-465).
В соответствии с настоящим изобретением пептидная метка рекомбинантного белка представляет собой пептид DED, характеризующийся аминокислотной последовательностью Gly-Pro-Ala-Pro-Gln-Pro-Asp-Glu-Asp-Leu-Lys-Arg-Gln.
Два содержащихся в пептиде DED высокогидрофильных трипептида определяют общее высокое значение гидрофильности С-конца пептида по широко известным шкалам (Норр Т.Р. and Woods K.R. (1981) Proc. Natl. Acad. Sci. USA 78:3824-8; Kyte J. and Doolittle R.F. (1982) J. Mol. Biol. 157(6):105-142). Было показано, что при трехмерной укладке белка такие гидрофильные последовательности занимают положение на поверхности белковой молекулы, что определяет, помимо прочего, высокую антигенность гибридной молекулы (Норр Т.Р. (1986) J. Immunol. Methods 88:1-18).
Олигопептид согласно настоящему изобретению может кодироваться большим множеством молекул нуклеиновых кислот, что является результатом хорошо известного в данной области техники явления вырожденности генетического кода. Суть феномена состоит в том, что любая аминокислота (за исключением триптофана и метионина), входящая в состав природных пептидов, может кодироваться более чем одним триплетным нуклеотидным кодоном (см. таблицу). Все эти вырожденные кодирующие молекулы нуклеиновых кислот подпадают под объем настоящего изобретения.
Одна из возможных молекул нуклеиновых кислот, кодирующих пептид DED согласно настоящему изобретению, представляет собой 5'- ggcccggctc cccagcctga cgaggatctg aagaggcaa-3' (SEQ ID NO:1).
В соответствии с настоящим изобретением вспомогательным вектором для получения нуклеотидных последовательностей, кодирующих интересующий рекомбинантный белок с пептидной меткой, является вектор pDED. Вектор pDED позволяет путем простого клонирования в него кДНК интересующего белка с сохранением одной рамки считывания получить химерную кДНК, кодирующую рекомбинантный белок с меткой. Далее эта кДНК может быть переклонирована в любой желаемый вектор для продукции рекомбинантного белка. При этом нет необходимости использовать набор векторов, кодирующих метку, для различных систем экспрессии, а также синтезировать олигонуклеотиды и использовать полимеразную цепную реакцию для введения метки в состав рекомбинантного белка.
Преимуществом плазмиды pDED является то, что клонирование в нее кДНК может сопровождаться использованием так называемой «бело-голубой» селекции, удобного инструмента отбора колоний клеток, содержащих клонированную кДНК. Селекция основана на том, что сайты узнавания рестриктаз для клонирования в вектор находятся внутри последовательности, кодирующей фермент β-галактозидазу. Если плазмида не содержит вставки, то в E.coli при определенных условиях продуцируется β-галактозидаза, которая способна конвертировать хромогенный субстрат (Х-gal), что приводит к сине-голубой окраске колоний. При наличии вставки рамка считывания кДНК β-галактозидазы нарушается, фермент не продуцируется, и колонии не синеют, а сохраняют белую окраску. При проведении клонирования это позволяет по цвету колоний определить, в клетках каких из них содержится требуемая плазмида, содержащая вставку. Вектор pDED создан на основе вектора для клонирования pBluescript II SK(+), который позволяет использовать бело-голубую селекцию.
Итак, вектор предназначен для клонирования интересующих кДНК с целью получения кДНК, кодирующих рекомбинантный белок с меткой Gly-Pro-Ala-Pro-Gln-Pro-Asp-Glu-Asp-Leu-Lys-Arg-Gln. При этом нуклеотидная последовательность, содержащаяся в векторе pDED и кодирущая метку, не сдвигает рамку считывания β-галактозидазы и не влияет на ее активность. Это позволяет при клонировании в вектор pDED интересующих кДНК проводить бело-голубую селекцию, что обеспечивает легкую идентификацию колонии с плазмидами, содержащими вставки кДНК.
Примеры
Пример 1
Конструирование вектора pDED
Нуклеотидная последовательность фрагмента вектора pDED для получения кДНК, кодирующих белки с идентификационным пептидом DED и позволяющего осуществлять цветную бело-голубую селекцию в клетках Е.coli для выявления клонов, содержащих плазмиды с кДНК, кодирующими рекомбинантные белки с идентификационным пептидом. Вектор pDED получен путем встраивания нуклеотидной последовательности SEQ ID NO:1, кодирующей пептид DED, дополненной на 5'-конце нуклеотидной последовательностью ggagga, кодирующей дипептид глицина, и фланкированной фрагментами сайтов узнавания рестрикционных эндонуклеаз XbaI на 5'-конце и NotI на 3'-конце, в вектор pBluescript SKII(+) (Stratagene, США) между сайтами узнавания рестрикционных эндонуклеаз XbaI и NotI. При этом сохраняется открытая рамка считывания гена lacZ, кодирующего β-галактозидазу, что позволяет, как и в исходном векторе pBluescript SKII(+), осуществлять идентификацию рекомбинантных ДНК, содержащих вставку в полилинкере, на основании цветной бело-голубой селекции колоний E.coli; кДНК, кодирующая белки с идентификационным пептидом DED, полученные в векторе pDED, может быть вырезана из вектора по любому присутствующему сайту в полилинкере между сайтами узнавания между XbaI и KpnI с 5'-конца и любому сайту между NotI и SacI с 3'-конца и вставлена в любой желаемый вектор для продукции рекомбинантного белка с идентификационным пептидом DED на его С-конце (см. Пример 5). Приведены нуклеотидная последовательность района полилинкера вектора pDED (фиг.1А) и для сравнения - аналогичного района вектора pBluescript SKII(+) (фиг.1Б). Структура вектора pDED подробно проиллюстрирована на фиг.1.
Пример 2
Демонстрация специфической детекции белка, имеющего в своем составе идентификационный пептид DED, методом Вестерн-блот анализа
Для демонстрации специфической детекции белков, содержащих последовательность, идентичную последовательности пептида DED, методом Вестерн-блот анализа был использован белок Рабаптин-5-гамма мыши (последовательность кДНК GeneBank Асc. No. AF248289), который содержит внутри полипептидной цепи последовательность, идентичную последовательности пептида DED, и белок Рабаптин-5 мыши (последовательность кДНК GeneBank Асc. № D86066), который содержит внутри полипептидной цепи последовательность, идентичную последовательности пептида DED, первые 7 аминокислотных остатков которого отделены от последующих 6 аминокислотных остатков 43 аминокислотными остатками (фиг.2). Белки Рабаптин-5-гамма и Рабаптин-5 были продуцированы в клетках линии COS-7 с С-концевыми FLAG-эпитопами для возможности мониторинга присутствия белков, для чего клетки линии COS-7 были транзиторно трансфицированы плазмидными векторами для экспрессии соответствующих белков. Через 48 часов после трансфекции клетки подвергали лизису в буфере для нанесения на ДСН-полиакриамидный гель и аликвоты лизатов анализировали методом Вестерн-блоттинга с анти-FLAG антителами (Sigma, США) или с моноклональными антителами, специфически узнающими пептид DED. Анти-FLAG антитела выявили белки Рабаптин-5 и Рабаптин-5-гамма с FLAG-эпитопами в лизатах трансфицированных, но не в лизате нетрансфицированных клеток (фиг.3Б). При этом моноклональные антитела, специфически узнающие пептид DED, уверенно детектировали только белок Рабаптин-5-гамма, но не белок Рабаптин-5, в лизатах трансфицированных клеток (фиг.3А). Незначительный сигнал в лизатах клеток, продуцирующих Рабаптин-5, обусловлен присутствием эндогенного белка Рабаптин-5-гамма в клетках линии COS-7, о чем позволяет сделать вывод присутствие сигнала аналогичной интенсивности в лизатах нетрансфицированных клеток.
Пример 3
Демонстрация возможности специфической детекции белка, имеющего в своем составе идентификационный пептид DED, иммуноцитохимическими методами
Для анализа использовали клетки линии ВНК-21, транзиторно продуцирующие в результате трансфекции соответствующими плазмидами белки Рабаптин-5-гамма или Рабаптин-5, экспрессирующимися как химеры с зеленым флуоресцентным белком на N-концах. Клетки фиксировали метанолом при -20°С и после фиксации окрашивали моноклональными антителами мыши, специфически узнающими пептид DED, с их последующей детекцией антителами к иммуноглобулинам мыши, меченными флуорофором AlexaFluor546 (Invitrogen, США). Клетки, продуцирующие белки-химеры Рабаптина-5-гамма или Рабаптина-5 с зеленым флуоресцентым белком, идентифицировали по зеленому свечению (фиг.4А, Б, В). Окраску моноклональными антителами к пептиду DED выявляли по красному свечению (фиг.4А', Б', В'). Моноклональные антитела к пептиду DED способны узнавать только химеру зеленого флуоресцентного белка с Рабаптином-5-гамма (фиг.4А, А'), но не с Рабаптином-5 (фиг.4Б, Б'). О специфичности окраски антителами к пептиду DED свидетельствует полное отсутствие окраски антителами химеры белка Рабаптин-5-гамма с зеленым флуоресцентным белком при инкубации антител перед детекцией с 10-кратным молярным избытком синтетического пептида DED (фиг.4В, В').
Пример 4
Демонстрация очистки белка, имеющего в своем составе пептид DED, с помощью моноклональных антител, специфически узнающих пептид DED
Клетки линии COS-7, продуцирующих белок Рабаптин-5-гамма мыши, который содержит внутри полипептидной цепи последовательность, идентичную последовательности пептида DED, или белок Рабаптин-5 мыши, который содержит внутри полипептидной цепи последовательность, идентичную последовательности пептида DED, первые 7 аминокислотных остатка которого отделены от последующих 6 аминокислотных остатков 43 аминокислотными остатками, с С-концевыми FLAG-эпитопами (описаны в Примере 2) использовали для приготовления лизатов. Для этого клетки лизировали в трис-солевом буфере, содержащем 1% Тритона Х-100, 0,5% Нонидента Р-40 и ингибиторы протеаз, и после центрифугирования лизаты инкубировали с Протеин G-Сефарозой (GE Healthcare) с иммобилизованными на ней моноклональными антителами, специфически узнающими пептид DED, в течение 3 часов при 4°С. После инкубации Сефарозу промывали 3 раза холодным трис-солевым буфером, добавляли буфер для нанесения на ДСН-полиакриламидный гель и кипятили 5 минут для элюции белков и последующего анализа методом Вестерн-блоттинга с поликлональными анти-FLAG антителами кролика или с моноклональными антителами мыши к пептиду DED. В результате анализа было установлено, что в то время как оба белка - Рабаптин-5-гамма и Рабаптин-5 с С-концевыми FLAG-эпитопами - присутствовали в лизатах (фиг.3Б), только белок Рабаптин-5-гамма, но не Рабаптин-5, оказывался связанным с Протеин G-Сефарозой с иммобилизованными антителами к пептиду DED (фиг.5).
Пример 5
Использование вектора pDED для получения плазмиды для продукции белка с идентифицирующим пептидом на С-конце
Фрагмент кДНК фактора роста эндотелия сосудов-165 человека (hVEGF-165), содержащий 16 пар оснований 5'-нетранслируемой области и всю открытую рамку считывания, был амплифицирован в полимеразной цепной раекции с праймерами 5'-agctgaattcgggcctccgaaacc-3' и 5'-taatctagaccgcctcggcttgtcacat-3', с помощью которых были введены уникальные сайты узнавания рестрикционных эндонуклеаз EcoRI и XbaI на 5'- и 3'-концах кДНК, соответственно, а также кодон терминации трансляции TGA заменен на ТСТ, кодирующий остаток серина. Положение сайта узнавания рестрикционной эндонуклеазы XbaI на 3'-конце кДНК было подобрано таким образом, чтобы рамка считывания кДНК hVEGF-165 и пептида DED в векторе pDED совпадали. Полученный продукт амплификации был обработан рестрикционными эндонуклеазами EcoRI и XbaI и лигирован в вектор pDED, обработанный теми же ферментами. После трансформации в клетки E.coli штамма XL1Blue MRF' (Stratagene) были высеяны на селективную среду, содержащую IPTG и X-gal. Колонии E.coli, содержащие клонированную в вектор pDED кДНК hVEGF-165, были идентифицированы по белой окраске, и наличие вставке в плазмиде было подтверждено рестрикционным анализом изолированных плазмидных ДНК. Затем кДНК, кодирующая hVEGF-165 с С-концвым идентификационным пептидом DED, была субклонирована по сайтам EcoRI и NotI в вектор для экспрессии белков в клетках эукариот (плазмида pVEGF-DED).
Пример 6
Продукция белка hVEGF-165 с идентификационным пептидом DED, расположенным на С-конце, и демонстрация его идентификации с помощью моноклональных антител, специфически узнающих идентификационный пептид DED
Белок hVEGF-165 является секреторным белком, и его секреция обуславливается наличием сигнального пептида на N-конце предшественника белка. Для продукции белка hVEGF-165-DED в культуральную среду клетки линии СНО были трансфицированы плазмидой pVEGF-DED. Через 24 часа после трансфекции среда заменялась на среду, не содержащую сыворотку (бессывороточная среда). Клетки инкубировались в бессывороточной среде в течение 48 часов, после чего среда собиралась и фильтровалась через мембрану с диаметром пор 0,22 мкм для удаления клеток и дебриса. Белки из полученной кондиционной среды концентрировались путем их осаждения метанолом. Для этого к аликвоте среды добавляли 3 объема метанола, и после инкубации в течение ночи при -20°С белки осаждались центрифугированием и растворялись в буфере для нанесения на ДСН-полиакриламидный гель. Аликвоты кондиционной среды и осажденных из нее белков анализировали методом Вестерн-блот анализа с моноклональными антителами, специфически узнающими идентификационный пептид DED. Для контроля использовали кондиционную среду и осажденные из нее белки нетрансфицированных клеток линии СНО. Антитела к пептиду DED детектировали белок с молекулярным весом около 25 кДа, что соответствует молекулярному весу белка hVEGF-165, продуцируемого эукариотическими клетками, только в кондиционной среде клеток, трансфицированных плазмидой pVEGF-DED и осажденных из нее белков, но не нетрансфицированных клеток (фиг.6А), что свидетельствует о продукции белка, содержащего идентификационный пептид. Для подтверждения того, что белок, содержащий идентификационный пептид, действительно является hVEGF-165, те же образцы были проанализировали методом Вестерн-блот анализа с антителами, специфически узнающими белок hVEGF-165. При использовании антител против белка hVEGF-165 был выявлен тот же характер окраски, что и с антителами к пептиду DED (фиг.6Б), что свидетельствует о том, что белок, содержащий идентификационный пептид DED, является белком hVEGF-165.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛИНИЯ КЛЕТОК CHO[V3D], СЕКРЕТИРУЮЩИХ РЕКОМБИНАНТНЫЙ ФАКТОР РОСТА ЭНДОТЕЛИЯ СОСУДОВ (VEGF) ЧЕЛОВЕКА, ИЗОФОРМА А165, С ЭКЗОГЕННЫМ 3×DED-ЭПИТОПОМ | 2012 |
|
RU2493251C1 |
| ВЫДЕЛЕННЫЙ ПЕПТИД, ОБЛАДАЮЩИЙ СПЕЦИФИЧЕСКОЙ СВЯЗЫВАЮЩЕЙ АНТИ-GDF-8 АНТИТЕЛО АКТИВНОСТЬЮ, ВЫДЕЛЕННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, ВЕКТОР ЭКСПРЕССИИ, КЛЕТКА-ХОЗЯИН, СПОСОБ ПОЛУЧЕНИЯ ПЕПТИДА, ВАКЦИННАЯ КОМПОЗИЦИЯ И СПОСОБ ВЫЗЫВАНИЯ ИММУННОГО ОТВЕТА АНТИ-GDF-8, СПОСОБ СКРИНИНГА ДЛЯ ОТБОРА АНТИ-GDF-8 АНТИТЕЛА И СПОСОБ ПОНИЖАЮЩЕГО РЕГУЛИРОВАНИЯ АКТИВНОСТИ GDF-8 У ЖИВОТНОГО | 2004 |
|
RU2422460C2 |
| АНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕСЯ С MUC1, И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2746413C1 |
| НАНОАНТИТЕЛО V9, СВЯЗЫВАЮЩЕЕ VEGF, И СПОСОБ ЕГО ПОЛУЧЕНИЯ, КОДИРУЮЩАЯ V9 НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ И СОДЕРЖАЩИЙ ЕЕ ВЕКТОР, СПОСОБ ИНГИБИРОВАНИЯ ПРОЛИФЕРАЦИИ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК | 2008 |
|
RU2395521C1 |
| НАНОАНТИТЕЛО V93, СВЯЗЫВАЮЩЕЕ VEGF, И СПОСОБ ЕГО ПОЛУЧЕНИЯ, КОДИРУЮЩАЯ V93 НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ И СОДЕРЖАЩИЙ ЕЕ ВЕКТОР, СПОСОБ ИНГИБИРОВАНИЯ ПРОЛИФЕРАЦИИ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК | 2008 |
|
RU2395522C1 |
| СОБАЧИЙ ТИМУСНЫЙ СТРОМАЛЬНЫЙ ЛИМФОПОЭТИЧЕСКИЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2007 |
|
RU2457217C2 |
| БЕЛОК СЛИЯНИЯ, СОДЕРЖАЩИЙ ТИМУСНЫЙ СТРОМАЛЬНЫЙ ЛИМФОПОЭТИЧЕСКИЙ БЕЛОК, ВАКЦИНА, СОДЕРЖАЩАЯ ЭКСПРЕССИОННЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ БЕЛКА, И ВАКЦИНА, СОДЕРЖАЩАЯ УКАЗАННЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ | 2012 |
|
RU2617957C2 |
| Антитела, связывающие опухолеассоциированный MUC1, и способы их получения | 2016 |
|
RU2652901C1 |
| СОСУДИСТЫЙ БЕЛОК - 1 АДГЕЗИИ, ОБЛАДАЮЩИЙ АМИНООКСИДАЗНОЙ АКТИВНОСТЬЮ | 1998 |
|
RU2204838C2 |
| АНТИТЕЛО ПРОТИВ ИНТЕРФЕРОН-АЛЬФА/БЕТА-СВЯЗЫВАЮЩЕГО БЕЛКА II (IFNAB-BPII) | 1995 |
|
RU2362781C2 |



Изобретение относится к области молекулярной биологии. Получают идентификационный пептид с аминокислотной последовательностью: Gly-Pro-Ala-Pro-Gln-Pro-Asp-Glu-Asp-Leu-Lys-Arg-Gln. Полученный пептид применяют для идентификации, очистки или выделения содержащих его рекомбинантных белков. Для получения рекомбинантных белков с полипептидной меткой применяют экспрессирующий вектор pDED. Вектор содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность идентификационного белка. Изобретение позволяет расширить спектр методов идентификации рекомбинантных белков. 5 н. и 9 з.п. ф-лы, 6 ил., 1 табл.
1. Идентификационный пептид, характеризующийся аминокислотной последовательностью Gly-Pro-Ala-Pro-Gln-Pro-Asp-Glu-Asp-Leu-Lys-Arg-Gln.
2. Нуклеиновая кислота, кодирующая аминокислотную последовательность идентификационного пептида по п.1.
3. Применение пептида по п.1 для идентификации рекомбинантных белков.
4. Применение по п.3, причем пептид находится на С-конце или N-конце рекомбинантного белка.
5. Применение по п.3, причем пептид находится в составе рекомбинантного белка в положении, отличном от С-конца или N-конца.
6. Применение по п.3, причем идентификация рекомбинантных белков осуществляется методом вестерн-блоттинга.
7. Применение по п.3, причем идентификация рекомбинантных белков осуществляется иммуноцитохимическими методами.
8. Применение пептида по п.1 для очистки рекомбинантных белков.
9. Применение по п.8, причем пептид находится на С-конце или N-конце рекомбинантного белка.
10. Применение по п.8, причем пептид находится в составе рекомбинантного белка в положении, отличном от С-конца или N-конца.
11. Применение по п.8, причем очистка рекомбинантных белков осуществляется с использованием метода вестерн-блоттинга.
12. Применение по п.8, причем очистка рекомбинантных белков осуществляется с использованием иммуноцитохимических методов.
13. Применение по п.8, причем при очистке рекомбинантных белков моноклональные антитела иммобилизованы на неподвижной фазе.
14. Экспрессирующий вектор pDED на основе вектора pBluescript SKII(+) для получения рекомбинантных белков, включающих в себя пептид по п.1, причем указанный вектор содержит нуклеотидную последовательность по п.2, дополненную нуклеотидной последовательностью ggagga, кодирующей дипептид глицина, на 5'-конце и фланкированную сайтами рестрикции вектора pBluescript SKII(+) ХbаI на 5'-конце и NotI на 3'-конце.
| ПОЛУЧЕНИЕ ОЧИЩЕННЫХ (ПОЛИ)ПЕПТИДОВ | 1997 |
|
RU2182155C2 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |

