Настоящее изобретение относится к продуцированию холестерина у организмов царства Fungi.
Холестерин (см. фиг.1) является наиважнейшим животным стерином. Он представляет собой фундаментальный компонент клеточных мембран, текучесть которых он регулирует, и присутствует во всех животных тканях и особенно в нервной ткани.
Холестерин является продуктом, представляющим большой промышленный интерес. Так, он используется обычно в косметической промышленности. Он также используется в фармацевтической промышленности, например, для введения лекарственных средств ("Drug delivery"), а также в клеточной культуре.
Холестерин используется также в промышленном синтезе витамина D3. Этот витамин используется затем для дополнения питания человека (например, в молочных продуктах) и животных. Холестерин также предпочтительно используют как добавку к пище животных, в частности к пище, предназначенной для искусственно выращиваемых креветок.
В настоящее время большинство выпускаемого на продажу холестерина экстрагируют из животной ткани (ничтожно малое количество получают химическим синтезом). Два больших источника, используемых для экстракции холестерина: спинной мозг крупного рогатого скота и ланолин, который является природным жировым материалом овечьей шерсти.
Использование животной ткани как исходного продукта связано с определенными проблемами. Таким образом, недавние проблемы, связанные с передачей приона, ответственного за овечью трясучку, крупному рогатому скоту (заболевание, названное ESB (губчатая энцефалопатия крупного рогатого скота) у крупного рогатого скота), напомнили о необходимости соблюдения осторожности в процессе использования животной ткани в качестве сырья. Между тем, несмотря на принятые меры, риск передачи патогенного фактора не может быть абсолютно исключенным, и было бы крайне желательно, таким образом, располагать источником холестерина, не происходящим из животной ткани.
Настоящее изобретение имеет целью предоставить источник холестерина, обильный и безопасный с санитарной точки зрения. Авторы изобретения неожиданно показали, что возможно изменить естественную продукцию эргостерола у Fungi таким образом, чтобы продуцировался холестерин.
Общее описание изобретения
Первый аспект изобретения касается организма царства Fungi, продуцирующий автономно холестерин.
Второй аспект изобретения касается организма царства Fungi, такого как определенный выше, отличающегося тем, что этот последний генетически модифицирован.
Третий аспект изобретения касается организма царства Fungi, такого как определенный выше, отличающегося тем, что этот последний продуцирует холестерин на основе простого источника углерода.
Изобретение касается также организма царства Fungi, такого как определенный выше, экспрессирующего ферменты 7-дегидрохолестерин редуктазу и 3β-гидроксистерин Δ24-редуктазу. В частности, изобретение касается организма, такого как определенный выше, в котором инактивирован фермент стерол 24-C-метилтрансфераза и/или инактивирован фермент C-22 стерол десатураза.
Другой аспект изобретения касается организма царства Fungi, такого как определенный выше, отличающегося тем, что экспрессию ферментов 7-дегидрохолестерин редуктазы и 3β-гидроксистерин Δ24-редуктазы осуществляют за счет трансформации организма.
Изобретение касается также организма царства Fungi, такого как определенный выше, отличающегося тем, что инактивацию фермента стерол 24-С-метилтрансферазы осуществляют за счет генетической инактивации, и/или инактивацию фермента C-22 стерол десатуразы осуществляют за счет генетической инактивации.
Другой аспект изобретения касается организма царства Fungi, такого как определенный выше, отличающегося тем, что его выбирают из типа Ascomycete, в частности из подтипа Saccharomycotina, предпочтительно, из класса Saccharomycetes или Schizosaccharomycetes, более предпочтительно, из порядка Saccharomycetales или Schizosaccharomycetales, еще более предпочтительно, из семейства Saccharomycetaceae или Schizosaccharomycetaceae, еще более предпочтительно, из рода Saccharomyces или Schizosaccharomyces.
Другой аспект изобретения касается организма царства Fungi, такого как определенный выше, отличающегося тем, что он представляет собой дрожжи вида Saccharomyces cerevisiae или Schizosaccharomyces pombe.
Изобретение касается также способа получения холестерина неживотного происхождения, включающего культивирование организма, такого как определенный выше.
В частности, этот способ отличается тем, что после стадии культивирования организма следует стадия экстракции холестерина. Предпочтительно, экстракцию холестерина осуществляют с использованием растворителя, не смешивающегося с водой.
В частности, способ, такой как определенный выше, отличается тем, что перед экстракцией холестерина осуществляют стадию омыления. Предпочтительно, способ, такой как определенный выше, отличается тем, что перед омылением или экстракцией холестерина осуществляют стадию механического дробления клеток.
Другой аспект изобретения касается применения организма царства Fungi, такого как определенный выше, для получения холестерина или одного из промежуточных продуктов его метаболизма, или смеси стеринов, меченых 13C или 14С.
Изобретение также относится к способу получения холестерина или одного из промежуточных продуктов его метаболизма, или смеси стеринов, меченых 13C или 14C, включающему следующие стадии:
- культивирование организма царства Fungi, такого как определено выше, на субстрате, меченом 13C или 14C, и
- экстракция указанного холестерина или одного из его метаболизма промежуточных продуктов, или из смеси стеринов.
Изобретение касается также способа получения изотопной смеси холестерина, промежуточных продуктов или метаболитов холестерина, меченых в различных положениях при помощи изотопных маркеров, включающего культивирование организма царства Fungi, такого как определено выше, на меченом субстрате, затем на немеченом субстрате, причем продолжительность культивирования на каждом из этих субстратов выбирают таким образом, чтобы получить определенный изотопный профиль. Изобретение касается также образца молекул холестерина, промежуточных продуктов или метаболитов холестерина, меченых в различных положениях при помощи изотопных маркеров, имеющего определенный изотопный профиль, и который может быть получен этим способом.
Изобретение касается также композиции, содержащей в качестве маркера для отслеживания происхождения изотопную смесь холестерина, промежуточных продуктов или метаболитов холестерина, меченых в различных положениях при помощи изотопных маркеров и имеющих определенный изотопный профиль. В частности, эта композиция предназначена для области питания или терапии человека или животных.
Подробное описание изобретения
Настоящее изобретение касается продукции холестерина у организмов царства Fungi. У Fungi холестерин не встречается в естественном состоянии, поскольку он является животным стерином. Основным стерином клеточных мембран этих организмов является эргостерол.
Настоящее изобретение позволяет осуществить синтез холестерина путем размножения Fungi в присутствии простого источника углерода. Метод, предложенный в настоящем изобретении, позволяет таким образом получить большое количество холестерина, при низкой стоимости, так как в способе осуществляют культивирование организмов царства Fungi и добавление простого источника углерода, легко доступного в продаже.
Под простым источником углерода согласно настоящему изобретению понимают источники углерода, используемые специалистом для нормального роста fungus, а в частности дрожжей. В частности, под такими источниками понимают различные усваиваемые сахара, такие как глюкоза, галактоза или сахароза, или патоки, или субпродукты этих сахаров. Особенно предпочтительным простым источником углерода является этанол и глицерин.
Тот факт, что продуцирование осуществляется автономно, означает, что не является необходимым добавлять субстраты для получения холестерина, но что организм может его продуцировать исходя только из первоначального простого источника углерода. Также ясно, что штамм может продуцировать холестерин путем использования субстрата, располагающегося выше в пути метаболизма, при условии, что штамм организма согласно настоящему изобретению содержит все гены, необходимые для полного осуществления метаболического пути получения холестерина.
Изобретение касается, в частности, генетически модифицированного организма царства Fungi (Fungus), автономно продуцирующего холестерин, исходя из простого источника углерода.
Некоторое количество генетических изменений у fungus могут быть осуществлены для того, чтобы изменить естественный метаболический путь получения эргостерола таким образом, чтобы продуцировался холестерин. Настоящее изобретение касается таким образом генетически модифицированного организма царства Fungi, экспрессирующего ферменты 7-дегидрохолестерин редуктазу и 3β-гидроксистерин Δ24-редуктазу. Измененный таким образом штамм организма царства Fungi продуцирует холестерин. Заявитель смог составить модель благодаря полученным результатам (см. экспериментальную часть настоящей заявки), пути метаболизма, приводящего к эргостеролу и к некоторым из его производных (см., фиг.2). Экспрессия ферментов 7-дегидрохолестерин-редуктазы и 3β-гидроксистерин Δ24-редуктазы у fungus S. cerevisiae может обеспечить продукцию холестерина, частично изменяя путь биосинтеза эргостерола.
Фермент 7-дегидрохолестерин редуктаза несет номер EC: 1.3.1.21 в международной классификации ферментов (Enzyme Classification). Ее также называют дельта 5,7-стерол-дельта 7-редуктазой, 7-DHC- редуктазой или Стерол-дельта 7-редуктазой, а также дельта 7-стерол-редуктазой, дельта 7Red, дельта 7-редуктазой или Δ7-редуктазой в следующей части этого документа. Этот фермент катализирует в естественном состоянии у растений, например НАДФН-зависимое восстановление 5,7-холестадиенола до дельта-5-холестенола или восстановление промежуточных стериновых продуктов, имеющих двойную связь в положении 7-8 (Taton and Rahier, 1991). Ген, кодирующий фермент 7-дегидрохолестерин-редуктазу, был впервые выделен у растения Arabidopsis thaliana, выделение соответствующего гена и экспрессия этого фермента у дрожжей Saccharomyces cerevisiae описаны в патенте EP 727489. Последовательности этого гена и белка доступны под следующим номером доступа в GenBank: U49398 (Lecain et al., 1996).
Некоторое число гомологов этого гена были описаны у других видов. Таким гомологом является, например, гомологичный ген у человека (нуклеотидная последовательность которого доступна под номером GenBank AF034544, белковая последовательность под номером GenBank: AAC05086) (Moebius et al., 1998). Гомологичный ген у крысы Rattus norvegicus (нуклеотидная последовательность которого доступна под номером GenBank: AB016800, белковая последовательность под номером GenBank: BAA34306). Гены-гомологи также были идентифицированы у курицы Gallus gallus с номером в Genbank BM490402, или у жабы Xenopus laevis с номером в Genbank B1315007, или у рыбы-зебры Danio rerio с номером в Genbank BQ132664. Ген, кодирующий активность дельта 7-стерол редуктазы, также обнаруживается у растений, таких как рис Oryza sativa с номером в Genbank CA753545, картофель Solanum tuberosum с номером в Genbank BF342071. Этот ген, кодирующий активность дельта 7-стерол редуктазы, может быть также обнаружен у простейшего Mastigomoeba balamuthi с номером в Genbank BE636562.
Специалист может легко выделить другие гомологичные гены, кодирующие фермент 7-дегидрохолестеринредуктазу у других организмов. Он может обратиться, в частности, к способу клонирования, описанному в примере 1 патента EP 727489, в котором описан метод клонирования, позволяющий выделить кДНК, кодирующую белок, обладающий активностью дельта 5,7-стерол-дельта-7-редуктазы. Специалист может также легко определить активность 7-дегидрохолестеринредуктазы соответствующих белков, в частности используя тест на активность, также описанный в примере 1 патента EP 727489.
Экспрессия фермента 7-дегидрохолестерин редуктазы у организма царства Fungi согласно изобретению может быть получена любым способом, известным специалисту. Это может быть, в частности, трансформация организма с помощью конструкции, включающей кассету экспрессии, состоящую из промотора транскрипции, преимущественно гомологичного, открытой рамки считывания, кодирующей фермент 7-дегидрохолестерин редуктазу, и терминатора транскрипции, адаптированного согласно обычным правилам, известным специалисту. В качестве гомологичного промотора обычно используют промотор, подходящий для обеспечения достаточной и функциональной экспрессии гетерологичного белка. Промотором может быть, например, промотор PGK, промотор ADH, промотор CYCl, промотор GALIO/CYCI, промотор TDH3 или промотор TPI. Терминатором может быть, например, терминатор гена фосфоглицерат киназы (PGK). Указанная кассета экспрессии может быть встроена в форме одной или некоторых копий в ядерный или митохондриальный геном хозяина, или находиться в составе искусственной структуры типа искусственной хромосомы дрожжей (YAC) или в составе эписомального генетического элемента, такого как плазмида. Для осуществления этого типа экспрессии могут быть использованы, например, дрожжи типа Yarrowia lipolitica, Kluyveromyces lactis или Pichia pastoris.
Предпочтительно, экспрессируемым ферментом 7-дегидрохолестерин редуктазой является фермент растения Arabidopsis thaliana (пример способа экспрессии этого фермента у дрожжей Saccharomyces cerevisiae описан в патенте EP 727489). Тем не менее, это может быть любой, гомологичный или нет, природный или искусственный фермент, обладающий той же ферментативной активностью.
Фермент 3β-гидроксистерин Δ24-редуктаза, также называемый DHCR24 или 24-дегидрохолестеринредуктазой, катализирует в естественном состоянии восстановление десмостерола (холеста 5, 24 диенола) или производных ланостерола, имеющих двойную связь в положении 24-25 на боковой цепи (например, 14 десметилланостерола, зимостерола или холеста 7,24 диенола), необходимое для биосинтеза холестерина, в частности у человека (Waterham HR. et al, 2001). Этот фермент в следующих частях этого документа называют дельта 24-(25) стерол редуктазой, дельта 24 стерол редуктазой или Δ24-редуктазой.
Ген, кодирующий фермент 3β-гидроксистерин Δ24-редуктазу, был впервые выделен у человека, выделение соответствующего гена и экспрессия этого фермента у дрожжей Saccharomyces cerevisiae описаны в публикации Waterham HR. et al., 2001. Последовательности этого гена и белка доступны под следующими номерами доступа GenBank: NM_014762.et NP_055577.
Некоторое число гомологов этого гена были описаны у других видов. Например, таким гомологом является гомологичный ген мыши (Mus musculus) (нуклеотидная последовательность которого доступна под номером GenBank: NM_053272, белковая последовательность - под номером GenBank: NP_444502). Гомологи были описаны у червя Caenorhabditis elegans, в частности комплементарная ДНК с номером в Genbank AF026214. Гомологичные последовательности были описаны также у растений, таких как хлопок Gossypium hirsutum с номером в Gehbank AAM 47602.1, рис Orysa sativa с номером в Genbank AAP53615, горох Pisuni satinum с номером в Genbank AAK15493.
Специалист может легко выделить другие гомологичные гены, кодирующие фермент 3β-гидроксистерин Δ24-редуктазу у других организмов. Он может, в частности, обратиться к способу клонирования, описанному в публикации Waterham HR. et al., 2001. Специалист может также легко определить активность 3β-гидроксистерин Δ24-редуктазы соответствующих белков, в частности, используя тест на активность, также описанный в этой публикации (Waterham et al., 2001).
Экспрессия фермента 3β-гидроксистерин Δ24-редуктазы у организма царства Fungi согласно изобретению может быть получена любым способом, известным специалисту. Это могут быть, в частности, средства, описанные выше в отношении экспрессии фермента 7-дегидрохолестерин редуктазы.
Предпочтительно, экспрессируемый фермент 3β-гидроксистерин Δ24-редуктаза представляет собой человеческий фермент. Пример выделения соответствующего гена и экспрессии этого фермента у дрожжей Saccharomyces cerevisiae описан в публикации Waterham HR. еt al, 2001. Тем не менее, это может быть любой гомологичный или природный, или искусственный фермент, обладающий той же ферментативной активностью.
Предпочтительно, организмы царства Fungi согласно настоящему изобретению экспрессируют ферменты 7-дегидрохолестерин редуктазу и 3β-гидроксистерин Δ24-редуктазу, и, кроме того, осуществляют инактивацию фермента стерол 24-C-метилтрансферазы.
Фермент стерол 24-C-метилтрансфераза несет номер EC-2.1.1.41 в международной классификации ферментов (Enzyme Classification). Его также называют ERG6p, дельта(24)-метилтрансферазой, дельта(24)-стерол метилтрансферазой, зимостерол-24-метилтрансферазой, S-аденозил-4-метионин:стерол дельта(24)-метилтрансферазой, SMT1, 24-стерол C-метилтрансферазой, S-аденозил-L-метионин:дельта(24(23))-стерол метилтрансферазой или фитостерол метилтрансферазой.
Этот фермент катализирует в естественном состоянии метилирование в положении C-24 зимостерола, что приводит к образованию фекостерола.
Ген, кодирующий фермент стерол 24-C-метилтрансферазу, был назван Erg6 у дрожжей Saccharomyces cerevisiae. Последовательность этого гена доступна под следующим номером доступа GenBank: NC_001145. Последовательность соответствующего белка доступна под следующим номером доступа GenBank: NP_013706 (Bowman et al., 1997), (Goffeau et al., 1996).
Некоторое число гомологов этого гена были описаны у других Fungi. Таким гомологом является, например, гомологичный ген у Schizosaccharomyces pombe (нуклеотидная последовательность которого доступна под номером GenBank Z99759, белковая последовательности - под номером GenBank: CABl 6897) (Wood et al., 2002). Гомологичный ген у Neurospora crassa (нуклеотидная последовательность которого доступна под номером GenBank: NCB24P7, белковая последовательность под номером GenBank: CAB97289). Гомологичный ген у Candida albicans (нуклеотидная последовательность которого доступна под номером GenBank: AF031941, белковая последовательность под номером GenBank: AAC26626) (Jensen-Pergakes et al., 1998). Гены, кодирующиеся фермент, гомологичный ERG6, были описаны также у Candida lusitaniae с номером Genbank AA021936.1, а также у Pneumocystis carinii (Kanesbiro et al., 2002) или у Kluveromyces lactis (Ozier-Kalogeropoulos et al., 1998).
Специалист может легко выделить другие гены, гомологичные гену ERG6, в организмах царства Fungi. Специалист может также легко определить активность стерол 24-C-метилтрансферазы соответствующих белков, в частности, используя в качестве теста на активность функциональную комплементацию штамма дрожжей, у которого отсутствуют эти гены. О комплементации свидетельствует образование стеринов, включенных в положении 24, в частности стеринов типа эргоста-, несущих метиленовую группу в положении 24-28. Присутствие биологической активности стерол 24-C-метилтрансферазы типа ERG6 также определяют in vitro благодаря методам, разработанным (McCammon et al., 1984) или Taylor и Parks (Тэйлор и Пакс, 1978). С другой стороны, полученные стерины и субстрат фермента ERG6 разделяют Хроматографией в газовой фазе согласно технологии, разработанной Nes в (Methods in Enzymology Steroids and Isoprenoids Volume 111 part B, 1985 «A comparison of Methods for the Identification of Sterols» pp3-37).
Штамм организма царства Fungi согласно настоящему изобретению, экспрессирующий ферменты 7-дегидрохолестерин редуктазу и 3β-гидроксистерин Δ24-редуктазу, и в котором, кроме того, инактивирован фермент стерол 24-C-метилтрансфераза, продуцирует холестерин. Заявитель неожиданно смог определить, что инактивация фермента стерол 24-C-метилтрансферазы блокирует путь биосинтеза эргостерола на более ранних стадиях и позволяет штамму fungus продуцировать повышенные количества холестерина (см. экспериментальную часть настоящей заявки).
Экспрессию ферментов 7-дегидрохолестерин редуктазы и 3β-гидроксистерин Δ24-редуктазы осуществляют, как описано выше.
Инактивация фермента стерол 24-C-метилтрансферазы может быть осуществлена любым способом, известным специалисту. Это может быть, в частности, введение путем мутагенеза нонсенс-мутации, вставки или делеции, приводящие к изменению рамки считывания в гене, кодирующем указанный белок.
Это также может быть экспрессия антисмысловой РНК, комплементарной по отношению к информационной РНК, кодирующий указанный белок, или системы генной экстинкции, известной специалисту под именем siРНК (small interfering RNA), и ассоциированных ферментных систем, если они отсутствуют в естественных условиях в хозяине. Мутагенез может быть осуществлен в кодирующей последовательности или в некодирующей последовательности таким образом, чтобы сделать кодируемый белок неактивным или воспрепятствовать его экспрессии или трансляции. Мутагенез может быть осуществлен in vitro или in situ путем супрессии, замены, делеции и/или добавления одного или нескольких оснований в соответствующий ген, или инактивацией гена.
В частности, может быть осуществлено введение экзогенной ДНК в кодирующую или промоторную последовательность (например, кассеты экспрессии с гомологичным промотором и/или терминатором и гетерологичной кодирующей частью). Кассета экспрессии предпочтительно обеспечивает экспрессию селективного маркера. Также возможно модифицировать промотор гена для понижения уровня экспрессии. Для fungi инактивация также может происходить за счет прерывания кодирующей последовательности последовательностью, кодирующей гетерологичный или гомологичный маркерный ген. Основные методы прерывания гена fungi описаны в статье Johnston et al (2002) (Methods in Enzymology Volume 350 Edited by Christine Guthrie and Gerry Fink ; «Gene Disruption»; M. Johnston,- L. Riles, J. Hegemann pp 290-315).
Предпочтительно, организмы царства Fungi согласно настоящему изобретению экспрессируют ферменты 7-дегидрохолестерин редуктазу и 3β-гидроксистерин Δ24-редуктазу и в них инактивирован фермент C-22 стерола десатураза.
Фермент C-22 стерол десатураза также называют ERG5p, Cyp61, цитохром p-45061, стерол дельта22-десатураза. Этот фермент катализирует в естественном состоянии конверсию эргоста-5,7,24(28)-триенола в эргоста-5,7,22,24(28)-тетраенол, добавляя двойную связь в положении C22 (см. фиг.2).
Ген, кодирующий фермент C-22 стерол десатуразу, был назван Erg5 у дрожжей Saccharomyces cerevisiae. Последовательность этого гена доступна под следующим номером доступа GenBank: U34636. Последовательность соответствующего белка доступна под следующими номерами доступа GenBank: AAB06217 (Skaggs et al., 1996) или P54781 (Bowman et al., 1997).
Некоторое число гомологов этого гена были описаны у других Fungi. Таким гомологом является, например, гомологичный ген у Schizosaccharomyces pombe (нуклеотидная последовательность которого доступна под номером GenBank Z98974, белковая последовательность - под номером GenBank: CAB11640) (Wood et al., 2002). Гомологичный ген у Symbiotаphrina buchneri (нуклеотидная последовательность которого доступна под номером GenBank: AB086896, белковая последовательность - под номером GenBank: BACOl 142) (Noda and Koizumi, 2003). Гомологичный ген у Symbiotaphrina kochii (нуклеотидная последовательность которго доступна под номером GenBank: AB086890, белковая последовательность - под номером GenBank: BAC01139) (Noda and Koizumi, 2003). Гомологичный ген у Candida albicans (нуклеотидная последовательность которого доступна под номером GenBank: AL033396, белковая последовательность - под номером GenBank: CAA21953) (Tait et al., 1997). Ген ERG5 был также описан у Candida lusitaniae с номером Genbank AAO48601.
Специалист может легко выделить другие гены, гомологичные гену Erg5, в организмах царства Fungi. Специалист может также легко определить активность C-22 стерол десатуразы соответствующих белков, в частности, используя тест на активность, описанный Skaggs, B.A., et al, 1996. Эта активность может также быть выявлена функциональной комплементацией дрожжей S. Cerevisiae, у которых предварительно был осуществлен разрыв на уровне гена erg5. Об этой комплементации будет свидетельствовать присутствие в комплементированном штамме эргоста 5,7,22 триенола. Активность C-22 стерол десатуразы может быть измерена in vitro с использованием метода, описанного Kelly and Baldwin et al JBC (1997) после лизиса дрожжей (Kelly et al., 1997).
Штамм организма царства Fungi согласно настоящему изобретению, экспрессирующий ферменты 7-дегидрохолестерин редуктазу и 3β-гидроксистерин Δ24-редуктазу, и у которого, кроме того, инактивирован фермент C-22 стерол десатураза, продуцирует холестерин. Заявитель смог определить, что инактивация фермента C-22 стерол десатуразы предпочтительно блокирует превращение холестерина в холеста 5,22 диенол и обеспечивает стабилизацию продукции холестерина (см. экспериментальную часть настоящей заявки). Эта блокировка происходит также на уровне превращения холеста 5,7 диенола, предшественника холестерина, в холеста 5,7,22 триенол, предшественник холеста 5,22 диенола. Неожиданно, фермент C-22 стерол десатураза использует в качестве субстрата холестерин, который он превращает в холеста 5,22 диенол. Эта паразитная реакция может быть устранена путем инактивации фермента-22 стерол-десатуразы, как удалось определить заявителю.
Экспрессию ферментов 7-дегидрохолестерин редуктазы и 3β-гидроксистерин Δ24-редуктазы осуществляют, как описано выше. Инактивация фермента C-22 стерол десатуразы может быть осуществлена любым известным специалисту способом. Это могут быть, в частности, методы, описанные выше относительно инактивации фермента стерол 24-C-метилтрансферазы.
Предпочтительно, организмы царства Fungi согласно настоящему изобретению экспрессируют ферменты 7-дегидрохолестерин редуктазу и 3β-гидроксистерин Δ24-редуктазу, и, кроме того, у них инактивирован фермент C-22 стерол десатураза и фермент стерол 24-C-метилтрансфераза. Эти штаммы действительно обладают кумулированными преимуществами, связанными с инактивацией обоих ферментов, и являются штаммами-продуцентами холестерина.
Экспрессию ферментов 7-дегидрохолестерин редуктазы и 3β-гидроксистерин Δ24-редуктазы и инактивацию ферментов C-22 стерол десатуразы и стерол 24-C-метилтрансферазы осуществляют, как описано выше.
В способе осуществления холестерин присутствует в штамме организма согласно настоящему изобретению в количестве более 20%, преимущественно 35%, наиболее предпочтительно 50% или более от общего количества стеринов, продуцируемых штаммом, согласно изобретению (в частности, промежуточных продуктов синтеза).
Предпочтительно, организмы царства Fungi согласно настоящему изобретению выбирают из типа Ascomycetes, более предпочтительно, их выбирают из подтипа Saccharomycotina, еще более предпочтительно, их выбирают из класса Saccharomycetes или Schizosaccharоmycetes, еще более предпочтительно, их выбирают из порядка Saccharomycetales или Schizosaccharomycetales, еще более предпочтительно, их выбирают из семейства Saccharomycetaceae или Schizasaccharomycetaceae, еще более предпочтительно, их выбирают из рода Saccharomyces или Schizosaccharomyces, наиболее предпочтительно, организмы царства Fungi согласно изобретению принадлежат виду Saccharomyces cerevisiae или Schizosaccharomyces pombe.
Настоящее изобретение касается также способа получения холестерина неживотного происхождения, включающего следующие стадии:
- культивируют организм царства Fungi такой, как описан выше,
- экстрагируют холестерин, продуцированный этим организмом.
Экстракция основана на обработке fungus растворителем холестерина, предпочтительно, не смешивающимся с водой. Эта обработка может быть предпочтительно скомбинирована с каким-либо методом механического дробления клеток. Более предпочтительно, fungus обрабатывают до экстракции растворителем омыляющей смесью, предназначенной для высвобождения холестерина, возможно связанного с другими клеточными компонентами, такими как, в частности, жирные кислоты. Эта омыляющая смесь может состоять из основания, например, гидроксида аммония, гидроксида натрия, гидроксида калия, растворенного в воде или, более предпочтительно, в органическом растворителе, поддающемся смешиванию с водой, таком как, например, метанол или этанол, или в смеси растворитель-вода. Омыление может быть осуществлено без или, предпочтительно, с нагреванием при температуре 60-120°C, при атмосферном давлении или при пониженном давлении. Экстракция растворителем, не смешивающимся с водой, может быть заменена экстракцией в твердой фазе на гидрофобной смоле. Метод экстракции стеринов описан L. Parks и сотрудниками (1985) (Methods in Enzymology 111 Edited by L Rilling, L. Parks, C. Bottema, R. Rodriguez and Thomas Lewis pp333-339).
Полученный таким образом сырой холестерин может быть очищен любыми способами, известными специалисту, в частности описанными Boselli E, Velazco V, Caboni Mf и Lercker G J Chromatogr A. 2001 May 1 1; 917(1-2):239-44.
Другие методы могут также быть использованы, например, описанный способ экстракции холестерина из овечьей шерсти. Специалист может, в частности, обратиться к способам, описанным в патентах США 2688623 или 2650929, или в английских патентах GB690879, GB646227 или GB613778.
Другой аспект изобретения касается применения штаммов согласно настоящему изобретению для получения холестерина или одного из промежуточных продуктов его метаболизма, или меченой смеси стеринов. Под промежуточным продуктом метаболизма холестерина понимают, в частности, стерины, указанные на фиг.2. Это может, в частности, быть холеста 8,24(25) диенол, холеста 7,24(25) диенол, холеста 5,7,24(25)триенол, холеста 5,24(25) диенол, холеста 5,22 диенол.
Принцип получения меченого холестерина описан на фиг.10. Эта манипуляция состоит в том, чтобы сначала вырастить штамм fungus на полностью меченом субстрате. Клетки затем культивируют на не меченом субстрате. Таким образом происходит изменение изотопной метки источника углерода, далее следует синтез de novo промежуточных продуктов метаболизма, затем стерина, включая холестерин, содержащего постепенное изменение метки. Речь идет, таким образом, о сложном, но вполне поддающемся экспериментальному определению профиле, который представляет уникальную изотопную подпись, которая зависит одновременно:
1) от протокола мечения и, в частности, сроков и условий культивирования на меченом и не меченом субстрате;
2) от точной генетической структуры использованного штамма;
3) точного времени остановки культивирования.
После остановки культивирования (например, лизисом клеток или путем остановки культивирования в присутствии суб-летальной концентрации цитотоксических или цитостатических противогрибковых продуктов) меченый холестерин или один из промежуточных продуктов его метаболизма, или меченую смесь стеринов экстрагируют и очищают, как описано выше.
Изотопный профиль меченого холестерина или одного из промежуточных продуктов его метаболизма, или меченой смеси стеринов обладает несколькими уникальными свойствами:
1) его можно при необходимости менять за счет изменений условий культивирования, используемого штамма и выбранного стерина, таким образом может быть получен уникальный регистр меток;
2) он является "комбинируемым", а именно некоторые изотопные подписи, соответствующие некоторым уникальным стеролам, меченым изотопными профилями, которые сами по себе являются модулируемыми, могут быть скомбинированы с получением «молекулярного алфавита»;
3) он является воспроизводимым и легко определимым экспериментально;
4) он соответствует трассирующей молекулярной смеси, легкой в выделении, стабильной, бесцветной и лишенной запаха, нелетучей, не токсичной и которая может быть внедрена в продукты питания, в лекарственное средство, добавки или другие продукты, потребляемые человеком;
5) он не поддается фальсификации, если неизвестны специфические рекомбинантные штаммы и очень точные условия мечения, культивирования и экстракции. К тому же знание изотопной подписи не позволяет выявить параметры, которые могли бы позволить осуществить его получение.
Таким образом, не поддающийся фальсификации «изотопный алфавит» общего назначения и который может быть включен в любой тип продуктов, в том числе употребляемых в пищу, может быть легко получен благодаря настоящему изобретению. Число «изотопных слов», которые можно построить исходя из такого алфавита, является практически неограниченным благодаря использованию одновременно профилей мечения и различных типов стеринов. Включение таких подписей в состав разнообразных продуктов представляет собой, таким образом, уникальный способ не поддающейся фальсификации маркировки, в отличие, например, от подписей ДНК, которые, став один раз известными, могут быть воспроизведены. Подпись может быть к тому же считана способами, не наносящими повреждений, например лазерной ионизацией с последующим анализом масс-спектометрией (MALDI-TOF и т.п.).
Использование субстрата, меченого 13C, вместо немеченых источников углерода для культивирования штаммов fungus согласно изобретению, позволяет осуществить синтез стеринов и, в частности, очень сильно меченого холестерина (включающего по меньшей мере 95% углерода 13C). Получение стеринов и радиоактивного холестерина 14C также возможно с использованием того же подхода. Способ может быть также использован в отношении штаммов дрожжей, продуцирующих стероиды, в частности, гидрокортизон (см. заявку на патент WO 02/061109) для получения стероидов, меченых 13C или 14C, например, для тестов RIA.
Описание чертежей
Фиг.1. Химическая формула холестерина, а также номенклатура, обычно используемая для нумерации различных атомов углерода и названия различных циклов. Четыре цикла молекулы холестерина названы соответственно A, B, C и D, и углероды пронумерованы от 1 до 27.
Фиг.2. Упрощенная схема поздней части пути биосинтеза стеринов типа эргоста- и холеста- в натуральных или модифицированных дрожжах. Схема не является исчерпывающей, но позволяет определить стадии, в которых участвуют ферменты, процитированные в этом документе. Белки ERG2p, ERG3p, ERG5p и ERG6p являются белками fungus или дрожжей, в то время как белки Дельта-7Red (Дельта 7 стерол редуктаза), Дельта 24-(25)Red (Дельта 24-(25) стерол редуктаза) являются гетерологичными белками, происходящими от млекопитающих или растений.
Фиг.3. Профиль, сравненный в ВЭЖХ с УФ-детекцией при 206 нм, свободных стеринов штаммов, производных от штамма BMA64, и идентификация этих стеринов. Были исследованы следующие штаммы: WGIF01 (штамм BMA64 с разрушенным геном erg6 (см. пример 1)), WGIF02 (штамм BMA64 с разрушенным геном erg6 и экспрессирующий Δ24-редуктазу, пример 12), WGIF03 (штамм BMA64 с разрушенным геном erg6 и экспрессирующий Δ7-редуктазу, пример 13), WGIF04 (штамм BMA64 с разрушенным геном erg6 и экспрессирующий Δ7-редуктазу и Δ24-редуктазу, пример 14). C5: холеста 5 енол (холестерин); C5,22: холеста 5,22 диенол; C5,24: холеста 5,24 диенол (десмостерол); C8,24: холеста 8,24 диенол (зимостерол); C5,7,22: холеста 5,7,22 триенол; C5,7,24: холеста 5,7,24 триенол; C5,22,24: холеста 5,22,24 триенол; C5,7,22,24: холеста 5,7,22,24 тетраенол; lan: ланостерол.
Фиг.4. Профиль, сравниваемый в ВЭЖХ с УФ-детекцией при 206 нм, свободных стеринов штамма WGIF04 (штамм BMA64 с разрушенным геном erg6 и экспрессирующий Δ7-редуктазу и Δ24-редуктазу, пример 14) после 0,2,4,8,24 часов индукции галактозой. Δ:штамм WGIF01 (пример 1). Для штамма WGIF04 отборы проб осуществляли через 0, 2, 4, 8 и 24 ч после замены источника углерода галактозой. Представленный профиль штамма BMA64, имеющего разрушение в гене erg6 (WGIF01), соответствует профилю, полученному сразу после замены на галактозу. Этот профиль остается практически неизменным в течение индукции (0-24 ч). Сигнал поглощения при 206 нм соответствует коэффициентам поглощения, меняющимся от одного стерина к другому. C5:10 холеста 5 енол (холестерин); C5,22: холеста 5,22 диенол; C5,24: холеста 5,24 диенол (десмостерол); C8,24: холеста 8,24 диенол (зимостерол); C5,7,22: холеста 5,7,22 триенол; C5,7,24: холеста 5,7,24 триенол; C5,22,24: холеста 5,22,24 триенол; C5,7,22,24: холеста 5,7,22,24 тетраенол; lan: ланостерол.
Фиг.5. Профиль, сравниваемый в ВЭЖХ с детекцией с электрораспылением и положительной ионизацией (масс-спектометрия) свободных стеринов штамма WGIF04 (пример 14) после 0, 2, 4, 8, 24 часов индукции галактозой. Δ:штамм WGIF01. C5:холеста 5 енол (холестерин); C5,22: холеста 5,22 диенол; C5,24: холеста 5,24 диенол (десмостерол); C8,24: холеста 8,24 диенол (зимостерол). Профили ВЭЖХ получены в том же тесте, что и в случае фиг.4.
Фиг.5A (слева): детекция при m/z=367, Фиг.5B (справа) m/z=3 69.
Ордината: число подсчитанных ионов/в секунду. Абсцисса: продолжительность элюирования в минутах.
Фиг.6. Деталь профиля при m/z=369 в ВЭЖХ для трех штаммов: WGIF01, CA10, несущего плазмиду для экспрессии дельта 24 стерол редуктазы и WGIF04, холестерин вводится в качестве внутреннего стандарта. Общее количество стеринов, введенных для всех трех штаммов, соответствует экстракции, осуществленной, исходя из идентичных количеств культуры, измеренных по поглощению при 600 нм.
Фиг.7. Сравниваемые профили общих стеринов (свободных и в форме сложных эфиров) в хроматографии в газовой фазе штаммов WGIF01 (делеция erg6), WGIF02 (делеция erg6 с экспрессией Δ24:редуктазы); WGIF03 (делеция erg6 с экспрессией Δ7-редуктазы), WGIF04 (делеция erg6 с экспрессией Δ24:редуктазы и Δ7-редуктазы) и CA10 pYES_Delta24 (генетическая основа FY1679, делеция erg5 с экспрессией Δ24:редуктазы, Δ-7-редуктазы, erg5). Масштабы ответа (токи ионизации пламени) являются произвольными. Профили разных штаммов должны сравниваться только в качественном отношении. Масштаб времени удержания является, тем не менее, одним и тем же для всех штаммов (времена удержания выражены в минутах). Идентификацию стеринов осуществляют согласно критериям, описанным в настоящей заявке.
Фиг.8. Количественное распределение главных свободных стеринов в штаммах дрожжей CBMA64 (фиг.8A), WGIF01 (фиг.8B), WGIF02 (фиг.8C) и WGIF03 (фиг.8D)), оцененное на основе спектров УФ. Распределение дано в % совокупности видов, представленных на чертеже, и которые являются единственными поддающимися обнаружению в ощутимых количествах. В отсутствие стандарта для нескольких промежуточных стеролов определение количества осуществляют на основе спектров УФ, ассоциированных с каждым из пиков хроматограммы ВЭЖХ с использованием коэффициентов поглощения, приведенных ниже (см. таблицу 1, коэффициенты поглощения выражены в мМ на 1 литр и на 1 см). С этой целью коэффициенты поглощения, соответствующие ненасыщенным структурным звеньям, присутствующим в структуре данного стерина, находят в таблице 1 и, в случае необходимости, складывают (если несколько звеньев присутствуют в одной и той же молекуле), чтобы получить оценку коэффициента погашения каждого типа стерина. Оценку делают, используя значения при 280 нм, если присутствует по меньшей мере одно звено, поглощающее при этой длине волны, в противном случае используют длину волны 235 нм, и при отсутствии поглощения при этой последней используют длину волны 206 нм для оценки концентрации каждого из стеринов исходя из соответствующих сигналов поглощения в ВЭЖХ.
Фиг.9. Количественное распределение главных свободных стеринов в штамме дрожжей WGIF4, определенное на основе спектров УФ. Определения количества осуществляют таким же образом, как и в случае фиг.8.
Фиг.10. Принцип изотопного мечения стеринов с заменой источников углерода.
Фиг.11. Оценка профилей изотопного мечения холестерина, продуцируемого в штамме WGIF04 после 4, 8, и 24 часов индукции. Свободные стерины экстрагируют и разделяют ВЭЖХ, как описано. Масс-спектр между значениями m/z 300 и m/z=450 получают каждые 0,2 секунды в течение элюирования. Эти спектры затем усредняют по окнам 1,8 секунды, затем подвергают мультилинейной регрессии, используя как основание регрессии все 24 вектора, представляющие теоретические распределения масс меченого холестерина, для произвольного включения независимыми тиражами углерода 13 в каждое из 27 положений молекулы с вероятностью мечения в каждом переменном углероде от 0 до 1, в зависимости от рассматриваемого вектора. Вероятности мечения различных векторов, используемых как основание, выбирают таким образом, чтобы коэффициент кросс-корреляции распределений двух последовательных векторов основания составлял 0,92, основание начинается вектором, соответствующим 100% вероятности присутствия во всех положениях для углерода 12. Мультилинейное урегулирование осуществляют по статистическому критерию наименьшего квадрата, аннулируя не-диагональные термины матрицы продуктов частичных производных метода Гаусса (максимальное цифровое фильтрование). После анализа осуществляют реконструкцию масс-спектров на оптимизированной базе. Кривые, изображенные на фигурах, представляют, таким образом, результат оптимальной фильтрованной реконструкции после нормализации максимальной амплитуды на значении 100.
Для каждого времени индукции обе кривые представляют два независимых профиля, соответствующих временам элюирования, отличным от 1,8 секунд, и соответствующим спектрам, расположенным в центральной зоне пика элюирования холестерина. Фигура доказывает высокую воспроизводимость анализа.
Фиг.12. Пример различных изотопных подписей на уровне различных стеринов или различного времени индукции. Тот же расчет и представление, как для фиг.11, но для различных стеринов и различного времени индукции.
Значение RT указывает гамму времени удержания, использованного для расчета (в минутах). Значения этой гаммы являются следующими:
Фиг.12A: RТ=12,25-12,42,
фиг.12B: RТ=12,2-12,7,
фиг.12C: RT=12,25-12,35,
фиг.12D: RT=13,3-13,6,
время индукции 8 или 24 часа.
Значения m/z указывают левую и правую границу m/z. Наиболее низкое значение m/z для каждой рамки соответствует m/z для стерина, полностью построенного из углерода 12.
Фиг.13. Сравниваемые профили общих стеринов (свободных и в форме сложных эфиров) в хроматографии в газовой фазе штаммов YIM59/pIM303 (часть A фигуры) и штамма YIM59/pIM331 (часть B фигуры) (см. пример 18). Масштаб ответа является произвольным. Масштаб времени удержания является одним и тем же для обоих штаммов (время удержания выражено в минутах). Идентификацию стеринов осуществляют согласно критериям, описанным в настоящей заявке.
Настоящее изобретение иллюстрируется при помощи следующих далее примеров, которые должны быть рассмотрены как иллюстративные и неограничительные.
Использованные методы молекулярной биологии описаны Ausubel et al, некоторые манипуляции над дрожжами описаны Adams et al (Adams and Holm, 1996).
ПРИМЕР 1
Конструирование штамма дрожжей S. cerevisiae с прерванным геном ERG6 (штамм WGIF01)
Штамм дрожжей S. cerevisiae WGIF01, ген ERG6 которого прерван геном TRPJ, был получен трансформацией штамма BM64 продуктом ПЦР, несущим функциональный ген TRP1, фланкированный краями, гомологичными гену ERG6.
Штамм BM64 (генотип MATa; uraЗ-52; TRP1AZ; leu2-3_112; his3-ll; ade2-1; canl-100) получают из штамма дрожжей S. cerevisiae, W303 МАТα путем полной делеции гена TRP1. Штамм BMA64 и штамм W303 МАТα описаны в публикации Baudin-Baillieu et al (Baudin- Baillieu et al., 1997).
Чтобы выделить ген TRP1, ген TRP1 плазмиды pFL44 (Bonneaud et al., 1991) амплифицировали, используя Z-TaqI (ДНК-зависимая ДНК-полимераза), предоставленный компанией Takara (Pan Vera LLC 501 Charmany Drive Madison, W1 53719 USA).
Использованная пара праймеров позволяет с помощью ДНК-полимеразы амплифицировать ген TRP1, фланкированный последовательностями, соответствующими гену ERG6.
Последовательность этих праймеров является следующей:
OERG6trp1: 5' (CCTAGCGACGAAAAGCATCATTGGAGTGAATAACTTGGACTTACCAttcttag cattttgacg) 3' (SEQ ID N 1).
OERG6trp2: 5' (GCATAAGAGTGAAACAGAATTGAGAAAAAGACAGGCCCAATTCAaattcggg tcgaaaaaagaaaagg) 3' (SEQ ID N 2).
Продукт ПЦР (Polymerase Chain Reaction), полученный таким образом, очищают электроэлюированием фрагмента, соответствующего ожидаемому размеру, и используют для трансформации штамма BM64 методикой хлорида лития, как описано (Gietz et al., 1995).
После трансформации обработанные дрожжи размещают на минимальной среде, не содержащей триптофан (Gietz et al., 1995). Получают таким образом 41 колонию трансформированного BM64, прототрофные в отношении триптофана. Эту 41 колонию затем тестируют в отношении трех свойств: чувствительность к нистатину, геномная структура вставки гена TRP1, профиль в хроматографии в газовой фазе общих стеринов, которые они продуцируют.
Для этого эту 41 колонию переносят на минимальную среду, содержащую соответственно 10, 20 или 50 мкг/мл нистатина, при этом десяток колоний способны расти на среде, содержащей дозу 50 мкг/мл нистатина. Эти резистентные колонии отбирают для проверки их генной структуры, а также состава их стеринов.
Включение гена TRP1 в ген ERG6 проверяют с помощью ПЦР, используя пару олигонуклеотидов, перекрывающих соединение между функциональным геном TRP1 и разорванным ERG6. Эта пара олигонуклеотидов является следующей: OERG6trp3: AGGGCCGAACAAAGCCCCGATCTTC (SEQ ID № 3) и OERG6trp4: GGCAAACCGAGGAACTCTTGG (SEQ ID № 4).
Некоторые штаммы представляют ожидаемый профиль ПЦР, то есть фрагмент из 800 пар основания, соответствующий размеру, ожидаемому для включения TRP1 в ERG6.
Для проверки того, что ген ERG6 действительно инактивирован в этих штаммах, проводили анализ состава стеринов этих штаммов хроматографией в газовой фазе и высокоэффективной жидкостной хроматографией (Duport et al., 2003; Szczebara et al., 2003).
Эти анализы подтверждают отсутствие синтеза эргостерола и аккумуляцию совместимых аномальных стеринов с ожидаемыми нарушениями пути биосинтеза в штамме с разорванным геном.
Один штамм был, в частности, отобран и назван WGIF01.
ПРИМЕР 2
Конструирование штаммов CA10, CA14 и CА23
Штаммы CA10 (генотип: МАТα, rho + , GAL2, ura3-52, trp1-Δ63, his3-Δ200, erg5::HYGRO R , ade2::GAL10/CYC1::ΔРедуктаза::PGK1, LEU2::GAL10/CYC1:: matADR:.PGK1), CA14 (генотип: МАТα, rho + , GAL2, ura3-52, trp1-Δ63, his3-Δ200, erg5::HYGRO R , atf2::G418 R , ade2::GAL10/CYC1::ΔРедуктаза::PGK1, LEU2::GAL10/CYC1::matADR::PGK1), и CA23 (генотип: МАТα, rho + , GAL2, ura3-52, trp1-Δ63, his3-Δ200, erg5::HYGRO R , are1::G418 R , are2::HIS3, ade2::GAL10/CYC1::ΔРедуктаза::PGK1, LEU2::GAL10/CYC1::matADR::PGK1), а также их строение описаны в ссылке Duport et al., техническое содержание которой относительно конструирования этих штаммов включено в настоящую заявку путем ссылки.
Эти штаммы продуцируют и содержат в своих мембранах неприродные стерины (как описано в заявке на европейский патент EP 0727489) и, в частности, эргоста-5-енол (кампестерол).
Эти три штамма не экспрессируют продукт гена ERG5, который является не функциональным за счет включения в его кодирующую последовательность гена резистентности к гигромицину. Кроме того, эти штаммы экспрессируют кДНК, кодирующую Δ7 редуктазу растений (в заявке на европейский патент EP 0727489 описано, в частности, клонирование Δ7 редуктазы растения Arabidopsis thaliana, эта заявка включена в настоящую заявку путем ссылки, номер доступа в GenBank этой последовательности - ATU49398).
Штамм СА14 получают из штамма CA10 за счет разрыва гена ATF2. Продукт этого гена приводит к ацетилированию прегненолона в положении 3 (как описано в заявке на патент WO 99/40203). Штамм CA23 представляет собой штамм, полученный от штамма CA10 за счет делеции генов ARE1 и ARE2, оба белка Are1p и Are2p являются ответственными за этерификацию эргостерола (Sturley, 2000) и, возможно, холестерина, так как они гомологичны ферменту, ответственному за этерификацию холестерина у млекопитающих (ACAT).
ПРИМЕР 3
Конструирование плазмиды экспрессии Δ24-25 редуктазы человеческого происхождения (плазмида pYES_Delta24)
Конструирование этой плазмиды было описано Waterham et al. 2001. Конструирование состоит в том, чтобы поместить кДНК, кодирующую Дельта 24 стерин-редуктазу, под контроль промотора pGAL1 и терминатора tCYCl в векторе pYES2 (Invitrogen SARL, Cergy Pontoise, Франция). Эта плазмида является челночной плазмидой E.coli/S. cerevisiae и содержит сайт начала репликации 2 микрон и ген URA3, что обеспечивает ее репликацию в дрожжах и облегчает отбор дрожжей, трансформированных этой плазмидой.
Промотор GAL1, кроме того, индуцируется галактозой.
ПРИМЕР 4
Конструирование плазмиды pAG1, экспрессирующей Δ7-редуктазу A. thaliana
Была сконструирована плазмида для специфической экспрессии Дельта 7 Редуктазы A. thaliana на однокопийном векторе. Для этого использовали плазмиду pAM1. Эта плазмида, конструирование которой описано в заявке PCT WO 02/061109 (см. пример 9.b указанной заявки, которая включена в настоящую заявку путем ссылки), является челночной плазмидой E.coli./S cerevisiae, основанной на последовательности автономной репликации и центромере (ARS CEN). Селекционным маркером является ген ADE2. Эта плазмида является совместимой и может таким образом реплицироваться одновременно с плазмидой, основанной на сайте начала репликации 2 микрон. Эта плазмида обладает, в частности, уникальным сайтом NotI, позволяющим клонировать кассеты экспрессии, как описано в указанной заявке PCT.
Этот сайт был использован, чтобы клонировать кассету экспрессии Дельта7-редуктазы A. thaliana, происходящую из штамма CA10. Действительно, эта кассета экспрессии является очень эффективной и позволяет штамму CA10, в котором к тому же разорван ERG5, продуцировать кампестерол (эргоста-5-енол) как мажоритарный стерин (Duport et al., 1998). Фрагмент геномной ДНК штамма CA10, содержащий ген дельта7-редуктазы, амплифицируют при помощи следующих праймеров:
OSA72 5' (TATATAGCGGCCGCTTTCGCTGATTAATTACCCCAG) 3' (SEQ ID № 5)
OSA77 5' (TATATAGCGGCCGCGAGAAGTGACGCAAGCATCA) 3' (SEQ ID № 6).
Амплификацию осуществляли геномной ДНК штамма СА10, полученной методом быстрой экстракции в смеси фенол/хлороформ, как описано Adams et al. (Adams and Holm, 1996).
Пятьдесят нанограмм геномной ДНК СА10 использовали в качестве матрицы для амплификации при помощи праймеров OSA72 и OSA77. Taq DNA полимераза и ферментативные условия были получены от компании Stratagene. Условия амплификации были следующими. Первоначальная денатурация 5 мин при 95°C, затем тридцать циклов, заключающихся в денатурации 30 с при 95°C, гибридизация в течение 30 с при 50°C, затем элонгация в течение 1 мин при 72°C. Реакция завершается окончательным растягиванием в течение 10 мин при 72°C.
Фрагмент ПЦР затем расщепляют ферментом NotI и очищают на агарозном геле, затем клонируют обычным образом на уровне уникального сайта NotI плазмиды pAM1. Плазмиду, полученную таким образом, назвали pAG1.
Она представляет собой однокопийный вектор, обеспечивающий экспрессию Дельта7 Редуктазы A. thaliana у дрожжей, в котором ген Дельта7 Редуктазы помещен под контроль промотора GALl0/CYCl (Lecain et al., 1996).
ПРИМЕР 5
Экстракция свободных и этерифицированных стеринов у дрожжей для анализов
1) Условия экстракции свободных и этерифицированных стеринов у дрожжей (процедура 1).
a) Условие экстракции свободных стеринов
Клеточный осадок промывают два раза с 500 мкл деионизованной и фильтрованной воды в стеклянной пробирке.
Клетки затем ресуспендируют в 500 мкл воды, содержащей стеклянные шарики диаметром 0,5 мм, что соответствует 150 мкл жидкости в пробирке.
Экстракцию осуществляют два раза с 2 мл 1,2-дихлорэтана при сильном взбалтывании на вортексе в течение 10 минут. После первой экстракции смесь клеток, стеклянных шариков, растворителя центрифугируют в течение 5 минут при 1500g для разделения двух фаз.
Обе органические фракции, происходящие из обеих последовательных экстракций, объединяют и высушивают в токе азота в течение нескольких часов. Стериновый экстракт суспендируют в 100 мкл ацетонитрила для анализа высокоэффективной жидкостной хроматографией (ВЭЖХ) (Szczebara et al., 2003) или в 100 мкл гексана для анализа хроматографией в газовой фазе (GC) (Duport et al., 2003).
b) Условия экстракции общих стеринов: Омыление и экстракция этерифицированных стеринов, качественный анализ, процедура 1
Клеточный осадок ресуспендируют в 500 мкл очищенной воды. К этой суспензии добавляют 2 мл гидроксида калия KOH, 10% в метаноле. Смесь нагревают в течение одного часа при 60°C в закрытых пробирках. Потом инкубации и сразу по достижении пробирками температуры окружающей среды смесь экстрагируют три раза с 2 мл гексана. Между каждой экстракцией обе фазы разделяют центрифугированием в течение 5 мин при 1500g. После каждой экстракции органическую фазу переносят в новую пробирку, затем три органические фазы объединяют, затем высушивают в токе азота.
Стериновый остаток ресуспендируют в 100 мкл ацетонитрила (100%) для анализа высокоэффективной жидкостной хроматографией (ВЭЖХ) (Szczebara et al., 2003) или в 100 мкл гексана для анализа хроматографией в газовой фазе (GC) (Duport et al., 2003).
2) Условия экстракции свободных и этерифицированных стеринов у дрожжей для качественного анализа (процедура 2).
Штаммы культивируют в богатой среде (10 г бактопептона на 1 л и 10 г экстрактов дрожжей на 1 л) с 2% глюкозы в качестве источника углерода, получая 500 мг лиофилизованных клеток. Эти высушенные клетки помещают в 3 мл метанола (100%), содержащего 1 г KOH, и следовые количества пирогаллола, затем смесь инкубируют в течение 45 минут при 90°C. После возвращения к температуре окружающей среды стерины экстрагируют с 5 мл гексана. Органическую фазу разделяют на три образца одного и того же объема и высушивают в токе воздуха. Два из образцов экстрагированных стеринов помещают в 100 мкл гексана для анализа хроматографией в газовой фазе (GC) и хроматографией в газовой фазе, объединенной с масс-спектометрией GC/MS, а третий образец помещают в 150 мкл метанола для исследований высокоэффективной жидкостной хроматографией (ВЭЖХ).
ПРИМЕР 6
Анализ свободных и этерифицированных стеринов у дрожжей хроматографией в газовой фазе (GС)
1) Хроматография в газовой фазе (GС) с детекцией FID (ионизация пламени)
Стериновый экстракт (свободный или общий), суспендированный в гексане, получают согласно процедуре 1 (см. пример 5 1) a) и b)). Контроль инъекции добавляют к смеси стерина, обычно холестерина, в концентрации от 10 до 50 нг/мкл.
Затем инъецируют от 1 до 3 мкл образца в аппарат для хроматографии в газовой фазе в следующих условиях. От 1 до 3 мкл впрыскивают на колонку Alltech типа SE30 (Данные колонки: 30 м×0,32 мм IDX 0,25 мкм). Используемым газом является гелий. Отношение Сплита составляет от 50 до 80. Давление в вершине колонки составляет 30 psi. Инжектор отрегулирован на 280°C. Первоначальная температура колонки составляет 130°C в течение 0,5 минуты. Ее повышают до 230°C из расчета 40°C/мин, затем с 230°C до 280°C из расчета 3°C/мин. Колонку поддерживают затем при 290°C. Температура детектора составляет 310°C.
2) Хроматография в газовой фазе (GC) с детекцией FID (ионизация пламени), объединенная с масс-спектометрией (GC/MS)
Экстракт общего стерина, суспендированный в гексане, получают согласно процедуре 2. Используемая GC оснащена классическим инжектором «split split less» с классической колонкой DB5, с длиной 30 метров и диаметром 0,25 мм.
Инъекцию осуществляют при 230°C с гелием в качестве газа-носителя с расходом 2 мл/мин. Температуру колонки повышают с 130 до 290°C в 4 стадии. Колонку поддерживают при 130°C до инъекции, затем повышают до 230°C со скоростью 40°C/мин, затем с 230°C до 280°C со скоростью 3°С/мин, затем с 280°C до 290°C со скоростью 30°C/мин. Колонку поддерживают в течение 5 минут при 290°C.
По выходе из колонки для хроматографии в газовой фазе молекулы анализируют масс-спектометрией с выпариванием в камере ионизации, такой как камера аппарата типа Turbo Mass от Perkin Elmer. Молекулы фрагментируют электронным пучком высокой энергии. Различные фрагменты затем разделяют на четырехполюсном фильтре, затем детектируют на детекторе ионов. Каждой массе, локализованной на графике тока ионов, соответствует масс-спектр, который объединяет массы всех продуктов фрагментации иона M+. Этот масс-спектр, полученный для заданного времени удержания на колонке, сравнивают с библиотеками фрагментированных продуктов, а также с библиотеками, описанными для стеринов Quail and Kelly. (Methods in Molecular Biology Vol53 Yeast Protocols Edited by Evans; M, Quail and S, Kelly «The Extraction and Analysis of Sterols from Yeast» pp 123-131 (1996)).
Таким образом, удалось выявить результат делеции гена ERG6 в штамме WGIF01, в частности отсутствие эргоста 8, 24(28) диенола и присутствие стерина типа холеста, имеющего двойную связь в 24(25).
ПРИМЕР 7
Анализ свободных и этерифицированных стеринов у дрожжей высокоэффективной жидкостной хроматографией (ВЭЖХ) с УФ-детекцией или детекцией масс-спектометрией
1) Анализ ВЭЖХ с УФ-детекцией
От десяти до 30 мкл стеринового экстракта (суспендированного в ацетонитриле или в метаноле и полученного согласно процедуре 1 или 2 (см. пример 5)) впрыскивают на колонку типа X terra RP18, 4,6×100 мм (Waters, Milford, MA01757 USA).
Разделение осуществляется на градиенте, состоящем из воды, содержащей 0,02% TFA (трифторуксусная кислота) (буфер A) и чистого ацетонитрила (буфер B). Колонку поддерживают при 60°C в течение анализа.
Использованный аппарат ВЭЖХ типа «Waters 600 E System Controller» (Waters, Milford, MA01757 USA). УФ-детекцию осуществляют на детекторе с диодной решеткой, покрывающей длину волны от 206 до 350 нм. Колонка была уравновешена с буфером, содержащим 20% (об./об.) буфера A (ацетонитрил) и 80% буфера B (вода, содержащая 0,02% TFA (трифторуксусная кислота)). Линейный градиент осуществляют исходя из раствора, содержащего 50% буфера A и 50% буфера B. По истечении 10 мин композиция буфера элюирования включает 25% буфера A на 75% буфера B. Новый линейный градиент применяют так, чтобы за 30 мин градиент достиг показателя 100% буфера B. Эту величину поддерживают в течение 5 минут, чтобы очистить колонку.
2) Анализ ВЭЖХ с детекцией масс-спектометрией (ВЭЖХ/MС)
В случае анализа масс-спектометрией образец поддерживают при 30°C, и колонку поддерживают при 60°C в ходе анализа. Использованный аппарат для ВЭЖХ является аппаратом типа «Alliance HT Waters 2790», соединенным с масс-детектором «Waters MicroMass ZQ». В отличие от предыдущего метода детекции, буфер элюирования A не содержит TFA, но оба буфера A и B содержат 0,01% (об./об.) муравьиной кислоты.
Колонка была уравновешена с буфером, содержащим 80% буфера A (вода, содержащая 0,01% (об./об.) муравьиной кислоты) и 20% буфера B (ацетонитрил, содержащий 0,01% (об./об.) муравьиной кислоты).
Инъекция начинается с буфером, содержащим 50% этих двух буферов. Линейный градиент с двумя скоростями роста осуществляют, исходя из раствора, содержащего 50% буфера A и 50% буфера B.
По истечении 10 минут композиция буфера элюирования включает 25% буфера A на 75% буфера B. Скорость роста градиента затем изменяют, чтобы получить 12,5% буфера A и 87,5% буфера B после 25 минут анализа, затем 100% буфера B за 30 минут. Эту величину поддерживают в течение 5 минут для регенерации колонки.
Масс-детектор «Waters MicroMass ZQ» настраивают на сканирование в положительной ионизации с электрораспылением. Значения m/z составляют от 295 до 450. Для сканирования выбирают режим “continum”. Кроме того, экстракцию сигнала в режиме "SIR" осуществляют параллельно всем массам, ожидаемым в естественном изотопном изобилии, для анализируемых стеринов. Детектор настраивают на полное разрешение без интерференции молекул, отличающихся на 1 единицу по m/z. Совокупность фиксаций параметризуют так, чтобы общая продолжительность фиксации, соответствующая сканированию, и по общему времени фиксации всех SIR составляла менее 2 секунд.
ПРИМЕР 8
Культивирование штаммов дрожжей для анализа содержания стерина с или без мечения 13 C
Анализируемые штаммы культивировали в объеме 50 мл среды Kappeli (Kappeli et al., 1985), содержащей 2% нормальной D-глюкозы или D-глюкозы-U-13С6 (в отношении мечения, см. фиг.10).
Оптическая плотность первоначальной культуры составляет 0,1 при 600 нм. Эту культуру инкубируют 72 часа при температуре 30°C при взбалтывании со скоростью 200 об/мин.
Клетки затем получают центрифугированием среды при 600 g в течение 10 минут. Клеточный осадок анализируют затем непосредственно методами анализа, представленными в примере 5 (для тестов, не требующих индукции в галактозе).
Тем не менее, для тестов кинетики индукции экспрессии дельта 7-редуктазы и дельта 24-редуктазы (штаммы, трансформированные плазмидой pYES_Delta24 и/или pAG1), осадок ресуспендируют в 50 мл свежей среды Kappeli, содержащей 2% галактозы (не меченой углеродом 13C).
Эту культуру инкубируют при температуре 30°C при перемешивании со скоростью 200 об/мин. Десять мл культуры отбирают после 0 часов, 2 часов, 4 часов, 8 часов и 24 часов культивирования.
Эти образцы культуры центрифугируют при 800 g в течение 10 минут, клеточный осадок замораживают и сохраняют при -20°C до экстракции стеринов методами, описанными в примере 5.
ПРИМЕР 9
Идентификация стеринов, присутствующих в анализируемых штаммах
Идентификация стеринов основана на сочетании следующих принципов.
- Сравнение поведения в GC, ВЭЖХ, GC/MS и ВЭЖХ/MS с подлинными стандартами в случае кампестерола (эргоста 5-енол), эргостерола (эргоста 5,7,22 триенол), холестерина (холеста 5-енол), десмостерола (холеста 5, 24 диенол), холеста 5,22 диенола и зимостерола (холеста 8,24 диенол).
- Анализ спектра поглощения в ВЭЖХ и в УФ-детекции с диодной решеткой (см. пример 7-1)).
Этот метод позволяет однозначно идентифицировать на базе спектров пять классов стеринов: 1) класс SA1: отсутствие сопряженной диентовой системы, 2) класс SA2: присутствие 5,7-диеновой системы; 3) класс SA3: присутствие 22,24(25) диеновой системы; 4) класс SA4: присутствие 8,14 диеновой системы; 5) класс SA5: присутствие 22, 24 (28) диеновой системы. Классы SA3 и SA5 не могут сосуществовать по структурным причинам. Классы SA2 и SA4 не могут сосуществовать по причинам биосинтеза, класс SA2 может быть скомбинирован со структурными звеньями классов SA1, SA3, SA5, образуя дополнительные суммарные спектры.
- Анализ времен удержания в GC и в ВЭЖХ на базе приблизительной аддитивности перемещений времени удержания, ассоциированных с каждым типом ненасыщенности и в присутствии скелета типа эргоста или холеста. Поскольку этот критерий не является абсолютным, его используют в качестве вспомогательного для идентификации и для устранения неоднозначности, но он представляет риск ошибки при индивидуальном использовании. Его используют таким образом только в сочетании с другими критериями.
- Анализ в GC/MS (см. пример 6-2)), который выдает массу молекулы и профиль фрагментации, который может быть сравнен с библиотеками спектров.
- Анализ в ВЭЖХ/MS (см. пример 7-2)) с электрораспылением, который выдает, в случае 3-гидроксистеринов, главный сигнал в молекулярной массе -17 (протонирование (+1) и потерю молекулы воды (-18)).
- Анализ со всеми вышеуказанными системами состава стеринов различных эталонных штаммов дрожжей в различных точках биосинтеза.
- Анализ изменений состава стеринов в течение комплементации различных ферментов биосинтеза и кинетики этой комплементации в течение индукции этой комплементации.
- Анализ профиля мечения различных стеринов изотопом 13 углерода.
- Анализ УФ-спектра стеринов, отделенных ВЭЖХ при данном времени удержания. Две сопряженные двойные связи 5,7 представляют типичный спектр с двумя пиками поглощения между 265 и 280 нм, в то время как две сопряженные двойные связи 22,24 представляют пик поглощения 235 нм. Последняя сопряженная двойная связь 8,14 распознается по пику поглощения 245 нм.
ПРИМЕР 10
Идентификация стеринов, присутствующих в штамме BMA64
Штамм BMA64 культивируют в объеме 50 мл среды Kappeli, содержащей 2% D-глюкозы, для количественного и сравнительного анализа стеринов.
Оптическая плотность первоначальной культуры составляет 0,1 при 600 нм. Эту культуру инкубируют 72 часа при температуре 30°C при взбалтывании со скоростью 200 об/мин.
Клетки затем получают центрифугированием среды при 600 g в течение 10 минут, клеточный осадок анализируют методами, представленными в примере 5. Различные описанные анализы позволили идентифицировать стерины, продуцируемые этим штаммом.
Определили таким образом, что этот штамм аккумулирует более 80% своих свободных стеринов в форме эргостерола (эргоста 5,7,22 триенол) (см. фиг.8). Два других поддающихся обнаружению миноритарных стерина продуцируются этим штаммом, речь идет об эргоста 5,7 диеноле (субстрат продукта гена ERG5) (12%) и зимостероле (эргоста 8,24 диенол) (5%). Холестерина не обнаруживается вообще (порог детекции этого метода составляет приблизительно 0,5% обнаруживаемых стеринов). Малые количества ланостерола также детектируются (только на уровне анализов общих стеринов).
ПРИМЕР 11
Идентификация стеринов, присутствующих в штамме WGEFOL
Штамм WGIFOL (см. пример 1) культивируют в объеме 50 мл среды Kappeli (Kappeli et al., 1985), содержащей 2% D-глюкозы, для количественного и сравнительного анализа стеринов.
Оптическая плотность первоначальной культуры составляет 0,1 при 600 нм. Эту культуру инкубируют 72 часа при температуре 30°C при взбалтывании со скоростью 200 об/мин. Клетки затем получают центрифугированием среды при 600 g в течение 10 минут, клеточный осадок анализируют методами, представленными в примере 5. Различные описанные анализы позволили идентифицировать стерины, продуцируемые этим штаммом.
Поиск в хроматограмме эргоста 5,7,22 триенола (эргостерол) или эргоста 5,7 диенола дает отрицательный результат (менее 0,5% величины, полученной у BMA64) в ВЭЖХ, соединенном с масс-спектометрией. На уровне свободных стеринов штамм аккумулирует 50% от общего количества зимостерола (холеста 8,24 диенол), субстрата продукта гена ERG6 и 30, и 20% соответственно, холеста 5,7,24 триенола и холеста 5,7,22,24 тетраенола, происходящих, вероятно, из механизма синтеза, идентичного приводящему к эргоста 5,7 диенолу и эргоста 5,7,22 триенолу у родительского штамма (см. фиг.3 и 8). Это ясно показывает, что путь биосинтеза заблокирован на уровне erg6, так как фермент ERG6р (S-аденозилметионин дельта 24 стерол C-метил трансфераза) превращает холеста 8,24(25) диенол в эргоста 8,24(28) диенол (см. фиг.2). Эта аккумуляция ясно указывает, что штамм WGIF01 не содержит функциональной копии гена erg6. Результаты указывают также, что путь нормального биосинтеза эргостерола у дрожжей и, в частности, стерол 8,7-изомераза, стерол 5-десатураза и стерол дельта 22-десатураза, способны превращать субстраты типа холеста с активностью, которая остается высокой.
ПРИМЕР 12
Конструирование штамма WGIF02 и идентификация стеринов, присутствующих в этом штамме
Штамм WGIF02 был получен трансформацией штамма WGIF01 плазмидой pYES2, несущей кассету экспрессии Δ24 Редуктазы (pYES_Delta24, см. пример 3). Клоны были отобраны на среде, не содержащей урацил, и присутствие и экспрессию кДНК Δ24 редуктазы проверяли, анализируя стерины этих трансформантов с использованием процедуры 1 (см. пример 5-1)).
Клон, названный WGIF02, был отобран, так как он обладал стериновым профилем, отличным от штамма WGIF01, к тому же дополнительный стерол имел время удержания, близкое к холестерину (см. фиг.7). Штамм WGIF02 культивировали в объеме 50 мл среды Kappeli (Kappeli et al., 1985), содержащей 2% D-глюкозы, для количественного и сравнительного анализа стеринов.
Оптическая плотность первоначальной культуры составляла 0,1 при 600 нм. Эту культуру инкубируют 72 часа при температуре 30°C при взбалтывании со скоростью 200 об/мин.
Клетки затем получают центрифугированием среды при 600 g в течение 10 минут, клеточный осадок анализируют методами, представленными в примере 5. Различные описанные анализы позволили идентифицировать стерины, продуцируемые этим штаммом.
Оба профиля стериновых экстрактов штамма WGIF01 и штамма WGIF02 подобны, за исключением нового пика, идентифицированного на основании его массы, его времени удержания и его сопряженных двойных связей как холеста 5,7,22 триенол (фиг.2, 3 и 7). Присутствие этого соединения указывает ожидаемое присутствие активности 24-25 стерол редуктазы на двойную связь в положении 24(25) холеста 5, 7, 25 22, 24 (25) тетраенола. Кроме того, количество холеста 5, 7 24 триенола снижается с появлением холеста 5,7,22 триенола в штамме WGIF02 (фиг.7 и 8). Активность фермента выявляют превращением холеста 5,7,24 триенола, представляющего 30% в штамме WGIF01, и только 12% в WGIF02, т.е. разница составляет 18% и встречается полностью в виде холеста 5,7,22 триенола в штамме WGIF02. Речь идет о неожиданном результате, поскольку продуктом превращения холеста 5,7,24 дельта 24-редуктазой является холеста 5,7, который отсутствует в WGIF02, таким образом он количественно превращен в холеста 5,7,22. Это выявляет другой неожиданный результат, а именно то, что холеста 5,7 является субстратом стерин 22-десатуразы, в то время как холеста 5,7,24, согласно профилю стеринов штамма WGIP01, является плохим субстратом.
ПРИМЕР 13
Конструирование штамма WGIF03 и идентификация стеринов, присутствующих в этом штамме
Штамм WGIF03 был получен трансформацией штамма WGIF01 плазмидой pAG1. Эта челночная плазмида между E. coli и S. cerevisiae несет кассету экспрессии Δ7 редуктазы, соответствующая кДНК, которой оказывается под контролем промотора GAL10/CYC1. Штамм WGIF01 был трансформирован техникой хлорида лития, и трансформанты были отобраны на среде, не содержащей аденин. Экспрессию Дельта 7-редуктазы проверяли на основании появления в стериновом профиле клонов холеста 5,24(25) диенола. Один клон, отвечающий этим критериям, был специфически отобран и назван WGIF03.
Штамм WGIP03 культивировали в объеме 50 мл среды Kappeli (Kappeli et al., 1985), содержащей 2% D-глюкозы, для количественного и сравнительного анализа стеринов.
Оптическая плотность первоначальной культуры составляла 0,1 при 600 нм. Эту культуру инкубируют 72 часа при температуре 30°C при взбалтывании со скоростью 200 об/мин.
Клетки затем получают центрифугированием среды при 600 g в течение 10 минут, клеточный осадок анализируют методами, представленными в примере 5. Различные описанные анализы позволили идентифицировать стерины, продуцируемые этим штаммом.
Экспрессия дельта 7 стерол редуктазы в штамме WGIF01 (для получения штамма WGIF03), в отличие от экспрессии дельта 24 стерол редуктазы, ведет к глубокому изменению стеринового профиля штамма с практически полным исчезновением холеста 5,7,22,24 тетраенола, холеста 7,24 диенола и холеста 8,24 диенола.
Эта активность также отмечена появлением мажоритарного пика, идентифицированного, как описано раньше, как холеста 5,24 диенол или десмостерол. Количество холеста 8,24 диенола составляет от 12 до 48%. Обнаруженный холеста 5,7,24 триенол составляет от 30 до 3% и обнаруженный холеста 5,7,22,24 тетраенол - от 23 до 4% соответственно для штаммов WGTF01 и WGIF03. Эти наблюдения указывают неожиданным образом, что стерол дельта 7-редуктазы восстанавливает холеста 5,7 диенол практически независимо от природы ненасыщенностей, находящихся на боковой цепи стеринов. Этот результат противоположен наблюдаемому со стерол дельта 24-редуктазой. Неожиданным образом, экспрессия стерол дельта 7-редуктазы приводит также к аккумуляции (12%) молекулы, со-мигрирующей с холеста 5,7 диенолом. Представляется, тем не менее, маловероятным, хотя и не исключается, чтобы эта молекула была молекулой холеста 5,7 диенола, чей теоретический уровень должен был бы уменьшаться, а не увеличиваться в этих условиях. Появление малого количества холеста 5,22,24 триенола (8%) также представляет интерес. Этот последний стерин представляет собой ожидаемый продукт действия стерол 22-десатуразы на холеста 5,24 диенол, мажоритарный стерин (60%) в штамме WGIF03 (происходящий из восстановления холеста 5,7,24 триенол стерол дельта 7-редуктазой). Малая аккумуляция холеста 5,22,24 триенола указывает, неожиданным образом, что холеста 5,24 диенол не является хорошим субстратом для стерол 22-десатуразы. Благодаря результатам, полученным со штаммом WGIF02 (см. пример 12), может быть сделан вывод, что присутствие ненасыщенности в положении 24 (холеста 5,24 диенол или холеста 5,7,24 триенол) делает стерины с трудом метаболизируемыми стерол 22-десатуразой. Действие фермента ERG6р, трансформирующего ненасыщенные в положении 24 холеста в эргоста, приводит таким образом к трансформации плохих субстратов гена ERG5 (22-десатуразы) в хорошие субстраты.
ПРИМЕР 14
Конструирование штамма WGIF04 и идентификация стеринов, присутствующих в этом штамме
Штамм WGIF04 был получен трансформацией штамма WGIP02 плазмидой pAG1 техникой хлорида лития, и трансформанты были отобраны на среде, не содержащей ни аденина, ни урацила. Трансформанты подтверждали на основании обнаружения аккумуляции холестерина. Клон, отвечающий этим критериям, был специфически отобран и назван WGIF04. Образец штамма WGTP04 был депонирован в Национальной Коллекция Культур Микроорганизмов (CNCM) в Институте Пастера), 25 rue du Docteur Roux, 75724 Paris Cedex 15, France, 22 апреля 2004 под регистрационным номером 1-3203.
Неразличимые штаммы WGIF04 могут быть получены также трансформацией штамма WGIF03 плазмидой pYES_Delta24 и с использованием такого же отбора.
Штамм WGTP04 культивировали в объеме 50 мл среды Kappeli, содержащей 2% D-глюкозы, для количественного и сравнительного анализа стеринов.
Оптическая плотность первоначальной культуры составляла 0,1 при 600 нм. Эту культуру инкубируют 72 часа при температуре 30°C при взбалтывании со скоростью 200 об/мин.
Клетки затем получают центрифугированием среды при 600 g в течение 10 минут, клеточный осадок анализируют методами, представленными в примере 5. Различные описанные анализы позволили идентифицировать стерины, продуцируемые этим штаммом.
Холестерин составляет 25% свободных стеринов штамма WGIF04 (см. фиг.9). Образование холестерина в этом штамме доказывают самостоятельно в GC и в ВЭЖХ со-миграцией с подлинным стандартом и подтверждают одновременно в GC/MC и в ВЭЖХ/MS. Холестерин не обнаруживается (<0,5% общих стеринов) во всех штаммах, которые не экспрессируют одновременно дельта 7-редуктазу и дельта 24-редуктазу.
Штаммы, в которых ген erg6 не является разорванным, могут продуцировать холестерин, однако он составляет менее 5% общих свободных стеринов. Таким образом, конструировали штамм BMA64-PYES_DELTA24-PAGL, полученный исходя из BMA64 совместной трансформацией pYES_Delta24 и pAG1. Этот штамм продуцирует холестерин, причем этот последний составляет несколько % от общих стеринов.
Штамм СА10 был трансформирован плазмидой pYES_Delta24. Этот штамм также продуцирует холестерин, причем этот последний составляет несколько % от общих стеринов.
Кроме того, удалось показать, что образование холестерина требует индукции промоторов дельта 7-редуктазы и дельта 24-редуктазы (см. фиг.4 и 5). Содержащие эти гены штаммы не продуцируют холестерин в отсутствие индукции, фиг.5 показывает, что максимального уровня холестерина достигают приблизительно после 24 ч индукции. Параллельно образованию холестерина (холеста 5-енола) также наблюдают образование холеста 5,22 диенола. Анализ фиг.4 и 5 показывает, что образование этого последнего соединения происходит быстрее после индукции, чем образование холестерина, и начинается даже до индукции (см. фиг.5A: m/z=367). Тем не менее, это соединение отсутствует абсолютно, если штамм не несет двух плазмид pAG1 и pYES_Delta24. Образование 22-дегидрохолестерина представляет собой таким образом процесс более быстрый, чем процесс образования холестерина, но в этом процессе участвует предшественник, который быстро исчезает после индукции, оставляя место для образования холестерина. Холестерин может быть образован исходя из холеста 5,24 через действие Δ24-редуктазы, или исходя из холеста 5,7 через действие Δ7~редуктазы. Так, было показано, что холеста 5,7 диенол не может аккумулироваться вследствие своей немедленной конверсии в холеста 5,7,22 триенол. Источником холестерина является таким образом холеста 5,24 диенол, который отсутствует во время индукции и накапливается к 4-8 часам индукции, после чего его количество снижается к 24 ч (фиг.5). Это объясняет запоздалое появление холестерина, так как требуется предварительный синтез холеста 5,24 диенола. Напротив, оба возможных предшественника холеста 5,22 диенола представляют собой холеста 5,7,22 триенол и холеста 5,22,24 триенол. Этот последний отсутствует в начале индукции (фиг.4), в то время как первый присутствует, затем его количество быстро сокращается параллельно со стабилизацией образования холеста 5,22 диенола. Исходя из этого, можно прийти к заключению, что источником холестерина является восстановление 5,24 диенол Δ24-редуктазой, в то время как образование холеста 5,22 диенол происходит в результате восстановления 5,7,22 триенола Δ7-редуктазой. Образование холеста 5,22 диенола под действием Δ22-десатуразы на холестерин не может быть полностью исключено, но представляется миноритарным процессом на основании предпочтительного накопления холестерина по отношению к холеста 5,22 диенолу в долговременной (24 ч) кинетике (фиг.4 и 5).
ПРИМЕР 15
Оптимизация пути биосинтеза холестерина, роль Δ22-десатуразы
При длительности индукции порядка 24 ч штамма WGIF04 аккумуляция холеста 5,22 диенола составляет приблизительно 50% от аккумуляции холестерина на уровне свободных стеринов (фиг.4). Разрушение гена Δ22-десатуразы является дополнительной опцией для оптимизации продукции холестерина. Можно осуществить конструирование штамма с двойным разрывом на уровне Δ22-десатуразы (ген erg5) и гена erg6 и экспрессирующего Δ7-редуктазу и Δ24-редуктазу. Получали штамм, несущий подмножество: разрыв Δ22-десатуразы, экспрессия Δ7-редуктазы и экспрессия Δ24-редуктазы.
Этот штамм был получен трансформацией штамма СА10 плазмидой pYES_Delta24 техникой хлорида лития и отбором на основании прототрофности по отношению к урацилу. Полученный штамм был назван CA10/Δ24.
Штамм СА10, экспрессирующий Δ24 стерол редуктазу, продуцирует относительно малое количество холестерина (см. фиг.6 и 7) и аккумулирует в основном эргоста 5-енол и в среднем количестве эргоста 5,7 диенол. Накопление холеста 5,7 диенола происходит очень слабо в таком штамме, что указывает на необходимость разрыва гена erg6 для значительного накопления производных серии холеста. Активность Δ24-редуктазы является таким образом неожиданно мало конкурентоспособной по сравнению с активностью продукта гена erg6. Таким образом, на основании этих результатов можно прийти к заключению, что синхронное разрушение генов erg5 и erg6 важно для оптимизации продукции холестерина.
Разрушение гена erg5 в штамме WGIP04 позволило бы специалисту увеличить существенно продукцию холестерина. Возможность получения такого штамма установлена благодаря конструированию штаммов WGIF04 и CAl0/Δ24 и тому факту, что этот штамм является всего лишь очевидной генетической комбинацией двух предыдущих штаммов. Данные, полученные с WGIF04, показывают быстрое исчезновение холеста 5,24 в ходе экспрессии Δ24-редуктазы (сравнивая результаты, полученные с WGIF03, и результаты, полученные с WGIF04). Это позволяет предсказать очень эффективный синтез холестерина в штамме, в котором одновременно разрушены гены erg5 и erg6, и который совместно экспрессирует Δ7-редуктазу и Δ24-редуктазу; в этом случае холестерин оказывается единственным конечным стерином.
В завершение минимум, необходимый для продукции холестерина в количестве, большем или равном 20% от общих стеринов, состоит в разрушении гена erg6, экспрессии Δ7-редуктазы и Δ24-редуктазы. Дополнительное разрушение Δ22-десатуразы, по-видимому, может позволить улучшить производительность в отношении холестерина и устранить паразитное образование холеста 5,22 диенола в качестве конечного стерина.
ПРИМЕР 16
Изотопное мечение холестерина и определение изотопных подписей
Принцип получения меченого холестерина описан на фиг.10. Эта манипуляция состоит в том, что сначала выращивают дрожжи на глюкозе, полностью меченой 13C, в течение 72 часов при 30°C.
Клетки затем получают центрифугированием среды при 600 g в течение 10 минут. Осадок клеток затем суспендируют в 50 мл свежей среды Kappeli, содержащей 2% галактозы, не меченой углеродом 13C. Культуры останавливают через 2 часа, 4 часа, 8 часов или 24 часа после переноса на галактозу, стерины экстрагируют, затем анализируют (см. пример 7). Переход от глюкозы к галактозе провоцирует индукцию промоторов GAL10/CYC1, контролирующих одновременно ген Δ7-редуктазы и ген Δ24-редуктазы. Одновременно происходит изменение изотопной метки источника углерода. Происходит синтез de novo промежуточных продуктов метаболизма, затем стерина, включая холестерин, что сопровождается постепенной заменой метки. Это изменение метки может быть охарактеризовано на основании профиля масс каждого промежуточного стерина. Действительно, включение каждого атома 13С приводит к изменению массы на одну атомную единицу массы (АЕМ). Так, например, холестерин проявляется с массой, варьирующей от 386 до 413 дальтон, в зависимости от уровня мечения. В анализе путем ВЭЖХ-масс с использованием электрораспыления с положительной ионизацией это соответствует величинам m/z (масса/заряд), варьирующим от 369 до 396 (ион М+Н+ - Н2О, т.е. М+1-18=385-17=369). Время удержания стеринов в ВЭЖХ не зависит в обнаружимой степени от уровня мечения, спектр масс пика ВЭЖХ, соответствующего единственному стерину «Х», соответствует массовому распределению, которое является таким образом наложением (суммой) массовых распределений стерина "X", синтезированного в различные моменты времени после мечения. Речь идет, таким образом, о сложном профиле (фиг.11), но вполне определимом экспериментально, который представляет единственную изотопную подпись, которая зависит одновременно:
1) от протокола мечения и, в частности, сроков и условий культивирования в 12C глюкозе и в 13C галактозе;
2) точной генетической структуры используемого штамма;
3) точного времени остановки культур.
У этого изотопного профиля имеются некоторые уникальные свойства:
1) его можно при необходимости менять за счет изменений условий культивирования, используемого штамма и выбранного стерина. Таким образом, может быть получен уникальный регистр меток;
2) он является "комбинируемым", а именно некоторые изотопные подписи, соответствующие некоторым уникальным стеролам, меченым изотопными профилями, которые сами по себе являются модулируемыми, могут быть скомбинированы с получением «молекулярного алфавита»;
3) он является воспроизводимым и легко определимым экспериментально (см. фиг.11 и 12), двойные и тройные линии, которые указывают на воспроизводимость профилей;
4) он соответствует трассирующей молекулярной смеси, легкой в выделении, стабильной, бесцветной и лишенной запаха, нелетучей, нетоксичной, и которая может быть внедрена в продукты питания, в лекарственное средство, добавки или другие продукты, потребляемые человеком;
5) он не поддается фальсификации, если не известны специфические рекомбинантные штаммы и очень точные условия мечения, культивирования и экстракции. К тому же знание изотопной подписи не позволяет выявить параметры, которые могли бы позволить осуществить его получение.
Таким образом, изобретен не поддающийся фальсификации «изотопный алфавит» общего назначения, который может быть включен в любой тип продуктов, в том числе употребляемых в пищу. Число «изотопных слов», которые можно построить, исходя из такого алфавита, является практически неограниченным благодаря использованию одновременно профилей мечения и различных типов стеринов. Включение таких подписей в состав разнообразных продуктов представляет собой, таким образом, уникальный способ не поддающейся фальсификации маркировки, в отличие, например, от подписей ДНК, которые, став один раз известными, могут быть воспроизведены. Подпись может быть к тому же считана способами, не наносящими повреждений, например лазерной ионизацией с последующим анализом масс-спектометрией (MALDI-TOF и т.п.).
ПРИМЕР 17
Продукция не животного холестерина, в высокой степени меченого 13 C
Использование 13C галактозы или 13C глюкозы и этанола вместо не меченых источников углерода для культуры штамма WGIP04 в условиях, описанных выше в отношении анализов стеринов, позволяет осуществить синтез стеринов, и, в частности, очень сильно меченого холестерина (включающего по меньшей мере 95% углерода 13C). Получение стеринов и радиоактивного холестерина 14C возможно также с использованием того же подхода. Способ также может быть использован в отношении штаммов дрожжей, продуцирующих стероиды, в частности гидрокортизон (см. заявку на патент WO 02/061109) для получения стероидов, меченых 13C или 14C, например, для тестов RIA.
ПРИМЕР 18
Конструирование штамма, продуцирующего в основном холестерин
Штамм CDR07 Мata описан в заявке на патент, опубликованной под номером WO 02061109, и депонирован в Национальной Коллекции Культур Микроорганизмов (CNCM), 25 rue du Docteur Roux, 75724 Paris Cedex 15, France, согласно положениям Будапештского Соглашения 24 января 2001 под порядковым номером 1-2616.
Штамм CDR07 МАТа (релевантные маркеры: ade2::GAL10/CYCl p ::Δ 7 ; erg5::PGKl p ::hygro R ; ERG6) был скрещен со штаммом WGIF01, описанным в примере 1 (релевантные маркеры: ERG5; erg6::TRP1).
После диплоидного спорообразования определяли стериновый состав спор, чтобы найти споры, продуцирующие десмостерол, который является предшественником холестерина, как описано в примере 6. Идентифицировали таким образом спору, продуцирующую десмостерол и обладающую функциональным геном TRP1, и несущую кДНК Δ7 редуктазы Arabidopsis thaliana, как показывает анализ ПЦР. Кроме того, этот штамм, названный YIM59, чувствителен к гигромицину, что показывает, что ген ERG5 является функциональным.
Получение стеринов, осуществленное, как описано в примерах 6 и 9, показало, что этот штамм YTM59 продуцирует стерины, имеющие то же время удержания, что и зимостерол и десмостерол. Штамм YTM59 также показал ауксотрофность в отношении аденина, лейцина, урацила и гистидина. Присутствие десмостерола доказывает, что этот штамм экспрессирует стерол Δ7-редуктазу, и что он несет не функциональный аллель erg6.
Для улучшения уровня экспрессии человеческого DHCR24 нуклеотидная последовательность кДНК DHCR24 была модифицирована; для улучшения трансляции на уровне инициатора ATG, промотор, контролирующий экспрессию DHCR24, также был модифицирован. Новый выбранный промотор представляет собой промотор CYC1 цитохрома c1 вместо индуцируемого промотора GAL1 плазмида pYES_Delta24 (см. пример 3).
Последовательность, соответствующая NH2-концу DHCR24 редуктазы, слитому с CYC1, была модифицирована следующим образом:
tagcgtggatggccaggcaactttagtgctgacacatacaggcatatatatatgtgtgcgacgacacatgatcatatggcatgcatgtgctdgfatgtatataaaactttgttttcttctWctctaaatattcttttacttctatagacacgcaaacacaaaggaattgacaagtttgtacaaaaaagcaggctaaaaaATGGAACCTGCC GTGTCGCTGGCCGTGTGCG (SEQ ID № 7).
Маленькими символами обозначена частичная нуклеотидная последовательность промотора CYC1, затем последовательности рекомбинации AttB1, затем последовательность AAAA, которая предшествует инициатору ATG. Последовательность первых двух кодонов была также модифицирована (последовательность GAA-CCT после кодона инициатора ATG).
Конечная плазмида, несущая кДНК DHCR24 под контролем промотора CYC1, а также сайт начала репликации 2µ S. cerevisiae и селективный маркер URA3-d, была названа pIM331. Ее эквивалент без кДНК DHCR24 был назван рIМ303.
Штамм YIM59 трансформировали независимо с помощью плазмид pIM303 и pIM331 и специфически отбирали два трансформанта, несущих плазмиду pIM303 (штамм YIM59/pIM303) или рIМЗЗ 1 (штамм YIM59/pIM331).
Эти штаммы культивировали в богатой реконструированной среде типа Kappeli в течение 72 часов при 28°C для достижения поглощения 40 при 600 нм. Экстракты общих стеринов (этерифицированные стерины и свободные стерины) штамма YIM59/pIM303 (не несущего кДНК DHCR24) и штамма YTM59/pIM331 (несущего кДНК DHCR24) получали в присутствии метанольного гидроксида калия (см. примеры 5 и 6). Эти два штамма тестировали в отношении их способности продуцировать холестерин. Часть результатов (GC) представлена на фиг.13. Представленные времена удержания даны в минутах на обеих хроматограммах.
Таким образом, удалось показать, что штамм, который не несет вектор экспрессии DHCR24 (штамм YIM59/pIM303), не продуцирует холестерин (часть A), но главным образом десмостерол (часть A). Наоборот, штамм, который несет кДНК DHCR24 (штамм YTM59/pIM331), продуцирует стерин, имеющий время удержания, характерное для холестерина (часть B). Методами хроматографии в газовой фазе, соединенной с масс-спектрометрией с электронным ударом (как описано на примере 6), удалось показать, что этот стерин является холестерином. С использованием площадей каждого из пиков стерина было показано, что 57% стеринов, продуцируемых штаммом YTM59/pIM331, составляет холестерин.
Депонирование биологического материала
Следующие организмы были депонированы 22 апреля 2004 в Национальной Коллекции Культур Микроорганизмов (CNCM), 25 rue du Docteur Roux, 75724 Paris Cedex 15, France согласно положениям Будапештского Соглашения.
Штамм WGIF04 депонирован под регистрационным номером 1-3203.
Все процитированные публикации и патенты включены в настоящую заявку путем ссылки.
(коэффициенты экстинкции мM-1см-1)
Список литературы
1. Adams, A.K., and C. Holm. 1996. Mol Cell Biol. 16:4614-20.
2. Ausubel Fred M., Brent Roger, Kingston Robert E., Moore David D., Seidman J.G., Smith John A., Struhl Kevin (Editeurs). Current Protocols in Molecular Biology, publie par John Wiley & Sons Inc. Current Protocols Customer Service, 605 Third Avenue, 9th Floor New York, NY 10158 USA (edition mise a jour de Mars 2002).
3. Baudin-Baillieu, A., E. Guillemet, C. Cullin, and F. Lacroute. 1997. Yeast. 13:353-6.
4. Bonneaud, N., O. Ozier-Kalogeropoulos, G.Y. Li, M. Labouesse, L. Minvielle-Sebastia, andF. Lacroute. 1991. Yeast. 7:609-15.
5. Bowman, S. et al. 1997. Nature. 387:90-3.
6. Duport, C, B. Schoepp, E. Chatelain, R. Spagnoli, B. Dumas, and D. Pompon. 2003. Eur J Biochem. 270:1502-14.
7. Gietz, R.D., R.H. Schiestl, A.R. Willems, and R.A. Woods. 1995. Yeast. 11:355-60.
8. Goffeau, A., et al. 1996. Science. 274:546, 563-7.
9. Jensen-Pergakes, K.L., M.A. Kennedy, N.D. Lees, R. Barbuch, C. Koegel, and M. Bard. 1998. Antimicrob Agents Chemother. 42:1160-7.
10. Kaneshiro, E.S., J.A. Rosenfeld, M. Basselin-Eiweida, J.R. Stringer, S.P. Keely, A.G. Smulian, and J.L. Giner. 2002. Mol Microbiol. 44:989-99.
11. Kappeli, O., M. Arreguin, and M. Rieger. 1985. J Gen Microbiol. 131:1411-6.
12. Kelly, S.L., D.C. Lamb, B.C. Baldwin, A.J. Corran, and D.E. Kelly. 1997. J Biol Chem. 272:9986-8.
13. Lecain, E., X. Chenivesse, R. Spagnoli, and D. Pompon. 1996. J Biol Chem. 271:10866-73.
14. McCammon, M.T., M.A. Hartmann, CD. Bottema, and L.W. Parks. 1984. J Bacteriol. 157:475-83.
15. Moebius, F.F., B.U. Fitzky, J.N. Lee, Y.K. Paik, and H. Glossmann. 1998. Proc Natl Acad Sci USA. 95:1899-902.
16. Noda, EL, and Y. Koizumi. 2003. Insect Biochem Mol Biol. 33:649-58.
17. Ozier-Kalogeropoulos, O., A. Malpertuy, J. Boyer, F. Tekaia, and B. Dujon. 1998. Nucleic Acids Res. 26:5511-24.
18. Skaggs, B.A., J.F. Alexander, C.A. Pierson, K.S. Schweitzer, K.T. Chun, C. Koegel, R. Barbuch, and M. Bard. 1996. Gene. 169:105-9.
19. Sturley, S.L. 2000. Biochim Biophys Acta. 1529:155-63.
20. Szczebara, F.M., et al. 2003. Nat Biotechnol 21:143-9.
21. Tait, E., M.C. Simon, S. King, A.J. Brown, N.A. Gow, and D.J. Shaw. 1997. Fungal Genet Biol. 21:308-14.
22. Taton, M., and A. Rahier. 1991. Biochem Biophys Res Commun. 181:465-73. Taylor, F.R., and T.W. Parks. 1978. J Bacteriol. 136:531-7.
23. Waterham, H.R., J. Koster, G.J. Romeijn, R.C. Hennekam, P. Vreken, H.C. Andersson, D.R. FitzPatrick, R.I. Kelley, and R.J. Wanders. 2001. Am J Hum Genet. 69:685-94.
24. Wood, V., et al. 2002. Nature. 415:871-80.


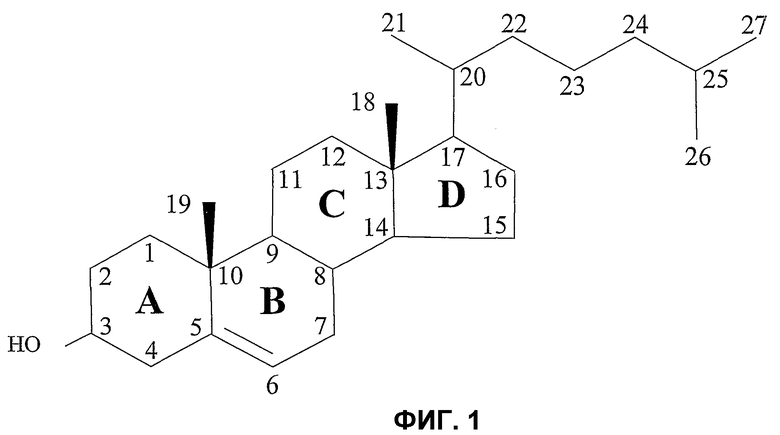
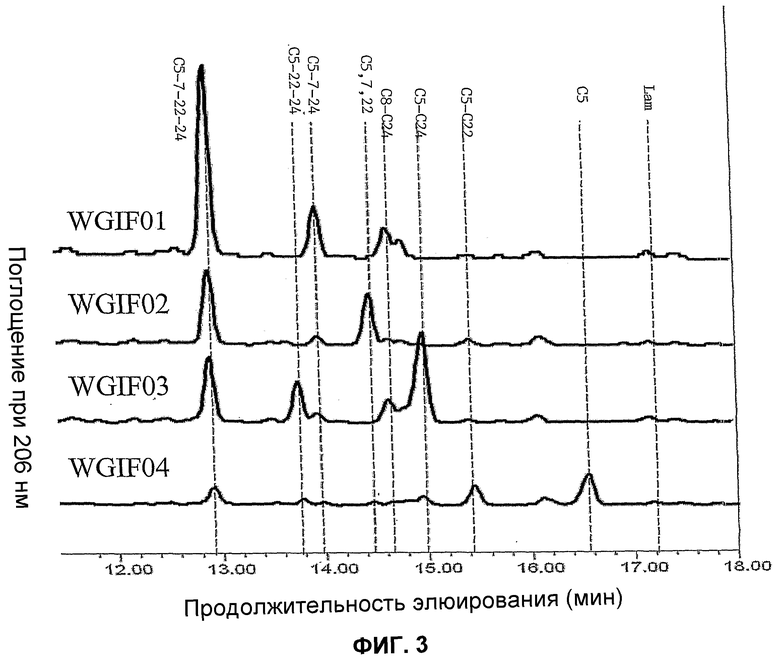
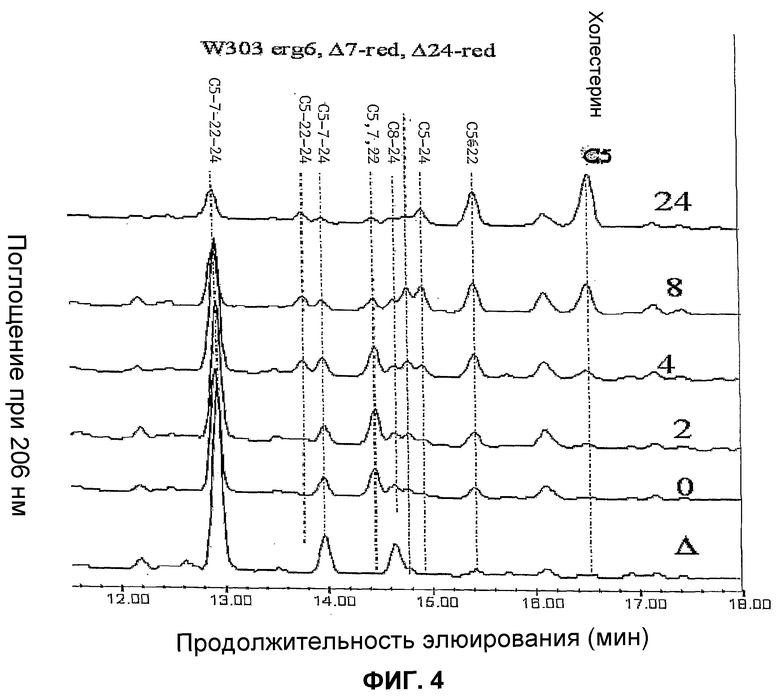
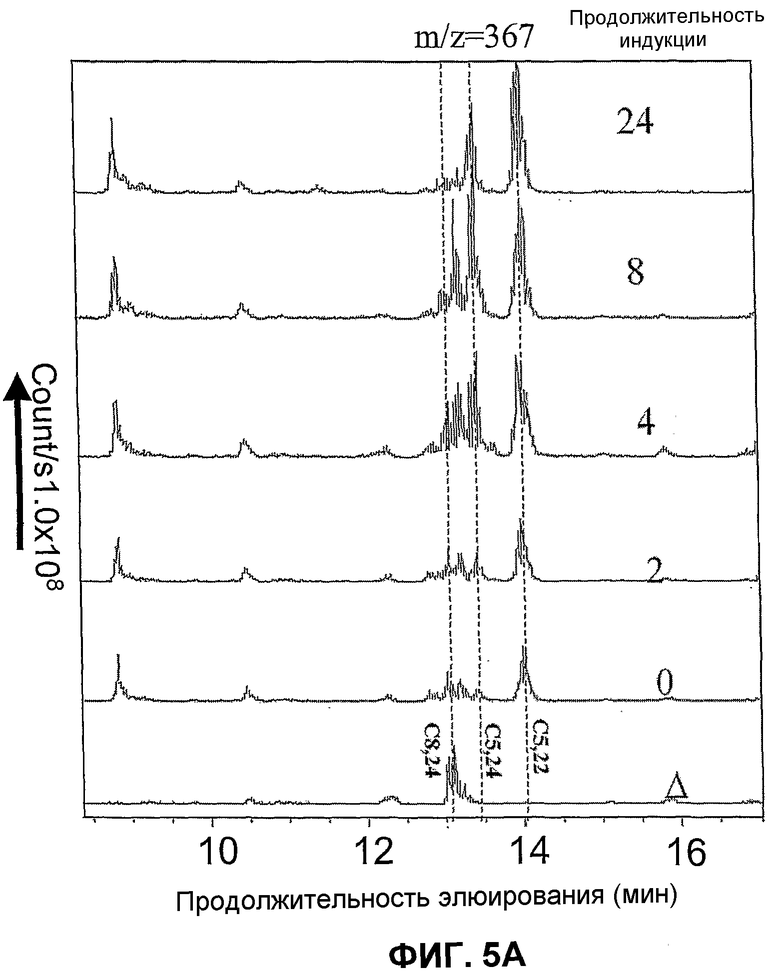
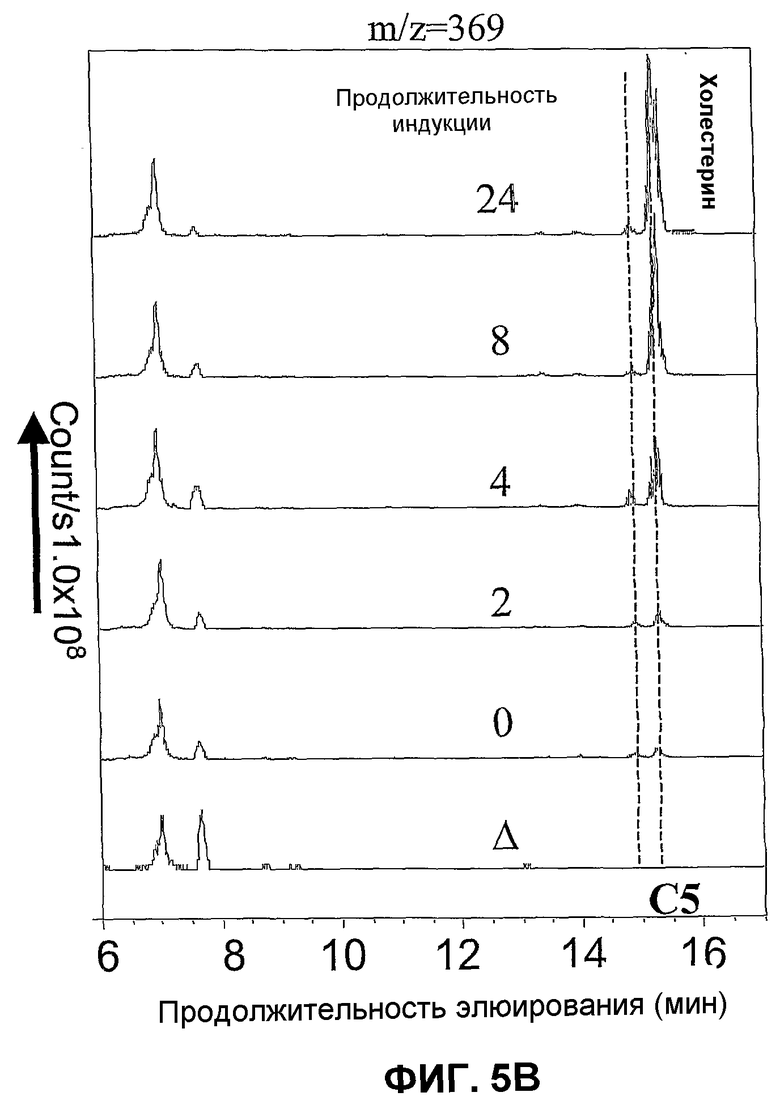
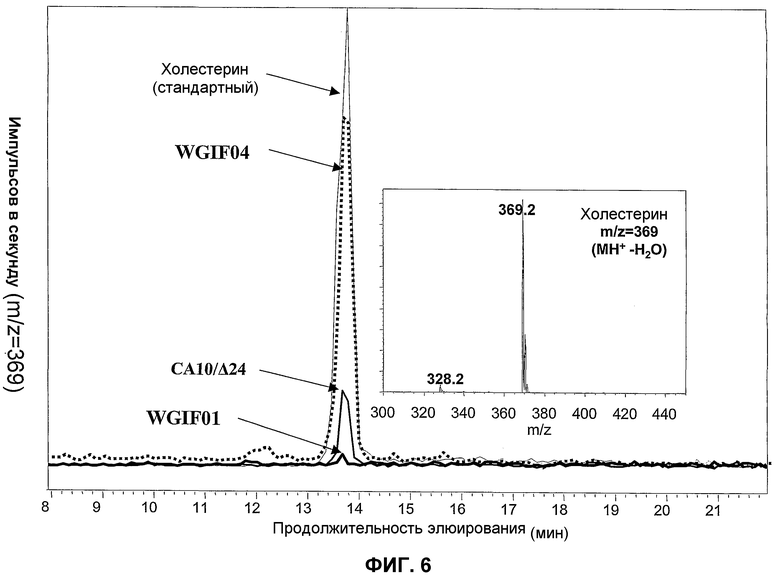

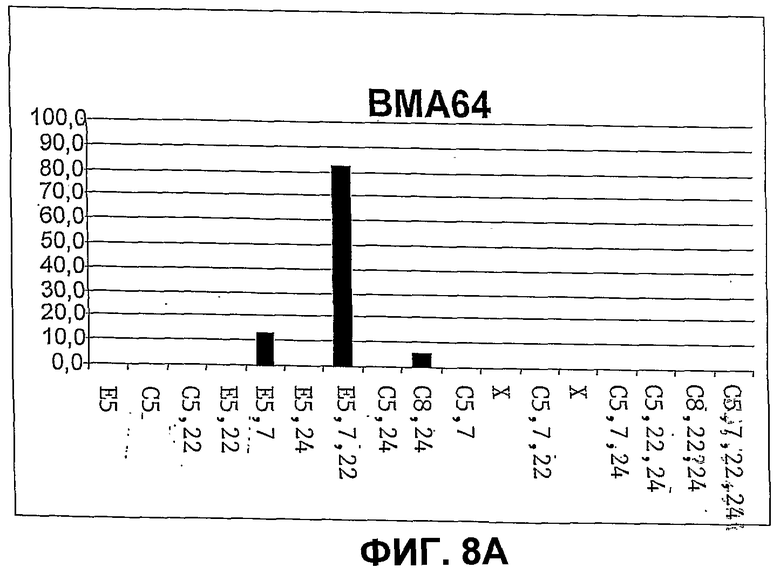
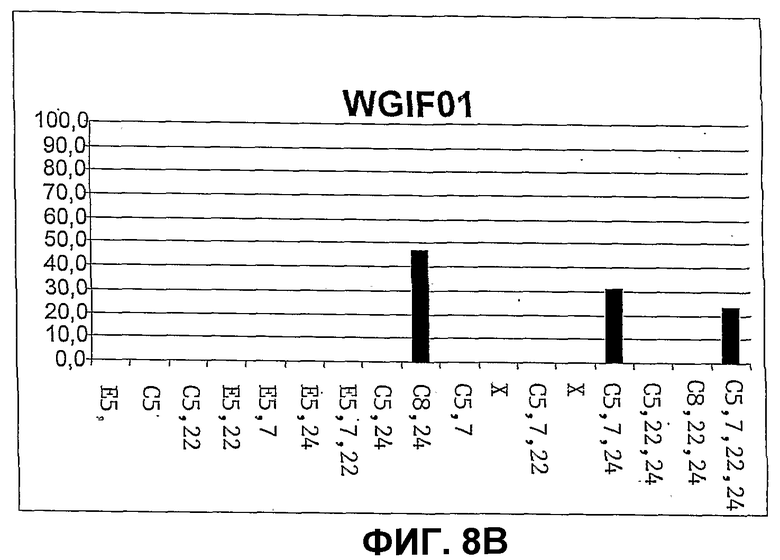
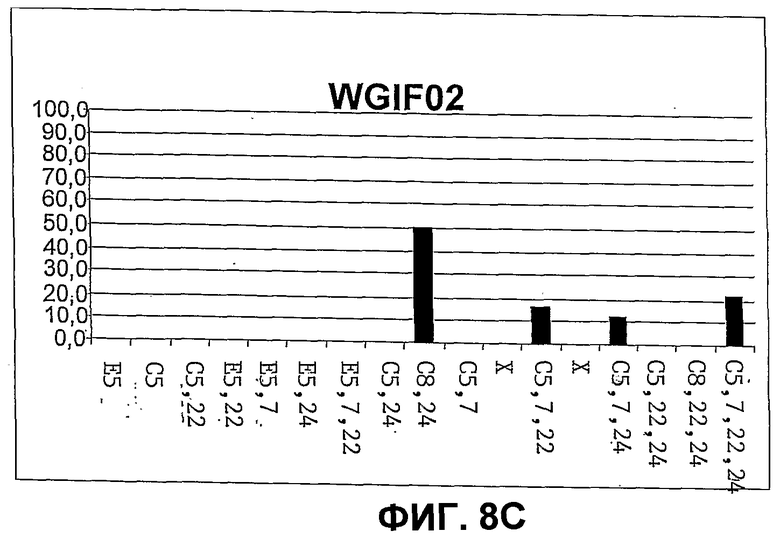
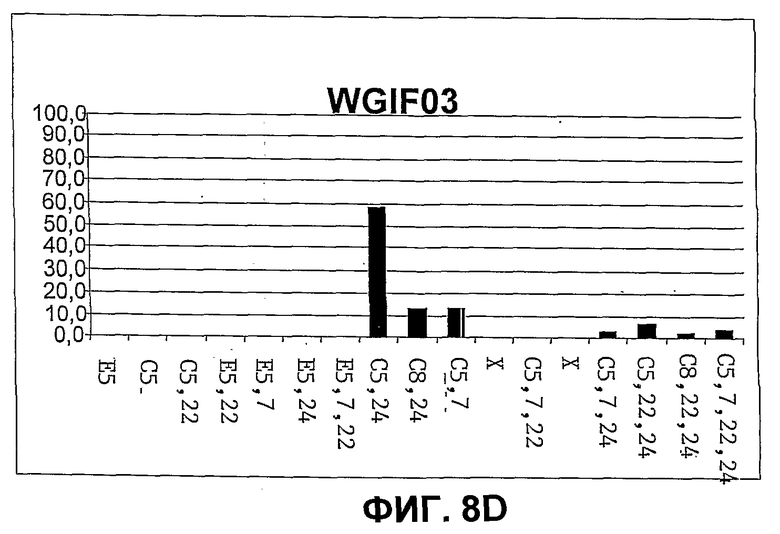
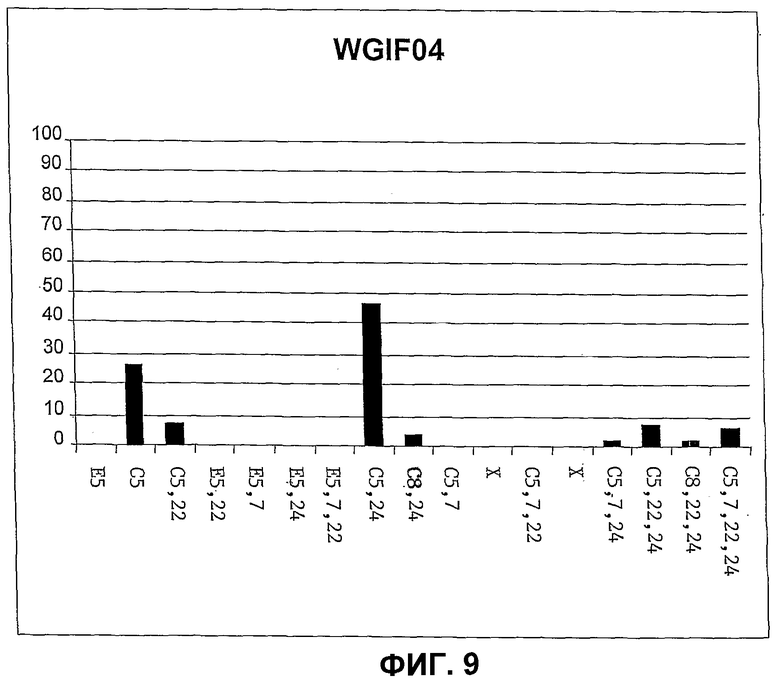
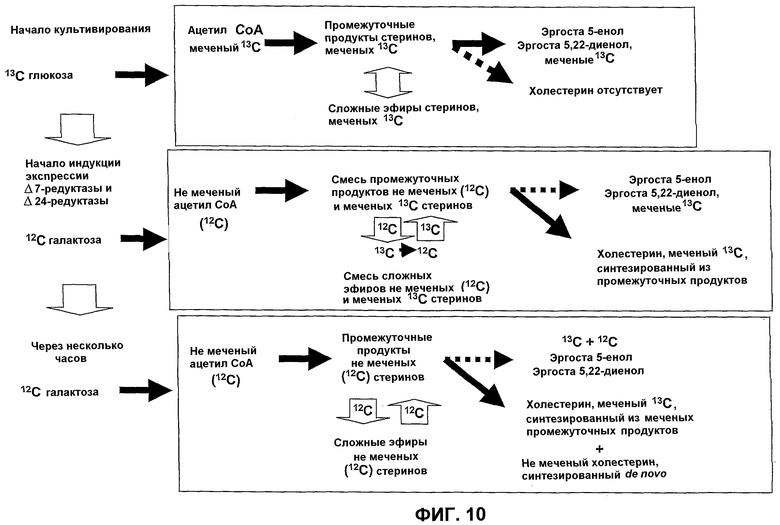
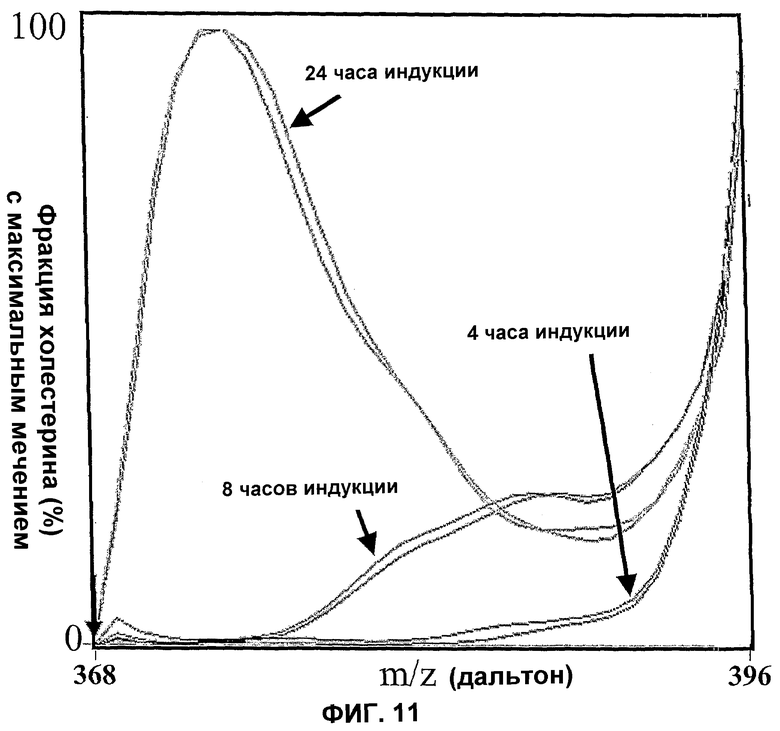
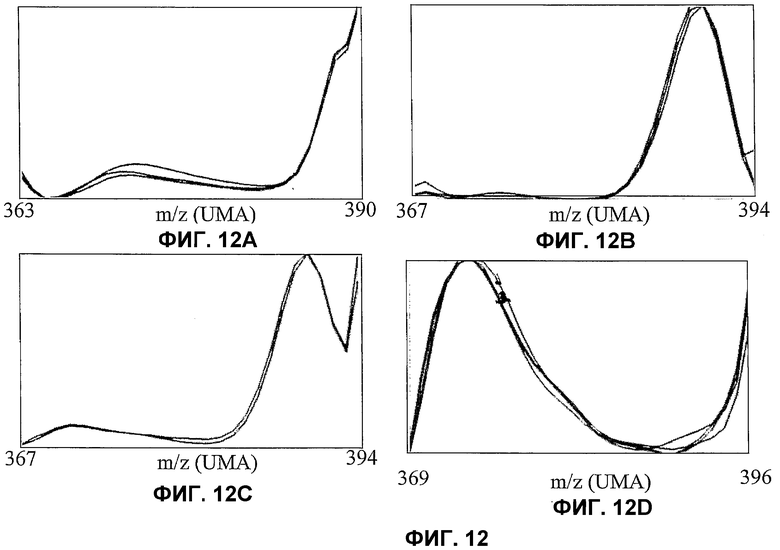
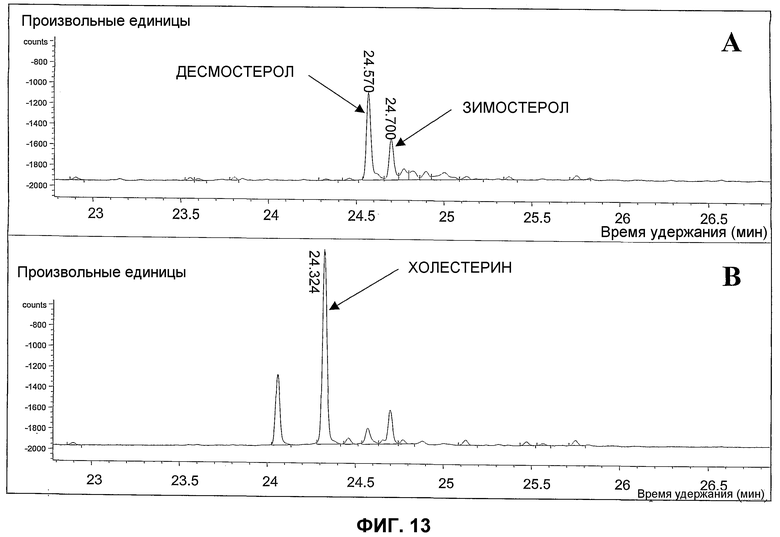
Настоящее изобретение относится к генетически модифицированным дрожжам, автономно продуцирующим холестерин исходя из простого источника углерода. Данные дрожжи экспрессируют ферменты 7-дегидрохолестерин редуктазу и 3β-гидроксистерин Δ24-редуктазу, а фермент стерол 24-С-метилтрансфераза у них инактивирован. Продукция холестерина у них равна или превышает 20% от всех продуцируемых стеринов. Кроме того, предлагается способ получения холестерина и применение штамма дрожжей по изобретению для получения не меченого и меченого холестерина. Данное изобретение позволяет получать безопасный с санитарной точки зрения холестерин в больших количествах. 5 н. и 7 з.п. ф-лы, 13 ил., 1 табл.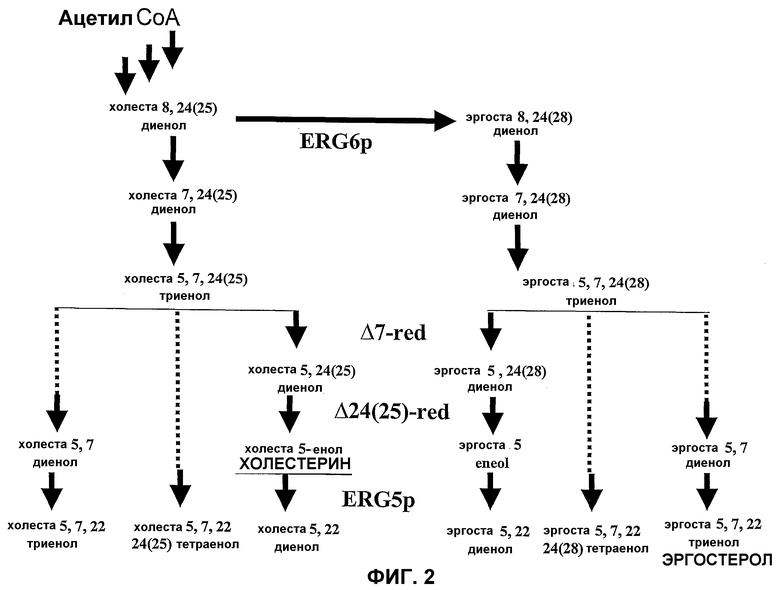
1. Генетически модифицированный штамм дрожжей, автономно продуцирующий холестерин, исходя из простого источника углерода, такого как глюкоза, галактоза, сахароза, этанол, глицерин или патоки, или субпродукты этих сахаров, отличающийся тем, что он экспрессирует ферменты 7-дегидрохолестерин редуктазу и 3β-гидроксистерин Δ24-редуктазу и тем, что фермент стерол 24-С-метилтрансфераза инактивирован, и тем, что продукция холестерина равна или превышает 20% от всех продуцируемых стеринов.
2. Штамм дрожжей по п.1, отличающийся тем, что экспрессию ферментов 7-дегидрохолестерин редуктазы и 3β-гидроксистерин Δ24-редуктазы получают трансформацией организма.
3. Штамм дрожжей по п.1, отличающийся тем, что инактивация фермента стерол 24-С-метилтрансферазы осуществлена генетической инактивацией.
4. Штамм дрожжей по п.1, отличающийся тем, что его выбирают из рода Saccharomyces или Schizosaccharomyces.
5. Штамм дрожжей Saccharomyces cerevisiae по п.4, отличающийся тем, что он представляет собой штамм WGIF04, депонированный в Национальной Коллекции Культур Микроорганизмов (CNCM) 22 апреля 2004 под регистрационным номером I-3203.
6. Способ получения холестерина неживотного происхождения, включающий культивирование штамма дрожжей по одному из пп.1-5 и экстракцию холестерина.
7. Способ по п.6, отличающийся тем, что экстракцию холестерина осуществляют с помощью растворителя, не смешивающегося с водой.
8. Способ по п.6, отличающийся тем, что перед экстракцией холестерина осуществляют стадию омыления.
9. Способ по п.6 или 8, отличающийся тем, что перед омылением или экстракцией холестерина осуществляют стадию механического дробления клеток.
10. Применение штамма дрожжей по одному из пп.1-5 для получения холестерина или одного из промежуточных продуктов его метаболизма, или смеси стеринов, меченой 13С или 14С.
11. Способ получения холестерина или одного из промежуточных продуктов его метаболизма, или смеси стеринов, меченой 13С или 14С, включающий следующие стадии:
культивирование штамма дрожжей по одному из пп.1-5 на галактозе или глюкозе, меченой или нет 13С или 14С, и
экстракция указанного холестерина или одного из промежуточных продуктов его метаболизма, или смеси стеринов.
12. Способ получения изотопной смеси холестерина, промежуточных продуктов или метаболитов холестерина, меченых в различных положениях при помощи изотопных меток 13С или 14С, включающий культивирование штамма дрожжей по одному из пп.1-5 на меченой галактозе или глюкозе, затем на немеченой галактозе или глюкозе, причем продолжительность культивирования на каждом из этих субстратов выбирают таким образом, чтобы получить определенный изотопный профиль, и экстракцию изотопной смеси холестерина, промежуточных продуктов или метаболитов холестерина.
| XU S | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| OSTLUND J.R.E | |||
| Насос | 1917 |
|
SU13A1 |
| Journal of lipid research, 1993, v.34, n.10, p.1825-1831 | |||
| Подвеска сиденья | 1977 |
|
SU727489A2 |

