Область техники, к которой относится изобретение
Данное изобретение относится к мечению биологических соединений стабильными изотопами. В способах данного изобретения микроорганизмы выращивают на минеральных средах, содержащих источники углерода и азота, которые содержат стабильные изотопы, для получения биомассы, тотально меченной стабильными изотопами. Биомассу экстрагируют органическим растворителем в целях получения липидов. Обезжиренную биомассу гидролизуют в целях получения меченых аминокислот и других питательных веществ, которые используют для приготовления питательной среды для выбранной клетки-хозяина в целях получения биологических соединений, тотально меченных стабильными изотопами. Тотальное мечение стабильными изотопами позволяет определять трехмерную структуру биологического соединения ЯМР-спектроскопией. Предпочтительно, биологическое соединение представляет собой биологическую макромолекулу, такую как, например, трансмембранный белок млекопитающего.
Уровень техники
Концепция «рационального конструирования лекарственных средств» требует детального знания трехмерной структуры (3-D) биологических макромолекул, в частности белков. Рациональное конструирование лекарственных средств включает в себя определение 3-D-структуры «активной части» конкретной биологической молекулы, которая, как предполагается, участвует в этиологии заболевания. Этой биологической молекулой может быть, например, рецептор, фермент, гормон или любая другая биологически активная молекула. После определения 3-D-структуры биологической молекулы или, по меньшей мере, ее активной части ученые могут использовать компьютерное моделирование для конструирования молекул, способных блокировать, имитировать или усиливать природную биологическую активность молекулы. Таким образом, определение 3-D-структуры биологических молекул имеет большое практическое и коммерческое значение.
Первым методом, доступным для определения 3-D-структур белков и других больших биомолекул, был рентгенофазовый анализ. Структуры гемоглобина и ДНК были среди первых структур, определенных этим способом. Однако требованием рентгенофазового анализа является доступность подлежащей исследованию молекулы в кристаллической форме, так как 3-D-структуру рассчитывают из рентгенограммы рентгеновских лучей, преломленных атомами упорядоченных молекул в этом кристалле. Вследствие этого применение рентгенофазового анализа для определения 3-D возможно лишь для молекул, которые могут быть кристаллизованы. В результате подавляющее большинство мембранных белков не может быть изучено рентгенофазовым анализом, также попытки кристаллизации многих растворимых белков потерпели неудачу, так как кристаллизация является скорее эмпирической областью, чем наукой. В настоящее время база данных белков насчитывает только 100 вводов (0,7%) интегральных мембранных доменов, проанализированных с помощью кристаллов рентгенофазовым анализом (из которых одиннадцать представляют собой не более, чем отдельные α-спиральные трансмембранные домены), в противоположность 13600 вводам для растворимых белков. Самые последние способы и стратегии, примененные для структурной геномики, обсуждаются подробно Essen (2002, Gene Funct. Dis. 3: 39; смотрите также Jack, 2002 DDT 7: 35).
Затем был разработан другой метод, спектроскопия ядерного магнитного резонанса («ЯМР»), для определения 3-D-структуры биологических молекул и, в частности, белков. ЯМР-спектроскопия не требует кристаллизации подлежащей анализу молекулы и, следовательно, может быть применима в принципе к любой представляющей интерес биологической молекуле, в том числе к трансмембранным белкам и другим молекулам, которые не поддаются кристаллизации. Следующим преимуществом метода ЯМР-спектроскопии является его способность получения детальной картины динамики взаимодействия белков с их лигандами (Roberts, G.C.K., (2000) DDT, 5: 230).
ЯМР-спектроскопия включает в себя помещение подлежащей анализу молекулы (обычно в подходящем растворителе) в сильное магнитное поле и облучение ее сильным радиосигналом. Ядра различных атомов переориентируются этим магнитным полем по мере воздействия радиосигналом. Затем они поглощают энергию магнитного поля и излучают ее (резонируют) при частоте, которая зависит от i) типа ядра и ii) химического окружения, в значительной степени определяемого образованием связей этого ядра. Кроме того, резонансы могут передаваться от одного ядра к другому либо через связи, либо через 3-D-пространство, предоставляя тем самым информацию об окружении конкретного ядра и ядер вблизи него.
Однако не все ядра являются ЯМР-активными, в частности не все изотопы одного и того же элемента являются активными. Наиболее часто используемыми стабильными изотопами для исследования макромолекул методом ЯМР являются 13С, 15N и 2H. Известны также природные ЯМР-активные ядра, такие как 31P, 19F и 1H. Для более крупных молекул, таких как белки, для получения достаточно сильного сигнала в ЯМР-спектрах необходимо обогащение ЯМР-активными стабильными изотопами. В зависимости от используемого ЯМР-зонда, например, «обычный» водород, 1Н, является ЯМР-активным, тогда как тяжелый водород (дейтерий), т.е. 2H, не является ЯМР-активным. Таким образом, любая содержащая водород молекула может стать «невидимой» в водородном ЯМР-спектре при замещении всех атомов водорода 1Н атомами 2H. В связи с этим ЯМР-спектры водорастворимых веществ определяют в растворе в 2H2O (тяжелой воде) во избежание сигнала воды. Напротив, «обычный» углерод, 12С, является ЯМР-неактивным, в то время как стабильный изотоп 13С (приблизительно 1% всего углерода в природе) является активным. Подобным образом, «обычный» азот, 14N, является ЯМР-неактивным, тогда как стабильный изотоп 15N (опять, приблизительно 1% всего азота в природе) является активным. Таким образом, наиболее часто используемыми стабильными изотопами для биомолекулярной спектроскопии ЯМР являются 13С, 15N и 2H.
Для небольших молекул, т.е. молекул с молекулярной массой менее приблизительно 1000 Дальтон, как было обнаружено, низкие природные уровни ЯМР-активных изотопов являются достаточными для получения требуемого сигнала в ЯМР-спектрах. Однако для больших молекул, таких как белки, для получения достаточно сильного сигнала в ЯМР-спектрах требуется обогащения ЯМР-активными стабильными изотопами. Изотопное обогащение приводит к увеличенной чувствительности и разрешению и к упрощению ЯМР-спектров. Изотопное обогащение позволило эффективно использовать гетероядерные многомерные эксперименты ЯМР-спектроскопии и обеспечило альтернативные подходы для отнесения резонансов спектров и дальнейших структурных определений на основе спин-спинового взаимодействия.
Способ изотопного обогащения или замещения в биомолекулах заключался в выращивании микроорганизмов, способных продуцировать белки, на питательных средах, меченных изотопами. Для этой цели бактерии или дрожжи, трансформированные генной инженерией для продуцирования конкретного выбранного белка, например, важного с точки зрения медицины белка млекопитающего, выращивали в среде, содержащей 13С- и/или 15N-меченные субстраты. Эти среды обычно содержат 13С-меченную глюкозу и/или 15N-меченные соли аммония (смотрите, например, Кау et al., 1990, Science, 249: 411 и ссылки в этой работе). WO 90/15525 описывает питательные среды для бактерий и дрожжей, содержащие меченые белковые гидролизаты, для улучшения роста и продуцирования белка.
Однако этот подход не является подходящим при рациональном конструировании лекарств для большинства белков, которые по определению являются белками млекопитающих и, предпочтительно, имеют человеческое происхождение. Многие белки млекопитающих, которые экспрессируются в микроорганизмах, таких как бактерии или дрожжи, не продуцируются в форме, идентичной форме нативного белка, или сходной с нативным белком. В частности, это относится к мембранным белкам млекопитающих и секретируемым белкам, которые могут подвергаться существенным посттрансляционным модификациям, в том числе соответствующему фолдингу, сшиванию меж- и внутримолекулярных цепей дисульфидными мостиками, гликозилированию, ацилированию, фосфорилированию и другим химическим модификациям, и протеолитическому расщеплению в активные формы. Многие из этих модификаций не могут надежным образом выполняться бактериальными и дрожжевыми клетками-хозяевами. Вследствие этого, продуцируемые бактериями или дрожжами белки не могут быть использованы для структурно-функциональных исследований, так как они не имеют и даже не напоминают соответствующую нативную структуру. Часто они не обладают биологической активностью нативного белка и, в некоторых случаях, белки млекопитающих вообще не могут продуцироваться в бактериях. По этим причинам были разработаны векторные системы клеток-хозяев, использующие как клетки млекопитающих, так и клетки насекомых. Было обнаружено, что линии клеток млекопитающих, такие как клетки яичника Китайского хомячка (СНО), клетки COS, и линии клеток насекомых, такие как линии клеток Spodoptera frugiperda Sf9 и Sf21 (Luckow and Summers, 1998, Biotechnol. 6: 47-55), продуцируют рекомбинантные белки млекопитающих с посттрансляционными модификациями, сходными с модификациями природного белка.
Однако проблема использования клеток млекопитающих или клеток насекомых для мечения белков стабильными изотопами, такими как 13С или 15N, заключается в том, что, в то время как бактерии могут расти на простых минеральных средах, клетки млекопитающих и насекомых требуют использования сложных смесей питательных веществ, включающих, по меньшей мере, аминокислоты. В данной области исследования эту проблему решали с помощью использования гидролизатов белок-содержащей меченой биомассы в качестве источника меченых аминокислот. Предпочтительно, для получения меченой биомассы использовали микроорганизмы, такие как зеленые или сине-зеленые водоросли, которые могут расти только на простых (и, следовательно, дешевых) источниках меченого углерода и азота, таких как 15NH3, 15NO3 -, 15NO2 -, Н13СО3 -, 15N2, 13СО2 или 2H2O, при этом фотосинтетически включая меченые атомы в сложные биомолекулы, такие как белки, углеводы, липиды и нуклеиновые кислоты. Практический способ тотального изотопного мечения рекомбинантных белков в клетках млекопитающих описан Hansen et al., (1992, Biochem. 31: 12713). Авторами описан гидролиз изотопно меченых белков водорослей и бактерий для получения изотопно-меченных белков в клетках млекопитающих в целях структурных исследований методом ЯМР. Гидролиз проведен с помощью метансульфоновой кислоты в присутствии триптамина и имидазола, и аминокислоты очищены ионообменной хроматографией. Однако при использованных условиях гидролиза аспарагин, глутамин и цистеин разрушаются полностью, и отмечаются лишь следовые количества триптофана. Поэтому состав питательной среды для культивирования клеток млекопитающих должен быть дополнен коммерчески доступным цистеином и ферментативно синтезированным глутамином, меченным по атомам N и С. В среду дополнительно добавляли 5% обработанную нагреванием сыворотку, содержащую, следовательно, немеченые вещества, которые могут использоваться для синтеза белка.
В WO 94/18339 описаны питательные среды для мечения белков, экспрессируемых в клетках млекопитающих и насекомых. Эти питательные среды состоят из меченых аминокислот, выделенных из гидролизованной биомассы зеленой водоросли Chlorella, с добавлением синтетически меченых аминокислот цистеина, глутамина и аспарагина и с добавлением меченой глюкозы и пирувата. Сходные способы для получения изотопно-меченных белков и макромолекул, таких как гликопротеины, в клетках млекопитающих и насекомых были описаны, например, в патентах США 5393669 и 5627044, Weller (1996, Biochem. 35: 8815-23) и Lustbader (1996, J. Biomol. NMR 7: 295-304).
В WO 99/11589 описаны способы мечения белков по пептидному скелету основной цепи, но не по боковым цепям аминокислот. Эти белки продуцируются в питательных средах, в состав которых входят меченные по основной цепи аминокислоты, химически синтезированные из аминокислот-производных глицина с двумя защитными группами.
При разработке питательной среды для получения меченых белков в клетках млекопитающих или насекомых исследователи в данной области сосредоточивались в основном на получении изотопно-меченных аминокислот из гидролизованной биомассы. Аминокислоты, которые не присутствовали в достаточных количествах в гидролизатах, а также другие питательные вещества, такие как глюкоза или пируват, просто получали из коммерческих источников для дополнения состава этих сред. Несмотря на ощутимую в течение продолжительного времени потребность в данной области для ЯМР-структурных исследований (трансмембранных) белков млекопитающих и несмотря на тот факт, что такие питательные среды были заявлены в 1994 году, до сих пор не было сообщений о ЯМР-структурных исследованиях изотопно-меченных белков, полученных в клетках млекопитающих или насекомых, выращенных в этих средах. Таким образом, целью данного изобретения является улучшение питательных сред для выращивания клеток насекомых и млекопитающих в целях получения изотопно-меченных белков для ЯМР-анализа.
Описание изобретения
В первом аспекте данное изобретение относится к способу получения питательной среды для выращивания клеток млекопитающих или насекомых в культуре. В этой питательной среде, по меньшей мере, для одного из Н, С или N, по существу все атомы в субстратах, которые используются клетками для синтеза биомолекул, являются изотопно-меченными. Этот способ предпочтительно предусматривает стадии: (а) выращивания организма на минеральной среде, поддерживающей рост этого организма, где по существу все из ассимилируемых атомов, по меньшей мере, для одного из Н, С или N, являются изотопно-меченными, в целях получения меченой биомассы; (b) автолиза биомассы организма, выращенного, как описанного в (а), с получением автолизата; и (с) приготовления питательной среды на основе комбинации автолизата, полученного в (b), с дополнительными компонентами, необходимыми для роста клеток млекопитающих или насекомых. Этим организмом является предпочтительно гриб, дрожжи или водоросли. Предпочтительные дрожжи включают в себя хлебопекарные дрожжи и метилотрофные дрожжи, а предпочтительные водоросли включают в себя красные водоросли. Предпочтительным организмом является организм, который принадлежит к роду, выбранному из Saccharomyces, Pichia, Hansenula, Kluyveromyces, Candida, Brettanomyces, Debaryomyces, Torulopsis, Yarrowia, Galdieria, Cyanidium, Porphyridium, Cystoclonium, Audouinella, Cyanidioschyzon.
Под автолизатом понимают композицию, содержащую автолитически солюбилизированные клеточные компоненты, такие как аминокислоты, полипептиды, нуклеотиды, белки, гликоген, сахара, В-витамины, органические кислоты, липиды и другие компоненты. Автолиз организма обычно включает в себя инкубирование клеток организма при повышенной температуре (30-50°С) в течение продолжительного периода времени (3-18 часов) в присутствии плазмолизирующего агента, такого как NaCl, этанол, этилацетат, хлороформ или декстроза. Во время инкубирования клеточные компоненты гидролизуются эндогенными гидролитическими ферментами клеток, клеточная стенка разрушается и дезинтегрируется и высвобождает белковое содержимое в водное окружение. Нерастворимые остатки клеток (дебрис) удаляют центрифугированием и/или фильтрованием и получают автолизат, содержащий вышеупомянутые растворимые компоненты. Хотя обычно это не является обязательным, автолизаты в данном изобретении не исключают возможности использования экзогенных гидролитических ферментов.
Этот способ предпочтительно предусматривает дополнительно стадии: (а) выращивания организма на минеральной среде, которая поддерживает рост этого организма, посредством чего в этой среде по существу все из ассимилируемых атомов, по меньшей мере, для одного из Н, С или N, являются изотопно-меченными, для получения меченой биомассы; (b) экстракции биомассы организма органическим растворителем для получения экстракта, содержащего липиды, причем этот организм выращивается, как описано в (а), или выращивается, как описано в (а), на среде без изотопного замещения; (с) гидролиза биомассы организма, выращенного, как описано в (а), при нещелочном рН для получения гидролизата, содержащего аминокислоты; и (а) приготовления питательной среды на основе автолизата, полученного, как описано выше, с добавлением липидов, полученных, как описано в (b), и/или с аминокислотами, полученными, как описано в (с), и с добавлением дополнительных компонентов, необходимых для роста клеток млекопитающих или насекомых. Предпочтительно, этой питательной средой является среда, которая пригодна для продуцирования белка клетками млекопитающих или насекомых в культуре. Предпочтительно, к этой питательной среде добавляют дополнительные компоненты, необходимые для продуцирования белка клетками млекопитающих или насекомых в этой питательной среде. Предпочтительно, по существу все из ассимилируемых атомов в этих дополнительных компонентах, по меньшей мере, для одного из Н, С или N, являются изотопно-меченными. Нещелочным рН является предпочтительно рН ниже или приблизительно равный рН 8,0.
В данном контексте, выражение (термин), что молекула является «по существу меченой» или что «по существу все» атомы конкретного элемента в молекуле или в композиции являются «изотопно-меченными» или являются атомами «конкретной изотопной формы», обозначает, что эта молекула или композиция, содержащая эту молекулу, является достаточно обогащенной желаемым изотопом, так что может быть получена содержательная ЯМР-спектральная информация. В случае ЯМР-активных изотопов, таких как 13С или 15N, степень обогащения будет такой, что информация о трехмерной структуре может быть получена из ЯМР-спектров. В контексте данного изобретения, это означает, что приблизительно 95% или более атомов конкретного элемента будет в желаемой изотопной форме, предпочтительно более чем приблизительно 98, 99, 99,5 или 99,9%. В случае обогащения одним 2H, степень обогащения будет такой, что меченая молекула не будет продуцировать ЯМР-сигнал, достаточный для интерференции с анализом ЯМР-активных частиц, находящихся в комплексе с ней или присутствующих в образце для анализа методом ЯМР. В этом случае, уровень обогащения предпочтительно является превышающим примерно 70%, более предпочтительно превышающим примерно 80, 90, 95 или 98%. Альтернативно, уровень обогащения 2H является таким, что сигналы ЯМР-активных ядер lH, 13С и 15N усиливаются или лучше разрешены. Обычно этот уровень обогащения будет находиться в диапазоне от приблизительно 20% - до приблизительно 40, 50, 60, 70, 80, 90 или 100%. Квалифицированному специалисту будет понятно, что хотя выражение "изотопно-меченный" или "конкретной изотопной формы" в контексте данного изобретения обычно относятся к изотопам, которые применимы для получения ЯМР-спектров, однако данное изобретение не является обязательно ограниченным ими и может также относиться к другим изотопам, таким как, например, нестабильные радиоактивные изотопы.
В данном изобретении молекулы являются обычно тотально мечеными, что означает, что по существу все атомы конкретного элемента в молекуле являются изотопно-меченными до указанной степени или являются атомами конкретной изотопной формы. Альтернативно, в данном изобретении молекулы могут быть специфически меченными, что означает, что только по существу все атомы конкретного элемента в одном или нескольких положениях в этой молекуле являются изотопно-меченными или являются атомами конкретной изотопной формы. Пример специфически меченых аминокислот и белков, состоящих из них, приведен в WO 99/11589, в котором описаны химически синтезированные аминокислоты, которые используют для получения белков, изотопно-меченных по пептидному скелету основной цепи, но не по боковым цепям аминокислот.
Биомолекула, синтезированная клетками млекопитающих или насекомых, может быть любой молекулой, которая синтезируется такими клетками, в том числе, например, белками, полипептидами, пептидами, нуклеиновыми кислотами, такими как полидезоксинуклеотиды и полирибонуклеотиды, углеводами, липидами, метаболитами и их комбинациями. Эта биомолекула может быть молекулой, которая природно синтезируется клетками млекопитающих или насекомых, или она может быть молекулой, которая синтезируется этими клетками в результате генной инженерии. В последнем случае, эта молекула будет обычно представлена полипептидом или нуклеиновой кислотой, однако метаболиты, которые продуцируются в результате генной инженерии, например, введением одного или нескольких генов, кодирующих конкретные ферментативные активности, также включены в данное изобретение.
Получение изотопно-меченной биомассы
Организмы, которые выращивают в стадии (а) способа данного изобретения, предпочтительно выращивают на минеральной среде или питательной среде с определенным химическим составом. В зависимости от подлежащего изотопному мечению элемента (С, N и/или Н) питательная среда предпочтительно содержит единственный источник углерода и/или источник азота, тотально меченный рассматриваемым изотопом, более предпочтительно по существу все атомы в единственном источнике углерода и/или источнике азота являются изотопно-меченными. Для этой цели могут быть использованы различные содержащие углерод и/или азот соединения, в том числе, например, глюкоза и другие сахара, малые органические кислоты, мочевина и т.п. Однако предпочтительно используются источники углерода и/или азота, которые содержат только один атом углерода или атом азота, в том числе неорганические субстраты CO2, N2, NH3, NO3 -, NO2 -, НСО3 - или органические C1-соединения, такие как метанол, метан, метиламин, формальдегид и муравьиная кислота.
Вследствие этого, организм, выращиваемый на стадии (а), является предпочтительно фотосинтетическим, хемолитотрофным или метилотрофным организмом. Таким образом, этот организм будет предпочтительно являться микроорганизмом, хотя не исключены и растения. Растения могут выращиваться в виде клеточной культуры растений in vitro или в виде растений, и в этом случае они могут выращиваться в замкнутом пространстве, содержащем меченый диоксид углерода, и на субстрате, обеспеченном минеральной средой. Эти организмы предпочтительно выращивают автотрофно. При выращивании в клеточной культуре эти организмы выращивают предпочтительно в водной культуре, предпочтительно в ферментере с перемешиванием с контролируемыми рН, температурой, светом и подачей кислорода, диоксида углерода, источника углерода/азота и других питательных веществ, которые могут быть использованы или которые общеизвестны в данной области.
Подходящие организмы, подлежащие выращиванию в стадии (а) данного способа, включают в себя цианобактерии (сине-зеленые водоросли) и другие фотосинтезирующие бактерии, в том числе неоксигенные пурпурные серные и несерные бактерии, азотфиксирующие бактерии, эукариотические водоросли, в том числе зеленые, красные, а также бурые водоросли, диатомовые и другие хризофиты, а также динофлагеллаты, криптомонады и эугленоиды, метилотрофные бактерии и грибы, в том числе дрожжи. Предпочтительные организмы включают в себя Rhodophyka, Cyanidiophyceae (в большинстве случаев они упоминаются как принадлежащие к Rhodophyta), Chlorophyta, Cyanophyta, Diatoms, Phaeophyceae и Din ofla gel ate как источники липидов, в том числе полиненасыщенных жирных кислот (PUFA, ПНЖК), липидрастворимых витаминов и стеролов; метилотрофные бактерии а качестве источника белков (для последующего получения аминокислот); метилотрофные дрожжи в качестве источника белков (для последующего получения аминокислот), и витаминов; Cyanophyta в качестве источника углеводов, таких как глюкоза, сахароза, фруктоза и витамины; Chlorophyta, Dinoflagelate в качестве источника глицерина и органических кислот, высвобождаемых в среду; Chlorobiaceae (зеленые серные) и Rhodospirillaceae (пурпурные несерные) бактерии в качестве источника органических кислот, высвобождаемых в темноте в среду.
Следующие примеры подходящих родов в указанных классах организмов приведены в качестве предпочтительных для применения в способе данного изобретения.
Cyanophyta (сине-зеленые водоросли: Cyanobacteria): Spirulina, Synechoccus, Anabaena или Microcystis. Chlorophyta (зеленые водоросли): Chlorella, Neochloris, Scenedesmus, Dunaliella, Haematococcus, Staurastrum. Rhodophyta (красные водоросли): Porphyridium, Cyanidium, Cystocloniurn или Audouinella. Cyanidiophyceae (термофильные Rhodophyceae): Cyanidium, Galdieria, Cyanidioschyzon. Phaeophyka (бурые водоросли): Ectocarpus или Streblonema. Водоросли из группы Heterokontophyta (гетероконт-хромофиты), Prymnesiophyta (Haptophyta), Bacillariophyceae (диатомовые), Chryptophyta, Dinophyta (Pyrrhophyta, Dinoflagelate), Eugenophyta (эугеноиды), жгутиковые и облигатные гетеротрофные жгутиковые. Heterokontophyta: Rhinomonas, Syncripta или Heteroccus, Primnesiophyta (Haptophyta): Pseudoisochrysis, Phaeocystis, Prymnestum или Emiliania. Группа Chrysophyta, включающая в себя Bacillariophyceae (диатомовые) класса Bacillariophyta: Phaeodactylum, Navicula, Nitzchia, Amphora, Centronella, Eucampia, Fragilaria, Chryophyceae класса Chrysophyta (золотые водоросли), Haptophyceae класса Haptophyta. Chryptophyka: Cryptomonas, Chromomonas или Rhodomonas. Dinophyta: Peridinium, Oxyrrhis или Crypthecodinium. Euglenophyta: Euglena или Astasia. Жгутиковые: Metopus, Cyclidium. Фототрофные бактерии (Rhodospirilleae) подразделены на два подпорядка, пурпурные бактерии (Rhodospirillineae) и зеленые бактерии (Chlorobineae}. Зеленые серные бактерии (Chlorobiaceae): Chlorobium или Pelodickyon. Зеленые несерные бактерии (Chloroflexaceae): Chloroflexus. Пурпурные серные бактерии (Chromatiaceae): Chromatium, Amoebobacter или Thiodictyon. Пурпурные несерные бактерии (Rhodospirillaceae): Rhodobacter, Rhodopseudomonas, Rhodophila, Rhodospirillum или Rhodomicrobium. Heliobacteria (Heliobacteriaceae): Heliobacterium или Heliobacillus. Аэробные фотосинтезирующие бактерии: Erythrobacter. Метилотрофные микроорганизмы включают в себя облигатные метилотрофные бактерии: Methylobacillus, Methylophilus, Methylomonas, Methylococcus или Methylobacter; факультативно метилотрофные бактерии: Brevibacterium; и дрожжи в целом, например Saccharomyces, Kluyveromyces (например, выращенные на глюкозе или этаноле) и предпочтительно метилотрофные дрожжи: Pichia или Hansenula. Хемолитотрофные бактерии: Ralstonia или Nocardia.
Способы получения меченой биомассы хорошо известны в данной области, что следует из публикаций, касающихся выращивания бактерий в присутствии меченого углевода и солей (Кау, et al., supra), выращивания бактерий на лизатах водорослей (Chubb, R.T., et al., Biochemistry, 30, 7718 (1991)), выращивания дрожжей на лизатах водорослей (Powers, R., et al., Biochemistry, 31, 4334 (1992)), выращивания бактерий и дрожжей на меченом метаноле (смотрите Moat, A.G. and Foster, J.W., Microbial Physiology, 2d Ed., John Wiley & Sons, New York (1988), стр.218) и фототрофного культивирования водорослей в присутствии изотопно-меченного СО2 и/или N-солей (Сох, J., et al., Stable Isotopes in Pediatric Nutritional and Metabolic Research, Capman, Т.Е. et al., Eds., Intercept Ltd., Andover House, England (1990), p.1615).
Экстракция липидов
В стадии (b) способа данного изобретения биомассу, полученную, как описано в (а), экстрагируют органическим растворителем для получения экстракта, содержащего липиды. Этот липидный экстракт может содержать (три-, ди- и/или моно-)глицериды, стеролы, фосфолипиды, сфинголипиды, растворимые в жирах витамины и/или свободные жирные кислоты. Липиды, жирные кислоты и т.д. определяются здесь со ссылкой на Christie (2003, In: "Lipid Analysis" 3-d Ed., The Oily Press) как жирные кислоты и их производные и вещества, которые являются родственными биосинтетически или функционально этим соединениям. В контексте данного изобретения термин липиды подразумевает группу природно встречающихся соединений, которые имеют общую растворимость в органических растворителях, таких как хлороформ, бензол, простые эфиры и спирты, и включают в себя, например, такие разные соединения, как жирные кислоты и их производные, стероиды, каротиноиды, терпены, желчные кислоты и растворимые в жирах витамины.
Перед экстракцией биомасса может быть отделена от культуральной среды методами, известными в данной области, в том числе, например, центрифугированием или другой формой фильтрования. Затем, при необходимости, биомасса может быть промыта с использованием, например, воды или забуференного и/или водного раствора конкретной осмотической силы. Перед экстракцией биомасса может быть высушена с использованием, например, распылительной сушки или сушки вымораживанием (лиофилизации). Высушенная биомасса может обычным образом храниться до экстракции. Отработанная культуральная среда, от которой была отделена биомасса, может содержать ценные меченые соединения, такие как, например, аминокислоты, секретированные белки и углеводы. Поэтому эту отработанную культуральную среду предпочтительно также сушат, например, распылительной сушкой или лиофилизацией. Высушенный остаток может быть экстрагирован и/или гидролизован, как описано ниже для биомассы. Альтернативно, биомассу и культуральную среду не разделяют. В таких случаях биомассу и культуральную среду предпочтительно лиофилизируют вместе для концентрирования и/или хранения и последующей экстракции и/или гидролиза, как описано ниже.
В предпочтительном способе экстракции клетки в биомассе будут измельчаться любым известным в данной области подходящим способом. Эти способы включают в себя, например, обработку ультразвуком, но, более предпочтительно, механическое воздействие, такое как дробление или размалывание биомассы. Альтернативно, клетки в биомассе могут быть также обработаны таким образом, чтобы ослабить их клеточные стенки, например, обработкой ферментами. Измельчение может применяться перед экстракцией, но, более предпочтительно, его применяют во время экстракции в среде растворителя. Это будет увеличивать эффективность экстракции. Различные типы известных устройств для экстракции или, скорее, для измельчения могут быть использованы с целью разрушения клеток в биомассе, в том числе установки для мокрого процесса измельчения, такие как шаровые мельницы, мельницы с фрикционными дисками, смесители Геншеля, Френч-прессы и т.п. Предпочтительно, клетки в биомассе по меньшей мере частично разрушаются или разрываются силой сжатия или механической силой трения в установке измельчения. Однако следует отметить, что эти клетки не должны быть разрушены до образования слишком мелких частиц, которые были бы слишком малы для их легкого отделения от смеси растворителей.
Основой любого способа экстракции является приведение биомассы в контакт с подходящей экстракционной средой в течение некоторого периода времени с последующим разделением смеси биомассы и экстракционной среды на (1) нерастворенные компоненты этой смеси (т.е. экстрагированную биомассу) и (2) растворенный экстракт (т.е. экстракционную среду с растворенными компонентами, экстрагированными из биомассы). При идеальных условиях экстракции твердое вещество должно растворяться в растворителе быстро и полностью в течение короткого периода времени. Однако этого нельзя ожидать в случае сложных смесей веществ, не говоря уже о веществах в клетках водорослей, связанных с мембранами и ассоциированных с белковыми комплексами. Для более эффективной экстракции биомассу и экстракционную среду смешивают, используя какую-либо форму встряхивания, перемешивания, смешивания или гомогенизации, в том числе механическими средствами, указанными выше. Разделение смеси может выполняться различными способами, обычно известными в данной области, в том числе осаждением, центрифугированием и фильтрованием. Затем отделенная экстрагированная биомасса может быть опять экстрагирована той же самой или другой экстракционной средой. Различные растворенные экстракты могут быть объединены и экстрагированные компоненты могут быть извлечены из растворенного экстракта различными средствами, известными в данной области, в том числе, например, дистилляцией или (быстрым) выпариванием растворителя (растворителей) экстракционной среды.
Подходящими растворителями для экстракции липидов из биомассы являются низшие алифатические спирты или низшие алифатические углеводороды, причем низшие алифатические означает C1-C8. Однако, в принципе, равным образом может быть использован любой другой органический растворитель, в том числе, например, ацетон, галогенированные низшие алифатические спирты или углеводороды, а также низшие алифатические простые эфиры. Подобным образом могут быть использованы смеси органических растворителей. Подходящие способы включают в себя использование подходящего растворителя (растворителей) при условиях, которые будут предотвращать окисление ненасыщенных липидов, например, при проведении экстракции под вакуумом или под инертным газом, таким как азот, аргон или т.п. Спирты являются предпочтительными растворителями для экстракции в сравнении с ацетоном. Однако поскольку известно, что спирты стимулируют образование алломеров хлорофилла, и поскольку хлорофиллы представляют коммерческую ценность, содержащую хлорофилл биомассу (например, биомассу водорослей) предпочтительно экстрагировать ацетоном в качестве растворителя.
Хотя это не является критическим, количество растворителя, используемое в данном изобретении, находится в диапазоне приблизительно 2-7 масс. частей, предпочтительно приблизительно 3-6 масс. частей на 1 масс. часть биомассы в расчете на сухую массу. Предпочтительно, чтобы во время экстракции растворителем присутствовала вода. Количество воды, которое должно быть использовано, должно находиться в диапазоне приблизительно 0,2-0,7 масс. частей или, предпочтительно, 0,3-0,6 масс. частей на 1 масс. часть растворителя на безводной основе. Такое количество воды может добавляться отдельно в виде рассчитанной массы, когда биомасса является полностью сухой, но это количество отдельно добавляемой воды должно быть уменьшено, когда биомасса содержит воду, как это имеет место при использовании сырого осадка остатков биомассы, получаемых центрифугированием или фильтрованием для удаления культуральной жидкости. В случае экстракции водной суспензии микроорганизмов отношение между количеством водной суспензии и используемым экстракционным растворителем само по себе не является критическим. Обычно используют отношения по массе между 1:1 и 1:100. Используют обычно отношения между 1:2 и 1:50 и, наконец, предпочтительно используют в работе отношения между 1:5 и 1:20.
Несколько способов экстракции липидов из (микробной) биомассы известны в данной области. Один общепринятый способ, например, включает в себя гомогенизацию биомассы в смеси растворителей, состоящей из хлороформа и метилового спирта. Другой подходящий способ для экстракции липидов из биомассы описан в патенте США №4857329, который включен здесь в качестве ссылки для этой цели. В этом патенте описана экстракция липидов из клеток гриба рода Mortierella сначала измельчением этих клеток в присутствии или в отсутствие горячего спиртового растворителя и затем экстракцией обработанных спиртом измельченных клеток растворителем, предпочтительно в надкритическом состоянии, или смесью растворителя в надкритическом состоянии с низшим алифатическим спиртом или низшим алифатическим углеводородом. Низшим алифатическим спиртом является предпочтительно спирт, имеющий точку кипения от приблизительно 40°С до приблизительно 120°С (например, этанол, пропанол, изопропанол, бутанол и изобутанол). Предпочтительные низшие алифатические углеводороды включают в себя бутан, пентан, гексан, гептан и циклогексан. Предпочтительные условия процесса включают в себя температуры приблизительно от 35°С до 90°С и давления приблизительно от 200 до 600 кг/см2. Другой подходящий способ описан в патенте США №4870011, который также включен здесь в качестве ссылки для этой цели. Этот способ предусматривает механическое разрушение из двух стадий, первой, со смесью спирт/вода с получением фракции, богатой полярными липидами, и затем с углеводородным растворителем, например, гексаном, с получением фракции, обедненной в отношении полярных липидов. Особенно предпочтительной системой растворителей является смесь гексана и изопропанола при отношении объем:объем приблизительно 3 к 2. Предпочтительные способы экстракции для экстракции липидов и жирных кислот используют смеси из хлороформа и метанола. Предпочтительно хлороформ заменяют менее токсичным дихлорметаном, причем последний легче удалять выпариванием.
Липиды могут быть чувствительными к окислению, свету, нагреванию, кислотам и щелочам. Таким образом, во время экстракции липиды могут подвергаться нежелательным структурным изменениям вследствие, например, окисления или изомеризации. Квалифицированному специалисту будет ясно, как принять необходимые меры предосторожности для уменьшения таких нежелательных изменений во время процесса экстракции.
Липиды обычно представляют собой обнаруженные в клетках водонерастворимые органические вещества, которые могут быть экстрагированы неполярными растворителями. Существует несколько основных классов и подклассов липидов, большинство из которых могут встречаться в виде различных молекулярных разновидностей, в зависимости от структуры их жирнокислотных компонентов, и каждые имеют предпочтительные способы экстракции. Ацилглицерины (также называемые нейтральными жирами или глицеридами) предпочтительно экстрагируют простым эфиром, хлороформом, бензолом или этанолом; фосфоглицериды (например, PEA (цефалин), PC (лецитин)) предпочтительно экстрагируют смесями хлороформ/метанол; стероиды, в частности стеролы, предпочтительно экстрагируют хлороформом, простым эфиром, бензолом или горячим спиртом; терпены (минорные липидные компоненты клетки), которые включают в себя каротиноиды, предпочтительно экстрагируют с использованием смешивающихся с водой растворителей, таких как ацетон, метанол или этанол; выделяемые жирорастворимые витамины в основном могут быть экстрагированы органическими растворителями; сфинголипиды (цереброзид), гликолипиды (гликозилдиацилглицерины, цереброзид, ганглиозид), простагландины и холестерин могут быть экстрагированы, как описано Christie (supra).
Пигменты, такие как хлорофиллы и каротиноиды, предпочтительно экстрагируют с использованием смешивающихся с водой органических растворителей (смотрите выше).
Организмы для получения липидного экстракта предпочтительно выбирают на основе высоких содержаний липидов и/или их специфического липидного состава. В обзоре Nagashima обсуждается распределение основных жирных кислот в различных классах водорослей (1994, в: "Evolutionary Pathways and Enigmatic algae: Cyanidium caldarium (Rhodophyta) and related Cells", J. Seckbach, ed., Kluwer Academic Publishers, стр.201-214). Большинство водорослей могут служить в качестве источника С16- и С10-ряда жирных кислот, а также C18-ненасыщенных жирных кислот. Rhodophyta, Chryptophyta, Chromophyta и Haptophyta могут служить в качестве источника C20:4 и С20:5.
Картина распределения ненасыщенных жирных кислот варьируется в рамках индивидуальных классов водорослей. Chlorophyceae является предпочтительным источником для типов C16:3 (n-3), C16:4 (n-3), Rhodophyceae является предпочтительным источником для типов С20:4 (n-6), С20:5 (n-3) и Phaeophyceae является предпочтительным источником для типов C18:3 (n-3), C18:4 (n-3), С20:4 (n-6), С20:5 (n-3) (Takagi, et al., (1995) Yukagaku, 34, 1008-1012, смотрите также Banaigs et al., (1984), Phytochemistry 23: 2951-2952).
Красные водоросли в целом (например, Cystoclonium purpureum) являются предпочтительными источниками для длинноцепочечных полиненасыщенных жирных кислот (т.е. C18 и более длинных) и их производных (Pohl and Zurheide, (1979) In Hoppe, H.A., Levring, Т., Tanaka, Y. Eds. Marine Algae in Pharmaceutical Science. Walter de Gruyter, Berlin, pp.437-523). Виды Porphyridium, такие как, например, Р. aerrugentium (пресная вода) и Р. ruentum (соленая вода), являются предпочтительными источниками для арахидоновой кислоты C20:4 (n-6) (36% всех жирных кислот) (Arad, 1986, Int. Indus. Biotechnol. 7: 281-283; Arad et al., 1985, In: "Biosalinity in Action: Bioproduction with saline water", Pasternak, D., San Pietro A. eds, Nijhoff Publishers, стр.117-128; Gudin, 2002, In: "Algal Biotechnology. A sea of opportunities". Book of Abstracts. Universidad de Almeria, Servicio de Publicaciones).
Dinophyta, такие как, например, Cripthecodintum cohnii, являются предпочтительными источниками длинноцепочечных полиненасыщенных жирных кислот (PUFA, ПНЖК), таких как DHA - C22:6 (n-3) (смотрите, например, Borowitzka, 1999, J. Biotechnology 70: 313; U.S. 5,711,983; U.S. 4,670,285; WO 91/14427; Kyle et al., 1992, In: "Biotecnology and nutrition". Bills, D.D., Kung, S.D., Eds., Butterworth-Heinemann, Boston, pages 451-468; Gudin, Sijtsma et al., and Mendoza, 2002, In: "Algal Biotechnology. A sea of opportunities". Book of Abstracts. Universidad de Almeria, Servicio de Publicaciones).
Гетеротрофные диатомовые водоросли, такие как Nitzschia, Cyclotella, Navicula, являются предпочтительным источником для ЕРА - С20:5 (n-3) (смотрите WO 91/14427). Другим источником для ЕРА является морская диатомовая водоросль Phaeodactylum (смотрите Belarbi, E.H. et al., Acien, F.G. et al., Sanchez, M. et al., and Cini Zitelli, G., et al., in Algal Biotechnology. A sea of opportunities. Book of Abstracts. Universidad de Almeria, Servicio de Publicaciones) и морская диатомовая водоросль Tetraselmis (Ceron, M.C., et al., 2002, в той же самой книге).
Chlorophyta, такие как Chlorella, Dunaliella, Haematococcus, Parietochloris, являются предпочтительным источником PUFA (смотрите Cohen, et al., Gudin, 2002, в: "Algal Biotechnology. A sea of opportunities". Book of Abstracts. Universidad de Almeria, Servicio de Publicaciones). Cyanophyta Spirulina может быть использована в качестве источника PUFA.
Зеленая водоросль Dunaliella является источником глицерина (20-40% от органической массы водоросли), и внутриклеточная концентрация глицерола прямо пропорциональна концентрации внеклеточных солей и зависит от условий культивирования. При температуре ниже 25°С в среде обнаруживали небольшое количество глицерола или его отсутствие. При температуре выше 25°С скорость высвобождения глицерола в среду постепенно увеличивалась и при температуре выше 40°С она разительно увеличивается, так что при 50°С водоросль выделяет свой глицерол в среду в течение нескольких минут (Wegmann, et al., 1980).
Таким образом, сине-зеленые водоросли являются предпочтительным источником для С18-жирных кислот, зеленые водоросли для C16-жирных кислот и красные водоросли и диатомовые для С16-С20-жирных кислот. Высшие водоросли в целом, т.е. эукариотические водоросли, являются предпочтительными источниками для фосфатидилхолина (лецитина).
Квалифицированный специалист знает, что изменением условий выращивания можно влиять на фактический состав липидов, продуцируемых конкретным организмом. Например, хорошо известно, что увеличение температуры вызывает соответствующее уменьшение количества ненасыщенных жирных кислот. Другим примером влияния температуры на состав липидов является Cyanldium caldarium (смотрите Kleinschmidt and McMahon, 1970, Plant Physiol. 46: 290-293). Клетки С. caldarium, выращенные при 20°С, содержали значимо большие количества гликолипидов (моно- и дигалактозилдиглицерида) и фосфолипидов (фосфатидилхолина и фосфатидилэтаноламина) по сравнению с клетками, выращенными при 55°С. Подобным образом, клетки, выращенные при 20°С, содержали полярные липидные компоненты, в которых степень ненасыщенности является в три раза более высокой, чем в клетках, выращенных при 55°С. С. caldarium культивируют обычно при 25°С.
В целом, в отношении получения и выделения природных продуктов из водорослей читатель отсылается к Cresswell, Rees и Shah (1989, Algal and Cyanobacterial Biotechnology, Longmann and Scientific technical), Cohen (1999, Chemicals from microalgae. Taylor and Fransis) и Book of Abstracts "Algal Biotechnology. A sea of opportunities" (2002, supra).
Водоросли обычно продуцируют широкий спектр стеролов. Распределение стеролов в водорослях обсуждается, например, в обзоре Patterson (1991, в: "Physiology and Biochemistry of Sterols", Eds. Patterson, G.W. and Nes, W.D., American Oil Chemists's Society, Champaign, Illinois, стр.351-354). Эргостерол и ситостерол могут быть, например, получены из большинства отделов водорослей, а также из высших растений. Красные водоросли, такие как, например, Rhodophyka, являются предпочтительными источниками для холестерола и, кроме того, могут служить источником С27-, С28-, C29- стеролов (причем преимущественно С28), а также в качестве источника ситостерола, метилен-24-холестерола и кампестерола (24-метилхолестерола). Chlorophyta (зеленые водоросли) являются предпочтительными источниками ситостерола, (Chlorella), кампестерола, пориферастерола (24-этилхолеста-5,22-диенола), эргостерола (24-метилхолеста-5,7,22-триенола) и других стеролов. Цианобактерии (сине-зеленые водоросли) являются предпочтительными источниками для большого спектра стеролов, в том числе холестерола, эргостерола, хондрилластерола, ситостерола и кампестерола и других 24-алкилстеролов. Бурые водоросли являются предпочтительными источниками для фукостерола (24(Е)-этилиденхолестерина). Cyanidiophyceae (Cyanidium) являются предпочтительными источниками эргостерола, β-ситостерола и кампестерола. Таким образом, водоросли могут рассматриваться как источник для получения тотально меченого стерола из липидных экстрактов. Стандартные процедуры для их выделения обычно известны в данной области. Два способа часто использовали для выделения стеролов из экстрактов растений или водорослей. Значительное количество стерола может быть осаждено из растворов в бензине, оставленных на ночь при -10°С, но более эффективная процедура включает в себя осаждение стеролов в виде их дигитонидов (смотрите также Voikman, J.K. Sterols in microorganisms. Appi. Microbiol. Biotechnol. (2003) 60: 495-506, Patterson, G.W. (1991) In: Physiology and Biochemistry of Sterols (Eds, Patterson, G.W. and Nes, W.D.) American Oil Chemists Society, Champaign, Ilinois, pp.118-157).
Гидролизаты аминокислот
В стадии (с) способа данного изобретения биомассу организма, выращенного, как описано в стадии (а), гидролизуют для получения гидролизата, содержащего аминокислоты. Биомассу предпочтительно гидролизуют при нещелочном рН, предпочтительно при рН ниже 8,0. Белковый материал в биомассе может быть гидролизован ферментативно, обработкой кислотой или комбинациями обоих способов. Опубликованы много процедур для гидролиза белков, в том числе гидролиз хлористоводородной кислотой, метансульфоновой кислотой и ферментативный гидролиз.
В случае ферментативного гидролиза, предпочтительно протеолитические ферменты иммобилизуют на твердом носителе таким образом, что они могут быть удобным образом удалены из гидролизата, предпочтительно с другими нерастворимыми компонентами, например, осаждением, центрифугированием или фильтрованием. Альтернативно, может быть удобным подкисление гидролизата после ферментативной реакции. Преимуществом является денатурация и осаждение фермента, который может быть затем удален из гидролизата, например, центрифугированием или фильтрованием. Способы ферментативного гидролиза белков или белкового материала в биомассе описаны Milligan и Holt (1977, Adv. Exp.Med. Biol. 86B: 277-284) и Bergmeyer (1984, в: "Methods of enzymatic analysis: Enzymes 3: peptidases, proteinases and their inhibitors", VCH Publishing, 3-d ed.). Коммерчески доступными смесями эндо- и экзопротеаз, которые могут быть использованы в данном изобретении для ферментативного гидролиза, являются, например, Sumizyme™, FP, Sumizyme™, LP-протеазы (обе из Shin Nihon, Japan), протеаза Flavourzyme™ (Novo Nordisk A/S, Denmark) и Protease M™ Amano (Amano, Japan). Также могут быть использованы другие ферменты со сходными свойствами. Ввиду кислых рН-оптимумов грибных ферментов смеси эндо- и экзопротеаз предпочтительно получают из видов Aspergillus, в частности из таких видов, как A. oryzae или A. soiae, хотя ферменты из других видов Asperglllus или фактически из других видов грибов могут быть использованы сходным образом. Смеси этих протеаз, например, с другими протеазами, такими как, например, Pescalase™ (DSM, The Netherlands) протеазой, которая является бактериальной протеазой, могут быть также использованы. Предпочтительным ферментом для ферментативного гидролиза белков и белкового материала в биомассе является проназа, получаемая от Fluka (Buchs SG, Switzerland).
Однако в данном способе, предпочтительным является кислотный гидролиз. Способы кислотного гидролиза биомассы для получения меченых аминокислот подробно описаны в WO 94/18339. В основе этих способов лежит использование сильной минеральной кислоты, такой как хлористоводородная кислота, азотная кислота или серная кислота. Однако более предпочтительными являются сульфоновые кислоты, такие как п-толуолсульфоновая кислота или метансульфоновая кислота, причем последняя является наиболее предпочтительной. Концентрация кислоты может изменяться, в зависимости от природы белкового субстрата, но обычно является достаточной для проведения полного гидролиза. Обычно концентрации кислоты находятся в диапазоне приблизительно от 1 н до приблизительно 8 н, предпочтительно приблизительно от 4 н до приблизительно 7 н, более предпочтительно приблизительно от 5 н до приблизительно 6 н. Кислотный гидролиз проводят предпочтительно при неокисляющих условиях. Эти условия могут быть достигнуты проведением этой реакции в вакууме или барботированием инертным газом, таким как азот, аргон или т.п. Гидролизуемый белок может быть добавлен в среду гидролиза при концентрации приблизительно от 50 г/л до 500 г/л, предпочтительно при концентрации приблизительно от 100 г/л до 250 г/л. Гидролиз проводят при температуре и в течение времени, достаточных для выполнения по существу полного гидролиза, в то же самое время при уменьшении рацемизации или потери лабильных аминокислот. Температура гидролиза обычно находится в диапазоне приблизительно от 90 до 140°С, но для минимизации рацемизации аминокислот температура находится предпочтительно в диапазоне от 100 до 130°С, более предпочтительно от 110 до 120°С, причем особенно предпочтительной является температура 115°С. Время гидролиза может быть в диапазоне от 10 до 72 часов, в зависимости от гидролизуемого белка. Предпочтительно, используют время гидролиза приблизительно 22 часа. Во время реакции гидролиза аминокислоты, которые являются чувствительными к окислению, предпочтительно защищены присутствием восстанавливающего агента. Предпочтительно, используют сильный сульфгидрилсодержащий восстанавливающий агент, такой как тиогликолевая кислота (Fasman, G.D., Ed., Practical Handbook of Biochemistry and Molecular Biology, CRC, New York (1989), стр.106). Задачей этого восстанавливающего агента является не только защита уязвимых остатков триптофана и гистидина. При использовании тиогликолевой кислоты она может легко удаляться в соответствии с процедурами данного изобретения. Восстанавливающий агент используют в гидролизуемой смеси в концентрации, достаточной для предотвращения разрушения триптофана и гистидина. Что касается тиогликолевой кислоты, ее концентрация обычно находится в диапазоне приблизительно от 1 до приблизительно 7% об./об., предпочтительно приблизительно от 3 до приблизительно 15% об./об. Более предпочтительно или дополнительно, разрушение триптофана и гистидина может быть уменьшено или предотвращено включением «саморазрушающихся оснований», подобных триптамину и имидазолу (смотрите, например, Hansen et al., 1992, Biochem. 31: 12713). Каждый из триптамина и имидазола предпочтительно добавляют при 10-15 г/л смеси для кислотного гидролиза. Предпочтительные условия гидролиза включают в себя, например, гидролиз 10 г биомассы, предпочтительно свободной от липидов и пигментов (смотрите ниже), в 150 мл 4 М метансульфоновой кислоты при 115°С в течение 22 часов в вакууме и в присутствии 2 г триптамина и 2 г имидазола.
Поскольку глутамин и аспарагин могут существенно деаминироваться при вышеописанных условиях гидролиза, предпочтительно их добавляют из других источников, например, полученными химическим или ферментативным синтезом, ферментативным гидролизом меченой биомассы или белков, которые богаты глутаминовой кислотой и/или аспарагиновой кислотой, или они могут быть получены непосредственно ферментацией сверхпродуцирующих штаммов (смотрите, например, R. Faurie., J. Thommel. 2003 Microbial production of L-amino acids. Springer Verlag).
В предпочтительном варианте, белковый материал биомассы сначала подвергают ферментативному гидролизу для частичного гидролиза белкового материала. После удаления протеолитических ферментов этот материал дополнительно гидролизуют кислотой до достижения полного гидролиза, как описано выше. При комбинировании ферментативного и кислотного гидролиза предпочтительно используют смесь протеолитических ферментов, содержащую, по меньшей мере, эндопротеазы, легкодоступные в виде относительно чистых и экономически выгодных продуктов. Предпочтительно, используют асептические эндопротеазы. Подходящие коммерчески доступные композиции, содержащие эндопротеазы, упомянуты выше.
Предпочтительно, в этом способе биомассу экстрагируют органическим растворителем для удаления липидов и пигментов из биомассы перед ее гидролизом. Преимущество экстракции липидов и особенно пигментов из биомассы перед гидролизом заключается в том, что эти соединения могут быть сами по себе токсичными для насекомого или млекопитающего или могут превращаться в токсичные соединения во время гидролиза или они могут мешать гидролизу. Липиды и пигменты могут быть экстрагированы из биомассы, как описано выше. Предпочтительно, экстракцию оптимизируют для, по меньшей мере, эффективной экстракции пигментов. Биомассу предпочтительно экстрагируют более чем один раз, посредством чего отдельные стадии экстракции могут быть оптимизированы для экстракции липидов и для экстракции пигментов. Предпочтительные условия для экстракции пигментов описаны выше.
Предпочтительно, нерастворимые вещества удаляют из гидролизата центрифугированием или фильтрованием. Аминокислоты могут быть выделены из гидролизата, например, обращенно-фазовой или ионообменной хроматографией, как описано inter alia в Egorova et al., (1995, J. Chrom. Biomed. Appl. 665: 53-62) и WO 94/18339.
Наконец, в способе данного изобретения питательную среду готовят на основе липидов, полученных, как описано в (b), с аминокислотами, полученными, как описано в (с), и с добавлением дополнительных компонентов, необходимых для роста клеток млекопитающих и насекомых. Эта питательная среда может быть приготовлена из липидов и аминокислот, полученных из одного и того же организма или из, по меньшей мере, двух различных организмов. Предпочтительно, каждый гидролизат из аминокислот и липидный экстракт получают, по меньшей мере, из двух различных организмов. Таким образом, один или несколько организмов могут быть выбраны для получения гидролизата из аминокислот и один или несколько организмов могут быть выбраны для получения липидного экстракта.
Организмы для получения аминокислотного гидролизата предпочтительно выбирают на основе высоких содержаний белка и/или его аминокислотного состава. Подходящие для этой цели организмы упомянуты выше и включают в себя, например, сине-зеленые водоросли, такие как Spirulina, с содержанием белка 70% сухой массы, или метилотрофные бактерии, такие как Methylobacillus. Патент России SU 989867 описывает бактериальный штамм вида Methylobacillus methylophilus (VSB-932) с содержанием белка 80% сухой массы.
Далее, предпочтительные организмы для получения гидролизатов аминокислот включают в себя дрожжи в целом, например Saccharomyces, Kluyveromyces, Candida, Hansenula, Pichia, Brettanomyces, Debaryomyces и Tolrulopsis. Предпочтительные роды включают в себя Pichia, Hansenula, Saccharomyces. Особенно предпочтительными являются такие метилотрофные дрожжи, как Pichia pastoris и Hansenula polymorpha, которые могут расти на целом ряде источников углерода, в том числе на глюкозе, сахарозе, трегалозе, мальтозе, глицерине, эритрите, ксилите, манните, метаноле и этаноле. Предпочтительный способ получения гидролизата аминокислот из дрожжей может включать в себя автогидролиз биомассы дрожжей, описанный, например, в патенте США US 4165391 и в Lukondeh et al., (2003, J. Ind. Microbiol. Biotechnol., 30: 52). Мечение стабильными изотопами дрожжей и приготовление из них экстрактов описано в патенте Японии JP 6161743 (1994).
Дрожжевой автолизат является концентрированным продуктом, который содержит автолитически солюбилизированные клеточные компоненты, такие как аминокислоты, полипептиды, нуклеотиды, белки, гликоген, сахара, В-витамины, органические кислоты и другие компоненты. Автолиз дрожжей обычно предусматривает инкубирование клеток дрожжей при повышенной температуре (30-50°С) в течение продолжительного периода времени (3-18 часов) в присутствии плазмолизирующего агента, такого как, например, NaCl, этанол, этилацетат, хлороформ или декстроза. NaCl используют в концентрации 5-20% масс./об., предпочтительно от 2 до 10% масс./об. и растворители используют в концентрации от 1 до 10% об./об. Во время этого инкубирования клеточные компоненты гидролизуются эндогенными гидролитическими ферментами дрожжей, клеточная стенка разрушается и дезинтегрируется и высвобождает белковое содержимое в водное окружение. Нерастворимые остатки клеток (дебрис) удаляют центрифугированием и/или фильтрованием и получают автолизат, содержащий вышеупомянутые растворимые компоненты.
В данном изобретении предпочтительный вид дрожжей, указанный выше, культивируют в питательной среде, содержащей источник углерода и источник азота (меченые стабильным 13С- и/или 15N-изотопом), неорганические соли и т.д., и клетки собирают центрифугированием и промывают водой подходящим образом и полученные живые клетки дрожжей используют для получения экстракта. Водным компонентом может быть вода, вода, содержащая разбавленные соли, или природная жидкость ферментера/культуры. Дрожжевые клетки суспендируют в воде при концентрации предпочтительно приблизительно 5%-15% в расчете на сухую массу и подвергают тепловому шоку 50-75°С, предпочтительно 65-70°С, в течение приблизительно 30 секунд - 1 минуты для индукции трещин в клеточных стенках и индукции автолиза. Для поддержания активности протеаз и других протеолитических ферментов, участвующих в автолизе, обработка в течение более продолжительного времени и при температуре выше приблизительно 65°С не является предпочтительной и предпочтительным является быстрое охлаждение после теплового шока. Затем клетки дрожжей автолизируют при рН 6,5-10,0, предпочтительно рН 7,5-8,0 и при 10% масса/объем NaCl. рН может доводиться до или после теплового шока. Для доведения рН может быть использован разбавленный 15N-меченый гидроксид аммония или гидроксид натрия, гидроксид калия. Затем суспензию подвергнутых тепловому шоку клеток дрожжей в воде хранят при приблизительно 30-50°С, предпочтительно 40-45°С в течение приблизительно 3-12 часов, чтобы сделать возможным прохождение ферментативного автолиза. Затем растворимые компоненты извлекают центрифугированием или фильтрованием с получением автолизата дрожжей, который может быть лиофилизирован в целях долгосрочного хранения.
В контексте данного изобретения, термины "дрожжевой экстракт", "yeastolate", «гидролизат дрожжей» и/или «гидролизат, содержащий аминокислоты» (полученный из дрожжей) включают в себя содержащий аминокислоты гидролизат, полученный способами гидролиза, которые могут включать в себя любой химический гидролиз, ферментативный гидролиз как экзогенными, так и эндогенными ферментами, причем последний называют также автолизом.
Дополнительные питательные вещества
Изотопно-меченная глюкоза может быть выделена из цианобактерий или может быть получена коммерчески. Условия солевого стресса (например, рост в 0,5 М NaCl) могут быть использованы для увеличения углеводного содержимого биомассы, например Spirulina.
Органические кислоты могут быть удобным образом получены из Dinoflagelate, которые продуцируют до 69% органических кислот. Dinoflagellate (род Symbiodinium) являются одноклеточными подвижными водорослями с особым признаком постоянно конденсированных хромосом. Dinoflagellate являются источником глицерола и органических кислот. В большинстве симбиотических ассоциаций динофлагеллаты локализованы внутриклеточно в гастродермальном слое и окружены мембраной, происходящей из хозяина. Основными высвобождающимися соединениями являются глицерин (21-95%), органические кислоты (0-69%), глюкоза (0,5-21%) и аланин (1-9%). Различия между Symbionidium, выделенными из различных хозяев, можно видеть на примере Zoanthus pacifica и Scyphozoan Rhyzostoma. Симбионты из Zoanthus высвобождают 42% их продукта фотосинтеза, из которых 95% составляет глицерол и 3% органические кислоты, тогда как симбионты Rhyzostoma высвобождают 20% их продукта фотосинтеза, из которых 21% составляет глицерол и 69% органические кислоты (Trench, 1971 Proc. R. Soc. Lond. B, 177, 251-264).
Зеленые серные бактерии Chlorobium, при инкубировании в темноте, экскретируют органические кислоты, такие как ацетат, пропионат, капроат и сукцинат (смотрите Sirevag, 1995, In: "Anoxigenic photosynthetic bacteria", Eds. Blankenship, R.E., et al., Kluwer Academic Publisher, стр.879). Органические кислоты могут быть также использованы в качестве источника углерода для гетеротрофного или миксотрофного роста микроорганизмов, и гетеротрофное культивирование, например, на ацетате или глюкозе в качестве источников углерода использовали в течение некоторого времени. Однако гетеротрофное культивирование не является возможным для всех микроводорослей, и химический состав водорослей часто изменяется при гетеротрофных условиях (Borowitzka, 1999, Biotechnol. 70: 313-321).
Пурпурные несерные бактерии являются наиболее метаболически разнообразными организмами, обнаруженными в природе. Ферментация органических кислот Rhodospirillum rub-rum обсуждается Gorell и Uffen (1977, J. Bacteriol. 131: 533-543) и Kohlmirrer и Gest (1951, J. Bacteriol. 61:269-292).
Органические кислоты, в частности кислоты цикла Кребса, такие как фумаровая, малеиновая, янтарная, щавелевая, яблочная кислоты, могут быть выделены экстракцией в системе жидкость-жидкость и экстракцией в системе жидкость-твердое вещество, очисткой экстракта, дериватизацией и предварительным фракционированием (Liebich, H.M. 1990. Analytica-Chimica-Acta. 236(1), 121-130, также патент США US 3875222). Они могут быть экстрагированы органическими растворителями из культурального бульона, если они секретируются, или они могут быть экстрагированы вместе с липидом из биомассы (Lian et al., 1999. J. Pharm. Biomed. Analysis. 19: 621-625). Органические кислоты могут быть, например, экстрагированы вместе со свободными аминокислотами с использованием 80% (об./об.) кипящего этанола.
Альтернативно, меченные стабильными изотопами растения могут быть использованы в качестве источника меченых компонентов среды. Тотальное (15N более 98%) мечение стабильным изотопом растений описано, например, Ippel et al., (2004, Proteomics 4: 226-234 и ссылки в ней). Меченные изотопами растения могут быть использованы в качестве источника для (содержащих аминокислоты) гидролизатов, автолизатов, липидов и углеводов, в основном, как описано ниже.
Состав питательной среды для выращивания клеток насекомых или млекопитающих
В следующем аспекте данное изобретение относится к питательной среде, которая поддерживает рост клеток насекомых или млекопитающих. Предпочтительно, эта питательная среда поддерживает продуцирование белков, предпочтительно рекомбинантных белков, и вирусных продуктов. Этой питательной средой является, предпочтительно, среда, в которой для, по меньшей мере, одного из Н, С или N по существу все атомы в субстратах, которые ассимилируются клетками насекомых для синтеза биомолекул, являются изотопно-меченными. Эта питательная среда может быть использована для изотопного мечения биомолекул, таких как белок, для анализа структуры при помощи ЯМР. Эта среда является, предпочтительно, свободной от сыворотки, компонентов из сыворотки или компонентов животного происхождения.
Эта питательная среда, предпочтительно, состоит из различных содержащих аминокислоты гидролизатов, экстрактов липидов, углеводов и органических кислот, которые могут быть получены из микробных культур, как описано выше. Эта среда может быть далее дополнена, в случае необходимости, химически синтезированными или коммерчески доступными компонентами. Различные гидролизаты, экстракты липидов и препараты углеводов и органических кислот были подвергнуты скринингу и тестированы для нахождения наиболее значительного действия этих компонентов среды и их концентраций на конечную концентрацию клеток и/или уровни продуцирования белка.
Предпочтительная питательная среда для мечения изотопами биомолекул, продуцируемых в клетках насекомых и млекопитающих, в соответствии с данным изобретением, содержит:
(a) смесь неорганических солей;
(b) источник изотопно-меченных аминокислот;
(c) изотопно-меченный источник энергии, обычно в форме углеводов, таких как глюкоза;
(d) источник липидов;
(e) защитный реагент;
(f) (при необходимости) витамины и/или органические соединения, требующиеся в низких концентрациях;
(g) (при необходимости) органические кислоты;
(h) (при необходимости) микроэлементы.
Смеси неорганических солей для включения в клеточные среды хорошо известны в данной области. Смесь неорганических солей предпочтительно обеспечивает физиологическую ионную силу и рН-буферную способность. Подходящей смесью неорганических солей для клеток насекомых является, например, смесь солей, известная в качестве среды Грейса или среды Шнайдера. Подходящей смесью неорганических солей для клеток млекопитающих является, например, смесь солей, известная как модифицированная по способу Дульбекко среда Игла (D-MEM) (Price, P.J., et al., 1995, Focus, 17: 75), ВМЕ (базальная среда Игла, смотрите Eagle, H. (1965) Proc. Soc. Exp.Bioi. Med. 89: 362), питательная смесь F-10, F-12 (Ham, 1963, Exp.Cell. Res. 29: 515) и их модификации или среда CHO-SSFM1 (Gibco). Предпочтительно, в случае, когда эта смесь содержит неорганическую соль, содержащую один из элементов H, С или N, по существу все эти атомы в этих солях являются изотопно-меченными в соответствии с типом изотопного мечения других компонентов, таких как аминокислоты. Например, NaHCO3 присутствует в ВМЕ при 2,2 г/л, а в D-MEM при 3,7 г/л. Кроме того, D-MEM содержит также Fe(NO3)×9Н2О при 0,05 мг/л или 0,1 мг/л. Большинство, если не все, культуральных сред для клеток насекомых основаны на композиции Грейса (Grace, 1962, Nature 195: 788) и также содержат NaHCO3 в концентрации 0,35-0,7 мг/л. IPL-41 содержит (NH4)6(Mo7O24×4H2O) при 0,04 мг/л.
Подобным образом, композиции микроэлементов (определяемых как неорганические соединения или встречающиеся в природе элементы, которые обычно требуются в очень низких концентрациях, обычно в микромолярном диапазоне) для оптимального включения в питательную среду и их конечные концентрации в среде хорошо известны в данной области как для клеток насекомых, так и для клеток млекопитающих (смотрите, например, Thilly, 1986, In: "Mammalian cell technology", p.109, Butterworths Publishers; Vlak et al., 1996, In: "Insect cell cultures: fundamental and applied aspects" Volume 2, Kluwer Academic Publishers; и Perelch and Vinoi, 2003, In: "Handbook of industrial cell culture: mammalian, microbial and plant cells", Humana Press).
Источник энергии для клеток насекомых или млекопитающих в культуре будет обычно содержать углеводы, предпочтительно глюкозу. Изотопно-меченные глюкоза или другие сахара, такие как моно- или дисахариды, могут быть получены коммерчески или выделены из биомассы водорослей. Предпочтительной биомассой в качестве источника углеводов, таких как глюкоза, является биомасса зеленых водорослей, предпочтительно выращенных в условиях солевого стресса.
Источник аминокислот
Источник изотопно-меченных аминокислот для питательной среды данного изобретения предпочтительно выбирают из большого разнообразия гидролизатов биомассы или белковых продуктов, гидролизаты которых могут быть использованы по отдельности или в комбинации. Предпочтительно, гидролизаты получают из обезжиренной или экстрагированной растворителем биомассы водорослей, бактерий или грибов (дрожжей), полученной, как описано выше. Эти гидролизаты заменяют дорогие изотопно-меченные аминокислоты и (в некоторой степени) витамины. Этот источник аминокислот может быть дополнен индивидуальными аминокислотами, которые присутствуют в недостаточных количествах в гидролизатах, полученными из других источников, таких как химический или ферментативный синтез или ферментация. Такими аминокислотами могут быть, в частности, глутамин, аргинин, цистеин, гистидин и триптофан. Однако следовые количества триптофана (после кислотного гидролиза) могут быть достаточными для роста клеток насекомых и млекопитающих (как показано Hansen et al., supra). Предпочтительно, источник аминокислот содержит изотопно-меченный гидролизат дрожжей, полученный из дрожжей в целом или, более конкретно, из метилотрофных дрожжей (таких как Pichia или Hansenula), изотопно-меченный гидролизат водорослей (Cyanidium, Spirulina) и/или изотопно-меченный гидролизат, полученный из биомассы метилотрофных бактерий (Methylobacillus).
Бессывороточная среда для крупномасштабной культуры клеток насекомых (Spodoptera frugiperda) описана в Maiorella et al., (1988, Biotechnology 6: 1406-1410). Кроме базальной среды, эта среда содержала дрожжевой экстракт, метиловые эфиры PUFA жира печени трески, холестерол и Твин. В данном изобретении жир печени трески может быть заменен экстрактами липидов водорослей и бактерий, которые являются предпочтительно изотопно-меченными.
Патент WO 92/05247 описывает бессывороточную среду для культуры клеток насекомых, которая содержит базальную среду, к которой добавлены гидролизат дрожжей и альбумин или декстран. Соответственно, в данном изобретении гидролизат дрожжей может быть добавлен в количестве от 1 до 10 г/л.
Концентрация источника аминокислот в среде данного изобретения может быть настолько высокой, насколько позволяют суммарные наивысшие допустимые концентрации индивидуальных гидролизатов, где наивысшая допустимая концентрация каждого гидролизата является концентрацией, при которой гидролизат не является токсичным или ингибиторным в отношении роста клеток. Наивысшая допустимая концентрация гидролизата может изменяться не только в зависимости от конкретного препарата гидролизата, но и в зависимости от конкретной клеточной линии, которую выращивают в этой среде. Квалифицированный специалист сможет легко определить наивысшую допустимую концентрацию, например, добавлением увеличивающегося количества конкретного препарата гидролизата к клеточной линии, растущей в обычной среде.
Обычно предпочтительные конечные концентрации источника аминокислот в среде данного изобретения могут находиться в диапазоне приблизительно 1-15 грамм источника аминокислот (сухая масса) на литр среды, более предпочтительно конечная концентрация находится в диапазоне приблизительно 2-10 г/л.
Предпочтительный источник аминокислот для питательной среды данного изобретения для клеток насекомых, таких как линия клеток Sf9 Spodoptera frugiperda, содержит 0-7 г/л изотопно-меченного гидролизата водорослей, предпочтительно 2,5-4 г/л, и/или от 0 до 7 г/л изотопно-меченного бактериального гидролизата, предпочтительно 1,5-3 г/л, и/или от 0 до 8 г/л изотопно-меченного грибного/дрожжевого гидролизата, предпочтительно 5-7 г/л.
Предпочтительный источник аминокислот для питательной среды данного изобретения для клеток млекопитающих, таких как, например, клетки СНО, содержит 0-10 г/л изотопно-меченного гидролизата водорослей, предпочтительно 4-6 г/л, и/или от 0 до 10 г/л изотопно-меченного бактериального гидролизата, предпочтительно 4-6 г/л, и/или от 0 до 12 г/л изотопно-меченного грибного/дрожжевого гидролизата, предпочтительно 7-9 г/л.
Предпочтительно, гидролизаты, которые являются источником аминокислот для питательной среды данного изобретения, очищают ультрафильтрацией для удаления оставшихся протеаз, используемых во время получения этого пептонного продукта, эндотоксинов и других высокомолекулярных компонентов, которые могли бы мешать получению и/или очистке (рекомбинантных) белков, экспрессируемых клетками-хозяевами. Пептонные фракции предпочтительно предварительно фильтруют и затем ультрафильтруют через мембрану с пределом отсечения молекулярной массы, выбранным таким образом, чтобы он был меньше, чем молекулярная масса рекомбинантного или вирусного продукта, для облегчения последующей очистки, предпочтительно через мембрану с пределом отсечения молекулярной массы 2000-15000, более предпочтительно через мембрану с пределом отсечения молекулярной массы 10000, такую как мембрана РМ10 (Amicon). Процесс ультрафильтрации проводят предпочтительно в аппарате для фильтрации с поперечным потоком, например, в находящейся под давлением перемешиваемой ячейке для малого масштаба или патроне с полыми волокнами или фильтрационном устройстве с планшетом или рамой для крупномасштабной фильтрации. Ультьрафильтрат может быть простерилизован с использованием мембранного фильтра перед добавлением базальной среды, к которой также добавляют другие компоненты сред этого изобретения.
Исходные растворы изотопно-меченного гидролизата готовят предпочтительно в концентрации 10-15% (сухая масса/объем) для гидролизатов водорослей, 15-20% для гидролизатов грибов/дрожжей и 15-18% для бактериальных гидролизатов.
Несколько партий каждого лизата тестируют на их способствующие росту свойства, например, в отношении клеток Sf9 или клеток СНО.
Понятно, что эти применяемые гидролизаты, в частности автолизаты, могут содержать олигопептиды, наряду со свободными аминокислотами. В некоторых гидролизатах присутствие аминокислот в форме олигопептидов достигает до примерно 20% от общего содержания аминокислот. Концентрации свободных аминокислот в культуральной среде могут изменяться во время культивирования клеток и могут, например, зависеть от пептидаз/протеаз, высвобождаемых этими клетками. Это следует принимать во внимание при определении потребления аминокислот (выражаемого в виде процента исходного содержания в культуральной среде). Концентрация источника аминокислот, подлежащих добавлению в среду, будет зависеть от таких факторов, как конкретные фракции источников аминокислот, которые используются, природа клеточной линии, подлежащей культивированию, уровень, при котором конкретный пептон становится токсичным или ингибиторным в отношении роста клеток. Оптимальная концентрация для каждой комбинации клеточной линии и источника аминокислот может быть определена эмпирически.
Обычно общее количество источника аминокислот в питательной среде должно находиться в диапазоне приблизительно от 8 до 30 г/л, более предпочтительно приблизительно от 12 до 24 г/л.
Относительные концентрации каждой индивидуальной аминокислоты и каждого витамина могут устанавливаться в соответствии с потребностями конкретной клеточной линии. Например, хотя глутамин является необходимым для некоторых линий клеток насекомых, в данной области известно, что другие клеточные линии растут без глутамина и могут быть способны синтезировать эту аминокислоту из предшественников (смотрите, например, Misuhashi, 1987, Appi. Entomol. Zool. 22: 533-536). В зависимости от предполагаемого применения среды концентрации индивидуальных свободных аминокислот и витаминов могут устанавливаться в соответствии с известными характеристиками различных клеточных линий насекомых или млекопитающих.
Источник липидов
Потребности в липидах клеток насекомых обычно сходны с потребностями в липидах млекопитающих позвоночных животных, за исключением того, что для клеток насекомых необходим источник холестерола. Источник липидов для питательной среды по данному изобретению предпочтительно содержит липиды, которые являются незаменимыми для роста клеток насекомых и млекопитающих. Предпочтительно, эти липиды являются изотопно-меченными липидами, но это не является обязательным для данного изобретения. Источник липидов предпочтительно содержит, по меньшей мере, (1) жирные кислоты, (2) стероиды и (3) растворимые в жирах витамины.
Жирные кислоты в источнике липидов могут присутствовать в различных формах, таких как, например, триглицериды, свободные жирные кислоты или алкиловые эфиры жирных кислот (предпочтительно С1-С4-алкильные), из которых предпочтительными являются метиловые эфиры. Жирные кислоты предпочтительно содержат полиненасыщенные жирные кислоты, имеющие длину цепи C12-C22, предпочтительно C13-C19.
Предпочтительная смесь (изотопно-меченных) полиненасыщенных жирных кислот для сред данного изобретения присутствует в экстракте липидов из изотопно-меченной биомассы Rhodophyta, Cyanidiophyceae (в большинстве случаев это семейство упоминается как принадлежащее к Rhodophyta), Chlorophyta, Cyanophyta, Diatoms, Phaeophyceae, Dinoflagelate, Dinophyta.
Конечные концентрации в питательной среде для (изотопно-меченных, полиненасыщенных) жирных кислот и/или их (метилового) эфиров находятся предпочтительно в диапазоне от 2 мг/л до приблизительно 100 мг/л, более предпочтительно от 10 до 30 мг/л и наиболее предпочтительно 18-22 мг/л.
Стероиды в источнике липидов являются предпочтительно стеролами, такими как ланостерол, стигмастерол, ситостерол и холестерол, причем холестерол является особенно предпочтительным. Стероиды могут быть получены из липидных экстрактов осаждением, как описано Young и Britton (1993, Carotenoids in Photosynthesis. Chapman and Hall) и Volkman 2003 Appl. Microbiol. Biotechnol., 60(5): 495-506. Конечная концентрация в питательной среде для (изотопно-меченных) стероидов равна предпочтительно от 2 до 10 мг/л, более предпочтительно от 5 до 7 мг/л.
Растворимые в жирах витамины в качестве источника липидов предпочтительно содержат, по меньшей мере, витамин Е (альфа-токоферол) и могут дополнительно содержать витамин А. Конечная концентрация в питательной среде для (изотопно-меченного) альфа-токоферола составляет от приблизительно от 0,5 мг/л до приблизительно 4 мг/л, более предпочтительно от 1,5 до 3 мг/л. В липидных экстрактах должны присутствовать достаточные количества витамина А.
Прямое добавление липидов к водным средам данного изобретения является непрактичным вследствие их низкой растворимости в воде. Таким образом, липиды предпочтительно обеспечивают в форме подходящей липидной эмульсии, такой как, например, микроэмульсии, описанные Maniella et al. (1988, BioTechnol. 6: 1406). Такая микроэмульсия содержит раствор липида/органического растворителя, содержащего небольшое количество эмульгатора. Предпочтительные эмульгаторы включают анионогенное поверхностно-активное вещество, обычно фосфолипиды, такие как лецитин, или нетоксичные, неионогенные детергенты, такие как, например, полисорбат 20 или 80. Дополнительной возможностью является добавление липидов, растворенных в смешивающемся с водой растворителе, таком как, например, диметилформамид и различные спирты (C1-С6-спирты), причем этанол является предпочтительным. Исходный раствор липидов в смешивающемся с водой органическом растворителе является предпочтительно таким, что конечная концентрация органического растворителя в питательной среде составляет менее чем 0,1% (об./об.), более предпочтительно составляет менее чем 0,05 или 0,01%, во избежание токсичности этого растворителя в отношении клеток насекомых или млекопитающих. Квалифицированный специалист эмпирически определит конечную концентрацию органического растворителя в зависимости от выбранного растворителя, типа выращиваемых клеток и т.п.
Предпочтительный изотопно-меченный источник липидов для питательной среды данного изобретения содержит на литр среды: 20 мг изотопно-меченного экстракта липидов, 6 мг изотопно-меченного холестерола, растворенных в 1 мл этанола.
При необходимости в питательную среду данного изобретения могут быть включены защитные агенты. Защитные агенты обычно известны в данной области и определяются как нетоксичные, водорастворимые компоненты, которые функционально действуют для защиты клеток насекомых или млекопитающих от повреждения и смерти в перемешиваемой и барботируемой культуре клеток насекомых. Поскольку их концентрация (масса/объем) является очень низкой (0,01 - приблизительно 1%) и они обычно не сразу метаболизируются клетками в культуре, они могут добавляться в немеченой форме. Защитные агенты могут добавляться отдельно или могут объединяться с источником липидов. Защитный компонент предпочтительно содержит блоксополимеры пропиленоксида и этиленоксида и более предпочтительно плурониковые полиолы, такие как плуроник F68 и F88 (BASF Wyandotte Corp.). Другие подходящие соединения, которые могут служить в качестве защитных агентов, включают, например, гидроксиэтилкрахмал, метилцеллюлозу, карбоксиметилцеллюлозу, декстрансульфат, полипропиленгликоль, альгиновую кислоту, Фиколл и поливинилпирролидон.
К при необходимости профильтрованному/простерилизованному 1 мл раствора липидного компонента (описанному выше) медленно добавляют 10 мл 10% Плуроника F68 в воде (при необходимости профильтрованного/простерилизованного) при перемешивании с использованием вортекса. Плурониковые полиолы коммерчески доступны от BASF Wyandotte Corp. (101 Cherry Hill road, P.O. Box 181, Parsipanny, N.J. 07054, U.S.A.). В результате этого образуется микроэмульсия липидного компонента. Успешность приготовления липидной эмульсии может зависеть от температуры, поэтому эмульсию готовят при температурах в диапазоне 35-40°С. Для облегчения приготовления эмульсии может быть использовано встряхивание на высокоскоростном вортексе. Предпочтительно, источник липидов фильтруют и стерилизуют фильтрованием через микропористую мембрану. Альтернативно, приготовленную питательную среду можно стерилизовать фильтрованием с использованием микропористой мембраны. Стерины обеспечиваются из органического экстракта вместе с другими классами липидов.
Подходящими источниками стеролов являются Cyanidiophyceae, красные водоросли, другие водоросли. Основная часть водорастворимых изотопно-меченных витаминов обеспечивается изотопно-меченным пептонным компонентом, обычно изотопно-меченным гидролизатом дрожжей и изотопно-меченными метилотрофами. Источником растворимых в липидах витаминов и других классов липидов являются органические экстракты.
Кроме того, кровь насекомых содержит необычайно высокий уровень свободных органических кислот, таких как цитрат, сукцинат, оксалат или малат, в диапазоне 0,1-35 ммоль на насекомое (Grace, (1962), Nature, 192, 788, Vaughn, J.L., (1968) Curr. Top.Microbiol. Immunol. 42, 108). Промежуточные продукты цикла Кребса являются хорошими хелатообразующими агентами и, следовательно, играют важную роль в катионном балансе гемолимфы. Питательные среды для культивирования клеток этого изобретения могут быть дополнены одним или несколькими промежуточными продуктами цикла Кребса и/или пируватом, которые предпочтительно добавляют до максимальной концентрации 50 мг/л индивидуальной органической кислоты. Некоторые среды, имеющие уменьшенные количества или вообще не имеющие этих компонентов, все еще поддерживают рост клеток насекомых. Например, (Gardiner and Stockdale, 1975), WO 01/985517 описывают, что большинство дорогих из свободных органических кислот, таких как фумарат, малат, сукцинат, кетоглутаровая кислота и гидроксипролин, может быть полностью исключено из сред для клеток насекомых без отрицательных последствий и заменены большими количествами (до 250 мг/л для индивидуального витамина) витаминов, таких как тиамин, рибофлавин, ниацин, витамин В6, фолиевая кислота, витамин В12, биотин, пантотеновая кислота, холин, пара-аминобензойная кислота, инозит, сахара, такие как глюкоза, и пептоны.
Подходящие источники органических кислот описаны выше и включают в себя Scyphozoan rhizostoma (высвобождает органические кислоты в культуральную среду), зеленую серную бактерию Chlorobium (при инкубировании в темноте высвобождает органические кислоты), пурпурную несерную бактерию Rhodo spirillum rubrum (высвобождает органические кислоты во время анаэробного роста в темноте). Органические кислоты обеспечиваются или органическим экстрактом биомассы, или отдельно в виде экстракта культурального бульона.
Эти среды могут быть дополнительно дополнены меченым пируватом при 60-90 мг/л, получаемым, например, от Isotec Inc.
При необходимости свободные, очищенные изотопно-меченные аминокислоты могут быть добавлены к средам данного изобретения (наряду с описанными выше источниками аминокислот). Препараты свободных аминокислот, органических кислот и витаминов могут быть или очищены из изотопно-меченных природных источников, или могут быть получены коммерчески и обычным образом добавлены к основной базальной среде независимо от любых аминокислот и/или витаминов, происходящих из гидролизатов или компонента дрожжевого гидролизата полной среды.
Способы получения изотопно-меченных биомолекул
В другом аспекте, данное изобретение относится к способам получения изотопно-меченных биомолекул. Предпочтительно, этим способом является способ получения биомолекулы, с использованием которого по существу все атомы в этой биомолекуле являются изотопно-меченными. Этот способ предпочтительно предусматривает стадии (а) выращивания культуры клеток млекопитающих или насекомых, способных производить эту биомолекулу, при условиях, способствующих получению этой биомолекулы, в питательной среде, получаемой в способе по данному изобретению, и (b) извлечение этой биомолекулы. Предпочтительно, этой биомолекулой является полипептид или белок (оба термина используются здесь взаимозаменяемо). Способы данного изобретения особенно пригодны для получения изотопно-меченных мембранных белков, в том числе трансмембранных белков и белков наружной мембраны. Однако способ данного изобретения равным образом пригоден для получения растворимых белков или других немембранных белков. Этим белком является предпочтительно белок млекопитающего, более предпочтительно белок человека.
В способе данного изобретения белок продуцируется культивированием клеток млекопитающих или насекомых, которые способны продуцировать этот белок, в питательной среде, в которой, по меньшей мере, для одного из Н, С или N по существу все атомы в субстратах, используемых клетками для синтеза этой биомолекулы, являются изотопно-меченными. Этот белок может быть эндогенным белком, который природно продуцируется этими клетками в культуре. Однако обычно этот белок будет получен рекомбинантным способом.
Для этой цели нуклеотидную последовательность, кодирующую представляющий интерес полипептид, экспрессируют в подходящих клетках млекопитающего или клетках-хозяевах, таких как описанные в руководствах Ausubel et al., (eds.), "Current Protocols in Molecular Biology", Greene Publishing and Wiley-Interscience, New York (1987) и в Sambrook and Russel (2001) "Molecular Cloning: A Laboratory Manual", 3rd edition. Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, New York; оба из которых включены здесь в качестве ссылки в их полном виде.
Нуклеотидная последовательность, кодирующая представляющий интерес полипептид, должна быть введена в клетки-хозяева посредством экспрессирующего вектора. Этот вектор может быть репликативным вектором, содержащим сайт инициации репликации (или последовательность автономной репликации), что гарантирует размножение этого вектора в подходящем хозяине для этого вектора. Альтернативно, этот вектор способен к интеграции в геном клетки-хозяина, например, посредством гомологичной рекомбинации или другим образом. Подходящие экспрессирующие векторы для клеток млекопитающих хорошо известны из Sambrook и Russel (2001, supra). Подходящие векторы для клеток насекомых основаны на хорошо известном бакуловирусе (Merrington et al., 1997, Mol. Biotechnol. 8: 283-97). Выражение «экспрессирующий вектор» обычно относится к нуклеотидным последовательностям, которые способны осуществлять экспрессию представляющей интерес кодирующей последовательности в хозяевах, совместимых с такими нуклеотидными последовательностями. Эти экспрессирующие векторы обычно включают в себя, по меньшей мере, подходящие транскрипционные (промотор) регуляторные последовательности и регуляторные последовательности инициации и терминации трансляции, функционально связанные с кодирующим этот полипептид сегментом. ДНК-сегмент является «функционально связанным», когда он помещен в функциональную связь с другим ДНК-сегментом. В экспрессирующем векторе нуклеотидная последовательность, кодирующая представляющий интерес полипептид, функционально связана с промотором, способным управлять экспрессией этой кодирующей последовательности в клетке-хозяине.
В данном контексте, термин «промотор» относится к фрагменту нуклеиновой кислоты, который функционирует для регуляции транскрипции одного или нескольких генов, расположенного слева относительно направления транскрипции от сайта инициации транскрипции этого гена, и структурно идентифицируется присутствием сайта связывания для ДНК-зависимой РНК-полимеразы, сайтов инициации транскрипции и любыми другими ДНК-последовательностями, в том числе, но не только, сайтов связывания факторов транскрипции, сайтов связывания репрессорных и активаторных белков и любых других последовательностей нуклеотидов, о которых квалифицированному специалисту известно, что они действуют прямо или опосредованно для регуляции количества транскрипции от этого промотора. «Конститутивным» промотором является промотор, который является активным при большинстве физиологических условий и связанных с развитием условий. «Индуцибельным» промотором является промотор, который регулируется в зависимости от физиологических и связанных с развитием условий. «Тканеспецифический» промотор является активным только в специфических типах дифференцированных клеток/тканей.
Выбор подходящих промоторных последовательностей обычно зависит от клетки-хозяина, выбранной для экспрессии конкретного ДНК-сегмента. Примеры подходящих промоторных последовательностей млекопитающих хорошо известны в данной области (смотрите, например, Sambrook и Russel, 2001, supra). Подобным образом, подходящие промоторные последовательности для использования в клетках насекомых, известны в данной области (Merrington et al., 1997, supra). Примеры применимых комбинаций клеточных линий и экспрессирующих векторов описаны в Sambrook и Russel (2001, supra), для клеток млекопитающих в Wurm и Barnard (1999, Curr. Opin. In Biotechnology 10: 156-159), для клеток насекомых в Massotte (2003, Biochim. Biophys. Acta, 1610: 77-89) и в Ikonomou et al., (2001, In Vitro Cell Dev. Biol.-Animal. 37: 549-559) и для дрожжей в Metzger et al. (1988, Nature 334: 31-36).
Таким образом, способ получения меченых полипептидов изобретения предусматривает стадию культивирования клетки-хозяина, определенной выше, при условиях, способствующих экспрессии этого полипептида. При необходимости, этот способ может предусматривать извлечение этого полипептида. Полипептид может быть, например, извлечен из культуральной среды стандартными способами очистки белков, в том числе различными хроматографическими способами, известными в данной области per se (смотрите ниже).
Очистка меченого белка из клеток/культуры
Очистка (меченых) белков из клеток или культуральной среды хорошо известна в данной области. В отношении растворимых белков смотрите, например, Deutscher (1990, Methods in Enzymology, Vol.182) и Yokoyama (2003, Current Opinin. Chemical Biology, 7: 39-43 и ссылки в этой работе). Способы получения больших количеств конкретного белка достаточной чистоты для анализа структуры доменов обычно известны квалифицированным специалистам. Хотя не все способы очистки белков применимы к представляющему интерес белку, понятно, что следующие способы являются предпочтительными: аффинная хроматография, осаждение сульфатом аммония, диализ, FPLC, ионообменная хроматография, ультрацентрифугирование и т.д. По поводу общего обзора очистки белков смотрите Burgess (1987, In: "Protein Purification", Oxender, et al., Eds., Protein Engineering, p.p.71-82, Liss), Jacoby, Ed. Methods Enzymol. 104: Part С (1984); Scopes, Protein Purification: Principles and practice (2-nd ed.), Springer-Verlag (1987).
Очистка мембранных белков описана, например, Bosman et al. (2003, In: "Methods in Molecular Biology", volume 228: 73, Ed. Selinsky, Humana Press Inc., Totowa, NJ), Reeves et al. (2002, Proc. Natl. Acad. Sci. USA 99: 13413-13418) и Eilers et al., (1999, Proc. Natl. Acad. Sci. USA 96: 487-492).
WO 99/22019 описывает приготовление образца белка для ЯМР. Условие растворителя, подходящее для определения биофизического свойства белка, описано в WO 99/30165. Приготовление образцов мембранных белков для ЯМР рассматривается Sanders и Oxenoid (2000, Biochim. Biophys. Acta 1508: 128-145 и ссылках). Восстановление мембранных белков в липидные бицеллы для ЯМР-исследований описано Sanders et al. (1995 Biochem. 34: 4030-4040). ЯМР-спектроскопия крупных белков описана в патенте США US 6198281.
В следующем аспекте данное изобретение относится к мембранному белку млекопитающих, причем по существу все атомы в этом белке являются изотопно-меченными изотопом, выбранным из 15N, 13C, 2H, 15N и 13C, 15N и 2Н, 13С и 2H, или 15N, 13С и 2Н. Изотопное мечение по существу всех атомов в белке означает, что, по меньшей мере, приблизительно 95% или более атомов конкретного элемента будут находиться в желаемой изотопной форме, предпочтительно более чем приблизительно 98, 99, 99,5 или 99,9%. Предпочтительно, этим мембранным белком млекопитающего является белок, в котором 20-100% атомов водорода тотально заменены изотопом Н. Предпочтительно этот мембранный белок является белком человека. Под мембранным белком подразумевают здесь белок, который содержит полипептидную цепь, имеющую длину, по меньшей мере, 30, 40, 50, 70 или 100 аминокислот, чтобы отличить этот белок от пептида, который может быть получен из полноразмерного белка.
Способ определения трехмерной структуры
В следующем аспекте данное изобретение относится к способу получения информации о структуре и/или информации о связи структура-функция биомолекулы. Предпочтительно, этот способ является способом определения трехмерной структуры (структуры 3-D) биомолекулы. Этот способ предусматривает получение изотопно-меченной биомолекулы в клетках насекомых или млекопитающих, которые выращивают в питательной среде данного изобретения, описанной выше. Затем эту изотопно-меченную биомолекулу, предпочтительно (трансмембранный) белок, извлекают и при необходимости очищают от культивируемых клеток и/или среды, при необходимости дополнительно очищают и подвергают спектроскопическому анализу. Таким спектроскопическим анализом может быть спектроскопический анализ ядерного магнитного резонанса («ЯМР»), инфракрасный спектроскопический анализ с Фурье-преобразованием и/или спектроскопический анализ комбинационного рассеяния (света), для определения информации относительно структуры, связи структура-функция и предпочтительно трехмерной структуры биомолекулы.
В предпочтительном варианте осуществления этой биомолекулой является белок, находящийся в комплексе со второй биомолекулой. Эта вторая биомолекула может быть также получена способом в соответствии с данным изобретением, причем предпочтительно 20-100% атомов водорода во второй биомолекуле являются тотально замененными изотопом 2H. Этой второй биомолекулой может быть белок.
Спектроскопия ядерного магнитного резонанса («ЯМР»), разработанная Felix Bloch и Edward Purcell в 1946 году, представляет важный метод с хорошо известными применениями в химии, естественных науках и молекулярной диагностике, в том числе структурном анализе биомолекул, таких как белки и нуклеиновые кислоты (Wutrich, К., (1986) NMR of proteins and nucleic acids, Wiley, New York). Дальнейшие технические успехи в ЯМР, в том числе, например, импульсные последовательности и методологические развития, увеличили диапазон применения для ЯМР в исследовании взаимодействий белок-лиганд, образуя основу для конструирования лекарственных средств на основе структуры (Marrasi and Opella, 1998, Curr. Opin. Struct. Biol. 8: 640; Watts et al., 1995, Mol. Membr. Biol. 12: 233). Разнообразие применений ЯМР к созданию лекарственных средств и возможный вклад ЯМР в процесс обнаружения лекарственных средств обсуждаются Roberts (2000; DDT, 5: 230). В настоящее время такие стратегии, как SAR by NMR (исследование зависимости активности от структуры с использованием ядерного магнитного резонанса) (Shuker et al., 1996, Science, 274: 1531), «скрининг конфигурации» (Bemis, 1996, J. Med. Chem. 39: 2887) и NMR-Solve (structurally oriented library valency engineering, ЯМР инжиниринг структурно ориентированной библиотеки валентностей), имеют широкое применение в разработке лекарственных средств (Pellecchia, et al., (2002, Nature, 1: 218).
Тотальное мечение изотопами, селективное мечение и сегментное мечение являются известными способами мечения, которые используются в ЯМР. Тотальное изотопное мечение белка позволяет отнесение резонансов посредством последовательного отнесения с помощью многомерных трехрезонансных экспериментов и подтверждает совокупность конформационных ограничений при de novo определении структуры белка (Кау, et al., 1997, Curr. Opin. Struct. Biol. 7: 722).
Селективное мечение индивидуальных аминокислот или определенных типов аминокислот в белках обычно приводит к явному упрощению спектра (Pellecia, et al., 2002, J. Biol. NMR 22: 165; Strauss, et al., 2003, J. Biomol. NMR 26: 367-372). Селективное мечение может выполняться с использованием культуральной среды с полным набором аминокислот и замены выбранных немеченых аминокислот аминокислотами, меченными желаемым стабильным изотопом. При обычных условиях количество гидролизата дрожжей и водорослей может быть уменьшено до 0,1% (масса/объем) без значимого влияния на рост клеток или продукцию белка, и это будет разбавлять включение метки максимально на 5%. Однако замена некоторых индивидуальных аминокислот, в частности Glu и Gin, будет приводить к введению изотопной метки в другие (заменимыми) аминокислоты.
Возможным подходом является «сегментное мечение», в котором дискретные сегменты полипептидной цепи метят тотально, тогда как другие не метят (Xu et al., 1999, Proc. Natl. Acad. Sci. USA 96: 388).
Для ЯМР-анализа мембранных белков наиболее подходящим способом является Solid State NMR (SS NMR ЯМР твердого тела), (Ernst, R.R., et al., 1987, in: Principles of Nuclear Magnetic Resonance in One and Two Dimensions, Oxford, Claredon press; Mehring, M., 1983, in: Principles of High Resolution NMR in Solids, Springer-Verlag Berlin). SS NMR inter alia позволяет выполнять рутинные анализы взаимодействий лиганд-белок для лигандов, когда они связаны с их рецептором-мишенью в мембране (de Groot, 2000, Curr. Opin. Struct. Biol., 10: 593). Этот способ представляет особый интерес для интегральных мембранных белков фармацевтической промышленности, поскольку 60% всех прописываемых лекарственных средств нацелено на мембранные белки, в первую очередь, семейство G-белок связанных рецепторов (Essen, L-O., (2002) Gene Funct. Dis., 2, 39). В способе SS NMR имеются два подхода, которые могут быть применены для анизотропных взаимодействий, которые приводят к ЯМР-спектральному расширению и позволяют извлечь молекулярную структурную информацию. Один из них заключается в использовании спектральной анизотропии в ориентированных образцах в целях придания молекулярной ориентации в статических образцах (Griffin, R.G., (1998) Nat. Struct. Biol., 5, 508, Opella, S.J. et al., (1999) Nat. Struct. Biol., 6, 374), а второй заключается в использовании случайных дисперсий в ЯМР с вращением под магическим углом (MAS) (Andrew, E.R., et al., (1958) Nature 182, 1659), в которых информация об ориентации может быть потеряна, но может быть восстановлена из анализа побочных полос вращения. Кроме того, диполярные связывания могут быть перефокусированы для получения точной информации о расстояниях, а также информации о химическом сдвиге для определения локальных характеристик окружения.
Способы на основе MAS могут успешно применяться для определения межъядерных расстояний или торсионных углов за пределами разрешения, получаемого при рентгенофазовом анализе (Thomson, L.K., (2002) Curr. Opin. Struct. Biol., 12, 661). Необходимыми условиями для определения структуры тотально меченых пептидов и белков являются гетеро- и гомоядерные способы отнесения резонансов. Для обоих аспектов в данной области доступны разнообразные импульсные схемы (Baldus, et al., (1998) Mol Phys., 95, 1197; и Bennet, 1994, In: "NMR Basic Principles and Progress", Springer-Verlag Berlin, 33: 1). Недавние примеры структур белков, определенных при помощи MAS NMR (Castellani, et al., (2002) Nature, 420 (7): 98; и Pauli., et al., (2000, J. Magn. Res., 143: 411), включают домен Src-гомологии 3 (SH3) α-спектрина, содержащий 62 аминокислотных остатка, где почти полное отнесение 13C- и 15N-резонанса для микрокристаллического препарата белка образовало основу для определения ряда расстояний. Были отнесены многие из резонансов пронизывающего мембрану светособирающего комплекса LH2 из пурпурной несерной фотосинтезирующей бактерии Rhodopseudomonas acidophila 10050, (Egorova-Zachernyuk, et al., (2001) J. Biomol. NMR, 19, 243). Частичные отнесения резонансов были сообщены также для убиквитина (Hong, (1999) J. Biomol. NMR, 15: 1; Pellegrini, M., (1999) Biopolymers, 51, 208) и BPTI (McDermott, et al., (2000) J. Biomol. NMR., 16, 209). Количество остатков в этих пептидах также вполне сравнимо с пронизывающими мембраны или связанными с поверхностями пептидами, исследованными недавно в ориентированных липидных бислоях (Opella, S.J., et al., (1999) Nat. Struct. Biol., 6, 374). Таким образом, можно сделать вывод, что методы корреляции на основе MAS могут быть использованы для исследования топологий целых мембранных белков или их частей.
В общем, с использованием дополнительной комбинации улучшенных схем мечения изотопами и дальнейших инструментальных ЯМР-усовершенствований будет расширен размер и сложность изучаемых ЯМР-систем рецептор-лиганд. Кроме того, наряду с успехами в экспрессии мембранных белков ЯМР представляет значительные перспективы для будущего. ЯМР твердого тела позволяет прямое исследование белка в некристаллическом окружении, прямое исследование белка в мембране, либо в виде отдельных молекул (реконструированных в специальных липидах), либо в гетерогенном окружении с другими белками и липидами (в природных мембранах), но, что важно, при ненарушаемых условиях; подробное описание ориентационных аспектов относительно исследуемой мембраны, идентификацию и разделение специфических частей белка, в особенности, при наличии необходимых сайтов связывания с лигандом, исследование белка без ограничения его молекулярной массы (в этом случае стратегии отнесения резонансов должны быть дополнительно разработаны), определение фармакофоров лигандов для рецепторов связанных с мембраной, определяемых из результатов химических сдвигов для связанных лигандов. Однако к настоящему времени только небольшое число мембранных белков или пептидов исследовали с использованием SS NMR. Оба структурных метода исследования, 3-D-кристаллизация и рентгенофазовый анализ, или, альтернативно, ЯМР-спектроскопия требуют, по меньшей мере, около 10 мг высокоочищенных мембранных белков.
Описание графического материала
Фиг.1А. 15N 1D MAS SS NMR-спектр меченного стабильным изотопом препарата мембран, очищенного из клеток Sf9, зарегистрированный при 750 МГц с MAS-частотой 10 кГц при температуре 200 К. Этот спектр был получен в результате 5000 сканирований. Во время получения использовали ТРРМ (Bennet, А.Е., et al., J. Chem. Phys., 1955, 103, 6951) протонную развязку при силе высокочастотного поля 80 кГц. Использовали время смешивания 1H-15N поперечной поляризацией 2 мс. Основной резонанс при 120 м.д. представляет атомы азота пептидного скелета основной цепи. Другие небольшие резонансы представляют резонансы аминокислотных остатков боковых цепей этого белка. В качестве стандарта использовали резонанс азота Gly при 34,3 м.д.
Фиг.1B. FT-IR - разностный спектр (Спектр инфракрасной спектроскопии с Фурье-преобразованием), демонстрирующий включение 15N в клеточные мембраны Sf9. Комбинация положительный/отрицательный пик при 1580/1535 (на наклонном крае пика 1620) представляет смещение колебания Амида II в результате мечения пептидного скелета основной цепи.
Фиг.2. Быстрый анализ состава биомассы с использованием трансмиссионной инфракрасной спектроскопии с Фурье-преобразованием (FT-IR). Колебательные полосы белков (1700-1500 см-1) и углеводов (1200-1000 см-1) легко детектируются при определении воспроизводимости состава (меченой) биомассы. Me - Methylobacterium extorquens; Cc - Cyanidium caldarium; Cv - Chlorella vulgaris; Sp - Spirulina platensis; Gs - Galdieria sulphuraria; So - Scenedesmus obliquus.
Фиг.3. Мониторинг мечения стабильным изотопом 15N с использованием FT-IR на основании смещения колебания Амида II (1542 см-1). На этом чертеже показана растянутая часть инфракрасного спектра биомассы дрожжей Hansenula polymorpha, культивируемой на немеченой среде (А) и на среде, меченной стабильным изотопом 15N (В). Эти спектры получали после коррекции базовой линии (фон) и масштабирования с использованием самого сильного пика (1658 см-1) в качестве стандарта. Включение метки 15N составляло 99%.
Примеры
Пример 1. Получение меченной стабильным изотопом биомассы, гидролизата и экстрактов из нее (дрожжи в качестве источника)
1.1. Экстракты дрожжей, содержащие источник аминокислот и сахаров, полученные из Saccharomyces cerevisiae
Saccharomyces cerevisiae (ATCC 13057) выращивали в среде, содержащей (г/л): тотально меченную 13С глюкозу - 15 г/л, меченый KH2PO4 - 2 г, 15NH4Cl - 1,5 г, MgSO4×7H2O - 0,5 г, CaCl2×6H2O - 0,25 г, FeSO4×6H2O - 0,036 г, ZnSO4×7H2O - 0,001 г, MnCl2×4H2O - 0,001 г, CoCl2×6H2O - 0,001 г. Витамины добавляли в тех же самых концентрациях, что описаны Heine, W., et al., в Stable Isotopes in Pediatric Nutritional and Metabolic Research (1990) Eds., Т.Е. Chapman, R. Berger, D.J. Reijngould and A. Okken, Intercept Ltd., p.84. Дрожжи культивировали при взбалтывании культуры в конических колбах на 10 л, содержащих 2,5 л стерилизованной при 120°С в течение 20 минут среды при рН 4,5, доведенном NaOH. В эту колбу переносили 100 мл посевной культуры. Посевную культуру получали при взбалтывании в колбе при 2 7°С в течение 18 часов. Дрожжи выращивали с перемешиванием при 800 об/мин при 27°С в течение 29 часов. Клетки дрожжей собирали центрифугированием при 5000 g в течение 10 минут при 4°С и промывали дважды водой. К промытым клеткам добавляли воду с получением 500 мл суспензии дрожжей, приблизительно 100 мг/мл в расчете на сухую массу.
Половину объема этой суспензии лиофилизировали и использовали для получения гидролизата дрожжей. Гидролизат дрожжей получали, как описано в примере 5.1.
Другую половину этой суспензии использовали для автолиза. Ее продавливали через изогнутый шприц при 65°С (внутренний диаметр 2-3 мм) с временем переноса менее 20 секунд, и затем собирали в колбу, охлажденную на льду. Затем эту суспензию выдерживали в масляном термостате при 45°С в течение 4 часов при доведении рН до 8,0, добавлением по каплям 20% NaOH. После автолиза рН доводили до 6,5 2N HCl и автолизат нагревали до 80°С. Затем автолизат центрифугировали и осадок промывали дважды водой. Объединенный супернатант 400 мл, содержащий автолизат, фильтровали через фильтр с пределом отсечения молекулярной массы 10000 и лиофилизовали с получением 15 г дрожжевого экстракта.
Химический анализ этого дрожжевого экстракта выполняли, как описано в Т. Hernawan, G. Fleet., J. Ind. Microbiology (1995) 14, 440. Обычно, в расчете на исходную сухую массу клеток этот растворимый автолизат Saccharomyces cerevisiae состоял из углеводов (4-8%), белка (12-15%), органических кислот (3-5%), свободных аминокислот (10-12%), олигопептидов (30-34%), продуктов нуклеиновых кислот (3-5%), липидов (2-3%) и других физиологически активных соединений.
1.2. Альтернативный протокол для автолиза дрожжей
К суспензии дрожжей, полученной, как описано в примере 1.1, добавляли 2 объемных процента этанола и автолиз проводили при 50°С в течение 12 часов. Затем дрожжевой автолизат обрабатывали с использованием ультразвукового дезинтегратора в течение 6 минут с последующим центрифугированием при 5000 g в течение 10 минут. Остаток промывали дважды дистиллированной водой и центрифугировали в течение 15 минут. Объединенные супернатанты фильтровали через мембранный фильтр с пределом молекулярной массы 10000 и лиофилизировали. В результате получали смеси аминокислот, пептидов, углеводов, компонентов нуклеиновых кислот, органических кислот и других компонентов. Эта процедура позволяет увеличить содержание свободных аминокислот до 80%.
1.3. Дрожжевой экстракт Pichia pastoris
Pichia pastorls NRRL-Y-11430 выращивали на меченном 13С глицерине и глюкозе в качестве углеродного энергетического субстрата и (15NH4)2SO4 в качестве источника азота, как описано в M.J. Wood and E.A. Komives, (1999) J. Biomol. NMR 13, 149, или на 13С-метаноле в качестве источника углерода и энергии. Дрожжевые экстракты (кислотный гидролизат и автолизаты) получали с использованием способов, описанных в примерах 1.1 и 1.2 и 5.1.
1.4. Дрожжевой экстракт Hansenula polymorpha
Hansenula polymorpha CBS 4732 (синонимы АТСС 34438, NRRL-Y-5445) выращивали в непрерывной культуре с ограниченным содержанием метанола и регулируемой подачей на среде, сходной со средой, описанной в Y. Laroche, et al., (1994) Biotechnology 12: 1119, Th. Egli, et al.. Arch. Microbiol. (1982) 131, 8, и Van Dijken, et al., (1976) Arch. Microbiol. Ill: 137, где меченную 15N соль использовали в качестве источника азота и тотально меченную 13С глюкозу и меченный 13С метанол использовали в качестве источника углерода и энергии. Культивирование выполняли при 35°С и при рН 5,0. Дрожжевой экстракт (кислотный гидролизат и автолизаты) получали с использованием тех же способов, что описаны в примерах 1.1 и 1.2 и 5.1, соответственно.
1.5. Крупномасштабное получение биомассы дрожжей
3-литровый ферментер (BioFlo3000, New Brunswick Scientific) использовали для культуры дрожжей в целях полного контроля над системой (температура, рН, подача кислорода, азота, воздуха, растворенный кислород). Оптимальный рост дрожжей достигался с использованием аэрации 25% кислородом и 75% азотом с уровнем растворенного кислорода 35%. Дрожжи росли при 30°С с использованием 2 литров модифицированной среды Heine или модифицированной среды Invitrogen, в которой глицерин был заменен глюкозой. Скорость перемешивания составляла 500 об/мин. В обоих случаях все источники углерода и азота заменяли изотопно-меченными аналогами. Глюкозу добавляли периодически, или использовали культуру с подпиткой.
Пример 2. Получение меченной стабильным изотопом биомассы, гидролизата и экстрактов из нее (водоросли в качестве источника)
2.1. Получение меченной стабильным изотопом биомассы Cyanidiurn
Cyanidium caldarium (SAG 16.91) и Galdieria sulphuraria (SAG 17.91) выращивали автотрофно при 25°С в колбах на 5 л с магнитными перемешивающими мешалками на водяной бане с постоянной температурой при постоянном рН 2 и собирали во время экспоненциальной фазы роста. Питательная среда для 13C, 15N меченных культур Cyanidium caldarium и Galdieria sulphuraria содержала на литр: 1,5 г (15NH4)2SO3, 0,3 г MgSO4×7H2O, 0,3 г KH2PO4, 0,02 г CaCl2×2H2O, 1,5 мл раствора Fe-ЭДТА (раствор Fe-ЭДТА готовили добавлением 0,690 г FeSO4 и 0,930 г ЭДТА к объему до 100 мл дистиллированной воды и кипячением раствора), и 2 мл раствора микроэлементов, который готовили отдельно (смотрите ниже). рН этой среды доводили до величины рН 1,8 1N H2SO4. Раствор микроэлементов содержит на литр: 2,86 г Н3ВО3, 1,82 г MnCl2, 0,22 г ZnSO4×7H2O, 0,130 г Na2MoO4×2H2O, 80 мг CuSO4×5H2O, 40 мг NaVO3×4H2O и 40 мг CoCl2×6H2O.
Источником непрерывного света 8000 люкс служили флуоресцентные лампы на 15 Ватт, и использовалась концентрация 13СО2 до 5%. Рост суспензии водорослей анализировали измерениями оптической плотности при 550 нм. Когда рост достигал экспоненциальной фазы (обычно 5 дней) (0,64 OD), клетки собирали с использованием центрифуги Бекмана при 5000 об/мин в течение 10 минут и осадки промывали дистиллированной водой для удаления следов растворимых питательных веществ. Полученную пастообразную массу замораживали и лиофилизировали. Лиофилизированные клетки использовали для приготовления липидных экстрактов и гидролизата.
2.3. Получение меченной стабильным изотопом биомассы зеленой водоросли (Scenedesmus obliquus)
13С-, 15N- меченную биомассу водоросли Scenedesmus obliquus получали, как описано Patzelt, H., et а1., 1999 Phytochemistry 50: 215-217. Штамм Scenedesmus obliquus 276/ЗС (полученный из Коллекции культур Водорослей и Простейших (ССАР), Institute of Freshwater ecology, Ambleside, UK) выращивали автотрофно при 30°C при рН 6,5 и собирали во время экспоненциальной фазы роста. Среда для меченных 13С и 15N культур штамма Scenedesmus obliquus содержит на литр: 0,18 г Na2HPO4×2H2O, 0,48 г NaH2PO4, 0,46 г NaCl, 0,25 г MgSO4×7H2O, 6 мг CaCl2×2H2O, 6,5 г FeSO4×7H2O, 8 мг ЭДТА, 2,9 мг Н3ВО3, 1,82 г MnCl2×4H2O, 0,22 г ZnSO4×7H2O, 0,25 мг Na2MoO4×2H2O, 0,08 мг CuSO4×5H2O, 0,08 мг CoCl2×6Н2О и 0,81 г K15NO3. рН среды составлял 6,5. Среду готовили в дистиллированной воде, которую интенсивно дегазировали для удаления растворенного CO2. 13CO2 подавали в эту питательную среду в виде NaH13CO3 в карбонатном буфере.
Для предварительного культивирования использовали два резервуара. Нижний резервуар содержал 2М карбонатный (Na2 13CO3/NaHl3CO3)-буфер, рН 7,5 (100 мл), верхняя колба содержала инокулят. После инокуляции эти колбы герметично закрывали и выращивали при перемешивании 60 об/мин при 30°С при постоянном освещении в течение 10 дней. Источником непрерывного света 2000 люкс являлись флуоресцентные лампы на 15 Ватт (холодные белые). Для приготовления буфера 1 л 1,2 М NaOH барботировали с помощью 16 л 13CO2, до достижения рН 9,3, добавляли 50 мг фенолфталеина для мониторинга рН.
Для препаративной ферментации использовали замкнутую систему, состоящую из 10 колб Фернбаха на 2,5 л (1 л среды в колбе с 5 мл инокулята), карбонатного буфера и насоса для газа. Воздух циркулирует по кругу таким образом, что клеточные культуры снабжаются 13CO2-обогащенным воздухом. Водоросли выращивали в течение 10-14 дней. Постоянный поток 5% 13CO2 в сжатом воздухе продували через колбы Фернбаха. Рост суспензии водоросли контролировали по измерениям оптической плотности при 685 нм. Когда рост достигал экспоненциальной фазы (17 дней) (1,0 OD), клетки собирали при температуре 4°С с использованием центрифуги Бекмана при 5000 об/мин в течение 10 минут и осадки промывали дистиллированной водой для удаления следов растворимых питательных веществ. Культуральную жидкость использовали для дополнительного цикла роста (после фильтрования). Отделенную от культуральной жидкости пасту замораживали и лиофилизировали. Процесс лиофилизации является важным, так как присутствие воды может вызвать разрушение хлорофиллов хлорофиллазой. Выход биомассы составил 2 г лиофилизированных клеток на 1 литр культуры. Лиофилизированные клетки использовали для получения липидных экстрактов и гидролизата.
Масштабирование роста водорослей до 20 л биореактора давало сходные результаты.
Пример 3. Получение меченной стабильным изотопом биомассы, гидролизата и экстрактов из нее (метилотрофная бактерия в качестве источника)
3.1. Получение биомассы из метилотрофной бактерии
Облигатный метилотроф Methylobacillus flagellatus (ATCC 51484, VKM B-1610, DSM 6875) выращивали при 30°С при рН 6,8 на 13С-меченном метаноле в качестве источника углерода и 15NH4Cl в качестве источника азота на АТСС-среде 784 AMS без агара. Концентрация 13С-меченного метанола составляла 1%. Клетки выращивали в ферментере на 1 л и собирали после 3 дней выращивания. Выход биомассы составлял 53% в расчете на 13С-метанол. Биомассу лиофилизировали. Выход белка составлял 75% в расчете на сухую биомассу. Лиофилизированные клетки использовали для получения липидных экстрактов и гидролизата.
Пример 4. Экстракия меченных стабильным изотопом липидов
4.1. Экстракция биомассы водорослей (процедура 1)
Лиофилизированные клетки (10 г) Cyanidium caldarium гомогенизировали в дихлорметане посредством обработки ультразвуком в течение 30 минут на льду (охлаждение достигалось льдом с солью), с чередующейся 50% мощностью в течение 30 секунд и переносили в аппарат Сокслета для экстракции. Экстракцию проводили в течение 30 часов и затем продолжали в течение еще 24 часов с 90% водным метанолом, пока экстракт не становился бесцветным. Объединенный липидный экстракт упаривали на роторном испарителе в атмосфере азота и разделяли на нелипидную и липидную фракции хроматографией на Сефадексе, Wuthier, R.E. (1966) J. Lipid. Res. 7: 557-561. Липидную фракцию упаривали и растворяли в смеси н-гексан-диэтиловый эфир с последующей хроматографией на колонке кремниевой кислоты, как описано Kleinschmidt, M.G. and McMahon, V.A. (1970, Plant Physiol. 46: 286-298). Типичными липидами были воски, углеводороды, сложные эфиры стерола, триглицериды, стеролы, свободные жирные кислоты, моноглицериды, диглицериды, гликолипиды, сульфолипиды и фосфолипиды. В целом из 1 г биомассы может быть выделено 150 мг липидов. Основными липидами Cyanidium caldarium являются моно- и дигалактозилглицерид и сульфолипид, лецитин, фосфатидилглицерин, фосфатидилинозит, фосфатидилэтаноламин.
Альтернативно, очистка жирных кислот и стеролов могла быть выполнена из объединенных экстрактов из аппарата Сокслета (общее количество 160 мг липидного экстракта), полученных из 1 г сухой биомассы Cyanidium caldarium, как описано Ikan, R., Seckbach, J. (1972) Phytochemistry, 11: 1077-1082. В результате получали 32 мг свободных жирных кислот и 39 мг жирных кислот омылением нейтральной фракции. Стерины выделяли из не подвергнутой омылению фракции (Ikah, supra) и их количество составляло 2 мг.
Эту процедуру использовали для экстракции липидов из сине-зеленых, красных водорослей и водорослей семейства Cyanidiophyceae.
4.2. Экстракция липидов из биомассы водорослей (процедура 2)
К лиофилизированной 13С-, 15N-меченной биомассе Scenedesmus obliquus в количестве 10 грамм добавляли 100 мл гексана и обрабатывали ультразвуком в ультразвуковом дезинтеграторе при -4°С (охлаждение достигалось льдом с солью) в течение, по меньшей мере, 60 минут. Растворитель удаляли фильтрованием, и остаток биомассы экстрагировали 100 мл ацетона с помощью процедур, описанных выше для экстракции гексаном. Наконец, остаток биомассы экстрагировали 100 мл смеси дихлорметан/метанол (2:1 об./об.). Фильтраты отдельно упаривали досуха в атмосфере азота и хранили при -80°С. Ацетоновый экстракт содержит источник тотально меченых пигментов, которые могут быть дополнительно выделены в чистом виде и использованы, например, для исследований зависимости функции от структуры пигмент-белкового взаимодействия в фотосинтезе. В случае сине-зеленых, красных водорослей и водорослей Cyanidiophyceae использовали дополнительную стадию, предусматривающую экстракцию 90% водным МеОН. Эти экстракты обрабатывали, как описано выше. Затем обезжиренную биомассу (8 г сухого веса) использовали для получения гидролизата.
Эту процедуру использовали для экстракции липидов из всех источников.
4.3. Экстракция биомассы красных водорослей (Porphyridium)
Porphyridium purpureum (= Porphyridium cruentum) (ATCC 50161) выращивали на среде ATCC 1495 ASW, но с K15NO3 и NaH13CO3 вместо KNO3 и NaHCO3 и с 54 г NaCl на литр среды при 25°С и рН 7,6. Культуры водорослей освещали непрерывно флуоресцентными лампами при интенсивности света 25 В/м2. Липиды экстрагировали в соответствии с процедурой 1 (пример 4.2) и определяли в соответствии со способом, описанным в Ohta, et al., 1993, Botanica marina, 36: 1043-107. С использованием таких условий для выращивания получали высокую концентрацию С20:4 (n-6) и C20:5 (n-3), а именно, 8,2 мг сухой массы и 14,3 мг сухой массы на 1 г сухой биомассы.
4.4. Экстракция биомассы метилотрофов (Methylobacillus)
Биомассу Methylobacillus flagellatus (10 г) экстрагировали, как описано в 4.3. Обезжиренную биомассу (8,5 г) использовали для получения гидролизата, как описано в примерах 5.1 и 5.2.
4.5. Переэтерификация липидов для анализа
Экстрагированные липиды из Galdieria sulphuraria переэтерифицировали для образования метиловых эфиров жирных кислот, позволяющих идентифицировать и определять количественно жирные кислоты с использованием способа газовой хроматографии, как описано Lewis et al., (2000, J. Microbiol. Meth. 43: 107-116). Лиофилизированные клетки взвешивали (5 г), добавляли к ним свежий раствор смеси для реакции переэтерификации (10/1/1 об./об./об.), как описано. Клетки суспендировали в этом растворе смешиванием на вортексе и немедленно помещали при 90°С для переэтерификации и выдерживали в течение 60 минут. Затем пробирки для переэтерификации охлаждали, в каждую пробирку добавляли воду (1 мл) и экстрагировали метиловые эфиры жирных кислот (гексан/хлороформ, 4/1 об./об. 3×4 мл). Химическую идентичность экстрактов в органическом растворителе подвергали мониторингу при помощи ТСХ (например, силикагель, гексан/EtOAc, 5:1) и ВЭЖХ (С18, градиент Ме2СО-H2O (1:4) - Ме2СО). Состав изотопов определяли с использованием масс-спектрометрии (EI-MS) (70 эВ) и/или инфракрасной спектроскопии.
Пример 5. Гидролиз биомассы
5.1. Кислотный гидролиз изотопно-меченной биомассы
К количеству биомассы, свободной от липидов и пигментов (пример 4.2), полученному таким образом и помещенному в колбу на 240 мл, добавляли 15 мл 6 н HCl и 0,6 мл тиогликолевой кислоты в качестве восстанавливающего агента (для предотвращения деструкции гистидина и триптофана). Гидролиз выполняли в атмосфере аргона при 110°С (на масляной бане) в течение 24 часов. 15 мл дистиллированной воды добавляли к гидролизату, который охлаждали на льду с последующим фильтрованием через стеклянный фильтр Р3. После этого гидролизат разделяли на две части и лиофилизировали в двух круглодонных колбах. Затем к каждому из гидролизатов добавляли количество 10 мл дистиллированной воды, один из них нейтрализовали 30% NaOH, a другой 30% 15NH4OH. Половину каждой из этих смесей пропускали через активированный уголь с грубыми порами 20-100 Å (колонка высотой 2 см), с использованием стеклянного фильтра Р3. В результате получали четыре гидролизата, которые затем лиофилизировали. Лиофилизация приводит к получению белого твердого порошка (смеси аминокислоты/сахара) для гидролизатов, пропущенных через уголь, и к получению порошка цвета лесного ореха для гидролизатов, не обработанных углем. Полученные гидролизаты (4 различных гидролизата) дополнительно измельчали с получением гомогенного порошка.
Альтернативно, гидролизаты получали без «стадии угля» и, например, в случае со Scenedesmus obliquus (а также Cyanidium caldarium) количество 0,6 г гидролизата, нейтрализованного 15NH4OH, и 0,7 г гидролизата, нейтрализованного NaOH, получали из 1 г обезжиренной биомассы. В большинстве случаев использовали масштабированную процедуру гидролиза для гидролиза 20 г обезжиренной биомассы. Гидролизаты, нейтрализованные 15NH4OH, использовали предпочтительно для приготовления питательной среды для клеток насекомых и млекопитающих.
Аминокислотный состав (определяемый с использованием Fmoc-дериватизации на основе обращенно-фазовой ВЭЖХ), например, гидролизата Cyanidium caldanum, нейтрализованного 15NH4OH, представлял собой: Ala - 8,8%, Arg - 5,1%, Asp - 5,2%, Glu - 9,5%, Gly - 7,3%, His - 0,8%, Ile - 6,7%, Leu - 6,5%, Lys - 5,6%, Met - 3,2%, Phe - 6,3%, Pro - 7,6%, Cys - 0,8%, Ser - 11%, Trp - 0,8%, Thr - 6,1%, Tyr - 1,2%, Val - 7,5%.
Гидролизаты использовали как для прямого приготовления питательных сред для клеток насекомых или млекопитающих, так и для получения очищенных аминокислот (с помощью ионообменной хроматографии (Crespi and Katz, 1972, Methods Enzymology 26, 627) или обращенно-фазовой хроматографии (Egorova, et al., T.A., (1995) J. Chromat., Biomedical Application 665: 53-62).
Альтернативно, гидролизат мог быть подвергнут ультрафильтрации через кассетную мембрану с пределом отсечения 8 К и фильтрованию с помощью стерильного фильтра Durapore 0,2 микрон. Стерилизованный гидролизат хранили в течение 6 месяцев при 4°С.
Усовершенствование способа кислотного гидролиза нацелено на получение гидролизатов, имеющих более низкое содержание солей. В этой усовершенствованной процедуре стадию лиофилизации выполняли дважды: сначала, немедленно после гидролиза (для удаления также большей части HCl) и, затем, после нейтрализации с использованием или гидроксида натрия, или меченного 15N гидроксида аммония.
5.2. Ферментативный гидролиз меченной стабильным изотопом биомассы
Обезжиренную биомассу Methylobacillus flagellakus ферментативно гидролизовали способом, подобным описанному Troxler, et al., (1975) 14(2): 268-274, иммобилизованную проназу (Fluka, Switzerland) получали в соответствии с инструкциями Pierce и использовали для ферментативного гидролиза. Продукт гидролиза лиофилизировали и использовали в качестве компонента питательной среды для клеток млекопитающих и насекомых.
5.3. Автолиз меченной стабильным изотопом биомассы водорослей
Процедуру для получения аутолитических экстрактов дрожжей, описанную в примерах 1.1 и 1.2, применяли для получения автолизатов из биомассы водорослей. Таким образом автолизовали меченную стабильным изотопом биомассу из различных водорослей, например, Cyanidium caldarium и Galdieria sulphurarla. Выход автолизатов составлял 35-45% в расчете на сухой вес биомассы. Оставшийся материал биомассы подвергали кислотному гидролизу (описанному в примере 5.1) для получения гидролизатов водорослей и для эффективного использования меченой биомассы.
Пример 6. Состав меченных стабильным изотопом питательных сред
6.1.1. Состав I меченной стабильным изотопом питательной среды для клеток насекомых
Состав среды в отношении микроэлементов, солей и витаминов был основан на композиции IPL-41 (Weiss, et al., In Vitro, 17: 495-502), за исключением того, что использовали NaH13CO3 и молибдат натрия. Другие компоненты этой среды добавляли в концентрации, приведенной в таблице ниже:
Липидный экстракт добавляли в среду в виде этанольного раствора, приготовленного в концентрации 50 мг липидного экстракта (пример) на 1 мл этанола. Этанольный липидный раствор добавляли в концентрации 5 мл/л культуральной среды.
Гидролизаты анализировали аминокислотным анализом (ОФ-ВЭЖХ в виде Fmoc-производных) и недостающие аминокислоты (в соответствии с IPL-композицией), такие как 13С-, 15N-меченный цистеин, добавляли отдельно в среду в концентрации 80 мг/л.
рН доводили до 6,2, осмолярность доводили до 340 мОсмоль на кг (NaCl). Полную среду стерилизовали пропусканием через фильтр 0,22 микрон и хранили при 4°С.
При помощи 5-10 пассажей линию клеток Sf9 адаптировали к росту в меченной изотопами среде в соответствии с (Bosman, et al., in Method Mol. Biology V. 228, Ed. Selinsky, Humana Press Inc.). Клетки насекомых культивировали при температуре 27°С, и добавление CO2 не требовалось. Время удвоения адаптированных клеток было 26-28 часов, что сравнимо с другими коммерчески доступными бессывороточными средами (S3777, Sigma, X-Cell 420 JRH Biosciences, HyQ SFX-Insect, Hyclone). Максимальная достигаемая плотность клеток составляла 4-6×106 клеток/мл.
6.1.2. Состав II меченной стабильным изотопом питательной среды для клеток насекомых
Состав среды в отношении микроэлементов, солей и витаминов был основан на композиции IPL-41 (Weiss, et al., In Vitro, 17: 495-502), за исключением того, что использовали NaH13CO3 и молибдат натрия. Другие компоненты этой среды добавляли в концентрации, приведенной в таблице ниже:
Липидный экстракт добавляли в среду в виде этанольного раствора, который готовили в концентрации 50 мг липидного экстракта (пример) на 1 мл этанола. Этанольный липидный раствор добавляли в концентрации 5 мл/л питательной среды. Следует обратить внимание на то, что липидный экстракт в этом примере является необязательным и используемая концентрация увеличивала рост клеток не более чем до 10%. Без липидного экстракта клетки также росли и продуцировали белок. Гидролизаты анализировали аминокислотным анализом (ОФ-ВЭЖХ в виде Fmoc-производных) и недостающие аминокислоты (в соответствии с IPL-композицией), такие как 13С-, 15N-меченные цистеин и глутамин, добавляли отдельно в среду в концентрации 80 мг/л. рН доводили до 6,2, осмолярность доводили до 340 мОсмоль на кг (NaCl). Полную среду стерилизовали пропусканием через фильтр 0,22 микрон и хранили при 4°С. При помощи 5-10 пассажей линию клеток Sf9 адаптировали к росту в меченной изотопами среде в соответствии с (Bosman, et al., in Method Mol. Biology V. 228, Ed. Selinsky, Humana Press Inc.). Клетки насекомых культивировали при температуре 27°С, и добавление CO2 не требовалось. Время удвоения адаптированных клеток было 26-28 часов, что сравнимо с другими коммерчески доступными бессывороточными средами (S3777, Sigma, X-Cell 420 JRH Biosciences, HyQ SFX-Insect, Hyclone). Максимальная достигаемая плотность клеток составляла 4-6×106 клеток/мл.
6.1.3. Влияние концентрации различных компонентов на среды для насекомых
В целом исследовали биомассы из более чем 10 различных микроорганизмов; более чем 100 различных гидролизатов и автолизатов готовили из этих биомасс и тестировали более чем 10 различных липидных экстрактов, приходящихся на одну биомассу микроорганизма. На основании этих тестов были сделаны следующие общие наблюдения.
Автолизаты дрожжей обычно включают в среды при концентрации в диапазоне 0,1-1,6% (масса/объем). Автолизаты на основе биомассы Pichia pastoris, выращенной на коммерческих средах Invitrogen, в которых глицерин заменяют глюкозой, показали наилучшую воспроизводимость и эффективность в отношении поддержания роста клеток насекомых с оптимальной концентрацией для этого автолизата в диапазоне 0,1-0,3%, причем концентрация 0,2% была оптимальной (масса/объем). Автолизаты на основе биомассы Hansenula polymorpha, выращенные на модифицированной среде Heine (без молочной кислоты), показали наилучшую воспроизводимость, эффективность в отношении поддержания роста клеток насекомых с оптимальной концентрацией для этого автолизата в диапазоне 0,4%-0,8% (масса/объем). Дрожжевые автолизаты обычно эффективны при добавлении небольших количеств гидролизатов (для дополнительных аминокислот) или даже только небольшого количества некоторых аминокислот. При использовании разработанных автором питательных сред на основе IPL-41 и клеточных линий в дополнение к дрожжевому автолизату требовалось добавление глутамина.
Автолизаты водорослей обычно включают при концентрации в диапазоне 0,1-0,8% (масса/объем). Оптимальная концентрация может зависеть от водоросли-источника, например, для Galdieria sulphuraria наиболее эффективная концентрация была 0,1%, но для Cyanidium caldarium она была 0,4% (масса/объем). Надо отметить, что автолизаты водорослей также обычно являются эффективными при добавлении небольших количеств гидролизатов (для дополнительных аминокислот) или даже только небольшого количества выбранных аминокислот. В испытаниях авторов изобретения только глутамин требовался в дополнение к автолизату водорослей. В противоположность этому, гидролизаты водорослей не были эффективными в виде единственного компонента (исследовали концентрацию в диапазоне 0,1-0,8% (масса/объем) сред, и при этом требовалось добавление более чем несколько индивидуальных аминокислот.
Бактериальные гидролизаты включают обычно в среды в концентрациях в диапазоне 0,1-0,4% (масса/объем). Бактериальные автолизаты включают обычно в диапазоне 0,1-0,6% (масса/объем).
Наиболее эффективной в поддержании роста насекомых и продуцирования белка является комбинация автолизатов на основе красных водорослей, предпочтительно Cyanidium caldarium или Galdieria sulphuraria, и дрожжевого автолизата, предпочтительно Hansenula polymorpha (смотрите пример 6.1.2). Эффективность, например, среды на основе автолизата, полученного из биомассы Galdieria sulphuraria, могла быть увеличена более чем на 70%, при добавлении к этой среде 0,8% автолизата Hansenula polymorpha, и/или эффективность среды на основе автолизатов, полученных из биомассы Hansenula polymorpha, могла быть увеличена более чем на 60%, при добавлении автолизата Galdieria sulphur aria.
6.2.1. Состав меченной стабильным изотопом питательной среды для клеток млекопитающих
Состав питательной среды в отношении микроэлементов, солей и витаминов был основан на композиции DMEM (смотрите, например, JRH Biosciences, Cell culture and services), за исключением того, что содержащие углерод и азот соли были заменены солями, меченными стабильными изотопами. Другие компоненты этой среды добавляли в концентрации, приведенной в таблице ниже:
Липидный экстракт добавляли в среду в виде этанольного раствора, приготовленного в концентрации 150 мг липидного экстракта (пример) на 1 мл этанола. Этанольный липидный раствор добавляли в концентрации 1 мл/л питательной среды. Гидролизаты анализировали с помощью аминокислотного анализа (ОФ-ВЭЖХ в виде Fmoc-производных) и недостающие аминокислоты (в соответствии с DMEM-композицией), такие как 13С-, 15N-меченный цистеин и 13С-, 15N-меченный глутамин, добавляли отдельно в среду в концентрации 70 мг/л и 1000 мг/л, соответственно. рН доводили до 7,2, осмолярность доводили до 310 мОсмоль на кг (NaCl). Полную среду стерилизовали пропусканием через фильтр 0,22 микрон и хранили при 4°С. При помощи 5-10 пассажей линию клеток СНО адаптировали к росту в меченной изотопами среде в соответствии с (Bosman, et al., in Method Mol. Biology V. 228, Ed. Selinsky, Humana Press Inc.). Клетки СНО культивировали при температуре 37°С, и добавление 13CO2 не требовалось. Время удвоения адаптированных клеток было 28-30 часов, что сравнимо с другими коммерчески доступными бессывороточными средами (С4726, Sigma, Ex-Cell 302 JRH, CD СНО AGT, Invitrogen). Максимальная достигаемая плотность клеток составляла 4-6×106 клеток/мл.
6.2.2. Состав меченной стабильным изотопом питательной среды, подходящей для клеток НЕК 293
Состав питательной среды в отношении микроэлементов, солей и витаминов был основан на композиции DMEM (смотрите, например, JRH Biosciences, Cell culture and services), за исключением того, что содержащие углерод и азот соли были заменены солями, меченными стабильными изотопами. Другие компоненты этой среды добавляли в концентрации, приведенной в таблице ниже:
Липидный экстракт добавляли в среду в виде этанольного раствора, который готовили в концентрации 150 мг липидного экстракта (пример) на 1 мл этанола. Этанольный липидный раствор добавляли в концентрации 1 мл/л культуральной среды. Гидролизаты анализировали аминокислотным анализом (ОФ-ВЭЖХ в виде Fmoc-производных) и недостающие аминокислоты (в соответствии с DMEM-композицией), такие как 13С-, 15N-меченный цистеин и 13С-, 15N-меченный глутамин, добавляли отдельно в среду в концентрации 70 мг/л и 1000 мг/л, соответственно. рН доводили до 7,2, осмолярность доводили до 310 мОсмоль на кг (NaCl). Питательную среду стерилизовали пропусканием через фильтр 0,22 микрон и хранили при 4°С. При помощи 5-10 пассажей линию клеток НЕК-293 адаптировали к росту в меченной изотопами среде в соответствии с Bosman, et al., in Method Mol. Biology V. 228, Ed. Selinsky, Humana Press Inc. Клетки НЕК-293 культивировали при температуре 37°С, и добавление 13CO2 не требовалось. Время удвоения адаптированных клеток было 26-30 часов, что сравнимо с другими коммерчески доступными бессывороточными средами (С4726, Sigma, Ex-Cell 302 JRH, CD CHO AGT, Invitrogen). Максимальная достигаемая плотность клеток была 3-5×106 клеток/мл.
6.2.3. Состав меченной стабильным изотопом питательной среды, подходящей для клеток CHO
Состав питательной среды в отношении микроэлементов, солей и витаминов был основан на композиции DMEM (смотрите, например, JRH Biosciences, Cell culture and services), за исключением того, что содержащие углерод и азот соли были заменены солями, мечеными стабильными изотопами. Другие компоненты этой среды добавляли в концентрации, приведенной в таблице ниже:
Липидный экстракт добавляли в среду в виде этанольного раствора, приготовленного в концентрации 150 мг липидного экстракта (пример) на 1 мл этанола. Этанольный липидный раствор добавляли в концентрации 1 мл/л питательной среды. Гидролизаты анализировали аминокислотным анализом (ОФ-ВЭЖХ в виде Fmoc-производных) и недостающие аминокислоты (в соответствии с DMEM-композицией), такие как 15N-меченный цистеин и 15N-меченный глутамин, добавляли отдельно в среду в концентрации 70 мг/л и 1000 мг/л, соответственно. рН доводили до 7,2, осмолярность доводили до 310 мОсмоль на кг (NaCl). Питательную среду стерилизовали пропусканием через фильтр 0,22 микрон и хранили при 4°С. При помощи 5-10 пассажей линию клеток СНО адаптировали к росту в меченной изотопами среде в соответствии с Bosman, et al., in Method Mol. Biology V. 228, Ed. Selinsky, Humana Press Inc. Клетки СНО культивировали при температуре 37°С, и они действительно требовали добавления CO2. Время удвоения адаптированных клеток было 26-30 часов, что сравнимо с другими коммерчески доступными бессывороточными средами (С4726, Sigma, Ex-Cell 302 JRH, CD СНО AGT, Invitrogen). Максимальная достигаемая плотность клеток была 4-6×106 клеток/мл.
Пример 7. Получение изотопно меченного рекомбинантного белка
7.1.1. Изотопно-меченный рекомбинантный аквапорин-2, продуцируемый в клетках насекомых
Клетки Sf9 выращивали во вращающихся 500-мл колбах, содержащих 100 мл питательной среды. Культуральную среду готовят, как показано в примере 6.1.2. Клетки Sf9 получали из АТСС (CRL-1711) и адаптировали к приготовленной среде ступенчатой адаптацией, как описано Bowman et al., supra. Клетки Sf9 инокулировали при плотности 3×105 клеток/мл и выращивали до плотности 2×106 клеток/мл. Затем клетки инфицировали рекомбинантным бакуловирусом, кодирующим аквапорин-2 (AQP2), с введенным на С-конец полигистидином (НТ), сконструированным, как описано в Werten et al., (2001, FEBS Lett. 504: 200). Бакуловирус добавляли при MOI 1,0. Ежедневно пробу 10 мкл из культуры брали для мониторинга экспрессии HT-AQP2 при помощи дот-блоттинга. Уровень экспрессии был максимальным при приблизительно 3 днях dpi (дни после инфекции). Клетки собирали 10-минутным центрифугированием при 5000 g при 4°С. Рекомбинантный HT-AQP2 очищали с использованием мочевины с последующей аффинной хроматографией с иммобилизованным металлом через гранулы Ni-NTA (Qiagen), как описано в Werten et al., (2001, supra). Очищенный HT-AQP2 элюировали 100 мМ L-гистидином и восстанавливали в протеолипосомы смешиванием с липидами Е. coli и диализом в течение ночи против буфера, содержащего 20 мМ фосфат, 50 мМ NaCl, pH 7,2, при 4°С. Наконец, получали очищенный белок в количестве 60 мкг. Включение метки стабильного изотопа определяли FT-IR-спектроскопией, подобно тому, как это описано в Talvenheimo et al., 2002, Biopolymers 67(1): 10-19, и было обнаружено включение 98% (13С) и 99% (15N).
7.1.2. Изотопно-меченный рекомбинантный рецептор гистамина H1, продуцируемый в клетках насекомых
кДНК, кодирующую гистаминовый рецептор одного человека с введенным на С-конец полигистидином (НТ-Н1), получали для экспрессии из рекомбинантного бакуловируса в клетках Sf9, как описано Ratnala et al. (2004, Eur. J. Biochem. 271: 2636-46). Рекомбинантный вирус очищали анализом с использованием бляшек и амплифицировали (Klaasen and de Grip, 2000, Methods Enzymology, 315: 12-29). Клетки Sf9 выращивали во вращающейся колбе на 500 мл, содержащей 100 мл культуральной среды, приготовленной, как показано в примере 6.1.2. Клетки Sf9 получали из АТСС (CRL-1711) и адаптировали к приготовленной среде ступенчатой адаптацией, как описано Bowman et al. (supra). Клетки Sf9 инокулировали при плотности 3×105 клеток/мл и выращивали до плотности 3×106 клеток/мл. Затем клетки инфицировали рекомбинантным бакуловирусом, кодирующим НТ-Н1, сконструированным, как описано выше. Бакуловирус добавляли при MOI 1,0. Ежедневно пробу 10 мкл из культуры брали для мониторинга экспрессии НТ-Н1 при помощи дот-блоттинга. Уровень экспрессии был максимальным при приблизительно 4 днях dpi (дни после инфекции). Клетки собирали 10-минутным центрифугированием при 5000 g при 4°С. Функциональность проверяли анализом связывания радиоактивного лиганда, как описано Ratnala et al. (2004, supra).
Рекомбинантный НТ-Н1 очищали с использованием аффинной хроматографии с иммобилизованным металлом через гранулы Ni-NTA (Qiagen), как описано в Werten et al., (2001, supra). Очищенный НТ-Н1 элюировали 150 мМ имидазолом и восстанавливали в протеолипосомы смешиванием с азолектином с последующей экстракцией циклодекстрина детергентом, как описано Grip et al. (1998, Biochem. J. 330: 667-674). В результате, получали очищенный функциональный белок в количестве 250 мкг.
Включение метки стабильного изотопа проверяли FT-IR-спектроскопией (FT-IR-спектрофотометр Mattson Cygnus, Madison, WI) посредством количественного определения колебательных смещений, индуцируемых мечением стабильными изотопами, и было обнаружено включение 95% для обоих ядер 13С и 15N (смотрите ниже в примере 8).
Кроме того, данные 15N-SSNMR регистрировали на спектрометре Bruker 750 при 200 К и скорости вращения 8-12 кГц в целях оптимизации условий приготовления образца для последующих структурных исследований (смотрите фиг.1). Процедуру масштабировали до биореактора (4 литра) с выходом по объему, по меньшей мере, 4 мг функционального меченного стабильным изотопом рецептора НТ-Н1 на литр среды.
7.1.3. Изотопно-меченный рекомбинантный рецептор гистамина H1R, продуцируемый в клетках СНО
кДНК, кодирующую рецептор гистамина 1 человека, клонировали в вектор pcDEF, полученный от профессора J. Langer, Robert Wood Johnson Medical School, Piscataway, NJ, USA, клетки СНО выращивали во вращающейся колбе на 500 мл, содержащей 100 мл культуральной среды, приготовленной, как показано в примере 6.2.3. Клетки СНО адаптировали к приготовленной среде ступенчатой адаптацией, как описано Bowman et al. (supra). Клетки СНО инокулировали при плотности 6×105 клеток/мл и выращивали до плотности 3×106 клеток/мл. Затем эти клетки трансфицировали плазмидой, кодирующей меченный His-меткой рецептор гистамина H1, сконструированный, как описано выше. Ежедневно пробу 10 мкл из культуры брали для мониторинга экспрессии НТ-Н1 при помощи дот-блоттинга. Уровень экспрессии был максимальным при приблизительно 3-4 днях dpi (дни после инфекции). Клетки собирали 10-минутным центрифугированием при 5000 g при 4°С. Функциональность проверяли анализом связывания радиоактивного лиганда, как описано Ratnala et al., supra. Рекомбинантный НТ-Н1 очищали и реконструировали, как это описано в примере 7.1.2, и получали очищенный функциональный белок в количестве 150 мкг. Включение метки стабильного изотопа проверяли FT-IR-спектроскопией (FT-IR-спектрофотометр Mattson Cygnus, Madison, WI), как описано ниже в примере 8, посредством количественного определения колебательных смещений, вызванных мечением стабильными изотопами, и было обнаружено включение метки, превышающее 95%, для ядер (15N). Процедуру можно масштабировать до масштаба биореактора со сходными выходами по объему, по меньшей мере, 2,5 мг функционального меченного стабильным изотопом рецептора на литр среды.
Пример 8. Инфракрасная спектроскопия с Фурье-преобразованием для анализа состава биомассы и для мониторинга мечения стабильными изотопами
8.1. Предыстория в отношении FT-IR-метода
Основные классы биомолекул (белки, липиды, углеводы) могут быть легко идентифицированы инфракрасной спектроскопией с Фурье-преобразованием (FT-IR), поскольку типичные классы биомолекул поглощают в различных частотных диапазонах: 1500-1700 см-1 для пептидных амидных групп белков, 1700-1750 см-1 и 2800-3000 см-1 для эфирных групп липидов и валентных колебаний связей С-Н, соответственно, и 1000-1200 см-1 для групп С-ОН и С-О-С углеводов. Поскольку молярная оптическая плотность этих колебательных переходов значительно различается, количественная оценка состава биомассы не может быть легко достигнута, но качественный анализ состава может быть проведен, что является очень полезным для сравнения разных партий биомассы, как показано на фиг.2.
Колебательная частота зависит от массы участвующих атомов, таким образом, FT-IR может быть очень хорошо применен также для определения включения изотопной метки. Введение 15N-метки будет смещать колебание амида II пептидной связи, которая поглощает при 1520-1550 см-1 и доминирует деформационным CN-H колебанием, на 10-20 см-1. Введение 13С-метки будет смещать колебание амида I пептидной связи, которая поглощает при 1620-1670 см-1 и доминирует валентным С=O колебанием, на 40-60 см-1 и, кроме того, будет смещать колебания С-ОН и С-О-С углеводов на 30-50 см-1. 2H-метка, наконец, будет смещать колебание С-Н липидных групп на приблизительно 700 см-1 и колебание амида II белков на приблизительно 100 см-1. С использованием стратегии разделения пиков (обращения свертки, второй производной) смещения пиков вследствие включения метки могут быть определены количественно с точностью приблизительно 5%. Таким образом, этот способ позволяет быстрый анализ состава биомассы и полученных из биомассы молекул, а также мониторинг мечения стабильными изотопами. Например, определение эффективности 15N-мечения может быть достигнуто мониторингом отношения оптических плотностей А1542/А1534 и А1518/1534 в районе амида II и показано на фиг.3. Полоса амида II, в основном представляющая деформационное колебание N-H в пептидном скелете основной цепи белка, смещается при 15N-мечении от приблизительно 1542 до 1530 см-1. Однако соседняя полоса амида I около 1658 см-1 (в основном валентное С=O колебание) и колебательная полоса боковой цепи Туr около 1518 см-1 не смещаются при введении изотопной метки. Известно, что поглощение около 1518 см-1 фактически смещается после введения изотопной метки (смещение другого амида II до приблизительно 1530 см-1).
8.2. Материалы и способы FTIR
FTIR-спектры определяют при температуре окружающей среды на спектрометре Mattson Cygnus 100 IR или в режиме ATR (нарушенного полного внутреннего отражения), или в режиме трансмиссии, при разрешении 8 см-1 и спектральном диапазоне 4000-800 см-1. Для ATR-анализа биомассу суспендировали в деионизованной воде (приблизительно 20 мг/мл) в ультразвуковой ванне и образец, содержащий приблизительно 5 мг биомассы, наносили равномерно на поверхности пластины из германия вспомогательной части Specac ATR. После дегидратации в течение ночи на воздухе тонкую пленку биомассы помещали на пластину из германия. Затем эту вспомогательную часть ATR устанавливали в спектрометр и пленку биомассы дополнительно дегидратировали в продувке газообразным азотом спектрометра, пока пары воды не становились недетектируемыми, или посредством спиновой сушки (dark, et al., (1980) Biophys. J. 31, 65-96). При анализе трансмиссии объем суспендированной биомассы, содержащий 0,5-1,0 мг, наносили на окно AgCl (Fisher Scientific, диаметр 1,6 см) и дегидратировали в течение ночи на воздухе. Затем это окно вставляли в изготовленное в лаборатории приспособление для смены проб, установленное в спектрометре и контролируемое компьютером, и дополнительно дегидратировали продувкой газообразного азота. Наконец, регистрировали спектры после того, как пары воды уже не были детектируемыми. Спектры второй производной рассчитывали из спектров оптической плотности с использованием программного алгоритма обработки данных Mattson, инсталлированного в компьютер, контролирующий спектрометр.
8.3. Анализ меченных стабильными изотопами компонентов
Воспроизводимость от партии к партии меченных стабильными изотопами компонентов сред контролировали с помощью FT-IR-спектроскопии. Содержание белка в биомассе определяли из элементного анализа посредством определения азота и аминокислотного анализа после полного гидролиза с использованием Fmoc-дериватизации. Креатинфосфокиназу из мышцы кролика (ЕС 2.7.3.2., Boehringer Mannheim) использовали в качестве стандарта для количественного определения белков в препаратах биомассы при помощи FTIR.
Дериватизацию тринитробензолсульфоновой кислотой (TNBS) использовали для определения общего количества содержащих аминогруппу компонентов в гидролизатах и автолизатах. Однако этот TNBS-способ не применим к гидролизатам, нейтрализованным 15N-гидроксидом аммония вследствие реакции с аммиаком.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ИЗОТОПНОМЕЧЕНЫХ C ПОЛИНЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ | 2007 |
|
RU2361922C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАВНОМЕРНО ИЗОТОПОМОДИФИЦИРОВАННЫХ ПРИРОДНЫХ L α АМИНОКИСЛОТ | 1993 |
|
RU2100434C1 |
| МИКРООРГАНИЗМЫ ДЛЯ ПОВЫШЕННОГО ПРОДУЦИРОВАНИЯ АМИНОКИСЛОТ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2015 |
|
RU2688486C2 |
| НЕ СОДЕРЖАЩАЯ ЖИВОТНЫХ БЕЛКОВ СРЕДА ДЛЯ КУЛЬТИВИРОВАНИЯ КЛЕТОК | 2005 |
|
RU2383616C2 |
| Тиазол-оксазол-модифицированные пептиды, обладающие способностью ингибировать бактериальную рибосому | 2018 |
|
RU2674581C1 |
| СРЕДА ДЛЯ РОСТА ГЕТЕРОТРОФНЫХ ВОДОРОСЛЕЙ ИЛИ ГЕТЕРОТРОФНЫХ БАКТЕРИЙ, СПОСОБ ИХ КУЛЬТИВИРОВАНИЯ И СУБСТРАТ ДЛЯ ИХ РОСТА | 2003 |
|
RU2343192C2 |
| Способы получения липидов | 2011 |
|
RU2636344C2 |
| ИММУНИЗАЦИЯ ПРОТИВ CHLAMYDIA TRACHOMATIS | 2002 |
|
RU2331435C2 |
| Генетическая конструкция на основе двух индуцибельных экспрессионных векторов для экспрессии тиазол/оксазол модифицированного пептида в клетках бактерий E. coli; способ получения рекомбинантного штамма бактерий E. coli и способ получения модифицированного пептида на его основе | 2017 |
|
RU2694044C2 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ VIP3AB И CRY1AB ДЛЯ РЕГУЛИРОВАНИЯ УСТОЙЧИВЫХ НАСЕКОМЫХ | 2011 |
|
RU2608500C2 |
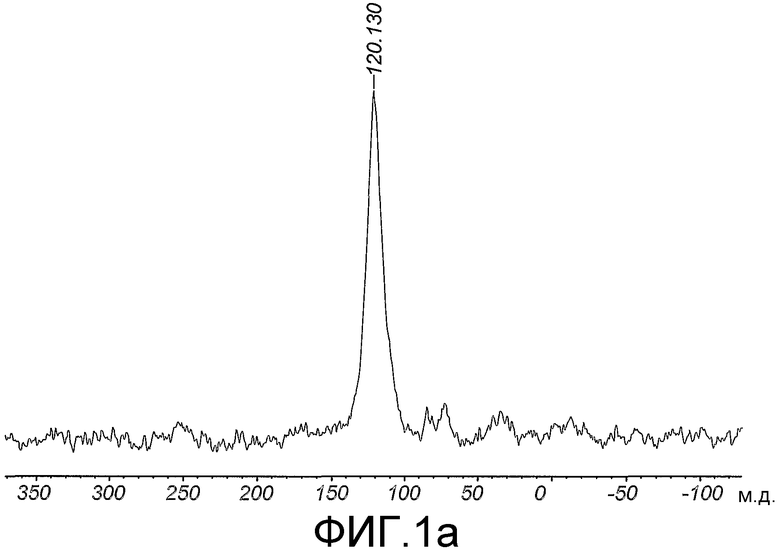
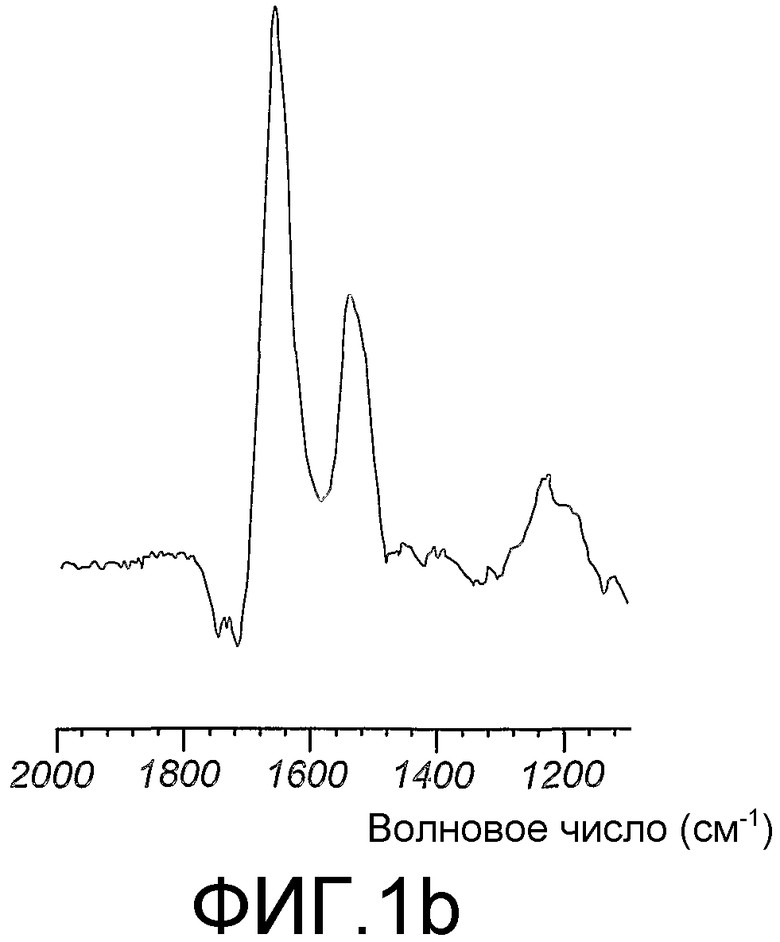
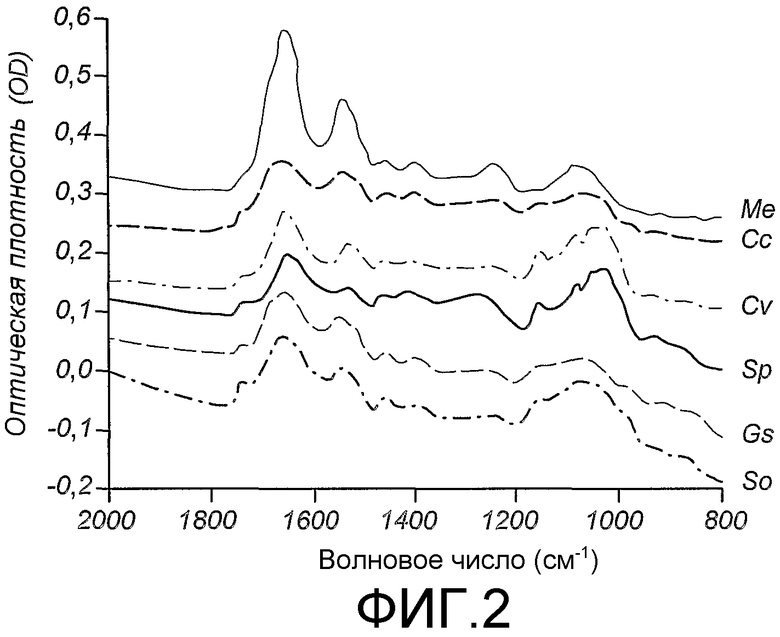
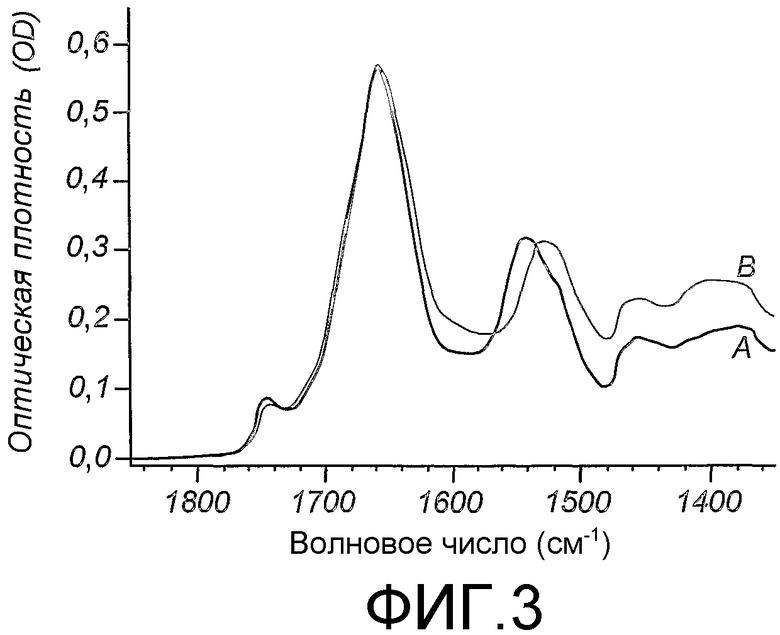
Для приготовления питательной среды используют микроорганизмы, которые выращивают на минеральной среде, поддерживающей рост этого организма, причем по существу все из ассимилируемых атомов являются изотопно-меченными, для получения меченой биомассы. Полученную биомассу подвергают автолизу для получения меченого автолизата. Эта биомасса может быть дополнительно экстрагирована органическим растворителем для получения липидов. Делипидированную биомассу гидролизуют для получения меченых аминокислот и других питательных веществ, которые используют вместе с автолизатом, экстрагированными липидами и дополнительными компонентами для приготовления питательной среды для выращивания клеток млекопитающих или насекомых в культуре для синтеза биомолекул. Полученная биомолекула является мембранным белком млекопитающего, в частности, в котором 20-100% атомов водорода в этом белке являются тотально замещенными изотопом 2Н. Однородное мечение стабильными изотопами позволяет определить трехмерную структуру биомолекулы, такой как мембранный белок млекопитающего, например, ЯМР-спектроскопией. 6 н. и 24 з.п. ф-лы, 4 ил.
1. Способ получения питательной среды для выращивания клеток млекопитающих или насекомых в культуре, где, по меньшей мере, для одного из Н, С или N, по существу, все атомы в субстратах, которые используются клетками для синтеза биомолекул в этой питательной среде, являются изотопно-мечеными, предусматривающий стадии:
(a) выращивания организма на минеральной среде, поддерживающей рост этого организма, причем в этой среде, по существу, все из ассимилируемых атомов, по меньшей мере, для одного из Н, С или N являются изотопно-мечеными, для получения меченой биомассы;
(b) автолиза полученной биомассы организма для получения меченого автолизата; и
(c) смешивания автолизата, полученного на стадии (b), с дополнительными компонентами, необходимыми для роста клеток млекопитающих или насекомых.
2. Способ по п.1, где этот организм является грибом, дрожжами или водорослями.
3. Способ по п.2, где этим организмом является организм, который принадлежит к роду, выбранному из Saccharomyces, Pichia, Hansenula, Kluyveromyces, Candida, Brettanomyces, Debaryomyces, Torulopsis, Yarrowia, Galdieria, Cyanidium, Porphyridium, Cystoclonium, Audouinella и Cyanidioschyzon.
4. Способ по любому из пп.1-3, где этот способ дополнительно предусматривает стадии:
(a) выращивания организма на минеральной среде, которая поддерживает рост этого организма, причем в этой среде, по существу, все из ассимилируемых атомов, по меньшей мере, для одного из Н, С или N являются изотопно-мечеными, для получения меченой биомассы;
(b) экстракции биомассы организма органическим растворителем для получения экстракта, содержащего липиды, причем этот организм выращивается, как описано в (а), или выращивается, как описано в (а), на среде без изотопного замещения;
(c) гидролиза биомассы организма, выращенного, как описано в (а), при нещелочном рН для получения гидролизата, содержащего аминокислоты;
(d) приготовления питательной среды на основе автолизата, полученного по любому из пп.1-3, с аминокислотами, полученными в (с), с добавлением дополнительных компонентов, необходимых для роста клеток млекопитающих или насекомых.
5. Способ по п.4, где в стадии (d) питательную среду готовят на основе автолизата, полученного по любому из пп.1-3, с аминокислотами, полученными в (с), и липидами, полученными в (b), с добавлением дополнительных компонентов, необходимых для роста клеток млекопитающих или насекомых.
6. Способ по п.4, где питательная среда состоит из автолизата, липидов и аминокислот, полученных, по меньшей мере, из двух различных организмов.
7. Способ по п.4, где перед гидролизом в (с) липиды и пигменты экстрагируют из биомассы с использованием органического растворителя.
8. Способ по п.4, где организм, из которого экстрагируют липиды, принадлежит к роду, выбранному из группы, состоящей из Rhodophyta, Cyanidiophyceae, Chlorophyta, Cyanophyta, Diatoms, Phaeophyceae, Dinoflagelate, Dinophyta и Galdieria.
9. Способ по п.4, где организм, из которого получают гидролизат, содержащий аминокислоты, является организмом, выбранным из водорослей, грибов, дрожжей и метилотрофных бактерий.
10. Способ по п.9, где этот организм принадлежит к роду, выбранному из группы, состоящей из Pichia, Saccharomyces, Hansenula, Cyanidium, Galdieria, Porphyridium, Spirulina и Methylobacillus.
11. Способ по п.l, где дополнительные компоненты, необходимые для роста клеток млекопитающих или насекомых, содержат один или несколько компонентов, таких как
(a) одна или несколько из глюкозы, фруктозы и сахарозы;
(b) один или несколько промежуточных продуктов цикла Кребса, выбранных из группы, состоящей из цитрата, сукцината, фумарата, малеиновой кислоты, щавелевой кислоты и малата;
(c) пируват; и
(d) один или несколько витаминов, выбранных из группы, состоящей из тиамина, рибофлавина, ниацина, витамина В6, фолиевой кислоты, витамина В12, биотина, пантотеновой кислоты, холина, пара-аминобензойной кислоты и альфа-токоферола.
12. Способ по п.1, где, по существу, все атомы в субстратах, которые используются клетками млекопитающих или насекомых для синтеза биомолекул в питательной среде, являются изотопно-меченым изотопом, выбранным из 15N; 13С; 2H; 15N и 13С; 15N и 2Н; 13С и 2H или 15N, 13С и 2H.
13. Способ получения биомолекулы, включающей белок или клеточный метаболит, причем, по существу, все атомы в этой биомолекуле являются изотопно-мечеными, предусматривающий стадии:
а) выращивания культуры клеток млекопитающих или насекомых, способных продуцировать эту биомолекулу при условиях, способствующих продукции этой биомолекулы, в питательной среде, содержащей изотопно-меченые атомы в субстратах, полученной в способе по любому из пп.1-12; и
(b) извлечения этой изотопно-меченой биомолекулы.
14. Способ по п.13, где этот белок является растворимым белком млекопитающего или мембранным белком.
15. Способ по п.13, где клетки млекопитающих или насекомых, способные продуцировать этот белок, содержат экспрессирующий вектор, содержащий нуклеотидную последовательность, кодирующую этот белок.
16. Способ получения информации о структуре белка или клеточного метаболита, предусматривающий стадии:
(a) получения белка или клеточного метаболита, причем, по существу, все атомы в этом белке или клеточном метаболите являются изотопно-мечеными, в способе по любому из пп.13-15;
(b) определения трехмерной структуры этого белка или клеточного метаболита путем проведения спектроскопического анализа.
17. Способ по п.16, где этот спектроскопический анализ включает в себя ЯМР-спектроскопию.
18. Способ по п.16, где этот белок или клеточный метаболит предварительно очищают.
19. Способ по п.16, где этот белок находится в комплексе со вторым белком или клеточным метаболитом.
20. Способ по п.19, где 20-100% атомов водорода во втором белке или клеточном метаболите являются тотально замещенными изотопом 2H.
21. Способ по п.20, где белок находится в комплексе со вторым белком.
22. Питательная среда для выращивания клеток млекопитающих или насекомых в культуре, где, по меньшей мере, для одного из Н, С или N, по существу, все атомы в субстратах, которые используются клетками для синтеза биомолекул в этой среде, являются изотопно-мечеными, характеризующаяся тем, что она получена способом по любому из пп.1-12, содержащая
(a) смесь неорганических солей;
(b) источник аминокислот;
(c) углеводный источник энергии;
(d) источник липидов;
(e) при необходимости, защитный агент;
(f) при необходимости, витамины и/или органические соединения;
(g) при необходимости, органические кислоты; и (h) при необходимости, микроэлементы;
причем, по существу, все атомы в (а), (b) и (с) и, при необходимости, в (d), (е), (f), (g) и (h), являются изотопно-мечеными, по меньшей мере, для одного из Н, С или N или причем 20-100% атомов водорода в (а), (b) и (с) и, при необходимости, в (d), (е), (f), (g) и (h) тотально замещены изотопом 2H.
23. Питательная среда по п.22, в которой источник аминокислот содержит гидролизат, содержащий аминокислоты, который получен из биомассы дрожжей, причем гидролиз этой биомассы предусматривает автогидролиз.
24. Питательная среда по п.22, в которой источник липидов содержит жирные кислоты, стероиды и растворимые в жирах витамины.
25. Питательная среда по п.22, в которой углеводным источником энергии является один или несколько компонентов из глюкозы, фруктозы и сахарозы; органическими кислотами являются один или несколько из пирувата и промежуточных продуктов цикла Кребса, выбранных из группы, состоящей из цитрата, сукцината, фумарата, малеиновой кислоты, оксалата и малата; витаминами являются один или несколько витаминов, выбранных из группы, состоящей из тиамина, рибофлавина, ниацина, витамина В6, фолиевой кислоты, витамина В12, биотина, пантотеновой кислоты, холина, пара-аминобензойной кислоты и альфа-токоферола.
26. Питательная среда по п.22, причем, по существу, все атомы в (а), (b) и (с) и, при необходимости, в (d), (e), (f), (g) и (h), являются изотопно-меченым изотопом, выбранным из 15N; 13С; 2H; 15N и 13С; 15N и 2H; 13С и 2H; или 15N, 13С и 2H.
27. Мембранный белок млекопитающего, полученный способом по любому из пп.13-15, причем, по существу, все атомы в этом белке являются изотопно-меченым изотопом, выбранным из 15N; 13С; 2H; 15N и 13С; 15N и 2H; 13С и 2H; или 15N, 13С и 2H.
28. Мембранный белок млекопитающего по п.27, причем этот белок является белком человека.
29. Мембранный белок млекопитающего, полученный способом по любому из пп.13-15, причем 20-100% атомов водорода в этом белке являются тотально замещенными изотопом 2H.
30. Мембранный белок млекопитающего по п.29, причем этот белок является белком человека.
| Экономайзер | 0 |
|
SU94A1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| US 5627044 A, 06.05.1997 | |||
| Штамм облигатно-метилотрофной бактерии @ @ ВСБ-792 | 1980 |
|
SU989867A1 |
| HANSEN ANDREW P | |||
| et al | |||
| A practical method for uniform isotopic labeling of recombinant protein in mammalian cells | |||
| Biochemistry, 1992 | |||
| v | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| Способ запрессовки не выдержавших гидравлической пробы отливок | 1923 |
|
SU51A1 |

