Область изобретения
Данное изобретение относится к способу получения вакцины против возбудителей туберкулеза и других внутриклеточных патогенов. Названная вакцина действует непосредственно на внутриклеточные возбудители заболеваний, в частности, в данном случае, против патогенных Mycobacterium tuberculosis и Salmonella.
Преимущество данного изобретения заключается в разработке вакцины против внутриклеточных возбудителей заболеваний, которые являются причиной туберкулеза, бруцеллеза, лейшманиоза, лейстериоза, лепры, малярии, брюшного тифа, трипаносомоза и стрептококковой инфекции и HIV-инфекции (ВИЧ). Патогенная Mycobacterium tuberculosis (M.tuberculosis), объект данного изобретения, является агентом, вызывающим туберкулез. В данном изобретении предусмотрено выращивание M. tuberculosis в аллогенных и сингенных макрофагах и в клеточных линиях макрофагов. Затем комплекс макрофаги-М.tuberculosis облучали, чтобы убить макрофаги, а также микобактерии.
Предпосылки изобретения
Туберкулез является хроническим инфекционным заболеванием, которое продолжает убивать около 3 миллионов человек в год. Каждый год возникает приблизительно 8 миллионов новых случаев и их количество продолжает увеличиваться. Приблизительно одна треть населения мира инфицирована М.tuberculosis. Появление AIDS (СПИДа) реактивировало туберкулез у миллионов индивидуумов со скрытой инфекцией, вызвав резкое увеличение числа случаев заболевания и смертей. Поэтому среди всех инфекционных агентов М.tuberculosis ответственны за очень высокий коэффициент заболеваемости. Единственная имеющаяся вакцина, BCG (БЦЖ), является как непредсказуемой, так и изменчивой. Сомнительная эффективность вакцинации BCG заставляет научное сообщество настойчиво разрабатывать эффективные средства вакцинации против М.tuberculosis (Bloom, B.R. et al., Annu. Rev. Immunol. 10:1992:453).
Во всем мире в течение последних многих десятилетий широко использовали BCG в качестве вакцины. Несколько сотен миллионов детей и новорожденных были реципиентами вакцины BCG. Однако, несмотря на широкое применение вакцины BCG, туберкулез все еще остается наиболее быстро распространяющимся заболеванием не только в развивающихся странах, но также в индустриальном мире. Кроме того, защитная эффективность имеющейся вакцины BCG оказалась как непредсказуемой, так и высоко изменчивой, и она остается наиболее спорной из всех используемых в настоящее время вакцин. Сомнительная эффективность BCG в контрольных испытаниях усилила беспокойство относительно ее применения в качестве вакцины (Bloom and Fine, Tuberculosis, B. Bloom (ed.), 1994:531; Bloom, B.R. et al., Annu. Rev. Immunol. 10:1992:453). Кроме того, проведенные в Мадрасе интенсивные клинические испытания показали одинаковую степень предупреждения заболевания у вакцинированных BCG и невакцинированных индивидуумов, указывая, что BCG индуцировала нулевую защиту (Ind. J. Med. Res. 1980:72 (Suppl.):1-74). Таким образом, очевидно, что вакцинация BCG не предотвращает трансмиссию.
В прошлом также уже возникали многочисленные вопросы относительно безопасного применения вакцины BCG.
Главная катастрофа, которая бросила тень на репутацию вакцинации BCG, произошла в 1929 г. В Любеке, Германия, 251 ребенок получил вакцину BCG, приготовленную в местном институте, и 72 из этих детей умерло. Последующее исследование выявило, что в институте также сохраняли культуры вирулентных туберкулезных бацилл и что партия вакцины BCG, данная детям, случайно была загрязнена одним из этих штаммов Mycobacterium tuberculosis (Lubeck. 1935. Die Sauglingstuberkulose in Lubeck. Springer, Berlin).
Новый вопрос возник относительно безопасности BCG для HIV-инфицированных индивидуумов. Описано небольшое количество случаев диссеминированного BCG-оза (BCG-туберкулеза) среди детей, которым вводили вакцину BCG и которые, как было впоследствии обнаружено, были HIV-сероположительными (Von Reyn, et al. Lancet 1987 : ii:669-672; Braun, et al., Pediatr. Infect. Dis. J. 1992:11:220-227; Weltman, et al., AIDS 7:1993:149). WHO (ВОЗ) рекомендовала прекратить применение вакцины BCG детям, проявляющим симптомы иммунодефицита (Всемирная Организация Здравоохранения. 1992. Expanded Program for Immunization. Program Report. World Health Organization, Geneva. World Health Organization. Weekly Epidemiol. Rec. 62:1987:53).
Недавно были завершены исследования большого количества добровольцев, проведенные Dahlstrom и Difts (Scand J. Respir Dis Suppl. 65:1968:35), и мета-анализ BCG для предупреждения туберкулеза, базировавшийся на 13 проспективных исследованиях и 10 контрольных исследованиях пациентов (Colditz et al., J. Amer. Med. Assoc. 271:1994:698-702). В то же время сделан вывод, что в среднем BCG защищает около 50% при предупреждении туберкулеза, биологическая и прикладная значимость усреднения, по существу, таких очень отклоняющихся результатов является сама по себе спорной.
До появления СПИДа в наиболее богатых странах, по крайней мере, в течение столетия происходило снижение заболеваемости туберкулезом. Это проиллюстрировано проведенными сравнениями между Нидерландами (которые никогда не применяли вакцинацию BCG) и Соединенным Королевством и Скандинавскими странами (которые ввели государственную вакцинацию BCG в 1950 г.). Уменьшение случаев туберкулеза, описанное в названных странах, было одинаковым (Styblo, K., Selected Papers R. Netherland Tuberc. Assoc. 24:1991:136; Sutherland, Bull. Int. Union. 57:1981:17). Таким образом, оказалось нецелесообразным считать, что снижение обусловлено только вакцинацией BCG.
Проведение вакцинации BCG базируется на гипотезе, что BCG эффективна против первичной инфекции у детей и эндогенной реактивации застарелых инфекций, но не против экзогенной инфекции (ten Dam, H.G. Adv. Tuberc. Res. 21:1984:79; ten Dam, H.G. and A. Pio. Tubercle 63:1988:226). Эпидемиологические данные позволяют предположить, что вакцинация BCG обеспечивает в большей степени или более стойкую защиту против системного заболевания, в частности милиарного туберкулеза и туберкулезного менингита у детей, чем против заболевания легких (Rodrigues, et al.. Int. J. Epidemiol. 22:1993:1154). Исследования Лурье (Lurie) продемонстрировали, что количество CFU (КОЕ) М.tuberculosis, выделенных из легких кроликов, иммунизированных BCG, против неиммунизированных, не выявило никаких различий в количестве организмов, извлеченных из легких и других тканей и способных культивироваться.
Другое представление возникает из внутриклеточной локализации микобактерий. Данные электронной микроскопии свидетельствуют о том, что по существу BCG полностью остается в фаголизосомах после инфицирования макрофагов in vitro, тогда как вирулентные М.tuberculosis (штамм H37Rv) могут выходить из фаголизосом и проникать в цитоплазму (McDonough, et al., Infect. Immun. 61:1993:276). Это может иметь определенное значение, поскольку они являются антигенами в эндосомальном компартменте антиген-представляющих клеток, которые присутствуют вместе с детерминантами МНС класса II к CD4+ Т-хелперным клеткам, тогда как цитоплазматические антигены присутствуют в ассоциации с детерминантами главного комплекса гистосовместимости (МНС) класса I к CD8+ цитотоксичным Т-клеткам (CTL). Если полученные in vitro показатели являются общими, они объясняют, почему М.tuberculosis, при элиминации, оказываются более зависимыми от МНС класса I-рестриктированных CTL, чем BCG, и позволяют предположить, что BCG не может быть очень эффективной в активации МНС класса I-рестриктированных CTL (Stover, et al., Nature 351:1991:456). В этом контексте Rich, 1951 (The Pathogenesis of Tuberculosis, 2nd, 1028р. Charles С Thomas, Publisher, Springfield, III), Canetti, 1955 (The Tubercule Bacilli in the Pulmonari Lesion of Man, 226p.Springer, New York) и Lurie, 1964 (Resistance to Tuberculosis. Experimental Studies in Native and Acquired Defense, 391p.Harvard University Press, Cambridge Press, Cambridge, Mass) поясняли, что выздоровление после инфекции М.tuberculosis обеспечивало более сильную защиту против будущего туберкулеза, чем может BCG.
Эффективная резистентность к инфекции М.tuberculosis требует участия как специфических CD8+ CTL для лизиса макрофагов или паренхиматозных клеток, неспособных ограничить их инфекцию, так и специфических CD4+ Т-клеток, способных продуцировать IL-2, IFN-γ, TNF-α и другие лимфокины, вовлеченные в активацию макрофагов.
Учитывая упомянутые недостатки вакцины BCG, в данном изобретении использовали тот факт, что вакцину следует применять в виде облученного препарата и без опасения заражения больных СПИДом или детей с ослабленным иммунитетом. BCG вводят в виде аттенуированного препарата и не рекомендуют введение упомянутым субъектам, так как она вызывает диссеминированный BCG-оз. В настоящее время WHO (ВОЗ) рекомендует прекратить применение вакцины BCG детям, проявляющим явные симптомы иммунодефицита (World Health Organization. 1992. Expanded Program for Immunization. Program Report. World Health Organization, Geneva. World Health Organization. Weekly Epidemiol. Rec. 1987:62:53-54).
Еще одно представление связано с внутриклеточной локализацией микобактерий. BCG по существу полностью остается в фаголизосомах макрофагов, тогда как вирулентные М.tuberculosis могут просачиваться из фаголизосом и проникать в цитоплазму (McDonough, К.A., Y. Kress, and B.R. Bloom. 1993. Infect. Immun. 61:2763:2773). В эндосомальном компартменте антиген-представляющих клеток антигены присутствуют вместе с детерминантами МНС класса II к CD4+ Т-хелперным клеткам, тогда как цитоплазматические антигены присутствуют в ассоциации с детерминантами главного комплекса гистосовместимости (МНС) класса I к CD8+ цитотоксическим Т-клеткам. М.tuberculosis для их элиминации, оказываются более зависимыми от МНС-класса I-рестриктированных CTL. BCG является не эффективной в активировании комплекса МНС-класса I-рестриктированные CTL (Stover, et al., Nature 351:1991:456). Данная вакцина содержит облученный препарат М.tuberculosis, выращенных в макрофагах. Описано, что макрофаги, инфицированные М.tuberculosis, эффективно генерируют CTL (Stover, et al., Nature 351:1991:456). Кроме того, также описано, что облученные клетки претерпевают апоптоз и могут быть фагоцитированы дендритными клетками (Albert, et al., Nature 392:1998:86), и это приводит к возникновению антиген-специфического ответа CD4+ и CD8+ Т-клеток. Описанный апоптоз-зависимый путь может иметь значение не только при изучении вакцинации, но также для терапевтического воздействия на иммуннную систему, чтобы индуцировать in vivo ответ Т-хелперов и CTL на целый ряд антигенов, включая опухоль, и возможность модулировать благоприятный иммунный ответ.
Rich (The pathogenesis of Tuberculosis, 2nd ed, 1028р., 1951. Charles С Thomas, Publisher, Springfield, III), Canetti (The tubercle Bacillus in the pulmonary Lesion of Man, 226p, 1955. Springer, New York), и Lurie (Resistance to Tuberculosis. Experimental Studies in Native and Acquired Defense, 391, 1964. Harvard University Press, Cambridge, Mass) отмечали, что выздоровление после инфекции М.tuberculosis обеспечивало более устойчивую защиту против будущего туберкулеза, чем в случае BCG. В контексте вышеприведенных утверждений предлагаемая вакцина имеет преимущество перед существующей вакциной BCG, так как она содержит М.tuberculosis, выращенные в естественной среде макрофагов, которые секретируют уникальные антигены, ответственные за индукцию защитного иммунного ответа, и могут генерировать образование CD4+ Т-хелперных клеток и CD8+ CTL. Эффективная резистентность к М.tuberculosis требует участия как специфических CD8+ CTL для лизиса макрофагов или паренхиматозных клеток, неспособных ограничивать их инфекционность, и специфических CD4+ Т-клеток, способных продуцировать IL-2, IFN-γ, TNF-α и другие лимфокины, вовлеченные в активацию макрофагов.
Основными логическими задачами данного способа являются следующие составляющие: разработка вакцины против туберкулеза и других внутриклеточных заболеваний, где МНС-совместимые (сингенные) и несовместимые (аллогенные) макрофаги, укрывающие М.tuberculosis при облучении претерпевают апоптоз, при этом дендритные клетки поглощают эти макрофаги и присутствующий на их поверхности антиген (микобактерии-белки и алломакрофагальные пептиды) и стимулируют наивные Т-клетки к дифференцировке в эффекторные CD4+ Th1 клетки. Упомянутые дендритные клетки также активируют CD8+ Т-клетки для опосредованного клеткой иммунитета. Алломакрофаги в системе генерируют аллореакцию, в результате которой образуются цитокины, подобные IL-2, IL-12, IFN-γ и так далее, которые усиливают ответ Th1 и иммунный ответ, опосредованный клеткой. Известно, что Th1-тип ответа обеспечивает защиту против туберкулеза. Следовательно, главное преимущество способа заключается в производстве мощной и специфической вакцины против М.tuberculosis.
Цели изобретения
Таким образом, главным аспектом данного изобретения является разработка вакцины против туберкулеза и других внутриклеточных заболеваний, подобных лепре, лейшманиозу, брюшному тифу, трипаносомозу, малярии, бруцеллезу, лейстериозу, СПИДу, стрептококковой инфекции и раку.
Другим аспектом данного изобретения является культивирование патогенного микроорганизма внутри сингенных и аллогенных макрофагов и предоставление им возможности секретировать антигены в клетках.
Еще один аспект заключается в разработке способа, посредством которого патогенные микроорганизмы уничтожаются уже известными лекарственными средствами и в дальнейшем перед применением подвергаются гамма-облучению; известно, что гамма-облученные клетки претерпевают апоптоз и поглощаются дендритными клетками. Дендритные клетки являются мощным активатором Тh1 клеток и CD8+ цитотоксических клеток.
Следующим аспектом является разработка вакцины, которая действует против как сингенных макрофагов, захвативших патогенные микроорганизмы (а именно, М.tuberculosis, М.leprae, leishmania, salmonella, trypanosoma, malaria, brucella, leisteria, HIV, streptococcus) (например, SMTV, S= сингенный, М= макрофаг, Т= туберкулез, V= вакцина), так и аллогенных макрофагов, захвативших патогенную вакцину (например, AMTV, А= алло, М= макрофаг, Т= туберкулез, V= вакцина), чтобы генерировать защитный иммунный ответ.
Еще одним аспектом данного изобретения является разработка вакцины, основанной на поглощении патогенного микроорганизма аллогенными клетками, что должно вызвать иммунный ответ независимо от генетического фона, то есть она будет действовать как смешанная вакцина и, следовательно, она будет действовать независимо от генетического разнообразия у субъектов.
Краткое изложение изобретения
Данное изобретение относится к способу получения вакцины против возбудителей туберкулеза и других внутриклеточных патогенов. Данная вакцина действует непосредственно на внутриклеточные патогенные микроорганизмы, в частности, в данном случае, патогенные Mycobacterium tuberculosis и Salmonella.
Подробное описание изобретения
Новизна данного изобретения заключается в том, что защитные антигены, секретируемые микобактериями внутри макрофагов, можно использовать в качестве вакцины без выделения их из макрофагов.
Вакцину использовали после облучения, а облученные клетки, как известно, подвергаются апоптоэу. Претерпевшие апоптоз клетки поглощаются дендритными клетками. Дендритные клетки активировали наивные Т-клетки к дифференцировке в Тh1 клетки и цитотоксические клетки. Известно, что клетки являются существенными в формировании защитного иммунитета против внутриклеточных инфекций и рака.
Алломакрофаги в системе генерировали аллореакцию, в результате которой продуцируется большое количество цитокинов, подобные IL-2, IL-12, IFN-γ и так далее, которые ускоряют Тh1-ответ и опосредованный клетками иммунный ответ. Используемые при создании вакцины аллогенные клетки должны вызывать иммунный ответ независимо от генетического фона, то есть вакцина будет действовать как смешанная вакцина. Следовательно, ее можно применять субъектам независимо от генетического разнообразия.
Целью данного изобретения является разработка вакцины против туберкулезной, сальмонеллезной и других внутриклеточных инфекций. М.tyberculosis и Salmonella typhimurium культивировали в аллогенных (AMTV) и сингенных (SMTV) макрофагах, убивали γ-облучением и использовали в качестве вакцины. In vivo AMTV предпочтительно поглощаются дендритными клетками (γ-облучение заставляет клетки претерпевать апоптоз, а дендритные клетки поглощают апоптотические клетки) и затем активируют микобактерия-реактивные наивные Т-клетки. Аллогенные макрофаги, которые использовали для иммунизации, действовали как адъювант и стимулировали аллогенные реактивные Т-клетки, которые продуцировали огромное количество IL-2, IFN-γ, IL-12. Названные цитокины являются существенными для роста и дифференцировки наивных Т-клеток в CD4+ и CD8+ эффекторные Т-клетки. Дендритные клетки являются предпочтительными антиген-представляющими клетками (АРС) для Th1 и цитотоксических Т-клеток (CTL). Тh1 и CD8+ CTL являются главными клетками в образовании эффективного и защитного иммунитета против М.tuberculosis. Резистентные и чувствительные к туберкулезу штаммы мышей прививали вакциной.
На чертеже схематически представлен способ действия алло-макрофагальной туберкулезной вакцины (AMTV).
Логическое представление способа действия алломакрофагальной туберкулезной вакцины (AMTV) схематически представлено на чертеже М.tuberculosis культивировали в МНС-несовместимых (аллогенных) и сингенных макрофагах. Полученный препарат подвергали γ-облучению и использовали в качестве вакцины. In vivo AMTV предпочтительно будет поглощаться дендритными клетками (известно, что γ-облучение заставляет клетки претерпевать апопотоз, а дендритные клетки поглощают апоптотические клетки) и затем стимулировать микобактерия-реактивные наивные Т-клетки. Однако макрофаги, нагруженные микобактериями, не могут непосредственно стимулировать наивные Т-клетки. Аллогенные макрофаги, которые используются для иммунизации, должны стимулировать аллогенные реактивные Т-клетки, которые продуцируют огромное количество IL-2, IFN-γ, IL-12. Названные цитокины являются существенными для роста и дифференцировки наивных Т-клеток в CD4+ и CD8+ эффекторные Т-клетки. Дендритные клетки являются предпочтительными антиген-представляющими клетками (АРС) для Th1 и цитотоксических Т-клеток (CTL). Они вызывают стимуляцию наивных Т-клеток к дифференцировке в антиген-реактивные Th1 клетки и цитотоксические Т-лимфоциты. Кроме того, дендритные клетки захватывают чужеродный антиген (в этом случае микобактериальный антиген) и действуют как резервуар, медленно высвобождая антиген в систему для активации Т-клеток, и для поддержания клеток памяти. IL-2, IFN-γ и IL-12, секретированные аллореактивными Т-клетками, будут способствовать клональной экспансии микобактерия-реактивных Th1 и цитотоксических Т-клеток. Th1 и CD8+ CTL являются главными клетками в образовании эффективного и защитного иммунитета против М.tuberculosis. Резистентные и чувствительные к туберкулезу штаммы мышей прививали вакциной. Логическое объяснение способа действия алло-макрофагальной туберкулезной вакцины (AMTV) продемонстрировано культивированием М.tuberculosis в МНС-несовместимых (аллогенных) и сингенных макрофагах. Полученный препарат подвергали γ-облучению и использовали в качестве вакцины. In vivo AMTV предпочтительно должна поглощаться дендритными клетками (известно, что γ-облучение заставляет клетки претерпевать апоптоз, а дендритные клетки поглощают апоптотические клетки) и затем стимулировать микобактерия-реактивные наивные Т-клетки. Однако макрофаги, нагруженные микобактериями, не могут непосредственно стимулировать наивные Т-клетки. Алломакрофаги, которые используются для иммунизации, должны стимулировать алло-реактивные Т-клетки, которые продуцируют огромное количество IL-2, IFN-γ, IL-12. Названные цитокины являются существенными для роста и дифференцировки наивных Т-клеток в CD4+ и CD8+ эффекторные Т-клетки. Дендритные клетки являются предпочтительными антиген-представляющими клетками (АРС) для Тh1 и цитотоксических Т-клеток (CTL). Они стимулируют наивные Т-клетки к дифференцировке в антиген-реактивные Тh1 клетки и цитотоксические Т-лимфоциты. Кроме того, дендритные клетки захватывают чужеродный антиген (в этом случае микобактериальный антиген) и действуют как резервуар, медленно высвобождая антиген в систему для активации Т-клеток и для поддержания клеток памяти. IL-2, IFN-γ и IL-12, секретированные аллореактивными Т-клетками, будут способствовать размножению клонов микобактерия-реактивных Тh1 и цитотоксических Т-клеток. Тh1 и CTL являются главными в образовании эффективного и защитного иммунитета против М.tuberculosis (Albert, M.L., et al., Nature 392:1998:86; Wang, B. et al., Proc. Natl. Acad. Sci. USA 90:1993:4156). Резистентные и чувствительные к туберкулезу штаммы мышей вакцинировали AMTV и SMTV. Эффективность вакцины контролировали посредством инфицирования мышей живыми М.tuberculosis и регистрации их гибели и определения количества оставшихся в живых бактерий в легких, селезенке и печени. Вакцинированным (4-12 недель) мышам проводили контрольное заражение 105-106 жизнеспособными М.tuberculosis H37Rv. После дополнительного периода 3-4 недель у инфицированных мышей удаляли легкие, селезенку и печень и серийные разведения гомогената органа помещали в чашки с агаром, чтобы установить количество жизнеспособных туберкулезных бацилл, оставшихся в перечисленных органах. У вакцинированных животных также следили за образованием Тh1 и Th2 клеток посредством измерения IFN-γ и IL-4. Вакцину инокулировали в подушечку стопы мыши и непрерывно контролировали индукцию аллергической реакции замедленного типа, измеряя толщину подушечки стопы.
Согласно данному изобретению предусмотрены новая вакцина против возбудителей туберкулеза и других внутриклеточных патогенов и способ ее получения. Туберкулезная вакцина (SMTV и AMTV) содержит М.tuberculosis, выращенные в МНС-совместимых и несовместимых макрофагах. Препараты облучали и использовали как индивидуальные вакцины.
Так как вакцина соответствует всем требованиям, необходимым для формирования благоприятного иммунного ответа против М.tuberculosis, полагают, что такие препараты должны действовать эффективно против туберкулеза.
Вакцина AMTV действует по смешанному способу, так как она не следует правилам МНС-рестрикции и основана на аллостимуляции и поглощении чужеродных - апоптотических клеток дендритными клетками. Тогда как вакцина SMTV действует по способу МНС-рестрикции.
Инфицированные клетки выращивали в достаточном количестве и хранили после обработки изониазидом и γ-облучения. Препарат тщательно проверяли в отношении жизнеспособных микобактерий, определяя количество живых. В вакцине ни одна из бактерий не была жизнеспособной. Мышей вакцинировали вакциной внутрибрюшинно или подкожно и проводили контрольное заражение жизнеспособными М.tuberculosis H37Rv. Проверяли жизнеспособность туберкулезных бацилл, оставшихся в легких, селезенке и печени. Животных иммунизировали вакциной и поглощение апоптотических клеток дендритными клетками регистрировали с помощью иммунофлуоресценции. Животных вакцинировали SMTV и AMTV и изучали пролиферацию и дифференцировку наивных CD4+ Th клеток в эффекторные Тh1 и Th2 подтипы. В качестве контроля также исследовали М.tuberculosis, поглощенные сингенными макрофагами. Способность SMTV и AMTV генерировать образование CD8+ цитотоксических Т-клеток контролировали с помощью стандартного анализа высвобождения Cr51.
Для проверки гипотезы аллостимуляции мышей линий Balb/c (IAd) и C57BL/6 (IAb) иммунизировали овальбумином, заключенным в аллогенные и сингенные АРС, обработанные митомицином С. Для исключения возможности превращения предпочтительно аллоответа во вторичный ответ изменяли гаплотип аллоАРС. Животным производили вторичную бустер-инъекцию овальбумина, заключенного в АРС мышей СВА (IAk). Наблюдали полную активацию CD4+ и CD88 Т-клеток. Отмечены антиген-специфическая Т-клеточная пролиферация и преимущественный Thl-ответ, о чем свидетельствует, главным образом, продукция IL-2 и IFN-γ и IgG2а-изотипа. Отмечена высокая продукция IL-2, которая указывает, что при иммунизации антигеном, заключенным в алло-АРС, обработанные митомицином С, имеет место апоптоз. Апоптотические клетки поглощаются дендритными клетками, которые затем индуцируют микобактерия-специфический и аллореактивный Т-клеточный ответ. Алло-Т-клетки составляют >10% всей Т-клеточной популяции и, как известно, индуцируют высокую секрецию IL-2. Затем IL-2, продуцируемые алло-Т-клетками, способствуют пролиферации антиген-специфических Т-клеток.
Поэтому в данном изобретении осуществлена разработка эффективной туберкулезной вакцины, основанной на новой системе доставки, направленной на дендритные клетки; М.tuberculosis культивировали в линии макрофагальных клеток, а именно J774 или в аллогенных и сингенных макрофагах. Инфицированные макрофаги обрабатывали изониазидом и облучали, а затем использовали для исследования при вакцинации против М.tuberculosis.
Рациональное объяснение способа действия алломакрофагальной туберкулезной вакцины (AMTV) продемонстрировано культивированием М.tuberculosis в МНС-несовместимых (аллогенных) и сингенных макрофагах. Полученный препарат подвергали γ-облучению и использовали в качестве вакцины. In vivo AMTV предпочтительно будет поглощаться дендритными клетками (известно, что γ-облучение заставляет клетки претерпевать апоптоз, а дендритные клетки поглощают апоптотические клетки) и затем стимулировать микобактерия-реактивные наивные Т-клетки. Однако макрофаги, нагруженные микобактериями, не могут непосредственно стимулировать наивные Т-клетки. Алломакрофаги, которые используются для иммунизации, будут стимулировать аллореактивные Т-клетки, которые продуцируют огромное количество IL-2, IFN-γ, IL-12. Названные цитокины являются существенными для роста и дифференцировки наивных Т-клеток в CD4+ и CD8+ эффекторные Т-клетки. Дендритные клетки являются предпочтительными антиген-представляющими клетками (АРС) для Тh1 и цитотоксических Т-клеток (CTL). Они стимулируют наивные Т-клетки к дифференцировке в антиген-реактивные Тh1 клетки и цитотоксические Т-лимфоциты. Кроме того, дендритные клетки захватывают чужеродный антиген (в этом случае микобактериальный антиген) и действуют как резервуар, медленно высвобождая антиген в систему для активации Т-клеток и для поддержания клеток памяти. IL-2, IFN-γ и IL-12, секретированные аллореактивными Т-клетками, будут способствовать размножению клонов микобактерия-реактивных Тh1 и цитотоксических Т-клеток. Тh1 и CTL являются главными в образовании эффективного и защитного иммунитета против М.tuberculosis (Albert, M.L., et al., Nature 392:1998:86; Wang, B. et al., Proc. Natl. Acad. Sci. USA 90:1993:4156). Резистентные и чувствительные к туберкулезу штаммы мышей вакцинировали AMTV и SMTV. Эффективность вакцины контролировали посредством инфицирования мышей живыми М.tuberculosis и регистрации их гибели и определения количества оставшихся в живых бактерий в легких, селезенке и печени. Вакцинированным (4-12 недель) мышам проводили контрольное заражение 105-106 жизнеспособными М.tuberculosis H37Rv. После дополнительного периода 3-4 недель у инфицированных мышей удаляли легкие, селезенку и печень и серийные разведения гомогената органа помещали в чашки с агаром, чтобы установить количество жизнеспособных туберкулезных бацилл, оставшихся в перечисленных органах. Также контролировали у вакцинированных мышей образование Тh1 и Th2 клеток посредством измерения IFN-γ и IL-4. Вакцину инокулировали в подушечку стопы мыши и индукцию гиперчувствительности замедленного типа непрерывно контролировали, измеряя толщину подушечки стопы.
Согласно данному изобретению предусмотрена вакцина против туберкулеза и других внутриклеточных возбудителей заболеваний, выбранных из группы, состоящей из Mycobacterium leprae, leishmania, salmonella, trypanosoma, plasmodium, brucella, leisteria, HIV, streptococcus, и рака. Изобретение также предусматривает способ получения названной вакцины, включающий в себя стадии:
i) культивирования патогенных микроорганизмов, выбранных из группы, включающей в себя Mycobacterium tuberculosis, Mycobacterium leprae, leishmania, salmonella, trypanosoma, plasmodium, brucella, leisteria, HIV, streptococcus,
ii) культивирования сингенных (тот же вид), аллогенных (другой вид) и ксеногенных (различные виды, как овца и коза) макрофагов и макрофагальных клеточных линий, выбранных из группы, состоящей из J774A, P388D1, RAW, BMC-2, ТНР-1 и так далее,
iii) инфицирования макрофагов и клеточных линий патогенным микроорганизмом,
iv) обработки инфицированных клеток известными лекарственными средствами с последующим гамма-облучением, чтобы получить вакцину,
v) иммунизации резистентных и чувствительных к заболеванию линий животных полученной выше вакциной,
vi) инфицирования животных живым патогенным микроорганизмом и регистрации их смертности и подсчета оставшихся в живых инфекционных агентов в легких, селезенке и печени, и
vii) мониторинга вакцинированных животных относительно пролиферации и образования CD4+ Тh1 и Th2 клеток и CD8+ цитотоксических Т-клеток, свидетельствующих о развитии опосредованного клетками иммунитета.
Кроме того, в изобретении предусмотрен способ получения вакцины против туберкулеза, причем названный способ включает в себя стадии:
i) культивирования Mycobacterium tuberculosis H37Rv,
ii) культивирования сингенных и аллогенных макрофагов и макрофагальных клеточных линий, выбранных из группы, состоящей из J774A, P388D1, RAW, BMC-2, ТНР-1 и так далее,
iii) инфицирования макрофагов и клеточных линий (J774, P388D1, RAW, BMC-2, ТНР-1) М.tuberculosis,
iv) обработки инфицированных клеток изониазидом и гамма-облучением для получения вакцины,
v) иммунизации резистентных и чувствительных к туберкулезу линий мышей аллогенной макрофагальной туберкулезной вакциной (AMTV) и сингенной макрофагальной туберкулезной вакциной (SMTV), полученными выше,
vi) инфицирования мышей живыми М.tuberculosis и регистрации их смертности и подсчета живых бактерий в легких, селезенке и печени,
vii) непрерывного контроля пролиферации и образования CD4+ Тh1 и Th2 клеток и CD8+ цитотоксических Т-клеток, свидетельствующих об образовании опосредованного клетками иммунитета, у вакцинированных животных,
viii) инокулирование вакцины в подушечку стопы мыши и исследования гиперчувствительности замедленного типа посредством измерения припухлости подушечки стопы, появляющейся из-за защитного иммунитета.
В изобретении также предусмотрен способ получения вакцины против salmonella, причем названный способ включает в себя стадии:
i) культивирования Salmonella typhimurium, ii) культивирования сингенных и аллогенных макрофагов и макрофагальных клеточных линий, выбранных из группы, состоящей из J774A, P388D1, RAW, BMC-2, ТНР-1 и так далее,
iii) инфицирования макрофагов и клеточных линий (J774, P388D1, RAW, BMC-2, ТНР-1) S.typhimurium,
iv) обработки инфицированных клеток митомицином С и гамма-облучением для получения вакцины,
v) иммунизации резистентных и чувствительных к туберкулезу линий мышей вакциной, полученной выше,
vi) инфицирования мышей живыми S.typhimurium и регистрации их смертности и подсчета живых бактерий в легких, селезенке и печени,
vii) непрерывного контроля у вакцинированных животных пролиферации и образования CD4+ Th1 и Th2 клеток и CD8+ цитотоксических Т-клеток, свидетельствующих об образовании опосредованного клетками иммунитета, и
viii) инокулирование вакцины в подушечку стопы мыши и исследования гиперчувствительности замедленного типа посредством измерения припухлости подушечки стопы, возникающей из-за защитного иммунитета.
В изобретении предусмотрено получение вакцины в результате поглощения М.tuberculosis, Salmonella и других внутриклеточных патогенных микроорганизмов аллогенными и сингенными макрофагами и ее использование для защиты против инфекционного агента.
Способ согласно данному изобретению проиллюстрирован примерами, представленными ниже, которые, однако, не следует рассматривать как ограничение объема притязаний данного изобретения.
Пример 1: Способ получения вакцины против возбудителей туберкулеза и других внутриклеточных патогенов
Внутриклеточные патогенные микроорганизмы, а именно Mycobacterium tuberculosis, Mycobacterium leprae, leishmania, salmonella, trypanosoma, plasmodium, brucella, leisteria, HIV, streptococcus, культивировали в макрофагах сингенных и аллогенных мышей, макрофагальных клеточных линиях J774, P388D1, RAW, ВМС-2, ТНР-1 (АТСС, Rockville). Инфицированные клетки обрабатывали изониазидом (20 мкг/мл) в течение 48 часов при 37°С/5% CO2 и облучали с дозой 0,05 кГй.
i. (а) Полученные инфицированные клетки обрабатывали лекарственным средством, специфическим в отношении патогенного микроорганизма, и потом облучали и использовали как вакцину, а их эффективность проверяли с помощью контрольного заражения вакцинированных мышей живыми бактериями. Эффективность вакцины оценивали, определяя через определенные интервалы времени жизнеспособность инфекционных микроорганизмов в легких, селезенке и печени инфицированных мышей в серийных разведениях гомогенатов органов, помещенных в чашки с агаром. Аналогично, не вакцинированным животным проводили контрольное заражение живыми бактериями и регистрировали их гибель и количество жизнеспособных бактерий определяли в легких, селезенке и печени.
(b) У вакцинированных животных осуществляли непрерывный контроль пролиферации и дифференцировки CD4+ Th клеток в бактерия-реактивные эффекторные цитотоксические Т-клетки, Th1 и Th2 клетки посредством определения IFN-γ и IL-4 с помощью ELISA-анализа.
(с) CD8+ цитотоксические клетки регистрировали, определяя высвобождение 51Cr.
Пример 2: Способ получения вакцины против туберкулеза
В другом примере Mycobacterium tuberculosis H37Rv, полученные из Центрального JALMA Института лепры, Агра, культивировали в макрофагах сингенных и аллогенных мышей, макрофагальных клеточных линиях J774, P388D1, RAW, ВМС-2, ТНР-1 (АТСС, Rockville). Инфицированные клетки обрабатывали изониазидом (20 мкг/мл) в течение 48 часов при 37°С/5% СО: и облучали с дозой 0,05 кГй.
i. (а) Полученные инфицированные клетки обрабатывали изониазидом и затем подвергали гамма-облучению, чтобы использовать как вакцину, а их эффективность проверяли контрольным заражением вакцинированных мышей живыми бактериями с дозой 105-106. В этом случае у инфицированных мышей легкие, селезенку и печень удалили через дополнительные 3-4 недели и серийные разведения гомогенатов органов помещали в чашки с агаром для определения количества жизнеспособных М.tuberculosis, остающихся в перечисленных органах. Аналогично, невакцинированным животным проводили контрольное заражение живыми бактериями и регистрировали их гибель, и количество жизнеспособных бактерий определяли в легких, селезенке и печени.
(b) У вакцинированных животных осуществляли непрерывный контроль пролиферации и дифференцировки Т-клеток в бактерия-реактивные эффекторные CD8+ цитотоксические Т-клетки и CD4+ Тh1 и Th2 клетки, определяя IFN-γ и IL-4 с помощью ELISA-анализа.
(c) CD8+ цитотоксические клетки регистрировали, определяя высвобождение 51Cr.
(d) Вакцину инокулировали в подушечку стопы мыши и определяли гиперчувствительность замедленного типа, измеряя толщину подушечки стопы.
Пример 3: Способ получения вакцины против сальмонелл
В следующем примере Salmonella typhimurium (MTCC98) культивировали в макрофагах, полученных от сингенных и аллогенных мышей, и макрофагальных клеточных линиях J774, ВМС-2 и RAW. Инфицированные клетки обрабатывали митомицином С (50 мкг/мл) и подвергали гамма-облучению (0,05 кГй).
i. (а) Полученные инфицированные клетки обрабатывали лекарственным средством и облучали и использовали как вакцину, а их эффективность проверяли контрольным заражением вакцинированных мышей 105-106 живых бактерий. Смертность животных регистрировали в течение 21 дня. У инфицированных животных удаляли легкие, селезенку и печень, а серийные разведения гомогенатов органов помещали в чашки с агаром, чтобы установить количество сальмонеллезных бацилл, оставшихся в перечисленных органах. Аналогично, невакцинированным животным проводили контрольное заражение живыми бактериями и регистрировали их гибель, и количество живых бактерий определяли в легких, селезенке и печени.
(b) У вакцинированных животных осуществляли непрерывный контроль пролиферации и дифференцировки CD4+ Th клеток в бактерия-реактивные эффекторные Тh1 и Th2 клетки, определяя IFN-γ и IL-4 с помощью ELISA-анализа.
(с) CD8+ цитотоксические Т клетки регистрировали, определяя высвобождение 51Cr.
(d) Вакцину инокулировали в подушечку стопы мыши и определяли гиперчувствительность замедленного типа, измеряя толщину подушечки стопы.
Результаты
Основные преимущества данного изобретения
(i) Приблизительно одна треть населения мира инфицирована М.tuberculosis. Приблизительно только у 5-10% развивается активный туберкулез и у 90% индивидуумов вырабатывается эффективный иммунитет против М.tuberculosis. М.tuberculosis, присутствующие в макрофагах хозяина, секретируют уникальные антигены, которые являются эффективными индукторами продолжительного защитного иммунитета. В противоположность этому, М.tuberculosis, когда культивировали in vitro на искусственной среде, секретировали антигены, которые не индуцировали оптимальный уровень защиты, а стимулированный иммунитет был кратковременным. Выдающейся особенностью способа является то, что защитные антигены микобактерий, секретированные внутри макрофагов, утилизировались без выделения их из макрофагов.
(ii) Описанные алломакрофаги, использованные в системе, функционируют как уникальная система доставки антигенов, секретированных живыми микобактериями, в дендритные клетки, так и как адъювант для усиления секреции аллореактивными Т-клетками цитокинов, а именно IL-2, IL-12, IFN-γ и так далее. Избыток секретированных IL-2 утилизировался микобактерия-реактивными защитными Т-клетками. Аллореактивные Т-клетки продуцируют, главным образом, IL-2, IL-12, IFN-γ, цитокины, ответственные за генерацию Th1-подобного иммунного ответа. Тh1 являются значимыми для индукции защитного иммунного ответа против М.tuberculosis.
(iii) Преимущество изобретения заключается в том, что γ-облученные макрофаги, инфицированные микобактериями, поглощаются дендритными клетками. Апоптотические клетки, поглощенные дендритными клетками, индуцируют активацию CD4+ Тh1 и CD8+ цитотоксических Т-клеток. Цитотоксические Т-клетки являются ответственными за уничтожение макрофагов, инфицированных микобактериями. Лизис мишени оказывается существенным в случае заболеваний, подобных туберкулезу, брюшному тифу, лепре, лейшманиозу, СПИДу и так далее, при которых возбудитель находится и размножается в макрофагах. Лизис этих клеток высвобождает возбудителей и дает возможность активированным макрофагам поглощать бактерии и элиминировать их. Дендритные клетки экспрессируют высокие уровни В7-1 и секретируют IL-12 и являются единственными мощными АРС, которые могут активировать наивные Т-клетки. Кроме того, дендритные клетки могут способствовать дифференцировке наивных Т-клеток в Тh1 и CD8+ цитотоксические Т-клетки. Тh1 и цитотоксические Т-клетки являются существенными для индукции защитного иммунитета против М.tuberculosis (Wakeham, et al., J. Immunol., 160:1998:6101).
(iv) Другим преимуществом и уникальностью изобретения является то, что дендриты на дендритных клетках захватывают чужеродные антигены и действуют как резервуар. Упомянутый антиген медленно высвобождается из дендритов и является ответственным за поддержание клеток памяти.
(v) AMTV-вакцина действует без рестрикции по антигенам МНС, так как она основана на аллостимуляции и поглощении апоптотических клеток дендритными клетками. Она действует у всех людей независимо от генетического разнообразия.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАКЦИНА MYCOBACTERIUM TUBERCULOSIS | 2009 |
|
RU2473365C1 |
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛИРОВАНИЯ СОЗРЕВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СОДЕРЖАЩАЯ ГИБРИДНЫЙ БЕЛОК Rv2299c/ESAT-6 | 2017 |
|
RU2733886C2 |
| ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИНА С УЛУЧШЕННОЙ ЭФФЕКТИВНОСТЬЮ | 2004 |
|
RU2342400C2 |
| ПРИМЕНЕНИЕ МОНОМИКОЛИЛГЛИЦЕРИНА (MMG) В КАЧЕСТВЕ АДЪЮВАНТА | 2008 |
|
RU2479317C2 |
| СОЧЕТАНИЕ РЕКОМБИНАНТНОЙ МИКРОБАКТЕРИИ И БИОЛОГИЧЕСКИ АКТИВНОГО СРЕДСТВА В КАЧЕСТВЕ ВАКЦИНЫ | 2005 |
|
RU2495677C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ВАКЦИНЫ | 2010 |
|
RU2444570C1 |
| ВАКЦИНА НА ОСНОВЕ В-КЛЕТОК, НАГРУЖЕННЫХ ЛИГАНДОМ Т-КЛЕТОК-ПРИРОДНЫХ КИЛЛЕРОВ И АНТИГЕНОМ | 2006 |
|
RU2420311C2 |
| СПОСОБ ПОЛУЧЕНИЯ АУТОЛОГИЧНОЙ ВАКЦИНЫ ДЛЯ ЛЕЧЕНИЯ ТУБЕРКУЛЕЗА | 2008 |
|
RU2372936C1 |
| КОМБИНИРОВАННЫЙ ПРЕПАРАТ ДЛЯ УВЕЛИЧЕНИЯ ИММУНОГЕННОСТИ ВАКЦИНЫ (ВАРИАНТЫ) | 2008 |
|
RU2491090C2 |
| ПОЛИАНТИГЕННАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ И ВСПОМОГАТЕЛЬНОГО ЛЕЧЕНИЯ ТУБЕРКУЛЕЗА | 2019 |
|
RU2724896C1 |
Изобретение относится к медицине и касается способа получения вакцины для лечения туберкулеза и других внутриклеточных инфекционных заболеваний и вакцины, полученной по этому способу. Сущность изобретения заключается в инфицировании макрофагов и клеток макрофагальных клеточных линий патогенными микроорганизмами, их обработки лекарственными средствами с последующим гамма-облучением, иммунизации резистентных и чувствительных к заболеванию линий животных и инфицировании вакцинированных животных живыми патогенными микроорганизмами. Преимущество изобретения заключается в повышении иммунитета к патогенным микроорганизмам. 4 н.п. ф-лы, 1 ил.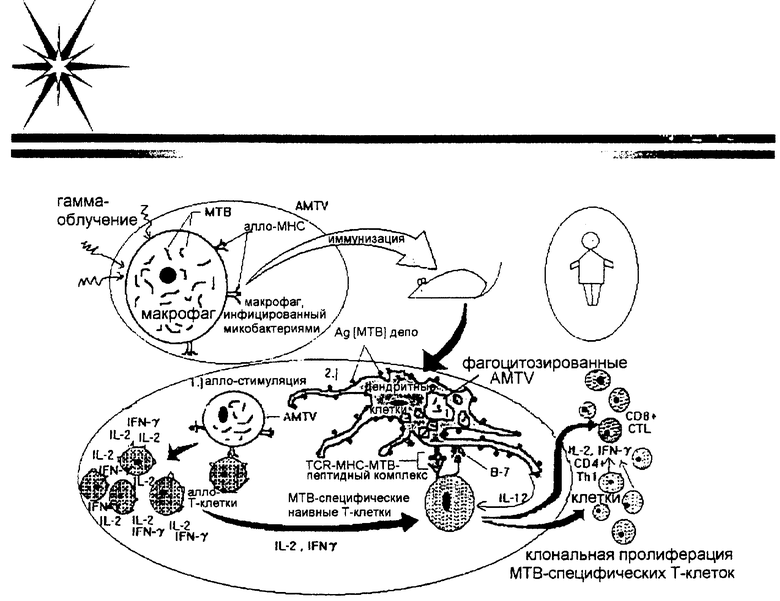
i) культивирования внутриклеточных патогенов;
ii) культивирования сингенных (тот же вид), аллогенных (другой вид) и ксеногенных (различные виды, как овца и коза) макрофагов и клеток макрофагальных клеточных линий, выбранных из группы, состоящей из J774A, P388D1, RAW, ВМС-2 и ТНР-1;
ii) инфицирования макрофагов и клеток макрофагальных клеточных линий патогенными микроорганизмами со стадии (i);
iv) обработки инфицированных макрофагов и клеток макрофагальных клеточных линий лекарственными средствами, вызывающими гибель патогена, с последующим гамма-облучением, вызывающим гибель макрофагов или макрофагальных клеточных линий и оставшихся патогенов, для получения композиции;
v) иммунизации резистентных и чувствительных к заболеванию линий животных указанной композицией;
vi) инфицирования вакцинированных животных выбранными живыми патогенными микроорганизмами и регистрации гибели животных и подсчета живых патогенов в легких, селезенке и печени,
vii) контроля у вакцинированных животных пролиферации и образования CD4+ Тh1 и Th2 клеток и CD8+ цитотоксических Т-клеток, свидетельствующих о создании опосредованного клетками иммунитета против выбранного патогена;
viii) где композиция представляет собой вакцину, если введение композиции приводит к понижению уровня гибели вакцинированных животных и/или понижению количества живых патогенов в легких, селезенке и печени инфицированных животных по сравнению с неиммунизированными животными.
i) культивирования М.tuberculosis H37Rv;
ii) культивирования сингенных, аллогенных и ксеногенных макрофагов и макрофагальных клеточных линий, выбранных из группы, состоящей из J774A, P388D1, RAW, ВМС-2 и ТНР-1;
iii) инфицирования макрофагов и клеток клеточных линий (J774, P388D1, RAW, ВМС-2, ТНР-1) М.tuberculosis;
iv) обработки инфицированных макрофагов и клеток клеточных линий изониазидом и гамма-облучением, вызывающим гибель макрофагов или макрофагальных клеточных линий и оставшихся М.tuberculosis, для получения композиции;
v) иммунизации резистентных и чувствительных к туберкулезу линий мышей аллогенной, сингенной или ксеногенной макрофагальной туберкулезной композицией;
vi) инфицирования вакцинированных групп мышей живыми М.tuberculosis, регистрации гибели мышей и подсчета оставшихся в живых М.tuberculosis в легких, селезенке и печени;
vii) контроля у вакцинированных животных пролиферации и образования CD4+ Th1 и Th2 клеток и CD8+ цитотоксических Т-клеток, свидетельствующих о создании опосредованного клетками иммунитета против M.tuberculosis;
viii) где композиция представляет собой вакцину, если введение композиции приводит к понижению уровня гибели вакцинированных животных и/или понижению количества М.tuberculosis в легких, селезенке и печени инфицированных животных по сравнению с неиммунизированными животными.
i) культивирования Salmonella typhimurium;
ii) культивирования сингенных, аллогенных и ксеногенных макрофагов и макрофагальных клеточных линий, выбранных из группы, состоящей из J774A, P388D1, RAW, ВМС-2 и ТНР-1;
iii) инфицирования макрофагов и клеток макрофагальных клеточных линий (J774, P388D1, RAW, ВМС-2, ТНР-1) S.typhimurium;
iv) обработки инфицированных макрофагов и клеток макрофагальных клеточных линий митомицином С и гамма-облучением, вызывающим гибель макрофагов или макрофагальных клеточных линий и оставшихся S.typhimurium, для получения композиции;
v) иммунизации резистентных и чувствительных к сальмонелле линий мышей указанной композицией;
vi) инфицирования вакцинированных групп мышей живыми S.typhimurium и регистрации гибели мышей и подсчета оставшихся в живых S.typhimurium в легких, селезенке и печени,
vii) контроля у вакцинированных животных пролиферации и образования CD4+ Th1 и Th2 клеток и CD8+ цитотоксических Т-клеток, свидетельствующих о создании опосредованного клетками иммунитета против S.typhimurium;
viii) где композиция представляет собой вакцину, если введение композиции приводит к понижению уровня гибели вакцинированных животных и/или понижению количества S.typhimurium в легких, селезенке и печени инфицированных животных по сравнению с неиммунизированными животными.
| ЕР 0223218 А, 27.05.1987 | |||
| US 4064007 А, 20.12.1997 | |||
| MOSMANN T.R | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Review immunology | |||
| today | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| WIEGESHAUS et al | |||
| Evaluation of the protective | |||
| potency of new tuberculosis vaccines | |||
| Rev | |||
| of Infect | |||
| Diseases | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

