Изобретение относится к медицине, и может быть использовано для получения стимулированных дендритных клеток для индукции иммунного ответа против туберкулеза человека.
Известен способ получения вакцин для внутривенного введения, основанный на иммунизации комбинированной сибиреязвенной вакциной, содержащей антитела к ПА сибиреязвенного микроба в титре не менее 1:400 по результатам ИФА, плазмы крови доноров [RU 2348429 С2, А61К 39/40, 2009.03.10].
Недостатком способа является не индивидуальность вакцины, поскольку получена из плазмы донора, а не пациента. Это снижает эффективность ее применения и может вызвать побочные заболевания.
Наиболее близким по своей сущности к предложенному является способ, основанный на извлечении из костного мозга незрелых дендритных клеток и выдерживании их в течение нескольких дней вместе с антигенами возбудителя пневмонии - клеточным лизатом или бактериальными липополисахаридами [www.cbio.ru].
Полученную вакцину - активированные дендритные клетки - внутрибрюшинно вводили мышам, а на следующий день заражали их летальной дозой Klebsiella pneumoniae. От этой инфекции погибла вся контрольная группа мышей. Все привитые мыши остались живы.
Недостатком наиболее близкого решения является относительно узкая область применения, поскольку полученная индивидуальная вакцина может быть использована для профилактики пневмонии, но не может быть использована в других областях медицины, в частности, для профилактики и лечения онкологических заболеваний с учетом особенностей, возникающих, например, при ее использовании для пациентов, больных туберкулезом.
Требуемый технический результат заключается в расширении области применения.
Требуемый результат достигается тем, что, по способу, основанному на заборе у пациента незрелых дендритных клеток, их культивировании ex vivo для созревания и формирования аллостимуляторной активности для последующей дополнительной обработки антигеном и обратном введении созревших дендритных клеток тому же пациенту с целью формирования адаптивного иммунитета, при культивировании ех vivo незрелых дендритных клеток в культуральную среду вводят фрагментированную аллогенную двуцепочечную геномную ДНК с фрагментами размером 200-6000 п.о. и составляющими полный геном физиологически и генетически здорового донора в количестве 5 мкг/мл среды.
Кроме того, требуемый результат достигается тем, что, в способе по п.1 препарат фрагментированной аллогенной двуцепочечной геномной ДНК с фрагментами размером 200-6000 п.о. и составляющими полный геном физиологически и генетически здорового донора вводят в культуральную среду, содержащую незрелые дендритные клетки больных туберкулезом в количестве 25 мкг/мл среды.
В более широком плане целью настоящего изобретения является разработка обоснованного и экспериментально подтвержденного способа воздействия на выделенные из организма пациента и культивируемые ех vivo незрелые дендритные клетки фрагментированной аллогенной двуцепочечной геномной ДНК с фрагментами размером 200-6000 п.н. и составляющими полный геном физиологически и генетически здорового донора, таким образом, что в результате воздействия на ДК экзогенной ДНК происходит их созревание и индуцируется их аллостиммуляторная активность. После добавления антигена и последующего введения этих ДК обратно в организм того же пациента должно происходить развитие специфического адаптивного иммунитета у этого больного, связанного с активированной дендритными клетками пролиферацией обученных против этих антигенов цитотоксических CD8+ Т лимфоцитов, то есть происходит специфическая вакцинация пациента.
В современной литературе отсутствуют указания на предлагаемый способ получения стимулированных дендритных клеток для индукции иммунного ответа против туберкулеза человека, основанный на заборе у пациента незрелых дендритных клеток, их культивирования ex vivo в присутствии фрагментированной экзогенной аллогенной двуцепочечной геномной ДНК, необходимой для созревания и формирования аллостимуляторной активности этих дендритных клеток. Последующая дополнительная обработка этих дендритных клеток специфическим антигеном и обратное введение их тому же пациенту необходимо для формирования адаптивного специфического иммунитета у таких пациентов.
Следовательно, предложение отвечает критериям новизны изобретательского уровня.
Ниже приводятся теоретические и экспериментальные данные, подтверждающие, что изобретение отвечает и критерию практической (промышленной) применяемости.
На чертеже представлены:
на фиг.1 - электронно-микроскопические фотографии препаратов ДНК, распластанных по методу Клейншмидта - а) фрагментированная ДНК плаценты человека, используемая в эксперименте, b) розеткоподобная структура, встречающаяся в препарате, с) нуклеосомы на ДНК, выделенной из клеток Hela;
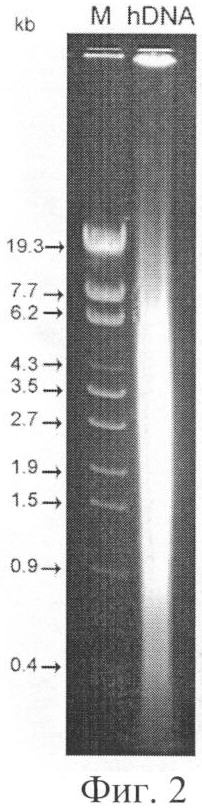
на фиг.2 - электрофоретическая характеристика экзогенной терапевтической ДНК плаценты человека (М - маркер молекулярного веса - lambda DNA BssT1I digest);
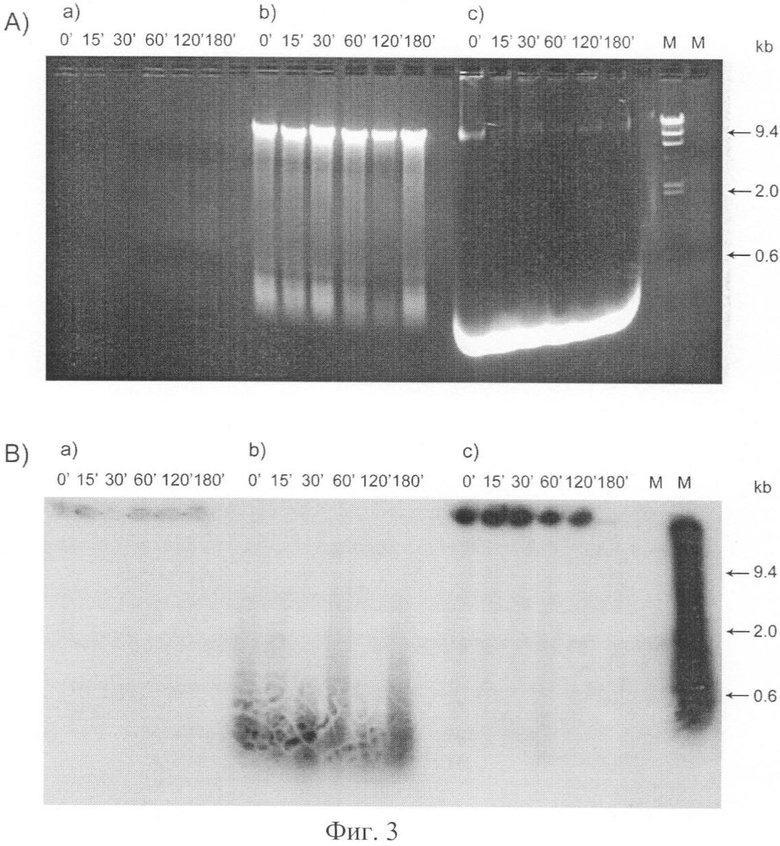
на фиг.3 - ДК человека инкубировали с α 32P меченой ДНК человека;
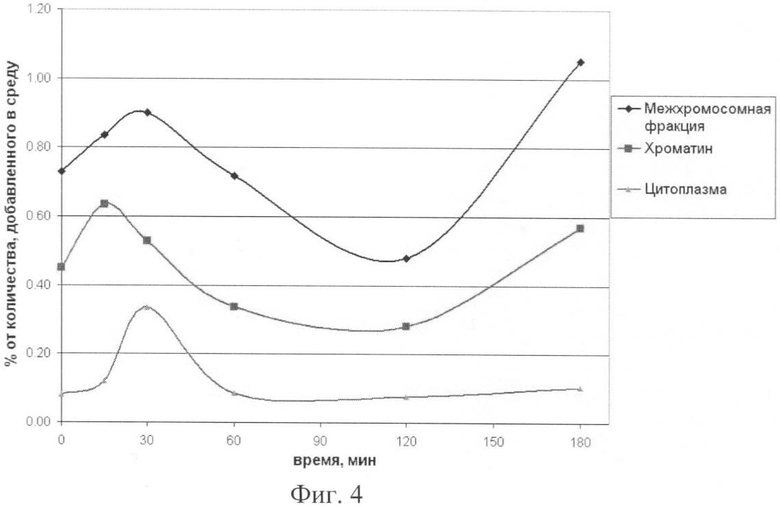
на фиг.4 - динамика доставки меченого материала во внутренние компартменты ДК человека в зависимости от времени инкубации культуры клеток и радиоактивно меченой ДНК;
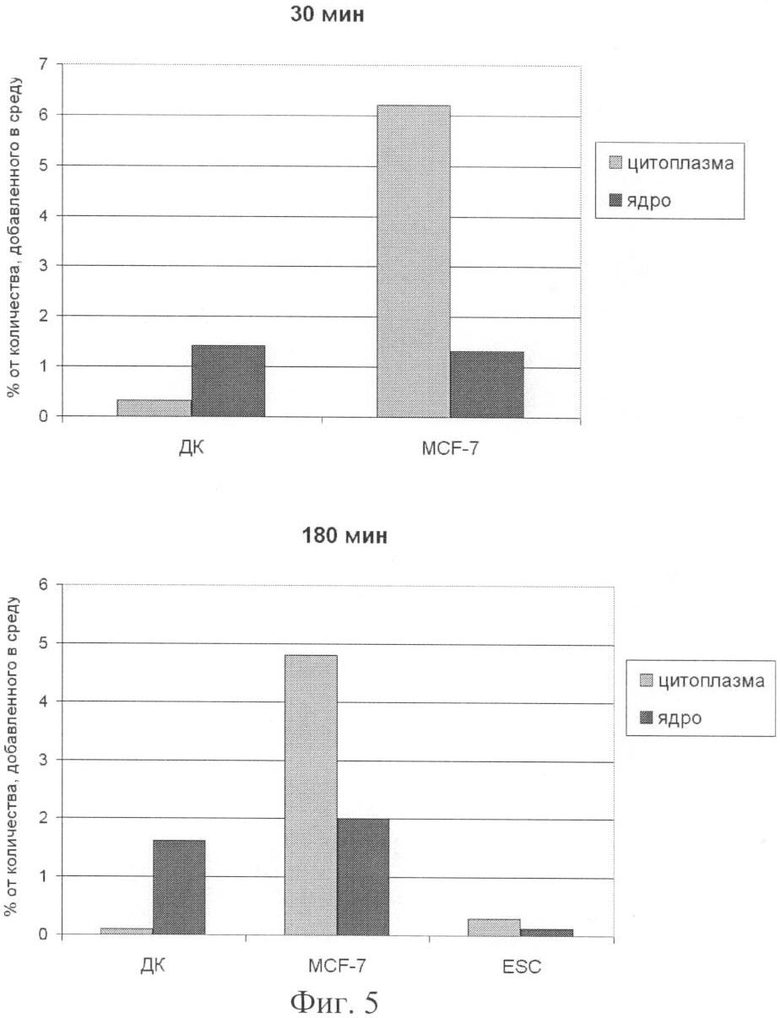
на фиг.5 - анализ соотношения экзогенной ДНК, доставляемой в цитоплазму и ядро трех типов клеток в точках ее максимального присутствия (пиках) [1, 2].
Относительно представленного на фиг.3 следует указать, что после инкубации клетки последовательно лигировали неионным детергентом Тритон X100 и SDS и фракционировали на (а) цитоплазматическую фракцию, (b) межхромосомную фракцию и (с) хроматин. ДНК, выделенную из каждой фракции, анализировали электрофорезом в агарозном геле. А) Блок 0,7%-ной агарозы, окрашенный бромистым этидием; В) Рентгенограмма этого же блока после высушивания. Цифры над блоками указывают время инкубации культуры клеток с меченой экстраклеточной ДНК. М - молекулярные маркеры - λHindIII ДНК и исходная меченая ДНК, добавляемая в культуральную среду. Цифры справа от блоков - маркеры молекулярных весов в т.п.о.
Предложенный способ получения стимулированных дендритных клеток для индукции иммунного ответа против туберкулеза человека реализуется следующим образом.
Способ получения стимулированных дендритных клеток для индукции иммунного ответа против туберкулеза человека основан на применении дозревающих стимулов при ex vivo культивировании. По предложенному способу в культуральную среду, содержащую незрелые дендритные клетки, добавляют фрагментированную аллогенную двуцепочечную геномную ДНК с фрагментами размером 200-6000 п.о. и составляющими полный геном физиологически и генетически здорового донора, что приводит к созреванию ДК и индукции у них аллостимуляторной активности, определяющей развитие специфического адаптивного иммунитета.
При этом фрагменты экзогенной ДНК добавляют в культуральную среду, содержащую выделенные из организма больного дендритные клетки в количестве 5 мкг/мл.
Фрагменты экзогенной ДНК добавляют в культуральную среду, содержащую выделенные из организма больного туберкулезом дендритные клетки в количестве 25 мкг/мл.
Созревшие и активированные ex vivo ДК дополнительно пульсированные специфическим антигеном(ами) и введенные обратно в организм того же самого пациента будут стимулировать развитие в организме этого пациента специфического адаптивного иммунитета.
Пример реализации способа.
Мононуклеарные клетки (МНК) выделяли из гепаринизированной, венозной крови стандартно в градиенте плотности фиколла-верографина. Дендритные клетки получали путем культивирования прилипающей фракции МНК в 24-луночных планшетах (Nunc, Дания) в течение 5 суток в среде RPMI-1640 (Sigma-Aldrich, США), дополненной 0,3 мг/мл L-глютамина, 5 мМ HEPES-буфера, 100 мкг/мл гентамицина и 5% инактивированной сыворотки крови AB(IV) группы, в присутствии GM-CSF (Leucomax, Шеринг-Плау, Швейцария) 1000 Ед/мл и INF-a (Роферон-А, Roche, Швейцария) 1000 Ед/мл. В качестве дозревающего стимула использовали аллогенную двуцепочечную геномную ДНК с фрагментами размером 200-6000 п.о. и составляющими полный геном физиологически и генетически здорового донора в количестве 5 мкг/мл, которую добавляли за 24 ч до окончания культивирования ДК и в количестве 25 мкг/мл для ДК, полученных от пациентов больных туберкулезом. Позитивным контролем служили ДК, созревание которых индуцировали ЛПС, негативным - ДК, генерируемые в отсутствие дозревающего стимула.
Аллостимуляторную активность ДК оценивали в реакции смешанной культуры лимфоцитов (СКЛ). В качестве отвечающих клеток использовались МНК доноров (0,1×106/лунку), которые культивировали в 96-луночных круглодонных планшетах для иммунологических исследований в среде RPMI-1640 и 10% инактивированной сыворотки крови AB(IV) группы. Стимуляторами служили ДК в соотношении МНК:ДК=10:1. Ответ оценивался на 5 сут радиометрически по включению 3H-тимидина (1 мкКю/лунку), который вносился за 18 ч до окончания культивирования. Индекс влияния (ИВ) экзогенной ДНК на аллостимуляторную активность ДК вычисляли как соотношение интенсивности включения 3H-тимидина в опыте к интенсивности включения 3Н-тимидина в контроле.
Эффект ДНК на фенотип (экспрессию CD 14 и CD83) дендритных клеток оценивали по влиянию различных доз ДНК, вносимой за 24 ч до окончания культивирования ДК. Позитивным контролем служили ДК, созревание которых индуцировали ЛПС, негативным - ДК, генерируемые в отсутствие дозревающего стимула. Поскольку дифференцировка моноцитов в ДК связана с потерей моноцитами CD 14 молекул, а созревание незрелых ДК в зрелые - с приобретением маркера терминальной дифференцировки CD83, уменьшение доли CD14+ клеток и увеличение CD83+ клеток расценивали как признак дифференцировки и созревания ДК. Фенотипирование клеток проводили методом проточной цитофлюориметрии (FACSCalibur, Becton Dickinson), с использованием РЕ-меченных моноклональных анти-CD14-антител и FITC-меченных анти-CD83 антител (BD Bioscience Pharmingem, США).
Для подтверждения того, что предложенный способ позволяет достигнуть требуемого результата, приведем его теоретическое и экспериментальное обоснование.
Рассмотрим концепцию активации специфического адаптивного иммунитета антигенпрезентирующими дендритными клетками, созревшими и активированными ex vivo, которые дополнительно могут быть пульсированы специфическим антигеном(ами), и введенными обратно в организм того же самого пациента на примере злокачественных онкологических заболеваний.
Известно, что течение многих злокачественных заболеваний сопряжено с развитием опухоль-ассоциированной иммуносупрессии, на фоне которой опухолевые клетки «ускользают» от иммунного надзора. Иммунорезистентность опухоли связывают с несколькими механизмами, которые реализуются на клеточном и гуморальном уровне, в первую очередь, в микроокружении развивающейся опухолевой ткани. Среди них выделяют: а) сдвиг цитокинового баланса в сторону иммуносупрессорных медиаторов, подавляющих функциональную активность иммунокомпетентных клеток, инфильтрирующих опухоль; б) связанный с этим дисбаланс эффекторных, цитотоксических и регуляторных Т-клеток; в) возникновение молекулярных дефектов в системах клеток, участвующих в распознавании и презентации антигена и передаче сигнала внутрь клетки [3]; г) формирование опухолью защитного барьера с повышенной концентрацией «опухоленосной» ДНК, поддерживающей генетическую стабильность клеток малигнизированной ткани и экспортирующей опухолевые гены в другие части организма [4-7].
Эффективность противоопухолевого иммунитета во многом зависит от активности иммунокомпетентных клеток - натуральных киллерных клеток, цитотоксических Т-лимфоцитов, макрофагов и дендритных клеток (ДК). Эти клетки участвуют в запуске, организации и реализации реакций врожденного и адаптивного иммунитета и через различные гуморальные и клеточно-опосредованные механизмы элиминируют раковые клетки. Первой линией противоопухолевой защиты является система неспецифической резистентности (innate immunity), которую составляют натуральные киллерные клетки и активированные макрофаги. В случае несостоятельности механизмов неспецифической защиты, связанной с нарушением экобиосистем жизнедеятельности, генетическими особенностями, возрастом индивида, тяжестью сопутствующей патологии, наличием исходной иммунокомпрометированности и т.д., в организме включаются механизмы адаптивного иммунитета [8, 9]. Во многом это зависит от уровня экспрессии на опухолевых клетках специфических опухолеассоциированных антигенов, которые распознаются иммунокомпетентными клетками и против которых индуцируются эффекторные реакции, направленные на элиминацию опухолевого клона. В эффективной реализации реакций адаптивного иммунитета большое значение имеют два этапа, связанные, во-первых, с процессингом/презентацией опухолеассоциированных антигенов антиген-презентирующими клетками (главным образом, ДК), и, во-вторых, со стимуляцией пролиферации и созревания цитотоксических Т-лимфоцитов.
В наших предыдущих работах [10, 11] был описан противоопухолевый эффект фрагментированной экзогенной ДНК. В частности, было установлено, что препараты фрагментированной геномной ДНК обладают способностью тормозить рост некоторых экспериментальных опухолей мышей. Одновременно они оказывают умеренный блокирующий эффект на развитие опухолевых метастазов.
Параллельно с этими исследованиями была проведена работа по анализу действия экзогенной ДНК на клетки аденокарциномы молочной железы человека MCF-7, растущие в культуре. Проведенный анализ свидетельствовал о том, что в этом случае происходит изменение неопластичных клеток на генетическом уровне. Известно, что основным фактором неконтролируемой пролиферации данного типа клеток является дефект в гене ключевого фермента апоптоза - каспазы 3. Ген экзекьюторной каспазы 3 в экзоне 4 имеет делецию 47 п.о., что приводит к сдвигу рамки считывания и невозможности синтеза функционально активного фермента [12]. Этот факт позволил выдвинуть гипотезу о том, что регрессия экспериментальных опухолей мыши связана с прямым воздействием фрагментов экзогенной ДНК на геном раковой клетки. Предполагалось, что такое воздействие может в статистическом режиме приводить к замещению последовательностей, мутации в которых привели к неотрансформации клетки на недефектные структуры ДНК [13]. Однако количественная оценка ДНК, доставляемой в клетки солидных опухолей in vivo, свидетельствовала о том, в используемых экспериментальных условиях менее одной копии фрагмента размером ~1000 п.о. интернализуется в межхромосомном пространстве ядра. Очевидно, что такое количество экзогенной ДНК не в состоянии индуцировать какие-либо значимые генетические изменения в клетке [14].
Мы предположили, что наиболее вероятным путем воздействие на развитие солидных опухолей в проведенных экспериментах in vivo [10, 11] является активация определенных звеньев иммунной системы, а именно адаптивного иммунитета, связанного с активностью ДК. Хотя нельзя исключить вероятность двойного противоракового действия, основанного как на прямом цитотоксическом эффекте иммунной системы в отношении опухоли, так и на реверсивном генетическом перерождении раковых клеток в зонах опухоли, доступных для интернализации фрагментов экзогенной ДНК в количестве, достаточном для модификации генома раковой клетки [15].
Для того чтобы ответить на вопрос, может ли наблюдаемая регрессия экспериментальных опухолей быть связанной с активацией иммунного ответа, индуцируемого ДК, мы провели анализ влияния фрагментированной геномной ДНК человека на ДК в системе ех vivo.
Было показано, что фрагментированная геномная ДНК человека эффективно захватывается дендритными клетками и интернализуется в основном в межхромосомном пространстве ядра, не подвергаясь конкатамеризации. Незначительное количество меченого материала обнаруживается в цитоплазме.
Было обнаружено, что экзогенная ДНК стимулирует эффективное созревание ДК, характеризующееся изменением соотношения их поверхностных маркеров зрелости и развитием аллостимуляторной активности. Способность ДК человека, дозревающих в присутствии экзогенной ДНК, стимулировать пролиферативный ответ в смешанной культуре лимфоцитов была сравнима по силе и в некоторых случаях превосходила стандартный активатор ДК - ЛПС из стенки бактерий. Внесение ДНК в культуры ДК человека сопровождалось увеличением относительного содержания CD83+ клеток и снижением доли CD14+ клеток.
Таким образом, мы показали, что экзогенная ДНК поглощается ДК и стимулирует их созревание и аллостимуляторную активность.
Используемые методики.
Анализ препарата ДНК методом электронной микроскопии
Препараты были приготовлены методом распластывания по Клейншмидту с формамидом [16, 17]. Образец ДНК (50 мкг) в растворе цитохрома С (Sigma, США) с поверхности воды снимали на угольную пленку, после чего распластанные молекулы оттеняли напылением платины-палладия.
Анализ препарата ДНК биохимическим методом
Биохимический анализ образца был проведен методом фракционирования белков в системе Лэммли, с последующим двойным окрашиванием геля Кумаси-К250-серебро.
Генерация дендритных клеток
Мононуклеарные клетки (МНК) выделяли из гепаринизированной, венозной крови стандартно в градиенте плотности фиколла-верографина. Дендритные клетки получали путем культивирования прилипающей фракции МНК в 24-луночных планшетах (Nunc, Дания) в течение 5 суток в среде RPMI-1640 (Sigma-Aldrich, США), дополненной 0,3 мг/мл L-глютамина, 5 мМ HEPES-буфера, 100 мкг/мл гентамицина и 5% инактивированной сыворотки крови AB(IV) группы, в присутствии GM-CSF (Leucomax, Шеринг-Плау, Швейцария) 1000 Ед/мл и INF-α (Роферон-А, Roche, Швейцария) 1000 Ед/мл. В качестве дозревающего стимула использовали ЛПС (10 мкг/мл, LPS E.coli 0111:В4, Sigma-Aldrich), который добавляли за 24 ч до окончания культивирования ДК.
Обработка культуры ДК меченой ДНК, анализ ДНК в клеточных компартментах
8 мкг меченой с помощью полимеразы Кленова α 32P ДНК плаценты человека добавляли в культуру ДК человека (3×106 клеток), через 0, 15, 30, 60, 120 и 180 мин после этого клетки обрабатывали трипсином и центрифугировали 10 мин при 4000 об/мин на К23 центрифуге. Осадок ресуспендировали в буфере А с 1% тритоном X100 и инкубировали 10 мин на льду. После этого суспензию наслаивали на столбик 12%-ной сахарозы (3 мл) и центрифугировали 20 мин при 2000 об/мин на К23 центрифуге. Осаждали ДНК из цитоплазматической фракции (супернатант). Осадок ядер ресуспендировали в буфере А, добавляли хлорид натрия до 2М, SDS до 1% и инкубировали 15 мин при 65°С. Центрифугировали 30 мин при 30°С на 21000 об/мин на центрифуге J2-21 Beckman. Осаждали ДНК из межхромосомной фракции, осадок хроматина просто растворяли в воде. Количество меченого материала ДНК в трех клеточных фракциях определяли стандартным способом в пробирках Eppendorf, опущенных в счетные флаконы, на счетчике Rac Betta. Образцы фракционировали в 0.7% агарозном геле.
Характеристика доноров крови
В работе были исследованы ДК здоровых доноров и больных туберкулезом легких (ТБ). Из 8 обследованных пациентов у 7 человек была выявлена инфильтративная и у одного пациента - фибрознокавернозная форма ТБ. У 6 пациентов было выявлено активное бацилловыделение, у 5 обнаружена резистентность к терапии стандартными туберкулостатиками.
Преинкубация ДК с экзогенной ДНК
ДНК плаценты человека и ДНК, обогащенную CpG последовательностями (CpG), добавляли в качестве дозревающего стимула в дозах 1, 5 и 25 мкг/мл за 24 ч до окончания культивирования ДК. Позитивным контролем служили ДК, созревание которых индуцировали ЛПС, негативным - ДК, генерируемые в отсутствие дозревающего стимула.
Оценка аллостимуляторной активности ДК
Аллостимуляторную активность ДК оценивали в реакции смешанной культуры лимфоцитов (СКЛ). В качестве отвечающих клеток использовались МНК доноров (0,1×106/лунку), которые культивировали в 96-луночных круглодонных планшетах для иммунологических исследований в среде RPMI-1640 и 10% инактивированной сыворотки крови AB(IV) группы. Стимуляторами служили ДК в соотношении МНК:ДК=10:1. Ответ оценивался на 5 сут радиометрически по включению 3Н-тимидина (1 мкКю/лунку), который вносился за 18 ч до окончания культивирования. Индекс влияния (ИВ) экзогенной ДНК на аллостимуляторную активность ДК вычисляли как соотношение интенсивности включения 3H-тимидина в опыте к интенсивности включения 3H-тимидина в контроле.
Оценка влияния препаратов ДНК на фенотип ДК
Эффект ДНК на фенотип (экспрессию CD14 и CD83) дендритных клеток оценивали по влиянию различных доз ДНК, вносимой за 24 ч до окончания культивирования ДК. Позитивным контролем служили ДК, созревание которых индуцировали ЛПС, негативным - ДК, генерируемые в отсутствие дозревающего стимула. Поскольку дифференцировка моноцитов в ДК связана с потерей моноцитами CD14 молекул, а созревание незрелых ДК в зрелые - с приобретением маркера терминальной дифференцировки CD83, уменьшение доли CD14+ клеток и увеличение CD83+ клеток расценивали как признак дифференцировки и созревания ДК. Фенотипирование клеток проводили методом проточной цитофлюориметрии (FACSCalibur, Becton Dickinson), с использованием РЕ-меченных моноклональных анти-CD14-антител и FITC-меченных анти-CD83 антител (BD Bioscience Pharmingem, США).
Остановимся на анализе молекулярных характеристик препарата экзогенной аллогенной геномной двуцепочечной фрагментированной ДНК.
Известно, что ДНК в форме нуклеосом, представленных в физиологической концентрации, активируют миелоидные и плазматические ДК [18, 19]. Препараты ДНК, используемые в настоящем исследовании, были получены безфенольным способом солевой экстракцией и последующим осаждением ацетоном. Для того чтобы определить минимальное загрязнение нуклеосомной фракцией препаратов используемой ДНК, мы провели биохимический и электронно-микроскопический анализ этих препаратов. В проведенных экспериментах мы не обнаружили ни следов гистонов при двойном окрашивании Кумаси-R250-серебро белков, фракционированных из препаратов ДНК, в системе Леммли, ни нуклеосом при электронно-микроскопическом исследовании. На микроскопическом уровне препарат ДНК представлял собой фрагменты различной длины и розетки, сформированные, по-видимому, остаточными белками ядерного матрикса (фиг.1).
Рассмотрим некоторые особенности интернализации фрагментированной экзогенной аллогенной двуцепочечной геномной ДНК в межхромосомном пространстве ядер ДК человека
Для того чтобы оценить эффективность доставки экзогенной ДНК во внутренние компартменты ДК, мы инкубировали сгенерированные, как описано в разделе материалы и методы, ДК с меченой ДНК человека, фрагментированной до размеров 1-20 нуклеосомных мономеров (170-6000 п.о.) (фиг.2).
Для количественной оценки доставляемых во внутренние компартменты ДК фрагментов экзогенной ДНК были определены следующие параметры.
В каждую экспериментальную точку бралось 8 мкг ДНК.
На каждую экспериментальную точку брали 3×106 клеток.
В каждой клетке 6 pg ДНК.
Общее количество ДНК, выделяемое из клеток каждой экспериментальной точки, составляло 18 мкг.
Геном человека 3×109 п.н.
В результате проведенного анализа были определены следующие особенности интернализации экзогенной ДНК в ДК (Таблица 1).
Как следует из электрофоретического анализа (фиг.3) и анализа счета радиоактивности, захваченная дендритными клетками ДНК сразу поступает в межхромосомное пространство, не задерживаясь в цитоплазме. В межхромосомной фракции ДНК находится в деградированном состоянии (200-1000 п.н.), не подвергаясь конкатамеризации, характерной для ДНК, поступившей в межхромосомную фракцию культуры клеток MCF-7 или эмбриональных стволовых клеток человека [1, 2].
Наличие меченого материала во фракции хроматина связано либо со встраиванием фрагментов экзогенной ДНК в хроматин вследствие рекомбинационных процессов либо с инкорпорацией при синтетических процессах, происходящих в клетке, меченых мономеров, образовавшихся в результате деградации экзогенной ДНК.
Количественная оценка полученных результатов свидетельствует о том, что в межхромосомном пространстве ядер ДК может находиться в свободном состоянии около 10 тыс. фрагментов длиной 1000 п.о. Анализ кинетики доставки меченого материала выявил два пика - 30 и 180 мин (фиг.4). Предполагается, что пики отражают время оборота рецепторов, связывающих ДНК. Первый пик связан с захватом и поступлением экзогенной ДНК во внутренние компартменты клетки, спад соответствует полной занятости рецепторов, затем рецепторы освобождаются и обеспечивают второй пик доставки меченой экзогенной ДНК. Как было показано для культуры клеток MCF-7, меченая ДНК также доставляется в клеточные компартменты двумя пиками (в точках 30 и 180 мин).
При сравнении количества меченого материала, доставляемого в ядерное пространство и находящегося при этом в цитоплазме, было обнаружено следующее различие между ДК и другими культурами клеток (MCF-7 и эмбриональные стволовые клетки человека), проанализированными в наших предыдущих работах. Оказалось, что для ДК человека количество находящейся меченой экзогенной ДНК в цитоплазматической фракции постоянно значительно ниже, чем во фракциях, представляющих ядерные компартменты (межхромосомная фракция и хроматин) (фиг.5). Эти факты могут говорить о том, что для ДК существует другой путь (по сравнению с указанными типами клеток) интернализации фрагментов экзогенной ДНК, опосредующийся специфическими рецепторами. Также, по-видимому, этот путь отличен от такового, определенного для CpG ДНК, для которой показана ее интернализация в эндосомы и последовательная транспортировка и концентрация в тубулярном лизосомальном компартменте цитоплазмы, где она связывается с TLR рецепторами [20].
Одной из наиболее интересных характеристик состояния фрагментов экзогенной ДНК, находящихся в межхромосомном пространстве ядра, явилось полное отсутствие их видимой конкатамеризации. Согласно литературным данным [21, 22] и нашим исследованиям [1, 2], выполненным на других типах клеток (MCF-7 и эмбриональные стволовые клетки), фрагменты ДНК, попавшие во внутриядерное пространство, лигируются один с другим, образуя конкатомеры. Электрофоретически это выглядит так: начиная с времени инкубации 15 мин меченый материал замедляет свою подвижность в агарозном геле, формируя иногда характерную лестницу [2]. В случае ДК меченый материал выглядит неизменным, независимо от времени инкубации (фиг.3). Проведенный анализ интернализации фрагментов экзогенной ДНК в ДК свидетельствует о том, что идет активное взаимодействие и проникновение экзогенной ДНК во внутренние компартменты указанного типа клеток. При этом в межхромосомном пространстве ядра, а также в цитоплазме, депонируется (или транзитно находится, предположительно для цитоплазмы) большое количество фрагментов экзогенной ДНК, которые с большой долей вероятности могут запускать многочисленные процессы, активирующие ДК.
Рассмотрим влияние препаратов ДНК на фенотип и функциональную активность дендритных клеток in vitro.
Иммуноактивные свойства ДНК млекопитающих, и в частности человеческого происхождения, особенно в отношении ДК, практически не исследованы, хотя имеются веские основания предполагать, что фрагментированная ДНК, источником которой могут быть клетки, разрушившиеся в результате апоптоза или некроза, способна взаимодействовать с антигенпрезентирующими клетками и оказывать влияние на процессы их дифференцировки и созревания [18, 19, 23-26].
В этой связи мы оценили степень влияния геномной человеческой ДНК из плаценты человека, представляющей смесь фрагментированной двуцепочечной ДНК здорового человека, на процессы дифференцировки/созревания, а также функциональную активность ДК. В качестве сравнения были использованы образцы Cotl человеческой ДНК, представляющей собой смесь повторяющихся последовательностей ДНК человека в двуцепочечной форме размером 100-300 п.о., не содержащих нуклеосом, и стандартный индуктор активации ДК ЛПС.
Для изучения влияния препаратов ДНК человека использовались ДК, генерированные по интерфероновому протоколу (GM-CSF+IFN-α). Данный тип ДК обладает свойствами как миелоидных, так и плазмоцитоидно-подобных клеток различной степени зрелости, в частности, экспрессирует TLR7 и TLR9 [27-29].
Проведенные нами ранее исследования показали, что ДК больных ТБ характеризуются задержкой дифференцировки/созревания и у значительной части больных - снижением аллостимуляторной активности [30, 31]. Поэтому наряду со здоровыми донорами оценка влияния ДНК на фенотип и функции ДК проводилась также в культурах ДК больных ТБ. Эти исследования представляли интерес как в плане анализа эффектов ДНК на ДК, так и в плане потенциального применения препарата ДНК для коррекции нарушений ДК при инфекционных заболеваниях.
Данные, характеризующие влияние ДНК на аллостимуляторную активность ДК здоровых доноров, представлены в Таблице 2. ЛПС усиливал аллостимуляторную активность ДК у 2-х из 3 здоровых доноров (доноры 2 и 3). В этом случае ДК, обработанные ЛПС, индуцировали более высокий уровень пролиферативного ответа в СКЛ, чем контрольные ДК, культивированные без дозревающего стимула (в отсутствие ЛПС). Использование ДНК в качестве дозревающего стимула ДК оказывало практически такой же эффект, как и ЛПС с наибольшим стимулирующим действием в дозе 5 и 25 мкг/мл. В одном случае (донор 1) ЛПС практически не усиливал аллостимуляторную активность ДК (ИВ=1,06), что, по-видимому, связано с индивидуальными (генетическими?) особенностями, обеспечивающими достаточно эффективное созревание ДК под влиянием GM-CSF+IFN-α в отсутствии дополнительной, митогенной стимуляции. Интересно отметить, что в этом случае и ДНК не оказывала стимулирующего эффекта ни в одной из анализируемых доз. Таким образом, в целом у здоровых доноров эффект ДНК на аллостимуляторную активность является схожим с эффектом ЛПС и проявляется в усилении способности ДК стимулировать пролиферативный ответ в СКЛ.
Примечание: представлены значения пролиферативного ответа (имп/мин) в аллогенной СКЛ. Отвечающие МНК доноров культивировали с ДК в соотношении 10:1. Позитивным контролем (ЛПС) служили ДК, созревание которых индуцировали ЛПС, негативным контролем (О) - ДК, генерируемые в отсутствие дозревающего стимула. ИВ - индекс влияния, отношение ответа в СКЛ при стимуляции ДК, генерированными с дозревающим стимулом, к ответу в СКЛ, при стимуляции ДК, генерированными без дозревающего стимула.
На следующем этапе был проведен анализ влияния ДНК на аллостимуляторную активность ДК, полученных от 5 больных ТБ (Таблица 3). Уровень пролиферативного ответа в СКЛ, индуцированной ДК больных, был ниже, чем в СКЛ, индуцированной ДК здоровых доноров. Так, например, при использовании ДК, обработанных ЛПС, пролиферативный ответ в СКЛ варьировал от 6938 до 18524 имп/мин, тогда как в группе доноров - от 36583 до 50089 имп/мин. По сравнению с негативным контролем (ДК, генерируемые в отсутствие дозревающего стимула) ЛПС усиливал аллостимуляторную активность ДК (ИВ>1,2) у 3-х из 5 пациентов. Усиление пролиферации в СКЛ после обработки ДК препаратом ДНК наблюдалось также у 3-х больных. В двух случаях (больной 1 и 2) стимулирующий эффект ДНК проявлялся в отношении ДК, которые отвечали на ЛПС-стимуляцию. В одном случае (больной 5) стимулирующий эффект ДНК проявлялся в культурах ДК, резистентных к действию ЛПС. При этом следует отметить, что по сравнению с донорами у больных ТБ не наблюдалось, во-первых, дозозависимого эффекта ДНК в отношении ДК, и, во-вторых, аллостимуляторная активность ДК, индуцированная ДНК, была ниже, чем при использовании ЛПС в качестве дозревающего стимула.
Эффект ДНК, обогащенной повторяющимися последовательностями (Cotl), на аллостимуляторную активность ДК был проанализирован в культурах ДК 4 больных ТБ (Таблица 3).
Примечание: в таблице представлены значения пролиферативного ответа в СКЛ (имп/мин) мононуклеаров доноров при стимуляции ДК больных ТБ. ИВ - индекс влияния, отношение ответа в СКЛ при стимуляции ДК, генерированными с дозревающим стимулом, к ответу в СКЛ, при стимуляции ДК, генерированными без дозревающего стимула.
Стимулирующий эффект обогащенной повторами ДНК (Cotl) регистрировался у 3 из 4-х больных. Причем, если у больного 2 аллостимуляторная активность ДК усиливалась как при обработке ЛПС, так и Cotl ДНК, то у больных 4 и 5 усиление аллостимуляторной активности ДК индуцировалось только Cotl, но не ЛПС. Это означает, что эффект ЛПС и обогащенной повторами ДНК, очевидно, реализуется через различные внутриклеточные сигнальные пути. Важно также подчеркнуть, что у больного 4 усиление аллостимуляторной активности ДК наблюдалось только при обработке ДК Cotl ДНК, и не выявлялось при обработке ЛПС или ДНК, что указывает на возможные различия механизмов влияния фрагментированной ДНК и обогащенной повторами ДНК.
Поскольку известно, что более зрелые миелоидные ДК обладают более выраженной аллостимуляторной активностью эффект ДНК может быть связан с индукцией созревания ДК. Чтобы убедиться в этом, был проведен анализ влияния ДНК человека и ДНК, обогащенной повторяющимися последовательностями, на процентное содержание CD83+и CD14+ ДК у здоровых доноров (Таблица 4). Внесение в культуры ДК ДНК сопровождалось снижением доли CD14+ клеток и увеличением относительного содержания CD83+ клеток (донор 4 и донор 5). Наиболее эффективным при этом было использование ДНК в дозе 5 мкг/мл. Причем, если у донора 5 эффект ДНК (5 мкг/мл) был сравним по выраженности и направленности с действием ЛПС, то у донора 4 ЛПС не оказывал заметного эффекта на созревание ДК, а обработка ДНК (все три исследуемые дозы) индуцировала их созревание (т.е. приводила к снижению доли CD14+ и увеличению доли CD83+ клеток).
Примечание: здесь и в таблице 5 ИВ - индекс влияния, отношение числа клеток, выращенных в присутствии дозревающего стимула, к числу клеток, выращенных в отсутствие дозревающего стимула.
Влияние обогащенной повторами ДНК было протестировано в культурах ДК донора 6, созревание которых эффективно индуцировалось ЛПС. Увеличение доли CD83+ клеток наблюдалось при обработке ДК Cotl в дозе 1 мкг/мл, тогда как снижение доли CD14+ клеток требовало больших концентраций обогащенной повторами ДНК.
В заключение был исследован эффект геномной ДНК и Cotl на созревание ДК больных ТБ (Таблица 5).
Как видно, у всех трех пациентов созревание ДК эффективно индуцировалось ЛПС, о чем свидетельствовало увеличение числа CD83-позитивных ДК. Аналогичный эффект, хотя и менее выраженный, регистрировался при использовании ДНК в качестве дозревающего стимула. Причем, если у доноров наиболее эффективна была доза 5 мкг/мл, то у больных ТБ - 25 мкг/мл. Обогащенная повторами ДНК также индуцировала созревание ДК. При этом оптимальный эффект наблюдался при обработке ДК Cotl в дозе 5 мкг/мл, тогда как у доноров - в дозе 1 мкг/мл.
Рассмотрим феномен активации ДК при воздействии на них экзогенной аллогенной фрагментированной двуцепочечной геномной ДНК.
В проведенной работе мы определили некоторые особенности взаимодействия ДК и экзогенной ДНК в форме статистических геномных фрагментов размером, кратным нуклеосомному мономеру (~200 п.о.) и составляющим 1-20 нуклеосом (200-6000 п.о.).
Известно, что в плазме крови и других интерстициальных жидкостях организма присутствует экстраклеточная ДНК, происходящая от клеток, подвергшихся апоптозу (или от любых других погибших клеток, например, раковых или клеток некротической ткани) [32-36]. До настоящего времени мало что известно о судьбе этой ДНК. Очевидно, что деградация до мономеров и их последующее участие в синтетических процессах клетки самый простой и легко объяснимый путь утилизации экзогенной экстраклеточной ДНК. Тем не менее, с одной стороны известно, что экстраклеточная ДНК может интернализоваться в клеточных компартментах и принимать участие в механизмах репаративной рекомбинации [7, 22, 37]. С другой стороны показано, что ДНК микроорганизмов или ДНК, выделяющаяся в кровь при повреждении и некрозе тканей, может активировать реакции врожденного и адаптивного иммунитета [24, 38]. Это во многом объясняет существующие многочисленные данные о том, что экзогенная ДНК может действовать и на опухоли, тормозя их прогрессию. Все эти свойства экзогенной ДНК имеют прямое отношение к функционированию иммунокомпетентных клеток, и в частности ДК [39, 40]. В этой связи мы провели серию экспериментов, направленных на анализ влияния экзогенной ДНК на активность ДК человека. Оказалось, что экзогенная ДНК проникает в ядерное пространство ДК человека, практически не задерживаясь в цитоплазме. В межхромосомном пространстве фрагменты экзогенной ДНК депонируются и как минимум в течение 6 часов не подвергаются процессингу (конкатамеризации) (Фиг.3), описанному для других типов клеток [2, 21]. Количество интернализованных в межхромосомном пространстве ядра фрагментов экзогенной ДНК достигает 10000 (если считать средний размер фрагмента 1000 п.о.). Характерной особенностью используемой ДНК являлось отсутствие гистонов и нуклеосомной организации, хотя розетки белков ядерного матрикса, ассоциированных с фрагментами ДНК, наблюдались в достаточном количестве при электронно-микроскопическом анализе (Фиг.1). Также важными характеристиками используемого препарата ДНК являлось его человеческое происхождение и размер, кратный 1-20 нуклеосомным мономерам (200-6000 п.о.), который соответствует физиологическому размеру ДНК циркулирующей в крови (фиг.2).
В то же самое время мы установили, что экзогенная ДНК в такой форме индуцирует созревание ДК человека и усиливает их аллостимуляторную активность с эффективностью, соответствующей ЛПС, часто используемого в качестве «стандартного» дозревающего стимула [41, 42]. Традиционно ДК генерируют путем культивирования прилипающей фракции мононуклеарных клеток в присутствии GM-CSF и IL-4 с последующим добавлением «коктейля» дозревающих цитокинов - IL-1, IL-6, PGE2, TNF-α [43, 44]. Однако в последнее время было показано, что частично зрелые ДК могут быстро генерироваться при культивировании моноцитов с GM-CSF и IFN-α, и такой путь генерации представляется более физиологичным [45]. Полученные в этом случае ДК, названные «интерфероновыми» ДК, имеют ряд преимуществ - генерируются быстрее по времени, характеризуются высокой способностью к захвату антигена, сохраняют стабильность в отсутствие цитокинов, имеют более высокую миграционную активность и эффективно стимулируют ТЫ ответ [45, 46]. В проведенных исследованиях мы использовали именно такой протокол генерации ДК. Анализ активации ДК экзогенной ДНК свидетельствует о следующих особенностях этого процесса. Как геномная фрагментированная ДНК, так и Cotl ДНК человека индуцируют созревание ДК и повышают их аллостимуляторную активность. К стимулирующему действию ДНК чувствительны ДК как здоровых доноров, так и больных ТБ. При этом в обеих анализируемых группах (доноров и больных) выявлены количественные отличия для геномной, и для Cotl ДНК в стимуляции ДК. Выраженность ДНК-опосредованной стимуляции оказалась сопоставимой с эффектом ЛПС. Тем не менее, были выявлены доноры, отвечающие на один индуктор (например, ДНК) и не отвечающие на ЛПС, и наоборот. Этот факт согласуется с тем, что индукция созревания ДК под влиянием ЛПС происходит TLR4 зависимым путем, тогда как для геномной и Cotl ДНК это путь полностью не изучен, и более того, как минимум для геномной ДНК, не связан с TLR9-CpG индукцией [42, 47].
Известно, что процесс созревания ДК из моноцитов крови человека сопровождается изменением экспрессии поверхностных антигенов - CD 14 и CD83 [48]. В популяции незрелых ДК доля CDH-позитивных клеток выше, чем CD83+, а при созревании соотношение меняется на противоположное. Оказалось, что эффективность созревания ДК, индуцированная как геномной, так и Cotl ДНК, сравнима с действием ЛПС и имеет дозозависимую выраженность. К стимулирующему действию геномной или Cotl ДНК были чувствительны ДК как здоровых доноров, так и больных ТБ, хотя в последнем случае реализация эффекта требует больших концентраций ДНК/Cotl. Полученные результаты свидетельствуют о способности препарата ДНК и ДНК, обогащенной повторяющимися последовательностями, стимулировать созревание ДК и усиливать их аллостимуляторную активность. При сходстве эффекта ДНК с ЛПС, действие этих индукторов опосредуется, по-видимому, разными сигнальными путями и/или различными субпопуляциями «интерфероновых» ДК, например, миелоидными или плазмоцитоидно-подобными клетками, отличающимися по степени зрелости.
Обобщенная картина полученных данных может свидетельствовать о следующем. Экзогенная ДНК в форме фрагментов геномной ДНК (или в виде Cotl повторов) поглощается ДК и интернализируется в межхромосомном пространстве клеточного ядра, не задерживаясь в цитоплазме. Взаимодействие с клеткой, продвижение в ядерное пространство и депонирование в нем фрагментов экзогенной ДНК приводит к активации ДК, что сопровождается их созреванием (изменением CD14/CD83 фенотипа) и усилением функциональной активности (способности стимулировать пролиферацию аллогенных Т-лимфоцитов в СКЛ).
ДК, являясь наиболее «профессиональными» антиген-презентирующими клетками, обеспечивают тесную взаимосвязь реакций врожденного и приобретенного иммунитета. Активация и развитие неспецифического и антиген-специфического иммунного ответа, во многом зависит от эффективности связывания соответствующих рецепторов с определенным типом агонистов. Агонистами для активации врожденного иммунитета являются так называемые PAMP (pathogen associated molecular patterns), которые связываясь с рецептором PRR (pattern-recognition receptor) индуцируют синтез и секрецию цитокинов и костимулирующих молекул. Адаптивный иммунный ответ связан с антиген-специфическими рецепторными комплексами Т- и В-лимфоцитов (TCR и BCR, соответственно). Соматическая перестройка генов TCR и BCR формирует в высшей степени разнообразный репертуар молекул с возможностью опознавать почти все антигенные детерминанты специфическим образом [38, 49]. Факты, полученные в исследованиях последних лет, предполагают, что нуклеиновые кислоты являются индукторами как врожденного, так и адаптивного иммунитета.
Наибольший интерес в плане рассматриваемой проблемы противоопухолевой активности иммунной системы представляет собой специфическое взаимодействие рецептора TLR9 и CpG ДНК. Этот рецептор располагается в эндосомальных компартментах в составе эндоплазматического ретикулюма [20, 42, 47]. Эффект CpG ДНК на развитие иммунного ответа может быть прямым или опосредоваться через активацию иммунных клеток, конститутивно экспрессирующих TLR9, таких как В-лимфоциты и ДК [50]. При этом ДК становятся компетентными индуцировать развитие CD8+ и CD4+ Т лимфоцитов в направлении преимущественной дифференцировки цитотоксических CD8+ Т клеток по сравнению с нормальным Т генезом [5 0-53]. TLR9 до настоящего времени является единственным хорошо изученным рецептором, который при взаимодействии с CpG ДНК формирует иммунный ответ [24, 39]. Предполагается, что TRL9 опосредованный путь активации иммунной системы связан как с присутствием неметилированных последовательностей CpG, так и с любой ненормальной структурой олигонуклеотида, содержащего, например, фосфоротиоатный остов. Все лиганды TRL9 являются полимерами с циклическими компонентами гидрофобной природы [24].
В последнее время стало очевидным, что двуцепочечная ДНК различного происхождения и в различной форме и не зависимо от последовательности может участвовать в регуляции обоих типов иммунного ответа. Причем эта регуляция не зависит от активации TLR9 и предполагает наличие альтернативных молекулярных механизмов активации иммунокомпетентных клеток [18, 25, 26, 54, 55]. Охарактеризованы различные индукторные формы ДНК и определены эффекторные свойства иммунокомпетентных клеток, индуцированных этой ДНК. Показано, что двуцепочечная ДНК в форме нуклеосом [18, 19], очищенная геномная ДНК млекопитающих, бактерий, вирусов [25, 55], двуцепочечная ДНК, не содержащая CpG мотивы [55, 56], одноцепочечные CpG олигонуклеотиды [57] приводят к созреванию и усилению функциональной активности ДК и макрофагов. Этот процесс характеризуется активацией синтеза костимуляторных молекул (МНС I и II класса, CD40, CD54, CD86, CD83) [18, 25, 56, 58-60], цитокинов (INF-α/β, TNF-α, IL-6, IL-8, IL-12p40/70) [18, 25, 55], нитрооксида (NO) [59].
До настоящего времени не известно, каким образом происходит такого рода активация, хотя известно, что для этого процесса необходимо функционирование определенного набора специфических генов [56].
Проведенные в настоящей работе исследования свидетельствуют о том, что фрагментированная экзогенная аллогенная ДНК активирует созревание ДК, что проявляется увеличением числа СD83+ клеток и снижения доли СD14-позитивных клеток. Экзогенная ДНК повышает аллостимуляторную активность ДК в СКЛ, что указывает на способность ДК под влиянием ДНК эффективно распознавать аллоантигены и в ответ на это секретировать в окружающую среду различные цитокины (IL-12, IFN-γ), стимулирующие пролиферацию отвечающих Т-клеток. Нахождение экзогенной ДНК длительное время в межхромосомном пространстве ядер ДК может быть одним из факторов, определяющих сдвиг молекулярных процессов ДК в сторону их активации. Известно, что экзогенная ДНК двуцепочечными концами активирует систему иерархических киназ и арестует клеточный цикл до полного удаления раздражителя [61-63]. Можно предположить, что в случае ДК происходит та же самая ситуация, что приводит к запуску репаративных молекулярных процессов и, в конечном итоге, к активации генов, определяющих иммунокомпетентность ДК.







| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2009 |
|
RU2429019C2 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2010 |
|
RU2489169C2 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2008 |
|
RU2387456C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2675233C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2007 |
|
RU2345792C2 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2006 |
|
RU2313349C2 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2012 |
|
RU2490028C1 |
| СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 2006 |
|
RU2322264C1 |
| СПОСОБ ТОРМОЖЕНИЯ РОСТА ПОДКОЖНОГО ТРАНСПЛАНТАТА ЭКСПЕРИМЕНТАЛЬНОЙ ГЛИОБЛАСТОМЫ ЧЕЛОВЕКА U-87, ПЕРЕВИТОГО ИММУНОДЕФИЦИТНЫМ МЫШАМ NU/J | 2019 |
|
RU2717218C1 |
| СПОСОБ ДЕТЕКЦИИ СТВОЛОВЫХ РАКОВЫХ КЛЕТОК | 2013 |
|
RU2530170C1 |
Изобретение относится к биотехнологии и может быть использовано при лечении заболеваний, в частности онкологических. Способ основан на заборе у пациента незрелых дендритных клеток, их культивирования ex vivo для созревания и формирования аллостимуляторной активности. Клетки дополнительно собирают антигеном и вводят их тому же пациенту для формирования адаптивного иммунитета. При культивировании ех vivo незрелых дендритных клеток в культуральную среду вводят фрагментированную аллогенную двуцепочечную геномную ДНК с фрагментами, размером 200-6000 п.о. в количестве 5 мкг/мл среды. Изобретение позволяет расширить область применения полученного продукта с повышением общего иммунного состояния пациентов. 1 з.п. ф-лы, 5 табл., 5 ил.
1. Способ получения стимулированных дендритных клеток для индукции иммунного ответа против туберкулеза человека, основанный на заборе у пациента незрелых дендритных клеток, их культивировании ex vivo для созревания и формирования аллостимуляторной активности для последующей дополнительной обработки антигеном и обратном введении созревших дендритных клеток тому же пациенту с целью формирования адаптивного иммунитета, отличающийся тем, что при культивировании ех vivo незрелых дендритных клеток в культуральную среду вводят фрагментированную аллогенную двуцепочечную геномную ДНК с фрагментами размером 200-6000 п.о., составляющими полный геном физиологически и генетически здорового донора, в количестве 5 мкг/мл среды.
2. Способ по п.1, отличающийся тем, что препарат фрагментированной аллогенной двуцепочечной геномной ДНК с фрагментами размером 200-6000 п.о., составляющими полный геном физиологически и генетически здорового донора, вводят в культуральную среду, содержащую незрелые дендритные клетки больных туберкулезом, в количестве 25 мкг/мл среды.
| ИММУНОГЛОБУЛИН ЧЕЛОВЕКА ПРОТИВОСИБИРЕЯЗВЕННЫЙ ДЛЯ ВНУТРИВЕННОГО ВВЕДЕНИЯ | 2006 |
|
RU2348429C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВАКЦИНЫ ДЛЯ ЛЕЧЕНИЯ ТУБЕРКУЛЕЗА И ДРУГИХ ВНУТРИКЛЕТОЧНЫХ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ И ВАКЦИНА, ПОЛУЧЕННАЯ ПО НАЗВАННОМУ СПОСОБУ | 2001 |
|
RU2262950C2 |

