Изобретение относится к медицине и может быть использовано при комплексном лечении пациентов препаратами цитостатической группы, формирующих двуцепочечные ковалентные сшивки между цепями молекулы ДНК с одновременным лечением препаратом, содержащим аллогенный генетический материал, в частности, для лечения пациентов с онкологическими заболеваниями методами заместительной терапии поврежденных и огомозигоченных аллелей генома фрагментами аллогенной ДНК, когда причиной онкологических заболеваний являются мутации в онкогенах, генах онкосупрессоров и тотальное огомозигочивание аллелей генов клетки, претерпевающей раковое перерождение.
Известен способ лечения, основанный на исправлении точечных мутаций в клетках [1].
Способ отличается относительно низкой эффективностью лечения и ограниченностью применения, поскольку согласно этому способу мутации должны быть точно определены еще до начала лечения.
Ввиду того что ни для одной мутации нет достоверного доказательства того, что именно эта мутация является причиной рака, развитие этого метода требует скрупулезных и длительных исследований по выявлению конкретных мутаций, требующих коррекции, а следовательно, существует необходимость в создании способов лечения, которые могли бы применяться на основе уже имеющихся знаний о причинах рака, не делая акцента на конкретных изменениях генома, приведших к злокачественному перерождению.
Известны также способы лечения, основанные на локальном применении ДНК фрагментов для лечения предраковых состояний в коже пациентов и стимуляции солнечного загара [2, 3].
Эти способы также имеют ограниченную применимость, поскольку ни конкретные последовательности, ни источники ДНК в указанных патентах не определены. В них предлагается использовать как природную, так и синтетическую ДНК из «любых подходящих источников», например ДНК лосося длиной от 200 мононуклеотидов и нуклеозидов, включая димеры, что в соответствии с тестами авторов является наиболее эффективным. Однако действие и последствия применения ДНК иной природы на человеческий организм пока не полностью изучены, что предопределяет определенную опасность применения способов.
Наиболее близким по своей сущности к предложенному является способ, основанный на использовании цитостатических препаратов, например препарата цисплатины для лечения больных с плоскоклеточным раком кожи туловища, путем использования цисплатины в дозе 100 мг/м2 (1 день), внутривенно капельно 30 мг/м2 1 раз в неделю, или 60-150 мг/м2 каждые 3-5 недель, или 20 мг/м2 ежедневно 5 дней с повторением каждые 4 недели, или 50 мг/м2 в 1-й и 8-й дни каждые 4 недели, препарата митомицин С для лечения больных раком желудка, кишечника, поджелудочной железы, мочевого пузыря, молочной железы, легкого, вульвы, предстательной железы (в составе лекарственных комбинаций), путем использования митомицина С внутривенно в дозе 10 мг/м2 1 раз в 3-4 недели (в составе лекарственных комбинаций) или также 2 мг/м2 с 1-го по 5-й и с 8-го по 12-й дни курса, суммарная доза 50 мг/м2, или препарата дактиномицина для лечения больных с хориокарциномой матки, саркомой мягких тканей, злокачественных опухолей яичка, меланомы, лимфосаркомы, нейробластом путем использования дактиномицина внутривенно 350 мкг/м2 ежедневно в течение 5 дней каждые 3-5 недель или 1-2 раза в неделю в течение 3-5 недель, суммарная доза 3000 мкг [4].
Действие цитостатических препаратов, таких как псорален, цисплатина, ЦФ, нитроген мустард, мельфолан митомицин С и др., используемых при терапии различных раков, основано на их свойстве образовывать двуцепочечные сшивки в произвольных местах генома.
При терапевтической дозе цитостатика в геноме клеток организма образуются до 2000 межцепочечных сшивок. Репаративная система клетки не в состоянии справиться с таким количеством одновременных повреждений, что приводит к гибели раковой клетки.
Однако наиболее близкий способ обладает относительно низкой эффективностью лечения, обусловленной тем, что раковая клетка гибнет не изменяя своего ракового статуса и распространяет в окружающее клетку пространство фрагменты ДНК несущие онкомутации, которые могут служить источником генометастазирования. При этом одновременно с раковыми гибнут все остальные активно пролиферирующие клетки организма, такие как СКК, клетки эпителиев, клетки волосяных фолликул и т.д., что приводит к крайне негативным и даже летальным последствиям.
Все это снижает эффективность лечения пациентов с онкологическими заболеваниями, причиной которых являются как мутации в онкогенах и генах онкосупрессоров, так же как и тотальное огомозигочивание генов.
Требуемый результат заключается в повышении эффективности лечения пациентов с онкологическими заболеваниями, причиной которых являются как мутации в онкогенах и генах онкосупрессоров, так и тотальное огомозигочивание генов.
В более широком плане задачей настоящего изобретения является разработка обоснованного и экспериментально подтвержденного способа совместного воздействия на раковые клетки и опухоль в целом цитостатических препаратов, индуцирующих сшивку цепей молекулы ДНК, и препарата фрагментированной гомологической ДНК, составляющей полный геном физиологически и генетически здорового донора таким образом, что в целом в результате индукции межцепочечных сшивок активируется вся рекомбинационная система клетки, приводящая к тому, что формируется открытая для рекомбинационной машины структура хроматина, создаются сайты репаративного гомологичного обмена в местах индукции кросс-линков и многократно усиливается рекомбинация, связанная с нахождением концевых гомологий экзогенных фрагментов по всему геному, включая ранее недоступный ввиду конформационных особенностей хроматин. При этом в местах индуцированных цитостатическим кросс-линком репаративных событий происходит гомологическое репаративное замещение поврежденных участков хроматина на гомологические фрагменты экзогенной терапевтической ДНК, депонированной в ядерном пространстве. Вследствие химической природы цитостатика сшивки молекулы ДНК происходят статистически по геному и не зависят от структуры хроматина и других ядерных факторов. Виду этого будет формироваться открытая для рекомбинационной машины структура хроматина. Поэтому онкомутации, содержащиеся в геноме, будут с определенной долей вероятности замещаться на недефектные последовательности за счет экзогенных терапевтических фрагментов, участвующих как в репаративной гомологической рекомбинации, так и в рекомбинации, связанной с нахождением концевых гомологий экзогенных фрагментов по всему геному, включая ранее недоступный, ввиду конформационной закрытости хроматина.
При совместном воздействии цитостатика и препарата фрагментированной гомологичной ДНК на протяжении нескольких курсов химиотерапии (с постоянным мониторингом биохимических маркеров онкотрансформации) происходит изменение генного гомеостаза, когда клетки претерпевают реверсивное генетическое перерождение, при котором исчезает основное свойство раковой клетки - ее неограниченная пролиферативная активность.
Требуемый результат достигается тем, что по способу лечения онкологических заболеваний, основанному на введении в организм пациента препарата цитостатической группы, механизм действия которого основан на формировании межцепочечных сшивок в молекуле ДНК раковых и других активно пролиферирующих клеток организма, в организм пациента дополнительно вводят препарат фрагментированной ДНК с фрагментами, имеющими биологически активный размер и составляющими полный геном физиологически и генетически здорового донора.
Кроме того, требуемый результат достигается тем, что препарат фрагментированной ДНК с фрагментами, имеющими биологически активный размер и составляющими полный геном физиологически и генетически здорового донора, вводят пациенту за один час и через час после введении препарата цитостатической группы и далее каждые два часа до завершения полного репаративного цикла в количестве, равном или превышающем количество собственно ДНК плазмы крови и тканевых жидкостей пациента, но не более максимально допустимого количества, равного 30 мкг/мл крови.
Кроме того, требуемый результат достигается тем, что полный реперативный цикл составляет 12 часов.
Кроме того, требуемый результат достигается тем, что препарат фрагментированной ДНК с фрагментами, имеющими биологически активный размер и составляющими полный геном физиологически и генетически здорового донора, используют в качестве субстрата для гомологической рекомбинации с однонитчатыми участками ДНК и областями двуцепочечных разрывов молекулы ДНК, возникающими при репарации межцепочечных сшивок, которые в силу химической природы цитостатика появляются в геноме статистически и место их локализации не зависит от структуры хроматина и связанных с этой структурой других факторов клетки.
Кроме того, требуемый результат достигается тем, что число процедур, основанных на введении в организм пациента препаратов цитостатической группы, механизм действия которых основан на формировании межцепочечных сшивок в молекуле ДНК раковых и других активно пролиферирующих клеток организма, и дополнительном введении в организм пациента препарата фрагментированной ДНК с фрагментами, имеющими биологически активный размер и составляющими полный геном физиологически и генетически здорового донора, определяется запуском нескольких механизмов, определяющих скорость и эффективность заместительного терапевтического действия фрагментов экзогенной ДНК. К таким механизмам относятся: а) освобождение молекулы ДНК от гистонов на протяжении нескольких мегабаз по обе стороны от возникшего двуцепочечного разрыва; б) происходящие при этом конформационные изменения хроматина в местах хромосом, недоступных для прямого замещения фрагментами экзогенной ДНК, и возникающая при этом возможность прямого замещения гомологичных участков хромосом экзогенными фрагментами ДНК; в) многократно усиливающийся, постоянно протекающий в ядре гомологический обмен между экзогенными фрагментами ДНК и теми участками хроматина, которые упакованы таким образом, что они постоянно доступны для гомологического обмена; г) гомологическая рекомбинация между фрагментами экзогенной ДНК и интермедиатами, возникающими в районе кросс-линка в ходе репарации.
Фактически число химиотерапевтических процедур совместно с введением препарата фрагментированной ДНК определяется и контролируется по совокупности всех маркерных тестов, сопровождающих лечение каждого конкретного рака.
Воздействие химиотерапевтического цитостатика приводит к возникновению поперечных сшивок в молекуле ДНК. Это приводит к вынужденной (аварийной) остановке репликативной вилки, включению механизма репарации. Механизм репарации состоит из нескольких событий, при которых первоначально образуется двуцепочечный разрыв. После этого эксцизионная система репарации делает серию разрезов и оголяет одоцепочечный участок размером около 1 килобазы. В результате формирования таких структур многократно активируется система репаративной гомологической рекомбинации. В результате этого крайне рекомбиногенные структуры - двуцепочечные разрывы и одноцепочечный гэп - ищут и находят гомологичные последовательности, расположенные в ядре (гомологичная хромосома, сестринская хроматида), и совершается акт гомологическокой рекомбинационной рапарации. В результате этого восстанавливается целостность хроматина, ликвидируется дефект и на участке хромосомы, участвующей в рекомбинации, происходит замещение участка хроматина в пределах 1-6 килобаз на аллельные варианты, находящиеся в составе экзогенных фрагментов ДНК из ядерного пространства. Если в ядерном пространстве будут находиться терапевтические фрагменты ДНК, то они будут являться субстратом для репаративной гомологической рекомбинации.
В современной литературе отсутствуют указания на предлагаемый способ лечения онкологических пациентов с использованием одновременного воздействия кросс-линкирующих цитостатиков и препарата фргаментированной генетически здоровой ДНК, естественного механизма ее доставки, депонирования в межхромосомном пространстве и активации всей рекомбинационной системы клетки, приводящей к тому, что формируется открытая для рекомбинационной машины структура хроматина, создаются сайты репаративного гомологичного обмена в местах индукции кросс-линков и многократно усиливается рекомбинация, связанная с нахождением концевых гомологий экзогенных фрагментов по всему геному, включая ранее недоступный ввиду конформационных особенностей хроматин.
Следовательно, предложение отвечает критериям новизны и изобретательского уровня.
Ниже приводятся теоретические и экспериментальные данные, подтверждающие, что изобретение отвечает и критерию практической (промышленной) применимости.
На чертежах представлены:
на фиг.1 - электрофоретические характеристики экзогенной терапевтической ДНК плаценты человека, используемой для инъекций в организм опытных мышей (М - маркер молекулярного веса - BssT1I гидролизат фага λ (lambda DNA - BssT1I digest));
на фиг.2.-2.6 - таблицы (1, 2, 3, 4) соотношений и динамика количества форменных элементов крови после введения циклофосфана и в комбинации с препаратом фрагментированной ДНК человека;
на фиг.3, А - элаймент консенсусов повторяющихся последовательностей Alu человека и В1 мыши;
на фиг.3, В - последовательность фрагмента X-Alu (стрелками и жирным шрифтом выделены используемые в ПНР реакции праймеры);
на фиг.4 - ПЦР анализ геномной ДНК экспериментальных животных на присутствие в ней последовательностей человеческой ДНК. А) ПЦР анализ геномной ДНК экспериментальных мышей на присутствие специфических ПЦР фрагментов по подвижности совпадающих с ПЦР продуктами, выявляемыми при аналогичном анализе в геноме человека. Левый блок - электрофоретически фракционированные ПЦР фрагменты экспериментальных и конрольных животных. Правый блок - Саузерн-блот гибридизация этого же геля с ДНК 32Р ПЦР меченного X-Alu фрагмента. В) Повторная гибридизация позитивных образцов, специфические фрагменты которых (280 п.о. для человека и обеих мышей) в дальнейшем использовались для определения нуклеотидной последовательности. Левый блок - электрофоретически фракционированные ПЦР фрагменты, правый блок - Саузерн-блот гибридизация этого же геля с ДНК 32Р ПЦР меченного X-Alu фрагмента. Цифры слева от блоков (280 и 310) указывают на фрагменты, соответствующие двум мажорным ПЦР продуктам, выявляемым в геноме человека. 1-15 - номера образцов, bp - пары оснований. Human - человек, СВА - реципиентная линия мышей, m №1 и m №8 - экспериментальные мыши, М - маркеры молекулярных весов (100 bp лестница). +hDNA - образцы ДНК мышей, обработанных только человеческой ДНК; +CP+hDNA образцы ДНК мышей, обработанных совместно циклофосфамидом и человеческой ДНК.
на фиг.5, А - саузерн-блот анализ геномной ДНК экспериментальных мышей №1 и №8 на присутствие в ней последовательностей человеческой ДНК. М - маркеры молекулярных весов (HindIII гидролизат фага λ); 1-4 - BamHI+HindIII гидролизаты геномной ДНК: m №1 - экспериментальное животное №1, m №8 - экспериментальное животное №8, СВА -реципиентная линия мышей, human - человек. Левый блок электрофоретически фракционированная ДНК, гидролизованная BamHI+HindIII. Правый блок - геномный блот этого же геля после гибридизации с 32Р меченым фрагментом X-Alu. Стрелками для левого блока указаны маркерные фрагменты. Стрелками для правого блока указаны гибридизующиеся фрагменты геномов экспериментальных животных.
На фиг.5, В - определение копийности человеческого фрагмента X-Alu в геноме человека, реципиентной линии СВА и экспериментальных мышей. Гибридизация проводилась с ДНК 32Р ПЦР меченного X-Alu фрагмента. Цифрами над всеми блоками указано титрованное количество ДНК, нанесенное на мембрану. SS - ДНК спермы лосося; Alu - ДНК X-Alu; hDNA - ДНК человека; mouse №1, mouse №8 - ДНК экспериментальных мышей.
Предложенный способ лечения онкологических заболеваний реализуется следующим образом.
Способ лечения онкологических заболеваний основан на введении в организм пациента препаратов цитостатической группы, механизм действия которых основан на формировании межцепочечных сшивок в молекуле ДНК раковых и других активно пролиферирующих клеток организма. При этом, вместе с вводимым цитостатическим препаратом в организм вводится препарат фрагментированной ДНК с фрагментами, имеющими биологически активный размер и составляющими полный геном физиологически и генетически здорового донора.
Фрагменты терапевтической ДНК вводят пациенту за один час и через час после введении препарата цитостатической группы и далее каждые два часа до завершения полного репаративного цикла в количестве, равном или превышающем количество собственно ДНК плазмы крови и тканевых жидкостей пациента, но не более максимально допустимого количества, равного 30 мкг/мл крови. Полный реперативный цикл составляет 12 часов.
Фрагменты терапевтической ДНК являются субстратом для гомологической рекомбинации с однонитчатыми участками ДНК и областями двуцепочечных разрывов молекулы ДНК, возникающими при репарации межцепочечных сшивок, которые в силу химической природы цитостатика появляются в геноме статистически и место их локализации не зависит от структуры хроматина и связанных с этой структурой других факторов клетки.
Кроме того, фрагменты терапевтической ДНК, являясь субстратом для гомологической рекомбинации и не имея по происхождению онкомутаций, замещают в процессе гомологического обмена онколокусы генома раковых клеток пациента, если в непосредственной близости от которых или в них самих была индуцирована поперечная сшивка молекулы ДНК, тем самым, изменяя раковый статус таких клеток на не раковый.
При этом, фрагменты терапевтической ДНК, являясь субстратом для гомологической рекомбинации, участвуют в завершающем этапе репарации, формируя новую неповрежденную цепь ДНК на участках хромосом, не содержащих онкомутаций, подвергшихся атаке цитостатика как в раковых, так и во всех других активно пролиферирующих клетках (СКК, эпителиях, клеток волосяных фолликул), тем самым спасая их от апоптоза и сохраняя их функции. Важнейшим моментом одновременной терапии препаратом фрагментированной ДНК при лечении цитостатиками является момент искусственного создания неестественной для клетки рекомбиногенной активности. При этом рекомбинация происходит как по всему геному (вследствие естественного механизма гомологического замещения в районах хроматина, доступного для таких событий), так и в тех местах, где был индуцирован кросс-линк (практически в любом месте генома). Кросс-линк, в силу химической природы индукторов сшивки, возникает в любом месте генома независимо от его организации любого уровня и независимо от любой конформационной связи ДНК с белками.
Кроме того, в результате искусственно созданной рекомбиногенной ситуации в ядре запускаются несколько механизмов, определяющих скорость и эффективность заместительного терапевтического действия фрагментов экзогенной ДНК. К таким механизмам относятся: а) освобождение двойной цепи ДНК от гистонов на протяжении нескольких мегабаз по обе стороны от возникшего двуцепочечного разрыва; б) происходящие при этом конформационные изменения хроматина в местах хромосом, ранее недоступных для прямого замещения фрагментами экзогенной ДНК, и возникающая, одновременно многократно усиленная возможность прямого замещения гомологичных участков хромосом экзогенными фрагментами ДНК; в) многократно усиливающийся, постоянно протекающий в ядре гомологический обмен между экзогенными фрагментами ДНК и теми участками хроматина, которые упакованы таким образом, что они постоянно доступны для гомологического обмена; г) гомологическая рекомбинация между фрагментами экзогенной ДНК и интермедиатами, возникающими в районе кросс-линка в ходе репарации.
Фактически число химиотерапевтических процедур совместно с введением препарата фрагментированной ДНК донора определяется и контролируется по совокупности всех маркерных тестов, сопровождающих лечение каждого конкретного рака.
Полное замещение всех онкомутаций в геноме зависит от количества кросс-линков на геном, которые попали в места, содержащие онкомутаций, и изменили конформацию хроматина и эффективности механизма прямого гомологичного обмена между фрагментами экзогенной ДНК и соответствующими локусами хроматина, многократно усиливающимся при активации рекомбиногенной ситуации в ядре.
Пример реализации способа
Вместе с вводимым цитостатическим препаратом цисплатины для лечения больных с плоскоклеточным раком кожи туловища путем использования цисплатины в дозе 100 мг/м2 (I день), внутривенно капельно 30 мг/м2 1 раз в неделю, или 60-150 мг/м2 каждые 3-5 недель, или 20 мг/м2 ежедневно 5 дней с повторением каждые 4 недели, или 50 мг/м2 в 1-й и 8-й дни каждые 4 недели, или препаратом митомицин С для лечения больных раком желудка, кишечника, поджелудочной железы, мочевого пузыря, молочной железы, легкого, вульвы, предстательной железы (в составе лекарственных комбинаций), путем использования митомицина С внутривенно в дозе 10 мг/м2 1 раз в 3-4 недели (в составе лекарственных комбинаций) или также 2 мг/м2 с 1-го по 5-й и с 8-го по 12-й дни курса, суммарная доза 50 мг/м2, или препаратом дактиномицина для лечения больных с хориокарциномой матки, саркомой мягких тканей, злокачественных опухолей яичка, меланомы, лимфосаркомы, нейробластом путем использования дактиномицина внутривенно 350 мкг/м2 ежедневно в течение 5 дней каждые 3-5 недель или 1-2 раза в неделю в течение 3-5 недель, суммарная доза 3000 мкг в организм пациента вводится препарат фрагментированной ДНК донора с фрагментами, имеющими биологически активный размер и составляющими полный геном физиологически и генетически здорового донора, в количестве, равном или превышающем количество собственно ДНК плазмы крови и тканевых жидкостей пациента, но не более максимально допустимого количества, равного 30 мкг/мл крови, за один час и через час после введения цитостатика и далее каждые два часа до завершения полного репаративного цикла (12 часов).
Для подтверждения того, что предложенный способ позволяет достигнуть требуемого результата, приведем его теоретическое и экспериментальное обоснование.
Рассмотрим концепцию заместительной терапии препаратом фрагментированной ДНК донора.
Главным моментом заместительной терапии с использованием алогенной ДНК в форме фрагментов с размером, соответствующим фрагментам ДНК, образующимся естественным путем в результате нормально протекающего апоптоза и всегда присутствующих в плазме крови и других инцесциальных жидкостях, является либо создание искусственной общеклеточной рекомбиногенной ситуации в момент нахождения терапевтических фрагментов ДНК во внутриядерном пространстве, либо доставка терапевтических фрагментов в клетки, находящиеся в таком рекомбинационно активном состоянии.
Искусственно индуцированная рекомбиногенная ситуация в клетке возникает при индукции межцепочечных сшивок цитостатическими препаратами.
Действие цитостатических препаратов, используемых при терапии различных раков, основано на их свойстве образовывать двуцепочечные сшивки в произвольных местах генома.
Известно, что одного такого повреждения на клетку при отсутствии системы репарации достаточно, чтобы клетка погибла, вступив на путь апоптотического самоуничтожения. Для клетки гибельным оказывается цитостатический удар, при котором образуется 122 кросс-линка на клетку. При терапевтической дозе цитостатика в геноме клеток организма образуются до 2000 межцепочечных сшивок [8].
Для низших эукариот показано, что в покоящихся клетках (G1/G2 фазы клеточного цикла) репарация возникших межцепочечных сшивок идет за счет активности полимеразы ζ (Rev3), которая способна проходить такое повреждение. Однако использование клеткой этого пути репарации приводит к появлению нуклеотидных замен (мутаций) в последовательности ДНК в непосредственной близости от точки кросс-линка (сшивки), которые затем закрепляются входе репликативного цикла [5]. Кроме этого, как было указано выше, такой путь репарации характерен и для не делящихся клеток высших экуариот и занимает незначительный удельный вес в общей системе репарации межцепочечных сшивок [6, 7].
В активно пролиферирующих клетках, к которым относятся раковые клетки, стволовые клетки разного генеза, клетки волосяных фолликул, клетки различных эпителиев, при индукции поперечных сшивок в молекуле ДНК возникает смертельная для клетки ситуация. Репликативная вилка, которая формируется с частотой порядка одна на 50 т.п.о. (тысяч пар оснований), наталкивается на стерическое препятствие, которое репликативный ферментативный комплекс не в состоянии преодолеть. Репликативная вилка сталируется. Именно остановка репликативной вилки запускает каскад репаративных событий. Немедленно в месте сталирования репликативной вилки формируется эксцизионный комплекс.
Процесс репарации имеет три стадии, разбитых во времени. Первоначально в непосредственной близости к повреждению на уже реплицированной цепи ДНК возникают двуцепочечные разрывы [8, 9, 5], механизм появления которых до настоящего времени остается не изученным. Сразу после этого события активируется эксцизионный репаративный комплекс. Специфические эндонуклеазы делают одноцепочечные надрезы в непосредственной близости от сшивки. Гетеродимер XPF-ERCC1 в присутствии RPA репликативного белка А своей 3'-5' экзонуклеазной активностью гидролизует одну цепь ДНК, продвигаясь на большое расстояние от места сшивки, проходя повреждение насквозь [11, 12]. После завершения этих двух стадий репарации, а именно индукции двуцепочечных разрывов и эксцизионных действий специфических эндонуклеаз, в месте кросс-линка формируется продолжительный одноцепочечный участок, фланкированный двуцепочечным разрывом [10, 9, 7], - структура в высшей степени рекомбиногенная.
Для репарации такого интермедиата клетка использует механизм гомологической рекомбинации. При этом в качестве субстрата для гомологической рекомбинации используется либо сестринская хроматида, либо гомологическая хромосома [9]. После восстановления целостности хромосомы в месте сшивки вновь реассоциируется репликативная ферментативная машина, и репликация возобновляется.
Другой возможный вариант репарации интермедиата, состоящего из двуцепочечного разрыва, расположенного в непосредственной близости к месту формирования репликативной вилки (небольшого участка двуцепочечной ДНК, расположенного перед сайтом кросс-линка, и продолжительного одноцепочечного участка, сформированного в результате экзонуклеазного движения XPF-ERCC1 комплекса), - это двойной гомологический обмен. Вначале по механизму генной конверсии в районе одноцепочечного участка происходит спаривание гомологической последовательности и одноцепочечного участка, содержащего нуклеотид аддукт. После этого эксцизионный репаративный комплекс вырезает место, содержащее кросс-линк на противоположной цепи. Происходит обмен цепями, и брешь достраивается по вновь интегрированной матрице. Далее двуцепочечный разрыв репарируется по механизму кроссинговера с гомологичной последовательностью ДНК, как это было описано выше.
Рассмотрим, что происходит при множественных межцепочечных сшивках, индуцируемых противораковыми препаратами (псорален, цисплатина, митомицин С, мельфолан, нитроген мустард, циклофосфамид и т.п.).
В этом случае немедленно активируется репаративная система клетки. Однако, во-первых, клетка не в состоянии одновременно сформировать достаточное количество репарационных комплексов и в этом случае часть повреждений остается не обслуженной, что приводит к апоптозу всех таких клеток. Во-вторых, при привлечении в качестве субстрата для гомологической рекомбинации гомологической хромосомы или сестринской хроматиды при множественных индуцированных межцепочечных сшивках возникают стерические препятствия к одновременному синапсису нескольких поврежденных участков с несколькими гомологическими участками. Это также препятствует полной репарации всех межцепочечных сшивок и с неизбежностью приводит к апоптозу и гибели клетки. И, в-третьих, при использовании в качестве субстрата для репаративной гомологической рекомбинации гомолога или сестринской хроматиды генетическая мода клетки остается точно такой же, как и до появления сшивки. То есть, если цитостатический диаддукт возник в области гена, мутация в котором привела к ораковлению клетки, то репарация с использованием эндогенного клеточного субстрата не приведет к генетическому изменению мутировавшего гена. Это означает, что если в геноме были раковые мутации, то при такой репарации они сохраняются.
Следовательно, единственный путь избавиться от такой онкомутации - это убить раковую клетку, а заодно и все остальные активно делящиеся клетки организма, что и делают цитостатические препараты.
Препарат фрагментированной ДНК донора с размером фрагментов, соответствующим фрагментам ДНК, образующимся естественным путем в результате нормально протекающего апоптоза и всегда присутствующим в плазме крови и других инцесциальных жидкостях [13, 14, 15], присутствуя в ядерном пространстве в момент возникновения сшивок, становится субстратом для гомологической репаративной рекомбинации, спасая клетки от апоптоза и изменяя раковый статус клетки, как это было описано выше. Фрагменты ДНК принимают участие в завершающей стадии репаративного процесса при восстановлении сшитых между собой цепей ДНК. Известно, что в межхромосомном пространстве ядра могут присутствовать до 2% от гаплоидного генома экстраклеточной ДНК в виде фрагментов с размером, соответствующим размеру фрагментов ДНК, образующимся естественным путем в результате нормально протекающего апоптоза [16]. В результате индукции сшивок и произошедшего сталирования репликативных вилок возникает крайне рекомбиногенная ситуация и активируется рекомбинационная система клетки. Если в этот момент в ядре присутствуют фрагменты ДНК, имеющие гомологию с репарируемым местом и являющиеся субстратом для гомологической рекомбинации, то таковая непременно произойдет и произойдет не с дефектным эндогенным гомологом, а с экзогенной терапевтической ДНК, не содержащей мутации. Вновь интегрированная последовательность закрепится в результате клеточного деления и онкомутация будет ликвидирована.
Важнейшим моментом одновременной терапии препаратом фрагментированной ДНК при лечении цитостатиками является момент искусственного создания неестественной для клетки рекомбиногенной активности. При этом рекомбинация происходит как по всему геному (вследствие естественного механизма гомологического замещения в районах хроматина, доступного для таких событий), так и в тех местах, где был индуцирован кросс-линк (практически в любом месте генома). Кросс-линк, в силу химической природы индукторов сшивки, возникает в любом месте генома независимо от его организации любого уровня и независимо от любой конформационной связи ДНК с белками. В результате искусственно созданной рекомбиногенной ситуации, как было сказано, в ядре запускаются несколько механизмов, определяющих скорость и эффективность заместительного терапевтического действия фрагментов экзогенной ДНК. К таким механизмам относятся: а) освобождение двойной цепи ДНК от гистонов на протяжении нескольких мегабаз по обе стороны от возникшего двуцепочечного разрыва; б) происходящие при этом конформационные изменения хроматина в местах хромосом, недоступных для прямого замещения фрагментами экзогенной ДНК, и возникающая, одновременно многократно усиленная возможность прямого замещения гомологичных участков хромосом экзогенными фрагментами ДНК; в) многократно усиливающийся, постоянно протекающий в ядре гомологический обмен экзогенными фрагментами ДНК и теми участками хроматина, которые упакованы таким образом, что они постоянно доступны для гомологического обмена; г) гомологическая рекомбинация между фрагментами экзогенной ДНК и интермедиатами, возникающими в районе кросс-линка в ходе репарации.
Фактически число химиотерапевтических процедур совместно с введением препарата фрагментированной ДНК донора определяется и контролируется по совокупности всех маркерных тестов, сопровождающих лечение каждого конкретного рака.
Полное замещение всех онкомутаций в геноме зависит от количества кросс-линков на геном, которые попали в места, содержащие онкомутации, и изменили конформацию хроматина, и эффективности механизма прямого гомологичного обмена между фрагментами экзогенной ДНК и соответствующими локусами хроматина, многократно усиливающимся при активации рекомбиногенной ситуации в ядре.
Для определения возможности использования предложенного способа лечения для широкого класса представителей животного мира последний этап репарации межцепочечной сшивки, а именно интеграция экзогенной ДНК в реципиентный геном после обработки цитостатиком, был изучен на модели мышь - человеческая фрагментированная ДНК. В результате проведенных экспериментов удалось интегрировать в геном экспериментальных мышей Alu-повторы человека в области, содержащие гомологические последовательности, относящиеся к классу умеренных повторов мыши В1.
Аналогичным образом экзогенная ДНК воздействует и на здоровые клетки организма, подвергшиеся воздействию цитостатика. В этом случае фрагменты экзогенной терапевтической ДНК, участвуя в репарационных событиях в здоровых клетках, спасают эти клетки от апоптоза, чем способствуют сохранению клеточных популяций различных тканей - СКК, эпителия и т.д., облегчая тем самым протекание химиотерапии и позволяя более четко планировать последующие циклы обработки цитостатиком.
В современной научной литературе подробно изучен вопрос репарации межцепочечных сшивок, индуцированных кросс-линкирующими агентами. Однако нигде не поднимался вопрос об использовании фрагментов экзогенной экстрахромосомальной ДНК для гомологического обмена при завершающей стадии репарации сшивок. Ниже приводится экспериментальное доказательство участия экзогенных фрагментов ДНК в репарации рекомбиногенных структур - одноцепочечного участка и двуцепочечных разрывов индуцированных возникшим кросс-линком между цепями хромосомной ДНК.
В наших ранних работах [16, 17] была высказана гипотеза о существовании естественного механизма, который может влиять на генетическую составляющую клетки многоклеточных организмов, используя внеклеточную геномную ДНК из биологических жидкостей, как внешний геномный стандарт. Согласно предложенному механизму ДНК связывающие рецепторы, располагающиеся на клеточной поверхности, доставляют фрагменты геномной ДНК из внешней среды (плазма крови, межтканевая жидкость, лимфа), образовавшиеся в результате естественно протекающего апоптоза в ядро, и этот оборот непрерывен. В ядерном пространстве интернализированные фрагменты ДНК могут участвовать во всех описанных системах репарации, в которых наличие неповрежденной гомологичной последовательности является необходимым условием протекания процесса.
Согласно анализу литературных данных и приведенной выше концепции, если в кровяном русле организма, подвергающегося воздействию сильного кросс-линкирующего мутагена, будут присутствовать фрагменты ДНК, имеющие гомологию с геномными последовательностями реципиентного организма, то эти фрагменты будут использоваться как субстрат для гомологической рекомбинации при репарации индуцированных цитостатиком повреждений и одновременно как субстрат для прямого гомологичного обмена с соответствующими локусами хроматина, многократно усиливающийся при активации рекомбиногенной ситуации в ядре.
В организм экспериментальных мышей внутримышечно были введены препарат фрагментированной ДНК человека и алкилирующий цитостатик циклофосфамид. В результате молекулярно-генетического анализа геномной ДНК экспериментальных животных были обнаружены фрагменты человеческой ДНК, интегрированные в реципиентный геном мыши.
Используемые методики.
Схема введения препаратов.
Циклофосфан вводился в/б самцам мышей линии СВА (n=20) из расчета 200 мг/кг веса. Одной группе мышей (n=10) за 30 мин до введения препарата в/б вводили 1 мг ДНК человека (фиг.1), через 30 мин после инъекции циклофосфана вводили еще по 0.5 мг ДНК и столько же на 2-й и 3-й день. Аналогично контрольным мышам (n=10) вводился физраствор.
Схема забора образцов крови для подсчета количества форменных элементов у экспериментальных животных.
Кровь для приготовления мазков брали из хвостовой вены индивидуально у каждой мыши непосредственно перед введением и спустя 4, 7, 10, 14 и 17 суток после введения препаратов. Опытная и экспериментальная группы состояли из 10 животных. Расчет лейкограммы проводили стандартными методами [18].
ПЦР амплификация.
Для амплификации специфических фрагментов человеческого генома использовался следующий температурный режим: 94°С - 2 мин - 1 цикл (94°С - 30 сек, 72°С - 1 мин) - 33 цикла, 4°С - хранение. Для выявления минорных фрагментов человеческого происхождения в геномах экспериментальных мышей использовали следующий подход. Первоначально минорные человеческие матрицы геномов экспериментальных мышей амплифицировали в присутствии одного специфического праймера. Далее полученный материал в новом раунде ПЦР амплифицировали в присутствии двух специфических праймеров при указанном температурном режиме. Такой подход позволил выявить в геномах двух экспериментальных животных №1 и №8 фрагменты человеческой ДНК.
Используемые специфические для человеческого генома праймеры:
Pr.9 CGAGGCGGGAGGATCACTTGAGCCC (25)
Pr.10 CGGCTCACTGCAGCCTCGACCTCCC (25)
Pr.11 GCGCGCGCCACCACGCCCGGC(21)
Амплификацию проводили на приборе Терцик ДНК-Технология (Россия)
Секвенирование ПЦР фрагментов.
Секвенирование экспериментально полученных фрагментов было проведено в Межинститутском центре секвенирования ДНК СО РАН (Россия). Для секвенирования фрагментов использовался краситель Big Dye 3,1 ("Applied Biosystems", США).
Саузерн-блот-дот гибридизация.
Гибридизация и отмывка проводились в жестких условиях при 68°С в 0,1% SDS и 0,1×SSC. Количество меченого материала (пятен и полос гибридизации, вырезанных из гибридизованного мембранного фильтра) определяли стандартным способом в счетных флаконах на счетчике RacBetta.
Расчет потенциального количества сайтов замещения фрагментами экзогенной экстрахромосомальной ДНК при репарации ДНК кросс-линков.
Геном мыши ˜2.5×109п.o.=2.5×106 т.п.о.
Геном человека 3,3×109п.о.=3,3×106 т.п.о.
Длина Alu повтора составляет ˜0.3 т.п.о. (kb) [19].
Количество Alu повторов в геноме человека ˜10% [20], что составляет ˜106 т.п.о. (копий) 0.3 kb мономера.
Длина В1 повтора составляет ˜0.15 т.п.о. (kb) [21].
Количество В1 повторов в геноме мыши ˜10%, что составляет ˜105 т.п.о. (копий) 0.15 kb мономера.
Alu элементы располагаются один через каждые 3-4 т.п.о. (kb).
В1 повторы располагаются один через каждые 3 т.п.о. (kb).
При терапевтических дозах кросс-линкирующего агента в клетке индуцируется 1000-2000 межцепочечных кросс-линков в молекуле ДНК [9, 22, 23].
Фрагменты экзогенной ДНК имеют размер, соответствующий 2-30 нуклеосомным единицам, что составляет ˜0.5-6.0 т.п.о. (в среднем 3.0 т.п.о.).
При введении в организм мыши (˜20 грамм) 1000-2000 мкг фрагментированной ДНК, что соответствует 50-100 мкг препарата на 1 г (мл) массы тела животного, в ядерном экстрахромосомальном пространстве активно пролиферирующих клеток организма может находиться до 2% (от гаплоидного генома) фрагментов экзогенной ДНК [16]. Таким образом, в ядерном пространстве одномоментно может находиться порядка 20000 Alu-мономеров, расположенных во фрагментах ДНК, размером в среднем 3.0 т.п.о.
В организм экспериментальных животных парентерально были введены ЦФ и препарат фрагментированной ДНК человека. ЦФ за счет своего метаболита, фосфорамид мустарда, образует поперечные сшивки в молекуле ДНК клеток экспериментальных мышей, при этом происходит блокирование репликативной вилки. При возникновении рекомбиногенной ситуации при сталировании репликативной вилки и активации инцизионной ферментативной системы для завершения репарации требуются гомологичные последовательности для осуществления репаративного гомологичного обмена. Мы предположили, что Alu-мономеры экзогенной ДНК человека вследствие высокой гомологии с В1 структурами генома мыши завершат репаративный гомологичный обмен за счет разделенных участков гомологии с В1 повторами. Следствием этого будет интеграция экзогенной ксеногенной ДНК в реципиентный геном мыши.
В1-повторы располагаются в геноме через каждые 3.0 т.п.о. В ядре индуцируется 1000-2000 кросс-линков. В межхромосомном пространстве присутствует около 20000 Alu-мономеров, размещенных во фрагментах, размером ˜3.0 т.п.о. То есть существует 10-кратное превосходство количества потенциальных замещающих последовательностей над количеством сайтов замещения.
Вероятность гомологичного замещения В1 повтора Alu-мономером рассчитывается следующим образом:
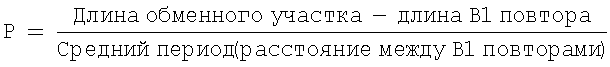
и составляет 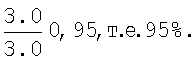
Оценка количества геномов клеток обработанных тканей, содержащих последовательности, гомологичные X-Alu, на основе абсолютных счетов меченого материала в дотах.
ДНК экспериментальных и контрольных организмов в количествах, указанных в таблице, наносилась на мембрану Hibond N (Amersham) и гибридизовалась с αР32 ПЦР меченым X-Alu фрагментом ДНК.
Для оценки счета уверенных зон гибридизации были выбраны пятна, где количество нанесенной ДНК не превышало удельную емкость мембраны (Hibond N). Количество импульсов в дотах двух экспериментальных мышей оценивалось после вычета усредненного специфического фона и составило для мыши №1 - на уровне фона, для мыши №8 - на несколько импульсов выше или равное усредненному специфическому фону. Чувствительность метода не позволяет определить точное соотношение нанесенного количество ДНК и количества импульсов в доте. По этой причине и следуя результатам Саузерн-блот гибридизации, мы провели оценочное определение количества копий для X-Alu мономеров в геноме экспериментальных животных. Для мыши №8 было определено 4 X-Alu мономера (мы определили как одна - несколько копий) и для мыши №1 было определено менее одной копии на гаплоидный геном.
1. Дот, содержащий 0.1 ng X-Alu, имеет одинаковый счет с дотом, содержащим 10 ng ДНК человека. Следовательно, в геноме человека содержится ˜1% фрагментов X-Alu человека.
2. Дот, содержащий 5000 ng ДНК экспериментальной мыши №8, практически не превышает уровень специфического фона и оценочно считает в ˜50 раз меньше, чем дот, содержащий 0.1 ng ДНК X-Alu или 10 ng ДНК человека. То есть в геноме экспериментальной мыши присутствует 0.00004% фрагментов X-Alu человека (0.1:250000×100%).
3. Гаплоидный геном человека содержит ориентировочно 3,3×109 п.о. ДНК. Соответственно 1% X-Alu размером ˜300 п.о. будет составлять 105 копий на гаплоидный геном.
4. Размер генома мыши составляет 2.5х109 п.о., что немногим меньше размера генома человека. Для удобства счета будем считать геномы обоих организмов одинакового размера. Тогда, если 1% от гаплоидного генома составляет 105 копий X-Alu мономера, то в геноме экспериментального животного №8 будет присутствовать ориентировочно 4 его копии. То есть в геноме одной клетки присутствует минимально 4 (одна - несколько) копий мономера Х-Alu. Для экспериментального животного №1 оцененное количество в пять-шесть раз меньше, чем для животного №8, и составляет менее одной копии на гаплоидный геном. При возможной кластеризации X-Alu мономеров в блоки и возможных нескольких сайтов интеграции менее чем одна клетка из составляющих три обработанных органа содержит мономеры X-Alu для обоих животных.
Таким образом, в реципиентных геномах экспериментальных мышей может присутствовать несколько т.п.о. чужеродной человеческой ДНК, гомологичной Alu-повторам. При этом, как следует из анализа геномного блота, X-Alu повторы ограничены рестриктазами BamHI, HindIII или BamHI-HindIII.
Некоторые цифры, необходимые для оценки возможных вариантов интеграции экзогенной ДНК.
Количество клеток в органах, из которых выделялась ДНК.
Печень 1.0 мг - 106 клеток. Средний вес печени 2 г. Общее количество клеток - 2×109.
Тимус содержит (5-6)×107 клеток.
Селезенка содержит ˜3×108 ядросодержащих клеток.
Селезеночная колония - потомок стволовой клетки крови - содержит 106 клеток.
И, таким образом, общее количество клеток составляет 23,5×108 клеток, где селезеночные колонии составляют 0.04%.
Рассмотрим влияние фрагментированной ДНК на восстановление кроветворной функции клеток костного мозга у мышей, формулу крови, выживаемость экспериментальных животных.
Характерной реакцией белой крови на введение циклофосфана является выраженная лейкопения на 4 сутки после введения. При использовании комбинации циклофосфана с фрагментированной ДНК человека картина практически не изменяется. Восстановление белой крови на 7 сутки происходит в двух группах - "циклофосфан" и "циклофосфан + ДНК человека" - несколько различно. Общим является резко выраженный лейкоцитоз с появлением в крови молодых форм гранулоцитарного ряда - юных и палочкоядерных нейтрофилов (в таблице обе формы объединены в графе "палочкоядерные нейтрофилы"). Класс клеток гранулоцитарного ряда становится преобладающим. На 7 сутки в опытной группе наблюдается появление зрелых форм гранулоцитов и восстановление исходного соотношения форменных элементов в крови происходит быстрее, чем у животных в контрольной группе, где этот процесс запаздывает до 14 суток. На 17 сутки картина крови в целом восстанавливается в обеих группах. В иллюстрациях приведены сравнительные данные отдельно по каждой группе гранулоцитов (фиг.2).
Обращают на себя внимание следующие факты. В группе "циклофосфан + ДНК человека" на 6 и 7 сутки пали мыши №5, №7 и №9 (отмечены в таблице зеленым). Причины не ясны - исходная картина крови у них была нормальной, без выраженной патологии. У животных №1 и №8 (отмечены синим) в картине крови отмечалось преобладание гранулоцитов даже спустя 14 суток после введения циклофосфана, в то время как у остальных животных этот показатель нормализовался. На 17-й день эти мыши - №1 и №8 из опытной группы - имели явные признаки каких-то болезней. На 18-й день их забили и выделили ДНК.
Еще две мыши из опытной группы (№2 и №10) пали на 24-е и на 45-е сутки от начала эксперимента. Оставшихся трех мышей - №3, №4 и №6 (опыт) забили на 56-й день для выделения и исследования их ДНК. При этом одна из них уже была больна.
За все это время из контрольной группы погибла только одна мышь. Таким образом, в экспериментальной группе выживаемость составила 20%, а в контрольной - 90%. Обнаруженное совместное действие препарата ДНК и цитостатика циклофосфана предполагало нарушение причинно-следственных механизмов в физиологических системах экспериментальных мышей. Мы предположили, что выявленные нарушения связаны с катастрофическими изменениями генетической информации, заложенной в хромосомах опытных животных, вследствие интеграции в них фрагментов чужеродной ДНК.
Остановимся на подборе праймеров для выявления фрагментов ДНК человека, интегрированных в геном мыши.
Для анализа интеграции ДНК человека в геном экспериментальных животных в качестве поисковой мишени была использована умеренно повторяющаяся последовательность человеческого генома Alu, которая составляет до 10% от общего количества ДНК человека. В геноме мыши присутствуют умеренные повторы В1, аналогичные человеческим Alu, - консенсусные последовательности Alu и В1 имеют высокую степень гомологии (˜65%). Большинство Alu-повторов человека имеют димерную структуру и составляют ˜290 п.о., тогда как В1 повторы, содержащиеся в мышином геноме, почти всегда мономеры (˜130 п.о.) [21, 24]. При выравнивании последовательностей Alu повтора и димера В1 (фиг.3,А) можно увидеть, что на фоне высокой гомологии имеются делетированные участки или участки, не имеющие между собой гомологии. Для конструирования специфичных для Alu повторов праймеров были использованы именно эти районы таким образом, чтобы 3' конец праймера приходился на участок, не имеющий гомологии в структуре повтора В1. Кроме того, праймеры (см. раздел материалы и методы и фиг.3,А) были подобраны так, чтобы в паре один праймер приходился на первую половину Alu, а другой - на вторую, т.е. чтобы ПЦР с указанными праймерами для отдельно взятого мономерного В1 повтора была невозможна.
Был проведен поисковый анализ по базе данных возможных ПЦР продуктов в обоих геномах. В результате проведенного поиска мы отобрали те праймеры, которые не образовывали теоретических ПЦР продуктов с мышиным геномом, при этом с человеческим геномом они давали определенные ПЦР продукты. Оказалось, что реальная картина ПЦР бэндов не соответствует картине, полученной на основе компьютерного анализа. При всех комбинациях всех специфических для человека праймеров в мышином геноме выявлялись четкие бэнды. ПЦР анализ ДНК экспериментальных животных с праймерами на Alu в его классическом виде не позволил достоверно определить наличие какой-либо человеческой последовательности, интегрированной в мышиный геном. Выявлялись яркие мажорные бэнды, характерные и для контрольных, и для экспериментальных мышей (фиг.4,А). По-видимому, последовательности выбранных человеческих праймеров могли спариваться с участками гомологий, существующих в мышином геноме, которые не были выявлены используемой программой. В условиях стандартной ПЦР реакции они имели количественное конкурентное преимущество по отношению к человеческим последовательностям, имеющим полную гомологию с используемыми праймерами, но находящимся в крайне незначительном количестве. Для исключения такого действия мы провели ПЦР амплификацию в жестких условиях с использованием единственного праймера (были проанализированы все синтезированные праймеры, смотри раздел Материалы и методы). Предполагалось, что праймер, обладающий полной гомологией с определенной человеческой последовательностью спарится именно с этой последовательностью и будет синтезирован ПЦР продукт, ограниченный с одного края специфическим для человека праймером. Таким образом, ПЦР материал будет обогащен человеческими последовательностями, исходно присутствующими в крайне малых количествах. Во втором раунде ПЦР амплификации уже с двумя праймерами эти фрагменты не будут подвержены конкурентному вытеснению. При таком экспериментальном подходе анализ ПЦР продуктов позволил четко выделить фрагменты, по подвижности соответствующие фрагментам, выявляемым в ПЦР реакции при использовании в качестве матрицы ДНК человеческого генома. Специфические фрагменты были выявлены в геномах двух экспериментальных мышей №1 и №8 (фиг.4) и их определенная электрофоретическая подвижность составляла около 300 п.о. (280 и 310 п.о.). Поскольку для синтеза были выбраны праймеры, локализованные в последовательности Alu повтора, то мы полагали, что два мажорных ПЦР фрагмента в образце, содержащем ДНК человека, являются двумя вариантами Alu повтора генома человека.
Таким образом, нами доказаны предпосылки использования предложенного способа для ширкого спектра представителей животного мира.
Остановимся на гибридизационном и нуклеотидном анализе последовательностей ПЦР продуктов, выделенных из генома экспериментальных мышей.
Обнаружив в паттерне ПЦР продуктов, полученных с тотальной ДНК экспериментальных мышей, фрагменты, по подвижности совпадающие с мажорными фрагментами, выявляемыми в паттерне ПЦР продуктов, полученных с тотальной ДНК человека, была проведена блот гибридизация с ПЦР меченым фрагментом, являющимся одним из мажорных бэндов в ПЦР реакции с тотальной ДНК человека размером 280 п.о. (фиг.4). Как показала гибридизация с электрофоретически фракционированными ПЦР продуктами всех экспериментальных мышей, только два ПЦР продукта размером 280 и 310 п.о. образов геномной ДНК экспериментальных животных под номерами 1 и 8 гибридизуются с фрагментом размером 280 п.о., полученным в ПЦР реакции с тотальной ДНК человека. Это означало, во-первых, что фрагменты 280 и 310 п.о. в высокой степени гомологичны друг другу и, во-вторых, что в геноме двух экспериментальных мышей появилась ДНК человека.
При анализе нуклеотидных последовательностей фрагментов, полученных в ПЦР реакции с тотальной ДНК человека и обеих мышей (280 п.о.), была установлена их полная гомология между собой (фиг.3,В). При этом они имели участок в ˜150 п.о., практически полностью совпадающий с частью консенсусного Alu-повтора. Это могло означать, что в геном мыши интегрировал(и) фрагмент(ы) большего размера, а выбранные праймеры ограничивают только ту часть, которая непосредственно относится к повторам типа Alu. Вновь описанные фрагменты были названы нами X-Alu. Скрининг экспериментально полученных последовательностей по существующим банкам данных выявил полную гомологию с районом хромосомы 16 человека (АС002400.1) и частичную гомологию с большим количеством фрагментов генома человека. Проведенная саузерн-блот гибридизация ПЦР меченного X-Alu фрагмента с ДНК человека и экспериментальных мышей также выявила большое количество гомологичных фрагментов в геноме человека, что еще раз свидетельствовало о родстве вновь найденного фрагмента ДНК с повторами Alu-семейства.
Остановимся на сравнительном Саузерн-блот анализе и определении количества копий X-AluI фрагмента в геноме человека и в геноме экспериментальных мышей.
В рамках настоящего исследования необходимо было определить, как интегрированная ДНК расположена в геноме и как по количеству интегрированных копий отличаются две экспериментальные мыши между собой. Мы провели саузерн-блот гибридизацию геномной ДНК экспериментальных мышей, геномной ДНК мышей линии СВА и геномной ДНК человека, каждая из которых была одновременно гидролизована рестриктазами BamHI и HindIII, с ПЦР меченым фрагментом, синтезированным с ДНК матрицы генома человека (280 п.о.). Гибридизационная картина представлена на фиг.5,А.
Отмечаются следующие особенности гибридизации.
Во-первых, несмотря на то что условия гибридизации были выбраны максимально жесткими, дорожка с ДНК человека имеет сильную засветку по всей длине, что свидетельствует о большом числе гомологичных используемому зонду последовательностей в геноме человека.
Во-вторых, ДНК мышей линии СВА не имеет гомологичных последовательностей, гибридизующихся в выбранных условиях с зондом X-Alu.
В-третьих, анализ паттерна гибридизации геномной ДНК экспериментальных мышей демонстрирует набор дискретных (BamHI, HindIII или BamHI-HindIII) бэндов в случае мыши №8 и один бэнд в случае мыши №1. Такая картина засветки говорит о следующем.
1) Экспериментальные мыши действительно содержат в своих геномах фрагмент(ы) ДНК человека, гомологичные Alu-повторам.
2) Выявление дискретных бэндов свидетельствуют о том, что полученный в ПЦР анализе и последующей гибридизации результат не является артефактом. В случае контаминации образцов человеческой ДНК можно было бы наблюдать картину засветки, аналогичную выявляемой при гибридизации с суммарной человеческой ДНК (шмер).
3) Количество интегрированной ДНК для двух трансгенных организмов различное.
Неравномерный, фокусированный перенос на мембрану мы объяснить не можем. Он может быть связан с тем, что не во всех клетках присутствуют интегрированные последовательности, что приводит к неравномерному распределению незначительного количества ДНК в полосе агарозного геля и, как следствие, к искажению переноса и фокусировке. Как следует из анализа геномного блота, X-Alu-повторы ограничены рестриктазами BamHI, HindIII или BamHI-HindIII и выявляются в виде дискретных фрагментов, количество которых различно для двух экспериментальных животных. Для животного №1 - это один фрагмент размером ˜2.2 т.п.о. Для животного №8 - это три сильно гибридизующихся фрагмента размером ˜2.2, 3.1 и 4.0 т.п.о. и два размером ˜2.0 и 6.0 т.п.о. (фиг.5,А).
Мы провели серию количественных дот-блот гибридизаций с тем, чтобы определить количество встроенной ДНК и тем самым определить, все ли клетки органов экспериментальных животных, из которых была выделена тотальная ДНК, имеют найденную человеческую ДНК. Оказалось, что определенное в гибридизации количество копий зонда X-Alu соответствует 1% для геномной ДНК человека (порядка 105 копий), для мышки №1 - менее одной копии, а для мыши №8 - ˜4 (одна - несколько) копий на гаплоидный геном (фиг.5,В, см. материалы и методы). Можно полагать, что если мономеры интегрировали блоками, по нескольку копий в блоке, и (или) если интеграция произошла в нескольких местах генома в одной клетке, то менее чем одна клетка из суммарного количества клеток содержит интегрированные фрагменты человеческой ДНК. (Будет выяснено после молекулярного анализа гомологичных X-Alu фрагментов, клонированных из геномов экспериментальных мышей.)
Следует отметить факт, что в дополнительных контрольных экспериментах с использованием только одной из двух субстанций (либо ЦФ, либо препарата фрагментированной ДНК человека) мы не обнаружили каких-либо отклонений от нормы. Все мыши были живы, интеграции анализируемой мишени X-Alu экспериментально не показано (фиг.4,А).
Рассмотрим феномен интеграции чужеродной ДНК в геном клеток соматических тканей взрослого организма.
В предлагаемом исследовании была предпринята попытка на основании наших ранних наблюдений обнаружить факт интеграции экзогенной ДНК человека в геном мыши. В наших предыдущих исследованиях было показано, что экзогенная ДНК воздействует на развитие предшественников эритро- и миелопоэза, что приводит к очень быстрому восстановлению белого и красного кровяных ростков после воздействия цитостатика [Yakubov et al., Николин и др., 2006]. При этом наблюдалась как стимуляция деления СКК, подвергшихся действию цитостатика, так и спасение (сохранение жизнеспособности) значительной доли популяции СКК при жесткой химиотерапии [17, 25, 26]. В результате проведенных исследований было сделано предположение, основывающееся на теоретических предпосылках, опубликованных в работе [16, 17], о том, что экзогенная ДНК, проникнув в ядра СКК (и любых других активно делящихся клеток) принимает участие в репарировании повреждений ДНК, вызванных действием цитостатика. Основываясь на данных о стимуляции кроветворения под воздействием как аллогенной, так и ксеногенной ДНК [17, 25] мы также предположили, что ДНК таксономически близких видов способна принимать участие в гомологичном обмене при репарации индуцированного цитостатиком повреждения и, следовательно, интегрировать в реципиентный геном.
Предполагалось, что моментом интеграции ДНК (как локализованной на хромосомах, так и экстрахромосомальной локализации аллогенного или ксеногенного происхождения) может служить рекомбиногенная ситуация, возникающая при блокировании движения репликативной вилки, формировании двуцепочечных разрывов в месте сшивки и активации системы эксцизионной репарации. В связи с этим главным условием дизайна проведенных экспериментов было обязательное присутствие экзогенной ДНК в ядрах соматических клеток тканей экспериментальных животных во время инъекции цитостатика.
Основными критериями произошедших событий, повлиявших на физиологическое состояние экспериментальных мышей, были скорость восстановления кроветворной функции, определяемой по изменению формулы крови, неожиданные изменения в формуле крови, связанные с одновременным воздействием цитостатика и экзогенной ДНК, и общее поведение подопытных животных.
Полная картина изменений формулы крови (фиг.2) подтвердила результаты аналогичных экспериментов, демонстрирующих ускорение восстановления угнетенного кроветворения при одновременной терапии цитостатиком и препаратом фрагментированной ДНК у всех экспериментальных животных. Однако у двух подопытных животных №1 и №8 необычно возросшее количество гранулоцитов не возвратилось к норме. Причем известно, что причинами нейтрофилеза могут являться заболевания, сопровождающиеся распадом тканей, или заболевания, сопровождающиеся напряжением иммунной системы [27].
К нашему удивлению на шестые сутки начался падеж экспериментальных животных, подвергшихся одновременной обработке двумя препаратами. Последняя мышка погибла на 56 сутки. Было отмечено, что погибали мышки с различной внешней симптоматикой. Было сделано предположение, что столь драматические последствия вызвала интеграция чужеродной ДНК в геном экспериментальных мышей.
Также, по-видимому, позитивное воздействие экзогенной ДНК на угнетенный белый кровяной росток не связано с гомологичным замещением участков хроматина, подвергшихся атаке цитостатика, поскольку, как следует из полученных фактов, интеграция чужеродной ДНК гибельна для реципиентного животного.
К сожалению, паталогоанатомического исследования, в связи с непредсказуемостью событий, сделано не было. Из всех погибших животных были забраны печень, селезенка и тимус для выделения ДНК и последующего ее анализа.
Мишенью для анализа возможной интеграции была выбрана последовательность Alu-повтора.
Некоторые молекулярные характеристики интегрированных последовательностей, имеющих гомологию с AluI повторами человека.
Выбор для анализа Alu-повтора диктовался следующими соображениями. Известно, что Alu-повторы составляют до 10% от гаплоидного генома, что соответствует порядка 106 копий 300 п.о. Alu мономера. Alu-повторы человека распределены в геноме неравномерно, образуя кластеры, и в среднем встречаются через каждые 3-4 т.п.о. Большинство Alu-повторов человека имеют димерную структуру. Найден также ряд тетрамерных последовательностей, мономерные же повторы были обнаружены в единичных случаях. Аналогичные в структурном отношении повторы, названные В1, содержатся в мышином геноме, которые, однако, почти всегда мономеры [21, 24]. Между повторами В1 и Alu существует высокая степень гомологии. Консенсусные последовательности этих структур имеют до 65% гомологии. Между реальными мономерами эта цифра может достигать 90%. В1 повторы мыши организованы в геноме сходным с Alu-повторами образом.
В ядерном пространстве клетки может скапливаться до 2% от гаплоидного генома фрагментов экзогенной экстрахромосомальной ДНК [16]. И, следовательно, в ядрах клеток мышей, подвергшихся обработке экзогенной ДНК, может присутствовать ˜20000 копий Alu мономеров в составе геномных фрагментов среднего размера порядка 3 т.п.о.
При терапевтических дозах цитостатиков, используемых в онкологической практике и научных исследованиях, индуцируется порядка 1000-2000 кросс-линков на ядро [22, 23]. Подобно Alu, B1 повтор встречается в геноме мыши один на каждые 3 т.п.о, и, как следует из расчетов (см. материалы и методы), вероятность гомологичного замещения между двумя родственными повторами составляет более 95%. То есть практически в каждой клетке может присутствовать кросс-линк, который находится в непосредственной близости или непосредственно в самом B1 повторе и может индуцировать гомологичный обмен с Alu последовательностью экстрахромосомального происхождения.
При возникновении повреждений, затрагивающих повторяющиеся последовательности генома, существует большое количество потенциальных гомологичных участков в самых различных его частях, которые могли бы принять участие в репарации повреждения за счет имеющейся гомологии, в случае, если такая репарация требует гомологичного обмена. Для гомологичных районов, в одном из которых присутствует двуцепочечный разрыв, вызванный сталированием репликативной вилки, возможны несколько вариантов репарации. Это генная конверсия, при которой происходит нереципрокный перенос ДНК от донора к реципиенту, при этом при репарации двуцепочечных разрывов поврежденная аллель является, как правило, реципиентом. Также возможно спаривание одиночной цепи (SSA) с последующим синтезом и миграцией цепи, при котором не происходит кроссинговер [28, 29, 30, 31]. Если экстрахромосомальный фрагмент достаточно большого размера и если в геноме находятся линейно расположенные последовательности, фланкирующие повреждение и имеющие гомологию с его концами, то он может полностью заместить поврежденный локус двойным реципрокным обменом по механизму, описанному в работах [32, 33, 28. 29, 17]. При этом центральная часть последовательности, которая образует концевыми участками гетеродуплексы с гомологичными участками хромосомы, интегрирует в реципиентный геном без каких-либо изменений, просто замещая участок хромосомы, ограниченный двумя спаренными концевыми участками. Когда в ядре при гомологичной репарации двуцепочечного разрыва присутствуют многочисленные гомологичные последовательности, например повторяющиеся последовательности генома, два важных фактора влияют на выбор гомологичного сегмента - это его близость к месту гомологичного обмена и степень гомологии [34].
Предполагалось, что большое количество интернализированных в ядерном пространстве мышиных клеток фрагментов, содержащих Alu-повторы, и высокая степень гомологии между Alu-повторами человека и В1 повторами мыши позволит этим повторам спариться друг с другом и вступить в гомологичный обмен.
В проведенных опытах для выделения ДНК использовались три органа экспериментальных животных - тимус, печень и селезенка. Проведенный анализ копийности X-Alu повтора у двух позитивных мышей составил для мыши №1 менее одной копии и для мыши №8 - ˜4 (одна - несколько) копий на гаплоидный геном. Это означает, что только часть (в случае животного №1 без допущений и в случае животного №8 с допущением, что X-Alu мономеры интегрировали блоками и (или) в несколько мест генома в одной клетке) из всех клеток выбранных тканей могут содержать интегрированные копии X-Alu. Если же клетка получила несколько копий мономера, или в случае кластеризации Х-Alu мономеров, еще меньшая часть клеток будет содержать ксеногенные последовательности, интегрированные в геном.
Анализ картины засветки геномных блотов говорит о следующих характеристиках интегрированных в геном мыши человеческих ДНК, содержащих X-Alu-повторы.
1) Соотношение определенной копийности и количества выявляемых бэндов, а также имеющиеся экспериментальные данные по организации Alu и В1 повторов в геноме, свидетельствует о том, что для обоих животных, по-видимому, только часть клеток, из которых выделялась ДНК, содержит гомологичные X-Alu последовательности. При этом обнаружено, что произошла интеграция нескольких различных фрагментов, содержащих X-Alu мономеры.
2) Маловероятно, что интеграция экзогенных фрагментов ДНК в разных клетках произошла в единственное уникальное место. Тогда выявление индивидуальных гибридизующихся фрагментов, содержащих последовательности, гомологичные X-Alu, можно объяснить следующим образом. Интеграция произошла в различные участки генома, имеющие гомологию с X-Alu последовательностью. Интегрировали фрагменты различной длины, но не короче ˜2 т.п.о. Внутри интегрированных фрагментов расположен единственный X-Alu повтор или кластер мономеров, которые ограниченны рестриктазами BamHI, HindIII или BamHI-HindIII. При гидролизе этими рестриктазами высвобождается один и тот же мономер или кластеры мономеров независимо от их локализации и количества в реципиентном геноме. Для мыши №1 это будет фрагмент размером ˜2.2 т.п.о., для мыши №8 - фрагменты размером ˜2.0, 2.2, 3.1, 4.0 и 6.0 т.п.о. Почему только эти фрагменты, содержащие гомологичные X-Alu последовательности, интегрировали в геном экспериментальных животных мы не знаем. Однако полученные различия в количестве и распределении по фрагментам X-Alu-повтора в геномах двух экспериментальных животных свидетельствует о существовании индивидуальной специфичности при интеграции в указанных экспериментальных условиях.
3) Размер интегрированных фрагментов, содержащих кластеры X-Alu, находится в пределах 2-6 т.п.о., что может говорить о необходимости протяженной области гомологии, требуемой для интеграции по типу генной конверсии, либо свидетельствовать о том, что интеграция произошла по механизму, в котором происходит двойной реципрокный обмен спаренных концевых участков фрагмента.
4) Все остальные погибшие мыши, так же как и животные из других серий экспериментов (циклофосфан моноинъекция, препарат человеческой ДНК моноинъекция, всего 30 животных), не содержали интегрированных X-Alu последовательностей. Однако их гибель на длительном экспериментальном отрезке времени дает основания предположить, что в геномы этих животных также произошла интеграция фрагментов экзогенной человеческой ДНК, последовательности которых представляют различные части генома человека, нашедшие гомологичные участки в геноме экспериментальных мышей.
Существует другая возможность, при которой интеграция произошла в нескольких СКК в костном мозге, выжившие потомки которых сформировали индивидуальные колонии в селезенке. Эти колонии и являются клонированными клетками, в которых регистрируется интеграция индивидуального фрагмента. При этом интегрированный фрагмент будет находиться в единственном месте генома, определенном родительской СКК. Оценка копийности X-Alu мономера в геномах экспериментальных животных при возможной кластеризации мономеров X-Alu не противоречит такому видению произошедших событий (оценочное количество: от менее одной до нескольких копий на гаплоидный геном; количество клеток селезеночной колонии составляет 0.04% от общего количества обработанных клеток, количество колоний в селезенке до 40 штук).
Возможные события при интеграции экзогенной ДНК в геном при одновременном воздействии цитостатика и препарата экзогенной ДНК.
Основным вопросом проведенного исследования является вопрос о том, как произошел трансгенез в клетках тканей у взрослых мышей при одновременной обработке цитостатиком, индуцирующим кросс-линки, и фрагментами экзогенной экстраклеточной ксеногенной ДНК. Согласно многочисленным исследованиям последних лет при появлении в геноме межцепочечных сшивок в активно делящихся клетках происходит блокирование репликативной вилки, наталкивающейся на стерическое препятствие. В результате этого события активируется репарационный процесс, в результате чего в области сшивки формируется крайне рекомбиногенная структура. А именно образуется двуцепочечный разрыв во вновь синтезированной цепи в непосредственной близости от места кросс-линка и остановки репликативной вилки и одноцепочечный участок ДНК с размером от 70 п.о. [16] до 700 п.о. [35], образующийся в результате активности XRCC 1-XPF-RPA комплекса. Обе структуры в месте индивидуального кросс-линка образуются последовательно. Однако, по-видимому, существует отрезок времени, когда они присутствуют одновременно. Описано несколько моделей репарации возникшего повреждения и все они включают эксцизионно-экзонуклеазную активность XRCC 1-XPF-RPA комплекса и следующую за этим гомологичную рекомбинацию [10, 9]. При возникновении такой структуры, по-видимому, появляется возможность интеграции в рекомбиногенный сайт экзогенной экстрахромосомальной ДНК с использованием того же самого механизма, как это описано у Kucherlapati [36], или как это происходит при репаративном гомологичном обмене с сестринской хроматидой [9]. Очевидно, что репарировать необходимо два повреждения. По-видимому, в предлагаемом варианте событий возможен любой из известных механизмов репарации разрывов в нити ДНК [37, 38, 39, 40, 41, 28, 29, 30, 31, 32, 33, 17]. Однако обязательным конечным условием такой репарации является репаративный гомологичный обмен между поврежденным участком и гомологичной последовательностью, интернализованной в ядерном пространстве.
Можно предположить, что сшивка цепей ДНК произошла между двумя В1 повторами. В результате поиска гомологичных последовательностей для репарации двуцепочечного разрыва, индуцированного сталированной репликативной вилкой, фрагмент, содержащий Alu-повтор или кластер Alu-повторов, который входит в состав выявленных по геномному блоту фрагментов размером 2-6 т.п.о., своими краевыми мономерами спарился с этими В1 последовательностями. Далее по механизму, описанному в работе [32], интермедиат реализуется с интеграцией всего фрагмента в реципиентный геном. Этим объясняется достаточно большой размер участков человеческой ДНК, интегрированных в геном мыши.
В недавней работе [42] было описано явление ретранспозиции интегрированных в мышиный геном Alu-повторов. Авторы использовали этопозид как генотоксический агент. Он блокирует действие топоизомеразы II, что приводит к возникновению двуцепочечных разрывов и к глобальной активации всех репарационных механизмов. Предполагается, что ретротранспозиция произошла в результате активации транскрипции и конкоминантной активации обратной транскриптазы мышиного генома, ответственной за транспозицию В1 элементов.
Мы не исключаем возможности включения механизма ретротранспозиции для фрагментов, несущих X-Alu последовательности и содержащих промоторную область для РНК полимеразы III. Тем более, что описанное явление транспозиции показано для практически идентичной с нашей биологической системы мышь-человек, а значит существует ферментативная система в мышиной клетке, способная транспозировать человеческие Alu последовательности.
Однако до настоящего времени неизвестен факт кластерной ретротранспозиции Alu-повторов, тем более с использованием в качестве исходной матрицы фрагментов экстрахромосомальной локализации. Единичные копии повторов интегрируют в сайт мишень по механизму обратной транспозиции, включающей синтез РНК, обратную транскрипцию РНК и транспозицию вновь синтезированной копии ДНК в сайт-мишень. Для такого процесса, во-первых, копия Alu элемента должна находиться в составе реципиентного генома. Кроме этого, для акта сайт-специфической интеграции требуется специфическая эндонуклеаза - обратная транскриптаза, фермент, индуцирующий разрывы и осуществляющий непосредственно встраивание и репарацию интегрированной копии Alu ДНК. Ни в одной из известных нам работ не указывается на присутствие этого фермента при репарационных событиях, связанных с репарацией кросс-линков. И последнее, при сайт-специфической ретротранспозиции Alu-повторов в ядерном пространстве клеток экспериментальных мышей, так же как и в случае контаминации образцов человеческой ДНК, невозможно выявить в геномной гибридизации четких бэндов. Это связано с тем, что интегрируемый Alu-повтор не содержит сайтов рестрикции для выбранных рестриктаз, которые при этом статистически распределены по геному. И, таким образом, при случайной транспозиции в разных клетках в различные участки генома не может появиться унифицированный рестрикционный фрагмент.
Таким образом, мы полагаем, что в проведенных экспериментах была осуществлена циклофосфанопосредованная in vivo трансгенная соматическая трансформация половозрелых мышей фрагментами экзогенной ксеногенной ДНК.
В настоящей работе мы показали, что экзогенная ДНК человека, введенная в организм взрослой мыши парентенрально совместно с цитостатиком циклофосфамидом, индуцирующим поперечные сшивки в молекуле ДНК, достигает ядерного пространства клеток, как минимум, трех изученных органов - печени, тимуса, селезенки, и интегрирует в геном. Интеграция экзогенной, ксеногенной ДНК приводит к изменению формулы крови и гибели животных.
Это означает, что
- для осуществления гомологичного обмена между экзогенной экстрохромосомальной ДНК достаточно уже той гомологии, которую имеют два родственных видовых повтора Alu и В1.
- ДНК чужеродного происхождения, будучи интегрированной в геном, приводит к нарушению причинно-следственных механизмов в физиологических системах экспериментальных мышей.
На основании ранее проведенных работ [43] следует, что фрагменты гомологичной ДНК человека, имеющие биологически активный размер и составляющие полный геном физиологически и генетически здорового донора, будут выполнять терапевтическую функцию, замещая различные локусы вследствие естественного механизма гомологической рекомбинации в районах хроматина, доступного для таких событий. В результате искусственно созданной рекомбиногенной ситуации в ядре запускаются несколько механизмов, определяющих скорость и эффективность заместительного терапевтического действия фрагментов экзогенной ДНК. К таким механизмам относятся: а) освобождение двойной цепи ДНК от гистонов на протяжении нескольких мегабаз по обе стороны от возникшего двуцепочечного разрыва; б) происходящие при этом конформационные изменения хроматина в местах хромосом, недоступных для прямого замещения фрагментами экзогенной ДНК, и возникающая, одновременно многократно усиленная возможность прямого замещения гомологичных участков хромосом экзогенными фрагментами ДНК; в) многократно усиливающийся, постоянно протекающий в ядре гомологический обмен экзогенными фрагментами ДНК и теми участками хроматина, которые упакованы таким образом, что они постоянно доступны для гомологического обмена; г) гомологическая рекомбинация между фрагментами экзогенной ДНК и интермедиатами, возникающими в районе кросс-линка в ходе репарации.
Важнейшим моментом одновременной терапии препаратом фрагментированной ДНК при лечении цитостатиками является момент искусственного создания неестественной для клетки рекомбиногенной активности. При этом формируется открытая для рекомбинационной машины структура хроматина, создаются сайты репаративного гомологичного обмена в местах индукции кросс-линков и многократно усиливается рекомбинация, связанная с нахождением концевых гомологий экзогенных фрагментов по всему геному, включая ранее недоступный, ввиду конформационных особенностей, хроматин.
Таким образом, теоретически обосновано и подтверждено экспериментально, что при совместном воздействии цитостатика и препарата фрагментированной гомологичной ДНК будет осуществляться интеграция экзогенной ДНК в гомологичные локусы на хромосомах, ДНК которых вследствие индукции двуцепочечных разрывов будет доступна для осуществления как репаративной гомологической рекомбинации, так и рекомбинации, связанной с нахождением концевых гомологий экзогенных фрагментов по всему геному, включая ранее недоступный, ввиду конформационной закрытости, хроматин.
Это существенно увеличивает эффективность лечения онкологических заболеваний.
При проведении указанных терапий на протяжении определенного времени, контролируемого по биохимическим маркерам онкотрансформации, будет происходить изменение генного гомеостаза, когда клетки претерпевают реверсивное генетическое перерождение, при котором исчезает основное свойство раковой клетки - ее неограниченная пролиферативная активность. Это подтверждает, что предложенный способ практически применим и будет эффективен для лечения раковых заболеваний у животных. При этом имеются все предпосылки проведения дальнейших исследований для определения возможности использования предложенного метода в практики лечения онкологических заблеваний у человека.
Источники информации
1. US, No. 5,795,972, 18.08.1998.
2. US, No. 5,955,059, 21.09.1999.
3. US, No. 5,470,577, 28.11.1995.
4. http://www.netoncology.ru.
5. McHugh PJ, Sones WR, Hartley JA. Repair of intermediate structures produced at DNA interstrand cross-links in Saccharomyces cerevisiae. Mol Cell Biol. 2000 May; 20(10): 3425-33.
6. Wang X., Peterson C.A., Zheng H., Nair R.S., Legerski R.J., Li L. Involvement ofnucleotide excision repair in a recombination-independent and error-prone pathway of DNA interstrand cross-link repair. Molecular and Cellular Biology, 2001, vol. 21, №3, p.713-720.
7. Barber LJ, Ward ТА, Hartley JA, McHugh PJ. DNA interstrand cross-link repair in the Saccharomyces cerevisiae cell cycle: overlapping roles for PS02 (SNM1) with MutS factors and EX01 during S phase. Mol Cell Biol. 2005 Mar; 25(6): 2297-309.
8. Magana-Schwencke N., Henriques J-A.P., Chanet R., Moustacchi E. The fate of 8-methoxypsoralen photoinduced crosslinks in nuclear and mitochondrial yeast DNA: comparison of wild-type and repair-deficient strains. Proc. Natl. Acad. Sci., 1982, vol 79, p.1722-1726.
9. Niedemhofer L.J., Odijk H., Budzowska M., van Drunen E., Maas A., Theil A.F., de Wit J., Jaspers N.G.J., Beverloo H.B., Hoeijmakers J.H.J. Kanaar R. The Structure-Specific Endonuclease Erccl-XpfIs Required To Resolve DNA Interstrand Cross-Link-Induced Double-Strand Breaks. Molecular and Cellular Biology, 2004, v.24, №13, p.5776-5787.
10. De Silva I.U., McHugh P.J., Clingen P.H., Hartley J.A. Defining the roles ofnucleotide excision repair and recombination in the repair of DNA interstrand cross-links in mammalian cells. Molecular and Cellular Biology, 2000, vol. 20, No 21, p.7980-7990.
11. Bessho T, Mu D., Sancar A. Initiation of DNA interstrand cross-link repair in humans: the nucleotide excision repair system makes dual incisions 5' to the cross-linked base and removes a 22- to 28-nucleotide-long damage-free strand. Molecular and Cellular Biology, 1997a, vol. 17, No 12, p.6822-6830.
12. Mu D., Bessho Т., Nechev L.V., Chen D.J., Harris T.M., Hearst J. E., Sancar A. DNA interstrand cross-links induce futile repair synthesis in mammalian cell extracts. Molecular and Cellular Biology. 2000, vol. 20, №7, p.2446-2454.
13. Anker P, Mulcahy H, Chen XQ, Stroun M. Detection of circulating tumour DNA in the blood (plasma/serum) of cancer patients. Cancer Metastasis Rev. 18,65-73(1999).
14. Anker Ph. Quantitative aspects of plasma/serum DNA in cancer patients // 2000. New York Acad. Sci. V.906. P.5-7.
15. Anker P., Mulcahy H., Stroun M. Circulating nucleic acids in plasma and serum as a noninvasive investigation for cancer: Time for large-scale clinical studies? // Int J Cancer, 2003, V. 103(2), P.149-152.
16. Rogachev V.A., Likhacheva A., Vratskikh 0., Mechetina L.V., Sebeleva Т.Е., Bogachev S.S., Yakubov L.A., Shurdov M.A. Qualitative and quantitative characteristics of the extracellular DNA delivered to the nucleus of a living cell. Cancer Cell Int., 2006, 6:23.
17. Yakubov L.A., Popova N.A., Nikolin V.P., Semenov D.V., Bogachev S.S., Os'kina I.N. Extracellular genomic DNA protects mice against radiation and chemical mutagens. Genome Biol. 2003. 5, р.3.
18. Лабораторные методы исследования в клинике. /Под. ред. Меньшикова В.В. - М, 1987.
19. Jurka J. Evolutionary impact of human Alu repetitive elements. Current. Opinion in Genetics and Development. 2004. 14:1-6.
20. Bogerd HP, Wiegand HL, Hulme AE, Garcia-Perez JL, O'Shea KS, Moran JV, Cullen BR. Cellular inhibitors of long interspersed element 1 and Alu retrotransposition. Proc Nati Acad Sci USA. 2006 Jim 6; 103(23): 8780-5.
21. Kolchanov N.A., Shahmuradov I.A., Kapitonov V.V., Omelyanchuk L.V. Evolutionare aspects of the mammalia Alu repeats. Mol Biol. 1988; 22(5): 1335-44.
22. Warren A.J., Mustra D.J., Hamilton J.W. Detection ofmitomycin C-DNA adducts in human breast cancer cells grown in culture, as xenografted tumors in nude mice, and biopsies of human breast cancer patient tumors as determind by "P-postlabeling. Clin. Cancer Res. 2001. 7(4): 1033-1042.
23. Palom Y., Kumar G.S., Tang L.Q., Paz M.M., Musser S.M., Rockwell S., Tomasz M. Relative toxicities ofDNA cross-links and monoadducts: new insights from studies of decarbamoyl mitomycin С and mitomycin C. Chem. Res. Toxicol. 2000; 15: 1398-1406.
24. Britten RJ, Baron WF, Stout DB, Davidson EH. Sources and evolution of human Alu repeated sequences. Proc Nati Acad Sci USA. 1988 Jul; 85(13):4770-4.
25. Nikolin V.P., PopovaN.A., Sebeleva Т.Е., Strounkin D.N., Rogachev V.A., Semenov D.V., Bogachev S.S., Yakubov L.A., Shurdov M.A. Effect of exogenous DNA injection on leukopoietic repair and antitumor action of cyclophosphamide. Voprosy onkologii, 2006, 52(3): 336-340.
26. Patent application №2006127134.
27. Ройтберг Г.Е., Струтынский А.В. Лабораторная и инструментальная диагностика заболеваний внутренних органов. 1999. Издательство: M.: ЗАО «Издательство БИНОМ».
28. Langston L.D., Symington L.S. Gene targeting in yeast is initiated by two independent strand invasions. Proc. Natl. Acad. Sci. U.S.A., 2004, vol. 101, p.15392-97.
29. Leung W.Y., Malkova A., Haber J.E. Gene targeting by linear duplex DNA frequently occurs by assimilation of a single strand that is subject to preferential mismatch correction. Proc. Natl. Acad. Sci. USA, 1997, vol. 94, p.6851-6.
30. Bartsch S., Kang L.E., Symington L.S. RAD51 is required for the repair of plasmid double-strand DNA Gaps from either plasmid or chromosomal templates. Molecular and Cellular Biology, 2000, vol. 20, No 4, p.1194-1205.
31. NickoloffJA. Recombination: mechanisms and roles in tumorigenesis. Encyclopedia of Cancer. 2002; 2nd ed., vol.4: 49-59. Elsevier Science, San Diego.
32. Li J., Read L.R., Baker M.D. The mechanism of mammalian gene replacement is consistent with the formation of long regions ofheteroduplex DNA associated with two crossing-over events. Molecular and Cellular Biology. 2001, vol.21, №2, p.501-510.
33. Hastings P.J., McGill C., Shafer В., Strathem J.N. Ends-In vs. ends-out recombination in Yeast. Genetics, 1993, vol. 135, p.973-980.
34. Schildkraut E., Miller C.A., Nickoloff J.A. Transcription of a donor enhances its use during double-strand break-induced gene conversion in human cells. Molecular and Cellular Biology, 2006, vol.26, №8, p.3098-3105.
35. Sinden R.R., Cole R.S. Repair of cross-linked DNA and survival of Escherichia coli treated with psoralen and ligyt: Effects of mutations influencing genetic recombination and DNA metabolism. J. Bacteriol. 1968; 136: 538-547.
36. Kucherlapati R.S., Eves E.M., Song K-Y., Morse B.S., Smithies O. Homologous recombination between plasmids in mammalian cells can be enhanced by treatment of input DNA. Proc. Natl. Acad. Sci. USA, 1984, vol. 81, p.3153-3157.
37. Thomas KR, Folger KR, Capecchi MR. High frequency targeting of genes to specific sites in the mammalian genome. Cell 1986 Feb 14; 44(3): 419-28.
38. Van Houten В., Gamper H., Holbrook S.R., Hearts J.E., Sancar A. Action mechanism of ABC excision nuclease on a DNA substrate containing a psoralen crosslink at a defined position. Proc. Natl. Acad. Sci. USA. 1986, vol. 83, p.8077-8081.
39. Rouet P., Smih F., Jasin M. Introduction of double-stand breaks into the genome of mouse cells by expression of a rare-cutting endonuclease. Molecular and Cellular Biology, 1994, vol. 14, №12, p.8096-8106.
40. Jasin M. Genetic manipulation ofgenomes with rare-cutting endonucleases. TIG June 1996, vol.12, №6, p.224-228.
41. Liang F., Han M., Romanienko P.J., Jasin M. Homology-directed repair is a major double-strand break repairpathway in mammalian cells. Proc. Natl. Acad. Sci. USA, 1998, vol. 95, p.5172-5177.
42. Hagan CR, Sheffield RF, Rudin CM. Human Alu element retrotransposition induced by genotoxic stress. Nat Genet. 2003 Nov; 35(3): 219-20.
43. Lihacheva et al., 2006.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 2006 |
|
RU2322264C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2006 |
|
RU2313349C2 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2008 |
|
RU2387456C1 |
| СПОСОБ ДЕТЕКЦИИ СТВОЛОВЫХ РАКОВЫХ КЛЕТОК | 2013 |
|
RU2530170C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2009 |
|
RU2429019C2 |
| СПОСОБ ЭРАДИКАЦИИ СТВОЛОВЫХ ИНИЦИИРУЮЩИХ РАКОВЫХ КЛЕТОК | 2013 |
|
RU2542410C1 |
| СПОСОБ ПОЛУЧЕНИЯ СТИМУЛИРОВАННЫХ ДЕНДРИТНЫХ КЛЕТОК ДЛЯ ИНДУКЦИИ ИММУННОГО ОТВЕТА ПРОТИВ ТУБЕРКУЛЕЗА ЧЕЛОВЕКА | 2009 |
|
RU2401664C1 |
| СПОСОБ ЛЕЧЕНИЯ АСЦИТНОЙ ФОРМЫ РАКА | 2015 |
|
RU2595864C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2662354C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2012 |
|
RU2490028C1 |

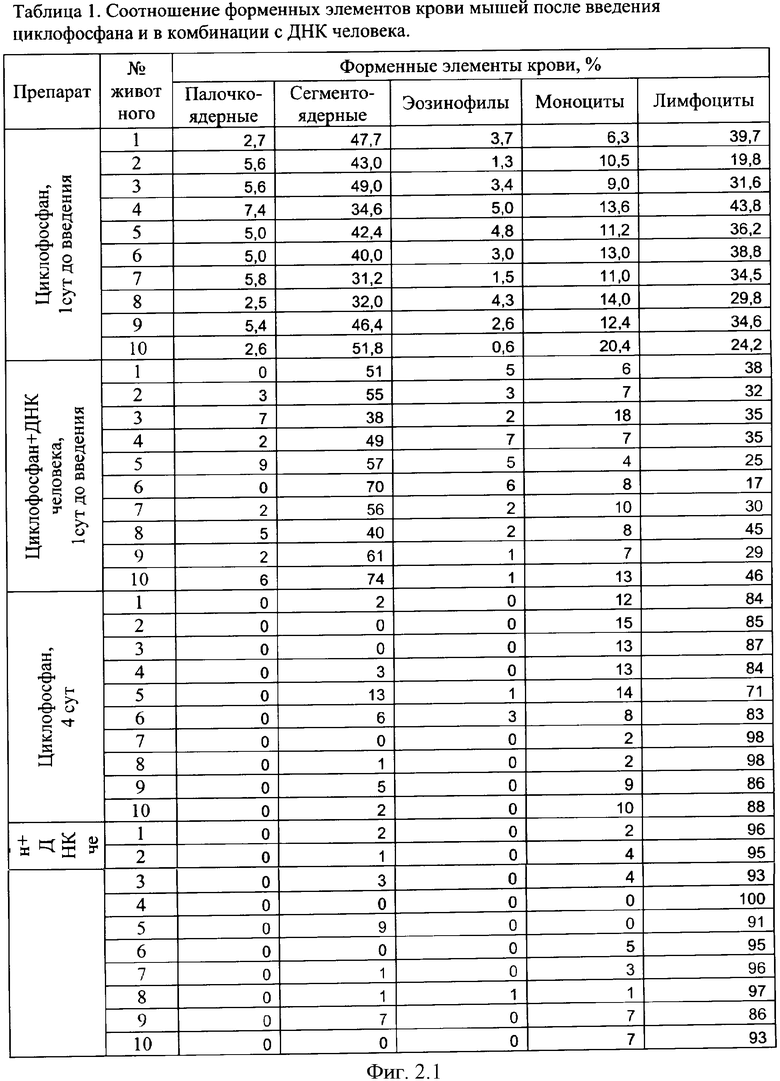
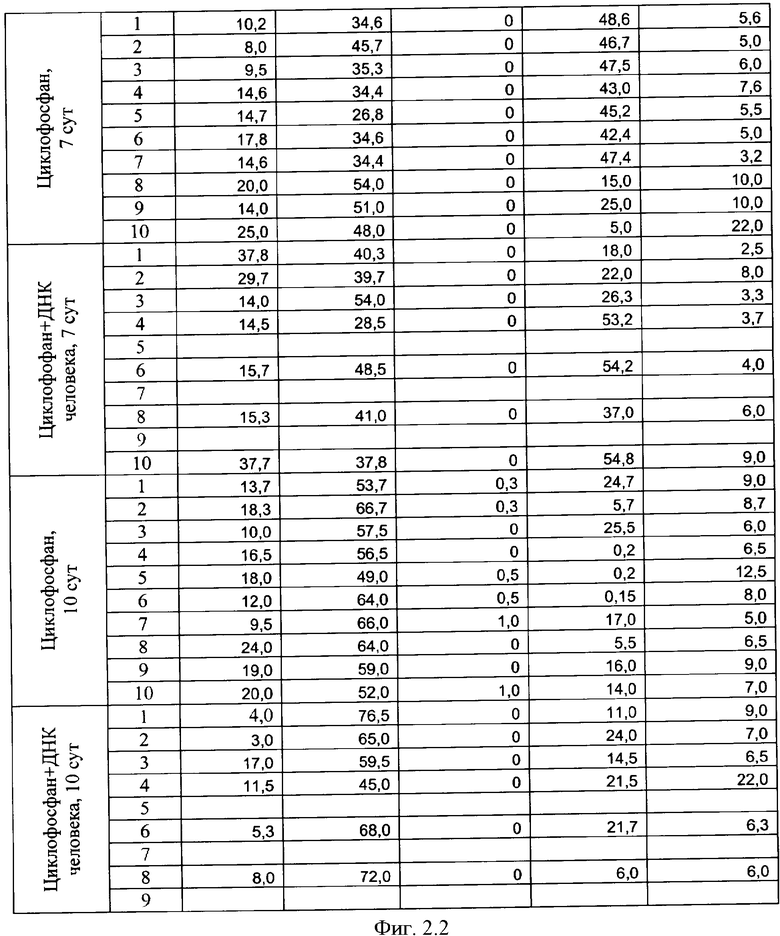
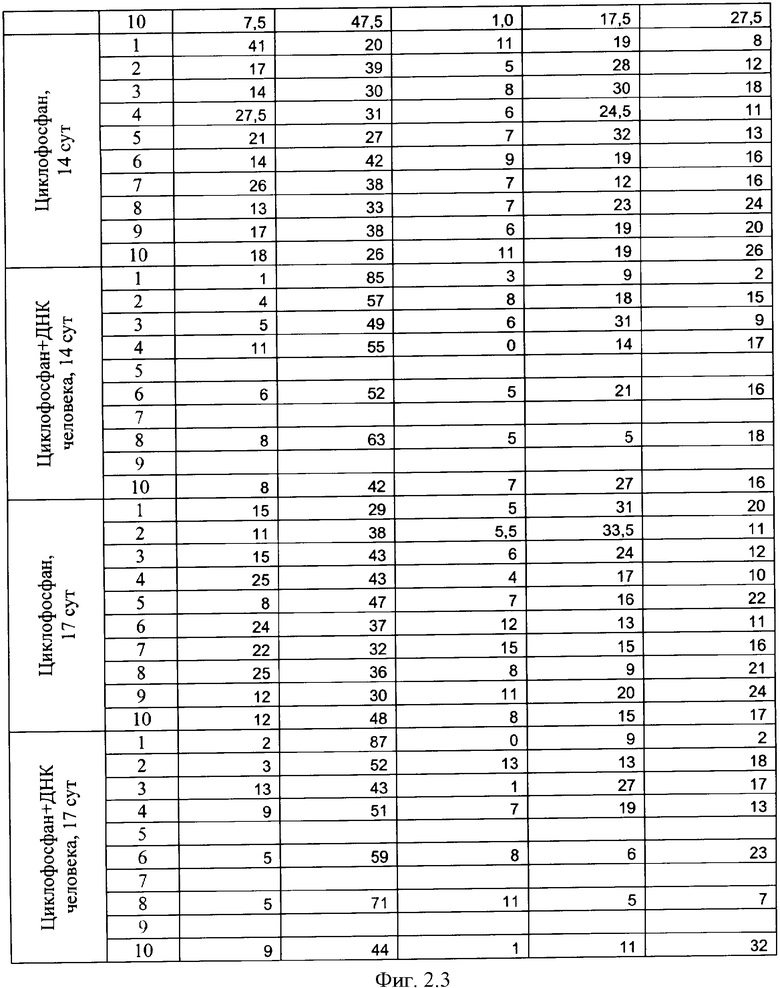
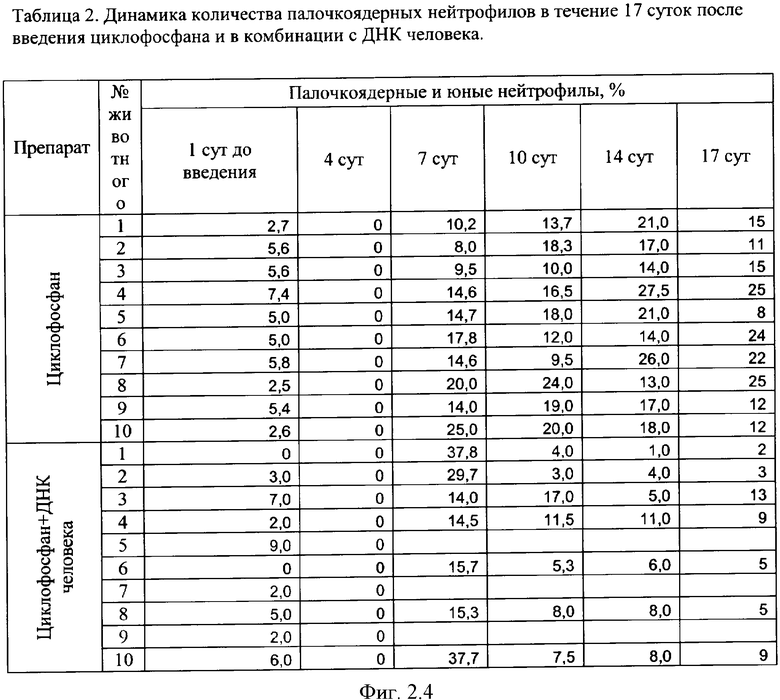
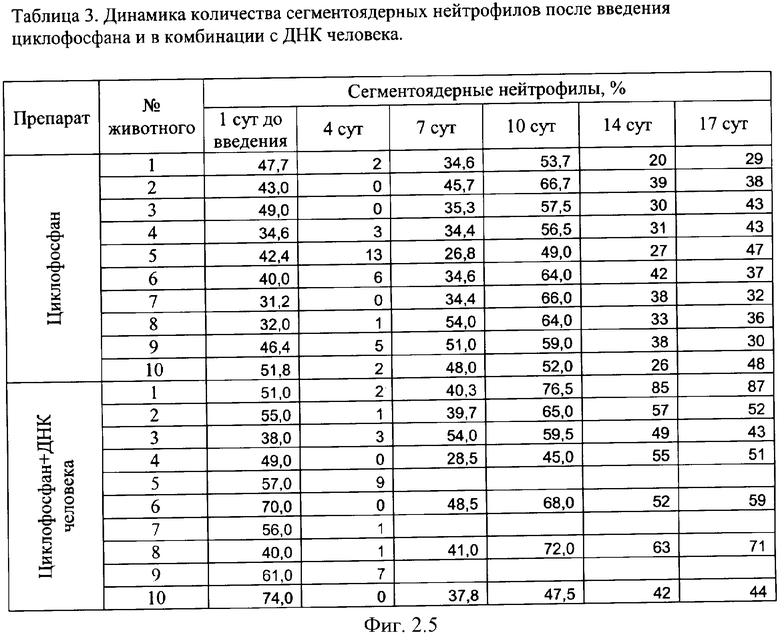
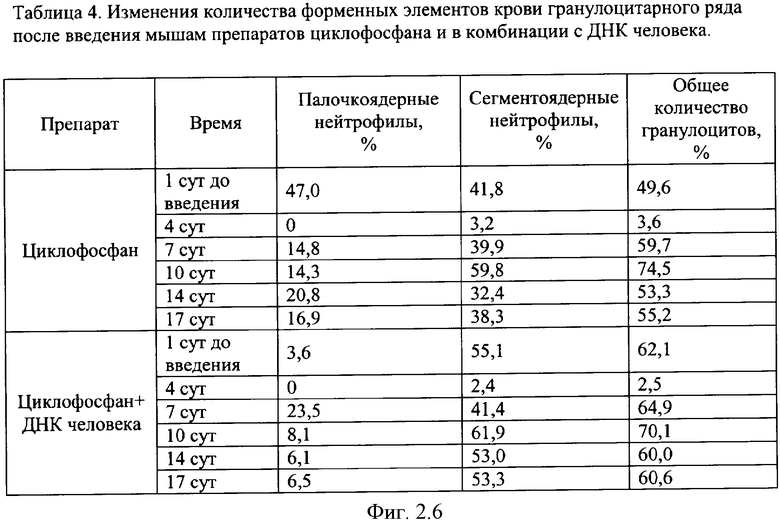
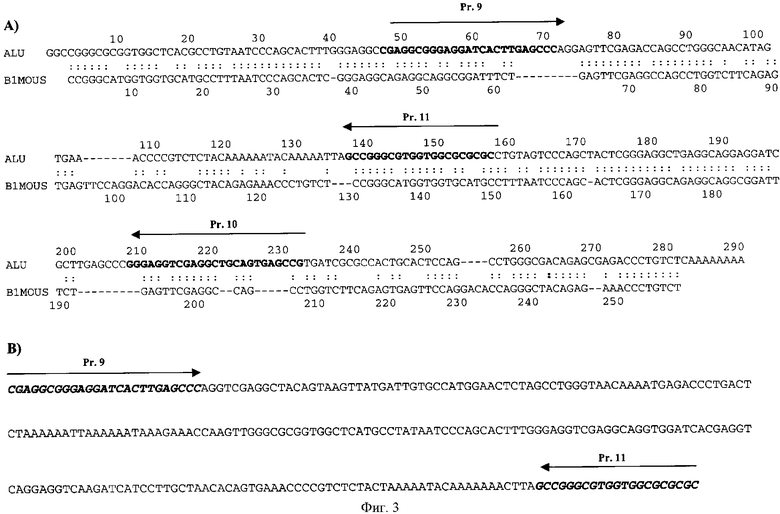
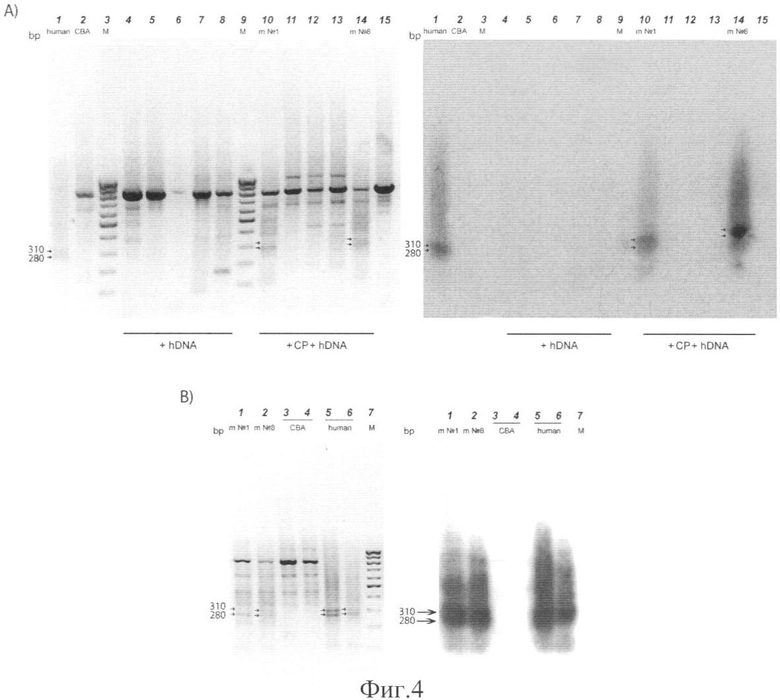
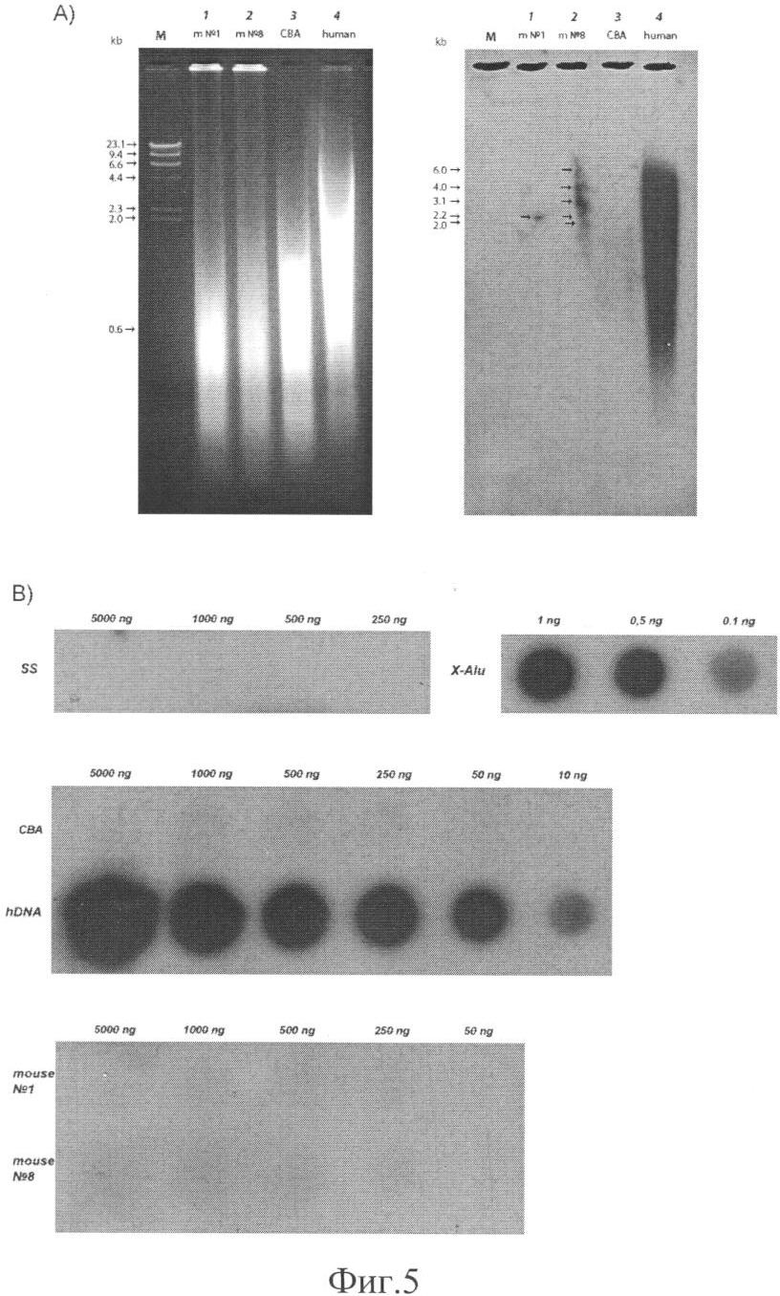
Изобретение относится к медицине, а именно к онкологии, и может быть использовано при комплексном лечении пациентов с онкологическими заболеваниями. Способ основан на введении в организм пациента препарата цитостатической группы, механизм действия которого основан на формировании межцепочечных сшивок в молекуле ДНК раковых клеток организма, совместно с препаратом фрагментированной аллогенной ДНК с фрагментами, имеющими биологически активный размер и составляющими полный геном физиологически и генетически здорового донора. Способ позволяет активировать рекомбинационную систему клетки и провести гомологичную интеграцию препарата ДНК в геном реципиента. 2 з.п. ф-лы, 5 ил., 1 табл.
| NIKOLIN V.P | |||
| et al | |||
| Effect of exogenous dna injection on leukopoietic repair and antitumor action of cyclophosphamide // Vopr Onkol | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| ПРЕПАРАТ, ОБЛАДАЮЩИЙ ПРОТИВООПУХОЛЕВЫМ, АНТИТОКСИЧЕСКИМ И РАДИОПРОТЕКТОРНЫМ ДЕЙСТВИЕМ | 2003 |
|
RU2234323C1 |
| НАБОР ПРОДУКТОВ НА НЕДЕЛЬНУЮ ДИЕТУ ДЛЯ ОНКОЛОГИЧЕСКИХ БОЛЬНЫХ И ДОЛГОЖИТЕЛЕЙ | 2005 |
|
RU2286679C1 |
| US 5795972, 18.08.1998 | |||
| US | |||

