Предшествующий уровень техники
Существует потребность в улучшенных вакцинах. Современные стратегии вакцинации против гриппа нацелены на формирование устойчивых гуморальных ответов (образования антител) против гемагглютининов. Ввиду высокой степени антигенного дрейфа между циркулирующими штаммами вируса гриппа в течение одного года, вакцинные штаммы следует заменять специально для каждого сезона гриппа. Хотя годовая (или сезонная) вакцинация против гриппа является успешной в разной степени для различных возрастных категорий, необходима, безусловно, более эффективная защита, особенно для молодых и пожилых людей. Кроме того, существует большой постоянный риск того, что будут развиваться реассортантные вирусы, которые приобрели самые различные гены гемагглютининов (ГА) в результате процесса, известного как «антигенный сдвиг». Это создало бы чрезвычайную ситуацию в здравоохранении, поскольку в настоящее время вакцины гриппа основаны в основном на гемагглютининовом антигене.
Вирус гриппа является оболочечным, одноцепочечным вирусом с отрицательной смысловой РНК из семейства вирусов Orthomyxoviridae, который разделяется на 3 основных типа: А, В и С. Вирусы гриппа А заражают самых различных животных, включая людей, птиц, свиней, лошадей, летучих мышей и многих других, хотя тропизм любого конкретного вируса гриппа, как правило, в высшей степени приспособлен к определенному хозяину. Вирусы гриппа В заражают меньшее количество видов, а именно, человека и тюленей, но также являются основной причиной ежегодных эпидемий гриппа. Большинство гриппозных инфекций человека вызваны вирусом гриппа А или В; вирусы гриппа С, которые заражают людей и свиней, реже бывают причиной серьезных инфекций или эпидемий у людей (Lamb, 2001).
Современные вакцины на основе инактивированного вируса гриппа вызывают образование антител, защищающих от близкородственных штаммов вируса. Вакцины, разрешенные к применению в настоящее время, вызывают, главным образом, образование штамм-специфических нейтрализующих антител против гемагглютинина (ГА), главной антигенной детерминанты на поверхности вируса, которые обладают высокой иммуногенностью и могут предотвратить заболевание, вызванное заражением соответствующим штаммом вируса. Тем не менее, ГА обладает существенной антигенной изменчивостью, что исключает его использование в виде однокомпонентной вакцины, предназначенной для обеспечения широкой защиты. По этой причине большой интерес представляют альтернативные стратегии вакцинации, которые формируют защитные ответы, направленные против менее изменчивых мишеней.
Естественное заражение вирусом гриппа А вызывает как гуморальный, так и клеточный иммунитет. Длительный клеточный иммунитет направлен преимущественно против консервативных внутренних вирусных белков, таких как нуклеопротеин (NP, от англ. nucleoprotein). Нуклеопротеиновый антиген (NP антиген) является иммуногенным для организма человека, перенесшего естественные инфекции, но образовавшиеся при этом цитотоксические Т-лимфоциты имеют короткий период существования (McMichael, 1983а, McMichael, 1983b).
Клеточный иммунитет против NP является важным, поскольку он направлен против различных вариантов эпитопов NP, а ДНК-вакцины, для которых мишенью служит NP, вызывают перекрестный иммунитет у животных (Schotsaert, 2012).
Нуклеопротеиновый антиген (NP антиген) уже давно признан в качестве высоко консервативного антигена: даже самые дивергентные штаммы вируса гриппа А обладают 90% идентичностью кодируемых в них белков NP (Gorman, Xu, 1991). Антигенные изменения NP редки и встречаются лишь в незначительной степени ( , 2010).
, 2010).
Применение нуклеопротеина в качестве антигена в вакцинах
Применение нуклеопротеина вируса гриппа в качестве антигена было описан в 1980-х годах (Wraith, 1987). Клеточные иммунные ответы у мышей против NP способны вызывать иммунитет, и в частности давать перекрестный иммунитет против дивергентных штаммов вируса типа А. Было показано, что иммунизация мышей NP, выделенным из вируса H3N2, может обеспечить прочную защиту (75%) при летальном заражении гетерологичным вирусом (H1N1), но не предотвращает инфекцию.
ДНК-вакцины, использующие ген NP, известны двадцать лет: их применяли в первых экспериментах «по доказательству правильности концепции» самой ДНК-вакцинации (Ulmer, 1993).
Экспрессия NP с помощью вирусного вектора была впервые продемонстрирована в 1980 (Yewdell, 1985), а иммунизация с помощью этого вектора была сопряжена с улучшенным образованием цитотоксических Т-лимфоцитов против разнообразных штаммов вируса гриппа типа А, но не типа В, по сравнению с ДНК-вакцинами.
Так, было показано, что иммунизация мышей с помощью вектора MVA, экспрессирующего нуклеопротеин PR8, защищает их от заражения низкой дозой гетеросубтипических вирусов гриппа (Altstein, 2006). Совсем недавно вирусный вектор, кодирующий белок NP, слитый с белком М1, был применен для иммунизации людей (Lillie, Berthoud, 2012; Antrobus, 2012). Эти исследования показали, в частности, что клеточные иммунные ответы на NP могут быть существенно увеличены у пожилых людей (Antrobus, 2012), когда гуморальный ответ снижается в связи со «старением» иммунной системы.
Секреция нуклеопротеина
Ряд исследований показал, что белок NP локализуется преимущественно в ядре, снижая иммуногенность таких ДНК-вакцин (, 2010).
Улучшенные клеточные иммунные ответы против NP могут быть получены путем стимуляции секреции NP, например, при помощи слияния сигнального пептида tPA с геном NP (Luo, 2008), с помощью ДНК-содержащих композиций (Greenland, 2007; Sullivan, 2010) и с помощью применения электропорации (Laddy, 2008) для улучшения доставки ДНК.
Мономерные нуклеопротеины гриппа
Предпочтительное применение мономерных антигенов в слиянии с доменами олигомеризации C4bp было описано в заявке на патент WO 2005/014654. Однако опасность мономерных антигенов заключается в их сниженной иммуногенности. Это было продемонстрировано Bachmann с коллегами на гликопротеине G вируса везикулярного стоматита (Bachmann, 1993) и Fiers с коллегами (Fiers, 2001) для антигена нейраминидазы, или NA, вируса гриппа. Следует ожидать, что уменьшение или разрушение структур более высокого порядка у нуклеопротеинов вируса гриппа будет снижать их иммуногенность.
Ряд мутаций, как было показано, преобразует нуклеопротеин вируса гриппа, который в естественном состоянии является олигомером, в мономерную форму (Ye, 2006). То, что мономерные варианты NP, описанные в этой статье 2006 года, действительно являются мономерными, подтверждается в более поздних работах (Tarus, 2012а; Ye, 2012). Две описанные в этих работах точечные мутации, которые делают мономерным нуклеопротеин вируса гриппа типа А, являются консервативными в нуклеопротеинах штаммов вируса гриппа типов В и С (Фигура 3 в Nakada, 1984). Поэтому те же самые точечные мутации могут быть введены в нуклеопротеины штаммов вируса гриппа типов В и С для того, чтобы сделать мономерными эти нуклеопротеины вируса гриппа. Однако никаких исследований иммуногенности мономерных нуклеопротеинов не проводилось.
Основной технической проблемой при получении вакцины против гриппа с NP антигеном является достижение сильных и прочных клеточных иммунных ответов. «Клеточный иммунный ответ» представляет собой иммунный ответ, который не вовлекает в участие антитела, а вызывает активацию антиген-специфических Т-лимфоцитов, и в частности цитотоксических Т-лимфоцитов, а также высвобождение различных цитокинов в ответ на антиген. Клетки CD4 или хелперные Т-клетки обеспечивают защиту против различных патогенов путем секреции цитокинов, которые активируют иммунный ответ. Цитотоксические Т-клетки (CD8) вызывают гибель патогенов путем апоптоза без использования цитокинов.
Хотя остаются споры о том, CD4 или CD8 ответы против NP являются более важными для защиты (Epstein, 2000), есть мнение, что клеточные, а не гуморальные ответы на нуклеопротеин являются ключом к защите, которую этот антиген может вызывать (Thomas, 2006). Вакцины, которые обеспечивают защиту, вызывая сильный цитотоксический Т-клеточный ответ, могут быть полезными, когда Т-клеточные эпитопы получены из высоко консервативного белка NP (Epstein, 2000; Roy, 2007). Клеточные иммунные ответы, опосредованные Т-лимфоцитами, функционируют, главным образом, путем распознавания инфицированных вирусом гриппа клеток, при помощи ингибирования репликации вируса и посредством ускорения клиренса вируса.
Специфические Т-клетки, участвующие в обеспечении иммунитета, включают как CD4+, так и CD8+ Т-клетки, и часто выполняют свои функции под действием секретируемых цитокинов и цитолитической активности, соответственно. В частности, CD8+ цитотоксические Т-лимфоциты (ЦТЛ) специфичные к NP вируса гриппа могут играть важную роль в гетеросубтипическом защитном иммунитете против летального заражения мышей вирусом гриппа (Gschoesser, 2002), обеспечивая клиренс вируса гриппа с поверхностей слизистой оболочки верхних дыхательных путей (Mbawuike, 2007) и способствуя выживанию и восстановлению после заражения (Epstein, 2000). Оптимальная вакцина на основе NP позволит улучшить как CD4, так и CD8 клеточные ответы.
Данная заявка на патент обеспечивает способы улучшения клеточного иммунного ответа на нуклеопротеины вируса гриппа.
Краткое описание изобретения
Настоящее изобретение относится к способу повышения иммуногенности и, в частности, клеточной иммуногенности NP антигенов из вирусов гриппа путем слияния, по меньшей мере, одного варианта NP антигена из штаммов вируса гриппа типа А, В или С с вариантом домена олигомеризации C4bp курицы в качестве белка-носителя.
Настоящее изобретение относится, в частности, к гибридному белку, содержащему, по меньшей мере, один мономерный вариант NP антигена из штамма вируса гриппа типа А, В или С, и белок-носитель IMX313, имеющий последовательность, показанную в SEQ ID NO: 1, например, такую, как описанная в заявке на патент WO 2007/062819.
Настоящее изобретение, в частности, относится к гибридному белку, включающему мономерный вариант NP антигена из штамма вируса гриппа типа А, содержащий точечные мутации Е339А и R416A, как показано в SEQ ID NO: 2, и вариант белка-носителя IMX313, содержащий С-концевую замену, по меньшей мере, одним положительно заряженным пептидом, имеющим последовательность ZXBBBBZ где (i) Ζ обозначает любую аминокислоту или отсутствует, (ii) X представляет собой любую аминокислоту и (iii) В является аргинином (R) или лизином (K), как показано в SEQ ID NO: 3, например, как описано в заявке на патент РСТ/ЕР 2013/076289, поданной 11 декабря 2013 года. Предпочтительный вариант белка-носителя IMX313 не вызывает образование антител, которые перекрестно реагируют с протамином.
Настоящее изобретение, в частности, относится к гибридному белку, включающему мономерный вариант NP антигена и модифицированный белок-носитель IMX313T или IMX313P, показанный соответственно в SEQ ID NO: 4 и SEQ ID NO: 5.
Настоящее изобретение также относится к иммуногенной композиции, содержащей последовательность ДНК в плазмиде или вирусном векторе, дополнительно содержащей сигнальный пептид, например, tPA, показанный в SEQ ID NO: 6.
Настоящее изобретение также относится к рекомбинантной ДНК, кодирующей указанные гибридные белки.
Настоящее изобретение также относится к иммуногенной композиции, содержащей последовательность ДНК, кодируемую плазмидой или вирусным вектором, или к гибридному белку, дополнительно содержащему вакцинные адъюванты или лиганды на основе нуклеиновых кислот для внутриклеточных ТПР, как описано в заявке на патент РСТ/ЕР 2013/076289, поданной 11 декабря 2013 года.
Настоящее изобретение также относится к плазмидной ДНК, вирусному вектору, гибридному белку или иммуногенной композиции для использования в качестве вакцины или для иммунотерапии в качестве способа профилактики или лечения гриппа.
Подробное описание изобретения
Перед подробным описанием настоящего изобретения следует понять, что данное изобретение не ограничивается, в частности, приведенными примерами способов и может, безусловно, варьироваться. В частности, настоящее изобретение относится к гибридным белкам, содержащим, по меньшей мере, один нуклеопротеиновый антиген вируса гриппа, и не ограничивается конкретным нуклеопротеином вируса гриппа.
Все публикации, патенты и патентные заявки, цитируемые здесь, независимо от того являются ли они supra или infra, включены в данное описание путем ссылки. Тем не менее, упомянутые в настоящем изобретении публикации цитируются с целью описания и раскрытия протоколов, реагентов и векторов, которые приведены в публикациях и могут быть использованы в связи с настоящим изобретением.
Кроме того, для осуществления настоящего изобретения используются, если не указано иное, обычная очистка белков и методы молекулярной биологии из данной области техники. Такие методы хорошо известны специалистам в данной области техники и полностью описаны в литературе. В нижеследующей формуле изобретения и в последовательном описании изобретения, если контекст не требует иного для того, чтобы выразить язык или необходимый смысл, слова «вмещать», «содержать», «заключать в себе» или «включать» или их варианты, такие как «содержит», «содержащий», «включающий», «включенный», «включает», «включающий» используются в включающем смысле, т.е. для определения присутствия заявленных признаков, что не исключает наличия или добавления дополнительных признаков в различные формы осуществления изобретения.
Следующие ниже термины определены для лучшего понимания изобретения.
Вирусы гриппа бывают трех типов А, В и С. Эта классификация была первоначально серологической: антисыворотки против нуклеопротеина вируса гриппа типа А перекрестно реагируют с нуклеопротеинами других вирусов гриппа типа А, но не реагируют с нуклеопротеинами вирусов гриппа типа В или типа С. Вирусы гриппа типа А классифицируются на подтипы на основе серологических перекрестных реакций их гликопротеинов гемагглютинина (Н) и нейраминидазы (N).
Под термином «нуклеопротеины вируса гриппа» подразумеваются нуклеопротеины вирусов гриппа всех трех типов (А, В и С).
Термин «белок-носитель», как правило, обозначает белок, с которым конъюгированы или слиты антигены, за счет чего они становятся более иммуногенными. В настоящем изобретении этот термин специально используется в значении «белок, несущий антиген». Функцией этого белка является повышение иммуногенности указанного антигена, с которым он конъюгирован или слит.
Термин «вариант нуклеопротеина» обозначает все белки с последовательностью, имеющей идентичность, по меньшей мере, 90% с версиями дикого типа нуклеопротеина вируса гриппа из штаммов вируса гриппа типа А, В и С.
Термин «вариант домена олигомеризации курицы» обозначает вариант домена C4bp с последовательностью SEQ ID NO: 1, описанный в заявках на патент WO 2007/062819 и РСТ/ЕР 2013/076289, поданных 11 декабря 2013 года, обе ссылки включены в настоящее изобретение путем ссылки; в частности, фрагмент, состоящий, по меньшей мере, из 48 смежных аминокислот и/или обладающий, по меньшей мере, 70% идентичностью с аминокислотной последовательностью SEQ ID NO: 1, описанный заявке на патент WO 2007/062819.
«Вариант белка-носителя IMX313» описан в заявке на патент РСТ/ЕР 2013/076289, поданной 11 декабря 2013 года.
Протамин состоит из группы гетерогенных пептидов со средней молекулярной массой 4500 дальтон, полученных из рыбы. Приблизительно 67% аминокислотного состава протамина составляет аргинин. Протамин уже давно используется для получения инсулина (нейтральный протамин Хагедорна), или для нейтрализации гепарина.
Термин «гибридный белок» обозначает рекомбинантный белок, не существующий в природе, содержащий два домена из различных источников, которые были подвергнуты слиянию. Точнее, в данном изобретении, гибридный белок включает нуклеопротеиновый антиген вируса гриппа, слитый с вариантом домена-носителя из домена олигомеризации C4bp курицы, в частности, «IMX313T» или «IMX313P». Процесс слияния обладает тем преимуществом, что образует гомогенный продукт.«Конъюгация» может быть более формально описана с генетической точки зрения таким образом: ДНК, кодирующую про-иммуногенный белок-носитель, сращивают с ДНК, кодирующей антиген. Антиген может быть слит с N- или С-концом белка-носителя.
Изобретение относится к иммуногенной композиции, содержащей, по меньшей мере, один вариант нуклеопротеинового антигена вируса гриппа и вариант домена олигомеризации C4bp, и вызывающей повышенный клеточный иммунный ответ против нуклеопротеинового антигена вируса гриппа.
В соответствии с настоящим изобретением, нуклеопротеин, слитый с вариантом белка-носителя домена олигомеризации C4bp курицы, в частности IMX313T или IMX313P, может представлять собой нуклеопротеин вирусов гриппа любого типа (А, В или С).
Антиген нуклеопротеина может быть слит с N- или С-концом белка-носителя, в частности IMX313T или IMX313P.
Согласно изобретению, по меньшей мере, один нуклеопротеин слит с одним белком-носителем, в частности IMX313T или IMX313P; тем не менее, с одним и тем же белком-носителем могут быть слиты два или более, одинаковых или различных, нуклеопротеина.
В соответствии с предпочтительным аспектом настоящего изобретения, антиген нуклеопротеина слитый с вариантом домена олигомеризации C4bp курицы, в частности IMX313T или IMX313P, представляет собой мономерный антиген. Использовать мономерные антигены, как описано в патентной заявке WO 2005/014654, действительно выгодно при условии, что мономеризация не уменьшает их иммуногенность. Кроме того, NP образует тример в кристаллах (Ye, 2006) и другие олигомеры in vivo (Arranz, 2012; Moeller, 2012). Слияние тримерного или олигомерных белков в гептамерный белок, такой как IMX313T или IMX313P, ведет к риску возникновения стерических конфликтов. С другой стороны, мономерные формы естественных олигомерных белков имеют уменьшенную иммуногенность (Fiers, 2001).
Для получения мономерного антигена нуклеопротеина специалистам в данной области техники известны различные точечные мутации, которые могут быть введены в белковую последовательность NP антигена для того, чтобы вызвать его мономеризацию. В частности, NP антиген имеет, по меньшей мере, одну из двух следующих точечных мутаций: Е339А и R416A.
В одной форме осуществления изобретения NP антиген представляет собой NP антиген из штамма вируса гриппа типа А.
В предпочтительной форме осуществления настоящего изобретения NP антиген содержит обе точечные мутации Е339А и R416A, и, следовательно, является мономерным.
В другой форме осуществления настоящего изобретения NP антиген имеет полипептидную последовательность, показанную в SEQ ID NO: 2.
Увеличение клеточных иммунных ответов на антигены, экспрессированные из ДНК-вакцин, было ранее получено путем слияния гена антигена с геном, кодирующим IMX313 (Spencer, 2012). Предпочтительно, если варианты этого домена имеют С-концевую замену, по меньшей мере, из одного положительно заряженного пептида, обладающего последовательностью ZXBBBBZ, где (i) Z обозначает любую аминокислоту или отсутствует, (ii) X представляет собой любую аминокислоту и (iii) В является аргинином (R) или лизином (K), как показано в SEQ ID NO: 3, которая позволяет улучшить иммунный ответ на антиген, слитый с данным антигеном, например, как описано в патентной заявке РСТ/ЕР 2013/076289, поданной 11-е декабря 2013 года. Предпочтительный вариант белка-носителя IMX313 не вызывает образования антител, которые перекрестно реагируют с протамином.
Конкретные улучшенные варианты, которые называются IMX313T и IMX313P, недавно описаны в заявке на патент РСТ/ЕР 2013/076289, поданной 11 декабря 2013 года. Их пептидные последовательности являются следующими:

В другой форме осуществления изобретения гибридный белок включает в себя NP антиген, который содержит сигнальный пептид. Некоторые исследования показали, что белок NP локализуется, прежде всего, в ядре, что потенциально могло бы уменьшить иммуногенность таких ДНК-вакцин. Таким образом, обеспечение возможности секреции NP антигена путем добавления сигнального пептида является целесообразным. В частности, этот сигнальный пептид представляет собой тканевый активатор плазминогена (tPA), секреторный сигнальный пептид, как описано в (Luo, 2008).
В конкретной форме осуществления настоящего изобретения NP антиген является мономерным и содержит сигнальный пептид.
В другой форме осуществления настоящего изобретения NP антиген включает обе мутации Е339А и R416A, а также сигнальный пептид tPA. Как показано в примерах, гибридный белок, содержащий IMX313T и мономерный NP антиген, включающий сигнальный пептид, вызывает у мышей при введении в форме ДНК-вакцины Th1-ответ (IgG2a) более сильный, чем Th2-ответа (Фиг. 13). По общему мнению иммунологов, Th1-ответы предпочтительнее Th2-ответов. Однако способы улучшения преимущественно Th1-ответов на антиген без использования вспомогательных средств, разработанных для этой цели, в данной области не известны. В приведенных ниже примерах показано, что слияние IMX313T или IMX313P с нуклеопротеиновыми антигенами вируса гриппа предпочтительно улучшает Th1-ответы.
Изобретение также относится к гибридному белку, включающему любой белок-носитель, который содержит биспиральный домен и, по меньшей мере, один нуклеопротеиновый (NP) антиген вируса гриппа. При этом данный нуклеопротеиновый антиген является мономерным.
Настоящее изобретение также относится к нуклеиновой кислоте, кодирующей гибридный белок, такой как описано выше, и, в частности:
- кодирующей гибридный белок, содержащий NP антиген и IMX313T или IMX313P;
- кодирующей гибридный белок, содержащий мономерный NP антиген и IMX313T или IMX313P;
- кодирующей гибридный белок, включающий NP антиген, который содержит сигнальный пептид и IMX313T или IMX313P;
- кодирующей гибридный белок, включающий мономерный NP антиген, который содержит сигнальный пептид и IMX313T или IMX313P.
В предпочтительных формах осуществления изобретения нуклеиновая кислота кодирует гибридный белок, содержащий мономерный NP антиген из вируса гриппа А. В предпочтительных формах осуществления изобретения нуклеиновая кислота кодирует гибридный белок, который не образует антитела, перекрестно реагирующие с протамином. В частности, эти нуклеиновые кислоты представляют собой последовательности, как показано в SEQ ID NO: 6 и SEQ ID NO: 7.
Настоящее изобретение также относится к вектору, содержащему нуклеиновую кислоту, представленную выше, и генетические элементы, такие как промоторы и энхансеры, для того, чтобы обеспечить экспрессию ДНК кассеты в клетках-хозяевах.
Настоящее изобретение также относится к иммуногенной композиции, содержащей:
- гибридный белок, или нуклеиновую кислоту, или вектор, упомянутые выше, и
- лиганды на основе нуклеиновых кислот для внутриклеточных ТПР и/или каких-либо других адъювантов.
Толл-подобные рецепторы (ТПР)
Клетки врожденной иммунной системы обнаруживают патогены через ограниченный набор рецепторов, кодируемых генами зародышевой линии. Эти рецепторы врожденного иммунитета распознают ряд экспрессируемых патогенами консервативных молекулярных структур, называемых патоген-ассоциированными молекулярными паттернами (ПАМП).
Эти производимые патогенами молекулы представляют собой сложные молекулы, которые высоко специфичны для набора патогенов. Толл-подобные рецепторы (ТПР) представляют собой набор рецепторов, распознающих иммунные паттерны, которые способны привести в состояние готовности иммунную систему немедленно после заражения патогеном. Они играют важную роль в качестве ключевых компонентов между врожденным и приобретенным иммунитетом и могут распознавать многие болезнетворные микроорганизмы, в диапазоне от вирусов до паразитов. Первый охарактеризованный ТПР, названный «толл», отвечает, как было показано, за противогрибковые ответы у взрослых мух дрозофил (Drosophila) и на сегодняшний день определены 10 его эквивалентов у человека, участвующих в распознавании патогена. ТПР могут быть классифицированы на различные группы в зависимости от их локализации и типа ПАМП, которые они распознают. ТПР 1, 2, 4, 5 и 6 экспрессируются, главным образом, на поверхности клетки, где они в основном распознают бактериальные продукты, тогда как ТПР 3, 7, 8 и 9 локализованы в внутриклеточных компартментах и распознают в основном вирусные продукты и нуклеиновые кислоты.
Внутриклеточные толл-подобные рецепторы
Кроме того, для совершенствования способов иммунизации также имеет большое значение ограничение передачи сигналов через ТПР рецепторы. Толл-подобные рецепторы (ТПР) представляют собой класс белков, которые играют ключевую роль в иммунной системе. После того, как микробы нарушили физические барьеры организмов, их распознают ТПР. Распознаваемые признаки микробов включают двухцепочечную РНК вирусов, неметилированные сайты CpG-островков бактерий и вирусов и некоторых РНК и ДНК молекулы.
Существует значительный интерес к таким нуклеиновым кислотам, поскольку они являются лигандами для класса толл-подобных рецепторов (далее ТПР) и, в частности для ТПР3, ТПР7, ТПР8, ТПР9 и ТПР13 (Blasius, 2010 и ссылки внутри статьи). Иногда они классифицируются как «внутриклеточные толл-подобные рецепторы», но, по меньшей мере, ТПРЗ также присутствует на некоторых поверхностях клеток. ТПР3 экспрессируется на множестве эпителиальных клеток, включая эпителиальные клетки дыхательных путей, матки, роговицы, влагалища, шейки матки, желчных протоков и кишечника, и при этом такие клетки экспрессируют ТПР3 на своей клеточной поверхности (Akira, 2006).
Важность ограничения сигналов через эти рецепторы, и в частности рецептор ТПР3, зависит от дозы. Таким образом, прочное связывание лигандов на основе нуклеиновой кислоты с антигеном важно для того, чтобы предотвратить их связывание с ТПР в отсутствие антигена. Поэтому прочно связанные внутриклеточные лиганды ТПР намного предпочтительнее композиций, в которых связывание является менее прочным. Вследствие этого специалисты в данной области заняты поиском антигенных композиций, способных эффективно связывать ТПР лиганды таким образом, чтобы они не отделялись от антигена прежде, чем антиген поступит в клетки, где он вызовет иммунный ответ, с целью уменьшения потенциальных побочных эффектов, опосредованных связыванием лигандов с ТПР рецепторами в других местах.
В настоящей заявке на патент и, в частности, в примерах, были использованы следующие внутриклеточные лиганды ТПР:
- для ТПР3: полиинозин-полицитидиловая кислота poly (I:C), представляющая собой дуплекс полинуклеотида полиинозиновой кислоты гибридизованный с полицитидиловой кислотой, аналог двухцепочечной РНК. Длина ее цепи составляла двадцать нуклеотидов для каждой нити.
- для ТПР7: олигонуклеотид, называемый ssRNA40, с последовательностью 5' GsCsCsCsGsUsCsUsGsUsUsGsUsGsUsGsAsCsUsC 3', где символ «s» представляет собой фосфотиоатную связь (SEQ ID NO: 8);
- для ТПР9: олигонуклеотид, называемый ODN1826 с последовательностью: 5' tccatgacgttcctgacgtt 3' (SEQ ID NO: 9).
В конкретном аспекте настоящего изобретения иммуногенная композиция включает:
- гибридный белок, или нуклеиновую кислоту, или вектор, представленные выше, и
- полиинозин-полицитидиловую кислоту poly (I:C).
Изобретение также относится к гибридному белку, такому, как описано выше, для применения в качестве вакцины с целью профилактики и лечения заболеваний гриппом. Эта вакцина против гриппа может применяться для нескольких направлений использования:
- профилактики сезонного гриппа;
- профилактики в ситуации пандемии;
- «универсальной» профилактики, т.е. иммунизации вакцинами против всех типов вируса гриппа;
- иммунотерапии всех видов гриппа.
Способы профилактики или лечения гриппа могут быть выполнены с помощью специфических вакцин в соответствии с настоящим изобретением, в организме человека или животных. Специалистам в данной области известно, как адаптировать композиции вакцин для каждого конкретного применения и конкретных пациентов.
Изобретение также относится к нуклеиновой кислоте, например, как описано выше, для применения в качестве ДНК-вакцины с целью профилактики заболеваний гриппом.
Изобретение также относится к вектору, например, как описано выше, для применения в качестве вирусной вакцины с целью профилактики заболеваний гриппом.
Изобретение также относится к иммуногенной композиции, как описано выше, для применения в качестве вакцины с целью предупреждения заболевания гриппом.
Изобретение также относится к способу повышения клеточного иммунного ответа на нуклеопротеиновый антиген вируса гриппа, включающему слияние этого антигена с белком-носителем IMX313T или IMX313P, имеющим последовательность, показанную в SEQ ID NO: 4 и SEQ ID NO: 5.
В другой форме осуществления изобретения гибридный белок, или нуклеиновая кислота, или вектор, или иммуногенная композиция, такие как описано выше, применяются в иммунотерапии заболеваний гриппом.
Краткое описание графических материалов
Фигура 1. Карта исходной плазмиды pcDNA3-NP. Эта плазмида и производные от нее плазмиды, сконструированные, как описано в примерах, были использованы для ДНК-вакцинации.
Фигура 2. Сравнение суммарных Т-клеток, секретирующих IFN-γ в ответ на иммунизацию плазмидами, кодирующими NP или NP, слитый с IMX313.
Фигура 3. Сравнение CD8 и CD4 Т-клеток, секретирующих IFN-γ в ответ на иммунизацию плазмидой, кодирующей NP или плазмидой, кодирующей NP, слитый CIMX313.
Фигура 4. Сравнение иммунных ответов в виде образования антител IgG против рекомбинантного NP, вызванных ДНК плазмидами, кодирующими либо NP, либо NP, слитый с IMX313.
Фигура 5. Сравнение иммунных ответов в виде образования антител подкласса IgG против рекомбинантного NP, вызванных ДНК плазмидами, кодирующими либо NP, либо NP, слитый с IMX313.
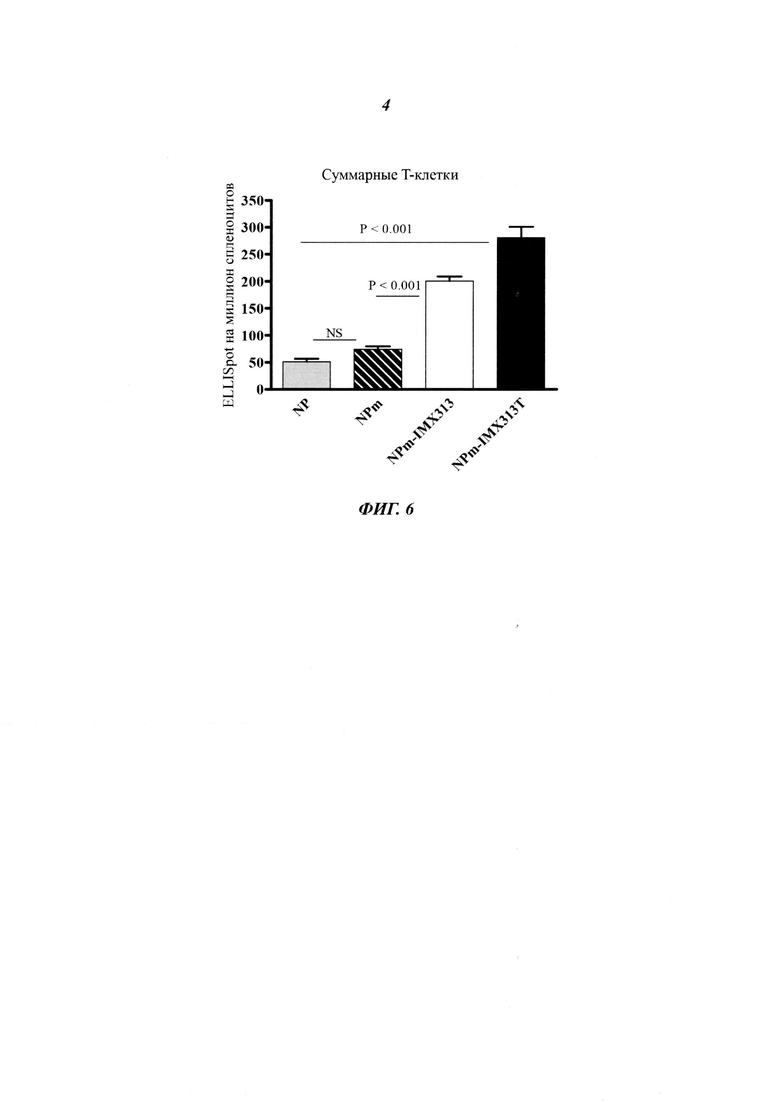
Фигура 6. Сравнение суммарных Т-клеточных ответов на плазмиды, кодирующие NP, мономерный NP (NPm), мономерный NP слитый с IMX313 (NPm-IMX313) и мономерный NP слитый с IMX313T (NPm-IMX313T).
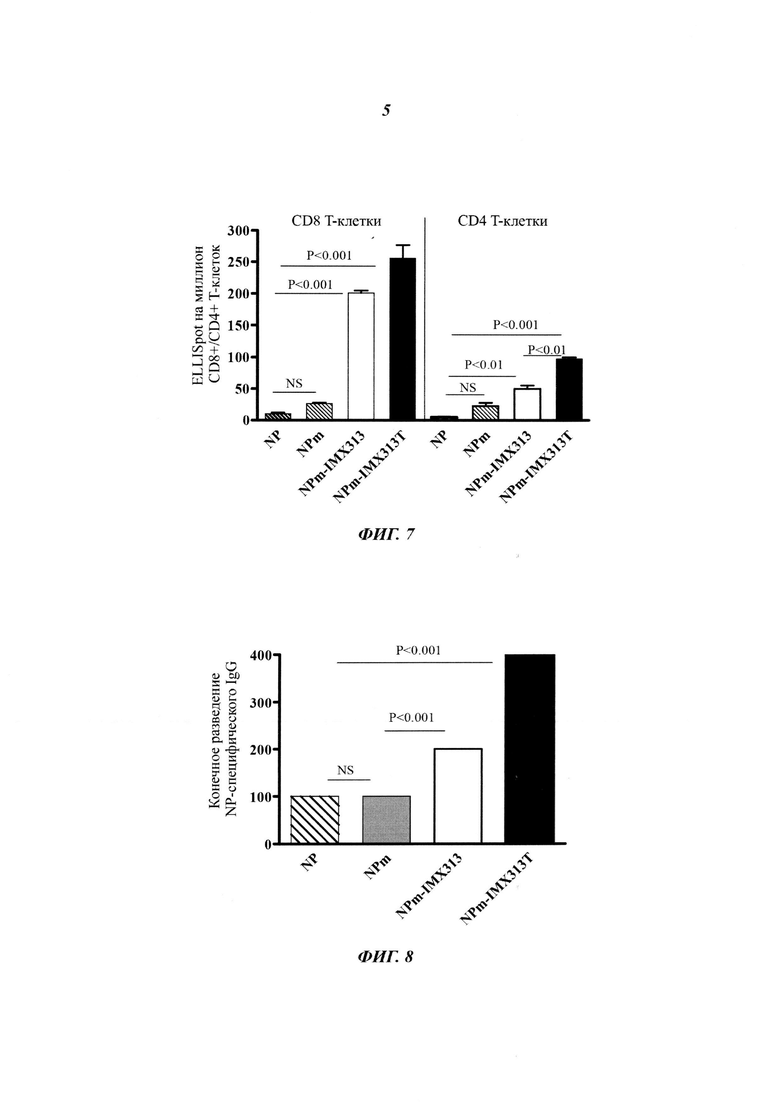
Фигура 7. Сравнение иммунных ответов в виде образования CD8+ и CD4+ Т-клеток на плазмиды, кодирующие NP, мономерный NP (NPm), мономерный NP слитый с IMX313 (NPm-IMX313) и мономерный NP слитый с IMX313T (NPm-IMX313T).
Фигура 8. Сравнение иммунных ответов в виде образования антител IgG, измеренных методом иммуносорбентного ферментного анализа (ELISA, от англ. enzyme-linked immunosorbent assay) с использованием рекомбинантного NP, на плазмиды, кодирующие NP, мономерный NP (NPm), мономерный NP слитый с IMX313 (NPm-IMX313) и мономерный NP слитый с IMX313T (NPm-IMX313T).
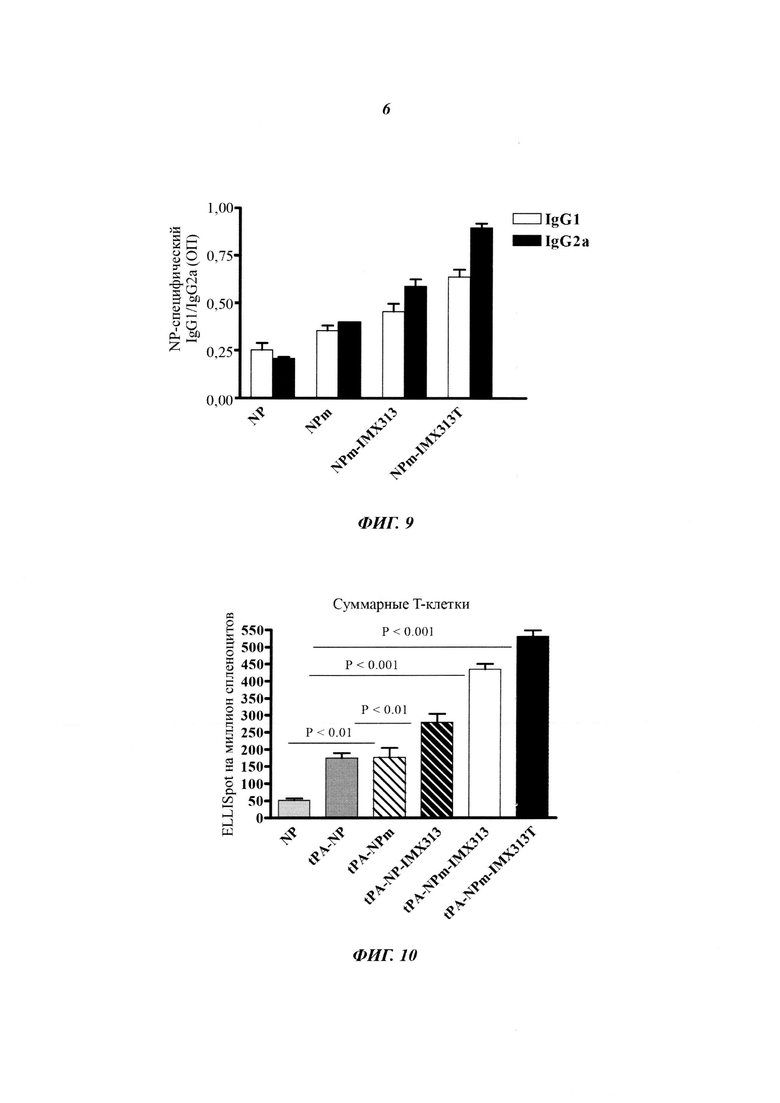
Фигура 9. Сравнение иммунных ответов в виде образования антител подкласса IgG, измеренных с использованием рекомбинантного NP, на плазмиды, кодирующие NP, мономерный NP (NPm), мономерный NP слитый с IMX313 (NPm-IMX313) и мономерный NP слитый с IMX313T (NPm-IMX313T).
Фигура 10. Влияние секреции, вызванной сигнальным пептидом tPA, на различные гибридные белки NP. Количество суммарных Т-клеток измеряли по секреции IFN-γ методом метод иммуноферментных пятен (ELISpot, от англ. enzyme-linked immunospot) по сравнению с NP, секретируемым NP (tPA-NP), секретируемым мономерным NP (tPA-NPm), секретируемым NP, слитым с IMX313 (tPA-NP-IMX313), секретируемым мономерным NP, слитым с IMX313 (tPA-NPm-IMX313) и секретируемым мономерным NP, слитым с IMX313T (tPA-NPm-IMX313T).
Фигура 11. Влияние секреции, вызванной сигнальным пептидом tPA, на CD8+ и CD4+ ответы против различных гибридных белков NP, измеренные по секреции IFN-γ, методом ELISpot сравнению с NP, секретируемым NP (tPA-NP), секретируемым мономерным NP (tPA-NPm), секретируемым NP, слитым с IMX313 (tPA-NP-IMX313), секретируемым мономерным NP, слитым с IMX313 (tPA-NPm-IMX313) и секретируемым мономерным NP, слитым с IMX313T (tPA-NPm-IMX313T).
Фигура 12. Влияние секреции, вызванной сигнальным пептидом tPA, на образование IgG против различных гибридных белков NP, измеренное по секреции IFN-γ, методом ELISpot по сравнению с NP, секретируемым NP (tPA-NP), секретируемым мономерным NP (tPA-NPm), секретируемым NP, слитым с IMX313 (tPA-NP-IMX313), секретируемым мономерным NP, слитым с IMX313 (tPA-NPm-IMX313) и секретируемым мономерным NP, слитым с IMX313T (tPA-NPm-IMX313T).
Фигура 13. Влияние секреции, вызванной сигнальным пептидом tPA, на образование антител подкласса IgG против различных гибридных белков NP, измеренное по секреции IFN-γ, методом ELISpot по сравнению с NP, секретируемым NP (tPA-NP), секретируемым мономерным NP (tPA-NPm), секретируемым NP, слитым с IMX313 (tPA-NP-IMX313), секретируемым мономерным NP, слитым с IMX313 (tPA-NPm-IMX313) и секретируемым мономерным NP, слитым с IMX313T (tPA-NPm-IMX313T).
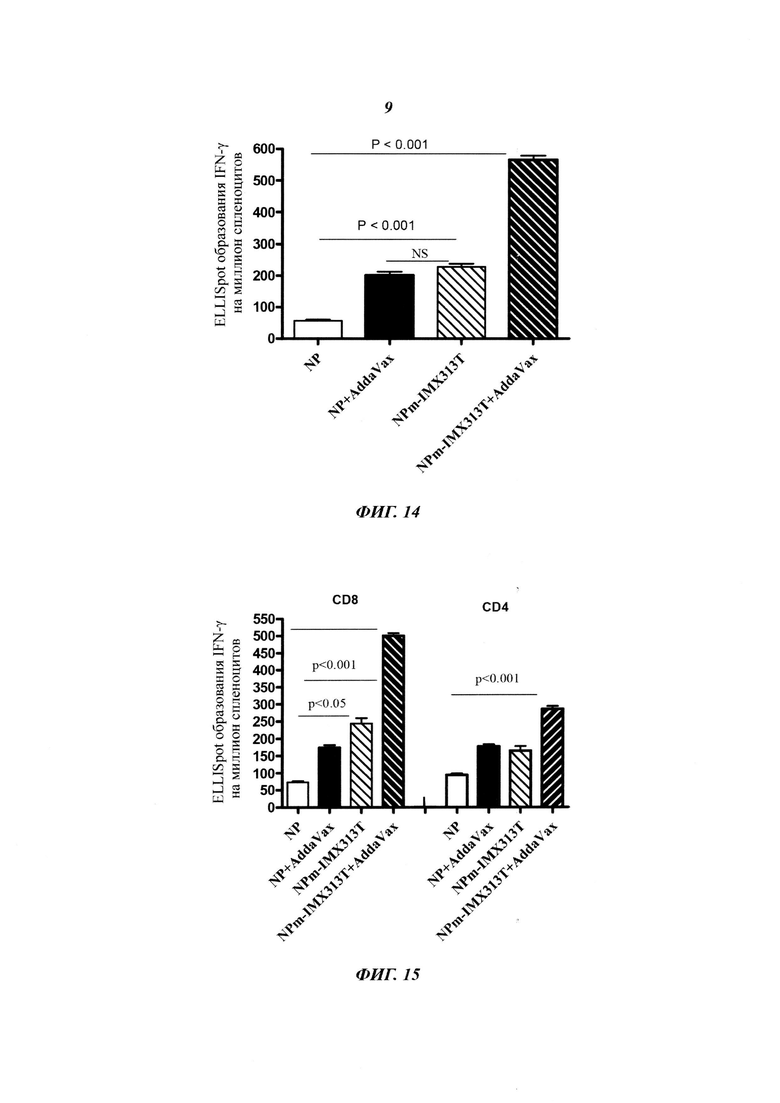
Фигура 14. Слияние нуклеопротеина с IMX313T увеличивает иммуногенность NP в такой же степени, как форма выпуска NP в адъюванте типа масло-в-воде AddaVax (Invivogen); при этом применение AddaVax совместно с гибридным белком NPM-IMX313T дает синергический эффект.
Фигура 15. Анализ результатов, представленных на Фигуре 14 после отделения CD4 и CD8 клеток. Синергетический эффект AddaVax с белком NPm-IMX313T виден как в CD4 ответах, так и в CD8 ответах.
Фигура 16. Образование IgG против нуклеопротеина. Слияние нуклеопротеина с IMX313T в отсутствие адъюванта AddaVax не показывало значительных изменений в титрах IgG по сравнению с NP. Но в присутствии AddaVax гибридный белок является значительно более иммуногенным, чем нуклеопротеин.
Фигура 17. Сравнение иммунных ответов в виде образования антител подкласса IgG, измеренное с использованием рекомбинантного NP, после иммунизации NP или NPm-IMX313T, совместно с AddaVax или без AddaVax. Как видно из Таблицы 4, NP совместно с AddaVax или без AddaVax вызывал Th1-ответ. Однако гибридный белок NPm-IMX313T, совместно с AddaVax или без AddaVax, дополнительно поляризовал иммунный ответ в виде образования IgG в сторону Th1-ответа.
Фигура 18. Анализ методом ДСН-ПААГ-электрофореза рекомбинантных белков, применяемых для иммунизации. Полоса 1: маркеры молекулярной массы (New England Biolabs); полоса 2: рекомбинантный NP (Imgenex); полоса 3: очищенный NP; полоса 4: очищенный NPm-IMX313T.
Фигура 19. Анализ методом ДСН-ПААГ-электрофореза рекомбинантных белков NPm-IMX313P. Полоса 1: очищенный NP; полоса 2: очищенный NPm-IMX313T; полоса 3: очищенный NPm-IMX313P; полоса 4: маркеры молекулярной массы (New England Biolabs).
Фигура 20. Образование IgG против протамина или IMX313P после иммунизации мышей белком IMX313P. Оно показывает, что, хотя мыши производят IgG антитела на IMX313P (и некоторые из них перекрестно реагируют с IMX313), никаких антител, не вступающих в перекрестную реакцию с протамином, найдено не было.
ПРИМЕРЫ
Для ДНК-вакцинации исходную плазмиду pcDNA3-NP, показанную на Фигуре 1, модифицировали так, как описано в приведенных ниже примерах. Плазмиды pIMX494 и pIMX497 описаны в заявке на патент РСТ/ЕР 2013/076289, поданной 11 декабря 2013 года.
Пример 1. Введение IMX313 в плазмиды, кодирующие нуклеопротеин Последовательность, кодирующую IMX313 амплифицировали из плазмиды pIMX494 с использованием олигонуклеотидных праймеров IMX1289 (5' caatgcagaggagtacgacaatggatccaagaagcaaggtgatgctgatg 3' - SEQ ID NO: 10) и IMX1290 (5' GTAGAAACAAGGGTATTTTTCTTtattactccttgctcagtccttgc 3' - SEQ ID NO: 11) и встраивали в плазмиду pcDNA3-NP, как описано Geiser.
Пример 2. Вставка сигнального пептида tPA
Сигнальный пептид tPA амплифицировали из вектора pSG2-85A (Spencer, 2012) с использованием олигонуклеотидов IMX1305 (5' cactgagtgacatcaaaatcatgGATGCAATGAAGAGAGGGC 3' - SEQ ID NO: 12) и IMX1306 (5' cgtaagaccgtttggtgccttggctagctcttctgaatcgggcatggatttcc 3' - SEQ ID NO: 13) и встраивали в рамку считывания с N-концом последовательности, кодирующей NP в ряде плазмид, как описано Geiser.
Пример 3. Создание двух точечных мутаций в NP для того, чтобы сделать его мономерным
Олигонуклеотидные праймеры IMX1287 (5' ccattctgccgcatttgCagatctaagag 3' -SEQ ID NO: 14) и IMX1288 (5' CAAAAGGGAGATTTGCCTGTACTGAGAAC 3' - SEQ ID NO: 15) использовали для амплификации внутреннего фрагмента гена NP, и полученный в результате этого продукт ПЦР встраивали в плазмиды, кодирующие NP, как описано Geiser. Поскольку оба олигонуклеотида не полностью соответствовали гену NP, вставка продукта ПЦР создавала две точечных мутации. Праймер IMX1287 создавал мутацию Е339А (замена GAA на GCA), тогда как праймер IMX1288 создавал мутацию R416A в гене NP (замена AGA на GCA).
Пример 4. Вставка IMX313T
Последовательность, кодирующую IMX313T, амплифицировали из плазмиды pIMX497 с использованием олигонуклеотидных праймеров IMX1289 (SEQ ID NO: 10) и IMX051 (5' GTAGAAACAAGGGTATTTTTCTTtattaggagcgacggcgacgc 3' - SEQ ID No: 16) и встраивали в различные плазмиды, являющиеся производными плазмиды pcDNA3-NP, как описано Geiser.
Пример 5. ДНК иммунизация нуклеиновыми кислотами согласно настоящему изобретению
5.1. Протокол
Группы из пяти самок мышей линии BALB/C иммунизировали внутримышечно дважды с интервалом в 14 дней различными ДНК-плазмидами, используя 20 мкг каждой плазмиды на инъекцию. Иммунные ответы измеряли на 28 день, определяя влияние различных модификаций: +/- IMX313 или IMX313T; +/- сигнальный пептид tPA; +/- мономеризующие мутации.
Антиген-специфические Т-клеточные ответы были измерены при помощи ELISpot с использованием спленоцитов на 28-й день. Очищенные CD4+, CD8+ и суммарные Т-клетки селезенки, выделенные из организма иммунизированных мышей, культивировали совместно с пептидом из NP вируса гриппа типа А (аминокислоты 366-374), приобретенным у компании Eurogentec.
Анализы методом ELISpot: нитроцеллюлозные 96-луночные планшеты с плоским дном (Millititer; Millipore) покрывали антителами IFN-γ mAb (15 мкг/мл; Mabtech, Стокгольм) и инкубировали в течение ночи при 4°С. После промывки фосфатно-солевым буфером (ФСБ), планшеты блокировали 10%-ной фетальной бычьей сывороткой в течение одного часа при 37°С. Клетки в концентрации 2×106 на лунку стимулировали соответствующими пептидами в конечной концентрации 2 мкг/мл (пептид из NP вируса гриппа типа А) на мембранах IPVH, покрытых античеловеческим интерфероном-γ в количестве 15 мкг/мл и инкубировали в течение 20 часов. После инкубации планшеты тщательно промывали ФСБ для удаления клеток и добавляли в каждую лунку антитела IFN-γ mAb (1 мкг на мл биотина, Mabtech). После инкубации в течение 2 ч при 37°С планшеты промывали и проводили обнаружение со стрептавидином, конъюгированным с пероксидазой хрена (1 мкг/мл; Mabtech) в течение одного часа при комнатной температуре: После промывки добавляли субстрат (3-амино-9-этилкарбазол (Sigma)) и инкубировали в течение 15 минут. После дополнительной промывки, пятна красного цвета подсчитывали под микроскопом.
Для изучения гуморального иммунного ответа авторы оценивали уровни антител с помощью ELISA конкретно для суммарных IgG, и раздельно для антител IgG1 и IgG2a, с целью определения относительных пропорций Th1 и Th2. Мыши линии BALB/C, как правило, реагируют на вакцины против гриппа путем иммунного ответа Th2-типа, который связан со стимуляцией IgG1 антител. Тем не менее, основным изотипом антител, присутствующим в сыворотках мышей, которые выживают при вирусных инфекциях, является IgG2a, который стимулируется в процессе иммунных ответов Thl-типа (Huber, 2006). Стимуляция IgG2a антител сопровождалась увеличением эффективности вакцинации против гриппа.
Для проведения ELISA антигены разбавляли до концентрации 5 мг/мл в 0,1 M растворе натрия карбоната/бикарбоната (рН 9,6), а затем использовали для покрытия лунок планшетов Maxisorb (Nunc-Immulon, Denmark). В лунки добавляли двукратные серийные разведения исследуемых сывороток и после промывания обнаруживали связанные антитела с помощью антимышиного IgG, или антимышиного IgG1, или антимышиного IgG2a (Sigma), конъюгированного с пероксидазой хрена. Поглощение при 490 нм определяли после добавления о-фенилендиамина (Sigma) и H2O2; реакции останавливали с помощью 1М серной кислоты.
Результаты показаны на Фигурах со 2 по 5.
5.2. В предварительных экспериментах авторы исследовали суммарные Т-клеточные ответы на NP, вызванные ДНК-вакцинами, кодирующими либо NP, либо NP, слитый с IMX313. Суммарные Т-клетки, выделенные из организма мышей, иммунизированных NP-IMX313, показывали значительно более высокое образование IFN-γ по сравнению с таковым у мышей, иммунизированных NP, что подтверждало способность IMX313 увеличивать Т-клеточные ответы.
Фигура 2 показывает, что слияние исходного гена NP антигена с геном IMX313 повышает Т-клеточные ответы на NP даже тогда, когда гибридный белок экспрессируется в цитозоле.
5.3. Чтобы определить, был ли IFN-γ, обнаруженный при помощи ELISpot, произведен CD4 или CD8 Т-клетками, авторы выделяли CD4+ и CD8+ Т-клетки селезенки из организма иммунизированных мышей и культивировали их совместно с пептидом из NP вируса гриппа типа А. В группе, иммунизированной NP-IMX313, обнаруживали значительное увеличение продукции IFN-γ CD8+ Т-клетками. Процентное отношение антиген-специфических CD8+ клеток, продуцирующих IFN-γ, было выше, чем процентное соотношение соответствующей популяции CD4+ Т-клеток (Фиг. 3).
Фигура 3 показывает, что слияние гена NP антигена с геном IMX313 улучшает как CD4+ ответы, так и CD8+ ответы на NP антиген.
5.4. Затем авторы исследовали образование антител на NP после иммунизации и спустя 14 дней после последней иммунизации; в сыворотке определяли образование NP-специфических антител IgG. Контрольные мыши и мыши, получавшие NP-IMX313, показывали умеренное образование NP-специфических антител IgG (Фиг. 4), которое было выше в группе, иммунизированной NP-IMX313.
Фигура 4 показывает, что слияние гена NP с геном IMX313 повышает образование антител IgG против NP антигена.
5.5. Сыворотки были также исследованы на присутствие NP-специфических IgG1 и IgG2a антител (отражающих соответственно Th2- и Th1-тип ответов у мышей линии BALB/C). NP-специфические антитела изотипов IgG1 и IgG2a были обнаружены в сыворотке мышей, иммунизированных NP-IMX313; однако образцы сыворотки мышей, получавших только NP, показывали очень низкие уровни антител IgG1 и IgG2a (Фиг. 5).
Фигура 5 показывает распределение подклассов антител, индуцированных против NP антигена. Слияние с геном IMX313 сильнее улучшало образование IgG2a, чем образование IgG1, преобразуя Th2-тип ответа против NP в Th1-тип ответа против NP-IMX313.
Результаты приведены ниже в таблице:
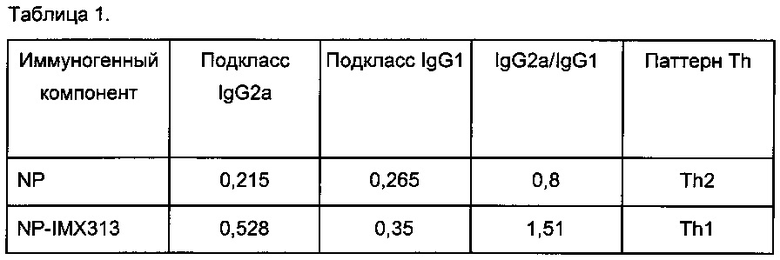
Фигура 6 показывает, что мономеризация NP (NPm) немного улучшает его иммуногенность (хотя улучшение не является статистически значимым: NS); что иммуногенность NPm дополнительно улучшается путем слияния с геном IMX313; и, наконец, что слияние мономерного NP с геном IMX313T еще больше повышает иммуногенность NP. Удивительно то, что мономеризация NP не уменьшает его иммуногенности.
Фигура 7 показывает, что для анализа ответов CD4+ и CD8+ наблюдается такой же уровень порядка как на Фигуре 6: мономеризация NP немного улучшает иммуногенность NP, но это не является статистически значимым (NS); иммуногенность NP дополнительно улучшается при помощи слияния с геном IMX313, но, наибольшее улучшение NP иммуногенности достигается с помощью слияния мономерного NP с геном IMX313T.
Фигура 8 показывает, что для В-клеточных ответов наблюдается такой же уровень порядка, как для Т-клеточных ответов на Фигурах 6 и 7 (как для CD4+, так и для CD8+). Суммарный IgG ответ против NP был выше с IMX313T, чем с IMX313.
На Фигуре 9 показано распределение подклассов антител, индуцированных мономерным NP антигеном. Слияние NP с геном IMX313 усиливает образование IgG2a больше, чем образование IgG1, трансформируя ответ Th2-типа против NP (0,8) в ответ Th1-типа против NP-IMX313 (1,51). Такое обращение Th2-типа ответа в Thl-тип более выражено поддерживается путем слияния с IMX313T, чем с IMX313 (1,5). Экспрессия IgG2a антител в вакцинах против гриппа коррелирует с клиренсом вируса и повышенной защитой от летального заражения гриппом. Увеличение индукции обоих изотипов антител, измеренное ELISA, имело лучшую корреляцию с эффективностью вакцины, чем с одной лишь нейтрализацией вируса (Huber, 2006).

Пример 6. Секреция NP антигена улучшала его иммуногенность
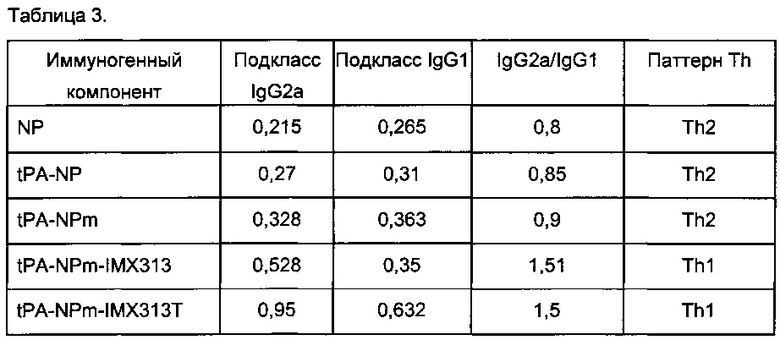
Авторы сделали серию конструктов ДНК-вакцин NP, содержащих секреторную сигнальную последовательность (tPA) тканевого активатора плазминогена: tPA-NP, tPA-NPm, tPA-NPm-IMX313 и tPA-NPm-IMX313T. Анализировали результаты слияния tPA и NP на гуморальные и клеточные иммунные ответы иммунизированных животных.
Мыши, иммунизированные конструктами, содержащими tPA, показывали значительно более высокое образование IFN-γ по сравнению с образованием IFN-γ мышей, иммунизированных NP, что подтверждало способность IMX313T и мономеризующих мутаций увеличивать Т-клеточные ответы.
Фигура 10 показывает, что стимуляция секреции NP антигена улучшала его иммуногенность (NP по сравнению с tPA-NP), независимо от того, был ли он мономерным или нет (tPA-NP по сравнению с tPA-NPm). Однако слияние с IMX313 показывало, что использование мономерной версии NP было более иммуногенным, чем использование немодифицированного антигена (TPA-NP-IMX313 по сравнению с tPA-NPm-IMX313). А замена IMX313 на IMX313T улучшала иммуногенность NP (tPA-NPm-IMX313 по сравнению с tPA-NPm-IMX313T).
Фигура 11 показывает CD8+ и CD4+ ответы на различные секретируемые версии NP. Наблюдается такой же уровень порядка, как на Фигуре 10, и при добавлении IMX313 снова четко выражена целесообразность мономеризации антигена. Как и на предыдущих Фигурах, более выраженные иммунные ответы наблюдаются тогда, когда IMX313T используют вместо IMX313.
Фигура 12 показывает суммарный IgG ответ на NP антиген и побуждает к таким же заключениям, как Фигура 11 для Т-клеточных ответов: наиболее выраженные ответы наблюдаются тогда, когда используется IMX313T, но секреция (NP по сравнению с tPA-NP) и мономеризация (tPA-NP-IMX313 по сравнению с tPA-NPm-IMX313) также вносят важный вклад.
Мыши, иммунизированные только NP (в виде NP, tPA-NP или tPA-NPm), не имели совсем антител IgG против NP или имели очень низкие уровни антител IgG против NP в их сыворотке (Фиг. 12). Мыши, иммунизированные tPA-NP-IMX313, tPA-NPm-IMX313 или tPA-NPm-IMX313T, напротив, показывали высокий уровень системных NP-специфических иммунных ответов в виде образования антител IgG; и снова мыши, иммунизированные tPA-NPm-IMX313T, имели значительно более высокие (р<0,001) иммунные ответы в виде образования антител IgG по сравнению со всеми другими группами иммунизированных мышей. Это показывает, что сочетание всех модификаций (мономеризующие мутации, tPA и IMX313T) придает существенно улучшенную иммуногенность антигену по сравнению с иммуногенностью исходной последовательности или другими комбинациями.
Фигура 13 показывает анализ подклассов В-клеточных реакций на NP и иллюстрирует, что начальный Th2-тип ответов при использовании одного лишь NP обращается при использовании IMX313 и IMX313T. В то время как секреция оказывает сама по себе мало влияния (NP по сравнению с tPA-NP), мономеризация (tPA-NP-IMX313 по сравнению с tPA-NPm-IMX313), а затем замена IMX313 на IMX313T (tPA-NPm-IMX313 по сравнению с tPA-NPm-IMX313T) способствуют улучшенным Th1-ответам (IgG2a) по сравнению с Th2-ответами (IgG1).
Очень важно, что tPA-NPm-IMX313T сам по себе почти эквивалентно улучшает Th1- и Th2-ответ. Слияние NP с IMX313 показывает, что как Th1-, так и Th2 ответ увеличиваются, и никаких значительных изменений в типе реакции не отмечается. Но при сочетании IMX313T с мономеризующими мутациями Th1-ответ (IgG2a) становится преобладающим. По общему мнению иммунологов, Th1-ответы предпочтительнее Th2-ответов (Фиг. 13).
Эти результаты приведены ниже в таблице:
Пример 7. Получение рекомбинантного белка NPm-IMX313T
Плазмиды производные от рЕТ22, экспрессирующие дикий тип белка H1N1 NP из штамма вируса гриппа A/WSN/33 (Tarus, 2012b) с С-концевой гистидиновой меткой 6-His-tag, экспрессировали в бактериальном штамме C43R. Этот штамм был создан путем преобразования C43(DE3) с помощью редкого кодона, экспрессирующего плазмиду pRARE2 (Novagen). Экспрессию индуцировали при помощи IPTG в среде ТВ (от англ. Terrific Broth). Суперэкспрессированный белок очищали сначала, как описано Ye и Tarus для стадий очистки от взвешенных примесей и ионного обмена, но на конечной стадии гибридный белок очищают при помощи аффинной хроматографии на гепарин-сефарозе и гель-фильтрации (Hi Prep 26/60 Sephacryl S-300), как описано в заявке на патент РСТ/ЕР 2013/076289, поданной 11 декабря 2013 года.
Для экспрессии белка NPm-IMX313T, плазмиду, экспрессирующую NP, модифицировали в две стадии. На первой стадии вводили мономеризующие мутации, как в Примере 3, с использованием олигонуклеотидных праймеров, IMX1287 (5' ccattctgccgcatttgCagatctaagag 3' - SEQ ID NO: 14) и IMX1288 (5' CAAAAGG G AG ATTTG CCTGTACTG AG AAC 3' - SEQ ID NO: 15). На второй стадии гистидиновая метка 6-His-tag была заменена последовательностью, кодирующей IMX313T, с использованием тех же олигонуклеотидных праймеров, как в Примере 4: IMX1289 (SEQ ID NO: 10) и IMX051 (5' GTAGAAACAAGGGTATTTTTCTTtattaggagcgacggcgacgc 3' - SEQ ID NO: 16). Затем вместо гистидиновой метки 6-His-tag проводили вставку продукта ПЦР, как описано Geiser.
Гибридный белок NPm-IMX313T экспрессировали таким же образом и в том же штамме, что и белок NP, и очищали с использованием тех же хроматографических стадий.
Пример 8. Иммунизация
Затем выполняли иммунизацию мышей для сравнения иммуногенности NPm-IMX313T, с применением или без применения препарата, содержащего адъювант AddaVax (Invivogen). Белок NP, с добавлением или без добавления препарата, содержащего адъювант AddaVax, был использован в качестве контроля.
С этой целью четыре группы по пять самок мышей BALB/C иммунизировали подкожно дважды с интервалом 14 дней, используя 20 мкг каждого белка на инъекцию. Индукцию антиген-специфических Т-клеточных ответов измеряли при помощи ELISpot с использованием спленоцитов, на 28-й день. Очищенные CD4+, CD8+ и суммарные Т-клетки селезенки, выделенные из иммунизированных мышей, культивировали совместно с белком NP или пептидом NP вируса гриппа типа А (366-374). Предварительный иммунный ответ и гуморальный иммунный ответ на 28 день были измерены при помощи ELISA с NP в качестве антигена.
Пример 9. IMX313T не разрушается под действием протеаз при прохождении через пути секреции
Результаты, полученные при помощи ДНК-иммунизации плазмидами, содержащими IMX313T, убедительно свидетельствуют о том, что концевой участок молекулы не расщепляется протеазами, когда она проходит через пути секреции, где протеазы в изобилии. Чтобы изучить этот вопрос более определенно, проводили трансфекцию клеток линии СНО-К1 плазмидой pcDNA3, используемой для экспрессии NPm-IMX313T in vivo. Трансфекцию проводили так, как описано в другом источнике (Krammer, 2010).
Спустя период от восемнадцати до двадцати четырех часов супернатанты трансфицированных клеток выделяли центрифугированием и фильтровали перед загрузкой на колонку с гепарин-сефарозой, как описано в заявке на патент РСТ/ЕР 2013/076289, поданной 11 декабря 2013 года.
Отмечали небольшой «пик С», который содержал белок NPm-IMX313T, что подтверждалось при помощи ДСН-ПААГ-электрофореза и Вестерн-блоттинга.
Пример 10. Получение рекомбинантного белка NPm-IMX313P
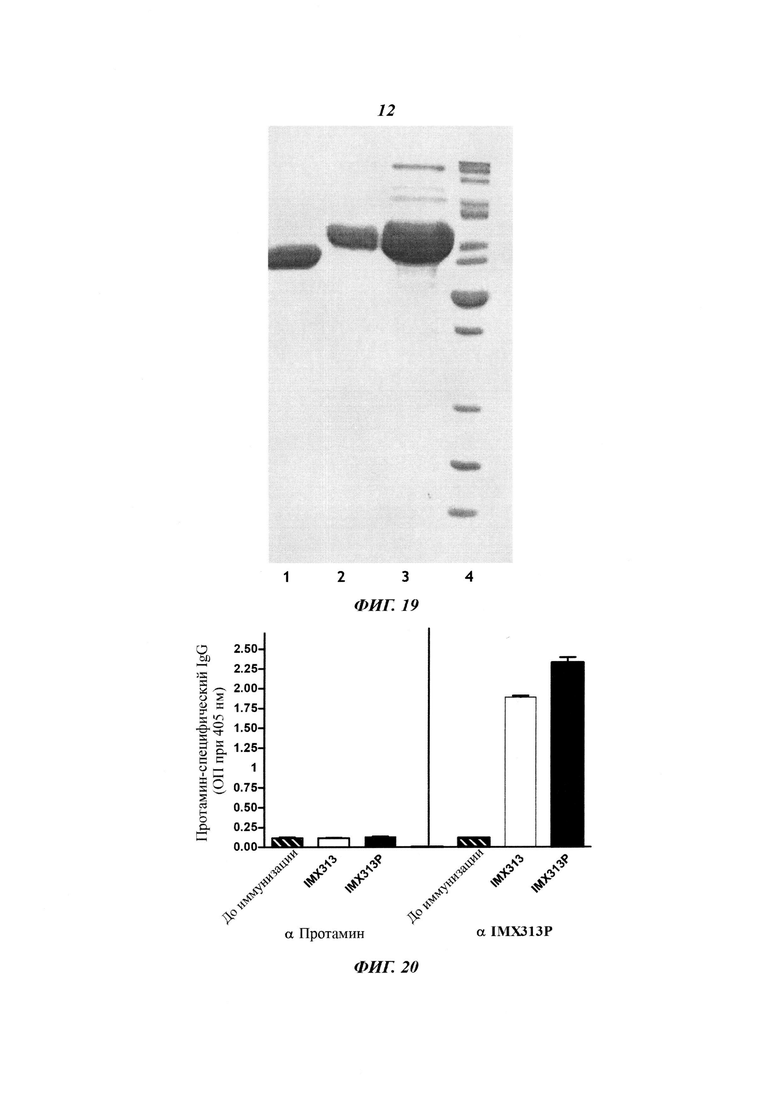
Для экспрессии белка NPm-IMX313P плазмиду, экспрессирующую NPm-IMX313T, модифицировали путем замены гена IMX313T на ген IMX313P при помощи замены соответствующего фрагмента в плазмиде, кодирующей белок NPm-IMX313T, рестрикционным фрагментом (Pml I-Hind III) плазмиды, кодирующей IMX313P. Затем гибридный белок экспрессировали и очищали, как описано в Примере 7. Фигура 19 показывает очищенный белок; основная полоса принадлежит мономеру, но выше основной полосы также заметны олигомерные формы (на сильно концентрированном геле).
Пример 11. Получение гипериммунной антисыворотки IMX313P
Группу из пяти самок мышей линии BALB/C иммунизировали внутримышечно шесть раз с 14-дневными интервалами белком IMX313P в дозе 50 мкг на инъекцию.
Сыворотки тестировали на содержание IgG антител с помощью модифицированного метода ELISA. Протаминсульфат Grade X (Sigma), IMX313 или IMX313P были использованы для покрытия лунок микропланшета с целью захвата антител. Идентифицирующие антитела представляли собой конъюгированный с пероксидазой хрена козий антимышиный иммуноглобулин IgG (Sigma), который вводили в реакцию с перекисью водорода для того, чтобы определить поглощение при 405 нм.
Все сыворотки, полученные из мышей, иммунизированных IMX313P, показывали высокие титры антител IgG против IMX313P и небольшое количество антител, перекрестно реагирующих с IMX313; но не давали никаких антител, перекрестно реагирующих с протамином (Фигура 20).
Ссылки:
Akira S, Uematsu S, Takeuchi O. 2006. Pathogen recognition and innate immunity. Cell 124:783-801.
Altstein AD, Gitelman AK, Smirnov YA, Piskareva LM, Zakharova LG, Pashvykina GV, Shmarov MM, Zhirnov OP, Varich NP, llyinskii PO, Shneider AM. (2006) Immunization with influenza A NP-expressing vaccinia virus recombinant protects mice against experimental infection with human and avian influenza viruses. Arch Virol. 151:921-931.
Antrobus RD, Lillie PJ, Berthoud TK, Spencer AJ, McLaren JE, Ladell K, Lambe T, Milicic A, Price DA, Hill AV, Gilbert SC. (2012) A T cell-inducing influenza vaccine for the elderly: safety and immunogenicity of MVA-NP+M1 in adults aged over 50 years. PLoS One. 7:e48322.
Arranz R, Coloma R, Chichon FJ, Conesa JJ, Carrascosa JL, Valpuesta JM,  J,
J,  -Benito J. (2012) The structure of native influenza virion ribonucleoproteins. Science. 338:1634-1637.
-Benito J. (2012) The structure of native influenza virion ribonucleoproteins. Science. 338:1634-1637.
Bachmann MF, Rohrer UH,  TM,
TM,  K, Hengartner H, Zinkernagel RM. (1993) The influence of antigen organization on В cell responsiveness. Science 262:1448-1451.
K, Hengartner H, Zinkernagel RM. (1993) The influence of antigen organization on В cell responsiveness. Science 262:1448-1451.
Berthoud TK, Hamill M, Lillie PJ, Hwenda L, Collins KA, Ewer KJ, Milicic A, Poyntz HC, Lambe T, Fletcher HA, Hill AV, Gilbert SC. (2011) Potent CD8+ T-cell immunogenicity in humans of a novel heterosubtypic influenza A vaccine, MVA-NP+M1. Clin Infect Dis. 52:1-7.
Blasius AL & Beutler B. 2010. Intracellular Toll-like Receptors. Immunity 32:305-315.
Couch RB, Atmar RL, Franco LM, Quarles JM,  D, Wells JM, Arden N, Cheung S, Belmont JW. (2012) Prior infections with seasonal influenza A/H1N1 virus reduced the illness severity and epidemic intensity of pandemic H1N1 influenza in healthy adults. Clin Infect Dis. 54:311-317.
D, Wells JM, Arden N, Cheung S, Belmont JW. (2012) Prior infections with seasonal influenza A/H1N1 virus reduced the illness severity and epidemic intensity of pandemic H1N1 influenza in healthy adults. Clin Infect Dis. 54:311-317.
Epstein SL, Stack A, Misplon JA, Lo CY, Mostowski H, Bennink J, Subbarao K. (2000) Vaccination with DNA encoding internal proteins of influenza virus does not require CD8(+) cytotoxic T lymphocytes: either CD4(+) or CD8(+) T cells can promote survival and recovery after challenge. Int Immunol. 12:91-101.
Epstein SL, Tumpey TM, Misplon JA, Lo CY, Cooper LA, Subbarao K, Renshaw M, Sambhara S, Katz JM. (2002). DNA vaccine expressing conserved influenza virus proteins protective against H5N1 challenge infection in mice. Emerg Infect Dis. 8:796-801. Fiers W, Neirynck S, Deroo T, Saelens X, Jou WM. (2001) Soluble recombinant influenza vaccines. Philos Trans R Soc Lond В Biol Sci. 356:1961-1963.
Fodor E, Devenish L, Engelhardt OG, Palese P, Brownlee GG,  -Sastre A. (1999) Rescue of influenza A virus from recombinant DNA. J Virol. 73:9679-9682.
-Sastre A. (1999) Rescue of influenza A virus from recombinant DNA. J Virol. 73:9679-9682.
Gammelin M,  A, Reinhardt U, Mandler J, Harley VR, Hudson PJ, Fitch WM, Scholtissek C. (1990) Phylogenetic analysis of nucleoproteins suggests that human influenza A viruses emerged from a 19th-century avian ancestor. Mol Biol Evol. 7:194-200. Geiser M,
A, Reinhardt U, Mandler J, Harley VR, Hudson PJ, Fitch WM, Scholtissek C. (1990) Phylogenetic analysis of nucleoproteins suggests that human influenza A viruses emerged from a 19th-century avian ancestor. Mol Biol Evol. 7:194-200. Geiser M,  R; Drewello D, Schmitz R. 2001. Integration of PCR fragments at any specific site within cloning vectors without the use of restriction enzymes and DNA ligase. Biotechniques 31:88-92.
R; Drewello D, Schmitz R. 2001. Integration of PCR fragments at any specific site within cloning vectors without the use of restriction enzymes and DNA ligase. Biotechniques 31:88-92.
Gorman ОТ, Bean WJ, Kawaoka Y, Donatelli I, Guo YJ, Webster RG. (1991) Evolution of influenza A virus nucleoprotein genes: implications for the origins of H1N1 human and classical swine viruses. J Virol. 65:3704-3714.
Greenland JR, Letvin NL. (2007) Chemical adjuvants for plasmid DNA vaccines. Vaccine 25:3731-3741.
Gschoesser C, Almanzar G, Hainz U, Ortin J, Schonitzer D, Schild H, Saurwein-Teissl M, Grubeck-Loebenstein B. (2002). CD4+ and CD8+ mediated cellular immune response to recombinant influenza nucleoprotein. Vaccine 20:3731-3738.
Huang B, Wang W, Li R, Wang X, Jiang T, Qi X, Gao Y, Tan W, Ruan L. (2012) Influenza A virus nucleoprotein derived from Escherichia coli or recombinant vaccinia (Tiantan) virus elicits robust cross-protection in mice. Virol J. 9:322.
Huber VC, McKeon RM, Brackin MN, Miller L, Keating R, Brown SA, Makarova N, Perez DR, MacDonald GH, McCullers JA. (2006) Distinct Contributions of Vaccine-Induced Immunoglobulin G1 (IgG1) and IgG2a Antibodies to Protective Immunity against Influenza. CLINICAL AND VACCINE IMMUNOLOGY 13: 981-990.
Krammer F, Pontiller J, Tauer C, Palmberger D, Maccani A, Baumann M, Grabherr R. (2010) Evaluation of the influenza A replicón for transient expression of recombinant proteins in mammalian cells. PLoS One. 5:e13265.
Laddy DJ, Yan J, Kutzler M, Kobasa D, Kobinger GP, Khan AS, Greenhouse J, Sardesai NY, Draghia-Akli R, Weiner DB. (2008) Heterosubtypic protection against pathogenic human and avian influenza viruses via in vivo electroporation of synthetic consensus DNA antigens. PLoS One. 3:e2517.
Lamb RA, Krug RM. Orthomyxoviridae: the viruses and their replication. In: Knipe DM, Howley PM, editors., eds. Fields Virology Vol 1 4th ed. Philadelphia, PA: Lippincott Williams & Wilkins; 2001:1487-1531.
Lillie PJ, Berthoud TK, Powell TJ, Lambe Τ, Mullarkey С, Spencer AJ, Hamill M, Peng Y, Biais ME, Duncan CJ, Sheehy SH, Havelock T, Faust SN, Williams RL, Gilbert A, Oxford J, Dong T, Hill AV, Gilbert SC. (2012) Preliminary assessment of the efficacy of a T-cell-based influenza vaccine, MVA-NP+M1, in humans. Clin Infect Dis. 55:19-25.
Luo M, Tao P, Li J, Zhou S, Guo D, Pan Z. (2008) Immunization with plasmid DNA encoding influenza A virus nucleoprotein fused to a tissue plasminogen activator signal sequence elicits strong immune responses and protection against H5N1 challenge in mice. J Virol Methods. 154:121-127.
Mbawuike I, Zang Y, Couch RB. (2007) Humoral and cell-mediated immune responses of humans to inactivated influenza vaccine with or without QS21 adjuvant. Vaccine 25:3263-3269.
McMichael AJ, Gotch FM, Dongworth DW, Clark A, Potter CW. (1983a) Declining T-cell immunity to influenza, 1977-82. Lancet 2(8353):762-724.
McMichael AJ, Gotch FM, Noble GR, Beare PA. (1983b) Cytotoxic T-cell immunity to influenza. N Engl J Med. 309:13-17.
McMichael AJ, Michie CA, Gotch FM, Smith GL, Moss B. (1986) Recognition of influenza A virus nucleoprotein by human cytotoxic Τ lymphocytes. J Gen Virol. 67:719-726.
Moeller A, Kirchdoerfer RN, Potter CS, Carragher B, Wilson IA. (2012) Organization of the influenza virus replication machinery. Science 338:1631-1634.
Nakada S, Creager RS, Krystal M, Palese P. (1984) Complete nucleotide sequence of the influenza C/California/78 virus nucleoprotein gene. Virus Res. 1:433-441.
Ogun SA, Dumon-Seignovert L, Marchand JB, Holder AA, Hill F.(2008). The oligomerization domain of C4-binding protein (C4bp) acts as an adjuvant, and the fusion protein comprised of the 19-kilodalton merozoite surface protein 1 fused with the murine C4bp domain protects mice against malaria. Infect Immun. 76:3817-3823.
Roy S, Kobingerb GP, Lina J, Figueredoa J, Calcedoa R, Kobasab D, Wilson JM. (2007) Partial protection against H5N1 influenza in mice with a single dose of a chimpanzee adenovirus vector expressing nucleoprotein. Vaccine 25:6845-6851.
Schotsaert M, Ysenbaert T, Neyt K,  LI, Bogaert P, Schepens B, Lambrecht BN, Fiers W, Saelens X. (2012) Natural and long-lasting cellular immune responses against influenza in the M2e-immune host. Mucosal Immunology 1-12.
LI, Bogaert P, Schepens B, Lambrecht BN, Fiers W, Saelens X. (2012) Natural and long-lasting cellular immune responses against influenza in the M2e-immune host. Mucosal Immunology 1-12.
Spencer AJ, Hill F, Honeycutt JD, Cottingham MG, Bregu M, Rollier CS, Furze J, Draper SJ,  KC, Gilbert SC, Wyllie DH, Hill AV. (2012) Fusion of the Mycobacterium tuberculosis antigen 85A to an oligomerization domain enhances its immunogenicity in both mice and non-human primates. PLoS One. 7:e33555.
KC, Gilbert SC, Wyllie DH, Hill AV. (2012) Fusion of the Mycobacterium tuberculosis antigen 85A to an oligomerization domain enhances its immunogenicity in both mice and non-human primates. PLoS One. 7:e33555.
 Ζ,
Ζ,  Ε. (2010) Conserved epitopes of influenza A virus inducing protective immunity and their prospects for universal vaccine development. Virology 7:351 Sullivan SM, Doukas J, Hartikka J, Smith L, Rolland A. (2010) Vaxfectin: a versatile adjuvant for plasmid DNA- and protein-based vaccines. Expert Opin Drug Deliv. 7:1433-1446.
Ε. (2010) Conserved epitopes of influenza A virus inducing protective immunity and their prospects for universal vaccine development. Virology 7:351 Sullivan SM, Doukas J, Hartikka J, Smith L, Rolland A. (2010) Vaxfectin: a versatile adjuvant for plasmid DNA- and protein-based vaccines. Expert Opin Drug Deliv. 7:1433-1446.
Sutter G, Wyatt LS, Foley PL, Bennink JR, Moss B. (1994) A recombinant vector derived from the host range-restricted and highly attenuated MVA strain of vaccinia virus stimulates protective immunity in mice to influenza virus. Vaccine 12:1032-1040.
Tarus В, Bakowiez О, Chenavas S, Duchemin L, Estrozi LF, Bourdieu C, Lejal N, Bernard J, Moudjou M, Chevalier C, Delmas B, Ruigrok RW, Di Primo С, Slama-Schwok A. (2012a) Oligomerization paths of the nucleoprotein of influenza A virus. Biochimie. 94:776-785.
Tarus B, Chevalier C, Richard CA, Delmas B, Di Primo С, Slama-Schwok A. (2012b) Molecular dynamics studies of the nucleoprotein of influenza A virus: role of the protein flexibility in RNA binding. PLoS One. 7:e30038.
Thomas PG, Keating R, Hulse-Post DJ, and Doherty PC. (2006) Cell-mediated Protection in Influenza Infection. Emerging Infectious Diseases 12: 48-54.
Ulmer JB, Donnelly JJ, Parker SE, Rhodes GH, Feigner PL, Dwarki VJ, Gromkowski SH, Deck RR, DeWitt CM, Friedman A, et al. (1993) Heterologous protection against influenza by injection of DNA encoding a viral protein. Science. 259:1745-1749.
Wraith DC, Vessey AE, Askonas BA. (1987) Purified influenza virus nucleoprotein protects mice from lethal infection. J Gen Virol. 68:433-440.
Xu J, Christman MC, Donis RO, Lu G. (2011) Evolutionary dynamics of influenza A nucleoprotein (NP) lineages revealed by large-scale sequence analyses. Infect Genet Evol. 11:2125-2132.
Ye Q, Guu TS, Mata DA, Kuo RL, Smith B, Krug RM, Tao YJ. (2012) Biochemical and structural evidence in support of a coherent model for the formation of the double-helical influenza A virus ribonucleoprotein. MBio. 4:e00467-12.
Ye Q, Krug RM, Tao YJ. (2006) The mechanism by which influenza A virus nucleoprotein forms oligomers and binds RNA. Nature.444:1078-1082.
Yewdell JW, Bennink JR, Smith GL, Moss B. (1985) Influenza A virus nucleoprotein is a major target antigen for cross-reactive anti-influenza A virus cytotoxic Τ lymphocytes. Proc Natl Acad Sci USA. 82:1785-1789.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЕ СУПЕРСПИРАЛЬНЫЕ БЕЛКИ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2013 |
|
RU2677799C2 |
| Универсальная противогриппозная вакцина | 2015 |
|
RU2618918C2 |
| РЕКОМБИНАНТНЫЙ ВИРУС ГРИППА, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ПРОФИЛАКТИКИ COVID-19 И ГРИППА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2022 |
|
RU2802058C1 |
| СЛИТЫЙ БЕЛОК ТЕЛОМЕРАЗНОЙ ОБРАТНОЙ ТРАНСКРИПТАЗЫ, КОДИРУЮЩИЕ ЕГО НУКЛЕОТИДЫ И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2473691C2 |
| ЭФФЕКТИВНЫЕ ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ (VLPS) ГРИППА | 2004 |
|
RU2369405C2 |
| КОНСТРУКЦИЯ ДНК (ВАРИАНТЫ), ДНК-ВЕКТОР, ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГРИППА, СПОСОБ ИНДУКЦИИ ИММУННОГО ОТВЕТА, ВАКЦИНА И СПОСОБ ВАКЦИНАЦИИ | 1994 |
|
RU2193065C2 |
| ОПТИМИЗИРОВАННЫЕ С ПОМОЩЬЮ ВЫЧИСЛИТЕЛЬНЫХ СРЕДСТВ АНТИГЕНЫ С ШИРОКИМ СПЕКТРОМ РЕАКТИВНОСТИ ДЛЯ ВИРУСОВ ГРИППА H5N1 И H1N1 | 2013 |
|
RU2639551C2 |
| ПОЛИВАЛЕНТНАЯ ВАКЦИНА ПРОТИВ ГРИППА НА ОСНОВЕ ГИБРИДНОГО БЕЛКА | 2013 |
|
RU2531235C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ВИРУСНОГО ВЕКТОРА APMV-8 | 2010 |
|
RU2592544C2 |
| СЛИТЫЕ БЕЛКИ ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ ИММУНОГЕННЫХ УСИЛИВАЮЩИХ АГЕНТОВ ДЛЯ ИНДУЦИРОВАНИЯ АНТИГЕНСПЕЦИФИЧЕСКОГО Т-КЛЕТОЧНОГО ОТВЕТА | 2013 |
|
RU2631002C2 |

















Представленные изобретения относятся, в частности, к гибридному белку, содержащему вариант нуклеопротеидного антигена из штамма вируса гриппа типа А, содержащий точечные мутации Е339А и R416A, слитый с вариантом домена олигомеризации С4bр курицы, в частности IMX313T и IMX313P. Изобретения также относятся к нуклеиновой кислоте, которая кодирует гибридный белок; вектору, содержащему указанную нуклеиновую кислоту; иммуногенной композиции, которая может использоваться в качестве вакцины или для иммунотерапии с целью профилактики заболеваний гриппом, которая содержит представленный гибридный белок. Изобретения позволяют улучшить иммуногенность NP антигена вируса гриппа А и могут быть использованы в медицинской промышленности. 6 н. и 8 з.п. ф-лы 20 ил., 3 табл., 11 пр.
1. Гибридный белок, индуцирующий клеточный иммунный ответ против штамма вируса гриппа типа А, содержащий по меньшей мере один вариант нуклеопротеинового (NP) антигена из штамма вируса гриппа типа А, слитый с вариантом домена олигомеризации C4bp курицы, который представляет собой последовательность SEQ ID NO: 4 или SEQ ID NO: 5.
2. Гибридный белок по п. 1, отличающийся тем, что нуклеопротеиновый антиген представляет собой мономерный антиген.
3. Гибридный белок по п. 2, в котором мономерный нуклеопротеиновый антиген из штамма вируса гриппа типа А содержит по меньшей мере одну из двух следующих точечных мутаций: Е339А и R416A.
4. Гибридный белок по п. 3, в котором мономерный нуклеопротеиновый антиген кодируется последовательностью SEQ ID NO: 2.
5. Гибридный белок по п. 1, в котором NP антиген содержит сигнальный пептид.
6. Нуклеиновая кислота, кодирующая гибридный белок по любому из пп. 1-5.
7. Нуклеиновая кислота по п. 6, представляющая собой последовательности SEQ ID NO: 6 или SEQ ID NO: 7.
8. Экспрессионный вектор, содержащий нуклеиновую кислоту по любому из пп. 6 и 7.
9. Иммуногенная композиция для профилактики заболевания штаммом вируса гриппа типа А, содержащая:
- в эффективном количестве гибридный белок по любому из пп. 1-5, или нуклеиновую кислоту по любому из пп. 6 и 7, или вектор по п. 8, и
- лиганды на основе нуклеиновых кислот для внутриклеточных ТПР (Толл-подобные рецепторы).
10. Иммуногенная композиция по п. 9, в которой лиганды на основе нуклеиновых кислот для внутриклеточных ТПР являются полиинозин-полицитидиловой кислотой poly(I:C).
11. Иммуногенная композиция по п. 9, дополнительно содержащая вакцинный адъювант.
12. Иммуногенная композиция по любому из пп. 9-11 для применения в качестве вакцины или для иммунотерапии с целью профилактики заболевания штаммом вируса гриппа типа А.
13. Способ повышения клеточного иммунного ответа на нуклеопротеиновый антиген штамма вируса гриппа типа А путем слияния нуклеопротеинового антигена, имеющего последовательность, показанную в SEQ ID NO: 2 с белком-носителем, имеющим последовательности, показанные в SEQ ID NO: 4 или SEQ ID NO: 5, и введения полученного гибридного белка в организм человека или животного.
14. Способ профилактики заболевания штаммом вируса гриппа типа А у нуждающегося в этом человека или животного, включающий введение в организм такого человека или животного иммуногенной композиции по любому из пп. 9-11.
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| WO 2007062819 A2, 07.06.2007 | |||
| Устройство для выпрямления опрокинувшихся на бок и затонувших у берега судов | 1922 |
|
SU85A1 |
| WO 2011045612 A1, 21.01.2011 | |||
| ДУПЛИК Н.В | |||
| и др | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |

