По настоящей заявке испрашивается приоритет предварительной заявки на патент США № 60/801172, поданной 17 мая 2006, полное содержание которой включено в настоящее описание посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области биотехнологии. Более конкретно, настоящее изобретение относится к идентификации фрагментов нуклеиновых кислот, кодирующих фермент дельта-5-десатуразу жирных кислот, и применению этой десатуразы для получения полиненасыщенных жирных кислот (PUFA) с длинными цепями.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Важность PUFA бесспорна. Например, некоторые PUFA являются важными биологическими компонентами здоровых клеток и признаются «существенными» жирными кислотами, которые не могут быть синтезированы de novo у млекопитающих, а вместо этого должны быть получены или вместе с пищей, или с помощью дальнейшего уменьшения насыщенности и элонгации линолевой кислоты (LA; 18:2 омега-6) или α-линолевой кислоты (ALA; 18:3 омега-3); составными частями плазматических мембран клеток, в которых их можно обнаружить в таких формах, как фосфолипиды или триацилглицерины необходимыми для правильного развития (особенно при развитии головного мозга детей младшего возраста) и для формирования и восстановления ткани; и предшественниками нескольких биологически активных эйкозаноидов, важных для млекопитающих (например, простациклинов, эйкозаноидов, лейкотриенов, простагландинов). Кроме того, прием в большом количестве PUFA омега-3 с длинными цепями вызывает эффекты защиты сердечно-сосудистой системы (Dyerberg, J. et al., Amer. J. Clin. Nutr., 28: 958-966 (1975); Dyerberg, J. et al., Lancet, 2(8081): 117-119 (July 15, 1978); Shimokawa, H., World Rev. Nutr. Diet., 88: 100-108 (2001); von Schacky, C. and Dyerberg, J., World Rev. Nutr. Diet., 88: 90-99 (2001)). И в многочисленных других исследованиях подтверждается польза для здоровья в широком диапазоне, предоставляемая введением PUFA омега-3 и/или омега-6, против множества симптомов и заболеваний (например, астмы, псориаза, экземы, диабета, рака).
Ряд различных хозяев, включающих растения, водоросли, грибы и дрожжи, исследуются в качестве средств для коммерческого получения PUFA. С помощью генной инженерии продемонстрировано, что природные способности некоторых хозяев (даже тех, которые природно ограничены в продукции жирных кислот LA и ALA) можно в значительной степени изменить для получения в результате продукции на высоком уровне различных PUFA омега-3/омега 6 с длинными цепями. Независимо от того, является ли это результатом естественных возможностей или рекомбинантной технологии, для продукции арахидоновой кислоты (ARA; 20:4 омега-6), эйкозапентаеновой кислоты (EPA; 20:5 омега-3) и докозагексаеновой кислоты (DHA; 22:6 омега-3) может быть необходима экспрессия дельта-5-десатуразы.
Большинство идентифицированных до настоящего времени ферментов дельта-5-десатураз имеют первичную способность превращать дигомо-гамма-линоленовую кислоту (DGLA; 20:3 омега-6) в ARA, при этом вторичной активностью является превращение эйкозатетраеновой кислоты (ЕТА; 20:4 омега-3) в ЕРА (причем из ЕРА после реакции с дополнительной С20/22-элонгазой и дельта-4-десатуразой впоследствии синтезируется DHA). Дельта-5-десатураза играет роль как в пути с участием дельта-6-десатуразы/дельта-6-элонгазы (который главным образом обнаружен в водорослях, мхах, грибах, нематодах и у людей и который характеризуется продукцией гамма-линоленовой кислоты (GLA; 18:3 омега-6) и/или стеаридоновой кислоты (STA; 18:4 омега-3)), так и в пути с участием дельта-9-элонгазы/дельта-8-десатуразы (который функционирует в некоторых организмах, таких как жгутиковые, и который характеризуется продукцией эйкозадиеновой кислоты (EDA; 20:2 омега-6) и/или эйкозатриеновой кислоты (ETrA; 20:3 омега-3)) (фиг.1).
Основываясь на том, какую роль ферменты дельта-5-десатуразы играют в синтезе, например, ARA, EPA и DHA, прилагалось значительное усилие для идентификации и характеристики этих ферментов из различных источников. В качестве таковых, многочисленные дельта-5-десатуразы были раскрыты как в открытых источниках информации (например, № доступа в GenBank - AF199596; № AF226273, № AF320509, № AB072976, № AF489588, № AJ510244, № AF419297, № AF07879, № AF067654 и № AB022097), так и в патентных источниках информации (например, патенты США № 5972664 и 6075183). Также в находящейся в совладении и в процессе одновременного рассмотрения заявке, имеющей № предварительной заявки 60/801119 (поданной 17 мая 2006), раскрываются аминокислотная и нуклеотидная последовательности фермента дельта-5-десатуразы из вида CCMP626 рода Peridium, в то время как в находящейся в совладении и в процессе одновременного рассмотрения заявке, имеющей № предварительной заявки 60/915733 (ВВ1614) (поданной 3 мая 2007), раскрываются аминокислотная и нуклеотидная последовательности фермента дельта-5-десатуразы из Euglena anabaena.
Настоящее изобретение относится к идентификации и выделению дополнительных генов, кодирующих дельта-5-десатуразы из Euglena gracilis, которые могли бы быть пригодны для гетерологичной экспрессии в ряде организмов-хозяев для применения в продукции жирных кислот омега-3/омега-6.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к выделенному полинуклеотиду, включающему
(а) нуклеотидную последовательность, кодирующую полипептид, обладающий активностью дельта-5-десатуразы, причем полипептид обладает по меньшей мере 70%, 75%, 80%, 85%, 90%, 95% или 100% идентичности аминокислотной последовательности, на основе способа выравнивания Clustal W, при сравнении с аминокислотной последовательностью, представленной в SEQ ID NO: 2;
(b) нуклеотидную последовательность, кодирующую полипептид, обладающий активностью дельта-5-десатуразы, причем нуклеотидная последовательность обладает по меньшей мере 70%, 75%, 80%, 85%, 90%, 95% или 100% идентичности последовательности, на основе способа выравнивания BLASTN, при сравнении с нуклеотидной последовательностью, представленной в SEQ ID NO: 1 или SEQ ID NO: 3;
(с) нуклеотидную последовательность, кодирующую полипептид, обладающий активностью дельта-5-десатуразы, причем нуклеотидная последовательность гибридизуется в жестких условиях с нуклеотидной последовательностью, представленной в SEQ ID NO: 1 или SEQ ID NO: 3; или
(d) комплемент нуклеотидной последовательности (а), (b) или (с), причем комплемент и нуклеотидная последовательность состоят из одинакового числа нуклеотидов и комплементарны на 100%.
Во втором варианте осуществления настоящее изобретение относится к рекомбинантной ДНК-конструкции, включающей любой из выделенных полинуклеотидов по настоящему изобретению, функционально связанный с по меньшей мере одной регуляторной последовательностью.
В третьем варианте осуществления настоящее изобретение относится к клетке, содержащей в своем геноме рекомбинантную ДНК-конструкцию по настоящему изобретению. Такие клетки могут быть клетками растений или дрожжевыми клетками.
В четвертом варианте осуществления настоящее изобретение относится к способу трансформации клетки, включающему трансформацию клетки рекомбинантной конструкцией по настоящему изобретению или выделенным полинуклеотидом по настоящему изобретению и отбор тех клеток, которые трансформированы рекомбинантной конструкцией или выделенным полинуклеотидом.
В пятом варианте осуществления настоящее изобретение относится к трансгенному семени, содержащему в своем геноме рекомбинантную конструкцию по настоящему изобретению, или трансгенному семени, полученному от растения, полученного способом по настоящему изобретению. Также представляет интерес масло или побочные продукты, получаемые из таких трансгенных семян.
В шестом варианте осуществления настоящее изобретение относится к способу получения полиненасыщенных жирных кислот с длинными цепями в клетке растения, включающему
(а) трансформацию клетки рекомбинантной конструкцией по настоящему изобретению; и
(b) отбор тех трансформированных клеток, которые продуцируют полиненасыщенные жирные кислоты с длинными цепями.
В седьмом варианте осуществления настоящее изобретение относится к способу получения по меньшей мере одной полиненасыщенной жирной кислоты в клетке растения с масличными семенами, включающему
(а) трансформацию клетки растения с масличными семенами первой рекомбинантной ДНК-конструкцией, включающей выделенный полинуклеотид, кодирующий по меньшей мере один полипептид дельта-5-десатуразы, функционально связанный с по меньшей мере одной регуляторной последовательностью, и по меньшей мере одной дополнительной рекомбинантной ДНК-конструкцией, включающей выделенный полинуклеотид, функционально связанный с по меньшей мере одной регуляторной последовательностью, кодирующий полипептид, выбираемый из группы, состоящей из дельта-4-десатуразы, дельта-5-десатуразы, дельта-6-десатуразы, дельта-8-десатуразы, дельта-12-десатуразы, дельта-15-десатуразы, дельта-17-десатуразы, дельта-9-десатуразы, дельта-9-элонгазы, С14/16-элонгазы, С16/18-элонгазы, С18/20-элонгазы и С20/22-элонгазы;
(b) регенерацию растения с масличными семенами из трансформированной клетки стадии (а); и
(с) отбор тех полученных от растений стадии (b) семян, которые имеют измененный уровень полиненасыщенных жирных кислот по сравнению с уровнем в семенах, полученных от не трансформированного растения с масличными семенами.
В восьмом варианте осуществления настоящее изобретение относится к растению с масличными семенами, содержащему в своем геноме рекомбинантную конструкцию по настоящему изобретению. Подходящие растения с масличными семенами включают, без ограничения, сою, виды Brassica, подсолнечник, кукурузу, хлопок, лен и сафлор.
В девятом варианте осуществления настоящее изобретение относится к растению с масличными семенами, содержащему
(а) первую рекомбинантную ДНК-конструкцию, включающую выделенный полинуклеотид, кодирующий по меньшей мере один полипептид дельта-5-десатуразы, функционально связанный с по меньшей мере одной регуляторной последовательностью, и
(b) по меньшей мере одну дополнительную рекомбинантную ДНК-конструкцию, включающую выделенный полинуклеотид, функционально связанный с по меньшей мере одной регуляторной последовательностью, кодирующий полипептид, выбираемый из группы, состоящей из дельта-4-десатуразы, дельта-5-десатуразы, дельта-6-десатуразы, дельта-8-десатуразы, дельта-12-десатуразы, дельта-15-десатуразы, дельта-17-десатуразы, дельта-9-десатуразы, дельта-9-элонгазы, С14/16-элонгазы, С16/18-элонгазы, С18/20-элонгазы и С20/22-элонгазы.
Также представляют интерес трансгенные семена, полученные из таких растений с масличными семенами, а также масло или побочные продукты, получаемые из этих трансгенных семян. Предпочтительным побочным продуктом является лецитин.
В десятом варианте осуществления настоящее изобретение относится к пищевому продукту или корму, включающему масло или семя по настоящему изобретению, или пищевому продукту или корму, включающему ингредиент, получаемый в процессе переработки семян.
В одиннадцатом варианте осуществления настоящее изобретение относится к потомству растений, полученному от растения, полученного способом по настоящему изобретению, или растения с масличными семенами по настоящему изобретению.
КРАТКОЕ ОПИСАНИЕ ФИГУР И СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
На фиг. 1 иллюстрируется путь биосинтеза жирных кислот омега-3/омега-6.
На фиг. 2 показана хроматограмма липидного профиля экстракта клеток Euglena gracilis, описанного в примере 1.
На фиг. 3 показана часть выравнивания белков дельта-5-десатураз и белков дельта-8-десатураз и между ними, используя анализ Clustal W (программа MegAlignTM программного обеспечения DNASTAR).
На фиг. 4 графически представлена взаимосвязь между SEQ ID NO: 1, 2, 4, 5, 6, 8 и 10, каждая из которых относится к дельта-5-десатуразе Euglena gracilis.
На фиг. 5А иллюстрируется стратегия клонирования, используемая для амплификации гена дельта-5-десатуразы Euglena gracilis (EgD5). На фиг. 5В представлена карта плазмиды pZUF17, в то время как на фиг. 5С представлена карта плазмиды pDMW367.
На фиг. 6 предоставляются карты следующих плазмид: (А) pKUNF12T6E; (B) pEgD5S и (C) pDMW369.
На фиг. 7 показано сравнение последовательности ДНК гена дельта-5-десатуразы Euglena gracilis (обозначенного «EgD5»; SEQ ID NO: 1) и синтетического гена (обозначенного «EgD5S»; SEQ ID NO: 3), оптимизированного в отношении кодонов для экспрессии в Yarrowia lipolytica.
На фиг. 8А и 8В показано выравнивание Clustal V (с использованием параметров по умолчанию) дельта-8-десатуразы Pavlova lutheri (SEQ ID NO: 18), дельта-8-десатуразы Pavlova salina (SEQ ID NO: 64), дельта-8-десатуразы Euglena gracilis (SEQ ID NO: 16) и двух различных дельта-6-десатураз жирных кислот Rhizopus stolonifer (SEQ ID NO: 51 и 63).
На фиг. 9 представлена карта плазмиды pY98.
На фиг. 10A представлены профили жирных кислот Yarrowia lipolytica, который экспрессирует pY98 (SEQ ID NO: 76; включающую ген дельта-5-десатуразы Mortierella alpina (обозначенный «MaD5»)), или pDMW367 (SEQ ID NO: 23; включающую ген дельта-5-десатуразы Euglena gracilis, обозначенный «EgD5») и которому обеспечивали различные субстраты. На фиг. 10В представлено сравнение специфичности MaD5 и EgD5 в отношении субстратов омега-3 и омега-6.
На фиг. 11 представлены карты следующих плазмид: (А) pKR916; (B) pKR1037 и (C) pKR328.
На фиг. 12А представлены средние профили жирных кислот для десяти случаев наличия наибольшей дельта-5-десатуразной активности, когда в зародыши сои трансформируют фермент Mortierella alpina (MaD5). На фиг. 12В представлены средние профили жирных кислот для десяти случаев наличия наибольшей дельта-5-десатуразной активности, когда в зародыши сои трансформируют фермент Euglena gracilis (EgD5). Жирные кислоты определяют как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), LA, ALA, EDA, SCI, DGLA, ARA, ERA, JUP, ЕТА и ЕРА. Жирные кислоты, перечисленные как «другие», включают 18:2 (5,9), GLA, STA, 20:0, 20:1(11), 20:2 (7,11) или 20:2 (8,11) и DPA. Относительное содержание каждой из этих «других» жирных кислот составляет менее 3% от жирных кислот в целом. Составы жирных кислот для отдельного зародыша выражали в виде весового процента (вес%) от жирных кислот в целом, а средний состав жирных кислот представляет собой среднее шести отдельных зародышей для каждого случая. На фиг. 12В показано, что активность EgD5 в зародышах сои является очень высокой, при этом среднее превращение (% правильной дельта-5-десатурации) составляет от 77% до 99% в десяти случаях наличия наибольшей дельта-5-десатуразной активности.
На фиг. 13 представлена активность дельта-5-десатуразы в отношении «правильных» субстратов («% правильной дельта-5-десатурации»), откладываемая по X-оси, в сравнении с активностью дельта-5-десатуразы в отношении «неправильных» субстратов («% неправильной дельта-5-десатурации»), откладываемой по Y-оси, для MaD5 (смотри фиг. 12А) и EgD5 (смотри фиг. 12В). По сравнению с MaD5 специфичность EgD5 в отношении субстратов больше в отношении «правильных» субстратов по сравнению с «неправильными» субстратами.
Полнее настоящее изобретение можно понять из следующего подробного описания и описаний сопроводительных последовательностей, составляющих часть настоящей заявки.
Следующие последовательности соответствуют 37 C.F.R. § 1.821-1.825 («требованиям к заявкам на патенты, содержащим сообщения о нуклеотидных последовательностях и/или аминокислотных последовательностях - правилам для последовательностей») и согласуются со стандартом ST.25 (1998) Всемирной организации по защите интеллектуальной собственности (WIPO) и требованиями к списку последовательностей Европейского патентного ведомства и РСТ (правилами 5.2 и 49.5 (a-bis), и разделом 208 и приложением С Административных инструкций). Для символов и формата, используемых для данных в виде нуклеотидных и аминокислотных последовательностей, соблюдаются правила, изложенные в 37 C.F.R. § 1.822.
SEQ ID NO: 1-26, 48, 49, 51-54, 61-64, 67-72 и 75-76 представляют собой ORF, кодирующие гены, или белки (или их части), или плазмиды, определенные в таблице 1.
Краткое изложение SEQ ID NO нуклеиновых кислот и белков
кислота
SEQ ID NO.
SEQ ID NO.
(«EgD5»)
(1350 п.о.)
(449 АК)
происходящая из Euglena gracilis,
оптимизированная в отношении кодонов для
экспрессии в Yarrowia lipolytica
(«EgD5S»)
(1350 п.о.)
(449 АК)
(590 п.о.)
(196 АК)
pT-EgD5-5'C2
(797 п.о.)
gracilis, родственная SEQ ID NO: 4
(559 п.о.)
pT-EgD5-5'2nd
(273 п.о.)
gracilis, родственная SEQ ID NO: 6
(20 п.о.)
pT-EgD5-3'
(728 п.о.)
gracilis, родственная SEQ ID NO: 4
(464 п.о.)
(№ доступа в GenBank - AAL13311)
(456 АК)
Phytophthora megasperma
(№ доступа в GenBank - CAD53323)
(477 АК)
Phaeodactylum tricornutum
(№ доступа в GenBank - AAL92562)
(469 АК)
Dictyostelium discoideum
(№ доступа в GenBank - XP_640331)
(467 АК)
(публикации РСТ-заявок № WO 2006/012325
и WO 2006/012326)
(421 АК)
(CCMP459)
(1269 п.о.)
(423 АК)
(7 АК)
(7 АК)
Thalassiosira pseudonana
(№ доступа в GenBank - AAX14502)
(476 АК)
(8165 п.о.)
(8438 п.о.)
(12649 п.о.)
происходящий из Thraustochytrium aureum
(патент США № 6677145),
оптимизированный в отношении кодонов для
экспрессии в Yarrowia lipolytica
(«EL2S»)
(819 п.о.)
(272 АК)
(4070 п.о.)
(8438 п.о.)
Rhizopus stolonifer
(NCBI № доступа - AAX22052)
(459 АК)
часть вставки кДНК из клона
eps1c.pk002.f22
(5'-конец вставки кДНК)
(695 п.о.)
полностью секвенированная EST
eps1c.pk002.f22:fis
(полная последовательность вставки)
(1106 п.о.)
трансляция нуклеотидов 1-864
полностью секвенированной EST
eps1c.pk002.f22:fis
(полной последовательности вставки;
SEQ ID NO:53)
(287 АК)
полная 5'-концевая последовательность
от прогулки по геному
(1294 п.о.)
собранная последовательность
(1927 п.о.)
Rhizopus stolonifer
(NCBI № доступа - ABB96724)
(459 АК)
(427 АК)
(1338 п.о.)
(446 АК)
(6473 п.о.)
(6970 п.о.)
фосфат-дегидрогеназы
Yarrowia lipolytica (GPD)
(968 п.о.)
(8630 п.о.)
(6339 п.о.)
(8319 п.о.)
(«EgD9е»)
(774 п.о.)
(«EgD8»)
(1263 п.о.)
(4311 п.о.)
(7085 п.о.)
(2540 п.о.)
(4359 п.о.)
(5147 п.о.)
(6559 п.о.)
(9014 п.о.)
(5561 п.о.)
(11889 п.о.)
(5661 п.о.)
(5578 п.о.)
(11907 п.о.)
(8671 п.о.)
(«SdD5»)
(1413 п.о.)
SEQ ID NO: 27-30 соответствуют вырожденным олигонуклеотидным праймерам 5-1А, 5-1В, 5-1С и 5-1D соответственно, которые кодируют консервативный район 1.
SEQ ID NO: 31-34 соответствуют вырожденным олигонуклеотидным праймерам 5-5AR, 5-5BR, 5-5СR и 5-5DR соответственно, которые кодируют консервативный район 2.
SEQ ID NO: 35-40 соответствуют праймерам ODMW480, CDSIII 5', ODMW479, DNR CDS 5', YL791 и YL792 соответственно, используемым для 5'-RAGE.
SEQ ID NO: 41-43 соответствуют праймерам ODMW469, AUAP и ODMW470, соответственно, используемым для 3'-RAGE.
SEQ ID NO: 44-47 соответствуют праймерам YL794, YL797, YL796 и YL795 соответственно, используемым для амплификации полноразмерной кДНК EgD5.
SEQ ID NO: 50 соответствует праймеру T7, используемому для секвенирования библиотеки кДНК Pavlova lutheri (CCMP459).
SEQ ID NO: 55 и 56 соответствуют праймерам SeqE и SeqW соответственно, используемым для секвенирования клонов Pavlova lutheri (CCMP459).
SEQ ID NO: 57 и 58 соответствуют универсальному праймеру AP1 и праймеру GSP PvDES соответственно, используемым для амплификации геномной ДНК Pavlova lutheri (CCMP459).
SEQ ID NO: 59 и 60 соответствуют праймерам M13-28Rev и PavDESseq соответственно, используемым для секвенирования геномных вставок Pavlova lutheri (CCMP459).
SEQ ID NO: 65 и 66 соответствуют праймеру AP и олигонуклеотидному праймеру Smart IV соответственно, используемым для синтеза кДНК Pavlova lutheri.
SEQ ID NO: 73 и 74 соответствуют праймерам GPDsense и GPDantisense соответственно, используемым для амплификации промотора GPD.
SEQ ID NO: 79 и 80 соответствуют праймерам oEugEL1-1 и oEugEL1-2 соответственно, используемым для амплификации дельта-9-элонгазы Euglena gracilis (EgD9e).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Все упоминаемые здесь патенты, заявки на патенты и публикации полностью включаются в настоящее описание посредством ссылки. Они, в частности, включают следующие находящиеся в процессе одновременного рассмотрения заявки правопреемников заявителей: патент США № 7125672, патент США № 7189559, патент США № 7192762, патент США № 7198937, патент США № 7202356, заявки на патенты США № 10/840579 и № 10/840325 (поданные 6 мая 2004), заявку на патент США № 10/869630 (поданную 16 июня 2004), заявку на патент США № 10/882760 (поданную 1 июля 2004), заявки на патенты США № 10/985254 и № 10/985691 (поданные 10 ноября 2004), заявку на патент США № 11/024544 (поданную 29 декабря 2004), заявку на патент США № 11/166993 (поданную 24 июня 2005), заявку на патент США № 11/183664 (поданную 18 июля 2005), заявку на патент США № 11/185301 (поданную 20 июля 2005), заявку на патент США № 11/190750 (поданную 27 июля 2005), заявку на патент США № 11/198975 (поданную 8 августа 2005), заявку на патент США № 11/225354 (поданную 13 сентября 2005), заявку на патент США № 11/253882 (поданную 19 октября 2005), заявки на патенты США № 11/264784 и № 11/264737 (поданные 1 ноября 2005), заявку на патент США № 11/265761 (поданную 2 ноября 2005), заявку на патент США № 11/737772 (поданную 20 апреля 2007), заявку на патент США № 11/787772 (поданную 17 апреля 2007), заявку на патент США № 11/740298 (поданную 26 апреля 2007), заявки на патенты США № 60/801172 и № 60/801119 (поданные 17 мая 2006), заявку на патент США № 60/853563 (поданную 23 октября 2006), заявку на патент США № 60/855177 (поданную 30 октября 2006), заявки на патенты США № 11/601563 и № 11/601564 (поданные 16 ноября 2006), заявку на патент США № 11/635258 (поданную 7 декабря 2006), заявку на патент США № 11/613420 (поданную 20 декабря 2006), заявку на патент США № 60/909790 (поданную 3 апреля 2007), заявку на патент США № 60/911925 (поданную 16 апреля 2007), заявку на патент США № 60/910831 (поданную 10 апреля 2007) и заявку на патент США № 60/915733 (ВВ1614) (поданную 3 мая 2007). Они, кроме того, включают следующие находящиеся в процессе одновременного рассмотрения заявки правопреемников заявителей: публикацию заявки на патент № 2005/0136519, касающуюся продукции PUFA в растениях, и патент США № 7129089, касающийся промотора аннексина и его применения для экспрессии трансгенов в растениях.
Как используются в настоящем описании и в прилагаемой формуле изобретения, формы единственного числа включают формы множественного числа, если только из контекста ясно следует иное. Таким образом, например, ссылка на «растение» включает множество таких растений, ссылка на «клетку» включает одну или несколько клеток и их эквивалентов, известных квалифицированным в данной области техники специалистам, и т.д.
В соответствии с данным изобретением авторы идентифицируют новый фермент дельта-5-десатуразу Euglena gracilis и ген, кодирующий этот фермент, который можно использовать для манипуляции биохимическими путями для получения целебных PUFA. Таким образом, данное изобретение находит множество применений.
Полученные с помощью раскрытой здесь методологии PUFA, или их производные, можно использовать в качестве диетических заместителей, или добавок, в частности, составов для детей младшего возраста, для пациентов, подвергаемых внутривенному кормлению, или для профилактики или лечения нарушения питания. В альтернативном случае очищенные PUFA (или их производные) можно включить в кулинарные масла, жиры или маргарины, приготовленные так, что при стандартном использовании реципиент будет получать желаемое количество для добавки к рациону. PUFA можно также включить в составы для детей младшего возраста, дополнения к питанию или другие пищевые продукты, и они могут найти применение в качестве противовоспалительных или понижающих холестерин агентов. Необязательно композиции могут использоваться для фармацевтического применения (для человека или ветеринарного).
Дополнение рационов людей или животных PUFA, получаемых рекомбинантным способом, может привести к увеличению уровней добавленных PUFA, а также продуктов их метаболизма. Например, лечение ЕРА может привести не только к увеличению уровней ЕРА, но также нижеследующих продуктов ЕРА, таких как эйкозаноиды (т.е. простагландины, лейкотриены, тромбоксаны). Сложные регуляторные механизмы могут сделать желательным объединение различных PUFA или добавление различных конъюгатов PUFA для того, чтобы предотвратить, контролировать или преодолеть такие механизмы для достижения желаемых уровней конкретных PUFA у индивидуума.
Определения
В настоящем описании используется ряд терминов и сокращений. Предоставляются следующие определения.
«Открытая рамка считывания» - сокращенно ORF.
«Полимеразная цепная реакция» - сокращенно ПЦР.
«Американская коллекция типовых культур» - сокращенно АТСС.
«Полиненасыщенная жирная кислота(ы)» - сокращенно PUFA.
«Триацилглицерины» - сокращенно TAG.
Используемый здесь термин «изобретение» или «настоящее изобретение», как предусматривается, не ограничивается каким-либо одним конкретным вариантом осуществления по настоящему изобретению, а применяется обычно к любому и всем вариантам осуществления по настоящему изобретению, охарактеризованным в формуле изобретения и описании.
Термин «жирные кислоты» относится к длинноцепочечным алифатическим кислотам (алкановым кислотам) с варьирующими длинами цепей, от приблизительно С12 до С22 (хотя известны кислоты как с более длинными, так и более короткими цепями). Преобладающие длины цепей находятся между С16 и С22. Структура жирных кислот представляется с помощью простой системы обозначений «Х:Y», где Х представляет собой общее число атомов углерода (С) в конкретной жирной кислоте, а Y представляет собой число двойных связей. Дополнительные детали, касающиеся дифференциации между «насыщенными жирными кислотами» и «ненасыщенными жирными кислотами», между «мононенасыщенными жирными кислотами» и «полиненасыщенными жирными кислотами» (или «PUFA») и между «жирными кислотами омега-6» (омега-6 или n-6) и «жирными кислотами омега-3» (омега-3 или n-3), предоставлены в публикации заявки на патент США № 2005/0136519.
Жирные кислоты описываются здесь с помощью простой системы обозначений «Х:Y», где Х представляет собой общее число атомов углерода (С) в конкретной жирной кислоте, а Y представляет собой число двойных связей. Число, следующее за обозначением жирной кислоты, указывает положение двойной связи от карбоксильного конца жирной кислоты с аффиксом «с» для цис-конфигурации двойной связи (например, пальмитиновая кислота (16:0), стеариновая кислота (18:0), олеиновая кислота (18:1, 9с), петроселиновая кислота (18:1, 6с), LA (18:2, 9c, 12c), GLA (18:3, 6c, 9c, 12c) и ALA (18:3, 9c, 12c, 15c). Если не указано иное, 18:1, 18:2 и 18:3 относятся к олеиновой кислоте, LА и ALA соответственно. Если специально не написано иное, предполагается, что двойные связи находятся в цис-конфигурации. Например, предполагается, что двойные связи в 18:2 (9,12) находятся в цис-конфигурации.
Номенклатура, используемая для описания PUFA в настоящем описании, продемонстрирована ниже в таблице 2. В колонке, озаглавленной «Сокращенное обозначение», используется ссылочная омега-система для указания числа атомов углерода, числа двойных связей и положение двойной связи, ближайшей к омега-углероду, считая от омега-углерода (которому присваивают номер 1 для этой цели). В остальной части таблицы суммируются общепринятые названия жирных кислот омега-3 и омега-6 и их предшественников, сокращения, которые будут использоваться на протяжении описания, и химическое название каждого соединения.
Номенклатура полиненасыщенных жирных кислот и предшественников
название
обозначение
октадекадиеновая
омега-6
октадекатриеновая
омега-6
эйкозадиеновая
омега-6
линолевая
эйкозатриеновая
омега-6
эйкозатетраеновая
омега-6
октадекатриеновая
омега-6
октадекатетраеновая
омега-3
ERA
эйкозатриеновая
омега-3
эйкозатриеновая
омега-6
эйкозатетраеновая
омега-3
эйкозатетраеновая
омега-3
эйкозапентаеновая
омега-3
докозапентаеновая
омега-3
докозагексаеновая
омега-3
Термины «триацилглицерин», «масло» или «TAG» относятся к нейтральным липидам, состоящим из трех ацильных остатков жирных кислот, этерифицированных до молекулы глицерина (и такие термины будут использоваться здесь взаимозаменяемо на протяжении описания по настоящему изобретению). Такие масла могут содержать длинноцепочечные PUFA, а также более короткие насыщенные и ненасыщенные жирные кислоты и насыщенные жирные кислоты с более длинными цепями. Таким образом, «биосинтез масла», как правило, относится к синтезу TAG в клетке.
«Процент (%) PUFA в общей липидной и масляной фракциях» относится к проценту PUFA относительно жирных кислот в целом в этих фракциях. Термин «общая липидная фракция» или «липидная фракция» относится к сумме всех липидов (т.е нейтральных и полярных) в содержащем или продуцирующем масло организме, включая, таким образом, те липиды, которые находятся в фосфатидилхолиновой (РС) фракции, фосфатидилэтаноламиновой (РЕ) фракции и триацилглицериновой (TAG или масляной) фракции. Однако термины «липид» и «масло» будут использоваться взаимозаменяемо на протяжении этого описания.
Метаболический путь или биосинтетический путь, в биохимическом смысле, можно рассматривать как ряд химических реакций, происходящих в клетке, катализируемых ферментами, с достижением или образованием метаболического продукта, используемого или сохраняемого клеткой, или инициации другого метаболического пути (затем называемой этапом создания потока). Многие из этих путей являются сложными и включают модификацию шаг за шагом первоначального вещества с формированием продукта, имеющего точную химическую структуру, которая является желаемой.
Термин «путь биосинтеза PUFA» относится к метаболическому процессу, который превращает олеиновую кислоту в LA, EDA, GLA, DGLA, ARA, ALA, STA, ETrA, ETA, EPA, DPA и DHA. Этот процесс хорошо описан в литературе (например, смотри публикацию РСТ-заявки № WO 2006/052870). Вкратце, в этот процесс вовлечена элонгация углеродной цепи благодаря добавлению атомов углерода и уменьшения насыщенности молекулы благодаря добавлению двойных связей, с помощью ряда специальных ферментов уменьшения насыщенности и элонгации (т.е. «ферментов пути биосинтеза PUFA»), присутствующих в мембране эндоплазматического ретикулума. Более конкретно «ферменты пути биосинтеза PUFA» относятся к любому из следующих ферментов (и генов, которые кодируют указанные ферменты), связанных с биосинтезом PUFA, включающих дельта-4-десатуразу, дельта-5-десатуразу, дельта-6-десатуразу, дельта-12-десатуразу, дельта-15-десатуразу, дельта-17-десатуразу, дельта-9-десатуразу, дельта-8-десатуразу, дельта-9-элонгазу, С14/16-элонгазу, С16/18-элонгазу, С18/20-элонгазу и С20/22-элонгазу.
Термин «путь биосинтеза жирных кислот омега-3/омега-6» относится к множеству генов, которые при экспрессии в соответствующих условиях кодируют ферменты, которые катализируют продукцию одной из двух или обеих жирных кислот омега-3 и омега-6. Как правило, гены, вовлеченные в путь биосинтеза жирных кислот омега-3/омега-6, кодируют ферменты пути биосинтеза PUFA. На фиг. 1 проиллюстрирован характерный путь, обеспечивающий превращение миристиновой кислоты через различные промежуточные химические соединения в DHA, который демонстрирует, каким образом обе жирные кислоты омега-3 и омега-6 могут быть продуцированы из общего источника. В природе путь делится на две части, причем одна часть будет создавать жирные кислоты омега-3, а другая часть - только жирные кислоты омега-6. Та часть, которая создает только жирные кислоты омега-3, будет упоминаться здесь как путь биосинтеза жирных кислот омега-3, в то время как та часть, которая создает только жирные кислоты омега-6, будет упоминаться здесь как путь биосинтеза жирных кислот омега-6.
Термин «функциональный», используемый здесь в контексте с путем биосинтеза жирных кислот омега-3/омега-6, означает, что некоторые (или все) гены в этом пути экспрессируют активные ферменты, приводящие к in vivo катализу или превращению субстрата. Следует понимать, что термин «путь биосинтеза жирных кислот омега-3/омега-6» или «функциональный путь биосинтеза жирных кислот омега-3/омега-6» не подразумевает, что необходимы все гены, перечисленные в вышеприведенном параграфе, поскольку для ряда продуктов жирных кислот будет только необходима экспрессия подмножества генов этого пути.
Термин «путь с участием дельта-6-десатуразы/дельта-6-элонгазы» относится к пути биосинтеза PUFA, который минимально включает по меньшей мере одну дельта-6-десатуразу и по меньшей мере одну С18/20-элонгазу, делая, таким образом, возможным биосинтез DGLA и/или ETA из LA и ALA соответственно с GLA и/или STA в качестве промежуточных жирных кислот. При экспрессии других десатураз и элонгаз могут также синтезироваться ARA, EPA, DPA и DHA.
Термин «путь с участием дельта-9-элонгазы/дельта-8-десатуразы» относится к пути биосинтеза PUFA, который минимально включает по меньшей мере одну дельта-9-элонгазу и по меньшей мере одну дельта-8-десатуразу, делая, таким образом, возможным биосинтез DGLA и/или ETA из LA и ALA соответственно с EDA и/или ETrA в качестве промежуточных жирных кислот. При экспрессии других десатураз и элонгаз могут также синтезироваться ARA, EPA, DPA и DHA.
Термин «промежуточная жирная кислота» относится к любой жирной кислоте, продуцируемой в пути метаболизма жирных кислот, которая может далее превратиться в намеченный продукт в виде жирной кислоты в этом пути под действием других ферментов метаболического пути. Например, когда продуцируется ЕРА с использованием пути с участием дельта-9-элонгазы/дельта-8-десатуразы, могут продуцироваться EDA, ETrA, DGLA, ETA и ARA, которые считаются «промежуточными жирными кислотами», поскольку эти жирные кислоты могут далее превратиться в ЕРА под действием других ферментов метаболического пути.
Термин «побочная жирная кислота» относится к любой жирной кислоте, продуцируемой в пути метаболизма жирных кислот, которая не является ни намеченным продуктом этого пути, ни «промежуточной жирной кислотой» этого пути. Например, когда продуцируется ЕРА с использованием пути с участием дельта-9-элонгазы/дельта-8-десатуразы, могут также продуцироваться сциадоновая кислота (SCI) и юнипероновая кислота (JUP) под действием дельта-5-десатуразы или на EDA, или на ETrA соответственно. Считается, что они являются «побочными жирными кислотами», поскольку ни одна из них не может далее превратиться в ЕРА под действием других ферментов метаболического пути.
Термин «десатураза» относится к полипептиду, который может десатурировать, т.е. вводить двойную связь в одну или несколько жирных кислот с получением представляющей интерес жирной кислоты или предшественника. Несмотря на использование ссылочной омега-системы на протяжении описания для ссылки на конкретные жирные кислоты, более удобно указывать активность десатуразы, ведя счет с карбоксильного конца субстрата с использованием дельта-системы. Особый интерес здесь представляют дельта-5-десатуразы, которые катализируют превращение DGLA в ARA и/или ETA в EPA. Другие десатуразы включают: 1) дельта-17-десатуразы, которые десатурируют жирную кислоту между 17 и 18 атомами углерода, пронумерованными с концевого карбоксильного конца молекулы, и которые, например, катализирует превращение ARA в EPA и/или DGLA в ETA; 2) дельта-6-десатуразы, которые катализирует превращение LA в GLA и/или ALA в STA; 3) дельта-12-десатуразы, которые катализирует превращение олеиновой кислоты в LA; 4) дельта-15-десатуразы, которые катализирует превращение LA в ALA и/или GLA в STA; 5) дельта-4-десатуразы, которые катализирует превращение DPA в DHA; 6) дельта-8-десатуразы, которые катализирует превращение EDA в DGLA и/или ETrA в ETA; и 7) дельта-9-десатуразы, которые катализирует превращение пальмитата в пальмитолеиновую кислоту (16:1) и/или стеарата в олеиновую кислоту. В данной области техники дельта-15- и дельта-17-десатуразы также иногда упоминаются как «омега-3-десатуразы», «w-3-десатуразы» и/или «омега-3-десатуразы» на основе их способности превращать жирные кислоты омега-6 в их аналоги омега-3 (т.е. превращать LA в ALA и ARA в EPA соответственно). В некоторых вариантах осуществления по настоящему изобретению особенно желательно эмпирически определить специфичность конкретной десатуразы жирной кислоты с помощью трансформации подходящего хозяина геном для десатуразы жирной кислоты и определения ее эффекта на профиль жирных кислот хозяина.
Термин «дельта-5-десатураза» относится к ферменту, который десатурирует жирную кислоту между пятым и шестым атомами углерода, пронумерованными с концевого карбоксильного конца молекулы. Предпочтительно дельта-5-десатураза превращает дигомо-гамма-линоленовую кислоту [20:3, DGLA] в арахидоновую кислоту [20:4, ARA] или превращает эйкозатетраеновую кислоту [20:4, ЕТА] в эйкозапентаеновую кислоту [20:5, ЕРА].
Для целей по настоящему изобретению термин «EgD5» относится к ферменту дельта-5-десатуразе (SEQ ID NO: 2), выделенной из Euglena gracilis, кодируемой SEQ ID NO: 1 здесь. Аналогично термин «EgD5S» относится к синтетической дельта-5-десатуразе, происходящей из Euglena gracilis, которая оптимизирована в отношении кодонов для экспрессии в Yarrowia lipolytica (т.е. SEQ ID NO: 3 и 2).
Термины «эффективность превращения» и «процент превращения субстрата» относятся к эффективности, с которой конкретный фермент (например, десатураза) может превращать субстрат в продукт. Эффективность превращения определяют по следующей формуле: ([продукт]/[субстрат + продукт])*100, где продукт включает непосредственный продукт и все продукты в пути, происходящие из него.
Термин «элонгаза» относится к полипептиду, который может удлинять углеродную цепь жирной кислоты с получением кислоты, которая длиннее на 2 углерода субстрата в виде жирной кислоты, на который действует элонгаза. Механизм этого процесса элонгации является многоступенчатым, связанным с синтазой жирных кислот, как описано в публикации заявки на патент США № 2005/0132442. Примерами реакций, катализируемых системами элонгаз, являются превращение GLA в DGLA, STA в ETA и EPA в DPA. Обычно субстратная избирательность элонгаз является довольно широкой, но разделяется как по длине цепи, так и по степени и типу ненасыщенности. Например, С14/16-элонгаза будет использовать С14 субстрат (например, миристиновую кислоту), С16/18-элонгаза будет использовать С16 субстрат (например, пальмитат), С18/20-элонгаза (также известная как дельта-6-элонгаза, поскольку термины могут использоваться взаимозаменяемо) будет использовать С18 субстрат (например, GLA, STA), и С20/22-элонгаза будет использовать С20 субстрат (например, EPA). Подобным образом, дельта-9-элонгаза способна катализировать превращение LA и ALA в EDA и ETrA соответственно. Важно отметить, что некоторые элонгазы имеют широкую специфичность, и, следовательно, один фермент может быть способен катализировать несколько реакций элонгазы (например, действуя как в качестве С16/18-элонгазы, так и в качестве С18/20-элонгазы).
Термин «содержащие или продуцирующие масло» относится к таким организмам, которые имеют тенденцию запасать свой энергетический источник в форме липида (Weete, In: Fungal Lipid Biochemistry, 2nd Ed., Plenum, 1980). Термин «содержащие или продуцирующие масло дрожжи» относится к таким организмам, отнесенным к дрожжам, которые могут продуцировать масло. Как правило, содержание масла или TAG в клетке содержащих или продуцирующих масло микроорганизмов следует сигмоидальной кривой, причем концентрация липида увеличивается до достижения максимума в поздней логарифмической или ранней стационарной фазе роста и затем постепенно снижается во время поздней стационарной фазы и фазы гибели (Yongmanitchai and Ward, Appl. Environ. Microbiol., 57: 419-25 (1991)). Для содержащих или продуцирующих масло микроорганизмов не является необычной аккумуляция избытка, составляющего приблизительно 25% их сухого клеточного веса, в виде масла. Примеры содержащих или продуцирующих масло дрожжей включают, но без ограничения, следующие роды: Yarrowia, Candida, Rhodotorula, Rhodosporidium, Cryptococcus, Trichosporon и Lipomyces.
Термин «Euglenophyceae» относится к группе одноклеточных бесцветных или фотосинтетических жгутиковых, обнаруживаемых живущими в пресноводном, морском, почвенном и паразитическом окружениях. Класс характеризуется одиночными клетками, причем большинство из них являются свободноплавающими и имеют два жгутика (один из которых может быть не аварийным), возникающих из передней инвагинации, известной как вместилище. Фотосинтетические жгутиковые содержат от одной до множества хлоропластов, которые варьируют от небольших дисков до увеличенных в объеме пластин или лент. Бесцветные жгутиковые зависят от осмотрофии или фаготрофиии для усвоения пищи. Приблизительно 1000 видов было идентифицировано или отнесено к приблизительно 40 родам и 6 порядкам. Примеры Euglenophyceae включают, но без ограничения, следующие роды: Euglena, Eutreptiella и Tetruetreptia.
Термин «консервативная замена аминокислоты» относится к замене аминокислотного остатка в данном белке другой аминокислотой без изменения химической или функциональной природы этого белка. Например, в данной области техники хорошо известно, что изменения в гене, которые приводят к продукции химически эквивалентной аминокислоты в данном сайте (но которые не оказывают влияние на структурные и функциональные свойства кодируемого, уложенного белка), являются общепринятыми. Для целей по настоящему изобретению «консервативные замены аминокислот» определяются как замены в пределах одной из следующих пяти групп:
1) небольших алифатических, неполярных или слабополярных остатков: Ala[A], Ser[S], Thr[T] (Pro[P], Gly[G]);
2) полярных, отрицательно заряженных остатков и их амидов: Asp[D], Asn[N], Glu[E], Gln[Q];
3) полярных, положительно заряженных остатков: His[H], Arg[R], Lys[K];
4) больших алифатических, неполярных остатков: Met[M], Leu[L], Ile[I], Val[V] (Cys[C]); и
5) больших ароматических остатков: Phe[F], Tyr[Y], Trp[W].
Консервативные аминокислотные замены обычно сохраняют 1) структуру остова полипептида в области замены; 2) заряд или гидрофобность молекулы в целевом сайте; или 3) большую часть боковой цепи. Кроме того, во многих случаях также не ожидается, что изменения N-концевой и С-концевой частей молекулы белка приведут к изменению активности белка.
Как здесь используется, «нуклеиновая кислота» означает полинуклеотид и включает одно- или двухцепочечный полимер из дезоксирибонуклеотидных или рибонуклеотидных оснований. Нуклеиновые кислоты могут также включать фрагменты и модифицированные нуклеотиды. Таким образом, термины «полинуклеотид», «последовательность нуклеиновой кислоты», «нуклеотидная последовательность» или «фрагмент нуклеиновой кислоты» используются взаимозаменяемо и означают полимер РНК или ДНК, который является одно- или двухцепочечным, необязательно содержащим синтетические, неприродные или измененные нуклеотидные основания. На нуклеотиды (обычно обнаруживаемые в их 5'-монофосфатной форме) ссылаются с помощью обозначения одной буквой следующим образом: «А» для аденилата или дезоксиаденилата (для РНК или ДНК соответственно), «С» для цитидилата или дезоксицитидилата, «G» для гуанилата или дезоксигуанилата, «U» для уридлата, «T» для дезокситимидилата, «R» для пуринов (А или G), «Y» для пиримидинов (С или Т), «К» для G или Т, «Н» для А или С или Т, «I» для инозина и «N» для любого нуклеотида.
Термины «субфрагмент, являющийся функционально эквивалентным» и «функционально эквивалентный субфрагмент» используются здесь взаимозаменяемо. Эти термины относятся к части или подпоследовательности выделенного фрагмента нуклеиновой кислоты, в которой способность изменять экспрессию гена или порождать определенный фенотип сохраняется независимо от того, кодирует ли фрагмент или подфрагмент активный фермент или нет. Например, фрагмент или субфрагмент можно использовать для конструирования химерных генов для получения желаемого фенотипа в трансформированном растении. Химерные гены можно создать для использования в супрессии с помощью связывания фрагмента нуклеиновой кислоты или его субфрагмента, независимо от того, кодирует ли он активный фермент или нет, в смысловой или антисмысловой ориентации относительно последовательности промотора растения.
Термин «консервативный домен» или «мотив» означает совокупность аминокислот, являющихся консервативными в специфических положениях вдоль выровненной последовательности эволюционно родственных белков. Хотя аминокислоты в других положениях могут варьировать между гомологичными белками, аминокислоты, являющиеся в высокой степени консервативными в специфических положениях, указывают аминокислоты, являющиеся существенными для структуры, стабильности или активности белка. Поскольку их идентифицируют по их высокой степени консервативности в выровненных последовательностях семейства белков-гомологов, их можно использовать в качестве идентификаторов или «характерных признаков» для определения того, принадлежит ли белок с недавно определенной последовательностью семейству ранее идентифицированных белков.
Термины «гомология», «гомологичный», «в значительной степени схожий» или «соответствующий в значительной степени» используются здесь взаимозаменяемо. Они относятся к фрагментам нуклеиновых кислот, в которых замены одного или нескольких нуклеотидных оснований не нарушают способность фрагмента нуклеиновой кислоты опосредовать экспрессию гена или порождать определенный фенотип. Эти термины также относятся к модификациям фрагментов нуклеиновых кислот по настоящему изобретению, таким как делеция или вставка одного или нескольких нуклеотидов, не изменяющих в значительной степени функциональных свойств результирующего фрагмента нуклеиновой кислоты относительно первоначального, немодифицированного фрагмента. Поэтому понятно, как это понятно квалифицированным в данной области техники специалистам, что настоящим изобретением охватываются не только конкретные, приведенные в качестве примеров последовательности.
Кроме того, квалифицированный специалист знает, что в значительной степени схожие последовательности нуклеиновых кислот, охватываемые настоящим изобретением, также определяются по их способности гибридизоваться (в умеренно жестких условиях (например, в 0,5Х SSC, 0,1% SDS при 60°С)) с последовательностями, приведенными здесь в качестве примеров, или с любой частью нуклеотидных последовательностей, раскрытых здесь, и функционально эквивалентны любой из последовательностей нуклеиновых кислот, раскрытых здесь. Для скринирования от умеренно схожих фрагментов, таких как гомологичные последовательности из отдаленно родственных организмов, до в высокой степени схожих фрагментов, таких как гены, дублирующие функциональные ферменты из близкородственных организмов, условия жесткости можно отрегулировать. Условия жесткости определяют промывки после гибридизации.
Термин «избирательно гибридизуется» включает ссылку на гибридизацию, в жестких условиях гибридизации, последовательности нуклеиновой кислоты с точно установленной последовательностью нуклеиновой кислоты-мишенью в явно большей степени (например, в по меньшей мере 2 раза выше фона), чем степень ее гибридизации с не являющейся мишенью последовательностью нуклеиновой кислоты, и на значительное исключение не являющихся мишенями нуклеиновых кислот. Избирательно гибридизующиеся последовательности, как правило, идентичны на по меньшей мере 80% или 90%, вплоть до и включительно 100% (т.е. полностью комплементарны) друг с другом.
Термин «жесткие условия» или «жесткие условия гибридизации» включают ссылку на условия, при которых зонд избирательно гибридизуется со своей последовательностью-мишенью. Жесткие условия зависят от последовательности и будут различными при различных обстоятельствах. Путем контролирования жесткости гибридизации и/или условий промывок, можно идентифицировать последовательности-мишени, которые комплементарны на 100% зонду (гомологичное зондирование). В альтернативном случае жесткие условия можно установить так, чтобы сделать возможной некоторое несоответствие последовательностей, так что определяется меньшая степень схожести (гетерологичное зондирование). Как правило, размер зонда составляет приблизительно 1000 нуклеотидов, необязательно менее 500 нуклеотидов.
Как правило, жесткие условия будут такими условиями, при которых концентрация соли составляет менее приблизительно 1,5 М ионов Na, обычно приблизительно 0,01-1,0 М концентрация ионов Na (или других солей), при рН 7,0-8,3, а температура составляет по меньшей мере приблизительно 30°С для коротких зондов (например, из 10-50 нуклеотидов) и по меньшей мере приблизительно 60οС для длинных зондов (например, из более 50 нуклеотидов). Жестких условий можно также достигнуть добавлением дестабилизирующих агентов, таких как формамид. Примеры условий низкой жесткости включают гибридизацию с использованием буферного раствора, включающего 30-35% формамида, 1М NaCl, 1% SDS (натрия додецилсульфата), при 30°С и промывку в 1Х-2Х SSC (20Х SSC = 3,0 М NaCl/0,3 М тринатрия цитрат) при 50-55°С. Примеры условий средней жесткости включают гибридизацию в 40-45% формамида, 1М NaCl, 1% SDS при 37°С и промывку в 0,5Х-1Х SSC при 55-60°С. Примеры условий высокой жесткости включают гибридизацию в 50% формамида, 1М NaCl, 1% SDS при 37°С и промывку в 0,1Х SSC при 60-65°С.
Специфичность, как правило, функционально зависит от промывок после гибридизации, при этом решающими факторами являются ионная сила и температура конечного промывочного раствора. Для гибридов ДНК:ДНК Тm можно аппроксимировать из уравнения Meinkoth и др. (Anal. Biochem. 138: 267-284 (1984)): Тm = 81,5°С + 16,6 (log M) + 0,41 (% G+C) - 0,61 (% формамида) - 500/L, где М представляет собой молярность одновалентных катионов, % G+C представляет собой процент нуклеотидов, включающих гуанозин и цитозин, в ДНК; % формамида представляет собой % формамида в растворе для гибридизации, и L представляет собой длину гибрида в парах оснований. Тm представляет собой температуру (при точно определенных ионной силе и рН), при которой 50% комплементарной последовательности-мишени гибридизуется с полностью соответствующим зондом. Тm уменьшают на приблизительно 1°С для каждого 1% несовпадений; таким образом, Тm, условия гибридизации и/или промывок можно установить для гибридизации с последовательностями желаемой идентичности. Например, если искомыми являются последовательности, идентичные на ≥90%, Тm можно уменьшить на 10°С. Как правило, жесткие условия выбирают так, чтобы они были на приблизительно 5°С ниже температуры (точки) плавления (Тm) для конкретной последовательности и ее комплемента при заданных ионной силе и рН. Однако в существенно жестких условиях могут использоваться гибридизация и/или промывка при температуре на 1, 2, 3 или 4°С ниже температуры (точки) плавления (Тm); в умеренно жестких условиях могут использоваться гибридизация и/или промывка при температуре на 6, 7, 8, 9 или 10°С ниже температуры (точки) плавления (Тm); в условиях низкой жесткости могут использоваться гибридизация и/или промывка при температуре на 11, 12, 13, 14, 15 или 20°С ниже температуры (точки) плавления (Тm). Используя уравнение, композиции для гибридизации и промывок и требуемую Тm, специалисты со средним уровнем компетентности будут понимать, что изменения жестокости гибридизации и/или растворов для промывок по существу описаны. Если желаемая степень несоответствий приводит к Тm, составляющей меньше 45°С (водный раствор) или 32°С (раствор формамида), предпочтительно увеличить концентрацию SSC с тем, чтобы можно было использовать более высокую температуру. Подробное руководство для гибридизации нуклеиновых кислот можно найти у Tijssen (Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, Part I, Chapter 2 “Overview of principles of hybridization and the strategy of nucleic acid probe assays”, Elsevier, New York (1993)) и в Current Protocols in Molecular Biology, Chapter 2, Ausubel et al., Eds., Greene Publishing and Wiley-Interscience, New York (1995). Условия гибридизации и/или промывок можно применять в течение по меньшей мере 10, 30, 60, 90, 120 или 240 минут.
«Существенная часть» аминокислотной или нуклеотидной последовательности является такой частью, которая включает достаточно аминокислотной последовательности полипептида или нуклеотидной последовательности гена для того, чтобы предположительно идентифицировать этот полипептид или ген, или с помощью немеханизированной оценки последовательности квалифицированным в данной области техники специалистом, или с помощью компьютерно-автоматизированного сравнения последовательностей и идентификации с использованием таких алгоритмов, как BLAST (Basic Local Alignment Search Tool (базисный, поисковый способ локального выравнивания); Altschul, S.F., et al., J. Mol. Biol., 215: 403-410 (1993)). Как правило, для предположительного установления гомологии полипептидной последовательности или последовательности нуклеиновой кислоты известному белку или гену необходима последовательность из десяти или более последовательных аминокислот или тридцати или более нуклеотидов. Кроме того, что касается нуклеотидных последовательностей, в зависимых от последовательностей способах идентификации (например, при гибридизации по Саузерну) и выделения генов (например, при in situ гибридизации бактериальных колоний или бактериофаговых бляшек) можно использовать специфичные для генов олигонуклеотидные зонды, включающие 20-30 последовательных нуклеотидов. Кроме того, короткие олигонуклеотиды из 12-15 оснований можно использовать в качестве праймеров для амплификации в ПЦР для того, чтобы получить конкретный фрагмент нуклеиновой кислоты, включающий праймеры. Соответственно, «существенная часть» нуклеотидной последовательности включает достаточно последовательности для того, чтобы специфически идентифицировать и/или выделить фрагмент нуклеиновой кислоты, включающий последовательность. В настоящей заявке сообщаются полные аминокислотные и нуклеотидные последовательности, кодирующие конкретные белки жгутиковых. Извлекая пользу из сообщенных здесь последовательностей, квалифицированный специалист может теперь использовать целиком или существенную часть раскрытых последовательностей для целей, известных квалифицированным в данной области техники специалистам. Соответственно, настоящее изобретение включает полные последовательности, представленные в сопроводительном списке последовательностей, а также существенные части этих последовательностей, определенные выше.
Термин «комплементарный» используется для описания взаимоотношения между нуклеотидными основаниями, которые способны гибридизоваться друг с другом. Например, что касается ДНК, аденозин комплементарен тимину, а цитозин комплементарен гуанину. Соответственно, настоящее изобретение также включает выделенные фрагменты нуклеиновых кислот, которые комплементарны полным последовательностям, представленным в сопроводительном списке последовательностей, а также в значительной степени схожие последовательности нуклеиновых кислот.
Термины «гомология» или «гомологичный» используются здесь взаимозаменяемо. Они относятся к фрагментам нуклеиновых кислот, в которых замены одного или нескольких нуклеотидных оснований не нарушают способность фрагмента нуклеиновой кислоты опосредовать экспрессию гена или порождать определенный фенотип. Эти термины также относятся к модификациям фрагментов нуклеиновых кислот по настоящему изобретению, таким как делеция или вставка одного или нескольких нуклеотидов, не изменяющих в значительной степени функциональных свойств результирующего фрагмента нуклеиновой кислоты относительно первоначального, немодифицированного фрагмента. Поэтому понятно, как это понятно квалифицированным в данной области техники специалистам, что настоящим изобретением охватываются не только конкретные, приведенные в качестве примеров последовательности.
Кроме того, квалифицированный специалист знает, что гомологичные последовательности нуклеиновых кислот, охватываемые настоящим изобретением, также определяются по их способности гибридизоваться, в умеренно жестких условиях (например, в 0,5Х SSC, 0,1% SDS при 60°С), с последовательностями, приведенными здесь в качестве примеров, или с любой частью нуклеотидных последовательностей, раскрытых здесь, и функционально эквивалентны любой из последовательностей нуклеиновых кислот, раскрытых здесь.
«Вырожденность кода» относится к природе генетического кода, делающей возможной вариацию нуклеотидной последовательности без изменения аминокислотной последовательности кодируемого полипептида. Соответственно, настоящее изобретение относится к любому фрагменту нуклеиновой кислоты, который кодирует всю или существенную часть аминокислотной последовательности полипептида жгутиковых по настоящему изобретению, приведенной в SEQ ID NO: 2. Квалифицированный специалист хорошо знает о «кодоновом предпочтении», проявляемом конкретной клеткой-хозяином при использовании нуклеотидных кодонов для определения данной аминокислоты. Следовательно, при синтезе гена для увеличенной экспрессии в клетке-хозяине желательно конструировать ген так, чтобы частота использования кодонов в нем приближалась к частоте использования предпочтительных кодонов клетки-хозяина.
Термин «химически синтезированная», относящийся к последовательности ДНК, означает, что компоненты-нуклеотиды были собраны in vitro. Немеханизированный химический синтез ДНК можно выполнить, используя хорошо известные процедуры, или автоматизированный химический синтез можно выполнить, используя одну из ряда коммерчески доступных машин. «Синтетические гены» можно собрать из олигонуклеотидных конструктивных блоков, которые химически синтезируют, используя известные квалифицированным в данной области техники специалистам процедуры. Эти конструктивные блоки подвергают лигированию и отжигу с образованием сегментов гена, которые затем ферментативно собирают с созданием целого гена. Соответственно, гены могут быть разработаны для оптимальной экспрессии гена на основе оптимизации нуклеотидной последовательности так, чтобы она отражала кодоновое предпочтение клетки-хозяина. Квалифицированный специалист оценивает вероятность успешной экспрессии гена, если использование кодонов смещено в сторону тех кодонов, которые пользуется преимуществом у хозяина. Определение предпочтительных кодонов может быть основано на исследовании генов, происходящих из клетки-хозяина, при доступности информации о последовательности.
Термин «ген» относится к фрагменту нуклеиновой кислоты, который экспрессирует конкретный белок, и он может относиться только к кодирующей последовательности или может включать регуляторные последовательности, предшествующие (5'-некодирующие последовательности) кодирующей последовательности и следующие (3'-некодирующие последовательности) за такой последовательностью. «Природный ген» относится к обнаруживаемому в природе гену со своими собственными регуляторными последовательностями. «Химерный ген» относится к любому гену, не являющемуся природным геном, включающему регуляторные и кодирующие последовательности, не обнаруживаемые вместе в природе. Соответственно, химерный ген может включать регуляторные последовательности и кодирующие последовательности, происходящие из различных источников, или регуляторные последовательности и кодирующие последовательности, происходящие из одного и того же источника, но расположенные отличным образом по сравнению с расположением, обнаруживаемым в природе. «Эндогенный ген» относится к природному гену в его природном местонахождении в геноме организма. «Чужеродный» ген относится к гену, введенному в организм-хозяина путем переноса генов. Чужеродные гены могут включать природные гены, встроенные в неприродный организм, природные гены, введенные в новое месторасположение в природном хозяине, или химерные гены. «Трансген» представляет собой ген, введенный в геном с помощью процедуры трансформации. «Оптимизированный в отношении кодонов ген» представляет собой ген, имеющий частоту использования кодонов, созданную для воспроизводства частоты использования предпочтительных кодонов клетки-хозяина.
«Кодирующая последовательность» относится к последовательности ДНК, кодирующей конкретную аминокислотную последовательность. «Подходящие регуляторные последовательности» относятся к нуклеотидным последовательностям, расположенным 5' (5'-некодирующие последовательности) от кодирующей последовательности, внутри кодирующей последовательности или 3' (3'-некодирующие последовательности) от кодирующей последовательности и оказывающим влияние на транскрипцию, процессинг или стабильность РНК или трансляцию ассоциируемой кодирующей последовательности. Регуляторные последовательности могут включать промоторы, направляющие трансляцию последовательности, интроны, последовательности идентификации полиаденилирования, сайты процессинга РНК, сайты связывания эффектора и шпилечные структуры.
Термин «аллель» относится к одной из нескольких альтернативных форм гена, занимающих данный локус на хромосоме. Если все присутствующие в данном локусе на хромосоме аллели являются одинаковыми, такое растение является гомозиготным по данному локусу. Если присутствующие в данном локусе на хромосоме аллели отличаются, такое растение является гетерозиготным по данному локусу.
Термин «промотор» относится к последовательности ДНК, способной контролировать экспрессию кодирующей последовательности или функциональной РНК. Как правило, кодирующая последовательность располагается 3' от последовательности промотора. Промоторы могут полностью происходить из природного гена, или они состоят из различных элементов, происходящих из различных промоторов, обнаруживаемых в природе, или даже включают синтетические сегменты ДНК. Квалифицированным в данной области техники специалистам понятно, что различные промоторы могут направлять экспрессию гена в различных типах тканей или клеток или на различных стадиях развития, или в ответ на различные условия окружающей среды или физиологические условия. Промоторы, которые вызывают экспрессию гена в большинстве типов клеток в большинстве периодов времени, обычно упоминаются как «конститутивные промоторы». Кроме того, понятно, что, поскольку в большинстве случаев точные границы регуляторных последовательностей не полностью определены, ДНК-фрагменты различных длин могут иметь идентичную промоторную активность.
Промоторная последовательность может состоять из проксимальных и более дистальных 5'-элементов, более дистальные 5'-элементы часто упоминаются как энхансеры. Соответственно, «энхансер» представляет собой последовательность ДНК, которая может стимулировать активность промотора, и он может быть природным элементом промотора или гетерологичным элементом, встроенным для увеличения уровня или тканеспецифичности промотора. Новые промоторы различных типов, применимые в клетках растений, постоянно обнаруживают; многочисленные примеры можно найти в сводке данных Okamuro, J.K. и Goldberg, R.B. (Biochemistry of Plants, 15: 1-82 (1989)).
«Направляющая трансляцию последовательность» относится к полинуклеотидной последовательности, расположенной между промоторной последовательностью гена и кодирующей последовательностью. Направляющая трансляцию последовательность присутствует в полностью процессированной мРНК 5' от последовательности начала трансляции. Направляющая трансляцию последовательность может оказывать влияние на процессинг первичного транскрипта в мРНК, стабильность мРНК или эффективность трансляции. Примеры направляющих трансляцию последовательностей описаны (Turner, R. and Foster, G.D., Mol. Biotechnol., 3: 225-236 (1995)).
Термины «3'-некодирующие последовательности» и «терминатор транскрипции» относятся к последовательностям ДНК, расположенным 3' от кодирующей последовательности. Они включают последовательности идентификации полиаденилирования и другие последовательности, кодирующие регуляторные сигналы, способные оказывать воздействие на процессинг мРНК или экспрессию гена. Сигнал полиаденилирования обычно характеризуется влиянием на добавление участков полиадениловой кислоты к 3'-концу предшественника мРНК. 3'-Район может оказывать влияние на транскрипцию, процессинг и стабильность РНК или трансляцию ассоциируемой кодирующей последовательности.
«РНК-транскрипт» относится к продукту, являющемуся результатом катализируемой РНК-полимеразой транскрипции последовательности ДНК. Когда РНК-транскрипт является идеальной комплементарной копией последовательности ДНК, он упоминается как первичный транскрипт, или он может быть последовательностью РНК, являющейся результатом посттранскрипционного процессинга первичного транскрипта, и упоминается как зрелая РНК. «Информационная РНК» или «мРНК» относится к РНК без интронов, которая может транслироваться клеткой в белок. «кДНК» относится к двухцепочечной ДНК, комплементарной мРНК и происходящей из нее. «Смысловая» РНК относится к РНК-транскрипту, который включает мРНК и значит может транслироваться клеткой в белок. «Антисмысловая» РНК относится к РНК-транскрипту, который комплементарен полностью или части целевого первичного транскрипта или мРНК и который блокирует экспрессию целевого гена (патент США № 5107065; публикация РСТ-заявки № WO 99/28508). Антисмысловая РНК может быть комплементарна любой части транскрипта конкретного гена, т.е. 5'-некодирующей последовательности, 3'-некодирующей последовательности или кодирующей последовательности. «Функциональная РНК» относится к антисмысловой РНК, рибозимной РНК или другой РНК, которая не транслируется, но все же оказывает эффект на клеточные процессы.
Термин «функционально связанный» относится к связи последовательностей нуклеиновых кислот в одном фрагменте нуклеиновой кислоты так, что на функционирование одной из них оказывает влияние другая. Например, промотор функционально связан с кодирующей последовательностью, если он способен влиять на экспрессию этой кодирующей последовательности (т.е. эта кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующие последовательности могут быть функционально связаны с регуляторными последовательностями в смысловой или антисмысловой ориентации.
Используемый здесь термин «экспрессия» относится к транскрипции и стабильной аккумуляции смысловой (мРНК) или антисмысловой РНК, происходящей из фрагментов нуклеиновых кислот по настоящему изобретению. Экспрессия может также относиться к трансляции мРНК в полипептид.
«Зрелый» белок относится к подвергшемуся посттрансляционному процессингу полипептиду, т.е. полипептиду, из которого были исключены любые пре- или пропептиды, присутствующие в первичном продукте трансляции. Белок-«предшественник» относится к первичному продукту трансляции мРНК, т.е. с пре- и пропептидами, все еще присутствующими. Пре- и пропептиды могут быть (но без ограничения) сигналами внутриклеточной локализации.
«Трансформация» относится к переносу молекулы нуклеиновой кислоты в организм-хозяина, что приводит к генетически стабильному наследованию. Молекула нуклеиновой кислоты может быть плазмидой, которая реплицируется автономно, например, или она может интегрироваться в геном организма-хозяина. Организмы-хозяева, содержащие трансформированные фрагменты нуклеиновых кислот, упоминаются как «трансгенные», или «рекомбинантные», или «трансформированные» организмы.
Термины «плазмида», «вектор» и «кассета» относятся к экстрахромосомному элементу, часто несущему гены, не являющиеся частью главного метаболизма клетки, и находящемуся обычно в форме кольцевых двухцепочечных ДНК-фрагментов. Такие элементы могут быть автономно реплицирующимися последовательностями, интегрирующимися в геном последовательностями, фаговыми или нуклеотидными последовательностями, линейными или кольцевыми, одно- или двухцепочечной ДНК или РНК, происходящими из любого источника, в которых ряд нуклеотидных последовательностей соединен или рекомбинирован в уникальную конструкцию, способную ввести в клетку фрагмент промотора и последовательность ДНК для выбранного генного продукта вместе с соответствующей 3'-нетранслируемой последовательностью. «Экспрессирующая кассета» относится к специфическому вектору, содержащему чужеродный ген и имеющему помимо чужеродного гена элементы, позволяющие увеличение экспрессии этого гена в чужеродном хозяине.
Термин «процент идентичности», известный в данной области техники, представляет собой взаимосвязь между двумя или более полипептидными последовательностями или двумя или более полипептидными последовательностями, определяемую сравнением последовательностей. В данной области техники «идентичность» также означает степень связанности между полипептидными или полинуклеотидными последовательностями, например, определяемой с помощью сопоставления последовательностей символов таких последовательностей. «Идентичность» и «схожесть» можно легко рассчитать с помощью известных способов, включающих, но без ограничения, способы, описанные в 1) Computational Molecular Biology (Lesk, A.M., Ed.) Oxford University: NY (1988); 2) Biocomputing: Informatics and Genome Projects (Smith, D.W., Ed.) Academic: NY (1993); 3) Computer Analysis of Sequence Data, Part I (Griffin, A.M., and Griffin, H.G., Eds.) Humania: NJ (1994); 4) Sequence Analysis in Molecular Biology (von Heinje, G., Ed.) Academic (1987); и 5) Sequence Analysis Primer (Gribskov, M. and Devereux, J., Eds.) Stockton: NY (1991).
Предпочтительные способы определения идентичности разработаны для получения наибольшего соответствия проверяемых последовательностей. Способы определения идентичности и схожести кодифицированы в публично доступных компьютерных программах. Выравнивания последовательностей и расчеты процентов идентичности можно проводить с использованием программы MegAlignTM комплекта вычислительных программ для биоинформатики LASERGENE (DNASTAR Inc., Madison, WI). Множественное выравнивание последовательностей проводят с использованием «способа выравнивания Clustal», который включает несколько выборов алгоритма, в том числе «способа выравнивания Clustal V», который соответствует способу выравнивания, обозначенному Clustal V (описанному Higgins and Sharp, CABIOS. 5: 151-153 (1989); Higgins, D.G. et al., Comput. Appl. Biosci., 8: 189-191 (1992)) и обнаруживаемому в программе MegAlignTM комплекта вычислительных программ для биоинформатики LASERGENE (DNASTAR Inc.). При множественных выравниваниях величины, используемые по умолчанию, соответствуют пенальти за промежуток = 10 и пенальти за длину промежутка = 10. Используемыми по умолчанию параметрами для попарных выравниваний и расчета процента идентичности белковых последовательностей с использованием способа Clustal V являются (оптимальный), используемый при выравнивании, = 1, пенальти за промежуток = 3, окно = 5 и сохраняемые диагонали = 5. Для нуклеиновых кислот этими параметрами являются (оптимальный) размер слова, используемый при выравнивании = 2, пенальти за промежуток = 5, окно = 4 и сохраняемые диагонали = 4. После выравнивания последовательностей с использованием программы Clustal V можно получить «процент идентичности» путем просмотра таблицы «несходства последовательностей» в этой же программе. Кроме того, доступен «способ выравнивания Clustal W», и он соответствует способу выравнивания, обозначенному Clustal W (описанному Higgins and Sharp, CABIOS. 5: 151-153 (1989); Higgins, D.G. et al., Comput. Appl. Biosci., 8: 189-191 (1992)) и обнаруживаемому в версии 6.1 программы MegAlignTM комплекта вычислительных программ для биоинформатики LASERGENE (DNASTAR Inc.). Используемые по умолчанию параметры для множественного выравнивания соответствуют пенальти за промежуток = 10, пенальти за длину промежутка = 0,2, обусловленная запаздыванием дивергенция последовательностей (%) = 30, переходный вес ДНК = 0,5, белковая весовая матрица = ряд Гоннета, ДНК весовая матрица = IUB. После выравнивания последовательностей с использованием программы Clustal W можно получить «процент идентичности» путем просмотра таблицы «несходства последовательностей» в этой же программе.
«Способ выравнивания BLASTN» представляет собой алгоритм, предоставленный National Center for Biotechnology Information (NCBI) для сравнения нуклеотидных последовательностей с использованием параметров по умолчанию.
Квалифицированному в данной области техники специалисту вполне понятно, что множество степеней идентичности последовательностей пригодны при идентификации полипептидов из других видов, причем такие полипептиды имеют одинаковую или схожую функцию или активность. Подходящие фрагменты нуклеиновых кислот (выделенные полинуклеотиды по настоящему изобретению) кодируют полипептиды, которые идентичны на по меньшей 70%, предпочтительно на по меньшей мере 75% и более предпочтительно на по меньшей мере 80% аминокислотным последовательностям, сообщаемым здесь. Предпочтительные фрагменты нуклеиновых кислот кодируют аминокислотные последовательности, которые идентичны на по меньшей 85% аминокислотным последовательностям, представленным здесь. Более предпочтительные фрагменты нуклеиновых кислот кодируют аминокислотные последовательности, которые идентичны на по меньшей 90% аминокислотным последовательностям, представленным здесь. Наиболее предпочтительными являются фрагменты нуклеиновых кислот, кодирующие аминокислотные последовательности, которые идентичны на по меньшей 95% аминокислотным последовательностям, сообщаемым здесь. Хотя предпочтительные диапазоны описаны выше, при описании по настоящему изобретению может использоваться аминокислотная идентичность на любое целое число от 39% до 100%, такая как 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% и 99%.
Подходящие фрагменты нуклеиновых кислот не только гомологичны на указанный выше процент, но, как правило, кодируют полипептид, имеющий по меньшей мере 50 аминокислот, предпочтительно по меньшей мере 100 аминокислот, более предпочтительно по меньшей мере 150 аминокислот, еще более предпочтительно по меньшей мере 200 аминокислот и наиболее предпочтительно по меньшей мере 250 аминокислот.
Термин «консервативный домен» или «мотив» означает совокупность аминокислот, являющихся консервативными в специфических положениях вдоль выровненной последовательности эволюционно родственных белков. Хотя аминокислоты в других положениях могут варьировать между гомологичными белками, аминокислоты, являющиеся в высокой степени консервативными в специфических положениях, указывают аминокислоты, являющиеся существенными для структуры, стабильности или активности белка. Поскольку их идентифицируют по их высокой степени консервативности в выровненных последовательностях семейства белков-гомологов, их можно использовать в качестве идентификаторов или «характерных признаков» для определения того, принадлежит ли белок с недавно определенной последовательностью семейству ранее идентифицированных белков.
Термин «программное обеспечение для анализа последовательностей» относится к любому алгоритму работы ЭВМ или программе, которые применимы для анализа нуклеотидных или аминокислотных последовательностей. «Программное обеспечение для анализа последовательностей» может быть коммерчески доступным или его можно независимо разработать. Типичное программное обеспечение для анализа последовательностей будет включать, но без ограничения, 1) комплект программ GCG (Wisconsin Package Version 9.0, Genetics Computer Group (GCG), Madison, WI); 2) BLASTP, BLASTN, BLASTX (Altschul et al., J. Mol. Biol., 215: 403-410 (1990); 3) DNASTAR (DNASTAR, Inc. Madison, WI); 4) Sequencher (Gene Codes Corporation, Ann Arbor, MI); и 5) программу FASTA, включающаяую алгоритм Смита-Уотермана (W.R. Pearson, Comput. Methods Genome Res., [Proc. Int. Symp.] (1994), Meeting Date 1992, 111-20. Editor(s): Suhai, Sandor. Plenum: New York, NY). В контексте этой заявки будет понятно, что если для анализа используется программное обеспечение для анализа последовательностей, результаты анализы будут основаны на «используемых по умолчанию величинах» программы, на которую приводится ссылка, если только не указано иное. Как здесь используются, «используемые по умолчанию величины» будут означать любую совокупность величин или параметров, первоначально загруженных с программным обеспечением при первой инициализации. Что касается используемого здесь алгоритма BLASTP, параметры по умолчанию будут включать повторяемости аминокислот Robinson и Robinson (Robinson A.B., Robinson L.R., Proc. Natl. Acad. Sci. U.S.A., 88: 8880-8884 (1991)), матрицу количественной оценки BLOSUM62 и счет за промежуток Δ(g)=11+g.
Термин «части растения» включает дифференцированные и недифференцированные ткани, включающие, но без ограничения, корни, стебли, ростки, листья, пыльцу, семена, ткани вздутий и различные формы клеток и культур (например, отдельные клетки, протопласты, зародыши и ткань каллюса). Ткани растения могут быть в растении или в органе, ткани растения или клеточной культуре.
Термин «орган растения» относится к ткани растения или группе тканей, которые создают морфологически и функционально отличную часть растения.
Термин «геном» относится к следующему: 1) полному комплементу генетического материала (генов и некодирующих последовательностей), присутствующего в каждой клетке организма, или вируса, или органелле; 2) полному набору хромосом, наследуемому как (гаплоидная) единица от одного родителя.
Используемые здесь стандартные методы рекомбинантных ДНК и молекулярного клонирования хорошо известны в данной области техники и описываются Sambrook, J., Fritsch, E.F. и Maniatis, T., (Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory: Cold Spring Harbor, NY (1989) (в дальнейшем «Maniatis»)) и Silhavy, T.J., Bennan, M.L. и Enquist, L.W. (Experiments with Gene Fusions, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1984)), и Ausubel, F.M. и др. (Current Protocols in Molecular Biology, опубликованные Greene Publishing Assoc. и Wiley-Interscience, Hoboken, NJ (1987)).
Термины «рекомбинантная конструкция», «экспрессирующая конструкция», «химерная конструкция», «конструкция» и «рекомбинантная ДНК-конструкция» используются здесь взаимозаменяемо. Рекомбинантная конструкция включает искусственную комбинацию фрагментов нуклеиновых кислот, т.е. регуляторных и кодирующих последовательностей, не обнаруживаемых вместе в природе. Например, химерная конструкция может включать регуляторные последовательности и кодирующие последовательности, происходящие из различных источников, или регуляторные последовательности и кодирующие последовательности, происходящие из одного и того же источника, но размещенные отличным от обнаруживаемого в природе образом. Такая конструкция может использоваться сама по себе или она может использоваться в объединении с вектором. Если используется вектор, его выбор зависит от способа, который будет использоваться для трансформации клеток-хозяев, как это хорошо известно квалифицированным в данной области техники специалистам. Например, можно использовать плазмидный вектор. Квалифицированному специалисту хорошо известны генетические элементы, которые должны присутствовать на векторе для успешной трансформации, отбора и размножения клеток-хозяев, содержащих любой из выделенных фрагментов нуклеиновых кислот по настоящему изобретению. Квалифицированному специалисту также понятно, что различные независимые случаи трансформации будут приводить к различным уровням и характерам экспрессии (Jones et al., EMBO J. 4: 2411-2418 (1985); De Almeida et al., Mol. Gen. Genetics 218: 78-86 (1989)), и, следовательно, необходимо скринировать множество случаев для того, чтобы получить линии, проявляющие желаемый уровень и характер экспрессии. Такое скринирование можно провести с помощью анализа ДНК по Саузерну, Норзерн-анализа экспрессии мРНК, анализа экспрессии белков иммуноблоттингом или фенотипического анализа, среди прочих.
Используемый здесь термин «экспрессия» относится к продукции функционального конечного продукта (например, мРНК или белка [или предшественника, или зрелого]).
Термин «введенный» означает предоставление нуклеиновой кислоты (например, экспрессирующей конструкции) или белка в клетку. Введенный включает ссылку на включение нуклеиновой кислоты в эукариотическую или прокариотическую клетку, где нуклеиновая кислота может включиться в геном клетки, и включает ссылку на временное предоставление нуклеиновой кислоты или белка клетке. Введенный включает ссылку на способы стабильной или временной трансформации, а также скрещивание половым путем. Таким образом, «введенный» в контексте включения фрагмента нуклеиновой кислоты (например, рекомбинатной ДНК-конструкции/экспрессирующей конструкции) в клетку означает «трансфекцию», или «трансформацию», или «трансдукцию» и включает ссылку на включение фрагмента нуклеиновой кислоты в эукариотическую или прокариотическую клетку, где фрагмент нуклеиновой кислоты может включиться в геном клетки (например, хромосому, плазмиду, пластиду или митохондриальную ДНК), превратиться в автономный репликон или временно экспрессироваться (например, трансфицированная мРНК).
«Зрелый» белок относится к подвергшемуся посттрансляционному процессингу полипептиду (т.е. полипептиду, из которого были исключены любые пре- или пропептиды, присутствующие в первичном продукте трансляции). Белок-«предшественник» относится к первичному продукту трансляции мРНК (т.е. с пре- и пропептидами, все еще присутствующими). Пре- и пропептиды могут быть, но без ограничения, сигналами внутриклеточной локализации.
«Стабильная трансформация» относится к переносу фрагмента нуклеиновой кислоты в геном организма-хозяина, включающий и ядерный, и органоидный геномы, что приводит к генетически стабильному наследованию. Напротив, «временная трансформация» относится к переносу фрагмента нуклеиновой кислоты в ядро, или ДНК-содержащую органеллу, организма-хозяина, что приводит к экспрессии гена без интеграции или стабильного наследования. Организмы-хозяева, содержащие трансформированные фрагменты нуклеиновых кислот, упоминаются как «трансгенные» организмы.
Как здесь используется, «трансгенное(ая)» относится к растению или клетке, которые содержат в своих геномах гетерологичный полинуклеотид. Предпочтительно гетерологичный полинуклеотид стабильно интегрируется в геном так, что полинуклеотид передается последующим поколениям. Гетерологичный полинуклеотид может интегрироваться в геном отдельно или в виде части экспрессирующей конструкции. Как здесь используется, трансгенное(ая) включает любую клетку, клеточную линию, каллюс, ткань, часть растения или растение, генотип которых был изменен присутствием гетерологичной нуклеиновой кислоты, в том числе те из них, которые были первоначально таким образом изменены, а также те, которые были созданы с помощью полового скрещивания или неполового размножения первоначальной трансгенной клетки или растения. Используемым здесь термином «трансгенное(ая)» не охватывается изменение генома (хромосомного или экстрахромосомного) с помощью стандартных способов селекции растений или с помощью происходящих в природе явлений, таких как случайное перекрестное оплодотворение, нерекомбинантная вирусная инфекция, нерекомбинантная бактериальная трансформация, нерекомбинантная транспозиция или спонтанная мутация.
«Антисмысловое ингибирование» относится к продукции антисмысловых РНК-транскриптов, способных подавлять экспрессию целевого белка. «Косупрессия» относится к продукции смысловых РНК-транскриптов, способных подавлять экспрессию идентичных или в значительной степени схожих чужеродных или эндогенных генов (патент США № 5231020). Косупрессирующие конструкции в растениях ранее конструировали, фокусируясь на сверхэкспрессии последовательности нуклеиновой кислоты, гомологичной эндогенной мРНК, в смысловой ориентации, что приводит к уменьшению всех РНК, гомологичных сверхэкспрессируемой последовательности (Vaucheret et al., Plant J. 16: 651-659 (1998); Gura, Nature 404: 804-808 (2000)). Общая эффективность этого явления очень низка, а степень уменьшения РНК широко варьирует. В более недавней работе описывается использование структур «шпилек», которые включают всю или часть кодирующей последовательности мРНК в комплементарной ориентации, которая приводит к возможной структуре «ножка-петля» экспрессированной РНК (публикация РСТ-заявки № WO 99/53050, опубликованной 21 октября 1999; публикация РСТ-заявки № WO 02/00904, опубликованной 3 января 2002). Это увеличивает частоту косупрессии в получаемых трансгенных растениях. В другой вариации описывается использование последовательностей вирусов растений для направления супрессии, или «глушения», проксимальных кодирующих последовательностей мРНК (публикация РСТ-заявки № WO 98/36083, опубликованной 20 августа 1998). Оба эти явления косупрессии были истолкованы механистически, хотя генетические данные начали объяснять эту сложную ситуацию (Elmayan et al., Plant Cell 10: 1747-1757 (1998)).
Обзор: биосинтез жирных кислот и триацилглицеринов в микроорганизмах
Обычно аккумуляция липидов в содержащих или продуцирующих масло микроорганизмах инициируется в ответ на отношение общего углерода к азоту в среде для роста. Этот процесс, приводящий к de novo синтезу свободного пальмитата (16:0) в содержащих или продуцирующих масло микроорганизмах, подробно описывается в публикации РСТ-заявки WO 2004/101757. Пальмитат является предшественником производных длинноцепочечных насыщенных и ненасыщенных жирных кислот, которые образуются под действием элонгаз и десатураз (фиг. 1)
TAG (первичные единицы запаса жирных кислот) образуются с помощью ряда реакций, которые включают: 1) этерификацию одной молекулы ацил-КоА с выходом глицерол-3-фосфата с помощью ацилтрансферазы для продуцирования лизофосфатидной кислоты; 2) этерификацию второй молекулы ацил-КоА с помощью ацилтрансферазы с выходом 1,2-диацилглицеролфосфата (обычно определяемого как фосфатидная кислота); 3) удаление фосфата с помощью фосфатазы фосфатидной кислоты с выходом 1,2-диацилглицерина (DAG); и 4) добавление третьей жирной кислоты под действием ацилтрансферазы с образованием TAG. В TAG может быть включен широкий диапазон жирных кислот, включающих насыщенные и ненасыщенные жирные кислоты и жирные кислоты с короткими и длинными цепями.
Биосинтез жирных кислот омега
Метаболический процесс, в котором олеиновая кислота превращается в жирные кислоты омега-3/омега-6, включает элонгацию углеродной цепи благодаря добавлению атомов углерода и десатурацию молекулы благодаря добавлению двойных связей. Для этого требуется ряд специальных ферментов десатураз и элонгаз, присутствующих в мембране эндоплазматического ретикулума. Однако, как видно на фиг. 1 и описывается ниже, часто существует множество альтернативных путей продукции конкретных жирных кислот омега-3/омега-6.
В частности, для всех путей требуется первоначальное превращение олеиновой кислоты в LA, первую из жирных кислот омега-6, под действием дельта-12-десатуразы. Затем, используя «путь с участием дельта-6-десатуразы/дельта-6-элонгазы», жирные кислоты омега-6 образуются следующим образом: (1) LA превращается в GLA под действием дельта-6-десатуразы; (2) GLA превращается в DGLA под действием С18/20-элонгазы; и (3) DGLA превращается в ARA под действием дельта-5-десатуразы. В альтернативном случае для образования жирных кислот омега-3 может использоваться «путь с участием дельта-6-десатуразы/дельта-6-элонгазы» следующим образом: 1) LA превращается в ALA, первую из жирных кислот омега-3, под действием дельта-15-десатуразы; (2) ALA превращается в STA под действием дельта-6-десатуразы; (3) STA превращается в ETA под действием С18/20-элонгазы; (4) ETA превращается в EPA под действием дельта-5-десатуразы; (5) EPA превращается в DPA под действием С20/22-элонгазы; и (6) DPA превращается в DHA под действием дельта-4-десатуразы. Необязательно жирные кислоты омега-6 могут превращаться в жирные кислоты омега-3; например, ЕТА и ЕРА продуцируются из DGLA и ARA, соответственно под действием активности дельта-17-десатуразы.
В альтернативных путях биосинтеза жирных кислот омега-3/омега-6 используются дельта-9-элонгаза и дельта-8-десатураза. Более конкретно, LA и ALA могут превратиться в EDA и ETrA соответственно, под действием дельта-9-элонгазы; затем дельта-8-десатураза превращает EDA в DGLA и/или ETrA в ETA.
Предполагается, что конкретные функциональные возможности, требуемые быть выраженными в конкретном организме-хозяине для продукции жирных кислот омега-3/омега-6, будут зависеть от клетки-хозяина (и его природного профиля PUFA и/или профиля десатураз/элонгаз), доступности субстрата и желаемого конечного продукта(ов). Квалифицированный в данной области техники специалист сможет идентифицировать различные возможные гены, кодирующие каждый из ферментов, требуемых для биосинтеза жирных кислот омега-3/омега-6. Последовательности пригодных десатураз и элонгаз могут быть получены из любого источника, например, выделены из природного источника (например, бактерий, водорослей, грибов, растений, животных и т.п.), продуцированы полусинтетическим путем или синтезированы de novo. Хотя конкретный источник генов десатураз и элонгаз, вводимых в хозяина, не является критическим, соображения в отношении выбора конкретного полипептида, обладающего активностью десатуразы или элонгазы, включают: 1) субстратную специфичность полипептида; 2) то, является ли полипептид или его компонент ограничивающим скорость ферментом; 3) то, является ли десатураза или элонгаза существенной для синтеза желаемой PUFA; и/или 4) кофакторы, требуемые для полипептида. Экспрессированный полипептид предпочтительно имеет характеристики, совместимые с биохимическим окружением его нахождения в клетке-хозяине (смотри публикацию РСТ-заявки WO 2004/101757 для дополнительных деталей).
В дополнительных вариантах осуществления также полезно принимать во внимание эффективность превращения для каждой конкретной десатуразы и/или элонгазы. Более конкретно, поскольку каждый фермент редко функционирует со 100% эффективностью превращения субстрата в продукт, конечный профиль липидов неочищенных масел, продуцируемых в клетке-хозяине, будет, как правило, представлять собой смесь различных PUFA, состоящих из желаемых жирных кислот омега-3/омега-6, а также различных верхних промежуточных PUFA. Таким образом, эффективность превращения для каждого фермента также является параметром для принятия во внимание при оптимизации биосинтеза желаемой жирной кислоты.
Имея в виду каждое из вышеотмеченных соображений, возможные гены, обладающие соответствующими активностями десатураз и элонгаз (например, дельта-6-десатураз, С18/20-элонгаз, дельта-5-десатураз, дельта-17-десатураз, дельта-15-десатураз, дельта-9-десатураз, дельта-12-десатураз, С14/16-элонгаз, С16/18-элонгаз, дельта-9-элонгаз, дельта-8-десатураз, дельта-4-десатураз и С20/22-элонгаз), можно идентифицировать сообразно с публично доступными источниками информации (например, GenBank), патентными источниками информации и экспериментальным анализом организма, обладающего способностью продуцировать PUFA. Эти гены будут пригодны для введения в конкретный организм-хозяина для того, чтобы сделать возможным или увеличить синтез организмом PUFA.
Идентификация последовательности новой дельта-5-десатуразы из Euglena gracilis
В настоящем изобретении из Euglena gracilis выделена нуклеотидная последовательность (SEQ ID NO: 1), кодирующая дельта-5-десатуразу (SEQ ID NO: 2), обозначенную здесь «EgD5».
Сравнение последовательности нуклеотидных оснований и выведенной аминокислотной последовательности EgD5 с общедоступными базами данных показывает, что большая часть схожих известных последовательностей идентична на приблизительно 39% аминокислотной последовательности EgD5, представленной здесь, на протяжении длины 449 аминокислот, используя способ выравнивания Clustal W. Более предпочтительные аминокислотные фрагменты идентичны на по меньшей мере приблизительно 70%-80% последовательностям по настоящему изобретению, причем те последовательности, которые идентичны на по меньшей мере приблизительно 80%-90%, являются особенно подходящими, а те последовательности, которые идентичны на по меньшей мере приблизительно 90%-95%, являются наиболее предпочтительными. Аналогично, предпочтительными кодирующими EgD5 последовательностями нуклеиновых кислот, соответствующими ORF по настоящему изобретению, являются последовательности, которые кодируют активные белки и которые идентичны на по меньшей мере приблизительно 70%-80% последовательностям нуклеиновых кислот для EgD5, представленным здесь, причем те последовательности, которые идентичны на по меньшей мере приблизительно 80%-90%, являются особенно подходящими, а те последовательности, которые идентичны на по меньшей мере приблизительно 90%-95%, являются наиболее предпочтительными.
В альтернативных вариантах осуществления последовательность десатуразы EgD5 по настоящему изобретению можно оптимизировать в отношении кодонов для экспрессии в конкретном организме-хозяине. Как хорошо известно в данной области техники, это может быть полезным способом дальнейшей оптимизации экспрессии фермента в альтернативном хозяине, поскольку использование предпочтительных для хозяина кодонов может значительно увеличить экспрессию чужеродного гена, кодирующего полипептид. Как правило, предпочтительные для хозяина кодоны можно определить в конкретном представляющем интерес виде хозяина с помощью исследования использования кодонов в белках (предпочтительно белках, экспрессируемых в больших количествах) и определения того, какие кодоны используются с наибольшей частотой. Затем последовательность, кодирующую представляющий интерес полипептид, обладающий, например, активностью десатуразы, можно синтезировать целиком или частично, используя кодоны, предпочтительные для вида хозяина.
В одном предпочтительном варианте осуществления по настоящему изобретению EgD5 оптимизировали в отношении кодонов для экспрессии в Yarrowia lipolytica. Это стало возможным благодаря, во-первых, определению профиля использования кодонов Y. lipolytica (смотри, публикацию РСТ-заявки № WO 04/101757 и патент США № 7125672) и идентификации тех кодонов, которые являлись предпочтительными. Затем, для дальнейшей оптимизации экспрессии генов в Y. lipolytica, была определена консенсусная последовательность вокруг кодона инициации «ATG». Эта оптимизация привела к модификации 196 п.о. кодирующей области размером 1350 п.о. (14,5%) и оптимизации 189 кодонов из 449 кодонов в целом (42%). Ни одна из модификаций в оптимизированном в отношении кодонов гене («EgD5S»; SEQ ID NO: 3) не изменяла аминокислотную последовательность кодируемого белка (SEQ ID NO: 2). Как описывается в примере 11, оптимизированный в отношении кодонов ген на 36% более эффективно десатурировал DGLA до ARA, чем ген дикого типа, при экспрессии в Y. lipolytica.
Квалифицированный в данной области техники специалист сможет использовать приведенные здесь учения для создания различных других оптимизированных в отношении кодонов белков дельта-5-десатураз, подходящих для оптимальной экспрессии в альтернативных хозяевах (т.е. отличных от Yarrowia lipolytica) на основе последовательности EgD5 дикого типа. Соответственно, настоящее изобретение относится к любому оптимизированному в отношении кодонов белку дельта-5-десатуразы, происходящему из EgD5 дикого типа (т.е. кодируемому SEQ ID NO: 2). Это включает, но без ограничения, нуклеотидную последовательность, представленную в SEQ ID NO: 3, которая кодирует синтетический белок дельта-5-десатуразы (т.е. EgD5S), которая была оптимизирована в отношении кодонов для экспрессии в Yarrowia lipolytica.
Идентификация и выделение гомологов
Любую из последовательностей десатураз по настоящему изобретению (т.е. EgD5, EgD5S) или их части можно использовать для поиска гомологов дельта-5-десатуразы в одних и тех же или разных видах бактерий, водорослей, грибов, жгутиковых или растений, используя программное обеспечение для анализа последовательностей. Как правило, такое компьютерное программное обеспечение сравнивает схожие последовательности по определению степеней гомологии к различным заменам, делециям и другим модификациям.
В альтернативном случае любую из последовательностей десатураз по настоящему изобретению или их части можно использовать в качестве реагентов для гибридизации для идентификации гомологов дельта-5-десатуразы. Основные компоненты теста на гибридизацию нуклеиновых кислот включают зонд, образец, предположительно содержащий представляющий интерес ген или фрагмент гена, и конкретный способ гибридизации. Зонды по настоящему изобретению представляют собой обычно одноцепочечные последовательности нуклеиновых кислот, комплементарные обнаруживаемым последовательностям нуклеиновых кислот. Зонды «способны гибридизоваться» с обнаруживаемыми последовательностями нуклеиновых кислот. Хотя размер зонда может варьировать от 5 оснований до десятков тысяч оснований, как правило, подходит размер зонда, составляющий приблизительно 15 оснований-приблизительно 30 оснований. Необходимо, чтобы одна часть молекулы зонда была комплементарна обнаруживаемой последовательности нуклеиновой кислоты. Кроме того, не требуется, чтобы комплементарность между зондом и последовательностью-мишенью была идеальной. В действительности гибридизация происходит между не вполне комплементарными молекулами, что является результатом того, что некоторая фракция оснований в гибридизующемся районе не спаривается с соответствующим комплементарным основанием.
Способы гибридизации достаточно описаны. Как правило, зонд и образец должны быть смешаны в условиях, которые сделают возможной гибридизацию нуклеиновых кислот. Это включает приведение в контакт зонда и образца в присутствии неорганической или органической соли в соответствующей концентрации и температурных условиях. Зонд и нуклеиновые кислоты в качестве образца должны быть приведены в контакт в течение периода времени, достаточно длинного для того, чтобы могла произойти какая-либо возможная гибридизация между зондом и нуклеиновой кислотой в качестве образца. Концентрация зонда или мишени в смеси будет определяться временем, необходимым для того, чтобы произошла гибридизация. Чем выше концентрация зонда или мишени, тем короче требуемое время инкубации для гибридизации. Необязательно можно добавить разобщающий агент (например, гуанидиния хлорид, гуанидиния тиоцианат, натрия тиоцианат, лития тетрахлорацетат, натрия перхлорат, рубидия тетрахлорацетат, калия йодид, цезия трифторацетат). Если желательно, в смесь для гибридизации можно добавить формамид, обычно 30-50% (вес./вес.).
Можно использовать различные растворы для гибридизации. Как правило, они включают от приблизительно 20 до 60% в объемном отношении, предпочтительно 30%, полярного органического растворителя. В общепринятых растворах для гибридизации используется приблизительно 30-50% в весовом отношении формамида, приблизительно 0,15-1 М натрия хлорид, приблизительно 0,05-0,1 М буферы (например, натрия цитрат, Tris-HCl, PIPES или HEPES (рН в диапазоне приблизительно 6-9)), приблизительно 0,05-0,2% детергента (например, натрия додецилсульфата) или 0,5-20 мМ EDTA, фиколл (Pharmacia Inc.) (приблизительно 300-500 килодальтон), поливинилпирролидон (приблизительно 250-500 килодальтон) и сывороточный альбумин. Также в типичный раствор для гибридизации будут включаться немеченые нуклеиновые кислоты-носители от приблизительно 0,1 до 5 мг/мл, фрагментированная ядерная ДНК (например, ДНК из тимуса теленка или молок лососевых, или РНК дрожжей) и необязательно от приблизительно 0,5 до 2% (вес/объем) глицина. Могут быть также включены другие добавки, такие как агенты объемного расширения, которые включают ряд полярных растворимых в воде или способных к набуханию агентов (например, полиэтиленгликоль), анионные полимеры (например, полиакрилат или полиметилакрилат) и анионные сахаридные полимеры (например, декстрана сульфат).
Гибридизацию нуклеиновых кислот можно адаптировать к множеству форматов анализа. Одним из наиболее подходящих форматов является формат сэндвич-анализа. Сэндвич-анализ можно, в частности, адаптировать к гибридизации в неденатурирующих условиях. Первичным компонентом сэндвич-анализа является твердая подложка. На твердой подложке адсорбирован иммобилизованный зонд в виде нуклеиновой кислоты, который является немеченым и комплементарен части последовательности, или такой зонд ковалентно связан с твердой подложкой.
В дополнительных вариантах осуществления любой из описанных здесь фрагментов нуклеиновых кислот для дельта-5-десатураз (или любой идентифицированный здесь гомолог) можно использовать для выделения генов, кодирующих гомологичные белки, из одного и того же и различных видов бактерий, водорослей, грибов, жгутиковых или растений. Выделение гомологичных генов с использованием зависимых от последовательностей протоколов хорошо известно в данной области техники. Примеры зависимых от последовательностей протоколов включают, но без ограничения, 1) способы гибридизации нуклеиновых кислот; 2) способы амплификации ДНК и РНК, иллюстрируемые различными применениями методов амплификации нуклеиновых кислот [например, полимеразная цепная реакция (ПЦР), Mullis et al., патент США № 4683202; лигазная цепная реакция (LCR), Tabor, S. et al., Proc. Natl. Acad. Sci. U.S.A., 82: 1074 (1985); или амплификации с замещением цепи (SDA), Walker et al., Proc. Natl. Acad. Sci. U.S.A., 89: 392 (1992)]; и 3) способы конструирования библиотек и скринирования на комплементарность.
Например, гены, кодирующие схожие с описанными здесь дельта-5-десатурами белки или полипептиды, можно выделить непосредственно с использованием всей или части фрагментов нуклеиновых кислот по настоящему изобретению в качестве зондов для ДНК-гибридизации для скринирования библиотек, например, из любых желаемых дрожжей или грибов, используя методологию, хорошо известную квалифицированным в данной области техники специалистам (причем предпочтительными будут такие организмы, которые продуцируют ARA [или ее производные]). Специфические олигонуклеотидные зонды, основанные на последовательностях нуклеиновых кислот по настоящему изобретению, можно сконструировать и синтезировать с помощью способов, известных в данной области техники (Maniatis, выше). Кроме того, последовательности целиком можно использовать непосредственно для синтеза ДНК-зондов с помощью способов, известных квалифицированному специалисту (например, произвольного мечения ДНК праймеров, ник-трансляции или способов концевого мечения), или РНК-зондов, используя доступные системы in vitro транскрипции. Кроме того, специфические праймеры можно сконструировать и использовать для амплификации части (или полного размера) последовательностей по настоящему изобретению. Получаемые в результате продукты амплификации можно пометить непосредственно во время реакций амплификации или пометить после реакций амплификации и использовать в качестве зондов для выделения полноразмерных фрагментов ДНК в условиях соответствующей жесткости.
Как правило, в способах амплификации типа ПЦР праймеры имеют различные последовательности и не комплементарны друг другу. В зависимости от желаемых условий проверки последовательности праймеров должны быть сконструированы так, чтобы обеспечить как эффективную, так и точную репликацию нуклеиновой кислоты-мишени. Способы конструирования ПЦР-праймеров являются общепринятыми и хорошо известными в данной области техники (Thein and Wallace, “The use of oligonucleotides as specific hybridization probes in the Diagnosis of Genetic Disorder”, in Human Genetic Diseases: A Practical Approach, K.E. Davis Ed., (1986) pp. 33-50, IRL: Herndon, VA; и Rychlik, W., In Methods in Molecular Biology, White, B.A. Ed., (1993) Vol. 15, pp. 31-39, PCR Protocols: Current Methods and Applications. Humania: Totowa, NJ).
Как правило, два коротких сегмента последовательностей по настоящему изобретению можно использовать в ПЦР-протоколах для амплификации более длинных фрагментов нуклеиновых кислот, кодирующих гомологичные гены, с ДНК или РНК. ПЦР можно также проводить с использованием библиотеки клонированных фрагментов нуклеиновых кислот, причем последовательность одного праймера происходит из фрагментов нуклеиновых кислот по настоящему изобретению, а в интересах последовательности другого праймера пользуются присутствием участков полиадениловой кислоты на 3'-конце предшественника мРНК, кодирующего эукариотические гены.
В альтернативном случае последовательность второго праймера может основываться на последовательностях, происходящих из вектора для клонирования. Например, квалифицированный специалист может следовать RACE-протоколу (Frohman et al., Proc. Natl. Acad. Sci. U.S.A., 85: 8998 (1988)) для создания кДНК с использованием ПЦР для амплификации копий района между одним местом в транскрипте и 3'- или 5'-концом. Праймеры, ориентированные в 3'- и 5'-направлениях, можно сконструировать из последовательностей по настоящему изобретению. Используя коммерчески доступные 3'RACE- или 5'RACE-системы (Gibco/BRL, Gaithersburg, MD), можно выделить специфичные 3' или 5' фрагменты кДНК (Ohara et al., Proc. Natl. Acad. Sci. U.S.A., 86: 5673 (1989); Loh et al., Science, 243: 217 (1989)).
В других вариантах осуществления любой из описанных здесь фрагментов нуклеиновых кислот для дельта-5-десатураз (или любой идентифицированный здесь гомолог) можно использовать для создания новых и улучшенных десатураз жирных кислот. Как хорошо известно в данной области техники, для получения мутаций во встречающихся в природе генах десатураз можно использовать in vitro мутагенез и отбор, химический мутагенез, способы «перестановки генов» или другие способы. Альтернативно, улучшенные жирные кислоты можно синтезировать с помощью замены домена, причем функциональный домен из любого из описанных здесь фрагментов нуклеиновых кислот для дельта-5-десатураз заменяется функциональным доменом в альтернативном гене десатуразы, что приводит, таким образом, к новому белку.
Способы получения различных жирных кислот омега-3 и/или омега-6
Ожидается, что введение химерных генов, кодирующих описанные здесь дельта-5-десатуразы (например, EgD5, EgD5S или другие мутантные ферменты, оптимизированные в отношении кодонов ферменты или их гомологи) под контролем соответствующих промоторов приведет к увеличенной продукции ARA и/или EPA в трансформированном организме-хозяине соответственно. Как таковым, настоящим изобретением охватывается способ прямого получения PUFA, включающий подвергание субстрата в виде жирной кислоты (т.е. DGLA или ETA) действию описанных здесь ферментов десатураз (например, EgD5, EgD5S), так что субстрат превращается в желаемый продукт в виде жирной кислоты (т.е. ARA или EPA соответственно).
Более конкретно, целью по настоящему изобретению является обеспечение способа получения ARA в клетке-хозяине (например, содержащих или продуцирующих масло дрожжах, сое), в котором клетка-хозяин содержит
(i) выделенную нуклеотидную молекулу, кодирующую полипептид дельта-5-десатуразы, обладающий по меньшей мере 39% идентичности при сравнении с полипептидом, имеющим аминокислотную последовательность, представленную в SEQ ID NO: 2, на основе алгоритмов BLASTP или способа выравнивания Clustal W; и
(ii) источник дигомо-γ-линолевой кислоты,
причем клетку-хозяина выращивают в таких условиях, что дельта-5-десатураза экспрессируется, и DGLA превращается в ARA, и ARA необязательно выделяют.
Для квалифицированного в данной области техники специалиста понятно, что широкий диапазон субстратов дельта-5-десатуразы может сделать возможным использование фермента для превращения ЕТА в ЕРА. Соответственно, настоящим изобретением обеспечивается способ получения ЕРА, в котором клетка-хозяин содержит
(i) выделенную нуклеотидную молекулу, кодирующую полипептид дельта-5-десатуразы, обладающий по меньшей мере 39% идентичности при сравнении с полипептидом, имеющим аминокислотную последовательность, представленную в SEQ ID NO: 2, на основе алгоритмов BLASTP или способа выравнивания Clustal W; и
(ii) источник эйкозатетраеновой кислоты,
причем клетку-хозяина выращивают в таких условиях, что дельта-5-десатураза экспрессируется, и ЕТА превращается в ЕРА, и ЕРА необязательно выделяют.
В альтернативном случае каждый ген дельта-5-десатуразы и его соответствующий ферментный продукт, описанный здесь, можно использовать опосредованно для продукции жирных кислот омега-3 (смотри публикацию заявки на патент США № 2005/0136519). Опосредованная продукция PUFA омега-3/омега-6 происходит, когда субстрат в виде жирной кислоты превращается опосредованно в желаемый продукт в виде жирной кислоты посредством промежуточной стадии(й) или через промежуточное соединение(я) пути. Таким образом, предусматривается, что описанные здесь дельта-5-десатуразы (например, EgD5, EgD5S или другие мутантные ферменты, оптимизированные в отношении кодонов ферменты или их гомологи) можно экспрессировать в соединении с дополнительными генами, кодирующими ферменты пути биосинтеза PUFA (например, дельта-6-десатуразы, С18/20-элонгазы, дельта-17-десатуразы, дельта-15-десатуразы, дельта-9-десатуразы, дельта-12-десатуразы, С14/16-элонгазы, С16/18-элонгазы, дельта-9-элонгазы, дельта-8-десатуразы, дельта-4-десатуразы, С20/22-элонгазы), для того, чтобы получить более высокие уровни продукции жирных кислот омега-3 с более длинными цепями (например, EPA, DPA и DHA). Конкретные гены, включаемые в конкретную экспрессирующую кассету, будут зависеть от клетки-хозяина (и его профиля PUFA и/или профиля десатураз/элонгаз), доступности субстрата и желаемого конечного продукта(ов).
В альтернативных вариантах осуществления может быть полезным разрушение природной дельта-5-десатуразы организма-хозяина основанное на полных последовательностях, описанных здесь, комплементе этих полных последовательностей, существенных частях этих последовательностей, оптимизированных в отношении кодонов десатуразах, происходящих из них, и таких последовательностях, которые в значительной степени гомологичны им.
Экспрессирующие системы, кассеты и векторы растений и трансформация растений
В одном варианте осуществления настоящее изобретение относится к рекомбинантной конструкции, включающей какой-либо один из полинуклеотидов дельта-5-десатураз по настоящему изобретению, функционально связанный с по меньшей мере одной регуляторной последовательностью, подходящей для экспрессии в растении. Промотор представляет собой последовательность ДНК, которая направляет клеточные механизмы растения на продукцию РНК с непрерывной кодирующей последовательности, находящейся 3' от промотора. Промоторная область оказывает влияние на скорость, стадию развития и тип клетки, в которой РНК-транскрипт гена продуцируется. РНК-транскрипт подвергается процессингу с продуцированием мРНК, которая служит в качестве матрицы для трансляции последовательности РНК в аминокислотную последовательность кодируемого полипептида. 5'-нетранслируемая лидерная последовательность представляет собой район мРНК, находящийся 5' от кодирующей белок области, который может играть роль в инициации и трансляции мРНК. 3'-сигнал терминации транскрипции/полиаденилирования представляет собой нетранслируемый район, находящийся 3' от кодирующей белок области, который функционирует в клетке растения с вызовом терминации РНК-транскрипта и добавления нуклеотидов полиаденилатов с 3'-конца РНК.
Происхождение промотора, выбираемого для управления экспрессией кодирующей дельта-5-десатуразу последовательности, не является важным, пока он обладает достаточной транскрипционной активностью для осуществления по настоящему изобретению с помощью экспрессии способной транслироваться мРНК для желаемых фрагментов нуклеиновых кислот в желаемых тканях хозяина в подходящее время. Или гетерологичные, или не гетерологичные (т.е. эндогенные) промоторы можно использовать для осуществления на практике по настоящему изобретению. Например, подходящие промоторы включают, но без ограничения, промотор альфа штрих-суъединицы бета-конглицинина, 3' промотор ингибитора трипсина Кунитца, промотор аннексина, промотор глицинина Gy1, промотор бета-суъединицы бета-конглицинина, промотор P34/Gly Bd m 30K, промотор альбумина, промотор Leg A1 и промотор Leg A2.
Промотор аннексина, или Р34, описывается в публикации РСТ-заявки № WO 2004/071178 (опубликованной 26 августа 2004). Уровень активности промотора аннексина сравним с уровнями активности многих известных сильных промоторов, таких как (1) промотор CaMV 35S (Atanassova et al., Plant Mol. Biol. 37: 275-285 (1998); Battraw and Hall, Plant Mol. Biol. 15: 527-538 (1990); Holtorf et al., Plant Mol. Biol. 29: 637-646 (1995); Jefferson et al., EMBO J. 6: 3901-3907 (1987); Wilmink et al., Plant Mol. Biol. 28: 949-955 (1995)); (2) промоторы олеозина Arabidopsis (Plant et al., Plant Mol. Biol. 25: 193-205 (1994); Li, Texas A&M University Ph.D. dissertation, pp. 107-128 (1997)); (3) промоторы белков, добавляющих убиквитин Arabidopsis (Callis et al., J. Biol. Chem. 265(21): 12486-93 (1990)); (4) промотор гена убиквитина томатов (Rollfinke et al., Gene 211(2): 267-76 (1998)); (5) промотор белка теплового шока сои (Schoffl et al., Mol. Gen. Genet. 217(2-3): 246-53 (1989)); и (6) промотор гена гистона Н3 кукурузы (Atanassova et al., Plant Mol. Biol. 37(2): 275-85 (1998)).
Другой полезной особенностью промотора аннексина является его профиль экспрессии в развивающихся семенах. Промотор аннексина наиболее активен в развивающихся семенах на ранних стадиях (раньше, чем 10 дней после опыления) и в большой степени неактивен на поздних стадиях. Профиль экспрессии промотора аннексина отличается от профиля экспрессии многих специфических для семян промоторов, например, промоторов накапливаемых в семенах белков, которые часто обеспечивают наибольшую активность на поздних стадиях развития (Chen et al., Dev. Genet. 10: 112-122 (1989); Ellerstrom et al., Plant Mol. Biol. 32: 1019-1027 (1996); Keddie et al., Plant Mol. Biol. 24: 327-340 (1994); Plant et al., (выше); Li (выше)). Промотор аннексина имеет более обычный профиль экспрессии, но остается отличным от других известных специфических для семян промоторов. Таким образом, промотор аннексина будет очень привлекательным кандидатом, когда желательна сверхэкспрессия, или супрессия, гена в зародышах на ранней стадии развития. Например, может быть желательной сверхэкспрессия гена, регулирующего раннее развитие зародыша, или гена, вовлеченного в метаболизм до созревания семян.
После идентификации соответствующего промотора, подходящего для экспрессии кодирующей конкретную дельта-5-десатуразу последовательности, промотор затем функционально связывают в смысловой ориентации, используя общепринятые способы, хорошо известные квалифицированным в данной области техники специалистам.
Используемые здесь стандартные методы рекомбинантных ДНК и молекулярного клонирования хорошо известны в данной области техники и описываются более полно Sambrook, J. и др. в Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, New York (1989) (в дальнейшем «Sambrook et al., 1989») или Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A. и Struhl, K., Eds., в Current Protocols in Molecular Biology; John Willey and Sons: New York, 1990 (в дальнейшем «Ausubel et al., 1990»).
После создания рекомбинантной конструкции ее затем можно ввести в предпочтительную клетку растения с помощью способов, хорошо известных специалистам со средним уровнем компетентности в данной области техники, (например, трансфекции, трансформации и электропорации). Клетки растения с масличными семенами являются предпочтительными клетками растений. Трансформированные клетки растения затем культивируют и регенерируют в подходящих условиях, делающих возможной экспрессию PUFA с длинной цепью, которую затем необязательно выделяют и очищают.
Рекомбинантные конструкции по настоящему изобретению можно ввести в одну клетку растения или, альтернативно, каждую конструкцию можно ввести в отдельные клетки растения.
Экспрессия в клетке растения может быть осуществлена временно или постоянно, как описано выше.
Желаемые длинноцепочечные PUFA можно экспрессировать в семени. Также в объем этого изобретения входят семена или части растения, полученные из таких трансформированных растений.
Части растения включают дифференцированные и недифференцированные ткани, включающие, но без ограничения, корни, стебли, ростки, листья, пыльцу, семена, ткани вздутий и различные формы клеток и культур (например, отдельные клетки, протопласты, зародыши и ткань каллюса). Ткани растения могут быть в растении или в органе, ткани растения или клеточной культуре.
Термин «орган растения» относится к ткани растения или группе тканей, которые создают морфологически и функционально отличную часть растения. Термин «геном» относится к следующему: 1) полному комплементу генетического материала (генов и некодирующих последовательностей), присутствующего в каждой клетке организма, или вируса, или органелле; и/или 2) полному набору хромосом, наследуемому как (гаплоидная) единица от одного родителя.
Таким образом, настоящее изобретение также относится к способу трансформации клетки, включающему трансформацию клетки рекомбинантной конструкцией по настоящему изобретению и отбор тех клеток, которые трансформированы рекомбинантной конструкцией по настоящему изобретению.
Также представляет интерес способ получения трансформированного растения, включающий трансформацию клетки растения полинуклеотидом дельта-5-десатуразы по настоящему изобретению и регенерацию растения из трансформированной клетки растения.
Способы трансформации двудольных растений (главным образом с помощью использования Agrobacterium tumefaciens) и получения трансгенных растений были опубликованы, среди прочих, для хлопка (патент США № 5004863; патент США № 5159135); сои (патент США № 5569834; патент США № 5416011); Brassica (патент США № 5463174); земляного ореха (Cheng et al., Plant Cell Rep. 15: 653-657 (1996); McKently et al. Plant Cell Rep. 14: 699-703 (1995)); папайи (Ling, K. et al. Bio/Technology 9: 752-758 (1991)) и гороха (Grant et al. Plant Cell Rep. 15: 254-258 (1995)). Для обзора других обычно используемых способов трансформации растений смотри Newell, C.A. (Mol. Biotechnol. 16: 53-65 (2000)). В одном из этих способов трансформации используют Agrobacterium rhizogenes (Tepfler, M. and Casse-Delbart, F. Microbiol. Sci. 4: 24-28 (1987)). Имеется публикация о трансформации сои с использованием прямой доставки ДНК, используя PEG-слияние (публикация РСТ-заявки № WO 92/17598), электропорацию (Chowrira, G.M. et al., Mol. Biotechnol. 3: 17-23 (1995); Christou, P. et al., Proc. Natl. Acad. Sci. U.S.A. 84: 3962-3966 (1987)), микроинъекцию и бомбардировку частицами (McCabe, D.E. et al., Bio/Technology 6: 923 (1988); Christou et al., Plant Physiol. 87: 671-674 (1988)).
Имеется множество способов регенерации растения из ткани растения. Конкретный способ регенерации будет зависеть от исходной ткани растения и конкретного вида растения, подвергаемого регенерации. Регенерация, развитие и выращивание растения из одиночных трансформантов-протопластов растения или из различных трансформированных эксплантов хорошо известны в данной области техники (Weissbach and Weissbach, In Methods for Plant Molecular Biology, (eds.), Academic: San Diego, CA (1988)). Этот процесс регенерации и роста обычно включает стадии отбора трансформированных клеток и выращивание этих индивидуализированных клеток через обычные стадии развития зародышей через стадию ростка с корнем. Аналогичным образом регенерируют трансгенные зародыши и семена. Получаемые в результате трансгенные ростки с корнями впоследствии высаживают в соответствующую среду для роста растений, такую как почва. Предпочтительно регенерируемые растения являются самоопыляемыми для обеспечения гомозиготных трансгенных растений. В противном случае пыльцу, полученную от регенерированных растений, переносят на выращиваемые из семян растения агрономически важных линий. В противоположном случае пыльцу от растений этих важных линий используют для опыления регенерированных растений. Трансгенное растение по настоящему изобретению, содержащее желаемый полипептид, выращивают с использованием способов, хорошо известных квалифицированному в данной области техники специалисту.
Помимо обсуждаемых выше процедур практикующие специалисты знают о стандартных ресурсных материалах, в которых описываются конкретные условия и процедуры для конструирования, манипуляции и выделения макромолекул (например, молекул ДНК, плазмид и т.п.), создания фрагментов рекомбинантных ДНК и рекомбинантных экспрессирующих конструкций и скринирования и выделения клонов. Смотри, например, Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor: NY (1989); Maliga et al., Methods in Plant Molecular Biology, Cold Spring Harbor: NY (1995); Birren et al., Genome Analysis: Detecting Genes, Vol. 1, Cold Spring Harbor: NY (1998); Birren et al., Genome Analysis: Analyzing DNA, Vol.2, Cold Spring Harbor: NY (1998); Plant Molecular Biology: A Laboratory Manual, eds. Clark, Springer: NY (1997).
Примеры растений с масличными семенами включают, но без ограничения, сою, виды Brassica, подсолнечник, кукурузу, хлопок, лен и сафлор.
Примеры PUFA, имеющих по меньшей мере двадцать атомов углерода и четыре или более углерод-углеродных двойных связей, включают, но без ограничения, жирные кислоты омега-3, такие как EPA, DPA и DHA, и жирную кислоту омега-6 ARA. В объем по настоящему изобретению входят семена, получаемые из таких растений, а также масло, полученное из таких семян.
Таким образом, в одном варианте осуществления настоящее изобретение относится к растению с масличными семенами, содержащему
(а) первую рекомбинантную ДНК-конструкцию, включающую выделенный полинуклеотид, кодирующий по меньшей мере один полипептид дельта-5-десатуразы, функционально связанный с по меньшей мере одной регуляторной последовательностью, и
(b) по меньшей мере одну дополнительную рекомбинантную ДНК-конструкцию, включающую выделенный полинуклеотид, функционально связанный с по меньшей мере одной регуляторной последовательностью, кодирующий полипептид, выбираемый из группы, состоящей из дельта-4-десатуразы, дельта-5-десатуразы, дельта-6-десатуразы, дельта-8-десатуразы, дельта-9-десатуразы, дельта-9-элонгазы, дельта-12-десатуразы, дельта-15-десатуразы, дельта-17-десатуразы, С14/16-элонгазы, С16/18-элонгазы, С18/20-элонгазы и С20/22-элонгазы.
Дополнительные десатуразы обсуждаются, например, в патентах США № 6075183, 5968809, 6136574, 5972664, 6051754, 6410288 и публикациях РСТ-заявок № WO 98/46763, WO 98/46764, WO 00/12720 и WO 00/40705.
Выбор комбинации используемых кассет зависит отчасти от профиля PUFA и/или профиля десатураз/элонгаз клеток растения с масличными семенами, подвергаемых трансформации, и длинноцепочечных PUFA, которые следует экспрессировать.
В другом аспекте настоящее изобретение относится к способу получения PUFA с длинными цепями в клетке растения, включающему
(а) трансформацию клетки рекомбинантной конструкцией по настоящему изобретению; и
(b) отбор тех трансформированных клеток, которые продуцируют PUFA с длинными цепями.
В еще одном аспекте настоящее изобретение относится к способу получения по меньшей мере одной PUFA в клетке сои, включающему
(а) трансформацию клетки сои первой рекомбинантной ДНК-конструкцией, включающей
(i) выделенный полинуклеотид, кодирующий полипептид дельта-5-десатуразы, функционально связанный с по меньшей мере одной регуляторной последовательностью, и
(ii) по меньшей мере одной дополнительной рекомбинантной ДНК-конструкцией, включающей выделенный полинуклеотид, функционально связанный с по меньшей мере одной регуляторной последовательностью, кодирующий полипептид, выбираемый из группы, состоящей из дельта-4-десатуразы, дельта-5-десатуразы, дельта-6-десатуразы, дельта-8-десатуразы, дельта-9-десатуразы, дельта-9-элонгазы, дельта-12-десатуразы, дельта-15-десатуразы, дельта-17-десатуразы, С14/16-элонгазы, С16/18-элонгазы, С18/20-элонгазы и С20/22-элонгазы;
(b) регенерацию растения сои из трансформированной клетки стадии (а); и
(с) отбор тех полученных от растений стадии (b) семян, которые имеют измененный уровень PUFA по сравнению с уровнем в семенах, полученных от не трансформированного растения сои.
В других предпочтительных вариантах осуществления по меньшей мере одна дополнительная рекомбинантная ДНК-конструкция кодирует полипептид, обладающий активностью дельта-9-элонгазы, например, дельта-9-элонгазы, выделенной или происходящей из Isochrysis galbana (№ доступа в GenBank - AF390174; IgD9e), или дельта-9-элонгазы, выделенной или происходящей из Euglena gracilis.
В других предпочтительных вариантах осуществления по меньшей мере одна дополнительная рекомбинантная ДНК-конструкция кодирует полипептид, обладающий активностью дельта-8-десатуразы. Например, в публикации РСТ-заявки № WO 2005/103253 (опубликованной 22 апреля 2005) раскрываются аминокислотная последовательность и последовательность нуклеиновой кислоты для фермента дельта-8-десатуразы из Pavlova salina (смотри также публикацию заявки на патент США № 2005/0273885). Sayanova и др. (FEBS Lett. 580: 1946-1952 (2006)) описывают выделение и характеристику кДНК из свободноживущей амебы почвы Acanthamoeba castellanii, которая при экспрессии в Arabidopsis кодирует С20-дельта-8-десатуразу. Также в находящейся в процессе одновременного рассмотрения заявке правопреемников заявителей, имеющей № предварительной заявки 60/795810, поданной 28 апреля 2006 (№ в реестре поверенного - ВВ-1566), раскрываются аминокислотная и нуклеотидная последовательности фермента дельта-8-десатуразы из Pavlova lutheri (CCMP459). В предварительной заявке США № 60/853563 (подданной 23 октября 2006; № в реестре поверенного - ВВ-1574) раскрываются аминокислотная и нуклеотидная последовательности фермента дельта-8-десатуразы из Tetruetreptia pomquetensis CCMP1491, вида CCMP389 рода Eutreptiella и Eutreptiella cf_gymnastica CCMP1594.
Экспрессирующие системы, кассеты и векторы микроорганизмов и трансформация микроорганизмов
Описанные в настоящем документе гены дельта-5-десатураз и продукты генов (т.е. EgD5 или другие мутантные ферменты, оптимизированные в отношении кодонов ферменты или их гомологи) можно также продуцировать в гетерологичных микробных клетках-хозяевах, в частности в клетках, содержащих или продуцирующих масло дрожжей (например, Yarrowia lipolytica).
Микробные экспрессирующие системы и экспрессирующие векторы, содержащие регуляторные последовательности, управляющие экспрессией на высоком уровне чужеродных белков, хорошо известны квалифицированным в данной области техники специалистам. Любые из них можно использовать для конструирования химерных генов для продукции любого из продуктов генов-последовательностей по настоящему изобретению. Эти химерные гены можно затем ввести в соответствующие микроорганизмы посредством трансформации с обеспечением экспрессии кодируемых ферментов на высоком уровне.
Векторы или ДНК-кассеты, применимые для трансформации подходящих микробных клеток-хозяев, хорошо известны в данной области техники. Конкретный выбор последовательностей, присутствующих в конструкции, зависит от желаемых продуктов экспрессии (выше), природы клетки-хозяина и предполагаемого способа отделения трансформированных клеток от нетрансформированных клеток. Как правило, однако, вектор или кассета содержит последовательности, направляющие транскрипцию и трансляцию релевантного гена(ов), селектируемый маркер и последовательность, делающую возможной автономную репликацию или хромосомную интеграцию. Подходящие векторы включают 5'-район гена, контролирующий инициацию транскрипции (например, промотор), и 3'-район фрагмента ДНК, контролирующий терминацию транскрипции (т.е. терминатор). Наиболее предпочтительно, когда оба контролирующих района происходят из генов трансформированной микробной клетки-хозяина, хотя понятно, что такие контролирующие районы могут также происходить из генов, не являющихся природными относительно конкретного вида, выбранного в качестве хозяина продукции.
Контролирующие инициацию районы или промоторы, пригодные для стимуляции экспрессии ORF дельта-5-десатуразы в желаемой микробной клетке-хозяине, многочисленны и хорошо известны квалифицированным в данной области техники специалистам. Для настоящего изобретения подходит практически любой промотор, способный стимулировать экспрессию этих генов в выбранной клетке-хозяине. Экспрессия в микробной клетке-хозяине может осуществляться временно или постоянно. Временную экспрессию можно осуществить с помощью индукции активности регулируемого промотора, функционально связанного с представляющим интерес геном. Постоянную экспрессию можно осуществить с помощью использования конститутивного промотора, функционально связанного с представляющим интерес геном. В качестве примера, когда клетка-хозяин является клеткой дрожжей, обеспечиваются функциональные в клетках дрожжей районы, контролирующие транскрипцию и трансляцию, в частности из вида хозяина (например, смотри, публикации РСТ-заявок № WO 2004/101757 и WO 2006/052870 в отношении предпочтительных регулирующих инициацию транскрипции районов для использования в Yarrowia lipolytica). Любую из ряда регуляторных последовательностей можно использовать в зависимости от того, является ли желательной конститутивная или индуцируемая транскрипция, эффективности промотора при экспрессии представляющей интерес ORF, простоты конструкции и т.п.
Обнаружено, что нуклеотидные последовательности, окружающие кодон инициации «ATG», оказывают влияние на экспрессию в клетках дрожжей. Если желаемый полипептид плохо экспрессируется в дрожжах, нуклеотидные последовательности экзогенных генов можно модифицировать с включением последовательности для эффективной инициации трансляции в дрожжах для получения оптимальной экспрессии генов. Для экспрессии в дрожжах это можно осуществить с помощью сайт-направленного мутагенеза неэффективно экспрессируемого гена путем слияния его в рамке считывания с эндогенных геном дрожжей, предпочтительно высоко экспрессируемым геном. Альтернативно можно определить консенсусную последовательность инициации трансляции в хозяине и создать эту последовательность в гетерологичных генах для их оптимальной экспрессии в представляющем интерес хозяине.
Контролирующий терминацию район может происходить из 3'-района гена, из которого был получен контролирующий инициацию район, или из отличного гена. Большое количество контролирующих терминацию районов известно и функционирует удовлетворительно в ряде хозяев (при использовании как в одном и том же роде и виде, так и в роде, и виде, отличном от рода и вида, из которого они были получены). Контролирующий терминацию район обычно выбирают больше из-за удобства, а не из-за какого-либо конкретного свойства. Предпочтительно, когда микробным хозяином является клетка дрожжей, контролирующий терминацию район происходит из гена дрожжей (в частности, Saccharomyces, Schizosaccharomyces, Candida, Yarrowia или Kluyveromyces). Также известно, что в дрожжах функционируют 3'-районы генов млекопитающих, кодирующих γ-интерферон и α-2-интерферон. Контролирующие терминацию районы можно также получить из различных генов, природных относительно предпочтительных хозяев. Необязательно сайт терминации может быть не нужен; однако наиболее предпочтительно, если его включают. Районы терминации, применимые в описании настоящего изобретения, включают, хотя, как подразумевается, без ограничения, ~100 п.о. 3'-района экстраклеточной протеазы Yarrowia lipolytica (XPR; № доступа в GenBank - M17741); терминаторы ацил-коА-оксидазы (Aco3: № доступа в GenBank - AJ001301 и № САА04661; Рох3: № доступа в GenBank - XP_503244); терминатор Pex20 (№ доступа в GenBank - AF054613); терминатор Pex16 (№ доступа в GenBank - U75433); терминатор Lip1 (№ доступа в GenBank - Z50020); терминатор Lip2 (№ доступа в GenBank - AJ012632); и терминатор 3-оксоацил-коА-тиолазы (OCT; № доступа в GenBank - X69988).
Как известно квалифицированному в данной области техники специалисту, только встраивание гена в вектор для клонирования не гарантирует то, что он будет успешно экспрессироваться на требуемом уровне. В ответ на потребность в высокой скорости экспрессии было создано много специализированных экспрессирующих векторов с помощью манипулирования рядом различных генетических элементов, которые контролируют аспекты транскрипции, трансляции, стабильности белка, недостаток кислорода и секрецию из микробной клетки-хозяина. Более конкретно, некоторые из молекулярных особенностей, которыми можно манипулировать для контролирования экспрессии генов, включают (1) природу релевантных последовательностей промотора и терминации транскрипции; (2) число копий клонированного гена и то, находится ли ген в плазмиде или он интегрирован в геном клетки-хозяина; (3) конечное расположение в клетке синтезированного чужеродного белка; (4) эффективность трансляции и правильную укладку белка в организме-хозяине; (5) внутреннюю стабильность мРНК и белка клонированного гена в клетке-хозяине; и (6) использование кодонов в клонированном гене, так чтобы частота использования кодонов в нем приближалась к частоте использования предпочтительных кодонов клетки-хозяина. В настоящее изобретение входит каждый из этих типов модификаций в качестве средств для дальнейшей оптимизации экспрессии дельта-5-десатуразы, описанной здесь.
После получения ДНК, кодирующей полипептид, подходящей для экспрессии в соответствующей микробной клетке-хозяине (например, в содержащих или продуцирующих масло дрожжах) (например, химерного гена, включающего промотор, ORF и терминатор), ее помещают в плазмидный вектор, способный автономно реплицироваться в клетке-хозяине, или она непосредственно интегрируется в геном клетки-хозяина. Интеграция экспрессирующих кассет может происходить произвольно в пределах генома хозяина или она может быть нацеленной благодаря использованию конструкций, содержащих районы гомологии с геномом хозяина, достаточной для нацеливания рекомбинации в локусе хозяина. Если конструкции нацелены на эндогенный локус, все или некоторые регулирующие транскрипцию и трансляцию районы могут быть обеспечены эндогенным локусом.
В настоящем изобретении предпочтительный способ экспрессии генов в Yarrowia lipolytica происходит с помощью интеграции линейной ДНК в геном хозяина; и интеграция во множество мест в пределах генома может быть особенно полезна, когда желательна экспрессия генов на высоком уровне [например, в локус Ura3 (№ доступа в GenBank - AJ306421), локус гена Leu2 (№ доступа в GenBank - AF260230), ген Lys5 (№ доступа в GenBank - M34929), локус гена Асо2 (№ доступа в GenBank - AJ001300), локус гена Рох3 (Рох3; № доступа в GenBank - XP_503244; или Aco3: № доступа в GenBank - AJ001301), локус гена дельта-12-десатуразы (публикация РСТ-заявки № WO 2004/104167), локус гена Lip1 (№ доступа в GenBank - Z50020) и/или локус гена Lip2 (№ доступа в GenBank - AJ012632)].
В преимущественном случае ген Ura3 можно использовать многократно в комбинации с отбором с использованием 5-фтороротовой кислоты (5-фторурацил-6-карбоновой кислоты моногидрата; «5-FOA») (ниже) для того, чтобы легко сделать возможным интеграцию генетических модификаций в геном Yarrowia.
При экспрессии двух или более генов с отдельных реплицирующихся векторов желательно, чтобы каждый вектор содержал различные средства отбора и не был гомологичен другой конструкции(ям) для поддержания стабильной экспрессии и предотвращения образования гибридов элементов среди конструкций. Экспериментально можно определить благоразумный выбор регуляторных районов, средств отбора и способа размножения введенной конструкции(й), вследствие чего все введенные гены будут экспрессироваться на необходимых уровнях с обеспечением синтеза желаемых продуктов.
Конструкции, содержащие представляющий интерес ген, можно вводить в микробную клетку-хозяина с помощью любого стандартного метода. Эти методы включают трансформацию например, трансформацию с использованием лития ацетата [Methods in Enzymology, 194: 186-187 (1991)], слияние протопластов, болистическое воздействие, электропорацию, микроинъекцию или любой другой метод, с помощью которого представляющий интерес ген вводится в клетку-хозяина. Более конкретные учения, применимые для содержащих или продуцирующих масло дрожжей (т.е. Yarrowia lipolytica), включают патенты США № 4880741 и 5071764 и Chen, D.C. и др. (Appl. Microbiol. Biotechnol., 48(2): 232-235 (1997)).
Для удобства, клетка-хозяин, которой манипулировали с помощью любого метода для приобретения ею последовательности ДНК (например, экспрессирующей кассеты), будет упоминаться здесь как «трансформированная» или «рекомбинантная». Таким образом, термин «трансформированная» и «рекомбинантная» используется здесь взаимозаменяемо. Трансформированный хозяин будет иметь по меньшей мере одну копию экспрессирующей конструкции и может иметь две или более копий, в зависимости от того, интегрирован ли ген в геном, амплифицирован ли или присутствует на экстрахромосомном элементе, имеющем множественное число копий.
Трансформированную клетку-хозяина можно идентифицировать с помощью различных методов отбора, описанных в публикациях РСТ-заявок № WO 2004/101757 и WO 2006/052870. Предпочтительными для использования здесь способами отбора являются устойчивость к канамицину, гигромицину и аминогликозиду G418, а также способность расти на среде без урацила, лейцина, лизина, триптофана или гистидина. В альтернативных вариантах осуществления для отбора дрожжевых мутантов Ura- используют 5-FOA. Соединение является токсичным для дрожжевых клеток, обладающих функционирующим геном URA3, кодирующим оротидин-5'-монофосфат-декарбоксилазу (ОМР-декарбоксилазу); таким образом, 5-FOA, на основе токсичности, особенно полезен для отбора и идентификации мутантных штаммов дрожжей Ura- (Bartel, P.L. and Fields, S., Yeast 2-Hybrid System, Oxford University: New York, v. 7, pp. 109-147, 1997). Более конкретно, сначала можно выключить природный ген Ura3 с получением штамма, имеющего фенотип Ura-, причем отбор происходит на основе устойчивости к 5-FOA. Затем кластер множества химерных генов и новый ген Ura3 можно интегрировать в отличный локус генома Yarrowia для получения, таким образом, нового штамма, имеющего фенотип Ura+. Последующая интеграция приводит к продуцированию нового штамма Ura3- (снова идентифицируемого с помощью отбора с использованием 5-FOA), когда введенный ген Ura3 выключается. Таким образом, ген Ura3 (в комбинации с отбором с использованием 5-FOA) можно использовать в качестве селектируемого маркера при множестве раундов трансформации.
После трансформации субстраты, подходящие для дельта-5-десатуразы по настоящему изобретению (и необязательно других ферментов PUFA, которые коэкспрессируются в клетке-хозяине), могут продуцироваться клеткой или природно, или трансгенно, или их можно обеспечить экзогенно.
Микробные клетки-хозяева для экспрессии генов и фрагментов нуклеиновых кислот по настоящему изобретению могут включать хозяев, которые растут на различном сырье, включающем простые или сложные углеводы, жирные кислоты, органические кислоты, масла и спирты, и/или углеводы, в широком диапазоне температур и величин рН. Основываясь на потребностях правопреемника заявителей, гены, описанные в настоящем изобретении, будут экспрессированы в продуцирующих или содержащих масло дрожжах (и, в частности, в Yarrowia lipolytica); однако предусматривается, что вследствие высокой консервативности транскрипции, трансляции и аппарата биосинтеза белков любая бактерия, дрожжевая клетка, водоросль и/или гриб будут подходящим микробным хозяином для экспрессии фрагментов нуклеиновых кислот по настоящему изобретению.
Предпочтительными микробными хозяевами, однако, являются продуцирующие или содержащие масло дрожжи. Эти организмы природно способны синтезировать и аккумулировать масло, причем масло может составлять более чем приблизительно 25% сухого веса клетки, более предпочтительно более чем приблизительно 30% сухого веса клетки и наиболее предпочтительно более чем приблизительно 40% сухого веса клетки. Роды, обычные определяемые как продуцирующие или содержащие масло дрожжи, включают, но без ограничения, Yarrowia, Candida, Rhodotorula, Rhodosporidium, Cryptococcus, Trichosporon и Lipomyces. Более конкретно, иллюстративные синтезирующие масло дрожжи включают Rhodosporidium toruloides, Lipomyces starkeyii, L. lipoferus, Candida revkaufi, C. pulcherrima, C. tropicalis, C.utilis, Trichosporon pullans, T. cutaneum, Rhodotorula glutinus, R. graminis и Yarrowia lipolytica (ранее определяемый как Candida lipolytica).
Наиболее предпочтительными являются продуцирующие или содержащие масло дрожжи Yarrowia lipolytica, и в дополнительном варианте осуществления наиболее предпочтительными являются штаммы Y. lipolytica, обозначенные ATCC #20362, ATCC #8862, ATCC #18944, ATCC #76982 и/или LGAM S(7)1 (Papanikolaou S., and Aggelis G., Bioresour. Technol. 82(1): 43-9 (2002)).
Исторически различные штаммы Y. lipolytica использовались для производства и продукции изоцитратлиазы, липаз, полигидроксиалканоатов, лимонной кислоты, эритрита, 2-оксоглутаровой кислоты, γ-декалактона, γ-додекалактона и пировиноградной кислоты. Конкретные учения, применимые для создания продукции ARA, EPA и DHA в Y. lipolytica, обеспечиваются в заявке на патент США № 11/264784 (WO 2006/055322), заявке на патент США № 11/265761 (WO 2006/052870) и заявке на патент США № 11/264737 (WO 2006/052871) соответственно.
Другие предпочтительные микробные хозяева включают продуцирующие или содержащие масло бактерии, водоросли и другие грибы, и в пределах этой широкой группы микробных хозяев представляющими особый интерес являются микроорганизмы, которые синтезируют жирные кислоты омега-3/омега-6 (или микроорганизмы, которые можно генетически сконструировать для этой цели [например, другие дрожжи, такие как Saccharomyces cerevisiae]). Таким образом, например, трансформация Mortierella alpina (который коммерчески используется для продукции ARA) любым геном дельта-5-десатуразы по настоящему изобретению под контролем индуцируемого или регулируемого промотора может привести к получению организмов-трансформантов, способных синтезировать увеличенные количества DGLA. Способ трансформации M. alpina описывается Mackenzie и др. (Appl. Environ. Microbiol., 66: 4655 (2000)). Аналогично способы трансформации микроорганизмов Thraustochytriales описываются в патенте США № 7001772.
Основываясь на приведенных выше учениях, в одном варианте осуществления настоящее изобретение относится к способу получения или ARA, или EPA соответственно, включающему
(а) получение содержащей или продуцирующей масло дрожжевой клетки, содержащей
(i) первую рекомбинантную ДНК-конструкцию, включающую выделенный полинуклеотид, кодирующий полипептид дельта-5-десатуразу, функционально связанный с по меньшей мере одной регуляторной последовательностью, и
(ii) источник субстрата для десатуразы, состоящий или из DGLA, или из ETA соответственно, и
(b) выращивание дрожжевой клетки стадии (а) в присутствии подходящего подвергаемого ферментации источника углерода, причем ген, кодирующий полипептид дельта-5-десатуразу, экспрессируется, а DGLA превращается в ARA или ЕТА превращается в ЕРА соответственно, и
(с) необязательно выделение ARA или ЕРА соответственно стадии (b).
Может быть необходимой подача субстрата.
Конечно, поскольку продуцируемые в природе PUFA в продуцирующих или содержащих масло дрожжах ограничиваются 18:2 жирными кислотами (т.е. LA) и менее обычно 18:3 жирными кислотами (т.е. ALA), в более предпочтительных вариантах осуществления по настоящему изобретению будут генетически конструировать содержащие или продуцирующие масло дрожжи для экспрессии множества ферментов, необходимых для биосинтеза длинноцепочечных PUFA (делая, таким образом, возможной продукцию, например, ARA, EPA, DPA и DHA), помимо дельта-5-десатураз, описанных здесь.
В частности, в одном варианте осуществления настоящее изобретение относится к содержащей или продуцирующей масло дрожжевой клетке, содержащей
(а) первую рекомбинантную ДНК-конструкцию, включающую выделенный полинуклеотид, кодирующий полипептид дельта-5-десатуразу, функционально связанный с по меньшей мере одной регуляторной последовательностью, и
(b) по меньшей мере одну дополнительную рекомбинантную ДНК-конструкцию, включающую выделенный полинуклеотид, функционально связанный с по меньшей мере одной регуляторной последовательностью, кодирующий полипептид, выбираемый из группы, состоящей из дельта-4-десатуразы, дельта-5-десатуразы, дельта-6-десатуразы, дельта-9-десатуразы, дельта-12-десатуразы, дельта-15-десатуразы, дельта-17-десатуразы, дельта-9-элонгазы, С14/16-элонгазы, С16/18-элонгазы, С18/20-элонгазы и С20/22-элонгазы.
В особенно предпочтительных вариантах осуществления по настоящему изобретению по меньшей мере одна дополнительная рекомбинантная ДНК-конструкция кодирует полипептид, обладающий активностью дельта-9-элонгазы, например, дельта-9-элонгазы, выделенной или происходящей из Isochrysis galbana (№ доступа в GenBank - AF390174; IgD9e или IgD9eS), или дельта-9-элонгазы, выделенной или происходящей из Euglena gracilis.
Конструирование метаболизма в виде биосинтеза жирных кислот омега-3 и/или омега-6 в микроорганизмах
Квалифицированным в данной области техники специалистам хорошо известны способы манипулирования биохимическими путями, и ожидают, что будут возможны многочисленные манипуляции для максимизирования биосинтеза жирных кислот омега-3 и/или омега-6 в содержащих или продуцирующих дрожжах и, в частности, в Yarrowia lipolytica. Для этой манипуляции может потребоваться создание метаболизма непосредственно в пределах пути биосинтеза PUFA или дополнительная скоординированная манипуляция различных других метаболических путей.
В случае манипуляций в пределах пути биосинтеза PUFA может быть желательным увеличение продукции LA для того, чтобы сделать возможной продукцию жирных кислот омега-6 и/или омега-3. Этого можно достичь введением и/или амплификацией генов, кодирующих дельта-9- и/или дельта-12-десатуразы. Для максимизирования продукции ненасыщенных жирных кислот омега-6, как хорошо известно квалифицированному в данной области техники специалисту, предпочтительна продукция в организме-хозяине, который по существу лишен ALA; таким образом, предпочтительно хозяина выбирают или получают с помощью исключения или ингибирования активности десатуразы типа дельта-15 или омега-3, делающей возможным превращение LA в ALA. Альтернативно может быть желательным максимизирование продукции жирных кислот омега-3 (и минимизирования синтеза жирных кислот омега-6). В этом случае можно использовать микроорганизм-хозяина, в котором активность дельта-12-десатуразы, делающей возможным превращение олеиновой кислоты в LA, исключена или ингибирована; впоследствии в хозяина вводят соответствующие экспрессирующие кассеты вместе с соответствующими субстратами (например, ALA) для превращения в жирные кислот омега-3 - производные ALA (например, STA, ETrA, ETA, EPA, DPA, DHA).
В альтернативных вариантах осуществления по настоящему изобретению биохимические пути, конкурирующие с путями биосинтеза жирных кислот омега-3 и/или омега-6 за энергию или углерод, или природные ферменты пути биосинтеза PUFA, мешающие продукции конкретного конечного продукта PUFA, можно элиминировать с помощью разрушения генов или супрессировать другим способом (например, с помощью антисмысловой мРНК).
В публикациях РСТ-заявок № WO 2006/055322, WO 2006/052870 и WO 2006/052871 представлено подробное обсуждение манипуляций в пределах пути биосинтеза PUFA в качестве способа увеличения ARA, EPA или DHA (и связанные с ними методы) соответственно в виде желаемых манипуляций в пути биосинтеза TAG и пути деградации TAG (и связанных с ними методов).
В контексте по настоящему изобретению может быть полезной модуляция экспрессии пути биосинтеза жирных кислот с помощью одной из описанных выше стратегий. Например, настоящим изобретением обеспечиваются способы, посредством которых гены, кодирующие ключевые ферменты в биосинтетическом пути с участием дельта-9-элонгазы/дельта-8-десатуразы, вводятся в содержащие или продуцирующие масло дрожжи для продукции жирных кислот омега-3 и/или омега-6. Особенно полезной будет экспрессия генов дельта-5-десатураз по настоящему изобретению в содержащих или продуцирующих масло дрожжах, которые не обладают природно путями биосинтеза жирных кислот омега-3 и/или омега-6, и координирование экспрессии этих генов для максимизирования продукции предпочтительных продуктов PUFA, используя различные средства создания метаболизма организма-хозяина.
Микробные процессы ферментации для получения PUFA
Трансформированную клетку-хозяина выращивают в условиях, которые оптимизируют экспрессию химерных генов десатураз и приводят к получению наибольшего и наиболее экономичного выхода желаемых PUFA. Как правило, условия сред, которые можно оптимизировать, включают тип и количество источника углерода, тип и количество источника азота, отношение углерода к азоту, количество различных минеральных ионов, уровень кислорода, температуру роста, рН, продолжительность фазы продукции биомассы, продолжительность фазы аккумуляции масла и время и способ сбора клеток. Yarrowia lipolytica обычно выращивают в сложной среде (например, среде дрожжевой экстракт-пептон-декстроза (YPD)) или в точно определенной минимальной среде, в которой отсутствует компонент, необходимый для роста, и которая, тем самым, форсирует отбор желаемых экспрессирующих кассет (например, в Основе азотного агара для классификации дрожжей (DIFCO Laboratories, Detroit, MI)).
Среда для ферментации в настоящем изобретении должна содержать подходящий источник углерода. О подходящих источниках углерода говорится в публикации РСТ-заявки № WO 2004/101757. Хотя предусматривается, что источник углерода, используемый в настоящем изобретении, может включать широкий выбор содержащих углерод источников, предпочтительными источниками углерода являются сахара, глицерин и/или жирные кислоты. Наиболее предпочтительными являются глюкоза и/или жирные кислоты, содержащие 10-22 атомов углерода.
Азот может обеспечиваться из неорганического (например, (NH4)2SO4) или органического (например, мочевина или глютамат) источника. Помимо подходящих источников углерода и азота среда для ферментации должна также содержать подходящие минеральные вещества, соли, кофакторы, буферы, витамины и другие компоненты, известные квалифицированным в данной области техники специалистам, подходящие для роста содержащих или продуцирующих масло хозяев и активации ферментативных путей, необходимых для продукции PUFA. Особое внимание уделяют нескольким ионам металлов (например, Mn+2, Co+2, Zn+2, Mg+2), которые активируют синтез липидов и PUFA (Nakahara, T. et al., Ind. Appl. Single Cell Oils, D.J. Kyle and R. Colin, eds. pp. 61-97 (1992)).
Предпочтительными в настоящем изобретении средами для роста являются обычные, коммерчески приготовленные среды, такие как Основа азотного агара для классификации дрожжей (DIFCO Laboratories, Detroit, MI). Можно также использовать другие точно определенные или синтетические среды для роста, и квалифицированному в области микробиологии или науке ферментации специалисту известна соответствующая среда для роста трансформированных клеток-хозяев. Подходящий диапазон рН для ферментации обычно составляет приблизительно 4,0-8,0, причем в качестве диапазона для условий первоначально роста предпочтительным является рН 5,5-рН 7,5. Ферментацию можно проводить в аэробных или анаэробных условиях, причем предпочтительными являются микроаэробные условия.
Как правило, для аккумуляции высоких уровней PUFA в содержащих или продуцирующих масло дрожжевых клетках требуется двустадийный процесс, поскольку метаболическое состояние должно быть «сбалансировано» между ростом и синтезом/запасом жиров. Таким образом, наиболее предпочтительно, когда двустадийный процесс ферментации необходим для продукции PUFA в содержащих или продуцирующих масло дрожжевых клетках (например, Yarrowia lipolytica). Этот подход, а также разработки для различных подходящих процессов ферментации (т.е. одноразовой загрузки, подпитываемой и непрерывной) и анализы во время роста описываются в публикации РСТ-заявки № WO 2004/101757.
Очистка и переработка масел, содержащих PUFA
PUFA можно обнаружить в хозяевах - микроорганизмах и растениях - в виде свободных жирных кислот или в этерифицированной форме, такой как ацилглицерины, фосфолипиды, сульфолипиды или гликолипиды, и их можно выделить из клеток-хозяев благодаря множеству средств, хорошо известных в данной области техники. Одним из обзоров методов выделения, анализа качества и стандартов-нормативов для липидов дрожжей является обзор Z. Jacobs (Critical Reviews in Biotechnology, 12(5/6): 463-491 (1992)). Краткий обзор последовательной переработки также доступен благодаря A. Singh и O. Ward (Adv. Appl. Microbiol., 45: 271-312 (1997)).
Вообще средства для очистки PUFA могут включать экстракцию органическими растворителями, обработку ультразвуком, селективную экстракцию с использованием газа в надкритическом состоянии (например, с использованием углекислого газа), омыление и физические средства, такие как сжатия, и их комбинации. Для дополнительных деталей приводится ссылка на учения публикации РСТ-заявки № WO 2004/101757. Способы выделения масел из семян хорошо известны в данной области техники (Young et al., Processing of Fats and Oils, In The Lipid Handbook, Gunstone et al., eds., Chapter 5 pp. 253-257; Chapman & Hall: London (1994)). Например, соевое масло получают используя ряд стадий, включающих экстракцию и очистку пищевых продуктов в виде масел из содержащих масла семян. Соевые масла и побочные продукты сои получают используя обобщенные стадии, показанные в таблице 3.
Обобщенные стадии для получения соевого масла
и побочных продуктов
процесса
получаемые побочные продукты
гидратацией
с использованием щелочи
или физическое
рафинирование
кислоты, пигменты
рафинированного масла
охлаждением)
токоферолы, стерины,
летучие вещества
Более конкретно, семена сои чистят, термически обрабатывают, подвергают обрушиванию и вальцуют, увеличивая, таким образом, эффективность экстракции масла. Экстракцию масла обычно выполняют с помощью экстракции растворителем (например, гексаном), но можно также выполнить с помощью комбинации физического сжатия и/или экстракции растворителем. Получаемое в результате масло называют неочищенным маслом. Неочищенное масло можно рафинировать гидратацией с помощью гидратации фосфолипидов и других полярных и нейтральных липидных комплексов, что облегчает их отделение от негидратируемой фракции триглицеридов (соевого масла). Получаемые в результате лецитиновые клеи можно далее переработать с получением коммерчески важных продуктов лецитина, используемых во множестве пищевых и промышленных продуктов в качестве агентов для образования эмульсий и разъединения (т.е. для предотвращения слипания или прилипания). Подвергнутое рафинированию гидратацией масло можно далее рафинировать для удаления примесей (главным образом свободных жирных кислот, пигментов и остающихся смол). Рафинирование выполняют путем добавления каустических агентов, которые реагируют со свободной жирной кислотой с образованием мыла и гидратов фосфатидов, и белков в неочищенном масле. Воду используют для вымывания остатков мыла, образованного во время рафинирования. Побочные продукты соапстока можно использовать непосредственно в кормах для животных или ацилировать для извлечения свободных жирных кислот. Краску удаляют благодаря адсорбции с использованием отбеливающей земли, которая удаляет большую часть соединений хлорофилла и каротеноидов. Рафинированное масло можно гидрировать, что приводит, таким образом, к жирам с различными свойствами плавления и строениями. Фракционирование рафинированного масла охлаждением (фракционирование) может использоваться для удаления стеарина из подвергнутого гидрированию масла благодаря кристаллизации в тщательно контролируемых условиях охлаждения. Дезодорация (главным образом через паровую дистилляцию под вакуумом) является последней стадией и предназначена для удаления соединений, которые придают маслу запах или вкус. Другие ценные побочные продукты, такие как токоферолы и стерины, можно удалить во время процесса дезодорации. Подвергнутый дезодорации дистиллят, содержащий эти побочные продукты, можно продать для производства природного витамина Е и других ценных фармацевтических продуктов. Рафинированные, обесцвеченные, (подвергнутые гидрированию, фракционированию) и подвергнутые дезодорации масла и жиры можно упаковать и продать непосредственно или далее переработать в более специализированные продукты. Более подробную ссылку на переработку семян сои, производство соевого масла и использование побочных продуктов можно найти у Erickson (Practical Handbook of Soybean Processing and Utilization, The American Oil Chemical' Society and United Soybean Board (1995)). При комнатной температуре соевое масло является жидкостью, поскольку в нем относительно мало насыщенных жирных кислот по сравнению с такими маслами, как кокосовое, пальмовое, косточковое пальмовое и какао-масло.
Содержащие PUFA масла из растений и микроорганизмов, которые были рафинированы и/или очищены, можно гидрировать для того, чтобы получить в результате жиры с различными свойствами плавления и структурами. Для многих подвергнутых переработке жиров (включающих бутербродные пасты, кондитерские жиры, твердые масла, маргарины, кулинарные жиры и т.п.) необходимы варьирующие степени твердости при комнатной температуре, и их можно получить только через изменение физических свойств исходного масла. Самым обычным образом этого можно достичь через каталитическое гидрирование.
Гидрирование представляет собой химическую реакцию, при которой водород добавляется к ненасыщенным двойным связям в жирных кислотах с помощью катализатора, такого как никель. Например, в высокой степени олеиновое соевое масло содержит ненасыщенные олеиновые кислоты, LA и линоленовые жирные кислоты, и каждую из этих кислот можно гидрировать. Гидрирование имеет два основных эффекта. Во-первых, окислительная стабильность масла увеличивается в результате восстановления доли ненасыщенных жирных кислот. Во-вторых, изменяются физические свойства масла, поскольку модификации жирных кислот увеличивают температуру плавления, приводя к полужидкому или твердому жиру при комнатной температуре.
Существует много параметров, оказывающих влияние на реакцию гидрирования, которая в свою очередь изменяет состав конечного продукта. Рабочие условия, включающие давление, температуру, тип и концентрацию катализатора, перемешивание и конструкцию реактора, находятся среди наиболее важных параметров, которые можно контролировать. Селективные условия гидрирования можно использовать для гидрирования более ненасыщенных жирных кислот, отдавая предпочтение менее ненасыщенным жирным кислотам. Очень легкое или ручное гидрирование часто используется для увеличения стабильности жидких масел. Дальнейшее гидрирование превращает жидкое масло в физически твердый жир. Степень гидрирования зависит от желаемых эксплуатационных показателей и свойств плавления, создаваемых для конкретного конечного продукта. Жидкие кулинарные жиры (используемые в производстве выпекаемых изделий, твердых жиров и кулинарных жиров, используемых для торговых операций жарения и обжаривания) и основы для производства маргарина находятся среди несчетного числа возможных масляных и жировых продуктов, получаемых благодаря гидрированию. Более подробное описание гидрирования и подвергнутых гидрированию продуктов можно найти у Patterson, H.B.W., Hydrogenation of Fats and Oils: Theory and Practice. The American Oil Chemists' Society (1994).
Подвергнутые гидрированию масла стали довольно спорными из-за присутствия трансизомеров жирных кислот, являющихся результатом процесса гидрирования. С потреблением больших количеств трансизомеров были связаны пагубные для здоровья эффекты, включающие увеличение отношений липопротеинов низкой плотности к липопротеинам высокой плотности в плазме крови и увеличение риска коронарной болезни сердца.
Содержащие PUFA масла для использования в пищевых продуктах
В настоящее время на рынке поддерживается большое множество пищевых продуктов и продуктов питания, включающих жирные кислоты омега-3 и/или омега-6 (в частности, ARA, EPA и DHA). Предусматривается, что масла из растений/семян, измененные семена и масла микроорганизмов по настоящему изобретению, содержащие PUFA, будут функционировать в пищевых продуктах и продуктах питания с приданием полезных для здоровья свойств настоящих композиций. По сравнению с другими растительными маслами масла по настоящему изобретению, как полагают, функционируют схоже с другими маслами при пищевых применениях с физической точки зрения (например, частично гидрированные масла, такие как соевое масло, широко используются в качестве ингредиентов для мягких бутербродных паст, маргарина и кулинарных жиров для жарения и обжаривания).
Масла из растений/семян, измененные семена и масла микроорганизмов, содержащие жирные кислоты омега-3 и/или омега-6, описанные здесь, будут подходить для использования во множестве пищевых продуктов и продуктов питания, включающих, но без ограничения, пищевые аналоги, мясные продукты, зерновые продукты, выпеченные продукты, продукты для легкой закуски, молочные продукты. Кроме того, масла из растений/семян, измененные семена и масла микроорганизмов по настоящему изобретению можно использовать в композициях для придания полезных для здоровья свойств медицинским продуктам, включающим медицинские питательные вещества, добавки к рациону, составы для детей младшего возраста, а также фармацевтические продукты. Квалифицированный в области пищевой переработки и пищевого приготовления специалист понимает, каким образом количество и композицию масел из растений и микроорганизмов можно добавить к пищевым продуктам или продуктам питания. Такое количество будет упоминаться здесь как «эффективное количество», и оно будет зависеть от пищевого продукта или продукта питания, рациона, который продукт, как планируется, дополняет, или медицинского состояния, которое медицинский продукт или медицинское питательное вещество, как планируется, корректирует или лечит.
Пищевые аналоги можно получить с использованием процессов, хорошо известных квалифицированным в данной области техники специалистам. Можно отметить аналоги мяса, аналоги сыра, аналоги молока и т.п. Аналоги мяса, полученные из сои, содержат соевый белок или соевый творог и другие ингредиенты, смешанные вместе для имитации различных видов мяса. Эти альтернативы мяса продают в виде замороженных, консервированных или сухих продуктов. Обычно их можно использовать так же, как и пищевые продукты, которые они заменяют. Альтернативы мясу, изготовленные из сои, являются превосходными источниками белка, железа и витаминов В. Примеры аналогов мяса включают, но без ограничения, аналоги окорока, колбасные аналоги, аналоги бекона и т.п.
Пищевые аналоги группируют на имитации или заменители в зависимости от их функциональных и композиционных характеристик. Например, сыр-имитация должен только напоминать сыр, который он, как планируется, заменяет. Однако обычно продукт можно назвать сыром-заменителем только в том случае, если он в пищевом отношении эквивалентен сыру, который он заменяет, и удовлетворяет минимальным композиционным требованиям для этого сыра. Следовательно, часто сыр-заменитель будет иметь более высокие уровни белка, чем сыр-имитация, и в нем будет увеличена концентрация витаминов и минеральных веществ.
Аналоги молока или немолочные пищевые продукты включают, но без ограничения, молоко-имитацию и немолочные замороженные десерты (например, те, которые изготовлены из сои и/или содержащие соевый белок продукты).
Мясные продукты включают широкий выбор продуктов. В Соединенных Штатах «мясо» включает «мясо», получаемое от крупного рогатого скота, свиней и овец. Помимо мяса существуют изделия из домашней птицы, включающей кур, индюшек, гусей, цесарок, уток, и рыба, и моллюски. Существует широкий ассортимент мясных продуктов с добавками обогатителей и продуктов обработки мяса: рыба, подвергнутая технической обработке и жаренная, и подвергнутая технической обработке и варке. Колбасы и горячие сосиски являются примерами продуктов обработки мяса. Таким образом, используемый здесь термин «мясные продукты» включает, но без ограничения, продукты обработки мяса.
Зерновой пищевой продукт представляет собой пищевой продукт, получаемый при переработке хлебных злаков. Хлебный злак включает любое растение семейства трав, которое дает пищевое зерно (семя). Наиболее популярными зерновыми являются ячмень, кукуруза, просо, овес, квиноа, рис, рожь, сорго, род Triticale, пшеница и дикий рис. Примеры зерновых пищевых продуктов включают, но без ограничения, целое зерно, дробленое зерно, крупы, муку, отруби, зародыш, блюда из зернового продукта для завтрака, продукты экструзии, пасты и т.п.
Выпеченные продукты-товары включают любой из зерновых пищевых продуктов, упоминаемых выше, и подвергнутый выпечке или обработке способом, сходным с выпеканием (т.е. высушиванию или затвердению с помощью подвергания воздействию тепла). Примеры выпеченных продуктов-товаров включают, но без ограничения, хлеб, торты, пончики, батоны, пасты, панировочные сухари, выпеченные легкие закуски, маленькие галеты, маленькие крекеры, маленькие пирожные и маленькие претцели. Как упоминалось выше, в качестве ингредиента можно использовать масла по настоящему изобретению.
Пищевой продукт для легкой закуски включает любой из описываемых выше или ниже пищевых продуктов.
Жареный пищевой продукт включает любой из описываемых выше или ниже пищевых продуктов, подвергнутый жарке.
Целебный пищевой продукт включает любой пищевой продукт, который приносит пользу здоровью.
Напиток может быть жидкостью или в любой порошковой форме.
Например, можно указать негазированные напитки, такие как фруктовые соки, свежие, замороженные, консервированные или в виде концентрата; ароматные или простые молочные напитки и т.п. Составы для питания взрослых и детей младшего возраста хорошо известны в данной области техники и коммерчески доступны (например, Similac®, Ensure®, Jevity® и Alimentum® от Ross Products Division, Abbort Laboratories).
Составы для детей младшего возраста являются жидкостями или восстанавливаемыми порошками, которыми кормят детей младшего возраста и новорожденных. «Составы для детей младшего возраста» определяют здесь как энтеральный продукт питания, которым можно заменить грудное молоко человека при кормлении детей младшего возраста и который состоит из желаемого процента жира, смешанного с желаемыми процентами углеводов и белков в водном растворе (смотри, например, патент США № 4670285). На основе всемирных исследований состава, а также уровней, установленных группами экспертов, в среднем грудном молоке человека обычно содержится приблизительно 0,20%-0,40% жирных кислот в целом (считая, что приблизительно 50% калорий от жира), и, как правило, отношение DHA к ARA находится в диапазоне от приблизительно 1:1 до 1:2 (смотри, например, составы Enfamil LIPILTM (Mead Johnson & Company) и Similac AdvanceTM (Ross Products Division, Abbott Laboratories)). Составы для детей младшего возраста должны играть особую роль в питании детей младшего возраста, поскольку они часто являются единственным источником питательных веществ для детей младшего возраста; и хотя кормление грудью является все еще самым лучшим питанием для детей младшего возраста, состав для детей младшего возраста является достаточно близким вторым питанием, которое позволяет детям не только выживать, но и процветать.
Молочный продукт представляет собой продукт, получаемый из молока. Аналог молока или немолочный продукт получают из источника, отличного от молока, например, соевого молока, как обсуждалось выше. Эти продукты включают, но без ограничения, цельное молоко, сепарированное молоко, кисломолочные продукты, такие как йогурт или простокваша, сливки, масло, сгущенное молоко, дегидрированное молоко, используемый в кофе порошковый заменитель молока, используемый в кофе порошковый заменитель сливок, мороженое, сыр и т.п.
Дополнительными пищевыми продуктами, в которые можно было бы включить содержащие PUFA масла, являются, например, жевательные резинки, кондитерские изделия и глазури, желатины и пудинги, твердые и мягкие конфеты, джемы и желе, белый сахарный песок, заместители сахара, сладкие колбасы, украшения десерта и сиропы, и смешанные сухими порошковые смеси.
Содержащие PUFA масла для использования в целебных пищевых продуктах и фармацевтических препаратах
Целебный пищевой продукт представляет собой любой пищевой продукт, который приносит пользу здоровью, и включает функциональные продукты питания, медицинские продукты питания, медицинские питательные вещества и добавки к рациону. Кроме того, масла из растений/семян, измененные семена и масла микроорганизмов по настоящему изобретению можно использовать в стандартных фармацевтических композициях (например, содержащие длинноцепочечные PUFA масла можно легко включить в любой из упоминаемых выше пищевых продуктов для получения, таким образом, функционального или медицинского продукта питания). Более концентрированные композиции, содержащие PUFA, включают капсулы, порошки, таблетки, мягкие желатиновые оболочки, окружающие жидкое заполнение, растворимый каркас из желатина, жидкие концентраты и эмульсии, которые можно использовать к качестве добавок к рациону людей или животных, отличных от людей.
Содержащие PUFA масла для использования в кормах для животных
Корма для животных в общем определяют здесь как продукты, предназначенные для использования в качестве корма или для вмешивания в корм для животных, отличных от людей. Масла из растений/семян, измененные семена и масла микроорганизмов по настоящему изобретению можно использовать в качестве ингредиентов в различных кормах для животных.
Более конкретно, хотя здесь без ограничения, ожидается, что масла по настоящему изобретению можно использовать в пищевых продуктах для домашних животных, пищевых продуктах для жвачных животных и домашней птицы и аквакультуральных пищевых продуктах. Пищевыми продуктами для домашних животных являются продукты, предназначенные для кормления домашнего животного (например, собаки, кошки, птицы, рептилии, грызуна). Эти продукты могут включать зерновые и лечебные пищевые продукты, упоминаемые выше, а также мясо и мясные субпродукты, содержащие соевой белок продукты, травяные и сенные продукты (например, люцерну, тимофеевку, овес или костер, овощи). Пищевые продукты для жвачных животных и домашней птицы являются продуктами, которые предназначены для кормления животного (например, индюшек, кур, крупного рогатого скота, свиней). Как и в случае упоминаемых выше продуктов для домашних животных, эти продукты могут включать зерновые и лечебные пищевые продукты, содержащие соевой белок продукты, мясо и мясные субпродукты и травяные и сенные продукты, перечисленные выше. Аквакультуральные пищевые продукты (или «аквакорма») являются продуктами, предназначенными для использования в аквафермерстве, т.е. которое относится к размножению, выращиванию или фермерству акватических животных и/или животных в пресной или морской воде.
ПРИМЕРЫ
Настоящее изобретение далее описывается в следующих примерах. Следует понимать, что эти примеры, хотя и показывающие предпочтительные варианты осуществления по настоящему изобретению, приведены только в качестве иллюстрации. Из приведенного выше обсуждения и этих примеров квалифицированный в данной области специалист может выяснить существенные характеристики настоящего изобретения и без отклонения от сущности и объема изобретения может осуществить различные изменения и модификации по настоящему изобретению для его адаптации к различным применениям и условиям.
ОБЩИЕ СПОСОБЫ
Стандартные методы рекомбинантных ДНК и молекулярного клонирования, описываемые в примерах, хорошо известны в данной области техники и описываются 1) Sambrook, J., Fritsch, E.F. и Maniatis, T. (Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory: Cold Spring Harbor, NY (1989) (Maniatis)); 2) T.J. Silhavy, M.L. Bennan и L.W. Enquist (Experiments with Gene Fusions, Cold Spring Harbor Laboratory: Cold Spring Harbor, NY (1984)), и 3) Ausubel, F.M. и др. (Current Protocols in Molecular Biology, опубликованные Greene Publishing Assoc. и Wiley-Interscience, Hoboken, NJ (1987)).
Материалы и методы, подходящие для поддержания и роста культур микроорганизмов, хорошо известны в данной области техники. Методы, подходящие для применения в следующих примерах, можно найти представленными в Manual of Methods for General Bacteriology (Phillipp Gerhardt, R.G.E. Murray, Ralph N. Costilow, Eugene W. Nester, Willis A. Wood, Noel R. Krieg and G. Briggs Phillips, Eds), American Society for Microbiology: Washington, DC. (1984)) или Thomas D. Brock в Biotechnology: A Textbook of Industrial Microbiology, 2nd ed., Sinauer Associates: Sunderland, MA (1989). Все реагенты, рестрикционные ферменты и материалы, используемые для роста и поддержания клеток микроорганизмов, получали от Aldrich Chemicals (Milwaukee, WI), DIFCO Laboratories (Detroit, MI), GIBCO/BRL (Gaithersburg, MD) или Sigma Chemical Company (St. Louis, MO), если не указано иное. Компетентные клетки E.coli (XL1-Blue) покупали у Stratagene Company (San Diego, CA). Штаммы E.coli обычно выращивали при 37°С на чашках со средой Лурия-Бертани (LB).
Обычное молекулярное клонирование проводили в соответствии со стандартными способами (Sambrook, выше). Последовательность ДНК создавали на автоматическом секвенсере ABI, используя технологию терминирующих красителей (патент США № 5366860; ЕР 272007) с использованием комбинации вектора и специфичных для вставки праймеров. Прочтение последовательностей выполняли на секвенсере (Gene Codes Corporation, Ann Arbor, MI). Все последовательности прочитывали по меньшей мере дважды в обоих направлениях. Сравнения генетических последовательностей выполняли с использованием программного обеспечения DNASTAR (DNASTAR Inc., Madison, WI).
Значение сокращений является следующим: «сек» означает секунду(ы), «мин» означает минуту(ы), «ч» означает час(ы), «д» означает день (дни), «мкл» означает микролитр(ы), «мл» означает миллилитр(ы), «л» означает литр(ы), «мкМ» означает микромолярный, «мМ» означает миллимолярный, «М» означает молярный, «ммоль» означает миллимоль(и), «мкмоль» означает микромоль(и), «г» означает грамм(ы), «мкг» означает микрограмм(ы), «нг» означает нанограмм(ы), «Е» означает единицу(ы), «п.о.» означает пару(ы) оснований, «т.о.» означает тысячу (тысячи) оснований.
Трансформация и культивирование Yarrowia lipolytica
Штамм АТСС #20362 Yarrowia lipolytica приобретали из Американской коллекции типовых культур (Rockville, MD). Штаммы Y. lipolytica обычно выращивали при 28°С на агаре YPD (1% дрожжевой экстракт, 2% бактопептона, 2% глюкозы, 2% агара).
Трансформацию Y. lipolytica осуществляли согласно способу Chen, D.C. и др. (Apl. Microbiol. Biotechnol., 48(2): 232-235 (1997)), если не отмечено иное. Вкратце, Yarrowia штриховали на чашки YPD и выращивали при 30°С в течение приблизительно 18 ч. С чашки соскребали несколько больших петель клеток, и ресуспендировали их в 1 мл буфера для трансформации, содержащего 2,25 мл 50% PEG, средняя М.м. 3350; 0,125 мл 2 М ацетата Li, pH 6,0; 0,125 мл 2 М DTT и 50 мкг фрагментированной ДНК из молок лососевых. Затем приблизительно 500 нг линеаризованной плазмидной ДНК инкубировали в 100 мкл ресуспендированных клеток и сохраняли при 39°С в течение 1 ч с вихревым перемешиванием с интервалами в 15 мин. Клетки помещали на чашки со средой для отбора и сохраняли при 30°С в течение 2-3 дней.
Для отбора трансформантов обычно использовали минимальную среду («ММ»); ММ имеет следующий состав: 0,17% основы азотного агара для классификации дрожжей (DIFCO Laboratories, Detroit, MI) без сульфата аммония или аминокислот, 2% глюкозы, 0,1% пролина, рН 6,1. Добавляли, если уместно, добавки урацила до конечной концентрации, составляющей 0,01% (получая, таким образом, среду для отбора «ММU», приготовленную с 20 г/л агара).
В альтернативном случае трансформанты отбирали на среде для отбора с использованием 5-фтороротовой кислоты («FOA»; также 5-фторурацил-6-карбоновой кислоты моногидрата), содержащей 0,17% основы азотного агара для классификации дрожжей (DIFCO Laboratories) без сульфата аммония или аминокислот, 2% глюкозы, 0,1% пролина, 75 мг/л урацила, 75 мг/л уридина, 900 мг/л FOA (Zymo Research Corp., Orange, CA) и 20 г/л агара.
Анализ жирных кислот Yarrowia lipolytica
Для анализа жирных кислот клетки собирали с помощью центрифугирования, и липиды экстрагировали, как описано Bligh, E.G. & Dyer, W.J. (Can. J. Biochem. Physiol., 37: 911-917 (1959)). Эфиры метилового спирта и жирных кислот готовили с помощью трансэтерификации экстракта липидов с использованием натрия метилата (Roughan, G., and Nishida I., Arch Biochem Biophys., 276(1): 38-46 (1990)) и впоследствии анализировали с использованием Hewlett-Packard 6890 GC, снабженного колонкой 30-м × 0,25 мм (внутренний диаметр) HP-INNOWAX (Hewlett-Packard). Температура термостата была от 170°С (удерживаясь на этом значении в течение 25 мин) до 185°С с подъемом на 3,5°С/мин.
Для прямой базовой трансэтерификации собирали культуру Yarrowia (3 мл), промывали ее один раз в дистиллированной воде и сушили под вакуумом в Speed-Vac в течение 5-10 мин. К образцу добавляли натрия метилат (100 мкл 1%), и затем образец встряхивали и качали в течение 20 мин. После добавления 3 капель 1 М NaCl и 400 мкл гексана образец встряхивали и центрифугировали. Верхний слой удаляли и анализировали с помощью GC, как описано выше.
ПРИМЕР 1
Условия роста Euglena gracilis, профиль липидов и выделение мРНК
Euglena gracilis получали из лаборатории Dr. Richard Trierner в Michigan State University (East Lansing, MI). Из 10 мл активно растущей культуры переносили 1 мл-аликвоту в 250 мл среды для Euglena gracilis (Eg) в 500-мл стеклянную колбу. Среду для Eg получали объединением 1 г натрия ацетата, 1 г бычьего экстракта (каталожный #U126-01, Difco Laboratories, Detroit, MI), 2 г Bacto®tryptone (каталожный #0123-17-3, Difco Laboratories) и 2 г дрожжевого экстракта Bacto® (каталожный #0127-17-9, Difco Laboratories) в 970 мл воды. После стерилизации фильтрованием асептически добавляли 30 мл супернатанта почвенной воды (каталожный #15-3790, Carolina Biological Supply Co., Burlington, NC) с получением конечной среды для Eg. Культуры Euglena gracilis выращивали при 23°С с использованием цикла, состоящего из 16 ч света и 8 ч тьмы, в течение 2 недель без перемешивания.
Через 2 недели отбирали 10 мл культуры для липидного анализа и центрифугировали при 1800×g в течение 5 мин. Осадок промывали один раз водой и повторно центрифугировали. Получаемый в результате осадок сушили в течение 5 мин под вакуумом, ресуспендировали в 100 мкл триметилсульфония гидроксида (TMSH) и инкубировали при комнатной температуре в течение 15 мин со встряхиванием. После этого добавляли 0,5 мл гексана, и пробирки инкубировали в течение 15 мин при комнатной температуре со встряхиванием. Эфиры метилового спирта и жирных кислот (5 мкл вводили из слоя гексана) отделяли и определяли количество с использованием газового хроматографа Hewlett-Packard 6890, снабженного капиллярной колонкой с кварцевым стеклом Omegawax 320 (каталожный #24152, Supelco Inc., Bellefonte, PA). Программировали, чтобы температура термостата удерживалась при 220°С в течение 2,5 мин, увеличивалась до 240°С с подъемом на 20°С/мин и затем удерживалась в течение дополнительных 2,3 мин. Газ-носитель поставлялся генератором водорода Whatman. Времена удержания сравнивали со временами удержания метиловых эфиров стандартов, коммерчески доступных (каталожный #U-99-F, Nu-Chek Prep, Inc., Elysian, MN), и результирующая хроматограмма показана на фиг. 2.
Остающуюся 2-недельную культуру (240 мл) осаждали центрифугированием при 1800×g в течение 10 мин, промывали один раз водой и повторно центрифугировали. Общую РНК экстрагировали из получаемого в результате остатка, используя реагент RNA STAT-60TM (TEL-TEST, Inc., Friendswood, TX) и следуя предоставленному производителем протоколу (использовать 5 мл реагента, растворить РНК в 0,5 мл воды). Таким образом из осадка было получен 1 мг общей РНК (2 мг/мл). Из 1 мг общей РНК выделяли мРНК с использованием набора для очистки мРНК (Amersham Biosciences, Piscataway, NJ), следуя предоставленному производителем протоколу. Таким образом было получено 85 мкг мРНК.
ПРИМЕР 2
Синтез кДНК Euglena gracilis
кДНК синтезировали непосредственно с мРНК Euglena gracilis следующим образом. В частности, мРНК праймировали адаптерным праймером АР (SEQ ID NO: 65) из набора для 3'-RACE Invitrogen (Carlsbad, CA) в присутствии олигонуклеотида Smart IV (SEQ ID NO: 66) из набора для библиотеки кДНК CreatorTM SmartTM BD-Clontech (Mississauga, ON, Канада). Обратную транскрипцию проводили с использованием обратной транскриптазы Superscript II из набора для 3'-RACE в соответствии с протоколом для набора для библиотеки кДНК CreatorTM SmartTM.
Смесь синтеза 1-й цепи кДНК использовали в качестве матрицы для амплификации с помощью ПЦР, используя АР в качестве 3' праймера и CDSIII 5' праймер (SEQ ID NO: 36) в качестве 5' праймера (поставляемого с набором для библиотеки кДНК CreatorTM SmartTM BD-Clontech). Амплификацию проводили с использованием смеси кДНК-полимеразы Clontech Advantage при 94°C в течение 30 сек с последующими 20 циклами, состоящими из 94°C в течение 10 сек и 68°C в течение 6 мин. Осуществляли конечное удлинение при 68°C в течение 7 мин.
ПРИМЕР 3
Выделение части кодирующей области гена дельта-5-десатуразы Euglena gracilis
В этом примере описывается идентификация части гена Euglena gracilis, кодирующего дельта-5-десатуразу (обозначенную здесь «EgD5» (SEQ ID NO: 1 и 2)), с помощью использования праймеров, происходящих из консервативных районов последовательностей других известных дельта-5- и дельта-8-десатураз.
При оценке того, на основе каких десатураз можно сконструировать вырожденные праймеры, подходящие для выделения дельта-5-десатуразы Euglena gracilis, принимались во внимание различные соображения. В частности, заявители знали, что только последовательности дельта-5-, дельта-6- и дельта-8-десатураз включают консервативный мотив «HPGG» на своих N-концах (причем домен «HPGG» является частью домена хорошо известного цитохрома В5); напротив, дельта-9-десатуразы обладают мотивом «HPGG» домена цитохрома В5 на своих С-концах, в то время как у дельта-17-, так и у дельта-12-десатураз отсутствует домен цитохрома В5. Предположили, что в Euglena gracilis функционирует путь с участием дельта-9-элонгазы/дельта-8-десатуразы; поэтому среди десатураз, разделяющих N-концевой консервативный мотив «HPGG», только дельта-5- и дельта-8-десатуразы ожидались в этом организме. Наконец, хотя известно только малое число последовательностей дельта-8-десатураз, публично доступны многочисленные дельта-5-десатуразы. Заявители отобрали те последовательности дельта-5-десатураз, которые менее гомологичны генам «традиционных» дельта-5-десатураз и которые также разделяют высокую гомологию друг к другу.
На основе отмеченного выше были подвергнуты выравниванию четыре дельта-5-десатуразы и две дельта-8-десатуразы, представленные ниже в таблице 3А, используя способ Clustal W (медленного, точного варианта Гоннета; Thompson et al., Nucleic Acids Res., 22: 4673-4680 (1994)) программы MegAlignTM программного обеспечения DNASTAR.
Выравнивание дельта-5- и дельта-8-десатураз для идентификации районов консервативных аминокислот
NO:
irregulare
AAL13311
megasperma
CAD53323
tricornutum
AAL92562
discoideum
XP_640331
№ WO 2006/012325 и
WO 2006/012326
На фиг. 3 демонстрируется часть результирующего выравнивания, содержащую несколько протяжений последовательности консервативных аминокислот среди 6 различных организмов. На основе этого выравнивания были сконструированы два набора вырожденных олигонуклеотидов для амплификации части кодирующей области гена дельта-5-десатуразы из Euglena gracilis, соответствующих районам фиг. 3, которые обозначены «Консервативный район 1» и «Консервативный район 2». В частности, была создана последовательность консервативных аминокислот GHH(I/V)YTN (SEQ ID NO: 19), соответствующая консервативному району 1, тогда как последовательность консервативных аминокислот N(Y/F)Q(V/I)EHH (SEQ ID NO: 20) была создана для соответствия консервативному району 2. Для уменьшения вырожденности олигонуклеотидов были сконструированы 4 набора олигонуклеотидов (т.е. 5-1А, 5-1В, 5-1С и 5-1D), кодирующие консервативный район 1, и были сконструированы 4 набора олигонуклеотидов (т.е. 5-5АR, 5-5ВR, 5-5СR и 5-5DR), кодирующие антисмысловую цепь консервативного района 2.
Таблица 4
Вырожденные олигонуклеотиды, используемые для амплификации гена дельта-5-десатуразы Euglena gracilis
 [Замечание: для SEQ ID NO: 27-34 использовался следующий код вырожденности нуклеиновых кислот: R=A/G; Y=C/T; W=A/T; B=G/T/C; V=G/A/C; и H=A/C/T.]
[Замечание: для SEQ ID NO: 27-34 использовался следующий код вырожденности нуклеиновых кислот: R=A/G; Y=C/T; W=A/T; B=G/T/C; V=G/A/C; и H=A/C/T.]
На основе полноразмерных последовательностей дельта-5-десатураз таблицы 3 выдвинули гипотезу, что размер фрагмента гена дельта-5-десатуразы Euglena gracilis, амплифицированный, как описано выше, должен был бы составлять приблизительно 600 п.о. (без приблизительно 210 аминокислот на его N-конце и 70 аминокислот на его С-конце).
В целом провели шестнадцать различных ПЦР-амплификаций, поскольку были проверены все комбинации праймеров (т.е. праймер 5-1А использовали с каждым из 5-5АR, 5-5ВR, 5-5СR и 5-5DR, индивидуально; аналогично праймер 5-1В использовали с каждым из 5-5АR, 5-5ВR, 5-5СR и 5-5DR, и т.д.). ПЦР-амлификации проводили в общем объеме 50 мкл, включающем ПЦР-буфер (содержащий 10 мМ KCl, 10 мМ (NH4)2SO4, 20 мМ Tris-HCl (pH 8,75), 2 мМ MgSO4, 0,1% Triton X-100), 100 мкг/мл BSA (конечная концентрация), 200 мкМ концентрацию каждого дезоксирибонуклеотидтрифосфата, 10 пмоль каждого праймера, 10 нг кДНК E. gracilis и 1 мкл Taq ДНК-полимеразы (Epicentre Technologies, Madison, WI). Условия термоциклера были заданы на 35 циклов при 95°С в течение 1 мин, при 56°С в течение 30 сек и при 72°С в течение 1 мин с последующим конечным удлинением при 72°С в течение 10 мин.
Продукты ПЦР очищали с использованием набора для очистки продуктов ПЦР Qiagen (Valencia, CA). Один фрагмент приблизительно ожидаемого размера затем очищали после гель-электрофореза в 1% (вес/объем) агарозе и затем клонировали в вектор pGEM-T-easy (Promega, Madison, WI). Подвергнутую лигированию ДНК использовали для трансформации клеток E.coli DH10B, и отбор трансформантов осуществляли на агаре LB (1% бактотриптона, 0,5% бакто-дрожжевого экстракта и 1% NaCl), содержащем ампициллин (100 мкг/мл). Анализ плазмидной ДНК из группы из 12 трансформантов подтвердил присутствие вставки ожидаемого размера (плазмиды были обозначены «pT-F10-1», «pT-F10-2», «pT-F10-3» и т.д. до «pT-F10-12»).
Анализ последовательностей показал, что плазмида pT-F10-1 содержала фрагмент размером 590 п.о. (SEQ ID NO: 4), который кодирует 196 аминокислот (SEQ ID NO: 5) (в том числе аминокислоты, которые соответствуют консервативному району 1 и 2). Подлинность последовательности Euglena определяли с помощью проведения поисков BLAST (Basic Local Alignment Search Tool (базисный, поисковый способ локального выравнивания); Altschul, S.F., et al., J. Mol. Biol., 215: 403-410 (1993)) на схожесть последовательностей, содержащихся в базе данных BLAST «nr» (включающей все не являющиеся лишними трансляции CDS GenBank, последовательности, происходящие из банка данных о 3-мерных структурах белков Brookhaven, базы данных о белковых последовательностях SWISS-PROT, базы данных EMBL и DDBJ). Последовательность анализировали на схожесть со всеми публично доступными последовательностями ДНК, содержащимися в базе данных «nr», используя алгоритм BLASTN, предоставленный National Center for Biotechnology Information (NCBI). SEQ ID NO: 4 анализировали на схожесть со всеми публично доступными белковыми последовательностями, содержащимися в базе данных «nr», используя алгоритм BLASTX (Gish, W. and States, D.J., Nature Genetics, 3: 266-272 (1993)), предоставленный NCBI.
Результаты сравнений BLASTX с выводом, касающимся последовательности, с которой наиболее схожа SEQ ID NO: 4, сообщаются в соответствии с % идентичности, % схожести и ожидаемым значением. «% Идентичности» определяют как процент аминокислот, являющихся идентичными между двумя белками. «% Схожести» определяют как процент аминокислот, являющихся идентичными или консервативными между двумя белками. «Ожидаемое значение» оценивает статистическую значимость соответствия, определяя число соответствий, с данной оценкой, которое ожидается при поиске в базе данных этого размера абсолютно случайно. Таким образом, транслированная аминокислотная последовательность SEQ ID NO: 4 (т.е. SEQ ID NO: 5) идентична на 38% и схожа на 53% с аминокислотной последовательностью дельта-8-десатуразы сфинголипидов Thalassiosira pseudonana (№ доступа в GenBank - AAX14502; SEQ ID NO: 21), с ожидаемым значением 5Е-28; кроме того, частичный фрагмент SEQ ID NO: 4 идентичен на 37% и схож на 52% с дельта-5-десатуразой жирных кислот Phaeodactylum tricornutum (№ доступа в GenBank - AAL92562, SEQ ID NO: 14), с ожидаемым значением 7Е-28.
ПРИМЕР 4
Выделение 5' кодирующей области гена дельта-5-десатуразы Euglena gracilis
Для выделения N-концевой части предполагаемой дельта-5-десатуразы, идентифицированной в примере 3, использовали модифицированный метод 5'-RACЕ, основанный на протоколах для RACЕ от двух различных компаний (т.е. Invitrogen и BD-Clontech).
Вкратце, двухцепочечную кДНК Euglena gracilis (пример 2) использовали в качестве матрицы в эксперименте 5' RACE, включающем два отдельных раунда ПЦР-амплификации. В первом раунде ПЦР-амплификации олигонуклеотидные праймеры состояли из специфического для гена одигонуклеотида (т.е. ODMW480; SEQ ID NO: 35) и общего олигонуклеотидного 5' праймера CDDIII (SEQ ID NO: 36) из набора для библиотеки кДНК CreatorTM SmartTM BD-Clontech. ПЦР-амлификации проводили в общем объеме 50 мкл, включающем 25 мкл предварительно подготовленной смеси LA TaqTM (TaKaRa Bio Inc., Otsu, Shiga, 520-2193, Япония), 10 пмоль каждого праймера и 1 мкл Taq ДНК-полимеразы (Epicentre Technologies, Madison, WI). Условия термоциклера были заданы на 35 циклов при 95°С в течение 1 мин, при 56°С в течение 30 сек и при 72°С в течение 1 мин с последующим конечным удлинением при 72°С в течение 10 мин.
Для второго раунда ПЦР-амплификации использовали 1 мкл продукта из ПЦР первого раунда в качестве матрицы. Праймеры состояли из специфического для гена олигонуклеотида (т.е. ODMW479; SEQ ID NO: 37) и общего 5' олигонуклеотида DNR CDS (SEQ ID NO: 38), поставляемого из набора для библиотеки кДНК CreatorTM SmartTM BD-Clontech. Амплификацию проводили, как описано выше.
Продукты ПЦР второго раунда подвергали электрофорезу в 1% (вес/объем) агарозе. Продукты между 400 п.о. и 800 п.о. затем очищали из геля и клонировали в вектор pGEM-T-easy (Promega, Madison, WI). Подвергнутую лигированию ДНК использовали для трансформации E.coli DH10B, и отбор трансформантов осуществляли на агаре LB, содержащем ампициллин (100 мкг/мл).
Анализ плазмидной ДНК из одного трансформанта, включающего 5'-район гена предполагаемой дельта-5-десатуразы, подтвердил присутствие ожидаемой плазмиды, обозначенной pT-EgD5-5'C2. Анализ последовательностей показал, что pT-EgD5-5'C2 содержала фрагмент размером 797 п.о. (SEQ ID NO: 6), которые перекрывались с 238 п.о. 5'-конца фрагмента размером 590 п.о. pT-F10-1 (пример 3, SEQ ID NO: 4) и дополнительно обеспечивали 559 п.о. 5' последовательности (SEQ ID NO: 7) (фиг. 4). Последовательность pT-EgD5-5'C2 также корректировала последовательность, соответствующую консервативному району 1, являющуюся результатом использования вырожденного олигонуклеотида для первоначальной ПЦР-амплификациии фрагмента размером 590 п.о. в pT-F10-1 (пример 3). Однако в удлиненном фрагменте размером 797 п.о. SEQ ID NO: 6 не было кодона инициации трансляции.
Второй раунд модифицированного метода 5'-RACE проводили, как описано выше, за исключением, что в качестве специфических для гена праймеров использовали олигонуклеотиды YL791 (SEQ ID NO: 39) и YL792 (SEQ ID NO: 40). Продукты между 200 п.о. и 400 п.о. затем очищали из геля и клонировали в вектор pGEM-T-easy (Promega, Madison, WI). Подвергнутую лигированию ДНК использовали для трансформации E.coli DH10B, и отбор трансформантов осуществляли на агаре LB, содержащем ампициллин (100 мкг/мл).
Анализ плазмидной ДНК из одного трансформанта, включающего 5'-район гена предполагаемой дельта-5-десатуразы, подтвердил присутствие ожидаемой плазмиды, обозначенной pT-EgD5-5'2nd. Анализ последовательностей показал, что pT-EgD5-5'2nd содержала фрагмент размером 273 п.о. (SEQ ID NO: 8), которые перекрывались с 253 п.о. 5'-конца ДНК-фрагмента в pT-EgD5-5'C2, описанной выше, и дополнительно обеспечивали 20 п.о. 5' последовательности (SEQ ID NO: 9). Семнадцать (17) п.о. из 20 п.о. кодировали N-концевую часть гена предполагаемой дельта-5-десатуразы, включающую кодон инициации трансляции, обеспечивая, таким образом, полную 5' последовательность гена.
ПРИМЕР 5
Выделение 3' кодирующей области гена дельта-5-десатуразы Euglena gracilis
Для выделения С-концевой части предполагаемой дельта-5-десатуразы, идентифицированной в примере 3, использовали метод 3'-RACЕ. Методология была описана выше в примере 4; однако праймерами, используемыми как в первом, так и во втором раунде ПЦР-амиплификации, были праймеры, показанные ниже в таблице 5.
Олигонуклеотидные праймеры, используемые для 3'-RACЕ
гена олигонуклеотид
олигонуклеотид
После выделения и очистки продуктов (т.е. 400-800 п.о. ) фрагменты клонировали в вектор pGEM-T-easy (Promega) и трансформировали в E.coli DH10B, как описано в примере 4.
Анализ плазмидной ДНК из одного трансформанта, включающего 3'-район гена дельта-5-десатуразы, подтвердил присутствие ожидаемой плазмиды, обозначенной pT-EgD5-3'. Анализ последовательностей показал, что pT-EgD5-3' содержала фрагмент размером 728 п.о. (SEQ ID NO: 10), которые перекрывались с 264 п.о. 3'-конца фрагмента размером 590 п.о. pT-F10-1 (пример 3, SEQ ID NO: 4) и обеспечивали 464 п.о. дополнительной 3' последовательности (SEQ ID NO: 11). Первые 184 п.о. фрагмента размером 464 п.о., включенные в pT-EgD5-3', кодировали С-концевую кодирующую область (в том числе стоп-кодон трансляции) гена предполагаемой дельта-5-десатуразы. Последовательность pT-EgD5-3' также корректировала последовательность, соответствующую консервативному району 2, являющуюся результатом использования вырожденного олигонуклеотида для первоначальной ПЦР-амплификациии фрагмента размером 590 п.о. pT-F10-1 (пример 3).
После 2 раундов 5'-RACE и одного раунда 3'-RACE была определена последовательность ДНК полной кодирующей предполагаемую дельта-5-десатуразу области Euglena gracilis (EgD5). Как показано на фиг. 4, размер CDS EgD5 составлял 1350 п.о. (SEQ ID NO: 1), и она кодировала полипептид с 449 аминокислотами (SEQ ID NO: 2), на основе выравнивания SEQ ID NO: 4, 6, 8 и 10. Результаты поисков BLASTP с использованием полноразмерного гена EgD5 в качестве запрашиваемой последовательности показал, что он идентичен на 39% и схож на 56% с дельта-5-десатуразой жирных кислот Phaeodactylum tricornutum (№ доступа в GenBank - AAL92562, SEQ ID NO: 14), с ожидаемым значением 1Е-80. Кроме того, полноразмерный ген EgD5 идентичен на 37% и схож на 55% с дельта-8-десатуразой сфинголипидов Thalassiosira pseudonana (№ доступа в GenBank - AAX14502, SEQ ID NO: 21), с ожидаемым значением 3Е-75.
ПРИМЕР 6
Создание конструкции pDMW367, включающей EgD5
В этом примере описывается создание конструкции pDMW367, включающей химерный ген FBAIN::EgD5::Pex20-3' (фиг. 5С). Она предназначена для интеграции химерного гена в геном Yarrowia lipolytica и затем исследования функционирования дельта-5-десатуразы Euglena gracilis в Yarrowia lipolytica.
Основываясь на полноразмерной кДНК EgD5 (SEQ ID NO: 1), в качестве праймеров для амплификации первой части EgD5 использовали олигонуклеотиды YL794 и YL797 (SEQ ID NO: 44 и 45 соответственно) (фиг. 5А). Праймер YL794 содержал сайт для NcoI, а праймер YL797 содержал сайт для HindIII. Затем в качестве праймеров для амплификации второй части EgD5 использовали праймеры YL796 и YL795 (SEQ ID NO: 46 и 47 соответственно). Праймер YL796 содержал сайт для HindIII, а праймер YL797 содержал сайт для NotI. ПЦР-реакции, используя пары праймеров YL794/YL797 или YL796/YL795, c кДНК Euglena gracilis (пример 2) в качестве матрицы проводили отдельно в общем объеме 50 мкл, включающем ПЦР-буфер (содержащий 10 мМ KCl, 10 мМ (NH4)2SO4, 20 мМ Tris-HCl (pH 8,75), 2 мМ MgSO4, 0,1% Triton X-100), 100 мкг/мл BSA (конечная концентрация), 200 мкМ концентрацию каждого дезоксирибонуклеотидтрифосфата, 10 пмоль каждого праймера и 1 мкл Pfu ДНК-полимеразы (Stratagene, San Diego, CA). Условия термоциклера были заданы на 35 циклов при 95°С в течение 1 мин, при 56°С в течение 30 сек и при 72°С в течение 1 мин с последующим конечным удлинением при 72°С в течение 10 мин.
Отдельные продукты ПЦР очищали с использованием набора для очистки продуктов ПЦР Qiagen. ПЦР-продукты из реакции, амплифицированные с помощью праймеров YL794/YL797, расщепляли NcoI и HindIII, в то время как ПЦР-продукты из реакции, амплифицированные с помощью праймеров YL796/YL795, расщепляли HindIII и NotI. NcoI/HindIII- и HindIII/NotI-расщепленные фрагменты ДНК очищали после гель-электрофореза в 1% (вес/объем) агарозе и затем направлено лигировали с NcoI/NotI-расщепленной конструкцией pZUF17 (фиг. 5В; SEQ ID NO: 22; содержащей синтетический ген дельта-17-десатуразы [«D17st»] из Saprolegnia diclina (публикация заявки на патент США № 2003/0196217 A1), оптимизированный в отношении кодонов для экспрессии в Yarrowia lipolytica (публикация РСТ-заявки № WO 2004/101757)). Продуктом этого лигирования была плазмида pDMW367 (фиг. 5С; SEQ ID NO: 23), которая содержала, таким образом, следующие компоненты:
Компоненты плазмиды pDMW367 (SEQ ID NO: 23)
нуклеотиды в пределах
SEQ ID NO: 23
химерного гена
(7416-1671)
• FBAIN: промотор FBAIN Yarrowia
lipolytica (публикация РСТ-заявки №
WO 2005/049805; патент США № 7202356)
• EgD5: дельта-5-десатураза Euglena
gracilis (SEQ ID NO: 1, описанная здесь; обозначенная «Euglena D5DS» на фиг.)
• Pex20: последовательность терминатора
Pex20 гена Pex20 Yarrowia (№ доступа в
GenBank - AF054613)
для отбора в E.coli
последовательность Yarrowia (ARS18;
№ доступа в GenBank - A17608)
GenBank - AJ306421)
Термин «промотор FBAIN» или «промоторный район FBAIN» относится к 5' нетранслируемому району перед кодоном инициации трансляции «ATG» фермента фруктозо-бифосфат-альдолазы (Е.С. 4.1.2.13), кодируемой геном fba1, который необходим для экспрессии, плюс часть 5' кодирующей области, которая включает интрон гена fba1.
ПРИМЕР 7
Создание штамма М4 Yarrowia lipolytica для продукции приблизительно 8% DGLA от липидов в целом
В этом примере описывается создание штамма М4, происходящего из Yarrowia lipolytica ATCC #20362, способного продуцировать 8% DGLA относительно липидов в целом. Этот штамм создан для экспрессии пути с участием дельта-6-десатуразы/дельта-6-элонгазы посредством введения конструкции pKUNF-12T6E (фиг. 6A; SEQ ID NO: 24). Эта конструкция сконструирована для интеграции четырех химерных генов (включающих дельта-12-десатуразу, дельта-6-десатуразу и две С18/20-элонгазы) в локус Ura3 штамма ATCC #20362 Yarrowia lipolytica дикого типа, делая, таким образом, возможной продукцию DGLA. Таким образом, pKUNF-12T6E содержала следующие компоненты:
Описание плазмиды pKUNF-12T6E (SEQ ID NO: 24)
нуклеотиды в пределах
SEQ ID NO: 24
химерных генов
(9420-8629)
Yarrowia (№ доступа в GenBank -
AJ306421)
(12128-1)
Yarrowia (№ доступа в GenBank -
AJ306421)
(6380-8629)
• FBAIN: промотор FBAIN Yarrowia
lipolytica (публикация РСТ-заявки №
WO 2005/049805; патент США № 7202356;
обозначенный «Fba1+ intron» на фиг.)
• EL1S: оптимизированный в отношении
кодонов ген элонгазы 1 (публикация
РСТ-заявки № WO 2004/101753),
происходящий из Mortierella alpina
(№ доступа в GenBank - AX464731)
• Pex20: последовательность терминатора
Pex20 из гена Pex20 Yarrowia (№ доступа
в GenBank - AF054613)
(4221-6380)
• TEF: промотор TEF Yarrowia
lipolytica (№ доступа в GenBank -
AF054508)
• delta-6S: оптимизированный в отношении кодонов ген дельта-6-десатуразы
(публикация РСТ-заявки № WO 2004/101753),
происходящий из Mortierella alpina
(№ доступа в GenBank - AF465281)
• Lip1: последовательность терминатора
Lip1 из гена Lip1 Yarrowia (№ доступа в
GenBank - Z50020)
(4207-1459)
• FBA: промотор FBA Yarrowia
lipolytica (публикация РСТ-заявки №
WO 2005/049805; патент США 7202356;
обозначенный «FBA1» на фиг.)
• F.delta-12: ген дельта-12-десатуразы
Fusarium moniliforme (публикация
РСТ-заявки № WO 2005/047485)
• Lip2: последовательность терминатора
Lip2 из гена Lip2 Yarrowia (№ доступа в
GenBank - AJ012632)
(1459-1)
• TEF: промотор TEF Yarrowia
lipolytica (№ доступа в GenBank -
AF054508)
• EL2Syn: оптимизированный в отношении
кодонов ген элонгазы (SEQ ID NO: 25),
происходящий из Thraustochytrium aureum
(патент США № 6677145)
• XPR2: ~100 п.о. 3'-района гена Xpr
Yarrowia (№ доступа в GenBank - M17741)
Плазмиду pKUNF-12T6E расщепляли AscI/SphI и затем использовали для трансформации Y. lipolytica ATCC #20362 дикого типа согласно разделу «Общие способы». Клетки-трансформанты помещали на чашки со средой для отбора с FOA и сохраняли при 30°С в течение 2-3 дней. Устойчивые к FOA колонии отбирали и штриховали на чашках для отбора ММ и ММU. Колонии, растущие на чашках ММU, но не чашках ММ, отбирали в качестве штаммов Ura-. Отдельные колонии штаммов Ura- затем инокулировали в жидкую ММU при 30°С и встряхивали при 250 оборотов/мин в течение 2 дней. Клетки собирали центрифугированием, экстрагировали липиды, и эфиры метилового спирта и жирных кислот готовили с помощью трансэтерификации и впоследствии анализировали с помощью Hewlett-Packard 6890 GC.
Анализ GC показал присутствие DGLA в трансформантах, содержащих 4 химерных гена pKUNF-12T6E, но не в контрольном штамме Yarrowia дикого типа. Большинство отобранных 32 штаммов Ura- продуцировали приблизительно 6% DGLA от липидов в целом. Существовало 2 штамма (т.е. штаммы М4 и 13-8), которые продуцировали приблизительно 8% DGLA от липидов в целом.
ПРИМЕР 8
Функциональный анализ гена EgD5 в штамме М4 Yarrowia lipolytica
Плазмиду pDMW367 (пример 6; включающую химерный ген FBAIN::EgD5::Pex20) трансформировали в штамм М4 (пример 7), как описано в разделе «Общие способы». Трансформанты отбирали на чашках ММ. Через 2 дня роста при 30°С 3 трансформанта, выросшие на чашках ММ, отбирали и повторно штриховали на свежих чашках ММ. После того, как они вырастали, эти штаммы индивидуально инокулировали в 3 мл жидкой ММ при 30°С и встряхивали при 250 оборотов/мин в течение 2 дней. Клетки собирали центрифугированием, экстрагировали липиды, и эфиры метилового спирта и жирных кислот готовили с помощью трансэтерификации и впоследствии анализировали с помощью Hewlett-Packard 6890 GC.
Анализ GC показал, что во всех трех трансформантах продуцировалось приблизительно 5,6% DGLA и 2,8% ARA от липидов в целом, причем эффективность превращения DGLA в ARA во всех трех штаммах, как определено, составляла приблизительно 33% (в среднем). Эффективность превращения определяли по следующей формуле: ([продукт]/[субстрат + продукт])∗100, где «продукт» включает непосредственный продукт и все продукты в пути, происходящие от него. Таким образом, эти экспериментальные данные продемонстрировали, что клонированная дельта-5-десатураза Euglena gracilis, описанная здесь как SEQ ID NO: 1 и 2, эффективно десатурирует DGLA в ARA.
ПРИМЕР 9
Синтез оптимизированного в отношении кодонов гена дельта-5-десатуразы («EgD5S») для экспрессии в Yarrowia lipolytica
Использование кодонов гена дельта-5-десатуразы Euglena gracilis (SEQ ID NO: 1 и 2; EgD5) оптимизировали для экспрессии в Yarrowia lipolytica аналогично тому, как это описано в публикации РСТ-заявки № WO 2004/101753 и патенте США № 7125672. В частности, оптимизированный в отношении кодонов ген дельта-5-десатуразы (обозначенный «EgD5S», SEQ ID NO: 3) конструировали на основе кодирующей последовательности гена дельта-5-десатуразы EgD5 в соответствии с характером использования кодонов Yarrowia (публикация РСТ-заявки № WO 2004/101753), консенсусной последовательностью вокруг кодона инициации трансляции «ATG» и общими правилами стабильности РНК (Guhaniyogi, G. and J. Brewer, Gene, 265(1-2): 11-23 (2001)). Помимо модификации сайта инициации трансляции модифицировали 196 п.о. кодирующей последовательности размером 1350 п.о. (14,5%; фиг. 7), и оптимизировали 189 кодонов (42%). Содержание G+C уменьшилось с 55,5% в гене дикого типа (т.е. EgD5) до 54,4% в синтетическом гене (т.е. EgD5S). Сайты для NcoI и NotI были включены вокруг кодона инициации трансляции и после стоп-кодона EgD5S соответственно. Ни одна из модификаций в оптимизированном в отношении кодонов гене не изменяла аминокислотную последовательность кодируемого белка (SEQ ID NO: 2). Сконструированный ген EgD5S (SEQ ID NO: 3) был синтезирован GenScript Corporation (Piscataway, NJ) и клонирован в pUC57 (№ доступа в GenBank - Y14837) с созданием pEgD5S (фиг. 6В, SEQ ID NO: 48).
ПРИМЕР 10
Создание конструкции pDMW369, включающей EgD5S
В этом примере описывается создание плазмиды pDMW369, включающей химерный ген FBAIN::EgD5S::Pex20. Плазмиду pDMW369 (фиг. 6С; SEQ ID NO: 49) создавали путем замены NcoI/NotI-фрагмента pZUF17 (фиг. 5В; SEQ ID NO: 22) NcoI/NotI-фрагментом EgD5S из pEgD5S (фиг. 6В; SEQ ID NO: 48). Продуктом этого лигирования была pDMW369, которая содержала, таким образом, следующие компоненты:
Компоненты плазмиды pDMW369 (SEQ ID NO: 49)
нуклеотиды в пределах
SEQ ID NO: 49
химерного гена
(6063-318)
• FBAIN: промотор FBAIN Yarrowia
lipolytica (публикация РСТ-заявки №
WO 2005/049805; патент США № 7202356;
обозначенный «FBA1+ intron» на фиг.)
• EgD5S: оптимизированная в отношении
кодонов дельта-5-десатураза
(SEQ ID NO: 3, описанная здесь как
EgD5S), происходящая из Euglena gracilis
• Pex20: последовательность терминатора
Pex20 гена Pex20 Yarrowia (№ доступа в
GenBank - AF054613)
для отбора в E.coli
последовательность Yarrowia (ARS18;
№ доступа в GenBank - A17608)
GenBank - AJ306421)
ПРИМЕР 11
Экспрессия оптимизированной в отношении кодонов дельта-5-десатуразы («EgD5S») в штамме М4 Yarrowia lipolytica
Плазмиду pDMW369 (пример 10; включающую химерный ген FBAIN::EgD5S::Pex20) трансформировали в штамм М4 (пример 7), как описано в разделе «Общие способы». Трансформанты отбирали на чашках ММ. Через 2 дня роста при 30°С 3 трансформанта, выросшие на чашках ММ, отбирали и повторно штриховали на свежих чашках ММ. После того как они вырастали, эти штаммы индивидуально инокулировали в 3 мл жидкой ММ при 30°С и встряхивали при 250 оборотов/мин в течение 2 дней. Клетки собирали центрифугированием, экстрагировали липиды, и эфиры метилового спирта и жирных кислот готовили с помощью трансэтерификации и впоследствии анализировали с помощью Hewlett-Packard 6890 GC.
Анализ GC показал, что во всех трех трансформантах продуцировалось приблизительно 3,3% DGLA и 2,7% ARA от липидов в целом, причем эффективность превращения DGLA в ARA в этих трех штаммах, как определено, составляла приблизительно 45% (в среднем, рассчитанная, как описано в примере 8). Таким образом, эти экспериментальные данные продемонстрировали, что синтетическая дельта-5-десатураза Euglena gracilis, оптимизированная в отношении кодонов для экспрессии в Yarrowia lipolytica (EgD5S, представленная в SEQ ID NO: 3), на приблизительно 36% более эффективно десатурирует DGLA в ARA, чем ген EgD5 дикого типа (SEQ ID NO: 1).
ПРИМЕР 12
Выделение дельта-8-десатуразы Pavlova lutheri (CCMP459)
В этом примере описывается выделение дельта-8-десатуразы Pavlova lutheri (CCMP459), используемой в примере 3 и на фиг. 3 (также описанной в заявке на патент США № 11/737772, поданной 20 апреля 2007). Для этого потребовались синтез кДНК Pavlova lutheri (CCMP459), конструирование библиотеки и секвенирование, идентификация гомологов дельта-8-десатуразы и клонирование полноразмерной дельта-8-десатуразы из геномной ДНК.
Синтез кДНК Pavlova lutheri (CCMP459), конструирование библиотеки и секвенирование
Библиотеку кДНК Pavlova lutheri (CCMP459) синтезировали, как описано в публикации РСТ-заявки № WO 2004/071467 (опубликованной 26 августа 2004). Вкратце, замороженные осадки Pav459 получали из Provasoli-Guillard National Center for Culture of Marine Phytoplankton (CCMP, West Boothbay Harbor, ME). Эти осадки разрушали в жидком азоте, и из Pav459 экстрагировали общую РНК с помощью использования набора Qiagen RNeasy® Maxi (Qiagen, Valencia, CA) по инструкциям производителя. Из этой общей РНК выделяли мРНК, используя целлюлозный полимер с олиго-dT, которую затем использовали для конструирования библиотеки кДНК, используя вектор pSport1 (Invitrogen, Carlsbad, CA). Полученную таким образом кДНК направленно клонировали (5' SalI/3' NotI) в вектор pSport1. Библиотека Pav459 содержала приблизительно 6,1×105 клонов в мл, при этом средний размер вставки в каждом клоне составлял приблизительно 1200 п.о. Библиотеку Pavlova lutheri назвали eps1c.
Для секвенирования клоны сначала отбирали из архивированных в глицерине культур, выращенных/замороженных в 384-луночных планшетах с замораживающей средой, и инокулировали с использованием автоматического отборщика колоний QPix® (Genetix) в 96-луночные планшеты с глубокими лунками, содержащие LB + 100 мг/мл ампициллина. После выращивания в течение 20 ч при 37°С клетки осаждали с помощью центрифугирования и хранили при -20°С. Плазмиды затем выделяли на робототехническом устройстве Eppendorf 5Prime, используя модифицированный мини-препаративный способ щелочного лизиса в 96-луночном формате (Eppendorf PerfectPrep®). Вкратце, после преципитации ацетатом для облегчения удаления клеточного дебриса использовали фильтр и вакуумный коллектор. Плазмидную ДНК затем связывали на втором планшете с фильтром непосредственно из фильтрата, промывали, сушили и извлекали.
Плазмиды подвергали секвенированию концевых последовательностей в 384-луночных планшетах, используя праймирующий вектор праймер Т7 (SEQ ID NO: 50) и набор для секвенирования ABI BigDye версии 3 Prism. Для реакции секвенирования использовали 100-200 нг матрицы и 6,4 пмоль праймера, и 25 раз повторяли следующие реакционные условия: 96°С в течение 10 сек, 50°С в течение 5 сек и 60°С в течение 4 мин. После основанной на этаноле очистки продукты циклических реакций секвенирования растворяли и определяли на автоматических секвенсерах Perkin-Elmer ABI 3700.
Идентификация гомологов дельта-8-десатуразы из библиотеки кДНК Pavlova lutheri eps1c
Клоны кДНК, кодирующие гомологи дельта-8-десатуразы Pavlova lutheri (называемые, таким образом, дельта-8-десатуразами), идентифицировали путем проведения поисков BLAST на схожесть последовательностей, содержащихся в базе данных BLAST «nr» (как описано в примере 3). Р-величины (вероятность) наблюдения соответствия последовательности кДНК последовательности, содержащейся в подвергаемых поиску базах данных, не более чем случайно, рассчитываемые BLAST, сообщали здесь в виде величин «pLog», которые представляют собой отрицательную величину логарифма сообщаемой Р-величины. Следовательно, чем больше величина pLog, тем выше вероятность того, что последовательность кДНК и «хит» BLAST представляют собой гомологичные белки.
С помощью поиска BLASTX с использованием нуклеотидной последовательности из клона eps1c.pk002.f22 обнаружена схожесть белка, кодируемого кДНК, с дельта-6-десатуразой из Rhizopus stolonifer (SEQ ID NO: 51) (№ доступа в NCBI - AAX22052 (GI 60499699), локус AAX22052, CDS AY795075; Lu et al., не опубликованное сообщение). Последовательность части вставки кДНК из клона eps1c.pk002.f22 показана в SEQ ID NO: 52 (5'-конец вставки кДНК). Впоследствии была получена последовательность всей вставки (eps1c.pk002.f22::fis), и она показана в SEQ ID NO: 53. Последовательность выведенной аминокислотной последовательности (от нуклеотида 1 SEQ ID NO: 53 до первого стоп-кодона в положении нуклеотида 864 SEQ ID NO: 53) показана в SEQ ID NO: 54. Секвенирование всей вставки осуществляли с использованием модифицированного протокола транспозиции. Клоны, определенные для секвенирования всей вставки, отбирали из архивированных в глицерине штоков в виде отдельных колоний, и выделяли плазмидную ДНК с помощью щелочного лизиса. Плазмидные матрицы подвергали транспозиции с помощью набора для транспозиции Template Generation System (TGS II) (Finnzymes Oy, Espoo, Финляндия), следуя протоколу производителя. Подвергнутую транспозиции ДНК трансформировали в электрокомпетентные клетки ЕН10В (Edge BioSystems, Gaithersburg, MD) с помощью электропорации. Из каждой реакции транспозиции произвольно отбирали множество трансформантов, готовили плазмидную ДНК, и матрицы секвенировали, как описано выше (ABI BigDye v.3.1), вне сайта транспозиционного события, используя уникальные праймеры SeqE (SEQ ID NO: 55) и SeqW (SEQ ID NO: 56).
Данные о последовательности собирали (программное обеспечение ABI Prism Collection) и компоновали с использованием программы для компоновки последовательности Phrap (P. Green, University of Washington, Seattle). Компоновки просматривались с помощью программы для редактирования последовательностей Consed (D. Gordon, University of Washington, Seattle) для окончательного прочтения.
Аминокислотную последовательность, представленную в SEQ ID NO: 54, оценивали с помощью BLASTP с получением величины pLog - 19,52 (Е-величина - 3е-20) в сравнении с дельта-6-десатуразой из Mortierella alpina (№ доступа в NCBI - BAC82361 (GI 34221934), локус ВАС82361, CDS AB070557; Sakuradani and Shimizu, Biosci. Biotechnol. Biochem., 67: 704-711 (2003)). Основываясь на результатах BLASTP-сравнения с десатуразой Mortierella alpina и другими десатуразами жирных кислот, дельта-8-десатураза Pavlova lutheri и не была полноразмерной, и в ней отсутствовала последовательность на 5'-конце.
Клонирование полноразмерной дельта-8-десатуразы из геномной ДНК Pavlova lutheri
Геномную ДНК выделяли из Pavlova lutheri (ССМР459), используя набор для максимального приготовления Qiagen DNeasy® Plant Maxi Prep Kit в соответствии с протоколом производителя. Используя 1 колонку maxi на 1 г замороженного клеточного осадка, из 4 г культуры Pavlova lutheri выделяли в целом 122 мкг геномной ДНК. Конечная концентрация геномной ДНК составляла 22,8 нг/мкл. Библиотеки GenomeWalker синтезировали с использованием набора Universal GenomeWalkerTM (BD Biosciences Clonetech, Palo Alto, CA), следуя протоколу производителя (протокол #PT3042-1, версия PRO3300). Вкратце, согласно протоколу проводилось четыре рестрикционных расщепления с использованием 300 нг геномной ДНК на реакцию. После очистки с использованием фенола осадки растворяли в 4 мкл воды, и согласно протоколу подвергались лигированию адаптеры.
Для первоначальной ПЦР использовали набор для ПЦР Advantage®-GC Genomic PCR (BD Biosciences Clonetech), следуя протоколу производителя (протокол #PT3090-1, версия PR1Х433). Для каждого рестрикционного расщепления 1 мкл библиотеки объединяли с 22,8 мкл воды ПЦР-степени, 10 мкл 5Х реакционного буфера для ПЦР GC Genomic PCR, 2,2 мкл 25 мМ Mg(CH3CO2)2, 10 мкл GC-Melt (5 M), 1 мкл 50X смеси dNTP (10 мМ каждого), 1 мкл смеси Advantage®-GC Genomic Pol. (50X), 1 мкл универсального праймера АР1 GenomeWalkerTM (10 мкМ, SEQ ID NO: 57) и 1 мкл GSP PvDES (10 мкМ, SEQ ID NO: 58). После денатурации при 95°С 35 раз повторяли следующие условия реакции: 94°С в течение 30 сек, 68°С в течение 6 мин. После этих условий реакции в течение 6 мин осуществляли конечное удлинение при 68°С с последующим охлаждением до 15°С до момента удаления.
Первоначальную ПЦР-реакцию для каждой библиотеки анализировали с помощью электрофореза в агарозном геле, и наблюдали полосы ДНК с молекулярными массами приблизительно 6 т.о., 3,5 т.о., 2,5 т.о. и 1,2 т.о. ДНК-полосы для каждой библиотеки очищали с использованием набора для очистки ДНК из геля ZymocleanTM (Zymo Research, Orange, CA), следуя протоколу производителя. Получаемую в результате ДНК клонировали в вектор pGEM®-T Easy (Promega), следуя протоколу производителя, и вставки секвенировали с использованием праймеров Т7 (SEQ ID NO: 50) и M13-28Rev (SEQ ID NO: 59), как описано выше. Дополнительная последовательность затем была получена с использованием специфичного для гена праймера для секвенирования PavDES seq (SEQ ID NO: 60), полученного из вновь появившихся данных о последовательности. Последовательность полного 5'-конца, полученная с помощью прогулки по геному, показана в SEQ ID NO: 61. Последовательность перекрывающихся областей геномной последовательности (SEQ ID NO: 61) и полностью секвенированную EST eps1c.pk002.f22:fis (SEQ ID NO: 53) выравнивали, используя SequencherTM (версия 4.2, Gene Codes Corporation, Ann Arbor, MI) с использованием алгоритма компоновки Large Gap. Интересно, что сравнение показало, что у EST, которая была первоначально секвенирована (SEQ ID NO: 53), отсутствовали 459 п.о. по сравнению с геномной последовательностью (SEQ ID NO: 61). По-видимому, эта пропущенная последовательность в EST является делецией, а не интроном, поскольку в геномной ДНК на 5'-конце гена не наблюдалось четких сайтов сплайсинга интронов. Геномную последовательность для 5'-конца (SEQ ID NO: 61) объединяли с 3'-концом последовательности EST (SEQ ID NO: 53) с получением SEQ ID NO: 62. Используя программное обеспечение для анализа последовательностей EditSeqTM 6.1 (DNASTAR Inc., Madison, WI), определили ORF (SEQ ID NO: 17). Аминокислотная последовательность, кодируемая SEQ ID NO: 17, показана в SEQ ID NO: 18.
Аминокислотную последовательность, представленную в SEQ ID NO: 18, оценивали с помощью BLASTP с получением величины pLog - 35,10 (Е-величина - 8е-36) в сравнении с дельта-6-десатуразой из Rhizopus stolonifer (SEQ ID NO: 63)(№ доступа в NCBI - ABB96724 (GI 83027409), локус ABB96724, CDS DQ291156; Zhang et al., не опубликованное сообщение). Кроме того, дельта-8-десатураза Pavlova lutheri идентична на 78,0% последовательности дельта-8-десатуразы Pavlova salina (SEQ ID NO: 64), раскрытой в публикации РСТ-заявки № WO 2005/103253 (опубликованной 22 апреля 2005), используя способ Jotun Hein. Расчеты процентов идентичности последовательностей, проведенные по способу Jotun Hein (Hein, J.J., Meth. Enz., 183: 626-645 (1990)), осуществляли с использованием версии 6.1 программы MegAlignTM комплекта вычислительных программ для биоинформатики LASERGENE (DNASTAR Inc., Madison, WI), используя параметры по умолчания для попарного выравнивая ((оптимальный) размер слова, используемый при выравнивании = 2). Дельта-8-десатураза Pavlova lutheri идентична на 76,4% последовательности дельта-8-десатуразы Pavlova salina, используя способ Clustal V. Расчеты процентов идентичности последовательностей, проведенные с использованием способа Clustal V (Higgins, D.G. and Sharp, P.M., Comput. Appl. Biosci, 5: 151-153 (1989); Higgins et al., Comput. Appl. Biosci., 8: 189-191 (1992)), осуществляли с использованием версии 6.1 программы MegAlignTM комплекта вычислительных программ для биоинформатики LASERGENE (DNASTAR Inc.), используя параметры по умолчания для попарного выравнивая ((оптимальный) размер слова, используемый при выравнивании = 1, пенальти за промежуток = 3, окно = 5, сохраняемые диагонали = 5 и пенальти за длину промежутка = 10). BLAST-оценки и вероятности указывают на то, что фрагмент SEQ ID NO: 17 кодирует целую дельта-8-десатуразу Pavlova lutheri.
На фиг. 8А и 8В демонстрируется выравнивание Clustal V (с использованием параметров по умолчанию) SEQ ID NO: 18 (аминокислотной последовательности дельта-8-десатуразы Pavlova lutheri), SEQ ID NO: 64 (аминокислотной последовательности дельта-8-десатуразы Pavlova salina, выше), SEQ ID NO: 16 (аминокислотной последовательности дельта-8-десатуразы Euglena gracilis, раскрытой как SEQ ID NO: 2 в публикации РСТ-заявки № WO 2006/012325, опубликованной 2 февраля 2006), SEQ ID NO: 63 (аминокислотной последовательности дельта-6-десатуразы жирных кислот Rhizopus stolonifer (№ доступа в NCBI - ABB96724, выше) и SEQ ID NO: 51 (аминокислотной последовательности дельта-6-десатуразы жирных кислот Rhizopus stolonifer (№ доступа в NCBI - AAХ22052, выше). Результаты выравнивания Clustal V показывают, что SEQ ID NO: 18 идентична на 76,4%, 22,6%, 22,2% и 22,2% SEQ ID NO: 64, SEQ ID NO: 16, SEQ ID NO: 63 и SEQ ID NO: 51 соответственно.
ПРИМЕР 13
Трансформация соматических культур зародышей сои
Условия культивирования
Образующую зародыши сои суспензионную культуру (распространенная версия Jack) поддерживали в 35 мл жидкой среды SB196 (ниже) на вращающейся качалке, 150 оборотов/мин, при 26°С с холодными белыми люминесцентными излучениями во время фотопериода 16:8 ч день/ночь при интенсивности света 60-85 мкЭрг/м2/сек. Культуры субкультивировали каждые 7 дней-2 недели путем инокуляции приблизительно 35 мг ткани в 35 мл свежей жидкой SB196 (предпочтительный интервал для субкультуры - каждые 7 дней).
Образующие зародыши сои суспензионные культуры трансформировали экспрессирующими плазмидами сои по способу пушечной бомбардировки частицами (Klein et al., Nature, 327: 70 (1987)) с использованием прибора DuPont Biolistic PDS1000/HE (гелиевая модификация) для всех трансформаций.
Инициирование образующей зародыши сои суспензионной культуры
Соевые культуры инициировали дважды в месяц с 5-7-днями между каждой инициацией. Стручки с незрелыми семенами от имеющихся растений сои через 45-55 дней после посадки собирают, отделяют от створок и помещают в подвергнутый стерилизации пурпурно-красный бокс. Семена сои стерилизуют с помощью встряхивания их в течение 15 мин в 5% растворе Клоракс с 1 каплей мыла «Айвори» (т.е. 95 мл подвергнутой автоклавированию дистиллированной воды плюс 5 мл Клоракса и 1 капля мыла, хорошо смешанные). Семена промывают двумя литровыми бутылками стерильной дистиллированной воды, и семена, которые меньше 4 мм, помещают на отдельные микроскопические предметные стекла. Маленький конец семени отрезают, и семядоли выдавливают из оболочки семени. Семядоли переносят в чашки, содержащие среду SB1 (25-30 семядолей на чашку). Чашки обертывают волокнистой лентой и хранят в течение 8 недель. Сразу после этого вторичные зародыши разрезают и помещают в жидкую среду SB196 на 7 дней.
Приготовление ДНК для бомбардировки
Для бомбардировки используют или интактную плазмиду, или ДНК-фрагмент плазмиды, содержащий представляющие интерес гены дельта-6-десатураз и селектируемый маркер. Фрагменты экспрессирующих плазмид сои, содержащие дельта-5-десатуразу по настоящему изобретению, получают с помощью выделения из геля расщепленных плазмид. Получаемые в результате ДНК-фрагменты разделяют с помощью гель-электрофореза в 1% агарозе SeaPlaque GTP (BioWhitaker Molecular Applications), и ДНК-фрагменты, содержащие кассеты генов, вырезают из агарозного геля. ДНК очищают от агарозы с использованием расщепляющего фермента GELase, следуя протоколу производителя.
50 мкл-аликвоту стерильной дистиллированной воды, содержащую 3 мг частиц золота, добавляют к 5 мкл 1 мкг/мкл раствора ДНК (или интактной плазмиды, или ДНК-фрагмента, приготовленного, как описано выше), 50 мкл 2,5 М CaCl2 и 20 мкл 0,1 М спермидина. Смесь встряхивают в течение 3 мин на уровне 3 встряхивателя с вихревым перемешиванием и центрифугируют в течение 10 сек в настольной микроцентрифуге. После промывки 400 мкл 100% этанола осадок суспендируют под воздействием ультразвука в 40 мкл 100% этанола. Суспензию ДНК (5 мкл) распределяют на каждый подвижный диск диска прибора Biolistic PDS1000/HE. Каждая 5 мкл-аликвота содержит приблизительно 0,375 мг частиц золота на бомбардировку (т.е. на диск).
Приготовление ткани и бомбардировка ДНК
Приблизительно 150-200 мг суспензионной культуры 7-дневных зародышей помещают в пустые, стерильные чашки Петри 60×15 мм, и чашку покрывают пластиковой сеткой. Ткань подвергают бомбардировке 1 или 2 выстрелами на чашку с разрывающим мембрану давлением, установленным на 1100 фунтов на квадратный дюйм, и камеру разрежают до вакуума 27-28 дюймов ртутного столба. Ткань помещают на расстоянии приблизительно 3,5 дюйма от удерживающего/останавливающего экрана.
Отбор трансформированных зародышей
Трансформированные зародыши отбирают с использованием гигромицина в качестве селектируемого маркера. В частности, после бомбардировки ткань помещают в свежую среду SB196 и культивируют, как описано выше. Через шесть дней после бомбардировки SB196 заменяют свежей SB196, содержащей 30 мг/л гигромицина. Среду для отбора освежают еженедельно. Через четыре-шесть недель после отбора наблюдают рост зеленой трансформированной ткани из нетрансформированных, некротических зародышевых скоплений. Выделенную зеленую ткань извлекают и инокулируют в многолуночные планшеты для регенерации новых, клонально размноженных, трансформированных, образующих зародыши суспензионных культур.
Созревание зародышей
Зародыши культивируют в течение 4-6 недель при 26°С в SB196 под лампами холодного белого люминесцентного излучения (Phillips cool white Econowatt F40/CW/RS/EW) и лампами Argo (Phillips F40 Argo) (40 Вт) во время фотопериода 16:8 ч при интенсивности света 90-120 мкЭрг/м2/сек. Сразу после этого зародышевые скопления перемещают на твердую агаровую среду SB166 на 1-2 недели. Скопления затем субкультивируют в среде SB103 в течение 3 недель. Во время этого периода отдельные зародыши извлекают из скоплений и скринируют на изменения в их составах жирных кислот, как описано выше.
Перечень ингредиентов сред
SB196 - жидкая среда для пролиферации FN Lite (на литр)
Твердая среда SB1 (на литр)
MS соли одной упаковки (Gibco/BRL - каталожный № - 11117-066)
1 мл 100Х маточного раствора витаминов В5
31,5 г сахарозы
2 мл 2,4-D (20 мг/л конечная концентрация)
рН 5,7
2 г ТС агара
Твердая среда SB 166 (на литр)
MS соли одной упаковки (Gibco/BRL - каталожный № - 11117-066)
1 мл 100Х маточного раствора витаминов В5
60 г мальтозы
750 мг MgCl2 гексагидрата
5 г активированного угля
рН 5,7
2 г гелрита
Твердая среда SB 103 (на литр)
MS соли одной упаковки (Gibco/BRL - каталожный № - 11117-066)
1 мл 1000Х маточного раствора витаминов В5
60 г мальтозы
750 мг MgCl2 гексагидрата
рН 5,7
2 г гелрита
Твердая среда SB 74-4 (на литр)
В5 соли Gamborg одного флакона (Gibco/BRL - каталожный № 21153-036)
рН 5,7
5 г ТС агара
Маточный раствор 2,4-D
Получают заранее приготовленным от Phytotech, каталожный № D 295 - концентрация 1 мг/мл.
Маточный раствор витаминов В5 (на 100 мл)
Хранить в аликвотах при -20°С
10 г миоинозита
100 мг никотиновой кислоты
100 мг пиридоксина HCl
1 г тиамина
Если раствор не растворяется достаточно быстро, применяют нагревание низкой степени через горячую плитку-перемешиватель.
ПРИМЕР 14
Функциональный анализ дельта-5-десатуразы (SEQ ID NO: 1 и 2) в соматических зародышах сои
Зрелые соматические зародыши сои являются хорошей моделью зиготных зародышей. При нахождении в глобулярном зародышевом состоянии в жидкой культуре соматические зародыши сои содержат очень низкие количества триацилглицерина (TAG) или запасаемых белков, типичных для созревающих, зиготных зародышей сои. На этой стадии развития отношение общего триацилглицерина к общему полярному липиду (фосфолипидам и гликолипидам) составляет приблизительно 1:4, что является типичным для зиготных зародышей сои на стадии развития, с которой инициировалась культура соматических зародышей. Также на глобулярной стадии мРНК для главных белков семени, α'-субъединицы β-конглицинина, ингибитора 3 трипсина Кунитца и лектина семян, по существу отсутствуют. При переносе безгормонной среды для того, чтобы сделать возможной дифференциацию до состояния созревающих соматических зародышей, TAG становится самым обильным классом липидов. Также мРНК для α'-субъединицы β-конглицина, ингибитора 3 трипсина Кунитца и лектина семян становятся очень обильными блоками информации в общей популяции мРНК. На этом основании система соматических зародышей сои ведет себя очень схоже с созревающими зиготными зародышами сои in vivo и является, следовательно, хорошей и быстрой модельной системой для анализа фенотипических эффектов модификации экспрессии генов в пути биосинтеза жирных кислот (смотри публикацию РСТ-заявки № WO 2002/00904, пример 3). Наиболее важным является то, что модельная система является также прогнозирующей в отношении состава жирных кислот семян от растений, происходящих от трансгенных зародышей.
Трансгенные соматические зародыши сои, содержащие дельта-5-десатуразу по настоящему изобретению, анализируют следующим образом. Из отдельных, созревших соматических зародышей сои с помощью трансэтерификации готовят эфиры метилового спирта и жирных кислот. Отдельные зародыши помещают в пробирку, содержащую 50 мкл триметилсульфония гидроксида (TMSH) и 0,5 мл гексана, и инкубируют в течение 30 мин при комнатной температуре со встряхиванием. Эфиры метилового спирта и жирной кислоты (5 мкл вводят из слоя гексана) отделяют и определяют количество с использованием газового хроматографа Hewlett-Packard 6890, снабженного капиллярной колонкой с кварцевым стеклом Omegawax 320 (каталожный #24152, Supelco Inc.). Программируют, чтобы температура термостата удерживалась при 220°С в течение 2,6 мин, увеличивалась до 240°С с подъемом на 20°С/мин и затем удерживалась в течение дополнительных 2,4 мин. Газ-носитель поставляется генератором водорода Whatman. Времена удержания сравнивают со временами удержания метиловых эфиров стандартов, коммерчески доступных (Nu-Chek Prep, Inc.). Обычно 5-10 зародышей на каждый случай анализируют с помощью GC, используя описанную выше методологию.
ПРИМЕР 15
Коэкспрессия других комбинаций кассет промотор/ген/терминатор в соматических зародышах сои
Квалифицированному в данной области техники специалисту понятно, что помимо описанных здесь кассет генов, промоторов и терминаторов можно синтезировать другие комбинации кассет промотор/ген/терминатор схожим с описанным здесь способом, но не ограничиваясь этим способом. Например, в публикациях РСТ-заявок № WO 2004/071467 и WO 2004/071178 описывается выделение ряда последовательностей промоторов и терминаторов транскрипции для использования в специфической для зародышей экспрессии в сое. Кроме того, в публикациях РСТ-заявок № WO 2004/071467, WO 2005/047479 и WO 2006/012325 описывается синтез множества комбинаций кассет промотор/ген/терминатор с помощью лигирования вместе отдельных промоторов, генов и терминаторов транскрипции в уникальных комбинациях. Как правило, для клонирования желаемого гена используют сайт для NotI, фланкированный подходящим промотором (например, промоторами, перечисленными, но без ограничения, в таблице 9) и терминатором транскрипции (например, терминаторами транскрипции, перечисленными, но без ограничения, в таблице 10). Сайты для NotI можно добавить к представляющему интерес гену, такому как гены, перечисленные, но без ограничения, в таблице 11, используя ПЦР-амплификацию с олигонуклеотидами, сконструированными для введения сайтов для NotI с 5' и 3' концов гена. Получаемый в результате продукт ПЦР затем расщепляют NotI и клонируют в подходящую кассету промотор/NotI/терминатор.
Кроме того, в публикациях РСТ-заявок № WO 2004/071467, WO 2005/047479 и WO 2006/012325 описывается дальнейшее связывание отдельных кассет генов в уникальных комбинациях вместе с подходящими кассетами селектируемых маркеров для того, чтобы получить желаемую фенотипическую экспрессию. Хотя это осуществляют, главным образом, с использованием сайтов для различных рестрикционных ферментов, квалифицированному в данной области техники специалисту понятно, что для получения желаемой комбинации промотор/ген/терминатор транскрипции можно использовать ряд методов. Действуя таким образом, можно получить любую комбинацию кассет специфическую для зародышей промотор/ген/терминатор транскрипции. Квалифицированному в данной области техники специалисту также понятно, что эти кассеты могут быть расположены на отдельных ДНК-фрагментах или множестве фрагментов, причем коэкспрессия генов является результатом котрансформации множества ДНК-фрагментов.
Специфические для семян промоторы
β-конглицинина
4: 3047-3053 (1985)
трипсина Кунитца
1: 1079-1093 (1989)
2004/071467
2004/071467
Genet., 225: 148-157 (1991)
β-конглицинина
2004/071467
называемого Р34)
2004/071467
Genet., 225: 148-157 (1991)
Терминаторы транскрипции
транскрипции
2004/071467
трипсина Кунитца 3'
2004/071467
называемого Р34) 3'
2004/071467
2004/071467
2004/071467
Гены пути биосинтеза PUFA
десатуразы
№ WO 2002/081668
десатуразы
№ WO 2000/12720;
патент США № 6403349
десатуразы
десатуразы
№ WO 2002/081668
десатуразы
№ WO 2005/047479
десатуразы
№ WO 2002/081668
№ WO 2002/08401;
патент США № 6677145
J., 384: 357-366 (2004)
десатуразы
№ WO 2002/090493
элонгазы
№ WO 2002/077213
элонгазы
№ 11/601563
десатуразы
№ WO 2000/34439;
патент США № 6825017;
публикация РСТ-заявки
№ WO 2004/057001;
публикация РСТ-заявки
№ WO 2006/012325
десатуразы
Lett., 580: 1946-1952
(2006)
десатуразы
№ WO 2005/103253
десатуразы
№ 11/737772
ПРИМЕР 16
Отбор с использованием хлорсульфурона (ALS) и регенерация растений
Отбор с использованием хлорсульфурона (ALS)
После бомбардировки ткань растения делят между 2 колбами со свежей средой SB196 и культивируют, как описано в примере 13. Через шесть-семь дней после бомбардировки SB196 заменяют свежей SB196, содержащей агент для отбора - 100 нг/л хлорсульфурона. Среду для отбора освежают еженедельно. Через четыре-шесть недель после отбора наблюдают рост зеленой трансформированной ткани из нетрансформированных, некротических зародышевых скоплений. Выделенную зеленую ткань извлекают и инокулируют в многолуночные планшеты, содержащие SB196, для регенерации новых, клонально размноженных, трансформированных, образующих зародыши суспензионных культур.
Регенерация соматических зародышей сои в растения
Для получения целых растений из образующих зародыши суспензионных культур необходимо подвергнуть регенерации ткань. Зародыши подвергают созреванию, как описано в примере 13. После субкультивирования в среде SB103 в течение 3 недель из скоплений отбирают отдельные зародыши, и скринируют на изменения в составе жирных кислот, как описано в примере 14. Следует отметить, что на этой стадии можно скринировать любой определяемый фенотип, являющийся результатом экспрессии представляющих интерес генов. Это включает, но без ограничения, изменения профиля жирных кислот, профиля и содержания белков, содержания углеводов, скорости роста, жизнеспособности или способности развиваться нормально в растение сои.
Зрелые индивидуальные зародыши освобождают от влаги путем размещения их в пустую, маленькую чашку Петри (35×10 мм) в течение приблизительно 4-7 дней. Чашки герметизируют волокнистой лентой (создавая камеру невысокой влажности). Освобожденные от влаги зародыши высаживают в среду SB71-4, в которой их оставляют прорастать в одинаковых с описанными выше культуральных условиях. Развившиеся ростки удаляют из среды для прорастания и промывают тщательно водой, и затем высаживают в среду Redi-Earth в 24-ячеечном лотке-таре, покрытом прозрачным пластиковым колпаком. Через 2 недели колпак снимают, и растения закаливают в течение еще недели. Если ростки выглядят стойкими, их пересаживают в 10'' горшки c Redi-Earth в количестве до 3 ростков на горшок. Через 10-16 недель собирают зрелые семена, откалывают края и анализируют на жирные кислоты, как описано в примере 14.
Рецепты сред можно найти в примере 13, и маточный раствор хлорсульфурона представляет собой 1 мг/мл в 0,01Н аммония гидроксида.
ПРИМЕР 17
Сравнение субстратной специфичности дельта-5-десатуразы Mortierella alpina (MaD5) с дельта-5-десатуразой Euglena gracilis (EgD5) в Yarrowia lipolytica
В данном примере описывается сравнение субстратной специфичности дельта-5-десатуразы Mortierella alpina (MaD5; SEQ ID NO: 67 и 68), которая описывается в патенте США № 6075183 и РСТ-публикациях № WO 2004/071467 и WO 2005/047479, с субстратной специфичностью EgD5 (SEQ ID NO: 2) в Yarrowia lipolytica.
Эта работа включала следующие стадии: (1) конструирование экспрессирующего вектора Yarrowia pY98, включающего MaD5; (2) трансформацию pY98 и pDMW367 в штамм Y2224 Yarrowia; и (3) сравнение профиля липидов в организмах-трансформантах, включающих pY98 или pDMW367, после подачи субстратов жирных кислот.
Конструирование экспрессирующего вектора Yarrowia pY98, включающего MaD5
Плазмида pY5-22 (SEQ ID NO: 69) является плазмидой-челноком, которая может реплицироваться как в E.coli, так и в Yarrowia lipolytica, содержащей следующие компоненты: автономно реплицируемую последовательность Yarrowia (ARS18; № доступа в GenBank - M91600); начало репликации плазмиды ColE1; ген устойчивости к ампициллину (AmpR) для отбора в E.coli; ген Ura3 Yarrowia (№ доступа в GenBank - AJ306421) для отбора в Yarrowia; и химерную кассету TEF::NcoI/NotI::XPR, где «XPR» был ~100 п.о. 3'-района гена XPR Yarrowia (№ доступа в GenBank - M17741). Хотя конструкция плазмиды pY5-22 не описывается здесь подробно, ее получали из pY5 (ранее описанной в публикации РСТ-заявки № WO 2004/101757).
Плазмиду pY5-22GPD (SEQ ID NO: 70) создавали из плазмиды pY5-22 (SEQ ID NO: 69) путем замены промотора TEF промотором GPD Yarrowia lipolytica (SEQ ID NO: 71), используя хорошо известные квалифицированному в данной области техники специалисту методы. «Промотор GPD» Yarrowia относится к 5' нетранслируемому району перед кодоном инициации трансляции «ATG» белка, кодируемого геном глицеральдегид-3-фосфат-дегидрогеназы (GPD) Yarrowia lipolytica, и необходим для экспрессии (публикация РСТ-заявки № WO 2005/003310). Более конкретно, промотор GPD Yarrowia lipolytica амплифицировали с плазмиды pYZDE2-S (SEQ ID NO: 72), которая была ранее описана в заявке на патент США № 11/737772 (содержание которой, таким образом, включено посредством ссылки), используя олигонуклеотиды GPDsense (SEQ ID NO: 73) и GPDantisense (SEQ ID NO: 74). Получаемый в результате фрагмент ДНК расщепляли SalI/NotI и клонировали в SalI/NotI-фрагмент pY5-22 (SEQ ID NO: 69), заменяя, таким образом, промотор TEF и сайт для NcoI/NotI промотором GPD и уникальным сайтом NotI и получая, таким образом, pY5-22GPD (SEQ ID NO: 70).
Ген дельта-5-десатуразы Mortierella alpina (SEQ ID NO: 67) высвобождали из pKR136 (SEQ ID NO: 75; которая была ранее описана в публикации РСТ-заявки № WO 2004/071467 (содержание которой, таким образом, включено посредством ссылки)) с помощью расщеплении NotI и клонировали в сайт для NotI pY5-22GPD с получением pY98 (SEQ ID NO: 76; фиг. 9).
Трансформация pY98 (включающей MaD5) и pDMW367 (включающей EgD5) в штамм Y2224 Yarrowia и сравнение профиля липидов
Штамм Y2224 выделяли следующим образом: клетки Yarrowia lipolytica АТСС #20362 с чашки с агаром YPD (1% дрожжевого экстракта, 2% бактопептона, 2% глюкозы, 2% агара) штриховали на чашку ММ (75 мг/л урацила и уридина, каждого, 6,7 г/л YNB с сульфатом аммония, без аминокислоты, и 20 г/л глюкозы), содержащую 250 мг/л 5-FOA (Zymo Research). Чашки инкубировали при 28°С, и четыре результирующие колонии распределяли по отдельности по чашкам ММ, содержащим 200 мг/л 5-FOA, и чашкам ММ без урацила и уридина для подтверждения ауксотрофии Ura3 в отношении урацила.
Штамм Y2224 трансформировали pY98 (SEQ ID NO: 76; фиг. 9) и pDMW367 (SEQ ID NO: 23; фиг. 5С; пример 6), как описано в разделе «Общие способы».
Одиночные колонии трансформанта-Yarrowia lipolytica, содержащего pY98 (SEQ ID NO: 76) или pDMW367 (SEQ ID NO: 23), выращивали в 3 мл среды ММ без урацила, дополненной 0,2% тергитола, при 30°С в течение 1 дня. После этого 1 мл переносили в 3 мл той же среды, или дополненной EDA, ETrA, DGLA, ETA, или не дополненной никакой жирной кислотой. Их инкубировали в течение 16 ч при 30°С, при 250 оборотах/мин, и затем получали с помощью центрифугирования осадки. Клетки промывали один раз водой, осаждали с помощью центрифугирования и сушили на воздухе. Осадки трансэтерифицировали (Roughan, G, and Nishida, I., Arch. Biochem. Biophys., 276(1): 38-46 (1990)) с использованием 500 мкл 1% натрия метилата в течение 30 мин при 50°С, после чего добавляли 500 мкл 1 М NaCl и 100 мкл гептана. После тщательного перемешивания и центрифугирования эфиры метилового спирта и жирных кислот (FAME) анализировали с помощью GC.
FAME (5 мкл вводили из слоя гексана) отделяли и определяли количество с использованием газового хроматографа Hewlett-Packard 6890, снабженного капиллярной колонкой с кварцевым стеклом Omegawax 320 (каталожный #24152, Supelco Inc.). Программировали, чтобы температура термостата удерживалась при 220°С в течение 2,6 мин, увеличивалась до 240°С с подъемом на 20°С/мин и затем удерживалась в течение дополнительных 2,4 мин. Газ-носитель поставлялся генератором водорода Whatman. Времена удержания сравнивали со временами удержания метиловых эфиров стандартов, коммерчески доступных (Nu-Chek Prep, Inc.).
Профили жирных кислот Yarrowia lipolytica, который экспрессирует pY98 (SEQ ID NO: 76) или pDMW367 (SEQ ID NO: 23) и которому подавались различные субстраты, показаны на фиг. 10А. На фиг. 10А затемнение указывает подаваемые субстраты и продуцируемые продукты; жирные кислоты определяют как 16:0 (пальмитат), 16:1, 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), LA, GLA, ALA, STA, EDA, SCI (сциадоновая кислота или цис-5,11,14-эйкозатриеновая кислота; 20:3 омега-6), DGLA, ARA, ETrA, JUP (юнипероновая кислота или цис-5,11,14,17-эйкозатриеновая кислота; 20:4 омега-3), ЕТА и ЕРА. Составы жирных кислот выражали в виде весового процента (вес.%) от жирных кислот в целом.
Процент дельта-5-десатурации («% дельта-5-десатурации») EgD5 и MaD5 для каждого субстрата показан на фиг. 10В и рассчитывали делением вес% для продукта (или SCI, JUP, ARA или ЕРА) на сумму вес.% для субстрата и продукта (или EDA и SCI, ETrA и JUP, DGLA и ARA, или ETA и EPA соответственно) и умножением на 100 для выражения в виде %, в зависимости от подаваемого субстрата.
Активности EgD5 и MaD5 сравнивали, используя отношение процентов дельта-5-десатурации («отношения десатураций Eg/Ma») на фиг. 10В, и рассчитывали делением процента дельта-5-десатурации для EgD5 в отношении конкретного субстрата на процент дельта-5-десатурациии для MaD5 в отношении того же субстрата.
Субстратная специфичность EgD5 и MaD5 в отношении правильного субстрата жирной кислоты омега-6 (т.е. DGLA) в сравнении с побочной жирной кислотой (т.е. SCI) или в отношении правильного субстрата жирной кислоты омега-3 (т.е. ETA) в сравнении с побочной жирной кислотой (т.е. JUP) также продемонстрирована на фиг. 10В. В частности, субстратную специфичность («отношение продукт/побочный продукт») для субстратов омега-6 рассчитывали делением процента дельта-5-десатурации (% дельта-5-десатурации) для DGLA на процент дельта-5-десатурации (% дельта-5-десатурации) для EDA, и она продемонстрирована на тех же строках, что и результаты для DGLA. Субстратную специфичность («отношение продукт/побочный продукт») для субстратов омега-3 рассчитывали делением процента дельта-5-десатурации (% дельта-5-десатурации) для ETA на процент дельта-5-десатурации (% дельта-5-десатурации) для ETrA, и она продемонстрирована на тех же строках, что и результаты для ETA. Кроме того, определяли отношение субстратных специфичностей («отношение продукт/побочный продукт Eg/Ma») для субстратов омега-6 делением субстратной специфичности для EgD5 в отношении субстратов омега-6 (т.е. DGLA/EDA) на субстратную специфичность для MaD5. Отношение субстратных специфичностей («отношение продукт/побочный продукт Eg/Ma») для субстратов омега-3 рассчитывали делением субстратной специфичности для EgD5 в отношении субстратов омега-3 (т.е. ETA/ETrA) на субстратную специфичность для Mad5.
Предпочтение EgD5 и MaD5 в отношении субстратов омега-6 или омега-3 сравнивают с использованием отношения процента дельта-5-десатурации («отношения n-6/n-3») на фиг. 10В, и его рассчитывают с помощью деления процента дельта-5-десатурации для EgD5 и MaD5 в отношении конкретного субстрата омега-6 (или DGLA, или EDA) на процент дельта-5-десатурации в отношении соответствующего субстрата омега-3 (или ETA, или ETrA соответственно).
Из результатов, представленных на фиг. 10В, ясно, что EgD5 приблизительно в 2,6-2,9 раз более активна в Yarrowia, чем MaD5, когда в качестве субстратов используются DGLA, EDA и ETA. Исключением является активность в отношении ETrA, которая является приблизительно одинаковой для обоих ферментов. Субстратная специфичность EgD5 и MaD5 в отношении правильного субстрата жирной кислоты омега-6 (т.е. DGLA в сравнении с EDA) является приблизительно одинаковой в Yarrowia, но существует приблизительно 2,5-кратное предпочтение EgD5 в отношении ЕТА (в сравнении с ETrA) по сравнению с MaD5. Высокая активность и предпочтительная субстратная специфичность EgD5 в отношении ЕТА в сравнении с ETrA могут быть полезны при продуцировании длинноцепочечных PUFA. EgD5 также имеет предпочтение в отношении субстратов омега-6 (т.е. EGA и DGLA) по сравнению с субстратами омега-3 (т.е. ETrA и ETA) соответственно.
ПРИМЕР 18
Конструирование экспрессирующего вектора сои pKR916 для коэкспрессии дельта-5-десатуразы Mortierella alpina (MaD5) с дельта-9-элонгазой, происходящей из Euglena gracilis, (EgD9е) и дельта-8-десатуразой, происходящей из Euglena gracilis, (EgD8)
В данном примере описывается конструирование вектора сои для коэкспрессии MaD5 (SEQ ID NO: 67; пример 17) с EgD9e (SEQ ID NO: 77; которая описывается в заявке на патент США № 11/601563 (поданной 16 ноября 2006; № в реестре поверенного - ВВ-1562) и EgD8 (SEQ ID NO: 78; описанной как Eg5 в публикации РСТ-заявки № WO 2006/012325).
Дельта-9-элонгаза Euglena gracilis (EgD9е)
Клон из библиотеки кДНК Euglena (eeg1c), названный eeg1c.pk001.n5f, содержащий дельта-9-элонгазу Euglena gracilis (EgD9е; SEQ ID NO: 77), использовали в качестве матрицы для амплификации EgD9е с использованием олигонуклеотидных праймеров oEugEL1-1 (SEQ ID NO: 79) и oEugEL1-2 (SEQ ID NO: 80) и ДНК-полимеразы VentR® (каталожный № M0254S, New England Biolabs Inc., Beverly, MA), следуя протоколу производителя. Получаемый в результате фрагмент ДНК клонировали в вектор для клонирования pCR-Blunt®, используя набор для клонирования продуктов ПЦР Zero Blunt® (Invitrogen Corporation), следуя протоколу производителя, с получением pKR906 (SEQ ID NO: 81).
Исходная плазмида pKR72 (№ доступа в АТСС - PTA-6019; SEQ ID NO: 82; последовательность размером 7085 п.о.), производная pKS123, которая ранее описана в публикации РСТ-заявки № WO 02/008269 (содержание которой, таким образом, включено посредством ссылки), содержит ген гигромицин В-фосфотрансферазы (НРТ) (Gritz, L. and Davies, J., Gene, 25: 179-188 (1983)), фланкированный промотором Т7 и терминатором транскрипции Т7 (т.е. кассету промотор Т7/НРТ/терминатор Т7) и бактериальное начало репликации для отбора и репликации в бактериях (например, E.coli). Кроме того, pKR72 также содержит НРТ, фланкированный промотором 35S (Odell et al., Nature, 313: 810-812 (1985)) и 3' терминатором транскрипции NOS (Depicker et al., J. Mol. Appl. Genet., 1: 561-570 (1982)) (т.е. кассету 35S/НРТ/NOS3'), для отбора в растениях, таких как соя. pKR72 также содержит рестрикционный сайт для NotI, фланкированный промотором для α'-субъединицы β-конглицинина (Beachy et al., EMBO J., 4: 3047-3053 (1985)) и 3'-районом терминации транскрипции гена фазеолина (Doyle et al., J. Biol. Chem., 261: 9228-9238 (1986)), что делает, таким образом, возможной сильную тканеспецифическую экспрессию в семенах сои генов, клонированных в сайт для NotI.
AscI-фрагмент плазмиды pKS102 (SEQ ID NO: 83), ранее описанной в публикации РСТ-заявки № WO 02/00905 (содержание которой, таким образом, включено посредством ссылки), содержащий кассету промотор Т7/hpt/терминатор Т7 и бактериальное начало репликации, объединяли с AscI-фрагментом плазмиды pKR72 (SEQ ID NO: 82), содержащим кассету промотор β-конглицинина/NotI/терминатор фазеолина, с получением pKR197 (SEQ ID NO: 84), ранее описанной в публикации РСТ-заявки № WO 04/071467 (содержание которой, таким образом, включено посредством ссылки).
Ген дельта-9-элонгазы Euglena gracilis высвобождали из pKR906 (SEQ ID NO: 81) с помощью расщепления NotI и клонировали в сайт для NotI pKR197 (SEQ ID NO: 84) с получением промежуточного вектора для клонирования pKR911 (SEQ ID NO: 85).
Дельта-8-десатураза Euglena gracilis (EgD8)
Плазмида pKR680 (SEQ ID NO: 86), которая ранее описана в публикации РСТ-заявки № WO 2006/012325 (содержание которой, таким образом, включено посредством ссылки), содержит ген дельта-8-десатуразы Euglena gracilis (EgD8; SEQ ID NO: 78, описываемой как Eg5 в WO 2006/012325), фланкированный промотором ингибитора трипсина сои Кунитца (KTi) (Jofuku et al., Plant Cell, 1: 1079-1093 (1989)) и 3'-районом терминации KTi, выделение которого описывается в патенте США № 6372965, за которым следует терминатор транскрипции альбумина сои (Salb3'), который был ранее описан в публикации РСТ-заявки № WO 2004/071467 (т.е. кассету Kti/NotI/Kti3'Salb3').
Плазмиду pKR680 (SEQ ID NO: 86) расщепляли BsiWI, и фрагмент, содержащий EgD8, клонировали в сайт для BsiWI pKR911 (SEQ ID NO: 85) с получением pKR913 (SEQ ID NO: 87).
Дельта-5-десатураза Mortierella alpina (MaD5)
Плазмида pKR767 (SEQ ID NO: 88), которая ранее описана в публикации РСТ-заявки № WO 2006/012325 (содержание которой, таким образом, включено посредством ссылки), содержит ген дельта-5-десатуразы Mortierella alpina (MaD5; SEQ ID NO: 67), фланкированный промотором гена Gy1 глицинина сои и 3'-районом терминации транскрипции легумина А2 гороха (т.е. кассету Gy1/MaD5/legA2; конструирование которой описывается в WO 2006/012325).
Кассету Gy1/MaD5/legA2 высвобождали из pKR767 (SEQ ID NO: 88) с помощью расщепления SbfI, и получаемый в результате фрагмент клонировали в сайт для SbfI pKR913 (SEQ ID NO: 87) с получением pKR916 (SEQ ID NO: 89). Схематическое изображение pKR916 представлено на фиг. 11А. Таким образом дельта-9-элонгаза Euglena gracilis (обозначенная «eug el1» на фиг. 11А) была коэкспрессирована с дельта-8-десатуразой Euglena gracilis (обозначенной «eug d8-sq5» на фиг. 11А) и дельта-5-десатуразой Mortierella alpina (обозначенной «DELTA 5 DESATURASE M ALPINA» на фиг. 11А) под контролем сильных, специфических для семени промоторов.
ПРИМЕР 19
Конструирование экспрессирующего вектора сои pKR1037 для коэкспрессии дельта-5-десатуразы Euglena gracilis (EgD5) с дельта-9-элонгазой, происходящей из Euglena gracilis, (EgD9е) и дельта-8-десатуразой, происходящей из Euglena gracilis, (EgD8)
В данном примере описывается конструирование вектора сои для коэкспрессии EgD5 (SEQ ID NO: 1; пример 5) с EgD9e (SEQ ID NO: 77; пример 18) и EgD8 (SEQ ID NO: 78; пример 18).
Исходная плазмида pKR974 (SEQ ID NO: 90) идентична pKR767 (SEQ ID NO: 88, пример 18), за исключением того, что NotI-фрагмент, содержащий MaD5, заменен NotI-фрагментом, содержащим дельта-5-десатуразу Saprolegnia diclina (SdD5; SEQ ID NO: 94; которая описывается в публикации РСТ-заявки № WO 2004/071467). Кроме того, сайт для MfeI в терминаторе legA2 pKR767 (SEQ ID NO: 88) удален с помощью расщепления MfeI, заполнения сайта для MfeI и повторного лигирования (т.е. CAATTG превращался в CAATTAATTG), и, следовательно, размер терминатора legA2 pKR974 (SEQ ID NO: 90) составляет 770 п.о. в сравнении с 766 п.о. для pKR767 (SEQ ID NO: 88).
Для того чтобы клонировать EgD5 в экспрессирующий вектор сои, было необходимо ввести в 5'-конец гена рестрикционный сайт для NotI. Квалифицированный в данной области техники специалист понимает, что существует множество путей введения рестрикционных сайтов в гены, такие как, но без ограничения, ПЦР или с помощью субклонирования в векторы, содержащие соответствующие сайты. В этом случае, для того чтобы ввести сайт для NotI в 5'-конец EgD5 (SEQ ID NO: 1), pDMW367 (SEQ ID NO: 23) расщепляли MfeI и затем частично расщепляли NcoI. NcoI/MfeI-фрагмент, содержащий полноразмерный EgD5 (SEQ ID NO: 1), клонировали в сайт для NcoI/MfeI промежуточного вектора для клонирования, имеющего сайт для NotI непосредственно выше сайта для NcoI (т.е. GCGGCCGCAAACCATGG). Получаемую в результате плазмиду расщепляли NotI, и фрагмент, содержащий EgD5 (SEQ ID NO: 1), клонировали в сайт для NotI pKR974 (SEQ ID NO: 90) с получением pKR1032 (SEQ ID NO: 91).
Кассету Gy1/EgD5/legA2 высвобождали из pKR1032 (SEQ ID NO: 91) с помощью расщепления SbfI, и получаемый в результате фрагмент клонировали в сайт для SbfI pKR913 (SEQ ID NO: 87) с получением pKR1037 (SEQ ID NO: 92). Схематическое изображение pKR1037 (SEQ ID NO: 92) представлено на фиг. 11В. Таким образом дельта-9-элонгаза Euglena gracilis (обозначенная «eug el1» на фиг. 11В) была коэкспрессирована с дельта-8-десатуразой Euglena gracilis (помеченной «eug d8-sq5» на фиг. 11В) и дельта-5-десатуразой Euglena gracilis (обозначенной «eug d5 DS» на фиг. 11B) под контролем сильных, специфических для семени промоторов.
ПРИМЕР 20
Коэкспрессия дельта-9-элонгазы Euglena gracilis, дельта-8-десатуразы Euglena gracilis и дельта-17-десатуразы Saprolegnia diclina или с дельта-5-десатуразой Mortierella alpina (pKR916 & pKR328), или с дельта-5-десатуразой Euglena gracilis (pKR1037 & pKR328) в соматических зародышах сои
В данном примере описываются трансформация и экспрессия в соматических зародышах сои pKR916 (SEQ ID NO: 89, пример 18; содержащей EgD9e, EgD8 и MaD5) с pKR328 (SEQ ID NO: 93, фиг. 11С, ранее описанной в публикации РСТ-заявки № WO 04/071467), содержащей ген дельта-17-десатуразы Saprolegnia diclina (SdD17) и ген гигромицин-фосфотрансферазы для отбора на гидромицин. В данном примере, кроме того, описывается трансформация и экспрессия в соматических зародышах сои pKR1037 (SEQ ID NO: 92, пример 19; содержащей EgD9e, EgD8 и EgD5) с pKR328 (SEQ ID NO: 93, фиг. 11С).
Образующую зародыши сои суспензионную культуру (распространенная версия Jack) трансформировали AscI-фрагментом, содержащим экспрессирующую кассету pKR916 (SEQ ID NO: 89), и интактной плазмидой pKR328 (SEQ ID NO: 93), или AscI-фрагментом, содержащим экспрессирующую кассету pKR1037 (SEQ ID NO: 92), и интактной плазмидой pKR328 (SEQ ID NO: 93), как описано в примере 13.
Созревание зародышей происходило в жидкой среде для гистодифференциации и созревания сои (жидкой среде SHaM; Schmidt et al., Cell Biology and Morphogenesis, 24: 393 (2005)), используя модифицированную процедуру. Вкратце, после 4 недель отбора в SB196, как описано в примере 13, зародышевые скопления перемещали в 35 мл SB228 (жидкой среды SHaM) в 250-мл конической колбе Эрленмейера. Ткань сохраняли в жидкой среде SHaM на вращающемся встряхивателе при 130 оборотах/мин и 26°С с холодными белыми люминесцентными излучениями во время фотопериода 16:8 ч день/ночь при интенсивности света 60-85 мкЭрг/м2/сек в течение 2 недель до созревания зародышей. Зародыши, выращиваемые в течение 2 недель в жидкой среде SHaM, были эквивалентны по размеру и содержанию жирных кислот зародышам, культивируемым на SB166/SB103 в течение 5-8 недель, как описывается в примере 13.
Перечень ингредиентов сред
SB 228 - среда для гистодифференциации и созревания сои (SHaM) (на литр)
Довести объем до 900 мл
рН 5,8
Автоклавировать
Добавлять к охлажденной среде (≤30°С):
∗Примечание: Конечный объем после добавления глютамина будет составлять 1010 мл.
Поскольку глютамин относительно быстро разрушается, его можно предпочтительно добавлять непосредственно перед использованием среды. По истечении 2 недель после добавления глютамина основную среду можно хранить дольше без глютамина.
FN-Lite макросоли для SHaM - 10x маточный раствор #1 (на литр)
Довести до объема
Автоклавировать
MS микросоли - 1000x маточный раствор #2 (на литр)
Довести до объема
Автоклавировать
FeEDTA - 100x маточный раствор #3 (на литр)
∗ EDTA необходимо полностью растворить до добавления железа.
Довести до объема
Раствор является светочувствительным. Флакон(ы) следует обернуть фольгой, не пропускающей свет.
Автоклавировать
Ca - 100x маточный раствор #4 (на литр)
Довести до объема
Автоклавировать
1000Х маточный раствор витаминов В5 #5 (на литр)
Довести до объема
Хранить замороженным
4% глютамин - маточный раствор #6 (на литр)
Постепенно добавлять при перемешивании и применении небольшого нагревания
Не превышать 35°С
Довести до объема
Стерилизовать фильтрованием
Хранить замороженным∗
∗Замечание: Нагреть оттаявший маточный раствор в бане на 31°С до полного растворения кристаллов.
После созревания в жидкой среде SHaM отдельные зародыши извлекали из скоплений, сушили и скринировали на изменения состава жирных кислот, как описано выше.
Подмножество соевых зародышей (т.е. шесть зародышей на каждый случай), трансформированных или pKR916 (SEQ ID NO: 89) и pKR328 (SEQ ID NO: 93), или pKR1037 (SEQ ID NO: 92) и pKR328 (SEQ ID NO: 93), собирали и отбирали в стеклянные пробирки для GC, и с помощью трансэтерификации готовили эфиры метилового спирта и жирных кислот. Для трансэтерификации 50 мкл триметилсульфония гидроксида (TMSH) и 0,5 мл гексана добавляли к зародышам в стеклянных пробирках и инкубировали в течение 30 мин при комнатной температуре со встряхиванием. Эфиры метилового спирта и жирных кислот (5 мкл вводили из слоя гексана) отделяли и определяли количество с использованием газового хроматографа Hewlett-Packard 6890, снабженного капиллярной колонкой с кварцевым стеклом Omegawax 320 (каталожный #24152, Supelco Inc.). Программировали, чтобы температура термостата удерживалась при 220°С в течение 2,6 мин, увеличивалась до 240°С с подъемом на 20°С/мин и затем удерживалась в течение дополнительных 2,4 мин. Газ-носитель поставлялся генератором водорода Whatman. Времена удержания сравнивали со временами удержания метиловых эфиров стандартов, коммерчески доступных (Nu-Chek Prep, Inc.).
Таким образом проанализировали 60 случаев трансформации pKR916 (SEQ ID NO: 89) и pKR328 (SEQ ID NO: 93) и 45 случаев трансформации pKR1037 (SEQ ID NO: 92) и pKR328 (SEQ ID NO: 93). Средние профили жирных кислот для десяти случаев наличия наибольшей дельта-5-десатуразной активности для каждой трансформации (pKR916 и pKR328, pKR1037 и pKR328) продемонстрированы на фиг. 12А и фиг. 12В соответственно.
На фиг. 12А и 12В жирные кислоты определяют как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), LA, ALA, EDA, SCI, DGLA, ARA, ERA, JUP, ЕТА и ЕРА. Жирные кислоты, перечисленные как «другие», включают 18:2 (5,9), GLA, STA, 20:0, 20:1(11), 20:2 (7,11) или 20:2 (8,11) и DPA. Относительное содержание каждой из этих «других» жирных кислот составляет менее 3% от жирных кислот в целом. Составы жирных кислот для отдельного зародыша выражали в виде весового процента (вес.%) от жирных кислот в целом, а средний состав жирных кислот представляет собой среднее шести отдельных зародышей для каждого случая.
Активность дельта-5-десатуразы в отношении «правильных» субстратов (т.е. DGLA и ETA) выражали в виде процента дельта-5-десатурации («процента правильной дельта-5-десатурации»), рассчитываемого согласно следующей формуле: ([продукт]/[субстрат + продукт])∗100. Более конкретно, процент дельта-5-десатурации для «правильных» субстратов определяли как ([ARA+EPA]/[DGLA+ETA+ARA+EPA])∗100.
Активность дельта-5-десатуразы в отношении «неправильных» субстратов (т.е. EDA и ERA) также выражают в виде процента дельта-5-десатурации («процента неправильной дельта-5-десатурации»), рассчитываемого как ([SCI+JUP]/[EDA+ERA+SCI+JUP])∗100.
Сравнивали субстратные специфичности MaD5 и EgD5 в отношении «правильных» субстратов (т.е. DGLA и ETA) в сравнении с «неправильными» субстратами (т.е. EDA и ERA), и сравнение продемонстрировано на фиг. 13. На фиг. 13 активность дельта-5-десатуразы в отношении «правильных» субстратов («% правильной дельта-5-десатурации») откладывают по X-оси, а активность дельта-5-десатуразы в отношении «неправильных» субстратов («% неправильной дельта-5-десатурации») откладывают по Y-оси для MaD5 (данные с фиг. 12А) и EgD5 (данные с фиг. 12В).
На фиг. 12В демонстрируется, что активность EgD5 в зародышах сои является очень высокой, при этом среднее превращение (% правильной дельта-5-десатурации) составляет от 77% до 99% в десяти случаях наличия наибольшей дельта-5-десатуразной активности. По сравнению с MaD5 специфичность EgD5 в отношении субстратов (фиг. 13) больше в отношении «правильных» субстратов по сравнению с «неправильными» субстратами. С учетом высокой активности и субстратной специфичности EgD5 может быть полезна при продуцировании PUFA, таких как, но без ограничения, EPA и DHA, в клетке-хозяине.
| название | год | авторы | номер документа |
|---|---|---|---|
| МУЛЬТИЗИМЫ И ИХ ИСПОЛЬЗОВАНИЕ В ПОЛУЧЕНИИ ПОЛИНЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ | 2008 |
|
RU2517608C2 |
| ГЕНЫ ДЕЛЬТА-8-ДЕСАТУРАЗЫ, ФЕРМЕНТЫ, КОДИРУЕМЫЕ ИМИ, И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2517615C2 |
| НОВАЯ ДЕЛЬТА-9-ЭЛОНГАЗА ДЛЯ ПОЛУЧЕНИЯ МАСЕЛ, ОБОГАЩЕННЫХ ПОЛИНЕНАСЫЩЕННЫМИ ЖИРНЫМИ КИСЛОТАМИ | 2010 |
|
RU2558302C2 |
| НОВАЯ ЛИЗОФОСФОЛИПИД-АЦИЛТРАНСФЕРАЗА | 2010 |
|
RU2534559C2 |
| ПРОМОТОР ЦЕЛОГО СЕМЕНИ | 2010 |
|
RU2561463C2 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И СПОСОБЫ ПОЛУЧЕНИЯ МАСЛА С ИЗМЕНЕННЫМ СОСТАВОМ ИЗ СЕМЯН | 2007 |
|
RU2392795C2 |
| ДЕЛЬТА 6-АЦЕТИЛЕНАЗА/ДЕЛЬТА-6-ДЕСАТУРАЗА И ДЕЛЬТА-6-ДЕСАТУРАЗА ИЗ Ceratodon purpureus | 2000 |
|
RU2268939C2 |
| Способы получения липидов | 2011 |
|
RU2636344C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЛИПИДОВ | 2012 |
|
RU2656996C2 |
| ГЕНЫ ДИАЦИЛГЛИЦЕРОЛ-АЦИЛТРАНСФЕРАЗЫ И ИХ ИСПОЛЬЗОВАНИЕ | 2010 |
|
RU2514655C2 |


















































































































































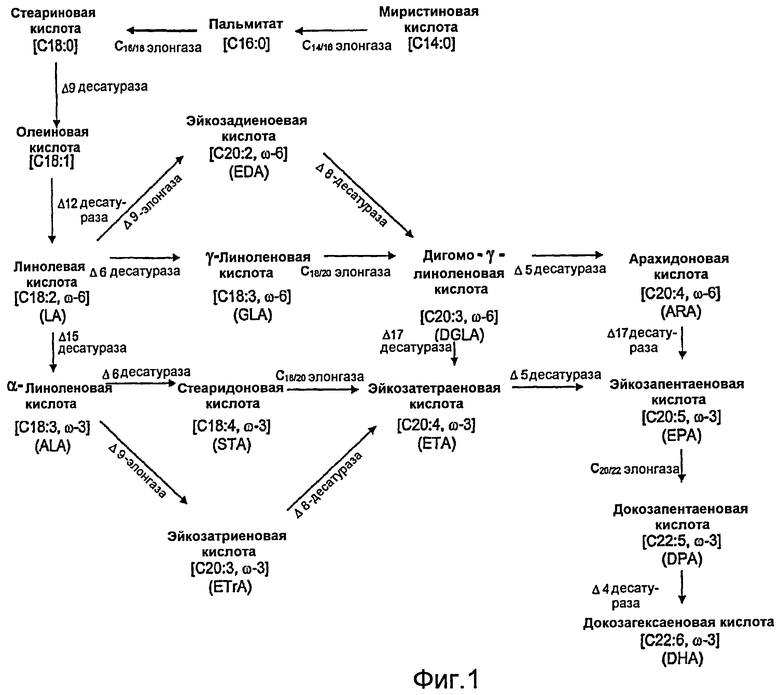
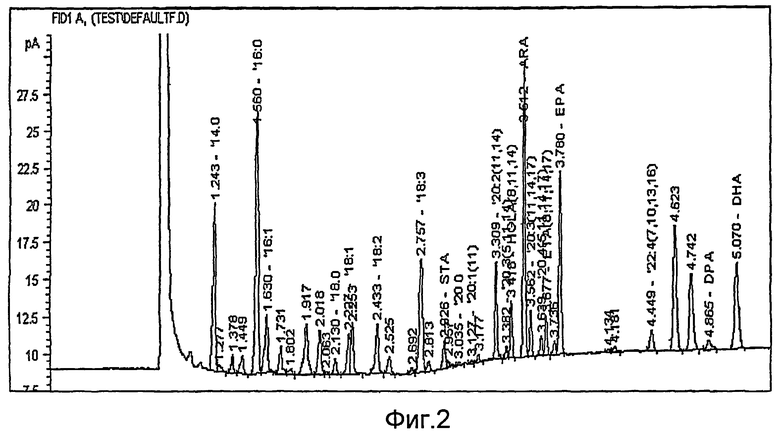

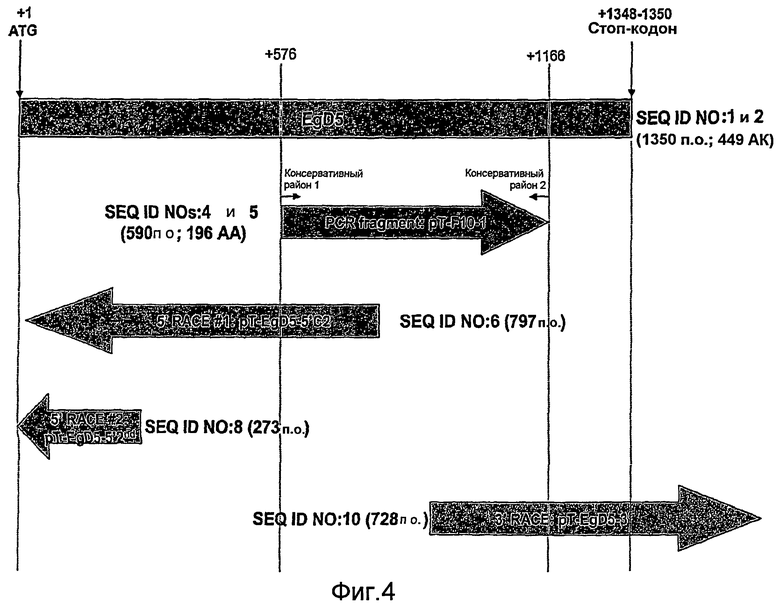
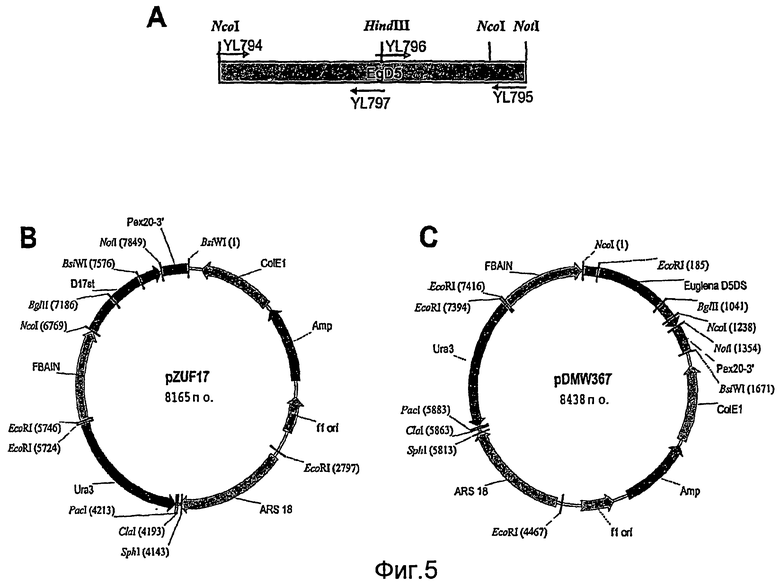



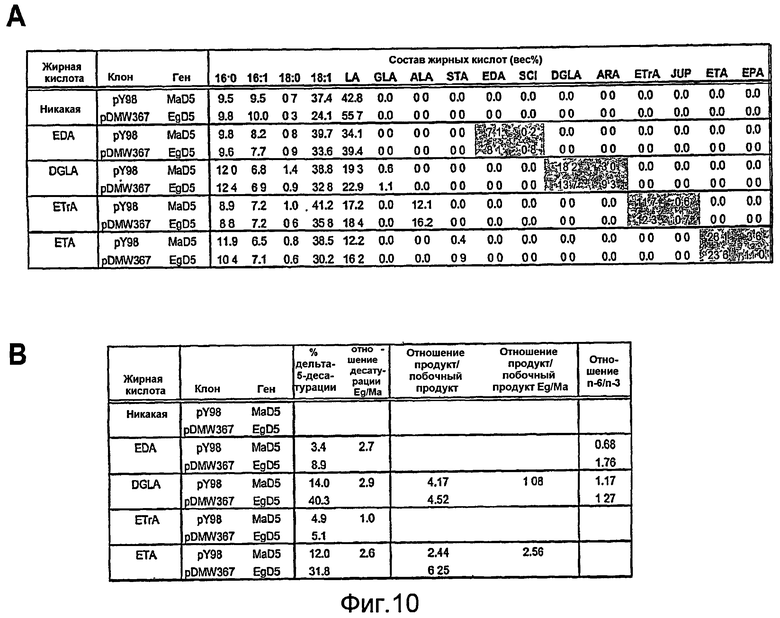
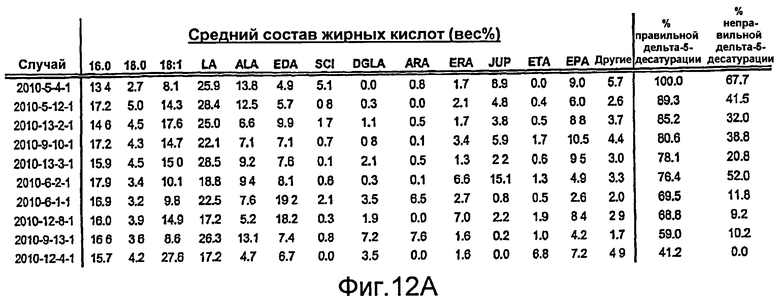
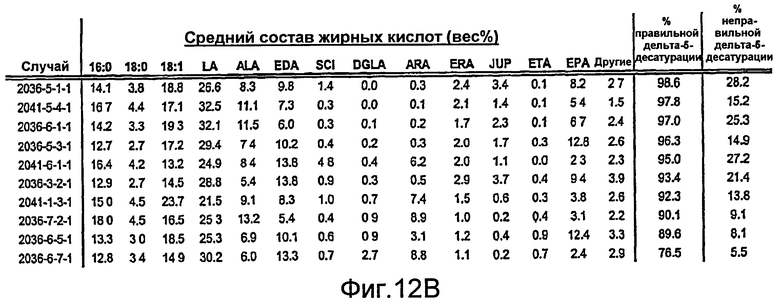
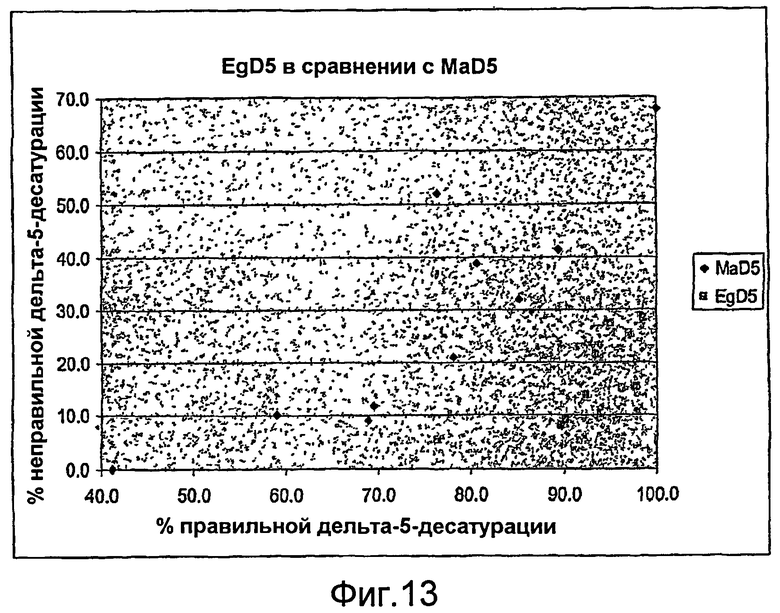
Изобретение относится к области биотехнологии. Представлен выделенный полинуклеотид для экспрессии дельта-5-десатуразы, включающий нуклеотидную последовательность: 1) кодирующую полипептид дельта-5-десатуразы, причем полипептид обладает по меньшей мере 80% идентичности SEQ ID NO:2; 2) кодирующую полипептид дельта-5-десатуразы, причем нуклеотидная последовательность обладает по меньшей мере 90% идентичности SEQ ID NO:1 или SEQ ID NO:3; 3) кодирующую полипептид дельта-5-десатуразы, причем нуклеотидная последовательность гибридизуется в жестких условиях с нуклеотидной последовательностью, представленной в SEQ ID NO:1 или SEQ ID NO:3; 4) комплемент нуклеотидной последовательности 1)-3). Описаны рекомбинанатная ДНК-конструкция, клетка-хозяин, трансгенное растение с масличными семенами, трансгенное семя, содержащие указанный полинуклеотид. Предложены способы: трансформации клетки-хозяина, экспрессии полиненасыщенных жирных кислот с длинными цепями в клетке растения, получения по меньшей мере одной полиненасыщенной жирной кислоты в растении, с использованием указанного полинуклеотида. Описана возможность применения указанных трансгенных семян для получения пищевых продуктов для питания людей, кормов для животных, получения масла и лецитина. Изобретение позволяет получить измененный профиль полиненасыщенных жирных кислот по сравнению с профилем полиненасыщенных жирных кислот нетрансформированного растения. 15 н. и 9 з.п. ф-лы, 12 табл., 15 ил., 20 пр.
1. Выделенный полинуклеотид для экспрессии дельта-5-десатуразы, включающий
(a) нуклеотидную последовательность, кодирующую полипептид, обладающий активностью дельта-5-десатуразы, причем полипептид обладает по меньшей мере 80% идентичности аминокислотной последовательности, на основе способа выравнивания Clustal W, предоставленного программой MegAlign™ V6.1 комплекта вычислительных программ для биоинформатики DNASTAR LASERGENE (параметры по умолчанию: пенальти за промежуток = 10, пенальти за длину промежутка = 0,2, обусловленная запаздыванием дивергенция последовательностей (%) = 30, переходный вес ДНК = 0,5, белковая весовая матрица = ряд Гоннета, ДНК весовая матрица = IUB), при сравнении с аминокислотной последовательностью, представленной в SEQ ID NO:2;
(b) нуклеотидную последовательность, кодирующую полипептид, обладающий активностью дельта-5-десатуразы, причем нуклеотидная последовательность обладает по меньшей мере 90% идентичности последовательности, на основе способа выравнивания Basic Local Alignment Search Tool (BLASTN), предоставленного National Center for Biotechnology Information (параметры по умолчанию: короткие запросы = автоматически устанавливаемые параметры для коротких последовательностей ввода, ожидаемый порог = 10, размер слова = 11, индексы соответствия/несоответствия = 2, -3, штрафы за пропуск = (наличие:5, удлинение:2) и фильтр = области низкой сложности), при сравнении с нуклеотидной последовательностью, представленной в SEQ ID NO:1 или SEQ ID NO:3;
(c) нуклеотидную последовательность, кодирующую полипептид, обладающий активностью дельта-5-десатуразы, причем нуклеотидная последовательность гибридизуется в жестких условиях с нуклеотидной последовательностью, представленной в SEQ ID NO:1 или SEQ ID NO:3, где жесткие условия включают гибридизацию в 50% формамида, 1М NaCl, 1% SDS при 37°С и промывку в 0,1Х SSC при 60°С; или
(d) комплемент нуклеотидной последовательности (а), (b) или (с), причем комплемент и нуклеотидная последовательность состоят из одинакового числа нуклеотидов и комплементарны на 100%.
2. Полинуклеотид по п.1, в котором нуклеотидная последовательность включает SEQ ID NO:1 или SEQ ID NO:3.
3. Полинуклеотид по п.1, где аминокислотная последовательность полипептида включает
(a) SEQ ID NO:2; или
(b) аминокислотную последовательность, которая отличается от аминокислотных последовательностей (а) по меньшей мере одной консервативной аминокислотной заменой.
4. Рекомбинантная ДНК-конструкция для экспрессии дельта-5-десатуразы, включающая полинуклеотид по пп.1, 2 или 3, функционально связанный с по меньшей мере одной регуляторной последовательностью.
5. Клетка-хозяин для экспрессии дельта-5-десатуразы, содержащая в своем геноме рекомбинантную ДНК-конструкцию по п.4.
6. Клетка-хозяин по п.5, которая выбрана из группы, состоящей из растений и дрожжей.
7. Способ трансформации клетки-хозяина, включающий трансформацию клетки-хозяина рекомбинантной конструкцией по п.4 и отбор тех клеток-хозяев, которые трансформированы рекомбинантной конструкцией по п.4.
8. Способ по п.7, в котором клетка-хозяин представляет собой клетку растения.
9. Способ по п.8, в котором клеткой растения является клетка растения сои.
10. Трансгенное семя для экспрессии дельта-5-десатуразы, содержащее в своем геноме рекомбинантную конструкцию по п.4.
11. Способ экспрессии полиненасыщенных жирных кислот с длинными цепями в клетке растения, включающий
(a) трансформацию клетки рекомбинантной конструкцией по п.4;
(b) отбор тех трансформированных клеток, которые экспрессируют дельта-5-десатуразу, кодируемую рекомбинантной конструкцией; и
(c) отбор тех трансформированных клеток со стадии (b), которые экспрессируют полиненасыщенные жирные кислоты с длинными цепями.
12. Способ получения по меньшей мере одной полиненасыщенной жирной кислоты в растении с масличными семенами, включающий (а) трансформацию клетки растения с масличными семенами рекомбинантной конструкцией по п.4 и по меньшей мере одной дополнительной рекомбинантной ДНК-конструкцией, включающей выделенный полинуклеотид, функционально связанный с по меньшей мере одной регуляторной последовательностью, кодирующий полипептид, выбранный из группы, состоящей из дельта-4-десатуразы, дельта-5-десатуразы, дельта-6-десатуразы, дельта-8-десатуразы, дельта-12-десатуразы, дельта-15-десатуразы, дельта-17-десатуразы, дельта-9-десатуразы, дельта-9-элонгазы, С14/16-элонгазы, С16/18-элонгазы, C18/20-элонгазы и С20/22-элонгазы;
(b) регенерацию трансгенного растения с масличными семенами из трансформированной клетки стадии (а); и
(c) отбор таких полученных от растений стадии (b) семян, которые имеют измененный уровень по меньшей мере одной из полиненасыщенных жирных кислот по сравнению с уровнем в семенах, полученных от нетрансформированного растения с масличными семенами.
13. Способ по п.12, в котором растение с масличными семенами выбрано из группы, состоящей из сои, видов Brassica, подсолнечника, кукурузы, хлопка, льна и сафлора.
14. Трансгенное растение с масличными семенами для экспрессии дельта-5-десатуразы, полученное способом по п.8.
15. Растение с масличными семенами для экспрессии дельта-5-десатуразы, содержащее
(a) рекомбинантную ДНК-конструкцию по п.4, и
(b) по меньшей мере одну дополнительную рекомбинантную ДНК-конструкцию, включающую выделенный полинуклеотид, функционально связанный с по меньшей мере одной регуляторной последовательностью, кодирующий полипептид, выбранный из группы, состоящей из дельта-4-десатуразы, дельта-5-десатуразы, дельта-6-десатуразы, дельта-8-десатуразы, дельта-12-десатуразы, дельта-15-десатуразы, дельта-17-десатуразы, дельта-9-десатуразы, дельта-9-элонгазы, С14/16-элонгазы, C16/18-элонгазы, C18/20-элонгазы и С20/22-элонгазы.
16. Растение с масличными семенами по п.14 или 15, которое выбрано из группы, состоящей из сои, видов Brassica, подсолнечника, кукурузы, хлопка, льна и сафлора.
17. Трансгенное семя для экспрессии дельта-5-десатуразы, полученное от трансгенного растения с масличными семенами по п.14, где трансгенное семя содержит рекомбинантную ДНК-конструкцию.
18. Трансгенное семя для экспрессии дельта-5-десатуразы, полученное от трансгенного растения с масличными семенами по п.15, где трансгенное семя содержит рекомбинантную ДНК-конструкцию.
19. Применение трансгенного семени по пп.10, 17 или 18 для получения пищевого продукта для питания людей.
20. Применение по п.19, в котором указанный пищевой продукт представляет собой крахмал, белок, крупу, муку или зерно.
21. Применение трансгенного семени по пп.10, 17 или 18 для получения корма для питания животных.
22. Применение трансгенного семени по п.17 или 18 для получения масла.
23. Применение по п.22, дополнительно включающее получение лецитина.
24. Выделенная молекула нуклеиновой кислоты, представленная в SEQ ID NO:3, где указанная молекула нуклеиновой кислоты кодирует полипептид дельта-5-десатуразу, и у которой по меньшей мере один кодон оптимизирован в отношении кодонов для экспрессии в видах Yarrowia.
| СПОСОБ РАЗДЕЛЕНИЯ СЛОЖНОГО РАСТВОРА | 1994 |
|
RU2081668C1 |
| WO 9846764, 22.10.1998 | |||
| СИНТЕЗ ГАММА-ЛИНОЛЕНОВОЙ КИСЛОТЫ ПОД ДЕЙСТВИЕМ ДЕЛЬТА-6-ДЕСАТУРАЗЫ | 1995 |
|
RU2181772C2 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| СПОСОБ ПОЛУЧЕНИЯ 3-АЦЕТИЛБЕНЗО[f]ХИНОЛИНА | 2014 |
|
RU2559360C1 |
| SHAMSUDIN L | |||
| "Lipid and fatty acid contents in red tides from tropical fish ponds of the coastal water of South China Sea", vol 104, no 1, 1996, pp.36-42 | |||
| CHEN CHIH-YU et al | |||
| "Screening of red algae filaments as a | |||

