ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Гранулоцитарно-макрофагальный колониестимулирующий фактор, GM-CSF, изначально был идентифицирован как гемопоэтический фактор роста. Он вырабатывается целым рядом типов клеток, включая лимфоциты, моноциты, эндотелиальные клетки, фибробласты и некоторые клетки злокачественных новообразований (Metcalf et al., 1986; Clark and Kamen, 1987; Hart et al, 1988; Metcalf et al., 1986). Кроме функции стимулирования роста и дифференциации гемопоэтических клеток-прекурсоров, также было обнаружено, что GM-CSF оказывает разнообразное воздействие на клетки иммунной системы, экспрессируя GM-CSF рецептор (обзоры см. в Hamilton, 2002; de Groot et al., 1998). Основная из этих функций состоит в активации моноцитов, макрофагов и гранулоцитов в нескольких иммунных и воспалительных процессах (Gasson et al., 1990b; Gasson et al., 1990a; Hart et al., 1988; Rapoport et al., 1992).
Зрелый GM-CSF представляет собой мономерный белок длиной 127 аминокислот с двумя сайтами гликозилирования. Варьирующая степень гликозилирования приводит к молекулярной массе в интервале от 14 кДа до 35 кДа. Негликозилированный и гликозилированный GM-CSF демонстрирует сходную активность in vitro (Cebon et al., 1990). Кристаллографический анализ GM-CSF выявил структуру в форме бочонка, состоящую из четырех коротких альфа-спиралей (Diederichs et al., 1991). Общий характер сворачивания сходен с другими факторами роста, такими как гормон роста, интерлейкин-2 и интерлейкин-4.
GM-CSF оказывает свое биологическое действие путем связывания со своим рецептором (Kastelein and Shanafelt, 1993; Sisson and Dinarello, 1988). Наиболее важные сайты экспрессии рецептора GM-CSF (GM-CSF-R) присутствуют на клеточной поверхности миелоидных клеток и эндотелиальных клеток, тогда как лимфоциты являются GM-CSF-R-негативными. Природный рецептор состоит как минимум из двух субъединиц, альфа и бета. Субъединица альфа влияет на специфичность лиганда и связывается с GM-CSF с наномолярным сродством (Gearing et al., 1989; Gasson et al., 1986). Субъединица бета также представляет собой часть комплексов рецептора интерлейкина-3 и интерлейкина-5 и, в сочетании с альфа-субъединицей рецептора GM-CSF и GM-CSF, ведет к образованию комплекса с пикомолярным сродством связывания (Hayashida et al., 1990). Построена карта доменов связывания на GM-CSF для рецептора: GM-CSF взаимодействует с субъединицей бета его рецептора через очень ограниченный участок в первой альфа-спирали GM-CSF (Shanafelt et al., 1991b; Shanafelt et al., 1991 a; Lopez et al., 1991). Связывание с субъединицей альфа могло бы проецироваться на третью альфа-спираль, спираль С, начальные остатки петли, соединяющей спирали С и D, а также на карбоксиконцевой хвост GM-CSF (Brown et al., 1994).
Образование тримерного комплекса рецептора GM-CSF ведет к активации комплексных каскадов проведения сигнала с участием молекул семейств JAK/STAT, Shc, Ras, Raf, киназ митогенактивируемого белка, фосфатидилинозитол-3-киназы и NFkB, что, в конце концов, ведет к транскрипции c-myc, c-fos и c-jun. Активация по большей части индуцируется субъединицей бета-рецептора (Hayashida et al., 1990; Kitamura et al., 1991; Sato et al., 1993). Общая субъединица бета также ответственна за функции частичного перекрытия, которые проявляют IL-3, IL-5 и GM-CSF (обзор см. в de Groot et al., 1998).
Помимо стимулирующего влияния на рост и дифференциацию гемопоэтических клеток GM-CSF конкретно функционирует как провоспалительный цитокин. Макрофаги и моноциты, а также нейтрофилы и эозинофилы активируются GM-CSF, что ведет к высвобождению других цитокинов и хемокинов, расщепляющих матрикс протеаз, повышенной экспрессии HLA и повышенной экспрессии молекул клеточной адгезии или рецепторов для СС-хемокинов. Последнее, в свою очередь, ведет к повышению хемотаксиса воспалительных клеток в воспаленную ткань (Chantry et al., 1990; Hamilton, 2002; Sisson and Dinarello, 1988; Zhang et al., 1998; Hamilton et al., 1993; Lopez et al., 1986; Cheng et al., 2001; Gomez-Cambronero et al., 2003). GM-CSF часто проявляет свою активность в синергии с другими воспалительными стимулирующими факторами, такими как другие цитокины или LPS, например нейтрофилы, обработанные GM-CSF, в комбинации с, например, LPS демонстрируют усиленную окислительную вспышку (Kaufman et al., 1989; Rapoport et al., 1992).
GM-CSF как цель для противовоспалительной терапии
Благодаря наличию разнообразных активирующих функций в иммунной системе GM-CSF может рассматриваться как цель для противовоспалительной терапии. Состояние пациентов при хронических и острых воспалительных заболеваниях, таких как ревматоидный артрит (RA), рассеянный склероз (МС), болезнь Крона, псориаз, астма, атопический дерматит или шок, может значительно улучшиться в результате блокирования активности GM-CSF и последующего снижения вредной активности реагирующих на GM-CSF клеток (Hamilton, 1993; Zhang et al., 1998; Hamilton, 2002).
Артрит
Несколько групп продемонстрировали, что GM-CSF, а также его рецептор, присутствуют в синовиальном суставе больных артритом (Alvaro-Gracia et al., 1991; Xu et al., 1989; Haworth et al., 1991). Кроме того, было показано, что GM-CSF вызывает обострение ревматоидного артрита у пациентов, которые получали лечение GM-CSF по поводу нейтропении при синдроме Фелти (Hazenberg et al., 1989) или после химиотерапии (de Vries et al., 1991).
Первые указания на пригодность антител, блокирующих GM-CSF, для лечения артрита были получены в результате исследований на мышах in vivo (Campbell et al., 1997; Campbell et al., 1998; Cook et al., 2001). Конкретно, Cook et al. показали, что нейтрализующие антитела к GM-CSF эффективны на модели индуцированного коллагеном артрита. Блокирование GM-CSF приводило к уменьшению тяжести заболевания и сопутствующего воспаления, разрушения хряща и прогресса заболевания в изначально пораженных конечностях или распространения на другие конечности.
Существует несколько эффектов анти-GM-CSF терапии, которые могли бы быть полезны для больных ревматоидным артритом или другими воспалительными заболеваниями.
Ожидается, что блокирование GM-CSF будет ингибировать или уменьшать:
a) активацию и количество зрелых моноцитов, макрофагов и нейтрофилов. Особенно нейтрофилы и макрофаги широко встречаются в синовиальной жидкости и мембране. Показано, что количество макрофагов в синовии коррелирует со степенью эрозии в пораженных ревматоидным артритом суставах (Mulherin et al., 1996; Burmester et at., 1997). Макрофаги представляют собой источник различных других провоспалительных цитокинов и расщепляющих матрикс протеаз. Выработка H2O2 нейтрофилами представляет собой часть разрушительных процессов, происходящих в пораженных артритом суставах (Babior, 2000);
b) дифференциацию миелоидных дендритных клеток (DC) и активацию синовиальных DC (=синовиоциты). GM-CSF регулирует вверх и поддерживает экспрессию класса II HLA на DC и синовиоцитах при ревматоидном артрите (Alvaro-Gracia JM et al., 1991). DCS активируются в суставе, чтобы приобрести функции, связанные с селективной активацией воспалительных Т-клеток. Конкретные аллели HLA-DR связаны с восприимчивостью к ревматоидному артриту, и активация Т-клеток через представление антигена DC может играть критическую роль при данном типе иммунного заболевания (Santiago-Schwarz et al., 2001).
Рассеянный склероз
При рассеянном склерозе повышенные уровни GM-CSF коррелируют с активной фазой рассеянного склероза (Carrieri et al., 1998; McQualter et al., 2001) и у GM-CSF-/- мышей не может развиться заболевание на модели рассеянного склероза - экспериментальном энцефаломиелите (ЕАЕ, McQualter et al., 2001).
Астма
Сообщалось об увеличении количества GM-CSF в легких при астме (Broide and Firestein, 1991). В то же время увеличивается количество эозинофилов, где GM-CSF синергично с интерлейкином-5 действует в трех направлениях: i) стимулирует дифференциацию клеток-прекурсоров до эозинофилов, ii) стимулирует их функциональную активацию, и iii) увеличивает продолжительность жизни эозинофилов в легких (Broide et al., 1992; Yamashita et al., 2002). Таким образом, уменьшение продолжительности жизни эозинофилов в пораженных астмой дыхательных путях путем блокирования GM-CSF, вероятно, облегчает тяжесть заболевания. Пригодность нейтрализующих анти-GM-CSF антител была дополнительно показана на модели астмы у мышей, где введение таких антител приводило к значительному уменьшению гиперреактивности дыхательных путей и воспалению дыхательных путей (Yamashita et al., 2002).
В другой модели на мышах LPS-зависимое воспаление легкого могло быть уменьшено применением анти-GM-CSF антитела 22Е9 у мышей (Bozinovski et al., 2003).
Токсическое действие
Мыши, гомозиготные по разрушенному гену гранулоцитарно/макрофагального колониестимулирующего фактора (GM-CSF), развиваются нормально и не демонстрируют никаких существенных нарушений гемопоэза до возраста 12 недель. Хотя большинство GM-CSF-дефицитных мышей внешне здоровы и способны давать потомство, у всех таких животных развивается дезорганизация сосудистого внеклеточного матрикса с разрушенными и уменьшенными пучками коллагена и аномальное легкое с нарушенным клиренсом легочного сурфактанта и сниженной резистентностью к микробным патогенам в легком. Характеристики последней патологии сходны с легочным альвеолярным протеинозом (РАР) у человека. По-видимому, GM-CSF не является существенным для поддержания нормальных уровней основных типов зрелых гемопоэтических клеток и их прекурсоров в крови, костном мозге и селезенке. Однако указывается на незаменимость GM-CSF для нормального развития сосудов, физиологии легких и сопротивления локальной инфекции (Stanley et al., 1994; Dranoff et al., 1994; Plenz et al., 2003; Shibata et al., 2001). Недавно получены дополнительные доказательства выраженной связи аутоантител к GM-CSF с РАР в виде аномалий проведения сигнала GM-CSF в патогенезе РАР у человека. Взятые вместе, эти наблюдения демонстрируют, что GM-CSF играет критическую роль в регулировании гомеостаза сурфактанта и врожденных иммунных функций альвеолярных макрофагов в легком (Bonfield et al., 2002; Trapnell and Whitsett, 2002; Uchida et al., 2004; Kitamura et al., 1999).
Высокие титры аутоантител с блокированием активности GM-CSF были описаны у больных миастенией гравис. Такие больные не проявляли каких-либо других аутоиммунных феноменов или дефицитов гемопоэза или "других очевидных клинических корреляций" (Meager et al., 1999).
Соединение E21R, модифицированная форма GM-CSF, которая осуществляет антагонизм в отношении функции GM-CSF, было оценено в фазе I исследования безопасности и продемонстрировало благоприятный профиль безопасности у больных раком (Olver et al., 2002).
Таким образом, помимо легочной функции, которую следует тщательно контролировать, не ожидается развития других побочных эффектов при применении анти-GM-CSF терапии.
До сих пор были получены только антитела, происходящие из нечеловеческих видов с функцией нейтрализации GM-CSF. Например, ЕР 0499161 А1 описывает антитело, полученное иммунизацией мышей олигопептидами, последовательность которого получена из GM-CSF. К тому же заявка раскрывает способ облегчения у млекопитающего, которое в этом нуждается, нежелательного влияния GM-CSF, который включает введение указанному млекопитающему GM-CSF-ингибирующего количества иммуноглобулина. Однако указанное антитело является мышиным и не подходит для введения человеку.
Кроме того, WO 03/068920 раскрывает ингибиторное химерное мышиное/человеческое антитело IgG1. Вероятно антитела, которые содержат нечеловеческие последовательности, вызывают иммунную реакцию у пациента-человека и не подходят для терапевтического введения. Например, при заболеваниях, где необходимо длительное лечение (например, хронические воспалительные заболевания, такие как ревматоидный артрит, астма и рассеянный склероз), длительное введение терапевтического агента нечеловеческого происхождения повышает вероятность выраженной воспалительной реакции и выработки человеческих антител, которые могут нейтрализовать терапевтический агент.
Соответственно, в свете большого потенциала терапии анти-GM-CSF антителом, существует большая потребность в человеческих анти-GM-CSF антителах с высоким сродством, которые эффективно блокируют взаимодействие рецептора GM-CSF/GM-CSF. Кроме того, было бы предпочтительным получить одно или больше антител, которые могут перекрестно реагировать с GM-CSF одного или больше нечеловеческих видов с целью проверки их эффективности в моделях in vivo на животных.
Настоящее изобретение удовлетворяет эти и другие потребности, предлагая высокоэффективные анти-GM-CSF антитела, которые описаны ниже.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Одним из объектом изобретения являются человеческие и гуманизированные антитела, способные эффективно блокировать взаимодействие рецептора GM-CSF/GM-CSF.
Другим объектом изобретения являются антитела, которые безопасны с точки зрения введения человеку.
Еще одним объектом настоящего изобретения являются способы лечения заболеваний или и/или состояний, связанных с присутствием GM-CSF, с применением одного или больше антител по изобретению. Эти и другие объекты изобретения более полно описаны в данном описании.
В одном аспекте изобретение предлагает антигенсвязывающий участок, специфичный для человеческого GM-CSF, где изолированное человеческое или гуманизированное антитело или его функциональный фрагмент способно (i) блокировать взаимодействие 0,5 мкг/мл человеческого GM-CSF с альфа-цепью человеческого рецептора GM-CSF, экспрессированного на приблизительно 2×105 клеток СНО-K1 как минимум на 50% в следующих условиях: (а) концентрация альфа-цепи человеческого рецептора GM-CSF, экспрессированного на клетках СНО-K1, сходна с концентрацией альфа-цепи человеческого рецептора GM-CSF, экспрессированного на приблизительно 2×105 клеток CHO-GMRa#11, и (b) концентрация изолированного человеческого или гуманизированного антитела или его функционального фрагмента составляет приблизительно 5 мкг/мл; и (ii) нейтрализовать 0,25 нг/мл человеческого GM-CSF в анализе пролиферации TF-1 со значением IC50 как минимум в 5 раз большим, чем у антитела сравнения BVD2-21C11 и/или антитела сравнения МАВ215. В данном описании "анализ пролиферации TF-1" определяется как анализ, в существенной мере аналогичный описанному в Примере 5В. Опытный специалист может получить клетки СНО-К1, экспрессирующие альфа-цепь человеческого рецептора GM-CSF в концентрации, сходной с той, которая экспрессируется на приблизительно 2×105 клеток CHO-GMRa#11 в соответствии с указаниями данного описания.
Изобретение дополнительно предлагает изолированное человеческое или гуманизированное антитело или функциональный фрагмент антитела, который содержит антигенсвязывающий участок, как раскрыто в данном описании. Такое антитело или его функциональный фрагмент может содержать антигенсвязывающий участок, который содержит участок H-CDR3, представленный в SEQ ID NO:11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 49, 50, 51 или 52; антигенсвязывающий участок также может содержать участок H-CDR2, представленный в SEQ ID NO:11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 49, 50, 51 или 52; и антигенсвязывающий участок также может содержать участок Н-CDRI, представленный в SEQ ID NO:11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 49, 50, 51 или 52. Такое антитело или его функциональный фрагмент может содержать антигенсвязывающий участок, который содержит вариабельную тяжелую цепь, представленную в SEQ ID NO:11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 49, 50, 51 или 52. Такое GM-CSF-специфическое антитело по изобретению может содержать антигенсвязывающий участок, который содержит участок L-CDR3, представленный в SEQ ID NO:31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 58, 59, 60 или 61; антигенсвязывающий участок дополнительно может содержать участок L-CDR2, представленный в SEQ ID NO:31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 58, 59, 60 или 61; и антигенсвязывающий участок также может содержать участок L-CDRI, представленный в SEQ ID NO:31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 58, 59, 60 или 61. Такое антитело или его функциональный фрагмент может содержать антигенсвязывающий участок, который содержит вариабельную легкую цепь, представленную в SEQ ID NO:31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 58, 59, 60 или 61.
Варианты пептидных последовательностей, раскрытых в данном описании, также включены в настоящее изобретение. Соответственно, изобретение включает анти-GM-CSF антитела, содержащие аминокислотную последовательность тяжелой цепи с идентичностью последовательности как минимум 60% в участках CDR, где последовательности участков CDR представлены в SEQ ID NO:11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 49, 50, 51 или 52; и/или гомологией последовательности как минимум 80% в участках CDR, где последовательности участков CDR представлены в SEQ ID NO:11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 49, 50, 51 или 52. Кроме того, включены анти-GM-CSF антитела, содержащие аминокислотную последовательность легкой цепи с идентичностью последовательности как минимум 60% в участках CDR, где последовательности участков CDR представлены в SEQ ID NO:31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 58, 59, 60 или 61; и/или гомологию последовательности как минимум 80% в участках CDR, где участки CDR представлены в SEQ ID NO:31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 58, 59, 60 или 61.
Антитело по изобретению может представлять собой IgG (например, IgG1), в то время как фрагмент антитела может представлять собой, например, Fab или scFv. Фрагмент антитела по изобретению, соответственно, может представлять собой или может содержать антигенсвязывающий участок, который ведет себя одним способом или больше, как описано в данном описании.
Изобретение также связано с изолированными последовательностями нуклеиновых кислот, каждая из которых может кодировать антигенсвязывающий участок человеческого или гуманизированного антитела или его функциональный фрагмент, который является специфичным в отношении GM-CSF. Такая последовательность нуклеиновой кислоты может кодировать вариабельную тяжелую цепь изолированного человеческого или гуманизированного антитела или его функционального фрагмента, содержащую SEQ ID NO:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 44, 45, 46, 47 или 48, или последовательность нуклеиновой кислоты, которая гибридизуется в условиях высокой строгости с комплементарной цепью SEQ ID NO:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 44, 45, 46, 47 или 48. Нуклеиновая кислота может кодировать вариабельную легкую цепь изолированного человеческого или гуманизированного антитела или его функционального фрагмента, содержащую SEQ ID NO:21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 53, 54, 55, 56 или 57, или последовательность нуклеиновой кислоты, которая гибридизуется в условиях высокой строгости с комплементарной цепью SEQ ID NO:21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 53, 54, 55, 56 или 57.
Последовательность нуклеиновой кислоты может кодировать антигенсвязывающий участок человеческого или гуманизированного антитела или функционального фрагмента антитела, который является специфичным в отношении человеческого GM-CSF, где антитело или его функциональный фрагмент может (i) блокировать взаимодействие 0,5 мкг/мл человеческого GM-CSF с альфа-цепью человеческого рецептора GM-CSF, экспрессированного на 2×105 клеток СНО-K1, как минимум на 50% в следующих условиях: (а) концентрация альфа-цепи человеческого рецептора GM-CSF, экспрессированного на клетках СНО-K1, сходна с концентрацией альфа-цепи человеческого рецептора GM-CSF, экспрессированного на приблизительно 2×105 клеток CHO-GMRa#11, и (b) концентрация изолированного человеческого или гуманизированного антитела или его функционального фрагмента составляет приблизительно 5 мкг/мл; и (ii) нейтрализовать 0,25 нг/мл человеческого GM-CSF в анализе пролиферации TF-1 со значением IC50, как минимум в 5 раз большим, чем у антитела сравнения BVD2-21C11 и/или антитела сравнения МАВ215.
Нуклеиновые кислоты по изобретению являются пригодными для рекомбинантного продуцирования. Таким образом, изобретение также относится к векторам и клеткам-хозяевам, содержащим последовательность нуклеиновой кислоты по изобретению. Такие клетки-хозяева могут быть бактериальными или эукариотными клетками.
Композиции по изобретению могут применяться для терапевтических или профилактических целей. Таким образом, изобретение включает фармацевтическую композицию, содержащую антитело по изобретению (или функциональный фрагмент антитела) и фармацевтически приемлемый носитель или вспомогательное вещество. В связанном аспекте изобретение предлагает способ лечения расстройства или состояния, связанного с нежелательным присутствием GM-CSF или экспрессирующих GM-CSF клеток. Такой способ включает стадии введения субъекту, который в этом нуждается, эффективного количества фармацевтической композиции, содержащей антитело по изобретению, как описано или предусмотрено в данном описании. Такое расстройство или состояние может представлять собой воспалительное заболевание, такое как ревматоидный артрит, рассеянный склероз, болезнь Крона, псориаз, астма, атопический дерматит и шок.
Человеческие или гуманизированные антитела (и их функциональные фрагменты) по настоящему изобретению могут быть перекрестно-реагирующими с GM-CSF крысы и/или макаки резус, что определяется равновесным титрованием раствора (SET) и/или анализом пролиферации TF1.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На Фигуре 1a представлены последовательности нуклеиновых кислот различных новых вариабельных участков тяжелой цепи антитела.
На Фигуре 1b представлены аминокислотные последовательности различных новых участков вариабельной тяжелой цепи антитела. CDR участки HCDR1, HCDR2 и HCDR3 обозначены от N-конца к С-концу полужирным начертанием шрифта.
На Фигуре 2а представлены последовательности нуклеиновых кислот различных новых вариабельных участков легкой цепи антитела.
На Фигуре 2b представлены аминокислотные последовательности различных новых вариабельных участков легкой цепи антитела. CDR участки LCDR1, LCDR2 и LCDR3 обозначены от N-конца к С-концу полужирным начертанием шрифта.
На Фигуре 3 представлены аминокислотные последовательности вариабельных участков тяжелой цепи основанных на консенсусе основных последовательностей гена антитела HuCAL®. CDR участки HCDR1, HCDR2 и HCDR3 обозначены от N-конца к С-концу полужирным начертанием шрифта (SEQ ID NO:41).
На Фигуре 4 представлены аминокислотные последовательности вариабельных участков легкой цепи основанных на консенсусе основных последовательностей гена антитела HuCAL®. CDR участки LCDR1, LCDR2 и LCDR3 обозначены от N-конца к С-концу полужирным начертанием шрифта (SEQ ID NO:42).
На Фигуре 5 представлен пример последовательности ДНК вектора экспрессии pMORPH®X9_MOR03929_FH (SEQ ID NO:43).
На Фигуре 6 представлен уровень экспрессии GM-CSF рецептора альфа, как определяется анализом FACS с использованием антитела МАВ1006, специфичного в отношении GM-CSF рецептора альфа. СНО-GMRa#11 (сплошная линия) показан в сравнении с СНО-K1 (пунктирная линия). Ось абсцисс представляет относительное значение интенсивности флуоресценции (RFL), измеренной в канале FL2; ось ординат представляет количество клеток.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано на открытии новых антител, которые являются специфичными или обладают высоким сродством в отношении GM-CSF, а также обладают одним или больше других новых свойств. Предпочтительно, антитело по изобретению может приносить терапевтическую пользу субъекту. Антитела по изобретению, которые могут быть человеческими или гуманизированными, могут использоваться во многих контекстах, которые более полно описаны в данном описании.
"Человеческое" антитело или функциональный фрагмент человеческого антитела, таким образом, определяется как такой, который не является химерным (например, не является "гуманизированным") и не происходит из (полностью или частично) нечеловеческого вида. Человеческое антитело или функциональный фрагмент антитела может происходить от человека или может представлять собой синтетическое человеческое антитело. "Синтетическое человеческое антитело" определяется в данном описании как антитело, содержащее полученную последовательность, в целом или частично, in silico из синтетических последовательностей, которые основаны на анализе известных последовательностей человеческого антитела. In silico дизайн последовательности человеческого антитела или его фрагмента может быть осуществлен, например, путем анализа базы данных последовательностей человеческого антитела или фрагмента антитела и изобретения полипептидной последовательности с использованием полученных данных. Другим примером человеческого антитела или функционального фрагмента антитела является такой, который кодируется нуклеиновой кислотой, выделенной из библиотеки последовательностей антител человеческого происхождения (т.е. такая библиотека базируется на антителах, полученных из человеческого природного источника).
"Гуманизированное антитело" или функциональный фрагмент гуманизированного антитела определяется в данном описании как такой, который (i) происходит из нечеловеческого источника (например, трансгенная мышь, которая несет гетерологичную иммунную систему), причем такое антитело основано на последовательности человеческой зародышевой линии; или (ii) является химерным, где вариабельный домен имеет нечеловеческое происхождение, и постоянный домен имеет человеческое происхождение или (iii) CDR-пересаженный, где участки CDR вариабельного домена имеют нечеловеческое происхождение, в то время как один или больше каркасов вариабельного домена имеют человеческое происхождение, и постоянная область (если она присутствует) имеет человеческое происхождение.
В данном описании антитело "специфично связывается с", "обладает специфичностью в отношении" или "специфично распознает" антиген (в данном случае GM-CSF), если такое антитело способно осуществлять различие между таким антигеном и одним или больше антигенов сравнения, поскольку специфичность связывания представляет собой не абсолютное, а относительное свойство. В самой общей форме (без упоминания какого-либо определенного источника), "специфическое связывание" означает способность антитела осуществлять различие между целевым антигеном и другим антигеном, что определяется, например, в соответствии с одним из нижеуказанных способов. Такие способы включают, не ограничиваясь ими, Вестерн-блоттинг, анализы ELISA, RIA, ЭХЛ, IRMA и сканирование пептидов. Например, может быть проведен стандартный анализ ELISA. Количественная оценка может быть произведена путем стандартного образования окрашивания (например, вторичное антитело с пероксидазой хрена и тетраметилбензидин с пероксидом водорода). Протекание реакции в определенных лунках измеряют по оптической плотности, например, на длине волны 450 нм. Типичный фон (=отрицательная реакция) может соответствовать 0,1 ед. оптической плотности; типичная положительная реакция может соответствовать 1 ед. оптической плотности. Это подразумевает, что различие позитивный/негативный может быть более чем 10-кратным. Обычно определение специфичности связывания выполняется с использованием не одного антигена сравнения, а набора из приблизительно 3-5 антигенов, не имеющих отношения к целевому антигену, например, молочного порошка, бычьего сывороточного альбумина, трасферрина и т.п.
Однако термин "специфическое связывание" также может означать способность антитела осуществлять различие между целевым антигеном и одним или больше близкородственным антигеном, которые используются в качестве ориентиров, например, между GM-CSF и IL3, IL5, IL-4, IL13 или М-CSF. Кроме того, "специфическое связывание" может означать способность антитела осуществлять различие между различными частями целевого антигена, например различными областями или участками GM-CSF, или между одним или больше ключевым аминокислотным остатком или участком аминокислотных остатков GM-CSF.
Также, в данном описании термин "иммуноглобулин" (Ig) определяется как белок, принадлежащий к классу IgG, IgM, IgE, IgA или IgD (или любому их подклассу), и включает все общеизвестные антитела и их функциональные фрагменты. Таким образом, "функциональный фрагмент" антитела/иммуноглобулина определяется как фрагмент антитела/иммуноглобулина (например, вариабельный участок IgG), который сохраняет антигенсвязывающий участок. "Антигенсвязывающий участок" антитела обычно содержится в одном или больше гипервариабельном участке антитела, т.е. участке CDR-1, -2 и/или -3; однако вариабельные "каркасные" области могут также играть важную роль в связывании с антигеном, например, обеспечивая каркас для участков CDR. Предпочтительно, "антигенсвязывающий участок" включает как минимум остатки аминокислоты 4-103 вариабельной легкой (VL) цепи и 5-109 вариабельной тяжелой (VH) цепи, более предпочтительно, остатки аминокислоты 3-107 VL и 4-111 VH, и особенно предпочтительно, полные цепи VL и VH (положения аминокислот 1-109 VL и 1-113 VH; нумерация в соответствии с WO 97/08320). Предпочтительным классом иммуноглобулинов для использования в настоящем изобретении является IgG.
"Функциональные фрагменты" по изобретению включают домен фрагмента F(ab')2, фрагмента Fab, scFv или конструкты, содержащие отдельные вариабельные домены иммуноглобулина или однодоменные полипептиды антитела, например отдельные вариабельные домены тяжелой цепи или отдельные вариабельные домены легкой цепи. F(ab')2 или Fab может быть сконструирован с целью минимизации или полного включения межмолекулярных дисульфидных взаимодействий, которые происходят между доменами CH1 и CL.
Антитело по изобретению может происходить из библиотеки рекомбинантных антител, которая базируется на аминокислотных последовательностях, созданных in silico и кодируемых нуклеиновыми кислотами, созданными синтетически. Дизайн последовательности антитела in silico осуществляют, например, путем анализа базы данных человеческих последовательностей и изобретения полипептидной последовательности с использованием полученных данных. Способы проектирования и получения созданных in silico последовательностей описаны, например, в Knappik et al., J. Mol. Biol. (2000) 296:57; Krebs et al., J. Immunol. Methods. (2001) 254:67; и Патенте США №6,300,064, выданном Knappik et al., которые, таким образом, включены в данное описание путем ссылки во всей их полноте.
Антитела по изобретению
В данном документе содержатся ссылки на следующие характерные антитела по изобретения: "антитело №" или "MOR" 03684, 04251, 03929, 04252, 04287, 04290, 04302, 04350, 04354, 04357, 03682, 04283, 04297 и 04342. MOR03684 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:1 (ДНК)/SEQ ID NO:11 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:21 (ДНК)/ SEQ ID NO:31 (белок). MOR04251 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:2 (ДНК)/SEQ ID NO:12 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:22 (ДНК) SEQ ID NO:32 (белок). MOR03929 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:3 (ДНК) SEQ ID NO:13 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:23 (ДНК) SEQ ID NO:33 (белок). MOR04252 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:4 (ДНК) SEQ ID NO:14 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:24 (ДНК) SEQ ID NO:34 (белок). MOR04287 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:5 (ДНК) SEQ ID NO:15 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:25 (ДНК) SEQ ID NO:35 (белок). MOR04290 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:6 (ДНК) SEQ ID NO:16 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:26 (ДНК) SEQ ID NO:36 (белок). MOR04302 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:7 (ДНК) SEQ ID NO:17 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:27 (ДНК) SEQ ID NO:37 (белок). MOR04350 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:8 (ДНК) SEQ ID NO:18 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:28 (ДНК) SEQ ID NO:38 (белок). MOR04354 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:9 (ДНК) SEQ ID NO:19 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:29 (ДНК) SEQ ID NO:39 (белок). MOR04357 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:10 или 48 (ДНК) SEQ ID NO:20 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:30 или 57 (ДНК) SEQ ID NO:40 (белок). MOR03682 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:44 (ДНК) SEQ ID NO:49 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO: 53 (ДНК) SEQ ID NO:58 (белок). MOR04283 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:45 (ДНК) SEQ ID NO:50 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:54 (ДНК) SEQ ID NO:59 (белок). MOR04297 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:46 (ДНК) SEQ ID NO:51 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:55 (ДНК) SEQ ID NO:60 (белок). MOR04342 представляет антитело, содержащее вариабельный тяжелый участок, соответствующий SEQ ID NO:47 (ДНК) SEQ ID NO:52 (белок) и вариабельный легкий участок, соответствующий SEQ ID NO:56 (ДНК) SEQ ID NO:61 (белок).
В одном аспекте изобретение предлагает антитела, содержащие антигенсвязывающий участок, который может специфично связаться или обладает высоким сродством к GM-CSF. В данном описании указывается, что антитело обладает "высоким сродством" к антигену, если измеренные показатели сродства составляют как минимум 100 нМ (одновалентное сродство Fab фрагмента). Антитело или антигенсвязывающий участок по изобретению предпочтительно может связываться с GM-CSF со сродством менее чем приблизительно 100 нМ, более предпочтительно, менее чем приблизительно 60 нМ, и все еще более предпочтительно, менее чем приблизительно 30 нМ. Кроме того, предпочтительными являются антитела, которые связываются с GM-CSF со сродством менее чем приблизительно 10 нМ, и более предпочтительно, менее чем приблизительно 3 нМ. Например, сродство антитела по изобретению к GM-CSF может составлять приблизительно 10,0 нМ или 1 пМ (одновалентное сродство Fab фрагмента).
В табл.1 представлено краткое описание сродства характерных антител по изобретению по данным резонанса поверхностного плазмона (Biacore) и анализа равновесного титрования в растворе (SET).
Приведенные в табл.1 данные сродства MOR03684, 04251, 03929, 04252, 04357, 04290, 04302, 04350 и 04354 были измерены по методу резонанса поверхностного плазмона (Biacore) на иммобилизированном рекомбинантном GM-CSF. Формат Fab MOR03684, 04251, 03929, 04252, 04357, 04290, 04302, 04350 и 04354 демонстрирует интервал одновалентного сродства приблизительно от 6420 до 7 пМ.
Формат Fab также использовали для определения сродства равновесным титрованием в растворе (SET). В правом столбике табл.1 указана прочность связывания в интервале приблизительно от 16000 до 0,4 пМ MOR в данном способе.
Антитело по изобретению предпочтительно относится к виду, обладающему перекрестной реактивностью с человеком и как минимум одним другим видом, который может представлять собой вид грызунов или негуманоидных приматов. Негуманоидный примат может быть макакой резус. Вид грызунов может представлять собой крысу. Антитело, которое обладает перекрестной реактивностью как минимум с одним видом грызунов, например, может обеспечить большую гибкость и преимущество по сравнению с известными анти-GM-CSF антителами для проведения исследований in vivo на различных видах с использованием одного и того же антитела.
Предпочтительно, антитело по изобретению не только может связываться с GM-CSF, но также может блокировать взаимодействие человеческого GM-CSF с альфа-цепью человеческого рецептора GM-CSF, экспрессированного на клетках СНО-K1 как минимум на 25%, предпочтительно как минимум на 50%, более предпочтительно как минимум на 60%, более предпочтительно как минимум на 70%, предпочтительно как минимум на 85% и наиболее предпочтительно, как минимум на 100%. В предпочтительном варианте антитело по изобретению может блокировать взаимодействие 0,5 мкг/мл человеческого GM-CSF с альфа-цепью человеческого рецептора GM-CSF, экспрессированного на приблизительно 2×105 клеток СНО-K1 как минимум на 50% в следующих условиях: концентрация альфа-цепи человеческого рецептора GM-CSF, экспрессированного на клетках СНО-K1, сходна с концентрацией альфа-цепи человеческого рецептора GM-CSF, экспрессированного на приблизительно 2×105 клеток CHO-GMRa#11, и концентрация антитела по изобретению составляет приблизительно 5 мкг/мл.
В этом отношении опытный специалист может получить клетки СНО-K1, экспрессирующие альфа-цепь человеческого рецептора GM-CSF в концентрации, сходной с той, которая экспрессируется на приблизительно 2×105 клеток CHO-GMRa#11, например, трансфекцией популяции клеток СНО-K1 подходящим вектором экспрессии, кодирующим GM-CSF рецептор альфа, чтобы генерировать различные стабильные линии клетки, экспрессирующие определенные уровни GM-CSF рецептора альфа; далее стабильные линии клеток анализируют с помощью анализа FACS, чтобы определить уровни экспрессии GM-CSF рецептора альфа в соответствии с протоколом, в существенной мере так, как описано в Примере 3С; линию клетки, которая экспрессирует альфа человеческого GM-CSF рецептора в концентрации, сходной с той, которая экспрессируется на приблизительно 2×105 клеток CHO-GMRa#11, идентифицируют, сравнивая медианное значение интенсивности флуоресценции (MFL) таких трансфицированных клеток со значением MFL, полученным в Примере 3С. В данном описании линия клетки определяется как экспрессирующая альфа GM-CSF рецептора в концентрации, "сходной" с той, которая экспрессируется приблизительно на 2×105 клеток CHO-GMRa#11, если значение MFL трансфицированной линии клеток отличается не более чем в 2 раза от значения MFL для клеток CHO-GMRa#11, полученных, как описано в Примере 3С.
Кроме того, антитело по изобретению может нейтрализовать человеческий GM-CSF в анализе пролиферации TF-1 с более низким значением IC50, чем для антитела сравнения BVD2-21C11 и/или МАВ215, предпочтительно как минимум в 5 раз более низкое значение IC50, более предпочтительно как минимум в 10 раз более низкое значение IC50, чем для антитела сравнения BVD2-21C11 и/или МАВ215, более предпочтительно, как минимум в 15 раз более низкое значение IC50, чем для антитела сравнения BVD2-21C11 и/или МАВ215, более предпочтительно, как минимум в 20 раз более низкое IC50, чем для антитела сравнения BVD2-21C11 и/или МАВ215, более предпочтительно, как минимум в 30 раз более низкое IC50, чем для антитела сравнения BVD2-21C11 и/или МАВ215, более предпочтительно, как минимум в 50 раз более низкое IC50, чем для антитела сравнения BVD2-21C11 и/или МАВ215, более предпочтительно, как минимум в 100 раз более низкое IC50, чем для антитела сравнения BVD2-21C11 и/или МАВ215 и наиболее предпочтительно, как минимум в 120 раз более низкое IC50, чем для антитела сравнения BVD2-21C11 и/или МАВ215.
Пептидные варианты
Антитела по изобретению не ограничены конкретными пептидными последовательностями, приведенными в данном описании. Скорее, изобретение также включает варианты этих полипептидов. Со ссылкой на данное раскрытие и общепризнанные доступные технологии и ссылки квалифицированный специалист сможет получить, исследовать и применять функциональные варианты антител, раскрытых в данном описании, при условии, что варианты, которые обладают способностью блокировать взаимодействие GM-CSF с альфа-цепью GM-CSF рецептора, находятся в пределах контекста настоящего изобретения. В данном контексте "способность блокировать взаимодействие GM-CSF с альфа-цепью GM-CSF рецептора" подразумевает функциональную характеристику, которая приписывается анти-GM-CSF антителу по изобретению.
Вариант может включать, например, антитело, которое содержит как минимум один измененный определяющий комплементарность участок (CDR) (гипервариабельный) и/или каркасный домен/положение (FR) (вариабельный), по отношению к пептидной последовательности, раскрытой в данном описании. Чтобы лучше проиллюстрировать данное понятие, далее приведено краткое описание структуры антитела.
Антитело состоит из двух пептидных цепей, каждая из которых содержит один (легкая цепь) или три (тяжелая цепь) постоянных домена и вариабельный участок (VL, VH), причем последний в каждом случае состоит из четырех участков FR и трех прерывистых участков CDR. Антигенсвязывающий сайт образован одним или больше участком CDR, кроме того, участки FR обеспечивают структурный каркас для участков CDR, а также могут играть важную роль в связывании с антигеном. Изменяя один или больше аминокислотных остатков в участке CDR или FR, квалифицированный специалист может шаблонно генерировать мутантные или разнообразные последовательности антител, для которых может быть проведен скрининг в отношении антигена, например, для поиска новых или улучшенных свойств.
В табл.2а (VH) и 2b (VL) показаны участки CDR и FR для некоторых антител по изобретению и сравниваются аминокислоты в данном положении по отношению друг к другу, а также к соответствующим консенсусным последовательностям или последовательностям "образца гена" (как описано в Патенте США №6,300,064):




Оригинальные образцы гена HuCAL® конструируются с их подлинными N-концами, например, VL лямбда 3 содержит аминокислоты "SY" в положениях 1 и 2; и VH3 содержит аминокислоту "Е" в положении 1. В ходе конструирования библиотек Fab HuCAL®, включая библиотеку HuCAL GOLD®, первые две аминокислоты заменены на "DI" в цепи VL лямбда 3; и первая аминокислота заменена на "Q" в цепи VH3.
Квалифицированный специалист может использовать данные табл.2а и 2b для разработки вариантов пептида, которые находятся в пределах контекста настоящего изобретения. Предпочтительно, варианты конструируются путем замены аминокислот в пределах одного или больше участков CDR; вариант также может содержать одну или больше измененных каркасных областей. Ссылаясь на сравнение новых антител друг с другом, остатки-кандидаты, которые могут быть изменены, включают, например, остатки 27 или 51 вариабельной легкой и, например, остатки 32 или 56 вариабельной тяжелой цепи MOR04251, поскольку указанные остатки варьируют друг относительно друга. Изменения также могут быть внесены в каркасные области. Например, область FR пептида могла бы быть изменена там, где присутствует расхождение остатка по сравнению с последовательностью зародышевой линии.
Ссылаясь на сравнение новых антител с соответствующей консенсусной последовательностью или последовательностью "образца гена", остатки-кандидаты, которые могут быть изменены, включают, например, остатки 27, 50 или 90 вариабельной легкой цепи MOR04251 по сравнению с VLλ3 и, например, остатки 33, 52 или 96 вариабельной тяжелой цепи MOR04251 по сравнению с VH3. Альтернативно, квалифицированный специалист мог бы провести такой же анализ, сравнивая аминокислотные последовательности, раскрытые в данном описании, с известными последовательностями аналогичного класса таких антител, с применением, например, методики, описанной Knappik et al. (2000) и в Патенте США №6,300,064, выданном Knappik et al.
Кроме того, варианты могут быть получены с использованием одного MOR в качестве исходной точки для оптимизации путем внесения разнообразия в один или больше аминокислотных остатков в MOR, предпочтительно аминокислотные остатки в одном или больше участках CDR, с последующим скринингом полученной коллекции вариантов антитела на предмет поиска вариантов с улучшенными свойствами. Особенно предпочтительным является разнообразие одного или больше аминокислотных остатков в CDR-3 VL, CDR-3 VH, CDR-1 VL и/или CDR-2 VH. Разнообразие может быть достигнуто путем синтеза коллекции молекул ДНК с использованием технологии мутагенеза тринуклеотида (TRIM) (Virnekas et al., 1994).
Консервативные аминокислотные варианты
Могут быть получены полипептидные варианты, где сохранена общая молекулярная структура последовательности пептида антитела, описанная в данном описании. С учетом свойств отдельных аминокислот, некоторые рациональные замены будут понятны квалифицированному специалисту. Замены аминокислот, т.е. "консервативные замены", могут быть осуществлены, например, на основании сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфифильной природы задействованных остатков.
Например, (а) неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; (b) полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; (с) положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; и (d) отрицательно заряженные (кислые) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту. Замены обычно могут быть осуществлены в пределах групп (a)-(d). Кроме того, глицин и пролин могут быть взаимозаменяемыми на основе их способности разрушать α-спирали. Подобным образом, некоторые аминокислоты, такие как аланин, цистеин, лейцин, метионин, глутаминовая кислота, глутамин, гистидин и лизин, чаще встречаются в α-спиралях, в то время как валин, изолейцин, фенилаланин, тирозин, триптофан и треонин чаще встречаются в β-складчатых листах. Глицин, серин, аспарагиновая кислота, аспарагин и пролин обычно встречаются в изгибах. Некоторые предпочтительные замены могут быть осуществлены среди следующих групп: (i) S и Т; (ii) Р и G; и (iii) А, V, L и I. С учетом известного генетического кода, а также методов рекомбинации и синтеза ДНК квалифицированный специалист легко может сконструировать ДНК, кодирующие консервативные варианты аминокислот.
В данном описании "идентичность последовательности" между двумя полипептидными последовательностями указывает на процент аминокислот, которые идентичны для двух последовательностей. "Гомология последовательности" указывает на процент аминокислот, которые является идентичными или представляют консервативные замены аминокислот. Предпочтительные полипептидные последовательности по изобретению демонстрируют идентичность последовательности в участках CDR как минимум 60%, более предпочтительно как минимум 70% или 80%, еще более предпочтительно как минимум 90% и наиболее предпочтительно как минимум 95%. Предпочтительные антитела также демонстрируют гомологию последовательности в участках CDR как минимум 80%, более предпочтительно 90% и наиболее предпочтительно 95%.
Молекулы ДНК по изобретению
Настоящее изобретение также относится к молекулам ДНК, которые кодируют антитело по изобретению. Такие последовательности включают, не ограничиваясь ими, молекулы ДНК, показанные на фиг.10 и 2а.
Молекулы ДНК по изобретению не ограничены последовательностями, раскрытыми в данном описании, но включают также их варианты. Варианты ДНК в пределах изобретения могут быть описаны со ссылкой на их физические свойства в гибридизации. Квалифицированному специалисту будет понятно, что ДНК может использоваться для идентификации комплементарной к ней цепи и, поскольку ДНК является двухцепочечной, ее эквивалента или гомолога с использованием методов гибридизации нуклеиновых кислот. Также будет понятно, что гибридизация может происходить при менее чем 100% комплементарности. Однако с учетом подходящего выбора условий методы гибридизации могут применяться для различия последовательностей ДНК на основе их структурной связи с конкретным зондом. Руководство относительно таких условий см. в Sambrook et al., 1989 (Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989) Molecular Cloning: A laboratory manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, USA) и Ausubel et al., 1995 (Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Sedman, J. G., Smith, J. A., & Struhl, K. eds. (1995). Current Protocols in Molecular Biology. New York: John Wiley and Sons).
Структурное сходство между двумя полинуклеотидными последовательностями может быть выражено как функция "строгости" условий, в которых две последовательности гибридизуются друг с другом. В данном описании термин "строгость" означает степень, до которой условия препятствуют гибридизации. Строгие условия значительно препятствуют гибридизации, и только наиболее тесно связанные со структурной точки зрения молекулы будут гибридизоваться друг с другом в таких условиях. С другой стороны, нестрогие условия способствуют гибридизации молекул, обладающих меньшей степенью структурного родства. Таким образом, строгость гибридизации непосредственно коррелирует со структурным сходством двух последовательностей нуклеиновых кислот. Следующие взаимоотношения являются пригодными с точки зрения корреляции гибридизации и сходства (где Tm - температура плавления спаренной нуклеиновой кислоты):
a. Tm=69,3+0,41(G+C)%,
b. Tm спаренной ДНК уменьшается на 1°C с каждым увеличением на 1% количества рассогласованных пар оснований,
с. (Тm)µ2 - (Тm)µ1=18,5 log10µ2/µl,
где µl и µ2 - ионная сила двух растворов.
Строгость гибридизации является функцией многих факторов, в том числе общей концентрации ДНК, ионной силы, температуры, размера зонда и присутствия агентов, которые нарушают образование водородных связей. Факторы, способствующие гибридизации, включают высокие концентрации ДНК, высокую ионную силу, низкую температуру, более длинный зонд и отсутствие агентов, которые нарушают образование водородных связей. Гибридизация обычно выполняется в две фазы: фаза "связывания" и фаза "промывания".
Сначала в ходе фазы связывания зонд связывается с мишенью в условиях, способствующих гибридизации. Строгостью обычно управляют на данной стадии, изменяя температуру. Для получения условий высокой строгости температура обычно составляет от 65°С до 70°С, если только не используются короткие (<20 нуклеотидов) олигонуклеотидные зонды. Типичный раствор для гибридизации включает 6×(натрия хлорид + натрия цитрат), 0,5% натрия лаурилсульфата, 5Х раствора Денхардта и 100 мкг неспецифичного носителя ДНК. См. Ausubel et al., section 2.9, Supplement 27 (1994). Конечно, известно множество различных и тем не менее эквивалентных с функциональной точки зрения, буферных условий. Если степень сходства ниже, может быть выбрана более низкая температура. Значения температуры для низкой строгости связывания составляют приблизительно от 25°С до 40°С. Температура для средней строгости составляет от как минимум 40°С до менее чем приблизительно 65°С. Температура для высокой строгости составляет как минимум приблизительно 65°С.
Во-вторых, избыток зонда удаляют промыванием. Именно в данной фазе обычно применяют более строгие условия. Таким образом, в определении сходства посредством гибридизации именно стадия "промывания" является наиболее важной. Растворы для промывания обычно содержат более низкие концентрации соли. Пример раствора для гибридизации средней строгости содержит 2×(натрия хлорид + натрия цитрат) и 0,1% натрия лаурилсульфата. Раствор для промывания гибридизации высокой строгости содержит эквивалент (по ионной силе) менее чем приблизительно 0,2×(натрия хлорид + натрия цитрат) с раствором предпочтительной строгости, содержащим приблизительно 0,1×(натрия хлорид + натрия цитрат). Значения температуры, связанные с различным уровнем строгости, являются такими же, как обсуждалось выше для стадии "связывания." Раствор для промывания обычно заменяют несколько раз в ходе промывания. Например, типичные условия промывания для высокой строгости включают промывание дважды в течение 30 минут при 55°С и трижды в течение 15 минут при 60°С.
Соответственно, настоящее изобретение включает молекулы нуклеиновой кислоты, которые гибридизуются с молекулами, определенными ранее на фиг.1а и 2а в условиях высокой строгости связывания и промывания, где такие нуклеиновые молекулы кодируют антитело или его функциональный фрагмент с такими свойствами, как описано в данном описании. Предпочтительными молекулами (с точки зрения перспектив мРНК) являются те, которые обладают как минимум 75% или 80% (предпочтительно как минимум 85%, более предпочтительно как минимум 90% и наиболее предпочтительно как минимум 95%) гомологией или идентичностью последовательности с одной из молекул ДНК, описанных в данном описании.
Варианты, равноценные с функциональной точки зрения
Понятно, что варианты молекул ДНК, предложенных в данном описании, могут быть сконструированы несколькими различными способами. Например, они могут быть сконструированы как полностью синтетические ДНК. Широко известны способы эффективного синтеза олигонуклеотидов в интервале от 20 до приблизительно 150 нуклеотиднов. См. Ausubel et al., section 2.11, Supplement 21 (1993). Частично перекрывающиеся олигонуклеотиды могут быть синтезированы и собраны способом, впервые описанным Khorana et al., J. Mol. Biol. 72:209-217 (1971); см. также Ausubel et al., section 8.2, выше. Синтетические ДНК предпочтительно конструируют с пригодными рестрикционными сайтами на 5' и 3' концах гена, чтобы облегчить клонирование в подходящий вектор.
Как показано, способ генерации вариантов состоит в том, чтобы начать с одной из ДНК, раскрытых в данном описании, а затем провести сайт-направленный мутагенез. См. Ausubel et al., section 8, Supplement 37 (1997), выше. В типичном способе целевую ДНК клонируют в одноцепочечную ДНК бактериофага-носителя. Одноцепочечную ДНК выделяют и гибридизуют с олигонуклеотидом, содержащим целевое изменение(я) нуклеотидов. Синтезируют комплементарную цепь, и двухцепочечный фаг вводят в хозяина. Некоторые из полученных потомков будут являться целевыми мутантами, что может быть подтверждено с использованием секвенирования ДНК. Кроме того, существуют различные способы, которые увеличивают вероятность того, что фаг потомка будет целевым мутантом. Такие способы хорошо известны специалистам в данной области, а соответствующие наборы для генерации таких мутантов являются коммерчески доступными.
Рекомбинантные конструкты ДНК и экспрессия
Настоящее изобретение дополнительно предлагает рекомбинантные конструкты ДНК, содержащие одну или больше нуклеотидных последовательностей по настоящему изобретению. Рекомбинантные конструкты по настоящему изобретению используются в сочетании с вектором, таким как плазмида, фагемида, фаг или вирусный вектор, в который вставлена молекула ДНК, кодирующая антитело по изобретению.
Кодируемый ген может быть продуцирован по методам, описанным в Sambrook et al., 1989, и Ausubel et al., 1989. Альтернативно, последовательности ДНК могут быть синтезированы химическим способом с использованием, например, устройств для синтеза. См., например, методы, описанные в Oligonucleotide Synthesis (1984, Gait, ed., IRL Press, Oxford), которая включена в данное описание путем ссылки во всей ее полноте. Рекомбинантные конструкты по изобретению содержатся в векторах экспрессии, которые способны экспрессировать РНК и/или белковые продукты кодирующей их ДНК. Кроме того, вектор может включать регулирующие последовательности, в том числе промотор, функционально связанный с открытой рамкой для считывания (ORF). Кроме того, вектор может включать маркерную последовательность с целью селекции. Конкретные сигналы инициации и бактериальные секреторные сигналы также могут потребоваться для эффективной трансляции вставленных последовательностей, кодирующих целевой ген.
Настоящее изобретение дополнительно предлагает клетки-хозяева, содержащие как минимум одну из ДНК по настоящему изобретению. Клетка-хозяин фактически может представлять собой любую клетку, для которой доступны векторы экспрессии. Она может являться, например, высшей эукариотной клеткой-хозяином, такой как клетка млекопитающего, низшей эукариотной клеткой-хозяином, такой как дрожжевая клетка, или прокариотной клеткой, такой как бактериальная клетка. Введение рекомбинантного конструкта в клетку-хозяин может производиться путем трансфекции фосфатом кальция, диэтиламиноэтилом, декстраном, опосредованной трансфекцией, элекропорацией или заражением фага.
Бактериальная экспрессия
Пригодные векторы экспрессии для использования в бактериях конструируют, вставляя структурную последовательность ДНК, кодирующую целевой белок, вместе с подходящими сигналами инициации и терминации трансляции в фазе функционального считывания с функциональным промотором. Вектор будет содержать один или больше фенотипических селекционных маркеров и источник репликации, чтобы обеспечить обслуживание вектора и, при желании, обеспечить амплификацию в хозяине. Подходящие прокариотные хозяева для трансформации включают Е.coli, Bacillus subtilis, Salmonella typhimurium и различные виды в пределах родов Pseudomonas, Streptomyces и Staphylococcus.
Бактериальные векторы могут базироваться, например, на бактериофаге, плазмиде или фагемиде. Эти векторы могут содержать селекционный маркер и бактериальный источник репликации, полученный из коммерчески доступных плазмид, обычно содержащих элементы хорошо известного вектора клонирования pBR322 (ATCC 37017). Последующее превращение подходящего штамма хозяина и культивирование штамма хозяина до подходящей плотности клеток для выбранного промотора устраняют подавление/индуцируют с помощью подходящих сред (например, сдвиг температуры или химическая индукция) и клетки культивируют на протяжении дополнительного периода времени. Клетки обычно собирают центрифугированием, разрушают физическими или химическими средами, и полученный сухой экстракт оставляют для дальнейшей очистки.
В бактериальных системах целый ряд векторов экспрессии может быть предпочтительно выбран в зависимости от предусмотренного применения экспрессированного белка. Например, если необходимо произвести большое количество такого белка, для генерации антител или для скриниговых библиотек пептидов, например, могут быть желательными векторы, которые направляют экспрессию высоких уровней продуктов в виде слитого белка, который легко подвергается очистке.
Терапевтические способы
Терапевтические способы включают введение субъекту, который нуждается в лечении, терапевтически эффективного количества антитела по изобретению. "Терапевтически эффективное количество", таким образом, определяется как количество антитела, которое является достаточным для эффективного блокирования взаимодействия между GM-CSF и рецептором на леченном участке субъекта - в виде единичной дозы или в соответствии с многодозовой схемой, отдельно или в комбинации с другими агентами, что ведет к облегчению неблагоприятного состояния, причем указанное количество является приемлемым с токсикологической точки зрения. Субъект может быть человеком или животным (например, крыса или макака резус).
Антитело по изобретению может быть введено совместно с известными лекарственными средами, и в некоторых случаях антитело само по себе может быть модифицировано. Например, антитело может быть конъюгировано с иммунотоксином или радиоизотопом для потенциального дальнейшего повышения эффективности.
Антитела по изобретению могут использоваться в качестве терапевтического или диагностического инструмента в различных ситуациях, где GM-CSF нежелательным образом экспрессируется или присутствует. Расстройства и состояния, особенно пригодные для лечения антителом по изобретению, представляют собой воспалительные заболевания, такие как ревматоидный артрит (РА), рассеянный склероз, болезнь Крона, псориаз, астма, атопический дерматит или шок.
Для лечения любого из вышеуказанных расстройств фармацевтические композиции для применения в соответствии с настоящим изобретением могут быть введены в обычную лекарственную форму с использованием одного или больше физиологически приемлемых носителей или вспомогательных веществ. Антитело по изобретению может быть введено любыми пригодными средствами, которые могут измениться в зависимости от вида расстройства, которое подлежит лечению. Возможные способы введения включают парентеральный (например, внутримышечный, внутривенный, внутриартериальный, внутрибрюшной или подкожный), внутрилегочный и интраназальный, и, если целью является местное иммуносупрессивное лечение, введение в очаги поражения. Кроме того, антитело по изобретению может быть введено импульсной инфузией, например, с уменьшением доз антитела. Предпочтительно, дозы вводят инъекционно, наиболее предпочтительно, с помощью внутривенных или подкожных инъекций, отчасти в зависимости от того, будет ли введение кратковременным или хроническим. Количество для введения будет зависеть от различных факторов, таких как клинические симптомы, масса тела индивидуума, вводят ли другие лекарственные среды. Квалифицированному специалисту будет понятно, что способ введения будет варьировать в зависимости от расстройства или состояния, которое подлежит лечению.
Определение терапевтически эффективного количества нового полипептида в соответствии с данным изобретением в значительной степени будет зависеть от характеристик конкретного пациента, способа введения и природы расстройства, которое подлежит лечению. Общее руководство может быть найдено, например, в публикациях International Conference on Harmonisation и Remington's Pharmaceutical Sciences, chapters 27 and 28, pp.484-528 (18th ed., Alfonso R. Gennaro, Ed., Easton, Pa.: Mack Pub. Co., 1990). Более конкретно, определение терапевтически эффективного количества будет зависеть от таких факторов, как токсичность и эффективность лекарственной среды. Токсичность может быть определена с использованием способов, хорошо известных в данной области, которые описаны в вышеуказанных источниках. Эффективность может быть определена с применением таких же принципов в сочетании со способами, описанными в Примерах ниже.
Диагностические способы
GM-CSF экспрессируется клетками различных типов, в том числе лимфоцитами, моноцитами, эндотелиальными клетками, фибробластами и некоторыми клетками злокачественных новообразований; таким образом, анти-GM-CSF антитело по изобретению может применяться для того, чтобы отображать или визуализировать локализацию возможной аккумуляции GM-CSF в различных тканях пациента. В этом отношении, антитело может содержать поддающуюся обнаружению метку, в том числе радиоизотопы, метки сродства (такие как биотин, авидин и т.п.), флуоресцентные метки, парамагнитные атомы и т.п. Способы введения такой метки хорошо известны из уровня техники. Клиническое применение антител в диагностической визуализации рассмотрено Grossman, Н.В., Urol. Clin. North Amer. 13:465-474 (1986)), Unger, E.С. et al., Invest. Radiol. 20:693-700 (1985)), и Khaw, В. et al., Science 209:295-297 (1980)).
Обнаружение очагов таких поддающихся обнаружению меченых антител могло бы указывать, например, на локализацию воспаления. В одном варианте такое обследование выполняют, извлекая образцы ткани или крови и инкубируя такие образцы в присутствии поддающихся обнаружению меченых антител. В предпочтительном варианте данная техника имеет неинвазивную форму путем применения магнитной визуализации, флюорографии и т.п. Такое диагностическое обследование может применяться в ходе мониторинга успешности лечения заболеваний, где присутствие или отсутствие GM-CSF является адекватным показателем. Изобретение также предусматривает использование анти-GM-CSF антитела, описанного в данном описании, для диагностики в условиях ex vivo.
Терапевтические и диагностические композиции
Антитела по настоящему изобретению могут быть введены в лекарственные формы в соответствии с известными способами получения фармацевтически приемлемых композиций, где антитело по изобретению (в том числе какой-либо его функциональный фрагмент) объединено в смеси с фармацевтически приемлемым носителем. Подходящие носители и их препараты описаны, например, в Remington's Pharmaceutical Sciences (18th ed., Alfonso R. Gennaro, Ed., Easton, Pa.: Mack Pub. Co., 1990). Для того чтобы получить фармацевтически приемлемую композицию, подходящую для эффективного введения, такие композиции будут содержать эффективное количество одного или больше антител по настоящему изобретению вместе с подходящим количеством носителя.
Рецептура препаратов может быть составлена таким образом, чтобы обеспечивать контролируемое высвобождение активного соединения. Препараты с контролируемым высвобождением могут быть получены путем использования полимеров для образования комплекса или абсорбирования анти-GM-CSF антитела. Контролируемая доставка может быть обеспечена выбором подходящих макромолекул (например, полиэфиры, полиаминокислоты, поливинилпирролидон, этиленвинилацетат, метилцеллюлоза, карбоксиметилцеллюлоза или протамина сульфат) и концентрации макромолекул, а также способов введения для того, чтобы управлять высвобождением. Другой возможный способ управления продолжительностью действия с помощью препаратов контролируемого высвобождения заключается в инкорпорации анти-GM-CSF антитела в частицы полимерного материала, например полиэстеры, полиаминокислоты, гидрогели, поли(молочная кислота) или этиленвинилацетатные сополимеры. Альтернативно, вместо инкорпорации этих агентов в полимерные частицы можно заключить указанные материалы в микрокапсулы, полученные, например, методами коацервации или полимеризации на границе фаз, например микрокапсулы из гидроксиметилцеллюлозы или желатина или микрокапсулы из поли(метилметакрилата), соответственно, или в коллоидных системах доставки лекарственных средств, таких как липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы, или в микроэмульсиях. Такие методы раскрыты в Remington's Pharmaceutical Sciences (1980).
Соединения могут быть введены в препараты для парентерального введения путем инъекции, например инъекции болюса или непрерывной инфузии. Препараты для инъекции могут быть представлены в однодозовой форме, например в ампулах, или в многодозовых контейнерах с добавлением консерванта. Композиции могут приобретать такие формы, как суспензии, растворы или эмульсии в масляном или водном носителе, и могут содержать вспомогательные вещества, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. Альтернативно, активный ингредиент может иметь форму порошка для разведения перед использованием подходящим растворителем, например стерильной, не содержащей пирогенов водой.
Композиции могут, при желании, быть представлены в упаковке или устройстве для отпуска, которое может содержать одну или больше единиц дозированной лекарственной формы, содержащих активный ингредиент. Упаковка, например, может включать металлическую или пластиковую фольгу, например пакет из фольги. Упаковка или устройство для отпуска может сопровождаться инструкцией по применению.
Изобретение далее будет более подробно описано со ссылкой на следующие рабочие примеры, которые приведены для иллюстрации и не должны ограничивать изобретение.
ПРИМЕРЫ
Пример 1: Генерация человеческих антител, специфичных в отношении GM-CSF из библиотеки HuCAL GOLD®
А. Извлечение фагемиды, амплификация и очистка фага
Библиотеку HuCAL GOLD® амплифицируют в среде 2×YT, содержащей 34 мкг/мл хлорамфеникола и 1% глюкозы (2×YT-CG). После инфицирования хелперным фагом (VCSM13) при OD600 0,5 (30 минут при 37°С без встряхивания; 30 минут при 37°С со встряхиванием при 250 об/мин), клетки осаждают (4120 g; 5 мин; 4°С), повторно суспендируют в 2×YT/34 мкг/мл хлорамфеникола/50 мкг/мл канамицина/0,25 мМ изопропилтиогалактозида и культивируют на протяжении ночи при 22°С. Фаги ПЭГ-осаждают из супернатанта, повторно суспендируют в фосфатном буферном растворе/20% глицерина и хранят при -80°С. Амплификацию фага между двумя циклами кругами пэннинга проводят, как указано ниже: середина логарифмического роста Е.coli TG1 клетки инфицируют элюированными фагами и покрывают LB-агар, содержащий 1% глюкозы и 34 мкг/мл хлорамфеникола. После инкубации на протяжении ночи при 30°С колонии соскабливают и используют для инокуляции 2×YT-CG, до достижения OD600 нм 0,5, после чего добавляют хелперный фаг, как описано выше.
В. Пэннинг с HuCAL GOLD®
Для селекции антител, распознающих человеческий GM-CSF, были применены несколько стратегий пэннинга. Если коротко, антитела-фаги HuCAL GOLD® были разделены на три динамические области, содержащие различные основные гены VH. Эти динамические области по отдельности подвергали одному из следующего: а) твердофазный пэннинг на биотинилированном человеческом GM-CSF белке (произведен на заказ R&D Systems, Minneapolis, MN), непосредственно нанесенном на покрытые нейтравидином 96-луночные планшеты (Pierce, Rockford, IL) в качестве твердой подложки для трех кругов, или b) пэннинг в растворе на биотинилированном человеческом GM-CSF белке, фиксированном на покрытых стрептавидином Dynabeads (Dynal, Осло, Норвегия) для трех кругов.
Если подробно, для пэннинга на иммобилизированном биотинилированном GM-CSF ячейки нейтравидинового планшета трижды промывают по 300 мкл ФБР. Антиген разбавляют до концентрации 3 мкг/мл (200 нМ) в ФБР и 0,1 мл покрывают лунку в течение 2 ч при комнатной температуре. После двух стадий промывания по 300 мкл ФБР лунки инкубируют с блокирующим буфером, содержащим 2×Chemiblocker (Chemicon, Temecula, CA), разведенный 1:1 в ФБР.
Перед селекцией 100 мкл фагов HuCAL GOLD® предварительно адсорбируют в 100 мкл блокирующего буфера, содержащего 0,4 мкл 25% Твина 20 в течение 0,5 ч при комнатной температуре. Блокированные фаги переносят аликвотами по 100 мкл в лунки нейтравидинового планшета на 0,5 ч при комнатной температуре. Эту стадию повторяют дважды для обеспечения предварительной абсорбции.
После промывания (2×300 мкл ФБР) покрытого и блокированного нейтравидинового планшета для микротитрования, 0,1 мл предварительно адсорбированных фагов добавляют в покрытые лунки и инкубируют в течение 1,5 ч при комнатной температуре и осторожном встряхивании. Инкубация завершают 10 циклами промывания с ФБР/0,05% Твина 20 при комнатной температуре.
Связанные фаги элюируют добавлением 120 мкл 20 мМ дитиотрейтола в 10 мМ Трис рН 8,0 на лунку и выдерживают в течение 10 минут при комнатной температуре. Элюат удаляют и добавляют 14 мл Е.coli TG1, культивированной до достижения OD600 нм 0,6-0,8. Лунки дополнительно промывают 200 мкл ФБР, и полученный раствор также добавляют к клеткам TG1. Инфицирование фага Е.coli осуществляют в течение 45 минут при 37°С без встряхивания. Кроме того, 200 мкл клеток TG1, выращенных до ОD600 нм 0,6-0,8, добавляют к селекционным лункам, выдерживая в течение 45 минут при 37°С без встряхивания. Полученные клетки TG-1 добавляют к 14 мл культуры, уже содержащей фаги с первой стадии элюирования. После центрифугирования в течение 10 минут при 5000 об/мин каждую гранулу бактерий повторно суспендируют в 500 мкл среды 2×YT, помещают на агаровые пластины 2×YT-CG и инкубируют в течение ночи при температуре 30°С. Далее колонии соскабливают и проводят амплификацию, как описано выше.
Второй и третий круги селекции осуществляют способом, аналогичным первому кругу селекции, с единственной разницей в том, что условия промывания после связывания фага являются более строгими. Кроме того, в третьем круге выделения фаги подвергают дополнительной стадии предварительной адсорбции на покрытых стрептавидином гранулах (Dynabeads M-280; Dynal). Пробирки Эппендорфа блокируют раствором Chemiblocker путем инкубации в течение 30 минут при комнатной температуре. Аликвоту 0,3 мл из каждого пула фага смешивают 1:1 с раствором 2×Chemiblocker, содержащим 0,05% Твина 20, и инкубируют в течение 1 ч при комнатной температуре в блокированных пробирках Эппендорфа на роторном устройстве. Блокированные фаги далее переносят в свежие блокированные пробирки Эппендорфа и добавляют 50 мкл Dynabeads M-280, выдерживая еще 30 минут для предварительной адсорбции.
Гранулы удаляют с помощью магнитного устройства (Dynal MPC-E). Далее аликвоты фагов по 150 мкл переносят на нейтравидиновые планшеты для дальнейшей предварительной адсорбции, как в круге 1 и 2 (см. выше).
Для раствора пэннинга с использованием биотинилированного GM-CSF, связанного с Dynabeads, применяют следующий протокол: пробирки Эппендорфа объемом 1,5 мл блокируют 1,5 мл 2×Chemiblocker, разведенного 1:1 фосфатным буферным раствором в течение ночи при 4°С. Покрытые 200 мкл стрептавидина магнитные гранулы (Dynabeads M-280; Dynal) промывают 1×200 мл фосфатного буферного раствора и повторно суспендируют в 200 мл 1×Chemiblocker (разбавленном в 1×фосфатного буферного раствора). Блокирование гранул выполняют в предварительно блокированных пробирках в течение ночи при 4°С. Фаги, разбавленные в 500 мл фосфатного буферного раствора для каждого состояния пэннинга, смешивают с 500 мл 2×Chemiblocker/0,1% Твина в течение 1 ч при комнатной температуре (роторное устройство). Предварительную адсорбцию фагов осуществляют дважды: 50 мкл блокированных магнитных гранул стрептавидина добавляют к блокированным фагам и инкубируют в течение 30 минут при комнатной температуре на ротаторе. После отделения гранул с помощью магнитного устройства (Dynal MPC-E) супернатант фага (~1 мл) переносят в новую блокированную пробирку и предварительную адсорбцию повторяют на 50 мкл блокированных гранул в течение 30 минут. Затем 200 нМ биотинилированного hGM-CSF добавляют к блокированным фагам в свежих блокированных пробирках объемом 1,5 мл и инкубируют в течение 1 ч при комнатной температуре на ротаторе. 100 мл блокированных магнитных гранул стрептавидина добавляют к каждому пулу фага пэннинга и инкубируют в течение 10 минут при комнатной температуре на ротаторе. Фаг, связанный с биотинилированным GM-CSF и, таким образом, иммобилизированный на магнитных гранулах, собирают с помощью магнитного разделителя частиц (Dynal MPC-E). Гранулы далее промывают 7×ФБР/0,05% Твина с использованием ротатора с последующим промыванием еще трижды фосфатным буферным раствором. Элюацию фага из Dynabeads осуществляют, добавляя 300 мл 20 мМ дитиотрейтола в 10 мМ Трис/НСl (рН 8) в каждую пробирку на 10 минут. Dynabeads удаляют с помощью магнитного разделителя частицы и к супернатанту добавляют 14 мл культуры Е.coli TG-1, культивированной до OD600 нм 0,6-0,8. Далее гранулы промывают один раз 200 мл фосфатного буферного раствора, и фосфатный буферный раствор, дополнительно содержащий удаленный фаг, добавляют к 14 мл культуры E.coli TG-1.
После центрифугирования в течение 10 минут при 5000 об/мин каждую гранулу бактерий повторно суспендируют в 500 мл среды 2×YT, наносят на пластины агара 2×YT-CG и инкубируют в течение ночи при 30°С. Затем колонии соскабливают с планшетов, фаги выделяют и амплифицируют, как изложено выше.
Вторые и третьи круги пэннинга в растворе на биотинилированном GM-CSF осуществляют в соответствии с протоколом первого круга за исключением увеличения строгости процедуры промывания.
С. Субклонирование выбранных фрагментов Fab и экспрессия растворимых фрагментов Fab
Кодирующие Fab вставки выбранных фагемид HuCAL GOLD® субклонируют в вектор экспрессии pMORPH®X9_Fab_FH (фиг.5), чтобы облегчить быструю экспрессию растворимого Fab. ДНК отобранных клонов расщепляют с помощью XbaI и EcoRI, таким образом, вырезая кодирующую Fab вставку (ompA-VLCL и phoA-Fd) и клонируют в расщепленный XbaI/EcoRI вектор pMORPH®X9_Fab_FH. Fab, которые экспрессируются в указанных векторах, несут две С-концевых метки (FLAG™ и 6xHis, соответственно), для обнаружения и очищения.
D. Микроэкспрессия антител Fab HuCAL GOLD® в Е.coli
Отдельные колонии, полученные после субклонирования в pMORPH®X9_Fab_FH, используют для инокуляции в лунки стерильного 96-луночного планшета для микротитрования, содержащего 100 мкл среды 2×TY/Cm/0,1% Glu на лунку и культивируют в течение ночи при 37°С. 5 мкл каждой культуры TG-1 Е.coli перемещают в новый стерильный 96-луночный планшет для микротитрования, содержащий 100 мл среды 2×TY/Cm/0,1% Glu на лунку. Планшеты для микротитрования инкубируют при 30°С и встряхивании при 400 об/мин на шейкере для микропланшетов пока культуры не становятся слегка мутными (~2-4 ч) с OD600 нм 0,5.
На полученные планшеты для экспрессии добавляют по 20 мкл 2×YT/Cm/3 мМ изопропилтиогалактозида на лунку (конечная концентрация 0,5 мМ IPTG), опечатывают проницаемой для газа лентой и инкубируют в течение ночи при 30°С и встряхивании при 400 об/мин.
Генерация лизатов цельных клеток (экстракты BEL)
В каждую лунку планшетов для экспрессии добавляют 40 мкл буфера BEL (2×буферизованный барбиталом солевой раствор/ЭДТА: 24,7 г/л борной кислоты, 18,7 г NaCl/л, 1,49 г ЭДТА/л, рН 8), содержащего 2,5 мг/мл лизоцима, и инкубируют в течение 1 ч при 22°С на шейкере для планшетов для микротитрования (400 об/мин). Экстракты BEL используют для анализа связывания ELISA или на анализаторе BioVeris M-series® 384 (см. Пример 2).
Е. Экспрессия HuCAL® GOLD Fab антител в Е.coli и очистка
Экспрессию фрагментов Fab, кодируемых pMORPH®X9_Fab_FH в клетках TG-1 осуществляют в культурах, находящихся в колбах для шейкера, с помощью 1 л среды 2×YT, содержащей 34 мкг/мл хлорамфеникола. После индукции с помощью 0,5 мМ IPTG клетки культивируют при 22°С в течение 16 час. Цельноклеточные экстракты клеточных гранул готовят с помощью французского пресса и фрагменты Fab выделяют хроматографией никель/NTA (Qiagen, Hilden, Германия). Концентрацию определяют УФ-спектрофотометрией (Krebs et al., 2001).
Пример 2: Идентификация специфичных в отношении hGM-CSF антител
Экстракты BEL отдельных клонов Е.coli, выбранных с помощью описанной выше стратегии пэннинга, анализируют с помощью ELISA или BioVeris (анализатор BioVeris M-series® 384) для идентификации клонов, кодирующих hGM-CSF специфические Fab.
А. Методы анализа на основе связанного с иммуносорбентом фермента (ELISA)
Человеческим рекомбинантным биотинилированным GM-CSF (R&D Systems) в фосфатном буферном растворе 1,5 мкг/мл покрывают планшеты нейтравидина для микротитрования в течение 2 ч при комнатной температуре.
После покрытия антигеном лунки блокируют фосфатным буферным раствором/0,05% Твина (ФБР-Т), содержащим 1% бычьего сывороточного альбумина в течение 1 ч при комнатной температуре. После промывания лунок ФБР-Т экстракт BEL, очищенный Hu-CAL® Fab или контрольные IgG разводят в фосфатном буферном растворе, добавляют в лунки и инкубируют в течение 1 ч при комнатной температуре. Для обнаружения первичных антител применяют следующие вторичные антитела: (АР)-конъюгированный фрагмент F(ab')2 AffiniPure щелочной фосфатазы, козий античеловеческий, антимышиный или антикрысиный IgG (Jackson Immuno Research). Для обнаружения АР-конъюгатов используют флуорогенные субстраты, такие как AttoPhos (Roche) в соответствии с инструкциями производителя. Между всеми стадиями инкубации лунки планшета для микротитрования трижды промывают ФБР-Т и трижды после заключительной инкубации с вторичным антителом. Флуоресценцию измеряют с помощью устройства для считывания планшетов TECAN Spectrafluor.
В. Основанный на электрохемилюминесценции (BioVeris) анализ связывания для обнаружения связывающихся с GM-CSF Fab в лизатах
В качестве альтернативы экспериментам ELISA для обнаружения связывающихся с GM-CSF антител Fab в лизатах Е.coli (экстракты BEL), связывание было проанализировано с помощью BioVeris M-SERIES® 384 AnalyzerBioVeris, Europe, Witney, Oxforfshire, UK).
Для этого экстракт BEL разбавляют как минимум 1:50, максимум 1:1000 в буфере анализа (ФБР/0,05% Твина 20/0,5% бычьего сывороточного альбумина) для использования в скрининге BioVeris. Биотинилированный GM-CSF (R&D Systems) связывают с покрытыми стрептавидином парамагнитными гранулами (Dynabeads, Dynal) в концентрации 0,1 мкг/мл. На каждую лунку 96- или 384-луночного планшета используют 25 или 15 мкл разведения 1:25 запасного раствора Dynabead. Гранулы промывают трижды буфером анализа перед добавлением биотинилированного GM-CSF на 30 минут при комнатной температуре на шейкере. Затем гранулы трижды промывают буфером анализа и в конце повторно суспендируют в свежем буфере анализа. Античеловеческий (Fab)'2 (Dianova) метят рутением с помощью BV-tag™ (BioVeris Europe, Witney, Oxfordshire, UK). Полученное вторичное антитело добавляют к связанным с GM-CSF гранулам в концентрации 6 мкг/мл непосредственно перед использованием. 100 мкл или 60 мкл разведенного экстракта BEL (см. выше) культуры Е.coli для экспрессии, содержащих антитела Fab, который вносят в лунки 96- или 384-луночного планшета и, соответственно, 25 или 15 мкл связанных с GM-CSF гранул плюс анти-Fab-BV-tag™ вторичное антитело добавляют в каждую лунку и инкубируют в течение 2 ч при комнатной температуре на устройстве для встряхивания планшетом. Планшеты анализируют на анализаторе BioVeris M-SERIES® 384.
После анализа последовательности были идентифицированы семьдесят четыре (74) уникальных клона, которые продемонстрировали достаточно прочное связывание (соотношение сигнал/шум больше 10:1 в ELISA или 50:1 в BioVeris). Данные клоны экспрессируют, очищают и изучают в функциональных анализах.
С. Определение молекулярной специфичности и перекрестной реактивности с другими видами для выбранных анти-hGM-CSF Fabs
Перекрестную реактивность анти-hGM-CSF антител определяли в отношении следующих анализируемых веществ: GM-CSF крысы и мыши, человеческий IL-3, человеческий IL-4, человеческий IL-5, человеческий IL-13, человеческий M-CSF (все от Peprotech, London, UK). Анализ проводили на фиксированных молекулах по методу резонанса поверхностного плазмона (Biacore 3000, Uppsala, Швеция).
Микросхемы СМ5 (Biacore, Швеция) покрывают 5000-6000 условных единиц анти-F(ab)2 (Dianova, Affinipure F(ab)2 фрагмент козьего античеловеческого IgG, F(ab)2 специфичный фрагмент); ацетатный буфер 80 мкг/мл 10 мМ, рН 4 во всех 4-х проточных кюветах, с помощью стандартной химии сочетания этил(диметиламинопропил)карбодиимид/N-гидроксисукцинимид с амином. На проточных кюветах 2-4 фиксируют специфичные в отношении GM-CSF Fab (20 мкл 500 нМ Fab со скоростью потока 5 мкл/мл, 300-400 условных единиц). После фиксации специфичных Fab осуществляют инъекцию буфера, чтобы определить диссоциацию анти-Fab/Fab взаимодействия. В следующем цикле вводят фактор роста анализируемого вещества (20 мкл, скорость потока 20 мл/мин) в интервале концентраций от 15 до 2000 нМ для определения специфического сигнала. Далее полученную буферную сенсограмму вручную вычитают из специфической. После каждого цикла проточные кюветы регенерируют с помощью 100 мМ HCl (5 мкл).
Семь HuCAL® анти-hGM-CSF антител, в том числе MOR03684 и MOR03682, были изучены, продемонстрировали специфичность в отношении человеческого GM-CSF и не связывались с любыми другими цитокинами или GM-CSF мыши или крысы. В противоположность этому, Fab MOR03929 продемонстрировал значительную перекрестную реактивность с крысиным GM-CSF.
Пример 3: Идентификация Fab кандидатов против человеческого GM-CSF, которые ингибируют взаимодействие между GM-CSF и GM-CSF рецептором альфа
74 различных антитела, специфичных в отношении hGM-CSF, выбранных из библиотеки HuCAL GOLD®, изучают на предмет активности с точки зрения ингибирования взаимодействия между hGM-CSF и его рецептором. Взаимодействие исследовали двумя путями: (i) анализ пролиферации с помощью GM-CSF зависимой клеточной линии TF-1 (Kitamura et al., 1989) и (ii) анализ FACS с использованием линии рекомбинантных клеток СНО, которые экспрессирует альфа-цепь GM-CSF рецептора. В анализе пролиферации TF-1 способность анти-GM-CSF антитела блокировать взаимодействие GM-CSF с эндогенным GM-CSF рецептором, состоящим из альфа- и бета-цепей, ведет к уменьшению пролиферации клеток. В анализе FACS определяют специфическое ингибирование взаимодействия между GM-CSF и альфа-цепью GM-CSF рецептора.
А. Клонирование и экспрессия GM-CSF macaca mulata и человека
кДНК полной длины GM-CSF macaca mulata (номер доступа GenBank AY007376) получают методом генного синтеза (geneART GmbH, Regensburg, Германия) гена и клонируют в вектор PCR-Script-Amp (Stratagene, LaJolla, CA, США). Далее кДНК клонируют в эукариотный вектор экспрессии pcDNA3.1 (+) (Invitrogen, Paisley, UK) с получением pcDNA-macGM-CSF. кДНК человеческого GM-CSF (номер доступа Genbank NP_000749) клонируют методом ПЦР с обратной транскрипцией из РНК, выделенной из 1×10е7 клеток TF-1 с помощью набора RNeasya (Qiagen, Hilden, Германия). Обратную транскрипцию выполняют с помощью набора Superscriptll с использованием случайных гексамеров (Gibco) и последующей амплификацией GM-CSF кДНК с помощью ПЦР. Полученный продукт ПЦР клонируют в вектор экспрессии pcDNA3.1 (+) с получением pcDNA-huGM-CSF.
Клетки НЕK293 временно трансфицируют полученными векторами экспрессии, соответственно, с помощью липофектамина (Stratagene, LaJolla, США). Среду, содержащую секретированный рекомбинантный GM-CSF макаки или человека, собирают через 4 дня после трансфекции.
В. Ингибирование GM-CSF зависимой пролиферации клеток TF-1 анти-hGM-CSF Fab с использованием GM-CSF макаки или человека
Клетки TF-1 (Kitamura et al., 1989) выращивают в соответствии с протоколом поставщика (DSMZ, Braunschweig, Германия; DSMZ, № АСС 334). Клетки TF-1 дважды промывают средой RPMI 1640 (10% сыворотки телячьего эмбриона), после чего инокулируют в концентрации 2×105 клеток/мл в 50 мкл на лунку 96-луночного планшета для культивирования клеток с плоским дном. Человеческий рекомбинантный GM-CSF ("Leucomax", ESSEX Pharma, Munchen) в концентрации 0,5 нг/мл и HuCAL® антитела Fab (200 нг/мл-200 мкг/мл, разведенные в среде RPMI 1640, 10% сыворотки телячьего эмбриона) перемешивают в течение 30 минут и 50 мкл смеси добавляют к клеткам TF-1 таким образом, что конечная концентрация GM-CSF составляет 0,25 нг/мл. Максимальную пролиферацию клеток (0% ингибирования) измеряют путем инкубирования клеток TF-1 с конечной концентрацией GM-CSF 0,25 нг/мл, без добавления антитела. 100% ингибирование пролиферации TF-1 измеряют путем исключения GM-CSF из анализа и содержания клеток только в среде RPMI 1640 (10% FCS). Далее клетки TF1 инкубируют в течение 72 часов при 37°C с 5% СO2 в увлажненной камере. Жизнеспособность клеток измеряют, добавляя реактив МТТ или ХТТ (Roche, Mannheim, Германия) в соответствии с рекомендациями производителя. Были идентифицированы всего 19 Fab, которые продемонстрировали значительное ингибирование пролиферации TF-1. Связующие агенты MOR03682, MOR03684 и MOR03929 продемонстрировали систематическое ингибирование пролиферации клеток TF1 более чем на 50% в концентрации 2 мкМ. Ингибирующая активность этих не оптимизированных Fab не была достаточно сильной, чтобы определить дозу IC50, поскольку полное ингибирование не могло быть достигнуто. Для сравнения, моноклональные антитела BVD2-21C11 (BD Biosciences Pharmingen; кат. №554503) и МАВ215 (R&D Systems; кат. № МАВ215) были способны полностью ингибировать пролиферацию TF-1.
Дополнительно, связывание MOR03682 и MOR03684 с природным человеческим GM-CSF изучали в анализе пролиферации TF-1. Вместо добавления очищенного человеческого рекомбинантного GM-CSF к клеткам TF-1 использовали супернатант клеток 5637 (DSMZ, № АСС 35), которые секретируют природный человеческий GM-CSF в среду. На основе кривой «доза-реакция», сравнивающей влияние рекомбинантного человеческого GM-CSF с различными разведениями супернатанта 5637, было определено, что среда содержит ~5 нг/мл природного человеческого GM-CSF. Путем предварительной инкубации супернатанта 5637 с античеловеческим GM-CSF Fab MOR03682 или MOR03684 связывание природного человеческого GM-CSF с клетками TF-1 было блокировано таким образом, что жизнеспособность клеток уменьшалась по сравнению с экспериментом с использованием рекомбинантного человеческого GM-CSF. Таким образом, MOR03684 и MOR03682 связываются с природным человеческим GM-CSF. Fab MOR03929 не изучали в данном анализе.
Дополнительно, перекрестную реактивность с GM-CSF макаки изучали в анализе пролиферации TF-1. Вместо добавления очищенного человеческого GM-CSF к клеткам TF-1 использовали супернатант трансфицированных клеток НЕК293, которые секретируют рекомбинантный GM-CSF макаки в среду.
Клетки TF-1 пролиферировали в присутствии GM-CSF макаки, содержащего супернатант, но не в присутствии супернатанта нетрансфицированных клеток НЕK293. На основе кривой «доза-реакция», сравнивающей влияние рекомбинантного человеческого GM-CSF с различными разведениями среды НЕK-293, было определено, что среда трансфицированных клеток содержит ~2 мкг/мл GM-CSF макаки. Путем предварительной инкубации супернатанта GM-CSF макаки с Fab против человеческого GM-CSF MOR03682 или MOR03684 связывание GM-CSF макаки с клетками TF-1 было блокировано таким образом, что жизнеспособность клеток существенно уменьшалась.
Таким образом, MOR03682 и MOR03684 обладают перекрестной реактивностью с GM-CSF макаки. Fab MOR03929 не изучали в данном анализе.
С. Блокирование анти-hGM-CSF Fab связывания GM-CSF с GM-CSF рецептором альфа
Для того чтобы изучить связывание GM-CSF с поверхностью клеток, экспрессирующих альфа-цепь рецептора GM-CSF, кДНК клонируют в вектор экспрессии и стабильно трансфицируют в клетки СНО-K1 (DSMZACC 110).
Клонирование стабильной линии клеток СНО-K1, экспрессирующей альфа-цепь GM-CSF рецептора
Клонируют кДНК альфа-цепи человеческого GM-CSF рецептора (номер доступа Genbank M64445) техникой ПЦР с обратной транскрипцией из РНК, выделенной из 1×10е7 клеток TF-1 с помощью набора RNeasy (Qiagen, Hilden, Германия). Обратную транскрипцию выполняют с помощью набора Superscriptll с использованием случайных гексамеров (Gibco). Далее кДНК альфа-цепи GM-CSF-рецептора амплифицируют с использованием следующих праймеров:
5': N-GCRa-plusSS: TTCTCTGGATCCGCCACCATGCTTCTCCTGGTGACAAGCC
и
3': C-flGCRa: ACCCTCCAATTGTCAGGTAATTTCCTTCACGGTC.
Реакция ПЦР дает продукт длиной ~1250 пар оснований, который расщепляют с помощью EcoRI и BamHI (New England BioLabs). Вектор экспрессии pcDNA3.1(+) (Invitrogen, Paisley, UK) расщепляют такими же ферментами. После очистки расщепленного вектора и продукта ПЦР фрагменты лигируют и трансформируют электропорацией в клетки Е.coli DH10B. Правильные клоны идентифицируют после приготовления плазмиды ДНК и секвенирования. Правильные клоны (pcDNA3.1(+)-GM-CSFRalpha) содержат кДНК человеческого GM-CSF рецептора альфа полной длины.
Клетки СНО-K1 культивируют в соответствии с протоколом поставщика (DSZM, Braunschweig, Германия; DSMZ, № АСС 110). Для трансфекции клетки выращивают до 80% слияния в 6-луночном планшете и инкубируют с 5 мкг ДНК pcDNA3.1(+)-GM-CSFRalpha смешивают с 10 мкл реактива Lipofectamine 2000 (Invitrogen). После прохождения 48 ч клетки подкармливают 1 мг/мл G418 (Gibco), и после прохождения еще 24 ч среду заменяют на среду, содержащую 2 мг/мл G418. После прохождения двух недель отдельные клетки инокулируют в лунки 96-луночного планшета для культивирования. Отдельные клоны выращивают и 5×105 клеток каждого клона исследуют на предмет экспрессии GM-CSFR-альфа с помощью анализа FACS с использованием мышиного IgG MAB1006 (Chemicon International, Temecula, CA) в качестве первичного антитела в концентрации 1 мкг/мл и (R-PE-AffiniPure (Fab')2 козьего антимышиного IgG (Dianova) в качестве вторичного антитела в разведении 1:200. Первичные и вторичные антитела инкубируют с клетками в течение 1 ч последовательно, пока клетки промывают в буфере FACS (фосфатный буферный раствор, 3% сыворотки телячьего эмбриона) между этими стадиями. Интенсивность флуоресценции окрашенных клеток определяли в канале FL2 с использованием системы FACSCalibur (Becton Dickinson).
Среди проанализированных клонов CHO-GMRa#11 продемонстрировали наиболее высокую медиану интенсивности флуоресценции. Медианное значение (значение MFL) 157 было определено для CHO-GMRa#11 (фиг.6).
Анализ FACS связывания GM-CSF с альфа GM-CSF рецептора. экспрессированной на CHO-GMRa#11
Перед добавлением клеток антитела с увеличивающейся концентрацией (0,1-100 мкг/мл) инкубируют вместе с биотинилированным GM-CSF (0,5 мкг/мл) в буфере (ФБР/3% FBS/NaN3 0,05%) FACS в течение 30 мин при комнатной температуре.
Окрашивание выполняют в круглодонных 96-луночных планшетах для культивирования (Nalge Nunc) с 1-5×105 клеток на лунку. 2×10Е5 клеток CHO-GMRa#11 переносят в 50 мкл буфера FACS, содержащего антитело/GM-CSF, и инкубируют при 4°С в течение 1 ч. Далее клетки промывают один раз 150 мкл буфера FACS/лунку и переносят в 100 мкл меченного фикоэритрином стрептавидина (BD Biosciences Pharmingen), разбавленного 1:400 в буфере FACS. После инкубации в течение 1 ч при 4°С клетках дважды промывают буфером FACS, повторно суспендируют в 100 мкл буфера FACS и связывание биотинилированного GM-CSF определяют путем измерения интенсивности свечения FL2 клеток с помощью FACScalibur (Becton Dickinson). Значения IC50 определяют на основе кривых «доза-реакция», полученных с помощью программного обеспечения GraphPad Prism v3.03 с применением нелинейной аппроксимации кривой регрессии.
Антитела Fab MOR03682, MOR03684 и MOR03929 продемонстрировали значительное ингибирование связывания GM-CSF с поверхностью клетки, экспрессирующей GM-CSF рецептор альфа.
Пример 4: Дозревание сродства выбранного Fab постадийным обменом кассет CDR
А. Генерация библиотек дозревания сродства Fab и пэннинг
Для того чтобы увеличить сродство и ингибирующую активность анти-GM-CSF Fab фрагментов, клоны MOR03682, MOR03684 и MOR03929 подвергают дозреванию сродства. Для этого участки CDR оптимизируют кассетным мутагенезом с применением тринуклеотид-направленного мутагенеза (Virnekas et al., 1994; Nagy et al., 2002). Анализ последовательности не выявил гомологии последовательности между CDR трех проанализированных материнских клонов. В табл.2а и 2b представлены шесть пептидных последовательностей CDR для материнских клонов MOR03682, MOR03684 и MOR03929.
Ниже кратко описан протокол, используемый для оптимизации Fab. Фрагменты Fab из вектора экспрессии pMORPH®X9Fab_FH клонируют в фагемидный вектор (США 6,753,136). Далее применяют две различные стратегии для оптимизации сродства и эффективности материнских Fab.
Сначала генерируют одну библиотеку антител Fab фага, где L-CDR3 каждого материнского заменен на репертуар отдельных последовательностей CDR3 легкой цепи лямбда. Во второй библиотеке участок H-CDR2 заменен на репертуар отдельных последовательностей CDR2 тяжелой цепи.
Библиотеки созревания сродства генерируют путем преобразования разнообразных клонов в Е.coli TOP10F' (Invitrogen). Фаги получают, как описано в Примере 1А. Библиотеки L-CDR3 MOR03684 и MOR03682 объединяют, а также объединяют библиотеки H-CDR2, полученные из MOR03684 и MOR03682, в то время как библиотеки L-CDR3 и H-CDR2, полученные из MOR03929, содержат отдельно в ходе процедуры селекции.
Пэннинг выполняют на биотинилированном GM-CSF в растворе для трех по способу, в существенной мере сходному с описанным в Примере 1В, с применением более строгих условий селекции.
В. Основанный на электрохемилюминесценции анализ связывания (BioVeris) для выявления Fab с улучшенным связыванием GM-CSF в лизатах
Для обнаружения антител Fab, связывающихся с GM-CSF в лизатах E.coli (экстракты BEL), связывание анализируют с помощью BioVeris M-384 SERIES® Workstation (Europe BioVeris, Witney, Oxforfshire, UK) по способу, в существенной мере сходному с описанным в Примере 2В.
Fabs с самыми высокими значениями ЭХЛ очищают и подвергают измерению сродства по методу равновесного титрования в растворе (SET; Haenel et al., 2005) и резонанса поверхностного плазмона (Biacore) (см. Пример 4D).
С. Х-Клонирование улучшенных VL (L-CDR3) с улучшенными VH (Н-CDR2)
Для дальнейшего улучшения сродства независимо оптимизированные H-CDR2 и L-CDR3 из зрелых Fab, полученных от одного материнского клона, объединяли, поскольку существовала высокая вероятность того, что данная комбинация приведет к дальнейшему повышению сродства (Yang et al., 1995; Schier et al., 1996; Chen et al., 1999). Данную процедуру под названием X-клонирования применяли к связывающимся веществам, которые происходят из материнского клона MOR03929, поскольку Fab с улучшенным сродством были идентифицированы как из H-CDR2, так и L-CDR3 библиотеки. Это было достигнуто переносом легких цепей целиком (фрагмент XbaI/SphI) из L-CDR3-оптимизированного донорского клона к H-CDR2-оптимизированному акцепторному клону.
MOR04290
MOR04252
MOR04354
MOR04357
Для 4-х полученных Fab было проведено секвенирование VL и VH, чтобы подтвердить перенос правильного VL на соответствующий векторный скелет с улучшенным H-CDR2. В табл.2а и 2b показаны белковые последовательности VH и VL для всех производных MOR03929 и 3682, которые перечислены в табл.3.
D. Определение пикомолярного сродства с использованием равновесного титрования в растворе (SET) и резонанса поверхностного плазмона (Biacore)
Для определения КD используют мономерные фракции (содержание мономера как минимум 90% по данным аналитической эксклюзионной хроматографии; Superdex75, Amersham Pharmacia) Fab. Основанное на электрохемилюминесценции (ЭХЛ) определение сродства в растворе и оценку данных в основном выполняют, как описано в Haenel et al., 2005. Постоянное количество Fab уравновешивают с различными концентрациями (серийные 5-кратные разведения) человеческого GM-CSF (Leucomax) в растворе. Добавляют биотинилированный человеческий GM-CSF (R&D Systems), связанный с парамагнитными гранулами (стрептавидин М-280, Dynal) и BV-tag™ (Europe BioVeris, Witney, Oxfordshire, UK), меченный античеловеческим (Fab)'2 (Dianova), и инкубируют в течение 15-30 минут. Далее производят количественное определение концентрации свободного Fab с помощью детектирования ЭХЛ с использованием анализатора M-SERIES® 384 (Europe BioVeris).
В соответствии с Friguet et al., 1985, предпринимают меры для избежания значительного сдвига равновесия в сторону твердой фазы в ходе обнаружения.
С использованием описанных выше условий анализа определяли сродство для Fab, которое показано в табл.4.
Дополнительно осуществляли кинетический анализ резонанса поверхностного плазмона на микросхеме F1 (Biacore, Швеция), покрытой с плотностью ~100 условных единиц рекомбинантным человеческим GM-CSF (Peprotech) в 10 мМ растворе Na-ацетата рН 4,5 с использованием стандартной химии сочетания этил(диметиламинопропил)карбодиимида/N-гидроксисукцинимида амина. Соответствующее количество человеческого сывороточного альбумина иммобилизируют на проточной кювете сравнения. Фосфатный буферный раствор (136 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,76 мМ KН2РO4 рН 7,4)+0,005% Твина 20 используют в качестве подвижного буфера. Fab используют в сериях концентраций 6,3-200 нМ со скоростью потока 20 мкл/мин. Фазу ассоциации устанавливают на отметке 60 сек, и фазу диссоциации - на отметке 120 сек (материнский) или до 600 сек (оптимизированное сродство). Для того чтобы контролировать фазу диссоциации в течение более длинного периода, используют следующие условия, в основном в соответствии с Drake et al., (2004): Fab используют в единой концентрации 200 нМ; скорость потока устанавливают на отметке 100 мкл/мин, и фазу диссоциации - на отметке 6000-18000 сек. На основе определенных в данных условиях значений скорости диссоциации оценивают значения сродства для Fab, которые показаны в табл.4.
Е. Определение сродства к крысиному GM-CSF с использованием равновесного титрования в растворе (SET)
Определение сродства к крысиному GM-CSF проводят по способу, в существенной мере сходному с описанным в Примере 4D, с использованием крысиного GM-CSF (Peprotech) в качестве анализируемого вещества в растворе вместо человеческого GM-CSF. Значения сродства вычисляют в соответствии с Haenel et al. (2005). В данном анализе сродство Fab MOR04357 к крысиному GM-CSF определено на уровне KD=1,0 нМ.
Пример 5: Характеристика оптимизированных античеловеческих GM-CSF Fab, которые ингибируют взаимодействие между GM-CSF и альфа-цепью GM-CSF рецептора
А. Анализ связывания с альфа GM-CSF рецептора
Анализ связывания с рецептором GM-CSF выполняют, как описано выше (Пример 3С) с использованием 0,5 мкг/мл (35 нМ) биотинилированного GM-CSF. Максимальное связывание GM-CSF с клетками CHO-GMRa#11 (0% ингибирования) измеряют путем инкубирования клеток с конечной концентрацией GM-CSF 0,5 мкг/мл биотинилированного GM-CSF, без добавления антитела. 100% ингибирование GM-CSF связывания измеряют, исключая из анализа GM-CSF. Значения IC50 определяют на основе кривых «доза-реакция», полученных с помощью программного обеспечения GraphPad Prism v3.03 с применением нелинейной аппроксимации кривой регрессии.
Проанализированы Fab с улучшенным сродством, материнские Fab и моноклональные IgG сравнения. В табл.5 приведены значения IC50, полученные в описанных анализах. % ингибирования, достигнутый в концентрации антитела 5 мкг/мл, также представлен в табл.5.
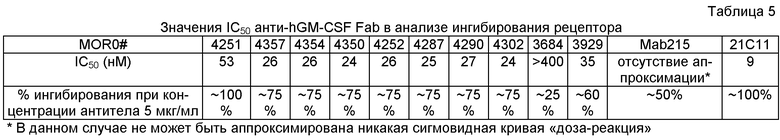
Данный анализ качественно показывает, что Fab, полученные в результате дозревания сродства и Х-клонирования, препятствуют связыванию GM-CSF с альфа-цепью GM-CSF рецептора и, таким образом, демонстрируют механизм блокирования, присущий их материнским Fab. Анализ необходимо выполнять с концентрацией 35 нМ (0,5 мкг/мл) биотинилированного GM-CSF, для того чтобы получить значительный сигнал в анализе FACS. Таким образом, 17,5 нМ Fab (или 8,75 нМ IgG) теоретически необходимо для блокирования 50% GM-CSF, таким образом, устанавливая предел для определения значений IC50.
В. Ингибирование GM-CSF зависимой пролиферации TF-1 с помощью анти-hGM-CSF Fab с использованием человеческого GM-CSF
Анализ пролиферации TF-1 выполняют, как описано в Примере 3В. Проанализированы Fab с улучшенным сродством и материнские Fab, а также моноклональные IgG сравнения. Значения IC50 определяют на основе кривых «доза-реакция», полученных с помощью программного обеспечения GraphPad Prism v3.03 с применением нелинейной аппроксимации кривой регрессии. В табл.6 приведены значения IC50, полученные в описанных анализах.
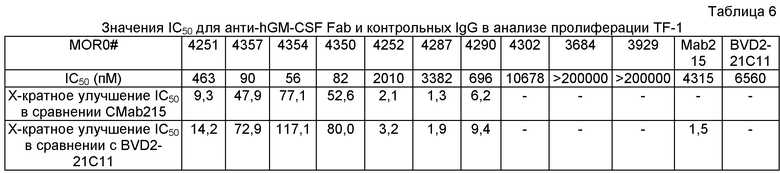
В другом наборе экспериментов значения IC50 в анализе пролиферации TF-1 определяли для материнского Fab MOR03682, его производных со зрелым сродством MOR04283, MOR04297 и Х-клонированного варианта MOR04342. В табл.7 приведены значения IC50, полученные в указанных анализах.
Эти эксперименты продемонстрировали значительное улучшение значений IC50 после дозревания сродства и Х-клонирования. Например, MOR04357, MOR04350, MOR04354 демонстрируют >2000-кратное улучшение значений IC50 по сравнению с материнским MOR03929, что превышает активность BVD2-21C11 и Маb215.
Пример 6: Превращение MOR04357 в формат человеческого IgG1
А. Генная оптимизация ДНК последовательностей Fab для экспрессии в системах экспрессии млекопитающих.
Для оптимизации ДНК VH и VL MOR04357 с целью экспрессии гена в млекопитающих (например, использование замененного кодона, содержание GC и т.п.) использовали программное обеспечение GeneOptimizer™ от Geneart (Regensburg, Германия) для определения таких оптимизированных последовательностей ДНК VH и VL, которые были синтезированы генным способом в Geneart (Regensburg, Германия), и клонировали в векторы pPCR-Script с получением 055906pPCR-Script и 055907pPCR-Script. SEQ ID NO:48 показывает соответствующую последовательность VH, тогда как SEQ ID NO:57 показывает соответствующую последовательность VL.
В. Клонирование Fab MOR04357 в формат человеческого IgG1 и экспрессия IgG1
Для того чтобы экспрессировать иммуноглобулин полной длины (Ig), фрагменты вариабельных доменов генно-оптимизированых тяжелой (VH) и легкой (VL) цепей субклонировали из векторов pPCR-Script (Пример 5а) в серию векторов pMORPH®2_h_lg для человеческого IgG1. Кодон-оптимизированную VH MOR04357 выделяли из 055906pPCR-Script путем расщепления NheI/BIpI и вставляли в главный вектор pMorph2_h_IgG1f, расщепленный такими же рестрикционными ферментами. Данный вектор уже содержал постоянный участок человеческого гамма 1. Полученная экспрессионная плазмида получила название pMorph2_h_IgG1f_MOR04357_co. Кодон-оптимизированную VL MOR04357 выделяли из 055907pPCR-Script путем расщепления NheI/HpaI и вставляли в главный вектор pMorph2_h_Iglambda2, расщепленный такими же рестрикционными ферментами. Данный вектор уже содержал человеческий постоянный участок лямбда. Полученная экспрессионная плазмида получила название pM2_h_Iglambda2_MOR04357_co.
С. Временная экспрессия и очистка человеческого IgG
Эукариотные клетки НKВ11 трансфицируют эквимолярным количеством вектора экспрессии ДНК тяжелой и легкой цепей IgG. Супернатант культуры клеток собирают через 3-7 дней после трансфекции. После доведения рН супернатанта до 8,0 и стерильной фильтрации раствор подвергают стандартной аффинной хроматографии с протеином A (rProteinA FF или MabSelect SURE, GE Healthcare). Осуществляют обмен буфера на 1×Дульбекко фосфатный буферный раствор (рН 7,2, Invitrogen), и образцы стерилизуют фильтрацией (0,2 мкм). MOR04357 IgG1 подвергают диализу против 1×Дульбекко фосфатного буферного раствора (рН 6,5, Invitrogen). Чистоту IgG изучают в денатурирующих восстанавливающих условиях электрофореза в полиакриламидном геле с додецилсульфатом натрия или с помощью Agilent BioAnalyzer, а также в природном состоянии с помощью эксклюзионной ВЭЖХ.
D. Определение пикомолярного сродства с использованием равновесного титрования в растворе (SET)
Для определения KD используют мономерные фракции (содержание мономера как минимум 90% по данным аналитической эксклюзионной хроматографии; Superdex75, Amersham Pharmacia) IgG1. Основанное на электрохемилюминесценции (ЭХЛ) определение сродства в растворе и оценку данных в основном выполняют, как описано в Haenel et al., 2005 и как описано в Примере 4В. Полученное значение KD для MOR04357 IgG1 против человеческого рекомбинантного GM-CSF составило 1,1 пМ.
Е. Определение сродства к крысиному GM-CSF с использованием равновесного титрования в растворе (SET)
Определение сродства к крысиному GM-CSF проводят по способу, в существенной мере сходному с описанным в Примере 4D, с использованием крысиного GM-CSF (Peprotech) в качестве анализируемого вещества в растворе вместо человеческого GM-CSF. Значения сродства вычисляют в соответствии с Haenel et at. (2005). Полученное значение KD для MOR04357 IgG1 против крысиного рекомбинантного GM-CSF составило 130 пМ.
Пример 7: Характеристика MOR04357 IgG1, полученного из оптимизированных античеловеческих GM-CSF Fab
А. Ингибирование GM-CSF-зависимой пролиферации TF-1 анти-hGM-CSF IgG с использованием GM-CSF человека и макаки резус
Анализ пролиферации TF-1 выполняют, как описано в Примере 3В. MOR04357 проанализирован в формате IgG1, и в качестве контрольных проанализированы моноклональные IgG сравнения. Значения IC50 определяют на основе кривых «доза-реакция», полученных с помощью программного обеспечения GraphPad Prism v3.03 с применением нелинейной аппроксимации кривой регрессии. В табл.8 приведены значения IC50, полученные в описанных анализах. В данном анализе использовали три различных варианта GM-CSF: во-первых, рекомбинантный человеческий GM-CSF в концентрации 0,25 нг/мл, продуцированный Е.coli, во-вторых, супернатант культуры НЕK293, которые были временно трансфицированы pcDNA-huGM-CSF (см. Пример 3А), содержащим рекомбинантный человеческий GM-CSF, и в-третьих, супернатант культуры клеток НЕК293, которые были временно трансфицированы pcDNA-macGM-CSF (см. Пример 3А), содержащим рекомбинантный GM-CSF mасаса mulata (макака резус). Для анализов пролиферации TF-1 использовали супернатанты культуры НЕK293 как источник соответствующего GM-CSF в таких разведениях, что клетки TF-1 демонстрируют пролиферацию, сходную с пролиферацией в определенной концентрации 0,25 нг/мл очищенного рекомбинантного человеческого GM-CSF, продуцированного Е.coli.
В данном эксперименте продемонстрировано значительное улучшение, достигнутое в значениях IC50 после дозревания сродства и Х-клонирования, если они сохранялись после превращения Fab в формат IgG1. IgG1 MOR04357 демонстрирует >2000-кратное улучшение значений IC50 по сравнению с Fab MOR03929, что превышает активность BVD2-21C11 и Маb215.
Литература:



























































Изобретение относится к биохимии и представляет собой изолированное человеческое или гуманизированное антитело или его функциональный фрагмент, которое является специфическим в отношении человеческого GM-CSF. Также предложены вариабельные тяжелая и легкая цепи антитела, изолированные последовательности нуклеиновой кислоты, вектор экспрессии, клетка-хозяин, фармацевтическая композиция и способ лечения связанного с GM-CSF состояния. Изобретение может использоваться для лечения заболеваний, связанных с GM-CSF, в диагностических целях, а также для дальнейшего исследования роли GM-CSF в прогрессировании различных расстройств. 11 н. и 91 з.п. ф-лы, 6 ил., 8 табл., 7 пр.
1. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент, содержащее антигенсвязывающий участок, который является специфичным в отношении человеческого GM-CSF, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 49, 50, 51 или 52, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 58, 59, 60 или 61, как показано на Фиг.2b.
2. Изолированный функциональный фрагмент антитела по п.1, отличающийся тем, что представляет собой Fab или scFv фрагмент антитела.
3. Изолированное человеческое или гуманизированное антитело по п.1, отличающееся тем, что представляет собой IgG.
4. Человеческое антитело по п.1, отличающееся тем, что человеческое антитело представляет собой синтетическое человеческое антитело.
5. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 11, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 31, как показано на Фиг.2b.
6. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 12, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 32, как показано на Фиг.2b.
7. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 13, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 33, как показано на Фиг.2b.
8. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 14, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 34, как показано на Фиг.2b.
9. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 15, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 35, как показано на Фиг.2b.
10. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 16, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 36, как показано на Фиг.2b.
11. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 17, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 37, как показано на Фиг.2b.
12. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 18, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 38, как показано на Фиг.2b.
13. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 19, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 39, как показано на Фиг.2b.
14. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 20, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 40, как показано на Фиг.2b.
15. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 49, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 58, как показано на Фиг.2b.
16. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 50, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 59, как показано на Фиг.2b.
17. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 51, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 60, как показано на Фиг.2b.
18. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.1, отличающееся тем, что содержит участки H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 52, как показано на Фиг.1b, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 61, как показано на Фиг.2b.
19. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.5, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 11, и вариабельную легкую цепь, представленную в SEQ ID NO: 31.
20. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.6, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 12, и вариабельную легкую цепь, представленную в SEQ ID NO: 32.
21. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.7, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 13, и вариабельную легкую цепь, представленную в SEQ ID NO: 33.
22. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.8, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 14, и вариабельную легкую цепь, представленную в SEQ ID NO: 34.
23. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.9, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 15, и вариабельную легкую цепь, представленную в SEQ ID NO: 35.
24. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.10, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 16, и вариабельную легкую цепь, представленную в SEQ ID NO: 36.
25. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.11, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 17, и вариабельную легкую цепь, представленную в SEQ ID NO: 37.
26. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.12, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 18, и вариабельную легкую цепь, представленную в SEQ ID NO: 38.
27. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.13, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 19, и вариабельную легкую цепь, представленную в SEQ ID NO: 39.
28. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.14, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 20, и вариабельную легкую цепь, представленную в SEQ ID NO: 40.
29. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.15, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 49, и вариабельную легкую цепь, представленную в SEQ ID NO: 58.
30. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.16, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 50, и вариабельную легкую цепь, представленную в SEQ ID NO: 59.
31. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.17, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 51, и вариабельную легкую цепь, представленную в SEQ ID NO: 60.
32. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.18, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 52, и вариабельную легкую цепь, представленную в SEQ ID NO: 61.
33. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент, содержащее антигенсвязывающий участок, который является специфичным в отношении человеческого GM-CSF, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 49, 50, 51 или 52, и участки L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 58, 59, 60 или 61, как показано на Фиг.2b.
34. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 11.
35. Изолированное антитело или его функциональный фрагмент по п.34, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 31, как показано на Фиг.2b.
36. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 12.
37. Изолированное антитело или его функциональный фрагмент по п.36, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 32, как показано на Фиг.2b.
38. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 13.
39. Изолированное антитело или его функциональный фрагмент по п.38, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 33, как показано на Фиг.2b.
40. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 14.
41. Изолированное антитело или его функциональный фрагмент по п.40, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 34, как показано на Фиг.2b.
42. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 15.
43. Изолированное антитело или его функциональный фрагмент по п.42, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 35, как показано на Фиг.2b.
44. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 16.
45. Изолированное антитело или его функциональный фрагмент по п.44, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 36, как показано на Фиг.2b.
46. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 17.
47. Изолированное антитело или его функциональный фрагмент по п.46, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 37, как показано на Фиг.2b.
48. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 18.
49. Изолированное антитело или его функциональный фрагмент по п.48, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 38, как показано на Фиг.2b.
50. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 19.
51. Изолированное антитело или его функциональный фрагмент по п.50, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 39, как показано на Фиг.2b.
52. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 20.
53. Изолированное антитело или его функциональный фрагмент по п.52, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 40, как показано на Фиг.2b.
54. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 49.
55. Изолированное антитело или его функциональный фрагмент по п.54, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 58, как показано на Фиг.2b.
56. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 50.
57. Изолированное антитело или его функциональный фрагмент по п.56, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 59, как показано на Фиг.2b.
58. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 51.
59. Изолированное антитело или его функциональный фрагмент по п.58, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 60, как показано на Фиг.2b.
60. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.33, отличающееся тем, что содержит вариабельную тяжелую цепь, представленную в SEQ ID NO: 52.
61. Изолированное антитело или его функциональный фрагмент по п.60, отличающееся тем, что содержит L-CDR1, L-CDR2 и L-CDR3, представленные в SEQ ID NO: 61, как показано на Фиг.2b.
62. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент, содержащее антигенсвязывающий участок, который является специфичным в отношении человеческого GM-CSF, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 58, 59, 60 или 61, и участки H-GDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 49, 50, 51 или 52, как показано на Фиг.1b.
63. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 31.
64. Изолированное антитело или его функциональный фрагмент по п.63, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 11, как показано на Фиг.1b.
65. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 32.
66. Изолированное антитело или его функциональный фрагмент по п.65, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 12, как показано на Фиг.1b.
67. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 33.
68. Изолированное антитело или его функциональный фрагмент по п.67, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 13, как показано на Фиг.1b.
69. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 34.
70. Изолированное антитело или его функциональный фрагмент по п.69, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 14, как показано на Фиг.1b.
71. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 35.
72. Изолированное антитело или его функциональный фрагмент по п.71, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 15, как показано на Фиг.1b.
73. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 36.
74. Изолированное антитело или его функциональный фрагмент по п.73, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 16, как показано на Фиг.1b.
75. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 37.
76. Изолированное антитело или его функциональный фрагмент по п.75, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 17, как показано на Фиг.1b.
77. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 38.
78. Изолированное антитело или его функциональный фрагмент по п.77, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 18, как показано на Фиг.1b.
79. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 39.
80. Изолированное антитело или его функциональный фрагмент по п.79, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 19, как показано на Фиг.1b.
81. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 40.
82. Изолированное антитело или его функциональный фрагмент по п.81, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 20, как показано на Фиг.1b.
83. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 58.
84. Изолированное антитело или его функциональный фрагмент по п.83, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 49, как показано на Фиг.1b.
85. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 59.
86. Изолированное антитело или его функциональный фрагмент по п.85, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 50, как показано на Фиг.1b.
87. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 60.
88. Изолированное антитело или его функциональный фрагмент по п.87, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 51, как показано на Фиг.1b.
89. Изолированное человеческое или гуманизированное антитело или его функциональный фрагмент по п.62, отличающееся тем, что содержит вариабельную легкую цепь, представленную в SEQ ID NO: 61.
90. Изолированное антитело или его функциональный фрагмент по п.89, отличающееся тем, что содержит H-CDR1, H-CDR2 и H-CDR3, представленные в SEQ ID NO: 52, как показано на Фиг.1b.
91. Вариабельная тяжелая цепь изолированного человеческого или гуманизированного антитела или функционального фрагмента антитела, отличающаяся тем, что кодируется (i) последовательностью нуклеиновой кислоты, содержащей SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 44, 45, 46, 47 или 48, или (ii) последовательностями нуклеиновой кислоты, которые гибридизуются в условиях высокой строгости с комплементарной цепью SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 44, 45, 46, 47 или 48, причем указанное антитело или его функциональный фрагмент обладает специфичностью в отношении человеческого GM-CSF.
92. Вариабельная легкая цепь изолированного человеческого или гуманизированного антитела или функционального фрагмента антитела, отличающаяся тем, что кодируется (i) последовательностью нуклеиновой кислоты, содержащей SEQ ID NO: 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 53, 54, 55, 56 или 57, или (ii) последовательностями нуклеиновой кислоты, которые гибридизуются в условиях высокой строгости с комплементарной цепью SEQ ID NO: 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 53, 54, 55, 56 или 57, причем указанное антитело или его функциональный фрагмент обладает специфичностью в отношении человеческого GM-CSF.
93. Изолированная последовательность нуклеиновой кислоты, отличающаяся тем, что кодирует антигенсвязывающий участок человеческого или гуманизированного антитела или функционального фрагмента антитела, содержащая (i) SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 44, 45, 46, 47 или 48 или (ii) последовательности нуклеиновой кислоты, которые гибридизуются в условиях высокой строгости с комплементарной цепью SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 44, 45, 46, 47 или 48, причем указанное антитело или его функциональный фрагмент обладает специфичностью в отношении человеческого GM-CSF.
94. Изолированная последовательность нуклеиновой кислоты, отличающаяся тем, что кодирует антигенсвязывающий участок человеческого или гуманизированного антитела или функционального фрагмента антитела, содержащая SEQ ID NO: 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 53, 54, 55, 56 или 57 или (ii) последовательности нуклеиновой кислоты, которые гибридизуются в условиях высокой строгости с комплементарной цепью SEQ ID NO: 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 53, 54, 55, 56 или 57, причем указанное антитело или его функциональный фрагмент обладает специфичностью в отношении человеческого GM-CSF.
95. Вектор экспрессии, отличающийся тем, что содержит последовательность нуклеиновой кислоты по любому из пп.93 и 94.
96. Изолированная клетка-хозяин, предназначенная для продуцирования антитела, специфичного в отношении человеческого GM-CSF, отличающаяся тем, что содержит вектор по п.95.
97. Изолированная клетка-хозяин по п.96, отличающаяся тем, что указанная клетка представляет собой бактериальную клетку.
98. Изолированная клетка-хозяин по п.96, отличающаяся тем, что указанная клетка представляет собой эукариотную клетку.
99. Фармацевтическая композиция, которая является специфичной в отношении человеческого GM-CSF, отличающаяся тем, что содержит эффективное количество человеческого или гуманизированного антитела или функционального фрагмента по любому из пп.1-90 и фармацевтически приемлемый носитель или вспомогательное вещество.
100. Способ лечения расстройства или состояния, связанного с нежелательным присутствием GM-CSF, отличающийся тем, что включает стадию введения субъекту, который нуждается в этом, эффективного количества фармацевтической композиции по п.99.
101. Способ по п.100, отличающийся тем, что указанное расстройство или состояние представляет собой воспалительное заболевание.
102. Способ по п.101, отличающийся тем, что указанное воспалительное заболевание выбрано из группы, состоящей из ревматоидного артрита, рассеянного склероза, болезни Крона, псориаза, астмы, атопического дерматита и шока.
| WO 03068920, 21.08.2003 | |||
| HERCUS TR, ET AL | |||
| Identification of residues in the first and fourth helices of human granulocyte-macrophage colony-stimulating factor involved in biologic activity and in binding to the alpha- and beta-chains of its receptor | |||
| Blood, 83(12), 15.06.1994, p.:3500-3508 | |||
| RAUCHENBERGER R, ET AL | |||
| Human combinatorial Fab library |

