Область, к которой относится изобретение
Настоящее изобретение относится к антагонистам нейропилина-2 (Nrp2), в частности к антителам против Nrp2, и к их использованию для профилактики и лечения метастазов опухоли.
Предшествующий уровень техники
В настоящее время точно установлено, что ангиогенез участвует в патогенезе ряда расстройств. Такими расстройствами являются солидные опухоли и метастазы, атеросклероз, ретролентальная фиброплазия, гемангиомы, хроническое воспаление, внутриглазные неоваскулярные заболевания, такие как пролиферирующие ретинопатии, например диабетическая ретинопатия, возрастная дегенерация желтого пятна (AMD), неоваскулярная глаукома, иммунное отторжение трансплантированной ткани роговицы и других тканей, ревматоидный артрит и псориаз. Folkman et al., J. Biol, Chem., 267:10931-10934 (1992); Klagsbrun et al., Annu. Rev. Physiol. 53:217-239 (1991); и Garner A., «Vascular diseases», In: Pathobiology of Ocular Disease. A Dynamic Approach, Garner A., Klintworth GK, eds., 2nd Edition (Marcel Dekker, NY, 1994), pp.1625-1710.
В случае роста опухоли ангиогенез, очевидно, является ключевым фактором, играющим важную роль в переходе гиперплазии в неоплазию и в обеспечении питательными веществами, необходимыми для роста и метастазирования опухоли. Folkman et al., Nature 339:58 (1989). В отличие от нормальных клеток, для роста опухолевых клеток и для их автономной пролиферации необходима неоваскуляризация. Опухоли обычно образуются из одной аберрантной клетки, которая может пролиферировать лишь до определенного размера, составляющего несколько кубических миллиметров, что определяется ее расстоянием от капиллярного русла, и такая клетка может оставаться «спящей», то есть может не подвергаться дальнейшему росту и диссеминации в течение длительного периода времени. Затем некоторые опухолевые клетки переключаются на ангиогенный фенотип с последующей активацией эндотелиальных клеток, которые пролиферируют и созревают, образуя новые капиллярные кровеносные сосуды. Эти вновь образованные кровеносные сосуды обеспечивают не только постоянный рост первичной опухоли, но также диссеминацию и реколонизацию метастатических опухолевых клеток. В соответствии с этим наблюдается связь между плотностью микрососудов в срезах опухоли и выживаемостью больных раком молочной железы, а также с опухолями некоторых других типов. Weidner et al., N. Engl. J. Med 324:1-6 (1991); Horak et al., Lancet 340:1120-1124 (1992); Macchiarini et al., Lancet 340:145-146 (1992). Точный механизм регуляции переключения ангиогенеза пока еще хорошо не изучен, однако очевидно, что неоваскуляризация опухолевой массы обусловлена суммарным балансом множества стимуляторов и ингибиторов ангиогенеза (Folkman Nat. Med. 1(l):27-31 (1995)).
В настоящее время считается, что в случае солидных опухолей подавляющее большинство, а по оценкам специалистов 90% летальных исходов связаны с метастазами (Gupta and Massague, Cell 127, 679-695 (2006)). Сложный процесс образования метастазов состоит из ряда отдельных стадий, включающих отделение опухолевых клеток от первичной опухоли, интравазацию опухолевых клеток в лимфатические или кровеносные сосуды и экстравазацию и рост опухолевых клеток во вторичных участках. Анализ региональных лимфоузлов при опухолях многих типов позволяет предположить, что лимфатическая сосудистая сеть является важным каналом диссеминации злокачественных опухолей человека. Кроме того, почти при всех карциномах присутствие опухолевых клеток в лимфоузлах является наиболее важным отрицательным прогностическим фактором. Хотя ранее считалось, что такие метастазы образуются в результате прохождения злокачественных клеток по лимфатическим сосудам, уже существующим возле опухолей, однако недавно проведенные экспериментальные исследования и отчеты о клинических патологиях (описанные в публикации Achen et al., Br. J. Cancer 94 (2006), 1355-1360 и Nathanson, Cancer 98, 413-423 (2003)) позволяют предположить, что лимфангиогенез может быть индуцирован солидными опухолями и тем самым может стимулировать распространение этих опухолей. Эти и другие недавно проведенные исследования позволяют предположить, что направленное воздействие на лимфатические сосуды и лимфангиогенез может служить терапевтической стратегией ограничения развития метастазов рака, и такая стратегия должна обеспечивать значительный благоприятный эффект у большого числа пациентов.
VEGFC, который является членом семейства факторов сосудистых эндотелиальных клеток (VEGF), представляет собой один из наиболее хорошо изученных медиаторов развития лимфатических сосудов. Было обнаружено, что сверхэкспрессия VEGFC в опухолевых клетках стимулирует опухолевый лимфангиогенез, приводящий к усилению развития метастазов в региональных лимфоузлах (Karpanen et al., Faseb. J. 20, 1462-1472 (2001); Mandriota et al. EMBO J. 20, 672-682 (2001); Skobe et al., Nat. Med. 7, 192-198 (2001); Stacker et al., Nat. Rev. Cancer 2, 573-583 (2002); Stacker et al., Faseb. J. 16, 922-934 (2002)). Экспрессия VEGFC также коррелирует с опухолевым лимфангиогенезом и метастазами в лимфоузлах при различных злокачественных заболеваниях человека (как описано в публикации Achen et al., 2006, см. выше). Кроме того, было обнаружено, что блокада VEGFC-опосредуемой передачи сигнала приводит к подавлению опухолевого лимфангиогенеза и к развитию метастазов в лимфоузлах у мышей (Chen et al., Cancer Res 65, 9004-9011 (2005); He et al., J. Natl Cancer Inst 94, 8190825 (2002); Krishnan et al., Cancer Res 63, 713-722 (2003); Lin et al., Cancer Res 65, 6901-6909 (2005)).
Известно, что VEGFC связывается по меньшей мере с двумя рецепторами, относящимися к семейству рецепторов клеточной поверхности, такими как рецепторы тирозинкиназы VEGF и рецепторы нейропилина (Nrp).
VEGFC, один из трех рецепторов VEGF, может связываться с VEGFR2 и VEGFR3, что приводит к димеризации этого рецептора (Shinkai et al., J Biol Chem 273, 31283-31288 (1998)), к активации киназы и к его аутофосфорилированию (Heldin, Cell 80, 213-223 (1995); Waltenberger et al., J. Biol Chem 269, 26988-26995 (1994)). Фосфорилированный рецептор индуцирует активацию множества субстратов, что приводит к ангиогенезу и лимфангиогенезу (Ferrara et al., Nat. Med. 9, 669-676 (2003)).
Семейство нейропилинов (Nrp) состоит из двух гомологичных белков, нейропилина-1 (Nrp1) и нейропилина-2 (Nrp2). С Nrp2, помимо рецепторов VEGF, также связывается VEGFC, который был сначала идентифицирован как рецептор семафорина класса 3 и медиатор регуляции действия аксонов (Favier et al., Blood 108, 1243-1250 (2006); Soker et al., J. Cell Biochem. 85, 357-368 (2002)). Существует множество данных, указывающих на участие Nrp2 в развитии сосудистой и лимфатической систем. У гомозиготных мутантов по Nrp2 было показано значительное снижение числа мелких лимфатических сосудов и капилляров в пренатальном периоде (Yuan et al., Development 129, 4797-4806 (2002)). Кроме того, серьезные нарушения и дефекты, приводящие к гибели эмбриона, наблюдаемые у мутантных мышей, гомозиготных по Nrp1, усиливаются в результате потери функции Nrp2, что приводит к раннему летальному исходу (Takashima et al., Proc. Natl. Acad. Sci. USA, 99, 3657-3662 (2002)). Однако роль Nrp2 в модуляции биологических процессов в кровеносных и лимфатических сосудах у взрослых и, в частности, его роль в развитии метастазов пока еще неясны.
Нейропилины (Nrp) имеют короткие внутриклеточные домены, однако неизвестно, обладают ли они какой-либо ферментативной активностью или активностью в передаче сигнала. Было высказано предположение, что функция Nrp заключается в усилении передачи сигнала VEGFR за счет повышения уровня связывания рецептора VEGF с лигандом (Favier et al., 2006, см. выше; Soker et al., 2002, см. выше). Кроме того, было обнаружено, что sema3F, семафориновый лиганд Nrp2, модулирует поведение эндотелиальных клеток in vitro и in vivo (Bielenberg et al., J. Clin. Invest. 114, 1260-1271 (2004); Favier et al., Blood 1243-12503 (2006)). Однако недавно появившиеся сообщения позволяют высказать другое предположение, а именно что Nrp могут функционировать независимо от рецепторов VEGF либо семафорин действует как модулятор миграции эндотелиальных клеток (EC) (Murga et al., Blood 105, 1992-1999 (2005); Pan et al., Cancer Cell 11, 53-67 (2007); Wang et al., J Biol Chem 278, 48848-48860 (2003)).
Антитела, нейтрализующие VEGF, подавляют рост различных опухолевых клеточных линий человека у «голых» мышей (Kim et al., Nature 362:841-844 (1993); Warren et al., J. Clin. Invest. 95: 1789-1797 (1995); Borgström et al., Cancer Res. 56:4032-4039 (1996); Melnyk et al., Cancer Res. 56:921-924 (1996)), а также ингибируют внутриглазной ангиогенез у моделей с ишемической ретинопатией (Adamis et al., Arch. Ophthalmol 114:66-71 (1996)). Поэтому моноклональные антитела против VEGF и другие ингибиторы действия VEGF являются перспективными кандидатами для лечения опухолей и различных внутриглазных неоваскулярных заболеваний. Такие антитела описаны, например, в EP 817648, опубликованном 14 января 1998; и в заявках WO 98/45331 и WO 98/45332, опубликованных 15 октября 1998. Одно из таких антител против VEGF, а именно бевацизумаб, было разрешено Управлением по контролю за качеством пищевых продуктов, медикаментов и косметических средств (FDA) для использования в комбинации с химиотерапией для лечения метастазирующего рака прямой и ободочной кишки (CRC) и немелкоклеточного рака легких (NSCLC). В настоящее время проводится большое число клинических испытаний по применению бевацизумаба для лечения различных злокачественных опухолей.
Специалистам известны также и другие антитела против VEGF и антитела против Nrp1, которые описаны, например, в публикациях Liang et al., J. Mol. Biol. 366, 815-829 (2007); Pan et al., Cancer Cell 11, 53-67 (2007); и Liang et al., J. Biol. Chem. 281, 951-961 (2006).
Описание сущности изобретения
Настоящее изобретение основано, по меньшей мере частично, на экспериментальных данных, полученных для высокоаффинного антитела, блокирующего функцию Nrp2. Результаты, полученные для этого антитела, показали, что Nrp2 играет определенную роль в модуляции миграции лимфатических эндотелиальных клеток (LEC) и что его функция выходит за пределы ранее приписываемой ей роли усилителя активации рецептора VEGF. Кроме того, полученные результаты продемонстрировали, что блокирование Nrp2 приводит к ингибированию лимфангиогенеза и значительному снижению метастазов в лимфоузлах и дистальных органах.
В одном из аспектов настоящее изобретение относится к способу ингибирования миграции эндотелиальных клеток лимфатических сосудов, включающему введение млекопитающему, при необходимости, эффективного количества антагониста нейропилина-2 (Nrp2).
В другом аспекте настоящее изобретение относится к способу ингибирования опухолевого лимфангиогенеза, включающему введение млекопитающему с опухолью эффективного количества антагониста нейропилина-2 (Nrp2).
В другом аспекте настоящее изобретение относится к способу ингибирования метастазов опухоли, включающему введение млекопитающему с опухолью эффективного количества антагониста нейропилина-2 (Nrp2).
Во всех вариантах изобретения предпочтительным млекопитающим является человек, такой как больной со злокачественной опухолью, у которого могут быть диагностированы метастазы или который может иметь риск развития метастазов.
В одном из вариантов изобретения злокачественная опухоль выбрана из группы, состоящей из карциномы, лимфомы, бластомы, саркомы и лейкоза.
В другом варианте изобретения злокачественная опухоль выбрана из группы, состоящей из плоскоклеточного рака, мелкоклеточного рака легких, немелкоклеточного рака легких, аденокарциномы легких, плоскоклеточной карциномы легких, перитонеального рака, гепатоцеллюлярного рака, рака желудка, рака желудочно-кишечного тракта, рака поджелудочной железы, глиобластомы, рака шейки матки, рака яичника, рака печени, рака мочевого пузыря, гепатомы, рака молочной железы, рака толстой кишки, рака прямой и ободочной кишки, карциномы эндометрия или матки, карциномы слюнных желез, рака почек или ренального рака, рака печени, рака предстательной железы, рака вульвы, рака щитовидной железы, карциномы печени и рака головы и шеи различных типов, В-клеточной лимфомы, хронического лимфоцитарного лейкоза (CLL); острого лимфобластного лейкоза (ALL);
волосато-клеточного лейкоза; хронического миелобластного лейкоза; лимфопролиферирующего расстройства после трансплантации (PTLD), патологической пролиферации сосудов, связанной с факоматозом, отеков, связанных с опухолями головного мозга, и синдрома Мейгса.
В еще одном варианте изобретения В-клеточная лимфома выбрана из группы, состоящей из низкозлокачественной/фолликулярной не-ходжкинской лимфомы (NHL); мелкоклеточной лимфоцитарной (SL) NHL; NHL средней стадии дифференцировки/фолликулярной NHL; диффузной NHL средней стадии дифференцировки; высокозлокачественной иммунобластной NHL; высокозлокачественной лимфобластной NHL; высокозлокачественной мелкоклеточной недифференцированной NHL; генерализованной NHL; лимфомы клеток коры головного мозга; лимфомы, связанной со СПИД; и макроглобулинемии Вальденстрема.
Антагонистами Nrp2 могут быть, но ими не ограничиваясь, антитело против Nrp2, включая антитела против Nrp2B и Nrp2A, такие как, например, антитела YW68.4.2, YW68.4.2.36, YW126.20, и их фрагменты и варианты, такие как аффинно-зрелые варианты.
В другом аспекте настоящее изобретение относится к антителу против Nrp2B, содержащему последовательность вариабельной области тяжелой и/или легкой цепи антитела, выбранного из группы, состоящей из YW68.4.2, YW68.4.2.36, и его фрагменту или варианту.
В еще одном аспекте настоящее изобретение относится к антителу против Nrp2A, содержащему последовательности вариабельных областей тяжелой и/или легкой цепи YW126.20, или его фрагменту или варианту.
Настоящее изобретение также относится к композиции, содержащей антитело по любому из пп.33-38 формулы изобретения в смеси с фармацевтически приемлемым носителем.
В другом аспекте настоящее изобретение относится к фармацевтической композиции для профилактики или лечения метастазов опухолей, содержащей эффективное количество антагониста Nrp2 в смеси с фармацевтически приемлемым носителем.
В других аспектах настоящее изобретение относится к антагонистам Nrp2, которые могут быть использованы для профилактики или лечения метастазов, и к применению антагонистов Nrp2, таких как антитела против Nrp2, для профилактики или лечения метастазов опухоли.
Краткое описание графического материала
Фигура 1. Характеристика моноклональных антител (mAb) против Nrp2B (анти-Nrp2B mAb). (A) Схематически представлена локализация Sema- и VEGF-связывающей области на Nrp2 по отношению к областям эпитопов для антитела против Nrp2B. (B) ELISA-анализ, демонстрирующий связывание антитела против Nrp2B с ECD hNrp2 (заштрихованные квадраты) и доменами B1-B2 hNrp2 (заштрихованные кружки), но не с ECD hNrp1 (незаштрихованные квадраты) или с доменами A1-A2 hNrp2 (незаштрихованные кружки). (C) Блокирование связывания VEGFC с Nrp2 под действием антитела против Nrp2B. mAb в возрастающих количествах предварительно инкубировали в планшетах, покрытых ECD Nrp2 человека (5 мкг/мл), в течение 1-2 часов, а затем добавляли предварительно оттитрованный биотинилированный VEGFC человека (1 нМ) в течение 15 минут. Процент связанного VEGFC определяли с использованием конъюгатов «стрептавидин-HRP (ПХ)». (D) Блокирование связывания VEGF165 с Nrp2 под действием антитела против Nrp2B. (Е) Блокирование связывания Sema3F с LEC. LEC инкубировали с кондиционированной средой, содержащей Sema3F, связанный со щелочной фосфатазой (AP) (REF), в присутствии или в отсутствие антитела против Nrp2B. AP-активность связанного с AP Sema3F детектировали колориметрически с одновременным проявлением. Какого-либо связывания с AP не наблюдалось (левая панель). Антитело против Nrp2B не блокировало связывания Sema3F с LEC (средние панели). ECD Nrp2 использовали в качестве положительного контроля для блокирования связывания (правая панель). Масштабная шкала.
Фигура 2. Антитело против Nrp2B подавляет VEGFC-индуцированную функцию in vitro и in vivo. (A) Примеры изображения окрашенных LEC, мигрирующих в ответ на действие 200 нг/мл VEGFC в течение 18 часов в присутствии или в отсутствие антитела против Nrp2B (50 мкг/мл) или ECD VEGFR3 (50 мкг/мл). (B) Количественная оценка миграции LEC в ответ на действие 200 нг/мл VEGFC (n=6 для каждого условия эксперимента). (C) Количественная оценка миграции LEC в ответ на действие 10 нг/мл VEGF165 в присутствии или в отсутствие антитела против Nrp2B (50 мкг/мл) или ECD VEGFR3 (50 мкг/мл). N=6 для каждого условия эксперимента. (D) Количественная оценка числа пикселей, проводимая с помощью анализа микрокармана роговицы и описанная в (E). *p<0,05. (E) Примеры изображения LYVE-1-окрашенной роговицы, иллюстрирующие действие 150 нг гранул VEGFC (P) внутри роговицы и системного введения антитела против Nrp2B (10 мг/кг, два раза в неделю) или ECD VEGFR3 (25 мг/кг, два раза в неделю). Для облегчения визуализации проводили LYVE-1-окрашивание, которое давало псевдоокраску красным, *p<0,05; величина ошибки представлена в виде стандартной ошибки среднего.
Фигура 3. Обработка Nrp2B приводит к снижению активации VEGF-рецептора и к ингибированию образования комплекса Nrp2/VEGF-рецептор. (A) FACS-анализ уровней Nrp2, VEGFR2 и VEGFR3 на поверхности LEC после обработки контрольным антителом (10 мкг/мл; зеленая линия) или антителом против Nrp2B (10 мкг/мл) в течение 5 минут (синяя линия) или в течение 20 часов (красная линия). (B) Количественная оценка миграции LEC в ответ на действие 20 нг/мл HGF в присутствии или в отсутствие антитела против Nrp2B (50 мкг/мл) или ECD VEGFR3 (50 мкг/мл). N=6 для каждого условия эксперимента. (C) Уровень фосфорилирования VEGFR2 в LEC, детектированный путем ELISA-анализа с использованием антител, которые распознают общий или фосфорилированный по тирозину VEGFR2. VEGFC (в указанной концентрации) добавляли в течение 10 минут в присутствии или в отсутствие антитела против Nrp2B (10 мкг/мл) или ECD VEGFR3 (10 мкг/мл) для индуцирования фосфорилирования VEGFR2, n=3 для каждого условия эксперимента. Уровень фосфорилирования VEGFR2 в клетках, обработанных антителом против Nrp2B (10 мкг/мл), значительно отличался от уровня фосфорилирования VEGFC, стимулированного при 200 нг/мл, и постоянно составлял в пределах от уровня фосфорилирования, индуцированного 175 нг/мл, до уровня фосфорилирования, индуцированного 150 нг/мл VEGFC. (D) Количественная оценка миграции LEC в ответ на VEGFC (в указанной концентрации) в присутствии или в отсутствие антитела против Nrp2B (10 мкг/мл) или ECD VEGFR3 (10 мкг/мл). Значительное снижение уровня миграции наблюдалось при 50 нг/мл VEGFC или при блокировании под действием ECD VEGFR3. *p<0,05; величина ошибки представляет собой стандартную ошибку среднего. Каждый эксперимент повторяли минимум три раза. (E) CO-IP.
Фигура 4. Nrp2 экспрессируется в лимфатических сосудах мышей с опухолью. (A-D) окрашивание LYVE-1 (левая колонка - красный) для мечения лимфатических сосудов, окрашивание Nrp2 (средняя колонка - зеленый) и перекрывание (A) в тонком кишечнике и (B) в лимфоузлах здоровой взрослой мыши. Сигнал Nrp2 не совпадает с локализацией LYVE-1-меченных лимфатических сосудов в любом органе. Окрашивание Nrp2 воспалительных клеток редко наблюдалось в фибростромальном ядре кишечных ворсинок и в зародышевых центрах лимфатических узлов. (C) В лимфатических узлах животных с опухолями сигнал Nrp2 не совпадает с локализацией LYVE-1-положительных лимфатических сосудов, выстилающих LN-синусы. Наблюдалось также дополнительное окрашивание Nrp2 воспалительных клеток. (D) Интенсивное окрашивание Nrp2 также наблюдалось в лимфатических сосудах опухолей 66cl4. На опухолевых клетках также можно было наблюдать слабое мембранное окрашивание. На второй день только окрашенный контроль не обнаруживал какого-либо сигнала. Области, обрамленные рамкой, показаны в виде вставки с большим увеличением. Масштабная шкала *** для A-C и ** для D.
Фигура 5. Обработка антителом против Nrp2B приводила к снижению числа метастазов в легких у модели с опухолью 66c14. (A) Ниже проанализирован график среднего объема опухоли 66c14, построенный при исследовании на модели с указанной опухолью. Животным два раза в неделю i.p. вводили дозу 10 мг/кг антитела против Nrp2B или контрольного антитела после достижения среднего размера опухоли 100 мм3, и такую дозу вводили на протяжении всего исследования. (B) Количественную оценку проводили путем визуального наблюдения числа узлов с метастазами на одно легкое у животных, обработанных контролем и антителом против Nrp2B. (C) Примеры изображения легких для животных, обработанных контролем (слева) и антителом против Nrp2B (справа). Перед фиксацией легкие раздували путем перфузии правого желудочка сердца. Для облегчения визуализации узлы помечали белым цветом. (D, E) Пример трехмерного изображения легких, сканированных с помощью микро-KT (компьютерной томографии), иллюстрирующего узлы с метастазами (красным) у животных, обработанных контролем (D) и антителом против Nrp2B (E). Положения продольного среза (верхняя вставка) и поперечного среза (нижняя вставка) показаны черными и красными пунктирными линиями соответственно. Этот анализ подтвердил, что большинство узлов находится на поверхности легких. (F) Количественная оценка числа узлов с метастазами на легкое, проводимая с помощью микро-KT-анализа легких. (G) FACS-анализ in vitro уровней Nrp2 на поверхности культивированных опухолевых клеток 66c14. (H) окрашивание H&E узлов легких, иллюстрирующих опухолевые клетки с метастазами. Величина ошибки представлена в виде стандартной ошибки среднего. Масштабная шкала *** для C и ** для H.
Фигура 6. Обработка антителом против Nrp2B приводила к снижению числа метастазов в легких у модели с опухолью С6. (A) Ниже проанализирован график среднего объема опухоли С6, построенный при исследовании на модели с указанной опухолью. Животным два раза в неделю i.p. вводили дозу антитела против Nrp2B (10 мг/кг), ECD VEGFR3 (25 мг/кг) или контрольного антитела (10 мг/кг) после достижения среднего размера опухоли 100 мм3, и такую дозу вводили на протяжении всего исследования. (B) Количественную оценку проводили путем визуального наблюдения числа узлов с метастазами на легкое у животных, обработанных контролем, ECD VEGFR3 и антителом против Nrp2B. (C) Примеры изображения легких животных, обработанных контролем (слева), ECD VEGFR3 (в середине) и антителом против Nrp2B (справа). Перед фиксацией легкие раздували путем перфузии правого желудочка сердца. Для облегчения визуализации узлы помечали белым цветом. (D) Пример трехмерного изображения легких, сканированных с помощью микро-KT (компьютерной томографии), иллюстрирующего узлы с метастазами (красным) у животных, обработанных контролем (слева) и антителом против Nrp2B (справа). Положения продольного среза (верхняя вставка) и поперечного среза (нижняя вставка) показаны черными и красными пунктирными линиями, соответственно. Этот анализ подтвердил, что большинство узлов находится на поверхности легких. (Е) FACS-анализ уровней Nrp2 на поверхности in vitro культивированных опухолевых клеток 66c14. (F) окрашивание H&E узлов легких, демонстрирующее опухолевые клетки с метастазами. Величина ошибки представлена в виде стандартной ошибки среднего. Масштабная шкала *** для C и ** для F.
Фигура 7. Обработка антителом против Nrp2B приводила к уменьшению числа опухолевых лимфатических сосудов. (A, B) Количественная оценка плотности сосудистой системы (А), детектируемой с помощью IHC PECAM-1, и плотности лимфатических сосудов (В), детектируемой с помощью IHC LYVE-1 в опухолях 66cl4, обработанных контрольным антителом или антителом против Nrp2B. Плотность сосудов определяли по 6 примерам изображений, полученных для каждой из 6 опухолей на группу и оцениваемых по среднему числу пикселей с помощью ImageJ. (C) Примеры изображений окрашенных PECAM-1 сосудов (верхний ряд) и окрашенных LYVE-1 лимфатических сосудов (средний и нижний ряды) в опухолях C6, обработанных контрольным антителом (левый столбец), ECD VEGFR3 (средний столбец) или антителом против Nrp2B (правый столбец). Области в рамке, показанные в среднем ряду, представлены с большим увеличением в нижнем ряду. Справа от этих изображений приводится количественная оценка плотности сосудистой системы (верхний график) и плотности лимфатических сосудов (нижний график). (D) окрашенные LYVE-1 опухоли, собранные у животных, обработанных антителом против Nrp2B (нижние панели) на день 4 (сбор 1) и на день 11 (сбор 2), демонстрируют разрушение лимфатических сосудов по сравнению с лимфатическими сосудами, собранными у животных, обработанных контролем (верхние панели). Слева представлены кривые роста в зависимости от времени сбора. Величина ошибки представлена в виде стандартной ошибки среднего. Масштабная шкала ***.
Фигура 8. Обработка антителом против Nrp2B приводила к уменьшению числа функциональных лимфатических сосудов опухоли и к замедлению развития метастазов в первичном лимфоузле. (A, B) После внутрикожной лимфангиографии полистирольные флуоресцентные микросферы (зеленые) наблюдались исключительно в лимфатических сосудах, меченных LYVE-1 IHC (красные). Области в рамке на фигуре A представлены на фигуре В с большим увеличением. (C-D) Обработка антителом против Nrp2B приводила к уменьшению интенсивности Эванса голубого в опухолях C6 (C) (P=0,035) и 66cl4 (D) (P=0,005), что указывает на уменьшение числа функциональных лимфатических сосудов в этих обработанных опухолях. (E) Процент животных с небольшими лимфоузлами (SLN), содержащими β-gal-экспрессирующие опухолевые клетки C6, в различные периоды времени после имплантации опухоли в уши мышей, обработанных контролем (черные) и антителом против Nrp2B (красные). Обработка антителом против Nrp2B приводила к замедлению поступления клеток в SLN (p=0,006). Число животных на каждое условие обработки в каждый период времени составляло 7 (N=7).
Фигура 9. Экспрессия Nrp2 в различных злокачественных опухолях человека. (A-F) Данные микромассива Affymetrix HG-U133A и B GeneChip® для оценки уровня экспрессии Nrp2 в здоровой толстой кишке и в аденокарциноме прямой и ободочной кишки (A), в здоровых тканях головы и шеи и в плоскоклеточной карциноме головы и шеи (B), в здоровой поджелудочной железе и в аденокарциноме поджелудочной железы (C), в здоровой коже и в злокачественной меланоме (D), в здоровой щитовидной железе и в папиллярной карциноме щитовидной железы (E), и в здоровой молочной железе и в Her2-инфильтрирующей аденокарциноме протоков (F). Каждый период времени представлен для одного пациента.
Фигура 10. Аминокислотная последовательность Fab-фрагментов антитела против Nrp2B YW68.4.2 и YW68.4.2.36.
Фигура 11. Аминокислотная последовательность Fab-фрагмента антитела против Nrp2A YW126.20.
Фигура 12. Выравнивание последовательностей вариабельного домена легкой цепи антитела против Nrp2A YW126.20 с последовательностью κ1 человека.
Фигура 13. Выравнивание последовательностей вариабельного домена легкой цепи антитела против Nrp2A YW126.20 с последовательностью III (hum III) человека.
Дополнительная фигура 1. FACS-анализ уровней рецепторов для VEGF всех типов на поверхности in vitro культивированных LEC. FACS-анализ Nrp1, Nrp2, VEGFR1, VEGFR2 и VEGFR3 на культивированных LEC.
Дополнительная фигура 2. Антитело против Nrp2B не блокирует индуцированную VEGF165 миграцию. (A-B) Количественная оценка клеток HUVEC (A) и LEC (B), мигрирующих в ответ на 200 нг/мл VEGF в течение 18 часов в присутствии или в отсутствие антитела против Nrp1B, антитела против Nrp2B (50 мкг/мл) или обоих антител (50 мкг/мл). *p<0,05. Величина ошибки представлена в виде стандартной ошибки среднего.
Дополнительная фигура 3. Влияние антитела против Nrp2B на опосредуемую VEGFC пролиферацию, сосудистую проницаемость и связанную VEGFC передачу внутриклеточного сигнала. (A) Количественная оценка пролиферации LEC, индуцированной 200 нг/мл VEGFC в присутствии или в отсутствие антитела против Nrp2B (50 мкг/мл) или ECD VEGFR3 (50 мкг/мл), как было определено по включению BrdU (n=6 на одно условие эксперимента). (B) Анализ сосудистой проницаемости кожи мышей. Изображения получали для кожи одного и того же животного. Синяя окраска указывает на утечку Эванса голубого из сосудистой системы в ответ на чрескожную доставку VEGFC после системной обработки антителом против Nrp2B (10 мг/кг) или ECD VEGFR3 (25 мг/кг). (C) Количественная оценка красителя Эванса голубого, экстрагированного из образцов кожи в анализе на проницаемость. Указанные величины представляют собой средние величины, полученные для 6 независимых экспериментов. *p<0,05; величина ошибки представлена в виде стандартной ошибки среднего.
Дополнительная фигура 4. Антитело против Nrp2B не блокирует индуцированный Sema3F коллапс конуса роста. (A) Изображения конусов роста гиппокампа E17.5, окрашенных фаллоидином, конъюгированным с родамином. В контрольных конусах роста обнаружены крупные, богатые актином структуры на окончании каждого аксона, которые уменьшались после обработки Sema3F. Антитело против Nrp2B (50 мкг/мл) не блокирует такой коллапс. В противоположность этому, ECD Nrp2 (10 мкг/мл) блокирует такой коллапс. (B) Контроль качества для иммуногистохимического анализа с использованием антитела против Nrp2. Изображение представляет фаговый клон, который распознает Nrp2, функционирующий в IHC на свежезамороженных срезах. Уровень экспрессии белка Nrp2 аналогичен уровню экспрессии Nrp2, наблюдаемому с использованием in situ гибридизации (Chen et al., Neuron 19, 547-559 (1997)). Это антитело было затем использовано для иммуногистологического анализа (IHC) на свежезамороженных срезах опухоли. Масштабная шкала для главного изображения = **мкм и для изображений вставки = **мкм.
Подробное описание изобретения
Определения
Термины «нейропилин», «NRP» или «Nrp» являются взаимозаменяемыми и означают нейропилин-1 (NRP1, Nrp1), нейропилин-2 (NRP2, Nrp2) и их изоформы и варианты, описанные в публикации Rossignol et al. (2000) Genomics 70:211-222. Нейропилины представляют собой не-тирозинкиназные рецепторы размером 120-130 кДа. Существует множество сплайсированных вариантов и растворимых изоформ NRP-1 и NRP-2. Основная структура нейропилинов включает пять доменов: три внеклеточных домена (a1a2, b1b2 и c), трансмембранный домен и цитоплазматический домен. Домен a1a2 гомологичен компонентам комплемента C1r и C1s (CUB), которые обычно содержат четыре цистеиновых остатка, образующие два дисульфидных мостика. Домен b1b2 гомологичен факторам свертывания крови V и VIII. Центральная часть домена с обозначена MAM, что обусловлено ее гомологией с меприном, A5 и рецепторными белками тирозинфосфатазы µ. Домены a1a2 и b1b2 ответственны за связывание с лигандом, а домен с играет решающую роль в гомодимеризации или гетеродимеризации. Gu et al. (2002) J. Biol. Chem. 277; 18069-76; He and Tessier-Lavigne (1997) Cell 90:739-51.
Термин «опосредуемая нейропилином биологическая активность» означает, в основном, физиологические или патологические события, в которых важную роль играют нейропилин-1 и/или нейропилин-2. Неограничивающими примерами таких активностей являются регуляция аксонов в процессе развития эмбриональной нервной системы или регенерации нейронов, ангиогенез (включая моделирование сосудов), онкогенез и образование метастазов опухоли.
Используемый в настоящем описании термин «опосредуемая нейропилином-2 биологическая активность» или «опосредуемая Nrp2 биологическая активность» означает, в основном, физиологические или патологические события, в которых важную роль играет Nrp2, например, такие как усиление активации рецептора VEGF и, в частности, способность модулировать миграцию эндотелиальных клеток (EC) лимфатических сосудов, лимфангиогенез у взрослых, в частности лимфангиогенез и образование метастазов опухоли.
Используемый в настоящем описании термин «антитело» используется в самом широком смысле, и, в частности, термин охватывает моноклональные антитела (в том числе полноразмерные моноклональные антитела), поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они обладают нужной биологической активностью.
Используемый в настоящем описании термин «моноклональное антитело» означает антитело, полученное из популяции, в основном, гомогенных антител, то есть отдельных антител, входящих в эту популяцию и являющихся идентичными, за исключением возможных природных мутаций, которые могут присутствовать в небольших количествах. Моноклональные антитела являются в высокой степени специфическими и нацелены на одну антигенную детерминанту. Кроме того, в отличие от стандартных препаратов (поликлональных) антител, которые обычно содержат различные антитела, нацеленные на различные детерминанты (эпитопы), каждое моноклональное антитело нацелено на одну детерминанту антигена. Термин «моноклональный» указывает на тип антитела, полученного по существу из гомогенной популяции антител, но этот термин не подразумевает, что это антитело должно быть получено каким-либо конкретным методом. Например, моноклональные антитела по изобретению могут быть получены с помощью гибридомной технологии, впервые описанной Kohler et al. (1975) Nature 256:495, или методами рекомбинантных ДНК (см., например, патент США № 4816567). «Моноклональные антитела» могут быть также выделены из фаговых библиотек антител методами, описанными, например, Clackson et al. (1991) Nature 352:624-628 и Marks et al. (1991) J. Mol. Biol. 222:581-597.
Представленные в настоящем описании моноклональные антитела, в частности, включают «химерные» антитела (иммуноглобулины), в которых часть тяжелой цепи и/или легкой цепи идентична или гомологична соответствующим последовательностям, присутствующим в антителах конкретного вида или принадлежащим конкретному классу или подклассу антител, а остальная часть этой(их) цепи(ей) идентична или гомологична соответствующим последовательностям, присутствующим в антителах другого вида или принадлежащим другому классу или подклассу антител; а также фрагменты этих антител, при условии, что они обладают нужной биологической активностью (см. патент США № 4816567; и Morrison et al. (1984) Proc. Natl. Acad. Sci., USA, 81:6851-6855).
«Гуманизованные» формы не-человеческих (например, мышиных) антител представляют собой химерные антитела, которые содержат минимальную последовательность не-человеческого иммуноглобулина. По большей части, гуманизованные антитела представляют собой иммуноглобулины человека (антитела-реципиенты), в которых остатки гипервариабельной области реципиента заменены остатками гипервариабельной области не-человеческого антитела (донорного антитела), такого как антитело мыши, антитело крысы, антитело кролика или антитело примата, не являющегося человеком, которое имеет нужную специфичность, аффинность или функцию. В некоторых случаях остатки каркасной области Fv (FR) иммуноглобулина человека могут быть заменены соответствующими не-человеческими остатками. Кроме того, гуманизованные антитела могут содержать остатки, которые отсутствуют в антителе-реципиенте или в донорном антителе. Такие модификации могут быть введены для дополнительного улучшения функции антитела. Как правило, гуманизованное антитело может содержать по существу весь по меньшей мере один, а обычно два вариабельных домена, в которых все или по существу все гипервариабельные петли, соответствующие петлям не-человеческих иммуноглобулинов, и все или по существу все области FR являются последовательностями иммуноглобулина человека. Гуманизованное антитело может также, но необязательно, содержать по меньшей мере часть константной области иммуноглобулина (Fc), обычно иммуноглобулина человека. Более подробное описание см. в публикации Jones et al. (1986) Nature 321:522-525; Riechmann et al. (1988) Nature 332:323-329; и Presta (1992) Curr. Op. Struct. Biol. 2:593-596.
Термин «видоспецифическое антитело» означает антитело, которое обладает более высокой аффинностью связывания с антигеном млекопитающего первого вида, чем с гомологом антигена млекопитающего другого вида. Обычно видоспецифическое антитело «специфически связывается» с антигеном человека (то есть имеет величину аффинности связывания (Kd), составляющую не более чем примерно 1×10-7 M, предпочтительно не более чем примерно 1×10-8 M и наиболее предпочтительно не более чем примерно 1×10-9 M), но имеет аффинность связывания с гомологом антигена млекопитающего, не являющегося человеком, другого вида, которая по меньшей мере примерно в 50 раз, или по меньшей мере примерно в 500 раз, или по меньшей мере примерно в 1000 раз меньше аффинности связывания с антигеном человека. Видоспецифическое антитело может представлять собой антитело любого типа, определенного выше, и предпочтительным является гуманизованное антитело или антитело человека.
Используемый в настоящем описании термин «мутантное антитело» или «вариант антитела» означает вариант аминокислотной последовательности видоспецифического антитела, в котором один или несколько аминокислотных остатков видоспецифического антитела модифицированы. Такие мутанты обязательно имеют последовательность, которая менее чем на 100% идентична или аналогична последовательности видоспецифического антитела. В предпочтительном варианте изобретения мутантное антитело имеет аминокислотную последовательность, которая по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, еще более предпочтительно по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90% и наиболее предпочтительно по меньшей мере на 95% идентична или аналогична аминокислотной последовательности вариабельного домена тяжелой или легкой цепи видоспецифического антитела. Идентичность или сходство с этой последовательностью определяются в настоящем описании как процент аминокислотных остатков в выбранной последовательности, которые идентичны (то есть являются одинаковыми) или аналогичны остаткам (то есть аминокислотным остаткам, принадлежащим одной и той же группе аминокислот, классифицированных по общим свойствам боковых цепей, см. ниже) видоспецифических антител, как было определено путем выравнивания последовательностей с введением пропусков, если это необходимо для достижения максимального процента идентичности последовательностей. При этом ни одна из N-концевых модификаций, С-концевых модификаций или модификаций типа внутренних добавлений, делеций или инсерций в последовательности антитела, находящихся за пределами вариабельного домена, не должна рассматриваться как модификация, влияющая на идентичность или сходство последовательностей.
«Выделенное» антитело представляет собой антитело, которое было идентифицировано и отделено от компонентов своего природного окружения и/или выделено из таких компонентов. Примесными компонентами его природного окружения являются вещества, которые могут препятствовать диагностическому или терапевтическому использованию указанного антитела, и такими веществами могут быть ферменты, гормоны и другие белковые или не-белковые растворенные вещества. В предпочтительных вариантах изобретения указанное антитело может быть очищено (1) более чем на 95% по массе антитела, как может быть определено методом Лаури, и наиболее предпочтительно более чем на 99% по массе антитела, (2) до уровня, достаточного для получения N-концевой или внутренней аминокислотной последовательности, состоящей по меньшей мере из 15 остатков, с использованием секвенатора с центрифужными чашками, или (3) до гомогенности, оцениваемой с помощью SDS-PAGE в восстанавливающих или невосстанавливающих условиях с использованием Кумасси синего или предпочтительно красителя с серебром. Выделенным антителом считается антитело in situ в рекомбинантных клетках, поскольку в них отсутствует по меньшей мере один компонент природного окружения антитела. Однако, как правило, выделенное антитело может быть получено по меньшей мере в одну стадию очистки.
Используемый в настоящем описании термин «вариабельный домен антитела» означает части легкой и тяжелой цепей молекул антител, которые включают аминокислотные последовательности гипервариабельных областей (областей, определяющих комплементарность, CDR; то есть CDR1, CDR2 и CDR3) и каркасных областей (FR). VH означает вариабельный домен тяжелой цепи. VL означает вариабельный домен легкой цепи. В соответствии со способами, используемыми в настоящем изобретении, положения аминокислот, присвоенные CDR и FR, могут быть определены в соответствии с нумерацией по Кабату (Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md., 1997 and 1991)). Нумерация аминокислот антител или антиген-связывающих фрагментов также дана по Кабату.
Используемый в настоящем описании термин «гипервариабельные области» (CDR, то есть CDR1, CDR2 и CDR3) означает аминокислотные остатки вариабельного домена антитела, присутствие которых необходимо для связывания с антигеном. Каждый вариабельный домен обычно имеет три области CDR, обозначенные как CDR1, CDR2 и CDR3. Каждая гипервариабельная область может содержать аминокислотные остатки «гипервариабельной области», определенной по Кабату (то есть примерно остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (H1), 50-65 (H2) и 95-102 (H3) в вариабельном домене тяжелой цепи; Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)), и/или остатки «гипервариабельной петли» (то есть примерно остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (Н1), 53-55 (Н2) и 96-101 (Н3) в вариабельном домене тяжелой цепи; Chothia and Lesk (1987) J. Mol. Biol., 196:901-917). В некоторых случаях гипервариабельная область может содержать аминокислоты обеих областей CDR, определенных по Кабату, и аминокислоты гипервариабельной петли. Например, CDRH1 тяжелой цепи антитела 4D5 содержит аминокислоты 26-35.
«Каркасными областями» (далее обозначаемыми FR) являются области, в которых остатки вариабельных доменов отличаются от остатков CDR. Каждый вариабельный домен обычно имеет четыре FR, обозначенных как FR1, FR2, FR3 и FR4. Если CDR определены по Кабату, то остатки FR легкой цепи имеют примерно следующие положения: 1-23 (LCFR1), 35-49 (LCFR2), 57-88 (LCFR3) и 98-107 (LCFR4), а остатки FR тяжелой цепи имеют примерно следующие положения: 1-30 (HCFR1), 36-49 (HCFR2), 66-94 (HCFR3) и 103-113 (HCFR4). Если CDR содержат аминокислотные остатки гипервариабельных петель, то остатки FR легкой цепи имеют примерно следующие положения: 1-25 (LCFR1), 33-49 (LCFR2), 53-90 (LCFR3) и 97-107 (LCFR4), а остатки FR тяжелой цепи имеют примерно следующие положения: 1-25 (HCFR1), 33-52 (HCFR2), 56-95 (HCFR3) и 102-113 (HCFR4). В некоторых случаях если CDR содержит аминокислоты как CDR, определенной по Кабату, так и гипервариабельной петли, то положения остатков FR могут быть соответственно скорректированы. Например, если CDRH1 содержит аминокислоты H26-H35, то остатки FR1 тяжелой цепи присутствуют в положениях 1-25, а остатки FR2 присутствуют в положениях 36-49.
Используемый в настоящем описании термин «набор кодонов» означает набор различных последовательностей нуклеотидных триплетов, кодирующих нужные варианты аминокислот. Набор олигонуклеотидов может быть синтезирован, например, путем твердофазного синтеза, включая последовательности, которые представляют все возможные комбинации нуклеотидных триплетов, обеспечиваемых указанным набором кодонов, и которые кодируют нужную группу аминокислот. Стандартной формой обозначения кодонов является известный код, который был принят Международным биохимическим союзом (IUB) и описан в настоящей заявке. Набор кодонов обычно представляют 3 заглавными буквами курсивом, например NNK, NNS, XYZ, DVK и т.п. Так, например, используемый в настоящем описании термин «неслучайный набор кодонов» означает набор кодонов, которые кодируют определенные аминокислоты, частично, а предпочтительно полностью удовлетворяющие критериям отбора аминокислот, описанным в настоящей заявке. Способ синтеза олигонуклеотидов с определенной степенью «вырожденности» в некоторых положениях хорошо известен специалистам, например способ TRIM (Knappek et al. (1999) J. Mol. Biol. 296:57-86); Garrard & Henner (1993) Gene 128:103). Такие наборы олигонуклеотидов, имеющие определенные наборы кодонов, могут быть синтезированы с помощью коммерчески доступных синтезаторов нуклеиновых кислот (поставляемых, например, Applied Biosystems, Foster City, CA), либо они могут быть закуплены (например, у компании Life Technologies, Rockville, MD). Следовательно, синтезированный набор олигонуклеотидов представляет собой конкретный набор кодонов, который обычно включает множество олигонуклеотидов с различными последовательностями, где указанные различия определяются конкретным набором кодонов по всей последовательности. Олигонуклеотиды, используемые в соответствии с настоящим изобретением, имеют последовательности, которые позволяют им гибридизоваться с матрицей нуклеиновой кислоты вариабельного домена, а также могут, но необязательно, содержать рестрикционные сайты, подходящие, например, для клонирования.
«Fv»-фрагмент представляет собой фрагмент антитела, содержащий полноразмерный антиген-распознающий сайт и антиген-связывающий сайт. Эта область состоит из димера одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, жестко связанных друг с другом связью, которая по своей природе может быть ковалентной, например, в scFv. Этот фрагмент имеет конфигурацию, при которой три CDR в каждом вариабельном домене взаимодействуют друг с другом таким образом, что они образуют антиген-связывающий сайт на поверхности димера VH-VL. В целом, шесть CDR или их подпоследовательность сообщают антителу специфичность связывания с антигеном. Однако даже один вариабельный домен (или половина Fv, содержащая только три гипервариабельных области, специфичные к антигену) обладает способностью распознавать антиген и связываться с ним, хотя и с меньшей аффинностью, чем весь сайт связывания.
«Fab»-фрагмент содержит вариабельный и константный домен легкой цепи и вариабельный домен и первый константный домен (СН1) тяжелой цепи. F(ab')2-фрагменты антитела содержат пару Fab-фрагментов, которые обычно ковалентно связаны возле их карбокси-концов шарнирными цистеинами, расположенными между ними. Специалистам также известны другие химические связи, соединяющие фрагменты антитела.
«Одноцепочечный Fv-фрагмент» или «scFv-фрагмент» антитела содержит домены VH и VL антитела, где указанные домены присутствуют в одной полипептидной цепи. Обычно полипептид Fv также содержит полипептидный линкер между доменами VH и VL, где указанный полипептид обеспечивает образование scFv со структурой, необходимой для связывания с антигеном. Описание scFv можно найти в публикации Pluckthun «The Pharmacology of Monoclonal Antibodies», vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994).
Термин «диатела» (diabodies) означает небольшие фрагменты антител с двумя антиген-связывающими сайтами, где указанные фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной и той же полипептидной цепи (VH-VL). Если линкер является слишком коротким для образования пары между двумя доменами на одной и той же цепи, то домены вынуждены спариваться с комплементарными доменами другой цепи и образовывать два антиген-связывающих сайта. Диатела более подробно описаны, например, в ЕР 404097; WO 93/11161 и у Hollinger et al. Proc. Natl. Acad. Sci., USA, 90:6444-6448 (1993).
Термин «линейные антитела» означает антитела, описанные в публикации Zapata et al. (1995 Protein Eng, 8(10):1057-1062). Коротко, такие антитела содержат пару тандемных сегментов (VH-CH1-VH-CH1), которые вместе с комплементарными полипептидами легкой цепи образуют антиген-связывающие области. Линейные антитела могут быть биспецифическими и моноспецифическими.
Используемый в настоящем описании термин «библиотека» означает множество последовательностей антител или фрагментов антител (например, полипептидов по изобретению) или нуклеиновых кислот, кодирующих эти последовательности, где указанные последовательности отличаются комбинациями различных аминокислот, которые вводят в эти последовательности способами по изобретению.
«Фаговый дисплей» означает метод, с помощью которого полипептидные варианты презентируются как слитые белки, связанные по меньшей мере с частью белка оболочки на поверхности фага, например, частиц нитчатого фага. Метод фагового дисплея основан на возможности сортировки крупных библиотек рандомизированных белковых вариантов быстро и эффективно по последовательностям, которые связываются с антигеном-мишенью с высокой аффинностью. Презентация пептидных и белковых библиотек на фаге используется для скрининга миллионов полипептидов с целью обнаружения полипептидов, обладающих специфическими связывающими свойствами. Методы поливалентного фагового дисплея используются для презентации небольших рандомизированных пептидов и небольших белков посредством соединения их с геном III или геном VIII нитчатого фага. См. публикации Wells and Lowman (1992) Curr. Opin. Struct. Biol. 3:355-362 и цитируемые в них ссылки. В методе одновалентного фагового дисплея белковую или пептидную библиотеку присоединяют к гену III или к его части и экспрессируют на низких уровнях в присутствии белка дикого типа, кодируемого геном III, так, чтобы фаговые частицы презентировали одну копию слитых белков или не презентировали ни один из слитых белков. Эффекты авидности, достигаемые в случае использования одновалентного фага, по сравнению с эффектами, достигаемыми в случае использования поливалентного фага, снижаются, и поэтому сортинг осуществляют на основе природной аффинности к лиганду и используют фагмидные векторы, облегчающие проведение манипуляций с ДНК. Lowman and Wells (1991) Methods: A companion to Methods in Enzymology 3:205-0216.
«Фагмида» представляет собой плазмидный вектор, имеющий точку начала репликации бактериальной природы, например ColEl, и копию межгенной области бактериофага. Фагмида может быть использована на любом известном бактериофаге, включая нитчатый бактериофаг и лямбдоидный бактериофаг. Плазмида также, как правило, содержит селективный маркер резистентности к антибиотикам. Сегменты ДНК, клонированные в эти векторы, могут быть размножены в виде плазмид. Если клетки, содержащие эти векторы, имеют все гены, необходимые для продукции фаговых частиц, то механизм репликации плазмиды переключают на репликацию типа «катящегося кольца» с образованием копий одной цепи плазмидной ДНК и упакованных фаговых частиц. Фагмида может образовывать инфекционные или неинфекционные фаговые частицы. Этот термин включает фагмиды, содержащие ген белка оболочки фага или его фрагмент, сцепленные с гетерологичным геном полипептида в виде слитого гена, который кодирует гетерологичный полипептид, представленный на поверхности фаговой частицы.
Термин «фаговый вектор» означает двухцепочечную репликативную форму бактериофага, содержащую гетерологичный ген и способную к репликации. Фаговый вектор имеет фаговую точку начала репликации, обеспечивающую репликацию фага и образование фаговых частиц. Указанным фагом предпочтительно является нитчатый бактериофаг, такой как фаг M13, fl, fd, Pf3 или их производное, или лямбдоидный фаг, такой как фаг лямбда, 21, phi80, phi81, 82, 424, 434 и т.д. или их производные.
Используемый в настоящем описании термин «положение, доступное для растворителя» означает положение аминокислотных остатков в вариабельных областях тяжелой и легкой цепей исходного антитела или антиген-связывающего фрагмента, где указанное положение определяют исходя из структуры, комбинации структур и/или моделированных структур антитела или антиген-связывающего фрагмента как доступное для растворителя и/или для контакта с молекулой, такой как антиген-специфическое антитело. Такие положения обычно находятся в CDR и за пределами белка. Положения антитела или антиген-связывающего фрагмента, доступные для растворителя, как определено в настоящем описании, могут быть определены с помощью ряда алгоритмов, известных специалистам в данной области. Положения, доступные для растворителя, предпочтительно определяют с использованием координат трехмерной модели антитела, предпочтительно с использованием компьютерной программы, такой как программа InsightII (Accelrys, San Diego, CA). Положения, доступные для растворителя, могут быть также определены с использованием алгоритмов, известных специалистам в данной области (например, Lee and Richards (1971) J. Mol. Biol. 55, 379 and Connolly (1983) J. Appl. Cryst. 16, 548). Определить положения, доступные для растворителя, можно с помощью компьютерной программы, подходящей для моделирования белков и поиска информации о 3-мерной структуре антитела. Компьютерной программой, которая может быть использована в этих целях, является программа SYBYL Biopolymer Module (Tripos Associates). Обычно и предпочтительно при работе с этим алгоритмом (программой) необходимо, чтобы пользователь вводил параметр размера, причем «размер» зонда, используемого в вычислениях, устанавливают приблизительно на 1,4 ангстрем или меньшего радиуса. Кроме того, методы определения областей и участков, доступных для растворителя, с помощью компьютерной программы, разработанной для персональных компьютеров, описаны в публикации Pacios (1994) Comput. Chem. 18(4):377-386.
«Ангиогенный фактор или средство» представляет собой фактор роста, который стимулирует развитие кровеносных сосудов, например стимулирует ангиогенез, стимулирует рост эндотелиальных клеток, повышает стабильность кровеносных сосудов и/или васкулогенез и т.д. Например, ангиогенными факторами являются, но ими не ограничиваясь, VEGF и члены семейства VEGF, PIGF, PDGF и семейства факторов роста фибробластов (FGF), лиганды TIE (ангиопоэтины), эфрины, Del-1, факторы роста фибробластов: кислотный (aFGF) и основный (bFGF), фоллистатин, гранулоцитарный колониестимулирующий фактор (G-CSF), фактор роста гепатоцитов (HGF)/фактор рассеивания (SF), интерлейкин-8 (IL-8), лептин, мидкин, нейропилины, плацентарный фактор роста, тромбоцитарный фактор роста эндотелиальных клеток (PD-ECGF), тромбоцитарный фактор роста, в частности PDGF-BB или PDGFR-бета, плейотропин (PTN), програнулин, пролиферин, трансформирующий фактор роста-альфа (TGF-альфа), трансформирующий фактор роста-бета (TGF-бета), фактор некроза опухоли-альфа (TNF-альфа) и т.д. Такими факторами также являются факторы, способствующие заживлению ран, такие как гормон роста, инсулиноподобный фактор роста-I (IGF-I), VIGF, эпидермальный фактор роста (EGF), CTGF и члены его семейства, и TGF-альфа, и TGF-бета. См., например, Klagsbrun and D'Amore (1991) Annu. Rev. Physiol. 53:217-39; Streit and Detmar (2003) Oncogene 22:3172-3179; Ferrara & Alitalo (1999) Nature Medicine 5(12):1359-1364; Tonini et al. (2003) Oncogene 22:6549-6556 (см., например, таблицу 1, где перечислены известные ангиогенные факторы); и Sato (2003) Int. J. Clin. Oncol. 8:200-206.
Термин «средство против ангиогенеза» или «ингибитор ангиогенеза» означает низкомолекулярное вещество, полинуклеотид, полипептид, выделенный белок, рекомбинантный белок, антитело или его конъюгаты или слитые белки, которые напрямую или опосредованно ингибируют ангиогенез, васкулогенез или нежелательную сосудистую проницаемость. Следует отметить, что средствами против ангиогенеза являются средства, которые связываются с ангиогенным фактором или его рецептором и блокируют ангиогенную активность указанного фактора или его рецептора. Например, средством против ангиогенеза являются антитело или другой антагонист ангиогенного средства, определенного выше, например антитела против VEGF-A или антитела против рецептора VEGF-A (например, рецептора KDR или рецептора Fit-1), анти-PDGFR ингибиторы, такие как Gleevec™ (мезилат иматиниба). Средствами против ангиогенеза также являются ингибиторы природного ангиогенеза, например ангиостатин, эндостатин и т.д. См., например, публикации Klagsbrun and D'Amore (1991) Annu. Rev. Physiol. 53:217-39; Streit and Detmar (2003) Oncogene 22:3172-3179 (см., например, таблицу 3, в которой приводится список схем антиангиогенной терапии злокачественной меланомы); Ferrara & Alitalo (1999) Nature Medicine 5(12): 1359-1364; Tonini et al. (2003) Oncogene 22:6549-6556 (см., например, таблицу 2, в которой приводится список известных антиангиогенных факторов); и Sato (2003) Int. J. Clin. Oncol. 8:200-206 (см., например, таблицу 1, в которой приводится список антиангиогенных средств, используемых в клинических испытаниях).
Используемый в настоящем описании термин «VEGF» или «VEGF-A» означает фактор роста сосудистых эндотелиальных клеток человека, состоящий из 165 аминокислот, и родственные факторы роста сосудистых эндотелиальных клеток человека, состоящие из 121, 189 и 206 аминокислот, как описано в публикации Leung et al. (1989) Science 246:1306 and Houck et al. (1991) Mol. Endocrin, 5:1806, вместе с их природными аллельными и процессированными формами. Термин «VEGF» также означает VEGF животных, не являющихся человеком, таких как мышь, крыса или примат. Иногда VEGF конкретных видов обозначается, например, hVEGF для VEGF человека, mVEGF для VEGF мыши и т.д. Термин «VEGF» также используется для обозначения усеченных форм полипептида, содержащего аминокислоты 8-109 или 1-109 фактора роста сосудистых эндотелиальных клеток человека, состоящего из 165 аминокислот. В этой связи любые формы VEGF могут быть идентифицированы в настоящей заявке, например, как «VEGF (8-109)», «VEGF (1-109)» или «VEGF165». Положения аминокислот для «усеченного» нативного VEGF пронумерованы, как указано в нативной последовательности VEGF. Например, положение аминокислоты 17 (метионин) в усеченном нативном VEGF также соответствует положению 17 (метионин) в нативном VEGF. Усеченный нативный VEGF по сравнению с нативным VEGF обладает аффинностью связывания с рецепторами KDR и Fit-1, сравнимой с аффинностью связывания с нативным VEGF.
«Антитело против VEGF» представляет собой антитело, которое связывается с VEGF с достаточной аффинностью и специфичностью. Антитело против VEGF по изобретению может быть предпочтительно использовано в качестве терапевтического средства для лечения и предотвращения заболеваний или состояний, которые связаны с активностью VEGF. Антитело против VEGF обычно не связывается ни с другими гомологами VEGF, такими как VEGF-B или VEGF-C, ни с другими факторами роста, такими как PIGF, PDGF или bFGF. Предпочтительным антителом против VEGF является моноклональное антитело, которое связывается с тем же самым эпитопом, с которым связывается антитело против VEGF A4.6.1, продуцируемое гибридомой ATCC HB 10709. Более предпочтительным антителом против VEGF является рекомбинантное гуманизованное моноклональное антитело против VEGF, полученное, как описано в публикации Presta et al. (1997) Cancer Res. 57:4593-4599, включая, но им не ограничиваясь, антитело, известное как бевацизумаб (BV; Avastin™, авастин).
Антитело против VEGF «бевацизумаб (BV)», также известное как «rhuMAb VEGF» или «авастин®», представляет собой рекомбинантное гуманизованное моноклональное антитело против VEGF, полученное, как описано в публикации Presta et al. (1997) Cancer Res. 57:4593-4599. Указанное антитело содержит мутированные каркасные области IgG1 человека и антиген-связывающие гипервариабельные области моноклонального анти-hVEGF антитела мыши A.4.6.1 и блокирует связывание VEGF человека с рецепторами. Приблизительно 93% аминокислотной последовательности бевацизумаба, включая большинство его каркасных областей, является IgG1 человека, а приблизительно 7% последовательности является антителом A4.6.1 мыши. Бевацизумаб имеет молекулярную массу приблизительно 149000 дальтон и является гликозилированным.
Термины «VEGFC» и «VEGF-C» являются взаимозаменяемыми и означают полипептид человека, состоящий из 419 аминокислот (SwissProt: VEGFC_HUMAN P49767), и его не-человеческие ортологи млекопитающих, впервые описанные в публикации Joukov et al., EMBO J. 15, 290-98 (1996) and EMBO J. 15, 1751 (1996).
Используемый в настоящем описании термин «антагонист Nrp2» означает молекулу, способную нейтрализовать, блокировать, ингибировать, отменять, снижать или подавлять способность Nrp2 модулировать миграцию эндотелиальных клеток (EC) лимфатических сосудов или лимфангиогенез у взрослых, в частности опухолевый лимфангиогенез и образование метастазов опухоли.
Термин «антагонист VEGF» означает молекулу, способную нейтрализовать, блокировать, ингибировать, отменять, снижать или подавлять активность VEGF, включая, но этим не ограничиваясь, связывание с одним или несколькими рецепторами VEGF. Антагонистами VEGF являются, но ими не ограничиваясь, антитела против VEGF и их антиген-связывающие фрагменты, молекулы рецепторов и производные, которые специфически связываются с VEGF и тем самым ограничивают его связывание с одним или несколькими рецепторами, антитела против рецептора VEGF и антагонисты рецептора VEGF, такие как низкомолекулярные ингибиторы VEGFR-тирозинкиназ. Используемый в настоящем описании термин «антагонист VEGF», в частности, включает молекулы, а именно антитела, фрагменты антител, другие связывающие полипептиды, пептиды и не-пептидные малые молекулы, которые связываются с нейропилином-1 и/или с нейропилином-2 (Nrp-1 и/или Nrp-2) и обладают способностью нейтрализовать, блокировать, ингибировать, отменять, снижать или подавлять активность VEGF, где указанными молекулами являются, но ими не ограничиваясь, антитела против Nrp1 и Nrp2 и антитела, перекрестно реагирующие с Nrp1 и Nrp2, при условии, что они обладают способностью нейтрализовать, блокировать, ингибировать, отменять, снижать или подавлять активность VEGF. Таким образом, термин «VEGF-активность» конкретно включает опосредованную нейропилином биологическую активность VEGF (как определено выше).
Термин «антагонисты семафорина» означает молекулу, обладающую способностью нейтрализовать, блокировать, ингибировать, отменять, снижать или предотвращать семафориновую активность, включая, но этим не ограничиваясь, способность связываться с одной или несколькими рецепторами семафорина. Антагонистами семафорина являются, но ими не ограничиваясь, антитела против семафорина и их антиген-связывающие фрагменты; рецепторные молекулы и их производные, которые специфически связываются с семафорином, ограничивая их связывание с одним или несколькими рецепторами; антитела против семафоринового рецептора и антагонисты семафоринового рецептора, такие как низкомолекулярные ингибиторы семафоринов. Используемый в настоящем описании термин «антагонист семафорина», в частности, означает молекулы, включая антитела, фрагменты антител, другие связывающие полипептиды, пептиды и не-пептидные малые молекулы, которые связываются с нейропилином-1 и/или нейропилином-2 (Nrp-1 и/или Nrp-2) и обладают способностью нейтрализовать, блокировать, ингибировать, отменять, снижать или предотвращать активность семафорина, где указанными молекулами являются, но ими не ограничиваясь, антитела против Nrp1 и Nrp2 и антитела, перекрестно реагирующие с Nrp1 и Nrp2, при условии, что они обладают способностью нейтрализовать, блокировать, ингибировать, отменять, снижать или предотвращать семафориновую активность. Таким образом, термин «семафориновая активность», в частности, включает опосредуемую нейропилином биологическую активность (определенную выше) семафоринов класса 3. Такой биологической активностью является, например, действие, ингибирующее рост нейритов в процессе развития эмбриональной нервной системы и регенерации нейронов.
Термин «лечение» означает как терапевтическое, так и профилактическое лечение заболевания или меры его профилактики. Нарушениями, на которые направлено лечение, являются уже существующие нарушения, а также нарушения, развитие которых необходимо предупредить.
«Нарушение» представляет собой любое состояние, симптомы которого могут быть ослаблены в результате лечения. В качестве примера может служить нарушение, которым страдает млекопитающее или которое требует профилактических мер по устранению патологического ангиогенеза (избыточного, несоответствующего или неконтролируемого ангиогенеза) или патологической проницаемости сосудов. Этот термин означает хронические и острые нарушения или заболевания, включая патологические состояния, которые определяют предрасположенность млекопитающего к данному нарушению. Неограничивающими примерами указанных в настоящем описании нарушений, на которые направлено лечение, являются злокачественные и доброкачественные опухоли; злокачественные опухоли, не являющиеся лейкозом, и лимфоидные злокачественные опухоли и, в частности, метастазирующие опухоли (рак).
Термин «злокачественная опухоль» означает или относится к физиологическому состоянию у млекопитающих, при котором обычно происходит нерегулируемый клеточный рост. Примерами злокачественных опухолей являются, но ими не ограничиваясь, карцинома, лимфома, бластома, саркома и лейкоз. Более конкретными примерами таких злокачественных опухолей являются плоскоклеточный рак, рак легких (включая мелкоклеточный рак легких, немелкоклеточный рак легких, аденокарциному легких и плоскоклеточную карциному легких), перитонеальный рак, гепатоцеллюлярный рак, рак кишечника или желудка (включая рак желудочно-кишечного тракта), рак поджелудочной железы, глиобластома, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатома, рак молочной железы, рак толстой кишки, рак прямой и ободочной кишки, карцинома эндометрия или матки, карциномы слюнных желез, рак почек или ренальный рак, рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, карцинома печени и рак головы и шеи различных типов, а также В-клеточная лимфома (включая низкозлокачественную/фолликулярную не-ходжкинскую лимфому (NHL); мелкоклеточную лимфоцитарную (SL) NHL; NHL средней стадии дифференцировки/фолликулярную NHL; диффузную NHL средней стадии дифференцировки; высокозлокачественную иммунобластную NHL; высокозлокачественную лимфобластную NHL; высокозлокачественную мелкоклеточную недифференцированную NHL; генерализованную NHL; лимфому клеток коры головного мозга; лимфому, связанную со СПИД, и макроглобулинемию Вальденстрема); хронический лимфоцитарный лейкоз (CLL); острый лимфобластный лейкоз (ALL); волосато-клеточный лейкоз; хронический миелобластный лейкоз и лимфопролиферирующее расстройство после трансплантации (PTLD), а также патологическая пролиферация сосудов, связанная с факоматозом, отек (такой как отек, связанный с опухолями головного мозга) и синдром Мейгса.
Термин «противоопухолевая композиция» означает композицию, которая может быть использована для лечения злокачественной опухоли и включает по меньшей мере одно активное терапевтическое средство, например «противоопухолевое средство». Примерами терапевтических средств (противоопухолевых средств) являются, но ими не ограничиваясь, химиотерапевтические средства, средства, ингибирующие рост опухоли, цитотоксические средства, средства, используемые в лучевой терапии, средства против ангиогенеза, апоптотические средства, антитубулиновые средства и другие средства, используемые для лечения злокачественных опухолей, такие как антитела против HER-2, антитела против CD20, антагонист рецептора эпидермального фактора роста (EGFR) (например, ингибитор тирозинкиназы), ингибитор HER1/EGFR (например, эрлотиниб (тарцева™)), ингибиторы тромбоцитарного фактора роста (например, гливек™ (мезилат иматиниба)), ингибитор COX-2 (например, целекоксиб), интерфероны, цитокины, антагонисты (например, нейтрализующие антитела), которые связываются с одним или несколькими из следующих мишеней, таких как рецептор(ы) ErbB2, ErbB3, ErbB4, PDGFR-бета, BlyS, APRIL, BCMA или VEGF, а также TRAIL/Apo2 и другие биологически активные и органические химические средства и т.д. Настоящее изобретение также включает их комбинации.
Используемый в настоящем описании термин «цитотоксическое средство» означает вещество, ингибирующее или блокирующее функцию клеток и/или вызывающее деструкцию клеток. Этот термин включает радиоактивные изотопы (например, 131I, 125I, 90Y и 186Re), химиотерапевтические средства и токсины, такие как ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, или их фрагменты.
«Химиотерапевтическое средство» представляет собой химическое соединение, используемое для лечения злокачественной опухоли. Примерами химиотерапевтических средств являются химические соединения, которые могут быть использованы для лечения злокачественной опухоли. Примерами химиотерапевтических средств являются алкилирующие средства, такие как тиотепа и цитоксан® (циклофосфамид); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метилмеламины, включая альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилолмеламин; ацетогенины (в частности, буллатацин и буллатацинон); камптотецин (включая синтетический аналог топотекан); бриостатин; каллистатин; СС-1065 (включая его синтетические аналоги, такие как адозелезин, карзелезин и бизелезин); криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги, KW-2189 и CB1-TM1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; производные азотистого иприта, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлоретамин, гидрохлорид оксида мехлоретамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урациловый иприт; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, в частности калихеамицин гамма II и калихеамицин омега II (см., например, Agnew (1994) Chem. Intl. Ed. Engl. 33:183-186); динемицин, включая динемицин А; бисфосфонаты, такие как клодронат; эсперамицин, а также неокарциностатиновый хромофор и родственные хромопротеиновые энедииновые антибиотики-хромофоры), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин ADRIAMYCIN® (адриамицин) (включая морфолино-доксорубицин, цианоморфолино-доксорубицин, 2-пирролино-доксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU);
аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, пропионат дромостанолона, эпитиостанол, мепитиостан, тестолактон; антиадренергические средства, такие как аминоглютетимид, митотан, трилостан; заменитель фолиевой кислоты, такой как фролиновая кислота; ацеглатон; альдофосфамидный гликозид; аминолевулиновая кислота; энилурацил; амсакрин; бестрабуцил; бизантрен; эдатраксат; дефофамин; демеколцин; диазиквон; элфорнитин; ацетат эллиптиния; эпотилон; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидамин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопидамол; нитракрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновая кислота; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновая кислота; триазиквон; 2,2',2”-трихлортриэтиламин; трихотецены (в частности, токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид (“Ara-C”); циклофосфамид; тиотепа; таксоиды, например паклитаксел TAXOL® (таксол) (Bristol-Myers Squibb Oncology, Princeton, N.J.), не содержащий кремофора и сконструированный на основе альбумина препарат паклитаксела в виде наночастиц, ABRAXANE™ (абраксан) (American Pharmaceutical Partners, Schaumberg, Illinois), и доксетаксел TAXOTERE® (таксотер) (Rhone-Poulenc Rorer, Antony, France); хлоранбуцил; гемцитабин GEMZAR® (гемзар); 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; винорелбин NAVELBINE® (навелбин); новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселода; ибандронат; иринотекан (Camptosar (камптосар), CPT-11) (включая курс лечения иринотеканом с 5-FU и лейковорином); ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; комбретастатин; лейковорин (LV); оксалиплатин, включая курс лечения оксалиплатином (FOLFOX (фолфокс)); ингибиторы PKC-альфа, Raf, H-Ras, EGFR (например, эрлотиниб (тарцева (Tarceva™)) и VEGF-A, который снижает уровень пролиферации клеток, и фармацевтически приемлемые соли, кислоты или производные всех вышеуказанных соединений.
В определение этого термина также входят противогормональные средства, регулирующие или ингибирующие действие гормонов на опухоли, такие как антиэстрогены и селективные модуляторы эстрогеновых рецепторов (SERM), включая, например, тамоксифен (в том числе тамоксифен NOLVADEX® (нольвадекс)), ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и FARESTON(фарестон)-торемифен; ингибиторы ароматазы, которые ингибируют фермент ароматазу и регулируют уровень продукции эстрогенов в коре надпочечника, такие как, например, 4(5)-имидазолы, аминоглютетимид, ацетат мегестрола MEGASE®, экземестан AROMASIN® (аромазин), форместан, фадрозол, ворозол RIVISOR®, летрозол FEMARA® (фемара) и анастрозол ARIMIDEX® (аримидекс); и антиандрогены, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; а также троксацитабин (нуклеозидный аналог 1,3-диоксоланцитозина); антисмысловые олигонуклеотиды, в частности олигонуклеотиды, которые ингибируют экспрессию генов путем передачи сигнала, участвующих в патологической пролиферации клеток, такие как, например, PKC-альфа, Raf и H-Ras; рибозимы, такие как ингибитор экспрессии VEGF (например, рибозим ANGIOZYME® (ангиозим)) и ингибитор экспрессии HER2; вакцины, такие как вакцина для генотерапии, например, вакцина ALLOVECTIN® (алловектин), вакцина LEUVECTIN® и вакцина VAXID®; rIL-2 PROLEUKIN® (пролейкин); ингибитор топоизомеразы 1 LURTOTECAN® (луртотекан); rmRH ABARELIX® (абареликс); винорелбин и эсперамицины (см. патент США № 4675187) и их фармацевтически приемлемые соли, кислоты или производные всех вышеуказанных соединений.
Используемый в настоящем описании термин «пролекарство» означает предшественник или производное фармацевтически активного вещества, которые, по сравнению с родительским лекарственным средством, являются менее цитотоксичными по отношению к опухолевым клеткам и способны ферментативно активироваться или превращаться в более активную зрелую форму. См., например, Wilman (1986) “Prodrugs in Cancer Chemotherapy” Biochemical Society Transactions, 14, pp. 375-382, 615th Meeting Belfast and Stella et al. (1985) “Prodrugs: A Chemical Approach to Targeted Drug Delivery”, Directed Drug Delivery, Borchardt et al. (ed.), pp. 247-267, Humana Press. Пролекарствами по изобретению являются, но ими не ограничиваясь, фосфат-содержащие пролекарства, тиофосфат-содержащие пролекарства, сульфат-содержащие пролекарства, пептид-содержащие пролекарства, пролекарства, модифицированные D-аминокислотой, гликозилированные пролекарства, β-лактам-содержащие пролекарства; пролекарства, содержащие необязательно замещенный феноксиацетамид, или пролекарства, содержащие необязательно замещенный фенилацетамид; 5-фторцитозиновые и другие 5-фторуридиновые пролекарства, которые могут быть преобразованы в более активное цитотоксическое свободное лекарственное средство. Примерами цитотоксических лекарственных средств, которые могут быть дериватизированы с получением пролекарственной формы для ее использования в настоящем изобретении, являются, но ими не ограничиваясь, химиотерапевтические средства, описанные выше.
Термин «низкомолекулярное соединение» определен в настоящем описании как соединение с молекулярной массой ниже примерно 500 дальтон.
«Выделенная» молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая идентифицирована и отделена по меньшей мере от одной примесной молекулы нуклеиновой кислоты, с которой она обычно связана в природном источнике нуклеиновой кислоты, кодирующей антитело. Выделенная молекула нуклеиновой кислоты отличается от другой молекулы по своей форме или природной локализации. Поэтому выделенные молекулы нуклеиновой кислоты отличаются от молекулы нуклеиновой кислоты, присутствующей в природных клетках. Однако выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно экспрессируют антитело, где, например, молекула нуклеиновой кислоты имеет хромосомную локализацию, отличающуюся от ее локализации в природных клетках.
Термин «регуляторные последовательности» означает последовательности ДНК, необходимые для экспрессии связанной кодирующей последовательности в конкретном организме-хозяине. Регуляторными последовательностями, подходящими для прокариотов, являются, например, промотор, необязательно последовательность оператора и сайт связывания с рибосомой. Известно, что в эукариотических клетках имеются промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является «функционально связанной», если она находится в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК для пре-последовательности или секреторной лидерной последовательности считается функционально связанной с ДНК полипептида, если она экспрессируется как пре-белок, который участвует в секреции полипептида; либо промотор или энхансер считаются функционально связанными с кодирующей последовательностью, если они влияют на транскрипцию последовательности; либо сайт связывания с рибосомой считается функционально связанным с кодирующей последовательностью, если его расположение способствует облегчению трансляции. Обычно термин «функционально связанный» означает, что связанные последовательности ДНК являются смежными, а в случае секреторной лидерной последовательности они являются смежными и находятся в одной и той же рамке считывания. Однако энхансеры не должны быть смежными. Связывание осуществляется посредством лигирования в подходящих рестрикционных сайтах. Если такие сайты отсутствуют, то синтетические олигонуклеотидные адапторы или линкеры используются в соответствии с общепринятой практикой.
Используемые в настоящем описании термины «клетка», «клеточная линия» и «клеточная культура» являются взаимозаменяемыми, и все эти термины включают также их потомство. Таким образом, термины «трансформанты» и «трансформированные клетки» включают первичную рассматриваемую клетку и ее культуры без учета числа пассажей. Следует также отметить, что все потомство не может быть абсолютно идентичным по содержанию ДНК, что обусловлено специально введенными или самопроизвольными мутациями. Этот термин включает мутированное потомство, которое имеет такие же функции или биологическую активность, которые, как показал скрининг, обнаруживаются в исходных трансформированных клетках. Если далее используются какие-либо другие термины, то они будут понятны из контекста описания изобретения.
Способы осуществления изобретения
В одном из аспектов настоящее изобретение основано на экспериментальных данных, указывающих на то, что блокирование функции Nrp2 приводит к ингибированию метастазов опухоли.
Основное событие в многостадийном процессе образования метастазов заключается в выходе опухолевых клеток из первичной опухолевой массы. В солидных опухолях лимфатическая система часто обеспечивает путь для отделения клеток. Известно, что VEGF является основным модулятором лимфангиогенеза и метастазов во многих опухолевых моделях, а ингибирование VEGF всех типов рассматривается как перспективная стратегия ингибирования развития метастазов. До создания настоящего изобретения Nrp2, корецептор VEGFC, не рассматривался как мишень для ингибирования метастазов опухоли, что, вероятно, обусловлено отсутствием повреждений в лимфатической системе у взрослых мышей, мутантных по Nrp2.
Исследования, лежащие в основе настоящего изобретения и описанные ниже в примерах, подтвердили важную роль Nrp2 в лимфангиогенезе и метастазах опухоли, отчасти только благодаря модулированию передачи VEGFR3-сигнала. Кроме того, данные, представленные в примерах, продемонстрировали присутствие функциональных лимфатических сосудов в опухолях и показали, что обработка антителом против Nrp2B приводит к уменьшению числа этих функциональных лимфатических сосудов.
Селективные функции VEGFC, регулируемые Nrp2 частично по механизму, независимому от активации рецептора VEGF
В настоящее время известно, что индуцирование миграции и пролиферации клеток осуществляется двумя центральными клеточными функциями VEGFC, описанными в литературе (Joukov et al., Embo J. 16, 3898-3911 (1997)). Таким образом, авторами настоящего изобретения было неожиданно обнаружено, что блокирование Nrp2 под действием антитела против Nrp2B приводит к блокированию миграции, но не пролиферации LEC (фигуры 2, 3). О такой селективности недавно сообщалось в экспериментах по ингибированию киРНК Nrp2, но этот факт был объяснен техническими недостатками эксперимента (Favier et al., Blood 108, 1243-1250 (2006)). Представленные в настоящем описании данные показали, что функциональная селективность Nrp2 также наблюдается in vivo, где обработка антителом против Nrp2B приводила к снижению уровня VEGFC-регулируемого лимфангиогенеза, а не сосудистой проницаемости (фигуры 2, 3). Такие наблюдения дали основание предположить, что ингибирование под действием антитела против Nrp2B осуществляется не только в результате нарушения передачи VEGFC-сигнала. Однако было установлено, что блокирование Nrp2 приводит к умеренному снижению уровня фосфорилирования рецептора VEGF (фигура 3), что подтверждает наличие механизма, в котором одна из ролей Nrp2 заключается в усилении функции рецептора VEGF. Это повышает вероятность того, что для осуществления различных VEGF-индуцированных физиологических событий могут потребоваться различные уровни активации рецепторов VEGF. Таким образом, снижение уровня активации рецептора может быть достаточным для подавления миграции, но не для пролиферации или сосудистой проницаемости.
Для выявления этого факта зависимость дозы VEGFC от ответа на фосфорилирование рецептора VEGF сравнивали с зависимостью дозы от ответа на миграцию LEC (фигура 3). Дозы VEGFC, которые приводят к определенному уровню фосфорилирования рецептора, эквивалентному уровню, наблюдаемому при обработке антителом против Nrp2B, не снижали или не ингибировали миграцию. Это указывало на то, что одно лишь снижение уровня активации рецептора не может служить объяснением эффектов блокирования функции под действием антитела против Nrp2B.
В связи с этим были исследованы другие механизмы, под действием которых блокирование Nrp2 может селективно влиять на миграцию, такие как модуляция адгезии или подвижности. Обработка антителом против Nrp2B не оказывала какого-либо воздействия на LEC-опосредованную адгезию или миграцию, индуцированную VEGF165 (фигура 2), HGF (фигура 3) или FGF-2, что указывало на то, что такая обработка по существу не оказывает какого-либо влияния на миграцию посредством нарушения подвижности. Кроме того, было высказано предположение, что sema3F, другой лиганд Nrp2, может модулировать миграцию LEC или EC, действуя как хеморепеллент (Bielenberg et al., J. Clin. Invest 114, 1260-1271 (2004); Favier et al., Blood 108, 1243-1250 (2006)). Однако антитело против Nrp2B не ингибировало или не усиливало связывание sema3F с Nrp2 (фигура 1) или функциональное действие sema3F на восприимчивые нейроны (дополнительная фигура 4). Таким образом, маловероятно, что снижение уровня VEGFC-индуцированной миграции под действием антитела против Nrp2B может быть объяснено модуляцией функции sema3F.
Было также оценено действие антитела против Nrp2B на образование комплекса Nrp2/VEGF-рецептор. В отличие от Nrp1, Nrp2 образует комплекс с VEGFR2 и VEGFR3 в отсутствие лиганда (Favier et al., 2006, см. выше; Karpanen et al., Faseb. J 20, 1462-1472 (2006)). Важно отметить, что антитело против Nrp2B в значительной степени ингибирует образование указанных комплексов. Кроме того, это наблюдение, помимо того факта, что Nrp2 играет важную роль не только в усилении функции VEGF-рецептора, позволяет создать модель, в которой Nrp2 будет обеспечивать дополнительную функциональность, заключающуюся в специфической модуляции миграции, возможно, посредством сообщения комплексу VEGF-рецептора дополнительной функции.
Nrp2 играет важную роль в модуляции лимфангиогенеза у взрослых
Анализ Nrp2 у KO-мышей демонстрирует, что Nrp2 представляет собой модулятор развивающегося лимфангиогенеза, вероятно, благодаря своей роли в качестве ко-рецептора VEGFC (Yuan et al., 2002, см. выше). Однако у этих мутантных мышей после их рождения образуются функциональные лимфатические сосуды, что указывает на то, что в данном случае либо происходит нарушение, которое заключается не в ингибировании, а в замедлении роста лимфатических сосудов, либо происходит функциональная компенсация за счет другого молекулярного медиатора. Поэтому роль Nrp2 в сохранении зрелых лимфатических сосудов и в модуляции лимфангиогенеза у взрослых пока еще неясна. Анализ экспрессии (фигура 4) не подтвердил роли Nrp2 в сохранении лимфатических сосудов. Интересно отметить, что Nrp2 в значительной степени экспрессируется в лимфатических сосудах, которые присутствуют в опухолях и в лимфоузлах (LN), находящихся рядом с опухолями, что дает основание предполагать, что Nrp2 может играть определенную роль в активации или росте лимфатических сосудов. Наблюдения in vitro демонстрируют, что антитело против Nrp2B является эффективным средством для оценки роли Nrp2 в этих процессах. Поэтому антитело против Nrp2B было протестировано in vivo в анализе микрокармана роговицы (фигура 2). Антитело против Nrp2B эффективно блокировало VEGFC-индуцированный лимфангиогенез, и было неожиданно обнаружено, что такое блокирование соразмерно блокированию ECD VEGFR3-индуцированного лимфангиогенеза. Интересно отметить, что антитело против Nrp2B обладает селективной ингибирующей функцией in vivo, а также не оказывает влияния на VEGFC-индуцированную сосудистую проницаемость. Эти данные соответствуют наблюдениям in vitro, указывающим на то, что Nrp2 специфически модулирует миграцию, то есть процесс, который играет важную роль в лимфангиогенезе, но маловероятно, что он играет какую-либо роль в сосудистой проницаемости. И наконец, у здоровых взрослых животных, получавших антитело против Nrp2B, не наблюдалось каких-либо изменений в лимфатических сосудах тонкого кишечника, что указывает на то, что Nrp2 не играет роли в сохранении зрелых лимфатических сосудов.
Ингибирование Nrp2 приводит к уменьшению числа функциональных лимфатических сосудов в опухоли и к уменьшению числа метастазов, что, вероятно, обусловлено ингибированием опухолевых клеток, покидающих основную опухолевую массу по лимфатическим путям
Ингибирование VEGFC всех типов, чаще всего с использованием ECD VEGFR3, является одной из наиболее распространенных стратегий сокращения числа метастазов (Chen et al., 2005, см. выше; He et al., 2002, см. выше; Krishnan et al., 2003, см. выше; Lin et al., 2005, см. выше). VEGFC может способствовать сокращению числа метастазов благодаря инициации лимфангиогенеза и тем самым увеличения площади поверхности опухолевых клеток при контактировании с LEC посредством модуляции адгезивных свойств LEC или экспрессии цитокинов или посредством повышения сосудистой проницаемости (Alitalo and Carmeliet, Cancer Cell 1, 219-227 (2002)). Поскольку антитело против Nrp2B модулирует селективные VEGFC-опосредуемые функции, включая ингибирование VEGFC-индуцированного лимфангиогенеза у взрослых, были проведены исследования влияния блокирования Nrp2 на метастазы. Для минимизации ненужных сопутствующих переменных и для однозначной оценки влияния антитела против Nrp2B на метастазы авторами настоящего изобретения были созданы модели, когда блокирование Nrp2 не влияет на рост первичной опухоли, а затем всех животных в этом исследовании анализировали через одинаковые промежутки времени (Withers and Lee, Semin Radiat Oncol. 16, 111-119 (2006)).
В обеих моделях с опухолями 66c14 и C6 обработка антителом против Nrp2B приводила к значительному снижению числа метастатических узлов в легких при визуальном наблюдении (фигуры 5, 6). Это подтверждается более чувствительными и количественными методами микро-КТ (Li et al., Technol. Cancer Res. Treat. 5, 147-155 (2006)). Сравнение данных обработки антителом против Nrp2B и ECD VEGFR3 в опухолях 66c14 было невозможно из-за уменьшения размера первичной опухоли после VEGFR3-обработки. Однако этот анализ проводили на опухолях C6, поскольку на их рост не влияла ECD VEGFR3-обработка. Как и в случае анализа, проводимого в микрокармане роговицы, обработка антителом против Nrp2B приводила к блокированию метастазов, соразмерному блокированию, наблюдаемому в случае ECD VEGFR3.
Проведенный авторами гистологический анализ показал отсутствие какого-либо влияния на первичные опухолевые клетки. Таким образом, авторами были оценены два потенциальных пути развития метастазов, которые являются доступными для опухолевых клеток, а именно метастазы в кровеносных и лимфатических сосудах (фигура 7). Введение антитела против Nrp2B не оказывало влияния на структуру или плотность кровеносных сосудов. Исходя из данных in vitro и данных in vivo, полученных с помощью анализа в микрокармане роговицы, была высказана гипотеза, что блокирование Nrp2 должно приводить к уменьшению количества опухолевых лимфатических сосудов. Введение антитела против Nrp2B приводило к резкому снижению плотности лимфатических сосудов, но и в этом случае степень такого снижения была эквивалента степени снижения, наблюдаемого при ECD VEGFR3-обработке. Однако эти два введения отличаются по своему воздействию на морфологию образующихся лимфатических сосудов. ECD VEGFR3-введение приводило к образованию редкой сети лимфатических сосудов, состоящих из пораженных на вид лимфатических клеток. С другой стороны, введение антитела против Nrp2B приводило к развитию коротких сосудов и карманов из отдельных здоровых на вид лимфатических клеток. Кроме того, эти различия подтверждались моделью, в которой Nrp2 не просто действует как усилитель активации рецептора VEGF, но также сообщает уникальные функциональные свойства, опосредующие биологическое действие VEGFC. Эти результаты также показали, что тестируемые экспериментальные парадигмы заключаются в том, что антитело против Nrp2B действует как ингибитор лимфангиогенеза (фигура 7). Однако нельзя исключить тот факт, что антитело против Nrp2B также может разрушать более развитые лимфатические сосуды в опухолях.
Авторами настоящего изобретения было также проведено исследование для определения, являются ли внутриопухолевые лимфатические сосуды функциональными и, следовательно, способствуют ли они образованию метастазов. Для идентификации редких функциональных внутриопухолевых лимфатических сосудов была проведена лимфангиография (фигура 8). Используемый метод не является адекватно аналитическим методом определения количества функциональных внутриопухолевых лимфатических сосудов. Однако они, очевидно, представляют собой небольшую часть от общего числа лимфатических сосудов (Padera et al., Mol. Imaging 1, 9-15 (2002)). Поскольку возможно, что авторам удалось уменьшить общую плотность лимфатических сосудов с малым числом функциональных сосудов (которые могут иметь отличающуюся чувствительность к антителу против Nrp2B), была проведена оценка влияния блокирования Nrp2 на образование функциональных лимфатических сосудов. Антитело против Nrp2B подавляло образование функциональных сосудов, что указывало на более прямую взаимосвязь влияния на опухолевые лимфатические сосуды с наблюдаемым сокращением числа метастазов.
Наконец, для подтверждения последствий уменьшения числа функциональных лимфатических сосудов было оценено действие антитела против Nrp2B на метастазы в SLN. SLN представляют собой первую ткань, куда попадают опухолевые клетки после их миграции из опухоли по лимфатическим сосудам. Таким образом, они представляют собой ткань, где происходит одна из самых ранних стадий образования метастазов в периферических органах (Stracke and Liotta, In vivo 6, 309-316 (1992)). Как было предсказано, обработка антителом против Nrp2B приводит к замедлению образования метастазов в SLN, что соответствует мнению о том, что из первичной опухолевой массы происходит отток меньшего числа клеток. Это совпадает с данными, свидетельствующими о том, что VEGFC способствует увеличению числа метастазов путем индуцирования лимфатической гиперплазии и увеличению доставки раковых клеток в лимфоузлы (Hoshida et al., Cancer Research 66, 8065-8075 (2006)). Таким образом, имеется множество данных, указывающих на механизм, посредством которого блокирование Nrp2 приводит к уменьшению числа функциональных опухолевых лимфатических сосудов и тем самым к предотвращению вовлечения опухолевых клеток в метастатический процесс в результате выхода этих клеток из первичной опухолевой массы.
Nrp2 как мишень для образования метастазов
Многочисленные клинико-патологические исследования показали, что экспрессия VEGFC и VEGFR3 напрямую связана с формированием метастазов в лимфоузлах и в дистальных органах при различных злокачественных опухолях человека (подробный обзор можно найти в публикациях Stacker et al., Nat. Rev. Cancer, 2002, см. выше; Stacker et al., Faseb J, 2002, см. выше, и He et al., 2004, см. выше). Однако информация, относящаяся к экспрессии Nrp2 и ее связи с развитием метастазов, очень ограничена. Действительно, была установлена лишь взаимосвязь между экспрессией Nrp2 и развитием злокачественных опухолей, в частности рака поджелудочной железы (Cohen et al., Biochem Biophys Res Commun 284, 395-403 (2001); Fukahi et al., Clin Cancer Res 10, 581-590 (2004)) и рака легких (Kawakami et al., Cancer 95, 2196-2201 (2002); Lantuejoul et al., J Pathol 200, 336-347 (2003)). Аналогичным образом было обнаружено, что экспрессия Nrp2, по сравнению с экспрессией в соответствующих контрольных тканях, наблюдается на более высоком уровне не только в клетках поджелудочной железы, но также и в клетках аденокарциномы толстой кишки, плоскоклеточной карциномы головы и шеи, меланомы, папиллярной карциномы щитовидной железы и инфильтрирующей аденокарциномы протоков молочной железы (фигура 9). Более важным является тот факт, что при разделении этих опухолей на подгруппы метастазирующих и не-метастазирующих опухолей статистически более высокий уровень экспрессии Nrp2 наблюдался в большинстве опухолей этого типа, относящихся к метастатической группе. Интересно отметить, что все опухоли этого типа обнаруживали внутриопухолевый лимфангиогенез, который, кроме того, напрямую связан с метастазами в лимфоузлах (Achen et al., 2006, см. выше; Achen and Stacker, Int. J. Cancer 119, 1755-1760 (2006)). Это указывает на то, что результаты, полученные в обсуждаемых экспериментах, позволяют предположить, что такие исследования могут быть продолжены с участием людей с опухолями различных типов.
В заключение следует отметить, что данные, обсуждаемые в настоящем описании и представленные в нижеследующих примерах, указывают на то, что Nrp2 играет определенную роль в модуляции VEGFC-индуцированной миграции клеток, и дают основание предположить, что Nrp2 может действовать по множеству механизмов, включая усиление активации рецептора VEGF, и по механизмам, которые не зависят от передачи сигнала рецептором VEGF. Кроме того, блокирование функции Nrp2 с использованием антитела против Nrp2B приводит к резкому снижению VEGFC-индуцированного лимфангиогенеза у взрослых мышей. Такая обработка также приводит к сокращению числа метастазов, вероятно, благодаря подавлению развития функциональных лимфатических сосудов. Эти данные, наряду с анализами на экспрессию Nrp2 в ряде опухолей у человека, позволяют предположить, что Nrp2 является истинной мишенью для модуляции метастазов.
Получение антител против Nrp2
В настоящем изобретении рассматривается получение и применение антител против Nrp2. Примеры способов получения антител более подробно описаны в разделах ниже.
Антитела против NRP2 выбирают, используя антиген NRP2 млекопитающего. Предпочтительным антигеном является NRP2 человека (hNRP2). Однако в качестве антигена-мишени могут быть также использованы NRP2 других млекопитающих, такие как NRP2 мыши (mNRP2). Антигены NRP2 млекопитающих различных видов могут быть выделены из природных источников. В других вариантах изобретения антиген получают рекомбинантными методами или с использованием других методов синтеза, известных специалистам в данной области.
Выбранное антитело обычно обладает достаточно высокой аффинностью связывания с антигеном NRP2. Например, антитело может связываться с hNRP2 с величиной Kd не более чем примерно 5 нМ, предпочтительно не более чем примерно 2 нМ и более предпочтительно не более чем примерно 500 пМ. Аффинность антител может быть определена с помощью анализа, например, на основе поверхностного плазмонного резонанса (такого как анализ BIAcore, описанный в примерах); твердофазного иммуноферментного анализа (ELISA); и анализа на конкурентное связывание (например, РИА).
Кроме того, антитело можно анализировать с помощью других анализов на биологическую активность, например, для оценки эффективности этого антитела в качестве терапевтического средства. Такие анализы известны специалистам в данной области, и их выбор зависит от антигена-мишени и от целей применения антитела. Примерами являются анализ на ингибирование HUVEC (описанный ниже в примерах); анализы на ингибирование роста опухолевых клеток (описанные, например, в WO 89/06692); анализы на антитело-зависимую клеточную цитотоксичность (ADCC) и комплемент-опосредуемую цитотоксичность (CDC) (патент США 5500362); и анализы на агонистическую активность или гемопоэз (см. WO 95/27062).
Для скрининга антител, которые связываются с конкретным эпитопом на представляющем интерес антигене, может быть проведен рутинный анализ на перекрестное блокирование, такой как анализ, описанный в публикации Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988). Альтернативно, чтобы определить, связывается ли антитело с представляющим интерес эпитопом, может быть осуществлено картирование эпитопов, например, как описано в публикации Champe et al. (1995) J. Biol. Chem. 270:1388-1394.
Получение антител против NRP2 из синтезированных фаговых библиотек антител
В предпочтительном варианте изобретения антитела против NRP2 отбирают с использованием уникального метода фагового дисплея. Такой метод предусматривает получение синтезированных фаговых библиотек антител на одной каркасной матрице, создание достаточного разнообразия вариабельных доменов, презентацию полипептидов, имеющих разнообразные вариабельные домены, отбор антител-кандидатов с высокой аффинностью по отношению к антигену-мишени NRP и выделение отобранных антител.
Подробное описание методов фагового дисплея можно найти, например, в заявке WO 03/102157, опубликованной 11 декабря 2003.
В одном из аспектов изобретения библиотеки антител могут быть получены путем мутаций, вводимых в положения, доступные для растворителя, и/или в положения с высоким разнообразием по меньшей мере одной CDR вариабельного домена антитела. Некоторые или все CDR могут быть мутированы с использованием представленных в настоящем описании методов. В некоторых вариантах изобретения может оказаться предпочтительным получение разнообразных библиотек антител путем мутаций, вводимых в положения CDRH1, CDRH2 и CDRH3 с образованием одной библиотеки, или путем мутаций, вводимых в положения CDRL3 и CDRH3 с образованием одной библиотеки, или путем мутаций, вводимых в положения CDRL3 и CDRH1, CDRH2 и CDRH3 с образованием одной библиотеки.
Библиотека вариабельных доменов антител может быть получена, например, путем введения мутаций в положения, доступные для растворителя, и/или в положения с высоким разнообразием CDRH1, CDRH2 и CDRH3. Другая библиотека может быть получена путем введения мутаций в CDRL1, CDRL2 и CDRL3. Такие библиотеки могут быть также использованы в комбинации друг с другом с получением связывающих агентов с нужной аффинностью. Например, после проведения одного или нескольких раундов отбора библиотеки тяжелых цепей для связывания с антигеном-мишенью, библиотека легких цепей может быть заменена на ряд связывающих агентов тяжелой цепи для проведения дополнительных раундов отбора в целях повышения аффинности связывающих агентов.
Предпочтительно библиотеку создают путем замены исходных аминокислот аминокислотными вариантами в CDRH3 вариабельной области последовательности тяжелой цепи. Полученная библиотека может содержать множество последовательностей антител, где разнообразие последовательностей достигается, главным образом, в области CDRH3 последовательности тяжелой цепи.
В одном из аспектов изобретения библиотеку создают на основе последовательности гуманизованного антитела 4D5 или последовательности каркасных аминокислот гуманизованного антитела 4D5. Предпочтительно библиотеку создают путем замены по меньшей мере 95-100 аминокислотных остатков тяжелой цепи аминокислотами, кодируемыми набором кодонов DVK, где указанный набор кодонов DVK используется для кодирования серии аминокислотных вариантов для каждого из этих положений. Примером набора олигонуклеотидов, который может быть использован для создания таких замен, является последовательность (DVK)7. В некоторых вариантах изобретения библиотеку создают путем замены по меньшей мере 95-100 аминокислотных остатков аминокислотами, кодируемыми обоими наборами кодонов DVK и NNK. Примером набора олигонуклеотидов, который может быть использован для создания таких замен, является последовательность (DVK)6(NNK). В другом варианте изобретения библиотеку создают путем замены по меньшей мере 95-100 аминокислотных остатков аминокислотами, кодируемыми обоими наборами кодонов DVK и NNK. Примером набора олигонуклеотидов, который может быть использован для создания таких замен, является последовательность (DVK)5(NNK). Другим примером набора олигонуклеотидов, который может быть использован для создания таких замен, является последовательность (NNK)6. Другие примеры подходящих олигонуклеотидных последовательностей могут быть определены специалистами в соответствии с описанными в данном документе критериями.
В другом варианте изобретения различные конструкции CDRH3 используют для выделения связывающих агентов с высокой аффинностью и для выделения связывающих агентов для ряда эпитопов. CDRH3, полученный в этой библиотеке, имеет длину от 11 до 13 аминокислот, хотя может быть получен CDRH3 с другой длиной. Разнообразие H3 может быть увеличено с использованием набора кодонов NNK, DVK и NVK, а также оно может быть ограничено у N- и/или C-концов.
Разнообразие может быть также создано в CDRH1 и CDRH2. Создание разнообразий CDR-H1 и H2 достигается в соответствии со стратегией, направленной на репертуар описанных природных антител-миметиков, при этом модификация направлена на получение разнообразия, которое больше соответствует природному разнообразию, чем разнообразию предыдущих конструкций.
Для получения разнообразия CDRH3 может быть отдельно сконструировано множество библиотек с различной длиной Н3, которые могут быть затем объединены для отбора агентов, связывающихся с антигенами-мишенями. Множество библиотек может быть объединено и отсортировано методами отбора на твердом носителе и методами сортинга в растворе, как было описано ранее и как описано ниже в настоящей заявке. Может быть использовано множество стратегий сортинга. Так, например, один вариант включает отбор по мишени, связанной с твердым веществом, с последующим сортингом по метке, которая может присутствовать на слитом полипептиде (например, анти-gD-метке), и последующим другим сортингом по мишени, связанной с твердым веществом. Альтернативно, библиотеки могут быть сначала отсортированы по мишени, связанной с твердой поверхностью, а затем элюированные связывающие агенты могут быть отсортированы путем связывания в растворе со снижающимися концентрациями антигена-мишени. Использование комбинаций различных методов сортинга позволяет минимизировать отбор только высокоэкспрессируемых последовательностей и проводить отбор ряда различных клонов с высокой аффинностью.
Агенты, связывающиеся с антигеном-мишенью NRP2 с высокой аффинностью, могут быть выделены из библиотек. Ограниченное разнообразие области H1/H2 снижает вырожденность кода примерно в 104-105 раз, а более высокое разнообразие области H3 требует более высокой аффинности связывающего агента. Для использования библиотек с различными типами разнообразия CDRH3 (например, с использованием кодонов DVK или NVT) требуется выделить связывающиеся агенты, которые могут связываться с различными эпитопами антигена-мишени.
После выделения связывающих агентов из объединенных библиотек, описанных выше, было обнаружено, что аффинность может быть дополнительно увеличена путем ограничения разнообразия в легкой цепи. В этом варианте разнообразие в легкой цепи может быть создано следующим образом: в CDRL1: аминокислота в положении 28 кодируется кодоном RDT; аминокислота в положении 29 кодируется кодоном RKT; аминокислота в положении 30 кодируется кодоном RVW; аминокислота в положении 31 кодируется кодоном ANW; аминокислота в положении 32 кодируется кодоном THT; необязательно, аминокислота в положении 33 кодируется кодоном CTG; в CDRL2: аминокислота в положении 50 кодируется кодоном KBG; аминокислота в положении 53 кодируется кодоном AVC; и, необязательно, аминокислота в положении 55 кодируется кодоном GMA; в CDRL3: аминокислота в положении 91 кодируется кодонами TMT или SRT или и тем и другим; аминокислота в положении 92 кодируется кодоном DMC; аминокислота в положении 93 кодируется кодоном RVT; аминокислота в положении 94 кодируется кодоном NHT; и аминокислота в положении 96 кодируется кодонами TWT или YKG или и тем и другим.
В другом варианте изобретения получают библиотеку или библиотеки с разнообразием областей CDRH1, CDRH2 и CDRH3. В этом варианте изобретения разнообразие CDRH3 достигается с использованием областей H3 различной длины и с использованием, главным образом, наборов кодонов XYZ и NNK или NNS. Библиотеки могут быть получены с использованием отдельных олигонуклеотидов и их комбинаций, либо олигонуклеотиды могут быть объединены с образованием подпоследовательности библиотек. В этом варианте изобретения библиотеки могут быть отсортированы по мишени, связанной с твердым веществом. Клоны, выделенные в результате проведения множества процедур сортинга, могут быть скринированы на специфичность и аффинность с помощью ELISA-анализов. Для определения специфичности клоны могут быть скринированы на нужные антигены-мишени, а также на другие антигены, не являющиеся мишенями. Агенты, связывающиеся с антигеном-мишенью NRP1, могут быть затем скринированы на аффинность в ELISA-анализе на конкурентное связывание в растворе или в точечном анализе на конкурентное связывание. Связывающие агенты с высокой аффинностью могут быть выделены из библиотеки с использованием наборов кодонов XYZ, полученных, как описано выше. Эти связывающие агенты могут быть легко получены в клеточной культуре в виде антител или антиген-связывающих фрагментов с высоким выходом.
В некоторых вариантах изобретения может оказаться желательным получение библиотек с большим разнообразием длин областей CDRH3. Например, может оказаться желательным получение библиотек с областями CDRH3, имеющими длину примерно от 7 до 19 аминокислот.
Высокоаффинные связывающие агенты, выделенные из библиотек этих вариантов изобретения, могут быть легко получены в культуре бактериальных и эукариотических клеток с высоким выходом. Эти векторы могут быть сконструированы так, чтобы последовательности, такие как метки gD и последовательность компонента белка вирусной оболочки, могли быть легко удалены и/или добавлены в последовательности константной области для получения полноразмерных антител или их антиген-связывающих фрагментов с высоким выходом.
Библиотека с мутациями в CDRH3 может быть объединена с библиотекой, содержащей модифицированные варианты других CDR, например CDRL1, CDRL2, CDRL3, CDRH1 и/или CDRH2. Например, в одном из вариантов изобретения библиотеку CDRH3 объединяют с библиотекой CDRL3, созданной на основе последовательности гуманизованного антитела 4D5 с вариантами аминокислот в положениях 28, 29, 30, 31 и/или 32, с использованием предварительно определенных наборов кодонов. В другом варианте изобретения библиотека с мутациями в CDRH3 может быть объединена с библиотекой, содержащей вариабельные домены тяжелой цепи вариантов CDRH1 и/или CDRH2. В одном из вариантов изобретения библиотеку CDRH1 создают с использованием последовательности гуманизованного антитела 4D5 с вариантами аминокислот в положениях 28, 30, 31, 32 и 33. Библиотека CDRH2 может быть создана с использованием последовательности гуманизованного антитела 4D5 с вариантами аминокислот в положениях 50, 52, 53, 54, 56 и 58 и с использованием предварительно определенных наборов кодонов.
Мутанты антител против NRP2
Антитело против NRP2, полученное из фаговых библиотек, может быть дополнительно модифицировано с получением мутантных антител, которые, по сравнению с родительским антителом, обладают улучшенными физическими, химическими или биологическими свойствами. Если используемым анализом является анализ на биологическую активность, то в выбранном анализе мутантное антитело предпочтительно должно обладать биологической активностью, которая по меньшей мере примерно в 10 раз, предпочтительно по меньшей мере примерно в 20 раз, более предпочтительно по меньшей мере примерно в 50 раз, а иногда по меньшей мере примерно в 100 или 200 раз превышает биологическую активность исходного антитела в данном анализе. Так, например, мутантное антитело против NRP1 предпочтительно имеет аффинность связывания с NRP, которая по меньшей мере примерно в 10 раз, предпочтительно по меньшей мере примерно в 20 раз, более предпочтительно по меньшей мере примерно в 50 раз, а иногда по меньшей мере примерно в 100 или 200 раз превышает аффинность связывания родительского антитела против NRP.
Для получения мутантного антитела в одну или несколько гипервариабельных областей родительского антитела вводят одну или несколько аминокислотных модификаций (например, замен). Альтернативно или дополнительно, в родительское антитело могут быть введены одна или несколько модификаций (например, замен) остатков каркасной области, что будет приводить к повышению аффинности связывания мутантного антитела с антигеном млекопитающего другого вида. Примерами модифицируемых остатков каркасной области являются остатки, которые непосредственно и нековалентно связываются с антигеном (Amit et al. (1986) Science 233:747-753); взаимодействуют с CDR/влияют на конформацию CDR (Chothia et al. (1987) J. Mol. Biol. 196:901-917); и/или участвуют в образовании пограничной области VL-VH (EP 239400 B1). В некоторых вариантах изобретения модификация одного или нескольких таких остатков каркасной области приводит к повышению аффинности связывания антитела с антигеном млекопитающего другого вида. Например, в этом варианте изобретения могут быть модифицированы примерно от одного до пяти каркасных остатков. Иногда может оказаться достаточным получения мутантного антитела с выходом, подходящим для его использования в доклинических испытаниях даже в том случае, если ни один из остатков гипервариабельной области не является модифицированным. Однако обычно мутантное антитело содержит дополнительную(ые) модификацию(и) в гипервариабельной области.
Остатки гипервариабельной области, которые являются модифицированными, могут быть заменены произвольно, особенно если родительское антитело имеет исходную аффинность связывания, при которой такие произвольно полученные мутанты антител могут быть легко скринированы.
Одной из методик, подходящих для получения таких мутантных антител, является методика, называемая «аланин-сканирующим мутагенезом» (Cunningham and Wells (1989) Science 244:1081-1085). В данном случае один или несколько остатков гипервариабельной области заменяют аланиновым(и) или полиаланиновым(и) остатком(ами) в целях обеспечения влияния на взаимодействие аминокислот с антигеном млекопитающего другого вида. Такой(ие) остаток(остатки) гипервариабельной области, обладающий(ие) функциональной чувствительностью к заменам, затем уточняют путем введения дополнительных или других мутаций в участки замен или рядом с этими участками. Таким образом, хотя участок для введения изменений в аминокислотную последовательность заранее определен, природа мутации per se необязательно должна быть определена заранее. Такие ala-мутантны, получаемые этим способом, скринируют на биологическую активность, как описано в настоящей заявке.
Обычно осуществление процедуры замены следует начинать с введения консервативных замен, таких как замены, представленные в нижеследующей таблице, озаглавленной “Предпочтительные замены”. Если такие замены приводят к изменению биологической активности (например, аффинности связывания), то могут быть введены более существенные замены, названные в нижеследующей таблице «характерные замены», или замены, дополнительно описанные ниже с указанием классов аминокислот, и полученные продукты могут быть скринированы.
Более значительные изменения биологических свойств антитела могут быть достигнуты путем выбора замен, которые оказывают значительное влияние на сохранение (а) структуры полипептидного остова в области замены, например, складчатой или спиральной конформации, (b) заряда или гидрофобности молекулы в нужном сайте или (с) объема боковой цепи. Природные остатки подразделяются на нижеследующие группы по общим свойствам боковых цепей:
(1) гидрофобные остатки: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные остатки: Cys, Ser, Thr; Asn; Gln;
(3) кислотные остатки: Asp, Glu;
(4) основные остатки: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro; и
(6) ароматические остатки: Trp, Tyr, Phe.
Не-консервативные замены приводят к заменам члена одного из этих классов членом другого класса.
В другом варианте изобретения сайты, отобранные для модификации, подвергают процедуре аффинного созревания методом фагового дисплея (см. выше).
Молекулы нуклеиновой кислоты, кодирующие мутантные аминокислотные последовательности, получают различными способами, известными специалистам. Такими способами являются, но ими не ограничиваясь, опосредуемый олигонуклеотидом (или сайт-направленный) мутагенез, ПЦР-мутагенез и кластерный мутагенез уже полученного мутанта или немутантного варианта родительского антитела. Предпочтительным способом получения мутантов является сайт-направленный мутагенез (см., например, Kunkel (1985) Proc. Natl. Acad. Sci. USA 82:488).
В некоторых вариантах изобретения мутантное антитело имеет только одну замену остатка в гипервариабельной области. В других вариантах изобретения были заменены два или более остатка гипервариабельной области родительского антитела, например примерно от двух до десяти остатков.
Обычно мутантное антитело с улучшенными биологическими свойствами имеет аминокислотную последовательность, которая по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, еще более предпочтительно по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90% и наиболее предпочтительно по меньшей мере на 95% идентична или аналогична аминокислотной последовательности вариабельного домена тяжелой или легкой цепи родительского антитела. Идентичность или сходство этих последовательностей определяют в настоящем описании как процент аминокислотных остатков в последовательности-кандидате, которые являются идентичными (то есть одни и те же остатки) или аналогичными (то есть аминокислотные остатки, принадлежащие к одной и той же группе исходя из общих свойств их боковых цепей) остаткам родительского антитела после выравнивания последовательностей и введения пропусков, если это необходимо, для достижения максимального процента идентичности последовательностей. Ни одна из N-концевых, С-концевых или внутренних модификаций, таких как удлинения, делеции или инсерции, вводимые в последовательность антитела за пределами вариабельного домена, не должна негативно влиять на идентичность или сходство последовательностей.
После получения мутантного антитела определяют биологическую активность полученной молекулы по сравнению с родительским антителом. Как указано выше, это может быть осуществлено путем определения аффинности связывания и/или других биологических активностей антитела. В предпочтительном варианте изобретения панель мутантных антител получают и скринируют на аффинность связывания с антигеном, таким как NRP1 или его фрагмент. Одно или несколько мутантных антител, выбранных в результате первоначального скрининга, подвергают, но необязательно, анализу на одну или несколько дополнительных биологических активностей для того, чтобы определить, может(гут) ли такое(ие) мутантное(ые) антитело(а) с повышенной аффинностью связывания быть использовано(ы), например, в доклинических исследованиях.
Выбранное(ые) таким образом мутантное(ые) антитело(а) можно подвергнуть дополнительным модификациям, часто в зависимости от целей использования антитела. Такие модификации могут включать дополнительную модификацию аминокислотной последовательности, слияние с гетерологичным(и) полипептидом(ами) и/или ковалентные модификации, такие как модификации, описанные ниже. Характерные модификации аминокислотной последовательности описаны выше. Так, например, любой цистеиновый остаток, не участвующий в сохранении соответствующей конформации мутантного антитела, может быть также заменен, обычно серином, для повышения стабильности молекулы к окислению и предотвращения нежелательного перекрестного связывания. И наоборот, в указанное антитело можно ввести цистеиновую(ые) связь(и) для повышения его стабильности (в частности, если указанным антителом является фрагмент антитела, такой как Fv-фрагмент). Аминокислотный мутант другого типа имеет измененный характер гликозилирования. Такой характер может быть достигнут путем делеции одной или нескольких углеводных групп, присутствующих в антителе, и/или добавления одного или нескольких сайтов гликозилирования, не присутствующих в антителе. Гликозилирование антител обычно является либо N-связанным, либо О-связанным. N-связанное гликозилирование означает присоединение углеводной группы к боковой цепи аспарагинового остатка. Трипептидные последовательности «аспарагин-Х-серин» и «аспарагин-Х-треонин», где Х обозначает любую аминокислоту, за исключением пролина, представляют собой последовательности распознавания для ферментативного присоединения углеводной группы к боковой цепи аспарагина. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде способствует созданию потенциального сайта гликозилирования. О-связанное гликозилирование означает присоединение одного из сахаров, таких как N-ацетилгалактозамин, галактоза или ксилоза, к гидроксиаминокислоте, главным образом, к серину или треонину, хотя ими могут быть также 5-гидроксипролин или 5-гидроксилизин. Присоединение сайтов гликозилирования к антителу обычно осуществляют путем такой модификации аминокислотной последовательности, в результате которой эта аминокислотная последовательность будет содержать одну или несколько вышеописанных трипептидных последовательностей (для сайтов N-связанного гликозилирования). Такая модификация может быть также осуществлена путем добавления одного или нескольких сериновых или треониновых остатков к последовательности или их замены в последовательности исходного антитела (для сайтов О-связанного гликозилирования).
Векторы, клетки-хозяева и рекомбинантные способы
Антитела против Nrp2 по изобретению могут быть получены рекомбинантно с использованием легко доступных способов и материалов.
Для рекомбинантного получения антитела против NRP2 нуклеиновую кислоту, кодирующую такое антитело, выделяют и встраивают в реплицируемый вектор для последующего клонирования (амплификации ДНК) или экспрессии. ДНК, кодирующая антитело, может быть легко выделена или синтезирована стандартными способами (например, с использованием олигонуклеотидных зондов, обладающих способностью специфически связываться с ДНК, кодирующими тяжелые и легкие цепи антитела). Многие векторы являются общедоступными. Векторными компонентами, как правило, являются, но ими не ограничиваясь, один или несколько из нижеследующих компонентов, таких как сигнальная последовательность, точка начала репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции.
(i) Компонент сигнальная последовательность
Антитело по изобретению может быть рекомбинантно получено не только прямым методом, но также и в виде слитого полипептида с гетерологичным полипептидом, который предпочтительно представляет собой сигнальную последовательность или другой полипептид, имеющий специфический сайт расщепления у N-конца зрелого белка или полипептида. Выбранной гетерологичной сигнальной последовательностью предпочтительно является последовательность, которая распознается и процессируется (то есть отщепляется сигнальной пептидазой) в клетке-хозяине. Если прокариотические клетки-хозяева не распознают и не процессируют нативную сигнальную последовательность антитела, то такая сигнальная последовательность может быть заменена прокариотической сигнальной последовательностью, выбранной, например, из группы, состоящей из лидерных последовательностей щелочной фосфатазы, пенициллиназы, lpp или термостабильного энтеротоксина II. Для секреции в дрожжах нативная сигнальная последовательность может быть заменена, например, лидерной последовательностью инвертазы дрожжей, лидерной последовательностью α-фактора (включая лидерные последовательности α-фактора Saccharomyces и Kluyveromyces) или лидерной последовательностью кислой фосфатазы, лидерной последовательностью глюкоамилазы C.albicans или сигнальной последовательностью, описанной в WO 90/13646. Для экспрессии в клетках млекопитающих используются сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например сигнальная последовательность gD вируса простого герпеса.
ДНК для такой области предшественника лигируют с ДНК, кодирующей антитело, с сохранением рамки считывания.
(ii) Компонент точка начала репликации
Экспрессионные и клонирующие векторы содержат последовательность нуклеиновой кислоты, которая способствует репликации данного вектора в одной или нескольких выбранных клетках-хозяевах. Как правило, в клонирующих векторах такой последовательностью является последовательность, которая позволяет данному вектору реплицироваться независимо от ДНК хромосомы хозяина и которая содержит точку начала репликации или автономно реплицирующиеся последовательности. Такие последовательности для различных бактерий, дрожжей и вирусов хорошо известны. Точка начала репликации плазмиды pBR322 является подходящей для большинства грамотрицательных бактерий, а именно точка начала репликации плазмиды 2 мкм является подходящей для дрожжей, а различные точки начала репликации вирусов (SV40, полиомавируса, аденовируса, VSV или BPV) могут быть использованы для клонирования векторов в клетках млекопитающих. Как правило, компонент точка начала репликации не является необходимым для векторов, экспрессирующихся в клетках млекопитающих (точка начала репликации SV40 обычно используется только в тех случаях, когда содержит ранний промотор).
(iii) Компонент селективный ген
Экспрессионный и клонирующий векторы могут содержать селективный ген, также называемый селективным маркером. Обычно селективные гены кодируют белки, которые (а) сообщают резистентность к антибиотиками или другим токсинам, например к ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементируют дефицит, обусловленный ауксотрофией, или (с) обеспечивают поступление важных питательных веществ, которые не поступают из комплексных сред, например, ген, кодирующий D-аланин-рацемазу Bacillus.
Одним из примеров схемы отбора является использование лекарственного средства, прекращающего рост клетки-хозяина. Клетки, которые были успешно трансформированы гетерологичным геном, продуцируют белок, сообщающий резистентность к лекарственному средству и тем самым способствующий выживанию этих клеток в селективной среде. Для такого доминантного отбора могут быть использованы, например, лекарственные средства, такие как неомицин, микофеноловая кислота и гигромицин.
Другими примерами подходящих селективных маркеров для клеток млекопитающих являются маркеры, которые позволяют идентифицировать клетки, способные встраивать в свой геном нуклеиновую кислоту, кодирующую антитело, такую как гены DHFR, тимидинкиназы, металлотионеина-I и II и предпочтительно гены, кодирующие металлотионеин приматов, аденозиндезаминазу, орнитиндекарбоксилазу и т.д.
Так, например, клетки, трансформированные селективным геном DHFR, сначала идентифицируют путем культивирования всех трансформантов в культуральной среде, содержащей метотрексат (Mtx), конкурентный антагонист DHFR. При использовании DHFR дикого типа подходящей клеткой-хозяином является клеточная линия яичника китайского хомячка (СНО), дефицитная по DHFR-активности.
Альтернативно, клетки-хозяева (в частности, хозяева дикого типа, содержащие эндогенный DHFR), трансформированные или ко-трансформированные ДНК-последовательностями, кодирующими антитело, белок DHFR дикого типа и другой селективный маркер, такой как аминогликозид-3'-фосфотрансфераза (АРН), могут быть отобраны путем культивирования клеток в среде, содержащей агент для отбора, проводимого с использованием селективного маркера, такого как аминогликозидный антибиотик, например канамицин, неомицин или G418. См. патент США № 4965199.
Подходящим селективным геном для использования в дрожжах является ген trp1, присутствующий в дрожжевой плазмиде YRp7 (Stinchcomb et al. (1979) Nature 282:39). Ген trp1 представляет собой селективный маркер для мутантного штамма дрожжей, который не способен расти в присутствии триптофана, например, штамма, депонированного в ATCC № 44076 или РЕР4-1. Jones (1977) Genetics, 85:12. Наличие мутации в гене trp1 в геноме дрожжевой клетки-хозяина позволяет эффективно осуществлять детектирование трансформации путем культивирования в отсутствие триптофана. Аналогичным образом, Leu2-дефицитные дрожжевые штаммы (ATCC 20622 или 38626) комплементировали известными плазмидами, несущими ген Leu2.
Кроме того, векторы кольцевой 1,6-мкм плазмиды pKD1 могут быть использованы для трансформации дрожжей Kluyveromyces. Альтернативно, экспрессионная система для крупномасштабного получения рекомбинантного химозина теленка была описана для K.lactis. Van den Berg (1990) Bio/Technology, 8:135. Были также описаны стабильные многокопийные экспрессионные векторы для секреции зрелого рекомбинантного сывороточного альбумина человека промышленными штаммами Kluyveromyces. Fleer et al. (1991) Bio/Technology, 9:968-975.
(iv) Компонент промотор
Экспрессионные и клонирующие векторы обычно содержат промотор, распознаваемый организмом-хозяином и функционально связанный с нуклеиновой кислотой, кодирующей антитело. Промоторами, подходящими для использования в прокариотических хозяевах, являются промотор phoA, промоторные системы β-лактамазы и лактозы, щелочная фосфатаза, промоторная система триптофана (trp) и гибридные промоторы, такие как промотор tac. Однако подходящими также являются и другие известные бактериальные промоторы. Промоторы, используемые в бактериальных системах, также содержат последовательность Шайна-Дальгарно (S.D.), функционально связанную с ДНК, кодирующей антитело.
Промоторные последовательности, подходящие для эукариотов, известны специалистам. Фактически все гены эукариотов имеют АТ-богатую область, локализованную приблизительно в области, расположенной на 25-30 нуклеотидов выше от сайта инициации транскрипции. Другой такой последовательностью, локализованной приблизительно в области, расположенной на 70-80 нуклеотидов выше от сайта инициации транскрипции многих генов, является область CNCAAT, где N может означать любой нуклеотид. У 3'-конца большинства эукариотических генов расположена последовательность ААТААА, которая может служить сигналом для присоединения poly A-хвоста к 3'-концу кодирующей последовательности. Все указанные последовательности могут быть соответствующим образом встроены в эукариотические экспрессионные векторы.
Примерами промоторных последовательностей, подходящих для использования в дрожжевых клетках-хозяевах, являются промоторы 3-фосфоглицераткиназы или других гликолитических ферментов, таких как энолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа.
Другими дрожжевыми промоторами, которые представляют собой индуцибельные промоторы, имеющие дополнительное преимущество, заключающееся в их способности регулировать транскрипцию в определенных условиях культивирования, являются промоторные области алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, деструктивных ферментов, связанных с метаболизмом азота, металлотионеина, глицеральдегид-3-фосфатдегидрогеназы и ферментов, ответственных за утилизацию мальтозы и галактозы. Векторы и промоторы, подходящие для экспрессии в дрожжах, также описаны в ЕР 73657. Наряду с дрожжевыми промоторами также преимущественно используются дрожжевые энхансеры.
Транскрипция антитела из векторов в клетках-хозяевах млекопитающих регулируется, например, промоторами, полученными из геномов таких вирусов, как вирус полиомы, вирус оспы домашней птицы, аденовирус (такой как аденовирус 2), вирус коровьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и наиболее предпочтительно обезьяний вирус 40 (SV40); гетерологичными промоторами млекопитающих, например промотором актина или промотором иммуноглобулина; и промоторами белков теплового шока, при условии, что такие промоторы являются совместимыми с системами клеток-хозяев.
Ранние и поздние промоторы вируса SV40 обычно получают в виде рестрикционного фрагмента SV40, который также содержит точку начала репликации вируса SV40. Предранний промотор цитомегаловируса человека обычно получают в виде рестрикционного HindIII-фрагмента Е. Система экспрессии ДНК в клетках-хозяевах млекопитающих, полученная на основе бычьего папилломавируса, используемого в качестве вектора, описана в патенте США № 4419446. Модификация такой системы описана в патенте США № 4601978. Экспрессия кДНК β-интерферона человека в мышиных клетках под контролем промотора тимидинкиназы вируса простого герпеса также описана в публикации Reyers et al. (1982) Nature 297:598-601. Альтернативно, в качестве промотора может быть использован вирус саркомы Рауса, имеющий длинный концевой повтор.
(v) Компонент энхансерный элемент
Транскрипция ДНК, кодирующей антитело по изобретению, в высших эукариотах часто усиливается при встраивании в вектор энхансерной последовательности. В настоящее время известно множество энхансерных последовательностей генов млекопитающих (генов глобина, эластазы, альбумина, α-фетопротеина и инсулина). Однако обычно используется энхансер от вируса эукариотической клетки. Примерами являются энхансер SV40, локализованный в поздней области точки начала репликации (100-270 п.н.), энхансер раннего промотора цитомегаловируса, энхансер полиомавируса, локализованный в поздней области точки начала репликации, и энхансеры адновируса. См. также публикацию Yaniv (1982) Nature 297:17-18, в которой описаны энхансерные элементы для активации эукариотических промоторов. Энхансер может быть сплайсирован в вектор в 5'- или 3'-положении по отношению к последовательности, кодирующей антитело, но предпочтительно он находится в 5'-положении от промотора.
(vi) Компонент сайт терминации транскрипции
Экспрессионные векторы, используемые в эукариотических клетках-хозяевах (в дрожжах, грибах, насекомых, растениях, у животных, у человека или в ядро-содержащих клетках других многоклеточных организмов), также содержат последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно расположены со стороны 5'-конца, а иногда 3'-конца от нетранслируемых областей эукариотических или вирусных ДНК или кДНК. Эти области содержат нуклеотидные сегменты, транскрибированные в виде полиаденилированных фрагментов в нетранслированной части мРНК, кодирующей антитело. Одним из подходящих компонентов терминации транскрипции является область полиаденилирования бычьего гормона роста. См. описание экспрессионных векторов в WO 94/11026 и в настоящей заявке.
(vii) Отбор и трансформация клеток-хозяев
Клетками-хозяевами, подходящими для клонирования или экспрессии ДНК в представленных в настоящем описании векторах, являются клетки прокариотов, дрожжей или высших эукариотов, описанные выше. Прокариотами, подходящими для этой цели, являются Eubacteria, такие как грамотрицательные или грамположительные микроорганизмы, например Enterobacteriaceae, такие как Escherichia, например E.coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans и Shigella, а также Bacilli, такие как B.subtilis и B.licheniformis (например, B.licheniformis 41Р, описанная в патенте DD 266710, опубликованном 12 апреля 1989), Pseudomonas, такие как P.aeruginosa, и Streptomyces. Предпочтительным клонирующим хозяином E.coli является E.coli 294 (АТСС 31446), хотя могут быть использованы и другие штаммы, такие как E.coli В, E.coli Х1776 (АТСС 31537) и E.coli W3110 (АТСС 27325). Эти примеры приводятся лишь для иллюстрации и не ограничивают объем изобретения.
Кроме прокариотов, подходящими хозяевами для клонирования или экспрессии векторов, кодирующих антитело, являются эукариотические микроорганизмы, такие как гифомицеты или дрожжи. Из низших эукариотических микроорганизмов-хозяев наиболее часто используются Saccharomyces cerevisiae или стандартные пекарские дрожжи. Однако в данном случае могут быть использованы и другие часто применяемые и доступные микроорганизмы другого рода, других видов и штаммов, таких как Schizosaccharomyces pombe; хозяева Kluyveromyces, такие как, например, K.lactis, K.fragilis (АТСС 12424), K.bulgaricus (АТСС 16045), K.wickeramii (АТСС 24178), K.waltii (АТСС 56500), K.drosophilarum (АТСС 36906), K.thermotolerans и K.marxianus; yarrowia (ЕР 402226); Pichia pastoris (ЕР 183070); Candida; Trichoderma reesia (ЕР 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и гифомицеты, такие как, например, Neurospora, Penicillium, Tolypocladium и Aspergillus, такие как A.nidulans и A.niger.
Клетками-хозяевами, подходящими для экспрессии гликозилированного антитела, являются клетки многоклеточных организмов. Примерами клеток беспозвоночных являются клетки растений и насекомых. Были идентифицированы различные бакуловирусные штаммы и варианты, а также соответствующие пермиссивные клетки-хозяева насекомых, таких как Spodoptera frugiperda (гусеница совки), Aedes aegypti (желтолихорадочный комар), Aedes albopictus (белопятнистый комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori (тутовый шелкопряд). В настоящее время имеется ряд доступных вирусных штаммов, которые могут быть использованы для трансфекции, например вариант L-1 Autographa californica NPV и штамм Bm-5 Bombyx mori NPV, и такие вирусы могут быть использованы в настоящем изобретении, в частности, для трансфекции клеток Spodoptera frugiperda. В качестве клеток-хозяев могут быть также использованы клетки растительных культур хлопчатника, кукурузы, картофеля, сои, петунии, томатов и табака.
Однако самый большой интерес представляют клетки позвоночных, и размножение клеток позвоночных в культуре (тканевой культуре) уже становится рутинной процедурой. Примерами обычно используемых линий клеток-хозяев млекопитающих являются клеточная линия CV1 почек обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); клеточная линия почек эмбриона человека (клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al. (1977) J. Gen. Virol. 36:59); клетки почек детеныша хомячка (ВНК, АТСС CCL 10); клетки яичника китайского хомячка/-DHFR (CHO, Urlaub et al. (1980) Proc. Natl. Acad. Sci. USA 77:4216); клетки Сертоли мыши (TM4, Mather (1980) Biol. Reprod. 23:243-251); клетки почек обезьяны (CV1 ATCC CCL 70); клетки почек африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почек собак (MDCK, ATCC CCL 34); клетки печени лабораторной крысы Buffalo (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухолевые клетки молочной железы мыши (MMT 060562, ATCC CCL 51); клетки TR1 (Mather et al. (1982) Annals N.Y. Acad. Sci. 383:44-68); клетки MRC 5; клетки FS4; клеточная линия гепатомы человека (Hep G2).
Клетки-хозяева трансформируют вышеописанными экспрессионными или клонирующими векторами для получения антитела и культивируют в подходящих питательных средах, модифицированных, при необходимости, для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих нужные последовательности.
(viii) Культивирование клеток-хозяев
Клетки-хозяева, используемые для получения антитела по изобретению, могут быть культивированы в различных средах. Средами, подходящими для культивирования клеток-хозяев, являются коммерчески доступные среды, такие как среда Хэмса F10 (Sigma), минимальная поддерживающая среда ((МЕМ), (Sigma), RPMI-1640 (Sigma) и модифицированная по Дульбекко среда Игла ((DMEM), Sigma). Кроме того, в качестве культуральной среды для культивирования клеток-хозяев может быть использована любая среда, описанная в публикации Ham et al. (1979) Meth. Enz. 58:44, Barnes et al. (1980) Anal. Biochem. 102:255, в патентах США №№ 4767704, 4657866, 4927762, 4560655 или 5122469; в WO 90/03430; WO 87/00195 или в патенте США Re. № 30985. В любую из этих сред могут быть добавлены, при необходимости, гормоны и/или другие факторы роста (такие как инсулин, трансферрин или эпидермальный фактор роста), соли (такие как хлорид натрия и фосфат кальция и магния), буферы (такие как HEPES), нуклеотиды (такие как аденозин и тимидин), антибиотики (такие как лекарственное средство гентамицин™), микроэлементы (определенные как неорганические соединения, обычно присутствующие в конечных концентрациях в микромолярных дозах) и глюкоза или эквивалентный источник энергии. Могут быть также включены любые другие необходимые добавки в соответствующих концентрациях, известные специалистам в данной области. Условия культивирования, такие как температура, рН и т.п., аналогичны условиям, ранее используемым для экспрессии выбранных клеток-хозяев, и известны среднему специалисту в данной области.
(ix) Очистка антитела
C использованием техники рекомбинантных ДНК антитело может быть получено внутри клеток или в периплазматическом пространстве, либо оно может быть непосредственно секретировано в среду. Если на первой стадии антитело получается внутри клеток, то клеточный дебрис, или клетки-хозяева, или их лизированные фрагменты удаляют, например, путем центрифугирования или ультрафильтрации. В публикации Carter et al. (1992) Bio/Technology 10:163-167 описана методика выделения антител, которые секретируются в периплазматическое пространство E.coli. Коротко, клеточный состав оттаивают в присутствии ацетата натрия (рН 3,5), EDTA и фенилметилсульфонилфторида (PMSF) на протяжении примерно 30 минут. Клеточный дебрис может быть удален путем центрифугирования. Если антитело секретируется в среду, то супернатанты от таких экспрессионных систем, как правило, сначала концентрируют с использованием коммерчески доступного фильтра для концентрирования белка, например устройства для ультрафильтрации Amicon или Millipore Pellicon. Ингибитор протеазы, такой как PMSF, может быть введен в любую из вышеуказанных стадий для ингибирования протеолиза, а антибиотики могут быть введены для предупреждения роста случайно внесенных примесных микроорганизмов.
Композиция антитела, полученная из клеток, может быть очищена, например, с помощью хроматографии на гидроксиапатитах, гель-электрофореза, диализа и аффинной хроматографии, причем предпочтительным способом очистки является аффинная хроматография. Подходит ли белок А в качестве аффинного лиганда, зависит от вида и изотипа Fc-области любого иммуноглобулина, присутствующего в антителах. Белок А может быть использован для очистки антител, содержащих тяжелые цепи γ1, γ2 или γ4 иммуноглобулина человека (Lindmark et al. (1983) J. Immunol. Meth. 62:1-13). Для всех изотипов мышей и для γ3 иммуноглобулина человека рекомендуется белок G (Guss et al. (1986) EMBO J. 5:1567-1575). В качестве матрицы, с которой связывается аффинный лиганд, наиболее часто используется агароза, но могут быть использованы и другие матрицы. Механически стабильные матрицы, такие как стекло с регулируемым размером пор или поли(стиролдивинил)бензол, обеспечивают более высокую скорость потока и более короткое время обработки, чем это может быть достигнуто с использованием агарозы. Если антитело содержит домен СН3, то для очистки может быть использована смола Bakerbond ABX™ (J.T. Baker, Phillipsburg, NJ). В зависимости от выделяемого антитела могут быть также использованы и другие способы очистки белка, такие как фракционирование на ионообменной колонке, преципитация этанолом, обращенно-фазовая ВЭЖХ, хроматография на диоксиде кремния, хроматография на гепарин-сефарозе™, хроматография на анионо- или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и преципитация сульфатом аммония.
После проведения любой(ых) стадии(й) предварительной очистки смесь, содержащая представляющее интерес антитело и примеси, может быть подвергнута гидрофобной хроматографии при низком рН с использованием элюирующего буфера при рН примерно 2,5-4,5, предпочтительно при низких концентрациях солей (например, примерно 0-0,25 M).
Фармацевтические композиции
Терапевтические композиции, содержащие антитело, получают в удобных для хранения формах путем смешивания антитела, имеющего нужную степень чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)) с получением лиофилизованных препаратов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы в используемых дозах и концентрациях должны быть нетоксичными для реципиентов, и ими являются буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные полипептиды (имеющие примерно менее чем 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатообразующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионогенные поверхностно-активные вещества, такие как твин™, плюроник™ или полиэтиленгликоль (ПЭГ).
Представленная в настоящем описании композиция может также содержать несколько активных соединений, необходимых для конкретного показания, предпочтительно имеющих дополнительную активность и не оказывающих отрицательного влияния друг на друга. Например, может быть желательным дополнительное введение иммуносупрессора, противоопухолевого средства, средства против ангиогенеза, антинеопластического средства, цитотоксического средства и/или химиотерапевтического средства. Такие молекулы могут присутствовать в комбинации в количествах, эффективных для использования в нужных целях.
Активные ингредиенты могут быть также введены в микрокапсулу, полученную, например, способами коацервации или путем межфазной полимеризации, например, в гидроксиметилцеллюлозную или желатиновую микрокапсулу и полиметилметакрилатную микрокапсулу, соответственно, в системы для доставки коллоидальных лекарственных средств (например, в липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы описаны в руководстве Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Композиции, используемые для введения in vivo, должны быть стерильными. Это может быть легко достигнуто путем фильтрации через стерильные фильтрующие мембраны.
Могут быть получены препараты пролонгированного высвобождения. Подходящими примерами препаратов пролонгированного высвобождения являются полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, где указанные матрицы имеют форму готовых изделий, например пленок или микрокапсул. Примерами матриц пролонгированного высвобождения являются полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поливиниловый спирт), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразлагаемый сополимер этилена-винилацетата, разлагаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOT™ (микросферы для инъекций, состоящие из сополимера молочной кислоты-гликолевой кислоты и ацетата лейпролида) и поли-D-(-)-3-гидроксимасляная кислота. Полимеры, такие как сополимер этилена-винилацетата и сополимер молочной кислоты-гликолевой кислоты, способны высвобождать молекулы в течение более 100 дней, а некоторые гидрогели высвобождают белки за более короткие периоды времени. Если инкапсулированные антитела сохраняются в организме в течение длительного периода времени, то они могут денатурироваться или агрегироваться под воздействием влаги при 37°C, что приводит к потере биологической активности и к возможным изменениям иммуногенности. Для стабилизации, в зависимости от конкретного механизма, могут быть разработаны рациональные стратегии. Так, например, если было обнаружено, что механизмом агрегации является образование межмолекулярной связи S-S посредством тиодисульфидного обмена, то стабилизация может быть достигнута путем модификации сульфгидрильных остатков, лиофилизации из кислотных растворов, регуляции содержания влаги с использованием соответствующих добавок и получения конкретных композиций на основе полимерной матрицы.
Терапевтическое использование
Считается, что антитела по изобретению могут быть использованы для лечения млекопитающего. В одном из вариантов изобретения антитело вводят млекопитающему, не являющемуся человеком, в целях получения, например, доклинических данных. Примерами млекопитающих, не являющихся человеком, которые могут получать лечение, являются приматы, не являющиеся человеком, собаки, кошки, грызуны и другие млекопитающие, участвующие в доклинических исследованиях. Такими млекопитающими могут быть животные с созданной моделью заболевания, получаемые лечение антителом, либо такие животные могут быть использованы для исследования токсичности представляющего интерес антитела. В каждом из этих вариантов изобретения на млекопитающих могут быть проведены исследования с увеличением дозы. Если указанным антителом является антитело против NRP2, то оно может быть введено, например, грызуну-хозяину с моделью солидной опухоли.
Дополнительно или альтернативно, антитело используют для лечения человека, например пациента, страдающего заболеванием или расстройством, симптомы которого могут быть уменьшены после введения указанного антитела.
Настоящее изобретение включает профилактику и лечение опухолевого лимфангиогенеза, профилактику и лечение метастазирующей опухоли и антиангиогенную терапию рака, новую стратегию лечения рака, направленную на ингибирование развития опухолевых кровеносных сосудов, которым для поддержания их роста необходимы питательные вещества. Настоящее изобретение, в частности, включает ингибирование неопластического роста опухоли в области ее первичного образования, а также профилактику и/или лечение метастазов опухоли в области образования вторичной опухоли и, следовательно, воздействие на опухоли другими терапевтическими средствами. Примерами описанных здесь злокачественных опухолей, подвергаемых лечению (включая профилактику этих опухолей), являются, но ими не ограничиваясь, карцинома, лимфома, бластома, саркома и лейкоз. Более конкретными примерами таких злокачественных опухолей являются плоскоклеточный рак, рак легких (включая мелкоклеточный рак легких, немелкоклеточный рак легких, аденокарциному легких и плоскоклеточную карциному легких), перитонеальный рак, гепатоцеллюлярный рак, рак кишечника или желудка (включая рак желудочно-кишечного тракта), рак поджелудочной железы, глиобластома, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатома, рак молочной железы, рак толстой кишки, рак прямой и ободочной кишки, карцинома эндометрия или матки, карцинома слюнных желез, рак почек или ренальный рак, рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, карцинома печени и рак головы и шеи различных типов, а также В-клеточная лимфома (включая низкозлокачественную/фолликулярную не-ходжкинскую лимфому (NHL); мелкоклеточную лимфоцитарную (SL) NHL; NHL средней стадии дифференцировки/фолликулярную NHL; диффузную NHL средней стадии дифференцировки; высокозлокачественную иммунобластную NHL; высокозлокачественную лимфобластную NHL; высокозлокачественную мелкоклеточную недифференцированную NHL; генерализованную NHL; лимфому клеток коры головного мозга; лимфому, связанную со СПИД, и макроглобулинемию Вальденстрема); хронический лимфоцитарный лейкоз (CLL); острый лимфобластный лейкоз (ALL); волосато-клеточный лейкоз; хронический миелобластный лейкоз и лимфопролиферирующее расстройство после трансплантации (PTLD), а также патологическая пролиферация сосудов, связанная с факоматозом, отек (такой как отек, связанный с опухолями головного мозга) и синдром Мейгса. Более конкретно, злокачественными опухолями, которые поддаются лечению антителами по изобретению, являются рак молочной железы, рак прямой и ободочной кишки, рак прямой кишки, немелкоклеточный рак легких, не-ходжкинская лимфома (NHL), почечно-клеточный рак, рак предстательной железы, рак печени, рак поджелудочной железы, саркома мягких тканей, саркома Капоши, карциноидная карцинома, рак головы и шеи, меланома, рак яичника, мезотелиома и множественная миелома.
В настоящем изобретении предусматривается, что антитела по изобретению, при их использовании для лечения различных заболеваний, могут быть объединены с другими терапевтическими средствами, подходящими для лечения тех же самых или аналогичных заболеваний. Антитела по изобретению, при их использовании для лечения рака, могут быть использованы в комбинации со стандартной терапией рака, такой как хирургическая операция, лучевая терапия, химиотерапия или их комбинации.
В некоторых аспектах изобретения другими терапевтическими средствами, используемыми в комбинированной терапии рака вместе с антителом по изобретению, являются другие средства против ангиогенеза. Многие средства против ангиогенеза идентифицированы и хорошо известны специалистам, включая средства, перечисленные в публикации Carmeliet and Jain (2000).
В одном из аспектов антитело по изобретению используют в комбинации с антагонистом VEGF или антагонистом рецептора VEGF, такими как антитела против VEGF, варианты VEGF, фрагменты растворимых рецепторов VEGF, аптамеры, способные блокировать VEGF или VEGFR, нейтрализующие антитела против VEGFR, ингибиторы VEGFR-тирозинкиназ и любые их комбинации. Альтернативно или дополнительно, два или более антитела против NRP1 могут быть совместно введены пациенту. В более предпочтительном варианте изобретения антитело против NRP1A или NRPB по изобретению используют в комбинации с антителом против VEGF для получения аддитивных или синергических эффектов. Предпочтительными антителами против VEGF являются антитела, которые связываются с тем же эпитопом, с которым связывается антитело против hVEGF A4.6.1. Более предпочтительным антителом против VEGF является бевацизумаб или ранибизумаб.
В некоторых других аспектах изобретения другими терапевтическими средствами, используемыми для комбинированной терапии опухоли вместе с антителом по изобретению, является антагонист других факторов, которые участвуют в росте опухоли, таких как EGFR, ErbB2 (также известный как Her2), ErbB3, ErbB4 или TNF. Предпочтительно антитело против NRP1 по изобретению может быть использовано в комбинации с низкомолекулярными ингибиторами тирозинкиназных рецепторов (RTKI), которые нацелены на один или несколько тирозинкиназных рецепторов, таких как рецепторы VEGF, рецепторы FGF, рецепторы EGF и рецепторы PDGF. Специалистам известны многие терапевтические низкомолекулярные RTKI, включая, но ими не ограничиваясь, ваталаниб (PTK787), эрлотиниб (тарцева, TARCEVA®), OSI-7904, ZD6474 (зактима, ZACTIMA®), ZD6126 (ANG453), ZD1839, сунитиниб (сутент, SUTENT®), семаксаниб (SU5416), AMG706, AG013736, иматиниб (гливек, GLEEVEC®), MLN-518, CEP-701, PKC-412, лапатиниб (GSK572016), VELCADE® (велкейд), AZD2171, сорафениб (нексавар, NEXAVAR®), XL880 и CHIR-265.
Антитело против Nrp по изобретению, взятое отдельно или в комбинации с другим терапевтическим средством (таким как антитело против VEGF), может быть также использовано в комбинации с одним или несколькими химиотерапевтическими средствами. Различные химиотерапевтические средства могут быть использованы в комбинированных методах лечения по изобретению. Примеры и неограничивающий список рассматриваемых химиотерапевтических средств приведены в настоящем описании в разделе «Определения».
При введении антитела против Nrp вместе со вторым терапевтическим средством сначала может быть введено второе терапевтическое средство, а затем антитело против Nrp. Однако также рассматривается вариант, в котором указанные средства вводят одновременно либо антитело против Nrp вводят первым. Подходящими дозами второго терапевтического средства являются дозы, используемые в настоящем изобретении, и такие дозы могут быть снижены благодаря комбинированному (синергическому) действию указанного средства и антитела против Nrp.
Соответствующая доза антитела, используемая для профилактики или лечения заболевания, зависит от типа заболевания, подвергаемого лечению, тяжести и стадии развития заболевания, независимо от того, вводится ли данное антитело в профилактических или терапевтических целях, от типа проводимой ранее терапии, от анамнеза пациента и ответа пациента на данное антитело, а также от назначения лечащего врача. Антитело может быть введено пациенту один раз или через некоторые периоды времени в процессе лечения.
В зависимости от типа и тяжести заболевания, примерно 1 мкг/кг-50 мг/кг (например, 0,1-20 мг/кг) антитела является возможной начальной дозой для введения пациенту, проводимого либо, например, путем одного или нескольких отдельных введений, либо путем непрерывного вливания. Типичная суточная доза может составлять примерно от 1 мкг/кг до 100 мг/кг или более, в зависимости от вышеупомянутых факторов. При повторном введении в течение нескольких дней или дольше, в зависимости от состояния пациента, лечение может быть продолжено до тех пор, пока не будет достигнуто желательное ослабление симптомов заболевания. Однако могут быть использованы другие схемы введения доз. В предпочтительном аспекте изобретения антитело по изобретению вводят каждые две-три недели в дозе, составляющей примерно от 5 мг/кг до 15 мг/кг. Более предпочтительно такая схема введения доз используется в комбинации с химиотерапией в качестве терапии первого ряда для лечения метастазирующего рака прямой и ободочной кишки. В некоторых аспектах изобретения химиотерапия включает традиционное периодическое введение высоких доз. В некоторых других аспектах изобретения химиотерапевтические средства вводят в меньших дозах и более часто, что позволяет проводить терапию без перерывов («метрономная химиотерапия»). Преимущество терапии по изобретению заключается в возможности легкого осуществления ее мониторинга с помощью стандартной техники и анализов.
Композицию антитела получают, дозируют и вводят в соответствии со стандартизированными международными требованиями к качеству лечебного процесса. Факторами, рассматриваемыми в этой связи, являются конкретное расстройство, подвергаемое лечению, конкретное млекопитающее, подвергаемое лечению, клиническое состояние отдельного пациента, этиология конкретного расстройства, участок доставки данного средства, способ введения, схема введения и другие факторы, известные практикующим врачам. «Терапевтически эффективное количество» вводимого антитела назначается исходя из оценки указанных факторов и представляет собой минимальное количество, необходимое для профилактики, ослабления симптомов или лечения заболевания или расстройства. Данное антитело не должно быть обязательно получено вместе с одним или несколькими средствами, используемыми в настоящее время для профилактики или лечения рассматриваемого расстройства. Эффективное количество таких других средств зависит от количества антитела, присутствующего в данной композиции, от типа расстройства или лечения и других обсуждаемых выше факторов. Обычно такое количество вводят в тех же самых дозах и теми же самыми способами введения, которые описаны выше, или в дозах, составляющих примерно от 1 до 99% от вышеупомянутых доз. Как правило, улучшение состояния или лечение заболевания или расстройства включает ослабление одного или нескольких симптомов или патологий, связанных с указанным заболеванием или расстройством. В случае рака, терапевтически эффективное количество лекарственного средства может быть введено отдельно или в комбинации с нижеследующей терапией, направленной на: снижение числа раковых клеток; уменьшение размера опухоли; ингибирование (то есть снижение до некоторой степени и/или прекращение) инфильтрации раковых клеток в периферические органы; ингибирование метастазов опухоли; ингибирование до некоторой степени роста опухоли; и/или ослабление до некоторой степени одного или нескольких симптомов, связанных с раком. Лекарственное средство, в зависимости от того, в какой степени оно способно предупреждать рост раковых клеток и/или уничтожать имеющиеся раковые клетки, может оказывать цитостатическое и/или цитотоксическое действие. В некоторых вариантах изобретения композиция по изобретению может быть использована для профилактики развития или рецидива заболевания или расстройства у индивидуума или млекопитающего.
Нижеследующие примеры приводятся лишь в целях иллюстрации и не должны рассматриваться как ограничение объема изобретения. Содержание всех цитируемых патентов и научных публикаций во всей своей полноте включено в настоящее описание посредством ссылки.
ПРИМЕРЫ
Пример 1
Получение и характеристика антител против Nrp2 B
Антитело против Nrp2B было выделено из синтезированных фаговых библиотек антител человека, как описано ранее (Lee et al., J Mol. Biol. 340, 1073-1093 (2004)). Кратко, синтезированные библиотеки антител для фагового дисплея конструировали на одной каркасной области человека путем искусственного внесения разнообразия в доступные для растворителя положения гипервариабельных областей тяжелой цепи (CDR). Для улучшения свойств библиотеки были сконструированы библиотеки одновалентных и двухвалентных антиген-связывающих фрагментов (Fab), которые анализировали на разнообразие различных CDR-H3 путем изменения аминокислотного состава и длины CDR. Затем эту библиотеку расширяли путем увеличения вариабельности длин CDR-H3 и использования специально сконструированных кодонов, которые имитируют аминокислотный состав природных последовательностей CDR-H3. Используя такие библиотеки с полностью синтезированными CDR, представленными на одном каркасе, получали высокоаффинные антитела. Более подробное описание стратегий и способов получения синтезированных библиотек антител с одной матрицей можно найти, например, в заявке WO 03/102157, опубликованной 11 декабря 2003, содержание которой во всей своей полноте включено в настоящее описание посредством ссылки.
Процедуры отбора анти-Nrp клонов состоят из различных комбинаций сортинга, известного специалистам и проводимого на основе связывания на твердом носителе и в растворе. При проведении сортинга на твердом носителе фаговую библиотеку антител подвергали пэннингу с использованием антигена-мишени, нанесенного на 96-луночный иммунопланшет NUNC Maxisorp в концентрации 5 мкг/мл. В способе сортинга на основе связывания в растворе фаговую библиотеку инкубировали со снижающейся концентрацией биотинилированного антигена в растворе, который затем захватывается нейтравидином, нанесенным на 96-луночный планшет Maxisorp (2-5 мкг/мл). Снижающиеся концентрации позволяют создать более жесткие условия при пэннинге, проводимом в целях поиска более тесно связывающихся агентов.
В результате объединения способов сортинга на твердом носителе и в растворе один клон библиотеки VH (YW68.4) и другой клон VHVL библиотеки (YW126.20) были идентифицированы как NRP-2-связывающие агенты. Для оценки свойств и активностей выбранных новых антител против NRP был проведен ряд in vitro анализов, включая анализы на аффинное связывание (такие как BIAcore) и анализы на блокирование (такие как анализ на индуцированный семафорином коллапс конуса роста и анализы HUVEC).
CDR «необученных» клонов конструировали для повышения их аффинности и стабильности, а затем были получены антитела против Nrp2 YW68.4.2 и YW68.4.2.36. Клетки CHO, экспрессирующие слитый белок mNrp2(a1a2b1b2)-His, hNrp2(a1a2b1b2)-Fc, и клетки насекомых, экспрессирующие hNrp2(b1b2), были использованы для скрининга и характеристики антител. Области VL и VH фагового антитела против Nrp2B (имеющие первоначальное обозначение YW68.4.2.36) клонировали в экспрессионный вектор млекопитающего соответственно. Человеческие антитела против Nrp2B IgG1 или mIgG2a экспрессировали в клетках CHO млекопитающих и очищали на аффинной колонке с белком А.
Аминокислотные последовательности антител против Nrp2B YW68.4.2 и YW68.4.2.36
представлены на фигуре 10. Аминокислотная последовательность антитела против Nrp2B YW126.20 представлена на фигуре 11. Выравнивание последовательности вариабельного домена легкой цепи антитела против Nrp2A YW126.20 с последовательностью κ1 человека показано на фигуре 12. Выравнивание последовательности вариабельного домена тяжелой цепи антитела против Nrp2A YW126.20 с последовательностью III человека (hum III) показано на фигуре 13.
В нижеследующих экспериментах использовали антитело против Nrp2B YW68.4.2.36,
которое далее обозначается «анти-Nrp2B». Это антитело нацелено на домены фактора свертывания крови V/VII (b1-b2) Nrp2 (фигура 1A), поскольку эти домены необходимы для связывания VEGFC с нейропилинами (Karpanen et al., Faseb. J. 20, 1462-1472 (2006)). Кроме того, это антитело связывается с аналогичной аффинностью с мышиным и человеческим Nrp2, но не связывается с Nrp1 (фигура 1B). Было подтверждено, что анти-Nrp2B связывается исключительно с доменами b1-b2 и не связывается с доменами CUB (a1-a2) Nrp2 человека, которые, главным образом, ответственны за связывание с семафорином (Chen et al., Neuron 21, 1283-1290 (1998); Giger et al., Neuron 21, 1079-1092 (1998)).
Для определения аффинностей связывания антител против Nrp2B IgG1 проводили измерения методом поверхностного плазмонного резонанса (SRP) на оборудовании BIAcore™-3000. Для получения приблизительно 200 единиц отклика (RU, ЕО), прежде всего, человеческие анти-Nrp2B IgG иммобилизовали на биосенсорных чипах CM5, покрытых антителом кролика против IgG человека. Для измерения кинетики двукратные серийные разведения мышиного или человеческого Nrp2 (a1a2b1b2) (0,5 нМ-250 нМ) отдельно инъецировали в PBT-буфер (PBS с 0,05% (об./об.) твина 20) при 25°С при скорости потока 30 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) вычисляли с использованием простой лангмюровской модели связывания 1:1 (BIAcore Evaluation Software version 3.2).
Константу равновесной диссоциации (KD) вычисляли как отношение koff/kon.
Анти-Nrp2B связывается с Nrp мыши с Kd 4,9 нМ и с Nrp2 человека с Kd 5,3 нМ, как было оценено путем измерения методом поверхностного плазмонного резонанса.
Способность анти-Nrp2B блокировать связывание VEGFC с Nrp2 тестировали в формате анализа ELISA и в клеточных анализах на связывание.
Для проведения ELISA-анализов на специфичность связывания трехкратные серийные разведения анти-Nrp2B IgG (0,002 нМ-500 нМ) в буфере PBST (в буфере PBT с 0,5% (мас./об.) BSA) инкубировали с 1 мкг/мл антигена, нанесенного на 96-луночные планшеты Maxisorp, по меньшей мере в течение 1 часа, а затем планшеты промывали буфером РВТ. Связанные антитела детектировали с использованием HRP(ПХ)-конъюгированного античеловеческого антитела, разведенного 1:2500 в буфере PBST, после чего проявляли с использованием субстрата TMB приблизительно в течение 5 минут, гасили добавлением 1M H3PO4 и считывали на спектрофотометре при 450 нм.
Для оценки блокирования связывания Nrp2 с VEGF трехкратные серийные разведения анти-Nrp2B IgG сначала инкубировали на 96-луночном планшете Maxisorp, сенсибилизированном NRP2-Fc (5 мкг/мл), в буфере PBST в течение 2 часов, а затем добавляли биотинилированный VEGF165 или VEGFC (полноразмерный) в течение 15 минут. Уровень связывания биотинилированного VEGF с Nrp2 определяли с использованием конъюгатов «стрептавидин-HRP(ПХ)».
Для оценки уровня клеточного связывания осуществляли связывание биотинилированного VEGF165 или VEGFC с LEC как описано ранее (Jia et al., J. Biol. Chem. 281, 13493-13502 (2006)), а затем связывание определяли с использованием конъюгатов «стрептавидин-щелочная фосфатаза». Связывание с Sema3F осуществляли, как описано ранее (Chen et al., 1998, см. выше).
Анти-Nrp2B в значительной степени блокировало связывание VEGFC с Nrp2 (фигура 1C) и с клетками HEK-293, трансфицированными полноразмерным Nrp2. Поскольку Nrp2 может также связываться с VEGF (Gluzman-Poltorak et al., J Biol Chem 275, 29922 (2000)), вероятно, с участием тех же самых доменов, то была также проанализирована способность анти-Nrp2B блокировать связывание VEGF165 с Nrp2. Анти-Nrp2B также в значительной степени блокирует связывание VEGF165 с Nrp2 (фигура 1D) с аналогичной IC50 (0,1 нМ). Однако анти-Nrp2B не обладало способностью блокировать связывание Sema3F с LEC (фигура 1Е), которые в значительной степени экспрессируют Nrp2 (дополнительная фигура 1). Полученные данные соответствуют данным предыдущих наблюдений, которые указывают на то, что домены a1-a2, главным образом, ответственны за связывание с семафорином, а домены b1-b2 ответственны за связывание с VEGF (фигура 1A).
Пример 2
Антитело против Nrp2 B блокирует селективные VEGFC-опосредуемые функции in vitro
Материалы и методы
Клеточные культуры
HMVEC-dLyAd - эндотелиальные клетки лимфатических микрососудов (LEC) кожи человека и HUVEC закупали у Cambrex и культивировали в среде EGM-2 (Cambrex). Клетки C6 LacZ закупали у ATCC. 66С14 были любезно предоставлены доктором Fred R. Miller. Опухолевые клетки культивировали в среде DMEM (Gibco), в которую был добавлен 10% FBS. Все клетки выдерживали при 37°С в 5% СО2 в инкубаторе с 95%-ной влажностью.
Анализ на пролиферацию клеток
96-Луночный планшет с черным прозрачным дном (VWR) сенсибилизировали 5 мкг/мл фибронектина (Invitrogen) при 37°С в течение 2 часов. LEC собирали и ресуспендировали в аналитической среде (0,1% BSA, EGM-2), содержащей 3000 клеток/100 мкл, а затем добавляли в лунки. Клетки инкубировали при 37°С в течение 16 часов. Затем добавляли раствор для BrdU-мечения (набор для анализа ELISA на пролиферацию клеток; Roche) и клетки инкубировали еще 24 часа при 37°С. Включение BrdU определяли с помощью хемилюминесцентного иммуноанализа (6 лунок на одно условие эксперимента).
Результаты
Оценивали роль Nrp2 в VEGFC-опосредуемой миграции и пролиферации. Известны основные клеточные активности, индуцированные VEGFC (Joukov et al., Embo J. 16, 3898-3911 (1997)). Ранее было показано, что LEC являются в высокой степени восприимчивыми к VEGFC (Makinen et al., Embo J. 20, 4762-4773 (2001); Veikkola et al., Faseb 17, 2006-2013 (2003); Whitehurst et al., Lymphat Res Biol 4, 119-142 (2006)). C использованием системы нескольких лунок LEC человека вводили в верхнюю часть камеры, а VEGFC добавляли в нижнюю часть камеры для стимуляции миграции. Затем LEC, которые мигрировали на дно, фиксировали, окрашивали (фигура 2A) и количественно оценивали (фигура 2B). Белок внеклеточного домена VEGFR3 (ECD), содержащий первые три (связывающиеся с лигандом) Ig-домена VEGFR3, использовали в качестве положительного контроля для блокирования VEGFC-опосредуемой миграции в этом и последующем экспериментах (Makinen et al., Nat Med 7, 199-205 (2001)). Чтобы определить, требуется ли Nrp2 для миграции LEC, анти-Nrp2B mAb добавляли в клетки, находящиеся в верхней части камеры, непосредственно перед добавлением VEGFC. Анти-Nrp2B существенно снижало уровень VEGFC-опосредуемой миграции LEC (фигура 2A, 2B; p=0,004). Уровень ингибирования был меньше уровня ингибирования, наблюдаемого в случае ECD VEGFR3, который полностью ингибировал VEGFC-опосредуемую миграцию LEC (фигура 2A, 2B; p<0,001 по сравнению с контролем; p=0,002 по сравнению с анти-Nrp2B). Аналогичные результаты были получены с использованием других восприимчивых к VEGFC первичных клеточных линий, HUVEC.
Поскольку анти-Nrp2B также блокировало связывание VEGF165 c Nrp2, то была оценена роль Nrp2 в модуляции VEGF165-опосредуемой миграции. VEGF165 в значительной степени индуцировал миграцию LEC, как описано ранее (Hirakawa et al., Am J Pathol 162, 575-586 (203); Hong et al., Faseb J 18, 1111-1113 (2004); Makinen et al., Embo J. 20, 4762-4773 (2001); Veikkola et al., Faseb J 17, 2006-2013 (2003)). Перекрестную реактивность антитела против VEGF (B20.4.1) у многих видов использовали в качестве положительного контроля для блокирования такой VEGF-индуцированной миграции (Liang et al., J Biol Chem 281, 951-961 (2006)). Интересно отметить, что анти-Nrp2B не оказывало какого-либо влияния на VEGF165-опосредуемую миграцию (фигура 2C), что, возможно, обусловлено присутствием Nrp1 (дополнительная фигура 1). Блокирование функции Nrp1 под действием анти-Nrp1 mAb, анти-Nrp1B (Pan et al., Cancer Cell 11, 53-67 (2007)) приводит к резкому снижению уровня VEGF-опосредуемой миграции, что подтверждает эту гипотезу (дополнительная фигура 2). Добавление анти-Nrp1B и анти-Nrp2B не приводило к какому-либо дополнительному ингибированию миграции в отличие от ингибирования, наблюдаемого в присутствии одного анти-Nrp1B (дополнительная фигура 2), что указывает на то, что Nrp2 не играет какой-либо роли в VEGF165-опосредуемой миграции.
Затем анализировали действие анти-Nrp2B на VEGFC-индуцированнную пролиферацию LEC. Примечательно, что анти-Nrp2B не влияет на пролиферацию LEC, тогда как ECD VEGFR3 в значительной степени блокирует пролиферацию LEC (дополнительная фигура 3), что соответствует полученным ранее данным, указывающим на то, что киРНК Nrp2 не ингибирует VEGFC-индуцированнную пролиферацию эндотелиальных клеток (Favier et al., Blood 108, 1243-1250 (2006)). Таким образом, очевидно, что Nrp2 играет важную роль в VEGFC-индуцированной миграции, но не пролиферации.
Затем тестировали способность анти-Nrp2B модулировать функцию семафорина. Авторами настоящего изобретения был проведен анализ на коллапс конуса роста нейронов гиппокампа, который ранее показал, что Nrp2 необходим для Sema3F-опосредуемого усечения богатых актином структур (Pozas et al., 2001). Добавление анти-Nrp2B не оказывало какого-либо влияния на индуцированный семафорином коллапс, а добавление либо рекомбинантного домена Nrp2 A1A2, либо ECD Nrp2 приводило к полному ингибированию такого коллапса (дополнительная фигура 4). Полученный результат соответствовал результатам, полученным в проведенных авторами предварительных исследованиях, которые показали, что анти-Nrp2B не связывается с семафорин-связывающей областью и не блокирует связывание Sema3F с Nrp2. Таким образом, анти-Nrp2B блокирует специфические аспекты функции Nrp2, ингибируя VEGFC-ответы, но не VEGF или Sema3F-опосредуемые клеточные ответы.
Пример 3
Антитело против Nrp2 B блокирует VEGFC-опосредуемый лимфангиогенез in vivo
Материалы и методы
Анализ, проводимый в микрокармане роговицы мышей
Взрослых мышей CD-1 (Charles-River) анастезировали и на расстоянии 1 мм от центра роговицы в эпителии создавали карман размером 2×3 мм путем микродиссекции (Polverini et al., Methods Enzymol 198, 440-450 (1991)). Агенты, тестируемые на лимфангиогенную активность, иммобилизовали на инертных гидроновых гранулах (2×2 мм). Затем эти гранулы имплантировали в основание кармана. Животным внутрибрюшинно (i.р.) вводили контрольное антитело (10 мг/кг), анти-Nrp2B (10 мг/кг) или ECD VEGFR3 (25 мг/кг) два раза в неделю в течение 2 недель. Затем животных умерщвляли и иссекали роговицы. Лимфатические узлы визуализировали с помощью полного гистологического анализа (IHC) на предметном стекле с использованием антитела против LYVE-1 (R&D Systems 1:500). Роговицы фотографировали и проводили количественную оценку LYVE-1-положительных лимфатических сосудов лимба.
Анализ на проницаемость кожных сосудов мышей
Спинки и бока взрослых самок мышей C57BL6J выбривали и делили на 4 зоны в области грудных сосков. Затем мышам i.v. инъецировали 150 мкл 0,5% раствора Эванса голубого. Через 1 час после инъекции Эванса голубого мышам чрескожно и произвольно в любую одну из четырех зон инъецировали 20 мкл PBS, содержащего BSA или hVEGF (7,5 мкг/мл), в присутствии или в отсутствие антитела (0,5 мг/мл). Через 1 час животных умерщвляли, а затем кожу иссекали и фотографировали. Образцы кожи для каждой зоны инъекции разрезали и инкубировали в формамидном растворе при 55°С в течение 48 часов для экстракции синего красителя. Затем измеряли оптическую плотность раствора на спектрометре при 600 нм.
Результаты
Принимая во внимание значительное снижение уровня миграции LEC путем блокирования Nrp2 in vitro, проверяли, требуется ли Nrp2 для модуляции функции VEGFC in vivo. Авторами настоящего изобретения были исследованы две хорошо охарактеризованные VEGFC-опосредуемые in vivo активности в отношении лимфангиогенеза у взрослых и сосудистой проницаемости (Cao et al., Circ Res 94, 664-670 (2004); Joukov et al., J Biol Chem 273, 6599-6602 (1998); Kubo et al., Proc Natl Acad Sci USA 99, 8868-8873 (2002); Saaristo et al., Faseb J 16, 1041-1049 (2002)). Для исследования лимфангиогенеза проводили анализ в микрокармане мышиной роговицы (Kubo et al., 2002, см. выше). В этом анализе гранулы VEGFC вводили в несосудистую часть роговицы взрослой мыши. Через 14 дней в ответ на VEGFC наблюдалось разрастание плотного сплетения лимфатических сосудов в роговицу из лимба (фигура 2E; 12000 пикселей2 для VEGFC-обработки по сравнению с 2284 пикселей2 для контроля). Эти сосуды метили LYVE-1 иммуногистохимическим методом (IHC), а затем количественно оценивали (фигура 2D). Системное введение ECD VEGFR3 почти полностью блокировало такой VEGFC-индуцированный лимфангиогенез (2671 пикселей2; p<0,001). Анти-Nrp2B также эффективно блокировало лимфангиогенный ответ в роговице (3281 пикселей2; p<0,001). Такое блокирование было аналогично блокированию, наблюдаемому с использованием ECD VEGFR3 (фигура 2E, 2D; p=0,67).
Для оценки сосудистой проницаемости проводили множество анализов (Brkovic and Sirois, J. Cell Biochem. 100, 727-37 (2007); Eriksson et al., Circulation 107, 532-1538 (2003)). Этот анализ проводили с использованием чрескожной инъекции VEGFC для индуцирования сосудистой проницаемости и внутрисосудистой инъекции красителя Эванса голубого, используемого в качестве индикатора для детектирования и количественной оценки проницаемости кожных сосудов (дополнительная фигура 3). Примечательно, что обработка антителом против Nrp2B не оказывала влияния на VEGFC-индуцированную проницаемость, в отличие от блокирования, наблюдаемого при обработке ECD VEGFR3 (p=0,038). Эти результаты показали, что в соответствии с данными, наблюдаемыми in vitro, Nrp2, очевидно, играет важную роль в селективных VEGFC-опосредуемых функциях in vivo.
Пример 4
Антитело против Nrp2 B модулирует функцию VEGFC путем ингибирования образования комплекса Nrp2/VEGF-рецептор
Материалы и методы
Анализ на миграцию клеток
Анализы на миграцию клеток проводили с использованием модифицированной камеры Бойдена, содержащей встроенную 24-луночную систему Falcon с размером пор 8 мкМ (BD Biosciences). Планшеты сенсибилизировали 5 мкг/мл фибронектина (Invitrogen) в течение 2 часов при 37°С. В верхнюю камеру добавляли клетки в 100 мкл аналитической среды (0,1% BSA, EGM-2) в присутствии/в отсутствие антител. В нижнюю камеру добавляли хемоаттрактант в 500 мкл аналитической среды и клетки инкубировали при 37°С в течение 16 часов. Клетки на верхней мембране удаляли губчатым тампоном, а клетки на нижней поверхности фиксировали в 70% этаноле и окрашивали зеленым красителем Sytox (Molecular Probes). Затем получали изображение всей нижней поверхности лунки и подсчитывали число мигрировавших клеток (6 лунок на одно условие эксперимента).
FACS-анализ
Конфлюэнтные LEC инкубировали с контрольными антителами против Nrp2B (10 мкг/мл) в течение 5 минут, 2 часов или 20 часов при 37°С. Клетки собирали с использованием буфера для диссоциации клеток, не содержащего фермента (Gibco), и инкубировали с биотинилированным антителом в отношении 1:100 в FACS-буфере (PBS, 2% FBS, 2 мМ EDTA, 0,1% азида натрия), содержащем 5% нормальной мышиной сыворотки, 2% нормальной крысиной сыворотки и 10% 10 мкг/мл IgG человека. Антитела биотинилировали с использованием мини-набора FluoReporter, содержащего биотин-хх-белок, для мечения (Molecular Probes). Затем клетки промывали FACS-буфером и окрашивали стрептавидином-ФЭ(PE) (BD Biosciences). Полученные данные анализировали с помощью системы FacsCalibur (BD Biosciences).
Анализ на клеточную адгезию
Субконфлюэнтные LEC предварительно инкубировали в 100 мкл среды 199, содержащей контроль или антитела против Nrp2B (10 мкг/мл), в течение 30 минут при 37°С, а затем их высевали на 96-луночные планшеты NUNC Maxisorp с плоским дном (eBioscience), покрытые 1 мкг/мл фибронектина (Roche) при плотности 10000 клеток на лунку. Планшеты центрифугировали в течение 1 минуты при 140g для синхронизации контакта клеток с субстратом и инкубировали при 37°С в течение 30 минут. Затем планшеты 3 раза промывали PBS и замораживали при -80°С. Плотность клеток определяли с помощью набора CyQuant (Molecular Probes).
Анализы на передачу сигнала VEGF-рецептором
Конфлюэнтные клетки HUVEC в течение 10 минут стимулировали 200 нг/мл VEGFC в присутствии или в отсутствие контрольного антитела или антитела против Nrp2B. Клетки подвергали лизису и анализировали на множество медиаторов, которые, как известно, играют определенную роль при передаче сигнала рецептором VEGF. Активацию VEGFR3 оценивали с помощью ELISA-анализов на общий VEGFR2 и фосфо-VEGFR2 (набор для ELISA, DuoSet IC ELISA kit, R&D). Активацию VEGFR3 оценивали с помощью анализа на активацию киназного рецептора (KIRA) с использованием клеточной линии VEGFR3-293, описанной в литературе (Sadick et al., J Pharm. Biomed Anal 19, 883-891 (1999)). Кратко, стабильные клеточные линии 293, экспрессирующие полноразмерный Flag-меченный hVEGFR3 человека, анализировали на фосфорилирование рецептора после стимуляции. Клетки в количестве 5×104 культивировали на минимальной среде в течение ночи (DMEM с 0,1% BSA), а затем стимулировали 40 нг/мл VEGFA (Genentech South San Francisco, CA) или 200 нг/мл VEGFC (Genentech South San Francisco, CA) в течение 8 минут. Клетки подвергали лизису в PBS, содержащем 1% тритон и ортованадат натрия. ELISA-планшеты покрывали иммобилизованным Flag-антителом (Sigma St Louis, MO). Затем планшеты сенсибилизировали (PBS + 1 мкг/мл антитела) в течение ночи и блокировали (PBS + 0,5% BSA) в течение 1 часа. После 3 промывок (PBS + 0,05% твин 20) добавляли лизаты в течение 2 часов, три раза промывали, а затем добавляли антитело 4G10 для детектирования фосфорилирования (Upstate Lake Placed, NY) в течение 2 часов. Детектирование осуществляли с использованием HRP(ПХ)-конъюгированного антитела (Amersham Piscataway, NJ) и субстрата TMB. Планшеты считывали на 450 нм. Затем проводили оценку на общий AKT, фосфо-AKT, общий Erk1/2, фосфо-Erk1/2, общий Src, фосфо-Src, общий p38 MAPK и фосфо-p38 MAPK с использованием наборов для «сэндвич»-ELISA от BioSource.
Результаты
Тот факт, что анти-Nrp2B подавляет функцию VEGFC, соответствует данным, свидетельствующим о том, что это антитело блокирует связывание VEGFC с Nrp2. Кроме того, в высшей степени неожиданным оказался тот факт, что это антитело блокирует только селективные функции как in vitro, так и in vivo. Одним из возможных объяснений того, почему наблюдается нарушение миграции LEC и лимфангиогенеза, а не пролиферации LEC или сосудистой проницаемости, является то, что анти-Nrp2B может по существу ингибировать миграцию, возможно, за счет нарушения адгезии LEC
к внеклеточному матриксу. Для проверки этого факта оценивали влияние анти-Nrp2B на миграцию LEC, индуцированную различными мотогенами. Анти-Nrp2B не оказывало никакого влияния на миграцию, индуцированную VEGF (фигура 2C), HGF (фигура 3B) или FGF. Поэтому анти-Nrp2B по существу не нарушало миграцию LEC. Кроме того, анти-Nrp2B не оказывало какого-либо действия на адгезию LEC к фибронектину или коллагену, то есть к двум субстратам внеклеточного матрикса.
Вторым возможным объяснением является то, что анти-Nrp2B mAb может вызывать интернализацию Nrp2 (Jaramillo et al., Exp. Cell Res. 312, 2778-2790 (2006)). Поскольку Nrp2 образует комплекс с VEGFR3 даже в отсутствие лиганда (Favier et al., 2006, см. выше; Karkkainen and Alitalo, Semin Cell Dev Biol 13, 9-18 (2002)), то это приводит к селективной совместной интернализации VEGFR3, влияющей на специфические VEGFC-опосредуемые функции. Эта возможность также подтверждается данными, указывающими на то, что VEGFC-индуцированная сосудистая проницаемость опосредуется VEGFR2, а не VEGFR3 (Joukov et al., 1998, см. выше). Для проверки этой возможности LEC инкубировали с анти-Nrp2B при 37°С в течение 5 минут, 2 часов или 20 часов, а затем проводили FACS-анализ с использованием антител против Nrp2, VEGFR2 и VEGFR3 для определения уровня рецепторов на клеточной поверхности. Какого-либо отличия между результатами этих обработок не наблюдалось, что дает основание предположить, что анти-Nrp2B не вызывает значительной интернализации Nrp2, VEGFR2 или VEGFR3 (фигура 3A).
Поскольку было высказано предположение, что Nrp2 усиливает передачу сигнала рецептором VEGF (Favier et al., 2006, см. выше), то было исследовано действие анти-Nrp2B на активацию VEGFR2 и VEGFR3, где стимуляция VEGFC приводит к димеризации и аутофосфорилированию рецептора. В соответствии с полученными ранее данными in vitro и in vivo можно утверждать, что ECD VEGFR3 полностью блокирует VEGFC-опосредуемое фосфорилирование VEGFR2 (фигура 3C; p<0,001) и VEGFR3. Обработка антителом против Nrp2B приводила к снижению активации VEGFR2 (фигура 3C; p<0,001) и VEGFR3, но в значительно меньшей степени, чем обработка ECD VEGFR3 (p<0,001). Это наблюдение повышает вероятность того, что селективная ингибирующая активность анти-Nrp2B может быть результатом различных условий активации рецептора VEGF, необходимых для миграции и пролиферации, причем для миграции требуются более высокие уровни активации рецептора, чем для пролиферации. Для проверки этого предположения оценивали зависимость «доза-ответ» для фосфорилирования VEGFR2, связанного со стимуляцией VEGFC (фигура 3C). При этом абсолютно очевидно, что снижение уровня фосфорилирования VEGFR2, вызываемое обработкой антителом против Nrp2B, было эквивалентно фосфорилированию VEGFR2, достигаемому при стимуляции 175 нг/мл или 150 нг/мл VEGFC. Затем проводили анализ зависимости «доза-ответ» для миграции, связанной со стимуляцией VEGFC (фигура 3D). Было отмечено, что LEC, стимулированные 175 нг/мл или 150 нг/мл VEGFC, мигрировали в такой же степени, как и клетки, стимулированные 200 нг/мл VEGFC. Действительно, значительного снижения уровня миграции не наблюдалось до тех пор, пока уровни VEGFC не снижались до 50 нг/мл. В связи с этим авторы настоящего изобретения утверждают, что снижение уровня фосфорилирования VEGFR2, индуцированного анти-Nrp2B, было само по себе недостаточным для снижения уровня миграции.
Авторами настоящего изобретения была проведена дополнительная оценка влияния анти-Nrp2B на последующие события передачи сигнала, опосредуемой рецепторами VEGF. Обработка антителом против Nrp2B или стимуляция 175 нг/мл или 150 нг/мл VEGFC не приводила к значительному снижению уровня активации Erk1/2, Akt или p38 MAPK, который обеспечивает модуляцию VEGFR2-опосредуемой пролиферации, проницаемости и подвижности соответственно, то есть к снижению, аналогичному тому, которое наблюдается в случае Nrp1 (Pan et al., 2007). Это указывает на то, что Nrp2 может регулировать миграцию LEC и лимфангиогенез по механизму, отличающемуся от механизма усиления активации рецептора VEGF или последующей передачи сигнала этим рецептором.
Наконец, авторами настоящего изобретения было протестировано действие анти-Nrp2B на образование комплекса Nrp2/VEGF-рецептор. Как сообщалось ранее, Nrp2 может быть подвергнут совместной иммунопреципитации VEGFR2 и VEGFR3 в присутствии или в отсутствие VEGFC (Favier et al., 2006, см. выше; Karpanen et al., 2006, см. выше) (фигура 3E). Такое взаимодействие резко ослаблялось под действием анти-Nrp2B (фигура 3E). Полученный результат дает основание предположить, что комплекс Nrp2/VEGF-рецептор играет важную роль в осуществлении специфических VEGFC-опосредуемых функций. Кроме того, роль Nrp2 заключается не только исключительно в усилении передачи сигнала рецептором VEGF в ответ на стимуляцию лиганда.
Пример 5
Nrp2 экспрессируется в лимфатических сосудах, связанных с опухолью
Материалы и методы
Иммуногистохимический анализ
Срезы тканей толщиной 18 мкм разрезали и помещали на предметные стекла. Эти срезы инкубировали в течение ночи с «первым» антителом (анти-NRP2B) (1:500, контрольное окрашивание осуществляли в мышином спинном мозге E12.5, где экспрессия была хорошо охарактеризована), с антителом против LYVE-1 (анти-R&D, 1:200), антителом против PECAM-1 (Benton Dickinson, 1:500), антителом против PROX-1 (Chemicon, 1:1000) или Ki67 (Neovision, 1:100) при 4°С. Затем образцы окрашивали «вторыми» антителами Alexa 488 или Alexa 568 (1:200; Molecular Probes) в течение 4 часов при комнатной температуре. Окрашивание только «вторым» антителом использовали в качестве контроля. TUNNEL-окрашивание осуществляли с помощью коммерчески доступного набора (Roche). Изображения фиксировали на флуоресцентном микроскопе Zeiss Axiophot. Площади кровеносных и лимфатических сосудов определяли по 6 репрезентативным изображениям для каждой из 6 опухолей на группу, которые были оценены на среднее число пикселей с помощью ImageJ.
Результаты
Чтобы определить, играет ли Nrp2 какую-либо роль в биологии лимфатических сосудов у взрослых животных, оценивали экспрессию Nrp2 в лимфатических сосудах у взрослых животных. Что касается сосудистой системы, то экспрессия Nrp2 в венах и лимфатических сосудах была описана ранее (Herzog et al., Mech Dev 109, 115-119 (2001); Moyon et al., Development 128, 3359-3370 (2001); Yuan et al., Development 129, 4797-4806 (2002)). Как описано выше, LEC в культуре экспрессируют Nrp2 на высоком уровне (дополнительная фигура 1). Однако авторам настоящего изобретения не удалось детектировать Nrp2 с помощью IHC в LYVE-I-положительных лимфатических сосудах толстой кишки здоровых взрослых мышей (фигура 4A; окрашивание положительного контроля см. на дополнительной фигуре 4). Проводили оценку толстой кишки, так как она богата сплетениями лимфатических сосудов с достаточно стереотипированной картиной. Авторам настоящего изобретения также не удалось детектировать экспрессию Nrp2 с помощью IHC в лимфатических сосудах лимфоузлов (фигура 4B) и в коже здоровых взрослых мышей. Эти результаты были подтверждены гибридизацией Nrp2 in situ (ISH). В противоположность этому, высокий уровень экспрессии Nrp2 наблюдался в LYVE-1-положительных лимфатических сосудах лимфоузлов, расположенных рядом с ортотопически или гетеротопически трансплантированными опухолями (фигура 4C). Такой уровень наблюдался для ряда опухолевых линий, включая ортотопически трансплантированную линию аденокарциномы молочной железы мыши, 66cl4 (Aslakson and Miller, Cancer Res 52, 1399-1405 (1992)), и гетеротопически трансплантированную линию глиобластомы крысы, C6 (данные не приводятся). Nrp2 также наблюдался в околоопухолевых и во внутриопухолевых лимфатических сосудах (фигура 4D) для ряда опухолевых линий, включая 66c14, C6 и PC3 (линию карциномы предстательной железы человека). Такая экспрессия подтверждалась ISH-анализом, проводимым для субпопуляции опухолей различных типов.
Пример 6
Антитело против Nrp2 B снижает число метастазов в легких у множества моделей с опухолью
Материалы и методы
Все исследования на животных проводили в соответствии с руководством по уходу за лабораторными животными и их использованию, опубликованным Национальным институтом здравоохранения (NIH Publication 85-23, revised 1985). Все протоколы по исследованиям на животных были одобрены Институциональным комитетом по уходу за животными и их использованию (IACUC).
Модели опухолей
В случае использования клеток 66C14, эти клетки собирали путем трипсинизации, промывали и ресуспендировали в PBS при концентрации 2×105 клеток в 10 мкл PBS. Мышей анестезировали с использованием 75 мг/кг кетамина и 7,5 мг/кг ксилазина и делали разрез под правой передней конечностью. Клетки в концентрации 2×105 клеток в 10 мкл PBS инъецировали непосредственно в доступное 4-е жировое тело молочной железы 6-8-недельных самок мышей balb-C. В случае использования клеток C6, 2×106 опухолевых клеток в 100 мкл PBS инъецировали подкожно в правый бок 6-8-недельных самок «голых» мышей balb-C. На обеих стадиях исследований проводили мониторинг роста опухоли 3 раза в неделю. Когда опухоль достигала среднего размера 80-120 мм3, мышей разделяли по среднему размеру опухоли на почти идентичные группы и начинали обработку. В каждом исследовании такая обработка рассматривалась как день 1. Животных обрабатывали контрольным антителом (10 мг/кг), анти-Nrp2B (10 мг/кг) или ECD VEGFR3 (25 мг/кг) i.р. два раза в неделю до завершения исследований. Для гарантии воспроизводимости все исследования повторяли 3 раза.
По окончании исследований животных анестезировали, а затем им вливали 4% PFA. Опухоли собирали, содержали в условиях криопротекции и замораживали в OCT (Tissue-Tek). Легкие раздували посредством правовентрикулярной перфузии 10 мл PBS,
а затем 4% PFA и перед проведением микро-КТ-анализа осуществляли визуальный подсчет метастатических поражений.
Метастазы в SLN оценивали путем гетеротопической имплантации опухолевых клеток C6 в ухо. Кратко, группу животных предварительно обрабатывали контрольным антителом (10 мг/кг) или анти-Nrp2B (10 мг/кг) за один день до имплантации опухолевых клеток и два раза в неделю после такой имплантации. Клетки C6 lacZ в количестве 1×105 подкожно инъецировали в ухо 70 самок «голых» мышей balb/c. Мышей умерщвляли на дни 3, 6, 9, 13 и 15, а затем «сторожевые» лимфоузлы идентифицировали с помощью подкожной лимфангиографии и иссекали. Лимфоузлы гомогенизировали и лизат анализировали на β-галактозидазную активность (Pierce).
Внутрикожную лимфангиографию осуществляли на контрольных мышах и мышах с опухолью следующим образом. Краситель Эванс голубой (2 мкл, 3% масс.), содержащий 1% 20 нм полистирольных флуоресцентных микрогранул (Molecular Probes), инъецировали чрескожно в верхушку опухоли. Животных оставляли на 2 часа для восстановления, а затем умерщвляли. Опухоли осторожно иссекали так, чтобы они не включали околоопухолевую ткань. Опухоли либо фиксировали, а затем подвергали гистохимическому анализу, либо инкубировали в формамиде для экстракции Эванса голубого и количественно оценивали путем измерения оптической плотности OD600 на спектрофотометре.
Микро-KT-анализ легких
Легкие погружали в 10% NBF на 24 часа, а затем на 24 часа погружали в 20%-ный раствор содержащего йод контрастного вещества для рентгеновской компьютерной томографии, Isovue370 (Bracco Diagnostics Inc, Princeton, NJ), разведенный в PBS. Затем легкие погружали и подвергали перфузии через трахеотомическую трубку 20 мл соевого масла (Sigma-Aldrich, St. Louis, MO) при скорости 0,25 мл/мин. Соевое масло использовали для удаления избытка контрастного вещества и получения фоновой среды для изображений.
Легкие мышей визуализировали ex vivo с помощью системы рентгеновской микрокомпьютерной томографии (микро-КТ) VivaCT (SCANCO Medical, Basserdorf, Switzerland). Сагиттальное обзорное изображение, сравнимое со стандартным плоским рентгеновским изображением, получали в целях определения начальных и конечных точек для аксиальной комплектации серии микро-КТ-изображений срезов. Положение и число аксиальных изображений выбирали с учетом полного охвата легких. Легкие погружали в соевое масло, используемое в качестве фоновой среды. Микро-КТ-изображения создавали с помощью рентгеновской трубки, работающей в следующем режиме: энергетический уровень 45 кВ, ток 160 мкА и время интеграции 450 миллисекунд. Аксиальные изображения получали при изотропном разрешении 21 мкм. Оценку опухолей легких (число и объем) проводили с помощью алгоритма полуавтоматизированного анализа изображений, который включает стадию осмотра с помощью специально сконструированного ридера. Опухоли легких выглядели как гиперинтенсивная твердая масса в отличие от пористой ситообразной структуры нормальных легких. Это обусловлено абсорбцией содержащего йод контрастного вещества твердыми структурами (стенками бронхов и альвеол, опухолями, трахеями и средостением), имеющимися в легких. Избыток контрастного вещества удаляли из заполненного воздухом пространства на стадии перфузии маслом. Потенциальную опухолевую массу экстрагировали путем проведения серии стадий обработки изображений. Программа для анализа изображений была разработана лабораторией. Эта программа была записана на языке программирования C++, и в ней использовались команды библиотеки функций программы для анализа изображений Analyze (AnalyzeDirect Inc., Lenexa, KS, USA). В этом алгоритме использовались следующие параметры: порог интенсивности, морфологическая фильтрация и образование опухолевых областей для экстракции всех потенциальных опухолевых масс. Порог интенсивности (1480 единиц Хаунсфилда) был определен с помощью анализа гистограмм 5 произвольно выбранных легких, используемых для разработки алгоритма, а оптимальный порог был выбран так, чтобы он включал элементы объемного изображения опухоли и исключал какой-либо фоновый сигнал. Морфологические параметры (эрозия, дилатация) и образование опухолевых областей были применимы для соединения элементов объемного изображения гиперинтенсивных областей и для удаления любых элементов объемного изображения с аналогичной плотностью, наблюдаемой в тонких стенках бронхиол и альвеол. Стадия образования опухолевых областей требует минимального объема из 2300 связанных элементов объемного изображения (более чем 0,0231 мм3), принимаемых за объект (массу). Идентифицированные объекты затем оценивали на специально сконструированном ридере с помощью программы для визуализации Analyze 3D. Отдельные объекты принимали или отбрасывали как возможные опухоли по их внешнему виду и локализации в легких. Объекты отбрасывали, если они находились за пределами легких (например, в средостении, в дебрисе других тканей) или были похожи на кровеносные сосуды. Для каждого легкого подсчитывали число опухолей, объем отдельной опухоли и общий объем опухолей. Такой аналитический метод подтверждался хорошо разработанной моделью метастазов опухоли. Одиннадцать животных с ортотопической трансплантацией опухолевых клеток аденокарциномы молочной железы 4T1 исследовали на метастазы в легких с помощью такого микро-КТ-метода с последующим серийным гистологическим анализом легких. Величины объема опухолей легких в высокой степени коррелировали (r=0,9, p=0,0002) с гистологической оценкой размера опухоли (подсчет пикселей; дополнительная фигура 5).
Результаты
Одним из главных способов ингибирования метастазов является ингибирование VEGFC всех типов (Chen et al., Cancer Res 65, 9004-9011 (2005); He et al., J Natl Cancer Inst 94, 819-825 (2002); Krishnan et al., Cancer Res 63, 713-722 (2003); Lin et al., Cancer Res 65, 6901-6909 (2005)). Чтобы определить, может ли блокирование функции Nrp2 также модулировать развитие метастазов, была проведена оценка влияния обработки антителом против Nrp2B на образование метастазов в легких у двух различных моделей с опухолями - модели 66c14, модели рака молочной железы и модели глиобластомы C6. 66cl4 представляет собой линию аденокарциномы молочной железы мыши, происходящую от спонтанно развивающейся опухоли молочной железы у мышей Balb/c (Aslakson and Miller, Cancer Res 52, 1399-1405 (1992)). Эти клетки экспрессируют VEGFC и образуют метастазы в легких под действием лимфатической системы (Aslakson and Miller, 1992, см. выше). Ортотопическая трансплантация этих клеток у мышей Balb/c приводила к репродуцируемому развитию опухолей и метастазов в легких. Обработка антителом против Nrp2B не оказывала влияния на скорость роста первичной опухоли (фигура 5A). Поскольку ECD VEGFR3 не вызывал резкого снижения скорости роста первичной опухоли, то он был исключен из любого последующего анализа метастазов. Группы животных (N=6 в каждой группе) с аналогичными размерами опухолей в обеих группах обработки одновременно умерщвляли, а затем легкие иссекали и раздували для облегчения анализа на метастатические узлы. Анти-Nrp2B вызывало резкое снижение среднего числа визуально детектируемых метастатических узлов на одно легкое по сравнению с числом метастатических узлов у животных, обработанных контрольным IgG (фигура 5B, C), составляющим в среднем 3,5-0,8 (P=0,03). Для подтверждения этого результата и использования этих способов для оценки метастазов в паренхиме легких, которая не была доступна для визуальной оценки, авторами настоящего изобретения был проведен микро-КТ-анализ (Li et al., Technol Cancer Res Treat 5, 147-155 (2006)) легких после аутопсии. Этот анализ подтвердил, что у животных, обработанных анти-Nrp2B, по сравнению с контрольными животными, наблюдалось уменьшение числа метастазов в легких (фигура 5D-F). Однако микро-КТ-анализ был более чувствительным, чем визуальный анализ, и позволял выявить большее абсолютное число метастатических узлов у обеих групп. Микро-КТ-анализ позволил авторам изобретения определить общую метастатическую нагрузку в легких. Обработка антителом против Nrp2B также приводила к сокращению общего объема метастазов (0,74 см3) по сравнению с обработкой контролем (1,78 см3). Кроме того, этот анализ подтвердил, что подавляющее большинство поражений присутствовало на поверхности легких как у контрольных животных, так и у животных, обработанных антителом против Nrp2B. Поэтому анти-Nrp2B не вызывало снижения числа метастазов, обусловленного перемещением узлов с поверхности в паренхиму легких.
FACS-анализ, осуществляемый, как описано в примере 4, показал, что на опухолевых клетках 66cl4 экспрессировался Nrp2, но не VEGFR2 или VEGFR3. Это повышает вероятность того, что обработка антителом против Nrp2B оказывает прямое воздействие на поведение опухолевых клеток, приводящее к образованию метастазов. Этот факт является маловероятным, если учесть отсутствие влияния такой обработки антителом против Nrp2B на рост первичной опухоли. Кроме того, анти-Nrp2B не оказывало какого-либо влияния на пролиферацию, апоптоз или миграцию опухолевых клеток in vitro. Однако, учитывая вероятность того, что снижение числа метастазов обусловлено влиянием анти-Nrp2B на опухолевые клетки, авторами настоящего изобретения была проведена оценка влияния анти-Nrp2B на модель глиобластомы С6 крысы. Эти клетки не экспрессировали Nrp2 на своей поверхности до какой-либо значительной степени (фигура 6E), но экспрессировали VEGFC и, вероятно, образовывали метастазы в легких через лимфатическую систему (Bernstein and Woodard, 1995). Кроме того, эти клетки были получены так, чтобы они экспрессировали β-галактозидазу, которая может быть использована в качестве маркера, облегчающего детектирование опухолевых клеток.
Подкожная трансплантация этих клеток «голым» мышам приводила к постоянному развитию опухолей и метастазов в легких. Обработка антителом против Nrp2B не влияла на скорость роста этих первичных опухолей (фигура 6A). Кроме того, ECD VEGFR3 не давал резкого снижения скорости роста первичной опухоли на этой модели опухоли. Это позволяет проводить сравнения антиметастатических эффектов ECD VEGFR3 и анти-Nrp2. И в этом случае группу животных (N=10) с аналогичными размерами опухолей во всех группах обработки умерщвляли, легкие иссекали и раздували для облегчения анализа на метастатические узлы. Обработка антителом против Nrp2B и ECD VEGFR3 вызывала уменьшение среднего числа визуально детектируемых метастатических узлов на одно легкое (фигура 6B, C). Снижение, наблюдаемое с использованием анти-Nrp2B, было сравнимо со снижением, наблюдаемым с использованием ECD VEGFR3. Микро-КТ-анализ легких подтвердил эти данные (фигура 6D), а также подтвердил, что подавляющее большинство метастазов локализуется на поверхности легких у всех групп обработки. Как показал гистологический анализ обеих моделей опухоли, узлы представляли собой метастатические поражения (фигуры 5H и 6G). Кроме того, общая аутопсия не выявила каких-либо узлов на поверхности других органов у любой модели с опухолью.
Хотя приведенное выше описание настоящего изобретения проиллюстрировано со ссылками на конкретные варианты его осуществления, однако настоящее изобретение не ограничивается этими вариантами. Действительно, различные модификации настоящего изобретения, помимо тех, которые указаны и описаны в настоящей заявке, будут очевидны для специалиста в данной области исходя из приведенного выше описания и входят в объем прилагаемой формулы изобретения.
Все публикации, цитируемые в настоящем описании, и работы, цитируемые в этих публикациях, точно и во всей своей полноте включены в настоящее описание посредством ссылки.









| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ panNrp (ВАРИАНТЫ) | 2007 |
|
RU2454427C2 |
| Способ прогнозирования рецидива у больных базальноклеточным раком кожи после хирургического удаления опухоли | 2024 |
|
RU2823211C1 |
| АКТИВНАЯ ИММУНОТЕРАПИЯ ПРОТИВ АНГИОГЕНЕЗА | 2003 |
|
RU2329824C2 |
| АНТАГОНИСТЫ ALK1 И ИХ ПРИМЕНЕНИЕ В ЛЕЧЕНИИ ПОЧЕЧНО-КЛЕТОЧНОЙ КАРЦИНОМЫ | 2013 |
|
RU2633638C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА СЛИТЫЙ БЕЛОК, В КОТОРОМ СЛИТЫ ПРОНИКАЮЩИЙ В ОПУХОЛЬ ПЕПТИД И АНТИАНГИОГЕННОЕ СРЕДСТВО, ДЛЯ ПРЕДУПРЕЖДЕНИЯ И ЛЕЧЕНИЯ РАКА ИЛИ СВЯЗАННЫХ С АНГИОГЕНЕЗОМ ЗАБОЛЕВАНИЙ | 2016 |
|
RU2727238C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2009 |
|
RU2547596C2 |
| ИНГИБИРОВАНИЕ МЕТАСТАЗИРОВАНИЯ ОПУХОЛИ ПРИ ПОМОЩИ АНТАГОНИСТОВ Bv8 ИЛИ G-CSF | 2010 |
|
RU2567803C2 |
| КОМПОЗИЦИИ ГУМАНИЗИРОВАННЫХ СЛИТЫХ БЕЛКОВ NOTCH И СПОСОБЫ ЛЕЧЕНИЯ | 2008 |
|
RU2532830C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ОПУХОЛЕЙ И МЕТАСТАЗОВ С ИСПОЛЬЗОВАНИЕМ КОМБИНАЦИИ АНТИАНГИОГЕННОЙ ТЕРАПИИ И ИММУНОТЕРАПИИ | 2000 |
|
RU2236251C2 |
| АНТИТЕЛА ПРОТИВ DLL4 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2415869C2 |
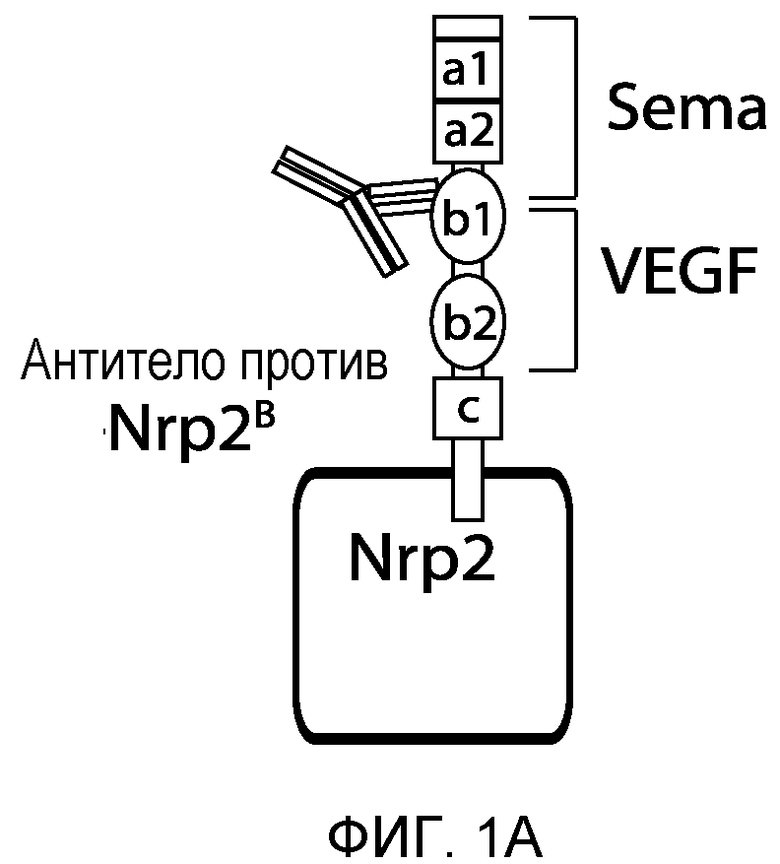


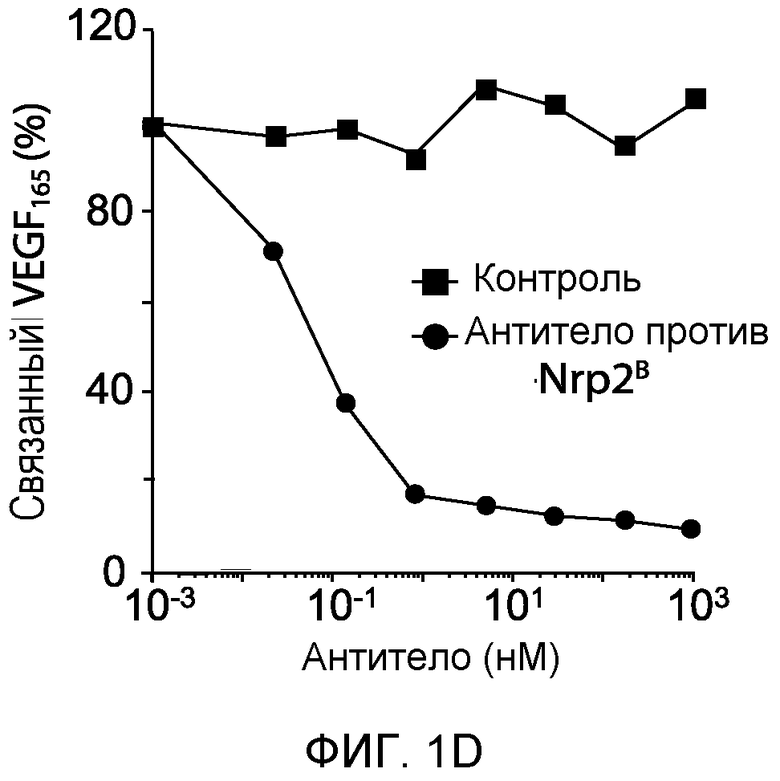
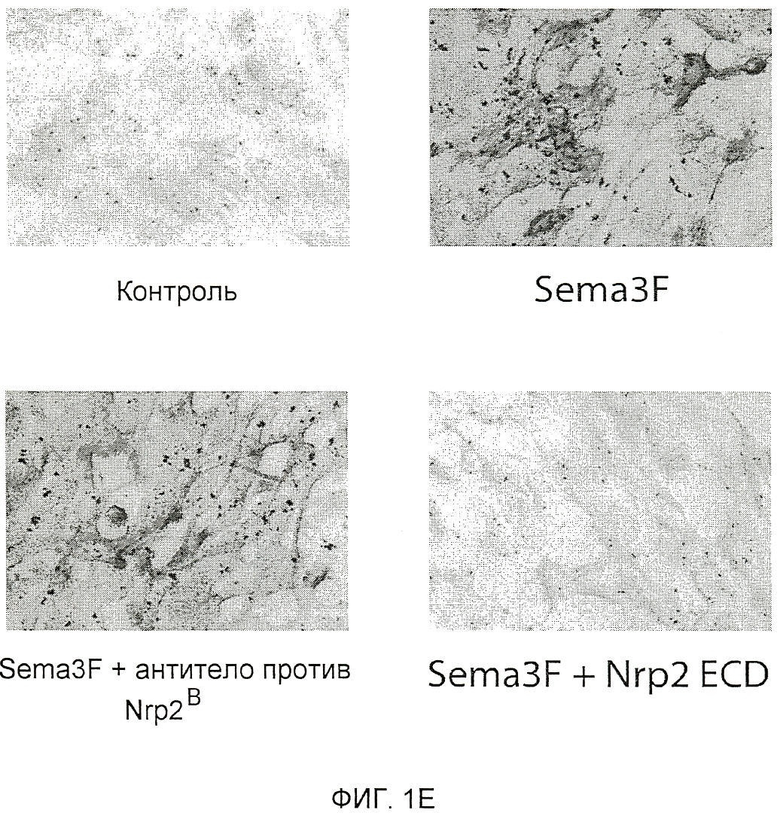
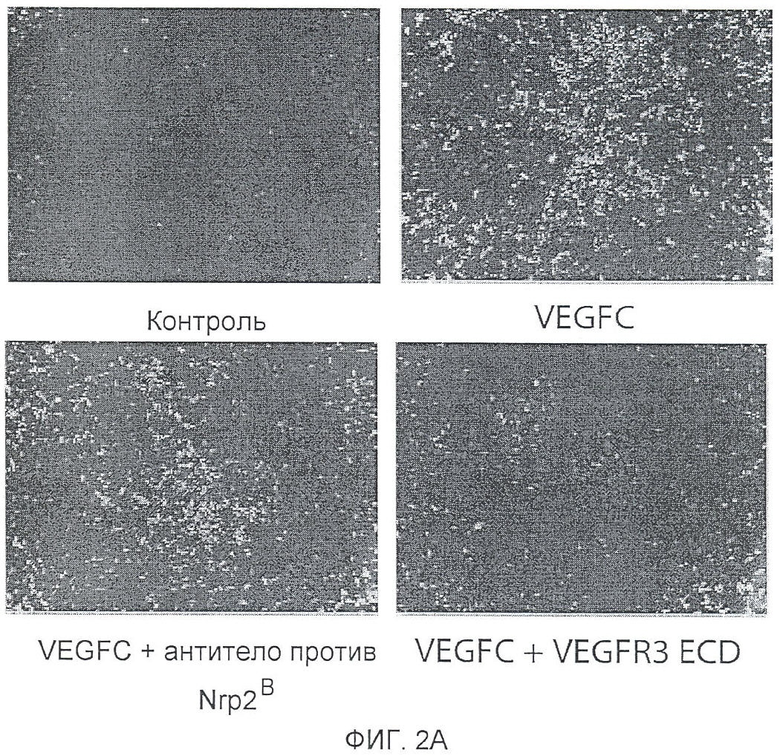
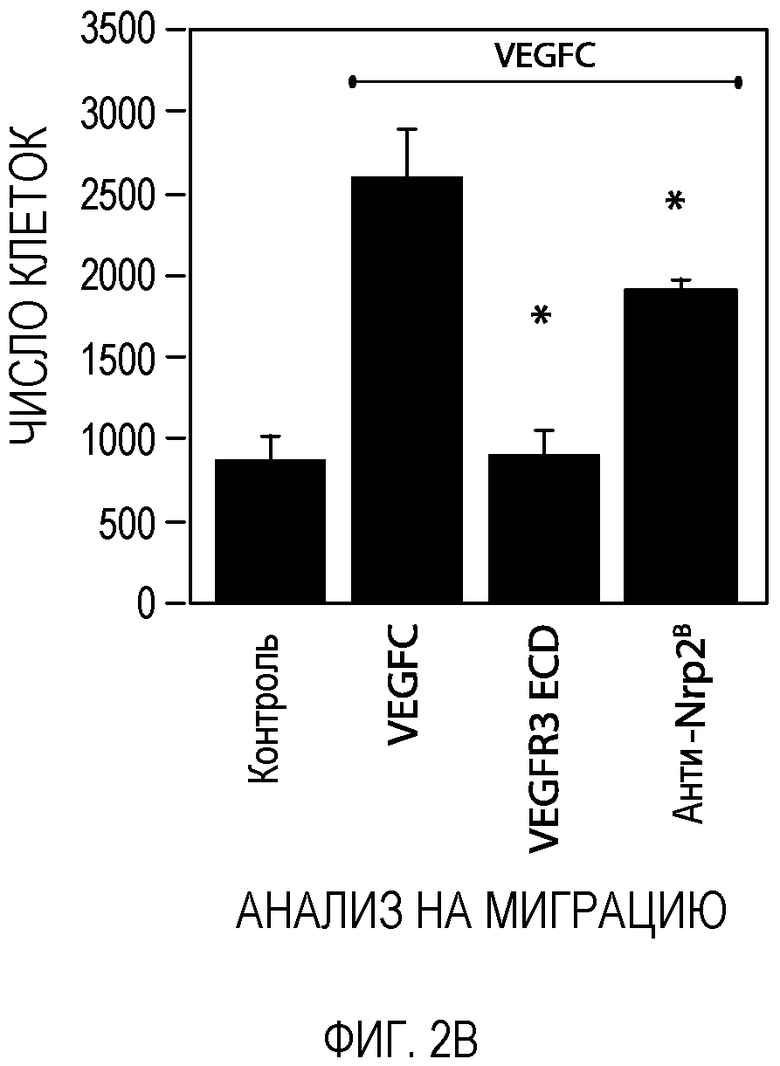

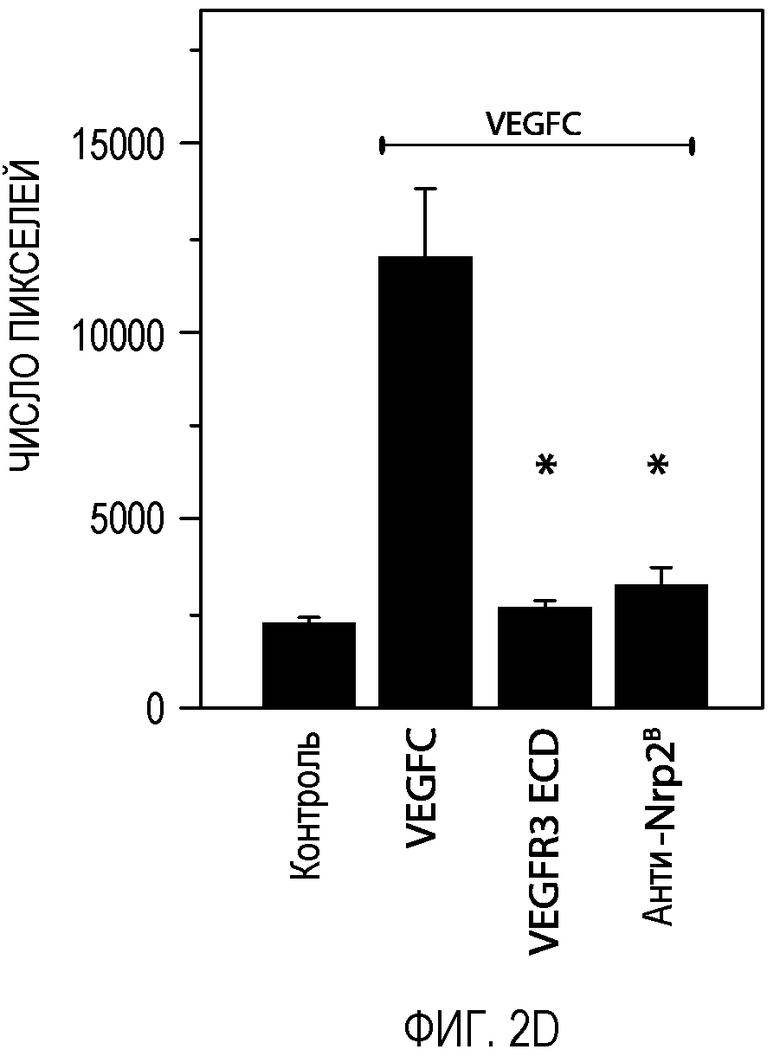
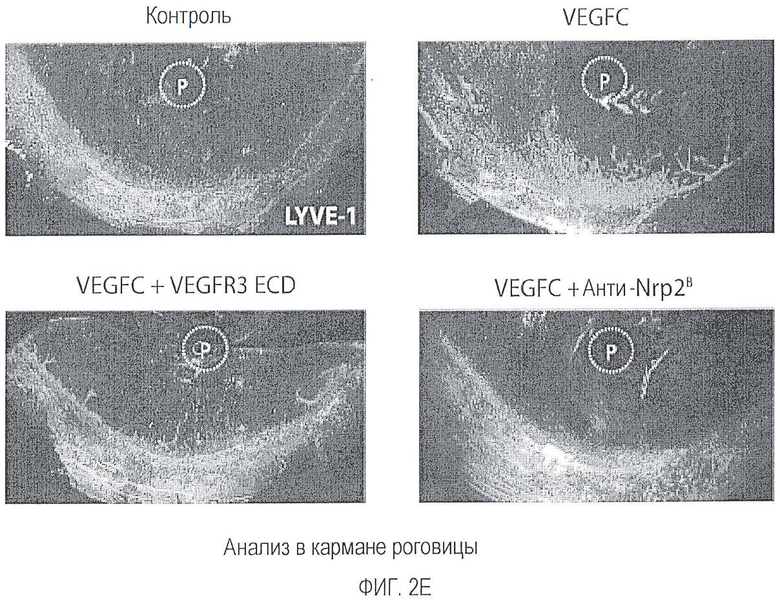
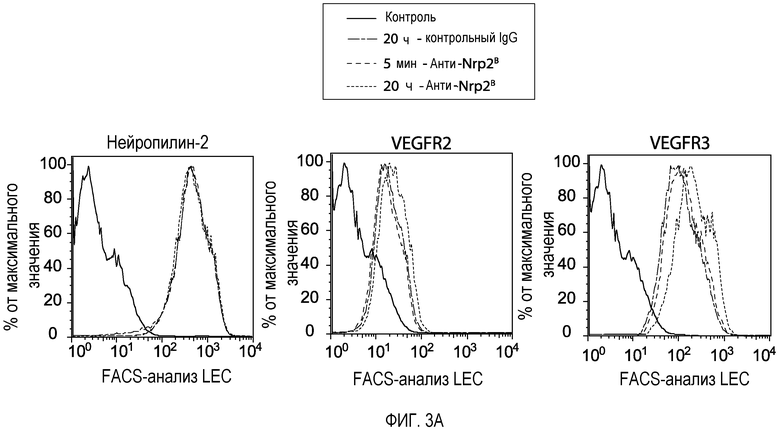







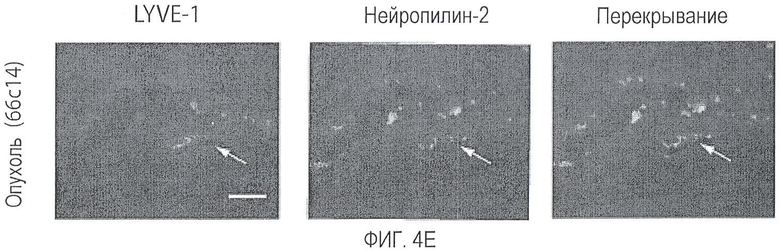
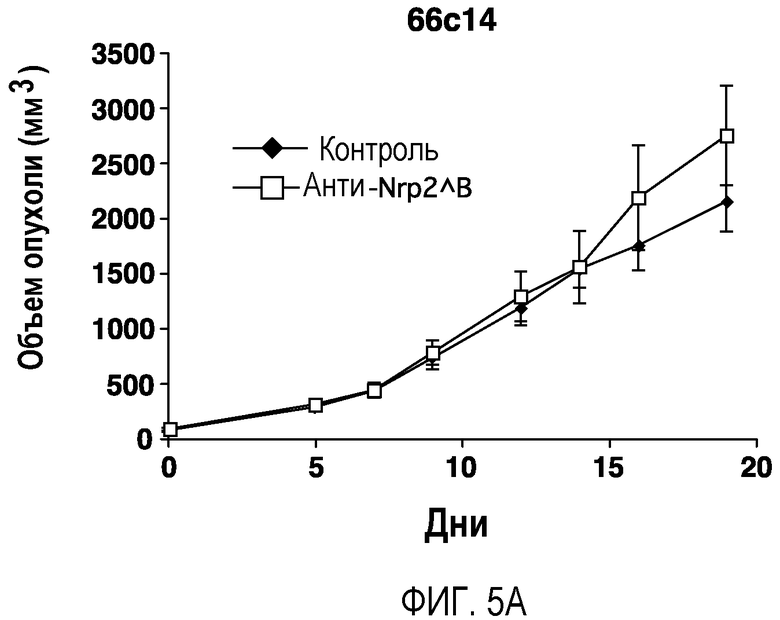
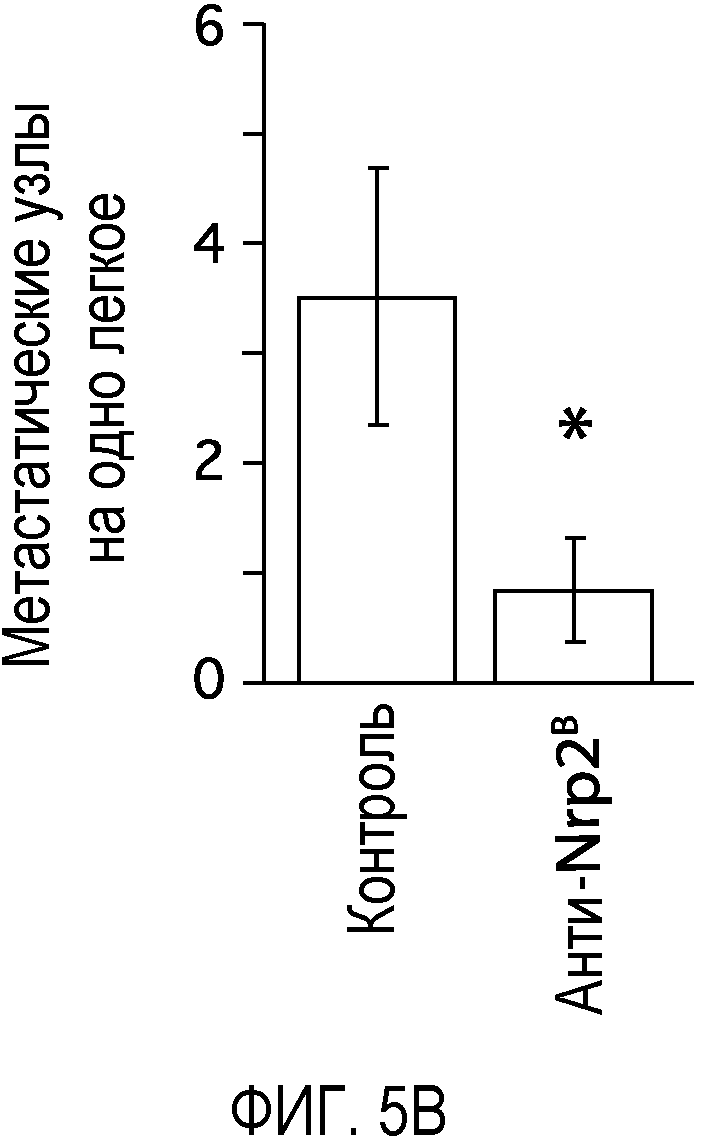
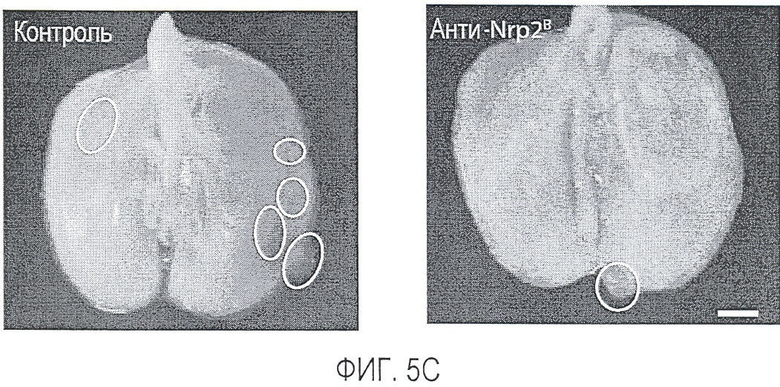
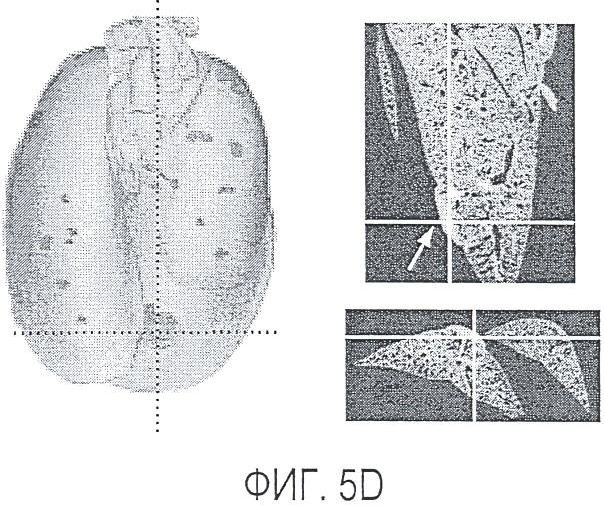
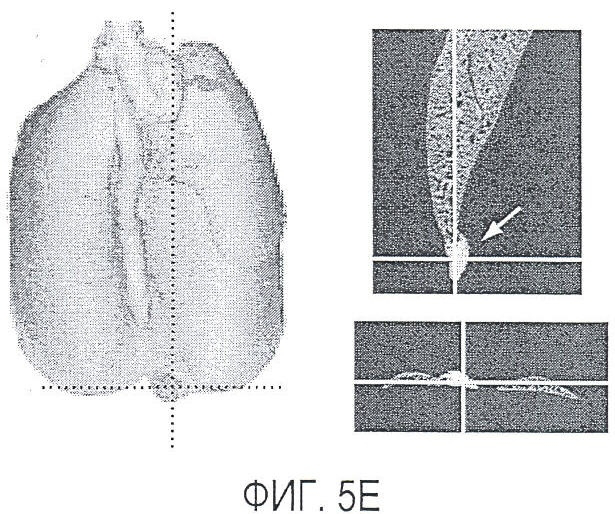


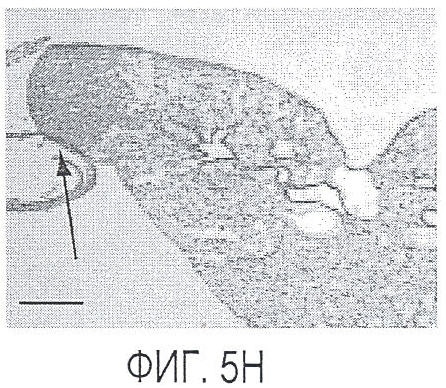
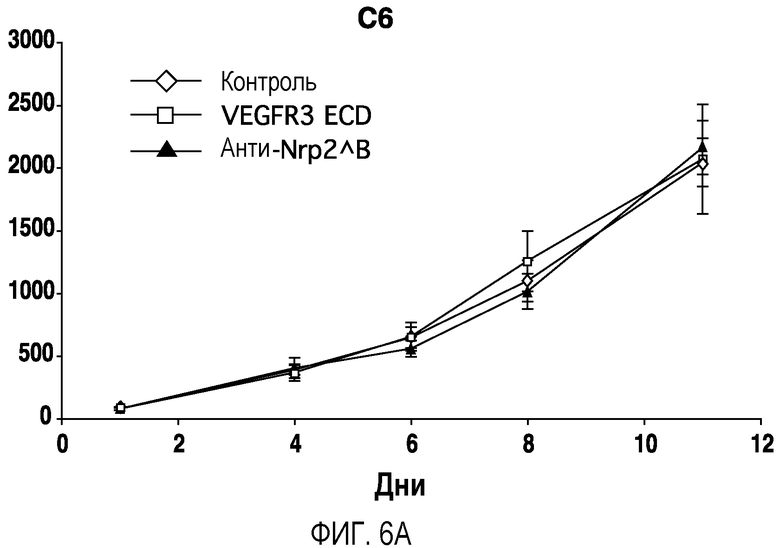
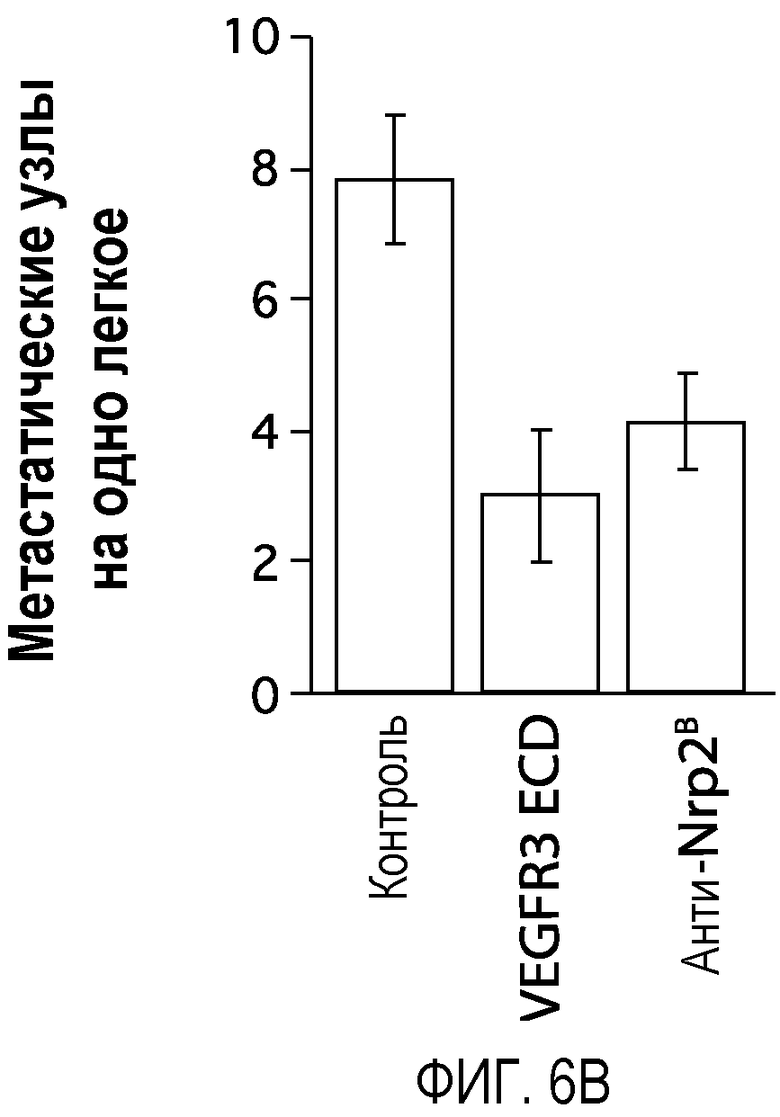
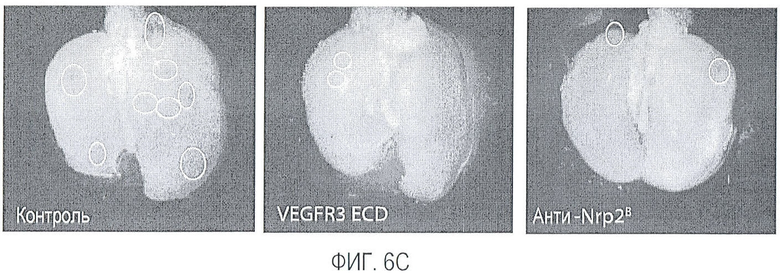
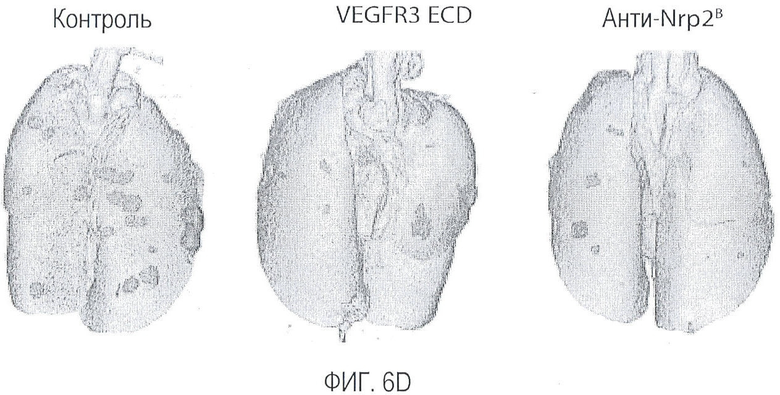
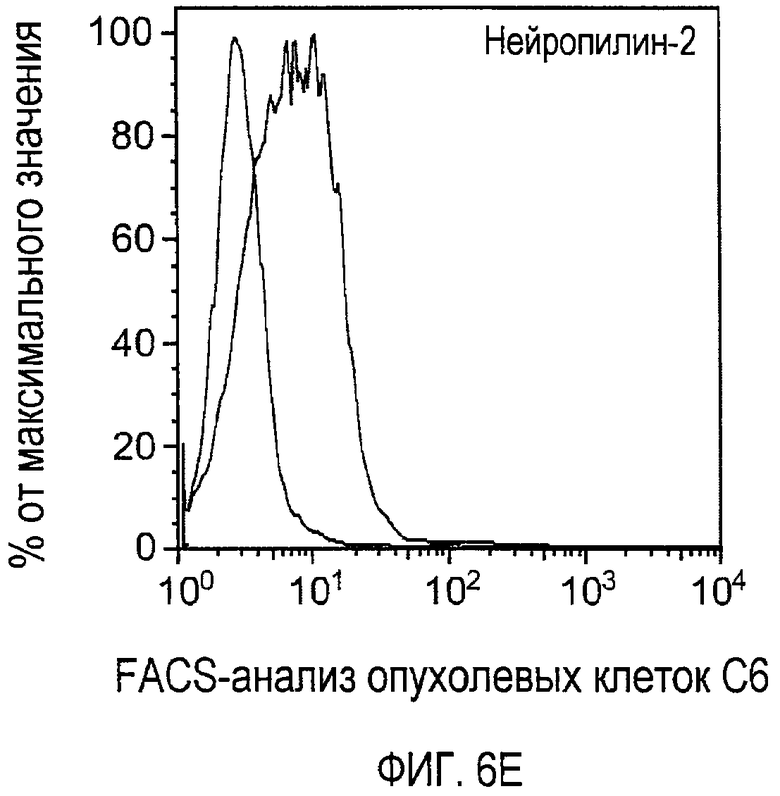

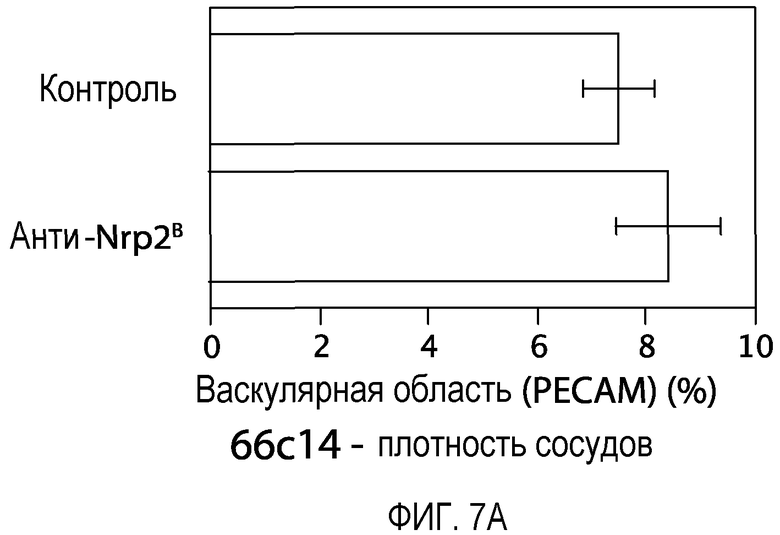

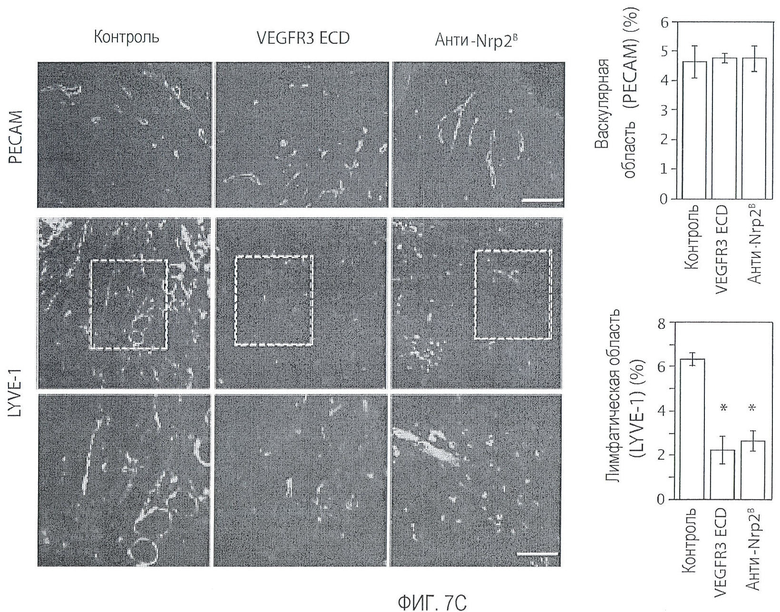
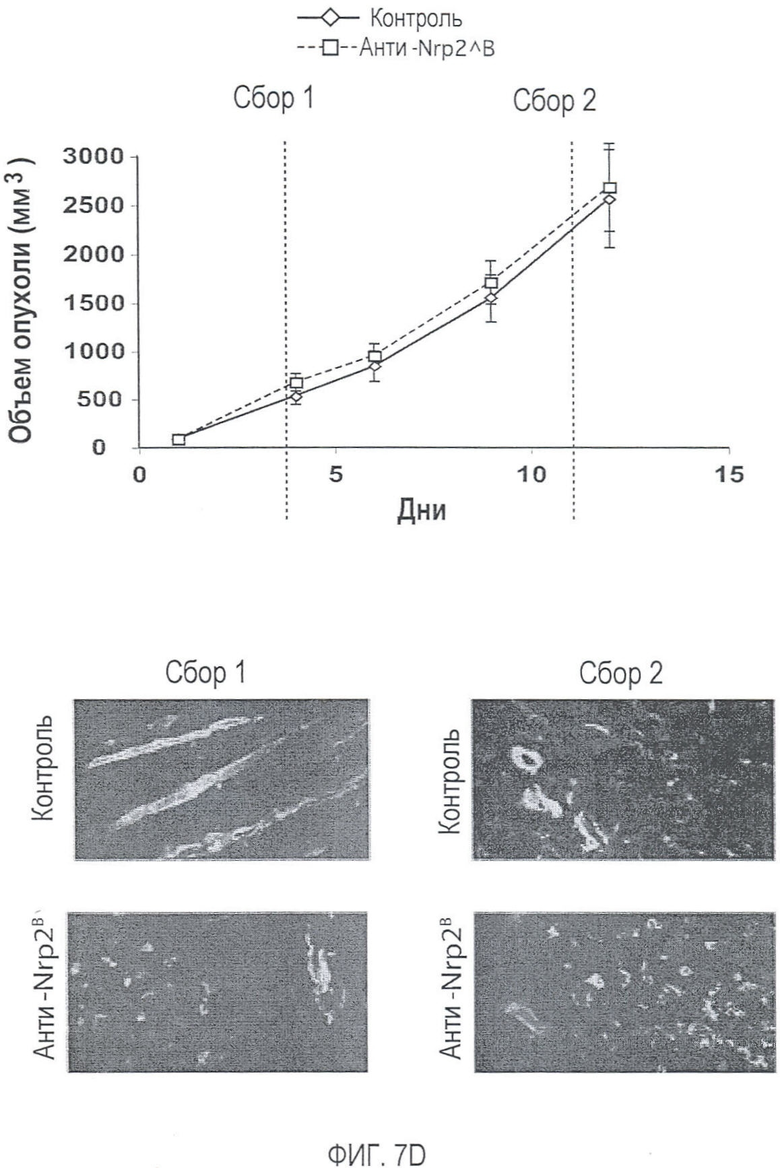


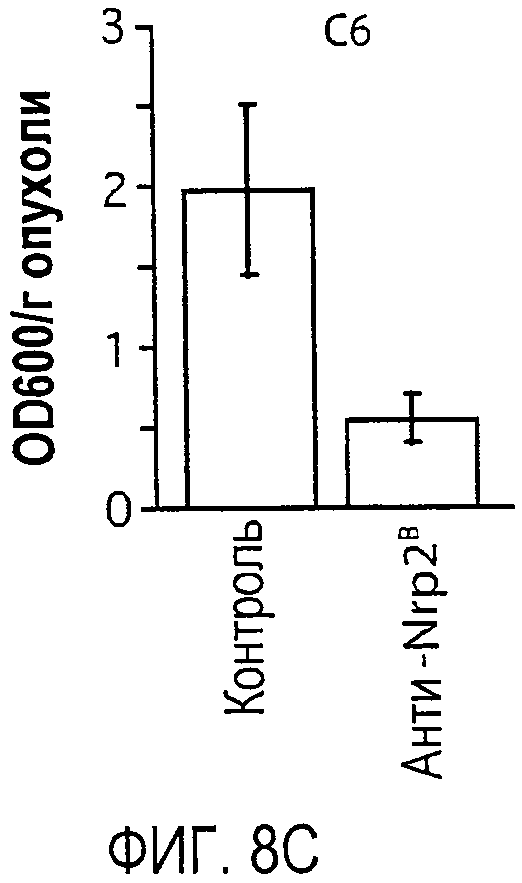


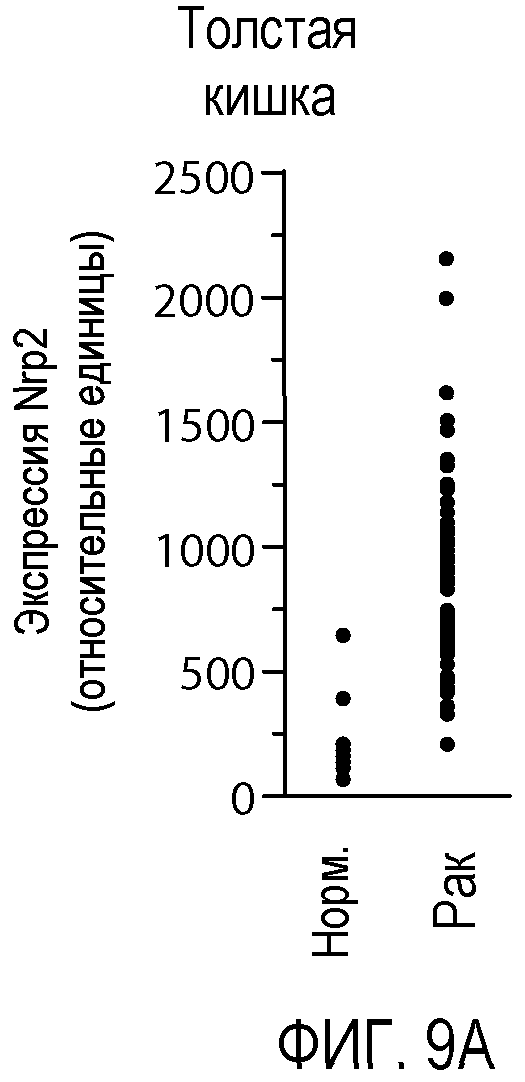
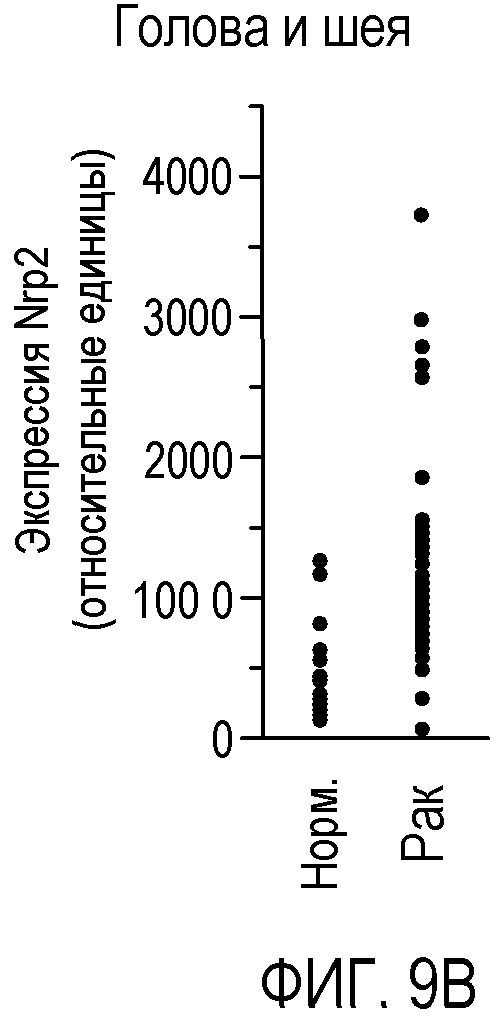
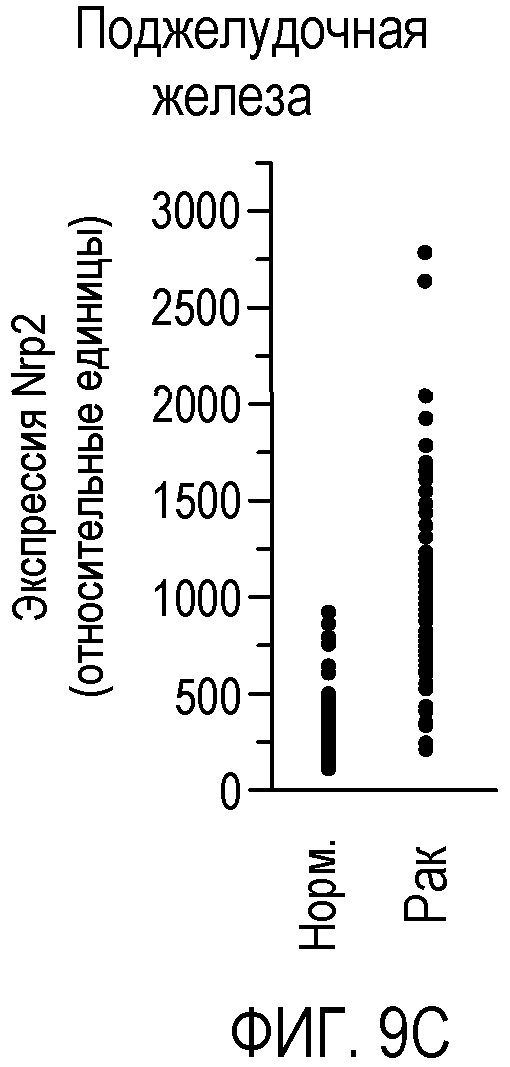
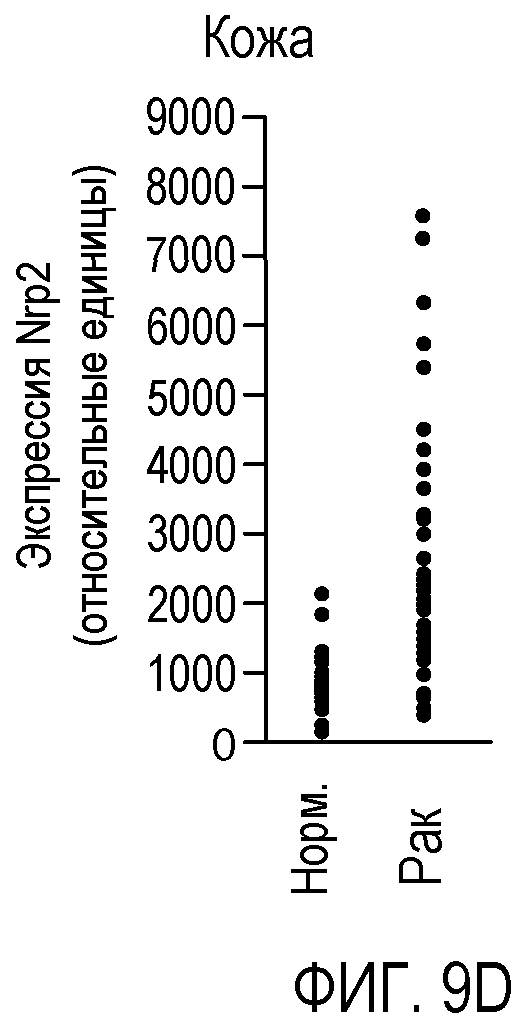

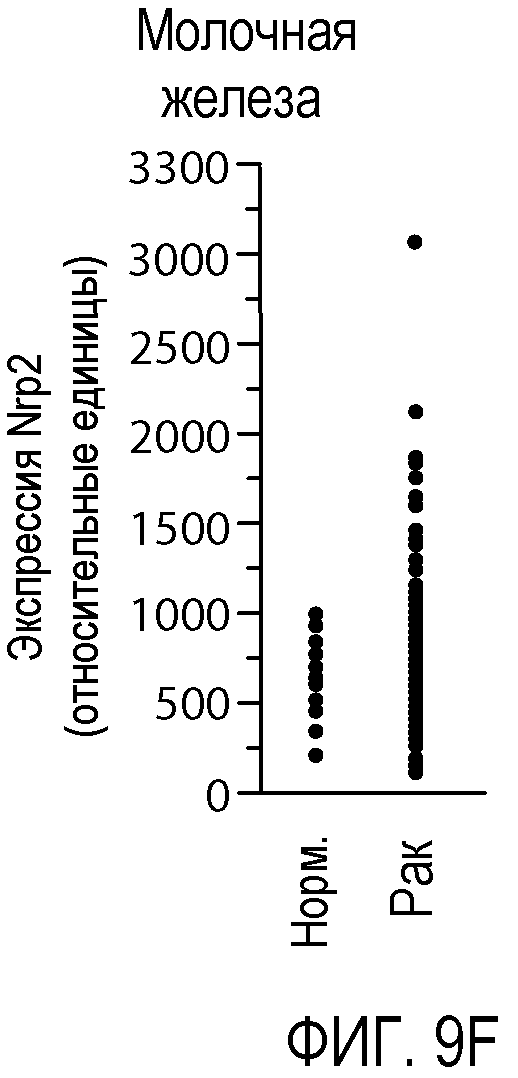


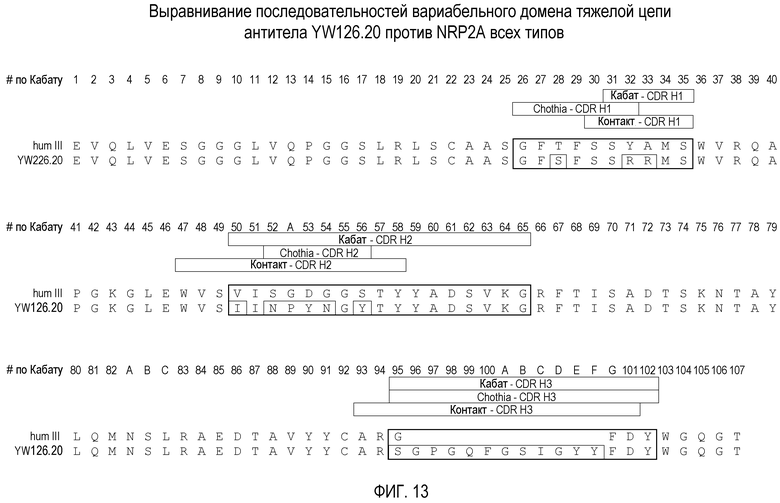






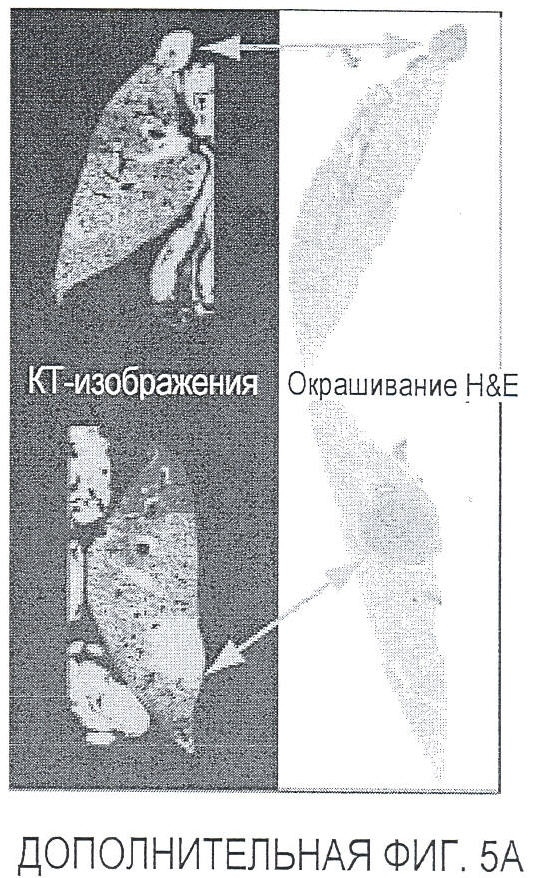

Группа изобретений относится к области медицины и фармакологии. В качестве антагониста нейропилина-2 используют антитело против Nrp2B, или его антиген-связывающий фрагмент, которое содержит последовательности областей, определяющих комплементарность, (CDR) вариабельных доменов легкой и/или тяжелой цепи антитела YW68.4.2 или антитела YW68.4.2.36, или вариант указанного антитела или указанного антиген-связывающего фрагмента, где аминокислотная последовательность Fab фрагмента антитела YW68.4.2 показана на фигуре 10А и аминокислотная последовательность Fab фрагмента антитела YW68.4.2.36 показана на фигуре 10В. Группа изобретений позволяет повысить эффективность профилактики и лечения метастазов опухоли. 4 н. и 17 з.п. ф-лы, 13 ил.
1. Антитело против Nrp2B или его антигенсвязывающий фрагмент, которое содержит последовательности областей, определяющих комплементарность (CDR) вариабельных доменов легкой и/или тяжелой цепи антитела YW68.4.2 или антитела YW68.4.2.36 или вариант указанного антитела или указанного антигенсвязывающего фрагмента, где аминокислотная последовательность Fab фрагмента антитела YW68.4.2 показана на фигуре 10А и аминокислотная последовательность Fab фрагмента антитела YW68.4.2.36 показана на фигуре 10В.
2. Антитело против Nrp2B или его антигенсвязывающий фрагмент по п.1, которое содержит последовательности CDR вариабельных доменов легкой и тяжелой цепи антитела YW68.4.2 или антитела YW68.4.2.36 или вариант указанного антитела или указанного антигенсвязывающего фрагмента.
3. Антитело против Nrp2B или его антигенсвязывающий фрагмент по п.1, которое содержит последовательности вариабельных областей легкой и/или тяжелой цепи антитела YW68.4.2 или антитела YW68.4.2.36 или вариант указанного антитела или указанного антигенсвязывающего фрагмента, имеющих одно или более изменений в каркасных областях, где аминокислотная последовательность фрагмента Fab антитела YW68.4.2 показана на фигуре 10А и аминокислотная последовательность Fab фрагмента антитела YW68.4.2.36 показана на фигуре 10В.
4. Антитело против Nrp2B или его антигенсвязывающий фрагмент по п.3, которое содержит последовательности вариабельных областей легкой и тяжелой цепи антитела YW68.4.2 или антитела YW68.4.2.36 или вариант указанного антитела или указанного антигенсвязывающего фрагмента, имеющих одно или более изменений в каркасных областях, где аминокислотная последовательность фрагмента Fab антитела YW68.4.2 показана на фигуре 10А и аминокислотная последовательность Fab фрагмента антитела YW68.4.2.36 показана на фигуре 10В.
5. Антитело против Nrp2B по п.1, которое содержит YW68.4.2 или YW68.4.2.36 или антигенсвязывающий фрагмент антитела YW68.4.2 или YW68.4.2.36.
6. Антитело против Nrp2B по п.1, которое содержит YW68.4.2.36, или его антигенсвязывающий фрагмент.
7. Антитело против Nrp2B по п.1, которое представляет собой YW68.4.2.36 или его антигенсвязывающий фрагмент.
8. Антитело против Nrp2B, или его антигенсвязывающий фрагмент, или вариант по любому из пп.1-7, которое является биспецифическим.
9. Антитело против Nrp2B, или антигенсвязывающий фрагмент, или вариант по п.8, которое дополнительно связывается с VEGF или с рецептором VEGF.
10. Антитело против Nrp2B, или антигенсвязывающий фрагмент, или вариант по любому из пп.1-7, где указанный фрагмент выбран из группы, состоящей из фрагментов Fab, F(ab')2 и scFv.
11. Антитело против Nrp2B, или антигенсвязывающий фрагмент, или вариант по любому из пп.1-7, где указанный вариант является аффинно-зрелым вариантом.
12. Антитело против Nrp2B по любому из пп.1-7, где указанное антитело, антигенсвязывающий фрагмент, или вариант является химерным или гуманизированным.
13. Композиция, содержащая антитело, антигенсвязывающий фрагмент, или вариант по любому из пп.1-7 в смеси с носителем.
14. Фармацевтическая композиция для профилактики или лечения опухолевых метастазов, содержащая эффективное количество антитела против Nrp2B, антигенсвязывающего фрагмента или варианта по любому из пп.1-7 в смеси с фармацевтически приемлемым носителем.
15. Способ ингибирования миграции лимфатических эндотелиальных клеток, включающий введение нуждающемуся млекопитающему эффективного количества антитела против Nrp2B, антигенсвязывающего фрагмента, или варианта по любому из пп.1-7.
16. Способ по п.15, где указанное млекопитающее является человеком.
17. Способ по п.16, где у указанного человека диагностирован рак.
18. Способ по п.17, где у указанного человека диагностированы опухолевые метастазы или имеется риск возникновения опухолевых метастазов.
19. Способ по п.18, где указанный метастаз находится в лимфатической системе или в отдаленном органе.
20. Способ по п.17, которые дополнительно включает введение антитела против VEGF.
21. Способ по п.21, где указанное антитело против VEGF является бевацизумабом.
| WO 9929729 А2, 17.06.1999 | |||
| WO 9929858 А, 17.06.1999. |

