Настоящая заявка испрашивает приоритет по предварительной заявке на патент США, серийный №61/474201, поданной 11 апреля 2011 года, которая включена в данный документ во всех отношениях и во всей своей полноте посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение в целом относится к композициям и способам улучшения признаков полевых и садовых сельскохозяйственных культур, включая повышение урожайности сельскохозяйственных культур. Более конкретно, настоящее изобретение относится к композициям и способам улучшения одного или нескольких признаков полевых или садовых культур путем нарушения одного или нескольких белков (KRP), родственных белку-ингибитору киназы (KIP), в однодольных и двудольных растениях.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Наиболее важным признаком в качестве объекта для улучшения сельскохозяйственных культур является урожайность. Исследования по повышению урожайности сельскохозяйственных культур путем получения новых сортов растений можно разделить на два подхода. Один из них заключается в уменьшении потерь урожая сельскохозяйственных культур путем селекции или создания сортов сельскохозяйственных культур с повышенной устойчивостью к абиотическим стрессовым условиям, таким как засуха, холод или содержание солей, или биотическим стрессовым условиям, связанным с вредителями или болезнетворными патогенами. Несмотря на то, что данный подход важен, он не обеспечивает существенного улучшения урожая сельскохозяйственных культур в отсутствие стрессовых условий и, фактически, на такую резистентность могут быть направлены те ресурсы растения, которые в ином случае были бы доступны для повышения его урожайности. Второй подход заключается в селекции или создании новых сортов сельскохозяйственных культур, обладающих повышенной основной урожайностью.
Классические программы селекции первоначально приводили к значительному повышению урожая различных сельскохозяйственных культур. Однако за обычно наблюдавшимся первоначально значительным приростом урожая следовали дополнительные поэтапные улучшения, которые становятся все менее масштабными и более сложными в получении. Разработанные в последнее время подходы, основанные на молекулярно-биологических методиках, в принципе, обладают потенциалом для достижения значительного улучшения урожая сельскохозяйственных культур путем изменения временного паттерна, локализации или уровня экспрессии генов растений или гетерологичных генов, которые играют роль в росте и/или развитии растений. За последние двадцать лет был достигнут существенный прогресс в идентификации генов растений и или гетерологичных генов, которые участвуют в росте и/или развитии растений. Несмотря на эти достижения в применении молекулярных подходов, по-прежнему существует большая потребность в улучшении полевых и садовых растений, полученных посредством более традиционной селекции растений. Из-за сложности регулирования роста растений и конечной связи такого регулирования с признаками урожайности, все еще не ясно, какие из определенных генов, если таковые имеются, могут быть явными кандидатами, улучшающими урожай сельскохозяйственных культур посредством селекции растений и/или применения молекулярных методик.
Белки KRP принадлежат к классу ингибиторов клеточного цикла, которые связывают и ингибируют комплексы циклин/циклин-зависимая киназа (CDK). Ожидается, что мутации консервативных остатков у представителей семейства KRP могут модифицировать способность KRP функционировать в качестве ингибитора комплексов циклин/CDK-киназа. В частности, некоторые мутации в генах KRP приводят к экспрессии нефункционального ингибитора клеточного цикла KRP или ингибитора клеточного цикла с пониженной активностью. Такая потеря или снижение ингибирующей активности циклин/CDK-киназы ведет к повышению активности циклин/CDK-киназы в клетках, тогда как в нормальных условиях данные клетки будут обладать пониженной активностью циклин/CDK. Такая потеря или пониженная ингибирующая активность комплексов циклин/CDK-киназа приведет к увеличению скорости деления клеток в тканях, в которых экспрессируется вариант нормального KRP дикого типа. Такое повышенное деление клеток приведет к возникновению положительных агротехнических признаков, таких как повышение выхода, увеличение веса, размера и/или числа одного или нескольких органов, например, увеличение размера семян, увеличение размера растения, увеличение листьев, увеличение корней и т.д. Для обоснования методик, связанных с KRP, смотри, например, WO/2007/016319 и US 20070056058, каждый из которых включен в данный документ во всей своей полноте посредством ссылки для всех целей. Настоящее изобретение идентифицирует новые гены и белки KRP и предлагает способы их применения для получения улучшенных полевых и садовых растений путем общепринятой селекции растений и/или с применением молекулярных методик.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения использовали способы целенаправленно индуцированных локальных повреждений геномов (TILLING®) растений для идентификации мутантных KRP, например, в однодольных растениях, таких как растения трибы Triticeae (например, растения рода Triticum) и растения трибы Oryzeae (например, растения рода Oryza), или в двудольных растениях, например, в Glycine spp. Идентифицированные мутантные KRP могут применяться для увеличения веса, размера и/или числа одного или нескольких органов растения. Органом может быть любая часть растения, например, органы, которые вносят вклад в урожайность растения. В некоторых вариантах осуществления органами являются семена, листья, ветки, корни, побеги, семяпочки, пыльца, семенные коробочки, головки семян или отростки. Например, в некоторых вариантах осуществления настоящее изобретение предлагает способы увеличения веса семян растений, размера семян, числа семян и/или урожая.
Данное изобретение описывает поиск мутаций в KRP растений, которые экспрессируются в клетках развивающихся семян. Стратегия заключалась в идентификации представителя(-ей) семейства KRP, которые экспрессируются в семенах, и применении TILL® в отношении мутаций в данных KRP. Определенные мутации, которые влияют на функцию KRP, затем могут быть охарактеризованы по положительным агротехническим и садоводческим признакам, таким как повышение урожая, ранняя всхожесть, ускоренный рост и т.д. Кроме того, эта же методика может быть использована в отношении KRP, которые экспрессируются в других тканях, таких как листья или стебли, поскольку повышение скорости деления клеток в данных других тканях, имеющих определенный характер экспрессии KRP, также может привести к положительным агротехническим признакам.
В настоящее время представители семейства KRP в тех видах растений, которые представляют интерес для сельского хозяйства, садоводства и/или промышленности, и их паттерн экспрессии в развивающихся семенах еще требует изучения. Стратегия авторов изобретения включала идентификацию KRP, которые экспрессируются в развивающихся семенах. Идентифицированы KRP, паттерн экспрессии которых в процессе развития является тканеспецифическим. Например, ген риса KRP4 (OsKRP4) продемонстрировал практически исключительную экспрессию в развивающихся семенах. Также были идентифицированы последовательности, эквивалентные этой, в рисе и других видах растений.
Настоящее изобретение предлагает гены KRP, мутированные в сравнении с KRP, имеющие последовательность нуклеиновой кислоты, которая содержит последовательность, выбранную из группы, включающей SEQ ID NO: 60-86, 100-107, 111-128, полученные на основании этих последовательностей фрагменты и варианты, которые кодируют ген KRP. В некоторых вариантах осуществления паттерн экспрессии KRP в ходе развития является тканеспецифическим. В некоторых вариантах осуществления экспрессия является специфичной для зародышей, специфичной для пыльцы или специфичной для семян.
В некоторых вариантах осуществления настоящее изобретение предлагает гены KRP, мутированные в сравнении с KRP дикого типа, содержащие последовательность, содержащую последовательность нуклеиновой кислоты, характеризующуюся по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% идентичностью с последовательностями SEQ ID NO: 60-86, 100-107, 111-128, 138-139, 141-142 и 144-145.
В некоторых вариантах осуществления настоящее изобретение предлагает гены KRP, мутированные по сравнению с KRP дикого типа, кодирующие аминокислотную последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% идентичностью с последовательностями SEQ ID NO: 87-99, 108-110, 129-137, 140, 143 и 146.
Например, настоящее изобретение предлагает мутанты пшеницы (Та) по KRP1A, KRP1B, KRP1D, KRP2A, KRP2B, KRP2D, KRP4B, KRP4D, KRP5A, KRP5D, KRP6A, KRP6B, KRP6D. Очевидно, что гексаплоидная пшеница подвергалась естественному отбору с тем, чтобы нести то, что является, по-видимому, нокаут-мутациями по KRP4A и KRP5B. Это было обнаружено в ходе проведения TILLING®. Что касается KRP4A, то этот ген похоже полностью отсутствует в геноме гексаплоидной пшеницы, хотя он все еще присутствует в тетраплоидном геноме. Что касается KRP5B, то в этом гене похоже отсутствует один нуклеотид, что ведет к сдвигу трансляционной рамки и в дальнейшем к предсказанному отсечению нескольких аминокислот.
Настоящее изобретение в другом аспекте предлагает клетки растений, части растений, культуру ткани или целые растения, содержащие один или несколько нарушенных генов KRP, как описано в данном документе. В некоторых вариантах осуществления настоящее изобретение предлагает клетки растений, части растений, культуру ткани или целые растения, содержащие по меньшей мере один ген KRP, где геном растения содержит одну или несколько копий гена и где функция одной или нескольких копий гена KRP нарушена.
В некоторых вариантах осуществления растение является однодольным. В некоторых вариантах осуществления однодольное растение является видом трибы Triticeae или трибы Oryzeae. В некоторых вариантах осуществления растение трибы Triticeae является растением рода Triticum, а растение трибы Oryzeae является растением рода Oryza.
В некоторых дополнительных вариантах осуществления растением рода Triticum является пшеница, а растением рода Oryza является рис.
В некоторых дополнительных вариантах осуществления растение пшеницы является тетраплоидным или гексаплоидным.
В некоторых вариантах осуществления растение является двудольным. В некоторых вариантах осуществления растение является видом семейства Fabaceae, например, Glycine spp., таким как соя.
В некоторых вариантах осуществления KRP в пшенице является TaKRP1, TaKRP2, TaKRP4, TaKRP5, TaKRP6, например, SEQ ID NO: 60-86, 138-139, 141-142, 144-145, или их функциональные варианты; KRP в рисе является OsKRP1, OsKRP2, OsKRP4 или OsKRP5, например, SEQ ID NO: 100-107, или их функциональные варианты; и KRP в Glycine spp. является Gm0003×00821, Gm0013×00399, Gm0043, Gm0053×00526, Gm0087×00306, Gm0102×00087, Gm0119×00131, Gm0151×00019, Gm0067×00001, например, SEQ ID NO: 111-128, или их функциональные варианты. Например, KRP в тетраплоидном растении пшеницы является TaKRP1A, TaKRP1B, TaKRP2A, TaKRP2B, TaKRP4A, TaKRP4B, TaKRP5A, TaKRP5B, TaKRP6A или TaKRP6B. В гексаплоидной пшенице KRP является TaKRP1A, TaKRP1B, TaKRP1D, TaKRP2A, TaKRP2B, TaKRP2D, TaKRP4A, TaKRP4B, TaKRP4D, TaKRP5A, TaKRP5B, TaKRP5D, TaKRP6A, TaKRP6B или TaKRP6D.
В некоторых вариантах осуществления функция генов KRP нарушена посредством замены нуклеотидов, делеции, вставки, гомологичной рекомбинации, Т-ДНК, транспозона, двухцепочечного олигонуклеотида, антисмыслового олигонуклеотида, инвертированного повтора или их комбинации.
В некоторых вариантах осуществления нарушенный KRP в клетках растения, части растения, культуре ткани или целом растении содержит одну или несколько мутаций, выбранных из любой мутации, изложенных в таблицах 2-12, 25, 28, 29-37 в графической части для определенного гена KRP.
В другом аспекте настоящее изобретение предлагает способы повышения веса, размера и/или числа одного или нескольких органов, например, повышение веса семян растений, размера семян, числа семян и/или урожайности растения, которые включают нарушение одного или нескольких KRP в растении. В одном варианте осуществления растение является двудольным растением или однодольным растением. В некоторых вариантах осуществления растение может быть растением, выбранным из трибы Triticeae, трибы Oryzeae или семейства Fabaceae, например, пшеницей, рисом и соей. В некоторых вариантах осуществления способы нарушения функции гена включают, но не ограничивают, мутагенез (например, химический мутагенез, радиационный мутагенез, транспозонный мутагенез, инсерционный мутагенез, мутагенез с использованием уникальных последовательностей в качестве меток (STM), сайт-специфический мутагенез и естественный мутагенез), создание антисмысловых последовательностей, нокауты и/или РНК-интерференцию. В некоторых вариантах осуществления растение с увеличенным весом, размером и/или числом одного или нескольких органов, например, растение с увеличенным весом семян растений, размером семян, числом семян и/или урожайностью, не является генетически модифицированным организмом или трансгенным растением. Например, нарушение KRP в геноме растения может быть следствием естественной мутации или мутаций, индуцированных химическим мутагенезом или радиационным мутагенезом.
В некоторых вариантах осуществления мутации, описанные в таблицах 2-12, 25, 28, 29-37, могут быть интегрированы в виды, близкородственные растениям трибы Triticeae, трибы Oryzeae, семейства Fabaceae, или в растения, близкородственные таким растениям, как пшеница, рис или соя. В некоторых вариантах осуществления в доменах или участках, которые являются консервативными по сравнению с ортологами KRP у других видов, может производиться замена или деления аминокислот с получением мутантов со сниженной или отсутствующей активностью и/или мутантов, которые приводят к потере функции (например, нестабильность белков). В некоторых вариантах осуществления один или несколько KRP в растении подвергнуты нокдауну или нокаутированы при помощи одного или нескольких способов, известных специалистам в данной области техники.
В некоторых вариантах осуществления нарушены одна или несколько копий одного или нескольких генов KRP. Например, в тетраплоидном растении пшеницы одна или две копии гена KRP нарушены (например, KRP1A, KRP1B; KRP2A, KRP2B; KRP4A, KRP4B; KRP5A, KRP5B и KRP6A, KRP6B); в гексаплоидном растении пшеницы нарушены одна или несколько копий одного, двух или трех копий гена KRP (например, KRP1A, KRP1B, KRP1D; KRP2A, KRP2B, KRP2D; KRP4A, KRP4B, KRP4D; KRP5A, KRP5B, KRP5D и KRP6A, KRP6B, KRP6D).
Настоящее изобретение также предлагает растение, характеризующееся увеличенным весом, размером и/или числом одного или нескольких органов, например, растение с увеличенным размером семян, числом семян и/или урожаем семян по сравнению с контрольным растением дикого типа, где растение имеет одну или несколько мутаций по одному или нескольким генам KRP. В некоторых вариантах осуществления вышеупомянутое растение является однодольным растением. В некоторых вариантах осуществления вышеупомянутое однодольное растение является растением трибы Triticeae или трибы Oryzeae. В некоторых вариантах осуществления вышеупомянутое растение является растением пшеницы или риса. В некоторых вариантах осуществления вышеупомянутое растение является двудольным растением. В некоторых вариантах осуществления, вышеупомянутое двудольное растение происходит из семейства Fabaceae, как, например, растение сои.
Настоящее изобретение дополнительно предлагает семена, фрукты, клетки растения или части трансгенного растения, как описано в данном документе. Например, настоящее изобретение предлагает пыльцу растения, семяпочку растения, генетически родственную популяцию растений, культуру ткани регенерируемых клеток растения. В некоторых вариантах осуществления регенерируемые клетки получают из зародышей, протопластов, клеток меристемы, каллуса, пыльцы, листьев, пыльников, стеблей, черешков, корней, кончик корня, фруктов, семян, цветов, семядолей и/или гипокотилей.
Настоящее изобретение также предлагает способы снижения активности одного или нескольких белков KRP в клетке растения, части растения, культуре ткани или в целом растении, включающие воздействие на клетку растения, часть растения, культуру ткани или целое растение ингибирующей нуклеиновой кислотой, характеризующейся комплементарностью к гену, кодирующему вышеупомянутый белок KRP. В некоторых вариантах осуществления растение является растением трибы Triticeae или трибы Oryzeae. В некоторых вариантах осуществления вышеупомянутое растение является растением пшеницы или риса. В некоторых вариантах осуществления вышеупомянутое растение является двудольным растением. В некоторых вариантах осуществления вышеупомянутое двудольное растение происходит из семейства Fabaceae, как например, растение сои.
Настоящее изобретение также предлагает способы селекции видов сельскохозяйственных культур, характеризующихся увеличенным весом, размером и/или числом одного или нескольких органов, например, видов сельскохозяйственных культур с увеличенным размером семян, числом семян, весом семян и/или урожаем семян по сравнению с контрольным растением дикого типа, включающие введение генетических материалов растения с нарушенным KRP в растение-реципиент.
В некоторых вариантах осуществления такие способы включают проведение скрещивания мутанта Triticum sp., имеющего одну или несколько мутаций, которые перечислены в таблицах 2-12, со вторым Triticum sp. для получения растения F1 или с видом трибы Triticeae, который может перекрестно скрещиваться с вышеупомянутым первым Triticum sp. Способ может дополнительно включать возвратное скрещивание растения F1 со вторым Triticum sp. или с видом трибы Triticeae и повторение этапа возвратного скрещивания для получения практически изогенной линии, где одна или несколько мутаций интегрированы в геном вышеупомянутого растения второго Triticum sp. или вида трибы Triticeae; где практически изогенная линия, полученная из второго Triticum sp. или вида трибы Triticeae с интегрированной мутацией, имеет измененный вес, размер и/или число одного или нескольких органов, например, измененный вес семян, размер семян, число семян и/или урожай семян. Необязательно, такие способы можно упростить при помощи молекулярных маркеров или TILLING®.
В некоторых вариантах осуществления такие способы включают проведение скрещивания мутанта Oryza sp., имеющего одну или несколько мутаций, которые перечислены в таблице 25, со вторым Oryza sp. для получения растения F1, или с видом трибы Oryzeae, который может перекрестие скрещиваться с вышеупомянутым Oryza sp. Способ может дополнительно включать возвратное скрещивание растения F1 со вторым Oryza sp. или с видом трибы Oryza; и повторение этапа возвратного скрещивания для получения практически изогенной линии, где одна или несколько мутаций интегрированы в геном вышеупомянутого растения второго Oryza sp. или вида трибы Oryza; где практически изогенная линия, полученная из второго Oryza sp. или вида трибы Oryza, с интегрированной мутацией, имеет измененный вес, размер и/или число одного или нескольких органов, например, измененный вес семян, размер семян, число семян и/или урожай семян. Необязательно, такие способы можно упростить при помощи молекулярных маркеров или TILLING®.
В некоторых вариантах осуществления такие способы включают проведение скрещивания мутанта Glycine sp., имеющего одну или несколько мутаций, изложенных в таблицах 29-37, со вторым Glycine sp. для получения растения F1, или с видом семейства Fabaceae, который может перекрестие скрещиваться с вышеупомянутым первым Glycine sp. Способ может дополнительно включать возвратное скрещивание растения F1 со вторым Glycine sp. или с видом семейства Fabaceae, и повторением этапа возвратного скрещивания для получения практически изогенной линии, где одна или несколько мутаций интегрированы в геном вышеупомянутого второго Glycine sp. или вида из семейства Fabaceae; где близкая изогенная линия, полученная из второго Glycine sp. или вида из семейства Fabaceae с интегрированной мутацией, имеет измененный вес, размер и/или число одного или нескольких органов, например, измененный вес семян, размер семян, число семян и/или урожай семян. Необязательно, такие способы можно упростить при помощи молекулярных маркеров или TILLING®.
Настоящее изобретение предлагает выделенную последовательность нуклеиновой кислоты, содержащую последовательность, выбранную из группы, включающей SEQ ID NO: 138, 139, 141, 142, 144 и 145, а также фрагменты и варианты, полученные из них, которые кодируют ген KRP пшеницы.
В одном варианте осуществления настоящее изобретение предлагает выделенный полинуклеотид, кодирующий белок KRP растения, содержащий последовательность нуклеиновой кислоты, которая характеризуется по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% идентичностью с SEQ ID NO: 138, 139, 141, 142, 144 и 145.
Настоящее изобретение дополнительно предлагает выделенную аминокислотную последовательность (например, пептид, полипептид и подобные), содержащую последовательность, выбранную из группы, включающей SEQ ID NO: 140, 143 и 146, и фрагменты и варианты, полученных из них, которые образуют белок KRP.
В некоторых вариантах осуществления настоящее изобретение предлагает выделенную аминокислотную последовательность, образующую белок, который имеет аминокислотную последовательность которого характеризуется по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% идентичностью с SEQ ID NO: 140, 143 и 146.
В одном варианте осуществления выделенные полинуклеотиды настоящего изобретения содержат последовательность, выбранную из группы, включающей (а) последовательности, представленные в SEQ ID NO: 138, 139, 141, 142, 144 и 145, или их части; (b) последовательности, комплементарные последовательностям, представленным в SEQ ID NO: 138, 139, 141, 142, 144 и 145, или их части; (с) последовательности, обратно комплементарные последовательностям, представленным в SEQ ID NO: 138, 139, 141, 142, 144 и 145, или их части; (d) обратные последовательности последовательностей, представленных в SEQ ID NO: 138, 139, 141, 142, 144 и 145, или их части; и (е) последовательности, характеризующиеся по меньшей мере 50%, 75%, 90%, 95% или 98% идентичностью, как указано в данном документе, с последовательностью (a)-(d) или специфическим участком последовательности (a)-(d).
Настоящее изобретение также предлагает химерный ген, содержащий выделенную последовательность нуклеиновой кислоты, относящуюся к любому из вышеописанных полинуклеотидов, функционально сцепленных с подходящими регуляторными последовательностями.
Настоящее изобретение также представляет рекомбинантные конструкты, содержащие химерный ген, который описан выше.
Настоящее изобретение дополнительно предлагает конструкты для интерферирующих РНК (RNAi) на основе последовательностей нуклеиновых кислот, настоящего изобретения. В некоторых вариантах осуществления конструкты для RNAi могут быть трансформированы в растение пшеницы для снижения экспрессии одного или нескольких KRP. RNAi-конструкт может быть, но без ограничений, конструктом антисмыслового олигонуклеотида, конструктом двухцепочечного олигонуклеотида, siRNA-конструктом или конструктом инвертированного повтора. В некоторых вариантах осуществления конструкты для RNAi содержат промотор растения, такой как конститутивный промотор, индуцируемый промотор или тканеспецифический промотор. В некоторых вариантах осуществления промотор является специфическим по отношению к зародышу или специфическим по отношению к семенам.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фигуре 1А изображен график амплификации сигнала флуоресценции в зависимости от числа циклов для геномного образца пшеницы гомозиготного по аллелю дикого типа KRP4B пшеницы. Верхняя кривая представляет амплификацию с зондом дикого типа, тогда как нижняя кривая представляет амплификацию с мутантным зондом. На фигуре 1В изображен график амплификации сигнала флуоресценции в зависимости от числа циклов для геномного образца пшеницы, гомозиготного по мутантному аллелю пшеницы KRP4B. Верхняя кривая представляет амплификацию с мутантным зондом, тогда как нижняя кривая представляет амплификацию с зондом дикого типа. На фигуре 1C изображен график амплификации сигнала флуоресценции в зависимости от числа циклов для геномного образца пшеницы, гетерозиготного по мутантному аллелю KRP4B пшеницы. На фигуре 1D изображена хроматограмма последовательности, демонстрирующая два нуклеотида, обнаруженные в геномном образце пшеницы, гетерозиготном по мутантному аллелю KRP4B пшеницы.
На фигуре 2 изображен график аллельной дискриминации для образцов в анализе аллельной дискриминации KRP4B. Образцы, гомозиготные по аллелю гена KRP4B дикого типа, выделены синим, образцы, гомозиготные по мутантному аллелю KRP4B P109L, выделены красным, а образцы, гетерозиготные по мутантному аллелю KRP4B Р109, выделены зеленым. Черные квадратики указывают на отсутствие контролей матрицы, а черные крестики обозначают неустановленные образцы.
На фигуре 3 изображены С-терминальные аминокислотные последовательности, которые содержат домены связывания циклина и циклин-зависимой киназы (CDK) для KRP1A, 2D, 4А и 5А. Нонсенс и тяжелые миссенс TILLING®-мутации типа I, протестированные в in vitro анализе киназ, помечены звездочками, или указаны мутантные аминокислоты, соответственно, под аминокислотами дикого типа.
На фигуре 4 изображен авторадиографически снимок анализов киназ, проведенных с применением ZmCyclinD4/CDKA; 1-киназного комплекса, указанного KRP дикого типа Triticum aestivum и указанного TILLING®-мутантного KRP Triticum aestivum. Гистон H1 (HH1) применяли в качестве субстрата для фосфорилирования. Полосы 1 и 18: киназный комплекс без KRP дикого типа или TILLING®-мутанта. Полосы 2 и 19: только киназный комплекс в буфере. Полосы 3, 4, 5: киназный комплекс и TaKRP1A дикого типа в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 6, 7, 8; киназный комплекс и мутант TaKRP1A P232L в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 9, 10, 11: киназный комплекс и мутантный TaKRP1A G236S в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 12, 13, 14: киназный комплекс и мутантный TaKRP1A W240* в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 15, 16, 17: киназный комплекс и TaKRP2D дикого типа в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 20, 21, 22: киназный комплекс и мутантный TaKRP2D P228S в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 23, 24, 25: киназный комплекс и мутантный TaKRP2D D254N в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 26, 27, 28: киназный комплекс и мутантный TaKRP2D R257C в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 29, 30: киназный комплекс и TaKRP4A дикого типа в количестве 0,25 и 0,1 мкг, соответственно. Полосы 31, 32, 33: киназный комплекс и мутантный TaKRP4A W186* в количестве 0,5; 0,25 и 0,1 мкг, соответственно.
На фигуре 5 изображен авторадиографический снимок анализа киназ, проведенного с применением ZmCyclinD4/CDKA; 1-киназного комплекса, указанного KRP дикого типа Triticum aestivum и указанного TILLING®-мутантного KRP Triticum aestivum. Гистон HI (НН1) применяли в качестве субстрата для фосфорилирования. Полоса 1: киназный комплекс без KRP дикого типа или TILLING®-мутанта. Полосы 2 и 18: только киназный комплекс в буфере. Полосы 3, 4, 5: киназный комплекс и TaKRP2D дикого типа в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 6, 7, 8: киназный комплекс и мутантный TaKRP2D A238V в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 9, 10, 11: киназный комплекс и мутантный TaKRP2D А239Т в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 12, 13, 14: киназный комплекс и TaKRP5D дикого типа в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 15, 16, 17: киназный комплекс и мутантный TaKRP5D W199* в количестве 0,5; 0,25 и 0,1 мкг, соответственно.
На фигуре 6А изображен авторадиографический снимок повторных анализов киназ, проведенных с применением ZmCyclinD4/CDKA; 1-киназного комплекса, указанного KRP дикого типа Triticum aestivum и указанного TILLING®-мутантного KRP Triticum aestivum. Гистон H1 (HH1) применялся в качестве субстрата для фосфорилирования. Полоса 1: киназный комплекс без KRP дикого типа или TILLING®-мутанта. Полоса 2: только киназный комплекс в буфере. Полосы 3, 4, 5: киназный комплекс и TaKRP1A дикого типа в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 6, 7, 8: киназный комплекс и мутантный TaKRP1A W240* в количестве 0,5; 0,25 и 0,1 мкг, соответственно. Полосы 9, 10: киназный комплекс и TaKRP4A дикого типа в количестве 0,25 и 0,1 мкг, соответственно. Полосы 11, 12, 13: киназный комплекс и мутантный TaKRP4A W186* в количестве 0,5; 0,25 и 0,1 мкг, соответственно. На фигуре 6 В изображен авторадиографический снимок анализа киназ с применением ZmCyclinD4/CDKA; 1-киназного комплекса и мутанта ZmKRP2. Мутант ZmKRP2 является мутантом белка кукурузы krp2 с эквивалентным преждевременным стоп-кодоном, расположенным очень близко к концу белка. Полосы 1 и 2: киназный комплекс и мутант ZmKRP2 в количестве 0,3 и 3 мкг, соответственно.
На фигуре 7 изображен средний урожай зерна (фунтов/акр) TILLING®-мутантов яровой пшеницы по KRP и контрольного сорта Express. Боземан, Монтана, Форт Коллинз, Колорадо. *, ** обозначают существенные различия между мутантным и контрольным сортом при уровнях вероятности 0,05 и 0,01, соответственно.
На фигуре 8 изображен средний урожай зерна (фунтов/акр) для дикого типа (WT) и гомозиготных (Homo) зиготных групп для TILLING®-мутантов по KRP яровой пшеницы и для контрольного сорта Express. Боземан, Монтана, Форт Коллинз, Колорадо. *, ** обозначают существенные различия между группами дикого типа (WT) и гомозиготными (Homo) зиготными группами при уровнях вероятности 0,05 и 0,01, соответственно.
На фигуре 9 изображен средний урожай зерна (фунтов/акр) в группах дикого типа (WT) и гомозиготных (Homo) зиготных группах в пределах сестринских линий F1 '148E04' и 149Е05' TILLING®-мутанта по KRP яровой пшеницы KRP4D Stop. Боземан, Монтана, Форт Коллинз, Колорадо.
На фигуре 10 изображено филогенетическое дерево KRP риса (Os), кукурузы (Zm) и пшеницы (Та).
На фигуре 11 изображено выравнивание последовательностей белков KRP риса (Os), кукурузы (Zm) и пшеницы (Та).
На фигуре 12 изображено выравнивание последовательностей белков KRP сои (Gm), Arabidopsis (At) и Brassica napus (Bn).
ПОСЛЕДОВАТЕЛЬНОСТИ
Перечень последовательностей SEQ ID NO: 1-SEQ ID NO: 152 является частью настоящей заявки и включен в данный документ посредством ссылки. Список последовательностей приведен в конце данного документа.
ПОДРОБНОЕ ОПИСАНИЕ
Все публикации, патенты и патентные заявки, включая все графические материалы и приложения, а также все последовательности нуклеиновых кислот и полипептидные последовательности, идентифицированные по номерам доступа в GenBank, включены в данный документ посредством ссылки в том же объеме, как если бы для каждой отдельной публикации или патентной заявки было специально и индивидуально указано их включение посредством ссылки.
Следующее описание включает информацию, которая может быть полезна для понимания настоящего изобретения. Это не является признанием того, что любая информация, представленная в данном документе, относится к уровню техники, известному до даты приоритета заявленного изобретения, или относится к изобретению настоящей заявки, или что любая публикация, на которую конкретно или неявно ссылаются, относится к известному уровню техники.
Определения
Используемые в данном документе в описании и формуле изобретения глаголы "включать/содержать" и производные от них формы употребляются в неограничивающем смысле, что означает, что элементы, следующие за данными словами, включены, но не исключены упомянутые конкретно элементы.
Используемое в данном документе выражение "растение" относится к любому живому организму, принадлежащему к царству растений (т.е., любой род/вид царства растений). К ним относятся такие известные организмы, которые включают, но не ограничивают, деревья, лекарственные травы, кусты, травы, вьющиеся растения, папоротники, мхи и зеленые водоросли. Выражение относится как к однодольным растениям, так и к двудольным растениям. Примеры конкретных растений включают, но не ограничивают, растения трибы Triticeae (например, растения рода Triticum), растения трибы Oryzeae (например, растения рода Oryza), растения трибы Andropogoneae (например, растения рода Zea, такие как кукуруза). Другие неограничивающие примеры растений включают, картофель, розы, яблони, подсолнечники, бананы, томаты, тыкву-горлянку, тыкву обыкновенную, тыкву крупноплодную, салат, капусту, дубы, гусманию, герань, гибискус, клематис, пуансеттию, сахарный тростник, колоказию, утиный сорняк, сосны, мятлик луговой, цойсию, кокосовые пальмы, капустные культуры (например, брокколи, итальянская брокколи, брюссельская капуста, белокочанная капуста, китайская капуста (бок-чой и напа), цветная капуста, черная капуста, капуста листовая, капуста, кольраби, горчица салатная, рапс и другие капустные культуры), луковичные культуры (например чеснок, лук-порей, репчатый лук (сухая луковица, зеленый лук и лук-батун), шалот и другие луковичные культуры), цитрусовые фрукты (например грейпфрут, лимон, лайм, апельсин, мандарин, цитрусовые гибриды, помело и другие цитрусовые фруктовые культуры), тыквенные культуры (например, огурец, арбуз кормовой, съедобные тыквы, корнишон, дыня мускусная (в том числе гибриды и/или культурные сорта растения дыни мускусной), арбуз, канталупа и другие тыквенные культуры), плодоносящие овощи (включая баклажан, вишню кустарниковую, пепино, перец, томат, физалис клейкоплодный и другие плодоносящие овощные культуры), виноград, листовые овощи (например, салат-ромен), корнеплоды/клубневые и клубнелуковицы (например, картофель) и орехи (миндаль, орех пекан, фисташка и грецкий орех), ягоды (например, томаты, барбарис, смородина, ягоды бузины, крыжовник, жимолость, подофилл щитовидный, калина канадская, орегонский виноград, облепиха крушинная, черемуха, толокнянка, брусника, клубника, морской виноград, черная смородина, морошка, логанова ягода, малина обыкновенная, малина превосходная, малина душистая и малина японская), зерновые культуры (например, кукуруза, рис, пшеница, ячмень, сорго, просо, овес, рожь, тритикале, гречиха, фонио, лебеда, масличная пальма), семечковые плоды (например, яблоки, груши), косточковые плоды (например, кофе, ююба, манго, маслины, кокосовые орехи, масличная пальма, фисташки, миндаль, абрикосы, вишни, терносливы, нектарины, персики и сливы), виноград (например, столовый виноград, технический виноград), прядильные культуры (например, конопля, хлопок), декоративные растения и подобные.
Используемое в данном документе выражение "часть растения" относится к любой части растения, включая, но без ограничений, побег, корень, стебель, семена, прилистники, листья, лепестки, цветы, семяпочки, прицветники, ветви, черешки, междоузлия, кору, пушок, отростки, корневища, листья, слоевища, пыльцу, тычинки и подобные. Две основные части растений, выращенных в определенной среде, такой как почва, часто называют "надземная" часть, которую также часто называют "побег", и "подземная" часть, которую также часто называют "корни".
Единственное число относится к одной или нескольким единицам; например, слово "ген" относится к одному или нескольким генам или по меньшей мере одному гену. Таким образом, выражения, указывающие на единственное число, "один или несколько" и "по меньшей мере один" используются в данном документе взаимозаменяемо. Кроме того, ссылка на "элемент" с использованием единственного числа не исключает возможности того, что присутствует более чем один элемент, если только контекст не указывает четко, что существует один и только один из элементов.
Используемое в данном документе выражение "нуклеиновая кислота" относится к полимерной форме нуклеотидов любой длины, рибонуклеотидов или дезоксирибонуклеотидов или их аналогов. Этот выражение относится к первичной структуре молекулы и таким образом, включает двух- и одноцепочечную ДНК, а также двух- и одноцепочечную РНК. Оно также включает модифицированные нуклеиновые кислоты, такие как метилированные и/или кэппированные нуклеиновые кислоты, нуклеиновые кислоты, содержащие модифицированные основания, модификации остова и подобные. Выражения "нуклеиновая кислота" и "нуклеотидная последовательность" используются как синонимы.
Используемые в данном документе выражения "полипептид", "пептид" и "белок" используются здесь взаимозаменяемо для обозначения полимеров из аминокислот любой длины. Эти выражения также включают белки, которые посттрансляционно модифицированы посредством реакций, которые включают гликозилирование, ацетилирование и фосфорилирование.
Используемое в данном документе выражение "гомологичный" или "гомолог" или "ортолог" известно в данной области техники и относится к родственным последовательностям, которые имеют общего предшественника или представителя семейства, и определяются на основании степени идентичности последовательности. Выражения "гомология", "гомологичный", "по сути, аналогичный" и "по сути, соответствующий" используются в данном документе взаимозаменяемо. Они относятся к фрагментам нуклеиновых кислот, где изменения в одном или нескольких нуклеотидных основаниях не влияют на способность фрагмента нуклеиновой кислоты опосредовать экспрессию гена или обеспечивать определенный фенотип. Данные выражения также относятся к модификациям фрагментов нуклеиновых кислот настоящего изобретения, таким как делеции или вставки одного или нескольких нуклеотидов, которые, по сути, не изменяют функциональные свойства полученного фрагмента нуклеиновой кислоты по отношению к исходному, немодифицированному фрагменту. Следовательно, необходимо понимать, как специалисты в данной области техники оценят то, что настоящее изобретение содержит более чем конкретные иллюстративные последовательности. Данные выражения описывают связь между геном, обнаруженном в одном виде, подвиде, разновидности, сорте или штамме, и соответствующим или эквивалентным геном, обнаруженном в другом виде, подвиде, разновидности, сорте или штамме. Для целей данного изобретения сравнивают гомологичные последовательности. "Гомологичные последовательности" или "гомологи" или "ортологи", предполагаются, считаются или известны функционально связанными. Функциональная взаимосвязь может быть указана любым из ряда способов, включая, но не ограничивая (а) степень идентичности последовательностей и/или (b) одинаковую или сходную биологическую функцию. Предпочтительно, чтобы были указаны как (а), так и (b). Степень идентичность последовательностей может варьировать, но в одном варианте осуществления она составляет по меньшей мере 50% (при использовании стандартных программ выравнивания последовательностей, известных в данной области техники), по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере приблизительно 91%, по меньшей мере приблизительно 92%, по меньшей мере приблизительно 93%, по меньшей мере приблизительно 94%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98% или по меньшей мере 98,5% или по меньшей мере приблизительно 99% или по меньшей мере 99,5% или по меньшей мере 99,8% или по меньшей мере 99,9%. Гомологию можно определять с использованием таких компьютерных программ, существующих и доступных в данной области техники, которые рассмотрены в Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987) Supplement 30, section 7.718, Table 7.71. Некоторые программы выравнивания последовательностей включают Mac Vector (Oxford Molecular Ltd, Oxford, U.K.), ALIGN Plus (Scientific and Educational Software, Pennsylvania) и AlignX (Vector NTI, Invitrogen, Carlsbad, CA). Другой программой выравнивания является Sequencher (Gene Codes, Ann-Arbor, Michigan) с использованием параметров по умолчанию.
Используемое в данном документе выражение "изменение нуклеотида" относится, например, к замене, делеции и/или вставке нуклеотида, что хорошо известно в данной области техники. Например, мутации характеризуются изменениями, которые приводят к молчащим заменам, дополнениям или делециям, но не изменяют свойства или активности кодируемых белков или структуры белков.
Используемое в данном документе выражение "модификация белка" относится, например, к замене аминокислот, модификации аминокислот, делеции и/или вставке, что хорошо известно в данной области техники.
Используемое в данном документе выражение "полученный из" относится к оригинальному источнику или источнику и может включать молекулы естественного происхождения, рекомбинантные, неочищенные или очищенные. Нуклеиновые кислоты или аминокислоты, полученные из оригинального источника или источника получения, могут содержать все виды нуклеотидных изменений или белковых модификаций, которые описаны в любой части данного документа.
Используемое в данном документе выражение "по меньшей мере часть" или "фрагмент" нуклеиновой кислоты или полипептида означает часть, которая характеризуется минимальными параметрами размера таких последовательностей или любого более крупного фрагмента молекулы полной длины, вплоть до и включая молекулу полной длины. Например, часть нуклеиновой кислоты может иметь 12 нуклеотидов, 13 нуклеотидов, 14 нуклеотидов, 15 нуклеотидов, 16 нуклеотидов, 17 нуклеотидов, 18 нуклеотидов, 19 нуклеотидов, 20 нуклеотидов, 22 нуклеотидов, 24 нуклеотидов, 26 нуклеотидов, 28 нуклеотидов, 30 нуклеотидов, 32 нуклеотидов, 34 нуклеотидов, 36 нуклеотидов, 38 нуклеотидов, 40 нуклеотидов, 45 нуклеотидов, 50 нуклеотидов, 55 нуклеотидов и так далее, до полной длины нуклеиновой кислоты. Аналогично, часть полипептида может иметь 4 аминокислоты, 5 аминокислот, 6 аминокислот, 7 аминокислот и так далее, до полной длины полипептида. Длина используемой части будет зависеть от конкретного применения. Часть нуклеиновой кислоты, используемой в качестве гибридизационного зонда, может иметь всего 12 нуклеотидов; в одном варианте осуществления имеет 20 нуклеотидов. Часть полипептида, используемого в качестве эпитопа, может иметь всего 4 аминокислоты. Часть полипептида, которая выполняет функцию полноразмерного полипептида, как правило, будет больше 4 аминокислот.
Используемое в данном документе выражение "идентичность последовательностей" или "идентичность" в контексте двух нуклеиновых кислот или полипептидных последовательностей относится к остаткам в двух последовательностях, которые одинаковы при выравнивании, выполняемом для определения максимального соответствия на протяжении указанного окна сравнения. В случае если процент идентичности последовательностей используется по отношению к белкам, считается, что положения остатков, которые не являются идентичными, часто отличаются по консервативным заменам аминокислот, где аминокислотные остатки заменяют на другие аминокислотные остатки со сходными химическими свойствами (например, такими как заряд или гидрофобность) и поэтому не изменяют функциональные свойства молекулы. В тех случаях, когда последовательности отличаются консервативными заменами, процент идентичности последовательностей может быть скорректирован в сторону повышения для учета консервативной природы замен. Последовательности, которые отличаются такими консервативными заменами, называют имеющими "сходные последовательности" или "сходство". Средства для такой коррекции хорошо известны специалистам в данной области техники. Как правило, они включают оценку консервативных замен в качестве частичного, а не полного несоответствия, тем самым повышая процент идентичности последовательностей. Таким образом, например, если идентичная аминокислота получает 1 балл, а неконсервативная замена получает ноль баллов, то консервативная замена получает балл в диапазоне между нулем и 1. Подсчет баллов по консервативным заменам рассчитывают, например, в соответствии с алгоритмом Meyers and Miller, Computer Applic. Biol. Sci., 4:11-17 (1988).
Используемое в данном документе выражение "по сути, комплементарный" означает, что комплементарность двух последовательностей нуклеиновых кислот составляет по меньшей мере приблизительно 65%, предпочтительно приблизительно 70% или 75%, более предпочтительно приблизительно 80% или 85%, еще более предпочтительно 90% или 95% и наиболее предпочтительно приблизительно 98% или 99%. Это означает, что праймеры и зонды должны обладать достаточной комплементарностью к их матрице и целевой нуклеиновой кислоте, соответственно, для гибридизации в жестких условиях. Таким образом, последовательности праймера и зонда не должны показывать точную комплементарную последовательность участка связывания на шаблоне и могут использоваться дегенерированные праймеры. Например, некомплементарный нуклеотидный фрагмент может быть присоединен к 5'-концу праймера, при этом остаток последовательности праймера является комплементарным цепи. Альтернативно, некомплементарные основания или более длинные последовательности могут быть встроены в праймер, при условии, что праймер характеризуется достаточной комплементарностью с последовательностью одной из цепей, которая должна быть амплифицирована для гибридизации с ней с образованием дуплексной структуры, которая может быть расширена путем полимеризации. Некомплементарные нуклеотидные последовательности праймеров могут содержать участки ферментов рестрикции. Добавление сайта фермента рестрикции к концу(концам) целевой последовательности будет особенно полезным для клонирования целевой последовательности. По сути, комплементарной последовательностью праймера является такая, которая характеризуется достаточной комплементарностью к матрице амплификации, чтобы привести к присоединению праймера и синтезу второй цепи. Специалисту в данной области техники известны требования к достаточной комплементарности последовательности праймеров по отношению к матрице амплификации.
Используемые в настоящем документе выражения "полинуклеотид", "полинуклеотидная последовательность", "последовательность нуклеиновой кислоты", "фрагмент нуклеиновой кислоты" и "выделенный фрагмент нуклеиновой кислоты" используются в данном документе взаимозаменяемо. Данные выражения охватывают нуклеотидные последовательности и подобные. Полинуклеотид может быть полимером РНК или ДНК, которая является одно- или двухцепочечной, необязательно содержит синтетические, неприродные или измененные нуклеотидные основания. Полинуклеотид в форме полимера ДНК может содержать один или несколько сегментов кДНК, геномной ДНК, синтетической ДНК или их смеси. Нуклеотиды (обычно находящиеся в форме 5'-монофосфата) обозначают одной буквой следующим образом: "А" для аденилата или дезоксиаденилата (для РНК или ДНК, соответственно), "С" для цитидилата или дезоксицитидилата, "G" для гуанилата или дезоксигуанилата, "U" для уридилата, "Т" для дезокситимидилата, "R" для пуринов (А или G), "Y" для пиримидинов (С или Т), "K" для G или T, "H" для A или C или T, "I" для инозина и "N" для любого нуклеотида.
Используемое в данном документе словосочетание "биологически активный вариант" или "функциональный вариант" по отношению к белку относится к аминокислотной последовательности, которая изменена в одной или нескольких аминокислотах относительно контрольной последовательности, но при этом сохраняет значительную биологическую активность контрольной последовательности. Вариант может иметь "консервативные" изменения, где замещенная аминокислота обладает сходными структурными или химическими свойствами, как например, при замене лейцина на изолейцин. Альтернативно, вариант может иметь "неконсервативные" изменения, например, замену глицина триптофаном. Аналогичные несущественные вариации также могут включать делеции или вставки аминокислот или обе. Руководство по определению того, какие аминокислотные остатки могут быть замещены, вставлены или удалены без подавления биологической или иммунологической активности, может быть получено с применением компьютерных программ, хорошо известных в данной области техники, например, программного обеспечения DNASTAR.
Используемое в данном документе выражение "праймер" относится к олигонуклеотиду, который может отжигаться в целевом участке амплификации, обеспечивая возможность прикрепления ДНК-полимеразы, тем самым выступая в качестве точки инициации синтеза ДНК при помещении в условия, в которых индуцируется синтез продукта элонгации праймера, то есть в присутствии нуклеотидов и средства для полимеризации, такого как ДНК-полимераза, и при соответствующих значениях температуры и pH. Праймер (для амплификации) является предпочтительно одноцепочечным для максимальной эффективности при амплификации. Предпочтительно, праймер является олигодезоксирибонуклеотидом. Праймер должен быть достаточно длинным, чтобы инициировать синтез продуктов достройки в присутствии средства для полимеризации. Точные длины праймеров будут зависеть от многих факторов, в том числе от температуры и композиции (содержания A/T по сравнению с G/C) праймера. Пара двунаправленных праймеров состоит из одного прямого и одного обратного праймера, которые обычно используются в такой области техники амплификации ДНК, как ПЦР-амплификация.
Выражения "жесткость" или "жесткие условия гибридизации" относятся к условиям гибридизации, которые влияют на стабильность гибридов, например, температура, концентрация солей, pH, концентрация формамида и подобные. Эти условия опытным путем оптимизируют для достижения максимального специфического связывания и минимального неспецифического связывания праймера или зонда со своей целевой последовательностью нуклеиновой кислоты. Используемые выражения включают ссылки на условия, при которых зонд или праймер гибридизируется со своей целевой последовательностью в большей степени, чем другие последовательности (например, по меньшей мере в 2-раза больше по сравнению с фоном), что поддается обнаружению. Строгие условия зависят от последовательности и будут отличаться при различных обстоятельствах. Более длинная последовательность гибридизируется исключительно при более высоких температурах. В целом, жесткие условия выбирают так, чтобы температура была приблизительно на 5°C ниже температуры точки плавления (Tm) для конкретной последовательности при определенных значениях ионной силы и pH. Tm - это температура (при определенной ионной силе и pH), при которой 50% комплементарной целевой последовательности гибридизируется с полностью соответствующим зондом или праймером. Как правило, жесткие условия будут означать такие, при которых концентрация солей составляет менее, чем приблизительно 1,0 М ионов Na+, как правило, с концентрацией ионов Na+ (или других солей) приблизительно от 0,01 до 1,0 М при pH от 7,0 до 8,3, а температура составляет по меньшей мере приблизительно 30°C для коротких зондов или праймеров (например, от 10 до 50 нуклеотидов) и по меньшей мере приблизительно 60°C для длинных зондов или праймеров (например, длиной более 50 нуклеотидов). Жесткие условия также могут быть достигнуты путем добавления дестабилизирующих средств, таких как формамид. Иллюстративные условия низкой жесткости или "условия пониженной жесткости" включают гибридизацию с буферным раствором 30% формамида, 1М NaCl, 1% SDS при 37°C и отмывание в 2× SSC при 40°C. Иллюстративные условия высокой жесткости включают гибридизацию в 50% формамиде, 1М NaCl, 1% SDS при 37°C и отмывание в 0,1× SSC при 60°C. Процедуры гибридизации хорошо известны в данной области техники и описаны, например, Ausubel et al., 1998 и Sambrook et al., 2001.
Используемое в данном документе выражение "кодирующая последовательность" относится к последовательности ДНК, которая кодирует специфическую аминокислотную последовательность. "Регуляторные последовательности" относятся к нуклеотидным последовательностям, которые расположены выше (5' некодирующие последовательности), в пределах или в ниже (3' некодирующие последовательности) относительно кодирующей последовательности, и которые влияют на транскрипцию, процессинг РНК или стабильность РНК или на трансляцию ассоциированной кодирующей последовательности.
Используемое в данном документе выражение "регуляторные последовательности" может включать, но не ограничивать, промоторы, лидерные последовательности для трансляции, интроны и распознаваемые последовательности для полиаденилирования.
Используемое в данном документе выражение "промотор" относится к последовательности ДНК, которая способна контролировать экспрессию кодирующей последовательности или функциональной РНК. Последовательность промотора состоит из проксимальных и более дистальных, расположенных выше, элементов, последние элементы часто называют энхансерами. Соответственно, "энхансер" представляет собой последовательность ДНК, которая может стимулировать активность промотора и может быть изначально присущим промотору элементом или гетерологичным элементом, встроенным в него в целях повышения активности или тканеспецифичности промотора. Промоторы могут быть получены в полном объеме из нативного гена или составлены из различных элементов, полученных из различных промоторов, встречающихся в природе, или даже могут включать сегменты синтетической ДНК. Специалистам в данной области техники понятно, что различные промоторы могут управлять экспрессией гена в различных тканях или типах клеток или на разных стадиях развития или в ответ на различные условия окружающей среды. Кроме того, считают, что поскольку в большинстве случаев точные границы регуляторных последовательностей не были полностью определены, фрагменты ДНК некоторых вариантов могут характеризоваться идентичной промоторной активностью.
Используемое в данном документе выражение "растительный промотор" является промотором, который обладает способностью инициировать транскрипцию в клетках растений, независимо от того, является ли его источником растительная клетка. Например, хорошо известно, что промоторы Agrobacterium функциональны в клетках растений. Таким образом, растительные промоторы охватывают промоторную ДНК, полученную из растений, растительных вирусов и бактерий, таких как Agrobacterium и Bradyrhizobium. Растительный промотор может быть конститутивным промотором или неконститутивным промотором.
Используемое в данном документе выражение "конститутивный промотор" является промотором, который проявляет активность в большинстве условий и/или на протяжении большинства стадий развития. Существует несколько преимуществ в применении конститутивных промоторов в векторах экспрессии, используемых в биотехнологии растений, такие как высокий уровень продуцирования белков, используемых для отбора трансгенных клеток или растений; высокий уровень экспрессии белков-репортеров или оцениваемых маркеров, которые обеспечивают легкое выявление и количественное определение; высокий уровень продуцирования транскрипционного фактора, который является частью системы регулирования транскрипции; продуцирование соединений, требующее активности во всех тканях растения; и производство соединений, которые необходимы на всех этапах развития растения. Неограничивающие примеры типичных конститутивных промоторов включают CAMV35S-промотор, промоторы опинов, промотор убиквитина, промотор актина, промотор алкогольдегидрогеназы и т.д.
Используемое в данном документе выражение "неконститутивный промотор" относится к промотору, который проявляет активность в определенных условиях, в определенных типах клеток и/или на протяжении определенных этапов развития. Например, тканеспецифичные, предпочтительные для определенной ткани, специфичные к определенному типу клеток, предпочтительные для определенного типа клеток, индуцируемые промоторы и промоторы, находящиеся под контролем развития, являются неконститутивными промоторами. Примеры промоторов, находящихся под контролем развития, включают промоторы, которые преимущественно инициируют транскрипцию в определенных тканях, таких как стебли, листья, корни или семена.
Используемое в данном документе выражение "индуцируемый" или "репрессируемый" промотор относится к промотору, который находится под контролем химических факторов или условий окружающей среды. Примеры условий окружающей среды, которые могут влиять на транскрипцию, опосредованную индуцируемыми промоторами, включают анаэробные условия или определенные химические вещества или присутствие света.
Используемое в данном документе выражение "тканеспецифический" промотор относится к промотору, который инициирует транскрипцию только в некоторых тканях. В отличие от конститутивной экспрессии генов тканеспецифичная экспрессия является результатом нескольких взаимодействующих уровней регуляции генов. Таким образом, в данной области техники иногда предпочтительнее применение промоторов из гомологичных или близкородственных видов растений для достижения эффективной и надежной экспрессии трансгенов в определенных тканях. Это одна из основных причин значительного количества тканеспецифичных промоторов, выделенных из определенных растений и тканей, что отражено в научной литературе и патентах. Неограничивающие примеры тканеспецифичных промоторов включают промоторы гена бета-амилазы или гена гордеина ячменя (для экспрессии генов в семенах), промоторы генов томата pz 7 и pz 130 (для экспрессии генов в завязи), промотор гена табака RD2 (для экспрессии генов в корнях), промотор гена банана TRX и промотор гена актина дыни (для экспрессии генов в фруктах) и эмбрионспецифические промоторы, например, промотор, связанный с геном пермеазы аминокислот (ААР1), геном олеат-12-гидроксилазьгдесатуразы из Lesquerella fendleri (LFAH12), геном альбумина 2S2 (2S2), геном элонгазы жирных кислот (FAE1) или геном семядольного листа (LEC2).
Используемое в данном документе выражение "предпочтительный для определенных тканей" промотор относится к промотору, который, в основном, инициирует транскрипцию, но не обязательно полностью или исключительно в определенных тканях.
Используемое в данном документе выражение "специфический к определенному типу клеток" промотор относится к промотору, который в первую очередь управляет экспрессией в определенных типах клеток в одном или нескольких органах, например, в сосудистых клетках в корнях, листьях, клетках стебля и стволовых клетках.
Используемое в данном документе выражение "предпочтительный для определенного типа клеток" промотор относится к промотору, который в первую очередь управляет экспрессией в определенных типах клеток в одном или нескольких органах, например, в сосудистых клетках в корнях, листьях, клетках стебля и стволовых клетках.
Используемое в данном документе выражение “3' некодирующие последовательности” или “3' нетранслируемые участки” относятся к последовательностям ДНК, расположенным ниже кодирующей последовательности, и содержащим распознаваемые последовательности для полиаденилирования и другие последовательности, кодирующие регуляторные сигналы, которые обладают способностью влиять на процессинг и РНК или экспрессию генов. Сигнал полиаденилирования обычно характеризуется влиянием на добавление трактов полиадениловой кислоты к 3' концу предшественника и РНК. Пример применения различных 3' некодирующих последовательностей приведен в Ingelbrecht, I.L., et al. (1989) Plant Cell 1:671-680.
Используемое в данном документе "РНК-транскрипт" относится к продукту, полученному в результате транскрипции последовательности ДНК, катализируемой РНК-полимеразой. Если РНК-транскрипт является полностью комплементарной копией последовательности ДНК, то он называется первичным транскриптом. РНК-транскриптом называют зрелую РНК, если эта последовательность РНК получена в результате посттранскрипционного процессинга первичного транскрипта. “Информационная РНК (иРНК)” относится к РНК, которая не содержит интроны и может транслироваться клеткой в белок. Выражение “кДНК” относится к ДНК, которая комплементарна и синтезирована на матрице иРНК с применением фермента обратной транскриптазы. КДНК может быть одноцепочечной или может быть преобразована в двухцепочечную форму с применением фрагмента Кленова ДНК-полимеразы I. “Смысловая” РНК означает РНК-транскрипт, который содержит иРНК и может быть транслирован в белок в клетке или в условиях in vitro. "Антисмысловая РНК" означает кРНКтранскрипт, который является комплементарным всему или части целевого первичного транскрипта или иРНК, и который блокирует экспрессию целевого гена (патент США №5107065). Антисмысловая РНК может быть комплементарна любой части специфического генного транскрипта, т.е. на 5' некодирующей последовательности, 3' некодирующей последовательности, интронам или кодирующей последовательности. Выражение "функциональная РНК" относится к антисмысловой РНК, рибосомальной РНК или другим типам РНК, которые не могут подвергаться трансляции, но оказывают влияние на процессы, проходящие в клетке. Выражения "комплементарная последовательность" и "обратная комплементарная последовательность" используются в данном документе взаимозаменяемо по отношению к иРНК-транскриптам, и предназначены для определения антисмысловой РНК транскрипта.
Используемое в данном документе выражение "функционально сцепленный" относится к такой взаимосвязи последовательностей нуклеиновых кислот в отдельном фрагменте нуклеиновой кислоты, при которой функция одной из последовательностей регулируется другой последовательностью. Например, промотор является функционально сцепленным с кодирующей последовательностью, когда он обладает способностью регулировать экспрессию этой кодирующей последовательности (то есть, кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующие последовательности могут быть функционально сцеплены с регуляторными последовательностями в смысловой или антисмысловой ориентации. В другом примере комплементарные участки РНК настоящего изобретения могут быть функционально сцепленными, прямо или опосредованно, 5' концом с целевой иРНК или 3' концом с целевой иРНК или внутри целевой иРНК, или первый комплементарный участок может быть связан 5' концом, а комплементарная ему последовательность связана 3' концом с целевой иРНК.
Используемое в данном документе выражение "рекомбинантный" относится к искусственной комбинации двух сегментов последовательности, разделенных другим способом, например, путем химического синтеза или с применением манипуляций выделенных сегментов нуклеиновых кислот методиками генной инженерии.
Используемые в данном документе словосочетания "рекомбинантный конструкт", "экспрессионный конструкт", "химерный конструкт", "конструкт" и "конструкт рекомбинантной ДНК" используются в данном документе взаимозаменяемо. Рекомбинантный конструкт содержит искусственную комбинацию фрагментов нуклеиновых кислот, например, регуляторных и кодирующих последовательностей, которые вместе отсутствуют в природе. Например, химерный конструкт может содержать регуляторные последовательности и кодирующие последовательности, которые получены из различных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного и того же источника, но расположенные таким образом, который отличается от встречающегося в природе. Такой конструкт может быть использован отдельно или может быть использован в сочетании с вектором. Если используют вектор, то выбор вектора зависит от способа, который будут использовать для трансформации клеток-хозяев, что хорошо известно специалистам в данной области техники. Например, можно использовать плазмидный вектор. Специалистам хорошо известно о генетических элементах, которые должны присутствовать в векторе для успешной трансформации, селекции и размножения клеток-хозяев, содержащих какие-либо из выделенных фрагментов нуклеиновых кислот настоящего изобретения. Специалисту в данной области также понятно, что различные независимые трансформационные события приведут к различным уровням и различному характеру экспрессии (Jones et al., (1985) EMBO J. 4:2411-2418; De Almeida et al., (1989) Mol. Gen. Genetics 218:78-86) и, таким образом, с целью получения линий, демонстрирующих нужный уровень и характер экспрессии, множественные события должны быть подвергнуты скринингу. Такой скрининг может быть выполнен с применением саузерн-блоттинга для анализа ДНК, нозерн-блоттинга для анализа экспрессии иРНК, иммуноблоттинга для анализа экспрессии белков или фенотипического анализа, помимо прочих. В качестве векторов могут быть использованы плазмиды, вирусы, бактериофаги, провирусы, фагмиды, транспозоны, искусственные хромосомы и подобные, которые автономно реплицируются или могут интегрироваться в хромосому клетки-хозяина. Вектор может также быть голым РНК-полинуклеотидом, голым ДНК-полинуклеотидом, полинуклеотидом, состоящим как из ДНК, так и из РНК в одной и той же цепи, полилизин-конъюгированной ДНК или РНК, пептид-конъюгированной ДНК или РНК, липосомно-конъюгированной ДНК или подобные, которые не подвергаются автономной репликации.
Используемое в данном документе выражение "экспрессия" относится к продуцированию функционального конечного продукта, например, иРНК или белка (предшественника или зрелой формы).
Используемое в данном документе словосочетание "растительный селектируемый или скринируемый маркер" относится к генетическому маркеру, который является функциональным в клетке растения. Селектируемый маркер позволяет клеткам, которые содержат и экспрессируют такой маркер, расти в условиях неблагоприятных для роста клеток, не экспрессирующих данный маркер. Скринируемый маркер облегчает идентификацию клеток, которые экспрессируют такой маркер.
Используемое в данном документе выражение "инбредный", "инбредное растение" используют в контексте настоящего изобретения. Оно также включает любые конверсии отдельных генов этого инбредного растения. Используемое в данном документе выражение "растение с единичным конвертированным аллелем" относится к тем растениям, которые получены с применением методики селекции растений, называемой обратным скрещиванием, где выделены по существу все желаемые морфологические и физиологические характеристики инбредного растения, в дополнение к единичному аллелю, перенесенному в инбредное растение методикой возвратного скрещивания.
Используемое в данном документе выражение "образец" включает образец из растения, части растения, клетки растения или из вектора, передающего генетическую информацию, или образец почвы, воды или воздуха.
Используемое в данном документе выражение "потомок" относится к любому растению, полученному в качестве потомка в результате вегетативного или полового размножения от одного или несколько родительских растений или происходящих от них растений. Например, растение-потомок может быть получено путем клонирования или самоопыления материнского растения или путем скрещивания двух родительских растений и включает самоопыляющиеся растения, а также F1 или F2 или следующие далее поколения. F1 представляет собой потомков первого поколения, полученных от родителей, одного из которых по меньшей мере в первый раз используют в качестве донора признака, в то время как потомки второго поколения (F2) или последующих поколений (F3, F4 и т.д.) являются растениями, полученными из самоопыляющихся растений поколений F1, F2 и т.д. Таким образом, потомок F1 может быть (и обычно является) гибридом, появившимся в результате скрещивания между родителями чистой линии (чистая линия является гомозиготной по признаку), тогда как F2 может быть (и обычно является) потомком, появившимся в результате самоопыления вышеупомянутых гибридов F1.
Используемое в данном документе выражение "скрещивание", "перекрестное опыление" или "кросс-бридинг" относятся к способу, посредством которого пыльца одного цветка с одного растения попадает (искусственно или естественно) на семяпочку (рыльце пестика) цветка другого растения.
Используемое в данном документе выражение "культурный сорт растения" относится к сорту, линии или расе растения, который был получен с применением методик садоводства или агротехники, и, как правило, не обнаруживается в диких популяциях.
Используемое в данном документе выражение "двудольное растение" или "двудольное" относится к цветковому растению, зародыш которого содержит две половины семени или семядоли. Двудольные растения по меньшей мере включают группы Eudicot, Magnoliid, Amborella, Nympheales, Austrobaileales, Chloranthales и Ceratophyllum. Eudicot включают следующие клады: Ranunculales, sabiales, Proteales, Trochodendrales, Buxales и Core Eudicots (например, Berberidopsidales, Dilleniales, Gunnerales, Caryophyllales, Santalales, Saxifragales, Vitales, Rosids и Asterids). Неограничивающие примеры двудольных растений включают табак, томат, горох, люцерну, клевер, фасоль, сою, арахис, представителей семейства Brassicaceae (например, рыжик, рапс, масличный рапс и т.д.), амарант, подсолнечник, сахарную свеклу, хлопок, дуб, клен, розу, мяту, тыкву, маргаритку, орехи; кактусы, фиалки и лютики.
Используемое в данном документе выражение "однодольное растение" или "однодольное" относится к любому цветковому растению подкласса (Monocotyledoneae), зародыш которого содержит только одну семядолю и обычно имеет листья с параллельным жилкованием, части цветка собранные по три, и у которых отсутствует вторичный рост ствола и корня. Неограничивающие примеры однодольных растений включают лилии, орхидеи, кукурузу, рис, пшеницу, ячмень, сорго, просо, овес, рожь, тритикале, гречиху, фонио, лебеду, травы, такие как овсяница, эгилопс и мятлик луговой; зерновые, такие как пшеница, овес и ячмень, ирисы, лук, пальмы.
Используемое в данном документе выражение "ген" относится к любому сегменту ДНК, связанному с биологической функцией. Таким образом, к генам относятся, но без ограничений, кодирующие последовательности и/или регуляторные последовательности, необходимые для их экспрессии. Гены могут также содержать неэкспрессирующиеся сегменты ДНК, которые, например, образуют последовательности узнавания для других белков. Гены могут быть получены из различных источников, в том числе путем клонирования из источника интереса, или синтеза на основании известной или предсказанной информации о последовательности, и могут содержать последовательности, созданные для получения требуемых параметров.
Используемое в данном документе выражение "генотип" относится к генетическому составу отдельной клетки, культуры клеток, ткани, организма (например, растения) или группы организмов.
Используемое в данном документе выражение "гемизиготный" относится к клетке, ткани или организму, в котором ген встречается в генотипе только один раз, как ген в гаплоидной клетке или организме, сцепленный с полом ген в гетерогаметном поле или ген, локализованный на сегменте хромосомы в диплоидной клетке или организме, где парный ему сегмент был удален.
Используемые в данном документе выражения "гетерологичный полинуклеотид" или "гетерологичная нуклеиновая кислота" или "экзогенный сегмент ДНК" относятся к полинуклеотиду, нуклеиновой кислоте или сегменту ДНК, которые происходят из источника, чужеродного по отношению к конкретной клетке-хозяину, или, если получены из того же источника, являются модифицированными по отношению к оригинальной форме. Таким образом, гетерологичный ген в клетке-хозяине содержит ген, который является эндогенным для конкретной клетки-хозяина, но не был модифицирован. Таким образом, выражения относятся к сегменту ДНК, который является чужеродным или гетерологичным для клетки или гомологичным для клетки, но находится в таком положении в нуклеиновой кислоте клетки-хозяина, в котором элемент обычно не обнаруживается. Экзогенные сегменты ДНК экспрессируются для получения экзогенных полипептидов.
Используемое в данном документе выражение "гетерологичный признак" относится к фенотипу, которым наделяет трансформированную клетку-хозяин или трансгенный организм экзогенный сегмент ДНК, гетерологичный полинуклеотид или гетерологичная нуклеиновая кислота.
Используемое в данном документе выражение "гетерозигота" относится к диплоидной или полиплоидной отдельной клетке или растению, которые имеют различные аллели (формы данного гена), присутствующие по меньшей мере в одном локусе.
Используемое в данном документе выражение "гетерозиготный" относится к наличию различных аллелей (форм данного гена) в определенном локусе гена.
Используемые в данном документе выражения "гомолог" или "гомологичный" относится к последовательности нуклеиновой кислоты или пептида, которая имеет общее происхождение и является функционально аналогичной последовательности нуклеиновой кислоты или пептида из других видов.
Используемое в данном документе выражение "гомозигота" относится к отдельной клетке или растению, которые имеют одинаковые аллели в одном или нескольких или всех локусах. Когда выражение используют в отношении конкретного локуса или гена, это по меньшей мере означает, что данный локус или ген имеет одинаковые аллели.
Используемые в данном документе выражения "гомозиготный" или "HOMO" относятся к наличию одинаковых аллелей в одном или нескольких или всех локусах в гомологичных сегментах хромосом. Когда выражение используется в отношении конкретного локуса или гена, это по меньшей мере означает, что данный локус или ген имеет одинаковые аллели
Используемое в данном документе выражение "гибрид" относится к любой отдельной клетке, ткани или растению, которые получены в результате скрещивания родительских организмов, которые отличаются по одному или нескольким генам.
Используемое в данном документе выражение "инбредный" или "инбредная линия" относится к чистой линии.
Используемое в данном документе выражение "линия" употребляется в широком смысле, включая, но не ограничивая, группу растений, произошедших от одного родительского растения путем вегетативного размножения с применением методик культивирования тканей, или группу инбредных растений, которые не являются очень сходными генетически в связи с их происхождением от общего родителя (родителей). Растение называют "относящимся" к определенной линии, если оно (а) является растением, представляющим собой первичный трансформант (Т0), воспроизведенным из материала этой линии, (b) имеет генеалогическую схему, включающую растение Т0 этой линии, или (с) является генетически очень сходным в связи с общим происхождением (например, путем инбридинга или самоопыления). В этом контексте выражение "генеалогическая схема" обозначает генеалогию растений, например, в отношении полового скрещивания, подвергающуюся изменениям таким образом, что ген или комбинация генов в гетерозиготном (гемизиготном) или гомозиготном состоянии, придает растению желаемый признак.
Используемые в данном документе выражения "мутантный" или "мутация" относятся к гену, клетке или организму с аномальной генетической конституцией, которая может привести к фенотипическому варианту.
Используемое в данном документе выражение "свободное опыление" относится к популяции растений, которая свободно подвергается определенному потоку генов в отличие от закрытой популяции, в которой есть эффективный барьер по отношению к потоку генов.
Используемые в данном документе выражения "свободноопыляемая популяция" или "свободноопыляемый сорт" относятся к растениям, которые обычно обладают способностью по меньшей мере к некоторому перекрестному оплодотворению, выбранному в качестве стандарта, которые могут демонстрировать различия, но которые обладают одной или несколькими генотипическими или фенотипическими характеристиками по которым популяция или сорт может быть дифференцирована от других. Гибрид, который не имеет барьеров по отношению к перекрестному опылению, относится к свободноопыляемой популяции или свободноопыляемому сорту.
Используемое в данном документе при обсуждении растений выражение "семяпочка" относится к женскому гаметофиту, тогда как выражение "пыльца" означает мужской гаметофит.
Используемое в данном документе выражение "фенотип" относится к наблюдаемым признакам отдельных клеток, культуры клеток, организма (например, растения) или группы организмов, что является результатом взаимодействия индивидуального набора генов (то есть, генотипа) и окружающей среды.
Используемое в данном документе выражение "растительная ткань" относится к любой части растения. Примеры органов растений включают, но не ограничивают, лист, стебель, корень, клубень, семя, ветку, опушенность, узелок, пазуху листьев, цветок, пыльцу, тычинку, пестик, лепесток, плодоножку, черешок, рыльце пестика, пестик, прицветник, фрукт, ствол, плодолистик, чашелистик, пыльник, семяпочку, цветоножку, иглу, шишку, корневище, столон, побег, околоплодник, эндосперм, плаценту, ягоду, тычинку и влагалище листа.
Используемое в данном документе выражение "самоскрещивание", "самоопыляющийся" или "самоопыление" означает, что пыльца из одного цветка попадает (искусственно или естественно) на семяпочку (рыльце пестика) того же или другого цветка на том же самом растении.
Используемое в данном документе выражение "трансформация" относится к переносу нуклеиновой кислоты (то есть, нуклеотидного полимера) в клетку. Используемое в данном документе выражение "генетическая трансформация" относится к переносу и встраиванию ДНК, особенно рекомбинантной ДНК в клетку.
Используемое в данном документе выражение "трансформант" относится к клетке, ткани или организму, которые подвергли трансформации. Оригинальный трансформант обозначается как "Т0" или "Т0". Самоопыление Т0 приводит к получению первого трансформированного поколения, которое обозначается как "Т1" или "T1".
Используемое в данном документе выражение "трансген" относится к нуклеиновой кислоте, которая встраивается в организм, клетку-хозяина или вектор таким образом, который обеспечивает ее функцию.
Используемое в данном документе выражение "трансгенный" относится к клеткам, клеточным культурам, организмам (например, растениям) и потомкам, которые получили чужеродный или модифицированный ген с применением одного из различных способов трансформации, где чужеродный или модифицированный ген происходит из того же вида или вида, отличного от вида организма, который получает чужеродный или модифицированный ген.
Используемое в данном документе выражение "транспозиционное событие" относится к перемещению транспозона от донорского сайта к целевому сайту.
Используемое в данном документе выражение "сорт" относится к подгруппе вида, состоящей из группы индивидуумов, принадлежащих к одному виду, которые отличаются по своей форме или функции от других подобных совокупностей индивидуумов.
Используемое в данном документе выражение "вектор", "плазмида" или "конструкт" относится в широком смысле к любой плазмиде или вирусу, кодирующему экзогенную нуклеиновую кислоту. Это выражение необходимо толковать как содержащий неплазмидные и невирусные соединения, которые обеспечивают перенос нуклеиновой кислоты в вирионы или клетки, такие как, например, соединения на основе полилизина и подобные. Вектором может быть вирусный вектор, который подходит в качестве средства для доставки нуклеиновой кислоты или ее мутантной формы в клетку, или вектором может быть невирусный вектор, который подходит для той же цели. Примеры вирусных и невирусных векторов для доставки ДНК в клетки и ткани хорошо известны в данной области техники и описаны, например, в Ма et al. (1997, Proc. Natl. Acad. Sci. U.S.A. 94:12744-12746).
Используемое в данном документе словосочетание "размер семени" относится к объему самого семенного материала, которое определяется как пространство, занимаемое составными частями семени.
Используемое в данном документе словосочетание "число семян" относится к среднему числу семян, получаемых из каждого плода, каждого растения или каждой заданной площади (например, 1 акр).
Используемое в данном документе словосочетание "натуральный вес" или "натуральный вес зерна" является определением объемной веса (масса/объем), которая измеряется для коммерческих целей при определенных условиях, установленных в США Федеральной инспекцией зерна Министерства сельского хозяйства США (USDA-GIPSA). Натуральный вес является общим показателем качества зерна, причем более высокий натуральный вес обычно означает более высокое качество зерна. Натуральный вес зерна в единицах фунтов на бушель указывает вес "объема", выраженного в бушелях, который составляет 32 кварты (30283 кубических сантиметров) зерна. При продаже зерна образцы, как правило, проверяют на качество, и оценка натурального веса является одним из проводимых испытаний. Показатели натурального веса были частью классов зерна в США с момента принятия Конгрессом в 1916 году закона США о стандартах на зерно. Классы США для большинства видов зерна указывают минимальные значения натурального веса для каждого класса. Например, официальная минимальная допустимая натурального веса в США для желтой кукурузы №1 составляет 56 фунтов/бушель, а для желтой кукурузы №2 составляет 54 фунта/бушель (USDA-GIPSA, 1996). По закону "весовой" бушель кукурузы составляет точно 56 фунтов, бушель сои составляет 60 фунтов, а бушель пшеницы составляет 60 фунтов, независимо от натурального веса. "Весовой" бушель используется в основе оплаты за зерно, но ценовые уступки часто связаны с поставками зерна низкого класса, обладающего низким натуральным весом.
Используемое в данном документе словосочетание "кажущаяся плотность зерна" относится к плотности зерна, которую определяют таким способом, при котором насыпную плотность (масса/объем) семян зерновых иногда измеряют с помощью газового пикнометра, который, как правило, использует гелий и измеряет объем образца. Ядра зерна содержат внутренние пустоты и межклеточные пространства и не являются полностью проницаемыми для гелия. Поскольку газ не может достигнуть всех внутренних пространств, то объем материала, образующего ядро, может быть завышен при применении газовой пикнометрии и определяемая плотность ниже, чем "истинная плотность" зернового материала (Chang, CS (1988) Cereal Chem: 65:13-15).
Используемое в данном документе словосочетание "истинная плотность зерна" относится к насыпной плотности зерна, которая выражается в виде отношения веса, разделенной на объем, при условии, что все пустое пространство, не содержащее твердых материалов семян, было уже исключено или вычтено при определении объема, используемого для расчета (Chang, CS (1988) Cereal Chem:65:13-15).
Используемое в данном документе выражение "циклин-зависимый ингибитор киназы" (который в данном документе также обозначается как "ингибитор CDK" или "CKI") относится к классу белков, которые отрицательно регулируют циклин-зависимые киназы (CDK). CDK, подходящими под описание настоящего изобретения, являются такие циклин-зависимые киназы, отдельные полипептидные участки которых обладают способностью независимо связывать циклин и CDK. Такие CKI включают, например, идентифицированные семейства растительных CKI (семь идентифицированных CKI Arabidopsis), обладающих гомологией к белка-ингибиторам киназ (KIP) у животных, называемых KIP-родственные белки (KRP) (также известные как ингибиторы "CDK" или "ICK").
Выражение "естественного происхождения" в контексте полипептидов и нуклеиновых кислот CKI означает полипептид или нуклеиновую кислоту, имеющие аминокислотную или нуклеотидную последовательность, которая встречается в природе, то есть, аминокислотную или нуклеотидную последовательность, которая может быть выделена из природного источника (организма) и которая не была намеренно модифицирована вмешательством человека. Используемые в данном документе лабораторные линии растений, которые возможно были избирательно выведены согласно классической генетике, считаются растениями естественного происхождения.
Используемое в данном документе выражение "ген CKI дикого типа" или "нуклеиновая кислота CKI дикого типа" относится к последовательности нуклеиновой кислоты, соответствующей генетическому локусу CKI в геноме организма, который кодирует генный продукт, выполняющий нормальную функцию белка CKI, кодируемого нуклеотидной последовательностью естественного происхождения, соответствующей генетическому локусу. Генетический локус может иметь более чем одну последовательность или аллель в популяции индивидуумов, следовательно, выражение "дикий тип" охватывает все такие аллели естественного происхождения, которые кодируют генный продукт, выполняющий нормальную функцию. Выражение "дикий тип" также охватывает последовательности генов, которые необязательно имеют естественное происхождение, но которые кодируют генный продукт с нормальной функцией (например, гены, имеющие молчащие мутации или кодирующий белки с консервативными заменами).
Используемое в данном документе выражение "полипептид CKI дикого типа" или "белок CKI дикого типа" относится к полипептиду CKI, который кодируется геном дикого типа. Генетический локус может иметь более чем одну последовательность или аллель в популяции индивидуумов, следовательно, выражение "дикий тип" охватывает все такие аллели естественного происхождения, которые кодируют генный продукт, выполняющий нормальную функцию.
Способы селекции
Классические способы селекции могут быть включены в настоящее изобретение для введения одного или нескольких рекомбинантных KRP по настоящему изобретению в другие сорта растения в или другие близкородственные виды, которые совместимы для того, чтобы подлежать скрещиванию с трансгенным растением настоящего изобретения.
Свободноопыляемые популяции. Улучшение свободноопыляемых популяций таких сельскохозяйственных культур как рожь, многие виды маиса и сахарной свеклы, кормовые культуры, бобовые, такие как люцерна и клевер, а также тропические древесные культуры, такие как какао, кокосовые орехи, масличная пальма и каучуконосные деревья, существенно зависит от изменения частот генов в отношении фиксации благоприятных аллелей при сохранении высокой (но далеко не максимальной) степени гетерозиготности. Однородность в таких популяциях невозможна и сортовая чистота в свободноопыляемом сорте является статистической особенностью популяции в целом, не являясь характеристикой отдельных растений. Таким образом, неоднородность свободноопыляемых популяций контрастирует с однородностью (или фактической однородностью) инбредных линий, клонов и гибридов.
Способы улучшения популяции по своей природе разделяют на две группы: те, которые основаны на чисто фенотипическом отборе, обычно называемом массовым отбором, и те, которые основаны на отборе с тестированием потомков. Внутрипопуляционное улучшение использует концепцию популяций с открытым скрещиванием; что позволяет генам циркулировать из одной популяции в другую. Растения в одной популяции (сорт культурного растения, линия, экотип или любой источник зародышевой плазмы) скрещиваются или естественным путем (например, ветром), или вручную, или пчелами (часто Apis mellifera L. или Megachile rotundata F.) с растениями из других популяций. Селекцию применяют для улучшения одной (или иногда обеих) популяции(-ий) путем выделения растений с желаемыми признаками из обоих источников.
Существует два принципиально основных способа улучшения свободноопыляемой популяции. Во-первых, это ситуация, при которой популяцию изменяют целиком с применением выбранного метода селекции. Результатом является улучшенная популяция, которая бесконечно размножается путем случайных скрещиваний внутри изолированной популяции. Во-вторых, при синтетическом сорте достигают того же конечного результата, как и при улучшении популяции, но сам по себе синтетический сорт не способен к размножению как таковому, поэтому он должен быть реконструирован из родительской линии или клонов. Данные способы селекции растений, направленные на улучшение свободноопыляемых популяций, хорошо известны специалистам в данной области техники, а полные обзоры селекционных способов, которые обычно используют для улучшения растений с перекрестным опылением, представлены в многочисленных текстах и статьях, включая Allard, Principles of Plant Breeding, John Wiley & Sons, Inc. (1960); Simmonds, Principles of Crop Improvement, Longman Group Limited (1979); Hallauer и Miranda, Quantitative Genetics in Maize Breeding, Iowa State University Press (1981); и Jensen, Plant Breeding Methodology, John Wiley & Sons, Inc. (1988).
Массовый отбор. При массовом отборе выбирают желаемые отдельные растения, собирают их и получают семена без тестирования потомков для получения следующего поколения. Поскольку отбор основан только на материнских растениях и контроль опыления отсутствует, массовый отбор является формой случайного скрещивания с селекцией. Как указано в данном описании, целью массового отбора является увеличение доли улучшенных генотипов в популяции.
Синтетические сорта. Синтетический сорт получают путем скрещивания между собой определенного количества генотипов, выбранных по их хорошей комбинационной способности во всех возможных гибридных комбинациях с последующим поддержанием сорта путем свободного опыления. Принадлежат ли родители (более или менее инбредные) к линиям, размножающимся семенами, как некоторые сорта сахарной свеклы и фасоли (горошка), или размножаются клонированием, как кормовые культуры, клевер и люцерна, не имеет никакого принципиального значения. Родителей выбирают по общей комбинационной способности, иногда с применением анализирующего скрещивания или линейного скрещивания или, в более общем смысле, с применением поликросс-методики. Родительские семенные линии могут намеренно подвергаться инбредному скрещиванию (например, путем самоопыления или скрещивания сибсов). Однако даже если родительские растения не подвергаются намеренно инбредному скрещиванию, отбор в линии во время поддержания линии будет обеспечивать некоторую степень инбридинга. Вегетативно размножающиеся родительские растения, конечно, остаются неизменными и с высокой степенью гетерозиготности.
Может ли синтетический сорт быть непосредственно отправлен с участка производства родительских семян в фермерское хозяйство или должен сначала пройти один или два цикла размножения, зависит от производства семян и масштабов спроса на семена. На практике, травы и клевер, как правило, проходят один или два цикла размножения и, таким образом, значительно отдаляются от первоначального синтетического сорта.
В то время как иногда используют массовый отбор, проверка по потомкам является обычно предпочтительнее для поликроссов из-за функциональной простоты и очевидного соответствия по отношению к цели, а именно использования общей комбинационной способности синтетического сорта.
Количество родительских линий или клонов, которые входят в синтетический сорт, колеблется в широких пределах. На практике число родительских линий варьирует в диапазоне от 10 до нескольких сотен, при этом в среднем составляет 100-200. Можно было бы ожидать, что синтетические сорта, основанные на большом количестве родительских линий, которые формируются из 100 или более клонов, будут более стабильными при размножении семенами, чем синтетические сорта, основанные на малом количестве родительских линий.
Сорта с родословной. Сорт с родословной имеет превосходный генотип, разработанным путем селекции отдельных растений из расщепляющейся популяции, с последующим размножением и увеличением числа семян самоопыляющихся потомков и тщательного тестирования генотипа в течение нескольких поколений. Данный способ свободного опыления хорошо работает у видов с естественным самоопылением. Этот способ может быть использован в комбинации с массовым отбором в разработке сорта. Варианты комбинаций генеалогической схемы и массового отбора являются наиболее распространенными способами для создания сортов сельскохозяйственных культур с самоопылением.
Гибриды. Гибрид является отдельным растением, полученным в результате скрещивания родителей с различными генотипами. Коммерческие гибриды в настоящее время широко используют во многих сельскохозяйственных культурах, включая кукурузу (маис), сорго, сахарную свеклу, подсолнечник и брокколи. Гибриды могут образовываться различными способами, в том числе путем прямого скрещивания двух родителей (простые гибриды), скрещивания простого гибрида с другим родителем (тройные гибриды) или скрещивания двух различных гибридов (двойные межлинейные гибриды).
Строго говоря, большинство индивидуумов в популяции с неродственным скрещиванием (то есть, свободноопыляемые) являются гибридами, но данное выражение, как правило, употребляют в тех случаях, в которых родители являются индивидуумами, геномы которых достаточно различны, чтобы их признали как различные виды или подвиды. Гибриды могут быть фертильными или стерильными, в зависимости от качественного и/или количественного различия в геномах двух родителей. Гетерозис или гибридная сила, как правило, связан с увеличением степени гетерозиготности, что приводит к увеличению силы роста, выживаемости и фертильности гибридов по сравнению с родительскими линиями, которые были использованы для получения гибридов. Максимальной степени гетерозиса обычно достигают путем скрещивания двух генетически различных, высоко инбредных линий.
Получение гибридов представляет собой хорошо развитую отрасль хозяйства, связанную с изолированным получением родительских линий и гибридов, которые являются результатом скрещивания этих линий. Подробное обсуждение способа получения гибридов представлено, например, в Wright, Commercial Hybrid Seed Production 8:161-176, In Hybridization of Crop Plants.
Целенаправленно индуцированные локальные повреждения в геномах (TILLING)
TILLING (целенаправленно индуцированные локальные повреждения в геномах) - это способ в молекулярной биологии, позволяющий осуществлять направленную идентификацию мутаций в определенном гене. Способ TILLING® был представлен в 2000 году с применением модельного растения Arabidopsis thaliana. С тех пор TILLING® используют как способ обратной генетики в других организмах, таких как рыбы данио, кукуруза, пшеница, рис, соя, томаты и салат.
Данный способ сочетает стандартную и эффективную методику мутагенеза с химическим мутагеном (например, этилметансульфонат (EMS)) с чувствительной скрининг-методикой ДНК, которая определяет мутации по единичным основаниям (также называемые точечные мутации) в гене-мишени. EcoTILLING является способом в котором используется методика TILLING® для поиска естественных мутаций у индивидуумов, как правило, для анализа генетики популяции. Смотри Comai, et al., 2003. Efficient discovery of DNA polymorphisms in natural populations by Ecotilling. The Plant Journal 37, 778-786. Gilchrist et al. 2006. Use of Ecotilling as an efficient SNP discovery tool to survey genetic variation in wild populations of Populus trichocarpa. Mol. Ecol. 15, 1367-1378. Mejlhede et al. 2006. EcoTILLING for the identification of allelic variation within the powdery mildew resistance genes mlo and Mla of barley. Plant Breeding 125, 461-467. Nieto et al. 2007, EcoTILLING for the identification of allelic variants of melon eIF4E, a factor that controls virus susceptibility. BMC Plant Biology 7, 34-42; каждая из которых включен в данный документ посредством ссылки для всех целей. DEcoTILLING является модификацией TILLING® и EcoTILLING, в котором используется недорогой способ идентификации фрагментов (Garvin et al., 2007, DEco-TILLING: An inexpensive method for SNP discovery that reduces ascertainment bias. Molecular Ecology Notes 7, 735-746).
Способ TILLING® основан на образовании гетеродуплексов, которые образуются при амплификации множественных аллелей (которые могут происходить от гетерозиготы или пула нескольких гомозигот и гетерозигот) в ПЦР, нагревании, а затем медленном охлаждении. В участке нарушения комплементарности двух цепей ДНК образуется "пузырь" (индуцированная мутация в TILLING® или спонтанная мутация или SNP в EcoTILLING), который затем расщепляют одноцепочечными нуклеазами. Эти продукты затем разделяют по размеру на различных платформах.
В мире существует несколько центров, использующих методику TILLING®, которые сосредоточили усилия на агротехнически важных видах: UC Davis (USA), который сосредоточил усилия на рисе; Purdue University (USA), который сосредоточил усилия на маисе; University of British Columbia (CA), который занимается Brassica napus; John Innes Centre (UK), который сосредоточил усилия на Brassica rapa; Fred Hutchinson Cancer Research, который сосредоточил усилия на Arabidopsis; Southern Illinois University (USA), который сосредоточил усилия на сое; John Innes Centre (UK), который сосредоточил усилия на лотосе и люцерне; и INRA (France), который сосредоточил усилия на горохе и томатах.
Более подробное описание способов и композиций, касающихся TILLING®, можно найти в ссылках №1-35b, US 5994075, US 2004/0053236 A1, WO 2005/055704 и WO 2005/048692, каждая из которых включена в данный документ посредством ссылки для всех целей.
Триба Triticeae
Интенсивное использование диких растений трибы Triticeae отмечалось в странах Леванта 23000 лет назад. Triticeae является трибой в подсемействе трав Pooideae, которое включает рода со многими одомашненными видами. В данную трибу входят основные рода сельскохозяйственных культур, которые, включают пшеницу (см. таксономию пшеницы), ячмень и рожь; сельскохозяйственные культуры других родов включают некоторые культуры, используемые в пищу человеком и другие, используемые для кормления животных или защиты пастбищных угодий. Среди культивируемых в мире видов данная триба имеет одну из наиболее сложных генетических историй. Примером может служить пшеница обыкновенная, которая содержит геномы трех видов, только один из которых первоначально относился к виду пшеница рода Triticum.
Рода, относящиеся к трибе Triticeae, включают, но не ограничивают, Aegilops (эгилопсы - эгилопс коленчатый, эгилопс Тауша и др.); Agropyron (пырей гребенчаты - пырей пустынный, пырей ползучий и др.); Amblyopyrum (тупочешуйник безостый - тупочешуйник и др.); Australopyrum (австралийский пырей - пшеница английская, пырей гребенчатый и др.); Cockaynea (смотри Stenotachys; Cockaynea - это более молодое и, следовательно, недействительное название для Stenotachys и др.); Crithopsis (Crithopsis delileana и др.); Dasypyrum (сорго лимонное; и др.); Elymus (Elymus (виды волоснеца - Elymus glaucus, Elymus interruptus и др.); Elytrigia; Eremium (Eremium erianthus и др.); Eremopyrum (плевел - Eremopyrum bonaepartis, Eremopyrum triticeum и др.); Festucopsis; Haynaldia; Henrardia; Heteranthelium; Hordelymus; Hordeum (ячмень - ячмень обыкновенный, ячмень гривастый и др.); Hystrix (miscanthus sinensus, шероховатка и др.); Kengyilia; Leymus (колосняк - американский колосняк мягкий, колосняк песчаный и др.); Lophopyrum (пырей удлиненный); Malacurus Pascopyrum (пырей Смита и др.); Peridictyon; Psathyrostachys (Psathyrostachys juncea и др.); Pseudoroegneria (Pseudoroegneria spicata и др.); Secale (виды ржи - Secale cereale, гималайская рожь и др.); Sitanion; Stenotachys (Stenostachys gracilisu др.); Taeniatherum (Taeniatherum caput-medusae и др.); Thinopyrum (Thinopyrum intermedium, Thinopyrum junceiforme, Thinopyrum pycnanthum и др.); Triticum (пшеница - пшеница обыкновенная, пшеница твердая и др.).
Triticeae и сестринская ей триба Bromeae (возможные культурные сорта: Bromus mango из Южной Америки) при объединении образуют сестринскую кладу с Роеае и Aveneae (дубы). С ранних стадий эти таксоны характеризует межродовой поток генов. Например, Роеае и Aveneae имеют общий генетический маркер с ячменем и 10 другими представителями трибы Triticeae, тогда как все 19 родов трибы Triticeae несут маркер пшеницы вместе с Bromeae. Рода в пределах Triticeae содержат диплоидный, аллотетраплоидный и/или аллогексаплоидный геномы, при этом способность образовывать аллополиплоиные геномы варьирует в пределах трибы. В данной трибе большинство испытанных диплоидных видов являются близко родственными по отношению к эгилопсу, а более отдаленные представители (ранние точки ветвления) включают Hordeum (ячмень), Eremian, Psathyrostachys.
Многие рода и виды из Triticeae являются типичными аллополиплоидами, имеющими большее число хромосом, чем то, которое наблюдается у типичных диплоидов. Как правило, аллополиплоиды являются тетраплоидными или гексаплоидными, ААВВ или AABBDD. Создание полиплоидных видов является результатом естественных случайных событий, которые происходят с растениями, способными к полиплоидии. Аналогично, природные аллополиплоидные растения могут иметь селективные преимущества и могут позволить рекомбинацию отдаленно связанного генетического материала, обеспечивая позднее возвращение обратно к диплоидному набору. Пшеница тургидум является примером стабильной аллотетраплоидной пшеницы.
Род Aegilops, по-видимому, лежит в основе нескольких таксонов, таких как Triticum, Ambylopyrum и Crithopsis. Некоторые виды, такие как Aegilops speltoides, могут потенциально представлять основные варианты таксона. Размещение рода может быть скорее вопросом номенклатуры. Рода Aegilops и Triticum являются очень близкородственными, так как виды рода Aegilops занимают большую часть базальных точек ветвления в эволюции пшеницы обыкновенной, указывая на то, что род Triticum образовался из рода Aegilops приблизительно 4 миллиона лет назад. За расхождением геномов следует аллотетраплоидизация гибридов Aegilops speltoid x основные виды пшеницы Triticum boeoticum, при этом линии в ближневосточном регионе являются источником культурной пшеницы двузернянки.
Triticum spp.
Triticum sp. является травянистым растением, которое культивируется по всему миру. В 2007 году мировой объем производства пшеницы составлял 607 миллионов тонн, делая ее третьей по производству зерновых культур после кукурузы (784 миллионов тонн) и риса (651 миллионов тонн). В мировом масштабе пшеница является основным источником растительного белка в пище человека, содержание которого в пшенице выше, чем содержание белка в кукурузе или рисе, других основных зерновых культурах. В отношении общей веса продукции, используемой в пищу, пшеница в настоящее время является второй после риса в составе основных продуктов питания человека.
Пшеница выращивается в ограниченном количестве в качестве корма для сельскохозяйственных животных, а ее солома может быть использована в качестве материала для конструкции соломенной кровли. Шелуха зерна, отделенная при перемалывании белой муки, называется отрубями. Зародыш пшеницы является зародышевой частью ядра пшеницы. Он представляет собой концентрированный источник витаминов, минералов и белка и поддерживается более крупным, крахмалсодержащим участком ядра - эндоспермом.
Неограничивающие примеры видов Triticum включают, Т. aestivum (например, пшеница обыкновенная или хлеб, также известный как Triticum aestivum L. subsp. Aestivum; пшеница карликовая, также известная как Triticum aestivum subsp. compactum (Host) MacKey; маха, также известная как Triticum aestivum subsp. macha (Dek. и Men) MacKey; пшеница Вавилова, также известная как Triticum aestivum subsp. vavilovi (Tuman) Sears; пшеница круглозерная, также известная как Triticum aestivum subsp. sphacrococcum (Perc.) MacKey), T. aethiopicum, T. araraticum, T. boeoticum (например, дикая пшеница однозернянка, также известная как Triticum boeotictim Boiss), T. carthlicum, T. compactum, T. dimitrium, T. dicoccoides (например, дикая пшеница двузернянка, также известная как Triticum dicoccoides (Koern. ex Ascb. & Graebn.) Aaronsohn.), T. dicoccum (например, двузернянка), T. durum (например, твердая пшеница), Т. ispahanicum, Т. karamyschevii, Т. macha, T. militinae, Т. monococcum (например, однозернянка, также известная как Triticum monococcum L.), Т. polonicum, T. spelta, Т. sphaerococcum, T. timopheevii (например, пшеница Тимофеева, также известная как Triticum timopheevii (Zbuk.) Zbuk.), T. turanicum (например, восточная пшеница, также известная как Triticum turanicum jakubz), T. turgidum (например, пшеница английская, также известная как Triticum turgidum L.), T. urartu, T. vavilovii и Т. zhukovskyi.
Генетика пшеницы является более сложной, чем у большинства других домашних видов. Некоторые виды пшеницы являются диплоидными с двумя наборами хромосом, но многие из них представляют собой стабильные полиплоиды с четырьмя наборами хромосом (тетраплоиды) или шестью наборами хромосом (гексаплоиды). Большинство тетраплоидных видов пшеницы (например, полба и твердая пшеница) происходят от дикой двузернянки, Т. dicoccoides. Дикая двузернянка является результатом гибридизации двух диплоидных диких травянистых растений, Т. urartu и дикого эгилопса, таких видов как Aegilops searsii или Ае. speltoides. Неизвестные травянистые растения никогда не были идентифицированы среди существующих в настоящее время диких трав, но ближайшим родственным видом, существующим в настоящее время, является Aegilops speltoides. Гибридизация, в результате которой образовалась дикая двузернянка (ААВВ), произошла в дикой природе задолго до одомашнивания под действием естественного отбора. Гексаплоидная пшеница появилась на фермерских полях. Пшеница обыкновенная (Triticum aestivum, 2n = 42, AABBDD) является одной из наиболее важных зерновых сельскохозяйственных культур в мире. Одомашненная двузернянка или твердая пшеница гибридизируется с еще одной дикой диплоидной травой (Aegilops cylindrica) для получения гексаплоидной пшеницы, пшеницы двузернянки и пшеницы обыкновенной. Они имеют три набора парных хромосом, что в три раза больше, чем у диплоидной пшеницы. В настоящее время используют синтетические гексаплоиды, полученные при скрещивании дикого предка эгилопса Aegilops tauschii и различных сортов твердой пшеницы, которые повышают генетическое разнообразие культивируемых сортов пшеницы.
Способы селекции растений для Triticum spp. хорошо известны. Неограничивающие способы для селекции Triticum spp. и сельскохозяйственно важные признаки (например, повышение урожайности пшеницы, устойчивость к биотическим стрессам и устойчивость к абиотическим стрессам и др.) описаны в ссылках №№36-51, US 7652204, US 6197518, US 7034208, US 7528297, US 6407311, US 20080040826, US 20090300783, US 20060223707, US 20110027233, US 20080028480, US 20090320152, US 20090320151, WO/2001/029237 A2, WO/2008/025097 A1 и WO/2003/057848 A2, каждая из которых включена в данный документ во всей своей полноте посредством ссылки.
Генетические материалы могут передаваться между Triticum spp. и другими видами, например, некоторыми видами растений, относящихся к трибе Triticeae. Xiang et al., описывают соматические гибриды пшеницы и Setaria italica (Genome 47: 680-688 (2004)); Ge et al. описывают электрослияние протопластов пшеницы обыкновенной и рейграса итальянского (In Vitro Cellular и Developmental Biology - Plant 42(2): 179-187. 2006); Yue et al. описывают асимметричную соматическую гибридизацию Aeleuropus littorulis sinensis и пшеницы (Plant Science, Volume 161, Issue 2, July 2001, Pages 259-266); Cai et al. описывают соматические гибриды между Festuca arundinacea Schreb. и пшеницей (Triticum aestivum L.); Xiang et al. описывают асимметричную соматическую гибридизацию между пшеницей и Avena sativa L. (Science in China, Vol 46(3), 243-252); Zhou et al. описывают асимметричную соматическую гибридизацию между пшеницей и асимметричную соматическую гибридизацию между пшеницей и Avena sativa Haynaldia villosa (Science in China, 44(3): 294-304); Xia et al. описывают асимметричную соматическую гибридизацию между пшеницей и Agropyron elongatum (Host) Nevishi (Theor Appl Genet. 2003 Jul; 107(2):299-305. Epub 2003 Mar 19); Li et al. описывают симметричную соматическую гибридизацию между пшеницей и Psathyrostachys juncea (Sheng Wu Gong Cheng Xue Bao. 2004 Jul; 20(4):610-4). Другие примеры гибридизации между Triticum spp. и другими видами описаны в ссылках №№77-86.
Триба Oryzeae
Триба Oryzeae (Poaceae), согласно условному делению, включает приблизительно 12 родов и более 70 видов, распространенных в тропических и умеренных регионах мира (Clayton and Renvoize, 1986; Vaughan, 1994). Как наиболее крупная триба в подсемействе Ehrhartoideae, Oryzeae включает более половины всех родов и видов подсемейства (Guo et al., 2005, Watson and Dallwitz, 1999; GPWG, 2001).
Рода трибы Oryzeae, включают, но не ограничивают, Chikusichloa Koidz, Hygroryza Nees, Leersia Sw., Luziola Juss, Maltebrunia Kunth, Oryza L., Porteresia Tateoka, Potamophila R. Br., Prosphytochloa Schweick., Rhynchoryza Baill., Zizania L. и Zizaniopsis Doll & Asch.
Oryza spp.
Oryza является родом, включающим от семи до двадцати видов трав в трибе Oryzeae, которые происходят из тропических и субтропических регионов Азии, Северной Австралии и Африки. Они являются высокими травами заболоченных земель, высотой до 1-2 м; род включает как однолетние, так и многолетние виды.
Род Oryza входит в трибу Oryzeae, которая морфологически характеризуется наличием колосков с единичными цветками, колосковые чешуи которого почти полностью отсутсвуют. У растений рода Oryza чешуи имитируют две стерильные нижние цветковые чешуи. Триба Oryzeae входит в состав подсемейства Bambusoideae группы триб Роасеае с определенными общими особенностями внутреннего строения листа. Bambusoideae входят в семейство злаковых, поскольку все они имеют волокнистую корневую систему, цилиндрические стебли, покровные листья с параллельным жилкованием и соцветия с колосками.
Неограничивающие примеры Oryza spp. включают О. sativa (например, азиатский рис), О. barthii, О. glaberrima (например, африканский рис), О. longistaminata, О. meridionalis, О. nivara, О. rufipogon (например, коричневый и красный рис), О. punctata, О. latifolia, О. alta, О. grandiglumis, О. eichingeri, О. officinalis, О. rhisomatis, О. minuta, О. australiensis, О. granulata, O. meyeriana и О. brachyantha.
Вид Oryza sativa включает два основных подвида: клейкая, короткозернистая разновидность japonica или sinica и нелипкая, длиннозернистая разновидность indica. Japonica, как правило, культивируют на сухих полях, в умеренном поясе Восточной Азии, горных и предгорных районах Юго-Восточной Азии и на большой высоте в районах Южной Азии, в то время как indica является затопляемым рисом, который в основном выращивают при погружении в воду в тропической Азии. Рис, как известно, может быть различных цветов, в том числе белого, коричневого, черного, фиолетового и красного цветов. Третий подвид, который является широкозернистым и произрастает в тропических условиях, был выделен на основе морфологии и первоначально назывался javanica, но теперь известен как tropical japonica. Примеры данной разновидности включают среднезерновые сорта 'Tinawon' и 'Unoy', которые выращивают на высокогорных рисовых террасах гор Кордильеры в Северном Лусоне, Филиппины. Glaszmann (1987) применял изоферменты для разделения Oryza sativa на шесть групп: japonica, aromatic, indica, aus, rayada и ashina; Garris et al. (2004) применяли SSR для разделения Oryza sativa на пять групп; temperate japonica, tropical japonica и aromatic входят в состав разновидности japonica, тогда как indica и aus входят в состав разновидности indica.
Способы селекции растений для Oryza spp. хорошо известны. Неограничивающие способы селекции Oryza spp. и агротехнически важные признаки (например, повышение урожайности пшеницы, устойчивость к биотическим стрессам и устойчивость к абиотическим стрессам и др.) описаны в ссылках №№56-76, US 20050097639, US 20040168232. US 20100287664, US 20080109919, US 5981842 и US 20050183173, WO/2003/000904 A2.
Между Oryza spp. и другими видами, например, некоторыми видами растений трибы Oryzeae может происходить обмен генетическим материалом. Yan et al. (Plant Cell Rep. 2004 Mar; 22(8):569-75. Epub 2003 Nov 1.) и Yu et al. (Phytochemistry. 2008 Jul; 69(10):1989-96) описывают асимметричную соматическую гибридизацию между О. meyeriana L. и О. sativa L; а Shan et al. описывает асимметричную соматическую гибридизацию между рисом (О. sativa) и диким рисом (Zizania latifolia Griseb.). Описаны соматические гибриды риса и ежовника обыкновенного (Terada et al., 1987), межвидовые соматические гибриды культурных и диких видов (Hayashi et al., 1988) и диплоидные гибридные растения, полученные в результате слияния гаплоидных клеток (Toriyama и Hinata 1988). Больше примеров гибридизации между Oryza spp. и другими видами описано в ссылках №№86-92.
Семейство Fabaceae и соя
Fabaceae или Leguminosae - это большое и экономически важное семейство цветковых растений, которое известно как семейство бобовых. Название 'Fabaceae' происходит от более не существующего рода Faba, который в настоящее время включен в род Vicia. Leguminosae - это более старое название, которое все еще признается действительным и относится к типичному плоду этих растений, который называется боб.
Fabaceae является третьим по величине семейством цветковых растений после Orchidaceae и Asteraceae, включающим 730 родов и более 19400 видов, по утверждению Королевского ботанического сада. Самые крупные рода включают Astragalus с более чем 2000 видами. Acacia с более чем 900 видами и Indigofera с приблизительно 700 видами. Другие крупные рода включают Crotalaria с 600 видами и Mimosa с 500 видами.
Виды данного семейства, произрастающие в различной среде и различных климатических условиях, обнаружены по всему миру. Ряд важных сельскохозяйственных растений, включая Glycine max (соя), Phaseolus (бобы), Pisum sativum (горох), Cicer arietinum (нут), Medicago sativa (люцерна), Arachis hypogaea (арахис), Ceratonia siliqua (рожковое дерево) и Glycyrrhiza glabra (лакрица) относятся к наиболее широко известным членам семейства Fabaceae. Ряд видов также включает сорняки, произрастающие в различных регионах мира, такие как Cytisus scoparius (ракитник) и Pueraria lobata (кудзу), а также и ряд видов рода Lupinus.
Соя (США) или соевые бобы (Великобритания) (Glycine max) является видом семейства бобовых родом из Восточной Азии, который широко выращивают из-за его съедобных бобов, имеющих множество применений. Растение классифицируют скорее как масличное, а не как бобовое. Обезжиренные соевые пищевые продукты являются первичным дешевым источником белка для животных кормов и большинства расфасованных продуктов питания; соевое растительное масло является еще одним ценным продуктом переработки сельскохозяйственной культуры сои. Например, соевые продукты, такие как текстурированный растительный белок (TVP) являются важными ингредиентами во многих заменителях мяса и молока.
Белок (KRP), родственный белку-ингибитору киназы (KIP)
Растения имеют циклин-зависимые киназы (CDK) которые регулируют переходы между различными фазами клеточного цикла (Verkest et al., 2005, Switching the Cell Cycle. Kip-Related proteins in Plant Cell Cycle Control, Plant Physiology, November 2005, Vol. 139, pp. 1099-1106, включена в данный документ во всей своей полноте посредством ссылки).
В арабидопсисе (Arabidopsis thaliana) по меньшей мере два класса CDK участвуют в регуляции клеточного цикла: CDK А-типа, которые представлены только одним геном в модельных видах арабидопсиса (обозначен как Arath; CDKA; 1), и семейством CDK В-типа, которое включает четырех представителей, сгруппированных в подклассы B1 (Arath; CDKB 1; 1 и Arath; CDKB 1; 2) и В2 (Arath; CDKB 2; 1 и Arath; CDKB 2; 2) (Vandepoele et al., 2002, Genome-wide analysis of core cell cycle genes in Arabidopsis. Plant Cell 14: 903-916). CDK А-типа проявляют киназную активность, начиная с конца фазы G1 до окончания митоза, что указывает на роль, которую играет данная конкретная CDK в точках перехода G1-в-S и G2-в-M (Magyar et al., 1997; Porceddu et al., 2001; Sorrell et al., 2001). Центральная роль, которую играет CDKA; 1 в контроле числа клеток была показана с применением трансгенного растения табака (Nicotiana tabacum) со сниженной активностью CDK А-типа (Hemerly et al., 1995). Необходимость в Arath; CKDA; 1 по меньшей мере для вступления в митоз была показана также на нуль-мутантах по cdka; 1, которые не способны проходить второе деление митоза в процессе развития мужского гаметофита (Nowack et al., 2005). Группа CDK В-типа проявляет пиковую активность только при переходе из фазы G2 в фазу М (Magyar et al., 1997; Porceddu et al., 2001; Sorrell et al., 2001), указывая на то, что данные киназы играют определенную роль в инициации или прогрессии митоза. Соответственно, клетки растений с пониженной активностью CDK В-типа блокируются в G2 фазе клеточного цикла (Porceddu et al., 2001; Boudolf et al., 2004).
CDK регулируются циклинами. Циклины растений очень сложные. В арабидопсисе существует по меньшей мере 49 различных циклинов, которые были разделены на семь подклассов (А, В, С, D, Н, P и Т) (Vandepoele et al., 2002; Wang et al., 2004). CDK также регулируются за счет присоединения небольших белков, широко известных как ингибиторы CDK (CDK). CDK были выявлены у многих организмов, например, у почкующихся дрожжей (Saccharomyces cerevisiae), делящихся дрожжей (Schizosaccharomyces pombe), у млекопитающих и растений, смотри Mendenhall, 1998; Kwon T.K. et al. 1998; Vlach J. et al. 1997; Russo et al., 1996; Wang et al., 1997, 1998 и 2000; Lui et al., 2000; De Veylder et al., 2001; Jasinski et al., 2002a, 2002b; Coelho et al., 2005; Jasinski S. et al., 2002, каждая из данных публикация включена в данный документ во всей своей полноте посредством ссылки).
Растительные CKI также известны как KIP-родственные белки (KRP). Они имеют циклин-связывающий и CDK-связывающий домены в С-терминальных концах, однако механизм, регулирующий стабильность и функцию этого белка остается неизвестным (Zhou et al., 2003a; Weinl et al. 2005). Активность KRP может регулироваться как на транскрипционном уровне, так и на посттрансляционном уровне (Wang et al., 1998; De Veylder et al., 2001; Jasinski et al., 2002b; Ormenese et al., 2004; Coqueret, 2003; Hengst, 2004; Verkest et al., 2005; Coelho et al., 2005, каждая из которых включена в данный документ во всей своей полноте посредством ссылки). KRP в растении обычно локализуется в ядре (Jasinski et al., 2002b; Zhou et al., 2003a; Weinl et al., 2005).
KRP может функционировать в качестве интегратора сигналов развития и контролировать начало эндоцикла, в различных программах клеточного цикла (например, в пролиферации, эндоредупликации и выходе из клеточного цикла). Смотри Wang et al., 1998; Richard et al., 2001; Himanen et al., 2002; Grafi и Larkins, 1995; Joube's et al., 1999; Verkest et al., 2005; Weinl et al., 2005; Boudolf et al., 2004b.
Мутации KRP
Настоящее изобретение дополнительно предлагает полинуклеотиды KRP и аминокислотные последовательности нарушенного KRP в сравнении с геном KRP дикого типа или белком KRP дикого типа, В некоторых вариантах осуществления настоящее изобретение предлагает мутации в одном или нескольких генах KRP, которые могут быть использованы для повышения веса, размера и/или числа одного или нескольких органов, например, увеличения размера семян, числа семян, веса семян и/или урожая семян в растении.
Мутации в мутантном гене KRP, представленные в настоящем изобретении, могут быть в кодирующем участке или некодирующем участке гена KRP. Мутации могут приводить или не приводить к изменениям аминокислот в кодированных полипептидах KRP. В некоторых вариантах осуществления мутации могут быть миссенс-мутациями, тяжелыми миссенс-мутациями, молчащими или нонсенс-мутациями. Например, мутация может представлять собой замены, вставки, делеции нуклеотидов или реорганизацию генома, которая в свою очередь может привести к сдвигу рамки считывания, изменению сплайсинга, замене, вставке, делеции аминокислот и/или отсечению полипептидов. В результате мутантный ген KRP кодирует полипептид KRP, обладающий меньшей ингибирующей активностью в отношении комплекса циклин/CDK по сравнению с полипептидом, который кодируется соответствующим геном KRP дикого типа.
Используемое в данном документе выражение нонсенс-мутация относится к точечной мутации, например, полиморфизму единичных нуклеотидов (SNP) в последовательности ДНК, которая приводит к образованию преждевременного стоп-кодона или нонсенс-кодона в транскрибируемой иРНК и отсеченного, неполного и, как правило, нефункционального белкового продукта. Миссенс-мутация (тип несинонимичных мутаций) является точечной мутацией, при которой изменяется один нуклеотид, приводя к образованию кодона, который кодирует другую аминокислоту (мутации, которые изменяют аминокислоту с образованием стоп-кодона, считаются нонсенс-мутациями, а не миссенс-мутациями). Это может привести к потере функциональности полученного белка. Молчащими являются мутации ДНК, которые не приводят к изменению аминокислотной последовательности белка. Они могут возникнуть в некодирующем участке (за пределам гена или внутри интронов) или могут возникнуть в экзоне таким образом, который не изменяет конечную аминокислотную последовательность. Тяжелая миссенс-мутация изменяет аминокислоту, которая приводит к критическим изменениям конформации, заряда и т.д.
Мутации могут быть локализованы в любой части гена KRP, например, в 5', в середине или в 3' гена KRP, приводя к мутации в любой части кодируемого белка KRP, например, в CDK-связывающем домене или в циклин-связывающем домене, при условии, что мутантный ген кодирует мутантный полипептид KRP частично или полностью теряет способность ингибировать один или несколько комплексов циклин/CDK, по сравнению с белком, кодирующим соответствующий дикий тип гена KRP. KRP и комплексы циклин/CDK могут принадлежать к одному и тому же виду растений, различным видам растений, принадлежащим к одному роду, или различным видам растений, принадлежащих к разным родам.
Настоящее изобретение предлагает эффективные системы для проверки того, теряет ли мутантный белок KRP ингибирующую активность в отношении комплекса циклин/CDK в сравнении с белком KRP дикого типа. Эффективные системы включают анализ киназ ("анализ KRP-циклин-CDK-киназ in vitro"), неограничивающий пример которого описан в данном изобретении.
В основе данного анализа киназ лежит in vitro анализ киназ. В данном анализе кандидат мутантный KRP, полученный из KRP дикого типа из вида растений А, белок KRP дикого типа из вида растений А, белок циклин дикого типа из вида растений В и белок CDK дикого типа из вида растений В, рекомбинантно экспрессируют и очищают. Затем рекомбинантный белок циклин дикого типа и белок CDK дикого типа смешивают с образованием комплекса (альтернативно, белок циклин и белок CDK можно совместно экспрессировать и совместно очищать в виде комплекса). В некоторых вариантах осуществления рекомбинантные белки экспрессируют в клетках насекомых. Вид растений А может быть таким же или отличаться от вида растений В. Данную активность киназ вышеупомянутого комплекса затем измеряют с применением стандартного анализа киназ, описанного ниже. Выбирают субстратный белок, который может быть активирован (то есть фосфорилирован) комплексом циклин-CDK. Такой субстратный белок может быть гистоном HI (HHI) или рекомбинантным белком ретинобластомы табака (Nt Rb). По меньшей мере три смеси могут быть получены путем добавления рекомбинантных белков в киназный буферный коктейль в соответствии с таблицей ниже:
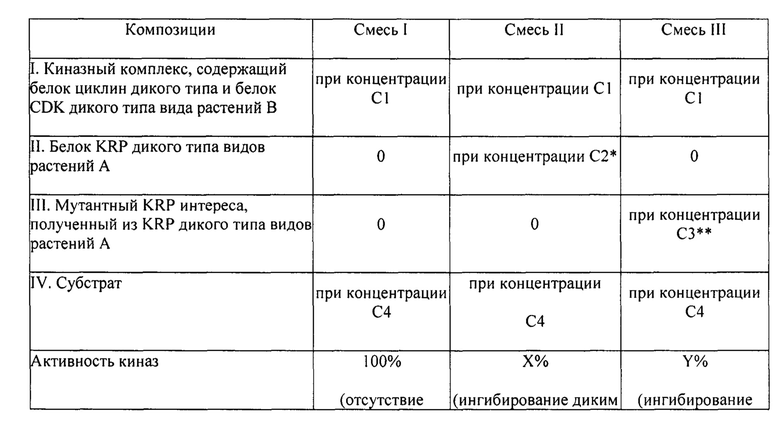

Неограничивающий пример киназного буферного коктейля включает KAB: 50 мМ Трис pH 8,0, 10 мМ MgCl2, 100 мкМ АТФ плюс 0,5 мкКи/мл 32РγАТФ и субстратный белок. Концентрации C1, C2 и С3 могут быть определены и оптимизированы специалистом в данной области техники в зависимости от условий эксперимента.
Для определения теряет ли кандидат мутантный KRP ингибирующую способность в отношении киназного комплекса, C2 должна быть приблизительно эквимолярна C1; а С3 должна составлять не более, чем 50Х от C2, или не более, чем 40Х от C2, или не более, чем 30Х от C2, или не более, чем 20Х от C2, или не больше, чем 10Х от C2, или не больше, чем 5Х от C2. Например, в некоторых случаях количество С3 составляет приблизительно 1X, или приблизительно 2Х, или приблизительно 3Х, или приблизительно 4Х, или приблизительно 5Х, или приблизительно 6Х, или приблизительно 7Х, или приблизительно 8Х, или приблизительно 9Х, или приблизительно 10Х, или приблизительно ИХ, или приблизительно 12Х, или приблизительно 13Х, или приблизительно 14Х, или приблизительно 15Х, или приблизительно 16Х, или приблизительно 17Х, или приблизительно 18Х, или приблизительно 19Х, или приблизительно 20Х от количества C2. В некоторых ситуациях, однако, количество С3 может составлять приблизительно 25Х, или приблизительно 30Х, или приблизительно 35Х, или приблизительно 40Х, или приблизительно 45Х, или приблизительно 50Х от количества C2. Как обсуждается в других частях данного документа, количество С3 которое используют в каждой конкретной ситуации должно быть физиологически достижимым в клетке растения, ткани растения или целом растении, чтобы оказывать доминирующий отрицательный эффект на KRP дикого типа.
Композицию I и/или композицию III инкубируют на льду в течение определенного количества времени (например, 30 минут). Затем Композицию II добавляют к смеси и инкубируют при 4°C в течение определенного количества времени (например, 30 минут) чтобы обеспечивать связывание с киназным комплексом. Затем киназную реакцию инициируют добавлением буферного коктейля (KAB) в смесь, содержащую киназный комплекс (I, II или III) и инкубируют при 27°C в течение определенного количества времени (например, 30 минут) для обеспечения завершения реакции. Киназную реакцию в каждой смеси останавливают путем добавления равного объема 2х буфера Лэммли и кипятят в течение 5 минут. Затем, в каждой смеси измеряют включение фосфата [32P] в субстратный белок с применением авторадиографии и/или системы Molecular Dynamics PhosphorImager после применения процедуры SDS-PAGE. Интенсивность сигнала включения фосфата [32P] в смеси I задают как 100%-ное восстановление функции киназы. Степень включения фосфата [32P] в смеси II по сравнению с таковой в смеси I рассчитывают как X%; Степень включения фосфата [32P] в смеси III по сравнению с таковой в смеси I рассчитывают как Y%. Например, если сила сигнала составляет половину от таковой, наблюдаемой для смеси I, расчетное процентное значение извлечения киназной активности составляет 50%.
X% сравнивают с Y% и эффект тестируемого мутантного KRP рассчитывают следующим образом: пусть Z%=(Y%-Х%), a Zmax% - это максимальное значение Z% в допустимом диапазоне С2 и С3; если значение Zmax% статистически не выше 0% (то есть Y% ≤ X%), то тестируемый мутантный KRP не теряет ингибирующую активность в отношении комплекса по сравнению с соответствующим диким типом KRP; если значение Zmax% статистически выше 0% (то есть Y%>X%), но менее 30%, то тестируемый мутантный KRP обладает слабой ингибирующей активностью по сравнению с тестируемым диким типом KRP; если Zmax% выше 30%, но менее 50%, то тестируемый мутантный KRP в значительной степени теряет ингибирующую активность по сравнению с тестируемым диким типом KRP; если Zmax% выше 50%, то тестируемый мутантный KRP полностью теряет ингибирующую активность по сравнению с тестируемым диким типом KRP. В некоторых вариантах осуществления мутантные KRP настоящего изобретения существенно не ингибируют киназную активность комплекса циклин/CDK, даже если присутствуют в значительно молярном избытке по отношению к комплексу циклин/CDK. Мутантные KRP со значением Zmax% выше 0% особенно пригодны для повышения веса, размера и/или числа одного или нескольких органов, например, для повышения урожая семян, размера семян, числа семян и/или урожая семян в растении.
Белок CDK и белок циклин в комплексе циклин/CDK могут быть получены из любого растения, например, любых двудольных растений или однодольных растений, особенно агротехнически или промышленно важных растений. Белок CDK и белок циклин могут быть получены из одного и того же вида растений или из различных видов. Белок KRP может быть получен из того же вида растений из которого получены белок CDK и белок циклин или из другого вида растений. В некоторых вариантах осуществления комплексы циклин/CDK содержат белок CDK, выбранный из группы, которая включает Zea mays CDK A; 1 (ZmCDKA; 1), Zea mays CDK A; 2 (ZmCDKA; 2), белки CDK пшеницы (например, номер доступа в GenBank № AAD10483) и белки CDK риса (например, номер доступа в GenBank № NP_001048772 и NP_001045731) и белки CDK из растения трибы Triticeae или трибы Oryzeae или семейства Fabaceae; белки циклины, выбранные из группы, которая включает циклин D1, D2, D3, D4, D5, D6, D7 Zea mays, белки циклины пшеницы (например, номер доступа в GenBank № AAQ08041 (циклин D2)), белки циклины риса (например, учетная запись в базе данных GenBank № Q67V81 (циклин-D1; 1), Q8H339 (циклин-D1; 2), Q0J233 (циклин-D2; 1), Q10K98 (циклин-D2; 3), Q69QB8 (циклин-D3; 1), Q0DQA9 (циклин-D5; 1), Q53MB7 (циклин-D7; 1)), белки циклины из растения трибы Triticeae или трибы Oryzeae или семейства Fabaceae и их комбинации, и белки KRP дикого типа, выбранные из группы, которая включает KRP1 пшеницы (TaKRP1), KRP2 пшеницы (TaKRP2), KRP3 пшеницы (TaKRP3), KRP4 пшеницы (TaKRP4), KRP5 пшеницы (TaKRP5), KRP6 пшеницы (TaKRP6), например, SEQ ID NO: 87-98, 140, 143 и 146, или их функциональные варианты, KRP1 риса, KRP2 риса, KRP3 риса, KRP4 риса, KRP5 риса, KRP6 риса, KRP7 риса, например, SEQ ID NO: 99, 108-110, или их функциональные варианты, KRP сои, например, SEQ ID NO: 129-137, или их функциональные варианты, KRP из растения трибы Triticeae или трибы Oryzeae или семейства Fabaceae и их комбинации. Например, KRP дикого типа представляет собой KRP1 пшеницы, KRP2 пшеницы, KRP4 пшеницы, KRP5 пшеницы или KRP6 пшеницы. Нумерация KRP пшеницы или риса не обязательно соответствует нумерации KRP других видов (например, арабидопсис или маис). Например, KRP1 пшеницы не обязательно является эквивалентным или ортологичным KRP1 арабидопсиса.
В некоторых вариантах осуществления циклин Zea mays выбирают из 59 циклинов, описанных в Hu et al., 2010, которая включена в данный документ во всей своей полноте посредством ссылки. В некоторых вариантах осуществления циклин Zea mays выбирают из 21 белка циклина D, которые описаны в Hu et al., 2010. Например, циклин выбирают из группы, которая включает циклин Zea mays D1; 1, D2; 1, D2; 2, D3; 1, D3; 2, D4; 1, D4; 2, D4; 3, D4; 4, D4; 5, D4; 6, D4; 7, D4; 8, D4; 9, D4; 10, D5; 1, D5; 2, D5; 3, D5; 4, D6; 1, D7; 1 и их комбинации.
В некоторых вариантах осуществления вышеупомянутый мутантный KRP получен из KRP пшеницы. В некоторых вариантах осуществления мутантный KRP получен из KRP1 пшеницы (А, В или D), KRP2 пшеницы (А, В или D), KRP4 пшеницы (А, В или D), KRP5 пшеницы (А, В или D) или KRP6 пшеницы (А, В или D). В некоторых вариантах осуществления вышеупомянутый мутантный KRP получен из KRP риса. В некоторых вариантах осуществления мутантный KRP получен из KRP1 риса, KRP2 риса, KRP3 риса, KRP4 риса или KRP5 риса. В некоторых вариантах осуществления вышеупомянутый мутантный KRP получен из KRP сои. В некоторых других вариантах осуществления вышеупомянутый мутантный KRP получен из биологически активного варианта KRP дикого типа пшеницы, риса, сои или его фрагмента. Мутантный KRP может быть результатом спонтанной мутации или мутации, искусственно индуцированной с применением способов, хорошо известных специалисту в данной области техники.
Мутантный белок KRP настоящего изобретения может иметь одну или несколько модификаций KRP дикого типа или биологически активного варианта или его фрагмента. Особенно подходящие модификации включают аминокислотные замены, вставки, делеции или отсечение. Например, аминокислотные замены могут быть получены как модификации в CDK или циклин-связывающем участке KRP, которые сокращают или блокируют связывание. Аналогично, аминокислотные замены могут быть получены как модификации в CDK или циклин-связывающем участке KRP, которые сокращают или блокируют ингибирующую активность KRP по отношению к комплексу циклин/CDK. В типичный вариантах осуществления проводят по меньшей мере одну неконсервативную аминокислотную замену, вставку или делецию в CDK-связывающем участке или циклин-связывающем участке для нарушения или модифицикации связывания полипептида CKI с белком CDK или циклином. Замены могут быть одиночными, при которых заменяется только одна аминокислота в молекуле, или могут быть множественными, при которых в одной и той же молекуле заменяются две или более аминокислоты. Инсерционные мутантные KRP являются такими, в которых одна или несколько аминокислот встроены в непосредственной близости к аминокислоте, занимающей специфическое положение в молекуле белка KRP дикого типа, биологически активном варианте или его фрагменте. Вставка может содержать одну или несколько аминокислот. Вставка может содержать, например, одну или две консервативные аминокислоты. Аминокислоты, сходные по своему заряду и/или структуре аминокислотам, прилегающим к участку вставки, определяют как консервативные. Альтернативно, мутантный белок KRP содержит вставку аминокислоты, заряд и/или структура которой существенно отличается от аминокислот, прилегающих к участку вставки. В некоторых других вариантах осуществления мутантный KRP представляет собой усеченный белок, теряющий один или несколько доменов по сравнению с соответствующим белком KRP дикого типа.
Способы повышения веса органов, размера органов, числа органов и/или урожая
Настоящее изобретение дополнительно предлагает способы повышения веса, размера и/или числа одного или нескольких органов, например, способы повышения веса семян, размера семян, числа семян и/или урожая растения. Растение может быть двудольным растением или однодольным растением, В некоторых вариантах осуществления растение является однодольным растением. В некоторых вариантах осуществления растение является видом растения трибы Triticeae или трибы Oryzeae, трибы семейства Fabaceae, к примеру, растением пшеницы или растением риса. В некоторых вариантах осуществления способы включают нарушение одного или нескольких KRP в растении. Нарушение может происходить на геномном уровне, транскрипционном уровне, посттранскрипционном уровне, трансляционном уровне и/или посттрансляционном уровне. В некоторых вариантах осуществления способы включают введение одной или нескольких мутаций в один или несколько генов KRP в растении. В некоторых вариантах осуществления способы включают нокдаун участков, отвечающих за стабильность иРНК KRP в растении. В некоторых вариантах осуществления способы включают подавление активности одного или нескольких белков KRP растения.
Например, в некоторых вариантах осуществления способы включают введение одного или нескольких мутантных KRP настоящего изобретения в геном растения. В некоторых вариантах осуществления способы включают гибридизацию первого растения, имеющего один или несколько мутантных KRP настоящего изобретения, со вторым растением. В некоторых вариантах осуществления этап гибридизации включает скрещивание первого растения со вторым растением. В некоторых вариантах осуществления этап гибридизации включает перенос генетического материала первого растения во второе растение путем селекции in vitro, например, соматической гибридизации.
Альтернативно, способы включают получения мутаций в одном или нескольких KRP растения. Способы получения мутации в целевом гене известны специалисту в данной области техники. Эти способы включают, но не ограничивают, мутагенез (например, химический мутагенез, радиационный мутагенез, транспозонный мутагенез, инсерционный мутагенез, мутагенез с использованием уникальных последовательностей в качестве меток (STM), сайт-специфический мутагенез и спонтанный мутагенез), TILLING®, гомологичную рекомбинацию, нокаут/нокин, создание антисмысловых последовательностей и РНК-интерференцию. Различные типы мутагенеза могут быть использованы для получения и/или выделения вариантов нуклеиновых кислот, которые кодируют молекулы белка, и/или дополнительного модифицирования/изменения белков настоящего изобретения. Они включают, но не ограничивают, сайт-специфический, случайный точечный мутагенез, гомологичную рекомбинацию, перестановку в ДНК, мутагенез с использованием урацилсодержащих матриц, олигонуклеотид-направленный мутагенез, мутагенез с использованием модифицированной фосфотиоатом ДНК, мутагенез с использованием гэпированного ДНК-дуплекса и т.п. Дополнительные подходящие способы включают мутагенез по механизму точечного ошибочного спаривания при репарации, мутагенез, связанный с недостаточностью функционирования системы репарации в штамме-хозяине, мутагенез на основе системы рестрикции-селекции и рестрикции-очищения, делеционный мутаганез, мутагенез, осуществляемый путем полного синтеза гена, репарация двухцепочечных разрывов и т.п. Мутагенез, в котором, например, используют химерные конструкты, также включен в настоящее изобретение. В одном варианте осуществления мутагенез может быть направлен известной информацией о молекуле с естественным происхождением или мутантной молекуле с естественным происхождением, например, такой как последовательность, сравнение последовательностей, физические свойства, кристаллическая структура и т.п. Более подробную информацию о мутагенезе растений, включая средства, протоколы, смотри в ссылках №№94-98, каждая из которых включена в данный документ во всей своей полноте посредством ссылки.
В некоторых вариантах осуществления случайные мутации в генах KRP создают in vitro. Например, может быть получена библиотека генов KRP с одной или несколькими случайными мутациями, при этом полученные мутантные гены KRP подвергают in vitro анализу киназ KRP-циклин-CDK, который описан в настоящем изобретении для определения может ли мутантный ген KRP быть использован для повышения веса, размера и/или числа одного или нескольких органов, например, для увеличения размера семян, числа семян, веса семян и/или урожая. Способы мутагенеза in vitro включают, но не ограничивают, ПЦР пониженной точности, ПЦР пониженной точности с использованием амплификации по механизму катящегося кольца, использование мутаторных штаммов, использование нестабильных мутаторных штаммов, инсерционный мутагенез, химический мутагенез (например, с применением этилметансульфоната (EMS), азотистой кислоты и др.), перестановка в ДНК и случайный сайт-направленный мутагенез. Дополнительные способы описаны в Chusacultanachai et al, Fujii et al., Braman и Trower. Доступны коммерческие наборы для случайного мутагенеза, такие как Random Mutagenesis Kits от Jena Bioscience, № по каталогу РР-101, Diversify® PCR random mutagenesis kit от Clontech.
В некоторых вариантах осуществления мутантные KRP настоящего изобретения получают in vivo с применением таких способов как TILLING®, сайт-специфический мутагенез, гомологичная рекомбинация и т.д. Полученные мутантные гены KRP подлежат скринингу и анализу киназ KRP-циклин-CDK in vitro, который описан в настоящем изобретении для определения, могут ли мутантные гены KRP быть использованы для повышения веса, размера и/или числа одного или нескольких органов, например, для повышения размера семян, числа семян, веса семян и/или урожая.
В некоторых вариантах осуществления способы включают нокдаун экспрессии одного или нескольких KRP в растении. Методики, которые могут быть использованы в соответствии с настоящим изобретением для нокдауна экспрессии генов, включают, но не ограничивают (1) нарушение транскрипта гена, такое как нарушение иРНК-транскрипта гена; (2) нарушение функции полипептида, кодируемого геном, или (3) нарушение самого гена.
Например, методики, использующие антисмысловую РНК, рибозим, dsRNAi, РНК-интерференцию (RNAi), могут применяться в настоящем изобретении для нацеливания на РНК-транскрипты одного или нескольких генов KRP. Методика, использующая антисмысловую РНК, включает экспрессию в клетке или введение в клетку молекулы РНК (или производного РНК), которая является комплементарной или антисмысловой к последовательностям, обнаруживаемым в определенной иРНК в клетке. Антисмысловая РНК может ингибировать трансляцию гена, кодирующего продукт, путем ассоциации с иРНК. Применение методики, использующей антисмысловую РНК для снижения или ингибирования экспрессии специфических генов растения, было описано, например, в Европейской патентной публикации №271988, Smith et al., Nature, 334:724-726 (1988); Smith et. al., Plant Mol. Biol, 14:369-379 (1990)).
Рибозим представляет собой молекулу РНК, которая имеет как каталитический домен, так и последовательность, комплементарную определенной иРНК. Рибозим функционирует за счет связывания с молекулой иРНК (посредством комплементарного домена рибозима) с последующим расщеплением (деградацией) транскрипта с применением каталитического домена.
РНК-интерференция (RNAi) представляет собой процесс последовательность-специфического посттранскрипционного сайленсинга генов или транскрипционного сайленсинга генов у животных и растений, инициированного двухцепочечной РНК (dsRNA), гомологичной в своей последовательности гену, подвергающемуся сайленсингу. Методика RNAi описана, например, в Elibashir, et al., Methods Enzymol. 26:199 (2002); McManus & Sharp, Nature Rev. Genetics 3:737 (2002); в заявке PCT WO 01/75164; Martinez et al., Cell 110:563 (2002); Elbashir et al., выше; Lagos-Quintana et al., Curr. Biol. 12:735 (2002); Tuschl et al., Nature Biotechnol. 20:446 (2002); Tuschl, Chembiochem. 2:239 (2001); Harborth et al., J. Cell Sci. 114:4557 (2001); et al., EMBO J. 20:6877 (2001); Lagos-Quintana et al., Science 294:8538 (2001); Hutvagner et al., в цитированном месте, 834; Elbashir et al., Nature 411:494 (2001).
Выражение "dsRNA" или "молекула dsRNA" или "двухцепочечная эффекторная молекула РНК" относится к по меньшей мере частично двухцепочечной молекуле рибонуклеиновой кислоты, содержащей участок из по меньшей мере приблизительно 19 или более нуклеотидов, которые находятся в составе двухцепочечной структуры. Двухцепочечная эффекторная молекула РНК может быть дуплексной двухцепочечной РНК, образованной двумя отдельными цепями РНК, или может представлять собой одноцепочечную РНК с участками самокомплементарности, которая обладает способностью по меньшей мере частично образовывать двухцепочечную структуру (то есть, dsRNA со шпилечной структурой или dsRNA со структурой типа стебель-петля). В различных вариантах осуществления dsRNA состоит полностью из рибонуклеотидов или состоит из смеси рибонуклеотидов и дезоксинуклеотидов, таких как гибриды РНК/ДНК. Молекула dsRNA может представлять собой отдельную молекулу с участками самокомплементарности, такими как нуклеотиды в одном сегменте пары оснований молекулы с нуклеотидами в другом сегменте молекулы. В одном аспекте участки самокомплементарности сцеплены между собой участком из по меньшей мере приблизительно 3-4 нуклеотидов или приблизительно из 5, 6, 7, 9-15 нуклеотидов или более, в котором отсутствует комплементарность к другой части молекулы и поэтому она остается одноцепочечной (то есть "участок петли"). Такая молекула принимает частично двухцепочечную структуру типа стебель-петля, необязательно, с короткими одноцепочечными 5' и/или 3' концами. В одном аспекте участки самокомплементарности шпильковой dsRNA или двухцепочечный участок дуплексной dsRNA будут содержать эффекторную последовательность и эффекторную комплементарную цепь (например, связанные между собой одноцепочечным петлевым участком в шпильковой dsRNA). Эффекторной последовательностью или эффекторной комплементарной цепью является такая цепь двухцепочечного дуплексного участка или дуплекса, которая встроена в или связана с RISC. В одном аспекте двухцепочечная эффекторная молекула РНК будет содержать эффекторную последовательность из по меньшей мере 19 смежных нуклеотидов, предпочтительно из 19-29, 19-27 или 19-21 нуклеотидов, которая является цепью, обратно комплементарной РНК KRP, или промежуточным звеном репликации противоположной цепи, или антигеномной плюс цепью РНК, не являющейся иРНК, последовательностей KRP. В одном варианте осуществления представлены вышеупомянутые двухцепочечные эффекторные молекулы РНК, путем обеспечения растения, ткани растения или клетки растения экспрессионным конструктом, содержащим одну или несколько двухцепочечных эффекторных молекул РНК. В одном варианте осуществления экспрессионный конструкт содержит двухцепочечную РНК, полученную из любой из SEQ ID NO: 60-86, 100-107, 111-128, 138-139, 141-142 и 144-145. Специалист в данной области сможет сконструировать подходящую двухцепочечную эффекторную молекулу РНК на основе нуклеотидных последовательностей KRP настоящего изобретения.
В некоторых вариантах осуществления эффекторная молекула dsRNA настоящего изобретения является "шпильковой dsRNA", "dsRNA-шпилькой", "короткой шпильковой РНК" или "shRNA", то есть молекулой РНК из менее, чем приблизительно 400-500 нуклеотидов (нт) или менее, чем 100-200 нт, в которой по меньшей мере один фрагмент из меньшей мере 15-100 нуклеотидов (например, 17-50 нт, 19-29 нт) спарен с основаниями комплементарной последовательности, локализованной в той же молекуле РНК (одноцепочечная РНК), и где вышеупомянутая последовательность и комплементарная последовательность разделены неспаренным участком из по меньшей мере приблизительно 4-7 нуклеотидов (или от приблизительно 9 до приблизительно 15 нт, от приблизительно 15 до приблизительно 100 нт, от приблизительно 100 до приблизительно 1000 нт), который формирует одноцепочечную петлю над стеблевой структурой, образованной двумя участками с комплементарными основаниями. Молекулы shRNA содержат по меньшей мере одну структуру стебель-петля, содержащую двухцепочечный стеблевой участок от приблизительно от 17 до приблизительно 100 п.о.; от приблизительно 17 до приблизительно 50 п.о.; от приблизительно 40 до приблизительно 100 п.о.; от приблизительно 18 до приблизительно 40 п.о.; или от приблизительно 19 до приблизительно 29 п.о.; гомологичный и комплементарный целевой последовательности, которая должна ингибироваться; и неспаренный петлевой участок из по меньшей мере приблизительно 4-7 нуклеотидов или от приблизительно 9 до приблизительно 15 нуклеотидов, от приблизительно 15 до приблизительно 100 нт, от приблизительно 100 до приблизительно 1000 нт, который формирует одноцепочечную петлю над стеблевой структурой, образованной двумя участками комплементарных оснований. Следует, однако, признать, что включение петлевого участка или петлевой последовательности не является строго обязательным, поскольку молекула РНК, содержащая последовательность, сразу за которой следует обратная ей комплементарная последовательность, будет иметь тенденцию к образованию структуры петля-стебель, даже если она не разделена несоответствующей спейсерной последовательностью.
Растения с одним или несколькими нарушенными KRP по настоящему изобретению могут быть использованы для многих целей. В одном варианте осуществления растение настоящего изобретения используют в качестве растения-донора генетического материала, который может быть перенесен в растение-реципиент для получения растения с желаемыми агротехническими признаками, которое содержит перенесенный генетический материал, характеризуется повышенной массой, размером и/или числом одного или нескольких органов, например, характеризующиеся повышенный массой семян, размером семян, числом семян и/или урожаем. Любой подходящий способ, известный в данной области техники, может быть применен для переноса генетического материала из растения-донора в растение-реципиент. В большинстве случаев такой генетический материал является геномным материалом.
Описания других способов селекции, часто используемых для различных признаков и сельскохозяйственных культур, можно найти в одной из нескольких справочных книг (например, R.W. Allard, 1960, Principles of Plant Breeding, John Wiley и Son, pp. 115-161; N.W. Simmonds, 1979, Principles of Crop Improvement, Longman Group Limited; W.R. Fehr, 1987, Principles of Crop Development, Macmillan Publishing Co.; N.F. Jensen, 1988, Plant Breeding Methodology, John Wiley & Sons).
В некоторых вариантах осуществления используют способ селекции возвратного скрещивания. Способ селекции возвратного скрещивания включает следующие этапы: (а) скрещивание первого растения пшеницы, содержащего один или несколько нарушенных генов KRP, со вторым растением, которое содержит желаемый признак (признаки); (b) отбор растений потомков F1, которые содержат желаемый признак (признаки); (с) скрещивание выбранных растений потомков F1 с первым растением пшеницы или вторым растением пшеницы для получения потомков растений путем возвратного скрещивания; (d) выбор потомков растений, полученных путем возвратного скрещивания, которые содержат желаемый признак (признаки) и один или несколько нарушенных генов KRP, для получения отобранных потомков растения, полученных путем возвратного скрещивания; и (е) повторение этапов (с) и (d) один, два, три, четыре, пять, шесть, семь, восемь, девять или более раз подряд для получения выбранных потомков растения, полученных путем возвратного скрещивания, потомков второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого или последующих поколений, которые содержат вышеупомянутые нарушенные гены KRP и/или желаемый признак (признаки).
Настоящее изобретение дополнительно предлагает способы получения сортов пшеницы в рамках селекционной программы для пшеницы с применением способов селекции растений, включая рекуррентную селекцию, возвратное скрещивание, селекцию на основе генеалогической схемы, улучшенную селекцию с применением молекулярных маркеров (изоферментный электрофорез, полиморфизм длин рестрикционных фрагментов (RFLP), случайно амплифицируемую полиморфную ДНК (RAPD), полимеразную цепную реакцию с произвольными праймерами (AP-PCR), ДНК-амплификационный фингерпринтинг (DAF), амплифицированные участки, охарактеризованные секвенированием (SCAR), полиморфизмы длин амплифицированных фрагментов (AFLP) и простые повторяющиеся последовательности (SSR), которые также называются микросателлитами и др.), улучшенная селекция с применением генетических маркеров и трансформация. Семена, растения и их часть(части), полученные с применением таких способов селекции, также являются частою настоящего изобретения.
В одном варианте осуществления целый геном растений по настоящему изобретению, содержащий нарушенный KRP, переносят в растение-реципиент. Это может быть осуществлено путем традиционной селекции, такой как скрещивание или соматическая гибридизация. В другом варианте осуществления переносят по меньшей мере части генома растения-донора, содержащие нарушенный KRP. Это может быть осуществлено путем скрещивания растения-донора с растением-реципиентом для получения растения F1, с проведением последующего одного или нескольких обратных скрещиваний с одним из родительских растений для того, чтобы получить растения с желаемым генетическим фоном. Селекция с применением молекулярных маркеров может быть использована для контроля переноса генетического материала. Полученные потомки могут быть выбраны на основе увеличенного веса, размера и/или числа одного или нескольких органов, например, увеличения веса семян, размера семян, числа семян и/или урожая.
В одном варианте осуществления растение-реципиент принадлежит к элитной линии с одним или несколькими определенными агротехнически важными признаками. Используемое в данном документе выражение "агротехнически важные признаки" включает любой фенотип в растении или части растения, который является полезным или предпочтительным для использования человеком. Примеры агротехнически важных признаков включают, но не ограничивают, такие, которые приводят к увеличению производства биовеса, увеличению производства пищевых продуктов, улучшению качества пищевых продуктов, уменьшению растрескивания, быстрому изменению цвета при созревании плода и т.д. Дополнительные примеры агротехнически важных признаков включают устойчивость к вредителям, мощность, время развития (время сбора урожая), повышенное содержание питательных веществ, увеличение содержания масла в семенах, новые модели роста, запахи или цвета, солеустойчивость, жароустойчивость, засухоустойчивость и холодоустойчивость и подобные.
Другие агротехнически важные признаки включают устойчивость к биотическому и/или абиотическому стрессам. Используемое в данном документе словосочетание "биотический стресс" или "биотическое давление" относится к ситуации, когда повреждение растения вызвано другими живыми организмами, такими как бактерии, вирусы, грибы, паразиты, насекомые, сорняки, животные и человек. Используемое в данном документе словосочетание "абиотический стресс" или "абиотическое давление" относится к негативному воздействию неживых факторов на растения в специфической окружающей среде. Переменный фактор, не относящийся к живой природе, должен влиять на окружающую среду, превышая нормальные пределы его вариации, чтобы неблагоприятно воздействовать на характеристики популяции или индивидуальную физиологию растений в существенной степени. Неограничивающими примерами источников стресса являются сильные ветры, экстремальные температуры, засуха, наводнение и другие природные бедствия, такие как торнадо и пожары.
В некоторых вариантах осуществления способ включает i) скрещивание растения, содержащего один или несколько нарушенных генов KRP, со вторым растением для получения растения F1, например, растения пшеницы, риса или сои, содержащего один или несколько нарушенных генов KRP. Необязательно, этот способ дополнительно включает ii) возвратное скрещивание растения F1 с первым или вторым растением; и iii) повторение этапа возвратного скрещивания для получения практически изогенной линии, в которой один или несколько нарушенных генов KRP интегрированы в геном второго растения.
В некоторых вариантах осуществления нарушенным геном KRP является KRP пшеницы, выбранный из группы, включающей TaKRP1A, TaKRP1B, TaKRP1D, TaKRP2A, TaKRP2B, TaKRP2D, TaKRP4A, TaKRP4B, TaKRP4D, TaKRP5A, TaKRP5B, TaKRP5D, TaKRP6A, TaKRP6B или TaKRP6D, например, SEQ ID NO: 60-86, 138-139, 141-142 и 144-145, или их функциональные варианты. В некоторых вариантах осуществления первое растение содержит одну или несколько мутаций, выбранных из списка мутаций, изложенных в таблицах 2-12 для определенного гена KRP.
В некоторых вариантах осуществления нарушенным геном KRP является KRP риса, выбранный из группы, включающей OsKRP1, OSKRP2, OsKRP4 и OsKRP5, например, SEQ ID NO: 100-107, или их функциональные варианты. В некоторых вариантах осуществления первое растение содержит одну или несколько мутаций, выбранных из списка мутаций, изложенных в таблице 25 для определенного гена KRP.
В некоторых вариантах осуществления нарушенным геном KRP является KRP сои. В некоторых вариантах осуществления KRP выбирают из группы, включающей Gm0003×00821, Gm0013×00399, Gm0043, Gm0053×00526, Gm0087×00306, Gm0102×00087, Gm0119×00131, Gm0151×00019, Gm0067×00001, например, SEQ ID NO: 111-128, или их функциональные элементы. В некоторых вариантах осуществления первое растение содержит одну или несколько мутаций, выбранных из списка мутаций, изложенных в таблицах 29-37 для определенного гена KRP.
В некоторых вариантах осуществления способы, представленные в настоящем изобретении, могут повышать средний вес, размер и/или число одного или нескольких органов, например, средняя масса семян растений, размер семян, число семян и/или урожай растений по меньшей мере на 5%, по меньшей мере на 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20%, 21%, 22%, 23%, 24%, 25%, 26%, 27%, 28%, 29%, 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%,79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 100%, 110%, 120%, 130%, 140%, 150%, 160%, 170%, 180%, 190%, 200%, 210%, 220%, 230%, 240%, 250%, 260%, 270%, 280%, 290%, 300%, 310%, 320%, 330%, 340%, 350%, 360%, 370%, 380%, 390%, 400% или более, по сравнению с контрольным растением, не содержащим один или несколько нарушенных KRP.
Мутантные KRP в геноме растения можно рассматривать как локусы количественных признаков (QTL), связанные с массой, размером и/или числом одного или нескольких органов, например, QTL, связанные с массой семян, размером семян, числом семян и/или урожайностью. QTL представляет собой участок ДНК, который ассоциирована с определенным фенотипическим признаком - данные QTL часто находятся на разных хромосомах. Информация о числе QTL, которое определяет вариацию по определенному фенотипическому признаку, сообщает о генетической структуре признака. Она может показать, что растение с предпочтительным фенотипом контролируется множественными генами, оказывающими незначительный эффект или несколькими генами, оказывающими значительный эффект. Таким образом, QTL-картирование может применяться для определения тех частей генома растения-донора, которые содержат мутантные KRP, и способствовать осуществлению способов селекции.
Один или несколько таких QTL мутантных KRP в растении-доноре могут быть перенесены в растение-реципиент, подтверждая фенотип с увеличенной массой, размером и/или числом одного или нескольких органов, например, с увеличенными массой семян, размером семян, числом семян и/или урожаем. В некоторых дополнительных вариантах осуществления QTL, связанные с мутантными KRP, могут быть объединены с одним или несколькими другими QTL, которые вносят вклад в формирование агротехнически важных фенотипов, таких как повышение урожайности, устойчивости к биотическому и абиотическому стрессам и др. Праймеры в настоящем изобретении, применяемые для генотипирования мутантных KRP, можно использовать в качестве молекулярных маркеров, указывающих на присутствие или отсутствие мутантных KRP. Кроме того, также можно применять молекулярные маркеры, тесно связанные с мутантными KRP. Способы разработки молекулярных маркеров и их применения описаны Avise (Molecular markers, natural history and evolution, Publisher: Sinauer Associates, 2004, ISBN0878930418, 9780878930418), Srivastava et al. (Plant biotechnology и molecular markers. Publisher: Springer, 2004, ISBN1402019114, 9781402019111) и Vienne (Molecular markers in plant genetics and biotechnology. Publisher: Science Publishers, 2003), каждая из этих публикация включена в данный документ в своей полноте посредством ссылки.
Без привязки к какой-либо теории, помимо увеличенного размера семян, числа семян, веса семян и/или урожая, растение, содержащее один или несколько нарушенных KRP, может иметь один или несколько других фенотипов, являющихся агротехнически или промышленно важными, которые включают, но не ограничивают, увеличение мощности растений, размера органов, улучшение адаптации к окружающей среде, повышение продукции масла, повышение продукции биовеса и признаки, которые обеспечивают лучший рост растения в определенных условиях с определенными температурами, состояниями почвы и уровнями солнечного света и осадков по сравнению с контрольным растением дикого типа.
Культивирование и прививка тканей
Современное культивирование тканей растений проводят в асептических условиях в атмосфере профильтрованного воздуха. Поверхности живых материалов растений из окружающей среды по своей природе (а иногда и ткани) контаминированы микроорганизмами, что требует стерилизации поверхности исходных материалов (эксплантатов) в химических растворах (обычно спирте или водном растворе гипохлорита натрия). Затем эксплантаты, как правило, помещают на поверхность плотной питательной среды, но иногда помещают непосредственно в жидкую среду, особенно, если требуется получение суспензионных культур клеток. Плотные и жидкие среды обычно имеют в составе неорганические соли, а также некоторые органические питательные вещества, витамины и растительные гормоны. Твердые среды готовят из жидких сред с добавлением гелеобразующего средства, как правило, очищенного агара.
Композиция среды, в частности растительные гормоны и источник азота (нитрат аммония по сравнению с солями или аминокислотами), оказывают сильное воздействие на морфологию тканей, которые растут из первоначального эксплантата. Например, избыток ауксинов часто приводит к пролиферации корней, а избыток цитокининов может повысить урожай побегов. Баланс ауксинов и цитокининов часто приводит к неорганизованному росту клеток или каллуса, но морфологический результат будет зависеть от вида растений, а также от композиции среды. В процессе роста культуры, куски ткани, как правило, отрезают и переносят в новую среду (пересев) для обеспечения роста или изменения морфологии культуры. Навыки и опыт специалиста, проводящего культивирование ткани, играют важную роль при оценке того, какие части оставить в культуре, а какие отсеять. При появлении в культуре всходов, их можно отрезать и укоренить с применением ауксина для получения саженцев, которые после созревания могут быть перенесены на почвенную смесь для дополнительно роста в теплице как нормальные растения.
Ткань, полученная из растения для культивирования, называется эксплантат. На основе работы с определенными модельными системами, в частности с табаком, неоднократно утверждалось, что тотипотентный эксплантат может быть выращен из любой части растения. Однако на практике эта идея оказалась недействительной. Во многих видах растений эксплантаты различных органов различаются по своим темпам роста и регенерации, а некоторые вообще не растут. Выбор материала эксплантата также определяет, будут ли саженцы, выращенные с применением культивирования ткани, гаплоидными или диплоидными. Также при использовании неподходящих эксплантатов повышается риск микробного загрязнения. Таким образом, очень важно перед культивированием ткани сделать соответствующий выбор эксплантата.
Характерные различия в потенциале регенерации различных органов и эксплантатов имеют различные объяснения. Значимые факторы включают различия в стадии клеток в клеточном цикле, возможность или способность транспортировать эндогенные регуляторы роста и метаболические возможности клеток. Наиболее часто применяемыми эксплантатами тканей являются меристематические ткани концевых участков растений, таких верхушка ствола, вспомогательная верхушка почки и кончик корня. Эти ткани обладают высокой скоростью деления клеток и либо концентрируют, либо продуцируют необходимые вещества, регулирующие рост, включая ауксины и цитокинины. Некоторые эксплантаты, такие как кончик корня, трудно выделить, и они контаминированы почвенной микрофлорой, что является проблемой в процессе культивирования ткани. Определенная почвенная микрофлора может образовывать тесные ассоциации с корневой системой или даже расти в тканях корня. Частицы почвы, связанные с корнями, трудно удалить без повреждения корней, которое затем позволяет проникновение микробов. Эта связанная микрофлора обычно проводит к зарастанию культуральной тканевой среды до достижения значительного роста растительной ткани. Воздушные (находящиеся над поверхностью почвы) эксплантаты также богаты нежелательной микрофлорой. Однако данную микрофлору легче удалить из эксплантата путем аккуратного промывания, и оставшуюся микрофлору обычно можно уничтожить путем стерилизации поверхности. Большая часть поверхностной микрофлоры не формирует тесных ассоциаций с растительной тканью. Подобные ассоциации обычно можно найти путем визуального осмотра в виде мозаичной болезни, обесцвечивания или локализованного некроза на поверхности эксплантата.
Альтернативой для получения неконтаминированных эксплантатов является получение эксплантатов из рассады, выращенной в асептических условиях из семян с простерилизованной поверхностью. Твердая поверхность семян является менее проницаемой для проникновения средств для стерилизации неровных поверхностей, таких как гипохлорит, поэтому приемлемые условия стерилизации, используемые для семян, могут быть гораздо более жесткими, чем для вегетативных тканей.
Растения, полученные из культуры тканей, являются клонами; если оригинальное материнское растение, используемое для получения первых эксплантатов, восприимчиво к патогену или условиям окружающей среды, тогда вся сельскохозяйственная культура будет восприимчива к этой же проблеме и, наоборот, в линии также сохранятся любые положительные признаки. Культура ткани растений широко используется в растениеводстве; она также имеет различные виды коммерческих применений. Применения включают:
1. Вегетативное размножение широко применяют в лесном хозяйстве и в цветоводстве. Вегетативное размножение также быть использовано для сохранения редких или находящихся под угрозой вымирания видов растений,
2. Растениевод-селекционер может применять культуру ткани для скрининга клеток, а не растений, в отношении полезных характеристик, например, устойчивости к патогенам/резистентности.
3. Масштабный рост клеток растений в жидкой культуре в биореакторах в качестве источника вторичных продуктов, таких как рекомбинантные белки, которые применяют в качестве биофармацевтических препаратов.
4. Для скрещивания дальнородственных видов путем слияния протопластов и регенерации нового гибрида.
5. Для перекрестного опыления дальнородственных видов и последующего культивирования ткани полученного зародыша, который в другом случае, как правило, погибает (сохранение зародыша).
6. Для получения растений с удвоенным моноплоидным генотипом (дигаплоидных) из гаплоидных культур для более быстрого получения гомозиготных линий в программах селекции, как правило, путем обработки колхицином, которая вызывает удвоение числа хромосом.
7. Как ткань для трансформации, за которой следует или краткосрочное тестирование генетических конструктов, или регенерация трансгенных растений.
8. Определенные методики, такие как культивирование апикальной меристемы, можно применять для получения чистого растительного материала из инфицированного биоматериала, такого как картофель и многие виды ягодных культур.
9. Вегетативное размножение с применением культуры меристемы и проростков для получения большого числа идентичных индивидуумов.
Неограничивающие иллюстративные способы культивирования тканей пшеницы, риса, кукурузы, описаны Trione et al., Dodig, et al., O'Hara et al., Zaidi et al., Wang et al., Ting et al., Hawes et al. и Sheridan, при этом каждая из данных публикаций включена в данный документ во своей полноте посредством ссылки.
Настоящее изобретение также предлагает черенок, корневой побег, привой или эксплантат, полученный от растения настоящего изобретения.
Прививка тканей - это способ бесполого размножения растений, который широко применяют в земледелии и садоводстве, при котором провоцируют объединение тканей одного растения с тканями другого растения. Этот способ чаще всего используют для размножения деревьев и кустарников, выращиваемых в коммерческих целях. В большинстве случаев одно растение выбирают по корневой системе и называют подвоем или корневым побегом. Другое растение выбирают по стеблям, листьям, цветам или фруктам и называется привоем. Привой содержит желаемые гены, которые должны быть продублированы в получаемом в будущем подвое/привое. При прививке к стеблю, которая является часто применяемым способом прививки, черенок выбранного, желаемого сорта культурного растения прививают на подвой другого типа. В другой распространенной форме, называемой окулировкой, спящую боковую почку прививают к стеблю другого растения-подвоя и после успешного объединения тканей ее рост стимулируют путем обрезания стебля над новой почкой.
Для того, чтобы прививка была успешной, ткани сосудистого камбия подвоя и привоя растений должны быть приведены в контакт друг с другом. Оба вида тканей должны поддерживаться в живом состоянии, пока не примется трансплантат, что, как правило, занимает несколько недель. Успешная прививка требует лишь того, чтобы между двумя тканями были установлены сосудистые связи. На привое может возникать физически слабое место, так как структурные ткани двух различных растений, такая как древесина, могут не объединиться.
Иллюстративные методики прививки включают прививку сближением, прививку окулировкой (окулировку пластинкой или четырехугольным кусочком коры, окулировку прорастающим глазком, окулировку с надрезом в виде буквы Т), прививку врасщеп, прививку в боковой отщеп, прививку копулировкой, прививку в пенек, прививка "шилом", прививку черенком, прививку за кору, прививку с язычком и др. Подробные неограничивающие способы прививки тканей пшеницы и кукурузы описаны Lacadena, 1968, и Katsumi et al., при этом каждая из данных публикаций включена в данный документ в своей полноте посредством ссылки.
Трансформация растений
Полинуклеотиды настоящего изобретения могут быть трансформированы в растение. Наиболее распространенный способ введения нового генетического материала в геном растения включает применение живых клеток патогенных бактерий Agrobacterium tumefaciens с тем, чтобы в буквальном смысле ввести кусок ДНК, называемый транспортной или Т-ДНК, в отдельные растительные клетки (обычно после скарификации ткани), где она направляется в ядро растения для интеграции в хромосомы. Существует множество патентов, в которых применяют опосредованную Agrobacterium трансформацию и конкретные плазмиды для доставки ДНК, сконструированные специально для применения с Agrobacterium - например, US 4536475, ЕР 0265556, ЕР 0270822, WO 8504899, WO 8603516, US 5591616, ЕР 0604662, ЕР 0672752, WO 8603776, WO 9209696, WO 9419930, WO 9967357, US 4399216, WO 8303259, US 5731179, ЕР 068730, WO 9516031, US 5693512, US 6051757 и ЕР 904362А1. Agrobacterium-опосредованная трансформация растений включает в качестве первого этапа введение фрагментов ДНК, клонированных в плазмиды, в живые клетки Agrobacterium, которые затем используют для трансформации в отдельные клетки растений. Agrobacterium-опосредованная трансформация растений, таким образом, является непрямым методом трансформации растений. Способы Agrobacterium-опосредованной трансформации растений, которые связаны с применением векторов, не содержащих Т-ДНК, также хорошо известны специалистам в данной области и могут быть применимы в настоящем изобретении. Смотри например, патент США №7250554, который использует Р-ДНК вместо Т-ДНК в векторе для трансформации.
Также были описаны способы прямой трансформации растений. Исторически первым из описанных способов была электропорация, которая использует электрический ток, применимый к раствору, содержащему клетки растений (М.Е. Fromm et al., Nature, 319, 791 (1986); H. Jones et al., Plant Mol. Biol., 13, 501 (1989) и H. Yang et al., Plant Cell Reports, 7, 421 (1988). Другой прямой способ, называемый "биолистической бомбардировкой", использует ультрамелкие частицы, как правило, из вольфрама или золота, покрытые ДНК, которые затем распыляют на поверхность ткани растения с силой, достаточной для того, чтобы частицы проникли в клетки растения, в том числе через толстую клеточную стенку и мембрану ядерной оболочки, не вызывая при этом гибели по меньшей мере некоторых из них (US 5204253, US 5015580). Третий прямой способ использует волокнистые формы металла или керамики, состоящие из острых, пористых или полых иглоподобных бомбардирующих частиц, которые буквально пронзают клетки, а также оболочки клеточного ядра. Нитевидные кристаллы как из карбида кремния, так и из бората алюминия применяли для трансформации растений (Mizuno et al., 2004; Petolino et al., 2000; US 5302523 US Application 20040197909), а также для трансформации бактериальных клеток и клеток животных (Kaepler et al., 1992; Raloff, 1990; Wang, 1995). Помимо опубликованных других способов несомненно будут разработаны дополнительные способы. Тем не менее, эффективность каждого из данных непрямых или прямых способов во введении чужеродной ДНК в клетки растений неизменно является крайне низкой, что делает необходимым применение определенного способа отбора только тех клеток, которые были трансформированы, и кроме этого, обеспечение роста и регенерации в растениях только тех клеток, которые были трансформированы.
Для эффективной трансформации растений, должен применяться такой способ селекции, при котором целые растения регенерируют из единственной трансформированной клетки и каждая клетка трансформированного растения несет ДНК интереса. Данные способы могут использовать позитивный отбор, при котором в клетку растения вводят чужеродный ген, который обеспечивает утилизацию субстрата, присутствующего в среде, который в ином случае не может быть утилизирован, такой как манноза или ксилоза (например, смотри US 5767378; US 5994629). Однако, в основном используют негативный отбор, поскольку он является более эффективным, используя такие средства отбора как гербициды или антибиотики, которые либо вызывают гибель, либо подавляют рост нетрансформированных клеток растений и снижают возможность образования химер. Гены устойчивости, которые эффективны против средств для негативного отбора, присутствуют в вводимой чужеродной ДНК, используемой для трансформации растений. Например, одно из наиболее популярных используемых средств для отбора антибиотик канамицин вместе с геном устойчивости, кодирующим неомицин-фосфотрансферазу (nptII), которая придает устойчивость к канамицину и родственным антибиотикам (смотри, например, Messing & Vierra, Gene 19: 259-268 (1982); Bevan et al., Nature 304:184-187 (1983)). Несмотря на это множество различных антибиотиков и генов устойчивости к антибиотикам может применяться в целях трансформация (смотри US 5034322, US 6174724 и US 6255560). В дополнение, в целях трансформация используются несколько гербицидов и генов устойчивости к гербицидам, в том числе ген bar, который придает устойчивость к гербициду фосфинотрицину (White et al., Nucl Acids Res 18: 1062 (1990), Spencer et al., Theor Appl Genet 79: 625-631(1990), US 4795855, US 5378824 и US 6107549). Кроме того, для селекции использовали ген dhfr, который придает устойчивость к противораковому средству метотрексату (Bourouis et al., EMBO J. 2(7): 1099-1104 (1983).
Элементы, контролирующие экспрессию, применяемые для регуляции экспрессии определенного белка, могут быть либо контролирующими экспрессию элементами, которые обычно находятся в связи с кодирующей последовательностью (гомологичный элемент для экспрессии) или могут быть гетерологичными контролирующими экспрессию элементами. В данной области техники известны различные гомологичные и гетерологичные контролирующие экспрессию элементы, которые можно легко применять для получения единиц экспрессии для применения в рамках настоящего изобретения. Участки инициации транскрипции, например, могут содержать участки инициации транскрипции опинов, таких как октопин, маннопин, нопалин и подобные, которые обнаружены в Ti-плазмидах Agrobacterium tumefaciens. Альтернативно, также можно применять промоторы от растительных вирусов, такие как промоторы 19S и 35S вируса мозаики цветной капусты (промоторы CaMV 19S и CaMV 35S, соответственно) для контроля экспрессии генов в растении (например, патенты США №№5352605; 5530196 и 5858742). Также можно применять энхансерные последовательности, полученные из CaMV (например, патенты США №№5164316; 5196525; 5322938; 5530196; 5352605; 5359142; и 5858742). Наконец, также можно применять промоторы растительных клеток, такие как промотор пролиферации, промоторы, специфичные для фруктов, промотор Ар3, промоторы теплового шока, промоторы, специфичные для семян, и др.
Как правило, гамето-специфический промотор, конститутивный промотор (такой как промотор CaMV или Nos), органо-специфический промотор (например, промотор, специфический для стволовых клеток) или индуцируемый промотор лигируют с белком или антисмысловой кодирующей областью с применением стандартных методик, известных в данной области техники. Единица экспрессии может быть дополнительно оптимизирована путем применения дополнительных элементов, таких как терминаторы транскрипции и/или энхансерные элементы. Кассета экспрессии может содержать, например, промотор, специфический для семян (например, промотор фазеолина (патент США №5504200). Выражение "промотор, специфический для семян" означает, что ген, который экспрессируется под контролем промотора, экспрессируется преимущественно в семенах растения, с отсутствием или незначительной экспрессией, составляющей, как правило, менее 10% от общего уровня экспрессии в других тканях растения. Промоторы, специфические для семян, известны в данной области техники, например, патент США №5623067, 5717129, 6403371, 6566584, 6642437, 6777591, 7081565, 7157629, 7192774, 7405345, 7554006, 7589252, 7595384, 7619135, 7642346, и публикация заявки на патент США №20030005485, 20030172403, 20040088754, 20040255350, 20050125861, 20050229273, 20060191044, 20070022502, 20070118933, 20070199098, 20080313771 и 20090100551.
Таким образом, для экспрессии в растениях, единицы экспрессии будут, как правило, содержать в дополнение к белковой последовательности область промотора растительных клеток, сайт инициации транскрипции и последовательность терминации транскрипции. Как правило, на 5'- и 3'-концах единицы экспрессии включают уникальные участки фермента рестрикции, которые способны обеспечить легкую вставку единицы экспрессии в существующий вектор.
В конструкции гетерологичного промотора/структурного гена или антисмысловых комбинаций промотор предпочтительно располагается приблизительно на таком же расстоянии от сайта начала гетерологичной транскрипции, как и от сайта начала транскрипции в естественных условиях. Однако, как известно в данной области техники, это расстояние может несколько изменяться без потери функции промотора.
В дополнение к последовательности промотора кассета экспрессии может также содержать участок терминации транскрипции ниже структурного гена для обеспечения эффективной терминации. Участок терминации может быть получен из того же гена, что и последовательность промотора, или может быть получен из других генов. Если иРНК, кодируемая структурным геном, должна подвергаться эффективному процессингу, то к векторному конструкту также часто добавляют последовательности ДНК, которые направляют полиаденилирование РНК. Последовательности для полиаденилирования включают, но не ограничивают, сигнальную последовательность октопин-синтазы Agrobacterium (Gielen et al., EMBO J 3:835-846 (1984)) или сигнальную последовательность нопалин-синтазы (Depicker et al., Mol. and Appl. Genet. 1:561-573 (1982)). Полученную в результате единицу экспрессии лигируют с вектором или конструируют таким образом, чтобы ввести в пригодный для трансформации высших растений вектор. Одна или несколько единиц экспрессии могут быть введены в один и тот же вектор. Вектор, как правило, будет содержать единицу экспрессии гена селектируемого маркера, по которой трансформированные клетки растений могут быть идентифицированы в культуре. Как правило, маркерный ген будет кодировать устойчивость к антибиотику, такому как G418, гигромицин, блеомицин, канамицин или гентамицин или к гербицидам, таким как глифосат (Round-Up) или глюфосинат (BASTA) или атразин. Обычно также вводят последовательности для репликации бактериального или вирусного происхождения для обеспечения клонирования вектора в бактерии-хозяине или фаге, при этом предпочтительно вводят последовательности для репликации, полученные из широкого спектра прокариот. Также может быть введен селектируемый маркер для бактерий, позволяющий проводить отбор бактериальных клеток, несущих желаемый конструкт. Подходящие селектируемые маркеры прокариот содержат устойчивость к антибиотикам, таким как ампициллин, канамицин или тетрациклин. Также в векторе могут присутствовать другие последовательности ДНК, кодирующие дополнительные функции, как известно в данной области техники. Например, в случае трансформаций Agrobacterium последовательности Т-ДНК также будут введены для последующего переноса в хромосомы растения.
Введение желаемого гена или набора генов стандартными способами требует проведения полового скрещивания между двумя линиями с последующим повторным обратным скрещиванием между гибридным потомством и одним из родителей до получения растения с желаемыми характеристиками. Данный процесс, однако, ограничен растениями, которые могут гибридизоваться половым путем, так как будут перенесены дополнительные гены, помимо требуемого гена.
Технологии рекомбинантной ДНК позволяют исследователям растений обойти эти ограничения, позволяя изучающим растения генетикам идентифицировать и клонировать специфические гены, отвечающие за желаемые признаки, такие как устойчивость к насекомым-вредителем, и ввести эти гены в уже полезные сорта растений. После того, как чужеродные гены были введены в растение, данное растение можно использовать в стандартных схемах селекции растений (например, селекция на основе генеалогии, схема селекции способом индивидуального отбора, реципрокный повторяющийся отбор) для получения потомков, которые также содержат ген интереса.
Гены могут быть введены сайт-направленным способом с применением гомологичной рекомбинации. Гомологичная рекомбинация обеспечивает сайт-специфичную модификацию эндогенных генов, и таким образом унаследованные или приобретенные мутации могут быть исправлены и/или новые изменения могут быть созданы в геноме. Гомологичную рекомбинацию и сайт-специфическую интеграцию в растениях рассматривают, например, в патентах США №№5451513; 5501967 и 5527695.
Способы получения трансгенных растений хорошо известны средним специалистам в данной области техники. В настоящее время трансгенные растения могут быть получены с применением различных способов трансформации, включая, но не ограничивая, электропорацию; микроинъекцию; баллистическую трансфекцию, также известную как ускорение частиц или биолистическую бомбардировку; вирус-опосредованную трансформацию; и Agrobacterium-опосредованную трансформацию. Смотри, например, патенты США №№5405765; 5472869; 5538877; 5538880; 5550318; 5641664; 5736369 и 5736369; публикации международных заявок на патент №№ WO 2002/038779 и WO/2009/117555; Lu et al., (Plant Cell Reports, 2008, 27:273-278); Watson et al., Recombinant DNA, Scientific American Books (1992); Hinchee et al., Bio/Tech. 6:915-922 (1988); McCabe et al., Bio/Tech. 6:923-926 (1988); Toriyama et al., Bio/Tech. 6:1072-1074 (1988); Fromm et al., Bio/Tech. 8:833-839 (1990); Mullins et al., Bio/Tech. 8:833-839 (1990); Hiei et al., Plant Molecular Biology 35:205-218 (1997); Ishida et al., Nature Biotechnology 14:745-750 (1996); Zhang et al., Molecular Biotechnology 8:223-231 (1997); Ku et al., Nature Biotechnology 17:76-80 (1999); и Raineri et al., Bio/Tech. 8:33-38 (1990)), каждая из которых специально включена во всей полноте в данный документ посредством ссылки.
Agrobacterium tumefaciens является встречающейся в природе бактерией, которая обладает способностью встраивать свою ДНК (генетическую информацию) в растения, что приводит к такому типу повреждения растения, которое известно как корончатый галл. Большинство видов растений в настоящее время могут быть трансформированы с применением данного способа, включая виды тыквенных.
Баллистическая трансфекция, также известна как ускорение частиц, биолистическая бомбардировка и генная пушка (Biolistic® Gene Gun). Генную пушку применяют для выстреливания частиц, несущих на своей поверхности гены (например, кодирующих желаемые признаки), в семена растения или ткани растения для того, чтобы получить клетки растений, в которых затем будут экспрессироваться новые гены. Генная пушка использует фактическую энергию взрыва (холостые пули калибра .22) для приведения в движение материала. Сжатый воздух или пар также могут быть использованы в качестве заряда. Генная пушка Biolistic® была изобретена в 1983-1984 в Корнельском университете John Sanford, Edward Wolf и Nelson Allen. Данная пушка и соответствующая зарегистрированная торговая марка в настоящее время являются собственностью Е.I. du Font de Nemours and Company. Большинство видов растений были трансформированы с применением этого способа.
Трансгенное растение, созданное с применением способов трансформации Agrobacterium, как правило, содержит единственный ген на одной хромосоме, хотя возможны множественные копии. Такие трансгенные растения могут называться гемизиготными за счет добавленного гена. Более точным названием такого растения является независимый сегрегант, поскольку каждое трансформированное растение представляет уникальное явление интеграции Т-ДНК (патент США №6156953). Трансгенные локусы обычно характеризуются по присутствию и/или отсутствию трансгена. Гетерозиготный генотип, в котором один аллель соответствует отсутствию трансгена, также называют гемизиготным (патент США №№6,008,437).
Общепринятые способы трансформации и специфические способы трансформации определенных видов растений (например, кукурузы, риса, пшеницы, ячменя, сои) описаны в патентах США №№4940838, 5464763, 5149645, 5501967, 6265638, 4693976, 5635381, 5731179, 5693512, 6162965, 5693512, 5981840, 6420630, 6919494, 6329571, 6215051, 6369298, 5169770, 5376543, 5416011, 5569834, 5824877, 5959179, 5563055 и 5968830, каждый из которых включен в данный документ во всей своей полноте посредством ссылки.
Настоящее изобретение дополнительно проиллюстрировано следующими примерами, которые не следует толковать как ограничивающие. Содержания всех ссылок, патентов и опубликованных патентных заявок, цитируемые в данной заявке, а также фигуры и перечень последовательностей, включены в данный документ посредством ссылки.
ПРИМЕРЫ
Материалы и Способы
Мутагенез
В одном варианте осуществления настоящего изобретения семена тетраплоидной пшеницы (Triticum turgidum) культурного сорта "Kronos" и гексаплоидной пшеницы (Triticum aestivum) культурного сорта "Express" (PVP # 9000012) пропитывали H2O под вакуумом (приблизительно 1000 семян/100 мл Н2О в течение приблизительно 4 минут). Затем семена помещали на вибрационное сито (45 оборотов в минуту) в вытяжном шкафу при комнатной температуре. Мутаген этилметансульфонат (EMS) добавляли к пропитывающимся семенам до конечных концентраций в диапазоне от приблизительно 0,75% до приблизительно 1,2% (объем/объем). По окончании 18 часов инкубационного период раствор EMS заменяли 4 раза свежей Н2О. Затем семена промывали под проточной водой в течение приблизительно 1 часа. Наконец, мутагенизированные семена высевали (96/лоток) в почву и выращивали в помещении. Растения в возрасте от четырех до шести недель переносили на поле для выращивания до полностью зрелых растений M1. Зрелые растения M1 самоопылялись, после чего семена от растений M1 собирали и высевали для получения растений М2.
Получение ДНК
ДНК из данных растений М2 экстрагировали и подготавливали с целью идентификации растений М2, которые несли мутацию в своем локусе KRP. ДНК растения М2 получали с применением способов и реагентов, содержащихся в Qiagen® (Валенсия, Калифорния, США) DNeasy® 96 Plant Kit. Приблизительно 50 мг замороженного образца растения помещали в пробирку для образца с вольфрамовым шариком, замораживали в жидком азоте и измельчали 2 раза в течение 1 минуты каждый раз при 20 Гц с применением Retsch® Mixer Mill MM 300. Затем к образцу добавляли 400 мкл раствора AP1 [буфер AP1, раствор DX и РНКазы (100 мг/мл)] при 80°C. Пробирку закрывали и встряхивали в течение 15 секунд. После добавления 130 мкл буфера АР2 пробирку встряхивали в течение 15 секунд. Образцы помещали в морозильную камеру при минус 20°C по меньшей мере на 1 час. Затем образцы центрифугировали в течение 20 минут при 5600 × g. Аликвоту надосадочной жидкости объемом 400 мкл переносили в другую пробирку для образца. После добавления 600 мкл буфера АР3/Е данную пробирку для образца закрывали и встряхивали в течение 15 секунд. Планшет с фильтрующим дном помещали на блок с квадратными лунками и в каждую лунку вносили 1 мл раствора образца и запечатывали планшет. Планшет и блок центрифугировали в течение 4 минут при 5600 × g. Затем 800 мкл буфера AW добавляли в каждую лунку планшета с фильтрующим дном, запечатывали и центрифугировали в течение 15 минут при 5600 × g в блоке с прямоугольными лунками. Планшет с фильтрующим дном затем помещали на новый набор пробирок с образцом и на фильтр вносили 80 мкл буфера АЕ. Планшет накрывали и инкубировали при комнатной температуре в течение 1 минуты, а затем центрифугировали в течение 2 минут при 5600 × g. Этот этап повторяли с добавлением 80 мкл буфера АЕ. Удаляли планшет с фильтрующим дном и накрывали пробирки, содержащие объединенные фильтраты. Отдельные образцы затем стандартизировали по концентрации ДНК 5-10 нг/мкл.
TILLING®
ДНК М2 объединяли в группы из двух отдельных растений. Концентрация ДНК для каждого индивидуума в пуле составляла приблизительно 1-2 нг/мкл с конечной концентрацией 2-4 нг/мкл для пула в целом. Затем 5 или 10 мкл объединенных образцов ДНК (или от 10-40 нг) наносили на титрационный микропланшет и подвергали ген-специфической ПЦР.
ПЦР-амплификацию проводили в объемах реакционной смеси 15 или 20 мкл, содержащих от 10-40 нг объединенной ДНК. Реакционная смесь включала 1,24 мкл буфера 10Х ExTaq (Takara®), 0,73 мкл 25 мМ MgCl2, 1,98 мкл 10 мМ дНТФ, 0,066 мкл смеси праймеров с концентрацией 100 мкМ и 0,11 мкл ДНК-полимеразы Ex-Taq (Takara®) с концентрацией 5 Ед/мкл с 6,87 мкл Н2О. Дополнительные компоненты для ПЦР, такие как диметилсульфоксид (DMSO), бетаин или Polymer-Aide PCR Enhancer (Sigma Aldrich®, Сент-Луис, Миссури, США), могут применяться для повышения эффективности ПЦР. ПЦР-амплификацию проводили с применением термоциклера MJ Research® следующим образом: 95°C в течение 2 минут; 8 циклов "ступенчатая ПЦР" (94°C в течение 20 секунд, за которым следует этап отжига, на котором начальная температура составляет 70-68°C в течение 30 секунд с последующим снижением на 1°C в каждом цикле, за этим следует ступенчатое повышение температуры на 0,5°C в секунду до 72°C с поддержанием температуры на уровне 72°C в течение 1 минуты); 25-45 циклов с температурой 94°C в течение 20 секунд, 63-67,5°C в течение 30 секунд, ступенчатое повышение температуры на 0,5°C в секунду до 72°C, 72°C в течение 1 минуты; 72°C в течение 8 минут; 98°C в течение 8 минут; 80°C в течение 20 секунд; 60 циклов при температуре 80°C в течение 7 секунд - 0,3 градуса/цикл.
ПЦР-праймеры (Eurofins MWG/Operon, Хантсвилл, Алабама, США) смешивали следующим образом:
12,5% 100 мкМ левого праймера, меченого IRD-700;
37,5% 100 мкМ левого праймер;
25% 100 мкМ правого праймера, меченого IRD-8700;
25% 100 мкМ правого праймер.
Метка может быть присоединена к каждому праймеру, как описано, или только к одному праймеру. Альтернативно, могут быть использованы Су5.5-модифицированные праймеры. Метку соединяли с олигонуклеотидом с применением методов традиционной фосфорамидитной химии.
Продукты ПЦР (15 или 20 мкл) расщепляли в 96-луночных планшетах. Затем добавляли 30 мкл раствора, содержащего 10 мМ HEPES [4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота] (pH 7,5), 10 мМ MgSO4, 0,002% (вес/объем) Triton® X-100, 20 нг/мл бычьего сывороточного альбумина, 5 мкл SURVEYOR® Nuclease (Transgenomic, Inc.) и 5 мкл SURVEYOR® Enhancer (Transgenomic, Inc.) с перемешиванием на льду, и планшет инкубировали при 45°C в течение 30 минут. Реакции останавливали путем добавления 10 мкл 2,5 М раствора NaCl с 0,5 мг/мл голубого декстрана и 75 мМ EDTA, после чего добавляли 80 мкл изопропилового спирта. Реакционные смеси осаждали при комнатной температуре центрифугированием при 4000 оборотов в минуту в течение 30 минут в Eppendorf Centrifuge 5810. Осадок ресуспендировали в 6 мкл 33% формамида с 0,017% красителя бромфенола синего, нагревали при 80°C в течение 7 минут, а затем при 95°C в течение 5 минут. Образцы переносили на мембранный гребень с применением гребнезагрузочного робота (MWG Biotech). Гребенчатую мембрану погружали в акриламидный гель (6,5%) и проводили электрофорезу в течение 4 ч 15 мин при 1500 В, 40 Вт и 40 мА при 50°C.
Во время электрофореза изображение геля получали с применением сканера LICOR® (Lincoln, NE), который настраивали на канал, способный обнаруживать метки IR Dye 700 и 800. Изображение геля демонстрировало последовательность-специфический паттерн фоновых бэндов, общий для всех 96 полос. Редкие события, такие как мутации, создают новые бэнды, которые располагаются выше фонового рисунка. Растения с бэндами, указывающими на наличие мутации интереса, оценивали по отдельным представителям, идентифицированным с применением TILLING®, из общего пула, смешанного с ДНК дикого типа, а затем секвенировали отдельные продукты ПЦР.
Пример 1.
TILLING для мутантных KRP Triticum
Сконструировали геном-специфические праймеры и применяли для TILL KRP пшеницы (см. Таблицу 1 в графической части).
Пример 2.
Мутации генов KRP Triticum, идентифицированных с применением TILLING®
Скрининг популяции TILLING®, в отношении мутантных KRP привел к обнаружению растений с молчащими мутациями, мутациями сайта сплайсинга, нонсенс-мутациями (преждевременные стоп-кодоны) и/или миссенс-мутациями (тяжелыми или нетяжелыми) в генах KRP1, KRP2, KRP4 и KRP5 (геномы А, В и Соположения и эффекты мутаций в генах KRP1, KRP2, KRP4 и KRP5 (геномы А, В и D) приведены в таблицах 2-12 в графической части (* обозначает мутацию, которая приводит к образованию стоп-кодона, = обозначает молчащую мутацию).
Пример 3.
Селекционная программа для пшеницы.
TILLING®-мутантные KRP пшеницы в селекционной программе располагали по приоритету следующем порядке: 1) нонсенс-мутанты и мутанты сайта сплайсинга; 2) тяжелые миссенс-мутации типа I; и 3) тяжелые миссенс-мутации типа II. Тяжелые миссенс-мутации типа I являются неконсервативными заменами аминокислот в тех участках белка KRP, которые известны как необходимые для связывания с циклином или циклин-зависимой киназой (CDK), и для которых, согласно анализу SIFT, ожидается вредное воздействие на функцию белка. Тяжелые миссенс-мутации типа II являются неконсервативными заменами аминокислот, которые расположены вне циклин- и CDK-связывающих доменов, но которые удовлетворяют двум дополнительным критериям. Во-первых, они располагаются в участках белка, которые согласно анализу BLOCKS (Henikoff, S. и Henikoff, J.G. (1991) Nucleic Acids Res., 19, 6565-6572) являются эволюционно консервативными и поэтому, вероятно, функционально значимы. Во-вторых, они имеют балл SIFT (Ng, P.С. и Henikoff, S. (2003) Nucleic Acids Res. July 1; 31(13): 3812-3814) менее 0,05 и поэтому предполагается их вредное воздействие на функцию белка.
В таблицах 13-22 в графической части перечислены TILLING®-мутантные KRP пшеницы, которые входят в селекционую программу и являются набором мутантов, перечисленных в таблицах 2-12. Указанная группа пшеницы (WH) является числом, которое идентифицирует конкретную мутацию в конкретном гене.
Выращивают зерно М3, гомозиготное или гетерозиготное по данной TILLING®-мутации KRP. Проводят возвратное скрещивание с гексаплоидным родительским яровым сортом Express, используемым в качестве фона, оптимально в несколько этапов (до уровня ВС3 или ВС4) для исключения вредных фоновых мутаций. Фоновые мутации могут приводить к нежелательным свойствам, таким как задержка созревания, преждевременное старение, повышенная восприимчивость к патогенам, поражающим пшеницу, медленное прорастание и/или бесплодие. Потомки каждого возвратного скрещивания (F1, BC1, ВС2 и др.) помимо этого самоопыляются с получением линии F2. Линии F2 генотипируют для выявления тех, которые являются гомозиготными по дикому типу или мутантному аллелю krp. Гомозигота по дикому типу и мутантные сибсы являются семенами поколения F3, полученными для полевых испытаний.
Мутантные аллели krp пшеницы интрогрессируют в другую яровую и озимую пшеницу чтобы передать свойство повышенной урожайности коммерческим сортам.
Скрещивания между мутантами проводятся для получения различных мутантов в пределах данного гена KRP (например, krp1A/1B, krp1B/krp1D, krp1A/1B/1D и др., все возможные комбинации) или по различным генам KRP (например, krp 1A/2A, krp2B/krp4B, krp4D/krp5A, krp1B/krp2A/krp5D и др., все возможные комбинации).
Определяют общий урожай зерна на единицу площади (например, фунт/акр), и оценивают такие компоненты урожая как число семян, размер семян/вес (вес тысячи зерен), число семян в колосе, число верхушек (колосьев) колоса, длина колоса, длина ости и/или число ростков. Также оценивают агронотехнические характеристики, такие как густота стояния растений, сроки созревания и параметры плодоножки.
Пример 4.
Генотипирование TILLING®-мутаций по KRP в пшенице
TILLING®-мутации, по KRP пшеницы генотипировали с применением наборов праймеров/зондов для аллельного дискриминантного анализа в ПЦР в режиме реального времени. Во-первых, геном-специфичные праймеры применяли для амплифицикации данного гена KRP пшеницы из образца геномной ДНК пшеницы. Продукт амплификации затем генотипировали с применением аллельного дискриминантного анализа Taqman® (AD Assay, Applied Biosystems). Приведенное ниже является специфичным примером анализа для генотипирования KRP4B2 P109L.
Прямой праймер ТТС СТТ АТТ ТТТ TAT GAC TAT TGA TAT GTG TTC ТТС (SEQ ID NO: 28) и обратный праймер GTG GTC АТТ ТСА GAA TGA GCT GCT ААС CGT T (SEQ ID NO: 29) применяли для амплифицикации KRP4B2 из геномной ДНК пшеницы. Реакционная смесь ПЦР содержит 2 мкл геномной ДНК, 2,6 мкл реакционного буфера 10Х Ex Taq, 2,0 мкл 2,5 мМ дНТФ, 0,75 каждого из 5 мМ растворов прямого и обратного праймеров, 0,1 мкл полимеразы Ех Taq (5 единиц/мкл, TaKaRa) и 17,8 мкл Н2О в 26 мкл общего объема реакционной смеси. Условиями ПЦР-амплификации были: первичная денатурация при 94°C, 2 минуты; 40 циклов при 94°C, 30 секунд, 58°C, 30 секунд, 72°C, 1 минута 30 секунд; окончательная элонгация при 72°C, 5 минут.
Продукт амплификации затем использовали в анализе AD с фланкирующим прямым праймером (TGTGTATGTATGTTTTGTGGCTAGCA, SEQ ID NO: 30), фланкирующим обратным праймером (CGTTCCCGAGTCCCTAATCAAG SEQ ID NO: 31), меченым зондом, специфичным для аллеля дикого типа (VIC - TGC AGG GCG TCG ТС - MGB - NFQ SEQ ID NO: 32) и меченым зондом, специфичным для мутантного аллеля (FAM - С TGC AGA GCG TCG ТС - MGB - NFQ SEQ ID NO: 33). Реакционная смесь ПЦР содержала 2 мкл продукта амплификации KRP4B2, 3 мкл ABI-мастер-микса для генотипирования в ПЦР в режиме реального времени и 1 мкл 6Х (по 5,4 мкмоль/мл каждого праймера на 1,2 мкмоль/мл каждого зонда) SNP-специфичных для анализа AD смеси праймер/зонд в общем объеме реакционной смеси 6 мкл. ПЦР в режиме реального времени проводили на приборе ABI для ПЦР в режиме реального времени в соответствии с условиями ПЦР для ABI.
На фигуре 1А-1С показано, что праймер/зонды могут быть созданы для различения аллелей дикого типа и мутантных аллелей KRP4B в анализе AD. Мутантные гетерозиготы по аллелю KRP4B P109L подтверждали путем секвенирования (фигура 1D). На фигуре 2 дополнительно показано, что популяции образцов гомозиготных по аллелю KRP4B дикого типа, гомозиготных по мутантному аллелю KRP4B P109L или гетерозиготных по мутантному аллелю KRP4B P109L можно легко различить в анализе AD.
In vitro анализы для тестирования ингибирования киназной активности TILLING®-мутантными белками KRP
Отобранные тяжелые миссенс типа I и TILLING®-нонсенс-мутантные KRP пшеницы тестировали в in vitro анализе для определения, обладают ли по-прежнему мутантные белки KRP способностью ингибировать киназную активность киназного комплекса ZmCyclinD4/ZmCDKA; 1 (фигура 3) (анализ описан в "Dominant Negative Mutant Kip-Related Proteins (KRP) in Zea Mays and Methods for their use", PCT/US2011/060598).
Рекомбинантные комплексы ZmCyclinD4 и ZmCDKA; 1 были получены и выделены из клеток насекомых S.frugiperda Sf9. Все гены TaKRP дикого типа были синтезированы с применением DNA2.0.
Мутагенез TaKRP.
TaKRP1A P232L (pTG1947)
TaKRP1A G236S (pTG1948)
TaKRP1A W240* (pTG1949)
TaKRP2D P228S (pTG1958)
TaKRP2D A238V (pTG1959)
TaKRP2D A239T (pTG1960)
TaKRP2D D254N (pTG1961)
TaKRP2D R257C (pTG1962)
TaKRP4A W186* (pTG1950)
TaKRP5D W199* (pTG1951)
Праймеры для получения мутантов, описанных выше, приведены в таблице 28 в графической части.
Общие способы экспрессии рекомбинантных белков в бактерии, очистки и детектирования описаны ниже:
I. Клетки насекомых и среда.
Система экспрессии бакуловирусов является универсальной эукариотической системой для экспрессии гетерологичных генов. Эта система обеспечивает корректную укладку белка, образование дисульфидного мостика и другие важные посттрансляционные модификации. Все способы были взяты из Baculovirus expression vector system: Procedures and methods manual. (BD Biosciences, Pharmingen, San Diego, Calif. 6th Ed.). Клетки насекомых Sf9 выращивали при 27°C в TNM-FH средах для клеток насекомых (BD Biosciences), для описываемых исследований. Нужно отметить, что альтернативные среды хорошо известны специалисту в данной области техники и также пригодны. Аналогично, альтернативные линии клеток насекомых, такие как клетки Sf21 и High Five™ также будут работать для получения вирусов и белков.
II. Вестерн-блоттинги и иммунопреципитации
Рекомбинантный белок, экспрессированный в клетках насекомых, контролировали при помощи вестерн-блоттинга. Экстракты белков (35 мкг) кипятили в присутствии буфера Лэммли, наносили на 10% или 12% гели SDS-PAGE и переносили на мембрану PVDF с помощью погружного устройства для переноса (BioRad). После переноса мембрану блокировали в TBS-T (25 мМ Tris pH 7,5; 75 мМ NaCl; 0,05% Tween), который содержит 5% порошка сухого обезжиренного молока. Первичное антитело применяли в разведении 1:1000 в блокирующем буфере TBS-T в течение ночи. Блоты промывали три раза по 15 минут при комнатной температуре. Соответствующее вторичное антитело, конъюгированное с пероксидазой хрена (HRP), применяли в разведении 1:10000 в блокирующем буфере TBS-T. Блоты инкубировали со вторичными антителами в течение 1 часа, а затем три раза промывали в TBS-T по 15 мин каждый. Блоты затем обрабатывали, как описано в протоколе для системы ECL (Amersham Biosciences). Обычно используемыми антителами были: моноклональные антитела к flag M2 (Sigma), моноклональные или поликлональные антитела к НА (Babco), антитела к PSTAIR (Sigma-Aldrich), моноклональные или поликлональные антитела к Мус 9Е10 (А-14) (Santa Cruz Biotechnology). Используемыми вторичными антителами были: антитело к IgG мыши, конъюгированное с пероксидазой хрена, антитело к IgG кролика, конъюигрованное с пероксидазой хрена (GE Healthcare).
III. Конструкция бакуловирусного вектора
Системой на основе бакуловируса была система Bac-to-bac (Invitrogen). Также можно применять альтернативные геномы бакуловируса. Все бакмиды, содержащие наши гены интереса, независимо трансфицировали в 293 клетки с применением трансфекционных реагентов на основе липидов, таких как Fugene или Lipofectamine. Клетки Sf9 S. frugiperda высевали в концентрации 9×106 клеток на 60 мм чашку и временно трансфицировали 1 мкг бакмиды с применением 3 мкл реагента для трансфекции Fugene 6 в соответствии с протоколом производителя (Roche Diagnostics). Через 4 часа после трансфекции раствор Fugene/ДНК удаляли и заменяли его 3 мл среды TNM-FH. Четыре (4) дня спустя собирали надосадочную жидкость и в дальнейшем использовали при инфицировании дополнительных клеток для амплификации вируса. Такую амплификацию повторяли до тех пор, пока титр вируса не составил по меньшей мере 109 вирусных частиц/мл. Вирус амплифицировали путем инфицирования клеток Sf9 при множественности заражения (moi)<1. Титр вируса контролировали с применением световой и флуоресцентной микроскопии.
IV. Экспрессия рекомбинантных белков в бактериях и очистка
Все бактериальные экспрессионные плазмиды, несущие вставки, были трансформированы в BL21 RosettaBlue (DE3) (Novagen). Бактериальные колонии от данной свежей трансформации использовали для инокуляции 400 мл LB, содержащей 100 мкг/мл ампициллина, и культивировали при 37°C. Когда культура достигала OD600 между 0,6 и 0,8, экспрессию рекомбинантного белка индуцировали добавлением 1 мМ изопропил-О-тиогалактопиранозида (IPTG). Клетки затем выращивали при 30°C в течение трех часов. Клетки собирали центрифугированием с применением ротора для центрифугирования Beckman JLA 10.500. Осажденные бактериальные клетки или хранили при -80°C или немедленно лизировали. Бактерии лизировали в 10 мл фосфатного буфера для лизиса (100 мМ фосфатного буфера pH 7,0, 150 мМ NaCl, 1% Triton X100), содержащего ингибиторы протеазы и не содержащим ЭДТА. Ресуспендированную бактериальную культуру лизировали посредством френч-пресса или повторяющейся ультразвуковой обработкой. Лизированные клетки центрифугировали при 14000 об/мин с применением ротора для центрифугирования Beckman JA20.1 в течение 15 минут при 4°C. Меченые молекулы KRP были в основном нерастворимыми. Нерастворимые меченые KRP растворяли в буфере мочевины (8М мочевины, 100 мМ фосфатного буфера, pH 7,0) вручную, с помощью пипетки. Белки, нерастворяющиеся в мочевине, удаляли центрифугированием при 14000 об./мин. в роторной центрифуге Beckman JA20.1 в течение 15 минут при 4°C. Меченые KRP очищали на установке периодического действия с применением металл-афинной смолы BD Talon Co2+, уравновешенной в буфере мочевины. Очистку на установке периодического действия проводили с инкубированием при 4°C в течение периода от 3 часов до всей ночи при медленном вращении. Суспензию наносили на колонку и смолу промывали 36 объемами слоя буфера мочевины, а затем 12 объемами слоя буфера мочевины, содержащего 5 мМ имидазола pH 7,0. Связанный меченый белок KRP элюировали с применением буфера мочевины, содержащего 300 мМ имидазола с рН 7,0. Фракции контролировали по меченным KRP с помощью SDS-PAGE и/или анализом белков по Брэдфорду (Bio-Rad). Рефолдинг денатурированного меченого KRP1 проводили с применением диализа со ступенчатым разведением. Фракции, содержащие большую часть меченого белка KRP, объединяли и диализировали в 1 мМ мочевины, 100 мМ фосфатного буфера с pH 7,0 и 1 мМ дитиотреитола в течение 20 часов при 4°C.
Диализный буфер затем заменяли на раствор 0,5 мМ мочевины, 100 мМ фосфатного буфера с рН 7,0 и 1 мМ дитиотреитола, и продолжали диализ в течение дополнительных 12 часов. Рекомбинантный белок собирали, оценивали количественно с применением анализа по Брэдфорду и хранили при 4°С.
В in vitro анализах киназ несколько мутантных белков TaKRP проявляли пониженную ингибирующую активность по отношению к циклин/CDK киназному комплексу, тогда как другие проявляли себя как TaKRP дикого типа. Примечательно, что TaKRP4A W186* (фигуры 4 и 6А) и TaKRP5D W199* (фигура 5) не ингибировал киназную активность ZmCyclinD4/ZmCDKA; 1 в количестве 0,1; 0,25 или 0,5 мкг. TaKRP1A W240* также не ингибировал киназную активность ZmCyclinD4/ZmCDKA; 1 в количестве 0,1; 0,25 или 0,5 мкг, хотя его ингибирующая активность не была такой, как в количестве 0,25 и 0,5 мкг (фигуры 4 и 6А). TaKRP1A G236S мог только сильно ингибировать ZmCyclinD4/ZmCDKA; 1 в количестве 0,5 мкг и слабо в количестве 0,25 мкг (фигура 4). Аналогично, TaKRP2D P228S мог только сильно ингибировать ZmCyclinD4/ZmCDKA; 1 в количестве 0,5 мкг и очень слабо в количестве 0,25 мкг (фигура 4). TaKRP4A W186*, TaKRP5D W199* и TaKRP1A W240* содержат преждевременные стоп-кодоны очень близко к концам белков KRP и, таким образом, все еще сохраняют циклин- и CDK-связывающие домены. Однако удаление последних 8 аминокислот TaKRP1A W240*, последних 6 аминокислот TaKRP5D W199* и последних 5 аминокислот TaKRP4A W186* нарушает способность мутантных KRP ингибировать киназный комплекс. Мутантный белок KRP2 кукурузы с эквивалентным преждевременным стоп-кодоном возле конца белка также был не способен ингибировать комплекс ZmCyclinD4/ZmCDKA; 1 в количествах 0,3 или 3 мкг (фигура 6В).
Другие мутанты, такие как TaKRP1A P232L (фигура 4), TaKRP2D A238V (фигура 5), TaKRP2D А239Т (фигура 5), TaKRP2D D254N (фигура 4) и TaKRP2D R257C (фигура 4) ингибируют комплекс ZmCyclinD4/ZmCDKA; 1 так же, как и их аналоги дикого типа при указанных концентрациях белков.
Ингибирующие активности различных TILLING®-мутантных KRP пшеницы по сравнению с их аналогами дикого типа приведены в таблице 23 в графической части.
Пример 5.
Предварительные результаты полевых исследований по М поколению TILLING®-мутантов по KRP пшеницы.
М5 поколение выбранных TILLING®-мутантов no KRP пшеницы выращивали в Юме, Аризона, в течение осень 2009-зима 2010 для определения урожайности. Шестьдесят объектов использовали в исследовании урожайности (таблица 24 в графической части) с тем, что каждый объект в исследовании был повторен три раза случайным образом. Каждый участок имел размеры 4 фута на 10 футов с 7 рядами на участок и расстоянием между рядами в 7 дюймов. Приблизительно 75 граммов семян высевали на участок, или приблизительно 2500 семян/участок или приблизительно 360 семян на ряд длиной в 10 футов. Высевание производили в октябре.
Там где возможно, сегрегант дикого типа данного гомозиготного или гетерозиготного TILLING®-мутанта по KRP высаживали для сравнения. Родительский сорт Express также включали в исследование. Густоту стояния определяли перед сбором урожая, уровень созревания определяли во время цветения, и параметры плодоножки, число верхушек, число зерен, длину колосьев и длину ости определяли после сбора урожая субвыборок. Сроки созревания и параметры плодоножки оценивали путем сравнения по балльной системе по сравнению с исходным родительским растением сорта Express. Срок созревания: 1 раннее, 3 аналогичное сорту Express, 5 позднее/вегетативное. Плодоножка: 1 очень тонкая, 3 аналогичная сорту Express, 5 очень толстая (таблица 24 в графической части).
Сбор урожая проводили в два этапа в мае. Участки повторности 1 скашивали вручную, растения связывали в пучки и оставляли для высыхания в поле в течение нескольких дней до стационарного обмолота. На втором этапе использовали комбайны для сбора урожая на оставшихся участках. Перед сбором второго урожая верхушки от пяти растений на участок повторностей 2 и 3 (то есть, всего 10 головок на объект) собирали для измерения верхушек. Надрез делали приблизительно на уровне на 5-6 дюймов ниже основания колоса.
Вес семян и количество семян определяли для каждой части выборки, включающей 5 верхушек, на основании которой мог быть рассчитан вес семени.
Массу урожая с участка определяли вручную. Число семян на участке рассчитывали по весу семени и массе урожая с участка. Вес тысячи зерен (TKW) определяется как вес семени, умноженный на 1000.
Ряд мутантов по KRP показал отличный урожай семян. Эти мутанты содержали WH68, WH3, WH91, WH124, WH4 и WH16 (таблица 24). WH68 (KRP4D, P146L) занимал первое место с урожаем 3584,90 фунтов/акр. Два мутанта, WH3 и WH4, с изменениями в смежных нуклеотидах, что приводит к мутации в сайте сплайсинга в одной и той же границе интрон/экзон продемонстрировали хороший урожай, занимая второе (WH3) и шестое место (WH4) из 60 объектов. Другой мутант по сайту сплайсинга в KRP5D, WH124, занял пятое место по урожаю. Интересно, что мутант WH16, который занял седьмое место по урожаю, имеет две мутации в гене KRP2A (V258M и G169S).
Пример 6. TILLING®-мутант no KRP риса
TILLING®-библиотеку риса подвергали скринингу на предмет мутаций в KRP 1, 2, 4 и 5 риса (SEQ ID NO: 100-107). Был идентифицирован мутант, содержащий преждевременный стоп-кодон в OsKRP4 (таблица 25 в графической части). Семена М3 из двух отдельных сиблинговых растений М2, гетерозиготных по мутации R167*, высевали в теплице и генотипировали для идентифицирования растений, гомозиготных по аллелю R167* или гомозиготных по аллелю дикого типа. Растения, гомозиготные по R167* и дикому типу, самоопылялись для получения гомозиготных мутантов и семян дикого типа М4. Дополнительно, гомозиготные по R167* растения подвергали возвратному скрещиванию с рекуррентным родительским растением сорта Cypress для получения семян F1.
Число семян и вес семян определяли для семян М4, гомозиготных по R167* и гомозиготных по дикому типу (таблица 26 в графической части). Существует указание на то, что гомозиготы по RI527 R167* имеют по меньшей мере общее увеличение в весе тысячи зерен (TK.W) по сравнению с их сиблингами дикого типа (таблица 27 в графической части) и гомозиготы по RI526 R167* могут быть лучше их контроля по числу семян. Эти же измерения будут снова проведены на семенах, полученных после успешного возвратного скрещивания поколений.
Семена F1 и М4 высевали в теплице в следующем цикле. Растения F1 генотипировали для подтверждения гетерозиготности по R167*. Растения, гетерозиготные по R167*, второй раз подвергали возвратному скрещиванию с рекуррентным родительским растением, чтобы сделать семя BC1F1. Одновременно, растения F1, гетерозиготные по R167*, самоопылялись, чтобы сделать сегрегирующее семя F2. Растения F2, гомозиготные по R167*, идентифицировали генотипированием и семена поколения F3, полученные для полевых исследований.
Семена BC1F1 выращивали в теплице и растения генотипировали для идентификации гетерозиготных по аллелю R167* или аллелю дикого типа. Гетерозиготные по R167* растения подвергали возвратному скрещиванию для получения BC2F1 и самоопыляли для получения BC1F2. Растения BC1F2, гомозиготные по R167*, идентифицировали путем генотипирования и получали семена поколения BC1F3 для полевых исследований. Обратное скрещивание может быть продолжено до уровня ВС3 или ВС4. В ходе возвратного скрещивания и самоопыления сиблинги дикого типа переносятся в следующее поколение, чтобы служить в качестве контроля. Обратные скрещивания также проводили для сорта Nipponbare для перемещения мутации в короткозерновой рис.
Пример 7
Мутации генов KRP Glycine max, идентифицированные при помощи TILLING®
Поиск генов-кандидатов KRP Glycine max (соя) проводили в Phytozome и были идентифицированы девять генов KRP (SEQ ID NO: 111-128). TILLING®-библиотеку сои скринировали на предмет мутаций в данных KRP сои.
Репрезентативные мутации в KRP Glycine max показаны в таблицах 29-37 в графической части (* указывает мутацию, которая приводит к образованию стоп-кодона). Это только репрезентативный перечень, который не должен быть интерпретирован как ограничивающий каким-либо образом.
Пример 8.
Предварительные полевые результаты оценки поколения F2:3 TILLING®-мутантов по KRP пшеницы
Цель состояла в том, чтобы оценить возможность получения достоверных данных полевых оценок KRP TILLING®-линий яровой пшеницы и определить характеристики гомозиготных (F2:3) мутантных линий.
Материалы и способы
Пять полевых экспериментов были проведены в течение вегетационного периода в Форт-Коллинз, Колорадо, и Боземан, Монтана (таблица 38 в графической части), в которых было оценено число мутантных линий (таблица 39 в графической части).
Основные результаты
Мутантные материалы произвели значительно меньшие урожай, чем контрольный сорт (фигура 7). Это не удивительно, так как мутантные линии скрещивали только один раз с родителем сорта Express и они могли все еще содержать вредные поражения, как результат мутагенеза при помощи EMS, по сравнению с не подвергавшимся мутагенезу сортом Express. Примечательно, что разница в урожае между мутантной линией и контролем существенно не отличалась для KRP4D Stop и KRP2A Splice WH4 в условиях высокой урожайности (Монтана). Влияние мутации оказалось меньше для одиночных мутантов, чем для двойных мутантов (фигура 7). Мутант KRP2AB Splice/Splice имел набольшее снижение урожайности (46% и 39% в Монтане и Колорадо), а мутант KRP4D Stop имел наименьшее снижение урожайности (4% и 11% в Монтане и Колорадо).
Между зиготными группами линии дикого типа (WT) имели тенденцию к более высокому урожаю, чем их мутантные аналоги (Homo), хотя в условиях высокой урожайности (Монтана) для двойных мутантов KRP4BD missense/stop и splice/splice KRP2AB было справедливо обратное (фигура 8). Среди групп Homo мутанты KRP4D Stop и KRP2A Splice WH11 демонстрировали самые высокие относительные показатели урожая. Гомозиготный мутант KRP2A Splice WH11 стабильно имел урожай близкий к среднему его аналога дикого типа в условиях выращивания в Колорадо и в Монтане.
Для KRP TILLING®-мутантов, в которые были включены сестринские линии F1, линия "149Е05" в рамках мутанта по стоп KRP4D показала дифференциальный ответ: гомозиготные линии давали урожай выше, или по меньшей мере такой же, как их аналоги дикого типа (фигура 9).
Зависимость, проявляющаяся в относительном урожае зерна между диким типом и гомозиготными зиготными группами, может выражаться в виде соотношения WT/Homo по урожаю зерна и различным компонентам урожая. Можно распознать предполагаемые механизмы, связанные с ответом в виде относительного урожая зерна (таблица 40 в графической части). В Монтане наибольшее положительное влияние на урожай (WT/Homo отношение = 0,92) наблюдали для двойного мутанта " KRP4BD Missense/Stop", что проявлялось в основном в числе остей, весе тысячи зерен (TKW) и высоте растения, тогда как наибольшее отрицательное влияние на урожай (WT/Homo отношение = 1,09) наблюдали для мутанта "KRP2A Splice WH4", линия 42D06, в основном за счет нарушений в числе зерен. В Колорадо наибольшее положительное влияние на урожай (WT/Homo отношение = 0,93) наблюдали для мутанта "KRP4D стоп", линия 149Е05, что проявлялось в основном в весе тысячи зерен (TKW) и высоте растения, тогда как наибольшее отрицательное влияние на урожай (WT/Homo отношение == 1,18) наблюдали для варианта мутанта "KRP2A Splice WH4", линия 42D06, что в основном проявлялось в числе остей и числе зерен.
Пример 9.
Идентификация и получение других KRP пшеницы
Рис служит в качестве диплоидного модельного вида злака для пшеницы. В рисе присутствуют и экспрессируются два дополнительных гена KRP: KRP3 и KRP6. KRP3 экспрессируется в очень специфичный момент времени в процессе развития семени, в то время как KRP6, по-видимому, экспрессируется в на протяжении развития семени риса (Mizutani et al. 2010). Для того чтобы завершить набор TILLING®-мутантов по KRP пшеницы, другие KRP пшеницы идентифицированы для применений TILLING® в будущем.
Последовательность ДНК KRP6 риса применяли для идентифицирования EST KRP6 пшеницы в ходе поиска по базе данных BLAST в NCBI. Тогда более длинные последовательности KRP6 пшеницы извлекали из базы данных геномных последовательностей пшеницы (WheatBP) в Университете Бристоля с применением EST KRP6 пшеницы из NCBI. База данных имела 5Х охват сорта пшеницы Chinese Spring. TILLING®-библиотека для пшеницы основана на сорте Express, но ожидается не так много полиморфизмов между этими двумя сортами.
Последовательности KRP6 пшеницы изменялись в диапазоне ~300-800 п.о. ContigExpress из Vector NTI (Invitrogen) применяли для сборки различных считываний в контиги. Считывания собирали в три контига, которые представляли TaKRP6A, TaKRP6B и TaKRP6D. Дополнительную характеристику проводили, чтобы определить, какой контиг соответствует какому геному. Границы экзон/интрон для генов KRP6 пшеницы предсказывали на основе последовательности KRP6 риса. Предсказанные кодирующие последовательности были транслированы в вектор для получения белковых последовательности. Таблица 41 в графической части представляет сравнение длин гена KRP6, кДНК и белка, для пшеницы, риса, Brachypodium, кукурузы и сорго.
Используя идентифицированные последовательности, были сконструированы три пары праймеров (таблица 42 в графической части). Не было получено никаких продуктов амплификации KRP6 пшеницы из геномной ДНК пшеницы с применением корректирующей Phusion-полимеразы при температурах отжига в диапазоне от 55°C до 65°C. С применением набора SureBand PCR Optimization (Bioline) при температуре отжига 58°C и с применением одного из двенадцати буферов из этого набора был получен бэнд небольшой интенсивности для TaKRP6 49F и TaKRP6 258R. При повторении амплификации с тем же буфером и парой праймеров и проведении тестирования в диапазоне температур отжига от 54°C до 66,5°C были получены уникальные бэнды сильной интенсивности при 60°C и выше. Бэнд вырезали из геля и ДНК, очищали и клонировали с применением наборов для клонирования ТА и Zero Blunt (Invitrogen). Скрининг колоний показал несколько положительных трансформантов.
Проводили минипрепы на культурах от 30 колоний и все они были секвенированы с обоих концов вставки с применением праймеров M13F и M13R.
Полученные последовательности сначала собирали в контиги (M13F и M13R последовательности от каждого из 30 клонов), и затем эти контиги собирали в более крупные контиги. После сборки и выравнивания, было установлено, что все три версии KRP6 пшеницы был клонированы: 14 клонов KRP6A, 1 клон KRP6B и 14 клонов KRP6D.
Последовательность кДНК KRP3 риса применяли в поиске BLAST в NCBI. Было идентифицировано несколько EST пшеницы. Идентифицированные выше последовательности KRP6 пшеницы применяли для поиска контигов KRP пшеницы по базе данных злаков. С применением данного подхода в базе данных для пшеницы был идентифицирован контиг, который выглядел сходным, но не идентичным KRP6 или другому известному KRP пшеницы.
Пример 10.
TILLING® других KRP пшеницы и характеристика мутантов Конструирование и тестирование геном-специфичных TILLING® праймеров и TILLING® для других KRP пшеницы, таких как KRP6 пшеницы, проводят, как описано выше в материалах и способах и в примере 1. Полученных KRP TILLING®-мутантов подвергали возвратному скрещиванию, интрогрессировали в другие сорта пшеницы и комбинировали с другими мутантами пшеницы по KRP как описано выше в примере 3. Определение урожая, компоненты урожая и агрономические характеристики максимально описаны выше в примерах 3, 5 и 8.
Если не определено иначе, все технические и научные термины, приведенные в настоящем документе, имеют то же значение, как часто понимается средним специалистом в данной области техники, к которой относится изобретение. Хотя любые способы и материалы, сходные или эквивалентные тем, которые описаны в данном документе, могут быть использованы в применении на практике или в тестировании настоящего изобретения, предпочтительные способы и материалы описаны в данном документе. Все публикации, патенты и патентные публикации, цитируемые в данном документе, включены в него посредством ссылки во всей их полноте для всех целей.
Обсуждаемые в данном документе публикации представлены исключительно для их раскрытия до даты подачи настоящей заявки. Ничто из представленного в данном документе не должно интерпретироваться как признание того, что настоящее изобретение не имеет права датировать такую публикацию более ранним числом посредством предшествующего изобретения.
Хотя настоящее изобретение было описано в связи с конкретными вариантами его осуществления, следует понимать, что оно может подвергаться дополнительным модификациям и, предполагается, что данная заявка охватывает любые вариации, применения или адаптации настоящего изобретения, следуя, в целом, принципам, представленным в настоящем изобретении, включая такие отклонения от настоящего раскрытия, которые входят в известную или общепринятую практику в рамках области техники, к которой относится настоящее изобретение, и которые могут быть применены к существенным признакам, изложенным выше в данном документе, и которые указаны в объеме прилагаемой формулы изобретения.
СПИСОК ЛИТЕРАТУРЫ
1. Bentley, А., В. MacLennan, et al. (2000). "Targeted Recovery of Mutations in Drosophila." Genetics 156: 1169-1173.
2. Comai, L. and S. Henikoff (2006). "TILLING: practical single-nucleotide mutation discovery." Plant J 45(4): 684-94.
3. Comai, L., K. Young, et al. (2004). "Efficient discovery of DNA polymorphisms in natural populations by Ecotilling." Plant J 37(5): 778-86.
4. Cooper, J.L., E.A. Greene, et al. (2008). "Retention of induced mutations in a Drosophila reverse-genetic resource." Genetics 180(1): 661-7.
5. Cooper, J.L., B.J. Till, et al. (2008). "Fly-TILL: reverse genetics using a living point mutation resource." Fly (Austin) 2(6); 300-2.
6. Cooper, J.L., B.J. Till, et al. (2008). "TILLING to detect induced mutations in soybean." BMC Plant Biol 8: 9.
7. Eddy, S.R. (2004). "Where did the BLOSUM62 alignment score matrix come from?" Nat Biotechnol 22(8): 1035-6.
8. Gilchrist, E. and G. Haughn "Reverse genetics techniques: engineering loss and gain of gene function in plants." Brief Funct Genomics 9(2): 103-10. 2010.
9. Gilchrist, E.J. and G.W. Haughn (2005). "TILLING without a plough: a new method with applications for reverse genetics." Curr Opin Plant Biol 8(2): 211-5.
10. Gilchrist, E.J., G.W. Haughn, et al. (2006). "Use of Ecotilling as an efficient SNP discovery tool to survey genetic variation in wild populations of Populus trichocarpa." Mol Ecol l5(5): 1367-78.
11. Gilchrist, E.J., N.J. O'Neil, et al. (2006). "TILLING is an effective reverse genetics technique for Caenorhabditis elegans." BMC Genomics 7: 262.
12. Greene, E.A., C.A. Codomo, et al. (2003). "Spectrum of chemically induced mutations from a large-scale reverse-genetic screen in Arabidopsis." Genetics 164(2): 731-40.
13. Henikoff, S., В.J. Till, et al. (2004). "TILLING. Traditional mutagenesis meets functional genomics." Plant Physiol 135(2): 630-6.
14. Himelblau, E., E.J. Gilchrist, et al. (2009). "Forward and reverse genetics of rapid-cycling Brassica oleracea." Theor Appl Genet 118(5): 953-61.
15. McCallum, С.М., L. Comai, et al. (2000). "Targeted screening for induced mutations." Nat Biotechnol 18(4); 455-7 (referenced in Anawah patents).
16. McCallum, C.М., L. Comai, et al. (2000). "Targeting induced local lesions IN genomes (TILLING) for plant functional genomics." Plant Physiol 123(2): 439-42 (referenced in Anawah patents).
17. Ng, P.C. and S. Henikoff (2003). "SIFT: Predicting amino acid changes that affect protein function." Nucleic Acids Res 31(13): 3812-4.
18. Slade, A.J., S.I. Fuerstenberg, et al. (2005). "A reverse genetic, nontransgenic approach to wheat crop improvement by TILLING®." Nat Biotechnol 23(1): 75-81.
19. Slade, A.J. and V.C. Knauf (2005). "TILLING moves beyond functional genomics into crop improvement." Transgenic Res 14(2): 109-15.
20. Stemple, D.L. (2004). "TILLING--a high-throughput harvest for functional genomics." Nat Rev Genet 5(2): 145-50.
21. Styczynski, M.Р., K.L. Jensen, et al. (2008). "BLOSUM62 miscalculations improve search performance." Nat Biotechnol 26(3): 274-5.
22. Talame, V., R. Bovina, et al. (2008). "TILLMore, a resource for the discovery of chemically induced mutants in barley." Plant Biotechnol J 6(5): 477-85.
23. Taylor, N.E. and E.A. Greene (2003). "PARSESNP: A tool for the analysis of nucleotide polymorphisms." Nucleic Acids Res 31(13): 3808-11.
24. Till, В.J., С. Burtner, et al. (2004). "Mismatch cleavage by single-strand specific nucleases." Nucleic Acids Res 32(8): 2632-41.
25. Till, В.J., Т. Colbert, et al. (2006). "High-throughput TILLING® for Arabidopsis." Methods Mol Biol 323: 127-35.
26. Till, В.J., Т. Colbert, et al. (2003). "High-throughput TILLING® for functional genomics." Methods Mol Biol 236: 205-20.
27. Till, В.J., J. Cooper, et al. (2007). "Discovery of chemically induced mutations in rice by TILLING®." BMC Plant Biol 7: 19.
28. Till, В.J., S.H. Reynolds, et al. (2003). "Large-scale discovery of induced point mutations with high-throughput TILLING®." Genome Res 13(3): 524-30.
29. Till, В.J., S.H. Reynolds, et al. (2004). "Discovery of induced point mutations in maize genes by TILLING®." BMC Plant Biol 4: 12.
30. Till, В.J., Т. Zerr, et al. (2006). "A protocol for TILLING® and Ecotilling in plants and animals." Nat Protoc 1(5): 2465-77.
31. Triques, K., E. Piednoir, et al. (2008). "Mutation detection using ENDO1: application to disease diagnostics in humans and TILLING® and Eco-TILLING in plants." BMC Mol Biol 9: 42.
32. Triques, K., В. Sturbois, et al. (2007). "Characterization of Arabidopsis thaliana mismatch specific endonucleases: application to mutation discovery by TILLING® in pea." Plant J 51 (6): 1116-25.
33. Uauy, C., F. Paraiso, et al. (2009). "A modified TILLING® approach to detect induced mutations in tetraploid and hexaploid wheat." BMC Plant Biol 9: 115.
34. Weil, С.F. and R. Monde (2007). "Getting the Point-Mutations in Maize." Crop Science 47 (S1)(No. 1): S-60-67.
35a. Zerr, T. and S. Henikoff (2005). "Automated band mapping in electrophoretic gel images using background information." Nucleic Acids Res 33(9): 2806-12.
35b. Tsai, H. et al. (2011), Discovery of Rare Mutations in Populations: TILLING by Sequencing, Plant Physiology, 156(3): 1257-1268.
36. Slafer and Araus (2007), Springer, "Physiological traits for improving wheat yield under a wide range of conditions", Scale and Complexity in Plant Systems Research: Gene-Plant-Crop Relations, 147-156.
37. Reynolds, "Physiological approaches to wheat breeding", Agriculture and Consumer Protection. Food and Agriculture Organization of the United Nations.
38. Richard et al., "Physiological Traits to Improve the Yield of Rainfed Wheat: Can Molecular Genetics Help", published by International Maize and Wheat Improvement Center.
39. Reynolds et al., "Evaluating Potential Genetic Gains in Wheat Associated with Stress-Adaptive Trait Expression in Elite Genetic Resources under Drought and Heat Stress Crop science". Crop Science 2007 47: Supplement_3: S-172-S-189.
40. Setter et al., Review of wheat improvement for waterlogging tolerance in Australia and India: the importance of anaerobiosis and element toxicities associated with different soils. Annals of Botany, Volume 103(2): 221-235.
41. M.J. Foulkes, N.D. Paveley, A. Worland, S.J. Welham, J. Thomas, J.W. Snape. Major Genetic Changes in Wheat with Potential to Affect Disease Tolerance. Phytopathology, July, Volume 96, Number 7, Pages 680-688 (doi: 10.1094/PHYTO-96-0680).
42. Rosyara, U.K., K. Pant, E. Duveiller and R.C. Sharma. 2007. Variation in chlorophyll content, anatomical traits and agronomic performance of wheat genotypes differing in spot blotch resistance under natural epiphytotic conditions. Australasian Plant Pathology 36: 245-251.
43. Rosyara, U.R., R.C. Sharma, and E. Duveiller. 2006. Variation of canopy temperature depression and chlorophyll content in spring wheat genotypes and association with foliar blight resistance. J. Plant Breed. Gr. 1: 45-52.
44. Rosyara, U.R., R.C. Sharma, S.M. Shrestha, and E. Duveiller. 2005. Canopy temperature depression and its association with helminthosporium leaf blight resistance in spring wheat. Journal of Institute of Agriculture and Animal Science 26: 25-28.
45. Rosyara, U.R., R.C. Sharma, S.M. Shrestha, and E. Duveiller. 2006. Yield and yield components response to defoliation of spring wheat genotypes with different level of resistance to Helminthosporium leaf blight. Journal of Institute of Agriculture and Animal Science 27. 42-48.
46. Rosyara, U.R. 2002. Physio-morphological traits associated with Helminthosporium leaf blight resistance in spring wheat. Masters' Thesis. Tribhuvan University, Institute of Agriculture and Animal Science, Rampur, Chitwan, Nepal. supported by CIMMYT International. Available at CIMMYT library.
47. Hayward, M.D., N.O. Bosemark, and I. Romangosa. 1993. Plant Breeding: Principle and Prospects. Chapman and Hall, London.
48. Wood, D.R., K.M. Rawal, and M.N. Wood (eds). 1983. Crop Breeding. American Society of Agronomy, Crop Science Society of America, Madison, Wisconsin.
49. Allard, R.W. 1960. Principles of Plant Breeding. John Wily and Sons Inc. New York.
50. Simmonds, N.W. 1979. Principles of Crop Improvement. Longman Group Limited, London.
51. Singh, B.D. 2000. Plant Breeding. Sixth ED. Kalyani Publishers, New Delhi.
52. Guo et al., 2005, American Journal of Botany 92(9); 1548-1558.
53. Watson et al. 1999. Grass genera of the world: descriptions, illustrations, identification, and information retrieval; including synonyms, morphology, anatomy, physiology, phytochemistry, cytology, classification, pathogens, world and local distribution, and references. Version: 18 August 1999.
54. GPWG. 2001. Phylogeny and subfamilial classification of the grasses (Poaceae). Annals of the Missouri Botanical Garden 88: 373-457.
55. Clayton et al., 1986. Genera Graminum. Kew Bulletin Additional Series XIII: 1-389.
56. Vaughan, 1994. The wild relative of rice: a genetic resources handbook. International Rice Research Institute, Manila, Philippines.
57. Donald C.M., 1968. The breeding for crop ideotypes. Euphytica. 17, 385-403.
58. Gao P.W., Wang B.L. et al., 1988. Studies on physiology and ecology for rice with high yield. Liaoning Agricultural Science. 1, 7-11.
59. International Rice research Institute, 1996. Rice research in Asia. IRRI. 1-70.
60. Shao G.J. et al., 1995. Summarization and discussion on rice breeding research and development in Liaoning Province. Liaoning Agrirultural Science. 6, 28-33.
61. Wang B.L. et al., 1989. Incident light distribution over the population with high yield in rice. Liaoning Agricultural Science. 6, 27-30.
62. Wang B.L., 1992. The Trail and method of rice breeding for super high yield. The Proceedings of Rice Research. Chinese Science and Technological Press. 97-104.
63. Wang В.L. et al., 1997. Studies on genetic activities of semi-dwarfism and erect-panicle in rice. Journal of Shenyang Agricultural University. 28(2), 83-87.
64. Wang B.L., 2000. Studies on rice breeding for high yield, good quality and multiple resistance. Prospects of Rice Genetics and Breeding for the 21 st Century. China Agricultural Scientech Press. 191-195.
65. Wang B.L. et al., 2002. Analysis of rice breeding in Liaoning Province in 1949-2000. Liaoning Agricultural Science. 5, 5-8.
66. Yang S.J., et al 1987. Research on rice breeding through crossing indica and japonica and its evolution in the past thirty six years. Journal of Shenyang Agricultural University. 18(3), 3-9.
67. Tan et al., The three important traits for cooking and eating quality of rice grains are controlled by a single locus in an elite rice hybrid, Shanyou 63, Thero. Appl. Genet (1999) 99: 642-648.
68. Yan et al., Comparative analyses of QTL for important agronomic traits between maize and rice, Yi Chuan Xue Bao, 2004, 31 (12); 1401-1407.
69. Khush et al., Rice Genetics IV, Int. Rice Res. Inst. 2001.
70. Yamamoto et al., "Towards the Understanding of Complex Traits in Rice: Substantially or Superficially?", DNA RESEARCH pp. 1-14, (2009).
71. Viraktamath et al., 1997, Hybrid rice breeding manual, ISBN 9712201031, 9789712201035.
72. International Rice Research Institute, 1972, Rice Breeding.
73. Rice breeding: Papers presented at the Symposium on Rice Breeding held at the International Rice Research Institute, 6-10 Sept., 1971.
74. Chakraborty, 2001, Rice Breeding and Genetics, ISBN 8170228743, 9788170228745.
75. International Rice Research Institute, 2003, Two-line hybrid rice breeding manual, ISBN 9712201856, 9789712201851.
76. Marshall and Wadsworth, Rice science and technology, vol. 59 of Food science and technology, CRC Press, 1994, ISBN 0824788877, 9780824788872.
77. Xia, Progress of chromosome engineering mediated by asymmetric somatic hybridization., J Genet Genomics. 2009 Sep; 36(9): 547-56. Review.
78. Liu et al., Generation of high frequency of novel alleles of the high molecular weight glutenin in somatic hybridization between bread wheat and tall wheatgrass. Theor Appl Genet. 2009 Apr; 118(6): 1193-8. Epub 2009 Feb 8.
79. Zhou et al., Comparative study of symmetric and asymmetric somatic hybridization between common wheat and Haynaldia villosa. Sci China С Life Sci. 2001 Jun; 44(3): 294-304.
80. Wang et al., Proteomic analysis on a high salt tolerance introgression strain of Triticum aestivum/Thinopyrum ponticum. Proteomics. 2008 Apr; 8(7): 1470-89.
81. Cai et al., Genotyping of somatic hybrids between Festuca arundinacea Schreb. and Triticum aestivum L., Plant Cell Rep. 2007 Oct; 26(10): 1809-19. Epub 2007 Jun 27.
82. Deng et al., Analysis of remote asymmetric somatic hybrids between common wheat and Arabidopsis thaliana., Plant Cell Rep. 2007 Aug; 26(8): 1233-41. Epub 2007 Apr 4.
83. Zhou et al., Genetic characterization of asymmetric somatic hybrids between Bupleurum scorzonerifblium Willd and Triticum aestivum L.: potential application to the study of the wheat genome. Planta. 2006 Mar; 223(4): 714-24. Epub 2005 Nov 4.
84. Li et al., Regeneration of asymmetric somatic hybrid plants from the fusion of two types of wheat with Russian wildrye. Plant Cell Rep. 2004 Dec; 23(7): 461-7. Epub 2004 Jul 24.
85. Zhou et al., Introgression of the Haynaldia villosa genome into gamma-ray-induced asymmetric somatic hybrids of wheat. Plant Cell Rep. 2005 Jul; 24(5): 289-96. Epub 2005 Jun 3.
86. Xia et al., RAPD method for the identification of intergeneric asymmetric somatic hybrid plants of wheat. Shi Yan Sheng Wu Xue Bao. 1999 Sep; 32(3): 265-70. Chinese.
87. Mostageer et al., Establishment of a salt tolerant somatic hybrid through protoplast fusion between rice and ditch reed Arab J. Biotech., Vol. 6, No. (1) Jan. (2003): 1-12.
88. Nakajo et al., Somatic cell hybridization in rice (Oryza sativa L.) and birdsfoot trefoil (Lotus comiculatus L.) Japanese Journal of Breeding (Mar 1994).
89. Niizeki et al., Somatic hybridization in rice x soybean, Bajaj YPS (ed) Biotechnology in agriculture and forestry vol 8, Plant protoplasts and genetic engineering. Springer, Berlin Heidelberg New Tork, pp. 410-434.
90. Kisaka, et al., Intergeneric somatic hybridization of rice (Oryza sativa L.) and barley (Hordeum vulgare L.) by protoplast fusion, Plant Cell Reports, Volume 17, Number 5, 362-367.
91. Cocking, Rice biotechnology: Somatic hybridisation for improved salinity tolerance and xylem colonisation by rhizobia for endophytic nitrogen fixation Cahiers Options, vol 40.
92. Ishikawa et al., Rice interspecies hybrids show precocious or delayed developmental transitions in the endosperm without change to the rate of syncytial nuclear division. Plant J. 2011 Mar; 65(5): 798-806.
93. X. Hu, X. Cheng, H. Jiang, S. Zhu, B. Cheng and Y. Xiang, (2010), Genome-wide analysis of cyclins in maize (Zea mays), Genet. Mol. Res. 9 (3): 1490-1503.
94. Acquaah et al. Principles of plant genetics and breeding, Wiley-Blackwell, 2007, ISBN 1405136464, 9781405136464.
95. Harten, Mutation Breeding, Cambridge University Press, 1998.
96. Roy Davies and Wall, "Artificial Mutagenesis in Plant Breeding", Nature 182, 955-956 (04 October 1958).
97. Grotewold, Plant Functional Genomics, Volume 236 of Methods in molecular biology, Humana Press, ISBN 1588291456, 9781588291455.
98. Braman, In vitro mutagenesis protocols, Volume 182 of Methods in molecular biology, Human Press, 2002, ISBN 0896039102, 9780896039100.
99. Chusacultanachai et al., "Random mutagenesis strategies for construction of large and diverse clone libraries of mutated DNA fragments." Methods Mol Biol. 2004; 270: 319-34.
100. Fujii et al., One-step random mutagenesis by error-prone rolling circle amplification, Nucl. Acids Res. (2004) 32 (19): el45.
101. Trower, In vitro mutagenesis protocols, Volume 57 of Methods in molecular biology, John M. Walker Methods in molecular biology (Clifton, N.J.) 57, ISBN 0896033325, 9780896033320.
102. Katsumi, M., Foard, D.E. and Phinney, B.O. (1983) Evidence for the translocation of gibberellin A3 and gibberellin-like substances in grafts between normal, dwarfl and dwarf5 seedlings of Zea mays L. Plant Cell Physiol. 24, 379-388.
103. Lacadena, J.-R. Hybrid wheat. VII. Tests on the transmission of cytoplasmic male sterility in wheat by embryo-endosperm grafting, Euphytica, 17(3), 439-444.
104. Trione et al., 1968, IN VITRO CULTURE OF SOMATIC WHEAT CALLUS TISSUE American Journal of Botany, Vol. 55, No. 5, May - Jun., 19.
105. Dodig, et al., tissue culture response of different wheat genotypes, environmental effect and association with plant traits, Options MEditerraneennes, Series A, No. 81, pages 129 to 132.
106. O'HARA et al., Wheat Callus Culture: the Initiation, Growth and Organogenesis of Callus Derived from Various Explant Sources Ann Bot (1978) 42(5): 1029-1978.
107. Zaidi et al., Optimizing tissue culture media for efficient transformation of different indica rice genotypes Agronomy Research 4(2): 563-575, 2006.
108. Wang et al., Tissue Culture Responses from Different Explants of Rice, Rice Science, 2005, 12(3): 229-232.
109. Ting Y, Boyer A, McSweeney G (1978) Maize tissue culture. MNL 52:6.
120. Martha С. Hawes, Diana Z. Sharpe, Maria-Ines Plata, Steven G. Pueppke, Prem S. Chourey, Auxin-independent growth of maize tissue culture cells, Plant Science, Volume 40, Issue 3, September 1985, Pages 197-202.
121. SHERIDAN Tissue Culture of Maize, Physiologia Plantarum, 41(3): 172-174, 1977.
122. Mizutani et al. The syncytium-specific expression of the Orysa; KRP3 CDK inhibitor: implication of its involvement in the cell cycle control in the rice (Oryza sativa L.) syncytial endosperm J Exp Bot. 2010 March; 61(3): 791-798. Published online 2009 November 20. doi: 10.1093/jxb/erp343.
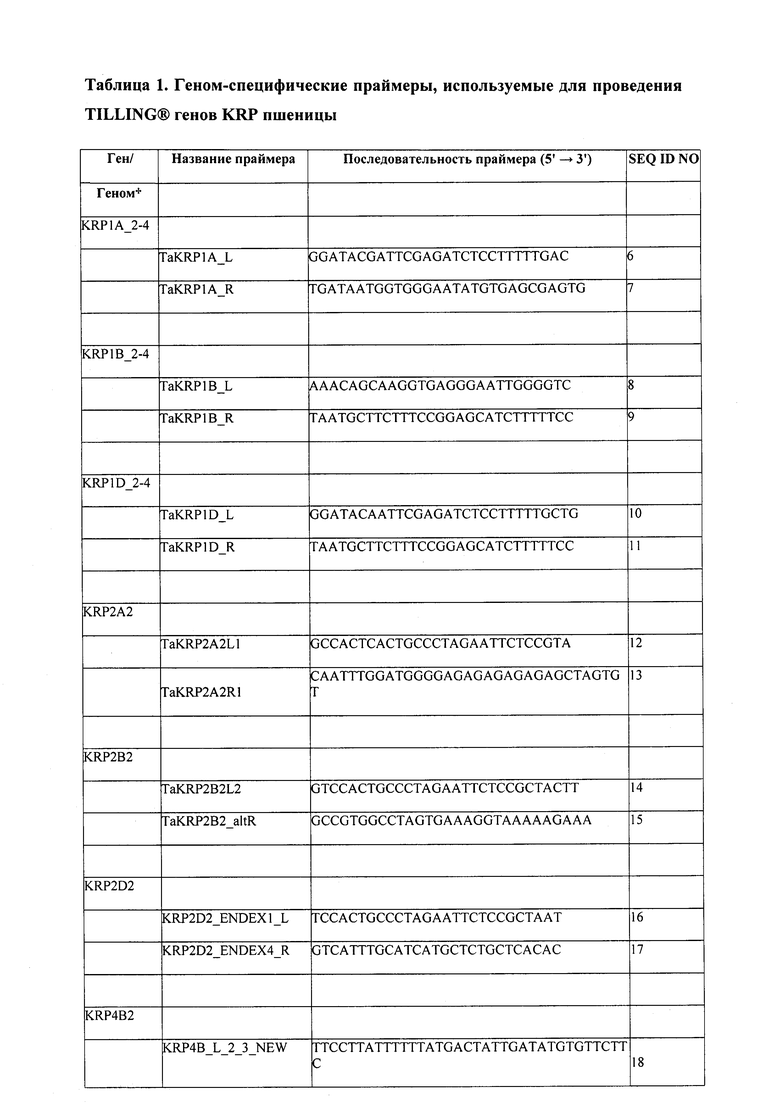
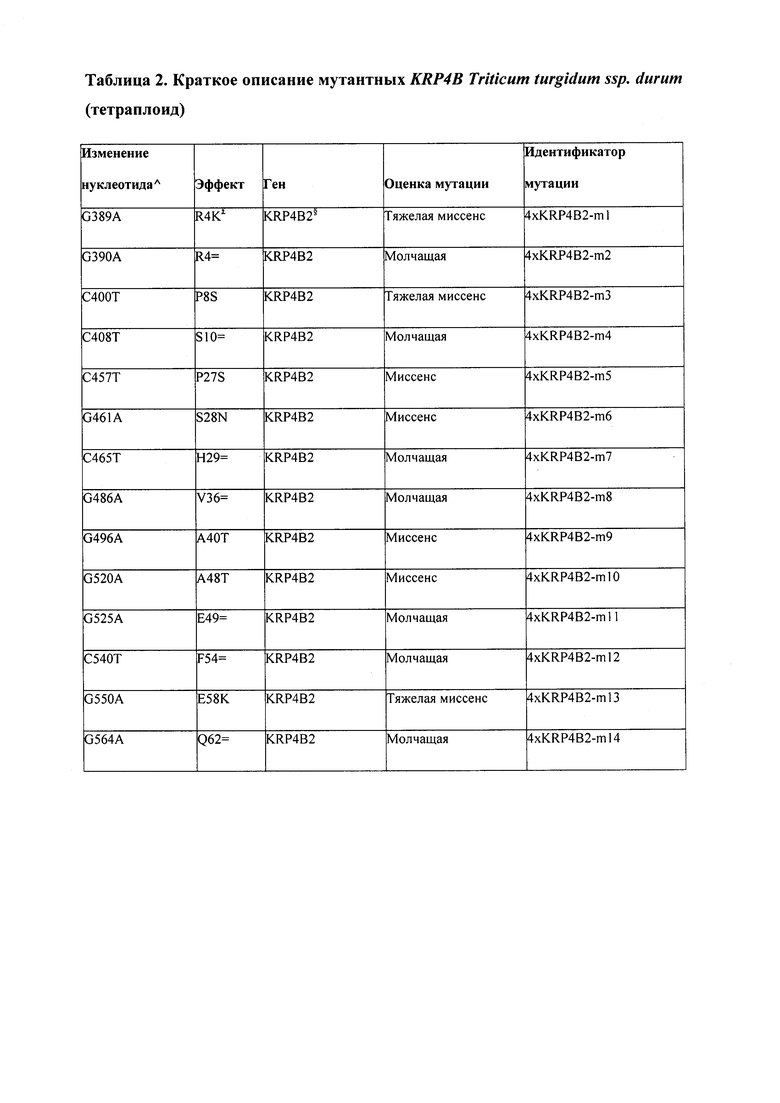
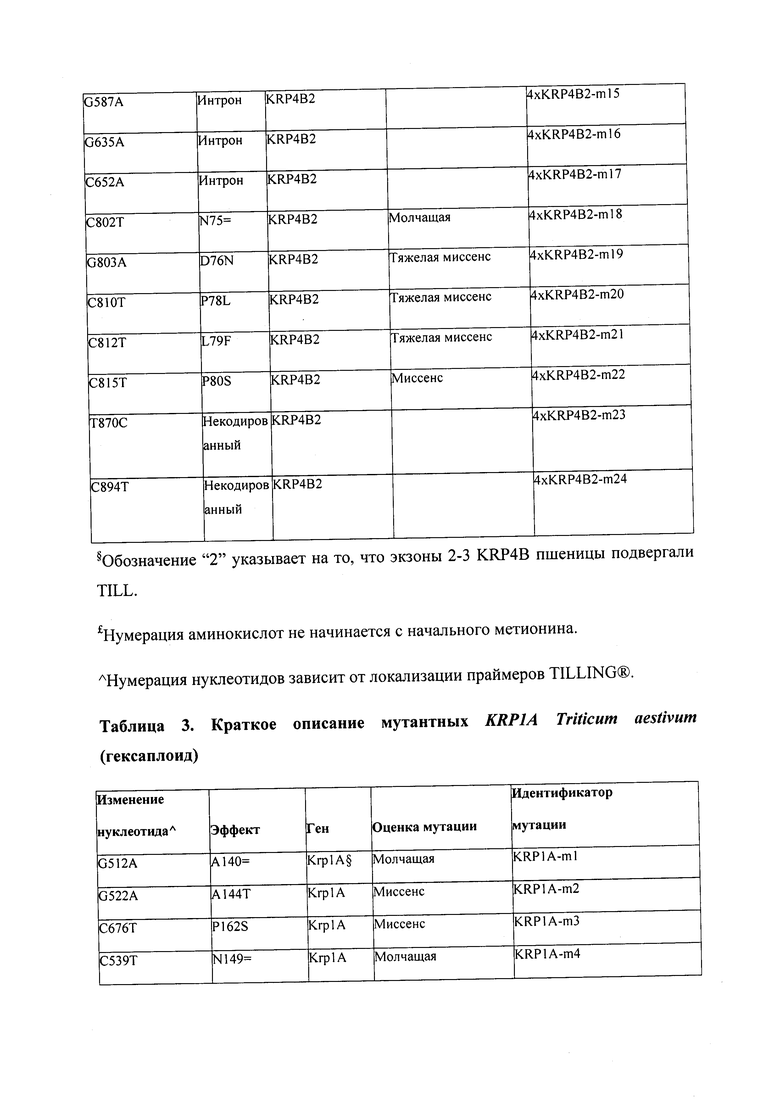
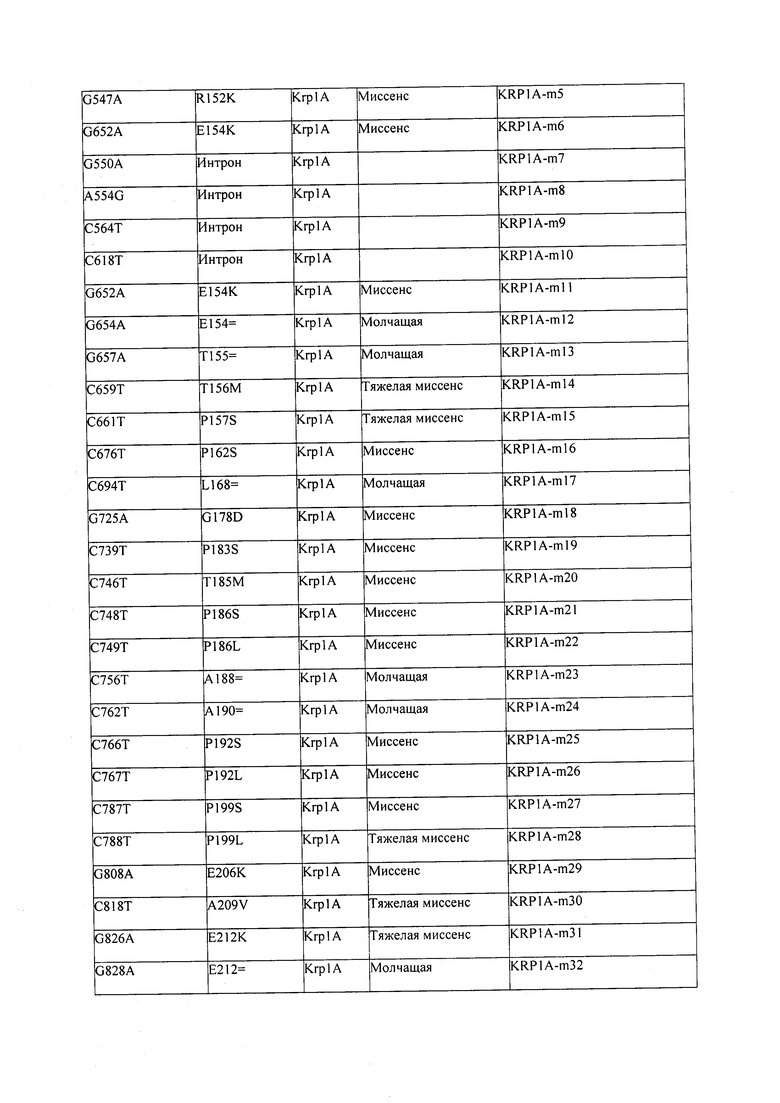
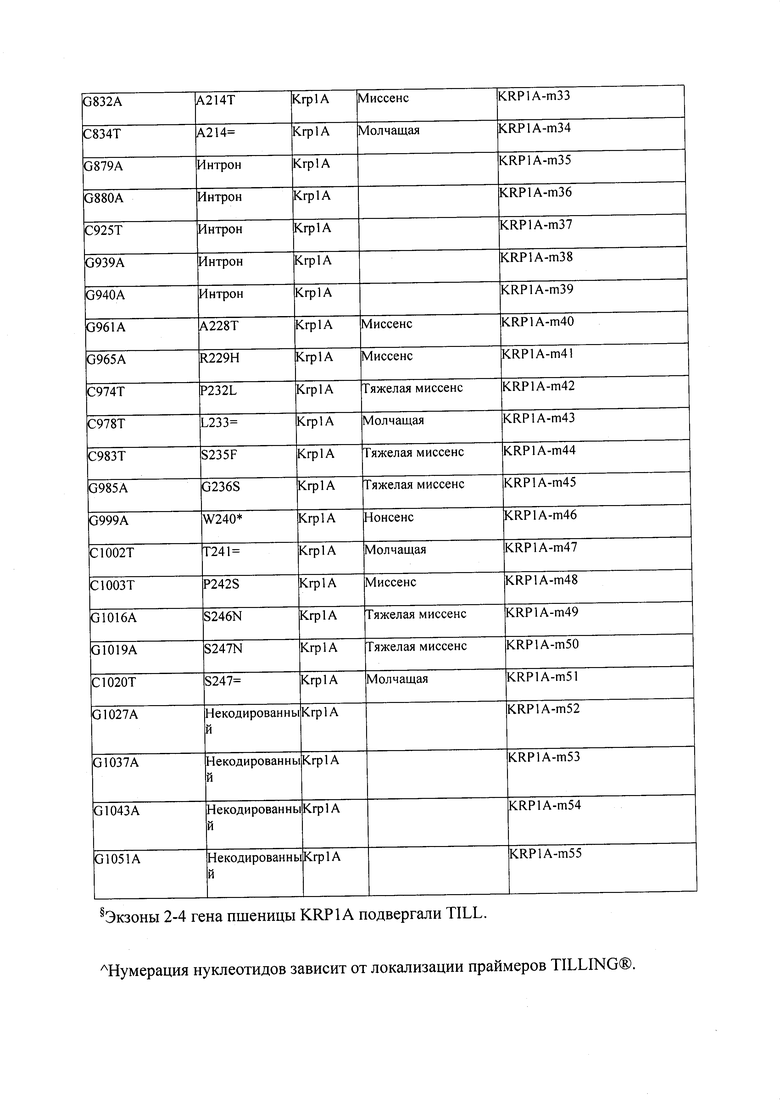
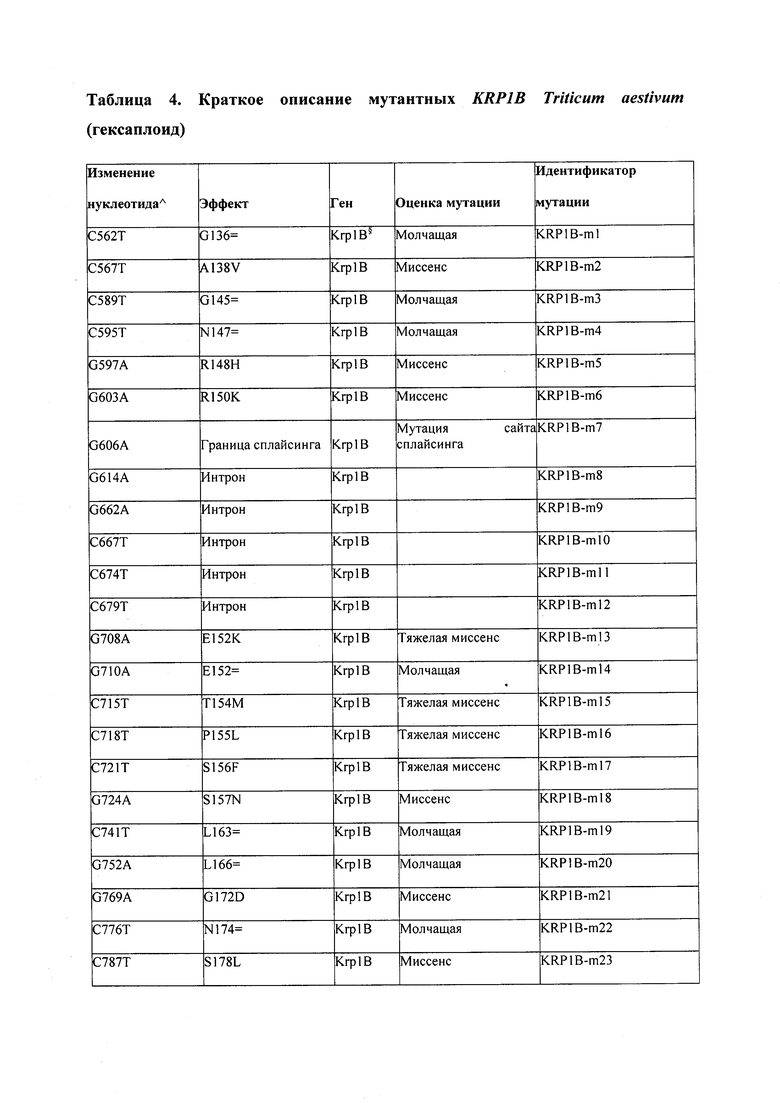
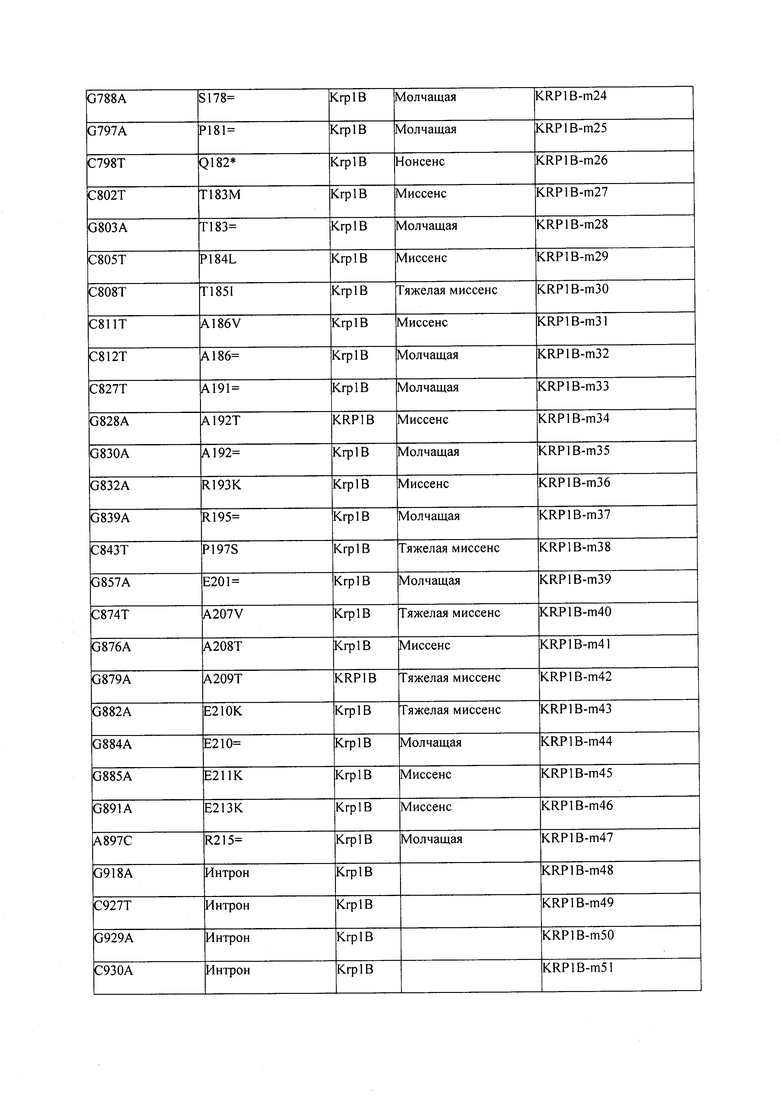
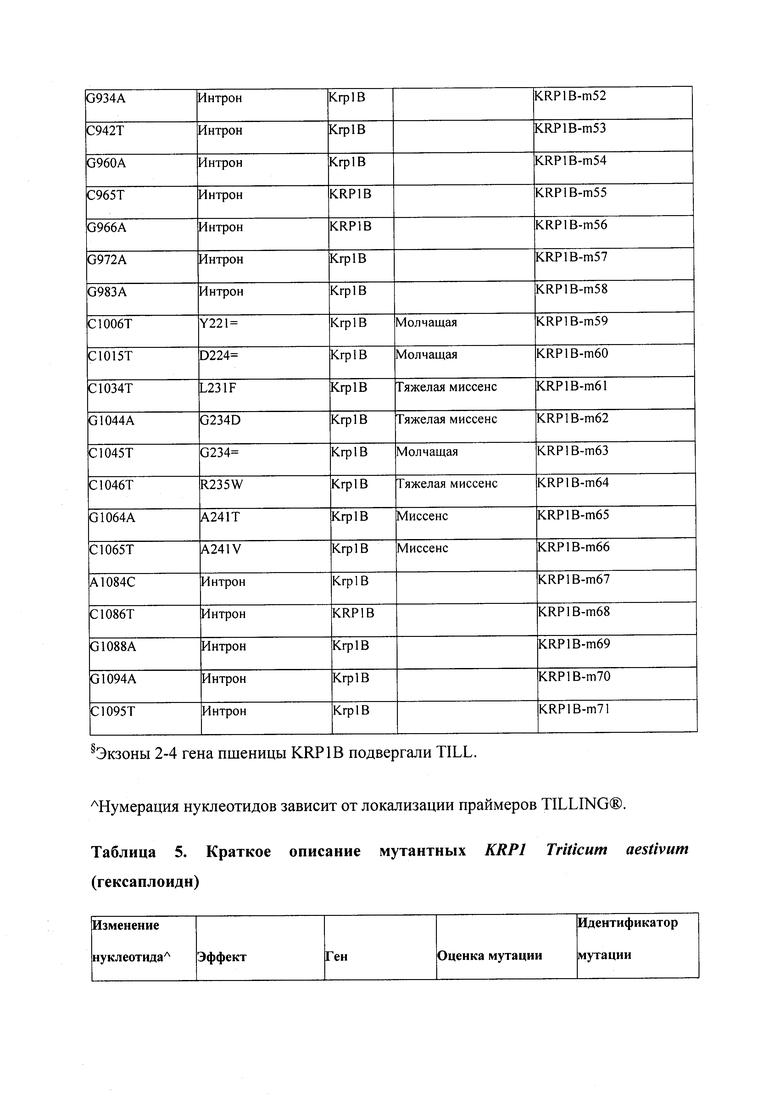
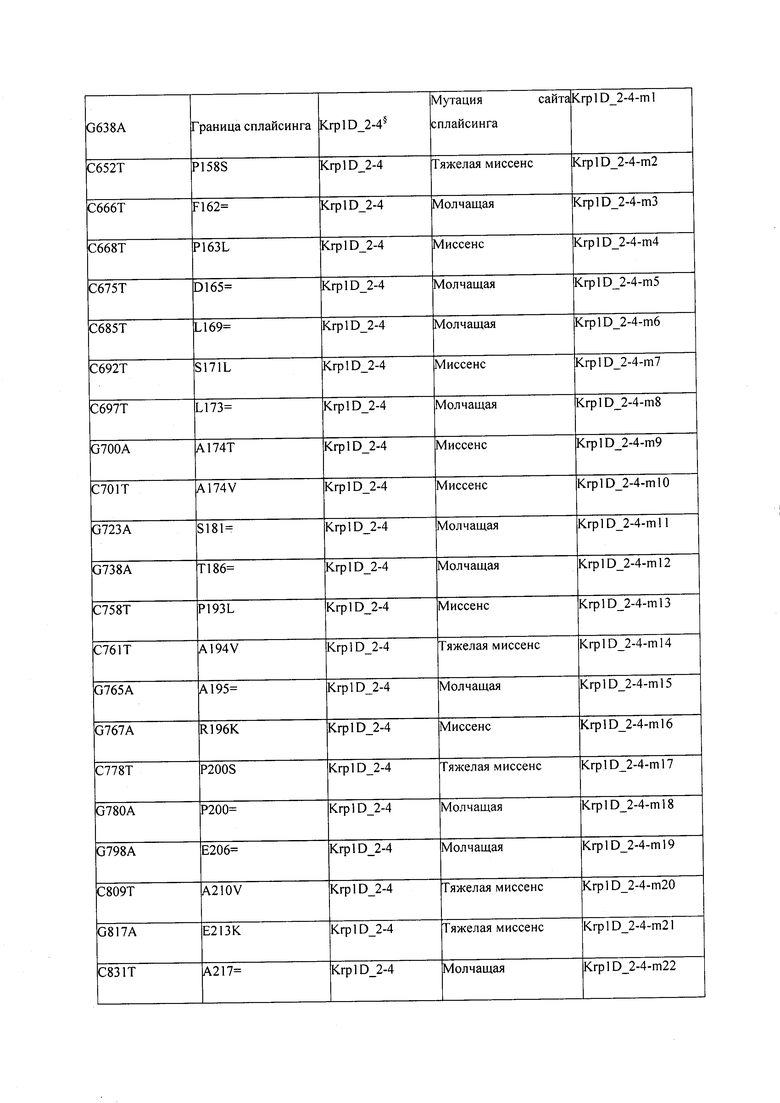
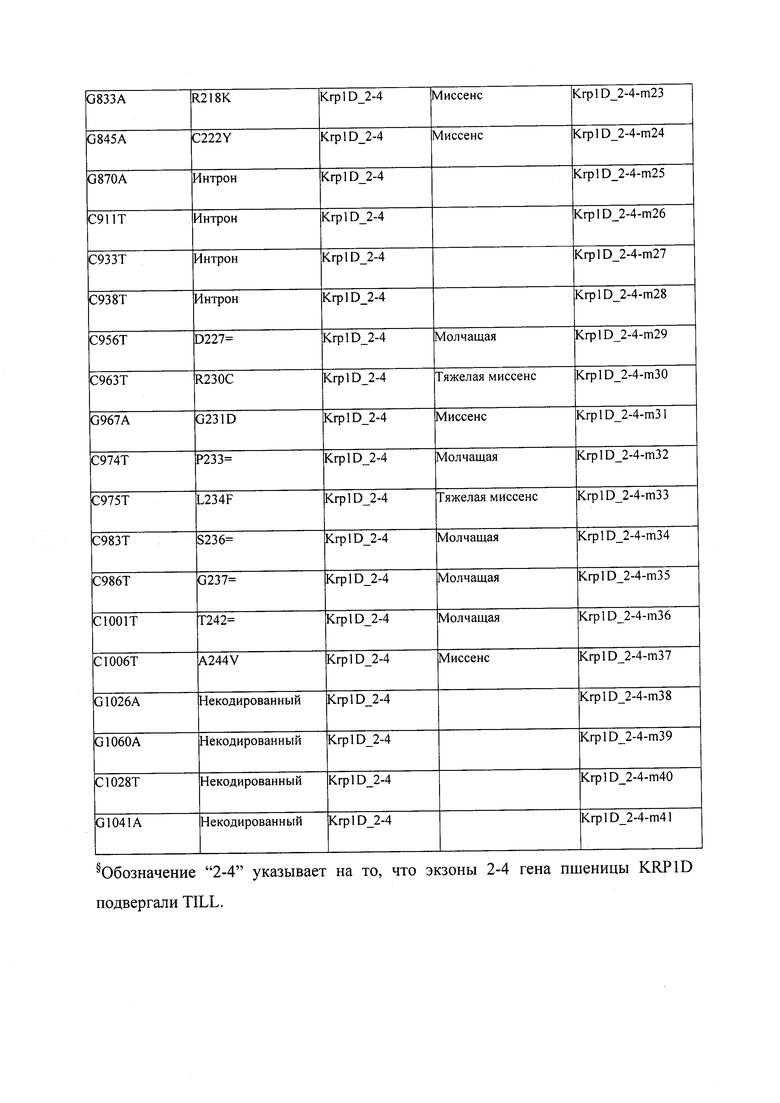
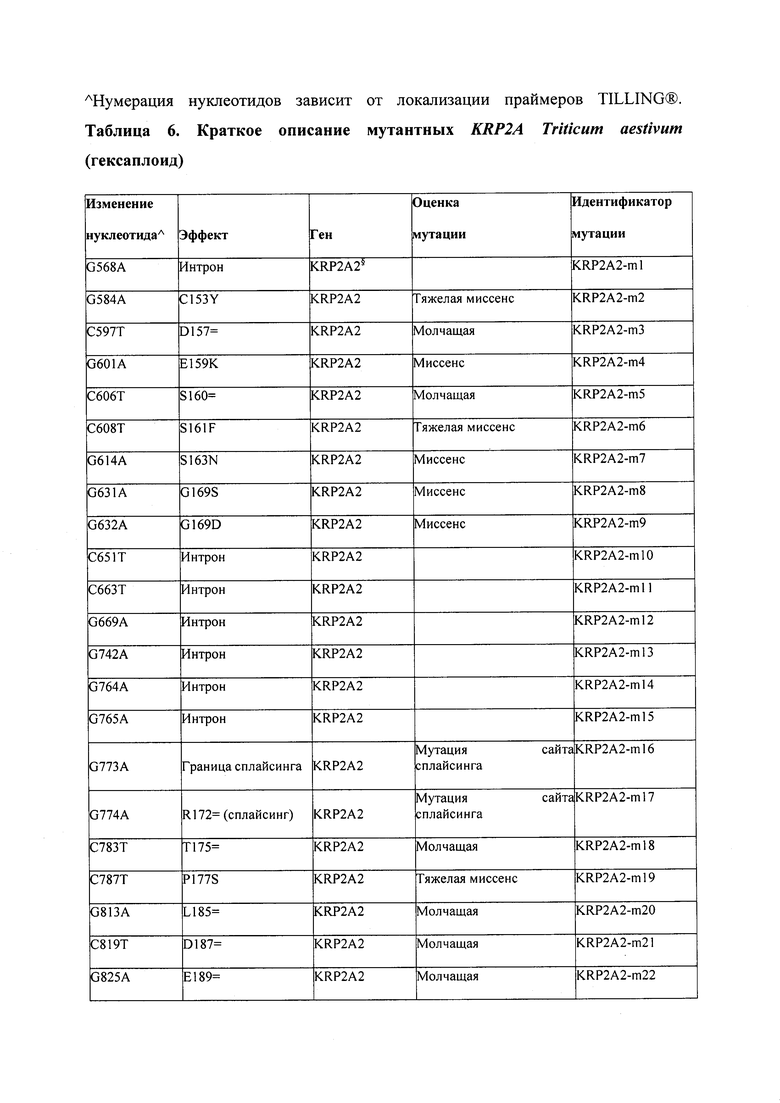
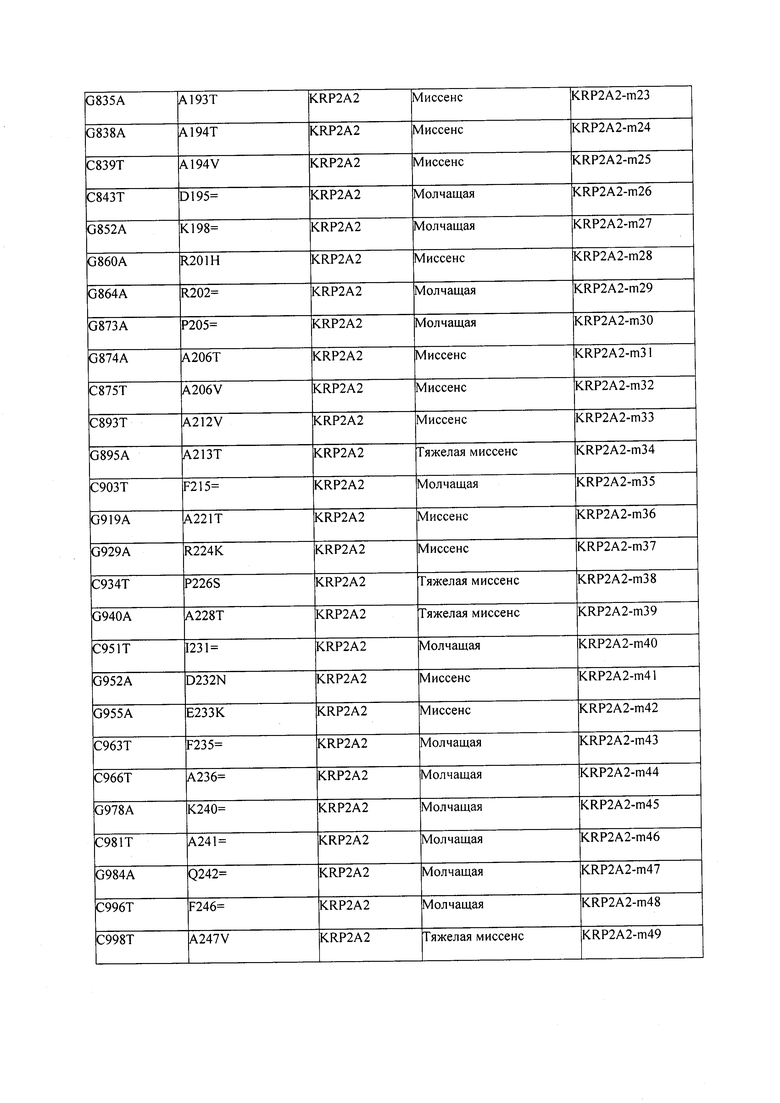
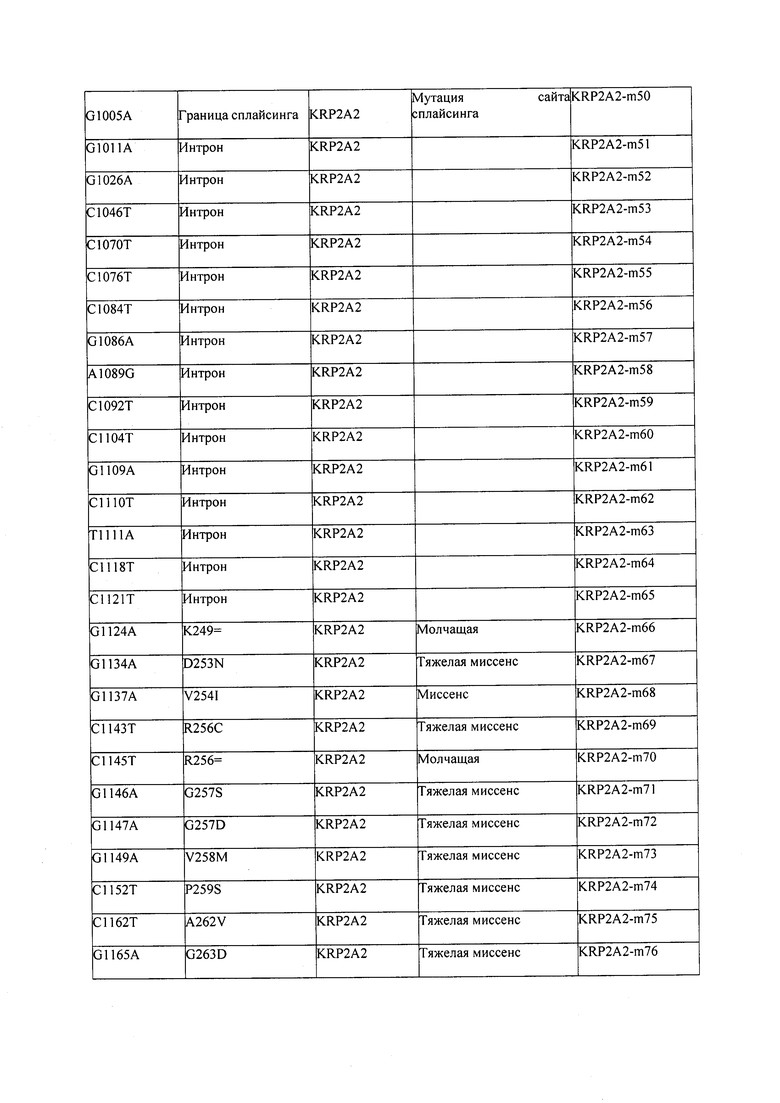
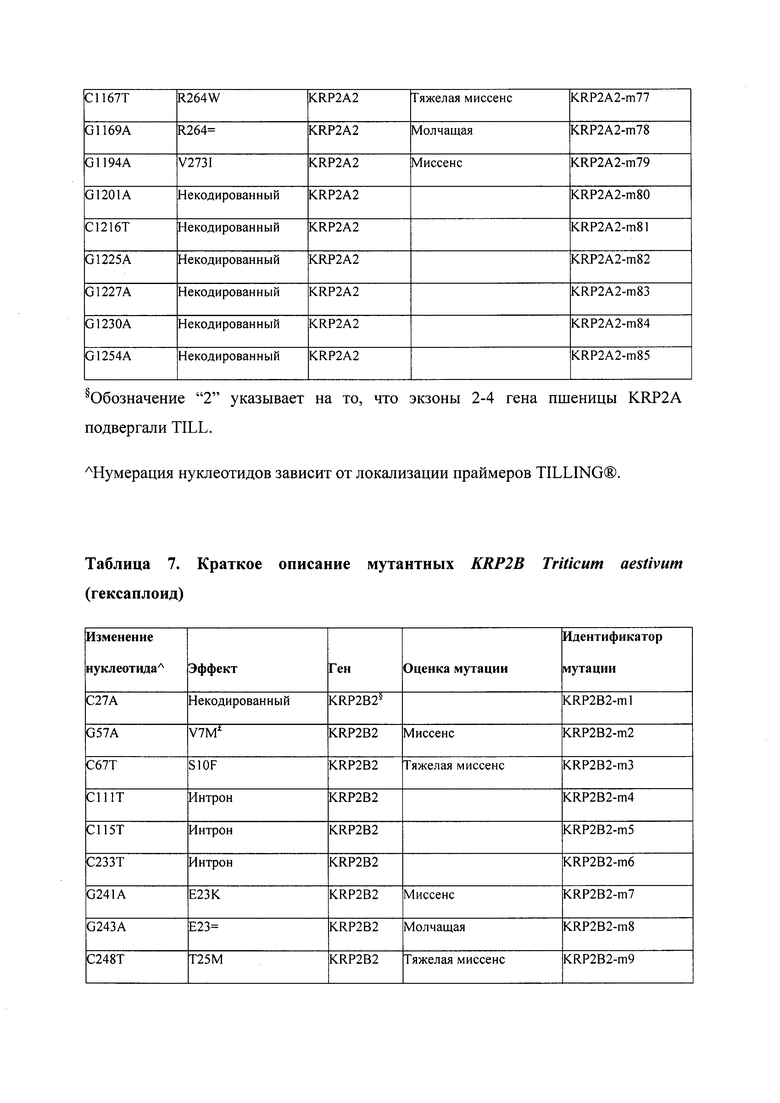
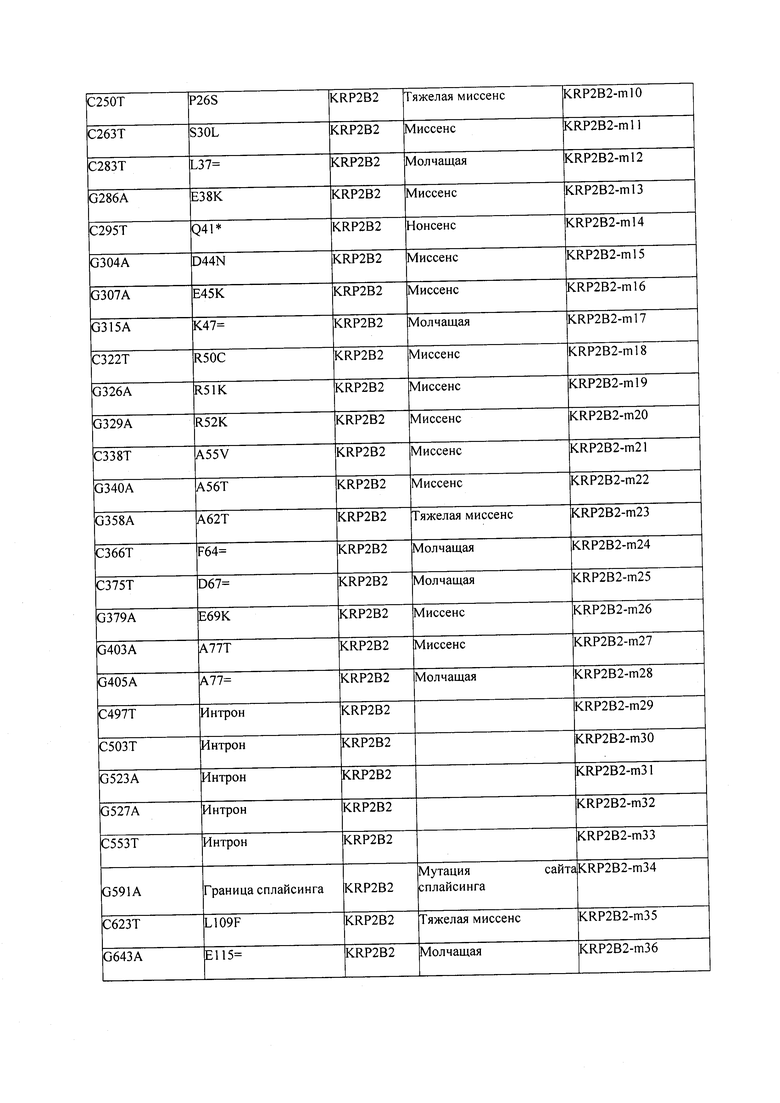
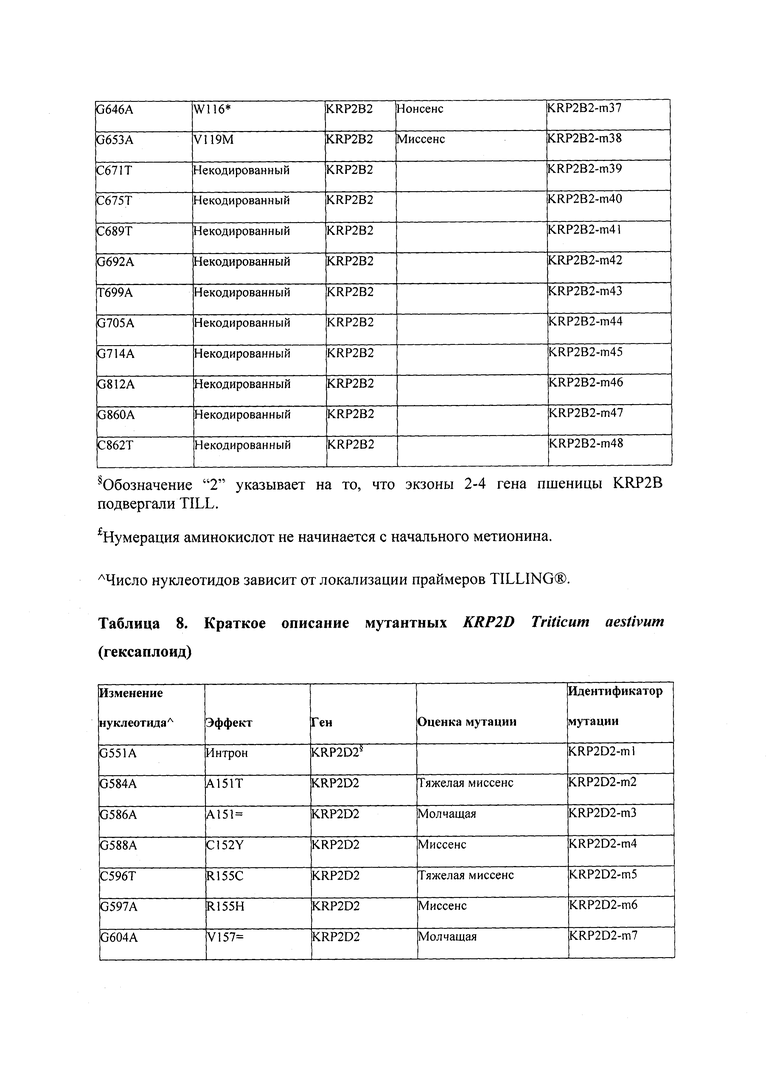
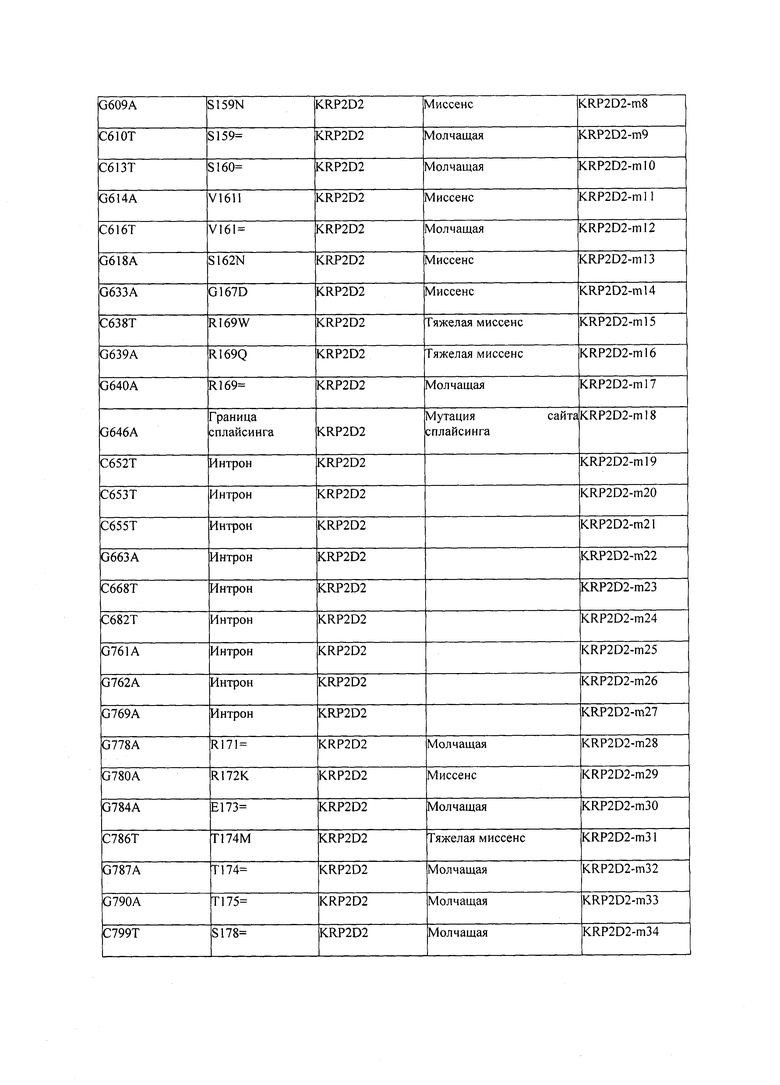
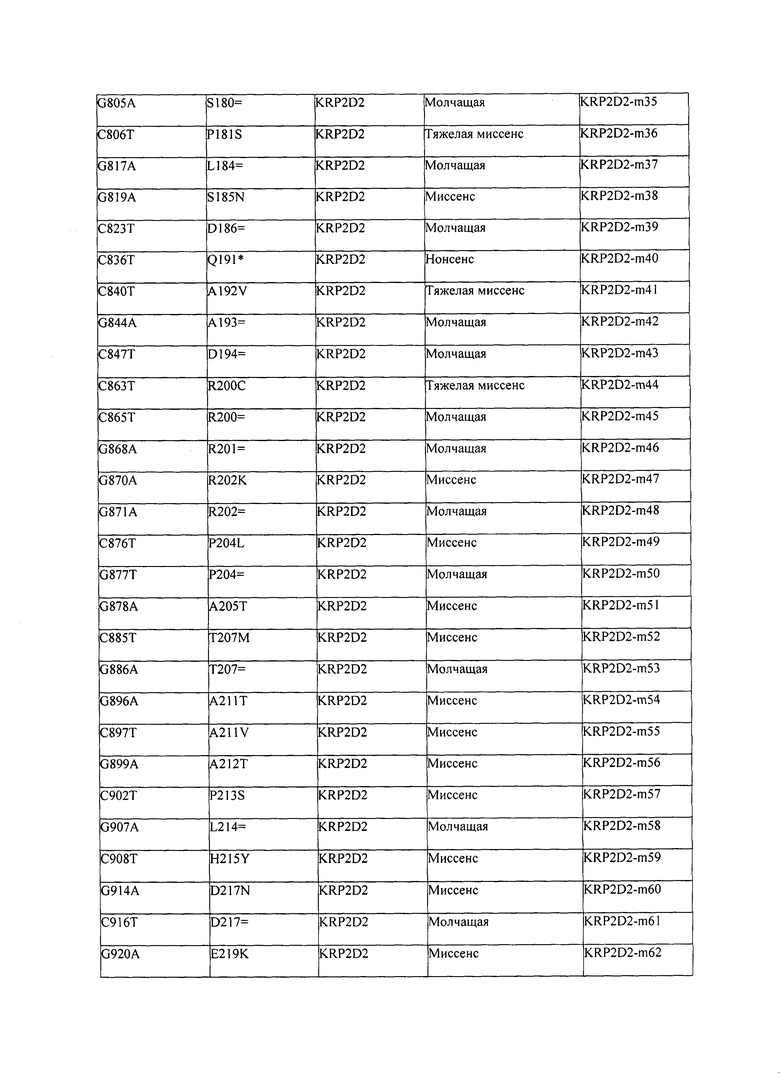
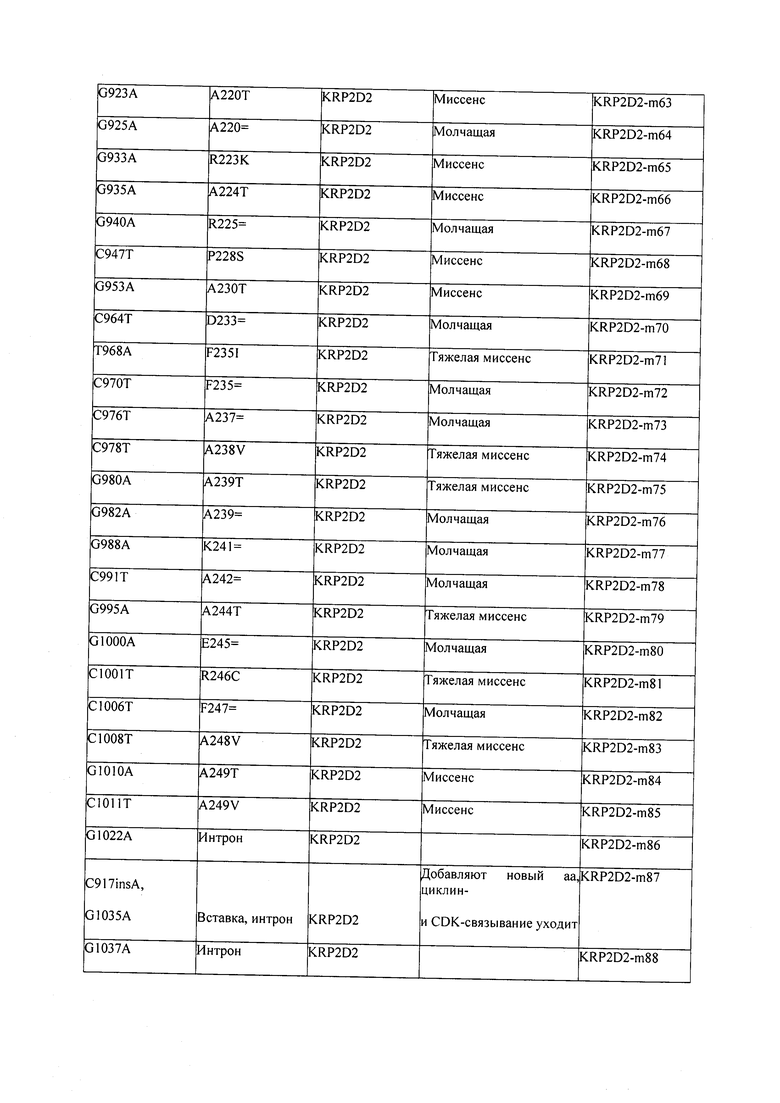
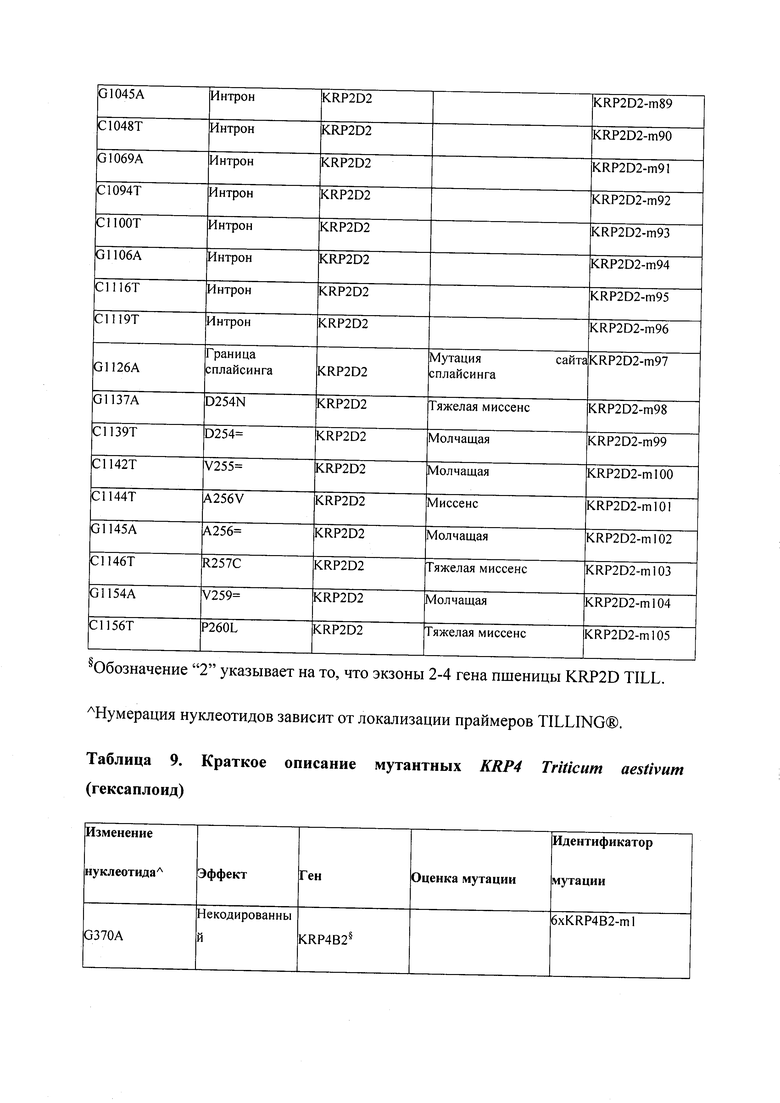
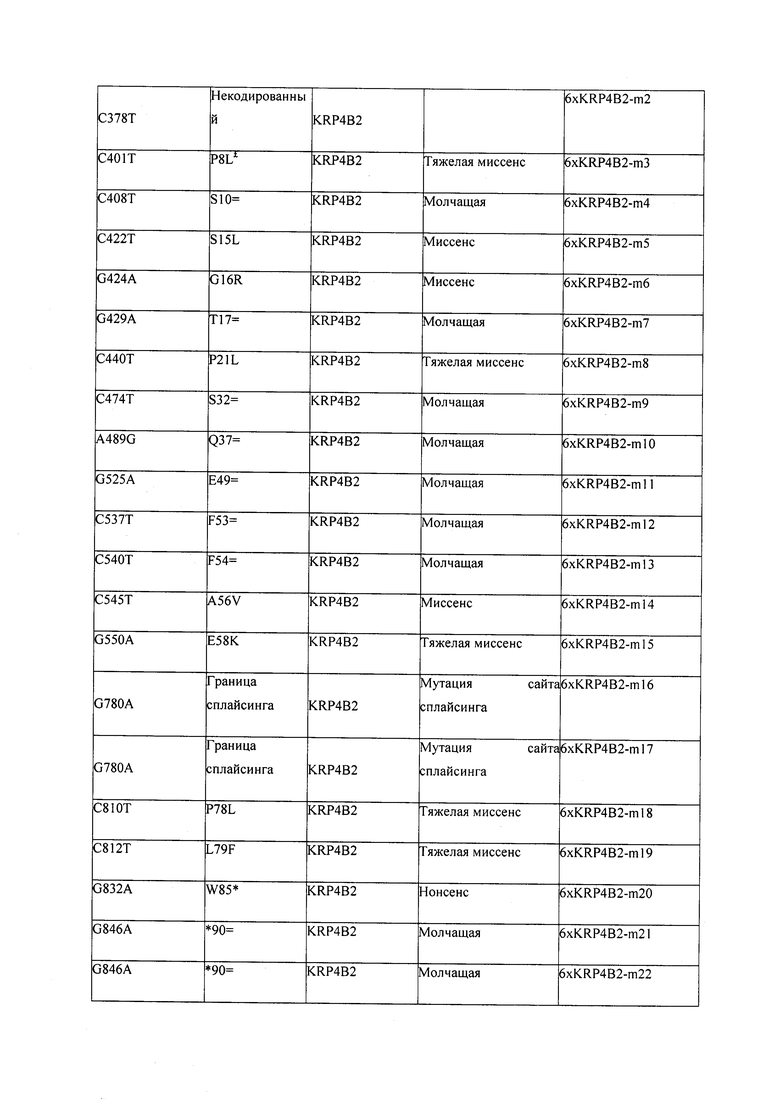
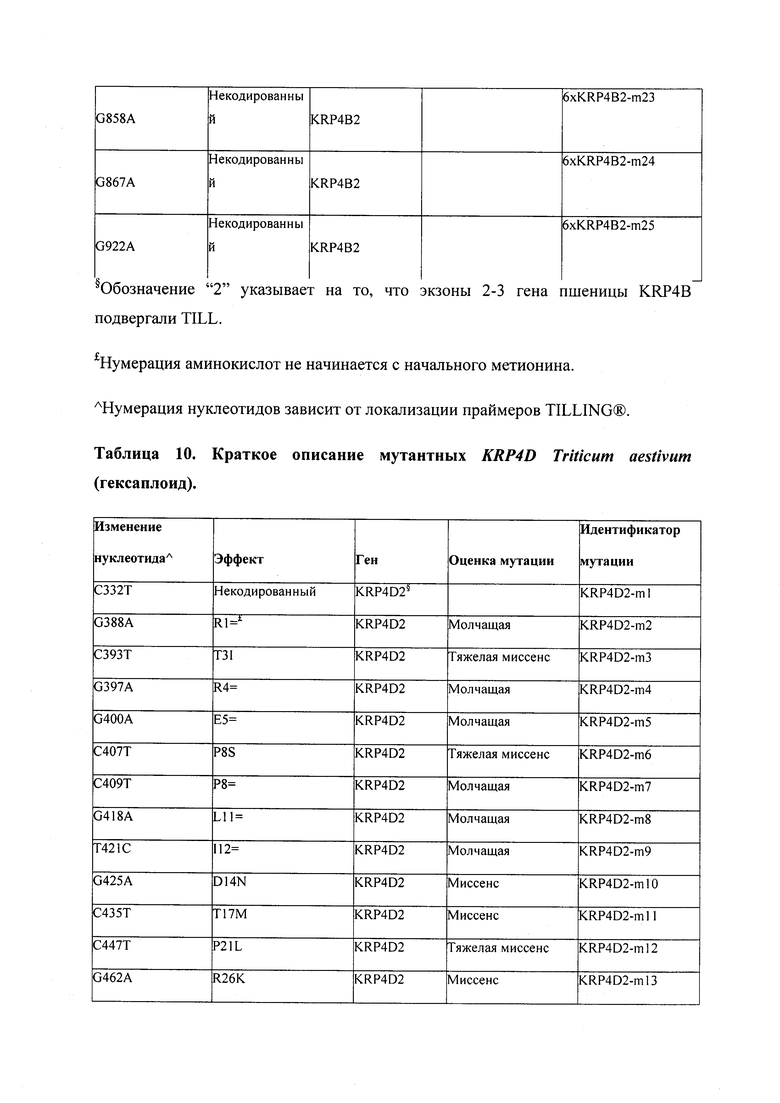
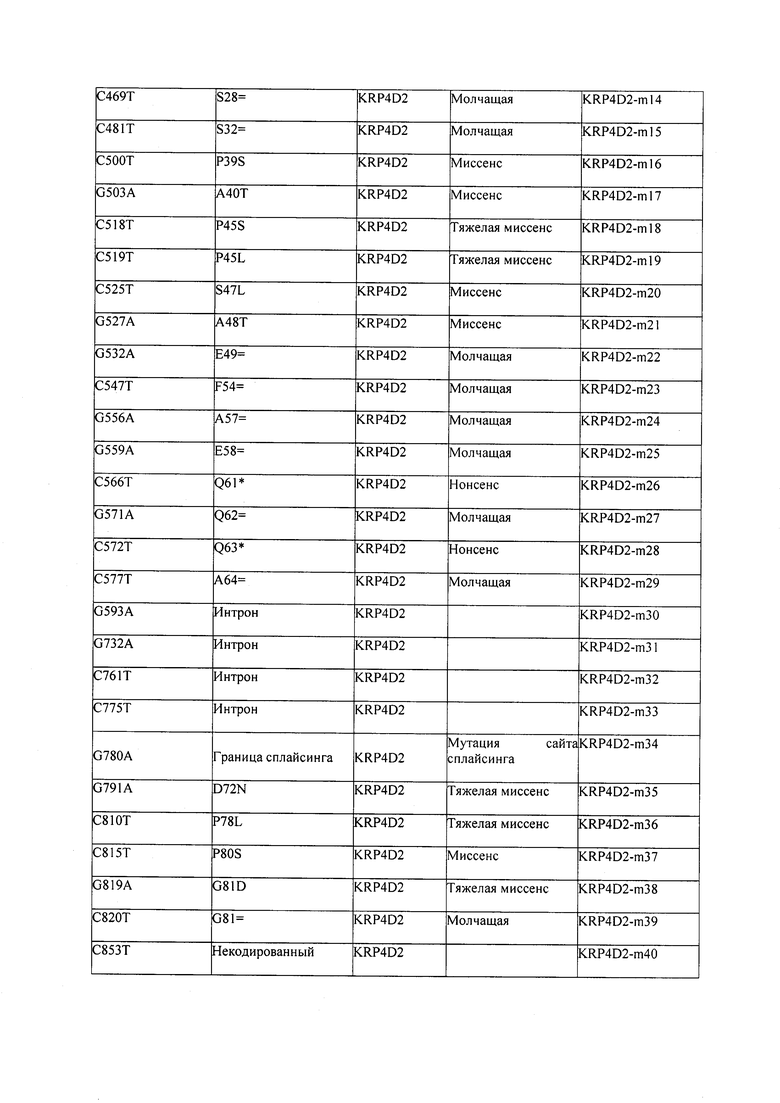
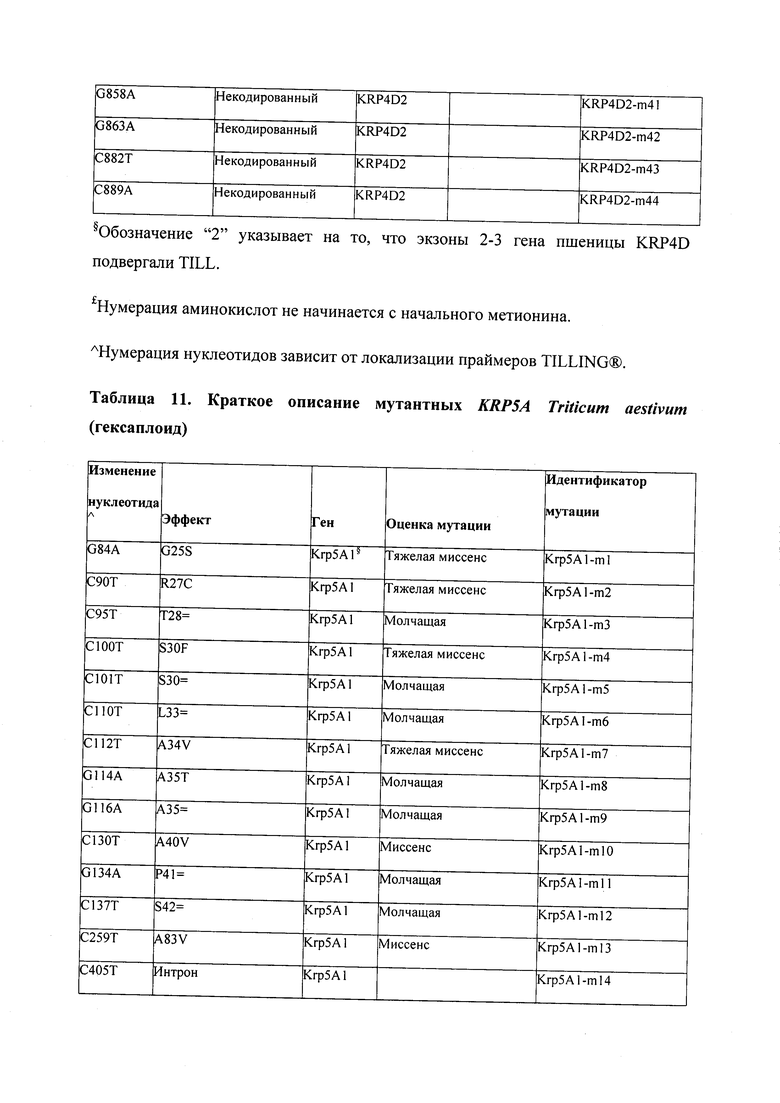
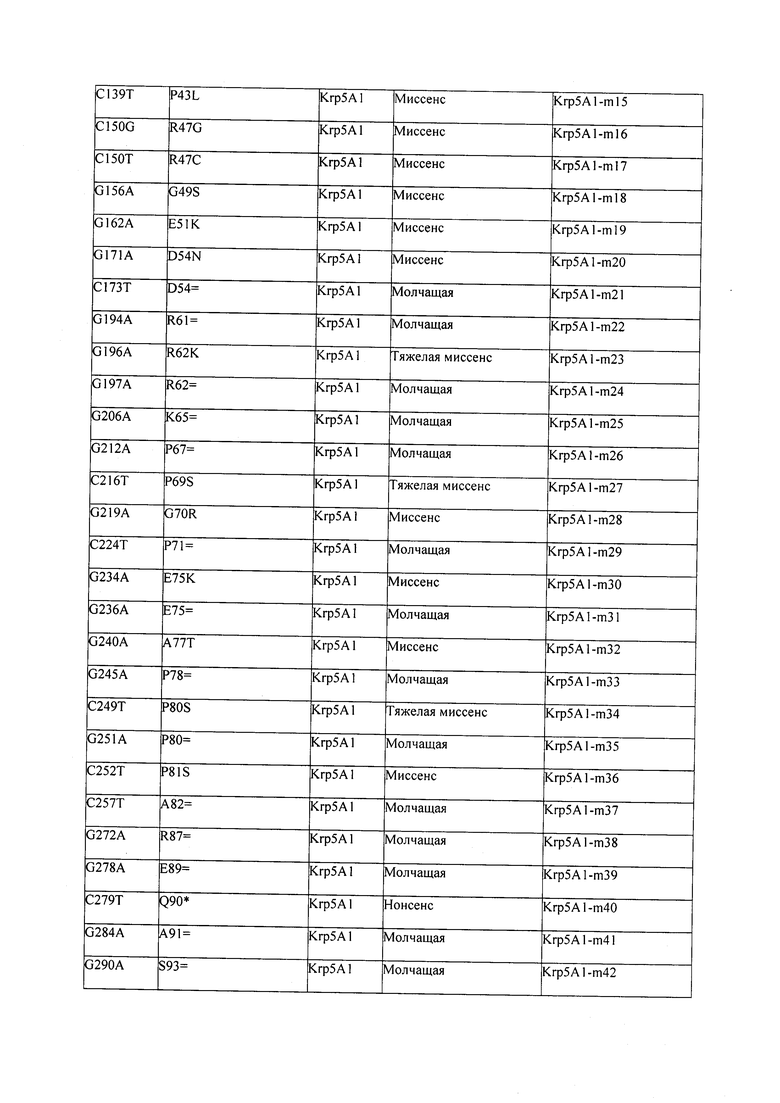
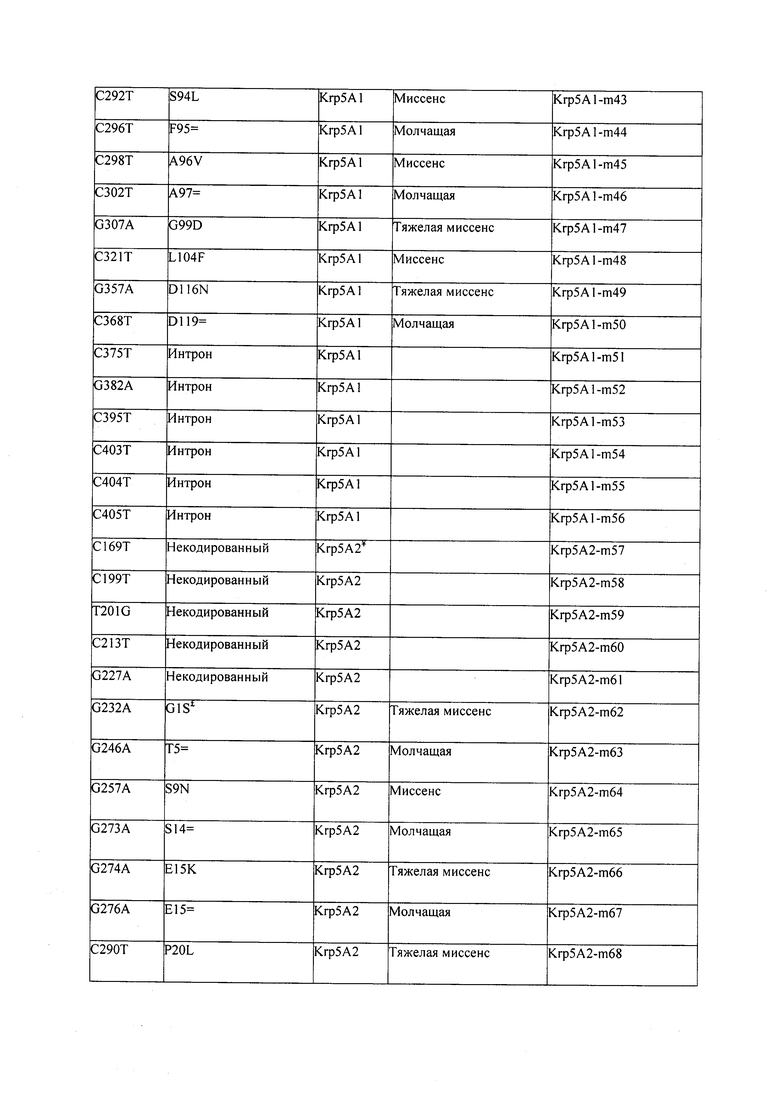
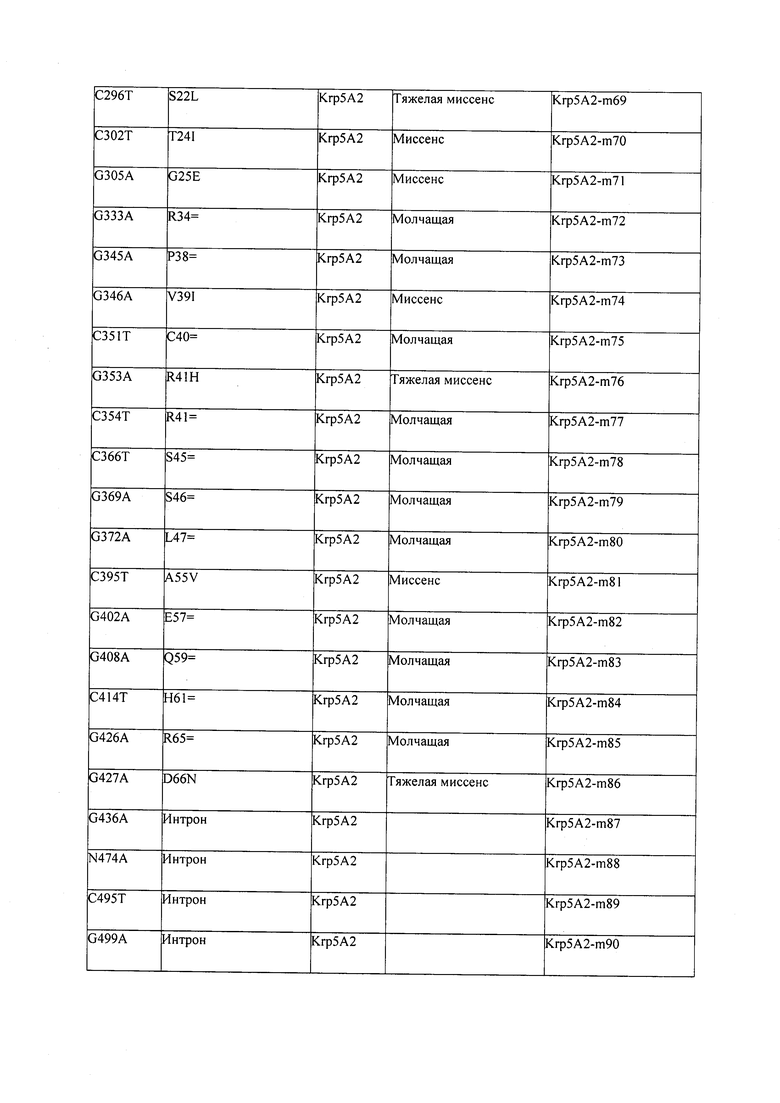
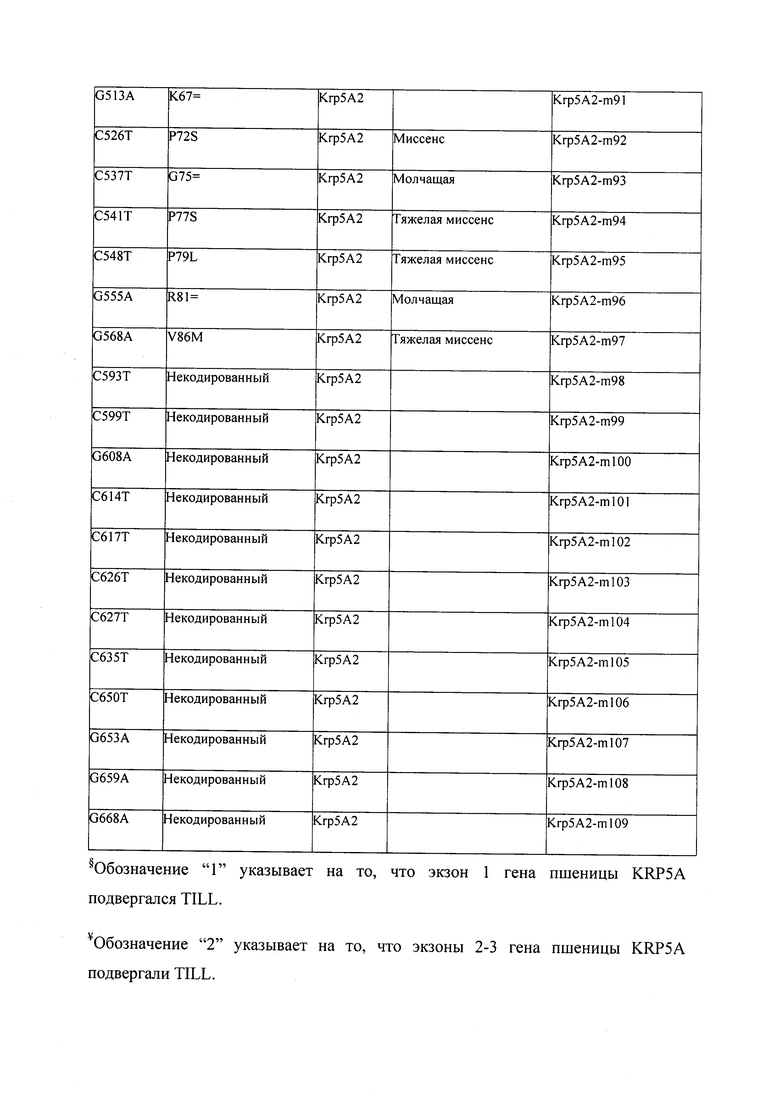
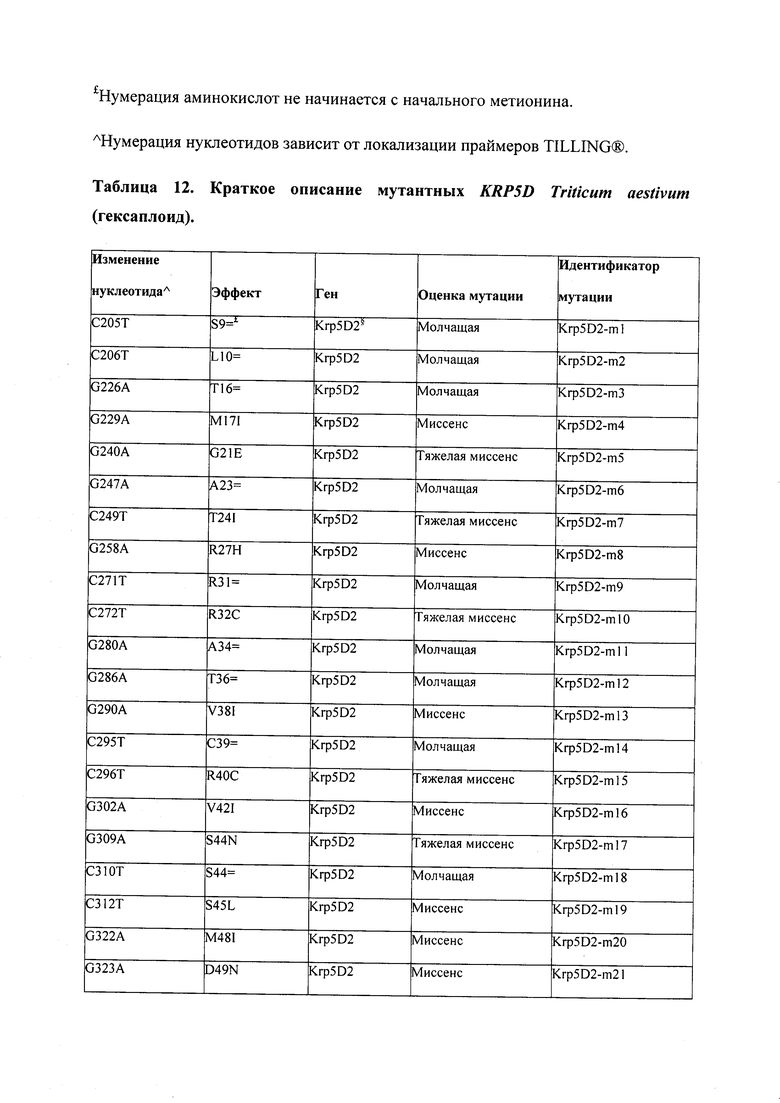
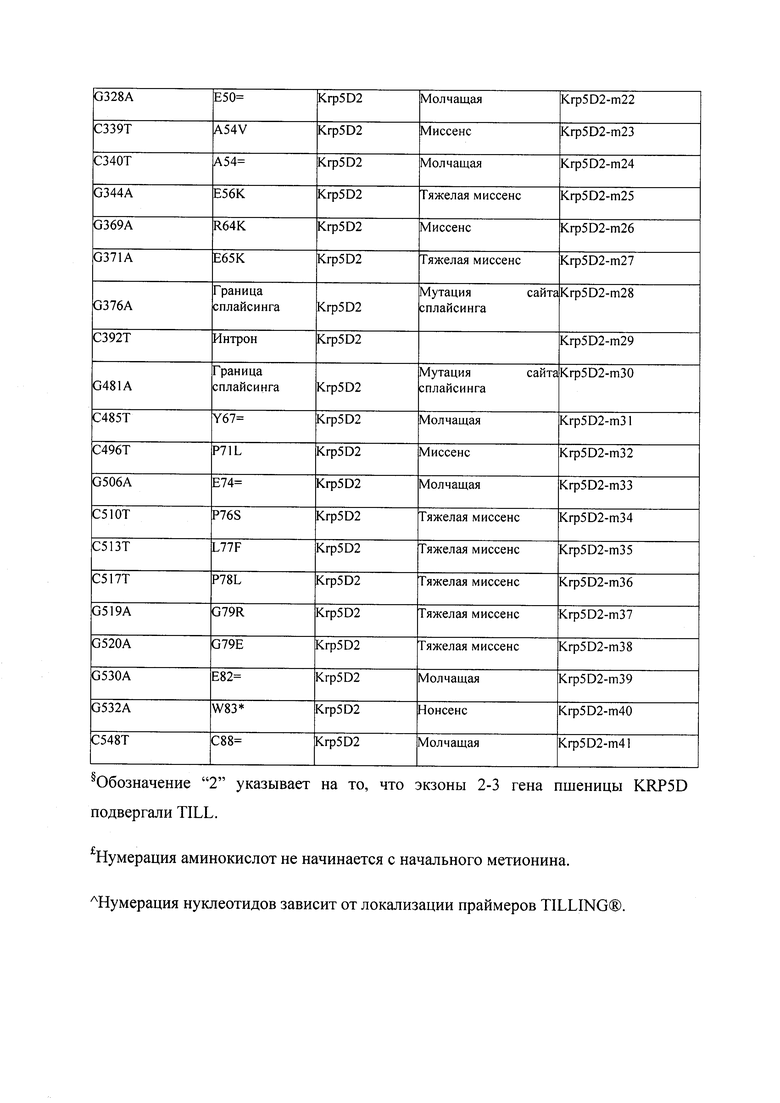
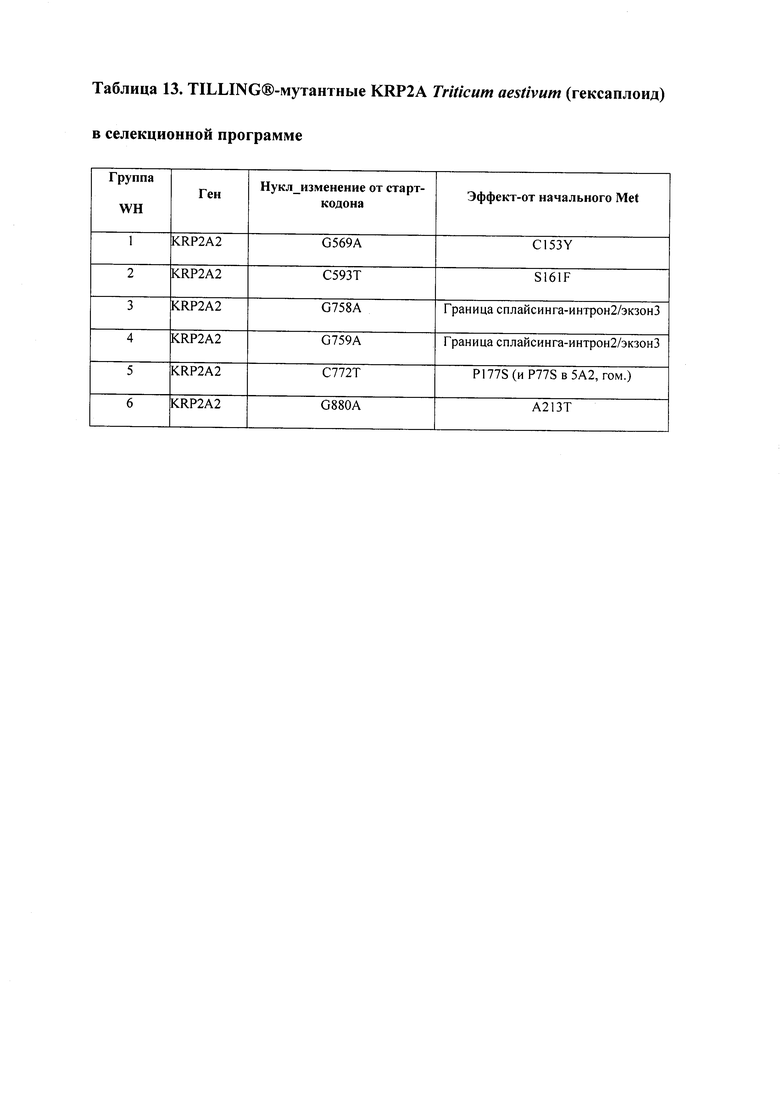
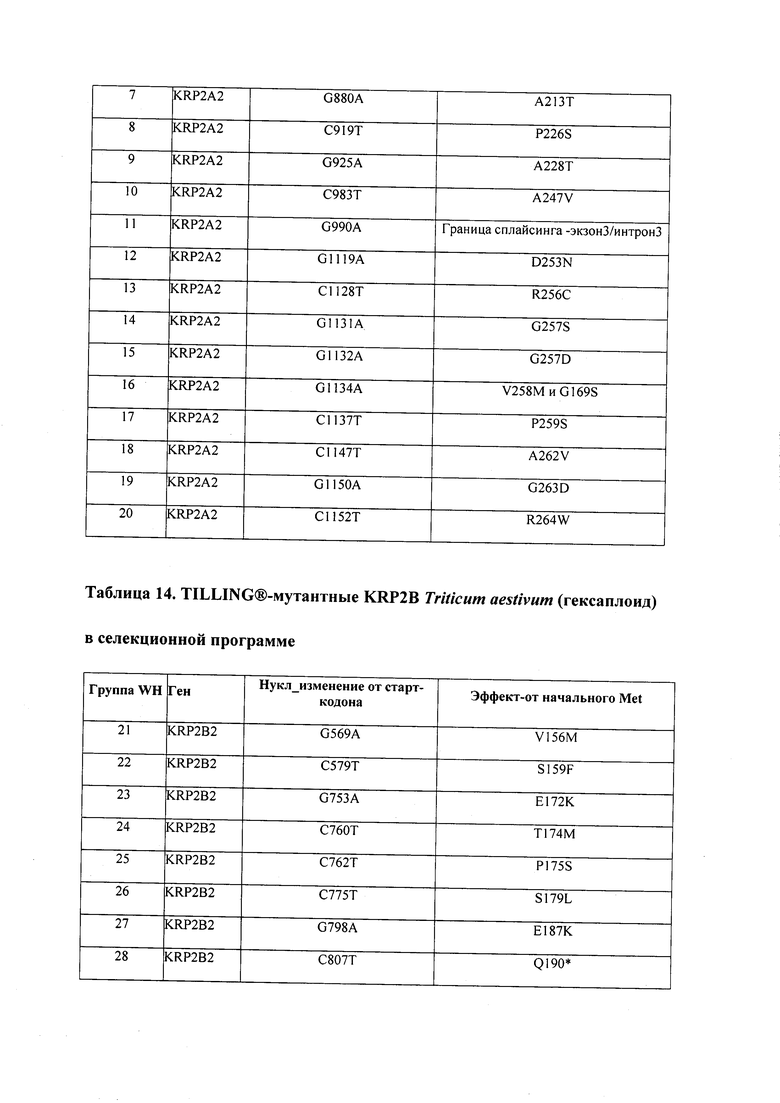
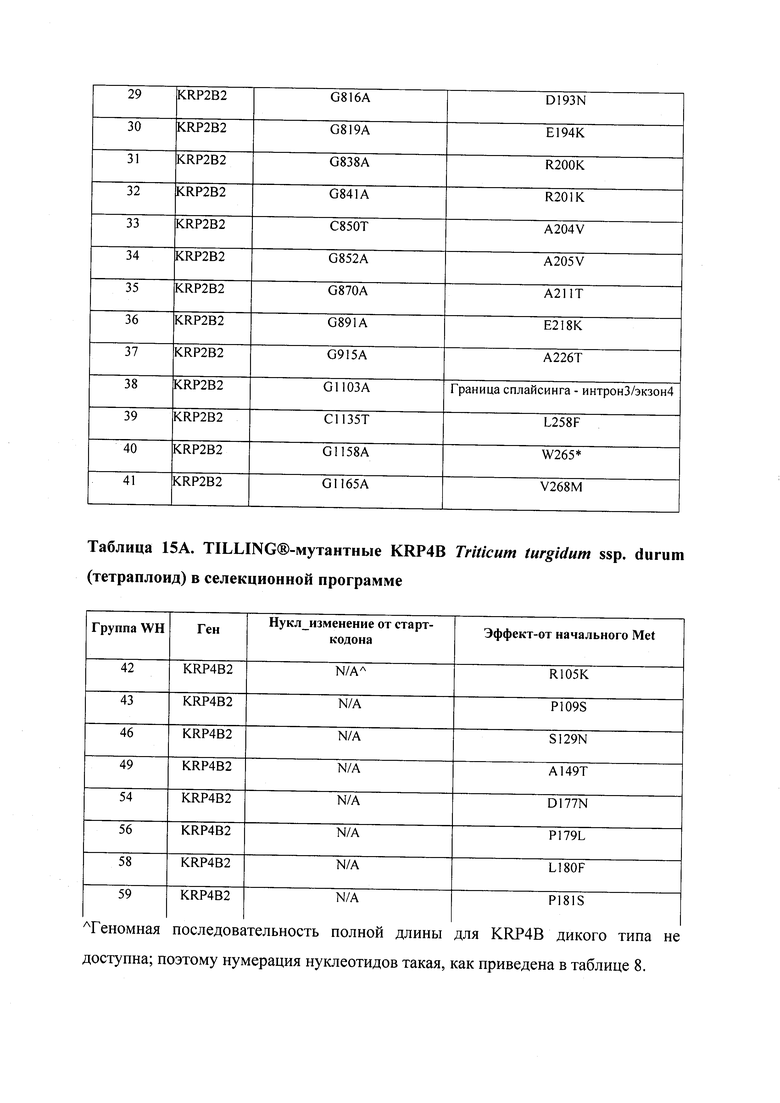
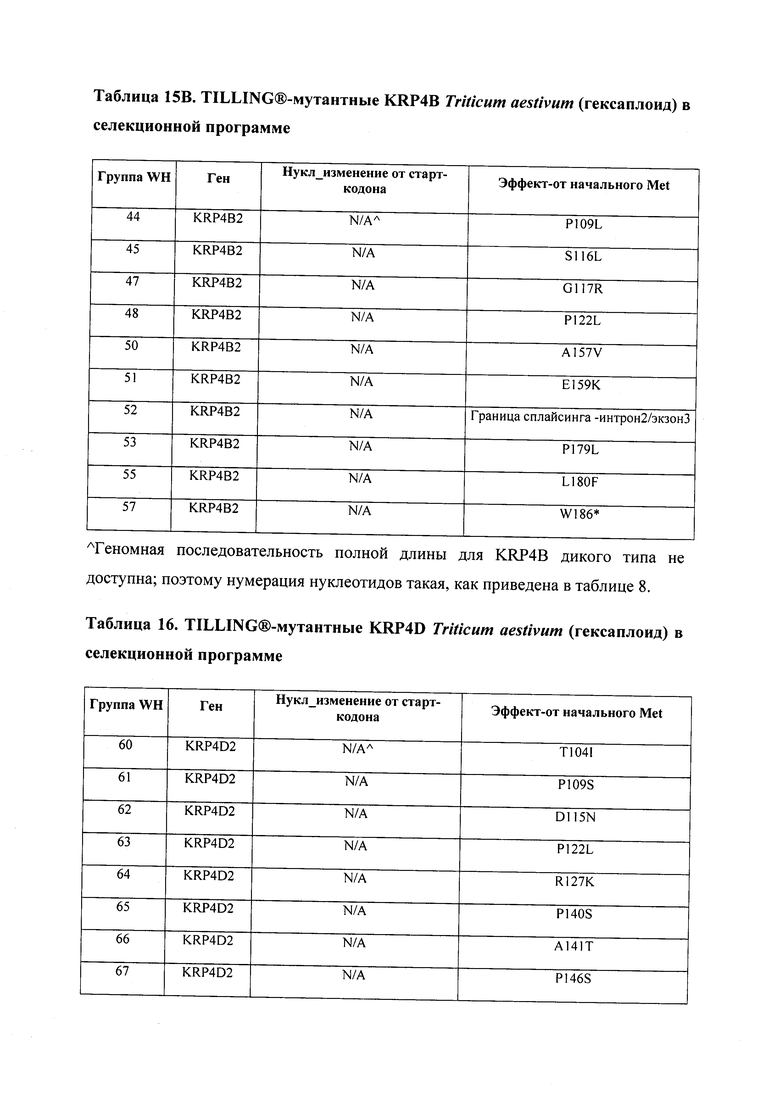
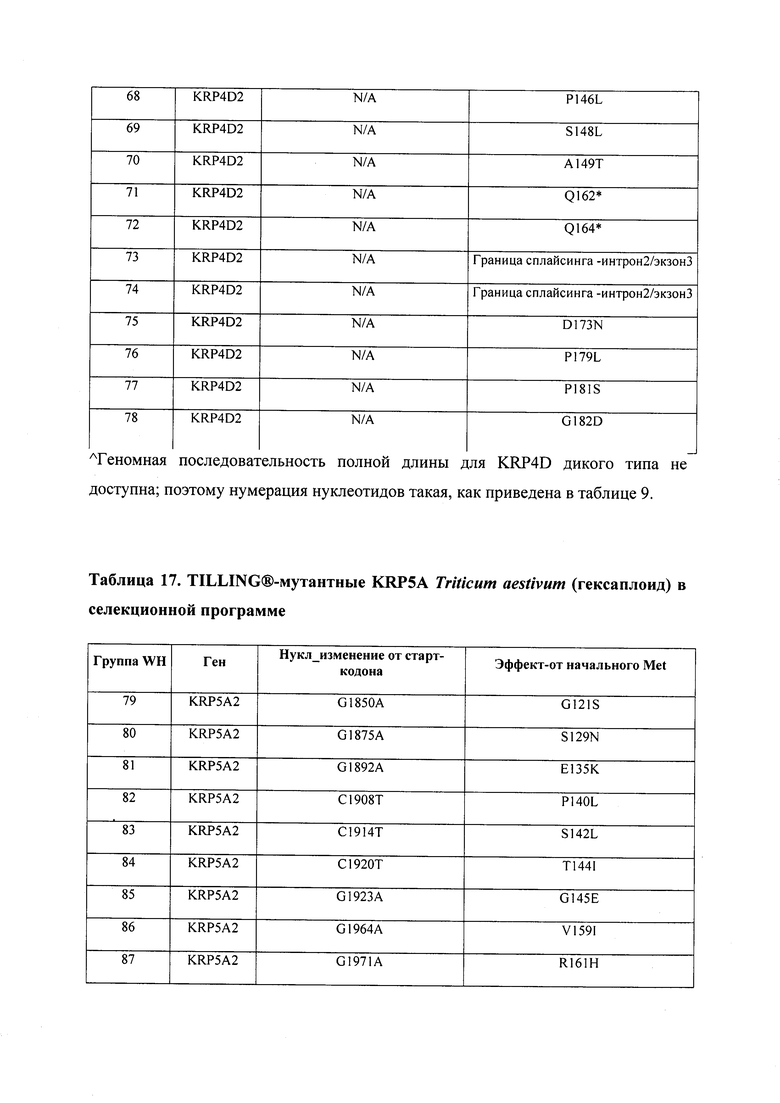
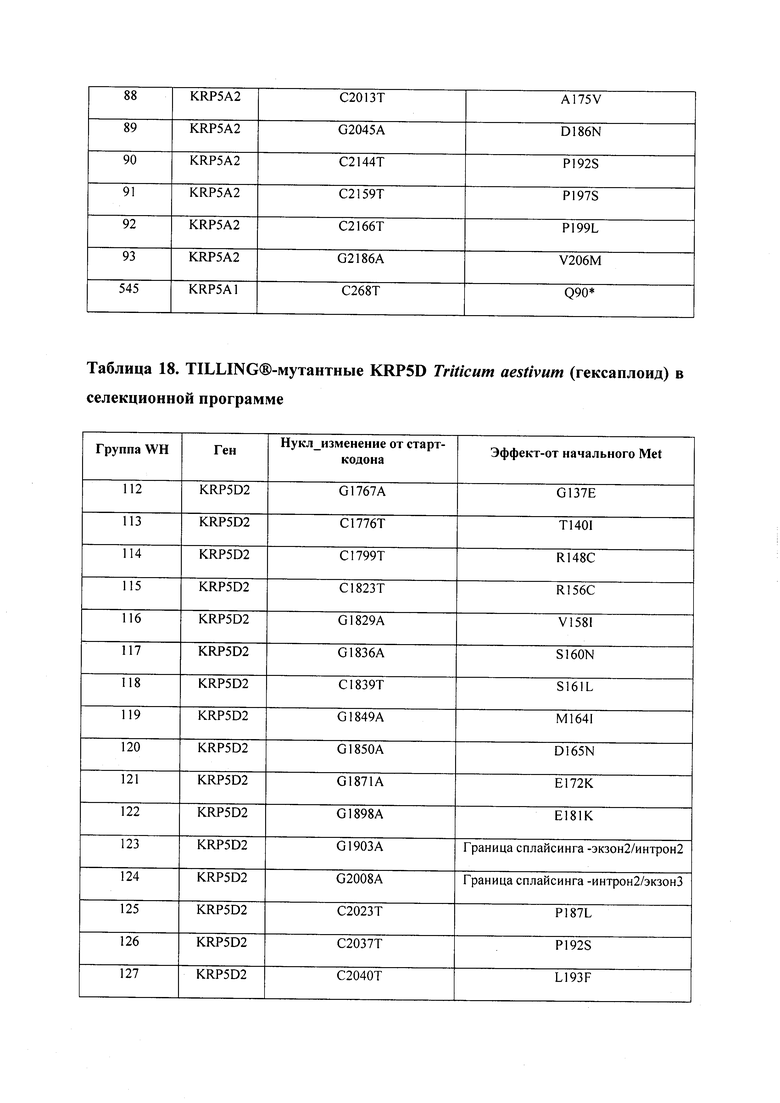
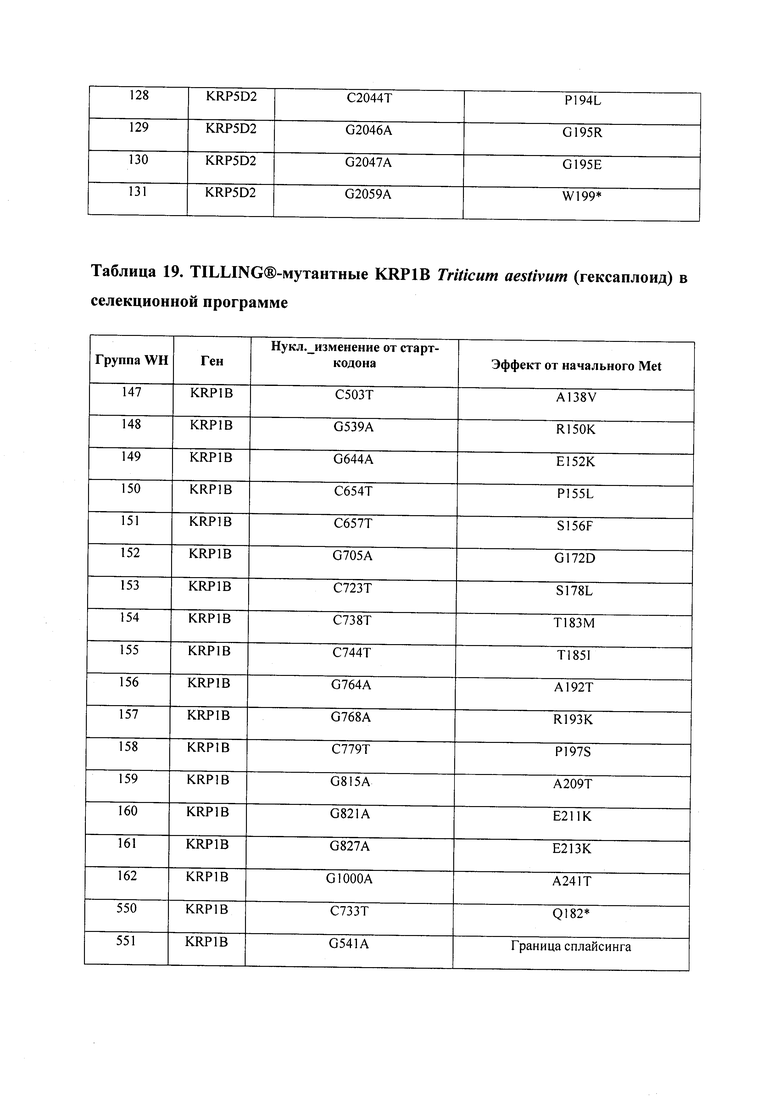
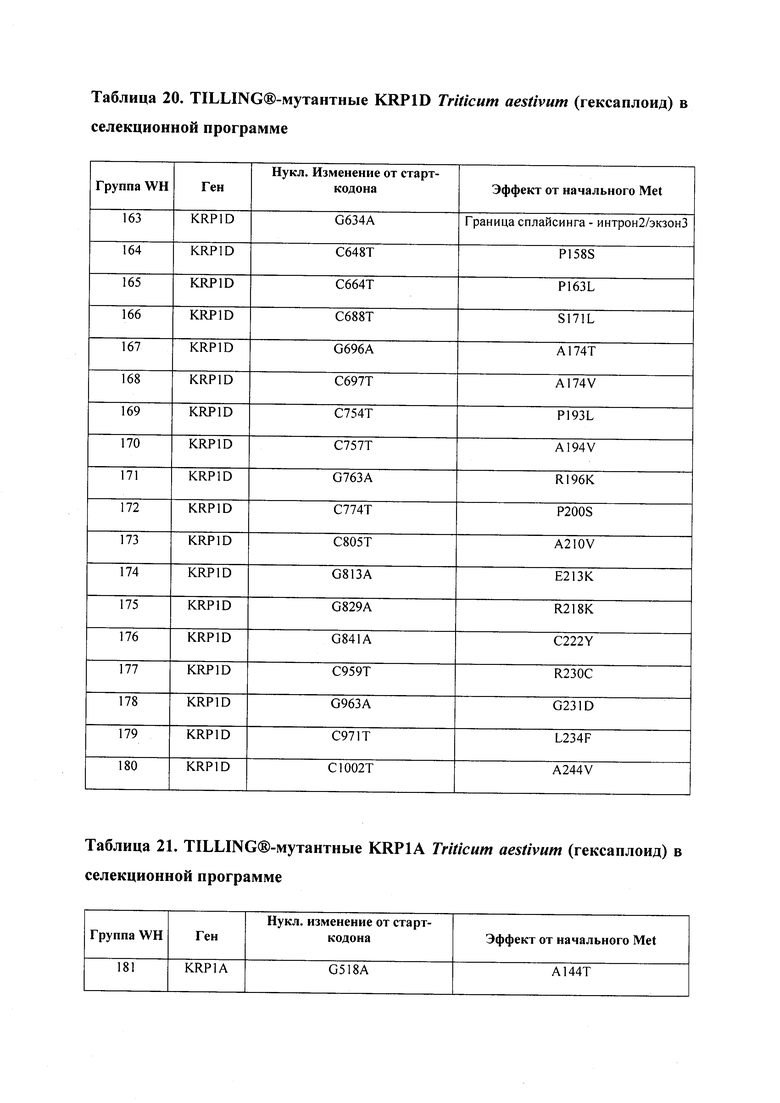
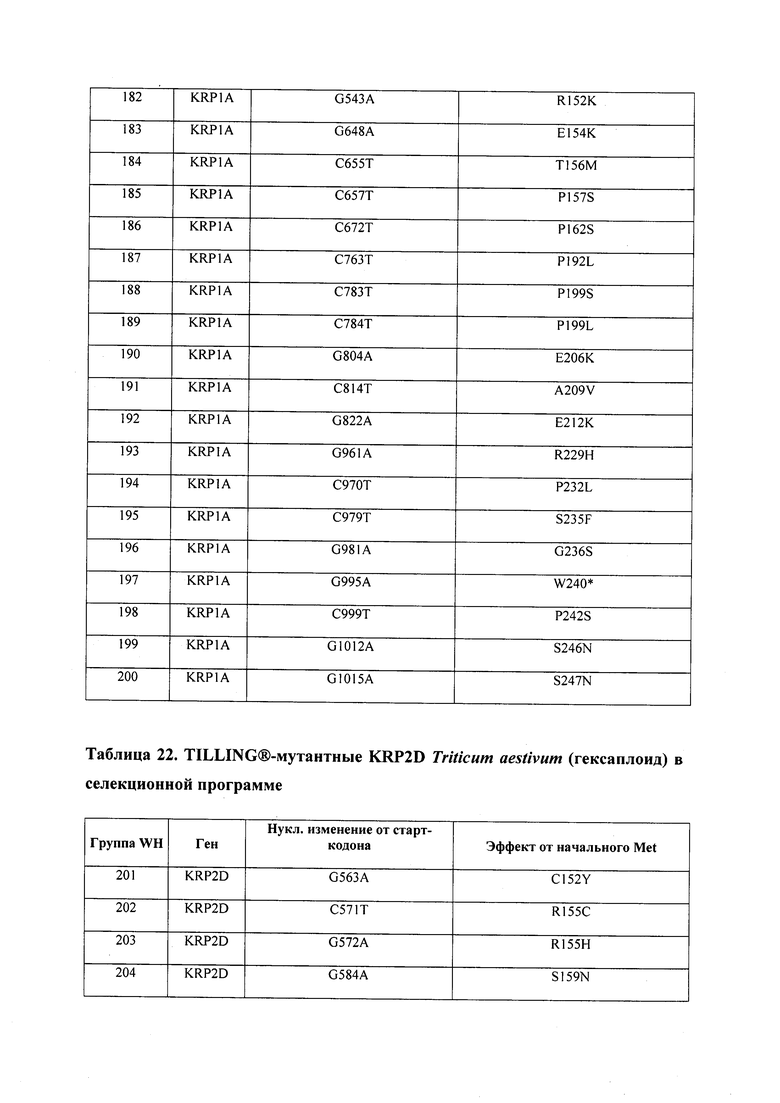
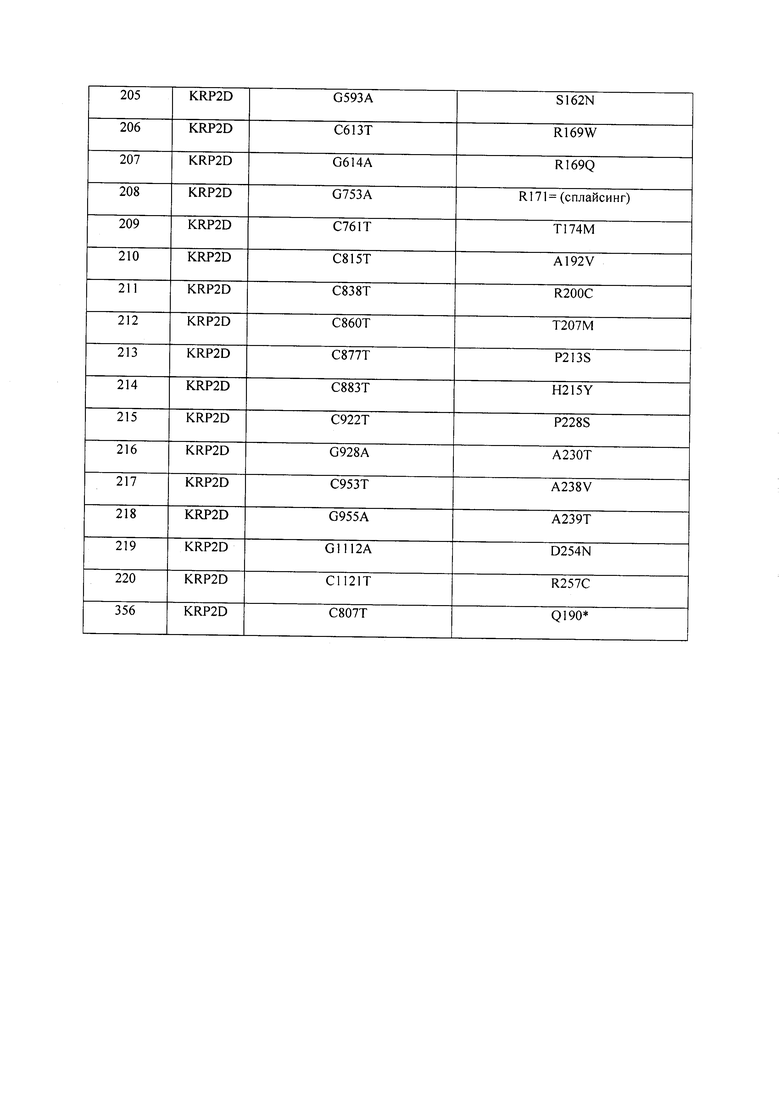

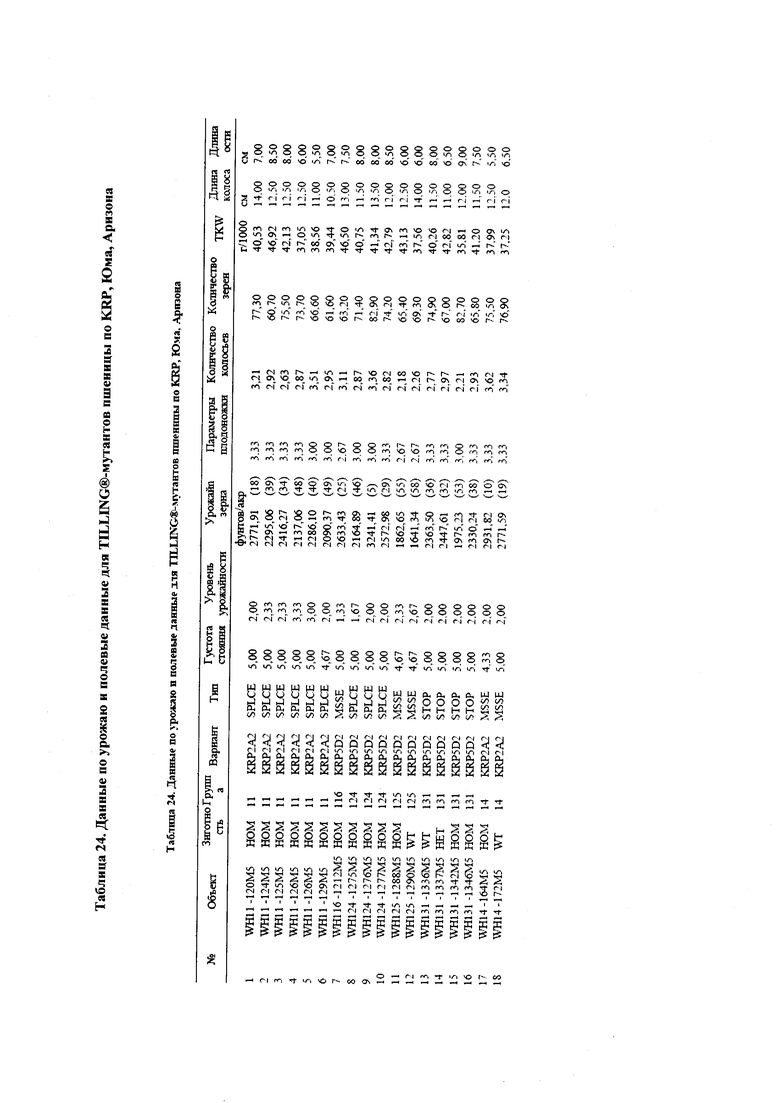


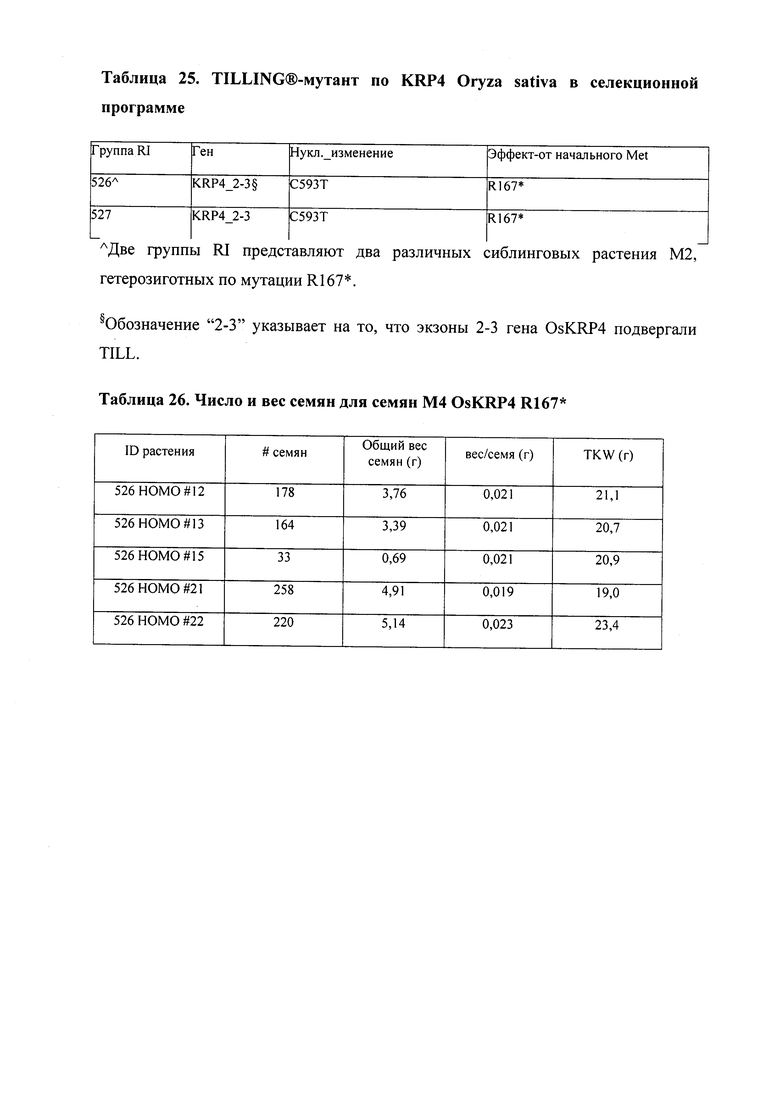
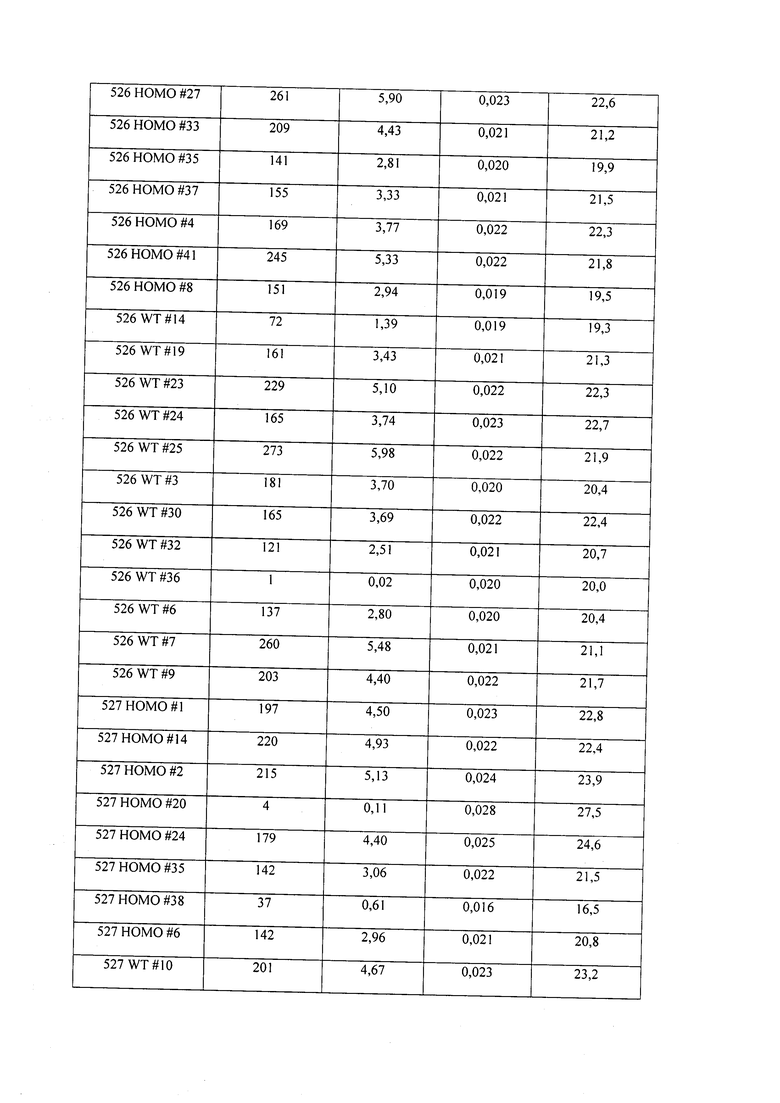
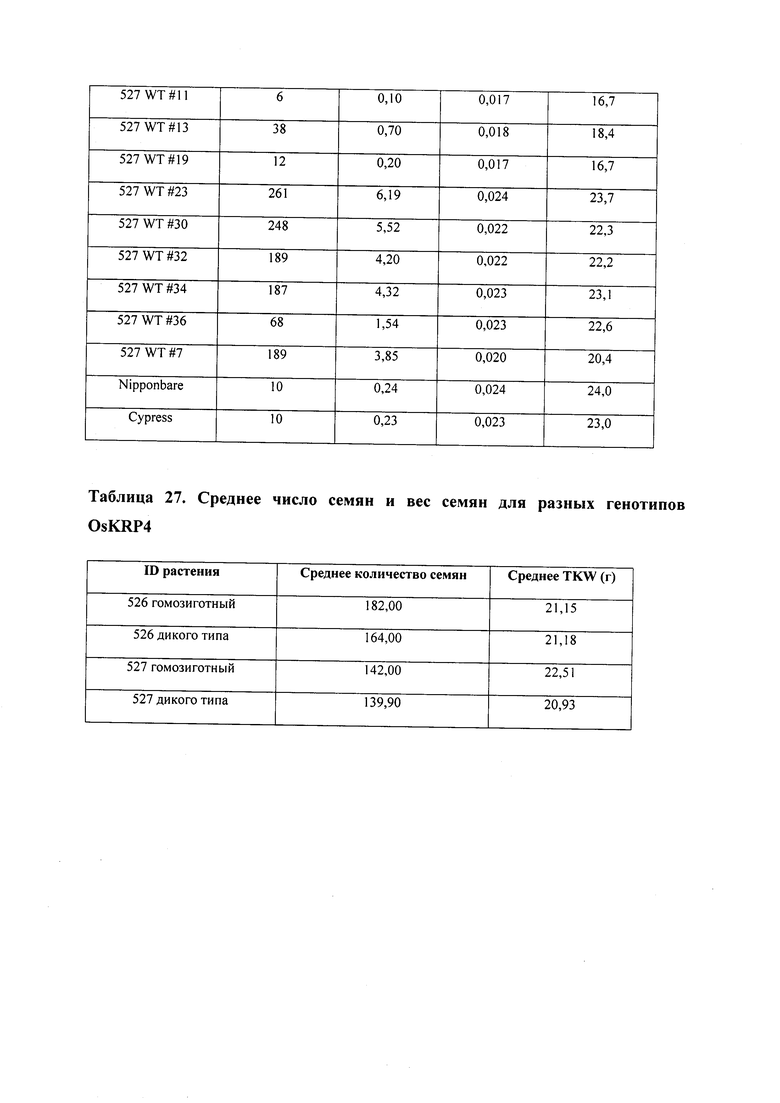
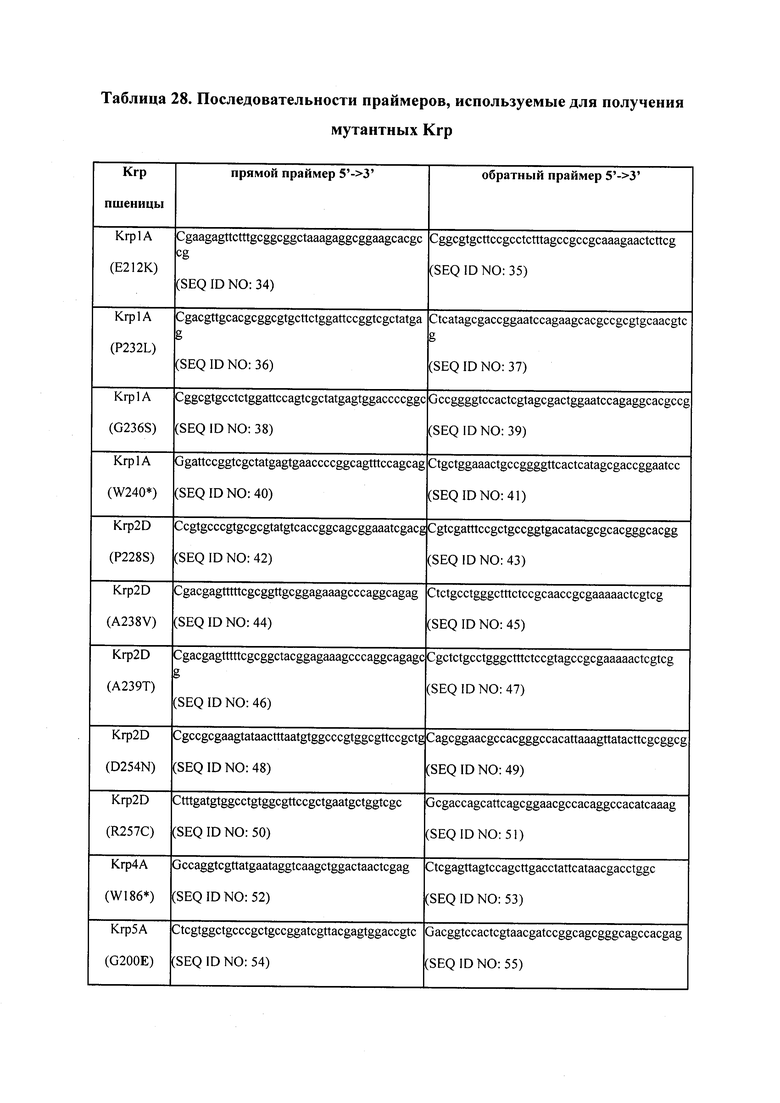
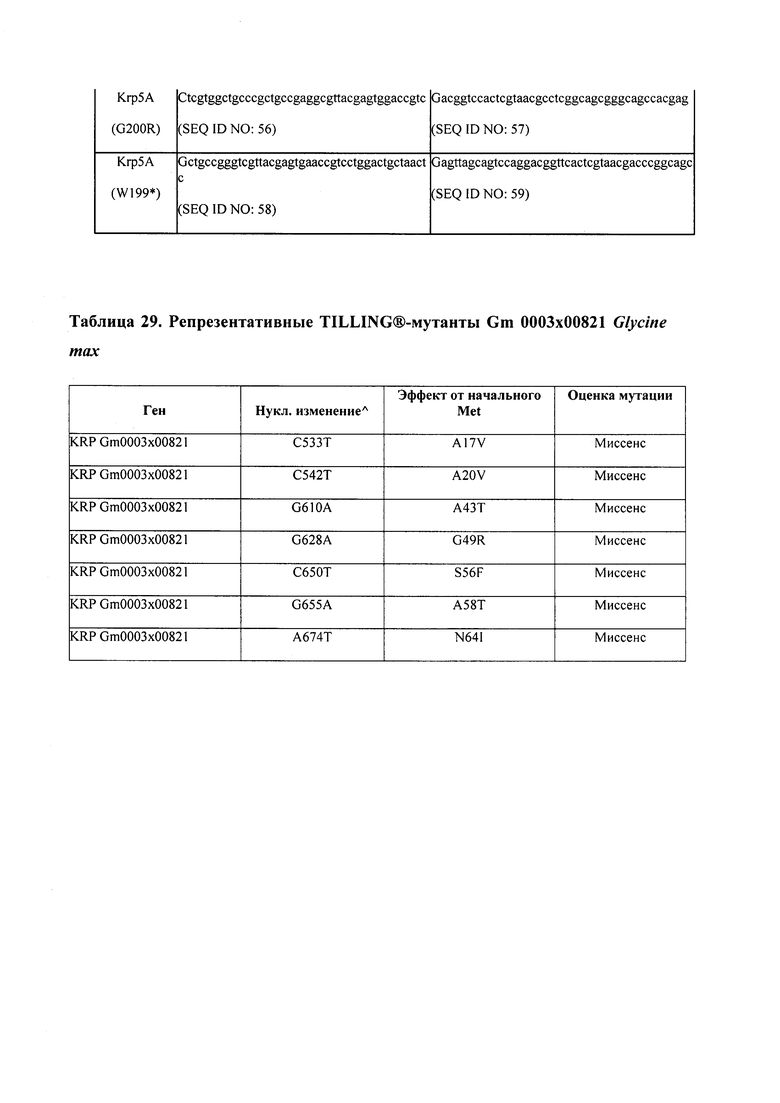
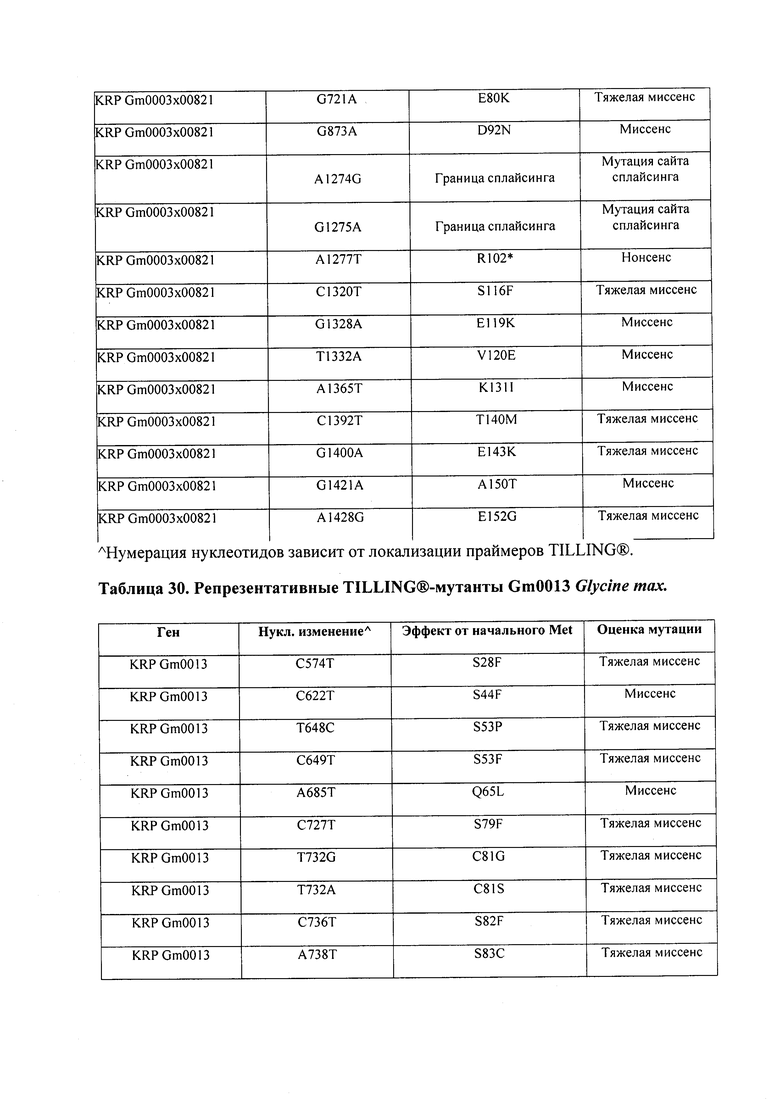
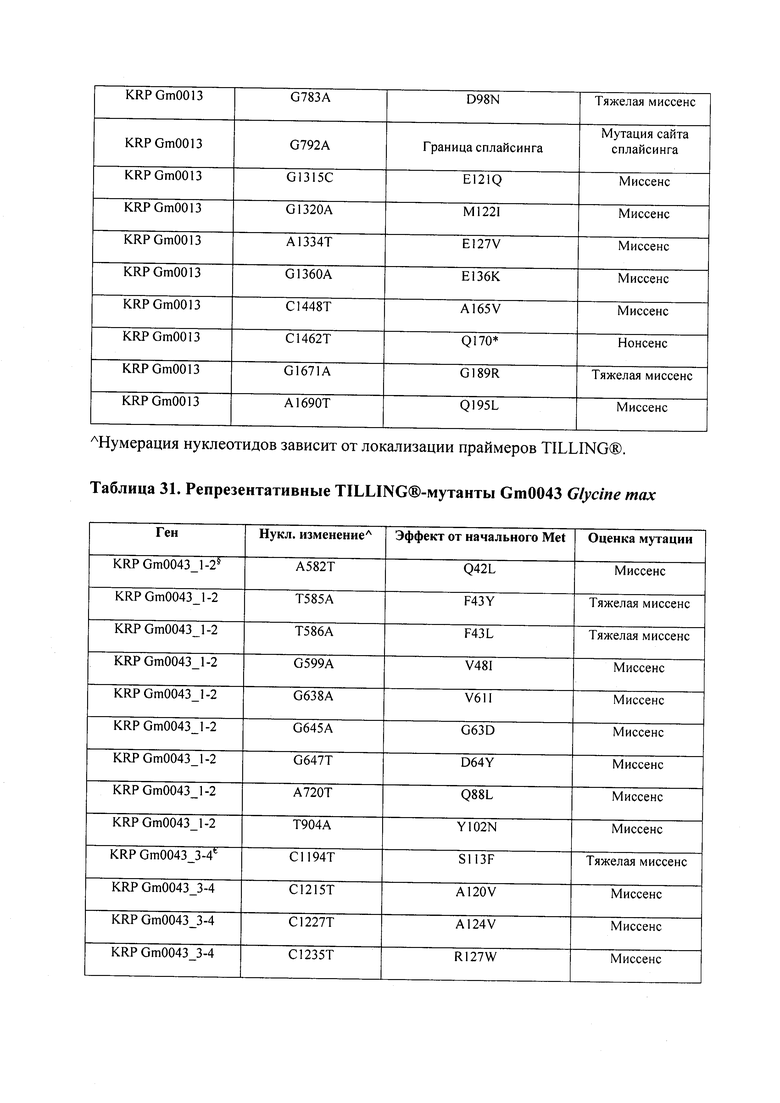
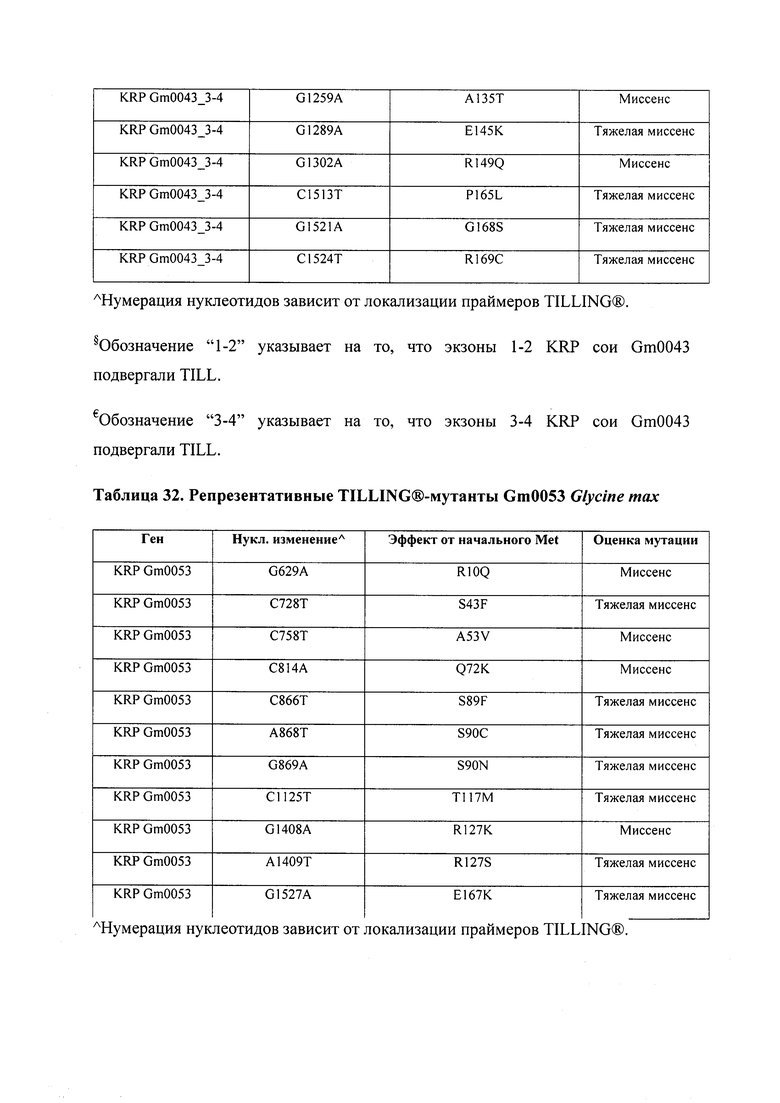
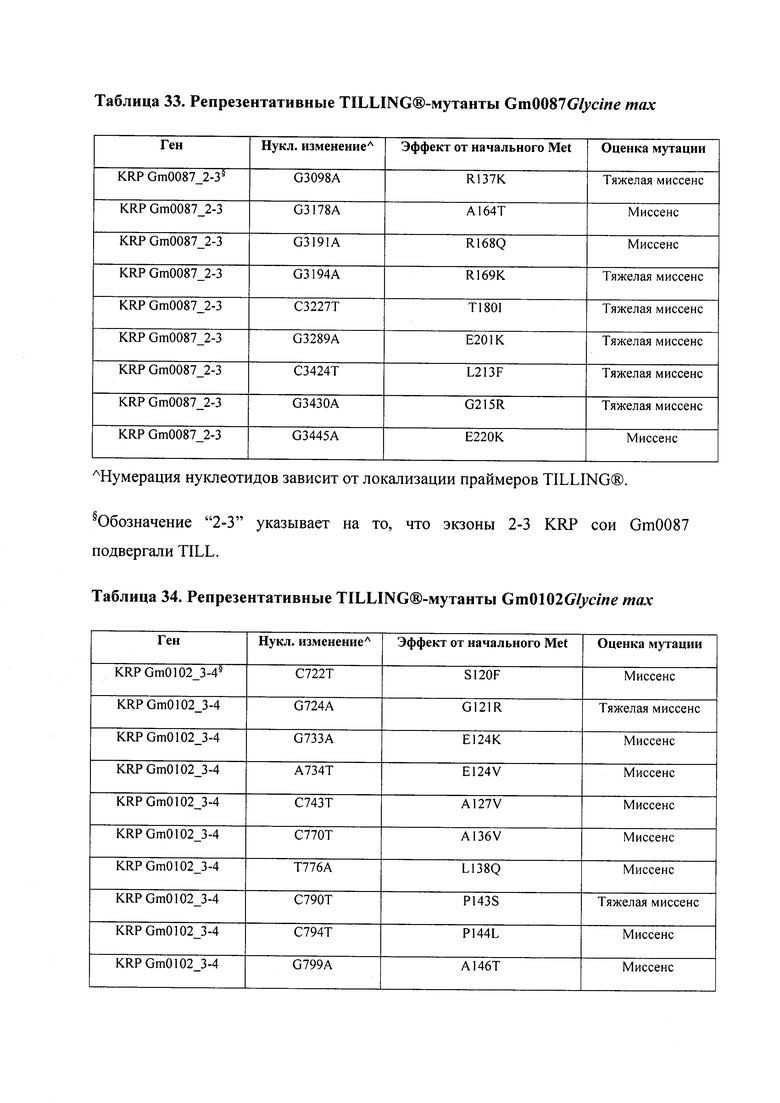
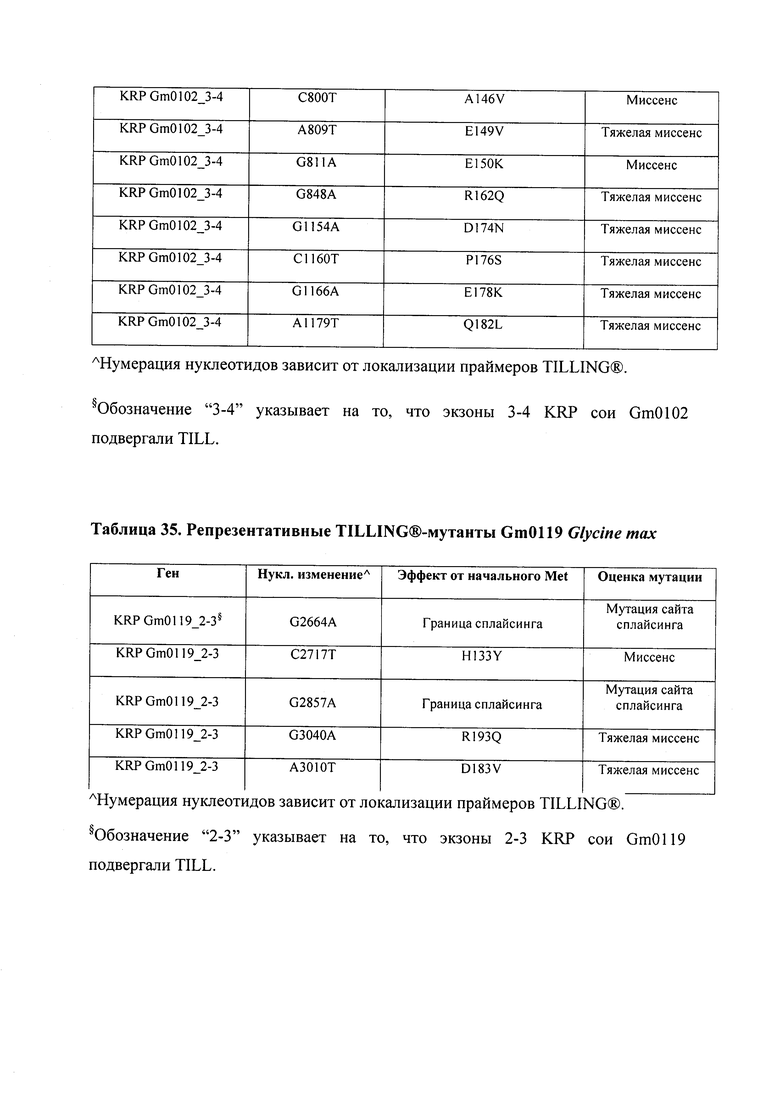
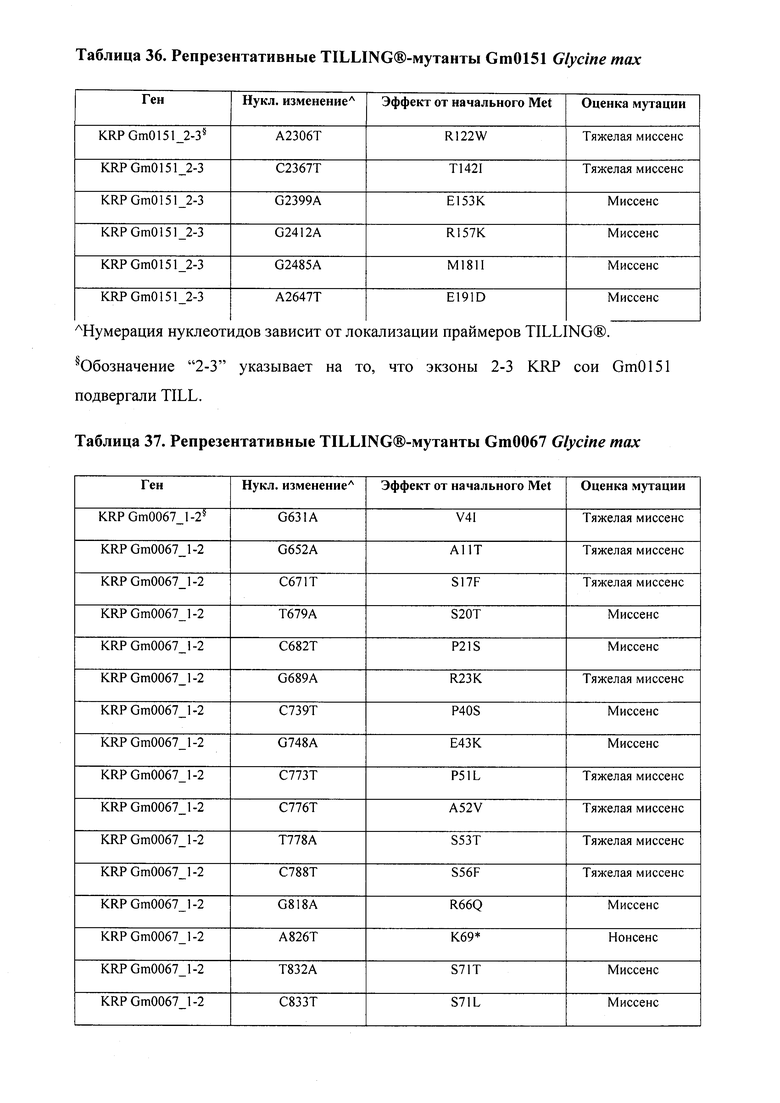


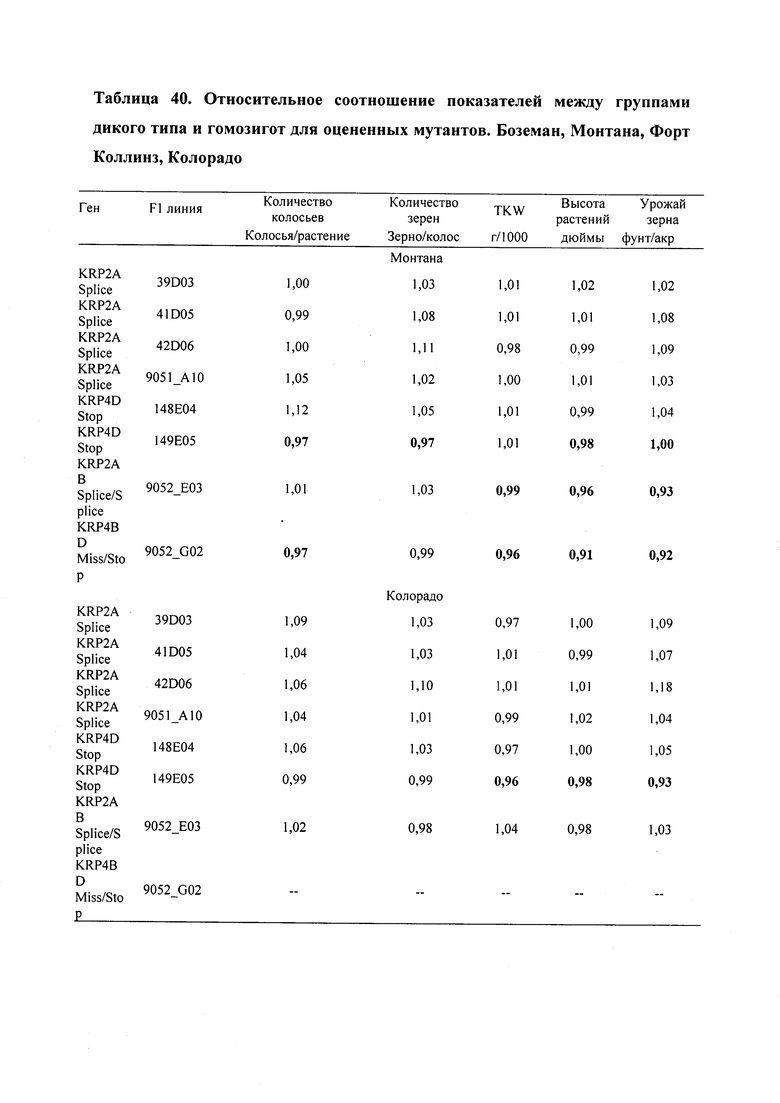
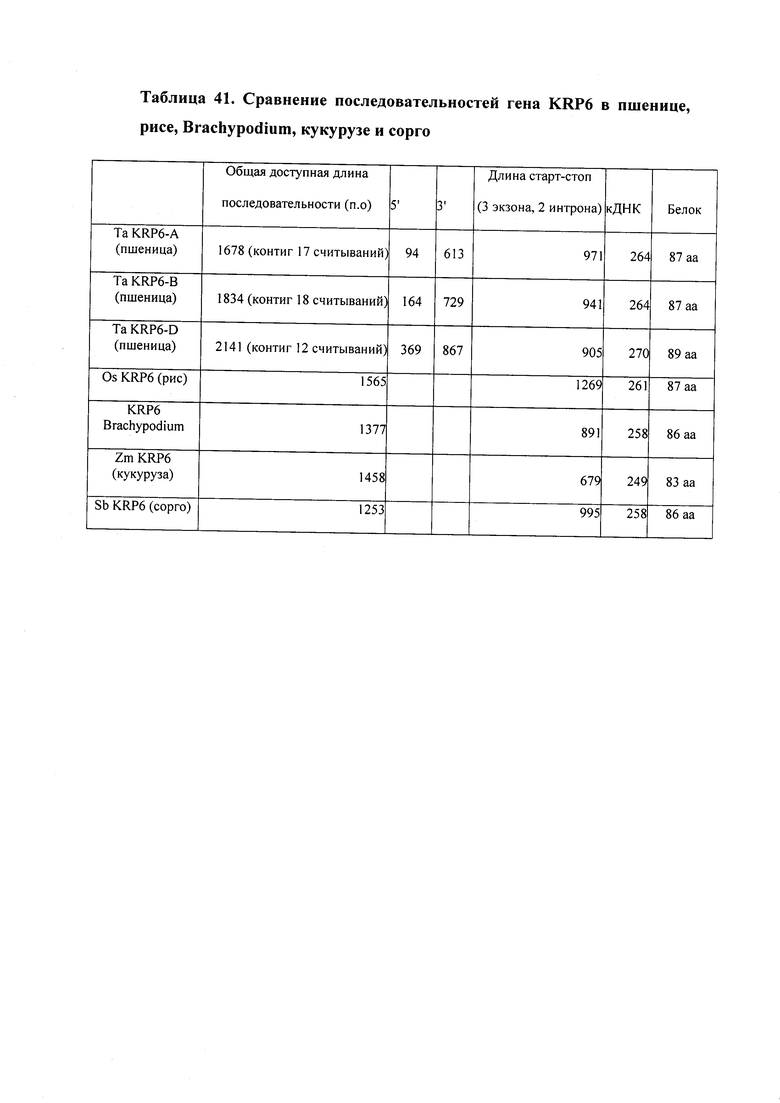
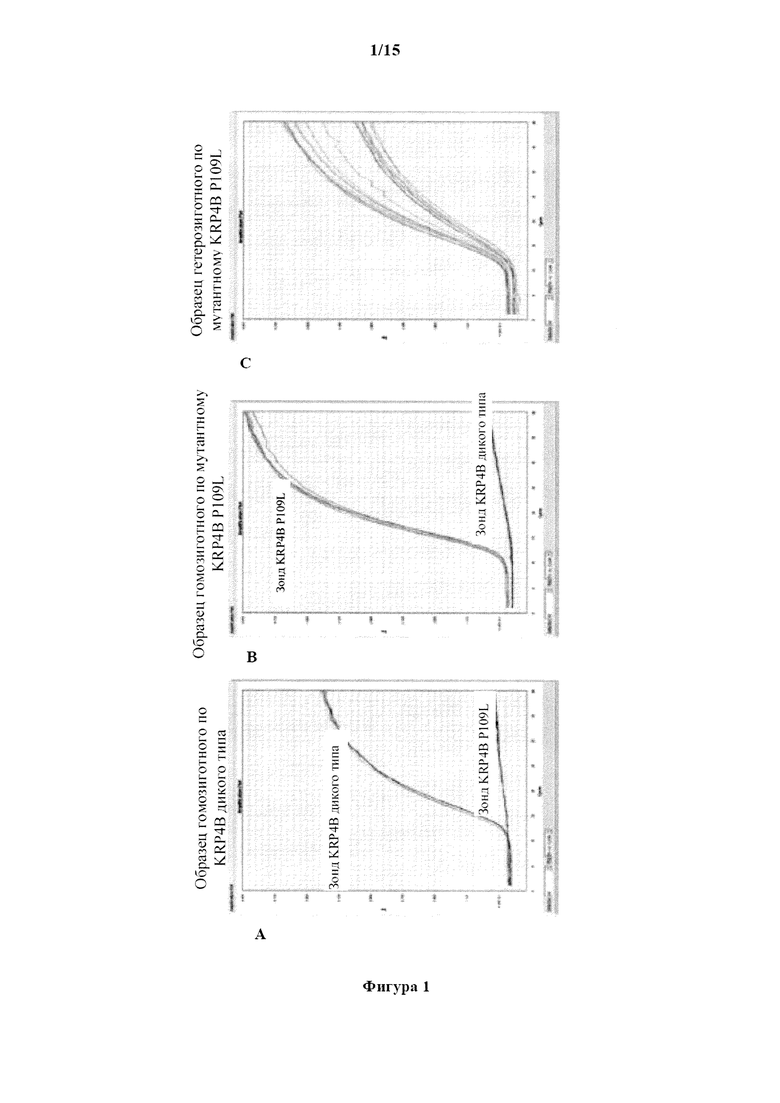
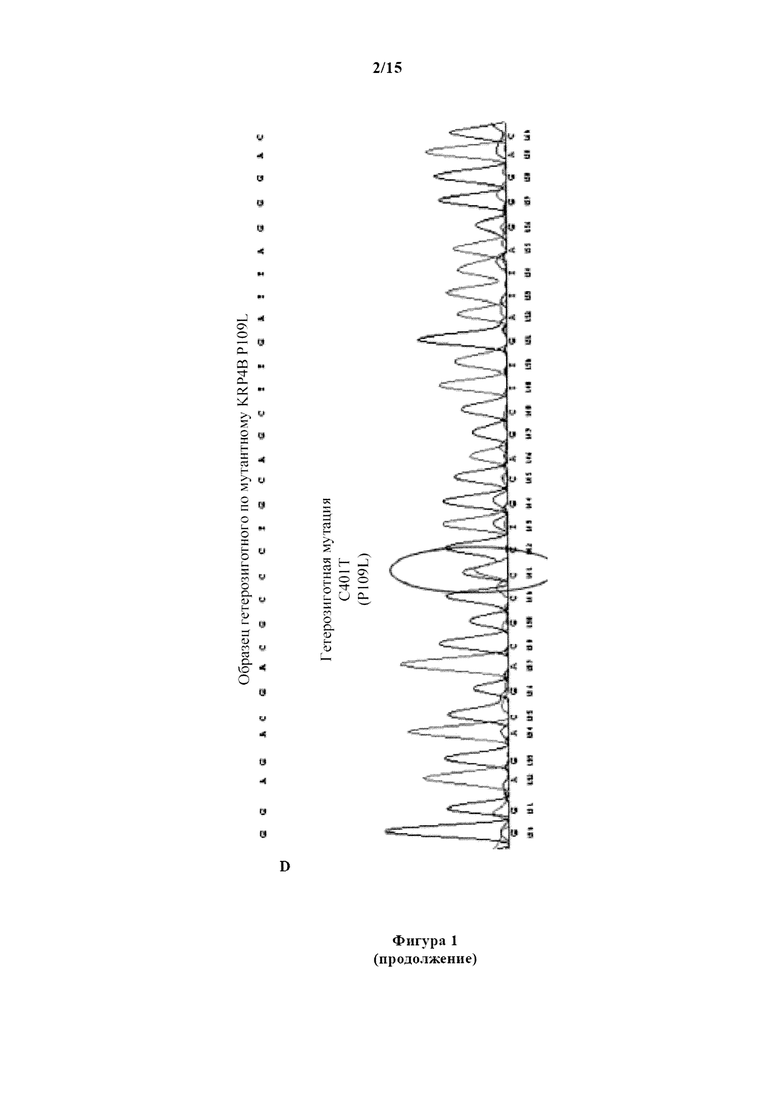
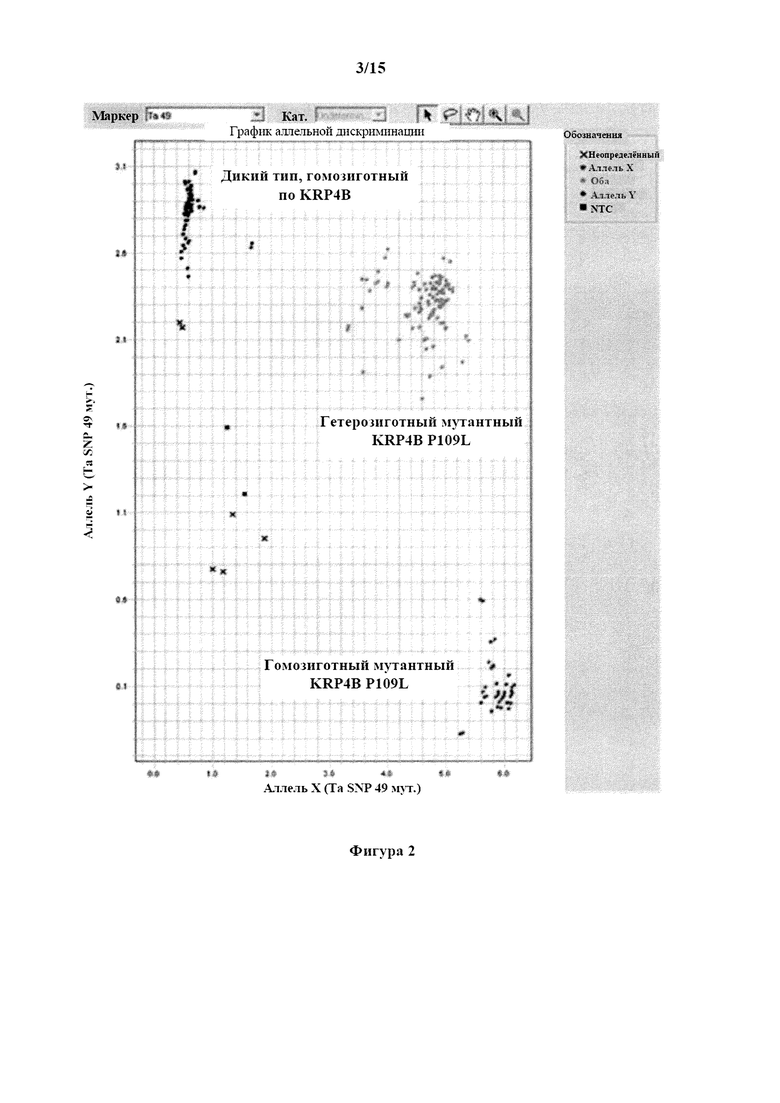

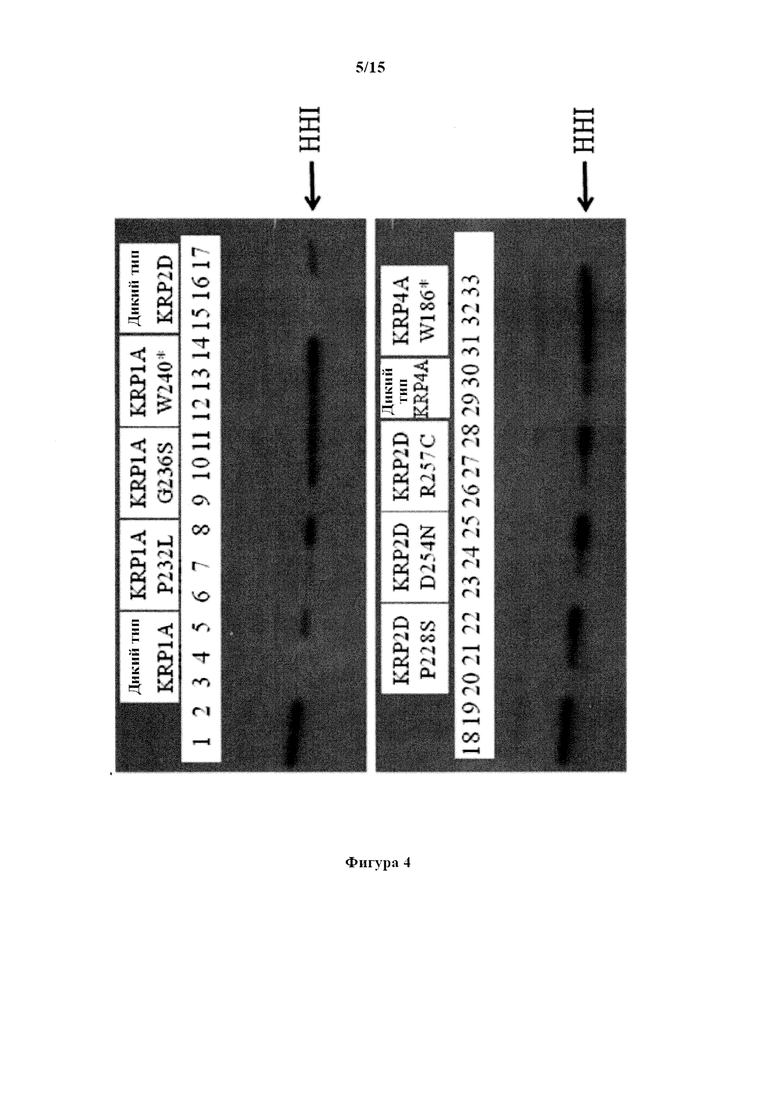

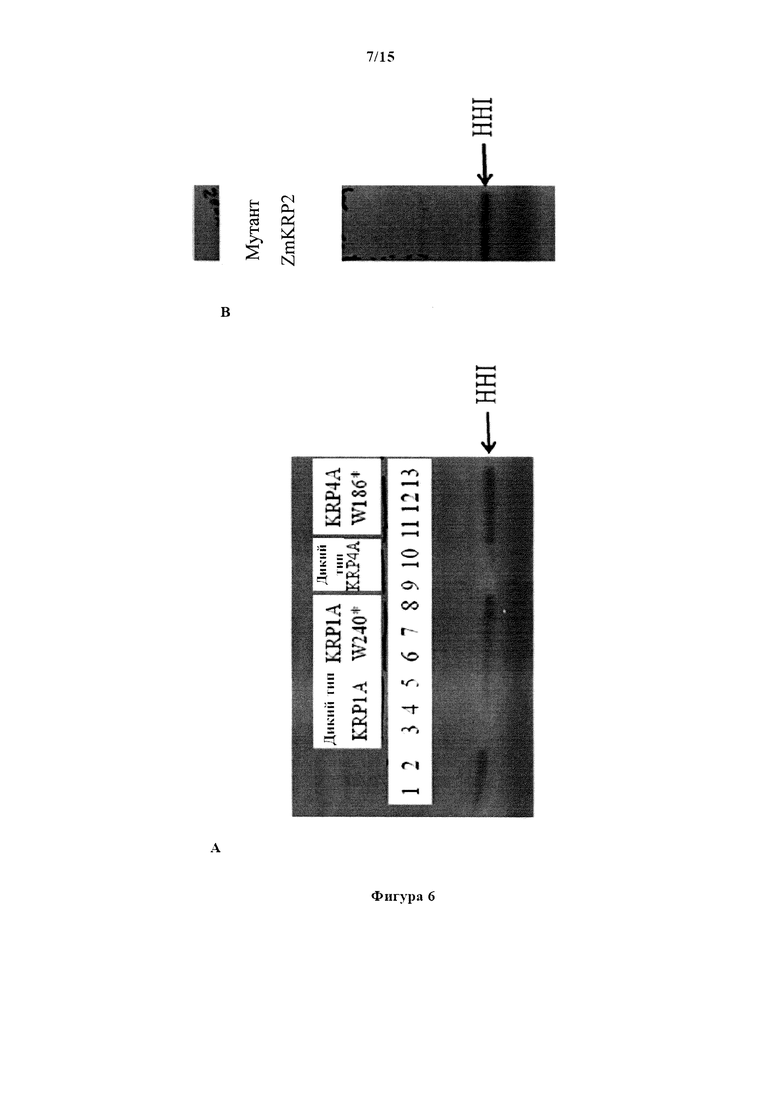
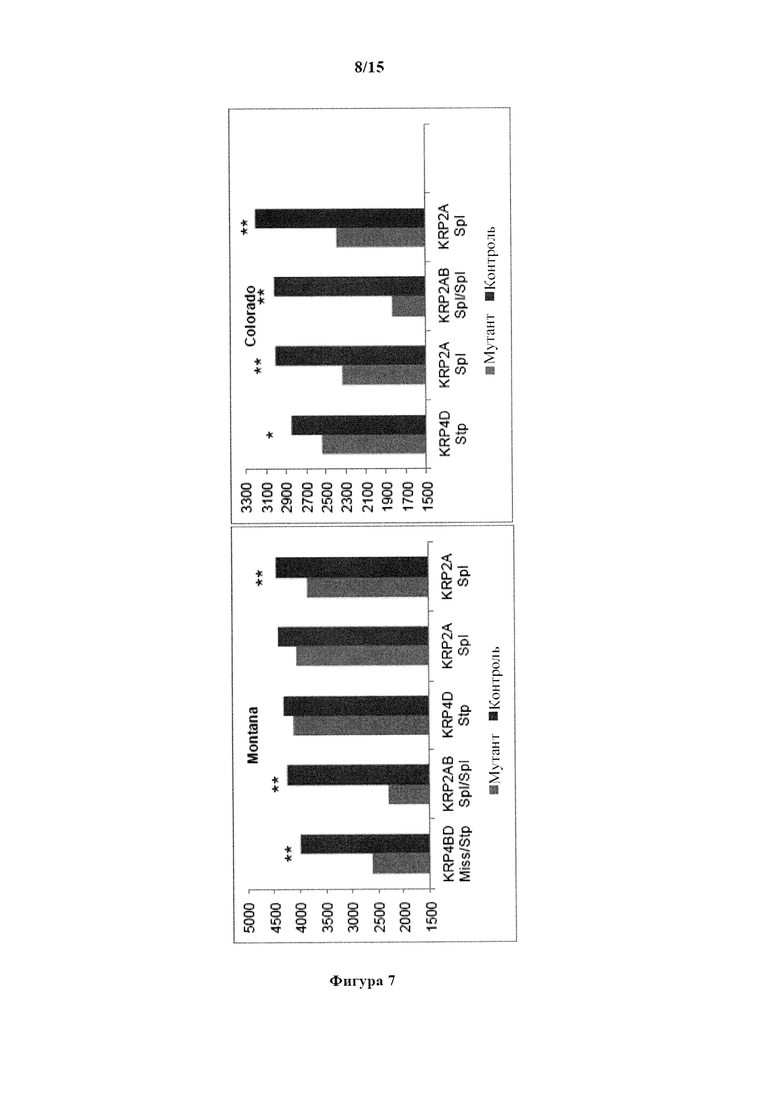
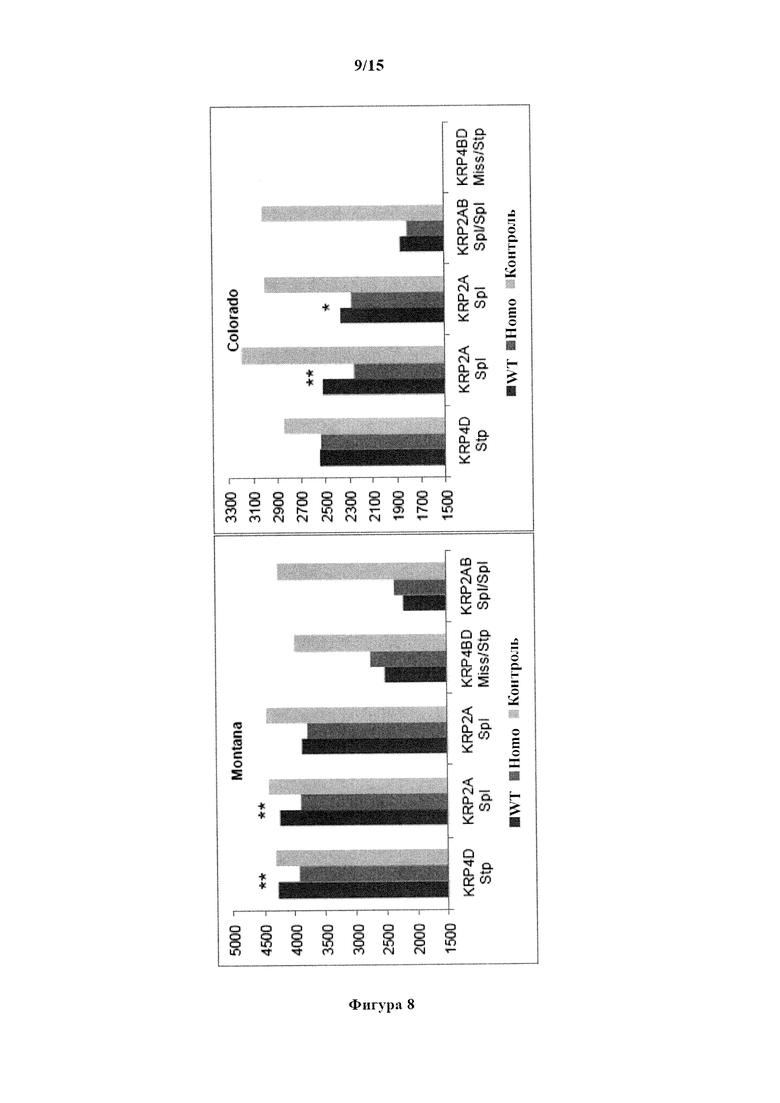
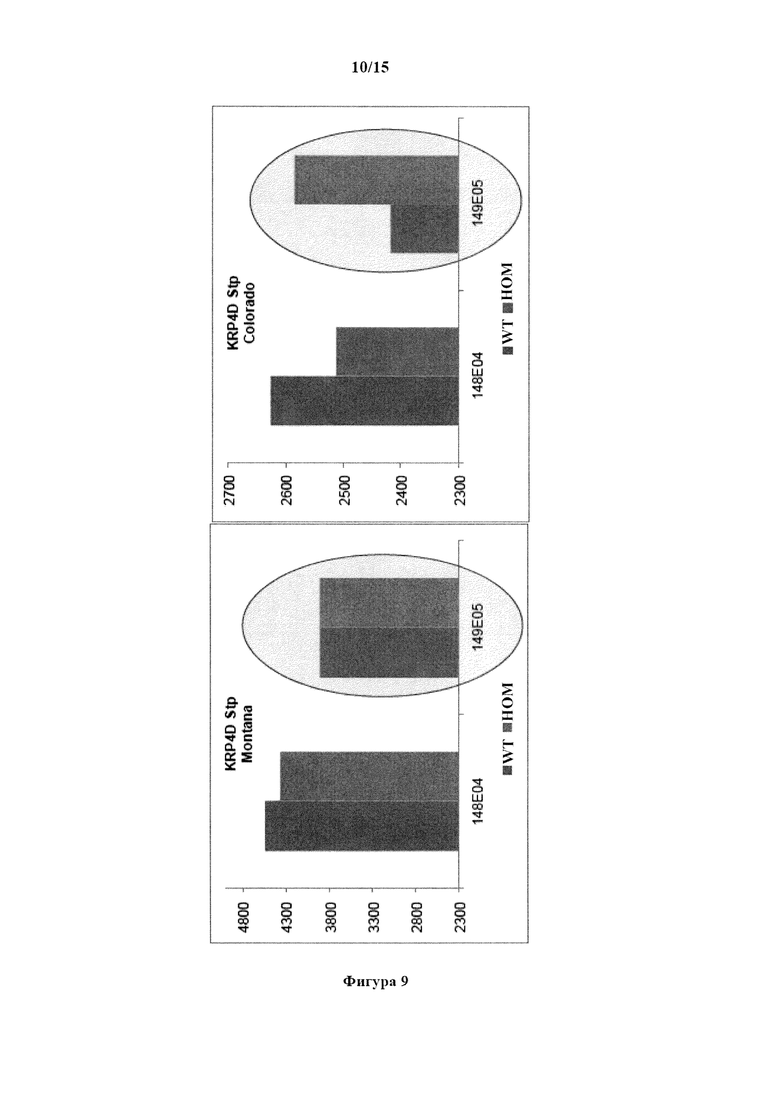





Изобретение относится к области биохимии, в частности к целому растению, имеющему увеличенный вес семени, размер семени и количество семян, и увеличенный урожай, путем введения определенных мутаций в указанные последовательности гена белка KRP, родственного белку-ингибитору киназы (KIP). Также раскрыто целое растение пшеницы, имеющее увеличенный вес семени, размер семени и количество семян, и увеличенный урожай, путем введения определенных мутаций в указанные последовательности гена белка KRP, родственного белку-ингибитору киназы (KIP). Раскрыт способ увеличения веса семени, размера семени и количества семян и урожая растения, путем введения определенных мутаций в указанные последовательности гена белка KRP, родственного белку-ингибитору киназы (KIP). Изобретение позволяет получать растения с увеличенным весом семени, размером семени и количеством семян, а также получать увеличенный урожай. 3 н. и 7 з.п. ф-лы, 15 ил., 42 табл., 10 пр.
1. Целое растение, имеющее увеличенный вес семени, размер семени и количество семян, и увеличенный урожай, содержащее по меньшей мере один мутантный ген неестественного происхождения белка (KRP), родственного белку-ингибитору киназы (KIP), где растение является видом трибы Triticeae, и где одна или несколько мутаций неестественного происхождения гена KRP содержат мутацию, выбранную из группы, состоящей из замены G на А в нуклеотиде 990 от старт-кодона нуклеотидной последовательности, приведенной под SEQ ID NO: 66, кодирующей ген KRP2A; замены С на Т в нуклеотиде 807 от старт-кодона нуклеотидной последовательности, приведенной под SEQ ID NO: 68, кодирующей ген KRP2B; и замены С на Т в нуклеотиде 572 нуклеотидной последовательности, приведенной под SEQ ID NO: 79, кодирующей ген KRP4D, или их комбинацию.
2. Целое растение по п. 1, где растение трибы Triticeae является растением рода Triticum.
3. Целое растение по п. 2, где растение рода Triticum является пшеницей.
4. Целое растение по п. 3, где растение пшеницы является тетраплоидным или гексаплоидным.
5. Целое растение пшеницы, имеющее увеличенный вес семени, размер семени и количество семян, и увеличенный урожай, содержащее по меньшей мере один мутантный ген неестественного происхождения белка (KRP), родственного белку-ингибитору киназы (KIP), и где одна или несколько мутаций неестественного происхождения гена KRP содержат мутацию, выбранную из группы, состоящей из замены G на А в нуклеотиде 990 от старт-кодона нуклеотидной последовательности, приведенной под SEQ ID NO: 66, кодирующей ген KRP2A; замены С на Т в нуклеотиде 807 от старт-кодона нуклеотидной последовательности, приведенной под SEQ ID NO: 68, кодирующей ген KRP2B; и замены С на Т в нуклеотиде 572 нуклеотидной последовательности, приведенной под SEQ ID NO: 79, кодирующей ген KRP4D, или их комбинацию.
6. Целое растение пшеницы по п. 5, где целое растение пшеницы является тетраплоидным или гексаплоидным.
7. Способ увеличения веса семени, размера семени, количества семян и урожая растения трибы Triticeae, включающий введения одной или нескольких мутаций неестественного происхождения в один или несколько генов KRP в растении, где одна или несколько мутаций неестественного происхождения содержит мутацию, выбранную из группы, состоящей из замены G на А в нуклеотиде 990 от старт-кодона нуклеотидной последовательности, приведенной под SEQ ID NO: 66, кодирующей ген KRP2A; замены С на Т в нуклеотиде 807 от старт-кодона нуклеотидной последовательности, приведенной под SEQ ID NO: 68, кодирующей ген KRP2B; и замены С на Т в нуклеотиде 572 нуклеотидной последовательности, приведенной под SEQ ID NO: 79, кодирующей ген KRP4D, или их комбинацию.
8. Способ по п. 7, где растение трибы Triticeae является растением рода Triticum.
9. Способ по п. 8, где растение рода Triticum является растением пшеницы.
10. Способ по п. 7, где растение является тетраплоидным растением пшеницы или гексаплоидным растением пшеницы.
| WO 2007016319 A2, от 08.02.2007 | |||
| US 20070056058 A1, от 08.03.2007 | |||
| WO 2000011175 A1, от 02.03.2000 | |||
| RU 2007107976 A, от 20.09.2008 | |||
| WO 2002057301 A2, от 25.07.2002. |

